ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ДАННОЕ ИЗОБРЕТЕНИЕ
Данное изобретение относится к способу отбора специфических пробиотических молочнокислых бактерий, продуцирующих гистамин, и к применению таких штаммов для доставки полезных эффектов для этого хозяина.
УРОВЕНЬ ТЕХНИКИ
Продовольственная и сельскохозяйственная организация Объединенных Наций (ФАО) определяет пробиотики как «живые микроорганизмы, которые после введения в достаточных количествах человеку производят пользу для здоровья (вызывают укрепление здоровья) этого человека». В настоящее время ряд различных бактерий используют в качестве пробиотиков, например, продуцирующие молочную кислоту бактерии, такие как штаммы Lactobacillus и Bifidobacteria.
Продуцирующие молочную кислоту бактерии не только используются вследствие их полезного действия на здоровье человека или животного, но они также широко используются в пищевой промышленности для процессов ферментации. Эффективность пробиотиков является штаммспецифической, и каждый штамм может способствовать здоровью хозяина через различные механизмы. Пробиотики могут предотвращать или ингибировать пролиферацию патогенов, подавлять продуцирование вирулентных факторов патогенами или модулировать иммунную реакцию в провоспалительном или противовоспалительном пути. Применение различных штаммов пробиотических, продуцирующих молочную кислоту бактерий Lactobacillus reuteri является многообещающей терапией для устранения пароксизмальной боли («детской колики»), ослабления экземы, уменьшения приступов профессиональных заболеваний и супрессии инфекции Helicobacter pylori. L. reuteri считается свойственным желудочно-кишечному тракту человека организмом и присутствует, например, на слизистой оболочке гастрального корпуса, гастральной полости, двенадцатиперстной кишки и подвздошной кишки. См., например, патенты США с номерами 5439678, 5458875, 5534253, 5837238 и 5849289.
При выращивании клеток L. reuteri в анаэробных условиях в присутствии глицерина они продуцируют антимикробное вещество, известное как реутерин (β-гидроксипропиональдегид).
Взаимосвязь между хозяином и его микробами является сложной, и для некоторых бактерий эта взаимосвязь хозяин/микроб развивалась на протяжении многих лет ко-эволюции. Это, по-видимому, является особенно верным для Lactobacillus reuteri. Наши знания мутуалистической взаимосвязи между микробами кишечника и человека-хозяина существуют в ее периоде новорожденности, но мы уже в высокой степени осознаем, что микробиом кишечника играет важную роль в развитии кишечника и иммунной системы, питании, и новые звенья устанавливаются между микробиомом кишечника и головным мозгом. Дисбиоз, пертурбация нормального микробиома кишечника, подразумевался в широком диапазоне связанных с заболеваниями процессов, включающих в себя процессы, влияющие на локальную среду кишечника, таких как воспалительная болезнь кишечника (IBD) и синдром раздраженной толстой кишки (слизистый колит) (IBS), и процессы в участках, удаленных от кишечника, такие как метаболический синдром. Существенный терапевтический потенциал лежит в микробиоме кишечника, и прилагается исследование в направлении дальнейшей задачи изменения микрофлоры для предотвращения и/или лечения различных, связанных с заболеванием процессов.
Таким образом, существует необходимость в понимании таких специфических взаимодействий между микробами и человеком, связанных с конкретным заболеванием или другими ситуациями, влияющими на здоровье хозяина таким образом, что большая часть подходящих пробиотических штаммов может быть отобрана и использована для противодействия таким развитиям.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Описанное здесь изобретение обеспечивает локальное продуцирование гистамина в млекопитающих, в частности, в человеке, причем это локальное продуцирование гистамина включает в себя, но не ограничивается ими, продуцирование в GI-тракте (гастроэнтерологическом тракте), мочеполовом тракте (GU), полости рта, в легких и дыхательных путях, на коже и т. д. тела человека посредством отбора определенных штаммов молочнокислых бактерий. Эти бактерии могут доставляться вместе с некоторыми аминокислотами и/или сахарами, вводиться отдельно или уже присутствовать в активном участке.
Первичной целью данного изобретения является отбор (селекция) штаммов, которые могут локально продуцировать гистамин в разнообразных местоположениях, включающих в себя GI-тракт, GU-тракт, полость рта, в легких и дыхательных путях, на коже и т. д. тела человека.
Следующей целью этого изобретения является обеспечение продуктов, содержащих указанные штаммы.
Следующей целью данного изобретения является комбинирование введения бактерий с введением гистидина или содержащих гистидин пищевых продуктов или композиций для гарантии локального генерирования гистамина.
Таким образом, данное изобретение относится к новому способу для отбора клеток молочнокислых бактерий, которые применимы в качестве пробиотиков и в терапии. Этот новый способ предусматривает скрининг и отбор на штаммы молочнокислых бактерий, которые имеют активный оперон гистидина и способны продуцировать гистамин. Неожиданно эти молочнокислые бактериальные штаммы, отобранные таким способом, применимы в качестве пробиотиков и в терапии, в частности, в продуцировании противовоспалительных эффектов, посредством локального продуцирования гистамина. Эти эффекты данных бактерий являются неожиданными, как обсуждается в другом месте здесь, и ранее присутствие в пищевых продуктах бактерий, продуцирующих гистамин, активно избегалось вследствие узнаваемого риска для здоровья, например, потенциальных токсичных эффектов. Так, введение млекопитающему молочнокислых бактерий, способных локально продуцировать гистамин, или фактически скрининг и селекция (отбор) молочнокислых бактерий на такую способность локального продуцирования гистамина на основе присутствия активного оперона гистамина является контринтуитивным относительно такой доктрины. Действительно, ранее никогда не сообщалось, что пробиотики продуцируют гистамин.
Таким образом, в самом широком смысле данное изобретение обеспечивает способ отбора продуцирующего молочную кислоту бактериального штамма для применения в локальном продуцировании гистамина в млекопитающем, где указанный способ предусматривает скрининг бактерий на присутствие активного оперона гистидина и отбор штамма, который имеет активный оперон гистидина и способен продуцировать гистамин.
Оперон гистидина содержит три гена (антипортера гистидина/гистамина, гистидин-декарбоксилазы-пирувоил типа А (HdcA) и гистидин-декарбоксилазы-пирувоил типа В (HdcB)). Считается, что активность каждого из этих генов является важной для данного изобретения. Так, в способах скрининга этого изобретения кандидатные бактерии оценивали на присутствие всех трех генов и отбирали штаммы, положительные в отношении всех трех генов. Любой подходящий способ может быть использован для детектирования присутствия всех трех генов, например, генетические способы, такие как ПЦР. Получение хороших уровней гистамина может быть также индикатором присутствия всех трех генов и присутствия активного оперона гистидина. Таким образом, этот способ отбора данного изобретения включает в себя также стадию отбора штамма, который способен продуцировать гистамин. Предпочтительными являются штаммы с высокими уровнями продуцирования гистамина. Таким образом, в предпочтительных вариантах осуществления штамм отбирают на его способность продуцировать гистамин на уровне, большем чем 200, предпочтительно большем чем 250, или более предпочтительно большем чем 300 пг/мл, например на уровне, большем чем 350, 400, 450 или 500 пг/мл. Такие величины обычно относятся к величинам гистамина, измеренным в супернатанте штаммов в культуре.
Подходящие способы измерения уровней продуцирования гистамина будут хорошо известны лицу с квалификацией в данной области. Здесь приводится в качестве примера и является предпочтительным способ масс-спектрометрии, более конкретно тройной квадрупольной масс-спектрометрии. Однако ELISA или иммуноанализы могут быть равным образом использованы для оценивания и количественного определения продуцирования гистамина. Так, в некоторых вариантах осуществления способ отбора будет включать в себя стадию детектирования количества или уровня гистамина, продуцируемого кандидатным штаммом. Вследствие дальнейших применений штаммов, которые отбирают способами этого изобретения, после отбора или выделения продуцирующих гистамин штаммов другие варианты осуществления будут включать в себя дополнительные способы культивирования или размножения таких штаммов или, возможно, хранения таких штаммов для последующих применений.
Такие дополнительные стадии (и фактически стадии отбора способов этого изобретения) будут обычно необходимыми для проведения в подходящей культуральной среде, которая поддерживает продуцирование гистамина. Предпочтительные культуральные среды будут содержать подходящий источник углерода, который будет поддерживать продуцирование гистамина указанным штаммом. В особенно предпочтительных вариантах осуществления эти среды будут содержать глюкозу в качестве источника углерода и предпочтительно не будут содержать сахарозы или по меньшей мере будут содержать только сахарозу при таком уровне, который не будет значимо ухудшать продуцирование гистамина этим штаммом. Гистидин или аналог гистидина могут быть также обеспечены, необязательно вместе с источниками других аминокислот.
В предпочтительных вариантах осуществления указанным штаммом является штамм Lactobacillus reuteri.
Сразу после отбора подходящего штамма с использованием способа данного изобретения он может быть затем использован для локального продуцирования гистамина в млекопитающем. Таким образом, указанные штаммы способны также к локальному продуцированию гистамина в млекопитающем.
Таким образом, один дополнительный аспект данного изобретения обеспечивает продукт, содержащий клетки штамма молочнокислых бактерий, получаемого способом отбора этого изобретения, где указанный штамм молочнокислых бактерий имеет активный оперон гистидина и способен продуцировать гистамин, для применения в локальном продуцировании гистамина в млекопитающем. Как будет описано в другом месте здесь, предпочтительными применениями являются применения в лечении и/или профилактике воспалительных состояний или в лечении и/или профилактике состояний или заболеваний, которые будут получать пользу от локального продуцирования гистамина. Например, такое локальное продуцирование гистамина может приводить к противовоспалительному эффекту.
Альтернативные варианты осуществления этого изобретения обеспечивают штамм молочнокислых бактерий, который способен продуцировать гистамин, для применения в локальном продуцировании гистамина в млекопитающем, где указанный штамм молочнокислых бактерий имеет активный оперон гистидина. Предпочтительные признаки этого штамма и его применения описаны в другом месте здесь.
Обеспечены также способы лечения или способы локального продуцирования гистамина в млекопитающем, причем указанные способы предусматривают введение продукта, содержащего клетки штамма молочнокислых бактерий, получаемые способом отбора этого изобретения, или введение штамма молочнокислых бактерий, где указанный штамм молочнокислых бактерий имеет активный оперон гистидина и способен продуцировать гистамин, указанному млекопитающему в количестве, эффективном для возможности локального продуцирования гистамина в указанном млекопитающем. Предпочтительные признаки этого штамма и его терапевтические применения описаны здесь в другом месте.
Данное изобретение обеспечивает также применение продукта, содержащего клетки молочнокислых бактерий, получаемые способом отбора этого изобретения, где указанный штамм молочнокислых бактерий имеет активный оперон гистидина и способен продуцировать гистамин, в приготовлении композиции или лекарственного средства для применения в локальном продуцировании гистамина в млекопитающем. Альтернативные варианты осуществления обеспечивают применение штамма молочнокислых бактерий, где указанный штамм молочнокислых бактерий имеет активный оперон гистидина и способен к локальному продуцированию гистамина в млекопитающем. Предпочтительные признаки этого штамма и его терапевтические применения описаны здесь в другом месте.
КРАТКОЕ ОПИСАНИЕ ФИГУР
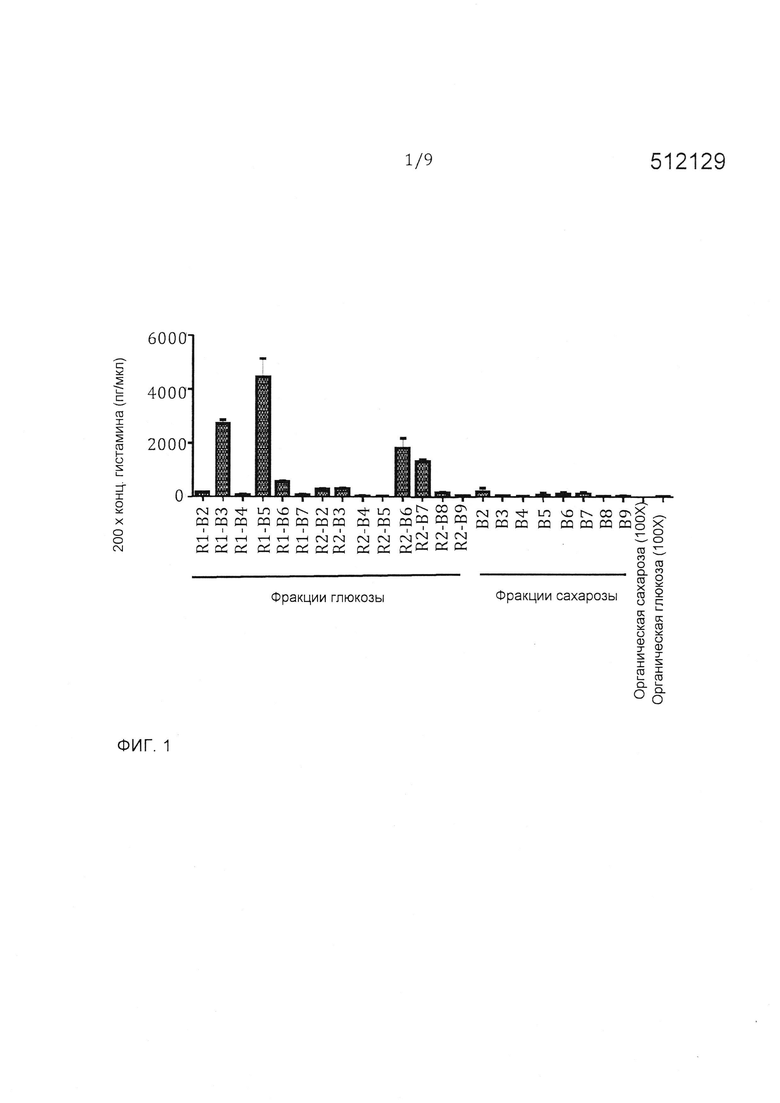
Фигура 1 - количественное определение гистамина во фракциях HILIC-HPLC. Использовали тройную квадрупольную масс-спектрометрии для количественного определения гистамина, присутствующего в выбранном диапазоне фракций HILIC-HPLC. TNF-ингибирующие фракции имели наивысшие количества гистамина из всех испытанных фракций.
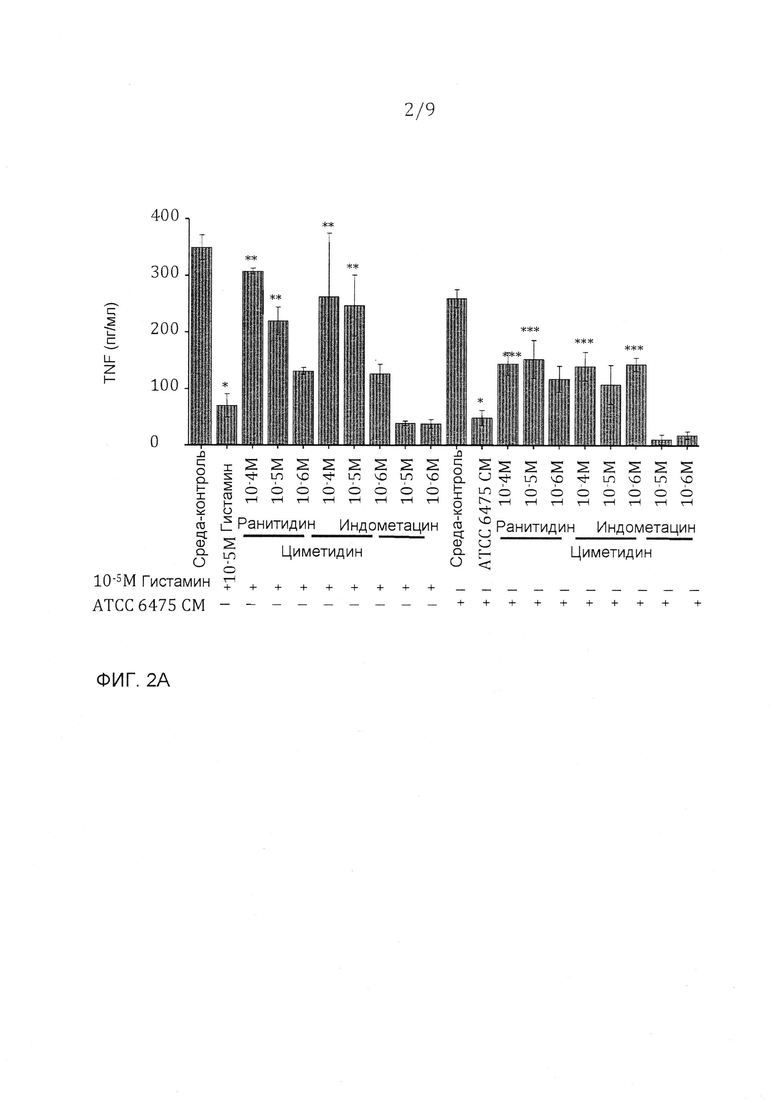
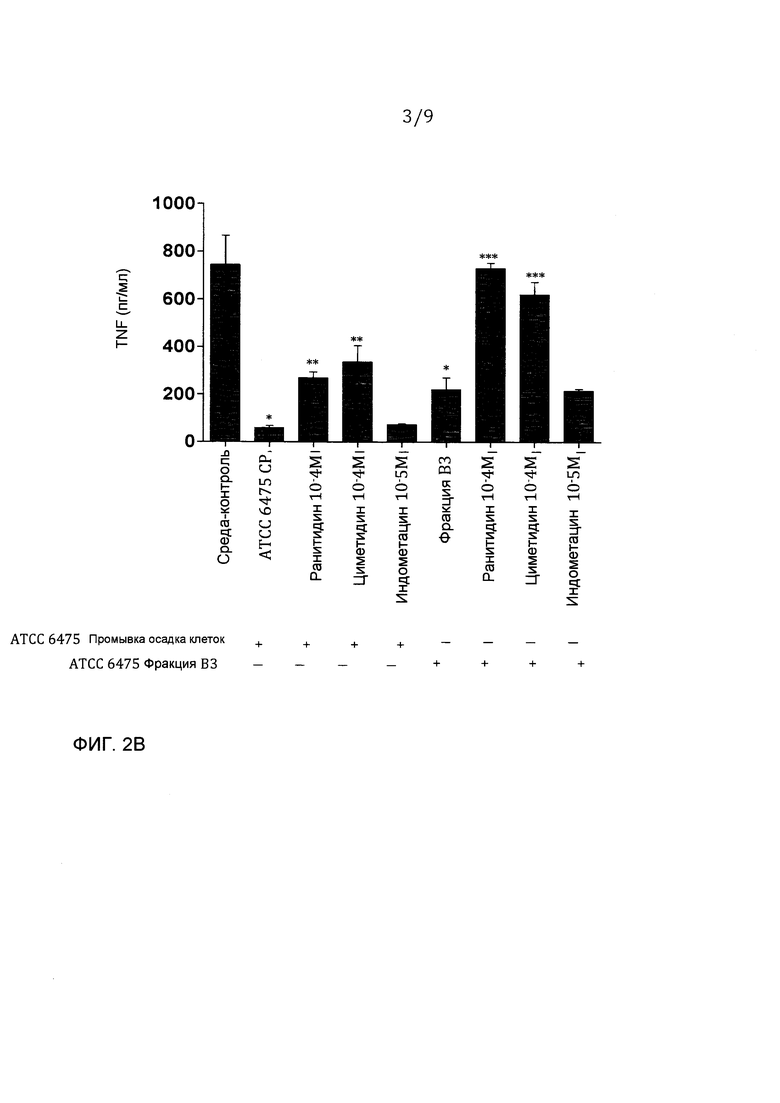
Фигура 2 - очищенный гистамин и гистамин из L. reuteri 6475 ингибирует продуцирование TNF через H2-рецептор гистамина. A. Очищенный гистамин значимо ингибировал продуцирование TNF, эффект, который блокируется специфическими антагонистами H2-рецепторов зависимым от дозы образом. Кондиционированные среды (или супернатант), содержащие секретированные факторы из штамма 6475 (включающие в себя гистамин), значимо ингибировали продуцирование TNF - эффект, который частично блокируется специфическими антагонистами H2-рецепторов. N=3, p-величина <0,05 в сравнении с контрольной средой, **p-величина <0,05 в сравнении с гистамином, ***p-величина <0,05 в сравнении с кондиционированной средой ATCC 6475 (CM). B. Промывка осадка клеток из штамма 6475, содержащая гистамин, подавляла продуцирование TNF - эффект, который частично блокировался специфическими антагонистами H2-рецепторов. Фракция В3, которая содержит относительно чистый гистамин, ингибировала продуцирование TNF - эффект, который полностью блокировался специфическими антагонистами H2-рецепторов. N=3, p-величина <0,05 в сравнении с контрольной средой, p-величина <0,05 в сравнении с промывкой клеток ATCC 6475 (CP), ***p-величина <0,05 в сравнении с фракцией B3.
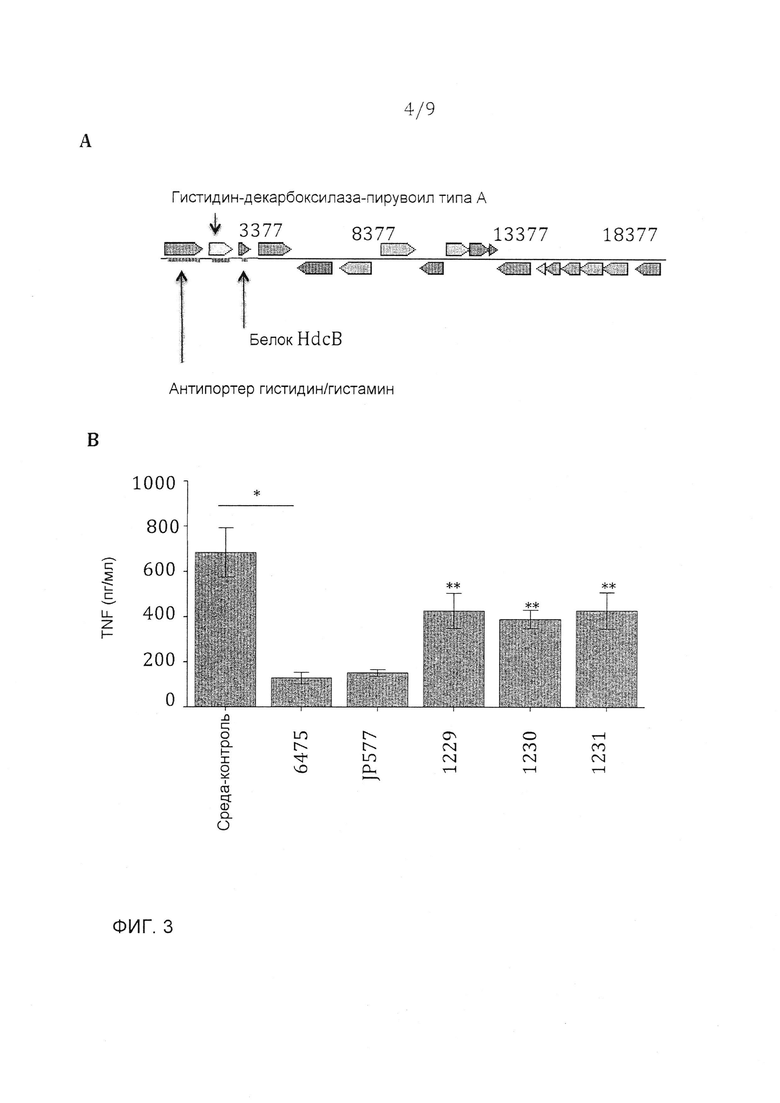
Фигура 3 - оперон гистидина является важным для TNF-ингибирующего фенотипа L. reuteri 6475. A. Этот оперон гистидина состоит из трех генов: антипортера гистидина/гистамина, hdcA и hdcB. B. Мутация в любом одном гене в этом опероне гистидина приводит к частичной потере супрессии TNF L. reuteri 6475. N=9, p-величина <0,05 в сравнении с контрольной средой, **p-величина <0,05 в сравнении с ATCC PTA 6475.
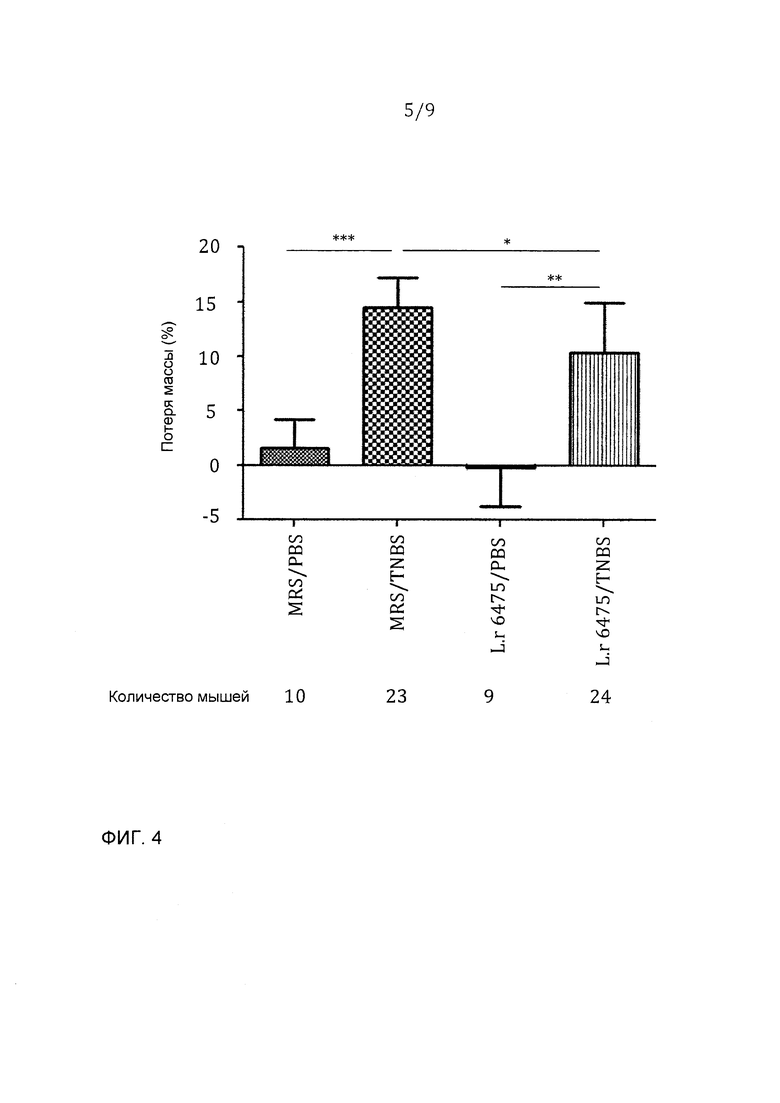
Фигура 4 - L. reuteri 6475 значимо уменьшал потерю массы, индуцированную введением TNBS; эта фигура представляет данные из двух независимых экспериментов: *p<0,05, **p<0,01, ***p<0,001.
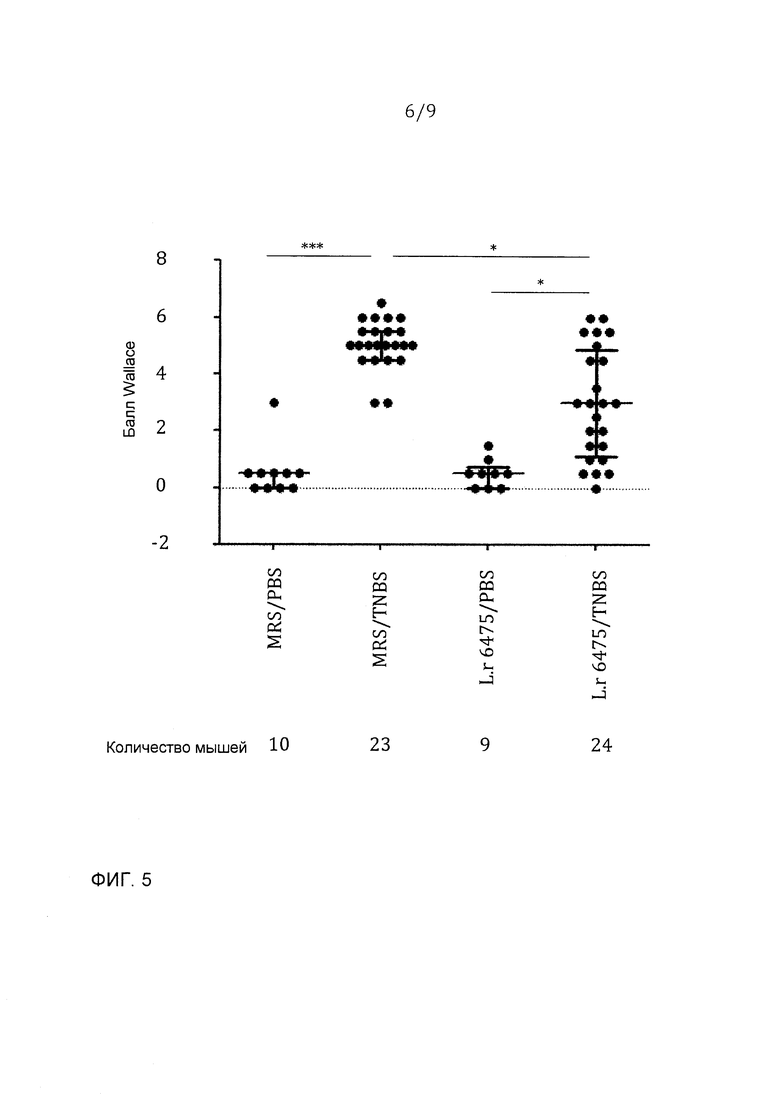
Фигура 5 - L. reuteri 6475 значимо уменьшал макроскопическое повреждение ободочной кишки, индуцированное введением TNBS; эта фигура представляет данные из двух независимых экспериментов: *p<0,05, ***p<0,001.
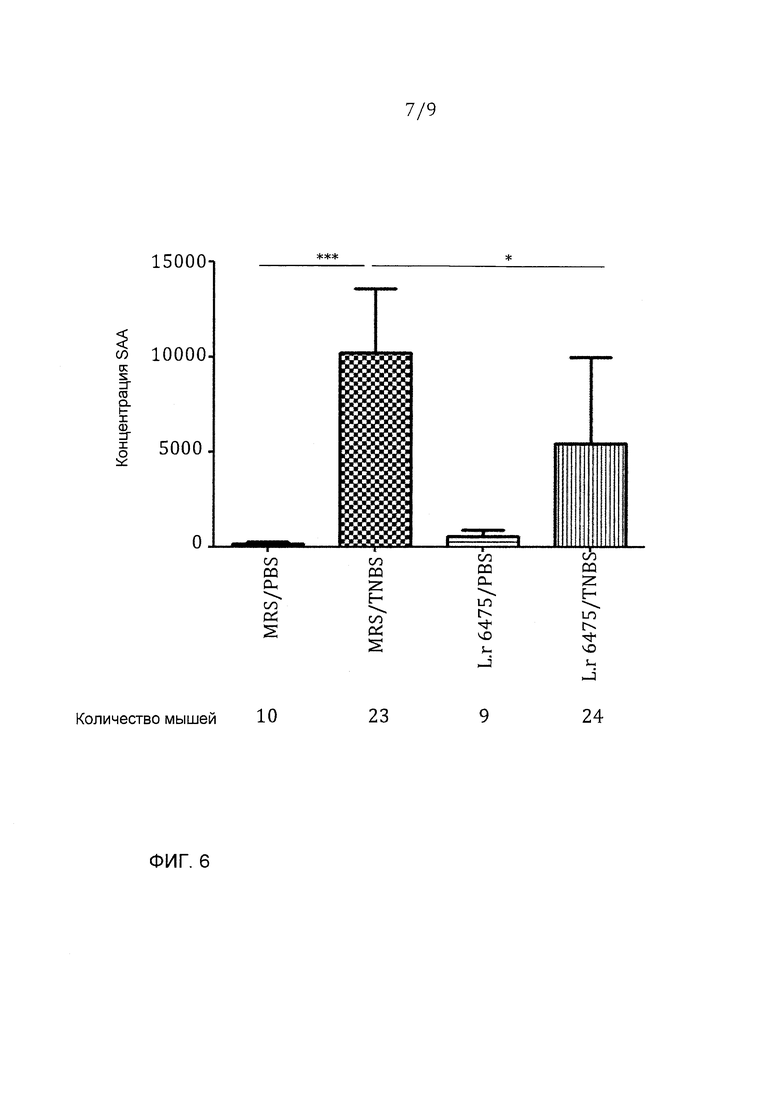
Фигура 6 - L. reuteri 6475 значимо уменьшал концентрацию SAA, индуцированную введением TNBS; эта фигура представляет данные из двух независимых экспериментов: *p<0,05, ***p<0,001.
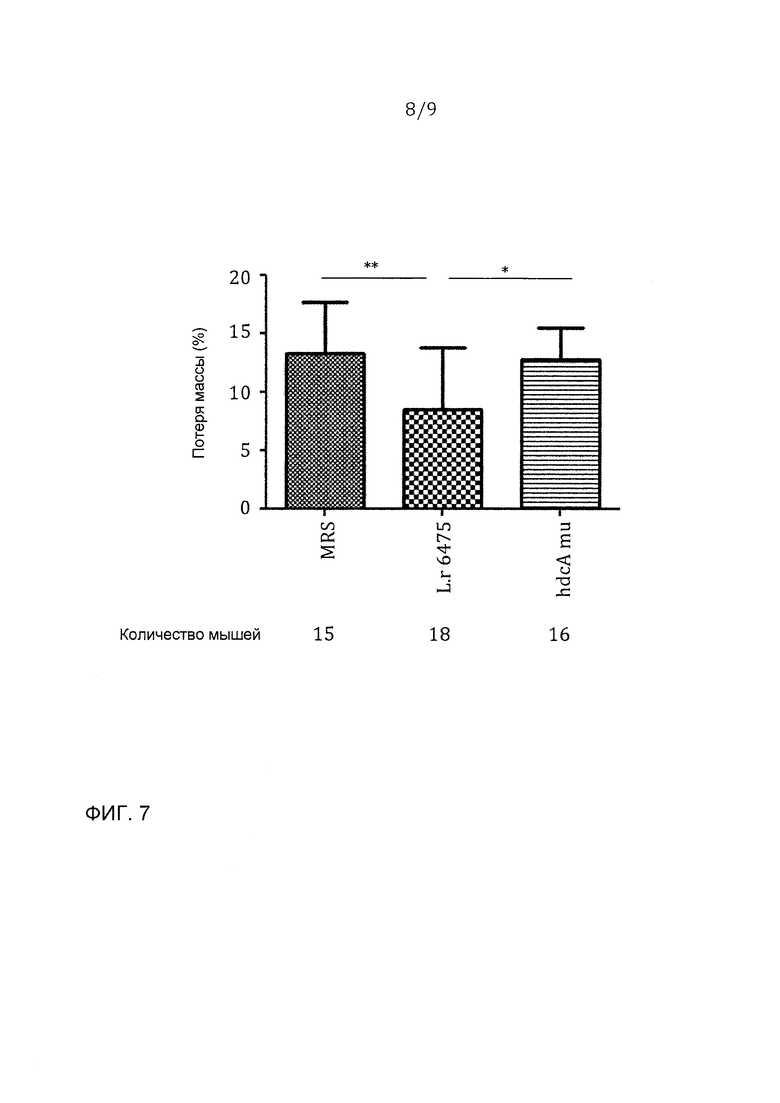
Фигура 7 - мутант hdcA давал уменьшенную способность аттенуированного колита; эта фигура представляет данные из двух независимых экспериментов: *p<0,05, ***p<0,001.
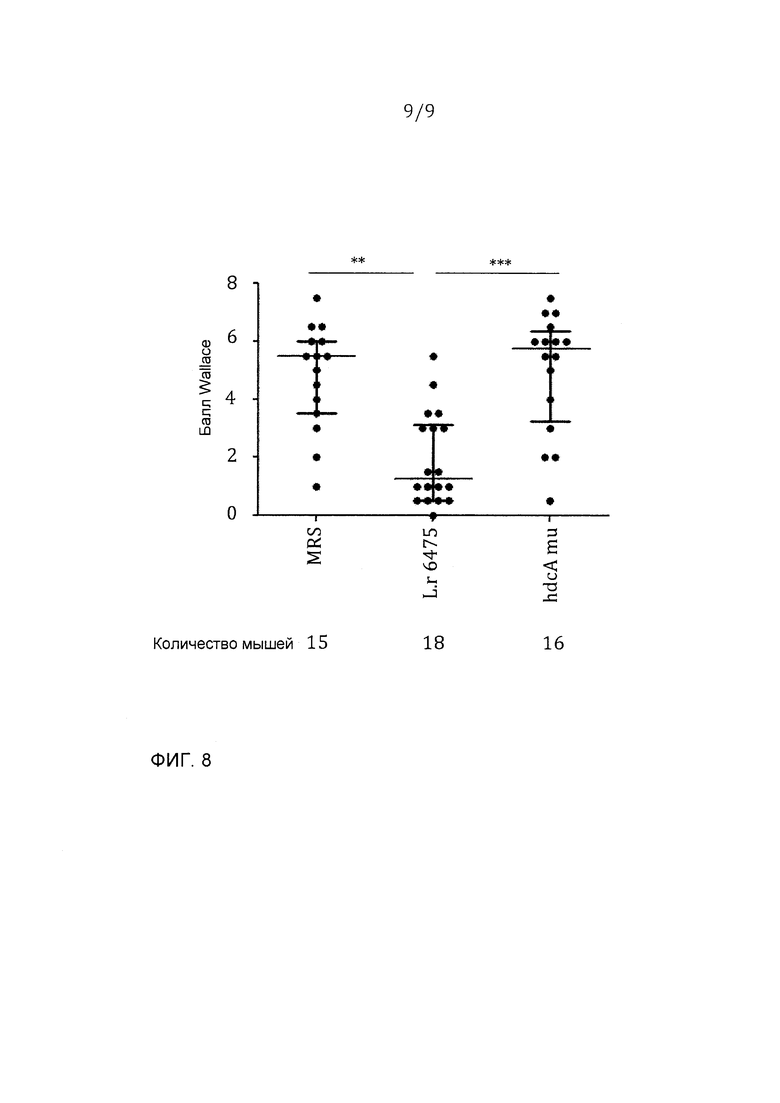
Фигура 8 - мутант hdcA давал уменьшенную способность аттенуированного колита; эта фигура представляет данные из двух независимых экспериментов: *p<0,01, ***p<0,001.
ПОДРОБНОЕ ОПИСАНИЕ ЭТОГО ИЗОБРЕТЕНИЯ И ЕГО ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Авторы данного изобретения обнаружили здесь, что отобранная группа лактобацилл, включающих в себя определенные штаммы Lactobacillus reuteri, локально продуцирует гистамин при конкретных условиях роста, и что такой гистамин будет давать пользу для хозяина, например, уменьшение воспаления, уменьшение некоторых типов рака и т. д.
Histamine
Гистамин является органическим азотистым соединением, участвующим в нескольких, ассоциированных со здоровьем процессов млекопитающего, включающих в себя локальные иммунные реакции, а также регулирующим физиологическую функцию в кишечнике и действующим в качестве нейротрансмиттера. В качестве части иммунной реакции на чужеродные патогены гистамин продуцируется базофилами и мастоцитами (тучными клетками). Гистамин может быть получен из декарбоксилирования аминокислоты гистидина - реакции, катализируемой ферментом L-гистидиндекарбоксилазой.
Бактерии способны продуцировать гистамин с использованием ферментов гистидиндекарбоксилаз, неродственных с ферментами, обнаруживаемыми в эукариотах. До сих пор такое продуцирование гистамина некоторыми бактериальными штаммами рассматривалось скорее как опасность для здоровья, чем как возможная польза для людей. Например, отравление Scombroid (форма неинфекционного заболевания новорожденных) обусловлено продуцированием гистамина бактериями в испортившейся пище, в частности, рыбы. Ферментированные пищевые продукты и напитки природно содержат малые количества гистамина вследствие сходного превращения, выполняемого ферментацией бактерий или дрожжей. Доставка некоторых контролируемых количеств гистамина из отобранных бактерий может неожиданно давать скорее полезные эффекты, чем вредные эффекты, как можно было бы ожидать из вышеупомянутых исследований.
Рецепторы гистамина являются классом G-белоксвязанных рецепторов с гистамином в качестве их эндогенного лиганда. Имеются четыре известных рецептора гистамина: Н1-рецептор (H1R), H2-рецептор (H2R), H3-рецептор (H3R) и H4-рецептор (H4R).
Vannier et al. (Histamine Suppresses Gene Expression and Synthesis of Tumor Necrosis Factor a via Histamine H2-Receptors; J Exp Med. 1991 July 1; 174(1):281-4) показали, что LPS-индуцируемый синтез TNF-α в мононуклеарных клетках периферической крови подавлялся гистамином, и авторы дополнительно предполагают, что высвобождение гистамина из тучных клеток может ограничивать степень воспалительных и иммунных реакций подавлением локального синтеза цитокинов в H2-рецепторнесущих клетках.
Противовоспалительная активность гистамина была ранее описана Wang et al. (Histamine Antagonizes Tumor Necrosis Factor (TNF) Signaling by Stimulating TNF Receptor Shedding from the Cell Surface and Golgi Storage Pool; J. Biol. Chem. 278(24): 21751-21760), которые показали, что гистамин вызывает транзиторную потерю поверхностного TNFR1, увеличивает выделение TNFR1 и мобилизацию молекул TNFR1 из аппарата Гольджи в культивируемых эндотелиальных клетках человека. Инъекция гистамина в кожу человека, трансплантированную на иммунонедостаточных мышах, вызывала выделение TNFR1 или уменьшала TNF-опосредованную индукцию эндотелиальных молекул адгезии.
Vannier et al. and Wang et al. не упоминали ничего ни об использовании бактериальных штаммов в качестве пробиотиков, ни о том, как отбирать эти штаммы на основе их гистамин-продуцирующих способностей для гарантии определенных, полезных для здоровья эффектов для хозяина, таких как противовоспалительные эффекты.
Цеплен, форму фармацевтической категории дигидрохлорида гистамина, используют для предотвращения рецидива в пациентах, диагностированных как имеющих острый миелоидный лейкоз (AML). Цеплен вводят вместе с низкими дозами иммуноактивирующего цитокина интерлейкина-2 (IL-2) в постремиссионной фазе AML, то есть когда пациенты завершили первоначальную химиотерапию. Исследования показали, что Цеплен/IL-2 может индуцировать иммуноопосредованное убивание лейкозных клеток. Это лечение (подкожные инъекции) предоставляется в виде 3-недельных циклов пациентами дома в течение 18 месяцев. Побочные эффекты Цеплена включают в себя транзиторные приливы и головную боль. Для пациентов было бы предпочтительным получение локально производимого гистамина по мере необходимости, вместо подкожных инъекций; эта стратегия доставки может быть достигнута введением бактериально-производимого гистамина пациенту с использованием штаммов, отобранных в соответствии с этим исследованием.
Было известно ранее, что грамотрицательные бактерии образуют гистамин, например, в сырой рыбе и сыром мясе после неприемлемой температуры, и что грамположительные бактерии вызывают порчу от гистамина ферментируемых продуктов, таких как сыр, колбаса, мисо, соевый соус, пиво и вино. Идентификация продуцирующих гистамин бактерий в пищевых продуктах была трудной.
Lactobacillus reuteri был ранее также ассоциирован с продуцированием гистамина, Casas et al. (Validation of the Probiotic Concept: Lactobacillus reuteri Confers Broad-spectrum Protection against Disease in Humans and Animals.; 2000, ISSN 0891-060X) сообщает, что было показано, что два штамма L. reuteri в руках Straub et.al. (Z Lebensm Unters Forsch (1995) 201: 79-82) декарбоксилируют L-гистидин с образованием гистамина, и эти авторы предостерегают против использования таких штаммов для ферментации пищевых продуктов и в качестве пробиотиков.
Trip et al. (HdcB, a novel enzyme catalyzing maturation of pyruvoyl-dependent histidine decarboxylase; Molecular Microbiology (2011) 79(4), 861-871) ссылаются на три типа генетической организации локусов декарбоксилирования гистидина среди гистамин-продуцирующих грамположительных бактерий. Наибольшая группа обнаружена в молочнокислых бактериях, включающих в себя L. hilgardii 0006, L. buchneri B301, L. reuteri F275 и T. halophilus. Было показано, что Lactobacillus hilgardii 0006 продуцирует гистамин; в исследовании, выполняемом Lucas et al. (Histamine-Producing Pathway Encoded on an Unstable Plasmid in Lactobacillus hilgardii 0006; APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Mar. 2005, Vol. 71, No. 3, p. 1417-1424), они дополнительно утверждают, что гистамин является примесью, которая появляется в некоторых продуктах во время роста нежелательных бактерий. Lucas et al. выполнили скрининг коллекции молочнокислых бактерий вина для идентификации генов, участвующих в гистамин-продуцирующем пути грамположительной бактерии вина.
Из швейцарского сыра был выделен гистамин-продуцирующий штамм Lactobacillus buchneri, который, по-видимому, участвовал во вспышке отравления гистамином. (Summer et al. Isolation of histamine-producing Lactobacillus buchneri from Swiss cheese implicated in a food poisoning outbreak.; Applied and Environmental Microbiology (1985), Vol. 50, Issue 4, p. 1094-1096).
Calles-Enriquez et al. (Sequencing and Transcriptional Analysis of the Streptococcus thermophiles Histamine Biosynthesis Gene Cluster: Factors That Affect Differential hdcA Expression; APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2010, Vol 76, No. 18, p. 6231-6238) описывают гистамин-продуцирующие штаммы Streptococcus thermophiles, термофильную закваску, используемую для приготовления йогурта и некоторых разновидностей сыра. Кроме того, они указывают на то, что присутствие штаммов со способностью декарбоксилирования гистидина могло бы приводить к продуктам, содержащим гистамин, продуцируемый во время приготовления или во время хранения, перед употреблением, и что это лежит в основе важности использования только гистаминотрицательных штаммов в приготовлении ферментированных молочных продуктов.
Даже если ранее было известно, что некоторый Lactobacillus может производить гистамин, определенно не известно, что способность продуцировать гистамин является ключевым фактором для гарантии определенных полезных эффектов для здоровья для его хозяина, например, противовоспалительных свойств некоторых штаммов Lactobacillus.
Из предыдущего уровня техники не известно и не является очевидным, что это может быть использовано для скрининга или отбора определенных пробиотических штаммов Lactobacillus.
Мастоциты (тучные клетки)
Тучная клетка (также известная как мастоцит и лаброцит) является резидентной тучной клеткой нескольких типов тканей и содержит многие гранулы, богатые гистомином и гепарином. Хотя они наиболее хорошо известны из-за их роли в аллергии и анафилаксии, тучные клетки также играют важную защитную роль, будучи, например, тесно связанными с заживлением ран и защитой против патогенов.
Тучные клетки присутствуют в большинстве тканей, обычно окружая кровеносные сосуды и нервы, и особенно заметны вблизи пограничных слоев между наружным миром и внутренней средой, такой как кожа, слизистая оболочка легких и пищеварительный тракт, а также в полости рта, конъюнктивы и носа.
В аллергических реакциях тучные клетки остаются неактивными, пока аллерген не связывается с IgE уже в ассоциации с этой клеткой. Другие события активации мембраны могут либо праймировать тучные клетки для последующей дегрануляции, либо могут действовать в синергии с трансдукцией сигнала FceRI. Гистамин из такой грануляции сужает посткапиллярные венулы, активирует эндотелий и увеличивает проницаемость кровеносных сосудов. Высвобождение гистамина приводит к локальному отеку (опуханию), теплу, покраснению и аттрагированию других воспалительных клеток к сайту высвобождения. Оно также раздражает нервные окончания (приводя к зуду или боли). Кожными признаками высвобождения гистамина является реакция «прилива крови к лицу и волдырей». Шишка и краснота сразу же после укуса комара являются хорошим примером этой реакции, которая осуществляется в течение секунд после обработки тучных клеток аллергеном. Другие физиологические активности тучных клеток являются гораздо менее хорошо понимаемыми. Несколько линий доказательства предполагают, что тучные клетки могут иметь явно фундаментальную роль в наследственном иммунитете - они способны усовершенствовать широкий ряд важных цитокинов и других воспалительных медиаторов, таких как TNFα, они экспрессируют множественные «рецепторы узнавания паттернов», которые, как считается, включены в распознавание широких классов патогенов, и мыши без тучных клеток считаются гораздо более чувствительными к различным инфекциям.
С учетом токсичности бактериального гистамина в пищевых продуктах и того факта, что рекомендуется избегание гистамин-продуцирующих штаммов в ферментированных продуктах (см. дополнительно примеры в Calles-Enriquez et al. Sequencing and Transcriptional Analysis of the Streptococcus thermophiles Histamine Biosynthesis Gene Cluster: Factors That Affect Differential hdcA Expression; APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2010, Vol. 76, No. 18, p. 6231-6238), нельзя считать очевидным применение определенных отобранных штаммов Lactobacillus для локального продуцирования гистамина в лечении и/или профилактике различных заболеваний.
Взаимоотношение между хозяином и его микробами является комплексным, так как имеются определенно собственные типы клеток млекопитающих, такие как тучные клетки. Это взаимоотношение хозяин-микроб развивалось на протяжении многих лет ко-эволюции, это включает в себя продуцирование микробами различных метаболитов, которые могут быть полезными для хозяина с точки зрения питания, иммунологически и т. д., действует в качестве всех или части антагонистов, агонистов, десенсибилизации и т. д. специфических рецепторов или других процессов. Таким образом, имеется необходимость в понимании таких специфических взаимодействий между микробами и человеком таким образом, чтобы большая часть подходящих пробиотических штаммов могла быть отобрана и использована для противодействия таким развитиям.
Авторы изобретения обнаружили здесь, что отобранная группа лактобацилл, включающая в себя некоторые штаммы Lactobacillus reuteri, локально продуцирует гистамин в специфических условиях роста. Такой локально продуцированный гистамин, в противоположность прежним мнениям, будет приносить пользу хозяину множественными путями, включающими в себя уменьшение воспаления, уменьшение определенных типов рака и т. д.
Одной целью этого изобретения является обеспечение продуктов, содержащих указанные штаммы вместе со специфическим источником углерода, для получения синбиотического продукта.
Другие цели и преимущества будут более полно очевидными из следующего описания и прилагаемой формулы изобретения.
Введение штаммов молочнокислых бактерий, отобранных в соответствии с данным способом, млекопитающему будет приводить к локально продуцируемому гистамину, который мог бы быть полезным по нескольким причинам.
Первичной целью данного изобретения является обеспечение способа отбора штаммов молочнокислых бактерий, гарантирующих хороший противовоспалительный эффект. Эти штаммы могли бы предпочтительно использоваться для лечения и/или профилактики воспалительных состояний, так как оперон гистидина и продуцирование гистамина является существенным для противовоспалительной способности определенной молочнокислой бактерии. Предпочтительно эти штаммы могут быть использованы для лечения и/или профилактики воспалительных процессов в GI-тракте, GU-тракте, полости рта, в легких и дыхательных путях, на коже и т. д. тела млекопитающего, в том числе, но не ограничивающихся ими, колита, IBD, IBS, дивертикулеза, гингивита, вагинита и т. д. Ранее было известно, что гистамин через H2-рецептор может уменьшать экспрессию гена TNF-альфа. Кроме того, тучные клетки способны вырабатывать широкий ряд важных цитокинов и других воспалительных медиаторов, таких как TNF-альфа. Однако ранее не было известно, что оперон гистидина и локальное продуцирование гистамина таких отобранных штаммов может быть полезным для этого хозяина и является, например, ключевым фактором в противовоспалительной способности отобранных штаммов L. reuteri. Ранее не было известно применение отобранного L. reuteri в лечении, требующем гистамина.
Предпочтительными продуктами и штаммами для лечения и/или профилактики воспалительных состояний являются Lactobacillus reuteri, в частности, Lactobacillus reuteri 6475 (ATCC PTA 6475). В других вариантах осуществления этого изобретения используемым штаммом не является Lactobacillus reuteri 6475 (ATCC PTA 6475).
Терапевтические применения этих штаммов, продуктов и композиций этого изобретения, определенных здесь, приводят к уменьшению или облегчению релевантных заболеваний или симптомов заболевания, например, могут приводить к значимому уменьшению уровней воспаления в млекопитающем. Например, локально производимый гистамин может быть активирующим H2-рецепторы на кишечных эпителиальных клетках, а также иммунных клетках для супрессии мукозного иммунитета хозяина, например, через ингибирование провоспалительных цитокинов. Таким образом, данное изобретение позволяет превращать диетический компонент (гистидин) в гистамин в сайте активности и локально модулировать иммунную реакцию хозяина (например, в кишечнике). Можно видеть, что такое локальное продуцирование гистамина, обеспечиваемое данным изобретением, может обеспечивать реальные преимущества относительно, например, перорального проглатывания или других форм введения гистамина, особенно в связи с тем фактом, что такое пероральное проглатывание не может пропагандироваться вследствие узнаваемых токсичных эффектов и рисков для здоровья.
В частности, когда речь идет о воспалительных заболеваниях кишечника, эти терапевтические применения штаммов, продуктов и композиций этого изобретения могут приводить к значимому уменьшению образования язв и повреждения кишечника (например, повреждения ободочной кишки), измеряемому стандартным способом, таким как показатель Wallace, значимому уменьшению потери массы или значимому уменьшению воспаления кишечника, например, ободочной кишки.
Такое уменьшение или облегчение заболевания или его симптомов может быть измерено любым подходящим анализом. Предпочтительно уменьшение или ослабление заболевания или симптомов является статистически значимым, предпочтительно с величиной вероятности <0,05. Такое уменьшение или ослабление заболевания или симптомов обычно определяют в сравнении с подходящим контрольным индивидуумом или контрольной популяцией, например, здоровым млекопитающим или не получающим лечения или получающим плацебо млекопитающим.
Один подходящий способ введения и приготовления этих штаммов и т. д. выбирают в зависимости от участка, в котором желательным является продуцирование гистамина. Одним предпочтительным способом введения является пероральный способ, однако равным образом для некоторых обработок будет подходящей местная или некая другая форма локального введения в кожу, прямую кишку, влагалище или десны или внутривенная или внутримышечная инъекция.
Хотя приведенные здесь примеры демонстрируют применение штаммов этого изобретения и их подходящие дозы для лечения колита, будет понятно, что это является только одним примером воспалительных состояний, которые могут лечиться в соответствии с данным изобретением, и что подходящие дозы этих штаммов, продуктов и композиций этого изобретения, описанного здесь, могут быть выбраны в зависимости от подлежащего лечению заболевания, способа введения и соответствующей композиции.
Диетические смеси, содержащие гистидин, могут быть использованы для гарантии присутствия гистидина и увеличения посредством этого эффективности этих бактерий. Гистидин может вводиться отдельно или вместе с этими бактериями.
Одной возможностью гарантии бактериальной доставки гистидина является употребление богатой гистидином пищи, в том числе, но не только, соевого белка, сыра, яиц, куриного мяса или свинины.
Было показано, что оперон гистидина в бактериях улучшает способность роста в условиях низкого рН или ограничения источника энергии (Calles-Enriquez et al.), но оперон гистидина не был ассоциирован с противовоспалительными признаками некоторых штаммов L. reuteri.
Другой целью данного изобретения является применение штаммов, отобранных в соответствии с данным способом, в раковой терапии. Гистамин в комбинации с IL-2 использовали для лечения AML. Применение штаммов данного изобретения будет приводить к локально продуцируемому гистамину, который в комбинации с IL-2 мог бы быть использован для лечения AML.
Другой целью данного изобретения является применение отобранных штаммов для уменьшения пищевой аллергии, других аллергических реакций или других аутоиммунных заболеваний. Системные увеличения гистамина являются, как было известно ранее, следствием аллергии, вызываемой грануляцией тучных клеток. При введении штаммов, отобранных в соответствии с этим изобретением, реципиенту этот локально продуцируемый гистамин будет приводить к эффекту десенсибилизации, который будет уменьшать аллергию или другие аутоиммунные заболевания.
Целью данного изобретения является также применение продуцирующих гистамин штаммов молочнокислых бактерий для уменьшения риска поноса путешественников. Пациенты, лечившиеся блокаторами гистамина, имеют увеличенный риск приобретения поноса путешественников, и этот увеличенный риск мог бы быть нейтрализован введением молочнокислых бактерий в соответствии с данным способом.
Еще одной целью данного изобретения является применение отобранных штаммов в лечении MS. Гистамин был предложен в качестве важной молекулы для развития новых способов лечения для MS, и штаммы, отобранные в соответствии с данным изобретением, будут обеспечивать пациента гистамином.
Еще одной целью данного изобретения является применение продуцирующих гистамин бактерий в качестве противовоспалительного лечения кожи с использованием доступных гистидина и аналогов гистидина в коже. Поскольку гистидин является субстратом уроканиновой кислоты, которую кожа производит при УФ-облучении, эта уроканиновая кислота проявляет противовоспалительные свойства на коже.
Другой целью является применение таких отобранных штаммов для ингибирования активации ERK1/2.
Другой целью этого изобретения является ингибирование TNF-альфа.
Другой целью этого изобретения является уменьшение воспаления, локально или системно.
Другой целью этого изобретения является усиление экзоцитоза синаптических пузырьков ингибированием ERK1/2.
Другой целью этого изобретения является стимуляция самообновления эмбриональных стволовых клеток ингибированием ERK1/2.
Другой целью этого изобретения является индуцирование экспрессии макрофага ABCA1 и отток холестерина ингибированием ERK1/2.
Другой целью этого изобретения является уменьшение сердечной гипертрофии и сердечной недостаточности ингибированием ERK1/2.
Другой целью этого изобретения является уменьшение пролиферации некоторых типов рака, включающих в себя лейкоз (например, AML) или злокачественную меланому. Таким образом, такие типы рака являются предпочтительными заболеваниями, которые могут лечиться с использованием этих штаммов, продуктов и композиций данного изобретения.
Другой целью этого изобретения является применение отобранных штаммов для продуцирования гистамина в определенных условиях в качестве нейротрансмиттера, например, во взаимодействиях GU-тракта с CNS, а также передаче невральных сигналов в случае локальной боли. Эта роль в качестве нейротрансмиттера может распространяться до действий на перистальтику кишечника (для лечения констипации или диареи) и для передачи болевых сигналов в кишечнике.
Другой целью этого изобретения является применение отобранных штаммов для воздействия на ось головной мозг - кишечник, когда отобранные LAB будут продуцировать гистамин и воздействовать на восприятие висцеральной боли и передачу сигнала в брюшную нервную систему. Таким образом, можно видеть, что данное изобретение может быть использовано для лечения и/или профилактики любого заболевания, которое будет иметь пользу от локального продуцирования гистамина, или для лечения и/или профилактики любого заболевания, которое может лечиться локальным введением гистамина.
Дополнительной целью этого изобретения является обеспечение продуктов, содержащих указанные штаммы.
Дополнительной целью этого изобретения является обеспечение продуктов, содержащих указанные штаммы, вместе со специфическим источником углерода, чтобы иметь синбиотический продукт, который будет, через специфическую стимуляцию этого гистамин-продуцирующего штамма, усиливать эти эффекты.
Дополнительной целью этого изобретения является обеспечение продуктов, содержащих указанные штаммы, вместе с гистидином, в том числе аналогами гистидина или гистидин-содержащими продуктами или композицией гистидина. Предпочтительно такую смесь вводят перорально в защитной капсуле для высвобождения ее содержимого в нижней части GI-тракта для гарантии выживания как гистидина, так и бактерий в участке действия.
Дополнительной целью этого изобретения является объединенное введение указанных штаммов с богатой гистидином диетой.
Еще один дополнительный аспект этого изобретения обеспечивает продукт для терапевтических применений, как определено здесь в другом месте, где указанное применение предусматривает введение по меньшей мере одного дополнительного терапевтического или пищевого агента. В таких вариантах осуществления этим дополнительным терапевтическим агентом может быть любой дополнительный агент, который применим в лечении рассматриваемого заболевания, например, дополнительный противовоспалительный агент или иммунотерапевтический агент, такой как, например, хемокин или цитокин (например, IL-2).
В предпочтительных вариантах осуществления указанный дополнительный агент содержит гистидин или аналог гистидина, подходящий источник углерода, который поддерживает продуцирование гистамина бактериальным штаммом, или их комбинацию.
Указанные дополнительные агенты могут вводиться вместе со штаммами этого изобретения или могут вводиться по отдельности. Кроме того, указанные дополнительные агенты могут вводиться в то же самое время, что и штаммы этого изобретения, или в различных временных точках.
Данное изобретение обеспечивает также композицию, содержащую:
(i) штамм молочнокислой бактерии, получаемый способом отбора этого изобретения (или штамм молочнокислой бактерии, способный продуцировать гистамин, определенный в других случаях здесь), где указанный штамм молочнокислой бактерии имеет оперон гистидина и способен продуцировать гистамин; и
(ii) по меньшей мере один дополнительный компонент, выбранный из группы, состоящей из подходящего источника углерода, который поддерживает продуцирование гистамина указанным штаммом, источника гистидина или аналога гистидина, и их комбинации.
В продуктах, композициях и применениях этого изобретения, описанного здесь, предпочтительно указанный гистидин или аналог гистидина находится в форме содержащего гистидин или аналог гистидина пищевого продукта или пищевой добавки, или указанный источник углерода содержит глюкозу. Предпочтительно указанный источник углерода не будет содержать сахарозы или по меньшей мере будет содержать сахарозу при таком уровне, который не будет значимо ухудшать продуцирование гистамина этим штаммом. Необязательно могут быть также обеспечены источники других аминокислот.
В альтернативных вариантах осуществления штаммы, определенные в части (i), могут быть объединены с дополнительным компонентом, который используется в лечении рассматриваемого заболевания, то есть дополнительным терапевтическим агентом, например, дополнительным противовоспалительным агентом или иммунотерапевтическим агентом, таким как, например, хемокин или цитокин (например, IL-2).
Lactobacillus reuteri является гетероферментативным видом молочнокислой бактерии, который природно заселяет кишечник людей и животных. Конкретные пробиотические штаммы L. reuteri сильно подавляют продуцирование TNFα человека, тогда как другие пробиотические штаммы L. reuteri усиливают продуцирование TNFα человека.
Описанное здесь изобретение стало возможным посредством механистических исследований пробиотического штамма L. reuteri 6475 и других штаммов, которые продемонстрировали их действие на активированных миелоидных клетках человека. Метаболиты L. reuteri выделяли с использованием HILIC-HPLC, и гистамин идентифицировали при помощи NMR-спектроскопии и масс-спектрометрии. Количественное определение гистамина тройной квадрупольной MS выявило, что штамм L. reuteri 6475 продуцирует относительно высокие концентрации гистамина при выращивании в минимальной среде на основе глюкозы. Прежние исследования транскриптомики предполагали, что два гена в опероне гистидина L. reuteri могут играть роль в ингибировании TNF штаммом 6475. Нацеленный мутагенез этих генов выявил, что каждый ген в опероне гистидина, антипортера гистидина-гистамина, HdcA и HdcB, является важным для TNF-ингибирующего фенотипа штамма 6475. Механистические исследования продемонстрировали, что гистамин ингибирует TNF через передачу сигнала через H2-, но не через H1-рецептор. Передача сигнала через H2-рецептор увеличивает внутриклеточную cAMP (цАМФ), которая активирует PKA. PKA-активность является необходимой для супрессии TNF гистамином. Гистамин блокирует активацию пути передачи сигнала MEK-ERK MAPK.
Гистамин является более известным в отношении его провоспалительных эффектов в аллергии и анафилаксии, но несколько исследований продемонстрировали противовоспалительные функции гистамина. Исследования in vitro показали, что гистамин может ингибировать продуцирование провоспалительных цитокинов, IL-1, IL-12 и TNF из LPS-стимулированных моноцитов и макрофагов человека, и этот эффект является обратимым антагонистами H2-рецептора. Кроме того, гистамин может стимулировать продуцирование противовоспалительного цитокина, IL-10, через H2-рецептор. Передача сигнала через H2-рецептор приводит к уменьшенной экспрессии CD14-рецептора, рецептора, участвующего в узнавании LPS, на поверхности моноцитов человека. Гистамин действует также на TNF-рецептор. Передача сигнала через H1-рецептор индуцирует выделение как TNFR1, так и TNFR2. Исследования in vivo также выявили противовоспалительную роль гистамина. Лечение димапритом, специфическим агонистом H2-рецептора, уменьшало уровни TNF в плазме в мышиных моделях эндотоксинового шока (LPS-индуцирование) и гепатита (индуцирование LPS плюс галактозамином).
Гистамин был протективным в мышиной модели LPS-индуцированного повреждения печени, и эти эффекты были атенуированными в мыши с нокаутом H2-рецептора. В этом кишечнике гистамин может способствовать защите против бактериальной инфекции. Передача сигнала через H2-рецептор в пейеровых бляшках способствует предотвращению инфекции Yersinia enterocolitica.
Это действие гистамина может быть определено по экспрессии рецепторов гистамина на клетке-мишени. В T-клетках это действие гистамина зависит от того, какой рецептор гистамина является активированным.
Посредством передачи сигнала через H1-рецептор гистамин усиливает реакции TH1-типа, но подавляет реакции как TH1, так и TH2 через H2-рецептор. Одно исследование выполняли с наблюдением экспрессии рецептора гистамина в желудочно-кишечном тракте человека. Многие из этих испытанных типов клеток экспрессировали множественные рецепторы гистамина. Например, иммунные клетки, включающие в себя макрофаги, высоко экспрессировали H1- и H2-рецептор и демонстрировали низкую экспрессию H4-рецептора. Увеличенные тучные клетки и гистамин участвовали в висцеральной гиперчувствительности, ассоциированной с IBS. Увеличенное число и увеличенная активность тучных клеток вблизи иннервации слизистой оболочки ободочной кишки может приводить к повышенному восприятию боли в животе, ассоциированной с IBS. Исследование с кетотифеном, стабилизирующим агентом тучных клеток, демонстрировали увеличенный порог боли в пациентах с IBS, уменьшенные симптомы IBS, но не изменение в числе или активности (определяемых по высвобождению гистамина и триптазы) тучных клеток в ткани биопсии тучных клеток. Эти эффекты кетотифена в улучшении IBS могут не быть результатом стабилизации тучных клеток, но могут быть отнесены к их другой роли в качестве антагониста H1-рецептора. Если активация H1-рецептора ассоциирована с провоспалительной реакцией, блокирование его активности кетотифеном может позволить гистамину, производимому либо тучными клетками, либо микробиотой кишечника, такой как L. reuteri, передавать сигнал только через H2-рецептор. Как уже было продемонстрировано авторами изобретения, передача сигнала через H2-рецептор может подавлять продуцирование TNF и вызывать противовоспалительный эффект. Этот механизм кетотифена может быть использован для новых терапий, объединяющих антагонисты H1-рецептора с общим пробиотическим эффектом штамма L. reuteri.
Дополнительное изменение источника углерода сред для выращивания с глюкозы на сахарозу является достаточным для супрессии TNF-ингибирующего фенотипа отобранного штамма, например, штамма L. reuteri 6475. Кроме того, значимую понижающую регуляцию (даун-регуляцию) всех трех генов в опероне гистидина наблюдали с сахарозой в условиях роста.
Эта идентификация гистамина как противовоспалительного соединения, продуцируемого отобранными пробиотическими штаммами Lactobacillus, будет способствовать определению терапевтических применений для таких штаммов. Механистические исследования связывали активацию H2-рецептора на клетках THP-1 с гистамином и супрессией ERK-активации. ERK-активация участвует во многих клеточных функциях наряду с продуцированием TNF. ERK-активация участвует в пролиферации, онкогенезе, дифференцировке и выживании клеток. Эти результаты предполагают роль отобранных штаммов, таких как L. reuteri 6475, в защите против рака посредством супрессии воспаления, пролиферации клеток и апоптоза через ингибирование ERK-активации. Кроме того, гистамин является известным нейротрансмиттером. Продуцирование гистамина отобранными штаммами может влиять на передачу сигнала в кишечной нервной системе, влиять на восприятие боли и перистальтику кишечника. Для гарантии продуцирования гистамина в месте действия может быть предпочтительным обеспечение этих бактерий гистидином. Гистидин может вводиться вместе с этими бактериями или отдельно, рацион, богатый гистидином, может также увеличивать продуцирование гистамина.
Данное изобретение обеспечивает некоторые штаммы молочнокислых бактерий и способ отбора таких штаммов и продуктов, содержащих такие штаммы. Эти бактерии выбирают с использованием скрининга на оперон гистидина; неожиданно было показано, что присутствие активного оперона гистидина является существенным для различных полезных эффектов, таких как иммуномодулирующие свойства штаммов молочнокислых бактерий.
Другие цели и преимущества данного изобретения станут очевидными для читателя, и предполагается, что эти цели и преимущества находятся в пределах объема данного изобретения.
Это изобретение будет описываться далее со ссылкой на следующие неограничивающие примеры:
ПРИМЕРЫ
Бактериальные штаммы, используемые в этом исследовании
Анализ транскриптомы оперона гистидина в мутантах штамма L. reuteri 6475
6475
глюкоза
§Дикий тип 6475, росший в LDMIIIS в сравнении с диким типом 6475, росшим в LDMIIIS, теряет способность ингибирования продуцирования TNF.
ПРИМЕР 1
Продуцирование гистамина выбранным Lactobacillus
Бактериальные штаммы и условия культивирования
Все бактериальные штаммы, использованные в этом исследовании, описаны в таблице 1. Lactobacillus reuteri ATCC PTA 6475 является изолятом молока финской матери (доступного из ATCC, Manassas, VA, USA). L. reuteri штаммы ATCC PTA 6475, ATCC 6475 JP577, ATCC 6475 1229, ATCC 6475 1230 и ATCC 6475 1231 будут называться штаммами 6475, JP577, 1229, 1230 и 1231 соответственно на протяжении этого описания. Штаммы L. reuteri культивировали при анаэробных условиях в течение 16-18 часов в средах deMan, Rogosa, Sharpe (Difco, Franklin Lakes, NJ), и инокулировали в 2 л полуопределенной среды, LDMIII (OD600, корректированной до 0,1), которая была описана ранее. Источником углерода были либо глюкоза, LDMIIIG, либо сахароза, LDMIIIS. Эту культуру выращивали в течение 24 часов при 37°С в анаэробной рабочей установке (MACS MG-500, Microbiology International, Frederick, MD), поставляемой со смесью 10% СО2, 10% H2 и 80% N2. Пробы брали в разных временных точках для прослеживания роста измерением OD600. В стационарной фазе (24 ч), эти клетки осаждали из 2-литровой культуры (4000×g, 10 мин). Осадки клеток и свободные от бактерий супернатанты хранили при -20°C перед дальнейшей обработкой для разделения ВЖХ и тестирования в биоанализе на ингибирование TNF.
Клеточная линия и реагенты
In vitro эксперименты выполняли с клетками THP-1 (моноцитоидной клеточной линией, ATCC, Manassas, VA), сохраняемой в среде RPMI (ATCC) и инактивированной нагреванием фетальной телячьей сыворотке (Invitrogen, Carlsbad, CA) при 37°C, 5% CO2. MEK1/2, фосфо-MEK1/2, ERK1/2 и фосфо-ERKl/2-антитела и MEK-ингибитор получали из Cell Signaling Technology (Dan vers, MA), и β-Актин-антитело получали из Abeam (Cambridge, MA). Все другие реагенты получали из Sigma (St. Louis, MO), если нет другого указания.
HILIC-HPLC-разделение связанных с клеточной стенкой факторов
Осадки клеток (7 г) из штамма 6475, выращенного либо в LDMIIIG, либо в LDMIIIS, промывали 30 мл, охлажденной на льду смесью 50% ацетонитрил/0,1% трифторуксусная кислота (TFA). Эту супензию клеток центрифугировали в течение 10 мин, 4000×g при 4°С. Супернатанты фильтровали через поливинилиденфторидные (PVDF) мембранные фильтры (размер пор 0,45 мкм, Millipore, Bedford, MA), лиофилизировали и ресуспендировали в 10 мл 0,1% муравьиной кислоты. Эту ресуспендированную пробу фракционировали по размеру при помощи центрифужных фильтровальных устройств Amicon Ultra-15, использующих ultracel-3-мембрану (Millipore, Bedford, MA). Этот фильтрат (9 мл) высушивали до 1 мл с использованием скоростного вакуума (speed-вакуума) и 0,75 мл использовали для HILIC-HPLC. Эту пробу растворяли с использованием 100% ацетонитрила перед пропусканием через колонку PolyLC Hydroxyethyl с градиентом 100-0% ацетонитрила, 0,1% муравьиной кислотой. Эта проба проходила в течение 25 минут и 25 фракций (A1-C1) собирали при 10 мл/мин/пробирка. Три миллилитра из каждой фракции лиофилизировали, ресуспендировали в 3 мл 0,1% уксусной кислоты и опять лиофилизировали для тестирования в биоанализе ингибирования TNF.
Биоанализ ингибирования TNF и TNF-ELISA
Бактериальные супернатанты (10 мл) из 24-часовой культуры LDMIII стерилизовали с использованием стерилизующих мембранных фильтров PVDF (размер пор 0,22 мкм, Millipore) и фракционировали по размеру, как описано выше. Один миллилитр фильтрата <3 кДа сушили при помощи speed-вакуума и ресуспендировали в среде RPMI. Эти пробы обработанного супернатанта называют кондиционированными средами. Все супернатанты нормализовали по объему относительно OD600=1-0. Лиофилизированные фракции из HILIC-HPLC-разделения ресуспендировали в 400 мкл 10 мг/мкл бикарбоната аммония, сушили в speed-вакууме и ресуспендировали в 400 мкл среды. Кондиционированные среды и промывочные фракции осадка клеток тестировали на их способность модулировать продуцирование TNF в моноцитоидных клетках. Вкратце THP-1-клетки (приблизительно 5×l04 клеток) стимулировали для получения TNF добавлением 100 нг/мл Pam3Cys-SKKKK×3 HC1 (EMC Microcollections, Tuebingen, Germany), как описано ранее. Ингибиторы - антагонисты H2-рецептора, ранитидин и циметидин (10-4-106 M), антагонист H1-рецептора, индометацин (10~5-106 M), ингибитор MEK, U0126 (10 мкМ) и ингибитор PKA, H89 (дигидрохлорид N-[2-(п-бромциннамиламино)этил]-5-изохинолинсульфонамида) (10-5 M) - добавляли к этим клеткам THP-1 с последующим добавлением кондиционированных сред L. reuteri или промывочных фракций осадка клеток (5% о/о), гистамина (105 M) или дибутирил cAMP (10~3-10-7 M). Планшеты инкубировали при 37°C и 5% CO2 в течение 3,5 ч. Клетки THP-1 осаждали (3000×g, 5 мин, 4°C) и использовали количественные ELISA для определения количеств TNF в супернатантах THP-1-клеток в соответствии с инструкциями изготовителя (R&D Systems, Minneapolis, MN).
TNF-ингибирующие соединения L. reuteri 6475 выделяли с использованием жидкостной хроматографии гидрофильного взаимодействия - высокоэффективной жидкостной хроматографии (HILIC-HPLC)
Осадки бактериальных клеток промывали для удаления соединений, рыхло связанных с клеточной поверхностью. Компоненты промывок осадков клеток разделяли на основе гидрофобности с использованием HILIC-HPLC и полученные 25 фракций тестировали на удерживание этого TNF-ингибирующего соединения. L. reuteri 6475, выращенный в минимальной среде с глюкозой в качестве единственного источника углерода, продуцирует TNF-ингибирующие факторы, которые удерживались в 3 отдельных фракциях HILIC-HPLC (B3, B5 и B6, данные не показаны). L. reuteri 6475, выращенный с сахарозой в качестве единственных источников углерода, теряет TNF-ингибирующий фенотип, и он служил в качестве отрицательного контроля. Ни одна из фракций HILIC-HPLC из промывок осадка клеток 6475 с сахарозой не демонстрировала значимого ингибирования TNF (данные не показаны).
Гистамин идентифицировали в TNF-ингибирующей фракции HILIC-HPLC с использованием NMR (ЯМР)-спектроскопии и масс-спектрометрии
TNF-ингибирующую фракцию HILIC-HPLC B3 анализировали при помощи 1H NMR и сравнивали с соседней, не ингибирующей TNF фракцией В4. Уникальный ряд пиков с химическим смещением между 7,0-7,5 м.д., которое является характеристикой ароматического соединения, наблюдали во фракции В3, но не во фракции В4 (данные не показаны). Этот кластер ароматических соединений дополнительно анализировали 2-мерным (2D) ЯМР с гетеронуклеарной одноквантовой когерентностью (HSQC) для идентификации его компонентов. Эти ароматические соединения состояли из триптофана, фенилаланина, гистамина и одного соединениия, которое было неидентифицируемым. Триптофан и фенилаланин являются компонентами среды для выращивания бактерий, тогда как гистамин не был компонентом среды. Эти результаты были подтвержены с использованием дополнительного 2D NMR-способа общей корреляционной спектроскопией (TOCSY). Гистамин является биогенным амином, который продуцируется из гистидина через гистидиндекарбоксилазу некоторыми ферментативными бактериями, в том числе молочнокислыми бактериями. Гистамин идентифицировали также во фракции В3 с использованием времяпролетной масс-спектрометрии с ионизацией в электроспрее (ESI TOF MS).
Гистамин не является ковалентно модифицированным на основе его картины фрагментации в анализе MS/MS. Анализ соответствующей фракции В3 L. reuteri 6475, выращенного в содержащей сахарозу среде, с использованием ESI TOF MS не выявил никакого гистамина. L. reuteri 6475, выращенный в содержащей глюкозу среде, продуцирует гистамин, который присутствует в TNF-ингибирующей фракции HILIC-HPLC.
Гистамин определяли количественно в отобранных фракциях HILIC-HPLC с использованием тройной квадрупольной масс-спектрометрии
Тройная квадрупольная масс-спектрометрия является установленным, высокочувствительным способом квантификации малых молекулярных соединений. Гистамин квантифицируют (определяют количественно) в выбранном диапазоне фракций HILIC-HPLC из L. reuteri 6475 глюкозы (B2-B7) и сахарозы (B2-B9), а также в супернатанте бактериальной культуры. Высокие уровни гистамина (>300 нг/мл) коррелировали со способностью фракций HILIC-HPLC ингибировать TNF (фигура 1). Низкие уровни гистамина измеряли в большинстве испытанных фракций, в том числе фракций из 6475 сахарозы (фигура 1). Эта способность гистамина ингибировать продуцирование TNF является, по-видимому, зависимой от концентрации.
Синтетический гистамин и гистамин, продуцируемый L. reuteri 6475, ингибируют продуцирование TNF через H2-рецептор
Гистамин может значимо ингибировать продуцирование TNF из TLR-2-активированных моноцитоидных клеток человека (THP-1) (фигура 4A). Гистамин может передавать сигнал через четыре различных рецептора гистамина, однако моноцитоидные клетки экспрессируют высокие уровни только H1- и H2-рецепторов. Прежние исследования показали эффекты гистамина на продуцировании TNF через H2-рецептор. H1- и H2-рецепторспецифические антагонисты использовали для определения, какой рецептор опосредовал эффект гистамина на клетках THP-1. H2-рецепторспецифические антагонисты, ранитидин и циметидин, могли блокировать TNF-ингибирование гистамином зависимым от концентрации образом. (фигура 2A). Анализ проточной цитометрии с H2-рецепторспецифическими антителами выявил, что THP-1-клетки высоко экспрессируют H2-рецептор (данные не показаны). Один H1-рецепторспецифический антагонист, индометацин, не оказывал действия на TNF-ингибирование гистамином (фигура 2А). Гистамин блокирует продуцирование TNF из TLR-2-активированных THP-1-клеток через передачу сигнала через H2-рецептор. Кондиционированные L. reuteri 6475 среды, содержащие гистамин, значимо ингибируют TNF в сравнении с контрольной средой, и этот эффект частично блокируется H2-рецептором, но не антагонистами H1-рецептора (фигура 2А). Частичное блокирование в супрессии TNF показывает, что гистамин, присутствующий в 6475-кондиционированной среде, передает сигнал через H2-рецептор, но что другие TNF-ингибирующие факторы, которые действуют через альтернативные механизмы, могут также присутствовать в этих кондиционированных средах. Промывка осадка клеток, содержащая гистамин штамма 6475, также подавляет продуцирование TNF (фигура 2B). Как видно с кондиционированными 6475 средами, антагонисты H2-рецептора частично блокируют эффект промывки осадка клеток 6475 (фигура 2B), что предполагает, что множественные иммуномодулины присутствуют в этой нефракционированной промывке остатка клеток. Эффекты TNF-ингибирующей фракции B3, которая содержит высокие количества очищенного гистамина, полностью блокировались добавлением антагонистов H2-рецептора (фигура 2B).
ПРИМЕР 2
Отбор штаммов, продуцирующих гистамин
Идентификация/отбор продуцирующих гистамин бактерий
Штаммы, подлежащие тестированию и, возможно, отбору, культивировали при анаэробных условиях в течение 16-18 ч в средах deMan, Rogosa, Sharpe (Difco, Franklin Lakes, NJ) и инокулировали в 2 л полуопределенной среды, LDMIII (ΟD600, доведенной до 0,1). Источником углерода была глюкоза, LDMIIIG. Каждую культуру выращивали в течение 24 ч при 37°С в анаэробной рабочей установке (MACS MG-500, Microbiology International, Frederick, MD), снабжаемой смесью 10% CO2, 10% H2 и 80% N2. Пробы брали в различных временных точках для наблюдения роста посредством измерения OD600. В стационарной фазе (24 ч) эти клетки брали для анализа с использованием ПЦР реального времени для тестирования на присутствие трех генов, антипортера гистидин/гистамин, HdcA и HdcB.
Для штаммов, положительных в отношении этих трех генов, уровни продуцируемого гистамина определяют тройной квадрупольной масс-спектрометрией. Штаммы с наивысшим продуцированием гистамина (>250 пг/мл) отбирают. Продуцирование гистамина может также оцениваться и квантифицироваться при помощи ELISA или иммуноанализов.
ПРИМЕР 3
Демонстрация иммуномодуляции
Оперон гистидина способствует TNF-ингибирующему фенотипу L. reuteri 6475
Три гена, которые, по-видимому, являются частью оперона, участвуют в продуцировании гистамина L. reuteri 6475. Этими генами являются антипортер гистидина/гистамина, гистидин-декарбоксилаза пирувоил типа А (HdcA) и HdcB (фигура 3A). Прежние исследования транскриптомики предполагали, что гены антипортера гистидина/гистамина и HdcA были потенциально важными для TNF-ингибирующего фенотипа штамма 6475. Все три гена являются в сильной степени даун-регулируемыми в 6475, растущем в глюкозной среде (таблица 2). Кроме того, по меньшей мере 1 ген в этом опероне является даун-регулируемым в 2 мутантах, которые теряют TNF-ингибирование (таблица 2). Эти мутанты исследовали ранее, и даже когда продукты генов не имели TNF-ингибирующих свойств, эти гены, по-видимому, являются важными для противовоспалительного фенотипа 6475. В отличие от этого 2 мутанта, которые не теряют TNF-ингибирования, демонстрировали отсутствие даун-регулирования любого из этих генов в опероне гистидина (таблица 2). Производили мутации в каждом из этих 3 генов инсертированием преждевременного стоп-кодона в эту генную последовательность (штаммы 1229, 1230 и 1231). Производили также мутацию в неродственном гене, гене устойчивости к рифампицину, для использования в качестве отрицательного контроля (штамм JP577). Мутации всего лишь в одном из этих генов в опероне гистидина было достаточно, чтобы вызвать частичную потерю TNF-ингибирования в сравнении со штаммом дикого типа (фигура 3B), что позволяет предположить, что каждый один из этих генов является важным для TNF-ингибирующего фенотипа L. reuteri 6475. Частичная потеря активности позволяет предположить, что другие активные иммуномодулины все еще продуцируются L. reuteri 6475.
Активация ERK1/2 является существенной для продуцирования TNF TLR2-стимулируемыми моноцитоидными клетками
ERK1/2 активируется фосфорилированием от расположенных слева MAPKK, MEK1/2, и, как было показано ранее, является важным для продуцирования TNF. THP-1-клетки обрабатывали специфическим ингибитором MEK1/2, U0126, для варьирования периодов времени до стимуляции агонистом TLR2 для супрессии активации ERK1/2. Обработка U0126 в течение 30 мин была достаточной для предотвращения продуцирования TNF (данные не показаны). ERK1/2 активируется после стимуляции TLR2 и является важным для стимуляции продуцирования TNF в модельной системе авторов этого изобретения.
Стимуляция H2-рецептора приводит к увеличению количества cAMP в этих клетках
H2-рецептор является G-белоксвязанным рецептором, который может активировать аденилатциклазу и увеличивать внутриклеточную cAMP (цАМФ). TNF может быть ингибирован на уровне транскрипции cAMP и аналогами cAMP. THP-1-клетки стимулировали агонистом TLR2 в присутствии контрольной среды, измеряли супернатант 6475 или гистамин с антагонистом или без антагониста H2-рецептора и внутриклеточные уровни cAMP. Супернатант L. reuteri 6475 вызывал малое, но значимое увеличение в cAMP (данные не показаны). Обработка H2-рецептором блокировала этот эффект. Увеличение в cAMP также наблюдали с обработкой гистамином, и этот эффект блокировался H2-антагонистом (данные не показаны). Синтетический аналог cAMP, дибутирил-cAMP (dcAMP), добавляли к TLR2-стимулированным THP-1-клеткам и действие на продуцировании TNF подвергали мониторингу. Добавление dcAMP (10-5-10-3 M) было достаточным для ингибирования для ингибирования продуцирования TNF (данные не показаны). Стимуляция H2-рецептора гистамина приводит к увеличенной cAMP, которая может блокировать последующее продуцирование TNF в активированных моноцитоидных клетках.
Активность протеинкиназы А (PKA) является важной для ингибирования TNF L. reuteri 6475, гистамином и dcAMP
Увеличенная концентрация cAMP может активировать PKA и затем ингибировать последующий путь передачи сигнала ERK MAPK. Для определения, является ли активность PKA важной для супрессии TNF гистамином, продуцируемым штаммом 6475, активированные клетки THP-1 обрабатывали специфическим ингибитором PKA, H89, в присутствии супернатанта 6475, фракции B3, гистамина или варьирующихся концентраций dcAMP. Добавление H89 частично блокировало ингибирование TNF всеми этими нормально TNF-ингибирующими соединениями (данные не показаны). Активность PKA является важной для супрессии TNF гистамином и dcAMP.
Передача сигнала через H2-рецептор блокирует активацию MEK1/2 и ERK1/2
Предыдущие исследования продемонстрировали, что PKA может ингибировать активацию Ras/Raf MEK и, следовательно, передачу сигнала ERK MAPK. Обработка активированных THP-1-клеток супернатантом 6475, гистамином или U0126 блокирует фосфорилирование как MEK1/2, так и затем ERK1/2 в сравнении со средой-контролем (данные не показаны). Обработка антагонистом H2-рецептора восстанавливает активацию как MEK1/2, так и ERK1/2 (данные не показаны). Не было различия в уровнях белка MEK1/2 и ERK1/2 с любым из способов этой обработки. Гистамин из штамма 6475 ингибирует активацию MEK и затем ERK с получением уменьшенного продуцирования TNF из TLR2-стимулируемых миелоидных клеток.
Таким образом, эти эксперименты показывают, что стимуляция H2-рецептора приводит к увеличенной cAMP, активации протеин киназы А (PKA) и ингибированию пути передачи сигнала MEK-ERK MAPK. Как описано выше, выполняли механистические исследования для определения действия гистамина на пути передачи сигнала активированной митогеном протеин киназы (MAPK). Ингибирование пути передачи сигнала MEK-ERK MEK-специфическим ингибитором является достаточным для блокирования продуцирования TNF. Обработка активированных THP-1-клеток супернатантом штамма 6475 или гистамином увеличивала внутриклеточную cAMP. Это увеличение в количестве cAMP блокировали ранитидином, специфическим антагонистом H2-рецептора. Обработка TLR2-стимулированных THP-1-клеток синтетическим аналогом cAMP, dcAMP, является достаточной для ингибирования продуцирования TNF. Ингибирование активности PKA частично блокирует супрессию TNF средой, ранее 6475-кондиционированной TNF-ингибирующими соединениями, фракцией B3, гистамином и dcAMP. Обработка активированных THP-1-клеток 6475-кондиционированной средой, гистамином или U0126 подавляла активацию MEK1/2, то есть производила эффект, который блокировался в присутствии ранитидина.
ПРИМЕР 4
Детектирование MEK1/2 и ERK1/2 с использованием вестерн-блоттинга
THP-1-клетки лизировали в охлажденном на льду лизирующем буфере, состоящем из 50 мМ Триса, pH 7,4, 250 мМ NaCl, 5 мМ ЭДТА, 50 мМ NaF, 1 мМ Na3VO4, 1% о/о Nonidet P40, 0,2% о/о NaN3 и ингибиторов протеазы и фосфатазы. Лизаты инкубировали на льду в течение 30 минут, встряхивали на вортексе каждые 10 минут и осветляли центрифугированием при 13000×g в течение 10 минут при 4°C. Концентрации белка измеряли с использованием набора Quant-iT™ Protein Assay (Invitrogen) и флуорометре Qubit в соответствии с инструкциями изготовителя. Равные количества белков загружали на гели электрофореза.
Анализ активации ERK1/2 выполняли с использованием специфических фосфо-ERKl/2-антител. Клеточные экстракты загружали на 10% ДСН-полиакриламидный гель и переносили на поливинилиден-дифторидные мембраны (Bio-Rad, Hercules, CA). Мембраны блокировали в течение ночи при 4°C в блокирующем буфере (Li-Cor Biosciences, Lincoln, NE). После нескольких промывок мембраны зондировали ERK1/2, фосфо-ERK1/2 или β-Актин-специфическими антителами, разведенными в блокирующем буфере (Li-Cor) в течение 1 часа при комнатной температуре. После промываний мембраны инкубировали с подходящим конъюгированным с пероксидазой хрена вторичным антителом в течение 1 часа при комнатной температуре и затем блоты проявляли с использованием хемилюминесцентного детектирования. Анализ активации MEK1/2 выполняли, как описано выше, за исключением того, что инкубирование первичных антител проводили на протяжении ночи при 4°C.
ПРИМЕР 5
hdcA-мутант дает уменьшенную способность ослабления колита
Бактериальные штаммы и культивирование
Мутанты генерировали с использованием RecT-опосредованной олигонуклеотидной рекомбинации. L. reuteri, экспрессирующий RecT (штамм RPRB0000), использовали для конструирования мутаций в rpoB (тэг локуса HMPREF0536_0828 (ZP_03961568)) и генах-мишенях, расположенных в кластере гистидиндекарбоксилазы HMPREF0536_1229 (ZP_03961969), HMPREF0536_1230 (ZP_03961970) и HMPREF0536_1231 (ZP_03961971) с получением штаммов RPRB3002, RPRB3004, RPRB3005 и RPRB3006 соответственно. Мутации проверяли при помощи ПЦК и целостность подтверждали анализом последовательностей.
L. reuteri ATCC PTA 6475 и мутант гена гистидиндекарбоксилазы (hdcA) культивировали в средах deMan, Rogosa, Sharpe (Difco, Franklin Lakes, NJ) при 37°C в анаэробной рабочей установке (MACS MG-500, Microbiology International, Frederick, MD), снабжаемой смесью 10% CO2, 10% H2 и 80% N2.
Приготовление клеток L. reuteri и введение мышам
Отдельную колонию каждого из штаммов L. reuteri инокулировали в 10 мл среды MRS и выращивали при 37°C в анаэробных условиях в течение 18-20 часов. Бактерии, доведенные до OD600=0,03, инокулировали в 40 мл MRS для начала ферментации и выращивали при 37°C при анаэробных условиях в течение 5,5 часов (OD600~2,5, бактерии находились в экспоненциальной фазе в этой временной точке). Эти клетки осторожно осаждали (2500×g, RT, 4 минуты) и суспендировали в MRS при концентрации 25×109 КОЕ/мл. В качестве среды-контроля использовали стерильную среду MRS. Каждая 8-недельная самка мыши Balb/c получала одну дозу свежеприготовленного L. reuteri 6475 дикого типа или hdcA-мутанта или MRS (0,2 мл в каждом случае) каждый день в течение семи дней принудительным кормлением с использованием зонда через полость рта и желудок после 10 дней акклиматизации. Все эксперименты на мышах выполняли в соответствии с одобренным протоколом (AN-4199; animal facility of Baylor College of Medicine). Мышей (45-дневных) получали из Harlan Laboratories (Houston, TX) и поддерживали при специфических, не содержащих патогенов условиях в клетках с фильтрующей крышкой (пять мышей на клетку), и они имели свободный отступ к дистиллированной воде и корму 2918 для грызунов Harlan. Мышей распределяли в различные группы случайным образом.
Индуцирование острого колита с использованием ректальной клизмы трибензолсульфоновой кислоты (TNBS)
Колит индуцировали за шесть часов перед шестым принудительным кормлением через зонд. Мышей анестезировали постоянной ингаляцией изофурана. 5% раствор TNBS в воде (Sigma-Aldrich, USA) разводили равным объемом абсолютного спирта и вводили в дозе 100 мг/кг массы тела внутриректально. Мышей держали вниз головой в вертикальном положении в течение 2 минут после клизмы для гарантии полного удерживания клизмы в ободочной кишке. Контрольные мыши процедуры получали 50% этанол в ЗФР. Мышей взвешивали перед введением TNBS и через два дня после введения TNBS. Затем мышей умерщвляли. Воспаление и повреждение ободочной кишки определяли по потере массы, макроскопическому баллу и концентрации SAA в сыворотке.
Макроскопическое оценивание колита
Ободочные кишки собирали, раскрывали продольно и изображения регистрировали цифровой камерой. Воспаление и повреждение ободочной кишки определяли согласно критериям Wallace (Morris et al., 1989). Каждую ободочную кишку оценивали вслепую. Статистику выполняли с использованием GraphPad Prism version 5.01 (GraphPad Software, La Jolla, CA). Тест Kruskal-Wallis использовали для детектирования значимого различия среди всех групп, включенных в этот анализ. Результаты суммировали в виде медианы и интерквартильного диапазона.
Измерение амилоидного белка А сыворотки (SAA) в качестве системного маркера воспаления
Пробы крови собирали сердечной пункцией, антикоагулировали и центрифугировали в течение 10 минут при 13000 об/мин для выделения плазмы. Концентрации сывороточного амилоида А (SAA) в пробах плазмы измеряли с использованием наборов ELISA из ALPCO (Salem, NH) в соответствии с инструкциями изготовителя. SAA является белком острой фазы, указывающим на системное воспаление в мышах, которое коррелирует с тяжестью колита.
Результаты
L. reuteri 6475 защищает мышей против TNBS-индуцированного острого колита
Противовоспалительные эффекты L. reuteri 6475 испытывали в TNBS-индуцированной модели острого колита мыши. Мышей, которые получали L. reuteri 6475 принудительным кормлением с использованием зонда через рот и желудок, сравнивали с мышами, которые получали среду-контроль. Мышей, которым вводили ЗФР вместо TNBS, также исследовали в качестве отрицательных контролей колита.
Фигуры 4-6 представляют данные из двух независимых экспериментов. Отрицательные контроли колита, которые получали ЗФР вместо TNBS интраректально, имели очень низкую потерю массы (или даже прибавляли массу), редкое повреждение ободочной кишки и низкие концентрации SAA в сыворотке. Колит-положительные мыши, которые получали среду MRS и TNBS/ETOH, развивали тяжелый колит, характеризующийся большой величиной потери массы, образованием язв с воспалением в ободочной кишке и основных участках повреждений, распространяющихся более чем на 1 см, и значимо увеличенными концентрациями SAA в сыворотке. Принудительное зондовое кормление L. reuteri 6475 через рот и желудок значимо уменьшало потерю массы, макроскопическое воспаление в ободочной кишке и концентрации SAA в сыворотке, показывая, что L. reuteri 6475 значимо аттенуирует колит.
Мутант hdcA дает уменьшенную способность аттенуирования колита
С использованием той же самой мышиной модели авторы изобретения испытывали, был ли ген hdcA, который кодирует гистидиндекарбоксилазу, необходим для противовоспалительных эффектов L. reuteri 6475. 8-недельных самок мышей BALB/c случайным образом распределяли в три группы, которые получали L. reuteri 6475 дикого типа, или hdcA-мутант, или среду MRS соответственно. Фигуры 7 и 8 представляют данные из двух независимых экспериментов. Опять зондовое введение L. reuteri 6475 через рот и желудок значимо уменьшало потерю массы и повреждение ободочной кишки в сравнении с группой среды-контроля. Мыши, получавшие hdcA-мутант, значимо увеличивали потерю массы и макроскопическое воспаление в ободочной кишке в сравнении с мышами, которые получали бактерии дикого типа, показывая, что hdcA-мутант дает уменьшенную способность аттенуирования колита.
Предложенная группа изобретений относится к области медицины. Предложен способ отбора пробиотического молочнокислого бактериального штамма для применения в локальном продуцировании гистамина в млекопитающем. Предложены продукт и композиция для локального продуцирования гистамина в млекопитающем, содержащие молочнокислый бактериальный штамм, имеющий активный оперон гистидина и способный продуцировать гистамин, для применения в лечении и/или профилактике воспалительных состояний. Предложенная группа изобретений обеспечивает локальное продуцирование гистамина в млекопитающем посредством отбора определенных штаммов молочнокислых бактерий. 3 н. и 10 з.п. ф-лы, 9 ил., 2 табл., 5 пр.
1. Способ отбора пробиотического молочнокислого бактериального штамма для применения в локальном продуцировании гистамина в млекопитающем, где указанный способ предусматривает скрининг бактерий на присутствие активного оперона гистидина и отбор штамма, который имеет активный оперон гистидина и способен продуцировать гистамин.
2. Способ по п. 1, где указанный штамм отбирают на его способность продуцировать гистамин на уровне, большем чем 250 пг/мл.
3. Способ по п. 1 или 2, где указанный штамм является Lactobacillus reuteri.
4. Продукт, содержащий клетки молочнокислого бактериального штамма, получаемый способом отбора по любому из пп. 1-3, где указанный бактериальный штамм имеет активный оперон гистидина и способен продуцировать гистамин, для применения в локальном продуцировании гистамина в млекопитающем, причем указанное применение является лечением и/или профилактикой воспалительных состояний.
5. Продукт по п. 4, где указанным млекопитающим является человек.
6. Продукт по п. 4 или 5, где локальное продуцирование гистамина происходит в желудочно-кишечном тракте (GI-тракте), мочеполовых путях (GU-тракте), полости рта, легких, дыхательных путях или на коже указанного млекопитающего.
7. Продукт по п. 4, где это воспалительное состояние выбрано из группы, состоящей из колита, воспалительной болезни кишечника, синдрома раздраженной толстой кишки (слизистого колита), дивертикулеза, гингивита и вагинита.
8. Продукт по п. 4 или 5, где указанным штаммом является Lactobacillus reuteri.
9. Продукт по п. 8, где указанным штаммом является Lactobacillus reuteri 6475.
10. Продукт по п. 4 или 5, где указанное применение дополнительно предусматривает введение по меньшей мере одного дополнительного терапевтического или пищевого агента.
11. Продукт по п. 10, где указанный дополнительный агент содержит гистидин или аналог гистидина, подходящий источник углерода, который поддерживает продуцирование гистамина указанным штаммом, или их комбинацию.
12. Композиция для локального продуцирования гистамина в млекопитающем, содержащая:
(i) молочнокислый бактериальный штамм, получаемый способом отбора по любому из пп. 1-3, где указанный молочнокислый бактериальный штамм имеет активный оперон гистидина и способен продуцировать гистамин; и
(ii) по меньшей мере один дополнительный компонент, выбранный из группы, состоящей из подходящего источника углерода, который поддерживает продуцирование гистамина указанным штаммом, источника гистидина или аналога гистидина, и их комбинации.
13. Продукт по п. 11 или композиция по п. 12, где указанный гистидин или аналог гистидина находится в форме содержащего гистидин или аналог гистидина пищевого продукта или пищевой добавки, или где указанный источник углерода содержит глюкозу.
| CALLES-ENRIQUEZ M | |||
| et al | |||
| Sequencing and transcriptional analysis of the Streptococcus thermophilus histamine biosynthesis gene cluster: factors that affect differential hdcA expression | |||
| Appl Environ Microbiol | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Найдено из Интернет: URL: http://www.ncbi.nlm.nih.gov/pubmed/20656875 | |||
| THOMAS C.M | |||
| et al | |||
| Probiotics-host communication: Modulation of signaling pathways in the intestine | |||
| Gut Microbes | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Найдено из Интернет: URL: http://www.ncbi.nlm.nih.gov/pubmed/20672012 | |||
| WO 2006110088 A1, 19.10.2006 | |||
| ПИЩЕВОЙ ПРОДУКТ С БИОЛОГИЧЕСКИ АКТИВНЫМИ ДОБАВКАМИ | 2000 |
|
RU2178975C1 |

