Область техники
Настоящее изобретение относится к области генной инженерии и биоинформатики, и, в частности, к способу создания нового гена в организме в отсутствии искусственной ДНК-матрицы, и к его применению.
Уровень техники
В общих словах, полная кассета экспрессии гена в организме содержит промотор, 5'-нетранслируемую область (5'-UTR), кодирующую область (CDS) или область некодирующей РНК (некодирующая РНК), 3'-нетранслируемую область (3'-UTR), терминатор и многие другие элементы. Некодирующая РНК может выполнять ее биологические функции на уровне РНК, включая рРНК, тРНК, мнРНК, мякРНК и микроРНК. Область CDS содержит экзоны и интроны. После трансляции транскрибированной РНК в белок аминокислоты различных сегментов обычно формируют различные домены. Конкретные домены определяют внутриклеточную локализацию и функцию белка (такие как сигнал ядерной локализации, направляющий в хлоропласты пептид, направляющий в митохондрии пептид, ДНК-связывающий домен, домен активации транскрипции, каталитический центр фермента и т.д.). В некодирующей РНК различные сегменты также имеют различные функции. Когда один или более элементов гена изменяются, образуется новый ген, который может иметь новые функции. Например, событие инверсии фрагмента хромосомы размером 1,7 м.п.н., произошедшее выше гена PpOFP1 плоского персика, может приводить к новому промотору, который значительно повышает экспрессию PpOFP1 в плоде персика плоской формы на стадии S2 развития плода по сравнению с экспрессией в плоде персика округлой формы, тем самым ингибируя вертикальное развитие плода персика и приводя к фенотипу плоской формы в плоском персике (Zhou et al. 2018. A 1,7-Mb chromosomal inversion downstream of a PpOFP1 gene is responsible for flat fruit shape in peach. Plant Biotechnol. J. DOI: 10.1111/pbi.13455).
Естественное образование новых генов в биологических геномах требует длительного эволюционного процесса. В ходе научной работы молекулярные механизмы получения новых генов включают перестройку экзонов, дупликацию генов, ретротранспозицию и встраивание подвижных элементов (транспозоны, ретротранспозоны), горизонтальный перенос генов, разделение слитых генов, формирование de novo и многие другие механизмы, и новые гены могут сохраняться в виде под действием естественного отбора путем преобразования и функциональной эволюции. Относительно молодые новые гены, которые идентифицированы в плодовых мушках, Arabidopsis thaliana и приматах, согласно вычислениям, имеют историю от сотен тысяч до миллионов лет (Long et al. 2012. The origin and evolution of new genes. Methods Mol Biol. DOI: 10,1007/978-1-61779-585-5_7). Таким образом, в области генной инженерии и биологической селекции, если взять растения в качестве примера, при желании внести новый ген в растение (даже если все генетические элементы нового гена происходят из различных генов самого этого вида) этого можно достигнуть только посредством трансгенной технологии. Иными словами, элементы из различных генов собирают вместе in vitro с образованием нового гена, который затем переносят в растение посредством трансгенной технологии. Это характеризуется тем, что сборка новых генов должна проводиться in vitro, что приводит к трансгенным культурам.
Инструменты редактирования генов, представителями которых являются CRISPR/Cas9 и т.п., могут эффективно и точно вносить двухцепочечные разрывы (DSB) в определенных участках генома организма, а затем двухцепочечные разрывы (DSB) репарируются посредством собственных механизмов негомологичной репарации концов или гомологичной рекомбинации, тем самым внося сайт-специфические мутации. Современные применения способа редактирования генов, главным образом, фокусируются на редактировании внутренних элементов одного гена, по большей части редактировании экзонной области CDS. Редактирование экзона обычно приводит к мутациям со сдвигом рамки считывания в гене, что приводит к потере функции гена. По этой причине инструменты редактирования генов, такие как CRISPR/Cas9, также известны как инструменты нокаута генов (т.е. разрушения генов). В дополнение к области CDS, для влияния на уровень экспрессии гена нокауту может быть подвергнут промотор, 5'UTR и другие области. Все эти способы вносят мутации в существующие гены без образования новых генов, так что трудно удовлетворить некоторым потребностям производства. Например, для большинства генов в существующей технологии редактирования генов является трудным достижение повышения экспрессии генов и также трудно достигнуть изменения субклеточной локализации белка или изменения функционального домена белка. Также в литературе существуют сообщения о встраивании промоторной или энхансерной последовательности выше существующего гена для изменения паттерна экспрессии гена для получения новых признаков (Lu et al. 2020. Targeted, efficient sequence insertion and replacement in rice. Nat Biotechnol. DOI: 10.1038/s41587-020-0581-5), однако этот способ требует предоставления чужеродных ДНК-матриц, таким образом применяются строгие регулятивные процедуры, сходные с генно-модифицированными культурами, и применение ограничено.
Сущность изобретения
Для решения вышеупомянутых проблем уровня техники настоящее изобретение относится к способу создания нового гена в организме в отсутствии искусственной ДНК-матрицы путем одновременного внесения двух или более двухцепочечных разрывов ДНК в наборе конкретных участков генома организма, и его применению.
В одном аспекте настоящее изобретение относится к способу создания нового гена в организме, включающему следующие стадии:
одновременное внесение разрывов ДНК в двух или более конкретных участках генома организма, где конкретные участки представляют собой геномные участки, которые могут разделять различные генетические элементы или различные белковые домены, лигирование разрывов ДНК друг с другом посредством негомологичного соединения концов (NHEJ) или гомологичной репарации, создавая новую комбинацию различных элементов генов или различных белковых доменов, которая отличается от исходной геномной последовательности, тем самым создавая новый ген.
В одном конкретном варианте осуществления "два или более различных конкретных участков" могут находиться на одной хромосоме или на различных хромосомах. Когда они находится на одной хромосоме, фрагмент хромосомы, образовавшийся в результате разрывов ДНК, одновременно произошедших в двух конкретных участках, может быть удален, инвертирован или репликационно удвоен после репарации; когда они находится на различных хромосомах, разрывы ДНК, внесенные в два конкретных участка, могут лигироваться друг с другом после репарации, приводя к событию кроссинговера плеч хромосом. Эти события могут быть идентифицированы и подвергнуты скринингу посредством секвенирования с использованием ПЦР со специально сконструированными праймерами.
В конкретном варианте осуществления "два или более различных конкретных участков" могут представлять собой конкретные участки на по меньшей мере двух различных генах или могут представлять собой по меньшей мере два различных конкретных участка на одном гене.
В конкретном варианте осуществления, направления транскрипции "по меньшей мере двух различных генов" могут быть одинаковыми или могут различаться (противоположные или друг к другу).
В конкретном варианте осуществления "разрывы ДНК" формируют посредством доставки нуклеазы со свойством нацеливания в клетку организма для контакта с конкретными участками геномной ДНК. Отсутствует существенное различие между этим типом разрывов ДНК и разрывами ДНК, внесенными традиционными способами (такими как радиационный или химический мутагенез).
В конкретном варианте осуществления "нуклеаза со свойством нацеливания" выбрана из мегануклеазы, нуклеазы с цинковыми пальцами (ZFN), TALEN и системы CRISPR/Cas.
Среди них, система CRISPR/Cas может образовывать два или более двухцепочечных разрывов ДНК в различных участках генома посредством двух или более направляющих РНК, нацеленных на различные последовательности; путем конструирования по отдельности белков ZFN или белков TALEN для двух или более последовательностей конкретных участков, системы нуклеазы с цинковыми пальцами и TALEN могут одновременно вносить двухцепочечные разрывы ДНК в два или более участка. Когда два разрыва находятся на одной хромосоме, могут быть достигнуты результаты репарации, такие как делеция, инверсия и удвоение; и, когда два разрыва находится на двух различных хромосомах, может происходить кроссинговер плеч хромосом. Делеция, инверсия, удвоение и обмен сегментами хромосом в двух разрывах ДНК может рекомбинировать различные элементы генов или белковые домены, тем самым создавая новый функциональный ген.
В конкретном варианте осуществления "нуклеаза со свойством нацеливания" существует в форме ДНК.
В другом конкретном варианте осуществления "нуклеаза со свойством нацеливания" существует в форме мРНК или белка, а не в форме ДНК.
В конкретном варианте осуществления способ доставки нуклеаз со свойством нацеливания в клетки выбран из группы, состоящей из: 1) ПЭГ-опосредуемой трансфекции клеток; 2) опосредуемой липосомами трансфекции клеток; 3) трансформации с использованием электрического шока; 4) микроинъекции; 5) бомбардировки из генной пушки; или 6) опосредуемой Agrobacterium трансформации.
"Элементы генов" включают промотор, 5'-нетранслируемую область (5'UTR), кодирующую область (CDS) или область некодирующй РНК (некодирующая РНК), 3'-нетранслируемую область (3'UTR) и терминатор гена.
В конкретном варианте осуществления комбинация различных элементов генов относится к комбинации промотора одного из двух генов с различными паттернами экспрессии и CDS или области некодирующей РНК другого гена.
В конкретном варианте осуществления одна из комбинаций различных элементов генов относится к сильному эндогенному промотору в организме, а другая представляет собой кодирующую область гена HPPD, EPSPS, PPO или GH1.
В другом конкретном варианте осуществления комбинация различных элементов генов относится к комбинации области из промотора с 5'UTR одного из двух генов с различными паттернами экспрессии и CDS или области некодирующей РНК другого гена.
В конкретном варианте осуществления "различные паттерны экспрессии" относятся к различным уровням экспрессии генов.
В другом конкретном варианте осуществления "различные паттерны экспрессии" относятся к различной тканевой специфичности экспрессии генов.
В другом конкретном варианте осуществления "различные паттерны экспрессии" относятся к различным специфичностям экспрессии генов к стадиям развития.
В другом конкретном варианте осуществления комбинация различных генных элементов представляет собой комбинацию соседних генных элементов в одном и том же гене.
"Белковые домены" относятся к фрагменту ДНК, соответствующему конкретному функциональному домену белка; он включает, но не ограничивается ими, сигнал ядерной локализации, направляющий в хлоропласты пептид, направляющий в митохондрии пептид, участок фосфорилирования, участок метилирования, трансмембранный домен, ДНК-связывающий домен, домен активации транскрипции, домен активации рецептора, каталитический центр фермента и т.д.
В конкретном варианте осуществления комбинация различных белковых доменов относится к комбинации области сигнала локализации одного из двух генов, кодирующих белки с различной субклеточной локализацией, и кодирующей зрелый белок области другого гена.
В конкретном варианте осуществления "различная субклеточная локализация" включает, но не ограничиваются ими, ядерную локализацию, цитоплазматическую локализацию, локализацию на клеточной мембране, локализацию в хлоропластах, локализацию в митохондриях и локализацию на мембране эндоплазматической сети.
В другом конкретном варианте осуществления комбинация различных белковых доменов относится к комбинации двух белковых доменов с различными биологическими функциями.
В конкретном варианте осуществления "различные биологические функции" включают, но не ограничиваются ими, распознавание конкретной консервативной последовательности ДНК или РНК, активацию экспрессии гена, связывание с белковым лигандом, связывание с низкомолекулярным сигнальным соединением, связывание иона или определенную ферментативную реакцию.
В другом конкретном варианте осуществления комбинация различных белковых доменов относится к комбинации соседних белковых доменов в одном и том же гене.
В другом конкретном варианте осуществления комбинация элементов генов и белковых доменов относится к комбинации белковых доменов и соседних промоторов, 5'UTR, 3'UTR или терминаторов в одном гене.
В частности, обмена промоторами различных генов можно достигать посредством встраивания фрагментов хромосом: когда два гена, находящиеся на одной хромосоме, имеют различные направления, могут быть внесены разрывы ДНК в определенных участках между промотором и CDS каждого из двух генов, область между разрывами может быть инвертирована, тем самым промоторы этих генов могут поменяться местами, и может быть получено два новых гена на обоих концах инвертированного сегмента хромосомы. Различные направления двух генов могут быть такими, чтобы их 5'-концы были внутренними, а именно, оба гена находились в противоположных направлениях, или их 5'-концы были внешними, а именно, оба гена были направлены друг к другу. Когда гены находятся в противоположных направлениях, промоторы генов могут быть инвертированы, как показано на схеме 1 фиг.2; когда гены направлены друг к другу, области CDS генов могут быть инвертированы, как показано на схеме 1 фиг.4. Инвертированная область может иметь длину только менее 10 т.п.н. без других генов в этой области; или инвертированная область может быть очень длинной, достигая вплоть до 300 т.п.н.-3 м.п.н., и может содержать сотни генов.
Также является возможным создание нового гена путем удвоения фрагмента хромосомы: когда два гена, находящиеся на одной хромосоме, имеют одно направление, могут быть внесены разрывы ДНК в конкретные участки между промотором и CDS каждого из двух генов, область между разрывами может быть удвоена посредством дупликации и может быть создан новый ген в точке соединения удвоенного сегмента путем слияния промотора нижележащего гена с областью CDS вышележащего гена, как показано на фиг.1, схема 1, и фиг.3. Длина удвоенной области может находиться в диапазоне от 500 п.н. до 5 м.п.н., и может быть очень короткой без других генов внутри нее, или может быть очень длинной и содержать сотни генов. Хотя этот способ индуцирует точковые мутации в областях между промоторами и областью CDS исходных двух генов, такие мелкомасштабные точковые мутации, как правило, имеют небольшой эффект на свойства экспрессии генов, в то время как новые гены, созданные посредством замены промоторов, могут иметь новые свойства экспрессии. Альтернативно, разрывы ДНК могут быть внесены в конкретные положения на обеих сторонах белкового домена одного и того же гена, и область между разрывами может быть удвоена путем дупликации, тем самым создавая новый ген с удвоенными определенными функциональными доменами.
В другом аспекте настоящее изобретение относится к способу создания нового гена в организме, включающему следующие стадии:
внесение разрывов ДНК в определенные участки по меньшей мере на двух различных генах на уровне генома или хромосомы организма, индуцируя перенос, удвоение, инверсию или делецию ДНК, так чтобы конкретный элемент гена одного эндогенного гена и элемент гена другого эндогенного гена лигировались посредством негомологичного соединения концов (NHEJ) или гомологичной репарации, тем самым создавая новый ген.
Настоящее изобретение также относится к новому гену, который может быть получен посредством настоящего способа.
По сравнению с исходными генами, новый ген может иметь отличающийся промотор и, таким образом, характеристики экспрессии, с точки зрения тканей или интенсивности, или стадий развития, или имеет новые аминокислотные последовательности.
"Новая аминокислотная последовательность" может представлять собой либо слияние целых или частичных кодирующих областей двух или более генов, либо удвоение частичной кодирующей белок области одного гена.
В конкретном варианте осуществления новый ген представляет собой эндогенный ген HPPD, EPSPS, PPO или GH1 с высокой экспрессией в организме.
Настоящее изобретение также относится к ДНК, содержащей данный ген.
Настоящее изобретение также относится к белку, кодируемому геном, или его биологически активному фрагменту.
Также настоящее изобретение относится к рекомбинантному экспрессирующему вектору, который содержит данный ген и промотор, функционально связанный с ним.
Настоящее изобретение также относится к экспрессирующей кассете, содержащей данный ген.
Настоящее изобретение также относится к клетке-хозяину, которая содержит экспрессирующую кассету. Предпочтительно клетка-хозяин представляет собой клетку растения, клетку животного или клетку гриба.
Кроме того, настоящее изобретение относится к организму, регенерировавшему из клетки-хозяина.
Кроме того, настоящее изобретение относится к применению гена для сообщения или улучшения признака резистентности/толерантности или признака преимущества роста в организме.
Кроме того, настоящее изобретение относится к композиции, которая содержит:
(a) промотор одного из двух генов с различными паттернами экспрессии и кодирующую область или область некодирующей РНК другого гена;
(b) область между промотором и 5'-нетранслируемой областью одного из двух генов с различными паттернами экспрессии и кодирующей областью или областью некодирующей РНК другого гена;
(c) соседние генные элементы одного гена;
(d) область сигнала локализации одного из двух генов, кодирующих белки с различной субклеточной локализацией, и кодирующую зрелый белок область другого гена;
(e) два белковых домена с различными биологическими функциями;
(f) соседние белковые домены одного и того же гена; или
(g) белковый домен и соседний промотор, 5'-нетранслируемую область, 3'-некодирующую область или терминатор одного и того же гена.
В конкретном варианте осуществления "различные паттерны экспрессии" относятся к различным уровням экспрессии генов.
В другом конкретном варианте осуществления "различные паттерны экспрессии" относятся к различным тканевым специфичностям экспрессии генов.
В другом конкретном варианте осуществления "различные паттерны экспрессии" относятся к различным специфичностям экспрессии генов к стадии развития.
В конкретном варианте осуществления "различная субклеточная локализация" включает, но не ограничиваются ими, ядерную локализацию, цитоплазматическую локализацию, локализацию на клеточной мембране, локализацию в хлоропластах, локализацию в митохондриях и локализацию на мембране эндоплазматической сети.
В конкретном варианте осуществления "различные биологические функции" включают, но не ограничиваются ими, распознавание конкретной консервативной последовательности ДНК или РНК, активацию экспрессии гена, связывание с белковым лигандом, связывание с низкомолекулярным сигнальным соединением, связывание иона или определенную ферментативную реакцию.
В конкретном варианте осуществления композиция является слитой in vivo.
В частности, настоящее изобретение также относится к способу редактирования для повышения уровня экспрессии эндогенного гена-мишени в организме независимо от экзогенного донорского фрагмента ДНК, который включает следующие стадии: одновременное внесение разрывов ДНК в конкретные участки между промотором и CDS каждого из эндогенного гена-мишени и необязательного эндогенного гена с высокой экспрессией; лигирование разрывов ДНК друг с другом посредством негомологичного соединения концов (NHEJ) или гомологичной репарации с образованием слитой конструкции in vivo между кодирующей областью эндогенного гена-мишени и необязательным сильным эндогенным промотором, тем самым создавая новый эндогенный ген с высокой экспрессией. Этот способ называется способом редактирования для повышения уровня экспрессии (knock-up) эндогенного гена.
В конкретном варианте осуществления эндогенный ген-мишень и необязательный эндогенный ген с высокой экспрессией находятся на одной хромосоме.
В другом конкретном варианте осуществления эндогенный ген-мишень и необязательный эндогенный ген с высокой экспрессии находятся на разных хромосомах.
В другом аспекте настоящее изобретение относится способу редактирования для повышения уровня экспрессии эндогенного гена HPPD в растении, включающему слияние кодирующей области гена HPPD с сильным эндогенным промотором растений in vivo, для получения нового эндогенного гена HPPD в растении с высокой экспрессией. Иными словами, способ включает одновременное внесение разрывов ДНК в конкретные участки между промотором и CDS каждого из гена HPPD и необязательного эндогенного гена с высокой экспрессией, лигирование разрывов ДНК друг с другом посредством внутриклеточного каскада репарации с образованием слитой конструкции in vivo между кодирующей областью гена HPPD и необязательным эндогенным сильным промотором, тем самым создавая новый ген HPPD с высокой экспрессией. В рисе сильный промотор предпочтительно представляет собой промотор гена убиквитина 2.
Настоящее изобретение также относится к эндогенному гену HPPD растения с высокой экспрессией, получаемому посредством описанного выше способа редактирования.
Настоящее изобретение также относится к эндогенному гену HPPD риса с высокой экспрессией, который имеет последовательность, выбранную из группы, состоящей из:
(1) последовательности нуклеиновой кислоты, как показано в SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 18 или SEQ ID NO: 19 или ее частичной последовательности, или комплементарной им последовательности;
(2) последовательности, обладающей идентичностью по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% с любой из последовательностей, как определено в (1); или
(3) последовательности нуклеиновой кислоты, способной гибридизоваться с последовательностью, как показано в (1) или (2) в жестких условиях.
В другом аспекте настоящее изобретение относится к способу редактирования для повышения экспрессии эндогенного гена EPSPS в растении, который включает слияние кодирующей области гена EPSPS с сильным эндогенным промотором растения in vivo с образованием нового эндогенного гена EPSPS растения с высокой экспрессией. Иными словами, способ включает одновременное внесение разрывов ДНК в конкретные участки между промотором и CDS каждого из гена EPSPS и необязательного эндогенного гена с высокой экспрессией, лигирование разрывов ДНК друг с другом посредством внутриклеточного каскада репарации с образованием слитой конструкции in vivo между кодирующей областью гена EPSPS и необязательным эндогенным сильным промотором, тем самым создавая новый ген EPSPS с высокой экспрессией. В рисе сильный промотор предпочтительно представляет собой промотор гена TKT.
Настоящее изобретение также относится к эндогенному гену EPSPS растения с высокой экспрессией, который может быть получен посредством описанного выше способа редактирования.
Настоящее изобретение также относится к эндогенному гену EPSPS риса с высокой экспрессией, который имеет последовательность, выбранную из группы, состоящей из:
(1) последовательности нуклеиновой кислоты, как показано в SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13 или SEQ ID NO: 14, или ее частичной последовательности, или комплементарной им последовательность;
(2) последовательности, обладающей идентичностью по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% с любой из последовательностей, как определено в (1); или
(3) последовательности нуклеиновой кислоты, способной гибридизоваться с последовательностью, как показано в (1) или (2), в жестких условиях.
В другом аспекте настоящее изобретение относится к способу редактирования для повышения экспрессии эндогенного гена PPO (PPOX) в растении, который включает слияние кодирующей области гена PPO с сильным эндогенным промотором растения in vivo с образованием нового эндогенного гена PPO растения с высокой экспрессией. Иными словами, способ включает одновременное внесение разрывов ДНК в конкретные участки между промотором и CDS каждого из гена PPO и необязательного эндогенного гена с высокой экспрессией, лигирование разрывов ДНК друг с другом посредством внутриклеточного каскада репарации с образованием слитой конструкции in vivo между кодирующей областью гена PPO и необязательным эндогенным сильным промотором, тем самым создавая новый ген PPO с высокой экспрессией. В рисе сильный промотор предпочтительно представляет собой промотор гена CP12. В Arabidopsis thaliana сильный промотор предпочтительно представляет собой промотор гена убиквитина 10.
Настоящее изобретение также относится к эндогенному гену PPO растения с высокой экспрессией, который может быть получен посредством описанного выше способа редактирования.
Настоящее изобретение также относится к эндогенному гену PPO риса с высокой экспрессией, имеющему последовательность, выбранную из группы, состоящей из:
(1) последовательности нуклеиновой кислоты, как показано в SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26, или ее частичной последовательности, или комплементарной им последовательности;
(2) последовательности, обладающей идентичностью по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% с любой из последовательностей, как определено в (1); или
(3) последовательности нуклеиновой кислоты, способной гибридизоваться с последовательностью, как показано в (1) или (2), в жестких условиях.
Настоящее изобретение также относится к ДНК, содержащей ген HPPD, EPSPS или PPO.
Настоящее изобретение также относится к белку, кодируемому геном HPPD, EPSPS или PPO, или его биологически активному фрагменту.
Настоящее изобретение также относится к рекомбинантному экспрессирующему вектору, который содержит ген HPPD, EPSPS или PPO и промотор, функционально связанный с ним.
Кроме того, настоящее изобретение относится к экспрессирующей кассете, содержащей ген HPPD, EPSPS или PPO.
Кроме того, настоящее изобретение относится к растительной клетке-хозяину, которая содержит экспрессирующую кассету.
Кроме того, настоящее изобретение относится к растению, регенерировавшему из растительной клетки-хозяина.
Настоящее изобретение также относится к способу получения растения с повышенной резистентностью или толерантностью к гербициду, который включает регенерацию растительной клетки-хозяина в растение и/или его потомка.
В конкретном варианте осуществления растение с повышенной резистентностью или толерантностью к гербициду представляет собой нетрансгенную линию, которая может быть получена путем скрещивания растения, регенерировавшего из растительной клетки-хозяина по изобретению, с растением дикого типа для удаления экзогенного трансгенного компонента посредством генетической сегрегации.
Настоящее изобретение также относится к резистентному к гербицидам рису, который содержит любой из вышеупомянутых эндогенного гена HPPD риса с высокой экспрессией, эндогенного гена EPSPS риса с высокой экспрессией, эндогенного гена PPO риса с высокой экспрессией или любую их комбинацию.
В конкретном варианте осуществления резистентный к гербицидам рис является нетрансгенным.
Кроме того, настоящее изобретение относится к применению эндогенного гена HPPD, EPSPS или PPO растения с высокой экспрессией для повышения резистентности или толерантности к ингибиторному гербициду в клетки растения, ткани растения, части растения или растении.
В другом аспекте настоящее изобретение относится к способу борьбы с сорняками на участке для возделывания растений, где растение включает вышеупомянутое растение или растение, полученное вышеупомянутым способом, который включает применение на участке для возделывания одного или более ингибирующих HPPD, EPSPS или PPO гербицидов в количестве для эффективной борьбы с сорняками.
В исследовательской работе, проведенной авторами изобретения, было обнаружено, что в клетках, одновременно подвергаемых двойному или множественному редактированию генов-мишеней, определенная доля концов двухцепочечных разрывов ДНК в различных мишенях одновременно лигировались друг с другом, что приводило к событиям делеции, инверсии или дупликации-удвоения фрагментов между мишенями на одной хромосоме, и/или обмену хромосомными фрагментами между мишенями на различных хромосомах. В литературе сообщалось, что это явление часто существует в растениях и животных (Puchta et al. 2020. Changing local recombination patterns in Arabidopsis by CRISPR/Cas mediated chromosome engineering. Nat Commun. DOI: 10.1038/s41467-020- 18277-z; Li et al. 2015. Efficient inversions and duplications of mammalian regulatory DNA elements and gene clusters by CRISPR/Cas9. J Mol Cell Biol. DOI: 10.1093/jmcb/mjv016).
Авторы настоящего изобретения неожиданно обнаружили, что посредством индукции двухцепочечных разрывов ДНК в комбинации редактирующих ген мишеней вблизи конкретных элементов представляющего интерес гена, вызывающей сампопроизвольную репарацию путем лигирования, может быть достигнута направленная комбинация различных элементов генов на геномном уровне без необходимости в предоставлении чужеродной ДНК-матрицы, и таким образом, является возможным образование из них нового функционального гена. Эта стратегия значительно ускоряет создание новых генов и имеет большой потенциал в разведении животных и растений, и изучении функции генов.
Подробное описание изобретения
В рамках настоящего изобретения, если нет иных указаний, научные и технические термины, используемые в настоящем описании, имеют значения, хорошо понятные специалистам в данной области. Кроме того, все термины и лабораторные методики, относящиеся к химии белков и нуклеиновых кислот, молекулярной биологии, культивированию клеток и тканей, микробиологии, иммунологии, используемые в настоящем описании, представляют собой термины и стандартные методики, широко используемые в соответствующих областях. Например, стандартные способы рекомбинантных ДНК и молекулярного клонирования, используемые в рамках настоящего изобретения, хорошо известны специалистам в данной области и полностью описаны в следующих документах: Sambrook, J., Fritsch, EF and Maniatis, T., Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, 1989. Для лучшего понимания настоящего изобретения ниже приводятся определения и пояснения соответствующих терминов.
Термин "геном", как используют в рамках изобретения, относится ко всем компонентам генетического материала (гены и некодирующие последовательности), присутствующим в каждой клетке, или вирусе, или органелле организма, и/или к полному геному, наследуемому от родителя в целом (гаплоид).
Термин "редактирование генов" относится к стратегиям и способам направленной специфической модификации любой генетической информации или генома живых организмов. Таким образом, данный термин включает редактирование кодирующих областей генов, а также включает редактирование областей, отличных от кодирующих областей генов генома. Также он включает редактирование или модификацию другой генетической информации ядра (при наличии) и клеток.
Термин " нуклеаза CRISPR/Cas" может представлять собой нуклеазу на основе CRISPR или последовательность нуклеиновой кислоты, кодирующую ее, включая, но не ограничиваясь ими: 1) Cas9, включая SpCas9, ScCas9, SaCas9, xCas9, VRER-Cas9, EQR-Cas9, SpG-Cas9, SpRY-Cas9, SpCas9-NG, NG-Cas9, NGA-Cas9 (VQR) и т.д.; 2) Cas12, включая LbCpf1, FnCpf1, AsCpf1, MAD7 и т.д., или любой вариант или производное вышеупомянутой нуклеазы на основе CRISPR; предпочтительно, где по меньшей мере одна нуклеаза на основе CRISPR содержит мутацию по сравнению с соответствующей последовательностью дикого типа, так что полученная нуклеаза на основе CRISPR распознает отличающуюся последовательность PAM. Как используют в рамках изобретения, "нуклеаза на основе CRISPR" представляет собой любую нуклеазу, которая идентифицирована во встречающейся в природе системе CRISPR, которая затем выделена из ее естественной среды и предпочтительно модифицирована или скомбинирована в представляющую интерес рекомбинантную конструкцию, пригодную в качестве инструмента для направленной инженерии генома. При условии, что исходная нуклеаза на основе CRISPR дикого типа обеспечивает распознавание ДНК, т.е. свойства связывания, можно использовать любую нуклеазу на основе CRISPR и необязательно перепрограммировать или иным образом подвергать мутации так, чтобы она была пригодной для различных вариантов осуществления изобретения.
Термин "CRISPR" относится к способу последовательность-специфической генетической манипуляции, который основан на коротких палиндромных повторах, регулярно расположенных кластерами, который отличается от РНК-интерференции, которая регулируют экспрессию генов на уровне транскрипции.
"Нуклеаза Cas9" и "Cas9" используются в настоящем описании взаимозаменяемо и относятся к РНК-направляемой нуклеазе, содержащей белок Cas9 или его фрагмент (например, белок, содержащий активный домен расщепления ДНК Cas9 и/или гРНК-связывающий домен Cas9). Cas9 является компонентом системы редактирования генома CRISPR/Cas (короткие палиндромные повторы, регулярно расположенные кластерами, и ассоциированные с ними системы). Она может нацеливаться на последовательности ДНК-мишени и расщеплять их, направляясь гидовой РНК, формируя двухцепочечные разрывы ДНК (DSB).
"Белок Cas" или "полипептид Cas" относится к полипептиду, кодируемому геном Cas (CRISPR-ассоциированный). Белок Cas включает эндонуклеазу Cas. Белок Cas может представлять собой белок бактерий или архей. Например, белки типов I-III Cas CRISPR, описанные в настоящем описании, как правило, происходят из прокариот; белки Cas типа I и типа III могут происходить из видов бактерий или архей, и белок Cas типа II (т.е. Cas9) может происходить из видов бактерий. "Белки Cas" включают белок Cas9, белок Cpf1, белок C2c1, белок C2c2, белок C2c3, Cas3, Cas3-HD, Cas5, Cas7, Cas8, Cas10, Cas12a, Cas12b, или их комбинацию или комплекс.
"Вариант Cas9" или "вариант эндонуклеазы Cas9" относится к варианту родительской эндонуклеазы Cas9, где, когда он ассоциирован с cr-РНК и trac-РНК или с sg-РНК, вариант эндонуклеазы Cas9 сохраняет способность распознавать, связывать всю или частью последовательности ДНК-мишени и необязательно расплетать всю или часть последовательности ДНК-мишени, вносить одноцепочечный разрыв во всей или части последовательности ДНК-мишени или разрезать всю или часть последовательности ДНК-мишени. Варианты эндонуклеазы Cas9 включают варианты эндонуклеазы Cas9, описанные в настоящем описании, где варианты эндонуклеазы Cas9 отличаются от родительской эндонуклеазы Cas9 следующим: варианты эндонуклеазы Cas9 (когда они в комплексе с гРНК с образованием полинуклеотид-направляемого эндонуклеазного комплекса, способного модифицировать участок-мишень) имеют по меньшей мере одно улучшенное свойство, такое как, но не ограничиваясь ими, повышенная эффективность трансформации, повышенная эффективность редактирования ДНК, сниженное разрезание вне мишени или любая их комбинации, по сравнению с исходной эндонуклеазой Cas9 (в комплексе с той же гРНК с образованием полинуклеотид-направляемого эндонуклеазного комплекса, способного модифицировать тот же участок-мишень).
Варианты эндонуклеазы Cas9, описанные в настоящем описании, включают варианты, которые могут связывать и вносить одноцепочечный разрыв в участка-мишенях двухцепочечной ДНК, когда они связаны с cr-РНК и tracr-РНК или с sg-РНК, в то время как исходная эндонуклеаза Cas может связываться с участком-мишенью и приводить к двухцепочечному разрыву (расщеплению), когда она ассоциирована с cr-РНК и tracr-РНК или с sg-РНК.
"Гидовая РНК" и "гРНК" используются в настоящем описании взаимозаменяемо и относятся к последовательности гидовой РНК, используемой для нацеливания на конкретный ген для коррекции с использованием технологии CRISPR, которая обычно состоит из молекул cr-РНК и tracr-РНК, которые являются частично комплементарными, формируя комплекс, где cr-РНК содержит последовательность, которая имеет достаточную комплементарность с последовательностью-мишенью, гибридизуясь таким образом с последовательностью-мишенью и направляя комплекс CRISPR (Cas9+cr-РНК+tracr-РНК) для специфического связывания с последовательностью-мишенью. Однако в данной области известно, что может быть сконструирована единая гидовая РНК (sg-РНК), которая имеет свойства как cr-РНК, так и tracr-РНК.
Термины "единая гидовая РНК" и "sg-РНК" используются в настоящем описании взаимозаменяемо, и они относятся к синтетической слитой конструкции двух молекул РНК, которая включает слитую конструкцию cr-РНК (CRISPR-РНК) вариабельного нацеливающего домена (связанного с образующей пару последовательностью tracr, гибридизованной с tracr-РНК) и tracr-РНК (трансактивирующая РНК CRISPR). Sg-РНК может содержать cr-РНК или фрагменты cr-РНК и tracr-РНК или фрагменты tracr-РНК системы CRISPR/Cas типа II, которые могут образовывать комплекс с эндонуклеазой Cas типа II, где комплекс гидовая РНК/эндонуклеаза Cas может направлять эндонуклеазу Cas к участку ДНК-мишени, так что эндонуклеаза Cas может распознавать, необязательно связываться с участком ДНК-мишенью, и необязательно вносить одноцепочечный разрыв в участок ДНК-мишень или разрезать (вносить одноцепочечный или двухцепочечный разрыв) участок ДНК, являющийся мишенью.
В определенных вариантах осуществления гидовая(ые) РНК и Cas9 могут быть доставлены в клетку в качестве рибонуклеопротеинового (RNP) комплекса. RNP состоит из очищенного белка Cas9 в комплексе с гРНК, и в данной области хорошо известно, что RNP может быть эффективно доставлен во многие типы клеток, включая, но не ограничиваясь ими, стволовые клетки и иммунные клетки (Addgene, Cambridge, MA, Mirus Bio LLC, Madison, WI).
Прилегающий к протоспейсеру мотив (PAM) в настоящем описании относится к короткой нуклеотидной последовательности, соседней с (нацеленной на) последовательностью-мишенью (преспейсер), распознаваемой эндонуклеазной системой гРНК/Cas. Если последовательность ДНК-мишени не является соседней с соответствующей последовательностью PAM, эндонуклеаза Cas может не быть способной успешно распознавать последовательность ДНК-мишень. Последовательность и длина PAM в рамках настоящего изобретения могут различаться в зависимости от используемого белка Cas или белкового комплекса Cas. Последовательность PAM может иметь любую длину, однако, как правило, она имеет длину 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидов.
Как используют в рамках изобретения, термин "организм" включает животные, растения, грибы, бактерии и т.п.
Как используют в рамках изобретения, термин "клетка-хозяин" включает клетки растений, клетки животных, клетки грибов, бактериальные клетки и т.п.
В раках настоящего изобретения под "растением" следует понимать любой дифференцированный многоклеточный организм, способный осуществлять фотосинтез, в частности, однодольные или двудольные растения, например, (1) пищевые культуры: Oryza spp., такие как Oryza sativa, Oryza latifolia, Oryza sativa, Oryza glaberrima; Triticum spp., такие как Triticum aestivum, T. Turgidumssp. durum; Hordeum spp., такие как Hordeum vulgare, Hordeum arizonicum; Secale cereale; Avena spp., такие как Avena sativa, Avena fatua, Avena byzantine, Avena fatua var.sativa, Avena hybrida; Echinochloa spp., такие как Pennisetum glaucum, сорго, Sorghum bicolor, Sorghum vulgare, тритикале, Zea mays или кукуруза, просо, рис, просо итальянское, просо обыкновенное, сорго двухцветное, паникум, Fagopyrum spp., Panicum miliaceum, Setaria italica, Zizania palustris, Eragrostis tef, Panicum miliaceum, Eleusine coracana; (2) бобовые культуры: Glycine spp., такие как Glycine max, Soja hispida, Soja max, Vicia spp., Vigna spp., Pisum spp., конские бобы, Lupinus spp., боб обыкновенный, Tamarindus indica, Lens culinaris, Lathyrus spp., бобы гиацинтовые, кормовые бобы, бобы мунг, красные бобы, турецкий горох; (3) масличные культуры: Arachis hypogaea, Arachis spp, Sesamum spp., Helianthus spp. такие как Helianthus annuus, Elaeis, такие как Eiaeis guineensis, Elaeis oleifera, соя, Brassicanapus, Brassica oleracea, Sesamum orientale, Brassica juncea, масличный рапс, Camellia oleifera, масличная пальма, олива, клещевина, Brassica napus L., канола; (4) волокнистые культуры: Agave sisalana, Gossypium spp., такие как Gossypium, Gossypium barbadense, Gossypium hirsutum, Hibiscus cannabinus, Agave sisalana, Musa textilis Nee, Linum usitatissimum, Corchorus capsularis L, Boehmeria nivea (L.), Cannabis sativa, Cannabis sativa; (5) плодовые культуры: Ziziphus spp., Cucumis spp., Passiflora edulis, Vitis spp., Vaccinium spp., Pyrus communis, Prunus spp., Psidium spp., Punica granatum, Malus spp., Citrullus lanatus, Citrus spp., Ficus carica, Fortunella spp., Fragaria spp., Crataegus spp., Diospyros spp., Eugenia unifora, Eriobotrya japonica, Dimocarpus longan, Carica papaya, Cocos spp., Averrhoa carambola, Actinidia spp., Prunus amygdalus, Musa spp. (musa acuminate), Persea spp. (Persea Americana), Psidium guajava, Mammea Americana, Mangifera indica, Canarium album (Oleaeuropaea), Caricapapaya, Cocos nucifera, Malpighia emarginata, Manilkara zapota, Ananas comosus, Annona spp., Citrus reticulate (Citrus spp.), Artocarpus spp., Litchi chinensis, Ribes spp., Rubus spp., груша, персик, абрикос, слива, восковница красная, лимон, кумкват, дуриан, апельсин, клубника, голубика, хамийская дыня, дыня мускатная, финиковая пальма, орех грецкий, вишня; (6) клубневые культуры: Manihot spp., Ipomoea batatas, Colocasia esculenta, клубневая горчица, Allium sulfur (лук), eleocharis tuberose (каштан водяной), Cyperus rotundus, Rhizoma dioscoreae; (7) овощные культуры: Spinacia spp., Phaseolus spp., Lactuca sativa, Momordica spp, Petroselinum crispum, Capsicum spp., Solanum spp. (такие как Solanum tuberosum, Solanum integrifolium, Solanum lycopersicum), Lycopersicon spp. (такие как Lycopersicon esculentum, Lycopersicon lycopersicum, Lycopersicon pyriforme), Macrotyloma spp., кудрявая капуста, Luffa acutangula, чечевица, окра, лук, картофель, артишок, спаржа, брокколи, брюссельская капуста, кочанная капуста, морковь, цветная капуста, сельдерей, капуста листовая, тыква, Benincasa hispida, Asparagus officinalis, Apium graveolens, Amaranthus spp., Allium spp., Abelmoschus spp., Cichorium endivia, Cucurbita spp., Coriandrum sativum, B.carinata, Rapbanus sativus, Brassica spp. (такие как Brassica napus, Brassica rapa ssp., канола, масличный рапс, репа масличная, горчица сарептская, капуста кочанная, горчица черная, канола (рапс), брюссельская капуста, Solanaceae (баклажан), Capsicum annuum (перец сладкий), огурец, люфа, китайская капуста, рапс, капуста кочанная, тыква-горлянка, лук душистый, лотус, корень лотуса, салат-латук; (8) цветочные культуры: Tropaeolum minus, Tropaeolum majus, Canna indica, Opuntia spp., Tagetes spp., Cymbidium (орхидея), Crinum asiaticum L., Clivia, Hippeastrum rutilum, Rosa rugosa, Rosa Chinensis, Jasminum sambac, Tulipa gesneriana L., Cerasus sp., Pharbitis nil (L.) Choisy, Calendula officinalis L., Nelumbo sp., Bellis perennis L., Dianthus caryophyllus, Petunia hybrida, Tulipa gesneriana L., Lilium brownie, Prunus mume, Narcissus tazetta L., Jasminum nudiflorum Lindl., Primula malacoides, Daphne odora, Camellia japonica, Michelia alba, Magnolia liliiflora, Viburnum macrocephalum, Clivia miniata, Malus spectabilis, Paeonia suffruticosa, Paeonia lactiflora, Syzygium aromaticum, Rhododendron simsii, Rhododendron hybridum, Michelia figo (Lour.) Spreng., Cercis chinensis, Kerria japonica, Weigela florida, Fructus forsythiae, Jasminum mesnyi, Parochetus communis, Cyclamen persicum Mill., Phalaenophsis hybrid, Dendrobium nobile, Hyacinthus orientalis, Iris tectorum Maxim, Zantedeschia aethiоpica, Calendula officinalis, Hippeastrum rutilum, Begonia semperflorenshybr, Fuchsia hybrida, Begonia maculataRaddi, Geranium, Epipremnum aureum; (9) медицинские культуры: Carthamus tinctorius, Mentha spp., Rheum rhabarbarum, Crocus sativus, Lycium chinense, Polygonatum odoratum, Polygonatum Kingianum, Anemarrhena asphodeloides Bunge, Radix ophiopogonis, Fritillaria cirrhosa, Curcuma Aromatica, Amomum villosum Lour., Polygonum multiflorum, Rheum officinale, Glycyrrhiza uralensis Fisch, Astragalus membranaceus, Panax ginseng, Panax notoginseng, Acanthopanax gracilistylus, Angelica sinensis, Ligusticum wallichii, Bupleurum sinenses DC., Datura stramonium Linn., Datura metel L., Mentha haplocalyx, Leonurus sibiricus L., Agastache rugosus, Scutellaria baicalensis, Prunella vulgaris L., Pyrethrum carneum, Ginkgo biloba L., Cinchona ledgeriana, Hevea brasiliensis (дикая), Medicago sativa Linn, Piper Nigrum L., Radix Isatidis, Atractylodes macrocephala Koidz; (10) сырьевые культуры: Hevea brasiliensis, Ricinus communis, Vernicia fordii, Morus alba L., Hops Humulus lupulus, Betula, Alnus cremastogyne Burk., Rhus verniciflua stokes; (11) пастбищные культуры: Agropyron spp., Trifolium spp., Miscanthus sinensis, Pennisetum sp., Phalaris arundinacea, Panicum virgatum, степные травы, индийская трава, бородач, Phleum pratense, дернистые растения, cyperaceae (Kobresia pygmaea, Carex pediformis, Carex humilis), Medicago sativa Linn, Phleum pratense L., Medicago sativa, Melilotus suavcolen, Astragalus sinicus, Crotalaria juncea, Sesbania cannabina, Azolla imbircata, Eichhornia crassipes, Amorpha fruticosa, Lupinus micranthus, Trifolium, Astragalus adsurgens pall, Pistia stratiotes linn, Alternanthera philoxeroides, Lolium; (12) сахарные культуры: Saccharum spp., Beta vulgaris; (13) культуры для производства напитков: Camellia sinensis, Camellia Sinensis, чай, кофе (Coffea spp.), Theobroma cacao, Humulus lupulus Linn.; (14) газонные травы: Ammophila arenaria, Poa spp.(Poa pratensis (bluegrass)), Agrostis spp. (Agrostis matsumurae, Agrostis palustris), Lolium spp. (плевел), Festuca spp. (Festuca ovina L.), Zoysia spp. (Zoysiajaponica), Cynodon spp. (Cynodon dactylon/бермудская трава) Stenotaphrum secunda tum (Stenotaphrum secundatum), Paspalum spp., Eremochloa ophiuroides (эремохлоя змеехвостая), Axonopus spp. (мутовчатка), Bouteloua dactyloides (бизонова трава), Bouteloua var. spp. (Bouteloua gracilis), Digitaria sanguinalis, Cyperusrotundus, Kyllingabrevifolia, Cyperusamuricus, Erigeron canadensis, Hydrocotylesibthorpioides, Kummerowiastriata, Euphorbia humifusa, Viola arvensis, Carex rigescens, Carex heterostachya, дернистые растения; (15) древесные культуры: Pinus spp., Salix spp., Acer spp., Hibiscus spp., Eucalyptus spp., Ginkgo biloba, Bambusa sp., Populus spp., Prosopis spp., Quercus spp., Phoenix spp., Fagus spp., Ceiba pentandra, Cinnamomum spp., Corchorus spp., Phragmites australis, Physalis spp., Desmodium spp., Populus, Hedera helix, Populus tomentosa Carr, Viburnum odoratissinum, Ginkgo biloba L., Quercus, Ailanthus altissima, Schima superba, Ilex pur-purea, Platanus acerifolia, ligustrum lucidum, Buxus megistophylla Levl., Dahurian larch, Acacia mearnsii, Pinus massoniana, Pinus khasys, Pinus yunnanensis, Pinus finlaysoniana, Pinus tabuliformis, Pinus koraiensis, Juglans nigra, Citrus limon, Platanus acerifolia, Syzygium jambos, Davidia involucrate, Bombax malabarica L., Ceiba pentandra (L.), Bauhinia blakeana, Albizia saman, Albizzia julibrissin, Erythrina corallodendron, Erythrina indica, Magnolia gradiflora, Cycas revolute, Lagerstroemia indica, хвойные растения, макрофанерофит, Frutex; (16) ореховые культуры: Bertholletia excelsea, Castanea spp., Corylus spp., Carya spp., Juglans spp., Pistacia vera, Anacardium occidentale, макадамия (Macadamia integrifolia), Carya illinoensis Koch, макадамия, фисташка, терминалия высокая, другие растения, которые продуцируют орехи; (17) прочие: arabidopsis thaliana, Bra chiaria eruciformis, Cenchrus echinatus, Setaria faberi, eleusine indica, Cadaba farinose, algae, Carex elata, ornamental plants, Carissa macrocarpa, Cynara spp., Daucus carota, Dioscorea spp., Erianthus sp., Festuca arundinacea, Hemerocallis fulva, Lotus spp., Luzula sylvatica, Medicago sativa, Melilotus spp., Morus nigra, Nicotiana spp., Olea spp., Ornithopus spp., Pastinaca sativa, Sambucus spp., Sinapis sp., Syzygium spp., Tripsacum dactyloides, Triticosecale rimpaui, Viola odorata и т.п.
В конкретном варианте осуществления, растение выбрано из риса, кукурузы, пшена, сои, подсолнуха, сорго, рапса, люцерны, хлопка, ячменя, проса, сахарного тростника, томата, табака, маниока, картофеля, сладкого картофеля, китайской капусты, капусты кочанной, огурца, китайской розы, Scindapsus aureus, арбуза, дыни, клубники, голубики, винограда, яблока, цитруса, персика, груши, банана и т.д.
Как используют в рамках изобретения, термин "растение" включает целое растение и любого потомка, клетку, ткань или часть растения. Термин "часть растения" включает любую часть растения, включая, например, но не ограничиваясь ими: семя (включая зрелое семя, незрелый зародыш без семенной оболочки и незрелое семя); побег растения; клетку растения; культуру клеток растения; орган растения (например, пыльца, зародыш, цветок, плод, почка, лист, корень, стебель и соответствующие эксплантаты). Ткань растения или орган растения могут представлять собой семя, ткань каллюса или любую другую популяцию клеток растения, организованную в структурную или функциональную единицу. Некоторые клетки растений или культуры тканей могут регенерировать в растение, которое имеет физиологические и морфологические характеристики растения, из которого происходит клетка или ткань, и могут регенерировать в растение, которое имеет по существу тот же генотип, что и указанное растение. Напротив, некоторые клетки растений не могут регенерировать в растения. Регенерирующие клетки в культурах клеток или тканей растений могут представлять собой клетки зародышей, протопластов, клетки меристемы, каллюса, пыльцы, листьев, пыльников, корней, верхушек корней, пестичных столбиков, цветков, зерен, початков, стержней початков, шелухи или стеблей.
Части растений включают пожинаемые части и части, которые могут использоваться для получения растений-потомков. Части растений, которые могут быть использованы для разведения, включают, например, но не ограничиваясь ими: семена; плоды; побеги; проростки; клубни и корневища. Пожинаемые части растений могут представлять собой любые подходящие части растений, включая, например, но не ограничиваясь ими: цветки; пыльцу; проростки; клубни; листья; стебли; плоды; семена и корни.
Клетки растения представляют собой структурные и физиологические элементы растений. Как используют в рамках изобретения, клетки растения включают протопласты и протопласты с частичными клеточными стенками. Клетки растений могут иметь форму выделенных единичных клеток или клеточных агрегатов (например, рыхлый каллюс и культивируемые клетки), и могут быть частью тканевых элементов более высокого порядка (например, ткани растений, органы растений и интактные растения). Таким образом, клетки растений могут представлять собой протопласты, гаметообразующие клетки, или клетки или совокупность клеток, способные регенерировать целое растение. Таким образом, в вариантах осуществления, описанных в настоящем описании, семя, содержащее множество клеток растения и способное регенерировать в целое растение, считается "частью растений".
Как используют в рамках изобретения, термин "протопласт" относится к клетке растения, чья клеточная стенка полностью или частично удалена и чья липидная бислойная мембрана обнажена. Как правило, протопласт представляет собой выделенную клетку растения без клеточной стенки, которая имеет потенциал к регенерации клеточной культуры или целого растения.
"Потомство" растения включает любые последующие поколения растения.
Термины "толерантность к ингибиторному гербициду" и "резистентность к ингибиторному гербициду" могут использоваться взаимозаменяемо и оба относятся к толерантности и резистентности к ингибиторному гербициду. "Повышение толерантности к ингибиторному гербициду" и "повышение резистентности к ингибиторному гербициду" означает, что толерантность или резистентность к ингибиторному гербициду повышается по сравнению с растением, содержащим ген дикого типа.
Термин "дикий тип" относится к молекуле нуклеиновой кислоты или белка, которые могут быть найдены в природе.
В рамках настоящего изобретения термин "участок для возделывания" включает участок, где растение по настоящему изобретению выращивают, такой как почва, и также он включает, например, семена растений, проростки растений и выросшие растения. Термин "эффективное количество для борьбы с сорняками" относится к количеству гербицида, которое является достаточным для влияния на рост или развитие сорняка, являющегося мишенью, например, для предупреждения или ингибирования роста или развития сорняка, являющегося мишенью, или для уничтожения сорняка. Преимущественно, эффективное количество для борьбы с сорняками не оказывает значительного влияния на рост и/или развитие семян растения, проростков растения или растений по настоящему изобретению. Специалисты в данной области могут определить эффективное количество для борьбы с сорняками посредством стандартных экспериментов.
Термин "ген" включает фрагмент нуклеиновой кислоты, экспрессирующий функциональную молекулу (такую как, но не ограничиваясь ими, конкретный белок), включая регуляторные последовательности до (5'-некодирующие последовательности) и после (3'-некодирующие последовательности) кодирующей последовательности.
Последовательность ДНК, которая "кодирует" конкретную РНК, представляет собой последовательность нуклеиновой кислоты ДНК, которая может транскрибироваться в РНК. Полинуклеотиды ДНК могут кодировать РНК (мРНК), которая транслируется в белок, или полинуклеотиды ДНК могут кодировать РНК, которая не может транслироваться в белок (например, тРНК, рРНК или ДНК-нацеливающая РНК; которые также известны как "некодирующая" РНК или "нкРНК").
Термины "полипептид", "пептид" и "белок" используют в рамках настоящего изобретения взаимозаменяемо, и они относятся к полимеру аминокислотных остатков. Термины применяются к полимерам аминокислот, в которых одна или более аминокислотных остатков представляют собой искусственные химические аналоги соответствующих и встречающихся в природе аминокислот, а также к встречающимся в природе полимерам аминокислот. Термины "полипептид", "пептид", "аминокислотная последовательность" и "белок" также могут включать их модифицированные формы, включая, но не ограничиваясь ими, гликозилирование, присоединение липида, сульфатацию, γ-карбоксилирование остатка глутаминовой кислоты, гидроксилирование и ADP-рибозилирование.
Термин "биологически активный фрагмент" относится к фрагменту, в котором один или более аминокислотных остатков делетированы с N и/или C-конца белка и одновременно сохранена его функциональная активность.
Термины "полинуклеотид" и "нуклеиновая кислота" используются взаимозаменяемо и включают ДНК, РНК или их гибриды, которые могут быть двухцепочечными или одноцепочечными.
Оба из терминов "нуклеотидная последовательность" и "последовательность нуклеиновой кислоты" относятся к последовательности оснований в ДНК или РНК.
Специалисты в данной области могут без труда использовать известные способы, такие как способы направленной эволюции и способы внесения точковой мутации для внесения мутаций во фрагменты ДНК, как показано в SEQ ID No: 9-SEQ ID No: 17 по настоящему изобретению. Искусственно модифицированные нуклеотидные последовательности, которые обладают по меньшей мере 75% идентичностью с любой из вышеуказанных последовательностей по настоящему изобретению и проявляют ту же функцию, считаются производными нуклеотидной последовательности по настоящему изобретению и эквивалентны последовательностям по настоящему изобретению.
Термин "идентичность" относится к сходству последовательностей с природной последовательностью нуклеиновой кислоты. Идентичность последовательностей можно оценивать путем наблюдения или с помощью компьютерного программного обеспечения. С использованием компьютерного программного обеспечения для выравнивания последовательностей, идентичность между двумя или более последовательностями может быть выражена в качестве процента (%), который может использоваться для оценки идентичности между родственными последовательностями. "Частичная последовательность" означает по меньшей мере 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 95% от данной последовательности.
Жесткие условия могут быть следующими: гибридизация при 50°С в смешанном растворе 7% додецилсульфата натрия (SDS), 0,5 M NaPO4 и 1 мМ EDTA, и промывание при 50°С в 2× SSC и 0,1% SDS; или альтернативно: гибридизация при 50°С в смешанном растворе 7% SDS, 0,5 M NaPO4 и 1 мМ EDTA, и промывание при 50°С в 1× SSC и 0,1% SDS; или альтернативно: гибридизация при 50°С в смешанном растворе 7% SDS, 0,5 M NaPO4 и 1 мМ EDTA, и промывание при 50°С в 0,5× SSC и 0,1% SDS; или альтернативно: гибридизация при 50°С в смешанном растворе 7% SDS, 0,5 M NaPO4 и 1 мМ EDTA, и промывание при 50°С в 0,1× SSC и 0,1% SDS; или альтернативно: гибридизация при 50°С в смешанном растворе 7% SDS, 0,5 M NaPO4 и 1 мМ EDTA, и промывание при 65°С в 0,1× SSC и 0,1% SDS; или альтернативно: гибридизация при 65°С в растворе 6× SSC, 0,5% SDS, а затем промывание мембраны 2× SSC, 0,1% SDS и 1× SSC, 0,1% SDS в каждом случае однократно; или альтернативно: гибридизация и промывание мембраны два раза в растворе 2× SSC, 0,1% SDS при 68°С, по 5 мин каждый раз, а затем гибридизация и промывание мембраны два раза в растворе 0,5× SSC, 0,1% SDS при 68°С, 15 мин каждый раз; или альтернативно: гибридизация и промывание мембраны в растворе 0,1× SSPE (или 0,1× SSC), 0,1% SDS при 65°С.
Как используют в рамках настоящего изобретения, "экспрессирующая кассета", "экспрессирующий вектор" и "экспрессирущая конструкция" относятся к вектору, таком как рекомбинантный вектор, пригодный для экспрессии представляющей интерес нуклеотидной последовательности в растении. Термин "экспрессия" относится к продуцированию функционального продукта. Например, экспрессия нуклеотидной последовательности может относиться к транскрипции нуклеотидной последовательности (такой как транскрипция для образования мРНК или функциональной РНК) и/или трансляции РНК в белок-предшественник или зрелый белок.
"Экспрессирующая конструкция" по настоящему изобретению может представлять собой линейный фрагмент нуклеиновой кислоты, кольцевую плазмиду, вирусный вектор, или в некоторых вариантах осуществления она может представлять собой РНК (такую как мРНК), которая может транслироваться.
"Экспрессирующая конструкция" по настоящему изобретению может содержать представляющие интерес регуляторные последовательности и нуклеотидные последовательности из различных источников, или представляющие интерес регуляторные последовательности и нуклеотидные последовательности из одного источника, но расположенные отличающимся образом от обычно встречающихся в природе.
"Ген с высокой экспрессией" в рамках настоящего изобретения относится к гену, уровень экспрессии которого превышает уровень экспрессии обычного гена в конкретной ткани.
Термины "рекомбинантный экспрессирующий вектор" или "конструкция ДНК" используются в настоящем описании взаимозаменяемо и относятся к молекуле ДНК, содержащей вектор и по меньшей мере одну вставку. Рекомбинантные экспрессирующие векторы обычно получают для экспрессии и/или увеличения в количестве вставки или для конструирования других рекомбинантных нуклеотидных последовательностей. Вставка может быть функционально или может быть нефункционально связана с промоторной последовательностью и может быть функционально или может быть нефункционально связана с регуляторной последовательностью ДНК.
Термины "регуляторная последовательность" и "регуляторный элемент" могут использоваться взаимозаменяемо, и они относятся к нуклеотидной последовательности, которая находится выше (5'-некодирующая последовательность), в середине или ниже (3'-некодирующая последовательность) кодирующей последовательности и влияет на транскрипцию, процессинг РНК, стабильность или трансляцию соответствующей кодирующей последовательности. Регуляторные элементы для экспрессии в растениях относятся к нуклеотидным последовательностям, которые могут контролировать транскрипцию, процессинг РНК, или стабильность или трансляцию представляющей интерес нуклеотидной последовательности в растениях.
Регуляторные последовательности могут включать, но не ограничиваться ими, промоторы, лидерные последовательности трансляции, интроны и последовательности распознавания поли-A.
Термин "промотор" относится к фрагменту нуклеиновой кислоты, способному контролировать транскрипцию другого фрагмента нуклеиновой кислоты. В некоторых вариантах осуществления настоящего изобретения промотор представляет собой промотор, способный контролировать транскрипцию гена в клетках растения, независимо от того, происходит ли он из клеток растения. Промотор может представлять собой конститутивный промотор или тканеспецифический промотор, или регулируемый стадией развития промотор или индуцибельный промотор.
Термин "сильный промотор" представляет собой хорошо известный и широко используемый в данной области термин. Многие сильные промоторы известны в данной области или могут быть идентифицированы посредством стандартных экспериментов. Активность сильного промотора превышает активность промотора, функционально связанного с молекулой нуклеиновой кислоты, подлежащей сверхэкспрессии в организме дикого типа, например, промотор обладает активностью, превышающей активность промотора эндогенного гена. Предпочтительно, активность сильного промотора является более высокой приблизительно на 2%, 5%, 8%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 150%, 200%, 250%, 300%, 350%, 400%, 450%, 500%, 600%, 700%, 800%, 900%, 1000% или более 1000%, чем активность промотора, функционально связанного с молекулой нуклеиновой кислоты, подлежащей сверхэкспрессии в организме дикого типа. Специалистам в данной области известно, как измерить активность промотора и сравнить активность различных промоторов.
Термин "конститутивный промотор" относится к промотору, который обычно вызывает экспрессию гена в большинстве типов клеток в большинстве случаев. "Тканеспецифический промотор" и "предпочтительный для ткани промотор" используют взаимозаменяемо, и они относятся к промотору, который в основном, но не обязательно исключительно, экспрессируется в ткани или органе, а также экспрессируется в конкретной клетке или типе клеток. "Регулируемый стадией развития промотор" относится к промотору, активность которого определяется обусловленным развитием событием. "Индуцибельный промотор" отвечает на эндогенный или экзогенный стимул (внешняя среда, гормон, химический сигнал и т.д.), селективно экспрессируя функционально связанную последовательность ДНК.
Как используют в рамках изобретения, термин "функционально связанный" относится к соединению регуляторного элемента (например, но не ограничиваясь ими, промоторная последовательность, последовательность терминации транскрипции и т.д.) с последовательностью нуклеиновой кислоты (например, кодирующая последовательность или открытая рамка считывания), так что транскрипция нуклеотидной последовательности контролируется и регулируется элементом регуляции транскрипции. Способы функционального связывания области регуляторного элемента с молекулой нуклеиновой кислоты известны в данной области.
"Введение" молекулы нуклеиновой кислоты (такой как плазмида, линейный фрагмент нуклеиновой кислоты, РНК и т.д.) или белка в растение относится к трансформации клетки растения нуклеиновой кислотой или белком, так чтобы нуклеиновая кислота или белок могли функционировать в клетке растения. Термин "трансформация", используемый в рамках настоящего изобретения, включает стабильную трансформацию и временную трансформацию.
Термин "стабильная трансформация" относится к тому, что введение экзогенной нуклеотидной последовательности в геном растения приводит к стабильному наследованию экзогенного гена. После стабильной трансформации экзогенная последовательность нуклеиновой кислоты стабильно встраивается в геном растения и любые его последующие поколения.
Термин "временная трансформация" относится к тому, что введение молекулы нуклеиновой кислоты или белка в клетку растения для выполнения функции не приводит к стабильному наследованию чужеродного гена. При временной трансформации экзогенная последовательность нуклеиновой кислоты не встраивается в геном растения.
Изменение экспрессии эндогенных генов в организмах включает два аспекта: интенсивность и пространственно-временные характеристики. Изменение интенсивности включает повышение (Knock-up), снижение (нокдаун) и/или выключение экспрессии гена (нокаут); пространственно-временная специфичность включает временную (рост и стадия развития) специфичность и пространственную (ткань) специфичность, а также способность к индукции. Кроме того, оно включает изменение нацеливания белка, например, изменение признака цитоплазматической локализации белка в признак локализации в хлоропластах или ядерной локализации.
Если не определено иначе, все технические и научные термины, используемые в настоящем описании, имеют те же значения, которые обычно понятны специалистам в области, к которой относится настоящее изобретение. Хотя любые способы и материалы, сходные или эквивалентные способам и материалам, описанным в настоящем описании, также можно использовать для применения на практике или тестирования настоящего изобретения, предпочтительные способы и материалы описаны в настоящем описании.
Все публикации и патенты, цитированные в настоящем описании, включены в настоящее описание в качестве ссылок, как если бы каждая индивидуальная публикация или патент были прямо и индивидуально указаны как включенные в качестве ссылки, и включены в настоящее описание в качестве ссылок для раскрытия и описания способов и/или материалов, связанных с цитированными публикациями. Цитирование любой публикации, которая была опубликована до даты подачи, не должно быть интерпретировано как допущение того, что настоящее изобретение не может быть способным превосходить публикацию существующего изобретения. Кроме того, приведенная дата публикации может отличаться от фактической даты публикации, что может потребовать независимой проверки.
Если конкретно не указано или не подразумевается иное, как используют в рамках изобретения, форма единственного числа означает "по меньшей мере один". Все патенты, патентные заявки и публикации, упоминаемые или цитируемые в настоящем описании, включены в настоящее описание в качестве ссылок в полном объеме с той же степенью цитирования, как если бы они были цитированы индивидуально.
Настоящее изобретение имеет следующие преимущественные технические эффекты:
настоящее изобретение всесторонне использует информацию следующих двух различных профессиональных областей для разработки способа для прямого создания новых генов в организмах, полностью изменяя общепринятое применение исходных инструментов для редактирования генов (т.е. нокаута генов), осуществляя их новое применение для создания новых генов, в частности, реализуя способ редактирования для повышения (knock-up) уровня экспрессии эндогенных генов с использованием технологии редактирования генов для повышения экспрессии генов-мишеней. Первой является информация в области редактирования генов, т.е., когда два или более различных участка-мишени и Cas9 одновременно нацелены на геном организма, могут происходить различные ситуации, такие как делеция, инверсия, удвоение или инверсия-удвоение. Второй является информация в области геномики, т.е. информация о положении и расстоянии различных генов в геноме, и конкретных положениях, направлениях и функциях различных элементов (промотор, 5'UTR, кодирующая область (CDS), различные доменные области, терминатор и т.д.) в генах, и специфичности экспрессии различных генов и т.д. Путем объединения информации в этих двух различных областях вносят разрывы ДНК в конкретные участки двух или более различных генов или в два или более конкретных участка в одном гене (конкретные участки могут быть определены в области геномики), новая комбинация различных элементов генов или функциональных доменов может быть образована путем делеции, инверсии, удвоения и инверсии-удвоения или обмена плечами хромосом, и т.д. (конкретные ситуации могут предусматриваться в области редактирования генов), тем самым специфически создавая новый ген в организме.
Новые гены, созданные посредством настоящего изобретения, образуются путем слияния или рекомбинации различных элементов двух или более генов под действием механизмов спонтанной репарации ДНК в организме, изменяя интенсивность экспрессии, пространственно-временную специфичность, специальные функциональные домены и т.п. исходного гена без экзогенного трансгена или синтетических элементов генов. Поскольку новый ген имеет слияние двух или более различных элементов генов, это значительно расширяет объем мутации генов и обеспечивает более обширные и разнообразные функции, таким образом, он имеет широкий диапазон перспектив применения. В то же время, эти новые гены не связаны с векторами для редактирования генов, так что элементы векторов могут быть удалены посредством генетической сегрегации, и, тем самым, приводить к нетрансгенным биологическим материалам, содержащим новые гены для разведения животных и растений. Альтернативно можно проводить временное редактирование без встраивания путем доставки мРНК или комплекса рибонуклеиновая кислота-белок (RNP) для создания не генно-модифицированных биологических материалов, содержащих новые гены. Этот процесс не является трансгенным и полученные отредактированные материалы также не содержат трансгена. В теории и фактически, эти новые гены также могут быть получены путем традиционных способов селекции (таких как радиационное облучение или химический мутагенез). Отличие состоит в том, что скрининг с использованием традиционных способов требует создания библиотек, содержащих большое количество случайных мутантов и, таким образом, скрининг новых функциональных генов является времязатратным и дорогостоящим. В то время как в рамках настоящего изобретения новые функциональные гены могут быть созданы посредством биоинформатического анализа в комбинации с технологией редактирования генов, длительность выведения может быть значительно укорочена. Способ по настоящему изобретению не подчиняется действующим нормам в отношении организмов с отредактированными генами в многих странах.
Кроме того, новая технология создания генов по настоящему изобретению может использоваться для изменения многих признаков в организмах, включая рост, развитие, резистентность, урожай и т.д., и имеет большую применимость. Созданные новые гены могут иметь новые регуляторные элементы (такие как промоторы), которые могут изменять интенсивность экспрессии и/или пространственно-временные характеристики исходных генов или могут иметь новые аминокислотные последовательности, и, таким образом, иметь новые функции. Если взять культуры в качестве примера, изменение экспрессии конкретных генов может повышать резистентность культур к вредоносным организмам, таким как паразиты и сорняки, и абиотическим стрессам, таким как засуха, заболачивание и соленость, и также может повышать урожай и улучшать качество. Если взять рыбу в качестве примера, изменение характеристик экспрессии гормона роста у рыб может значительно изменять их скорость роста и развития.
Краткое описание чертежей
На фиг.1 представлена схематическая диаграмма создания нового гена HPPD в рисе.
На фиг.2 представлена схематическая диаграмма создания нового гена EPSPS в рисе.
На фиг.3 представлена схематическая диаграмма создания нового гена PPOX в Arabidopsis thaliana.
На фиг.4 представлена схематическая диаграмма создания нового гена PPOX в рисе.
На фиг.5 представлены результаты секвенирования для схемы дупликации HPPD, протестированной с использованием протопласта риса.
На фиг.6 представлена карта вектора для трансформации риса посредством pQY2091 с использованием Agrobacterium.
На фиг.7 представлены результаты электрофореза продуктов ПЦР для детекции новых фрагментов генов в трансформированном посредством pQY2091 резистентном к гигромицину каллюсе риса. Стрелкой указана полоса ПЦР, соответствующая новому гену, созданному путем слияния промотора гена UBI2 с кодирующей областью HPPD. Числа представляют собой номера различных образцов каллюса. M обозначает ДНК-маркер и размеры полос представляют собой 100 п.н., 250 п.н., 500 п.н., 750 п.н., 1000 п.н., 2000 п.н., 2500 п.н., 5000 п.н., 7500 п.н. в указанном порядке.
На фиг.8 представлены результаты электрофореза продуктов ПЦР для детекции новых фрагментов генов в трансформированных посредством pQY2091 проростках риса T0. Стрелкой указана полоса ПЦР нового гена, созданного путем слияния промотора гена UBI2 с кодирующей областью HPPD. Числа представляют собой серийные номера различных проростков T0. M обозначает ДНК-маркер, и размеры полос последовательно представляют собой 100 п.н., 250 п.н., 500 п.н., 750 п.н., 1000 п.н., 2000 п.н., 2500 п.н., 5000 п.н., 7500 п.н.
На фиг.9 представлены результаты теста резистентности к Shuangzuocaotong поколения T0 QY2091 линии с удвоением HPPD. В одном и том же цветочном горшке Jinjing 818 дикого типа находится слева и линия с удвоением HPPD находится справа.
На фиг.10 представлены относительные уровни экспрессии генов HPPD и UBI2 в поколении T0 линии QY2091 с удвоением гена HPPD. 818CK1 и 818CK3 обозначают два контрольных растения Jinjing 818 дикого типа; 13M и 20M обозначают образцы первичных листьев побегов растений T0 QY2091-13 и QY2091-20; 13L и 20L обозначают образцы вторичных листьев побегов растений T0 QY2091-13 и QY2091-20, используемых в тесте резистентности к гербициду.
На фиг.11 представлена схематическая диаграмма возможных генотипов поколения T1 QY2091 и участки связывания праймеров для молекулярной детекции.
На фиг.12 представлено сравнение результатов секвенирования, выявляющего удвоение HPPD, и спрогнозированные удвоенные последовательности для QY2091-13 и QY2091-20.
На фиг.13 представлены результаты теста резистентности к гербицидам для поколения T1 линии QY2091 с удвоением HPPD на стадии проростка.
На фиг.14 представлена схематическая диаграмма типов возможного события редактирования, представляющего собой инверсию фрагмента хромосомы с геном PPO1 риса, и участки связывания праймеров для молекулярной детекции.
На фиг.15 представлены результаты секвенирования для детекции инверсии EPSPS.
На фиг.16 представлена карта вектора Agrobacterium pQY2234 для трансформации риса.
На фиг.17 представлены результаты электрофореза продуктов ПЦР для детекции новых фрагментов генов резистентного к гигромицину каллюса риса, трансформированного pQY2234. Стрелкой указана полоса ПЦР нового гена, созданного путем слияния промотора гена CP12 с кодирующей областью PPO1. Числа соответствуют серийным номерам различных образцов каллюса. M обозначает ДНК-маркер, и размеры полос последовательно представляют собой 100 п.н., 250 п.н., 500 п.н., 750 п.н., 1000 п.н., 2000 п.н., 2500 п.н., 5000 п.н., 7500 п.н.
На фиг.18 представлены результаты теста резистентности к 2081 линии QY2234 с инверсией гена PPO1 поколения T0. При одной и той же дозе обработки цветочный горшок слева представляет собой контроль Huaidao No.5 дикого типа, и горшок справа представляет собой линию с инверсией PPO1.
На фиг.19 представлены относительные уровни экспрессии генов PPO1 и CP12 в поколении T0 линии QY2234 с инверсией PPO1. H5CK1 и H5CK2 соответствуют двумя контрольным растениям Huaidao No.5 дикого типа; 252M, 304M и 329M соответствуют образцам первичных листьев побегов растений T0 QY2234-252, QY2234-304 и QY2234-329; 252L, 304L и 329L соответствуют образцам вторичных листьев побегов.
На фиг.20 представлено сравнение результата секвенирования инверсии PPO1 со спрогнозированной последовательностью инверсии на фоне Huaidao 5.
На фиг.21 представлено сравнение результата секвенирования инверсии PPO1 со спрогнозированной последовательностью инверсии на фоне Jinjing 818.
На фиг.22 представлены результаты теста резистентности к гербицидам для поколения T1 линии QY2234 с инверсией PPO1 на стадии проростка.
Конкретные способы осуществления изобретения
Настоящее изобретение далее описано с помощью следующих примеров. Приведенное ниже описание является только иллюстративным, и объем защиты настоящего изобретения не ограничивается им.
Пример 1: Способ редактирования для повышения (knock-up) экспрессии эндогенного гена HPPD путем индукции удвоения хромосомного фрагмента в растении - тест на протопластах риса
HPPD является ключевым ферментом каскада синтеза хлорофилла в растениях, и ингибирование активности HPPD может в конечном итоге приводить к белому хлорозу и гибели растений. Многие гербициды, такие как мезотрион и топрамезон, представляют собой ингибиторы с HPPD в качестве белка-мишени, и, таким образом, повышение уровня экспрессии эндогенного гена HPPD в растениях могло повысить толерантность растений к этим гербицидам. Ген HPPD риса (как показано в SEQ ID NO: 6, где 1-1067 п.н. представляют собой промотор, а остальные представляют собой экспрессируемую область) находится на хромосоме 2 риса. Посредством биоинформатического анализа было обнаружено, что ген убиквитина 2 риса (далее обозначаемый как UBI2) (как показано на SEQ ID NO: 5, где 1-2107 п.н. представляют собой промотор, а остальные представляют собой область экспрессии) находится приблизительно на 338 п.н. ниже гена HPPD, и ген UBI2 и ген HPPD имеют одно направление на хромосоме. В соответствии с данными о профиле экспрессии генов риса, приведенными International Rice Genome Sequencing Project (http://rice.plantbiology.msu.edu/index.shtml), интенсивность экспрессии гена UBI2 в листьях риса была в 3-10 раз более высокой, чем у гена HPPD, и промотор гена UBI2 представлял собой сильный промотор с конститутивной экспрессией.
Как показано на фиг.1, на схеме 1 показано, что двухцепочечные разрывы одновременно вносились в участках между промоторами и областью CDS генов HPPD и UBI2, соответственно, после скрининга и идентификации было выявлено событие удвоения области между двумя разрывами, и мог сформироваться новый ген путем слияния промотора UBI2 и кодирующей области HPPD вместе. Кроме того, в соответствии со схемой 2, как показано на фиг.1, новый ген, в котором были слиты промотор UBI2 и кодирующая область HPPD, также мог образовываться посредством двух последовательных инверсий. Сначала, схемы, как показано на фиг.1, тестировали в системе протопластов риса следующим образом:
1. Сначала последовательности геномной ДНК генов HPPD и UBI2 риса вводили в онлайн-инструмент CRISPOR (http://crispor.tefor.net/) для поиска доступных участков-мишеней для редактирования. После онлайн-оценки были отобраны следующие участки-мишени между промоторами и областями CDS генов HPPD и UBI2 для тестирования:
Гидовая РНК1 и гидовая РНК2 находились между промотором и областью CDS гена HPPD, вблизи инициирующего кодона белка HPPD, и гидовая РНК3 и гидовая РНК4 находились между промотором и областью CDS гена UBI2 вблизи кодона инициации белка UBI2.
Вектор pHUE411 (https://www.addgene.org/62203/) использовали в качестве остова, и следующие праймеры конструировали для вышеупомянутых участков-мишеней для проведения конструирования вектора, как описано в "Xing HL, Dong L, Wang ZP, Zhang HY, Han CY, Liu B, Wang XC, Chen QJ. A CRISPR/Cas9 Toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014 Nov 29; 14(1): 327".
Векторы для редактирования генов для следующей комбинации двойных мишеней конструировали в соответствии со способом, приведенном в вышеупомянутой литературе. В частности, в случае плазмиды pCBC-MT1T2 (https://www.addgene.org/50593/) в качестве матрицы, фрагменты двойных мишеней для sg-РНК1+3, sg-РНК1+4, sg-РНК2+3 и sg-РНК2+4 амплифицировали, соответственно, для конструирования экспрессирующих кассет sg-РНК. Остов вектора pHUE411 расщепляли BsaI и выделяли из геля, и фрагмент-мишень расщепляли и прямо использовали для реакции лигирования. ДНК-лигазу T4 использовали для лигирования остова вектора и фрагмента-мишени, и продуктом лигирования трансформировали компетентные клетки Trans5α. Различные моноклоны отбирали и секвенировали. Для экстракции плазмид из клонов с правильными последовательностями использовали Sparkjade High Purity Plasmid Mini Extraction Kit, тем самым получая рекомбинантные плазмиды, соответственно, названные pQY002065, pQY002066, pQY002067 и pQY002068, следующим образом:
pQY002065 pHUE411-HPPD-sgRNA1+3, комбинация OsHPPD-гидовой РНК1, гидовой RNA3
pQY002066 pHUE411-HPPD-sgRNA1+4, комбинация OsHPPD-гидовой РНК1, гидовой РНК4
pQY002067 pHUE411-HPPD-sgRNA2+3, комбинация OsHPPD-гидовой РНК2, гидовой РНК3
pQY002068 pHUE411-HPPD-sgRNA2+4, комбинация OsHPPD-гидовой РНК2, гидовой РНК4
2. Получали плазмиды с высокой чистотой и высокой концентрацией для вышеупомянутых векторов pQY002065-002068 следующим образом:
Плазмиды экстрагировали с использованием набора Promega Medium Plasmid Extraction Kit (Midipreps DNA Purification System, Promega, A7640) в соответствии с инструкциями. Конкретные стадии представляли собой:
(1) Добавление 5 мл Escherichia coli к 300 мл жидкой среде LB, содержащей канамицин, и качание при 200 об/мин, 37°C в течение от 12 до 16 часов;
(2) Помещение указанного выше раствора бактерий в 500-мл центрифужную пробирку и центрифугирование при 5000g в течение 10 минут, удаление супернатанта;
(3) добавление 3 мл раствора для ресуспендирования клеток (CRS), чтобы ресуспендировать клеточный осадок, и встряхивание для тщательного перемешивания;
(4) добавление 3 мл раствора для лизиса клеток (CLS) и перемешивание вверх и вниз медленно в течение не более 5 минут;
(5) добавление 3 мл раствора для нейтрализации и тщательное перемешивание путем переворачивания до достижения светлого цвета и прозрачности;
(6) центрифугирование при 14000g в течение 15 минут и дальнейшее центрифугирование в течение 15 минут, если не образовался плотный осадок;
(7) перенос супернатанта в новую 50-мл центрифужную пробирку, избегая засасывания белого осадка в центрифужную пробирку;
(8) добавление 10 мл смолы для очистки ДНК (смола для очистки, перед применением энергично встряхнуть) и тщательное перемешивание;
(9) переливание смеси смола/ДНК в колонку для фильтрования и обработка посредством способа отрицательного давления на вакуумном насосе (0,05 МПа);
(10) добавление 15 мл раствора для промывания колонки (CWS) с колонку для фильтрации, и применение вакуума.
(11) добавление 15 мл CWS и повторное применение вакуума один раз; вакуум продолжают в течение 30 с после пропускания раствора через колонку для фильтрации;
(12) отрезание колонки для фильтрации, перенос в 1,5-мл центрифужную пробирку, центрифугирование при 12000g в течение 2 минут, удаление остаточной жидкости и перенос колонки для фильтрации в новую 1,5-мл центрифужную пробирку;
(13) добавление 200 мкл стерилизованной воды, предварительно нагретой до 70°C и выдерживание в покое в течение 2 минут;
(14) центрифугирование при 12000g в течение 2 минут для элюирования плазмидной ДНК; и концентрация составляла, главным образом, приблизительно 1 мкг/мкл.
3. Получение протопластов риса и проведение ПЭГ-опосредуемой трансформации:
Сначала получали проростки риса для протопластов сорта Nipponbare. Семена были предоставлены Weeds Department of the School of Plant Protection, China Agricultural University, и были получены в теплице. Семена риса сначала очищали от шелухи, и очищенные от шелухи семена ополаскивали 75% этанолом в течение 1 минуты, обрабатывали 5% (об./об.) гипохлоритом натрия в течение 20 минут, затем промывали стерильной водой более 5 раз. После сушки воздухом на сверхчистом столе, их помещали в бутылку для культивирования тканей, содержавшую 1/2 среды MS, по 20 семян на каждую бутылку. Протопласты получали путем инкубации при 26°C в течение приблизительно 10 суток с 12 часами на свету.
Способы получения протопластов риса и ПЭГ-опосредуемой трансформации проводили в соответствии с "Lin et al., 2018 Application of protoplast technology to CRISPR/Cas9 mutagenesis: from single-cell mutation detection to mutant plant regeneration. Plant Biotechnology Journal https://doi.org /10.1111/pbi.12870". Стадии были следующими:
(1) влагалище листа проростков собирали, нарезали на фрагменты размером приблизительно 1 мм с использованием острого лезвия бритвы Geely и помещали в 0,6 M маннит и культуральную среду MES (состав: 0,6 M маннит, 0,4 M MES, pH 5,7) для последующего применения. Все материалы разрезали и переносили в 20 мл раствора для ферментного гидролиза (состав: 1,5% целлюлаза R10/RS (YaKult Honsha), 0,5% Mecerozyme R10 (YaKult Honsha), 0,5 M маннит, 20 мМ KCl, 20 мМ MES, pH 5,7, 10 мМ CaCl2, 0,1% BSA, 5 мМ β-меркаптоэтанол), заворачивали в оловянную фольгу и помещали в устройство для качания при 28°C, подвергали ферментативному гидролизу при 50 об/мин в темноте в течение приблизительно 4 часов и скорость увеличивали до 100 об/мин за последние 2 минуты;
(2) после ферментного лизиса добавляли равный объем раствора W5 (состав: 154 мМ NaCl, 125 мМ CaCl2, 5 мМ KCl, 15 мМ MES), качали горизонтально в течение 10 секунд для высвобождения протопластов. Клетки после ферментного лизиса фильтровали через сито калибра 300 и центрифугировали при 150 g в течение 5 минут для сбора протопластов;
(3) клетки ополаскивали два раза раствором W5 и протопласты собирали центрифугированием при 150 g в течение 5 минут;
(4) протопласты ресуспендировали с соответствующим количеством раствора MMG (состав: 3,05 г/л MgCl2, 1 г/л MES, 91,2 г/л маннита), и концентрация протопластов составляла приблизительно 2×106 клеток/мл.
Трансформацию протопластов проводили следующим образом:
(1) к 200 мкл вышеупомянутых ресуспендированных в MMG протопластов добавляли свободную от эндотоксинов плазмидную ДНК высокого качества (10-20 мкг) и постукивали для тщательного перемешивания;
(2) добавляли равный объем 40% (масс./об.) раствора ПЭГ (состав: 40% (масс./об.) PEG, 0,5 M маннит, 100 мМ CaCl2), постукивали для тщательного перемешивания и оставляли в покое при 28°C в темноте в течение 15 минут;
(3) после индукции трансформации медленно добавляли 1,5 мл раствора W5, постукивали для тщательного перемешивания клеток. Клетки собирали центрифугированием при 150g в течение 3 минут. Эту стадию повторяли один раз;
(4) добавляли 1,5 мл раствора W5 для ресуспендирования клеток и помещали в инкубатор при 28°C и культивировали в темноте в течение 12-16 часов. Для экстракции геномной ДНК протопластов культивирование необходимо проводить в течение 48-60 часов.
4. Нацеливание на геном и детекция нового гена:
(1) Сначала ДНК протопластов экстрагировали способом CTAB с некоторыми модификациями. Конкретный способ был следующим: протопласты центрифугировали, затем супернатант удаляли. Добавляли 500 мкл раствора для экстракции ДНК (состав: CTAB 20 г/л, NaCl 81,82 г/л, 100 мМ Tris-HCl (pH 8,0), 20 мМ EDTA, 0,2% β-меркаптоэтанол), встряхивали для тщательного перемешивания и инкубировали на водяной бане при 65°C в течение 1 часа; когда инкубированный образец остывал, добавляли 500 мкл хлороформа и перемешивали сверху вниз и центрифугировали при 10000 об/мин в течение 10 минут; 400 мкл супернатанта переносили в новую 1,5-мл центрифужную пробирку, добавляли 1 мл 70% (об./об.) этанола и смесь держали при -20°C для преципитации в течение 20 минут; смесь центрифугировали при 12000 об/мин в течение 15 минут для преципитации ДНК; после высушивания преципитата воздухом добавляли 50 мкл сверхчистой воды и хранили при -20°C для последующего применения.
(2) Праймеры для детекции в приведенной ниже таблице использовали для амплификации фрагментов, содержавших участки-мишени по обеим сторонам или спрогнозированные фрагменты в результате слияния промотора UBI2 и кодирующей области HPPD. Длина продуктов ПЦР составляла 300-1000 п.н., где комбинацию праймер 8-F+праймер 6-R использовали для детекции слитого фрагмента в средней точке соединения после удвоения фрагмента хромосомы, и ожидалось, что длина продукта будет составлять 630 п.н.
Реакционная система ПЦР была следующей:
(3) Реакцию ПЦР проводили в следующих общих условиях реакции:
(4) Детекцию продуктов реакции ПЦР проводили посредством 1% агарозного гель-электрофореза. Результаты показали, что положительная полоса размером 630 п.н. для спрогнозированного слитого фрагмента промотора UBI2 и кодирующей области HPPD может обнаруживаться образцах, трансформированных pQY002066 и pQY002068.
5. Положительные образцы слитого фрагмента промотора UBI2 и кодирующей области HPPD секвенировали для подтверждения и праймеры OsHPPDduplicated-primer8-F и OsHPPDduplicated-primer6-R использовали для секвенирования с обоих концов. Как показано на фиг.5, промотор гена UBI2 и экспрессируемая область гена HPPD могли лигироваться прямо, и событие редактирования слитой конструкции промотора гена UBI2 риса и экспрессируемой области гена HPPD могло обнаруживаться в геномной ДНК протопластов риса, трансформированных плазмидами pQY002066 и pQY002068, что указывает на то, что схема удвоения фрагментов хромосом для формирования нового гена HPPD была осуществимой, мог быть получен новый ген HPPD, экспрессия которого контролировалась сильным промотором, и это было определено как событие удвоения HPPD. Результат секвенирования протопластов, трансформированных вектором pQY002066, для тестирования события удвоения HPPD представлен в SEQ ID NO: 9; и результат секвенирования протопластов, трансформированных вектором pQY002068, для тестирования события удвоения HPPD, представлен в SEQ ID NO: 10.
Пример 2: Создание резистентного к гербициду риса с повышением (knock-up) экспрессии эндогенного гена HPPD посредством удвоения фрагмента хромосомы с использованием опосредуемой Agrobacterium трансформации
1. Конструирование редактирующего вектора для повышения экспрессии: на основе результатов теста протопластов в примере 1 отбирали комбинацию с двойной мишенью OsHPPD-гидовая РНК1: 5'GTGCTGGTTGCCTTGGCTGC3' и OsHPPD-гидовая РНК4: 5'GAAATAATCACCAAACAGAT3' с высокой эффективностью редактирования. Вектор pQY2091 для трансформации с использованием Agrobacterium конструировали в соответствии с примером 1. В качестве остова вектора использовали pHUE411, и его подвергали оптимизации кодонов для риса. Карта вектора представлена на фиг.6.
2. Трансформация каллюса риса с использованием Agrobacterium:
1) Трансформация с использованием Agrobacterium: 1 мкг плазмиды редактирующего вектора pQY2091 для повышения экспрессии добавляли к 10 мкл клеток Agrobacterium EHA105, компетентных посредством теплового шока (Angyu Biotech, каталожный номер № G6040), помещали на лед на 5 минут, погружали в жидкий азот для быстрого замораживания на 5 минут, затем удаляли и нагревали при 37°C в течение 5 минут, и наконец помещали на лед на 5 минут. Добавляли 500 мкл жидкой среды YEB (состав: дрожжевой экстракт 1 г/л, пептон 5 г/л, говяжий экстракт 5 г/л, сахароза 5 г/л, сульфат магния 0,5 г/л). Смесь помещали в устройство для качания и инкубировали при 28°C, 200 об/мин в течение 2~3 часов; бактерии собирали центрифугированием при 3500 об/мин в течение 30 секунд, собранные бактерии распределяли на чашке с YEB (канамицин 50 мг/л+рифампицин 25 мг/л) и инкубировали в течение 2 суток в инкубаторе при 28°C; единичные колонии отбирали и помещали в жидкую культуральную среду, и бактерии хранили при -80°С.
2) Культивирование Agrobacterium: единичные колонии трансформированных Agrobacterium на чашке с YEB отбирали, добавляли в 20 мл жидкой среды YEB (канамицин 50 мг/л+рифампицин 25 мг/л) и культивировали при перемешивании при 28°C до тех пор, пока OD600 не составила 0,5, затем клетки бактерий собирали центрифугированием при 5000 об/мин в течение 10 минут, добавляли 20-40 мл AAM (Solarbio, партия номер LA8580) жидкой среды для ресуспендирования клеток бактерий до достижения OD600 0,2-0,3, а затем добавляли ацетосирингон (Solarbio, номер артикля A8110) до достижения конечной концентрации 200 мкМ для инфицирования каллюса.
3) Индукция каллюса риса: сорта риса, являющиеся реципиентами трансформации, представляли собой Huaidao 5 и Jinjing 818, приобретенные на рынке семян в Huai’an, Jiangsu, и они были получены в теплице. 800-2000 чистых семян риса очищали от шелухи, затем промывали стерильной водой до тех пор, пока вода не становилась прозрачной после промывания. Затем семена дезинфицировали 70% спиртом в течение 30 секунд, затем добавляли 30 мл 5% гипохлорита натрия, и смесь помещали на горизонтальное устройство для качания и качали при 50 об/мин в течение 20 минут, а затем промывали стерильной водой 5 раз. Семена помещали на стерильную впитывающую бумагу, сушили воздухом для удаления воды на поверхности семян, инокулировали на индукционную среду и культивировали при 28°C с получением каллюса.
Состав для индуцирующей среды: 4,1 г/л порошка N6+0,3 г/л гидролизованного казеина+2,878 г/л пролина+2 мг/л 2,4-D+3% сахароза+0,1 г/л инозитола+0,5 г глутамина+0,45% фитагель, pH 5,8.
4) Инфицирование каллюса риса Agrobacterium: каллюс Huaidao No. 5 или Jinjing 818, субкультивированный в течение 10 суток, диаметром 3 мм, отбирали и собирали в 50-мл центрифужную пробирку; в центрифужную пробирку, содержавшую каллюс, переливали раствор для ресуспендирования Agrobacterium AAM с OD600, доведенной до 0,2-0,3, помещали ее на устройство для качания при 28°C при скорости 200 об/мин для проведения инфицирования в течение 20 минут; когда инфицирование завершалось, раствор с бактериями удаляли, каллюс помещали на стерильную фильтровальную бумагу и сушили воздухом в течение приблизительно 20 минут, затем помещали на чашку, содержавшую среду для сокультивирования, для проведения сокультивирования, и эту чашку накрывали стерильной фильтровальной бумагой, пропитанной жидкой средой AAM, содержавшей 100 мкМ ацетосирингон; после сокультивирования в течение 3 суток Agrobacterium удаляли путем промывания (сначала промывание стерильной водой 5 раз, затем промывание 500 мг/л антибиотика цефалоспорина в течение 20 минут) и подвергали селективному культивированию на селективной среде с 50 мг/л гигромицина.
Состав среды для сокультивирования: 4,1 г/л порошка N6+0,3 г/л гидролизованного казеина+0,5 г/л пролина+2 мг/л 2,4-D+200 мкМ AS+10 г/л глюкозы+3% сахароза+0,45% фитагель, pH 5,5.
3. Молекулярная идентификация и дифференцировка в проростки резистентного к гигромицину каллюса:
В отличие от процесса селекции при общепринятой трансформации риса, с использованием специфических праймеров для слитых фрагментов, образовавшихся после удвоения фрагмента хромосомы, резистентный к гигромицину каллюс мог быть молекулярно идентифицирован в ходе стадии селекции каллюса и культивирования в рамках настоящего изобретения, положительные события удвоения могли быть определены, и каллюс, содержавший новые гены, полученные в результате слияния различных генетических элементов, отбирали для культивирования с целью дифференцировки и индуцировали к формированию проростков. Конкретные стадии были следующими:
1) Сокультивированный каллюс переносили в селективную среду для первого раунда селекции (2 недели). Состав селективной среды представлял собой: 4,1 г/л порошка N6+0,3 г/л гидролизованного казеина+2,878 г/л пролина+2 мг/л 2,4-D+3% сахароза+0,5 г глутамина+30 мг/л гигромицина (HYG) + 500 мг/л цефалоспорина (cef) + 0,1 г/л инозитола+0,45% фитагель, pH 5,8.
2) После завершения первого раунда селекции вновь выращенный каллюс переносили в новую селективную среду для второго раунда селекции (2 недели). На этой стадии вновь выращенный каллюс диаметром более 3 мм зажимали пинцетом для взятия небольшого количества образца, его ДНК экстрагировали способом с CTAB, описанным в примере 1 для первого раунда молекулярной идентификации. В этом примере отбирали пару праймеров OsHPPDduplicated-primer8-F (8F) и OsHPPDduplicated-primer6-R (6R) для проведения идентификации способом ПЦР для каллюса, трансформированного вектором pQY2091, в случае которого реакционная система и условия реакции были сходными с реакционной схемой и условиями реакции примера 1. Среди всего 350 протестированных каллюсов, не было обнаружено положительного образца в каллюсах Huaidao 5, в то время как в каллюсах Jinjing 818 было обнаружено 28 положительных образцов. Результаты детекции с использованием ПЦР для некоторых каллюсов представлены на фиг.7.
3) Каллюсы, идентифицированные в качестве положительных способом ПЦР, переносили в новую селективную среду для третьего раунда селекции и экспансивного культивирования; после превышения каллюсом диаметра 5 мм каллюс, подвергаемый экспансивному культивированию, подвергали второму раунду молекулярной идентификации с использованием пары праймеров 8F+6R, желто-белый каллюс с хорошим статусом роста, идентифицированный в качестве положительного на втором раунде, переносили в среду для дифференцировки для проведения дифференцировки, и через 3-4 недели могли быть получены проростки размером приблизительно 1 см; дифференцированные проростки переносили в среду для укоренения культивирования; после укрепления проростков в ходе ускорения культивирования, их переносили в цветочный горшок с почвой и помещали в теплицу для культивирования. Состав среды для дифференцировки представлял собой: 4,42 г/л порошка MS+0,5 г/л гидролизованного казеина+0,2 мг/л NAA+2 мг/л KT+3% сахароза+ 3% сорбит+30 мг/л гигромицина+0,1 г/л инозитола+0,45% фитагель, pH 5,8. Состав среды для укоренения представляет собой: 2,3 г/л порошка MS+3% сахароза+0,45% фитагель.
4. Молекулярная детекция проростков с удвоением HPPD (поколение T0):
После второго раунда молекулярной идентификации 29 положительных по событию удвоения каллюсов подвергали совместной дифференцировке с получением 403 проростков поколения T0, и пару праймеров 8F+6R использовали для третьего раунда молекулярной идентификации 403 проростков, среди которых 56 имели положительные полосы. Положительные проростки переносили в теплицу для выращивания. Результаты детекции способом ПЦР для некоторых проростков T0 представлены на фиг.8.
5. Тест резистентности к ингибирующему HPPD гербициду для проростков с удвоенным HPPD (поколение T0):
Трансформированные проростки поколения T0, идентифицированные в качестве положительных по событию удвоения, пересаживали в пластмассовые ведра в теплицы для экспансивного выращивания с получением семян поколения T1. После того, как проростки начинали пускать побеги, собирали побеги энергично растущих линий и сеяли в те же горшки, что и побеги контрольных сортов дикого типа в тот же период роста. После достижения высоты растения приблизительно 20 см проводили тест резистентности к гербициду. Использованный гербицид представлял собой Shuangzuocaotong (CAS №. 1622908-18-2), производимый компанией авторов изобретения, и его дозировка для применения в полевых условиях составляет приблизительно 4 грамма активных ингредиентов на mu (4 г a.i./mu). В этом эксперименте Shuangzuocaotong применяли в градиенте дозировки 2 г a.i./mu, 4 г a.i./mu, 8 г a.i./mu и 32 г a.i./mu с использованием переносной оросительной колонны.
Результаты теста резистентности представлены на фиг.9. Через 5-7 суток после применения контрольные проростки риса дикого типа начинали демонстрировать альбинизм, в то время как все линии с событиями удвоения HPPD оставались нормально зелеными. Через 4 недели после применения проростки риса дикого типа практически погибли, в то время как все линии с событиями удвоения продолжали быть зелеными и росли нормально. Результаты теста показали, что линии с удвоением гена HPPD имели значительно повышенную толерантность к Shuangzuocaotong.
6. Количественная детекция относительной экспрессии гена HPPD в проростках с удвоением HPPD (поколение T0):
Было предположено, что повышенная резистентность линии с удвоением гена HPPD к Shuangzuocaotong была следствием слияния сильного промотора UBI2 и CDS гена HPPD, которое повышало экспрессию HPPD, так что линии поколения T0 QY2091-13 и QY2091-20 использовали для получения образцов из первичных побегов и вторичных побегов, использованных для теста резистентности гербициду для детекции уровней экспрессии генов HPPD и UBI2, соответственно, с Jinjing 818 дикого типа в качестве контроля. Конкретные стадии были следующими:
1) Экстракция тотальной РНК (способ с Trizol):
0,1-0,3 г свежих листьев собирали и растирали в порошок в жидком азоте. Добавляли 1 мл реагента Trizol для каждых 50-100 мг ткани для лизиса; лизат с Trizol вышеуказанной ткани переносили в 1,5-мл центрифужную пробирку, оставляли стоять при комнатной температуре (15-30°C) в течение 5 минут; добавляли хлороформ в количестве 0,2 мл на 1 мл Trizol; центрифужную пробирку закрывали крышкой, энергично встряхивали вручную в течение 15 секунд, оставляли стоять при комнатной температуре (15-30°C) в течение 2-3 минут, затем центрифугировали при 12000g (4°С) в течение 15 минут; верхнюю водную фазу удаляли и помещали в новую центрифужную пробирку, добавляли изопропанол в количестве 0,5 мл на 1 мл Trizol, смесь держали при комнатной температуре (15-30°C) в течение 10 минут, затем центрифугировали при 12000g (2-8°C) в течение 10 минут; супернатант удаляли и к осадку добавляли 75% этанол в количестве 1 мл на 1 мл Trizol для промывания. Смесь встряхивали и центрифугировали при 7500g (2-8°C) в течение 5 минут. Супернатант удаляли; преципитированную РНК сушили естественным путем при комнатной температуре в течение 30 минут; преципитат РНК растворяли 50 мкл свободной от РНКаз воды и хранили в холодильнике при -80°C после анализа с использованием электрофореза и определения концентрации.
2) Анализ с использованием электрофореза РНК:
Получали агарозный гель в концентрации 1%, затем отбирали 1 мкл РНК и смешивали с 1 мкл 2X буфера для внесения. Смесь наносили на гель. Напряжение было установлено на 180 В и время электрофореза составляло 12 минут. После завершения электрофореза агарозный гель отбирали, и положения и яркость фрагментов наблюдали с использованием УФ-системы для визуализации гелей.
3) Детекция чистоты РНК:
Концентрацию РНК измеряли с использованием анализатора нуклеиновых кислот Microprotein. РНК с высокой чистотой имела величину OD260/OD280 1,8-2,1. Величина ниже 1,8 указывала на серьезную белковую контаминацию и величина более 2,1 указывала на серьезную деградацию РНК.
4) Флуоресцентная количественная ПЦР в реальном времени
Экстрагированную тотальную РНК подвергали обратной транскрипции в кДНК с использованием специального набора для обратной транскрипции. Основная методика включала: определение сначала концентрации экстрагированной тотальной РНК и использование части РНК массой 1-4 мкг для синтеза кДНК посредством синтеза с использованием обратной транскриптазы. Полученную кДНК хранили при -20°C.
1. Раствор РНК-матрицы получали на льду, как указано в таблице ниже, и подвергали реакции денатурации и отжига на устройстве для ПЦР. Этот процесс способствовал денатурации РНК-матрицы и специфическому отжигу праймеров и матриц тем самым повышая эффективность обратной транскрипции.
Условия реакции для денатурации и отжига:
2. Реакционную систему для обратной транскрипции получали, как указано в таблице 2, для синтеза кДНК:
Evo M-MLV Plus RTase (200 Е/мкл)
1 мкл
Условия реакции для синтеза кДНК:
3. Ген UBQ5 риса был выбран в качестве гена внутреннего стандарта, и синтезированную кДНК использовали в качестве матрицы для проведения флуоресцентной количественной ПЦР. Праймеры, приведенные в таблице 3, использовали для получения реакционного раствора согласно таблице 4.
Rox II
0,2 мкл
4. Реакцию проводили в соответствии со стадиями количественной реакции ПЦР в реальном времени, приведенными в таблице 5. Реакцию проводили в течение 40 циклов.
5) Обработка данных и экспериментальные результаты
Как показано в таблице 6, в качестве внутреннего стандарта использовали UBQ5, ΔCt вычисляли путем вычитания величины Ct для UBQ5 из величины Ct для гена-мишени, а затем вычисляли 2-ΔCt, что соответствовало относительному уровню экспрессии гена-мишени. 818CK1 и 818CK3 представляли собой два контрольных растения Jinjing 818 дикого типа; 13M и 20M представляли собой образцы первичных листьев побегов растений T0 QY2091-13 и QY2091-20; 13L и 20L представляли собой образцы вторичных листьев побегов растений T0 QY2091-13 и QY2091-20, использованных для тестирования резистентности к гербицидам.
значение
значение
значение
Результаты представлены на фиг.10. UBQ5 риса использовали в качестве гена внутреннего стандарта для вычисления относительных уровней экспрессии генов OsHPPD и UBI2. Результаты показали, что уровень экспрессии HPPD в линии с удвоением HPPD был значительно более высоким, чем у дикого типа, что указывает на то, что слитый сильный промотор UBI2 повышал уровень экспрессии HPPD, тем самым создавая ген HPPD с высокой экспрессией, с повышением экспрессии (knock-up) гена HPPD. Небольшое снижение уровня экспрессии UBI2 могло быть следствием мелкомасштабных мутаций в результате редактирования промоторной области, и авторы изобретения действительно выявили инсерции, делеции оснований или делеции небольших фрагментов в участке-мишени UBI2. По сравнению с диким типом, уровни экспрессии UBI2 и HPPD имели выраженную тенденцию к соответствию и удовлетворению теоретических ожиданий; среди них, уровень экспрессии HPPD в образце 20M приблизительно в 6 раз превышал уровень экспрессии в группе CK3 дикого типа.
Указанные выше результаты показали, что в соответствии с эффективной программой удвоения фрагментов хромосом, протестированной в протопластах, каллюсах и трансформированных проростках, события удвоения могут быть отобраны посредством многократных раундов молекулярной идентификации в ходе трансформации с использованием Agrobacterium и культивирования тканей, и сильный промотор UBI2 в новой слитой конструкции гена HPPD, образовавшейся в трансформированных проростках, повышал уровень экспрессии гена HPPD, обеспечивая приобретение растениями резистентности к ингибирующему HPPD гербициду Shuangzuocaotong, вплоть до 8-кратной дозы в полевых условиях, и, таким образом, был получен резистентный к гербициду рис с повышением экспрессии эндогенного гена HPPD. Принимая это в качестве примера, с использованием технического решения удвоения хромосомного фрагмента примера 1 и примера 2, желаемый промотор также может быть введен в эндогенный ген, паттерн генной экспрессии которого должен быть изменен, для создания нового гена, и может быть создан новый сорт растений с желаемым паттерном экспрессии генов посредством опосредуемой Agrobacterium трансформации.
Пример 3: Молекулярная детекция и тест резистентности к гербициду поколения T1 резистентной к гербициду линии риса с повышением экспрессии эндогенного гена HPPD, вызванным удвоением фрагмента хромосомы
Физическое расстояние между геном HPPD и геном UBI2 в геноме риса дикого типа составляло 338 т.п.н., как показано на схеме 1 на фиг.1. Длина хромосомы была увеличена на 338 т.п.н. после удвоения фрагмента хромосомы между ними посредством дупликации, и был получен новый ген HPPD с высокой экспрессией с промотором UBI2 в точке соединения дуплицированного фрагмента для запуска экспрессии области CDS HPPD. Для определения того, мог ли новый ген стабильно наследоваться, и эффекта удвоения фрагмента хромосомы на генетическую стабильность, проводили молекулярную детекцию и тест резистентности к гербициду для поколения T1 линий с удвоением HPPD.
Во-первых, было обнаружено, что событие удвоения не имело значительного эффекта на фертильность растений поколения T0, поскольку все положительные линии T0 были способны продуцировать нормальные семена. Для линий QY2091-13 и QY2091-20 дополнительно проводили тест с посевом проростков поколения T1.
1. Получение образца:
Для QY2091-13 проводили посев всего 36 проростков T1, среди которых 27 росли нормально и 9 имели альбинизм. 32 было отобрано для экстракции и детекции ДНК, где № 1-24 представляли собой нормальные проростки и № 25-32 представляли собой проростки с альбинизмом.
Для QY2091-20 проводили посев всего 44 проростков T1, среди которых 33 росли нормально и 11 имели альбинизм. 40 было отобрано для экстракции и детекции ДНК, где № 1-32 представляли собой нормальные проростки и № 33-40 представляли собой проростки с альбинизмом.
В растениях поколения T1 наблюдались проростки с альбинизмом. Было предположено, что, поскольку HPPD представлял собой ключевой фермент в каскаде синтеза хлорофилла в растениях и растения поколения T0, являющиеся результатом двойного редактирования мишеней, возможно могли быть химерами многих генотипов, таких как удвоение, делеция, инверсия фрагментов хромосом или небольшая мутация фрагмента в отредактированном участке-мишени. Фенотип альбиноса мог присутствовать в растениях, где ген HPPD был разрушен, например, область CDS HPPD делетирована. Для определения возможных генотипов были сконструированы различные пары праймеров для ПЦР.
2. Молекулярная идентификация с использованием ПЦР:
1) Последовательности праймеров для детекции: последовательность 5’-3’
Праймер 8F: TCTGTGTGAAGATTATTGCCACTAGTTC
Праймер 6R: GAGTTCCCCGTGGAGAGGT
Test 141-F: CCCCTTCCCTCTAAAAATCAGAACAG
Праймер 4R: GGGATGCCCTCCTTATCTTGGATC
Праймер 3F: CCTCCATTACTACTCTCCCCGATTC
Праймер 7R: GTGTGGGGGAGTGGATGACAG
pg-Hyg-R1: TCGTCCATCACAGTTTGCCA
pg-35S-F: TGACGTAAGGGATGACGCAC
2) Участки связывания указанных выше праймеров представлены на фиг.11. Среди них, праймер 8F+праймер 6R использовали для детекции слитого фрагмента промотора UBI2 и CDS HPPD после удвоения фрагмента хромосомы, и длины продукта составляла 630 п.н.; Test 141-F+праймер 4R использовали для детекции события делеции фрагмента хромосомы, и длина продукта составляла 222 п.н.; и pg-Hyg-R1+pg-35S-F использовали для детекции фрагмента T-ДНК редактирующего вектора, и длина продукта составляла 660 п.н.
3) Реакционная система ПЦР, методика реакции и детекция с использованием гель-электрофореза были такими, как в примере 1.
3. Результаты молекулярной детекции:
Результаты детекции событий удвоения и делеции представлены в таблице 7. Можно было отметить, что события удвоения и события делеции фрагмента хромосомы были выявлены в растениях поколения T1 с различными соотношениями между различными линиями. События удвоения в QY2091-13 (29/32) были более частыми, чем в QY2091-20 (21/40), возможно вследствие различных химерных соотношений растений поколения T0. Результаты теста показали, что слитый ген, полученный путем удвоения, был наследуемым.
Праймеры pg-Hyg-R1+ pg-35S-F использовали для детекции фрагмента T-ДНК вектора для редактирования в описанных выше проростках T1. Результаты электрофореза продуктов ПЦР QY2091-20-17 и QY2091-13-7 были отрицательными для фрагмента T-ДНК, указывая на то, что они были гомозиготными по удвоению. Можно было наблюдать, что гомозиготные по удвоению нетрансгенные линии могли сегрегироваться из поколения T1 событий удвоения.
4. Детекция событий редактирования посредством секвенирования:
Слитые фрагменты с удвоением секвенировали в отношении гомозиготных по удвоению положительных образцов 1, 5, 7, 11, 18 и 19 поколения T1 QY2091-20 и в отношении гомозиготных по удвоению положительных образцов 1, 3, 7, 9, 10 и 12 поколения T1 QY2091-13. Одновременно амплифицировали левый участок-мишень гена HPPD и правый участок-мишень UBI2 для секвенирования с целью обнаружения событий редактирования в участка-мишенях. Среди них праймер 3F+праймер 7R использовали для детекции события редактирования левого участка-мишени HPPD, где контрольный продукт дикого типа имел длину 481 п.н.; праймер 8F+ праймер 4R использовали для детекции события редактирования правого участка-мишени UBI2, где контрольный продукт дикого типа имел длину 329 п.н.
1) Генотип событий удвоения:
Результат секвенирования для удвоения HPPD в QY2091-13 представлен в SEQ ID NO: 18, и результат секвенирования для удвоения HPPD в QY2091-20 представлен в SEQ ID NO: 19, см. фиг.12. По сравнению со спрогнозированными линкерными последовательностями удвоения, одно основание T было встроено в линкер в QY2091-13, 19 оснований были удалены из линкера в QY2091-20, и как вставка, так и делеция произошли в промоторной области UBI2 и не имели эффекта на кодирующую область белка HPPD. Из результатов детекции уровней экспрессии гена HPPD в примере 2 можно видеть, что уровни экспрессии этих новых генов HPPD, где промоторы UBI2 были слиты с областью CDS HPPD, были значительно увеличены.
2) События редактирования в исходных участках-мишенях HPPD и UBI2 на обеих сторонах:
Существовало больше типов событий редактирования в участках-мишенях с обеих сторон. В двух линиях три типа редактирования произошли в промоторной области HPPD, а именно, встраивание единичного основания, делеция 17 оснований и делеция 16 оснований; и два типа редактирования происходили в промоторной области UBI2, а именно, встраивание 7 оснований и делеция 3 оснований. Все растения T1, использованные для тестирования и взятия образцов, представляли собой зеленые проростки и росли нормально, что указывало на то, что мелкомасштабные мутации в этих промоторных областях не имели значительного эффекта на функцию гена, и из их потомков могли быть отобраны резистентные к гербицидам сорта риса.
5. Тест резистентности к гербициду на проростках поколения T1:
Резистентность к гербициду поколения T1 линии QY2091 с удвоением HPPD тестировали на стадии проростков. После проведения поверхностной дезинфекции семян поколения T1 их проращивали на среде 1/2 MS, содержавшей 1,2 мкМ Shuangzuocaotong, и культивировали при 28°C, 16 часов на свету/8 часов в темноте, где Jinjing 818 дикого типа использовали в качестве контроля.
Результаты теста резистентности представлены на фиг.13. После культивирования на свету в течение 10 суток контрольные проростки риса дикого типа продемонстрировали фенотип альбинизма, и практически все были альбиносами, в то время как линии с событиями удвоения HPPD QY2091-7, 13, 20, 22 продемонстрировали сегрегацию фенотипа хлороза и зеленых проростков. В соответствии с вышеупомянутыми результатами молекулярной детекции происходила сегрегация генотипов в поколении T1. Проростки-альбиносы появлялись в отсутствии обработки гербицидом, в то время как зеленые проростки продолжали оставаться зелеными и росли нормально после добавления 1,2 мкМ Shuangzuocaotong. Результаты теста показали, что высокая резистентность к Shuangzuocaotong линий с удвоением гена HPPD могла стабильно наследоваться поколением T1.
Пример 4: Способ редактирования для повышения (knock-up) экспрессии эндогенного гена PPO путем индукции инверсии фрагмента хромосомы - тест с протопластами риса
Ген PPO1 риса (также известный как PPOX1) (как показано в SEQ ID NO: 7, в котором 1-1065 п.н. представляли собой промотор, остальные представляли собой кодирующую область) находится на хромосоме 1, и ген белка цикла Кальвина CP12 (как показано в SEQ ID NO: ID NO: 8, в котором 1-2088 п.н. представляли собой промотор, а остальная часть представляла собой кодирующую область) находится на 911 т.п.н. ниже гена PPO1 с противоположным направлением. В соответствии с данными о профили экспрессии генов, приведенными International Rice Genome Sequencing Project (http://rice.plantbiology.msu.edu/index.shtml), интенсивность экспрессии гена CP12 в листьях риса в 50 раз превышала интенсивность экспрессии гена PPO1, и промотор гена CP12 представлял собой сильный промотор с высокой экспрессией в листьях.
Как показано на схеме 1 фиг.4, посредством одновременной индукции двухцепочечных разрывов между соответствующими промоторами и областью CDS двух генов и скрининга область между двумя разрывами можно было перевернуть с заменой промотора гена PPO1 промотором гена CP12, повышая уровень экспрессии гена PPO1 и достигая резистентности к ингибирующим PPO гербицидам, тем самым можно было отобрать резистентные к гербициду линии. Кроме того, как показано на схеме 2 фиг.4, новый ген PPO1, контролируемый промотором гена CP12, также мог быть создан путем сначала инверсии, а затем удвоения.
1. Сначала последовательности геномной ДНК PPO1 и CP12 риса вводили в онлайн-инструмент CRISPOR (http://crispor.tefor.net/) для поиска доступных участков-мишеней для редактирования. После онлайн-оценки были отобраны следующие участки-мишени между промоторами и областями CDS генов PPO1 и CP12 для тестирования:
Гидовая РНК1 и гидовая РНК2 находились между промотором и областью CDS гена PPO1, вблизи инициирующего кодона PPO1, и гидовая РНК3 и гидовая РНК4 находились между промотором и областью CDS гена CP12 вблизи кодона инициации CP12.
Как описано в примере 1, конструировали праймеры для указанных выше участков-мишеней для конструирования векторов с двойными мишенями с pHUE411 в качестве остова:
В частности, плазмиду pCBC-MT1T2 (https://www.addgene.org/50593/) использовали в качестве матрицы для амплификации фрагментов с двойной мишенью sg-РНК1+3, sg-РНК1+4, sg-РНК2+3, sg-РНК2+4 и конструирования экспрессирующих кассет для sg-РНК, соответственно. Остов вектора pHUE411 расщепляли BsaI и выделяли из геля, и фрагмент-мишень прямо использовали для реакции лигирования после расщепления. ДНК-лигазу T4 использовали для лигирования остова вектора и фрагмента-мишени, продуктом лигирования трансформировали компетентные клетки Trans5α, различные моноклоны отбирали и секвенировали. Для экстракции плазмид из клонов с правильными последовательностями использовали Sparkjade High Purity Plasmid Mini Extraction Kit, тем самым получая рекомбинантные плазмиды, соответственно, названные pQY002095, pQY002096, pQY002097, pQY002098, как показано ниже:
pQY002095 pHUE411-PPO-sgRNA1+3, содержащая комбинацию OsPPO-гидовой РНК1, гидовой РНК3
pQY002096 pHUE411-PPO-sgRNA2+3, содержащая комбинацию OsPPO-гидовой РНК2, гидовой РНК3
pQY002097 pHUE411-PPO-sgRNA1+4, содержащая комбинацию OsPPO-гидовой РНК1, гидовой РНК4
pQY002098 pHUE411-PPO-sgRNA2+4, содержащая комбинацию OsPPO-гидовой РНК2, гидовой РНК4
2. Получали плазмиды с высокой чистотой и высокой концентрацией для вышеупомянутых векторов pQY002065-002098, как описано для стадии 2 примера 1.
3. Протопласты риса получали и подвергали ПЭГ-опосредуемой трансформации с использованием вышеупомянутых векторов, как описано для стадии 3 примера 1.
4. Проводили нацеливание на геном и детекцию нового гена с использованием праймеров для детекции, представленных в таблице ниже для детекции с использованием ПЦР, как описано для стадии 4 примера 1.
Среди них, комбинацию PPO-R2 и CP-R2 использовали для амплификации контролируемого промотором CP12 нового фрагмента CDS гена PPO1, который находился справа после инверсии фрагмента хромосомы, и комбинацию PPO-F2 и CP-F2 использовали для амплификации контролируемого промотором PPO1 нового фрагмента CDS гена CP12, который находился слева после инверсии. Возможные генотипы в результате двойного редактирования мишени и участков связывания праймеров для молекулярной детекции представлены на фиг.14.
5. Результаты ПЦР и секвенирования показали, что в результате трансформации протопластов риса был получен ожидаемый новый ген, в котором промотор CP12 контролировал экспрессию PPO1. Событие редактирования, где промотор гена CP12 риса был слит с экспрессирующей областью гена PPO1, могло быть обнаружено в геномной ДНК трансформированных протопластов риса. Это показало, что схема образования нового гена PPO посредством инверсии фрагмента хромосомы была осуществимой, и мог быть получен новый ген PPO, контролируемый сильным промотором, который был определен как событие инверсии PPO1. Результаты секвенирования для инверсии фрагмента хромосомы в протопластах, трансформированных вектором pQY002095, представлены в SEQ ID NO: 15; результаты секвенирования для делеции фрагмента хромосомы в протопластах, трансформированных вектором pQY002095, представлены в SEQ ID NO: 16; и результаты секвенирования для инверсии фрагмента хромосомы в протопластах, трансформированных вектором pQY002098, представлены в SEQ ID NO: 17.
Пример 5: Получение резистентного к гербициду риса с повышением (knock-up) экспрессии эндогенного гена PPO, вызванным инверсией фрагмента хромосомы посредством опосредуемой Agrobacterium трансформации
1. Конструирование редактирующего вектора для повышения экспрессии: на основе результатов теста протопластов отбирали комбинацию с двойной мишенью OsHPPD-гидовая РНК1: 5'CCATGTCCGTCGCTGACGAG3' и OsHPPD-гидовая РНК4: 5'CGGATTTCTGCGTGTGATGT3' с высокой эффективностью редактирования для конструирования вектора pQY2234 для трансформации с использованием Agrobacterium. В качестве остова вектора использовали pHUE411, и его подвергали оптимизации кодонов для риса. Карта вектора представлена на фиг.16.
2. Трансформация каллюса риса с использованием Agrobacterium и два раунда молекулярной идентификации:
Плазмиду pQY2234 использовали для трансформации каллюса риса в соответствии со способом, описанным на стадии 2 примера 2. Реципиентными сортами были Huaidao No.5 и Jinjing 818. На стадии селекции каллюса проводили два раунда молекулярной идентификации на резистентном к гигромицину каллюсе, и каллюсы, положительные по событию инверсии, подвергали дифференцировке. В ходе молекулярной детекции каллюса амплификация контролируемого промотором CP12 нового фрагмента CDS гена PPO1, находившегося с правой стороны после инверсии фрагмента хромосомы посредством комбинации PPO-R2 и CP-R2, считалась положительным стандартом события инверсии, в то время как новый ген CP12, находившийся на левой стороне после инверсии, рассматривался после дифференцировки и появления проростков каллюса. Было протестировано всего 734 каллюса Huaidao No.5, из которых 24 каллюса были положительными по событию инверсии, и тестировали 259 каллюсов из Jinjing 818, из которых 29 каллюсов были положительными по событию инверсии. На фиг.17 представлены результаты детекции с использованием ПЦР каллюсов № 192-259 Jinjing 818.
3. Всего 53 положительных по событию инверсии каллюса подвергали двум раундам молекулярной идентификации, а затем совместно дифференцировали, и было идентифицированных 9 положительных по событию удвоения каллюса, которые подвергали двум раундам молекулярной идентификации, а затем совместно дифференцировали с получением 1875 проростков T0, в которых 768 линий имели фон Huaidao No.5, и 1107 линий имели фон Jinjing 818. Эти 1875 проростков далее подвергали третьему раунду молекулярной идентификации с парой праймеров PPO-R2 и CP-R2, из которых 184 линии фона Huaidao No.5 продемонстрировали положительные по инверсии полосы, 350 линий фона Jinjing 818 продемонстрировали положительные по инверсии полосы. Положительные проростки переносили в теплицу для культивирования.
4. Тест резистентности и ингибирующему PPO гербициду для проростков с инверсией PPO1 (поколение T0):
Проростки с трансформацией QY2234 поколения T0, идентифицированные в качестве положительных по инверсии, пересаживали в пластмассовые ведра в теплице для проращивания семян поколения T1. Существовало большое количество положительных проростков, так что отбирали некоторые проростки T0 и контрольные линии дикого типа со сходным периодом роста и статусом. Когда высота растения достигала приблизительно 20 см прямо проводили тест резистентности к гербициду. Использованный гербицид представлял собой высокоэффективный ингибирующий PPO гербицид  , производимый данной компанией (код 2081, см. катайскую патентную заявку на изобретение № 202010281666.4). В этом эксперименте гербицид применяли с градиентом из трех уровней, а именно 0,18, 0,4 и 0,6 г ai/mu, с использованием оросительной колонны переносного типа.
, производимый данной компанией (код 2081, см. катайскую патентную заявку на изобретение № 202010281666.4). В этом эксперименте гербицид применяли с градиентом из трех уровней, а именно 0,18, 0,4 и 0,6 г ai/mu, с использованием оросительной колонны переносного типа.
Результаты теста резистентности представлены на фиг.18. Через 3-5 суток после применения контрольные проростки риса дикого типа начинали вянуть с вершины листа, на листьях появлялись некротические пятна и растения постепенно увядали, в то время как большинство линий с событием инверсии PPO1 сохраняли нормальный рост, листья не имели заметной фитотоксичности. Кроме того, некоторые линии продемонстрировали фитотоксичность, возможно вследствие полигенотипического мозаицизма событий редактирования и низкого уровня экспрессии PPO1 в линиях поколения T0. Через две недели после применения проростки риса дикого типа погибли, и большинство из линий с инверсией продолжали оставаться зелеными и росли нормально. Результаты теста показали, что линии с инверсией PPO1 могли значительно повысить толерантность растений к 2081.
5. Количественная детекция относительного уровня экспрессии гена PPO1 в проростках с инверсией PPO1 (поколение T0):
Было предположено, что повышенная резистентность линий с инверсией гена PPO1 к 2081 была следствием слияния сильного промотора CP12 и CDS гена PPO1, которое повышало уровень экспрессии PPO1. Таким образом, были отобраны линии поколения T0 QY2234-252, QY2234-304 и QY2234-329 из фона Huaidao No.5, и получали образцы их первичных побегов и вторичных побегов и подвергали их детекции уровней экспрессии генов PPO1 и CP12. В качестве контроля использовали Huaidao No.5 дикого типа. Конкретные протоколы соответствовали стадии 6 примера 2 с геном UBQ5 риса в качестве внутреннего эталонного гена. Праймеры для количественного флуоресцентного определения были следующими: 5'-3'.
В качестве внутреннего стандарта использовали UBQ5. ΔCt вычисляли путем вычитания величины Ct для UBQ5 из величины Ct для гена-мишени. Затем вычисляли 2-ΔCt, что соответствовало относительному уровню экспрессии гена-мишени. H5CK1 и H5CK2 представляли собой два контрольных растения Huaidao No.5 дикого типа; 252M, 304M и 329M представляли собой образцы первичных листьев побегов растений T0 QY2234-252, QY2234-304 и QY2234-329; 252L, 304L и 329L представляли собой образцы их вторичных листьев побегов. Результаты представлены в таблице 8 ниже:
значение
значение
значение
Относительные уровни экспрессии PPO1 и CP12 в различных линиях представлены на фиг.19. Как показали результаты, в отличие от события удвоения в примере 2, уровни экспрессии генов этих линий с событием инверсии значительно различались. Уровни экспрессии CP12 значительно различались между двумя группами CK Huaidao No.5, возможно вследствие различных скоростей роста проростков. По сравнению с контрольной группой H5CK2, все уровни экспрессии CP12 в экспериментальных группах продемонстрировали тенденцию к снижению, в то время как уровни экспрессии PPO1 для 252L и 329M значительно возрастали, и уровни экспрессии PPO1 для 304L и 329L умеренно возрастали, и уровни экспрессии PPO1 для 252M и 304M снижались. В отличие от удвоения фрагментов хромосом, которое в основном повышало уровень экспрессии гена, инверсия фрагментов хромосом обеспечивала новые гены на обеих сторонах, таким образом, различные события редактирования могли происходить на мишенях с обеих сторон, и в то же время изменения направления транскрипции также влияют на уровень экспрессии генов. Иными словами, растения поколения T0 представляли собой комплексные химеры. Также могли существовать значительные различия уровней экспрессии генов между первичными и вторичными побегами одного и того же растения. Из результатов количественной ПЦР можно видеть, что события инверсии PPO1 продемонстрировали более высокую вероятность увеличения уровня экспрессии гена PPO1, и, таким образом, резистентные к гербициду линии с высоким уровнем экспрессии PPO1 могли быть отобраны по посредством селекции событий инверсии на основе резистентности к гербициду.
Описанные выше результаты показали, что, согласно схеме детекции эффективной инверсии фрагмента хромосомы в протопластах, можно было отобрать каллюсы и трансформированные проростки с событиями инверсии посредством нескольких раундов молекулярной идентификации в ходе трансформации Agrobacterium и культивирования тканей, и сильный промотор CP12, слитый с новым геном PPO1, образовавшихся в трансформированных проростках, действительно мог повышать уровень экспрессии гена PPO1, что могло сообщать растениям резистентность к ингибирующему PPO гербициду 2081, тем самым был получен резистентный к гербициду рис с повышением экспрессии эндогенного гена PPO. Принимая это в качестве примера, протокол инверсии фрагмента хромосомы примера 4 и примера 5 также применялся к другим эндогенным генам, паттерн экспрессии которых должен быть изменен путем введения и слияния с требуемым промотором, тем самым мог быть создан новый ген, и могли быть созданы новые сорта с желаемым паттерном экспрессии генов посредством опосредуемой Agrobacterium трансформации в растениях.
Пример 6: Молекулярная детекция и тест резистентности к гербициду растений поколения T1 резистентных к гербициду линий риса с повышением экспрессии (knock-up) эндогенного гена PPO1 посредством инверсии фрагмента хромосомы
Физическое расстояние между геном PPO1 и геном CP12 генома риса дикого типа составляет 911 т.п.н. Как показано на фиг.14 ген PPO1 с высокой экспрессией с контролируемой промотором CP12 областью CDS PPO1 получали с правой стороны после инверсии фрагмента хромосомы между двумя генами. Также могла происходить делеция фрагмента хромосомы. Для тестирования, может ли новый ген наследоваться стабильно, и влияния инверсии фрагмента хромосомы на генетическую стабильность проводили молекулярную детекцию и тест резистентности к гербициду на поколении T1 линии с инверсией PPO1.
В первую очередь, было обнаружено, что событие инверсии не имело значительного эффекта на фертильность растений поколения T0, поскольку все положительные линии T0 были способны нормально продуцировать семена. Для детекции были отобраны поколения T1 линий QY2234/H5-851 с фоном Huaidao No.5.
1. Получение образца:
Для QY2234/H5-851 было посеяно всего 48 проростков T1. Все растения росли нормально.
2. Молекулярная идентификация с использованием ПЦР:
1) Последовательность праймеров для детекции: 5’-3’
PPO-R2: AAGGCTGGAAGCTGTTGGG
CP-R2: CTGAGGAGGCGATAAGAAACGA
PPO-F2: CGGACTTATTTCCCACCAGAA
CP-F2: AGGTCTCCTTGAGCTTGTCG
pg-Hyg-R1: TCGTCCATCACAGTTTGCCA
pg-35S-F: TGACGTAAGGGATGACGCAC
2) Участки связывания указанных выше праймеров представлены на фиг.14, где PPO-R2+CP-R2 использовали для детекции слитого фрагмента правого промотора CP12 и кодирующей области PPO1 после инверсии фрагмента хромосомы, и длина продукта составляла 507 п.н.; PPO-F2+CP-F2 использовали для детекции слитого фрагмента левого промотора PPO1 и кодирующей области CP12 после инверсии фрагмента хромосомы, и длина продукта составляла 560 п.н.; PPO-F2+PPO-R2 использовали для детекции левого участка-мишени PPO перед инверсией, и длина продукта в контроле дикого типа составляла 586 п.н.; CP-F2+CP-R2 использовали для детекции правого участка-мишени CP12 перед инверсией, и длина продукта в контроле дикого типа составляла 481 п.н. Для детекции фрагмента T-ДНК редактирующего вектора использовали pg-Hyg-R1+pg-35S-F и длина продукта составляла 660 п.н.
3) Реакционная система для ПЦР и условия реакции:
Реакционная система (система 10 мкл):
Условия реакции:
Продукты ПЦР подвергали электрофорезу на 1% агарозном геле с напряжением 180 В в течение 10 минут.
3. Результаты молекулярной детекции:
Результаты детекции представлены в таблице 9. Было выявлено всего 48 растений, из которых 12 растений (2/7/11/16/26/36/37/40/41/44/46/47) были гомозиготными по инверсии, 21 растение (1/3/4/5/6/8/9/15/17/20/22/23/24/27/30/31/33/34/39/42/43) было гетерозиготным по инверсии и 15 растений (10/12/13/14/18/19/21/25/28/29/32/35/38/45/48) были гомозиготными по отсутствию инверсии. Соотношение гомозиготные по инверсии: гетерозиготные по инверсии: гомозиготные по отсутствию инверсии составляло 1:1,75:1,25, приблизительно 1:2:1. Таким образом, результаты детекции удовлетворяли закону наследования Менделя, что указывает на то, что новый ген PPO1, полученный путем инверсии, был наследуемым.
Для указанных выше проростков T1 праймеры Pg-Hyg-R1+pg-35S-F использовали для детекции фрагмента T-ДНК вектора для редактирования. Результаты электрофореза 16 и 41 были негативными в отношении фрагмента T-ДНК, указывая на гомозиготную инверсию. Можно было видеть, что нетрансгенные линии гомозиготной инверсии могли сегрегировать из поколения T1 события инверсии.
4. Детекция событий редактирования с использованием секвенирования:
Детекция генотипов для событий инверсии фокусировалась на событиях редактирования нового гена PPO на правой стороне. События мутаций с полной кодирующей рамкой белка гена PPO1 сохранялись. События редактирования участка CP12 на левой стороне, которые не влияли на нормальный рост растений при определении фенотипов в теплице и в полевых условиях, сохранялись. Генотипы событий редактирования, выявленные в положительных по событию инверсии линиях, приведены ниже, где "цельный" указывает на идентичный заданной последовательности слитого фрагмента после инверсии. Генотипы успешных событий инверсии QY2234 в фоне Huaidao No.5 были следующими:
Некоторые карты пиков секвенирования и результаты сравнения последовательностей представлены на фиг.20.
Генотипы успешной инверсии QY2234 на фоне Jinjing818 были следующими:
Некоторые карты пиков секвенирования и результаты сравнения последовательностей представлены на фиг.21.
Результаты секвенирования описанных выше различных новых генов PPO1 с промотором CP12, слитым с кодирующей областью PPO1, представлены в SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
5. Тест резистентности к гербициду проростков поколения T1:
Тест резистентности к гербициду проводили на поколении T1 линий QY2234/H5-851 с инверсией PPO1 на стадии проростков. В качестве контроля использовали Huaidao No.5 дикого типа, и их высевали одновременно с семенами поколения T1 линий с инверсией. Когда проростки достигали высоты растения 15 см, применяли 2081 путем опрыскивания в четырех уровнях дозы 0,3, 0,6, 0,9 и 1,2 г a.i./mu. Условия культивирования представляли 28°C с 16 часами на свету и 8 часами в темноте.
Результаты теста резистентности представлены на фиг.22. Через 5 суток после применения контрольные проростки риса дикого типа продемонстрировали заметную фитотоксичность в дозе 0,3 г a.i./mu. Они начали вянуть на вершинах листьев и на листьях появились некротические пятна; в дозе 0,6 г a.i./mu растения быстро погибали. Однако проростки T1 QY2234/H5-851 могли сохранять нормальный рост при дозе 0,3 г a.i./mu, и нельзя было наблюдать заметную фитотоксичность на листьях; при дозах 0,6 и 0,9 г ai/mu, некоторые проростки T1 продемонстрировали сухие концы листьев, однако большинство проростков T1 могли оставаться зелеными и продолжать расти, в то время как контроли по существу погибали. При дозе 1,2 г a.i./mu все контрольные растения погибли, в то время как некоторые из проростков T1 могли оставаться зелеными и продолжали расти. Результаты теста показали, что резистентность линий с инверсией гена PPO1 к 2081 могла стабильно наследоваться их поколением T1.
Пример 7: Способ редактирования для повышения (knock-up) экспрессии эндогенного гена EPSPS в растении
EPSPS представлял собой ключевой фермент в каскаде синтеза ароматических аминокислот в растениях и мишень биоцидного гербицида глифосата. Высокий уровень экспрессии гена EPSPS может обеспечить растениям резистентность к глифосату. Ген EPSPS (как показано в SEQ ID NO: 4, в котором 1-1897 п.н. представляли собой промотор, а остальная часть представляла собой область экспрессии) находится на хромосоме 6 в рисе. Вышележащий ген представляет собой транскетолазу (TKT, как показано в SEQ ID NO: 3, в которой 1-2091 п.н. представляли собой промотор, а остальные представляли собой область экспрессии) с противоположным направлением. Интенсивность экспрессии гена TKT в листьях в 20-50 раз превышала интенсивность экспрессии гена EPSPS. Как показано на фиг.2, посредством одновременной индукции двухцепочечных разрывов между промотором и областью CDS двух генов, соответственно, после скрининга может быть достигнута инверсия (схема 1) или удвоение с инверсией (схема 2) области между двумя разрывами. В обоих случаях промотор гена EPSPS может быть заменен промотором гена TKT, тем самым повышая уровень экспрессии гена EPSPS и достигая резистентности к глифосату. Кроме того, схемы 3, 4 и 5, как показано на фиг.2, также могли создавать новые гены EPSPS, контролируемые промотором гена TKT. Структура гена EPSPS, являющаяся соседней и противоположной по направлению относительно TKT, была консервативной в однодольных растениях (таблица 10). В то же время в двудольных растениях оба гена был соседними, но в то же время имели одно направление; таким образом, этот способ был универсальным в растениях.
Для этого в качестве остова использовали pHUE411, и следующие последовательности в качестве мишеней:
Конструировали несколько различных векторов с двойной мишенью:
pQY002061 pHUE411-EPSPS-sgRNA1+3
pQY002062 pHUE411-EPSPS-sgRNA2+3
pQY002063 pHUE411-EPSPS-sgRNA1+4
pQY002064 pHUE411-EPSPS-sgRNA2+4
pQY002093 pHUE411-EPSPS-sgRNA2+5
pQY002094 pHUE411-EPSPS-sgRNA2+6
(2) С использованием соответствующих праймеров для детекции, представленных в приведенной ниже таблице, амплифицировали фрагменты, содержавшие участки-мишени по обеим сторонам или спрогнозированные фрагменты, полученные путем слияния промотора UBI2 и кодирующей области HPPD, и длина продуктов составляет 300-1000 п.н.
После трансформации протопластов результаты детекции показали, что были достигнуты ожидаемые события инверсии. Как показано на фиг.15, результат секвенирования для детекции инверсии протопласта, трансформированного вектором pQY002062, представлен в SEQ ID NO: 11; результат секвенирования для детекции делеции в протопластах, трансформированных вектором pQY002062, представлен в SEQ ID No: 12; результат секвенирования для детекции инверсии в протопластах, трансформированных вектором pQY002093, представлен в SEQ ID NO: 13; и результат секвенирования для детекции делеции в протопластах, трансформированных вектором pQY002093, представлен в SEQ ID NO: 14.
Эти векторы переносили в Agrobacterium для трансформации каллюсов риса. Получали растения, содержавшие новый ген EPSPS. Результаты биоанализа с гербицидами показали, что растения имели очевидную резистентность к гербициду глифосату.
Пример 8: Способ редактирования для повышения экспрессии эндогенного гена PPO в Arabidopsis
Протопорфириногеноксидаза (PPO) является одной из основных мишеней гербицидов. Посредством высокой экспрессии эндогенной PPO растений резистентность к ингибирующим PPO гербицидам могла быть значительно увеличена. Ген PPO Arabidopsis (как показано в SEQ ID NO: 1, где 1-2058 п.н. представляют собой промотор, и остальные представляют собой область экспрессии) находится на хромосоме 4, и ген убиквитина 10 (как показано в SEQ ID NO: 2, где 1-2078 п.н. представляют собой промотор, а остальные представляют собой область экспрессии) находится на 1,9 м.п.н. ниже с тем же направлением, что и ген PPO.
Как показано на схеме, приведенной на фиг.3, происходило одновременное внесение двухцепочечных разрывов в участках между промотором и областью CDS генов PPO и убиквитина 10, соответственно. События удвоения области между двумя разрывами могли быть достигнуты путем скрининга, а именно, новый ген был получен путем слияния промотора убиквитина 10 и кодирующей области PPO. Кроме того, в соответствии со схемой 2, как показано на фиг.1, также мог быть создан новый ген, в котором промотор убиквитина 10 и кодирующая область PPO были слиты вместе.
Для этого использовали pHEE401E в качестве остова (https://www.addgene.org/71287/) и следующие положения использовали в качестве участков-мишеней:
Векторы с двойной мишенью конструировали способом, описанным "Wang ZP, Xing HL, Dong L, Zhang HY, Han CY, Wang XC, Chen QJ. Egg cell-specific promoter-controlled CRISPR/Cas9 efficiently generates homozygous mutants for multiple target genes in Arabidopsis in a single generation. Genome Biol. 2015 Jul 21;16:144.":
Arabidopsis трансформировали в соответствии со следующим способом:
(1) Трансформация с использованием Agrobacterium
Компетентные в отношении Agrobacterium GV3101 клетки трансформировали рекомбинантными плазмидами с получением рекомбинантных Agrobacterium.
(2) Получение раствора для инфицирования Agrobacterium
1) Активированные Agrobacterium инокулировали в 30 мл жидкой среды YEP (содержавшей 25 мг/л Rif и 50 мг/л Kan), культивировали при 28°C при качании при 200 об/мин в течение ночи до тех пор, пока величина OD600 не составляла приблизительно 1,0-1,5.
2) Бактерии собирали центрифугированием при 6000 об/мин в течение 10 минут и супернатант удаляли.
3) Бактерии ресуспендировали в растворе для инфицирования (не было необходимости корректировать pH) до достижения OD600=0,8 для последующего применения.
(3) Трансформация Arabidopsis
1) Перед трансформацией растений растения должны хорошо вырасти с густым соцветием и без стрессового ответа. Сначала трансформацию можно было проводить при условии, что высота растения достигала 20 см. Когда почва была сухой, проводили полив соответствующим образом. За сутки перед трансформацией выросшие стручки отрезали ножницами.
2) Соцветия растения, подлежащего трансформации, погружали в описанный выше раствор на от 30 секунд до 1 минуты при осторожном перемешивании. Растение после инфильтрации должно иметь слой жидкой пленки на нем.
3) После трансформации растение культивировали в темноте в течение 24 часов, а затем переносили в условия нормального освежения для роста.
4) Через одну неделю проводили вторую трансформацию аналогичным образом.
(4) Сбор семян
Семена собирали, когда они созревали. Собранные семена сушили в печи при 37°C в течение одной недели.
(5) Селекция трансгенных растений
Семена обрабатывали дезинфицирующим средством в течение 5 минут, промывали ddH2O 5 раз, а затем равномерно распределяли на селективную среду MS (содержавшую 30 мкг/мл Hyg, 100 мкг/мл Cef). Затем среду помещали в инкубатор с освещением (при температуре 22°С, 16 часов на свету и 8 часов в темноте, интенсивность освещения 100-150 мкмоль/м2/с и влажность 75%) для культивирования. Положительные проростки отбирали и пересаживали в почву через одну неделю.
(6) Детекция мутантных растений T1
(6.1) Экстракция геномной ДНК
1) Приблизительно 200 мг листьев Arabidopsis нарезали и помещали в 2-мл центрифужную пробирку. Добавляли стальные шарики, и листья растирали с использованием высокопроизводительного дезинтегратора тканей.
2) После тщательного растирания добавляли 400 мкл буфера для экстракции SDS и перемешивали сверху вниз. Смесь инкубировали на водяной бане при 65°С в течение 15 минут и перемешивали сверху вниз каждые 5 минут в ходе этого периода.
3) Смесь центрифугировали при 13000 об/мин в течение 5 минут.
4) 300 мкл супернатанта удаляли и переносили в новую 1,5-мл центрифужную пробирку, в центрифужную пробирку добавляли равный объем изопропанола, предварительно охлажденного до -20°C, а затем центрифужную пробирку держали при -20°C в течение 1 часа или в течение ночи.
5) Смесь центрифугировали при 13000 об/мин в течение 10 минут и супернатант удаляли.
6) в центрифужную пробирку добавляли 500 мкл 70% этанола для промывания осадка, промывочный раствор удаляли после центрифугирования (осторожно, не удаляя осадок). Осадок сушили при комнатной температуре, а затем добавляли 30 мкл ddH2O для растворения ДНК, а затем хранили при -20°С.
(6.2) Амплификация способом ПЦР
С использованием экстрагированного генома растения T1 в качестве матрицы амплифицировали фрагмент-мишень с использованием праймеров для детекции. 5 мкл продукта амплификации отбирали и подвергали детекции с использованием 1% агарозного гель-электрофореза, а затем визуализировали посредством устройства для визуализации гелей. Оставшийся продукт прямо секвенировали в компании, проводящей секвенирование.
Результаты секвенирования показали, что удвоение гена AtPPO1 было успешно достигнуто в Arabidopsis, и тест резистентности к гербициду показал, что растение с удвоением обладало резистентностью к PPO-гербицидам.
Пример 9: Создание гена GH1 с новыми характеристиками экспрессии у данио-рерио
Гены гормона роста (GH) у рыб контролируют их скорость роста и развития. В настоящее время высокая экспрессия гена GH у атлантических лососей посредством трансгенной технологии может значительно повысить их скорость роста. Способ имеет высокую экономическую ценность, однако был одобрен для выпуска на рынок только через десятилетия. Ген GH1 является геном гормона роста у данио-рерио. В рамках настоящего изобретения подходящие промоторы в данио-рерио (подходящие с точки зрения непрерывной экспрессии, силы и тканевой специфичности) подвергали слиянию вместе in vivo посредством делеции, инверсии, удвоения, удвоения с инверсией, хромосомного переноса и т.д. с получением быстрорастующей линии рыб.
Все публикации и патентные заявки, упомянутые в описании, включены в настоящее описание в качестве ссылок, как если бы каждая публикация или патентная заявка была индивидуально и конкретно указана как включенная в настоящее описание в качестве ссылки.
Хотя описанное выше изобретение описано более подробно посредством примеров и вариантов осуществления для простоты понимания, очевидно, что определенные изменения и модификации могут быть осуществлены в пределах объема прилагаемой формулы изобретения, и все такие изменения и модификации входят в объем настоящего изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> QINGDAO KINGAGROOT CHEMICAL COMPOUND CO., LTD.
<120> СПОСОБ СОЗДАНИЯ НОВОГО ГЕНА В ОРГАНИЗМЕ И ЕГО ПРИМЕНЕНИЕ
<130> 2
<160> 26
<170> SIPOSequenceListing 1.0
<210> 1
<211> 4439
<212> ДНК
<213> Ген PPO Arabidopsis thaliana (последовательность Genbank номер
AT4G01690)
<400> 1
gatattttga tgtcgtatgt atatcaatgt tgttagctgt aaactgtcca atgaaaatcc 60
atttaatatt tggacaaatt attattaggg aaattaatgt aatgtggagt attttatatg 120
aaatgtgtat taatgacgac gaccttactt gatcccggaa gtgtgtatta atttaatcgc 180
ggtgcatcgc tgtccacacg gttgggttca tgcgtacgtg gtatttgctg gcaagtggca 240
gtgaagtttc ttgtttgata ctgtatgttc cattcatgtc tttccctaga tgcatgtttc 300
ttctattttt atatattaat atttttttgt ttttcttatt atgttttctt tccctttgtg 360
aggagaataa ttgcgatacg attatttgtt ttaacagaaa aaagtgaagc gaaaaagatg 420
attcgaagtt tgaagtacga ttaaaatttt tctttttccg gatccgagtt tgatcttctt 480
tagaaacaaa aattatattt gtttagaaag tctgtcagtt gggttttggc ccaattgatt 540
tacctggtag tcagaaatgt aagtgacctg actacccttg acattagtca gaaaacattc 600
taaactcgag ataaacactg tggtagttct ttagaaagaa gtcgactcta aagtactttg 660
gcgtaaaaca aaaaaatttc ccttgaaatt atactttatt tgtttgacat aaatgtttca 720
aatataatct tcttttaatc aatgtagtta aatttttttg atagaagcac gtcagaattt 780
cacactagta atgtataagt ggtaatcatc attgcctaaa acaatgatta attaaaattt 840
tagaattcgt cgtcgacaaa gaatttttct tatgatgaag gttttgtgca taattatgaa 900
cggatccatt tatttcactc taactaggaa tagatgtttg atgggataca agtgaacacg 960
acacaaagta acaaaaattc atttaacgtc acaaattatt ctcaatcgaa tttcgaagta 1020
tcaagaaaaa gggcaattaa aaacgtaatc tcattatgtc tttgaaatct aaattaaaag 1080
ttgagtttat tttaattatt aatttccttt ttgatgatgt aggggatgac aattgacaac 1140
caatttgatt tgaaagatta acataaatca ttttgagcat acatatattg cttaaatcac 1200
acccgtttat ctctaaaaac ctccataatt tatatcagga aaataaatct agttattgat 1260
atcgccatca aatatttttt tcttaaattg tttttttttg ttgttgttga aaatttaatt 1320
agaaagataa gtggacaacg acgcgagaca aagacacaat ctttagggcc ttgagtaaat 1380
tagggacagc ataaacacat gcactagtcc tgtctttatt taattatgtg gtgtttactc 1440
taatgacgtc gctttgatta accaagtggt cataattaat atatcgatcg aattatataa 1500
ttatcataaa tttgaataag catgttgcct tttattaaag aggtttaata aagtttggta 1560
ataatggact ttgacttcaa actcgattct catgtaatta attaatattt acatcaaaat 1620
ttggtcacta atattaccaa attaatatac taaaatgtta attcgcaaat aaaacactaa 1680
ttccaaataa agggtcatta tgataaacac gtattgaact tgataaagca aagcaaaaat 1740
aatgggtttc aaggtttggt tatatatgac aaaaaaaaaa aaaggtttgg ttatatatct 1800
attgggccta taaccatgtt ataacaaatt tgggcctaac taaaataata aaataaacgt 1860
aatggtcctt tttatatttg ggtcaaaccc aactctaaac ccaaaccaaa gaaaaagtat 1920
acggtacggt acacagactt atggtgtgtg tgattgcagg tgaatatttc tcgtcgtctt 1980
ctcctttctt ctgaagaaga ttacccaatc tgaaaaaaac caagaagctg acaaaattcc 2040
gaattctctg cgatttccat ggagttatct cttctccgtc cgacgactca atcgcttctt 2100
ccgtcgtttt cgaagcccaa tctccgatta aatgtttata agcctcttag actccgttgt 2160
tcagtggccg gtggaccaac cgtcggatct tcaaaaatcg aaggcggagg aggcaccacc 2220
atcacgacgg attgtgtgat tgtcggcgga ggtattagtg gtctttgcat cgctcaggcg 2280
cttgctacta agcatcctga tgctgctccg aatttaattg tgaccgaggc taaggatcgt 2340
gttggaggca acattatcac tcgtgaagag aatggttttc tctgggaaga aggtcccaat 2400
agttttcaac cgtctgatcc tatgctcact atggtggtaa gttcttgaaa caatctgatt 2460
cgttaatact gagaacaatg tatttgtgaa ttgtttgatt tgtggattta gccttctctg 2520
tttatggtta gagtaggtag atagtggttt gaaggatgat ttggtgttgg gagatcctac 2580
tgcgccaagg tttgtgttgt ggaatgggaa attgaggccg gttccatcga agctaacaga 2640
cttaccgttc tttgatttga tgagtattgg tgggaagatt agagctggtt ttggtgcact 2700
tggcattcga ccgtcacctc cagtgtgtat tcttacgacc ttttagatca atatgatttt 2760
tagggcttta tgttggtaga atctgctcat gttcttagag tttggcaaag gtgacagggt 2820
cgtgaagaat ctgtggagga gtttgtacgg cgtaacctcg gtgatgaggt ttttgagcgc 2880
ctgattgaac cgttttgttc aggtagagtt atagataaaa cttcatgtga accatttatt 2940
catttagatg tcagcgaatc tgattatcta gcagttttca tttttgttct agtgcaccct 3000
taatgcttta gattttatgc ttttaggtgt ttatgctggt gatccttcaa aactgagcat 3060
gaaagcagcg tttgggaagg tttggaaact agagcaaaat ggtggaagca taataggtgg 3120
tacttttaag gcaattcagg agaggaaaaa cgctcccaag gcagaacgag acccgtgagt 3180
aacatcaaac ttttctgttg cttgggtctt ttgtcccttc atatatatat ttgtaaaaca 3240
actcaacctg cttcttcagg cgcctgccaa aaccacaggg ccaaacagtt ggttctttca 3300
ggaagggact tcgaatgttg ccagaagcaa tatctgcaag gtatccagta cttgtctctc 3360
tctattgttg gaagggaaaa gggaaaaact ttcaaatgtt ttctgaaatc ttttgcactt 3420
tgacagatta ggtagcaaag ttaagttgtc ttggaagctc tcaggtatca ctaagctgga 3480
gagcggagga tacaacttaa catatgagac tccagatggt ttagtttccg tgcagagcaa 3540
aagtgttgta atgacggtgc catctcatgt tgcaagtggt ctcttgcgcc ctctttctgt 3600
aagtttctca ttttgcgaac taggaatttg ctcaaaattc atagattcga caaatatggg 3660
ttcttaaatc gtgtgctgca actgttatct tctcaggaat ctgctgcaaa tgcactctca 3720
aaactatatt acccaccagt tgcagcagta tctatctcgt acccgaaaga agcaatccga 3780
acagaatgtt tgatagatgg tgaactaaag ggttttgggc aattgcatcc acgcacgcaa 3840
ggagttgaaa cattaggtat atatcttgta gttataatcg tcaattatgt caaaatgttc 3900
atagaatctt catgctgttg ctcgtatttc ttcaggaact atctacagct cctcactctt 3960
tccaaatcgc gcaccgcccg gaagaatttt gctgttgaac tacattggcg ggtctacaaa 4020
caccggaatt ctgtccaagg taaaaaacag caaacacttg taacacatct ttattcaacc 4080
aagtaaacct aagaactgat agttttcttc tctctctctc ttttgttttg attccgcagt 4140
ctgaaggtga gttagtggaa gcagttgaca gagatttgag gaaaatgcta attaagccta 4200
attcgaccga tccacttaaa ttaggagtta gggtatggcc tcaagccatt cctcagtttc 4260
tagttggtca ctttgatatc cttgacacgg ctaaatcatc tctaacgtct tcgggctacg 4320
aagggctatt tttgggtggc aattacgtcg ctggtgtagc cttaggccgg tgtgtagaag 4380
gcgcatatga aaccgcgatt gaggtcaaca acttcatgtc acggtacgct tacaagtaa 4439
<210> 2
<211> 3452
<212> ДНК
<213> ген убиквитина 10 Arabidopsis thaliana (последовательность Genbank
номер AT4G05320)
<400> 2
aaacaagact accaatatga tgaactgatg attttttttt tttttttttt ggctcaacaa 60
caaactttcc attcacccat ttatggatac acagtctagc tcaacagagc ttttaaccca 120
aattggtaca atagaataca actttagatc ataattctca aaagaaagag attccttagc 180
tattctatct gccactccat ttccttctcg gcttgtatgc acaagcataa aatcctcaaa 240
cttgctaagt agatacttta tgtcttggat aattggattg agacttgaca agcataactt 300
tcatgtaacc aaagacacaa gttgctgaga atccacctca aaaatgatct tcctataatt 360
gaatcgggat aatgacagca cagcccatct aagagcctcc acttctactt ccagcacgct 420
tcttactttt accacagctc ttgcacctaa ccataacacc ttccctgtat gatcgcgaag 480
cacccaccct aagccacatt ttaatccttc tgttggccat gccccatcaa agttgcactt 540
aacccaagat tgtggtggag cttcccatgt ttctcgtctg tcccgacggt gttgtggttg 600
gtgctttcct tacattctga gcctctttcc ttctaatcca ctcatctgca tcttcttgtg 660
tccttactaa tacctcattg gttccaaatt ccctcccttt aagcaccagc tcgtttctgt 720
tcttccacag cctcccaagt atccaaggga ctaaagcctc cacattcttc agatcaggat 780
attcttgttt aagatgttga actctatgga ggtttgtatg aactgatgat ctaggaccgg 840
ataagttccc ttcttcatag cgaacttatt caaagaatgt tttgtgtatc attcttgtta 900
cattgttatt aatgaaaaaa tattattggt cattggactg aacacgagtg ttaaatatgg 960
accaggcccc aaataagatc cattgatata tgaattaaat aacaagaata aatcgagtca 1020
ccaaaccact tgcctttttt aacgagactt gttcaccaac ttgatacaaa agtcattatc 1080
ctatgcaaat caataatcat acaaaaatat ccaataacac taaaaaatta aaagaaatgg 1140
ataatttcac aatatgttat acgataaaga agttactttt ccaagaaatt cactgatttt 1200
ataagcccac ttgcattaga taaatggcaa aaaaaaacaa aaaggaaaag aaataaagca 1260
cgaagaattc tagaaaatac gaaatacgct tcaatgcagt gggacccacg gttcaattat 1320
tgccaatttt cagctccacc gtatatttaa aaaataaaac gataatgcta aaaaaatata 1380
aatcgtaacg atcgttaaat ctcaacggct ggatcttatg acgaccgtta gaaattgtgg 1440
ttgtcgacga gtcagtaata aacggcgtca aagtggttgc agccggcaca cacgagtcgt 1500
gtttatcaac tcaaagcaca aatacttttc ctcaacctaa aaataaggca attagccaaa 1560
aacaactttg cgtgtaaaca acgctcaata cacgtgtcat tttattatta gctattgctt 1620
caccgcctta gctttctcgt gacctagtcg tcctcgtctt ttcttcttct tcttctataa 1680
aacaataccc aaagagctct tcttcttcac aattcagatt tcaatttctc aaaatcttaa 1740
aaactttctc tcaattctct ctaccgtgat caaggtaaat ttctgtgttc cttattctct 1800
caaaatcttc gattttgttt tcgttcgatc ccaatttcgt atatgttctt tggtttagat 1860
tctgttaatc ttagatcgaa gacgattttc tgggtttgat cgttagatat catcttaatt 1920
ctcgattagg gtttcataga tatcatccga tttgttcaaa taatttgagt tttgtcgaat 1980
aattactctt cgatttgtga tttctatcta gatctggtgt tagtttctag tttgtgcgat 2040
cgaatttgtc gattaatctg agtttttctg attaacagat gcagatcttt gttaagactc 2100
tcaccggaaa gacaatcacc ctcgaggtgg aaagctccga caccatcgac aacgttaagg 2160
ccaagatcca ggataaggag ggcattcctc cggatcagca gaggcttatt ttcgccggca 2220
agcagctaga ggatggccgt acgttggctg attacaatat ccagaaggaa tccaccctcc 2280
acttggtcct caggctccgt ggtggtatgc agattttcgt taaaacccta acgggaaaga 2340
cgattactct tgaggtggag agttctgaca ccatcgacaa cgtcaaggcc aagatccaag 2400
acaaagaggg tattcctccg gaccagcaga ggctgatctt cgccggaaag cagttggagg 2460
atggcagaac tcttgctgac tacaatatcc agaaggagtc cacccttcat cttgttctca 2520
ggctccgtgg tggtatgcag attttcgtta agacgttgac tgggaaaact atcactttgg 2580
aggtggagag ttctgacacc attgataacg tgaaagccaa gatccaagac aaagagggta 2640
ttcctccgga ccagcagaga ttgatcttcg ccggaaaaca acttgaagat ggcagaactt 2700
tggccgacta caacattcag aaggagtcca cactccactt ggtcttgcgt ctgcgtggag 2760
gtatgcagat cttcgtgaag actctcaccg gaaagaccat cactttggag gtggagagtt 2820
ctgacaccat tgataacgtg aaagccaaga tccaggacaa agagggtatc ccaccggacc 2880
agcagagatt gatcttcgcc ggaaagcaac ttgaagatgg aagaactttg gctgactaca 2940
acattcagaa ggagtccaca cttcacttgg tcttgcgtct gcgtggaggt atgcagatct 3000
tcgtgaagac tctcaccgga aagactatca ctttggaggt agagagctct gacaccattg 3060
acaacgtgaa ggccaagatc caggataagg aaggaatccc tccggaccag cagaggttga 3120
tctttgccgg aaaacaattg gaggatggtc gtactttggc ggattacaac atccagaagg 3180
agtcgaccct tcacttggtg ttgcgtctgc gtggaggtat gcagatcttc gtcaagactt 3240
tgaccggaaa gaccatcacc cttgaagtgg aaagctccga caccattgac aacgtcaagg 3300
ccaagatcca ggacaaggaa ggtattcctc cggaccagca gcgtctcatc ttcgctggaa 3360
agcagcttga ggatggacgt actttggccg actacaacat ccagaaggag tctactcttc 3420
acttggtcct gcgtcttcgt ggtggtttct aa 3452
<210> 3
<211> 5159
<212> ДНК
<213> Последовательность гена транскетолазы (TKT) риса (GenBank номер LOC4340025)
<400> 3
catgaaccaa agagtcaaat tcaagatggc gtgctgctct tggtcttttg aatgtggcag 60
cagccaggat caatgcatcc taatatatct atgttggctt agtgaatgac taattaaaca 120
caattaaccc cagggatggt cacatgacta ctgatctctt caaactgttt taatttccca 180
tgtcagaatc tacatctgta cgtacaattt aagctctcct tttttttttt ttaaaaaaaa 240
aagacgaggt taaactaact aactaagctt gagcatcttc taatgtagta aggtatataa 300
atcagaagca aaaacaacat caaaactgtt acaaaaacaa gcaaaccata taaataagaa 360
tattccacaa aggtagattc ctgctgaata gattctatgc ccatctgggt ctatcttgtt 420
atttattctc gtgtggctcg gtttcatcat ccatcttggc gaatcacaga aaagagcggt 480
ttgcaattta tatttggttg aaaagaatgg caattttaca tgccaggatc atatacctct 540
gcatctaaaa agttgatttt tttgacccca aattttcaac ataaacatca caatcaacca 600
tggaatgacg ccagaaaaca actgtgacgg cgacgacccc aacatacaca aggagccagg 660
acaatcttta cattattttt atctcttaga aaaaaatatg tcccataaca attttttttt 720
caaaaggctc accagaccac taacaaatag gcgaaaatat ctttaataat atccaatcgt 780
ttcctaccct cttccaacgg ccaggatcat cttgtttgcc atatcaacgg tcatggctga 840
tatcctcatc caccaaccca cctcaccaca cccatcccca cataaaccca gtccatacac 900
gcaaatctaa tcaaaaccac atgtatagct gcagaatata tataacaatt ctatcatata 960
tacatactac aaacttgcac tttattattc caagaattga taccatttcc agcatctttt 1020
ttaaacctaa ctgtagaatt ttgtgaaagc tacaaaatca gtcaggtcaa tgttcatgta 1080
aaacgaacta attctcatca gattagtgtg ctataaacat aagcaaaata tataaagaaa 1140
ttagtaagat cactcctcta tgtgaatatt ttcacacctt tgccaaatat gcataaattt 1200
tttatccaag atgttgcata tacttcctcc gtttcacaat gtaagtcact ctagcatttt 1260
ccacatttat attgatgtta acgaatctag acagtctaga ttcgtgtcta gcttcattaa 1320
catcaatatg aatgtgggaa atgctagaat gacttacatt gtgaaacgga gggagtactt 1380
acctaggtta acatttgtta agtttttttt attgtgtcat attttcatca tttgttgata 1440
atgctatact ttcttttcag tatagactat gtaagtaagt agctatagct tctgttaaaa 1500
caaatgtcag gttttatcta ttatctaaaa acatccaagt taccccctct gcttcatatt 1560
ataagtcgtt tatttttttc ctaatcaaat attgttaatt tcgacaaaat ttatagaaaa 1620
aattaacaac atctaagata ttaaattagt tttattaaat ctaagttgta tatattttga 1680
taatatgatt gttgtgttga aaatactaat atatttttct acatatttag ttaaacttaa 1740
aaaaagtttg ataaaatttt tttaaaggat ttataatata aaaggaggga gtaacatttg 1800
ttaactacaa tataaaaact aatttaaaga tcaaataagt acaataacta ctttataaag 1860
attataggtt caataattgc ggcgaagaaa gtgctcctaa tctcaccagc catggcggct 1920
taggatgctg gtccatgcac catgtccacg tccacaccat agccccctcc tttttattgg 1980
ctacgagtcc ccccaaaacg cacccccaag tctatacata acccatccca tctcccaccc 2040
cacaacctcg ccatctccgc ctctcctcgc caccacacca ctcctctcgc catggccgcg 2100
cactccgtcg ccgccgcgca cgccaccatc gccgcgcgcg cgggtgccgc cgcgccagcg 2160
cccgcgccgc cggagcgcct cgggttccgc ctcagcgcgc tcgccggccg cggcctccgc 2220
tccccgctcc cgcctcgccg cggcgcgcca tcggcgtccg cgtcgcgccg ccgccacaac 2280
aaccgcgtgc gcgcggcggc ggtcgagacg ctcgaggggc aggcggcgac gggggcgctg 2340
ctcgagaagt cggtgaacac gatccggttc ctcgccatcg acgccgtcga gaaggccaac 2400
tcggggcacc ccggcctgcc catggggtgc gcgcccatgg ggcacatcct ctacgacgag 2460
gtcatgcggt acaaccccaa gaacccctac tggttcaacc gcgaccgctt catcctctcc 2520
gccggccacg gctgcatgct ccagtacgcc ctgctccacc tcgccggcta cgacgccgtc 2580
ttggtaagcg cttcccatat ccctttgctc cttgcatctc tctttcgctt ctcgagttgg 2640
ttggtttggg gcggataaat cgggagtagg ttcgtgcaag tttcggatct tgttaggtga 2700
agcagtggaa agtgtggatc ttgactgcta tttttgtgtg gcggcttcgt ttttttccta 2760
gtaaaagtgt gcaccttttc cagtcgtcag agatctgttt agtggaattt aatcttatgt 2820
aatacttcct aggaagatta gttggaagat gctattaagt cgagcaattg ttcactgtaa 2880
attctggagt gatcggtgaa gtgtttaata tggttgactt cagtttcaac tgatttgttc 2940
atcgagcttg ctgtaactat ggccgcatat tgctgcagga ggaggacttg aagcaattca 3000
ggcaatgggg aagcaagact ccaggccatc ccgagaactt cgagacgccc ggagttgaag 3060
ttaccactgg tctgtcatct caactgaagc tcatagcacc tttaagtttc tatgcttgaa 3120
tacatacaga actgaatgtt tcggtttctt gtttggtttc gttctgtagg acctcttggt 3180
cagggtattg caaatgcagt tgggctggcc cttgctgaga agcacctggc tgctcgtttc 3240
aacaagcccg acagcgagat tgttgatcac tacacgtaat ttttttttta tcatgaatac 3300
ctttagtttt cactttagtt atggtggtgc aaccttggta acatgattca tggattttgt 3360
gcagctactg tattttggga gatgggtgcc agatggaggg tatctccaat gaagcttgct 3420
cgttggctgg ccattggggt cttggcaagc tgattgcatt ctatgatgac aaccacattt 3480
ccattgacgg agacacagag attgcattca ctgaggatgt gagtgctcgc tttgaggctc 3540
ttgggtggca tacaatctgg gttaagaatg gaaacgatgg ctatgatgag attcgtgccg 3600
ccattaagga agcaaaggcg gtaactgaca agccaacact aatcaaggtt agattcctgt 3660
acattctgtt atgctgtgtt ttgttataga gcttatcata tttagccaaa atgtactctt 3720
tttaggtgac caccacaatc ggttttggat cacccaacaa ggccaactca tacagcgtcc 3780
acggaagtgc tttgggtacc aaagaggttg aagcaaccag agagaacctt ggatggccct 3840
atgagccatt ctttgtgcct gaggatgtga agaggtttgt ctattgttgt tttggttatc 3900
tgaatgagat gttgaaattt catagcactc accttatggt ttatttgcat gggctgttac 3960
agccactgga gccgccatgt gcctcaaggt gctgcttttg aggctgactg gaatgctaag 4020
ttcgccgagt atgagaagaa gtacccagaa gatgcagcga ccttgaagag cattgtctca 4080
ggggagttgc ctgctggctg ggctgacgct cttcctgtaa gtcttttacc atttctactc 4140
ataacatttg agctggtatg tgaatgatga cattctaatg gtagattact tgatttgtac 4200
agaaatacac tccagagagc cctgcagatg ccaccaggaa tctgtcacag cagtgcttaa 4260
acgcacttgc taaagttgtt cctggtcttc ttggaggaag tgctgatctt gcatcctcca 4320
acatgacatt gcttaagatg ttcggtgact tccagaagga tacgcctgag gagcgcaatg 4380
tccgatttgg agtcagggag catggaatgg gcgccatttg caacggcatt gctctgcaca 4440
gcccaggact cattccatac tgtgctactt tctttgtttt cactgattac atgagagctg 4500
ccatgaggat ctcagccttg tgtgaagccg gagttatcta tgttatgacc catgactcta 4560
ttggtcttgg agaagatggt ccaacccatc agcccattga gcacttggtg agcttccgtg 4620
cgatgcccaa cattctgatg cttcgtcctg ctgatggtaa cgagactgct ggggcataca 4680
aaatcgcggt cctcaacagg aagaggccat ccgtccttgc tctctccagg caaaagcttg 4740
ctcagctgcc tggtacctcg attgagggtg ttgagaaggg tgggtacatc gtctctgaca 4800
actcaactgg caacaagcct gacttcattg tgatgagcac tggctctgaa ctagagattg 4860
tcgccaaggc tgctgatgag ttgaggaagg aggggaagac tgtccgtgtc gtgtcatttg 4920
tttgctggga gcttttcgat gaacagtcgg ctgagtacaa ggagagtgtt ctccctgagg 4980
ctgttactgc aagagtcagc cttgaagcag ggtctactct tggatggcag aagtacgtcg 5040
gaagcaaagg caaggctatt ggcatcgaca aattcggtgc aagtgctcct gctggaaaga 5100
tctaccagga gtatggcatc accgcggaga acgtcatcgc aacagcaaag agcctgtaa 5159
<210> 4
<211> 5164
<212> ДНК
<213> Ген EPSPS риса (GenBank номер LOC4340026)
<400> 4
gctgggtagc ctttcaaagc cccaaaagat tgggtcgggt ctctctttcg gctctcgtcg 60
gtggctgctt cccacaaaga ccgccatcag acgtgaatga actgcaagtc tgcaactacg 120
actccagttg ctatcccctt aaattacttt actactacct ttattctaag ggccggttcg 180
gataattgcc aaaatcaatc gtaccaaatt ttaataatat taggatgaaa taaatatgtt 240
gcgccaaaat tttctactgt tactgaaatt tggtaacaaa ctaaacacaa ccgtatattt 300
atcaatttta ccaaatagtg gtatggttta aaatgacatt aatctaaata ggtcctgaaa 360
tataaaaatg caccaaccat caatgcatgc agttacattc caaagttctc ctaggacaca 420
tatgccatat attttcgatg gatagcctta atgtctgact actctcttca tcgcaacaaa 480
tctaatataa tacaaatctg aataggagct tatataaata tatagtagta gaatatgtca 540
taatattaca gtggtattcc atattggatt tgttgattat agaacgacga aagtatgcgt 600
taacttacaa ggttgtttac attatgacga aatgaagaga aggaacgcac ctactgcgaa 660
gagctcaaca actacttcct ccaaatttcc actcaccaac tgatcaccac caattcactt 720
gtgcagtgcc acctacccaa tttcattcca gtggcgtggt acgtaggttc cgcatgtcag 780
tgtgacaatc gtacggactg ccatgtgtat tgtgttcagg taggtgcact ggtcgtggtc 840
tcaccatctc tcctccagtc gtacggcgcc acgtaggcag tacacgtaga cggccttctt 900
cctcctccct cgagttgagt tggttggtga gagtgagaca ccgacggaac ggaaggagaa 960
ccacgccgct tggatttttc ttttttacct tttcaaattt taatttaaaa aataaaacca 1020
ttttaaaaac ttatcttcaa atacaaatct tttaaaaaca ctaacacgtg acacacagcg 1080
ggcacgtcac ccaaacgggc gtgacaatat tgttttgcca caccaatcca gctggtgtgg 1140
acaaaatgtt catatattga aaataaaatt taaaacaatt tatatttttt atctatatca 1200
ttataaaaat tgaagatgtt tttaccggta ttttgttact catttgtgca tgagtcggtt 1260
tttaagtttg ttcgcttttg gaaatacata tccgtatttg agtatgtttt taagttcgtt 1320
cgttttttga aatacaaaag gaatcgtaaa ataaatctat tttaaaaaac tcgcatgcta 1380
acttgagacg atcgaactgc taattgcagc tcataatttt ccaaaaaaaa atatatccaa 1440
acgagttctt atagtagatt tcaccttaat taaaacatat aaatgttcac ccggtacaac 1500
gcacgagtat ttttataagt aaaattaaaa gtttaaaata aataaaaatc ccgccaccac 1560
ggcgcgatgg taaaaggggg acgcttctaa acgggccggg cacgggacga tcggccccga 1620
acccggccca tctaaccgct gtaggcccac cgcccaccaa tccaactccg tactacgtga 1680
agcgctggat ccgcaacccg ttaagcagtc cacacgactc gactcgactc gcgcactcgc 1740
cgtggtaggt ggcaaccctt cttcctcctc tatttcttct tcttcctccc ttctccgcct 1800
caccacacca accgcaccaa ccccaacccc gcgcgcgctc tcccctctcc cctcccacca 1860
accccacccc atcctcccga cctccacgcc gccggcaatg gcggcgacca tggcgtccaa 1920
cgccgcggct gcggcggcgg tgtccctgga ccaggccgtg gcggcgtcgg cggcgttctc 1980
gtcgcggaag cagctgcggc tgcccgccgc ggcgcgcggg gggatgcggg tgcgggtgcg 2040
ggcgcggggg cggcgggagg cggtggtggt ggcgtccgcg tcgtcgtcgt cggtggcagc 2100
gccggcggcg aaggcggagg agatcgtgct ccagcccatc agggagatct ccggggcggt 2160
tcagctgcca gggtccaagt cgctctccaa caggatcctc ctcctctccg ccctctccga 2220
ggtgagacgc ggatcccttc ctcttgcgtg aattccattt ctggagatga gattttaggg 2280
ggtttattag gtgaggtggc tgtgtttgtg aaatcctagg aattatctct caagtcaatc 2340
taacgatgag atataactga ggttctggtt ttaatcacac actcatataa ccaatttatt 2400
gaaacatttt ggtttggcat aagaaactgc ttacgaaggt atgatatcct cctacatgtc 2460
aggctactaa attttcacga cggtatgatc cactcaaaac aagtttctta acgagtctgg 2520
tgaggtctgt tatgaaattt gtgtaaacta aggcaacttt ggaggtttcg cactgtacca 2580
atgttatgtt tgaacatttt gcaagcagtg ctttctccca aaattatgca attttgaggc 2640
tcctctacat cattataatt ccccaataca ttgctcttta ttcttaatag ctttgatcgc 2700
gaaatttaac attttaattc ttgagctgtt attttgtagc atcagtttat catgagccat 2760
gtttggtact aaatatacaa tcccttgggt ttatttgttt ccaagcatgt cattaactta 2820
tcttaatgtg gacaagaaac tgatgcctgc ttacattgct attatttcaa gcgggtattg 2880
atcctttgac atgtgattga tcattttttt ttctctggtt attagggcac aacagtggtg 2940
gacaacttgc tgaacagtga ggatgttcac tacatgcttg aggccctgaa agccctcggg 3000
ctctctgtgg aagcagataa agttgcaaaa agagctgtag tcgttggctg tggtggcaag 3060
tttcctgttg agaaggatgc gaaagaggaa gtgcaactct tcttggggaa cgctggaact 3120
gcaatgcgac cattgacagc agccgtgact gctgctggtg gaaatgcaac gtatgttttt 3180
ttttttaatg tttatgaaaa tatgtatgga attcatgggg tatgttttat gacctttttc 3240
tttaccatca gttatgtgct tgatggagtg ccacgaatga gggagagacc gattggtgac 3300
ttggttgtcg ggttgaaaca acttggtgcg gatgtcgact gtttccttgg cactgaatgc 3360
ccacctgttc gtgtcaaggg aattggagga cttcctggtg gcaaggttag ttactcctaa 3420
actgcatcct ttgtacttct gtatgcacct caattctttg tcaaccttct gcatttataa 3480
ggaacattct atgatgcaat tcgaccttac actgcacagt aacttgaaat gtttcatgct 3540
taatcaatat gccatattcc tgccaagctc aagcgagcaa tatttgtttg aatttggtac 3600
catatttttg tatatttggg cattcctttt tggtcttgat gtcttctttt gaattagcat 3660
ttaactgaat tacactcaac aggttaagct ctctggttcc atcagcagtc agtacttgag 3720
tgccttgctg atggctgctc ctttggccct tggggatgtg gagatcgaaa tcattgacaa 3780
actaatctcc attccttacg ttgaaatgac attgagattg atggagcgtt ttggtgtgaa 3840
ggcagagcat tctgatagtt gggacagatt ctatattaag ggagggcaga agtacaagta 3900
agcttctacc tgccttactg agctgaatta ttcgggtgtt tatgattaac tccctaaact 3960
aacccttttt cttttctttg gcattgacag atctcctgga aatgcctatg ttgaaggtga 4020
tgcctcaagc gcgagctatt tcttggctgg tgctgcaatc actggaggca ctgtgacagt 4080
tcaaggttgt ggtacgacca gtttgcaggt ataactgtag tgcctgtttt gacattctac 4140
cgtttagtca agtttagtca gtagtcacat attcagaata tagcacaatc tgtattatgc 4200
cactgttaat caaatacgct tgacctagag agtgctatat accctagctt aatcttcaaa 4260
ctaaacagtt ctcttgtggc ttgctgtgct gttatgttcc ctgacctaca tgttaatatt 4320
acagggtgat gtcaaatttg ctgaggtact tgagatgatg ggagcaaagg ttacatggac 4380
tgacaccagt gtaaccgtaa ctggtccacc acgtgagcct tatgggaaga aacacctgaa 4440
agctgttgat gtcaacatga acaaaatgcc tgatgttgcc atgacccttg ccgttgttgc 4500
actcttcgct gatggtccaa ctgctatcag agatggtaaa cattaaggcc tattatacct 4560
gttctatcat actagcaatt actgcttagc attgtgacaa aacaaataac caaactttct 4620
tcaaaataac ttagaaatat aagaaaggtt cgttttgtgt ggtaaacagt actactgtag 4680
tttcagctat gaagtttgct gctggcaatt ttctgaacgg tttcagctaa attgcatgtt 4740
tgttcatcat acttatccat tgtcttccac agtggcttcc tggagagtaa aggaaaccga 4800
aaggatggtt gcaattcgga ccgagctaac aaaggtaaat tcattaggtc ccgtgtcctt 4860
tcattcttca agtagtttgt tcataagttg aattctcctt caatgatgtt taaattcatc 4920
atcttctttt ttggtgttgt gccagctggg agcatcggtt gaagaaggtc ctgactactg 4980
catcatcacc ccaccggaga agctgaacat cacggcaatc gacacctacg atgatcacag 5040
gatggccatg gccttctccc tcgctgcctg cgccgacgtg cccgtgacga tcagggaccc 5100
tggttgcacc cgcaagacct tccccaacta cttcgacgtt ctaagcactt tcgtcaggaa 5160
ctga 5164
<210> 5
<211> 3481
<212> ДНК
<213> Ген убиквитина 2 риса (GenBank номер LOC4328390)
<400> 5
gaaatcaaaa gggaaatgca ccatcttata tctccagttt atatgaacag attggataag 60
atcataagat caagtggttt atattatttt gaggaatata acatggattc atcctaatca 120
ctcgtctagg cagtatgtgt attcatgatg gatatggtac tatactacgg agttttttct 180
tcacaaaata acctgttatt ttgacctcca accaaacacg aattatacca aaaattgggt 240
tatttcatct atagtacaac tctattataa acatgcagta aattatccta cacatatacc 300
aaaattcaag tgtaataatc ctaatacaca gacttaaaaa acaaactatt tcctttttaa 360
gaaaaggaaa accatttttt taacggaagg aaaacaaatt cgggtcaagg cggaagccag 420
cgcgccaccc cacgtcagca aatacggagg cgcggggttg acggcgtcac ccggtcctaa 480
cggcgaccaa caaaccagcc agaagaaatt acagtaaaaa aaaagtaaat tgcactttga 540
tccacctttt attacctaag tctcaatttg gatcaccctt aaacctatct tttcaatttg 600
ggccgggttg tggtttggac taccatgaac aacttttcgt catgtctaac ttccctttca 660
gcaaacatat gaaccatata tagaggagat cggccgtata ctagagctga tgtgtttaag 720
gtcgttgatt gcacgagaaa aaaaaatcca aatcgcaaca atagcaaatt tatctggttc 780
aaagtgaaaa gatatgttta aaggtagtcc aaagtaaaac ttatagataa taaaatgtgg 840
tccaaagcgt aattcactca aaaaaaatca acgagacgtg taccaaacgg agacaaacgg 900
catcttctcg aaatttccca accgctcgct cgcccgcctc gtcttcccgg aaaccgcggt 960
ggtttcagcg tggcggattc tccaagcaga cggagacgtc acggcacggg actcctccca 1020
ccacccaacc gccataaata ccagccccct catctcctct cctcgcatca gctccacccc 1080
cgaaaaattt ctccccaatc tcgcgaggct ctcgtcgtcg aatcgaatcc tctcgcgtcc 1140
tcaaggtacg ctgcttctcc tctcctcgct tcgtttcgat tcgatttcgg acgggtgagg 1200
ttgttttgtt gctagatccg attggtggtt agggttgtcg atgtgattat cgtgagatgt 1260
ttaggggttg tagatctgat ggttgtgatt tgggcacggt tggttcgata ggtggaatcg 1320
tggttaggtt ttgggattgg atgttggttc tgatgattgg ggggaatttt tacggttaga 1380
tgaattgttg gatgattcga ttggggaaat cggtgtagat ctgttgggga attgtggaac 1440
tagtcatgcc tgagtgattg gtgcgatttg tagcgtgttc catcttgtag gccttgttgc 1500
gagcatgttc agatctactg ttccgctctt gattgagtta ttggtgccat gggttggtgc 1560
aaacacaggc tttaatatgt tatatctgtt ttgtgtttga tgtagatctg tagggtagtt 1620
cttcttagac atggttcaat tatgtagctt gtgcgtttcg atttgatttc atatgttcac 1680
agattagata atgatgaact cttttaatta attgtcaatg gtaaatagga agtcttgtcg 1740
ctatatctgt cataatgatc tcatgttact atctgccagt aatttatgct aagaactata 1800
ttagaatatc atgttacaat ctgtagtaat atcatgttac aatctgtagt tcatctatat 1860
aatctattgt ggtaatttct ttttactatc tgtgtgaaga ttattgccac tagttcattc 1920
tacttatttc tgaagttcag gatacgtgtg ctgttactac ctatctgaat acatgtgtga 1980
tgtgcctgtt actatctttt tgaatacatg tatgttctgt tggaatatgt ttgctgtttg 2040
atccgttgtt gtgtccttaa tcttgtgcta gttcttaccc tatctgtttg gtgattattt 2100
cttgcagatg cagatctttg tgaagacatt gaccggcaag actatcaccc tcgaggtgga 2160
gtcctctgac accatcgata atgtcaaggc taagatccaa gataaggagg gcatcccccc 2220
ggaccagcag cgtctcatct tcgctggcaa gcagctggag gatggcagga cccttgctga 2280
ctacaacatc cagaaggagt cgacccttca ccttgtcctc cgcctccgtg gtggcatgca 2340
gatctttgtc aagactctga ccggcaagac tatcaccctt gaggtggagt cttctgacac 2400
catcgacaac gtcaaggcca agatccagga caaagagggc atccccccag accagcagcg 2460
tctcatcttc gccggcaagc agctggagga tggcaggacc cttgctgact acaacatcca 2520
gaaggagtcc accctccacc ttgtcctccg cctccgtggt ggcatgcaga tctttgtcaa 2580
gacactgacc ggcaagacca tcaccctcga ggtggaatct tctgacacca tcgacaacgt 2640
caaggccaag atccaggaca aggagggcat tcccccggac cagcagcgtc tcatctttgc 2700
cggcaagcag cttgaggacg gcaggaccct tgctgactac aacatccaga aggagtcaac 2760
gcttcacctt gtcctccgtc tcaggggagg catgcaaatc ttcgtgaaga ctctgaccgg 2820
caagaccatc accctcgagg tggagtcttc tgataccatc gacaatgtca aggccaagat 2880
ccaggacaag gagggcattc ccccggacca gcagcgcctc atctttgctg gcaagcagct 2940
ggaggatggc aggacccttg ctgactacaa catccagaag gagtccaccc tccaccttgt 3000
gctccgcctt cgtggtggta tgcagatctt tgtcaagacc ctcacaggca agaccatcac 3060
cctggaggtt gagagctcgg acaccatcga caacgtcaag gccaagatcc aggacaagga 3120
gggcatcccc ccagaccagc agcgtctcat cttcgccggc aagcagctcg aggatggccg 3180
caccctcgcc gactacaaca tccagaagga gtctaccctc cacctggtgc ttcgtctccg 3240
tggtggtatg cagatcttcg tgaagacctt gactgggaag accatcactt tggaggttga 3300
gagctccgac accattgata atgtgaaggc caagatccag gacaaggagg ggattccccc 3360
agaccagcag cgtctgatct tcgctggcaa gcagctggag gatggacgca ccctcgccga 3420
ctacaacatc cagaaggagt ccaccctcca cctggtgctc cgcctccgtg gtggtcagta 3480
a 3481
<210> 6
<211> 3166
<212> ДНК
<213> Ген HPPD риса (GenBank номер LOC4328425 )
<400> 6
aaatggtatt gtttagaagt ttagaaaacg ttctgccgga cggaatggtt ccatccctta 60
aaaaaggacg ctgtttcaaa ttcgaaaaaa agaagagcca tcacgataac cagacggtta 120
ctgacgattt tgccgaaggt gattatcgat ggtcagtcaa caaagtataa accctgctta 180
gcggtaataa acacttgatg ttttacgtaa aatcttttga aacttagacg gccaacgact 240
atcaaaatat ttacatcgaa atcatgaaaa aatgttgtgt tcatttgtct ttgcaaataa 300
tttcaaggcc tcataaactt attttaatac aaaataatgg ctaacgtcac actaaatcaa 360
aatccaatat caaatgcttt taaggaagca gtagagatat cttcaattga gattttgact 420
gtttgagaca gctcattcta ccgcttttca tatgtctatc gagaccgagg taatatatgt 480
ggcacaaaac gttttagaga agtgatgtga aaaacactac catccatcca tttgcaataa 540
gaaaatcctc cattactact ctccccgatt cttaatagat aacactatca atttttaaaa 600
atatatattt atcattcatc taattaagga atataattat cttttatttt tttaacttgg 660
tttatcatta aaaacacttt taaatatgac ttatattttc ttatgtttac acatgttttt 720
aaaataaaac aaatgattga acatataatc acctagtaaa aacaagagct ttactccaag 780
ttaccacaaa atttgtttcc ccttccctct aaaaatcaga acagtcacca acttttctca 840
aaaaaaaaaa aagtaacaaa ctcatcagct tcacaaattc accagcagcc aaggcaacca 900
gcaccagagg acaaatcccg tccgcaacca ctagacttag cgtccacgtg gcgccatctc 960
ctccctttat ttaacccccc accaccaact cctcccccca cgccgccact gtcatccact 1020
cccccacacc ccacgacgcg ccacgccacg ccgcgccgcg ccgcgccatg cctcccactc 1080
ccacccccac cgccaccacc ggcgccgtct cggccgctgc ggcggcgggg gagaacgcgg 1140
ggttccgcct cgtcgggcac cgccgcttcg tccgcgccaa cccgcggagc gaccggttcc 1200
aggcgctcgc gttccaccac gtcgagctct ggtgcgccga cgccgcgtcc gccgcgggcc 1260
ggttcgcctt cgccctgggc gcgccgctcg ccgccaggtc cgacctctcc acggggaact 1320
ccgcgcacgc ctccctcctc ctccgctccg cctccgtcgc gttcctcttc accgccccct 1380
acggcggcga ccacggcgtc ggcgcggacg cggccaccac cgcctccatc ccttccttct 1440
ccccaggcgc cgcgcggagg ttcgccgcgg accacggcct cgcggtgcac gccgtggcgc 1500
tgcgcgtcgc cgacgcggcc gacgccttcc gcgccagcgt cgcggccggt gcgcgcccgg 1560
cgttccagcc cgccgacctc ggcggtggct tcggcctcgc ggaggtggag ctctacggcg 1620
acgtcgtgct ccgcttcgtc agccacccgg acggcgccga cgcgcccttc ctcccgggtt 1680
tcgagggcgt cagcaacccg ggcgccgtgg actacggcct ccgccggttc gaccacgtcg 1740
tcggcaacgt gccggagctc gctccggtag ccgcgtacat ctccgggttc accgggttcc 1800
acgagttcgc cgagttcacc gccgaggacg tgggcaccgc cgagagcggc ctcaactcgg 1860
tggtgctcgc caacaacgcg gagaccgtgc tgctgccgct caacgagccg gtgcacggca 1920
ccaagcggcg gagccagata cagacgtacc tggaccacca cggcggcccg ggggtgcagc 1980
acatcgcgct ggccagcgac gacgtgctcg ggacgctgag ggagatgcgg gcgcgctccg 2040
ccatgggcgg cttcgagttc ttggcgccgc cgccgcccaa ctactacgac ggcgtgcggc 2100
ggcgcgccgg ggacgtgctc tcggaggagc agatcaacga gtgccaggag ctcggggtgc 2160
tcgtggacag ggatgaccag ggggtgttgc tccagatctt caccaagcca gtaggagaca 2220
ggtaaaatcc tcacctcttt catgatgaaa atggcttatg aattcagatt tgcagttatt 2280
tgttggcaca tagcatcgat taggcgcaga aaggtgtcaa gcattatgaa attaatccag 2340
aatgcttgaa taatacagta taatatatga tagtgagctc tgtgatactc catggatact 2400
ctttatgtgt ctccatgaat ccatgatgcg cctttctgaa gattgtgaca ctagaaaggg 2460
aataaagctg aatgtgcata ggaaaaaaat gaaaagccaa tgtgtgtctg tttatgcctt 2520
cttgcaagca tatcccagtt cctttttgcc ggcatgttgt aatgcagata gccagccaca 2580
tatagctact taattagtga gtactccctc tcacaatgta agtcattcta gtattttcca 2640
cattcatatt gatgctaatc tatctagatt cattagcatc aatatgaata tgggaaatac 2700
tagaatgact tacattgtga aacggaggaa gtattactta ctacatctaa ggtccatgga 2760
ttcctttttt tacaaaagaa agaaagaatc ttatggcaac tccatcagca taaaccagca 2820
atgctgctgg gaacaactta aactttaggt tcaggaggtt gtaattgtct ttaagcttaa 2880
tagtctgatt cagtcagtat tctaatttct gctgcatctt tgctattgtt atttcctctc 2940
tgtgactcca aatctaactg gatcagctat ttcactcagg ccaacctttt tcttggagat 3000
gatacaaagg attgggtgca tggagaagga tgagagtggg caggagtacc agaagggcgg 3060
ctgcggcggg tttgggaagg gcaacttctc ggagctgttc aagtccattg aggagtatga 3120
gaaatccctt gaagccaagc aagcccctac agttcaagga tcctag 3166
<210> 7
<211> 4311
<212> ДНК
<213> Ген PPO риса (GenBank номер LOC4327918 )
<400> 7
caaaaaaaat tattcaaaag gaagcaggca gggcagcaca ttgtgaggga atcaggcggc 60
ggcgcacagt tctctccctt ggcaccctcc tttccctcat atgcaatgga ctcaaaaggg 120
cggctgcagc gtatgccggt ggattaggtt tcgagggtgg atccagtgag agcaaaggtg 180
gtggggtgac ccaaggacgc tgcttcgctc aagtccacat gtgcttcgcc tcaaggccag 240
gatgccgccg ccctactcga gactaggtag ctgacttgag actaggccgt tgtcgcgcgt 300
cggggccgcc cagccgctgc tccaccttgg gtcacgctgt cactctatct caaggctggg 360
ccatcgccac tatgcttgaa gccgcactac cgtcccacct caaggccggc accctgtcac 420
tccacctcga ggctacctag acgttatttt gttccgctgc tacgccttag agacacccca 480
ctgccgctct accttgggct atgccgtcgc tctttctcca aggcgcgctg ctacaactcc 540
gccctgagtc tccatgtgcc tgtgtggaga aaggagaggg aagaaagaaa gaaaggagaa 600
agaagtggga ggaatcacta acatgtgagc tctgtatatt gatttggtca gttggattag 660
gtcaacgagc catccaagag aaaacaaccc ttaaaatcat tgagaaaatc atttttaact 720
gttttaatag ttaagagatg aagatatatt tttttttata gaaagagatg aagatatccg 780
atatccaata tgataggaat cagaaaacgt atatttttga gggagtcaat gcggacttat 840
tcccaccaga aactgcagca tattggacgc agtggttcct tccgcggtag ctgcgcccgc 900
gcccacggga atgcgattcc cagcccattc cacccccttc ccattccggt gggccattcc 960
gaatccaacc aaccaaccaa ccatcgaacc cgcacccgca tcgtatccac tcctctccag 1020
tctccccgcc gctccgcatc ccgcagccgc tcgtcagcga cggacatggc cgccgccgcc 1080
gcagccatgg ccaccgccac ctccgccacg gcagcgccgc cgctccgcat tcgcgacgcc 1140
gcgaggagga cccgccgacg cggccacgtt cgctgcgccg tcgccagcgg cgcggccgag 1200
gcgcccgcgg cgcccggggc gcgggtgtcg gcggactgcg tcgtggtggg cggcggcatc 1260
agcgggctct gcaccgcgca ggcgctggcc acaaagcacg gcgtcggcga cgtgctcgtc 1320
acggaggccc gcgcccgccc cggcggcaac atcaccaccg ccgagcgcgc cggcgagggc 1380
tacctctggg aggaggggcc caacagcttc cagccttccg accccgtcct caccatggcc 1440
gtacgtcttc ttgctcccct tctcttctga ttctctcgcg gcggagacga catgaatggg 1500
aatggtggtg catggattgg ggcgcgcagg tggacagcgg gctcaaggac gatctcgtgt 1560
tcggggaccc caacgcgccg cggttcgtgc tgtgggaggg gaagctaagg ccggtgccgt 1620
ccaagcccgg cgacctgccg ttcttcgacc tcatgagcat ccccggcaag ctcagggccg 1680
gccttggcgc gctcggcgtt cgagcgccac ctccagtttg tgtgctctcc ccgctgtgca 1740
ttcttgattc acttgtgaaa ttcgattgtg ctgagcgttt ccggcgaagg tttcaggggc 1800
gtgaggagtc ggtggaggac ttcgtgcggc gcaacctcgg cgcggaggtc tttgagcgcc 1860
tcattgagcc tttctgctca ggtgtttatt gtagtgtgca attgctgttt tgtttttgat 1920
gattcagata agaatacggt gatttcggtg cttaggtgtg tatgctggtg atccttcaaa 1980
gctcagtatg aaggctgcat ttgggaaggt gtggaggctg gaggatactg gaggtagcat 2040
tattggtgga accatcaaaa caatccagga gagggggaaa aaccccaaac cgccgaggga 2100
tccgtgagtg agaaattgcc ttctttgttg gattaattgt ccattgtgtt acactgatat 2160
gccttcacca tttttagccg ccttccaacg ccaaaggggc agacagttgc atctttcagg 2220
aagggtctga ctatgctccc ggatgctatt acatctaggt ttgttatcat tgtctttgta 2280
atttacctag ttcttcaact atggatatta ggtgctgtag attgttcaga tagatgcaca 2340
ttgtacaaca atctaggtag attgattgct atggcttgtt gaattaactg tttcacttgc 2400
atcattgcct cagcacatat gaagcatatg gatagattct tcaatcattt atccctcaat 2460
aacacaattt tgacaccagt gcttctcctt tttttcatcc tgcttcatct ggctatccac 2520
aatataatta agcatacaaa agaggcactc tttgatggac aattcatagt gttgtgggtt 2580
aatattcatt tgcattcttt gagggacaat tcgttacacc ctaacatgaa ctagtaatga 2640
ttggggtgct taaccatttg ttctgcattt cctccatttt caggttgggt agcaaagtca 2700
aactttcatg gaagttgaca agcattacaa agtcagacaa caaaggatat gcattagtgt 2760
atgaaacacc agaaggggtg gtctcggtgc aagctaaaac tgttgtcatg accatcccat 2820
catatgttgc tagtgatatc ttgcggccac tttcagtaag ttatatatat ttaattaact 2880
ttctgttccc aaaatacact gcagcacttc attgcttcct gaggtcctcg attcattttt 2940
cggtagacag gaagtagtat tcatttgcac tttttaaggg attaattcaa catatccact 3000
ggaaatatac atatcctaca catcctgtca acatacttgc taaacagcat tttgtttgag 3060
ttgactggca tctcagcagc caattactat ctttagggga caagccacat tcttaataaa 3120
tcctgtcgga aatcactttt tgatttttat agattagtgt tgtcatagaa tttggcttgg 3180
tgatctactt ggtaaggtta actgattcac aagtcggaca atttcttcac caatctagca 3240
gtatgtttaa gtgtgttggt acttaattct aaatgtcctg cgcatggtaa catatcatat 3300
gcaaaaattc ctcagtaacc gaaatttata ctgtaagttt taactgtctt tacactgtta 3360
attttagaca tacttcttcc ttgcttgttc attgaacttg tttccccctt ccacagagtg 3420
atgcagcaga tgctctgtca atattctatt atccaccagt tgctgctgta actgtttcat 3480
atccaaaaga agcaattaga aaagaatgct taattgacgg agagctccag ggtttcggcc 3540
agctgcatcc gcgtagtcag ggagttgaga ctttaggtac ttataggaat tcaaccttat 3600
tattcttcta acatataaat gaactaatct ttcttgtcta gtttgcattt attgtggatt 3660
aagtttggtt atattgttct tacaagtttg tggtattatt ttgtatagga acaatatata 3720
gctcatcact ctttccaaat cgtgctccag ctggaagggt gttacttctg aactacatag 3780
gaggttctac aaatacaggg attgtttcca aggtatcgct gtcaagttgt ttattttgcg 3840
actatatgat tacagtatcc tgtttttcaa ctccagctgc tgttagactg tcataataaa 3900
tctgctacta catgtttgca cactatttga ctgcatttaa aaactcagat agcctatatt 3960
tttagttgcc tgctactggg tgtatttcta atgatcccat catgtttgca gactgaaagt 4020
gagctggtag aagcagttga ccgtgacctc aggaagatgc tgataaatcc taaagcagtg 4080
gaccctttgg tccttggcgt ccgggtatgg ccacaagcca taccacagtt cctcattggc 4140
catcttgatc atcttgaggc tgcaaaatct gccctgggca aaggtggtta tgatggattg 4200
ttcctcggag ggaactatgt tgcaggagtt gccctgggcc gatgcgttga aggtgcatat 4260
gagagtgcct cacaaatatc tgactacttg accaagtacg cctacaagtg a 4311
<210> 8
<211> 2463
<212> ДНК
<213> Ген CP12 риса (GenBank номер LOC4325791)
<400> 8
tttaaatcgt atttctactt ggactctcct ttccttttct aattttggat tttttttttg 60
aattttgatt aatctcgtat tgggttctta tatggaaact tctttcaata ttgcttattt 120
tcaattccga atttcagcta tttttaaatc gtatttctac ttggactctc cttttctttt 180
tttctttttc ttcgattaat gtgggaattt ctagccccca cagcgaacgt ggtgactatt 240
ttcaaagctg ttttaataat ataatagatt ttattttagg gcctgttcag attgtagcca 300
aaataaacct tatcaaaatt tgacaatacc aaaattttgt caagttgaca atattgccaa 360
aattttggca ggatttctta cgtatttatc aaatttggca acaaactaaa catagatatt 420
tttttggcaa ctttaccaaa aaagttgtat gattgaaaat gacatcaatc tgaacagccc 480
cttagaactt agaactacaa tctgaacagc cccttagaac ttagaactac agtggaatcc 540
atcttggcat atgtacttgg cagggaattt tttttacatt tttattatta aaaaattaca 600
aaattttttt tcgaaaattt acaaatctag acgcctaaca tcctttagga gggcgttttt 660
ttaaatcgcc cttctaaagg gcgttaaggg acctaaatgc aaaattttta tttgtattcg 720
aaccctttcc accagtttta attgcttaaa agctaataat gcttatttga cactccaaat 780
aattttaaat gaaaaggtaa taaactacaa agttgtagat ctcatcgaga tctatatctt 840
ttttataaag tttatctcca tccaatgtcg tttgaaatgt agttctgaga atttttttaa 900
ataggtttta gattttgtaa cgaatatttg gacatctaaa cgatcttaaa tgaaagttgt 960
taattacaaa cttgtagatc ttctccagct ctacaatttt gatataaact ttaatttcat 1020
atggctttat atgatatagt tttaaattgt aaaatcatag aggtagcaag ctatgatgga 1080
aaatttgtat ttaggtccct aacgccctct ggaagggtga tttgaaaaaa gccctctagg 1140
aggacgtgag gcacgactag atttgtaaat tttcggaatg aaaaattatt tttaaatatt 1200
ttaatcaaaa aatgtaaaaa taaaaaaaat tctcggcagg gtggcagcat gggcctaagg 1260
cccagtcaac tgtgggccta taagcgacta atccggctgt aactgggcct tgcaagaggc 1320
ttgtcttgtt ggtccgaact caggaagtcc aggttgcggg gacaacttca aggccatctg 1380
gtttccactt ctcttaccac ctcaattccg ctcttgatcc gagctagctt agtcccaatc 1440
taaaaacttt acaaagaaag aaccatacgc acctattggg caaaatgaaa aataatttgc 1500
tactcaccaa ataatttgag cacctctgca cctgtacact aaataactct gttccaccaa 1560
aatagttgag atatctagga cgtttcattt tgtccgttct tcaccaaact tttccatagt 1620
atctcagata ttttcgagac cgaaagtgat ctttctggcc ttagaccgag ttcacttccc 1680
tacaagccat tctttgctgg cacaacacga acctctacat caatttcgta tccaacctga 1740
acttctgcat acatgtacac acccacagtc atctgctcat gttttcacgg tcaaattaaa 1800
actgcttctc tcaccttaga ttcacccaag ggaaaagaaa aagatctcct ttgccaagtc 1860
cccatttcgc atgaaatatc tcaaaataca gcccacgtgg cacacgacga ttggctgagg 1920
aggcgataag aaacgagtgc acgtcgtcga atcctctctc cccttctccc ccaccccacg 1980
gagctatata tatataaacc ccatctcttc aatccgtgca acgaacgcct cgtcgcaaca 2040
gctacaaacg cccacatcac acgcagaaat ccgcatcaac agccagccat ggcgtccacg 2100
ctgaccaacg tcggcctgtc taccccggcg gcggcggcgt cgtccctcgt taggccggtc 2160
gccggagctg gacgcgtggt gtttccccgt gttggccgcg gcgggttcgc ggcggtgagg 2220
gcgagcgggc cggcgacgcc gccggacatc tcggacaaga tgtcggagag catcgacaag 2280
gcgaaggagg cgtgcgcgga ggacacggcg agcggcgagt gcgcggcggc gtgggacgag 2340
gtggaggagc tgagcgcggc ggcgagccac gcgcgcgaca agctcaagga gacctccgac 2400
ccgctcgagg cctactgcaa ggacaacccg gagaccgacg agtgccgcac ctacgacaac 2460
tga 2463
<210> 9
<211> 744
<212> ДНК
<213> Последовательность удвоения в результате трансформации протопласта
посредством pQY002066 (искусственная последовательность)
<400> 9
ttcaagtcca taggcgaggc gaccggcccg cggcggacgc ggcgtcggcg caccagagct 60
cgacgtggtg gaacgcgagc gcctggaacc ggtcgctccg cgggttggcg cggacgaagc 120
ggcggtgccc gacgaggcgg aaccccgcgt tctcccccgc cgccgcagcg gccgagacgg 180
cgccggtggt ggcggtgggg gtgggagtgg gaggcatggc gcggcgcggc gcggcgtggc 240
gtggcgcgtc gtggggtgtg ggggagtgga tgacagtggc ggcgtggggg gaggagttgg 300
tggtgggggg ttaaataaag ggaggagatg gcgccacgtg gacgctaagt ctagtggttg 360
cggacgggat ttgtcctctg gtgctggttg ccttggcaga tagggtaaga actagcacaa 420
gattaaggac acaacaacgg atcaaacagc aaacatattc caacagaaca tacatgtatt 480
caaaaagata gtaacaggca catcacacat gtattcagat aggtagtaac agcacacgta 540
tcctgaactt cataaataag tagaatgaac tagtggcaat aacttttcac acagaacatc 600
tcccttgttt tcaaatatgt agtaacagca cacgtatcct gtacttcaaa acttagtaga 660
atgaactagt ggcagtaatc tttcacacag aaagaatgaa ctagtggcaa taatctttca 720
cacagaatgt tggttgatgg atag 744
<210> 10
<211> 603
<212> ДНК
<213> Последовательность удвоения в результате трансформации протопласта
посредством pQY002068 (искусственная последовательность)
<400> 10
caacgtgcct taggcgaggc gaccggcccg cggcggacgc ggcgtcggcg caccagagct 60
cgacgtggtg gaacgcgagc gcctggaacc ggtcgctccg cgggttggcg cggacgaagc 120
ggcggtgccc gacgaggcgg aaccccgcgt tctcccccgc cgccgcagcg gccgagacgg 180
cgccggtggt ggcggtgggg gtgggagtgg gaggcatggc gcggcgcggc gcggcgtggc 240
gtggcgcgtc gtggggtgtg ggggagtgga tgacagtggc ggcgtggggg gaggagttgg 300
tggtgggggg ttaaataaag ggaggagatg gcgccacgtg gacgctaagt ctagtggttg 360
cggacgggat ttgtcctctg gtgctggttg ccttggagat agggtaagaa ctagcacaag 420
attaaggaca caacaacgga tcaaacagca aacatattcc aacagaacat acatgtattc 480
aaaaagatag taacaggcac atcacacatg tattcagata ggtagtaaca gcacacgtat 540
cctgaacttc agaaataagt agaatgaact agtggcaata acttttcccc ccaaaaacct 600
gtc 603
<210> 11
<211> 477
<212> ДНК
<213> Последовательно реверсии в результате трансформации протопласта
посредством pQY002062 (искусственная последовательность)
<400> 11
gggcggattt cgacggaggg gcggagagga ggaggatcct gttggagagc gacttggacc 60
ctggcagctg aaccgccccg gagatctccc tgatgggctg gagcacgatc tcctccgcct 120
tcgccgccgg cgctgccacc gacgacgacg acgcggacgc caccaccacc gcctcccgcc 180
gcccccgcgc ccgcacccgc acccgcatcc ccccgcgcgc cgcggcgggc agccgcagct 240
gcttccgcga cgagaacgcc gccgacgccg ccacggcctg gtccagggac accgccgccg 300
cagccgcggc gttggacgcc atggtcgccg ccattgcggt gtggtggcga ggagaggcgg 360
agatggcgag gttgtggggt gggagatggg atgggttatg tatagacttg ggggtgcgtt 420
ttggggggac tcgtagccaa taaaaaggag ggggctatgg tgtggccgtt tggacat 477
<210> 12
<211> 670
<212> ДНК
<213> Последовательность делеции в результате трансформации протопласта
посредством pQY002062 (искусственная последовательность)
<400> 12
ttttcgttgg aggggcggag aggaggagga tcctgttgga gagcgacttg gaccctggca 60
gctgaaccgc cccggagatc tccctgatgg gctggagcac gatctcctcc gccttcgccg 120
ccggcgctgc caccgacgac gacgacgcgg acgccaccac caccgcctcc cgccgccccc 180
gcgcccgcac ccgcacccgc atccccccgc gcgccgcggc gggcagccgc agctgcttcc 240
gcgacgagaa cgccgccgac gccgccacgg cctggtccag ggacaccgcc gccgcagccg 300
cggcgttgga cgccatggtc gccgccattg gacgagtcgg aatcgcagac cgataccagg 360
atcttgccat cctatggaac tgcctcggtc catggccgcg cactccgtcg ccgccgcgca 420
cgccaccatc gccgcgcgcg cgggtgccgc cgcgccagcg cccgcgccgc cggagcgcct 480
cgggttccgc ctcagcgcgc tcgccggccg cggcctccgc tccccgctcc cgcctcgccg 540
cggcgcgcca tcggcgtccg cgtcgcgccg ccgccacaac aaccgcgtgc gcgcggcggc 600
ggtcgagacg ctcgaggggc aggcggcgac gggggcgctg ctcgagaagt cggaaacccc 660
cgaatccaaa 670
<210> 13
<211> 777
<212> ДНК
<213> Последовательность реверсии в результате трансформации протопласта
посредством pQY002093 (искусственная последовательность)
<400> 13
gaacgatgcg agccccacgc tcgaccgccg ccgcgcgcac gcggttgttg tggcggcggc 60
gcgacgcgga cgccgatggc gcgccgcggc gaggcgggag cggggagcgg aggccgcggc 120
cggcgagcgc gctgaggcgg aacccgaggc gctccggcgg cgcgggcgct ggcgcggcgg 180
cacccgcgcg cgcggcgatg gtggcgtgcg cggcggcgac ggagtgcgcg gccatggcga 240
gaggagtgga tggtcacctg actaccggtc ccctcaaact ggtttaattt cccctggcaa 300
aatccacctc cggacctaca atttaagctc cccttttttt ttttaaaaaa aaaaaaaaag 360
agggtaaacc aactaaccta gcttggacct tttttaatgg agtaagggat ttaaaacaaa 420
aacaaaaaaa aaatccaaac tggtaaaaaa acaaacaaac catttaaata aaaattttcc 480
ccaaagggaa attcctggcg aaaaaaatct atgcccctct gggtctatct tggtattttt 540
tccccggggc tccgtttcat ccttcatttt ggcgaataca aaaaaaaccg tttgaatttt 600
ttttggttga aagaatggca atttactggc caggatcatg tactctgcat ctaagaattg 660
attttttgac cccaaatttc aacttagcct ccatcgcagt agtgcgcaca cacaggctga 720
aggtgactct tagacccaat gtcactatct cagcaatatg cagagagaat gacccaa 777
<210> 14
<211> 635
<212> ДНК
<213> Последовательность делеции в результате трансформации протопласта
посредством pQY002093 (искусственная последовательность)
<400> 14
agaacgatgc gcagcctcgg cgtctcgacc gccgccgcgc gcacgcggtt gttgtggcgg 60
cggcgcgacg cggacgccga tggcgcgccg cggcgaggcg ggagcgggga gcggaggccg 120
cggccggcga gcgcgctgag gcggaacccg aggcgctccg gcggcgcggg cgctggcgcg 180
gcggcacccg cgcgcgcggc gatggtggcg tgcgcggcgg cgacggagtg cgcggccatg 240
gcgagaggag tgtggtgggt accctttcaa acccccaaaa gagtgggtcg ggtctctctt 300
tcggctctcg gcgggggctg cttcccacaa agaccgccat cagacgtgag tgaactgcaa 360
gtctgcaact accactccag gtgctctccc cttaaattac tttactacta cctttattct 420
aggggccggt tcagataatt gccaaaatca ctcgcaccat tttttaataa tattaggatg 480
aaatatatat gtcgcgccaa atttttctac tgttactgaa atttggacac aaactacaca 540
ccaccgtata tttatctttt ttaccaaata ggggtatggt ttaaaatgac tttaatctaa 600
ataggccctg aaatataaaa gcgccccacc cctca 635
<210> 15
<211> 652
<212> ДНК
<213> Последовательность реверсии в результате трансформации протопласта
посредством pQY002095 (искусственная последовательность)
<400> 15
tttgagtaga ctcgccggcg cgctcggcgg tggtgatgtt gccgccgggg cgggcgcggg 60
cctccgtgac gagcacgtcg ccgacgccgt gctttgtggc cagcgcctgc gcggtgcaga 120
gcccgctgat gccgccgccc accacgacgc agtccgccga cacccgcgcc ccgggcgccg 180
cgggcgcctc ggccgcgccg ctggcgacgg cgcagcgaac gtggccgcgt cggcgggtcc 240
tcctcgcggc gtcgcgaatg cggagcggcg gcgctgccgt ggcggaggtg gcggtggcca 300
tggctgcggc ggcggcggcc atgtccgtcg ctgacatgcg gatttctgcg tgtgatgtgg 360
gcgtttgtag ctgttgcgac gaggcgttcg ttgcacggat tgaagagatg gggtttatat 420
atatatagct ccgtggggtg ggggagaagg ggagagagga ttcgacgacg tgcactcgtt 480
tcttatcgcc tcctcagcca atcgtcgtgt gccacgtggg ctgtattttg agatatttca 540
tgcgaaatgg ggacttggca aaggagatct ttttcttttc ccttgggtga atctaaggtg 600
agagaagcag ttttaatttg accgtgaaaa catgagccaa aaggaaaaaa gg 652
<210> 16
<211> 506
<212> ДНК
<213> Последовательность делеции в результате трансформации протопласта
посредством pQY002095 (искусственная последовательность)
<400> 16
acgcgtggtt cttccggtag ctgcgcccgc gcccacggga atgcgattcc cagcccattc 60
cacccccttc ccattccggt gggccattcc gaatccaacc aaccaaccaa ccatcgaacc 120
cgcacccgca tcgtatccac tcctctccag tctccccgcc gctccgcatc ccgcagccgc 180
tcgatgcgga tttctgcgtg tgatgtgggc gtttgtagct gttgcgacga ggcgttcgtt 240
gcacggattg aagagatggg gtttatatat atatagctcc gtggggtggg ggagaagggg 300
agagaggatt cgacgacgtg cactcgtttc ttatcgcctc ctcagccaat cgtcgtgtgc 360
cacgtgggct gtattttgag atatttcatg cgaaatgggg acttggcaaa ggagatcttt 420
ttcttttccc ttgggtgaat ctaaggtgag agaagcagtt ttaatttgac cgtgaaaaca 480
tagcccagaa tgacaaaata accttt 506
<210> 17
<211> 614
<212> ДНК
<213> Последовательность реверсии в результате трансформации протопласта
посредством pQY002098 (искусственная последовательность)
<400> 17
gcatcgtccg gccgtcggcg gtggtgatgt tgccgccggg gcgggcgcgg gcctccgtga 60
cgagcacgtc gccgacgccg tgctttgtgg ccagcgcctg cgcggtgcag agcccgctga 120
tgccgccgcc caccacgacg cagtccgccg acacccgcgc cccgggcgcc gcgggcgcct 180
cggccgcgcc gctggcgacg gcgcagcgaa cgtggccgcg tcggcgggtc ctcctcgcgg 240
cgtcgcgaat gcggagcggc ggcgctgccg tggcggaggt ggcggtggcc atggctgcgg 300
cggcggcggc catgttgtgg gcgtttgtag ctgttgcgac gaggcgttcg ttgcacggat 360
tgaagagatg gggtttatat atatatagct ccgtggggtg ggggagaagg ggaaagagga 420
ttcgacgacg tgcactcgtt tcttatcgcc tcctcagcca atcgtcgtgt gccacgtggg 480
ctgtattttg agatatttca tgcgaaatgg ggacttggca aaggagatct ttttcttttc 540
ccttgggtga atctaaggtg agagaagcag ttttaatttg accgtgaaaa catgagccca 600
aattgacaaa tccc 614
<210> 18
<211> 574
<212> ДНК
<213> Последовательность удвоения HPPD в QY2091-13 (искусственная
последовательность)
<400> 18
ggatacgtgt gctgttacta cctatctgaa tacatgtgtg atgtgcctgt tactatcttt 60
ttgaatacat gtatgttctg ttggaatatg tttgctgttt gatccgttgt tgtgtcctta 120
atcttgtgct agttcttacc ctatctgcca aggcaaccag caccagagga caaatcccgt 180
ccgcaaccac tagacttagc gtccacgtgg cgccatctcc tccctttatt taacccccca 240
ccaccaactc ctccccccac gccgccactg tcatccactc ccccacaccc cacgacgcgc 300
cacgccacgc cgcgccgcgc cgcgccatgc ctcccactcc cacccccacc gccaccaccg 360
gcgccgtctc ggccgctgcg gcggcggggg agaacgcggg gttccgcctc gtcgggcacc 420
gccgcttcgt ccgcgccaac ccgcggagcg accggttcca ggcgctcgcg ttccaccacg 480
tcgagctctg gtgcgccgac gccgcgtccg ccgcgggccg gttcgccttc gccctgggcg 540
cgccgctcgc cgccaggtcc gacctctcca cggg 574
<210> 19
<211> 551
<212> ДНК
<213> Последовательность удвоения HPPD в QY2091-20 (искусственная
последовательность)
<400> 19
aggatacgtg tgctgttact acctatctga atacatgtgt gatgtgcctg ttactatctt 60
tttgaataca tgtatgttct gttggaatat gtttgctgtt tgatccgttg ttgtgtcctt 120
aatcttgtcc aaggcaacca gcaccagagg acaaatcccg tccgcaacca ctagacttag 180
cgtccacgtg gcgccatctc ctccctttat ttaacccccc accaccaact cctcccccca 240
cgccgccact gtcatccact cccccacacc ccacgacgcg ccacgccacg ccgcgccgcg 300
ccgcgccatg cctcccactc ccacccccac cgccaccacc ggcgccgtct cggccgctgc 360
ggcggcgggg gagaacgcgg ggttccgcct cgtcgggcac cgccgcttcg tccgcgccaa 420
cccgcggagc gaccggttcc aggcgctcgc gttccaccac gtcgagctct ggtgcgccga 480
cgccgcgtcc gccgcgggcc ggttcgcctt cgccctgggc gcgccgctcg ccgccaggtc 540
cgacctctcc a 551
<210> 20
<211> 155
<212> ДНК
<213> Последовательность реверсии PPO1 в QY2234-20 (искусственная
последовательность)
<400> 20
tctctctccc ttctccccca ccccacggag ctatatatat ataaacccca tctcttcaat 60
ccgtgcaacg aacgcctcgt cgcaacagct acaaacgccc acaagcaacg aaggaaatgg 120
gcggccgccg ccgaaccctg ggccccggcc cctcc 155
<210> 21
<211> 447
<212> ДНК
<213> Последовательность реверсии PPO1 в QY2234-205 (искусственная
последовательность)
<400> 21
cccttctccc ccaccccacg gagctatata tatataaacc ccatctcttc aatccgtgca 60
acgaacgcct cgtcgcaaca gctacaaacg gacatggccg ccgccgccgc agccatggcc 120
accgccacct ccgccacggc agcgccgccg ctccgcattc gcgacgccgc gaggaggacc 180
cgccgacgcg gccacgttcg ctgcgccgtc gccagcggcg cggccgaggc gcccgcggcg 240
cccggggcgc gggtgtcggc ggactgcgtc gtggtgggcg gcggcatcgg cgggctctgc 300
accgcgcagg cgctggccac aaagcacggc gtcggcgacg tgctcgtcac ggaggcccgc 360
ccccgccccg gcggcaacat caccaccgcc gagcgcgccg gcgagggcta cctctgggag 420
gaggggccca acagcttcca gccttcc 447
<210> 22
<211> 463
<212> ДНК
<213> Последовательность реверсии PPO1 в QY2234-207 (искусственная
последовательность)
<400> 22
cccttctccc ccaccccacg gagctatata tatataaacc ccatctcttc aatccgtgca 60
acgaacgcct cgtcgcaaca gctacaaacg cccacagtca gcgacggaca tggccgccgc 120
cgccgcagcc atggccaccg ccacctccgc cacggcagcg ccgccgctcc gcattcgcga 180
cgccgcgagg aggacccgcc gacgcggcca cgttcgctgc gccgtcgcca gcggcgcggc 240
cgaggcgccc gcggcgcccg gggcgcgggt gtcggcggac tgcgtcgtgg tgggcggcgg 300
catcagcggg ctctgcaccg cgcaggcgct ggccacaaag cacggcgtcg gcgacgtgct 360
cgtcacggag gcccgcgccc gccccggcgg caacatcacc accgccgagc gcgccggcga 420
gggctacctc tgggaggagg ggcccaacag cttccagcct tcc 463
<210> 23
<211> 388
<212> ДНК
<213> Последовательность реверсии PPO1 в QY2234-580 (искусственная
последовательность)
<400> 23
cccttctccc ccaccccacg gagctatata tatataaacc ccatctcttc aatccgtgca 60
acgaacgcct cgtcgcaaca ggtacatggc cgccgccgcc gcagccatgg ccaccgccac 120
ctccgccacg gcagcgccgc cgctccgcat tcgcgacgcc gcgaggagga cccgccgacg 180
cggccacgtt cgctgcgccg tcgccagcgg cgcggccgag gcgcccgcgg cgcccggggc 240
gcgggtgtcg gcggactgcg tcgtggtggg cggcggcatc agcgggctct gcaccgcgca 300
ggcgctggcc acaaagcacg gcgtcggcga cgtgctcgtc acggaggccc gcgcccgccc 360
cggcggcaac atcaccaccg ccgagcgc 388
<210> 24
<211> 417
<212> ДНК
<213> Последовательность реверсии PPO1 в QY2234-H5-159 (искусственная
последовательность)
<400> 24
cccttctccc ccaccccacg gagctatata tatataaacc ccatctcttc aatccgtgca 60
acgaacgcct cgtcgcaaca gctacaaacg gacatggccg ccgccgccgc agccatggcc 120
accgccacct ccgccacggc agcgccgccg ctccgcattc gcgacgccgc gaggaggacc 180
cgccgacgcg gccacgttcg ctgcgccgtc gccagcggcg cggccgaggc gcccgcggcg 240
cccggggcgc gggtgtcggc ggactgcgtc gtggtgggcg gcggcatcag cgggctctgc 300
accgcgcagg cgctggccac aaagcacggc gtcggcgacg tgctcgtcac ggaggcccgc 360
gcccgccccg gcggcaacat caccaccgcc gagcgcgccg gcgagggcta cctctgg 417
<210> 25
<211> 430
<212> ДНК
<213> Последовательность реверсии PPO1 в QY2234-H5-232 (искусственная
последовательность)
<400> 25
ccttctcccc caccccacgg agctatatat atataaaccc catctcttca atccgtgcaa 60
cgaacgcctc gtcgcaacag ctacaaacgc ccgtcagcga cggacatggc cgccgccgcc 120
gcagccatgg ccaccgccac ctccgccacg gcagcgccgc cgctccgcat tcgcgacgcc 180
gcgaggagga cccgccgacg cggccacgtt cgctgcgccg tcgccagcgg cgcggccgag 240
gcgcccgcgg cgcccggggc gcgggtgtcg gcggactgcg tcgtggtggg cggcggcatc 300
agcgggctct gcaccgcgca ggcgctggcc acaaagcacg gcgtcggcga cgtgctcgtc 360
acggaggccc gcgcccgccc cggcggcaac atcaccaccg ccgagcgcgc cggcgagggc 420
tacctctggg 430
<210> 26
<211> 424
<212> ДНК
<213> Последовательность реверсии PPO1 в QY2234-H5-263 (искусственная
последовательность)
<400> 26
ataaacccca tctcttcaat ccgtgcaacg aacgcctcgt cgcaacagct acaaacgccc 60
acagtcagcg acggacatgg ccgccgccgc cgcagccatg gccaccgcca cctccgccac 120
ggcagcgccg ccgctccgca ttcgcgacgc cgcgaggagg acccgccgac gcggccacgt 180
tcgctgcgcc gtcgccagcg gcgcggccga ggcgcccgcg gcgcccgggg cgcgggtgtc 240
ggcggactgc gtcgtggtgg gcggcggcat cagcgggctc tgcaccgcgc aggcgctggc 300
cacaaagcac ggcgtcggcg acgtgctcgt cacggaggcc cgcgcccgcc ccggcggcaa 360
catcaccacc gccgagcgcg ccggcgaggg ctacctctgg gaggaggggc ccaacagctt 420
ccag 424
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СОЗДАНИЯ НОВЫХ МУТАЦИЙ В ОРГАНИЗМАХ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2833881C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ВСТРАИВАНИЯ ЭКЗОГЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ В ГЕНОМ РАСТЕНИЙ | 2014 |
|
RU2723130C2 |
| РАСТЕНИЕ, ОБЛАДАЮЩЕЕ ПОВЫШЕННОЙ УСТОЙЧИВОСТЬЮ ИЛИ ЧУВСТВИТЕЛЬНОСТЬЮ К ИНГИБИТОРУ 4-HPPD | 2011 |
|
RU2604793C2 |
| ВЫДЕЛЕННЫЙ ПОЛИНУКЛЕОТИД, СПОСОБНЫЙ ПРИДАВАТЬ РАСТЕНИЮ УСТОЙЧИВОСТЬ ИЛИ ТОЛЕРАНТНОСТЬ К ГЛИФОСАТНОМУ ГЕРБИЦИДУ, ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ, ТОЛЕРАНТНЫХ ИЛИ УСТОЙЧИВЫХ К ГЛИФОСАТНОМУ ГЕРБИЦИДУ, СПОСОБ РЕГЕНЕРАЦИИ ТРАНСФОРМИРОВАННОГО РАСТЕНИЯ И СПОСОБ СЕЛЕКТИВНОЙ БОРЬБЫ С СОРНЯКАМИ | 2000 |
|
RU2235778C2 |
| ЛОКУСЫ ФУНКЦИОНАЛЬНОСТИ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ К УЧАСТКУ-МИШЕНИ СВЯЗЫВАЮЩИЕ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656159C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОПОСРЕДОВАННОЙ НУКЛЕАЗОЙ НАПРАВЛЕННОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ | 2013 |
|
RU2650819C2 |
| СКОНСТРУИРОВАННАЯ СПОСОБАМИ ИНЖЕНЕРИИ ПЛАТФОРМА ДЛЯ ВСТРАИВАНИЯ ТРАНСГЕНА (ETIP) ДЛЯ НАЦЕЛИВАНИЯ ГЕНОВ И СТЭКИНГА ПРИЗНАКОВ | 2013 |
|
RU2666916C2 |
| НОВЫЙ КЛАСС ГЕНОВ УСТОЙЧИВОСТИ К ГЛИФОСАТУ | 2013 |
|
RU2634411C2 |
| ВЫДЕЛЕННАЯ ДНК, ПРЕДСТАВЛЯЮЩАЯ СОБОЙ ПРОМОТОР (ВАРИАНТЫ), ВЕКТОР, СОДЕРЖАЩИЙ ЭТУ ДНК, И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО РАСТЕНИЯ С ПОМОЩЬЮ ЭТОЙ ДНК (ВАРИАНТЫ) | 1999 |
|
RU2245921C2 |
| ЛОКУСЫ FAD3 ДЛЯ ВЫПОЛНЕНИЯ ОПЕРАЦИЙ И СООТВЕТСТВУЮЩИЕ СВЯЗЫВАЮЩИЕСЯ СО СПЕЦИФИЧЕСКИМИ САЙТАМИ-МИШЕНЯМИ БЕЛКИ, СПОСОБНЫЕ К ВЫЗОВУ НАПРАВЛЕННЫХ РАЗРЫВОВ | 2013 |
|
RU2665811C2 |
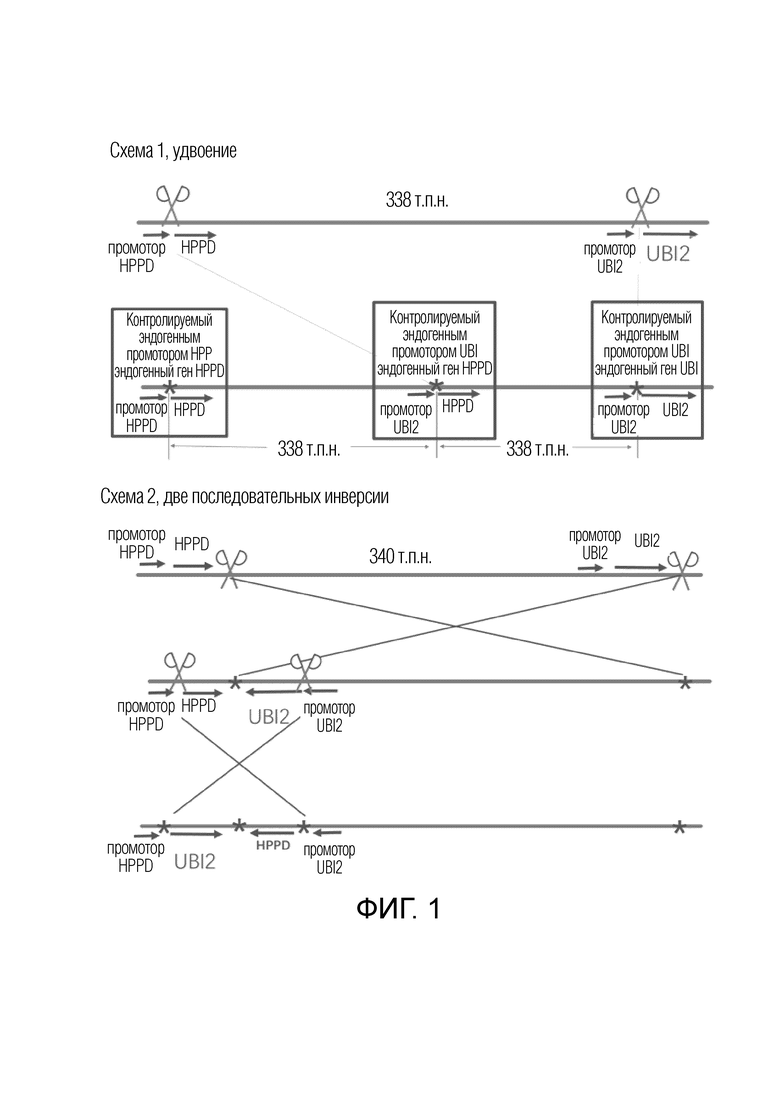
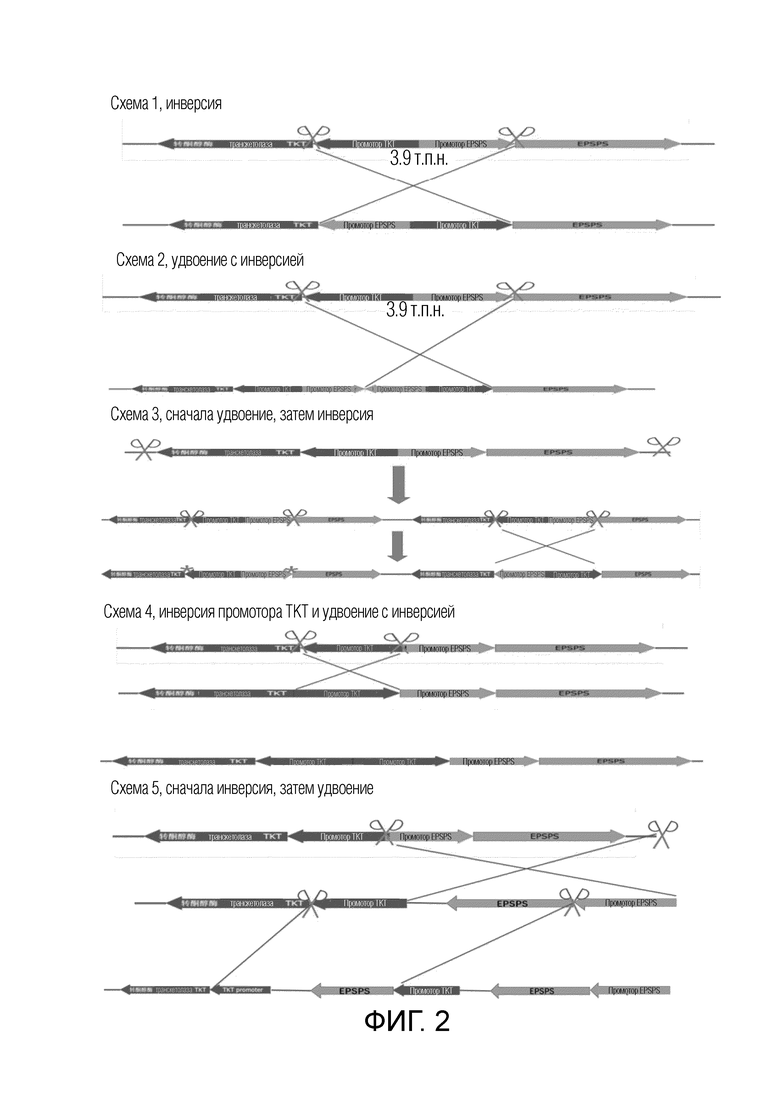
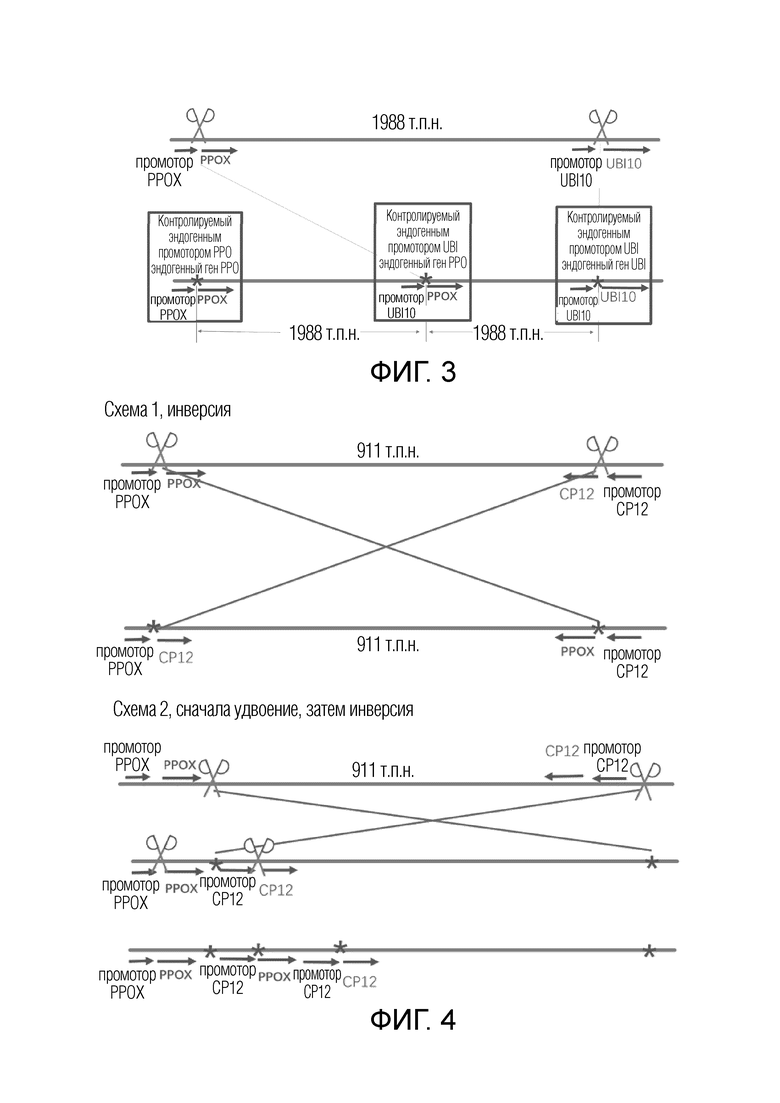

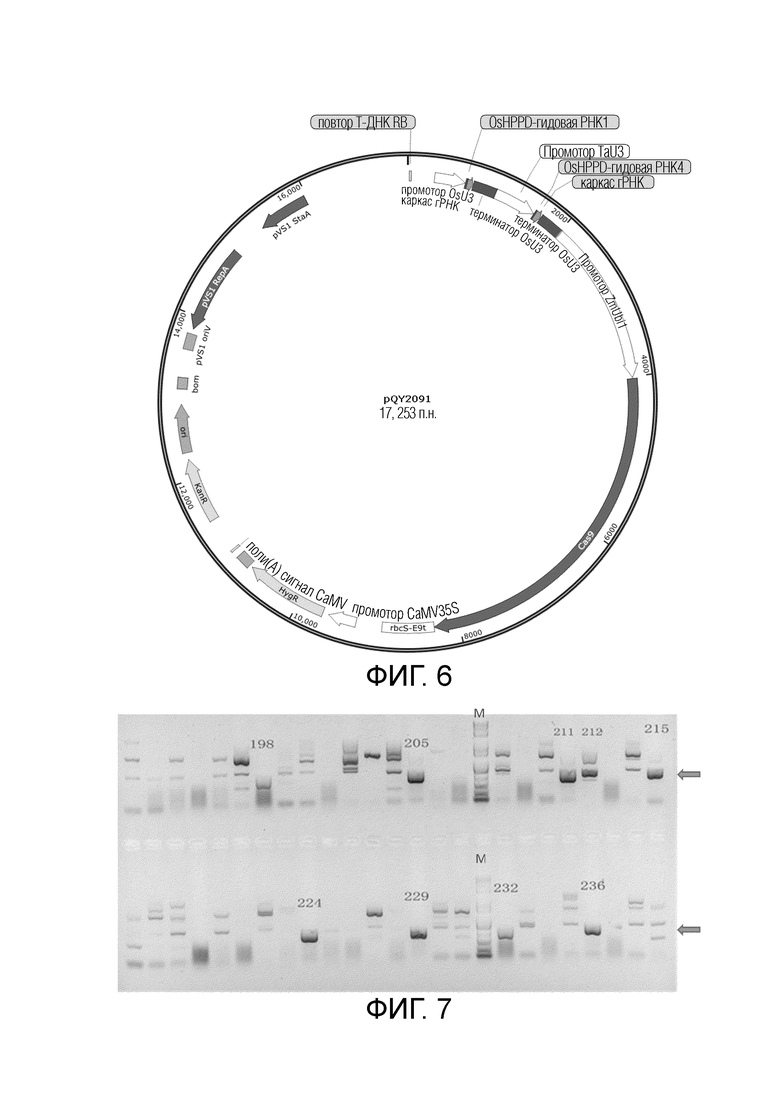
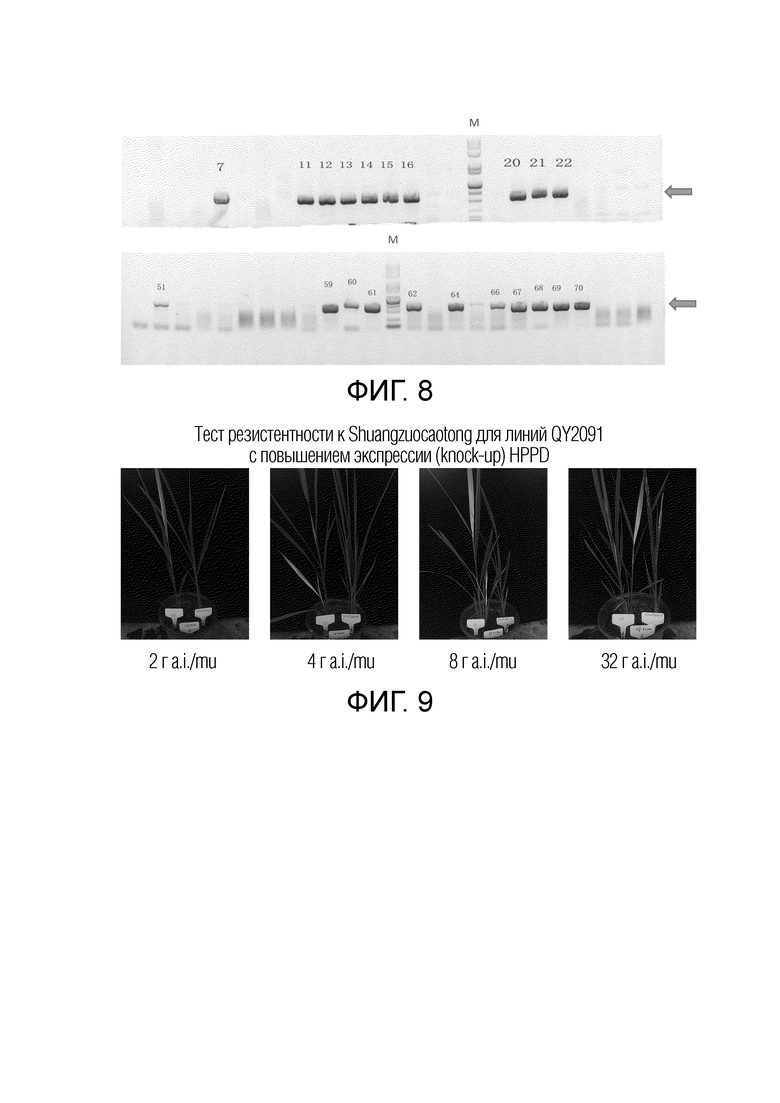
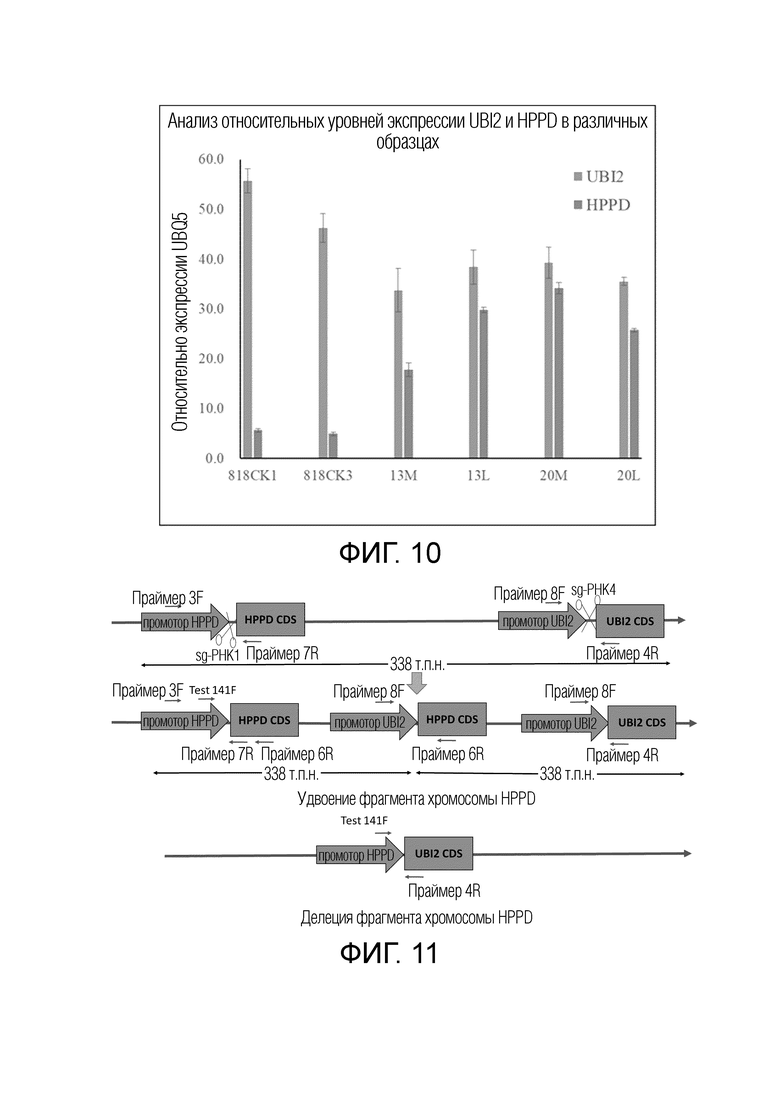
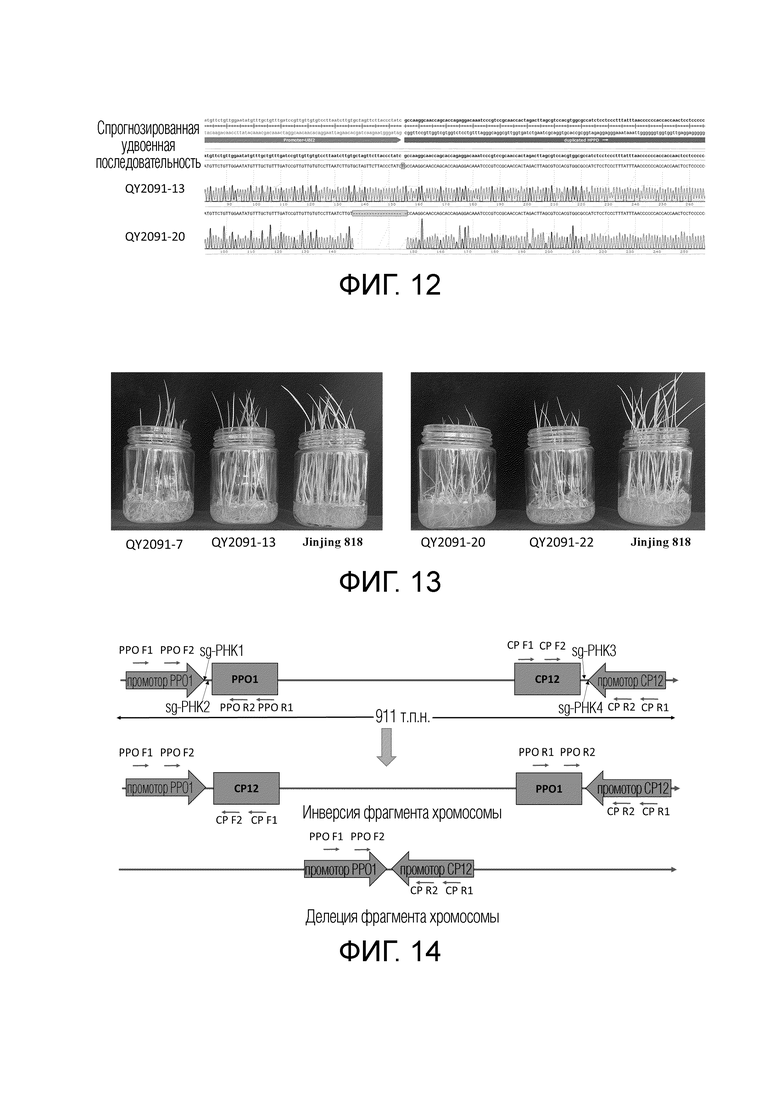

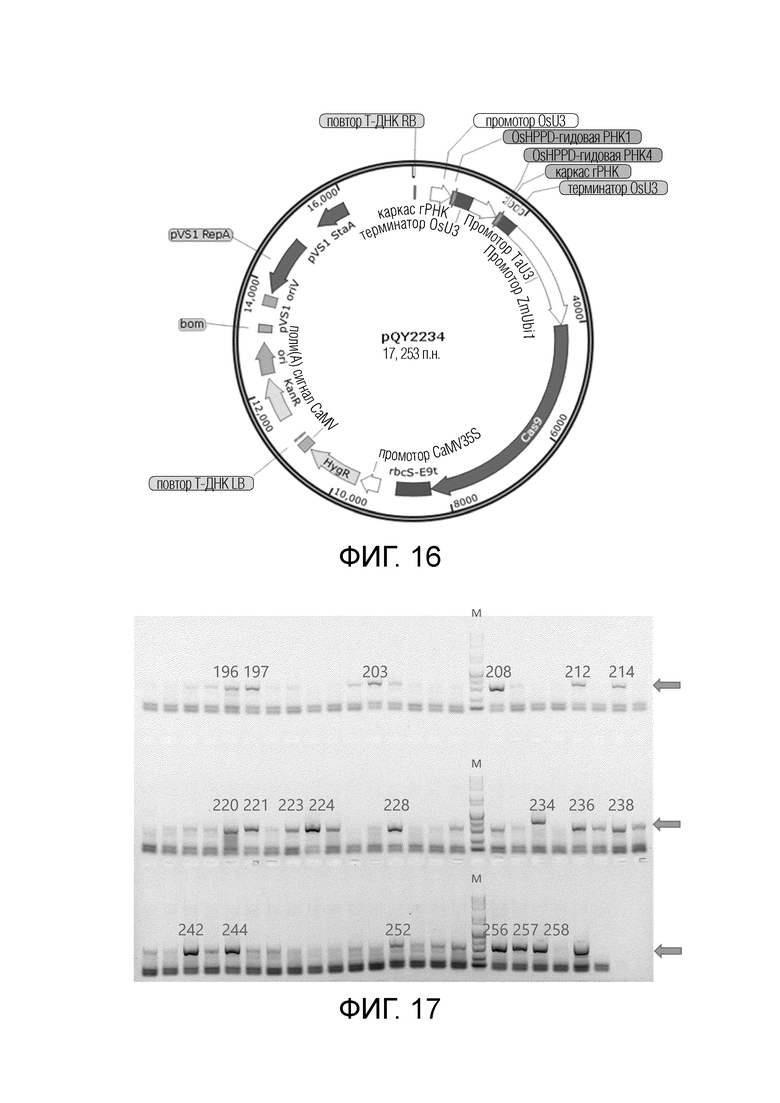
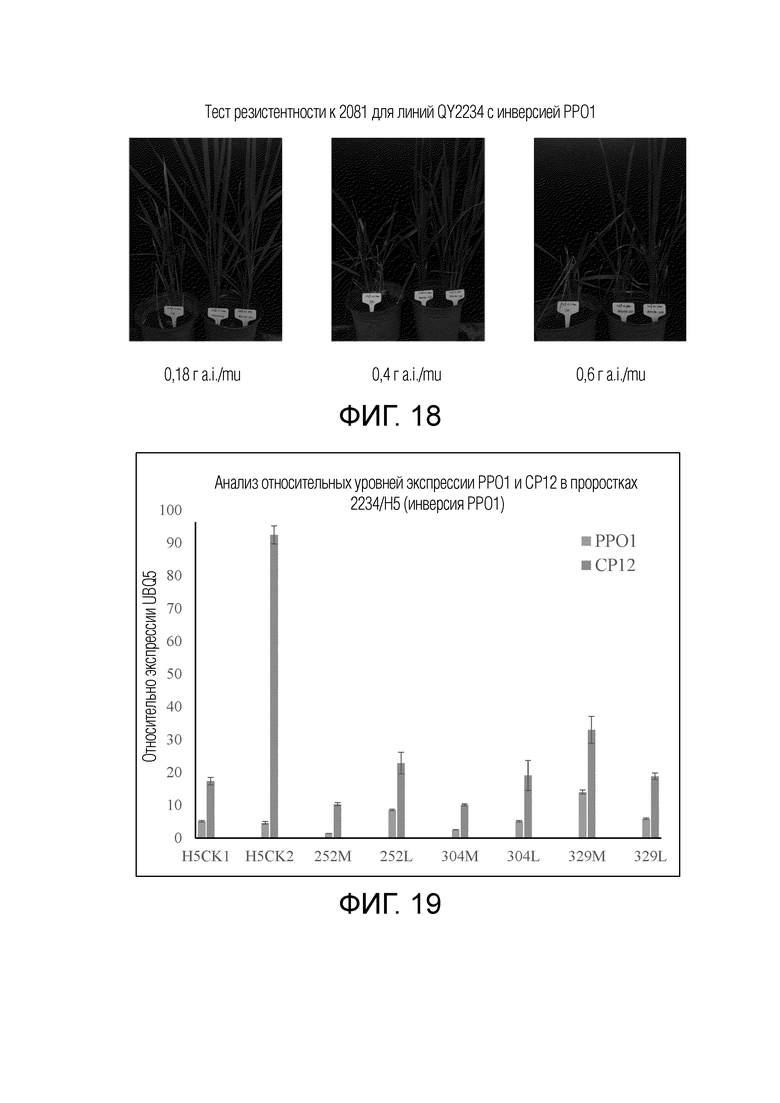
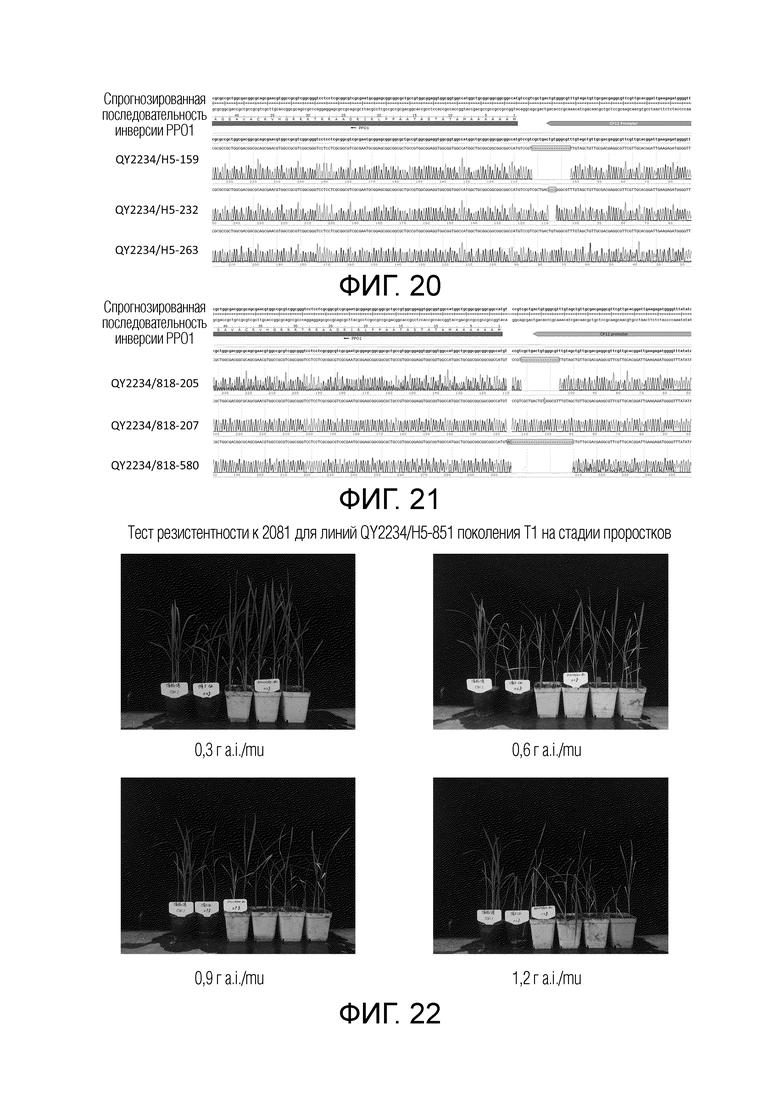
Изобретение относится к области биохимии, в частности к способу редактирования гена-мишени в организме. Также раскрыты отредактированные гены HPPD, EPSPS, PPO, поученные указанным способом; экспрессирующая кассета, содержащая указанные гены; клетка-хозяин, содержащая указанную кассету; организм, содержащий указанный ген. Изобретение позволяет эффективно редактировать ген-мишень. 13 н. и 21 з.п. ф-лы, 22 ил., 10 табл., 9 пр.
1. Способ редактирования гена-мишени в организме, отличающийся тем, что он включает:
(a) одновременное внесение разрывов ДНК в двух или более конкретных участках в гене-мишени или в двух или более различных специфических участках гена-мишени и одного или нескольких других генов генома организма, где конкретные участки представляют собой геномные участки, которые могут разделять различные генетические элементы или различные белковые домены, и
(b) лигирование разрывов ДНК друг с другом посредством негомологичного соединения концов или гомологичной репарации с созданием комбинации различных генных элементов или различных белковых доменов, которая отличается от исходной геномной последовательности, тем самым редактируя указанный ген.
2. Способ по п.1, отличающийся тем, что указанные два или более различных конкретных участков находятся на одной хромосоме или на различных хромосомах или где указанные по меньшей мере два различных гена могут иметь одинаковые или различные направления транскрипции.
3. Способ по п.1 или 2, отличающийся тем, что указанных разрывов ДНК достигают путем доставки нуклеазы со свойством нацеливания в клетку организма для контакта с конкретными участками геномной ДНК.
4. Способ по п.3, отличающийся тем, что указанная нуклеаза со свойством нацеливания выбрана из группы, состоящей из мегануклеазы, нуклеазы с цинковыми пальцами, TALEN и системы CRISPR/Cas.
5. Способ по п.3 или 4, отличающийся тем, что нуклеаза со свойством нацеливания имеет форму ДНК или существует в форме мРНК или белка, но не ДНК.
6. Способ по любому из пп.3-5, отличающийся тем, что нуклеазы со свойством нацеливания доставляют в клетку посредством: 1) ПЭГ-опосредуемой трансфекции клеток; 2) опосредуемой липосомами трансфекции клеток; 3) трансформации с использованием электрического шока; 4) микроинъекции; 5) бомбардировки из генной пушки или 6) опосредуемой Agrobacterium трансформации.
7. Способ по любому из пп.1-6, отличающийся тем, что указанные генные элементы выбраны из группы, состоящей из промотора, 5'-нетранслируемой области, кодирующей области или области некодирующей РНК, 3'-нетранслируемой области, терминатора гена или любой их комбинации.
8. Способ по любому из пп.1-7, отличающийся тем, что комбинация различных генных элементов представляет собой комбинацию промотора одного из двух генов с различными паттернами экспрессии и кодирующей области или области некодирующей РНК другого гена,
комбинацию области из промотора с 5'UTR одного из двух генов с различными паттернами экспрессии и CDS или области некодирующей РНК другого гена или
комбинацию соседних генных элементов одного и того же гена.
9. Способ по п.8, отличающийся тем, что в комбинации различных элементов генов один элемент представляет собой сильный эндогенный промотор организма, а другой представляет собой кодирующую область гена HPPD, EPSPS, PPO или GH1.
10. Способ по п.8 или 9, отличающийся тем, что различные паттерны экспрессии представляют собой различные уровни экспрессии генов, различную тканевую специфичность экспрессии генов или различную специфичность экспрессии генов к стадии развития.
11. Способ по любому из пп. 1-10, отличающийся тем, что белковый домен представляет собой фрагмент ДНК, соответствующий конкретному функциональному домену белка; предпочтительно включая, но не ограничиваясь ими, сигнал ядерной локализации, направляющий в хлоропласты пептид, направляющий в митохондрии пептид, участок фосфорилирования, участок метилирования, трансмембранный домен, ДНК-связывающий домен, домен активации транскрипции, домен активации рецептора или каталитический центр фермента.
12. Способ по любому из пп.1-11, отличающийся тем, что комбинация различных белковых доменов представляет собой комбинацию области сигнала локализации одного из двух белков с различной субклеточной локализацией и кодирующей зрелый белок области другого гена,
комбинацию двух белковых доменов с различными биологическими функциями или
комбинацию соседних белковых доменов одного гена.
13. Способ по п.12, отличающийся тем, что различные субклеточные положения выбраны из группы, состоящей из ядерной локализации, цитоплазматической локализации, локализации на клеточной мембране, локализации в хлоропластах, локализации в митохондриях и локализации на мембране эндоплазматической сети, или различные биологические функции выбраны из группы, состоящей из распознавания конкретной консервативной последовательности ДНК или РНК, активации экспрессии гена, связывания с белковым лигандом, связывания с низкомолекулярным сигнальным соединением, связывания иона или определенной ферментативной реакции и любой их комбинации.
14. Способ по любому из пп.1-7 и 11, отличающийся тем, что комбинация генных элементов и белковых доменов представляет собой комбинацию белковых доменов и соседних промоторов, 5'UTR, 3'UTR или терминаторов одного гена.
15. Способ по п.1, где указанные различные генные элементы или различные белковые домены включают промоторы и кодирующие области эндогенного гена-мишени и эндогенного гена с высокой экспрессией, и разрывы ДНК одновременно вносятся по отдельности в выбранных участках между промотором и кодирующей областью каждого из эндогенного гена-мишени и необязательного эндогенного гена с высокой экспрессией; разрывы ДНК лигируются друг с другом, что приводит к слиянию in vivo кодирующей области эндогенного гена-мишени и необязательного сильного эндогенного промотора, посредством чего повышается уровень генной экспрессии эндогенного гена-мишени в организме,
где эндогенный ген-мишень и необязательный эндогенный ген с высокой экспрессией находятся на одной хромосоме или на различных хромосомах.
16. Способ по п.15, где указанный эндогенный ген-мишень выбран из группы, состоящей из эндогенных генов HPPD, EPSPS и PPO, и указанный организм представляет собой растение.
17. Способ по п.16, где разрывы ДНК лигируются друг с другом посредством внутриклеточного каскада репарации.
18. Способ по п.16 или 17, где кодирующая область гена HPPD слита с сильным эндогенным промотором риса, где указанный сильный промотор представляет собой промотор гена убиквитина 2.
19. Способ по п.16 или 17, где кодирующая область гена EPSPS слита с сильным эндогенным промотором риса, где указанный сильный промотор представляет собой промотор гена TKT.
20. Способ по п.16 или 17, где кодирующая область гена PPO слита с сильным эндогенным промотором риса или Arabidopsis, где в рисе сильный промотор представляет собой промотор гена CP12; и в Arabidopsis сильный промотор представляет собой промотор гена убиквитина 10.
21. Отредактированный ген HPPD, получаемый способом по любому из пп.1-20, который содержит последовательность нуклеиновой кислоты, выбранную из группы, состоящей из:
последовательности нуклеиновой кислоты SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 18, SEQ ID NO: 19 и комплементарной им последовательности,
где уровень экспрессии указанного гена является повышенным по сравнению с неотредактированным геном HPPD.
22. Отредактированный ген EPSPS, получаемый способом по любому из пп.1-20, который содержит последовательность нуклеиновой кислоты, выбранную из группы, состоящей из:
последовательности нуклеиновой кислоты SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14 и комплементарной им последовательности,
где уровень экспрессии указанного гена является повышенным по сравнению с неотредактированным геном EPSPS.
23. Отредактированный ген PPO, получаемый способом по любому из пп.1-20, который содержит последовательность нуклеиновой кислоты, выбранную из группы, состоящей из:
последовательности нуклеиновой кислоты SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26 и комплементарной им последовательности,
где уровень экспрессии указанного гена является повышенным по сравнению с неотредактированным геном PPO.
24. Экспрессирующая кассета, содержащая ген HPPD по п.21.
25. Экспрессирующая кассета, содержащая ген EPSPS по п.22.
26. Экспрессирующая кассета, содержащая ген PPO по п.23.
27. Клетка-хозяин для экспрессии HPPD, которая содержит экспрессирующую кассету по п.24.
28. Клетка-хозяин для экспрессии EPSPS, которая содержит экспрессирующую кассету по п.25.
29. Клетка-хозяин для экспрессии PPO, которая содержит экспрессирующую кассету по п.26.
30. Клетка-хозяин по любому из пп.27-29, которая представляет собой бактериальную клетку, клетку гриба, клетку животного или клетку растения.
31. Организм с повышенной экспрессией гена HPPD по сравнению с организмом дикого типа, регенерировавший из клетки-хозяина по п.27.
32. Организм с повышенной экспрессией гена EPSPS по сравнению с организмом дикого типа, регенерировавший из клетки-хозяина по п.28.
33. Организм с повышенной экспрессией гена PPO по сравнению с организмом дикого типа, регенерировавший из клетки-хозяина по п.29.
34. Организм по любому из пп.31-33, где указанный организм представляет собой растение.
| WO 2016116032 A1, 28.07.2016 | |||
| CN 105916987 A, 31.08.2016 | |||
| RU 2018107701 A, 05.09.2019. |

