Изобретение относится к рекомбинантному штамму вируса осповакцины, обладающему противоопухолевой активностью, и может быть использовано в биотехнологии, в частности, в генетической инженерии для диагностики и разработки лекарственных средств нового поколения для борьбы с онкологическими заболеваниями (тераностики).
Тераностика (англ. theranostics) - новый подход к созданию фармацевтических композиций, заключающийся в комплексном решении терапевтических и диагностических проблем путем создания препаратов, которые являются одновременно и средством ранней диагностики, и терапевтическим агентом.
Известны радиоактивные препараты, которые могут быть использованы в рамках тераностики: например, препараты на основе изотопов галлия-68 - для диагностики, а с лютецием-177 - для терапии. Тераностика - это наиболее эффективный путь для лечения многих заболеваний, в том числе онкологических. Цель ее - сначала увидеть, диагностировать мишень, а потом с помощью того же самого инструмента - поразить (https://scientificrussia.ru/articles/radionuklidy-protiv-raka-v-mire-nauki-no-9-10).
Онколитические вирусы могут использоваться против рака способами, которые дополняют лизис инфицированных клеток. Добавление гена симпортера йодида натрия (NIS) к вирусному геному заставляет инфицированные опухолевые клетки экспрессировать NIS и накапливать йод. В сочетании с радиойодтерапией он позволяет проводить местную лучевую терапию опухоли, как это используется для лечения рака щитовидной железы. Радиоактивный йод также может быть использован для визуализации репликации вируса в организме с помощью гамма-камеры. Этот подход успешно использовался доклинически с аденовирусом, вирусом кори и вирусом осповакцины.
Известно изобретение, относящееся к новым методам лечения раковых заболеваний путем модуляции SAMHD1 для лечения устойчивости к терапии рака (международная заявка WO 2017076880, опубл. 11.05.2017). Лечение рака аналогами нуклеозидов (НА), которые специфически ингибируют быстро делящиеся клетки, может столкнуться с ранее существовавшей резистентностью к НК или развитием резистентных раковых клеток, что приводит к неблагоприятному клиническому прогнозу. Лечение рака онколитическими вирусами простого герпеса (ВПГ) может столкнуться с ранее существовавшей резистентностью или развитием резистентных раковых клеток, что приводит к плохому клиническому прогнозу. Изобретение преодолевает резистентность к химиотерапии или резистентность к онколитическому HSV, предоставляя способы обнаружения резистентности при раковом заболевании, основанные на экспрессии белка 1, содержащего домен SAM и домен HD (SAMHD1), в раковых клетках. Кроме того, предлагаются варианты лечения, направленные на устранение резистентности к химиотерапии, такие как комбинация ингибитора SAMHD1 с NA. Кроме того, предусмотрены варианты лечения, направленные на устранение устойчивости к онколитическому ВПГ, такие как комбинация ингибитора SAMHD1 или истощение SAMHD1 с онколитическим ВПГ. Изобретение обеспечивает новые лекарственные средства и сопутствующие диагностические средства, поддерживающие решения о клиническом лечении.
Наиболее близким аналогом (прототипом) является сконструированный ранее авторами настоящей заявки на изобретение рекомбинантный вариант вируса осповакцины VV-NS1-dGF, содержащий делению фрагмента гена вирусного ростового фактора, в район которой встроен синтетический ген NS1 парвовируса крыс Н-1 (патент РФ №2692628 - прототип) [1]. Белок NS1 парвовируса крыс является онкотоксическим белком и индуцирует апоптоз раковых клеток человека, что способствует повышению природной онколитической активности вируса осповакцины в отношении, в частности, опухолей мозга человека.
Однако такой штамм не позволяет одновременно проводить терапию и диагностику онкозаболевания, что ограничивает его функциональные возможности.
Техническим результатом заявляемого изобретения является создание более эффективного средства в онкологии на основе рекомбинантного штамма вируса осповакцины для обеспечения одновременной терапии и диагностики (тераностики) онкологических заболеваний человека.
Указанный технический результат достигается получением рекомбинантного штамма вируса осповакцины VV-mNIS-NS1, используемого для терапии и диагностики (тераностики) опухолей человека за счет адресной литической активности в отношении раковых клеток человека и способности аккумулировать в них рентгеноскопический контраст в виде йодида за счет экспрессии трансгена mNIS, содержащий синтетический ген мышиного симпортера йодида натрия mNIS, встроенного в район гена вирусной тимидинкиназы, с длиной генома 190181 п.н., в котором встройка трансгена NS1 расположена в позиции 7706-9877 п.н., а встройка трансгена mNIS - в позиции 83163-85076 п.н. Штамм депонирован в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером V-1206 (справка прилагается).
Рекомбинантный штамм VV-NS1-dGF был использован в качестве штамма-реципиента для встройки дополнительного трансгена симпортера йодида натрия мыши mNIS в центральную часть гена вирусной тимидинкиназы с одновременной делецией его фрагмента. Таким образом, полученный нами двойной рекомбинант имеет делеции двух генов - вирусного ростового фактора (virus growth factor, VGF) и тимидинкиназы (ТК). Гены VGF и ТК являются факторами вирулентности вируса осповакцины, поэтому их инактивация приводит к аттенуации вируса, что особенно важно при лечении иммунодефицитных раковых больных для предотвращения нежелательных побочных эффектов. Ранее нами был оценен уровень аттенуации рекомбинантных вариантов вируса осповакцины с делециями генов ТК и VGF по сравнению с используемым нами исходным не модифицированным штаммом Л-ИВП. Оценку уровня аттенуации проводили по величине 50%-ной летальной дозы (ЛД50) для 10-дневных куриных эмбрионов (КЭ). Оказалось, что ЛД50 (вирулентность) рекомбинантных вариантов Л-ИВП с фенотипом ТК-VGF- в 100 и более раз ниже, чем исходного штамма Л-ИВП (Koval, 2017) [2]. Инактивация генов VGF и ТК также способствует повышению онкоселективности вируса осповакцины поскольку приводит к торможению репликации вируса в неделящихся или слабо делящихся нормальных клетках, при этом эффективность разрушения раковых клеток такими двойными мутантами не отличается от исходного вируса (McCart, 2007) [3].
NIS представляет собой трансмембранный гликопротеин, который обеспечивает поглощение и концентрацию йодида внутри клеток. NIS играет центральную роль в метаболизме гормонов щитовидной железы и используется для выявления и лечения заболеваний щитовидной железы (Ravera, 2022) [4]. Встройка трансгена NIS в геном онколитических вирусов обеспечивает концентрацию йодида в тканях опухолей, что теоретически позволяет использовать его в качестве контраста при рентгеноскопической диагностике и направленном лечении опухолей с помощью радиотерапии (Семенова, 2022) [5]. NIS является консервативным белком позвоночных с высокой межвидовой гомологией (Thorne, 2007; Concilio, 2020) [6-7]. Для встройки в геном онколитического вируса нами была выбрана последовательность мышиного гена NIS (mNIS), соответствующая GenBank AF235001. Выбор обусловлен тем, что мышиный NIS более эффективно катализирует транспорт йодида в трансфицированные клетки, чем NIS человека, вследствие того, что мышиный белок преимущественно обнаруживался на плазматической мембране, а его человеческий ортолог был внутриклеточным примерно в 40% экспрессирующих клеток (Dayem, 2008) [8]. Последовательность, включающая трансген mNIS, последовательность Козака и сайты узнавания рестриктаз HindIII-EcoRI (1872 п.н.) была синтезирована ЗАО «Евроген» (г. Москва). В процессе синтеза последовательность была оптимизирована для усиления транскрипции, финальный вариант последовательности представлен на фиг. 1.
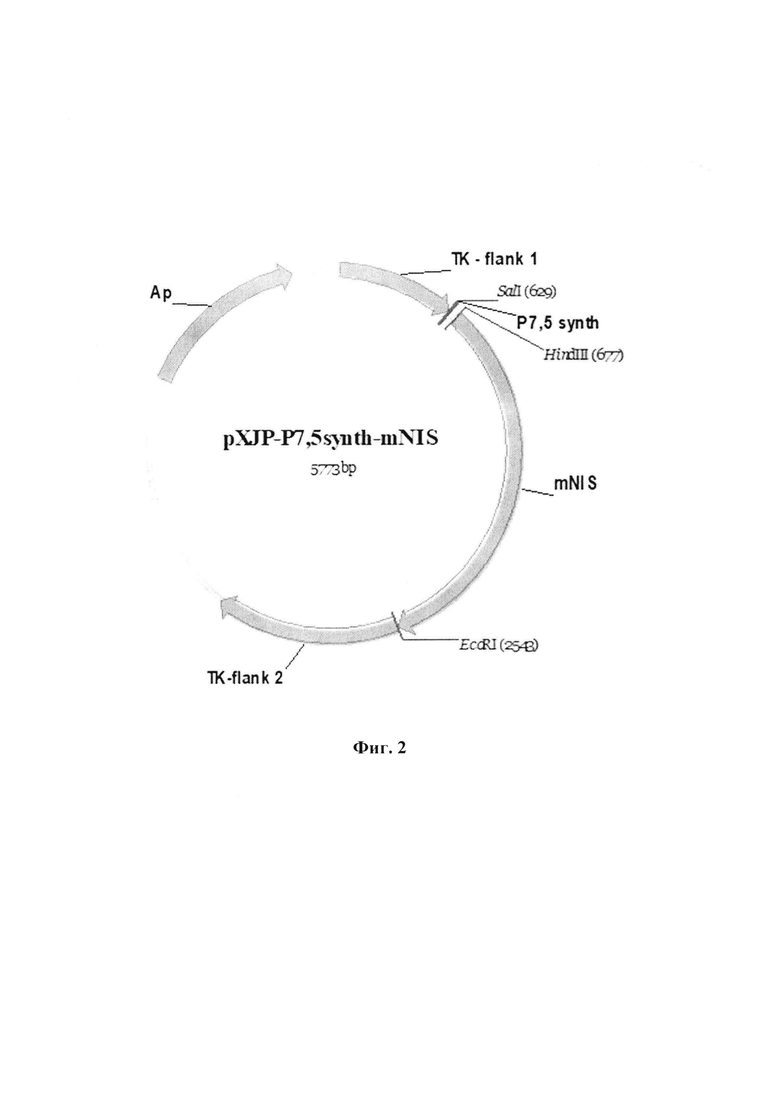
Для создания рекомбинантного варианта вируса осповакцины со встройкой трансгена mNIS нами была сконструирована инсерционная плазмида pXJP-P7,5 synth-mNIS (фиг. 2). Эта плазмида позволяет встраивать трансген mNIS в район гена тимидинкиназы вируса осповакцины с одновременной делецией фрагмента этого гена (фенотип ТК-). Трансген mNIS экспрессируются под контролем ранне-позднего синтетического промотора вируса осповакцины Р7,5 synth. Рекомбинантный штамм VV-mNIS-NS1 со встройкой трансгена mNIS был получен в результате двойного кроссинговера по гомологичным участкам плазмидной ДНК pXJP-P7,5 synth-mNIS и ДНК штамма-реципиента VV-NS1-dGF с последующей селекцией рекомбинантов в клетках Н143ТК- с добавлением в культуральную среду 5-бромдезоксиуридина. Для рекомбинантного варианта VV-mNIS-NS1 проведено полногеномное секвенирование и депонирование в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером V1206.
Штамм VV-mNIS-NS1 характеризуется следующими признаками: Морфологические признаки. Штамм обладает свойствами типичного представителя вируса осповакцины (vaccinia virus, VACV), но в отличие от исходного штамма Л-ИВП имеет фенотип TK-VGF-/NIS+NS1+. Показано, что при заражении раковых клеток человека U87MG (глиобластома), Mel8 (меланома), РС3 (карцинома простаты) и MDA-MB-231 (аденокарцинома молочной железы) штамм VV-mNIS-NS1 за счет экспрессии трансгена mNIS обеспечивает накопление йодида в количествах, значимо превышающих количество йодида в неинфицированных клетках.
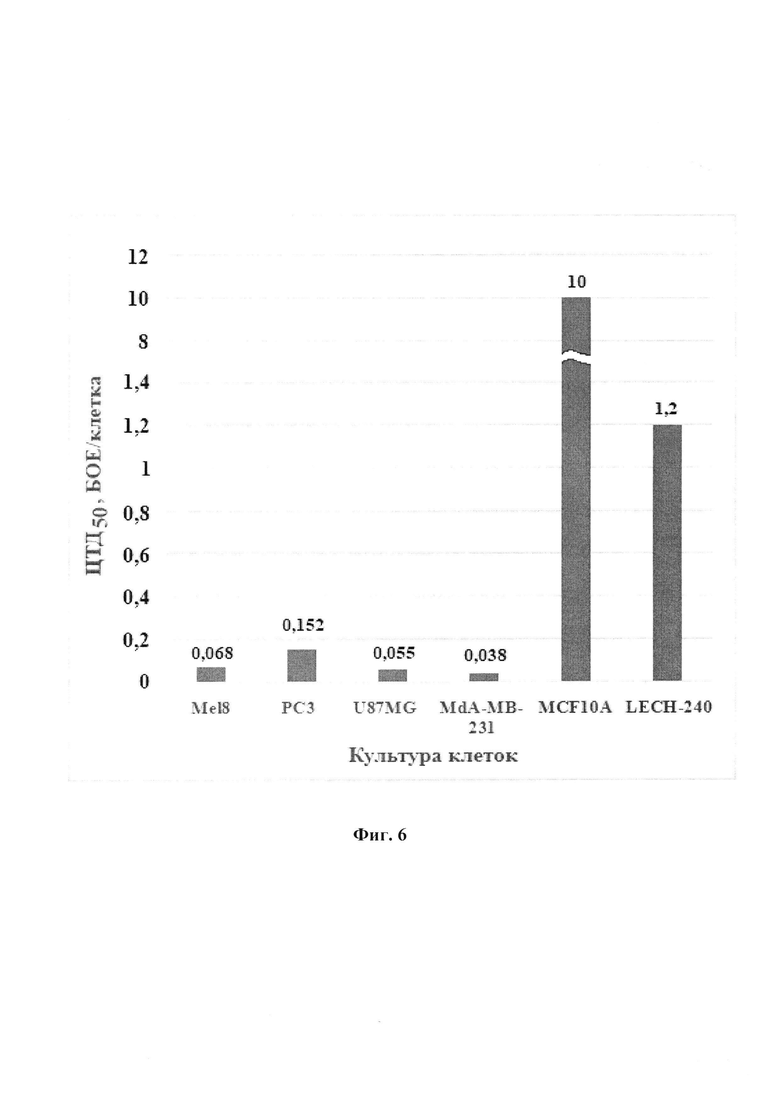
Физиолого-биохимические характеристики и культуральные свойства штамма. Секвенирование генома рекомбинанта выполнено с помощью технологии секвенирования нового поколения (NGS) на платформе MiSEQ. Длина генома рекомбинантного штамма VV-mNIS-NS1 составляет 190181 п.н., встройка трансгена NS1 расположена в позиции 7706-9877 п.н., а встройка трансгена mNIS - в позиции 83163-85076 п.н. Экспрессия мРНК mNIS в клетках опухолей человека, инфицированных VV-mNIS-NS1, подтверждена методом ПЦР в режиме реального времени (ПЦР-РВ) (Фиг. 4). Показано накопление йодида в раковых клетках человека в процессе инфекции штаммом VV-mNIS-NS1 (Фиг. 5). Рекомбинантный штамм VV-mNIS-NS1 селективно реплицируется в опухолевых клетках человека и обладает высокой онколитической активностью (Фиг. 6).
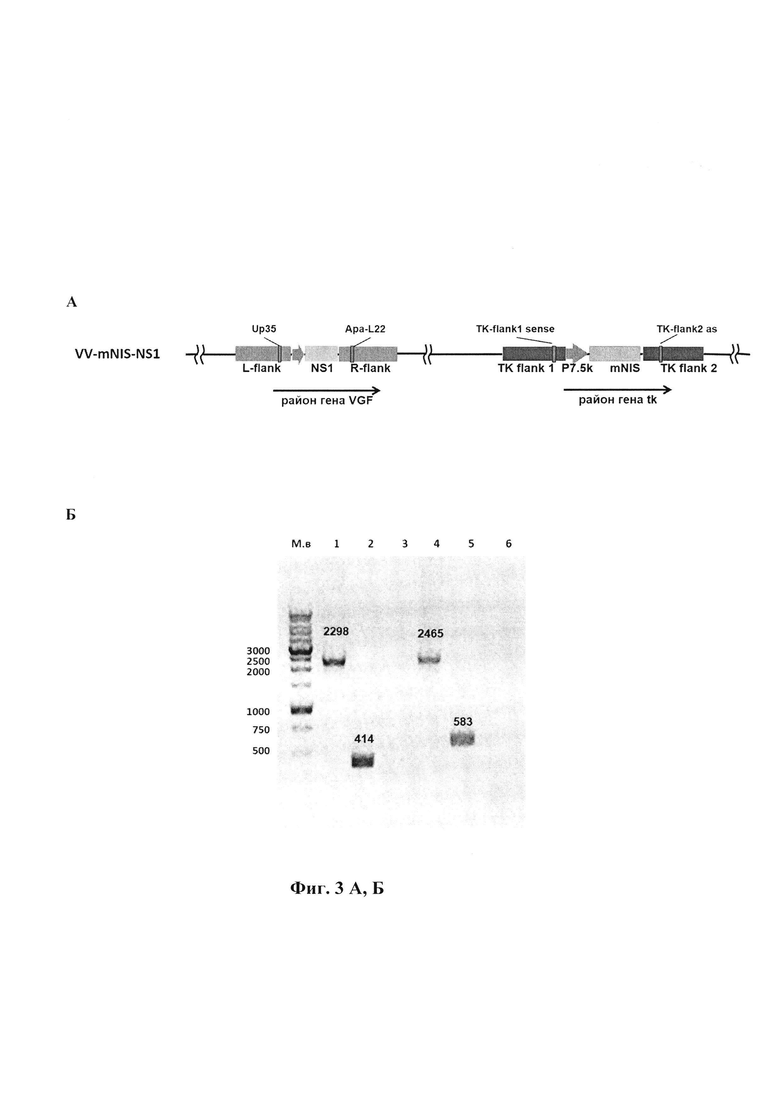
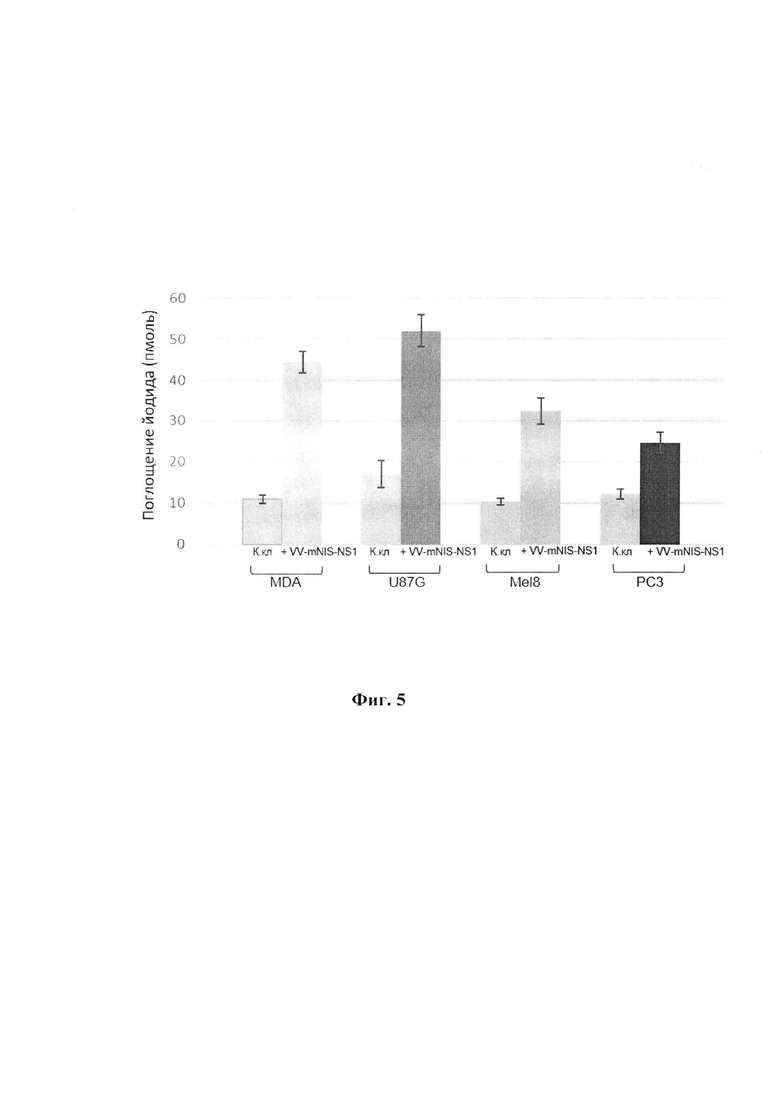
Изобретение иллюстрируется следующими фигурами графических изображений. На фиг. 1 приведена оптимизированная синтетическая нуклеотидная последовательность трансгена mNIS с заложенными по краям последовательности сайтами рестрикции HindIII и EcoRI (1872 п.н.). Жирным шрифтом помечены стартовый и стоп - кодоны. На фиг. 2 представлена физическая и генетическая карта плазмиды pXJP-P7,5 synth-mNIS. TK-flank 1 - фрагмент генома VACV штамма Л-ИВП длиной 595 п.н. (GenBank KP233807, позиции 80682-81277), расположенный слева от гена TK; TK-flank 2 - фрагмент генома VACV длиной 933 п.н., (GenBank КР233807, позиции 81307-82251), расположенный справа от гена TK; Р7,5 synth (36 п.н.) - синтетический ранний промотор VACV (Merchlinsky, 1997) [9]; mNIS - модифицированный синтетический ген NIS мыши, встроенный в плазмидную ДНК по сайтам рестрикции HindIII и EcoRI; Ар - ген β-лактамазы, определяющий устойчивость к ампициллину. На фиг. 3 приведено схематичное изображение структуры генома рекомбинантного варианта VV-mNIS-NS1 с указанием позиций праймеров (А). ПЦР анализ ДНК VV-mNIS-NS1 на наличие встроек трансгенов (Б). ПЦР проводили с использованием пар праймеров TK-flank 1 sense - ТК-flank 2 as (дорожки 1-3) и Up35-ApaL22 (дорожки 4-6). Дорожки 1, 4 - рекомбинант VV-mNIS-NS1; дорожки 2, 5 - исходный немодифицированный штамм Л-ИВП; дорожки 3, 6 - вода. На фиг. 4 представлена экспрессия мРНК mNIS в культурах раковых клеток человека, инфицированных рекомбинантным штаммом VV-mNIS-NS1 в множественности 0,1, 1 и 10 БОЕ/кл. Клетки инкубировали с вирусом в течение 24 ч. Уровень транскриптов анализировали методом qПЦР со специфическими праймерами после этапа обратной транскрипции. Уровень мРНК GAPDH и АСТВ использовали в качестве внутреннего стандарта. В качестве контроля использовали клетки, не инфицированные вирусом. Соотношение мРНК mNIS к нормировочным генам в контрольных клетках принимали за единицу. Данные представлены как среднее значение соотношения mNIS/GAPDH(ACTB)±SD. Статистические различия между контрольной и экспериментальными группами р<0.001. На фиг. 5 приведены данные по накоплению йодида в культурах раковых клеток человека, инфицированных рекомбинантным штаммом VV-mNIS-NS1. MDA, U87G, Mel8, РС3 - раковые клетки человека, инфицированные рекомбинантным штаммом VV-mNIS-NS1 при множественности 1 БОЕ/кл поглощают йодид в количестве 44 пмоль, 52 пмоль, 32 пмоль, 25 пмоль, соответственно, что значимо превышает поглощение ионов йода неинфицированными клетками (10,3-17,0 пмоль). На фиг. 6 представлена онколитическая активность рекомбинантного штамма VV-mNIS-NS1 в раковых и нормальных клетках человека. Клетки выращивали в 96-ти луночных планшетах, инфицировали 10-кратными разведениями вируса в диапазоне доз 0,001 - 10 БОЕ на клетку и рассчитывали 50%-ную цитотоксическую дозу для каждого вируса (ЦТД50 - значения указаны в вершинах столбиков) в фотометрическом тесте с использованием субстрата для митохондриальных дегидрогеназ 2,3-бис-(2-метокси-4-нитро-5-сульфо-фенил)-2Н-тетразолий-5-карбоксанилида (реагент ХТТ).
Для лучшего понимания сущности изобретения ниже следуют примеры его осуществления.
Пример 1. Конструирование рекомбинантного штамма VV-mNIS-NS1.
Для получения рекомбинантного варианта VV-mNIS-NS1 была сконструирована инсерционная плазмида pXJP-P7,5 synth-mNIS (фиг. 2), которая содержит фрагмент ДНК VACV, соответствующий позициям нуклеотидов 80682-82251 депонированного нами в GenBank штамма Л-ИВП (Accession number KP233807) (Shvalov, 2016) [10]. Фрагмент вирусной ДНК в плазмиде разделен на левый (595 п.н., позиции 80682-81277) и правый (933 п.н., позиции 81307-82251) фланки с делецией 30 п.н. в центре ТК-гена VACV, в область которой по сайтам SalI - EcoRI встроен синтетический фрагмент:
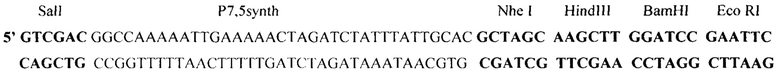
Фрагмент содержит синтетический промотор VACV Р7,5 synth (36 п.н.) и полилинкер (Nhe1, HindIII, BamH1, EcoRI), обеспечивающий встройку и экспрессию трансгенов под контролем синтетического ранне-позднего промотора вируса осповакцины (Р7,5 synth) (Merchlinsky, 1997) [9]. Оптимизированная последовательность трансгена mNIS (фиг. 1) была встроена в полилинкер плазмиды по HindIII - EcoRI сайтам рестрикции.
Для проведения трансфекции ДНК плазмиды pXJP-P7,5 synth-mNIS была наработана в препаративных количествах из 250 мл среды Лурия-Бертани и выделена с использованием набора лабораторных реагентов для выделения плазмидной ДНК, очищенной от эндотоксинов, EndoFree Plasmid Maxi Kit (Qiagen, США). Для трансфекции использовали реагент Lipofectamine™ LTX и реагент Plus (Invitrogen, США). Трансфекцию проводили на 90%-ном монослое дефектных по тимидинкиназе клеток остеосаркомы человека Н143ТК-, выращенном в шестилуночных планшетах (Greiner, США). Клетки инфицировали рекомбинантным штаммом-реципиентом VV-NS1-dGF с множественностью 0,05 БОЕ/клетка, и через 1 час инкубации при 37°С добавляли смесь плазмидной ДНК (5 мгк)+липофектамин (20 мкл)+Реагент Plus в соответствии с рекомендацией производителя в 1 мл среды Opti-MEM (Invitrogen, США). Через 1 час инкубации при 37°С в лунки добавляли 2 мл среды Opti-MEM и инкубировали при 37°С в атмосфере 5% СО2 еще 24-36 часов до развития цитопатического эффекта. Материал трижды замораживали-оттаивали и обрабатывали ультразвуком для получения гомогенной вирусной суспензии. Далее проводили селекцию рекомбинантов путем двукратного пассирования на монослое клеток Н143ТК- с добавлением бромдезоксиуридина (Sigma, США) в концентрации 25 мкг/мл среды DMEM (ООО «Биолот», Россия). Вирус клонировали методом бляшек под твердым агаровым покрытием и анализировали на наличие встройки трансгена mNIS методом ПЦР с использованием пары праймеров: TK-flank1 sense 5'-CAGAATTAATTAGACGAGTTAGACG и TK-flank2 as 5'-ATTGGGTGAGGAAACCGAGA (фиг. 3). Вирусную ДНК для проведения ПЦР выделяли с использованием наборов «ДНК-сорб» (ЗАО «Интерлабсервис», Россия). Как следует из фиг. 3Б, рекомбинант VV-mNIS-NS1 содержит встройку трансгена mNIS расчетной длины 2298 п.н.
Сохранность встройки трансгена NS1 в рекомбинантном штамме VV-mNIS-NS1 подтверждали методом ПЦР с использованием пары праймеров: Up35 5'-GTAAGCAAAGAATATAAGAATGAAGCGGTAATG и Apa-L22 5'-CCATCTCCCGGTATTGTGCTCG. Как видно на фиг. 3Б, рекомбинант VV-mNIS-NS1 несет в районе делеции гена VGF протяженную встройку трансгена NS1 (2465 п.н.) в отличие от исходного немодифицированного штамма Л-ИВП, с ДНК которого амплифицируется фрагмент размером 583 п.н.
Рекомбинантный вариант VV-mNIS-NS1 нарабатывали на монослое клеток почки африканской зеленой мартышки 4647 и очищали центрифугированием в градиенте плотности сахарозы (25-40%). Титр вируса определяли методом бляшек на монослое клеток 4647, окрашенном фиксирующим раствором кристаллического фиолетового (2 г/л кристаллический фиолетовый, 50 мл/л формальдегид, 100 мл/л этанол, вода). Очищенный рекомбинантный штамм VV-mNIS-NS1 с титром 108-9 БОЕ/мл хранится в расфасованном виде при -40°С и используется для дальнейшего анализа.
Пример 2. Оценка экспрессии трансгена mNIS рекомбинантным штаммом VV-mNIS-NS1.
Для оценки экспрессии трансгена mNIS в раковых клетках человека использовали метод ПЦР в реальном времени (ПЦР-РВ). На первом этапе проводили выделение суммарной клеточной РНК. С этой целью монослой клеток MDA, U87G, Mel8, РС3, выращенный в лунках 6-луночного планшета, был инфицирован рекомбинантными вирусами с множественностью 0,1, 1,0 и 10 БОЕ/кл. Через 24 часа лунки планшета промывали добавлением раствора PBS, снимали клетки с поверхности лунок с помощью культурального скребка в 1 мл PBS, переносили в пробирки и осаждали центрифугированием (1000xg, 10 мин, 4°С). Дальнейшее выделение суммарной клеточной РНК проводили с использованием набора для выделения РНК на колонках RU-250 (ООО «Биолабмикс», Россия) по протоколу производителя, как описано ниже.
Осадок клеток ресуспендировали в 50 мкл PBS, добавляли 350 мкл буфера для лизиса LB (предварительно добавляли 10 мкл 2-меркаптоэтанола на 1 мл буфера LB), перемешивали пипетированием, инкубировали 5-10 мин. К лизату добавляли равный объем этанола, переносили лизат на колонку, центрифугировали 30 сек, 10000xg, удаляли фильтрат. Наносили на колонку 500 мкл буфера для промывки WB1 (для получения буфера WB1 к 150 мкл концентрата буфера WB1 добавляли 350 мкл этанола), центрифугировали 30 сек, 10000xg, удаляли фильтрат. Наносили на колонку 500 мкл буфера для промывки WB2 (для получения буфера WB2 к 100 мкл концентрата буфера WB2 добавляли 400 мкл этанола), центрифугировали 30 сек, 10000xg, удаляли фильтрат. Дополнительно центрифугировали 3 минуты, 10000xg, для полного удаления буфера WB2. Переносили колонку в новую микроцентрифужную пробирку, осаждали в 60 мкл буфера для элюции ЕВ - инкубировали 1 минуту, центрифугировали 1 мин, 10000xg. Сохранность выделенной РНК анализировали электрофорезом в 1% агарозном геле.
Для удаления загрязнения геномной ДНК и исключения контаминаций при ПЦР проводили обработку полученных образцов РНК ДНКазой I (RNase-Free DNase, Ambion). В пробирку добавляли 1 мкл 10х DNase I Buffer (100 mM Tris, рН 7.5, 25 mM MgCl2, 5 mM CaCl2), 0,5 е.а. фермента DNase I, раствор РНК в воде до суммарного объема смеси 10 мкл. Реакцию проводили в течении 30 мин при 37°С в микротермостате. Для полной инактивации ДНКазы I пробы инкубировали 10 мин при 75°С.Сохранность выделенной РНК и отсутствие геномной ДНК анализировали электрофорезом в 1% агарозном геле.
Реакцию обратной транскрипции проводили с помощью набора «Обратная транскриптаза М-MuLV-RH» (ООО «Биолабмикс», Россия) по протоколу производителя, как описано далее. В пробирки для ПЦР объемом 200 мкл вносили: 2 мкл 20 мкМ Олиго-d(Т)18 праймеров, 2 мкл 20 мкМ гексапраймеров, 8 мкл тестируемого образца РНК. Смесь инкубировали при 70°С 2 мин, затем охлаждали на льду 1 мин. В пробирки добавляли: 4 мкл 5Х буфера, 1 мкл ревертазы MMLV (100 ед/мкл), 3 мкл воды. Смесь тщательно перемешивали, инкубировали 10 мин при 25°С, затем инкубировали 1 час при 42°С.Реакцию останавливали путем прогревания смеси при 70°С в течение 10 мин.
Для определения уровня экспрессии гена mNIS проводили количественный ПЦР в реальном времени в присутствии красителя SYBR Green I с использованием набора БиоМастер HS-qPCR SYBR Blue (2х) (ООО «Биолабмикс», Россия). Амплификацию проводили в ПЦР-амплификаторе CFX96 (BioRad, США), используя следующую программу: 1) денатурация при 95°С в течение 3 мин, 2) денатурация при 95°С в течение 10 сек, 3) отжиг при 58°С в течение 10 сек, 4) элонгация и режим чтения плашки при 72°С в течение 30 сек, 5) режим чтения плашки при 82°С в течение 3 сек, 6) этапы 2-5 повторяли 40 раз, 6) кривая плавления - нагрев от 65°С до 95°С с шагом 0,5°С по 5 сек. Каждая реакционная смесь содержала 2 мкл матрицы кДНК, 300 нМ каждого праймера и смесь БиоМастер HS-qPCR SYBR Blue (2х). В качестве внутренних контролей, относительно которых проводилось нормирование продуктов амплификации исследуемого гена, были использованы гены «домашнего хозяйства» глицеральдегид 3-фосфат-дегидрогеназы - GAPDH и β-актин - АСТВ. Праймеры к мРНК гена mNIS и к мРНК генов GAPDH и АСТВ подбирали с помощью программы "Primer Blast".
Структуры праймеров:
GAPDH-F 5'-GAAATCCCATCACCATCTTCCAGG-3'
GAPDH-R 5'-GAGCCCCAGCCTTCTCCATG-3'
ACTB-F 5'-CCTGGCACCCAGCACAAT-3'
ACTB-R 5'-GGGCCGGACTCGTCATAC-3'
NIS-for 5'-GATGCACCAATGCCTCTGTC-3'
NIS-rev 5'-TACCCAGAGCCCCGTAGTAG-3'
В каждом эксперименте на один планшет помещали образцы исследуемых кДНК с праймерами на целевой ген mNIS (по 2 повтора на образец кДНК); аналогичные образцы с праймерами на 2 гена сравнения GAPDH и АСТВ (также по 2 повтора), также в каждом планшете присутствовал отрицательный контроль обратной транскрипции, положительный контроль ПЦР реакции, отрицательный контроль наличия геномной ДНК. Порог детекции (Ct) для каждой реакции определялся автоматически программой Bio-Rad CFX Manager 3.1 (BioRad, США). Относительный уровень экспрессии (ΔCt) каждого гена интереса рассчитывали по разнице его Ct и среднего геометрического значения Ct генов домашнего хозяйства. Из ΔCt в опытных образцах вычитали ΔCt контрольных образцов, получая ΔΔCt для каждого гена. Изменение экспрессии рассчитывали как 2ΔΔCt (Schmittgen, 2008) [11]. Графический анализ данных и оценка экспрессии генов проводилась с помощью программного обеспечения амплификатора Bio-Rad CFX Manager 3.1 (Bio-Rad, США).
Результаты экспериментов представлены на фиг. 4, из которого следует, что обнаруженные уровни мРНК трансгена mNIS положительно коррелируют с увеличением множественности инфекции клеток рекомбинантным штаммом VV-mNIS-NS1. В неинфицированных клетках был обнаружен только низкий фоновый уровень экспрессии мРНК mNIS, который был принят за 1 условную единицу. Полученные данные свидетельствуют об эффективной экспрессии трансгена mNIS в инфицированных рекомбинантными штаммами клетках опухолей человека.
Пример 3. Оценка биологической активности трансгена mNIS в раковых клетках человека.
Биологическую активность трансгена mNIS в раковых клетках оценивали по их способности аккумулировать йодид в процессе инфекции рекомбинантным штаммом VV-mNIS-NS1. Измерение количества йодида, захваченного клетками, проводили спектрофотометрически в формате 96-луночного планшета с использованием реакции Санделла-Колтоффа (Dong, 2019) [12]. Реакция Санделла-Колтоффа обеспечивает колориметрический индикатор концентрации ионов йода посредством реакции между четырехвалентным церием и трехвалентным мышьяком, катализируемой свободным I- в растворе серной кислоты. Восстановление желтого церия(IV) до бесцветного церия(III) под действием мышьяка(III) протекает очень медленно, но следы йодида сильно ускоряют эту реакцию, причем скорость прямо пропорциональна концентрации йодида.
Выращивали 100%-ный монослой клеток (MDA, U87G, Mel8, РС3) в прозрачных плоскодонных 96-луночных микропланшетах, оставляя 1-ый и 12-ый столбик планшета без клеток для последующего нанесения стандартных растворов. Инфицировали клетки десятикратными разведениями вирусной суспензии с множественностью инфекции от 10 до 0,01 БОЕ/кл с десятичным шагом разведения по 8 лунок на каждую точку. Спустя 24 часа промывали планшет буфером HHBS (Hank's буфер с добавлением 10 мМ Hepes) 4 раза, после каждого цикла промывки оставляя в лунке 95 мкл. После заключительной промывки в каждую лунку добавляли 5 мкл раствора 100 мкМ NaI (конечная концентрация 5 мкМ). Клетки инкубировали 2 часа при комнатной температуре, после чего промывали 4 раза буфером HHBS. Переворачивали и высушивали планшет в течение 10 мин, чтобы удалить остаточный внеклеточный I-. Стандарты NaI (10, 20, 30, 40, 50, 60, 70 пмоль) и воду распределяли в двух повторах по 100 мкл в свободные лунки, оставленные для стандартных растворов. Во все лунки, кроме лунок со стандартными растворами, вносили по 100 мкл деионизованной воды. Затем во все лунки добавляли 100 мкл 10,5 мМ раствора сульфата аммония церия(IV) и 100 мкл 24 мМ раствора арсенита натрия(III). Планшет инкубировали в течение 30 минут при комнатной температуре. Поглощение при 405 нм регистрировали на планшетном спектрофотометре SpectraCount (Packard). Для каждого планшета строили кривую зависимости значений поглощения от стандартных концентраций йодида и использовали линейную регрессию для построения калибровочной кривой. Концентрации йодида в инфицированных клетках определяли на основании калибровочной кривой и оптической плотности испытуемого раствора. Значения для каждой точки определяли в восьми повторностях, рассчитывали среднее значение и дисперсию.
Результаты представлены на фиг. 5, из которой следует, что раковые клетки человека MDA, U87G, Mel8, РС3, инфицированные рекомбинантным штаммом VV-mNIS-NS1 при множественности 1 БОЕ/кл поглощают йодид в количестве 44 пмоль, 52 пмоль, 32 пмоль, 25 пмоль, соответственно. Это значимо (Р<0,01) превышает поглощение ионов йода неинфицированными клетками (10,3-17 пмоль). При другой множественности инфекции в диапазоне от 0,1 до 10 БОЕ/кл отличия в поглощении йодида в сравнении с контрольными неинфицированными клетками были также достоверны (данные не представлены).
Пример 4. Оценка цитотоксической активности рекомбинантного штамма VV-mNIS-NS1 в отношении раковых и нормальных клеток человека.
Исследование цитотоксической активности рекомбинантного варианта VV-mNIS-NS1 проводили в отношении раковых клеток человека U87MG, Ме18, РС3 и MDA-MB-231. Для оценки онкоспецифичности использовали две культуры клеток нормальных тканей человека: MCF10A - культура клеток нормального эпителия молочной железы человека и LECH-240 - диплоидная культура клеток легкого эмбриона человека. Клетки опухолей человека выращивали на среде DMEM (Invitrogen, США) с добавлением 10% эмбриональной телячьей сыворотки (HyClone, США). Клетки MCF10A растили на среде MEGM Bullet Kit (Lonza, Швейцария). Клетки LECH-240 растили на среде F-12 (Invitrogen, США).
Исследование цитотоксической активности проводили микрометодом на 96-луночных культуральных планшетах (Greiner, США) с использованием субстрата для митохондриальных дегидрогеназ 2,3-бис-(2-метокси-4-нитро-5-сульфо-фенил)-2Н-тетразолий-5-карбоксанилида (реагент ХТТ) (Sigma, США). Цитотоксическую активность рекомбинантного варианта VV-mNIS-NS1 выражали как ЦТД50 - эффективная концентрация вируса, выраженная в количестве бляшкообразующих единиц на клетку (БОЕ/кл.), необходимая для гибели 50% клеток. Для определения ЦТД50 в лунки планшета с 50%-ным монослоем клеток вносили десятикратные разведения вирусной суспензии с множественностью инфекции от 10 до 0,001 БОЕ/кл. (multiplicity of infection, MOI) в 100 мкл среды 199 (ООО «Биолот», Россия) с добавлением 2% фетальной сыворотки коров (ООО «Биолот», Россия). Планшеты помещали в термостат при температуре 37°С, 5% СО2, влажности 85% и инкубировали 72 часа. После инкубации в каждую тестируемую лунку добавляли по 50 мкл реагента ХТТ и феназин-метасульфата (PMS) (Sigma, США), который получали добавлением к рабочему раствору с содержанием ХТТ 1 мг/мл раствора PMS 1,25 мМ из расчета: на каждый 1 мл ХТТ - 20 мкл PMS. Планшет инкубировали еще 3 часа и определяли оптическую плотность ОП490/620 на планшетном спектрофотометре SpectraCount (Packard, США). Строили график зависимости ОП от MOI и определяли ЦТД50 - концентрацию вируса, при которой величина ОП490/620, измеренная в зараженных лунках, составляет 50% от величины ОП490/620, измеренной в лунках с незараженной культурой. Сравнивали ЦТД50 для разных культур клеток. Чем больше ЦТД50, тем меньше цитотоксическая активность исследуемого вирусного препарата для данной культуры клеток. Значения для каждой точки определяли в пяти повторностях, рассчитывали среднее значение и дисперсию. Все расчеты проводили с использованием программного обеспечения LabView. Результаты представлены на фиг. 6, из которой следует, что рекомбинантный штамм VV-mNIS-NS1 обладает значимо большей цитотоксической активностью в отношении раковых клеток по сравнению с нормальными (Р<0,01). Индекс онкоселективности, рассчитанный как отношение ЦТД50 в нормальных клетках к ЦТД50 в раковых клетках, составляет более 7 для клеток LECH-240 и более 65 для клеток MCF10A. Различия в индексах селективности связаны с тем, что клетки эмбриона обладают достаточно высокой пролиферативной активностью, что, в определенной степени, роднит их с раковыми и повышает репликативную активность штамма VV-mNIS-NS1.
Таким образом, выше изложенные результаты (примеры 1-4) подтверждают достижение заявляемого технического результата, а именно, создан рекомбинантный штамм VV-mNIS-NS1 вируса осповакцины, несущий встройки трансгенов онкотоксического белка NS1 парвовируса Н-1 и симпортера йодида натрия мыши mNIS. Рекомбинантный штамм может использоваться для терапии и диагностики (тераностики) опухолей и метастазов человека за счет следующих свойств: штамм VV-mNIS-NS1 обладает высокой онколитической активностью в отношении раковых клеток человека и способностью накапливать в них йодид, что может использоваться для рентгеноскопической диагностики опухолей и метастазов, а при использовании радиоактивного субстрата и для радиотерапии.
Источники научно-технической и патентной информации
1. Ткачева А.В., Сиволобова Г.Ф., Локтев В.Б., Гражданцева А.А., Кочнева Г.В. Рекомбинантный штамм VV-NS1-dGF вируса осповакцины, продуцирующий белок NS1 парвовируса Н-1 и обладающий онколитической активностью в отношении глиобластомы человека. // Патент РФ №2692628, опубл. 25.06.2019, Бюл. №18 (прототип).
2. Koval О., Kochneva G., Tkachenko A., Troitskaya О., Sivolobova G., Grazhdantseva A., Nushtaeva A., Kuligina E., Richter V. Recombinant vaccinia viruses coding transgenes of apoptosis-inducing proteins enhance apoptosis but not immunogenicity of infected tumor cells // BioMed Research International. - 2017. - Article ID 3620510. https://doi.org/10.1155/2017/3620510
3. McCart A., Bartlett D., Moss B. Combined growth factor-deleted and thymidine kinase-deleted vaccinia virus vector. // US Patent. №7208313. - 2007.
4. Ravera S., Nicola J.P., Salazar-De Simone G., Sigworth F.J., Karakas E., Amzel L.M., Bianchet M.A., Carrasco N. Structural insights into the mechanism of the sodium/iodide symporter. // Nature. - 2022. https://doi.org/10.1038/s41586-022-05530-2.
5. Семенова A.B., Сиволобова Г.Ф., Гражданцева A.A., Агафонов А.П., Кочнева Г.В. Репортерные трансгены для мониторинга противоопухолевой эффективности рекомбинантных онколитических вирусов. // Acta Naturae. - 2022. - Т. 14. - №3(54). - С. 46-56. https://doi.org/10.32607/actanaturae.11719.
6. Thorne S., Hwang Т., O'Gorman W., Bartlett D.L., Sei S., Kanji F., Brown C., Werier J., Cho J.-H., Lee D.-E., Wang Y., Bell J., Kirn D.H. Rational strain selection and engineering creates a broad-spectrum, systemically effective oncolytic poxvirus, JX-963. // J. Clin. Invest. - 2007. - V. 117. - P.3350-3358. https://doi.org/10.1172/JCI32727.
7. Concilio S.C, Zhekova H.R., Noskov S.Y., Russell S.J. Inter-species variation in monovalent anion substrate selectivity and inhibitor sensitivity in the sodium iodide symporter (NIS). // PLoS ONE. - 2020. - V. 15(2). - P. 1-25. https://doi.org/10.1371/journal.pone.0229085.
8. Dayem M., Basquin С., Navarro V., Carrier P., Marsault R., Chang P., Huc S., Darrouzet E., Lindenthal S., Pourcher T. Comparison of expressed human and mouse sodium/iodide symporters reveals differences in transport properties and subcellular localization. // Journal of Endocrinology. - 2008. - V. 197(1). - P. 95-109. https://doi.org/10.1677/JOE-07-0455.
9. Merchlinsky M., Eckert D., Smith E., Zauderer M. Construction and characterization of vaccinia direct ligation vectors. // Virology. - 1997. - V. 238. - P. 444-451. https://doi.org/10.1006/viro.1997.8828.
10. Shvalov A.N., Sivolobova G.F., Kuligina E.V., Kochneva G.V. Complete Genome Sequence of Vaccinia Virus Strain L-IVP. // Genome Announcements. - 2016. - V. 4 (3). - e00372-16. https://doi.org/10.1128/genomeA.00372-16.
11. Schmittgen T.D., Livak, K.J. Analyzing real-time PCR data by the comparative CT method. // Nat. Protoc. - 2008. - V. 3. - №6. - P. 1101-1108. https://doi.org/10.1038/nprot.2008.73.
12. Dong H., Atlas E., Wade M.G. Development of a non-radioactive screening assay to detect chemicals disrupting the human sodium iodide symporter activity. // Toxicology in Vitro. - 2019. - V. 57. - P. 39-47. https://doi.org/10.1016/j.tiv.2019.01.021.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Штамм VV-mNIS-NS1
Кочнева_20.07.2023.xml" softwareName="WIPO Sequence"
softwareVersion="2.3.0" productionDate="2023-07-20">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1234567</ApplicationNumberText>
<FilingDate>2023-07-20</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>123456</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>12345</ApplicationNumberText>
<FilingDate>2023-07-20</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное бюджетное учреждение
науки «Государственный научный центр вирусологии и биотехнологии
«Вектор» Федеральной службы по надзору в сфере защиты прав
потребителей и благополучия человека (ФБУН ГНЦ ВБ «Вектор»
Роспотребнадзора)</ApplicantName>
<ApplicantNameLatin>Federalnoe byudzhetnoe uchrezhdenie nauki
"Gosudarstvennyj nauchnyj tsentr virusologii i biotekhnologii
"Vektor" Federalnoj sluzhby po nadzoru v sfere zashchity
prav potrebitelej i blagopoluchiya cheloveka (FBUN GNTS VB
"Vektor" Rospotrebnadzora) (RU)</ApplicantNameLatin>
<InventionTitle languageCode="ru">Рекомбинантный штамм VV-mNIS-NS1
вируса осповакцины, продуцирующий симпортер йодида натрия мышей mNIS
и белок NS1 парвовируса крыс H-1 для тераностики злокачественных
опухолей</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>1872</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1872</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aagcttaccatggaaggtgctgaagcaggtgcaagagccacctttggtc
cctgggactatggagtgtttgccaccatgttgcttgtgtccactggcattggtctgtgggttggccttgc
tagaggaggccaaagaagtgcagatgacttcttcaccggtggtcggcagcttgctgcagtgcctgtgggg
ttgtcacttgcagctagcttcatgtctgctgttcaggtgcttggagtcccagcggaagcagcgcgctatg
gactcaagtttctgtggatgtgtgttggccagttgctcaactcgctgctcacagccttgcttttccttcc
catcttctatagacttggccttaccagcacatatcagtacctagaacttaggttcagtagagctgtcaga
ctctgtgggactctgcagtacttggttgccacgatgctgtacactggcattgtgatctacgcacctgctc
tcatcctgaaccaagtgacagggttggacatctgggcatcactactgtccacagggatcatctgcacctt
gtacacgaccgttggtggcatgaaagccgtggtctggacagatgtgttccaagtggttgtcatgctcgtc
ggcttctgggtgatccttgctcgaggtgtcatgctgatgggtggaccctggaacgtgctcagtctcgctc
agaaccactctcggatcaacctgatggactttgatccagatcctaggagtaggtacaccttctggacctt
tgtcgttggtggctcacttgtgtggctctccatgtatggtgtgaaccaagctcaggtgcaacggtatgtg
gcctgtcacacggagaggaaagccaagctggcactgcttgtcaatcagcttggactcttcttgattgttg
ctagtgcagcctgctgtggcattgtcatgtttgtctactacaaggactgcgatccactcttgacaggccg
catcgcagcccccgatcagtacatgcccttgcttgtgttggacatctttgaggatcttcctggagttcca
ggactcttccttgcctgtgcctacagtggcaccttgagcactgcctccaccagcatcaatgccatggctg
ctgtcactgtggaagacctcatcaaacccaggatgcccagtctggcaccacggaagctggtgttcatctc
taaaggactctccttcatctatggctcaacctgtctcacagttgctgcactgtccagtctgctaggaggt
ggtgtcctccaaggctccttcacagtgatgggtgtcatcagtggtcctctacttggagccttcacacttg
ggatgctactcccagcctgcaacactccaggtgttctctccggactgacagcaggcttagctgtgtccct
gtgggttgccgttggggccacactgtacccacctggagaacagaccatgggtgtgttacccacctctgct
gcaggatgcaccaatgcctcagttcttccgagtccacctggagctgccaacacttccagagggattccca
gctcaggaatggactcgggcagacctgcctttgccgacaccttctatgctgtctcctatctctactatgg
ggctctgggcacattgaccactatgctttgtggtgctctcatcagctatctgactggccccaccaaaagg
agctcacttggtcctggattgctgtggtgggatctcgctagacagacagcatctgttgcacccaaggagg
acaccaccactcttgaagacagtctggtcaaaggaccggaagacattcctgctgcgaccaagaaaccccc
tggcttcagaccagaagctgagactcatcctctgtatcttggtcatgatgttgaaaccaacctctgagaa
ttc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---


Изобретение относится к биотехнологии. Предложен рекомбинантный штамм вируса осповакцины VV-mNIS-NS1, используемый для терапии и диагностики опухолей человека, включающий трансген онкотоксического белка NS1 парвовируса Н-1 и трансген mNIS, имеющий искусственную нуклеотидную последовательность SEQ ID NO: 1 и содержащую синтетический ген мышиного симпортера йодида натрия mNIS. Штамм депонирован в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером V-1206. Изобретение обеспечивает эффективную терапию и диагностику опухолей человека. 6 ил., 4 пр.
Рекомбинантный штамм вируса осповакцины VV-mNIS-NS1, используемый для терапии и диагностики опухолей человека, депонированный в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером V-1206 и включающий трансген онкотоксического белка NS1 парвовируса Н-1 и трансген mNIS, имеющий искусственную нуклеотидную последовательность SEQ ID NO: 1 и содержащую синтетический ген мышиного симпортера йодида натрия mNIS.
| Рекомбинантный штамм VV-NS1-dGF вируса осповакцины, продуцирующий белок NS1 парвовируса H-1 и обладающий онколитической активностью в отношении глиобластомы человека | 2018 |
|
RU2692628C1 |
| база данных GenBank: AF235001.1, 01.02.2001 Mus musculus sodium iodide symporter NIS mRNA, complete cds | |||
| Найдено онлайн: https://www.ncbi.nlm.nih.gov/nucleotide/AF235001.1?report=genbank&log$=nucltop&blast_rank=1&RID=10T68JEN01N Дата обращения 16.01.2024 | |||
| HADDAD D | |||
| et al | |||
| Imaging characteristics, tissue distribution, | |||

