Предлагаемое изобретение относится к области медицины и биохимии, касается рекомбинантного таргетного токсина, специфичного к клеткам, экспрессирующим рецептор HER2, который может быть использован в качестве терапевтического агента для таргетной терапии опухолей, гиперэкспрессирующих рецептор HER2, характеризующихся повышенным риском метастазирования и устойчивостью к химиотерапии.
Хорошо изученными онкомаркерами являются белки семейства рецептора эпидермального фактора роста, HER1 и HER2/neu, гиперэкспрессия которых характерна для ряда карцином (молочной железы, простаты, легких, яичника, мочевого пузыря и др.). Разница в уровне экспрессии этих белков в нормальных и опухолевых клетках делает их перспективными мишенями для направленной доставки терапевтических и/или диагностических агентов [Поляновский О.Л., Лебеденко Е.Н., Деев С.М. // Биохимия. 2011. Т. 77. С. 289-311].
Разработка новых методов и подходов для терапии онкологических заболеваний на сегодняшний день - одна из наиболее актуальных и активно развивающихся областей биологии и медицины. Ведущим трендом в этой области является персонализированный подход, основанный на определении молекулярно-генетических особенностей опухоли для каждого конкретного случая и выборе оптимального метода лечения с учетом полученной информации. Хотя основными методами лечения солидных опухолей в клинике по-прежнему является хирургическое воздействие с последующей химио- или лучевой терапией, благодаря прогрессу в изучении молекулярных основ канцерогенеза в последние десятилетия все большую роль начинает играть таргетная (направленная) терапия. Краеугольным камнем этого подхода являются моноклональные антитела, связывающиеся преимущественно с опухолевыми клетками, экспрессирующими определенный, характерный только для них маркер, и вызывающих их гибель.
Поскольку такие антитела недостаточно эффективны как самостоятельные препараты, их конъюгируют с высокоэффективными токсичными молекулами; такие соединения получили название «иммунотоксины». Несмотря на то, что в доклинических испытаниях иммунотоксины показывают ярко выраженное специфическое цитотоксическое действие на опухолевые клетки, на сегодняшний день для клинического применения одобрены только два таких препарата trastuzumab emtansine (Kadcyla = TD-M1) и brentuximab vedotin (Adcetris).
За последние десятилетия были созданы и изучены в предклинических исследованиях сотни моноклональных антител к различным антигенам, десятки из них дошли до стадии клинических испытаний, отдельные моноклональные антитела получили разрешение на применение в качестве лекарственных средств [Деев С.М. Современные технологии создания неприродных антител для клинического применения. / Деев С.М., Лебеденко Е.Н. // Acta Naturae 2009. Т. 1. С.32-50; Bronte G. Monoclonal antibodies for the treatment of non-haematological tumours: update of an expanding scenario. / Bronte G., Sortino G., Passiglia F., Rizzo S., Lo Vullo F., Galvano A., Bazan V., Rolfo C., Russo A. // Expert Opin Biol Ther 2014. P. 1-15].
Недостаточная эффективность «ненагруженных» антител по отношению к мишеням побудили исследователей к конъюгированию антител с другими эффекторными молекулами. В этом случае моноклональные антитела могут использоваться как средства направленной доставки токсических агентов в опухоль. Такие соединения получили название «иммуноконъюгаты». Иммуноконъюгаты составляются, как правило, из трех основных компонентов: моноклональное антитело, терапевтический агент и линкер, связывающий антитело с терапевтическим агентом [Cao Y. Single-chain antibody-based immunotoxins targeting Her2/neu: design optimization and impact of affinity on antitumor efficacy and off-target toxicity. / Cao Y., Marks J.D., Huang Q., Rudnick S.I., Xiong C., Hittelman W.N., Wen X., Marks J.W., Cheung L.H., Boland K., Li C., Adams G.P., Rosenblum M.G. // Mol Cancer Ther 2012. V. 11, №1. P. 143-53; Kreitman R.J. Immunotoxins for targeted cancer therapy. [текст] / Kreitman R.J. // The AAPS Journal 2006. V. 8, №3. P. Е532-Е551]. Построенный таким образом иммуноконъюгат может служить для направленной доставки к опухолевым клеткам различных терапевтических агентов, в том числе радиоизотопов, токсинов, интерлейкинов, ферментов, активирующих лекарственных препаратов и т.д. [Деев С.М. Современные технологии создания неприродных антител для клинического применения. / Деев С.М., Лебеденко Е.Н. // Acta Naturae 2009. Т. 1. С. 32-50].
В качестве направляющего модуля чаще всего используют иммуноглобулины или их фрагменты, обладающие высокой специфичностью и аффинностью связывания с соответствующими антигенами. Однако за последние несколько лет появилось несколько альтернативных вариантов направляющих молекул неиммуноглобулиновой природы -лиганды специфических рецепторов, узнающие пептиды, аффибоди, DARPin'ы и т.д.
В роли терапевтического агента чаще всего выступают химически синтезированные или природные (бактериальные, растительные или животные) токсины. В арсенале исследователей на сегодняшний день - это ауристатины, мейтанзиноиды, калихеамицины и белковые токсины: рицин, псевдомонадный и дифтерийный токсины [Oldham R.K. Principles of Cancer Biotherapy. NY: Springer Science. 2009. 780 p.].
В настоящее время для клинического применения одобрены только два препарата на основе иммунотоксинов - это Брентуксимаб ведотин (brentuximab vedotin) и Трастузумаб Эмтансин (trastuzumab emtansine), применяющиеся для лечения некоторых лимфом и рака молочной железы [Dosio F. Immunotoxins and Anticancer Drug Conjugate Assemblies: The Role of the Linkage between Components. / Dosio F., Brusa P., Cartel L. // Toxins 2011. V. 3, №7. P. 848-883]. Во многом это обусловлено тем, что несмотря на то, что в доклинических испытаниях иммунотоксины показывают ярковыраженное специфическое цитотоксическое действие на опухолевые клетки, прогресс в области таргетной терапии опухолей тормозится тремя основными проблемами. Во-первых, опухоль обладает генетической гетерогенностью, связанной с особенностями генотипа каждого отдельного пациента и чрезвычайной нестабильностью генома опухолевых клеток, что приводит к возникновению резистентности опухолей к этим препаратам [Al-Lazikani В. Combinatorial drug therapy for cancer in the post-genomic era. / Al-Lazikani В., Banerji U., Workman P. // Nat Biotech 2012. V. 30, №7. P. 679-692]. Во-вторых, иммунотоксины демонстрируют значительную иммуногенность, обусловленную их происхождением, как правило, бактериальным, растительным или грибным. Кроме того, препараты на основе иммунотоксинов, как и другие противоопухолевые препараты, недостаточно глубоко проникают в опухолевую ткань. Это принято связывать с рядом причин, среди которых выделяют: (1) аномальное строение сосудистого русла опухоли и отсутствие нормального лимфодренажа, что приводит к повышению интерстициального давления в паренхиме опухоли и препятствует конвекционной транспортировке наноразмерньгх молекул и частиц из сосудов, расположенных, в основном, на периферии опухоли, к более глубоко лежащим клеткам паренхимы опухоли; (2) плотные межклеточные контакты опухолевых клеток; (3) фиброз опухолевой ткани. При этом важно понимать, что при использовании иммунотоксинов увеличенное однократное или многократное дозирование терапевтических препаратов, хотя и приводит к их более равномерному распределению в опухолевом объеме, мало приемлемо в силу их иммуногенности [Marcucci F. How to improve exposure of tumor cells to drugs - Promoter drugs increase tumor uptake and penetration of effector drugs. / Marcucci F., Corti A. // Advanced Drug Delivery Reviews 2012. V. 64, №1. P. 53-68]. Последовательное или комплексное решение перечисленных проблем лежит в основе всех современных разработок в направлении развития таргетной онкотерапии.
Так, например, известен иммунотоксин, содержащий направляющий модуль в виде кодирующей последовательности белка класса дарпин (DARPin), специфичный к белку ЕрСАМ, и токсический модуль в виде фрагмента псевдомонадного экзотоксина А ЕТА, соединенные между собой GS-линкером («The Promise of DARPins for Site-Specific Drug Conjugation & Pharmacokinetic Optimization)), Fabian Brande, 15.02.2016 г.).
Аминокислотные последовательности участков молекулы ЕрСАМ-специфичного дарпина, обеспечивающих аффинность к белку ЕрСАМ, отличаются от последовательностей соответствующих участков молекулы HER2-cпeцифичнoгo дарпина. Использование в качестве направляющего модуля ЕрСАМ-специфичного дарпина, не обладающего специфичностью к рецептору HER2/neu, исключает возможность его применения для таргетной терапии опухолей, гиперэкспрессирующих рецептор HER2.
Наиболее близким по технической сущности и достигаемому результату к предлагаемому изобретению является рекомбинантный иммунотоксин, специфичный к клеткам, экспрессирующим рецептор HER2, защищенный патентом RU №2576232 С1, кл. А61K 39/395, А61P 35/00, опубл. 27.02.2016 г., принятый за ближайший аналог (прототип).
Рекомбинантный иммунотоксин по прототипу содержит направляющий модуль в виде антитела формата scFv, включающего вариабельные домены легкой и тяжелой цепей моноклонального гуманизированного НЕR2-специфичного антитела, соединенные между собой линкером, и токсический модуль в виде фрагмента псевдомонадного экзотоксина А ЕТА. При этом направляющий модуль представлен антителом 4D5scFv, который соединен с токсическим модулем ЕТА гибким гидрофильным hinge-подобным линкером. На С-конце молекулы выполнена последовательность KDEL. Использование данного рекомбинантного иммунотоксина позволяет повысить токсичность для клеток, экспрессирующих рецептор HER2, и, следовательно, терапевтическую эффективность.
В задачу изобретения положено создание нового рекомбинантного таргетного токсина, специфичного к клеткам, экспрессирующим рецептор HER2.
Техническим результатом от использования предлагаемого изобретения является усиление токсического действия на клетки, экспрессирующие рецептор HER2, и повышение эффективности таргетной терапии.
Поставленная задача достигается тем, что рекомбинантный таргетный токсин, специфичный к клеткам, экспрессирующим рецептор HER2, содержит HER2-специфичный направляющий модуль, представленный кодирующей последовательностью HER2-специфичного белка класса дарпин DARPin, и токсический модуль в виде фрагмента псевдомонадного экзотоксина А ЕТА, соединенные между собой гибким гидрофильным 16-аминокислотным линкером, представленным последовательностью SEQ ID NO: 1, олигогистидиновую последовательность His6 и последовательность KDEL на С-конце молекулы, при этом рекомбинантный таргетный токсин представлен последовательностью SEQ ID NO: 2; ген, кодирующий рекомбинантный таргетный токсин, находится под контролем Т7-промотора.
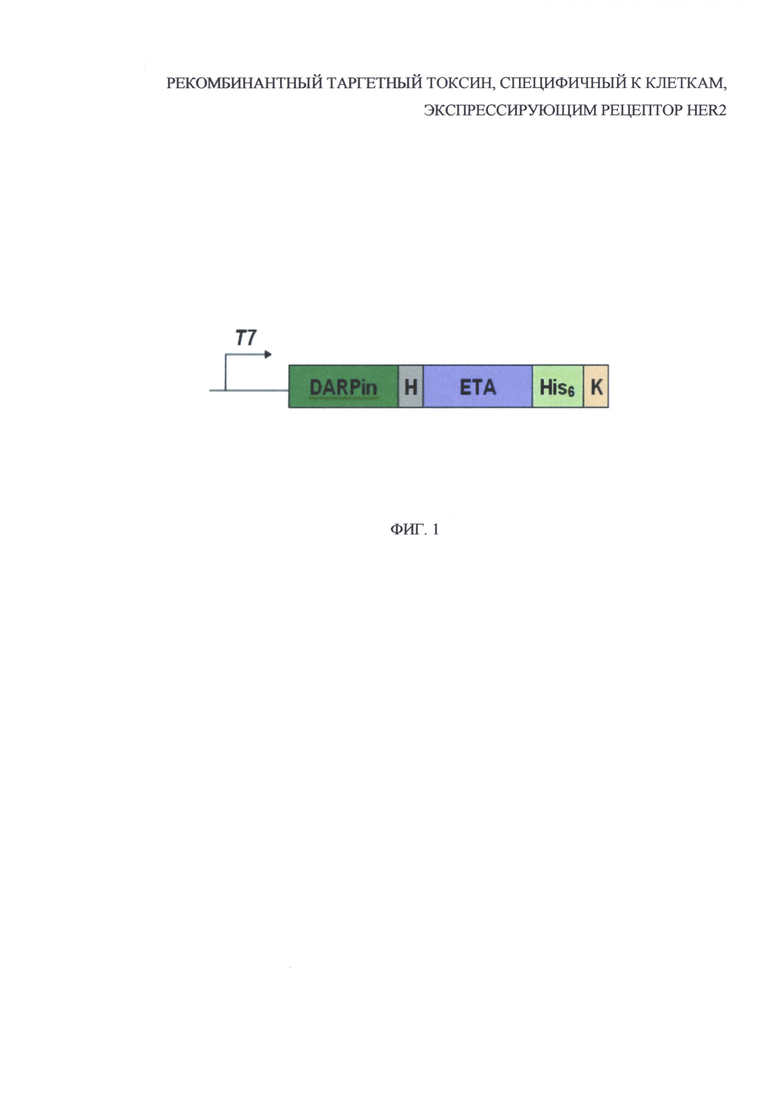
На фиг. 1 представлена схема генетической конструкции рекомбинантного таргетного токсина, содержащего HER2-специфичный дарпин и фрагмент псевомонадного экзотоксина A (DARPin-ETA), где: Т7 - регуляторный элемент плазмиды - промотор Т7, DARPin - кодирующая последовательность HER2-специфичного дарпина, Н - кодирующая последовательность гидрофильного линкера, ЕТА - кодирующая последовательность фрагмента псевдомонадного экзотоксина А, His6 - кодирующая последовательность полигистидинового олигопептида, К - последовательность KDEL.
На фиг. 2 представлены фотографии, отражающие результаты электрофоретического анализа полученного очищенного препарата рекомбинантного таргетного токсина, содержащего HER2-специфичный дарпин и фрагмент псевомонадного экзотоксина А, где: А - электрофорез в 12% ПААГ; Б - вестерн-блот анализ; М - белковый маркер молекулярного веса.
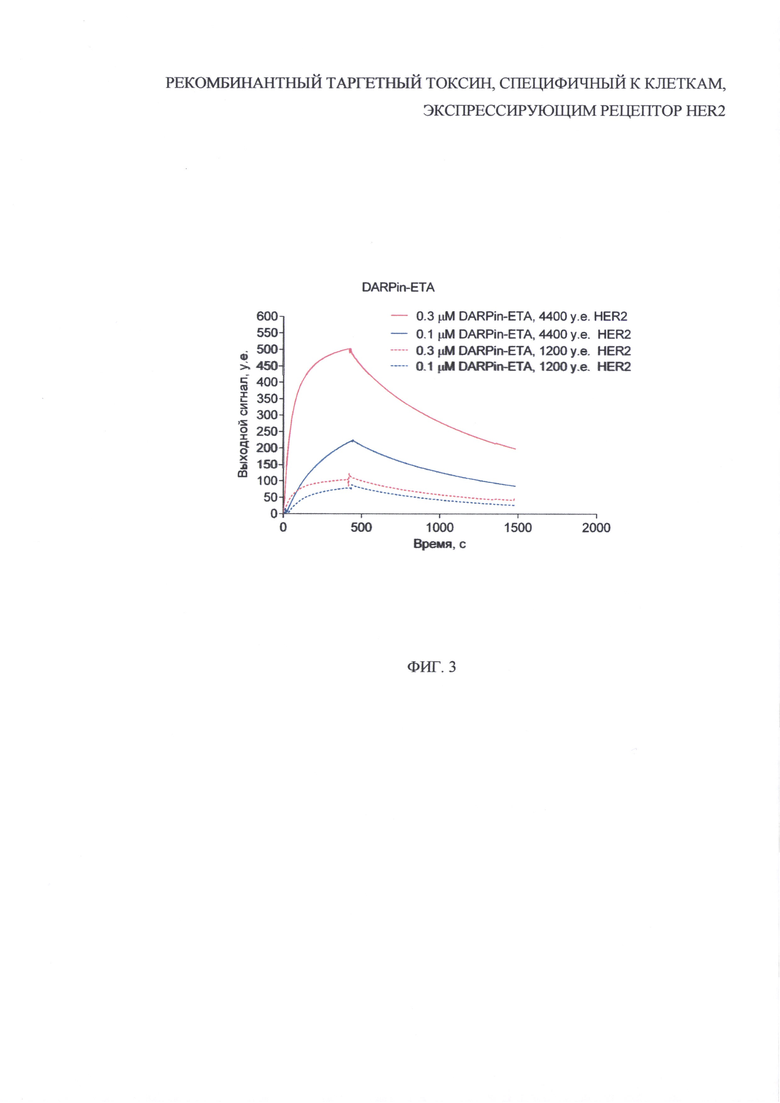
На фиг. 3 представлены графики, отражающие результаты анализа аффинности рекомбинантного таргетного токсина, содержащего HER2-специфичный дарпин и фрагмент псевомонадного экзотоксина А. Кинетика взаимодействия DARPin-ETA с внеклеточным доменом рецептора HER2, полученная с помощью метода поверхностного плазмонного резонанса при различной плотности рецептора на чипе и различной концентрации DARPin-ETA.
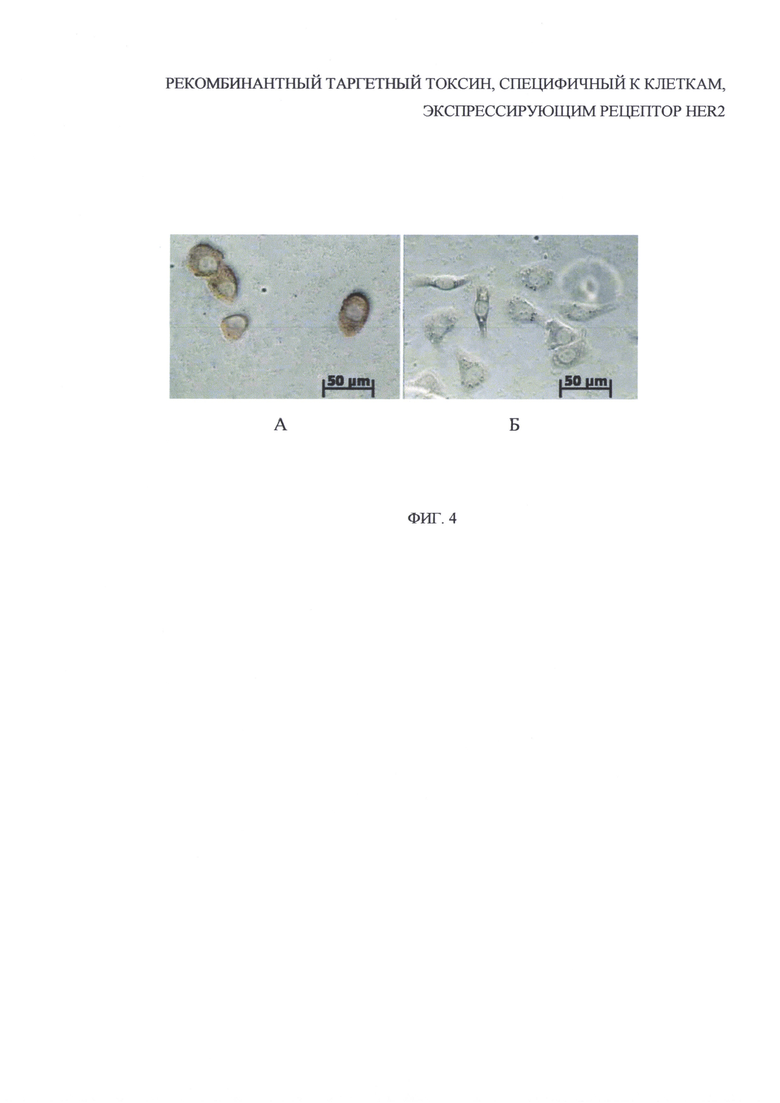
На фиг. 4 представлены результаты иммуноцитохимического анализа связывания белка DARPin-ETA с клетками SKOV3ip, где: А - клетки, инкубированные с
иммунотоксином; Б - клетки, инкубированные с фосфатно-солевым буфером в качестве контроля.
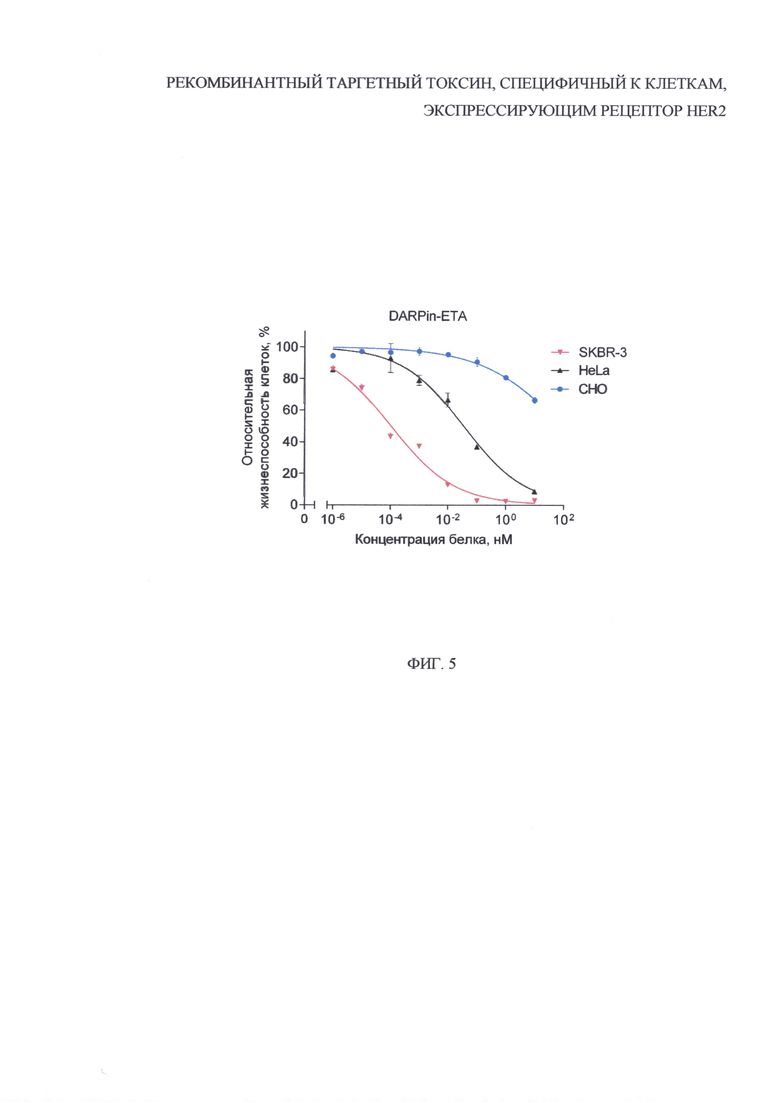
На фиг. 5 представлены графики, отражающие результаты анализа относительной жизнеспособности HER2-положительных клеток SKBR-3 (красная линия и перевернутый треугольник) и HeLa (черная линия и треугольник), и HER2-отрицательных клеток СНО (синяя линия и кружок) после их обработки рекомбинантным белком DARPin-ETA в течение 72 ч. Планки погрешностей представлены стандартной ошибкой среднего.
На фиг. 6 представлена диаграмма динамики роста HER2-гиперэкспрессирующей ксенографтной опухоли SKBR-3, где: серые столбцы - в группах контрольных животных, получивших инъекции фосфатно-солевого буфера, синие столбцы - в группах животных, получивших дозу DARPin-ETA 5×5 мкг, зеленые столбцы - в группах животных, получивших дозу DARPin-ETA 5×10 мкг, красные столбцы - в группах животных, получивших дозу DARPin-ETA 4×20 мкг. Дни инъекций DARPin-ETA обозначены стрелками. Планки погрешностей представлены стандартной ошибкой среднего. "*" - p<0.05 (сравнение с контрольной группой по критерию Даннета, n=4-6).
Генетически кодируемый HER2-специфичный таргетный (направленный) токсин, DARPin-ETA представляет собой рекомбинантный белок, в котором два независимых функциональных модуля белковой природы - направляющий и токсический - объединены в единую полипептидную цепь с помощью гибкого гидрофильного линкера (16 а.о.).
Ген, кодирующий таргетный токсин, под контролем Т7-промотора, представляющего собой промотор РНК-полимеразы бактериофага Т7. После трансформации клеток штамма-продуцента вектором, содержащими ген целевого белка, в культуральную среду добавляют индуктор lac-оперона - изопропилтиогалактозид (ИПТГ), синтетический аналог лактозы, который не метаболизируется клетками. В этих условиях происходит синтез РНК-полимеразы фага Т7, после чего запускается экспрессия целевого гена.
Направляющий модуль, DARPin, представляет собой HER2-специфичный каркасный белок класса дарпинов. DARPin характеризуется высокой аффинностью к рецептору HER2 (равновесная константа диссоциации 3.8 нМ), высокой стабильностью в широком диапазоне условий, малыми размерами (18 кДа), легкостью наработки в больших количествах в бактериальных продуцентах.
Токсический модуль, ЕТА, представляет собой фрагмент природного экзотоксина А из Pseudomonas aeruginosa (синегнойная палочка), содержащий домены II, Ib и III природного токсина, обеспечивающие эффективный внутриклеточный транспорт токсина в цитоплазму и токсический эффект - АДФ-рибозилирование фактора элонгации 2 эукариот (еЕТ2). Природный направляющий домен I экзотоксина А был заменен на необходимый для наших целей направляющий белок DARPin.
Направляющий и токсичный модули были соединены гибким гидрофильным 16-аминокислотным линкером, представленным последовательностью SEQ ID NO: 1, представляющим собой производное шарнирной области мышиного иммуноглобулина IgG3. Этот линкер может образовывать расстояние 2,5-2,7 нм между соединяемыми им участками белковой молекулы, позволяя двум доменам белка не испытывать стерических затруднений и сохранять свои функциональные свойства.
Рекомбинантный белок DARPin-ETA содержит полигистидновую последовательность («тэг») из 6 остатков гистидина, необходимую для очистки целевого белка из бактериального лизата методом металл-хелатной аффинной хроматографии на Ni2+-NTA-сефарозе. На С-конец полученной молекулы также введена последовательность KDEL, необходимая для проявления токсического действия псевдомонадного экзотоксина на клетку-мишень.
В экспериментах in vitro и in vivo было подтверждено, что полученный рекомбинантный белок с высокой аффинностью связывается с рекомбинантным внеклеточным доменом рецептора HER2, избирательно ингибирует рост HER2-положительных клеток в культуре, а также ингибирует рост ксенографтных HER2-гиперэкспрессирующих опухолей in vivo. Таким образом, DARPin-ETA является HER2-специфичным таргетным токсином.
Была создана генно-инженерная конструкция, кодирующая рекомбинантный белок DARPin-ETA, позволяющая нарабатывать его в высоких количествах в клетках кишечной палочки E. coli (фиг. 1).
Была разработана и оптимизирована методика выделения и очистки соответствующего рекомбинантного белка. Белок нарабатывали в клетках E.coli штамма BL21 и выделяли методами металл-хелатной аффинной и ионообменной хроматографии.
Электрофоретический анализ полученного рекомбинантного белка DARPin-ETA в 12% ПААГ в денатурирующих условиях показал, что полученный препарат очищенного белка был гомогенен и не содержал примесей, детектируемых с помощью окрашивания Coomassi blue G-250 при нанесении образца с перегрузом (фиг. 2А). Аутентичность полученного рекомбинантного белка была подтверждена методом вестерн-блота с использованием антигистидиновых моноклональных антител (фиг 2Б). Молекулярная масса полученного белка DARPin-ETA по результатам электрофореза и вестерн-блота совпадала с расчетной (58.1 кДа).
Была показана высокая аффинность полученного белка DARPin-ETA к внеклеточному домену рецептора HER2 методом поверхностного плазмонного резонанса с использованием оптического биосенсора BIAcore 3000 (GE Healthcare, США). Рекомбинантный внеклеточный домен HER2 ковалентно иммобилизовали на поверхности карбоксиметилдекстранового чипа СМ5 с плотностью 1200 и 2200 у.е. Таргетный токсин DARPin-ETA был использован в концентрации 0.3 и 0.1 мкМ. Значение равновесной константы диссоциации (KD), являющейся количественной мерой аффинности, рассчитывали с помощью программы «BIAevaluation 3.0 Software» на основе полученных сенсограмм (фиг. 3).
Было показано, что KD белка DARPin-ETA составила 10.8 нМ, что согласуется с ранее опубликованными значениями для свободного белка DARPin (3.8 нМ), являющегося направляющим модулем созданного таргетного токсина. Таким образом, направляющий белок DARPin сохраняет высокую аффинность к рецептору HER2 в составе сконструированного рекомбинантного белка DARPin-ETA.
Была показана способность полученного белка DARPin-ETA связываться с опухолевыми клетками, экспрессирующими рецептор HER2.
Для подтверждения специфичности связывания полученного иммунотоксина с рецептором HER2 была использована культура клеток аденокарциномы яичника человека SKOVip, гиперэкспрессирующая на поверхности клеток соответствующий рецептор. Суспензию клеток с концентрацией 2×104 кл/мл высевали на 96-луночные планшеты (Corning) и культивировали в течение ночи. Клетки фиксировали формальдегидом и инкубировали с раствором белка DARPin-ETA (10 мкг/мл). Далее клетки последовательно инкубировали с моноклональными антителами 6x-His Epitope Tag Antibody (HIS.H8) (Thermo Scientific, cat. # MA1-21315), специфичными к олигогистидиновой последовательности на С-конце белка DARPin-ETA и поликлональными антимышиными антителами с пероксидазой хрена HRP Goat Anti-Mouse Ig (BD Pharmingen, cat. # 554002). Клетки окрашивали с использованием в качестве субстрата для пероксидазы диаминобензидин.
Детекцию клеток проводили с помощью инвертированного микроскопа Axiovert 200 (Zeiss) c использованием объектива с увеличением 40х. После инкубации клеток SKOVip, гиперэкспрессирующих HER2/neu на наружной поверхности цитоплазматической мембраны, с белком DARPin-ETA на микрофотографиях заметно характерное коричневое окрашивание по краю клеток (фиг. 4А), значительно отличающееся от фона (фиг. 4Б).
Было показано специфическое цитотоксическое действие полученного таргетного токсина на опухолевые клетки, экспрессирующие рецептор HER2.
Были использованы клетки аденокарциномы молочной железы человека SKBR-3 (номер по каталогу АТСС - НТВ-30), гиперэкспрессирующие рецептор HER2, клетки карциномы шейки матки человека HeLa (номер по каталогу АТСС - CCL-2) с низким уровнем экспрессии HER2/neu (~ 1×104 молекул на клетку) и контрольные клетки яичника китайского хомячка СНО (номер по каталогу АТСС - CCL-61), не экспрессирующие данный рецептор. Клетки инкубировали в течение 72 ч при 37°С и 5% CO2 в ростовой среде, содержащей белок DARPin-ETA в различной концентрации. По окончании инкубации жизнеспособность клеток оценивали с помощью стандартного МТТ-теста.
Было показано, что полученный HER2-специфичный белок DARPin-ETA значительно снижает жизнеспособность HER2-положительных клеток SKBR-3 и HeLa, практически не влияя на HER2-отрицательные клеток СНО (фиг. 5). При этом выявленный цитотоксический эффект коррелирует с уровнем экспрессии рецептора HER2: значение IC50 (концентрации DARPin-ETA, вызывающей снижение жизнеспособности клеток на 50% относительно контроля) для HER2-гиперэкспрессирующих клеток SKBR-3 и клеток HeLa с низким уровнем экспрессии составило 0.1 пМ и 37 пМ соответственно.
Был показан противоопухолевый эффект белка DARPin-ETA в отношении HER2-гиперэкспрессирующей ксенографтной опухоли.
Для получения экспериментальной модели опухоли человека иммунодефицитным мышам линии nude была подкожно привита суспензия клеток SKBR-3 в количестве 10 млн клеток на животное. Мониторинг роста опухолей был проведен на основе данных об объеме опухолевых узлов. По достижении размера опухолей 100 мм3 животные были разделены на группы в соответствии с вводимой дозой DARPin-ETA: пятикратное введение по 5 мкг, пятикратное введение по 10 мкг или четырехкратное введение по 20 мкг DARPin-ETA (внутривенно).
Было выявлено выраженное замедление роста опухолей в ответ на введение DARPin-ETA, при этом данный эффект носил дозо-зависимый характер (фиг. 6). При введении белка DARPin-ETA в дозе 4×20 мкг не наблюдалось роста опухолей вплоть до окончания наблюдения, при этом рассчитанное значение коэффициента торможения роста опухоли составило 94%.
Высокая аффинность белка DARPin-ETA к рецептору HER2 обеспечивает более сильный цитотоксический эффект рекомбинантного таргетного токсина на опухолевые клетки, экспрессирующие рецептор HER2, а также больший противоопухолевый эффект в отношении HER2-гиперэкспрессирующей ксенографтной опухоли по сравнению с рекомбинантным иммунотоксином по прототипу.
Таким образом, использование HER2-специфичного дарпина в качестве направляющего модуля обеспечивает усиление токсического действия на клетки, экспрессирующие рецептор HER2, и повышение эффективности таргетной терапии.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ИММУНОТОКСИН, СПЕЦИФИЧНЫЙ К КЛЕТКАМ, ЭКСПРЕССИРУЮЩИМ РЕЦЕПТОР HER2 | 2014 |
|
RU2576232C1 |
| Способ получения рекомбинантного противоопухолевого токсина на основе белков барназа-барстар и адресного полипептида дарпина с эффектом моментальной отмены цитотоксического действия | 2015 |
|
RU2610179C1 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКОВ В ДВУХЦЕПОЧЕЧНОЙ ФОРМЕ С ДИСУЛЬФИДНЫМ МОСТИКОМ | 2006 |
|
RU2412253C2 |
| СПОСОБ КОНЪЮГАЦИИ КОНСТАНТНОГО ФРАГМЕНТА ТЯЖЕЛОЙ ЦЕПИ АНТИТЕЛА ЧЕЛОВЕКА И ПЕПТОИДНОГО АНАЛОГА АУТОАНТИГЕНА MOG35-55 ДЛЯ ТЕРАПИИ РАССЕЯННОГО СКЛЕРОЗА | 2018 |
|
RU2724714C1 |
| Конъюгат люминесцентных наночастиц диоксида кремния с рекомбинантными однодоменными нано - моноантителами C7b ламы, способными специфически взаимодействовать с рекомбинантным белком HER2/neu - аналогом природного рецептора эпидермального фактора роста EGFR/ERBB клеток человека | 2017 |
|
RU2679075C1 |
| ХИМЕРНЫЙ ИММУНОТОКСИН НА ОСНОВЕ НЕСТРУКТУРНОГО ГЛИКОПРОТЕИНА NSP4 РОТАВИРУСА ЧЕЛОВЕКА, ПРОЯВЛЯЮЩИЙ ПОВЫШЕННОЕ ЦИТОТОКСИЧЕСКОЕ ВОЗДЕЙСТВИЕ НА ОПУХОЛЕАССОЦИИРОВАННЫЙ MUC1 РАКОВЫХ КЛЕТОК | 2024 |
|
RU2837117C1 |
| РЕКОМБИНАНТНЫЙ ИММУНОТОКСИН, НАЦЕЛЕННЫЙ НА МЕЗОТЕЛИН | 2012 |
|
RU2600067C2 |
| Способ радионуклидной диагностики рака молочной железы с гиперэкспрессией Her2/neu | 2019 |
|
RU2720801C1 |
| КОМПЛЕКС ТЕХНЕЦИЯ-99М С РЕКОМБИНАНТНЫМИ АДРЕСНЫМИ МОЛЕКУЛАМИ БЕЛКОВОЙ ПРИРОДЫ С АНКИРИНОВЫМИ ПОВТОРАМИ ДЛЯ РАДИОНУКЛИДНОЙ ДИАГНОСТИКИ ЗЛОКАЧЕСТВЕННЫХ ОБРАЗОВАНИЙ С ГИПЕРЭКСПРЕССИЕЙ HER2/NEU И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2023 |
|
RU2812633C1 |
| МОДИФИЦИРОВАННЫЕ БЕЛКИ, СКОНСТРУИРОВАННЫЕ ТОКСИНЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2002 |
|
RU2305684C2 |



Изобретение относится к биотехнологии. Предложен рекомбинантный таргетный токсин, специфичный к клеткам, экспрессирующим рецептор HER2. Рекомбинантный таргетный токсин содержит HER2-специфичный направляющий модуль, представленный кодирующей последовательностью HER2-специфичного белка класса дарпин DARPin, и токсический модуль в виде фрагмента псевдомонадного экзотоксина А ЕТА, соединенные между собой гибким гидрофильным 16-аминокислотным линкером, представленным последовательностью SEQ ID NO: 1, олигогистидиновую последовательность His6 и последовательность KDEL на С-конце молекулы. При этом рекомбинантный таргетный токсин представлен последовательностью SEQ ID NO: 2. Изобретение обеспечивает усиление токсического действия на клетки, экспрессирующие рецептор HER2, и повышение эффективности таргетной терапии. 1 з.п. ф-лы, 6 ил.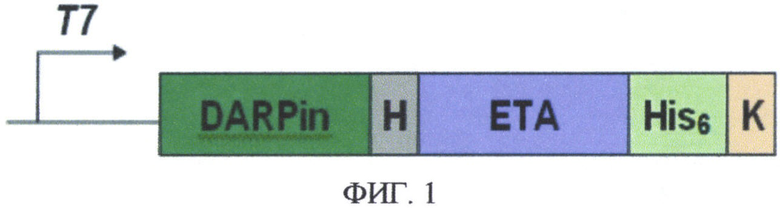
1. Рекомбинантный таргетный токсин, специфичный к клеткам, экспрессирующим рецептор HER2, содержит HER2-специфичный направляющий модуль, представленный кодирующей последовательностью HER2-специфичного белка класса дарпин DARPin, и токсический модуль в виде фрагмента псевдомонадного экзотоксина А ЕТА, соединенные между собой гибким гидрофильным 16-аминокислотным линкером, представленным последовательностью SEQ ID NO: 1, олигогистидиновую последовательность His6 и последовательность KDEL на С-конце молекулы, при этом рекомбинантный таргетный токсин представлен последовательностью SEQ ID NO: 2.
2. Рекомбинантный таргетный токсин по п. 1, отличающийся тем, что ген, кодирующий рекомбинантный таргетный токсин, находится под контролем Т7-промотора.
| РЕКОМБИНАНТНЫЙ ИММУНОТОКСИН, СПЕЦИФИЧНЫЙ К КЛЕТКАМ, ЭКСПРЕССИРУЮЩИМ РЕЦЕПТОР HER2 | 2014 |
|
RU2576232C1 |
| WO2014083208 A1, 05.06.2014 | |||
| PROSHKINA G.M | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| MARTIN-KILLIAS P | |||
| ET AL | |||
| A Novel Fusion Toxin Derived from an EpCAM-Specific Designed Ankyrin Repeat Protein Has Potent Antitumor Activity //Clin Cancer Res, 2011,17:100-110.. | |||

