1. Область изобретения
Данное изобретение относится в общем к областям молекулярной биологии и токсикологии. Более конкретно, оно относится к способам получения модифицированных белков, которые являются более короткими и/или менее антигенными полипептидами, а также к композициям, содержащим такие полипептиды. В данном описании описаны более короткие и менее антигенные варианты растительного токсина гелонина. Такие модифицированные белки имеют терапевтические и диагностические применения, например, в качестве иммунотоксинов.
2. Описание уровня техники
Пептиды, полипептиды и белки имеют многочисленные профилактические, диагностические и терапевтические полезные эффекты. Однако одним недостатком является то, что такие белковые соединения могут индуцировать иммунный ответ на эти соединения у субъекта, который надеется получить пользу от них. Иммунный ответ на эти соединения может уменьшать или полностью элиминировать полезные эффекты, которые могут быть достигнуты их применением. Таким образом, главной задачей является уменьшение антигенности или иммуногенности соединения, эффективность которого может уменьшаться индуцированием им иммунного ответа у хозяина.
Один особый тип белка, моноклональные антитела, были в центре большой исследовательской работы и разработок для достижения профилактических, диагностических и терапевтических полезных эффектов. На протяжении двух последних десятилетий были разработаны и испытаны высокоспецифические иммунотоксины, узнающие различные антигены клеточной поверхности. Привлекательной чертой иммунотоксинов является то, что эти сильнодействующие агенты требуют успешной доставки очень малого количества молекул к нужному внутриклеточному компартменту для индукции цитотоксического эффекта. Были сконструированы иммунотоксины, содержащие различные токсины, такие как сапонин, абрин, А-цепь рицина (RTA), экзотоксин Pseudomonas (РЕ), дифтерийный токсин (DT) и гелонин.
Проблемы, связанные с применением иммунотоксинов in vivo, обычно включают в себя повреждение сосудов, ведущее к синдрому подтекания капилляров, ошибочное нацеливание вследствие узнавания части токсина ретикулоэндотелиальной системой, гетерогенность экспрессии антигена-мишени и развитие антител против токсина, ведущее к суженному окну применения терапии приблизительно 14 дней. Индукция антител против токсинов и против конъюгатов может также препятствовать повторному лечению пациентов, несмотря на наличие противоопухолевого действия. Продолжительное применение иммунотоксинов у пациентов также вызывает проблемы. Было обнаружено, что иммуноконъюгаты, содержащие RTA и РЕ, являются высокоиммуногенными у пациентов. Кроме того, существует подозрение, что размер этих белков в конструкциях иммунотоксинов (приблизительно 30 кДа) препятствует эффективному проникновению иммуноконъюгатов в солидные опухоли. Структурная модификация белков типа I, таких как RTA, была большей частью безуспешной (Munishkin et al., 1995). Были получены многочисленные мутанты RTA, модифицирующие несколько аминокислот. В 1995 году Wool et al. описали 45 делеций отдельных аминокислот RTA. Было показано, что из них только 8 делеций отдельных аминокислот имели биологическую активность, хотя относительные биологические активности этих делеционных мутантов в сравнении с нативным RTA не испытывались. Эти исследования с испытанием RTA, хотя и являются интересными, имеют ограниченную ценность, так как, например, RTA имеет только 30% гомологию последовательности с другими токсинами, такими как гелонин.
Конкретные применения процедур на основе моноклональных антител (mAb) традиционно использовались в диагностике и терапии злокачественных опухолей человека. Однако клиническое применение этих агентов имело ограниченный успех вследствие недостатков, связанных с этим подходом, например, гетерогенности экспрессии антигена, слабого проникновения в опухоль в случае солидных опухолей частично из-за размера антител и антигенности этих антител (Roselli et al., 1993; Berkower, 1996; Pullibland et al., 1997; Panchagnula et al., 1997; Panchal, 1998). Для преодоления этих проблем был применен ряд молекулярных подходов для изменения конфигурации обычной структуры антитела в химеры мышь:человек, полностью человеческие антитела или фрагменты антител с измененной формой, содержащие антигенсвязывающие части исходной структуры в меньшем и более простом (одноцепочечном) формате (Bird et al., 1988; Kipriyanov et al., 1994; Owens et al., 1994; McCartney et al., 1995; Worn et al., 1998). Одноцепочечные антитела (scfv, sfv), сохраняющие связывающие свойства исходного иммуноглобулина (IgG) состоят из доменов VL и VH антитела, связанных сконструированным гибким пептидным линкером (Wels et al., 1992; Kurucz et al., 1993). Кроме того, scFv могут быть предпочтительными в клинических и диагностических приложениях, в настоящее время включающих общепринятые mAb или их Fab-фрагменты, так как их меньший размер может обеспечить лучшее проникновение в опухолевую ткань, улучшенные фармакокинетические свойства и уменьшение иммуногенности, наблюдаемой при внутривенном введении мышиных антител.
Среди нескольких антигенов-мишеней, которые высокопродуктивно экспрессируются в клетках меланомы в сравнении с нормальной тканью, находится поверхностный домен высокомолекулярного гликопротеина (gp240), обнаруживаемого на большинстве клеточных линий меланомы и свежих образцах опухолей (Kantor et al., 1982). Ранее были выделены и описаны два мышиных антитела (названные 9.2.27 и ZME-018), узнающих различные эпитопы на этом антигене (Morgan et al., 1981; Wilson et al., 1981). Мышиное моноклональное антитело ZME-018 обладает высокой специфичностью в отношении меланомы и является минимально реактивным с различными здоровыми тканями, что делает его перспективным кандидатом для дальнейшего исследования. Клинические испытания, исследующие способность этого антитела локализоваться в очагах меланомы, показали селективное концентрирование в метастатических опухолях (Macey et al., 1988; Koizumi et al., 1988).
Успешное развитие нацеленных на опухоль лекарственных средств зависит, отчасти, от сайт-специфической доставки лекарственных средств, а также от биологической активности доставляемого агента. Моноклональные антитела были использованы для придания селективности в иных условиях неселективным цитотоксическим агентам, таким как токсины, радионуклиды и факторы роста (Williams et al., 1990; Rowlinson-Busza et al., 1992; Wahl, 1994). Одной из таких молекул является гелонин, инактивирующий рибосомы растительный токсин 29 кДа, обладающий силой и механизмом действия, сходными с А-цепью рицина (RTA), но имеющий улучшенную стабильность и уменьшенную токсичность (Stirpe et al., 1992; Rosenblum et al., 1995). Предыдущие исследования в лаборатории авторов изобретения позволили идентифицировать и исследовать биологические свойства многочисленных химических конъюгатов растительного токсина гелонина и различных антител (Boyle et al., 1995; Xu et al., 1996; Rosenblum et al., 1999). В предыдущих исследованиях антитело ZME-018 химически связывали с очищенным гелонином, и этот иммуноконъюгат продемонстрировал специфическую цитотоксичность против антиген-положительных клеток меланомы как в культуре ткани, так и в моделях ксенотрансплантатов опухоли человека (Rosenblum et al., 1991; Mujoo et al., 1995). Однако эта конструкция, как и иммунотоксины вообще, имела присущие ей проблемы антигенности у пациентов-людей.
При наличии побочных эффектов иммунотоксинов и ограниченного прогресса, достигнутого в уменьшении этих проблем, продолжает существовать потребность в разработке менее антигенных белков, полипептидов и пептидов для применения в лечении, профилактике и диагностике заболеваний и состояний. Замена антигенных последовательностей в молекуле токсина является концепцией в отношении не являющихся антителами полипептидов, таких как токсины. Хотя эта концепция была успешно использована с заменой каркасных доменов мышиного иммуноглобулина каркасными доменами человеческого иммуноглобулина с образованием химерной молекулы человек/мышь, та же самая концепция никогда не применялась с успехом к другим молекулам, в частности токсинам или ферментам из растительных источников, или с использованием описанных в данном описании способов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к способам создания и получения белковых соединений, которые модифицируют с образованием модифицированного белка, обладающего преимуществом над немодифицированным или нативным белком. Данное изобретение относится также к композициям, которые получают этими способами.
Некоторые варианты осуществления данного изобретения представляют собой рекомбинантный токсин гелонин, который является измененным относительно последовательности нативного гелонина. Рекомбинантный токсин гелонин может иметь замененные или удаленные аминокислоты в сравнении с нативной последовательностью белка гелонина (показанной в SEQ ID NO:1), которая описана в патенте США № 5631348, включенном в данное описание в качестве ссылки, и которая имеет номер доступа GenBank L12243. Рекомбинантный токсин гелонин, или данное изобретение, не содержит всех аминокислот SEQ ID NO:1, но в некоторых вариантах осуществления содержит коровую область токсина, определяемую аминокислотными остатками 110-210 SEQ ID NO:1. Другие соединения данного изобретения включают в себя рекомбинантный токсин гелонин, который содержит коровую область токсина, наряду с наличием по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 или более непрерывных аминокислотных остатков SEQ ID NO:1 в дополнение к коровой области токсина. Предполагается, что соединения данного изобретения включают в себя также множественные области, которые включают в себя смежные аминокислотные остатки SEQ ID NO:1. Например, соединение может включать в себя коровую область токсина, наряду с 10 смежными аминокислотными остатками SEQ ID NO:1 перед коровой областью токсина и 20 смежными аминокислотными остатками SEQ ID NO:1 после коровой области токсина.
Рекомбинантный токсин гелонин данного изобретения также включает в себя токсин гелонин, который является укороченным относительно нативной последовательности, так что этот токсин не имеет по меньшей мере 5, 10, 20, 30, 40, 50 или более аминокислотных остатков SEQ ID NO:1. В некоторых вариантах осуществления данного изобретения этот токсин содержит коровую область токсина, но не содержит аминокислот, находящихся где-либо вне коровой области токсина. Кроме делеций, рекомбинантный токсин гелонин может содержать аминокислоту вместо удаленной аминокислоты. Например, остаток глицина в положении 7 в последовательности белка гелонина может быть заменен аминокислотным остатком, не являющимся глицином, или модифицированной аминокислотой. Если остаток глицина в положении 7 просто удален, аланин в положении 8 в SEQ ID NO:1 становится положением 7, но не считается заменой, так как положения аминокислот просто смещаются на одно положение. Предполагается, что могут быть заменены по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 и более аминокислотных остатков.
В следующих вариантах осуществления данного изобретения рекомбинантный токсин гелонин может быть присоединен ко второму полипептиду. В некоторых случаях второй полипептид служит для нацеливания токсина гелонина на определенный тип клеток (в том числе на клетки, имеющие определенный генотип или фенотип, например злокачественные клетки или клетку, инфицированную патогеном), часть тела или другое конкретное местоположение. Таким образом, белковые соединения данного изобретения включают в себя соединение, которое содержит как рекомбинантный токсин гелонин, так и модифицированный токсин гелонин и второй полипептид. В некоторых вариантах осуществления эти два полипептида конъюгированы друг с другом, в то время как в других вариантах осуществления эти полипептиды сконструированы рекомбинантно для получения слитого белка. Конъюгированные соединения могут быть соединены друг с другом линкером. Предполагается, что модифицированные белки данного изобретения могут включать в себя дополнительные полипептидные композиции, все или некоторые из которых могут быть ковалентно связаны друг с другом.
Данное изобретение относится к мультиполипептидным композициям, в которых более чем одна полипептидная частица присутствуют в виде единого соединения. Таким образом, модифицированный белок может быть присоединен ко второму, третьему, четвертому, пятому, шестому полипептидам или к большему количеству полипептидов. Альтернативно, два или более модифицированных белков могут присутствовать в виде единого белкового соединения. В некоторых вариантах осуществления данного изобретения второй полипептид является антителом, например антителом с антигенсвязывающей областью. Обсуждается, что антитело может быть направлено против опухолевого антигена, продукта онкогена, клеточного рецептора или любого другого соединения, которое определяет местоположение этой мультипептидной композиции. Как описано в данном описании, второй полипептид может быть ферментом, цитокином, цитотоксической молекулой, фактором роста, лигандом или рецептором или любой молекулой, которая способна модифицировать характеристики роста клеток.
Другие композиции данного изобретения включают в себя модифицированный фермент, получаемый способом, который предусматривает: а) идентификацию одной или нескольких антигенных областей в этом ферменте с использованием антитела; b) удаление одной или нескольких антигенных областей из этого фермента с получением модифицированного фермента и с) определение ферментативной активности модифицированного фермента. Фермент является биологической молекулой, которая катализирует конкретную химическую реакцию в клетке; он может быть молекулой белка или молекулой нуклеиновой кислоты. Однако предполагается, что любые способы, обсуждаемые в связи с ферментами, могут быть применены к полипептидам вообще. Антигенная область является областью полипептида, которая специфически узнается антителом или Т-клеточным рецептором конкретного организма. Понятно, что область может быть антигенной в одном виде, но не антигенной в другом виде, и, следовательно, антигенность соединения является характеристикой, которая относится к конкретному организму. Предполагается, что, кроме удаления аминокислот, которые являются частью антигенной области, аминокислоты из нескольких (не только из одного) антигенных областей могут быть удалены из фермента данного изобретения. Аминокислоты из всех или из части 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более антигенных областей могут быть удалены из этого полипептида. В некоторых случаях удаленную область заменяют областью, которая является менее антигенной, чем удаленная область. Конечно, должно быть понятно, что аминокислота, фланкирующая антигенную область, может быть также удалена, например, для удобства. Таким образом, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более аминокислот, фланкирующих одну или обе стороны антигенной области, могут быть удалены или заменены.
Менее антигенная область или области могут быть идентифицированы поиском в базе данных белков областей, которые являются гомологичными антигенной области или имеют несколько остатков, общих с антигенной областью. Антигенная область может быть идентифицирована, и эту последовательность используют для идентификации известных белковых последовательностей организма, в котором является желательной меньшая антигенность в отношении модифицированного белка. Таким образом, база данных белков человека может быть использована для нахождения белковых последовательностей человека, которые имеют множественные остатки, которые являются идентичными или сравнимыми с остатками антигенной области белка, которая должна быть менее антигенной у людей. Остаток является сравнимым с другим остатком, если эти остатки не являются идентичными, но имеют общие сходные химические свойства. Такие взаимосвязи хорошо известны специалистам с квалификацией в данной области.
В некоторых вариантах осуществления для идентификации антигенной области используют антитело. Предполагается, что антитело может быть поликлональным. Организмом-источником этого антитела является тот же самый вид организма, в котором модифицированный белок, как это желательно, должен быть менее антигенным. Таким образом, если желательно, чтобы фермент или белок был менее антигенным у человека, желательно в некоторых вариантах осуществления, чтобы были использованы антитела человека либо для идентификации антигенной области, либо для определения, является ли модифицированный белок менее антигенным, чем немодифицированный белок (нативный или рекомбинантный полноразмерный белок). В предпочтительном варианте осуществления модифицированный фермент или белок оценивают на уменьшенную или более низкую антигенность сравнением антигенности модифицированного фермента или белка с немодифицированным ферментом или белком; это может быть выполнено i) получением образца у субъекта перед подверганием действию или введением модифицированного белка и применением этого образца для сравнения антигенности модифицированного белка и немодифицированного варианта того же самого белка, или ii) получением образца из субъекта после подвергания действию или введения модифицированного белка и применением этого образца для сравнения антигенности модифицированного белка и немодифицированного варианта того же самого белка. Образцом может быть любая композиция, которая содержит антитела или иммунные клетки, в том числе жидкости тела, такие как кровь (сыворотка). Затем этот образец может быть использован для выполнения способа иммунодетектирования, такого как ELISA. Предполагается, что субъект может быть нативным в отношении немодифицированного белка, хотя предпочтительно, чтобы субъект давал образец, который ранее подвергался действию немодифицированного белка. В некоторых вариантах осуществления может быть удобным, чтобы образцом была культуральная среда из гибридомы моноклональных антител.
Хотя в других аспектах данного изобретения определение, обладает ли модифицированный белок или фермент активностью, может выполняться анализом модифицированного соединения на активность, например ферментативную активность.
Любой фермент может быть модифицирован в соответствии со способами данного изобретения. Ферментом может быть гидролаза (например, дезаминаза, эстераза, гликозидаза, липаза, нуклеаза, пептидаза, фосфатаза, фосфодиэстераза и протеиназа); изомераза (например, эпимераза, мутаза и рацемаза); лигаза или синтетаза (например, ацил-CoA-синтетаза, аминоацил-тРНК-синтетаза и карбоксилаза); лиаза (например, альдолаза, декарбоксилаза, дегидратаза и нуклеотидциклаза); оксидоредуктаза (например, дегидрогеназа, диоксигеназа, гидрогеназа, монооксигеназа, нитрогеназа, оксидаза и редуктаза) и трансфераза (например, ацилтрансфераза, аминотрансфераза, гликозилтрансфераза, киназа, метилтрансфераза, нуклеотидилтрансфераза, фосфорилаза и сульфотрансфераза). В конкретных вариантах осуществления этот фермент классифицируется как токсин, что означает, что он является токсичным для клетки, ткани или организма. В качестве части данного изобретения конкретно обсуждаются токсины, продуцируемые растениями, такие как гелонин. Как обсуждалось ранее, модифицированный фермент, такой как модифицированные полипептиды гелонина данного изобретения, может быть присоединен к дополнительным полипептидам. Понятно, что любой из вариантов, касающихся модифицированного гелонина, может быть применен к модифицированным ферментам, и наоборот.
Данное изобретение относится также к способам генерирования модифицированных белков, которые имеют уменьшенную антигенность, и в некоторых случаях, в частности, в отношении субъекта. В некоторых вариантах осуществления этот способ предусматривает: а) выбор белка, который желательно ввести первому субъекту; b) идентификацию области этого белка, которая является антигенной для первого субъекта, с использованием антисыворотки либо из первого субъекта, либо из второго субъекта того же самого вида, что и первый субъект; с) генерирование модифицированного белка, в котором отсутствует идентифицированная область, и d) подтверждение, что этот модифицированный белок имеет уменьшенную антигенность. Как обсуждалось ранее, эта последняя стадия может быть выполнена с использованием образца, такого как сыворотка, у индивидуума, который был ранее подвергнут действию немодифицированного варианта модифицированного белка, или у индивидуума, у которого является желательным уменьшенный иммунный ответ против модифицированного белка.
Далее предполагается, что способы генерирования модифицированного белка включают в себя скрининг базы данных белков человека для идентификации менее антигенной области, которая имеет гомологию в отношении антигенной области этого белка, и замену этой антигенной области всей идентифицированной областью или частью идентифицированной области, которая является менее антигенной, для образования модифицированного белка. Скрининг большой базы данных белков человека не является необходимым, но является желательным. Таким образом, если последовательность конкретного белка человека, которая имеет гомологию или идентичные остатки с антигенной областью, является известной независимо от скрининга базы данных белков человека, этот способ будет включен в объем данного изобретения. Например, может быть известной последовательность человеческого гомолога мышиного фермента, уменьшенная антигенность которого является желательной; замена областей в мышином белке остатками из последовательности человека относится к данному изобретению. Способы и композиции данного изобретения включают в себя замену, делецию и/или модификацию аминокислотных остатков полипептида. Остаток, который заменяют, делает как порядок, так и число остальных аминокислот такими же, какие были в полипептиде перед заменой остатка. Остаток может быть заменен консервативным или неконсервативным остатком. Остаток, который делетируется, не нарушает порядка остальных аминокислот, но уменьшает число остатков этого полипептида на один остаток. Остаток, который модифицируют, является остатком, который химически изменен; это изменение не изменяет порядка или числа остальных аминокислот в данном полипептиде.
В некоторых вариантах осуществления способы включают в себя использование технологии рекомбинантных нуклеиновых кислот для получения модифицированного белка или фермента. Так, кДНК-последовательность для фермента, который желательно модифицировать, может быть подвергнута манипуляции таким образом, что последовательность нуклеиновой кислоты, которая кодирует антигенную область, заменяется последовательностью нуклеиновой кислоты, которая кодирует менее антигенную область. Альтернативно, модифицированный белок может быть генерирован удалением идентифицированной области. Область, которую удаляют, считается отсутствующей. Отсутствующая область может содержать по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 или более аминокислотных остатков. Кроме того, модифицированный белок может иметь более чем одну удаленную или замененную антигенную область, и могут быть также удалены или заменены аминокислоты, фланкирующие эту область. Предполагается, что отсутствующая антигенная область может быть заменена тем же самым количеством аминокислотных остатков, какое было удалено.
В способах данного изобретения антигенная область может быть идентифицирована или модифицированная область может быть оценена при помощи анализа ELISA. Субъектом может быть млекопитающее, например человек.
Другие композиции данного изобретения включают в себя гуманизированный рекомбинантный токсин гелонин, имеющий по меньшей мере 3 аминокислоты из одного или нескольких из антигенных доменов 1, 2, 3 или 4, замененных аминокислотами, менее антигенными у человека, чем рекомбинантный токсин гелонин с этими замененными аминокислотами. Антигенные домены токсина гелонина описаны в другом месте. Предполагается, что по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50, 75, 100 или более аминокислотных остатков из антигенных доменов 1, 2, 3 и/или 4 являются замененными, делетированными или модифицированными. Аминокислоты из по меньшей мере 2, 3 или 4 антигенных доменов могут быть подвергнуты манипуляции.
Дополнительные варианты данного изобретения обеспечивают рекомбинантный токсин гелонин, получаемый способом, предусматривающим: а) идентификацию по меньшей мере одной области в токсине гелонине, которая является антигенной у млекопитающего, и b) замену по меньшей мере части этой антигенной области областью, менее антигенной у этого млекопитающего. Предполагается, что токсин гелонин может быть рекомбинантным, т.е. произведенным из последовательности нуклеиновой кислоты, которая была подвергнута манипуляции in vitro. Этот способ может также предусматривать сравнение идентифицированной антигенной области с аминокислотными последовательностями млекопитающего, посредством чего идентифицируют область, менее антигенную у этого млекопитающего, или идентификацию области, которая является менее антигенной у этого млекопитающего. В некоторых вариантах осуществления этим млекопитающим является человек. Как упоминалось ранее, любые из способов и композиций, описанных в данном описании, могут быть применены к любым другим способам и композициям, описанным в данном описании.
Данное изобретение относится также к способам лечения с использованием композиций данного изобретения. Они могут использоваться в лечении любого заболевания, в котором лечение имеет форму лизиса или элиминации определенных клеток или организмов, на которые действуют токсины данного изобретения. Предполагается, что варианты осуществления, обсуждаемые в отношении одной композиции или одного способа, могут быть применены к любой другой композиции или любому другому способу данного изобретения.
В некоторых вариантах осуществления описан способ лизиса злокачественных или опухолевых клеток предоставлением этим клеткам эффективного количества иммунотоксина, который включает в себя весь токсин гелонин или часть токсина гелонина, такую как коровую область токсина, или все антитело или часть антитела, которая используется для направления этого иммунотоксина к конкретной клетке. "Эффективным количеством" называют количество, которое выполняет предполагаемую задачу. В случае способа для лизиса злокачественной или опухолевой клетки оно является количеством, достаточным для лизиса злокачественной или опухолевой клетки. Другие способы данного изобретения включают в себя способы для лечения злокачественных опухолей у пациента введением этому пациенту эффективного количества композиции, содержащей иммунотоксин, содержащий коровую область токсина гелонина и одноцепочечное антитело, которое специфически нацелено на злокачественную клетку. "Эффективным количеством" в отношении лечения называют количество, которое является достаточным для обеспечения терапевтической пользы субъекту. Термин "терапевтическая польза", используемый во всей этой заявке, обозначает все, что стимулирует или усиливает хорошее самочувствие субъекта в связи с лекарственным лечением его состояния. В контексте злокачественных опухолей (хотя этот термин может применяться также к другим состояниям) терапевтическая польза, которая относится к лечению предрака, злокачественных опухолей и гиперпролиферативных заболеваний, включает в себя следующие неисчерпывающие примеры: продление жизни субъекта на любой период времени, ослабление или задержку развития заболевания, ослабление гиперпролиферации, ослабление роста опухоли, задержку метастазирования, ослабление скорости пролиферации злокачественных клеток или опухолевых клеток и ослабление боли у субъекта, которая может быть связана с этим состоянием субъекта.
В некоторых вариантах осуществления данного изобретения токсином является гелонин. В других вариантах этот иммунотоксин включает в себя всю аминокислотную последовательность или часть аминокислотной последовательности SEQ ID NO:1. Предполагается, что этот иммунотоксин может включать в себя меньше аминокислот, чем полноразмерная (непроцессированная) белковая последовательность гелонина, хотя в некоторых вариантах осуществления он включает в себя полноразмерную последовательность. Далее обсуждается, что этот токсин может быть гуманизирован, и он может представлять собой любой токсин или конструкцию, заявленные или описанные в данном описании.
В следующих вариантах осуществления антитело иммунотоксина является гуманизированным и/или является одноцепочечным антителом. В способах данного изобретения антитело нацеливает иммунотоксин на злокачественную клетку-мишень, хотя оно может быть не полноразмерным, пока оно обеспечивает возможность специфического нацеливания. В некоторых вариантах это антитело (которое может быть и фрагментами антител) специфически нацелено на антиген (т.е. связывается с антигеном) на поверхности клетки-мишени. В более специфических вариантах осуществления это антитело нацелено на опухолевый антиген. Антитело может быть любым антителом млекопитающих, хотя конкретно обсуждается, что антитело является антителом мыши, кролика, крысы, козы или обезьяны. Антитело, хотя и происходящее от различных видов, может быть гуманизировано в соответствии с данным изобретением или другими способами, известными специалистам с обычной квалификацией в данной области. В случаях, в которых антитело является одноцепочечным антителом, оно может включать в себя 9.2.27 или ZME-018, которые являются антителами, направленными на клетки меланомы. В конкретных примерах иммунотоксин представляет собой scfvMEL-2018 или scfvMEL-2025 (SEQ ID NO:11), описанные в данном описании.
Злокачественная клетка, которая является мишенью, может быть злокачественной клеткой из предстательной железы, легкого, головного мозга, кожи, печени, молочной железы, лимфоидного органа, желудка, яичек, яичника, поджелудочной железы, костной ткани, костного мозга, головы и шеи, шейки матки, пищевода, глаза, желчного пузыря, почки, надпочечников, сердца, ободочной кишки или крови. Альтернативно, больной злокачественной опухолью может иметь злокачественную опухоль органов/тканей, указанных выше. В некоторых вариантах осуществления данного изобретения злокачественной клеткой является клетка меланомы. Предполагается, что злокачественная или опухолевая клетка может находиться у пациента. В некоторых вариантах осуществления пациенту вводят эффективное количество терапевтической композиции, что подразумевает количество, необходимое для получения конкретного желаемого результата, например, лечения. В контексте злокачественной опухоли, например, желаемым результатом может быть лизис злокачественной или опухолевой клетки.
Иммунотоксин может быть включен в фармацевтически или фармакологически приемлемую композицию. В качестве части схемы лечения пациент может также получать другую противораковую терапию, такую как химиотерапия, лучевая терапия, генотерапия, хирургическое вмешательство или другая иммунотерапия.
В следующих вариантах осуществления обсуждается, что иммунотоксин может быть доставлен к клетке или пациенту с помощью экспрессирующей конструкции, которая содержит последовательность нуклеиновой кислоты, кодирующую иммунотоксин, и способна экспрессировать этот иммунотоксин. В некоторых вариантах осуществления этой экспрессирующей конструкцией является вирусный вектор, в том числе, но не только, аденовирусный вектор, аденоассоциированный вирусный вектор, вирус гепатита, герпес-вирус, лентивирус, ретровирус или вирус коровьей оспы.
Применение неопределенного артикля "а" или "an" вместе с термином «содержащий» в формуле изобретения и/или в описании может означать «один», но может также совпадать со значением «один или более», «по меньшей мере один» и «один или более чем один».
Другие цели, признаки и преимущества данного изобретения станут очевидными из следующего подробного описания. Однако должно быть понятно, что это подробное описание и конкретные примеры, хотя они представляют характерные варианты осуществления данного изобретения, приводятся только в качестве иллюстрации, так как различные изменения и модификации в идее и объеме данного изобретения будут очевидными специалистам с квалификацией в данной области из этого подробного описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Следующие чертежи образуют часть данного описания и включены для дополнительной демонстрации определенных аспектов данного изобретения. Данное изобретение может быть лучше понято со ссылкой на 1 или несколько из этих чертежей вместе с подробным описанием представленных в данном описании конкретных вариантов осуществления.
Фиг. 1. ELISA, выполненный с использованием антитела против рекомбинантного гелонина (rGelonin) на образцах сыворотки человека.
Фиг. 2. Эпитопы рекомбинантного гелонина, узнаваемые антителами человека против гелонина.
Фиг. 3А-3В. Делеционные конструкции гелонина. Показаны структуры делеционных конструкций гелонина.
Фиг. 4. Схематическое конструирование на основе ПЦР слитого токсина sfvMEL/rGel и лигирование в произведенные из рЕТ-32а векторы.
Фиг. 5. Полный анализ последовательности ДНК («сиквенс») слитой конструкции sfvMEL/rGel (SEQ ID NO:10).
Фиг. 6. Сравнительное связывание исходного химического конъюгата ZME-rGel и слитой конструкции sfvMEL-rGel (то же самое, что и sfvMEL/rGel). Связывание с клетками А-375 оценивали с использованием ELISA и поликлонального кроличьего антитела против гелонина. Связывание обеих конструкций с клетками-мишенями было сходным, хотя для рекомбинантной слитой конструкции наблюдали более высокое связывание.
Фиг. 7. Сравнительная цитотоксичность in vitro исходного химического конъюгата ZME-rGel и слитой конструкции sfvMEL-rGel на антиген-положительных клетках А375 меланомы человека. Клетки высевали и затем обрабатывали в течение 72 часов различными дозами слитой конструкции sfvMEL/rGel, химического конъюгата ZME-rGel или свободного рекомбинантного гелонина. Величины IC50 для обоих иммуноконъюгатов были приблизительно 8 нМ, хотя IC50 для рекомбинантного гелонина была на несколько порядков величин более высокой при приблизительно 2·103 нМ.
Фиг. 8. Конкурентное ингибирование иммунотоксина sfvMEL/rGel антителом ZME. Различные концентрации рекомбинантного иммунотоксина добавляли к клеткам А-375 меланомы человека в культуре log-фазы роста в четырех повторностях. Для другого ряда лунок фиксированную концентрацию антитела ZME (50 мкг/мл) смешивали с различными дозами иммунотоксина sfvMEL/rGel и инкубировали в течение 72 часов. Добавление свободного антитела ZME приводило к приблизительно 3-кратному уменьшению цитотоксичности иммунотоксина.
Фиг. 9. Голых мышей, несущих хорошо развившиеся опухоли меланомы (А-375), растущие в правом боку, обрабатывали (i.v.) либо солевым раствором (контроли), либо sfvMEL/rGel при 2 или 20 мг/кг (общая доза) в течение 4 последовательных дней (стрелки). Площади опухолей измеряли в течение 30 дней. Обработанные солевым раствором контрольные опухоли увеличивались от 30 до 150 мм2 на протяжении этого периода. Опухоли, обработанные самой низкой дозой иммунотоксина, увеличивались от 30 до 60 мм2. Животные, обработанные самой высокой дозой иммунотоксина, не показали общего увеличения в размере опухоли в сравнении с исходным размером 30 мм2.
Фиг. 10. Цитотоксичность scfvMEL-CFR2018 (также известного как sfvMEL-CFR2018) сравнивали с цитотоксичностью scfvMEL-CFR2025 на клетках меланомы А375-М в анализе цитотоксичности in vitro.
ОПИСАНИЕ ИЛЛЮСТРАТИВНЫХ ВАРИАНТОВ
Белки и полипептиды с уменьшенной антигенностью могут обеспечивать огромную пользу в составе композиций, вводимых организму с иммунной системой. В данном описании описаны способы конструирования и получения таких белков и полипептидов, а также полученные в результате молекулы. Ферменты являются особенно интересными кандидатами для этих способов, так как может быть желательным сохранение ферментативной активности конкретного фермента, а также уменьшение его антигенности у субъекта, который может иметь пользу от ферментативной активности этого белка. Примером такого белка являются инактивирующие рибосомы белки (RIP). Таким образом, в некоторых вариантах этого изобретения обеспечены композиции нуклеиновых кислот и полипептидов, которые включают в себя растительные токсины, такие как гелонин. Могут быть сконструированы белки для обеспечения токсической функции одного полипептида в комбинации с другим полипептидом, таким как нацеливающая молекула. Сконструированные токсины имеют большое разнообразие применений.
I. Белковые соединения
В некоторых вариантах осуществления данное изобретение относится к новым композициям, содержащим белковую молекулу, которая была модифицирована относительно нативного белка или белка дикого типа. В некоторых вариантах осуществления в этом белковом соединении были делетированы аминокислотные остатки; в других вариантах осуществления аминокислотные остатки белкового соединения были заменены, тогда как в иных вариантах осуществления были произведены как делеции, так и замены аминокислотных остатков в этом белковом соединении. Кроме того, белковое соединение может быть аминокислотной молекулой, содержащей более чем одну полипептидную часть. Применяемые в данном описании термины "белковая молекула", "белковая композиция", "белковое соединение", "белковая цепь" или "белковый материал" обычно относятся, но не только, к белку, содержащему более чем приблизительно 200 аминокислот, или полноразмерной эндогенной последовательности (первичному продукту трансляции), транслируемой из гена; полипептиду, имеющему более чем приблизительно 100 аминокислот, и/или пептиду, состоящему из приблизительно 3 - приблизительно 100 аминокислот. Все термины "белковый", описанные выше, могут использоваться в данном описании взаимозаменяемо. Кроме того, эти термины могут использоваться также в отношении слитых белков или белковых конъюгатов.
В некоторых вариантах осуществления размер по меньшей мере одной белковой молекулы может содержать без ограничения приблизительно или по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240, 250, 275, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1100, 1200, 1300, 1400, 1500, 1750, 3000, 2250, 2500 или больше аминокислотных остатков, и любой диапазон, который может быть получен из них. Соединения данного изобретения могут включать в себя вышеупомянутый ряд смежных аминокислот из SEQ ID NO:1 и/или SEQ ID NO:11. Предполагается, что варианты осуществления относительно SEQ ID NO:1 могут быть использованы в отношении любых других аминокислотных последовательностей, описанных в данном описании, в том числе SEQ ID NO:11, и наоборот, если это требуется.
Таким образом, термин «белковая композиция» включает в себя последовательности молекул аминокислот, содержащие по меньшей мере одну из 20 обычных аминокислот в природно синтезируемых белках или по меньшей мере одну модифицированную или необычную аминокислоту, в том числе, но не только, аминокислоты, приведенные в таблице 1 ниже.
Модифицированные и необычные аминокислоты
Применяемый в данном описании термин «аминокислота» относится к любой аминокислоте, любому производному аминокислоты или любому миметику аминокислоты, как известно специалисту с обычной квалификацией в данной области. В некоторых вариантах осуществления остатки белковой молекулы являются последовательными, без какой-либо не содержащей аминогруппы молекулы, прерывающей последовательность аминокислотных остатков. В других вариантах осуществления эта последовательность может содержать одну или несколько не являющихся аминокислотами частей молекулы. В конкретных вариантах осуществления последовательность остатков белковой молекулы может быть прервана частями, не являющимися аминокислотами.
1. Функциональные аспекты
Данное изобретение относится к модифицированным белкам, в частности к белкам, которые приносят терапевтическую пользу субъекту, поскольку этот модифицированный белок проявляет функциональную активность, которая сравнима с активностью немодифицированного белка, но все же этот модифицированный белок обладает дополнительным преимуществом у субъекта в сравнении с немодифицированным белком, таким как меньшая антигенность и/или индуцирование меньших побочных эффектов и/или обладание лучшей или более длительной эффективностью. Таким образом, когда в данной заявке ссылаются на функцию или активность "модифицированного белка", специалист с обычной квалификацией в данной области должен понимать, что речь идет, например, о белке, который 1) проявляет ту же самую активность или имеет ту же самую специфичность, что и немодифицированный белок, но который может иметь отличающийся уровень активности, и 2) проявляет дополнительное преимущество в сравнении с немодифицированным белком. Определение активности может быть выполнено с использованием анализов, известных специалистам с квалификацией в данной области, в частности, в отношении активности белка, и может включать в себя, для целей сравнения, например, применение нативного и/или рекомбинантного вариантов либо модифицированного, либо немодифицированного белка.
2. Модифицированные белки
Модифицированные белки данного изобретения могут содержать делеции и/или замены аминокислот; таким образом, белок с делецией, белок с заменой и белок с делецией и заменой являются модифицированными белками. В некоторых вариантах осуществления эти модифицированные белки могут дополнительно содержать инсерции или добавленные аминокислоты, например, как в случае слитых белков или белков с линкерами. "Модифицированный делетированный белок" лишен одного или нескольких остатков нативного белка, но обладает специфичностью и/или активностью нативного белка. "Модифицированный делетированный белок" может также иметь уменьшенную иммуногенность или антигенность. Примером модифицированного делетированного белка является белок, который имеет делетированный аминокислотный остаток из по меньшей мере одной антигенной области, т.е. области этого белка, определенной как антигенная область в конкретном организме, например в типе организма, которому может быть введен этот модифицированный белок.
Замещенные или замененные варианты обычно содержат замену одной аминокислоты другой аминокислотой в одном или нескольких сайтах в этом белке и могут быть сконструированы таким образом, чтобы модулировать одно или несколько свойств полипептида, в частности уменьшать его иммуногенность/антигенность, уменьшать любые побочные эффекты у субъекта или увеличивать его эффективность. Замены этого рода предпочтительно являются консервативными, т.е. одна аминокислота заменена другой аминокислотой сходной формы и со сходным зарядом. Консервативные замены хорошо известны в данной области и включают в себя, например, замену аланина на серин; аргинина на лизин; аспарагина на глутамин или гистидин; аспартата на глутамат; цистеина на серин; глутамина на аспарагин; глутамата на аспартат; глицина на пролин; гистидина на аспарагин или глутамин; изолейцина на лейцин или валин; лейцина на валин или изолейцин; лизина на аргинин; метионина на лейцин или изолейцин; фенилаланина на тирозин; лейцин или метионин; серина на треонин; треонина на серин; триптофана на тирозин; тирозина на триптофан или фенилаланин и валина на изолейцин или лейцин. Антигенная область полипептида может быть заменена менее антигенной областью; эта менее антигенная область может содержать остатки, которые являются идентичными соответствующим остаткам в нативном белке, но может также содержать некоторые консервативные замены и/или неконсервативные замены.
Кроме делеции или замены, модифицированный белок может иметь инсерцию остатков, которая обычно является добавлением по меньшей мере одного остатка в данный полипептид. Инсерцией может быть инсерция нацеливающего пептида или полипептида или просто единственного остатка. Терминальные (концевые) добавления, называемые слитыми белками, обсуждаются ниже.
Термин «биологически функциональный эквивалент» хорошо известен в данной области и дополнительно определяется подробно в данном описании. Таким образом, последовательности, которые имеют между приблизительно 70% и приблизительно 80%, или между приблизительно 81% и приблизительно 90%, или даже между приблизительно 91% и приблизительно 99% аминокислот, которые являются идентичными или функционально эквивалентными аминокислотам нативного полипептида, включены в этот термин, при условии, что сохраняется биологическая активность этого белка. Модифицированный белок может быть биологически функционально эквивалентным его нативной копии.
Должно быть также понятно, что аминокислотные последовательности и последовательности нуклеиновых кислот могут включать в себя дополнительные остатки, такие как дополнительные N- или С-концевые аминокислоты или 5'- или 3'-последовательности, и все еще быть по существу такими же, как приведенные в одной из описанных в данном описании последовательностей, пока эта последовательность удовлетворяет приведенным выше критериям, в том числе сохранению биологической активности белка, если идет речь об экспрессии белка. Добавление терминальных (концевых) последовательностей касается, в частности, последовательностей нуклеиновых кислот, которые могут, например, включать в себя различные некодирующие последовательности, фланкирующие либо 5'-, либо 3'-части кодирующей области, или могут включать в себя различные внутренние последовательности, т.е. интроны, которые, как известно, встречаются в генах.
Далее следует обсуждение, основывающееся на изменении аминокислот в белке для создания эквивалентной или даже улучшенной молекулы второго поколения. Например, определенные аминокислоты могут заменять другие аминокислоты в структуре белка без ощутимой потери интерактивной связывающей способности с такими структурами, как, например, сайты связывания в молекулах субстрата. Поскольку именно интерактивная способность и природа белка определяют биологическую функциональную активность этого белка, в последовательности белка и в определяющей его кодирующей последовательности ДНК могут быть произведены определенные аминокислотные замены с получением тем не менее белка со сходными свойствами. Таким образом, авторы изобретения полагают, что различные изменения могут быть произведены в последовательностях ДНК генов без ощутимой потери их биологической применимости или активности, как обсуждается ниже. Таблица 1 показывает кодоны, которые кодируют конкретные аминокислоты. Белковая молекула имеет "гомологию" или считается "гомологичной" со второй белковой молекулой, если удовлетворяется один из следующих "критериев гомологии": 1) по меньшей мере 30% этой белковой молекулы имеет идентичность последовательности в одних и тех же положениях со второй белковой молекулой; 2) имеется некоторая идентичность последовательности в одних и тех же положениях со второй белковой молекулой и в неидентичных остатках, по меньшей мере 30% из них являются консервативными различиями, как описано в данном описании, относительно второй белковой молекулы, или 3) по меньшей мере 30% белковой последовательности имеет идентичность последовательности со второй белковой молекулой, но с возможными пропусками неидентичных остатков между идентичными остатками. Применяемый в данном описании термин "гомологичные" может быть равным образом применен к району белковой молекулы, вместо всей молекулы. Если термин "гомология" или "гомологичная" определяется числом, например, "50% гомология" или "на 50% гомологичная", то критерии гомологии, относящиеся к 1), 2) и 3), корректируются изменением "по меньшей мере 30%" на "по меньшей мере 50%". Таким образом, предполагается, что может быть гомология по меньшей мере 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более между двумя белковыми молекулами или частями белковых молекул.
При введении таких изменений может учитываться гидропатический индекс аминокислот. Важность гидропатического индекса аминокислот в придании интерактивной биологической функции белку обычно является понятной в данной области (Kyte & Doolittle, 1982). Общепринятым является то, что относительный гидропатический характер аминокислоты вносит вклад во вторичную структуру полученного белка, которая, в свою очередь, определяет взаимодействие этого белка с другими молекулами, например ферментами, субстратами, рецепторами, ДНК, антителами, антигенами и т.п.
Также общепризнанным в данной области является то, что замена подобных аминокислот может эффективно производиться на основе гидрофильности. Патент США 4554101, включенный в данное описание в качестве ссылки, утверждает, что наивысшая локальная средняя гидрофильность белка, определяемая гидрофильностью его примыкающих друг к другу аминокислот, коррелирует с биологическим свойством данного белка. Как подробно описано в патенте США 4554101, следующие величины гидрофильности были приписаны аминокислотным остаткам: аргинин (+3,0); лизин (+3,0); аспартат (+3,0±1); глутамат (+3,0±1); серин (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5±1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5); триптофан (-3,4).
Должно быть понятно, что аминокислота может быть заменена другой аминокислотой, имеющей сходную величину гидрофильности, и все еще продуцировать биологически эквивалентный и иммунологически эквивалентный белок. В таких изменениях замена аминокислот, величины гидрофильности которых находятся в пределах ±2, является предпочтительной, аминокислоты, которые находятся в пределах ±1, являются особенно предпочтительными и аминокислоты, которые находятся в пределах ±0,5, являются даже еще более предпочтительными.
Как описано в общих чертах выше, замены аминокислот обычно основаны на относительном сходстве заместителей боковых цепей аминокислот, например, на их гидрофобности, гидрофильности, заряде, размере и т.п. Примеры замен, которые учитывают различные предыдущие характеристики, хорошо известны специалистам с квалификацией в данной области и включают в себя аргинин и лизин; глутамат и аспартат; серин и треонин; глутамин и аспарагин; и валин, лейцин и изолейцин.
В таблице 2 приведен перечень белков и полипептидов, которые могут быть модифицированы в соответствии со способами данного изобретения, описанными в данном описании. Полипептиды не человека особо обсуждаются в качестве мишеней способов данного изобретения для уменьшения их антигенности у человека. Предполагается, что белки не человека с терапевтической ценностью находятся в рамках данного изобретения. Любые другие белки или полипептиды, обсуждаемые в этом описании, могут быть модифицированы в соответствии со способами данного изобретения.
факторы роста
IL-11, IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IL-19
серосодержащие группы
селенсодержащие группы
Другим вариантом для получения модифицированных полипептидов в соответствии с данным изобретением является применение пептидных миметиков. Миметики являются пептидсодержащими молекулами, которые имитируют элементы вторичной структуры белков. См., например, Johnson (1993). Лежащим в основе обоснованием применения пептидных миметиков является то, что пептидный скелет белков существует главным образом для ориентации боковых цепей аминокислот таким образом, чтобы облегчить молекулярные взаимодействия, такие как взаимодействия антитела и антигена. Ожидается, что пептидный миметик обеспечит возможность молекулярных взаимодействий, сходных с взаимодействиями природной молекулы. Эти принципы могут быть использованы, вместе с описанными выше принципами, для конструирования модифицированных белковых молекул второго поколения, имеющих многие из природных свойств нативного белка, но с измененными и, в некоторых случаях, даже улучшенными характеристиками.
3. Мультиполипептидные белковые соединения
Данное изобретение относится к белковому соединению, которое может включать аминокислотные последовательности из более чем одного полипептида. Например, белковое соединение или белковая молекула могли бы включать в себя модифицированный токсин с антигенсвязывающей областью антитела. Мультиполипептидная белковая молекула может быть двумя или более белками, химически конъюгированными друг с другом, или она может быть слитым белком двух или более полипептидов, кодируемых одной и той же молекулой нуклеиновой кислоты. Слитый или конъюгированный белок, содержащий токсин и второй полипептид с активностью, может быть назван "двойным токсином". Таким образом, мультипептидное белковое соединение может состоять из всего первого полипептида или части первого полипептида и всего второго полипептида или части второго полипептида, третьего полипептида, четвертого полипептида, пятого полипептида, шестого полипептида, седьмого полипептида, восьмого полипептида, девятого полипептида, десятого полипептида или большего числа полипептидов.
Сами сконструированные токсины обычно не способны связываться с поверхностью клеток или интернализоваться в специфических клетках. Таким образом, эти агенты требуют либо химической конъюгации, либо слияния с агентами/белками, которые способны связываться со специфическими клетками-мишенями и эффективно интернализоваться в клетку после связывания. Таблица 3 обеспечивает перечень белков и полипептидов, которые могут быть конъюгированы или слиты с токсинами данного изобретения, в частности, в вариантах, предусматривающих нацеливание сконструированных белковых соединений к конкретно расположенным, например, специфическим типам клеток или частям тела. Далее, данное изобретение предусматривает присоединение всей молекулы токсина или ее части ко всему белку или его части из белков, перечисленных в таблице 2. Подразумевается, что данное изобретение включает в себя, но не ограничиваются ими, примеры, приведенные в таблицах 2 и 3.
факторы роста
а. Слитые белки
Особым типом инсерционного варианта является слитый белок. Эта молекула обычно содержит целую нативную молекулу или существенную часть нативной молекулы, связанную на N- или С-конце, с полным вторым полипептидом или частью второго полипептида. Например, слитые белки обычно используют лидерные последовательности из других видов для обеспечения возможности экспрессии рекомбинантного белка у гетерологичного хозяина. Другое применимое слияние включает в себя добавление иммунологически активного домена, такого как эпитоп антитела или другая метка, для облегчения нацеливания или очистки слитого белка. Применение 6хHis и GST (глутатион-S-трансферазы) в качестве меток хорошо известно. Встраивание сайта расщепления в области соединения или вблизи области соединения слитого белка будет облегчать удаление чужеродного полипептида после очистки. Другие применимые слияния включают в себя связывание функциональных доменов, таких как активные центры из ферментов, таких как гидролаза, домены гликозилирования, сигналы клеточного нацеливания или трансмембранные области.
Особо обсуждаются иммунотоксины в качестве варианта осуществления данного изобретения. Иммунотоксин представляет собой цитотоксическое соединение, содержащее по меньшей мере часть антитела и часть молекулы токсина. Антитело и токсин могут быть слиты или конъюгированы друг с другом. Более подробно иммунотоксины описаны ниже.
b. Конъюгированные белки
Данное изобретение относится также к конъюгированным полипептидам, таким как транслированные белки, полипептиды и пептиды, обычно моноклонального типа, которые связаны по меньшей мере с одним агентом с образованием конъюгата с антителом. Для увеличения эффективности молекул антител в качестве диагностических или лекарственных средств удобно присоединять или ковалентно связывать или связывать в виде комплекса по меньшей мере одну желательную молекулу или часть молекулы. Такой молекулой или частью молекулы может быть, но не только, по меньшей мере одна эффекторная или репортерная молекула. Эффекторные молекулы включают в себя молекулы, имеющие желательную активность, например, цитотоксическую активность. Неограничивающие примеры эффекторных молекул, которые были присоединены к антителам, включают в себя токсины, противоопухолевые агенты, терапевтические ферменты, радиоактивно меченные нуклеотиды, антивирусные агенты, хелатообразователи, цитокины, факторы роста и олиго- или полинуклеотиды. В противоположность этому, репортерная молекула определяется как любая часть молекулы, которая может быть определена с использованием анализа. Неограничивающие примеры репортерных молекул, которые были конъюгированы с антителами, включают в себя ферменты, радиоактивные метки, гаптены, флуоресцентные метки, фосфоресцентные молекулы, хемилюминесцентные молекулы, хромофоры, люминесцентные молекулы, фотоаффинные молекулы, окрашенные частицы или лиганды, такие как биотин.
В качестве основы для конъюгата антитела может быть использовано любое антитело достаточной селективности, специфичности или аффинности. Такие свойства могут оцениваться с использованием общепринятой методологии иммунологического скрининга, известной специалистам с квалификацией в данной области. Сайты для связывания с биологически активными молекулами в молекуле антитела, наряду с каноническими антигенсвязывающими сайтами, включают в себя сайты, которые находятся в вариабельном домене, которые могут связывать патогены, В-клеточные суперантигены, Т-клеточный корецептор CD4 и оболочку ВИЧ-1 (Sasso et al., 1989; Shorki et al., 1991; Silvermann et al., 1995; Cleary et al., 1994; Lenert et al., 1990; Berberian et al., 1993; Kreier et al., 1991). Кроме того, вариабельный домен участвует в аутосвязывании антител (Kang et al., 1988) и содержит эпитопы (идиотопы), узнаваемые анти-антителами (антителами против антител) (Kohler et al., 1989).
Определенными примерами конъюгатов антител являются конъюгаты, в которых антитело связано с определяемой меткой. «Определяемыми метками» являются соединения и/или элементы, которые могут быть определены вследствие их специфических функциональных свойств и/или химических характеристик, применение которых позволяет определять антитело, к которому они присоединены, и/или затем определить количественно, если желательно. Другим таким примером является образование конъюгата, содержащего антитело, связанное с цитотоксическим или антиклеточным агентом, причем такой конъюгат может называться "иммунотоксином".
Конъюгаты антител могут использоваться в качестве диагностических агентов. Содержащие антитело диагностические агенты обычно подразделяются на два класса: диагностические агенты для применения в диагностике in vitro, например, в различных иммуноанализах, и/или диагностические агенты для применения в диагностических протоколах in vivo, обычно известных как "визуализация на основе антител".
В данной области известны многие подходящие агенты визуализации, а также способы для их присоединения к антителам (см., например, патенты США с номерами 5021236; 4938948 и 4472509, каждый из которых включен в данное описание в качестве ссылки). Визуализирующими частями молекул могут быть парамагнитные ионы; радиоактивные изотопы; флуорохромы; ЯМР-определяемые вещества; рентгеновская визуализация.
В случае парамагнитных ионов можно упомянуть в качестве примера такие ионы, как хром (III), марганец (II), железо (III), железо (II), кобальт (II), никель (II), медь (II), неодим (III), самарий (III), иттербий (III), гадолиний (III), ванадий (II), тербий (III), диспрозий (III), гольмий (III) и/или эрбий (III), причем особенно предпочтительным является гадолиний. Ионы, применимые в других контекстах, таких как рентгеновская визуализация, включают в себя, но не ограничиваются ими, лантан (III), золото (III), свинец (II) и особенно висмут (III).
В случае радиоактивных изотопов для терапевтического и/или диагностического применений можно упомянуть 211астатин, 14углерод, 51хром, 36хлор, 57кобальт, 58кобальт, 67медь, 152европий,67галлий, 3водород, 123йод, 125йод, 131йод, 111индий, 59железо, 32фосфор, 186рений, 188рений, 75селен, 35сера, 99mтехнеций и/или 90иттрий. 125I является часто предпочтительным для использования в некоторых вариантах, а 99mтехнеций и/или 111индий являются также часто предпочтительными вследствие их низкой энергии и пригодности для определения большого диапазона. Радиоактивно меченные моноклональные антитела данного изобретения могут быть получены в соответствии с хорошо известными в данной области способами. Например, моноклональные антитела могут быть йодированы контактированием с йодидом натрия и/или калия и химическим окислителем, таким как гипохлорит натрия, или ферментативным окислителем, таким как лактопероксидаза. Моноклональные антитела данного изобретения могут быть помечены 99mтехнецием посредством способа обмена лигандами, например восстановлением пертехнетата (ТсО4 -) раствором ионов олова (II), хелатированием восстановленного технеция на колонке Сефадекса и нанесением антитела на эту колонку. Альтернативно, могут быть использованы способы прямого мечения, например, с использованием инкубирования пертехнетата, восстановителя, такого как SnCl2, буферного раствора, такого как раствор фталата натрия-калия, и антитела. Промежуточными функциональными группами, которые часто используют для связывания радиоактивных изотопов, которые существуют в виде ионов металлов, с антителом, являются диэтилентриаминпентауксусная кислота (ДТПУ) или этилендиаминтетрауксусная кислота (ЭДТУ).
Среди флуоресцентных меток, рассматриваемых для применения в качестве конъюгатов, находятся Alexa 350, Alexa 430, AMCA, BODIPY 630/650, BODIPY 650/665, BODIPY-FL, BODIPY-R6G, BODIPY-TMR, BODIPY-TRX, Cascade Blue, Cy3, Cy5,6-FAM, изотиоцианат флуоресцеина, HEX, 6-JOE, Oregon Green 488, Oregon Green 500, Oregon Green 514, Pacific Blue, REG, Rhodamin Green, Rhodamin Red, Renographin, ROX, TAMRA, TET, тетраметилродамин и/или Texas Red.
Другим типом конъюгатов антител, рассматриваемых в данном изобретении, являются конъюгаты, предназначенные прежде всего для применения in vitro, где антитело связано со вторичным связывающим лигандом и/или ферментом (ферментной меткой), который будет генерировать окрашенный продукт при контакте с хромогенным субстратом. Примеры подходящих ферментов включают в себя уреазу, щелочную фосфатазу, пероксидазу (хрена) или глюкозооксидазу. Предпочтительными лигандами вторичного связывания являются соединения биотин и/или авидин и стрептавидин. Применение таких меток хорошо известно специалистам с квалификацией в данной области, и они описаны, например, в патентах США 3817837; 3850752; 3939350; 3996345; 4277437; 4275149 и 4366241, каждый из которых включен в данное описание в качестве ссылки.
Другой известный способ сайт-специфического присоединения молекул к антителам предусматривает реакцию антител с аффинными метками на основе гаптенов. По существу, аффинные метки на основе гаптенов реагируют с аминокислотами в антигенсвязывающем сайте, разрушая посредством этого этот сайт и блокируя специфическую реакцию антигена. Однако это может быть невыгодным, так как это приводит к потере связывания антигена конъюгатом антитела.
Молекулы, содержащие азидогруппы, могут быть также использованы для образования ковалентных связей с белками посредством реактивных нитреновых промежуточных продуктов, которые образуются под действием ультрафиолетового света низкой интенсивности (Potter & Haley, 1983). В частности, 2- и 8-Азидоаналоги пуриновых нуклеотидов использовали в качестве сайт-направленных фотозондов для идентификации нуклеотидсвязывающих белков в неочищенных клеточных экстрактах (Owen & Haley, 1987; Atherton et al., 1985). 2- и 8-азидонуклеотиды использовали также для картирования нуклеотидсвязывающих доменов очищенных белков (Khatoon et al., 1989; King et al., 1989, и Dholakia et al., 1989), и они могут быть использованы в качестве антителосвязывающих агентов.
В данной области известно несколько способов для присоединения или конъюгации антитела с его конъюгированной частью. Некоторые способы присоединения предусматривают применение хелатного комплекса с металлом с использованием, например, органического хелатообразователя, такого как ангидрид диэтилентриаминпентауксусной кислоты (ДТПУ); этилентриаминтетрауксусная кислота; N-хлор-п-толуолсульфонамид и/или тетрахлор-3α-6α-дифенилглюкоурил-3, присоединенного к антителу (патенты США с номерами 4472509 и 4938948, каждый из которых включен в данное описание в качестве ссылки). Моноклональные антитела могут также взаимодействовать с ферментом в присутствии сопрягающего агента, такого как глутаровый альдегид или периодат. Конъюгаты с флуоресцеиновыми маркерами получают в присутствии этих сопрягающих агентов или реакцией с изотиоцианатом. В патенте США № 4938948 визуализация опухолей молочной железы достигается с использованием моноклональных антител, и определяемые визуализирующие части молекулы связывают с антителом с использованием линкеров, таких как метил-п-гидроксибензимидат или N-сукциниламидил-3-(4-гидроксифенил)пропионат.
В других вариантах осуществления обсуждается дериватизация иммуноглобулинов селективным введением сульфгидрильных групп в Fc-область иммуноглобулина с использованием условий реакции, которые не изменяют антигенсвязывающий центр антитела. Описано, что конъюгаты антител, получаемые в соответствии с этой методологией, проявляют улучшенную долговечность, специфичность и чувствительность (патент США № 5196066, включенный в данное описание в качестве ссылки). Сайт-специфическое присоединение эффекторных или репортерных молекул, где репортерную или эффекторную молекулу конъюгируют с углеводным остатком в Fc-области, также было описано в литературе (O'Shannessy et al., 1987). Сообщалось, что этот подход давал диагностически и терапевтически перспективные антитела, которые в настоящее время находятся в процессе клинического испытания.
i. Линкеры/сопрягающие агенты
Множественные пептиды или полипептиды, такие как конъюгированный иммунотоксин, могут быть соединены через биологически расщепляемую связь, такую как селективно расщепляемый линкер или аминокислотная последовательность. Например, обсуждаются пептидные линкеры, которые содержат сайт расщепления для фермента, преимущественно локализованного или активного в среде опухоли. Примерными формами таких пептидных линкеров являются формы, которые расщепляются урокиназой, плазмином, тромбином, фактором IXa, фактором Ха или металлопротеиназой, такой как коллагеназа, желатиназа или стромелизин. Альтернативно, пептиды или полипептиды могут быть присоединены к адъюванту.
Для отделения белковых частей молекул могут быть использованы аминокислоты, такие как селективно расщепляемые линкеры, синтетические линкеры или другие аминокислотные последовательности. Кроме того, хотя известны многочисленные типы содержащих дисульфидную связь линкеров, которые могут быть успешно использованы для конъюгирования токсиновой части с нацеливающим агентом, некоторые линкеры обычно будут предпочтительными в сравнении с другими линкерами, на основе отличающихся фармакологических характеристик и способностей. Например, должны быть предпочтительными линкеры, которые содержат дисульфидную связь, которая является стерически затрудненной, вследствие их более высокой стабильности in vivo, и, следовательно, предотвращающей высвобождение токсиновой части перед связыванием в сайте действия. Кроме того, определенные преимущества в соответствии с данным изобретением будут реализоваться посредством применения любой из ряда токсиновых частей молекул, в том числе гелонина и дегликозилированной А-цепи рицина.
В качестве общего руководства следует считать, что любой биохимический сшивающий линкер, который является подходящим для применения в иммунотоксине, будет также применим в данном контексте, и могут также рассматриваться дополнительные линкеры.
Сшивающие реагенты используют в форме молекулярных мостиков, которые связывают вместе функциональные группы двух различных молекул, например, стабилизирующего и коагулирующего агентов. Для связывания двух разных белков одноступенчатым образом могут быть использованы гетеробифункциональные сшивающие линкеры, которые элиминируют нежелательное образование гомополимера.
Предполагается, что сшивающие линкеры могут быть обеспечены модифицированными белковыми молекулами данного изобретения. Бифункциональные сшивающие реагенты интенсивно использовались для разных целей, в том числе получения аффинных матриксов, модификации и стабилизации разнообразных структур, идентификации сайтов связывания и исследований в области структуры соединений. В контексте данного изобретения такой сшивающий линкер может быть использован для стабилизации полипептида или превращения его в более применимый полипептид в качестве лекарственного средства, например, улучшением нацеливающей способности или общей эффективности модифицированного белка. Сшивающие линкеры могут быть также расщепляемыми, такими как дисульфиды, кислоточувствительные линкеры и другие. Было показано, что гомобифункциональные реагенты, которые несут две идентичные функциональные группы, являются высокоэффективными в индукции сшивания между идентичными и различными макромолекулами или субъединицами макромолекулы и связывании полипептидов со специфическими сайтами связывания на партнерах связывания. Гетеробифункциональные реагенты содержат две различные функциональные группы. Используя преимущество дифференциальных реактивностей двух различных функциональных групп, сшивания можно контролировать как в отношении селективности, так и в отношении последовательности событий. Бифункциональные сшивающие реагенты могут быть классифицированы в соответствии со специфичностью их функциональных групп, например амино-, сульфгидрил-, гуанидино-, индол-, карбоксилспецифических групп. Из них особенно популярными стали реагенты, направленные на свободные аминогруппы, вследствие их коммерческой доступности, легкости синтеза и мягких условий реакций, при которых они могут применяться. Большинство гетеробифункциональных сшивающих реагентов содержат первичную аминореактивную группу и тиолреактивную группу.
Примеры способов для сшивания лигандов с липосомами описаны в патенте США 5603872 и патенте США 5401511, каждый из которых особо включен в данное описание в качестве ссылки в его полном виде. Различные лиганды могут быть ковалентно связаны с поверхностями липосом посредством сшивания аминовых остатков. Липосомы, в частности многослойные везикулы (MLV) или однослойные везикулы, такие как микроэмульгированные липосомы (MEL) и большие однослойные липосомы (LUVET), каждая из которых содержит фосфатидилэтаноламин (PE), получали с использованием установленных процедур. Включение РЕ в липосомы обеспечивает активный функциональный остаток, первичный амин, на поверхностии липосом для целей сшивания. Такие лиганды, как эпидермальный фактор роста (EGF), были успешно связаны с РЕ-липосомами. Лиганды связываются ковалентно с дискретными сайтами на поверхностях липосом. Число и поверхностная плотность этих сайтов диктуется составом липосом и типом липосом. Поверхности липосом могут также иметь сайты для нековалентного связывания. Для образования ковалентных конъюгатов лигандов и липосом сшивающие реагенты исследовались на эффективность и биосовместимость. Сшивающие реагенты включают в себя глутаровый альдегид (GAD), бифункциональный оксиран (OXR), диглицидиловый простой эфир этиленгликоля (EGDE) и водорастворимый карбодиимид, предпочтительно 1-этил-3-(3-диметиламинопропил)карбодиимид (EDC). Посредством химического сшивания комплексов установлено связывание аминных остатков узнающего вещества и липосом.
В другом примере описаны гетеробифункциональные сшивающие реагенты и способы применения этих сшивающих реагентов (патент США 5889155, специально включенный в данное описание в качестве ссылки в его полном виде). Сшивающие реагенты комбинируют нуклеофильный гидразидный остаток с электрофильным малеимидным остатком, обеспечивая возможность связывания в одном примере альдегидов со свободными тиолами. Этот сшивающий реагент может быть модифицирован для сшивания различных функциональных групп и, следовательно, применим для сшивания полипептидов и сахаров. Таблица 4 показывает подробно некоторые гетеробифункциональные сшивающие линкеры, считающиеся применимыми в данном изобретении.
ГЕТЕРОБИФУНКЦИОНАЛЬНЫЕ СШИВАЮЩИЕ ЛИНКЕРЫ
В случаях, когда конкретный полипептид, например гелонин, не содержит остатка, доступного для конкретного сшивающего реагента в его нативной последовательности, могут быть использованы консервативные генетические или синтетические замены аминокислот в первичной последовательности.
4. Очистка белка
Хотя некоторые из вариантов осуществления данного изобретения включают в себя рекомбинантные белки, данное изобретение относится также к способам и процессам очистки белков, в том числе модифицированных белков и рекомбинантных белков. Обычно эти способы включают в себя, на одном уровне, грубое фракционирование клеточной среды на полипептидную и неполипептидную фракции. После отделения полипептида от других белков представляющий интерес полипептид может быть дополнительно очищен при помощи хроматографических и электрофоретических способов для достижения полной или частичной очистки (или очистки до гомогенности). Аналитическими способами, особенно пригодными для получения чистого пептида, являются ионообменная хроматография, гель-фильтрационная хроматография, электрофорез в полиакриламидном геле, изоэлектрическое фокусирование. Особенно эффективным способом очистки пептидов является жидкостная экспресс-хроматография белков или даже ВЭЖХ. Кроме того, условия, при которых выполняются такие способы, могут влиять на характеристики очищенных молекул, такие как функциональная активность.
Некоторые аспекты данного изобретения относятся к очистке и в конкретных вариантах осуществления к существенной очистке кодируемого белка или пептида. Термин "очищенный белок или пептид" в данном контексте относится к композиции, изолируемой от других компонентов, где этот белок или пептид очищен до любой степени относительно природно получаемого состояния. Таким образом, очищенным белком или пептидом называют также белок или пептид, не содержащий окружающей среды, в которой он может встречаться в природе.
"По существу очищенный" белок или пептид.
Обычно термин "очищенная" будет относиться к белковой или пептидной композиции, которая была подвергнута фракционированию для удаления различных других компонентов и которая по существу сохраняет ее экспрессируемую биологическую активность. При использовании термина "по существу очищенная" этот термин будет относиться к композиции, в которой белок или пептид образует основной компонент этой композиции, например, составляет приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 90%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99%, приблизительно 99,2%, приблизительно 99,4%, приблизительно 99,6%, приблизительно 99,8%, приблизительно 99,9% или более белков данной композиции.
Различные способы для количественного определения степени очистки белка или пептида будут известны специалистам с квалификацией в данной области в свете данного описания. Они включают в себя, например, определение удельной активности активной фракции или оценку количества полипептидов во фракции анализом при помощи электрофореза в ДСН/ПААГ. Предпочтительным способом оценки чистоты фракции является расчет удельной активности этой фракции для сравнения ее с удельной активностью исходного экстракта и, следовательно, расчет степени чистоты, оцениваемой в данном описании как "очистка в указанное число раз". Фактические единицы, используемые для представления величины активности, будут, конечно, зависеть от конкретного способа анализа, выбранного для прослеживания очистки, и от того, проявляет ли экспрессируемый белок или пептид определяемую активность.
Различные способы, подходящие для применения в очистке белка, хорошо известны специалистам с квалификацией в данной области. Они включают в себя, например, осаждение сульфатом аммония, ПЭГ, антителами и т.п. или денатурацией нагреванием с последующим центрифугированием; стадии хроматографии, например ионообменной хроматографии, гель-фильтрационной хроматографии, обращенно-фазовой, гидроксиапатитной и аффинной хроматографии; изоэлектрическое фокусирование; гель-электрофорез и комбинации таких и других способов. Как обычно известно в данной области, считается, что порядок проведения различных стадий очистки может изменяться или некоторые стадии могут опускаться и все еще обеспечивать получение подходящего способа для получения по существу очищенного белка или пептида.
Не существует общего требования, что данный белок или пептид должен быть всегда представлен в его наиболее очищенном состоянии. В самом деле, предполагается, что менее существенно очищенные продукты смогут находить применение в некоторых вариантах осуществления. Частичная очистка может выполняться с использованием меньшего числа стадий очистки в комбинации стадий или с использованием отличающихся форм той же самой общей схемы очистки. Например, понятно, что катионообменная колоночная хроматография, выполняемая на приборе ВЭЖХ, будет обычно приводить к очистке в большее число раз, чем тот же самый способ с использованием системы хроматографии низкого давления. Способы, обнаруживающие более низкую степень относительной очистки, могут иметь преимущество в общем выходе белкового продукта или в сохранении активности экспрессируемого белка.
Известно, что миграция полипептида может варьироваться, иногда значительно, в зависимости от различных условий электрофореза в ДСН/ПААГ (Capaldi et al., 1977). Таким образом, будет понятно, что при различающихся условиях электрофореза могут варьироваться видимые молекулярные массы очищенных или частично очищенных продуктов экспрессии.
Обсуждается также применение пептидной метки в комбинации со способами и композициями данного изобретения. Метка использует преимущество взаимодействия между двумя полипептидами. Часть одного из этих полипептидов, которая участвует во взаимодействии, может быть использована в качестве метки. Например, связывающая область глутатион-S-трансферазы (GST) может быть использована в качестве метки, так что могут быть использованы гранулы глутатиона для обогащения в отношении соединения, содержащего метку GST. Может быть использована эпитопная метка, которая является областью аминокислот, узнаваемой антителом или Т-клеточным рецептором. Эта метка может кодироваться сегментом нуклеиновой кислоты, который функционально связан с сегментом нуклеиновой кислоты, кодирующим модифицированный белок, так что этой молекулой нуклеиновой кислоты кодируется слитый белок. Другими подходящими слитыми белками являются слитые белки с β-галактозидазой, убиквитином, гексагистидином (6хHis) или т.п.
5. Антитела
В некоторых вариантах осуществления данное изобретение включает в себя антитела. Например, целое моноклональное, одноцепочечное или гуманизированное антитело или его часть могут быть химически конъюгированы или рекомбинантно слиты с другим белковым соединением, таким как модифицированный токсин гелонин. Альтернативно, другие аспекты данного изобретения включают в себя определение иммунного ответа, т.е. гуморального ответа, на конкретный антиген или область антигена, для конструирования и/или получения белкового соединения с меньшей иммуногенностью, чем иммуногенность нативной формы этого белкового соединения. Как подробно описано выше, кроме антител, генерируемых против полноразмерных белков, антитела могут быть также образованы в ответ на меньшие конструкции, содержащие коровые области эпитопа, в том числе эпитопа дикого типа и мутантного эпитопа. Эпитоп представляет собой антигенную детерминанту. Антиген является любым веществом, которое специфически узнается антителом или Т-клеточным рецептором. Иммуноген представляет собой антиген, который индуцирует специфический иммунный ответ.
В данном контексте термин "антитело" относится в широком смысле к любому иммунному связывающему агенту, такому как IgG, IgM, IgA, IgD и IgE. Обычно предпочтительными являются IgG и/или IgM, так как они являются наиболее обычными антителами в физиологической ситуации и так как они являются наиболее легко получаемыми в лабораторных условиях.
Считается, что моноклональные антитела (mAb) имеют определенные преимущества, например воспроизводимость и возможность крупномасштабного получения, и их применение обычно является предпочтительным. Таким образом, данное изобретение относится к моноклональным антителам человека, мышей, обезьяны, крысы, хомячка, кролика и даже курицы.
Термин "антитело" в данном контексте относится к любой подобной антителу молекуле, которая имеет антигенсвязывающую область, и включает в себя фрагменты антитела, такие как Fab', Fab, F(ab')2, антитела с единственным доменом (DAB), Fv, scFv (одноцепочечный Fv) и т.п. Способы получения и применения различных конструкций на основе антител и фрагментов хорошо известны в данной области. Средства для получения и характеристики антител также хорошо известны в данной области (См., например, справочник Harlow and Lane, "Antibodies: A Laboratory Manual", Cold Spring Harbor Laboratory, 1988, включенный в данное описание в качестве ссылки).
Способы для генерирования моноклональных антител (mAb) обычно начинаются так же, как и способы для получения поликлональных антител. Вкратце, поликлональные антитела могут быть получены иммунизацией животного иммуногенной полипептидной композицией в соответствии с данным изобретением и сбором антисывороток у иммунизированного животного. Альтернативно, в некоторых вариантах осуществления данного изобретения сыворотку собирают у лиц, которые, возможно, подвергались действию конкретного антигена. Взаимодействие с конкретным антигеном может происходить в условиях труда, так что эти лица профессионально подвергались действию конкретного антигена и развили поликлональные антитела к пептиду, полипептиду или белку. В некоторых вариантах осуществления данного изобретения поликлональную сыворотку у профессионально подвергаемых действию антигена лиц используют для идентификации антигенных областей в токсине гелонине посредством применения способов иммуноопределения.
В получении антисывороток может быть использован большой диапазон видов животных. Обычно животным, используемым для получения антисывороток, является кролик, мышь, крыса, хомячок, морская свинка или коза. Вследствие относительно большого объема крови у кроликов кролик является предпочтительным для получения поликлональных антител.
Как хорошо известно в данной области, получаемая композиция может варьироваться по ее иммуногенности. Таким образом, часто бывает необходимо проводить бустинг иммунной системы хозяина, что может быть достигнуто связыванием пептидного или полипептидного иммуногена с носителем. Примерами предпочтительных носителей являются гемоцианин фиссуреллы (KLH) и бычий сывороточный альбумин (БСА). В качестве носителей могут быть также использованы другие альбумины, такие как овальбумин, альбумин мышиной сыворотки или альбумин кроличьей сыворотки. Средства для конъюгирования полипептида с белком-носителем хорошо известны в данной области и включают в себя глутаровый альдегид, сложный эфир м-малеимидобензоил-N-гидроксисукцинимида, карбодиимид и бис-диазотизированный бензидин.
Как также хорошо известно в данной области, иммуногенность конкретной иммуногенной композиции может быть усилена использованием неспецифических стимуляторов иммунного ответа, известных как адъюванты. Подходящие молекулы адъювантов включают в себя все приемлемые иммуностимуляторные соединения, такие как цитокины, токсины или синтетические композиции.
Адъюванты, которые могут быть использованы, включают в себя IL-1, IL-2, IL-4, IL-7, IL-12, γ-интерферон, GM-CSF, BCG, гидроксид алюминия, соединения MDP, такие как thur-MDP и nor-MDP, CGP (MTP-PE), липид А и монофосфориллипид А (MPL). Обсуждаются также RIBI, который содержит три компонента, экстрагированных из бактерий, MPL, трегалозодимиколат (TDM) и скелет клеточной стенки (CWS) в эмульсии 2% сквален/Твин 80. Могут быть использованы даже антигены МНС. Например, часто предпочтительные адъюванты включают в себя полный адъювант Фрейнда (неспецифический стимулятор иммунного ответа, содержащий убитую бактерию Mycobacterium tuberculosis), неполные адъюванты Фрейнда и содержащий гидроксид алюминия адъювант.
Кроме адъювантов, может быть желательно одновременное введение модификаторов биологического ответа (BRM), которые, как было показано, положительно регулируют Т-клеточный иммунитет или отрицательно регулируют активность супрессорных клеток. Такие BRM включают в себя, но не ограничиваются ими, Циметидин (CIM; 1200 мг/д) (Smith/Kline, PA); Циклофосфамид в низкой дозе (CYP; 300 мг/м2) (Johnson/Mead, NJ), цитокины, такие как γ-интерферон, IL-2 или IL-12, или гены, кодирующие белки, участвующие в иммунных хелперных функциях, такие как В-7.
Количество иммуногенной композиции, используемой в получении поликлональных антител, варьируется в зависимости от природы иммуногена, а также животного, используемого для иммунизации. Для введения иммуногена могут быть использованы различные пути (подкожный, внутримышечный, внутрикожный, внутривенный и внутрибрюшинный). Продуцирование поликлональных антител может быть подвергнуто мониторингу взятием крови иммунизированного животного через различные промежутки времени после иммунизации.
Может также производиться вторая, бустерная инъекция. Процесс бустинга и определения титра повторяют, пока не будет достигнут подходящий титр. После получения желательного уровня иммуногенности у иммунного животного может быть собрана кровь, и сыворотку выделяют и хранят, и/или это животное может быть использовано для получения mAb.
mAb могут быть легко получены с использованием хорошо известных способов, таких как описанные в патенте США 4196265, включенном в данное описание в качестве ссылки. Обычно этот способ предусматривает иммунизацию подходящего животного выбранной композицией иммуногена, например, очищенного или частично очищенного полипептида, пептида или домена, как в случае композиции дикого типа, так и в случае мутантного белка. Композицию для иммунизации вводят таким способом, который является эффективным для стимуляции продуцирующих антитела клеток.
mAb могут быть, если желательно, очищены дополнительно с использованием фильтрации, центрифугирования и различных хроматографических способов, таких как ВЭЖХ или аффинная хроматография. Фрагменты моноклональных антител данного изобретения могут быть получены из полученных таким образом моноклональных антител способами, которые включают в себя расщепление ферментами, такими как пепсин или папаин, и/или расщепление дисульфидных связей химическим восстановлением. Альтернативно, фрагменты моноклональных антител, включенные в данное изобретение, могут быть синтезированы с использованием автоматического пептидного синтезатора.
Предполагается также, что для генерирования mAb может быть использован подход молекулярного клонирования. Для этого получают комбинаторные библиотеки иммуноглобулинсодержащих фагмид из РНК, выделенной из селезенки иммунизированного животного, и фагмиды, экспрессирующие подходящие антитела, отбирают пэннингом с использованием клеток, экспрессирующих этот антиген, и контрольных клеток. Преимущества этого подхода в сравнении с общепринятыми гибридомными способами заключаются в том, что может быть получено и подвергнуто скринингу в единственном цикле приблизительно в 104 раз больше антител и что новые специфичности генерируются посредством комбинирования Н- и L-цепей, что дополнительно увеличивает шанс нахождения подходящих антител.
Гуманизированные моноклональные антитела являются антителами животного происхождения, которые были модифицированы с использованием способов генетической инженерии для замены константной области и/или каркасных последовательностей вариабельной области человеческими последовательностями при сохранении исходной специфичности в отношении исходного антигена. Такие антитела обычно получают из антител грызунов, специфических в отношении антигенов человека. Такие антитела обычно применимы в терапевтических применениях in vivo. Эта стратегия уменьшает реакцию хозяина на чужеродное антитело и обеспечивает возможность отбора на эффекторные функции человека.
Обсуждаются также "гуманизированные" антитела, а также химерные антитела мыши, крысы или других видов, несущие домены константной и/или вариабельной области человека, биспецифические антитела, рекомбинантные и сконструированные антитела и их фрагменты. Способы получения гуманизированных иммуноглобулинов хорошо известны специалистам с квалификацией в данной области. Например, в патенте США № 5693762 описаны способы для получения и композиции гуманизированных иммуноглобулинов, имеющих одну или более определяющих комплементарность областей (CDR). При комбинировании введением в интактное антитело эти гуманизированные иммуноглобулины являются по существу неиммуногенными для человека и сохраняют по существу ту же самую аффинность, что и иммуноглобулин донора, в отношении антигена, такого как белок или другое соединение, содержащее эпитоп. Примеры других сообщений в этой области включают в себя патенты США с номерами 6054297, 5861155 и 6020192, которые специально включены в качестве ссылки. Способы генерирования антител, которые являются "изготовленными на заказ" относительно заболевания пациента, также известны, и такие "изготовленные на заказ" антитела также обсуждаются в данном изобретении.
6. Способы иммуноанализа
Как обсуждалось выше, в некоторых вариантах осуществления данное изобретение относится к способам иммуноанализа в отношении связывания, очистки, удаления, количественного определения и/или иного определения биологических компонентов, таких как антигенные области на полипептидах и пептидах. Способы иммуноанализа данного изобретения могут быть использованы для идентификации антигенных областей пептида, полипептида или белка, которые имеют терапевтические применения, в частности, в уменьшении иммуногенности или антигенности этого пептида, полипептида или белка у субъекта-мишени.
Способы иммуноанализа включают в себя твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (РИА), иммунорадиометрический анализ, флуороиммуноанализ, хемилюминесцентный анализ, биолюминесцентный анализ и Вестерн-блоттинг, хотя некоторые другие способы хорошо известны специалистам с обычной квалификацией в данной области. Стадии различных применимых способов иммуноанализа были описаны в научной литературе, например, Doolittle MH and Ben-Zeev O, 1999; Gulbis В. et al., 1993; De Jager R. et al., 1993 и Nakamura et al., 1987, которые включены в данное описание в качестве ссылки.
В общем, способы иммуносвязывания включают в себя получение образца, в котором предполагают присутствие белка, полипептида и/или пептида, и контактирование этого образца с первым антителом, моноклональным или поликлональным в соответствии с данным изобретением, например, в условиях, эффективных для обеспечения образования иммунных комплексов.
Эти способы включают в себя способы очистки белка, полипептида и/или пептида из образцов органелл, клеток, ткани или организма. В этих случаях антитело извлекает антигенный белок, полипептид и/или пептид из образца. Антитело предпочтительно связано с твердым носителем, например, в форме колоночного матрикса, и образец, в котором предполагают присутствие антигенного компонента белка, полипептида или пептида, наносят на это иммобилизованное антитело. Нежелательные компоненты будут вымываться из колонки, оставляя антиген, находящийся в иммунном комплексе с иммобилизованным антителом, который подлежит элюции.
Способы иммуносвязывания также включают в себя способы определения и количественного определения антигенного компонента в образце и определения и количественного определения любых иммунных комплексов, образованных во время процесса связывания. В данном описании получают образец, в котором предполагают присутствие антигена или антигенного домена, и контактируют этот образец с антителом против этого антигена или антигенного домена и затем детектируют и определяют количество иммунных комплексов, образованных в этих специфических условиях.
Что касается определения антигена, анализируемым биологическим образцом может быть любой образец, в котором предполагают присутствие антигена или антигенного домена, такой как, например, срез или образец ткани, экстракт гомогенизированной ткани, клетка, органелла, отделенные и/или очищенные формы любой из вышеупомянутых композиций или даже любая биологическая жидкость, которая приходит в контакт с этой клеткой или тканью, в том числе кровь и/или сыворотка.
Контактирование выбранного биологического образца с антителом в эффективных условиях и в течение периода времени, достаточных для образования иммунных комплексов (первичных иммунных комплексов), обычно является простым добавлением композиции антител к этому образцу и инкубированием этой смеси в течение периода времени, достаточно длительного для образования антителами иммунных комплексов, т.е. связывания любых присутствующих антигенов. После этого времени композицию образец-антитело, такую как срез ткани, планшет ELISA, дот-блот или Вестерн-блот, обычно промывают для удаления неспецифически связанных молекул антитела с оставлением для определения только тех антител, которые специфически связаны в первичных иммунных комплексах.
В общем, определение образования иммунных комплексов хорошо известно в данной области и может достигаться посредством применения многочисленных подходов. Эти способы обычно основаны на определении метки или маркера, таких как любой из радиоактивных, флуоресцентных, биологических и ферментных меток. Патенты США, относящиеся к применению таких меток, включают в себя 3817837; 3850752; 3939350; 3996345; 4277437; 4275149 и 4366241, каждый из которых включен в данное описание в качестве ссылки. Конечно, можно найти дополнительные преимущества посредством применения вторичного связывающего лиганда, такого как второе антитело, и/или использования биотин/авидин-связывания лиганда, как известно в данной области.
Антитело, используемое в этом определении, само может быть связано с определяемой меткой, где остается затем просто определять эту метку для определения количества первичных иммунных комплексов в определяемой композиции. Альтернативно, первое антитело, которое становится связанным в первичных иммунных комплексах, может быть определено с использованием второго лиганда связывания, который имеет связывающую аффинность в отношении этого антитела. В этих случаях второй лиганд связывания может быть связан с определяемой меткой. Второй лиганд связывания часто сам является антителом, которое, следовательно, может быть названо "вторичным" антителом. Первичные иммунные комплексы контактируют с меченым вторичным лигандом связывания, или антителом, в эффективных условиях и в течение периода времени, достаточных для образования вторичных иммунных комплексов. Затем вторичные иммунные комплексы обычно промывают для удаления любых неспецифически связанных вторичных антител или лигандов и затем определяют остающуюся метку во вторичных иммунных комплексах.
Следующие способы включают в себя определение первичных иммунных комплексов с использованием двустадийного подхода. Второй лиганд связывания, такой как антитело, которое имеет связывающую аффинность в отношении первого антитела, используют для образования вторичных иммунных комплексов, как описано выше. После промывания вторичные иммунные комплексы контактируют с третьим лигандом связывания или антителом, которое имеет связывающую аффинность в отношении второго антитела, опять в эффективных условиях и в течение периода времени, достаточных для образования иммунных комплексов (третичных иммунных комплексов). Третий лиганд (или антитело) связан с определяемой меткой, что позволяет определять эти образованные таким образом третичные иммунные комплексы.
Один способ иммуноанализа, разработанный Charles Cantor, использует два различных антитела. В первой стадии биотинилированное, моноклональное или поликлональное антитело первой стадии используют для определения антигена-мишени (антигенов-мишеней), а затем антитело второй стадии используют для определения биотина, присоединенного к этому находящемуся в комплексе биотину. В этом способе пробу, подлежащую тестированию, сначала инкубируют в растворе, содержащем антитело первой стадии. Если антиген-мишень присутствует, некоторая часть этого антитела связывается с антигеном с образованием комплекса биотинилированное антитело/антиген. Затем комплекс антитело/антиген амплифицируют инкубированием в последовательных растворах стрептавидина (или авидина), биотинилированной ДНК и/или комплементарно биотинилированной ДНК, причем каждая стадия добавляет дополнительные сайты биотина к комплексу антитело/антиген. Стадии амплификации повторяют, пока не будет достигнут подходящий уровень амплификации, и в этот момент этот образец инкубируют в растворе, содержащем антитело второй стадии против биотина. Это антитело второй стадии является меченым, например, ферментом, который может быть использован для определения присутствия комплекса антитело/антиген гистоэнзимологическими методами с использованием хромогенного субстрата. При подходящей амплификации может быть получен конъюгат, который является макроскопически видимым.
Другой известный способ иммуноанализа использует преимущество методологии иммуно-ПЦР (полимеразной цепной реакции). Способ ПЦР сходен со способом Cantor до инкубирования с биотинилированной ДНК, однако, вместо использования множественных циклов инкубирования стрептавидина и биотинилированной ДНК, комплекс ДНК/биотин/стрептавидин/антитело вымывают буфером с низким рН или высокой концентрацией соли, который высвобождает это антитело. Затем полученный промывной раствор используют для проведения реакции ПЦР с удобными праймерами с подходящими контролями. По меньшей мере теоретически огромная способность амплификации и специфичность ПЦР могут быть использованы для определения единственной молекулы антигена.
а. ELISA
Как подробно описано выше, иммуноанализы, в их наиболее простом и/или прямом смысле, являются анализами связывания. Некоторые предпочтительные иммуноанализы являются различными типами твердофазных иммуноферментных анализов (ELISA) и/или радиоиммуноанализов (РИА), известных в данной области. Иммуногистохимическое определение с использованием срезов тканей также является особенно применимым. Однако будет легко понятно, что определение не ограничивается такими способами, и могут также использоваться Вестерн-блоттинг, дот-блоттинг, FACS-анализы и/или т.п.
В одном примере ELISA антитела иммобилизуют на выбранной поверхности, проявляющей аффинность к белку, такой как лунка в полистироловом микротитрационном планшете. Затем тестируемую композицию, в которой предполагают присутствие антигена, такую как клинический образец, добавляют в эти лунки. После связывания и/или промывания для удаления неспецифически связанных иммунных комплексов может быть определен связанный антиген. Определение обычно достигается добавлением другого антитела, которое связано с определяемой меткой. Этот тип ELISA является простым "сэндвич-ELISA". Определение может также достигаться добавлением второго антитела с последующим добавлением третьего антитела, которое имеет связывающую аффинность в отношении второго антитела, причем это третье антитело связано с определяемой меткой.
В другом примере ELISA образцы, в которых предполагают присутствие антигена, иммобилизуют на поверхности лунок и/или затем контактируют с антителами. После связывания и/или промывания для удаления неспецифически связанных иммунных комплексов определяют связанные антитела. Если исходные антитела связаны с определяемой меткой, иммунные комплексы могут быть определены сразу же. Опять же иммунные комплексы могут быть определены с использованием второго антитела, которое имеет связывающую аффинность в отношении первого антитела, причем второе антитело связано с определяемой меткой.
Другой тип ELISA, в котором иммобилизованными являются антигены, предусматривает применение конкуренции антител в определении. В этом типе ELISA меченые антитела против антигена добавляют в лунки, дают им связаться и/или определяют посредством их метки. Затем количество антигена в неизвестном образце определяют смешиванием этого образца с мечеными антителами против этого антигена во время инкубирования с имеющими покрытие лунками. Присутствие антигена в этом образце приводит к уменьшению количества антитела против антигена, доступного для связывания с лункой, и таким образом к уменьшению конечного сигнала. Это также можно использовать для определения антител против антигена в неизвестном образце, где немеченые антитела связываются с покрытыми антигеном лунками и также уменьшают количество антигена, доступного для связывания меченых антител.
Независимо от используемого формата, ELISA имеют определенные общие признаки, такие как покрытие лунок, инкубирование и связывание, промывание для удаления неспецифически связанных молекул и определение связанных иммунных комплексов. Они описываются ниже.
В покрытии планшета либо антигеном, либо антителами обычно инкубируют лунки планшета с раствором антигена или антитела либо в течение ночи, либо в течение указанного периода времени. Затем лунки планшета промывают для удаления неполностью адсорбированного вещества. Затем любые оставшиеся доступные поверхности лунок "покрывают" неспецифическим белком, который является антигенно нейтральным в отношении тестируемых антисывороток. Эти белки включают в себя сывороточный альбумин (БСА), казеин или растворы сухого молока. Это покрытие обеспечивает блокирование сайтов неспецифической адсорбции на иммобилизующей поверхности и, следовательно, уменьшает фон, обусловленный неспецифическим связыванием антисывороток на этой поверхности.
В ELISA, возможно, более обычным является применение вторичных или третичных средств определения, чем прямой процедуры. Таким образом, после связывания белка или антитела с лункой, покрытия нереактивным материалом для уменьшения фона и промывания для удаления несвязанного материала иммобилизующую поверхность контактируют с биологическим образцом, подлежащим тестированию, в условиях, эффективных для обеспечения возможности образования иммунного комплекса (антиген/антитело). Затем определение этого иммунного комплекса требует меченого вторичного лиганда связывания или антитела и вторичного лиганда связывания или антитела вместе с меченым третичным антителом или третьим лигандом связывания.
Под «условиями, эффективными для обеспечения возможности образования иммунного комплекса (антиген/антитело)» подразумевают, что эти условия предпочтительно включают в себя разведение антигенов и/или антител растворами, такими как БСА, бычий гамма-глобулин (BGG) или фосфатно-буферный солевой раствор (ФБР)/Твин. Эти добавляемые агенты также склонны способствовать уменьшению неспецифического фона.
«Подходящие условия» означают также, что инкубирование выполняют при температуре или в течение периода времени, достаточных для обеспечения возможности эффективного связывания. Стадии инкубирования обычно проводят в течение приблизительно 1-2-4 часов или около этого периода времени при температурах предпочтительно порядка 25-27°С или в течение ночи при приблизительно 4°С или около этого.
После всех стадий инкубирования в ELISA контактированную поверхность промывают таким образом, чтобы удалить не образовавшее комплекса вещество. Пример процедуры промывания включает в себя промывание раствором, таким как ФБР/Твин или боратный буфер. После образования специфических иммунных комплексов между тестируемым образцом и первоначально связанным веществом и последующего промывания может быть определено наличие даже мельчайших количеств иммунных комплексов.
Для обеспечения определяющих средств второе или третье антитело будет иметь связанную с ними метку для возможности определения. Этой меткой может быть фермент, который будет генерировать развитие окраски при инкубировании с подходящим хромогенным агентом. Так, например, может быть желательным контактирование или инкубирование первого и второго иммунного комплекса с конъюгированным с уреазой, глюкозооксидазой, щелочной фосфатазой или пероксидазой водорода антителом в течение периода времени и в условиях, которые благоприятствуют развитию дополнительного образования иммунного комплекса (например, инкубирование в течение 2 часов при комнатной температуре в ФБР-содержащем растворе, таком как ФБР/Твин).
После инкубирования с меченым антителом и последующего промывания для удаления несвязанного вещества определяют количественно количество метки, например, инкубированием с хромогенным субстратом, таким как мочевина, или бромкрезоловый красный, или 2,2'-азиноди-(3-этилбензтиазолин-6-сульфоновая кислота (ABTS), или Н2О2, в случае пероксидазы в качестве ферментной метки. Затем определение количества достигается измерением степени генерируемой окраски, например, с использованием спектрофотометра с видимой частью спектра.
b. Иммуногистохимические методы
Антитела данного изобретения могут быть также использованы вместе как со свежезамороженными, так и/или фиксированными формалином, залитыми в парафин блоками тканей, приготовленными для исследования с использованием иммуногистохимических методов (IHC). Например, иммуногистохимические методы могут быть использованы для оценки конкретного иммунотоксина данного изобретения. Способ приготовления блоков тканей из этих состоящих из частиц образцов был успешено использован в предыдущих IHC-исследованиях различных прогностических факторов и/или хорошо известен специалистам с квалификацией в данной области (Brown et al., 1990; Abbondanzo et al., 1990; Allred et al., 1990).
Вкратце, замороженные срезы могут быть получены регидратацией 50 нг замороженной «измельченной (пульверизованной)» ткани при комнатной температуре в фосфатно-буферном солевом растворе (ФБР) в небольших пластиковых капсулах; осаждением этих частиц центрифугированием; ресуспендированием их в различных вязких средах для заливки (ОСТ); перевертыванием капсулы и/или повторным осаждением центрифугированием; быстрым (мгновенным) замораживанием в изопентане при температуре -70°С; разрезанием пластиковой капсулы и/или удалением замороженного цилиндра ткани; закреплением цилиндра ткани на кулачке зажимного патрона криостатного микротома и/или приготовлением 25-50 последовательных срезов.
Перманентные срезы могут быть приготовлены сходным способом, предусматривающим повторную гидратацию образца 50 мг в пластиковой микрофужной пробирке; осаждение центрифугированием; ресуспендирование в 10% формалине в течение 4 часов фиксации; промывание/осаждение центрифугированием; ресуспендирование в теплом 2,5% агаре; осаждение центрифугированием; охлаждение в ледяной воде для отверждения агара; удаление блока ткани/агара из пробирки; инфильтрацию и/или заливку этого блока в парафин и/или приготовление 50 последовательных перманентных срезов.
II. Молекулы нуклеиновых кислот
А. Полинуклеотиды, кодирующие природные белки или модифицированные белки
Данное изобретение относится к полинуклеотидам, выделенным из клеток, которые свободны от общей геномной ДНК и которые способны экспрессировать весь белок или полипептид или часть белка или полипептида. Этот полинуклеотид может кодировать нативный белок, который может быть подвергнут манипуляции для получения модифицированного белка. Альтернативно, этот полинуклеотид может кодировать модифицированный белок или он может кодировать полинуклеотид, который может быть использован для получения слитого белка с модифицированным белком. Например, полинуклеотид может кодировать множественные части молекулы, такие как модифицированный полипептид гелонина, который ковалентно связан с нацеливающим полипептидом, например опухолевым антигеном. Предполагается, что одна молекула полинуклеотида может кодировать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более различных полипептидов (целых или их частей). Любые из полипептидов, белков или пептидов, описанных в таблице 2, могут быть получены рекомбинантно в виде части описанного изобретения. Кроме того, любые из белковых соединений в таблице 2 могут кодироваться с одним или несколькими другими полипептидами, приведенными в таблице 2 или описанными в данном описании, на одной и той же молекуле нуклеиновой кислоты, так что создается слитый белок. Рекомбинантные белки могут быть очищены из экспрессирующих клеток с получением активных белков. Таким образом, варианты данного изобретения включают в себя применение нуклеиновых кислот, кодирующих всю последовательность SEQ ID NO:1 или ее часть. Такие нуклеиновые кислоты включают в себя всю последовательность SEQ ID NO:2 или ее часть, которая соответствует кДНК-последовательности, кодирующей полипептид гелонина (номер доступа L12243 GenBank). Таким образом, предполагается, что любые из способов и композиций, обсуждаемых в данном описании в отношении нуклеиновых кислот, могут применяться в отношении SEQ ID NO:2.
В применении в данном описании термин «сегмент ДНК» относится к молекуле ДНК, которая была выделена из общей геномной ДНК конкретного вида. Таким образом, сегмент ДНК, кодирующий полипептид, обозначает сегмент ДНК, который содержит кодирующие полипептид дикого типа, полиморфный или мутантный полипептид последовательности, но выделенный или очищенный из общей (тотальной) геномной ДНК млекопитающего или человека. В термин «сегмент ДНК» включены сегменты ДНК, кодирующие полипептид или полипептиды, сегменты ДНК, кодирующие меньшие молекулы, чем полипептид, и рекомбинантные векторы, в том числе, например, плазмиды, космиды, фаговые векторы, вирусы и т.п.
В применении в этой заявке термин «полинуклеотид» относится к молекуле нуклеиновой кислоты, которая была выделена из общей геномной нуклеиновой кислоты. Таким образом, термин «полинуклеотид, кодирующий нативный полипептид» относится к сегменту ДНК, который содержит последовательности, кодирующие полипептид дикого типа или полиморфный полипептид, выделенные или очищенные из общей геномной ДНК млекопитающего или человека. Таким образом, например, когда в данной заявке говорят о функции или активности гелонина, «нативного полипептида гелонина» или «модифицированного полипептида гелонина», который кодируется полинуклеотидом гелонина, это означает, что этот полинуклеотид кодирует молекулу, которая имеет ферментативную активность RIP.
Термин «кДНК» относится к ДНК, полученной с использованием мессенджер-РНК (мРНК) в качестве матрицы. Преимущество применения кДНК, в противоположность геномной ДНК или ДНК, полимеризованной из геномной, непроцессированной или частично процессированной РНК-матрицы, заключается в том, что эта кДНК прежде всего содержит кодирующие последовательности соответствующего белка. Могут быть случаи, когда полная или частичная геномная последовательность является предпочтительной, например, когда для оптимальной экспрессии требуются некодирующие последовательности или когда нацеливание происходит на некодирующие области, такие как интроны, в антисмысловой стратегии.
Предполагается также, что конкретный полипептид из конкретного вида может быть представлен природными вариантами, которые имеют слегка отличающиеся последовательности нуклеиновых кислот, но тем не менее кодируют тот же самый белок (см. таблицу 1 выше).
Подобным образом, полинуклеотид, содержащий выделенный или очищенный ген полипептида дикого типа, полиморфного или мутантного полипептида, обозначает сегмент ДНК, содержащий кодирующие последовательности полипептида дикого типа, полиморфного или мутантного полипептида и, в некоторых аспектах, регуляторные последовательности, выделенный по существу из других природно встречающихся генов или кодирующих белок последовательностей. В этом отношении термин «ген» используется для простоты для обозначения единицы, кодирующей функциональный белок, полипептид или пептид. Как должно быть понятно специалистам с квалификацией в данной области, этот функциональный термин включает в себя геномные последовательности, кДНК-последовательности и меньшие сконструированные сегменты гена, которые экспрессируют или могут быть адаптированы для такой экспрессии, белки, полипептиды, домены, пептиды, слитые белки и мутанты. Нуклеиновая кислота, кодирующая весь нативный или модифицированный полипептид или его часть, может содержать непрерывную последовательность нуклеиновой кислоты, кодирующую весь полипептид или часть такого полипептида следующей длины: приблизительно, по меньшей мере или самое большее, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240, 250, 260, 270, 280, 290, 300, 310, 320, 330, 340, 350, 360, 370, 380, 390, 400, 410, 420, 430, 440, 441, 450, 460, 470, 480, 490, 500, 510, 520, 530, 540, 550, 560, 570, 580, 590, 600, 610, 620, 630, 640, 650, 660, 670, 680, 690, 700, 710, 720, 730, 740, 750, 760, 770, 780, 790, 800, 810, 820, 830, 840, 850, 860, 870, 880, 890, 900, 910, 920, 930, 940, 950, 960, 970, 980, 990, 1000, 1010, 1020, 1030, 1040, 1050, 1060, 1070, 1080, 1090, 1095, 1100, 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000, 5500, 6000, 6500, 7000, 7500, 8000, 9000, 10000 или более нуклеотидов, нуклеозидов или пар оснований. Такие длины непрерывных остатков могут применяться к любой последовательности нуклеиновой кислоты, описанной или обсуждаемой в данном описании, в том числе SEQ ID NO:2-10.
В конкретных вариантах осуществления данное изобретение относится к выделенным сегментам ДНК и рекомбинантным векторам, включающим в себя последовательности ДНК, которые кодируют полипептид или пептид дикого типа, полиморфный или модифицированный полипептид или пептид, который содержит в его аминокислотной последовательности последовательность смежных аминокислот, соответствующую или по существу соответствующую нативному полипептиду. Таким образом, выделенный сегмент ДНК или вектор, содержащий сегмент ДНК, может кодировать, например, модифицированный полипептид гелонина, который имеет инактивирующую рибосомы активность и специфичность нативного полипептида гелонина, но имеет отличающиеся аминокислоты. Термин "рекомбинантный" может быть использован вместе с полипептидом или названием конкретного полипептида, и он обычно относится к полипептиду, продуцируемому из молекулы нуклеиновой кислоты, которая была подвергнута манипуляции in vitro, или к полипептиду, который является реплицируемым продуктом такой молекулы.
В других вариантах осуществления данное изобретение относится к выделенным сегментам ДНК и рекомбинантным векторам, включающим последовательности ДНК, которые кодируют полипептид или пептид, который содержит в его аминокислотной последовательности последовательность смежных аминокислот, соответствующую или по существу соответствующую этому полипептиду.
Сегменты нуклеиновых кислот, используемые в данном изобретении, независимо от длины самой кодирующей последовательности, могут быть объединены с другими последовательностями нуклеиновых кислот, такими как промоторы, сигналы полиаденилирования, дополнительные сайты рестриктаз, сайты множественного клонирования, другие кодирующие сегменты и т.п., так что их общая длина может в значительной степени варьироваться. Таким образом, предполагается, что может быть использован фрагмент нуклеиновой кислоты почти любой длины, причем общая длина предпочтительно лимитируется легкостью приготовления и применения в предполагаемом протоколе рекомбинантных ДНК.
Предполагается, что конструкции нуклеиновых кислот данного изобретения могут кодировать полноразмерный полипептид из любого источника или могут кодировать укороченный вариант этого полипептида, например укороченный полипептид гелонина, так что транскрипт кодирующей области представляет укороченный вариант. Затем укороченный транскрипт может транслироваться в укороченный белок. Альтернативно, последовательность нуклеиновой кислоты может кодировать полноразмерную полипептидную последовательность с дополнительными гетерологическими кодирующими последовательностями, например, для обеспечения возможности очистки этого полипептида, транспорта, секреции, посттрансляционной модификации или для терапевтических выгод, таких как нацеливание или эффективность. Как обсуждалось выше, к кодирующей модифицированный полипептид последовательности могут быть добавлены метка или другой гетерологичный полипептид, где термин "гетерологичный" относится к полипептиду, который не является тем же самым, что и модифицированный полипептид.
В неограничивающем примере могут быть приготовлены одна или несколько конструкций нуклеиновых кислот, которые включают в себя непрерывный отрезок нуклеотидов, идентичный или комплементарный конкретному гену, такому как ген токсина гелонина. Конструкция нуклеиновой кислоты может иметь длину по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 250, 300, 400, 500, 600, 700, 800, 900, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 15000, 20000, 30000, 50000, 100000, 250000, 500000, 750000, по меньшей мере 1000000 нуклеотидов, а специалистам с обычной квалификацией в данной области известны также конструкции большего размера до хромосомных размеров включительно (в том числе все промежуточные длины и промежуточные диапазоны), при условии создания таких конструкций нуклеиновых кислот, как искусственная хромосома дрожжей. Будет вполне понятно, что термины "промежуточные длины" и "промежуточные диапазоны" в данном контексте означают любую длину или любой диапазон, включающие в себя приведенные величины или находящиеся между приведенными величинами (т.е. все целые числа, включающие эти величины или находящиеся между этими величинами).
Сегменты ДНК, используемые в данном изобретении, включают в себя биологически функциональные эквивалентные модифицированные полипептиды и пептиды, например, модифицированный токсин гелонин. Такие последовательности могут возникать как следствие избыточности кодонов и функциональной эквивалентности, которые, как известно, встречаются природно в последовательностях нуклеиновых кислот и, следовательно, в кодируемых белках. Альтернативно, функционально эквивалентные белки и пептиды могут быть созданы применением технологии рекомбинантных ДНК, в которой изменения в структуре белка могут быть сконструированы на основе учета свойств заменяемых аминокислот. Изменения, конструируемые человеком, могут быть введены посредством применения способов сайт-направленного мутагенеза, например, для введения улучшений в антигенность белка, для уменьшения токсичных действий этого белка in vivo в отношении субъекта, которому вводят этот белок, или для увеличения эффективности любого лечения с использованием этого белка.
1. Векторы
Нативные и модифицированные полипептиды могут кодироваться молекулами нуклеиновых кислот, содержащимися в векторе. Термин "вектор" обозначает молекулу нуклеиновой кислоты-носителя, в которую может быть встроена последовательность нуклеиновой кислоты для введения в клетку, где она может реплицироваться. Последовательность нуклеиновой кислоты может быть "экзогенной", что означает, что она является чужеродной для клетки, в которую вводится этот вектор, или что эта последовательность является гомологичной относительно последовательности в клетке, но находится в положении в нуклеиновой кислоте клетки-хозяина, в котором эта последовательность обычно не обнаруживается. Векторы включают в себя плазмиды, космиды, вирусы (бактериофаги, вирусы животных и вирусы растений) и искусственные хромосомы (например, YAC). Специалист с квалификацией в данной области мог бы получить конструкцию вектора с использованием стандартных рекомбинантных способов, которые описаны в Sambrook et al., 1989, и Ausubel et al., 1996, обе включены в данное описание в качестве ссылки. Кроме кодирования модифицированного полипептида, такого как модифицированный гелонин, вектор может кодировать немодифицированные полипептидные последовательности, такие как метка или нацеливающая последовательность. Применимые векторы, кодирующие такие слитые белки, включают в себя векторы pIN (Inouye et al., 1985), векторы, кодирующие отрезок из гистидинов, и векторы pGEX, для применения в генерировании растворимых слитых белков с глутатион-S-трансферазой (GST) для последующей очистки и отделения или отщепления. Нацеливающей молекулой является молекула, которая направляет этот модифицированный полипептид в конкретные орган, ткань, клетку или другое местоположение в теле субъекта.
Термин «экспрессирующий вектор» относится к вектору, содержащему последовательность нуклеиновой кислоты, кодирующую по меньшей мере часть продукта гена, способного транскрибироваться. В некоторых случаях молекулы РНК затем транслируются в белок, полипептид или пептид. В других случаях эти последовательности не транслируются, например, при продуцировании антисмысловых молекул или рибозимов. Экспрессирующие векторы могут содержать различные «регуляторные последовательности», которые являются последовательностями нуклеиновых кислот, необходимыми для транскрипции и, возможно, трансляции функционально связанной кодирующей последовательности в конкретном организме-хозяине. Кроме регуляторных последовательностей, которые управляют транскрипцией и трансляцией, векторы и экспрессирующие векторы могут содержать последовательности нуклеиновых кислот, которые выполняют другие функции, а также описаны ниже.
а. Промоторы и энхансеры
«Промотор» является регуляторной последовательностью, которая является областью последовательности нуклеиновой кислоты, которой регулируется инициация и скорость транскрипции. Он может содержать генетические элементы, при которых могут связываться регуляторные белки и молекулы, такие как РНК-полимераза и другие факторы транскрипции. Фразы «функционально размещенные», «функционально связанные», «под контролем» и «под транскрипционным контролем» означают, что промотор находится в правильном функциональном местоположении и/или правильной ориентации в отношении последовательности нуклеиновой кислоты для контроля инициации транскрипции и/или экспрессии этой последовательности. Промотор может использоваться или может не использоваться вместе с "энхансером", который является цис-действующей регуляторной последовательностью, участвующей в активации транскрипции последовательности нуклеиновой кислоты.
Промотор может быть промотором, природно связанным с геном или последовательностью, как это может быть получено выделением 5'-некодирующих последовательностей, локализованных слева от кодирующего сегмента и/или экзона. Такой промотор может называться "эндогенным". Подобным образом, энхансер может быть энхансером, природно связанным с последовательностью нуклеиновой кислоты, расположенным либо справа, либо слева от этой последовательности. Альтернативно, некоторые преимущества могли бы достигаться помещением сегмента кодирующей последовательности нуклеиновой кислоты под контроль рекомбинантного или гетерологичного промотора, который является промотором, обычно не связанным с последовательностью нуклеиновой кислоты в ее природном окружении. Рекомбинантный или гетерологичный энхансер обозначает также энхансер, не связанный обычно с последовательностью нуклеиновой кислоты в ее природном окружении. Такие промоторы и энхансеры могут включать в себя промоторы или энхансеры других генов и промоторы или энхансеры, выделенные из любых других прокариотических, вирусных или эукариотических клеток, и промоторы или энхансеры, "не встречающиеся в природе", т.е. содержащие отличающиеся элементы отличающихся регуляторных областей транскрипции, и/или мутации, которые изменяют экспрессию. Кроме синтетического получения последовательностей нуклеиновых кислот промоторов и энхансеров, эти последовательности могут быть получены с использованием технологии рекомбинантного клонирования и/или амплификации нуклеиновых кислот, в том числе ПЦР™, вместе с описанными в данном описании композициями (см. патент США US 4683202, патент США US 5928906, каждый из которых включен в данное описание в качестве ссылки). Кроме того, предполагается, что могут быть использованы также регуляторные последовательности, которые управляют транскрипцией и/или экспрессией последовательностей в неядерных органеллах, таких как митохондрии, хлоропласты и т.п.
Само собой разумеется, что может быть важным использование промотора и/или энхансера, который эффективно управляет экспрессией сегмента ДНК в определенном типе клеток, органелле и организме, выбранных для экспрессии. Специалистам с квалификацией в области молекулярной биологии обычно известно применение промоторов, энхансеров и комбинаций клеточных типов для экспрессии белков, например, см. руководство Sambrook et al. (1989), включенное в данное описание в качестве ссылки. Используемые промоторы могут быть конститутивными, тканеспецифическими, индуцируемыми и/или применимыми при подходящих условиях для управления экспрессией высокого уровня введенного сегмента ДНК, например, выгодной в крупномасштабном производстве рекомбинантных белков и/или пептидов. Промотор может быть гетерологичным или эндогенным.
Таблица 5 дает перечень нескольких элементов/промоторов, которые могут быть использованы в связи с данным изобретением для регуляции экспрессии гена. Этот перечень не рассматривается в качестве исчерпывающего для всех возможных элементов, участвующих в стимуляции экспрессии, а приводится лишь в качестве примерного перечня. Таблица 6 дает примеры индуцируемых элементов, которые являются областями последовательности нуклеиновой кислоты, которые могут активироваться в ответ на специфический стимул.
Индуцируемые элементы
Тяжелые металлы
et al., 1989
1984; Ponta et al., 1985; Sakai
et al., 1988
поли(rc)
Вирус Ньюкаслской болезни крупного рогатого скота
Большой Т-антиген SV40
Идентичность тканеспецифических промоторов или элементов, а также анализы для характеристики их активности хорошо известны специалистам с квалификацией в данной области. Примеры таких областей включают в себя ген LIMK2 человека (Nomoto et al., 1999), ген рецептора 2 соматостатина (Kraus et al., 1998), мышиный эпидидимальный связывающий ретиноевую кислоту ген (Lareyre et al., 1999), CD4 человека (Zhao-Emonet et al., 1998), мышиный альфа2 (XI)-коллаген (Tsumaki et al., 1998), ген рецептора допамина D1A (Lee et al., 1997), инсулиноподобный фактор роста II (Wu et al., 1997), молекула адгезии-1 эндотелиальных клеток тромбоцитов человека (Almendro et al., 1996).
b. Сигналы инициации и внутренние сайты связывания рибосомы
Для эффективной трансляции кодирующих последовательностей может также требоваться специфический сигнал инициации. Эти сигналы включают в себя кодон инициации ATG или смежные последовательности. Может быть необходимым обеспечение экзогенных сигналов регуляции трансляции. Специалист с обычной квалификацией в данной области способен с легкостью определить это и обеспечить необходимые сигналы. Хорошо известно, что инициирующий кодон должен находиться «в рамке считывания» с рамкой считывания желательной кодирующей последовательности для обеспечения трансляции всего инсерта. Экзогенные сигналы регуляции трансляции и инициирующие кодоны могут быть либо природными, либо синтетическими. Эффективность экспрессии может быть повышена включением подходящих энхансерных элементов.
В некоторых вариантах осуществления данного изобретения элементы внутренних сайтов связывания рибосомы (IRES) используют для создания мультигенных, или полицистронных, матриц. Элементы IRES способны обходить модель рибосомного сканирования зависимой от 5'-метилированного Cap трансляции и начинать трансляцию при внутренних сайтах (Pelletier and Sonenberg, 1988). Были описаны элементы IRES из двух членов семейства пикорнавирусов (полиомиелита и энцефаломиокардита) (Pelletier and Sonenberg, 1988), а также IRES из матрицы млекопитающего (Macejak and Sarnow, 1991). Элементы IRES могут быть связаны с гетерологичными открытыми рамками считывания. Множественные открытые рамки считывания могут транскрибироваться вместе, каждая отделяемая посредством IRES, с образованием полицистронных матриц. Благодаря элементу IRES каждая открытая рамка считывания доступна рибосомам для эффективной трансляции. Множественные гены могут эффективно экспрессироваться с использованием единственного промотора/энхансера для транскрипции единой матрицы (см. патенты США US 5925565 и 5935819, включенные в данное описание в качестве ссылки).
с. Сайты множественного клонирования
Векторы могут включать в себя сайт множественного клонирования (MCS), который является областью нуклеиновой кислоты, которая содержит множественные сайты рестрикционных ферментов, любой из которых может быть использован вместе со стандартной рекомбинантной технологией для расщепления этого вектора (см. Carbonelli et al., 1999, Levenson et al., 1998, и Cocea, 1997, включенные в данное описание в качестве ссылки). "Расщеплением рестрикционными ферментами" называют каталитическое расщепление молекулы нуклеиновой кислоты ферментом, который функционирует только в специфических местоположениях в молекуле нуклеиновой кислоты. Многие из из этих рестрикционных ферментов являются коммерчески доступными. Применение таких ферментов хорошо понятно специалистам с квалификацией в данной области. Часто вектор линеаризуют или фрагментируют с использованием рестрикционного фермента, который разрезает в MCS, для лигирования экзогенных последовательностей в этот вектор. «Лигирование» обозначает процесс образования фосфодиэфирных связей между двумя фрагментами нуклеиновых кислот, которые могут быть или могут не быть смежными друг с другом. Способы с использованием рестриктаз и реакций лигирования хорошо известны специалистам с квалификацией в области рекомбинантной технологии.
d. Сайты сплайсинга
Большинство транскрибируемых эукариотических молекул РНК будет подвергаться сплайсингу РНК для удаления интронов из первичных транскриптов. Векторы, содержащие геномные эукариотические последовательности, могут требовать донорных и/или акцепторных сайтов сплайсинга для гарантии правильного процессинга транскрипта для экспрессии белка (см. работу Chandler et al., 1997, включенную в данное описание в качестве ссылки).
е. Сигналы терминации
Векторы или конструкции данного изобретения обычно будут содержать по меньшей мере один сигнал терминации. «Сигнал терминации» или «терминатор» состоит из последовательностей ДНК, участвующих в специфической терминации РНК-транскрипта РНК-полимеразой. Таким образом, в некоторых вариантах обсуждается сигнал терминации, который заканчивает продуцирование РНК-транскрипта. Терминатор может быть необходимым in vivo для достижения желаемых уровней матрицы.
В эукариотических системах область терминатора может также содержать специфические последовательности ДНК, которые обеспечивают возможность сайт-специфического отщепления нового транскрипта таким образом, чтобы открыть сайт полиаденилирования. Он сигнализирует специальной эндогенной полимеразе о необходимости добавления отрезка из приблизительно 200 остатков А (полиА) к 3'-концу этого транскрипта. Молекулы РНК, модифицированные этим полиА-хвостом, являются, по-видимому, более стабильными и транслируются более эффективно. Таким образом, в других вариантах осуществления, включающих в себя эукариоты, предпочтительно, чтобы терминатор содержал сигнал расщепления РНК и более предпочтительно, чтобы сигнал терминатора стимулировал полиаденилирование матрицы. Элементы сайта терминатора и/или сайта полиаденилирования могут служить для увеличения уровней матрицы и/или минимизации сквозного считывания из этой кассеты в другие последовательности.
Терминаторы, обсуждаемые для применения в этом изобретении, включают в себя любой известный терминатор транскрипции, описанный в данном описании или известный специалистам с обычной квалификацией в данной области, в том числе, но не только, например, последовательности терминации генов, такие как, например, последовательности терминатора бычьего гормона роста или последовательности терминации вируса, такого как, например, терминатора SV40. В некоторых вариантах осуществления сигналом терминации может быть отсутствие транскрибируемой или транслируемой последовательности, например, вследствие укорочения последовательности.
f. Сигналы полиаденилирования
При экспрессии, в частности, эукариотической экспрессии, обычно включают сигнал полиаденилирования для выполнения правильного полиаденилирования транскрипта. Считается, что природа сигнала полиаденилирования не является принципиально важной для успешного применения на практике данного изобретения, и/или может использоваться любая такая последовательность. Предпочтительные варианты включают в себя сигнал полиаденилирования SV40 и/или сигнал полиаденилирования бычьего гормона роста, которые являются удобными и/или о которых известно, что они действует хорошо в различных клетках-мишенях. Полиаденилирование может увеличивать стабильность транскрипта или может облегчать цитоплазматический транспорт.
g. Сайты инициации репликации
Для размножения вектора в клетке-хозяине вектор может содержать один или более сайтов инициации репликации (часто называемых «ori»), причем этот сайт является специфической последовательностью нуклеиновой кислоты, в которой инициируется репликация. Альтернативно, если клеткой-хозяином являются дрожжи, может быть использована автономно реплицирующаяся последовательность (ARS).
h. Отбираемые и поддающиеся скринингу маркеры
В некоторых вариантах осуществления данного изобретения клетки, содержащие конструкцию нуклеиновой кислоты данного изобретения, могут быть идентифицированы in vitro или in vivo включением в экспрессирующий вектор маркера. Такие маркеры будут придавать идентифицируемое изменение клетке, делая возможной легкую идентификацию клеток, содержащих этот экспрессирующий вектор. Обычно отбираемым маркером является маркер, который придает свойство, позволяющее производить отбор. Позитивным отбираемым маркером является маркер, когда присутствие этого маркера делает возможным его отбор, тогда как негативным отбираемым маркером является маркер, присутствие которого предотвращает его отбор. Примером позитивного селективного маркера является маркер устойчивости к лекарственным средствам.
Обычно включение маркера для отбора с лекарственным средством помогает клонированию и идентификации трансформантов, например, гены, которые придают устойчивость к неомицину, пуромицину, гигромицину, DHFR, GPT, зеоцину и гистидинолу, являются применимыми отбираемыми маркерами. Кроме маркеров, придающих фенотип, который позволяет дискриминацию трансформантов на основе создания условий, обсуждаются также другие типы маркеров, в том числе поддающиеся скринингу маркеры, такие как GFP, основой обнаружения которого является колориметрический анализ. Альтернативно, могут использоваться такие поддающиеся скринингу ферменты, как тимидинкиназа вируса простого герпеса (tk) или хлорамфениколацетилтрансфераза (САТ). Специалист с квалификацией в данной области будет также знать, как использовать иммунологические маркеры, возможно, в сочетании с FACS-анализом. Считается, что используемый маркер не является решающим, пока он способен экспрессироваться одновременно с нуклеиновой кислотой, кодирующей продукт гена. Дополнительные примеры отбираемых и поддающихся скринингу маркеров хорошо известны специалисту с квалификацией в данной области.
2. Клетки-хозяева
В применении в данном описании термины «клетка», «клеточная линия» и «культура клеток» могут быть использованы взаимозаменяемо. Все эти термины включают в себя также их потомство, которое является любыми или всеми последующими поколениями. Должно быть понятно, что все потомство не может быть идентичным вследствие произвольных или случайных мутаций. В контексте экспрессии гетерологичной последовательности нуклеиновой кислоты «клеткой-хозяином» называют прокариотическую или эукариотическую клетку, и она включает в себя любой трансформируемый организм, который способен реплицировать вектор и/или экспрессировать гетерологичный ген, кодируемый вектором. Клетка-хозяин может быть использована и была использована в качестве реципиента для векторов. Клетка-хозяин может быть «трансфицирована» или «трансформирована», что относится к процессу, при помощи которого экзогенная нуклеиновая кислота, например, кодирующая модифицированный белок последовательность, переносится или вводится в клетку-хозяина. Трансформированная клетка включает в себя первичную рассматриваемую клетку и ее потомство.
Клетки-хозяева могут происходить из прокариот или эукариот, в том числе дрожжевых клеток, клеток насекомых и клеток млекопитающих, в зависимости от того, является ли желаемым результатом репликация вектора или экспрессия части или всех кодируемых вектором последовательностей нуклеиновых кислот. Многочисленные клеточные линии и культуры являются доступными в качестве клетки-хозяина, и они могут быть получены через Американскую Коллекцию Типовых Культур (АТСС), которая является организацией, служащей в качестве архива для живых культур и генетических материалов (www.atcc.org). Подходящий хозяин может быть определен специалистом с квалификацией в данной области на основе скелета вектора и желаемого результата. Например, плазмида или космида может быть введена в прокариотическую клетку-хозяина для репликации многих векторов. Бактериальные клетки, используемые в качестве клеток-хозяев для репликации и/или экспрессии, включают в себя клетки DH5α, JM109 и КС8, а также ряд коммерчески доступных бактериальных хозяев, таких как SURE® Competent Cells и SOLOPACK™ Gold Cells (Stratagene®, La Jolla). Альтернативно, такие бактериальные клетки, как E. coli LE392, могли бы применяться в качестве клеток-хозяев для фаговых вирусов. Подходящие дрожжевые клетки включают в себя Saccharomyces cerevisiae, Saccharomyces pombe и Pichia pastoris.
Примеры эукариотических клеток-хозяев для репликации и/или экспрессии вектора включают в себя клетки HeLa, NIH3T3, Jurkat, 293, Cos, CHO, Saos и РС12. Многие клетки-хозяева из различных типов клеток и организмов являются доступными и должны быть известными специалистам с квалификацией в данной области. Подобным образом, вместе либо с эукариотической, либо с прокариотической клеткой-хозяином может быть использован вирусный вектор, в частности вектор, который является пермиссивным для репликации или экспрессии этого вектора.
Некоторые векторы могут использовать регуляторные последовательности, которые позволяют им реплицироваться и/или экспрессироваться как в прокариотических, так и в эукариотических клетках. Далее, специалисту с квалификацией в данной области будет понятно, какими являются условия для инкубирования всех вышеупомянутых клеток-хозяев для их поддержания и обеспечения возможности репликации вектора. Также понятными и известными являются способы и условия, которые обеспечат возможность крупномасштабного получения векторов, а также получения нуклеиновых кислот, кодируемых векторами, и их соответствующих полипептидов, белков или пептидов.
3. Экспрессирующие системы
Существуют многочисленные экспрессирующие системы, которые содержат по меньшей мере часть композиций или все композици, обсуждаемые выше. Системы на основе прокариот и/или на основе эукариот могут быть использованы с данным изобретением для получения последовательностей нуклеиновых кислот или соответствующих им полипептидов, белков и пептидов. Многие такие системы являются широко доступными коммерчески.
Система клетка насекомого/бакуловирус может продуцировать высокий уровень экспрессии белка с использованием сегмента гетерологичной нуклеиновой кислоты, как описано, например, в патентах США с номерами 5871986, 4879236, оба из которых включены в данное описание в качестве ссылки, и эта система может быть куплена, например, под названием MaxBac® 2.0 из Invitrogen® и BacPack™ Baculovirus Expression System из Clontech®.
Кроме описанных экспрессирующих систем данного изобретения, другие примеры экспрессирующих систем включают в себя индуцируемую систему экспрессии в млекопитающих Stratagene®'s Complete Control™, которая включает в себя синтетический индуцируемый экдизоном рецептор или его систему экспрессии рЕТ, систему экспрессии E. coli. Другой пример индуцируемой экспрессирующей системы доступен из Invitrogen®, причем эта система является T-Rex™ (регулируемой тетрациклином) системой, индуцируемой системой экспрессии млекопитающих, которая использует полноразмерный промотор CMV. Invitrogen™ обеспечивает также систему экспрессии дрожжей, называемую системой экспрессии Pichia methanolica, которая предназначена для получения на высоком уровне рекомбинантных белков в метилотрофных дрожжах Pichia methanolica. Специалисту с квалификацией в данной области будет ясно, как экспрессировать вектор, такой как экспрессирующая конструкция, для получения последовательности нуклеиновой кислоты или ее соответствующих полипептида, белка или пептида.
4. Вирусные векторы
Имеется ряд способов, в которых экспрессирующие векторы могут быть введены в клетки. В некоторых вариантах осуществления данного изобретения экспрессирующий вектор содержит вирус или сконструированный вектор на основе вирусного генома. Способность некоторых вирусов входить в клетки посредством опосредованного рецептором эндоцитоза для интеграции в геном клетки-хозяина и стабильной и эффективной экспрессии вирусных генов сделала их привлекательными кандидатами для переноса чужеродных генов в клетки млекопитающих (Ridgeway, 1988; Nicolas and Rubenstein, 1988; Baichwal and Sugden, 1986; Temin, 1986). Первыми вирусами, использованными в качестве генных векторов, были ДНК-вирусы, в том числе паповавирусы (вирус обезьян 40, бычий папилломавирус и вирус полиомы) (Ridgeway, 1988; Baichwal and Sugden, 1986) и аденовирусы (Ridgeway, 1988; Baichwal and Sugden, 1986). Они имеют относительно низкую емкость для чужеродных последовательностей ДНК и имеют ограниченный спектр хозяев. Кроме того, их онкогенный потенциал и цитопатические действия в пермиссивных клетках вызывают беспокойство в отношении безопасности. Они могут вмещать только 8 т.п.н. чужеродного генетического вещества, но могут быть легко введены в различные клеточные линии и в различных лабораторных животных (Nicolas and Rubenstein, 1988; Temin, 1986).
Ретровирусы являются группой одноцепочечных РНК-вирусов, отличающихся способностью превращать их РНК в двухцепочечную ДНК в инфицированных клетках; они могут быть также использованы в качестве векторов. Другие вирусные векторы могут также использоваться в качестве экспрессирующих конструкций в данном изобретении. Могут быть использованы векторы, произведенные из таких вирусов, как вирус коровьей оспы (Ridgeway, 1988; Baichwal and Sugden, 1986; Coupar et al., 1988), аденоассоциированный вирус (AAV) (Ridgeway, 1988; Baichwal and Sugden, 1986; Hermonat and Muzycska, 1984) и герпес-вирусы. Они предоставляют некоторые привлекательные признаки для различных клеток млекопитающих (Friedman, 1989; Ridgeway, 1988; Baichwal and Sugden, 1986; Coupar et al., 1988; Horwich et al., 1990).
В. Определение нуклеиновых кислот
Кроме их использования в управлении экспрессией сконструированного токсина и модифицированных белков, полипептидов и/или пептидов, последовательности нуклеиновых кислот, описанные в данном описании, имеют различные другие применения. Например, они находят применение в качестве зондов или праймеров для вариантов осуществления, включающих в себя гибридизацию нуклеиновых кислот. Определение нуклеиновых кислот, кодирующих модифицированные белки или сконструированные токсины, включено также в данное изобретение.
1. Гибридизация
Применение зонда или праймера с длиной 13-100 нуклеотидов, предпочтительно 17-100 нуклеотидов или в некоторых аспектах с длиной до 1-2 т.п.н. или более, позволяет образовать дуплексную молекулу, которая является как стабильной, так и селективной. Для увеличения стабильности и/или селективности полученных гибридных молекул обычно являются предпочтительными молекулы, имеющие комплементарные последовательности на протяжении непрерывных отрезков с длиной более 20 оснований. Обычно предпочтительно конструирование молекул нуклеиновых кислот для гибридизации, имеющих одну или несколько комплементарных последовательностей из 20-30 нуклеотидов или даже более длинных, если желательно. Такие фрагменты могут быть легко получены, например, прямым синтезом фрагмента химическими средствами или введением выбранных последовательностей в рекомбинантные векторы для получения рекомбинантов.
Таким образом, нуклеотидные последовательности данного изобретения могут быть использованы для их способности селективно образовывать дуплексные молекулы с комплементарными отрезками ДНК и/или РНК или для обеспечения праймеров для амплификации ДНК или РНК из образцов. В зависимости от предполагаемого применения желательным является применение варьирующихся условий гибридизации для достижения варьирующихся степеней селективности зонда или праймеров для последовательности-мишени.
Для применений, требующих высокой селективности, обычно будет желательно использовать условия относительно высокой жесткости для образования гибридов, например условия относительно низкой концентрации соли и/или высокой температуры, такие как обеспечиваемые приблизительно 0,02 М - приблизительно 0,10 М NaCl при температурах приблизительно 50°С - приблизительно 70°С. Такие условия высокой жесткости позволяют лишь мало, если вообще позволяют, ошибочных спариваний между зондом или праймерами и матрицей или цепью-мишенью и были бы особенно подходящими для выделения специфических генов или для определения специфических мРНК-транскриптов. Обычно удобно делать условия более строгими добавлением увеличивающихся количеств формамида.
Для некоторых применений, например для сайт-направленного мутагенеза, предпочтительны условия более низкой жесткости. В этих условиях гибридизация может происходить, даже если последовательности гибридизующихся цепей не являются точно комплементарными, а являются ошибочно спаренными в одном или нескольких положениях. Условия могут быть сделаны менее строгими увеличением концентрации соли и/или понижением температуры. Например, условия средней жесткости могли бы быть обеспечены приблизительно 0,1-0,25 М NaCl при температурах приблизительно 37°С - приблизительно 55°С, тогда как условия низкой жесткости могли бы быть обеспечены приблизительно 0,15 - приблизительно 0,9 М солью при температурах в диапазоне от приблизительно 20°С до приблизительно 55°С. Условиями гибридизации можно легко манипулировать в зависимости от желаемых результатов.
В других вариантах осуществления гибридизация может достигаться, например, при условиях 50 мМ Трис-HCl (рН 8,3), 75 мМ KCl, 3 мМ MgCl2, 1,0 мМ дитиотреитол, при температурах между приблизительно 20°С - приблизительно 37°С. Другие используемые условия гибридизации могли бы включать в себя приблизительно 10 мМ Трис-HCl (рН 8,3), 50 мМ KCl, 1,5 мМ MgCl2 при температурах в диапазоне от приблизительно 40°С до приблизительно 72°С.
В некоторых вариантах осуществления предпочтительно использовать нуклеиновые кислоты определенных последовательностей данного изобретения в комбинации с подходящими средствами, такими как метка, для определения гибридизации. В данной области известно большое разнообразие подходящих индикаторных средств, в том числе флуоресцентных, радиоактивных, ферментных или других лигандов, таких как авидин/биотин, которые могут быть определены. В предпочтительных вариантах осуществления может быть желательным применение флуоресцентной метки или ферментной метки, такой как уреаза, щелочная фосфатаза или пероксидаза, вместо радиоактивной или других нежелательных для окружающей среды реагентов. В случае ферментных меток, известны колориметрические индикаторные субстраты, которые могут быть использованы для обеспечения средства определения, которое является видимым или спектрофотометрически определяемым, для идентификации специфической гибридизации с содержащими комплементарные нуклеиновые кислоты образцами.
Обычно предполагается, что зонды или праймеры, описанные в данном описании, будут применимы в качестве реагентов в гибридизации в растворе, например, в ПЦР™, для обнаружения экспрессии соответствующих генов, а также в вариантах, использующих твердую фазу. В вариантах осуществления, использующих твердую фазу, тест-ДНК (или РНК) адсорбируют или иным образом прикрепляют к выбранному матриксу или к выбранной поверхности. Затем эту фиксированную, одноцепочечную нуклеиновую кислоту подвергают гибридизации с выбранными зондами в желательных условиях. Выбранные условия будут зависеть от конкретных обстоятельств (например, от содержания G+C, типа нуклеиновой кислоты-мишени, источника нуклеиновой кислоты, размера гибридизационного зонда и т.д.). Оптимизация условий гибридизации для представляющего интерес конкретного применения хорошо известна специалистам с квалификацией в данной области. После промывания гибридизованных молекул для удаления неспецифически связанных молекул зонда гибридизацию определяют и/или количественно определяют посредством определения количества связанной метки. Репрезентативные способы гибридизации на твердой фазе описаны в патентах США с номерами 5843663, 5900481 и 5919626. Другие способы гибридизации, которые могут быть использованы в практике данного изобретения, описаны в патентах США с номерами 5849481, 5849486 и 5851772. Релевантные части этих и других ссылок, цитированных в этом разделе описания, включены в данное описание в качестве ссылок.
2. Амплификация нуклеиновых кислот
Нуклеиновые кислоты, используемые в качестве матрицы для амплификации, могут быть выделены из клеток, тканей или других образцов в соответствии со стандартными методологиями (Sambrook et al., 1989). В некоторых вариантах осуществления анализ выполняют на целых клетках или на гомогенатах ткани или на образцах биологических жидкостей без существенной очистки нуклеиновой кислоты-матрицы. Нуклеиновая кислота может быть геномной ДНК или фракционированной или общей клеточной РНК. При использовании РНК может быть желательным предварительное превращение этой РНК в комплементарную ДНК.
Термин «праймер» в данном контексте включает в себя любую нуклеиновую кислоту, которая способна праймировать синтез образующейся нуклеиновой кислоты в зависимом от матрицы процессе. Обычно праймерами являются олигонуклеотиды длиной десять-двадцать и/или тридцать п.н., но могут использоваться и более длинные последовательности. Праймеры могут быть обеспечены в двухцепочечной и/или одноцепочечной форме, хотя предпочтительной является одноцепочечная форма.
Пары праймеров, сконструированных для селективной гибридизации с нуклеиновами кислотами, соответствующими SEQ ID NO:1 или любым другим последовательностям, контактируют с нуклеиновой кислотой-матрицей в условиях, позволяющих селективную гибридизацию. В зависимости от желаемого применения могут быть выбраны условия гибридизации высокой жесткости, которые будут допускать только гибридизацию с последовательностями, которые полностью комплементарны этим праймерам. В других вариантах гибридизация может происходить при пониженной жесткости для допущения амплификации нуклеиновых кислот, содержащих одно или несколько ошибочных спариваний с последовательностями праймеров. После гибридизации комплекс матрица-праймер контактируют с одним или несколькими ферментами, которые облегчают зависимый от матрицы синтез нуклеиновой кислоты. Множественные циклы амплификации, также называемые "циклами", проводят, пока не образуется достаточное количество амплифицируемого продукта.
Продукт амплификации может быть определен или количественно определен. В некоторых приложениях определение может быть выполнено визуально. Альтернативно, определение может включать в себя непрямую идентификацию продукта с использованием хемилюминесценции, радиоактивной сцинтиграфии включенной радиоактивной или флуоресцентной метки или даже с использованием системы, использующей электрические сигналы и/или сигналы термических импульсов (Affymax technology; Bellus, 1994).
Доступен ряд зависимых от матрицы процессов для амплификации олигонуклеотидных последовательностей, присутствующих в конкретном образце-матрице. Одним из наиболее известных способов амплификации является полимеразная цепная реакция (называемая ПЦР™), которая подробно описана в патентах США с номерами 4683195, 4683202 и 4800159 и в Innis et al., 1988, включенных в данное описание в качестве ссылки в их полном виде.
Процедура амплификации с использованием обратной транскриптазы-ПЦР™ может быть выполнена для количественного определения амплифицированной мРНК. Способы обратной транскрипции РНК с получением кДНК хорошо известны (см. Sambrook et al., 1989). Альтернативные способы для обратной транскрипции используют термостабильные ДНК-полимеразы. Эти способы описаны в WO 90/07641. Методологии полимеразной цепной реакции хорошо известны в данной области. Репрезентативные способы ОТ-ПЦР описаны в патенте США № 5882864.
Другим способом амплификации является лигазная цепная реакция (LCR), описанная в европейской заявке № 320308, включенной в данное описание в качестве ссылки в ее полном виде. В патенте США 4883750 описан способ, сходный с LCR, для связывания пары зондов с последовательностью-мишенью. Могут быть также использованы способ на основе ПЦР™ и олигонуклеотид-лигазный анализ (OLA), описанный в патенте США 5912148.
Альтернативные способы для амплификации последовательностей нуклеиновых кислот-мишеней, которые могут быть использованы в практике данного изобретения, описаны в патентах США с номерами 5843650, 5846709, 5846783, 5849546, 5849497, 5849547, 5858652, 5866366, 5916776, 5922574, 5928905, 5928906, 5932451, 5935825, 5939291 и 5942391, заявке Великобритании № 2202328 и РСТ заявке № РСТ/US89/01025, включенных в данное описание в качестве ссылок в их полном виде.
Q-бета-репликаза, описанная в заявке РСТ № РСТ/US87/00880, может быть также использована в качестве способа амплификации в данном изобретении. В этом способе к образцу добавляют репликативную последовательность РНК, которая имеет область, комплементарную области мишени, в присутствии РНК-полимеразы. Эта полимераза будет копировать репликативную последовательность, которая может быть затем определена.
В амплификации нуклеиновых кислот данного изобретения может быть также использован способ изотермической амплификации, в котором используют рестриктазы и лигазы, для получения амплификации молекул-мишеней, которые содержат 5'-[альфа-тио]трифосфаты в одной цепи сайта рестрикции (Walker et al., 1992). Амплификация с вытеснением цепи (SDA), описанная в патенте США № 5916779, является другим способом проведения изотермической амплификации нуклеиновых кислот, который использует множественные циклы вытеснения и синтеза цепи, т.е. ник-трансляцию.
Другие процедуры амплификации нуклеиновых кислот включают в себя системы амплификации на основе транскрипции (TAS), в том числе амплификацию на основе последовательности нуклеиновой кислоты (NASBA) и 3SR (Kwoh et al., 1989; РСТ заявка WO 88/10315, включенные в данное описание в качестве ссылки в их полном виде). Европейская заявка № 329822 описывает способ амплификации нуклеиновых кислот, включающий в себя циклический синтез одноцепочечной РНК (ssRNA), одноцепочечной ДНК (ssDNA) и двухцепочечной ДНК (dsDNA), который может быть использован в соответствии с данным изобретением.
Заявка РСТ WO 89/06700 (включенная в данное описание в качестве ссылки в ее полном виде) описывает схему амплификации последовательности нуклеиновой кислоты, основанную на гибридизации последовательности область промотора/область праймера с одноцепочечной ДНК-мишенью (ssDNA) с последующей транскрипцией многочисленных РНК-копий этой последовательности. Эта схема не является циклической, т.е. из полученных РНК-транскриптов не образуются новые матрицы. Другие способы амплификации включают в себя RACE и «одностороннюю ПЦР» (Frohman, 1990; Ohara et al., 1989).
3. Определение нуклеиновых кислот
После любой амплификации может быть желательным отделение продукта амплификации от матрицы и/или избыточного праймера. В одном варианте осуществления продукты амплификации разделяют электрофорезом в агарозном, агароза-акриламидном или полиакриламидном геле (Sambrook et al., 1989). Разделенные продукты амплификации могут быть вырезаны и элюированы из геля для дополнительной манипуляции. При помощи агарозных гелей с низкой точкой плавления отделенную полосу можно удалить нагреванием геля с последующей экстракцией нуклеиновой кислоты.
Разделение нуклеиновых кислот может быть также выполнено хроматографическими способами, известными в данной области. Имеются многочисленные типы хроматографии, которые могут быть использованы в практике данного изобретения, в том числе адсорбционная, распределительная, ионообменная, гидроксиапатитная, использующая молекулярные сита, обращенно-фазовая, колоночная, бумажная, тонкослойная и газовая хроматография, а также ВЭЖХ.
В некоторых вариантах продукты амплификации визуализируют. Обычный способ визуализации включает в себя окрашивание геля бромидом этидия и визуализацию под УФ-светом. Альтернативно, если продукты амплификации являются интегрально меченными, радиоактивно или флуорометрически меченными нуклеотидами, разделенные продукты амплификации могут быть экспонированы на рентгеновской пленке или визуализированы под подходящими спектрами возбуждения.
В одном варианте осуществления после разделения продуктов амплификации меченый зонд нуклеиновой кислоты приводят в контакт с амплифицированной маркерной последовательностью. Этот зонд предпочтительно конъюгирован с хромофором, но может быть радиоактивно меченным. В другом варианте зонд конъюгирован с партнером связывания, таким как антитело или биотин, или другим партнером связывания, несущим определяемую часть молекулы.
В конкретных вариантах осуществления определением является Саузерн-блоттинг и гибридизация с меченым зондом. Способы, используемые в Саузерн-блоттинге, хорошо известны специалистам с квалификацией в данной области (см. Sambrook et al., 1989). Один пример из предыдущего описания приведен в патенте США № 5279721, включенном в данное описание в качестве ссылки, который описывает устройство и способ для автоматического электрофореза и переноса нуклеиновых кислот. Устройство делает возможным электрофорез и блоттинг без внешней манипуляции с гелем и идеально пригодно для проведения способов данного изобретения.
Другие способы определения нуклеиновых кислот, которые могут быть использованы в практике данного изобретения, описаны в патентах США с номерами 5840873, 5843640, 5843651, 5846708, 5846717, 5846726, 5846729, 5849487, 5853990, 5853992, 5853993, 5856092, 5861244, 5863732, 5863753, 5866331, 5905024, 5910407, 5912124, 5912145, 2919630, 5925517, 5928862, 5928869,5929227, 5932413 и 5935791, каждый из которых включен в данное описание в качестве ссылки.
4. Другие анализы
Другие способы генетического скрининга могут быть использованы в рамках данного изобретения, например, для определения мутаций в образцах геномной ДНК, кДНК и/или РНК. Способы, используемые для обнаружения точковых мутаций, включают в себя электрофорез в денатурирующем градиентном геле (DGGE), анализ полиморфизмов длин рестрикционных фрагментов (RFLP), химические или ферментативные способы расщепления, прямое секвенирование областей мишени, амплифицированных при помощи ПЦР™ (см. выше), анализ одноцепочечных конформационных полиморфизмов (SSCP) и другие способы, хорошо известные в данной области.
Один способ скрининга на точковые мутации основан на расщеплении РНКазой ошибочных спариваний пар оснований в гетеродуплексах РНК/ДНК или РНК/РНК. В данном контексте термин «ошибочное спаривание» определяется как область одного или нескольких неспаренных или ошибочно спаренных нуклеотидов в двухцепочечной молекуле РНК/РНК, РНК/ДНК или ДНК/ДНК. Таким образом, это определение включает в себя ошибочные спаривания, обусловленные инсерционными/делеционными мутациями, а также точковыми мутациями одного или множественных нуклеотидов.
Патент США № 4946773 описывает анализ расщепления ошибочного спаривания РНКазой А, который предусматривает отжиг тестируемых образцов одноцепочечной ДНК или РНК с РНК-зондом и последующую обработку дуплексов нуклеиновых кислот РНКазой А. Для определения ошибочных спариваний одноцепочечные продукты обработки РНКазой А, электрофоретически разделенные в соответствии с размером, сравнивают с обработанными таким же образом контрольными дуплексами. Образцы, содержащие меньшие фрагменты (продукты расщепления), не обнаруживаемые в контрольном дуплексе, оцениваются как положительные.
Другие исследователи описали применение РНКазы I в анализах ошибочного спаривания. Применение РНКазы I для обнаружения ошибочного спаривания оснований описано в литературе из Promega Biotech. Promega продает набор, содержащий РНКазу I, которая, как сообщается, расщепляет три из четырех известных ошибочных спариваний. Другие исследователи описали применение белка MutS или других ферментов репарации ДНК для обнаружения ошибочных спариваний одиночных нуклеотидов.
Альтернативные способы для обнаружения делеционных, инсерционных или заменяющих мутаций, которые могут быть использованы в практике данного изобретения, описаны в патентах США с номерами 5849483, 5851770, 5866337, 5925525 и 5928870, каждый из которых включен в данное описание в качестве ссылки в его полном виде.
С. Способы переноса генов
Считается, что подходящие способы для доставки нуклеиновых кислот для выполнения экспрессии композициий данного изобретения включают в себя фактически любой способ, при помощи которого нуклеиновая кислота (например, ДНК, в том числе вирусные и невирусные векторы) может быть введена в органеллу, клетку, ткань или организм, как описано в данном описании или как должно быть известно специалисту с обычной квалификацией в данной области. Такие способы включают в себя, но не ограничиваются ими, прямую доставку ДНК, например, посредством инъекции (патенты США с номерами 5994624, 5981274, 5945100, 5780448, 5736524, 5702932, 5656610, 5589466 и 5580859, каждый из которых включен в данное описание в качестве ссылки), в том числе микроинъекции (Harlan and Weintraub, 1985; патент США № 5789215, включенные в данное описание в качестве ссылки); электропорацией (патент США № 5384253, включенный в данное описание в качестве ссылки); осаждением фосфатом кальция (Graham and Van Der Eb, 1973; Chen and Okayama, 1987; Rippe et al., 1990); с использованием ДЭАЭ-декстрана с последующим добавлением полиэтиленгликоля (Gopal, 1985); посредством прямой обработки ультразвуком (Frechheimer et al., 1987); опосредованной липосомами трансфекцией (Nicolau and Sene, 1982; Fraley et al., 1979; Nicolau et al., 1987; Wong et al., 1980; Kaneda et al., 1989; Kato et al., 1991); бомбардировкой микроснарядами (заявки РСТ с номерами WO 94/09699 и 95/06128; патенты США с номерами 5610042; 5322783; 5563055; 5550318; 5538877 и 5538880, каждый включен в данное описание в качестве ссылки); встряхиванием с волокнами карбида кремния (Kaeppler et al., 1990; патенты США с номерами 5302523 и 5464765, каждый из которых включен в данное описание в качестве ссылки); опосредованной Agrobacterium трансформацией (патенты США с номерами 5591616 и 5563055, каждый из которых включен в данное описание в качестве ссылки) или ПЭГ-опосредованной трансформацией протопластов (Omirulleh et al., 1993; патенты США с номерами 4684611 и 4952500, каждый из которых включен в данное описание в качестве ссылки); опосредованным обезвоживанием/ингибированием поглощением ДНК (Potrykus et al., 1985). С использованием таких способов органелла (органеллы), клетка (клетки), ткань (ткани) или организм (организмы) могут быть стабильно или транзиторно (временно) трансформированы.
III. Инактивирующие рибосомы белки
Инактивирующие рибосомы токсины (RIT) являются сильнодействующими ингибиторами белкового синтеза в эукариотах. Ферментативный домен этих белков действует как цитотоксическая н-гликозидаза, которая способна инактивировать каталитически рибосомы, как только они входят во внутриклеточное пространство. Это достигается расщеплением н-гликозидной связи аденина в положении 4324 рРНК 28S, которое необратимо инактивирует рибосому разрушением сайта связывания для факторов элонгации. RIT, которые были выделены из бактерий, являются преобладающими у высших растений. В растениях имеются два типа: токсины Типа I имеют единственную полипептидную цепь, которая имеет ингибирующую рибосомы активность, а токсины Типа II имеют А-цепь, сравнимую с белком Типа I, которая связана дисульфидной связью с В-цепью, обладающей связывающими клетки свойствами. Примерами RIT Типа I являются гелонин, додекандрин, трикосантин, трикокирин, бриодин, антивирусный белок Mirabilis, инактивирующий рибосомы белок ячменя (BRIP), антивирусные белки фитолакки американской (РАР), сапорины, луффины и момордины. Токсины Типа II включают в себя рицин и абрин. Токсины могут быть конъюгированы или экспрессированы в виде слитого белка с любым из обсуждаемых в данном описании полипептидов. Альтернативно, модифицированные токсины данного изобретения могут быть конъюгированы с малой молекулой, такой как химиотерапевтический или нацеливающий агент.
А. Иммунотоксины
Токсины данного изобретения особенно пригодны для применения в качестве компонентов цитотоксических лекарственных средств. Эти цитотоксические средства могут быть использованы in vivo для селективной элиминации конкретного типа клеток, на которые токсиновый компонент нацеливается посредством специфической связывающей способности второго компонента. Для образования цитотоксических агентов модифицированные токсины данного изобретения могут быть конъюгированы с моноклональными антителами, в том числе химерными и CDR-привитыми антителами, и доменами/фрагментами антител (например, Fab, Fab' и F(ab')2, одноцепочечными антителами и Fv или одиночными вариабельными доменами). Иммуноконъюгаты, включающие в себя токсины, могут быть описаны как иммунотоксины. Иммунотоксин может также состоять из слитого белка, а не иммуноконъюгата.
Модифицированные токсины, конъюгированные с моноклональными антителами, генетически сконструированные таким образом, что они содержат свободные остатки цистеина, также находятся в рамках данного изобретения. Примеры Fab'- и F(ab')2-фрагментов, применимых в данном изобретении, описаны в WO 89/00999, который включен в данное описание в качестве ссылки.
Альтернативно, модифицированные токсины могут быть конъюгированы или слиты с гуманизированными или сконструированными антителами человека. Такие гуманизированные антитела могут быть сконструированы из вариабельных доменов мышиных антител.
1. Области антител
Области из различных членов семейства иммуноглобулинов включены в данное изобретение. Данное изобретение включает в себя обе вариабельные области из специфических антител, включающие в себя определяющие комплементарность области (CDR) и нейтрализующие области антител, в том числе области, которые связывают эффекторные молекулы, такие как Fc-области. Антиген-специфические кодирующие области из антител, такие как вариабельные области из IgG, IgM или IgA, могут быть использованы с другой молекулой, такой как токсин, в комбинации с нейтрализующей областью антитела или с одним из описанных выше терапевтических соединений.
Еще в одном варианте осуществления один ген может содержать одноцепочечное антитело. Способы получения одноцепочечных антител хорошо известны специалистам с квалификацией в данной области. Специалист в данной области отсылается в отношении таких способов к патенту США № 5359046 (включенному в данное описание в качестве ссылки). Одноцепочечное антитело создается слиянием вместе вариабельных доменов тяжелой и легкой цепей с использованием короткого пептидного линкера, с восстановлением посредством этого антигенсвязывающего сайта на единой молекуле.
Вариабельные фрагменты одноцепочечных антител (scFv), в которых С-конец одного вариабельного домена соединен с N-концом другого через 15-25-аминокислотный пептид или линкер, были разработаны без существенного нарушения связывания антигена или специфичности этого связывания (Bedzyk et al., 1990; Chaudhary et al., 1990). Эти Fv лишены константных областей (Fc), присутствующих в тяжелой и легкой цепях нативного антитела. Иммунотоксины, использующие одноцепочечные антитела, описаны в патенте США № 6099842, специально включенном в данное описание в качестве ссылки.
Обсуждаются антитела к большому разнообразию молекул, таких как онкогены, опухолеассоциированные антигены, цитокины, факторы роста, гормоны, ферменты, факторы транскрипции или рецепторы. Также обсуждаются секретируемые антитела, нацеленные против сыворотки, ангиогенных факторов (VEGF/VPF; βFGF; αFGF и другие), факторов свертывания и эндотелиальных антигенов, необходимых для ангиогенеза (т.е. V3-интегрин). Особо рассматриваются факторы роста, такие как трансформирующий фактор роста, фибробластный фактор роста и тромбоцитарный фактор роста (PDGF) и члены семейства PDGF.
Кроме того, данное изобретение относится к композиции, нацеливающей на специфические патогены с использованием антиген-специфических последовательностей или нацеливающей на специфические типы клеток, например на клетки, экспрессирующие маркеры клеточной поверхности, для идентификации этой клетки. Примеры таких маркеров клеточной поверхности включают в себя ассоциированные с опухолями антигены или специфические маркеры типов клеток, такие как CD4 или CD8.
Антитела, используемые в данном изобретении в качестве части иммунотоксина, могут быть нацелены на любой антиген. Этот антиген может быть специфическим для организма, для типа клеток, для заболевания или состояния или для патогена. Примеры антигенов включают в себя белки клеточной поверхности, например ассоциированные с опухолями антигены, вирусные белки, микробные белки, посттрансляционные модификации или углеводы и рецепторы. Обычные опухолевые маркеры включают в себя карциноэмбриональный антиген, специфический антиген предстательной железы, антиген связанный с опухолью мочевых путей, фетальный антиген, тирозиназу (р97), gp68, TAG-72, HMFG, Sialyl Lewis Antigen, MucA, MucB, PLAP, рецептор эстрогена, рецептор ламинина, erb B и р155. Другие антигены, которые могут быть мишенями нацеливания, включают в себя рецепторы для EGF и VEGF, TIE-1 и -2, CD-33, CD-38, CD-20, CD-52, GP-240, Lym-1, MMO-2 и ММР-9.
В. Другие нацеливающие части молекулы
Применение области белка, которая опосредует белок-белковые взаимодействия, в том числе взаимодействия лиганд-рецептор, также обсуждается данным изобретением. Эта область могла бы использоваться в качестве ингибитора или конкурента белок-белкового взаимодействия или в качестве специфического мотива нацеливания. Таким образом, данное изобретение включает в себя применение нацеливающей части молекулы для рекрутинга токсина или другого терапевтического или диагностического полипептида к конкретной части тела, органу, ткани или клетке. После достижения композициями данного изобретения этой конкретной зоны с использованием мотива нацеливания токсин или другой полипептид может функционировать.
Нацеливающие части молекул могут использовать преимущество белок-белковых взаимодействий. Они включают в себя взаимодействия между белками и среди белков, таких как рецепторы и лиганды; рецепторы и рецепторы; полимерные комплексы; факторы транскрипции; киназы и мишени ниже по ходу процесса; ферменты и субстраты и т.д. Например, домен связывания лиганда опосредует взаимодействие белок:белок между лигандом и соответствующим ему рецептором. Таким образом, этот домен мог бы быть использован либо для ингибирования связывания, либо для конкуренции со связыванием эндогенного лиганда, либо для более специфического нацеливания на типы клеток, которые экспрессируют рецептор, узнающий домен связывания лиганда, функционально связанный с терапевтическим полипептидом, таким как токсин гелонин.
Примеры лигандсвязывающих доменов включают в себя такие лиганды, как VEGF/VPF; βFGF; αFGF; факторы свертывания и эндотелиальные антигены, необходимые для ангиогенеза (т.е. V3-интегрин); факторы роста, такие как трансформирующий фактор роста, фибробластный фактор роста, колониестимулирующий фактор, Kit-лиганд (KL), flk-2/flt-3 и тромбоцитарный фактор роста (PDGF) и члены семейства PDGF; лиганды, которые связываются с рецепторами клеточной поверхности, такие как молекулы МНС, среди прочих.
Наиболее широко охарактеризованными лигандами являются асиалоорозомукоид (ASOR) (Wu and Wu, 1987) и трансферрин (Wagner et al., 1990). Недавно в качестве носителя для доставки генов использовали синтетический неогликопротеин, который узнает тот же самый рецептор, что и ASOR (Ferkol et al., 1993; Perales et al., 1994), а эпидермальный фактор роста (EGF) использовали также для доставки генов в клетки плоскоклеточной карциномы (Myers, EPO 0273085).
В других вариантах Nicolau et al. (1987) использовали лактозилцерамид, галактозатерминальный асиалганглиозид, включенный в липосомы, и наблюдали увеличение поглощения гена инсулина гепатоцитами. В качестве рецептора для опосредованной доставки в ткань предстательной железы может быть также использован специфический для предстательной железы человека антиген (Watt et al., 1986).
В дополнительных вариантах молекулу лектина можно использовать для нацеливания соединения на клетку, экспрессирующую конкретный углевод на ее поверхности.
1. Цитокины
Другой класс соединений, который обсуждается для функционального связывания с терапевтическим полипептидом, таким как токсин, включает в себя интерлейкины и цитокины, такие как интерлейкин 1 (IL-1), IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14, IL-15, β-интерферон, α-интерферон, γ-интерферон, ангиостатин, тромбоспондин, эндостатин, МЕТН-1, МЕТН-2, лиганд Flk2/Flt3, GM-CSF, G-CSF, M-CSF и фактор некроза опухолей (TNF).
2. Факторы роста
В других вариантах осуществления данного изобретения факторы роста или лиганды могут находиться в комплексе с лекарственным средством. Примеры включают в себя VEGF/VPE, FGF, TGFβ, лиганды, которые связываются с TIE, опухолеассоциированные изоформы фибронектина, фактор подвижности, фактор роста гепатоцитов, фактор роста фибробластов, тромбоцитарный фактор (PF4), PDGF, KIT-лиганд (KL), колониестимулирующие факторы (CSF), LIF и TIMP (тканевый ингибитор металлопротеиназ).
3. Индукторы клеточной пролиферации
Другая группа белков, которые могут быть использованы вместе с модифицированными белками данного изобретения, такими как модифицированный токсин гелонин, содержит белки, индуцирующие клеточную пролиферацию. В некоторых вариантах осуществления этот токсин функционально связан с рибозимом, который может инактивировать индуктор клеточной пролиферации, тогда как в других вариантах осуществления токсин связан с самим индуктором. Альтернативно, токсин может быть присоединен к антителу, которое узнает индуктор пролиферации клеток.
Общей чертой всех этих белков является их способность регулировать клеточную пролиферацию. Например, форма PDGF, sis-онкоген, является секретируемым фактором роста. Онкогены редко возникают из генов, кодирующих факторы роста, и в настоящее время sis является единственным природно встречающимся онкогенным фактором роста. В одном варианте осуществления данного изобретения предполагается, что антисмысловая мРНК, нацеленная на конкретный индуктор клеточной пролиферации, применима для предотвращения экспрессии этого индуктора клеточной пролиферации.
Белки FMS, ErbA, ErbB и neu являются рецепторами фактора роста. Мутации в этих рецепторах приводят к потере регуляторной функции. Например, точковая мутация в трансмембранном домене рецепторного белка Neu приводит к онкогену neu. Онкоген erbA происходит из внутриклеточного рецептора тиреоидного гормона. Считается, что модифицированный онкогенный рецептор ErbA конкурирует с эндогенным рецептором тиреоидного гормона, вызывая неконтролируемый рост.
Самый большой класс онкогенов включает в себя трансдуцирующие сигнал белки (например, Src, Abl и Ras). Белок Src является цитоплазматической протеинтирозинкиназой, и его трансформация из протоонкогена в онкоген в некоторых случаях происходит через мутации при остатке тирозина в положении 527. В противоположность этому, трансформация белка ГТФ-азы ras из протоонкогена в онкоген, в одном примере, происходит из мутации валина в глицин при аминокислоте 12 в этой последовательности, уменьшая ГТФ-азную активность ras.
Белки Jun, Fos и Myc являются белками, которые непосредственно проявляют их действие на ядерные функции в качестве факторов транскрипции.
4. Ингибиторы клеточной пролиферации
Супрессоры опухолей функционируют, ингибируя избыточную пролиферацию клеток. Инактивация этих генов разрушает их ингибиторную активность, приводя к нерегулируемой пролиферации. Предполагается, что токсины могут быть присоединены к антителам, которые узнают мутантные опухолевые супрессоры или опухолевые супрессоры дикого типа. Альтернативно, токсин может быть связан с всем опухолевым супрессором или его частью. Опухолевые супрессоры р53, р16 и С-САМ описаны ниже.
Высокие уровни мутантного р53 были обнаружены во многих клетках, малигнизированных химическим канцерогенезом, облучением ультрафиолетом и несколькими вирусами. Ген р53 является частой мишенью мутационной инактивации в широком разнообразии опухолей человека, и уже документировано, что он является наиболее часто мутируемым геном в обычных злокачественных опухолях человека. Он является мутированным в более чем 50% NSCLC человека (Hollstein et al., 1991) и в широком спектре других опухолей.
Ген р53 кодирует фосфопротеин из 393 аминокислот, который может образовывать комплексы с белками хозяина, такими как большой Т-антиген и E1B. Этот белок обнаружен в нормальных тканях и клетках, но в концентрациях, которые являются незначительными в сравнении с малигнизированными клетками или тканью опухоли.
р53 дикого типа признается в качестве важного регулятора роста во многих типах клеток. Миссенс-мутации являются обычными для гена р53 и являются существенными для трансформирующей способности этого онкогена. Единственная генетическая замена, провоцируемая точковыми мутациями, может создать канцерогенный р53. Однако известно, что в отличие от других онкогенов точковые мутации р53 встречаются по меньшей мере в 30 различных кодонах, часто создавая доминантные аллели, которые производят смещения в клеточном фенотипе без уменьшения гомозиготности. Кроме того, многие из этих доминантных негативных аллелей, являются, по-видимому, переносимыми в организме и переходят в зародышевую линию. Различные мутантные аллели, по-видимому, находятся в диапазоне от аллелей с минимально нарушенной функцией до сильно пенетрантных, доминантных негативных аллелей (Weinberg, 1991).
Другим ингибитором клеточной пролиферации является р16. Основные транзиции клеточного цикла эукариот запускаются циклин-зависимыми киназами или CDK. Одна CDK, циклин-зависимая киназа 4 (CDK4) регулирует прохождение через фазу G1. Активность этого фермента может заключаться в фосфорилировании Rb в конце фазы G1. Активность CDK4 регулируется активирующей субъединицей, циклином D-типа, и ингибиторной субъединицей, причем ингибиторная субъединица, р16INK4, была биохимически охарактеризована как белок, который специфически связывается с CDK4 и ингибирует CDK4, и, следовательно, может регулировать фосфорилирование Rb (Serrano et al., 1993; Serrano et al., 1995). Поскольку белок р16INK4 является ингибитором CDK4 (Serrano, 1993), делеция этого гена может увеличивать активность CDK4, приводя к гиперфосфорилированию белка Rb. Известно также, что р16 регулирует функцию CDK6.
р16INK4 принадлежит к недавно описанному классу CDK-ингибиторных белков, который включает в себя также р16В, р19, р21WAF1 и р27KIP1. Ген р16INK4 картируется в 9р21, области хромосомы, часто делетированной во многих типах опухолей. Гомозиготные делеции и мутации гена р16INK4 являются частыми в опухолевых клеточных линиях человека. Этот факт предполагает, что ген р16INK4 является геном супрессора опухолей. Однако эта интерпретация была поставлена под сомнение наблюдением, что частота изменений гена р16INK4 является гораздо более низкой в первичных некультивированных опухолях, чем в культивируемых клеточных линиях (Caldas et al., 1994; Cheng et al., 1994; Hussissian et al., 1994; Kamb et al., 1994; Kamb et al., 1994; Mori et al., 1994; Okamoto et al., 1994; Nobori et al., 1995; Orlow et al., 1994; Arap et al., 1995). Восстановление функции р16INK4 дикого типа путем трансфекции плазмидным экспрессирующим вектором уменьшало образование колоний некоторыми клеточными линиями злокачественных опухолей человека (Okamoto, 1994; Arap, 1995).
Другие гены, которые могут быть использованы в соответствии с данным изобретением, включают в себя Rb, APC, mda-7, DCC, NF-1, NF-2, WT-1, MEN-I, MEN-II, zac1, p73, VHL, MMAC1/PTEN, DBCCR-1, FCC, rsk-3, p27, слияния р27/р16, слияния p27/p27, антитромботические гены (например, COX-1, TFPI), PGS, Dp, E2F, ras, myc, neu, raf, erb, fms, trk, ret, gsp, hst, abl, E1A, p300, гены, участвующие в ангиогенезе (например, VEGF, FGF, тромбоспондин, BAI-1, GDAIF или их рецепторы) и МСС.
5. Регуляторы запрограммированной гибели клеток
Апоптоз, или запрограммированная гибель клеток, является существенным процессом для нормального эмбрионального развития, поддержания гомеостаза в тканях взрослых и супрессии канцерогенеза (Kerr et al., 1972). Было показано, что Bcl-2-семейство белков и ICE-подобные протеазы являются важными регуляторами и эффекторами апоптоза в других системах. Белок Bcl-2, обнаруженный в связи с фолликулярной лимфомой, играет выдающуюся роль в регуляции апоптоза и усилении выживания клеток в ответ на различные апоптотические стимулы (Bakhshi et al., 1985; Cleary and Sklar, 1985; Cleary et al., 1986; Tsujimoto et al., 1985; Tsujimoto and Croce, 1986). В настоящее время признается, что эволюционно консервативный белок Bcl-2 является членом семейства родственных белков, которые могут быть разделены на агонистов или антагонистов гибели клеток.
Лиганд Аро2 (Apo2L, также называемый TRAIL) является членом семейства цитокинов фактора некроза опухолей (TNF). TRAIL активирует апоптоз во многих типах злокачественных клеток, но не является токсичным для здоровых клеток. мРНК TRAIL встречается в большом разнообразии тканей. Большинство нормальных клеток, по-видимому, являются устойчивыми к цитотоксическому действию TRAIL, что предполагает существование механизмов, которые могут защищать против индукции апоптоза TRAIL. Первый рецептор, описанный для TRAIL, называемый рецептором 4 гибели клеток (DR4), содержит цитоплазматический «домен гибели»; DR4 передает сигнал апоптоза, который несет TRAIL. Были идентифицированы дополнительные рецепторы, которые связываются с TRAIL. Один рецептор, названный DR5, содержит цитоплазматический домен гибели и передает сигнал апоптоза, очень сходно с DR4. мРНК DR4 и DR5 экспрессируются во многих нормальных тканях и линиях опухолевых клеток. Недавно были идентифицированы рецепторы-ловушки, такие как DcR1 и DcR2, которые препятствуют индукции апоптоза посредством TRAIL через DR4 и DR5. Таким образом, эти рецепторы-ловушки представляют новый механизм для регуляции чувствительности к проапоптотическому цитокину непосредственно на поверхности клетки. Предпочтительная экспрессия этих ингибиторных рецепторов в нормальных тканях предполагает, что TRAIL может быть применим в качестве противоракового агента, который индуцирует апоптоз в злокачественных клетках при сохранении нормальных клеток (Marsters et al., 1999).
После его открытия было показано, что Bcl-2 действует в качестве супрессора гибели клеток, запускаемой различными стимулами. В настоящее время является очевидным, что существует семейство Bcl-2 регуляторных белков гибели клеток, которые имеют общие структурные гомологии и гомологии последовательности. Было показано, что эти различные члены семейства обладают функциями, сходными с функцией Bcl-2 (например, BclXL, BclW, BclS, Mcl-1, Al, Bfl-1), или противодействуют функции Bcl-2 и стимулируют гибель клеток (например, Bax, Bak, Bik, Bim, Bid, Bad, Harakiri). Предполагается, что любой из этих полипептидов, в том числе TRAIL, или любые другие полипептиды, которые индуцируют или стимулируют апоптоз, могут быть функционально связаны с токсином или что антитело, узнающее любой из этих полипептидов, может быть также присоединено к токсину.
IV. Способы получения модифицированных белков и сконструированных токсинов
Данное изобретение включает в себя способы идентификации антигенных областей на белке, способы идентификации областей, которые являются менее антигенными, способы создания менее антигенного белка, который имеет активность, сравнимую с активностью нативного белка, и способы анализа и определения антигенности и активности.
А. Антигенные области
Общее обсуждение антител и способов определения антител может быть найдено в предыдущих разделах. Термин «антигенная область» относится к части белка, которая специфически узнается антителом или Т-клеточным рецептором. Термин «менее антигенный» означает, что белок или область белка индуцирует меньшую реакцию антител или узнается меньшим количеством антител (поликлональных) или уменьшается связывающая ассоциация с антителом.
Антигенность связана с конкретным организмом. Во многих вариантах осуществления данного изобретения организм является человеком, но антигенность может обсуждаться в отношении также других организмов, таких как другие млекопитающие: обезьяны, гориллы, коровы, кролики, мыши, овцы, кошки, собаки, свиньи, козы и т.д., а также птицы и любой другой организм, который может индуцировать иммунный ответ.
В некоторых вариантах осуществления данного изобретения применяют поликлональные сыворотки со способами иммуноанализа, обсужденными ранее, для идентификации антигенных областей в конкретном белке. Поликлональные сыворотки могут собираться из различных источников, в том числе у работников, в отношении которых существует предположение, что они профессионально подвергались действию конкретного белка; у пациентов, в отношении которых существует предположение, что они диагностированы как имеющие состояние или заболевание, которое сопровождается или обусловлено присутствием антител к конкретному белку или организму; у пациентов, которые в дальнейшем не лечились в отношении состояния или заболевания, которое сопровождалось присутствием антител к конкретному белку или организму, и у случайных субъектов.
В. Базы данных
В некоторых способах осуществления данного изобретения после идентификации предположительных антигенных областей в конкретном белке используют базы данных белков. Затем область сравнивают с базой данных, содержащей белковые последовательности у организма, в котором является желательным более низкий иммунный ответ против этой области. Ряд таких баз данных существуют как на коммерческой, так и на публичной основе, в том числе GenBank, GenPept, SwissProt, PIR, PRF, PDB, все из которых являются доступными из вебсайта Национального Центра Информации по Биотехнологии (http://www.ncbi.nlm.nih.gov/).
C. Удаление и/или замена антигенных областей
После идентификации антигенной области она может быть удалена созданием укороченного белка. Альтернативно, эта область может быть заменена областью, которая считается менее антигенной.
Для удаления этой области полипептид может быть расщеплен протеиназами или полинуклеотид, кодирующий этот полипептид, может быть подвергнут манипуляции для удаления антигенной области. Область может быть удалена из полинуклеотида с использованием обычной технологии рекомбинантных ДНК, например, рестриктазами или ДНКазами.
Эта область может быть также заменена введением аминокислотных замен. «Заменена» означает, что аминокислота в конкретном положении была заменена отличающимся аминокислотным остатком или модифицированной аминокислотой. Это может быть выполнено рядом путей. Эта область может быть сначала удалена, а затем область замены может быть включена в полинуклеотид или в полипептид. Для включения конкретной кодирующей области в полинуклеотид может быть использована технология рекомбинантных ДНК. Альтернативно, антигенная область может быть подвергнута мутагенезу с использованием способов сайт-специфического мутагенеза, которые хорошо известны специалистам с обычной квалификацией в данной области.
Обсуждается, что аминокислоты, фланкирующие каждую сторону антигенной области, могут быть также удалены или заменены либо для облегчения создания модифицированного белка, либо для улучшения этого белка любым путем, например, для уменьшения его антигенности, увеличения стабильности этого белка, увеличения активности этого белка, уменьшения активности этого белка и т.д. Кроме того, множественные аминокислоты могут быть заменены или удалены либо из антигенной области, либо из фланкирующей области, либо из обоих; таким образом могут быть удалены или заменены точно или по меньшей мере

или более аминокислот.
Анализы для определения антигенности или активности модифицированного белка описаны в данном описании, например, в разделе, описывающем способы иммуноанализа, или они хорошо известны специалистам с квалификацией в данной области. Подходящие анализы для конкретного белка будут варьироваться в зависимости от этого белка. Ферментативные анализы могут быть пригодными, например, для оценки активности фермента. Специалист с квалификацией в данной области способен оценить активность модифицированного белка относительно активности нативного белка. Как обсуждалось выше, модифицированный белок может быть соединен (конъюгирован или слит) с другим полипептидом, пептидом или белком. Специалист с квалификацией в данной области способен оценить любой модифицированный конъюгированный или слитый белок данного изобретения в зависимости от активности или активностей полипептидных компонентов.
V. Комбинаторные терапии
Для увеличения эффективности любой из терапевтических композиций данного изобретения может быть желательным комбинирование этих композиций с другими агентами, эффективными в лечении конкретного заболевания или состояния. Предполагается, что может лечиться большое разнообразие состояний или заболеваний, таких как состояния или заболевания микробной этиологии, СПИД, аутоиммунные заболевания, гиперпролиферативные нарушения, в том числе злокачественные опухоли, лейкозы, артрит, воспалительные заболевания, сердечно-сосудистые заболевания и состояния, патогенные заболевания и состояния и диабет. Особо обсуждается лечение СПИДа, злокачественных опухолей и других гиперпролиферативных нарушений. Могут использоваться различные комбинации лекарственных средств, например, в которых композиция, содержащая модифицированный белок, является «А», а второй агент является «В»:
А. Лечение гиперпролиферативных заболеваний
Гиперпролиферативные заболевания включают в себя злокачественную опухоль, для которой имеется большое разнообразие схем лечения, например противораковые агенты или хирургическое вмешательство. «Противораковый» агент способен отрицательно воздействовать на злокачественную опухоль у субъекта, например, лизисом злокачественных клеток, индукцией апоптоза в злокачественных клетках, уменьшением скорости роста злокачественных клеток, уменьшением встречаемости или числа метастазов, уменьшением размера опухоли, ингибированием роста опухоли, ослаблением кровоснабжения опухоли или злокачественных клеток, стимуляцией иммунного ответа против злокачественных клеток или опухоли, предотвращением или ингибированием прогрессирования злокачественных опухолей или увеличением продолжительности жизни субъекта, имеющего злокачественную опухоль.
Противораковые агенты включают в себя биологические агенты (биотерапевтические агенты), химиотерапевтические агенты и радиотерапевтические агенты. Более часто эти другие композиции могли бы обеспечиваться в комбинированном количестве, эффективном для лизиса клеток или ингибирования пролиферации клеток. Этот способ может включать в себя контактирование этих клеток с экспрессирующей конструкцией и агентом (агентами) или множественными факторами одновременно. Это может достигаться контактированием клетки с одной композицией или фармакологической формой, которая включает в себя оба агента, или контактированием клетки с двумя отдельными композициями или формами одновременно, где одна композиция включает в себя экспрессирующую конструкцию, а другая включает в себя второй агент (агенты).
Устойчивость опухолевых клеток к химиотерапевтическим агентам и радиотерапевтическим агентам представляет главную проблему в клинической онкологии. Одной из задач исследования злокачественных опухолей в настоящее время является нахождение путей для улучшения эффективности химиотерапии и лучевой терапии комбинированием их с генотерапией. Например, ген тимидинкиназы простого герпеса (HS-tK) при доставке к опухолям мозга ретровирусной векторной системой успешно индуцировал чувствительность к антивирусному агенту ганцикловиру (Culver, et al., 1992). В контексте данного изобретения предполагается, что терапия модифицированными белками могла бы быть использована подобным образом вместе с химиотерапевтическим, радиотерапевтическим, иммунотерапевтическим или другим биологическим вмешательством, в дополнение к другим про-апоптотическим или регулирующим клеточный цикл агентам.
Альтернативно, генотерапия или введение модифицированных белков может предшествовать лечению другим агентом или следовать после другого агента с интервалами в диапазоне от минут до недель. В вариантах осуществления, где другой агент и экспрессирующая конструкция применяются к клетке по отдельности, следовало бы бы гарантировать, чтобы значительный период времени не проходил между моментом каждой доставки, так чтобы этот агент и экспрессирующая конструкция все еще были способны проявлять преимущественно комбинированное действие на клетку. В таких случаях предполагается, что клетку можно контактировать с обоими компонентами комбинаторной терапии в пределах приблизительно 12-24 часов один от другого и более предпочтительно в пределах 6-12 часов один от другого. В некоторых ситуациях может быть желательным значительное пролонгирование временного периода для лечения, однако, чтобы между соответствующими введениями проходило от нескольких дней (2, 3, 4, 5, 6 или 7) до нескольких недель (1, 2, 3, 4, 5, 6, 7 или 8).
Введение терапевтических экспрессирующих конструкций данного изобретения пациенту должно проводиться в соответствии с общим протоколом для введения химиотерапевтических средств, с учетом токсичности, если она имеется, этого вектора. Ожидается также, что различные стандартные терапии, а также хирургическое вмешательство, могут применяться в комбинации с описанной терапией гиперпролиферативных клеток.
а. Химиотерапия
Терапии злокачественных опухолей также включают в себя различные комбинаторные терапии с лечением на основе как химических средств, так и облучения. Комбинаторные химиотерапии используют, например, цисплатин (CDDP), карбоплатин, прокарбазин, меклоретамин, циклофосфамид, камптотецин, ифосфамид, мелфалан, хлорамбуцил, бусульфан, нитромочевину, дактиномицин, даунорубицин, доксорубицин, блеомицин, пликомицин, митомицин, этопозид (VP16), тамоксифен, ралоксифен, агенты связывания рецептора эстрогена, таксол, гемцитабиен, навелбин, ингибиторы фарнезил-протеинтрансферазы, трансплатину, 5-фторурацил, винкристин, винбластин и метотрексат, Темазоломид (водную форму DTIC) или вариант любого аналога или производного предыдущих агентов. Комбинирование химиотерапии с биологической терапией известно как биохимиотерапия.
В некоторых вариантах осуществления данного изобретения предполагается, что химиотерапевтический агент функционально присоединен к модифицированному белку, такому как молекула токсина.
b. Лучевая терапия
Другие факторы, которые вызывают повреждение ДНК и были широко использованы, включают в себя факторы, которые обычно известны как γ-лучи, рентгеновские лучи и/или прямая доставка радиоактивных изотопов в опухолевые клетки. Другие формы факторов повреждения ДНК также рассматриваются, например микроволны и УФ-облучение. Наиболее вероятно, что все эти факторы осуществляют большой диапазон повреждения в отношении ДНК, предшественников ДНК, репликации и репарации ДНК и сборки и поддержания хромосом. Диапазоны доз для рентгеновских лучей находятся в диапазоне от доз один раз в день 50-200 рентген в течение пролонгированных периодов времени (3-4 недели) до отдельных доз 2000-6000 рентген. Диапазоны доз для радиоактивных изотопов варьируются очень широко и зависят от периода полураспада изотопа, силы и типа излучаемой радиации и поглощения неопластическими клетками.
Термины «контактированная» и «экспонированная» при применении к клетке используются в данном описании для описания процесса, при помощи которого терапевтическая конструкция и химиотерапевтический агент или агент лучевой терапии доставляется в клетку-мишень или помещается в непосредственной близости от клетки-мишени. Для получения лизиса клетки или стаза оба агента доставляются к клетке в комбинированном количестве, эффективном для лизиса клетки или предотвращения ее деления.
с. Иммунотерапия
Иммунотерапевтические средства обычно основаны на применении иммунных эффекторных клеток и молекул для нацеливания на злокачественные клетки и разрушения этих клеток. Иммунным эффектором может быть, например, антитело, специфическое для какого-либо маркера на поверхности опухолевой клетки. Антитело может само служить в качестве эффектора терапии или оно может рекрутировать другие клетки для фактического осуществления лизиса клеток. Как обсуждалось выше в связи с заявленными композициями, антитело может быть также конъюгировано с лекарственным средством или токсином (химиотерапевтическим агентом, радионуклидом, А-цепью рицина, холерным токсином, коклюшным токсином и т.д.) и служить лишь в качестве нацеливающего агента. Альтернативно, эффектором может быть лимфоцит, несущий поверхностную молекулу, которая взаимодействует, прямо или опосредованно, с опухолевой клеткой-мишенью. Различные эффекторные клетки включают в себя цитотоксические Т-клетки и NK-клетки. Комбинирование терапевтических способов воздействия, т.е. нацеливания цитотоксической активности и иммуннной активации, может обеспечивать терапевтическую пользу в лечении злокачественных опухолей, и, следовательно, ожидается, что иммунотерапевтические агенты могут быть использованы вместе с любой терапевтической композицией данного изобретения. Например, субъекту могут быть введены два различных иммунотоксина или иммунотоксин может быть введен в комбинации с другим иммунотерапевтическим соединением в лечении заболевания, такого как злокачественная опухоль.
В одном аспекте иммунотерапии опухолевая клетка должна нести какой-либо маркер, который доступен для нацеливания, т.е. не присутствует на большинстве других клеток. Существуют многие опухолевые маркеры, и любые из них могут быть пригодными для нацеливания в контексте данного изобретения, как обсуждалось выше. Альтернативным аспектом иммунотерапии является комбинирование про-апоптотического действия с иммунными стимуляторными эффектами. Однако существуют также другие иммуностимулирующие молекулы, в том числе цитокины, такие как IL-2, IL-4, IL-12, GM-CSF, гамма-интерферон (IFN), хемокины, такие как MIP-1, MCP-1, IL-8 и факторы роста, такие как FLT3-лиганд. Комбинирование иммуностимулирующих молекул либо в виде белков, либо с использованием доставки генов в комбинации с иммунотоксином, нацеленным на опухоль, может усиливать противоопухолевые действия.
Как обсуждалось ранее, примерами иммунотерапий, исследующихся или применяющихся в настоящее время, являются иммуноадъюванты (например, Mycobacterium bovis, Plasmodium falciparum, динитрохлорбензол и ароматические соединения) (патент США 5801005, патент США 5739169, Hui and Hashimoto, 1998; Christodoulides et al., 1998), терапия с использованием цитокинов (например, интерферонов α, β и γ; IL-1, GM-CSF и TNF) (Bukowski et al., 1998; Davidson et al., 1998; Hellstrand et al., 1998), генотерапия (например, TNF, IL-1, IL-2, р53) (Qin et al., 1998; Austin-Ward and Villaseca, 1998; патент США 5830880 и патент США 5846945) и моноклональные антитела (например, анти-ганглиозид GM2, анти-HER-2, анти-р185) (Pietras et al., 1998; Hanibuchi et al., 1998; патент США 5824311). Герцептин (транстузумаб) является химерным (мышь-человек) моноклональным антителом, которое блокирует HER2-neu-рецептор. Он обладает противоопухолевой активностью и был одобрен для применения в лечении злокачественных опухолей (Dillman, 1999).
i. Пассивная иммунотерапия
Существует ряд различных подходов для пассивной иммунотерапии злокачественных опухолей. Они могут быть в широком смысле разделены на следующие категории: инъекция только антител; инъекция антител, связанных с токсинами или химиотерапевтическими агентами; инъекция антител, связанных с радиоактивными изотопами; инъекция антиидиотипических антител и, наконец, очистка от опухолевых клеток костного мозга.
Предпочтительно в пассивной иммунотерапии используют моноклональные антитела человека, так как они производят мало побочных эффектов или не производят побочных эффектов у пациента. Однако их применение является в некоторой степени ограниченным их недостаточным количеством, и до сих пор их вводили только внутрь повреждений. Моноклональные антитела человека против ганглиозидных антигенов вводили внутрь повреждений пациентам, страдающим от кожной рецидивирующей меланомы (Irie and Morton, 1986). У шести из десяти пациентов наблюдали регресс (обратное развитие) после инъекций внутрь повреждений один раз в день или один раз в неделю. В другом исследовании умеренный успех достигался от инъекций двух моноклональных антител человека внутрь повреждений (Irie et al., 1989).
Может быть полезным введение более чем одного моноклонального антитела против двух различных антигенов или даже антител со множественной антигенной специфичностью. Протоколы введения могут также включать в себя лимфокины или другие иммунологические стимуляторы, как описано Bajorin et al. (1988). Получение моноклональных антител человека описано более подробно в другом месте в данном описании.
ii. Активная иммунотерапия
В активной иммунотерапии вводят антигенный пептид, полипептид или белок или аутологичную или аллогенную композицию опухолевых клеток или «вакцину», обычно с отличающимся бактериальным адъювантом (Ravindranath and Morton, 1991; Morton and Ravindranath, 1996; Morton et al., 1992; Mitchell et al., 1990; Mitchell et al., 1993). В иммунотерапии меланомы пациенты, которые индуцировали высокую IgM-реакцию, часто обнаруживали лучшее выживание, чем пациенты, которые не образовывали IgM-антитела или образовывали мало IgM-антител (Morton et al., 1992). IgM-антитела часто являются транзиторными (временными) антителами и в качестве исключения из правил являются, по-видимому, антиганглиозидными или противоуглеводными антителами.
iii. Адоптивная иммунотерапия
В адоптивной иммунотерапии циркулирующие в кровотоке лимфоциты или инфильтруирующие опухоль лимфоциты пациента выделяют in vitro, активируют лимфокинами, такими как IL-2, или трансдуцируют генами для некроза опухолей и вводят повторно (Rosenberg et al., 1988; 1989). Для достижения этого следует ввести животному или пациенту-человеку иммунологически эффективное количество активированных лимфоцитов в комбинации с содержащей адъювант композицией антигенного пептида, как описано в данном описании. Активированные лимфоциты должны быть, наиболее предпочтительно, собственными клетками пациента, которые были выделены ранее из образца крови или опухоли и активированы (или "размножены") in vitro. Эта форма иммунотерапии дала несколько случаев регресса меланомы и почечной карциномы, но проценты респондеров были низкими в сравнении с процентами пациентов, которые не отвечали на эту терапию.
d. Гены
Еще в одном варианте вторичной терапией является генотерапия, в которой терапевтический полинуклеотид вводят до, после или одновременно с модифицированным полипептидом или полинуклеотидом, кодирующим модифицированный полипептид. Альтернативно, может быть использован единственный вектор, кодирующий две различные терапевтические полипептидные молекулы. Различные белки включены в данном изобретении, некоторые из которых описаны ранее. Например, генотерапия может быть использована в связи с доставкой гена супрессора опухоли дикого типа в злокачественную клетку.
е. Хирургическое вмешательство
Приблизительно 60% лиц, имеющих злокачественную опухоль, должны подвергаться хирургическому вмешательству того или иного типа, которое включает в себя превентивное, диагностическое или определяющее стадию, лечебную и паллиативную хирургического вмешательства. Лечебное хирургическое вмешательство является лечением злокачественных опухолей, которое может быть использовано вместе с другими терапиями, такими как лечение по способу данного изобретения, химитотерапия, лучевая терапия, гормональная терапия, генотерапия, иммунотерапия и/или альтернативные терапии.
Лечебное хирургическое вмешательство включает в себя резекцию, в которой физически удаляется, вырезается и/или разрушается вся злокачественная ткань или часть злокачественной ткани. Резекцией опухоли называют физическое удаление по меньшей мере части опухоли. Кроме резекции опухоли, хирургическое лечение включает в себя лазерную хирургию, криохирургию, электрохирургию и контролируемую микроскопически хирургию (хирургию Mohs). Далее, ожидается, что данное изобретение может быть использовано вместе с удалением поверхностных злокачественных опухолей, предраков или несущественных количеств нормальной ткани.
При вырезании части всех злокачественных клеток, ткани или опухоли в теле может образоваться пустое пространство (полость). Лечение может выполняться перфузией, прямой инъекцией или местным применением к этой зоне дополнительной противораковой терапии. Такая обработка может повторяться, например, каждые 1, 2, 3, 4, 5, 6 или 7 дней или каждые 1, 2, 3, 4 и 5 недель или каждые 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев. Эти обработки могут проводиться также с варьирующимися дозами.
f. Другие агенты
Ожидается, что в комбинации с данным изобретением могут быть использованы другие агенты для улучшения терапевтической эффективности лечения. Эти дополнительные агенты включают в себя иммуномодуляторные агенты, агенты, которые влияют на повышающую регуляцию рецепторов клеточной поверхности и гэп-соединений, цитостатические агенты и агенты дифференцировки, ингибиторы адгезии клеток, агенты, увеличивающие чувствительность гиперпролиферативных клеток к апоптотическим индукторам или другие биологические агенты. Иммуномодуляторные агенты включают в себя фактор некроза опухолей, интерферон альфа, бета и гамма; IL-2 и другие цитокины; F42K и другие аналоги цитокинов или MIP-1, MIP-1бета, MCP-1, RANTES и другие хемокины. Предполагается также, что повышающая регуляция рецепторов поверхности клеток или их лигандов, таких как Fas/Fas-лиганд, DR4 или DR5/TRAIL (Apo-2-лиганд), могла бы потенциировать антираковые возможности данного изобретения установлением аутокринного или паракринного действия на гиперпролиферативные клетки. Увеличения межклеточной передачи сигнала повышением числа гэп-соединений могли бы увеличивать антигиперпролиферативные эффекты на соседнюю популяцию гиперпролиферативных клеток. В других вариантах осуществления цитостатические агенты или агенты дифференцировки могут быть использованы в комбинации с данным изобретением для улучшения антигиперпролиферативной эффективности лечения. Предполагается, что ингибиторы адгезии клеток могут улучшать эффективность данного изобретения. Примерами ингибиторов адгезии клеток являются ингибиторы киназы очаговой адгезии (FAK) и ловастатин. Кроме того, предполагается, что другие агенты, которые увеличивают чувствительность гиперпролиферативной клетки к апоптозу, такие как антитело с225, могли бы использоваться в комбинации с данным изобретением для улучшения эффективности лечения.
Многие успехи в терапии злокачественных опухолей были получены после введения цитотоксических химиотерапевтических лекарственных средств. Однако одним из последствий химиотерапии является развитие/приобретение устойчивых к лекарственным средствам фенотипов и развитие устойчивости ко множественным лекарственным средствам. Развитие устойчивости к лекарственным средствам остается главным препятствием в лечении таких опухолей, и, следовательно, существует явная потребность в альтернативных подходах, таких как генотерапия.
Исследования ряда исследователей продемонстрировали, что опухолевые клетки, которые являются устойчивыми к TRAIL, могут быть сенсибилизированы субтоксическими концентрациями лекарственных средств/цитокинов, и эти сенсибилизированные опухолевые клетки в значительной степени убиваются TRAIL (Bonavida et al., 1999; Bonavida et al., 2000; Gliniak et al., 1999; Keane et al., 1999). Кроме того, комбинация химиотерапевтических агентов, таких как СРТ-11 или доксорубицин, с TRAIL также приводит к повышенной противоопухолевой активности и к усилению апоптоза. Некоторые из этих эффектов могут быть опосредованы повышающей регуляцией TRAIL или соответствующих рецепторов, тогда как другие могут не быть опосредованы этим.
Другая форма терапии для применения вместе с химиотерапией, лучевой терапией или биологической терапией включает в себя гипертермию, которая является процедурой, в которой ткань пациента подвергают действию высоких температур (до 106оF). Внешние или внутренние нагревательные устройства могут использоваться в применении местной, регионарной гипертермии или гипертермии всего тела. Местная гипертермия включает в себя применение нагревания к небольшой зоне, такой как опухоль. Нагревание может генерироваться наружно высокочастотными волнами с нацеливанием на опухоль из устройства, находящегося вне тела. Внутреннее нагревание может предусматривать применение стерильного зонда, в том числе тонких нагреваемых проволок или полых трубок, заполненных теплой водой, имплантированной микроволновой антенны или радиочастотных электродов.
Орган или конечность пациента нагревают в случае регионарной терапии, что осуществляется с использованием устройств, которые производят высокую энергию, таких как магниты. Альтернативно, часть крови пациента может быть извлечена и нагрета перед перфузией в участок, который будет нагреваться изнутри. Нагревание всего тела может также осуществляться в случаях, когда злокачественная опухоль распространена по всему телу. Для этой цели могут быть использованы одеяла с теплой водой, горячий воск, индукционные катушки и термические камеры.
Гормональная терапия может быть также использована с данным изобретением или в комбинации с любой другой ранее описанной терапией. Применение гормонов может быть использовано в лечении некоторых злокачественных опухолей, таких как злокачественная опухоль молочной железы, предстательной железы или злокачественная опухоль шейки матки, для понижения уровня или ингибирования действий некоторых гормонов, таких как тестостерон или эстроген. Это лечение часто используют в комбинации по меньшей мере с одной другой раковой терапией в качестве необязательного лечения или для уменьшения риска метастазирования.
В. Вирусный патогенез
Конечно, понятно, что композиции и способы данного изобретения имеют отношение к лечению или диагностике вирусного патогенеза. Например, предполагается, что данное изобретение может быть использовано для лечения СПИДа, который вызывается ВИЧ-инфекцией. Таким образом, данное изобретение может быть использовано в комбинации с введением традиционных лекарственных средств. Некоторые из таких лекарственных средств описаны ниже.
1. AZT
Хорошо известным лекарственным средством для лечения СПИДа является зовидовудин (AZT™, доступный из Burroughs Wellcome). Он представляет класс аналогов нуклеозидов, известных как дидезоксинуклеозиды, которые блокируют репликацию ВИЧ ингибированием обратной транскриптазы ВИЧ. Лекарственное средство против СПИДа зидовудин (также известное как AZT) может быть также использовано в ограниченных обстоятельствах, главным образом в комбинации с рифампином, как описано Burger et al. (1993).
Описанные в данном описании композиции и способы будут особенно эффективными вместе с другими формами терапии, такими как AZT и/или ингибиторы протеаз, которые сконструированы для ингибирования вирусной репликации, посредством поддержания желательных уровней лейкоцитов. Это в результате дает пациенту время для проявления действия антивирусных лекарственных средств.
2. HAART
Новая комбинаторная лекарственная терапия обнаружила многообещающие результаты в лечении ВИЧ-инфицированных пациентов. Лечение комбинациями сильнодействующего анти-ВИЧ лекарственного средства называют "высокоактивной антиретровирусной терапией" (HAART), и она обеспечила клиническое улучшение, более длительное выживание и улучшенное качество жизни для людей, инфицированных ВИЧ, во время всех четырех стадий ВИЧ-заболевания. Примеры HAART включают в себя ингибитор протеаз (индинавир, нелфинавир, ритонавир, ритонавир/саквинавир или саквинавир), комбинированный с двумя нуклеозидными аналогами (AZT/ddI, d4T/ddI, AZT/ddC, AZT/3TC или d4T/3TC).
V. Фармацевтические композиции и пути введения
Данное изобретение рассматривает молекулы нуклеиновых кислот, кодирующие модифицированные белки (в том числе слитые белки), а также модифицированные белки, которые могут быть конъюгированы с другим белковым соединением или с малой молекулой. В некоторых вариантах осуществления фармацевтические композиции вводят субъекту. Различные аспекты данного изобретения включают в себя введение эффективного количества водной композиции. В другом варианте осуществления данного изобретения обсуждается конкретно гелонин в качестве иммунотоксина. Такие композиции обычно будут растворены или диспергированы в фармацевтически приемлемом носителе или водной среде. Кроме того, такие соединения могут вводиться в комбинации с другим лечением в зависимости от подлежащего лечению заболевания или состояния. Лечение СПИД могло бы включать в себя введение HAART или AZT, или обоих, тогда как лечение злокачественных опухолей могло бы включать в себя хирургическое вмешательство или назначение химиотерапии, лучевой терапии, иммунотерапии или гормонов.
А. Пути введения
Специалистам в данной области хорошо известно, как использовать доставку генов в ситуациях in vivo и ex vivo. Для вирусных векторов обычно готовят исходный материал вирусного вектора. В зависимости от типа вируса и получаемого титра доставляют 1-100, 10-50, 100-1000 или до 1·104, 1·105, 1·106, 1·107, 1·108, 1·109, 1·1010, 1·1011 или 1·1012 инфекционных вирусных частиц пациенту. Сходные цифры могут быть экстраполированы для липосомных или других невирусных форм доставки сравнением относительных эффективностей поглощения. Приготовление в качестве фармацевтически приемлемой композиции обсуждается ниже.
Фразы "фармацевтически приемлемые" или "фармакологически приемлемые" относятся к молекулярным частицам и композициям, которые не производят вредной, аллергической или другой нежелательной реакции при введении животному или человеку соответственно. В данном контексте "фармацевтически приемлемый носитель" включает в себя любой и все растворители, дисперсионные среды, покрытия, антибактериальные и антивирусные агенты, изотонические и задерживающие абсорбцию агенты и т.п. Применение таких сред и агентов для фармацевтически активных веществ хорошо известно в данной области. За исключением случаев, когда любые общепринятые среда или агент несовместимы с активными ингредиентами, предполагается их применение в этих терапевтических композициях. В эти композиции могут быть включены также другие противораковые агенты.
Кроме соединений, приготовляемых для парентерального введения, например для внутривенной или внутримышечной инъекции, другие фармацевтически приемлемые формы включают в себя, например, таблетки или другие твердые формы для перорального введения; капсулы для пролонгированного высвобождения и любая другая форма, используемая в настоящее время, в том числе кремы, лосьоны, жидкости для полоскания рта, ингалируемые агенты и т.п.
Активные соединения данного изобретения могут быть приготовлены для парентерального введения, например приготовлены для инъекции через внутривенный, внутримышечный, интраторакальный, подкожный или даже внутрибрюшинный пути. Приготовление водной композиции, которая содержит соединение или соединения, которые увеличивают экспрессию молекул МНС класса I, будет понятно специалистам с квалификацией в данной области в свете данного описания. Обычно такие композиции могут быть приготовлены в виде инъекционных растворов, либо в виде водных растворов, либо в виде суспензий; могут быть также приготовлены твердые формы, пригодные для применения в приготовлении растворов или суспензий при добавлении жидкости перед инъекцией; эти препараты могут быть также эмульгированы.
Растворы активных соединений в виде свободного основания или фармакологически приемлемых солей могут быть приготовлены в воде, подходящим образом смешанной с поверхностно-активным веществом, таким как гидроксипропилцеллюлоза. Дисперсии могут быть также приготовлены в глицерине, жидких полиэтиленгликолях и их смесях и в маслах. В обычных условиях хранения и применения эти препараты содержат консервант для предупреждения роста микроорганизмов.
Фармацевтические формы, пригодные для применения в виде инъекций, включают в себя стерильные водные растворы или дисперсии, формы, включающие в себя кунжутное масло, арахисовое масло или водный полипропиленгликоль, и стерильные порошки для импровизированного приготовления стерильных инъекционных растворов или дисперсий. Во всех случаях эта форма должна быть стерильной и должна быть жидкой до такой степени, при которой она может быть легко инъецирована. Она должна быть стабильной в условиях приготовления и хранения и должна содержать консервант против загрязняющего действия микроорганизмов, таких как бактерии и грибы.
Активные соединения могут быть приготовлены в виде композиции в нейтральной форме или в форме соли. Фармацевтически приемлемые соли включают в себя кислотно-аддитивные соли (образованные со свободными аминогруппами белка), которые образованы с неорганическими кислотами, такими как, например, хлористоводородная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная, миндальная и т.п. Соли, образованные со свободными карбоксильными группами, могут быть также образованы из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или трехвалентного железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и т.п.
Носителем может быть растворитель или дисперсионная среда, содержащая, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль, и т.п.), их подходящие смеси и растительные масла. Может поддерживаться подходящая текучесть, например, с использованием покрытия, такого как лецитин, посредством поддержания требуемого размера частиц в случае дисперсии и с использованием поверхностно-активных веществ. Предохранение от действия микроорганизмов может осуществляться различными антибактериальными и противогрибковыми агентами, например парабенами, хлорбутанолом, фенолом, сорбиновой кислотой, тимерозалом и т.п. Во многих случаях будет предпочтительным включение изотонических агентов, например сахаров или хлорида натрия. Пролонгированная абсорбция инъецируемых композиций может осуществляться использованием в композициях агентов, задерживающих абсорбцию, например моностеарата алюминия и желатина.
Стерильные инъекционные растворы готовят включением активных соединений в необходимом количестве в подходящем растворителе с различными другими перечисленными выше ингредиентами, по мере необходимости, с последующей фильтр-стерилизацией. Обычно дисперсии готовят включением различных простерилизованных активных ингредиентов в твердый носитель, который содержит основную дисперсионную среду и требуемые другие ингредиенты из перечисленных выше. В случае стерильных порошков для приготовления стерильных инъекционных растворов предпочтительными способами приготовления являются способы сушки в вакууме и лиофилизации, которые дают порошок активного ингредиента плюс любого дополнительного желаемого ингредиента из их заранее стерильно профильтрованного раствора.
В некоторых случаях терапевтические композиции данного изобретения могут быть также приготовлены в формах, пригодных для местного введения, например в виде кремов или лосьонов. Эти формы могут быть использованы для лечения связанных с кожей заболеваний, таких как различные саркомы.
Введение терапевтических композиций в соответствии с данным изобретением будет производиться обычным путем, пока ткань-мишень является доступной с использованием такого пути. В случаях, когда данное изобретение используется в виде вирусного вектора, следует прежде всего учитывать желательное местоположение для гетерологичных последовательностей, которые несет этот вектор. Способы введения включают в себя пероральный, назальный, буккальный, ректальный, вагинальный или локальный. Например, местное введение было бы, в частности, предпочтительным для лечения меланомы или связанных со СПИДом кожных состояний или в тех случаях, когда вирусный вектор несет гетерологичный ген, применимый в лечении кожного состояния. Альтернативно, введение может быть ортотопической, интрадермальной, подкожной, внутримышечной, внутрибрюшинной или внутривенной инъекцией. Такие композиции обычно будут вводиться в виде фармацевтически приемлемых композиций, которые включают в себя физиологически приемлемые носители, буферы или другие наполнители. Для лечения состояний легких предполагается аэрозольная доставка. Объем аэрозоля находится между приблизительно 0,01 и 0,5 мл. Подобным образом, предпочтительным способом для лечения связанного с ободочной кишкой заболевания могла бы быть клизма. Объем клизмы равен приблизительно 1 мл - приблизительно 100 мл. Предпочтительным способом является прямая внутриопухолевая инъекция, причем более конкретным вариантом осуществления является внутриопухолевая перфузия.
В некоторых вариантах осуществления может быть желательным обеспечение непрерывной подачи терапевтических композиций пациенту. Для внутривенного или внутриартериального путей это выполняется системой капельного вливания. Для местных применений можно использовать периодическое нанесение. Для различных подходов можно было бы использовать формы задержанного высвобождения, которые обеспечивают ограниченные, но постоянные количества лекарственного средства на протяжении продолжительного периода времени. Для внутреннего применения может быть предпочтительной непрерывная перфузия, например, вирусного вектора, несущего сегмент нуклеиновой кислоты представляющей интерес области. Это могло бы достигаться катетеризацией, в некоторых случаях послеоперационно, с последующим непрерывным введением лекарственного средства. Период времени для перфузии может быть выбран клиницистом для конкретных пациента и ситуации, но время могло бы быть в диапазоне от приблизительно 1-2 часов до 2-6 часов, до приблизительно 6-10 часов, до приблизительно 10-24 часов, до приблизительно 1-2 дней, до приблизительно 1-2 недель или более длительным. Обычно доза терапевтической композиции, вводимой непрерывной перфузией, будет эквивалентна дозе, предоставляемой единственной или множественными инъекциями, корректированной на период времени, на протяжении которого вводят эти инъекции. Однако считается, что перфузией могут вводиться более высокие дозы.
Для парентерального введения, например, в водном растворе этот раствор должен быть подходящим образом забуференным, если необходимо, и жидкий растворитель сначала должен быть сделан изотоническим с использованием достаточного количества солевого раствора или глюкозы. Эти конкретные водные растворы особенно пригодны для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В этой связи стерильные водные среды, которые могут использоваться, будут известны специалистам в данной области в свете данного описания. Например, одна доза может быть растворена в 1 мл изотонического раствора NaCl и либо добавлена к 1000 мл вводимой в подкожную клетчатку жидкости, либо инъецирована в предлагаемом месте инфузии (см., например, Remington's Pharmaceutical Sciences, 1990). Некоторое варьирование дозы будет обязательно производиться в зависимости от состояния проходящего лечение субъекта. Лицо, ответственное за введение, в любом случае должно определить подходящую дозу для индивидуального субъекта.
Эффективное количество терапевтической композиции определяют на основании предполагаемой задачи. Термин "унифицированная (стандартная) доза" или "доза" относится к физически дискретным единицам, пригодным для применения у субъекта, причем каждая единица содержит заранее определенное количество терапевтической композиции, рассчитанное для вызывания желаемых реакций, обсужденных выше, в связи с ее введением, т.е. подходящим путем введения и схемой лечения. Вводимое количество, в соответствии как с числом введений, так и с унифицированной дозой, зависит от желательной защиты.
Точные количества терапевтической композиции зависят также от мнения практикующего врача и являются специфическими для каждого индивидуума. Факторы, от которых зависит доза, включают в себя физическое и клиническое состояние пациента, способ введения, предполагаемую цель лечения (ослабление симптомов или излечение) и эффективность, стабильность и токсичность конкретного терапевтического вещества.
После приготовления растворы должны вводиться способом, совместимым с дозированной формой, и в таком количестве, которое является терапевтически эффективным. Эти готовые формы могут легко вводиться в различных дозированных формах, например, в описанных выше типах инъекционных растворов, но можно применять также капсулы пролонгированного высвобождения и т.п.
Применяемый в данном описании термин «введение in vitro» относится к манипуляциям, выполняемым на клетках, извлеченных у животного, в том числе, но не только, клетках в культуре. Термин «введение ex vivo» относится к клеткам, которые были манипулированы in vitro и затем введены живому животному. Термин «введение in vivo» включает в себя все манипуляции, выполняемые на клетках, находящихся внутри животного.
В определенных аспектах данного изобретения композиции могут вводиться либо in vitro, ex vivo, либо in vivo. В некоторых вариантах осуществления in vitro экспрессирующая конструкция, кодирующая модифицированный белок, может быть трансдуцирована в клетку-хозяина. Затем эти трансдуцированные клетки могут быть использованы для анализов in vitro или альтернативно для введения in vivo.
В патентах США с номерами 4690915 и 5199942, оба из которых включены в данное описание в качестве ссылки, описаны способы для манипуляции ex vivo с мононуклеарными клетками крови и клетками костного мозга для применения в терапевтических приложениях.
Предполагается также введение in vivo композиций данного изобретения. Примеры включают в себя, но не ограничиваются ими, трансдукцию эпителия мочевого пузыря введением трансдуцирующих композиций данного изобретения посредством внутрипузырной катетеризации в мочевой пузырь (Bass, 1995) и трансдукцию клеток печени инфузией подходящих трансдуцирующих композиций через воротную вену посредством катетера (Bao, 1996). Дополнительные примеры включают в себя прямую инъекцию опухолей с использованием не требующих длительного приготовления трансдуцирующих композиций и либо интраназальную, либо внутритрахеальную (Dong, 1996) инстилляцию трансдуцирующих композиций для осуществления трансдукции клеток легких.
Данное изобретение может вводиться внутривенно, интрадермально, внутриартериально, внутрибрюшинно, внутрь повреждения, интракраниально, внутрисуставно, внутрь предстательной железы, интраплеврально, интратрахеально, интраназально, интравитреально, интравагинально, ректально, местным введением, внутриопухолевым введением, внутримышечно, внутрибрюшинно, подкожно, внутрипузырно, через слизистую оболочку, интраперикардиально, перорально, местным введением, локально и/или с использованием аэрозоля, инъекции, инфузии, непрерывной инфузии, локализованной перфузии, омывающей клетки-мишени, непосредственно или посредством катетера и/или лаважа.
В. Липидные композиции
В некоторых вариантах осуществления данное изобретение относится к новой композиции, содержащей один или несколько липидов, связанных с полинуклеотидом или полипептидом заявленного изобретения. Липид является веществом, которое характерным образом нерастворимо в воде и может экстрагироваться органическим растворителем. Соединения, иные, чем соединения, конкретно описанные в данном описании, известны специалисту с квалификацией в данной области как липиды, и они также включены в композиции и способы данного изобретения.
Липид может быть природно встречающимся или синтетическим (например, сконструированным или полученным человеком). Однако обычно липид является биологическим веществом. Биологические липиды хорошо известны в данной области и включают в себя, например, нейтральные жиры, фосфолипиды, фосфоглицериды, стероиды, терпены, лизолипиды, гликосфинголипиды, глюколипиды, сульфатиды, липиды со связанными простыми эфирными или сложноэфирными связями жирными кислотами и полимеризуемые липиды или их комбинации.
1. Типы липидов
Нейтральный жир может содержать глицерин и жирную кислоту. Обычный глицерин является трехатомным спиртом. Жирная кислота обычно является молекулой, содержащей углеродную цепь с кислотной группой (например, карбоновую кислоту) на конце этой цепи. Углеродная цепь жирной кислоты может иметь любую длину, однако предпочтительно, чтобы длина углеродной цепи была равна приблизительно от 2, приблизительно 3, приблизительно 4, приблизительно 5, приблизительно 6, приблизительно 7, приблизительно 8, приблизительно 9, приблизительно 10, приблизительно 11, приблизительно 12, приблизительно 13, приблизительно 14, приблизительно 15, приблизительно 16, приблизительно 17, приблизительно 18, приблизительно 19, приблизительно 20, приблизительно 21, приблизительно 22, приблизительно 23, приблизительно 24, приблизительно 25, приблизительно 26, приблизительно 27, приблизительно 28, приблизительно 29 до приблизительно 30 или более атомов углерода и находиться в любом производимом из этих чисел диапазоне. Однако предпочтительным является диапазон от приблизительно 14 до приблизительно 24 атомов углерода в части цепи жирной кислоты, причем в некоторых вариантах осуществления предпочтительным является диапазон от приблизительно 16 до приблизительно 18 атомов углерода. В некоторых вариантах осуществления углеродная цепь жирной кислоты может содержать нечетное число атомов углерода, однако в определенных вариантах четное число атомов углерода в этой цепи может быть предпочтительным. Жирная кислота, содержащая только простые связи в ее углеродной цепи, называется насыщенной, тогда как жирная кислота, содержащая по меньшей мере одну двойную связь в ее цепи, называется ненасыщенной.
Конкретные жирные кислоты включают в себя, но не ограничиваются ими, линолевую кислоту, олеиновую кислоту, пальмитиновую кислоту, линоленовую кислоту, стеариновую кислоту, лауриновую кислоту, миристиновую кислоту, арахиновую кислоту, пальмитолеиновую кислоту, арахидоновую кислоту, рицинолеиновую кислоту, туберкулостериновую кислоту, лактобацилловую кислоту. Кислотная группа одной или более жирных кислот ковалентно связана с одной или более гидроксильными группами глицерина. Таким образом, моноглицерид содержит глицерин и одну жирную кислоту, диглицерид содержит глицерин и две жирные кислоты и триглицерид содержит глицерин и три жирные кислоты.
Фосфолипид обычно содержит глицериновую или сфингозиновую часть, ионную фосфатную группу для образования амфипатического соединения и одну или более жирных кислот. Типы фосфолипидов включают в себя, например, фосфоглицериды, где фосфатная группа связана с первым углеродом глицерина диглицерида, и сфинголипиды (например, сфингомиелин), где фосфатная группа этерифицирована с аминоспиртом сфингозином. Другим примером сфингофосфолипида является сульфатид, который содержит ионную сульфатную группу, которая делает эту молекулу амфипатической. Фосфолипид может, конечно, содержать дополнительные химические группы, такие как, например, спирт, присоединенный к фосфатной группе. Примеры таких спиртовых групп включают в себя серин, этаноламин, холин, глицерин и инозит. Таким образом, характерные фосфолипиды включают в себя фосфатидилсерин, фосфатидилэтаноламин, фосфатидилхолин, фосфатидилглицерин или фосфатидилинозит. Другие фосфолипиды включают в себя фосфатидиловую кислоту или диацетилфосфат. В одном аспекте фосфатидилхолин включает в себя диолеилфосфатидилхолин (также известный как кардиолипин), яичный фосфатидилхолин, дипальмитоилфосфатидилхолин, мономиристоилфосфатидилхолин, монопальмитоилфосфатидилхолин, моностеарилфосфатидилхолин, моноолеоилфосфатидилхолин, дибутиролфосфатидилхолин, дивалероилфосфатидилхолин, дикапроилфосфатидилхолин, дигептаноилфосфатидилхолин, дикаприлоилфосфатидилхолин или дистеароилфосфатидилхолин.
Гликолипид является сфингофосфолипидом, но содержит углеводную группу, а не фосфатную группу, присоединенную к первичной гидроксильной группе сфингозина. Тип гликолипида, называемый цереброзидом, содержит одну сахарную группу (например, глюкозу или галактозу), присоединенную к первичной гидроксильной группе. Другим примером гликолипида является ганглиозид (например, моносиалоганглиозид, GM1), который содержит от приблизительно 2, приблизительно 3, приблизительно 4, приблизительно 5, приблизительно 6 до приблизительно 7 или около этого сахарных групп, которые могут находиться в разветвленной цепи, присоединенной к первичной гидроксильной группе. В другом варианте осуществления этот гликолипид является церамидом (например, лактозилцерамидом).
Стероид является четырехчленной циклической системой, произведенной из фенантрена. Стероиды часто обладают регуляторными функциями в клетках, тканях и организмах и включают в себя, например, гормоны и родственные соединения в семействах прогестагенов (например, прогестерон), глюкокортикоидов (например, кортизол), минералокортикоидов (например, альдостерон), андрогенов (например, тестостерон) и эстрогенов (например, эстрон). Холестерин является другим примером стероида и обычно выполняет структурную функцию, а не регуляторные функции. Витамин D является другим примером стерина и участвует в абсорбции кальция из кишечника.
Терпен является липидом, содержащим одну или более пятиуглеродных изопреновых групп. Терпены имеют различные биологические функции и включают в себя, например, витамин А, кофермент Q и каротиноиды (например, ликопин и β-каротин).
2. Заряженные и нейтральные липидные композиции
В некоторых вариантах осуществления липидный компонент композиции является незаряженным или первично незаряженным. В одном варианте осуществления липидный компонент композиции содержит один или несколько нейтральных липидов. В другом аспекте липидный компонент композиции может быть по существу свободным от анионных и катионных липидов, таких как некоторые фосфолипиды и холестерин. В определенных вариантах липидный компонент незаряженной или первично незаряженной липидной композиции содержит приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или 100% липидов без заряда, по существу незаряженный липид (липиды) и/или смесь липидов с равными количествами положительных и отрицательных зарядов.
В других аспектах липидная композиция может быть заряженной. Например, заряженные фосфолипиды могут быть использованы для приготовления липидной композиции в соответствии с данным изобретением и могут нести в целом положительный заряд или в целом отрицательный заряд. В неограничивающем примере может быть использован диацетилфосфат для придания отрицательного заряда липидной композиции, а стеариламин может быть использован для придания положительного заряда липидной композиции.
3. Получение липидов
Липиды могут быть получены из природных источников, коммерческих источников или синтезированы химически, как будет известно специалисту с обычной квалификацией в данной области. Например, фосфолипиды могут быть из природных источников, такие как яичный или соевый фосфатидилхолин, фосфатидиловая кислота мозга, фосфатидилинозит из мозга или из растений, сердечный кардиолипин и растительный или бактериальный фосфатидилэтаноламин. В другом примере липиды, пригодные для применения в соответствии с данным изобретением, могут быть получены из коммерческих источников. Например, димиристилфосфатидилхолин (DMPC) может быть получен из Sigma Chemical Co., дицетилфосфат (DCP) получают из К & К Laboratories (Plainview, NY); холестерин (Chol) получают из Calbiochem-Behring; димиристилфосфатидилглицерин (DMPG) и другие липиды могут быть получены из Avanti Polar Lipids, Inc. (Birmingham, Ala.). В некоторых вариантах осуществления исходные растворы липидов в хлороформе или смеси хлороформ/метанол могут храниться при приблизительно -20°С. Предпочтительно хлороформ используют в качестве единственного растворителя, так как он более легко выпаривается, чем метанол.
4. Структуры липидных композиций
Соединение, связанное с липидом, может быть диспергировано в растворе, содержащем липид, растворено липидом, эмульгировано с липидом, смешано с липидом, комбинировано с липидом, ковалентно связано с липидом, содержащимся в виде суспензии в липиде или другим образом связано с липидом. Липидная или связанная с липидом композиция данного изобретения не ограничивается какой-либо конкретной структурой. Например, эти композиции могут также быть просто разбросаны в растворе, возможно, с образованием агрегатов, которые не являются однородными по размеру или форме. В другом примере они могут присутствовать в бислойной структуре, в виде мицелл, или иметь «спавшуюся» (в виде коллапса) структуру. В другом неограничивающем примере обсуждается также липофектамин (Gibco BRL) или комплекс Superfect (Qiagen).
В некоторых вариантах липидная композиция может содержать приблизительно 1%, приблизительно 2%, приблизительно 3%, приблизительно 4%, приблизительно 5%, приблизительно 6%, приблизительно 7%, приблизительно 8%, приблизительно 9%, приблизительно 10%, приблизительно 11%, приблизительно 12%, приблизительно 13%, приблизительно 14%, приблизительно 15%, приблизительно 16%, приблизительно 17%, приблизительно 18%, приблизительно 19%, приблизительно 20%, приблизительно 21%, приблизительно 22%, приблизительно 23%, приблизительно 24%, приблизительно 25%, приблизительно 26%, приблизительно 27%, приблизительно 28%, приблизительно 29%, приблизительно 30%, приблизительно 31%, приблизительно 32%, приблизительно 33%, приблизительно 34%, приблизительно 35%, приблизительно 36%, приблизительно 37%, приблизительно 38%, приблизительно 39%, приблизительно 40%, приблизительно 41%, приблизительно 42%, приблизительно 43%, приблизительно 44%, приблизительно 45%, приблизительно 46%, приблизительно 47%, приблизительно 48%, приблизительно 49%, приблизительно 50%, приблизительно 51%, приблизительно 52%, приблизительно 53%, приблизительно 54%, приблизительно 55%, приблизительно 56%, приблизительно 57%, приблизительно 58%, приблизительно 59%, приблизительно 60%, приблизительно 61%, приблизительно 62%, приблизительно 63%, приблизительно 64%, приблизительно 65%, приблизительно 66%, приблизительно 67%, приблизительно 68%, приблизительно 69%, приблизительно 70%, приблизительно 71%, приблизительно 72%, приблизительно 73%, приблизительно 74%, приблизительно 75%, приблизительно 76%, приблизительно 77%, приблизительно 78%, приблизительно 79%, приблизительно 80%, приблизительно 81%, приблизительно 82%, приблизительно 83%, приблизительно 84%, приблизительно 85%, приблизительно 86%, приблизительно 87%, приблизительно 88%, приблизительно 89%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99%, приблизительно 100% или любой произведенный из этих цифр диапазон конкретного липида, компонента липидного типа или нелипидного компонента, такого как лекарственное средство, белок, сахар, нуклеиновые кислоты или другое вещество, описанное в данном описании или известное специалисту с квалификацией в данной области. В неограничивающем примере липидная композиция может содержать приблизительно 10% - приблизительно 20% нейтральных липидов и приблизительно 33% - приблизительно 34% цереброзида и приблизительно 1% холестерина. В другом неограничивающем примере липосома может содержать приблизительно 4% - приблизительно 12% терпенов, где приблизительно 1% мицеллы является конкретно ликопином, оставляя приблизительно 3%-11% этой липосомы, содержащих другие терпены; и приблизительно 10% - приблизительно 35% фосфатидилхолина и приблизительно 1% лекарственного средства. Таким образом, предполагается, что липидные композиции данного изобретения могут содержать любые из липидов, типов липидов или других компонентов в любой комбинации или в любом процентном диапазоне.
а. Эмульсии
Липид может содержаться в эмульсии. Липидная эмульсия представляет собой по существу перманентную гетерогенную жидкую смесь двух или более жидкостей, которые обычно не растворяются друг в друге, получаемую механическим встряхиванием или добавлением малых количеств дополнительных веществ, известных как эмульгаторы. Способы приготовления липидных эмульсий и добавления дополнительных компонентов хорошо известны в данной области (например, Modern Pharmaceutics, 1990, включенная в данное описание в качестве ссылки).
Например, один или несколько липидов добавляют к этанолу или хлороформу или любому другому подходящему растворителю и встряхивают вручную или механическими способами. Затем растворитель выпаривают из этой смеси, оставляя высушенную глазурь липида. Эти липиды ресуспендируют в водной среде, такой как фосфатно-буферный солевой раствор, с получением эмульсии. Для достижения более гомогенного распределения размеров эмульгированных липидов эта смесь может быть обработана ультразвуком с использованием общепринятых способов обработки ультразвуком, дополнительно эмульгирована с использованием микрофлюидизации (с применением, например, Микрофлюидизатора, Newton, Mass.) и/или экструдирована при высоком давлении (например, 600 фунт/кв. дюйм) с применением экструдерного устройства (Lipex Biomembranes, Vancouver, Canada).
b. Мицеллы
Липид может содержаться в мицелле. Мицелла является кластером или агрегатом липидных соединений, обычно в форме липидного монослоя, и может быть получена с использованием любого протокола получения мицелл, известного специалистам с квалификацией в данной области (например, ссылки Canfield et al., 1990; El-Gorab et al., 1973; Colloidal Surfactant, 1963 и Catalys in Micellar and Macromolecular Systems, 1975, каждая из которых включена в данное описание в качестве ссылки). Например, один или более липидов обычно готовят в виде суспензии в органическом растворителе, растворитель выпаривают, липид ресуспендируют в водной среде, обрабатывают ультразвуком и затем центрифугируют.
5. Липосомы
В конкретных вариантах осуществления липид является липосомой. "Липосома" является общим термином, включающим в себя разнообразные однослойные и многослойные липидные носители, образованные генерированием замкнутых липидных бислоев или агрегатов. Липосомы могут характеризоваться как имеющие везикулярные структуры с бислойной мембраной, обычно содержащей фосфолипид, и внутренней средой, которая обычно содержит водную композицию.
Многослойная липосома имеет мультилипидные слои, разделенные водной средой. Они образуются самопроизвольно при суспендировании липидов, содержащих фосфолипиды, в избытке водного раствора. Липидные компоненты подвергаются самореаранжировке перед образованием замкнутых структур и заключают воду и растворенные в ней вещества между липидными бислоями (Ghosh and Bachhawat, 1991). Липофильные молекулы или молекулы с липофильными областями могут также растворяться в этом липидном бислое или связываться с этим липидным бислоем.
В специфических аспектах липид и/или модифицированный белок или полинуклеотид, кодирующий модифицированный белок, могут быть, например, инкапсулированы в водном внутреннем пространстве липосомы, разбросаны в липидном бислое липосомы, присоединены к липосоме через линкерную молекулу, которая связана как с липосомой, так и композицией, заключены в липосоме, находиться в комплексе с липосомой и т.д.
а. Получение липосом
Липосомы в соответствии с данным изобретением могут быть получены различными способами, как будет известно специалисту с обычной квалификацией в данной области.
Например, фосфолипид (Avanti Polar Lipids, Alabaster, AL), такой как, например, нейтральный фосфолипид диолеоилфосфатидилхолин (DOPC) растворяют в трет-бутаноле. Затем этот липид (липиды) смешивают с полинуклеотидом или полипептидом и/или другим компонентом (другими компонентами). К смеси липидов добавляют Твин 20, так что Твин 20 составляет около 5% веса этой композиции. К этой смеси добавляют избыток трет-бутанола, так что объем трет-бутанола составляет по меньшей мере 95%. Смесь встряхивают на вортексе, замораживают на бане со смесью сухой лед/ацетон и лиофилизируют в течение ночи. Лиофилизированный препарат хранят при -20°С, и он может быть использован в пределах трех месяцев. При необходимости лиофилизированные липосомы восстанавливают в 0,9% солевом растворе. Средний диаметр частиц, полученных с использованием Твина 20 для инкапсулирования соединения, равен приблизительно 0,7 - приблизительно 1,0 мкм.
Альтернативно, липосома может быть получена смешиванием липидов в растворителе в контейнере, например в стеклянной колбе в форме груши. Этот контейнер должен иметь объем, в 10 раз больший, чем объем ожидаемой суспензии липосом. С использованием роторного испарителя растворитель удаляют при приблизительно 40°С при отрицательном давлении. Обычно растворитель удаляется в пределах приблизительно 5 минут - 2 часов, в зависимости от желаемого объема липосом. Композиция может быть высушена дополнительно в эксикаторе под вакуумом. Высушенные липиды обычно выбрасывают после приблизительно 1 недели из-за тенденции разрушаться со временем.
Высушенные липиды могут быть гидратированы при приблизительно 25-50 мМ фосфолипида в стерильной апирогенной воде встряхиванием, пока вся липидная пленка не будет ресуспендирована. Затем водные липосомы разделяют на аликвоты, каждую из них помещают во флакон, лиофилизируют и герметизируют под вакуумом.
В других альтернативных способах липосомы могут быть приготовлены в соответствии с другими известными лабораторными процедурами (например, см. ссылки Bangham et al., 1965; Gregoriadis, 1979; Deamer and Uster, 1983; Szoka and Papahadjopoulos, 1978, каждая из которых включена в данное описание в качестве ссылки в релевантной части). Эти способы различаются по их способности заключать водное вещество и по их соответствующим соотношениям водное пространство - липид.
Высушенные липиды или лиофилизированные липосомы, полученные, как описано выше, могут быть повторно гидратированы и восстановлены в растворе ингибиторного пептида и разведены до подходящей концентрации подходящим растворителем, например DPBS. Затем эту смесь интенсивно встряхивают в вортексе (вихревом смесителе). Неинкапсулированные дополнительные материалы, такие как агенты, включающие в себя, но не ограничивающиеся ими, гормоны, лекарственные средства, конструкции нуклеиновых кислот и т.п., удаляют центрифугированием при 20000 g и липосомные остатки промывают. Промытые липосомы ресуспендируют при подходящей концентрации фосфолипида, например около 50-200 мМ. Количество дополнительного вещества или активного инкапсулированного агента может быть определено в соответствии со стандартными способами. После определения количества дополнительного вещества или активного агента, инкапсулированного в препарате липосом, липосомы могут быть разбавлены до подходящих концентраций и храниться при 4°С до использования. Фармацевтическая композиция, содержащая липосомы, обычно содержит также стерильный, фармацевтически приемлемый носитель или разбавитель, такой как вода или солевой раствор.
Размер липосомы варьируется в зависимости от способа синтеза. Липосомы в данном изобретении могут быть различных размеров. В некоторых вариантах осуществления липосомы являются небольшими, менее чем приблизительно 100 нм, приблизительно 90 нм, приблизительно 80 нм, приблизительно 70 нм, приблизительно 60 нм или менее приблизительно 50 нм в наружном диаметре. В получении таких липосом может быть использован любой протокол, описанный в данном описании или известный специалисту с обычной квалификацией в данной области. Дополнительные неограничивающие примеры получения липосом описаны в патентах США с номерами 4728578, 5728575, 4737323, 4533254, 4162282, 4310505 и 4921706; международных заявках РСТ/US85/01161 и РСТ/US89/05040, патентной заявке Великобритании GB 213095 A; Mayer et al., 1986; Hope et al., 1985; Mayhew et al., 1987; Mayhew et al., 1984; Cheng et al., 1987, и Liposome Technology, 1984, причем каждая из этих работ и каждый патент включены в данное описание в качестве ссылки.
Липосома, суспендированная в водном растворе, обычно имеет форму сферического пузырька (везикула), имеющего один или несколько концентрических слоев липидных бислойных молекул. Каждый слой состоит из параллельного ряда молекул, представленных формулой XY, где Х обозначает гидрофильную часть молекулы, а Y обозначает гидрофобную часть. В водной суспензии эти концентрические слои размещаются таким образом, что гидрофильные молекулы стремятся оставаться в контакте с водной фазой, а гидрофобные области стремятся к самоассоциации. Например, когда водные фазы присутствуют как внутри, так и вне липосомы, липидные молекулы могут образовать бислой, известный как ламелла, с размещением XY-YX. Агрегаты липидов могут образовываться, когда гидрофильная и гидрофобная части более чем одной липидной молекулы становятся связанными друг с другом. Размер и форма этих агрегатов будет зависеть от многих различных переменных, таких как природа растворителя и присутствие других соединений в растворе.
Получение липидных форм часто выполняют обработкой ультразвуком или последовательной экструзией липосомных смесей после (I) обращенно-фазового выпаривания, (II) дегидратации-регидратации, (III) детергентного диализа и (IV) гидратации тонкой пленки. В одном аспекте предполагаемым способом получения липосом в некоторых вариантах является нагревание, обработка ультразвуком и последовательная экструзия липидов через фильтры или мембраны с порами уменьшающегося размера с получением в результате небольших стабильных липосомных структур. Этот способ получения дает также липосомное/терапевтическое соединение или липосомы только подходящего и однородного размера, которые являются структурно стабильными и производят максимальную активность. Такие способы хорошо известны специалистам с квалификацией в данной области (см., например, Martin, 1990).
После приготовления липидные структуры могут быть использованы для инкапсулирования соединений, которые являются токсичными (например, химиотерапевтические средства) или лабильными (например, нуклеиновые кислоты), когда они находятся в кровотоке. Липосомное инкапсулирование приводило к меньшей токсичности и более продолжительному полупериоду существования в организме для таких соединений (Gabizon et al., 1990).
Многочисленные способы лечения заболевания используют стратегии переноса генов на основе липидов для усиления общепринятых или установления новых лекарственных средств, в частности лекарственных средств для лечения гиперпролиферативных заболеваний. Успехи в приготовлении липосом увеличили эффективность переноса генов in vivo (Templeton et al., 1997), и предполагается, что липосомы получают этими способами. Описаны другие способы получения форм на основе липидов для доставки нуклеиновых кислот (WO 99/18933).
В другой липосомной форме амфипатический носитель, называемый микроносителем разведения растворителя (SDMC), обеспечивает возможность интеграции конкретных молекул в бислой липидного носителя (патент США 5879703). SDMC может быть использован для доставки липополисахаридов, полипептидов, нуклеиновых кислот и т.д. Конечно, любые другие способы получения липосом могут быть использованы специалистом с квалификацией в данной области для получения желаемой липосомной формы в данном изобретении.
b. Нацеливание липосом
Связывание композиций данного изобретения с липосомой может улучшать ее биораспределение и другие свойства. Например, опосредованная липосомами доставка нуклеиновых кислот и экспрессия чужеродной ДНК in vitro были очень успешными (Nicolau and Sene, 1982; Fraley et al., 1979; Nicolau et al., 1987). Была также продемонстрирована возможность опосредованной липосомами доставки и экспрессии чужеродной ДНК в культивируемых куриных эмбрионах, клетках HeLa и гепатомы (Wong et al., 1980). Был осуществлен также успешный опосредованный липосомами перенос генов в крысах после внутривенной инъекции (Nicolau et al., 1987).
Предполагается, что липосомная композиция может содержать дополнительные вещества для доставки к ткани. Например, в некоторых вариантах осуществления данного изобретения липид или липосома могут быть связаны с гемагглютинирующим вирусом (HVJ). Было показано, что это облегчает слияние с клеточной мембраной и стимулирует вхождение в клетку инкапсулированной в липосоме ДНК (Kaneda et al., 1989). В другом примере липид или липосома могут быть связаны или использованы вместе с ядерными негистоновыми хромосомными белками (HMG-1) (Kato et al., 1991). В других вариантах липид может находиться в комплексе или использоваться вместе как с HVJ, так и HMG-1.
Нацеленная доставка достигается добавлением лигандов без нарушения способности этих липосом доставлять большие количества любого описанного соединения данного изобретения. Предполагается, что это будет обеспечивать доставку к специфическим клеткам, тканям и органам. Нацеливающая специфичность систем доставки на основе лигандов основана на распределении рецепторов этих лигандов на различных типах клеток. Нацеливающий лиганд может быть нековалентно или ковалентно связан с липидным комплексом и может быть конъюгирован с липосомами различными способами.
Примеры способов сшивания лигандов (некоторые обсуждались выше) с липосомами описаны в патенте США 5603872 и патенте США 5401511, каждый из которых включен в данное описание в качестве ссылки в его полном виде. Различные лиганды могут быть ковалентно связаны с поверхностями липосом посредством сшивания с аминными остатками. Липосомы, в частности, многоламеллярные везикулы (MLV) или одноламеллярные везикулы, такие как микроэмульгированные липосомы (MEL) и большие одноламеллярные липосомы (LUVET), каждые из которых содержат фосфатидилэтаноламин (РЕ), получали с использованием установленных процедур. Включение РЕ в липосому обеспечивает активный функциональный остаток, первичный амин, на липосомной поверхности для целей сшивания. Лиганды, такие как эпидермальный фактор роста (EGF), были успешно связаны с РЕ-липосомами. Лиганды связывались ковалентно с дискретными сайтами на поверхностях липосом. Число и поверхностная плотность этих сайтов будет диктоваться формой липосомы и типом липосомы. Липосомные поверхности могут быть также сайтами для нековалентной связи. Для образования ковалентных конъюгатов лигандов и липосом сшивающие реагенты исследовались на эффективность и биосовместимость. Сшивающие агенты включают в себя глутаровый альдегид (GAD), бифункциональный оксиран (OXR), диглицидиловый простой эфир этиленгликоля (EGDE) и водорастворимый карбодиимид, предпочтительно 1-этил-3-(3-диметиламинопропил)карбодиимид (EDC). Посредством химического способа образования комплексов сшиванием установлено образование связи аминных остатков узнающего вещества и липосом.
i. Нацеливающие лиганды
Нацеливающий лиганд может быть либо «заякорен» в гидрофобной части этого комплекса, либо присоединен к реакционноспособным терминальным группам гидрофильной части этого комплекса. Нацеливающий лиганд может быть присоединен к липосоме посредством связи с реакционноспособной группой, например, на дистальном конце гидрофильного полимера. Предпочтительные реакционноспособные группы включают в себя аминогруппы, карбоксильные группы, гидразидные группы и тиоловые группы. Связывание нацеливающего лиганда с гидрофильным полимером может быть выполнено стандартными способами органической химии, которые известны специалистам с квалификацией в данной области. В некоторых вариантах общая концентрация нацеливающего лиганда может быть от приблизительно 0,01 до приблизительно 10 мол.%.
Нацеливающими лигандами являются любые лиганды, специфические в отношении характерного компонента области-мишени. Предпочтительные нацеливающие лиганды включают в себя белки, такие как поликлональные или моноклональные антитела, фрагменты антител или химерные антитела, ферменты или гормоны или сахара, такие как моно-, олиго- и полисахариды (см. Heath et al. (1986)). Например, дисиалоганглиозид GD2 является опухолевым антигеном, который был идентифицирован в опухолях нейроэктодермального происхождения, таких как нейробластома, меланома, мелкоклеточная злокачественная опухоль легкого, глиома и некоторые саркомы (Cheresh et al., 1986; Schulz et al., 1984). Липосомы, содержащие моноклональные антитела против дисиалоганглиозида GD2, использовали для содействия нацеливанию этих липосом на клетки, экспрессирующие этот опухолевый антиген (Montaldo et al., 1999; Pagnan et al., 1999). В другом неограничивающем примере специфические антитела против антигена злокачественной опухоли молочной железы и гинекологических злокачественных опухолей описаны в патенте США 5939277, включенном в данное описание в качестве ссылки. В следующем неограничивающем примере специфические антитела против злокачественной опухоли предстательной железы описаны в патенте США с номером 6107090, включенном в данное описание в качестве ссылки. Таким образом, предполагается, что описанные в данном описании антитела или антитела, известные специалисту с обычной квалификацией в данной области, могут быть использованы для нацеливания на специфические ткани и типы клеток в комбинации с композициями и способами данного изобретения. В некоторых вариантах осуществления данного изобретения обсуждаемые нацеливающие лиганды взаимодействуют с интегринами, протеогликанами, гликопротеинами, рецепторами или транспортерами. Подходящие лиганды включают в себя любые лиганды, которые являются специфическими для клеток, происходящих из органа-мишени, или для структур органа-мишени, подвергающихся циркуляции в кровотоке в результате локальной патологии, такой как опухоли.
В некоторых вариантах осуществления данного изобретения для усиления трансдукции клеток, для увеличения трансдукции клеток-мишеней или для лимитирования трансдукции нежелательных клеток, антитело или нацеливающие на циклический пептид части (лиганды) связывают с липидным комплексом. Такие способы известны в данной области. Например, были описаны дополнительно липосомы, которые специфически нацеливают на клетки-мишени центральной нервной системы млекопитающих (патент США № 5786214, включенный в данное описание в качестве ссылки). Эти липосомы состоят по существу из N-глутарилфосфатидилэтаноламина, холестерина и олеиновой кислоты, где с этими липосомами конъюгировано моноклональное антитело, специфическое в отношении нейроглии. Предполагается, что моноклональное антитело или фрагмент антитела могут быть использованы для нацеливания доставки к специфическим клеткам, тканям или органам животного, таким как, например, мозг, сердце, легкое, печень и т.д.
Далее, соединение может быть доставлено к клетке-мишени посредством опосредованной рецептором доставки и/или нацеливающими носителями, содержащими липид или липосому. Они используют преимущество селективного поглощения макромолекул опосредованным рецептором эндоцитозом, который будет иметь место в клетке-мишени. В связи со специфическим в отношении типов клеток распределением различных рецепторов этот способ доставки добавляет следующую степень специфичности данному изобретению.
Таким образом, в некоторых аспектах данного изобретения будет выбираться лиганд для соответствия рецептору, специфически экспрессируемому на клеточной популяции-мишени. Клеточно-специфическая доставка соединений данного изобретения и/или нацеливающий носитель может содержать лиганд специфического связывания в комбинации с липосомой. Доставляемые соединения содержатся в липосоме, а лиганд специфического связывания функционально включен в мембрану липосомы. Таким образом, липосома будет специфически связываться с рецептором (рецепторами) клетки-мишени и доставлять содержимое в эту клетку. Было показано, что такие системы являются системами функционального использования, в которых, например, используют эпидермальный фактор роста (EGF) в опосредованной рецептором доставке нуклеиновой кислоты к клеткам, которые проявляют повышающую регуляцию рецептора EGF.
В некоторых вариантах опосредованная рецептором доставка и/или нацеливающие носители содержат рецептор-специфический лиганд и агент связывания. Другие содержат специфический в отношении клеточного рецептора лиганд, к которому функционально присоединены подлежащий доставке модифицированный белок или полинуклеотид, кодирующий модифицированный белок. Например, несколько лигандов использовали для опосредованного рецептором переноса генов (Wu and Wu, 1987; Wagner et al., 1990; Perales et al., 1994; Myers, EPO 0273085), который устанавливает функциональность этого способа. В другом примере была описана специфическая доставка в контексте другого типа клеток млекопитающих (Wu and Wu, 1993, включено в данное описание в качестве ссылки).
В следующих вариантах осуществления лиганд специфического связывания может содержать один или более липидов или гликопротеинов, которые направляют клеткоспецифическое связывание. Например, лактозилцерамид, галактоза-терминальный асиалоганглиозид, включали в липосомы и наблюдали увеличение поглощения гена инсулина гепатоцитами (Nicolau et al., 1987). Было также показано, что асиалогликопротеин, асиалофетуин, который содержит терминальные галактозильные остатки, нацеливает липосомы на печень (Spanjer and Scherphof, 1983; Hara et al., 1996). Сахара маннозил-, фукозил- или N-ацетилглюкозамин, при связывании со скелетом полипептида, связывают высокоаффинный рецептор маннозы (патент США 5432260, специально включенный в данное описание в качестве ссылки в его полном виде). Предполагается, что клетко- или тканеспецифические трансформирующие конструкции данного изобретения могут специфически доставляться в клетку-мишень или ткань-мишень сходным образом.
В другом примере лактозилцерамид и пептиды, которые нацеливают на белки, родственные рецептору LDL, такие как аполипопротеин Е3 (Apo E), были применимы в нацеливании липосом на печень (Spanjer and Scherphof, 1983; WO 98/0748).
Фолат и рецептор фолата также были описаны как применимые для клеточного нацеливания (патент США 5871727). В этом примере витамин фолат связан в этот комплекс. Рецептор фолата имеет высокую аффинность в отношении его лиганда и сверхэкспрессируется на поверхности нескольких линий злокачественных клеток, в том числе опухолях легкого, молочной железы и мозга. Антифолат, такой как метотрексат, может быть также использован в качестве нацеливающих лигандов. Опосредованные трансферрином системы доставки нацеливают на большой диапазон реплицирующихся клеток, которые экспрессируют рецептор трансферрина (Gilliand et al., 1980).
с. Комбинации липосома/нуклеиновая кислота
Предполагается, что когда липосомная композиция содержит клетко- или тканеспецифическую нуклеиновую кислоту, этот способ может иметь применимость в данном изобретении. В некоторых вариантах осуществления невирусные формы на основе липидов обеспечивают альтернативу способам вирусной генотерапии. Хотя многие исследования на культуре клеток документировали невирусный перенос генов на основе липидов, системная доставка генов последством форм на основе липидов была ограниченной. Основным недостатком невирусной доставки генов на основе липидов является токсичность катионных липидов, которые составляют невирусный доставляющий носитель. Токсичность липосом in vivo отчасти объясняет противоречия между результатами переноса генов in vitro и in vivo. Другим фактором, вносящим вклад в эту противоречивую информацию, является различие стабильности липосом в присутствии и в отсутствие сывороточных белков. Взаимодействие между липосомами и сывороточными белками оказывает разительное действие на характеристики стабильности липосом (Yang and Huang, 1997). Катионные липосомы притягивают и связывают отрицательно заряженные сывороточные белки. Липосомы, покрытые сывороточными белками, либо растворяются, либо поглощаются макрофагами, что приводит к их удалению из кровотока. Существующие способы липосомной доставки in vivo используют аэролизацию, подкожную, интрадермальную, внутриопухолевую или интракраниальную инъекцию для элиминации проблем токсичности и стабильности, связанных с катионными липидами в кровотоке. Взаимодействие липосом и белков плазмы в значительной степени ответственно за несоответствие между эффективностью in vitro (Felgner et al., 1987) и переносом генов in vivo (Zhu et al., 1993; Philip et al., 1993; Solodin et al., 1995; Liu et al., 1995; Thierry et al., 1995; Tsukamoto et al., 1995; Aksentijevich et al., 1996).
Был описан пример способа для нацеливания вирусных частиц на клетки, которые не имеют единственного клеткоспецифического маркера (патент США 5849718). В этом способе, например, антитело А имеет специфичность в отношении опухоли, но также в отношении нормальной ткани сердца и легкого, тогда как антитело В имеет специфичность в отношении опухоли, но также в отношении нормальных клеток печени. Применение антитела А или антитела В по отдельности для доставки антипролиферативной нуклеиновой кислоты в эту опухоль могло приводить к нежелательному повреждению клеток сердца и легкого или печени. Однако антитело А и антитело В могут быть использованы вместе для улучшенного нацеливания на клетки. Так, антитело А связывают с геном, кодирующим антипролиферативную нуклеиновую кислоту, и доставляют, через опосредованную рецептором систему поглощения, к опухоли, а также к тканям сердца и легкого. Однако этот ген не транскрибируется в этих клетках, так как они лишены необходимого фактора транскрипции. Антитело В связывают с универсально активным геном, кодирующим фактор транскрипции, необходимый для транскрипции антипролиферативной нуклеиновой кислоты, и доставляют к клеткам опухоли и клеткам печени. Таким образом, в клетки сердца и легкого доставляется только неактивная антипролиферативная нуклеиновая кислота, где она не транскрибируется, что приводит к отсутствию вредных побочных действий. В клетках печени ген, кодирующий фактор транскрипции, доставляется и транскрибируется, но не оказывает действия, поскольку ген антипролиферативной нуклеиновой кислоты отсутствует. Однако в опухолевых клетках оба гена доставляются и фактор транскрипции может активировать транскрипцию антипролиферативной нуклеиновой кислоты, приводя к опухолеспецифическим токсическим эффектам.
Добавление нацеливающих лигандов для доставки генов для лечения гиперпролиферативных заболеваний позволяет доставлять гены, генные продукты которых являются более токсичными, чем генные продукты, доставляемые ненацеленными системами. Примеры более токсичных генов, которые могут доставляться, включают в себя про-апоптотические гены, такие как Вах и Bak плюс гены, произведенные из вирусов и других патогенов, такие как аденовирусный ген Е4orf4 и ген пуриннуклеозидфосфорилазы E. coli, так называемый «суицидный ген», который превращает пролекарство дезоксирибозид 6-метилпурина в токсичный пурин 6-метилпурин. Другими примерами суицидных генов, используемых с пролекарствами, являются ген цитозинаминазы E. coli и ген тимидинкиназы HSV.
Можно также использовать ненацеленные или нацеленные липидные комплексы для получения рекомбинантных или модифицированных вирусов in vivo. Например, две или большее число плазмид могут быть использованы для введения ретровирусных последовательностей плюс терапевтического гена в гиперпролиферативную клетку. Ретровирусные белки, обеспеченные in trans из одной из этих плазмид, будут обеспечивать возможность упаковки второй несущей терапевтический ген плазмиды. Таким образом, трансдуцированные клетки получат сайт для продуцирования нерепликативных ретровирусов, несущих терапевтический ген. После этого ретровирусы будут способны инфицировать ближайшие клетки. Промотор для терапевтического гена может быть или может не быть индуцируемым или тканеспецифическим.
Подобным образом, переносимая нуклеиновая кислота может предоставлять ДНК для репликации компетентного или реплицирующегося только в определенных условиях вирусного генома, такого как аденовирусный геном, который лишен всей аденовирусной области E1a или E2b или который имеет один или несколько тканеспецифических промоторов или индуцируемых промоторов, запускающих транскрипцию от областей E1a и/или E1b. Эта реплицирующаяся или реплицирующаяся только в определенных условиях нуклеиновая кислота может содержать или может не содержать дополнительный терапевтический ген, такой как супрессорный ген опухолей или антионкоген.
d. Введение липидов
Фактическое дозовое количество липидной композиции (например, модифицированного липосомой белка или полинуклеотида, кодирующего модифицированный белок), вводимое пациенту, может определяться физическими и физиологическими факторами, такими как вес тела, тяжесть состояния, идиопатия пациента и способ введения. С учетом этого доза липидной композиции для конкретного субъекта и/или схема введения могут быть легко определены.
V. Примеры
Следующие примеры приведены для демонстрации конкретных вариантов данного изобретения. Специалистам с квалификацией в данной области должно быть понятно, что способы, описанные в следующих примерах, представляют способы, которые, как показал автор изобретения, работают хорошо в практике данного изобретения и, следовательно, могут рассматриваться как способы его применения на практике. Однако специалисты с квалификацией в данной области должны понимать, в свете данного описания, что могут быть произведены многие изменения в этих конкретных вариантах осуществления, которые описаны, и при этом может быть получен такой же или сходный результат, без отхода от идеи и объема данного изобретения.
ПРИМЕР 1
Последовательные делеционные исследования
Нуклеотидную последовательность, кодирующую рекомбинантный гелонин (SEQ ID NO:2), использовали в качестве матрицы для создания этих токсинов. Анализ последовательности и структурное моделирование rGel показали значительную укладку этой молекулы в β-складки, β-спирали и «шпильки», как показано. В соответствии с этими исследованиями аминокислоты 200-277 (С-концевые), по-видимому, укладываются в карман связывания, сходный с карманом связывания маркирующего порта RTA для его В-цепи. Поскольку rGel не имеет В-цепи, теоретически предполагалось, что этот «маркирующий порт» является рудиментарной частью этого токсина, которая может быть необязательной для биологической активности этого белка.
Были созданы последовательные делеционные мутанты кДНК-молекулы, кодирующей rGel, из С-конца и из N-конца, как показано на фиг. 3А и 3В и в таблице 7, и были названы CFR1901-CFR1905. В предварительных исследованиях конструкции CFR 1904, 1905, 2001, 2007 и 2024 продемонстрировали определяемую активность в анализах с лизатами кроличьих ретикулоцитов (RRLA). Эти конструкции были в 105-103 раз менее активными, чем CFR 1888, который находится в диапазоне «активных» молекул токсина (Munishkin et al., 1995).
CFR 1901
Для создания CFR 1901, который имеет делетированные аминокислоты 1-46, 1 мкг очищенной кДНК CFR 1888, содержащейся в векторе рХ2 (исходно основывающемся на рЕТ-22b, Novagen, Inc.), расщепляли 50 единицами рестрикционных эндонуклеаз NcoI и SmaI (Boehringer Mannheim). Затем выступающий фрагмент на сайте 5' NcoI затупляли добавлением 1 единицы нуклеазы фасоли золотистой (New England Biolabs) и инкубировали при 30°С в течение 0,5 ч. Набор для ПЦР-очистки Qiagen использовали для удаления нуклеазы фасоли золотистой. Полученную ДНК циркуляризовали лигированием с 3' затупленным сайтом SmaI.
CFR 1902
CFR 1888 расщепляли рестрикционными эндонуклеазами NcoI и ClaI. Концы достраивали для сохранения правильной рамки считывания с использованием фермента Кленова (New England Biolabs) на 3'-выступе для его затупления перед повторным лигированием.
Для экспрессии белка 50 нг плазмидной ДНК трансформировали в 50 мкл BL21(DE3)pLysS-компетентных клеток-хозяев E. coli (Novagen, Inc.). Отдельные колонии выскребали и выращивали при 37°С со встряхиванием в 100 мл бульона Luria, содержащего 200 мкг/мл ампициллина (Sigma, Chemical Co.) до OD600 между 0,6 и 0,8. Затем к культуре добавляли IPTG (Boehringer Mannheim) для индукции рекомбинантного белка в конечной концентрации 0,1 мМ. Культуре давали инкубироваться еще в течение 2 часов при 37°С перед сбором центрифугированием.

ПРИМЕР 2
Картирование антигенных линейных пептидных доменов
Антигенные домены на молекуле rGel не были ранее описаны в литературе. Антигенные домены молекулы rGel могут зависеть либо от углеводных, либо от пептидных последовательностей этой молекулы. Рекомбинантный rGel, продуцируемый в бактериях, не имеет гликозилирования белка, и, следовательно, антитела, направленные против rGel, должны узнавать пептидные домены на этой молекуле.
Для идентификации антигенных доменов на молекуле rGel сначала выделяли поликлональные антитела человека из сыворотки работников лаборатории, профессионально подвергающихся действию рекомбинантного гелонина. Сыворотки, полученные у трех работников лаборатории, добавляли в 96-луночные планшеты ELISA, покрытые rGel. Затем эти планшеты проявляли с использованием антител против антител человека для идентификации присутствия человеческих антител против гелонина. Сыворотка у двух из этих трех работников обнаружила значимые титры антител, сравнимые с титрами контрольной сыворотки человека (фиг. 1). Затем получали двадцать мл сыворотки у этих двух работников и получали поликлональные человеческие антитела против гелонина аффинной хроматографией с использованием аффинной смолы Affi-gel, содержащей rGel.
Были синтезированы десять пептидов, охватывающих всю длину молекулы rGel, и затем их использовали для покрытия 96-луночных планшетов. Раствор человеческих антител против гелонина добавляли в эти планшеты, давали взаимодействовать и присутствие человеческих антител, прикрепившихся к покрытым пептидом лункам, оценивали с использованием ELISA. Как показано на фиг. 2, значимую реактивность поликлональных антител получали с пептидами, простирающимися на 23-53 (домен 1), 72-89 (домен 2), 181-198 (домен 3) и 223-252 (домен 4). Были сконструированы токсины CFR-2001-2024 для делеции антигенных доменов, специфически узнаваемых человеческими поликлональными антителами к гелонину.
ПРИМЕР 3
Замена антигенных последовательностей последовательностями человека
Химерная молекула человек/растение была сконструирована с использованием информации, касающейся антигенных доменов, полученной с использованием человеческих антител против гелонина, как описано выше, для идентификации четырех специфических антигенных доменов в молекуле гелонина (аминокислоты 205-257, 23-42, 71-88 и 189-204). Эти последовательности анализировали дополнительно с использованием базы данных GenQuest/BLAST для поиска на гомологии с известными белками человека. Дополнительной задачей в этом исследовании было не только идентифицировать гомологичную «человеческую» последовательность, но также выстроить такую последовательность в сконструированной молекуле токсина таким образом, чтобы могла сохраняться ферментативная (н-гликозидная) функциональная группа полученной гибридной молекулы.
Четыре последовательности-кандидата для встраивания в сконструированный токсин идентифицировали из этой базы данных, которые и использовали в качестве основы для замен аминокислот (таблица 8). Человеческие гомологичные последовательности для домена 1 показали высокую гомологию (40%) с белком KELL человека, который является белком группы крови с цинксвязывающим доменом. Интересно, что ранние исследования природного гелонина предполагают, что эта молекула может связывать цинк (Sperti et al., 1986), но эти исследования не были подтверждены. Анализ домена 4 (аминокислоты 189-204) показал идентичность (40%) белку CFAH человека, который, по-видимому, играет роль в качестве кофактора в функции печени человека. Поиск на гомологию домену 2 (аминокислоты 23-42) показал 44% гомологию с белком UTRO человека, который, по-видимому, играет роль в цитоскелетном прикреплении клеточных плазматических мембран.

Сконструированные токсины, содержащие модификации антигенного сайта и делеции, дополнительно модифицировали в домене 3 для придания 100% идентичности с белком KELL человека. Аминокислоты в молекуле-дизайнере, фланкирующие последовательность KELL, также корректировали для близкой имитации выстраивания последовательностей в полноразмерном (непроцессированном) белке KELL. Конечные последовательности, сконструированные для замены домена 3, показаны в таблице 8. Таблица 9 показывает результат поисков гомологии последовательностей в GenBank на полноразмерные (непроцессированные) белки, содержащие гомологичные белки человека.
Результаты исследований авторов изобретения ясно показали, что с использованием рекомбинантного гелонина в качестве исходной «матрицы» могут быть спроектированы, сконструированы и испытаны уникальные делеции.
ПРИМЕР 4
Гелониновые сконструированные токсины
В следующем разделе описаны гелониновые сконструированные токсины, которые были сконструированы с использованием способов, описанных в предыдущих примерах.
Делеционные токсины
CFR 1888
На основе исходной «матрицы» рекомбинантного гелонина авторов данного изобретения С-конец из исходного KDPE был модифицирован с получением KDEL для облегчения внутриклеточного прохождения этого белка к внутриклеточному рибосомному компартменту.
CFR 1901-1905 (см. таблицу 7 и фиг. 3А)
С использованием CFR 1888 в качестве «матрицы» генерировали серию последовательных делеционных мутантов от N- и/или С-конца для определения участков этой молекулы, которые могли бы быть делетированы без влияния на биологическую активность. Как показано на фиг. 3А, было обнаружено, что индивидуальные делеции 1901, 1902 и 1903 были неактивными. Однако при объединении отдельных делеций (CFR 1904 и CFR 1905), биологическая активность повторно устанавливалась (фиг. 3А).
CFR 2018
С использованием CFR 1888 в качестве «матрицы» этот белок дополнительно модифицировали, как показано в таблице 7, для получения слегка меньшей молекулы и добавления остатка аланина на С-конце для обеспечения улучшенной стабильности in vivo во время продуцирования в бактериальном хозяине.
Токсины на основе исследований антигенности
Антигенные домены на этой молекуле картировали с использованием линейных пептидов, охватывающих молекулу гелонина. Как показано на фиг. 1, поликлональные антисыворотки человека против гелонина выявили четыре различных антигенных домена на этой молекуле. Домен 1 простирается на аминокислоты 205-257, домен 2 состоит из аминокислот 23-42, домен 3 состоит из аминокислот 71-88 и домен 4 состоит из аминокислот 89-204.
CFR-2024
Как показано на фиг. 3В, были созданы шесть делеционных мутантов на основе наблюдаемых антигенных доменов. Три белка обнаруживали биологическую активность, тогда как остальные три белка были неактивными.
CFR 2019-2042
Заместительные исследования по созданию химерных молекул человек/растение. Четыре антигенных домена были переданы в GenBank для анализа последовательности. Поиск по программе Swissprot protein sequence проводили для обнаружения гомологичных последовательностей человека на основе описанных выше четырех антигенных доменов, описанных выше.
Гомологичные последовательности человека
Было обнаружено, что домен 1 (аминокислоты 205-257) картируется относительно последовательности в белке группы крови человека KELL (P23276). 40% идентичность и 65% положительность была обнаружена относительно последовательности на этом белке.
Было обнаружено, что домен 2 (аминокислоты 23-42) имеет совпадение с 44% идентичностью с белком человека UTRO (Р46939).
Для домена 3 (аминокислоты 71-88) эта последовательность показала 37% идентичность и 68% положительность относительно белка человека МАОМ (Р23368). Этот белок описан в таблице 8.
Для последовательности домена 4 (189-204) 40% идентичность была обнаружена в белке человека CFAH (P08603).
На основании этих данных были созданы химерные последовательности человека, соответствующие этим четырем антигенным доменам (таблица 8). Эти последовательности представляют человеческие неантигенные замены антигенных доменов в растительном белке.
CFR 2019-2024
С использованием CFR 2019 и 2024 в качестве «матриц» генерировали многочисленные новые белки-дизайнеры CFR 2019-2042 (таблица 7). Эти белки представляют замену 1, 2, 3 или 4 доменов на этой молекуле человеческими химерными гомологами. Некоторые из этих сконструированных токсинов (CFR 2018, 2019, 2024 и 2025) экспрессировали в бактериях, содержащих метку 50 кДа, и очищали до гомогенности. Вестерн-анализ выполняли с использованием поликлональных антител к этой метке.
Молекулы 2019, 2024 и 2025 были уменьшены в размере в сравнении с исходным «матричным» белком 2018. Вестерн-анализ также показал приблизительно эквивалентную реактивность с антителами против метки, свидетельствующую об однородном нанесении молекул каждого токсина на ДСН-ПААГ. Вестерн-блот повторно зондировали с использованием антител к нативной молекуле CFR 1888. Была хорошая реактивность с белком 2018, как показал Вестерн-блот, однако, фактически не было реактивности этой поликлональной антисыворотки со сконструированными токсинами 2019 и 2025 и лишь слабая реактивность была со сконструированным токсином 2024. Это свидетельствует о том, что посредством специфической делеции (CFR 2024) или замены антигенных доменов (CFR 2019) или комбинации делеции и замены антигенных доменов (CFR 2025) могут быть созданы новые молекулы токсина, которые становятся фактически неузнаваемыми для антител к исходной молекуле и, следовательно, должны иметь уменьшенный антигенный профиль.
CFR 2143-2146
Эту серию гибридных молекул конструировали для включения оптимальных функциональных свойств белков CFR 1888 и CFR 2018.
CFR 2247-2458
Была получена серия молекул, объединяющих как н-гликозидные функции токсинов Типа I, так и функции выбранных про-апоптотических молекул человека, таких как ВАХ и Granzyme B. Эти молекулы анализировали на функциональную активность гелонинового компонента и на активность ВАХ и Granzyme B. Их также оценивали на ингибирование бесклеточного белкового синтеза.
Сначала может быть сделана оценка на этой серии молекул относительно экспрессии ВАХ в клетках. Это может быть выполнено с использованием антител к ВАХ, таких как универсальные антитела против ВАХ 6А7, в иммуноанализах, таких как иммунопреципитации или Вестерн-блоттинг. После подтверждения экспрессии ВАХ клетки анализируют на жизнеспособность клеток. Это может быть выполнено различными путями, в том числе с использованием конструкции люциферазы светляка. Для этого экспрессирующий вектор pGL3 млекопитающих (Promega), несущий структурный ген люциферазы светляка (Luc), может быть трансфицирован в клеточную линию млекопитающего вместе с плазмидами, кодирующими ВАХ и слитые с ВАХ белки. Активность люциферазы может быть измерена жидкостным сцинтилляционным счетом с использованием 20 мл клеточного экстракта. Жизнеспособность клеток измеряют в виде относительной активности люциферазы тестируемой конструкции в сравнении с указанной контрольной плазмидой.
Гибриды, которые включают в себя весь полипептид granzyme B или его часть, оценивают на их ферментативную активность с использованием флуориметрического измерения 2-нафтиламина после гидролиза L-глутамил-2-нафтиламида (Bachem, Philadelphia, PA). Активность амидазы измеряют при 21°С с 1,00 мМ L-глутамил-2-нафтиламидом в буфере А (0,3 М NaCl, 0,1 М HEPES, доведенный до рН 7,0 1 М NaOH, 1 мМ Na2-ЭДТА, 0,05 М (об./об.) Тритон Х-100) на спектрофлуориметре Perkin-Elmer 650-10М с возбуждением при 340 нм и испусканием флуоресценции, наблюдаемым при 415 нм (оба с полосным пропусканием 5 нм). Небольшие аликвоты растворов фермента добавляют к раствору субстрата и увеличение испускания флуоресценции подвергают мониторингу в течение 10-40 минут. Альтернативно, активность granzyme B определяют в непрерывном колориметрическом анализе с BAADT (N-α-трет-бутоксикарбонил-L-аланил-L-аспартилтиобензиловым эфиром) в качестве субстрата. Для анализа колоночных фракций 1-50 мкл добавляют к буферу А с 1 М (об./об.) 10 мМ BAADT в (СН3)250 и 1 М (об./об.) 11 нМ дитиобис(2-нитробензойной кислоты) (Sigma) в (СН3)250 при 21°С и скорость увеличения оптической плотности измеряют при 405 нм на планшет-ридере Thermomax (Molecular Devices Inc., Palo Alto, Ca). Увеличения оптической плотности преобразуют в ферментативные скорости.
ПРИМЕР 4
Материалы и способы для примера 5
Материалы
кДНК, кодирующую антитело ZME-018, амплифицировали из гибридомной РНК, полученной из гибридомных клеток, экспрессирующих мышиное антитело, с использованием наборов из Novagen (Madison, WI) и Invitrogen Corp. (Carlsbad, CA). Реагенты ПЦР получали от Fisher Scientific (Pittsburg, PA) и молекулярно-биологические ферменты покупали либо у Boehringer Mannheim (Indianapolis, IN) или New England Biolabs (Beverly, MA). Бактериальные штаммы и бактериальные экспрессирующие плазмиды pEt получали от Novagen (Madison, WI) и среду для выращивания покупали у Difco Laboratories (Detroit, MI). Все другие химикалии и реагенты были либо из Fisher Scientific, либо из Sigma Chemicals Co. (St. Louis, MO). Аффинную смолу с металлом (Talon) получали от Clontech Laboratories (Palo Alto, CA). Другие хроматографические смолы и материалы были от Pharmacia Biotech (Piscataway, NJ). Реагенты для культуры ткани были от Gibco BRL (Gaithersberg, MD).
Клонирование VH- и VL-доменов антитела ZME-018
Мессенджер-РНК из мышиной гибридомы FMT 112 P2, экспрессирующей антитело ZME-018 (IgG2А), выделяли с использованием набора Invitrogen Fast Track и транскрибировали с получением кДНК с использованием набора Invitrogen Copy с применением указанных условий. Амплификацию вариабельных областей легкой и тяжелой цепи антитела проводили с использованием набора Novagen Ig-Prime с набором мышиных Ig-праймеров. ПЦР-профиль для амплификации легкой цепи был следующим: 30 циклов 94°С·1 мин, 60°С·1 мин и 72°С·1 мин, с конечным 5-минутным инкубированием при 72°С. Для реакций тяжелой цепи использовали идентичные условия, за исключением того, что температура отжига была 50°С вместо 60°С. Затем амплифицированную при помощи этой процедуры ДНК клонировали в клонирующий вектор pCR II Invitrogen T/A без дополнительной очистки, трансформировали в E. coli XL1-Blue и идентифицировали с использованием процедур скрининга на основе отбора по цвету колоний (синие-белые). Положительные клоны (по пять из библиотек тяжелых цепей и библиотек легких цепей) секвенировали с использованием праймеров промоторов Т7 и SP6 и домены антител идентифицировали по гомологии с последовательностями других иммуноглобулинов.
Конструирование генов, кодирующих одноцепочечное антитело scfvMEL и иммунотоксин scfvMEL/rGel
Двухстадийный ПЦР-способ со сплайсинговым перекрывающимся удлинением цепи (Sambrook et al., 1989) использовали для конструирования одноцепочечного антитела ZME-018 с применением ДНК-клонов легкой и тяжелой цепи в качестве матриц. Последовательности легкой цепи амплифицировали с использованием праймеров А (5'-GCTGCCCAACCAGCC ATGGCGGACATTGTGATG-3') и С (5'-GCCGGAGCCTGGCTTGC(A/C)GCTGCCGCTGGTGGAGCCTTTGATC(A/T)CCAG-3'), тогда как ДНК тяжелой цепи амплифицировали с праймерами В (5'-AAGCCAGGCTCCGGCGAAGGCAGCACCAAAGG CGAAGTGAAGGTT-3') и D (5'-GCCACCGCCACCACTAGTTGAGGAGACTGT-3'). ПЦР-профили для каждой серии реакций были следующими: 30 циклов 1-минутной денатурации при 94°С, 1-минутный отжиг при 50°С и 1-минутное удлинение при 72°С, с последующим конечным 5-минутным инкубированием при 72°С. Одну десятую объема каждой из этих реакций объединяли и использовали непосредственно во второй ПЦР только с праймерами А и D с тем же самым профилем реакции, описанным выше. Конечный продукт очищали с использованием Geneclean II (Bio 101, Vista, CA), расщепляли рестриктазами NcoI и SpeI и клонировали в плазмидный вектор рЕТ-22b на основе Т7. Гены, кодирующие scfvMEL и рекомбинантный гелонин, сливали вместе с использованием ПЦР способа со сплайсинговым перекрывающимся удлинением с ДНК антитела и гелонина в качестве матриц и праймером NbsphZME (5'-GGCGGTGGCTCCGTGATGACGGACATTGTGATGACC CAGTCTCAAAAATTC-3'), праймером NTXOM (5'-GGTGGCGGTGGCTCCGGTCTAG ACACCGTGACG-3') и праймером XOMBAC (5'-AAGGCTCGTGTCGACCTCG AGTCATTAAGC TTTAGGATCTTTATC-3') (фиг. 4). Очищенные продукты ПЦР очищали и расщепляли затем, как описано выше, и клонировали в вектор рЕТ-32а. Затем секвенированные ДНК-клоны трансформировали в штамм E. coli AD494(DE3) pLys S, полученный из Novagen, для экспрессии слитого токсина.
Экспрессия белка в E. coli
Для экспрессии иммунотоксина бактериальные культуры инкубировали при 37°С в среде для выращивания 2хYT с отбором с сильными антибиотиками (200 мкг/мл ампициллина, 70 мкг/мл хлорамфеникола и 15 мкг/мл канамицина) и выращивали до ранней log-фазы (А600=0,4-0,8). Затем эти культуры разбавляли 1:1 свежей средой 2хYT, содержащей те же самые концентрации антибиотиков, и экспрессию белка-мишени индуцировали при 23°С добавлением 0,1 мМ IPTG и выдерживанием с ним в течение 16-23 часов. Затем индуцированные бактериальные культуры центрифугировали и хранили в замороженном виде при -80°С для более поздней очистки.
Очистка комплекса иммунотоксин/белок
Замороженные бактериальные осадки из индуцированных культур, экспрессирующих иммунотоксин scFvZME-Gel, оттаивали при комнатной температуре и лизировали добавлением 1 мг/мл лизоцима в 10 мМ Трис-HCl, рН 8,0, в течение 30 мин при 4°С. Затем эти бактериальные лизаты обрабатывали ультразвуком три раза в течение 10 сек каждый раз в приборе для разрушения клеток и центрифугировали при 14000 об/мин в течение 30 мин при 4°С. Супернатант переносили и сохраняли на льду и процедуру обработки ультразвуком повторяли с осадком клеток. Затем супернатанты из этих двух лизатов объединяли и ультрацентрифугировали при 40000 об/мин в роторе SS-34 в течение 45 мин при 4°С. Затем образцы, содержащие только растворимый белок, фильтровали (поры 0,22 мкм), доводили до 40 мМ Трис-HCl с использованием 1 М Трис-HCl (рН 8,0) и затем наносили при комнатной температуре на аффинную колонку с металлом Talon, предварительно уравновешенную некоторым количеством буфера. После нанесения колонку промывали 3 колоночными объемами буфера для нанесения с последующей промывкой 5 колоночными объемами 40 мМ Трис-HCl, рН 8,0, 500 мМ NaCl и 5 мМ имидазола. Затем связанный белок элюировали 5 колоночными объемами буфера, содержащего 40 мМ Трис-HCl (рН 8,0), 500 мМ NaCl и 100-200 мМ имидазол. Фракции, содержащие иммунотоксин, объединяли, определяли количественно и диализовали в 20 мМ Трис-HCl (рН 7,2), 50 мМ NaCl перед расщеплением энтерокиназой для удаления метки 6хHis с применением процедуры, установленной Novagen (Madison, WI).
ELISA и Вестерн-анализ
Все стадии инкубации ELISA проводили при комнатной температуре в течение 1 ч, если нет других указаний, и между инкубациями все лунки промывали промывочным буфером ELISA (10 мМ Трис-HCl, рН 8,0, 150 мМ NaCl, 0,2% Твин-20). Лунки 96-луночного микротитрационного планшета покрывали, каждую, 50000 gp240-антиген-положительными клетками меланомы А375М и сушили. Затем их регидратировали и блокировали 3% БСА в промывочном буфере. Планшеты инкубировали и добавляли образцы очищенного иммунотоксина, кроличье поликлональное антитело против гелонина (при 100 нг/мл в буфере для разведения [промывочный буфер ELISA, содержащий БСА в концентрации 1 мг/мл]) и конъюгированные с пероксидазой козьи антитела против кроличьего IgG (Sigma, используемые при разведении 1:5000 в буфере для разведения). Индивидуальные лунки промывали тщательно промывочным буфером и затем проявляли в течение 30 мин ABTS (2,2'-азино-бис[3-этилбензтиазолин-6-сульфоновой кислотой]) в 0,1 М цитратном буфере (рН 4,2) и сигнал измеряли при 405 нм.
Для Вестерн-блотов все инкубации выполняли при комнатной температуре в течение 1 ч, если нет других указаний. Вкратце, белки разделяли электрофорезом в ДСН-ПААГ и переносили на нитроцеллюлозу в течение ночи при 4°С в буфере для переноса (25 мМ Трис-HCl, рН 7,5, 190 мМ глицин, 20% (об./об.) метанол категории ВЭЖХ) при 40 В. Фильтры блокировали 5% БСА в блокирующем буфере для Вестерн-блотов (50 мМ Трис-HCl, рН 7,5, 150 мМ NaCl) и затем давали им взаимодействовать последовательно с кроличьим поликлональным антителом против гелонина (при концентрации 100 нг/мл в промывочном буфере для Вестерн-блотов TBS, рН 7,6, 0,5% Твин-20) и конъюгированными с пероксидазой козьими антителами против кроличьего IgG (Sigma, при разведении 1:10000 в промывочном буфере). Сигнал измеряли с использованием системы определения Amersham ECL.
Анализ трансляции in vitro с лизатами ретикулоцитов
Индуцируемое гелонином ингибирование включения радиоактивно меченного (3Н) лейцина в белок в бесклеточной системе синтеза белка после введения различных доз иммунотоксина проводили, как рекомендовано изготовителем (Promega) и как описано ранее (Press et al., 1986).
Иммунофлуоресцентное окрашивание
Антиген-положительные (меланома А375) клетки добавляли к покрытым полилизином 16-луночным предметным стеклам с камерами (Nunc) при 104 клеток на камеру и инкубировали при 37°С в течение ночи в атмосфере 5% СО2. Клетки обрабатывали слитой конструкцией scfvMEL/rGel в концентрации 50 мкг/мл в различных временных точках. Затем клетки промывали кратковременно ФБР и затем белки, связанные с поверхностью клеток, отделяли инкубированием в течение 10 мин с глициновым буфером (500 мМ NaCl, 0,1 М глицин, рН 2,5), нейтрализовали 5 мин 0,5 М Трис, рН 7,4, промывали кратковременно ФБР и затем фиксировали в 3,7% формальдегиде (Sigma) в течение 15 мин при комнатной температуре с последующим кратковременным промыванием ФБР. Затем клетки делали проницаемыми в течение 10 мин в ФБР, содержащем 0,2% Тритон Х-100m, промывали три раза ФБР и затем инкубировали с ФБР, содержащим 3% БСА, в течение 1 ч при комнатной температуре. После кратковременного промывания ФБР клетки инкубировали либо с кроличьими антителами против scFvMEL, либо с кроличьими поликлональными антителами против rGel, разведенными 1:500 в ФБР, содержащем 0,1% Твин-20 и 0,2% БСА, в течение 1 ч при комнатной температуре. Клетки промывали три раза в ФБР, содержащем 0,1% Твин-20 (PBST), и затем добавляли разведенное 1:100 связанное с изотиоцианатом флуоресцеина (FITC) антитело против кроличьего IgG (Sigma). Контрольные клетки инкубировали только со вторичным FITC-связанным антителом против кроличьего IgG (1:100). После трех конечных промывок PBST клетки промывали один раз ФБР в течение 10 мин и готовили препарат в среде для приготовления препаратов. Предметные стекла анализировали с использованием флуоресцентного микроскопа, и каждая фотография представляла по меньшей мере 10 полей зрения для каждого эксперимента при 400х увеличении.
Анализ цитотоксичности in vitro
Образцы анализировали с использованием стандартного 72-часового анализа пролиферации клеток с монослоями клеток log-фазы (5000 клеток на лунку) антиген-положительных А375М и антиген-отрицательных Ме-180 или SK-OV-3 клеток и с использованием процедур окрашивания красителем кристаллическим фиолетовым, как описано ранее (Nishikawa et al., 1992).
Исследования цитотоксичности in vivo
Бестимусных (голых) мышей в возрасте 4-6 недель делили на группы из 5 мышей на клетку. Клетки меланомы человека А-375 log-фазы (5·107 клеток на мышь) инъецировали подкожно в правый бок и давали развиться опухолям. Как только опухоли становились измеримыми (˜30-50 мм2), животных обрабатывали (i.v. в хвостовую вену) либо солевым раствором (контроль), либо различными концентрациями слитого токсина scfvMel/rGel в течение 4 последовательных дней. Животных подвергали мониторингу и опухоли измеряли в течение дополнительных 30 дней.
ПРИМЕР 5
Одноцепочечное рекомбинантное антитело против меланомы,
слитое с гелонином
Конструирование слитого белка scFvMEL/rGel
Гены вариабельной области для антитела ZME-018 и ген гелонина (Rosenblum et al., 1999) были матрицами для конструирования гена слитого белка антитело против меланомы-иммунотоксин. В качестве первой стадии авторы собирали иммунотоксин в одной ориентации и оценивали его связывание и цитотоксичность в отношении антиген-положительных клеток меланомы А375М. Гены, кодирующие фрагменты этого антитела и гелонина, связывали вместе с использованием способа на основе ПЦР для конструирования слияния в ориентациях антитело-гелонин. Ген иммунотоксина метили также на С-конце гексагистидиновой последовательностью и экспрессировали в E. coli AD494(DE3) pLysS с использованием экспрессирующего вектора на основе Т7 pET-32b Novagen.
Фиг. 4 иллюстрирует ориентацию экспрессируемого иммунотоксина и также показывает последовательности аминокислотных линкеров в местах соединений белковых доменов. Конструировали антитело для кодирования вариабельной области легкой цепи (VL) при N-конце этого белка с состоящим из 18 аминокислот гибким пептидным линкером (Alfthan et al., 1995) с С-концом VH. Гелонин (CFR 2018) (называемый rGel в этом примере) помещали по ходу транскрипции (справа) от VH после другого линкера. Авторы выбирают эту конфигурацию по причине присутствия в ней незатрудненной гибкости сайта связывания антитела. При помещении токсина при N-конце этого слитого белка мог бы требоваться более длинный пептид для обеспечения оптимальной пространственной ориентации этих двух белковых молекул, и конструирование такого варианта в настоящее время проводится. Исследования по секвенированию ДНК конечного слитого гена (фиг. 5) подтвердили эту последовательность конечного продукта и показали, что не были введены ошибки с использованием этого ПЦР-способа. Кроме того, секвенирование подтвердило также, что ген-мишень был встроен в правильной рамке считывания в векторе рЕТ-32b.
Активность ингибирования белкового синтеза в бесклеточных системах этого рекомбинантного слитого токсина, сравниваемая с активностью свободного рекомбинантного гелонина, предполагает, что не было значимого пространственного препятствия щели активного сайта гелонина из-за близости фрагмента антитела в сконструированной молекуле авторов данного изобретения. Кроме того, поскольку не было сайтов расщепления белка в этой слитой конструкции, эти данные предполагают также, что гелонин не требует обязательного отщепления от этой конструкции для сохранения биологической активности. Это находится в резком противоречии с исследованиями с А-цепью рицина (RTA), которая требует высвобождения из белка-носителя для восстановления биологической активности (Kim et al., 1988; O'Hare et al., 1990). Это является удивительным, так как гелонин и RTA имеют идентичные механизмы действия (Stirpe et al., 1992), а также имеют 30% гомологию последовательности (Rosenblum et al., 1999).
Экспрессия и очистка слитых белков
Плазмидный вектор рЕТ-32b, содержащий этот слитый ген, трансформировали в E. coli AD494(DE3) pLysS и экспрессию белка-мишени индуцировали добавлением IPTG. Как показано окрашенным Кумасси гелем, был индуцирован белок ожидаемой молекулярной массы (68 кДа). Этот белок очищали с использованием смолы IMAC и элюат подвергали действию рекомбинантной энтерокиназы (ЕК) для получения конечной нативной слитой конструкции, мигрирующей в геле в виде одной полосы при 56 кДа. Эту слитую конструкцию испытывали также при помощи Вестерн-блота с использованием как антитела против гелонина, так и антитела против одноцепочечного антитела. Слитая конструкция sfvMEL/rGel, мигрирующая в геле при 56 кДа, реагировала с обоими антителами, демонстрируя тем самым присутствие компонентов иммунореактивного антитела и токсина в этой слитой конструкции. Оцененные приближенно выходы растворимого иммунотоксина sfvMEL/rGel из индуцированных бактериальных культур были приблизительно 700 мкг/л; однако выход конечного, очищенного слитого токсина был приблизительно 200 мкг/л. Было обнаружено, что первичной причиной этого уменьшенного выхода является неспособность смолы IMAC улавливать доступный растворимый белок-мишень. Изменения, произведенные в отношении буферов связывания и условий, а также изменения сортов улавливающей смолы IMAC не улучшили эти результаты.
Связывание иммунотоксинов с использованием ELISA
Для гарантии того, что очищенный слитый белок сохранял антигенсвязывающую способность, связывание этого материала сравнивали со связыванием интактного химического конъюгата IgG ZME-018-гелонин и IgG ZME-018 в конкурентном анализе связывания на основе ELISA (фиг. 6) с использованием интактных антиген-положительных клеток меланомы человека в качестве источника антигена. Было обнаружено, что слитая конструкция scfvMEL/rGel сохраняет связывающие аффинности, сравнимые с химическим конъюгатом. Этот белок показал специфическую и значимую активность связывания в ELISA в отношении клеток-мишеней меланомы А375М с фоновыми уровнями связывания с клетками SK-OV-3 или ME-180.
Активность слитого с sfvMEL токсина в ингибировании бесклеточного белкового синтеза
Биологическая активность токсинов может нарушаться в значительной степени при включении в слитые конструкции. Для испытания н-гликозидной активности rGel-компонента этой слитой конструкции это вещество добавляли в реакционную смесь для трансляции белка in vitro с использованием включения 3Н-лейцина выделенными ретикулоцитами кролика. Кривые ингибирования для слитой конструкции и нативного rGel сравнивали, и было обнаружено, что величины IC50 для этих двух молекул были фактически идентичными (100 пМ против 104 пМ соответственно).
Связывание и интернализация scfvMEL/rGel, измеряемые по иммунофлуоресценции
Иммунофлуоресцентное окрашивание выполняли на клетках А375-М, обработанных scFvMEL/rGel, в разных временных точках после введения. Интернализованную конструкцию определяли с использованием либо кроличьего антитела против rGel, либо кроличьего антитела против антитела scFvMEL с последующим добавлением FITC-связанного антитела против кроличьего IgG. Компонент rGel слитого белка scFvMEL/rGel наблюдали прежде всего в цитозоле после обработки, и количество rGel в цитозоле увеличивалось со временем. Кроме того, scFvMel-компонент scFvMEL/rGel также наблюдали в цитозоле. Это свидетельствует о том, что слитая конструкция способна эффективно связываться с клетками и интернализовать токсин rGel после предоставления клеткам log-фазы.
Цитотоксическая активность иммунотоксинов in vitro
Очищенный слитый белок sfvMEL/rGel и исходную химическую конструкцию ZME/rGel испытывали на специфическую цитотоксичность против антиген-положительной (А375М) и антиген-отрицательной (SK-OV-3) клеточных линий. Как показано на фиг. 7, как полученная химически, так и слитая конструкция демонстрировали величины IC50 приблизительно 10 нМ. В противоположность этому, величины IC50 для токсина rGel были приблизительно в 200 раз более высокими (приблизительно 2000 нМ). Цитотоксические эффекты иммунотоксинов против антиген-отрицательных клеток SK-OV-3 были сходными с цитотоксическим эффектом одного гелонина. Совместное введение свободного антитела ZME с иммунотоксином sfvMEL/rGel (фиг. 8), как и ожидалось, показало умеренное смещение кривой доза-ответ, демонстрируя зависимость от узнавания поверхностного антигена для развития клеточной токсичности этой слитой конструкции.
Противоопухолевая активность sfvMEL/rGel в моделях ксенотрансплантатов
Мышей, несущих хорошо развитые ксенотрансплантаты меланомы А-375, обрабатывали либо солевым раствором (контроли), либо sfvMEL/rGel при 2 или 20 мг/кг в течение 4 дней. Как показано на фиг. 9, размер опухоли в контрольной группе увеличивался с 30 до 150 мм2 (500%-ное увеличение) на протяжении 28 дней этого эксперимента. В противоположность этому, мыши, обработанные слитым токсином при 2 мг/кг, показали небольшое уменьшение размера опухоли с последующим увеличением до приблизительно 60 мм2 (100% увеличение). Мыши, обработанные 20 мг/кг слитого токсина, показали 50% уменьшение размера опухоли во время обработки с последующим медленным восстановлением размера опухоли до исходного размера опухоли на протяжении 28 дней (не было увеличения в общем росте). Не наблюдали видимых токсичных эффектов иммунотоксина на мышах при этих дозах, что предполагает, что при этой схеме введения не достигалась максимальная переносимая доза (MTD).
ПРИМЕР 6
Анализ цитотоксичности in vitro
Способы культивирования клеток
Опухолевые клетки меланомы человека А375М поддерживали в культуре с использованием минимальной поддерживающей среды (МЕМ), дополненной 10% инактивированной нагреванием фетальной телячьей сывороткой и 100 мкМ не-незаменимыми аминокислотами, 2 мМ L-глутамином, 1 мМ пируватом натрия, витаминами и антибиотиками. Культивируемые клетки подвергали скринингу рутинным образом, и было показано, что они не были инфицированы микоплазмой.
Анализ пролиферации клеток
Клеточные линии поддерживали в культуре в полной среде при 37°С в термостате с 5% СО2-увлажненым воздухом. Для анализов с рекомбинантными токсинами и иммунотоксинами культуры промывали, клетки отделяли с использованием версена и ресуспендировали в полной среде при плотности 25·103 клеток на 1 мл. Аликвоты по 200 мкл распределяли в 96-луночные микротитрационные планшеты и затем клеткам давали прикрепляться к лункам. Это приводит к редко посеянной популяции клеток. Спустя 24 часа эту среду заменяли средой, содержащей различные концентрации либо иммунотоксина, либо гелонина. Клетки инкубировали в течение 72 часов и анализировали на относительную пролиферацию клеток с использованием окрашивания кристаллическим фиолетовым.
Окрашивание кристаллическим фиолетовым
Клетки промывали три раза ФБР, содержащим кальций и магний, фиксировали и окрашивали 20% (об./об.) метанолом, содержащим 0,5% (масса/объем) краситель кристаллический фиолетовый. Связавшийся краситель элюировали 150 мкл цитратного буфера Серенсона (0,1 М цитрат натрия, рН 4,2, 50% (об./об.) этанол) в течение 1 часа при комнатной температуре. Оптическую плотность измеряли при 600 нм с использованием микропланшет-ридера Bio-Tek. Относительную пролиферацию (RCP) рассчитывали следующим образом: 
[уравнение 1]
Образцы очищенных scfvMEL-CFR2018, scfvMEL-CFR2025 и CFR2018 анализировали с использованием обычного 72-часового анализа пролиферации с монослоями клеток log-фазы (5000 клеток на лунку) антиген-положительных клеток А375М и с использованием процедур окрашивания красителем кристаллическим фиолетовым, как описано ранее.
Результаты
Фиг. 10 показывает цитотоксичность каждой из рекомбинантных молекул, испытанных на 72-часовом росте линии клеток А-375 меланомы человека, как описано выше. Как можно видеть, слитые конструкции как scfvMEL-CFR2018, так и scfvMEL-CFR2025 ингибировали рост клеток меланомы в культуре. Концентрация каждого агента, требующаяся для ингибирования роста клеток до 50% контрольных величин (IC50) была 100 нМ. В противоположность этому, ингибирование роста клеток свободным токсином (CFR2018) происходило при концентрации более 800 нМ или почти в 8 раз более высокой в сравнении со слитыми с антителами конструкциями. Нацеливание антителами токсина CFR2018 на опухолевые клетки посредством слияния с антителом scfvMEL увеличивает токсичность в 8 раз. Кроме того, в сравнении с токсином CFR2018, сконструированный токсин, названный CFR2025, имеет цитотоксическую активность, сравнимую с цитотоксической активностью токсина CFR2018, когда они оба доставляются в опухолевые клетки антителом-носителем.
* * * * * * * *
Все композиции и способы, описанные и заявленные в данном описании, могут быть получены и проведены без чрезмерного экспериментирования в свете данного описания. Хотя композиции и способы данного изобретения были описаны в виде вариантов осуществления, специалистам с квалификацией в данной области будет очевидно, что к композициям и способам, и в стадиях и очередности стадий этих способов, описанных в данном описании, могут быть применены изменения без отхода от концепции, идеи и объема данного изобретения. Более конкретно, должно быть очевидным, что агенты, описанные в данном описании, могут быть заменены определенными агентами, химически и физиологически родственными им, с получением таких же или сходных результатов. Предполагается, что все такие сходные замены и модификации, очевидные для специалистов с квалификацией в данной области, находятся в рамках идеи, объема и концепции данного изобретения, определяемых прилагаемой формулой изобретения.
ССЫЛКИ
Следующие цитируемые работы, в той степени, в какой они обеспечивают примеры процедур или другие подробности, дополняющие приведенные в данном описании, специально включены в данное описание в качестве ссылок.





















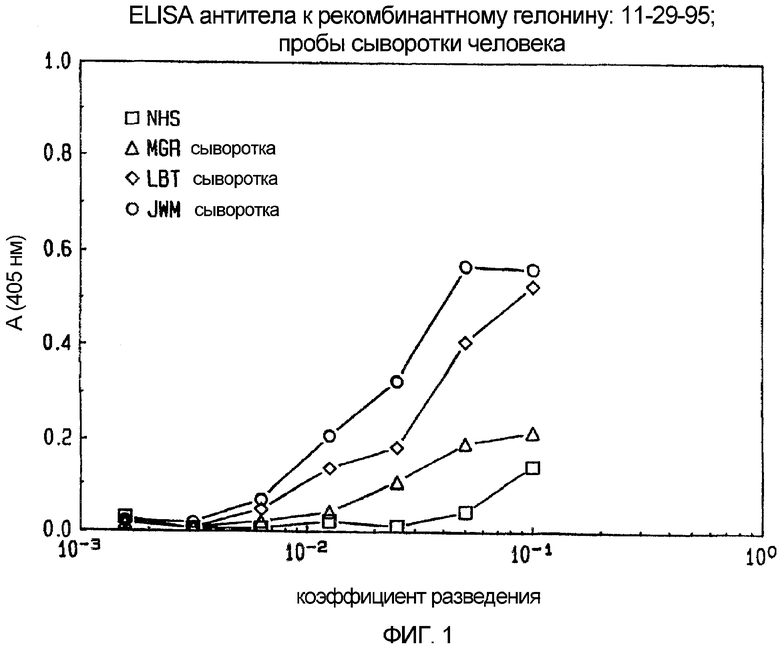
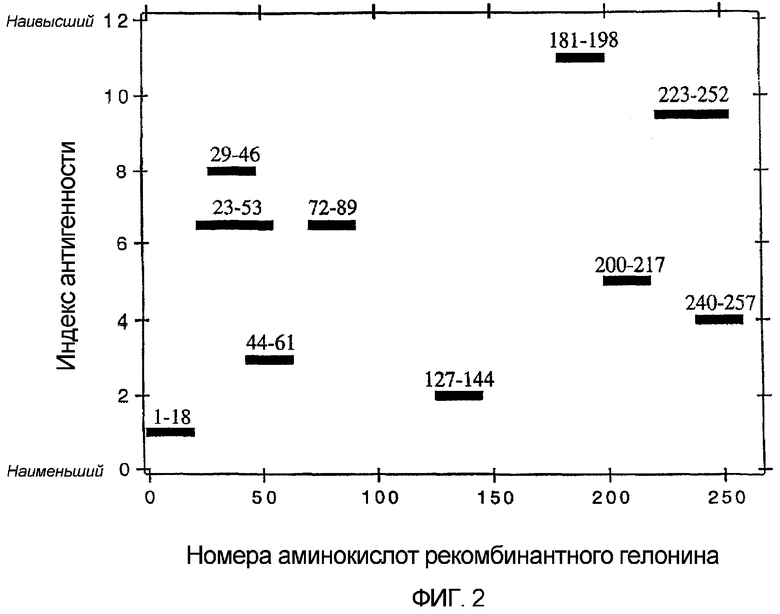
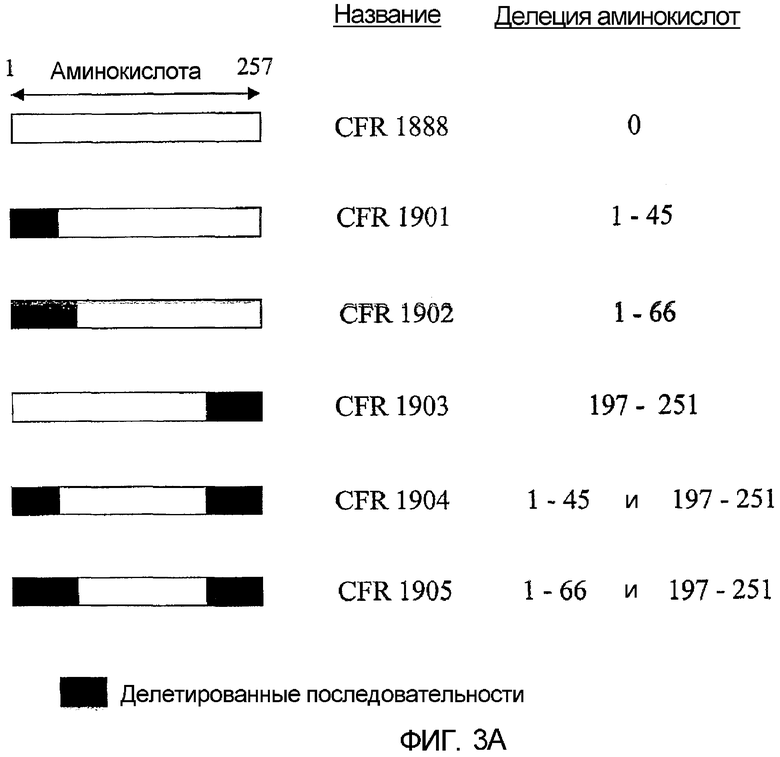

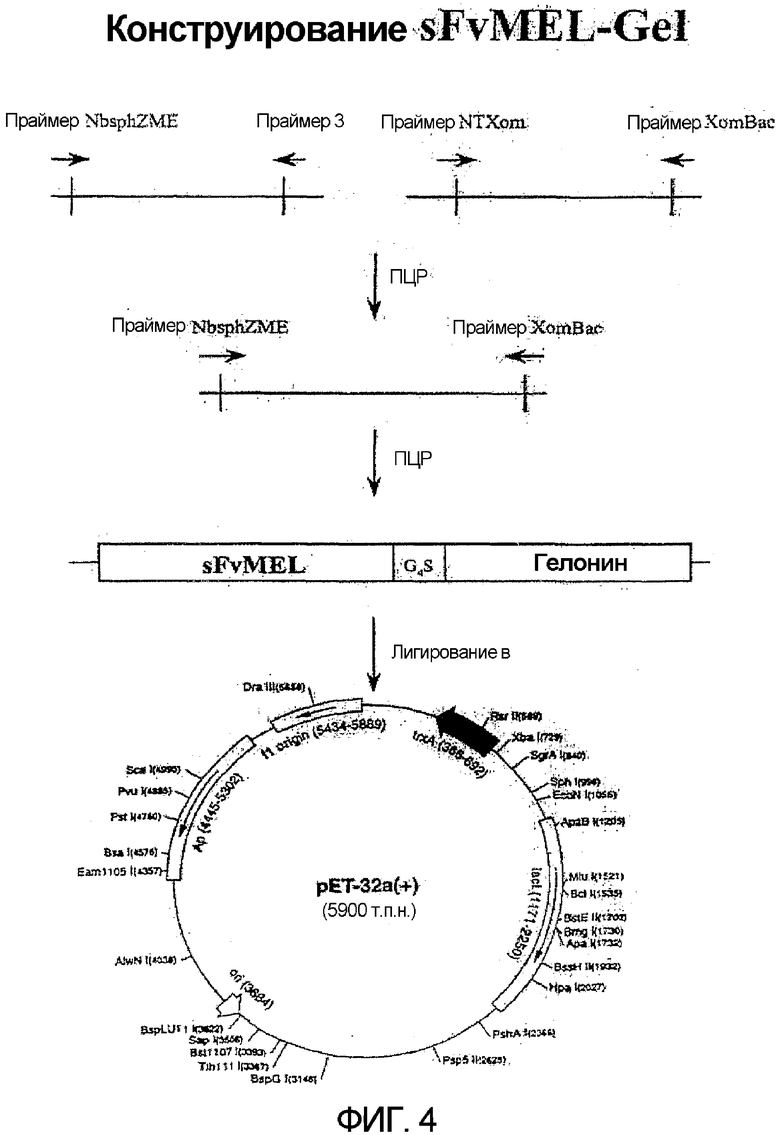

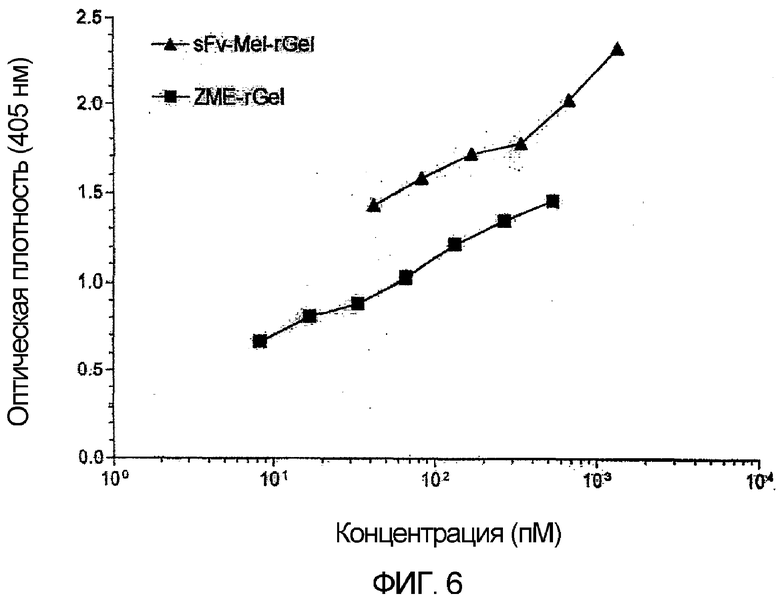
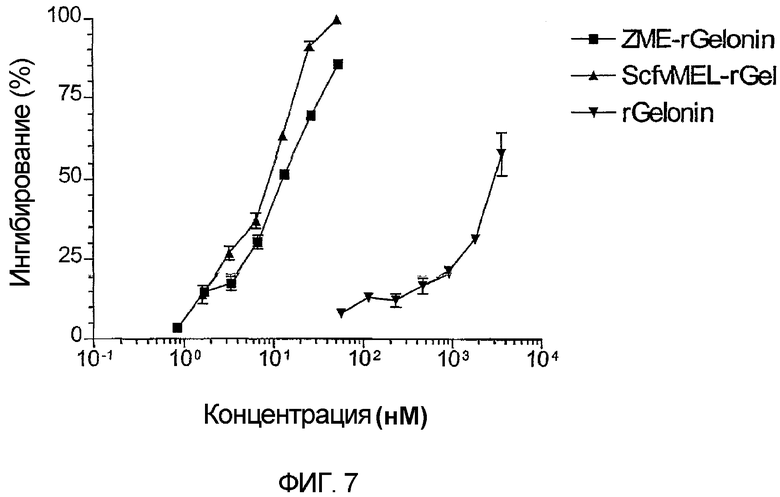
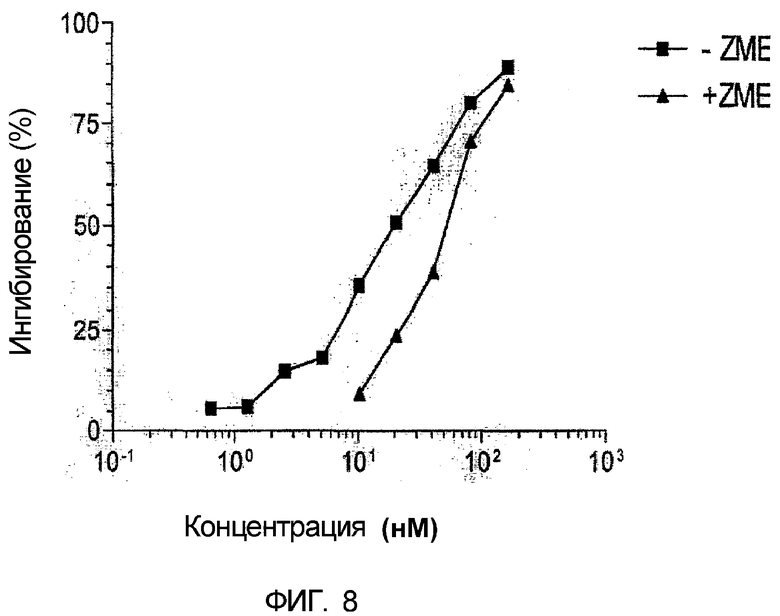
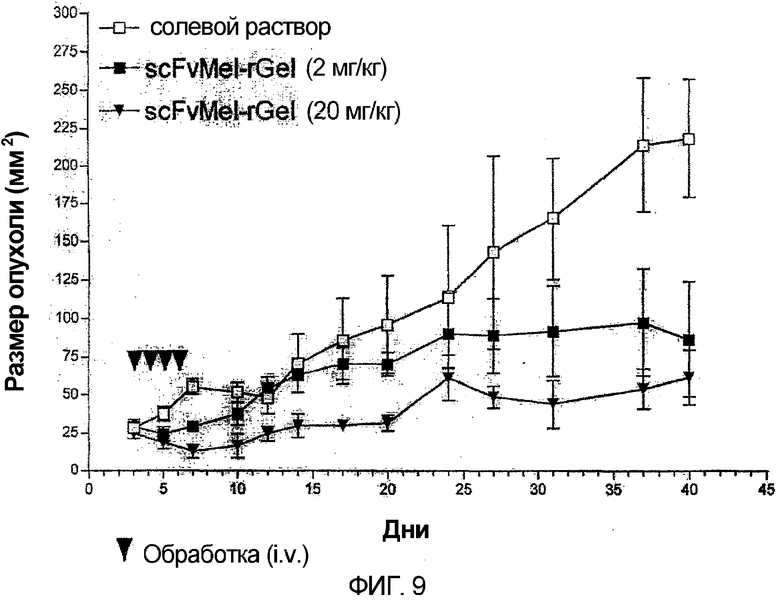
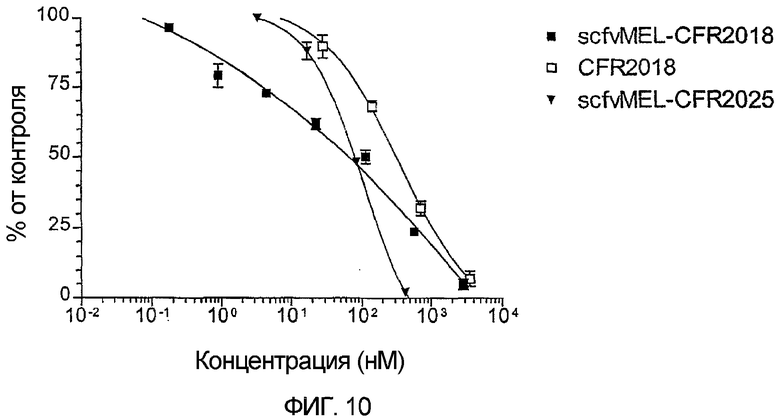
Изобретение относится к модифицированному ферменту гелонину, обладающему пониженной антигенностью, охарактеризованному способом его получения. Сущность способа заключается в идентификации антигенных областей в ферменте с помощью антитела, удалении по меньшей мере одной антигенной области и определении ферментативной активности полученного модифицированного фермента гелонина. Изобретение также касается мультипептидного белкового соединения на основе модифицированного фермента гелонина, который присоединен к второму полипептиду - антителу, токсину или другому ферменту, и обладает пониженной антигенностью. Изобретение раскрывает также гуманизированный рекомбинантный токсин гелонин (ГРТГ) с пониженной антигенностью, в аминокислотной последовательности которого по меньшей мере три аминокислотных остатка из одного или более антигенных доменов 1, 2, 3 или 4 заменены другими аминокислотными остатками. ГРТГ, модифицированный фермент гелонин и мультиполипептидное белковое соединения могут быть применены как лекарственные средства для лечения рака. Изобретение обеспечивает получение модифицированных белков с пониженной антигенностью и высокой цитотоксичностью в отношении опухолевых клеток, в частности клеток миеломы. 5 н. и 22 з.п. ф-лы, 11 ил., 9 табл.
a) идентификацию одной или нескольких антигенных областей в этом ферменте с использованием антитела;
b) удаление одной или нескольких антигенных областей из этого фермента для образования модифицированного фермента и
c) определение ферментативной активности этого модифицированного фермента.
| US 5621083 А, 15.04.1997 | |||
| US 6146850 А, 14.11.2000 | |||
| Глубиннонасосная установка для добычи нефти | 1981 |
|
SU1006727A1 |
| US 5631348 А, 20.05.1997 | |||
| WO 9426910 А, 24.11.1994. | |||

