ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет предварительной заявки на патент США №61/483531, поданной 6 мая 2011 г., которая включена в настоящий документ посредством ссылки во всей полноте.
ЗАЯВЛЕНИЕ О ПРАВАХ НА ИЗОБРЕТЕНИЯ, СДЕЛАННОЕ В РАМКАХ НИОКР, ФИНАНСИРУЕМЫХ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА
НЕТ ДАННЫХ
ССЫЛКА НА ПРИЛОЖЕНИЕ «ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ», ТАБЛИЦУ ИЛИ СПИСОК КОМПЬЮТЕРНЫХ ПРОГРАММ, ПРЕДСТАВЛЕННЫЕ НА КОМПАКТ-ДИСКЕ
НЕТ ДАННЫХ
ОБЛАСТЬ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Рекомбинантные иммунотоксины (RIT) - это искусственно созданные лечебные белки, в которых соединены фрагмент антитела и цитотоксический белок, полученный из бактериального или растительного источника. RIT созданы в качестве селективных агентов для целенаправленного уничтожения клеток без проявления множественной вторичной токсичности, характерной для химиотерапевтических стратегий. RIT для лечения раковых заболеваний могут быть сконструированы путем слияния вариабельного фрагмента (Fv) антител против опухолевых антигенов клеточной поверхности с фрагментом экзотоксина А Pseudomonas (РЕ). RIT с использованием 38-кДа фрагмента экзотоксина А Pseudomonas (PE38) имели определенный успех в клинических испытаниях, но имели также ограничения, которые включали плохое проникновение в твердые опухоли, высокую иммуногенность и неспецифическую токсичность (Kreitman RJ et al., Clin Cancer Res., 15(16):5274-9 (2009; Hassan R et al., Clin Cancer Res., 13(17):5144-9 (2007); Wayne AS et al., Clin Cancer Res., 16(6):1894-903 (2010); Kreitman RJ et al., J Clin Oncol., 27(18):2983-90 (2009); Sampson JH et al., Neuro Oncol., 10(3):320-9 (2008); Powell DJ Jr et al., J Immunol., 179(7):4919-28 (2007); Kreitman RJ, J Clin Oncol., 23(27):6719-29 (2005); Pal LH et al., Nat Med., 2(3):350-3 (1996)).
В стремлении улучшить результат лечения с использованием RIT важную роль для понимания строения этих белков играет знание пути токсического действия РЕ. RIT проникают в клетки путем рецептор-опосредованного эндоцитоза и транспортируются через эндолизосомальную систему в аппарат Гольджи, где они подвергаются обратному транспорту в эндоплазматический ретикулум (ER). Во время этого этапа токсин активируется за счет восстановления дисульфидной связи и расщепления протеазой фурином в определенном участке РЕ38, который отделяет Fv от фрагмента РЕ. Впоследствии активированный РЕ должен перемещаться в цитозоль, где он осуществляет АДФ-рибозилирование и инактивацию фактора элонгации 2, необходимого элемента трансляционного аппарата. Это останавливает синтез белка и в конечном итоге приводит к гибели клетки (обзор о пути токсического действия РЕ см. в 9). Предыдущие стратегии, направленные на улучшение цитотоксической активности RIT на основе РЕ, включали замену C-концевых остатков РЕ, REDLK (SEQ ID NO:15), на канонический сигнал возвращения в ER KDEL (SEQ ID NO:16) (Seetharam S et al., J Biol Chem., 266(26):17376-81 (1991); Du X, Но M, and Pastan I, J Immunother., 30(6):607-13 (2007); Rozemuller H. et al., Int J Cancer., 92(6):861-70 (2001); Kreitman RJ and Pastan I., Biochem J., 307 (Pt 1):29-37 (1995)). Известно, что это изменение повышает цитотоксичность РЕ, предположительно, за счет повышения эффективности ретроградного транспорта в ER из аппарата Гольджи. Эта стратегия эффективна, но, как правило, повышает также и неспецифическую токсичность RIT. Еще одна стратегия заключается в повышении эффективной интернализации комплекса RIT-рецептор, и увеличении тем самым количества токсина в клетке, за счет улучшения сродства между Fv и его мишенью (Salvatore G et al., Chn Cancer Res., 8(4):995-1002 (2002); Decker Т et al., Blood., 103(7):2718-26 (2004)).
Недавно был разработан протеазоустойчивый RIT, выдерживающий деградацию в эндолизосомальной системе, потенциальный барьер для эффективного лечения иммунотоксинами (Johannes L and Decaudin D, Gene Ther., 12(18):1360-8 (2005); Fitzgerald D. Why toxins Semin Cancer Biol., 7(2):87-95 (1996)). Этот «устойчивый к лизосомальной деградации» (LR) вариант RIT был получен путем удаления протеазочувствительных районов РЕ38 и направлением их на B-клеточный рецептор CD22 с использованием высокоаффинного Fv против CD22, полученного из RIT HA22 (Weldon JE, Blood., 113(16):3792-800 (2009)). Мутация LR практически не влияла на активность in vitro на клеточных линиях, но значительно понизила неспецифическую токсичность на мышах и резко повысила активность in vitro на клетках, полученных от пациента с хроническим лимфолейкозом (CLL). Кроме того, в варианте LR устранены две основные группы B-клеточных эпитопов у мышей (Onda M et al., J Immunol., 177(12):8822-34 (2006)) и участки процессинга антигена из РЕ38, что помогает уменьшить его иммуногенность у мышей (Hansen JK et al., J Immunother., 33(3):297-304 (2010)). Благодаря модульной природе RIT, вариант LR из РЕ может быть направлен на другие ассоциированные с опухолью антигены путем замены одного Fv на другой. Соответственно, предшествующий уровень техники, описывающий уменьшение доменов II и Ib РЕ, в общем показывает преимущества удаления протеазочувствительных и антигенных сайтов из молекулы. Уровень техники также демонстрирует фармакокинетические преимущества меньших RIT, что является следствием этих изменений.
Клинически значимым кандидатом-мишенью является ассоциированный с опухолью антиген мезотелин, который часто экспрессируется на высоком уровне при раковых заболеваниях, которые включают мезотелиомы и рак легких, яичников и поджелудочной железы. Соответственно, существует потребность в усовершенствованных RIT, которые специфически поражают раковые клетки, которые экспрессируют мезотелин на их поверхности. Данное изобретение обеспечивает эти и другие потребности, предоставляя RIT, фармацевтические композиции и способы лечения раковых опухолей, которые экспрессируют или сверхэкспрессируют мезотелин.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение предлагает усовершенствованный экзотоксин А Pseudomonas («РЕ») с уменьшенной иммуногенностью, улучшенной устойчивостью к лизосомальным протеазам и повышенной цитотоксичностью для клеток, экспрессирующих мезотелин. Структурно у усовершенствованного РЕ изобретения удален домен I РЕ, удалена большая часть домена II РЕ, и имеется функциональный домен III РЕ необязательно с замещением а) аминокислотных остатков домена III РЕ в позициях D406, R432, R467, R490, R513, Е548, K590 и/или Q592 на глицин, аланин или серин, и/или б) аминокислотных остатков домена III РЕ в позициях D403, R412, R427, Е431, R458, D461, R505, Е522, R538, R551, R576 и/или L597 на глицин, серин или аланин. Усовершенствование заключается во введении короткого и гибкого пептидного линкера («FL») от 3 до 8 аминокислот в длину, состоящего из остатков глицина и/или серина, введенного между сайтом расщепления фурином и функциональным доменом III РЕ улучшенного РЕ. Соответственно, короткий линкер состоит из остатков глицина и/или серина. В некоторых вариантах осуществления изобретения линкер представляет собой пептид формулы (Хаа1)n, где каждый Хаа1 независимо выбирают из глицина и серина, и n составляет от 3 до 8. Усовершенствованные молекулы РЕ изобретения сохраняют высокую цитотоксическую активность с удалением B-клеточных эпитопов. Неожиданно оказалось, что включение короткого гибкого линкера повышает цитотоксичность молекулы без существенного изменения расщепления молекулы фурином. Усовершенствованные молекулы РЕ проиллюстрированы частными вариантами осуществления изобретения, представленными здесь как (LR/FL/8X, SEQ ID NO:4) и LR/FL/8M (SEQ ID NO:5). Кроме того, существуют варианты осуществления, в которых молекулы РЕ имеют одну или более мутаций в их функциональном домене III, как показано в SEQ ID NO:4 или 5, по сравнению с функциональным доменом РЕ из SEQ ID NO:1. В других вариантах осуществления имеются дополнительные замены в одном или более остатках, соответствующих 609-613 из SEQ ID NO:1, которые, как предполагают, также функционируют для удержания в эндоплазматическом ретикулуме нативной последовательности. В других вариантах осуществления сайт расщепления фурином домена II РЕ модифицирован или заменен другим сайтом расщепления фурина.
В некоторых вариантах осуществления также в соответствии со всем вышеописанным РЕ содержит функциональный домен III, который имеет одну или более мутаций в функциональной области токсина РЕ из SEQ ID NO:2 (позиции с 12 до 230), или выбранных из следующей таблицы, которые удаляют один или несколько эпитопов домена III РЕ:
В близком аспекте изобретения предложены рекомбинантные иммунотоксины («RIT изобретения»), которые являются химерными молекулами, включающими (а) группу, направленную на мезотелин, конъюгированную или слившуюся с (б) модифицированным экзотоксином A Pseudomonas («РЕ»), как указано выше. В некоторых вариантах осуществления группа представляет собой антитело, выбранное из группы, состоящей из scFv, dsFv, Fab, однодоменного антитела и F(ab′)2, или полипептид, включающий CDR антитела. SS1 и MORab-009 являются предпочтительными направляющими группами. Кроме того, антитело к мезотелину может содержать вариабельную тяжелую цепь («VH») и вариабельную легкую цепь («VL»), причем каждая из цепей VH и VL содержит первую, вторую и третью области, определяющие комплементарность («CDR»), в которых первая CDR («CDR1»), вторая CDR («CDR2»), и третья CDR («CDR3») указанной тяжелой цепи соответственно имеют последовательность аминокислотных остатков, показанную для CDR1 (GYTMN, SEQ ID NO:51), CDR2 (LITPYNGASSYNQKFRG; SEQ ID NO:52), и CDR3 (GGYDGRGFDY; SEQ ID NO:53), и в которых CDR1, CDR2 и CDR3 указанной VL цепи соответственно имеют последовательность аминокислотных остатков, показанную для CDR1 (SASSSVSYMH; SEQ ID NO:54), CDR2 (DTSKLAS; SEQ ID NO:55), и CDR3 (QQWSGYPLT; SEQ ID NO:56). В некоторых вариантах осуществления CDR3 легкой цепи модифицирована и имеет последовательность QQWSKHPLT (SEQ ID NO:57), QQWSGHPLT (SEQ ID NO:58), QQWSAHPLT (SEQ ID NO:59), QQWSQIPLT (SEQ ID NO:60), QQWGFNPLT (SEQ ID NO:61), QQWGTNPLT (SEQ ID NO:62), QQWGSHPLT (SEQ ID NO:63), QQWGDFPLT (SEQ ID NO:64), QQWGDHPLT (SEQ ID NO:65), QQWSAHPLT (SEQ ID NO:66) или QQWSGYPTT (SEQ ID NO:67). В некоторых других вариантах осуществления антитело к мезотелину представляет собой scFv, dsFv, Fab или F(ab′)2. В некоторых других вариантах осуществления изобретения антитело к мезотелину для использования в RIT включает аминокислотную замену по меньшей мере одной аминокислоты в CDR, выбранной из группы, состоящей из VL CDR1, VL CDR2, VH CDR1 и VH CDR2, причем указанная аминокислота кодируется кодоном, который содержит нуклеотид, принадлежащий мотиву «горячей точки», выбранный из группы AGY или RGYW, где R представляет собой А или G, Y представляет собой С или Т, и W представляет собой А или Т.
В другом аспекте настоящее изобретение относится к фармацевтическим композициям, содержащим
(а) RIT изобретения, как указано выше, и (б) фармацевтически приемлемый носитель.
В другом аспекте настоящее изобретение относится к выделенным нуклеиновым кислотам, кодирующим модифицированный экзотоксин А Pseudomonas («РЕ»), FL или RIT, описанным выше. В некоторых вариантах осуществления нуклеиновая кислота далее кодирует все антитело, направленное на мезотелин, или его фрагмент (вариабельную легкую или тяжелую цепь или CDR).
Соответственно, первая группа вариантов осуществления изобретения относится к выделенным модифицированным экзотоксинам A Pseudomonas ("PE"s), содержащим непрерывную полипептидную последовательность со следующей формулой: FCS-FL-функциональный домен III РЕ или L1-FCS-FL-функциональный домен III РЕ, где L1 состоит из непрерывной последовательности пептида от 1 до 10 аминокислотных остатков в длину; FCS представляет собой сайт или последовательность расщепления фурином (например, RHRQPRGWEQL; SEQ ID NO:17), или другую последовательность, которая расщепляется фурином; FL представляет собой последовательность пептида гибкого линкера, содержащую от 3 до 8 аминокислотных остатков, независимо выбранных из глицина и серина; функциональный домен III РЕ включает остатки 395-613 из SEQ ID NO:1, необязательно содержащие (i) замещения в одном или более остатках, соответствующих 609-613 из SEQ ID NO:1, (ii) замещение глицина, аланина, валина, лейцина или изолейцина на аргинин в положении, соответствующем положению 490 в SEQ ID NO:1, (iii) замещение одного или более остатков, соответствующих остаткам SEQ ID NO:1, которые поддерживают иммуногенность эпитопа или субэпитопа домена III РЕ или (iv) комбинацию любого из (i) - (iii). В предпочтительных вариантах осуществления функциональный домен РЕ - это функциональный домен РЕ LR/FL/8X (SEQ ID NO:4) или LR/GGS/8M (SEQ ID NO:3).
В еще одной группе вариантов осуществления изобретения предложены химерные молекулы или RIT, включающие (а) направляющую группу или лиганд, который специфически связывается с мезотелином на поверхности клетки, конъюгированный или слившийся с (б) модифицированным экзотоксином А Pseudomonas (РЕ), как описано выше. В некоторых вариантах осуществления изобретения лиганд представляет собой антитело или его фрагмент, который сохраняет способность распознавать антиген. В предпочтительных вариантах осуществления антитела являются производными исходного антитела SS1. Предпочтительно, RIT является SS1-LR/GGS/8X, который имеет GGS FL, вставленный между FCS и функциональным доменом III. Эти RIT, соответственно, могут содержать вариабельную легкую цепь SS1 с SEQ ID NO:6 и последовательность вариабельной тяжелой цепи SS1 рекомбинантного иммунотоксина SEQ ID NO:8 или SEQ ID NO:9, где вариабельные легкая и тяжелая цепи SS1 образуют стабилизированное дисульфидными связями антитело.
В еще одной группе вариантов осуществления изобретения предложены терапевтические способы уничтожения клеток-мишеней или ингибирования роста клеток-мишеней, которые экспрессируют или сверхэкспрессируют мезотелин на поверхности клеток. Способы включают контактирование клеток с RIT изобретения. Мезотелин - это дифференцировочный антиген, присутствующий на поверхности клеток рака яичника, мезотелиом и некоторых других видов злокачественных новообразований человека. RIT согласно изобретению могут использоваться, например, in vitro или in vivo, чтобы подавить или замедлить рост рака яичников, желудка, клеток плоского эпителия, мезотелиом и других злокачественных клеток, экспрессирующих мезотелин. Предусмотрены способы лечения пациентов, имеющих эти состояния и нуждающихся в лечении с помощью RIT изобретения.
В еще одной группе вариантов осуществления изобретения предложены нуклеиновые кислоты, кодирующие мутировавшие РЕ и RIT, описанные выше.
В некоторых вариантах осуществления любого из вышеописанного гибкий линкер представляет собой GGS или GGSGGS (SEQ ID NO:18).
В других вариантах осуществления антитело выбрано из группы, состоящей из scFv, dsFv, Fab, однодоменного антитела и F(ab′)2. В некоторых других вариантах осуществления вышеописанного антитело представляет собой SS1 или модифицированное SS1 (scFv, dsFv, Fab, однодоменное антитело или F(ab′)2 антитела SS1 или фрагмент(ы), представляющий(ие) части CDR антитела SS1). В некоторых вариантах осуществления CDR антитела используют в качестве направляющей группы. В некоторых вариантах осуществления антитело является человеческим или гуманизированным. В некоторых вариантах осуществления изобретения модифицированный РЕ представляет собой LR/GGS/8M (SEQ ID NO:3) или LR/(Xaa1)n/8X (SEQ ID NO:4) или LR/(Xaa1)n/8M (SEQ ID NO:5). В некоторых других вариантах осуществления химерная молекула представляет собой SS1-LR/GGS/8X (SEQ ID NO:6 и 7) или SS1-LR/GGS/8M (SEQ ID NO:6 и 8), где их соответствующие направляющие группы включают VL и VH части антитела SS1.
Дополнительные варианты осуществления станут очевидными для специалистов и описаны в данном документе.
Соответственно, в некоторых вариантах осуществления изобретения предложена химерная молекула, содержащая фрагмент антитела к мезотелину, напрямую последовательно соединенный с первым пептидным линкером, содержащим от 3 до 8 аминокислот в длину, который непосредственно соединен в последовательности с сайтом расщепления полипептида фурином RHRQPRGWEQL (SEQ ID NO:17), который непосредственно соединен в последовательности со вторым пептидным линкером, содержащим от 3 до 6 аминокислот, выбранных из Gly и Ser, и который непосредственно соединен в последовательности с N-концевой аминокислотой функционального домена III экзотоксина A Pseudomonas. В некоторых последующих вариантах осуществления функциональный домен - это домен III LR или LR/8M, и фрагментом антитела является dsFv из SS1-LR. В некоторых вариантах осуществления первый пептидный линкер (L1) непосредственно соединен в последовательности с карбоксильным концом участка VH из dsFv. Также предложены фармацевтические композиции, содержащие химерные молекулы, а также их применение в способе лечения рака, который сверхэкспрессирует мезотелин, у субъекта, нуждающегося в этом. В последующих вариантах осуществления рак представляет собой аденокарциному легкого, рак яичника, мезотелиому или плоскоклеточный рак.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1. Рекомбинантные иммунотоксины. (А) Рекомбинантный иммунотоксин SS1P состоит из стабилизированных дисульфидными связями (ds) тяжелой (VH) и легкой (VL) полипептидных цепей вариабельного фрагмента (Fv) моноклонального антитела к мезотелину SS1, связанных с 38-кДа фрагментом экзотоксина A Pseudomonas (PE38) через короткий пептидный линкер (ASGG; SEQ ID NO:19) из тяжелой цепи. PE38 состоит из домена II, домена III и фрагмента домена Ib из нативного экзотоксина A Pseudomonas. Домен II включает гидрофильную петлю, ограниченную цистеинами, которые образуют дисульфидную связь, которая содержит сайт расщепления протеазой фурин (RHRQPRGWEQL; SEQ ID NO:17). (В) Вариант SS1P, устойчивый к лизосомальной деградации, SS1-LR, не имеет домена Ib и домена II РЕ, за исключением фрагмента из 11 остатков, содержащего сайт расщепления фурином из домена II. Созданы различные конструкции с мутациями (подчеркнуто) вокруг сайта расщепления фурином в SS1-LR (SEQ ID NO:20-24).
Фигура 2. Цитотоксичность SS1-LR на мезотелин-позитивных клеточных линиях. Клеточные линии L55, NCI-H322M, HAY, KB31, М30, А431/K5, OVCAR-8 и А1847 инкубировали с возрастающими концентрациями SS1P (незакрашенные кружки, сплошная линия) или SS1-LR (незакрашенные квадраты, пунктирная линия). Через 3 дня жизнеспособность клеток оценивали методом колориметрического анализа WST-8 и нормализовали между контрольными пробами, не обработанной и обработанной циклогексамидом. Показаны средние значения и стандартные ошибки для шести повторных измерений. SS1-LR содержит стабилизированную дисульфидными связями полипептидную цепь SS1 VL c SEQ ID NO:6 и полипептидную цепь SS1 VH-PE с SEQ ID NO:74.
Фигура 3. Высокие дозы SS1-LR обладают сильной противоопухолевой активностью. Мышам, лишенным волосяного покрова (линии nude), с ксенотрансплантатами опухоли А431/K5 вводили внутривенно на 5, 7 и 9 сутки после имплантации буферный раствор для RIT (0,2% HSA в PBS; кресты, сплошная линия), 0,3 мг/кг SS1P (незакрашенные кружки, сплошная линия) или SS1-LR с дозой 6 мг/кг (незакрашенные квадраты, пунктирная линия) или 15 мг/кг (закрашенные квадраты, пунктирная линия). Стрелки указывают дни, когда проводилось лечение. Размер опухоли измеряли в течение 22 дней. Точки представляют средние размеры опухоли у всех мышей в группе обработки (n=6). Планки погрешностей показывают стандартное отклонение каждого среднего значения.
Фигура 4. Процессинг интернализированного иммунотоксина. Клетки А431/K5 инкубировали непрерывно с (А) SS1P или (В) SS1-LR, лизировали в различные моменты времени от 0 до 24 часов и анализировали методом невосстанавливающего электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и вестерн-блоттинга с антителом к РЕ. Отмечены полосы, соответствующие полноразмерным, восстановленным и расщепленным фурином формам. (С) Интенсивность полосы, соответствующей форме, расщепленной фурином, относительно суммарной интенсивности всех полос в каждый момент времени показана для SS1P (незакрашенные кружки, сплошная линия) и SS1-LR (незакрашенные квадраты, пунктирная линия).
Фигура 5. Добавление гибкого линкера повышает цитотоксичность SS1-LR. Клеточные линии (А) KB31 и (В) NCI-H322M инкубировали с возрастающими концентрациями SS1P (незакрашенные кружки, сплошная линия), SS1-LR (незакрашенные квадраты, пунктирная линия), SS1-LR/GGS (незакрашенные ромбы, сплошная линия) или SS1-LR/GGS R279G (закрашенные шестиугольники, без линии). Через 3 дня жизнеспособность клеток оценивали методом колориметрического анализа WST-8 и нормализовали между контрольными пробами, не обработанной и обработанной циклогексамидом. Представлены средние значения и стандартные ошибки для шести повторных измерений.
Фигура 6. Цитотоксичность SS1-LR/GGS/8M на клетках пациентов. Клетки, полученные из плевральной жидкости или асцита пациентов с мезотелиомой, высевали с увеличивающимися концентрациями RIT SS1P (белая полоса) или SS1-LR/GGS/8M (серая полоса). После 4 дней клетки фиксировали и окрашивали кристаллическим фиолетовым, чтобы обнаружить интактные клетки. Полученное поглощение при 595 нм нормализовали относительно необработанной контрольной пробы. Представлены средние значения и стандартные ошибки для трех повторных измерений. Без звездочки = p>0,05; *=p<0,05; **=p<0,01; ***=p<0,001.
Фигура 7. Действие SS1-LR/GGS/8M in vivo. А) Противоопухолевая активность SS1-LR/GGS/8M. Мышам, лишенным волосяного покрова, с ксенотрансплантатом опухоли L55 внутривенно вводили на 7, 9 и 12 сутки после имплантации буферный раствор для RIT (0,2% HSA в D-PBS; кресты, сплошная линия), 0,4 мг/кг SS1P (незакрашенные кружки, сплошная линия), или SS1-LR/GGS/8M в дозе 0,4 мг/кг (незакрашенные квадраты, пунктирная линия) или 2,5 мг/кг (закрашенные квадраты, пунктирная линия). Стрелки указывают дни, когда проводилось лечение. Размер опухоли измеряли в течение 30 дней. Точки представляют средние размеры опухоли у всех мышей в группе обработки (n=7). Планки погрешностей указывают стандартное отклонение каждого среднего значения. В) Модель синдрома повышенной проницаемости капилляров на крысах. Крысам внутривенно вводили PBS, SS1P или SS1-LR/GGS/8M, наблюдали в течение 24 часов, а затем умерщвляли. Торакальную жидкость из эвтанизированных животных собирали и измеряли. Легкие нескольких крыс фиксировали, делали срезы и окрашивали гематоксилином и эозином. С) Показаны репрезентативные фотографии при 200-кратном увеличении. D) Фармакокинетика SS1-LR/GGS/8M. Мышам линии BalbC внутривенно вводили 10 мкг либо SS1P, либо SS1-LR/GGS/8M и брали кровь через определенные промежутки времени, от 2 до 60 минут после инъекции. Концентрацию иммунотоксина в сыворотке через различные промежутки времени определяли с использованием ELISA и аппроксимировали одноэкспоненциальной функцией распада. Рассчитывали соответствующий период полураспада (t1/2). Каждая точка представляет собой концентрацию иммунотоксина в сыворотке одной мыши, и концентрацию в каждый промежуток времени определяли по меньшей мере у двух различных мышей.
Фигура 8. Антигенность SS1-LR/GGS/8M для человека. Реакционную способность SS1P и SS1-LR/GGS/8M к существовавшим ранее антителам в сыворотке человека сравнивали, используя метод вытеснения, чтобы определить концентрацию, при которой два RIT уменьшали сигнал ELISA для выявления антител сыворотки на 50% (IC50). Здесь приведены относительные значения IC50 SS1P к SS1-LR/GGS/8M. Антигенность SS1-LR/GGS/8M резко понизилась по отношению к SS1P для всех сывороток.
Фигура 9. Заключение о цитотоксичности SS1-LR/GGS/8M для клеток пациентов. Зависимость относительной жизнеспособности от лечения. Клетки, полученные из плевральной жидкости или асцита у пациентов с мезотелиомой, высевали с увеличивающейся концентрацией SS1P (белая полоса) или SS1-LR/GGS/8M (серая полоса). Через 4 дня клетки фиксировали и окрашивали кристаллическим фиолетовым, чтобы обнаружить интактные клетки. Полученное поглощение при 595 нм нормализовали относительно необработанной контрольной пробы. Представлены средние значения и стандартные ошибки для трех повторных измерений. Звездочки указывают существенные различия p<0,01 (**) или p<0,001 (***).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложен менее токсичный и менее иммуногенный вариант RIT, направленных на мезотелин, на основе RIT SS1P, направленных на мезотелин, который создан на основе РЕ. Наша первоначальная оценка SS1-LR, произведенная на основе предыдущей работы с анти-CD22 RIT HA22-LR на основе РЕ (Weldon et al., Blood 113(6):3792-3800)(2009)), показала высоковариабельную активность в отношении экспрессирующих мезотелин клеточных линий in vitro. При исследовании ксенотрансплантатов опухоли А431/K5 на мышах SS1-LR (SEQ ID NO:6 и 7) был менее активен, чем SS1P, но SS1-LR мог быть введен в намного более высоких дозах для достижения значительной регрессии опухоли. При исследовании причин его высоковариабельной активности по отношению к SS1P мы изучали интернализацию и процессинг SS1-LR и обнаружили, что доля SS1-LR, расщепленного фурином, была гораздо ниже, чем у SS1P. Это позволило предположить, что пониженное расщепление фурином может ограничивать активность SS1-LR, и мы разработали и создали несколько мутантов, чтобы проверить эту гипотезу. Добавление короткого Gly-Gly-Ser линкера после сайта расщепления фурином повысило активность SS1-LR на клеточных линиях, но, на удивление, увеличенная цитотоксичность не соответствовала увеличенному расщеплению фурином. Настоящее изобретение относится к этому удивительному открытию значимости короткого гибкого линкера для цитотоксичность конструкции RIT, направленного на мезотелин, независимо от какого-либо влияния на расщепление РЕ фурином. В дальнейшей работе в SS1-LR/GGS были включены 8 точечных мутаций, на которых было показано уменьшение иммуногенности РЕ, а затем молекулу тестировали на первичных злокачественных клетках от пациентов с мезотелиомой. Конечная молекула, SS1-LR/GGS/8M (SEQ ID NO:6 и 8) демонстрировала цитотоксичность, аналогичную цитотоксичности SS1P. Кроме того, RIT в соответствии с изобретением могут обеспечить значительно сниженную неспецифическую токсичность (например, синдром повышенной проницаемости капилляров) у млекопитающих.
При использовании in vivo мышиной модели с ксенотрансплантатной опухолью A4311K5 наблюдали примерно 20-кратное различие в противоопухолевом эффекте между SS1-LR и SS1P. Это различие нельзя полностью отнести к цитотоксичности, поскольку данные in vitro указывают на 4-кратное снижение цитотоксичности на А431/K5 клетках. Видимо, остальная часть этой разницы связана с фармакокинетическими свойствами SS1-LR у мышей. Ранее нами было показано, что HA22-LR имеет почти в 2 раза более короткий период полураспада в сыворотке у мышей, чем НА22 (7,8 против 14,6 минут соответственно), и мы предположили, что разница связана с повышенной почечной фильтрации меньшей молекулы LR (Weldon JE, Blood., 113(16):3792-800 (2009)). Изучая площадь под кривой распада, мы обнаружили, что эта разница в полураспаде предполагает приблизительно 4-кратное различие в количестве доступного белка в течение одного часа. Таким образом, различие в активности in vivo можно отнести как к пониженной цитотоксичности, так и к более короткому периоду полураспада.
Хотя SS1-LR демонстрировал in vivo более низкую противоопухолевую активность, чем SS1P, его неспецифическая токсичность была также значительно снижена у мышей. Мы воспользовались этим свойством, чтобы резко увеличить дозу SS1-LR по сравнению с SS1P (в 50 раз) при анализе ксенотрансплантатной опухоли, что привело к значительному увеличению противоопухолевого эффекта. Предыдущие эксперименты показали, что LD50 для однократного внутривенного введения SS1P составляет 1,0 мг/кг для мышей линии Balb/C (Filpula D et al., Bioconjug Chem., 18(3):773-84 (2007)) и 0,75 мг/кг для мышей линии NIH Swiss (Onda M et al., Cancer Res., 61(13):5070-7 (2001)). При использовании графика дозирования QODx3, похожего на клинический график, мыши выносили максимальную дозу 0,3 мг/кг SS1P (неопубликованные наблюдения). SS1-LR, однако, вводили в режиме QODx3 мышам с ксенотрансплантатом опухоли А431/K5 в дозе 15 мг/кг без вредного воздействия. Ранее однократное внутривенное введение дозы HA22-LR 20 мг/кг не показало никакого токсичного эффекта у мышей (Weldon JE, Blood., 113(16):3792-800 (2009)), и мы вводили однократные дозы HA22-LR вплоть до 45 мг/кг мышам, не вызывая смерти (неопубликованные наблюдения). Хотя ни одна LR молекула не была испытана клинически, этот эффект означает, что LR вариант RIT может иметь пониженную токсичность у человека, что позволило бы избежать дозолимитирующей токсичности и вводить более высокие дозы.
Хотя SS1-LR была эффективна in vitro и in vivo, мы были обеспокоены пониженной в большинстве случаев активностью по сравнению с SS1P. Одним из возможных объяснений этого несоответствия является разница во внутриклеточном механизме интоксикации. LR вариант РЕ38 содержит обширные делеции в домене II и Ib РЕ, и эти делеции могли негативно отразиться на способности РЕ к перемещению в цитозоль. Интересно, что наши первоначальные эксперименты по обнаружению полноразмерного и процессированного РЕ в лизатах клеток, обработанных SS1P и SS1-LR, показали существенное различие в количестве обработанного фурином RIT. Большая часть от общего количества RIT в клетках, обработанных SS1P, была процессирована, но только небольшая часть от общего количества RIT была процессирована в обработанных SS1-LR клетках. Этот результат означал, что плохое расщепление фурином могло ограничивать активность SS1-LR, и мы намеревались улучшить этот этап токсического действия РЕ.
Наши усилия по повышению цитотоксичности SS1-LR за счет увеличения доступности сайта расщепления фурином привели к созданию более активного RIT, но мы не смогли продемонстрировать улучшенное расщепление фурином. Добавление короткого Gly-Gly-Ser линкера (SS1-LR/GGS, Фиг.1В), более длинного линкера (SS1-LR/GGSx2, Фиг.1В) или повторение сайта расщепления фурином в окружении коротких Gly-Gly-Ser линкеров (SS1-LR/2хФурин, Фиг.1В) привело к среднему повышению цитотоксичности. Ни у одной из этих молекул, однако, не повышалась доля расщепленного фурином SS1-LR в обработанных клетках А431/K5, а также не увеличивалась скорость расщепления фурином in vitro. Мы пришли к выводу, что добавление линкера должно повышать цитотоксичность через другой механизм, возможно, связанный с внутриклеточной миграцией молекулы в тестируемых клетках.
Эти эксперименты также показали абсолютную необходимость расщепления фурином для сохранения цитотоксичности SS1P. Точечная мутация в SS1-LR/GGS, которая заменяла аргинин, необходимый для расщепления, на глицин (SS1-LR/GGS R279G, Фиг.1В), приводила к получению белка, который не расщеплялся фурином. Такой RIT не проявлял никакой активности на обеих клеточных линиях - NCI-H322M и KB31. Необходимость расщепления фурином в пути интоксикации РЕ недавно была поставлена под сомнение (Morlon-Guyot J et al., Infect Immun., 77(7):3090-9 (2009)), но существует множество доказательств того, что фурин выполняет важную роль во время интоксикации (Ornatowski W et al., J Clin Invest, 117(11):3489-97 (2007); Shiryaev SA et al., J Biol Chem., 282(29):20847-53 (2007); Sarac MS et al., Infect Immun., 70(12):7136-9 (2002); Chiron MF, Fryling CM, and FitzGerald D, J Bid Chem., 272(50):31707-11 (1997); Gu M et al., Infect Immun., 64(2):524-7 (1996); Inocencio NM, Moehring JM, and Moehring TJ, J Biol Chem., 269(50):31831-5 (1994); Moehring JM et al., J Biol Chem., 268(4):2590-4 (1993)). В случае, представленном здесь, интоксикация РЕ нарушается при отсутствии сайта, пригодного для процессинга под действием фурина. Исследования по изучению связи между расщепления фурином и цитотоксичностью продолжаются.
Отдельное направление исследований в нашей лаборатории недавно привело к созданию варианта НА22, HA22-LR-8M, который имеет чрезвычайно низкую иммуногенность из-за ликвидации эпитопов В-клеток (Onda M et al., направлено для публикации в PNAS). HA22-LR-8M содержит те же делеции, что и LR вариант РЕ, но также включает восемь точечных мутаций в домене III РЕ. Эти мутации внесли в SS1P для создания SS1-LR/GGS/8M. Единственными различиями между HA22-LR-8M и SS1-LR/GGS/8M являются антитело Fv и линкер GGS после сайта расщепления фурином. Поскольку подавляющая часть иммунного ответа на RIT направлена на РЕ, SS1-LR/GGS/8M должен обладать аналогично сниженной иммуногенностью.
Цитотоксичность SS1-LR/GGS/8M сравнивали с SS1P на первичных злокачественных клетках от пациентов с мезотелиомой, и результаты показали, что цитотоксичность SS1-LR/GGS/8M была сравнима или лучше, чем SS1P. В дополнение к хорошей активности, SS1-LR/GGS/8M имеет потенциальные преимущества перед SS1P, которые включают пониженную неспецифическую токсичность и низкую иммуногенность. Эксперименты, описанные здесь, показывают, что SS1-LR/GGS/8M был бы превосходным кандидатом для применения в клинике из-за его низкой иммуногенности, низкой неспецифической токсичности и хорошей цитотоксичности.
Определения
Единицы, приставки и символы приведены в форме, принятой в Международной системе единиц (СИ). Числовые диапазоны включают числа, определяющие диапазон. Если не указано иное, нуклеиновые кислоты записаны слева направо в ориентации от 5′ к 3′; аминокислотные последовательности записаны слева направо от N-конца к С-концу. Заголовки, приведенные здесь, не ограничивают различные аспекты или варианты осуществления изобретения, которые необходимо воспринимать со ссылкой на описание в целом. Соответственно, термины, определенные непосредственно ниже, более полно определяются в соответствии с описанием в полном объеме.
Нативный экзотоксин A Pseudomonas («РЕ») является чрезвычайно активным мономерным белком (молекулярная масса 66 кДа), выделяемым синегнойной палочкой, который ингибирует синтез белков в эукариотических клетках. Нативная последовательность РЕ приведена в SEQ ID NO:1 патента США №5602095, включенного в данный документ посредством ссылки. Способ действия заключается в инактивации фактора элонгации 2 (EF-2) путем АДФ-рибозилирования. Экзотоксин состоит из трех структурных доменов, которые действуют совместно, проявляя цитотоксичность. Домен Ia (аминокислоты 1-252) опосредует связывание с клеткой. Домен II (аминокислоты 253-364) отвечает за транслокацию в цитозоль, и домен III (аминокислоты 400-613) опосредует АДФ-рибозилирование фактора элонгации 2. В исходной структуре РЕ домен III классифицировали как остатки 405-613, а не 400-613. Allured VS, Collier RJ, Carroll SF & McKay DB, Proc Natl Acad Sci USA 83, 1320-1324 (1986). Функция домена Ib (аминокислоты 365-399) остается неопределенной, хотя большая его часть, аминокислоты 365-380, могут быть удалены без потери цитотоксичности. См. Siegall, et al., J Biol Chem 264:14256-61 (1989). Многочисленные такие модификации известны в данной области техники и включают, без ограничения, удаление домена la, различные аминокислотные делеции в доменах Ib, II и III, единичные аминокислотные замены и добавление одной или более последовательностей к карбоксильному концу, таких как KDEL (SEQ ID NO:16) и REDL (SEQ ID NO:26). См. Siegall, et al., J. Biol. Chem. 264:14256-14261 (1989). Иммунотоксины данного изобретения способны к транслокации и рибозилированию EF-2 в клетке-мишени.
Мутации РЕ описаны в данном документе с указанием аминокислотного остатка, присутствующего в конкретном положении в 613-аминокислотной последовательности нативного РЕ (SEQ ID NO:1), с последующей аминокислотой, которой этот остаток был замещен в конкретной рассматриваемой мутации. Так, например, термин «R490A» указывает, что «R» (аргинин, стандартный однобуквенный код) в положении 490 в указанной молекуле замещен на «А» (аланин, стандартный однобуквенный код), a «K590Q» означает, что лизин, обычно присутствующий в положении 590, был замещен на глутамин. Стандартный однобуквенный код для распространенных аминокислот представлен ниже.
Термин «функциональный домен III РЕ» или «функциональный домен РЕ III» относится к остаткам 395-613 нативного РЕ (нативной последовательностью является SEQ ID NO:1). Хотя структурные границы домена III установлены как остатки 405-613, функциональный анализ показал, что домен III требует сегмент домена Ib для сохранения активности АДФ-рибозилирования (Hwang, J. et al., Cell, 48:129-136 (1987); Siegall, C.B. et al., J Biol Chem, 264:14256-14261 (1989)). Функциональный домен III РЕ, таким образом, определяют остатками 395-613 из РЕ (Kihara, A. and Pastan, I., Bioconjug Chem, 5:532-538 (1994)). В данном документе последовательность функционального домена III РЕ включает в себя необязательные модификации для снижения антигенности и необязательные альтернативные последовательности удержания в эндоплазматическом ретикулуме.
Концевые остатки домена III РЕ, REDLK (SEQ ID NO:15), могут меняться так, чтобы повысить цитотоксичность RIT, получающихся в соответствии с данным изобретением. Например, иммунотоксины, полученные с мутированным окончанием РЕ в виде последовательностей KDEL (SEQ ID NO:16), REEL (SEQ ID NO:27) или RDEL (SEQ ID NO:28), могут обладать гораздо более сильным цитотоксическим действием на клетки-мишени, чем иммунотоксины, полученные из РЕ38 с нативной концевой последовательностью. См. Kreitman and Pastan, Biochem J, 307(Pt 1):29-37 (1995). Повторы этих последовательностей также могут быть использованы в данных RIT. См., например, патенты США 5854044; 5821238 и 5602095, и международную публикацию WO 99/51643. В то время как РЕ, заканчивающиеся на KDEL (SEQ ID NO:16), полезны для исследований in vitro, они могут иметь большую неспецифическую токсичность у животных и являются менее предпочтительными для применения in vivo.
Термин «мезотелин» относится к белку и его фрагментам, присутствующим на поверхности некоторых клеток человека и связываемым, например, антителом K1. Нуклеотидные и аминокислотные последовательности мезотелина приведены, например, в опубликованной заявке РСТ WO 97/25068 и в патентах США 6083502 и 6153430. См. также Chang, K. & Pastan, I., Int. J. Cancer 57:90 (1994); Chang, K. & Pastan, I., Proc. Nat′l Acad. Sci. USA 93:136 (1996); Brinkmann U., et al., Int. J. Cancer 71:638 (1997); Chowdhury, P.S., et al., Mol. Immunol. 34:9 (1997), и патент США №6809184. Мезотелин экспрессируется в виде белка-предшественника примерно 69 кДа, который затем подвергается процессингу с выделением белка 30 кДа, оставляя прикрепленным к клеточной поверхности описанный в уровне техники гликопротеин клеточной поверхности, связанный с 40 кДа гликозилфосфатидилинозитолом. 40 кДа гликопротеин является гликопротеином, который в данном документе называется термином «мезотелин». Нуклеотидные и аминокислотные последовательности мезотелина известны для нескольких биологических видов, например, человека (NM_005823.4→NP_005814.2 и NM_013404.3→NP_037536.2), мыши (NM_018857.1→NP_061345.1), крысы (NM_031658.1→NP_113846.1), коровы (NM_001100374.1→NP_001093844).
Для удобства пользования ссылками используемый в данном документе термин «антитело» включает целые антитела (иногда называемые здесь «интактные»), фрагменты антител, которые сохраняют способность к распознаванию антигена и связыванию, будучи полученными либо путем модификации целых антител либо синтезированными de novo с использованием методик на основе рекомбинантной ДНК, моноклональные антитела, поликлональные антитела и имитаторы антител, если иное не следует из контекста. Антитело может быть IgM, IgG (например, IgG1, IgG2, IgG3 или IgG4), IgD, IgA или IgE.
Последовательности константных областей подклассов IgG хорошо известны в этой области техники в течение многих лет (например, Honjo et al., Cell, 18:559-68 (1979); Tucker et al., Science, 206:1303-6 (1979); Yamawaki et al., Nature 283:786-9 (1980); Ellison et al., Nucl Acids Res 10:4071-9 (1982); Ellison et al., DNA 1:11-8 (1981); Ellison and Hood, Proc Natl Acad Sci USA 79:1984-8 (1982)). Поскольку CDR вариабельных районов определяют специфичность антител, CDR или Fv антител против поверхностного антигена клетки-мишени могут быть привиты или встроены в выбранное антитело для придания специфичности в отношении поверхностного антигена клетки-мишени этому антителу. Например, CDR антитела против поверхностного антигена клетки-мишени могут быть соединены с каркасными районами человеческого антитела с известной трехмерной структурой (см., например, W098/45322, WO 87/02671; патенты США №5859205, 5585089 и 4816567; заявка ЕР 0173494; Jones, et al. Nature 321:522 (1986); Verhoeyen, et al., Science 239:1534 (1988), Riechmann, et al. Nature 332:323 (1988); и Winter & Milstein, Nature 349:293 (1991)) с образованием антитела, которое вызовет незначительный иммуногенный ответ или вообще его не вызовет при введении в организм человека. Альтернативно, константные области антител можно сконструировать, заменив остатки, обнаруженные у животных (не у человека), таких как мыши, на остатки, обычно встречающиеся у людей. Антитела, полученные таким образом, называют «гуманизированными антителами», и они являются предпочтительными, так как характеризуются меньшим риском появления побочных эффектов и могут дольше оставаться в кровотоке. Методы гуманизации антител известны в данной области техники и изложены, например, в патентах США 6180377, 6407213, 5693762, 5585089 и 5530101.
Термин «фрагменты антител» означает молекулы, которые включают часть интактного антитела, как правило, область, связывающую антиген, или вариабельную область интактного антитела. Примеры фрагментов антител включают Fab, Fab′, F(ab′)2, и Fv фрагменты, однодоменные антитела (см., например, Wesolowski, Med Microbiol Immunol. (2009) 198(3):157-74; Saerens, et al., Curr Opin Pharmacol. (2008) 8(5):600-8; Harmsen and de Haard, Appl Microbiol Biotechnol. (2007) 77(1):13-22); антитела, стабилизированные спиралями (см., например, Arndt et al., J Mol Biol 312:221-228 (2001); диатела (см. ниже); молекулы одноцепочечных антител («scFv», см., например, патент США №5888773); антитела, стабилизированные дисульфидными связями («dsFv», см., например, патент США №5747654 и 6558672) и доменные антитела («dAb», см., например, Holt et al., Trends Biotech 21(11):484-490 (2003), Ghahroudi et al., FEBS Lett. 414:521-526 (1997), Lauwereys et al., EMBO J 17:3512-3520 (1998), Reiter et al., J. Mol. Biol. 290:685-698 (1999), Davies and Riechmann, Biotechnology, 13:475-479 (2001)).
Термин «диатела» относится к небольшим фрагментам антител с двумя антигенсвязывающими сайтами, где фрагменты содержат вариабельный домен тяжелой цепи («VH» или «VH»), соединенный с вариабельным доменом легкой цепи («VL» или «VL») в той же полипептидной цепи (VH-VL). При использовании линкера, который является слишком коротким, чтобы позволить спаривание между этими двумя доменами одной и той же цепи, домены вынуждены спариваться с комплементарными доменами другой цепи и создавать два антигенсвязывающих сайта. Диатела и их получение описано более подробно, например, в ЕР 404,097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993).
Термин «исходное антитело» означает любое антитело, представляющее интерес, которое должно быть мутировано или изменено для получения антител или их фрагментов, которые связываются с тем же эпитопом, что и исходное антитело, но с более высоким сродством.
«Направляющей группой» является часть иммуноконъюгата, предназначенная для нацеливания иммуноконъюгата на клетку, представляющую интерес. Как правило, направляющей группой является антитело или фрагмент антитела, которые сохраняют способность распознавать антиген, такие как scFv, dsFv, Fab, или F(ab′)2.
«Токсический фрагмент» - это часть иммунотоксина, которая делает иммунотоксин цитотоксичным для клеток, представляющих интерес. Что касается иммунотоксинов, которые являются предметом настоящего изобретения, токсический фрагмент представляет собой экзотоксин A Pseudomonas, который был модифицирован/мутирован для уменьшения его неспецифической цитотоксичности, как более подробно описано ниже.
Как правило, иммуноглобулин имеет тяжелую и легкую цепи. Каждая тяжелая и легкая цепь содержит константную область и вариабельную область (области также известны как «домены»). Вариабельные области легких и тяжелых цепей содержат «каркасные» участки, прерывающиеся тремя гипервариабельными областями, называемыми также «областями, определяющими комплементарность» или «CDR». Определена протяженность каркасного участка и CDR. Последовательности каркасных участков различных легких и тяжелых цепей относительно стабильны в пределах биологического вида. Каркасные участки антитела, то есть объединенные каркасные участки образующих легких и тяжелых цепей, служат для размещения и выравнивания CDR в трехмерном пространстве.
CDR в первую очередь ответственны за связывание с эпитопом антигена. CDR каждой цепи обычно называют CDR1, CDR2 и CDR3, с последовательной нумерацией, начиная с N-конца, а также обычно идентифицируют по цепи, в которой конкретная CDR находится. Так, VH CDR3 находится в вариабельном домене тяжелой цепи антитела, в котором она обнаружена, в то время как VL CDR1 является CDR1 из вариабельного домена легкой цепи антитела, в котором она обнаружена.
Ссылки на «VH» или «VH» относятся к вариабельной области тяжелой цепи иммуноглобулина, включая Fv, scFv, dsFv или Fab. Ссылки на «VL» или «VL» относятся к вариабельной области легкой цепи иммуноглобулина, включая Fv, scFv, dsFv или Fab.
Фраза «одноцепочечный Fv» или «scFv» относится к антителу, в котором вариабельные домены тяжелой цепи и легкой цепи традиционного двуцепочечного антитела объединены с образованием одной цепи. Как правило, линкерный пептид вставляют между двумя цепями для обеспечения надлежащей укладки и создания активного сайта связывания.
Фразы «дисульфидная связь» или «цистеин-цистеин дисульфидная связь» относятся к ковалентному взаимодействию между двумя остатками цистеина, в которых атомы серы цистеина окисляются с образованием дисульфидной связи. Средняя энергия дисульфидной связи составляет около 60 ккал/моль по сравнению с 1-2 ккал/моль для водородной связи.
Фраза «стабилизированный дисульфидными связями Fv» или «dsFv» относится к вариабельному фрагменту иммуноглобулина, в котором есть дисульфидная связь между легкой и тяжелой цепями. В контексте настоящего изобретения цистеины, которые образуют дисульфидную связь, находятся в каркасных участках цепей антитела и служат для стабилизации конформации антитела. Как правило, антитело сконструировано таким образом, чтобы вводить в каркасный участок цистеиновые остатки в такое положение, где замена не будет мешать связыванию антигена.
Под термином «линкерный пептид» подразумевается пептид, находящийся в пределах связывающего фрагмента антитела (например, фрагмента Fv), который служит для непрямого связывания вариабельного домена тяжелой цепи с вариабельным доменом легкой цепи.
Термин «горячая точка» означает часть нуклеотидной последовательности CDR или каркасного участка вариабельного домена, которая является местом особенно высокой естественной изменчивости. Хотя CDR сами по себе считаются участками гипервариабельности, было обнаружено, что мутации распределены неравномерно по всем CDR. Особые сайты, или горячие точки, были определены как места, в которых сконцентрированы мутации. Горячие точки характеризуются рядом структурных особенностей и последовательностей. Такие «мотивы горячих точек» могут быть использованы для идентификации горячих точек. Двумя мотивами консенсусных последовательностей, которые особенно хорошо охарактеризованы, являются последовательность тетрануклеотида RGYW и последовательность серина AGY, где R представляет собой А или G, Y представляет собой С или Т, и W представляет собой А или Т.
Антитело, иммунологически активное к конкретному антигену, может быть получено рекомбинантными методами, такими как отбор из библиотек рекомбинантных антител в фаговых или подобных векторах, см., например, Huse, et al., Science 246:1275-1281 (1989); Ward, et al., Nature 341:544-546 (1989); and Vaughan, et al., Nature Biotech. 14:309-314 (1996), или путем иммунизации животного антигеном или ДНК, кодирующей антиген.
Термин «эффекторная группа» означает часть иммуноконъюгата, предназначенную оказывать эффект на клетку-мишень, определенную направляющей группой, или идентифицировать присутствие иммуноконъюгата. В контексте настоящего изобретения эффекторная группа представляет собой модифицированный или мутированный экзотоксин A Pseudomonas.
Под термином «иммуноконъюгат» подразумевается ковалентная связь эффекторной молекулы с антителом.
Термины «эффективное количество» или «количество, эффективное для» или «терапевтически эффективное количество» относятся к дозе терапевтического агента, достаточной для получения желаемого результата, такого как ингибирование синтеза клеточного белка, по меньшей мере, на 50% или распад клетки.
В контексте настоящего изобретения токсин является мутированным экзотоксином A Pseudomonas.
Термин «контактирование» означает расположение в непосредственной физической близости.
«Экспрессионная плазмида» включает нуклеотидную последовательность, кодирующую молекулу, представляющую интерес, которая функционально связана с промотором.
Используемые в данном документе термины «полипептид», «пептид» и «белок» используются взаимозаменяемо и относятся к полимеру из аминокислотных остатков. Эти термины применимы к полимерам аминокислот, в которых один или более аминокислотных остатков представляет собой искусственный химический аналог соответствующей аминокислоты природного происхождения, а также к полимерам аминокислот, встречающихся в природе. Эти термины распространяются также на полимеры, содержащие консервативные аминокислотные замены, в результате которых белок остается функциональным.
Термин «остаток» или «аминокислотный остаток» или «аминокислота» означает аминокислоту, которая включена в белок, полипептид или пептид (вместе обозначаемым «пептид»). Аминокислота может быть встречающейся в природе аминокислотой и, если не ограничено иначе, может включать известные аналоги природных аминокислот, которые могут функционировать таким же образом, как природные аминокислоты.
Аминокислоты и аналоги, упоминаемые в данном документе, описываются сокращенными обозначениями, как следует из Таблицы А:
«Консервативная замена» при описании белка относится к изменению в аминокислотном составе белка, которое по существу не изменяет активность белка. Так, «консервативно модифицированные варианты» конкретной аминокислотной последовательности относятся к аминокислотным заменам тех аминокислот, которые не являются особо важными для активности белка, или замене аминокислот другими аминокислотами, имеющими аналогичные свойства (например, кислые, основные, положительно или отрицательно заряженные, полярные или неполярные и т.д.), так что замещение даже ключевых аминокислот существенно не отразится на активности. Таблицы консервативных замен, представляющие функционально похожие аминокислоты, хорошо известны в данной области техники. Следующие шесть групп в Таблице В содержат аминокислоты, которые являются консервативными заменами по отношению друг к другу:
Таблица В
1) Аланин (А), Серин (S), Треонин (Т);
2) Аспарагиновая кислота (D), Глутаминовая кислота (Е);
3) Аспарагин (N), Глутамин (Q);
4) Аргинин (R), Лизин (K);
5) Изолейцин (I), Лейцин (L), Метионин (М), Валин (V); и
6) Фенилаланин (F), Тирозин (Y), Триптофан (W).
Смотри также Creighton, Proteins: Structures and Molecular Properties, W.H. Freeman and Company, New York (2nd Ed., 1992).
Термины «конъюгация», «соединение», «связь» или «связывание» относятся к объединению двух полипептидов в одну непрерывную полипептидную молекулы. В контексте настоящего изобретения эти термины означают присоединение фрагмента антитела к эффекторной молекуле (ЭМ). Связь может быть создана химическими или рекомбинантными средствами. Химические средства относятся к реакции между фрагментом антитела и эффекторной молекулой, приводящей к образованию между двух молекул ковалентной связи, формирующей одну молекулу.
Используемый в данном документе термин «рекомбинантный» относится к белку, полученному с использованием клеток, которые не имеют в своем нативном состоянии эндогенной копии ДНК, способной к экспрессии белка. Клетки продуцируют рекомбинантный белок, потому что они были генетически изменены путем введения соответствующей выделенной последовательности нуклеиновых кислот. Этот термин также относится к клетке, или нуклеиновой кислоте, или вектору, которые были модифицированы путем введения гетерологичной нуклеиновой кислоты или изменения нативной нуклеиновой кислоты до формы, не являющейся нативной для этой клетки, или к тому, что клетка получена из клетки, модифицированной таким образом. Так, например, рекомбинантные клетки экспрессируют гены, которые не встречаются в нативной (нерекомбинантной) форме клетки, экспрессируют мутантные гены, которые находятся в нативной форме, или экспрессируют нативные гены, которые в других обстоятельствах эскпрессируются аномально, не в полной мере или вообще не экспрессируются.
Используемые в данном документе термины «нуклеиновая кислота» или «последовательность нуклеиновых кислот» относятся к полимерам дезоксирибонуклеотидов или рибонуклеотидов либо в одноцепочечной, либо в двухцепочечной форме, и, если не ограничено иначе, охватывают известные аналоги природных нуклеотидов, которые создают гибриды с нуклеиновыми кислотами аналогично нуклеотидам природного происхождения. Если не указано иное, конкретная последовательность нуклеиновых кислот включает комплементарную последовательность нуклеиновых кислот, а также консервативные варианты, т.е. нуклеиновые кислоты присутствуют в неоднозначных позициях кодонов и вариантов, что при трансляции белка приводит к консервативной замене аминокислоты.
Используемый в данном документе термин «кодирование» применительно к определенной нуклеиновой кислоте относится к нуклеиновым кислотам, которые содержат информацию для трансляции в специфический белок. Информация задается использованием кодонов. Как правило, аминокислотная последовательность кодируется нуклеиновой кислотой с использованием «универсального» генетического кода. Однако могут быть использованы варианты универсального кода, такие как присутствующие в митохондриях некоторых растений, животных и грибов бактерии Mycoplasma capricolum (Proc. Nat′l Acad. Sci. USA 82:2306-2309 (1985) или инфузории Macronucleus, когда нуклеиновая кислота экспрессируется с использованием трансляционного аппарата этих организмов.
Фраза «слияние в рамке» относится к соединению двух или более последовательностей нуклеиновых кислот, которые кодируют полипептиды, так что соединенная последовательность нуклеиновых кислот транслируется в одноцепочечный белок («слитый белок»), который включает первоначальные полипептидные цепи.
Используемый в данном документе термин «экспрессируемый» относится к трансляции нуклеиновой кислоты в белок. Белки могут быть экспрессированы и оставаться внутри клетки, стать составной частью поверхностной мембраны клетки или секретироваться во внеклеточный матрикс или среду.
Термин «клетка-хозяин» означает клетку, которая может поддерживать репликацию или экспрессию экспрессионного вектора. Клетки-хозяева могут быть прокариотическими клетками, такими как E.coli, или эукариотическими клетками, такими как клетки дрожжей, насекомых, амфибий или млекопитающих.
Термины «идентичный» или процент «идентичности» в контексте двух или более последовательностей нуклеиновых кислот или полипептидов относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или имеют определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми при сравнении и выравнивании для максимального соответствия, как можно измерить с помощью одного из следующих алгоритмов сравнения последовательностей или путем визуальной проверки.
Фраза «по существу идентичные» в контексте двух нуклеиновых кислот или полипептидов относится к двум или более последовательностям или подпоследовательностям, которые имеют по меньшей мере 60%, более предпочтительно 65%, еще более предпочтительно 70%, еще более предпочтительно 75%, еще более предпочтительно 80% и наиболее предпочтительно 90-95% идентичных нуклеотидных или аминокислотных остатков при сравнении и выравнивании для максимального соответствия, как измерено с помощью одного из следующих алгоритмов сравнения последовательностей или путем визуальной проверки. Предпочтительно, идентичность по существу имеет место на протяжении области последовательностей, по меньшей мере, из приблизительно 50 остатков в длину, более предпочтительно, области, по меньшей мере, из приблизительно 100 остатков, и наиболее предпочтительно, области, в которой последовательности по существу идентичны, по меньшей мере, из приблизительно 150 остатков. В наиболее предпочтительном варианте осуществления изобретения последовательности являются по существу идентичными по всей длине пептида сравнения или кодирующих областей.
Для сравнения последовательностей обычно одна последовательность выступает в качестве эталонной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и эталонную последовательности вводят в компьютер, задают координаты подпоследовательности, если необходимо, и выбирают программные параметры алгоритма. Затем, на основании заданных программных параметров алгоритм сравнения последовательностей вычисляет процент идентичности для тестируемой(ых) последовательности(ей) относительно эталонной последовательности.
Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма локальной гомологии Smith & Waterman, Adv. Appl. Math. 2:482 (1981), алгоритма гомологичного выравнивания Needleman & Wunsch, J. Mol. Biol. 48:443 (1970), методом поиска сходства Pearson & Lipman, Proc. Nat′l. Acad. Sci. USA 85:2444 (1988), с помощью компьютеризированных воплощений этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в комплекте программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wl), либо путем визуального осмотра (см. Current Protocols in Molecular Biology, F.M. Ausubel et at., eds., Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (1995 Supplement) (Ausubel)).
Примерами алгоритмов, которые пригодны для определения процента идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al. (1990) J. Mol. Biol. 215:403-410 и Altschuel et al. (1977) Nucleic Acids Res. 25:3389-3402, соответственно. Программное обеспечение для осуществления анализов BLAST является общедоступным через Национальный центр биотехнологической информации (National Center for Biotechnology Information, http://www.ncbi.nlm.nih.gov/). Этот алгоритм предполагает в первую очередь выявление сильно совпадающих пар последовательностей (HSP) путем идентификации коротких слов длины W в запрашиваемой последовательности, которые соответствуют или удовлетворяют некоторому положительному пороговому значению Т при выравнивании со словом той же длины в последовательности из базы данных. Т называют пороговым значением соседних слов (Altschul et al, см. выше). Эти первые соседние совпадающие слова выступают в качестве стартовых точек для начала поисков, чтобы найти более длинные HSP, содержащие их. Совпадение слов затем распространяется в обоих направлениях вдоль каждой последовательности до тех пор, пока общий счет выравнивания может увеличиваться. Общий счет выравнивания рассчитывают для нуклеотидных последовательностей с использованием параметров М (поощрительный балл за пару совпадающих остатков; всегда >0) и N (штрафной балл за несовпадающие остатки; всегда <0). Для последовательностей аминокислот используют матрицу замен для расчета общего балла. Расширение слов в каждом направлении прекращают, когда: общий счет выравнивания снижается на величину Х от его максимального достигнутого значения; общий счет выравнивания падает до нуля или ниже вследствие накопления одного или более отрицательных баллов при выравнивании остатков; или при достижении конца любой последовательности. Параметры алгоритма BLAST W, Т и Х определяют чувствительность и скорость выравнивания. Программа BLASTN (для нуклеотидных последовательностей) использует по умолчанию длину слова (W) 11, ожидание (Е) 10, М=5, N=-4 и сравнение обеих цепей. Для аминокислотных последовательностей программа BLASTP по умолчанию использует длину слова (W) 3, ожидание (Е) 10 и матрицу замен BLOSUM62 (см. Henikoff & Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1989)).
В дополнение к расчету процента идентичности последовательностей, алгоритм BLAST также выполняет статистический анализ сходства между двумя последовательностями (см., например, Karlin & Altschul, Proc. Nat′l. Acad. Sci. USA 90:5873-5787 (1993)). Одной мерой сходства, оцениваемой алгоритмом BLAST, является наименьшая суммарная вероятность (Р(N)), которая указывает на вероятность, с которой совпадение между двумя нуклеотидными или аминокислотными последовательностями может произойти случайно. Например, нуклеиновая кислота считается сходной с эталонной последовательностью, если наименьшая суммарная вероятность при сравнении тестируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой составляет менее чем приблизительно 0,1, более предпочтительно - менее чем приблизительно 0,01 и наиболее предпочтительно - менее чем приблизительно 0,001.
Еще один признак того, что две последовательности нуклеиновых кислот или полипептиды по существу идентичны, заключается в том, что полипептид, кодируемый первой нуклеиновой кислотой, иммунологически перекрестно реагирует с полипептидом, кодируемым второй нуклеиновой кислотой, как описано ниже. Таким образом, полипептид, как правило, по существу идентичен второму полипептиду, например, когда два пептида отличаются только консервативными заменами. Другим признаком того, что две последовательности нуклеиновых кислот по существу идентичны, является то, что две молекулы гибридизуются друг с другом в жестких условиях, как описано ниже.
Термин «in vivo» означает внутри тела организма, из которого клетка была получена. «Ex vivo» и «in vitro» означает вне тела организма, из которого клетка была получена.
Фразы «злокачественная клетка» или «злокачественность» относятся к опухолям или опухолевым клеткам, которые являются инвазивными и/или способными подвергаться метастазированию, то есть раковым клеткам.
Используемый в данном документе термин «клетки млекопитающих» включает клетки, полученные из млекопитающих, включая человека, крыс, мышей, морских свинок, шимпанзе или макак. Эти клетки могут быть культивированы in vivo или in vitro.
Термин «селективно реагирующий» по отношению к антигену означает преимущественное связывание антитела, целого или части, с несущей этот антиген клеткой или тканью, а не с клетками или тканями, у которых этот антиген отсутствует. Разумеется, определенная степень неспецифического взаимодействия может иметь место между молекулой и нецелевой клеткой или тканью. Тем не менее, селективная активность может быть охарактеризована как опосредованная через специфическое распознавание антигена. Хотя селективно реагирующие антитела связываются с антигеном, они могут делать это с низким сродством. С другой стороны, специфическое связывание приводит к гораздо более сильной ассоциации между антителом и клетками, несущими антиген, чем между связанным антителом и клетками, не содержащими антиген. Специфическое связывание обычно приводит к более чем 5-кратному, предпочтительно - более чем 10-кратному, и наиболее предпочтительно - более чем 100-кратному увеличению количества связанного антитела (в единицу времени) с клеткой или тканью, несущей антиген-мишень, по сравнению с клеткой или тканью, не имеющей антигена-мишени. Специфическое связывание с белком в таких условиях требует антитела, которое выбрано по его специфичности в отношении конкретного белка. Различные иммунологические анализы подходят для отбора антител, специфически иммунореактивных к конкретному белку. Например, твердофазный иммунологический анализ ELISA, который обычно используют для выбора моноклональных антител, специфически иммунореактивных к белку. См. Harlow & Lane, ANTIBODIES, A LABORATORY MANUAL, Cold Spring Harbor Publications, New York (1988), для описания форматов иммунологического анализа и условий, которые могут использоваться для определения специфической иммунореактивности.
Термин «иммунологически активные условия» относится к условиям, которые позволяют антителу, созданному для конкретного эпитопа, связываться с этим эпитопом в заметно большей степени, чем практически со всеми другими эпитопами, и/или по существу исключая связывание практически со всеми другими эпитопами. Иммунологически активные условия зависят от формата реакции связывания антитела и обычно являются такими условиями, что используются в протоколах иммунологических анализов, или такими условиями, что встречаются in vivo. См. Harlow & Lane, выше, для описания форматов и условий иммунологических анализов. Предпочтительно, иммунологически активными условиями, используемыми в способах по настоящему изобретению, являются «физиологические условия», которые относятся к условиям (например, температуре, осмолярности, рН), которые являются типичными в организме млекопитающего или внутри живой клетки млекопитающего. Хотя общепризнанно, что некоторые органы могут иметь экстремальные условия, среда внутри организма и внутри клеток обычно имеет рН около 7 (т.е. от рН 6,0 до рН 8,0, более типично от рН 6,5 до 7,5), содержит воду в качестве основного растворителя и имеет температуру выше 0°С и ниже 50°С. Осмолярность находится в пределах, способствующих жизнеспособности клеток и пролиферации.
Термины «пациент», «субъект», «индивидуум» взаимозаменяемы и относятся к млекопитающим, например, человеку или отличным от человека приматам, домашним млекопитающим (например, собакам или кошкам), сельскохозяйственным млекопитающим (например, крупному рогатому скоту, свиньям, овцам, лошадям), лабораторным млекопитающим (мышам, крысам, хомякам, кроликам).
Термин «совместно вводимый» относится к одновременному присутствию двух активных агентов в крови индивидуума. Активные агенты, которые вводятся совместно, могут доставляться одновременно или последовательно.
Используемые в данном документе термины «лечение» и «обработка» относятся к задержке начала, замедлению или реверсии развития, или облегчению, или предупреждению либо заболевания, либо состояния, к которому эти термины применяются, или одного или нескольких симптомов такого заболевания или состояния.
Термины «ингибирование», «снижение», «уменьшение» в отношении опухоли или роста или развития рака относятся к ингибированию роста, распространения, метастазирования опухоли или рака у субъекта, измеренному с использованием любого способа, известного в этой области техники. Рост, развитие или распространение опухоли или рака ингибируется, снижается или уменьшается, если опухолевая масса уменьшается по меньшей мере приблизительно на 10%, 20%, 30%, 50%, 80% или 100% по сравнению с опухолевой массой до введения РЕ настоящего изобретения, например, в составе химерной молекулы. В некоторых вариантах осуществления рост, развитие или распространение опухоли или рака ингибируется, снижается или уменьшается по меньшей мере примерно в 1 раз, 2 раза, 3 раза, 4 раза или более по сравнению с опухолевой массой до введения РЕ.
Компоненты рекомбинантных иммунотоксинов
А. Сайты расщепления фурином (FCS)
Сайтом расщепления фурином может быть любой полипептидный сайт, расщепляемый фурином. Как сообщается в статье Duckert et al., Protein Engineering, Design & Selection 17(1):107-112 (2004) (далее - «Duckert et al.», которая включена в данный документ посредством ссылки в полном объеме и особенно в отношении расщепляемых фурином последовательностей и мотивов, раскрытых в ней), фурин является ферментом из «семейства эволюционно стабильных специфичных к одиночным и парным основным остаткам Са2+-зависимых сериновых протеаз, называемых субтилизин/кексин-подобными конвертазами пропротеинов». Там же, с.107. Фурин, также известный как «фермент, расщепляющий белок у парных основных аминокислот» или «РАСЕ», является одним из семи белков млекопитающих из этого семейства и участвует в процессинге нескольких эндогенных белков человека. См. в широком плане, например, Thomas G, Nat Rev Mol Cell Biol, (10):753-66 (2002). Это ассоциированный с мембраной белок, содержащийся в основном в транс-Гольджи сети. Последовательность человеческого фурина известна с начала 1990-х годов. Смотри, например, Hatsuzawa, K. et al., J. Biol Chem., 267:16094-16099 (1992); Molloy, S. et al., J. Biol. Chem., 267:16396-16402 (1992).
Минимальный сайт расщепления, записанный с помощью однобуквенного кода для аминокислотных остатков, - это обычно R-X-X-R, с расщеплением после второго «R». Duckert et al. обобщили информацию о последовательностях 38 белков, описанных в литературе как имеющих сайты расщепления фурином, в том числе белков млекопитающих, белков патогенных бактерий и вирусных белков. В статье сообщается, что 31 из рассмотренных мотивов расщепления, или 81%, имели консенсусную последовательность R-X-[R/K]-R, из которых 11, или 29%, имели последовательность R-X-R-R, и 20, или 52%, были R-X-K-R. Три из мотивов расщепления содержали только минимальную последовательность расщепления. Duckert et al. затем выровняли мотивы и идентифицировали остатки, находящиеся в каждой позиции в каждом фурине, как для самого мотива расщепления, так и в окружающих остатках. Фиг.1А в статье Duckert et al. показывает в относительных величинах остатки, наиболее часто встречающиеся в каждой позиции. По традиции остатки, окружающие сайт расщепления фурином, нумеруют от неустойчивой химической связи (которая обычно обозначается символом «↓»). Считая в направлении к N-концу, остатки субстрата обозначают Р1, Р2 и так далее, в то время как, считая в направлении к C-концу, остатки обозначают Р1′, Р2′ и так далее. См., например, Rockwell, N.C., and J.W. Thorner, Trends Biochem. Sci., 29:80-87 (2004); Thomas G., Nat. Rev. Mol. Cell Biol., 3:753-766 (2002). Таким образом, следуя традиции, можно использовать следующую последовательность для выравнивания и нумерации остатков минимальной последовательности расщепления и окружающих остатков:
Р6-Р5-Р4-Р3-Р2-Р1-Р1′-Р2′-Р3′-Р4′-Р5′,
где минимальная последовательность расщепления фурином пронумерована как Р4-Р1. Согласно Duckert et al. выравнивание 38 последовательностей, расщепляемых фурином, определяет вариации, допускаемые в зависимости от остатков, присутствующих в различных положениях. Например, если остаток в Р4 не является R, это может быть компенсировано наличием остатков аргинина или лизина в Р2 и Р6. Там же, с.109.
В нативном РЕ расщепление фурином происходит между аргинином 279 и глицином 280 в богатой аргинином петле, расположенной в домене II токсина. Нативная последовательность расщепления фурином в домене II РЕ приведена ниже (с числами, указывающими положения остатков в 613-аминокислотной нативной последовательности РЕ) и выровнена, чтобы показать ее нумерацию в соответствии с приведенной выше традицией:
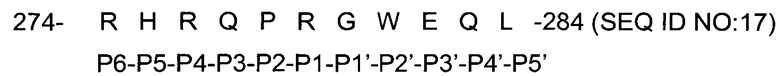
В исследованиях, лежащих в основе настоящего изобретения, были сделаны замены в положениях РЗ и Р2 для формирования следующей последовательности, с подчеркнутыми заменами:
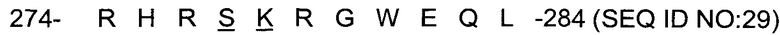 .
.
Эта последовательность показала более быстрое расщепление, чем нативная последовательность, и ее использование в образце иммунотоксина вызвало приблизительно такую же цитотоксичность в клетках-мишенях, как и использование нативной последовательности.
На основе этого и наших предыдущих исследований можно заключить, что последовательность расщепления фурином, используемая для прикрепления направляющей молекулы к домену III РЕ, может быть минимальной последовательностью расщепления фурином, R-X-X-R, или любой другой последовательностью расщепления фурином, известной в данной области или допускаемой Фиг.1А статьи Duckert et al., при условии, что, если есть остаток, присутствующий в положении, определенном как Р2′, то это должен быть триптофан или, если этот остаток не является триптофаном, он не должен быть валином или аланином. Например, в некоторых вариантах осуществления последовательность может представлять собой RKKR (SEQ ID NO:30), RRRR (SEQ ID NO:31), RKAR (SEQ ID NO:32), SRVARS (SEQ ID NO:33), TSSRKRRFW (SEQ ID NO:34) или ASRRKARSW (SEQ ID NO:35).
Как отмечено в статье Duckert et al., в положении Р4 может быть использован остаток менее предпочтительный, чем R (прежде всего валин), если это скомпенсировано остатками аргинина или лизина в положениях Р2 и Р6, так что по меньшей мере два из трех остатков в Р2, Р4 и Р6 являются основными. Так, в некоторых вариантах осуществления последовательность расщепления фурином представляет собой RRVKKRFW (SEQ ID NO:36), RNWRRDW (SEQ ID NO:37), или TRAVRRRSW (SEQ ID NO:38). Остаток в положении Р1 может быть аргинином, присутствующим в нативной последовательности, или лизином. Таким образом, лизин может быть заменен на аргинин в положении Р1, например, в любой из последовательностей, приведенных выше.
В некоторых вариантах осуществления за последовательностью расщепления фурином следует последовательность расщепления фурином РЕ: R-H-R-Q-P-R-G-W-E-Q-L (SEQ ID NO:15), или укороченный вариант нативной последовательности, при условии, что он содержит минимальную последовательность расщепления фурином и расщепляется фурином. Так, в некоторых вариантах осуществления последовательность расщепления фурином может представлять собой R-Q-P-R (SEQ ID NO:39), R-H-R-Q-P-R-G-W (SEQ ID NO:40), R-H-R-Q-P-R-G-W-E (SEQ ID NO:41), H-R-Q-P-R-G-W-E-Q (SEQ ID NO:42) или R-Q-P-R-G-W-E (SEQ ID NO:43). В некоторых вариантах осуществления последовательность представляет собой R-H-R-S-K-R-G-W-E-Q-L (SEQ ID NO:29) или укороченный вариант этой последовательности, при условии, что он содержит минимальную последовательность расщепления фурином и расщепляется фурином. Так, в некоторых вариантах осуществления последовательность расщепления фурином может представлять собой R-S-K-R (SEQ ID NO:44), R-H-R-S-K-R-G-W (SEQ ID NO:45), H-R-S-K-R-G-W-E (SEQ ID NO:46), R-S-K-R-G-W-E-Q-L (SEQ ID NO:47), H-R-S-K-R-G-W-E-Q-L (SEQ ID NO:48) или R-H-R-S-K-R (SEQ ID NO:49). Любая конкретная последовательность расщепления фурином может быть легко протестирована путем превращения ее в иммунотоксин с антителом, используемым в SS1-LR, и исследования полученного иммунотоксина in vitro на экспрессирующей мезотелин клеточной линии.
Подвержена ли какая-либо конкретная последовательность расщеплению фурином или нет, может быть определено способами, известными в данной области. Например, расщепление последовательности фурином может быть проверено путем инкубации последовательности с фурином в буфере для фурина (0,2 М NaOAc (pH 5,5), 5 мМ CaCl2) в молярном соотношении фермент: субстрат 1:10 при 25°С в течение 16 часов. Эти условия ранее были признаны оптимальными для расщепления фурином РЕ. Предпочтительно в качестве фурина использовать фурин человека. Рекомбинантный укороченный фурин человека является коммерчески доступным, например, от New England Biolabs (Beverly, MA). См. также Bravo et al., J Biol Chem, 269(14):25830-25837 (1994). Подходящие FCS также приведены в публикации патента РСТ WO 2009/032954, опубликованной 12 марта 2009 г., которая включена в настоящий документ посредством ссылки, особенно в отношении последовательностей расщепления фурином, раскрытых в ней.
В. Функциональный домен III
Подразумевается, что в структуру домена Ib включены остатки 365-399. Как обсуждается в данном документе далее, в то время как считается, что структурная граница домена III РЕ начинается с остатка 405, функциональные анализы показали, что домену III нужен сегмент структурного домена 1b, чтобы сохранить АДФ-рибозилирующую активность. Соответственно, функциональный домен III определяют как остатки 395-613 РЕ, и, таким образом, предпочтительно, чтобы токсины настоящего изобретения содержали остатки 395-613 РЕ, с некоторыми допустимыми вариантами, описанными ниже. Желательно удаление остатков 365-394, кроме тех, что содержатся в последовательности расщепления фурином, так как делеции устраняют любые иммуногенные эпитопы, присутствующие в этих частях молекулы РЕ. В РЕ настоящего изобретения последовательность расщепления фурином (или ее укороченный или модифицированный вариант) прикреплена своим карбоксильным концом к домену III, с внедренным между ними гибким линкером от 3 до 8 аминокислот, независимо выбранных из глицина и серина.
В предпочтительных вариантах осуществления функциональный домен молекул РЕ модифицирован так, чтобы заменить на аланин, глицин, серин или глутамин аминокислотные остатки, обычно присутствующие в положениях D406 и Q592 в пределах домена III. Замены в положениях D406 и Q592 можно совместить с заменами на аланин, глицин, серин или глутамин в положениях R432, R467, R490, R513, Е548 и K590 в пределах домена III. В некоторых вариантах осуществления, кроме того, по меньшей мере один аминокислотный остаток, соответствующий аминокислотному остатку в положении, выбранному из D403, R412, R427, Е431, R458, D461, R505, Е522, R538, R551, R576 и L597, заменен на аланин, глицин, серин или глутамин. Замены на эти остатки в положениях D406, R432, R467, R490, R513, Е548, K590 и Q592 домена III. В некоторых вариантах осуществления функциональный домен III РЕ является по существу идентичным или идентичным аминокислотной последовательности функционального домена РЕ SS1-LR/GGS/8M. В некоторых вариантах осуществления функциональный домен III РЕ по существу идентичен или идентичен аминокислотной последовательности функционального домена РЕ SS1-LR/GGS/8X.
Подразумевается, что последовательность нативного РЕ и варианты, описанные выше, могут иметь консервативные замены и сохранять цитотоксическую способность и, желательно, пониженную антигенность по сравнению с нативной последовательностью РЕ. В предпочтительных вариантах осуществления модифицированные варианты РЕ или их цитотоксические фрагменты имеют по меньшей мере 80% сходства последовательностей, предпочтительно - по меньшей мере 85% сходства последовательностей, более предпочтительно - по меньшей мере 90% сходства последовательностей, и наиболее предпочтительно - по меньшей мере 95% сходства последовательностей на уровне аминокислот, с представляющим интерес функциональным доменом III РЕ SS1-LR/GGS/8M или SS1-LR/GGS/8M. Публикация РСТ WO/2011/032022, опубликованная 17 марта 2011 г. и соответствующая PCT/US2010/048504, поданной 10 сентября 2010 г., раскрывает подходящие мутации, которые снижают антигенность функционального домена II РЕ. Эта опубликованная заявка включена посредством ссылки в полном объеме в отношении мутаций, замен и молекул, описанных в ней, которые обеспечивают пониженную иммуногенность функционального домена III.
Термин «консервативно модифицированные варианты» применяется как к аминокислотным, так и нуклеотидным последовательностям. Что касается конкретных нуклеотидных последовательностей, консервативно модифицированные варианты относятся к тем последовательностям нуклеиновых кислот, которые кодируют идентичные или по существу идентичные аминокислотные последовательности, или, если нуклеиновая кислота не кодирует аминокислотную последовательность, - к по существу идентичным последовательностям нуклеиновых кислот. Из-за вырожденности генетического кода любой взятый полипептид кодируется большим количеством функционально идентичных нуклеиновых кислот. Например, кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждой позиции, где находится кодон, задающий аланин, кодон может быть изменен на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие варианты нуклеиновых кислот являются «молчащими вариантами», которые являются одним из видов консервативно модифицированных вариантов. Каждая нуклеотидная последовательность, кодирующая полипептид, упомянутая здесь, также описывает каждый возможный молчащий вариант нуклеиновой кислоты. Специалисту будет понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который является обычно единственным кодоном для метионина) может быть модифицирован с образованием функционально идентичной молекулы. Соответственно, каждый молчащий вариант нуклеиновой кислоты, кодирующий полипептид, предполагается в каждой описанной последовательности.
Что касается аминокислотных последовательностей, специалисту в данной области будет понятно, что отдельные замены, делеции или добавления к последовательности нуклеиновых кислот, пептидов, полипептидов или белков, которые изменяют, добавляют или удаляют одну аминокислоту или небольшой процент аминокислот в закодированной последовательности, являются «консервативно модифицированными вариантами», в которых изменение приводит к замене аминокислоты на химически сходную аминокислоту.
Анализ цитотоксичности или антигенности РЕ
Экзотоксины Pseudomonas, используемые в изобретении, могут быть исследованы на желаемый уровень цитотоксичности с помощью анализов, хорошо известных специалистам в данной области техники. Так, цитотоксические фрагменты РЕ и консервативно модифицированные варианты таких фрагментов могут быть легко проанализированы на цитотоксичность. Большое количество молекул-кандидатов РЕ может быть одновременно проанализировано на цитотоксичность способами, хорошо известными в данной области. Например, подгруппы молекул-кандидатов могут быть проанализированы на цитотоксичность. Положительно реагирующие подгруппы молекул-кандидатов можно далее разделить и снова проанализировать, до тех пор пока желаемый(ые) цитотоксический(ие) фрагмент(ы) не будет(будут) идентифицирован(ы). Такие способы позволяют провести быстрый скрининг большого числа цитотоксических фрагментов или консервативных вариантов РЕ. Антигенность может быть проанализирована любым способом, известным в данной области, включая анализы, приведенные в WO 2007/016150.
С. Антитела к мезотелину
Направляющий компонент химерной молекулы специфически связывается с маркером клеточной поверхности мезотелином. Антигены на поверхности клеток, которые являются мишенями для химерных молекул, хорошо известны в данной области техники и приведены, например, в работах Mufson, Front Biosci (2006) 11:337-43; Frankel, Clin Cancer Res (2000) 6:326-334 и Kreitman, AAPS Journal (2006) 8(3):Е532-Е551. Злокачественные новообразования, приводимые в качестве примера, рост, распространение и/или развитие которых могут быть уменьшены или подавлены при использовании мезотелина в качестве мишени, включают рак яичников, мезотелиомы, немелкоклеточный рак легкого, аденокарциному легких, рак фаллопиевых труб, рак головы и шеи, рак шейки матки и рак поджелудочной железы. В другом предпочтительном варианте осуществления изобретения направляющая группа представляет собой фрагмент антитела, предпочтительно фрагмент антитела, специфически связывающийся с поверхностным маркером на клетке. Предпочтительный фрагмент антитела представляет собой одноцепочечный Fv. В данном документе описаны конструкции и характеристики иммунотоксинов на основе цитотоксина, в которых цитотоксин связан с scFv. Другие предпочтительные фрагменты антител, с которыми может быть соединен токсин или цитотоксический фрагмент, включают Fab, Fab′, F(ab′)2, Fv фрагмент, стабилизированное спиралями антитело, диатело, стабилизированное дисульфидными связями антитело и однодоменное антитело (например, верблюжье антитело). Антитела к мезотелину включают SS1, SSP1, HN1, HN2, MN, K1 и их варианты. MORAb-009 (гуманизированная версия SS1) является особенно подходящим антителом.
Показано, что SS1P специфически убивает экспрессирующие мезотелин клеточные линии и вызывает регрессию экспрессирующих мезотелин опухолей у мышей (Hassan, R. et al., Clin Cancer Res 8:3520-6 (2002); Onda, M. et al., Cancer Res 61:5070-7 (2001)). На основе этих исследований и соответствующих данных по безопасности в Национальном институте рака проводили 2 клинических испытания I фазы с SS1P у пациентов со злокачественными новообразованиями, экспрессирующими мезотелин (Chowdhury, P. S. et al., Proc Natl Acad Sci USA 95:669-74 (1998); Hassan, R. et al., Proc Am Soc Clin Oncol 21:29a (2002)), каждое из которых включено в данный документ посредством ссылки относительно объекта SS1P, раскрытого в них). Кроме того, другие виды терапии, направленные на мезотелин, находятся в стадии доклинической разработки (Thomas, A.M. et al., J Exp Med 200:297-306 (2004)). HN1 и HN2 являются человеческими антителами к мезотелину, описанными, например, в работе Feng, et al., Mol Cancer Ther (2009) 8(5):1113-8. Описаны иммунотоксины SS1P, в которых удалены кластеры расщепления для лизосомальных протеаз. Эти варианты описаны, например, в работе Weldon, et al., Blood, (2009) 113(16):3792-800 и в WO 2009/032954, которые включены в данный документ в полном объеме в отношении антител, FCS и функциональных доменов III, описанных в них.
RIT изобретения включают, без ограничения, молекулы, в которых имеется ковалентная связь молекулы РЕ с антителом или другим направляющим агентом. Цитотоксин соединяют с антителом или фрагментом антитела, как правило, через С-конец антитела или фрагмента антитела. Такое слияние обычно достигается с использованием методик рекомбинантной ДНК. Выбор конкретного направляющего агента зависит от конкретной клетки-мишени. Антитела, которые направляют иммунотоксин, могут быть поликлональными, моноклональными или рекомбинантными антителами, такими как химерные антитела или фрагменты вариабельных областей. Если антитело не является рекомбинантным, иммунотоксин должен быть образован путем химической конъюгации антитела с токсическим фрагментом. Если антитело получают рекомбинантным способом, антитело может быть присоединено к токсину посредством химического связывания или через рекомбинантное слияние. При рекомбинантном слиянии кДНК, кодирующая антитело, вставлена в рамке в плазмиду, которая уже содержит кДНК, которая кодирует токсин. Конечно, также может быть сделано обратное;
кДНК токсина может быть вставлена в плазмиду, несущую кДНК, которая кодирует антитело. Из-за возможного большого размера иммунотоксина иногда желательно присоединять только фрагмент антитела к токсическому фрагменту. Фрагменты Fab, Fab′ и F(ab)2 могут быть получены из поликлональных, моноклональных или химерных антител, а затем присоединены к токсину путем химического связывания. Альтернативно, может быть получена кДНК, в которой вариабельные области антитела соединены с необходимыми каркасными участками. Эти меньшие антитела затем секретируются в виде двухцепочечных антител Fv, или, если участки тяжелой и легкой цепей соединены непосредственно или через пептидный линкер, - как одноцепочечные антитела Fv (scFv). Особенно предпочтительны антитела к мезотелину и их фрагменты, в которых удалены кластеры расщепления для лизосомальных протеаз, как раскрыто в патентной публикации РСТ WO/2000/073346, опубликованной 12 июля 2000 г., которая соответствует PCT/US2009/014829, поданной 26 мая 2000 г., принадлежащей тому же заявителю, что и настоящее изобретение, и которая включена в настоящее изобретение посредством ссылки в полном объеме, в частности, в отношении объекта антитела, раскрытого в ней.
Один из способов создания scFv - это использование библиотек фагового отображения, полученных из мРНК селезенки мышей, иммунизированных иммуногеном (Chowdhury, et al., Mol. Immunol. 34:9-20 (1997)). Если белковый иммуноген в естественных условиях находится в млекопитающих, но его рекомбинантно экспрессировали в прокариотах, белок не будет иметь правильный профиль гликозилирования и может иметь неправильную конформацию. Антитела, вырабатываемые мышью в ответ на этот иммуноген, могут не распознать белок в его нативном состоянии. Одним из решений этой проблемы является иммунизация животных нативным белком, полученным в клетках млекопитающих, но очистка из клеток млекопитающих достаточного количества некоторых белков, в частности, белков клеточной поверхности, может оказаться невозможной. Другим решением, хотя и более редким, является иммунизация животных кДНК, которая кодирует иммуноген. Животному вводят кДНК под контролем соответствующего промотора. После повторных инъекций, когда титр антител достигает максимума, животных умерщвляют и, чтобы создать библиотеку фагового отображения, удаляют селезенку. Путем иммунизации мышей плазмидами, содержащими ДНК, кодирующую мезотелин, мы можем добиться высоких титров антител к мезотелину. Используя РНК из селезенки и технологию фагового отображения, можно выделить одноцепочечные Fv («scFv»), названные нами SS scFv, которые с высоким сродством связываются с мезотелином.
Антитела к мезотелину для применения в настоящем изобретении могут быть связаны с FCS через амино-конец FCS. Аналогично, FCS может быть непосредственно связан с тяжелым, легким, Fc (константной областью) или каркасным участками антитела. Связь может осуществляться через амино- или карбоксильные концы антитела, или через внутренний аминокислотный остаток. Антитела, используемые в композиции поливалентных иммуноконъюгатов настоящего изобретения, могут быть направлены на одинаковые или различные эпитопы мезотелина.
В предпочтительных вариантах осуществления настоящего изобретения антитело к мезотелину представляет собой рекомбинантное антитело, такое как scFv или стабилизированное дисульфидными связями антитело Fv. Антитела Fv обычно имеют массу около 25 кДа и содержат полный антигенсвязывающий сайт с тремя CDR в каждой тяжелой и легкой цепи. Если цепи VH и VL экспрессируют отдельно, цепи антитела Fv, как правило, соединены за счет нековалентных взаимодействий. Тем не менее, эти цепи, как правило, диссоциируют при разбавлении, поэтому были разработаны методы для сшивания цепей с помощью глутаральдегида, межмолекулярных дисульфидных связей или пептидного линкера.
В наиболее предпочтительном варианте осуществления антитело представляет собой одноцепочечный Fv (scFv). Области VH и VL антитела scFv составляют одну цепь, которая уложена таким образом, что образует антигенсвязывающий сайт, подобный тем, что имеются в двухцепочечных антителах. После укладки нековалентные взаимодействия стабилизируют одноцепочечное антитело. В более предпочтительном варианте осуществления scFv получают рекомбинантным способом. Специалисту будет понятно, что могут быть получены консервативные варианты антител настоящего изобретения. Такие консервативные варианты применительно к фрагментам scFv будут сохранять критические аминокислотные остатки, необходимые для правильной укладки и стабилизации между областями VH и VL. В некоторых вариантах осуществления настоящего изобретения антитело scFv непосредственно связано с FCS через легкую цепь.
Хотя области VH и VL антитела в некоторых вариантах осуществления могут быть непосредственно соединены вместе, специалисту будет понятно, что эти области могут быть отделены друг от друга пептидным линкером, состоящим из одной или более аминокислот. Пептидные линкеры и их применение хорошо известны в данной области. Смотри, например, работы Huston, et al., Proc. Nat′l Acad. Sci. USA 8:5879 (1988); Bird, et al., Science 242:4236 (1988); Glockshuber, et al., Biochemistry 29:1362 (1990), патент США №4946778, патент США №5132405 и Stemmer, et al., Biotechniques 14:256-265 (1993), каждый из которых включен в данный документ посредством ссылки. Как правило, пептидный линкер не имеет специфической биологической активности, кроме соединения областей или сохранения некоторого минимального расстояния или других пространственных отношений между ними. Тем не менее, составляющие пептидный линкер аминокислоты могут быть выбраны, чтобы воздействовать на некоторые свойства молекулы, такие как укладка, суммарный заряд или гидрофобность. Одноцепочечные антитела Fv (scFv) необязательно включают пептидный линкер из не более чем 50 аминокислот, обычно не более чем 40 аминокислот, предпочтительно не более чем 30 аминокислот и более предпочтительно не более чем 20 аминокислот в длину. В некоторых вариантах осуществления пептидный линкер является конкатемером последовательности Gly-Gly-Gly-Ser (SEQ ID NO:50), предпочтительно 2, 3, 4, 5 или 6 таких последовательностей. Тем не менее, следует понимать, что в пределах линкера могут быть сделаны некоторые аминокислотные замены. Например, вместо глицина может быть использован валин.
Предпочтительно, когда антитело или его фрагмент включает мутированную вариабельную область тяжелой цепи или вариабельную область легкой цепи антитела, причем полипептид имеет аффинность связывания с антигеном по меньшей мере в 5 раз выше, чем исходное антитело, полипептид имеет последовательность, которая отличается от исходного антитела заменой по меньшей мере одной аминокислоты в области, определяющей комплементарность (CDR), где аминокислота закодирована кодоном, который включает нуклеотид, принадлежащий мотиву горячей точки, выбранному из AGY или RGYW, в котором R представляет собой А или G, Y представляет собой С или Т, и W представляет собой А или Т. Замена может иметь место в CDR3 вариабельной области легкой или тяжелой цепи. Замена может быть в CDR1 или CDR2 вариабельной области легкой или тяжелой цепи. В некоторых вариантах осуществления антитело к мезотелину представляет собой антитело, описанное в патенте США №7081518, выданном 25 июля 2006 г., который включен посредством ссылки в отношении таких антител, их нуклеотидных последовательностей, применения и способов получения.
Антитело к мезотелину может содержать вариабельную тяжелую цепь («VH») и вариабельную легкую цепь («VL»), причем каждая из цепей VH и VL имеет первую, вторую и третью область, определяющую комплементарность («CDR»), где первая CDR («CDR1»), вторая CDR («CDR2») и третья CDR («CDR3») тяжелой цепи имеют соответственно последовательности аминокислотных остатков, указанную для CDR1 (GYTMN; SEQ ID NO:51), CDR2 (LITPYNGASSYNQKFRG; SEQ ID NO:52) и CDR3 (GGYDGRGFDY; SEQ ID NO:53), и где области CDR 1, 2 и 3 цепи VL имеют соответственно последовательности аминокислотные остатков, указанные для CDR1 (SASSSVSYMH; SEQ ID NO:54), CDR2 (DTSKLAS; SEQ ID NO:55) и CDR3 (QQWSGYPLT; SEQ ID NO:56). В некоторых вариантах осуществления CDR3 легкой цепи модифицирована и имеет последовательность QQWSKHPLT (SEQ ID NO:57), QQWSGHPLT (SEQ ID NO:58), QQWSAHPLT (SEQ ID NO:59), QQWSQIPLT (SEQ ID NO:60), QQWGFNPLT (SEQ ID NO:61), QQWGTNPLT (SEQ ID NO:62), QQWGSHPLT (SEQ ID NO:63), QQWGDFPLT (SEQ ID NO:64), QQWGDHPLT (SEQ ID NO:65), QQWSAHPLT (SEQ ID NO:66) или QQWSGYPTT (SEQ ID NO:67). В некоторых вариантах осуществления VH связана с VL линкерным пептидом GVGGSG4SG4S (SEQ ID NO:25). В некоторых других вариантах осуществления антитело к мезотелину представляет собой scFv, dsFv, Fab или F(ab′)2. В некоторых других вариантах осуществления изобретения антитело к мезотелину для использования в RIT содержит замену по меньшей мере одной аминокислоты в CDR, выбранной из группы, состоящей из CDR1 VL, CDR2 VL, CDR1 VH и CDR2 VH, причем эта аминокислота кодируется кодоном, который содержит нуклеотид, принадлежащий мотиву горячей точки, выбранному из AGY или RGYW, где R представляет собой А или G, Y представляет собой С или Т, и W представляет собой А или Т.
D. L1
Антитело связывается с FCS дополнительным линкером, который предпочтительно представляет собой связь или полипептид от 1 до 10 аминокислот в длину. В некоторых вариантах осуществления этот линкер имеет 1, 2, 3, 4, 5, 6, 7, 8 или 9 аминокислот в длину. В некоторых предпочтительных вариантах осуществления линкер состоит из остатков глицина и серина. В некоторых других вариантах осуществления линкер имеет последовательность ASGG (SEQ ID NO:19) или ASGGSGGG (SEQ ID NO:68). В предпочтительных вариантах осуществления линкер образует непрерывную полипептидную цепь, которая непосредственно соединяет карбоксильный конец антитела с N-концом FCS.
Е. Гибкий линкер
Гибкий линкер непосредственно соединяет FCS и функциональный домен III РЕ. Гибкий линкер является непрерывным пептидом формулы (Хаа1)n, где каждый Хаа1 независимо выбран из глицина и серина, и n составляет от 3 до 8. В предпочтительных вариантах осуществления n равно 3. В наиболее предпочтительном варианте осуществления линкер является GGS. В других вариантах осуществления n составляет 4, 5, 6 или 7. В других вариантах осуществления гибкий линкер представляет собой последовательность GGGS (SEQ ID NO:50), GGGSG (SEQ ID NO:69), GGGGSG (SEQ ID NO:70) или GGSGGS (SEQ ID NO:18).
Гибкий линкер слит в последовательности с С-концом FCS и непосредственно слит в последовательности с функциональным доменом III РЕ и, соответственно, образует одну непрерывную полипептидную цепь с FCS и функциональным доменом III.
Получение иммуноконъюгатов
I. Нерекомбинантные методы
В нерекомбинантном варианте осуществления изобретения направляющую молекулу, такую как антитело, связывают с молекулой РЕ настоящего изобретения, используя любое количество средств, известных специалистам в данной области. Могут быть применены как ковалентное, так и нековалентное связывание с молекулами РЕ настоящего изобретения.
Процедура связывания молекулы РЕ с антителом или другой направляющей молекулой («ТМ») будет изменяться в зависимости от химической структуры ТМ. Полипептиды обычно содержат разнообразные функциональные группы, например, карбоксильную группу (СООН), свободную аминогруппу (-NH2) или сульфгидрильную группу (-SH), которые доступны для реакции с подходящей функциональной группой на антителе, что приводит, например, к связыванию молекулы РЕ.
Альтернативно, антитело или другие ТМ подвергают дериватизации или к ним прикрепляют дополнительные реакционноспособные функциональные группы. Дериватизация может включать присоединение любого количества линкерных молекул, таких как те, что поставляет Pierce Chemical Company, Rockford Illinois.
«Линкер», используемый здесь, представляет собой молекулу, которая используется, чтобы присоединить ТМ к молекуле РЕ. Линкер способен образовывать ковалентные связи как с антителом, так и с эффекторной молекулой. Подходящие линкеры хорошо известны специалистам в данной области и включают, без ограничения, линкеры с прямой или разветвленной углеродной цепью, гетероциклические линкеры или пептидные линкеры. Если антитело и эффекторная молекула представляют собой полипептиды, линкеры могут быть присоединены к входящим в их состав аминокислотам через их боковые группы (например, посредством дисульфидной связи с цистеином). Однако, в предпочтительном варианте осуществления линкеры будут присоединены к амино- и карбоксильным группам концевых аминокислот у альфа-атомов углерода.
В некоторых случаях желательно освободить молекулу РЕ от ТМ, когда иммуноконъюгат достигнет своего сайта-мишени. Тогда в этих условиях иммуноконъюгаты будут содержать связи, которые расщепляются в непосредственной близости от сайта-мишени. Расщепление линкера, чтобы освободить молекулу РЕ от ТМ, может быть вызвано ферментативной активностью или условиями, которым подвергается иммуноконъюгат либо внутри клетки-мишени, либо в непосредственной близости от сайта-мишени. Когда сайтом-мишенью является опухоль, может быть использован линкер, который расщепляется в условиях, существующих в месте локализации опухоли (например, при воздействии опухолеассоциированных ферментов или кислых значений рН).
II. Рекомбинантные методы
Последовательности нуклеиновых кислот настоящего изобретения могут быть получены любым подходящим способом, включая, например, клонирование соответствующих последовательностей или прямой химический синтез с помощью таких методов, как фосфотриэфирный метод, описанный Narang, et al., Meth. Enzymol. 68:90-99 (1979); фосфодиэфирный метод, описанный Brown, et al., Meth. Enzymol. 68:109-151 (1979); диэтилфосфорамидитный метод, описанный Beaucage, et al., Tetra. Lett. 22:1859-1862 (1981), твердофазный фосфорамидиттриэфирный метод, описанный Beaucage & Caruthers, Tetra. Letts. 22(20):1859-1862 (1981), например, с использованием автоматизированного синтезатора, как описано, например, Needham-VanDevanter, et al. Nucl. Acids Res. 12:6159-6168 (1984); и метод с твердой подложкой, описанный в патенте США №4458066. Химический синтез дает одноцепочечный олигонуклеотид. Он может быть преобразован в двухцепочечную ДНК путем гибридизации с комплементарной последовательностью или путем полимеризации с участием ДНК-полимеразы при использовании одной цепи в качестве матрицы. Специалисту будет понятно, что, так как химический синтез ДНК ограничен последовательностью из приблизительно 100 оснований, более длинные последовательности могут быть получены путем лигирования коротких последовательностей.
В предпочтительном варианте осуществления последовательности нуклеиновых кислот по настоящему изобретению получают с помощью методов клонирования. Примеры подходящих методов клонирования и секвенирования, а также достаточные инструкции для специалистов в данной области, осуществляющих эксперименты по клонированию, имеются в Sambrook, et al., Molecular Cloning: A Laboratory Manual (2nd Ed.), Vol.1-3, Cold Spring Harbor Laboratory (1989)), Berger and Kimmel (eds.), Guide to Molecular Cloning Techniques, Academic Press, Inc., San Diego CA (1987)), или Ausubel, et al. (eds.), Current Protocols in Molecular Biology, Greene Publishing and Wiley-Interscience, NY (1987). Данные о продукте от производителей биологических реагентов и экспериментального оборудования также предоставляют полезную информацию. К таким производителям относятся химическая компания SIGMA (Сент-Луис, Миссури), R&D systems (Миннеаполис, Миннесота), Pharmacia LKB Biotechnology (Пискатауэй, Нью-Джерси), CLONTECH Laboratories, Inc. (Пало-Альто, Калифорния), Chem Genes Corp., химическая компания Aldrich (Милуоки, Висконсин), Glen Research, Inc., GIBCO BRL Life Technologies, Inc. (Гейтерсберг, Мэриленд), Fluka Chemica-Biochemika Analytika (Fluka Chemie AG, Букс, Швейцария), Invitrogen, Сан-Диего, Калифорния, и Applied Biosystems (Фостер-Сити, Калифорния), а также многие другие коммерческие источники, известные специалистам.
Нуклеиновые кислоты, кодирующие нативный РЕ, также могут быть модифицированы с образованием иммуноконъюгатов согласно настоящему изобретению. Модификация путем сайт-направленного мутагенеза хорошо известна в данной области. Нуклеиновые кислоты, кодирующие РЕ, могут быть амплифицированы с помощью методов in vitro. Методы амплификации включают полимеразную цепную реакцию (PCR), лигазную цепную реакцию (LCR), транскрипционную систему амплификации (TAS), самоподдерживаемую систему репликации последовательности (3SR). Широкое разнообразие методов клонирования, клеток-хозяев и методов амплификации in vitro хорошо известно специалистам в данной области.
В предпочтительном варианте осуществления иммуноконъюгаты получают путем вставки кДНК, которая кодирует антитело или другую ТМ по выбору, в вектор, который содержит кДНК, кодирующую желаемый РЕ настоящего изобретения. Вставку выполняют таким образом, чтобы направляющий агент (для простоты обсуждения в этом документе примем, что направляющий агент представляет собой Fv, хотя другие направляющие агенты могут быть использованы с равным эффектом) и РЕ считывались в рамке, то есть в виде одного непрерывного полипептида, который содержит функциональную область Fv и функциональную область РЕ. В особенно предпочтительном варианте осуществления кДНК, кодирующую РЕ изобретения, лигируют с scFv так, чтобы токсин находился на карбоксильном конце scFv. В других предпочтительных вариантах осуществления кДНК, кодирующую РЕ изобретения, лигируют с scFv так, чтобы токсин находился на аминоконце scFv.
Как только нуклеиновые кислоты, кодирующие РЕ, антитело или иммуноконъюгат согласно настоящему изобретению, выделяют и клонируют, можно экспрессировать желаемый белок в рекомбинантно сконструированные клетки, такие как клетки бактерий, растений, дрожжей, насекомых и клетки млекопитающих. Предполагается, что специалисты в данной области осведомлены о многочисленных системах экспрессии, пригодных для экспрессии белков, в том числе в E.coli, других бактериях, дрожжах и различных высших эукариотических клетках, таких как COS, CHO, HeLa и линии клеток миеломы. Не будет попыток подробно описывать различные известные методы экспрессии белков в клетках прокариот или эукариот. Если коротко, экспрессия природных или синтетических нуклеиновых кислот, кодирующих выделенные белки настоящего изобретения, обычно достигается путем функционального связывания ДНК или кДНК с промотором (который является либо конститутивным, либо индуцибельным), с последующим включением конструкции в состав экспрессионной кассеты. Кассеты могут быть пригодны для репликации и интеграции в клетки прокариот или эукариот. Типичные кассеты экспрессии содержат терминаторы транскрипции и трансляции, инициирующие последовательности и промоторы, нужные для регуляции экспрессии ДНК, кодирующей белок. Для получения высокого уровня экспрессии клонированного гена желательно создать кассеты экспрессии, которые содержат, как минимум, сильный промотор для направления транскрипции, сайт связывания рибосомы для инициации трансляции и терминатор транскрипции/трансляции. Для E.coli кассета включает промотор, такой как Т7, trp, lac или лямбда-промоторы, сайт связывания рибосомы и предпочтительно сигнал терминации транскрипции. Для эукариотических клеток контролирующие последовательности могут включать промотор и предпочтительно энхансер, полученный из генов иммуноглобулинов, SV40, цитомегаловируса, и последовательность полиаденилирования, и могут включать последовательности донора и акцептора сплайсинга. Кассеты согласно изобретению могут быть перенесены в выбранную клетку-хозяина хорошо известными методами, такими как трансформация с помощью хлорида кальция или электропорация для E.coli и обработка с фосфатом кальция, электропорация или липофекция для клеток млекопитающих. Клетки, трансформированные кассетами, могут быть отсортированы по резистентности к антибиотикам, предоставляемой генами, содержащимися в кассетах, такими как гены amp, gpt, neo и hyg.
Специалисту будет понятно, что могут быть сделаны модификации нуклеиновой кислоты, кодирующей полипептид согласно настоящему изобретению (т.е. РЕ или иммуноконъюгат, образованный из РЕ по изобретению), без снижения его биологической активности. Некоторые модификации могут быть сделаны, чтобы облегчить клонирование, экспрессию или включение направляющей молекулы в белок слияния. Такие модификации хорошо известны специалистам в данной области и включают, например, кодоны терминации, метионин, добавленный на аминоконце, чтобы обеспечить сайт инициации, дополнительные аминокислоты, размещенные по обоим концам последовательности, чтобы создать удобно расположенные сайты рестрикции, или дополнительные аминокислоты (такие как поли-His), чтобы содействовать стадиям очистки.
В дополнение к рекомбинантным методам иммуноконъюгаты и РЕ по настоящему изобретению также могут быть сконструированы полностью или частично с помощью стандартного пептидного синтеза. Твердофазный синтез полипептидов по настоящему изобретению из менее чем приблизительно 50 аминокислот в длину может быть осуществлен путем присоединения C-концевой аминокислоты последовательности к нерастворимой подложке с последующим последовательным добавлением остальных аминокислот в последовательность. Методы твердофазного синтеза описаны Barany & Merrifield, THE PEPTIDES: ANALYSIS, SYNTHESIS, BIOLOGY. VOL. 2: SPECIAL METHODS IN PEPTIDE SYNTHESIS, PART A. pp.3-284; Merrifield, et al. J. Am. Chem. Soc. 85:2149-2156 (1963), and Stewart, et al., SOLID PHASE PEPTIDE SYNTHESIS, 2ND ED., Pierce Chem. Co., Rockford, III. (1984). Белки большей длины могут быть синтезированы путем конденсации амино- и карбоксильного концов коротких фрагментов. Способы образования пептидных связей активацией карбоксильной концевой группы (например, путем использования связующего реагента N,N′-дициклогексилкарбодиимида) известны специалистам в данной области.
III. Очистка
Сразу после экспрессии рекомбинантные иммуноконъюгаты и РЕ по настоящему изобретению могут быть очищены в соответствии со стандартными процедурами, известными в данной области, в том числе осаждением сульфатом аммония, аффинной колоночной хроматографией, колоночной хроматографией и т.п. (см., в общем, R. Scopes, PROTEIN PURIFICATION, Springer-Verlag, N.Y. (1982)). Предпочтительны по существу чистые композиции с гомогенностью по меньшей мере приблизительно от 90 до 95%, и для фармацевтических целей наиболее предпочтительны композиции с гомогенностью от 98 до 99% или более. Затем полипептиды, очищенные частично или до гомогенности, по желанию, в случае терапевтического использования, должны фактически быть освобождены от эндотоксина.
Методы экспрессии одноцепочечных антител и/или рефолдинга в соответствующую активную форму, в том числе одноцепочечных антител, из бактерий, таких как E.coli, описаны и хорошо известны и применимы к антителам настоящего изобретения. См. Buchner, et al., Anal. Biochem. 205:263-270 (1992); Pluckthun, Biotechnology 9:545 (1991); Huse, et al., Science 246:1275 (1989) и Ward, et al., Nature 341:544 (1989), которые включены в данный документ посредством ссылки.
Зачастую функциональные гетерологичные белки из E.coli или других бактерий выделяют из телец включения, после чего они требуют растворения с использованием сильных денатурирующих агентов и последующего рефолдинга. На этапе солюбилизации, как хорошо известно в данной области, для разрушения дисульфидных связей должен присутствовать восстанавливающий агент. Буфер с восстанавливающим агентом, приводимый в качестве примера, содержит 0,1 М Трис с рН 8, 6 М гуанидина, 2 мМ ЭДТА, 0,3 М DTE (дитиоэритритол). Повторное окисление дисульфидных связей может происходить в присутствии низкомолекулярных тиоловых реагентов в восстановленной и окисленной форме, как описано в Saxena, et al., Biochemistry 9:5015-5021 (1970), включенной в данный документ посредством ссылки, и особенно, как описано в Buchner, et al., см. выше.
Ренатурацию обычно осуществляют путем разбавления (например, в 100 раз) денатурированного и восстановленного белка в буфере для рефолдинга. Буфер, приводимый в качестве примера, содержит 0,1 М Трис, рН 8,0, 0,5 М L-аргинина, 8 мМ окисленный глутатиона и 2 мМ EDTA.
В качестве модификации к протоколу очистки двухцепочечного антитела области тяжелой и легкой цепей отдельно растворяют и восстанавливают, а затем объединяют в растворе для рефолдинга. Предпочтительный выход получают, когда эти два белка смешивают в молярном отношении таким образом, чтобы не был превышен 5-кратный молярный избыток одного белка над другим. Желательно добавить в раствор для рефолдинга избыток окисленного глутатиона или других окисляющих низкомолекулярных соединений после завершения окислительно-восстановительной перестановки.
2. Фармацевтические композиции и введение
В одном аспекте настоящего изобретения предложена фармацевтическая композиция или лекарственный препарат, содержащие по меньшей мере один химерный белок по настоящему изобретению, предпочтительно токсин направленного действия, и, необязательно, фармацевтически приемлемый носитель. Фармацевтическую композицию или лекарственный препарат можно вводить пациенту для лечения состояния, включающего, без ограничения, злокачественное заболевание или рак.
а. Получение композиций
Фармацевтические композиции или лекарственные препараты для использования в настоящем изобретении могут быть получены с помощью стандартных методик с использованием одного или более физиологически приемлемых носителей или наполнителей. Подходящие фармацевтические носители описаны в данном документе и в Remington: The Science and Practice of Pharmacy, 21st Ed., University of the Sciences in Philadelphia, Lippencott Williams & Wilkins (2005). Химерные белки по настоящему изобретению могут быть получены для введения любым подходящим способом, в том числе посредством ингаляции, местно, интраназально, перорально, парентерально или ректально. Так, введение фармацевтической композиции может быть осуществлено путем интрадермального, субдермального, внутривенного, внутримышечного, интраназального, ингаляционного, интрацеребрального, внутритрахеального, внутриартериального, внутрибрюшинного, внутрипузырного, внутриплеврального, интракоронарного, подкожного введения или инъекции в опухоль, с помощью шприца или других устройств. Также предполагается трансдермальное введение, как и ингаляционное или аэрозольное введение. Таблетки и капсулы могут быть введены перорально, ректально или вагинально.
Композиции для введения обычно содержат раствор химерного белка, предпочтительно токсина направленного действия, растворенного в фармацевтически приемлемом носителе, предпочтительно в водном носителе. Могут быть использованы различные водные носители, например, буферный солевой раствор и тому подобное. Эти растворы являются стерильными и обычно не содержат нежелательных примесей. Эти композиции могут быть стерилизованы с помощью обычных, хорошо известных способов стерилизации. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, необходимые для создания физиологических условий, такие как регулирующие рН и буферные вещества, агенты, регулирующие токсичность и т.д., например, ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция, лактат натрия и тому подобное. Концентрация слитого белка в этих препаратах может широко варьировать, и будет выбрана в первую очередь на основе объемов жидкости, вязкостей, массы тела и т.д. в соответствии с конкретным выбранным способом введения и потребностями пациента.
Композиции токсинов направленного действия по данному изобретению пригодны для парентерального введения, в том числе внутривенного введения или введения в полость тела.
Химерные белки, предпочтительно токсины направленного действия, по настоящему изобретению могут быть введены в композиции для парентерального введения путем инъекции, например, болюсной инъекции или непрерывной инфузии. Композиции для инъекций могут быть представлены в единичной дозированной форме, например, в ампулах или в многодозовых контейнерах, с добавлением консерванта. Композиции для инъекций предпочтительно представляют собой водные изотонические растворы или суспензии, а суппозитории предпочтительно получают из жирных эмульсий или суспензий. Композиции могут быть стерилизованы и/или содержать вспомогательные вещества, такие как консерванты, стабилизаторы, смачивающие или эмульгирующие агенты, вещества, способствующие растворению, соли для регулирования осмотического давления и/или буферы. Альтернативно, активный ингредиент может быть в форме порошка для разведения перед применением подходящим носителем, например, стерильной апирогенной водой. Кроме того, композиции также могут содержать другие терапевтически полезные вещества. Композиции получают обычным смешиванием, гранулированием или нанесением покрытия, соответственно, и они содержат приблизительно от 0,1 до 75% активного ингредиента, предпочтительно приблизительно от 1 до 50% активного ингредиента.
Парентеральные композиции токсина направленного действия по настоящему изобретению с контролируемым высвобождением могут быть получены в виде имплантатов, масляных инъекций или систем, состоящих из частиц. Для широкого представления о системах доставки белков см. Banga, A.J., THERAPEUTIC PEPTIDES AND PROTEINS: FORMULATION, PROCESSING, AND DELIVERY SYSTEMS, Technomic Publishing Company, Inc., Lancaster, PA (1995), приведенную в данном документе посредством ссылки. Системы, состоящие из частиц, включают микросферы, микрочастицы, микрокапсулы, нанокапсулы, наносферы и наночастицы. Микрокапсулы содержат терапевтический белок в качестве центрального ядра. В микросферах терапевтический агент диспергирован по всему объему частицы. Частицы, микросферы и микрокапсулы размером меньше чем приблизительно 1 мкм обычно называют наночастицами, наносферами и нанокапсулами, соответственно. Капилляры имеют диаметр приблизительно 5 мкм, так что только наночастицы вводят внутривенно. Микрочастицы, как правило, имеют около 100 мкм в диаметре, и их вводят подкожно или внутримышечно. См., например Kreuter J., COLLOIDAL DRUG DELIVERY SYSTEMS, J. Kreuter, ed., Marcel Dekker, Inc., New York, NY, pp.219-342 (1994); и Tice & Tabibi, TREATISE ON CONTROLLED DRUG DELIVERY, A. Kydonieus, ed., Marcel Dekker, Inc. New York, NY, pp.315-339 (1992), обе публикации включены в данный документ посредством ссылки.
Для ионно-контролируемого высвобождения композиций с токсином направленного действия по настоящему изобретению могут быть использованы полимеры. В данной области известны различные разлагаемые и неразлагающиеся полимерные матрицы, применяемые для контролируемой доставки лекарственных средств (Langer R., Accounts Chem. Res., 26:537-542 (1993)). Например, блок-сополимер полаксамер 407 существует в виде вязкой, но еще подвижной жидкости при низких температурах, но образует полутвердый гель при температуре тела. Показано, что он является эффективным носителем для получения композиций и устойчивой доставки рекомбинантного интерлейкина-2 и уреазы (Johnston et al., Pharm. Res., 9:425-434 (1992); and Pec et al., J. Parent. Sci. Tech., 44(2):58 65 (1990)). В качестве микроносителя для контролируемого высвобождения белков альтернативно был использован гидроксиапатит (Ijntema et al., Int. J. Pharm., 112:215-224 (1994)). В еще одном аспекте для контролируемого высвобождения, а также направленной доставки инкапсулированного в липиды лекарственного средства используют липосомы (Betageri et al., LIPOSOME DRUG DELIVERY SYSTEMS, Technomic Publishing Co., Inc., Lancaster, PA (1993)). Известны многочисленные дополнительные системы для контролируемой доставки терапевтических белков. См., например, патенты США №5055303, 5188837, 4235871, 4501728, 4837028, 4957735, 5019369, 5055303, 5514670, 5413797 5268164, 5004697, 4902505, 5506206, 5271961, 5254342 и 5534496, каждый из которых включен в данный документ посредством ссылки.
Подходящие для трансдермального введения составы содержат эффективное количество композиции по настоящему изобретению с носителем. Предпочтительные носители включают абсорбируемые фармакологически приемлемые растворители, способствующие проникновению через кожу хозяина. Например, трансдермальные устройства в виде бандажа включают опорный элемент, резервуар, содержащий композицию, необязательно, с носителями, необязательно, контролирующий скорость барьер для доставки композиции на кожу хозяина с контролируемой и предварительно определенной скоростью в течение продолжительного периода времени, и средства для закрепления устройства на коже. Также могут быть использованы матричные трансдермальные рецептуры.
Составы, подходящие для местного применения, например, для нанесения на кожу и глаза, предпочтительно представляют собой водные растворы, мази, кремы или гели, хорошо известные в данной области. Они могут содержать солюбилизаторы, стабилизаторы, агенты, повышающие тоничность, буферы и консерванты.
Для перорального введения фармацевтическая композиция или лекарственный препарат может иметь форму, например, таблетки или капсулы, полученных известными способами с фармацевтически приемлемым наполнителем. Предпочтительными являются таблетки и желатиновые капсулы, включающие активный ингредиент, т.е. композицию по настоящему изобретению, вместе с (а) разбавителями или наполнителями, такими как лактоза, декстроза, сахароза, маннит, сорбит, целлюлоза (например, этилцеллюлоза, микрокристаллическая целлюлоза), глицин, пектин, полиакрилаты и/или гидрофосфат кальция, сульфат кальция, (b) смазывающими материалами, такими как диоксид кремния, тальк, стеариновая кислота, ее магниевая или кальциевая соль, стеараты металлов, коллоидный диоксид кремния, гидрогенизированное растительное масло, кукурузный крахмал, бензоат натрия, ацетат натрия и/или полиэтиленгликоль; для таблеток также (с) связующими веществами, такими как алюмосиликат магния, крахмальная паста, желатин, трагакант, метилцеллюлоза, натрий-карбоксиметилцеллюлоза, поливинилпирролидон и/или гидроксипропилметилцеллюлоза; при желании (d) разрыхляющими веществами, такими как крахмалы (например, картофельный крахмал или натрия карбоксиметилкрахмал), агар, альгиновая кислота или ее натриевая соль, или шипучие смеси; (е) смачивающими агентами, такими как лаурилсульфат натрия, и/или (f) абсорбентами, красителями, ароматизаторами и подсластителями.
Таблетки могут быть покрыты либо пленкой, либо энтеросолюбильной оболочкой способами, известными в данной области. Жидкие препараты для перорального введения могут быть в форме, например, растворов, сиропов или суспензий, или они могут быть представлены в виде сухого продукта для смешивания с водой или другим подходящим носителем перед использованием. Такие жидкие препараты могут быть получены обычными способами с фармацевтически приемлемыми добавками, например, суспендирующими агентами, например, сиропом сорбита, производными целлюлозы или гидрогенизированных пищевых жиров; эмульгирующими агентами например, лецитином или гуаровой камедью; неводными носителями, например, миндальным маслом, жирными сложными эфирами, этиловым спиртом или фракционированными растительными маслами; и консервантами, например, метил- или пропил-п-гидроксибензоатами или сорбиновой кислотой. Препараты также могут содержать буферные соли, ароматизирующие, красящие и/или подслащивающие агенты при необходимости. Если желательно, препараты для перорального введения могут быть составлены таким образом, чтобы обеспечивать контролируемое высвобождение активной субстанции.
Для введения путем ингаляции химерный белок, предпочтительно антитело и/или токсин направленного действия, может доставляться в виде аэрозольного спрея из упаковок под давлением или распылителя с применением подходящего пропеллента, например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, 1,1,1,2-тетрафторэтана, двуокиси углерода или другого подходящего газа. В случае аэрозоля под давлением единичная доза может быть определена с помощью клапана для доставки отмеренного количества. Капсулы и картриджи, например, из желатина, для использования в ингаляторе или инсуффляторе могут содержать порошкообразную смесь химерного белка, предпочтительно антитела и/или токсина направленного действия, и подходящей порошковой основы, например, лактозы или крахмала.
Композиции также могут быть составлены в виде ректальных композиций, например, суппозиториев или удерживающих клизм, например, содержащих обычные основы для суппозиториев, например, масло какао или другие глицериды.
Кроме того, композиции могут быть составлены в виде депо-препарата. Такие длительно действующие композиции можно вводить имплантацией (например, подкожно или внутримышечно) или путем внутримышечной инъекции. Так, например, композиция может быть составлена с подходящими полимерными или гидрофобными материалами (например, в виде эмульсии в приемлемом масле) или ионообменными смолами, или в виде умеренно растворимых производных, например, в виде умеренно растворимой соли.
При желании композиции могут быть представлены в упаковке или дозирующем устройстве, которое может содержать одну или более единичных дозированных форм, содержащих активный ингредиент. Упаковка может, например, содержать металлическую или пластиковую фольгу, например, блистерную упаковку. Упаковка или дозирующее устройство может сопровождаться инструкциями по применению.
b. Дозировка
В одном варианте осуществления настоящего изобретения фармацевтическую композицию или лекарственный препарат вводят пациенту в терапевтически эффективной дозе для профилактики, лечения или контроля заболевания или злокачественного состояния, такого как рак. Фармацевтическую композицию или лекарственный препарат вводят пациенту в количестве, достаточном, чтобы вызвать эффективный терапевтический или диагностический ответ у пациента. Эффективный терапевтический или диагностический ответ - это ответ, который по меньшей мере частично подавляет или замедляет симптомы или осложнения болезни или злокачественного состояния. Количество, соответствующее достижению этой цели, определяется как «терапевтически эффективная доза».
Дозировка химерных белков, предпочтительно вводимых токсинов направленного действия или композиций, зависит от вида теплокровного животного (млекопитающего), массы тела, возраста, индивидуального состояния, площади поверхности зоны, которая подлежит обработке, и от формы введения. Размер дозы также будет определяться наличием, природой и степенью любых негативных последствий, которые сопровождают введение конкретной композиции определенному субъекту. Стандартная лекарственная доза для введения млекопитающему массой приблизительно от 50 до 70 кг может содержать приблизительно от 5 до 500 мг активного ингредиента. Как правило, дозой композиции по настоящему изобретению является доза, достаточная для достижения желаемого эффекта.
Оптимальные схемы дозирования могут быть рассчитаны из измерений накопления химерного белка, предпочтительно токсина направленного действия, в организме субъекта. В большинстве случаев доза составляет от 1 нг до 1000 мг на кг массы тела и может вводиться один или несколько раз в день, неделю, месяц или год. Специалисты в данной области могут легко определить оптимальные дозировки, методики дозирования и частоту повторения. Специалист в данной области сможет определить оптимальную дозировку для введения химерного белка, предпочтительно токсина направленного действия, человеку в соответствии с установленными протоколами, известными в данной области, и описанием, приведенным в данном документе.
Оптимальные дозировки, токсичность и терапевтическая эффективность композиций могут меняться в зависимости от относительной эффективности отдельных композиций и могут быть определены с помощью стандартных фармацевтических процедур на клеточных культурах или на экспериментальных животных, например, путем определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной для 50% популяции). Соотношение между токсическим и терапевтическим эффектами представляет собой терапевтический индекс и может быть выражено как отношение LD50/ED50. Предпочтительными являются композиции, которые обладают большими терапевтическими индексами. В тех случаях, когда могут быть использованы композиции, демонстрирующие токсические побочные эффекты, следует уделить внимание разработке системы доставки, которая бы направляла такие композиции точно в область локализации пораженной ткани, чтобы минимизировать возможность повреждения нормальных клеток и тем самым уменьшить побочные эффекты.
Данные, полученные в результате, например, исследований на животных (например, грызунах и обезьянах) могут использоваться для определения диапазона доз, назначаемых людям. Дозировка композиций по настоящему изобретению лежит предпочтительно в пределах диапазона концентраций в кровотоке, который включает ED50 при небольшой токсичности или отсутствии токсичности. Дозировка может меняться в пределах этого диапазона в зависимости от применяемой дозированной формы и способа введения. Для применения любой композиции в способах согласно данному изобретению терапевтически эффективная доза может быть оценена первоначально из анализов на клеточных культурах. Доза может быть определена на моделях животных, чтобы получить диапазон концентраций в циркулирующей плазме, который включает IC50 (концентрацию тестируемого соединения, которая обеспечивает полумаксимальное ингибирование симптомов), как определено в клеточной культуре. Такая информация может быть использована для более точного определения полезных доз для человека. Уровни в плазме могут быть измерены, например, с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). Как правило, эквивалент дозы химерного белка, предпочтительно токсина направленного действия, составляет приблизительно от 1 нг/кг до 100 мг/кг для среднестатистического субъекта.
Стандартная доза композиции токсина направленного действия согласно настоящему изобретению для внутривенного введения составляет приблизительно от 0,1 до 10 мг на пациента в день. Могут быть использованы дозы от 0,1 до приблизительно 100 мг на пациента в день. Существующие в настоящее время методы получения вводимых композиций будут известны или очевидны специалистам в данной области, и описаны более подробно в таких публикациях как Remington: The Science and Practice of Pharmacy, 21st Ed., University of the Sciences in Philadelphia, Lippencott Williams & Wilkins (2005).
Приводимые в данном документе в качестве примеров дозы композиций включают миллиграммовые или микрограммовые количества композиции на килограмм массы субъекта или образца (например, от приблизительно 1 мкг на килограмм до приблизительно 500 миллиграммов на килограмм, от приблизительно 100 микрограммов на килограмм до приблизительно 5 мг на килограмм, или от приблизительно 1 микрограмма на килограмм до приблизительно 50 микрограммов на килограмм). Кроме того, понятно, что соответствующие дозы композиции зависят от активности композиции по отношению к желаемому эффекту, который должен быть достигнут. Когда одну или более из этих композиций должна быть введена млекопитающему, врач, ветеринар или исследователь может, например, предписать относительно низкую дозу в начале, затем увеличить дозу, пока не будет получен соответствующий ответ. Кроме того, следует понимать, что специфическая величина дозы для какой-либо отдельной особи млекопитающего будет зависеть от множества факторов, в том числе активности конкретной применяемой композиции, возраста, массы тела, общего состояния здоровья, пола и диеты субъекта, времени введения, пути введения, скорости экскреции, комбинации лекарственных средств и степени экспрессии или активности, которые необходимо модулировать.
В одном варианте осуществления настоящего изобретения фармацевтическую композицию или лекарственный препарат, содержащие химерный белок, предпочтительно токсин направленного действия, по настоящему изобретению вводят, например, в суточной дозе в интервале от приблизительно 1 мг композиции на килограмм массы субъекта (1 мг/кг) до приблизительно 1 г/кг. В другом варианте осуществления доза представляет собой дозу в диапазоне от приблизительно 5 мг/кг до приблизительно 500 мг/кг. В еще одном варианте осуществления доза составляет от приблизительно 10 мг/кг до приблизительно 250 мг/кг. В другом варианте осуществления доза составляет от приблизительно 25 мг/кг до приблизительно 150 мг/кг. Предпочтительная доза составляет приблизительно 10 мг/кг. Суточную дозу можно вводить один раз в день или разделить на меньшие дозы и вводить в виде многократных доз, например, два, три или четыре раза в день. Однако, как будет понятно специалисту в данной области, композиции, описанные в данном документе, могут быть введены в разных количествах и в разное время. Специалисту будет также понятно, что определенные факторы могут оказывать влияние на дозировку и время, необходимое для эффективного лечения субъекта, включая, без ограничения, тяжесть заболевания или злокачественного состояния, предшествующие виды лечения, общее состояние здоровья и/или возраст субъекта, и другие присутствующие заболевания. Кроме того, лечение субъекта терапевтически эффективным количеством композиции может включать одну лечебную процедуру или, предпочтительно, серию лечебных процедур.
После успешного лечения субъекту желательно пройти поддерживающую терапию для предотвращения рецидива излеченного заболевания или злокачественного состояния.
c. Введение
Композиции согласно настоящему изобретению могут быть введены для терапевтического лечения. При применении в терапевтических целях композиции вводят пациенту, страдающему от заболевания или злокачественного состояния, такого как рак, в количестве, достаточном для лечения или по меньшей мере частичной остановки развития заболевания и его осложнений. Количество, подходящее для достижения этой цели, определяется как «терапевтически эффективная доза». Количества, эффективные для такого использования, зависят от тяжести заболевания и общего состояния здоровья пациента. Эффективным количеством соединения является количество, которое обеспечивает либо субъективное облегчение симптома(ов), либо объективно определяемое улучшение, отмечаемое врачом или другим квалифицированным наблюдателем.
Определение эффективного количества находится в рамках возможностей специалистов в данной области, особенно в свете приведенного здесь подробного описания. Как правило, действующее или эффективное количество иммуноконъюгата определяют при введении сначала низкой дозы или небольшого количества иммуноконъюгата, затем постепенно увеличивая вводимую дозу или дозировку, добавляя, если необходимо, второй или третий лекарственный препарат, до тех пор, пока не будет наблюдаться желаемый эффект у подвергаемого лечению субъекта с минимальными или нулевыми токсическими побочными эффектами.
Введение композиций производят однократно или многократно в зависимости от дозы и частоты, которые требуются и переносимы пациентом. В любом случае композиция должна обеспечивать достаточное количество белков по настоящему изобретению, чтобы эффективно лечить пациента. Предпочтительно, дозу вводят один раз, но она может применяться периодически до тех пор, пока либо не будет достигнут терапевтический результат, либо побочные эффекты не станут основанием для прекращения терапии. Обычно доза является достаточной для лечения или облегчения симптомов или признаков болезни, не вызывая неприемлемой токсичности для пациента.
Для достижения желаемого терапевтического эффекта композиции могут вводиться в течение нескольких дней в терапевтически эффективных суточных дозах. Так, терапевтически эффективное введение композиции для лечения заболевания или злокачественного состояния, описанного в данном документе, у субъекта, может потребовать периодического (например, ежедневного) введения, которое продолжается от трех дней до двух недель или дольше. Как правило, композиции могут вводиться в течение по меньшей мере трех дней подряд, часто - по меньшей мере пяти дней подряд, чаще - по меньшей мере десяти дней, а иногда и в течение 20, 30, 40 и более дней подряд. Хотя последовательные ежедневные дозы предпочтительны для достижения терапевтически эффективной дозы, терапевтически полезный эффект можно достигнуть, даже если соединения или композиции не вводить ежедневно, при условии, что введение повторяется достаточно часто, чтобы поддерживать терапевтически эффективную концентрацию композиции у субъекта. Например, можно вводить композицию через день, каждый третий день или один раз в неделю, если применяются более высокие дозы и они хорошо переносятся субъектом.
3. Способы ингибирования роста опухоли
Композиции согласно настоящему изобретению находят применение для различных задач. Например, молекулы РЕ настоящего изобретения, например, как часть химерной молекулы, находят применение для (i) индуцирования апоптоза в клетке, несущей один или более поверхностный маркер, (ii) ингибирования нежелательного роста, гиперпролиферации или выживания клетки, несущей один или несколько маркеров клеточной поверхности, (iii) лечения состояний, таких как рак, и (iv) обеспечения терапии млекопитающего с развившимся заболеванием, вызванным наличием клеток, несущих один или несколько маркеров клеточной поверхности.
Любая клетка или опухолевая клетка, экспрессирующая мезотелин как маркер клеточной поверхности, может быть использована для практической реализации способа по настоящему изобретению. Способы настоящего изобретения могут быть осуществлены in vitro или in vivo. При упоминании клетки следует понимать, что этот термин также включает в себя популяцию клеток, то есть более чем одну клетку.
Применение композиций для индуцирования апоптоза в клетке. несущей один или несколько маркеров клеточной поверхности
Апоптоз играет центральную роль как в развитии, так и в гомеостазе многоклеточных организмов. «Апоптоз» относится к программируемой смерти клеток и характеризуется определенными клеточными особенностями, такими как блеббинг мембраны, конденсация и фрагментация хроматина, образование апоптотических тел и положительным окрашиванием «TUNEL» (опосредованное терминальной дезоксинуклеотидилтрансферазой мечение UTP концов однонитевых разрывов). Более поздней стадией в апоптозе является деградация плазматической мембраны, что делает апоптотические клетки чувствительными к различным красителям (например, пропидийиодиду).
Апоптоз может быть вызван несколькими независимыми сигнальными путями, которые сходятся на конечном эффекторном механизме, состоящем из множественных взаимодействий между несколькими рецепторами смерти и их лигандами, которые принадлежат к надсемейству рецепторов/лигандов фактора некроза опухоли (TNF). Наиболее охарактеризованными рецепторами смерти являются CD95 («Fas»), TNFR1 (р55), рецептор гибели 3 (DR3 или Аро3/TRAMO), DR4 и DR5 (apo2-TRAIL-R2). Конечным эффекторным механизмом апоптоза является активация серии протеиназ, обозначаемых как каспазы. Активация этих каспаз приводит к расщеплению серии жизненно важных клеточных белков и гибели клеток.
Настоящее изобретение относится к способам индуцирования апоптоза в клетке, экспрессирующей мезотелин. В одном аспекте способ индуцирования апоптоза в клетке включает стадию воздействия или контакта клетки, экспрессирующей мезотелин как маркер клеточной поверхности, с RIT настоящего изобретения. Как правило, клетки подвергают воздействию или вводят в контакт с эффективными количествами иммуноконъюгата, в результате чего происходит индуцирование апоптоза.
В другом аспекте настоящего изобретения предложен способ индуцирования апоптоза у опухолевых клеток, экспрессирующих мезотелин на их поверхности, включающий стадию введения субъекту RIT настоящего изобретения.
Применение композиций для ингибирования роста, гиперпролиферации или выживания клетки, несущей один или несколько маркеров клеточной поверхности
Целью настоящего изобретения является обеспечение улучшенной терапевтической стратегии для лечения злокачественных образований с применением композиций согласно изобретению. В одном аспекте настоящего изобретения предложен способ ингибирования по меньшей мере одного из следующих явлений: нежелательного роста, гиперпролиферации или выживания клетки. Этот способ включает стадию контакта клетки, экспрессирующей мезотелин в качестве поверхностного маркера, с эффективным количеством РЕ настоящего изобретения, например, в качестве части химерной молекулы, как описано в данном документе, отличающийся тем, что стадия контакта приводит к ингибированию по меньшей мере одного из следующих явлений: нежелательного роста, гиперпролиферации или выживания клетки. В одном варианте этот способ включает стадию определения, экспрессирует ли клетка один или несколько маркеров клеточной поверхности, например, рецептор клеточной поверхности. Как правило, клетки подвергают воздействию или вводят в контакт с эффективным количеством иммуноконъюгата, в результате чего происходит ингибирование по меньшей мере одного из следующих явлений: нежелательного роста, гиперпролиферации или выживания клетки.
Так, в одном аспекте настоящего изобретения предложены способы ингибирования роста популяции клеток, несущих мезотелин. В предпочтительном варианте осуществления этот способ включает следующие стадии: (а) контакт популяции клеток с химерным белком согласно изобретению. Многие опухоли образуют метастазы. Так, в другом аспекте настоящего изобретения используют композиции настоящего изобретения, чтобы предотвратить образование метастазов. Этот способ включает стадию введения композиции по настоящему изобретению к опухолевой клетке, в которой введение композиции ведет к предотвращению образования метастаз. В предпочтительном варианте осуществления композиция содержит токсин направленного действия, содержащий антитело против антигена клеточной поверхности, и РЕ настоящего изобретения. Как правило, клетки подвергают воздействию или вводят в контакт с эффективными количествами иммуноконъюгата, следствием которого является предотвращение образования метастаз. Приводимые в качестве примеров злокачественные образования, чей рост, распространение и/или развитие могут быть ингибированы, включают рак яичников, мезотелиомы, немелкоклеточный рак легких, аденокарциному легких и рак поджелудочной железы.
Применение композиций для лечения рака
Способы настоящего изобретения могут быть осуществлены in vitro и in vivo. Так, в еще одном аспекте настоящего изобретения предложен способ лечения субъекта, страдающего от злокачественного состояния. Этот способ включает в себя стадию введения субъекту с диагнозом рак терапевтически эффективных количеств RIT изобретения, как описано в данном документе, в котором злокачественное состояние характеризуется нежелательным ростом или пролиферацией клетки, экспрессирующей один или более маркеров клеточной поверхности, и в котором стадия введения приводит к излечению субъекта. Как правило, клетки подвергают воздействию или вводят в контакт с эффективными количествами иммунотоксина, результатом чего является излечение субъекта.
В одном варианте осуществления настоящего изобретения иммунотоксин, содержащий РЕ по настоящему изобретению, используют для лечения субъекта, страдающего от рака, опосредованного взаимодействием мезотелин-СА125. Приводимые в качестве примеров злокачественные образования, чей рост, распространение и/или развитие, по меньшей мере, частично опосредованы связыванием СА125/мезотелин, включают рак яичников, мезотелиомы, немелкоклеточный рак легких, аденокарциному легких и рак поджелудочной железы.
Способы лечения рака могут необязательно содержать одну или более из следующих стадий: получение биологического образца ткани или жидкости от индивидуума; скрининг биологического образца на экспрессию мезотелина путем приведения в контакт биологического образца с антителом к поверхностному маркеру или скрининг биологического образца на экспрессию полинуклеотида поверхностного маркера посредством обнаружения мРНК поверхностного маркера. Это может быть сделано с помощью стандартных методик, известных в данной области, например, Вестерн-блоттинга, Нозерн-блоттинга или ПЦР.
Применение композиций для лечения субъекта, имеющего развившееся заболевание, вызванное наличием клеток, несущих один или несколько маркеров клеточной поверхности
Также предусмотрен способ терапии млекопитающего с развившимся заболеванием, вызванным наличием или аберрантной пролиферацией клеток, преимущественно несущих мезотелин или свехрэкспрессирующих мезотелин. В предпочтительном варианте осуществления этот способ включает стадию введения указанному млекопитающему RIT изобретения. Как правило, клетки подвергают воздействию или вводят в контакт с эффективными количествами иммунотоксина, результатом чего является излечение субъекта.
В еще одном варианте осуществления настоящее изобретение относится к устранению клеток-мишеней in vitro или ex vivo с помощью RIT настоящего изобретения. Например, химерные молекулы, содержащие RIT по изобретению, могут быть использованы для удаления клеток-мишеней из популяции клеток в культуре. Так, например, клетки, взятые от пациента, имеющего рак, экспрессирующий мезотелин, могут быть очищены от раковых клеток путем контакта культуры с химерными молекулами, направленными против мезотелина, как описано в данном документе.
В некоторых случаях клетки-мишени могут содержаться в биологическом образце. «Биологический образец» в настоящем документе представляет собой образец биологической ткани или жидкости, содержащий клетки-мишени и клетки, не являющиеся мишенями. Такие образцы включают, без ограничения, ткани из биопсии, кровь и клетки крови (например, лейкоциты). Биологический образец обычно получают из многоклеточных эукариот, предпочтительно млекопитающих, таких как крысы, мыши, коровы, собаки, морские свинки, кролики или, более предпочтительно, приматы, например, макаки, шимпанзе или человек. Наиболее предпочтительны образцы от человека.
Способы мониторинга реакции на лечение с помощью RIT изобретения
Изобретение предусматривает способы обнаружения ингибирования роста опухоли у пациента, страдающего от рака или предрасположенного к раку, который можно лечить с помощью RIT изобретения. Эти способы особенно полезны для мониторинга курса лечения, который проводят пациенту с использованием RIT настоящего изобретения. Способы могут быть использованы для мониторинга как терапевтического лечения пациентов с клиническими проявлениями заболевания, так и профилактического лечения пациентов без симптомов заболевания.
Способы мониторинга включают определение базового уровня опухолевой массы у пациента до введения дозы RIT настоящего изобретения и сравнение этого уровня с уровнем опухолевой массой после лечения или с опухолевой массой у пациента, не получающего лечение.
Значительное снижение (то есть, больше, чем типичная граница экспериментальной ошибки в повторных измерениях одного и того же образца, выраженная в виде одного стандартного отклонения от среднего значения таких измерений) уровня опухолевой массы сигнализирует о положительном результате лечения (то есть, что введение RIT настоящего изобретения остановило прогрессирование роста опухоли и/или метастазов).
В других способах контрольное значение (т.е. среднее значение и стандартное отклонение) опухолевой массы определяют для контрольной популяции или нормальной популяции (например, масса = ноль). Как правило, индивидуумы в контрольной популяции ранее не получали лечение. Измеренные значения опухолевой массы у пациента после введения RIT по настоящему изобретению затем сравнивают с контрольным значением. Значительное снижение опухолевой массы по отношению к контрольной величине (например, более чем на одно стандартное отклонение от среднего) сигнализирует о положительном результате лечения. Отсутствие значительного снижения или увеличение сигнализирует о негативном результате лечения.
В других способах контрольное значение опухолевой массы (например, среднее и стандартное отклонение) определяют в контрольной популяции индивидуумов, которые подверглись лечению, принимая по схеме RIT по настоящему изобретению, например, как части химерной молекулы, как описано в данном документе. Измеренные значения опухолевой массы у пациента сравнивают с контрольным значением. Если измеренный уровень у пациента существенно не отличается (например, более чем на одно стандартное отклонение) от контрольного значения, лечение может быть прекращено. Если уровень опухолевой массы у пациента значительно выше контрольного значения, продолжение введения агента является оправданным.
В других способах пациента, который в настоящее время не получает лечение, но прошел курс лечения ранее, наблюдают на предмет опухолевой массы, чтобы определить, требуется ли возобновление лечения. Измеренное значение опухолевой массы у пациента можно сравнить со значением опухолевой массы, ранее достигнутым у пациента после предыдущего курса лечения. Значительное увеличение опухолевой массы по сравнению с предыдущим измерением (т.е. больше, чем типичная погрешность повторных измерений одного и того же образца) является свидетельством того, что лечение может быть возобновлено. С другой стороны, значение, измеренное у пациента, можно сравнить с контрольным значением (среднее плюс стандартное отклонение), определенным в популяции пациентов после прохождения курса лечения. Альтернативно, измеренное значение у пациента можно сравнить с контрольным значением в популяциях пациентов, подвергшихся профилактическому лечению, которые остаются без симптомов заболевания, или в популяциях пациентов, подвергшихся терапевтическому лечению, которые демонстрируют улучшение характеристик заболевания. Во всех этих случаях увеличение опухолевой массы по отношению к контрольному уровню (т.е. больше, чем стандартное отклонение) является показателем того, что лечение пациента должно быть возобновлено.
Образец ткани для анализа, как правило, представляет собой кровь, плазму, сыворотку, слизь, биопсию тканей, опухоль, асцитную или спинномозговую жидкость пациента. Образец можно проанализировать для обнаружения неоплазии. Неоплазия или опухолевая масса могут быть обнаружены с помощью любого способа, известного в данной области, например, визуального наблюдения биопсии квалифицированным патологоанатомом или других способов визуализации, например, рентгенографией, УЗИ, магнитно-резонансной томографией (МРТ).
НАБОРЫ, КОНТЕЙНЕРЫ, ПРИБОРЫ И СИСТЕМЫ
Для применения в диагностических, исследовательских и терапевтических целях, описанных выше, в изобретении также предусмотрены наборы и системы. Наборы по настоящему изобретению включают RIT по настоящему изобретению, например, как часть химерной молекулы. Кроме того, наборы и системы могут включать инструкции, содержащие указания (т.е. протоколы) для осуществления на практике способов настоящего изобретения. Эти инструкции могут присутствовать в наборах в различных формах, одна или более из которых могут присутствовать в наборе. Хотя инструкции обычно содержат письменные или печатные материалы, они не ограничиваются таковыми. Любой носитель, пригодный для хранения таких инструкций и передачи их конечному пользователю, предусмотрен в настоящем изобретении. К таким носителям относятся, без ограничения, электронные носители информации (например, магнитные диски, ленты, кассеты, чипы), оптические носители (например, CD-ROM) и тому подобное. Такие носители могут включать адреса сайтов в сети Интернет, на которых предоставляются такие инструкции.
В зависимости от предполагаемого пользователя набора и системы и особых потребностей этого пользователя могут быть подготовлены разнообразные наборы, системы и композиции в соответствии с настоящим изобретением.
Предусмотрены наборы с единичными дозами активной композиции, например, в оральной, вагинальной, ректальной, трансдермальной или инъекционной дозированной форме (например, для внутримышечной, внутривенной или подкожной инъекции). В такие наборы в дополнение к контейнерам, содержащим стандартные дозы, будет включен информационный вкладыш в упаковку, описывающий применение и сопутствующие преимущества композиции для лечения заболевания или злокачественного состояния. Подходящие активные композиции и единичные дозы описаны в данном документе выше.
Хотя вышеизложенное изобретение подробно описано с помощью иллюстраций и примеров для ясности и понимания, обычному специалисту в данной области в свете идей настоящего изобретения будет очевидно, что могут быть осуществлены определенные вариации, изменения, модификации и замещения эквивалентов без отхода от сущности и объема настоящего изобретения. В результате варианты осуществления, описанные здесь, могут быть предметом различных модификаций, изменений и т.п., причем объем настоящего изобретения определяется исключительно по формуле изобретения, приложенной к данному документу. Специалисты в данной области легко определят множество некритических параметров, которые могут быть заменены, изменены или модифицированы с получением по существу аналогичных результатов. Кроме того, следует понимать, что терминология, использованная в данном документе, служит только для описания отдельных вариантов осуществления и не предназначена для ограничения, поскольку объем настоящего изобретения будет ограничен только приложенной формулой изобретения.
Хотя каждый из элементов настоящего изобретения описан здесь как содержащий множество вариантов осуществления, следует понимать, что, если не указано иное, каждый из вариантов осуществления рассматриваемого элемента согласно настоящему изобретению может быть использован с каждым из вариантов осуществления других элементов по настоящему изобретению, и каждое такое применение предназначено для формирования отдельного варианта осуществления настоящего изобретения.
Приведенные патенты, патентные заявки и научная литература, в том числе номера доступа к базе данных последовательностей GenBank, о которых говорится здесь, включены в настоящий документ посредством ссылки в полном объеме, как если бы каждая отдельная публикация, патент или патентная заявка были особо и индивидуально указаны для включения посредством ссылки. Любое противоречие между любой из упомянутых в данном документе ссылок и приведенным раскрытием этого описания должно быть решено в пользу последнего. Точно так же, любое противоречие между известным из уровня техники определением слова или фразы и определением этого слова или фразы, представленным в данном описании, должно быть решено в пользу последнего.
Как можно понять из приведенного выше описания, настоящее изобретение имеет широкий спектр применений.
Далее изобретение иллюстрируется последующими примерами, которые являются только иллюстративными и не предназначены для ограничения определения и объема изобретения каким-либо образом.
ПРИМЕРЫ
Следующие примеры приведены в иллюстративных целях, но не для ограничения заявленного изобретения.
Пример 1
Белок
SS1P (SS1 (dsFv)-PE38), и все мутанты, полученные из SS1P, были экспрессированы в Escherichia coli BL21 (λDE3) с векторов для SS1 (VH)-PE и SS1 (VL), и впоследствии подвергнуты рефолдингу и очищены, как описано (Pastan I, Beers R, and Bera TK, Methods Mol Biol., 248:503-18 (2004)). Исходные растворы SS1P были приготовлены Advanced BioScience Laboratories, Inc. (Кенсингтон, Мэриленд). Все остальные RIT были приготовлены в Лаборатории молекулярной биологии Национального института рака (Бетесда, Мэриленд). Мутации в SS1P производились с применением системы сайт-направленного мутагенеза QuikChange (Stratagene, Ла-Хойя, Калифорния) с праймерами из Lofstrand Labs Limited (Гейтерсберг, Мэриленд).
Клеточные линии
Разнообразные мезотелин-положительные клеточные линии человека были использованы в этом исследовании. Линии клеток аденокарциномы легких L55 и мезотелиомы М30 были предоставлены Dr. Steven Albelda, Университет Пенсильвании (Филадельфия, Пенсильвания). Линия клеток мезотелиомы HAY была предоставлена Stehlin Foundation for Cancer Research (Хьюстон, Техас). Линии раковых клеток яичников OVCAR-8 и А1847 были предоставлены Dr. Hisataka Kobayashi и Dr. S. Aaronson, соответственно, из Национального института рака (Бетесда, Мэриленд). Линия клеток аденокарциномы легких NCI-H322M была получена от Dr. Mitchell Но из Национального института рака (Бетесда, Мэриленд). Клеточная линия KB31 является субклоном человеческой клеточной линии эпидермальной карциномы KB (Akiyama S et al., Somat Cell Mol Genet., 11(2):117-26 (1985)). Клеточная линия А431/K5 является производной от клеточной линии плоскоклеточного рака А431, трансфицированной мезотелином человека (Chowdhury PS et al., Proc. Nat. Acad. Sci. USA, 95:669-674 (1998)) и выращенной в модифицированной питательной среде Дульбекко (DMEM), дополненной 10% FBS, 2 мМ L-глутамином, 1 мМ пируватом натрия, 100 ед. пенициллина, 100 мкг стрептомицина и 750 мкг/мл G-418 (генетицина). Если не указано иное, все клеточные линии выращивали при 37°C с 5% СО2 в среде RPMI-1640, дополненной 10% FBS, 2 мМ L-глутамином, 1 мМ пируватом натрия, 100 ед. пенициллина и 100 мкг стрептомицина (Invitrogen Corporation, Карлсбад, Калифорния).
Анализ цитотоксичности
Жизнеспособность клеточных линий, обработанных иммунотоксинами, измеряли посредством Cell Counting Kit-8 WST-8 (Dojindo Molecular Technologies, Inc., Гейтерсберг, Мэриленд) по существу, как описано в техническом руководстве. Вкратце, от 5000 до 10000 клеток/лунку высевали в 96-луночные планшеты, позволяли прикрепиться и инкубировали с различными концентрациями RIT в течение 72 часов в конечном объеме 0,2 мл, после чего добавляли 10 мкл реагента CCK-8 в каждую лунку. Планшеты инкубировали при 37°С до тех пор, пока максимум поглощения при 450 нм не достигал значений OD около 1. Значения нормализовали между контрольными показателями циклогексамида (10 мкг/мл) и буфера (0,2% человеческий сывороточный альбумин в PBS) и аппроксимировали стандартным 4-параметрическим сигмоидальным уравнением с переменным наклоном с помощью программы GraphPad PRISM, чтобы получить концентрацию иммунотоксина, при которой достигалась гибель 50% клеток (EC50).
Анализ жизнеспособности клеток от пациентов с мезотелиомой проводили, как описано ниже. Вкратце, клетки были получены из плевральной жидкости или асцита у пациентов с мезотелиомой и высеяны при плотности 5×104 клеток/лунку в 24-луночный планшет с различными концентрациями RIT SS1P, SS1-LR, BL22 (анти-CD22/РЕ38) в качестве отрицательного контроля, и НВ21 (анти-рецептор трансферрина/РЕ38) в качестве положительного контроля. Клетки инкубировали в течение 96 часов, фиксировали и окрашивали красителем кристаллическим фиолетовым. Интенсивность окраски каждой лунки определяли с помощью планшетного спектрофотометра Versamax (Molecular Devices, Inc., Саннивейл, Калифорния) при длине волны 595 нм. Каждое значение определяли в трех параллельных анализах. Статистический анализ полученных данных по ANOVA проводили с применением программного обеспечения GraphPad Prism (GraphPad Software, Inc., Ла-Хойя, Калифорния).
Анализ противоопухолевой активности на ксенотрансплантатах у мышей
Двадцати четырем женским особям мышей, лишенных волосяного покрова, в день 0 вводили подкожно в бок клетки А431/K5, как описано ранее (Zhang Y, Clin Cancer Res., 12(15):4695-701 (2006)). Объем опухоли регулярно измеряли штангенциркулем в течение следующих 6 недель. Когда средний размер опухоли достигал -100 мм3, через 5 дней после имплантации мышей разделили на четыре группы по шесть особей и вводили согласно графику QODX3 0,2 мл 0,2% MSA в PBS (только носитель) или носитель, содержащий SS1P (0,3 мг/кг) или SS1-LR (6,0 или 15 мг/кг). Мышей умерщвляли, если их опухоль превышала 1000 мм3, или в конце эксперимента. С животными обращались в соответствии с руководством Национального института здравоохранения, утвержденным Комитетом по уходу и использованию животных Национального института рака.
Анализ интернализации RIT
Клетки А431/K5 (106/лунка) высевали в 6-луночный планшет (10 см2/лунка) и инкубировали в течение ночи. На следующий день среду в каждой лунке меняли на 1 мл свежей среды, содержащей 1 мкг RIT, подлежащего оценке. Клетки инкубировали при 37°С в течение различных интервалов времени, после чего лунку быстро ополаскивали 2 мл холодного PBS, 1 мл холодного буфера для очистки (1 мг/мл BSA в 0,2 М глицине, рН 2,5), и снова 2 мл холодного PBS. Клетки затем лизировали 200 мкл буфера RIPA (150 мМ NaCl, 1 мМ ЭДТА, 1% NP-40, 0,5% дезоксихолата натрия, 0,1% SDS, 50 мМ Трис-Cl, рН 8,0), содержащего коктейль ингибиторов протеаз (Sigma, Сент-Луис, Миссури). Образцы анализировали методом невосстанавливающего Трис-глицин SDS-PAGE Вестерн-блоттинга с применением кроличьих анти-РЕ38 поликлональных антител.
Вариабельная цитотоксичность SS1-LR
Первоначальные эксперименты с SS1-LR показали, что он имел высоковариабельную цитотоксичность относительно SS1P на выбранных мезотелин-экспрессирующих клеточных линиях, от более чем в 4 раза большей активности до в 20 раз меньшей активности. Общая тенденция, однако, свидетельствовала о меньшей активности молекулы. SS1-LR был более чем на 20% менее активен по сравнению с SS1P на 5 клеточных линиях из 8 испытанных, и более чем на 20% более активен только на одной клеточной линии. Эта тенденция заметно отличается от версии анти-CD22 молекулы LR, HA22-LR, которая, как правило, демонстрировала аналогичную или повышенную цитотоксичность как на клеточных линиях, так и на клетках пациента (Weldon JE, Blood., 113(16):3792-800 (2009)). Эти наблюдения показывают, что существует значительная разница между путем токсического действия РЕ в эпителиальных клетках, экспрессирующих мезотелин, и в B-клетках, экспрессирующих CD22.
Активность SS1-LR in vitro
Иммунотоксин SS1P (Фиг.1А) состоит из стабилизированной дисульфидными связями двойной цепи антимезотелинового фрагмента Fv SS1, присоединенного к РЕ38 (Chowdhury PS et al., Proc. Nat. Acad. Sci. USA, 95:669-674 (1998); Chowdhury PS and Pastan I., Nat Biotechnol., 17:568-572 (1999); Reiter Y and Pastan I., Clin. Cancer Res., 2:245-252 (1996)). Мы ввели мутацию LR (Weldon JE, Blood., 113(16):3792-800 (2009); РЕ Δ251-273 & Δ285-394) в SS1P, чтобы создать вариант SS1-LR (Фиг.1В), и оценили его активность по сравнению с SS1P против нескольких мезотелин-экспрессирующих клеточных линий in vitro. Фигура 2 показывает репрезентативную цитотоксичность по отношению к восьми различным клеточным линиям.
Линия клеток рака легких L55 (А) показала сходную чувствительность к обоим RIT. В противоположность этому, линия рака легких NCI-H322M (Pal LH et al., Nat Med., 2(3):350-3 (1996)) была приблизительно в 3 раза менее чувствительна к SS1-LR, чем к SS1P. Клеточная линия мезотелиомы HAY (G) была более чем в 4 раза более чувствительна к SS1-LR, чем к SS1P, но линия мезотелиомы М30 (D) была приблизительно на 20% менее чувствительна к SS1P, чем к SS1-LR. Клеточная линия А431/K5 (Н), линия эпителиальных клеток, стабильно трансфицированная мезотелином (Chowdhury PS et al., Proc. Nat. Acad. Sci. USA, 95:669-674 (1998); Chang K and Pastan I., Proc. Natl. Acad. Sci. USA, 93, 136-140 (1996)), была приблизительно в 4 раза менее чувствительна к SS1-LR по сравнению с SS1P. Кроме того, линия цервикального рака KB31 (С) чувствительна к уничтожению SS1P, но его EC50 в 20 раз меньше по сравнению с SS1-LR. Также были оценены чувствительности клеточных линий рака яичников А1847 (F) и OVCAR-8 (Е) к SS1-LR. SS1-LR показал EC50, аналогичную SS1P, на линии А1847, но в 2 раза сниженную EC50 на линии OVCAR-8. Таблица 1 суммирует эти данные и представляет относительные значения ЕС50, сравнивающие SS1P и SS1-LR. Активность SS1-LR относительно SS1P меняется в широких пределах среди различных клеточных линий.
Анализируя данные, мы заметили, что SS1-LR не удалось полностью уменьшить жизнеспособность многих клеточных линий, которые мы оценивали (NCI-H322M, KB31, М30, OVCAR-8 и А1847), до контрольных уровней, как определено в клетках, обработанных 10 мкг/мл циклогексамида. Это позволяет предположить, что субпопуляция клеток может быть чувствительна к SS1P, но устойчива к SS1-LR. Мы оценивали эту возможность, инкубируя клетки OVCAR-8 и А1847 в течение 4 дней с 10 нг/мл SS1-LR и пытаясь культивировать любые выжившие клетки. Никакого дальнейшего роста клеток не наблюдалось после обработки RIT. Мы пришли к выводу, что, хотя в нашем анализе не было достигнуто полное метаболическое подавление, в этих клеточных линиях отсутствует субпопуляция, резистентная к SS1-LR.
Активность SS1-LR in vivo
Затем мы оценивали эффективность SS1-LR, используя модель ксенотрансплантатной опухоли на мышах. Из клеточных линий, что мы тестировали in vitro, только А431/K5 стабильно росла в нашей ксенотрансплантатной модели. Мышам, лишенным волосяного покрова, с ксенотрансплантатными опухолями А431/K5 размером в среднем -100 мм3 вводили внутривенно согласно графику QODX3 (через день, три дозы) SS1-LR в дозе 6,0 или 15 мг/кг. Для сравнения дополнительным группам вводили внутривенно согласно графику QODX3 либо буфер (0,2% HSA в PBS), либо 0,3 мг/кг SS1P, максимально переносимую дозу SS1P в соответствии с приведенным графиком дозирования. Размер опухоли каждой мыши регулярно измеряли в течение 22 дней после имплантации (Фиг.3).
Опухоли PBS-обработанных мышей быстро выросли до среднего размера более чем 1000 мм3 на 14 день после имплантации. Мыши, получавшие в дни 5, 7 и 9 0,3 мг/кг SS1P продемонстрировали регрессию опухоли, которая привела средний размер опухоли к минимуму около 53 мм3 на 12 день. На 20 день возобновился быстрый рост всех опухолей. Доза SS1-LR 6,0 мг/кг была немного менее активна, чем 0,3 мг/кг SS1P. Опухоли у мышей, получавших 6,0 мг/кг SS1-LR, достигли среднего минимального размера 92 мм3 в дни 7 и 9, и все возобновили быстрый рост к 18 дню. Доза 15 мг/кг SS1-LR продемонстрировала значительно лучшую противоопухолевую активность, чем 0,3 мг/кг SS1P. Опухоли у мышей, которым вводили 15 мг/кг SS1-LR, регрессировали в среднем до минимального размера 17 мм3. На 22 день четыре из шести опухолей возобновили рост, одна опухоль была неизменной на уровне 10 мм3 с 18 дня, и одна опухоль не обнаруживалась после 15 дня. Хотя SS1-LR приблизительно в 20 раз менее активен, чем SS1P в этой модели, его низкая неспецифическая токсичность позволяет вводить мышам высокие дозы и достичь лучшего противоопухолевого эффекта.
Процессинг интернализованных RIT
Различия во внутриклеточном транспорте и процессинге между SS1P и SS1-LR лежат в основе вариативности активности, наблюдаемой между этими двумя RIT. Чтобы проверить эту гипотезу, мы исследовали интернализацию и процессинг SS1P и SS1-LR в клетках А431/K5 методом Вестерн-блоттинга (Фиг.4). Клетки А431/K5 были выбраны потому, что они экспрессируют мезотелин на высоком уровне (около 106 сайтов/клетка) и интернализуют большие количества RIT для визуализации. Мы провели невосстанавливающий SDS-PAGE Вестерн-блоттинг на клеточных лизатах целых клеток А431/K5, обработанных непрерывной инкубацией с 1 мкг/мл либо SS1P, либо SS1-LR при различных временных интервалах. На Фигуре 4А показана динамика обработки SS1P, в то время как на Фигуре 4В показана динамика обработки SS1-LR.
Как и ожидалось, клетки А431/K5, обработанные либо SS1P, либо SS1-LR, имеют характерные полосы в положениях, указывающих на полноразмерную (62 кДа и 50 кДа, соответственно) и фрагментированную (51 кДа и 39 кДа, соответственно) формы этих двух RIT. Оба RIT содержат три дисульфидные связи в области Fv: одна внутрицепочечная связь в каждом из фрагментов VH и VL, и одна межцепочечная связь между VH и VL (Фиг.1). SS1P содержит дополнительную дисульфидную связь в домене II между остатками цистеина, фланкирующими сайт расщепления фурином (Cys265 и Cys287). Степень окисления дополнительной дисульфидной связи в SS1P проявляется в изменении миграции SS1P в SDS-PAGE, вызывая появление наблюдаемого дублета (Фиг.4А, восстановленная). Эти полосы сливаются в одну полосу при обработке восстановителем (данные не показаны).
Мы также ожидали увидеть полосы, соответствующие C-концевым фрагментам расщепления фурином SS1P и SS1-LR (34 кДа и 24 кДа, соответственно). Лизаты обработанных SS1P клеток показывают заметную полосу вблизи 34 кДа, свидетельствующую о фрагменте расщепления фурином. В противоположность этому, лизаты клеток, обработанных SS1-LR, показывают только слабую полосу, соответствующую 24 кДа. Интенсивность каждой полосы оценивали количественно с помощью программного обеспечения ImageQuant (GE Healthcare). На Фиг.4С показан процент интенсивности полосы фрагмента расщепления фурином к полной интенсивности в каждый момент времени. Расщепление SS1P фурином достигает максимума немного более 30% от суммы интенсивностей всех полос, в то время как расщепление SS1-LR достигает максимума 6% от общей интенсивности. Существует более чем 5-кратное различие между долями расщепленного SS1P и расщепленного SS1-LR в каждый момент времени, что позволяет предположить, что SS1-LR может процессироваться фурином менее эффективно, чем SS1P.
Мутанты с гибким линкером
Так как наши данные позволяют предположить, что SS1-LR процессируется фурином менее эффективно, чем SS1P, мы попытались повысить активность SS1-LR за счет повышения эффективности его расщепления фурином. Мы создали несколько мутантов, чтобы исследовать, могут ли изменения сайта расщепления фурином повысить активность SS1-LR за счет повышения эффективности внутриклеточного расщепления фурином. Наша стратегия состояла в том, чтобы увеличить гибкость сайта расщепления фурином, и, таким образом, увеличить его доступность для фурина, путем добавления коротких неструктурированных Gly/Ser линкеров. SS1-LR/GGS (Фиг.1В) включает линкер Gly-Gly-Ser на С-конце сайта расщепления фурином из 11 остатков и имеет повышенную активность на обоих клеточных линиях NCIH322M и KB31 (Фиг.5). Увеличение длины линкера (SS1-LR/GGSx2, Фиг.1B) не повышает активность далее (данные не показаны). Дублирование сайта расщепления фурином с дополнительными областями линкеров (SS1-2 x фурин; Фиг.1B) также не повышает активность относительно SS1-LR/GGS (данные не показаны). В отличие от SS1-LR, все мутанты с удлиненным линкером демонстрируют полное подавление метаболизма, эквивалентное контролю циклогексамидом. Кроме того, для подтверждения важности расщепления фурином мы получили мутантный SS1-LR/GGS R279G, который включает мутацию R279G, которая делает сайт нерасщепляемым для фурина. Этот мутант был совсем неактивным в отношении как клеток NCI-H322M, так и KB31 (Фиг.5), что свидетельствует о необходимости расщепления фурином для активности.
Затем мы проверили нашу гипотезу, что линкер GGS повысил эффективность расщепления SS1-LR фурином. Вестерн-блоттинг клеток А431/K5, обработанных SS1-LR/GGS, однако, не показал улучшения в соотношении расщепленного RIT по отношению к общему RIT (данные не показаны). Расщепление SS1-LR/GGS фурином in vitro также не показало улучшения в скорости расщепления фурином (данные не показаны). Эти результаты показывают, что улучшенная цитотоксичность SS1-LR/GGS не связана с повышенным расщеплением фурином, но вместо этого является результатом другого механизма. Возможные объяснения включают повышенную транслокацию токсина во время внутриклеточного транспорта.
Активность на клетках пациентов
В продолжение этого исследования была проведена серия из 8 точечных мутаций в домене III, чтобы уменьшить иммуногенность РЕ у пациентов, подвергающихся лечению RIT, путем устранения известных эпитопов В-клеток (Onda M et al., направлена для публикации в PNAS.). Эти мутации (D406A, R432G, R467, R490A, R513A, E548S, Q592A и K590S) не влияют на активность RIT, но вместо этого снижают иммуногенность белка. Мы включили эти восемь мутаций в SS1-LR/GGS, создав новый вариант, называемый SS1-LR/GGS/8M, и испытали его цитотоксичность против первичных клеток, взятых из плевральной жидкости или асцита у пациентов с мезотелиомой. Жидкость, взятая у пациентов, содержит смесь клеток, не все из которых являются злокачественными, и они не экспрессируют мезотелин равномерно. Тем самым анализ обеспечивает хорошую оценку относительной активности, но это лишь грубая оценка абсолютной цитотоксичности. Клетки обрабатывали на раннем пассаже различными концентрациями SS1P или SS1-LR/GGS/8M и через четыре дня оценивали жизнеспособность посредством анализа с кристаллическим фиолетовым, как описано. Из нескольких клеток пациентов, которые оценивали на ответ на SS1P, клетки двух линий (NCI-M-02 и NCI-M-03) показали хорошую реакцию на лечение (>65% снижение жизнеспособности при уровне дозы 100 нг/мл). Мы оценивали активность SS1P-LR/GGS/8M на эти две популяции и сравнивали ее с активностью SS1P. Данные представлены на Фигуре 6 в качестве дробных значений относительно необработанного контроля.
Клетки пациентов NCI-M-02 и NCI-M-03 были чувствительны к лечению SS1P или SS1-LR/GGS/8M, демонстрируя более чем 60% снижение жизнеспособности по сравнению с необработанным контролем в дозах 100 нг/мл или ниже. NCI-M-02 и NCI-M-03 были особенно чувствительны к SS1-LR/GGS/8M в дозах 1,0 и 10 нг/мл, что свидетельствует о значительно более высокой цитотоксичности по сравнению с SS1P при этих концентрациях (p<0,05). В качестве контроля клетки пациентов были также обработаны RIT BL22 и НВ21 (данные не показаны). BL22, который направлен на специфический маркер CD22 В-клеток, не влиял на жизнеспособность каждой клеточной популяции в дозе 100 нг/мл. НВ21, который направлен на повсеместный рецептор трансферрина, и, как известно, очень активен по отношению к почти всем клеткам, снижал жизнеспособность обеих клеточных линий почти на 90% при дозе 10 нг/мл. В целом, данные показывают, что SS1-LR/GGS/8M обладал цитотоксичностью, аналогичной или лучшей, чем SS1P, в двух различных клеточных популяциях пациентов. Мы пришли к выводу, что эти два антимезотелиновых иммунотоксина, ведущих себя неодинаково, имеют сравнимую цитотоксическую активность.
Пример 2
Далее представлен отчет о создании, оценке и селективности SS1-LR/GGS/8M, варианта SS1P с улучшенными терапевтическими свойствами. Как уже говорилось выше, SS1-LR/GGS/8M включает мутации, которые, как ранее показано, улучшают RIT, а также новую мутацию, которая повышает его активность. SS1-LR представляет собой укороченный вариант SS1P, который содержит каталитический фрагмент РЕ, присоединенный к Fv SS1 посредством сайта расщепления фурином РЕ из 11 остатков. Цитотоксичность SS1-LR оценивали на семи клеточных линиях и установили, что она существенно ниже, чем у SS1P. Дальнейший анализ SS1-LR в культивируемых клетках показал, что он плохо процессируется фурином в пути токсического действия. Для улучшения расщепления фурином мы ввели линкер из 3 остатков в SS1-LR, создав SS1-LR7GGS. SS1-LR/GGS был значительно более активен на клеточных линиях, но не показал повышенное расщепление фурином. Мы ввели восемь точечных мутаций в каталитический фрагмент SS1-LR/GGS, которые, как было показано, уменьшают иммуногенность RIT, устраняя B-клеточные эпитопы. Этот новый RIT, SS1-LR/GGS/8M, демонстрирует отличную противоопухолевую активность, низкую неспецифическую токсичность у грызунов, а также сниженную реактивность с анти-SS1P сывороткой крови человека. Кроме того, первичные клетки от пациентов с мезотелиомой показывают повышенные ответы на SS1-LR/GGS/8M относительно SS1P. SS1-LR/GGS/8M является превосходным терапевтическим кандидатом для клинической разработки за счет своей низкой антигенности, низкой неспецифической токсичности и высокой активности.
В этом исследовании были использованы несколько мезотелин-положительных клеточных линий человека. Эти клетки получали и культивировали в большинстве случаев, как описано в Примере 1.
Анализ цитотоксичности
Жизнеспособность клеточных линий, обработанных иммунотоксинами, измеряли с помощью Cell Counting Kit-8 WST-8 (Dojindo Molecular Technologies, Inc., Гейтерсберг, Мэриленд). Клетки (2000 клеток/лунку) высевали в 96-луночные планшеты, оставляли на ночь для прикрепления и инкубировали с различными концентрациями RIT в течение 72 часов при конечном объеме 0,2 мл. В конце инкубационного периода в каждую лунку добавляли 10 мкл реагента CCK-8 и планшеты инкубировали при 37°С, пока в лунках максимум поглощения при 450 нм не достигал значений ~1 OD. Значения нормализовали между контрольными показателями с циклогексамидом (10 мкг/мл) и буфером (фосфатный буферный солевой раствор Дульбекко без Са и Mg (D-PBS; Quality Biological, Inc., Гейтерсберг, Мэриленд), содержащий 0,2% человеческого сывороточного альбумина (HSA)), а затем аппроксимировали сигмоидальным уравнением с переменным углом наклона для плато, базовой линии и наклона Хилла с помощью программного обеспечения GraphPad PRISM (GraphPad Software, Inc., Ла-Хойя, Калифорния). Уравнение затем использовали для интерполяции концентрации RIT, которая уменьшала жизнеспособность клеток на 50% (ЕС50).
Клетки от пациентов с мезотелиомой культивировали и оценивали на их ответ на SS1P и SS1-LR/GGS/8M по существу, как описано (Xiang X, et al., PLoS One 2011; 6: e14640). D-PBS и 10 нг/мл НВ21 (анти-рецептор трансферрина/РЕ40) использовали в качестве отрицательного и положительного контроля гибели клеток. Каждое условие оценивали в трех параллельных анализах. Статистический анализ полученных данных по ANOVA проводили, используя программное обеспечение GraphPad Prism.
Анализ противоопухолевой активности на ксенотрансплантатах у мышей
Двадцати восьми женским особям мышей, лишенных волосяного покрова, вводили подкожно в бок 5×106 клеток L55 в 0,2 мл RPMI с 4 мг/мл матригеля (BD Biosciences, Сан-Хосе, Калифорния) в день 0. Объем опухоли регулярно измеряли штангенциркулем в течение следующих 30 дней. Через семь дней после имплантации (средний размер опухоли ~100 мм3) мыши были разделены на четыре равные группы, и им вводили внутривенно в дни 7, 9 и 12 по 0,2 мл 0,2% HSA в D-PBS (носитель) или носитель, содержащий либо SS1P (0,4 мг/кг), либо SS1-LR/GGS/8M (0,4 или 2,5 мг/кг). Этот эксперимент и все последующие эксперименты на животных проводились в соответствии с руководством Национального института здравоохранения, утвержденным Комитетом по уходу и использованию животных Национального института рака.
Фармакокинетика в мышиной сыворотке
Группам из девяти женских особей мышей линии BALB/c вводили внутривенно 10 мкг SS1P или SS1-LR/GGS/8M в 0,2 мл D-PBS с 0,2% HSA. У групп из трех мышей брали кровь через интервалы времени 2 и 20, 5 и 30, или 10 и 60 минут. Анализировали сыворотки с помощью твердофазного иммуноферментного анализа (ELISA), как описано ранее (Bang S, et al., Clin Cancer Res 2005; 11:1545-50.25)).
Анализ повышения проницаемости капилляров у крыс
Описанную ранее модель RIT-индуцированного синдрома повышенной проницаемости капилляров у крыс (Siegall CB, et al., Proc Natl Acad Sci USA 1994; 91:9514-8) использовали для оценки неспецифической токсичности SS1-LR/GGS/8M. Вкратце, самкам Wistar Furthand Rowett, nu/nu (бестимусных) крыс (Харлан-Спраг-Доули) возраста шести-восьми недель вводили внутривенно D-PBS, SS1P (0,2 или 0,3 мг/кг) или SS1-LR/GGS/8M (6 или 12 мг/кг). Через 24 часа крыс умерщвляли под воздействием CO2. Жидкость гидроторакса собирали от эвтаназированных животных, помещая тело в спинное положение, удаляя вентральную грудную стенку и аспирируя жидкость с помощью шприца на 3 мл и иглы калибра 27,5. Легкие у нескольких крыс были удалены, фиксированы в течение 3 дней в 10% формалине, из них делали срезы и окрашивали.
Анализ интернализации RIT проводили по существу, как описано выше, образцы были проанализированы с помощью невосстанавливающего вестерн-блоттинга с применением кроличьих анти-РЕ38 поликлональных антител.
Анализ антигенности
Связывание SS1P или SS1-LR/GGS/8M с антителами в сыворотках пациентов анализировали по существу, как описано (Onda M, et al., Proc Natl Acad Sci USA 2011; 108:5742-7), за исключением того, что для выявления РЕ-специфических антител с помощью El-ISA были использованы CD22-rFc и НА22.
Результаты
Активность SS1-LR in vitro
Иммунотоксин SS1P (Фиг.1) является стабилизированным дисульфидной связью двухцепочечным антимезотелиновым фрагментом Fv SS1, присоединенным к РЕ38. Мы ввели мутацию LR ((Weldon JE, et al. Blood 2009; 113:3792-800); РЕ Δ251-273 & Δ285-394)) в SS1P для создания SS1-LR и оценили его активность на семи экспрессирующих мезотелин клеточных линиях in vitro. В таблице 2 приведены данные, по меньшей мере, 3 отдельных экспериментов по оценке цитотоксичности, в виде средних значений ЕС50 и стандартной ошибки среднего. По сравнению с SS1P, SS1-LR был более активным на клеточной линии HAY, но менее активным на остальных линиях. Активность SS1-LR широко варьировала среди различных клеточных линий, но он был наименее активным на линиях клеток рака яичников.
SS1-LR/GGS/8M
Иммуногенность остается серьезной проблемой для RIT на основе РЕ (Weldon JE, et al., FEBS J. 2011 Dec; 278(23):4683-700). Хотя работа по сопоставлению HA22-LR с НА22 (Hansen JK, et al., J Immunother 2010; 33:297-304)) заставляет предполагать, что SS1-LR будет менее иммуногенен, чем SS1P, оставшиеся элементы РЕ будут, тем не менее, быстро вызывать нейтрализующие антитела. Для удаления иммуногенных эпитопов В-клеток в РЕ недавно была разработана серия из восьми точечных мутаций в домене III (Onda M, et al., Proc Natl Acad Sci USA 2011; 108:5742-7). Эти мутации (D406A, R432G, R467A, R490A, R513A, E548S, Q592A и K590S) резко снизили иммуногенность HA22-LR у мышей, но незначительно снизили цитотоксичность. Мы включили мутации в SS1-LR/GGS, создав новый вариант под названием SS1-LR/GGS/8M, и протестировали RIT против семи экспрессирующих мезотелин клеточных линий. Значения ЕС50 этих экспериментов представлены в таблице 2. SS1-LR/GGS/8M более активен, чем SS1-LR, а иногда и более активен, чем SS1P (HAY, L55), но менее активен, чем SS1-LR/GGS. Снижение активности особенно заметно на линиях рака яичников.
Неспецифическая активность
В предыдущей работе с анти-CD22 RIT HA22 мы обнаружили, что одна внутривенная доза 2 мг/кг HA22 была смертельной для мышей, тогда как доза 20 мг/кг HA22-LR не показала токсичности (Weldon JE, et al., Blood 2009; 113:3792-800). Затем мы давали мышам единичные дозы HA22-LR вплоть до 45 мг/кг, не вызывая их смерть (неопубликованные наблюдения, данные не показаны), и мы ожидали подобное поведение от SS1-LR и его вариантов. Предыдущие эксперименты установили LD50 SS1P при одной дозе внутривенно на уровне 1,0 мг/кг для Balb/C мышей (Filpula D, et al., Bioconjug Chem 2007; 18:773-84) и 0,75 мг/кг для NIH Swiss мышей (Onda M., et al., Cancer Res 2001; 61:5070-7). С применением графика дозирования QODx3 (введения через день три раза), подобного клиническому графику, мыши переносили максимальную дозу 0,4 мг/кг SS1P (неопубликованные наблюдения, данные не показаны), но мыши, которым давали SS1-LR, получали дозы до 15 мг/кг QODx3 без токсичности (данные не показаны).
Хотя сниженная неспецифическая токсичность RIT на основе LR у мышей интригует, она не может иметь отношения к основной токсичности, наблюдаемой в клинических испытаниях. Неспецифическая токсичность RIT у мышей является результатом повреждения печени (Weldon JE, et al., Blood 2009; 113:3792-800; Onda M, et al., J Immunol. 2000:165:7150-6), которую обычно не наблюдают у пациентов. Более релевантной моделью неспецифической токсичности у животных является индуцированный RIT синдром повышенной проницаемости капилляров у крыс (Siegall CB, et al., Proc Natl Acad Sci USA 1994; 91:9514-8, Siegall CB, et al., Clin Cancer Res 1997; 3:339-45). Синдром повышенной проницаемости капилляров, при котором жидкость вытекает из кровеносных сосудов в интерстициальное пространство, является проявлением общей токсичности, наблюдаемой в клинических испытаниях RIT на основе РЕ. Используя эту модель, мы заметили, что крысы, получавшие внутривенно 2 мг/кг SS1P, становились больны после 24 часов, у них затруднялось дыхание, и накапливалась жидкость в их грудной полости (Фиг.7А). Увеличение дозы SS1P до 3 мг/кг увеличивало объем жидкости в грудной полости. В противоположность этому, крысы, получавшие либо D-PBS, либо SS1-LR/GGS/8M, не проявляли никаких признаков болезни и не накапливали жидкость. Дозы 6 мг/кг и 12 мг/кг SS1-LR/GGS/8M вводили без наблюдаемого токсического эффекта. Когда легкие крыс, получавших D-PBS, SS1P и SS1-LR/GGS/8M, фиксировали и окрашивали, у животных, получавших D-PBS или SS1-LR/GGS/8M, они оказывались нормальными, в то время как у крыс, получавших SS1P, появлялись признаки серьезного повреждения (Фиг.7B). Хотя ни одна молекула на основе LR не была протестирована клинически, это наблюдение усиливает предположение, что RIT на основе LR могут иметь сниженную токсичность у пациентов.
Активность SS1-LR/GGS/8M in vivo
Затем мы оценивали эффективность SS1-LR/GGS/8M in vivo с помощью модели ксенотрансплантата опухоли у мышей, используя клетки рака легких линии L55. Группам из семи мышей, лишенных волосяного покрова, с опухолями в среднем ~100 мм3 вводили внутривенно на 7, 9 и 12 день SS1-LR/GGS/8M в дозах 0,4 и 2,5 мг/кг. Для сравнения дополнительным группам вводили внутривенно по тому же графику носитель (0,2% HSA в D-PBS) или 0,4 мг/кг SS1P, максимально переносимую дозу SS1P в соответствии с указанным графиком дозирования. Размер опухоли у каждой мыши измеряли регулярно в течение 30 дней после имплантации (Фиг.7С).
Опухоли у мышей, которым вводили носитель, выросли до среднего размера приблизительно 500 мм3 на 16-й день после имплантации. У мышей, которым вводили 0,4 мг/кг SS1P, была небольшая задержка в росте опухоли: для достижения размера опухоли приблизительно 500 мм3 потребовалось 23 дня после имплантации. Мы наблюдали почти идентичный ответ у мышей, получавших 0,4 мг/кг SS1-LR/GGS/8M, предполагая паритет между SS1P и SS1-LR/GGS/8M в этой модели. Хотя SS1P из-за его неспецифической токсичности не может вводиться мышам при этом графике в дозах более высоких, чем 0,4 мг/кг, SS1-LR/GGS/8M может быть введен мышам при значительно более высоких дозах без вредных последствий. В этой модели опухоли была испытана примерно в 6 раз более высокая доза SS1-LR/GGS/8M (2,5 мг/кг). Мы наблюдали достоверную (p<0,01 с применением парного двустороннего t-теста) регрессию опухоли в этой группе мышей, чьи опухоли достигали минимального размера ~73 мм3 на 9 день. Эта группа мышей также испытывала повышенное подавление роста опухоли, которая достигала размера приблизительно 500 мм3 на 30 день после имплантации.
Хотя SS1-LR/GGS/8M имел активность, эквивалентную SS1P в модели ксенотрансплантата опухоли L55 у мышей, он показал повышенную активность по сравнению с SS1P на клетках L55 in vitro. Это расхождение в активности может быть обусловлено разницей во времени полужизни двух молекул в мышиной сыворотке. В предыдущей работе сравнивали время полужизни HA22-LR (~51 кДа) с НА22 (~63 кДа) у мышей (Weldon JE, et al., Blood 2009; 113:3792-800) и показали, что время полужизни у HA22-LR почти в 2 раза короче, чем у НА22 (7,8 и 14,6 минут соответственно). Мы предположили, что эта разница была связана с увеличением почечной фильтрации меньшей молекулы, и ожидали подобного результата при сравнении SS1-LR/GGS/8M (~50 кДа) с SS1P (~63 кДа). Анализ образцов сыворотки, взятых у мышей, которым вводили SS1-LR/GGS/8M, показал время полужизни 13 мин по сравнению с временем полужизни 19 минут для SS1P (Фиг.7D). Мы пришли к выводу, что разница во времени полужизни между SS1-LR/GGS/8M и SS1P лежит в основе расхождения относительной активности между этими двумя молекулами in vitro и in vivo.
Антигенность SS1-LR/GGS/8M
Основываясь на предыдущих исследованиях с RIT НА22 (Onda М, et al., Proc Natl Acad Sci USA 2011; 108:5742-7), мы ожидали, что мутации в SS1-LR/GGS/8M удалят B-клеточные эпитопы из SS1P. Для того чтобы оценить это предположение, мы сравнивали реактивность SS1P и SS1-LR/GGS/8M с сывороткой пяти пациентов, которые наработали нейтрализующие антитела в ответ на введение SS1P. Сыворотка пациентов сначала смешивалась с SS1P или SS1-LR/GGS/8M. Впоследствии несвязанные РЕ38-специфические антитела в сыворотке были выявлены с помощью ICC-ELISA (Onda М, et al., Proc Natl Acad Sci USA 2011; 108:5742-7). Из этих данных были определены концентрации SS1P и SS1-LR/GGS/8M, при которых сигнал ELISA был уменьшен на 50% (IC50). Как сообщалось ранее (Onda М., et al., J Immunol 2006; 177:8822-34), величины IC50 коррелируют со сродством взаимодействия антитело-антиген. Значения IC50 для SS1P относительно SS1-LR/GGS/8M отложены в процентах на Фигуре 8. Для всех сывороток пациентов соотношения величин IC50 SS1P к SS1-LR/GGS/8M были значительно ниже 10%, указывая, что основная фракция реагирующих с SS1P антител в сыворотке была инертна по отношению к SS1-LR/GGS/8M.
Активность на клетках пациентов
В дополнение к нашей оценке SS1-LR/GGS/8M на клеточных линиях in vitro и in vivo, мы также проверили его активность в отношении первичных клеток, полученных из плевральной жидкости или асцита пациентов с мезотелиомой и поддерживаемых в культуре в течение нескольких пассажей. Так как жидкость, отбираемая у пациентов, содержит смесь клеток, анализ обеспечивает хорошую оценку относительной активности, но это лишь грубая оценка абсолютной цитотоксичности. Клетки от двух дополнительных пациентов обрабатывали различными концентрациями SS1P, и их жизнеспособность оценивали через четыре дня, используя анализ с кристаллическим фиолетовым. Клетки мезотелиомы пациентов на раннем пассаже NCI-M-16 и NCI-M-19 показали четкие реакции на обработку SS1P (>75% снижение жизнеспособности при уровне дозы 100 нг/мл). Мы оценили активность SS1P-LR/GGS/8M на этих двух клеточных популяциях, полученных от дополнительных пациентов. Данные представлены на Фигуре 9 в виде дробных значений, нормализованных между контрольными обработками D-PBS (100% жизнеспособности) и 10 нг/мл антитрансферринового рецептора/РЕ40 RIT HB21 (0% жизнеспособности). Все клеточные популяции пациентов были крайне чувствительны к обработке SS1P-LR/GGS/8M, демонстрируя значительно повышенную цитотоксичность по сравнению с SS1P.
SS1P является антимезотелиновым рекомбинантным иммунотоксином на основе экзотоксина A Pseudomonas, который в настоящее время находится в клинической разработке для лечения мезотелиомы, но имеет потенциал для лечения различных твердых опухолей, которые экспрессируют мезотелин. В клинических испытаниях SS1P добился скромных, но обнадеживающих результатов. Его эффективность, однако, сведена к определенным пределам из-за ограничивающей дозу токсичности и быстрой выработки нейтрализующих антител у пациентов. Здесь мы сообщаем, что SS1-LR/GGS/8M, вариант SS1P с низкой антигенностью, имеет превосходную активность и заметно сниженную неспецифическую токсичность у грызунов.
SS1-LR/GGS/8M является высокоактивным, менее токсичным и менее антигенным вариантом антимезотелинового RIT SS1P на основе РЕ. Наша первоначальная оценка SS1-LR показала сильно варьирующую, но в целом низкую активность на отобранных клеточных линиях, экспрессирующих мезотелин, in vitro (Таблицы 1 и 2). Выясняя причины его низкой активности по сравнению с SS1P, мы изучали интернализацию и процессинг SS1-LR и обнаружили, что доля SS1-LR, расщепленного фурином, было намного ниже, чем у SS1P в клетках, обработанных этими двумя RIT. Это позволило предположить, что пониженное расщепление фурином может ограничивать активность SS1-LR, и для проверки этой гипотезы нами были разработаны и произведены несколько мутантов. Добавление короткого Gly-Gly-Ser линкера после сайта расщепления фурином не усиливало расщепление фурином, но повышало активность SS1-LR на клеточных линиях. Объединив SS1-LR с линкером GGS и еще восемью точечными мутациями, которые, как было показано, уменьшают иммуногенность РЕ, мы получили нашу конечную молекулу, SS1-LR/GGS/8M. По сравнению с SS1P, SS1-LR/GGS/8M демонстрировала значительно сниженную неспецифическую токсичность в модели повышенной проницаемости капилляров у крыс, повышенную цитотоксичность против клеток пациента, а также снижение реактивности с антителами в сыворотках пациентов. Первоначальные эксперименты с SS1-LR демонстрировали вариабельную цитотоксичность по отношению к SS1P на клеточных линиях. Первичное направление, однако, было к менее активной молекуле. SS1-LR была более активна на линии HAY и менее активна на остальных шести линиях. Эта тенденция заметно отличается от анти-CD22 версии молекулы LR, HA22-LR, которая демонстрировала похожую или повышенную цитотоксичность в большинстве клеточных линий и клеток пациентов (Weldon JE, et al., Blood 2009; 113:3792-800). Это расхождение говорит о том, что есть существенная разница между путем токсического действия РЕ, направленного на мезотелин на эпителиальных клетках, и РЕ, направленного на CD22 на В-клетках.
Что касается общего снижения активности SS1-LR относительно SS1P, одним из возможных объяснений для такого несоответствия является разница во внутриклеточном пути токсического действия. Вариант РЕ38 LR содержит обширные делеции в доменах II и Ib РЕ, и эти делеции могут иметь негативное влияние на способность РЕ перемешаться в цитозоле. Наши эксперименты по обнаружению полноразмерных и расщепленных РЕ в лизатах клеток А431/K5, обработанных SS1P и SS1-LR, показали резкое различие в количестве процессированных фурином RIT. Большая часть от общего количества RIT в клетках, обработанных SS1P, была процессирована, но только небольшая часть от общего количества RIT расщеплялась в клетках, обработанных SS1-LR. Этот результат позволил предположить, что слабое расщепление фурином может ограничивать активность SS1-LR, и мы приступили к улучшению этого этапа пути токсического действия РЕ.
Добавляя гибкий линкер к сайту расщепления фурином в SS1-LR, мы получили более активный RIT, но не смогли продемонстрировать повышенное расщепление фурином. Добавление короткого линкера Gly-Gly-Ser (SS1-LR/GGS, Фиг.1), более длинного линкера (SS1-LR/GGSx2, Фиг.1), или повтора сайта расщепления фурином, фланкированного короткими Gly-Gly-Ser линкерами (SS1-LR/2x фурин, Фиг.1) привело к умеренному повышению цитотоксичности. Ни одна из этих молекул, однако, не повышала долю SS1-LR, расщепленного фурином, в обработанных клетках А431/K5 и не увеличивала скорость расщепления фурином in vitro (данные не показаны). Мы пришли к заключению, что добавление линкера должно повышать цитотоксичность через другой механизм, возможно, связанный с внутриклеточным транспортом или повышенной стабильностью токсина, и продолжаем исследовать эти возможности. Наши эксперименты также показали необходимость расщепления фурином для цитотоксичности SS1P. Точечная мутация в SS1-LR/GGS, которая привела к замене аргинина, необходимого для расщепления, на глицин (SS1-LR/GGS R279G, Фиг.1) произвела белок, который не расщеплялся фурином. Этот RIT показал очень слабую активность на всех клетках, с незначительной активностью при концентрации 120 мкг/мл на шести из семи протестированных линий клеток. Нерасщепляемый мутант демонстрирует EC50 ниже 1 мкг/мл только на клетках А431/K5 (Таблица 2), но его активность, тем не менее, была существенно снижена. Кроме того, искусственно высокая экспрессия мезотелина в этой клеточной линии не может быть репрезентативной для тех линий, которые экспрессируют мезотелин на естественном уровне. Необходимость расщепления фурином в пути токсического действия РЕ недавно была поставлена под сомнение (Morlon-Guyot J, et al., Infect Immun 2009; 77:3090-9), но существует гораздо больше доказательств того, что фурин выполняет важную роль во время токсического действия (Ornatowski W, et al.; J Clin Invest 2007; 117:3489-97; Shiryaev SA, et al., J Biol Chem 2007; 282:20847-53; Sarac MS, et al., Infect Immun 2002; 70:7136-9; Chiron MF, et al., J Biol Chem 1997; 272:31707-11; Gu M, et al., Infect Immun 1996; 64:524-7; Inocencio NM, et al., J Biol Chem 1994; 269:31831-5; and Moehring JM, et al., J Biol Chem 1993; 18:2590-4). В случае, представленном здесь, токсическое действие РЕ обычно нарушается без сайта, пригодного для процессинга фурином.
Наша лаборатория недавно получила RIT, HA22-LR-8M, который имеет крайне низкую иммуногенность из-за удаления B-клеточных эпитопов (Onda M, et al., Proc Natl Acad Sci USA 2011; 108:5742-7). HA22-LR-8M содержит делеции варианта LR РЕ и дополнительные восемь точечных мутаций в домене III. Эти мутации был и в ведены в SS1P, образовав SS1-LR/GGS/8M. Поскольку основная часть иммунного ответа на RIT направлена на РЕ, мы ожидаем, что SS1-LR/GGS/8M продемонстрирует аналогичное снижение иммуногенности. Чтобы подтвердить, что SS1-LR/GGS/8M действительно удаляет эпитопы B-клеток человека из SS1P, мы исследовали реактивность SS1-LR/GGS/8M с сыворотками от пациентов, которые выработали нейтрализующие антитела во время лечения SS1P. В пяти случаях, что мы тестировали, SS1-LR/GGS/8M показал резко сниженную антигенность по сравнению с SS1P. Этот результат согласуется с наблюдениями НА22 и HA22-LR/8M (Onda M, et al., Proc Natl Acad Sci USA 2011; 108:5742-7), и указывает, что мы идентифицировали и удалили многие из иммуногенных эпитопов в RIT на основе РЕ.
Цитотоксичность SS1-LR/GGS/8M оценивали на нескольких клеточных линиях, в модели опухоли у мышей и на первичных клетках от пациентов с мезотелиомой. SS1-LR/GGS/8M демонстрировал отличную Цитотоксичность по отношению к SS1P в линиях клеток мезотелиомы и рака легких, но был слабо активен на линиях клеток рака яичников. Этот результат является неожиданным, так как линии клеток рака яичников были чувствительны к SS1-LR/GGS, и указывает, что восемь точечных мутаций в каталитическом домене РЕ могут мешать активности SS1-LR/GGS/8M. В модели опухоли у мышей ответы ксенотрансплантатов опухоли L55 на SS1-LR/GGS/8M и SS1P были аналогичными, хотя для усиления противоопухолевого эффекта можно было вводить более высокие дозы SS1-LR/GGS/8M. Наконец, при испытании на первичных злокачественных клетках от пациентов с мезотелиомой SS1-LR/GGS/8M демонстрировал удивительно повышенную Цитотоксичность по сравнению с SS1P. В целом, наша оценка показала отличную цитотоксическую активность SS1-LR/GGS/8M.
В дополнение к высокой активности и низкой антигенности SS1-LR/GGS/8M показал снижение неспецифической токсичности по сравнению с SS1P. Модель индуцированного RIT синдрома повышенной проницаемости капилляров у крыс эффективно демонстрирует эту разницу. Не было никаких существенных различий между крысами, получавшими PBS, и крысами, получавшими SS1-LR/GGS/8M, в то время как у крыс, получавших SS1P, наблюдалось вызывающее слабость накопление жидкости в легких. Синдром повышенной проницаемости капилляров (также называемый синдромом повышенной проницаемости сосудов) появляется, когда жидкость вытекает из капилляров, приводя к падению уровня сывороточного альбумина, задержке жидкости, отекам и набору веса. Эта токсичность часто наблюдалась у пациентов, получавших различные иммунотоксины, в том числе иммунотоксины на основе РЕ, и, предположительно, происходила в результате ненаправленного повреждения эндотелиальных клеток. Ограничение ненаправленной токсичности RIT потенциально может повысить их эффективность, позволяя безопасно вводить более высокие дозы. Этот эксперимент, наряду с другими, описанными здесь, говорит о том, что SS1-LR/GGS/8M был бы отличным кандидатом для клинического применения. Низкая антигенность, низкая неспецифическая токсичность и высокая цитотоксичность SS1-LR/GGS/8M являются многообещающими показателями для дальнейшей разработки антимезотелиновых RIT.
Подразумевается, что примеры и варианты осуществления, описанные в данном документе, приведены только для иллюстративных целей, и что различные модификации или изменения в свете этого предлагаются специалистам в данной области и должны быть включены в сущность и объем настоящей заявки и прилагаемой формулы изобретения. Все номера доступа, публикации, патенты и патентные заявки, цитируемые здесь, включены в данный документ посредством ссылки в полном объеме для любых целей.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЭКЗОТОКСИН А PSEUDOMONAS С МЕНЕЕ ИММУНОГЕННЫМИ Т-И/ИЛИ В-КЛЕТОЧНЫМИ ЭПИТОПАМИ | 2012 |
|
RU2627216C2 |
| МОДИФИЦИРОВАННЫЙ ЭКЗОТОКСИН А ПСЕВДОМОНАД | 2014 |
|
RU2687143C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| АКТИВИРУЮЩИЙ NK-КЛЕТКИ СЛИТЫЙ БЕЛОК, NK-КЛЕТКИ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ВКЛЮЧАЮЩАЯ ИХ | 2018 |
|
RU2740438C1 |
| ОДНОЦЕПОЧЕЧНЫЕ АНТИТЕЛА И ДРУГИЕ ГЕТЕРОМУЛЬТИМЕРЫ | 2013 |
|
RU2644341C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2831418C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2708032C2 |
| ПРОЛЕКАРСТВО ИНТЕРФЕРОНА ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2788736C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ TLR2-ОПОСРЕДОВАННЫХ ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2019 |
|
RU2791022C2 |


































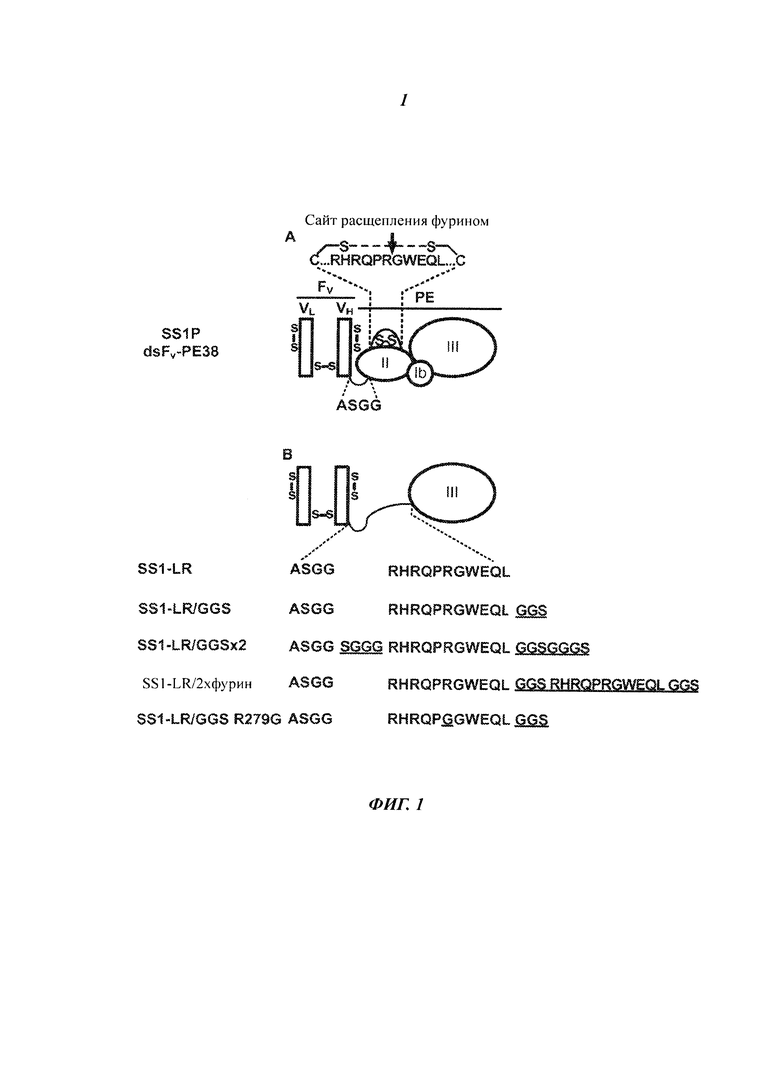
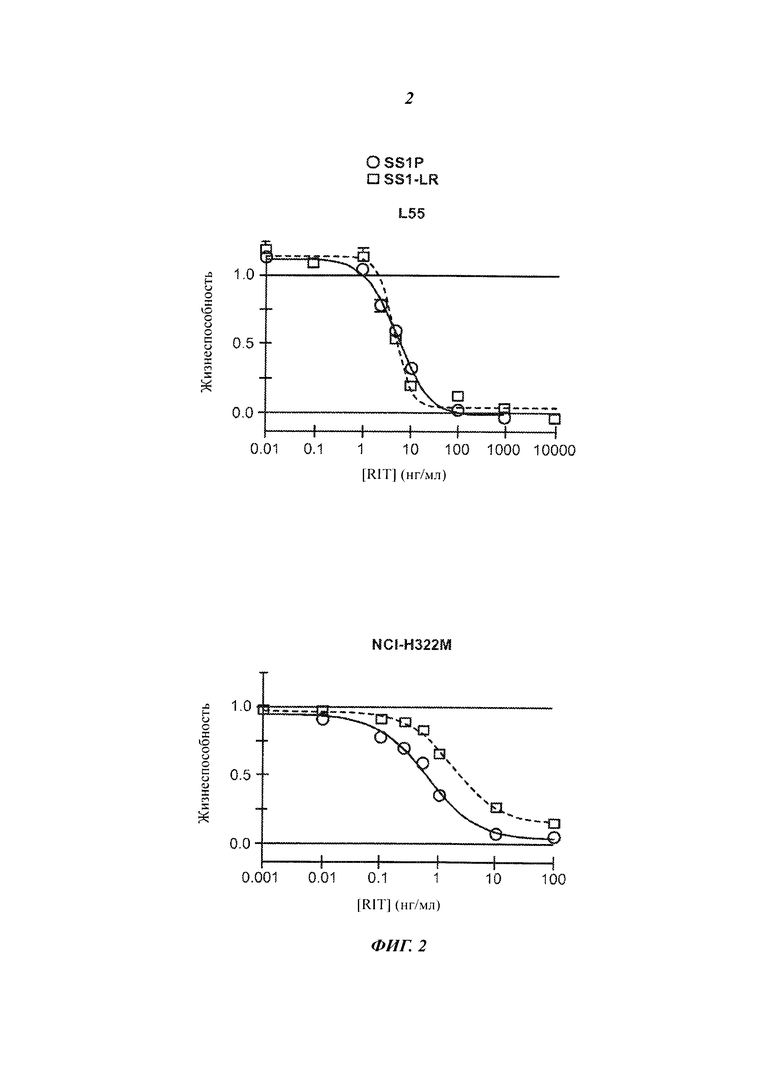
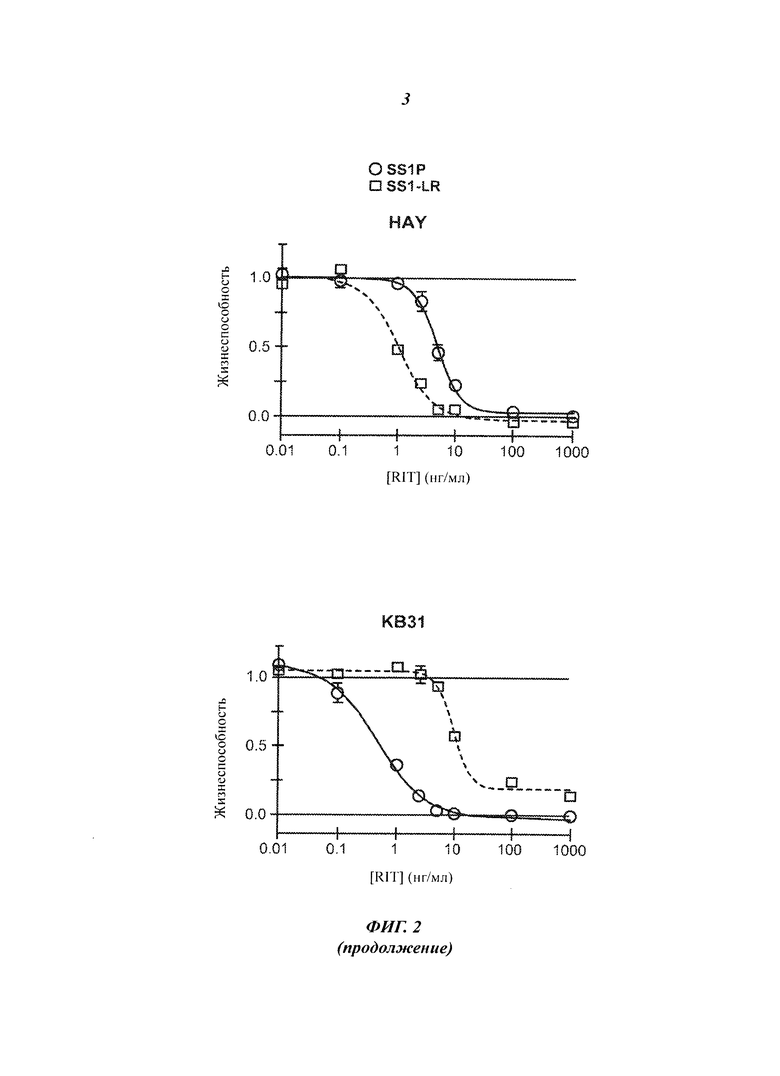
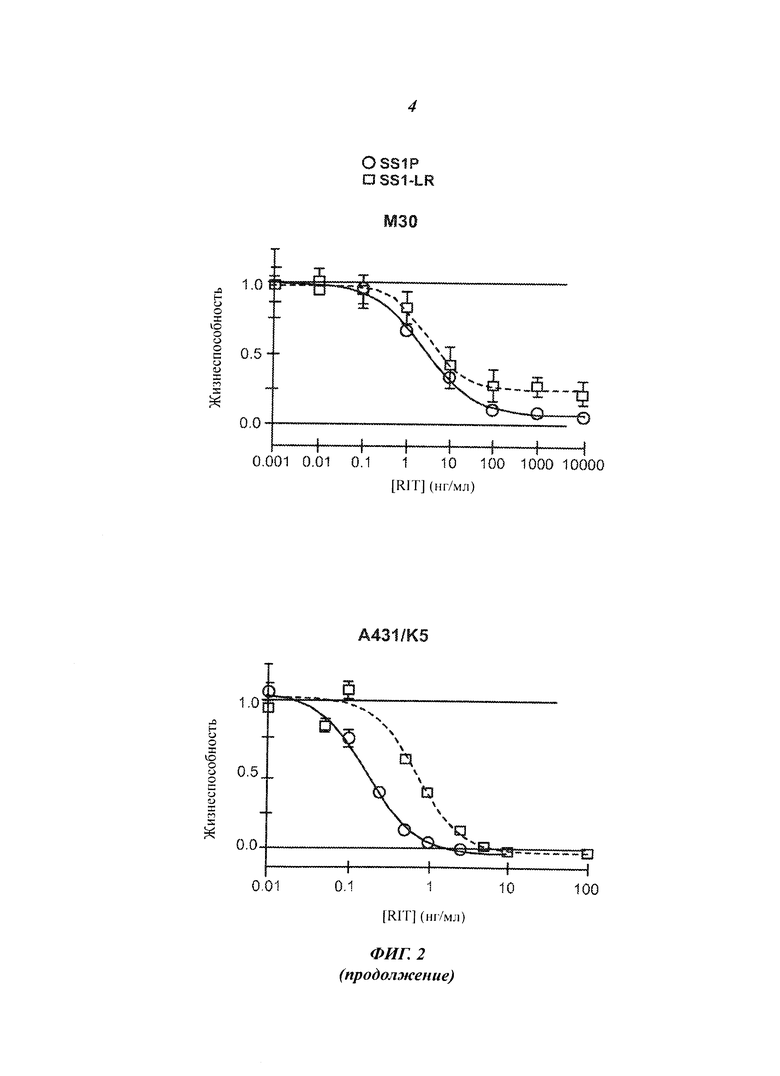
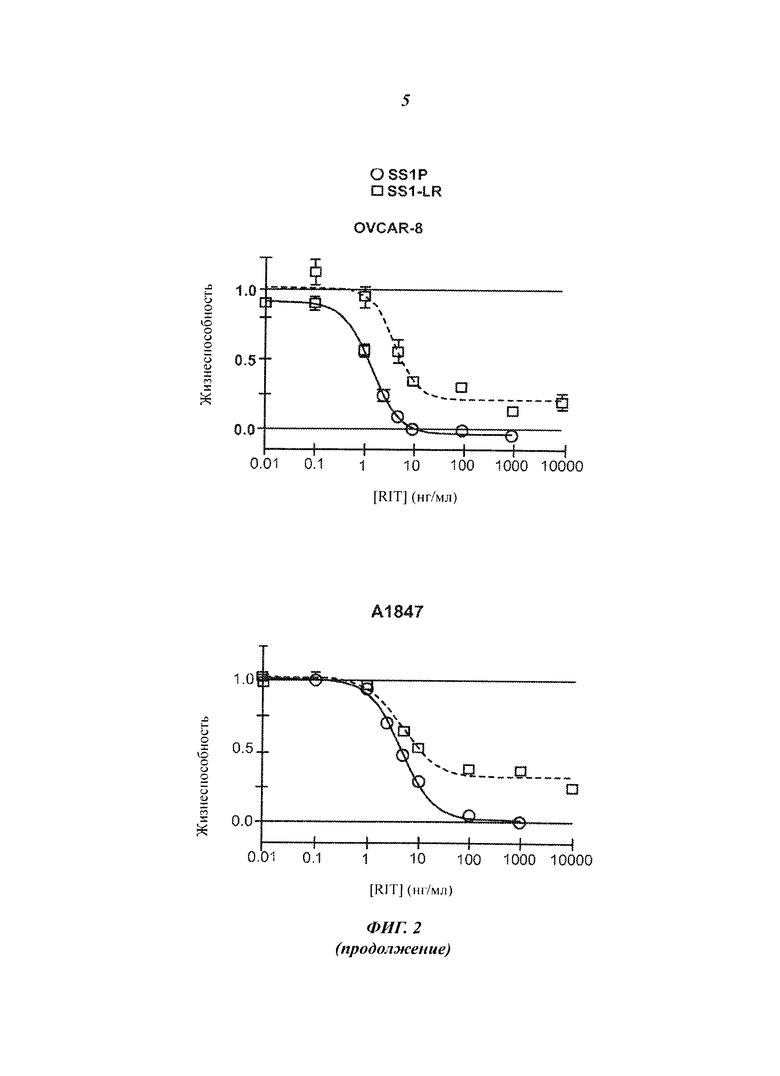
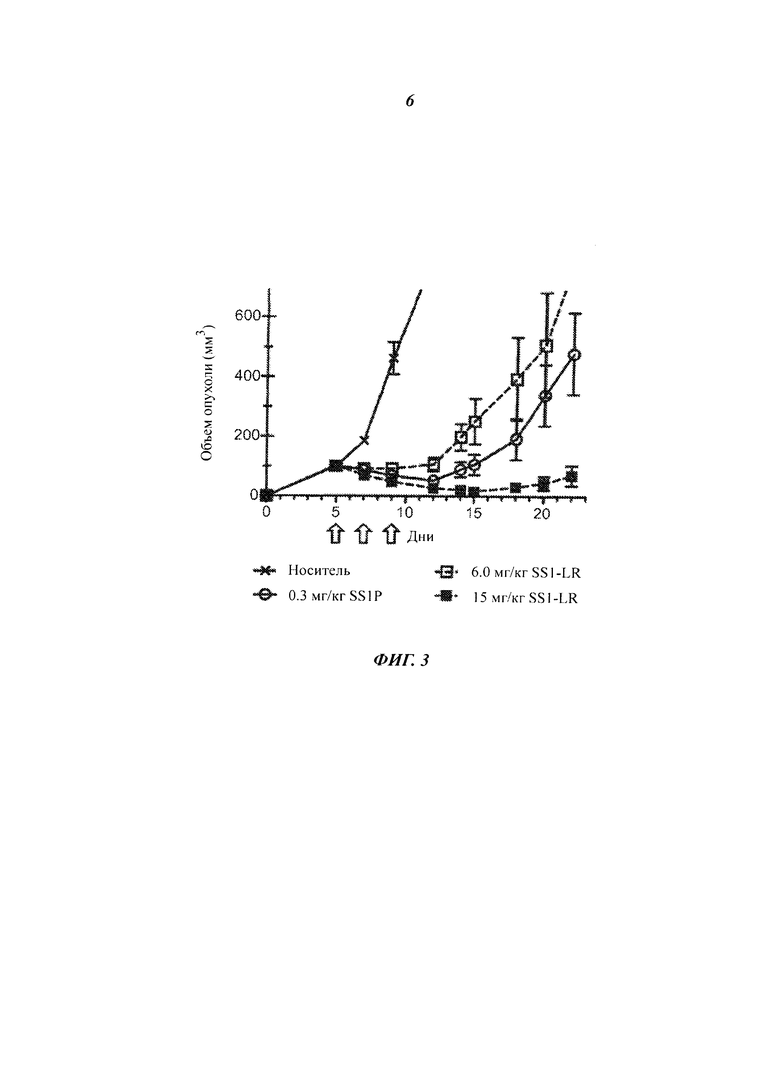
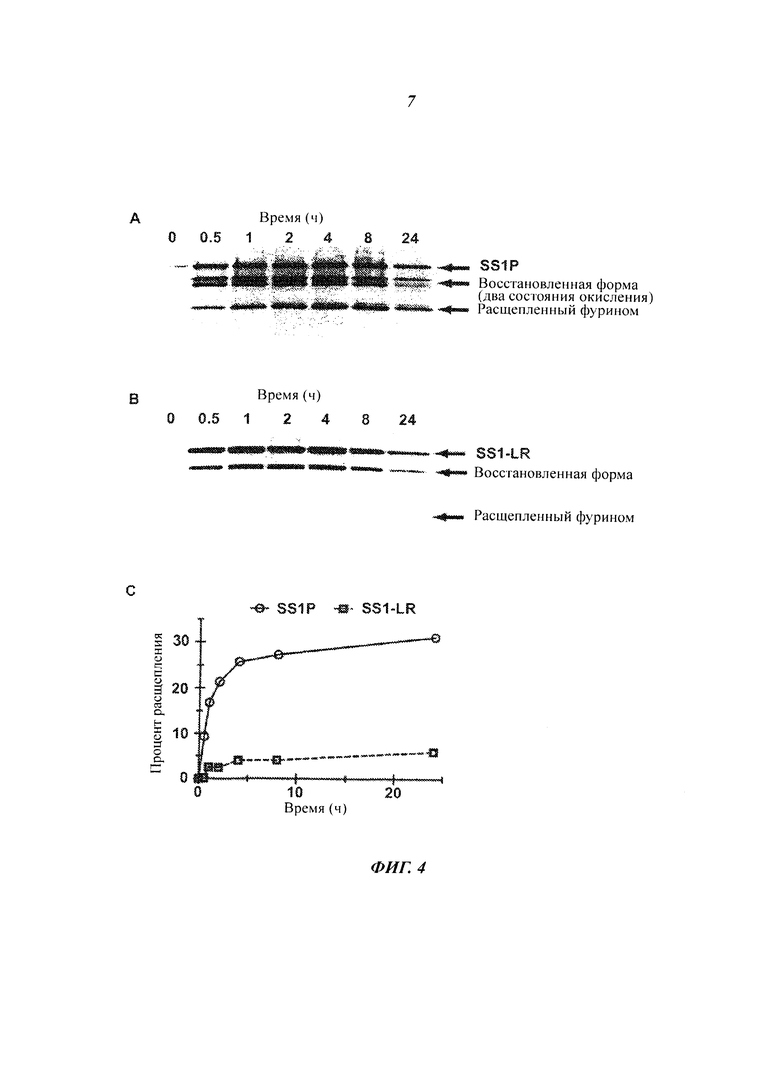

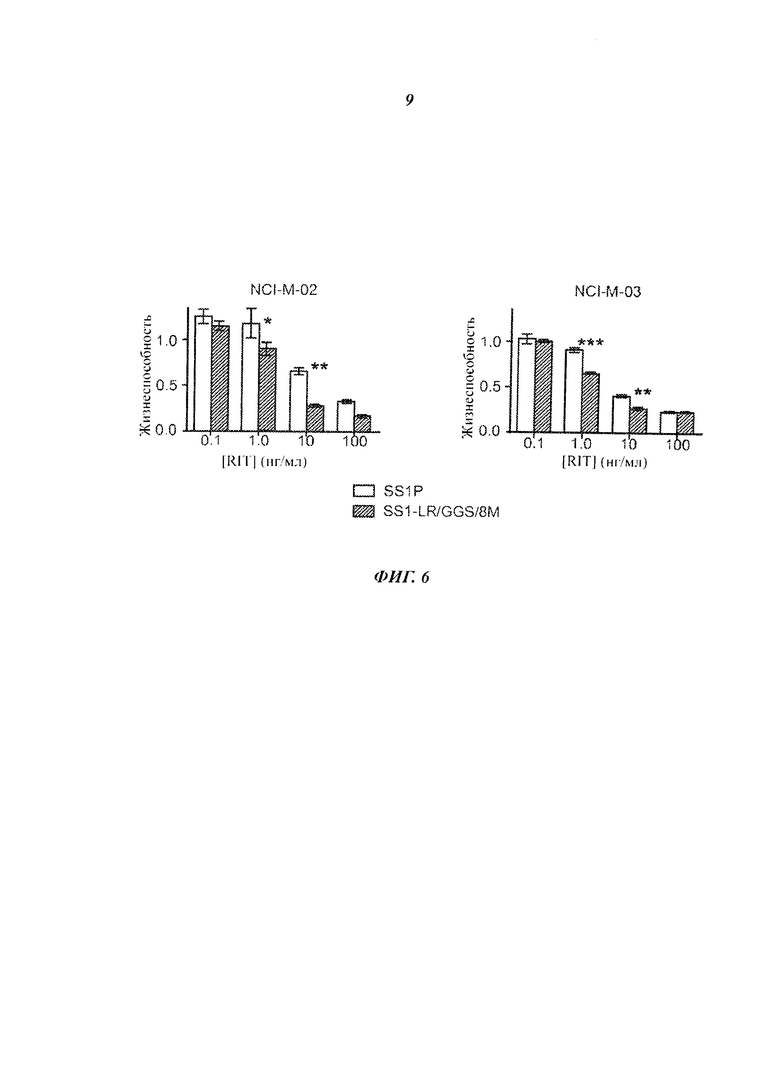
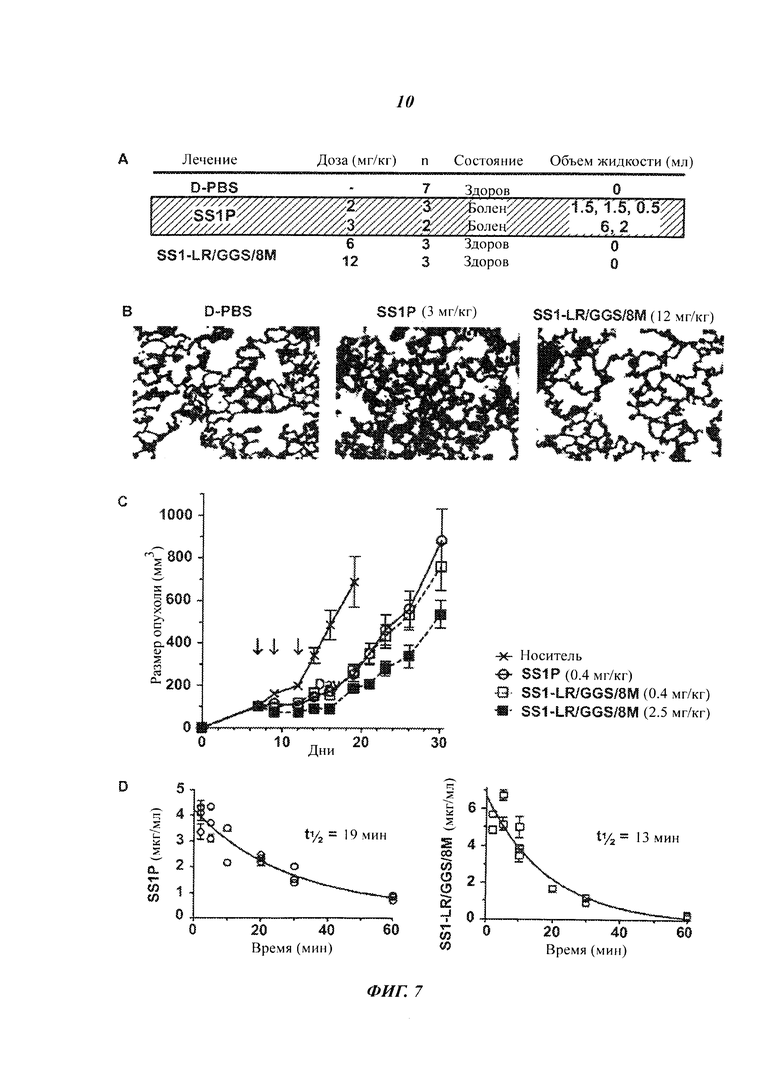
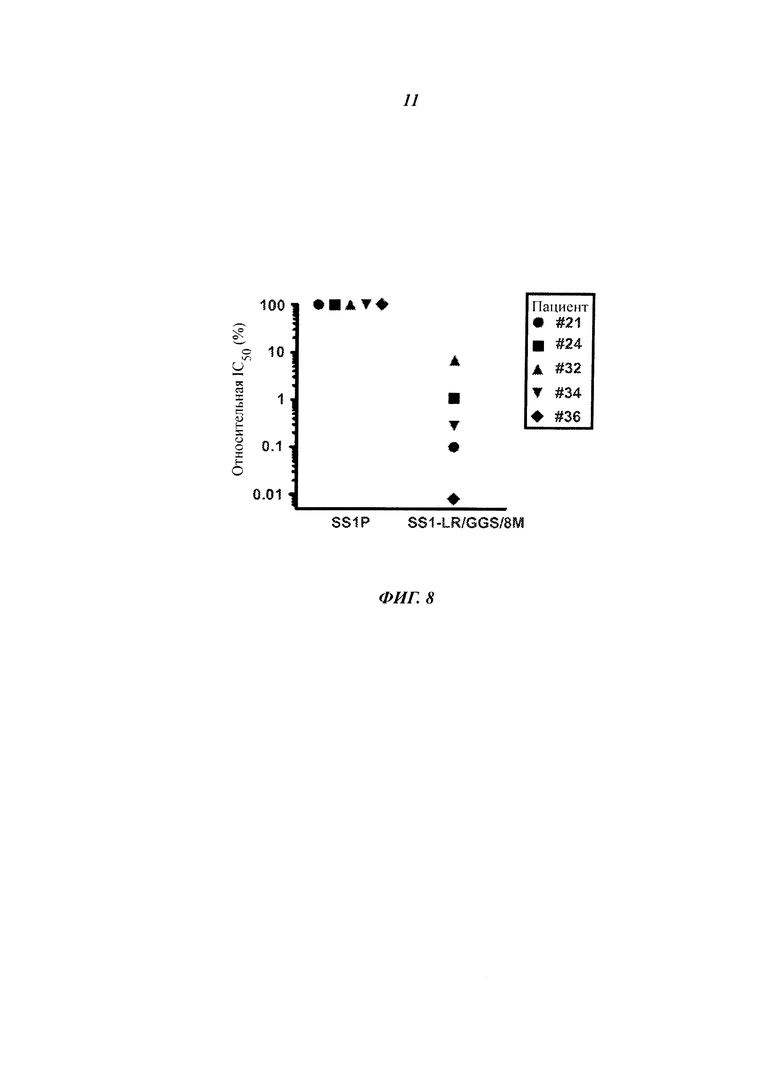
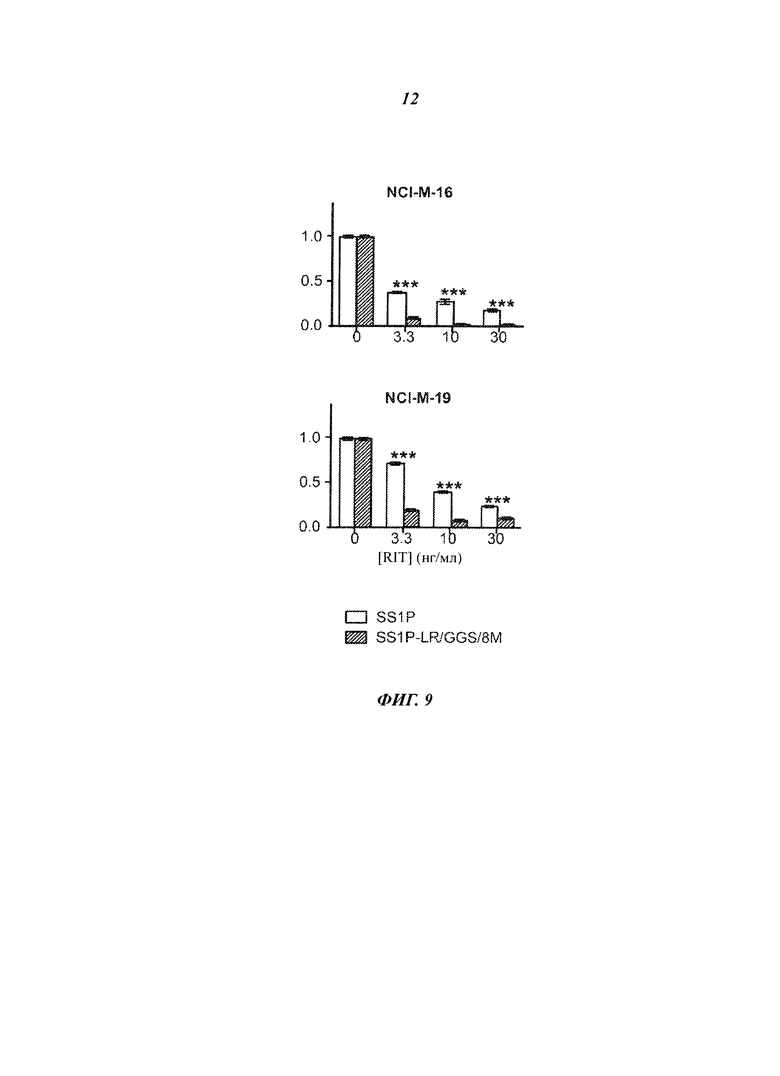
Изобретение относится к области биохимии, в частности к усовершенствованному рекомбинантному иммунотоксину для клеток, экспрессирующих мезотелин. Заявленный иммунотоксин представляет собой слитый белок, включающий антитело к мезотелину и фрагмент экзотоксина Pseudomonas, который модифицирован так, чтобы снизить его иммуногенность и чувствительность к протеазам. Иммунотоксин может быть использован в способе лечения рака, сверхэкспрессирующего мезотелин. Изобретение обеспечивает повышенную цитотоксичность иммунотоксина в отношении клеток, которые экспрессируют мезотелин. 6 н. и 15 з.п. ф-лы, 9 ил., 4 табл., 2 пр.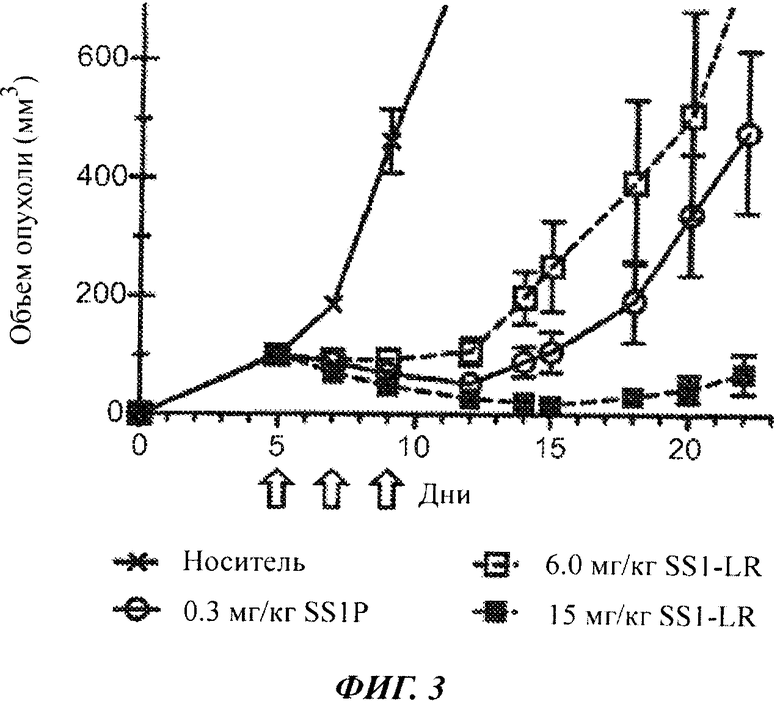
1. Химерная молекула, представляющая собой иммунотоксин для клеток, экспрессирующих мезотелин, содержащая слитый полипептид, имеющий формулу M-L1-FCS-FL-функциональный домен III РЕ, в которой
М представляет собой антитело, которое специфически связывается с мезотелином, или его фрагмент, который специфически связывается с мезотелином, где антитело или фрагмент антитела содержит:
цепь VH, содержащую аминокислотную последовательность CDR1 GYTMN (SEQ ID NO: 51), аминокислотную последовательность CDR2 LITPYNGASSYNQKFRG (SEQ ID NO: 52) и аминокислотную последовательность CDR3 GGYDGRGFDY (SEQ ID NO: 53), и
цепь VL, содержащую аминокислотную последовательность CDR1 SASSSVSYMH (SEQ ID NO: 54), аминокислотную последовательность CDR2 DTSKLAS (SEQ ID NO: 55) и аминокислотную последовательность CDR3, выбранную из группы, состоящей из QQWSGYPLT (SEQ ID NO: 56), QQWSKHPLT (SEQ ID NO: 57), QQWSGHPLT (SEQ ID NO: 58), QQWSAHPLT (SEQ ID NO: 59), QQWSQIPLT (SEQ ID NO: 60), QQWGFNPLT (SEQ ID NO: 61), QQWGTNPLT (SEQ ID NO: 62), QQWGSHPLT (SEQ ID NO: 63), QQWGDFPLT (SEQ ID NO: 64), QQWGDHPLT (SEQ ID NO: 65), QQWSAHPLT (SEQ ID NO: 66) и QQWSGYPTT (SEQ ID NO: 67);
L1 представляет собой линкер, состоящий из непрерывной последовательности от 1 до 10 аминокислотных остатков;
FCS представляет собой сайт расщепления фурином, состоящий из непрерывной последовательности аминокислотных остатков, способных расщепляться фурином;
FL состоит из непрерывной последовательности от 3 до 8 аминокислотных остатков, независимо выбранных из глицина и серина;
функциональный домен III РЕ состоит из непрерывной последовательности аминокислотных остатков, идентичной последовательности SEQ ID NO: 1 от положения 395 до положения 613, необязательно содержащей (i) замену непрерывной последовательности аминокислотных остатков в положениях 609-613 из SEQ ID NO: 1 на KDEL (SEQ ID NO: 16), REDL (SEQ ID NO: 26), REEL (SEQ ID NO: 27) или RDEL (SEQ ID NO: 28), (ii) замену аргинина на глицин, аланин, валин, лейцин или изолейцин в положении, соответствующем положению 490 в SEQ ID NO: 1, (iii) замену, независимо друг от друга, на аланин, глицин, серии или глутамин одного или более остатков, соответствующих остаткам D403, D406, R412, R427, Е431, R432, R458, D461, R467, R490, R505, R513, Е522, R538, Е548, R551, R576, K590, Q592 и L597 последовательности SEQ ID NO: 1, где остатки SEQ ID NO: 1 поддерживают иммуногенность эпитопа или субэпитопа домена III РЕ, или (iv) комбинацию любого из (i)-(iii).
2. Молекула по п.1, в которой антитело или фрагмент антитела представляет собой мини-антитело, диатело, триатело, фрагмент Fab′, фрагмент F(ab)′2, одноцепочечный белок Fv («scFv») или стабилизированный дисульфидными связями фрагмент белка Fv («dsFv»).
3. Молекула по п.1, в которой антитело или фрагмент антитела содержит стабилизированные дисульфидными связями вариабельные участки легкой и тяжелой цепей иммуноглобулина, слитые с остальной частью молекулы посредством связывания тяжелой цепи с L1.
4. Молекула по п.1, в которой аминокислотная последовательность CDR3 VL представляет собой QQWSKHPLT (SEQ ID NO: 57).
5. Молекула по п.1, в которой L1 составляет от 3 до 5 аминокислот в длину.
6. Молекула по п.1, в которой L1 представляет собой ASGG (SEQ ID NO: 19).
7. Молекула по п.1, в которой домен III РЕ имеет замену на аланин, глицин или серии по меньшей мере одного аминокислотного остатка, соответствующего аминокислотному остатку из SEQ ID NO: 1, выбранному из группы, состоящей из D403, R412, R427, Е431, R458, D461, R505, Е522, R538, R551, R576 и L597.
8. Молекула по п.1, в которой домен III РЕ имеет замену на аланин, глицин или серии по меньшей мере одного аминокислотного остатка, соответствующего аминокислотному остатку из SEQ ID NO: 1, выбранному из группы, состоящей из D406, R432, R467, R490, R513, Е548, K590 и Q592.
9. Молекула по п.1, в которой FL представляет собой GGS, (GGS)2 (SEQ ID NO: 18), GSGG (SEQ ID NO: 71), GGSGG (SEQ ID NO: 72), GGSG (SEQ ID NO: 73), GSG или GGG.
10. Молекула по п.1, в которой FL представляет собой GGS.
11. Молекула по п.1, в которой последовательность расщепления фурином представляет собой RHRQPRGWEQL (SEQ ID NO: 17).
12. Молекула по п.1, в которой функциональный домен III РЕ представляет собой функциональный домен III РЕ варианта LR, имеющий аминокислотную последовательность SEQ ID NO: 4 от положения 20 до положения 237 или SEQ ID NO: 5 от положения 20 до положения 237.
13. Молекула по п.1, в которой антитело или фрагмент антитела содержит стабилизированную дисульфидными связями вариабельную легкую цепь SS1 последовательности SEQ ID NO: 6 и вариабельную тяжелую цепь SS1 последовательности SEQ ID NO: 7.
14. Молекула по п.1, в которой молекула содержит вариабельную легкую цепь SS1 последовательности SEQ ID NO: 6 и вариабельную тяжелую цепь SS1 из слитого с РЕ полипептида последовательности SEQ ID NO: 8, где вариабельные легкие и тяжелые цепи образуют стабилизированное дисульфидными связями антитело.
15. Молекула по любому из пп. 1-14, в которой FCS идентичен последовательности SEQ ID NO: 1 от положения 274 до положения 284 или представляет собой аминокислотную последовательность RHRSKRGWEQL (SEQ ID NO: 29).
16. Фармацевтическая композиция для лечения рака, который сверхэкспрессирует мезотелин, у субъекта, нуждающегося в этом, содержащая молекулу по любому из пп.1-14 и фармацевтически приемлемый наполнитель.
17. Нуклеиновая кислота, кодирующая молекулу по любому из пп.1-14.
18. Экспрессионный вектор, содержащий нуклеиновую кислоту по п.17.
19. Клетка-хозяин, продуцирующая химерную молекулу по любому из пп.1-14, содержащая нуклеиновую кислоту по п.17.
20. Способ лечения рака, который сверхэкспрессирует мезотелин, у субъекта, нуждающегося в этом, включающий введение субъекту химерной молекулы по любому из пп.1-14.
21. Способ по п.20, где рак представляет собой аденокарциному легкого, рак яичников, мезотелиому и/или плоскоклеточный рак.
| WO 2009032954 A1, 12.03.2009 | |||
| Судовая гребная установка | 1980 |
|
SU954329A1 |
| US 20100215656 A1, 26.08.2010 | |||
| Устройство для резки полимерного полотна | 1979 |
|
SU861091A1 |
| US 20020151689 A1, 17.10.2002 | |||
| US 6074644 A1, 13.06.2000 | |||
| ПЕРЕКРЫВАЮЩЕЕ КРОВОТОК ТВЕРДОФАЗНОЕ СРЕДСТВО С ПОКРЫТИЕМ ИЗ ИММОБИЛИЗОВАННОГО СВЯЗЫВАЮЩЕГО СРЕДСТВА | 2002 |
|
RU2295329C2 |

