ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к областям биоинженерии и генетической инженерии растений для усовершенствования растения. Точнее говоря, настоящее изобретение относится к применению новых генов белков-факторов транскрипции с доменом «цинковые пальцы» риса и кодируемых ими белков или полипептидов для того, чтобы увеличить устойчивость к засухе и к действию солей у растений, к способам увеличения устойчивости к действию солей и/или засухе растений при ингибировании генов, описанных выше, или их экспрессирующихся белков и к трансгенным растениям.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Увеличивающийся мировой спрос на продукты питания и постоянное сокращение сельскохозяйственных земель оказывают постоянное давление на национальную продовольственную безопасность. При производстве пищевых продуктов засуха и действие солей являются основными небиологическими (абиотическими) стрессовыми факторами, вызывающими значительное сокращение количества и ухудшения качества сельскохозяйственных культур каждый год. Кроме того, засуха часто сопровождается воздействием солей. Например, засоление почвы представляет собой общее явление ухудшения состояния почвы в областях засухи.
Доступные данные показывают, что ежегодные потери риса из-за засухи в Китае составляют по стоимости, по меньшей мере, 2 миллиардов $ и засоление сельскохозяйственных земель также является основной причиной для пониженного производства и низкой урожайности. Засуха и действие солей представляют собой две серьезные проблемы, с которыми сталкивается сельское хозяйство Китая.
Следовательно, вопрос как увеличить устойчивость к засухе и/или к действию солей сельскохозяйственных культур для увеличения урожайности, решающего продовольственные проблемы в Китае и в мире, имеет большое значение. Исследование абиотических стрессовых факторов является одной из самых актуальных и перспективных областей в исследовании растений.
На сегодняшний день клонированы некоторые гены, связанные с устойчивость к засухе и к действию солей, включающие несколько факторов транскрипции, и получены некоторые устойчивые к стрессу штаммы растений или сельскохозяйственных культур с применением методик генетической инженерии. Цель генетической инженерии в отношении устойчивости к стрессовым воздействиям заключается в том, чтобы увеличить устойчивость растений к стрессам, модулируя транскрипцию и экспрессию гена, в которых факторы транскрипции играют ключевую роль.
В настоящее время клонированы некоторые факторы транскрипции, которые участвуют в экспрессии генов реакции на стресс. Тем не менее, из-за очень большого количества транскрипционных факторов, участвующих в таких сложных процессах реакции растений на стресс, данные транскрипционные факторы представляют собой только верхушку айсберга. Намного больше транскрипционных факторов предстоит обнаружить, исследовать и применить при селекции сельскохозяйственных культур с устойчивостью к стрессу.
Следовательно, существует насущная необходимость в области исследования факторов транскрипции, которые участвуют в экспрессии гена реакции на стресс для того, чтобы разработать новые способы селекции устойчивых к стрессу сельскохозяйственных культур и создать такие новые сельскохозяйственные культуры, тем самым улучшая качество и увеличивая количество сельскохозяйственных культур.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Одна цель настоящего изобретения заключается в том, чтобы получить новый ген, который тесно связан с устойчивостью к действию солей и засухе растений (особенно, сельскохозяйственных культур), ген белка-фактора транскрипции DST с доменом «цинковые пальцы» риса, и чтобы подтвердить, что данный фактор транскрипции является негативным регулятором устойчивости к действию солей и к засухе у растений. Другая цель настоящего изобретения заключается в том, чтобы создать новый способ повышения устойчивости к стрессовому воздействию солей и/или засухи. Другая цель настоящего изобретения заключается в том, чтобы предоставить способы создания трансгенных растений с повышенной устойчивостью к засухе и к действию солей и трансгенные растения, получаемые данными способами. Другая цель настоящего изобретения заключается в том, чтобы предоставить способы отбора растений, обладающих повышенной устойчивостью к засухе и к действию солей, и растения, получаемые данными способами отбора.
В первом аспекте настоящее изобретение относится к выделенным белкам-факторам транскрипции с доменом «цинковые пальцы», включающим полипептиды, содержащие аминокислотную последовательность SEQ ID NO: 2, полипептиды, содержащие консервативные мутации в предшествующих полипептидах, или гомологи предшествующих полипептидов.
В одном из предпочтительных вариантов осуществления указанные выше полипептиды включают структурный домен типа «цинковый палец» Cys-2/His-2.
В другом предпочтительном варианте осуществления указанные полипептиды участвуют в регуляции генов, связанных с ферментом, который воздействует на пероксиды, контролируя накопление перекиси водорода, и/или в регуляции устьичной поры, тем самым влияя на устойчивость риса к засухе и к действию солей.
В одном из вариантов осуществления настоящего изобретения, указанный полипептид выбран из следующей ниже группы:
(a) полипептид, обладающий аминокислотной последовательностью SEQ ID NO: 2;
(b) полипептид, полученный из (a) при помощи одной или нескольких замен аминокислотных остатков, делеций или вставок и способный увеличивать чувствительность к засухе и действию солей у растения; или
(c) полипептидный гомолог полипептида (a) или (b), содержащий структурный домен типа «цинковый палец» Cys2/His-2 и способный увеличивать чувствительность к засухе и действию соли у растения.
В предпочтительном варианте осуществления указанные растения представляют собой двудольные растения или однодольные растения, предпочтительно сельскохозяйственные культуры.
В другом предпочтительном варианте осуществления указанные растения выбраны из: Gramineae, Malvaceae gossypium, Cruciferae brassica, Compositae, Solanaceae, Labiatae или Umbelliferae, предпочтительно Gramineae.
В другом предпочтительном варианте осуществления указанные растения выбраны из: риса, кукурузы, пшеницы, ячменя, сахарного тростника, сорго, Arabidopsis, хлопка или канола, более предпочтительно, риса, кукурузы, пшеницы, ячменя, сахарного тростника или сорго.
В другом предпочтительном варианте осуществления указанные соли относятся к хлориду натрия, сульфату натрия, карбонату натрия или бикарбонату натрия.
Во втором аспекте настоящее изобретение относится к выделенным полинуклеотидам, которые включают нуклеотидные последовательности, кодирующие полипептиды по настоящему изобретению.
В одном из предпочтительных вариантов осуществления указанный полинуклеотид кодирует аминокислотную последовательность SEQ ID NO: 2 или его полипептидный гомолог.
В одном из вариантов осуществления настоящего изобретения, указанная полинуклеотидная последовательность представляет собой последовательность, выбранную из следующего ниже:
(a) нуклеотидная последовательность, содержащая последовательность SEQ ID NO: 1;
(b) нуклеотидная последовательность, содержащая нуклеотиды 1-435 в SEQ ID NO: 1;
или
(c) полинуклеотидная последовательность, комплементарная одной из нуклеотидных последовательностей (a)-(b).
В третьем аспекте настоящее изобретение относится к вектору, который содержит полинуклеотид по настоящему изобретению.
В предпочтительном варианте осуществления указанный вектор выбран из: бактериальной плазмиды, фага, дрожжевой плазмиды, вируса растений или вируса млекопитающих; предпочтительно, pCAMBIA1301, pEGFP-1, pBI121, pCAMBIA1300, pCAMBIA2301 или pHB, и, более предпочтительно, pCAMBIA1301.
В четвертом аспекте настоящее изобретение относится к генетически сконструированным клеткам-хозяевам, которые содержат вектор по настоящему изобретению или содержат полинуклеотид по настоящему изобретению, встроенный в геном.
В предпочтительном варианте осуществления указанная клетка-хозяин выбрана из прокариотической клетки, клетки низших эукариот или клетки высших эукариот, предпочтительно, бактериальной клетки, дрожжевой клетки или растительной клетки, более предпочтительно E. coli, Streptomyces, Agrobacterium, дрожжей, наиболее предпочтительно Agrobacterium, где указанный Agrobacterium включает в качестве неограничивающих примеров: EHA105, SOUP1301 или C58, предпочтительно, EHA105.
В пятом аспекте настоящее изобретение относится к цис-действующему элементу, который включает последовательность SEQ ID NO: 3, способный связываться с фактором транскрипции по настоящему изобретению.
В предпочтительном варианте осуществления указанный цис-действующий элемент содержит последовательность TGCTANN(A/T)TTG, в которой N выбран из A, C, G или T.
В другом предпочтительном варианте осуществления указанный цис-действующий элемент связывается с фактором транскрипции со структурным доменом «цинковый палец» по настоящему изобретению.
В другом предпочтительном варианте осуществления связывание указанного цис-действующего элемента с фактором транскрипции по настоящему изобретению может увеличить чувствительность к засухе и к действию солей.
В шестом аспекте настоящее изобретение относится к антагонистам белков или полинуклеотидов факторов транскрипции с доменом «цинковые пальцы».
В предпочтительном варианте осуществления антагонисты представляют собой малую интерферирующую РНК, антитела или антисмысловые олигонуклеотиды.
В седьмом аспекте настоящее изобретение относится к способам увеличения устойчивости растений к засухе и к действию солей, указанные способы включают ингибирование факторов транскрипции с доменом «цинковые пальцы» по настоящему изобретению, ингибирование экспрессии полинуклеотидов по настоящему изобретению или ингибирование связывания цис-действующего элемента с белком-фактором транскрипции с доменом «цинковые пальцы» по настоящему изобретению.
В одном из предпочтительных вариантов осуществления указанное ингибирование осуществляют способами делеции, мутации, РНКи, антисмысловой или доминантной негативной регуляции.
В другом предпочтительном варианте осуществления указанное ингибирование включает замену одной или нескольких аминокислот или нуклеотидов, делецию или вставку в факторах транскрипции по настоящему изобретению или полинуклеотидах по настоящему изобретению, приводящую к появлению указанных растений, обладающих повышенной устойчивостью к засухе и к действию солей.
В другом предпочтительном варианте осуществления согласно аминокислотной последовательности SEQ ID NO: 2 указанное ингибирование предполагает мутирование аспарагина в аспарагиновую кислоту в аминокислоте 69, мутирование аланина в треонин в аминокислоте 162, приводящее к появлению растений с данными мутантными последовательностями для увеличения устойчивости к засухе и к действию солей.
В другом предпочтительном варианте осуществления указанные способы включают применение антагонистов по настоящему изобретению в отношении растения.
В другом предпочтительном варианте осуществления указанное ингибирование включает: трансформацию растений при помощи вектора, содержащего малую интерферирующую РНК, нацеленную на белок-фактор транскрипции с доменом «цинковые пальцы» по настоящему изобретению или трансформацию растений с использованием клетки-хозяина, содержащей указанные векторы.
В другом предпочтительном варианте осуществления указанные способы дополнительно включают кроссбридинг растений, обладающих повышенной устойчивостью к засухе и к действию солей, получаемых способами, описанными выше, с нетрансгенными растениями или другими трансгенными растениями.
В другом предпочтительном варианте осуществления указанные соли относятся к хлориду натрия, сульфату натрия, карбонату натрия или бикарбонату натрия.
В восьмом аспекте настоящее изобретение относится к способам отбора растений с устойчивостью к засухе и к действию солей, указанные способы включают:
(i) определение в растении-кандидате уровня белка-фактора транскрипции с доменом «цинковые пальцы» по настоящему изобретению, уровня экспрессии полинуклеотида по настоящему изобретению и/или уровней связывания цис-действующего элемента по настоящему изобретению с белком-фактором транскрипции с доменом «цинковые пальцы» по настоящему изобретению; и
(ii) сравнение уровня в растении-кандидате, определенном на стадии (i), с уровнем в контрольном растении, и если уровень в растении-кандидате ниже, чем уровень в контрольном растении, то указанное растение-кандидат представляет собой устойчивое к засухе и действию солей растение.
В предпочтительном варианте осуществления указанные соли относятся к хлориду натрия, сульфату натрия, карбонату натрия или бикарбонату натрия.
В девятом аспекте настоящее изобретение относится к способам получения белка-фактора транскрипции с доменом «цинковые пальцы», отличающимся тем, что указанные способы включают:
(a) культивирование клетки-хозяина по настоящему изобретению в условиях, подходящих для экспрессии; и
(b) выделение белка-фактора транскрипции с доменом «цинковые пальцы» из среды для культивирования.
В другом аспекте настоящее изобретение относится к применению ингибитора или неконсервативной мутантной последовательности белка-фактора транскрипции с доменом «цинковые пальцы» или нуклеотидной последовательности по настоящему изобретению для увеличения устойчивости к засухе и к действию солей у растений.
В одном из предпочтительных вариантов осуществления указанный ингибитор представляет собой малую интерферирующую РНК, антитело или антисмысловой олигонуклеотид, действие которого направлено на указанный фактор транскрипции или нуклеотидную последовательность.
В другом предпочтительном варианте осуществления указанная неконсервативная мутантная последовательность ингибирует трансляцию или экспрессию белка-фактора транскрипции с доменом «цинковые пальцы» или нуклеотидной последовательности по настоящему изобретению у растений, содержащих указанную неконсервативную мутантную последовательность, приводя к возникновению повышенной устойчивости к засухе и к действию солей, по сравнению с устойчивостью к засухе и к действию солей у растений дикого типа, которые не содержат неконсервативной мутантной последовательности.
В другом предпочтительном варианте осуществления указанная неконсервативная мутантная последовательность представляет собой полинуклеотидную последовательность SEQ ID NO: 1 с двумя мутациями: A в позиции 205 мутирован на G, и G в позиции 484 мутирован на A; или аминокислотную последовательность SEQ ID NO: 2 с двумя мутациями: аспарагин в позиции 69 мутирован на аспарагиновую кислоту, и аланин в позиции 162 мутирован на треонин.
В одном из вариантов осуществления настоящего изобретения, указанное увеличение устойчивости растений к засухе и к действию солей включает:
(i) непосредственное нанесение ингибитора (антагониста), описанного выше на растение;
(ii) введение неконсервативной мутантной последовательности, описанное выше, в растение; или
(iii) конструирование молекулярного маркера, специфичного для неконсервативной мутантной последовательности, и применение указанного молекулярного маркера для отбора потомства, получаемого при кроссбридинге мутантного растения, содержащего неконсервативную мутантную последовательность с другими видами риса, чтобы выбрать отдельное потомство, содержащее неконсервативную мутантную последовательность.
В одном из предпочтительных вариантов осуществления настоящего изобретения указанный молекулярный маркер содержит пару праймеров, имеющих последовательности, показанные в SEQ ID NO: 10 и SEQ ID NO: 11, и/или пару праймеров, имеющих последовательности, показанные в SEQ ID NO: 12 и SEQ ID NO: 13.
В другом аспекте настоящее изобретение относится к способам увеличения устойчивости растений к засухе и к действию солей, где указанный способ включает: (A) получение ингибитора или неконсервативной мутантной последовательности для белка-фактора транскрипции с доменом «цинковые пальцы» или полинуклеотидной последовательности по настоящему изобретению; (B) воздействие на растение одной или несколькими обработками, выбранными из следующего ниже: (i) нанесение указанного ингибитора непосредственно на растение; (ii) введение в растение неконсервативной мутантной последовательности; или (iii) конструирование молекулярного маркера, специфичного для неконсервативной мутантной последовательности, и применение указанного молекулярного маркера для отбора потомства при кроссбридинге мутантного растения, содержащего неконсервативную мутантную последовательность, и другого штамма риса, чтобы выбрать отдельное потомство, которое содержит неконсервативную мутантную последовательность.
В одном из предпочтительных вариантов осуществления настоящего изобретения указанный молекулярный маркер представляет собой пару праймеров, имеющих последовательности, показанные в SEQ ID NO: 10 и SEQ ID NO: 11, и/или пару праймеров, имеющих последовательности, показанные в SEQ ID NO: 12 и SEQ ID NO: 13.
В другом аспекте настоящее изобретение относится к способам получения трансгенного растения, где указанные способы включают:
(1) трансфекцию растительной клетки, ткани растения или части растения конструкцией, содержащей неконсервативную мутантную последовательность белка-фактора транскрипции с доменом «цинковые пальцы» по настоящему изобретению, или конструкцией, содержащей неконсервативную мутантную последовательность полинуклеотида по настоящему изобретению;
(2) выбор растительной клетки, ткани растения или части растения, содержащей неконсервативную мутантную последовательность; и
(3) восстановление растения из растительной клетки, ткани растения или части растения, получаемой на стадии (2),
где получаемое трансгенное растение обладает более высокой устойчивостью к засухе и действию солей, чем нетрансгенное растение.
В другом предпочтительном варианте осуществления указанные способы также включают кроссбридинг получаемого трансгенного растения с нетрансгенным растением или другим трансгенным растением, и получением гибридного потомства, содержащего неконсервативную мутантную последовательность, причем указанное гибридное потомство обладает более высокой устойчивостью к засухе и действию солей, чем нетрансгенное растение, предпочтительно, указанное гибридное потомство обладает стабильным генетическим признаком.
В другом предпочтительном варианте осуществления указанные способы также включают конструирование молекулярного маркера, специфичного для неконсервативной мутантной последовательности, для отбора потомства, получаемого при кроссбридинге трансгенного растения, чтобы получить растение, обладающее повышенной устойчивостью к засухе и к действию солей.
На основе настоящего описания специалисту в данной области были бы очевидны другие аспекты по изобретению.
ОПИСАНИЕ ФИГУР
ФИГ. 1: Последовательность гена DST риса (ФИГ. 1A) и кодируемая им аминокислотная последовательность (ФИГ. 1 B).
ФИГ. 2: Сравнение фенотипов мутанта dst гена DST риса и фенотипов риса дикого типа в условиях засухи и действия солей. На каждом фотоснимке дикий тип (Zhonghua 11, ZH11) находится слева и мутант dst находится справа.
ФИГ. 3: Сравнение фенотипов в условиях засухи и действия солей дикого типа, мутанта dst, получаемого при комплементации гена DST, и растений с редуцированной функцией DST при помощи РНКи.
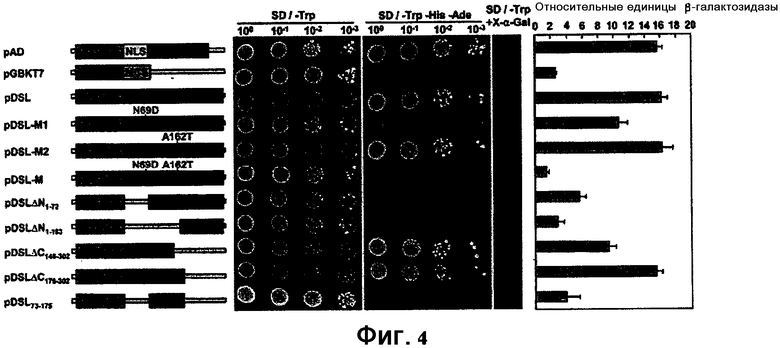
ФИГ. 4: Анализ транскрипционной активации DST с использованием дрожжевой двухгибридной системы 3 с GAL4 Matchmaker™ (Clontech).
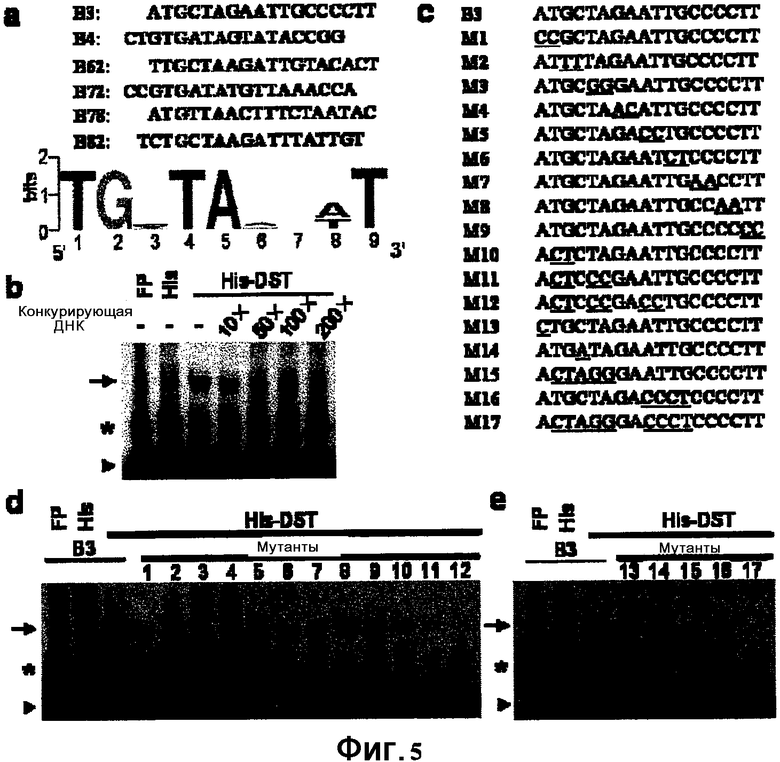
ФИГ. 5: Результаты анализа изменения электрофоретической подвижности (EMSA).
ФИГ. 6: Анализ выравнивания последовательности гомологичных доменов «цинковые пальцы» белка DST среди сельскохозяйственных растений Gramineae.
ПОДРОБНОЕ ОПИСАНИЕ
После продолжительного и интенсивного исследования авторы настоящего изобретения обнаружили новый ген белка-фактора транскрипции DST (ген устойчивости к засухе и действию солей) с доменом «цинковые пальцы» риса и доказали, что данный ген является фактором негативной регуляции в отношении устойчивости к засухе и действию солей, способным регулировать устойчивость к засухе и действию солей у растений, и что ингибирование экспрессии данного гена может увеличивать устойчивость к стрессовым воздействиям соли или к засухе у растений. Таким образом, данный ген играет важную роль при селекции растений, которые устойчивы к засухе и действию солей. На основании этого авторы изобретения осуществили изобретение на практике.
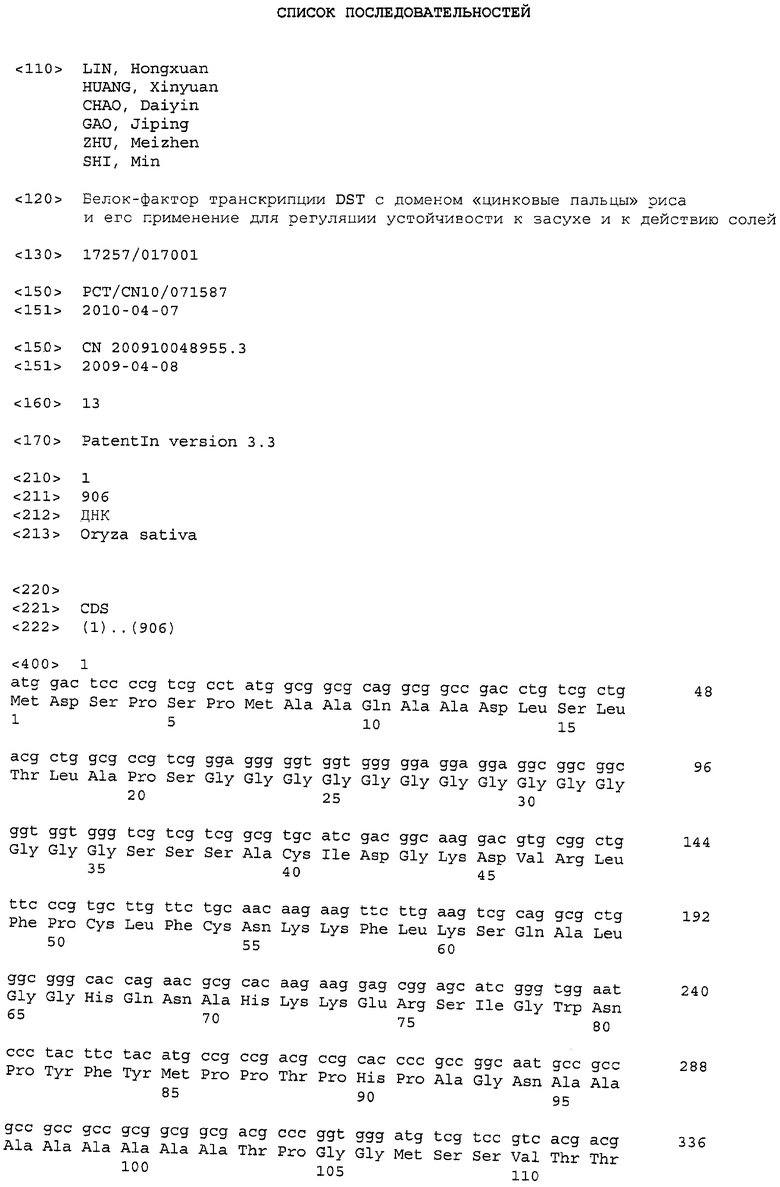
Точнее говоря, применяя библиотеку мутантов риса (мутагенез EMS) и методики клонирования на основе генетических карт, авторы изобретения провели масштабный скрининг в условиях стрессового воздействия солей, чтобы получить новый ген DST, который регулирует устойчивость к засухе и действию солей у риса. Длина геномного гена DST составляет 906 п.н., которые не включают никаких интронов. Следовательно, полноразмерная ORF (открытая рамка считывания) составляет 906 п.н. Данный ген кодирует 301 аминокислоту, белок приблизительно 29 кДа, который включает консервативный домен «цинковые пальцы». Данный белок является фактором транскрипции.
Результаты определения фенотипа показали, что мутанты данного гена (например, ген DST с 2 нуклеотидными мутациями, приводящими к возникновению 2 аминокислотных замен) демонстрируют устойчивость и к засухе, и к действию солей. Использование РНКи для понижающей регуляции экспрессии данного гена также приводит к появлению повышенной устойчивости к засухе и действию солей.
Результаты биохимических исследований показывают, что DST является фактором транскрипции, который включает не только домен активации транскрипции, но также и ДНК-связывающий домен. Анализ на генном чипе показал, что DST действует в качестве фактора транскрипции, который регулирует ряд генов последующих звеньев сигнальных каскадов.
Функциональные исследования показали, что по сравнению с диким типом мутанты содержат больше перекиси водорода (H2O2), накапливаемой вокруг устьица, обладают меньшим размером устьичных пор и позволяют листьям сохранять относительно более высокое содержание воды при стрессовом воздействии засухи. Следовательно, мутанты обладают большей устойчивостью к засухе. Кроме того, вследствие меньшего размера устьичных пор у мутантов устьичная проводимость ниже, и скорость испарения воды меньше. В результате транспорт ионов Na+ от корней к частям над землей (листья и так далее) уменьшен, и, следовательно, ниже токсичность Na+, тем самым повышается устойчивость к действию солей. Данное исследование показало, что DST участвует в регуляции относящихся к пероксидазе генов, регулирует накопление перекиси водорода (H2O2), регулирует размер устьичной поры, тем самым влияя на устойчивость к засухе и действию солей у риса.
Описанные выше исследования показали, что ген DST является фактором негативной регуляции в отношении устойчивости к засухе и к действию солей, ингибирование его экспрессии может увеличить устойчивость растений к стрессовым воздействиям солей или засухи. Данное свойство можно использовать для получения трансгенных растений со значительно более высокой устойчивостью к стрессовым воздействиям солей или засухи. Таким образом, ген DST обладает большим потенциалом в усовершенствовании способности сельскохозяйственных культур быть устойчивыми к неблагоприятным стрессовым воздействиям, таким как стресс, вызванный засолением, и засуха.
Кроме того, поиск по базе данных выявил: один гомологичный DST ген в геноме сорго (Sorghum bicolor) с белковой схожестью 54,3%; три гомологичных DST гена в геноме кукурузы (Zea mays) с белковой схожестью 51,7%, 36,1%, и 33,5%; один гомологичный ген DST в геноме ячменя (Hordeum vulgare), с белковой схожестью 38,4%; и три гомологичных DST гена в геноме сахарного тростника (Saccharum officinarum) с белковой схожестью 38,2%, 38,2% и 34,5%. Все данные гомологичные гены содержат консервативный структурный домен «цинковый палец» типа C2H2 с высокой степенью схожести на N-конце, предполагающей, что гены, гомологичные DST, у других растений (предпочтительно, Gramineae) будут обладать такими же функциями, как и функции DST гена риса.
БЕЛКИ ИЛИ ПОЛИПЕПТИДЫ DST И ИХ КОДИРУЮЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ
В настоящем изобретении, термины «белки или полипептиды DST», «белки или полипептиды, кодируемые геном DST» или «белки-факторы транскрипции с доменом «цинковый палец» относятся к белкам или полипептидам, кодируемым генами DST по настоящему изобретению. Данные определения включают в себя мутанты описанных выше белков или полипептидов с консервативными мутациями или их гомологичные полипептиды. Все они содержат структурные домены «цинковые пальцы» типа Cys-2/His-2 и, когда ингибируют экспрессию указанных белков или полипептидов, у растений может увеличиваться устойчивость к стрессовым воздействиям засухи или солей.
В одном из вариантов осуществления настоящего изобретения, указанные факторы транскрипции участвуют в регуляции родственных пероксидазе генов, регулируют накопление перекиси водорода и/или регулируют размер устьичной поры, тем самым влияя на устойчивость риса к засухе и действию солей.
Указанные последовательности белка или полипептида DST выбраны из: (a) полипептидов, имеющих аминокислотную последовательность SEQ ID NO: 2; (b) полипептидов, полученных из (a), содержащих одну или несколько замен аминокислотных остатков, делеций или вставок в аминокислотной последовательности SEQ ID NO: 2, и способных увеличивать восприимчивость растений к засухе и действию соли; или (c) полипептидные гомологи полипептидов (a) или (b), содержащие структурные домены «цинковые пальцы» типа Cys-2/His-2 и способные увеличивать восприимчивость растений к засухе и действию соли. Предпочтительно, указанные белки или полипептиды могут связываться с TGCTANN(A/T)TTG, где N представляет собой A, C, G или T.
Белки и полипептиды по настоящему изобретению могут представлять собой очищенные природные продукты или химически синтезированные продукты или продукты, получаемые с использованием рекомбинантной технологии из прокариотических или эукариотических клеткок-хозяев (например, клетки бактерии, дрожжей, высших растений, насекомых и млекопитающих). Белки и полипептиды DST по настоящему изобретению предпочтительно кодируются геном DST Gramineae (предпочтительно, риса) или его гомологичными генами или генами семейства.
Типы мутаций в белках или полипептидах по настоящему изобретению включают в качестве неограничивающих примеров: делецию, вставку и/или замену одной или нескольких (обычно, 1-50, предпочтительно, 1-30, более предпочтительно, 1-20, наиболее предпочтительно, 1-10, например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) аминокислот и добавление на C-конце и/или N-конце одной или нескольких (обычно, 20 или менее, предпочтительно, менее чем 10, наиболее предпочтительно, менее чем 5) аминокислот. Например, известно в данной области, что при замещении аминокислотами, обладающими родственными или схожими свойствами, функции белков или полипептидов обычно не изменяются. В другом примере, добавление одной или нескольких аминокислот на C-конце и/или N-конце обычно не изменяет функции белков или полипептидов. Например, белки или полипептиды DST по настоящему изобретению могут содержать или могут не содержать начальный остаток метионин и все же обладать активностью в отношении увеличения устойчивости растений к стрессовому воздействию тяжелых металлов и солей. Специалист в данной области, основываясь на общих знаниях в данной области и/или обычном экспериментировании, может с легкостью определить данные различные типы мутаций, которые бы не влияли на активность белков и полипептидов.
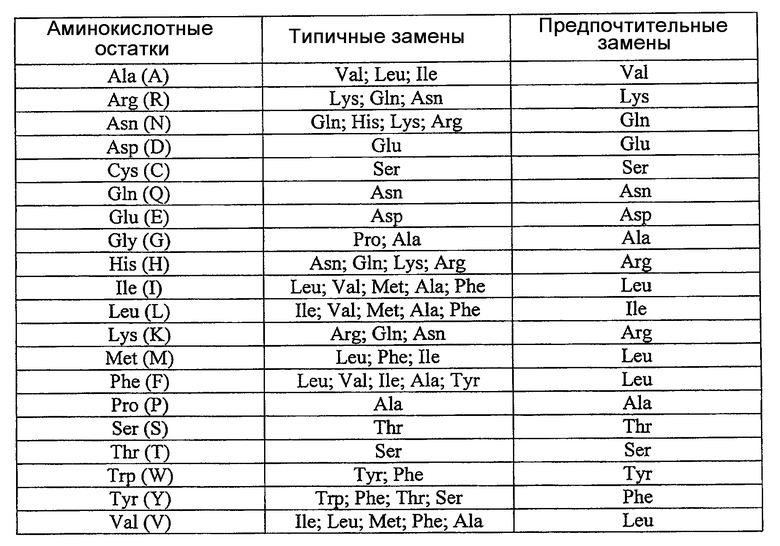
В настоящем изобретении термин «консервативные мутантные полипептиды» относится к полипептидам, содержащим по сравнению с аминокислотной последовательностью SEQ ID NO: 2, вплоть до 20, предпочтительно, вплоть до 10, более предпочтительно, вплоть до 5, наиболее предпочтительно, вплоть до 3 аминокислот, замещенных аминокислотами, обладающими родственными или схожими свойствами. Такие консервативные мутантные полипептиды можно лучше всего получить согласно следующей ниже таблице для аминокислотных замен:
Указанные белки и полипептиды из (b) можно получать при воздействии радиацией или мутагенами, чтобы вызвать случайный мутагенез, или посредством сайт-специфического мутагенеза или других известных методик молекулярной биологии. Последовательности, кодирующие белки или полипептиды можно использовать для конструирования трансгенных растений для скрининга и идентификации белков или полипептидов на основе того, обладают ли трансгенные растения измененными характеристиками.
Мутантные формы указанных полипептидов включают: гомологичные последовательности, консервативные мутанты, аллельные мутанты, природные мутанты, индуцированные мутанты, белки, кодируемые последовательностями, которые можно гибридизовать с кодирующими последовательностями белка DST при высокой и низкой степени строгости условий, и полипептиды или белки, получаемые с использованием антисыворотки против белка DST. Также в настоящем изобретении можно использовать другие полипептиды, такие как слитные белки, содержащие белок DST или его фрагмент. В дополнение к почти полноразмерным полипептидам настоящее изобретение также относится к растворимым фрагментам белков DST. Как правило, указанные растворимые фрагменты содержат, по меньшей мере, приблизительно 10 следующих друг за другом аминокислот в последовательности белка DST, обычно, по меньшей мере, приблизительно 30 следующих друг за другом аминокислот, предпочтительно, по меньшей мере, приблизительно 50 следующих друг за другом аминокислот, более предпочтительно, по меньшей мере, приблизительно 80 следующих друг за другом аминокислот, наиболее предпочтительно, по меньшей мере, приблизительно 100 следующих друг за другом аминокислот.
В зависимости от хозяев, используемых для получения рекомбинант, белки или полипептиды по настоящему изобретению могут быть гликозилированы, или могут быть негликозилированы. Термин также включает активные фрагменты и активные производные белков DST.
В настоящем описании термины «ген DST», «ген DST растений» или «кодирующие последовательности факторов транскрипции по настоящему изобретению» являются взаимозаменяемыми. Все они относятся к последовательностям, кодирующим белки или полипептиды DST по настоящему изобретению. Они высоко гомологичны последовательности гена DST риса (смотри SEQ ID NO: 1); они являются молекулами, которые могут гибридизоваться с указанной последовательностью гена в строгих условиях; или они являются молекулами генов семейства, высоко гомологичными указанным молекулам. Ингибирование экспрессии указанных генов приводит к значительному увеличению устойчивости растений к засухе или стрессу, вызванному засолением.
В одном из вариантов осуществления настоящего изобретения, указанный полинуклеотид включает: (a) нуклеотидные последовательности SEQ ID NO: 1; (b) нуклеотидную последовательность, содержащую нуклеотиды 1-435 из SEQ ID NO: 1; или (c) полинуклеотиды, комплементарные одной из нуклеотидных последовательностей в (a)-(b).
В настоящем описании термин «строгие условия» относится к: (1) гибридизации и промывке при низкой ионной силе и высоких температурах, таких как 0,2×SSC, 0,1% SDS, 60°C; или (2) гибридизация в присутствии денатурирующего средства, такого как 50% (об./об.) формамид, 0,1% сыворотка теленка/0,1% Ficoll, 42°C и так далее; или (3) гибридизация, которая имеет место, только когда гомология между двумя последовательностями достигает, по меньшей мере, 50%, предпочтительно 55% или более, 60% или более, 65% или более, 70% или более, 75% или более, 80% или более, 85% или более или 90% или более, более предпочтительно 95% или более. Например, указанные последовательности могут представлять собой последовательности, комплементарные последовательностям, определенным в (a).
Полноразмерные нуклеотидные последовательности или фрагменты нуклеотидных последовательностей гена DST по настоящему изобретению обычно можно получить ПЦР-амплификацией, рекомбинантными или синтетическими способами. Для ПЦР-амплификации близкородственные последовательности можно получать, конструируя праймеры, основанные на близкородственных нуклеотидных последовательностях, описываемых в настоящем изобретении, в особенности, на открытой рамке считывания, и используя коммерчески доступные библиотеки кДНК или библиотеки кДНК, созданные общепринятыми способами, известными специалистам в данной области в качестве матриц. Когда имеют дело с длинными последовательностями, обычно необходимо две или более ПЦР-амплификации и затем объединение фрагментов, получаемых при амплификации согласно правильному порядку.
Следует понимать, что ген DST по настоящему изобретению предпочтительно происходит из риса. Другие гены, получаемые из других растений, которые обладают высокой гомологией с геном DST риса (такой как 50% или более, предпочтительно 55% или более, 60% или более, 65% или более, 70% или более, 75% или более, 80% или более, более предпочтительно, 85% или более, такой как 85%, 90%, 95% или даже 98% идентичностью последовательности), также, как полагают, находятся в объеме настоящего изобретения. Способы и средства для сравнения идентичности последовательности, такие как BLAST, также хорошо известны в данной области.
РАСТЕНИЯ И ИХ УСТОЙЧИВОСТЬ К СТРЕССОВОМУ ВОЗДЕЙСТВИЮ СОЛИ И/ИЛИ ЗАСУХИ
Применяемые в настоящем описании, указанные «растения» включают (в качестве неограничивающих примеров): Gramineae, растения Malvaceae Gossypium, крестоцветные Brassica, Compositae, Solanaceae, растения Labiatae или Umbelliferae и так далее. Предпочтительно, указанные растения представляют собой растения Gramineae, более предпочтительно сельскохозяйственные культуры Gramineae. Например, указанные растения можно выбрать из: риса, кукурузы, пшеницы, ячменя, сахарного тростника, сорго, Arabidopsis, хлопка или канолы, более предпочтительно, риса, кукурузы, пшеницы, ячменя, сахарного тростника или сорго.
Применяемый в настоящем описании термин «сельскохозяйственные культуры» относится к растениям с экономической значимостью в отношении зерна, хлопка, масла и так далее, в сельском хозяйстве и промышленности. Экономическая значимость может проявляться в виде семян растений, фруктов, корней, стеблей, листьев и других полезных частей. Сельскохозяйственные культуры включают в качестве неограничивающих примеров: двудольные или однодольные растения. Предпочтительными однодольными растениями являются растения Gramineae, более предпочтительно, рис, пшеница, ячмень, кукуруза, сорго и так далее. Предпочтительные двудольные включают в качестве неограничивающих примеров: хлопчатник Malvaceae, крестоцветные растения, такие как Brassica, более предпочтительно, хлопок и канола.
Применяемый в настоящем описании термин «стрессовое воздействие солей» относится к явлению что, когда растения растут в почве или воде, содержащей высокую концентрацию солей, их рост будет замедляться или даже они могут погибнуть. Соли, которые вызывают стресс при засолении, включают (в качестве неограничивающих примеров) хлорид натрия, сульфат натрия, карбонат натрия или бикарбонат натрия. Гены DST по настоящему изобретению или кодируемые ими белки или полипептиды могут увеличивать устойчивость растений к стрессовому воздействию солей. Повышенную устойчивость можно наблюдать при сравнении с контрольными растениями, которые не обрабатывали указанными генами, белками или полипептидами. На рост и развитие указанных растений не влияет или влияет в меньшей степени высокая концентрация соли или указанные растения могут выживать при высоких концентрациях солей.
Применяемый в настоящем описании термин «стрессовое воздействие засухи» относится к явлению что, когда растения растут в сухой почве или другой засушливой окружающей среде, их рост будет замедляться или даже они могут погибнуть. Гены DST по настоящему изобретению или кодируемые ими белки или полипептиды могут увеличивать устойчивость растений к стрессовому воздействию засухи, повышенную устойчивость можно наблюдать при сравнении с контрольными растениями, которые не обрабатывали указанными генами, белками или полипептидами. На рост и развитие указанных растений не влияет или влияет в меньшей степени отсутствие воды, или указанные растения могут выживать при более тяжелых условиях засухи.
ВЕКТОРЫ, ХОЗЯЕВА И ТРАНСГЕННЫЕ РАСТЕНИЯ
Настоящее изобретение также относится к векторам, содержащим гены DST, и к клеткам-хозяевам, содержащим указанные вектора, получаемые посредством генетической инженерии, и к трансгенным растениям, получаемым трансфекцией гена и экспрессирующим высокие уровни DST.
Применяя общепринятую технологию рекомбинантных ДНК (Science, 1984; 224:1431), кодирующие последовательности по настоящему изобретению можно использовать для экспрессии или получения рекомбинантных белков DST. В основном, данные процессы включают следующие ниже стадии:
(1) трансфекции или трансформации подходящих клеток-хозяев полинуклеотидами (или мутантами), кодирующими белки DST по настоящему изобретению, или рекомбинантными экспрессирующими векторами, содержащими указанные полинуклеотиды;
(2) культивирования клеток-хозяев в подходящих средах для культивирования; и
(3) выделения и очистки белков или полипептидов от сред для культивирования или из клеток.
В настоящем изобретении можно использовать взаимозаменяемо термины «векторы» и «рекомбинантные экспрессирующие векторы», относящиеся к бактериальным плазмидам, бактериофагам, дрожжевым плазмидам, вирусам растительных клеток, вирусам клеток млекопитающих или другим векторам, которые хорошо известны в данной области. Вкратце, можно использовать любые плазмиды и векторы при условии, что они могут реплицироваться, и что они стабильны внутри клеток-хозяев. Одна важная характеристика экспрессирующих векторов заключается в том, что они обычно содержат точку начала репликации, промотор, маркерный ген и элемент регуляции трансляции.
Способы, хорошо известные специалисту в данной области, можно использовать для конструкции экспрессирующих векторов, содержащих кодирующие последовательности DST и подходящий транскрпционный/траснсляционный регулирующий сигнал. Такие способы включают технологию рекомбинантных ДНК in vitro, технологию синтеза ДНК и рекомбинантную технологию in vivo и так далее. Указанные последовательности ДНК можно эффективно связывать с подходящими промоторами в экспрессирующих векторах, направляя синтез мРНК. Экспрессирующие векторы также включают участки связывания рибосом для инициации трансляции и участки терминации транскрипции. В настоящем изобретении предпочтительно используют pEGFP-1, pBI121, pCAMBIA1300, pCAMBIA1301, pCAMBIA2301 или pHB.
Кроме того, экспрессирующие векторы предпочтительно включают один или несколько маркерных генов для отбора, обеспечивающих выявление фенотипа для выбора трансфицированных клеток-хозяев, таких как дигидрофолатредуктаза, устойчивость к неомицину и зеленый флуоресцентный белок (GFP) для применения в культуре эукариотических клеток или устойчивость к тетрациклину или ампицилину для применения в E. coli.
Векторы, содержащие описанные выше последовательности ДНК и подходящие промоторы или регулирующие элементы, можно использовать для трансформации подходящих клеток-хозяев, придавая им способность экспрессировать белки или полипептиды. Клетки-хозяева могут представлять собой прокариотические клетки, такие как бактериальные клетки; или клетки низших эукариот, такие как дрожжевые клетки; или клетки высших эукариот, такие как растительные клетки. Типичные примеры включают: E. coli, Streptomyces, Agrobacterium; относящиеся к грибам клетки, такие как дрожжи; растительные клетки и так далее. В настоящем изобретении клетки-хозяева предпочтительно представляют собой Agrobacterium.
Для экспрессии полинуклеотидов по настоящему изобретению в клетках высших эукариот транскрипцию можно увеличить, если в векторы встроить энхансерные последовательности. Энхансеры представляют собой цис-действующие факторы ДНК, обычно приблизительно от 10 до 300 п.н., действующие на промоторы для увеличения транскрипции гена. Специалист в данной области знает, как выбрать подходящие векторы, промоторы, энхансеры и клетки-хозяева.
Получаемые трансформанты, экспрессирующие полипептиды, кодируемые генами по настоящему изобретению, можно культивировать общепринятыми способами. В зависимости от клеток-хозяев среды для культивирования, применяемые при культивировании, можно выбрать из любых общепринятых сред. Культивирование можно проводить в условиях, подходящих для роста клетки-хозяина. Когда клетки-хозяева вырастают до соответствующей плотности клеток, подходящие способы (такие как температурный сдвиг или химическая индукция) применяют для индукции выбранных промоторов, и затем культивирование продолжают в течение другого периода времени.
Рекомбинантные полипептиды, получаемые описанными выше способами, можно экспрессировать в клетках, на клеточной мембране или секретировать из клеток. При необходимости рекомбинантные белки можно выделить и очистить, используя различные способы выделения, основанные на их физических, химических или других свойствах. Такие способы хорошо известны специалисту в данной области. Примеры таких способов включают в качестве неограничивающих примеров: общепринятую ренатурирующую обработку, обработку осаждающими белок веществами (способ высаливания), центрифугирование, осмотический лизис бактерий, обработку ультразвуком, ультрацентрифугирование, хроматографию на молекулярных ситах (гель-фильтрация), адсорбционную хроматографию, ионообменную хроматографию, высокоэффективную жидкостную хроматографию (ВЭЖХ) и различные другие методы жидкостной хроматографии и их сочетание.
Трансформацию растений можно провести, используя трансформацию Agrobacterium или трансформацию при помощи генной пушки и так далее, такой как опосредованная Agrobacterium трансформация листовых дисков. Трансформированные растительные клетки, ткани или части можно восстановить до растений, применяя общепринятые способы, приводящие к появлению повышенной устойчивости заболеваниям.
ЦИС -ДЕЙСТВУЮЩИЕ ЭЛЕМЕНТЫ
Применяемый в настоящем описании термин «цис-действующие элементы» относится к последовательностям, которые локализованы во фланкирующих областях генов и могут влиять на экспрессию гена. Их функции заключаются в том, чтобы принимать участие в регуляции экспрессии гена. Как таковой цис-действующий элемент не кодирует какой-либо белок, он только предоставляет участок взаимодействия. Он взаимодействует с транс-действующим фактором, чтобы привести к реализации его функций.
В ходе исследования авторы настоящего изобретения обнаружили, что белки DST по настоящему изобретению обладают связывающими ДНК способностями. Их коровые связывающие элементы представляют собой цис-действующие элементы, TGCTANN(A/T)TTG, где N представляет собой A, C, G или T. Цис-действующие элементы по настоящему изобретению связываются с факторами транскрипции DST, увеличивающими чувствительность к засухе и действию солей у растений, тем самым снижая устойчивость растений к засухе и действию солей. Указанные цис-действующие элементы предпочтительно взаимодействуют с доменом «цинковый палец» DST.
И наоборот, если ингибируют взаимодействия между цис-действующими элементами и факторами транскрипции DST по настоящему изобретению, это приводит к повышению устойчивости растений к засухе и действию солей.
СПОСОБЫ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ РАСТЕНИЙ К ЗАСУХЕ И ДЕЙСТВИЮ СОЛЕЙ
Как описано в настоящем описании белки DST по настоящему изобретению, их кодирующие последовательности или связывание белков DST с цис-действующими элементами тесно связаны с устойчивостью растений к засухе и действию солей. Ингибирование белков DST, их кодирующих последовательностей или связывания белков DST с цис-действующими элементами приведет к повышению устойчивости растений к засухе и действию солей.
Следовательно, настоящее изобретение также относится к способам повышения устойчивости растений к засухе и действию солей посредством ингибирования белков DST, их кодирующих последовательностей или связывания белков DST с цис-действующими элементами.
В одном из вариантов осуществления настоящего изобретения антагонисты белков DST или их кодирующих последовательностей можно использовать для ингибирования их экспрессии. Указанные антагонисты включают в качестве неограничивающих примеров: низкомолекулярную интерферирующую РНК, антитела, доминантные негативные регуляторы или антисмысловые олигонуклеотиды. Специалист в данной области, имеющий известные белки DST или их кодирующие последовательности, знает, как применить общепринятые способы и тесты для того, чтобы отобрать и получить указанные антагонисты.
Применяемый в настоящем изобретении термин «не сохраняющаяся мутация» или «неконсервативная мутация» относится к одной или нескольким заменам аминокислоты или нуклеотида, делеции или вставки (предпочтительно, неконсервативной) в белке DST или его кодирующей последовательности, приводящее к повышению устойчивости растений к засухе и действию солей.
В другом варианте осуществления настоящего изобретения известные в данной области способы можно использовать для встраивания неконсервативных мутаций в белки DST по настоящему изобретению или их кодирующие последовательности. Например, мутации нуклеотидов в кодирующих последовательностях белка DST можно использовать для того, чтобы внести неконсервативные мутации в аминокислотные последовательности белков DST, которые содержат SEQ ID NO: 2, обеспечивая появление растений, содержащих мутантные последовательности с повышенной устойчивостью к засухе и действию солей. Например, мутация аминокислоты 69 с аспарагина на аспарагиновую кислоту или мутация аминокислоты 162 с аланина на треонин.
В другом варианте осуществления настоящего изобретения можно получить трансгенные растения с заингибированной экспрессией генов DST или белков, и такие трансгенные растения можно необязательно скрещивать с нетрансгенными растениями или другими трансгенными растениями. Например, растения можно трансформировать векторами, содержащими низкомолекулярную интерферирующую РНК, антисмысловые векторы, векторы доминантной негативной регуляции, специфически направленные на белки DST, или их кодирующие белки, или клетками-хозяевами, несущими указанные векторы.
СПОСОБЫ ОТБОРА РАСТЕНИЙ, УСТОЙЧИВЫХ К ЗАСУХЕ И ДЕЙСТВИЮ СОЛЕЙ
В соответствии с уникальными свойствами генов DST по настоящему изобретению и кодируемых ими белков настоящее изобретение дополнительно включает способы отбора растений, устойчивых к засухе и действию солей.
В одном из вариантов осуществления способ отбора по настоящему изобретению включает:
(i) оценку в растении-кандидате уровня белка фактора транскрипции DST с доменом «цинковые пальцы» по настоящему изобретению, уровень экспрессии кодирующего его полинуклеотида и/или уровень связывания цис-действующего элемента по настоящему изобретению с белком-фактором транскрипции DST с доменом цинковые пальцы; (ii) сравнение уровня, определенного в растении-кандидате на стадии (i), с соответствующим уровнем в контрольном растении, если уровень в растении-кандидате ниже, чем уровень в контрольном растении, то растение-кандидат представляет собой устойчивое к засухе и к действию солей растение.
В другом варианте осуществления методику отбора с использованием молекулярного маркера, известную в данной области, можно использовать для введения гена устойчивости к засухе и к действию солей DST в другие варианты, чтобы отобрать и культивировать новые варианты, которые устойчивые к засухе и к действию солей. Указанные способы могут применять общепринятые способы кроссбридинга. Его преимущества заключаются в том, что не требуется никакого переноса генов, избегая проблем безопасности при переносе генов. Указанные способы могут включать: конструирование молекулярных маркеров, специфичных для неконсервативной мутантной последовательности, применение указанных молекулярных маркеров для отбора потомства в результате кроссбридинга мутантов, содержащих неконсервативные мутантные последовательности, и других вариантов риса, тем самым отбирая отдельные растения, несущие указанные неконсервативные мутантные последовательности.
ОСНОВНЫЕ ПРЕИМУЩЕСТВА НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Основные преимущества настоящего изобретения включают:
(1) идентификацию генов DST и кодируемых ими белков или полипептидов и подтверждение их взаимосвязи с устойчивостью к засухе и действию солей у растений, тем самым предлагая новые способы изучения устойчивости к засухе и действию солей у растений;
(2) получение трансгенных растений, обладающих повышенной устойчивостью к стрессовому воздействию солей и засухи, тем самым предлагая превосходные исходные материалы и продукты для получения и обработки зерен, хлопка и масел; и
(3) предоставление способов для отбора устойчивого к засухе и действию солей потомства с использованием молекулярных маркеров, которые можно реализовать, применяя общепринятые способы кроссбридинга без необходимости переноса генов, тем самым избегая проблем безопасности, относящихся к переносу генов.
Настоящее изобретение относится к новым подходам увеличения устойчивости к стрессовому воздействию солей и засухи у растений с огромными возможностями при применении.
ПРИМЕРЫ
Следующее ниже описание совместно с конкретными примерами дополнительно иллюстрирует настоящее изобретение. Следует понимать, что данные примеры используют для того, чтобы объяснить настоящее изобретение и их не следует использовать для ограничения объема настоящего изобретения.
В следующих ниже примерах, когда условия не определены в экспериментальных способах, они обычно основаны на общепринятых условиях (например, смотри, пожалуйста, Sambrook et al, "Molecular Cloning: A Laboratory Manual," third edition, 2001, Cold Spring Harbor Laboratory Press) или условиях в соответствии с рекомендациями производителя. Если не указано иначе, процентные соотношения и соотношения вычисляют на основе веса.
Если не определено иначе, все профессиональные и научные термины, применяемые в настоящем описании, имеют то же самое значение, что и термины, хорошо известные специалисту в данной области. Кроме того, любые способы и материалы, схожие или эквивалентные способам и материалам, описанным в настоящем описании, можно использовать в настоящем изобретении. Предпочтительные способы и материалы, описываемые в настоящем описании, используют только для иллюстрации.
Различные среды, используемые в примерах (жидкая среда для культивирования YEB, жидкая среда для культивирования AB, жидкая среда для культивирования AAM, среда для культивирования N6D2, среда для культивирования N6D2C, среда для совместного культивирования, среда культивирования для отбора N6D2S1, N6D2S2, среда культивирования для предварительной дифференцировки, среда культивирования для дифференцировки, 1/2 MS0H среда для культивирования, среда для культивирования риса, среда для культивирования SD и так далее), приготавливают согласно описаниям в соответствующей литературе (Molecular Cloning: Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989; Hiei, Y., etc., Plant J., 1994, 6, 271-282).
ПРИМЕР 1: ЭКСПЕРИМЕНТЫ ПО ПЕРЕНОСУ ГЕНА DST РИСА
1. Получение мутантов DST, обладающих высокой устойчивостью к засухе и действию солей, их характеристики и субклеточная локализация
Зерна риса обрабатывали 0,6% EMS (этилметансульфонатом) для создания библиотеки мутантов риса, содержащей приблизительно 9000 мутантных линий риса. Масштабный скрининг библиотеки мутантов риса проводили при стрессе, вызванным засолением 140 мМ хлоридом натрия. Фенотипы, устойчивые к засухе и действию соли, верифицировали, подвергая мутанты-кандидаты повторяющемуся стрессовому воздействию солью 140 мМ хлорида натрия и 20% PEG4000, имитирующему стрессовое воздействие засухи. Получают высокоустойчивые к засухе и действию солей мутанты (dst).
С использованием молекулярных маркеров ген DST предварительно локализуют на хромосоме 3 риса. Посредством кроссбридинга мутанта dst с чувствительными к действию солей штаммами создавали большое потомство F2. Используя молекулярные маркеры, проводили клонирование на основе генных карт для отбора смешанного потомства из группы, объединяющей в себе генотипы и фенотипы смешанного потомства. Это приводило к успешному клонированию гена DST. Указанные ген DST кодирует белок с доменом «цинковый палец» (фактор транскрипции) с неизвестной функцией, содержащий консервативные домен «цинковый палец» типа C2H2. В геноме риса не обнаружено никакой другой гомологичной копии DST, и никакого гомологичного гена не обнаружено в геноме Arabidopsis. Длина указанного геномного гена составляет 906 п.н. без интронов. Полноразмерная ORF (открытая рамка считывания) составляет 906 п.н. в длину, кодирующая 301 аминокислоту. Молекулярная масса белкового продукта, как оценивают, составляет 29 кДа (ФИГ. 1). Анализ сравнения последовательности показал, что ген DST в данном мутанте содержит 2 мутации нуклеотидов, которые приводят к появлению 2 аминокислотных замен (аминокислота 69 мутирована с аспарагина на аспарагиновую кислоту, и аминокислота 162 мутирована с аланина на треонин) и приводят к появлению фенотипа, устойчивого к засухе и действию солей. Данное наблюдение указывает на то, что DST является негативным регулятором устойчивости к засухе и действию солей.
Для того чтобы определить субклеточную локализацию DST, слитую конструкцию DST и GFP (зеленый флуоресцентный белок) получают и трансфицируют в эпидермальные клетки лука, используя способ генной пушки для транзиторной экспрессии. Распределение флуоресценции внутри клеток исследуют с использованием флуоресцентного конфокального микроскопа. Исходя из данного исследования субклеточной локализации, DST, как обнаружено, специфически локализован в ядре.
2. Конструирование трансгенных плазмид, содержащих геномные фрагменты DST
Клоны риса дикого типа BAC расщепляли с использованием рестрикционного фермента ApaLI, после чего следовала обработка ДНК-полимеразой T4 для того, чтобы получить тупые концы, которые затем расщепляли рестрикционным ферментом SalI. Геномный фрагмент 4,6 т.п.н. дикого типа (содержащий полноразмерную ORF DST, промоторную область и стоп-кодон с областью, расположенной по ходу транскрипции) таким образом восстанавливали. Растительный экспрессирующий бинарный вектор pCAMBIA1301 (приобретенный в CAMBIA) расщепляли EcoRI, после чего следовала обработка ДНК-полимеразой T4 для того, чтобы получить тупые концы, которые расщепляли при помощи SalI и затем лигировали с восстановленными фрагментами, упомянутыми выше, для успешного конструирования плазмиды p-DST, которую используют для трансформации мутантов и проведения экспериментов по комплементации. Все ферменты приобретены в New England Biolabs.
3. Конструирование экспрессирующих плазмид DST-RNAi
Использовали олигонуклеотиды с 5'- и 3'-концов в качестве праймеров (SEQ ID NO: 4 и 5), чтобы амплифицировать фрагмент DST, содержащий уникальную кодирующую область (535-п.н.) посредством ПЦР. Лигировали данный фрагмент с вектором p1300RNAi (получаемым при модификации pCAMBIA1300 посредством вставки интрона каталазы в качестве линкера, фланкируемого поли-A и поли-T с обоих концов), чтобы создать плазмиду DST-RNAi.
5'-последовательность олигонуклеотидного праймера представляет собой:
5'-AAGCTTTCCTTGCGAAGCCAAATAGC-3' (SEQ ID NO: 4)
3'- последовательность праймера представляет собой:
5'-GGATCCCGAGGCTCAAGTTGAGGTCGA-3' (SEQ ID NO: 5)
4. Трансгенный рис с DST:
Две рекомбинантные плазмиды, описанные выше, переносили в штамм Agrobacterium EHA105, применяя способ замораживания и оттаивания. Добавляли 0,5-1 мкг (приблизительно 10 мкл) ДНК плазмиды в каждые 200 мкл компетентных клеток EHA105, перемешивали и затем последовательно помещали на лед, в жидкий азот и в водяную баню при 37°C в течение 5 минут. Реакционную смесь разбавляли 1 мл свежей жидкой средой для культивирования YEB и затем инкубировали при перемешивании при 28°C в течение 2-4 часов. 200 аликвот были распределены в планшете с YEB, содержащем антибиотик канамицин (Kan) (50 мкг/мл). Инкубировали планшет при 28°C в течение 2-3 дней. Наносили полученные колонии в виде полосок три раза на планшеты с YEB, содержащие Kan (50 мкг/мл) для того, чтобы отобрать единичные колонии.
Отбирали единичную колонию Agrobacterium из планшета с YEB и инокулировали ее в 3 мл жидкой среды для культивирования YEB, содержащей 50 мкг/мл антибиотиков Kan и инкубировали при перемешивании при 28°C в течение ночи. На день 2 переносили 1% инокулят в 50 мл жидкой среды AB, содержащей 50 мкг/мл антибиотиков Kan, и продолжали инкубацию при перемешивании при 200 об./мин. до тех пор, пока OD600 не достигал значения приблизительно от 0,6 до 0,8. Центрифугировали свежую культуру Agrobacterium при 5000 об./мин. и 4°C в течение 5 минут. Собирали и ресуспендировали осадок в 1/3 объема жидкой среды для культивирования AAM. Данную суспензию можно было использовать для трансформации различных материалов-реципиентов риса.
В данном эксперименте использовали общепринятый опосредуемый Agrobacterium способ трансформации для того, чтобы трансформировать каллюс зародыша риса Zhonghua 11 (или его мутантов). Погружали незрелые зерна Zhonghua 11 (через 12-15 дней после опыления) в 70% этанол в течение 1 минуты, стерилизовали их в растворе NaClO (смешанном с водой в соотношении 1:3, с добавлением 2-3 капель Tween 20) в течение 90 минут или более и промывали зерна стерильной водой 4-5 раз. Затем извлекали зародыши из зерен, используя скальпель и микропинцет, и помещали на среды для культивирования N6D2, чтобы индуцировать образование ткани каллюса при культивировании при 26±1°C в темноте. Через 4 дня они были готовы к трансформации.
Погружали полученные ткани каллюса зародыша в свежие жидкие среды AAM для Agrobacterium при частом перемешивании. Удаляли части риса через 20 минут, используя стерильную фильтровальную бумагу для того, чтобы удалить излишек раствора бактерий, затем переносили их на среды для культивирования N6D2C, накрытые стерильной фильтровальной бумагой, и совместно культивировали при 26°C в течение 3 дней. Добавяли Acetosyringone к среде для совместного культивирования в качестве активатора гена Vir Agrobacterium в концентрации 100 мкмоль/л.
Через 3 дня удаляли ткани каллюса из среды для совместного культивирования, срезали завязи и переносили их на среду для отбора N6D2S1 (среда N6D2, содержащая 25 мг/л Hyg) для селекции. Через 7-12 дней переносили полученные ткани каллюса в среду для отбора N6D2S2 (среда N6D2, содержащая 50 мг/л Hyg) и продолжали отбор.
Через 10-12 дней переносили интенсивно растущие устойчивые ткани каллюса в среды культивирования для предварительной дифференцировки и инкубировали в течение приблизительно недели. Затем переносили их в среды культивирования для предварительной дифференцировки, чтобы позволить им дифференцироваться (12 ч свет/день). Как только у восстановленной рассады вырастали корни в среде для культивирования 1/2 MSOH, их переносили в горшок с почвой и выращивали в камере фитотрона.
После того, как восстановленные растения переживали трансплантацию, идентифицировали положительные трансгенные растения, используя известные в данной области способы, посредством определения β-глюкозидазы (беты-глюкуронидазы, GUS, смотри Jefferson et al. EMBO J. 6, 3901-3907, 1987) или смазывая листья 0,1% гербицидом. Экстрагировали полную ДНК из листьев положительных трансгенных растений и применяли ПЦР для дополнительной верификации данных для трансгенных растений.
В последующих экспериментах трансгенные растения в поколении T2, полученные упомянутыми выше способами, использовали, для исследования устойчивых к засухе и действию солей фенотипов, под воздействием засухи и стресса, вызванного засолением (140 мМ NaCl), чтобы подтвердить функции гена DST.
ПРИМЕР 2: КУЛЬТИВИРОВАНИЕ ТРАНСГЕННОГО РАСТЕНИЯ И ТЕСТИРОВАНИЕ УСТОЙЧИВОСТИ К ЗАСУХЕ И ДЕЙСТВИЮ СОЛЕЙ
Брали зерна трансгенного риса, полученного в ПРИМЕРЕ 1, и инкубировали их в печи при 45°C в течение недели, чтобы нарушить состояние покоя. Затем замачивали их в воде из-под крана при комнатной температуре в течение 3 дней, и подготавливали их для проращивания при 37°C в течение 2 дней. После прорастания их засевали в виде пятен в 96-луночные планшеты. Затем переносили их в легкие инкубаторы, инкубировали их при 30°C и подвергали их воздействию светом в течение 13 часов в день. Через один день постепенно уменьшали температуру до 28°C и 26°C и инкубировали их в течение одного дня каждый и культивировали их при 20°C ночью. После того, как вся рассада была выращена, заменили воду из-под крана на среды для культивирования риса и продолжали культивирование.
Приблизительно через 14 дней после культивирования рассада вырастала до состояния двух листьев и одной верхушечной почки. Их подвергали обработке солью в средах для культивирования риса, содержащих 140 мМ NaCl, в течение 12 дней или обработке PEG в средах для культивирования риса, содержащей 20% (масс./об.) PEG-4000, в течение 7 дней, чтобы имитировать стрессовое воздействие засухи.
Что касается воздействия засухи в трубах PVP (поливинилпирролидоновая труба, высотой 1,2 м, 20 см в диаметре, два сливных отверстия у основания трубы), выращивали рассаду в водяном инкубаторе в течение 25 дней в трубах PVP, содержащих почву и культивировали рассаду в камере фитотрона. Температура составляла 24-30°C, и влажность составляла 50-60%. Сливали воду через 30 дней после трансплантации, открыв нижние сливные отверстия, чтобы слить воду, и, выполняли воздействие засухой в течение 12 дней.
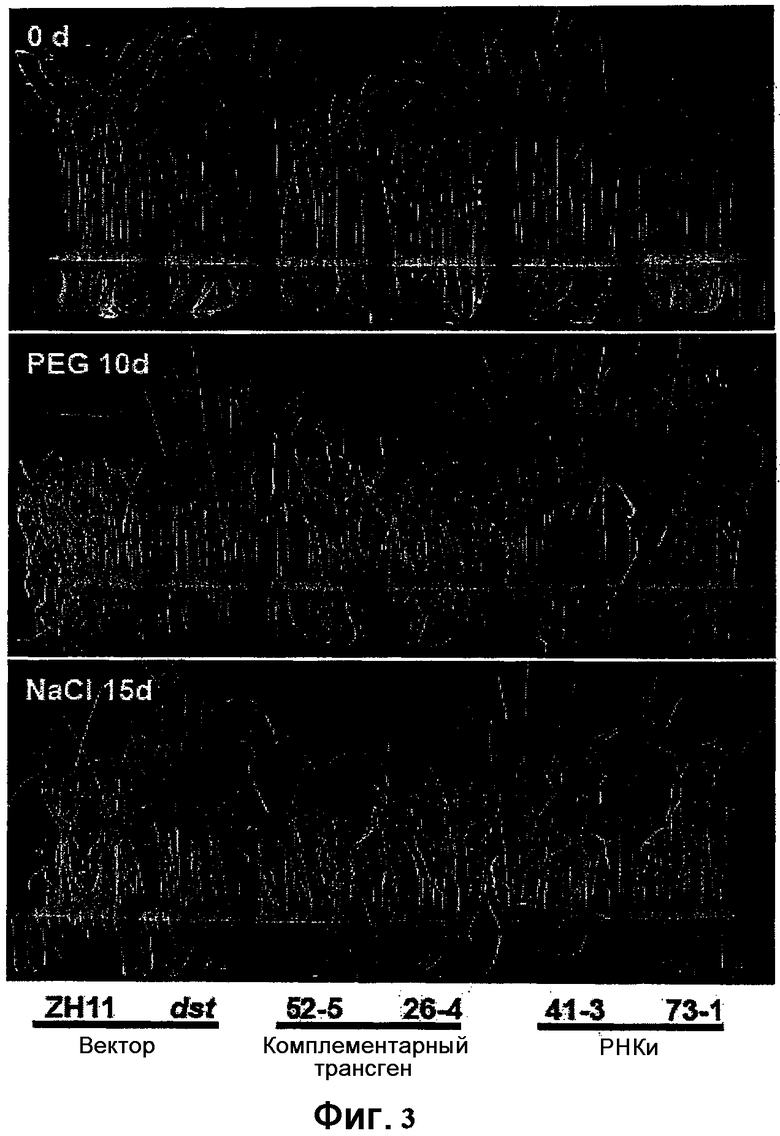
На ФИГ. 2 и ФИГ. 3 показаны экспериментальные результаты. Как показано на ФИГ. 2, мутантный рис dst обладает значительно более высокой устойчивостью к засухе и действию солей, чем дикий тип (Zhonghua 11, ZH11). В ходе наблюдения и сравнения также обнаружено: по сравнению с диким типом у мутантов большее количество перекиси водорода (H202), накопленной вокруг устьица, меньший размер устьичной поры, относительно более высокое содержание воды в листьях при стрессовом воздействии засухи. Таким образом, мутанты обладают более высокой устойчивостью к засухе. Кроме того, поскольку мутанты обладают меньшим размером устьичной поры, меньшей устьичной проводимостью и более медленными скоростями испарения воды, тем самым снижая транспорт ионов Na+ от корня к расположенным выше частям (листьям и так далее) и понижая токсичность Na+, и таким образом, обладают повышенной устойчивостью к действию солей.
Как показано на ФИГ. 3, трансфекция геномного фрагмента DST из риса дикого типа (Zhonghua 11, ZH 11) в мутант dst восстанавливает фенотип, чувствительный к засухе и действию солей дикого типа в трансгенных комплементарных растениях; тогда как, если уровень экспрессии DST уменьшают при помощи РНКи (трансформация Zhonghua 11), устойчивость к засухе и действию солей у Zhonghua 11 значительно возрастает.
Данные результаты показывают, что ген DST успешно клонируется и при помощи встраивания DST генетической инженерией устойчивость к стрессовому воздействию у риса может значительно увеличиваться.
ПРИМЕР 3: АНАЛИЗ АКТИВНОСТИ ТРАНСКРИПЦИОННОЙ АКТИВАЦИИ DST
Набор Matchmaker дрожжевой двухгибридной системы 3 с GAL4 (Clontech) применяли для анализа транскрипционной активации DST. Для конструирования положительного контрольного вектора pAD, последовательности доменов активации (AD) NLS и GAL4 амплифицировали при помощи ПЦР и встраивали в участки разрезания BamHI/SalI (праймеры представляют собой SEQ ID NO: 6 и 7) в pGBKT7 (приобретенный в Clontech), чтобы слить с доменом связывания (BD) ДНК GAL4 в pGBKT7.
Затем использовали ПЦР, чтобы амплифицировать полноразмерную ORF DST (праймеры представляют собой SEQ ID NO: 8 и 9). После подтверждения последовательности продукт ПЦР встраивали в вектор pGBKT7 в участки BamHI и Sall, чтобы слить с ДНК-связывающим доменом GAL4 для того, чтобы получить вектор pGBKT7-DST. Различные векторы затем трансформировали в дрожжи AH109. После выращивания в течение ночи культуру разбавляли и помещали на среды для культивирования SD без Trp или без трех аминокислот (-Trp/-His/-Ade). Затем наблюдали за ростом дрожжей и определяли активность DST в отношении транскрипционной активации.
Последовательности олигонуклеотидных праймеров представляют собой:
5'-AAAGGATCCAAGCGGAATTAATTCCCGAG-3' (SEQ ID NO: 6);
5'-AAAGTCGACCCTCTTTTTTTGGGTTTGGTGG-3' (SEQ ID NO: 7);
5'-AAAGGATCCTGATGGACTCCCCGTCGCCT-3' (SEQ ID NO: 8);
5'-AAAGTCGACCGAGGCTCAAGTTGAGGTCGAG-3' (SEQ ID NO: 9).
Результаты показаны на ФИГ. 4. Как показано на фигуре, белок DST риса обладает активностью в отношении транскрипционной активации, тогда как мутантный белок DST и белки с N-концевыми делециями утратили активность в отношении транскрипционной активации.
Результаты указывают на то, что pGBKT7-DST обладает более высокой активностью в отношении транскрипционной активации и домен активации транскрипции локализован на N-конце, указывая на то, что DST представляет собой фактор транскрипции с активностью транскрипционной активации.
ПРИМЕР 4: АНАЛИЗ БЕЛКОВ DST И АНАЛИЗ ИЗМЕНЕНИЯ ЭЛЕКТРОФОРЕТИЧЕСКОЙ ПОДВИЖНОСТИ
1. Прокариотическая экспрессия белков:
Расщепляли вектор pGBKT7-DST при помощи EcoRI и SalI, чтобы получить полноразмерную кДНК DST, которую затем перевстраивали в pET32a(+). Трансфицировали прокариотический экспрессирующий вектор pET32a(+), содержащий репкомбинантный DST в BL21, используя IPTG для индукции прокариотической экспрессии, и затем очищали белки, используя колонки с His-метками (бусы).
2. Получение антитела:
Иммунизировали кроликов очищенными белками, описанными выше, общепринятыми способами получения антитела против DST.
3. Синтезировали зонды, метили их биотином, очищали их в гелях PAGE и восстанавливали меченые зонды электроэлюцией.
4. Проводили реакцию меченых зондов с очищенными прокариотическими экспрессированными белками DST. Затем комплексы подвергали нативному электрофорезу PAGE и переносили на нейлоновые мембраны, используя полусухие способы переноса на мембрану. Нейлоновые мембраны затем помещали на рентгеновские пленки для авторадиографии, чтобы наблюдать сдвиг полос.
На ФИГ. 5 показаны экспериментальные результаты. Как показано на ФИГ. 5, DST обладает ДНК-связывающей способностью. Коровый элемент для связывания DST представляет собой цис-действующий элемент TGCTANN(A/T)TTG (SEQ ID NO: 3).
Настоящее исследование показывает, что связывание DST с указанным цис-действующим элементом может регулировать экспрессию генов, расположенных по ходу транскрипции, тем самым влияя на устойчивость растений к засухе и действию солей. Следовательно, связывание DST с цис-действующим элементом играет важную роль в негативной регуляции устойчивости к засухе и к действию солей.
ПРИМЕР 5: НАЛИЧИЕ ГОМОЛОГОВ ГЕНА DST В РАЗЛИЧНЫХ РАСТЕНИЯХ
Поиск по базе данных (http://plantta.jcvi.org/index.shtml) выявил один гомолог гена DST в геноме сорго (Sorghum bicolor) с белковой схожестью 54,3%; три гомолога гена в геноме кукурузы (Zea mays) с белковой схожестью 51,7%, 36,1% и 33,5%; один гомолог гена DST в геноме ячменя (Hordeum vulgare) с белковой схожестью 38,4%; три гомолога гена в геноме сахарного тростника (Saccharum officinarum) с белковой схожестью 38,2%, 38,2% и 34,5%.
Все данные гомологи гена содержат консервативные домены «цинковые пальцы» типа C2H2. Они обладают идентичным доменом «цинковые пальцы». В то же самое время схожесть выше в N-концевых доменах (на ФИГ. 6 показана общая последовательность данных гомологов гена. Общая последовательность представляет собой DGKDVRLFPCLFCNKKFLKSQALGGHQNAHKKERSIGWNPYFYM, то есть, позиции 42-85 в SEQ ID NO: 2). Белки с доменом «цинковые пальцы» типа C2H2 связываются с цис-действующими элементами через домены «цинковые пальцы». Следовательно, существует соответствующая взаимосвязь между данными гомологами генов и цис-действующими элементами, указывая на то, что гомологи гена DST других сельскохозяйственных культур Gramineae могут обладать схожими функциями, что и функции гена DST риса.
ПРИМЕР 6: ПРИМЕНЕНИЕ МЕТОДИК ОТБОРА С ПОМОЩЬЮ МОЛЕКУЛЯРНОГО МАРКЕРА ГЕНА DST РИСА И ДРУГИХ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР ДЛЯ СЕЛЕКЦИИ СЕЛЬСКОХОЗЯЙСТВЕННОЙ КУЛЬТУРЫ ДЛЯ УВЕЛИЧЕНИЯ УСТОЙЧИВОСТИ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР
В ходе химического мутагенеза (EMS) получили две мутации нуклеотидов в гене DST (A мутируют до G в нуклеотиде 205, и G мутируют до A в нуклеотиде 484), вызывающие появление двух замен аминокислот (аспарагин мутирует до аспарагиновой кислоты в положении 69, и аланин мутирует до треонина в положении 162), приводящие к появлению фенотипа, устойчивого к засухе и устойчивого к действию солей. Конструировали следующие ниже две пары праймеров, SNP5 и SNP3, в указанном гене, так чтобы продукты амплификации включали первую точечную мутацию и вторую точечную мутацию, соответственно.
SNP-5S: ATGGACTCCCCGTCGCCT (SEQ ID NO: 10)
SNP-5A: GTGCGCCGGGAGAAGCCC (SEQ ID NO: 11)
SNP-3S: GCGGTGCCGACGTCGTTCCC (SEQ ID NO: 12)
SNP-3A: GCCGCCGTCGTCGTCGTCTTC (SEQ ID NO: 13)
Первая точечная мутация создает участок разрезания для рестрикционного фермента ScrFI, тогда как вторая точечная мутация разрушает участок разрезания для рестрикционного фермента BstUI. Расщепление амплифицированных продуктов, получаемых при помощи праймеров SNP5 с ScrFI, приводило к получению фрагментов из 311 п.н., 85 п.н. и 31 п.н. в диком типе, тогда как мутантные амплифицированные продукты приводили к появлению фрагментов из 202 п.н., 109 п.н., 85 п.н., и 31 п.н., тем самым обеспечивая полиморфизм. Расщепление амплифицированных продуктов, получаемых при помощи праймеров SNP3 с BstUI, приводило к получению фрагментов из 66 п.н., 40 п.н. и 25 п.н. в диком типе, тогда как мутантные амплифицированные продукты приводили к появлению фрагментов из 91 п.н. и 40 п.н., тем самым также обеспечивая полиморфизм. Следовательно, такие две пары праймеров SNP5 и SNP3 можно использовать в качестве молекулярных маркеров для применения в селекционном разведении с помощью молекулярного маркера.
Устойчивые к засухе и к действию солей мутанты dst использовали для скрещивания с вариантами риса и для отбора потомства по одному молекулярному маркеру или по двум молекулярным маркерам, чтобы выбрать отдельные особи, несущие мутантный ген DST. Затем культивировали новые варианты (линии), обладающие повышенной устойчивостью к засухе и действию солей.
В указанном способе применяли общепринятые способы кроссбридинга без переноса генов, тем самым избегая проблем безопасности, связанные с переносом генов. Следовательно, такие способы являются преимущественными.
Всю литературу, цитируемую в настоящем изобретении, используют в качестве ссылок в настоящей заявке, как будто бы на каждый литературный источник ссылаются отдельно. Кроме того, следует понимать, что специалист в данной области, прочитавший описание настоящего изобретения, описанного выше, может изменять или модифицировать различные аспекты по настоящему изобретению. Данные эквиваленты находятся в объеме прилагаемой формулы изобретения в настоящей заявке.








Изобретение относится к области биохимии, в частности к способам получения растения с повышенной устойчивостью к засухе и действию солей по сравнению с диким видом растения путем снижения экспрессии/функции белка-фактора транскрипции у растения. Также изобретение относится к растению с повышенной устойчивостью к засухе и действию солей, полученное вышеуказанным способом. Изобретение позволяет эффективно получать растения с повышенной устойчивостью к засухе и действию солей по сравнению с диким видом растения. 4 н. и 2 з.п. ф-лы, 6 ил., 1 табл., 6 пр.
1. Способ получения растения с повышенной устойчивостью к засухе и действию солей по сравнению с диким видом растения, где способ включает:
(1) трансформацию растительной клетки, растительной ткани или растительного органа экспрессионным вектором, содержащим полинуклеотид, кодирующий неконсервативный мутант белка-фактора транскрипции, где белок-фактор транскрипции содержит последовательность SEQ ID NO: 2, где неконсервативный мутант белка-фактора транскрипции выбран из группы, состоящей из белка, имеющего последовательность, которая включает Asn69Asp (N69D) мутацию и/или Ala162Thr (А162Т) мутацию в последовательности SEQ ID NO: 2;
(2) отбор растительной клетки, растительной ткани или растительного органа, трансформированных полинуклеотидом, где функция белка-фактора транскрипции у растения снижена по сравнению с функцией белка-фактора дикого вида растения, и
(3) получение растения из растительной клетки, растительной ткани или растительного органа, полученных на стадии (2),
где растение является сельскохозяйственной культурой, выбранной из группы, состоящей из риса, кукурузы, ячменя, сахарного тростника и сорго.
2. Способ получения растения с повышенной устойчивостью к засухе и действию солей по сравнению с диким видом растения п. 1, где неконсервативный мутант белка-фактора транскрипции является белок, имеющий последовательность, которая включает Asn69Asp (N69D) мутацию и/или Ala162Thr (А162Т) мутацию в последовательности SEQ ID NO: 2.
3. Способ получения растения с повышенной устойчивостью к засухе и действию солей по сравнению с диким видом растения, где способ включает
(1) трансформацию растительной клетки, растительной ткани или растительного органа конструкцией, экспрессирующей олигонуклеотид РНКи, направленный против последовательности, кодирующей белок-фактор транскрипции, где белок-фактор транскрипции содержит последовательность SEQ ID NO: 2, где конструкция содержит последовательность, полученную с помощью ПЦР с использованием праймеров, содержащих последовательности SEQ ID NO: 4 и SEQ ID NO: 5;
(2) отбор растительной клетки, растительной ткани или растительного органа, трансформированных олигонуклеотидом, где экспрессия белка-фактора транскрипции у растения снижена по сравнению с экспрессией белка-фактора дикого вида растения, и
(3) получение растения из растительной клетки, растительной ткани или растительного органа, полученных на стадии (2),
где растение является сельскохозяйственной культурой, выбранной из группы, состоящей из риса, кукурузы, ячменя, сахарного тростника и сорго.
4. Способ получения растения с повышенной устойчивостью к засухе и действию солей по сравнению с диким видом растения по п. 3, где РНКи олигонуклеотид направлен против последовательности SEQ ID NO: 1.
5. Растение с повышенной устойчивостью к засухе и действию солей, полученное способом по п. 1, где растение содержит полинуклеотид, кодирующий неконсервативный мутант белка-фактора транскрипции, где белок-фактор транскрипции содержит последовательность SEQ ID NO: 2, и где неконсервативный мутант белка-фактора транскрипции выбран из группы, состоящей из белка, имеющего последовательность, которая включает Asn69Asp (N69D) мутацию и/или Ala162Thr (А162Т) мутацию в последовательности SEQ ID NO: 2.
6. Растение с повышенной устойчивостью к засухе и действию солей, полученное способом по п. 3, где растение содержит полинуклеотид, кодирующий олигонуклеотид РНКи, где РНКи олигонуклеотид направлен против последовательности SEQ ID NO: 1.
| SAKAMOTO H | |||
| Et al., Arabidopsis Cys2/His2-Type Zinc-Finger Proteins Function as Transcription Repressors under Drought, Cold, and High-Salinity Stress Conditions, Plant Physiology, 2004, vol | |||
| Регулятор для ветряного двигателя в ветроэлектрических установках | 1921 |
|
SU136A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Корпус судна | 1924 |
|
SU2734A1 |
| HU H | |||
| et al., Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in | |||

