Изобретение относится к области биологии и медицины.
Респираторно-синцитиальный вирус (РСВ) является широко распространенным вирусом простудных заболеваний, принадлежащим к семейству парамиксовирусов. РСВ является вирулентным, легко передающимся вирусом и наиболее распространенной причиной заболеваний нижних дыхательных путей у детей в возрасте до 2 лет. Обычно, до 98% детей, получающих дневную медицинскую помощь, инфицированы РСВ в течение одного сезона заболевания. От 0,5% до 3,2% детей с инфекцией РСВ требуют госпитализации. В Соединенных Штатах в год регистрируется приблизительно 90000 случаев госпитализации и 4500 случаев смерти. Основными факторами риска для госпитализации в связи с РСВ являются преждевременные роды, хронические заболевания легких, врожденные пороки сердца, ослабленный иммунитет и возраст менее 6 недель у здоровых в остальном детей. Кроме поддерживающей терапии в виде адекватного питания и кислородной терапии не существует эффективного лечения РСВ положительного бронхиолита (капиллярного бронхита). Противовирусная терапия, такая как терапия Рибавирином, не доказала свою эффективность в отношение РСВ инфекции. Для профилактики РСВ инфекции зарегистрированы в качестве лекарственного средства только моноклональные антитела Паливизумаб (также называемые Синагис (Synagis)). Паливизумаб представляет собой генно-инженерное (гуманизированное) моноклональное антитело к гибридным белкам РСВ. Однако Паливизумаб не всегда является эффективным. Следовательно, существует потребность в альтернативных способах лечения и антителах против РСВ.
Предметом настоящего изобретения является создание средств и методов противодействия и/или предотвращения связанных с РСВ заболеваний. Еще одним предметом изобретения является получение альтернативных и/или улучшенных антител против РСВ, или функциональных эквивалентов таких антител, а также обеспечение стабильных клеток, способных производить антитела или их функциональные эквиваленты против РСВ.
Настоящее изобретение обеспечивает антитела и их функциональные эквиваленты, которые способны специфически связывать РСВ. Такие антитела и/или функциональные эквиваленты, также называемые в настоящем документе "анти-РСВ антитела" или "РСВ-специфические антитела", способны специфически связывать по меньшей мере один из компонентов РСВ, такой как, например, эпитоп белка РСВ. Неспецифическое связывание не подпадает под определение "специфическое связывание". Анти-РСВ антитела и функциональные эквиваленты по настоящему изобретению особенно подходят для противодействия и/или по меньшей мере частичного предотвращения РСВ-инфекции и/или неблагоприятных последствий РСВ инфекции. Одним из наиболее предпочтительных анти-РСВ антител по настоящему изобретению является антитело, обозначенное "D25", обладающее областью тяжелой цепи и областью легкой цепи, как показано на Фигуре 11А-Г. CDR последовательность D25, которая, в частности, способствует антигенсвязывающим свойствам D25, показана на Фигуре 11Г. Антитело D25 обладает гораздо лучшими по сравнению с зарегистрированным анти-РСВ антителом Паливизумаб характеристиками (Фиг. 8). Например, D25 обладает значением IC50 около 0,4-1,5 нг/мл при in vitro анализе на обезвреживание, в котором клетки НЕр-2 инфицировали РСВ, а Паливизумаб обладает значением IC50 около 453 нг/мл.
Функциональный эквивалент антитела в настоящем документе определен как функциональная часть, производное или аналог антитела.
Функциональная часть антитела определяется как его часть, которая обладает, качественно, не обязательно количественно по меньшей мере одним одинаковым свойством указанного антитела. Указанная функциональная часть способна связывать тот же антиген, что и указанное антитело, хотя и не обязательно в той же степени. Функциональная часть антитела предпочтительно представляет собой однодоменное антитело, одноцепочечное антитело, одноцепочечный вариабельный фрагмент (ScFv), фрагмент Fab или F(ab')2 фрагмент.
Функциональное производное антитела определяется как антитело, которое было изменено таким образом, что по меньшей мере одно свойство, предпочтительно, способность связывать антиген, у полученного соединения в основном остается качественно, не обязательно количественно, неизменным. Производное антитела может быть получено различными способами, например, посредством консервативных аминокислотных замен, где аминокислотный остаток заменен другим остатком в целом с аналогичными свойствами (размером, гидрофобностью, и т.д.), так что функционирование, в целом, вероятно, не будет серьезно затронуто.
Специалист в данной области техники вполне способен получить аналоги антитела. Это, например, может быть осуществлено посредством скрининга пептидной библиотеки или фаговой дисплейной библиотеки. Такие аналоги обладают по меньшей мере качественно, не обязательно количественно, одним одинаковым свойством с указанным антителом.
Как хорошо известно специалисту в данной области техники, тяжелая цепь антитела является большей из двух типов цепей, составляющих молекулу иммуноглобулина. Тяжелая цепь включает константный домен и вариабельный домен, где вариабельный домен участвует в связывания антигена. Легкая цепь антитела является меньшей из двух типов цепей, составляющих молекулу иммуноглобулина. Легкая цепь включает константный домен и вариабельный домен. Вариабельный домен легкой цепи вместе с вариабельным доменом тяжелой цепи участвует в связывании антигена.
Определяющие комплементарность районы (CDR) являются гипервариабельными районами, присутствующими в вариабельных доменах тяжелых цепей и вариабельных доменах легких цепей. CDR тяжелой цепи и связанной с ней легкой цепи антитела вместе образуют антигенсвязывающей сайт.
CDR последовательности, приведенные на Фигуре 11, обеспечивают желаемые характеристики связывания РСВ, и специалист в данной области техники способен создать варианты, включающие по меньшей мере одну измененную последовательность CDR, например, с применением консервативной аминокислотной замены. Консервативная аминокислотная замена предполагает замещение одной аминокислоты другой в целом с аналогичными свойствами (размером, гидрофобностью, и т.д.), так что общее функционирование, вероятно, не будет серьезно затронуто.
Также возможно изменить по меньшей мере одну CDR последовательность, приведенную на Фигуре 11, с целью получения варианта антитела, или его функционального эквивалента по меньшей мере с одним измененным свойством по сравнению с D25. Предпочтительно, в настоящем документе предусмотрены антитело или его функциональный эквивалент, включающие последовательность CDR, обладающую по меньшей мере 70% идентичностью последовательности CDR, приведенной на Фигуре 11, так что предпочтительные характеристики связывания D25 по меньшей мере частично сохранены и даже улучшены. CDR последовательность, приведенная на Фигуре 11, предпочтительно изменена таким образом, что полученное антитело или его функциональный эквивалент обладает по меньшей мере одним улучшенным свойством, таким как, например, улучшенная аффинность связывания, избирательность и/или стабильность по сравнению с D25. Варианты антител или их функциональные эквиваленты, включающие аминокислотную последовательность, обладающую по меньшей мере 70% идентичностью последовательности CDR, приведенной на Фигуре 11, находятся в объеме настоящего изобретения. Для изменения аминокислотной последовательности в уровне техники доступны различные методы. Например, последовательности тяжелой цепи или легкой цепи с желаемой последовательностью CDR искусственно синтезированы. Предпочтительно, последовательность нуклеиновой кислоты, кодирующую последовательность CDR, мутировали, например, с использованием случайного или сайт-направленного мутагенеза.
Таким образом, в первом аспекте изобретение обеспечивает выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, способное специфически связывать респираторно-синцитиальный вирус, которое включает:
- последовательность CDR1 тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности NYIIN, и/или
- последовательность CDR2 тяжелой цепи, включающую последовательность, которая по меньшей мере на 75% идентична последовательности GIIPVLGTVHYAPKFQG, и/или
- последовательность CDR3 тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ETALVVSTTYLPHYFDN, и/или
- последовательность CDR1 легкой цепи, включающую последовательность, которая по меньшей мере на 85% идентична последовательности QASQDIVNYLN, и/или
- последовательность CDR2 легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности VASNLET.
Предпочтительно, указанное антитело также включает последовательность CDR3 легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QQYDNLP.
Предпочтительно, антитело или функциональный эквивалент по настоящему изобретению включают последовательность CDR, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% идентична по меньшей мере одной из последовательностей CDR, приведенных на Фигуре 11Г. Наиболее предпочтительно, антитело или функциональный эквивалент по настоящему изобретению включает CDR последовательность, которая по меньшей мере на 95% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 11Г.
Особенно предпочтительно антитело D25, описанное выше, включающее CDR последовательности, которые представляют собой CDR последовательности, приведенные па Фигуре 11Г. Таким образом, особенно предпочтителен вариант по настоящему изобретению, который включает выделенные, искусственные или рекомбинантные антитела или их функциональные эквиваленты, способные специфически связывать респираторно-синцитиальный вирус, которые включают:
- последовательность CDR1 тяжелой цепи, включающую последовательность NYIIN, и/или
- последовательность CDR2 тяжелой цепи, включающую последовательность GIIPVLGTVHYAPKFQG, и/или
- последовательность CDR3 тяжелой цепи, включающую последовательность ETALVVSTTYLPHYFDN, и/или
- последовательность CDR1 легкой цепи, включающую последовательность QASQDIVNYLN, и/или
- последовательность CDR2 легкой цепи, включающую последовательность VASNLET.
Предпочтительно, указанные антитела включают также CDR3 последовательность легкой цепи, включающую последовательность QQYDNLP.
В одном варианте предусмотрены антитело или его функциональный эквивалент, включающие три CDR последовательности тяжелой цепи и три CDR последовательности легкой цепи, приведенные на фигуре 11Г, или последовательности по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85% идентичные приведенным последовательностям. Кроме того, предусмотрены выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, включающие CDR1 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности NYIIN, и CDR2 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GIIPVLGTVHYAPKFQG, и CDR3 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ETALVVSTTYLPHYFDN, и CDR1 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QASQDIVNYLN, и CDR2 последовательности легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности VASNLET, и CDR3 последовательность легкой цепи, включающей последовательность, которая, по меньшей мере на 70% идентична последовательности QQYDNLP. Указанные антитела или их функциональные эквиваленты предпочтительно содержат CDR последовательности, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны CDR последовательностям тяжелой цепи и CDR последовательностям легкой цепи, приведенным на фигуре 11Г. Также предусмотрены антитело или его функциональный эквивалент, включающие вышеупомянутые CDR1, CDR2 и CDR3 последовательности тяжелой цепи, а также вышеупомянутые CDR1, CDR2 и CDR3 последовательности легкой цепи.
Также предусмотрены антитела или их функциональные эквиваленты, включающие вариабельную аминокислотную последовательность тяжелой цепи, которая по меньшей мере на 70% идентична последовательности тяжелой цепи, приведенной на Фигуре 11. Такие последовательности тяжелой цепи обеспечивают желаемые свойства связывания РСВ, о чем свидетельствует антитело D25. Кроме того, предусмотрены антитело или его функциональный эквивалент, обладающие последовательностью тяжелой цепи, включающей последовательность, которая по меньшей мере на 70% идентична последовательности

 Кроме того, аминокислотная вариабельная последовательность легкой цепи, которая по меньшей мере на 70% идентична последовательности легкой цепи, приведенной на Фигуре 11, обеспечивает желаемые свойства связывания РСВ, о чем свидетельствуют антитело D25. Поэтому также предусмотрены антитело или его функциональный эквивалент с последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности
Кроме того, аминокислотная вариабельная последовательность легкой цепи, которая по меньшей мере на 70% идентична последовательности легкой цепи, приведенной на Фигуре 11, обеспечивает желаемые свойства связывания РСВ, о чем свидетельствуют антитело D25. Поэтому также предусмотрены антитело или его функциональный эквивалент с последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности
 . Антитело или его функциональная часть по настоящему изобретению предпочтительно включают вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 11. Чем выше гомология, тем ближе указанное антитело или его функциональная часть к антителу D25. Антитело или его функциональная часть по настоящему изобретению предпочтительно состоит из тяжелой цепи, а также легкой цепи, которые похожи на тяжелую и легкую цепи D25. Поэтому, кроме того, предусмотрены антитело или его функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 11.
. Антитело или его функциональная часть по настоящему изобретению предпочтительно включают вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 11. Чем выше гомология, тем ближе указанное антитело или его функциональная часть к антителу D25. Антитело или его функциональная часть по настоящему изобретению предпочтительно состоит из тяжелой цепи, а также легкой цепи, которые похожи на тяжелую и легкую цепи D25. Поэтому, кроме того, предусмотрены антитело или его функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 11.
В одном варианте предусмотрены антитело или его функциональный эквивалент, включающие последовательность тяжелой цепи, состоящую из последовательности тяжелой цепи, приведенной на Фигуре 11, и последовательность легкой цепи, состоящую из последовательности легкой цепи, приведенной на Фигуре 11. Кроме того, как хорошо известно специалисту в данной области, возможно создать сокращенную последовательность тяжелой цепи или легкой цепи при сохранении желаемого свойства связывать антиген. Предпочтительно, такие полученные сокращенные тяжелые цепи или легкие цепи обладали сокращенным константным районом по сравнению с первоначальными тяжелыми или легкими цепями. Вариабельный домен предпочтительно сохранить. Например, получены фрагмент Fab или фрагмент F(ab')2 на основе последовательности тяжелой цепи или последовательности легкой цепи, приведенных на Фигуре 11. Поэтому также предусмотрен функциональный эквивалент антитела, включающий по меньшей мере функциональную часть последовательности, приведенной на Фигуре 11. Указанная функциональная часть имеет в длину не менее 20 аминокислот и включает последовательность, которая по меньшей мере на 70% идентична CDR1 последовательности тяжелой цепи, приведенной на фигуре 11Г, и/или последовательность, которая по меньшей мере на 75% идентична CDR2 последовательности тяжелой цепи, приведенной на Фигуре 11Г, и/или последовательность, которая по меньшей мере на 70% идентична CDR3 последовательности тяжелой цепи, приведенной на Фигуре 11Г, и/или последовательность, которая по меньшей мере на 85% идентична CDR1 последовательности легкой цепи, приведенной на Фигуре 11D, и/или последовательность, которая по меньшей мере на 70% идентична CDR2 последовательности легкой цепи, приведенной на Фигуре 11Г. Предпочтительно, указанная функциональная часть также включает последовательность, которая по меньшей мере на 70% идентична CDR3 последовательности легкой цепи, приведенной на Фигуре 11Г.
Другое особенно предпочтительное анти-РСВ антитело по настоящему изобретению представляет собой антитело, обозначаемое "АМ14", которое обладает тяжелой цепью и легкой цепью, приведенными на Фигуре 14А. CDR последовательности AM14, которые, в частности ответственны за антигенсвязывающую способность AM14, также представлены на Фигуре 14А.
В настоящем изобретении предусмотрено, что CDR последовательности, приведенные на Фигуре 14А, обеспечивают желаемые характеристики связывания РСВ, и специалист в области техники способен создать варианты, включающие по меньшей мере одну измененную последовательность CDR, например, применяя консервативные замены аминокислот. Консервативная замена аминокислоты предполагает замещение одной аминокислоты другой аминокислотой в целом с аналогичными свойствами (размером, гидрофобностью, и т.д.), так что общее функционирование, вероятно, не будет серьезно затронуто.
Также возможно изменить по меньшей мере одну CDR последовательность, приведенную на Фигуре 14А, с целью получения варианта антитела, или его функционального эквивалента по меньшей мере с одним измененным свойством по сравнению с AM14. Предпочтительно, предусмотрены антитело или его функциональный эквивалент, включающие последовательность CDR, которая, по меньшей мере на 70% идентична последовательности CDR, приведенной на Фигуре 14А, так что предпочтительные характеристики связывания антигена АМ14 по меньшей мере частично сохранены и даже улучшены. CDR последовательности, приведенные на Фигуре 14А, предпочтительно изменены таким образом, что полученное антитело или его функциональный эквивалент обладают, по меньшей мере, одним улучшенным свойством, таким как, например, улучшенная аффинность связывания, избирательность и/или стабильность, по сравнению с AM14. Варианты антител или их функциональные эквиваленты, включающие аминокислотные последовательности, которые по меньшей мере на 70% идентичны последовательности CDR, приведенной на Фигуре 14А, находятся в рамках настоящего изобретения. В уровне техники доступны различные методы для изменения последовательности аминокислот. Например, последовательности тяжелой цепи или легкой цепи с желаемыми последовательностями CDR искусственно синтезированы. Предпочтительно, последовательности нуклеиновой кислоты, кодирующие последовательности CDR, мутировали, например, с использованием случайного или сайт-специфического мутагенеза.
Таким образом, в одном аспекте изобретение обеспечивает выделенные, искусственные или рекомбинантные антитела или их функциональные части, их производные и/или их аналоги, способные специфически связывать респираторно-синцитиальный вирус и включающие:
- CDR1 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GFSFSHYA, и/или
- CDR2 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ISYDGENT, и/или
- CDR3 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ARDRIVDDYYYYGMDV, и/или
- CDR1 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QDIKKY, и/или
- CDR2 последовательности легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности DAS, и/или
- CDR3 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QQYDNLPPLT.
Предпочтительно, антитело или его функциональный эквивалент по настоящему изобретению включает CDR последовательность, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 14А. Наиболее предпочтительно, антитело или его функциональный эквивалент по настоящему изобретению включает CDR последовательность, которая по меньшей мере на 95% идентична, по меньшей мере, одной из CDR последовательностей, приведенных на Фигуре 14А. Особенно предпочтительное антитело AM14, описанное выше, включает CDR последовательности, которые состоят из CDR последовательностей, приведенных на Фигуре 14А. Таким образом, в особенно предпочтительном варианте по настоящему изобретению предусмотрены выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, способные специфически связывать респираторно-синцитиальный вирус, которые включают:
- CDR1 последовательность тяжелой цепи, включающую последовательность GFSFSHYA, и/или
- CDR2 последовательность тяжелой цепи, включающую последовательность ISYDGENT, и/или
- CDR3 последовательность тяжелой цепи, включающую последовательность ARDRIVDDYYYYGMDV, и/или
- CDR1 последовательность легкой цепи, включающую последовательность QDIKKY, и/или
- CDR2 последовательности легкой цепи, включающую последовательность DAS, и/или
- CDR3 последовательность легкой цепи, включающую последовательность QQYDNLPPLT.
В одном варианте предусмотрены антитело или его функциональный эквивалент, включающие три CDR последовательности тяжелой цепи и три CDR последовательности легкой цепи, приведенные на Фигуре 14А, или последовательности, которые по меньшей на 70% идентичны им. Поэтому дополнительно предусмотрены выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, включающие CDR1 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GFSFSHYA, и CDR2 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ISYDGENT, и CDR3 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ARDRIVDDYYYYGMDV, и CDR1 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QDIKKY, и CDR2 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности DAS, и CDR3 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QQYDNLPPLT. Указанные антитела или их функциональные эквиваленты предпочтительно содержат CDR последовательности, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны CDR последовательностям тяжелой цепи и CDR последовательностям легкой цепи, представленным на Фигуре 14А. Также предусмотрены антитело или его функциональный эквивалент, включающие вышеупомянутые CDR1, CDR2 и CDR3 последовательности тяжелой цепи, приведенные на Фигуре 14А, а также вышеупомянутые CDR1, CDR2 и CDR3 последовательности легкой цепи, приведенные на Фигуре 14А.
Также предусмотрены антитела или их функциональные эквиваленты, включающие тяжелые цепи, аминокислотные последовательности которых по меньшей мере на 70% идентичны последовательностям тяжелых цепей, приведенных на Фигуре 14А. Такие последовательности тяжелых цепей обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело АМ14. Поэтому, кроме того, предусмотрены антитело или его функциональные эквиваленты, обладающие последовательностью тяжелой цепи, включающей последовательность, которая по меньшей мере на 70% идентична последовательности

 Кроме того, аминокислотные последовательности легкой цепи, которые по меньшей мере на 70% идентичны последовательности легкой цепи, приведенной на Фигуре НА, также обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело АМ14. Поэтому также предусмотрены антитело или его функциональный эквивалент с последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности
Кроме того, аминокислотные последовательности легкой цепи, которые по меньшей мере на 70% идентичны последовательности легкой цепи, приведенной на Фигуре НА, также обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело АМ14. Поэтому также предусмотрены антитело или его функциональный эквивалент с последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности

 Антитело или его функциональная часть по настоящему изобретению предпочтительно включает вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентична последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 14А. Чем выше гомология, тем больше указанное антитело или его функциональная части напоминает антитело АМ14. Антитело или его функциональная часть по настоящему изобретению предпочтительно состоит из тяжелой цепи, а также легкой цепи, которые похожи на тяжелую и легкую цепи АМ14. Поэтому, кроме того предусмотрены антитело или его функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 14А.
Антитело или его функциональная часть по настоящему изобретению предпочтительно включает вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентична последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 14А. Чем выше гомология, тем больше указанное антитело или его функциональная части напоминает антитело АМ14. Антитело или его функциональная часть по настоящему изобретению предпочтительно состоит из тяжелой цепи, а также легкой цепи, которые похожи на тяжелую и легкую цепи АМ14. Поэтому, кроме того предусмотрены антитело или его функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 14А.
В одном варианте предусмотрено антитело или его функциональный эквивалент, включающие последовательность тяжелой цепи, состоящую из последовательности тяжелой цепи, приведенной на Фигуре 14А, и включающие последовательность легкой цепи, состоящую из последовательности легкой цепи, приведенной на Фигуре 14А. Кроме того, как хорошо известно специалисту в данной области, возможно создать сокращенную последовательность тяжелой цепи или легкой цепи при сохранении интересующей способности связывать антиген. Предпочтительно, создаются сокращенная тяжелая цепь или легкая цепь с сокращенным константным районом по сравнению с первоначальными тяжелой или легкой цепями. Вариабельный домен желательно сохранить. Например, получены Fab фрагмент или F(ab')2 фрагмент на основе последовательности тяжелой цепи или последовательности легкой цепи, приведенных на Фигуре 14А. Поэтому также предусмотрен функциональный эквивалент антитела, включающий по меньшей мере функциональную часть последовательности, приведенной на Фигуре 14А. Указанная функциональная часть имеет длину не менее 20 аминокислот и включает последовательность, которая по меньшей мере на 70% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 14А.
Другое особенно предпочтительное анти-РСВ антитело по настоящему изобретению представляет собой антитело, обозначенное "AM16ʺ, которое обладает районом тяжелой цепи и районом легкой цепи, приведенными на Фигуре 14Б. CDR последовательности AM16, которые, в частности, способствуют антигенсвязывающим свойствам AM16, также приведены на Фигуре 14Б.
В настоящем изобретении предусмотрено, что CDR последовательности, приведенные на Фигуре 14Б, обеспечивают желаемые характеристики связывания РСВ, и специалист в данной области способен создать варианты, включающие по меньшей мере одну измененную последовательность CDR, например, применяя консервативную замену аминокислот. Консервативная замена аминокислоты предполагает замещение одной аминокислоты другой аминокислотой в целом с аналогичными свойствами (размером, гидрофобностью, и т.д.), так что общее функционирование, вероятно, не будет серьезно затронуто.
Также возможно изменить по меньшей мере одну CDR последовательность, приведенную на Фигуре 14Б, с целью получения варианта антитела, или его функционального эквивалента по меньшей мере с одним измененным свойством по сравнению с AM16. Предпочтительно, предусмотрены антитело или его функциональный эквивалент, включающие последовательность CDR, которая по меньшей мере на 70% идентична последовательности CDR, приведенной на Фигуре 14Б, так что предпочтительные характеристики связывания AM16 по меньшей мере частично сохранены и даже улучшены. CDR последовательность, приведенная на Фигуре 14Б, предпочтительно изменена таким образом, что полученное антитело или функциональный эквивалент обладает по меньшей мере одним улучшенным свойством, таким как, например, улучшенная аффинность связывания, избирательность и/или стабильность по сравнению с AM16. Поэтому варианты антител или их функциональные эквиваленты, включающие аминокислотные последовательности, которые по меньшей мере на 70% идентичны последовательности CDR, приведенной на Фигуре 14Б, находятся в рамках настоящего изобретения. В уровне техники доступны различные методы для изменения последовательности аминокислот. Например, последовательности тяжелой цепи или легкой цепи с желаемой последовательностью CDR искусственно синтезированы. Предпочтительно, последовательность нуклеиновой кислоты, кодирующую последовательность CDR, мутировали, например, с использованием случайного или сайт-специфического мутагенеза.
Таким образом, в одном аспекте изобретение предусматривает выделенное, искусственное или рекомбинантное антитело или его функциональную часть, производное и/или его аналог, способные специфически связывать респираторно-синцитиальный вирус, которые включают:
- CDR1 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GFTFSSYN, и/или
- CDR2 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ISAGSSYI, и/или
- CDR3 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности AREDYGPGNYYSPNWFDP, и/или
- CDR1 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности SSNIGAGYD, и/или
- CDR2 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GNT, и/или
- CDR3 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности HSYDRSLSG.
Предпочтительно, антитело или функциональный эквивалент по настоящему изобретению включает CDR последовательность, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно, по меньшей мере, на 90% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 14Б. Наиболее предпочтительно, антитело или функциональный эквивалент по настоящему изобретению включает CDR последовательность, которая по меньшей мере на 95% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 14Б. Особенно предпочтительно антитело AM16, описанное выше, включающее CDR последовательности, которые состоят из CDR последовательностей, приведенных на Фигуре 14Б. Таким образом, в особенно предпочтительном варианте по настоящему изобретению предусмотрено выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, способные специфически связывать респираторно-синцитиальный вирус, которые включают:
- CDR1 последовательность тяжелой цепи, включающую последовательность GFTFSSYN, и/или
- CDR2 последовательность тяжелой цепи, включающую последовательность ISAGSSYI, и/или
- CDR3 последовательность тяжелой цепи, включающую последовательность AREDYGPGNYYSPNWFDP, и/или
- CDR1 последовательность легкой цепи, включающую последовательность SSNIGAGYD, и/или
- CDR2 последовательность легкой цепи, включающую последовательность GNT, и/или
- CDR3 последовательность легкой цепи, включающую последовательность HSYDRSLSGB.
В одном варианте предусмотрены антитело или его функциональный эквивалент, включающие три CDR последовательности тяжелой цепи и три CDR последовательности легкой цепи, приведенные на Фигуре 14Б, или включающие последовательности, которые по меньшей мере на 70% идентичны указанным последовательностям. Поэтому, кроме того, предусмотрены выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, включающие CDR1 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GFTFSSYN, и включающие CDR2 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности ISAGSSYI, и включающие CDR3 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере 70% идентична последовательности AREDYGPGNYYSPNWFDP, и включающие CDR1 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности SSNIGAGYD, и CDR2 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GNT, и CDR3 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности HSYDRSLSG. Указанное антитело или функциональный эквивалент предпочтительно содержат CDR последовательности, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны вышеупомянутым CDR последовательностям тяжелой цепи и вышеупомянутым CDR последовательностям легкой цепи приведенным на Фигуре 14Б. Также предусмотрены антитело или функциональный эквивалент, включающие вышеупомянутые CDR1, CDR2 и CDR3 последовательности тяжелой цепи, приведенные на Фигуре 14Б, а также вышеупомянутые CDR1, CDR2 и CDR3 последовательности легкой цепи, приведенные на Фигуре 14Б.
Также предусмотрены антитела или их функциональные эквиваленты, включающие аминокислотную последовательность тяжелой цепи, которая по меньшей мере на 70% идентична последовательности тяжелой цепи, приведенной на Фигуре 14Б. Такие последовательности тяжелой цепи обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело AM16. Поэтому, кроме того, предусмотрены антитело или его функциональные эквиваленты, обладающие последовательностью тяжелой цепи, включающей последовательность, которая по меньшей мере на 70% идентична последовательности
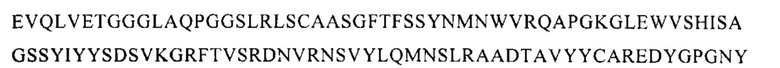
 Кроме того, аминокислотные последовательности легкой цепи, которые по меньшей мере на 70% идентичны последовательности легкой цепи, приведенной на Фигуре 14Б, также обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело AM16. Поэтому также предусмотрены антитело или его функциональный эквивалент, обладающие последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности
Кроме того, аминокислотные последовательности легкой цепи, которые по меньшей мере на 70% идентичны последовательности легкой цепи, приведенной на Фигуре 14Б, также обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело AM16. Поэтому также предусмотрены антитело или его функциональный эквивалент, обладающие последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности

 Антитело или функциональная часть по настоящему изобретению предпочтительно включают вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 14Б. Чем выше гомология, тем больше указанное антитело или функциональная часть похожи на антитело AM16. Антитело или функциональная часть по настоящему изобретению предпочтительно включают тяжелую цепь, а также легкую цепь, которые похожи на тяжелую и легкую цепи AM16. Поэтому, кроме того предусмотрены антитело или функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 14Б.
Антитело или функциональная часть по настоящему изобретению предпочтительно включают вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 14Б. Чем выше гомология, тем больше указанное антитело или функциональная часть похожи на антитело AM16. Антитело или функциональная часть по настоящему изобретению предпочтительно включают тяжелую цепь, а также легкую цепь, которые похожи на тяжелую и легкую цепи AM16. Поэтому, кроме того предусмотрены антитело или функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 14Б.
В одном варианте предусмотрены антитело или его функциональный эквивалент, включающие последовательность тяжелой цепи, состоящую из последовательности тяжелой цепи, приведенной на Фигуре 14Б, и включающие последовательность легкой цепи, состоящую из последовательности легкой цепи, приведенной на Фигуре 14Б. Альтернативно, как хорошо известно специалисту в данной области, возможно создать сокращенные последовательности тяжелой цепи или легкой цепи при сохранении интересующей способности связывать антиген. Предпочтительно, создаются сокращенные тяжелая цепь или легкая цепь, которые обладают сокращенным константным районом по сравнению с первоначальными тяжелой или легкой цепями. Вариабельный домен желательно сохранить. Например, получены фрагмент Fab или фрагмент F(ab')2 на основе последовательности тяжелой цепи или последовательности легкой цепи, приведенных на Фигуре 14Б. Поэтому также предусмотрен функциональный эквивалент антитела, включающий по меньшей мере функциональную часть последовательности, приведенной на Фигуре 14Б. Указанная функциональная часть имеет длину не менее 20 аминокислот и включает последовательность, которая по меньшей мере на 70% идентична по меньшей мере одной из CDR последовательностей, приведенной на Фигуре 14Б.
Другое особенно предпочтительное анти-РСВ антитело по настоящему изобретению представляет собой антитело, обозначенное "АМ23", которое обладает районом тяжелой цепи и районом легкой цепи, приведенными на Фигуре 14В. CDR последовательности АМ23, которые, в частности, способствуют антигенсвязывающим свойствам АМ23, также приведены на Фигуре 14В.
В настоящем изобретении предусмотрено, что CDR последовательности, приведенные на Фигуре 14В, обеспечивают желаемые характеристики связывания РСВ, и специалист в данной области способен создать варианты, включающие по меньшей мере одну измененную последовательность CDR, например, применяя консервативную замену аминокислот. Консервативная замена аминокислоты предполагает замещение одной аминокислоты другой аминокислотой в целом с аналогичными свойствами (размером, гидрофобностью, и т.д.), так что общее функционирование, вероятно, не будет серьезно затронуто.
Также возможно изменить по меньшей мере одну CDR последовательность, приведенную на Фигуре 14В, с целью получения варианта антитела, или его функционального эквивалента по меньшей мере с одним измененным свойством по сравнению с АМ23. Предпочтительно, предусмотрены антитело или его функциональный эквивалент, включающие последовательность CDR, которая по меньшей мере на 70% идентична последовательности CDR, приведенной на Фигуре 14В, так что предпочтительные характеристики связывания АМ23 по меньшей мере частично сохранены и даже улучшены. CDR последовательность, приведенная на Фигуре 14В, предпочтительно изменена таким образом, что полученное антитело или функциональный эквивалент обладает по меньшей мере одним улучшенным свойством, таким как, например, улучшенная аффинность связывания, избирательность и/или стабильность по сравнению с АМ23. Поэтому варианты антител или их функциональные эквиваленты, включающие аминокислотные последовательности, которые по меньшей мере на 70% идентичны последовательности CDR, приведенной на Фигуре 14В, находятся в рамках настоящего изобретения. В уровне техники доступны различные методы для изменения последовательности аминокислот. Например, последовательности тяжелой цепи или легкой цепи с желаемой последовательностью CDR искусственно синтезированы. Предпочтительно, последовательность нуклеиновой кислоты, кодирующую последовательность CDR, мутировали, например, с использованием случайного или сайт-специфического мутагенеза
Таким образом, в одном аспекте изобретение предусматривает выделенное, искусственное или рекомбинантное антитело или его функциональную часть, производное и/или его аналог, способные специфически связывать респираторно-синцитиальный вирус, которые включают:
- CDR1 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GFNFHNYG, и/или
- CDR2 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности VWYDGSKK, и/или
- CDR3 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности VRDKVGPTPYFDS, и/или
- CDR1 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности NIGSET, и/или
- CDR2 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности DDD, и/или
- CDR3 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QVWDRSNYHQV.
Предпочтительно, антитело или функциональный эквивалент по настоящему изобретению включает CDR последовательность, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 14В. Наиболее предпочтительно, антитело или функциональный эквивалент по настоящему изобретению включает CDR последовательность, которая по меньшей мере на 95% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 14В. Особенно предпочтительно антитело АМ23, описанное выше, включающее CDR последовательности, которые состоят из CDR последовательностей, приведенных на Фигуре 14В. Таким образом, в особенно предпочтительном варианте по настоящему изобретению предусмотрено выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, способные специфически связывать респираторно-синцитиальный вирус, которые включают:
- CDR1 последовательность тяжелой цепи, включающую последовательность GFNFHNYG, и/или
- CDR2 последовательность тяжелой цепи, включающую последовательность VWYDGSKK, и/или
- CDR3 последовательность тяжелой цепи, включающую последовательность VRDKVGPTPYFDS, и/или
- CDR1 последовательность легкой цепи, включающую последовательность NIGSET, и/или
- CDR2 последовательность легкой цепи, включающую последовательность DDD, и/или
- CDR3 последовательность легкой цепи, включающую последовательность QVWDRSNYHQV.
В одном варианте предусмотрены антитело или его функциональный эквивалент, включающий три CDR последовательности тяжелой цепи и три CDR последовательности легкой цепи, приведенные на Фигуре 14В, или включающий последовательности, которые по меньшей мере на 70% идентичны указанным последовательностям. Поэтому, кроме того, предусмотрены выделенное, искусственное или рекомбинантное антитело или его функциональный эквивалент, включающие CDR1 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности GFNFHNYG, и включающие CDR2 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности VWYDGSKK, и включающие CDR3 последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности VRDKVGPTPYFDS, и включающие CDR1 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности NIGSET, и CDR2 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности DDD, и CDR3 последовательность легкой цепи, включающую последовательность, которая по меньшей мере на 70% идентична последовательности QVWDRSNYHQV. Указанное антитело или функциональный эквивалент предпочтительно содержат CDR последовательности, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны вышеупомянутым CDR последовательностям тяжелой цепи и вышеупомянутым CDR последовательностям легкой цепи, приведенным на Фигуре 14 В. Также предусмотрены антитело или функциональный эквивалент, включающие вышеупомянутые CDR1, CDR2 и CDR3 последовательности тяжелой цепи, приведенные на Фигуре 14В, а также вышеупомянутые CDR1, CDR2 и CDR3 последовательности легкой цепи, приведенные на Фигуре 14В.
Также предусмотрены антитела или их функциональные эквиваленты, включающие аминокислотную последовательность тяжелой цепи, которая по меньшей мере на 70% идентична последовательности тяжелой цепи, приведенной на Фигуре 14В. Такие последовательности тяжелой цепи обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело АМ23. Поэтому, кроме того предусмотрены антитело или его функциональные эквиваленты, обладающие последовательностью тяжелой цепи, включающей последовательность, которая по меньшей мере на 70% идентична последовательности

 Кроме того, аминокислотные последовательности легкой цепи, которые по меньшей мере на 70% идентичны последовательности легкой цепи, приведенной на Фигуре 14В, также обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело АМ23. Поэтому также предусмотрены антитело или его функциональный эквивалент, обладающие последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности
Кроме того, аминокислотные последовательности легкой цепи, которые по меньшей мере на 70% идентичны последовательности легкой цепи, приведенной на Фигуре 14В, также обеспечивают желаемое свойство связывать РСВ, о чем свидетельствует антитело АМ23. Поэтому также предусмотрены антитело или его функциональный эквивалент, обладающие последовательностью легкой цепи, которая по меньшей мере на 70% идентична последовательности
 . Антитело или функциональная часть по настоящему изобретению предпочтительно включают вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 14В. Чем выше гомология, тем больше указанное антитело или функциональная часть похожи на антитело АМ23. Антитело или функциональная часть по настоящему изобретению предпочтительно включают тяжелую цепь, а также легкую цепь, которые похожи на тяжелую и легкую цепи АМ23. Поэтому, кроме того предусмотрены антитело или функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 14В.
. Антитело или функциональная часть по настоящему изобретению предпочтительно включают вариабельную последовательность тяжелой цепи и/или вариабельную последовательность легкой цепи, которые по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и/или последовательности легкой цепи, приведенным на Фигуре 14В. Чем выше гомология, тем больше указанное антитело или функциональная часть похожи на антитело АМ23. Антитело или функциональная часть по настоящему изобретению предпочтительно включают тяжелую цепь, а также легкую цепь, которые похожи на тяжелую и легкую цепи АМ23. Поэтому, кроме того предусмотрены антитело или функциональная часть, включающие последовательность тяжелой цепи и последовательность легкой цепи, которые по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичны последовательности тяжелой цепи и последовательности легкой цепи, приведенным на Фигуре 14В.
В одном варианте предусмотрены антитело или его функциональный эквивалент, включающие последовательность тяжелой цепи, состоящую из последовательности тяжелой цепи, приведенной на Фигуре 14В, и включающие последовательность легкой цепи, состоящую из последовательности легкой цепи, приведенной на Фигуре 14В. Альтернативно, как хорошо известно специалисту в данной области, возможно создать сокращенные последовательности тяжелой цепи или легкой цепи при сохранении интересующей способности связывать антиген. Предпочтительно, создаются сокращенные тяжелая цепь или легкая цепь, которые обладают сокращенным константным районом по сравнению с первоначальными тяжелой или легкой цепями. Вариабельный домен желательно сохранить. Например, получены фрагмент Fab или фрагмент F(ab')2 на основе последовательности тяжелой цепи или последовательности легкой цепи, приведенных на Фигуре 14В. Поэтому также предусмотрен функциональный эквивалент антитела, включающий по меньшей мере функциональную часть последовательности, приведенной на Фигуре 14С. Указанная функциональная часть имеет длину не менее 20 аминокислот и включает последовательность, которая по меньшей мере на 70% идентична по меньшей мере одной из CDR последовательностей, приведенных на Фигуре 14В.
Настоящее изобретение предусматривает РСВ-специфические антитела или их функциональные эквиваленты с улучшенными свойствами по сравнению с антителами, известными в уровне техники. Изобретатели добились успеха в создании РСВ-специфических антител с низкими значениями IC50. Такие антитела обладают высоким или сильным сродством к РСВ и поэтому особенно подходят для противодействия и/или по меньшей мере частичного предотвращения РСВ-инфекции и/или неблагоприятных последствий РСВ инфекции. В одном варианте предусмотрено антитело и функциональный эквивалент указанного антитела, которые обладают значением IC50 менее 10 нг/мл в in vitro анализе на обезвреживание, где клетки НЕр-2 инфицировали РСВ. Указанное антитело или функциональный эквивалент предпочтительно обладают значением IC50 менее 5 нг/мл, более предпочтительно менее 2 нг/мл. Предпочтительное D25 антитело обладает значением IC50 около 0,5-1,5 нг/мл в in vitro анализе на обезвреживание, описанном в Примерах (см. Фигуру 8).
Антитело по настоящему изобретению предпочтительно представляет собой человеческое антитело. Использование человеческих антител для лечения человека уменьшает вероятность побочных эффектов, вызванных иммунологической реакцией в человеческом организме против нечеловеческих последовательностей. В другом предпочтительном варианте антитело или его функциональная часть, его производное или аналог по настоящему изобретению представляют собой химерное антитело. Таким образом, интересующие последовательности, такие как, например, интересующий сайт связывания, могут быть включены в антитело или функциональный эквивалент по настоящему изобретению.
Изобретение также предусматривает выделенную, искусственную или рекомбинантную последовательность нуклеиновой кислоты, или ее функциональную часть, производное или ее аналог, кодирующие антитело или функциональный эквивалент по настоящему изобретению. Такие нуклеиновые кислоты, например, выделены из В-клеток, которые способны производить антитела по настоящему изобретению, как описано более подробно ниже. В предпочтительном варианте предусмотрена последовательность нуклеиновой кислоты, включающая последовательность, которая по меньшей мере на 70% гомологична по меньшей мере функциональной части последовательности нуклеиновой кислоты, приведенной на Фигуре 11, Фигуре 12, Фигуре 14А, Фигуре 14Б и/или Фигуре 14Б. Указанная последовательность нуклеиновой кислоты предпочтительно включает последовательность, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% гомологична, по меньшей мере, функциональной части последовательности нуклеиновой кислоты, приведенной на Фигуре 11, Фигуре 12, Фигуре 14А, Фигуре 14Б и/или Фигуре 14Б. Указанная функциональная часть имеет длину, по меньшей мере, 30 нуклеотидов, предпочтительно по меньшей мере 50 нуклеотидов, более предпочтительно по меньшей мере 75 нуклеотидов. Предпочтительно, указанная функциональная часть кодирует по меньшей мере одну последовательность нуклеиновой кислоты, приведенную на Фигуре 11Г, Фигуре 12, Фигуре 14А, Фигуре 14Б и/или Фигуре 14Б. Указанная последовательность предпочтительно представляет собой CDR последовательность.
Антитело или функциональный эквивалент по настоящему изобретению особенно подходит для использования в качестве профилактического лекарственного средства или профилактического агента. Поэтому в настоящем документе антитело по настоящему изобретению или его функциональная часть, производное или аналог, также предусмотрены для использования в качестве лекарственного средства и/или профилактического агента. В особенно предпочтительном варианте указанное антитело включает антитело D25, АМ14, AM16 и/или АМ23 или их функциональные части, производные или аналоги. Указанное лекарственное средство или профилактический агент предпочтительно используются для противодействия или по меньшей мере частичного предотвращения РСВ-инфекции или для противодействия или по меньшей мере частичного предотвращения неблагоприятных последствий РСВ-инфекции. Поэтому также предусмотрено использование антитела, его функциональной части, производного или аналога по настоящему изобретению для изготовления лекарственного средства и/или профилактического агента для по меньшей мере частичного лечения и/или предотвращения связанного с РСВ расстройства, а также предусмотрен способ по меньшей мере частичного лечения или профилактики связанного с РСВ расстройства, где способ предусматривает введение нуждающемуся в этом индивидууму терапевтически эффективного количество антитела или его функционального эквивалента по настоящему изобретению. Указанное антитело предпочтительно включает антитело D25, AM14, AM16 и/или АМ23 или их функциональные части, производные или их аналоги.
В целях противодействия РСВ, антитело или функциональный эквивалент по настоящему изобретению предпочтительно вводят индивидууму до появления РСВ-инфекции. Альтернативно, антитело или его функциональный эквивалент по настоящему изобретению вводят, когда индивидуум уже инфицирован РСВ. Указанное антитело или его функциональный эквивалент предпочтительно вводят индивидууму с повышенным риском РСВ-расстройства, такому как, например, недоношенный ребенок, индивидуум с хроническим заболеванием легких, врожденным пороком сердца и/или ослабленным иммунитетом и ребенок младше 6 недель. Кроме того, пожилые люди имеют повышенный риск связанного с РСВ расстройства. Антитела или функциональные эквиваленты по настоящему изобретению предпочтительно вводят перорально или с помощью одной или более инъекций. Диапазоны доз антитела и/или его функциональных эквивалентов по настоящему изобретению, которые используются в терапевтических целях, как описано здесь ранее, должны быть разработаны на основе исследования дозировки в клинике, в ходе клинических испытаний, для которых существуют строгие протокольные требования. Типичная доза составляет от 0,1 до 10 мг/кг веса тела. Для терапевтического применения антитела или функциональные эквиваленты по настоящему изобретению, как правило, совмещают с фармацевтически приемлемым носителем, адъювантом, растворителем и/или наполнителем. Примеры подходящих носителей, например, включают гемоцианин моллюска фиссуреллы (KLH), сывороточный альбумин (например, BSA или RSA) и яичный альбумин. Специалисту в данной области известно много подходящих адъювантов на масляной и на водной основе. В одном варианте указанный адъювант включает Specol. В другом варианте, указанный подходящий носитель включает раствор, например, физиологический раствор.
В еще одном варианте используется нуклеиновая кислота, кодирующая антитело или его функциональную часть по настоящему изобретению. При введении такой нуклеиновой кислоты, антитела или функциональные эквиваленты производятся с помощью механизмов хозяина. Производимые антитела или функциональные эквиваленты способны предотвратить и/или противодействовать РСВ-инфекции и/или негативным последствиям РСВ-инфекции. Поэтому в настоящем изобретении также предусмотрены последовательность нуклеиновой кислоты, функциональная часть, производное и/или аналог антитела по настоящему изобретению для использования в качестве лекарственного средства и/или профилактического агента. Указанная нуклеиновая кислота предпочтительно используется для противодействия РСВ. Поэтому, кроме того предусмотрено применение последовательности нуклеиновой кислоты, функциональной части, производного и/или аналога антитела по настоящему изобретению для изготовления лекарственного средства и/или профилактического агента по меньшей мере для частичного лечения и/или предотвращения связанного с РСВ расстройства.
По меньшей мере функциональная часть нуклеиновой кислоты по настоящему изобретению означает часть указанной нуклеиновой кислоты по меньшей мере составляющую 30 пар нуклеотидов в длину, предпочтительно по меньшей мере 50 пар нуклеотидов в длину, более предпочтительно по меньшей мере 100 пар нуклеотидов в длину, включающую, по меньшей мере, одну характеристику экспрессии (качественно, не обязательно количественно) нуклеиновой кислоты по настоящему изобретению. Указанная функциональная часть, по меньшей мере, кодирует последовательность аминокислот, включающую последовательность, которая по меньшей мере на 70% идентична последовательности CDR, приведенной на Фигуре 11Г, Фигуре 14А, Фигуре 14Б и/или Фигуре 14В.
Кроме того, изобретение предусматривает выделенные продуцирующие антитела клетки, способные производить антитело, функциональную часть, производное или аналог антитела по настоящему изобретению. Возможные (но не ограничивающие) способы получения таких продуцирующих антитела клеток подробно описаны в Примерах. Изобретатели разработали и использовали новые способы для улучшения стабильности продуцирующих РСВ-специфические антитела клеток. С применением этого способа созданы продуцирующие РСВ-специфические антитела клетки, которые являются стабильными по меньшей мере в течение шесть месяцев. В настоящем изобретении также предусмотрены клетки, продуцирующие РСВ-специфические антитела по настоящему изобретению, которые являются стабильными в течение девяти недель, предпочтительно по меньшей мере в течение трех месяцев, более предпочтительно по меньшей мере в течение шести месяцев.
Изобретатели использовали свое открытие, заключающееся в том, что стабильность продуцирующих РСВ-специфические антитела клеток находится под влиянием количества продукта экспрессии BCL6 и/или Blimp-1 в указанных продуцирующих антитела клетках. Количество продукта экспрессии BCL6 и/или Blimp-1 оказывает прямое или косвенное влияние. Предпочтительно регулировать количество продуктов экспрессии и BCL6, и Blimp-1 в указанных продуцирующих антитела клетках, так как оба продукта экспрессии влияют на стабильность продуцирующей антитела клетки. Стабильность продуцирующей антитела клетки определяется, как способность указанной продуцирующей антитела клетки оставаться в определенной стадии развития (предпочтительно после того, как указанная клетка была переведена в указанную стадию). Различные этапы развития клетки вызывают по меньшей мере различия одной характеристики указанной клетки. Например, В клетка памяти, как известно, дифференцируется при стимуляции в секретирующий антитела плазмоцит через этап, который некоторые ученые называют плазмабласт. В клетка памяти, плазмабласт и плазмацит представляют собой различные стадии развития В клетки, в которых В клетка обладает различными характеристиками. В клетка памяти проявляет низкую способность к пролиферации и секреции антител. Плазмабласт проявляет и более высокую способность к пролиферации, и более высокий уровень секреции антител по сравнению с В клеткой памяти, в то время как плазмацит проявляет высокий уровень секреции антител, но не способен пролиферировать. С применением способа, открытого авторами настоящего изобретения, стало возможным регулировать репликативную продолжительность жизни продуцирующей антитела клетки. Репликативная продолжительность жизни продуцирующей антитела клетки определяется здесь как промежуток времени, в течение которого В клетка и ее дочерние клетки способны к репликации при сохранении их способности производить антитела и/или развиваться в клетку, которая производит антитела. Предпочтительно, репликативная продолжительность жизни продуцирующей антитела клетки продлена, а это означает, что указанная продуцирующая антитела клетка не будет терминально дифференцировать, и продолжит пролиферировать in vitro, или будет терминально дифференцировать только после длительного периода по сравнению с такой же продуцирующей антитела клеткой, которая используется в настоящее время. По мнению изобретателей, возможно регулировать количество продукта экспрессии BCL6 и/или Blimp-1 в продуцирующей антитела клетке таким образом, что продуцирующая антитела клетка может быть приведена в заданное состояние и/или поддерживаться в заданном состоянии, в котором клетка продолжает пролиферировать. Таким образом с применением разработанного авторами способа становится возможным увеличить репликативную продолжительность жизни продуцирующей антитела клетки, поскольку возможно поддерживать В-клетку в определенной стадии развития, в которой происходит репликация, см. PCT/NL2006/000625, поданную тем же заявителем. Настоящее изобретение предусматривает средства и способы получения стабильных клеток, продуцирующих РСВ-специфические антитела.
Продуцирующая антитело клетка определяется как клетка, которая способна продуцировать и/или секретировать антитело или его функциональный эквивалент, и/или как клетка, способная превращаться в клетку, способную производить и/или секретировать антитело или его функциональный эквивалент. Продуцирующая РСВ-специфическое антитело клетка определяется в настоящем документе как клетка, способная производить и/или секретировать антитело или его функциональный эквивалент, которые способны специфически связывать РСВ и/или компоненты РСВ, такие как, например, эпитоп РСВ F (гибридный) белка, РСВ G (присоединение) белка или РСВ SH (небольшой гидрофобный) белка. Предпочтительно, указанная продуцирующая РСВ-специфическое антитело клетка включает В клетку и/или плазматическую клетку, производное В клетки. В клетка в настоящем документе называется продуцирующей антитело клеткой, даже если В клетка находится в стадии, когда продукция антител низка или отсутствует вовсе, такой как стадия наивной В клетки или В клетки памяти, активированной или неактивированной, потому что такие клетки способны превращаться в клетки, которые производят антитела, такие как плазмабласт и/или плазматическая клетка.
Продуцирующая РСВ-специфическое антитело клетка по настоящему изобретению предпочтительно включает клетки млекопитающих. Неограничивающие примеры включают продуцирующие антитело клетки, полученные из организма человека, грызуна, кролика, ламы, свиньи, коровы, козы, лошади, обезьяны, гориллы. Предпочтительно, указанная продуцирующая антитела клетка включает клетки человека, клетки мыши, клетки кролика и/или клетки ламы.
BCL6 кодирует репрессор транскрипции, который необходим для нормального развития и созревания В-клеток и Т-клеток и для формирования герминативных центров. (Ye, 1997). BCL6 интенсивно экспрессируется в В-клетках герминативного центра, в то же время слабо экспрессируется в плазматических клетках. BCL6 ингибирует дифференциацию активированных клеток в плазматические клетки. Индуцирующий В-лимфоциты репрессор транскрипции, белок созревания 1 (Blimp-1) требуется для развития В клетки в плазматическую клетку. Человеческий вариант Blimp-1 называется Prdm1. В настоящем документе любые ссылки на Blimp-1 включают ссылку на Prdm1. Blimp-1 направляет дифференцировку плазматической клетки. BCL6 и Blimp-1 подавляют экспрессию друг друга, и потому в естественной ситуации, когда один белок достигает более высокого уровня экспрессии, чем другой, достигается стадии дифференцировки. В человеческом организме, дифференциация плазматических клеток из активированных наивных клеток или В-клеток памяти предполагает снижение экспрессии BCL6 и увеличение экспрессии Blimp-1. В клетках герминативного центра экспрессия BCL6 находится на высоком уровне, а экспрессия Blimp-1 является низкой. В покоящихся клетках памяти экспрессия BCL6 и Blimp-1 является низкой. Сигналы, вызывающие дифференциацию, приводят к увеличению экспрессии Blimp-1, и образованный Blimp-1 противодействует экспрессии BCL6. Стадия, на которой экспрессируются как BCL6, так и Blimp-1, является кратковременной и называется плазмабласт. С постепенным увеличением уровня Blimp-1, экспрессия BCL6 затухает, что приводит к плазматической клетке.
В одном из вариантов по настоящему изобретению предусмотрена продуцирующая РСВ-специфические антитела клетка, в которой коэкспрессируются BCL6 и Blimp-1 (имеется в виду, что BCL6 и Blimp-1 экспрессируются в указанной продуцирующей антитела клетке по меньшей мере в течение 1 дня, предпочтительно по меньшей мере в течение одной недели, более предпочтительно по меньшей мере в течение шести недель, наиболее предпочтительно по меньшей мере в течение трех месяцев). Указанная продуцирующая РСВ-специфические антитела клетка способна пролиферировать при условии наличия соответствующего сигнала. Было установлено, что коэкспрессия BCL6 и Blimp-1 приводит к продуцирующей антитела клетке, которая способна как пролиферировать, так и производить антитела. BCL6 и Blimp-1 предпочтительно коэкспрессируются в В-клетке, предпочтительно человеческой В-клетке. Коэкспрессия BCL6 и Blimp-1 в В-клетке приводит к стабилизации указанной В-клетки на плазмабласт-подобной стадии. Плазмабласты, как плазматические клетки, способны секретировать антитела. Однако плазмабласты все еще способны пролиферировать, а плазматические клетки потеряли возможность пролиферировать. Поэтому плазматические клетки непригодны для культивирования антителопродуцирующих клеточных линий.
В одном предпочтительном варианте предусмотрена продуцирующая РСВ-специфические антитела клетка, включающая экзогенные последовательности нуклеиновой кислоты, кодирующие BCL6 или его функциональную часть, производное и/или его аналог. Экзогенная нуклеиновая кислота определена в настоящем документе как последовательность нуклеиновой кислоты, которая не относится к естественному геному клетки. С помощью таких экзогенных молекул нуклеиновой кислоты можно регулировать концентрацию BCL6 в продуцирующей антитело клетке независимо от экспрессии эндогенного BCL6. Таким образом, даже если экспрессия эндогенного BCL6 низка или отсутствует, что вызвано, например, Blimp-1, экзогенная последовательность нуклеиновой кислоты, кодирующая BCL6 или его функциональную часть, производное и/или его аналог, по-прежнему способна производить BCL6 в концентрации, которой достаточно для влияния на стабильность продуцирующей антитела клетки. Предпочтительно, указанная последовательности нуклеиновой кислоты, кодирующая BCL6 или его функциональную часть, производное и/или его аналог, конститутивно активна, так что экспрессия BCL6 сохраняется, даже когда экспрессия эндогенного BCL6 в указанной клетке подавлена эндогенным репрессором, таким как Blimp-1. Наиболее предпочтительно, экспрессия указанной последовательности нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или его аналог, регулируется экзогенным индуктором или репрессором, так что степень экспрессии BCL6 регулируется по желанию.
Предпочтительно, продуцирующая РСВ-специфическое антитело клетка по настоящему изобретению включает экзогенную последовательность нуклеиновой кислоты, кодирующую Bcl-xL или его функциональную часть, производное и/или его аналог, как указано более подробно ниже. Если присутствует Bcl-xL или его функциональная часть, производное и/или его аналог, возможно выращивать плазмабласты в условиях низкой плотности клеток. Экспрессия указанной последовательности нуклеиновой кислоты, кодирующей Bcl-xL или его функциональную часть, производное и/или его аналог, предпочтительно регулируется экзогенным индуктором или репрессором, так что степень экспрессии Bcl-xL регулируется по желанию. Поэтому в предпочтительном варианте предусмотрена продуцирующая РСВ-специфическое антитело клетка, включающая:
- экзогенную последовательность нуклеиновой кислоты, кодирующую BCL6 или его функциональную часть, производное и/или его аналог, и/или
- экзогенную последовательность нуклеиновой кислоты, кодирующую Bcl-xL или его функциональную часть, производное и/или его аналог. Указанная продуцирующая РСВ-специфическое антитело клетка предпочтительно включает как экзогенную последовательность нуклеиновой кислоты, кодирующую BCL6 или его функциональную часть, производное и/или его аналог, так и экзогенную последовательность нуклеиновой кислоты, кодирующую Bcl-xL или его функциональную часть, производное и/или его аналог. Предпочтительно, экспрессия указанной последовательности нуклеиновой кислоты, кодирующей BCL6, Bcl-xL или функциональную часть, производное и/или аналог BCL6 или Bcl-xL, регулируется активатором и/или репрессором, который индуцируется экзогенным соединением. Например, может быть использована индуцибельная промотерная система, такая как Тет(тетрациклин)-on или Tet-off системы.
Стабильная продуцирующая РСВ-специфическое антитело клетка по настоящему изобретению предпочтительно получена за счет коэкспрессии BCL6 и Blimp-1 в продуцирующей РСВ-специфическое антитело клетке. Продуцирующая РСВ-специфическое антитело клетка предпочтительно получена из субъекта, который был подвержен воздействию РСВ. Способы выделения продуцирующих антитело клеток хорошо известны в уровне техники. Например, полученные из РСВ соединения, снабженные меткой и/или тэгом, инкубируют с образцом из субъекта, который подвергался воздействию РСВ, где образец включает продуцирующие антитела клетки. Продуцирующие РСВ-специфическое антитело клетки, которые распознают меченые полученные из РСВ соединения, выделяют, в то время как несвязанные клетки смывают. Полученные продуцирующие РСВ-специфическое антитело клетки впоследствии стабилизируют коэкспресией BCL6 и Blimp-1.
В одном варианте предполагается в первую очередь общая стабилизация антитело-продуцирующих клеток из подвергнутого воздействию РСВ донора, а затем выделение клеток, которые распознают меченые полученные из РСВ соединения. В другом варианте продуцирующие антитела клетки снабжены маркером (флуоресцентным) В-клеточного рецептора (BCR, мембранная форма антитела), который подает сигнал, когда продуцирующая антитело клетка связывает не снабженный тэгом/меткой антиген через BCR. Продуцирующие антитела клетки с измененным маркером отбираются и впоследствии стабилизируются коэкспрессией BCL6 и Blimp-1. В другом варианте, когда отсутствуют доступные соединения из антигена, но при наличии доступных анализов для выявления уникальных антител, все/большая часть продуцирующих антитела клеток стабилизируются коэкспрессией BCL6, Blimp 1 и, по желанию, Bcl-xL. Согласно этому варианту клетки культивируют при низкой плотности, предпочтительно от 10 до 100 клеток в 96-ячеечном планшете, в присутствии L-клеток (общие мини-культуры, МВС). Культуральный супернатант может быть использован непосредственно для анализов, таких как ИФА, Вестерн-блот или для функциональных анализов, таких как ELISPOT, анализы на обезвреживание или для анализов клеточной миграции.
В одном варианте для получения клеточных линий интересующих продуцирующих моноклональные антитела клеток отбирают МВС и готовят культуры предельного разведения и, предпочтительно через 2-3 недели, супернатант указанных культур снова проверяют с помощью предпочтительного анализа.
Как хорошо известно специалисту в данной области, в уровне техники имеются многие альтернативные способы. Вышеупомянутые варианты не являются ограничивающими.
Поэтому, кроме того, предусмотрен способ получения продуцирующей антитела клетки, которая является стабильной по меньшей мере в течение трех месяцев и которая способна производить РСВ-специфические антитела или их функциональные эквиваленты, где способ включает:
- повышение уровня экспрессии Blimp-1 в клетке, которая способна производить РСВ-специфические антитела или их функциональные эквиваленты, а также
- увеличение и/или поддержание уровня экспрессии BCL6 в указанной клетке.
С использованием способа по настоящему изобретению стало возможным преобразовать РСВ-специфичные В-клетки памяти в плазмабласт-подобные клетки и стабилизировать указанные клетки, так что быстрая дифференциация в плазматические клетки не происходит. Это идет вразрез с естественным развитием плазматических клеток, где экспрессия Blimp-1 в В-клетках памяти приводит к быстрому развитию в плазматические клетки, тем самым, препятствуя экспрессии BCL6, так что в образовавшихся плазматических клетках почти не экспрессируется BCL6. В одном варианте по настоящему изобретению, таким образом, предполагается коэкспрессия BCL6 и Blimp-1 в РСВ-специфичной В-клетке, что приводит к клетке, способной как пролиферировать, так и производить антитела. Уровень экспрессии BCL6 в указанной РСВ-специфичной В-клетке предпочтительно находится и поддерживается, по существу, на том же уровне или на более высоком уровне по сравнению с уровнем экспрессии в плазмабласте. Таким образом, создается стабильная культура РСВ-специфичных клеток, которые способны производить РСВ-специфичные антитела. Эти РСВ-специфичные клетки, которые коэкспрессируют BCL6 и Blimp-1 предпочтительно в дальнейшем стабилизируют путем добавления антиапоптотического гена Bcl-xL. С введением Bcl-xL возможно культивировать плазмабласты в условиях низкой плотности клеток. Таким образом, изобретение также предусматривает способ культивирования плазмабластов в условиях низкой плотности клеток, включающий создание продуцирующих РСВ-специфические антитела клеток с уровнями экспрессии BCL6, Blimp-1 и Bcl-xL по любому из описанных здесь способов.
Количество продукта экспрессии BCL6 (предпочтительно BCL6 белка) в продуцирующей РСВ-специфические антитела клетке регулируется множеством способов.
В одном варианте предусмотрена продуцирующая антитела клетка с соединением, способным прямо или косвенно влиять на экспрессию BCL6. Продуцирующая антитела клетка предпочтительно предусмотрена с соединением, способным повышать экспрессию BCL6 для противодействия снижению экспрессии BCL6 во время экспрессия Blimp-1. Такие соединения предпочтительно включают воспринимающий сигнал активации и транскрипции белок 5 (STAT5) или его функциональную часть, производное и/или его аналог, и/или последовательность нуклеиновой кислоты, кодирующую его. STAT5 представляет собой датчик сигнала, способный повысить экспрессию BCL6. Существуют две известных формы STAT5, STAT5a и STAT5b, которые закодированы двумя различными, тандемно сцепленными генами. Введение и/или активация STAT5 приводит к повышению уровня BCL6. Следовательно, снижение экспрессии BCL6 под действием Blimp-1 по меньшей мере частично скомпенсировано повышением экспрессии BCL6 под действием STAT5 или его функциональной части, производного и/или аналога. Следовательно, STAT5 или его функциональная часть, производное и/или аналог способны непосредственно влиять на экспрессию BCL6. Также возможность косвенно влиять на экспрессию BCL6. Это может быть осуществлено, например, регулированием количества соединения, в свою очередь способного прямо или косвенно активировать STAT5 и/или регулировать экспрессию STAT5. Таким образом, в одном из вариантов изобретения увеличена экспрессия и/или активность эндогенного и/или экзогенного STAT5. Речь идет о возможности косвенного повышения экспрессии BCL6 при культивировании продуцирующих антитела клеток в присутствии интерлейкина (IL) 2 и/или IL 4, которые способны активировать STAT5.
В одном варианте предусмотрена продуцирующая РСВ-специфические антитела клетка с последовательностью нуклеиновой кислоты, кодирующей STAT5 или его функциональную часть, производное и/или его аналог, где указанная последовательность нуклеиновой кислоты конститутивно активна, что означает, что экспрессия STAT5 конститутивна независимо от присутствия (эндогенных) регуляторов. В том случае, если эндогенная экспрессия STAT5 низка или отсутствует, предпочтительно применить экзогенную конститутивно активную последовательность нуклеиновой кислоты, кодирующую STAT5 или его функциональную часть, производное и/или его аналог, что приведет к концентрации STAT5 или его функциональной части, производного и/или аналога, достаточной для повышения экспрессии BCL6. Наиболее предпочтительно, предусмотрена продуцирующая РСВ-специфические антитела клетка с последовательностью нуклеиновой кислоты, кодирующей соединение, включающее STAT5 или его функциональную часть, производное и/или его аналог, предпочтительно гибридный белок, активность которого регулируется экзогенным индуктором или репрессором, так что степень активации экспрессии BCL6 регулируется по желанию. Предусмотрена еще одна, Tet-on система, позволяющая индуцировать BCL-6, в которой добавление тетрациклина и/или производных тетрациклина вызывает активность трансактиватора, который индуцирует транскрипцию гена BCL6 и последующий синтез белка BCL. В одном из предпочтительных вариантов, продуцирующая антитело клетка снабжена последовательностью нуклеиновой кислоты, кодирующий рецептор эстрогена (ER) и STAT5 как гибридный белок ER-STAT5. Данный гибридный белок неактивен, поскольку образует комплекс с белками теплового шока в цитозоле. Таким образом, STAT5 не в состоянии добраться до ядра, и экспрессия BCL6 не увеличена. При введении экзогенного индуктора 4-гидрокси-тамоксифена (4НТ), гибридный белок ER-STAT5 диссоциирует от белков теплового шока, так что STAT5 может войти в ядро и активировать экспрессию BCL6.
Дополнительно или альтернативно, экспрессия BCL6 в продуцирующей РСВ-специфические антитела клетке повышается при культивировании указанной продуцирующей антитела клетки в присутствии соединения, способного прямо или косвенно повышать экспрессию BCL6.
Поэтому в одном варианте предусмотрен способ получения продуцирующих РСВ-специфические антитела клеток, включающий:
- обработку продуцирующей РСВ-специфические антитела клетки соединением, способным прямо или косвенно повышать экспрессию BCL6 и/или;
- культивирование продуцирующей РСВ-специфические антитела клетки в присутствии соединения, способного прямо или косвенно повышать экспрессию BCL6. Указанное соединение, способное прямо или косвенно повышать экспрессию BCL6, предпочтительно включает STAT5 или его функциональную часть, производное и/или его аналог. Поэтому предусмотрен способ по настоящему изобретению, включающий обработку указанной продуцирующей РСВ-специфические антитела клетки STAT5 или его функциональной частью, производным и/или его аналогом, либо последовательностью нуклеиновой кислоты, кодирующей STAT5 или его функциональную часть, производное и/или аналог. В одном варианте указанная продуцирующая антитела клетка культивируется после введения последовательности нуклеиновой кислоты, кодирующей STAT5 или его функциональную часть, производное и/или аналог в указанную клетку. Указанная последовательность нуклеиновой кислоты, например, вводится в указанную клетку путем трансфекции и/или вирус-опосредованного переноса генов. В уровне техники описано множество альтернативных способов введения последовательности нуклеиновой кислоты в клетку, которые нецелесообразно подробно приводить в настоящем документе.
С помощью соединения, способного прямо или косвенно повышать экспрессию BCL6, возможно усилить экспрессию эндогенного BCL6. В одном из предпочтительных вариантов продуцирующая антитела клетка снабжена последовательностью нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или его аналог. Как пояснено в настоящем документе выше, экзогенная нуклеиновая кислота, кодирующая BCL6, является предпочтительной, поскольку позволяет регулировать концентрацию BCL6 внутри клетки независимо от экспрессия эндогенного BCL6. Таким образом, даже если экспрессия эндогенного BCL6 низка или отсутствует, например, в результате активности Blimp-1, экзогенная последовательность нуклеиновой кислоты, кодирующая BCL6 или его функциональную часть, производное и/или аналог, по-прежнему способна производить концентрацию BCL6, достаточную для оказания влияния на стабильность продуцирующей антитела клетки. Поэтому, кроме того предусмотрен способ по настоящему изобретению, включающий снабжение продуцирующей РСВ-специфические антитела клетки последовательностью нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или его аналог. Предпочтительно, указанная продуцирующая антитела клетка снабжена конститутивно активной последовательностью нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или аналог, так что экспрессия BCL6 сохраняется, даже когда эндогенная экспрессия BCL6 в указанной клетке ингибируется таким эндогенным репрессором, как Blimp-1. Наиболее предпочтительно, экспрессия указанной последовательности нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или его аналог, регулируется экзогенным индуктором или репрессором, так что степень экспрессии BCL6 регулируется по желанию. Например, используется система индуцибильного промотера, такая как Tet-on Tet-off система, как было описано выше.
В другом предпочтительном варианте изобретение предусматривает способ, в котором количество BCL6 косвенно регулируется снабжением продуцирующей РСВ-специфические антитела клетки последовательностью нуклеиновой кислоты, кодирующей Е47 или его функциональную часть, производное и/или его аналог. Е47 кодирует фактор транскрипции, который принадлежит к семейству спираль-петля-спираль белков, называемых Е-белками. Существуют четыре Е-белка, Е12, Е47, Е2-2 и НЕВ, которые участвуют в развитии лимфоцитов. Е12 и Е47 кодируются одним геном, называемым Е2А, который подвергается альтернативному сплайсингу. Е-белки могут ингибироваться ингибиторами Е белков Id2, Id3 и ABF-1 (Матас С., 2006). Е белки были описаны как супрессоры опухолей, и было показано, что их гиперэкспрессия вызывает апоптоз. Одной из специфических мишеней Е47 являются Socs1 и Socs3 гены. Известно, что данные гены Socs представляют собой негативный регулятор STAT5b, и, таким образом, косвенный негативный регулятор BCL6. Иными словами, экспрессия Е47 в В-клетке повышает экспрессию Blimp-1, который приводит к дифференцировке В-клетки к фенотипу продукции антител (плазматической клетке).
Уровень экспрессии Blimp-1 в продуцирующей РСВ-специфические антитела клетке также регулируется разными способами. В одном варианте предусмотрена продуцирующая РСВ-специфические антитела клетка с соединением, способным прямо или косвенно влиять на экспрессию Blimp-1. Дополнительно или альтернативно продуцирующая антитела клетка культивируется в присутствии соединения, способного прямо или косвенно влиять на экспрессию Blimp-1. Поэтому, кроме того предусмотрен способ по настоящему изобретению, включающий снабжение продуцирующей РСВ-специфические антитела клетки соединением, способным прямо или косвенно влиять на экспрессию Blimp-1. Кроме того, предусмотрен способ по настоящему изобретению, включающий культивирование указанной продуцирующей антитела клетки в присутствии соединения, способного прямо или косвенно влиять на экспрессию Blimp-1. Предпочтительно использовать соединение, которое способно повышать экспрессию Blimp-1 для противодействия снижению экспрессии Blimp-1 во время экспрессии BCL6. Указанное соединение наиболее предпочтительно включает IL-21.
В одном из предпочтительных вариантов указанное соединение, способное прямо или косвенно влиять на экспрессию Blimp-1, включает воспринимающий сигнал активации и транскрипции белок 3 (STAT3) или его функциональную часть, производное и/или его аналог, и/или последовательность нуклеиновой кислоты, кодирующую его. STAT3 представляет собой датчик сигнала, который участвует в развитии и дифференцировке В-клеток. STAT3 способен увеличивать экспрессию Blimp-1. Кроме того в связи с этим предусмотрен способ по настоящему изобретению, в котором указанное соединение, включающее STAT3 или его функциональную часть, производное и/или его аналог, или последовательность нуклеиновой кислоты, кодирующую STAT3 или его функциональную часть, производное и/или его аналог, способно прямо или косвенно влиять на экспрессию Blimp-1. Наиболее предпочтительно, экспрессия указанной последовательности нуклеиновой кислоты, кодирующей STAT3 или его функциональную часть, производное и/или его аналог, регулируется экзогенным индуктором или репрессором, так что степень экспрессии STAT3 регулируется по желанию. Например, возможно использовать систему индуцибильного промотера, такую как, например, Tet-on Tet-off система. В одном варианте гибридный белок, включающий STAT3, его производное или аналог, и ER, вводится в указанную клетку, что позволяет регулировать экспрессию STAT3 с помощью гидрокситамоксифена.
Так как STAT3 способен влиять на экспрессию Blimp-1, также возможно косвенное регулирование экспрессии Blimp-1 введением соединения, способного прямо или косвенно регулировать активность и/или экспрессию STAT3. В одном варианте предусмотрена продуцирующая антитело клетка с соединением, которое способно повышать активность STAT3, так что экспрессия Blimp-1 также косвенно увеличивается. Кроме того, в связи с этим предусмотрен способ по настоящему изобретению, в котором предусмотрена продуцирующая антитела клетка с соединением, способным прямо или косвенно повышать активность STAT3.
Таким образом, в одном варианте предусмотрена продуцирующая антитела клетка с соединением, способным прямо или косвенно активировать STAT3, для повышения экспрессии Blimp-1.
STAT3 активируется различными способами. Предпочтительно, STAT3 активируется путем обеспечения продуцирующей антитела клетки цитокином. Цитокины естественно участвуют в дифференцировке В клеток, являясь очень эффективными в регулировании STAT белков. Очень эффективными активаторами STAT3 являются IL-21 и IL-6, известно, что IL-2, IL-7, IL-10, IL-15 и IL-27 также активируют STAT3. Кроме того, Toll-подобные рецепторы (TLRs), которые участвуют во врожденном иммунитете, также способны активировать STAT3. Поэтому в одном варианте предусмотрен способ по изобретению, в котором указанное соединение, способное прямо или косвенно влиять на экспрессию Blimp-1, включает IL-21, IL-2, IL-6, IL 7, IL-10, IL-15 и/или IL-27. Наиболее предпочтительно использовать IL-21, поскольку IL-21 особенно хорошо подходит для оказания влияния на стабильность продуцирующей антитела клетки. IL-21 способен увеличивать экспрессию Blimp-1, даже когда экспрессии Blimp-1 противодействует BCL6.
Дополнительно или альтернативно для активации STAT3 используется мутированная Янус-киназа (JAK). В естественных условиях JAK способен фосфорилировать STAT3 после того, как сам был активирован по меньшей мере одним цитокином. Мутированная Янус-киназа, способная активировать STAT3 независимо от наличия цитокинов, особенно подходит для способа по настоящему изобретению.
Как уже говорилось ранее, соединение, способное повышать экспрессию Blimp-1, в одном варианте включает последовательность нуклеиновой кислоты, кодирующую STAT3 или его функциональную часть, производное и/или его аналог. Присутствие экзогенной последовательности нуклеиновой кислоты, кодирующей STAT3 или его функциональную часть, производное и/или его аналог, обусловливает постоянное присутствие STAT3 или функциональной части, производного и/или его аналога, даже если экспрессия эндогенного STAT3 очень низка или отсутствует.
Также возможно уменьшить экспрессию и/или активность STAT5 для увеличения экспрессии Blimp-1. Если уменьшены количество и/или активность STAT5, также уменьшена активность экспрессии BCL6, что приводит к уменьшению количества продукта экспрессии BCL6. BCL6 и Blimp-1 противодействуют экспрессии друг друга, уменьшение количества продукта экспрессии BCL6 приводит к повышению количества продукта экспрессии Blimp-1. Соединения, способные понижать активность STAT5, таким образом, способны косвенно увеличивать экспрессию Blimp-1. Такие соединения, например, включают белки-супрессоры цитокиновой сигнализации (SOCS). В одном варианте количество продукта экспрессии Blimp-1 в продуцирующей РСВ-специфическое антитело клетке повышено посредством обеспечения указанной клетки SOCS белком и/или путем активации SOCS белка в указанной клетке.
В одном из предпочтительных вариантов уменьшена экспрессия и/или активность STAT5 в продуцирующей РСВ-специфические антитела клетке, снабженной последовательностью нуклеиновой кислоты, кодирующей Е47 или его функциональную часть, производное и/или его аналог. Экспрессия Е47 в В-клетке, экспрессирующей STAT5b на высоком уровне, предотвращает дифференциацию и пролиферацию, то есть блокировка STAT5 через Е47 и SOCS приводит к снижению уровня BCL6, а затем к увеличению уровня Blimp-1. Увеличение уровня Blimp-1 приводит к уменьшению пролиферации и дифференциации вовлеченной клетки к антитело-продуцирующей клетке. Иными словами, экспрессия Е47 в В-клетке повышает экспрессию Blimp-1, что приводит к В-клеточной дифференцировке к фенотипу продукции антител (плазматической клетке).
По меньшей мере функциональная часть белка STAT5, STAT3, белка Bcl-xL и/или BCL6 означает белковую молекулу, которая обладает, качественно, не обязательно количественно, таким же свойством влиять на стабильность продуцирующей антитела клетки по сравнению с белком STAT5, STAT3 белком, Bcl-xL и/или BCL6, соответственно. Функциональные части белка STAT5 и белка STAT3, например, лишены аминокислот, которые не участвуют или очень мало участвуют в указанном свойстве белка. Производное белка STAT5, белка STAT3, Bcl-xL и/или BCL6 определяется как белок, который был изменен таким образом, что свойство указанного белка влиять на стабильность продуцирующей антитело клетки по существу остается таким же, качественно, не обязательно количественно. Производное может быть получено различными способами, например, посредством консервативных аминокислотных замен, где одна аминокислота заменена другой аминокислотой, в целом, с аналогичными свойствами (размером, гидрофобностью, и т.д.), так что общее функционирование белка, скорее всего, не будет серьезно затронуто. Производное, например, включает гибридный белок, такой как STAT5-ER и STAT3-ER гибридные белки, активность которых зависит от наличия 4-гидрокситамоксифена (4НТ). Аналог белка STAT5, белка STAT3, Bcl-xL и/или BCL6 определяется как молекула, обладающая той же возможность влиять на стабильность продуцирующей антитело клетки, качественно, не обязательно количественно. Указанный аналог не обязательно является производным указанного STAT5 белка, STAT3 белка, Bcl-xL и/или BCL6.
В одном из предпочтительных вариантов указанные продуцирующие РСВ-специфические антитела клетки культивируются в присутствии IL-21 до обеспечения указанной продуцирующей антитела клетки последовательностью нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или его аналог. Культивирование продуцирующих РСВ-специфические антитела клеток, предпочтительно В-клеток, в присутствии IL-21 до обеспечения указанной клетки последовательностью нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или его аналог, является предпочтительным, поскольку в данном варианте стабильность, пролиферация и/или продукция антител особенно улучшены.
В предпочтительном варианте в настоящем изобретении предусмотрен способ влияния на стабильность продуцирующей РСВ-специфическое антитело клетки, как описано в настоящем документе, дополнительно включающий прямое или косвенное увеличение количества продукта экспрессии Bcl-xL в указанной продуцирующей антитела клетке. Это осуществляется, например, путем обеспечения указанной продуцирующей антитело клетки последовательностью нуклеиновой кислоты, кодирующей Bcl-xL или его функциональную часть, производное и/или его аналог, или последовательностью нуклеиновой кислоты, кодирующей другие антиапоптотические гены, включая, но, не ограничиваясь, Всl-2. В еще одном варианте это осуществляется обеспечением указанной продуцирующей антитело клетки соединением, способным прямо или косвенно повышать экспрессию Bcl-xL, предпочтительно указанное соединение включает APRIL, BAFF, CD40, BCR стимуляцию, цитокины, факторы роста или расположенные ниже в каскаде эффекторы, такие как JNK и АКТ (РКВ).
Bcl-xL является членом антиапоптотического семейства Всl-2, Всl2-белки взаимодействуют и противодействуют только так называемым членам семейства Всl-2 гомологичного домена 3 (ВН3), таким как Вах, Bak, Bim, и Bad, которые вызывают высвобождение цитохрома с и последующие внутренние стимулы смерти (Бойзе, Л.Г., 1993). Таким образом, защита целостности митохондриальных мембран через такие белки, как Bcl-xL, имеет решающее значение для выживания клеток.
Было показано, что активация STAT5 защищает клетки от гибели. Было показано, что STAT5 регулирует экспрессию Bcl-xL, что поддерживает антиапоптотическую роль STAT5. STAT5 положительно регулирует экспрессию Bcl-xL через STAT связывающие элементы в Bcl-xL промотере. In vivo экспрессия Bcl-xL отсутствует в костном мозге STAT5A/B дважды дефектных мышей. Кроме того, SТАТ5-опосредованное выживание эритробластов зависит от повышения Bcl-xL. Недавно было показано, что трансгенная гиперэкспрессия Bcl-xL в В-клетках мыши способствует выживанию В-клеток и доброкачественных очагов плазматических клеток.
Способ по изобретению особенно подходит для производства клеточных культур, включающих продуцирующие РСВ-специфические антитела клетки, которые способны пролиферировать и секретировать антитела. В одном варианте, для получения ex vivo клеточной культуры используется РСВ-специфичная В-клетка памяти. Указанная В-клетка памяти, предпочтительно, является человеческой и продуцирует человеческое антитело. Указанная В-клетка, предпочтительно, происходит из индивидуума, который ранее подвергался воздействию респираторного синцитиального вируса. В одном варианте, РСВ-специфичные В-клетки выделяют из образцов периферической крови и/или образцов миндалин с использованием способов, известных в данной области техники. В-клетки памяти, например, выделяют селекцией (сортировка с помощью магнитных бусин) по маркеру CD19 и/или CD22 и (последующий) селекцией на IgG клеточной поверхности и/или CD27 и/или негативной селекцией IgM, IgD и/или IgA. В В-клетках экспрессия BCL6 является высокой, в то время как экспрессия Blimp-1 является низкой. Естественное развитие в секретирующую антитела клетку включает повышение экспрессии Blimp-1. Так как Blimp-1 подавляет экспрессию BCL6, повышение экспрессии Blimp-1 в естественном положении приводит к снижению экспрессии BCL6. Однако в предпочтительном варианте по настоящему изобретению экспрессия Blimp-1 увеличена по меньшей мере при частично сохраненной экспрессии BCL6. Это приводит к продуцирующей РСВ-специфическое антитело клетке, в которой BCL6 и Blimp-1 коэкспрессируются. Указанная продуцирующая РСВ-специфическое антитело клетка способна пролиферировать и секретировать анти-РСВ антитела, и поэтому пригодна для использования в ex vivo культуре В-клеток. В еще одном предпочтительном варианте указанная продуцирующая антитела клетка защищена от апоптоза с помощью Bcl-xL. Продуцирующая РСВ-специфическое антитело клетка по настоящему изобретению обладает преимуществом, состоящим в том, что она является стабильной и не подвергается терминальной дифференцировке в течение длительного периода. Указанная продуцирующая антитела клетка по настоящему изобретению является стабильной по меньшей мере в течение одной недели, предпочтительно по меньшей мере в течение одного месяца, более предпочтительно по меньшей мере в течение трех месяцев, наиболее предпочтительно по меньшей мере в течение шести месяцев. В-клетки по настоящему изобретению предпочтительно культивировать в присутствии CD40L, так как репликация большинства В клеток поддерживается CD40L.
В одном варианте экспрессия BCL6 поддерживается на том же уровне или на более высоком уровне по сравнению с В клетками зародышевого (герминативного) центра, поскольку значительная экспрессия BCL6 вместе с экспрессией Blimp-1 приводит к предпочтительным пролиферативным свойствам, продукции антител и/или стабильности продуцирующей антитело клетки. В предпочтительном варианте, указанная экспрессия BCL6 и/или экспрессия Blimp-1 сопровождается экспрессией Bcl-xL, что приводит даже к более предпочтительным пролиферативным свойствам, продукции антител и/или стабильности.
Поэтому в одном варианте предусмотрен способ получения продуцирующей РСВ-специфическое антитело клетки, которая является стабильной по меньшей мере в течение одной недели, предпочтительно по меньшей мере в течение одного месяца, более предпочтительно по меньшей мере в течение трех месяцев, наиболее предпочтительно по меньшей мере в течение шести месяцев, где способ включает:
- получение РСВ-специфичной В-клетки памяти;
- повышение уровня экспрессии Blimp-1 в указанной клетке и
- увеличение и/или поддержание уровня экспрессии BCL6 в указанной клетке. Также предусмотрен способ для получения продуцирующей РСВ-специфическое антитело клетки ex vivo, который включает повышение уровня экспрессии Blimp-1 в РСВ-специфичной В-клетке памяти и увеличение и/или сохранение уровня экспрессии BCL6 в указанной клетке. Указанные уровни экспрессии BCL6 и Blimp-1 предпочтительно установлены и/или поддерживаются на одинаковом уровне, либо на более высоком уровне по сравнению с плазмабластом. В предпочтительном варианте указанная В клетка трансдуцирована BCL6 и Bcl-xL. Кроме того, поэтому предусмотрен способ получения продуцирующей РСВ-специфическое антитело клетки, которая стабильна по меньшей мере в течение трех месяцев, включающий:
- получение способной производить РСВ-специфическое антитело В-клетки с BCL6 или его функциональной частью, производным и/или его аналогом, а также
- обеспечение указанной В-клетки Bcl-xL или его функциональной частью, производным и/или его аналогом, а также
- культивирование указанной В-клетки.
Указанная В-клетка предпочтительно снабжена последовательностью нуклеиновой кислоты, кодирующей BCL6 или его функциональную часть, производное и/или его аналог, а также последовательностью нуклеиновой кислоты, кодирующей Bcl-xL или его функциональную часть, производное и/или его аналог.
Указанную В-клетку предпочтительно культивировать в присутствии соединения, способного повышать экспрессию Blimp-1, такого как, например, IL-21, IL-2, IL-6, IL-7, IL-10, IL-15, IL-27 или мутированная Янус-киназа. Предпочтительно использовать IL-21, потому что данный цитокин особенно подходит для повышения экспрессии Blimp-1 и стабилизации продуцирующей антитело клетки в соответствии со способом по настоящему изобретению. Кроме того, в целях повышения эффективности трансдукции указанную В-клетку предпочтительно культивировать в присутствии IL-21 до трансдукции указанной В-клетки последовательностью нуклеиновой кислоты, кодирующей BCL6 и/или Bcl-xL, или функциональную часть, производное и/или аналог.
В одном варианте указанная В-клетка обеспечивается SOCS белком или его функциональной частью, производным и/или его аналогом, или нуклеиновой кислотой, кодирующей их, так как белок SOCS или его функциональная часть, производное и/или его аналог способны косвенно повышать экспрессию Blimp-1. В другом альтернативном или дополнительном варианте указанная В-клетка обеспечивается Е47 или его функциональной частью, производным и/или его аналогом, или нуклеиновой кислотой, кодирующей их. Как уже отмечалось ранее, в результате повышения уровня Е47 или его функциональной части, производного и/или его аналога усиливается функция SOCS белка и косвенно увеличивается экспрессия Blimp-1.
В Примерах показаны особенно предпочтительные варианты. В одном из наиболее предпочтительных вариантов, РСВ-специфичные В-клетки, во-первых, культивируют в присутствии IL-21. Во-вторых, В-клетки подвергаются реакции трансдукции с использованием нуклеиновой кислоты, кодирующей BCL6, и нуклеиновой кислоты, кодирующей Bcl-xL. Предпочтительно используется трансдукция с вращением. Наиболее предпочтительно, В-клетки и вирус, включающий по меньшей мере одну желаемую нуклеиновую кислоту, смешивают и затем смесь подвергают вращению для достижения высокой эффективности трансдукции. После трансдукции В-клетки культивируют в отсутствие IL-21 и в присутствии IL-4 и L-клеток в течение 3-5 дней для осуществления экспрессии BCL6. Впоследствии, согласно этому предпочтительному варианту В-клетки вновь подвергаются реакции трансдукции с использованием нуклеиновой кислоты, кодирующей BCL6, и нуклеиновой кислоты, кодирующей Bcl-xL. Впоследствии В-клетки вновь культивируют в отсутствие IL-21 и в присутствии IL-4 и L-клеток в течение 3-5 дней для осуществления экспрессии BCL6. Впоследствии клетки, экспрессирующие BCL6 и Bcl-xL отделяют и в культуру вновь вводят IL-21 для повышения репликации и продукции антител. Антитела, выделяемые экспрессирующими Bcl-6, Blimp-1 и Bcl-xL клетками в культуральный супернатант, предпочтительно исследовали на in vitro способность /активность/ реактивность обезвреживать РСВ. Продуцирующие антитела клетки, которые производят указанные антитела, предпочтительно в дальнейшем отбирали, например, с использованием культуры предельных разведений. Таким образом получали стабильные РСВ-специфичные клетки, в которых коэкспрессировались BCL6 и Blimp-1. Указанные В-клетки способны к репликации и продукции антител в in vitro культуре в течение не менее шести месяцев.
В одном варианте предусмотрен способ по настоящему изобретению, дополнительно включающий отбор и/или выделение РСВ-специфических антител или их функциональных эквивалентов. В одном варианте отбирают и/или выделяют IgM-продуцирующие клетки и IgG-продуцирующие клетки. Предпочтительно отбирают и/или выделяют IgG-продуцирующие клетки.
Продуцирующие РСВ-специфические антитела клетки, полученные по способу по настоящему изобретению, пригодны для продукции антител против РСВ. Тем не менее, в одном из предпочтительных вариантов гены, кодирующие тяжелую и/или легкую цепи Ig, выделяют из указанных клеток и экспрессируют в других клетках, таких как, например, клетки яичника китайского хомяка (СНО) или клетки 293 (Т). Указанные другие клетки, которая также называются в настоящем документе продуцирующими клетками, предпочтительно адаптированы к коммерческому производству антител. Пролиферация указанной продуцирующей клетки приводит к линии продуцирующих клеток, способных производить РСВ-специфические антитела. Предпочтительно, указанная линия продуцирующих клеток подходит для получения соединений для применения на людях. Следовательно, указанная линия продуцирующих клеток предпочтительно свободна от патогенных агентов, таких как патогенные микроорганизмы.
Способ по изобретению предпочтительно использовать для создания продуцирующей антитело клетки, которая является стабильной по меньшей мере в течение одной недели, предпочтительно по меньшей мере в течение одного месяца, более предпочтительно по меньшей мере в течение трех месяцев, более предпочтительно по меньшей мере в течение шести месяцев, что делает возможным промышленное производство антител. Наиболее предпочтительно получение стабильной линии клеток, способных производить моноклональные антитела. Это предпочтительно осуществить с помощью В-клеток памяти, которые, например, были выделены из образца путем отбора CD19 и/или CD22 (маркеры В-клеток) и IgG на клеточной поверхности и/или CD27 (для маркирования клеток памяти) и/или негативного отбора IgM, IgD и/или IgA. Кроме того, продуцирующая РСВ-специфическое антитело клетка, например, может быть отобрана с помощью анализа на связывание с использованием РСВ или компонентов, полученных из РСВ, таких как, например, белок РСВ F, G белок и/или SH белок. Впоследствии, согласно этому предпочтительному варианту, в указанной продуцирующей РСВ-специфическое антитело клетке коэкспрессируются Blimp-1 и BCL6, что приводит к культуре клеток, способных специфически связывать РСВ (компонент РСВ). В еще одном предпочтительном варианте указанная В-клетка дополнительно снабжена Bcl-xL или его функциональной частью, производным и/или его аналогом.
Если использовать только одну клетку памяти, то будет получена клеточная линия по настоящему изобретению, производящая моноклональные антитела. Также возможно создать продуцирующую моноклональное антитело клеточную линию, начиная с В-клеток, способных производить антитела против РСВ. После получения стабильной клеточной культуры по способу согласно настоящему изобретению выделяют В-клетку, способную производить антитело против конкретного антигена РСВ, и, предпочтительно по меньшей мере функциональную часть гена из указанной В-клетки, кодирующего тяжелую цепь и/или легкую цепь Ig, экспрессируют в другой клеточной линии. Предпочтительно по меньшей мере функциональную часть гена, кодирующего тяжелую цепь Ig, и по меньшей мере функциональную часть гена, кодирующего легкую цепь Ig из указанной В-клетки, экспрессируют в другой клеточной линии.
В одном варианте продуцирующая антитело клетка, предпочтительно, но необязательно представляющая собой В-клетку памяти, полученную от индивидуума, который ранее был подвержен воздействию РСВ, используется в способе по настоящему изобретению. Таким образом, стало возможным производить желаемые человеческие антитела ex vivo.
Кроме того, предусмотрен способ получения антител, которые способны специфически связывать и/или обезвреживать респираторно-синцитиальный вирус, где способ включает:
- выработку продуцирующей антитела клетки, способной производить РСВ-специфическое антитело по способу согласно настоящему изобретению, и
- получение антител, вырабатываемых указанной продуцирующей антитела клеткой.
Также предусмотрены выделенное или рекомбинантное антитело, а также выделенная или рекомбинантная продуцирующая антитело клетка, получаемые по способу согласно настоящему изобретению, или их функциональные эквиваленты. Указанное антитело предпочтительно включает антитело D25, АМН, AM16 и/или АМ23 или его функциональную часть, производное или его аналог.
После получения продуцирующей РСВ-специфическое антитело клетки согласно изобретению, предпочтительно выделяют и/или получают искусственно по меньшей мере функциональную часть гена, кодирующего тяжелую цепь и/или легкую цепь Ig указанной клетки. В одном варианте предусмотрена последовательность нуклеиновой кислоты, включающая по меньшей мере функциональную часть последовательности нуклеиновой кислоты, приведенной на Фигуре 11, Фигуре 12, Фигуре 14А, Фигуре 14Б и/или Фигуре 14В. Указанная функциональная часть предпочтительно содержит по меньшей мере одну последовательность нуклеиновой кислоты, приведенную на Фигуре 11Г, Фигуре 12, Фигуре 14А, Фигуре 14Б и/или Фигуре 14 В. Указанная функциональная часть кодирует предпочтительно по меньшей мере один CDR, приведенный на Фигуре 11Г, Фигуре 12, Фигуре 14А, Фигуре 14Б и/или Фигуре 14В.
Кроме того, предусмотрены выделенная, искусственная или рекомбинантная последовательности нуклеиновой кислоты, включающие последовательность тяжелой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности 
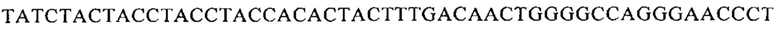
 где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела D25. Указанная последовательность тяжелой цепи предпочтительно включает последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична последовательности, приведенной на Фигуре 11Г. Выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи, также предусмотрена в настоящем документе.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела D25. Указанная последовательность тяжелой цепи предпочтительно включает последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична последовательности, приведенной на Фигуре 11Г. Выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи, также предусмотрена в настоящем документе.
Также предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность легкой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности

 где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела D25.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела D25.
Указанная последовательность легкой цепи предпочтительно включает последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична последовательности, приведенной на Фигуре 11Г. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей легкой цепи.
Кроме того, предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности

 где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела АМ14. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела АМ14. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
Также предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность легкой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности

 где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела AM14. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей легкой цепи.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела AM14. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей легкой цепи.
Кроме того предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности 

 где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела AM16. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела AM16. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
Также предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность легкой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности 

 где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела AM16. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей легкой цепи.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела AM16. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей легкой цепи.
Также предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности  где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела АМ23. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность тяжелой цепи предпочтительно получена из антитела АМ23. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
Также предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность легкой цепи, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% гомологична по меньшей мере части последовательности

 где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела АМ23. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
где указанная часть включает не менее 15 нуклеотидов. Указанная последовательность легкой цепи предпочтительно получена из антитела АМ23. Также в настоящем изобретении предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность тяжелой цепи, состоящую из любой из вышеупомянутых последовательностей тяжелой цепи.
Также предусмотрена последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% идентична по меньшей мере функциональной части аминокислотной последовательности, приведенной на Фигуре 11, Фигуре 14A, Фигуре 14Б и/или Фигуре 14В, где указанная часть включает не менее 5 аминокислотных остатков. Указанная последовательность нуклеиновой кислоты предпочтительно кодирует аминокислотную последовательность, которая по меньшей мере на 80% идентична CDR последовательности 1, 2 и/или 3 тяжелой цепи и/или CDR последовательности 1 или 2 легкой цепи, приведенным на Фигуре 11Г. В другом предпочтительном варианте указанная последовательность нуклеиновой кислоты кодирует аминокислотную последовательность, которая по меньшей мере на 80% идентична по меньшей мере одной из CDR последовательностей, приведенной на Фигуре 14А, на Фигуре 14Б и/или Фигуре 14В. В одном из предпочтительных вариантов указанная последовательность нуклеиновой кислоты кодирует аминокислотную последовательность, которая по меньшей мере на 70% идентична последовательности тяжелой цепи, приведенной на Фигуре 11А, последовательности тяжелой цепи, приведенной на Фигуре 14А, последовательности тяжелой цепи, приведенной на Фигуре 11Б, последовательности тяжелой цепи, приведенной на Фигуре 14В, последовательности легкой цепи, приведенной на Фигуре 11А, последовательности легкой цепи, приведенной на Фигуре 14А, последовательности легкой цепи, приведенной на Фигуре 14Б, и/или последовательности легкой цепи, приведенной на Фигуре 14В.
Кроме того предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность, кодирующую аминокислотную последовательность которая, на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85% идентична аминокислотной последовательности, приведенной Фигурах 11А-Г. Указанная последовательность нуклеиновой кислоты предпочтительно кодирует аминокислотную последовательность, которая по меньшей мере на 80% идентична CDR последовательности 1, 2 и/или 3 тяжелой цепи и/или CDR последовательности 1 или 2 легкой цепи, приведенным на Фигурах 11А-Г. В одном варианте предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность, кодирующую аминокислотную последовательность по меньшей мере на 70% идентичную аминокислотной последовательности NYIIN, и/или по меньшей мере на 75% идентичную последовательности GIIPVLGTVHYAPKFQG, и/или по меньшей мере на 70% идентичную последовательности ETALVVSTTYLPHYFDN, и/или по меньшей мере на 85% идентичную последовательности QASQDIVNYLN, и/или по меньшей мере на 70% идентичную последовательности VASNLET, и/или по меньшей мере на 70% идентичную последовательности

 и/или по меньшей мере на 70% идентичную последовательности
и/или по меньшей мере на 70% идентичную последовательности 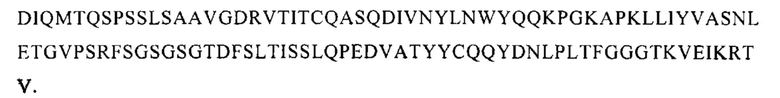
Последовательность нуклеиновой кислоты по настоящему изобретению предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% гомологична любой из вышеуказанных последовательностей.
Кроме того предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность, кодирующую аминокислотную последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85% идентична аминокислотной последовательности, приведенной Фигурах 14А-В. Указанная последовательность нуклеиновой кислоты предпочтительно кодирует аминокислотную последовательность, которая по меньшей мере на 70% идентична CDR последовательности, приведенной на Фигуре 14А, 14Б и/или 14В. В одном варианте предусмотрена выделенная, искусственная или рекомбинантная последовательность нуклеиновой кислоты, включающая последовательность, кодирующую аминокислотную последовательность, которая по меньшей мере на 70% идентична аминокислотной последовательности, выбранной из группы, состоящей из: 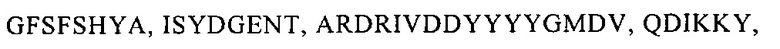

Последовательность нуклеиновой кислоты по настоящему изобретению предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% гомологична любой из вышеуказанных последовательностей.
Как уже указывалось в настоящем документе ранее, последовательность нуклеиновой кислоты по настоящему изобретению особенно подходит для экспрессии антитела или его функциональной части, производного или его аналога по настоящему изобретению, предпочтительно для экспрессии D25, AM14, AM16, АМ23 или его функциональной части, производного и/или его аналога в системе экспрессии нуклеиновых кислот. Последовательность нуклеиновой кислоты по настоящему изобретению предпочтительно экспрессируется в клетке, более предпочтительно, в продуцирующей клетке, адаптированной для производства антител.
Изобретение поясняется следующими примерами. Эти примеры не ограничивают сферу применения изобретения, а служат лишь для пояснения изобретения.
Примеры
Материалы и методы
Поддержание и выделение В клеток человека
CD19 положительные В-клетки человека были выделены из полученной из банка крови лейкоцитной пленки с использованием стандартных процедур (другие источники могут представлять собой свежую кровь с анти-коагулянтным фактором или лимфоидный орган, например, миндалины и селезенку). Были выделены все мононуклеарные клетки периферической крови (РВМС) с помощью разделения по плотности в фиколле (Амершам, Бакингемшир, Великобритания). Для положительного отбора В-клеток с помощью MACS техники сортировки клеток были использованы меченые CD22 бусины, согласно описанию завода-изготовителя (Miltenyi, Утрехт, Нидерланды). Клетки были впоследствии окрашены соответствующей комбинацией моноклональных антител (mAbs) к CD19, CD27, IgD, IgM и IgA (Becton Dickinson (BD), Франклин Лейке, Нью-Джерси, США). Позитивные по CD 19 и CD27 и негативные по IgM, IgA и IgD В-клетки памяти затем были отсортированы помощью FACSAria (BD) (Фигура 1). Кроме В-клеток памяти с помощью соответствующих маркеров могут быть выделены другие подтипы В-клеток, такие как наивные, фолликулярные В-клетки, В-клетки памяти, антитело-продуцирующие В-клетки, центробласты, центроциты, В-клетки зародышевого центра, плазмабласты, плазматические клетки, клетки маргинальной зоны, перисинусоидальные или переходные В-клетки (многие клетки из этих подтипов определены только у мышей).
Культура клеток
Сортированные клетки отмыли и культивировали в 24-луночных планшетах (от 1,5 до 2×105 клеток/мл) на облученных дозой 80 Грэй CD40L-экспрессирующих L-клетках (5×104 клеток/мл; предоставлены доктором J. Banchereau, Schering Plough France, Dardilly France) в полной среде (Модифицированная D минимальная среда Искова, содержащая 8% эмбриональной телячьей сыворотки (FCS) и пенициллин/стрептомицин). Если не указано иное, указанные СО40L-экспрессирующие L-клетки всегда присутствуют в культуре в сочетании с 8% FCS. Для подготовки В-клеток для ретровирусной трансдукции клетки культивировали в течение 36 часов в присутствии IL-21 мыши (50 нг/мл, R&D, Миннеаполис, Миннесота, США). После трансдукции клетки преимущественно культивировали в присутствии IL-21, хотя клетки реагируют на IL-4, IL-15 и IL-10 (не исключая другие цитокины). Например, индуцированный IL-4 рост В-клеток слабее по сравнению с индуцированным IL-21, а более низкий уровень деления клеток может потребоваться для некоторых экспериментов.
Ретровирусные конструкции и получение рекомбинантных ретровирусов
Конститутивно активные мутанты STAT5a и b были описаны ранее. Молекулы ДНК, кодирующие эти мутанты и STAT5b дикого типа, были получены от Т. Китамуры (IMSUT, Токио, Япония). Всl-6 был идентифицирован в исследовании на противодействие старению на мышиных фибробластах как ингибитор антипролиферативной сигнализации p19ARF-p53. Bcl-xL был идентифицирован как анти-апоптотический фактор, был любезно предоставлен доктором Корсмейером (Медицинский Институт Говарда Хьюза, Бостон, США). Данные молекулы ДНК лигировали в LZRS-линкер-IRES-GFP (или IRES-YFP, или IRES-NGFR) векторе, который был описан ранее (Хемскерк и др., 1997; Хемскерк и др., 1999). Вместо IRES-GFP маркера (зеленый флуоресцентный белок) также были использованы IRES-YFP (желтый флуоресцентный белок) или IRES-NGFR (рецептор фактора роста нервов). NGFR представляет собой сигналинг-некомпетентный мутант NGFR, любезно предоставленный доктором К. Бонини. Для визуализации NGFR-экспрессирующих клеток были использованы моноклональные антитела против NGFR (Chromaprobe, Маунтин Вью, Калифорния, США или Miltenyi).
Для получения рекомбинантного ретровируса ретровирусной плазмидой трансфецировали свободную от вируса-помощника амфотропную линию продуцирующих клеток Феникс-А, производное линии человеческих эмбриональных клеток почки 293 (Кинселла и Нолан, 1996) (предоставлены доктором Г. Нолан, Стэнфордский университет, Пало-Альто, Калифорния), с использованием Fugene-6 (Roche Diagnostics Netherlands, Алмере, Нидерланды) по протоколу производителя. Через два дня селекцию трансфецированных клеток начали добавлением 2 мкг/мл пуромицина (Becton Dickinson Clontech Laboratories, Пало-Альто, Калифорния). От десяти до 14 дней после трансфекции 6×106 клеток были высажены на 10 см чашку Петри (Becton Dickinson Discovery Labware, Бедфорд, Массачусетс) в 10 мл полной среды без пуромицина. На следующий день среду обновили, и на следующий день ретровирусный супернатант собрали, центрифугировали и заморозили в свободных от клеток аликвотах при температуре -70°С. Такой подход обеспечивает быстро воспроизводимую продукцию ретровирусов в большом масштабе с высоким титром свыше 3×106 инфекционных вирусных частиц/мл.
Ретровирусная трансдукция
Была проведена процедура трансдукции на фрагментах рекомбинантного человеческого фибронектина СН-296 (RetroNectinТМ; Такара, Оцу, Япония), как описано ранее (Хемскерк и др., 1997; Хемскерк и др., 1999). Не покрытые тканью обработанные для культивирования 24-луночные планшеты (Costar, Бадхеведорп, Нидерланды) были покрыты 0,3 мл 30 мг/мл раствора фрагментов рекомбинантного человеческого фибронектина СН-296 при комнатной температуре в течение 2 часов или в течение ночи при 4°С. При использовании не покрытых тканью культуральных планшетов разных размеров, реагенты используют в соответствующих пропорциях. Раствор СН-296 убрали, затем планшеты инкубировали с 2% человеческим сывороточным альбумином (ЧСА) в фосфатном буферном растворе (PBS) в течение 30 минут при комнатной температуре с последующей промывкой PBS. 5×105 В-клеток, которые были подготовлены для ретровирусной трансдукции, посеяли в 0,25 мл RPMI без FCS и L-клеток и смешали с 0,25 мл размороженного ретровирусного супернатанта. Для двойной трансдукции Bcl-6 и Bcl-xL 125 мкл Bcl-6-IRES-NGFR (или IRES-YFP) (А. Шварц и др., Genes Dev., 2002) и 125 мкл Bcl-XL-IRES-GFP (предоставлено К. Корсмейер, Медицинский Институт Говарда Хьюза, Детский госпиталь, Бостон, США) смешали и добавили к клеткам. Затем культуру центрифугировали при 1800 об/мин при 25°C в течение 60 минут и инкубировали в течение 6 часов при температуре 37°С. Затем были удалены 0,25 мл супернатанта, и было добавлено 0,25 мл свежего ретровирусного супернатанта. Культуру вновь центрифугировали при 1800 об/мин при 25°C в течение 60 минут и инкубировали при 37°C в течение ночи. Следующим утром клетки перенесли на 24-луночные покрытые тканью обработанные для культивирования планшеты (Costar) и культивировали в течение 3-5 дней при нормальных условиях в присутствии IL-4 человека (50 нг/мл) или IL-21 мыши (50 нг/мл, R & D, Миннеаполис, Миннесота, США). Эффективность трансдукции определяли с помощью окрашивания антителами против усеченного сигналинг-некомпетентного мутанта рецептора фактора роста нервов (NGFR, предоставлен К. Бонини, больница Сен-Рафаэле, Милан, Италия) или (ко)экспрессии GFP и/или YFP. Клетки, содержащие представляющий интерес трансген (трансгены), затем отбирали для дальнейших экспериментов.
Проточная цитометрия
Для анализа методом проточной цитометрии были использованы антитела против человеческих молекул IgD, IgG, CD3, CD19, CD20, CD27, CD38, CD40, CD45, CD56, CD70, CD80, CD86, HLA-DR (BD), непосредственно меченые FITC (флуоресцеин изотиоцианат), РЕ, PERCP (пиридинин-хлорофилл-белковый комплекс), РЕ-Су5 (фикоэритрин-цианин-5.18), АРС (аллофикоцианин) или АРС-Су7 и IgM, легкая цепь каппа, легкая цепь лямбда, CD138, прямо меченые РЕ (DAKO). Окрашенные клетки анализировали с помощью LSRII (BD) и данные FACS были обработаны с помощью FlowJo компьютерного программного обеспечения (Tree Star, Inc).
Исследование пролиферации
Наивные В-клетки и В-клетки памяти были выделены из свежих РВМС (мононуклеарных клеток периферической крови) на FACSAria: наивные В-клетки: CD19-PE-Cy7 положительные, CD27-APC отрицательные, IgD-PE положительные
В-клетки памяти: CD19-PE-Cy7 положительные, CD27-APC положительные, IgD-РЕ отрицательные, IgA-FlTC отрицательные
Клетки промыли PBS и ресуспендировали в 0,5 мл среды RPMI (37°С) без FCS. К смеси с клетками добавили равное количество IMDM (среда Дульбекко, модифицированная по способу Исков), содержащей 2 мкМ карбоксифлуоресцеинсукцинимидиловый эфир (CFSE), и инкубировали в течение 7 мин при температуре 37°С. Мечение клеток было остановлено промывкой клеток холодной FCS. Клетки ресуспендировали в 500 мкл IMDM-8% FCS и культивировали с L-клетками в отсутствие или в присутствии IL-21. Немеченые клетки были использованы в качестве контроля.
После 36 часов (непосредственно перед трансдукцией) часть клеток была проанализирована на содержание CFSE. Оставшиеся клетки трансдуцировали при вращении Bcl-6-IRES-NGFR, культивировали в течение 3 дней и проанализировали на содержание CFSE с использованием LSRII. Данные были проанализированы с помощью программного обеспечения FlowJo (Treestar)
Выделение антиген-специфичных В-клеток человека с помощью высокоскоростной сортировки по одной клетке
В дополнение к способу выделения В-клеток памяти, описанному выше, начинающемуся с МВС (т.е. культуры 100 клеток/лунку), человеческие В-клетки памяти могут также инкубироваться с флуоресцентно меченым антигеном и сортироваться по распознаванию антигена. Примером является выделение В-клеток, которые связывают меченый фикоэритрином (РЕ) столбнячный токсин (предоставлен А. Радбрух, Берлин, Германия) (Фигура 4). Клетки культивировали по 1 клетке/лунку и проверяли на связывание столбнячного токсина. Однако может быть использован любой другой меченый антиген.
Определение изменения экспрессии рецепторов В-клеток (BCR) в долгосрочной культуре Всl-6 и Bcl-xL трансдуцированных клеток
Известно, что В-клетки, дифференцирующиеся в ходе культивирования in vitro, теряют экспрессию мембранного BCR, которая наблюдается и в EBV (вирус Эпштейна-Барр) трансформированных лимфоцитах. Поэтому В-клетки, трансдуцированные Всl-6 и Bcl-xL и культивируемые в присутствии IL-21, были окрашены на GFP, NGFR, CD 19, каппа и/или лямбда, или IgG, или окрашены меченым столбнячным токсином. Чтобы показать полезность экспрессии BCR авторы отсортировали связывающие столбнячный токсин-РЕ (Радбрух) клетки с использованием системы для клеточного сортинга FACSAria (BD) по 1 клетке/лунку в 96-луночные планшеты, которые были засеяны L-клетками и заполнены содержащей IL-21 питательной средой. Через три недели связывание столбнячного токсина выросшими клонами было проверено с помощью FACS Canto (BD). Для этого клетки собрали и окрасили в 96-луночном планшете GFP, NGFR, CD 19 и столбнячным токсином-РЕ.
Развитие линий Всl-6 и Bcl-xL положительных В-клеток, которые выделяют антитела
Были созданы линии В-клеток, которые продуцируют моноклональные антитела, и которые являются на 100% Всl-6 и Bcl-xL положительными. Сначала пролиферацию и дифференцировку индуцировали с использованием IL-21. Указанные клетки трансдуцировали Bcl-6-IRES-NGFR и Bcl-xL-IRES-GFP ретровирусами. Клетки поддерживали на IL-4 в течение 3-4 дней. Клетки, которые трансдуцировали одним или обоими ретровирусами, затем экспрессировали трансген и поэтому экспрессировали NGFR или GFP белок. Экспрессия NGFR и/или GFP может быть визуализирована с помощью LSRII (BD). Если необходимо, клетки могут быть снова трансдуцированы для получения большего число клеток, экспрессирующих оба трансгена. Независимо от второй трансдукции клетки, которые экспрессируют оба трансгена, отсортировали с помощью FASC Aria (BD) и культивировали при плотности 10-500 клеток/96-ячеечный планшет в присутствии IL-21 и от 2500 до 5000 L-клеток/ячейку. Указанные общие мини-культуры (МВС) выделяют относительно большое количество антител в культуральный супернатант уже на 5-й день, указанный супернатант затем может быть использован для исследовательских целей. Исследование может быть основано на доступных для интересующего антигена способах, например ИФА/EIA/RIA, Вестерн-блоте, либо прямых функциональных анализах, таких как эксперименты на нейтрализацию блокирования цитокина. После исследования и селекции МВС, которые распознают интересующий антиген (столбнячный токсин и РСВ в настоящих экспериментах), клетки субклонируют при 0,5-1 клетке/ячейку в 96 ячейках в присутствии IL-21. Субклонирование обычно занимает 2-3 недели и может быть выполнено с помощью культур предельного разведения (LD) или сортировки по одной клетке с помощью проточной цитометрии (FACSAria).
Исходный раствор РСВ А-2 вируса и НЕр2 линия клеток
Вирус РСВ А-2 (любезно предоставлен Г. ван Бликом, WKZ, Утрехт) и НЕр2 линию клеток (клиническая лаборатория, АМС, Амстердам) культивировали в больших количествах и заморозили в жидком азоте.
Прикрепленные к субстрату клетки линии Нер2 культивировали в нормальной среде в Т175 Falcon флаконах и заморозили порциями.
Для получения исходного раствора РСВ с высоким титром НЕр2 клетки посеяли и культивировали до 50-60% скопления. К НЕр2 клеткам добавили исходный раствор РСВ (1/20 разбавление с общим объемом 5 мл) на 45 минут при комнатной температуре. Добавили 15 мл свежей среды и клетки оставили на ночь при 37°C с 5% CO2 с открытой крышкой. Следующим утром культуральный супернатант осторожно удалили и добавили 15 мл среды, содержащей 1% FCS. Клетки оставили на срок от 24 до 36 часов при температуре 37°C, 5% CO2 с закрытой крышкой. Когда индуцированный РСВ синцитий был четко виден, и большая часть синцития была еще цела, среда была собрана, отфильтрована (0,22 мкм) и подвергнута вращению при 1450 об/мин при комнатной температуре, после чего образцы быстро заморозили и хранили замороженными в жидком азоте. Среда может быть собрана второй раз при немедленном добавлении новой среды, содержащей 1% FCS, и замораживания этой партии 4-6 часов спустя.
РСВ лизат для ИФА
НЕр2 клетки, которые были инфицированы РСВ А-2 для получения запаса вируса, были использованы для выделения РСВ белков. Сначала клетки были тщательно промыты PBS и трипсинизированы. Трипсин (Gibco) был отмыт, и клеточный осадок лизировали с помощью 1% октилглюкозида (клеточный осадок из одного Т175 флакона обработали 2 мл октилглюкозида). Суспензию гомогенизировали с помощью шприца и иглы (10 движений поршня вверх и вниз), инкубировали в течение 1 часа на льду, а затем диализовали против 2 л TBS буфера с pH 7,4 в течение ночи при 4°С. После осаждения разрушенных клеток был получен супернатант. Содержание белка было определено на уровне 3,6 мг/мл, и белок был использован в концентрации 20 мкг/мл (50 мкл) для ИФА.
Определение ТСID50 и количества БОЕ исходного раствора РСВ
Для определения TCID50 104 НЕр2 посеяли в 96-луночные планшеты и инфицировали серийными разведениями (2 или 10 разведений) РСВ вируса в 4 раза. 2-3 дня спустя культуральный супернатант удалили и клетки фиксировали 80% ацетоном в течение 10 минут при комнатной температуре. После удаления ацетона слой фиксированных клеток высушили и хранили при 4°C или заморозили при температуре -20°С. Для окрашивания РСВ НЕр2 клеток планшеты сначала блокировали 5% сухим молоком в PBS с 0,1% Твин-20. Затем планшеты промыли 3 раза до инкубации в течение 3-5 часов при температуре 37°C с козьими поликлональными анти-PCB-HRP антителами (1:500, Biodesign, Saco, ME, США), после чего планшеты были тщательно промыты. Затем лунки инкубировали с АЕС субстратом в течение 30 минут при комнатной температуре. Зараженные очаги окрасились в красный цвет, их можно было наблюдать с помощью светового микроскопа и сосчитать. Для определения TCID50 было использовано стандартное программное обеспечение Excel.
Для определения количества бляшкообразующих единиц (БОЕ) вируса, 1×105 мл НЕр2 клеток инкубировали в 24-луночных планшетах с 10-кратными серийными разведениями (10-3-10-7) исходного раствора РСВ вируса в среде с 1% FCS при температуре 37°C в течение 45 минут (200 мкл), затем клетки и вирус были покрыты 0,5 мл теплого на ощупь 0,25% агара для выявления бляшек (Biozyme). Агарозный слой предотвращает распространение вируса на неинфицированные клетки через культуральную среду. Таким образом, вирус может заразить только соседние клетки, которые в конечном итоге погибают от вируса и образуют бляшки в монослое НЕр2 клеток. Эти бляшки могут быть наилучшим образом визуализированы окрашиванием фиксированных клеток (96% этанол-100%-ная уксусная кислота-10% формалин 6:2:1) 1%-ным раствором кристаллического фиолетового. Бляшки сосчитали (по меньшей мере, два разных человека), и определили количество БОЕ.
Выбор обезвреживающих респираторно-синцитиальный вирус (РСВ) антител
Чтобы получить клоны анти-респираторно-синцитиальный вирус (РСВ) В-клеток, клетки периферической крови (РВМС) от двух доноров были выделены из полученных из банка крови лейкоцитных пленок (доноры В62 и В63). До сортировки CD19полIgMотрIgDотрIgAотрСD27пол клеток с помощью FACSAria (BD) (фигура 1) с помощью MACS бусин и колонок (Miltenyi) были выделены CD22+ клетки. Только если не указано иное, клетки инкубировали с L-клетками. Клетки культивировали в течение 36 часов в присутствии IL-21 прежде чем трансдуцировать Bcl-6-IRES-NGFR. Через 12 часов клетки собрали и культивировали в течение 3 дней в присутствии IL-4, после чего экспрессирующие NGFR клетки отсортировали с помощью MACS бусин (Miltenyi) и сразу трансдуцировали Bcl-xL-IRES-GFP. В-клетки, которые не связались с MACS бусинами, были смыты и трансдуцированы Всl-6 и Bcl-xL одновременно. Через 12 часов клетки собрали, объединили и культивировали в течение 3 дней в присутствии IL-4, после чего отсортировали по GFP и NGFR экспрессии на FACSAria. Клетки промыли и культивировали при плотности 100 клеток/лунку в 96-луночном планшете (Costar) в присутствии IL-21.
Трансдуцированные Всl-6 и Bcl-xL культуры В-клеток были проверены на связывание РСВ с использованием ИФА лизата РСВ-инфицированных НЕр2 клеток и параллельно с использованием эксперимента на микрообезвреживание РСВ. 104 НЕр2 клеток высеяли на плоскодонные 96-луночные планшеты (Costar) в полной среде. На следующий день среду заменили на 1 час при комнатной температуре смесью РСВ вируса и супернатанта клеточной культуры, которую предварительно инкубировали в течение 30 минут при 37°C. Общий объем составил 25 мкл с конечной концентрацией РСВ 0,1 MOI (множественность заражения). Через 1 час смесь супернатанта с вирусом разбавили в 9 раз PBS и заменили 100 мкл IMDM/5% FCS. Через 2 дня клетки фиксировали 80% ацетоном и окрасили поликлональными анти-PCB-HRP антителами (Biodesign). При обработке Н2О2 и АЕС клетки, зараженные РСВ, проявили красное окрашивание. С использованием световой микроскопии в случае необходимости инфицированные клетки могут быть увидены и сосчитаны. В качестве контроля для обезвреживание РСВ использовали поликлональные козьи анти-РСВ антитела (Abcam, Кембридж, Массачусетс).
ПЦР в реальном времени и клонирование VH и VL районов
Из ~5×105 В-клеток была выделена тотальная РНК с помощью набора RNeasy® mini Kit (Qiagen, Венло, Нидерланды). 250 нг тотальной РНК было подвергнуто обратной транскрипции в объеме 20 мкл, содержащем IX буфер для синтеза первой цепи, 500 мкм дНТФ, 250 нг случайных гексамеров, 5 мМ DTT, 40 единиц RNasin (Promega) и 200 единиц Superscript III РТ (Invitrogen). кДНК разбавили в 10 раз в сверхчистой воде, 2,5 мкл кДНК были подвергнуты ПЦР в 50 мкл раствора, содержащих 20 мМ трис-HCl, 50 мМ KCl, 2,5 мМ MgCl2, 250 мкм дНТФ, 1 единицу AmpliTaq Gold ДНК-полимеразы (Applied Biosystems Inc.), и 25 пмоль каждого праймера. Условия ПЦР были следующими: стадия денатурирования составляла 8 мин при 96°C, затем следовали 35 циклов по 30 секунд при 96°C, 30 секунд при 60°C, 1 минуте при 72°C и по 10 минут удлинения при 72°С.
Продукты ПЦР разделили на агарозном геле, очистили и клонировали в pCR2.1 ТА векторе клонирования по рекомендациям производителя. Анализ последовательности провели с использованием BigDye Terminator chemistry (Applied Biosystems Inc.) и программного обеспечения Vector-NTI (Invitrogen).
Чтобы исключить мутации, индуцированные обратной транскриптазой и/или ДНК-полимеразой, были проведены ряд независимых превращений кДНК и ПЦР-реакций, их продукты были индивидуально клонированы и их последовательности были проанализированы. Консенсусные последовательности были определены с помощью программного обеспечения Vector-NTI Contig Express.
Для экспрессии рекомбинантных антител в клетках 293Т были созданы конструкции с тяжелой и легкой цепями полной длины в pcDNA3.1(+)Zeo (Invitrogen). Вектор экспрессии с тяжелой цепью был создан с помощью амплификации последовательности лидера и VH-района тяжелой цепи клона D25 полимеразной цепной реакцией и введения 5'-NheI сайта и 3'-Xhol сайта. Константную область IgG1 (CH1-шарнирная область-СН2-СН3) амплифицировали из той же кДНК с введением 5'-XhoI и 3'-Notl сайтов. Вектор экспрессии с тяжелой цепью полной длины был получен посредством лигирования в трех точках Nhel/NotI расщепленного pcDNA3.1(+)Zeo. Конструкция для экспрессии с легкой цепью полной длины была создана с помощью амплификации последовательности лидерной последовательности легкой цепи, VL района и константного района легкой цепи с праймерами, вводящими 5'-NheI и 3'-NotI сайты. Последний продукт был клонирован в Nhel/NotI расщепленном pcDNA3.1(+)Zeo с получением вектора экспрессии с легкой цепью полной длины.
Для подтверждения правильности конструкций для экспрессии был проведен анализ последовательности.
Для получения рекомбинантных моноклональных антител была проведена транзиентная двойная трансфекция (Fugene-6, Roche, Германия или липофектамин LTX, Invitrogen) 293Т клеток векторами экспрессии с тяжелой и легкой цепями. Для демонстрирования функционального связывания антител с F-белком РСВ было проведено FACS окрашивание инфицированных РСВ клеток Нер2 полученным культуральным супернатантом (48 часов).
Олигонуклеотиды, использованные для амплификации с помощью ПЦР:
VH района:

Vкаппа района:

Vлямбда района:


Олигонуклеотиды, использованные для получения векторов экспрессии: Вектор экспрессии с тяжелой цепью:

Вектор экспрессии с легкой цепью:

EBV ПЦР в реальном времени
Чтобы проверить, связана ли сильная пролиферативная реакция с присутствием EBV, была выполнена EBV ПЦР в реальном времени. Процедура в реальном времени описана выше. Условия ПЦР были следующими: 7-минутная стадия денатурирования при 94°C, затем 30 циклов по 30 секунд при 94°C, по 30 секунд при 62°C (HPRT1), 52°C (LMP-1) и 58°C (EBNA1/2) и по 30 секунд при 72°C, и наконец по 7 минут удлинения при 72°С.Для ПЦР в реальном времени были использованы следующие олигонуклеотиды: HPRT1 прямой 
 и HPRT1 обратный
и HPRT1 обратный 
 LMP-1 прямой:
LMP-1 прямой: 
 и LMP-1 обратный
и LMP-1 обратный 
 EBNA1/2 прямой
EBNA1/2 прямой 
 и EBNA1/2 обратный
и EBNA1/2 обратный 

В дополнение к ПЦР в реальном времени была проведена ПЦР непосредственно на ДНК клеточного осадка и супернатанта, которые были выделены с помощью комплекта для выделения QIAmp (Qiagen).
Пример 1
Результаты
Фенотип В-клеток
Использование человеческих В-клеток памяти в качестве основы для выделения лекарственных средств опирается на способность выращивать и тестировать эти клетки в течение относительно длительного периода времени. В-клетки человека можно выращивать и поддерживать в лабораторных условиях, однако, не достаточно долго для размножения, выбора и клонирования одной клеточной линии в зависимости от интересующего антигена. Мы разработали технику иммортализации, основанную на генетических изменениях В-клеток человека. Мы изучили нижестоящие мишени STAT5. Одной из мишеней является Bcl-6. Всl-6 подавляет дифференцировку В-клеток в плазматические клетки, которые не могут пролиферировать. Гиперэкспрессия Всl-6 сохраняет баланс BLIMP1, фактора транскрипции, экспрессия которого сильно возрастает при стимуляции В-клеток IL-21 (работает через STAT3). BLIMP1 необходим для развития Ig продуцирующих клеток (CD20-CD38+), в то время как Всl-6 может предотвратить это развитие (клетки, сохраняющие экспрессию CD20, так называемые клетки с фенотипом зародышевого центра).
Чтобы изучить возможные отклонения определенных популяций клеток в компартменте В-клеток, было осуществлено мечение CFSE до стимуляции свежих клеток памяти и наивных клеток человека, которое показало, что все клетки начинают делиться, и что все популяции клеток одинаково трансдуцированны (Фигура 2). В-клетки памяти, трансдуцированые Всl-6, культивировали в присутствии IL-21 и IL-4. Наивные В-клетки были трансдуцированны на более низком уровне и после 36 часов уровень их деления был ниже, но после 3 дней культивации был идентичен уровню деления В-клеток памяти (данные не представлены).
Затем мы показали, что Всl-6 вместе с Bcl-xL (антиапоптотическая нижерасположенная мишень STAT5), CD40L сигнализацией и в присутствии IL-21 поддерживают человеческие IgG В-клетки памяти в CD20+CD38низкий фенотипе в течение длительного периода времени (более 3 месяцев) (Фигура 3). Кроме того, Bcl-6 Bcl-xL-клетки имеют фенотип, соответствующий активированным В-клеткам (см. таблицу 1, проиллюстрированную FACS окрашиванием 3 ТТ+ клонов В-клеток), поскольку эти клетки имеют высокий уровень экспрессии CD80, CD86 и HLA-DR.

Экспрессия антител на мембране
Bcl-6 Bcl-xL трансдуцированные EBV отрицательные клетки поддерживали экспрессию BCR, что определяли по связыванию антигена или Каппа и Лямбда окрашиванию (Фигуры 3 и 4). Следовательно, такие клетки особенно подходят для выделения и/или отбора на желаемую специфичность после долгого периода культивирования, например, с использованием меченого антигена, потому что такие клетки будут связывать указанный меченый антиген с помощью их BCR. Это было подтверждено с помощью сортировки по одной клетке с использованием FACSAria Bcl-6 и Bcl-xL трансдуцированных В-клеток, которые связывают меченый РЕ столбнячный токсин. Через три недели отсортированные по одной клетке клоны были окрашены соответствующими маркерами и меченым РЕ столбнячным токсином в 96-луночных планшетах и измерены на связывание в FACS Canto (BD) (Фигура 4). В тех случаях, когда предпочтительно наличие В-клеточных рецепторов на В-клетках, например, в исследованиях по отбору, В-клетки предпочтительно трансдуцируют Bcl-6 и Bcl-xL, но не инфицируют EBV.
Клеточное деление и кривые роста
Bcl-6 Bcl-xL трансдуцированные В-клетки делятся, в среднем, 0,6 раз в день. Уровень деления варьирует в зависимости от донора и плотности клеток в культуре (Фигура 5а). Анти-РСВ клон D25 обладает уровнем деления 0,47 раз в день (Фигура 5б). Клетки могут быть выращены при плотности ниже 1 клетки/96 лунок для целей клонирования.
Секреция антител Bcl-6 Bcl-xL В-клетками
Bcl-6 Bcl-xL трансдуцированные В-клетки выделяют в среднем один мкг/мл антител, что достаточно для образования количества, необходимого для доклинических испытаний (Фигура 6). D25 анти-РСВ клон продуцирует в три раза больше антител по сравнению с другими исследованными клеточными линиями.
Определение содержания EBV
EBV ПЦР в реальном времени на мРНК Bcl-6 Bcl-xL клеточных линий, которые культивировали с IL-21 и CD40L сигнализацией. В клеточных линиях, полученных с помощью данной техники иммортализации не было обнаружено транскрипта гена EBV (данные не представлены).
Процедура отбора
Благодаря стабильности в росте и экспрессии BCR, данные клетки хорошо подходят для выделения антиген-специфичных клеток. Это дало нам возможность использовать несколько различных методов отбора и процедур клонирования. Один метод позволяет сразу получить антигенспецифичные клетки после введения Всl-6 и Bcl-xL с помощью FACS или сортировки с магнитными бусинами с использованием меченого интересующего антигена, тем самым, повышая вероятность получения нескольких антигенспецифичных клонов В-клеток. Другая возможность состоит в выращивании всех очищенных Bcl-6 Bcl-xL трансдуцированных В-клеток памяти (или любых других В-клеток) при низкой концентрации клеток (например, 100 клеток на лунку). Супернатанты из этих 100 клеток/лунку культур могут быть собраны и протестированы на их специфичность. 100 клеток/лунку культуры, которые распознают антигены, затем субклонируют с помощью культур предельного разведения для получения моноклональных клеточных линий. С использованием обоих способов можно выделить свыше 40 распознающих столбнячный токсин (ТТ) клонов В-клеток. Таким образом, эти клоны либо были отобраны по связыванию ТТ с BCR с помощью FACSAria, либо были отобраны с помощью ИФА скрининга серии культур, пока единственная анти-ТТ моноклональная клеточная линия ни была выделена (данные не представлены).
Выбор обезвреживающих РСВ антител
25 100 клеток/лунку культур от донора В63 полностью блокировали инфекцию и репликацию РСВ. Одна из обезвреживающих 100 клеток/лунку культур, D10, которая производила сильное анти-РСВ антитело, была клонирована с помощью культур предельного разведения. Одно из моноклональных антител, D25, было использовано для продолжения исследований. D25 представляет собой моноклональное антитело с IgGl тяжелой цепью, как это определено с помощью коммерческого ИФА (Sanquin, Амстердам, не показана), и каппа легкой цепью (Фигура 7), которое очень эффективно блокирует РСВ инфекцию со значением IC50 от 0,5 до 1,5 нг/мл (±10 пМ), причем IC50 стандартного анти-РСВ антитела, используемого в клинике (паливизумаб, разработан MeIlmmune), составляет 0,453 мкг/мл (3,02 нМ) (Н. Wu и др., 2005 J. Mol. Biol. и Mejias и др., 2005 Antimicrob. Agents Chemother.) (Фигура 8).
Распознавание антигена
В дополнение к экспериментам на обезвреживание, было определено связывание D25 с зараженными РСВ клетками НЕр2. НЕр2 клетки были инфицированы с использованием обычного протокола получения вирусов. НЕр2 клетки, зараженные РСВ, трипсинизировали и инкубировали с 25-50 мкл культурального супернатанта. Клетки промыли и окрасили мышь-анти-человек IgG-PE антителами (BD или Jackson) для обнаружения связывание антитела D25 с зараженными клетками. R-Biopharm ИФА контрольные антитела были использованы в качестве внутреннего контроля. На Фигуре 9А приведено связывание D25 с интактными, зараженными РСВ НЕр2 клетками.
Поскольку белки оболочки РСВ (мембранные) представляют собой два белка, а именно G и F белки, связывание D25 было исследовано на клетках, инфицированных псевдотипом VSV вируса, включающим РСВ F или РСВ G белок, или не включающим указанных белков (любезно предоставлены Джон К. Роз). Как показано на Фигуре 9Б, D25 сильно связывает в EL-4 клетки, зараженные VSV вирусом с F-белком. В попытке установить эпитоп, распознаваемый D25 против паливизумаб, инфицированные VSV вирусом с F-белком EL-4 клетки инкубировали с увеличивающимися количествами D25 или паливизумаб. Клетки промывали и окрашивали смесью 3 мышь-анти-PCB-F антител (Dako). В отличие от Паливизумаб, для которого показано конкуренция за связывание с инфицированными VSV вирусом с F клетками с мышь-анти-PCB-F антителами, D25 связывание не было затронуто (данные не представлены).
На Фигуре 9В показано связывания Паливизумаб (Synagis) и D25 с инфицированными клетками НЕр2 в зависимости от концентрации. Так как оба антитела связывают их целевой белок в отношении 1 к 1, не наблюдается какого-либо различия в связывании с инфицированными НЕр2 клетками.
Встречаемость связывающих РСВ антиген и обезвреживающих клонов
Мы подсчитали, что встречаемость специфичных к антигену В-клеткок памяти, которые связывают РСВ, составляет 17%, а встречаемость специфических к антигену клеток, которые нейтрализуют РСВ, составляет 6%, как это определено для донора В63. D25 связывается с конформационным эпитопом, который отличается то эпитопа, распознаваемого паливизумаб. Это проиллюстрировано на Фигуре 10, где показано, что D25 не связывается с денатурированными, линейными эпитопами, представленными нанесенным на планшеты ИФА лизатом инфицированных РСВ клеток, причем паливизумаб связывает денатурированный (F) белок.
Выделение и очистка фрагментов антител
Для нескольких линий клеток, в том числе высоко обезвреживающего РСВ клона D25, мы смогли получить объемы до 500 мл. Указанные культуральные супернатанты содержали по меньшей мере 2 мкг/мл антител, поэтому необходимо иметь возможность получить достаточное количество очищенных антител для осуществления доклинических исследований (на животных). Очистку осуществляли с помощью набора для очистки Montage Antigen Purification Kit (Millipore, Billerica, Массачусетс, США) и колонок HiTrap Protein A HP (GE Healthcare, Diegem, Бельгия).
Кроме того, 293T клетки трансфецировали тяжелой и легкой цепями D25, которые были субклонированы в pCDA3.1 векторе экспрессии с помощью липофектамина LTX (Invitrogen). Количество IgG, которое присутствовало в супернатанте, составляло, примерно, 22 мкг/мл (общий объем 50 мл). Указанное антитело, полученное из клонированной нуклеотидной последовательности антитела, экспрессируемого в линии В-клеток D25, также распознает НЕр2 инфицированные клетки (данные не представлены).
Последовательность антитела
На Фигуре 11А приведены нуклеотидные и аминокислотные последовательности тяжелой и легкой цепей B63D10-D25 клона. С использованием стандартной ПЦР в реальном времени и специфичных к антителам праймеров, были определены последовательности тяжелой (VH1-69) и легкой (Vkl 08/018) цепей. Вся последовательность антитела была клонирована с помощью ТОРО векторов, и после контроля последовательности, субклонирована в pcDNA3.1 векторе экспрессии млекопитающих (Invitrogen). На Фигурах 11Б и 11 в приведены VH и VL4 цепи клона, звездочки указывают мутации по сравнению с последовательностью в зародышевой линии клеток Vh1-69, которые должны были произойти во время созревания аффинности и дальнейшего отбора В-клеток.
В настоящем документе приведены методы выделения, характеристика и долгосрочная культура В-клеток памяти человека, полученная с использованием трансгенов Всl-6 и Bcl-xL. Указанная культура предоставляет необходимые инструменты для выделения антител с уникальными свойствами, таких как анти-РСВ моноклональное антитело B63D10-B25. Поскольку указанные В-клетки являются клетками человеческого происхождения, они могут быть легко использованы в качестве лекарственного средства.
Пример 2
Тяжелую и легкую цепи D25 клонировали в стандартных векторах экспрессии, как описано ранее (р44 'последовательность антитела'). Для создания конструкции для экспрессии, которая обусловливает максимальную экспрессию последовательностей тяжелой и легкой цепей D25, с помощью GENEART (Регенсбург, Германия) были оптимизированы кодоны. В ходе этой процедуры были созданы дополнительные сайты рестрикции для упрощения процедуры клонирования в будущем, но самое главное, нуклеотидные кодоны, которые транслируются в аминокислотные последовательности, были оптимизированы для достижения максимальной трансляции в белок. Таким образом, нуклеотидная последовательность была оптимизирована, но аминокислотная последовательность осталась неизменной. В Примере 4 показан потенциал обезвреживания D25, полученного из очищенного супернатанта В-клеток, рекомбинантного D25 и D25, оптимизированного с помощью GENEART. Все эффективно обезвреживают РСВ.
Изменения, введенные с помощью GENEART, по сравнению с исходной последовательностью D25 изображены на Фигуре 12.
Пример 3
После in vitro экспериментов на обезвреживание РСВ авторы тестировали моноклональное антитело D25 на in vivo моделях. Модели, которые были описаны для in vivo анти-РСВ исследований, включают BALB/c мышей и хлопкового хомяка (Sigmodon hispidus) (Mejias и др., Antimicrobial Agents and chemotherapy 2004; p 1811, Johnson S и др., JID 1997; стр. 1215 и Wu H и др., JMB 2007: стр. 652). BALB/c мышиная модель явно является неэффективной моделью, но так как хлопковых хомяков трудно получить и содержать, сначала D25 протестировали на BALB/c мышах.
Протокол: РСВ специфические антитела в BALB/c, 5 дней
План эксперимента:
День -1. внутрибрюшинная инъекция 100 мкл антител
День 0. интраназальное инфицирование 1×107 БОЕ РСВ А2 в 50 мкл
День с 1 до 5 - проверка общего состояния и веса мыши
День 5, вскрытие, изъятие крови и легких
Изъятие крови через прокол вены
Сбор 2,0 мл жидкости при лаваже легких через канюлю в трахее
Извлечение легких
Немедленно осуществили определение TCID50 в жидкости, полученной при лаваже легких (1 мл)
Заморозили 1 мл жидкости, полученной при лаваже легких (ИФА цитокинов / ПЦР в реальном времени), при -80°C
Выполнили определение TCID50 на подготовленном материале (1 мл)
Заморозили 1 мл материала (ИФА цитокинов/ПЦР в реальном времени) при -80°C
Собрали/центрифугировали кровь для hIgG ИФА сыворотки и хранили при -80°C
Результаты показаны на Фигуре 13:
(А) За один день до заражения РСВ (1×107 РСВ-А2 частиц) посредством интраназального распыления, животным внутрибрюшинно вводили различные количества Synagis (Medlmmune), очищенных D25 или IgG1 антител в качестве контроля (Eureka) (табл. 3). (Фигура 13Б) Уровень человеческого IgG определяли в мышиной сыворотке, начиная с 5 дня, и снижение уровня антител в сыворотке наблюдали в течение 5 дней; в Таблице 4 приведен обзор значений времени полураспада. На Фигуре 13В приведены титры вируса из жидкости, полученной при лаваже легких на 5-й день у обработанных и необработанных животных, в то время как на Фигуре 13Д приведено количество Т-клеток и В-клеток в периферической крови обработанных и необработанных мышей. На Фигуре 13Е приведены гистологические препараты легких и бронхов, инфильтрированные (как правило, в основном, эозинофилами) у необработанных и обработанных животных.
Заключение/результаты:
Оценка времени полураспада D25 составляет от 5 до 9 дней на основании расчета (линейного), учитывающего, что 60 мкг и 30 мкг антитела вводили в 0 день (2 и 1 мг/кг соответственно), а на 5-й день было обнаружено 33 мкг или 16 мкг (общий объем мыши 1,5). При введении в 0 день 0,5 мг/кг антитела животному уровень Ig затем снизился с 15 мкг до 11 мкг на 5-й день, что свидетельствует о времени полураспада 9 дней (табл.4).

Был определен титр вируса по данным TCID50 анализа, показано, что у контрольных животных может быть обнаружено 1×104 БОЕ, в то время как у обработанных Synagis (2 мг/кг) или D25 (2, 1 и 0,5 мг/кг) животных ни один вирус не был обнаружен.
Животные, получавшие Synagis или D25, поддерживают высокий % периферических CD4 Т-клеток и В220-лимфоцитов. Животные, получавшие Synagis (2 мг/кг), обладают более низким % CD4 Т-клеток по сравнению с получавшими D25 животными. Хотя это может не иметь значительных последствий, важно отметить, что животные, получавшие низкую дозу D25 (1 и 0,5 мг/кг), поддерживали высокий уровень В-клеток и Т-клеток по сравнению с контрольными животными.
Хотя данные гистологических исследований (Фигура 13Е) не являются количественными, ясно, что Synagis и D25 уменьшают приток иммунных клеток в легкие и вокруг бронхов по сравнению с контролем. При сравнении Synagis и D25, обработанные D25 животные меньше инфильтрированы клетками в легких и вокруг бронхов.
Для проверки D25 на хлопковых хомяках были проведены эксперименты для сравнения животных, обработанных перед заражением РСВ-Х вирусом, NVI (Институт вакцин Нидерландов) (Билтховен, Нидерланды) антителами Synagis и D25.
Пример 4
В дополнение к B63-D10-D25, мы выделили три новых мощных РСВ-нейтрализующих антитела (AM14, AM16 и АМ23) от того же донора (В63). Культуры всех В-клеток, содержащие 100 клеток на одну лунку, которые были изначально выбраны для обезвреживания РСВ, заморожены и хранились в жидком азоте, были разморожены, и их культуральный супернатант был исследован на связывание зараженных РСВ НЕр2 клеток. Мы протестировали связывание с зараженными Нер2 клетками, поскольку это является признаком распознавания антителами нативных, олигомерных мембранных белков РСВ, таких как F и G белки, и может служить хорошим показателем обезвреживания. Когда связывание было обнаружено, клетки культивировали по одной и проверили на связывание для получения клонов. Все три антитела были клонированы в векторе GENEART, который первоначально был сконструирован для D25. Кроме того, все указанные антитела, как D25, распознают F-белок РСВ (данные не приведены). После клонирования и экспрессии в 293Т клетках был очищен рекомбинантный белок (нуклеотидные и аминокислотные последовательности приведены на Фигурах 14А, Б и В). Антитела были испытаны на нейтрализацию против нескольких выделенных РСВ на Vero и НЕр2 клетках (Фигура 15). Все три антитела имеют IgG1 изотип. АМ14 обладает легкой цепью каппа, a AM16 и АМ23 обладают легкой цепью лямбда. Все три антитела, как и D25, содержат соматические гипермутации в их вариабельных районах, что предполагает, что они in vivo претерпели созревание аффинности во время реакции в зародышевом центре, процесса, при котором образуются уникальные последовательности антител.
Результаты представлены на фигурах 15-1 и 15-II: исследование на обезвреживание PC вируса с D25 (sD25), полученными из очищенного супернатанта линии В-клеток, с очищенными рекомбинантными D25 (rD25), с рекомбинантными D25 с GENEART оптимизированными кодонами (rD25 GA), с АМ14, AM16, АМ23 (все представляют собой очищенные рекомбинантные белки) и с Synagis. Обезвреживание вируса антителами было исследовано на двух различных клеточных линиях, (Фигура 15-I) Vero и (Фигура 15-II) Нер2 клетках, с различными антителами: А2 (А), X (Б) и 2006/1 (В) относятся к подтипу А РСВ, в то время как вирус Z (Г) и 2007-2 (Д) относятся к 100 TCID50 каждого вируса добавили к серийным разведениям антител в DMEM (среда Игла, модифицированная по способу Дульбекко) /1% FCS и инкубировали 1 час при 37 градусах до добавления 100 мкл Vero или НЕр2 клеток (1×106/мл). Смесь антител с вирусом не смывали. Через три дня супернатант удалили и клетки фиксировали 80% ацетоном в течение 10 минут при комнатной температуре. После удаления ацетона, слой фиксированных клеток высушили и хранили при 4°C или замороженными при температуре -20°C. Для окрашивания инфицированных РСВ клеток НЕр2, планшеты сначала были заблокированы 5% обезжиренным молоком в PBS с 0,1% Твин-20, затем планшеты промыли 3 раза, затем инкубировали в течение 3-5 часов при температуре 37°C с козьим поликлональными анти-PCB-HRP антителами (1: 500, Biodesign, Сако, Мэн, США) и интенсивно промыли. Впоследствии все лунки инкубировали с субстратом АЕС в течение 30 минут при комнатной температуре. Инфицированные очаги окрасились в красный цвет, и их можно было наблюдать с помощью светового микроскопа и сосчитать. Результат / выводы
Все антитела обезвреживали штаммы РСВ А и B (Таблица 5). В целом различные антитела D25 нейтрализовали РСВ вирусы эффективно, хотя были заметны небольшие экспериментальные вариации. АМ14 столь же эффективно, как D25, a AM16 столь же эффективно, как Synagis. АМ23 нейтрализует А-штамм РСВ очень эффективно, и менее эффективен в обезвреживании РСВ В-штамма, хотя его эффективность сопоставима с Synagis.

sD25 = очищенный полученный из В-клеток культуральный супернатант
rD25 = очищенный рекомбинантный D25
rD25 GA = супернатант 293Т клеток с рекомбинантным D25 с GENEART оптимизированными кодонами
Пример 5
Взаимодополняющие и блокирующие эффекты анти-РСВ антител.
Для исследования D25, Synagis или новых AM антител на предмет конкуренции между ними за распознавание белка F РСВ, авторы предварительно инкубировали РСВ-инфицированные клетки НЕр2 с увеличивающимися концентрациями немеченых антител, пока не достигли плато максимального связывания. Для каждого антитела авторы определили фазу плато, в которой при увеличении количества Ig не наблюдалось увеличение связывания (данные не приведены). После отмывания образцы инкубировали со стандартной дозой (3 пмоль) РЕ-меченых D25 или АРС-меченых Synagis. Эта доза также дает максимальное связывание.
Результаты
Как показано на Фигуре 16, меченые Synagis и D25 показывают уменьшение связывания с инфицированными РСВ клетками НЕр2, если эти клетки предварительно инкубировали с немечеными Synagis или D25. Кроме того, Synagis показывает незначительное уменьшение связывания, индуцированное AM16. Связывание D25 сильно блокируется АМ23, но после предварительной инкубации с AM14 связывание D25, напротив, сильно возрастает. Это свидетельствует о том, что распознаваемый D25 эпитоп в естественных условиях не полностью экспонирован, но экспонирование усиливается после связывания AM14 со своим нативным эпитопом. Это свидетельствует о том, что указанные два антитела могут работать вместе для повышения обезвреживания.
Краткое описание фигур
Фигура 1.
Выделение человеческих IgG положительных В-клеток памяти.
РВМС, выделенные из лейкоцитарной пленки с помощью разделения по плотности в фиколле (Amersham), инкубировали с анти-СБ22 магнитными бусинами перед выделением с помощью MACS колонок (Miltenyi). Затем CD22 положительные клетки инкубировали с антителами против человеческих CD19, CD27, IgM, IgA и IgD (BD). IgM, IgA и IgD отрицательные и CD19 и CD27 положительные клетки были отобраны с помощью высокоскоростной сортировки по одной клетке (FACSAria, BD).
Фигура 2
CFSE окрашивание.
Свежие человеческие В-клетки памяти были выделены, мечены CSFE и стимулированы IL-21 в течение 36 часов, затем трансдуцированы Bcl-6-IRES-NGFR. Клетки дополнительно выдерживали в течение 3 дней с IL-21 до определения содержания CFSE. Краситель CFSE разбавлялся с каждым клеточным делением.
Фигура 3
Пример В-клеток человека, трансдуцированных Всl-6 и Bcl-xL, и трансдуцированных только Bcl-xL.
Клетки поддерживали на облученных L-клетках, экспрессирующих CD40L, и цитокине IL-21. Слева показана экспрессия BCR, определенная по окрашиванию Каппа и Лямбда цепей (93% каппа-лямбда-положительных клеток имеют изотип IgG, данные не приведены). Справа на Х-оси показана экспрессия CD38, а на Y-оси показана CD20 экспрессия. CD38низкийCD20+ окрашивание указывает на В-клетки памяти или на В-клетки зародышевого центра; CD38+CD20- окрашивание указывает на плазмабласты.
Фигура 4
Выделение иммортализованных, антиген-специфичных В-клеток человека.
Человеческие В-клетки памяти выделили, как описано на Фигуре 1, а затем трансдуцировали Bcl-6-IRES-NGFR и Bcl-XL-IRES-GFP. Клетки, экспрессирующие NGFR, GFP и связывающие РЕ-меченый столбнячный токсин (ТТ), были выделены с помощью FACSAria. Клетки культивировали по одной в 96-луночных плоскодонных планшетах в присутствии облученных L-клеток и IL-21, затем их отбирали на основе связывания РЕ-меченого столбнячного токсина с использованием FACS Canto (BD).
Фигура 5
Совокупный рост и скорость деления клона В-клеток 6XL.
В-клетки из (А), два анти-ТТ клона и один (В) анти-РСВ клон (B63D10-D25) культивировали в присутствии IL-21 и облученных L-клеток.
Фигура 6
Свежие культуры начали с 200000 клеток/24 лунки в 1,0 мл IMDM с 8% FCS и пенициллином/стрептомицином. FCS была использована или в нормальном количестве (HyClone) или в очень малом количестве Ultralow Bovine IgG FCS (Gibco). Через 3 дня культуральный супернатант заменили и количество клеток было скорректировано до 200000 клеток/мл. Показана средняя продукция IgG в измеренных в течение 3 дней 3 последовательных точках, различие не было значимым (значение p 0,2).
Фигура 7
Для определения фенотипа легкой цепи D25 анти-РСВ клона, В-клетки D25 линии окрашивали каппа-фикоэритрин или лямбда-фикоэритрин (BD) антителами. Только каппа-фикоэритрин антитела связывали клеточную линию, показывая этим, что антитело имеет каппа легкую цепь.
Фигура 8
Культуры, содержащие 100 клеток/лунку, от донора В63 выращивали с использованием Bcl-6 Bcl-xL положительных В-клеток памяти человека. Одна из указанных культур, D10, продемонстрировала сильное обезвреживание. С помощью предельных разведений были получены моноклональные клеточные линии, одна из них, D25, эффективно обезвреживала РСВ-2 вирус. На фигуре показано действие D25 по сравнению с паливизумаб (synagis) и поликлональными козьими анти-РСВ антителами. На фигуре не показаны данные по культуральным супернатантам Bcl-6 Bcl-xL трансдуцированных клонов В-клеток, культивируемых с IL-21 и при CD40L сигнализации, которые производят высокие уровни антител, но не блокируют РСВ инфекцию. Клон D25 был использован для дальнейших исследований.
Фигура 9
Фигура 9А: НЕр2 клетки были высеяны по 10-12е6 клеток во флакон Т175 (Nunc) в IMDM/5% FCS. На следующий день среду заменили 5 мл среды с РСВ вирусом (1,0 MOI) и инкубировали 45 минут при комнатной температуре, затем добавили 20 мл свежей среды и клетки культивировали в течение ночи при 37°С.
На следующий день среду заменили IMDM/1% FCS и культивировали в течение ночи с закрытой крышкой при 37°С. На следующий день клетки промыли PBS и обработали трипсином. Для окрашивания инфицированных клеток сначала провели инкубацию с культуральным супернатантом. Последующая инкубация осуществлялась с анти-IgG человека-РЕ антителами (BD). Клетки проанализировали с помощью LSRII (BD). В качестве положительного контроля был использован положительный контроль коммерческого набора для ИФА от r-Biopharm.
Фигура 9Б: EL-4 клетки инфицировали псевдотипом VSV вируса с F или G белком РСВ (любезно предоставлен Джоном Роуз) и инкубировали с культуральным супернатантом D25. Клетки промывали и инкубировали с анти-IgG человека-РЕ антителами (Jackson) для обнаружения связывания D25 с зараженными клетками. Было обнаружено связывание D25 только с клетками, инфицированными псевдотипом вируса VSV с F-белком РСВ.
На Фигуре 9В показано связывание Паливизумаб и D25 с инфицированными клетками НЕр2 (Synagis) в зависимости от концентрации. Показана средняя интенсивность флуоресценции (MFI).
Фигура 10
Связывание поликлональных козьих анти-РСВ (положительный контроль), паливизумаб (synagis) и D25 с адсорбированным лизатом НЕр2 инфицированных клеток.
Фигура 11
Анализ последовательности D25 клона.
На Фигуре 11А приведена нуклеотидная последовательность и предуказанная аминокислотная последовательность вариабельных доменов тяжелой и легкой цепей. На Фигуре 11Б/В показана последовательность тяжелой и легкой цепи D25 по сравнению с предсказанной последовательностью исходной линии клеток. Звездочки указывают мутации, которые, вероятно, произошли во время отбора и созревания аффинности клонов В-клеток in vivo.
Фигура 12
Клонирование и экспрессия рекомбинантных человеческих антител из BCL6 BCL-xL трансдуцированной линии клеток.
Указанные процедуры уже описаны для антитела D25 (Фигура 11). Здесь приведены нуклеотидные модификации GENEART по сравнению с исходной последовательностью D25, необходимо отметить, что эти мутации не изменяют аминокислотной последовательности D25 антитела.
Фигура 13
Введение BALB/c мышам очищенных, полученных из супернатанта В-клеток, D25 и Synagis антител.
(А) За один день до заражения РСВ (1x107 РСВ-А2 частиц) посредством интраназального распыления животным интраперитонеально вводили различные количества Synagis (MedImmune), очищенных D25 или контрольных IgG1 антител (Eureka) (Таблица 3). (Б) Уровень человеческого IgG определяли в мышиной сыворотке, начиная с 5 дня, и снижение уровня антител в сыворотке наблюдали в течение 5 дней (В); в Таблице 4 приведен обзор значений времен полураспада. На Фигуре 13Г приведены титры вируса в жидкости, полученной при лаваже легких на 5-й день у обработанных и необработанных животных, в то время как на Фигуре 13Д приведено количество Т-клеток и В-клеток в периферической крови обработанных и необработанных мышей. (Е) Показаны гистологические препараты легких и бронхов и инфильтрация (как правило, в основном, эозинофилами) у необработанных и обработанных животных.
Фигура 14
Нуклеотидные и аминокислотные последовательности трех новых мощных РСВ-нейтрализующих антител (А) АМ14, (Б) и АМ16 (В) АМ23.
Фигура 15
Исследование обезвреживания PC вируса очищенным D25, полученным из супернатанта линии В-клеток (sD25), очищенным рекомбинантным D25 (rD25), рекомбинантным D25 с GENEART оптимизированными кодонами (rD25 GA), AM14, AM16, АМ23 (все представляют собой очищенные рекомбинантные белки) и Synagis антителом. Обезвреживание вируса антителами было протестировано на двух различных клеточных линиях, (Фигура 15-I) Vero и (Фигура 15-II) Нер2 клетках, с различными антителами: А2 (А), X (Б) и 2006/1 (В) относятся к подтипу А РСВ, в то время как вирус Z (Г) и 2007-2 (Д) относятся к подтипу В. 100 TCID50 каждого вируса добавили к серийным разведениям антител в DMEM/1% FCS и инкубировали 1 час при 37 градусах до добавления 100 мкл Vero или НЕр2 клеток (1×107 мл).
Фигура 16
Относительное связывание фиксированного количества (3 пмоль) меченых АРС Synagis и меченых РЕ rD25 с зараженными РСВ НЕр2 клетками, которые предварительно инкубировали с увеличивающимися концентрациями указанных немеченых антител.
Источники информации
Banchereau, J., de Paoli, P., Valle, A., Garcia, E., Rousset, F., (1991). Long term human В cell lines dependent on interleukin-4 and antibody to CD40, Science 251, 70-2.
Boise, L.H., M. Gonzalez-Garcia, С.E. Postema, L. Ding, T. Lindsten, L.A. Turka, X. Mao, G. Nunez, and С.B. Thompson. (1993). Bcl-x, a bcl-2-related gene that functions as a dominant regulator of apoptotic cell death. Cell 74: 597.
Dadgostar, H., Zarnegar, B., Hoffmann, A., Qin, X.F., Truong, U., Rao, G., Baltimore, D., и Cheng, G. (2002). Cooperation of multiple signaling pathways in CD40-regulated gene expression in В lymphocytes. Proc. Natl. Acad. Sci USA 99, 1497-1502.
Heemskerk и др., 1997: J. Exp. Med. том 186, стр. 1597-1602.
Heemskerk и др., 1999: Cell Immunol, том 195, стр. 10-17.
Kinsella и Nolan, 1996: Hum. Gene Ther. том 7 стр. 1405-1413.
Malisan, F., Briere, F., Bridon, J.M., Harindranath, N., Mills, F.C., Max, E.E., Banchereau, J., Martinez-Valdez, H. (1996). Interleukin-10 induces immunoglobulin G isotype switch recombination in human CD40-activated naive В lymphocytes, J. Exp. Med. 183, 937-47.
Mathas S, Janz M, Hummel F, Hummel M, Wollert-Wulf B, Lusatis S, Anagnostopoulos I, Lietz A, Sigvardsson M, Jundt F, Johrens K, Bommert K, Stein H, Dorken В (2006). Intrinsic inhibition of transcription factor E2A by HLH proteins ABF-1 and Id2 mediates reprogramming of neoplastic В cells in Hodgkin lymphoma. Nat Immunol. 7, 207-215.
Mejias А и др., Antimicrobial Agents and chemotherapy 2004; стр. 1811, Johnson S и др., JID 1997; стр. 1215
Wu H и др., JMB 2007: стр. 652.
Shvarts А. и др, 2002: Genes Dev. том 16, стр. 681-686.
Traggiai, Е., Becker, S., Subbarao, К., Kolesnikova, L., Uematsu, Y., Gismondo, M.R., Murphy, B.R., Rappuoli, R., Lanzavecchia, A. (2004). An efficient method to make human monoclonal antibodies from memory В cells: potent neutralization of SARS coronavirus. Nature Medicine, том 10, №8, 871-875.
Ye, В.H., Cattoretti, G., Shen, Q., Zhang, J., Hawe, N., de Waard, R., Leung, C., Nouri-Shirazi, M., Orazi, A., Chaganti, R.S., и др. (1997). The BCL-6 proto-oncogene controls germinal-centre formation and Th2-type inflammation. Nat Genet 16, 161-170.
















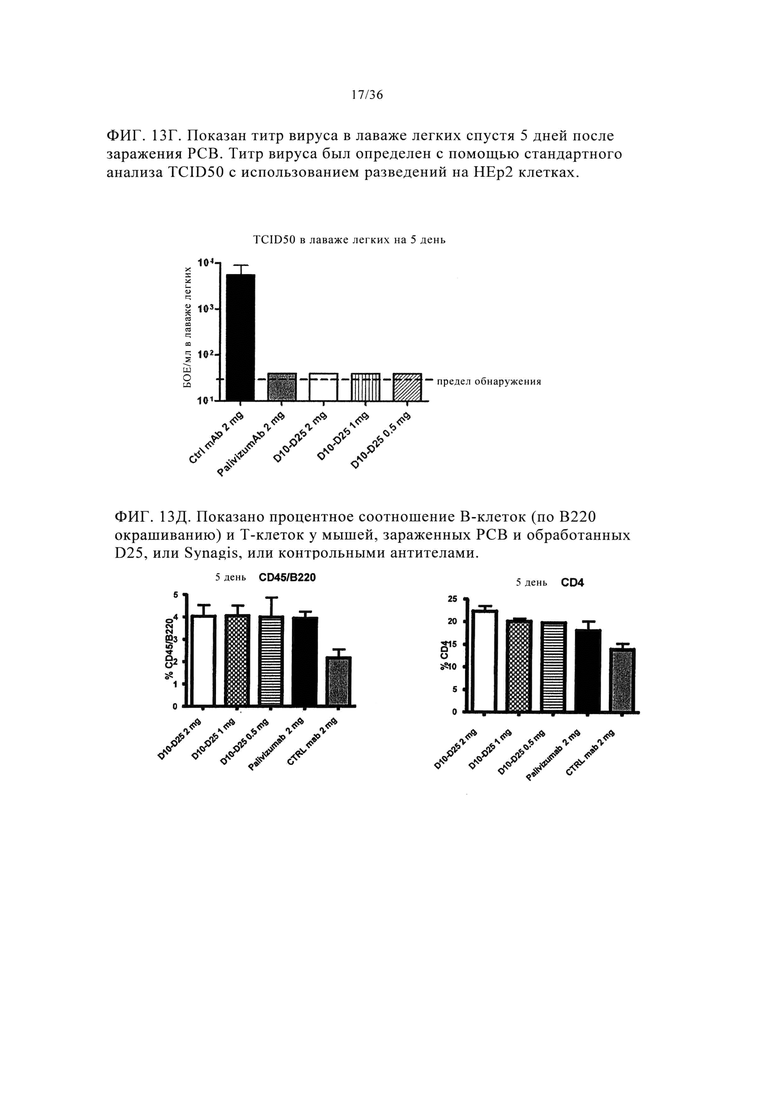



















Изобретение относится к области биохимии. Предложено выделенное антитело или его функциональная часть, способное специфически связывать респираторно-синцитиальный вирус (РСВ). Также предложена молекула нуклеиновой кислоты, кодирующая указанное антитело. Описаны вектор экспрессии и выделенные клетки, включающие указанную молекулу нуклеиновой кислоты и предназначенные для получения указанного антитела. Кроме того, изобретение относится к способу получения указанного антитела, а также к применению указанного антитела в способах лечения или предотвращения РСВ-связанного расстройства. Изобретение позволяет эффективно связывать РСВ и может быть использовано в терапии РСВ-связанных расстройств. 9 н. и 18 з.п. ф-лы, 16 ил., 2 табл., 5 пр.
1. Выделенное антитело или его функциональная часть, способное специфически связывать респираторно-синцитиальный вирус, где антитело или его функциональная часть включает:
- CDR1 последовательность тяжелой цепи, включающую последовательность GFSFSHYA (SEQ ID NO: 73), и
- CDR2 последовательность тяжелой цепи, включающую последовательность ISYDGENT (SEQ ID NO: 74), и
- CDR3 последовательность тяжелой цепи, включающую последовательность ARDRIVDDYYYYGMDV (SEQ ID NO: 75), и
- CDR1 последовательность легкой цепи, включающую последовательность QDIKKY (SEQ ID NO: 76), и
- CDR2 последовательность легкой цепи, включающую последовательность DAS, и
- CDR3 последовательность легкой цепи, включающую последовательность QQYDNLPPL (SEQ ID NO: 77).
2. Антитело или его функциональная часть по п.1, которое обладает значением IC50 менее 10 нг/мл, предпочтительно менее 2 нг/мл, в in vitro анализе обезвреживания, в котором НЕр-2 клетки инфицированы РСВ.
3. Антитело или его функциональная часть по п.1, имеющее последовательность тяжелой цепи, включающую последовательность, которая по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентична последовательности EVQLVESGGGVVQPGRSLRLSCAASGFSFSHYAMHWVRQAPGKGLEWVAVISYDGENTYYADSVKGRFSISRDNSKNTVSLQMNSLRPEDTALYYCARDRIVDDYYYYGMDVWGQGATVTVSS (SEQ ID NO: 78), и/или имеющее последовательность легкой цепи, которая по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентична последовательности DIQMTQSPSSLSASVGDRVTITCQASQDIKKYLNWYHQKPGKVPELLMHDASNLETGVPSRFSGRGSGTDFTLTISSLQPEDIGTYYCQQYDNLPPLTFGGGTKVEIKRTV (SEQ ID NO: 79).
4. Антитело или его функциональная часть по п.1, где антитело или его функциональная часть включает:
вариабельную последовательность тяжелой цепи, содержащую определяющую комплементарность область тяжелой цепи CDR1, включающую аминокислотную последовательность GFSFSHYA (SEQ ID NO: 73), последовательность CDR2 тяжелой цепи, включающую аминокислотную последовательность ISYDGENT (SEQ ID NO: 74), последовательность CDR3 тяжелой цепи, включающую аминокислотную последовательность ARDRIVDDYYYYGMDV (SEQ ID NO: 75), и
вариабельную последовательность легкой цепи, содержащую определяющую комплементарность область легкой цепи CDR1, включающую аминокислотную последовательность QDIKKY (SEQ ID NO: 76), последовательность CDR2 легкой цепи, включающую аминокислотную последовательность DAS, и последовательность CDR3 легкой цепи, включающую аминокислотную последовательность QQYDNLPPL (SEQ ID NO: 77).
5. Антитело или его функциональная часть по п.1, где антитело или его функциональная часть включает вариабельную последовательность тяжелой цепи, содержащую аминокислотную последовательность EVQLVESGGGVVQPGRSLRLSCAASGFSFSHYAMHWVRQAPGKGLEWVAVISYDGENTYYADSVKGRFSISRDNSKNTVSLQMNSLRPEDTALYYCARDRIVDDYYYYGMDVWGQGATVTVSS (SEQ ID NO: 78).
6. Антитело или его функциональная часть по п.1, где антитело или его функциональная часть включает вариабельную последовательность легкой цепи, содержащую аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCQASQDIKKYLNWYHQKPGKVPELLMHDASNLETGVPSRFSGRGSGTDFTLTISSLQPEDIGTYYCQQYDNLPPLTFGGGTKVEIKRTV (SEQ ID NO: 79).
7. Антитело или его функциональная часть по п.1, где антитело или его функциональная часть включает вариабельную последовательность тяжелой цепи, содержащую аминокислотную последовательность EVQLVESGGGVVQPGRSLRLSCAASGFSFSHYAMHWVRQAPGKGLEWVAVISYDGENTYYADSVKGRFSISRDNSKNTVSLQMNSLRPEDTALYYCARDRIVDDYYYYGMDVWGQGATVTVSS (SEQ ID NO: 78), и включает вариабельную последовательность легкой цепи, содержащую аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCQASQDIKKYLNWYHQKPGKVPELLMHDASNLETGVPSRFSGRGSGTDFTLTISSLQPEDIGTYYCQQYDNLPPLTFGGGTKVEIKRTV (SEQ ID NO: 79).
8. Антитело или его функциональная часть по любому из пп.1-7, причем антитело представляет собой человеческое антитело и/или химерное антитело.
9. Антитело или его функциональная часть по любому из пп.1-7, где антитело или его функциональная часть представляет собой однодоменное антитело, одноцепочечное антитело, одноцепочечный вариабельный фрагмент (scFv), Fab фрагмент или F(ab') фрагмент.
10. Антитело или его функциональная часть по любому из пп.1-7, где антитело или его функциональная часть является моноклональным антителом.
11. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его функциональную часть по любому из пп.1-10.
12. Вектор экспрессии, включающий молекулу нуклеиновой кислоты по п.11.
13. Выделенная клетка млекопитающего для получения антитела или его функциональной части по пп.1-10, включающая молекулу нуклеиновой кислоты по п.11.
14. Способ получения антитела или его функциональной части по пп.1-10, который включает культивирование клетки по п.13 in vitro и получение антител или их функциональных частей, которые продуцируются клеткой.
15. Выделенная продуцирующая антитело клетка, способная производить антитело или его функциональную часть по пп.1-10, включающая молекулу нуклеиновой кислоты по п.11 и коэкспрессирующая BCL6 и Blimp-1.
16. Продуцирующая антитело клетка по п.15, которая является стабильной в течение девяти недель, предпочтительно по меньшей мере в течение трех месяцев, более предпочтительно по меньшей мере в течение шести месяцев.
17. Продуцирующая антитело клетка по п.15 или п.16, включающая:
- последовательность экзогенной нуклеиновой кислоты, кодирующую BCL6, или ее функциональную часть, и/или
- последовательность экзогенной нуклеиновой кислоты, кодирующую Bcl-xL, или ее функциональную часть.
18. Продуцирующая антитело клетка по п.17, в которой экспрессия последовательности нуклеиновой кислоты, кодирующей BCL6, Bcl-xL или функциональную часть BCL6 или Bcl-xL, регулируется активатором и/или репрессором, который индуцируется экзогенным соединением.
19. Выделенная молекула нуклеиновой кислоты по п.11, где указанная последовательность нуклеиновой кислоты включает последовательность, которая выбрана из SEQ ID NO: 95, SEQ ID NO: 97, SEQ ID NO: 99, SEQ ID NO: 103, SEQ ID NO: 106, нуклеотидной последовательности GATGCATCC, SEQ ID NO: 107, SEQ ID NO 101 или SEQ ID NO: 108
20. Выделенная молекула нуклеиновой кислоты п.11, где последовательность нуклеиновой кислоты включает
а. нуклеотидную последовательность вариабельной области тяжелой цепи, которая содержит нуклеотидную последовательность SEQ ID NO: 101;
б. нуклеотидную последовательность вариабельной области легкой цепи, которая содержит нуклеотидную последовательность SEQ ID NO: 108; или
в. нуклеотидную последовательность вариабельной области тяжелой цепи, которая содержит нуклеотидную последовательность SEQ ID NO: 101, и нуклеотидную последовательность вариабельной области легкой цепи, которая содержит нуклеотидную последовательность SEQ ID NO: 108.
21. Способ по меньшей мере частичного излечения или предотвращения РСВ-связанного расстройства, где способ предусматривает введение индивидууму, нуждающемуся в этом, терапевтически эффективного количества антитела или его функциональной части по любому из пп.1-10.
22. Композиция для лечения или предотвращения РСВ-связанного расстройства, или предотвращения, или противодействия неблагоприятных последствий РСВ инфекции у субъекта, которым является человек, где композиция включает терапевтически эффективное количество антитела или его функциональной части по любому из пп.1-10, и фармацевтически приемлемый носитель, разбавитель или эксципиет.
23. Композиция по п.22, дополнительно включающая второе антитело или его функциональную часть, которое способно специфично связывать F антиген респираторного синовиального вируса РСВ и где второе антитело включает
вариабельную последовательность тяжелой цепи, содержащую определяющую комплементарность область тяжелой цепи CDR1, включающую аминокислотную последовательность NYIIN (SEQ ID NO: 1), последовательность CDR2 тяжелой цепи, включающую аминокислотную последовательность GIIPVLGTVHYAPKFQG (SEQ ID NO: 2), последовательность CDR3 тяжелой цепи, включающую аминокислотную последовательность ET-ALVVSTTYLPHYFDN (SEQ ID NO: 3), и
вариабельную последовательность легкой цепи, содержащую определяющую комплементарность область легкой цепи CDR1, включающую аминокислотную последовательность QASQDIVNYLN (SEQ ID NO: 4), последовательность CDR2 легкой цепи, включающую аминокислотную последовательность VASNLET (SEQ ID NO: 5), и последовательность CDR3 легкой цепи, включающую аминокислотную последовательность QQYDNLP (SEQ ID NO: 6).
24. Композиция по п.22 или 23, где человек является субъектом с хроническим заболеванием легких, врожденным пороком сердца, или ослабленным иммунитетом, или ребенком младше 6 недель, или пожилым субъектом.
25. Применение антитела или его функциональной части по пп.1-10 для получения лекарственного средства для лечения или предотвращения РСВ-связанного расстройства, или предотвращения, или противодействия неблагоприятных последствий РСВ инфекции у субъекта, которым является человек.
26. Применение по п.25, где лекарственное средство создано для введения в дозе от 0,1 до 10 мг/кг веса тела субъекта, которым является человек.
27. Применение по п.25, где человек является субъектом с хроническим заболеванием легких, врожденным пороком сердца, или ослабленным иммунитетом, или ребенком младше 6 недель, или пожилым субъектом.
| US 2005288491 A1, 29.12.2005 | |||
| JOHNSON S | |||
| et al., Development of a Humanized Monoclonal Antibody (MEDI-493) with Potent In Vitro and In Vivo Activity against Respiratory Syncytial Virus, J Infect Dis, 1997, vol.176 (5), pp.1215-1224 | |||
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ БЕЛОК РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА (ВАРИАНТЫ) | 1994 |
|
RU2160119C2 |
| АВЕРЬЯНОВ А.В., Острые респираторные вирусные инфекции, Атмосфера | |||
| Пульмонология и аллергология, 2003, N 4, с.32-34. | |||

