Область техники, к которой относится данное изобретение
Настоящее изобретение относится к клеткам растений, которые экспрессируют белок-мишень, к способу их получения и способу получения белка-мишени с использованием этих клеток. Кроме того, данное изобретение относится к клеткам растений, в которые был введен вектор, содержащий ген (гены), кодирующие белок-мишень, благодаря чему они экспрессируют данный белок-мишень, способу их получения, а также способу крупномасштабного производства белка-мишени с помощью клеток растений.
Уровень техники
В данном контексте термин "биофармацевтические препараты" относится к лекарственным средствам, продуцируемым из биологических материалов. В более широком смысле этот термин может относиться к лекарственным средствам, получаемым с помощью биоинженерных технологий, включающих генетическую рекомбинацию, слияние клеток и культивирование клеток. Такие биофармацевтические препараты классифицируют как белковые лекарственные средства, терапевтические антитела, вакцины, агенты для генотерапии и агенты для клеточной терапии.
В последнее время большинство рекомбинантных белков получают либо с использованием эукариотических клеток-хозяев, таких как клетки животных или клетки насекомых, либо с использованием микроорганизмов, таких как дрожжи или бактерии. Однако такие культуры клеток животных имеют ряд недостатков, заключающихся в том, что используемые среды являются дорогими, высока возможность загрязнения вирусами, способными инфицировать человека, и требуется дополнительный отдельный процесс очистки для удаления происходящих из бычьей сыворотки белков, которые могут находиться в ней (Huang and McDonald 2009).
По этой причине недавно исследователи начали уделять внимание культурам клеток растений в качестве альтернативной системы для получения рекомбинантных белков. Клетки растений считаются безопасной системой продуцирования, так как эти клетки растений не инфицируются вирусами или патогенами животного происхождения, и нет возможности попадания в них постороннего материала животного происхождения.
Однако культуры клеток растений характеризуется относительно низким уровнем экспрессии белка и низкой скоростью роста в сравнении с культурами других клеток-хозяев, в том числе культурами клеток животных. Таким образом, для коммерциализации полученных из растений биофармацевтических средств, необходимо создание новых систем получения рекомбинантных белков из новых культур растительных клеток.
Информация, описанная в разделе уровень техники, предназначена только для лучшего понимания предпосылок создания данного изобретения, и, следовательно, может не содержать информации, которая образует известный уровень техники, который уже известен среднему специалисту в этой области.
Техническая задача
Задачей данного изобретения является обеспечение способа получения белка-мишени с превосходным выходом продукции, что позволяет применять его в массовом производстве.
Техническое решение
Для решения вышеуказанной задачи, в настоящем изобретении предлагаются клетки растений для экспрессии белка-мишени, где эти клетки несут рекомбинантный вектор, содержащий встроенный в него ген (гены), кодирующий белок-мишень. Данные клетки растений включают клетки камбиальной меристемы (CMC) или каллюса, где клетки камбиальной меристемы или каллюса (CMC) являются типом полученной из растений клеточной линии, содержащей выделенные из растения по природе недифференцированные клетки, где клеточная линия выделена из камбиальной ткани растения и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс.
В данном изобретении также предложен способ получения клеток растений для экспрессии белка-мишени, где способ включает стадию трансфекции или трансформации популяции клеток растений, включающей клетки камбиальной меристемы (CMC) или каллюса, геном (генами), кодирующими белок-мишень; сокультивирования популяции клеток растений, включающих клетки камбиальной меристемы (CMC) или каллюса с Agrobacteria, содержащими вектор, несущий встроенный ген, кодирующий белок-мишень, где эти клетки камбиальной меристемы (CMC) являются клеточной линией, содержащей по природе недифференцированные клетки, выделенные из растения, причем эта клеточная линия выделена из камбиальной ткани растения и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс.
В данном изобретении также предложен способ получения белка-мишени клетками растений, предназначенными для экспрессии белка-мишени. Данный способ включает стадии: (а) стабильной трансформации популяции клеток растений, включающей клетки камбиальной меристемы (CMC) или каллюса, геном (генами), кодирующими белок-мишень, или временная экспрессия в них гена (генов), кодирующих белок-мишень, посредством сокультивирования этой популяции клеток растений, включающей клетки камбиальной меристемы (CMC) или каллюса, с Agrobacteria, содержащими вектор, несущим встроенный ген, кодирующий белок-мишень, где эти клетки камбиальной меристемы (CMC) являются клеточной линией, содержащей по природе недифференцированные клетки, выделенные из растения, где эта клеточная линия выделена из камбиальной ткани растения и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс; и (b) извлечение белка-мишени, экспрессированного в культуре клеток растения, которые инфицировали Agrobacteria посредством сокультивирования.
В настоящем изобретении также предлагается способ получения белка-мишени из трансгенного растения, предназначенного для экспрессии белка-мишени, где способ включает стадии:
(a) выращивания растения, трансформированного геном (генами), кодирующими белок-мишень;
(b) выделения трансгенных клеток камбиальной меристемы (ТСМС) из трансгенного растения;
(c) культивирования выделенных трансгенных клеток камбиальной меристемы (ТСМС) в среде; и
(d) выделения белка-мишени, экспрессированного в культивированных трансгенных клетках камбиальной меристемы (ТСМС).
Краткое описание фигур
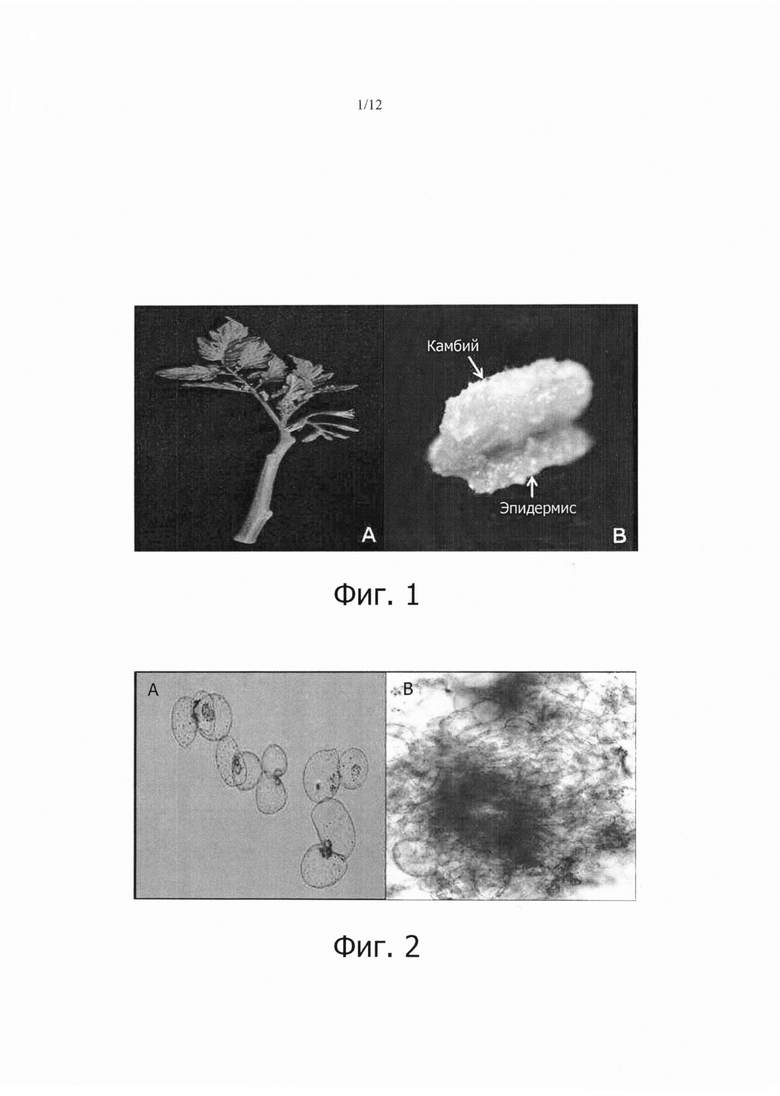
Фиг. 1А является фотографией растительного материала (стебля томата), а фиг. 1В является фотографией, показывающей, что клетки камбиальной меристемы (CMC) индуцировались и начали отделяться от каллюсного слоя, полученного из другой ткани.
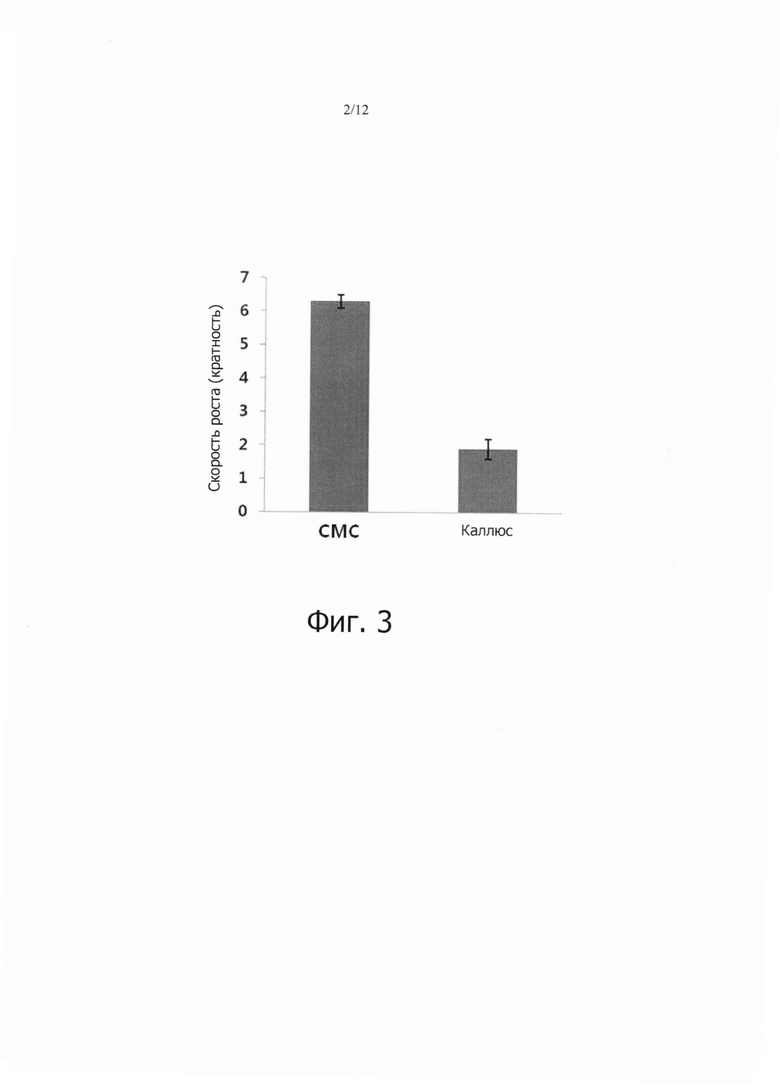
Фиг. 2 является набором микрофотографий, показывающим результаты наблюдения степени агрегации клеток у клеток камбиальной меристемы томата (CMC; фиг. 2А) в соответствии с данным изобретением и клеток каллюса томата (фиг. 2В).
Фиг. 3 является графиком, показывающим скорость роста клеток камбиальной меристемы томата (CMC) по данному изобретению и клеток каллюса томата.
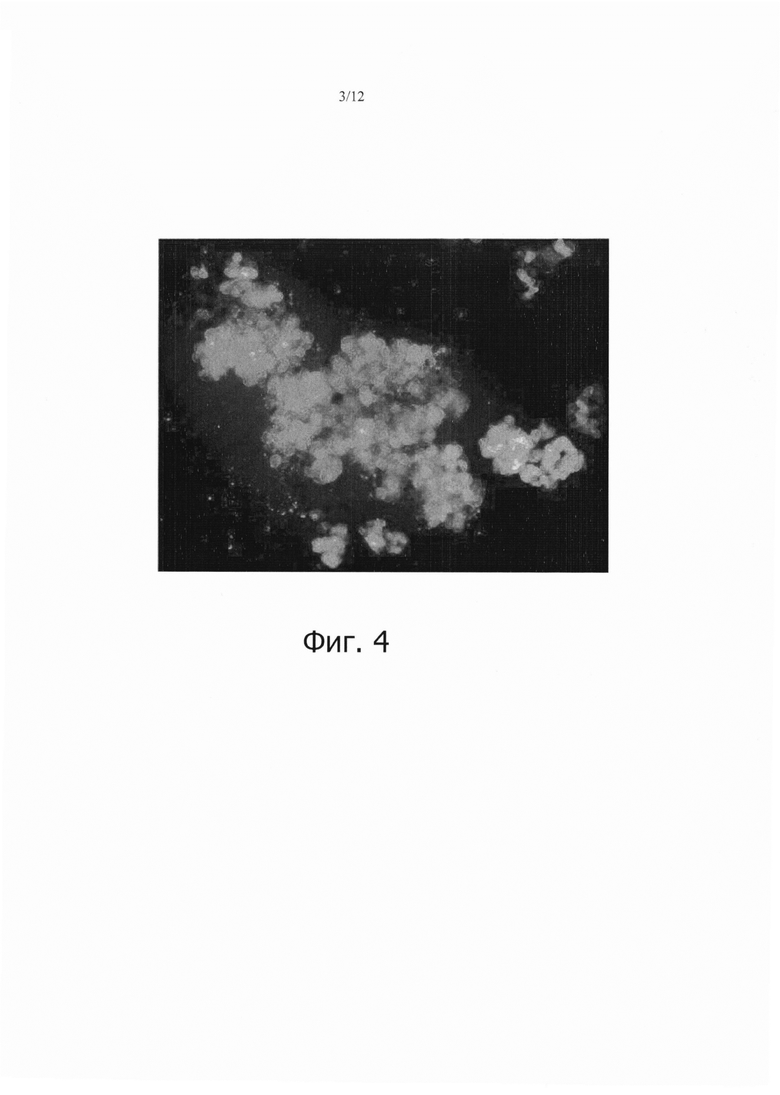
Фиг. 4 является фотографией, показывающей временную экспрессию зеленого флуоресцентного белка (GFP), что указывает на то, что 90% или более клеток камбиальной меристемы (CMC) томата были инфицированы Agrobacteria после 5 дней сокультивирования.
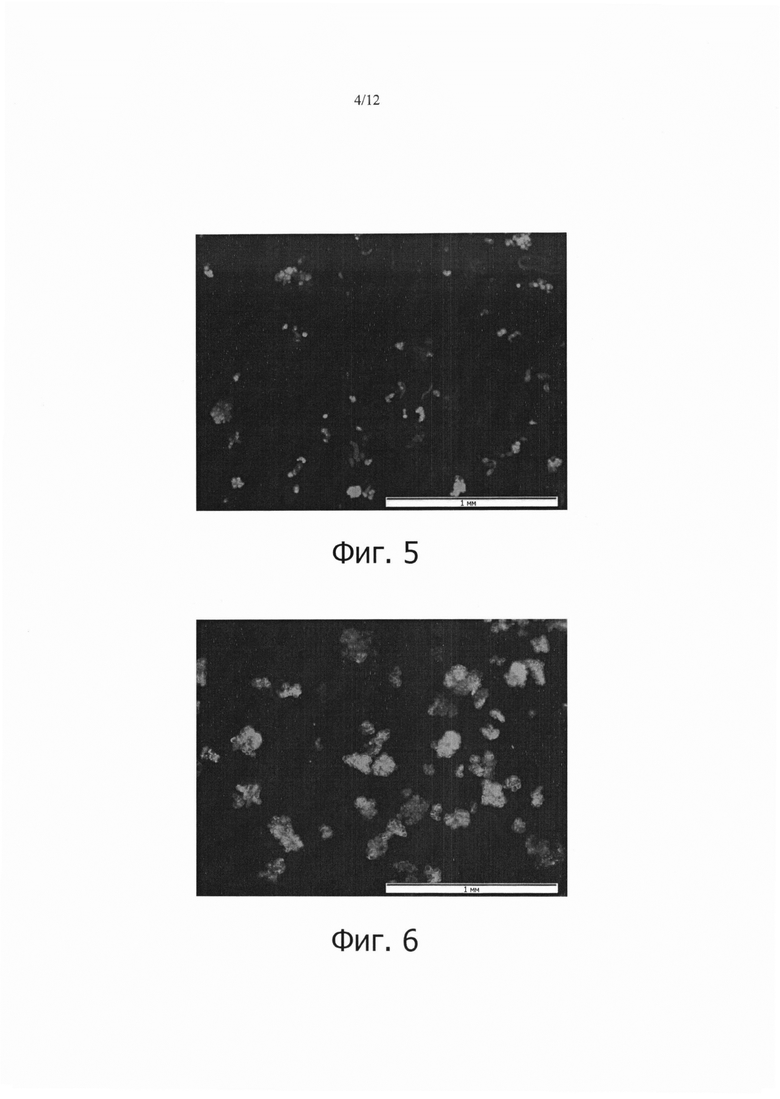
Фиг. 5 является фотографией, показывающей временную экспрессию GFP в клетках камбиальной меристемы (CMC) дикого женьшеня после 10 дней сокультивирования с Agrobacteria.
Фиг. 6 является фотографией показывающей временную экспрессию GFP в клетках камбиальной меристемы (CMC) моркови после 10 дней сокультивирования с Agrobacteria.
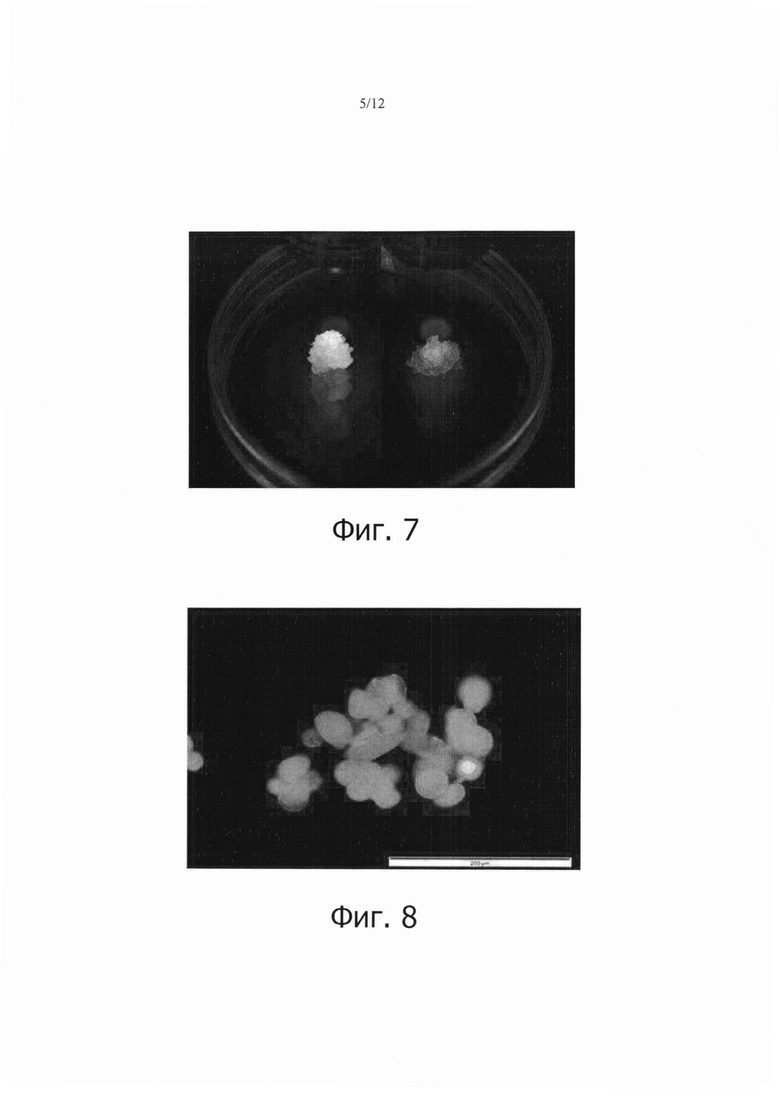
Фиг. 7 является фотографией, показывающей результаты наблюдения кластера, который экспрессирует GFP в результате стабильной трансформации клеток камбиальной меристемы (CMCs) томата, и нетрансформированного кластера, под УФ-светом.
Фиг. 8 является флуоресцентной микрофотографией клеток камбиальной меристемы (CMC), которые стабильно трансформированы GFP.
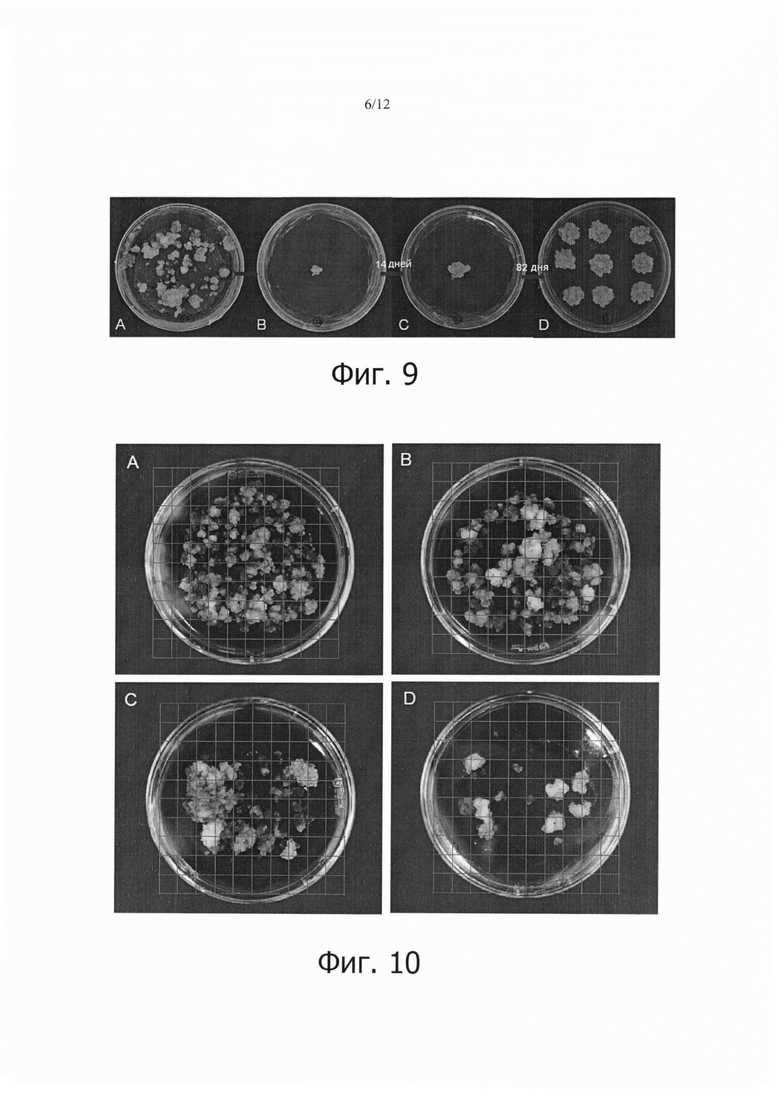
Фиг. 9 является набором микрофотографий, показывающих результаты пролиферации GFP-экспрессирующих клеток при непрерывном субкультивировании выбранных кластеров, для которых подтверждено, что они испускают флуоресценцию.
Фиг. 10 является набором фотографий, показывающих сравнение стабильной трансформации клеток камбиальной меристемы (CMC) томата или каллюса в случае, когда выполнялась седиментация, и в случае, когда не выполнялась седиментация.
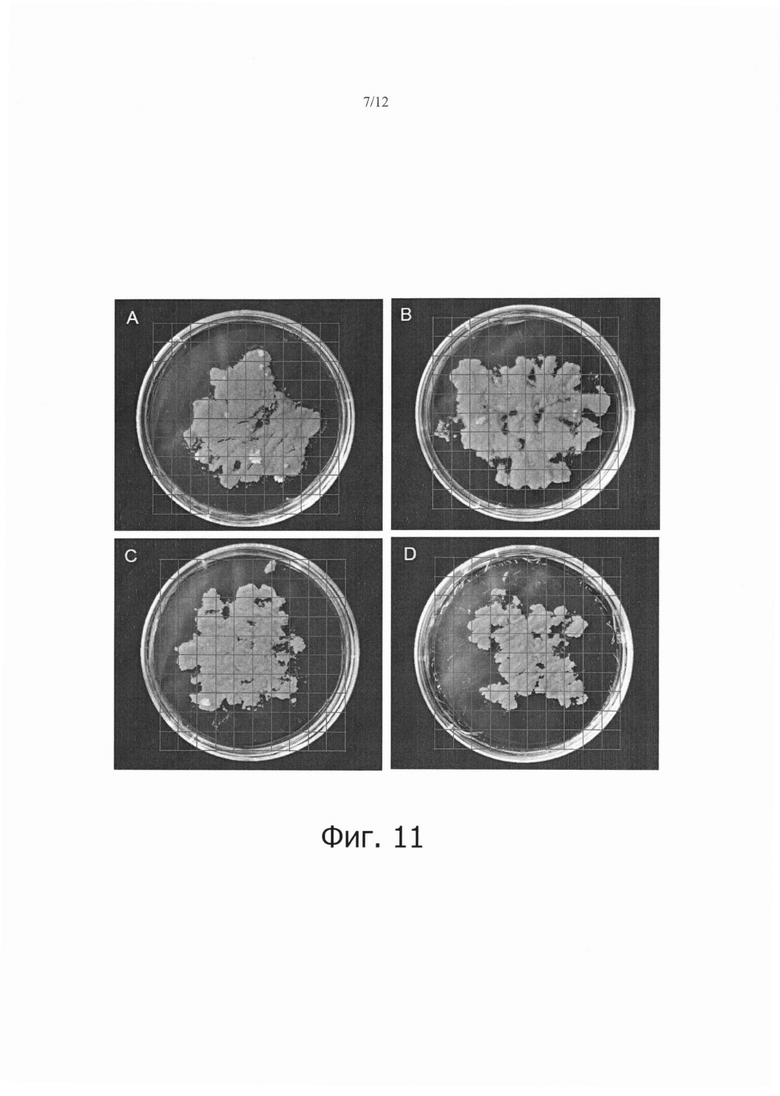
Фиг. 11 является набором фотографий, показывающих сравнение стабильной трансформации клеток камбиальной меристемы (CMC) или каллюса моркови в случае, когда выполнялась седиментация, и в случае, когда не выполнялась седиментация.
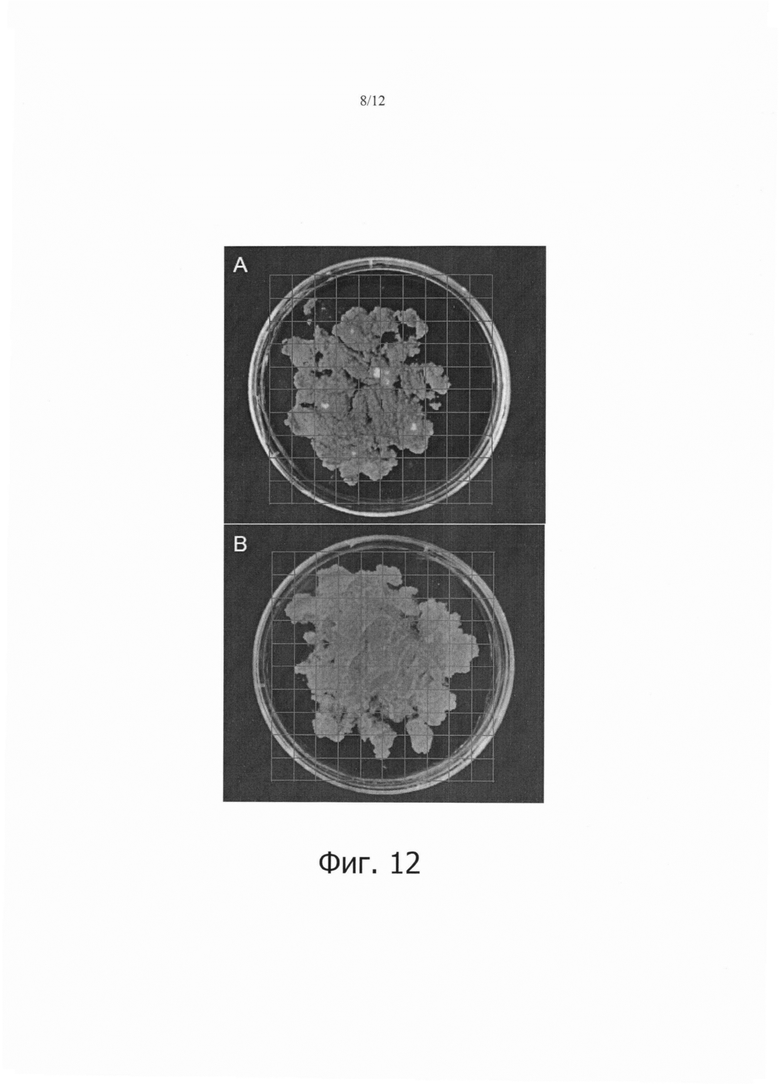
Фиг. 12 является набором фотографий, показывающих сравнение стабильной трансформации клеток камбиальной меристемы (CMC) дикого женьшеня в случае, когда выполнялась седиментация, и в случае, когда не выполнялась седиментация.
Фиг. 13 является фотографией, показывающей результаты экспрессии GFP после культивирования выделенных трансгенных по GFP клеток камбиальной меристемы томата (ТСМС) в 3-литровом биореакторе и облучении их УФ-светом.
Фиг. 14 является фотографией, показывающей результаты акклиматизации трансгенного Nicotiana benthamiana и выращивания акклиматизированного растения в цветочном горшке.
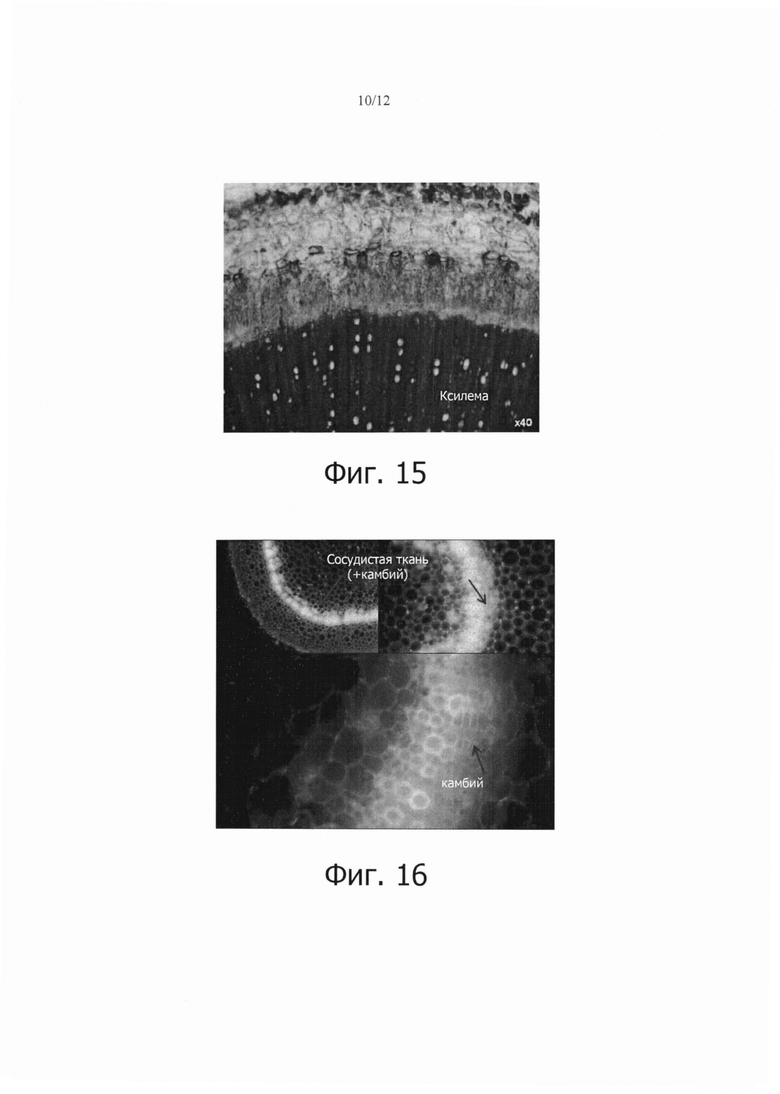
Фиг. 15 является фотографией, показывающей морфологию собранного стебля табака.
Фиг. 16 является фотографией, на которой представлено поперечное сечение растения табака, трансформированного GFP.
Фиг. 17 является фотографией, показывающей результаты выделения трансформированных клеток камбиальной меристемы (ТСМС) из камбиальной зоны трансгенного Nicotiana benthamiana.
Фиг. 18 является фотографией, показывающей результаты выделения клеток камбиальной меристемы (ТСМС) из камбиальной зоны трансгенного Nicotiana tabacum cv. xanthi.
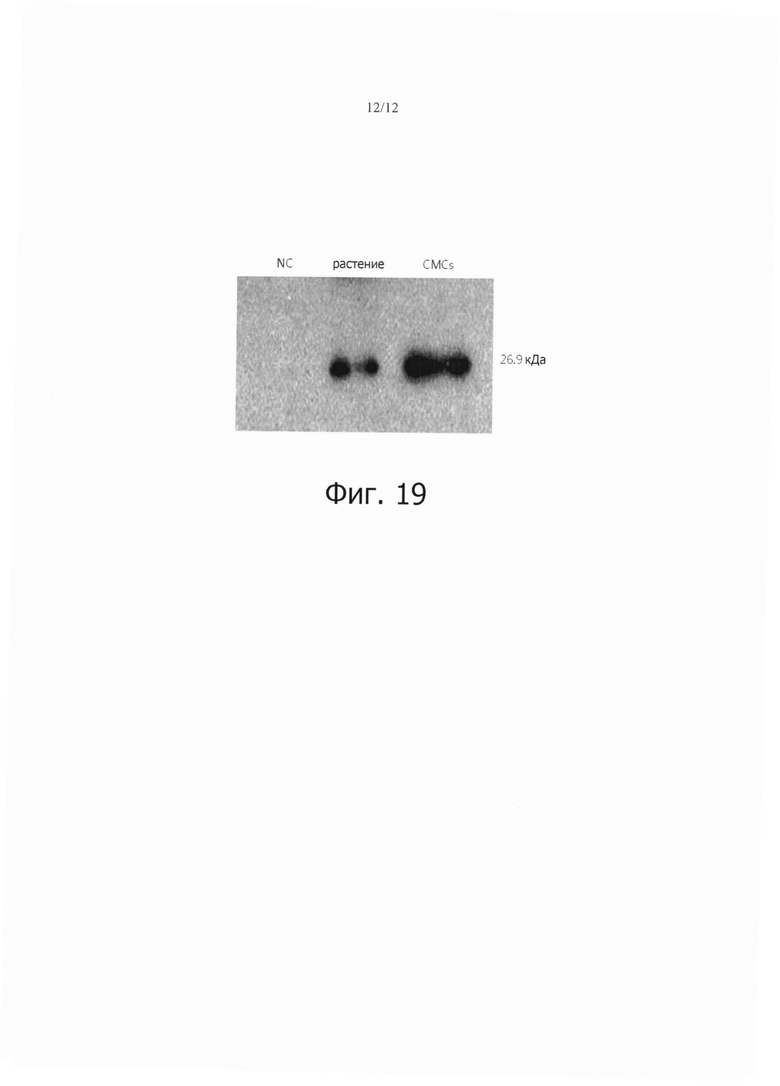
Фиг. 19 является фотографией, показывающей результаты вестерн-блот анализа общего растворимого белка, экстрагированном как из трансгенного по GFP растения, так и из клеток камбиальной меристемы (ТСМС), выделенных из растения.
Лучший способ осуществления изобретения
Если не указано другое, все технические и научные термины, используемые здесь, имеют то же самое значение, какое обычно понимается специалистом в области, к которой относится это изобретение. В общем используемая здесь номенклатура хорошо известна и широко используется в данной области.
Клетки растений для экспрессии белка-мишени содержат введенный в них рекомбинантный вектор, несущий ген (гены), кодирующие этот белок-мишень, и эти клетки растений включают клетки камбиальной меристемы (CMC) или каллюса.
В одном варианте осуществления, эти клетки растений могут быть трансгенными клетками камбиальной меристемы (ТСМС), в которые ввели рекомбинантный вектор, несущий ген (гены), кодирующие белок-мишень.
Авторы данного изобретения обнаружили, что введение гена, кодирующего белок-мишень в клетки камбиальной меристемы (CMC), позволяет преодолеть проблемы, связанные с медленной скоростью роста и низким уровнем экспрессии белка, которые возникают в культуре каллюса из предшествующего уровня техники. Кроме того, авторы этого изобретения обнаружили, что когда рекомбинантный белок получают трансформацией клеток камбиальной меристемы (CMC), то эффективность трансформации этих клеток значительно выше, чем эффективность трансформации ранее известных клеток растений, благодаря чему стало возможным значительно увеличить продукцию за счет применения технологии временной экспрессии белка-мишени, а также можно использовать стабильную трансформацию.
Здесь, к клеткам камбиальной меристемы (CMC) относят выделенную из растения клеточную линию, содержащую по природе недифференцированные клетки. Эту клеточную линию выделяют из камбиальной ткани растения, и она имеет меристематическую преемственность без прохождение через дедифференцировку в каллюс.
В данном изобретении каллюс является аморфной клеточной массой, которая образуется посредством дедифференцировки дифференцированной ткани вследствие повреждения. После дедифференцировки, каллюс теряет свои первоначальные характеристики и присутствует в недифференцированном состоянии. Клетки камбиальной меристемы растений (CMC), которые используют в данном изобретении, отличаются от каллюса тем, что они по природе поддерживаются в недифференцированном состоянии без прохождения через дедифференцировку.
Некоторые из авторов данного изобретения сначала выделили из камбия растений клетки камбиальной меристемы (CMC), которые являются по природе недифференцированными клетками, отличающимися от недифференцированного каллюса (KR 10-1064519 В1). Такие клетки камбиальной меристемы растений могут быть выделены способом, включающем стадии: (а) сбора камбий-содержащей ткани из растения; (b) культивирования собранной камбий-содержащей ткани в среде; и (с) выделения из культивируемой камбий-содержащей ткани камбиальных клеток, которые не включают ни части, отличные от камбия, ни каллюс, происходящий из частей, отличных от камбия. Данная камбий-содержащая ткань на стадии (а) может быть дополнительно стерилизована.
В данном контексте, термин "вектор" обозначает ДНК конструкцию, содержащую последовательность ДНК, функционально связанную с подходящей регуляторной последовательностью, способной влиять на экспрессию этой ДНК в подходящем хозяине. Вектором может быть плазмида, фаговая частица или просто последовательность, способная встраиваться в геном. После включения в подходящего хозяина вектор может реплицироваться и функционировать независимо от генома хозяина, или в некоторых случаях может интегрироваться непосредственно в геном. В данном описании, "плазмида" и "вектор" иногда используются взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора.
Для целей данного изобретения предпочтительно используют плазмидный вектор. Типичный плазмидный вектор, который может быть использован, содержит: (а) ориждин репликации, посредством которого осуществляется эффективная репликация, что создает несколько сотен плазмидных векторов на клетку-хозяина; (b) ген устойчивости к антибиотику, посредством которого могут быть отобраны клетки-хозяева, трансформированные плазмидным вектором; и (с) сайты узнавания рестрикционных ферментов, по которым могут быть встроены чужеродные ДНК-фрагменты. Даже если подходящие сайты узнавания рестрикционных ферментов не присутствуют в векторе, использование стандартных синтетических олигонуклеотидных адапторов или линкеров делает возможным легкое лигирование вектора и чужеродных ДНК-фрагментов. После лигирования, вектор должен быть трансформирован в подходящие клетки-хозяева. Трансформация может легко достигаться посредством кальций-хлоридного способа или электропорации (Neumann, et al., EMBO J., 1: 841, 1982).
В качестве вектора, используемого для сверхэкспрессии гена по данному изобретению, можно взять экспрессионный вектор, известный в данной области. В данном описании применялся бинарный вектор, часто используемый для трансформации растения.
Как также хорошо известно в данной области, для увеличения уровня экспрессии вводимого гена в клетке-хозяине, соответствующий ген должен быть функционально связан с регуляторными последовательностями, контролирующими процессы транскрипции и трансляции. Предпочтительно эти регуляторные последовательности, контролирующие экспрессию, и соответствующий ген включены в один экспрессионный вектор вместе с бактериальным селективным маркером и сайтом инициации репликации. Рекомбинантный вектор предпочтительно включает в себя также дополнительный маркер экспрессии, который эффективен в клетках растений.
Трансгенные клетки камбиальной меристемы (ТСМС) с вышеупомянутым экспрессионным вектором составляют другой аспект данного изобретения. В данном контексте, термин "трансформация" означает, что введенная в хозяина ДНК может быть реплицирована в виде фактора вне хромосомы или в составе хромосомы хозяина. Между тем, в данном контексте, термин "трансфекция" обозначает, что ДНК введена в клетки-хозяева таким образом, что она может реплицироваться в этих клетках-хозяевах.
Конечно должно быть понятно, что все векторы и последовательности, регулирующие экспрессию, не равнозначно функционируют в системе клеток камбиальной меристемы (CMC) по данному изобретению. Однако квалифицированный в данной области специалист может без дополнительного экспериментирования выбрать наиболее подходящий вектор и регуляторные последовательности, не отступая от сущности настоящего изобретения. Если говорить более конкретно, то при выборе должны учитываться число копий вектора, способность к регуляции числа копий и экспрессия этим вектором другого белка (например, маркера устойчивости к антибиотикам).
Как указывалось выше, ген (гены), кодирующие белок-мишень, могут временно экспрессироваться в клетках камбиальной меристемы (CMC) растения или клетки могут быть стабильно трансформированы этим геном (генами) с помощью вектора.
Однако ген (гены), кодирующие белок-мишень, могут быть не только введены в клетки камбиальной меристемы растений (CMC) и временно экспрессироваться в них, но также ген (гены), кодирующие белок-мишень, могут встраиваться в геном клеток камбиальной меристемы растения (CMC) так, что он будет присутствовать на хромосоме, то есть данные клетки могут быть стабильно трансформированы им. Квалифицированному в данной области специалисту будет очевидно, что в контексте настоящего изобретения встраивание гена (генов) белка-мишени в геном (хромосому) клеток камбиальной меристемы (CMC) растений будет такой же эффект, что и введение рекомбинантного вектора в клетки камбиальной меристемы (CMC), как описано выше.
Таким образом, в другом аспекте, данное изобретение относится к трансгенным клеткам камбиальной меристемы (ТСМС) для экспрессии белка-мишени, которые содержат ген(гены), кодирующий белок-мишень (белки-мишени), встроенный в хромосому клеток камбиальной меристемы (CMC).
В данном изобретении введение вектора, несущего ген (гены), кодирующие белок-мишень, или встраивание этого гена (генов), кодирующего белок-мишень, в хромосому клеток камбиальной меристемы (CMC) может выполняться путем добавления Agrobacteria, содержащих вектор, несущий ген (гены), кодирующие белок-мишень, к популяции растительных клеток, включающей клетки камбиальной меристемы (CMC) или каллюса, и сокультивированием Agrobacteria с этими растительными клетками.
В одном варианте осуществления сокультивирование может выполняться в условиях темноты. Сокультивирование может осуществляться путем культивирования клеток растений, включающих клетки камбиальной меристемы или каллюса, с культурой Agrobacteria, содержащей вектор, несущий ген (гены), кодирующий белок-мишень, с встряхиванием, и затем следует стационарное культивирование.
Термин "стационарное культивирование" обозначает в данном контексте способ культивирования клеток в контейнере в стационарном состоянии без встряхивания культуральной среды, который может быть использован взаимозаменяемо со способом седиментации клеток без встряхивания.
Стационарное культивирование может выполняться один раз или выполняться периодически. Когда стационарное культивирование выполняют один раз, растительные клетки и культуру Agrobacteria могут, например, сокультивировать со встряхиванием, затем стационарно культивировать и затем опять культивировать со встряхиванием. Когда стационарное культивирование выполняют периодически (периодически выполняемое стационарное культивирование), процесс состоит из повторяющихся от нескольких раз до нескольких десятков раз культивирований, осуществляемых следующим образом: сокультивирование растительных клеток и культуры Agrobacteria с встряхиванием, стационарное культивирование растительных клеток и культуры Agrobacteria и затем опять сокультивирование растительных клеток и культуры Agrobacteria со встряхиванием.
Более конкретно, этот процесс культивирования может выполняться сокультивированием растительных клеток и культуры Agrobacteria, содержащей вектор, несущий ген(ы), кодирующий белок-мишень, со встряхиванием в течение 1 минуты - 48 часов, затем стационарное культивирование растительных клеток и культуры Agrobacteria в течение 1 минуты - 96 часов, и затем культивирование со встряхиванием в течение 1-10 дней.
Agrobacteria, которые добавляют для сокультивирования, могут иметь OD600 0,00001-2,0. Если OD600 Agrobacteria является слишком низким, возникнет проблема, заключающаяся в том, что снижается частота трансфекции для временной экспрессии, а если OD600 Agrobacteria является слишком высоким, то возникнет проблема, заключающаяся в том, что жизнеспособность клеток-хозяев быстро уменьшается. Таким образом, предпочтительно добавлять для сокультивирования Agrobacteria, имеющие OD600 в определенном выше диапазоне.
Используемые в данном изобретении Agrobacteria могут быть которые наиболее часто используемые для трансформации растений Agrobacteria. Например, этими Agrobacteria могут быть Agrobacterium tumefaciens или Agrobacterium rhizogenes.
Как описано выше, белок-мишень может быть получен культивированием трансгенных клеток камбиальной меристемы (ТСМС) для экспрессии в них белка-мишени и извлечения этого экспрессированного белка-мишени.
В другом варианте осуществления, трансгенные клетки камбиальной меристемы (ТСМС) могут также быть выделены из трансгенного растения, содержащего ген (гены), кодирующие белок-мишень.
В данном изобретении, растение табака стабильно трансформировали геном (генами), кодирующим белок-мишень, и выращивали в цветочном горшке. В результате, было обнаружено, что уровень экспрессии гена (генов), кодирующего белок-мишень в камбии был выше, чем уровни экспрессии в других тканях.
Белок-мишень, который используют в данном изобретении, может, например, быть одним или несколькими белками, выбранными из группы, состоящей из антигенов, антител, фрагментов антител, структурных белков, регуляторных белков, факторов транскрипции, белков токсинов, гормонов, аналогов гормонов, цитокинов, ферментов, ингибиторов ферментов, транспортных белков, рецепторов, фрагментов рецепторов, индукторов иммунной защиты хозяина, запасных белков, транспортных белков (вирусов растений), белков защиты (exploitive proteins) и репортерных белков, но не ограничивающихся ими.
В одном примере настоящего изобретения, меристематические клетки выделяли из камбия растений томата, моркови, тиса и дикого женьшеня и затем трансформировали с помощью Agrobacteria, которые содержали ген зеленого флуоресцентного белка (GFP), после чего анализировали экспрессию GFP в этих клетках. В результате, было показано, что и временная экспрессия GFP и стабильная трансформация с GFP была успешной. Кроме того, можно было видеть, что даже когда клетки субкультивировали с интервалами в 2 недели, они клетки стабильно пролиферировались и белок-мишень GFP стабильно экспрессировался в них.
В одном примере данного изобретения, в результате временной экспрессии гена, кодирующего белок-мишень, путем трансформации клеток камбиальной меристемы томата (CMC) Agrobacteria, можно было видеть, что при 1-9 днях сокультивирования, и наиболее предпочтительно 5 днях сокультивирования, 90% или более жизнеспособных клеток инфицировались Agrobacteria и экспрессировали GFP. Во время периода сокультивирования клетки камбиальной меристемы томата демонстрировали уменьшение жизнеспособности, меньшее, чем 10%, и ригидность клеточной стенки сохранялась интактной. Ранее сообщалось, что трансформация на уровне культуры клеток имеет достаточно низкую эффективность, такую как 10% или ниже; однако как это можно видеть, при использовании клеток камбиальной меристемы растения (CMC) в соответствии с данным изобретением эффективность трансформации достаточно высокая, такая как 90% или выше.
Эта высокая эффективность трансформации указывает на то, что возможно крупномасштабное получение рекомбинантных белков посредством временной экспрессии. Это получение возможно без выполнения отдельного процесса отбора, и, следовательно, из вектора может быть элиминирована кассета маркеров селекции. Кроме того, может быть экспрессирован только белок-мишень и, следовательно, данное изобретение является лучшим с точки зрения эффективности.
Между тем каллюс томата культивировали при тех же самых условиях, которые описаны выше, и в результате для каллюса томата была выявлена низкая эффективность трансформации, примерно в 26,4%, ниже по сравнению с меристематическими клетками растения данного изобретения. Однако эта величина по меньшей мере в два раза выше, чем 10%, что является ранее сообщенной эффективностью трансформации каллюса. Это указывает на то, что способ продуцирования рекомбинантного белка в соответствии с данным изобретением может быть применен не только к клеткам камбиальной меристемы растений (CMC), но также к каллюсу.
В другом аспекте, данное изобретение направлено на способ продуцирования белка-мишени клетками растений, предназначенными для экспрессии белка-мишени, причем этот способ включает стадии:
(a) стабильной трансформации популяции клеток растений, включающей клетки камбиальной меристемы (CMC) или каллюса, геном (генами), кодирующими белок-мишень, или временная экспрессия гена (генов), кодирующих белок-мишень, сокультивированием этой популяции клеток растений, включающих клетки камбиальной меристемы (CMC) или каллюса, с Agrobacteria, содержащими вектор со встроенным в него геном (генами), кодирующими белок-мишень,
где эти клетки камбиальной меристемы (CMC) являются клеточной линией, содержащей по природе недифференцированные клетки, выделенные из растения, причем эта клеточная линия выделена из камбиальной ткани растения и имеет меристематическую преемственность без прохождения через дедифферентировку в каллюс; и
(b) извлечения белка-мишени, экспрессируемого в культуре клеток-мишеней, которую инфицировали Agrobacteria посредством сокультивирования.
В другом аспекте, данное изобретение направлено на способ продуцирования белка-мишени трансгенным растением с геном (генами), кодирующими белок-мишень, где способ предусматривает стадии:
(a) выращивания трансгенного растения, содержащего ген (гены), кодирующие белок-мишень;
(b) выделения трансгенных клеток камбиальной меристемы (ТСМС) из трансгенного растения;
(c) культивирования выделенных трансгенных клеток камбиальной меристемы (ТСМС) в среде; и
(d) извлечения белка-мишени, экспрессированного в культивированных трансгенных клетках камбиальной меристемы (ТСМС).
Примеры
В дальнейшем, данное изобретение будет описываться более подробно со ссылкой на примеры. Квалифицированному в данной области специалисту будет очевидно, что эти примеры представлен только в целях иллюстрации и не должны истолковываться как ограничивающие или изменяющие объем данного изобретения.
Пример 1. Продуцирование, пролиферация и характеристика клеток камбиальной меристемы (CMC) растений семейства Solanaceae
1-1: Приготовление растительного материала томата
Для выделения клеток камбиальной меристемы (CMC) из томата (Lycopericum esculentum, Sejong Seed Co., Ltd., Korea), у представителя рода Lycopersicon семейства Solanaceae, собрали стебель и побеги (фиг. 1А) и затем немедленно замочили их в 100 мг/л антиоксиданта - аскорбиновой кислоте (L-аскорбиновая кислота, DUCHEFA, The Netherlands), после чего их транспортировали и хранили.
Затем растение предварительно обрабатывали смешанным раствором 0,1% беномила (Dongbu Hannong Chemical, Korea), 0,1% даконила (Dongbu Hannong Chemical, Korea), 0,1% сульфатом стрептомицина (DUCHEFA, The Netherlands) и 0,01% натрий-цефотаксима (DUCHEFA, The Netherlands) в течение 10 минут, и промывали водопроводной водой в течение 5 минут для удаления фенольных соединений и оставшихся химикалиев. Потом поверхность растения стерилизовали 70% этанолом (DC Chemical, Korea) в течение 1 минуты, 1,5% перекисью водорода (LG Chemical, Korea) в течение 3 минут, 0,5% раствором CLOROX в течение 5 минут и 0,1% раствором CLOROX в течение 5 минут, и промывали 3-4 раза водой.
1-2: Приготовление камбий-содержащего эксплантата из стебля растения томата и отделение ткани
Стебель, который стерилизовали в Примере 1-1, разрезали и от ксилемы отделяли ткани флоэмы, кортекса и эпидермиса, содержащие камбий, имеющий превосходную способность делиться.
1-3: Индуцирование полученных из камбия томата стволовых клеток
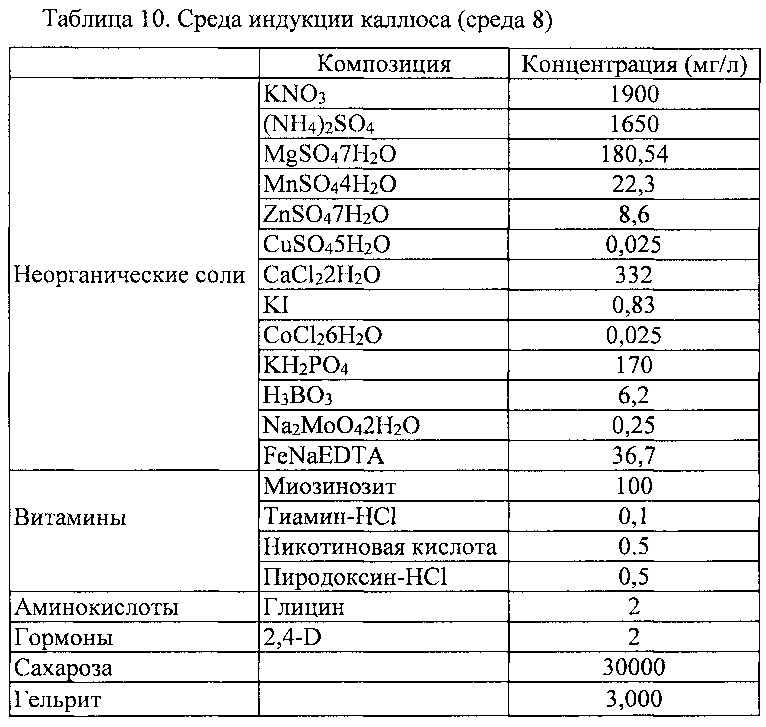
Камбий-содержащий эксплантат, приготовленный в Примере 1-2 выше, сажали и культивировали в среде для индукции клеток камбиальной меристемы (CMC) (среде 1), показанной в таблице 1 ниже.
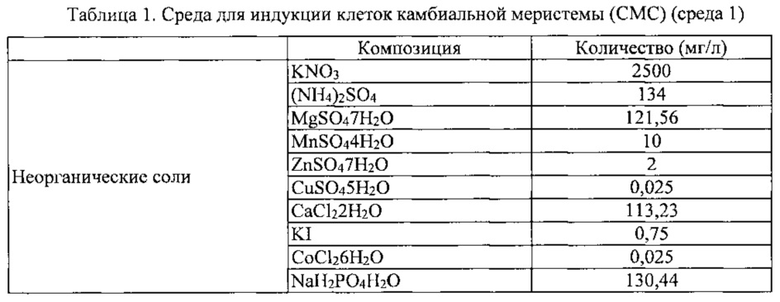
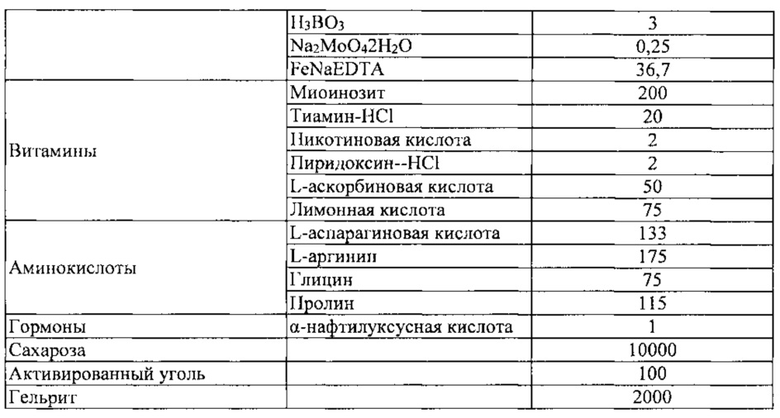
К этой среде может быть добавлен регулятор роста ауксин, такой как NAA, IAA, IBA, 2,4-D или пиклорам, в концентрации 0,5-5 мг/л. В этом Примере, NAA добавляют в концентрации 1 мг/мл. Культивирование проводили в темной комнате, в которой поддерживается температура 25±1°C.
В течение 7-10 дней первоначального культивирования визуально наблюдали деление клеток из камбия, а через 3 недели (21 день) культивирования из слоя, составленного из флоэмы, кортекса и эпидермиса, начинал индуцироваться аморфный каллюс, образованный дедифференцировкой. После 30 дней культивирования, эта ткань начала разделяться в культивируемый слой камбия и верхний содержащий флоэму слой, который является аморфным слоем каллуса (фиг. 1В). После того, как эта ткань естественным образом полностью разделялась на два слоя, от нее отделяли и культивировали только ту часть, которая является камбием. После отделения этой ткани, ее белую и мягкую часть субкультивировали в свежей среде того же состава с интервалами 14 дней.
Эти клетки камбиальной меристемы томата стабильно поддерживались без вариаций в их скорости роста, характере роста и степени агрегации в течение продолжительного периода их культивирования, что свидетельствует о том, что их можно использовать для крупномасштабного культивирования. Однако каллюс, полученный из стебля томата, обнаруживал варьирование в характере роста и скорости роста и высокую степень агрегации при культивировании в течение продолжительного периода времени. После этого клетки каллюса окрасились в коричневый цвет, и подвернулись некрозу. Таким образом эти клетки не подходят для стабильного крупномасштабного культивирования.
1-4: Культура каллюса томата
Каллюс томата (PC 10623), приобретенный из Biological Resource Center (Korea), субкультивировали с интервалами в 3 недели.
1-5: Пролиферация и характеристика клеток камбиальной меристемы (CMC) растений семейства Solanaceae
Клетки камбиальной меристемы томата (CMCs), выделенные в Примере 1-3 выше, поместили в колбу, содержащую жидкую среду, показанную в таблице 2 ниже. Затем клетки в данной колбе культивировали в ротационном шейкере в темноте при 100 об/мин и 25±1°C. Для непрерывного культивирования клетки камбиальной меристемы томата (CMC), культивированные для пролиферации, культивировали в виде суспензии при объемном соотношении клетка : среда 1:10 в течение 7 дней.
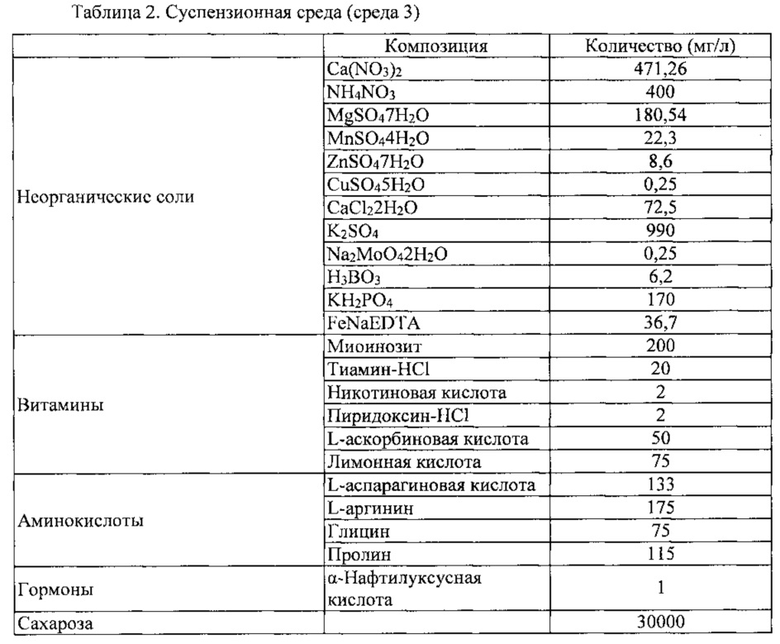
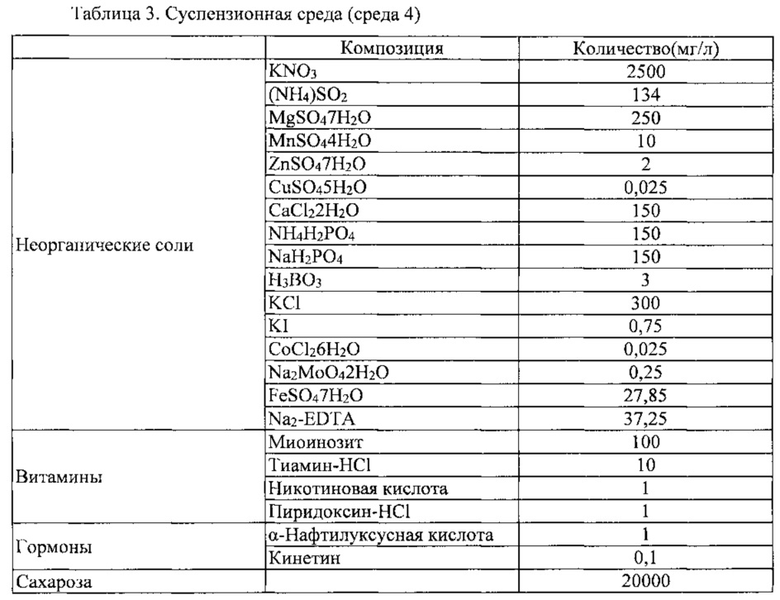
Каллюс томата (PC 10623) также высевали при том же самом соотношении, что описано выше, и жидкая среда, используемая для культуры каллюса томата, показана в таблице 3 ниже.
Степень агрегации этих клеток наблюдали с помощью микроскопа СХ31 (Olympus, Japan). Как показано на фиг. 2А, в результате наблюдалось, что культура клеток камбиальной меристемы (CMC) по данному изобретению во время суспензионного культивирования состояла из большого количества одиночных клеток, но некоторые клетки образовывали агрегаты клеток, имеющие очень малый размер. Конкретно, при культивировании клеток камбиальной меристемы (CMC) по данному изобретению, максимальный размер клеточных агрегатов составлял только 500 мкм. Напротив, при изучении каллюса томата (PC10623) каллюсные клетки образовывали множество агрегатов, что показано на фиг. 2В, и максимальный размер клеточных агрегатов был равен 10 мм. Кроме того, клетки камбиальной меристемы (CMC) по данному изобретению и клетки каллюса (PC10623) собирали после пролиферативного культивирования, но перед субкультивированием, и жизнеспособность этих клеток (в %) рассчитывали с использованием способа окрашивания 2%-ым Evan’s blue (5 мин). Как показано в Таблице 4, в результате клетки камбиальной меристемы (CMC) по данному изобретению обнаруживали жизнеспособность 96,33%, тогда как клетки каллюса обнаруживали жизнеспособность только 65,2%.

Пример 2. Получение клеток камбиальной меристемы (CMC) из запасающего корня растения
2-1: Получение клеток камбиальной меристемы дикого женьшеня (CMC) Был выбран здоровый дикий женьшень, не имеющий дефектов, и у него полностью удалили сосущие корни. Затем поверхность оставшихся тканей стерилизовали двумя стадиями. Для предотвращения побурения стерилизованной ткани, стерилизованный основной корень помещали в содержащую антиоксидант среду (BIM), показанную в таблице 5 ниже, и культивировали со встряхиванием в течение приблизительно 30 минут - 1 часа, после чего его помещали на стерилизованную фильтровальную бумагу для удаления влаги.

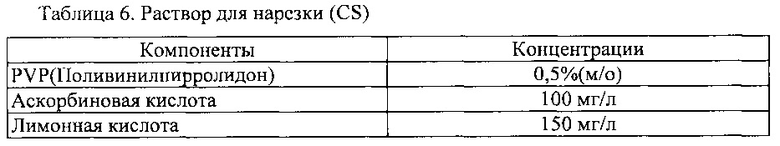
После завершения процесса стерилизации, для предотвращения побурения этого материала, его нарезали на кусочки размера 0,5-0,7 см (В) × 0,5-0,7 см (Д) × 0,2-0,5 мм (III) в антиоксидант-содержащем растворе для нарезки (CS), показанного в таблице 6 ниже, так что они включали бы в себя часть камбия, имеющего превосходную способность делиться.
Для индукции только камбия в приготовленном эксплантате, этот эксплантат помещали в колбу, содержащую раствор 1 М сахарозы (Duchefa, Netherlands) и подвергли осмотическому стрессу в течение 16-24 часов в холодном состоянии. Затем эксплантат обрабатывали в растворе 0,05 М сахарозы в течение 5 минут и в растворе 0,1 М сахарозы в течение 5 минут, тем самым снимая стресс, вызываемый сахарозой с высокой концентрацией. Полученный в результате эксплантат, помещали на преинокуляционную среду (среду 6) с лежащей на ней фильтровальной бумагой для удаления влаги.
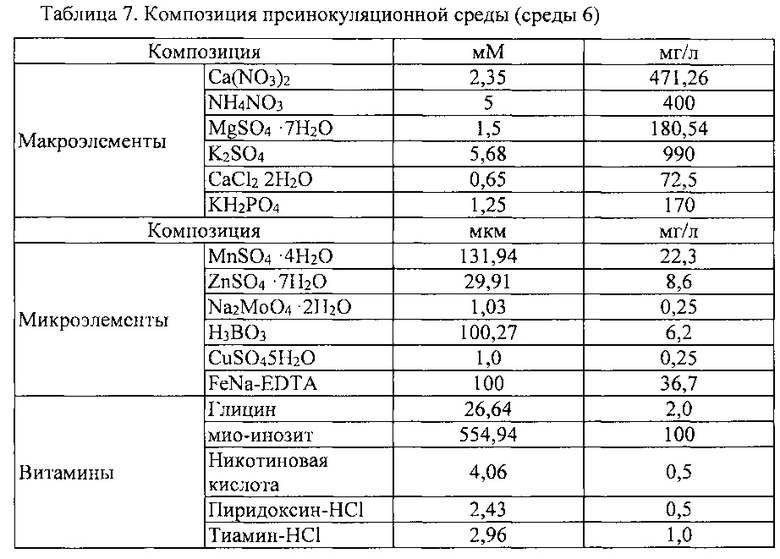
Для индукции клеток камбиальной меристемы дикого женьшеня (CMC), этот подвернутый осмотическому стрессу эксплантат помещали на среду для индукции клеточной линии (среду 7). Эта среда, используемая для индукции клеток камбиальной меристемы (CMC), показана в таблице 8 ниже.
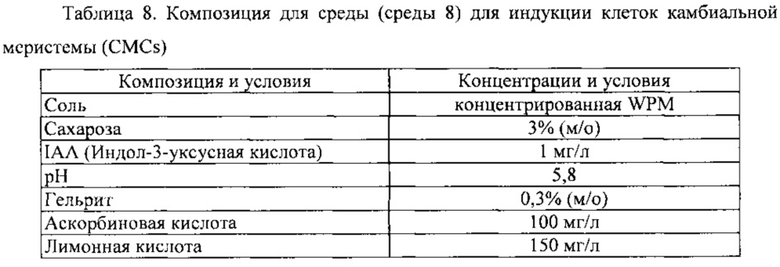
В этом эксплантате, помещенном на среду для индукции клеточной линии (среду 7) после описанной выше обработки, клетки камбиальной меристемы (CMC) индуцировались именно в камбии, и не индуцировались в других тканях.
После того как ткани, отличные от камбия, были некротизированы культивированием в среде 7, этот эксплантат культивировали в среде 3-1 для пролиферации только камбиальных клеток.
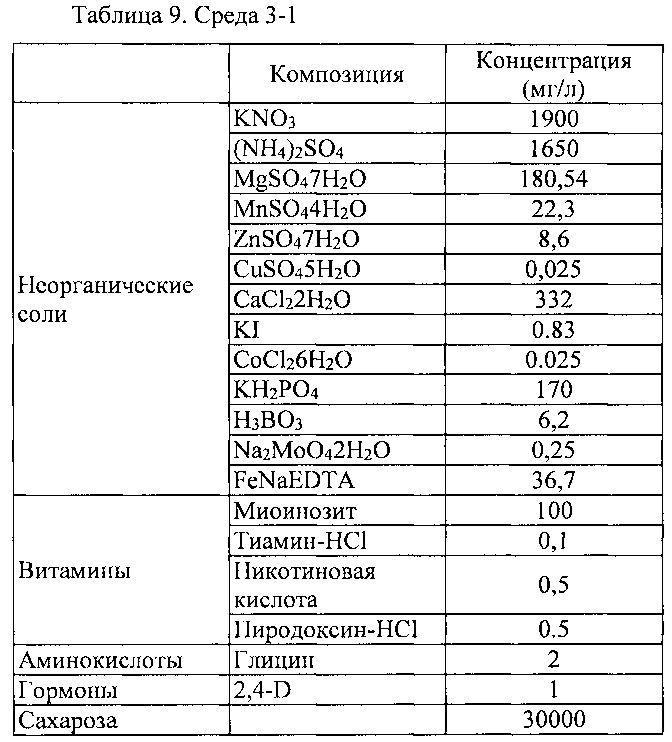
Между тем, в Biological Resource Center (Korea) приобретали полученный из котиледона каллюс (КСТС 10224) и субкультивировали с интервалами равными 3 неделям.
2-2: Получение клеток камбиальной меристемы моркови (CMC)
Получали растение моркови (Daucus carota L.) и стерилизовали поверхность таким же образом, как описано в Примере 2-1. Затем, эту полученный образец подвергли стрессу таким же образом, как описано в Примере 2-1, с последующей индукцией камбиальных клеток в нем.
В результате было показано, что отличные от камбия ткани были некротизированы, индуцировались клетки камбиальной меристемы (CMC), имеющие способность отделения, подобно тому, что было получено в Примере 2-1.
Между тем для культивирования каллюса моркови в качестве контроля взяли корень моркови и стерилизовали поверхность таким же образом, как описано в Примере 2-1, затем из этого корня моркови готовили эксплантат. Полученный эксплантат помещали на среду для индукции каллюса, показанную в таблице 10 ниже, и культивировали в темной комнате, с поддержанием температуры 21+1°C. Собирали аморфный каллюс и затем субкультивировали с интервалами, равными 14 дням.
2-3: Пролиферация и характеристика клеток камбиальной меристемы (CMC) из запасающего корня
Клетки камбиальной меристемы женьшеня (CMC), выделенные в Примере 1 выше, помещали в колбу, содержащую жидкую среду, показанную в таблице 2 ниже. Затем клетки в колбе культивировали в ротационном шейкере в темноте при 100 об/мин и 21+1°C. Для непрерывного культивирования клетки камбиальной меристемы женьшеня (CMC), культивируемые для пролиферации, культивировали в суспензии при объемном отношении клетка : среда 1:10 в течение 14 дней. Кроме того, каллюс дикого женьшеня, выделенный в Примере 2-1, культивировали в тех же самых условиях, и жидкая среда, используемая для культуры каллюса, была такая же, что и жидкая среда, используемая для культивирования клеток камбиальной меристемы дикого женьшеня (CMC).
Степень агрегации клеток наблюдали с помощью микроскопа СХ31 (Olympus, Japan). Как показано на фиг. 2А во время суспензионного культуривирования культура клеток камбиальной меристемы (CMC) по данному изобретению содержала большое количество отдельных клеток и некоторое количество агрегатов из клеток, имеющих очень малый размер. Конкретно, при культивировании клеток камбиальной меристемы (CMC) максимальный размер агрегатов клеток был только 200 мкм. Напротив когда изучали контроль, что показано на фиг. 2В, каллюсные клетки оказались высоко агрегированными, и максимальный размер клеточных агрегатов был 500 мкм. Кроме того, клетки камбиальной меристемы (CMC) по данному изобретению и клетки каллюса собирали после пролиферативного культивирования, но перед субкультивированием, и рассчитывали жизнеспособность клеток (%) из этих проб с использованием окрашивания 2% Evan’s blue (5 мин). В результате, клетки камбиальной меристемы (CMC) по этому изобретению обнаруживали жизнеспособность 94,3%, тогда как клетки каллюса обнаруживали жизнеспособность только 61%.
Между тем, клетки камбиальной меристемы моркови (CMC), выделенные в Примере 2-2 выше, помещали в колбу, содержащую жидкую среду (среду 3-1). Затем, клетки в колбе культивировали в ротационном шейкере в темноте при 100 об/мин и 25±1°С. Для непрерывного культивирования клетки камбиальной меристемы моркови (CMC), культивированные для пролиферации, культивировали в суспензии при соотношении объемов клетка : среда 1:10 в течение 14 дней. Кроме того, культивировали каллюс моркови, выделенный в Примере 2-2, в тех же самых условиях, и жидкая среда, используемая для культуривирования каллюса, была такой же, что и жидкая среда, используемая для культивирования клеток камбиальной меристемы (CMC) моркови.
Пример 3. Получение экспрессирующего вектора для трансформации клеток камбиальной меристемы растений (CMC) и для культуры Agrobacteria
Эксперимент выполняли с использованием бинарного экспрессирующего вектора растения, содержащего ген GFP и Agrobacterium tumefaciens LBA4404, приобретенные из Takara Korea Biomedical (LBA4404 Electro cells, cat no. 9115, Korea).
Введение бинарного GFP-содержащего вектора в агробактерии выполняли с использованием Bio-Rad Cuvette и Gene Pulser II в соответствии с инструкциями изготовителя (для Agrobacterium tumefaciens LBA4404).
Полученный pBINmGFP5ER/LBA4404 инокулировали с пулом платины в 15% растворе глицерина и выполняли посев штрихом на твердой среде YEP (среде 9), с добавлением 100 мг/л рифампицина (TCI, Japan) и 100 мг/л канамицина с последующим культивированием при 28°С в течение 3 дней в темноте.

Agrobacteria (pBINmGFP5ER7LBA4404) наносили штрихом на свежую среду с интервалами в 3 дня и субкультивировали при 28°С в темноте.
Agrobacteria подвергали суспензионному культивированию для трансформации клеток камбиальной меристемы растений (CMC) этими Agrobacteria.
Единичную колонии Agrobacteria, культивируемых на твердой среде, добавляли к 5 мл жидкой среды YEP (Таблица 12; среда 10) и культивировали при 28°С и 200 об/мин в течение 6-18 часов в темноте, после чего 1-5 мл культуры Agrobacteria добавляли к 100 мл среды YEP и культивировали при 28°С и 200 об/мин в течение 6-24 часов.

В качестве контроля использовали 1 мл каждой из приготовленных суспензий Agrobacteria и жидкой среды YEP (содержащей 100 мг/л рифампицина и 100 мг/л канамицина), которые отбирали пипеткой, помещали в кювету и измеряли оптическую плотность (OD600) при длине волны 600 нм с использованием УФ/видимого спектрофотометра. Этот используемый Уф-спектрофотометр был произведен в Amersham Bioscience.
Для индукции вирулентности Agrobacteria, суспензию Agrobacteria, имеющую величину OD600 0,4-2,0, помещали в коническую пробирку (BD FALCON, USA) и центрифугировали (Hanil Science Industrial Co., Ltd., Korea) при 4°С и 6000g в течение 3-10 минут. Собирали осадки Agrobacteria, образовавшиеся на стенке пробирки, и ресуспендировали их в 10 мл суспензионной среды (среды 2). Когда величина OD600 Agrobacteria достигала 0,00001-2,0, к суспензии Agrobacteria добавляли 10-200 мкМ ацетосирингона (Aldrich, USA) и затем инкубировали со встряхиванием при 28°C и 200 об/мин в течение 1 минуты - 24 часов.
Agrobacteria могут быть обработаны ацетосирингоном, как описано а данном изобретении, или клетки камбиальной меристемы растений (CMC) могут быть обработаны непосредственно ацетосирингоном. Альтернативно, и Agrobacteria и клетки камбиальной меристемы растений (CMC) могут быть одновременно обработаны ацетосирингоном.
Пример 4. Временная экспрессия белка-мишени в клетках камбиальной меристемы растений (CMC) с вектора для трансформации растений
Agrobacteria, предназначенные для трансформации клеток камбиальной меристемы растений (CMC), получали как описано в Примере 3 выше. Клетки растений на экспоненциальной фазе роста из примеров 1 и 2 получали в виде камбиальных меристематических клеток (CMC) и культивировали при соотношении клетка : среда 1:10.
10 мл вирулентно-индуцированной суспензии Agrobacteria помещали в колбу на 250 мл, содержащую клетки камбиальной меристемы томата (CMC), выделенные в Примере 1, и сокультивировали с этими клетками при 25°C и 100 об/мин. Для максимизации эффективности трансформации CMC томата в этом процессе, культивирование осуществляли в ротационном шейкере (Sejong, Korea) при 100 об/мин в течение 1 минуты - 48 часов в темноте. Затем проводили осаждение в течение 1 минуты - 48 часов без встряхивания. Потом полученную культуру повторно культивировали в ротационном шейкере при 100 об/мин в течение 1-9 дней.
1 мл культуры, сокультивированной в течение 1-9 дней, отбирали пипеткой и помещали в микропробирку на 1,5 мл. 10 мкл из этой 1 мл пробы помещали в гемацитометр (Marienfeld) и с использованием инвертированного фазово-контрастного микроскопа IX71 (источник флуоресцентного света: U-RFL-T) наблюдали в ней экспрессию GFP. В этом исследовании, использовали зеленый свет и длину волны 460-490/520 нм (возбуждение/барьер). Затем считали процент GFP-экспрессирующих клеток относительно общего числа живых клеток на одном и том же микроскопическом препарате (предметном стекле) с использованием оптического микроскопа.
GFP временно экспрессировался в клетках камбиальной меристемы томата (CMC) за счет Agrobacteria, и в результате, как можно видеть на фиг. 4, GFP временно экспрессировался в 90% живых клеток при 1-9 днях сокультивирования, и наиболее предпочтительно после 5 дней сокультивирования. Было показано, что во время периода сокультивирования, клетки камбиальной меристемы томата (CMC) демонстрировали уменьшение жизнеспособности, меньшее, чем 10%, и ригидность клеточной стенки оставалась интактной.
Другими словами, известно, что трансформация на уровне культуры клеток затруднительна, и что она не может быть рассчитана в процентах, но можно было видеть, что применение клеток камбиальной меристемы томата (CMC) по данному изобретению показывало значительную эффективность трансформации равную 90% или больше.
Между тем, каллюс томата сокультивировали в тех же самых условиях, которые описаны выше, и, в результате, каллюс томата из Примера 1 обнаруживал низкую эффективность трансформации равную 26,4%, в отличие от клеток камбиальной меристемы томата (CMC) по данному изобретению. Однако эта величина была по меньшей мере в два раза выше, чем 10%, что считалось ранее степенью эффективности трансформации каллюса.

Кроме того, таким же образом как описан выше был выполнен эксперимент с клетками камбиальной меристемы дикого женьшеня и моркови (CMC) из Примера 2, и результаты этого эксперимента показаны на фиг. 7 и 8.
Обращаясь к фиг. 5, можно видеть, что когда клетки камбиальной меристемы дикого женьшеня (CMC) временно сокультивировали с Agrobacteria, 13% или более живых клеток инфицировались Agrobacteria и экспрессировали GFP.
Обращаясь к фиг.6, можно видеть, что когда клетки камбиальной меристемы моркови (CMC) временно сокультивировали с Agrobacteria, 17% или более живых клеток инфицировались Agrobacteria и экспрессировали GFP. Уровни временной экспрессии GFP в клетках камбиальной меристемы женьшеня и моркови (CMC) были меньшими, чем уровни в клетках камбиальной меристемы томата (CMC), но ожидается, что клетки камбиальной меристемы женьшеня и моркови (CMC) могут обнаруживать уровни временной экспрессии, сравнимые с уровнями в клетках камбиальной меристемы томата, если проконтролировать время сокультивирования и стационарного культивирования.
Пример 5. Испытание стабильности трансформации клеток камбиальной меристемы (CMC) экспрессирующим вектором растений, кодирующим белок-мишень
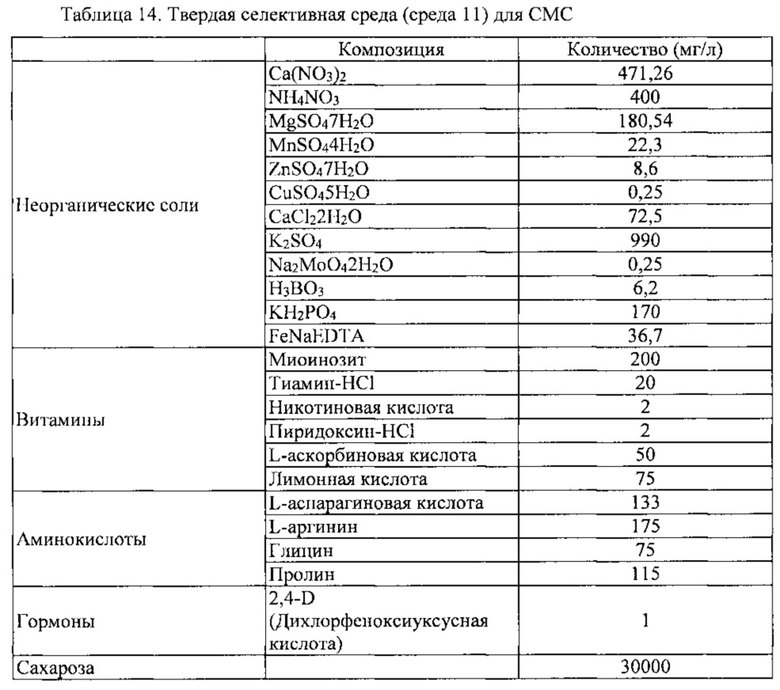
После выполнения процедуры, описанной в Примере 4, клетки камбиальной меристемы томата (CMC) сокультивировали с Agrobacteria в колбе на 250 мл при 25°C и 100 об/мин в течение 3-21 дней. В той временной точке, когда экспрессия GFP была наивысшей, культуру промывали суспензионной средой (средой 3), показанной в таблице 2 в течение 5-20 минут для удаления Agrobacteria. Затем культуру обрабатывали 300 мг/л канамицина (TCI, Japan) и 500 мг/л цефотаксима (TCI, Japan) и культивировали в суспензии со встряхиванием при 25°C и 100 об/мин в течение 1 недели. После этого, клетки камбиальной меристемы (CMC) осаждали, и максимально возможное количество среды удаляли декантацией, после чего оставшуюся среду обычно абсорбировали фильтровальной бумагой (70 мм, Toyo Roshi Kaisha, Japan), и клетки камбиальной меристемы томата (CMC) помещали на твердую среду отбора (таблица 14; среда 11). Помещенные клетки культивировали при 25±1°C в темноте, с получением тем самым трансгенных клеток камбиальной меристемы (ТСМС), которые экспрессируют GFP. Клетки камбиальной меристемы моркови (CMC) обрабатывали таким же образом, какой описан выше, а клетки камбиальной меристемы дикого женьшеня (CMC) высевали на среду, аналогичную среде 11 из которой исключили гормоны.

Полученную клеточную линию исследовали под УФ-светом, и, в результате, стабильно трансформированная клеточная линия испускала флуоресценцию GFP, как показано в левом скоплении (кластере) на фиг. 7, тогда как нетрансформированная клеточная линия не испускала флуоресценцию, что можно видеть в правом скоплении (кластере). Для испытания эмиссии зеленой флуоресценции из каждой клеточной линии в этих кластерах, степень эмиссии флуоресценции анализировали с использованием флуоресцентного микроскопа (Olympus IX71 инвертированный микроскоп, источник флуоресцентного света: U-RFL-T).
В результате, как показано на фиг. 8, GFP экспрессировался во всех клетках образцов, показывая, что стабильная трансформация клеток камбиальной меристемы растения (CMC) по данному изобретению оказалась успешной.
Кроме того, отбирали и субкультивировали кластеры, которые испускали флуоресценцию, как описано выше (фиг. 9А), и в них анализировали экспрессию GFP (фиг. 9В). Затем, эти кластеры субкультивировали для пролиферации с интервалами в 14 дней (фиг. 9С: после 14 дней; фиг. 9D: после 82 дней непрерывного субкультивирования), и можно было видеть, что эти кластеры стабильно экспрессируют GFP.
Это предполагает, что трансгенные клетки камбиальной меристемы (ТСМС), для которых выявили, что они стабильно трансформированные, могут непрерывно пролиферироваться посредством субкультивирования. Следует отметить, что эти трансгенные клетки камбиальной меристемы (ТСМС), трансформированные GFP, демонстрируют очень сильную зеленую флуоресценцию. Это количественно указывает на то, что GFP сильно экспрессируется в клетках камбиальной меристемы растений (CMC). Не только общий уровень экспрессии GFP, но также уровень экспрессии этого белка индивидуальными клетками влияет на общий выход. Таким образом, степень выраженности зеленой флуоресценции позволяет предположить, что белок-мишень может экспрессироваться на очень высоком уровне в трансгенных клетках камбиальной меристемы (ТСМС).
Кроме того, результаты стабильной трансформации клеток камбиальной меристемы томата, моркови и дикого женьшеня сравнивали в случае, когда выполняли седиментацию, и случае, когда седиментацию не выполняли. Кроме того, результаты для клеток камбиальной меристемы (CMC) сравнивали с соответствующими результатами для каллюса.
Как можно видеть на фиг. 10, количество образованных кластеров было большим в клетках камбиальной меристемы томата (ТСМС; фиг. 10А) или каллюсе томата (фиг. 10С), которые подвергали седиментации, чем в клетках камбиальной меристемы томата (ТСМС; фиг. 10В) или каллюсе томата (фиг. 10D), которые не подвергали седиментации. Кроме того, количество образованных кластеров было большим в клетках камбиальной меристемы томата (ТСМС; фиг. 10А и 10В), чем в каллюсе томата (фиг. 10С и 10D).
Как показано на фиг. 11, количество образованных кластеров (указанных частях на фиг. 13А и 13С) было большим в клетках камбиальной меристемы моркови (ТСМС; фиг. 11А или каллюсе моркови (фиг. 11С), которые подвергали седиментации, чем в клетках камбиальной меристемы моркови (ТСМС; фиг. 11В) или каллюсе моркови (фиг.11D FIG. 11D), которые не подвергали седиментации. Кроме того, количество образованных кластеров было большим в клетках камбиальной меристемы моркови (ТСМС; фиг. 11А и 11В), чем в каллюсе моркови (фиг. 11С и 11D).
Как показано на фиг. 12, количество образованных кластеров было большим в клетках камбиальной меристемы дикого женьшеня (ТСМС; фиг. 12А), которые подвергали седиментации, чем в клетках камбиальной меристемы дикого женьшеня (ТСМС; фиг. 12В).
Образование кластеров означает, что Т-ДНК агробактерий выстроилась в геном растительных клеток. Из результатов на фиг. 10-12, можно видеть, что седиментация является важным фактором в увеличении эффективности трансформации.
Кроме того, наблюдали, что количество образованных кластеров было большим в трансгенных клетках камбиальной меристемы (ТСМС), чем в каллюсе при одних и тех же условиях, позволяя предполагать, что применение клеток камбиальной меристемы (ТСМС) в качестве материала является важным фактором в увеличении эффективности трансформации.
Пример 6. Испытание возможности более масштабного культивирования
70 мл клеток, осажденных в колбе на 250 мл для которых подтверждено, что они имеют уровень экспрессии GFP 90% или выше в Примере 4, засевали в 3-литровый биореактор с воздушным потоком в соотношении клетка: среда 1:30. Сухая масса клеток была 0,6 г/л. Рабочий объем этого 3-литрового биореактора был равен 2100 мл и объем утилизации этого биореактора был 70% общего объема. Используемая среда была такой же, какую использовали в колбе на 250 мл. Добавляли антибиотик, после чего эти клетки культивировали при скорости аэрации 0,1-0,15 объем/объем/минута при 25°C в течение 7-10 дней в темноте. Субкультивирование в этом 3-литровом биореакторе выполняли с интервалами 7-10 дней и предпочтительно 7 дней.
Трансгенные клетки камбиальной меристемы томата (ТСМС) культивировали в 3-литровом биореакторе. В результате, как показано на фиг. 13, при облучении УФ-светом (350 нм) можно было видеть тот же уровень экспрессии GFP, что и в культуре из колбы на 250 мл при 7-10 днях пролиферативного культивирования. Скорость роста трансформированных клеток камбиальной меристемы томата (ТСМС) была по меньшей мере в 10 раз более высокой после завершения пролиферативного культивирования в сравнении с количеством клеток, посеянных в первоначальной стадии. После завершения этого пролиферативного культивирования, трансгенные клетки камбиальной меристемы томата (ТСМС) демонстрировали уменьшение жизнеспособности, меньшее, чем 10%, и результаты наблюдения с использованием оптического микроскопа показали, что ригидность клеточной стенки сохранялась без изменений.

Пример 7. Конструирование трансгенного растения табака
7-1: Культура Agrobacteria
Единичную колонию Agrobacteria pBINmGFP5ER/LBA4404 добавляли к 5 мл среды YEP и культивировали при 28°C в течение 6 часов. Затем, эту культуру добавляли к 50 мл среды YEP при отношении 1:50-1:100 и культивировали в течение 18-24 часов, и затем измеряли OD культуры Agrobacteria. Эту культуру Agrobacteria помещали в коническую пробирку и клетки осаждали с использованием центрифуги при 4°C. Среду культуры табака (MS + 2 мг/л ВА + 0,1 мг/л NAA + рН 5,8) добавляли к собранным после центрифугирования осадкам Agrobacteria, и основывались на измерении OD Agrobacteria, эту среду культуры табака смешивали со средой Agrobacteria, и добавляли к ней 10-200 мкМ ацетосирингона, с последующим культивированием при 250 об/мин при 28°C в течение 2 часов.
7-2: Приготовление эксплантатов листьев табака
Листья отделяли, держа черешки этого растения пинцетом, и помещали в чашку Петри. Эксплантат 1,0 см × 1,0 см (~0,5 см × 0,5 см), включающий в себя основную часть жилки вблизи черешка, делали с использованием ножа.
7-3: Сокультивирование
Эксплантат, представляющий собой положительный контроль (PC), отделяли, помещали на среду сокультивирования (MS + 2 мг/л ВА + 0,1 мг/л NAA + рН 5,8+0,8% агар), и затем культивировали при 25°C в течение 7 дней в темноте. Эксплантаты (за исключением положительного контроля), приготовленные в Примере 7-2, добавляли к культурам, приготовленным в Примере 7-1, после иммерсии в течение 20 минут при мягком встряхивании с интервалами 5 минут, агро-инокулируя тем самым этот эксплантат. Затем эксплантат вынимали и помещали на фильтровальную бумагу для удаления влаги, и впоследствии помещали на среду для сокультивирования (MS + 2 мг/л ВА + 0,1 мг/л NAA + рН 5,8 + 0,8% агар + 100 мкМ AS) и культивировали при 25°C в течение 3 дней в темноте.
7-4: Отбор
Сокультивированный эксплантат промывали три раза стерильной водой и сушили фильтровальной бумагой. Один эксплантат, выбранный в качестве отрицательного контроля (NC), отделяли от PC. PC помещали на среду регенерации (MS + 2 мг/л ВА + 0,1 мг/л NAA + рН 5,8 + 0,8% агар), NC помещали на селективную среду (MS + 2 мг/л ВА + 0,1 мг/л NAA + рН 5,8 + 0,8% агаровая среда + кан 100 мг/л + цеф 500 мг/л), и оставшийся эксплантат размещали на селективной среде.
После культивирования в течение 3 недель в темноте, каждый из эксплантатов культивировали с 16-часовыми светлыми/8-часовыми темными циклами для образования побегов. Когда побеги образовывались, только один побег отщепляли и переносили на среду для образования корней (MS + кан 100 мг/л + цеф 500 мг/л рН 5,8 + 0,8% агар) для образования корней. На фиг. 14 представлены результаты акклимации трансгенного N. benthamiana и выращивания акклиматированного растения в цветочном горшке.
7-5: Изучение морфологии
3-6-месячные стебли собирали и изучали их морфологию. Для исследования стебли разрезали поперечно и радиально и для различения тканей их окрашивали ксилема-специфическим реагентом-красителем флороглюцинол-HCl. Как можно видеть на фиг. 15, ксилема окрашивалась в красный цвет и 2-4 камбиальных слоя (3-6 камбиальных слоя для Nicotiana tabacum cv. Xanthi) присутствовали сразу же выше ксилемы.
Кроме того, растение табака, трансформированное GFP, разрезали поперечно и изучали с использованием фильтра GFP (возбуждение/барьер: 460-490/520 нм). В результате, как показано на фиг. 16, флуоресценция была более интенсивной в камбиальной зоне, чем в других тканях.
7-6: Отделение трансгенных клеток камбиальной меристемы (ТСМС) табака
На 4 день культивирования, в камбиальной зоне N. benthamiana наблюдали деление клеток. Через 2 недель культивирования, как показано на фиг.17(A), ткань флоэмы, отличную от камбия, отделяли пинцетом. Как показано на фиг. 17(B), разделение клеток или повреждение тканей, отличных от флоэмы, после отделения не наблюдали. В случае Nicotiana tabacum cv. Xanthi, клетки камбиальной меристемы отделяли таким же самым образом, какой описан для N. benthamiana, и, как показано на фиг. 18, те же самые результаты получали также для Nicotiana tabacum cv. Xanthi.
7-7: Анализ экспрессии белка в клетках камбиальной меристемы табака (ТСМС) и растении табака
Общий растворимый белок выделяли из каждой из клеток камбиальной меристемы (ТСМС) и растения табака трансформированных геном GFP и нетрансформированного растения табака.
Эти выделенные общие растворимые белки подвергали SDS-PAGE (электрофорезу в полиакриламидном геле) на двух полиакриламидных гелях и переносили на нитроцеллюлозную бумагу с использованием блоттера semi-dry transfer cell (BIO-RAD).
Бумагу блокировали в течение ночи 5% обезжиренным молоком и промывали TBST, после чего она реагировала последовательно с первичным и вторичным анти-GFP-антителом. Затем, ее промывали с использованием TBST/TNM и проявляли проявителем (проявляющим раствором BCIP/NBT), после чего анализировали полосы белков.
В результате, как показано на фиг. 19, ни одна полоса не была детектирована в нетрансгенном растении табака. В трансгенном растении табака и клетках камбиальной меристемы табака (TCMCs), выделенных из трансгенного растения табака, детектировали полосу в месте, сходном с размером GFP.
Кроме того, можно было видеть, что белок GFP сильнее экспрессировался в клетках камбиальной меристемы табака (ТСМС), выделенных из трансгенного растения табака, чем в трансгенном растении табака. Такие результаты согласуются с результатами фиг. 16, что указывает на то, что результаты для камбиальных меристематических клеток табака (ТСМС) были более высокими.
Хотя данное изобретение было описано подробно со ссылкой на конкретные признаки, квалифицированным в данной области специалистам будет очевидно, что это описание относится только к предпочтительному варианту и не ограничивает объем данного изобретения. Таким образом, значительный объем данного изобретения будет определен прилагаемыми пунктами формулы изобретения и их эквивалентами.
Промышленная применимость
Как описано выше, система экспрессии белка-мишени с использованием рекомбинантных растительных клеток в соответствии с этим изобретением может преодолеть проблемы традиционной культуры клеток растений. Кроме этого, она показывает значительно более высокую эффективность трансформации и, следовательно, может продуцировать высокий уровень белков-мишеней, в том числе биофармацевтических белков. Таким образом, это дает возможность коммерциализации биофармацевтических лекарственных средств, включающих в себя белковые продукты растительного происхождения.


Данное изобретение относится к области биохимии и биотехнологии, а именно к растительной клетке, которая экспрессирует рекомбинантный белок, способу ее получения и способу получения рекомбинантного белка. Эта растительная клетка имеет введенный в рекомбинантный вектор, содержащий ген, кодирующий рекомбинантный белок, а также представляет собой клетку камбиальной меристемы (CMC), которая является клеточной линией, содержащей по природе недифференцированные клетки, выделенные из растения, где клеточная линия выделена из камбиальной ткани растения, и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс. Система экспрессии рекомбинантного белка с использованием вышеуказанных растительных клеток в соответствии с данным изобретением показывает значимо высокую эффективность трансформации и, следовательно, может продуцировать большие количества рекомбинантных белков, в том числе биофармацевтических белков. Таким образом, она делает возможной коммерциализацию биофармацевтических лекарственных средств, включающих в себя белковые продукты растительного происхождения. 4 н. и 9 з.п. ф-лы, 19 ил., 15 табл., 7 пр.
1. Растительная клетка для экспрессии рекомбинантного белка,
где растительная клетка содержит рекомбинантный вектор, содержащий встроенный в него ген, кодирующий рекомбинантный белок,
где растительная клетка представляет собой клетку камбиальной меристемы (CMC),
где клетка камбиальной меристемы (CMC) является типом полученной из растения клеточной линии, содержащей по природе недифференцированные клетки, выделенные из растения, где клеточная линия выделена из камбиальной ткани растения и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс.
2. Растительная клетка для экспрессии рекомбинантного белка по п. 1, где ген, кодирующий рекомбинантный белок, введен в растительную клетку камбиальной меристемы так, что рекомбинантный белок временно экспрессируется в растительной клетке камбиальной меристемы.
3. Растительная клетка для экспрессии рекомбинантного белка по п. 1, где растительная клетка камбиальной меристемы (CMC) стабильно трансформированы геном, кодирующим рекомбинантный белок.
4. Растительная клетка для экспрессии рекомбинантного белка по п. 1, где клетка камбиальной меристемы (CMC) выделена из трансгенного растения с геном, кодирующим рекомбинантный белок.
5. Растительная клетка для экспрессии рекомбинантного белка по п. 1, где рекомбинантный белок является одним или более белками, выбранными из группы, состоящей из антигенов, антител, фрагментов антител, структурных белков, регуляторных белков, факторов транскрипции, белков токсинов, гормонов, аналогов гормонов, цитокинов, ферментов, ингибиторов ферментов, транспортных белков, рецепторов, фрагментов рецепторов, индукторов защитных сил хозяина, запасных белков, транспортных белков вирусов растений, белков защиты и репортерных белков.
6. Растительная клетка для экспрессии рекомбинантного белка по п. 1, где растение выбрано из группы, состоящей из томата, табака, моркови, тиса и дикого женьшеня.
7. Способ получения растительной клетки для экспрессии рекомбинантного белка, где способ включает стадию трансфекции или трансформации популяции растительных клеток, содержащей клетки камбиальной меристемы (CMC) с геном, кодирующим рекомбинантный белок, сокультивированием популяции клетки камбиальной меристемы (СМС) с Agrobacteria, содержащей вектор, содержащий встроенный в него ген, кодирующий рекомбинантный белок,
где клетка камбиальной меристемы (CMC) является клеточной линией, содержащей по природе недифференцированную клетку, выделенную из растения, причем клеточная линия выделена из камбиальной ткани растения и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс.
8. Способ по п. 7, где стадия трансфекции или трансформации дополнительно содержит стационарное культивирование.
9. Способ по п. 7, где стадия трансфекции или трансформации дополнительно содержит выполняемое однократное стационарное культивирование или периодически выполняемое стационарное культивирование.
10. Способ по п. 7, где сокультивирование включает культивирование растительной клетки и культуры Agrobacteria, содержащей вектор, содержащий ген, кодирующий рекомбинантный белок, со встряхиванием в течение от 1 минуты до 48 часов;
подвергание данной культуры стационарному культивированию в течение от 1 минуты до 96 часов; и
подвергание данной культуры культивированию со встряхиванием в течение 1-14 дней.
11. Способ по п. 7, где Arobacteria имеет OD600 0,00001-2,0.
12. Способ получения рекомбинантного белка из трансгенного растения для экспрессии белка-мишени, причем этот способ предусматривает стадии:
(a) выращивания трансгенного растения с геном, кодирующим белок-мишень;
(b) выделения трансгенной клетки камбиальной меристемы (ТСМС) из этого трансформированного растения;
(c) культивирования выделенной трансгенной клетки камбиальной меристемы (ТСМС) в среде; и
(d) извлечения белка-мишени, экспрессированного в культивированной трансгенной клетке камбиальной меристемы (ТСМС), где трансгенная клетка камбиальной меристемы (ТCMC) является клеточной линией, содержащей по природе недифференцированную клетку, выделенную из трансгенного растения, причем клеточная линия выделена из камбиальной ткани трансгенного растения и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс.
13. Способ производства рекомбинантного белка растительной клеткой для экспрессии этого рекомбинантного белка, причем этот способ включает стадии:
(a) стабильной трансформации популяции растительной клетки, представляющей собой клетку камбиальной меристемы (CMC) с геном, кодирующим рекомбинантный белок, или временно экспрессирующую ген, кодирующий рекомбинантный белок, сокультивированием популяции растительной клетки, представляющей собой клетку камбиальной меристемы (CMC) с Agrobacteria, содержащими вектор, содержащий встроенный в него ген, кодирующий рекомбинантный белок,
где клетки камбиальной меристемы (CMC) являются клеточной линией, содержащей по природе недифференцированную клетку, выделенную из растения, причем эта клеточная линия выделена из камбиальной ткани растения и имеет меристематическую преемственность без прохождения через дедифференцировку в каллюс; и
(b) извлечения рекомбинантного белка, экспрессируемого в культуре клетки растения, которая инфицирована Agrobacteria посредством сокультивирования.
| ДОИЛЬНЫЙ СТАКАН | 1999 |
|
RU2151498C1 |
| WO 2008062049 A1, 29.05.2008 | |||
| WO 2004048595 A2, 10.06.2004 | |||
| WO 2011025516 A1, 3.03.2011 | |||
| RU 97118361 A, 10.08.1999. | |||

