Изобретение относится к области молекулярной иммунологии, биотехнологии и медицины, касается химерного иммунотоксина на основе неструктурного гликопротеина NSP4 ротавируса человека и может быть использовано для получения антител, блокирующих опухолеассоциированный муцин 1, при разработке мониторинговых и терапевтических подходов к лечению больных злокачественными новообразованиями.
Химерные токсины (XT) - молекулярные конструкции, состоящие из двух генетически слитых субъединиц, предназначенные для направленного воздействия на клетки мишени. Одна субъединица обеспечивает направленную доставку XT к клеткам. Для этого используются моноклональные антитела или их фрагменты, лиганды белков поверхности клетки, антимикробные пептиды или искусственно созданные аффинные молекулы [М. Heiat, Н. Hashemi Yeganeh, S.M.Alavian, E. Rezaie. Immunotoxins. Immunotherapy against hepatocellular carcinoma: a promising prospect / VToxins (Basel). - 2021. Vol. 13. №10. - P. 719]. Другая субъединица представлена токсином, который воздействует на клетку. Известно применение в составе XT белков бактерий [S. Khoshnood, Н. Fathizadeh, F. Neamati, В. Baindara, М.А. Abdullah. Bacteria-derived chimeric toxins as potential anticancer agents//Front.Oncol. - 2022. Vol.12.], растений [R. Mazor, I. Pastan. Immunogenicity of immunotoxins containing Pseudomonas exotoxin A: causes, consequences, and mitigation // Front.Oncol. - 2020. Vol. 11] и насекомых [H. Liu, Y. Han, H. Fu, M. Liu, J. Wu et al. Costruction and expression of s-TRAIL - mellitin combining enhanced anticancer activity with antibacterial activity in Escherichia colillApplied microbiology and biotechnology. - 2013. Vol. 97. - P. 2877-2884; A.B.H. Choo, R.D.Dunn, K.W. Broady, R.L. Raison. Soluble expression of a functional recombinant cytolytic immunotoxins in insect cells//Protein. Expr. Purif. - 2002.Vol. 24. №3. - P. 338-347; M. Moreno, E. Giralt. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: mellitin, apamin and mastoporan // Toxins(Basel) - 2015. Vol. 7. №4. - P. 11-26-1150]. Рассматривается возможность использования токсинов морских полипов [СЕ. Carrison, W. Guan, М. Kato, Th.Tamsett, Т. Patel, Y. Sun, Т.P. Pathak. Structure-activity relationship evaluation of wasp toxin β-PMTX leads to analogs with superior activity for human neuronal sodium channels//ACS medicinal chemistry letters. - publ. 2019.11.011 и моллюсков [M.L. Giglio, S. Ituate, A.E. Ibanoz, M.S. Dreon et al. Novel role for animal innate immune molecules: enterotoxic activity of a snail egg MACPF-toxin // Front. Immunol. - 2020. Vol. 11].
Особый интерес представляет такая область применения XT, как направленная терапия онкологических заболеваний. На культурах раковых клеток и моделях опухолей у животных показана их эффективность в качестве терапевтического средства. В настоящее время разрабатывается множество XT, а препараты, основанные на дифтерийном токсине (Ontak -и Elzonris) и на псевдомонадном экзотоксине (Lumoxiti), прошли клинические испытания и одобрены в США для практического применения: Ontak - для лечения лейкемии и лимфом, включая Т-клеточную лимфому кожи; Elzonris - для лечения бластных плазмоцитоидных дендритоклеточных новообразований; Lumoxiti - для лечения рецидивирующего или рефракторного волосатоклеточного лейкоза [S. Hamamichi, Т. Fukuhara, N. Hattari. Immunotoxin screening system: a rapid and direct approach to obtain functional antibodies with internalization capacities / VToxins. - 2020. Vol. 12. №10. - P. 658.; F. Shafiee, M.G. Aucoin, Ali-Ja. Najafabadi. Targeted diphtheria toxin-based therapy: a review article // Front.Microbiol. - 2019. Vol. 19]. Еще одной областью применения XT является модуляция иммунологических реакций. Так, известно применение XT в трансплантологии для предотвращения реакции организма на трансплантат [С.A. Pennell, М.Е. Panza. CD7-specific single chain Fv mmunotoxins. Design and expressions // Methods Mol. Biol. - 2001. Vol. 166. №17. - P. 29] или удаления Т-клеток из трансплантатов [J.S. Bromberg, E.J. Alfrey, С.F. Barker, K.D. Chavin et al. Adrenal suppression and steroid supplementation in renal transplant recipients // Transplantation-1991.Vol. 51. №2. - P. 385-390], а также для снижения количества Т-регуляторных клеток у больных раком [M.J. Besser, R. Shapiro-Frommer, A.J. Tresei, D. Zippel et al. Minimally cultured or selected autologous tumor-infiltrating lymphocytes after a lymphodeleting chemotherapy regimen in metastatic melanoma patients // J.Immunother. - 2009. Vol. 32. №4. - P. 415 - 423]. Показана возможность применения XT в противовирусной [J.С.Umotoy, S.W. deTaeye. Antibody conjugates for targeted therapy against HIV-1 as an emerging tool for HIV-1 cure // Front.Immunol. - 2021. Vol. 12.] и антипаразитарной терапии [Н. Li, Ch. Gu., Yo. Ren, Ya. Dai, X.Zhu et al. The efficacy of NP11-4-derived immunotoxin scFv-artesunate in reducing hepatic fibrosis induced by Schistosoma japonicum in mice // Boimed. Res. - 2011.Vol. 25. №2. - P. 48-54.; Zh. Tian, Xi-ya Wang, Yun-fei Zhou, Qi-mei Feng et al. Schistosoma japonicum scFv-IL18 fusion DNA ameliorates hepatic fibrosis in schistosomiasis-infected mice via improving local concentration of IL-18 in liver // Exp. Parasitol.-2013. Vol. 34. №4. - P. 447-454.; Sh. Yoshida, D. Ioko, H. Matsuoka, H. Endo, A. Ishii. Bacteria expressing single-chain immunotoxin inhibit malaria parasite development in mosquitoes // Molecular and biochemical parasitology. - 2001. Vol. 1 13. №1. - P.89-96]. Однако разработка и испытание новых XT часто сталкиваются с вызванной токсином проблемой возникновения негативных побочных эффектов [S. Khoshnood, H. Fathizadhe, F. Neamoti et al. Dacteria-derived chimeric toxins as potential anticancer agents // Front.Oncol. - 2022. Vol. 12]. Например, рицин в составе XT может вызывать синдром повышенной проницаемости капилляров (болезнь Кларксона) [L. Polito, A. Djemil, M. Bortolotti Plant toxin-based immunotoxins for cancer therapy: a short overview // Biomedicines. - 2016. Vol. 4. - P.12], a псевдомонадный экзотоксин вызывает развитие гемолитико-уремического синдрома (болезнь Гассера) [R.J. Kreitman, M.D. Wyndham H. Wilson, K. Bergern et al. Efficacy of the anti-CD22 recombinant immunotoxin BL22 in chemotherapy-resistant hairy-cell leukemia // N.Engl.J.Med. - 2001. Vol. 345. №241. - P.247] или плеврита [R. Hassan, S. Bullock, R.J. Kreitman et al. Phase 1 study of SS1P, a recombinant anti-mesothelin immunotoxin given as a bolus I.V. infusion to patients with mesothelin-expressing mesothelioma, ovarian, and pancreatic cancers // Clin.Cancer Res. - 2007. Vol. l3. №7. - P. 5144 - 5149]. Наличие побочных эффектов обосновывает проведение исследований, направленных на поиск альтернативных токсинов.
Известно, что вирусы также кодируют белки, обладающие токсическими свойствами. Наиболее изученными являются токсины вирусов, вызывающих острые кишечные инфекции. Вирусные энтеротоксины, вовлеченные в патогенез гастроэнтерита, проявляют уникальные свойства, используя при этом механизмы, во многом отличающиеся от действия бактериальных токсинов [R.J. Ваша, Sh. Islam, Z. Soddik et al. Comparison of diagnostic tests for detection of bovine rotavirus A in calf feces // MacVetRev. - 2021. Vol. 44. №1. - P. 37-45.]. Действие некоторых вирусных энтеротоксинов не приводит к драматическим повреждениям тканей [M.D. Koci, L.A. Moser, L.A. Kelley, D. Larsen. Astrovirus induces diarrhea in the absence of inflammation and cell death // J.Virol. - 2003. Vol. 77. №21. - P. 11798-11808.; M. Zhang, C.Q.-Zeng, A.P. Morris, M.K. Estes. A functional NSP4 enterotoxin peptide secreted from rotavirus-infected cells / VJ.Virol-2000. Vol. 74. №24. - P.1663-1670.;C.L. Swaggerty, A.A. Frolov, M.J. McArthur, V.W. Cox, S.Tong, R.W. Compans, J.M. Ball. The envelope glycoprotein of simian immunodeficiency virus contains an enterotoxin domain // Virology. - 2000. Vol. 277. №2. - P. 250-261], в связи с чем использование вирусных белков в составе XT потенциально способно снизить негативные эффекты его применения.
В качестве мишени для разработки новых мониторинговых и терапевтических подходов в лечении больных со злокачественными новообразованиями известно применение опухолеассоциированного муцина 1.
Муцин 1 (MUC1) представляет собой гликопротеин, экспрессируется на высоком уровне в ряде эпителиальных злокачественных опухолей человека, в том числе при раке молочной железы, толстой кишки, яичников, а также на злокачественных клетках множественной миеломы. В раковых клетках регистрируются изменение экспрессии гена MUC1, отклонения от нормального гликозилированного белка и изменения внутриклеточной локализации [Joshi S., Kumar S., Choudhury A. et al. Altered mucins trafficking in benign and malignant conditions / VOncotarget. - 2014. Vol. 5. №17. - P. 7272-7284]. Изменение свойств MUC1 приводит к метаболическому перепрограммированию клеток, приобретению новых функций и играет важную роль в развитии опухоли [Nath S., Mukherjee P. MUC1: a multifaceted oncoprotein with a key role in cancer progression // Trends Mol.Med. - 2014. Vol. 20. №6. - P. 332- 342]. Известны способы индукции у больного иммунного ответа против MUC1 с использованием иммуногенного гликопептида в качестве вакцины против рака, а также использование экзогенных антител в терапии и диагностике рака [US 20140005366 А1, 02.01.2014]. Еще известны способы лечения больных немелкоклеточным раком легкого или раком простаты путем введения синтетических пептидов на основе MUC1 в составе липосомальной вакцины [US 20080131495 А1,05.04.2005]. Известно также использование пептидов, соответствующих цитоплазматическому домену, для терапии MUC1 позитивных опухолей. Эти пептиды способны ингибировать олигомеризацию белка MUC1, ингибировать взаимодействие MUC1 с NF-kappaB или STAT, тем самым блокировать воспалительную реакцию [US 20110015138 А1, 20.01.2011]. Известно антитело и его антиген-связывающий фрагмент, специфичность которых в отношении опухолеассоциированного MUC1 выше в 100 раз или более по сравнению с MUC1, экспрессирующимся на поверхности нормальных эпителиальных клеток [US 20120040375 A1, 16.02.2012]. Еще известны моноклональные антитела, направленные против эпитопов, расположенных в регионе тандемных повторов MUC1, используемые в качестве иммунотерапевтических средств для лечения и профилактики рака [US 20130034557 А1,07.02.2011]. Известно также использование антител, которые одновременно связывают место объединения альфа- и бета-субъединиц белка MUC1, в диагностике и иммунотерапии онкозаболеваний [US 20120207772 A1, 16.08.2012].
В качестве аналога выбрано моноклональное антитело ИК025, специфичное к антигену MUC1, характеризующееся высокой константной ассоциацией, определенной на модели клеток культуры рака легкого человека ОАТ75, и высоким числом сайтов связывания на клетку, а также высокой избирательностью внутривенного введения препарата [RU 2198681 C2, 2003]. Однако известное моноклональное антитело не обладает непосредственным цитологическим действием, а большой размер молекулы препятствует распознаванию некоторых скрытых эпитопов антигена MUC1 из-за структурных особенностей белка.
В качестве прототипа рассматриваются одноцепочные антитела (scFv), представляющие собой вариабельные фрагменты тяжелой и легкой цепей антител ИК025, связанные в одну молекулу линкером, подавляющие цитотоксическое действие в отношении линий раковых клеток Со1о205 (рак толстой кишки) и MCF7 (рак молочной железы) [RU 2652901 C1, 2018].
Разработки в данной области направлены на получение молекул, специфически блокирующих опухолеассоциированный MUC1.
Задача заявленного изобретения - химерный иммунотоксин на основе неструктурного гликопротеина NSP4 ротавируса человека, обладающий повышенным биологическим действием за счет инициации задержки клеточного цикла и апоптоза раковых клеток.
Технический результат - медико-биологический белок, проявляющий при использовании повышенное цитотоксическое воздействие на опухолеассоциированный MUC1 раковых клеток.
Технический результат достигается химерным иммунотоксином (scFv-NSP4), в котором структурные варианты рекомбинантных одноцепочных антител, связывающих опухолеассоциированный MUC1, сливаются в одну молекулу с пептидом, состоящим из последовательности SEQ ID No 112-174 аминокислот белка NSP4 ротавируса. Для сохранения функциональной активности связывающих участков антител и пептида ротавируса при объединении в одну молекулу между ними располагают линкер, состоящий из нуклеотидной последовательности, кодирующей аминокислотную последовательность SGGGGSGGGGS.
Изобретение осуществляется тем, что методами молекулярного клонирования проводится изготовление генетической конструкции, кодирующей рекомбинантный химерный иммунотоксин, которая в составе плазмидной ДНК содержит последовательности, кодирующие рекомбинантное одноцепочное антитело, объединенное в одну молекулу с пептидом, состоящим из последовательности SEQ ID No 112-174 аминокислот неструктурного гликопотеина NSP4 ротавируса (SEQ ID NO: 1), и находящейся под контролем промотора и терминатора бактериофага Т7 с последующим получением штамма бактерий Escherichia coli Rosetta2 (DE3) -продуцента рекомбинантного химерного иммунотоксина, связывающего опухолеассоциированный MUC1. Полученный штамм культивируют и индуцируют синтез химерного иммунотоксина. Клетки собирают центрифугированием, лизируют и очищают химерный иммунотоксин от примесей. Взаимодействие химерного иммунотоксина с опухолеассоциированным MUC1, изменения в клеточном цикле и апоптоз раковых клеток определяют методом проточной цитофлуореметрии. Цитотоксичную концентрацию химерного иммунотоксина определяют с использованием МТТ теста.
Изобретение поясняется графическими материалами, представленными на фиг.1-5.
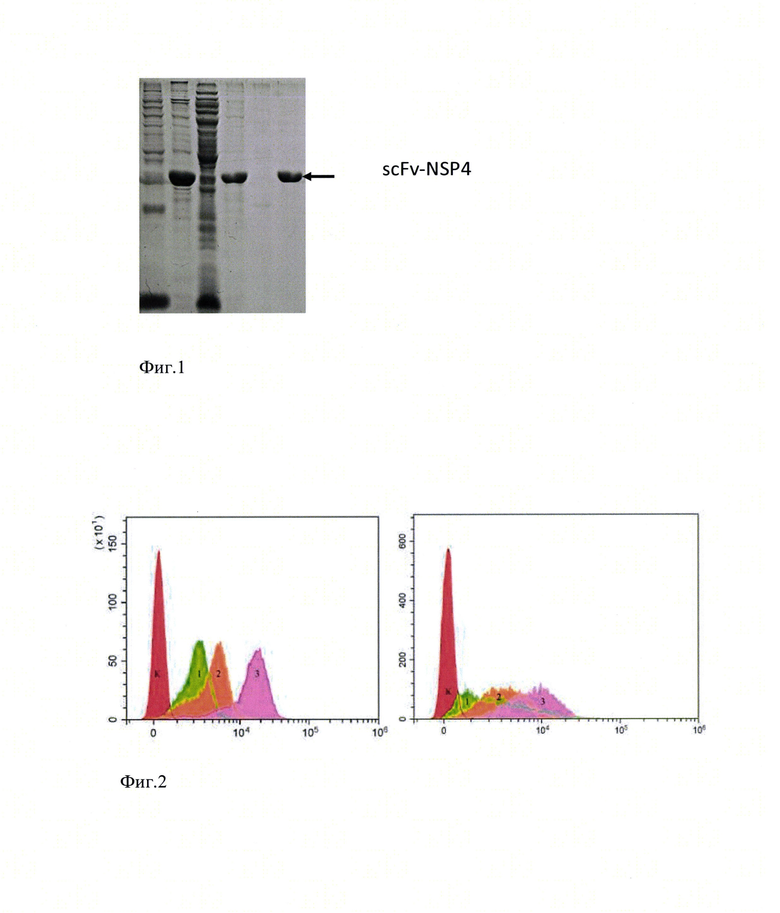
На фиг.1 представлена оценка степени очистки рекомбинантного химерного иммунотоксина методом электрофореза в ПААГ.
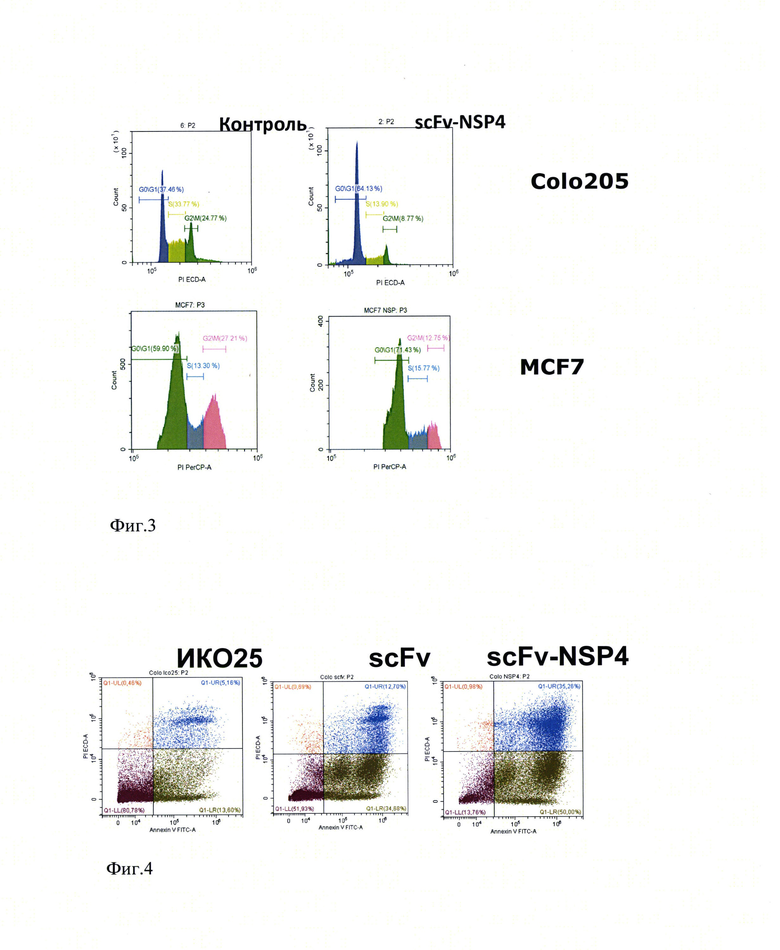
На фиг.2 представлены уровни флуоресценции клеток Colo 205 (А) и MCF-7 (Б), окрашенных с использованием антител ИК025 (1), scFv-NSP4 (2) и scFv (3), меченных Су5. Ось ординат соответствует количеству клеток. Ось абсцисс отражает уровень флуоресценции при длине волны 650 нм.
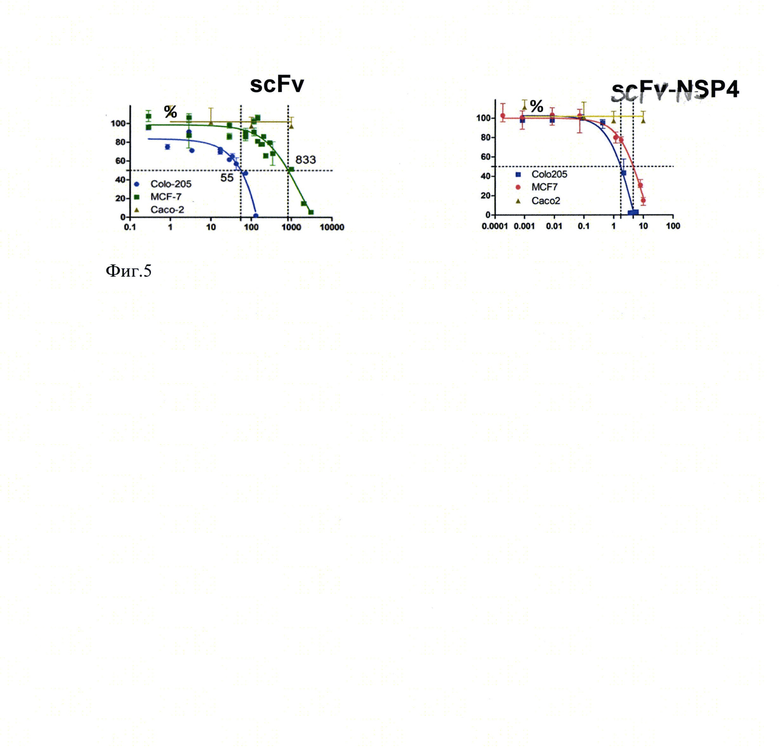
На фиг.3 представлены гистограммы количества ДНК в клетках Colo 205 (А) и MCF-7 (Б), окрашенных с использованием йодида пропидия. Ось ординат соответствует количеству клеток. Ось абсцисс отражает уровень флуоресценции при длине волны 610 нм.
На фиг.4 представлено распределение клеток Colo-205 по уровню окрашивания йодидом пропидия и аннексином V. Ось ординат отражает уровень флуоресценции при длине волны 610 нм. Ось абсцисс отражает уровень флуоресценции при длине волны 488 нм.
На фиг.5 представлена метаболическая активность клеток Colo-205, Сасо-2 и MCF-7 в присутствии различных концентраций рекомбинантных одноцепочных антител (scFv) и рекомбинантного химерного иммунотоксина (scFv-NSP4).
Сущность изобретения поясняется примерами.
Пример 1. Конструирование химерного иммунотоксина
Синтез комплементарной ДНК, кодирующей 112-174 аминокислот неструктурного гликоппротеина NSP4 ротавируса человека, проводили на матрице мРНК, выделенной из образца стула больного ротавирусным гастроэнтеритом методом обратной транскрипции - полимеразной цепной реакции с использованием олигонуклеотида 5'-CGAGTGCGGCCGCTCAATGATGATGGTGATGATGCATGGATGCAGT-3' и олигонуклеотида 5'-GAGCTCTGGCGGCGGCGGCTCCGGTGGTGGTGGTTCTATTGAT AAACTAACTACTCG-3'. Проводили 40 циклов полимеразной цепной реакции при следующих температурных условиях: 94°С - 30'', 50°С - 30'', 72°С - Г. Полученный фрагмент кДНК очищали с использованием агарозного геля и набора N-Gel (Биолабмих, РФ). Полученный фрагмент кДНК NSP4 ротавируса челоека и плазмиду pET22b+LH обрабатывали эндонуклеазами рестрикции Sac I и Not I, лигировали между собой и методами молекулярного клонирования отбирали плазмиду, содержащую генетическую конструкцию, кодирующую химерный иммунотоксин, состоящий из нуклеотидной последовательности одноцепочного антитела для связывания опухолеассоциированного MUC1, объединенного в одну молекулу с пептидом, состоящим из последовательности SEQ ID No 112-174 аминокислот неструктурного гликоппротеина NSP4 ротавируса человека, и находящуюся под контролем промотора и терминатора бактериофага Т7. Аминокислотную последовательность региона, кодирующего химерный иммунотоксин (SEQ ID NO: 1), предсказывали с использованием компьютерной программы «EditSEQ» производства Lasergen, США.
Пример 2. Экспрессия и очистка рекомбинантного одноцепочного антитела (SEQ ID NO: 1)
Для экспрессии рекомбинантного одноцепочного антитела (SEQ ID NO: 1) использовали штамм Escherichia coli Rosetta2 (DE3) (Novagen, США). Клетки Escherichia coli трансформировали плазмидной ДНК, в структуру которой была встроена генетическая конструкция, кодирующая аминокислотную последовательность SEQ ID NO 1: MEIVLTQSPASLAVSLGQRATISYRASKSVSTSGYSYMHWNQQKPGQPPRLLIYLVSNLESGVPARFSGSGSGTDFTLNIHPVEEEDAATYYCQHIRELTRSEGGPSWKSNSGGGGSGGGGSGGGGSEVQLVESGGGLVKPGGSLKLSCAASGFTFSSYAMSWVRQTPEKRLEWVATISSGGSYTYYPDSVKGRFTISRDNAKNTEYLQMSNLRSEDTAIYYCARQISLWYAMAYWGQGTSVTVSSGGGGSGGGGSIDKLTTREIEQVELLKRIHDNLITRPVDVIDMSKEFNQKNIKTLDEWESGKNPYEPSEVTASMHHHHHH с последующей очисткой препаратом плазмиды, высевали на твердую питательную среду 2YT (1.6% триптона, 1% дрожжевого экстракта, 0,5% NaCl, 1,5% агар-агара, ампицилина - 100 мкг/мл) и инкубировали в течение 18 часов при температуре 37°С. Клетки выращивали в жидкой среде LB (1% триптона, 0,5% дрожжевого экстракта, 1% NaCl, хлорамфеникола - 100 мкг/мл) при температуре 37°С и перемешивании до OD600=0,5 - 0,7. Индукцию синтеза рекомбинантного иммунотоксина проводили путем добавления изопропил-P-D-1-тиогалактопиранозида до конечной концентрации 0,1, 0,25 или 0,5 мМ. Биомассу инкубировали в течение 6 часов при температуре 37°С при перемешивании 150 об/мин. Клетки осаждали центрифугированием и лизировали. Рекомбинантный химерный иммунотоксин очищали от примесей с использованием сефарозы Ni-NTA Superflow (GE Healthcare). Проводили диализ против буфера, содержащего 100 мМ натрий карбоната рН 8.5, 10% глюкозы, 150 мМ NaCl, и оценивали степень очистки методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия.
Пример 3. Оценка взаимодействия химерного иммунотоксина с опухолеассоциированым MUC1
Взаимодействие химерного иммунотоксина с опухолеассоциированным MUC1 оценивали методом иммунофлуоресценции с использованием линий клеток, имеющих происхождение от опухолей больных раком молочной железы (MCF7) и толстой кишки (Colo 205). В качестве отрицательного контроля использовали раковые клетки Сасо 2, в которых экспрессия MUC1 подавлена. Клетки Colo205, MCF7 и Сасо 2 культивировали до образования монослоя с использованием среды DMEM (TermoFisher, США), содержащей 10% эмбриональной телячьей сыворотки, по 50 мкг пенициллина и стрептомицина, 5 мМ L-глютамина в термостате при 37°С и содержании СО2 5%. Очищенные рекомбинантный иммунотоксин, моноклональное антитело ИК025 и одноцепочечное антитело scFv метили активированным эфиром флуоресцентного красителя Су5 (Lumiprobe, РФ) согласно рекомендациям производителя и наносили на монослой раковых клеток. Инкубировали с меченными белками в течение 1 часа и отмывали несвязавшийся компонент с использованием фосфатно-солевого буфера. Клетки суспендировали с использованием раствора этилендиаминтетрауксусной кислоты. Методом проточной цитометрии регистрировали окрашивание клеток, что свидетельствовало о связывании рекомбинантного химерного иммунотоксина с MUC1, представленным на поверхности клеток.
Пример 4. Оценка влияния химерного иммунотоксина на пролиферацию
Для оценки влияния химерного иммунотоксина на пролиферацию получали монослой клеток линий Со1о205 или MCF7, как описано в примере 3. Клетки инкубировали в течение 24 часов в присутствии рекомбинантного химерного иммунотоксина. Клетки суспендировали с использованием раствора этилендиаминтетрауксусной кислоты, фиксировали этанолом и окрашивали йодидом пропидия. Методом проточной цитометрии получали гистограммы распределения клеток по фазам клеточного цикла. Между контрольными клетками и клетками, культивируемыми в присутствии рекомбинантного химерного иммунотоксина, сравнивали процентное содержание клеток, находящихся в G0\G1, S и G2\M фазах клеточного цикла. Под воздействием иммунотоксина регистрировали увеличение процентного содержания клеток, находящихся в G0\G1 фазе, и снижение процентного содержания клеток, находящихся в G2\M фазе.
Пример 5. Оценка влияния химерного иммунотоксина на апоптоз раковых клеток
Для оценки влияния химерного иммунотоксина на апоптоз раковых клеток получали монослой клеток линий Со1о205, как описано в примере 3. Клетки инкубировали в течение 24 часов в присутствии рекомбинантного химерного иммунотоксина, моноклонального антитела ИК025 и одноцепочечного антитела scFv. Клетки суспендировали с использованием раствора этилендиаминтетрауксусной кислоты и окрашивали с использованием йодида пропидия и Annexin V. Методом проточной цитофлуореметрии определяли количество клеток, имеющих наиболее высокий уровень флуоресценции, что свидетельствовало об апоптозе. В клетках, инкубированных в присутствии рекомбинантного химерного иммунотоксина, регистрировали наибольшее процентное содержание апоптотических клеток.
Пример 6. Определение цитотоксической концентрации химерного иммунотоксина
Цитотоксическую концентрацию химерного иммунотоксина определяли с использованием МТТ теста. Клетки инкубировали в течение 48 часов в питательной среде DMEM (TermoFisher, США), содержащей 10% эмбриональной телячьей сыворотки, по 50 мкг пенициллина и стрептомицина, 5 мМ L-глютамина, при температуре 37°С во влажной атмосфере, содержащей 5% СО2, в присутствии различных концентраций рекомбинантных одноцепочных антител и рекомбинантного химерного иммунотоксина. Добавляли МТТ (3-[4,5-диметилтиазолил-2-ел]-2,5-дифенилтетразолиум бромида) и инкубировали в течение 4 часов при температуре 37°С. Добавляли диметилсульфоксид и измеряли оптическую плотность фотометрически при длине волны 540 нм. На основе полученных результатов строили графики, отражающие метаболическую активность клеток. При сравнении полученных результатов установлено, что рекомбинантный химерный иммунотоксин проявлял цитотоксическую активность в меньших концентрациях.
Проведенные исследования подтвердили получение химерного иммунотоксина, содержащего неструктурный гликопротеин NSP4 ротавируса человека, который инициирует задержку клеточного цикла и апоптоза раковых клеток, что обеспечивает цитотоксическую активность в меньших концентрациях.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="НСП-4.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2024-02-27">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1</ApplicationNumberText>
<FilingDate>2024-02-27</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>1</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное бюджетное учреждение
науки «Нижегородский научно-исследовательский институт эпидемиологии
и микробиологии им. академика И.Н. Блохиной» Федеральной службы по
надзору в сфере защиты прав потребителей и благополучия человека
(ФБУН ННИИЭМ им. академика И.Н. Блохиной
Роспотребнадзора)</ApplicantName>
<ApplicantNameLatin>Tsyganova Maria</ApplicantNameLatin>
<InventorName languageCode="ru">Новиков Дмитрий
Викторович</InventorName>
<InventorNameLatin>Novikov Dmitriy</InventorNameLatin>
<InventionTitle languageCode="ru">ХИМЕРНЫЙ ИММУНОТОКСИН НА ОСНОВЕ
НЕСТРУКТУРНОГО ГЛИКОПРОТЕИНА NSP4 РОТАВИРУСА
ЧЕЛОВЕКА</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>325</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..325</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Rotavirus A</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MEIVLTQSPASLAVSLGQRATISYRASKSVSTSGYSYMHWNQQKPGQPP
RLLIYLVSNLESGVPARFSGSGSGTDFTLNIHPVEEEDAATYYCQHIRELTRSEGGPSWKSNSGGGGSGG
GGSGGGGSEVQLVESGGGLVKPGGSLKLSCAASGFTFSSYAMSWVRQTPEKRLEWVATISSGGSYTYYPD
SVKGRFTISRDNAKNTLYLQMSNLRSEDTAIYYCARQISLWYAMAYWGQGTSVTVSSGGGGSGGGGSIDK
LTTREIEQVELLKRIHDNLITRPVDVIDMSKEFNQKNIKTLDEWESGKNPYEPSEVTASMHHHHHH</IN
SDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Антитела, связывающие опухолеассоциированный MUC1, и способы их получения | 2016 |
|
RU2652901C1 |
| АНТИТЕЛО ПРОТИВ CSF-1R | 2009 |
|
RU2547586C2 |
| АНТИТЕЛО ПРОТИВ КСФ-1R | 2012 |
|
RU2621859C2 |
| СПЕЦИФИЧЕСКИЕ К ОПУХОЛИ АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2595403C2 |
| РЕКОМБИНАНТНЫЙ ИММУНОТОКСИН, НАЦЕЛЕННЫЙ НА МЕЗОТЕЛИН | 2012 |
|
RU2600067C2 |
| КОНСТРУИРОВАНИЕ ОБЛИГАТНОГО ВЕКТОРА НА ОСНОВЕ ОНКОЛИТИЧЕСКИХ ВИРУСОВ ПРОСТОГО ГЕРПЕСА (oHSV) И КОНСТРУКЦИИ ДЛЯ ТЕРАПИИ РАКА | 2016 |
|
RU2831084C2 |
| ЭКЗОТОКСИН А PSEUDOMONAS С МЕНЕЕ ИММУНОГЕННЫМИ Т-И/ИЛИ В-КЛЕТОЧНЫМИ ЭПИТОПАМИ | 2012 |
|
RU2627216C2 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА К CD3 И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2831840C2 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВО-ЭМБРИОНАЛЬНОМУ АНТИГЕНУ | 2015 |
|
RU2650858C2 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВО-ЭМБРИОНАЛЬНОМУ АНТИГЕНУ | 2015 |
|
RU2652955C1 |
Изобретение относится к области биотехнологии, молекулярной иммунологии и медицины. Предложен химерный иммунотоксин на основе неструктурного гликопротеина NSP4 ротавируса человека, проявляющий повышенное цитотоксическое воздействие на опухолеассоциированный MUC1 раковых клеток. Иммунотоксин содержит одноцепочное антитело, связывающее опухолеассоциированный MUC1, и пептид, состоящий из аминокислот 112-174 неструктурного гликопротеина NSP4 ротавируса человека, соединенные пептидным линкером. Химерный иммунотоксин инициирует задержку клеточного цикла и апоптоз раковых клеток, что обеспечивает цитотоксическую активность в меньших концентрациях. Изобретение может найти дальнейшее применение в разработке мониторинговых и терапевтических подходов в лечении больных злокачественными новообразованиями. 5 ил., 6 пр.
Химерный иммунотоксин на основе неструктурного гликопротеина NSP4 ротавируса человека, проявляющий повышенное цитотоксическое воздействие на опухолеассоциированный MUC1 раковых клеток, отличающийся тем, что содержит аминокислотную последовательность, представленную в SEQ ID NО: 1.
| ВАСИЛЬЧИКОВ П.И | |||
| и др | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Устройство для отыскания металлических предметов | 1920 |
|
SU165A1 |
| Антитела, связывающие опухолеассоциированный MUC1, и способы их получения | 2016 |
|
RU2652901C1 |
| МЕТОД УНИЧТОЖЕНИЯ НЕЖЕЛАТЕЛЬНЫХ КЛЕТОК-МИШЕНЕЙ | 1997 |
|
RU2182493C2 |
| US 8648172 B2, 11.02.2014. | |||

