Респираторно-синцитиальный вирус (PCB) является основной причиной заболеваний нижних отделов дыхательной системы у детей младенческого и раннего возраста /Meintosh and Chanock, 1985, Virology, Fields, B. (ed), Raven, HY, pp. 1285-1304/. Во всех географических зонах он является основной причиной бронхиолита у младенцев и маленьких детей. В период детства агент часто вызывает повторную инфекцию, но болезнь, вызванная повторной инфекцией, обычно слабее, чем болезнь, связанная с начальной инфекцией, и редко вызывает большие проблемы.
PC-вирус является содержащим РНК-оболочку вирусом семейства Poramyxo Viridal и рода пневмовируса. Два главных белка оболочки представляют собой C-белок, который ответственен за прикрепление вируса к мембране клетки-хозяина, и слитый белок /F-белок/, который ответственен за слияние вируса и клеточных мембран. Слияние вируса с клеткой является необходимой стадией инфекции. Слитый белок также требуется для слияния клетки с клеткой, которое является другим путем распространения инфекции от инфицированной клетки к неинфицированной клетке.
Антитела, направленные против слитого белка или против C-белка, могут нейтрализовать вирус. Однако только антитела к слитому белку будут блокировать распространение вируса между клетками, т.е. обладают активностью против слияния. Таким образом, антитела к слитому белку будут защищать от циркулирующего вируса так же, как и ингибировать распространение - между клетками - внедрившейся инфекции. Обнаружено, что антитела к слитому белку /как поликлональные сыворотки против очищенного слитого белка, так и моноклональные антитела, которые обладают как нейтрализующей активностью, так и активностью против слияния/ защищают животных-моделей от инфекции /Walsh et al., 1984, Infeсt Immun. 43: 756-758/.
Практическим способом защиты новорожденных и маленьких детей от респираторных заболеваний верхних дыхательных путей и нижних отделов дыхательной системы могла бы быть профилактическая вакцинация против PC-вируса. Вакцинация будущих матерей /активная иммунизация/ будет защищать новорожденных за счет пассивной передачи иммунитета либо трансплацентарно, либо через материнское молоко. Возможно несколько подходов к вакцине против PC-вируса, но некоторые из них в прошлом оказались безуспешными.
Предпринята попытка вакцинации вакциной с убитым PC-вирусом, и обнаружено, что она является неэффективной /Kim et al., 1969, Am. T. Epid 89:422/. Дети не только не защищались, но в некоторых случаях последующие инфекции с PC-вирусом приводили к нетипичному и более серьезному заболеванию, чем в контрольной невакционированной группе. Это явление не является свойственным только для PC-вируса и обнаруживается также в случае вакцин с убитым парамиксовирусом, такие как против кори. Предполагается, что причина неудач в прошлом с вакцинами с инактивированным PC-вирусом является следствием инактивации биологически функциональных эпитопов либо на одном, либо на обоих гликопротеинах оболочки вируса. То есть в вакцине с убитым вирусом вируснейтрализующая и связующая антигенные детерминанты "денатурируются". В результате вакцинированный субъект не испытывает действия биологически функциональных вируснейтрализующей и связующей антигенных детерминант. Поэтому, когда вакцинированный субъект сталкивается с живым вирусом, получающийся в результате антителогенез не приводит к защитному иммунитету. Вместо этого антитело посредничает в воспалительной реакции, которая часто дает в результате более тяжелое заболевание /Choppin and Sсheid, 1980, Rev. Inf. Dis. 2:40-61/.
Другим подходом к вакцине против PC-вируса стало ослабление живого вируса. Оказалось, что чувствительные к температуре мутанты /Wright et al., 1982, Infeet Immun. 37:397-400/ и пассажные ослабленные вирусы /Belshe et al. , 1982, I.Inf. Dis. 145:311-319/ являются малозаразными и неэффективны для предупреждения заболевания, когда используются в качестве иммуногенов в вакцинах против PC-вируса. Однако в этих случаях нетипичное заболевание, как результат вакцинации, не имеет места.
Из наших сегодняшних знаний о строении PC-вируса и иммунном ответе на заражение понятно, что полезная против такого вируса вакцина должна быть эффективной при индуцировании образования антител к слитому белку и/или C-белку. Особое значение для защитного иммунитета имеет продуцирование антител, которые ингибируют слияние и, следовательно, могут остановить распространение вируса между клетками в дыхательных путях. Кроме того, полезно индуцировать клеточно-опосредованную иммунную реакцию, включая стимуляцию цитотоксических Т-клеток /ЦТЛ/, которые пригодны против инфицированных PC-вирусом клеток. Различные вакцинные формулировки настоящего изобретения направлены на то, чтобы удовлетворить этим двум целям.
Краткое изложение сущности изобретения
Настоящее изобретение имеет отношение к открытию некоторых адъювантов, которые способны усилить иммунологическую реакцию на белки оболочки респираторно-синцитиального вируса, точнее на гликопротеин F РСВ. В частности, здесь показано, что адъювант QS-21, или, с другой стороны, 3D-монофосфориллипид A /MPL/ с алюмом гидроксид алюминия существенно увеличивают способность антител, индуцированных против гликопротеинов РСВ F и/или G-, нейтрализовать вирус, а также обеспечивать иммунологическую защиту, через клеточно-опосредованную реакцию, против вируса. Кроме того, эти адъюванты, как показано, предотвращают образование синцитий в зараженных вирусом клетках. На основе этих сведений могут быть изготовлены вакцинные композиции, содержащие белок /белки/ оболочки РСВ и адъюванты, которые выбирают среди QS-21, MPL, 3D-MPL и их сочетаний. Композиция может содержать, необязательно, алюм. Добавление алюма может еще усилить иммунологическую реакцию к антигену /антигенам/ РСВ, когда они вводятся с такими адъювантами. Присутствие таких адъювантов обеспечивает усиленную иммуногенность к антигену за счет усиления антителогенеза. В частности, опосредованное комплементом подавление бляшкообразования по сравнению с алюмом. Кроме того, присутствие адъюванта создает возможность изготовления вакцины с уменьшенным количеством антигена /антигенов/.
Краткое описание рисунков
На фиг. 1-3 изображаются результаты по клеточно-опосредованной цитотоксичности, полученные в экспериментах примера 5, который описан ниже.
Подробное описание изобретения
Настоящее изобретение относится к новым вакцинным формулировкам и применению их для предупреждения заражения РСВ. Вакцинная формулировка настоящего изобретения включает белок РСВ или его иммунологический фрагмент и адъювант, который, как было показано, усиливает иммунологическую реакцию к белку РСВ. Адъювант выбирают среди QS-21 и монофосфориллипида A и их сочетаний и, необязательно, алюма. Присутствие алюма в вакцине действует синергически с MPL, чтобы добиться реакции нейтрализации РСВ.
В одном из предпочтительных вариантов осуществления изобретения QS-21 формулируют с сапонином, который очищают из сырого экстракта Quillaja suponaria и который описан Kensil и Marciani в патенте США 5057540. Антитела, вызванные против формулировок, содержащих QS-21 и белок РСВ F или белки РСВ G и F, могут нейтрализовать PC-вирус. Иммуногенность белков РСВ F и G значительно возрастает при использовании в качестве адъюванта QS-21 по сравнению с формулировками, которые не содержат адъювантов, или которые содержат другие известные адъюванты, такие как алюм, когда они используются одни в качестве адъюванта.
Другим аспектом настоящего изобретения является то обстоятельство, что адъюванты могут использоваться в вакцине с белком РСВ G или белком F, чтобы добиться иммунного ответа, такого как антителогенез, который нейтрализует вирус как подгруппы A, так и подгруппы B. Это является существенным открытием, так как найдено, что другие адъюванты, в особенности алюм, с белком G нейтрализуют только подгруппу, из которой очищают белок.
В другом предпочтительном варианте осуществления изобретения для получения вакцинной формулировки MPL, конкретно - 3D-MPL, может быть использован в сочетании с алюмом, чтобы получить формулировку вакцины, которая может усилить стимуляцию комплемент-зависящих нейтрализующих антител к РСВ. Иммуногенность компонентов субъединиц РСВ значительно возрастает с таким адъювантом по сравнению с формулировками, которые не содержат адъювантов, или которые содержат алюм в качестве единственного адъюванта.
Белки и полипептиды, отнесенные к эпитопу /эпитопам/ нейтрализации и/или слияния слитого белка и/или G-белка PC-вируса, пригодны в качестве иммуногенов в субъединичной вакцине для защиты от заболеваний нижних отделов дыхательной системы и других болезненных симптомов PC-вирусной инфекции, и могут быть введены в состав вакцин настоящего изобретения. Субъединичные вакцины содержат относящийся к делу иммуногенный материал, необходимый для иммунизации хозяина, и адъюванты, идентифицируемые здесь как сильные иммуномодуляторы. Вакцины, приготовленные из иммуногенов, полученных генной инженерией, химически синтезированных иммуногенов и/или иммуногенов, содержащих аутеничный, по существу, чистый слитый белок PC-вируса или его фрагменты - одни или в сочетании с подобным образом полученным G-белком PC-вируса или его фрагментами, которые способны достигать защитной иммунной реакции, являются особенно выгодными, поскольку отсутствует риск заражения реципиентов. Химерные полипептиды, содержащие по крайней мере один иммуногенный фрагмент из обоих гликопротеинов PC-вируса - F и G - также могут использоваться в вакцинных формулировках настоящего изобретения. Такие химерные полипептиды РСВ описаны в патенте США 5194595, который включен в настоящее в качестве ссылки.
Слитый белок и/или G-белок PC-вируса и полипептиды могут быть очищены из рекомбинантов, которые экспрессируют связующую и вируснейтрализующую антигенные детерминанты. Такие рекомбинанты включают любые бактериальные трансформанты, трансформанты дрожжей, культивируемые клетки насекомых, зараженные рекомбинантными бакуловирусами, или культивируемые клетки млекопитающих, какие известны в технике, например, такие как клетки яичника китайского хомяка, которые экспрессируют эпитопы слитого белка PC-вируса. Рекомбинантный белок или полипептиды могут содержать многочисленные копии представляющего интерес эпитопа.
Родственные слитому белку и/или G-белку PC-вируса белок или полипептид могут быть синтезированы химически. С другой стороны, родственные слитому белку PC-вируса белок или полипептид, или родственные G-белку, могут быть выделены, по существу, в чистом виде из PC-вируса или культур клеток, зараженных PC-вирусом, и сформулированы с новыми адъювантами как вакцина против РСВ.
Независимо от способа получения слитый белок или C-белок PC-вируса, родственный белок или полипептид доводят до соответствующей концентрации, и его можно ввести в состав с адъювантом, выбираемым среди QS-21 или MPL с алюмом. MPL и его производное - 3-деацилированный MPL-/3D-MPL/, могут быть включены в состав с TDM и сквалентом и использованы в вакцинных формулировках настоящего изобретения. 3-D-MPL может быть получен в соответствии со способами, описанными в патенте Великобритании N 2220211 /Ribi Immunochem/.
Количество белка в каждой дозе вакцины выбирают как количество, которое индуцирует иммунозащитный отклик без существенных вредных побочных эффектов. Такое количество будет изменяться в зависимости от используемого иммуногена. Обычно каждая доза будет включать от 0,1 до 100 мкг белка, причем предпочтительным является количество от 5 до 50 мкг и, с другой стороны, предпочтительным является количество от 5 до 25 мкг на дозу. Количество адъюванта будет таким количеством, которое будет индуцировать иммуномодулирующий отклик без существенного вредного побочного действия. Оптимальное количество для конкретной вакцины может быть установлено стандартными исследованиями, включая исследования титров антител вакцины и их вируснейтрализующие способности. Количество адъюванта будет составлять от 1 до 100 мкг на дозу, причем предпочтительнее количество от 5 до 50 мкг на дозу, и, с другой стороны, предпочтительнее количество от 20 до 50 мкг на дозу.
Иммуногенные потенции вакцин, содержащих слитый белок или G-белок PC-вируса, или их иммунологические фрагменты, и их генетические или физические смеси, могут быть определены как мониторингом иммунного ответа испытуемых животных после иммунизации очищенным белком, синтетическим пептидом или рекомбинантным белком. К испытуемым животным могут относиться, но не ограничиваться этим перечислением, мыши, крысы, кролики, приматы и, со временем, люди. Способы введения иммуногена могут включать внутрикожный, внутримышечный, интраперитонеальный, внутривенный, чрескожный, интраназальный или другие любые обычные способы иммунизации. Иммунный отклик подопытных объектов может анализироваться многими способами:
/а/ по реактивности полученной иммунной сыворотки и аутеничным PC-вирусным антигенам при испытаниях с применением известных технических приемов, например, твердофазным иммуноферментным анализом /ELISA/, иммуноблотами, радиоиммунопреципитацией и т.п.;
/b/ по способности иммунной сыворотки нейтрализовать инфекционность PC-вируса in vitro;
/c/ по способности иммунной сыворотки ингибировать слияние вируса in vitro;
/d/ по способности иммунизировать животных генерировать активность зависящих от антигена цитотоксических T-лимфоцитов /ЦТЛ/; и
/e/ по защите от PC-вируса.
Многие способы могут использоваться, чтобы вводить описанные здесь вакцинные формулировки людям в целях профилактики. Эти способы включают, но не ограничиваются ими, внутрикожный, внутримышечный, интраперитонеальный, внутривенный, чрескожный и интраназальный способы. Секреторные IqA-антитела, продуцируемые ассоциированной со слизистой лимфоидной тканью, могут играть главную роль в защите от заражения PC-вирусом путем предотвращения первоначального взаимодействия патогенов с поверхностью слизистой, или путем нейтрализации важных эпитопов патогенов, включаются при заражении или распространении заболевания. Стимуляция иммунных ответов слизистых, в том числе образование секреторных IgA-антител, может иметь большое значение для приобретения защиты от инфекции верхних дыхательных путей и нижних отделов дыхательной системы.
Полипептиды и белки могут вводиться в формулировки, в основном, при концентрации в интервале от 0,1 мкг до 100 мкг на дозу.
В качестве носителей могут использоваться физиологически приемлемые среды. Такие среды включают, но не ограничиваются, стерильную воду, физиологический раствор, забуференный фосфатом физиологический раствор и т.п. К новым вакцинным формулировкам настоящего изобретения могут добавляться другие подходящие адъюванты, такие как минеральные гели, например гидроксид алюминия, фосфат алюминия и т.п. Иммуноген может также включаться в липосомы или конъюгироваться с полисахаридами и/или другими полимерами для применения в вакцинной формулировке.
Полипептиды и белки, которые могут вводиться в вакцинные формулировки настоящего изобретения, могут быть связаны с высокомолекулярными носителями. Предпочтительно, носитель, и полипептиды и белки превышают пять тысяч дальтон после связывания, и, предпочтительнее, носитель превышает пять килодальтон. Носитель, предпочтительно, представляет собой полиаминокислоту, либо природную, либо синтетическую, которая является иммуногенной в организме животного, включая человека. Способ связывания обычен. Многие технические приемы соединения раскрываются в патенте США N 4629783, который включается в настоящее изобретение в качестве ссылки. Многие сшивающие агенты раскрываются в Handbook and General Catalog 1986-87 и Pierce Chemical Company /Rockojord, Illinois/, pages 311-340.
Получают рекомбинантные вирусы, которые экспрессируют эпитопы, родственные слитому белку и/или C-белку PC-вируса. Такие вирусы могут использоваться для получения инактивированных рекомбинантных вирусных вакцин для защиты от инфекций нижних отделов дыхательной системы и от других признаков заболеваний от PC-вируса.
Инактивированные вакцины являются "мертвыми" в том смысле, что их инфекционность разрушена обычно путем химической обработки /например, формальдегидом/. В идеальном случае инфекционность вируса разрушается без воздействия на белки, которые связаны с иммуногенностью вируса. Для получения инактивированных вакцин необходимо вырастить в культуре большое количество рекомбинантного вируса, экспрессирующего слитый белок и/или G-белок PC-вируса, родственные белки или полипептиды, чтобы обеспечить необходимое количество нужных антигенов. Смесь инактивированных вирусов, которые экспрессируют различные эпитопы, может быть использована для составления "поливалентных" вакцин. В некоторых случаях такие "поливалентные" инактивированные вакцины можно предпочесть формулировке живой вакцины из-за возможных трудностей вследствие взаимной интерференции живых вирусов, вводимых вместе. В любом случае, инактивированный рекомбинантный вирус или смесь вирусов могут быть сформулированы с адъювантом по настоящему изобретению для усиления иммунологической реакции к антигенам.
Вакцины настоящего изобретения могут быть введены особи для предупреждения инфекции или симптомов заболевания, связанных с РСВ. Такое введение может быть осуществлено посредством одной дозы или многих доз с целью добиться у особи первичного иммунного отклика. Обычно многоразовая вакцинация людей будет осуществляться три раза с двухмесячными интервалами. Для стимуляции существующего иммунного отклика от предыдущей вакцинации или природной инфекции могут даваться бустер-дозы. Следующие далее примеры предлагаются в целях иллюстрации настоящего изобретения и не должны истолковываться как ограничение объема настоящего изобретения
Примеры
Пример 1.
Получение белка РСВ
A. Иммуноаффинный слитый белок-1 /РFР-1/
PFP-1 получают по методике Walsh et al. , I. Gen. Virol 66:409-415 (1985), со следующими изменениями. Иммуноаффинный элюированный материал пропускают через колонку DEAE и прошедший сквозь поток собирают, проводят диализ против ЗФР с 0,1% тритона Х-100, и стерилизуют фильтрацией через фильтр 0,2 мкм.
B. Ионообменный слитый белок-2 /1F/
1F получают, пропуская осветленный лизат инфицированных РСВ клеток через колонку с EAE. Собирают прошедший через нее поток и пропускают через колонку с гидроксиапатитом /НА/. После элюирования из НА элюированный F-белок диализуют против ЭФР с 0,1% тритона Х-100 и стерилизуют фильтрацией через фильтр 0,2 мкм.
C. Иммуноаффинный G-белок /G/
G-белок получают по методике Walsh et al., I. Gen. Virol 66:761-767 (1984) со следующими изменениями. После элюирования G-белок пропускают через иммуноаффинную колонку, специфическую для F-белка РСВ. Собирают вышедший поток, диализуют против ЗФР с 0,1% тритона Х-100 и стерильно фильтруют через фильтр 0,2 мкм.
D. Химерный белок F/G
Химерный белок F/G получают по патенту США N 5194595, и он поставляется Upjohn Corporation.
Пример 2.
Иммуноферментный анализ /EIA/
Определяют титр антител в образцах сыворотки, используя иммуноферментный анализ /EIA/, который выполняют следующим образом.
Слитый белок PC-вируса разбавляют до 200 нг/мл карбонат-бикарбонатным буфером, pH 9,6. Сто мкл разбавленного антигена вносят в каждую лунку рядов B-C 96-луночного аналитического планшета NUNCTM с лунками с плоским дном. В рядах A и H в каждую лунку добавляют только 100 мкл одного карбонат-бикарбонатного буфера. Планшет накрывают, инкубируют в течение 2 часов при 37oC при встряхивании и хранят при 4oC в течение ночи, чтобы иммобилизовать антиген.
Удаляют супернатанты из аналитического планшета NUNCTM, и планшет промывают 0,1% твина в ЗФР, pH 7,4, и планшет сушат подходящим способом.
На каждом планшете анализируют три образца антител. Каждый образец сначала разбавляют до первого разбавления в 0,2% твина, 0,01 М ЭДТК с ЗФР, pH 7,5 /0,2% TWN/. Первые разбавления разбавляют серийно далее в 96-луночном планшете FAL CONTM с лунками с U-образным дном следующим образом.
/a/ Первые разбавления образцов инокулируют в ряд 2 в количестве 200 мкл на лунку. Образец 1 вносят три раза, например в лунки A2, B2 и C2; образец 2 вносят два раза, например в лунки D2, E2; образец 3 вносят три раза, например, в лунки F2, C2 и H2.
/b/ В каждую лунку в рядах 3-12 вносят по 100 мкл 0,2% TWN.
/c/ Серийное разведение выполняют затем, перенося 100 мкл из лунки ряда 2 в соответствующую лунку ряда 3 /например, из B2 в B3; из C2 в C3/, из лунки в ряду 3 - в соответствующую лунку ряда 4 и т.д., пока не достигнут 12 ряда.
/d/ В ряду 1 в каждую лунку вводят по 100 мкл 0,2% ТWN как в контрольные лунки.
По 100 мкл первичного разбавления переносят из каждой лунки FAL CONTM в соответствующие лунки планшета NUNCTМ, например из A2 (FAL CONTM) в A2 (NUNCTM). Аналитический планшет NUNCTM накрывают и инкубируют в течение одного часа при 37oC при встряхивании. Удаляют супернатанты из аналитического планшета, и планшет промывают 0,1% твина в ЗФР и сушат подходящим способом.
Конъюгат козьего антимышиного IgC с щелочной фосфотазой /TAGOTM/ разводят 0,3% твина в ЗФР, pH 7,0 /0,3 TWM/ до разведения, с которым можно работать, например 1: 1500. Разведенный конъюгат /100 мкл/ добавляют в каждую лунку в рядах 2-12. В ряду 1 в качестве контроля в каждую лунку добавляют по 100 мкл 0,3% TWN. Планшет накрывают и инкубируют в течение 1 часа при 37oC при встряхивании. Инокулянты затем удаляют, и планшет промывают 0,1% твина в ЗФР, pH 7,4, и сушат подходящим образом.
Во все без исключения лунки добавляют по 100 мкл раствора субстрата - 1 мг/мл в диэтаноламинном буфере, pH 9,8 /SIG - MATM/. Позволяют протекать ферментативной реакции при комнатной температуре в течение 1 часа. Реакцию останавливают, добавляя в каждую лунку 100 мкл 3N NaOH. Степень ферментативной реакции определяют, считывая оптическую плотность при 410 нм.
Ряды A и H служат для негативного контроля, поскольку в них не присутствует антиген; ряд 1 служит также в качестве негативного контроля, поскольку в нем не присутствуют антитела.
Пример 3
Проверка нейтрализации вируса (реакция нейтрализации по редукции бляшек, PRNT)
Образцы испытуемых сывороток, которые разведены серийно, и позитивную контрольную сыворотку инактивируют нагреванием при 56oC в течение 30 минут. Все сыворотки затем разбавляют равными объемами, содержащими 30 бляшкообразующих единиц /PFU/ PC-вируса, и инкубируют при 37oC в течение одного часа, с /C' плюс PRNT/ или без /PRNT/ добавления 5% кроличьего комплемента. Для позитивного контроля используют сыворотку взрослого человека, которая предварительно охарактеризована иммуноферментным анализом, проверена на нейтрализацию и блокирование слияния. Сыворотки, которые предварительно проверены и о которых известно, что они неиммунные, используют для негативного контроля.
Каждую инкубированную смесь сыворотки с вирусом инокулируют в клетки НЕр-2 /ATCC N CCL 23/ в отдельных лунках 96-луночных планшетов, и позволяют проходить поглощению вирусов в течение 2 часов при 37oC. Инокуляты удаляют. Клеточные монослои промывают и покрывают модифицированной средой Игла с 5% фетальной коровьей сыворотки и 1% сефадекса® , и инкубируют при 37oC в течение 3 суток. Покрывающую среду удаляют, и клетки промывают забуференным фосфатом физиологическим раствором /ЗФР/.
В каждую лунку добавляют по 200 мкл охлажденного раствора ЗФР-метанол /1: 5/, и клетки фиксируют в течение 30 мин при комнатной температуре, ЭФР-метанольный фиксатор удаляют, и добавляют 200 мкл на лунку 5% растворимого молока CARNATION® в ЗФР, pH 6,8 /BL OTTO/. Планшет инкубируют в течение 30 минут при 37oC.
BL OTTO удаляют. Добавляют 50 мкл на лунку моноклональных антител против PC-вируса /предварительно оттированных и разветвленных BL OTTO до рабочей концентрации/, и планшет инкубируют при 37oC в течение 1 часа. Антитела удаляют, и фиксированные клетки дважды промывают BL OTTO, каждый раз по 30 минут.
Добавляют по 50 мкл на лунку пероксидазы из хрена, конъюгированной с козьим антимышиным IgG /разведенной 1:250 в BL OTTO/, и планшет инкубируют при 37oC в течение 1 часа. Козьи антитела удаляют, и фиксированные клетки дважды промывают BL OTTO, каждый раз по 30 минут.
Добавляют по 50 мкл на лунку раствора субстрата пероксидазы /0,05% 4-хлор-1-нафтол, 0,09% H2О2 в ЗФР, pH 6,8/, и оставляют для проявления окраски на 15-30 минут при комнатной температуре. Раствор субстрата удаляют, и лунки промывают водой и сушат на воздухе. Определяют число бляшек в каждой лунке.
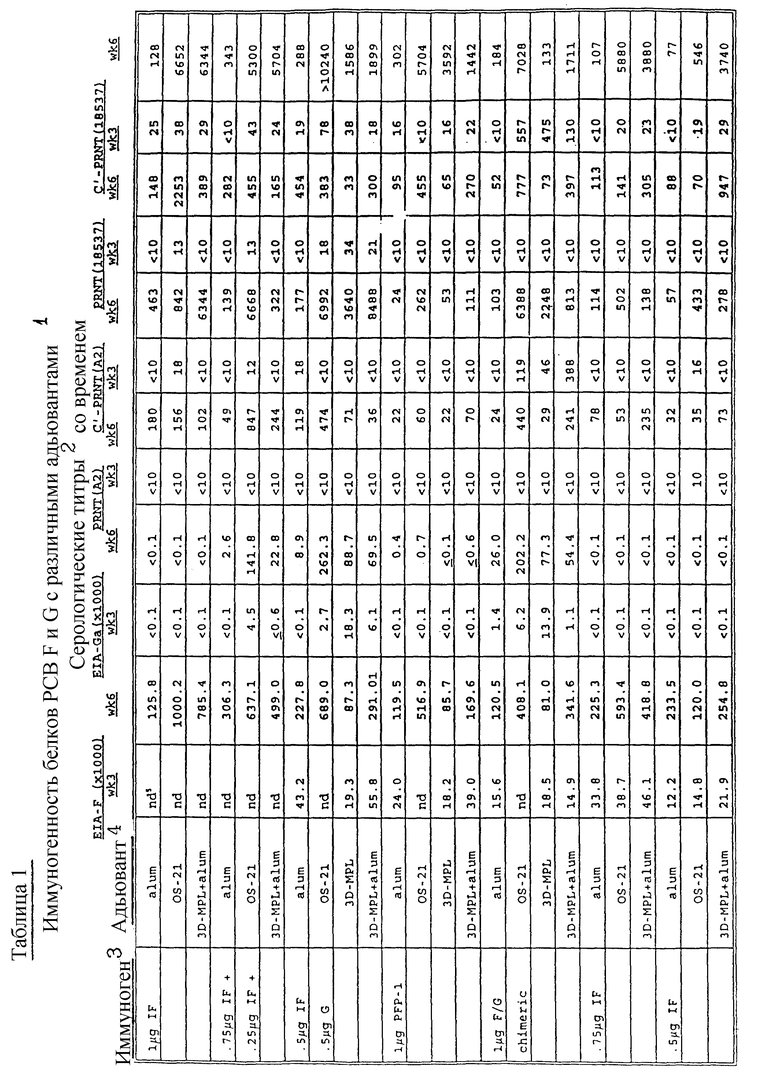
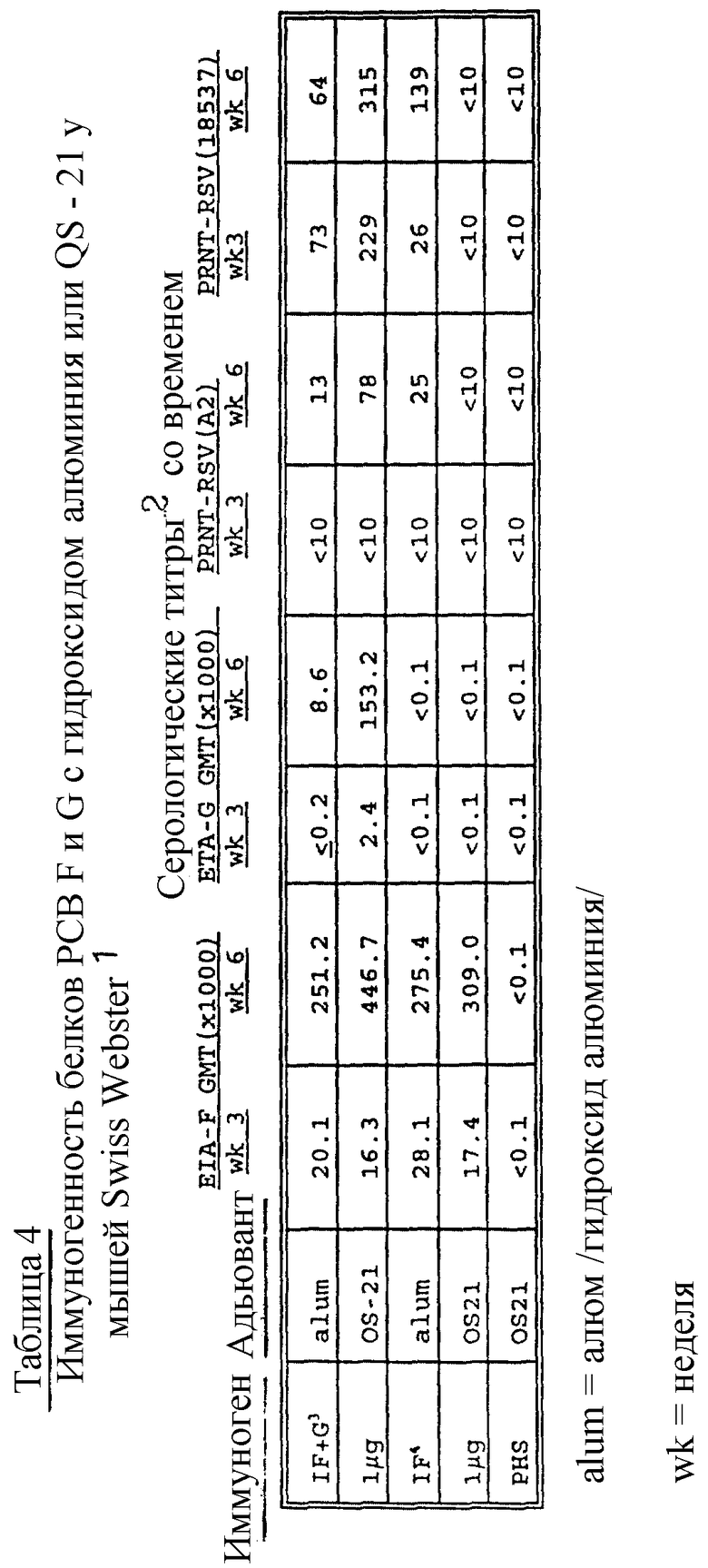
Нейтрализующую способность проверяемого образца сыворотки выражают как разбавление, которое дает в результате 60% уменьшение бляшкообразования при сравнении с контрольной неиммунной сывороткой. Результаты приводятся в таблицах 1 - 4.
Данные, приведенные в таблицах 1 - 4, представляющие результаты E1A и реакции подавления бляшкообразования, показывают улучшение биологического иммунного отклика при применении настоящих новых адъювантов при сравнении с одним алюмом. Вакцинные формулировки со слитым белком PC-вируса, G-белком, их смесями и химерным F/G-белком с новыми адъювантами /с или без добавления алюма/ являются значительно усиленными при сравнении с формулировками, содержащими один алюм.
Пример 4
Вирус и клеточные линии
Используют штаммы РСВ A2 и 18537, и штаммы вирусов выращивают либо в клетках Vero [American Type Culture Collection (ATCC) N CCL 81], или в клетках НЕр-2 /АТСС N CCL 23/, с последующими стандартными процедурами, очищают на сорбите с градиентом плотности и хранят при -70oC до использования. Линия клеток BCH4, устойчиво инфицированных давно существующим штаммом РСВ, и линия неинфицированных клеток BAL B/c /об обеих клеточных линиях см. Fernie et al., Proc. Soc. Exp. Biol. Med., 1981, 167:83-86/, подарены Dr. Bruce F. Fernie Последние клеточные линии хранятся в модифицированной по Дульбекко среде Игла /DMEM, Gibco. BRL Gaithersburg M.D/ с 10% /по объему/ инактивированного нагревания FBS /Hyclone Laboratories Inc., Logan, UT/.
Пример 5
Определение подкласса антитела против F-белка.
Титр подкласса антител против F-белка мышей, примированных 5 мкг F-белка, смешанного с QS-21, AL ОН или природной инфекцией, определяют EL ISA. Вкратце, 96-луночные планшеты с 20 нг F-белка или 5 мкг РСВ A2 готовят следующим образом. Очищенный F-белок /200 нг/мл/ или РСВ F2 /50 мкг/мл/ в карбонат-бикарбонатном буфере /pH 9,6/ наносят на 96-луночные планшеты /Nune, Roskilde Дания/ на 2 часа при 37oC и хранят их в течение ночи при 4oC. Затем планшеты промывают 5 раз ЗФР с 0,05% твина 20 /Sigma/, после чего два раза ополаскивают еще одним ЗФР. Затем в лунки добавляют серийно трехкратноразведенную сыворотку, приготовленную в ЗФР с 0,3% твина 20 и 0,01 М ЭДТК-буфером /pH 7,0/, и инкубируют в течение 1 часа при комнатной температуре. После 5-кратного промывания ЗФР с 0,1% твина 20 добавляют 100 мкл биотинилированного козьего антимышиного IgG /1:4000 Kirkegaard and Perry Laboratories/IgG1 /1: 3000, Zymed/ или IgG2a /1:5000 Zymed/, и инкубируют в течение 1 часа при комнатной температуре. После еще одной серии промывок в лунки добавляют 100 мкл стрептавидина, конъюгированного с пероксидазой из хрена /Разведение 1: 10000 в ЗФР с 0,3% твина 20; Zymed/, и инкубируют при комнатной температуре еще в течение 30 минут. После промывания в лунки добавляют перoксидазный субстрат /2,2'-азино-ди/3-этилбензтиазолин/сульфонат (6)/, и инкубируют при комнатной температуре в течение 20 минут, и в это время реакцию останавливают 100 мкл 1% додецилсульфата натрия /Pierce Rockjord IL/. Конечные точки титрования определяют при 410 нм.
Испытания на нейтрализацию вируса /PRNT/ выполняют так же, как в примере 3.
Титры нейтрализующих антител усиленных вспомогательным комплементом сывороток, установленные F/QS-21, коррелируют с индукцией антител против F-белка подкласса IgG2a /табл. 5/. Через три недели после первичной иммунизации существует связанное с дозой QS-21 увеличение протеин-специфического IgG2a, как и антител IqG1. Для сравнения, однократная инъекция F-белка, смешанного с одним физиологическим раствором или F/AL ОН, выделяет, в первую очередь, протеин-специфические антитела подкласса IgG1 /табл. 5/. Данные указывают, что F/QS-21 индуцирует гуморальные иммунные ответы, которые подобны ответам, генерированным экспериментальным заражением, и состоят из обоих комплементфиксирующих IqG2a, как и из IgG1 антител.
Пример 6
Определение титров перекрестно-нейтрализующих антител и инфекционности РСВ
Титрование сывороточных нейтрализующих антител выполняют дважды на монослоях клеток НЕр-2 в 96-луночных культуральных планшетах, как описано в примере 3.
В этом примере, как показано в таблице 5, наблюдают, что адъювант может давать возможность РСВ-белку индуцировать комплемент-зависимый IgG-антительный ответ, который нейтрализует вирусы как подгруппы A, так и подгруппы B /причем эти подгруппы в приведенной таблице 5 идентифицируются как подгруппа A2 и 18537 соответственно/. Такой перекрестный нейтрализующий иммунный ответ гетерологичного субтипа PC-вируса не достигался прежде при использовании одного очищенного G-белка. Вакцина, составленная с адъювантом (QS-21 и G-белком PC-вируса, генерирует желательный гетеротипический ответ нейтрализующих антител, который существенно больше, чем ответ, который достигается одним алюмом или естественным заражением.
Пример 7.
Сравнение QS-21 с AL ОН по способности выявлять локальную, зависящую от F-белка, киллерную активность
Проверяют также способность QS-21 выявлять локальную, зависящую от F-белка, киллерную активность и сравнивают с клеточно-опосредованной цитотоксичностью, генерируемой иммунизацией F/AL OH или экспериментальным заражением.
Выделение легочных мононуклеарных клеток /РМС/
РМС выделяют из легких коллагеназным перевариванием /см. Hancock et al., Vaccine 12: 267-274, 1994 и Anderson et al., I. Gen. Virol., 71:1561-1570, 1990/. Вкратце, иссеченные легкие помещают в холодную DMEM и отмывают от периферической крови. Затем легкие измельчают в свежей DMEM, переносят в 50-мл пробирку для центрифугирования и покачивают при 37oC в присутствии коллагеназы /коллагеназа типа 1V, Sigma Chemical Co., St, Louis, MO/ при конечной концентрации 2 мг/мл, 10 мМ HEPES-буфера и 1% /по объему/ инактивированного нагреванием FBS. После 90-минутной инкубации фрагменты пропускают через сито из нержавеющей стали 100 меш для культуры тканей /Sigma/.
Получающуюся в результате суспензию осаждают центрифугированием /400 г/, ресуспендируют в метризамиде /16%, в/о Accurate Chemical and Scientific Corp. Westbury NY/ покрывают слоем RPMI 1640 /Gibco BRL/, содержащей 10% инактивированного нагреванием FBS, и вращают /150 g/ в течение 20 минут при 5oC. Затем собирают слои РМС, отмывают от градиента и проверяют ex viro их цитотоксическую способность.
Определение процентной цитотоксичности
Антигензависимую клеточную цитотоксичность определяют в 4-часовых испытаниях с выделением 51Cr /Amersham Corp. Arlington Heights, IL/. Вкратце, инкубируют 50 мкл /5000 клеток/ сингенных, помеченных 51Cr контрольной клеточной линии или инфицированной РСВ клеточной линии- мишени /37oC, 5% CO2/, при 3-кратном повторении, в микролунках с V-образным дном /Costar, Cambridge, MA/ со 100 мкл мононуклеарных клеток селезенки или легких /разведенных серийно 2 раза в RPMI 1640, содержащей 10%, по объему, инактивированного нагреванием PBS/. Конечный объем составляет 150 мкл на лунку. После инкубации собирают супернатанты /Skatron Harvester, Skatron Inc., Sterling VA/, измеряют выделение 51Cr счетчиком Clin Gamma /Pharmacia LKB/ и сравнивают со спонтанным выделением /мишени, инкубированные с одной средой, 20 - 25%/ и общим выделением /мишени, инкубированные в культуральной среде с 1,0% /по объему/ тритона Х-100 в ЗФР/. Вычисляют удельное выделение, в процентах: 100 х [(среднее cpm экспериментальное) - (среднее cpm при спонтанном выделении)] / [(среднее cpm при общем выделении) - (среднее cpm при спонтанном выделении)] /cpm = число импульсов в минуту/.
Исследование ингибирования антителами
Очищенные моноклональные антитела, направленные против антигенов главного комплекса гистосовместимости /MHC H2Кd /клон SF-1.1, IqG 2a/, H-2Dd /клон AF4-62.4, IgG 2b/ и Н-2Kb /клон AF6-88.5, IgG 2a/ закупают в Pharmingen, San Diego, CA. Моноклональное антитело /E37-10, IgG 2b/, направленное против антигена дифтерийного токсоида, служит в качестве контроля. Моноклональное антитело, направленное против мышиных CD 8 поверхностных молекул /53-6.72, АТСС N TIB 105/ очищают от супернатанта гибридомной культуры в колонке с рекомбинантным белком G /Pharmаcia/. Очищенный крысиный IqG закупают в Calbiochem /San Diego, CA/. Чтобы блокировать клеточно-опосредованный цитолиз, 50 мкл антитела добавляют к 50 мкл клеток-эффекторов перед добавлением 50 мкл клеток-мишеней. Конечное соотношение эффектора и мишени составляет 60:1.
Мышей Balb/c вакцинируют в 0 и 3 недели 5 мкг F-белка, смешанного либо с 20 мкг QS-21 (), либо со 100 мкг AL ОН (Δ), и сравнивают с результатами, полученными на мышах, иммунизированных посредством экспериментального заражения (•). Через две недели после второй иммунизации проводят контрольное заражение мышей вирусом. Через четыре дня после заражения РМС из BAL B/c мышей, вакцинированных F/QS-21, способны убивать инфицированные РСВ мишени (сплошные линии) антигензависимым способом (см. фиг. 1). Наиболее заслуживает внимания то, что такая цитотоксическая активность по силе такая же, как активность РМС от мышей, предварительно инфицированных РСВ, и почти в три раза выше, чем активность, индуцированная в РМС мышей, вакцинированных А/AL ОН. Контрольные сингенные мишени (пунктирные линии), неинфицированные РСВ, не являются киллерными (фиг. 1). Активность является локальной, поскольку клетки селезенки от той же мыши не являются цитолитическими.
Результаты также наводят на мысль, что локальная активность киллерных клеток, индуцированная вакциной F/QS-21, опосредуется T-клетками фенотипа CD 8. Цитолиз ингибируется, когда в испытываемые смеси добавляют увеличивающиеся дозы моноклонального антигена, направленного против клеток, порождающих CD-8 поверхностные детерминанты (символ заполнен) (фиг. 2). Подобным образом, увеличивающиеся концентрации моноклональных антител против H2Dd и H2Kd (зачерненные обозначения) блокируют цитолиз. Контрольный иммуноглобулин (пустые обозначения) не является ингибирующим (фиг. 2 и 3).
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННАЯ ВАКЦИНА, СПОСОБ ОДНОВРЕМЕННОЙ КОМБИНИРОВАННОЙ ВАКЦИНАЦИИ И СПОСОБ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ | 1993 |
|
RU2121365C1 |
| Рекомбинантная вакцина против вируса ветряной оспы (VZV) | 2020 |
|
RU2772902C1 |
| F-БЕЛОК РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2464316C2 |
| Рекомбинантный химерный полипептид-иммуноген nTBI, обладающий способностью индуцировать антитела, нейтрализующие вирус иммунодефицита человека 1 типа, и предназначенный для использования в качестве компонента вакцины против ВИЧ-1 | 2016 |
|
RU2642258C1 |
| ВАКЦИНЫ ПРОТИВ ВИРУСА ПРОСТОГО ГЕРПЕСА 2 ТИПА: КОМПОЗИЦИИ И СПОСОБЫ ЗАПУСКА ИММУННОГО ОТВЕТА | 2010 |
|
RU2585961C9 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ ГЛИКОПРОТЕИНА G ВИРУСОВ HENDRA И NIPAH | 2012 |
|
RU2681529C2 |
| СПОСОБ БЫСТРОГО ОТБОРА ВАРИАНТОВ GP-120 ВИЧ | 2012 |
|
RU2603732C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ АДЪЮВАНТНЫХ ВИРОСОМ И АДЪЮВАНТНЫЕ ВИРОСОМЫ, ПОЛУЧАЕМЫЕ УКАЗАННЫМИ СПОСОБАМИ | 2014 |
|
RU2694367C2 |
| ВАКЦИНА ПРОТИВ РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА (РСВ) | 2014 |
|
RU2723328C2 |
| СЛИТЫЕ БЕЛКИ КАРЦИНОЭМБРИОНАЛЬНОГО АНТИГЕНА | 2005 |
|
RU2380375C2 |











Изобретение относится к области медицины. Изобретение характеризуется тем, что описываются вакцинные композиции, содержащие белки респираторно-синцитиального вируса или их иммунологические фрагменты, и адъюванты, выбираемые из группы, состоящей из QS-21, 3-деацилированного монофосфориллипида А, монофосфориллипида А и их сочетаний. Технический результат заключается в том, что вакцинная композиция с присутствием адъюванта (адъювантов) значительно увеличивает гуморальную и клеточно-опосредованную иммуногенность белков РСВ. 4 с. и 7 з.п.ф-лы, 3 ил., 5 табл.
| СПОСОБ ВЫПЛАВКИ СТАЛИ В ЭЛЕКТРОДУГОВОЙ ПЕЧИ | 2001 |
|
RU2220211C2 |
| ШТАММ РОТАВИРУСА ЧЕЛОВЕКА ДЛЯ ПОЛУЧЕНИЯ ДИАГНОСТИЧЕСКИХ И ПРОФИЛАКТИЧЕСКИХ СЫВОРОТОК | 1991 |
|
RU2026345C1 |
| СПОСОБ ИНАКТИВАЦИИ ВИРУСА СПИДА | 1992 |
|
RU2031945C1 |
| US 5194595 А, 16.03.93 | |||
| US 5057540 А, 15.10.91 | |||
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Экономайзер | 0 |
|
SU94A1 |

