Изобретение относится к рекомбинантному штамму вируса осповакцины (VACV), продуцирующему секретируемый химерный белок, состоящий из гранулоцитарно-макрофагального колониестимулирующего фактора человека (ГМ-КСФ) и онкотоксического белка лактаптина. Оба белка в химере сохраняют свои биологические свойства, а рекомбинантный штамм обладает онколитической активностью в отношении опухолей человека и может быть использован в биотехнологии, в частности, в генетической инженерии для разработки лекарственных средств нового поколения для борьбы с онкологическими заболеваниями, в частности раком молочной железы.
Адьювантные свойства ГМ-КСФ хорошо известны и широко используются при создании различных протективных и терапевтических вакцин (Weisbart, 1985; Fleischmann, 1986; Gaudernack, 1999; Warren, 2000; Morrissey, 1987; Yoon, 2006; Somasundaram, 2015 [1-7]). ГМ-КСФ эффективно стимулирует противоопухолевый иммунный ответ в комбинации с клеточными, вирусными и ДНК-вакцинными препаратами (Somasundaram, 2015; Lai, 2011; Hellerstein, 2012; Zhang, 2015; Chen, 2014; Fowler, 2012; Zheng, 2011 [7-13]). Показано, что ГМ-КСФ усиливает индукцию первичного иммунного ответа за счет активации и рекрутирования (chemo-attraction) антиген-презентирующих клеток (Weisbart, 1985; Fleischmann, 1986 [1-2]). ГМ-КСФ усиливает пролиферацию Т-клеток и индуцирует продукцию как Th1, так и Th2 (Morrissey, 1987; Yoon, 2006 [5-6]). ГМ-КСФ работает как генетический адьювант при совместном введении в клетку ДНК цитокина и антигена и увеличивает протективный иммунный ответ на 25-71% (Somasundaram, 2015; Wang, 2009; Hartoonian, 2009; Encke, 2006 [7, 14-16]).
Ген ГМ-КСФ широко используется в качестве трансгена при конструировании рекомбинантных онколитических вирусов (Кочнева, 2012 [17]). В частности, ген ГМ-КСФ человека был встроен в геном двух штаммов VACV: WR (JX-963) (Thorne, 2007 [18]) и Wyeth (JX-594) (Kirn, 2010 [19]) в район гена вирусной тимидинкиназы (tk-ген) под контролем синтетического ранне-позднего промотора VACV (Merchlinsky, 1997 [20]). Эти штаммы в настоящее время успешно проходят клинические испытания в качестве противоопухолевых препаратов (Breitbach, 2011 [21]).
На основе российского VACV штамма Л-ИВП (GenBank Acc. KP233807) нами сконструирован рекомбинантный штамм VV-GMCSF-S1/3, содержащий встройку гена ГМ-КСФ человека в центральной части tk-гена с одновременной его инактивацией (Кочнева, 2015 [22]). Ген ГМ-КСФ в составе рекомбинанта экспрессируется под контролем природного промотора VACV P7.5k и продуцирует секретируемую форму биологически активного ГМ-КСФ человека в клетках млекопитающих на уровне 1-40 мкг на мл культуральной среды (Гражданцева, 2015 [23]).
Представляется целесообразным использовать адьювантные свойства ГМ-КСФ с целью стабилизации структуры онкотоксических белков и усиления их противоопухолевой активности за счет индукции специфического иммунного ответа против раковых клеток. При этом наиболее перспективным является создание химерных конструкций, в которых ГМ-КСФ стоит в N-концевой части белка и через гибкий линкер соединяется с онкотоксическим белком. Такая конструкция обеспечит секрецию химерного белка в межклеточной пространство и, так называемый, «эффект соседа» (bystander effect) (Алексеенко, 2011 [24]) - проникновение онкотоксического белка в соседние раковые клетки и индукцию их гибели, что многократно усиливает терапевтический эффект.Кроме того, гибкий линкер позволяет сохранить биологические свойства обоих белков в химерной конструкции и, соответственно, индукцию специфического противоопухолевого иммунитета за счет экспрессии ГМ-КСФ.
В качестве онкотоксического белка нами использован лактаптин, который является фрагментом каппа-казеина молока человека (23-134 а.о.) и специфически индуцирует гибель клеток рака молочной железы человека in vitro и in vivo (Koval, 2014 [25]).
Для создания эффективного рекомбинантного онколитического вируса необходимо использовать репликативно-компетентные вектора, однако для повышения безопасности необходимо также удалять гены вирулентности, не существенные для репродукции вируса в быстро делящихся раковых клетках. Показано, что сочетанное подавление генов тимидинкиназы (tk-ген) и ростового фактора (virus growth factor, VGF) VACV приводит к практически полному отсутствию репликации вируса в неделящихся клетках (McCart, 2007 [26]). Таким образом, делеции в этих двух генах приводят не только к аттенуации вируса, но и к повышению его онкоселективности.
В качестве штамма-реципиента для создания онколитического вируса нами был использован сконструированный ранее на основе российского штамма VACV Л-ИВП рекомбинантный вариант VVdGF2/6 с делецией гена VGF (Кочнева, 2013 [27]). Химерная конструкция была введена в структурную часть tk-гена штамма VVdGF2/6 с одновременным удалением фрагмента tk-гена и, соответственно, его инактивацией. Химерная конструкция состоит из генов двух белков, ГМ-КСФ человека и лактаптина, соединенных гибким линкером GlyGlyGlySer.
Наиболее близким аналогом заявляемого изобретения (прототипом) является рекомбинантный штамм VV-GMCSF-S1/3 (Патент РФ №2565544, МПК C12N 15/27, опубл. 20.10.2015 г.[22]), сконструированный на основе штамма Л-ИВП вируса осповакцины, содержащий встройку гена ГМ-КСФ человека (GenBank Acc. M11220.1) в районе делеции фрагмента tk-гена между позициями 81277 и 81308 п.н. (GenBank Acc. KP233807.1); ген ГМ-КСФ человека экспрессируется под контролем природного промотора Р7.5k вируса осповакцины и продуцирует секретируемую форму биологически активного ГМ-КСФ человека в клетках млекопитающих на уровне 1-40 мкг на мл культуральной среды; штамм VV-GMCSF-S1/3 депонирован в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» под номером V-631.
Однако рекомбинантный штамм-прототип не обеспечивает дополнительную аттенуацию (ослабление) вируса в отношении нормальных клеток вследствие отсутствия в его геноме делеции гена VGF и дополнительное усиление литической активности в отношении раковых клеток вследствие отсутствия в его конструкции цитотоксических генов, обеспечивающих специфическую гибель раковых клеток.
Техническим результатом заявляемого изобретения является обеспечение дополнительной аттенуации (ослабления) рекомбинантного онколитического вируса VACV в отношении нормальных клеток и усиление его литической активности в отношении раковых клеток.
Указанный технический результат достигается созданием двойного рекомбинантного штамма VV-GMCSF/lact-dGF, сконструированного на основе штамма Л-ИВП вируса осповакцины и содержащего делецию фрагмента tk-гена, в район которой между позициями 81277 и 81307 п.н. (GenBank Acc. KP233807.1) встроен оперон, состоящий из химерного транскрипта GMCSF/lact [см. Приложение], экспрессируемый под контролем природного промотора P7.5k VACV, включающий 276 п.н. и состоящий из генов ГМ-КСФ человека (GenBank Acc. M11220.1) и лактаптина - фрагмента гена каппа-казеина человека, кодирующего пептид размером 23-134 а.о., соединенных гибким линкером ggc gga ggc gga tca (GlyGlyGlySer). Штамм также содержит делецию в левом концевом районе вирусного генома, соответствующую позициям 7770-8071 п.н. (GenBank Acc. KP233807.1), включающую фрагмент гена VGF размером 170 п.н., соответствующую позициям 7801-8071 п.н. Штамм VV-GMCSF/lact-dGF имеет сниженную более, чем в 100 раз вирулентность по сравнению с исходным штаммом Л-ИВП (вирулентность оценивали по величине 50%-ной летальной дозы вирусов для 10-дневных куриных эмбрионов), а также более высокую онколитическую активность в отношении клеток опухолей человека различного генеза. Штамм VV-GMCSF/lact-dGF депонирован в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» под номером V-711.
Штамм VV-GMCSF/lact-dGF образуется за счет одиночного кроссинговера между ДНК штамма VVdGF2/6 (Кочнева, 2013 [27]) и плазмидной ДНК pXJP-GMCSF/lact-PE/L-Pat (фиг. 1) в цитоплазме клеток CV-1, последующей селекции полученной нестабильной конструкции с добавлением в культуральную среду пуромицина за счет наличия в ней гена устойчивости к этому антибиотику (Pat) (Кочнева, 2013 [27]) и внутримолекулярной рекомбинации после снятия селективных условий.
Штамм VV-GMCSF/lact-dGF характеризуется следующими признаками. Морфологические признаки. Штамм обладает свойствами типичного представителя VACV, но в отличие от векторного вируса VVdGF2/6 с фенотипом TK+VGF- имеет фенотип ТК-ГМКСФ+VGF-Lact+ (фиг. 2А). Показано, что при заражении клеток CV-1 штамм VV-GMCSF/lact-dGF продуцирует секретируемый химерный белок GMCSF/lact (фиг. 3).
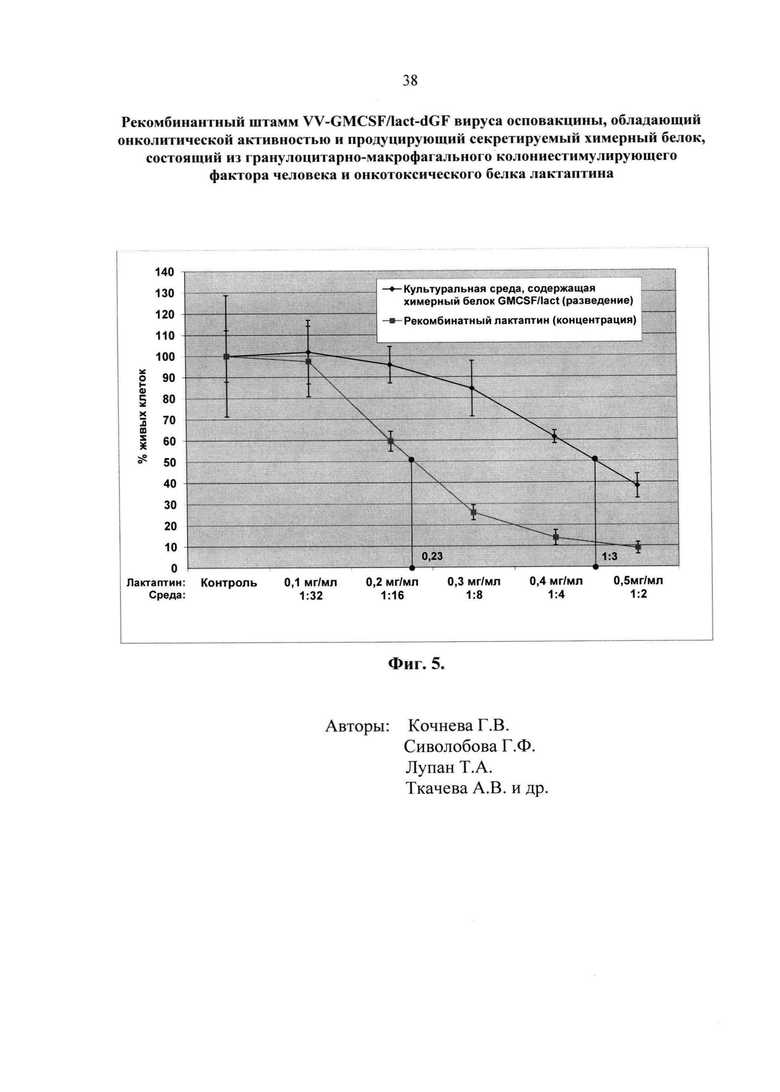
Физиолого-биохимические характеристики и культуральные свойства штамма. ДНК рекомбинантного штамма VV-GMCSF/lact-dGF имеет длину около 200000 п.н. Наличие в его геноме встройки химерного гена GMCSF/lact и делеции гена VGF подтверждено методом ПЦР (фиг. 2Б) с использованием пар праймеров на область tk-гена, трансгенов ГМ-КСФ и лактаптина и гена VGF. Позиции праймеров указаны на фиг. 2А. Экспрессия химерного гена GMCSF/lact и секреция белка в культуральную среду подтверждена Вестерн-блот анализом лизатов и культуральной среды клеток CV-1, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF с использованием коммерческих антител против ГМ-КСФ как описано в (Гражданцева, 2015 [23]) (фиг. 3A). Для выявления лактаптина в составе химерного белка использовали моноклональные антитела anti-RL2 (Koval, 2014 [25]) (фиг. 3Б). Биологическая активность ГМ-КСФ в составе секретируемого химерного белка была оценена в культуре клеток эритролейкоза человека TF-1, которая является цитокин-зависимой и пролиферирует только в присутствии ГМ-КСФ и ряда других цитокинов человека как описано в (Гражданцева, 2015 [23]) (фиг. 4). Как следует из фиг. 4, 2-кратный пролиферативный эффект (200%) достигается при разведении культуральной среды, полученной при инфицировании клеток CV-1, в 1100 раз (1:1100), что соответствует концентрации ГМ-КСФ 1,65 мкг/мл по калибровочной кривой. Активность ГМ-КСФ в составе химерного белка несколько ниже, чем индивидуально экспрессированного ГМ-КСФ (4 мкг/мл среды) в культуральной среде тех же клеток CV-1 (Кочнева, 2015 [22]), однако эти различия недостоверны при Р<0,05. Биологическую активность лактаптина в составе химерного белка GMCSF/lact оценивали по его способности индуцировать гибель раковых клеток молочной железы человека ВТ-549 (фиг. 5). Процент гибели клеток рассчитывали в фотометрическом тесте с использованием субстрата для митохондриальных дегидрогеназ 2,3-бис-(2-метокси-4-нитро-5-сульфо-фенил)-2H-тетразолий-5-карбоксанилида (реагент XTT). Как следует из фиг. 5, 50%-ный цитотоксический эффект достигается при разведении культуральной среды, содержащей химерный белок GMCSF/lact, 1:3. Аналогичный эффект достигается при концентрации 0,23 мг/мл рекомбинантного аналога лактаптина, экспрессированного в системе E.coli, что соответствует ранее опубликованным данным для этого белка (Koval, 2014 [25]). Учитывая разведение культуральной среды 1:3, концентрация химерного белка должна составлять 0,69 мг/мл, что существенно превосходит концентрацию, измеренную по активности ГМ-КСФ (1,65 мкг/мл). Такое несогласование связано, по всей видимости, как с существенным различием систем оценки биологической активности и свойств самих белок, так и с усилением онкотоксичности лактаптина в химерной молекуле за счет стабилизирующей и адьювантной активности ГМ-КСФ. Эти данные подтверждаются дальнейшими экспериментами по оценке онколитической активности рекомбинантного штамма VV-GMCSF/lact-dGF.
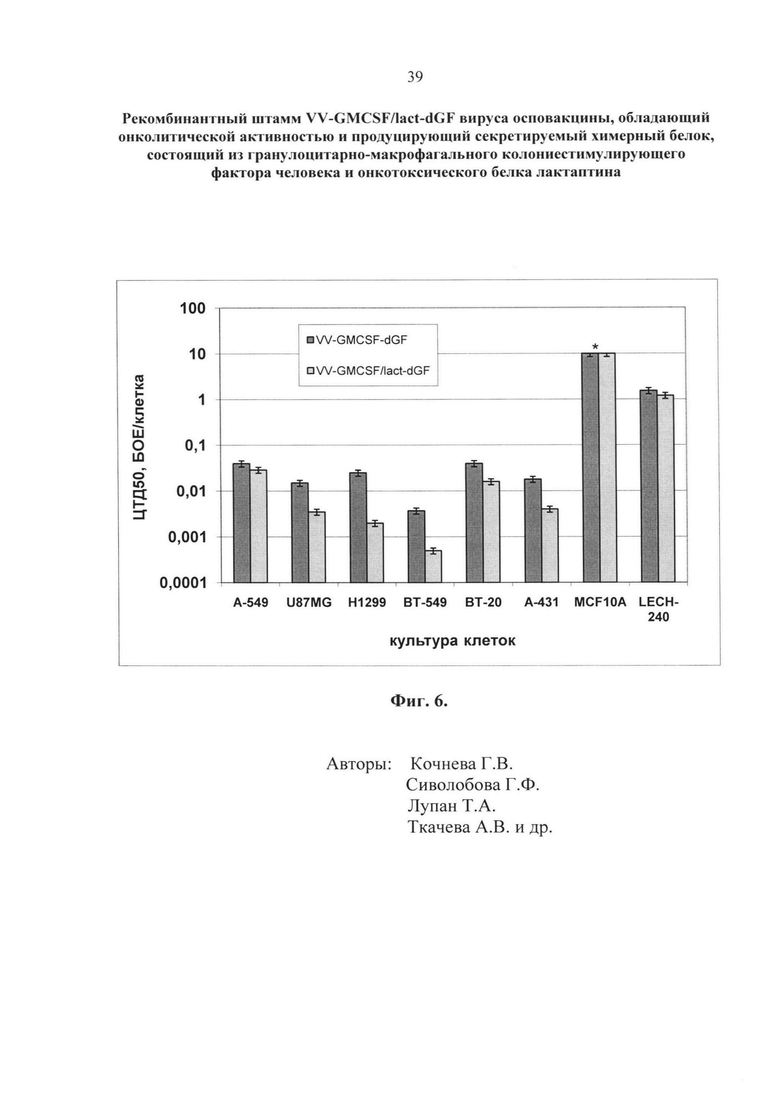
Рекомбинантный штамм VV-GMCSF/lact-dGF обладает повышенной цитотоксической активностью не только в отношении клеток опухолей молочной железы человека, но также и в отношении опухолей другого генеза (фиг. 6), в частности:
А-549 - клетки карциномы легкого человека;
U87MG - клетки эпителиальной глиобластомы/астроцитомы человека;
Н1299 - клетки немелкоклеточного рака легкого человека;
ВТ-549 - клетки эстроген-независимой инвазивной дуктальной карциномы молочной железы, первичный узел;
ВТ-20 - клетки эстроген-независимой карциномы молочной железы, первичный узел;
А-431 - клетки эпидермоидной карциномы шейки матки.
Для оценки вклада лактаптина специально сконструировали рекомбинант VV-GMCSF-dGF, в котором имеется встройка гена ГМ-КСФ человека в районе tk-гена (аналогично VV-GMCSF-1/3) и делеция гена VGF (аналогично ранее описанному варианту VVdGF2/6 (Кочнева, 2013 [27]), но отсутствует встройка гена лактаптина. Штамм VV-GMCSF-dGF депонирован в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» под номером V-712.
Как следует из фиг. 6, цитотоксическая активность в отношении клеток опухолей человека штамма VV-GMCSF/lact-dGF в 1,4-12,5 раз выше (ЦТД50 0,029-0,0005 БОЕ/кл.), чем штамма VV-GMCSF-dGF (ЦТД50 0,04-0,0037 БОЕ/кл.). Цифровые значения ЦТД50, соответствующие данным фиг.6, представлены в таблице 1.
Таблица 1. Цитотоксическая активность рекомбинантных штаммов VACV.
Для оценки онкоспецифичности мы использовали две культуры клеток из нормальных тканей человека: MCF10A – культура клеток нормального эпителия молочной железы человека и LECH-240 – диплоидная культура клеток легкого эмбриона человека. Как следует из данных, представленных в таблице 1 и на фиг. 6, индекс онкоселективности, рассчитанный как отношение ЦТД50 в нормальных клетках к ЦТД50 в раковых клетках для каждого вирусного штамма, составляет более 30 для клеток LECH-240 и более 250 для клеток MCF10A. Различия в индексах селективности связаны с тем, что клетки эмбриона обладают достаточно высокой пролиферативной активностью, что, в определенной степени, роднит их с раковыми и повышает репликативную активность рекомбинантов вируса осповакцины. Таким образом, созданный нами рекомбинантный штамм VV-GMCSF/lact-dGF обладает онкоспецифичностью и лизирует опухолевые клетки в 30-250 раз эффективнее, чем нормальные клетки человека. Штамм VV-GMCSF/lact-dGF обладает повышенной литической активностью в отношении опухолевых клеток человека за счет введения лактаптина в состав химерного белка.
Инактивация генов тимидинкиназы и вирусного ростового фактора обеспечивает существенное снижение вирулентности рекомбинантного штамма VV-GMCSF/lact-dGF по сравнению с немодифицированным вакцинным штаммом Л-ИВП. Оценку вирулентности проводили путем определения величины 50%-ной летальной дозы (ЛД50) вирусов для 10-дневных куриных эмбрионов (КЭ). КЭ заражали 10-кратными разведениями вирусной суспензии с заранее определенным титром и оценивали гибель через 48 часов. Использовали одинаковые дозы заражения, также как и их диапазон, при сравнении вирулентности исходного (Л-ИВП) и рекомбинантного (VV-GMCSF/lact-dGF) вирусных штаммов. Исходный титр обоих вирусных штаммов составил 1,0*109 БОЕ/мл.
В качестве отрицательного контроля использовали буферный раствор 0,005 М Трис-HCl буфер, рН-8,0. Этот раствор применялся для получения очищенных препаратов вируса и подготовки разведений.
Для расчета ЛД50 диапазон разведений вирусных образцов подбирается так, чтобы в разведении с наибольшей концентрацией произошла гибель 100% КЭ, а в разведении с наименьшей концентрацией - 0% КЭ. Мы использовали диапазон разведений от 1:10 (-1) до 1:10000000 (-7) для рекомбинантного штамма и Л-ИВП, что соответствует диапазону доз 107-10 БОЕ/КЭ. Каждое разведение делали в 5-ти повторностях (5 КЭ), время инкубации зараженных КЭ составило 48 часов. Определение ЛД50 проводили по методу Кербера с использованием следующей схемы расчета:
1. Для каждого разведения вируса определяем количество мертвых эмбрионов (есть эффект, +) и количество живых (нет эффекта, –).
2. Считаем доли эмбрионов (Li), где зарегистрирован эффект, то есть мертвых.
3. Считаем сумму (Σ Li) этих долей от максимального разведения с Li=1 до минимального разведения с Li=0 (в таблицах выделены жирным шрифтом).
4. Разведение, вызывающее 50%-ную гибель эмбрионов, считается по формуле: ЛД50=Cmax⋅100,5–Σ Li
Результаты экспериментов представлены в таблицах 2 и 3.
В таблице 2 приведены данные о вирулентности штамма VV-GMCSF/lact-dGF для 10-дневных КЭ.
Таблица 2. Вирулентность штамма VV-GMCSF/lact-dGF
для 10-дневных КЭ.
=10-1⋅10-1,7=10-2,7
103,7 ЛД50/мл
В таблице 3 представлены данные о вирулентности штамма Л-ИВП VACV для 10-дневных КЭ.
Таблица 3. Вирулентность штамма Л-ИВП VACV
для 10-дневных КЭ.
=10-4⋅10-0,9=10-4,9
105,9 ЛД50/мл
Введение буферного раствора гибели КЭ не вызывало.
Уровень аттенуации рассчитывали как разность десятичных логарифмов титра в единицах ЛД50/мл для штамма Л-ИВП и рекомбинантного штамма VV-GMCSF/lact-dGF. В данном случае она составила 158 (105,9–3,7=2,2), то есть рекомбинантный штамм в 158 раз менее вирулентен, чем Л-ИВП.
Изобретение иллюстрируется следующими фигурами графических изображений:
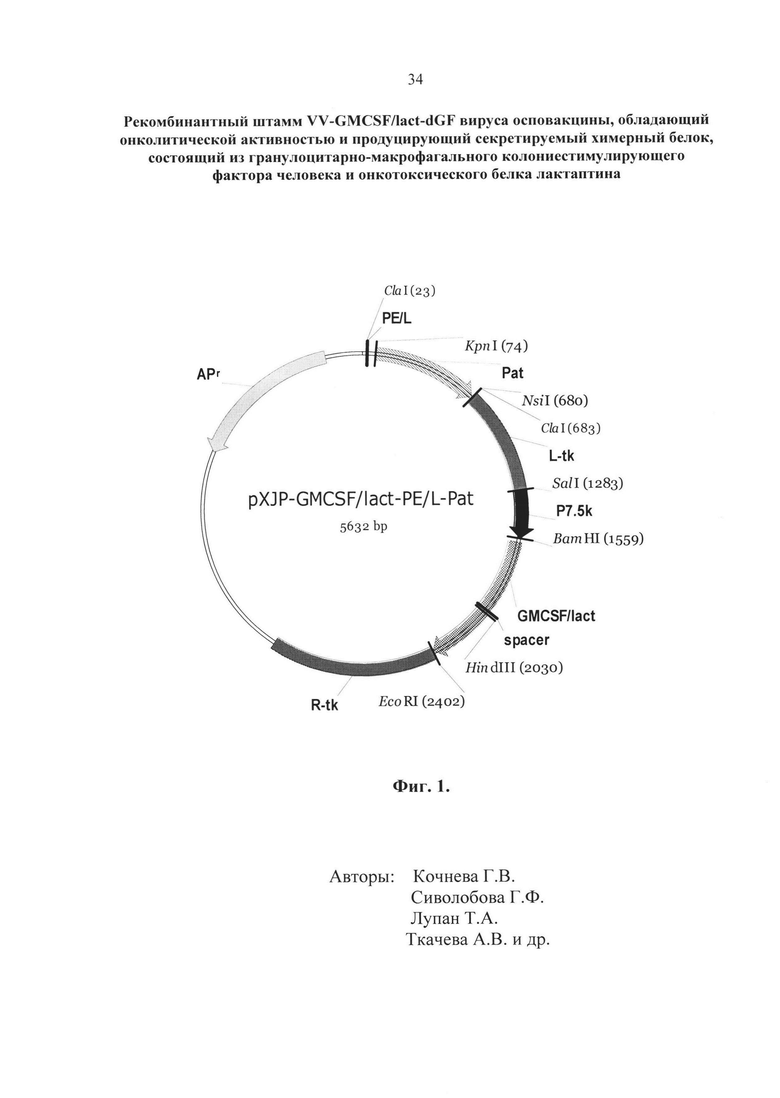
- Фиг. 1. Физическая и генетическая карта плазмиды pXJP-GMCSF/lact- PE/L-Pat. L-tk - фрагмент генома VACV штамма Л-ИВП размером 595 п.н., расположен в левой части гена тимидинкиназы и соответствует позициям 80682-81277 (GenBank, Accession KP233807); R-tk - фрагмент генома VACV штамма Л-ИВП размером 933 п.н., расположен в правой части tk-гена и соответствует позициям 81307-82251(GenBank, Accession KP233807); фланки расположены так, что образуется делеция размером 30 п.н. в центре tk-гена. В район делеции встроен оперон, состоящий из химерного гена GMCSF/lact под контролем природного промотора VACV P7.5k (276 п.н.). Spacer - гибкий линкер GlyGlyGlySer, связывающий ГМ-КСФ и лактаптин в химерной молекуле. Наличие фланков обеспечивает встройку оперона в район tk-гена. Наличие гена Pat под контролем синтетического промотора PE/L (Merchlinsky, 1997) обеспечивает селекцию рекомбинантных клонов.
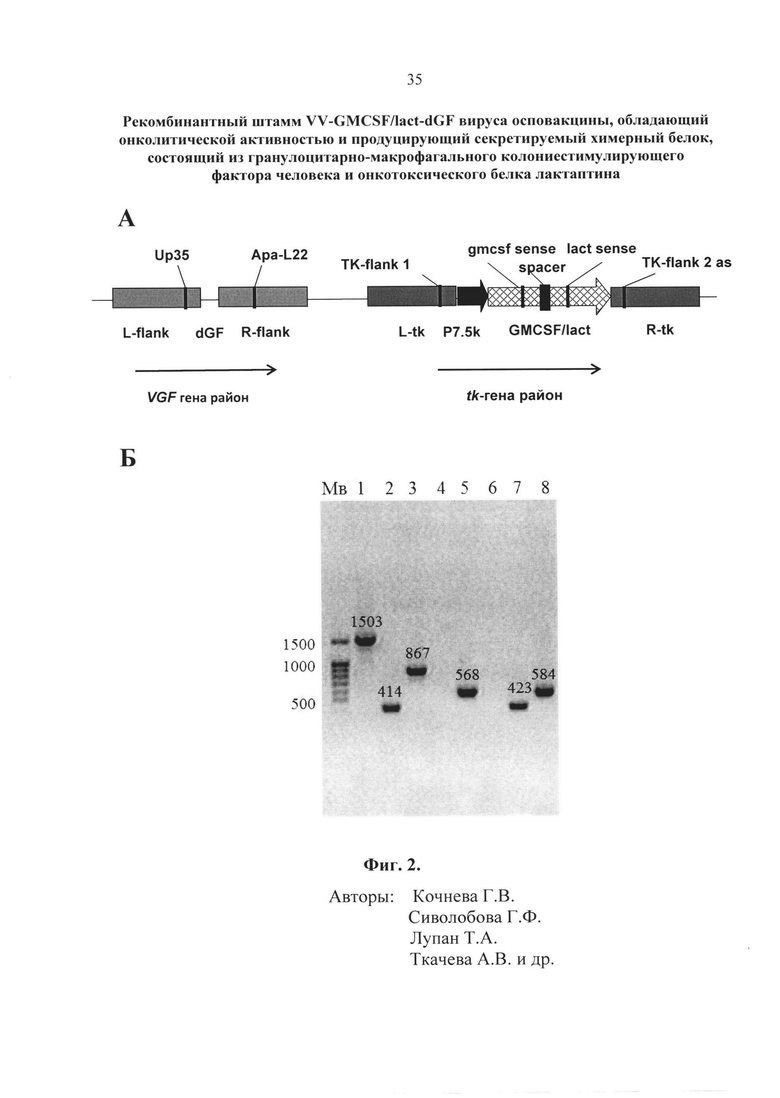
- Фиг. 2. Структура рекомбинантного штамма VV-GMCSF/lact-dGF. A – схема генома с указанием позиций праймеров, dGF - делеция VGF-гена, остальные подписи как на фиг. 1. Б – результаты ПЦР-анализа рекомбинантного штамма VV-GMCSF/lact-dGF в сравнении со штаммом-реципиентом VVdGF2/6 и исходным штаммом Л-ИВП с использованием трех пар праймеров (ТК-flank1 sense/ gmcsf sense/ lact sense × TK-flank 2 as) для подтверждения встройки химерного гена GMCSF/lact (дорожки 1-6) и пары праймеров (Up35×Apa-L22) для подтверждения делеции гена VGF (дорожки 7-8). Мв - маркеры молекулярных весов. 1 - рекомбинант VV-GMCSF/lact-dGF, праймеры ТК-flank1 sense×TK-flank 2 as (фрагмент 1503 п.н.), 2 - штамм Л-ИВП, праймеры ТК-flank1 sense×TK-flank 2 as (фрагмент 414 п.н.); 3 - рекомбинант VV-GMCSF/lact-dGF, праймеры gmcsf sense×TK-flank 2 as (фрагмент 867 п.н.), 4 – штамм Л-ИВП, праймеры gmcsf sense×TK-flank 2 as (нет фрагмента, поскольку праймер gmcsf sense лежит внутри встроенного гена ГМ-КСФ); 5 - рекомбинант VV-GMCSF/lact-dGF, праймеры lact sense ×TK-flank 2 as (фрагмент 568 п.н.), 6 – штамм Л-ИВП, праймеры lact sense×TK-flank 2 as (нет фрагмента, поскольку праймер lact sense лежит внутри встроенного гена Lact); 7 - рекомбинант VV-GMCSF/lact-dGF, праймеры Up35×Apa-L22 (фрагмент 423 п.н.), 8 - штамм Л-ИВП, праймеры Up35×Apa-L22 (фрагмент 584 п.н.).
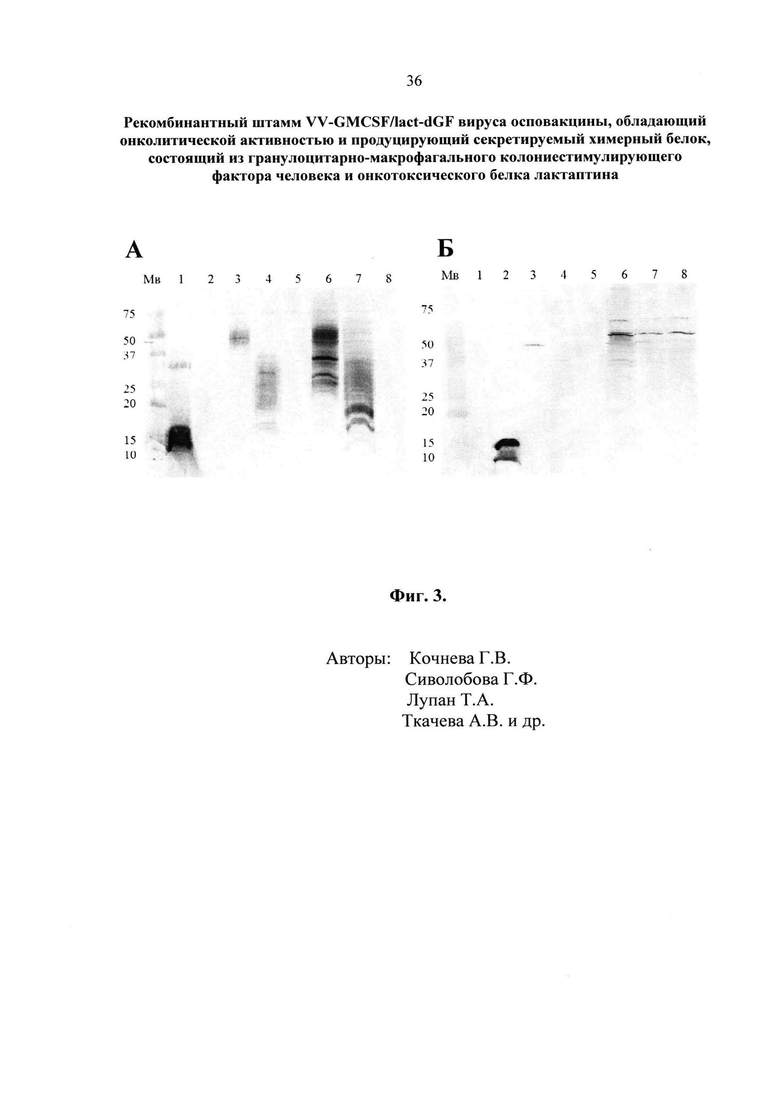
- Фиг. 3. Вестерн-блот анализ экспрессии химерного гена GMCSF/lact в составе рекомбинантного штамма VV-GMCSF/lact-dGF в культуре клеток почки африканской зеленой мартышки CV-1. A – в качестве первичных антител использовали коммерческие поликлональные антитела кролика против ГМ-КСФ человека (PerroTech), в качестве вторичных антител - антикроличьи IgG, коньюгированные в фосфатазой (Sigma). Б – для визуализации использовали моноклональные антитела к лактаптину, коньюгированные с пероксидазой хрена F14-HRP. Мв – контроль молекулярных весов. 1 – контроль, негликозилированный ГМ-КСФ человека, продуцированный в клетках E.coli (14.4 кДа); 2 - контроль, лактаптин, продуцированный в клетках E.coli (14 кДа); 3 – культуральная среда клеток, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF (полностью гликозилированная секретируемая форма химерного белка GMCSF/lact, 46 кДа); 4 – культуральная среда клеток, инфицированных рекомбинантным штаммом VV-GMCSF-S1/3 (Кочнева, 2015 [22]), продуцирующим ГМ-КСФ (гликозилированная секретируемая форма ГМ-КСФ, 25-32 кДа); 5 – отрицательный контроль, культуральная среда клеток, инфицированных исходным нерекомбинантным исходным штаммом Л-ИВП; 6 – лизат клеток, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF (внутриклеточная форма химерного белка GMCSF/lact с разной степенью гликозилирования, 28-46 кДа); 7-лизат клеток, инфицированных рекомбинантным штаммом VV-GMCSF-S1/3 (разная степень гликозилирования ГМ-КСФ, 18-32 кДа), 8 – отрицательный контроль, лизат клеток, инфицированных Л-ИВП.
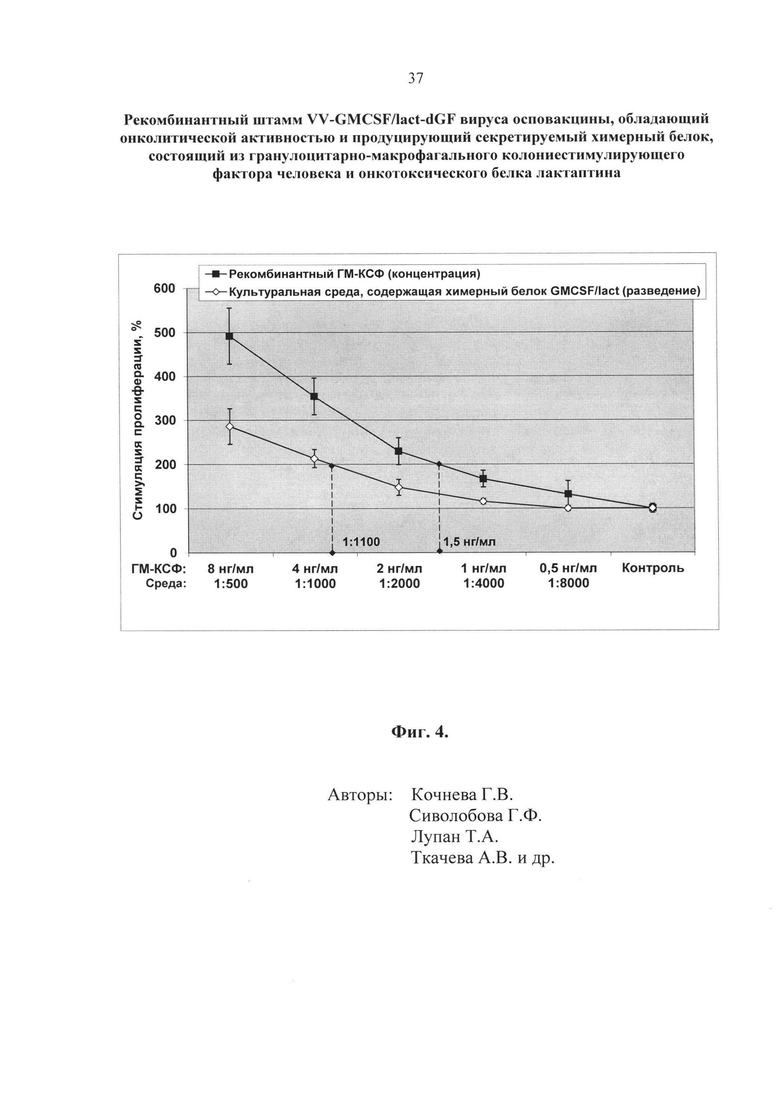
- Фиг. 4. Анализ биологической активности ГМ-КСФ человека в составе химерного белка GMCSF/lact. Биологическую активность ГМ-КСФ человека в составе химерного белка GMCSF/lact, секретируемого из клеток CV-1, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF, оценивали по выраженному в процентах уровню стимуляции пролиферации ГМКСФ-зависимых клеток эритролейкоза человека TF-1 (ось Y). За 100% принимали количество живых клеток, выращенных с добавлением неразведенной культуральной среды клеток CV-1, инфицированных исходным штаммом VACV Л-ИВП (отрицательный контроль). На оси Х представлены разведения культуральной среды клеток CV-1, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF, и концентрации рекомбинантного очищенного препарата ГМ-КСФ, полученного в клетках E. coli (калибровочная кривая).
- Фиг. 5. Анализ биологической активности лактаптина, экспрессированного в составе химерного белка GMCSF/lact. Биологическую активность лактаптина в культуральной среде клеток CV-1, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF, оценивали по выраженному в процентах уровню цитотоксичности различных разведений культуральной среды для клеток рака молочной железы человека ВТ-549 (ось Y). За 100% принимали количество живых клеток, инкубированных с добавлением неразведенной культуральной среды клеток CV-1, инфицированных исходным штаммом VACV Л-ИВП (положительный контроль). На оси Х представлены разведения культуральной среды клеток CV-1, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF, и концентрации рекомбинантного очищенного препарата лактаптина, полученного в клетках E. coli (калибровочная кривая).
- Фиг. 6. Сравнительная онколитическая активность рекомбинантных штаммов VACV в раковых клетках человека различного генеза. На оси Y представлены значения 50%-ной цитотоксической дозы в единицах БОЕ/клетка, логарифмическая шкала. На оси Х – культуры клеток. Использовали шесть раковых культур клеток: A-549; U87MG; H1299; BT-549, ВТ-20, А-431 и две нормальных культуры клеток человека: MCF10A и LECH-240. Клетки выращивали в 96-ти луночных планшетах, инфицировали 10-кратными разведениями вируса в диапазоне доз 0,0001–10 БОЕ на клетку и рассчитывали 50%-ную цитотоксическую дозу для каждого рекомбинантного штамма (ЦТД50) в фотометрическом тесте с использованием субстрата для митохондриальных дегидрогеназ 2,3-бис-(2-метокси-4-нитро-5-сульфо-фенил)-2H-тетразолий-5-карбоксанилида (реагент XTT).
Для лучшего понимания сущности изобретения ниже следуют примеры его осуществления.
Пример 1. Конструирование рекомбинантной плазмидной ДНК pXJP-GMCSF/lact-PE/L-Pat. Плазмида pXJP-GMCSF/lact-PE/L-Pat была получена на основе pXJP5.2 (Кочнева, 2015 [22]). На первом этапе в плазмиду pXJP5.2 по ClaI-сайту был встроен ген Pat под контролем синтетического промотора PE/L (Merchlinsky, 1997 [20]) для селекции рекомбинантных клонов VACV. Для этой цели были синтезированы два олигонуклеотида, которые содержали последовательность промотора PE/L и сайты узнавания ряда рестриктаз. При отжиге олигонуклеотиды образуют дуплекс следующей структуры:
ClaI KpnI SpeI Nsil ClaI
CGATGGCCAAAAATTGAAATTTTATTTTTTTTTTTTGGAATATAAAGGTACCACTAGTATGCATAT
TACCGGTTTTTAACTTTAAAATAAAAAAAAAAAACCTTATATTTCCATGGTGATCATACGTATAGC
После встройки этого дуплекса по ClaI-сайту в полученную плазмиду по KpnI – Nsil сайтам был встроен ген Pat. Фрагмент ДНК, содержащий ген Pat получали методом ПЦР на матрице pGEM-Puro-DS-Apo (Чумаков, 2013 [28]) с использованием пары праймеров, в состав которых введены сайты узнавания рестриктаз KpnI и Nsil:
KpnI
Puro Up 5´- GCATCGGTACCATGACCGAGTACAAGCCCACGG
Nsil
Puro Low 5´- GCATCATGCATTCAGGCACCGGGCTTGCGGGTCA
Полученная в результате лигирования плазмидная ДНК, обозначенная как pXJP5.2-PE/L-Pat, содержит ген Pat под контролем синтетического ранне-позднего промотора PE/L (для селекции рекомбинантных клонов VACV).
Для конструирования гена химерного белка GMCSF/lact в плазмиду pXJP5.2-PE/L-Pat были последовательно встроены ген GMCSF, лишенный стоп-кодона, но содержащий на С-конце нуклеотидную последовательность, кодирующую гибкий линкер Gly-Gly-Gly-Gly-Ser, и ген лактаптина со стоп-кодоном. На первом этапе в плазмиду pXJP5.2-PE/L-Pat был встроен ген лактаптина, полученный методом ПЦР на матрице плазмидной ДНК pGSDI/RL2 (Semenov, 2010 [29]) с использованием пары праймеров, содержащих сайты эндонуклеаз рестрикции HindIII, EcoRI:
HindIII
FR2 For: 5’ – AAATAAAGCTTATGAACCAGAAACAACCAGCA
EcoRI
FR2 Rev: 5’ –CTATCGAATTCTTAGTGATGGTGATGGTGATGTG
Полученный ПЦР-фрагмент встроен в плазмиду pXJP5.2-PE/L-Pat по HindIII - EcoRI и промежуточная плазмида была названа pXJP5.2-Lact-PE/L-Pat. На втором этапе в эту плазмиду по сайтам BamHI – HindIII, расположенным перед геном лактаптина, был встроен ген ГМ-КСФ, лишенный стоп-кодона, но содержащий на С-конце нуклеотидную последовательность, кодирующую гибкий линкер Gly-Gly-Gly-Ser. Ген GMCSF был получен из плазмиды pXJP5.2-GMCSF (Кочнева, 2015 [22]) методом ПЦР с использованием пары праймеров:
BamHI
ChimUp 5’- GCATCGGATCC GCTAAAGTTCTCTGGAGGATGTGGCTGCA
HindIII Ser Gly Gly Gly Gly
ChimLow 5’ - AAATAAAGCTTTGATCCGCCTCCGCCCTCCTGGACTGGCTCCCAGCAGT
Таким образом, полученная плазмидная ДНК pXJP-GMCSF/lact-PE/L-Pat (фиг. 1) содержит фрагмент ДНК VACV, соответствующий позициям нуклеотидов 80682-81277 генома штамма Л-ИВП (GenBank, Accession KP233807), разделенного на левый (595 п.н., позиции 80682-81277) и правый (933 п.н., позиции 81307-82251) фланки с делецией 30 п.н. в центре tk-гена. В район делеции встроен оперон, состоящий из химерного гена GMCSF/lact под контролем природного промотора VACV P7.5k (276 п.н.). Наличие фланков обеспечивает встройку оперона в район tk-гена. Наличие гена Pat под контролем синтетического промотора PE/L обеспечивает селекцию рекомбинантных клонов. Структура полученной плазмидной ДНК подтверждена секвенированием.
Пример 2. Трансфекция клеток CV-1 рекомбинантной плазмидой pXJP-GMCSF/lact-PE/L-Pat и получение рекомбинантного штамма VV-GMCSF/lact-dGF. Для проведения трансфекции ДНК плазмиды pXJP-GMCSF/lact-PE/L-Pat была наработана в препаративных количествах из 1000 мл среды Лурия-Бертани и выделена с использованием набора лабораторных реагентов для выделения плазмидной ДНК, очищенной от эндотоксинов «EndoFree Plasmid Maxi Kit» (Qiagen).
Рекомбинантные VACV получали с использованием реагента для трансфекции Lipofectamine™ LTX, 1 мл и реагент Plus (Invitrogen). Трансфекцию проводили на 90%-ном монослое клеток CV-1, выращенном в шести-луночных планшетах (Greiner). Клетки инфицировали штаммом-реципиентом VVdGF2/6 (Кочнева, 2013 [27]) с множественностью 0,05 БОЕ/клетка, и через 1 час инкубации при 37ºС добавляли смесь плазмидной ДНК (5 мгк) + липофектамин (20 мкл) + Реагент Plus в соответствии с рекомендацией производителя в 1 мл среды Opti-MEM (Invitrogen). Через 1 час инкубации при 37ºС в атмосфере 5% СО2 в лунки добавляли еще 2 мл среды Opti-MEM и инкубировали в тех же условиях еще 24-36 часов до развития цитопатического действия (ЦПД). Материал трижды замораживали-оттаивали и обрабатывали ультразвуком для получения гомогенной вирусной суспензии. Далее проводили селекцию рекомбинантов путем трех-кратного пассирования на монослое клеток CV-1 с добавлением пуромицина (Sigma) в концентрации 10 мкг/мл среды DMEM (Invitrogen). Вирус клонировали методом бляшек под твердым агаровым покрытием и анализировали на наличие встройки химерного гена GMCSF/lact методом ПЦР с использованием праймеров
TK-flank 1 sense 5´- CAGAATTAATTAGACGAGTTAGACG
TK-flank 2 as 5´- TCTCGGTTTCCTCACCCAAT.
Позиции праймеров указаны на фиг. 2А. Вирусную ДНК для проведения ПЦР выделяли с использованием наборов «ДНК-сорб-В» (ООО «АмплиПрайм»). Размер амплифицированного фрагмента ДНК рекомбинантного вируса со встройкой химерного гена GMCSF/lact составляет 1503 п.н. (дорожка 1, фиг. 2Б), а исходного не рекомбинантного штамма Л-ИВП (также как и штамма-реципиента VVdGF2/6) - 414 п.н. (дорожка 2, фиг. 2Б). Отобранные рекомбинантные клоны вируса далее анализировали методом ПЦР на правильность встроенной конструкции химерного гена с использованием внутренних праймеров на участок ГМ-КСФ (gmcsf sense 5´- CTGCTGAGATGAATGAAACAGTAGA) и участок лактаптина (lact sense 5´- AAGCTTATGAACCAGAAACAACCAGCA). Размер амплифицированного фрагмента ДНК рекомбинантного вируса со встройкой химерного гена GMCSF/lact с использованием в ПЦР пары праймеров gmcsf sense х TK-flank 2 as составляет 867 п.н. (дорожка 3, фиг. 2Б), а с использованием пары праймеров lact sense х TK-flank 2 as – 568 п.н. (дорожка 5, фиг. 2Б). В качестве отрицательного контроля в ПЦР использовали ДНК не рекомбинантного штамма Л-ИВП, в составе которой нет последовательностей, соответствующих праймерам gmcsf sense и lact sense, поэтому фрагменты в ПЦР с использованием этих пар праймеров не образуются (дорожки 4 и 6, фиг. 2Б).
Наличие делеции в гене VGF в составе полученного рекомбинанта подтверждали методом ПЦР вирусной ДНК с использованием пары праймеров на область гена VGF
Apa-L22 - 5’- CGAGCACAATACCGGGAGATGG
Up35 5’- GTAAGCAAAGAATATAAGAATGAAGCGGTAATG AT.
Позиции праймеров указаны на фиг.2А. Размер амплифицированного фрагмента ДНК рекомбинантного штамма с делецией фрагмента гена VGF составляет 423 п.н. (дорожка 7, фиг. 2Б), а штамма Л-ИВП без делеции – 584 п.н. (дорожка 8, фиг. 2Б). Таким образом, полученный рекомбинантный штамм VV-GMCSF/lact-dGF содержит встройку химерного гена GMCSF/lact в tk-гене вируса и делецию – в VGF гене. Отобранный рекомбинантный вариант VV-GMCSF/lact-dGF дважды реклонировали, чтобы избежать следовых примесей исходного вируса, нарабатывали на монослое клеток CV-1 и очищали центрифугированием в градиенте плотности сахарозы (25-40%). Титр вируса определяли методом бляшек на монослое клеток CV-1, окрашенном фиксирующим раствором кристаллического фиолетового (2 г/л кристаллический фиолетовый, 50 мл/л формальдегид, 100 мл/л этанол, вода). Очищенный рекомбинантный штамм VV-GMCSF/lact-dGF с титром 109 БОЕ/мл хранится в расфасованном виде при -80°С.
Пример 3. Оценка экспрессии химерного гена GMCSF/lact рекомбинантным штаммом VV-GMCSF/lact-dGF. ГМ-КСФ человека синтезируется в виде белка-предшественника (144 а.о.) с последующим отщеплением сигнального пептида (17 а.о.), таким образом, зрелый полипептид содержит 127 а.о. (14.4 кДа). Показано существование 16-ти изоформ ГМ-КСФ человека, продуцируемого в клетках эукариот. Эти изоформы, имеющие различный характер гликозилирования, обуславливают гетерогенность природного ГМ-КСФ и разброс молекулярных масс от 14.4 до 32 кДа. Лактаптин является фрагментом каппа-казеина человека с молекулярной массой 14 кДа, белок не гликозилируется в клетках эукариот. Таким образом, максимальная молекулярная масса химерного белка GMCSF/lact, в котором ГМ-КСФ полностью гликозилирован, должна составлять 46 кДа, а минимальная – 28,4 кДа.
Монослой клеток CV-1 (90% поверхности), выращенный в культуральном матрасе объемом 650 мл (Greiner), инфицировали рекомбинантными штаммами VV-GMCSF/lact-dGF, VV-GMCSF-S1/3 (положительный контроль, прототип) или исходным штаммом VACV Л-ИВП (отрицательный контроль) с множественностью 1 БОЕ/кл. Инкубировали 24 часа при 37°С в атмосфере 5% СО2, культуральную среду (DMEM) собирали, клетки разрушали лизирующим буфером (50mM TRIS-HClpH 7.5, 150mM NaCl, 1% Triton X-100, 5 mM MgCl2, proteases inhibitor cocktail) в объеме 7 мл, проводили 3 раунда замораживания-оттаивания, затем трехкратную обработку ультразвуком (20 сек при 200-300 W с 10-ти сек охлаждением после каждой обработки). Культуральную среду и лизаты клеток центрифугировали 14000 rpm, 30 мин, 4°C, супернатанты анализировали в Western blot анализе отдельно на присутствие ГМ-КСФ (фиг. 3А) и лактаптина (фиг. 3Б). Электрофоретическое разделение белков проводили в камере «BioRad» в 5% концентрирующем и 14% разделяющем акриламидном геле при V=100. Перенос белков с геля осуществляли в камере MiniTrans-Blot cell «BioRad» на мембраны Immun-Blot TMPVDF Membrane for Protein Blotting 0,2 μm при V=100 1 час 20 минут. Затем мембраны промывали буфером для переноса и помещали в блокирующий раствор - TBS pH 7,4 с 5% молоком (Skim Milk Powder, Biochemika, Fluka) на 1 час при комнатной температуре (КТ) на качалке. Отмывали мембраны 3 раза по 5 минут TBS pH 7,4 на качалке и анализировали со специфическими антителами.
При анализе ГМ-КСФ (фиг. 3А) в качестве первичных антител использовали кроличьи поликлональные антитела против ГМ-КСФ человека (PerroTech). В качестве вторичных антител – коньюгат антикроличьих антител с фосфатазой (Sigma). Мембрану инкубировали 16 часов, при +4ºС с первичными антителами в рабочей концентрации 0,2 мкг/мл, в TBS pH 7,4 с 0,1% Tween-20 и 5% молока. После связывания с первичными антителами мембрану отмывали 3 раза по 5 минут TBS pH 7,4 с 0,1% Tween-20 на качалке, затем проводили связывание с вторичными антителами в рабочем разведении 1:5000 в TBS pH 7,4 с 0,1% Tween-20 и 5% молока в течение 1 часа при КТ на качалке. После связывания с конъюгатом отмывали 3 раза по 5 минут TBS pH 7,4 с 0,1% Tween-20 на качалке и 1 раз буфером для субстрата (АР – буфер: 100 mM Tris, 100mM NaCl, pH 9,5 с добавлением 50 mM MgCl2). В качестве субстрата использовали BCIP (5-bromo-4-chloro-3-indolyl phosphate) и NBT (NitroBluetetrazolium). Останавливали реакцию промыванием мембраны в дистиллированной воде.
Полученные результаты показывают, что химерный белок GMCSF/lact выявляется как в лизатах, так и в культуральной среде клеток CV-1, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF (фиг.3A). В культуральной среде представлена секретированная форма химерного белка GMCSF/lact с большим молекулярным весом 46 кДа (фиг. 3А, дорожка 3) вследствие его полного гликозилирования, в отличие от внутриклеточного химерного белка GMCSF/lact, выявленного в лизатах клеток, где представлен широкий диапазон изоформ от 28 до 46 кДа (фиг. 3А, дорожка 6). Молекулярная масса 46 кДа складывается из массы полностью гликозилированного ГМ-КСФ человека 32 кДа (Гражданцева, 2015 [23]) и лактаптина 14 кДа, который не гликозилируется в клетках млекопитающих. В качестве положительного контроля использовали негликозилированную форму ГМ-КСФ с молекулярной массой 14.4 кДа, полученную в клетках E.coli (фиг. 3А, дорожка 1). В культуральной среде и лизатах клеток, инфицированных штаммом Л-ИВП, ГМ-КСФ не выявляется (фиг. 3А, дорожки 5 и 8). Для сравнения с экспрессией химерного белка GMCSF/lact нами был использован рекомбинант VV-GMCSF-S1/3 (Патент РФ №2565544 Опубликован 20.10.2015 Бюл. №29 [22]), продуцирующий ГМ-КСФ. Как следует из фиг. 3А (дорожки 4 и 7) в культуральной среде и лизатах клеток, инфицированных рекомбинантным штаммом VV-GMCSF-S1/3, выявляются разные изоформы ГМ-КСФ с молекулярной массой 18-32 кДа, соответствующие секретируемой и внутриклеточной формам белка.
При анализе лактаптина (фиг. 3Б) использовали моноклональные антитела к лактаптину, коньюгированные с пероксидазой хрена F14-HRP в концентрации 0,2 мкг/мл в TBS pH 7,4 с 0,1% Tween-20 и 0,1% BSA. Инкубировали мембрану с антителами в течение ночи, ~16 часов, при +4ºС. Затем после 3-х кратной промывки TBS pH 7,4 с 0,1% Tween-20 на качалке проявляли раствором DAB (ЗАО «Вектор-Бест»). Останавливали реакцию промыванием в дистиллированной воде. Как следует из фиг.3Б антитела на лактаптин в культуральной среде выявляют белок с молекулярной массой 46 кДа (фиг. 3Б, дорожка 3), то есть в той же позиции, что и антитела на ГМ-КСФ. Это подтверждает экспрессию и секрецию химерного белка GMCSF/lact в составе рекомбинантного штамма VV-GMCSF/lact-dGF, а также факт присутствия в этом белке иммунологически адекватных компонентов: ГМ-КСФ человека и лактаптина. В качестве положительного контроля использовали рекомбинантный аналог лактаптина с молекулярной массой 14 кДа, полученный в клетках E.coli (фиг. 3Б, дорожка 2). В лизатах клеток, аналогично фиг. 3А, представлен широкий диапазон изоформ химерного белка GMCSF/lact от 28 до 46 кДа (фиг. 3Б, дорожка 6) в зависимости от степени гликозирования его компоненты, соответствующей ГМ-КСФ.
Таким образом, химерный белок GMCSF/lact выявляется в среде и лизатах клеток, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF, что свидетельствует о его экспрессии и секреции в составе рекомбинантного вируса.
Пример 4. Анализ биологической активности ГМ-КСФ человека, экспрессированного в составе химерного белка GMCSF/lact. Биологическую активность ГМ-КСФ человека в составе химерного белка GMCSF/lact, секретируемого из клеток CV-1, инфицированных рекомбинантным штаммом VV-GMCSF/lact-dGF, оценивали по уровню стимуляции пролиферации клеток эритролейкоза человека TF-1. Данный тест основан на способности ГМ-КСФ-зависимых клеток линии TF-1 пролиферировать только в присутствии ГМ-КСФ. Пролиферативную активность оценивали микрометодом на 96-луночных культуральных планшетах с использованием реагента XTT (2,3-бис-(2-метокси-4-нитро-5-сульфо-фенил)-2H-тетразолий-5-карбоксанилид). Метод основан на способности митохондриальных дегидрогеназ конвертировать водорастворимый ХТТ в формазан, который кристаллизуется внутри клетки. Перевод формазана в раствор с помощью феназин-метасульфата (PMS) и последующая фотометрия позволяют точно соотнести изменение оптической плотности раствора с изменением количества жизнеспособных клеток. Мы оценивали стимулирующую пролиферативную активность культуральной среды, полученной через 48 часов после заражения клеток CV-1 рекомбинантным или контрольным (штамм Л-ИВП) VACV с множественностью 0,5 БОЕ/клетка. Перед использованием культуральную среду осветляли центрифугированием 10 минут при 5000 об/мин для осаждения клеточного дебриса и фильтровали через ячейку Amicon Ultra – 100K (Millipore) для удаления вируса. Концентрацию ГМ-КСФ в культуральной среде оценивали с использованием очищенного и охарактеризованного препарата рекомбинантного ГМ-КСФ, полученного в клетках E.coli (Гилева, 1997; Топоркова, 2009 [30-31]). Для проведения эксперимента в среде RPMI (Invitrogen) с добавлением 10% эмбриональной телячьей сыворотки готовили 2-кратные разведения культуральной среды от инфицированных рекомбинантом VV-GMCSF/lact-dGF клеток в диапазоне 1:500 - 8000 или рекомбинантного белка ГМ-КСФ до конечных концентраций 0,5; 1; 2; 4; 8 нг/мл (калибровка). В качестве контроля использовали неразведенную культуральную среду клеток, инфицированных исходным штаммом VACV Л-ИВП (отрицательный контроль). В лунки 96-ти луночного планшета вносили 50 мкл вышеуказанных разведений или контрольных образцов и добавляли 50 мкл суспензии клеток TF-1 в среде RPMI с добавлением 10% эмбриональной телячьей сыворотки по 2×104 клеток/лунка. Планшеты помещали в термостат при температуре 37°С, 5% СО2, влажности 85% и инкубировали 72 часа. После инкубации в каждую тестируемую лунку добавляли 50 мкл реагента ХТТ (Sigma) и PMS (Fluka), который получали добавлением к рабочему раствору с содержанием XTT 1 мг/мл раствора PMS 1,25 мМ из расчета: на каждый мл ХТТ – 20 мкл PMS. Планшет инкубировали еще 4 часа и определяли оптическую плотность ОП490/620 на планшетном спектрофотометре SpectraCount (Packard). Каждую точку делали в пяти повторностях, определяли среднее значение и дисперсию для различных концентраций культуральной среды и очищенного рекомбинантного белка ГМ-КСФ, строили график зависимости ОП от разведений и концентрации. Стимуляцию пролиферации TF-1 клеток рассчитывали в процентах по отношению к контролю. За 100% принимали количество живых клеток в отрицательном контроле. Все расчеты проводили с использованием программного обеспечения LabView. Как следует из результатов эксперимента (фиг. 4), ГМ-КСФ в составе химерного белка сохраняет свою биологическую активность на уровне 1,65 мкг/мл в пересчете на активность рекомбинантного ГМ-КСФ. Расчет произведен следующим образом: 2-кратный пролиферативный эффект (200%) достигается при разведении культуральной среды в 1100 раз (1:1100), что соответствует концентрации ГМ-КСФ 1,5 нг/мл по калибровочной кривой. Таким образом получаем 1,5 нг/мл × 1100=1650 нг/мл=1,65 мкг/мл.
Пример 5. Анализ биологической активности лактаптина, экспрессированного в составе химерного белка GMCSF/lact.
Биологическую активность лактаптина в составе химерного белка GMCSF/lact оценивали по его способности индуцировать гибель раковых клеток молочной железы человека ВТ-549 (фиг. 5). Подготовку культуральной среды, контрольной и содержащей белок GMCSF/lact, и оценку процента гибели клеток в фотометрическом тесте с использованием реагента XTT проводили аналогично тому, как описано в примере 4. В лунки 96-ти луночного планшета вносили по 1×104 клеток ВТ-549 в 200 мкл ростовой среды F-12 (Invitrogene) с добавлением 10% фетальной сыворотки коров (HyClone). Планшеты помещали в термостат при температуре 37°С, 5% СО2, влажности 85% и инкубировали 24 часа. Затем ростовую среду удаляли и в лунки планшета вносили 2-кратные разведения культуральной среды, содержащей белок GMCSF/lact, в диапазоне 1:2 - 32 или рекомбинантного белка лактаптина до конечных концентраций 0,1-0,5 мг/мл (калибровка) в 100 мкл среды 199 (Биолот) с добавлением 2% фетальной сыворотки коров (HyClone). Планшеты помещали в термостат при температуре 37°С, 5% СО2, влажности 85% и инкубировали 72 часа. После инкубации в каждую тестируемую лунку добавляли по 50 мкл реагента ХТТ с добавлением PMS как описано в примере 4. Планшет инкубировали еще 3 часа и определяли оптическую плотность ОП490/620 на планшетном спектрофотометре SpectraCount (Packard). Каждую точку делали в пяти повторностях, определяли среднее значение и дисперсию для различных концентраций культуральной среды и очищенного рекомбинантного белка лактаптина, строили график зависимости ОП от разведений и концентрации. Гибель клеток ВТ-549 рассчитывали в процентах по отношению к контролю. За 100% принимали количество живых клеток ВТ-549, инкубированных с добавлением неразведенной культуральной среды клеток CV-1, инфицированных исходным штаммом VACV Л-ИВП (положительный контроль в данном случае). Все расчеты проводили с использованием программного обеспечения LabView. Как следует из результатов, представленных на фиг. 5, 50%-ная гибель клеток достигается при разведении культуральной среды, содержащей химерный белок GMCSF/lact, 1:3. Аналогичный эффект достигается при концентрации рекомбинантного аналога лактаптина, экспрессированного в системе E.coli, 0,23 мг/мл.
Пример 6. Оценка цитолитической активности рекомбинантного штамма ВОВ VV-GMCSF/lact-dGF.
Исследование цитолитической активности рекомбинантного штамма VV-GMCSF/lact-dGF на культурах раковых и нормальных клеток человека проводили микрометодом на 96-луночных культуральных планшетах (Greiner) с использованием реагента XTT аналогично тому, как описано в примере 4. В качестве отрицательного контроля использовали рекомбинантный штамм VV-GMCSF-dGF, который также был сконструирован на основе VV-GMCSF-S1/3, но содержал только делецию гена ростового фактора и не содержал встройки гена лактаптина.
Цитотоксическую активность вирусов для соответствующих типов клеток выражали как ЦТД50 - эффективная концентрация вируса, выраженная в количестве бляшкообразующих единиц на клетку (БОЕ/кл.), необходимая для гибели 50% клеток. Для определения ЦТД50 в лунки планшета с монослоем соответствующих клеток вносили десятикратные разведения вирусной суспензии с множественностью инфекции от 10 до 0,0001 БОЕ/кл. (multiplicity of infection, MOI) в 100 мкл среды 199 (Биолот) с добавлением 2% фетальной сыворотки коров (HyClone). Планшеты помещали в термостат при температуре 37°С, 5% СО2, влажности 85% и инкубировали 72 часа. После инкубации в каждую тестируемую лунку добавляли по 50 мкл реагента ХТТ с добавлением PMS как описано в примере 4. Планшет инкубировали еще 3 часа и определяли оптическую плотность ОП490/620 на планшетном спектрофотометре SpectraCount (Packard). Строили график зависимости ОП от MOI и определяли ЦТД50 - концентрацию вируса, при которой величина ОП490/620, измеренная в зараженных лунках, составляет 50% от величины ОП490/620, измеренной в лунках с незараженной культурой. Сравнивали ЦТД50 для разных культур клеток. Чем больше ЦТД50, тем меньшей цитотоксической активностью обладает исследуемый вирусный препарат для данной культуры клеток. Все расчеты проводили с использованием программного обеспечения LabView. Проведенные эксперименты показали (фиг. 6), что ЦТД50 рекомбинантного штамма VV-GMCSF/lact-dGF для всех тестированных раковых культур за исключением А-549 достоверно ниже (Р<0,05) аналогичных показателей для контрольного вируса без встройки лактаптина VV-GMCSF-dGF. При этом цитотоксическая активность обоих рекомбинантов в отношении нормальных клеток достоверно ниже, чем в отношении раковых (Р<0,05).
Таким образом, выше изложенные результаты (примеры 1-6) подтверждают достижение заявляемого технического результата, а именно: создан аттенуированный рекомбинантный штамм VV-GMCSF/lact-dGF, обладающий адресной противоопухолевой активностью в отношении клеток рака человека.
Источники научно-технической и патентной информации
1. Weisbart R.H., Golde D.W., Cark S.C., Wong G.G., Gassan J.C. Human granulocyte-macrophage colony-stimulating factor is a neutrophil activator. // Nature. - 1985. - V. 314. - P. 361-363.
2. Fleischmann J., Golde D. W., Weisbart R. H., Gasson J. C. Granulocyte-macrophage colony-stimulating factor enhances phagocytosis of bacteria by human neutrophils. // Blood. - 1986. - V. 68(3). - P. 708-711.
3. Gaudernack G., Gjertsen M.K. Combination of GM-CSF with antitumor vaccine strategies. // Eur. J. Cancer. - 1999. - V. 35 (Suppl. 3). P. 33–35.
4. Warren T.L., Weiner G.J. Uses of granulocyte-macrophage colony-stimulating factor in vaccine development. // Curr. Opin. Hematol. - 2000. - V. 7. - P. 168–173.
5. Morrissey P.J., Bressler L., Park L.S., Alpert A., Gillis S. Granulocyte-macrophage colony-stimulating factor augments the primary antibody response by enhancing the function of antigen-presenting cells. // J. Immunol. - 1987. - V. 15. - P. 1113-1119.
6. Yoon H.A., Aleyas A.G., George J.A., Park S.O., Han Y.W., Cho J.-G., Eo S.K. Cytokine GM-CSF genetic adjuvant facilitates prophylactic DNA vaccine against pseudorabies virus through enhanced immune responses. // Microbiol. Immunol. - 2006. - V. 50(2). - P. 83-92.
7. Somasundaram C. Recent advances and current status of GM-CSF as an adjuvant in DNA vaccines for viral diseases. // J. Investig. Genomics. - 2015. - V. 2(3). - 00025. DOI: 10.15406/jig.2015.02.00025.
8. Lai L., Kwa S., Kozlowski P.A., Montefiori D.C., Ferrari G. et al. Prevention of infection by a granulocyte-macrophage colony-stimulating factor co-expressing DNA/modified vaccinia Ankara simian immunodeficiency virus vaccine. // J. Infect. Dis. - 2011. - V. 204(1). - P. 164-173.
9. Hellerstein M., Xu Y., Marino T., Lu S., Yi H. et al. Co-expression of HIV-1 virus-like particles and granulocyte-macrophage colony stimulating factor by GEO-D03 DNA vaccine. // Hum. Vaccin. Immunother. - 2012. - V. 8(11). - P. 1654-1658.
10. Zhang H., Qian P., Peng B., Shi L., Chen H. et al. A novel subunit vaccine co-expressing GM-CSF and PCV2b Cap protein enhances protective immunity against porcine circovirus type 2 in piglets. // Vaccine. - 2015. - V. 33(21). - P. 2449-2456.
11. Chen H., Gao N., Wu J., Zheng X., Li J. et al. Variable effects of the co-administration of a GM-CSF-expressing plasmid on the immune response to flavivirus DNA vaccines in mice. // Immunol. Lett. - 2014. - V. 162(1 Pt A). - P. 140-148.
12. Fowler V., Robinson L., Bankowski B., Cox S., Parida S. et al. A DNA vaccination regime including protein boost and electroporation protects cattle against foot-and-mouth disease. // Antiviral. Res. - 2012. - V. 94(1). - P. 25-34.
13. Zheng Q., Fan D., Gao N., Chen H., Wang J. et al. Evaluation of a DNA vaccine candidate expressing prM-E-NS1 antigens of dengue virus serotype 1 with or without granulocyte-macrophage colony-stimulating factor (GM-CSF) in immunogenicity and protection. // Vaccine. - 2011. - V. 29(4). - P. 763-771.
14. Wang X., Li J., Jiang P., Li Y., Zeshan B. et al. GM-CSF fused with GP3 and GP5 of porcine reproductive and respiratory syndrome virus increased the immune responses and protective efficacy against virulent PRRSV challenge. // Virus Res. - 2009. - V. 143(1). - P. 24-32.
15. Hartoonian C., Ebtekar M., Soleimanjahi H., Karami A., Mahdavi M. et al. Effect of immunological adjuvants: GM-CSF (granulocyte-monocyte colony stimulatingfactor) and IL-23 (interleukin-23) on immune responses generated against hepatitis C virus core DNA vaccine. // Cytokine. - 2009. - V. 46(1). - P. 43-50.
16. Encke J., Bernardin J., Geib J., Barbakadze G., Bujdoso R. et al. Genetic vaccination with Flt3-L and GM-CSF as adjuvants: Enhancement of cellular and humoral immune responses that results in protective immunity in a murine model of hepatitis C virus infection. // World J. Gastroenterol. - 2006. - V. 12(44). - P. 7118-7125.
17. Кочнева Г.В., Сиволобова Г.Ф., Юдина К.В., Бабкин И.В., Чумаков П.М., Нетесов С.В. Онколитические поксвирусы. //Молекулярная генетика, микробиология и вирусология. - 2012. - №1. - С.8–15.
18. Thorne S.H., Hwang T.H., O’Gorman W.E., Bartlett D.L., Sei S., Kanji F., Brown C., Werier J., Cho J., Lee D., Wang Y., Bell J., Kirn D.H. Rational strain selection and engineering creates a broad-spectrum, systemically effective oncolytic poxvirus, JX-963. // J. Clin. Invest. - 2007. - V. 117. - P. 3350-3358.
19. Kirn D. Oncolytic vaccinia virus cancer therapy. // U.S. Patent. - 2010. - №2010/0303714.
20. Merchlinsky M., Eckert D., Smith E., Zauderer M. Construction and characterization of vaccinia direct ligation vectors. // Virology. - 1997. - V. 238 - P. 444-451.
21. Breitbach C.J., Burke J., Jonker D., Stephenson J., Haas A.R., Chow L.Q. M., Nieva J., Hwang T., Moon A., Patt R., Pelusio A., Le Boeuf F., Burns J., Evgin L., De Silva N., Cvancic S., Robertson T., Je J., Lee Y., Parato K., Diallo J., Fenster A., Daneshmand M., Bell J.C., Kirn D.H. Intravenous delivery of a multi-mechanistic cancer-targeted oncolytic poxvirus in humans. // Nature. - 2011. - V. 477. - P. 99-102.
22. Кочнева Г.В., Сиволобова Г.Ф., Лупан Т.А., Гражданцева А.А., Чумаков П.М., Нетесов С.В. Рекомбинантный штамм VV-GMCSF-S1/3 вируса осповакцины, продуцирующий секретируемый гранулоцитарно-макрофагальный колониестимулирующий фактор человека. // Патент РФ. - опубл. 20.10.2015. - №2565544. - Бюл. №29 (прототип).
23. Гражданцева А.А., Сиволобова Г.Ф., Ткачева А.В., Гилева И.П., Кулигина Е.В., Рихтер В.А., Кочнева Г.В. Высокоэффективная продукция биологически активного секретируемого гранулоцитарно-макрофагального колониестимулирующего фактора человека рекомбинантным вирусом осповакцины. // Биотехнология, 2015, №5, C. 1-9.
24. Алексеенко И.В., Копанцев Е.П., Виноградова Т.В., Свердлов Е.Д. Бицистронный вектор для совместной экспрессии гена-убийцы HSVtk и цитокина GM-CSF в раковых клетках. // Доклады академии наук. - 2011. - Т. 439. - №4. - С.551-554.
25. Koval O.A. et al. Lactaptin induces p53-independent cell death associated with features of apoptosis and autophagy and delays growth of breast cancer cells in mouse xenografts. // PLoS one. - 2014. - V. 9 (4). - e93921.
26. McCart A., Bartlett D., Moss B. Combined growth factor-deleted and thymidine kinase-deleted vaccinia virus vector. // US Patent. №7208313. - 2007.
27. Кочнева Г.В., Бабкина И.Н., Лупан Т.А., Гражданцева А.А., Юдин П.В., Сиволобова Г.Ф., Швалов А.Н., Попов Е.Г., Бабкин И.В., Нетесов С. В., Чумаков П.М. Апоптин усиливает онколитическую активность вируса осповакцины in vitro. // Молекулярная биология. 2013. Т. 47. №5. С. 842-852.
28. Чумаков П.М., Кочнева Г.В., Бабкина И.Н., Лупан Т.А., Гражданцева А.А., Юдин П.В., Сиволобова Г.Ф., Попов Е.Г., Нетесов С. В. Рекомбинантная плазмидная ДНК pGEM-Puro-DS-Apo, содержащая синтетический ген апоптина, фланкированный последовательностями генома вируса осповакцины из района C10L-C12L, и рекомбинантный штамм VVdGF-ApoS24/2 вируса осповакцины, продуцирующий апоптин. // Патент РФ. - 2013. - №2492238. - Бюл. №25.
29. Semenov D.V., Fomin A.S., Kuligina E.V., Koval O.A., Matveeva V.A., Babkina I.N., Tikunova N.V., Richter V.A.. Recombinant analogs of a novel milk pro-apoptotic peptide, lactaptin, and their effect on cultured human cells. // Protein. J. - 2010. - V. 29. - P. 174-180.
30. Гилева И.П., Бондарь Т.С., Коробко В.Г., Кравченко В.В., Сандахчиев Л.С. Рекомбинантная плазмидная ДНК p280GM, кодирующая полипептид со свойствами гранулоцитарно-макрофагального колониестимулирующего фактора человека, и штамм E.coli SG20050/p280GM – продуцент полипептида со свойствами гранулоцитарно-макрофагального колониестимулирующего фактора человека. // Патент РФ №2091488. 1997.
31. Топоркова Л.Б., Орловская И.А., Сенников С. В., Сахно Л.В., Козлова Ю.Н., Лебедев Л.Р., Гилева И.П. Изучение in vitro биологических свойств рекомбинантного гранулоцитарно-макрофагального колониестимулирующего фактора. // Иммунология. - 2009. - №4. - С. 203-205.
Приложение
<110> Федеральное государственное бюджетное учреждение науки Институт химической биологии и фундаментальной медицины Сибирского отделения Российской академии наук (ИХБФМ СО РАН)
<120> Рекомбинантный штамм VV-GMCSF/lact-dGF вируса осповакцины, обладающий онколитической активностью и продуцирующий секретируемый химерный белок, состоящий из гранулоцитарно-макрофагального колониестимулирующего фактора человека и онкотоксического белка лактаптина
<160> 1
<210> 1
<211> 819
<212> DNA
<213> chimeric gene coding human GM-CSF and human milk kappa-casein peptide (residues 23-134) joined by flexible spacer GlyGlyGlySer (химерный ген, кодирующий ГМ-КСФ человека и фрагмент каппа-казеина человека (остатки 23-134), соединенные гибким линкером GlyGlyGlySer)
<400> 1
atg tgg ctg cag agc ctg ctg ctc ttg ggc act gtg gcc tgc agc atc tct 51
gca ccc gcc cgc tcg ccc agc ccc agc acg cag ccc tgg gag cat gtg aat 102
gcc atc cag gag gcc cgg cgt ctc ctg aac ctg agt aga gac act gct gct 153
gag atg aat gaa aca gta gaa gtc atc tca gaa atg ttt gac ctc cag gag 204
ccg acc tgc cta cag acc cgc ctg gag ctg tac aag cag ggc ctg cgg ggc 255
agc ctc acc aag ctc aag ggc ccc ttg acc atg atg gcc agc cac tac aag 306
cag cac tgc cct cca acc ccg gaa act tcc tgt gca acc cag att atc acc 357
ttt gaa agt ttc aaa gag aac ctg aag gac ttt ctg ctt gtc atc ccc ttt 408
gac tgc tgg gag cca gtc cag gag ggc gga ggc gga tca aag ctt atg aac 459
cag aaa caa cca gca tgc cat gag aat gat gaa aga cca ttc tat cag aaa 510
aca gct cca tat gtc cca atg tat tat gtg cca aat agc tat cct tat tat 561
gga acc aat ttg tac caa cgt aga cca gct ata gca att aat aat cca tat 612
gtg cct cgc aca tat tat gca aac cca gct gta gtt agg cca cat gcc caa 663
att cct cag cgg caa tac ctg cca aat agc cac cca ccc act gtg gta cgt 714
cgc cca aac ctg cat cca tca ttt att gcc atc ccc cca aag aaa att cag 765
gat aaa ata atc atc cct acc atc ggc gga tca cat cac cat cac cat cac taa 819
Представленный рекомбинантный штамм обладает адресной онколитической активностью и продуцирует секретируемый химерный белок, состоящий из ГМ-КСФ человека и лактаптина (GMCSF/lact). Штамм VV-GMCSF/lact-dGF сконструирован на основе штамма Л-ИВП вируса осповакцины, содержащего делеции фрагментов генов вирусной тимидинкиназы и ростового фактора. В район делеции гена тимидинкиназы встроен трансген химерного белка GMCSF/lact, в состав которого входит генный адьювант ГМ-КСФ, соединенный через гибкий линкер GlyGlyGlySer с онкотоксическим пептидом лактаптином. Заявленный штамм имеет сниженную более чем в 100 раз вирулентность по сравнению с исходным штаммом Л-ИВП, а также высокую адресную онколитическую активность в отношении клеток опухолей человека различного генеза. Штамм депонирован в Государственную коллекцию возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» под регистрационным номером V-711 (18.04.2016). Изобретение обеспечивает дополнительную аттенуацию вируса VACV в отношении нормальных клеток и усиливает его литическую активность в отношении раковых клеток, а также может быть использовано в биотехнологии. 8 ил., 3 табл., 6 пр.
Рекомбинантный штамм VV-GMCSF/lact-dGF, сконструированный на основе штамма Л-ИВП вируса осповакцины и содержащий делецию фрагмента гена вирусной тимидинкиназы, в район которой между позициями 81277 и 81307 п.н. (GenBank Acc. KP233807.1) встроен оперон, состоящий из химерного транскрипта GMCSF/lact, экспрессируемого под контролем природного промотора вируса осповакцины P7.5k, включающий 276 п.н. и состоящий из генов ГМ-КСФ человека (GenBank Acc. M11220.1) и лактаптина - фрагмента гена каппа-казеина человека, кодирующего пептид размером 23-134 а.о., соединенных гибким линкером ggc gga ggc gga tca (GlyGlyGlySer); содержащий делецию в левом концевом районе вирусного генома, соответствующую позициям 7770-8071 п.н. (GenBank Acc. KP233807.1), включающую фрагмент гена VGF размером 170 п.н., соответствующую позициям 7801-8071 п.н.; продуцирующий секретируемую форму химерного белка GMCSF/lact, обладающего биологическими активностями ГМ-КСФ, и лактаптина; депонированный в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ "Вектор" под номером V-711.
| РЕКОМБИНАНТНЫЙ ШТАММ VV-GMCSF-S1/3 ВИРУСА ОСПОВАКЦИНЫ, ПРОДУЦИРУЮЩИЙ СЕКРЕТИРУЕМЫЙ ГРАНУЛОЦИТАРНО-МАКРОФАГАЛЬНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР ЧЕЛОВЕКА | 2013 |
|
RU2565544C2 |
| ШТАММ ГИБРИДНЫХ КУЛЬТИВИРУЕМЫХ КЛЕТОК ЖИВОТНЫХ Mus. Musculus, ПРОДУЦИРУЮЩИЙ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К ПЕПТИДУ, ОБЛАДАЮЩЕМУ АПОПТОТИЧЕСКОЙ АКТИВНОСТЬЮ ПО ОТНОШЕНИЮ К РАКОВЫМ КЛЕТКАМ ЧЕЛОВЕКА | 2009 |
|
RU2402605C1 |
| R | |||
| CHAVAN et al., Expression of CCL20 and Granulocyte-Macrophage Colony-Stimulating Factor, but Not Flt3-L, from Modified Vaccinia Virus Ankara Enhances Antiviral Cellular and Humoral Immune Responses, JOURNAL OF VIROLOGY, Aug | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Капельная масленка с постоянным уровнем масла | 0 |
|
SU80A1 |
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Двухсторонний перекрестный стрелочный перевод | 1926 |
|
SU7676A1 |

