ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
[0001] Эта заявка испрашивает приоритет по предварительной заявке США №61/660,232, поданной 15 июня 2012 г., содержание которой включено в данное описание путем ссылки в полном объеме.
ССЫЛКА НА СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Список последовательностей, представленный вместе с этой заявкой под номером 37 CFR §1.821 в машиночитаемой форме (CRF) через EFS-Web в виде файла PC071914_SEQLIST_ST25.txt, включается в это описание путем ссылки. Электронная копия списка последовательностей была создана 14 мая 2013 г., с размером файла 71 килобайт.
ДЕПОНИРОВАНИЕ БИОЛОГИЧЕСКОГО МАТЕРИАЛА
[0003] Репрезентативные материалы согласно настоящему изобретению были депонированы в Американской коллекции типовых культур ("АТСС"), 10801 University Boulevard, Manassas, VA 20110-2209, USA, 14 июня 2012 г. Вектор OGD1.0.0-HC, имеющий учетный номер АТСС РТА-12980, представляет собой полинуклеотид, кодирующий вариабельный участок тяжелой цепи OGD 1.0.0, и вектор OGD1.0.0-LC, имеющий учетный номер АТСС РТА-12981, представляет собой полинуклеотид, кодирующий вариабельный участок легкой цепи OGD 1.0.0.
[0004] Депонирование осуществляют в соответствии с положениями Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры и его подзаконными актами ("Будапештский договор"). Это обеспечивает поддержание жизнеспособности депонируемой культуры в течение 30 лет со дня депонирования. Депонированный образец может быть предоставлен со стороны АТСС согласно условиям Будапештского договора и в соответствии с договором между Pfizer Inc. и АТСС, который гарантирует постоянный и неограниченный открытый доступ к потомству депонированной культуры после выдачи соответствующего патента США или после публикации любой патентной заявки США или иностранной патентной заявки, то есть более ранней з них, и гарантирует доступ к потомству лицам, определяемым Комиссаром США по патентам и товарным знакам, которые получают на это право в соответствии с Разделом 35 Кодекса законов США, § 122, и установленными Комиссаром соответствующими правилами (включая 37 CFR § 1.14 с конкретной ссылкой на 886 OG 638).
[0005] Правообладатель настоящей заявки согласился, что в случае гибели или потери культуры депонируемых материалов или ее уничтожения при культивировании в соответствующих условиях, материалы должны быть после уведомления безотлагательно заменены на другие такие же. Доступ к депонированному материалу не следует рассматривать как разрешение на практическое воплощение изобретения в нарушение прав, предоставляемых правительством любой страны в соответствии с действующими в ней патентными законами.
УРОВЕНЬ ТЕХНИКИ
[0006] Фактор роста и дифференцировки 8 (GDF-8), также известный как миостатин, представляет собой секретируемый белок и относится к надсемейству трансформирующих ростовых факторов бета (TGF-) структурно связанных факторов роста. Представители этого надсемейства обладают свойствами, регулирующими рост, и морфогенетическими свойствами (Kingsley et al. (1994) Genes Dev. 8:133-46; Hoodless et al. (1998) Curr. Topics Microbial. Immunol. 228:235-72). Человеческий GDF-8 синтезируют как состоящий из 375 аминокислот белок-предшественник, образующий гомодимерный комплекс. Во время обработки аминоконцевой пропептид, известный как "латентно-ассоциированный пептид" (LAP), расщепляется и может оставаться нековалентно связанным с гомодимером, образуя неактивный комплекс, называемый как "малый латентный комплекс" (Miyazono et al. (1988) J. Biol. Chem. 263:6407-15; Wakefield et al. (1988) J. Biol. Chem. 263:7646-54; Brown et al. (1999) Growth Factors 3:35-43; Thies et al. (2001) Growth Factors 18:251-59; Gentry et al. (1990) Biochemistry 29:6851-57; Derynck et al. (1995) Nature 316:701-05; Massague (1990) Ann. Rev. Cell Biol 12:597-641). Белки, такие как фоллистатин и родственные с ним белки, также связываются со зрелыми гомодимерами GDF-8 и ингибируют биологическую активность GDF-8 (Gamer et al. (1999) Dev. Biol. 208:222-32).
[0007] Выравнивание аминокислотной последовательности GDF-8 из разных видов демонстрирует, что GDF-8 является высококонсервативным (McPherron et al. (1997) Proc. Natl. Acad. Sci. U.S.A. 94:12457-61). Последовательности GDF-8 человека, мыши, крысы, свиньи и курицы являются на 100% идентичными на С-концевом участке, a GDF-8 бабуина, крупного рогатого скота и овцы отличаются всего на 3 аминокислоты на С-конце. Высокая степень консервативности GDF-8 в разных видах свидетельствует о том, что GDF-8 выполняет существенную физиологическую функцию.
[0008] Было продемонстрировано, что GDF-8 играет главную роль в регуляции развития и гомеостаза мышц путем подавления пролиферации и дифференциации миобластов и сателлитоцитов (Lee and McPherron (1999) Curr. Opin. Genet. Dev. 9:604- 07; McCroskery et al. (2003) J. Cell. Biol. 162:1135-47). Он демонстрирует раннюю экспрессию в развивающихся скелетных мышцах и продолжает экспрессироваться в развитых скелетных мышцах, преимущественно быстросокращающегося типа. Кроме того, GDF-8, который демонстрирует сверхэкспрессию у взрослых мышей, приводит к значительной атрофии мышечной ткани (Zimmers et al. (2002) Science 296:1486-88). К тому же, было продемонстрировано, что природные мутации, в результате которых ген GDF-8 становится неактивным, вызывают гипертрофию и гиперплазию у животных и у человека (Lee and McPherron (1997) supra). Например, трансгенные мыши с выключением гена GDF-8 характеризуются выраженной гипертрофией и гиперплазией скелетных мышц и измененной структурой кортикальной кости (McPherron et al. (1997) Nature 387:83-90; Hamrick et al. (2000) Bone 27:343-49). Подобное повышение массы скелетных мышц является очевидным при природных мутациях GDF-8 у крупного рогатого скота (Ashmore et al. (1974) Growth 38:501-07; Swatland et al. (1994) J. Anim. Sci. 38:752-57; McPherron et al, supra; Kambadur et al. (1997) Genome Res. 7:910-15). Кроме того, различные исследования показывают, что повышенная экспрессия GDF-8 связана с ВИЧ-индуцированной мышечной атрофией (Gonzalez-Cadavid et al. (1998) Proc. Natl. Acad Sci. U.S.A. 95:14938-43). GDF-8 также участвует в продуцировании мышечно-специфичных ферментов (например, креатинкиназы) и пролиферации миобластов (WO 00/43781).
[0009] Считается, что, помимо его свойств, регулирующих рост, и морфогенетических свойств, GDF-8 участвует во многих других физиологических процессах, включая гомеостаз глюкозы во время развития диабета 2 типа, нарушение толерантности к глюкозе, метаболические синдромы (т.е., такой синдром, как, например, синдром X, связанный с одновременным наличием группы состояний, к которым могут относиться резистентность к инсулину, абдоминальное ожирение, дислипидемия, гипертензия, хроническое воспаление, протромботическое состояние и т.п., что связано с возникновением высокого риска диабета 2 типа и/или сердечного заболевания у пациента), резистентность к инсулину (например, резистентность, вызванная травмой, такой как ожог, или дисбалансом азота) и расстройства жировой ткани (например, ожирение, дислипидемия, неалкогольная жировая болезнь печени и т.п.) (Kim et al. (2000) Biochem. Biophys. Res. Comm. 281:902-06).
[00010] Многие расстройства у человека и животных связаны с функциональными расстройствами мышечной ткани, например, амиотрофический латеральный склероз ("ALS"), мышечная дистрофия ("MD"; включая мышечную дистрофию Дюшенна), мышечная атрофия, атрофия органов, слабость, застойное обструктивное заболевание легких (COPD), саркопения, кахексия и синдромы мышечной атрофии, вызванные другими болезнями и состояниями. В настоящее время существует мало надежных и эффективных способов терапии для лечения этих нарушений. Патология этих заболеваний указывает на возможную роль сигнала GDF-8 в качестве объекта в лечении этих заболеваний.
[00011] ALS является нейродегенеративным заболеванием с поздним началом и смертельным исходом, характеризующимся дегенерацией центральной нервной системы и мышечной атрофией. ALS, как правило, вызывает отклонения в походке и потерю физической ловкости, а затем прогрессирует до паралича конечностей и диафрагмы. Хотя большинство случаев ALS являются спорадическими и имеют невыясненную этиологию, было обнаружено 5-10% случаев являются результатом доминантной семейной (FALS) наследственности. Приблизительно 10-20% случаев FALS объясняются мутациями в ген Cu/Zn супероксиддисмутазы (SOD1) (см. обзор в Bruijn et al. (2004) Ann. Rev. Neurosci. 27:723-49). SOD1 является гетеродимерным металлопротеином, катализирующим реакцию супероксида до перекиси водорода и двухатомного кислорода, и поскольку потеря SOD1 не приводит к болезни двигательного нейрона (Reaume et al. (1996) Nat. Genet. 13:43- 47), но считается, что он вызывает болезнь через токсическое усиление функции (см. обзор в Bruijn et al., supra). Конкретные механизмы вызываемой SOD1 гибели нейронных клеток остаются неизвестными и могут быть связаны с изменениями в аксональном транспорте, клеточными реакциями та неправильно свернутый белок, митохондриальной дисфункцией и усилением токсичности (Bruijn et al., supra).
[00012] Дегенерация двигательных нейронов, наблюдаемая при ALS, может осуществляться через различные механизмы, включая прерывание поглощения или транспорта трофических факторов двигательными нейронами (см. обзор в Holzbaur (2004) Trends Cell Biol. 14:233-40). Таким образом, ALS может поддаваться лечению с применением способов терапии, которые омолаживают дегенерирующий нейрон путем обеспечения оптимальной среды для выживания. Среда для нерва включает ненейронные клетки, такие как глиальные и мышечные клетки, иннервированные двигательным нейроном. Эта среда предусматривает трофические факторы и факторы роста, которые эндоцитозируются нейроном и переносятся через ретроградный аксональный транспорт в клеточное тело (Chao (2003) Neuron 39:1-2; Holzbaur, supra).
[00013] FALS моделировали у мышей и крыс путем сверхэкспрессии мутантного SOD1 (Howland et al. (2002) Proc. Natl. Acad. Sci. U.S.A.99:1604-09). Трансгенные мыши, сверхэкспрессирующие форму G93A мутантного SOD1, демонстрируют мышечную слабость и атрофию к 90-100-дневному возрасту и обычно умирают в возрасте приблизительно 130 дней (Gurney et al. (1994) Science 264:1772-75). Однако лежащая в основе 800093А-индуцируемая патология, включающая слабость захвата и потерю нервно-мышечного соединения, является значительной уже к 50-дневному возрасту (Frey et al. (2000) J. Neurosci. 20:2534-42; Fisher et al. (2004) Exp. Neuro. 185:232-40; Ligon et al. (2005) NeuroReport 16:533-36; Wooley et al. (2005) Muscle Nerve 32:43-50). Трансгенные крысы, экспрессирующие мутацию SODG93A, демонстрируют ту же динамику дегенерации (Howland et al., supra). Работы последнего времени показывают, что развитие патологии не является клеточно-автономным, что согласуется с гипотезой, согласно которой дегенерация двигательных нейронов, наблюдаемая при ALS, происходит путем различных механизмов, включая прерывание поглощения или транспорта трофических факторов двигательным нейроном (см. выше). Clement и сотрудники использовали химерных мышей, чтобы продемонстрировать, что ненейронные клетки дикого типа могут продлевать выживание двигательных нейронов, экспрессирующих мутантный SOD1 (Clement et al. (2003) Science 302:113-17). Эти наблюдения привели к исследованию средств терапии, которые могли бы замедлить дегенерацию нейронов, обеспечивая оптимальную микросреду для выживания. Например, лечение мышей SODG93A путем прямой внутримышечной инъекции вирусно-экспрессируемых факторов роста (включая IGF-1, GDNF и VEGF) продлевает выживаемость животных (Kaspar et al. (2003) Science 301:839-42; Azzouz et al. (2004) Nature 429:413-17; Wang et al. (2002) J. Neurosci. 22:6920-28). Кроме того, мышечно-специфичная экспрессия местной IGF-1-специфичной изоформы (mlGF-1) стабилизирует нервно-мышечные соединения, повышает выживаемость двигательных нейронов и задерживает начало и прогрессирование болезни в модели трансгенных мышей SODG93A, указывая на то, что прямое воздействие на мышцы может влиять на начало и прогрессирование заболевания у SOD 1-трансгенных животных (Dobrowolny et al. (2005) J. Cell Biol. 168:193-99). Связь между мышечным гиперметаболизмом и чувствительностью двигательных нейронов также отмечалась у мышей с ALS, что подтверждало гипотезу о том, что дефекты в мышцах могут быть задействованы в этиологии болезни (Dupois et al. (2004) Proc. Natl. Acad Sci. U.S.A. 101:11159-64). Таким образом, усиление роста мышц должно обеспечивать местную поддержку для двигательных нейронов, а значит, обеспечивать благоприятный терапевтический эффект.
[00014] Ингибирование экспрессии миостатина ведет к гипертрофии и гиперплазии мышц (Lee and McPherron, supra; McPherron et al., supra). Миостатин отрицательно регулирует регенерацию мышц после повреждения, и недостаток миостатина у мышей с нулевым GDF-8 ведет к ускоренной регенерации мышц (McCroskery et al., (2005) J. Cell. Sci. 118:3531-41). Нейтрализующие миостатин антитела повышают массу тела, массу скелетных мышц и размер и силу скелетных мышц мышей дикого типа (Whittemore et al. (2003) Biochem. Biophys. Res. Commun. 300:965-71) и mdx-мышей, модели мышечной дистрофии (Bogdanovich et al. (2002) Nature 420:418-21; Wagner et al. (2002) Ann. Neural. 52:832-36). Кроме того, антитело против миостатина у этих мышей снижало повреждение диафрагмы, мышцы, которая также является объектом во время патогенеза ALS. Была высказана гипотеза, что действие на мышцы факторов роста, таких как HGF, может быть обусловлено ингибированием экспрессии миостатина (McCroskery et al. (2005), supra), таким образом, способствуя сдвигу "push and pull" или баланса между регенерацией и дегенерацией в положительном направлении. Таким образом, ингибирование GDF-8 представляет потенциальное фармакологическое действие для лечения ALS, мышечной дистрофии (MD) и других GDF-8-ассоциированных нарушений, например, нервно-мышечных нарушений, при которых желательно повышение массы, силы, размера мышц и т.п. При наличии животных моделей (мышиных и крысиных) ALS существует возможность испытания терапевтических средств в двух разных видах, таким образом, повышая достоверность терапевтического эффекта для человека in vivo.
[00015] Помимо нервно-мышечных нарушений у человека, также существуют состояния, зависящие от фактора роста, связанные с потерей костной ткани, такие как остеопороз и остеоартрит, которые преимущественно поражают пожилых людей и/или женщин в период постменопаузы. Кроме того, к метаболическим болезням или костным расстройствам относятся низкая костная масса вследствие хронической глюкокортикоидной терапии, преждевременной гонадной дисфункции, подавления андрогена, дефицита витамина D, вторичного гиперпаратиреоза, пищевой недостаточности и нервной анорексии. Хотя многие применяемые в настоящее время средства терапии действуют путем ингибирования резорбции костной ткани, терапия, способствующая образованию костной ткани, была бы полезной в качестве альтернативного лечения. Поскольку GDF-8 играет роль в развитии костей, а также развитии мышц, регуляция GDF-8 также является отличным фармакологическим способом для лечения костно-дегенеративных нарушений.
[00016] Мышиное моноклональное антитело, специфично антагонизирующее GDF-8, было ранее описано как повышающее мышечную массу и силу в моделировании ALS на грызунах, помимо других биологических эффектов (Holzbaur, EL, et al, Myostatin inhibition slows muscle atrophy in rodent models of amyotrophic lateral sclerosis, Neurobiology of Disease (2006) 23(3):697-707). Таким образом, ожидается, что мышиное антитело и его гуманизированный аналог должны быть эффективными для увеличения мышечной массы и силы у пациентов с ALS, а также у пациентов, страдающих от других болезней и нарушений, характеризуемых или опосредованных избыточным количеством GDF-8, таких как описанные выше.
[00017] Гуманизированный вариант вышеупомянутого антитела мыши против GDF-8, как и многие моноклональные антитела и другие терапевтические средства на белковой основе, требует немалых усилий и материальных затрат для производства, поскольку для этого обычно требуется продуцирование в живых клетках млекопитающих. Таким образом, улучшение выработки этого антитела или других антител с подобной специфичностью позволяло бы получать такое же количество активного препарата с меньшими затратами. Это обеспечивает двойное преимущество снижения стоимости производства с одновременным освобождением ограниченных производственных мощностей для производства других биологических медикаментов. Оба преимущества способствуют повышению доступности для пациентов терапевтических антител против GDF-8, а также других биологических средств. Соответственно, в данной отрасли существует потребность в улучшенных вариантах антител против GDF-8, обеспечивающих больший выход продукции в клетках млекопитающих.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00018] Настоящее изобретение обеспечивает гуманизированные антитела против GDF-8 или их антигенсвязывающие фрагменты, которые могут экспрессироваться на более высоком уровне в клетках-хозяевах по сравнению с более ранними вариантами таких антител, имеющих общие определяющие комплементарность области (CDR). Также обеспечиваются композиции, включающие такие антител, для применения в соответствии со способами согласно настоящему изобретению.
[00019] В некоторых вариантах воплощения эти антитела имеют вариабельную тяжелую (VH) область, в которой CDR1 определяется аминокислотной последовательностью SEQ ID NO: 10 или SEQ ID NO: 20, CDR2 определяется аминокислотной последовательностью SEQ ID NO: 11 или SEQ ID NO: 21, CDR3 определяется аминокислотной последовательностью SEQ ID NO: 12, и в которой VH область является модифицированной таким образом, что аминокислота в позиции 108 по Kabat является лейцином вместо метионина. В других вариантах воплощения эти антитела имеют VH область, которая включает одинаковые CDR, и в которой четвертая каркасная область VH области включает аминокислоты 106-116 SEQ ID NO: 44. В других вариантах воплощения этих антител CDR прививают к сегменту VH-гена зародышевой линии человека DP-47, а затем соединяют с геном тяжелого J-сегмента человека JH4. В других вариантах воплощения VH область этих антител включает аминокислотную последовательность SEQ ID NO: 44.
[00020] Антитела согласно настоящему изобретению, имеющие лейцин в позиции 108 по Kabat, такие как приведенные для примера выше, характеризуются повышенным уровнем экспрессии по сравнению с вариантами, в которых метионин находится в той же позиции. В некоторых вариантах воплощения более ранний вариант экспрессируется на более высоком уровне в подобных условиях по сравнению с последним более, чем приблизительно на 50%, 100%, 150%, 200%, 250%, 300%, 400%, 500%, 600%, 700%, 800%, 900%, 1000%, 1200%, 1400%, 1600%, 1800% или даже 2000%.
[00021] В соответствии с другими вариантами воплощения для антител согласно изобретению, VH области, описанные в предыдущем абзаце, могут быть соединены с вариабельными легкими (VL) областями, в которых CDR1 определяется аминокислотной последовательностью SEQ ID NO: 13, CDR2 определяется аминокислотной последовательностью SEQ ID NO: 14 и CDR3 определяется аминокислотной последовательностью SEQ ID NO: 15, и в которых аминокислота в позиции 100 по Kabat VL области является глицином или глутамином. В других вариантах воплощения VH область соединяют с VL областью, в которой CDR легкой цепи привиты к сегменту VL-гена зародышевой линии человека DPK-9, а затем соединяют с геном легкого J-сегмента человека JH4. Согласно некоторым другим вариантам воплощения этих антител, ранее описанные VH области соединяют с VL областями, которые имеют вышеописанные CDR VL области и в которых четвертая каркасная область VL области включает аминокислоты 98-107 SEQ ID NO: 9 или SEQ ID NO: 46. В других вариантах воплощения ранее описанные VH области соединяют с VL областями, включающими аминокислотную последовательность SEQ ID NO: 9 или SEQ ID NO: 46.
[00022] В некоторых других вариантах воплощения вышеописанные VH области соединяют с константными областями тяжелой цепи подтипов человеческого антитела, включая IgA, IgG, IgD, IgE или IgM. Если константная область тяжелой цепи взята из IgG, в других вариантах воплощения константные области тяжелой цепи антитела взяты из подтипов IgG: IgG1, IgG2, IgG3 или IgG4. Константные области тяжелой цепи могут быть модифицированы, например, для устранения одной или нескольких эффекторных функций Fe домена, как показано на примере SEQ ID NO: 57. В других вариантах воплощения описанные выше VL области связывают с константными областями легкой цепи, которые могут относиться к лямбда или каппа подтипам.
[00023] В соответствии с другими вариантами воплощения VH область SEQ ID NO: 44 связывают с модифицированной тяжелой областью аминокислотной последовательности SEQ ID NO: 57 для создания тяжелой цепи антитела полной длины аминокислотной последовательности SEQ ID NO: 58. И наоборот, в других вариантах воплощения VL область SEQ ID NO: 46 может быть связана с каппа-константной легкой цепью аминокислотной последовательности SEQ ID NO: 17 для создания легкой цепи антитела полной длины аминокислотной последовательности SEQ ID NO: 59. В других вариантах воплощения каждая из тяжелой и легкой цепей антитела полной длины может комбинироваться для создания антитела против GDF-8, имеющего два антигенсвязывающих центра. В других вариантах воплощения антитела могут включать фрагменты или производные таких интактных антител полного размера, включая, например Fab, F(ab')2, scFv, scFv-Fc, scFv-CH, scFab, scFv-"молнию", диатело, триатело, тетратело, миниантитело, Fv и биспецифическое антитело.
[00024] Антитела согласно изобретению могут иметь различную аффинность связывания к GDF-8, например, приблизительно 5000 нМ или даже выше, например, как минимум примерно 4000 нМ, 3000 нМ, 2000 нМ, 1000 нМ, 900 нМ, 800 нМ, 700 нМ, 600 нМ, 500 нМ, 400 нМ, 300 нМ, 200 нМ, 100 нМ, 90 нМ, 80 нМ, 70 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 25 нМ, 20 нМ, 15 нМ, 10 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,1 нМ, 0,01 нМ или 0,001 нМ.
[00025] Изобретение также обеспечивает нуклеиновые кислоты, кодирующие антитела против GDF-8, а также векторы экспрессии, включающие такие нуклеиново-кислотные последовательности, и клетки-хозяева для экспрессии антител.
[00026] Настоящее изобретение также обеспечивает способы, применимые для лечения или профилактики у пациентов мышечных нарушений, характеризуемых сниженной мышечной массой или силой. Такие способы применяются при введении пациентам, нуждающимся в лечении или профилактике таких нарушений, терапевтически или профилактически эффективного количества композиции, включающей антитело или его антигенсвязывающий фрагмент, которые специфически связываются с GDF-8. К антителам, применимым для этих способов, относятся антитела, описанные выше и по всему тексту этой заявки.
[00027] В соответствии с некоторыми вариантами воплощения, композиции антитела согласно настоящему изобретению вводят в терапевтически или профилактически эффективном количестве пациенту, нуждающемуся в лечении или профилактике мышечного расстройства, включая выбранные из группы, к которой относятся мышечная дистрофия, мышечная атрофия, саркопения, кахексия, синдром мышечной атрофии, возрастная потеря мышечной массы или силы и слабость. В других вариантах воплощения мышечным расстройством является кахексия, вызванная раком. В других вариантах воплощения мышечным расстройством является мышечная дистрофия Дюшенна. В некоторых из последних вариантов воплощения введение антител против GDF-8 является эффективным для улучшения показателей пациента в тесте с шестиминутной ходьбой.
[00028] В некоторых вариантах воплощения изобретение также применяют для лечения пациента, страдающего от мышечной дистрофии, например, мышечной дистрофии Дюшенна, путем введения композиции, включающей антитело согласно настоящему изобретению, до, одновременно или после применения другого средства, эффективного для лечения мышечной дистрофии. В некоторых вариантах воплощения такое средство может быть глюкокортикоидом, таким как преднизон.
[00029] Также обеспечиваются способы увеличения массы или силы мышц млекопитающих путем введения млекопитающему композиции, включающей антитело против GDF-8 согласно изобретению в количестве, эффективном для увеличения массы или силы мышц млекопитающего. Во многих вариантах воплощения мышцы являются скелетными мышцами, включая одну или несколько мышц, активных при дыхании, или сердечную мышцу.
[00030] Также обеспечиваются способы лечения или профилактики нервно-мышечных нарушений путем введения субъекту, нуждающемуся в лечении или профилактике нервно-мышечного расстройства, терапевтически или профилактически эффективного количества антитела против GDF-8 согласно настоящему изобретению. В некоторых вариантах воплощения нервно-мышечным расстройством, подлежащим лечению или профилактике, является ALS.
[00031] Также обеспечиваются способы лечения или профилактики нарушений обмена веществ путем введения субъекту, нуждающемуся в лечении или профилактике нарушений обмена веществ, терапевтически или профилактически эффективного количества антитела против GDF-8 согласно настоящему изобретению. Во многих вариантах воплощения к расстройствам обмена веществ, подлежащим лечению или профилактике, относятся сахарный диабет 2 типа, метаболический синдром, синдром X, резистентность к инсулину и нарушение толерантности к глюкозе.
[00032] Также обеспечиваются способы лечения или профилактики нарушений жировой ткани путем введения субъекту, нуждающемуся в лечении или профилактике нарушений жировой ткани, терапевтически или профилактически эффективного количества антитела против GDF-8 согласно настоящему изобретению. Во многих вариантах воплощения к расстройствам жировой ткани, подлежащим лечению или профилактике, относятся ожирение и чрезмерно высокий индекс массы тела.
[00033] Также обеспечиваются способы лечения или профилактики нарушений, связанных с потерей костной массы, путем введения субъекту, нуждающемуся в лечении или профилактике нарушений, связанных с потерей костной массы, терапевтически или профилактически эффективного количества антитела против GDF-8 согласно настоящему изобретению. Во многих вариантах воплощения к расстройствам, связанным с потерей костной массы, подлежащим лечению или профилактике, относятся остеопороз, остеопения, остеоартрит и связанные с остеопорозом переломы костей.
[00034] В некоторых других вариантах воплощения к антителам против GDF-8, применимым для осуществления способов согласно изобретению, относятся антитела, конъюгированные с компонентами, которые благоприятно изменяют их функцию или характеристики, например, помимо прочего, увеличивают период полувыведения из сыворотки. В других вариантах воплощения аминокислотные изменения могут осуществляться с подобной целью или с другими целями.
[00035] Композиции антител для применения в соответствии со способами согласно изобретению могут быть получены в форме различных композиций, включая, помимо прочих, водную суспензию для введения различными путями, включая, помимо прочих, подкожное введение, внутривенное введение, внутримышечное введение, внутрибрюшинное введение, инфузионное введение или болюсное введение.
[00036] В некоторых вариантах воплощения эффективная доза антител против GDF-8 согласно изобретению составляет от 0,001 мг/кг до приблизительно 250 мг/кг, которая может быть введена за один раз или за несколько раз с интервалами.
[00037] Изобретение также обеспечивает фармацевтические наборы для применения клиническими врачами и другими специалистами для облегчения введения композиций антитела против GDF-8 пациентам. В некоторых вариантах воплощения комплекты включают антитело против GDF-8, согласно изобретению, в лиофилизированной форме или в форме водного раствора, разбавитель, такой как вода или буфер фармацевтической степени чистоты, и устройство для введения антитела против прогастрина, такое как шприц и игла. В других вариантах воплощения наборы дополнительно могут включать второе терапевтическое средство.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[00038] На ФИГ. 1А представлено выравнивание аминокислотных последовательностей VH областей некоторых антител против GDF-8 согласно изобретению, включая VH область мышиного антитела и две VH области гуманизированного антитела, созданные путем прививки мышиных CDR тяжелой цепи к VH области зародышевой линии человека DP-47 (т.е., VH0 и VH1). Дополнительно представлено выравнивание аминокислотных последовательностей других гуманизированных VH областей VH2-VH5. Аминокислотная последовательность CDR тяжелой цепи выделена жирным подчеркнутым шрифтом. Аминокислота в позиции 108 по Kabat выделена жирным шрифтом под звездочкой.
[00039] На ФИГ. 1 В представлено выравнивание аминокислотных последовательностей VL областей некоторых антител против GDF-8 согласно изобретению, включая VL область мышиного антитела и две VL области гуманизированного антитела, созданные путем прививки мышиных CDR легкой цепи к VL области зародышевой линии человека DPK-9 (т.е., VL0 и VL1). Дополнительно представлено выравнивание аминокислотных последовательностей других гуманизированных VL областей VL2-VL5. Аминокислотная последовательность CDR легкой цепи выделена жирным подчеркнутым шрифтом. Аминокислота в позиции 100 по Kabat выделена жирным шрифтом под звездочкой.
[00040] На ФИГ. 2А представлен график, на котором показано увеличение массы икроножной мышцы (GAS) и четырехглавой мышцы (QUAD) мышей С57 В1/6, получавших в течение двух недель 10 мг/кг антитела OGD 1.0.0, по сравнению с контрольной группой, получавшей носитель. На ФИГ. 2 В представлены те же данные, что и на ФИГ. 2А, как процент увеличения массы мышц у мышей получавших антитело OGD 1.0.0, относительно контрольной группы. На ФИГ. 2 В дополнительно показано процент увеличения мышечной массы длинного разгибателя пальцев (EDL) из одних и тех же групп мышей, получавших антитело и контрольный носитель.
[00041] На ФИГ. 3 представлен график, на котором показано увеличение общего тетанического усилия, создаваемого EDL-мышцей мышей С57 В1/6, получавших в течение двух недель 10 мг/кг антитела OGD 1.0.0 по сравнению с контрольной группой, получавшей носитель.
[00042] На ФИГ. 4А представлен график, на котором показано зависящее от дозы увеличение массы икроножной мышцы (Gastroc) и четырехглавой мышцы (Quad) мышей С57 В1/6, получавших еженедельно в течение четырех недель PBS-носитель и 0,3, 1,3, 10 и 30 мг/кг антитела OGD1.0.0. Данные представляют мышечную массу, измеряемую по окончании четвертой недели. На ФИГ. 4 В представлены те же данные, что и на ФИГ. 4А как процентное увеличение массы икроножной мышцы и четырехглавой мышцы у мышей получавших антитело OGD 1.0.0, относительно контрольной группы.
[00043] На ФИГ. 5А представлен график, на котором показано зависящее от дозы увеличение общей безжировой массы тела мышей С57 В1/6, получавших еженедельно в течение четырех недель PBS-носитель и 0,3, 1, 3, 10 и 30 мг/кг антитела OGD1.0.0. Данные представляют безжировую массу, измеряемую в конце каждой недели в течение четырех недель. На ФИГ. 5 В представлен график, на котором показано увеличение общей безжировой массы тела мышей С57 В1/6, получавших еженедельно в течение четырех недель PBS-носитель и 0,3, 1, 3, 10 и 30 мг/кг антитела OGD1.0.0, в конце четырехнедельного исследования.
[00044] На ФИГ. 6А представлен график, на котором показано увеличение общей безжировой массы тела мышей mdx, получавших еженедельно в течение восьми недель 10 мг/кг антитела OGD 1.0.0, относительно контрольной группы мышей mdx, получавших PBS-носитель. На ФИГ. 6 В представлен график, на котором показано увеличение максимального пикового усилия захвата мышей mdx, получавших еженедельно в течение восьми недель 10 мг/кг антитела OGD 1.0.0, относительно контрольной группы мышей mdx, получавших PBS-носитель.
[00045] На ФИГ. 7А представлен график, на котором показано увеличение массы икроножной мышцы и четырехглавой мышцы мышей mdx и мышей С57В1/6, получавших еженедельно в течение восьми недель 10 мг/кг антитела OGD1.0.0, относительно контрольной группы мышей, получавших PBS-носитель. Данные представляют мышечную массу, измеряемую по окончании восьмой недели. На ФИГ. 7В представлены те же данные, что и на ФИГ. 7А, как процентное увеличение массы икроножной мышцы и четырехглавой мышцы у мышей тех, получавших антитело OGD 1.0.0, относительно контрольной группы.
[00046] На ФИГ. 8 представлен график, на котором показано зависящее от дозы увеличение общей безжировой массы тела и безжировой массы конечности у яванских макак, получавших 0, 3, 10 и 30 мг/кг антитела OGD1.0.0.
[00047] На ФИГ. 9 представлен график, на котором показано, что увеличение общей безжировой массы тела яванских макак, получавших 10 мг/кг и 30 мг/кг антитела OGD1.0.0, сохранялось в течение нескольких недель после прекращения лечения с применением антитела.
[00048] На ФИГ. 10А представлен график, на котором показано, что объем эпаксиальных мышц, расположенных над позвонками L3-L5 яванских макак, получавших 10 мг/кг и 30 мг/кг антитела OGD1.0.0 в течение 8 недель, увеличился относительно контрольной группы животных, получавших PBS-носитель.
[00049] На ФИГ. 11 представлено трехмерное изображение эпаксиальной мышцы типичного подопытного животного до и после 4 недель лечения с применением антитела OGD1.0.0.
[00050] На ФИГ. 12А представлена аминокислотная последовательность типичной тяжелой цепи антитела против GDF-8, содержащей вариабельную тяжелую область, указанную в данном описании как VH0.
[00051] На ФИГ. 12В представлена аминокислотная последовательность типичной легкой цепи антитела против GDF-8, содержащей вариабельную легкую область, указанную в данном описании как VL0.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00052] Настоящее изобретение обеспечивает улучшенные варианты гуманизированного антитела против GDF-8, которые могут экспрессироваться в клетках на гораздо более высоком уровне по сравнению с предыдущими вариантами антитела. Соответственно, ожидается получение описываемых авторами улучшенных вариантов антитела против GDF-8 в большем количестве и с меньшими материальными затратами при таких же исходных материалах, как и в предыдущих вариантах. Настоящее изобретение также раскрывает различные способы лечения или профилактики с применением улучшенных антител. Таким образом, в некоторых типичных не ограничительных вариантах воплощения могут применяться улучшенные антитела против GDF-8 для лечения мышечной дистрофии, кахексии и других нарушений, при которых увеличение мышечной массы или силы субъекта может обеспечивать терапевтический эффект.
Структура и разнообразие антител
[00053] В контексте данного описания термин "антитело" означает интактный иммуноглобулин (Ig) или любой его антигенсвязывающий фрагмент, часть или составляющую и охватывает, помимо прочего, любой полипептид, включающий полный или частичный антигенсвязывающий центр, сохраняющий как минимум некоторую антигенсвязывающую специфичность. Антитело также может означать комбинацию антигенсвязывающих фрагментов, частей или составляющих, взятых из интактного антитела, с другой молекулой, включая другое антитело, белок из надсемейства Ig или белки или другие типы молекул, не относящиеся к иммунной системе. Такие производные антител могут включать небелковые составляющие или компоненты.
[00054] Антиген означает вещество, белковое или иное, способное специфически связываться антителом. Антиген может иметь более одной антигенной детерминанты или более одного эпитопа, представляющих часть антигена, связываемую антителом.
[00055] Иммуноглобулины (Ig) представляют собой гетеротетрамерные белки, включающие две тяжелые цепи приблизительно по 50 кДа каждая, и две легкие цепи приблизительно по 25 кДа каждая. Каждая цепь включает несколько Ig-доменов. Начиная с амино-конца, тяжелая цепь содержит один вариабельный участок (VH), за которым следуют, в зависимости подтипа Ig, три или четыре константных области, которые называются CH1, СН2, СН3 и, в случае наличия, СН4. Подобным образом, в легкой цепи один вариабельный участок (VL) находится на амино-конце полипептида, а за ним следует одна константная область (CL). Между областями СН1 и СН2 находится шарнирная область переменной длины, в зависимости от изотипа, которая придает молекуле гибкость. Карбокси-конец тяжелой цепи СН1, включая шарнир, СН2, СН3 и, в случае наличия, СН4, составляет Fe-область. Каждая вариабельная или константная область включает один Ig домен.
[00056] Легкие цепи Ig связываются с тяжелыми цепями Ig, и пары тяжелых цепей Ig связываются между собой через дисульфидные связи. Нековалентная взаимосвязь также может способствовать стабилизации межцепочечной четвертичной структуры между тяжелыми и легкими цепями и между спаренными тяжелыми цепями. В интактных молекулах Ig области VH и VL спаренных тяжелых и легких цепей располагаются по соседству одна с другой и взаимодействуют для образования антиген-связывающего центра. Поскольку интактные молекулы Ig содержат две пары спаренных тяжелых и легких цепей, т.е., всего две тяжелые и две легкие цепи, молекулы Ig содержат два антигенсвязывающих центра. Наличие шарнирной области обеспечивает гибкость между антигенсвязывающими центрами и остальной частью молекулы.
[00057] Константные области тяжелой и легкой цепей не принимают непосредственного участия в распознавании антигена. Однако тяжелая цепь, в частности, Fe область, содержит последовательности, способные взаимодействовать с эффекторными молекулами и клетками иммунной системы, что позволяет тяжелым константным областям опосредовать важные биологические функции молекул Ig.
[00058] Вариабельные участки тяжелых и легких цепей содержат три расположенных с промежутками участка повышенной вариабельности аминокислот, называемых гипервариабельными участками или определяющими комплементарность областями (CDR), которые имеют широкие различия в разных молекулах Ig по сравнению с более консервативными каркасными областями (FR), окружающими CDR. От амино-конца вариабельного участка последовательный порядок и нумерация FR и CDR таковы: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Каркасные области принципиально отвечают за определение третичной структуры Ig домена вариабельного участка. Для сравнения, CDR образуют петли, простирающиеся наружу от каждого вариабельного участка. CDR соседних VH и VL областей взаимодействуют для образования антигенсвязывающей поверхности, которая принципиально отвечает за определение антигенсвязывающей специфичности конкретных молекул Ig.
[00059] Исследователи, изучавшие структуру и функцию антитела, разработали различные схемы распознавания CDR тяжелой и легкой цепей, существующих в пределах аминокислотной последовательности любой конкретной VH или VL области. Многие из этих схем позволяют распознавать CDR в соответствии с неизменными или почти неизменными структурами, связанными с окружающим каркасом вариабельных тяжелой и легкой областей. Затем CDR определяют, применяя диапазоны чисел, соответствующие позиции их составляющих остатков в пределах VH и VL областей. Поскольку CDR, в частности, третья CDR, могут отличаться по длине, в схемах иногда также используются буквы для обозначения составляющих остатков. Одна из первых таких схем известна как система нумерации Kabat, которая основывается на выравнивании известных на данный момент последовательностей VH и VL для определения позиции вариабельных подпоследовательностей CDR в пределах более консервативных каркасных областей. К другим схемам определения CDR относятся система нумерации АЬМ и система нумерации Chothia. Также возможны и другие схемы. Например, CDR может определяться как остатки вариабельного участка, контактирующие с антигеном, даже если эти остатки точно не совпадают с более формализованными определениями для CDR, такими, как схемы нумерации Kabat или Chothia. См., например, публикацию Y. Ofran, et al., Automated identification of complementarity determining regions (CDRs) reveals peculiar characteristics of CDRs and В cell epitopes, J Immunol. 2008 Nov l;181(9):6230-5, включенную путем ссылки. Схема нумерации Kabat и некоторые другие схемы нумерации антител более детально описываются, например, в публикации Handbook of Therapeutic Antibodies (2007), ed. Stefan Dubel, Wiley-VCH Verlag GmbH & Co. KgaA, Weinheim, включенной путем ссылки.
[00060] Аминокислоты в пределах CDR вариабельных тяжелой и легкой областей контактируют с остатками в антигене и принципиально отвечают за определение специфичности связывания антитела к антигену. В зависимости от исследуемой пары антитело-антиген, все или менее, чем все остатки CDR могут прямо контактировать с антигеном. Кроме того, некоторые контакты могут больше, чем другие, способствовать определению специфичности и/или аффинности.
[00061] Конкретные контактные остатки, как в антителе, так и в антигене, могут определяться с применением рентгеновской кристаллографии или других способов, известных специалистам в данной области. Часто, хотя и не всегда, мутация таких контактных остатков отрицательно влияет на антигенсвязывающую специфичность и/или аффинность. С другой стороны, существует возможность мутации неконтактирующих остатков CDR, а также остатков FR, без существенного воздействия на антигенсвязывающую специфичность или аффинность. Хотя считается, что консервативные аминокислотные изменения с большей вероятностью сохраняют антигенсвязывающую специфичность и аффинность, фактический эффект любой конкретной мутации CDR или каркаса может определяться эмпирически с применением способов, известных специалистам в данной области.
[00062] Хотя CDR областей VH и VL, поддерживаемые их соответствующими каркасными областями, как правило, отвечают за установление антигенсвязывающей специфичности и аффинности, они могут быть исключениями. Например, в некоторых молекулах Ig остатки FR также могут способствовать связыванию с антигеном, в то время как в некоторых других молекулах Ig одна или несколько CDR могут прямо не контактировать с антигеном. Кроме того, в других молекулах Ig CDR отдельных областей VH и VL могут обладать значительной антигенсвязывающей специфичностью даже в отсутствие соответствующего вариабельного участка, с которым они обычно соединяются. Способность некоторых отдельных вариабельных участков молекулы Ig к специфичному связыванию с антигеном аналогична антигенсвязывающей специфичности антител акул или представителей семейства верблюдовых, которые включают спаренные тяжелые цепи, но без легких цепей.
[00063] Молекулы Ig некоторых видов могут классифицироваться в соответствии с различными изотипами. Например, у человека к изотипам Ig относятся IgA, IgG, IgD, IgE и IgM. Кроме того, изотипы IgA и IgG могут разбиваться на подтипы, называемые, соответственно, IgA1 и IgA2 и IgG1, IgG2, IgG3 и IgG4. Изотипы и подтипы определяются различиями в аминокислотных последовательностях константных областей тяжелой цепи. В результате различные изотипы и подтипы способны взаимодействовать с различными эффекторными молекулами на различных иммунных клетках, таким образом, обеспечивая различные эффекторные функции. Например, молекулы IgA способствуют иммунитету слизистой оболочки, в то время как молекулы IgE способствуют иммунитету против некоторых паразитов. Тяжелые цепи IgM и IgE содержат четыре тандемно расположенных СН Ig домена, пронумерованных, начиная с амино-концевого СН участка, как CH1, СН2, СН3 и СН4. Однако IgA, IgD и IgG содержат лишь три тандемно расположенных СН участка. Константные области легкой цепи включают два изотипа, называемые каппа и лямбда, которые не выполняют известных функций биологического эффектора. Природная молекула Ig имеет только один изотип константной области легкой цепи.
[00064] Гены, экспрессирующие тяжелые и легкие цепи антитела, составляют in vivo путем нескольких перестроек гена, известных как V(D)J рекомбинация. Этот процесс отвечает за создание большого репертуара антигенсвязывающих белков из сравнительно ограниченного репертуара последовательностей генов, находящихся в геноме. Больше информации об этом процессе содержится в публикации Abbas, А.К., Lichtman, А.Н. and Pillai, S., 2010, Cellular and Molecular Immunology, 5th Ed., Chapter 8, Saunders, Philadelphia, PA, включенной путем ссылки в полном объеме.
[00065] В ДНК зародышевой линии человека три отдельных генных локуса кодируют экзоны, необходимые для построения тяжелых цепей иммуноглобулина, каппа-легких цепей и лямбда-легких цепей. Локус тяжелой цепи находится на хромосоме 14, локус каппа-цепи находится на хромосоме 2, и локус лямбда-цепи находится на хромосоме 22. На 5'-конце каждого локуса находятся несколько вариабельных (V) сегментов гена, каждый длиной приблизительно 300 пар оснований, кодирующих большинство аминокислот, составляющих вариабельный участок тяжелых и легких цепей антитела, включая первую и вторую области, определяющие комплементарность (CDR1 и CDR2). У человека существует приблизительно 100 V генов в локусе тяжелой цепи, приблизительно 35 V генов в локусе каппа-цепи и приблизительно 30 V генов в локусе лямбда-цепи. Сегменты V гена разделены между собой нитронами.
[00066] После V-сегментов и перед константными (С) сегментами гена в локусе тяжелой цепи и локусе каппа-легкой цепи человека находятся кластеры соединительных (J) сегментов, как правило, имеющие длину приблизительно 30-50 пар оснований и отделенные друг от друга и соседних V и С генов некодирующей последовательностью. Локус тяжелой цепи содержит кластер из шести функциональных J-сегментов перед девятью функциональными С генами, связанными с различными изотипами Ig, а локус каппа-легкой цепи содержит кластер из пяти J-сегментов перед единственным Сκ геном. Локус лямбда-легкой цепи человека также содержит четыре функциональных J-сегмента, однако каждый располагается на 5'-стороне одного из четырех соответствующих функциональных Сλ генов- Локус тяжелой цепи человека также содержит кластер из более, чем 20 сегментов гена разнообразия (D), расположенных после V генов и пред кластером J-сегментов. Ни один из локусов легкой цепи не содержит сегментов D-гена.
[00067] В гене легкой цепи зрелого Ig V область кодируется V- и J-сегментами гена, в то время как в тяжелой цепи Ig V область кодируется V-, J- и D-сегментами гена. CDR1 и CDR2 в тяжелых и легких цепях кодируются V-сегментом гена. Однако построение CDR3 является более сложным. Для тяжелой цепи CDR3 кодируется соединением VDJ, включая D- и J-сегменты и соединительные остатки. Подобным образом CDR3 легкой цепи кодируется соединением VJ, включая J-сегмент и соединительные остатки.
[00068] В незрелых В-клетках все V-, D- и J-сегменты генов располагаются отдельно в зародышевой линии и не могут использоваться для экспрессии функциональных белков Ig. Вместо этого, с созреванием В-клеток сегменты генов подвергаются сложному процессу перестройки ДНК, известному как V(D)J рекомбинация, в результате которой случайно выбранные V-, D- и J-сегменты генов тяжелой цепи или V-и J-сегменты генов легкой цепи становятся смежными. Во время соединения V-, D- и J-сегментов генов молекулы, отвечающие за осуществление V(D)J рекомбинации случайно добавляют или удаляют нуклеотиды между сегментами. Таким образом, создаются экзоны полного вариабельного участка в геноме зрелых В-клеток, которые затем комбинируются с другими экзонами, включая кодирующие С области, в мРНК, кодирующих белки тяжелой и легкой цепи функционального Ig.
[00069] Случайная комбинация различных V-, D- и J-сегментов генов для построения экзонов V области и случайное добавление или удаление нуклеотидов между связываемыми сегментами генов, являются важными механизмами, через которые иммунная система создает большое разнообразие антигенсвязывающих центров. Эти явления, соответственно, называются комбинаторным разнообразием и соединительным разнообразием. Поскольку CDR3 образуется из последовательностей, состоящих из V-, D-и J-сегментов в случае тяжелых цепей или V и J-сегментов в случае легких цепей, соединительное разнообразие объясняет причину, по которой CDR3 является наиболее вариабельной из трех CDR и, как правило, образует наиболее широкий контакт с антигеном.
[00070] Поскольку структура молекул Ig по сути является модульной, в которое различные области выполняют различные функции, существует возможность получения фрагментов или производных антител против GDF-8, которые сохраняют способность к связыванию с GDF-8. Такие фрагменты или производные охватываются термином "антитело" в контексте этого описания. Неограничивающими примерами антигенсвязывающих фрагментов или производных, получаемых из молекул Ig, могут быть фрагменты Fab, представляющие собой моновалентные фрагменты, включающие VH, СН1, VL и CL области; F(ab')2, двухвалентный фрагмент, включающий два Fab, связанные между собой через шарнирную область; Fd фрагмент, включающий VH и СН1 области; Fv фрагмент, включающий VL и VH области; dAb фрагмент, включающий VH или VL область. Другим примером может быть одноцепочечная FV область (scFv), которая включает VH и VL области, тандемно расположенные в одной полипептидной цепи и разделенные полипептидным линкером, обеспечивающим возможность ассоциации вариабельных участков и образования моновалентного антигенсвязывающего центра. Могут быть построены одноцепочечные FV области, в которых VH область предшествует VL области, или, в альтернативном варианте, в которых VL область предшествует VH области. Неограничивающим примером линкера может быть состоящий из 15 остатков (Gly4Ser)3 пептид (SEQ ID NO: 34). Возможны также другие линкеры. К другим фрагментам или производным относятся Fab', суррогатные антитела, дисульфидно-стабилизированные Fv антитела (dsFv), диатела, триатела и однодоменные антитела, такие как антитело акулы или камелизированное антитело или нанотело. Также возможны другие фрагменты или производные. Антигенсвязывающие фрагменты, части или составляющие, такие как представленные в данном описании, могут быть получены рекомбинантным или способом или путем ферментного или химического расщепления интактных антител.
Примеры антител против GDF-8
[00071] GDF-8 означает фактор роста и дифференцировки-8, относящийся к надсемейству TGF-13. Аминокислотная последовательность зрелого человеческого GDF-8 представлена в SEQ ID NO: 1.
[00072] В предыдущих исследованиях было распознано мышиное моноклональное антитело, способное специфически связываться с GDF-8 и нейтрализовать его биологическую активность. Было продемонстрировано, что это антитело повышает мышечную массу и силу у мышей, включая моделирование амиотрофического латерального склероза (ALS) на мышах. См. заявку WO 2007/024535, включенную путем ссылки в полном объеме. VH область антитела мыши имеет аминокислотную последовательность SEQ ID NO: 3 и ее VL область имеет аминокислотную последовательность SEQ ID NO: 4. Эти VH и VL области показаны на ФИГ. 1А и ФИГ. 1 В, соответственно, на которых аминокислотная последовательность каждой CDR VH и VL областей обозначена жирным шрифтом. SEQ ID NO, ассоциированные с каждой CDR VH и VL в системах нумерации Kabat и АЬМ перечислены в Таблице 1, представленной ниже. CDR H1, Н2 и Н3, определяемые согласно системе нумерации Kabat, указываются как SEQ ID NO: 10, 11 и 12, соответственно, a CDR LI, L2 и L3 указываются как SEQ ID NO: 13, 14 и 15, соответственно. Согласно системе нумерации АЬМ, CDR H1, Н2 и Н3 указываются как SEQ ID NO: 20, 21 и 22, соответственно, a CDR LI, L2 и L3 указываются как SEQ ID NO: 23, 24 и 25, соответственно.
ТАБЛИЦА 1: Нуклеиново-кислотные и аминокислотные последовательности согласно изобретению, определяемые по идентификационным номерам последовательностей

[00073] Как объясняется детальнее в WO 2007/024535, мышиное антитело было гуманизировано путем прививки CDR. То есть, VH область мыши гуманизировали путем применения вариабельного тяжелого (VH) гена зародышевой линии человека DP47 (VH3-23; номер доступа в Genbank АВ019439) в качестве акцепторного каркаса человека с привитыми CDR VH мыши. Аминокислотная последовательность DP47 (SEQ ID NO: 33) показана на ФИГ. 1А. VL область мыши гуманизировали путем применения каппа-вариабельного легкого (VL) гена зародышевой линии человека DPK9 (012m Vkl; номер доступа в Genbank Х59315) в качестве акцепторного каркаса человека с привитыми CDR VI мыши. Аминокислотная последовательность DPK9 (SEQ ID NO: 32) показана на ФИГ. 1В.
[00074] Поскольку последовательности V области DP47 и DPK9 были взяты из зародышевой линии, а не из рекомбинированных генов V области, процесс гуманизации также требовал выбора J-сегмента гена человека для кодирования аминокислотной последовательности каждой частично гуманизированной VH и VL области на карбокси-конце относительно CDR3. Как описывается в WO 2007/024535, гуманизацию выполняли с применением J-сегмента тяжелой цепи JH3 (SEQ ID NO: 35) для VH области (т.е., DP47/JH3) и с применением J-сегмента легкой цепи JK1 (SEQ ID NO: 39) для VL области (т.е., DPK9/JK1). Аминокислотные последовательности, кодируемые этими генами J-сегмента, находятся непосредственно после VH CDR3 и CDR3 VL при выравнивании последовательностей, показанном на ФИГ. 1А и ФИГ. 1В, соответственно.
[00075] В контексте данного описания VH область гуманизированного антитела против GDF-8, построенная с применением DP47 и JH3, как описывается в WO 2007/024535, называется VH1 (SEQ ID NO: 7), a VL область гуманизированного антитела против GDF-8, построенная с применением DPK9 и JK1, называется VL1 (SEQ ID NO: 9). Также в контексте данного описания гуманизированное антитело против GDF-8, включающее VH1 и VL1, называется OGD 1.1.1. Согласно этой номенклатуре, номер VH область следует непосредственно после названия антитела "OGD1", а номер VL области следует непосредственно после номера VH области. Таким образом, например, номер антитела OGD 1.0.1 должен относиться к антителу, имеющему VH0 область и VL1 область, а название антитела OGD1.1.0 должно относиться к антителу, имеющему VH1 область и VL0 область. Выравнивание между VH и VL областями мыши, гуманизированными VH1 и VL1 областями и аминокислотами, кодируемыми генными последовательностями DPK9 и DP47 показано на ФИГ. 1А и ФИГ. 1 В.
[00076] Авторами описываются новые варианты гуманизированного антитела против GDF-8, обладающие удивительным свойством - способностью экспрессироваться клетками на значительно более высоком уровне по сравнению с OGD1.1.1, при этом сохраняя способность к специфичному связыванию GDF-8 с высокой аффинностью и к нейтрализации активности GDF-8.
[00077] В некоторых вариантах воплощения этих новых антител использовали другой тяжелый J-сегмент, т.е., JH4 (номер доступа в Genbank J00256) (SEQ ID NO: 37) после CDR3 в VH области. В результате этого изменения Leu (L) заменяет Met (M) в позиции 108 VH области согласно схеме нумерации Kabat по сравнению с VH1. В контексте данного описания эта новая гуманизированная VH область, в которой L находится в позиции 108 по Kabat, называется VH0. Аминокислотная последовательность VH0 (SEQ ID NO: 44) показана в выравнивании последовательности на ФИГ. 1А.
[00078] Поскольку в схеме нумерации Kabat используются буквы, добавляемые к некоторым из одинаковых номеров для обозначения CDR переменной длины, не обязательно существует точное соответствие между номером остатка по Kabat и его физическим расположением в последовательности остатков в полипептиде. Поэтому позиция 108 по Kabat VH области эквивалентна номеру аминокислоты 111 в SEQ ID NO: 44 (т.е., VH0) и описанных авторами аминокислотных последовательностях других гуманизированных VH областей.
[00079] В других вариантах воплощения антител согласно настоящему изобретению применяли другой легкий J-сегмент, т.е., JK4 (номер доступа в Genbank J00242) (SEQ ID NO: 41) после CDR3 в VL области. В результате этого изменения Gly (G) заменяет Gin (Q) в позиции 100 VL области согласно схеме нумерации Kabat по сравнению с VL 1. В контексте данного описания эта новая гуманизированная VL область, в которой G находится в позиции 100 по Kabat, называется VL0. Аминокислотная последовательность VL0 (SEQ ID NO: 46) показана при выравнивании последовательности ФИГ. 1В.
[00080] В данном описании гуманизированное антитело против GDF-8, включающее тяжелые цепи, включающие VH0, и легкую цепь, включающую VL0, называется OGD1.0.0.
[00081] Сегменты генов, последовательности и терминология, связанные с описанными выше вариантами воплощения VH и VL, представлены в Таблице 2 ниже.

[00082] Как описывается подробнее в Примерах, неожиданно было обнаружено, что антитела против GDF-8, включающие VH0, экспрессируются клетками на гораздо более высоком уровне по сравнению с антителами против GDF-8, включающими VH1. Например, в одном неограничивающем варианте воплощения, описанном в Примере 1, неожиданно было продемонстрировано, что интактный иммуноглобулин, включающий VH0 и VL0 (т.е., OGD1.0.0) временно экспрессируется на уровне, более, чем в 12 раз превышающем уровень подобного антитела, включающего VH1 и VL1 (т.е., OGD1.1.1). Устойчивый уровень экспрессии также был гораздо выше, как обсуждается в Примере 2. В Примере 3 было установлено, что повышенная экспрессия объясняется наличием VH0, поскольку повышенная экспрессия имела место независимо от того, соединялась ли VH0 с VL0 или VL1, и только тогда, когда VL0 соединялась с VH0, но не с VH1.
[00083] В некоторых вариантах воплощения антитела согласно настоящему изобретению являются интактными гетеротетрамерными молекулами Ig, включающими тяжелые и легкие цепи полной длины, в которых вариабельной тяжелой областью является VH0, и вариабельной легкой областью является VL0 (OGD1.0.0) или VL1 (OGD1.0.1), а в других вариантах воплощения антитела представляют собой специфично связывающиеся фрагменты GDF-8 или производные таких антител полной длины.
[00084] В соответствии с некоторыми вариантами воплощения, VH области антител согласно настоящему изобретению включают три CDR тяжелой цепи, т.е., CDRH1, CDRH2 и CDRH3, которые присутствуют в аминокислотной последовательности SEQ ID NO: 44 или ее мышином аналоге, SEQ ID NO: 3, и в которых аминокислотой в позиции 108 по Kabat является лейцин. В других вариантах воплощения VH область включает аминокислотную последовательность SEQ ID NO: 44 (т.е., VH0). В других вариантах воплощения VL области антител согласно настоящему изобретению включают три CDR легкой цепи, т.е., CORL1, CDRL2 и CDRL3, которые присутствуют в аминокислотной последовательности SEQ ID NO: 46 или ее мышином аналоге, SEQ ID NO: 5, и в которых аминокислотой в позиции 100 по Kabat является глицин или глутамин. В других вариантах воплощения VL область включает аминокислотную последовательность SEQ ID NO: 46 (т.е., VL0) или SEQ ID NO: 48 (т.е., VL1).
[00085] В антителах согласно настоящему изобретению, изотип тяжелой цепи антитела может относиться к любому из изотипов или подтипов Ig человека, т.е., IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4 или IgM. Изотипом легкой цепи антитела может быть каппа или лямбда. В конкретных неограничивающих вариантах воплощения константной тяжелой цепью антитела является аминокислотная последовательность SEQ ID NO: 19 или SEQ ID NO: 57, обе из которых являются подтипом IgG1. SEQ ID NO: 19 содержит две заместительных мутации в шарнирной области, которые препятствуют связыванию с Fe рецепторами на иммунных клетках, a SEQ ID NO: 57 содержит дополнительную мутацию шарнирной области, в общей сложности три, которые имеют подобный фенотип. В другом конкретном неограничивающем варианте воплощения СН область легкой цепи представляет собой аминокислотную последовательность SEQ ID NO: 17, которая относится к каппа-изотипу.
[00086] В конкретном неограничивающем варианте воплощения настоящего изобретения антитело против GDF-8 включает тяжелую цепь антитела полной длины в соответствии с аминокислотной последовательностью SEQ ID NO: 58 и легкую цепь антитела полной длины в соответствии с аминокислотной последовательностью SEQ ID NO: 59. Первая последовательность включает VH0 и константные области тяжелой цепи аминокислотной последовательности SEQ ID NO: 57, в то время как последняя последовательность включает VL0 и каппа-константную область легкой цепи аминокислотной последовательности SEQ ID NO: 17. В соответствии с другим типичным неограничивающим вариантом воплощения, антитело против GDF-8 согласно изобретению включает интактное гетеротетрамерное антитело, состоящее из двух тяжелых цепей антитела и двух легких цепей антитела согласно аминокислотным последовательностям SEQ ID NO: 58 и SEQ ID NO:59 (т.е., OGD1.0.0).
[00087] Как было указано выше, в других вариантах воплощения антитела согласно настоящему изобретению включают антигенсвязывающие фрагменты или производные иммуноглобулинов против GDF-8, которые включают VH0. В некоторых вариантах воплощения фрагментов или производных VH0 может попарно связываться с VL0 или VL1. Неограничивающими примерами фрагментов или производных согласно настоящему изобретению являются Fab', F(ab')2, Fab, Fv, scFv, dsFv, диатела, триатела и однодоменные антитела, такие как антитело акулы или камелизированное антитело или нанотело, включающие VH0. Возможны также и другие фрагменты или производные. Конкретный неограничивающий пример производной Ig согласно настоящему изобретению включает SEQ ID NO: 63, scFv, в которой VL0 является тандемно расположенной аминоконцевой областью относительно VH0. Другим неограничивающим примером является SEQ ID NO: 65, scFv, в которой V-области являются обращенной, с VH0, которая является тандемно расположенной аминоконцевой областью относительно VL0.
[00088] Хотя антитела согласно настоящему изобретению представлены на примере иммуноглобулина, в котором CDR тяжелой цепи мышиного антитела против GDF-8 были привиты к VH области зародышевой линии человека DP47, гуманизированные антитела против GDF-8 согласно изобретению не ограничиваются применением только этого вариабельного участка. Таким образом, например, антитела также включают интактные иммуноглобулины и их фрагменты или производные, в которых мышиные CDR тяжелой цепи (т.е., SEQ ID NO: 10-12 или 20-22) привиты к VH областям человека, отличающимся от DP47, и подвергаются дальнейшей модификации, таким образом, чтобы образовавшийся в результате полипептид VH области включал Leu (L) в позиции 108 по Kabat. Последовательность других VH областей зародышевой линии человека можно найти путем поиска в Genbank или различных открытых для публичного доступа базах данных в Интернет, включая VBASE (http://vbase.mrc-cpe.cam.ac.uk/) или VBAS Е2 (http://www.vbase2.org/).
[00089] Как описывается подробнее в Примере 10, совместную кристаллическую структуру OGD1.0.0 и химерного антитела против GDF-8, включающую мышиные VH и VL области SEQ ID NO: 3 и SEQ ID NO: 5, соответственно, связанные с GDF-8, определяли и использовали для распознавания контактных остатков в антителе, отвечающих за связывание с антигеном. На основе этой информации, и как подробнее описывается в Примере 11, VH и VL области подвергали дальнейшей гуманизации путем мутации неконтактных остатков в CDR для соответствия остаткам, присутствующим в той же позиции в вариабельной последовательности зародышевой линии человека. Как показано на ФИГ. 1А, другие гуманизированные вариабельные тяжелые области называются VH2, VH3, VH4 и VH5. И, как показано на ФИГ. 1В, другие гуманизированные вариабельные легкие области называются VL2, VL3, VL4 и VL5.
[00090] В некоторых вариантах воплощения антител согласно настоящему изобретению любая из гуманизированных VH областей может попарно связываться с любой из гуманизированных VL областей для создания интактных антител против GDF-8 или их антигенсвязывающих фрагментов или производных. Например, в некоторых вариантах воплощения VH0 может попарно связываться с любой из VL областей VL0, VL1, VL2, VL3, VL4 или VL5. В других вариантах воплощения VH1 может попарно связываться с любой из VL областей VL0, VL1, VL2, VL3, VL4 или VL5. В других вариантах воплощения VH2 может попарно связываться с любой из VL областей VL0, VL1, VL2, VL3, VL4 или VL5. В других вариантах воплощения VH3 может попарно связываться с любой из VL областей VL0, VL1, VL2, VL3, VL4 или VL5. В других вариантах воплощения VH4 может попарно связываться с любой из VL областей VL0, VL1, VL2, VL3, VL4 или VL5. В некоторых других вариантах воплощения VH5 может попарно связываться с любой из VL областей VL0, VL1, VL2, VL3, VL4 или VL5.
[00091] Как объяснялось выше, ожидается, что мутация неконтактных остатков в пределах CDR и каркасных областей должна минимально влиять на специфичность и/или аффинность связывания GDF-8, а мутация контактных остатков должна иметь большее влияние. Хотя мутации, в частности; контактных остатков, могут снижать специфичность и/или аффинность связывания, в некоторых случаях наблюдается, что мутации повышают специфичность и/или аффинность к GDF-8. Фактическое влияние на специфичность или аффинность любой конкретной мутации может определяться с применением способов, известных специалистам в данной области, например, поверхностного плазмонного резонанса или других способов.
[00092] С учетом вышеизложенных принципов в некоторых вариантах воплощения один, два, три или более неконтактных остатков в пределах одной или нескольких CDR VH и/или VL или каркасных областей антител согласно изобретению могут быть консервативно или неконсервативно замещены другим аминокислотным остатком и сохранять значительную специфичность аффинность связывания к GDF-8. В других вариантах воплощения один, два, три или более контактных остатков в пределах одной или нескольких CDR VH и/или VL могут быть консервативно замещены и могут сохранять значительную специфичность и аффинность связывания к GDF-8. В других вариантах воплощения мутации неконтактных остатков или контактных остатков в результате улучшают специфичность и/или аффинность к GDF-8.
[00093] В других вариантах воплощения антител согласно изобретению аминокислотные последовательности VH и/или VL области могут отличаться различным процентом конкретно указанных авторами последовательностей и сохранять существенную, или даже улучшенную, специфичность и/или аффинность к GDF-8. Таким образом, в некоторых вариантах воплощения VH область антитела против GDF-8 согласно изобретению может отличаться на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% от аминокислотной последовательности VH0, VH1, VH2, VH3, VH4 или VH5. В других вариантах воплощения VL область антитела против GDF-8 согласно изобретению может отличаться на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% от аминокислотной последовательности VL0, VH1, VH2, VH3, VH4 или VH5. В других вариантах воплощения VH и VL области антител против GDF-8 могут отличаться от конкретно приведенных авторами с подобными процентными отличиями при сохранении существенной или даже улучшенной GDF-8 специфичности и/или аффинности связывания.
[00094] Антитела согласно настоящему изобретению также могут быть дериватизированы, ковалентно модифицированы или конъюгированы с другими молекулами для изменения их свойств или улучшения их функции. Например, помимо прочих, к дериватизированным антителам относятся антитела, которые были модифицированы, например, путем гликозилирования, фукозилировани, ацетилирование, пегилирования, фосфорилирования, амидирования, формилирования, дериватизации известными защитными/блокирующими группами, сцепление с клеточным лигандом или другим белком и т.п.
[00095] В некоторых вариантах воплощения С-концевой лизин тяжелой цепи антитела против GDF-8 согласно настоящему изобретению может быть расщеплен и удален. Таким образом, например, в некоторых вариантах воплощения настоящего изобретения антитело против GDF-8 включает константные области тяжелой цепи SEQ ID NO: 19 или SEQ ID NO: 57, в которых отсутствует С-концевой лизин, или может включать тяжелую цепь антитела с последовательностью SEQ ID NO: 58, в которой отсутствует С-концевой лизин.
[00096] Некоторые модификации структуры антител против GDF-8 могут существовать в природе в результате типа клетки, в которой они вырабатываются. В неограничивающем примере синтез антител в клетках млекопитающих, таких как клетки СНО, может вести к гликозилированию в одной или нескольких аминокислотах в цепях антител. В типичном неограничивающем варианте воплощения антитела против GDF-8 аминокислота N296 в тяжелой цепи является гликозилированной. Также возможно гликозилирование в других центрах. Как станет понятно специалистам в данной области, в результате выработки антител в некоторых других типах клеток, например, бактериальных клеток, могут образовываться цепи антител, которые не являются гликозилированными. Другие типы модификации антител могут происходить естественным или искусственным путем химических или ферментных модификаций, осуществляемых во время или после очистки антитела.
[00097] В альтернативном варианте конкретные аминокислоты в вариабельных или константных областях могут быть изменены для изменения или улучшения функции. В одном неограничивающем примере аминокислотные остатки в Fe области антитела могут быть изменены для продления периода полувыведения антитела из сыворотки путем повышения его связывания с FcRn. См., например, заявку WO 2000/009560, включенную в это описание путем ссылки. В других неограничивающих примерах аминокислоты антитело могут изменяться для уменьшения связывания с одним или несколькими Fe рецепторами, комплементом или другими иммунными рецепторами, которые опосредуют биологические эффекторные функции Ig. В другом неограничивающем примере аминокислоты в CDR, каркасных областях или константных областях могут изменяться для повышения аффинности связывания GDF-8 или для уменьшения иммуногенности. В конкретном неограничивающем примере некоторые человеческие каркасные остатки VH или VL областей антител согласно настоящему изобретению могут быть обратно изменены на их мышиные аналоги, как в аминокислотных последовательностях SEQ ID NO: 26 и SEQ ID NO: 27, соответственно.
[00098] В других вариантах воплощения антитела метят обнаружимым веществом, и они могут быть обнаружены в соответствии со способами, известными специалистам в данной области. Такие метки могут быть конъюгированы, прямо или опосредованно, с антителом согласно изобретению. Метка сама может быть непосредственно обнаружимой (например, радионуклидной или флуоресцентной меткой), или может быть опосредованно обнаружимой по ее способности к образованию обнаружимых молекул (например, ферментной меткой, катализирующей субстрат для образования продукта, который является прямо обнаружимым). Примерами обнаружимых меток могут быть ферменты (например, пероксидаза хрена, 13-галактозидаза, люцифераза, щелочная фосфатаза и т.п.), простетические группы (например, биотин и т.п.), флуоресцентные красители или компоненты (например, FITC, родамин, люминофоры на основе комплексов лантанидов), люминесцентные компоненты, биолюминесцентные компоненты, радионуклиды (например, 3Н, 14С, 15N, 35S, 90Y, 99Тс, 111In, 125I, 131I и т.п.), позитронно-активные атомы или ионы, магнитные атомы или ионы, атомы или ионы парамагнитных металлов или пептидные эпитопы, которые могут специфически связываться с другими антителами. В некоторых вариантах воплощения метки могут прикрепляться с использованием спейсеров различной длины для уменьшения или предотвращения возможного стерического несоответствия с антигенсвязывающим центром.
[00099] Антитела согласно настоящему изобретению могут экспрессироваться в культуре или в организме животных из клеток любого типа, способных поддерживать экспрессию белков млекопитающих. Неограничивающими примерами могут быть клетки человека, мыши, крысы или других грызунов, клетки других млекопитающих, клетки СНО, дрожжевые клетки или клетки других грибов, растительные клетки или бактериальные клетки. Способы, применяемые для клонирования ДНК, кодирующих молекулы Ig, или их фрагменты или производные, в векторы экспрессии, с последующей временной или устойчивой трансфекцией клеток такими векторами, хорошо известны специалистам в данной области. Условия культивирования могут изменяться для обеспечения максимального уровня экспрессии антитела. Антитела также могут экспрессироваться в организме животных с применением способов, известных специалистам в данной области, с последующей очисткой от молока или других жидкостей организма, антитела также могут быть полностью или частично синтетическими.
[000100] Антитела согласно настоящему изобретению связываются с GDF-8 с высокой аффинностью, например, с равновесной константой диссоциации (KD) как минимум приблизительно 1×10-6 Μ, 1×10-7 M, 1×10-8, 1×10-9, 1×10-10, 1×10-11 Μ или выше. Ко антитела против GDF-8 для GDF-8 определяют в соответствии с различными способами, известными специалистам в данной области. Неограничивающими примерами таких способов могут быть поверхностный плазмонный резонанс (SPR) и ELISA. Как известно специалистам в данной области, из-за эффекта авидности видимая аффинность связывания антитела против GDF-8, имеющего два или более антигенсвязывающих центров, может быть большей, чем у фрагмента антитела, имеющего моновалентный антигенсвязывающий центр.
[000101] Хотя антитела согласно настоящему изобретению являются специфичными к GDF-8, такие антитела, в зависимости от распознаваемого эпитопа или эпитопов, также могут быть способны связываться с высокой аффинностью с близкородственным фактором роста и дифференцировки, известным как GDF-11. Таким образом, антитело, специфичное к GDF-8 не обязательно исключает антитела, способные связываться с молекулами GDF-11.
[000102] В контексте данного описания нейтрализующим антителом против GDF-8 является антитело, снижающее биологическую активность GDF-8 по сравнению с неспецифичным контрольным антитело или другим подходящим контролем. Не желая быть связанными соответствием конкретной теории, авторы полагают, что как минимум один способ, которым антитело против GDF-8 может нейтрализовать биологическую функцию, опосредованную GDF-8, состоит в предотвращении связывания зрелого GDF-8 с его высокоаффинным рецептором, например, ActRJlB, или одним или несколькими из его низкоаффинными рецепторами. Однако возможны и другие механизмы, благодаря которым нейтрализующее антитело против GDF-8 может препятствовать биологической активности GDF-8.
[000103] Специалистам в данной области известны многие виды биологической активности, опосредованной GDF-8, которые могут быть снижены нейтрализующим антителом согласно изобретению. Неограничивающими примерами могут быть связывание GDF-8 с ActRllB, которое может быть измерено, например, с применением анализа на основе ELISA. Другой пример включает активацию с помощью GDF-8 его пути клеточного сигнала, которая может быть обнаружена, например, с применением трансфицированного гена-репортера, включающего так называемые GAGA-элементы. См., например, публикации Lee, et al., Regulation of muscle growth by multiple ligands signaling through activin type II receptors, PNAS (2005) 102:18117-18122, и Thies, et al., GDF-8 Propeptide Binds to GDF-8 and Antagonizes Biological Activity by Inhibiting GDF-8 Receptor Binding, Growth Factors (2001) 18:251-59, которые включены путем ссылки. Еще один пример включает фосфорилирование белков SMAD, отвечающих за передачу опосредованного GDF-8 сигнала от рецептора GDF-8 на поверхности клетки в ядро. См., например, публикацию Philip, et al., Regulation of GDF-8 signaling by the p38 МАРК, Cellular Signalling (2005) 17:365-375, включенную путем ссылки. Фосфорилирование белков SMAD может быть обнаружено, например, при помощи количественного вестерн-блоттинга с применением антител против фосфо-SMAD. Также может быть обнаружена модуляция дальнейшей генной экспрессии генов, обычно активируемых или подавляемых GDF-8. Еще одним примером опосредованной GDF-8 активности, которая может быть снижена с применением нейтрализирующих антител согласно настоящему изобретению, является отрицательная регуляция мышечной массы или силы. Возможны также другие виды активности.
[000104] Нейтрализующие антитела согласно настоящему изобретению могут снижать биологическую активность, опосредованную GDF-8, в разной степени в зависимости от переменных, известных специалистам в данной области, таких как концентрация антитела и антигена и аффинность связывания, а также другие. Типичные неограничивающие проценты снижения опосредованной GDF-8 биологической активности, вызываемого связыванием антитела с GDF-8, означают снижение как минимум приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99% или более по сравнению с соответствующими контрольными образцами.
[000105] Ингибирование антителом против GDF-8 опосредованной GDF-8 биологической активности для удобства может быть выражено как концентрация такого антитела, которая способна ингибировать 50% биологической активности при любых выбранных условиях анализа. Эта концентрация также называется IC50. В некоторых вариантах воплощения антитела против GDF-8 согласно настоящему изобретению имеют значения IC50, равные или меньшие, чем приблизительно 500 нМ, 250 нМ, 100 нМ, 75 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 5 нМ, 1 нМ, 0,5 нМ, 0,1 нМ или менее. Секреторные лидерные последовательности
[000106] Согласно некоторым вариантам воплощения, гены, кодирующие тяжелые и легкие цепи антитела, могут иметь последовательность, кодирующую аминоконцевой секреторный лидерный пептид, направляющий новые синтезированные белки в секреторный компартмент. Посттрансляционная обработка затем удаляет лидерные пептиды до секреции зрелого антитела из клетки. В конкретных неограничивающих вариантах воплощения антител согласно настоящему изобретению обеспечиваются области VH0, VH1, VL0 и VL1 с секреторным лидерным пептидом длиной 19 аминокислот. Этим V областям, включающим лидерную последовательность, присвоены следующие идентификационные номера последовательностей: VH0 (SEQ ID NO: 50); VL0 (SEQ ID NO: 52); VH1 (SEQ ID NO: 54); и VL1(SEQ ID NO: 56). Также могут применяться другие секреторные лидерные последовательности. Неограничивающими примерами могут быть первые 19 аминокислот мышиной VH области и ее лидерная последовательность (SEQ ID NO: 29) и первые 20 аминокислот мышиной VL области и ее лидерная последовательность (SEQ ID NO:31).
Экспрессия антитела против GDF-8
[000107] Как объясняется более детально в Примерах, OGD1.0.0 экспрессируется на значительно более высоком уровне по сравнению с OGD1.1.1 в клетках млекопитающих. Например, при экспрессии OGD1.0.0 и OGD 1.1.1 во временно трансфицированных клетках COS OGD1.0.0 экспрессировался на приблизительно в 12 раз более высоком уровне по сравнению с OGD1.1.1. Подобным образом при экспрессии этих антител в устойчиво трансфицированных клетках СНО OGD1.0.0 экспрессировался на приблизительно в 6 раз более высоком уровне по сравнению с OGD 1.1.1. Также, как объясняется в Примерах, разница в уровне экспрессии объясняется, главным образом, наличием в антителе VH0 вместо VH1.
[000108] Таким образом, антитела согласно настоящему изобретению, включающие VH0, демонстрируют большую экспрессию по сравнению с подобными антителами, включающими VH1 при экспрессии в подобных условиях. Например, в некоторых вариантах воплощения уровень экспрессии антител, включающих VH0, выше, чем у подобного антитела, включающего VH1, экспрессируемого в подобных условиях, как минимум приблизительно в 1,5 раза, 2 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 20 раз, 30 раз или более. Степень различий в уровне экспрессии между антителами может зависеть, например, от типа клетки-хозяина, используемой для экспрессии антител, например, клеток COS или клеток СНО, или от временной либо устойчивой трансфекции клеток-хозяев. Сравнительные уровни экспрессии антител, содержащих вариабельные тяжелые области VH0 или VH1, могут меняться с изменением других условий роста и могут определяться с применением способов, известных специалистам в данной области.
[000109] В других вариантах воплощения уровень экспрессии антитела с VH0 выше уровня подобного антитела, содержащего VH1, которое экспрессируется в подобных условиях, как минимум приблизительно на 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 75%, 100%, 150%, 200%, 250%, 300%, 400%, 500%, 600%, 700%, 800%, 900%, 1000%, 1100%, 1200%, 1300%, 1400%, 1500%, 1600%, 1700%, 1800%, 1900%, 2000%, 3000%, 4000%, 5000% или более. Также возможны другие различия в уровне экспрессии, например, между указанными процентными значениями.
[000110] Уровень экспрессии антител измеряют, применяя способы, известные специалистам в данной области. В одном неограничивающем примере уровень экспрессии антител измеряют, применяя количественный анализ ELISA. Также могут применяться другие количественные анализы в соответствии с информацией, которой располагают специалисты в данной области.
Молекулы нуклеиновых кислот, кодирующих антитела против GDF-8
[000111] Настоящее изобретение также обеспечивает молекулы нуклеиновых кислот или полинуклеотидов, кодирующих антитела против GDF-8. Нуклеиновые кислоты могут включать ДНК или РНК, в которых U заменяет Τ в последовательности нуклеотидных оснований ДНК. Нуклеиновые кислоты также могут содержать модификации, такие как нестандартные нуклеотидные основания (например, 5-метилцитозин) или модифицированный остов (например, фосфотиоат). Возможны и другие модификации. Нуклеиновые кислоты могут быть одноцепочечными или двухцепочечными. Нуклеиновые кислоты могут быть получены из природного источника, такого, как клетка или целый организм. Неограничивающими примерами природных нуклеиновых кислот могут быть геномная ДНК, амплифицированная плазмидная ДНК или мРНК. В альтернативном варианте нуклеиновые кислоты могут быть синтезированы. Неограничивающими примерами синтетических нуклеиновых кислот могут быть кДНК, продукт ПЦР или нуклеиновая кислота, синтезированная в устройстве для синтеза нуклеиновых кислот.
[000112] В некоторых вариантах воплощения нуклеиновые кислоты согласно настоящему изобретению кодируют аминокислотную последовательность тяжелой цепи антитела или ее фрагмент или производную, включающую VH0. В других вариантах воплощения нуклеиновые кислоты кодируют аминокислотную последовательность легкой цепи антитела или ее фрагмент или производную, включающую VL0. В других вариантах воплощения нуклеиново-кислотные последовательности, кодирующие VH0 и VL области, такие как VL0 или VL1, присутствуют в разных или одних и тех же выделенных полинуклеотидах.
[000113] Хотя настоящее изобретение обеспечивает конкретные нуклеиново-кислотные последовательности, кодирующие антитела против GDF-8, или их фрагменты или производные, специалистам в данной области станет понятно, что из-за дегенерации генетического кода такие последовательности являются лишь примерами и не должны рассматриваться как ограничивающие. Таким образом, типичными нуклеиново-кислотными последовательностями, кодирующими VH и VL области мышиного антитела против GDF-8, являются SEQ ID NO: 2 и SEQ ID NO: 4, соответственно. Типичными нуклеиново-кислотными последовательностями, кодирующими VH1, являются SEQ ID NO:6 и SEQ ID NO: 47. Типичными нуклеиново-кислотными последовательностями, кодирующими VL1, являются SEQ ID NO: 8 и SEQ ID NO: 48. Типичной нуклеотидной последовательностью, кодирующей VH0, является SEQ ID NO:43. Типичной нуклеотидной последовательностью, кодирующей VL0, является SEQ ID NO:45. Типичной нуклеотидной последовательностью, кодирующей аминокислотную последовательность SEQ ID NO: 19, которая включает СН области IgG1, включающие две мутации шарнирной области, является SEQ ID NO: 18. Типичной нуклеотидной последовательностью, кодирующей аминокислотную последовательность SEQ ID NO: 17, которая включает каппа-область CL человека, является SEQ ID NO: 16. Типичными нуклеиново-кислотными последовательностями, кодирующими VH и VL области мышиного антитела против GDF-8, перед которыми находятся лидерные последовательности, являются SEQ ID NO: 28 и SEQ ID NO: 30, соответственно. Типичными нуклеиново-кислотными последовательностями, кодирующими аминокислотные последовательности VH0, VL0, VH1 и VL1 областей, перед которыми находятся лидерные последовательности, являются SEQ ID NO: 49; SEQ ID NO: 51; SEQ ID NO: 53; и SEQ ID NO: 55, соответственно.
[000114] Согласно некоторым вариантам воплощения, молекула нуклеиновой кислоты включает нуклеотидную последовательность, кодирующую аминокислоты любой из следующих последовательностей SEQ ID NO: 7, 9, 10, 11, 12, 13, 14, 15, 17, 19, 20, 21, 22, 23, 24, 25, 26, 27, 36, 38, 40, 42, 44, 46, 50, 52, 54, 56, 57, 58, 59, 63 или 65. В других вариантах воплощения молекулы нуклеиновых кислот включают нуклеотидную последовательность, кодирующую аминокислотные последовательности, которые как минимум на 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичны SEQ ID NO: 7, 9, 10, 11, 12, 13, 14, 15, 17, 19, 20, 21, 22, 23, 24, 25, 26, 27, 36, 38, 40, 42, 44, 46, 50, 52, 54, 56, 57, 58, 59, 63 или 65. В других вариантах воплощения молекулы нуклеиновых кислот включают нуклеотидную последовательность, кодирующую аминокислотные последовательности, которые как минимум на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичны VH0 (SEQ ID NO: 44), и в которых в позиции 108 по Kabat находится лейцин.
[000115] Согласно некоторым вариантам воплощения, молекула нуклеиновой кислоты включает нуклеотидную последовательность любой из следующих SEQ ID NO: 6, 8, 16, 18, 35, 37, 39, 41, 43, 45, 47, 48, 49, 51, 53, 55, 62 или 64. В других вариантах воплощения молекулы нуклеиновых кислот включают нуклеотидную последовательность, которая как минимум на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична нуклеиново-кислотным последовательностям SEQ ID NO: 6, 8, 16, 18, 35, 37, 39, 41, 43, 45, 47, 48, 49, 51, 53, 55, 62 или 64. В других вариантах воплощения молекулы нуклеиновых кислот включают нуклеотидную последовательность, которая гибридизируется в очень строгих условиях с нуклеиново-кислотными последовательностями SEQ ID NO: 6, 8, 16, 18, 35, 37, 39, 41, 43, 45, 47, 48, 49, 51, 53, 55, 62 или 64.
[000116] Неограничивающим примером очень жестких условий гибридизации может быть инкубация нуклеиновых кислот, которые гибридизируются в 1x SSC при 65°С, или 1x SSC и 50% формамида при 42°С, с последующим промыванием в 0,3х SSC при 65°С. Дополнительные примеры жестких условий представлены в публикации Sambrook et al., Molecular Cloning: A Laboratory Manual, Chs. 9 & 11, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989), включенной в данное описание путем ссылки.
[000117] Как станет понятно специалистам в данной области, некоторые из нуклеиновых кислот согласно настоящему изобретению могут быть лигированы в пределах рамки для создания сложных нуклеиново-кислотных последовательностей. Например, нуклеиновая кислота, кодирующая VH0, может быть лигирована в пределах рамки с нуклеиновой кислотой, кодирующей СН области, для создания сложной нуклеиновой кислоты, кодирующей полную тяжелую цепь. В неограничивающем примере нуклеотидная последовательность SEQ ID NO: 43, кодирующая VH0, может быть лигирована в пределах рамки с нуклеотидной последовательностью, кодирующей аминокислотную последовательность SEQ ID NO: 57, константной частью тяжелой цепи человеческого IgG1, содержащей три мутации, влияющие на эффекторную функцию. Возможны подобные лигирования для создания легкой цепи, так же, как и другие лигирования, для создания композиции описанных авторами нуклеиновых кислот.
Векторы
[000118] Нуклеиновые кислоты согласно настоящему изобретению могут быть включены в векторы с применением способов, хорошо известных специалистам в данной области. Векторы в некоторых вариантах воплощения включают плазмиды в целом, бактериальные плазмиды, эукариотные эписомы, дрожжевые искусственные хромосомы и вирусные геномы. К типичным неограничивающим вирусам относятся ретровирусы, аденовирусы, аденоассоциированные вирусы (AAV) и растительные вирусы, такие как вирус мозаики цветной капусты и вирус табачной мозаики. Возможны и другие типы векторов. В некоторых вариантах воплощения векторы способны к автономной репликации в соответствующих хозяевах. В других вариантах воплощения векторы поддерживаются в хозяевах внехромосомно или могут быть интегрированы в геном хозяина, что позволяет реплицировать вектор с геномом хозяина. Векторы, включающие ген и контрольные последовательности, достаточные для поддержания транскрипции и трансляции гена, называются векторами экспрессии. Векторы согласно настоящему изобретению могут быть выбраны или построены в соответствии с данными, известными специалистам в данной области, для функционирования в любом типе клеток, способном поддерживать экспрессию Ig генов, включая бактериальные клетки, другие прокариотные клетки, дрожжевые клетки, другие грибковые клетки, растительные клетки, животные клетки, клетки насекомых, клетки млекопитающих, клетки СНО и клетки человека или другие.
[000119] Векторы необязательно могут содержать одну или несколько контрольных последовательностей. Некоторые контрольные последовательности обеспечивают возможность репликации, такие как точки начала репликации. Другие контрольные последовательности регулируют или модулируют транскрипцию, такие как промоторы, энхансеры и центры терминации транскрипции. Неограничивающими примерами промоторов или энхансеров могут быть те, которые взяты из ретровирусных длинных концевых повторов, цитомегаловируса (CMV), вируса обезьян 40 (SV40), аденовируса (например, главного позднего промотора аденовируса (AdMLP)) или вируса полиомы. Дополнительными примерами могут быть ткане-специфические промоторы и энхансеры, конститутивно активные промоторы и энхансеры, индуцируемые промоторы и энхансеры, промоторы и энхансеры Ig-генов и промоторы и энхансеры актина. Возможны также другие промоторы и энхансеры.
[000120] Некоторые контрольные последовательности регулируют или модулируют посттранскрипционную обработку РНК, такие как сигналы сплайсинга и полиаденилирования или сигналы, повышающие или снижающие устойчивость мРНК. Другие контрольные последовательности регулируют или модулируют трансляцию белка, такие как последовательности инициации трансляции (например, консенсусная последовательность Козака), посттрансляционную обработку, такие как сигнальные пептидные последовательности, направляющие секрецию генного продукта из клетки-хозяина, или устойчивость белка. Сигнальные пептидные последовательности могут быть взяты из иммуноглобулинов или из секретируемых белков, которые не относятся к надсемейству Ig. Возможны также другие контрольные последовательности.
[000121] Векторы также могут включать селектируемые маркерные гены, позволяющие выбирать клетки-хозяева, принявшие векторы. Неограничивающими примерами могут быть селектируемые маркерные гены, обеспечивающие резистентный к медикаментам фенотип, такие как ген дигидрофолатредуктазы (DHFR) (для применения в клетках-хозяевах dhfr, которые позволяют производить селекцию с использованием метотрексата), нео-ген (позволяющий производить селекцию с использованием G418 или подобных медикаментов), hph-ген (позволяющий производить селекцию с использованием гигромицина В) и ген глутаматсинтетазы (позволяющий производить селекцию с использованием метионинсульфоксимина).
[000122] В некоторых вариантах воплощения векторы могут включать нуклеотидную последовательность, кодирующую одну тяжелую или легкую цепь Ig или ее антигенсвязывающие фрагменты, но не обе цепи в одном векторе. Как правило, экспрессия интактных антител из таких векторов включает введение отдельных векторов, включающих тяжелую цепь и легкую цепь, в одну и ту же клетку. В других вариантах воплощения векторы могут включать нуклеиново-кислотные последовательности, кодирующие тяжелые и легкие цепи Ig или их антигенсвязывающие фрагменты, в одном векторе.
[000123] Молекулы нуклеиновых кислот согласно настоящему изобретению или векторы, включающие такие нуклеиновые кислоты, могут быть введены в один или несколько типов клеток-хозяев, способных поддерживать экспрессию антитела. Способы введения нуклеиновых кислот или векторов в соответствующие клетки-хозяева хорошо известны специалистам в данной области. Неограничивающими примерами могут быть временная и устойчивая трансфекция, трансформация, трансдукция и вирусная инфекция заданных клеток-хозяев. Другими примерами могут быть опосредованная декстраном трансфекция, осаждение фосфатом кальция, опосредованная полибреном трансфекция, слияние протопластов, электропорация, инкапсуляция полинуклеотида(ов) в липосомах и прямая микроинъекция ДНК в ядра. Типичные неограничивающие способы обсуждаются, например, в Патентах США №№4,399,216, 4,912,040, 4,740,461 и 4,959,455, которые включены путем ссылки. Способы трансформации растительных клеток также хорошо известны специалистам в данной области, включая, например, опосредованную агробактериями трансформацию, биолистическую трансформацию, прямую инъекцию, электропорацию и вирусную трансформацию. Способы трансформации бактериальных и дрожжевых клеток также хорошо известны специалистам в данной области.
Клетки-хозяева
[000124] В некоторых вариантах воплощения нуклеиновые кислоты, кодирующие антитела согласно настоящему изобретению или их фрагменты или производные, вводят в соответствующие клетки-хозяева с целью экспрессии. К клеткам, способным эскпрессировать антитела, относятся бактериальные, грибковые, растительные, животные клетки и клетки млекопитающих. Также могут использоваться другие типы клеток в соответствии с информацией, которой располагают специалисты в данной обрасти.
[000125] Линии клеток млекопитающих, подходящие в качестве хозяев для экспрессии антител известны специалистам в данной области. Типичными неограничивающими примерами могут быть некоторые иммортализованные линии клеток, которые можно получить в Американской коллекции типовых культур (АТСС) или из других источников, и к которым относятся клетки яичников китайского хомячка (СНО), клетки NSO, клетки SP2, клетки НЕК-293Т, клетки NIH-3T3, клетки Hela, клетки почки новорожденного хомяка (ВНК), клетки почки африканской зеленой мартышки (например, клетки COS, CV-1 или Vero), клетки злокачественной гепатомы человека (например, HepG2), клетки А549, клетки А431, клетки Hela, L-клетки, клетки ВНК21, клетки HL-60, клетки U937, клетки НаК, клетки Jurkat и другие. Возможны и другие клетки животных, насекомых или клетки млекопитающих, подходящие в качестве хозяев для экспрессии антител.
[000126] В других вариантах воплощения используют линии клеток насекомых, растений, бактерий или грибов. Неограничивающими примерами клеток насекомых могут быть клетки Sf9 или SÍ21, которые часто используют вместе с экспрессирующей бакуловирусной векторной системой. Неограничивающими примерами растительных клеток могут быть клетки никоцианы, арабидопсиса, ряски, кукурузы, пшеницы и картофеля. Неограничивающими примерами бактерий могут быть штаммы Escherichia coli, Bacillus subtilis, Salmonella typhimurium и Streptomyces. Неограничивающими примерами грибов могут быть дрожжевые штаммы Schizosaccharomyces pombe, Saccharomyces cerevisiae, Pichia pastoris, Kluyveromyces и Candida. Возможны и другие клетки насекомых, растений, бактерий и грибов.
[000127] Способы выращивания и поддержания различных типов клеток-хозяев в условиях, ведущих к экспрессии антител, хорошо известны специалистам в данной области. После экспрессии антител экспрессированные таким образом антитела могут быть очищены от клеток-хозяев в соответствии с информацией, которой располагают специалисты в данной обрасти. Например, секретированные антитела могут быть очищены от сред, в которых выращивают клетки-хозяева. В некоторых альтернативных вариантах воплощения, в частности, в которых клетки-хозяева имеют стенки, клетки-хозяева могут подвергаться механическому, химическому или ферментному разрыванию для высвобождения экспрессированного антитела секвестрированного в клетках. Неограничивающими примерами способов очистки антител могут быть ионообменная хроматография, осаждение солями и гель-фильтрация. В других вариантах воплощения применяют аффинную хроматографию. Например, антитела мыши, распознающие последовательности константной области человека, могут быть зафиксированы на очистительных колонках. В альтернативном варианте могут быть экспрессированы антитела, слитые с эпитопными метками или большими аффинными метками, такими, как мальтозосвязывающий белок, глутатион-8-трансфераза и тиоредоксин, для очистки специфическими антителами или другими молекулами, которые тесно связываются с аффинной меткой. После этого эпитопы или аффинные метки расщепляют, применяя способы, известные специалистам в данной области, и антитела, очищенные с применением других способов, таких как описанные авторами. Также возможны другие способы очистки антител от сред и клеток-хозяев. Антитела также могут подвергаться дополнительным этапам обработки для дальнейшей очистки антител в соответствии с надлежащей производственной практикой или другими нормативными требованиями, в зависимости от конкретного случая. Подходящие способы очистки и этапы их осуществления могут быть определены специалистами в данной области.
Трансгенные животные и растения
[000128] Антитела согласно настоящему изобретению также могут вырабатываться в генетически модифицированных отличных от человека животных или растениях. Экспрессия антител в таких организмах может быть конститутивной или индуцируемой. Антитела, которые экспрессируются в таких организмах, затем могут быть выделены с применением способов, известных специалистам в данной области. Способы экспрессии антител и их антигенсвязывающих фрагментов в трансгенных отличных от человека организмах хорошо известны специалистам в данной области. В неограничивающем примере антитела согласно настоящему изобретению могут вырабатываться и извлекаться из молока коз, коров или других отличных от человека млекопитающих. См., например, Патенты США №№5,827,690, 5,756,687, 5,750,172 и 5,741,957, включенные в это описание путем ссылки. Другими неограничивающими примерами трансгенных млекопитающих, у которых могут вырабатываться антитела, могут быть мыши, крысы, овцы, свиньи или лошади. Дополнительным неограничивающим примером жидкости организма, из которой могут быть выделены антитела, является кровь. Возможны также другие жидкости организма. Антитела согласно настоящему изобретению также могут вырабатываться в растениях и извлекаться из них. См., например, Патенты США №№6,417,429, 6,046,037 и 5,959, 177, включенные в это описание путем ссылки.
Фармацевтические композиции
[000129] Для применения в соответствии со способами терапии и профилактики согласно настоящему изобретению описанные авторами антитела рецептируют в форме композиций. Необязательно композиции могут включать один или несколько дополнительных агентов, включая антитела, связывающиеся с эпитопами GDF-8, отличающимися от описываемых авторами, которые являются терапевтически или профилактически эффективными против опосредованных GDF-8 нарушений. Композиции, как правило, поставляются в составе стерильной фармацевтической композиции, которая обычно включает фармацевтически приемлемый носитель. Эта композиция может существовать в любой приемлемой форме в зависимости от нужного способа ее введения пациенту.
[000130] Антитела согласно настоящему изобретению вводят субъекту различными путями, как правило, парентерально, например, путем подкожной, внутривенной, внутрибрюшинной или внутримышечной инъекции. Введение может выполняться одной или несколькими болюсными инъекциями или одной или несколькими инфузиями. Также возможны другие способы введения в соответствии с информацией, которой располагают специалисты в данной области. Наиболее подходящий способ введения в любом конкретном случае может зависеть от конкретной вводимой композиции и характеристик субъекта, таких как нарушение, подлежащее лечению, возраст или пол.
[000131] Фармацевтические композиции для удобства могут быть представлены в форме стандартных доз, содержащих заданное количество антитела на дозу. Такая доза может содержать, например, от 5 мг до 5 г, от 10 мг до 1 г или от 20 до 50 мг. Фармацевтически приемлемые носители для применения согласно изобретению могут быть предусмотрены в различных формах, в зависимости, например, от способа введения.
[000132] Фармацевтические композиции согласно изобретению могут быть приготовлены для хранения в форме лиофилизированных композиций или водных растворов путем смешивания антитела, имеющего достаточную степень чистоты, с необязательными фармацевтически приемлемыми носителями, вспомогательными веществами или стабилизаторами, которые обычно применяются в данной области (под общим названием "носители"), т.е., буферными агентами, стабилизаторами, консервантами, изотонирующими добавками, неионными детергентами, антиоксидантами и разными другими добавками. См. Remington's Pharmaceutical Sciences, 16th edition (Osol, ed. 1980). Такие добавки должны быть нетоксичными для потребителя в применяемых дозах и концентрациях.
[000133] Буферные агенты способствуют поддержанию уровня рН в диапазоне, приближенном к физиологическим условиям. Они могут присутствовать в концентрации от приблизительно 2 мМ до приблизительно 50 мМ. К подходящим буферным агентам для применения согласно настоящему изобретению относятся органические и неорганические кислоты и их соли, такие как цитратные буферы (например, смесь мононатрийцитрата -динатрийцитрата, смесь лимонной кислоты - тринатрийцитрата, смесь лимонной кислоты - мононатрийцитрата и т.п.), сукцинатные буферы (например, смесь янтарной кислоты - мононатрийсукцината, смесь янтарной кислоты - гидроксида натрия, смесь янтарной кислоты - динатрийсукцината и т.п.), тартратные буферы (например, смесь винной кислоты - тартрат натрия, смесь винной кислоты - тартрата калия, смесь винной кислоты -гидроксида натрия и т.п.), фумаратные буферы (например, смесь фумаровой кислоты - мононатрийфумарата, смесь фумаровой кислоты - динатрийфумарата, смесь мононатрийфумарата - динатрийфумарата и т.п.), глюконатные буферы (например, смесь глюконовой кислоты - глюконата натрия, смесь глюконовой кислоты - гидроксида натрия, смесь глюконовой кислоты - глюконата калия и т.п.), оксалатный буфер (например, смесь оксалатной кислоты - оксалата натрия, смесь оксалатной кислоты - гидроксида натрия, смесь оксалатной кислоты - оксалата калия и т.п.), лактатные буферы (например, смесь молочной кислоты - лактата натрия, смесь молочной кислоты - гидроксида натрия, смесь молочной кислоты - лактата калия и т.п.) ацетатные буферы (например, смесь уксусной кислоты - ацетата натрия, смесь уксусной кислоты - гидроксида натрия и т.п.). Дополнительно могут применяться фосфатные буферы, гистидиновые буферы и триметиламиновые соли, такие как Tris.
[000134] Консерванты добавляют для сдерживания роста микробов, и обычно их добавляют в количестве 0,2%-4% (масса/объем). К подходящим консервантам для применения согласно настоящему изобретению относятся фенол, бензиловый спирт, мета-крезол, метилпарабен, пропилпарабен, хлорид октадецилдиметилбензиламмония, галогениды бензалкония (например, хлорид, бромид и иодид), хлорид гексаметония и алкилпарабены, такие как метил- или пропилпарабен, катехол, резорцин, циклогексанол и 3-пентанол. Вещества, поддерживающие изотоничность, которые иногда называют "стабилизаторами", могут добавляться для обеспечения изотоничности жидких композиций согласно настоящему изобретению, и к ним относятся многоатомные сахароспирты, например, трех- или более атомные сахароспирты, такие как глицерин, эритрит, арабит, ксилит, сорбит и маннит. Под стабилизаторами подразумевается широкая категория вспомогательных веществ с разнообразными функциями, от объемообразующего агента до добавки, которая солюбилизирует терапевтическое средство или предотвращает денатурацию или прилипание к стенкам сосуда. Типичными стабилизаторами являются многоатомные сахароспирты (перечисленные выше); аминокислоты, такие как аргинин, лизин, глицин, глутамин, аспарагин, гистидин, аланин, орнитин, L-лейцин, 2-фенилаланин, глутаминовая кислота, треонин и т.п., органические сахара или сахароспирты, такие как лактоза, трегалоза, стахиоза, маннит, сорбит, ксилит, рибит, миоинозит, галактит, глицерин и т.п., включая циклиты, такие как инозит; полиэтиленгликоль; аминокислотные полимеры; серосодержащие восстановители, такие как мочевина, глутатион, липоевая кислота, тиогликолят натрия, тиоглицерин, α-монотиоглицерин и тиосульфат натрия; низкомолекулярные полипептиды (например, пептиды из 10 или менее остатков); белки, такие как альбумин сыворотки человека, альбумин сыворотки крупного рогатого скота, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; моносахариды, такие как ксилоза, манноза, фруктоза, глюкоза; дисахариды, такие как лактоза, мальтоза, сахароза, и трисахариды, такие как раффиноза; и полисахариды, такие как декстран. Стабилизаторы могут присутствовать в количестве от 0,1 до 10000 массовых частей от массы активного белка.
[000135] Неионные поверхностно-активные вещества или детергенты (также известные как "смачивающие вещества") добавляют для обеспечения солюбилизации антител и защиты любых других терапевтических средств, которые могут быть включены, от агрегации, вызванной взбалтыванием, что также позволяет подвергать композицию поверхностному напряжению сдвига, не вызывая денатурации белка. К подходящим неионным поверхностно-активным веществам относятся полисорбаты (20, 80 и т.п.), полиоксамеры (184, 188 и т.п.), полиолы Pluronic®, моноэфиры полиоксиэтиленсорбита (lWEEN®-20, lWEEN®-80 и т.п.). Неионные поверхностно-активные вещества могут присутствовать в количестве от приблизительно 0,05 мг/мл до приблизительно 1,0 мг/мл, например, от приблизительно 0,07 мг/мл до приблизительно 0,2 мг/мл.
[000136] К разным дополнительным вспомогательным веществам относятся хелатирующие вещества (например, EDTA), антиоксиданты (например, аскорбиновая кислота, метионин, витамин Е) и сорастворители.
[000137] В неограничивающих примерах воплощения антитела согласно изобретению рецептируют в растворе, содержащем 20 мМ L-гистидина, 85 мг/мл сахарозы, 0,2 мг/мл PS-80, 0,05 мг/мл EDTA при рН 5,8. В других вариантах воплощения концентрация антитело в этой композиции составляет 100 мг/мл. В других вариантах воплощения антитело в этой композиции является лиофилизированным.
Фармацевтические комплекты
[000138] В некоторых вариантах воплощения изобретение обеспечивает фармацевтические комплекты для применения клиническими врачами или другими специалистами. Фармацевтический комплект представляет собой упаковку, включающую антитело против GDF-8 согласно изобретению (например, в лиофилизированной форме или в форме водного раствора) и один или несколько следующих компонентов: как минимум второе терапевтическое средство, как описывается в других разделах этого описания; устройство для введения антитела, например, иглу и/или шприц; и вода или буфер фармацевтической степени чистоты для повторного суспендирования или разведения антитела, если антитело предусмотрено в лиофилизированной или концентрированной форме. Комплекты также могут включать инструкции по приготовлению композиции антитела и/или введению композиции пациенту.
[000139] Каждая стандартная доза композиции антитела против GDF-8 может быть упакована отдельно, и комплект может содержать одну или несколько стандартных доз (например, две стандартных дозы, три стандартных дозы, четыре стандартных дозы, пять стандартных доз, семь стандартных доз, восемь стандартных доз, десять стандартных доз или более). В одном варианте воплощения одну или несколько стандартных доз помещают в шприц, а в другом варианте воплощения одна или несколько стандартных доз по отдельности содержатся в пакете или другом подобном вместилище, подходящем для подсоединения к системе для внутривенного введения.
Способы лечения и профилактики
[000140] Настоящее изобретение обеспечивает способы лечения и профилактики состояний и нарушений, при которых снижение активности GDF-8 прямо или косвенно приносит терапевтический эффект.Такие способы включают введение субъекту эффективного количества композиции, включающей антитело против GDF-8. В некоторых из этих вариантов воплощения антитело представляет собой OGD1.0.0 или его связывающиеся с GDF-8 фрагменты, части, составляющие или производные.
[000141] Субъектом, которому могут вводиться композиции антитела против GDF-8, может быть млекопитающее, например, отличное от примата (например, корова, свинья, лошадь, кошка, собака, крыса и т.п.), или примат (например, макаки, шимпанзе, другие человекообразные обезьяны или человек). Субъектом может быть человек, например, взрослый пациент или ребенок.
[000142] Состояния и расстройства, которые поддаются лечению с применением композиций антител согласно настоящему изобретению, являются те, которые опосредуются, как минимум частично, GDF-8, или при которых существуют научные основания предполагать, что снижение активности GDF-8 у субъекта должно обеспечить терапевтический эффект.
[000143] Хотя терапевтический эффект частично зависит от конкретного состояния или расстройства, терапевтический эффект возникает тогда, когда снижение активности GDF-8 у субъекта приводит к ослаблению симптомов, признаков или тяжести состояния или расстройства или прекращению или замедлению прогрессирующего ухудшения таких симптомов, признаков или тяжести. Терапевтический эффект также возникает тогда, когда снижение активности GDF-8 повышает продолжительность жизни, комфорт или качество жизни субъекта. Терапевтический эффект также возникает тогда, когда снижение активности GDF-8 улучшает или прекращает или замедляет ухудшение одной или нескольких физических или физиологических функций субъекта или показателей субъекта при тестах, отражающих такие функции.
[000144] Вывод о терапевтическом эффекте может быть сделан по результатам наблюдения субъекта, выполняющего определенные задачи, ответов субъекта на вопросы о том, что он (она) ощущает, или выполнения одного или нескольких тестов на субъекте в клинике или в лаборатории на образцах, полученных от субъекта. Терапевтический эффект также может подтверждаться маркерами ингибирования GDF-8. В качестве неограничивающего примера, мышцы субъекта, подлежащего лечению, могут быть подвергнуты биопсии и анализу на наличие или отсутствие маркеров, связанных с понижающей регуляцией пути трансдукции сигнала, стимулируемого GDF-8, например, снижением уровня фосфорилированного белка SMAD2 или SMAD3. Другие тесты, применимые для обнаружения терапевтического эффекта у субъекта, подвергающегося лечению композициями, включающими антитела согласно настоящему изобретению, известны специалистам в данной области. Полное излечение или реверсия состояния или расстройства, поддающегося лечению или профилактике, хотя и желательно, однако не является необходимым для возникновения терапевтического эффекта.
[000145] В некоторых вариантах воплощения композиции, включающие антитела согласно настоящему изобретению, применяют для лечения или профилактики состояний или нарушений, характеризующихся потерей массы и/или силы скелетных мышц, или при которых увеличение такой мышечной массы и/или силы обеспечивает терапевтический эффект. В некоторых из этих вариантов воплощения антитело представляет собой OGD1.0.0 или его связывающиеся с GDF-8 фрагменты или производные.
[000146] К состояниям или расстройствам, которые приводят к уменьшению мышечной массы и/или силы, и которые поддаются лечению или профилактике путем введения описанных авторами антител, помимо прочих, относятся, возрастная потеря мышечной массы или силы, слабость, саркопения и потеря мышечной массы или силы, вызываемая мышечной атрофией, неподвижностью или бездействием, например, после повреждения, денервации или длительного пребывания в среде с нулевой гравитацией. В других вариантах воплощения к состояниям или расстройствам, которые поддаются лечению или профилактике, относятся переломы костей, в частности, у пожилых людей и других людей, подверженных переломам костей, например, перелому шейки бедра, или переломам других костей, или стабилизации замены сустава. В некоторых других вариантах воплощения состояниями или расстройствами, которые поддаются лечению или профилактике, являются синдромы мышечной атрофии, включая те, которые обусловлены процессом первичного заболевания. Неограничивающими примерами синдромов мышечной атрофии могут быть кахексия, например, вызываемая раком, анорексия или другие типы неполноценного питания и мышечная атрофия, вызываемая СПИД, сепсисом, ожогами, хронической почечной недостаточностью, застойной сердечной недостаточностью (CHF) и хроническим обструктивным заболеванием легких (COPD).
[000147] В качестве неограничивающего примера, терапевтический эффект у субъектов, получающих композицию, включающую антитела согласно настоящему изобретению, может проявляться через увеличение массы или силы мышц в целом или конкретных мышц. Неограничивающими примерами мышц, масса и/или сила которых может увеличиваться благодаря лечению антителами против GDF-8, могут быть скелетные мышцы и сердечная мышца. Другими примерами могут быть мышцы, контролирующие дыхание, включая диафрагму и межреберные мышцы, а также вспомогательные дыхательные мышцы, включая грудино-ключично-сосцевидные лестничные мышцы и другие. Другими примерами скелетных мышц могут быть икроножная мышца, задняя большеберцовая мышца, камбаловидная мышца, передняя большеберцовая мышца, длинная, короткая, большая ягодичная мышца, бицепс бедра, полусухожильная мышца, полуперепончатая мышца, подвздошно-поясничная мышца, четырехглавая мышца бедра, приводящие мышцы бедра, мышца, поднимающая лопатку, трапециевидная мышца, прямая мышца живота, поперечная мышца живота, наружная косая мышца живота, внутренняя косая мышца живота, распрямляющая мышца спины, большая грудная мышца, двуглавая мышца плеча, трехглавая мышца плеча, брахиалис, круглый пронатор, плечелучевая мышца, ромбовидная мышца, дельтовидная мышца и широчайшая мышца спины. Также возможны и другие скелетные мышцы, масса и/или сила которых может быть увеличена благодаря лечению антителами против GDF-8 согласно настоящему изобретению.
[000148] Увеличение мышечной массы или силы определяют непосредственно, например, путем наблюдения способности субъекта к сопротивлению силе или поднятию веса, или косвенно, например, путем сканирования тела субъекта с применением MPT, КТ или двухэнергетической рентгеновской абсорбциометрии (DEXA). Возможны также и другие способы.
[000149] В альтернативном варианте вывод о терапевтическом эффекте может быть сделан по уменьшению тяжести симптомов, которые иначе бы неуклонно ухудшались. Улучшение также может быть продемонстрировано путем применения физиологических тестов функции мышц, таких как электромиография, гистологические тесты взятой путем биопсии мышечной структуры и биохимические тесты, например, на наличие в сыворотке креатинкиназы, фермента, высвобождаемого поврежденной мышцей. Возможны также и другие тесты мышечной структуры и функции, применимые для обнаружения терапевтического эффекта.
[000150] В других вариантах воплощения, касающихся мышечной массы и/или силы, настоящее изобретение обеспечивает способы лечения и профилактики мышечной дистрофии ("MD") путем введения пациентам, нуждающимся в таком лечении или профилактике, композиции, включающей антитела против GDF-8. В некоторых вариантах воплощения способов антитело представляет собой OGD 1.0.0 или его связывающиеся с GDF-8 фрагменты или производные. Согласно некоторым вариантам воплощения, субъектами являются дети, страдающие от мышечной дистрофии, а в других вариантах воплощения субъектами являются взрослые пациенты с мышечной дистрофией.
[000151] Как известно специалистам в данной области, существуют разные типы мышечной дистрофии, отличающиеся по характеру генетического повреждения или повреждений, отвечающих за болезнь, и фенотипу, который является результатом лежащих в основе генетических дефектов. Неограничивающими примерами типов мышечной дистрофии, которая поддается лечению или профилактике путем введения композиций, включающих антитела согласно настоящему изобретению, могут быть мышечная дистрофия Дюшенна (DMD) (также известная как мышечная псевдогипертрофическая дистрофия), мышечная дистрофия Беккера (BMD), мышечная дистрофия Эмери-Дрейфуса (EDMD), мышечная дистрофия Лейдена-Мебиуса (LGMD), плече-лопаточно-лицевая мышечная дистрофия (FSH или FSHD) (также известная как мышечная дистрофия Ландузи-Дежерина), миотоническая дистрофия (MMD) (также известная как DM или болезнь Штейнерта), окулофарингеальная мышечная дистрофия (OPMD), дистальная мышечная дистрофия (DD) (также известная как мышечная дистрофия Миоши) и врожденная мышечная дистрофия (CMD).
[000152] Помимо описанных выше способов оценки улучшений мышечной массы и/или силы, терапевтический эффект введения композиций, включающих антитела согласно настоящему изобретению, субъектам с MD, поддается количественному определению с применением теста с шестиминутной ходьбой ("6MWT"). См., например, публикацию McDonald, et al., The 6-minute walk test as a new outcome measure in Duchenne muscular dystrophy, Muscle Nerve (2010) 41:500-510, включенную в это описание путем ссылки.
[000153] При 6MWT субъектов тестируют, чтобы определить, сколько они способны пройти за 6 минут по заданному курсу. Как правило, субъекта тестируют перед началом лечения для определения исходного уровня, а затем с интервалами в процессе лечения. Терапевтический эффект наблюдается тогда, когда показатели субъекта с MD при 6MWT остаются неизменными или фактически улучшаются благодаря лечению, или, в альтернативном варианте, когда показатели субъекта не снижаются настолько быстро, как у среднего субъекта, не подвергаемого лечению. Неограничивающими примерами улучшений в показателях при 6MWT могут быть процентные улучшения приблизительно на 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 60%, 70%, 80%, 90%, 100% или более по сравнению с контрольными субъектами, не подвергаемыми лечению или получающими плацебо. 6MWT также применяют для обнаружения терапевтического эффекта у субъектов, получающих лечение от состояний или нарушений, влияющих на способность передвигаться, отличных от MD.
[000154] В других вариантах воплощения настоящее изобретение обеспечивает способы лечения и профилактики заболеваний двигательных нейронов путем введения пациентам, нуждающимся в таком лечении или профилактике, композиции, включающей антитела против GDF-8. В некоторых вариантах воплощения способов антитело представляет собой OGD1.0.0 или его связывающиеся с GDF-8 фрагменты или производные. Неограничивающими примерами типов заболеваний двигательных нейронов, которые поддаются лечению или профилактике путем введения композиций, включающих антитела согласно настоящему изобретению, могут быть амиотрофический латеральный склероз (ALS) (также известный как болезнь Лу Герига), спинальная мышечная атрофия детского возраста I типа (SMA1) (также известная как синдром Верднига-Гоффманна), спинальная мышечная атрофия II типа (SMA2), спинальная мышечная атрофия III типа (SMA3) (также известная как синдром Кугельберга-Веландера) и спинальная бульбарная мышечная атрофия (SBMA) (также известная как синдром Кеннеди).
[000155] В предыдущих исследованиях было продемонстрировано, что мышиное антитело против GDF-8 является эффективным для увеличения мышечной массы и силы у мышей и крыс с SOD1, которые являются малыми животными моделями человеческого ALS. См. WO 2007/024535 и публикацию Holzbauer, et al., Myostatin inhibition slows muscle atrophy in rodent models of amyotrophic lateral sclerosis, Neurobiology of Disease (2006) 23:697-707, включенные в это описание путем ссылки. Как сообщалось, лечение мышиным антителом увеличивает мышечную массу в диафрагмальных и скелетных мышцах мышей и крыс с SOD1 по сравнению с контрольными животными, получавшими PBS. Подобным образом лечение антителом снижает мышечную атрофию икроножной мышцы и диафрагмы у мышей с SOD1 по сравнению с контрольными животными, получавшими PBS. Трофические эффекты лечения антителом в принципе были очевидны во время ранних стадий развития болезни в отличие от конечной стадии, хотя сдерживание атрофии диафрагмы было очевидным в оба периода. Также наблюдалось, что лечение антителом увеличивает силу мышц конечностей, а также общую массу тела у мышей и крыс с SOD1, хотя антитело не продлевало срок выживания по сравнению с контрольными животными, получавшими лишь носитель. Поскольку гуманизированные антитела против GDF-8 согласно настоящему изобретению, такие как OGD1.0.0, имеют общие антигенсвязывающие детерминанты с обсуждавшимся выше мышиным антителом, ожидается, что они также должны быть эффективны для лечения или профилактики ALS у человека.
[000156] Другие врожденные или приобретенные болезни и расстройства мышц, центральной нервной системы и периферической нервной системы, влияющие на мышечную массу, функцию и/или силу, также поддаются лечению или профилактике путем введения субъектам, которые в этом нуждаются, композиций, включающих антитела согласно настоящему изобретению.
[000157] Настоящее изобретение также обеспечивает способы лечения и профилактики нарушений обмена веществ путем введения пациентам, нуждающимся в лечении от нарушений обмена веществ, композиции, включающей антитела против GDF-8. В некоторых из этих вариантов воплощения антитело представляет собой OGD1.0.0 или его связывающиеся с GDF-8 фрагменты или производные.
[000158] Неограничивающими примерами нарушений обмена веществ, которые поддаются лечению или профилактике путем введения антител согласно настоящему изобретению, могут быть сахарный диабет 2 типа, метаболические синдромы, такие как синдром X, резистентность к инсулину т нарушение толерантности к глюкозе.
[000159] В других вариантах воплощения настоящее изобретение обеспечивает способы лечения и профилактики расстройства жировой ткани путем введения пациентам, нуждающимся в лечении от таких нарушений, композиции, включающей антитела против GDF-8. В некоторых из этих вариантов воплощения антитело представляет собой OGD1.0.0 или его связывающиеся с GDF-8 фрагменты или производные.
[000160] Неограничивающими примерами расстройства жировой ткани, которые поддаются лечению или профилактике путем введения антител согласно настоящему изобретению, могут быть ожирение и более высокий по сравнению с нормой индекса массы тела (ΒΜΙ) для пола, возраста и роста конкретного субъекта.
[000161] В других вариантах воплощения настоящее изобретение обеспечивает способы лечения и профилактики нарушений, связанных с потерей костной массы, путем введения пациентам, нуждающимся в лечении от таких нарушений, композиции, включающей антитела против GDF-8. В некоторых из этих вариантов воплощения антитело представляет собой OGD 1.0.0, или его связывающиеся с GDF-8 фрагменты или производные.
[000162] Неограничивающими примерами связанных с потерей костной массы нарушений, которые поддаются лечению или профилактике путем введения антител согласно настоящему изобретению, могут быть остеопороз, гормональный остеопороз, остеопения, остеоартрит и связанные с остеопорозом переломы.
Комбинированная терапия
[000163] Согласно некоторым вариантам воплощения способов согласно настоящему изобретению, антитела против GDF-8 могут вводиться в композиции в качестве монотерапии или комбинированной терапии с как минимум вторым терапевтическим средством. Как правило, хотя и не во всех случаях, второе терапевтическое средство выбирают для лечения или профилактики того же состояния или расстройства, против которого было направлено антитело против GDF-8. Однако в других вариантах воплощения второе средство может быть выбрано для лечения или профилактики другого состояния или расстройства. Дозы антитела и второго терапевтического средства для применения в комбинированной терапии выбирают в соответствии с данными, известными специалистам в данной области, для обеспечения максимальной эффективности и минимальных побочных эффектов.
[000164] Композиции антитела против GDF-8 согласно настоящему изобретению вводят субъекту с применением такого же или другого режима введения по сравнению со вторым терапевтическим средством. В зависимости от химических и физических характеристик антител против GDF-8 и второго терапевтического средства, они могут комбинироваться в одной композиции. В альтернативных вариантах воплощения их вводят в виде отдельных композиций. Композиции антител и вторых терапевтических средств для удобства могут быть включены в комплекты согласно настоящему изобретению.
[000165] При введении в рамках комбинированной терапии антитело и второе терапевтическое средство могут вводиться одновременно, последовательно или отдельно.
[000166] Одновременное введение осуществляют в случаях, когда два или более средств вводят в одно и то же время, даже при частичном совпадении времени введения соответствующих средств, но с разным временем начала и конца их введения. Последовательное введение осуществляют в случаях, когда два или более средств вводят субъекту в один день, например, во время одного визита в клинику, но не одновременно.
[000167] Последовательное введение может производиться с интервалами в 1, 2, 3, 4, 5, 6, 7, 8 или более часов. Композиция антитела против GDF-8 может быть введена первой, с последующим введением второго средства, или наоборот.
[000168] Отдельное введение осуществляют в случаях, когда средства вводят субъекту в разные дни. Типичные интервалы между отдельными введениями агентов могут составлять 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, одну неделю, 2 недели, 3 недели или месяц или более. Как и при последовательном введении, введение композиции антитела против GDF-8 может предшествовать или следовать за отдельным введением второго средства.
[000169] В некоторых других вариантах воплощения настоящего изобретения композиция антитела против GDF-8 и второе терапевтическое средство могут вводиться многократно в поочередном порядке, независимо от того, вводятся ли они последовательно или отдельно.
[000170] В соответствии со способами лечения или профилактики расстройства обмена веществ антитела против GDF-8 согласно настоящему изобретению могут сочетаться со вторым средством, эффективным для лечения или профилактики таких нарушений. Неограничивающими примерами вторых средств, эффективных для этой цели, могут быть метформин, сульфонилмочевины, инсулин, прамлинтид, тиазолидиндионы, такие как розиглитазон и пиоглитазон, аналоги GLP-1, такие как эксенитид и ингибиторы OPP-IV, такие как вилдаглиптин.
[000171] В соответствии со способами лечения или профилактики потери костной массы, антитела против GDF-8 согласно настоящему изобретению могут сочетаться со вторым средством, эффективным для лечения или профилактики таких нарушений. Неограничивающими примерами вторых средств, эффективных для этой цели, могут быть бисфосфонаты, такие как алендронат и ризедронат, кальцитонин, ралоксифен и гормональные средства, такие как эстроген или паратиреоидный гормон (РТН).
[000172] В соответствии со способами лечения или профилактики мышечной дистрофии, антитела против GDF-8 согласно настоящему изобретению могут сочетаться со вторым средством, эффективным для лечения или профилактики мышечной дистрофии, таким, как кортикостероид. Другие средства, эффективные для лечения или профилактики мышечной дистрофии известны специалистам в данной области. Неограничивающими примерами кортикостероидов, эффективных для лечения мышечной дистрофии, могут быть метилпреднизолон, дефлазакорт, бетаметазон, преднизолон, гидрокортизон, кортизон, беклометазон, будесонид, кортизол, дексаметезон, флутиказон, преднизон, мометазон, триамцинолон и их производные. В других вариантах воплощения антитела против GDF-8 могут водиться со средствами для лечения кардиомиопатии, которая часто случается у пациентов с DMD, в частности, пожилых пациентов с DMD. К таким средствам, помимо прочих, относятся бета-адреноблокаторы и ингибиторы ангиотензин-преобразующих ферментов.
[000173] В соответствии со способами лечения или профилактики ALS, антитела против GDF-8 согласно настоящему изобретению могут сочетаться со вторым средством, эффективным для лечения или профилактики ALS, включая, помимо прочих, рилузол, талампанел, гликопирролат, бензотропин, скополамин, атропин, тригексифенидил гидрохлорид, амитриптилин, флувоксамин, баклофен, тизанидин, дантролен, диазепам, хинин, фенитоин, бензодиазепины, габапентин, спазмолитические средства, антидепрессанты и морфин или другие болеутоляющие средства.
[000174] В соответствии с другими вариантами воплощения, композиции, включающие антитела против GDF-8 согласно настоящему изобретению, вводят в сочетании с немедикаментозными видами терапии, включая, в качестве неограничивающего примера, упражнения, физиотерапию, респираторную терапию, вспомогательное дыхание, кардиотерапию и питательные добавки.
Эффективные дозы
[000175] Как было описано выше, композиции, включающие антитела против GDF-8 согласно настоящему изобретению, вводят субъекту, нуждающемуся в лечении или профилактике некоторых состояний или нарушений, в дозе, эффективной для достижения, как минимум частичного, желаемого терапевтического эффекта.
[000176] Связывание всех GDF-8 не обязательно требуется для достижения терапевтической эффективности. Также эффективным может быть снижение концентрации зрелого, активного GDF-8 в жидкости организма, такой, как кровь или сыворотка, или в ткани организма, такой, как мышцы или другие ткани или органы организма.
[000177] В соответствии с информацией, которой располагают специалисты в данной области, доза композиции антитела против GDF-8 может титроваться для пациента таким образом, чтобы снизить концентрацию активного GDF-8 в соответствующей ткани или жидкости организма в заданное время после введения как минимум приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%, или приблизительно 5%-10%, приблизительно 10%-45%, приблизительно 15%-20%, приблизительно 20%-25%, приблизительно 25%-30%, приблизительно 30%-35%, приблизительно 35%-40%, приблизительно 40%-45%, приблизительно 45%-50%, приблизительно 50%-55%, приблизительно 55%-60%, приблизительно 60%-65%, приблизительно 65%-70%, приблизительно 70%-75%, приблизительно 75%-80%, приблизительно 80%-85%, приблизительно 85%-90%, приблизительно 90%-95%, приблизительно 95%-99%, или с процентным снижением концентрации активного GDF-8 между любыми из вышеуказанных значений.
[000178] Количество антитела против GDF-8, вводимого субъекту, зависит от различных факторов, включая состояние или нарушение, подлежащее лечению или профилактике, размер и массу субъекта, форму, путь и место введения, лечебный режим (например, наличие или отсутствие применения второго терапевтического средства), возраст и состояние конкретного субъекта, уровень активного GDF-8, обнаруженного в соответствующей ткани или жидкости организма вышеупомянутого субъекта перед началом лечения и реакции или чувствительности субъекта к воздействию композиции антитела. Соответствующая доза может легко определяться специалистом в данной области. Наконец, соответствующие дозы определяются клиническим врачом или другим специалистом, оказывающим медицинскую помощь. Частота введения дозы зависит от потребности. При возникновении побочных эффектов количество и/или частота введения доз может меняться или снижаться в соответствии с нормальной клинической практикой. Надлежащую дозу и режим лечения определяют путем наблюдения за ходом терапии с применением способов, известных специалистам в данной области.
[000179] Эффективные дозы сначала определяют по результатам анализов in vitro. Например, первоначальная доза для применения на животных может быть составлена для достижения концентрации антитела против GDF-8 в циркулирующей крови или сыворотке на уровне или выше уровня аффинности связывания антитела к GDF-8, измеряемой in vitro. Расчет доз для достижения такой концентрации антитела против GDF-8 в циркулирующей крови или сыворотке с учетом биодоступности конкретного антитела может быть произведен специалистом в данной области средней квалификации. С наставлениями можно ознакомиться в Части 1 "Общие принципы" публикации "Goodman and Gilman's The Pharmacological Basis of Therapeutics," 11th Ed., Hardman, J. G., et al., Eds., McGraw-Hill Professional, и в приведенных в ней источниках. Первоначальные дозы также определяют по in vivo данным, например, по животным моделям. Специалисты в данной области смогут традиционным способом использовать эту информацию для определения доз, подходящих для введения человеку.
[000180] В некоторых вариантах воплощения дозу определяют для конкретного субъекта путем измерения концентрации активного GDF-8 в сыворотке, мышцах или другой соответствующей жидкости или ткани организма определенное количество раз в течение нескольких дней или недель, предшествующих введению композиции антитела, для расчета количества антитела против GDF-8, которое было бы насыщающим, т.е., количества, которое было бы достаточным для связывания по сути всего активного GDF-8. Как станет понятно специалистам в данной области, количество любого специфичного антитела, необходимое для достижения насыщения для данного количества GDF-8 в сыворотке, мышцах или где-либо еще, частично зависит от аффинности конкретного антитела к GDF-8. Способы расчета насыщающего количества для специфичных антител против GDF-8 с учетом фармакокинетических свойств и биодоступности конкретного антитела в случае необходимости, хорошо известны специалистам в данной области. Для обеспечения насыщения вводят количество, превышающее расчетное насыщающее количество, например, как минимум в 2, 3, 4, 5, 6, 7, 8, 9 или даже 10 превышающее расчетное насыщающее количество.
[000181] Эффективная доза композиции антитела против GDF-8 может составлять от приблизительно 0,01 мг/кг до приблизительно 250 мг/кг для одного (например, болюсного) введения, нескольких введений или непрерывного (например, инфузионного) введения или соответствует любому эффективному диапазону или значению в зависимости от таких факторов, как, например, состояние или нарушение, подлежащее лечению или профилактике, и т.п., как обсуждалось выше.
[000182] В некоторых вариантах воплощения каждая доза может составлять от приблизительно 0,1 мг/кг до приблизительно 0,5 мг/кг; от приблизительно 0,25 мг/кг до приблизительно 0,75 мг/кг; от приблизительно 0,5 мг/кг до приблизительно 1 мг/кг; приблизительно 2 мг/кг; от приблизительно 1,5 мг/кг до приблизительно 2,5 мг/кг; от приблизительно 2 мг/кг до приблизительно 3 мг/кг; от приблизительно 2,5 мг/кг до приблизительно 3,5 мг/кг; от приблизительно 3 мг/кг до приблизительно 4 мг/кг; от приблизительно 3,5 мг/кг до приблизительно 4,5 мг/кг; от приблизительно 4 мг/кг до приблизительно 5 мг/кг; от приблизительно 5 мг/кг до приблизительно 7 мг/кг; от приблизительно 6 мг/кг до приблизительно 8 мг/кг; от приблизительно 7 мг/кг до приблизительно 9 мг/кг; от приблизительно 8 мг/кг до приблизительно 10 мг/кг; от приблизительно 10 мг/кг до приблизительно 15 мг/кг; от приблизительно 12,5 мг/кг до приблизительно 17,5 мг/кг; от приблизительно 15 мг/кг до приблизительно 20 мг/кг; от приблизительно 17,5 мг/кг до приблизительно 22,5 мг/кг; от приблизительно 20 мг/кг до приблизительно 25 мг/кг; от приблизительно 22,5 мг/кг до приблизительно 27,5 мг/кг; от приблизительно 25 мг/кг до приблизительно 30 мг/кг; от приблизительно 30 мг/кг до приблизительно 40 мг/кг; от приблизительно 35 мг/кг до приблизительно 45 мг/кг; от приблизительно 40 мг/кг до приблизительно 50 мг/кг; от приблизительно 45 мг/кг до приблизительно 55 мг/кг; от приблизительно 50 мг/кг до приблизительно 60 мг/кг; от приблизительно 55 мг/кг до приблизительно 65 мг/кг; от приблизительно 60 мг/кг до приблизительно 70 мг/кг; от приблизительно 65 мг/кг до приблизительно 75 мг/кг; от приблизительно 70 мг/кг до приблизительно 80 мг/кг; от приблизительно 75 мг/кг до приблизительно 85 мг/кг; от приблизительно 80 мг/кг до приблизительно 90 мг/кг; от приблизительно 85 мг/кг до приблизительно 95 мг/кг; от приблизительно 90 мг/кг до приблизительно 100 мг/кг; от приблизительно 95 мг/кг до приблизительно 105 мг/кг; от приблизительно 100 мг/кг до приблизительно 150 мг/кг; от приблизительно 125 мг/кг до приблизительно 175 мг/кг; от приблизительно 150 мг/кг до приблизительно 200 мг/кг; от приблизительно 175 мг/кг до приблизительно 225 мг/кг; от приблизительно 200 мг/кг до приблизительно 250 мг/кг. Возможны также другие диапазоны доз.
[000183] Количество, частота и длительность введения зависят от различных факторов, таких как возраст, вес и состояние субъекта. Таким образом, в неограничивающих примерах терапевтический режим введения может охватывать 1 день или более, 2 дня или более, 3 дня или более, 4 дня или более, 5 дней или более, 6 дней или более, 1 неделю или более, от 2 недели до неопределенного времени, от 2 недель до 6 месяцев, от 3 месяцев до 5 лет, от 6 месяцев до 1 или 2 лет, от 8 месяцев до 18 месяцев или другие подобные периоды. Необязательно терапевтический режим предусматривает многократное введение, например, дважды в день, раз в день, раз в два дня, три дня, четыре дня, пять дней, шесть дней, раз в неделю, раз в две недели или раз в месяц. Многократное введение может производиться одинаковыми или разными дозами. Введение может повторяться один раз, два раза, три раза, четыре раза, пять раз, шесть раз, семь раз, восемь раз, девять раз, десять раз или более. Терапевтически эффективное количество композиции антитела против GDF-8 вводят единой дозой или курсом в терапевтическом режиме, например, в течение недели, двух недель, трех недель, одного месяца, трех месяцев, шести месяцев, одного года или дольше. Эффективная доза композиции антитела против GDF-8 для конкретного субъекта может со временем меняться с изменениями в состоянии субъекта или с возникновением других проблем со здоровьем.
ПРИМЕРЫ
ПРИМЕР 1
Анализ временной экспрессии OGD1.0.0 и OGD1.1.1
[000184] Временную экспрессию интактных гетеротетрамерных OGD1.1.1 и OGD1.0.0 испытывали в клетках COS-1 М6, и было продемонстрировано, что OGD1.0.0 экспрессируется на значительно более высоком уровне.
[000185] В общих чертах, ДНК, кодирующую VH0 и VH1 (SEQ ID NO: 49 и 53, соответственно) клонировали в векторы экспрессии IgG млекопитающих, таким образом, чтобы каждая из VH областей соединялась в рамке с нуклеотидной последовательностью, кодирующей константные тяжелые области человеческого IgG1, включая три мутации, устраняющие эффекторную функцию (SEQ ID NO: 57), таким образом, чтобы экспрессировалась тяжелая цепь антитела полной длины, включающая VH0 или VH1. Подобным образом ДНК, кодирующую VL0 и VL1 (SEQ ID NO: 51 и 55, соответственно) клонировали в векторы экспрессии IgG млекопитающих, таким образом, чтобы каждая из VL областей соединялась в рамке с нуклеотидной последовательностью, кодирующей каппа-константную легкую область человека SEQ ID NO: 17, таким образом, чтобы экспрессировалась легкая цепь антитела полной длины, включающая VL0 или VL1.
[000186] После создания векторов экспрессии приготавливали максипреп-ДНК с применением стандартных способов. Клетки помещали в 100 мм чашки Петри, а затем подвергали временной котрансфекции векторами экспрессии тяжелой и легкой цепей (т.е., VH0 и VL0, комбинированными в одной чашке, и VH1 и VL1, комбинированными в другой чашке). Реагент для трансфекции TranslT (Minis MIR2306) (40 мкл) добавляли к 2 мл среды для роста OptiMEM и глутамину (конечная концентрация 2 мМ) при комнатной температуре, перемешивали вихревым способом, а затем инкубировали при комнатной температуре в течение 15 минут. Максипреп-ДНК (по 8 мкг ДНК тяжелой и легкой цепей) добавляли к смеси и инкубировали при комнатной температуре в течение 15 минут. Раствор для трансфекции затем добавляли в чашки Петри, содержащие 8 мл среды для роста (DMEM, HIFBS, пенициллин, стрептомицин, глутамин). После инкубации в течение 24 часов при 37°С в 10% СО2 клетки промывали бессывороточной средой для роста R1CD1, а затем выращивали в течение 48 часов при 37°С в 10% СО2 в 10 мл R1CD1 (с добавлением пенициллина, стрептомицина, глутамина). Кондиционированную среду удаляли из клеток, центрифугировали для осаждения любого дебриса, и супернатант переносили в новую пробирку.
[000187] Концентрацию антитела, выработанного временно трансфицированными клетками COS-1, определяли с применением специфичного к полностью человеческому IgG-Fc ELISA-анализа. В общих чертах, на планшет с плоским дном для ELISA-анализа наносили IgG козы против человека (Pierce 31125) путем добавления 100 мкл антитела в PBS (1 мкг/мл) в каждую лунку и инкубировали в течение суток при комнатной температуре. Планшеты блокировали с использованием 100 мкл/лунку 0,02% раствора казеина в PBS в течение периода от 3 до 24 часов при комнатной температуре, а затем промывали. Стандарт и образцы последовательно разбавляли в аналитическом буфере (0,5% BSA, 0,02% Tween-20 в PBS), распределяли по планшету для ELISA (100 мкл/лунку) и инкубировали в течение периода от 3 до 24 часов при комнатной температуре. После промывания IgG козы против человека (Pierce 31413), разбавленное 1:5000 в аналитическом буфере, распределяли (100 мкл/лунку) и планшет инкубировали в течение 15 минут при комнатной температуре. После промывания планшет проявляли путем добавления BioFX ТМВ (TMBW-0100-01) (100 мкл/лунку). После прекращения реакции добавлением 0,18 N H2S04 (100 мкл/лунку) планшет считывали при 450 нМ с применением планшет-ридера Molecular Devices vMax. Концентрацию образцов рассчитывали с использованием линейного участка кривой, определяемого по серии разбавлений стандарта.
[000188] Результаты эксперимента с временной трансфекцией показаны ниже в таблице, в которой POI означает соответствующий пик при эксклюзионной хроматографии (SEC) после очистки белком Α. POI представляет пропорцию интактного антитела полного размера, экспрессируемого клетками, в отличие от высокомолекулярных агрегатов или продуктов распада.
[000189] Неожиданно было обнаружено, что антитело OGD1.0.0 экспрессировалось на гораздо более высоком уровне (т.е., с более, чем 10-кратным повышением) по сравнению с антителом OGD1.1.1 в тех же условиях после временной трансфекции. Важно отметить, что, как указывает значение POI, сильно увеличенные наблюдаемые показатели экспрессии почти полностью связаны с интактным антителом полной длины в отличие от высокомолекулярных комплексов или продуктов распада. Эта разница в экспрессии еще более удивительна, если учитывать, что между OGD1.0.0 и OGD1.1.1 существует разница лишь в одну аминокислоту в позиции 108 по Kabat VH областей (т.е., номер остатка 111 SEQ ID NO: 44 и 7) и разница в одну аминокислоту в позиции 100 по Kabat VL области (т.е., номер остатка 100 SEQ ID NO: 46 и 9). См. ФИГ. 1А и ФИГ. 1В.
[000190] Как объяснялось выше, эти структурные и функциональные различия объясняются использованием различных J-сегментов в VH0 и VL0 по сравнению с VH1 и VL1, соответственно. Как объясняется ниже, наиболее значимым различием, очевидно, является изменение в VH области. Следует отметить, что этот факт считается первой демонстрацией того, что выбор J-сегмента, используемого для построения гуманизированного антитела может влиять на уровень экспрессии антител в целом, не говоря уже о наблюдаемом существенном повышении степени. Это открытие особенно важно, поскольку ожидается, что оно должно значительно снизить стоимость товаров, необходимых для выработки OGD1.0.0. Без этого открытия не было бы экономии средств для выработки этого антитела в количестве, требуемом для его выпуска на рынок, от чего страдали бы пациенты, для которых лечение с его применением является благоприятным.

ПРИМЕР 2
Анализ устойчивой экспрессии OGD1.0.0 и OGD1.1.1
[000191] Устойчивую экспрессию OGD1.0.0 и OGD1.1.1 испытывали в клетках CHO-DUKX. В общих чертах, клетки выращивали до 80% конфлюэнтности, а затем контрансфицировали с применением реагента для трансфекции липофектамина с 25 мкг каждого из векторов экспрессии тяжелой и легкой цепи (в целом 50 мкг), как описано в предыдущем примере (т.е., VH0 и VL0 для одного набора клеток и VH1 и VL1 для другого набора клеток). После трансфекции использованную среду заменяли свежей средой R1CD1 плюс 10% FBS каждые три-четыре дня при создании устойчивых пулов.
[000192] После образования устойчивых трансфектантов испытывали способность клеток к экспрессии антитела против GDF-8 при выращивании в форме закрепленных клеток в бессывороточной среде R5CD1. В этих условиях клетки, экспрессирующие OGD1.0.0, экспрессировали 47,3 мг/л антитела после 96 часов выращивания, в то время как клетки, экспрессирующие OGD1.1.1, экспрессировали 41 мг/л антитела после 72 часов выращивания. Антитела очищали, применяя колонку с 1 мл белка А, а затем определяли концентрацию с применением HPLC.
[000193] После адаптации закрепленных клеток к росту суспензии в бессывороточной среде снова определяли экспрессию антител OGD1.0.0 и OGD1.1.1. Бессывороточную среду ASI высевали с 3,0×105 жизнеспособных клеток / мл и инкубировали при 37°С. На четвертый день уровень рН доводили до 7,3, добавляли питательный концентрат и температуру инкубации снижали до 31°С в течение дополнительных 3 дней выращивания. Экспрессирующие OGD1.0.0 клетки выращивали в объеме 100 л культуры, а экспрессирующие OGD1.1.1 клетки выращивали в объеме 50 мл культуры. Все остальные условия роста были одинаковыми для разных клеток. На седьмой день клетки экспрессирующие OGD1.0.0, экспрессировали 66,12 мг/л антитела, а клетки, экспрессирующие OGD1.1.1, экспрессировали 10,6 мг/л антитела, как было определено путем очистки белком А и количественного определения с применением HPLC. Когда эксперимент с использованием клеток OGD1.0.0 повторяли путем выращивания клеток в 100 л культуры в течение 9 дней, включая 5 дней при 31°С, концентрация антитела увеличилась до 207,2 мг/л. В другом эксперименте, в котором экспрессирующие OGD1.0.0 клетки выращивали в 25 л культуры в течение 11 дней, включая 7 дней при 31°С, концентрация антитела составляла 145 мг/л. В отдельном эксперименте, в котором экспрессирующие OGD1.1.1 клетки выращивали в 50 мл культуры в бессывороточной среде R5CD1 в течение 7 дней, включая 3 дня при 31°С, концентрация антитела составляла 39,3 мг/л.
[000194] Результаты эксперимента с устойчивой трансфекцией показаны ниже в таблице, в которой ΡΟΙ означает соответствующий пик при эксклюзионной хроматографии (SEC) после очистки белком А. По результатам, сопоставимым с полученными при экспрессии OGD1.0.0 и OGD1.1.1 во временно трансфицированных клетках COS-1, уровень экспрессии антитела OGD1.0.0 был значительно выше в устойчиво трансфицированных клетках СНО по сравнению с экспрессией OGD1.1.1 в подобных условиях. Этот неожиданный результат сопоставим с повышением уровня экспрессии, наблюдаемым в эксперименте с временной трансфекцией, описанном выше. Этот результат также указывает, что способ культивирования экспрессирующих клеток, либо прилипающих, либо в суспензии, и тип клеток существенно не влияют на повышенную экспрессию антитела OGD1.0.0 по сравнению с OGD1.1.1.
Таблица 4: Сравнение уровня экспрессии OGD1.0.0 и OGD1.1.1, устойчиво экспрессируемых в клетках СНО.

ПРИМЕР 3
Анализ временной экспрессии OGD1.0.1 и OGD1.1.0
[000195] Поскольку J-сегменты в каждом из вариабельных участков тяжелой и легкой цепей были изменены, не было выяснено, может ли отдельное изменение быть достаточным для заметного повышения наблюдаемого уровня экспрессии OGD1.0.0, или же оба изменения участвуют в повышенной экспрессии антитела.
[000196] Для исследования этого явления заявителями был повторен вышеописанный эксперимент с временной трансфекцией, однако дополнительно комбинировали последовательности VH0 и VL1 в одном планшете и комбинировали последовательности VH1 и VL0 в другом планшете, а затем определяли уровень экспрессии антител с применением ELISA. Результаты эксперимента, показанные ниже в таблице, демонстрируют, что замещение JH4 J-сегментом JH3 в VH области является достаточным для обеспечения значительно повышенной экспрессии антитела, наблюдаемой заявителями. И наоборот, изменение Kanna-J-сегментов (т.е., JK4 на JK1) не обнаружило существенного влияния на уровень экспрессии.
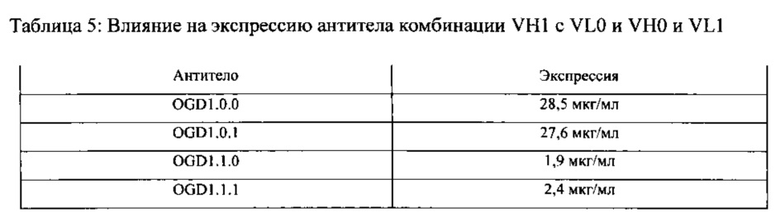
ПРИМЕР 4
Связывание GDF-8 антителами против GDF-8
[000197] Связывание GDF-8 родительским мышиным антителом, химерным мышиным-человеческим антителом (мышиные вариабельные домены и человеческие константные домены) и гуманизированными антителами OGD1.0.0 и OGD1.1.1 анализировали с применением количественного анализа ELISA и поверхностного плазмонного резонанса (SPR). В экспериментах с ELISA способность антител к ингибированию связывания GDF-8 с его родственным высокоаффинным рецептором ActRllB определяли путем вычисления значений IC50. SPR-анализ применяли для вычисления видимых значений Ко. Результаты показаны в Таблице 6.
[000198] Для ELISA-анализа слитый белок ActRllB-Fc (1 мкг/мл в 0,2 M буфера из карбоната натрия) наносили на 96-луночные аналитические планшеты с плоским дном в течение суток при 4°С. Эти планшеты затем блокировали 1 мг/мл BSA в PBS 0,1% Tween (200 мкл/лунку) в течение 1 часа при комнатной температуре или в течение суток при 4°С, а затем промывали. Антитела в разной концентрации комбинировали с 10 нг/мл GDF-8, конъюгированного с биотином, и инкубировали в течение 45 минут при комнатной температуре. После инкубации испытываемый раствор добавляли в блокированный планшет для ELISA (100 мкл/лунку) и дополнительно инкубировали в течение 1 часа при комнатной температуре. После промывания лунок количество GDF-8, связанного с иммобилизованным ActRllB-Fc, относительно контрольной группы обнаруживали с применением стрептавидина - пероксидазы хрена (30-минктная инкубация) и ТМВ. Колориметрические измерения при 450 нМ записывали в считывающем устройстве для микропланшетов. Эксперименты с использованием мышиного и химерного антител повторяли по четыре раза и выводили среднее значение.
[000199] SPR выполняли при 25°С с применением устройства BIACORE 3000 (GE Healthcare). Мышиное антитело захватывали, применяя антитела против IgG мыши, а гуманизированные антитела захватывали, применяя белок А. Белок А иммобилизовали на всех четырех проточных ячейках сенсорного чипа СМ5 с применением аминного связывания. Поверхность активировали путем введения раствора 0,2 M N-этил-N-диметил-амино-пропил-карбодиимида (EDC) и 50 мМ N-гидроксисукцинимида (NHS) в течение 7 минут. Белок А разбавляли до 50 мкг/мл в 10 мМ буфера из ацетата натрия при рН 5,0 и вводили в течение 3 минут при скорости потока 10 мкл в минуту. Затем поверхность блокировали 1М этаноламина (ΕΤΗ) в течение 7 минут. Конечный уровень иммобилизации белка А составлял 1000-1200 единиц ответа (RU). После процедуры иммобилизаци осуществляли несколько промываний подвижным буфером (0,01 M HEPES, рН 7,4, 0,15 M NaCl, 3 мМ EDTA, 0,005% Р20) для уравновешивания поверхности. Антитела разбавляли до 0,25 мкг/мл в буфере HBS-EP. Растворы каждого антитела (5 мкл) вводили над покрытыми белком А проточными ячейками 2, 3 или 4 со скоростью 10 мкл/мин, с получением на выходе приблизительно 200 RU захваченного антитела. Серии титрования GDF-8 (2-кратные разбавления от 4,0 нМ до 0,125 нМ) приготавливали в 0,01 M ацетата натрия при рН 5,0, 0,15М NaCl, 3 мМ EDTA, 0,005% Р20. Последний раствор также использовали в качестве подвижного буфера. Раствор GDF-8 вводили над захваченным антителом в течение 2 минут при скорости потока 50 мкл/мин и оставляли для диссоциации на 30 мин. После каждого цикла введения и захвата поверхность сенсорного чипа восстанавливали с использованием 30 мкл 10 мМ NaP04, 0,5 М NaCl при рН 2,5 при скорости потока 50 мкл/мин. Для анализа данных использовали программу BIAevaluation (вер. 4.1.1, GE Healthcare). Данные подвергали двойной привязке путем вычитания сигнала, который давали буфер и контрольная поверхность. Применяли модель Ленгмюра 1:1 для общего согласования данных сенсограммы и расчета значений KD. Эксперименты с применением OGD1.0.0 повторяли по три раза и выводили среднее значение.
[000200] Данные 1С50, определенные с применением ELISA, являются сравнимыми между разными вариантами испытанных антител. В более точном анализе SPR OGD1.0.0 обладает значительно большей аффинностью связывания к GDF-8 по сравнению с OGD1.1.1.
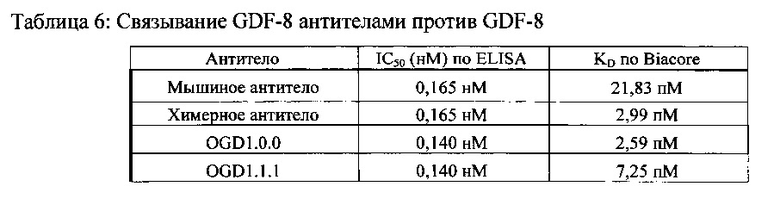
ПРИМЕР 5
Нейтрализующая GDF-8 способность антитела OGD1.0.0
[000201] Способность антител против GDF-8 к нейтрализации опосредованного GDF-8 сигнала подтверждали с применением анализа гена-репортера. Последовательность репортера, называемую pGL3(CAGA)12, составляли путем помещения 12 CAGA-боксов перед ТАТА-боксом и сайтом инициации транскрипции из главного позднего промотора аденовируса в репортерном векторе люциферазы pGL3 (Promega). CAGA-бокс, который находится в промоторе гена РА1-1, представляет собой реагирующий на TGF элемент, который также реагирует на GDF-8. Линию клеток рабдомиосаркомы человека А204 (АТСС НТВ-82) временно трансфицировали pGL3(CAGA)12 и культивировали в 96-луночных планшетах в среде McCoy 5 А с добавлением 2 мМ глутамина, 100 U/мл стрептомицина, 100 мкг/мл пенициллина и 10% сыворотки эмбрионов крупного рогатого скота в течение 16 часов. Антитела предварительно инкубировали с GDF-8 (10 нг/мл) в среде с добавлением 1 мг/мл BSA в течение 1 ч при комнатной температуре. Затем клетки обрабатывали в течение 6 ч при 37°С испытываемыми и контрольными образцами, включая образцы без GDF-8 и с GDF-8 (10 нг/мл) без добавления антитела. Активность люциферазы измеряли с применением системы анализа люциферазы (Promega). Эксперименты с применением мышиных и химерных антител повторяли дважды каждый и выводили среднее значение, а эксперименты с применением OGD 1.0.0 повторяли трижды и выводили среднее значение. Значения ЕС50, определяемые с применением анализа гена-репортера, являются сравнимыми между разными вариантами антител.
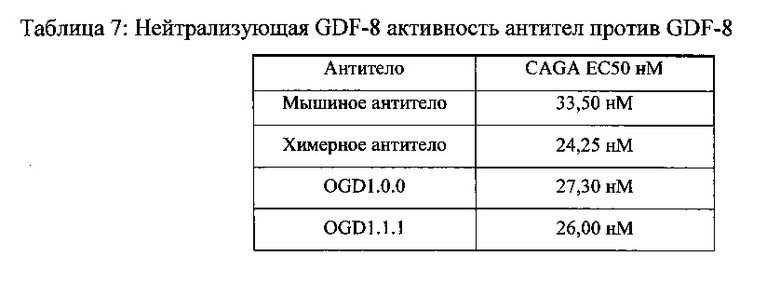
ПРИМЕР 6
OGD1.0.0 Антитело увеличивает мышечную массу, мышечную силу и безжировую массу у мышей
[000202] Восьминедельным самцам мышей C57Bl/6 внутрибрюшинно (IP) раз в неделю в течение двух недель вводили дозу OGD1.0.0 (10 мг/кг) или контрольный носитель (PBS). Всего в каждой группе использовали восемь мышей. На 14-й день полную безжировую массу тела определяли с применением ЯМР-визуализации мелких животных. После определения безжировой массы животных усыпляли и икроножную мышцу, четырехглавую мышцу и длинный разгибатель пальцев (EDL) вырезали и взвешивали. EDL-мышцу также испытывали на способность к созданию усилия ex vivo.
[000203] После двух недель лечения безжировая масса контрольных животных увеличивалась на 1,66±0,56 г, а безжировая масса животных, получавших OGD1.0.0, увеличивалась на 3,36±0,62 г, что составляет повышение на 102% по сравнению с контролем.
[000204] Как показано на ФИГ. 2А и ФИГ. 28, масса четырехглавой мышцы увеличивалась на 14,8% по сравнению с контролем, масса икроножной мышцы увеличивалась на 10,3% по сравнению с контролем, а мышечная масса EDL увеличивалась на 10,8% по сравнению с контролем у животных, получавших антитело OGD 1.0.0. Как показано на ФИГ. 3, у животных, получавших антитело OGD1.0.0, общее тетаническое усилие, прилагаемое EDL-мышцей, увеличивалось на 14,8% относительно усилия, создаваемого EDL-мышцей мышей получавших контрольный носитель. Данные показаны как среднее значение ± стандартная погрешность среднего.
[000205] Также определяли реакцию общей безжировой массы тела и мышечной массы четырехглавой мышцы и икроножной мышцы в ответ на лечение с применением OGD1.0.0. В этих экспериментах 12-недельных самок мышей C57Bl/6 разделяли на группы (n=6) и еженедельно вводили носитель или OGD1.0.0 в дозе 0,3, 1, 3, 10 или 30 мг/кг в течение 4 недель. После 7, 14, 21 и 28 дней периода лечения безжировую массу определяли при помощи ЯМР-визуализаци. В конце периода исследования четырехглавую мышцу и икроножную мышцу вырезали и взвешивали после усыпления испытуемых животных. Как показано на ФИГ. 4А и ФИГ. 48, мышечная масса четырехглавой мышцы и икроножной мышцы увеличивалась с повышением дозы антитела до приблизительно 10 мг/кг. Подобным образом, как показано на ФИГ. 5А и ФИГ. 5В, общая безжировая масса тела увеличивалась с повышением дозы антитела до приблизительно 10 мг/кг. Данные показаны как среднее значение ± стандартная погрешность среднего.
ПРИМЕР 7
Антитело OGD1.0.0 увеличивает мышечную массу и безжировую массу у mdx-мышей
[000206] Mdx-мутация Х-сцепленного гена дистрофина (Dmd) спонтанно возникает у мышей C57BL/1 OScSn и вызывает точечную мутацию в пределах экзона в позиции гена 3185, преобразуя глутаминовый кодон в терминирующий кодон, приводя к преждевременной терминации белка дистрофина. В результате у mdx-мышей отсутствует функциональный дистрофии, и они служат в качестве малой модели мышечной дистрофии Дюшенна. Начиная с 3-й недели, развивается некроз мышц с явными проявлениями мышечной слабости. Хотя скелетные мышцы конечностей характеризуются устойчивой и прогрессирующей дегенерацией и некрозом, это явление компенсируется регенеративной реакцией, активируемой сателлитоцитами, и мышечной гипертрофией. Мышцы mdx-мутантов в целом характеризуются снижением упругости, что делает их более восприимчивыми к повреждению из-за активации растяжения. Мышцы задних конечностей мутантных мышей сначала развивались нормально, однако дифференциация регенерированных мышечных трубочек в быстрые и медленные типы волокон в значительной мере подавляется. Сравнительно мягкий фенотип mdx-мышей частично может объясняться компенсирующей функцией родственного с дистрофином белка атрофина, который подвергается сильной повышающей регуляции в регенерирующих мышечных волокнах у взрослых mdx-мутантов. В отличие от мышц конечностей, мышцы диафрагмы mdx-мышей не подвергаются фазе значительной регенерации, и, таким образом непрерывная дистрофия с возрастом ослабляет эти мышцы. Удельное сокращающее усилие, удельное тетаническое усилие и максимальная сила снижаются в диафрагме mdx-мутантов.
[000207] Восьминедельным самцам mdx-мышей и контрольным мышам C57Bl/6 внутрибрюшинно (IP) раз в неделю в течение восьми недель вводили дозу OGD 1.0.0 (10 мг/кг) или контрольного носителя (PBS). В этих экспериментах десять mdx-мышей получали антитело, восьми вводили контрольный носитель, и шесть мышей C57Bl/6 получали антитело или PBS. В конце периода лечения измеряли полную безжировую массу тела, силу захвата и мышечную массу. Полную безжировую массу тела определяли с применением ЯМР-визуализации мелких животных. Силу захвата испытывали, помещая испытуемое животное на проволочную сетку, давая ему возможность ухватиться за сетку всеми конечностями, а затем оттягивая его за хвост и измеряя максимальное пиковое усилие при отпускании животным захвата. Усредняли данные 3-5 попыток для каждого животного. После измерения безжировой массы тела и силы захвата мышей усыпляли и четырехглавую мышцу и икроножную мышцу вырезали и взвешивали.
[000208] Как показано на ФИГ. 6А, лечение антителом OGD1.0.0 увеличивало безжировую массу у mdx-мышей в среднем на 7,28±0,4 г по сравнению со средним показателем 4,83±0,4 г у mdx-мышей, получавших PBS. Разница была статистически значимой при p<0,05. Таким образом, безжировая маса в восьминедельном исследовании увеличивалась на 50% ± 8,2% у mdx-мышей по сравнению с получавшей носитель контрольной группой у. Как показано на ФИГ. 6В, лечение антителом также увеличивало силу захвата у mdx-мышей по сравнению с получавшими носитель контрольными животными. Разница была статистически значимой при p<0,05.
[000209] Как показано на ФИГ. 7А, лечение антителом увеличивало массу икроножной мышцы и четырехглавой мышцы у mdx-мышей и C57Bl/6-мышей по сравнению с тем же типом мышей, получавшим PBS. Увеличение было статистически значимым (р=0,005 и р=0,002 для четырехглавой мышцы и икроножной мышцы mdx-мышей, соответственно, и р=0,001 и р=0,003 для четырехглавой мышцы и икроножной мышцы C57Bl/6-мышей, соответственно). Как показано на ФИГ. 7В, масса икроножной мышцы и четырехглавой мышцы получавших антитело mdx-мышей увеличивалась на 12,2% и 12,1%, соответственно, по сравнению с получавшими носитель контрольными мышами. Масса тех же типов мышц мышей C57Bl/6 также увеличивалась на 15,2% и 12,8% после лечения антителом по сравнению с получавшими носитель контрольными животными (не показано).
ПРИМЕР 8
Антитело OGD1.0.0 увеличивает объем мышц и безжировую массу у отличных от человека приматов
[000210] Планировали и осуществляли два исследования с изучением влияния введения антитела OGD1.0.0 у яванских макак на безжировую массу тела и объем мышц.
[000211] Каждое исследование длилось восемь недель, в которые животным еженедельно внутривенно вводили дозы антитела и предоставляли еду в избыточном количестве для обеспечения положительный баланс азота. В первом исследовании каждому из трех самцов и каждой из трех самок вводили PBS-носитель или антитело OGD 1.0.0 в дозах 3,0 мг/кг, 10 мг/кг и 30 мг/кг. Перед первым введением, а затем на четвертой неделе и восьмой неделе животных подвергали анестезии, а затем визуализировали с применением двухэнергетической рентгеновской абсорбциометрии (DEXA), компьютеризированной рентгеновской томографии (СТ) и магнитно-резонансной визуализации (MRI) для обнаружения и измерения состава тела, включая безжировую массу и содержание жира. Животных из Исследования 1 подвергали усыплению и вскрытию. Во втором исследовании использовали только самцов, которые получали только носитель (n=5) или антитело OGD1.0.0 в дозах 10 мг/кг и 30 мг/кг (n=5 и n=3 субъектов, соответственно). Животных подвергали визуализации через восемь недель в первом исследовании. После этого животных поддерживали на обогащенном рационе и снова подвергали визуализации через 12, 17 и 26 недель.
[000212] Восьминедельные данные для безжировой массы тела, измеряемой с применением DEXA в обоих исследованиях, комбинировали и анализировали. Результаты показаны на ФИГ. 8, на которой продемонстрировано, что после восьми недель лечения антителом OGD 1.0.0 существует зависящее от дозы увеличение общей безжировой массы и безжировой массы конечности. Данные выражены как средние значения ± стандартная погрешность среднего. Количество включенных в исследование субъектов, получавших только носитель, составляло n=11, получавших 3 мг/кг антитела - n=6, 10 мг/кг - n=10 и 30 мг/кг - n=8. Увеличение общей безжировой массы тела и безжировой массы конечности по сравнению с получавшими носитель контрольными животными было статистически значимым (р<0,05) для всех испытываемых доз антитела. Кроме того, увеличение безжировой массы задней конечности у субъектов, получавших 30 мг/кг, по сравнению с получавшими 10 мг/кг, также достигало статистической значимости (р<0,05).
[000213] Интересно, что, как показано на ФИГ. 9, увеличение безжировой массы тела среди субъектов второго исследования, получавших 10 мг/кг и 30 мг/кг антитела OGD1.0.0, длилось неделями после введения последней дозы антитела на седьмой неделе. Данные показаны как разница по сравнению с PBS-носителем в каждый момент времени. Увеличение относительно контрольной группы для более высокой дозы было статистически значимым при p<0,05 на всех показанных неделях. Увеличение для более низкой дозы было статистически значимым на 4-й и 8-й неделях при p<0,09.
[000214] Влияние лечение антителом OGD1.0.0 на объем эпаксиальных мышц, находящихся сзади от позвоночного столба над поясничными позвонками L3-L5, измеряли путем КТ-сканирования. Как показано на ФИГ. 10А и ФИГ. 10В, объем эпаксиальных мышц значительно увеличивался у животных, получавших 10 мг/кг (n=5) и 30 мг/кг (n=3) антитела OGD 1.0.0 в течение 8 недель по сравнению с контрольными животными, которым вводили PBS. Увеличение в объеме мышц было статистически значимым при p<0,05.
[000215] ФИГ. 11 представляет трехмерный рендеринг эпаксиальной мышцы типичного испытуемого субъекта после 4 недель лечения с применением 30 мг/кг антитела OGD1.0.0. По сравнению с исходным уровнем (слева) достигаемое в результате увеличение объема мышц, которое является визуально очевидным (справа), составляло 22%.
ПРИМЕР 9
Антитело OGD1.0.0 не обладает эффекторной функцией Fe домена
[000216] Связывание антителом OGD1.0.0, включающим три мутации в Fe области, которое, как известно, устраняет связывание Fcγ рецептора (FcγR) с панелью Fcγ рецепторов, испытывали с применением поверхностного плазмонного резонанса. Все эксперименты выполняли с применением устройства BIACORE Т200 (GE HealthCare). В общих чертах, 100 RU GDF-8 захватывали на сенсорном чипе-SA через биотиновую метку, на которой захватывали приблизительно 100 RU антитела OGD 1.0.0, с последующим потоком FcγR в диапазоне концентрации 0-21 мкМ для CD32a-131H, CD16a-158V, CD32b и 0-270 нМ для CD64. Для каждого эксперимента связывания FcγR осуществляли серию инъекций в одноцикловом кинетическом режиме. Фазы ассоциации и диссоциации длились по 120 сек каждая. В конце фазы диссоциации после последней инъекции поверхность, содержащую GDF-8, регенерировали, применяя 20 сек импульс 0,1% раствора TFA.
[000217] Связывание с CD16, CD32a и CD64 не наблюдалось. В то же время, связывание с CD32b наблюдалось, но только при наивысшей испытываемой концентрации (21 мкМ). Из-за отсутствия информационных точек свыше 21 мкМ точное значение Kd не было определено, однако предполагалось, что оно составляет более 21 мкМ, что считается очень слабым показателем по сравнению с Kd молекул IgGl дикого типа (т.е., 2-4 мкМ). Эти результаты указывают, что антитело OGD 1.0.0 не обладает или обладает значительно сниженной способностью к вызыванию эффекторных функций.
ПРИМЕР 10
Кристаллическая структура антител против GDF-8, связанных с GDF-8
[000218] Как объясняется в этом примере, кристаллическую структуру химерного мышиного и гуманизированного антител против GDF-8, связанных с человеческим GDF-8, разрешали и использовали для определения конкретных аминокислот в пределах антител и GDF-8, контактирующих между собой.
[000219] Fab фрагменты получали из химерного антитела против GDF-8, содержащего мышиные VH и VL области (SEQ ID NO: 3 and SEQ ID NO:5, соответственно), связанные с константными областями IgG1 человека. Fab фрагменты затем смешивали с белком GDF-8 человека для образования связанных комплексов. Белковые комплексы концентрировали до 10,75 мг/мл в 50 мМ трис гидрохлорида при рН 7,5 и 100 мМ хлорид натрия. Кристаллы формировали, применяя метод висячей капли с уравновешиванием при 18°С относительно раствора, содержащего 20% PEG ММЕ 5000 и 100 мМ бис-трис, рН 6,5. Кристаллы, содержащие Fab, полученный из гуманизированного антитела против GDF-8 OGD 1.0.0 и GDF-8 получали подобным образом, за исключением того, что белковый раствор уравновешивали относительно небуферированного раствора, содержащего 20% PEG 3350 и 200 иМ хлорида натрия.
[000220] Одночастотные (1,0 А) данные для каждого кристалла собирали на ID-канале пучка в SER-CAT, Advanced Photon Source, Argonne National Laboratory. Отдельный кристалл, охлажденный до -180°С, использовали для каждого набора данных. Данные обрабатывали, применяя DENZO and Scalepack (Z. Otwinowski and W. Minor, "Processing of X-ray Diffraction Data Collected in Oscillation Mode", Methods in Enzymology, Volume 276: Macromolecular Crystallography, part A, p. 307-326, 1997, C.W. Carter, Jr. & R.M. Sweet, Eds., Academic Press (New York)) (включено путем ссылки). Структуру химерного антитела в комплексе с GDF-8 разлагали путем молекулярного замещения с применением программы AMORE (Navaza, J. (2001). Implementation of molecular replacement in AMoRe. Acta Crystallogr. 57, 1367-1372) (включено путем ссылки). Зондом, который использовали при поиске молекулярного замещения, был белок под номером 1HZH в PDB. Перед уточнением 5% данных случайно отбирали и определяли как тестовый набор R^ee ДЛЯ отслеживания процесса уточнения. Структуру каждого комплекса затем восстанавливали в пределах Coot с использованием разностных карт (Emsley, Р. & Cowtan, К. (2004) Coot: инструменты построения модели для молекулярных графиков. Acta Crystallogr., Sect. D: Biol. Crystallogr. 60, 2126-2132) (включено путем ссылки). Статистические данные уточнения перечислены в Таблице 8. Структуру гуманизированного OGD 1.0.0 в комплексе с GDF-8 разлагали подобным образом, за исключением того, что используемым зондом была структура химерного антитела.

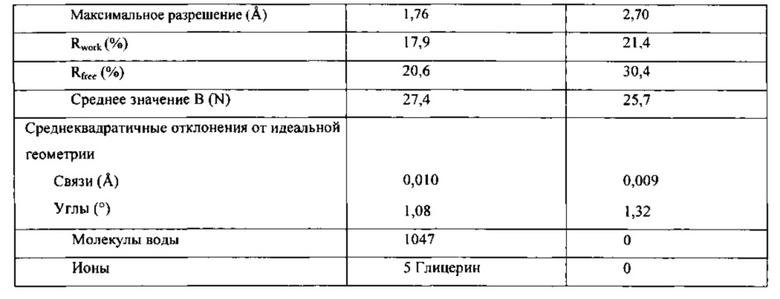
[000221] Остатки в антителах и GDF-8, которые предположительно контактируют между собой на основе совместной кристаллической структуры, перечислены в Таблице 9, в которой перед остатками антитела из VH цепи указывается буква "Н", и нумерация представлена относительно SEQ ID NO: 3. Перед остатками из VL цепи указывается буква "L", и нумерация представлена относительно SEQ ID NO: 5. Номера зрелых человеческих GDF-8 указаны относительно SEQ ID NO: 1. Остатки определяли как пребывающие в контакте между собой, если они содержали как минимум одну пару контактирующих атомов. Атомы определяли как контактирующие, если они имели контактный коэффициент С<1,3, где D12÷(R1+R2), D12 является расстоянием между атомами, R1 является vdW радиусом атома 1, и R2 является vdW радиусом атома 2. На практике среднее расстояние между контактными атомами составляло приблизительно 4,7 Å, хотя фактическое расстояние в любом конкретном случае колеблется в зависимости от соответствующих типов атомов.
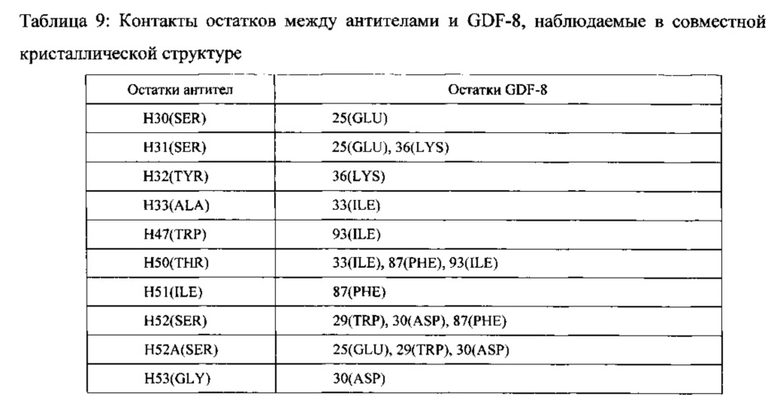
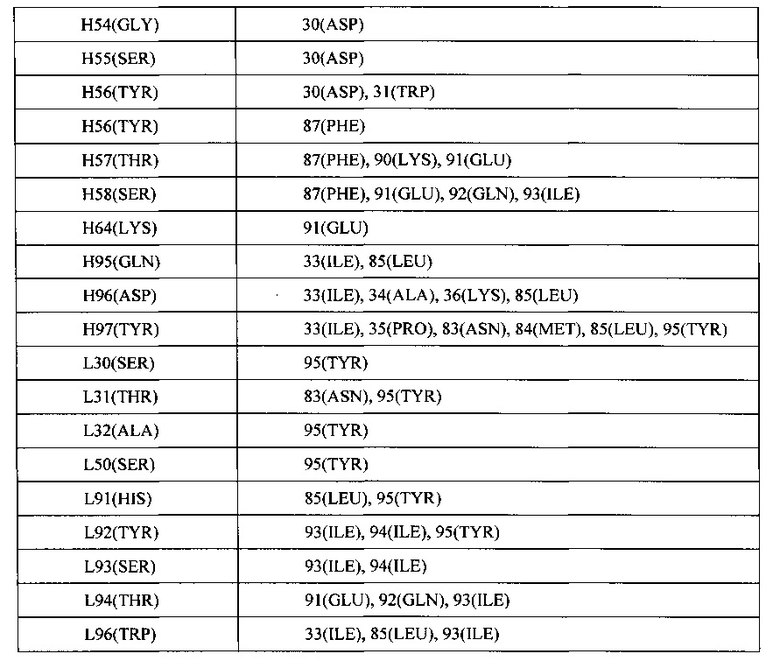
ПРИМЕР 11
Дальнейшая гуманизация VH и VL областей антетела
[000222] На основе результатов анализа последовательности и структуры антител против GDF-8, совместно кристаллизированных с GDF-8, VH и VL области антитела модифицировали с целью дальнейшей гуманизации их последовательностей. Выравнивание последовательности других гуманизированных VH областей показано на ФИГ. 1А. Выравнивание последовательности других гуманизированных VL областей показано на ФИГ. 1В. После создания экспрессионных последовательностей, содержащих новые VH и VL области, антитела вырабатывались во временно трансфицированных клетках COS-1, и их очищали с применением стандартных способов. Аффинность связывания к GDF-8 и нейтрализующую активность антител затем испытывали, как описывается авторами. Результаты указываются в Таблице 10.
[000223] Аминокислотную последовательность CDR2 из гуманизированных VH0 и VL0 областей (взятых из мышиного антитела) сравнивали с последовательностью CDR2 из VH области DP-47 и VL области DPK-9 зародышевой линии человека, соответственно. Все остатки в последовательностях CDR2 VH0 и VL0, отличающиеся от человеческой последовательности, меняли на человеческие. Новые VH и VL области обозначали как VH2 (SEQ ID NO: 66) и VL2 (SEQ ID NO: 67), соответственно. Интактные антитела, полученные с использованием VH2 и VL0 областей и VH0 и VL2 областей, испытывали на связывание с GDF-8 в эксперименте с конкурентным ELISA-анализом. Результаты показали, что полная гуманизация CDR2 VH значительно снижает связывание GDF-8, тогда как полная гуманизация CDR2 VL не приводила к потере связывания антигена.
[000224] Дальнейшая гуманизация VH и VL областей основывалась на совместной кристаллической структуре. В данном случае остатки мышиного происхождения в CDR VH и VL областей оставляли только в случаях, когда они контактировали с остатками GDF-8 в совместной кристаллической структуре. В других случаях все остатки CDR VH и VL меняли на соответствующие человеческие остатки в DP-47 и DPK9, соответственно, для образования VH3 (SEQ ID NO: 68) и VL3 (SEQ ID NO: 69). В эксперименте с конкурентным ELISA-анализом антитело, включающее VH3 и VL0 (т.е., OGD1.3.0), демонстрировало значительную потерю активности, а антитело, включающее VH0 и VL3, (т.е., OGD 1.0.3) полностью сохраняло активность.
[000225] На основе последовательности CDR2 в VL2 позицию 50 VL3 замещали аланином (т.е. S50A) для создания VL4 (SEQ ID NO: 71). Антитело, включающее VH0 и VL4 (т.е., OGD1.0.4), выработанное с использованием этой VL области, сохраняло существенную активность. Другую мутацию, W96L, включали в CDR3 VL3 для создания VL5 (SEQ ID NO: 73). Однако антитело, включающее VH0 и VL5 (т.е., OGD1.0.5), выработанное с использованием этой VL области, демонстрировало сниженную активность по сравнению с OGD1.0.0.
[000226] Новые мутации также включали тяжелый вариабельный участок. Два замещения включали в CDR3, т.е., M99F и N101D, для образования VH4 (SEQ ID NO: 70). Антитело, включающее VH4 и VL0 (т.е., OGD1.4.0), выработанное с использованием этой VH области, имело существенно сниженную активность, которая коррелировала со сниженной аффинностью связывания к GDF-8. В CDR2 замещение G53S осуществляли с целью создания VH5 (SEQ ID NO: 72). Антитело, включающее VH5 и VL0 (т.е., OGD1.5.0), выработанное с использованием этой VH области, сохраняло существенную активность.
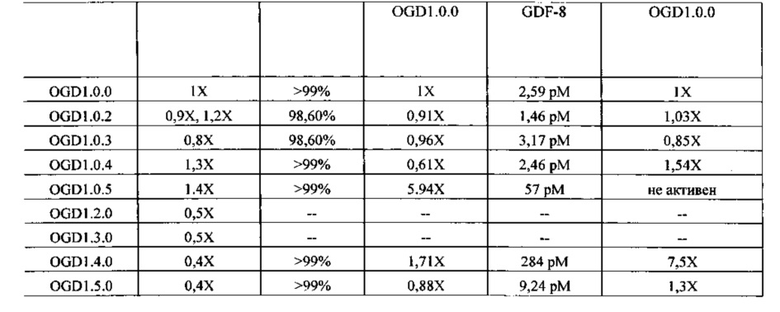
[000227] Все публикации, патенты, патентные заявки и другие документы, приведенные в этой заявке, таким образом, включены путем ссылки в их полном объеме во всех отношениях в той же мере, в которой каждая отдельная публикация, патент, патентная заявка или другой документ были бы отдельно указанные как включенные путем ссылки во всех отношениях.
[000228] Хотя были показаны и описаны различные конкретные варианты воплощения, специалистам в данной области станет понятно, что возможны различные изменения без отклонения от сущности и объема изобретения(й).
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА ПРОТИВ GDF-8 И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2360925C2 |
| Терапевтические антитела и их применения | 2016 |
|
RU2722381C2 |
| АНТИТЕЛА ПРОТИВ TFR И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ ПРОЛИФЕРАТИВНЫХ И ВОСПАЛИТЕЛЬНЫХ РАССТРОЙСТВ | 2016 |
|
RU2737637C2 |
| АНТИТЕЛА ПРОТИВ VEGF И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2567639C2 |
| АНТИТЕЛА ПРОТИВ TNF-α И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2595379C2 |
| Молекулы, связывающиеся с 4-1ВВ | 2011 |
|
RU2710717C2 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА К CD3 И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2831840C2 |
| Конъюгаты "антитело-лекарственное средство" | 2012 |
|
RU2624141C2 |
| АНТИТЕЛА ПРОТИВ PD-1 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2788616C2 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЯ ИЛИ НАРУШЕНИЯ СО СТОРОНЫ ПЕЧЕНИ, ПРЕДУСМАТРИВАЮЩЕЕ ПРИМЕНЕНИЕ АНТАГОНИСТОВ РЕЦЕПТОРОВ ACTRII | 2020 |
|
RU2838150C1 |










































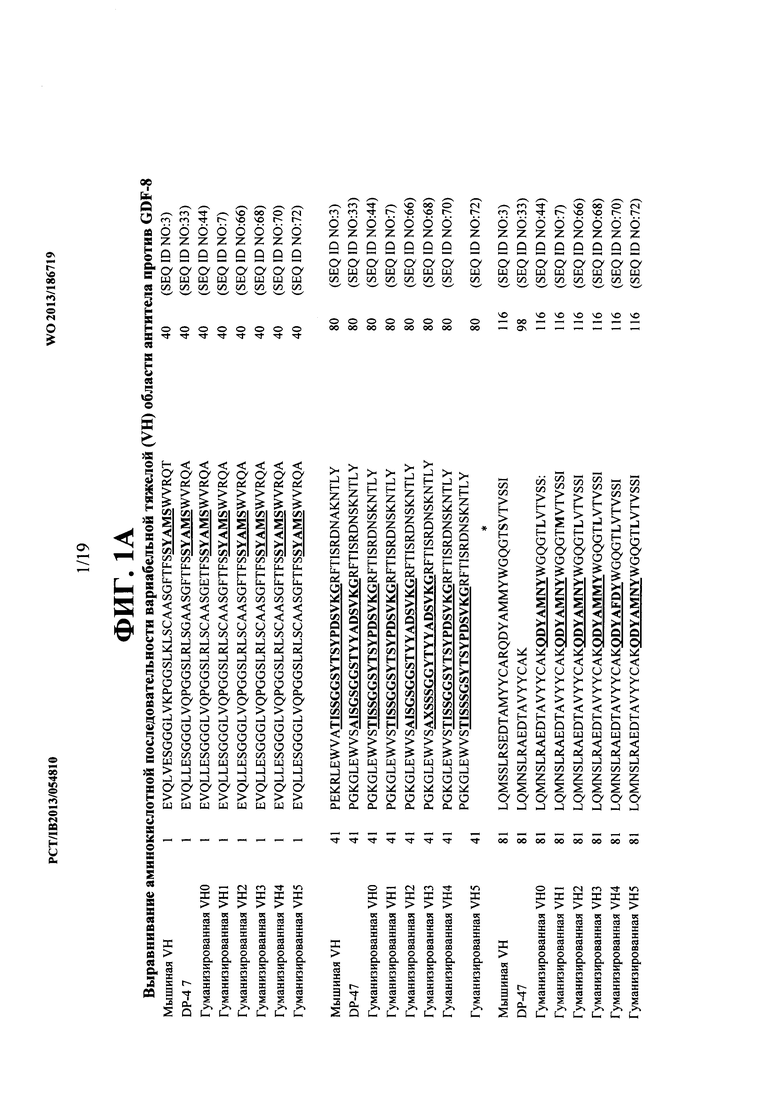

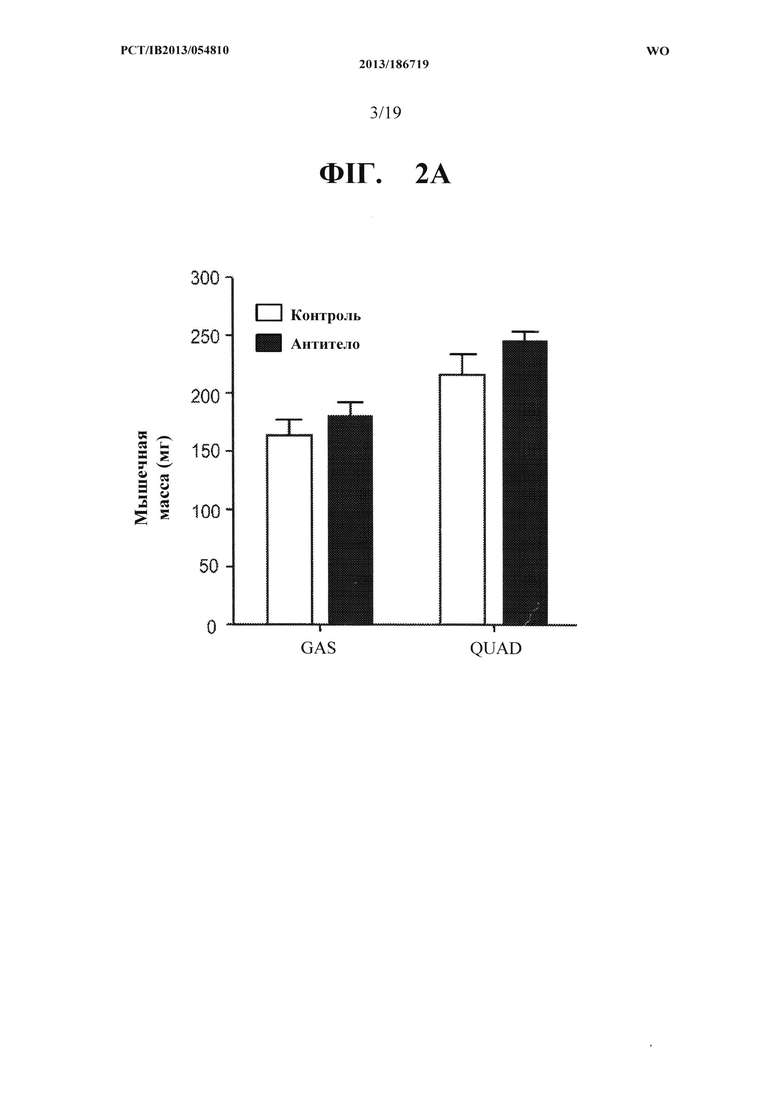
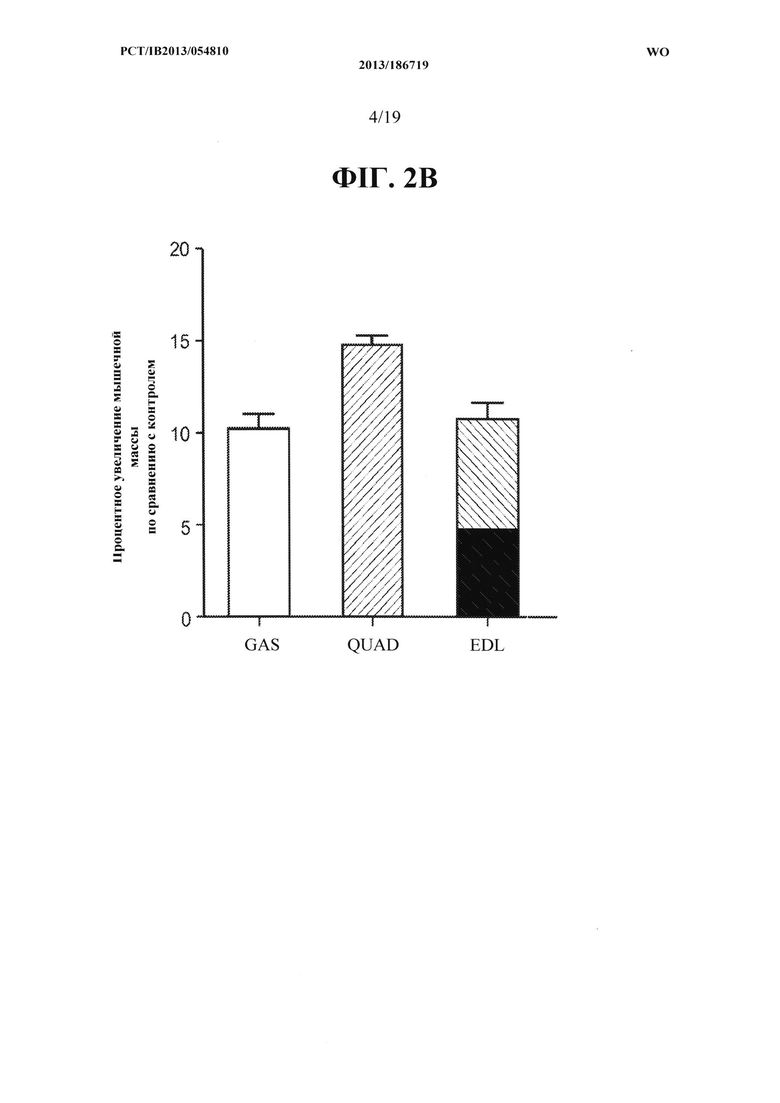
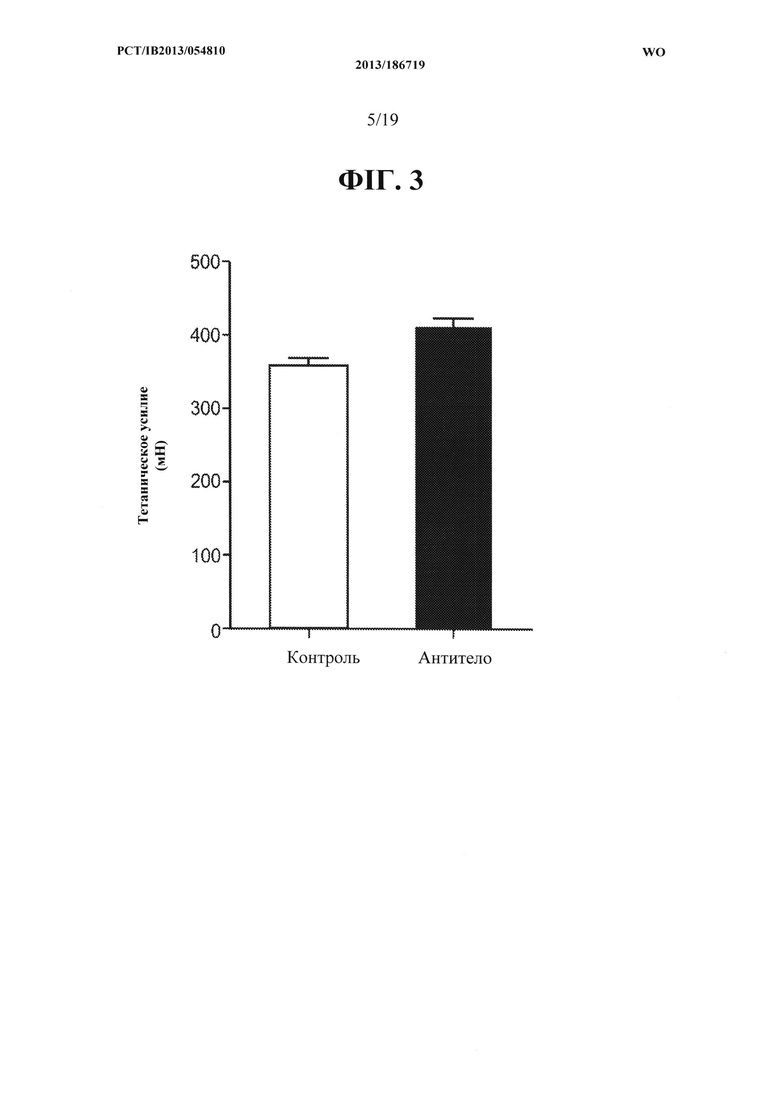
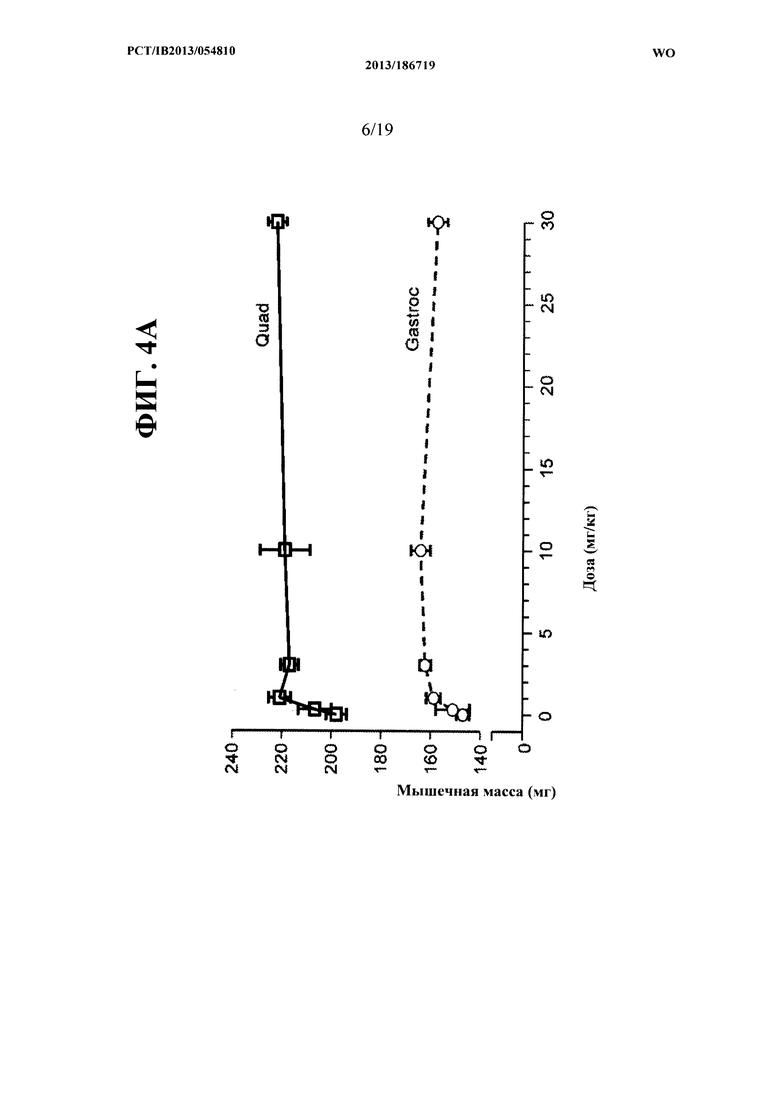
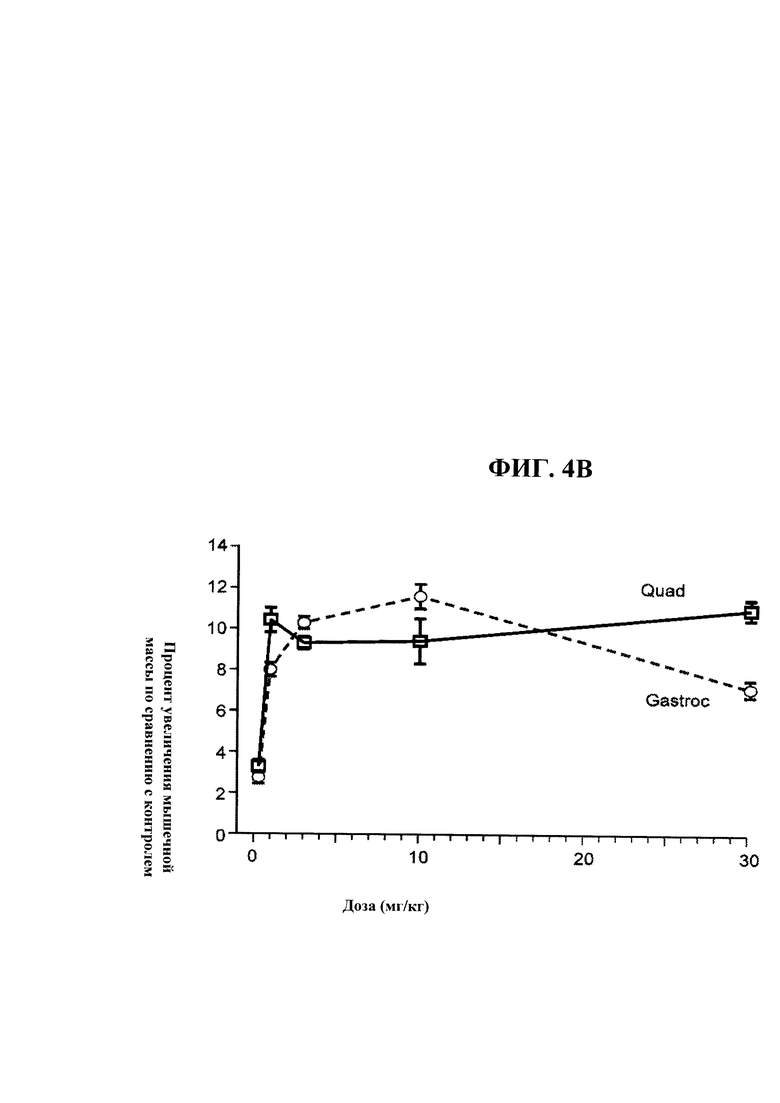
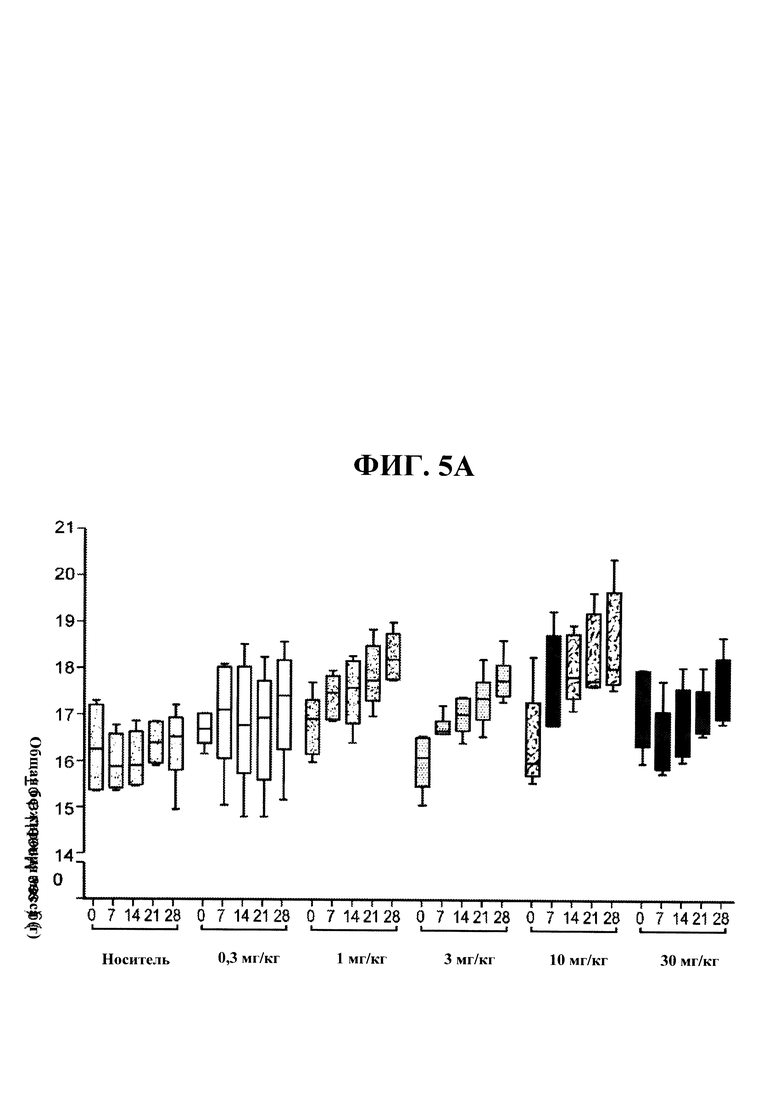
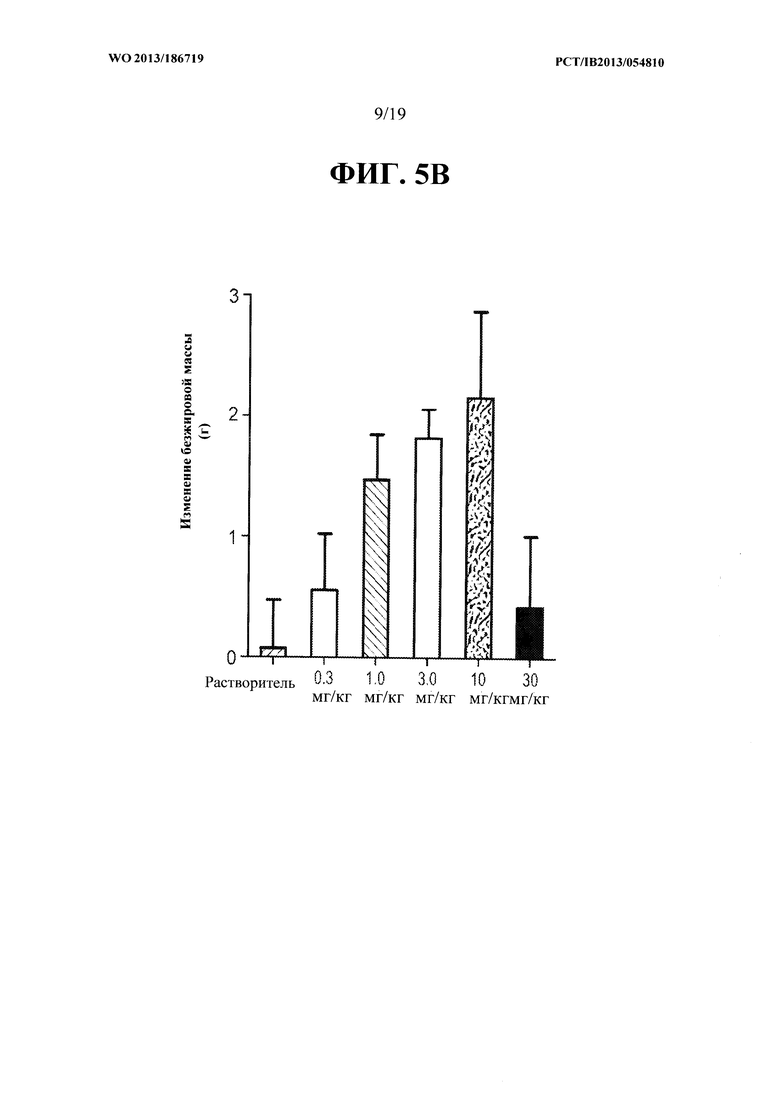
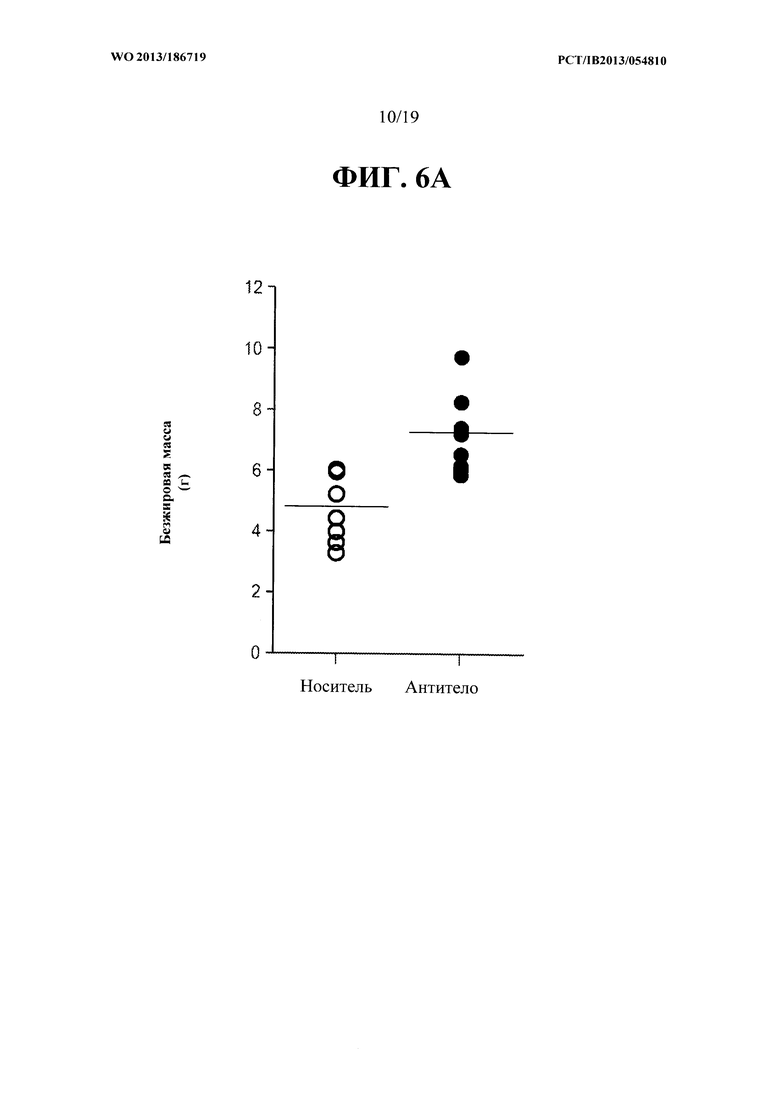
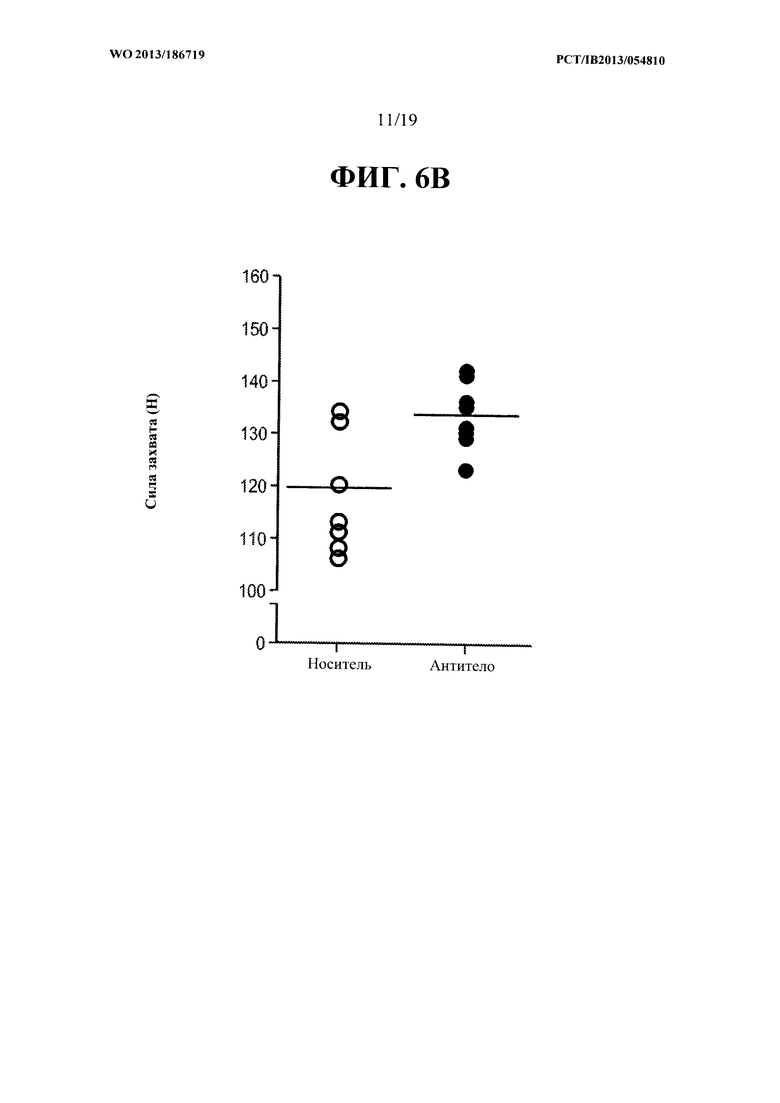
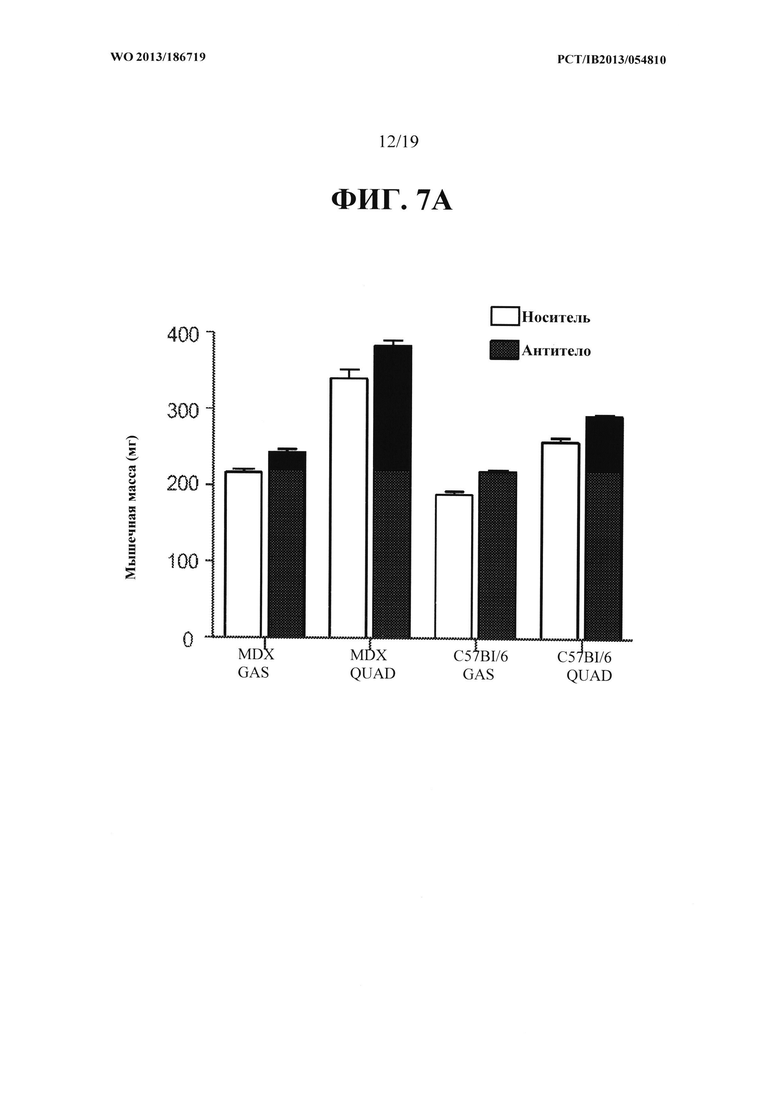
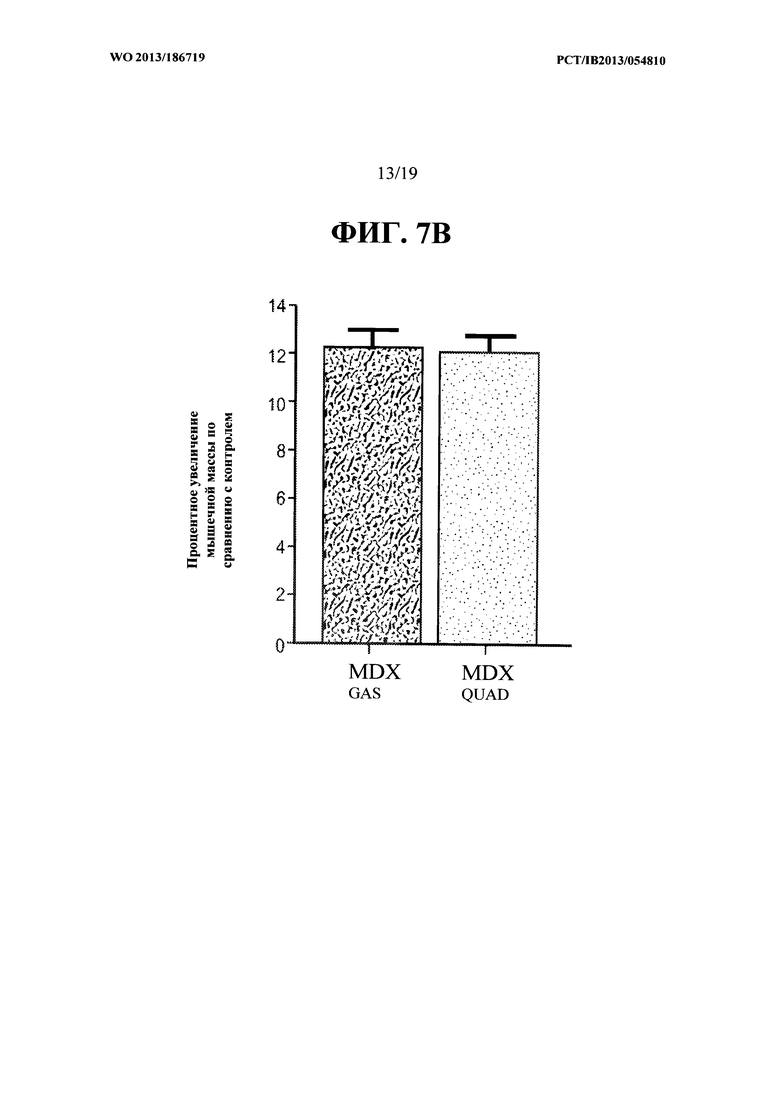
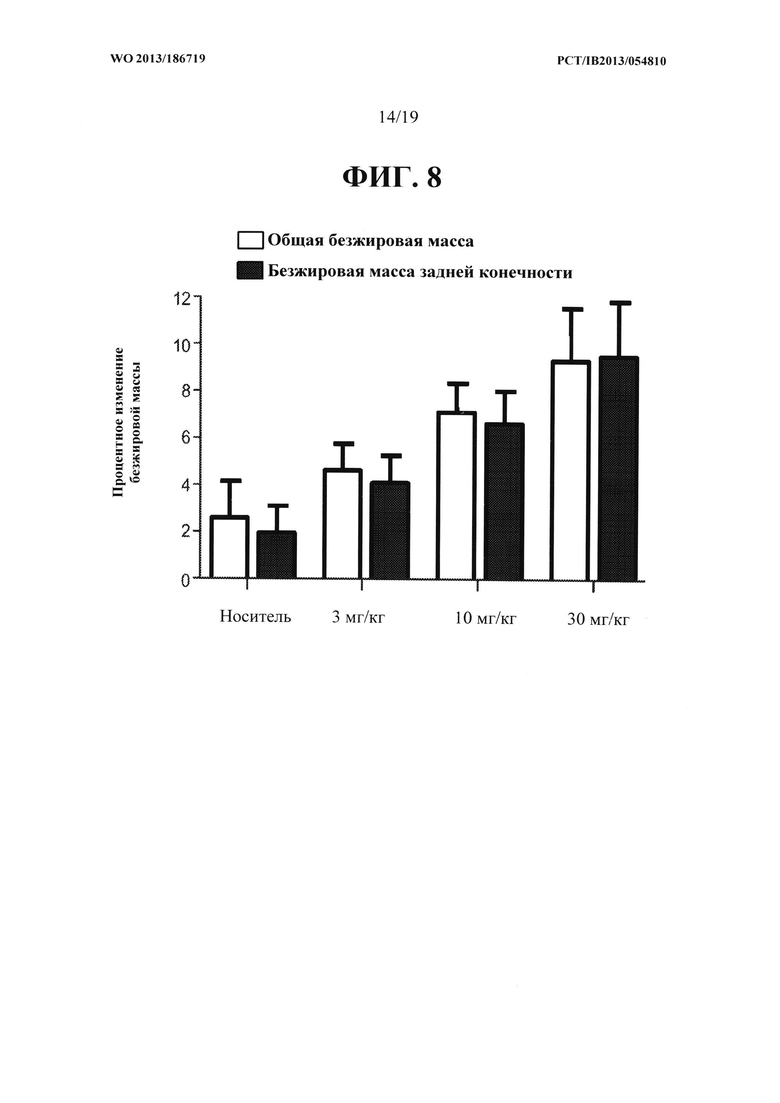
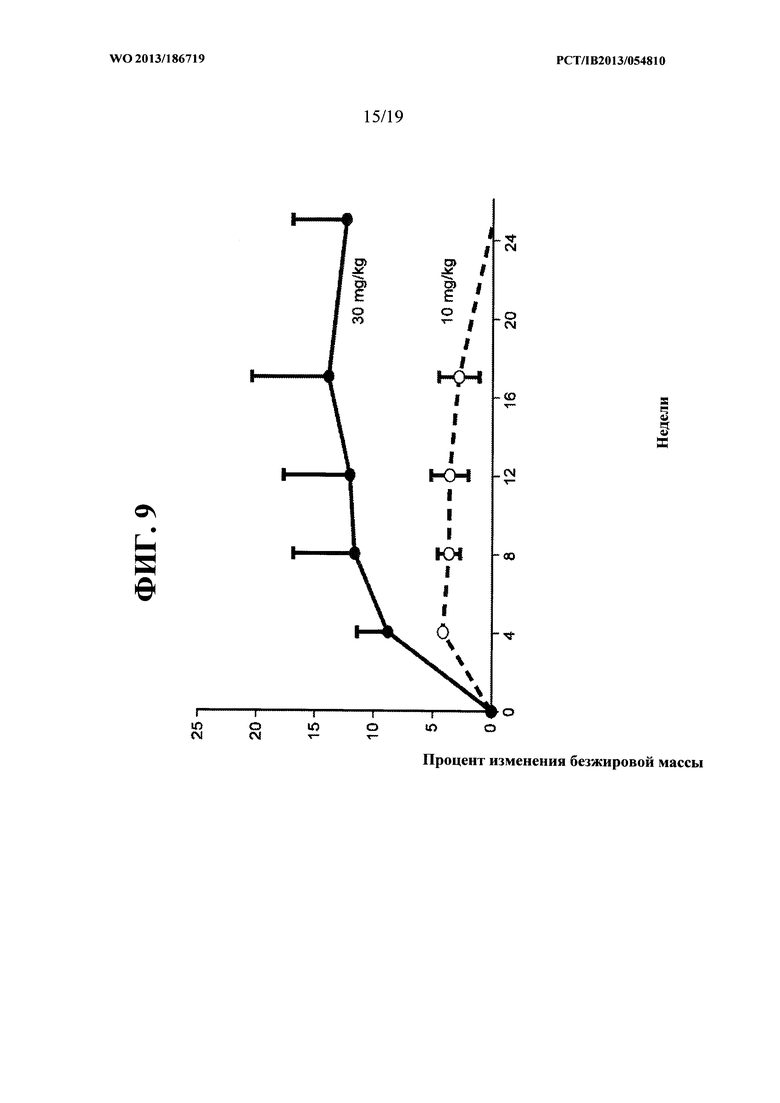

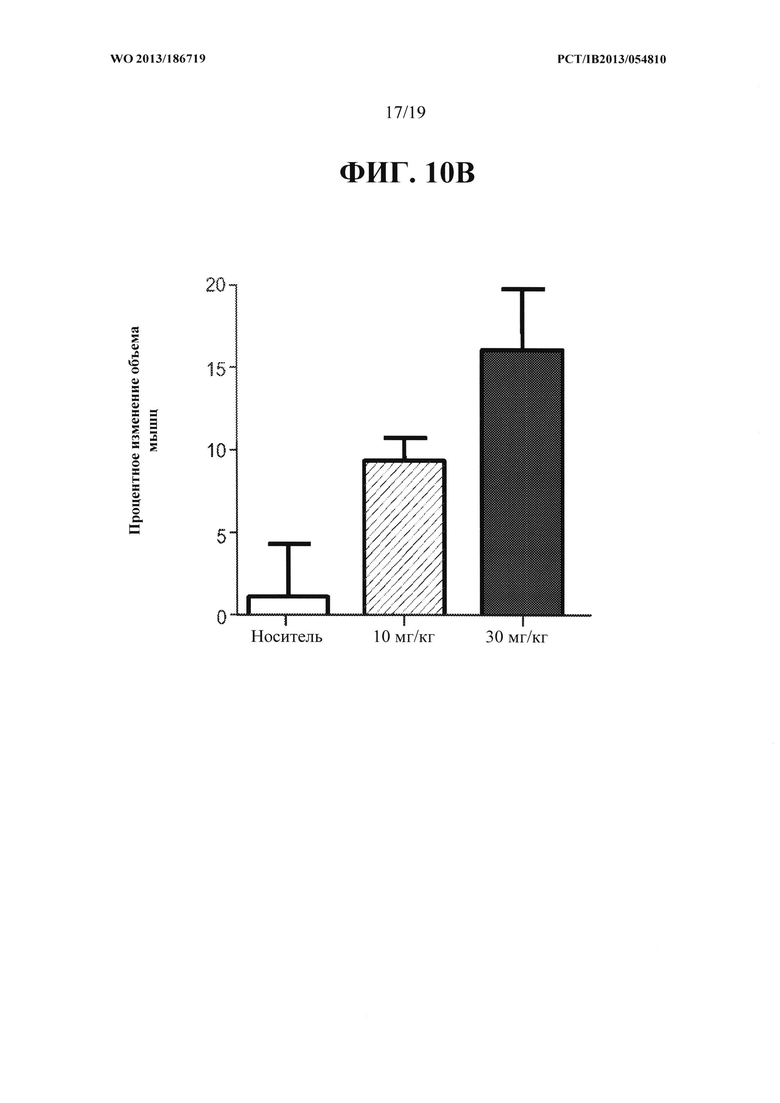
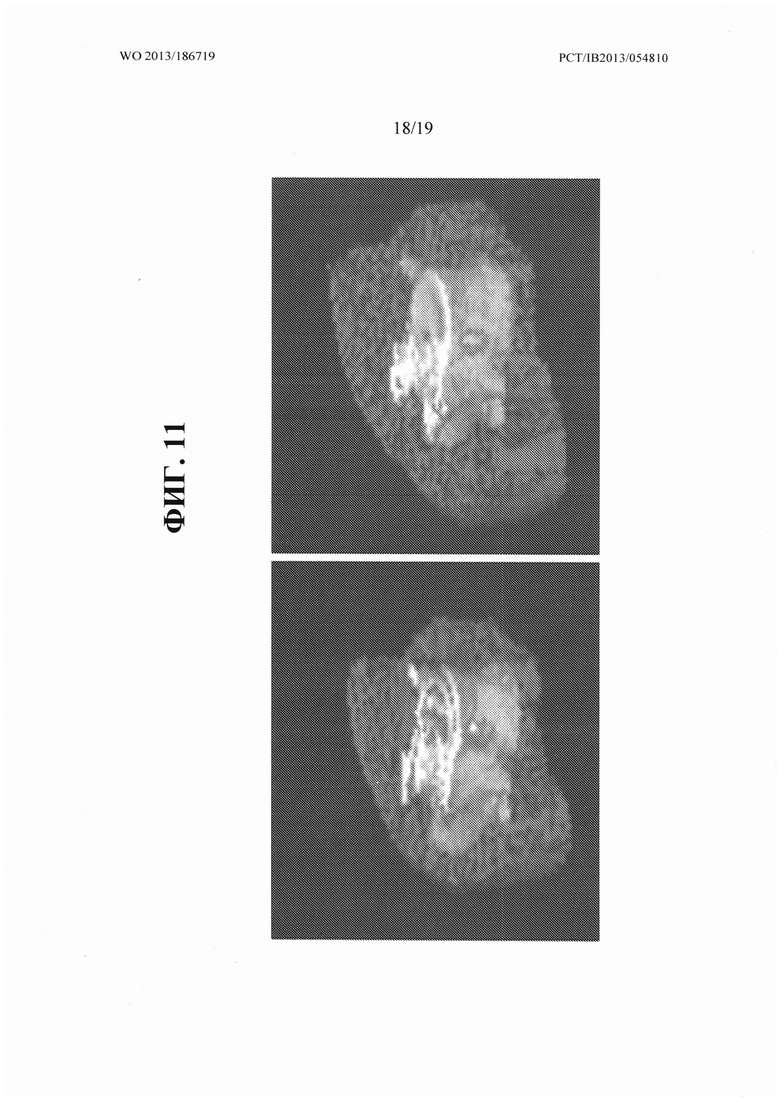
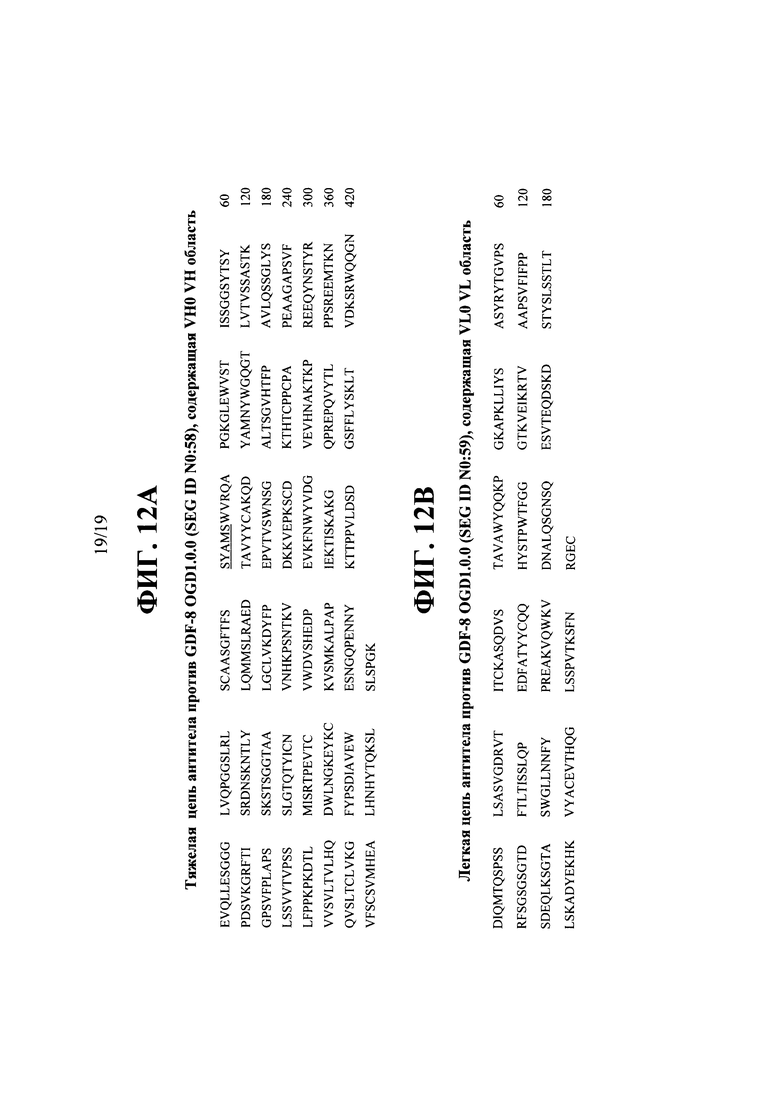
Изобретение относится к биохимии. Описано антитело или его антигенсвязывающий фрагмент, которые специфически связываются с GDF-8, причем антитело или фрагмент включают: вариабельную тяжелую (VH) область антитела, включающую первую, вторую и третью определяющие комплементарность области (CDR) из VH области, определяемой аминокислотной последовательностью SEQ ID NO: 44; и вариабельную легкую (VL) область антитела, включающую первую, вторую и третью определяющие комплементарность области (CDR) из VL области, определяемой аминокислотной последовательностью SEQ ID NO: 46; причем вышеупомянутая VH область включает лейцин в аминокислотной позиции, соответствующей номеру остатка 111 SEQ ID NO: 44 (позиция 108 по Kabat). Представлена фармацевтическая композиция, содержащая описанное антитело. Также представлены нуклеиновые кислоты, кодирующие лёгкую и тяжёлую цепь описанного антитела. Изобретение позволяет получать улучшенные нейтрализующие антитела против GDF-8, способные на существенно более высоком уровне экспрессироваться в клетках-хозяевах по сравнению с ранее применявшимися антителами против GDF-8. Изобретение также расширяет арсенал средств и способов для увеличения мышечной массы или силы и лечения или профилактики мышечных нарушений, нервно-мышечных нарушений, нарушений обмена веществ, расстройств жировой ткани или костных нарушений. 19 н. и 46 з.п. ф-лы, 12 ил., 10 табл., 11 пр.
1. Антитело или его антигенсвязывающий фрагмент, которые специфически связываются с GDF-8, причем антитело или фрагмент включают:
вариабельную тяжелую (VH) область антитела, включающую первую, вторую и третью определяющие комплементарность области (CDR) из VH области, определяемой аминокислотной последовательностью SEQ ID NO: 44; и
вариабельную легкую (VL) область антитела, включающую первую, вторую и третью определяющие комплементарность области (CDR) из VL области, определяемой аминокислотной последовательностью SEQ ID NO: 46;
причем вышеупомянутая VH область включает лейцин в аминокислотной позиции, соответствующей номеру остатка 111 SEQ ID NO: 44 (позиция 108 по Kabat).
2. Антитело или фрагмент по п. 1, отличающиеся тем, что CDR1 VH включает SEQ ID NO: 10, CDR2 VH включает SEQ ID NO: 11, CDR3 VH включает SEQ ID NO: 12, CDR1 VL включает SEQ ID NO: 13, CDR2 VL включает SEQ ID NO: 14 и CDR3 VL включает SEQ ID NO: 15.
3. Антитело или фрагмент по п. 1, отличающиеся тем, что четвертая каркасная область (FR4) вышеупомянутой VH области включает аминокислоты 106-116 SEQ ID NO: 44.
4. Антитело или фрагмент по п. 1, отличающиеся тем, что вышеупомянутая VL область включает глицин в аминокислотной позиции, соответствующей номеру остатка 100 SEQ ID NO: 46 (позиция по Kabat 100).
5. Антитело или фрагмент по п. 1, отличающиеся тем, что вышеупомянутая VL область включает глутамин в аминокислотной позиции, соответствующей номеру остатка 100 SEQ ID NO: 46 (позиция по Kabat 100).
6. Антитело или фрагмент по п. 4, отличающиеся тем, что четвертая каркасная область вышеупомянутой VL области включает аминокислоты 98-107 SEQ ID NO: 46.
7. Антитело или фрагмент по п. 5, отличающиеся тем, что четвертая каркасная область вышеупомянутой VL области включает аминокислоты 98-107 SEQ ID NO: 9.
8. Антитело или фрагмент по п. 2, отличающиеся тем, что VH область включает аминокислотную последовательность SEQ ID NO: 44.
9. Антитело или фрагмент по п. 8, отличающиеся тем, что VL область включает аминокислотную последовательность SEQ ID NO: 46.
10. Антитело или фрагмент по п. 8, отличающиеся тем, что VL область включает аминокислотную последовательность SEQ ID NO: 9.
11. Антитело или фрагмент по п. 2, отличающиеся тем, что также включают константную область (СН) тяжелой цепи антитела, производную из подтипа антитела, выбранного из группы, состоящей из IgA, IgG, IgD, IgE или IgM.
12. Антитело или фрагмент по п. 11, отличающиеся тем, что вышеупомянутый подтип IgG также выбран из группы, состоящей из IgG1, IgG2, IgG3 и IgG4.
13. Антитело или фрагмент по п. 12, отличающиеся тем, что вышеупомянутым подтипом IgG является IgG1.
14. Антитело или фрагмент по п. 12, отличающиеся тем, что вышеупомянутый подтип IgG включает как минимум одну мутацию, меняющую функцию Fc домена.
15. Антитело или фрагмент по п. 11, отличающиеся тем, что тяжелая цепь вышеупомянутого антитела или фрагмента включает аминокислотную последовательность SEQ ID NO: 57.
16. Антитело или фрагмент по п. 2, отличающиеся тем, что также включают константную область (CL) легкой цепи антитела.
17. Антитело или фрагмент по п. 16, отличающиеся тем, что вышеупомянутая CL область является каппа или лямбда CL областью.
18. Антитело или фрагмент по п. 16, отличающиеся тем, что легкая цепь вышеупомянутого антитела или фрагмента включает аминокислотную последовательность SEQ ID NO: 17.
19. Антитело или фрагмент по п. 8, отличающиеся тем, что вышеупомянутая VH область кодируется нуклеотидной последовательностью SEQ ID NO: 43.
20. Антитело или фрагмент по п. 1, отличающиеся тем, что вышеупомянутые антитело или фрагмент связывают GDF-8 с Kd как минимум 10-6 М, 10-7 М, 10-8 М, 10-9 М, 10-10 М или 10-11 М.
21. Антитело или фрагмент по п. 1, отличающиеся тем, что вышеупомянутое антитело или фрагмент выбраны из группы, состоящей из Fab, F(ab')2, scFv, scFv-Fc, scFv-CH, scFab, scFv-''молния", диатела, триатела, тетратела, миниантитела, Fv и биспецифического антитела.
22. Антитело по п. 1, отличающееся тем, что количество вышеупомянутого антитела, вырабатываемого трансфицированными клетками, больше вырабатываемого в подобных условиях количества идентичного в других отношениях антитела, в котором VH область включает метионин в позиции 108 по Kabat.
23. Антитело по п. 22, отличающееся тем, что количество вышеупомянутого антитела, вырабатываемого трансфицированными клетками, превышает количество вышеупомянутого идентичного в других отношениях антитела, вырабатываемого в подобных условиях, как минимум приблизительно в 1,5 раза, 2 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 20 раз и 30 раз.
24. Антитело по п. 1, в котором тяжелая цепь указанного антитела содержит аминокислотную последовательность SEQ ID NO: 58 и легкая цепь указанного антитела содержит аминокислотную последовательность SEQ ID NO: 59.
25. Антитело по п. 24, отличающееся тем, что вышеупомянутое антитело, по сути, состоит из двух тяжелых цепей антитела, каждая из которых определяется аминокислотной последовательностью SEQ ID NO: 58; и двух легких цепей антитела, каждая из которых определяется аминокислотной последовательностью SEQ ID NO: 59.
26. Фармацевтическая композиция для лечения и профилактики состояний и нарушений, при которых снижение активности GDF-8 прямо или косвенно приносит терапевтический эффект, включающая терапевтически эффективное количество антитела по п. 1 и фармацевтически приемлемый носитель.
27. Выделенный полинуклеотид, кодирующий вариабельную тяжелую область антитела или фрагмента по п. 1, включающий нуклеотидную последовательность, кодирующую вариабельную область (VH) тяжелой цепи антитела, включающую первую, вторую и третью определяющие комплементарность области (CDR) из VH области, определяемой аминокислотной последовательностью SEQ ID NO: 44, где вышеупомянутая VH область включает лейцин в аминокислотной позиции, соответствующей номеру остатка 111 SEQ ID NO: 44 (позиция 108 по Kabat).
28. Выделенный полинуклеотид по п. 27, отличающийся тем, что нуклеотидной последовательностью, кодирующей VH область, является нуклеотидная последовательность SEQ ID NO: 43.
29. Выделенный полинуклеотид по п. 27, отличающийся тем, что также включает нуклеотидную последовательность, кодирующую СН область человеческого антитела.
30. Выделенный полинуклеотид, кодирующий вариабельную легкую область антитела или фрагмента по п. 1, включающий нуклеотидную последовательность, кодирующую вариабельную область (VL) легкой цепи антитела, включающую первую, вторую и третью определяющие комплементарность области (CDR) из VL области, определяемой аминокислотной последовательностью SEQ ID NO: 46, где вышеупомянутая VL область включает глицин в аминокислотной позиции, соответствующей номеру остатка 100 SEQ ID NO: 46 (позиция по Kabat 100).
31. Выделенный полинуклеотид, включающий нуклеотидную последовательность, кодирующую вариабельную область (VL) легкой цепи антитела, включающую первую, вторую и третью определяющие комплементарность области (CDR) из VL области, определяемой аминокислотной последовательностью SEQ ID NO: 46, где вышеупомянутая VL область включает глицин в аминокислотной позиции, соответствующей номеру остатка 100 SEQ ID NO: 46 (позиция по Kabat 100).
32. Выделенный полинуклеотид по п. 30, отличающийся тем, что дополнительно включает нуклеотидную последовательность, кодирующую CL область человеческого антитела.
33. Выделенный полинуклеотид по п. 31, отличающийся тем, что CL область кодируется нуклеотидной последовательностью SEQ ID NO: 16.
34. Выделенный полинуклеотид по п. 30, отличающийся тем, что нуклеотидная последовательность, кодирующая VL область, является нуклеотидной последовательностью SEQ ID NO: 45.
35. Вектор экспрессии, включающий полинуклеотид по п. 27, функционально связанный с конститутивно активным промотером.
36. Вектор экспрессии, включающий полинуклеотид по п. 30, функционально связанный с конститутивно активным промотером.
37. Вектор экспрессии, включающий полинуклеотид по п. 27 и полинуклеотид по п. 30, функционально связанные с конститутивно активным промотером.
38. Клетка-хозяин для экспрессирования антител, включающая вектор экспрессии по пп. 34 и 35.
39. Клетка-хозяин для экспрессирования антител, включающая вектор экспрессии по п. 36.
40. Способ выработки антитела или его фрагмента, которые специфически связываются с GDF-8, включающий стадии культивирования клетки-хозяина по пп. 37, 38 и выделения вырабатываемых ею антитела или фрагмента.
41. Способ увеличения массы мышц у млекопитающего, включающий введение млекопитающему эффективного для увеличения массы или силы мышц млекопитающего количества антитела или его антигенсвязывающего фрагмента по п. 1.
42. Способ по п. 40, отличающийся тем, что мышца является скелетной мышцей или сердечной мышцей.
43. Способ по п. 40, отличающийся тем, что скелетная мышца действует при дыхании.
44. Способ лечения мышечного расстройства, включающий введение субъекту, нуждающемуся в лечении мышечного расстройства, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по п. 1.
45. Способ по п. 43, отличающийся тем, что мышечное расстройство выбрано из группы, состоящей из мышечной дистрофии, мышечной атрофии, саркопении, кахексии, синдрома мышечной атрофии, возрастной потери мышечной массы или силы и слабости.
46. Способ по п. 44, отличающийся тем, что мышечным расстройством является мышечная дистрофия.
47. Способ по п. 45, отличающийся тем, что мышечной дистрофией является мышечная дистрофия Дюшенна.
48. Способ по п. 46, отличающийся тем, что лечение является эффективным для повышения показателей субъекта в тесте с шестиминутной ходьбой.
49. Способ по п. 47, отличающийся тем, что вышеупомянутое антитело вводят субъекту, подвергаемому лечению глюкокортикоидом.
50. Способ профилактики мышечного расстройства, включающий введение субъекту, нуждающемуся в профилактике, количества антитела или его антигенсвязывающего фрагмента по п. 1, эффективного для профилактики мышечного расстройства.
51. Способ по п. 49, отличающийся тем, что мышечное расстройство выбрано из группы, состоящей из мышечной дистрофии, мышечной атрофии, саркопении, кахексии, синдрома мышечной атрофии, возрастной потери мышечной массы или силы и слабости.
52. Способ по п. 50, отличающийся тем, что мышечным расстройством является мышечная дистрофия.
53. Способ по п. 51, отличающийся тем, что мышечной дистрофией является мышечная дистрофия Дюшенна.
54. Способ профилактики или лечения нервно-мышечного расстройства, включающий введение субъекту, нуждающемуся в лечении нервно-мышечного расстройства, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по п. 1.
55. Способ по п. 53, отличающийся тем, что нервно-мышечным расстройством является ALS.
56. Способ профилактики или лечения расстройства обмена веществ, включающий введение субъекту, нуждающемуся в лечении расстройства обмена веществ, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по п. 1.
57. Способ по п. 55, отличающийся тем, что нарушение обмена веществ выбрано из группы, состоящей из: сахарного диабета 2 типа, метаболического синдрома, синдрома X, резистентности к инсулину и нарушения толерантности к глюкозе.
58. Способ профилактики или лечения расстройства жировой ткани, включающий введение субъекту, нуждающемуся в лечении расстройства жировой ткани, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по п. 1.
59. Способ по п. 57, отличающийся тем, что расстройством жировой ткани является ожирение.
60. Способ профилактики или лечения расстройства, связанного с потерей костной массы, включающий введение субъекту, нуждающемуся в лечении расстройства связанного с потерей костной массы, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по п. 1.
61. Способ по п. 59, отличающийся тем, что связанное с потерей костной массы нарушение выбрано из группы, состоящей из: остеопороза, остеопении, остеоартрита и связанных с остеопорозом переломов костей.
62. Антитело, специфически связывающееся с GDF-8, включающее:
VH область антитела, состоящую из SEQ ID NO: 44, и VL область антитела, состоящую из SEQ ID NO: 46,
причем вышеупомянутое антитело экспрессируется на более высоком уровне в подобных условиях по сравнению с идентичным в других отношениях вторым антителом, включающим VH область, состоящую из SEQ ID NO: 7, и VL область, состоящую из SEQ ID NO: 9.
63. Антитело по п. 61, отличающееся тем, что вышеупомянутое антитело экспрессируется на более высоком уровне по сравнению с вышеупомянутым вторым антителом как минимум приблизительно в 1,5 раза, 2 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 20 раз или 30 раз.
64. Антитело по п. 62, отличающееся тем, что вышеупомянутое антитело экспрессируется приблизительно в 12 раз выше, чем вышеупомянутое второе антитело, при экспрессии в клетках COS.
65. Антитело по п. 62, отличающееся тем, что вышеупомянутое антитело экспрессируется приблизительно в 6 раз выше, чем вышеупомянутое второе антитело, при экспрессии в клетках СНО.
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Гнездовая сеялка | 1928 |
|
SU14112A1 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА ПРОТИВ GDF-8 И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2360925C2 |

