Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к трансгенному растению и способу его получения. В частности, настоящее изобретение относится к введению молекулы нуклеиновой кислоты в растение для повышения биомассы и/или урожайности растения.
Предшествующий уровень техники настоящего изобретения
Из-за растущего населения в мире площадь доступной земли для сельского хозяйства уменьшается. Для повышения эффективности сельского хозяйства и увеличения разнообразия садоводческих растений остается основным объектом исследований. Традиционные методы улучшения сельскохозяйственных культур и садоводческих растений используют селекционные технологии разведения для идентификации растений с желаемыми свойствами. Однако этот вид селекционных технологий разведения имеет несколько недостатков: эти технологии, как правило, трудоемки и дают в результате растения, которые обычно содержат гетерогенные генетические комплементарные последовательности, которые не всегда передают желаемые признаки оригинальных растений.
Развитие молекулярной биологии позволяет людям манипулировать животными и растениями. Генная инженерия растения требует выделения генетических материалов и манипулирования ими (как правило, в форме ДНК или РНК), а затем введения генетических материалов в растения. Такая технология дает в результате растения с рядом улучшенных экономических, сельскохозяйственных или садоводческих признаков. Признаком, обладающим особым экономическим значением, является ростовая характеристика, например, высокая урожайность.
Урожайность семян является очень важным признаком, поскольку семена многих растений очень важны для питания человека и животных. Вне зависимости от того, потребляются ли семена как таковые, или потребляются мясные продукты на основе обработанных семян, сельскохозяйственные культуры, такие как кукуруза, рис, пшеница и соя, составляют более половины от общего количества калорий, потребляемых человеком. Они также являются источником Сахаров, масел и многих видов метаболитов, используемых для промышленной переработки. С учетом постоянной потребности в поиске генов для увеличения урожайности семян в уровне техники раскрыты способы манипулирования уровнями гормонов растений (WO 03/050287), манипулирования клеточными циклами (WO 2005/061702) и манипулирования генами, участвующими в реакциях на стресс, вызванный засолением (WO 2004/058980). В дополнение к урожайности семян, размер тысячи семян, масса одного растения, число корневых отростков и/или высота растения также являются важными признаками для измерения урожайности растений. Более того, следует уточнить, что скорость роста растения не обязательно связана с его урожайностью. Например, если удобрения достаточно, рис имеет тенденцию расти слишком быстро и слишком высоким на ранней стадии, а также демонстрирует полегание на поздней стадии. Это приводит к снижению урожайности.
С помощью трансгенных технологий вводят искусственно выделенные и модифицированные гены в геном организма и получают наследственные модификации признаков организма за счет экспрессии введенных генов. Исследования растений с использованием трансгенных технологий главным образом сосредоточены на областях генной инженерии по борьбе с насекомыми, генной инженерии по борьбе с болезнями, генной инженерии стрессе-устойчивости, генной инженерии качества и т.д. Трансгенные растения, которые были коммерциализированы, в основном представляют собой противоинсектицидные и противогербицидные сорта. Выращивание таких сортов снижает применение химических пестицидов на 37%, повышает урожайность сельскохозяйственных культур на 22% и увеличивает прибыль фермеров на 68%. Однако существующие трансгенные технологии повышают урожайность опосредованно за счет противоинсектицидных и противогербицидных свойств и т.д.
Предыдущие исследования эпигенетики в основном фокусируются на обратимых модификациях ДНК и гистонов. В последнее время интерес исследователей перемещается к области модификации РНК. На данный момент ученые обнаружили сотни модификаций РНК. N6-метиладенозин (m6A) является наиболее распространенной модификацией РНК в мРНК у всех эукариот. Модификация m6A была обнаружена более сорока лет назад, но ее функция не была известна до тех пор, пока автор настоящей заявки впервые не обнаружил m6A-деметилазу - белок FTO (Jia et al, N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol, 2011, 7 (12): 885-887), впервые сообщил об обратимости модификации РНК и начал «РНК-эпигенетическое (или эпитранскриптомное)» исследование в 2011 году. В 2012 году исследователи разработали антитело против m6A с помощью технологии высокопроизводительного секвенирования целого транскриптома m6A - m6A-seq (или MeRIP). Результаты секвенирования показывают, что имеется приблизительно 12000 сайтов m6A в клетках человека и мыши, главным образом распределенных в 7000 mRNA, транскрибированных из кодирующих генов, и в 300 некодирующих РНК (ncRNA), транскрибированных из некодирующих генов (Dominissini et al, Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature, 2012, 485 (7397): 201-206; Meyer et al, Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons. Cell, 2012, 149 (7): 1635-1646). На данный момент обнаружили, что у млекопитающих основными компонентами метилтрансферазы являются METTL3, METTL14 и WTAP (Liu et al, A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem Biol, 2014, 10 (2): 93-95; Ping et al, Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res, 2014, 24 (2): 177-189). Существуют два вида m6A-деметилаз, которые представляют собой FTO (ассоциированный с жировой массой и ожирением) и ALKBH5 (Jia et al, N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol, 2011, 7 (12): 885-887; Zheng et al, ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell, 2013, 49 (1): 18-29). m6A регулирует метаболическое процессирование mRNA, в том числе сплайсинг, ядерный экспорт, стабильность и трансляцию белка посредством m6A-связывающих белков.
Исследователи растений обнаружили, что mRNA всех растений в значительной степени содержат m6A. Он продуцируется модифицирующими m6A ферментами (найденные на данный момент субъединицы модифицирующего фермента классифицируют как МТА (гомологичный ген METTL3) и FIP37 (гомологичный ген WTAP)) и оказывает важное регуляторное влияние на рост и развитие растений. Исследователи обнаружили, что если МТА или FIP37 были нокаутированы у Arabidopsis thaliana, то семя не могло нормально прорастать. Если МТА или FIP37 был частично комплементарным при стадии прорастания, то после прорастания утрата m6A у Arabidopsis thaliana сильно ингибировала нормальный рост и развитие растения (Zhong, S., Li, Н., Bodi, Z., Button, J., Vespa, L., Herzog, M., and Fray, R.G (2008). MTA is an Arabidopsis messenger RNA adenosine methylase and interacts with a homolog of a sex-specific splicing factor. Plant Cell 20, 1278-1288; Shen, L., Liang, Z., Gu, X., Chen, Y., Teo, Z.W., Hou, X., Cai, W.M., Dedon, P.C., Liu, L., and Yu, H. (2016). N6-Methyladenosine RNA Modification Regulates Shoot Stem Cell Fate in Arabidopsis. Dev Cell 38,186-200).
Автор настоящей заявки ранее обнаружил, что в ходе процесса деметилирования m6A в РНК FTO продуцировал две относительно стабильные новые модификации - hm6A (N6-гадроксиметиладенозин) и f6A (N6-формиладенозин), которые обладали потенциальным регуляторным эффектом в отношении процессирования РНК (Fu et al, FTO-mediated formation of N6-hydroxymethyladenosine and N6-formyladenosine in mammalian RNA. Nat Commun, 2013, 4:1798). Новые экспериментальные данные автора настоящего изобретения показали, что FTO может удалить модификации метилирования tRNA (данные не были опубликованы).
Задача исследований всех ученых в области сельского хозяйства заключалась в том, как повысить урожайность сельскохозяйственных культур на ограниченном участке земли для обеспечения пищей растущего населения, и как непосредственно и эффективно повысить биомассу и урожайность растений. Для сельскохозяйственных культур (таких как рис, пшеница, кукуруза) традиционные трансгенные технологии и технологии гибридизационного скрещивания могут оптимизировать один определенный ген и повысить урожайность на 10% ~ 30%. Для достижения чрезвычайно высокой урожайности необходимы синергетические эффекты нескольких генов. Регулирование метаболического уровня mRNA путем модификации метилирования РНК дает возможность способа регулирования одного гена для достижения высокой урожайности или увеличения биомассы.
Краткое раскрытие настоящего изобретения
Автор настоящего изобретения неожиданно обнаружил, что с помощью введения m6A-деметилазы FTO в растения можно регулировать метаболический уровень mRNA и обеспечивать возможность способа регулирования одного гена для достижения высокой урожайности и/или увеличения биомассы. Для эффективного повышения урожайности и/или биомассы растений автор настоящего изобретения вводил гетерогенную m6A-деметилазу с помощью трансгенной технологии и динамически регулировал содержание m6A в mRNA растений с тем, чтобы регулировать сплайсинг, ядерный экспорт, стабильность и белковую трансляцию mRNA. По сравнению с традиционным способом гибридизационного скрещивания, который повышает урожайность на 20-30%, способ в соответствии с настоящим изобретением повышает урожайность растений в 4 раза и увеличивает биомассу в 4 раза за счет регулирования одного гена. Действительно достигалась высокая урожайность и высокая биомасса растений за счет регулирования одного гена.
В частности, настоящее изобретение относится к следующим аспектам.
В одном аспекте настоящее изобретение относится к трансгенному растению или клетке растения, в которых введена молекула нуклеиновой кислоты, кодирующая m6A-деметилазу, при этом указанная m6A-деметилаза имеет следующих два домена:
i) N-концевой домен (NTD), обладающий функцией окислительной деметилазы AlkB;и
ii) С-концевой домен (CTD).
В другом аспекте настоящее изобретение относится к способу получения трансгенного растения, демонстрирующего увеличенную биомассу, повышенную урожайность (например, повышенную урожайность семян/зерна, повышенную урожайность клубней, повышенную урожайность листьев, повышенную урожайность стеблей, повышенную урожайность корней, повышенную урожайность хлопкового семени) или их комбинацию, при этом указанный способ предусматривает:
a) введение молекулы нуклеиновой кислоты, кодирующей m6A-деметилазу, в способную к регенерации клетку растения, при этом указанная m6A-деметилаза имеет следующих два домена:
i) N-концевой домен (NTD), обладающий функцией окислительной деметилазы AlkB;и
ii) С-концевой домен (CTD); и
b) регенерацию трансгенного растения из способной к регенерации клетки растения, при этом трансгенное растение содержит в своем геноме указанную молекулу нуклеиновой кислоты, кодирующую m6A-деметилазу, и демонстрирует увеличенную биомассу, повышенную урожайность или их комбинацию по сравнению с контрольным растением, которое не содержит молекулу нуклеиновой кислоты, кодирующую m6A-деметилазу.
В соответствии с одним вариантом осуществления указанный способ дополнительно предусматривает:
c) получение растения-потомка, получаемого из трансгенного растения стадии b), при этом указанное растение-потомок содержит в своем геноме указанную молекулу нуклеиновой кислоты, кодирующую m6A-деметилазу, и демонстрирует увеличенную биомассу, повышенную урожайность или их комбинацию по сравнению с контрольным растением, которое не содержит молекулу нуклеиновой кислоты, кодирующую m6A-деметилазу.
В соответствии с одним вариантом осуществления указанной выше m6A-деметилазой является белок FTO. Указанный белком FTO является белок, который походит от позвоночных или морских водорослей.
В соответствии с одним вариантом осуществления указанный белок FTO обладает по меньшей мере 40%, предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 99%, наиболее предпочтительно 100% идентичностью по отношению к любой из SEQ ID NO: 1-4.
В соответствии с одним вариантом осуществления указанная молекула нуклеиновой кислоты, кодирующая m6A-деметилазу, обладает по меньшей мере 90%, предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 99%, наиболее предпочтительно 100% идентичностью по отношению к любой из SEQ ID NO: 5-12.
Трансгенное растение в соответствии с настоящим изобретением демонстрирует увеличенную биомассу, повышенную урожайность или их комбинацию по сравнению с контрольным растением, которое не содержит молекулу нуклеиновой кислоты, кодирующую m6A-деметилазу.
В соответствии с одним вариантом осуществления растение в соответствии с настоящим изобретением выбрано из группы, состоящей из риса, кукурузы (Zea mays), сои, табака, картофеля, люцерны (Medicago sativa), рапса (Brassica), кок-сагыза (Taraxacum Kok-saghyz), хлопчатника, пшеницы, проса (Panicum miliaceum), льна, подсолнечника и ложного льна (Camelina sativa).
Настоящее изобретение также относится к ткани, органу, пыльце, семени, зерну, плоду и растению-потомку указанного выше трансгенного растения.
Описание графических материалов и последовательностей
На фиг. 1 показана карта плазмиды pCAMBIA1307, в которую введена нуклеиновая кислота, кодирующая FTO.
SEQ ID NO: 1 представляет собой последовательность белка FTO человека (Homo sapiens).
SEQ ID NO: 2 представляет собой последовательность белка FTO свиньи (Sus scrofa).
SEQ ID NO: 3 представляет собой последовательность белка FTO быка (Bos taurus). SEQ ID NO: 4 представляет собой последовательность белка FTO Ostreococcus lucimarinus, из типа морских водорослей Ostreococcus lucimarinus.
SEQ ID NO: 5 и 6 представляют собой последовательности нуклеиновой кислоты, кодирующие человеческий белок FTO. SEQ ID NO: 5 представляет собой натуральную последовательность, выделенную из человека, a SEQ ID NO: 6 представляет собой последовательность, которая была кодон-оптимизирована для экспрессии в растениях.
SEQ ID NO: 7 и 8 представляют собой последовательности нуклеиновой кислоты, кодирующие белок FTO свиньи. SEQ ID NO: 7 представляет собой натуральную последовательность, выделенную из свиньи, a SEQ ID NO: 8 представляет собой последовательность, которая была кодон-оптимизирована для экспрессии в растениях.
SEQ ID NO: 9 и 10 представляют собой последовательности нуклеиновой кислоты, кодирующие белок FTO быка. SEQ ID NO: 9 представляет собой натуральную последовательность, выделенную из быка, a SEQ ID NO: 10 представляет собой последовательность, которая была кодон-оптимизирована для экспрессии в растениях.
SEQ ID NO: 11 и 12 представляют собой последовательности нуклеиновой кислоты, кодирующие белок FTO Ostreococcus lucimarinus. SEQ ID NO: 11 представляет собой натуральную последовательность, выделенную из Ostreococcus lucimarinus, a SEQ ID NO: 12 представляет собой последовательность, которая была кодон-оптимизирована для экспрессии в растениях.
Подробное раскрытие настоящего изобретения
m6A-деметилаза или ее гомологи и нуклеиновые кислоты, кодирующие указанную деметилазу или гомологов, могут быть использованы для получения трансгенного растения в соответствии с настоящим изобретением. m6A-деметилаза, используемая в соответствии с настоящим изобретением, может присутствовать у любых позвоночных и морских водорослей. Указанный фермент состоит из последовательности ядерной локализации (NLS) и следующих двух доменов: i) N-концевого домена (NTD), обладающего функцией окислительной деметилазы AlkB; и ii) С-концевого домена (CTD).
Используемый в настоящем документе термин «гомолог» означает белок из группы белков, которые выполняют одну и ту же биологическую функцию. Гомологи экспрессируются гомологичными генами. Гомологичные гены включают в себя встречающиеся в природе аллели и искусственно созданные варианты. Вырожденность генетического кода обеспечивает возможность замены по меньшей мере одного основания в кодирующей белок последовательности гена другим основанием без изменения аминокислотной последовательности полипептида, получаемого из гена. Гомологи представляют собой белки, которые при оптимальном выравнивании обладают по меньшей мере 40% идентичностью, более предпочтительно приблизительно 50% или выше, более предпочтительно приблизительно 60% или выше, более предпочтительно приблизительно 70% или выше, более предпочтительно по меньшей мере 80% и еще более предпочтительно по меньшей мере 90% идентичностью по всей длине с белком, идентифицированным как повышающий урожайность и/или биомассу растений при экспрессии в клетках растения.
Гомологи подлежат идентификации путем сравнения аминокислотной последовательности, например, вручную или с помощью применения компьютерного инструментария, с использованием известных алгоритмов поиска на основе гомологов, таких как алгоритмы, которые широко известны и называются BLAST, FASTA и алгоритм Смита-Уотермана. Программу локального выравнивания последовательностей, например, BLAST, можно использовать для поиска в базе данных последовательностей с целью выявления подобных последовательностей, и суммарное ожидаемое значение (Е-значение) используется для измерения подобия оснований последовательности. Поскольку белок, совпадающий с наилучшим Е-значением для конкретного организма, необязательно может быть ортологом или единственным ортологом, в настоящем изобретении используется обратный запрос для фильтрации совпадения последовательностей со значительными Е-значениями для идентификации ортолога. Обратный запрос предусматривает поиск значительных совпадений по базе данных аминокислотных последовательностей из основного организма, которые подобны последовательности запрашиваемого белка. Совпадение является вероятным ортологом, если наилучшим совпадением обратного запроса является сам запрашиваемый белок или белок, кодируемый дуплицированным геном, после идентификации. Следующий аспект настоящего изобретения предусматривает функциональные гомологичные белки, которые отличаются одной или несколькими аминокислотами от таковых в раскрываемом белке, в результате консервативных аминокислотных замен, например, заменами являются кислотные (отрицательно заряженные) аминокислоты, такие как аспарагиновая кислота и глютаминовая кислота; основные (положительно заряженные) аминокислоты, такие как аргинин, гистидин и лизин; нейтральные полярные аминокислоты, такие как глицин, серии, треонин, цистеин, тирозин, аспарагин и глутамин; нейтральные неполярные (гидрофобные) аминокислоты, такие как аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; аминокислоты, имеющие алифатические боковые цепи, такие как глицин, аланин, валин, лейцин и изолейцин; аминокислоты, имеющие алифатически-гидроксильные боковые цепи, такие как серии и треонин; аминокислоты, содержащие амидсодержащие боковые цепи, такие как аспарагин и глутамин; аминокислоты, содержащие ароматические боковые цепи, такие как фенилаланин, тирозин и триптофан; аминокислоты, имеющие основные боковые цепи, такие как лизин, аргинин и гистидин; аминокислоты, содержащие серосодержащие боковые цепи, такие как цистеин и метионин; естественно консервативные аминокислоты, такие как валин-лейцин, валин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, аспарагиновая кислота-глутаминовая кислота и аспарагин-глутамин. В следующем аспекте гомологов, кодируемых ДНК, применимыми в трансгенных растениях в соответствии с настоящим изобретением, являются те белки, которые отличаются от раскрываемого белка в результате делеции или вставки одной или нескольких аминокислот в нативной последовательности.
Используемый в настоящем документе термин «процент идентичности» означает степень, с которой два оптимально выровненных сегмента ДНК или белка являются инвариантными во всем окне выравнивания компонентов, например, нуклеотидной последовательности или аминокислотной последовательности. «Доля идентичности» для выравниваемых сегментов тестируемой последовательности и эталонной последовательности представляет собой число идентичных компонентов, которые являются общими для последовательностей двух выравниваемых сегментов, поделенное на общее число компонентов последовательности в эталонном сегменте по всему окну выравнивания, которым является меньшая из полной тестируемой последовательности или полной эталонной последовательности. «Процент идентичности» («% идентичности») представляет собой долю идентичности, умноженную на 100.
m6A-деметилазой, используемой в соответствии с настоящим изобретением, может быть SEQ ID NO: 1, 2, 3 или 4, или ее гомолог. Указанный гомолог обладает по меньшей мере 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательностей в отношении любой из SEQ ID NO: 1-4.
Нуклеиновой кислотой, кодирующей m6A-деметилазу, используемую в соответствии с настоящим изобретением, может быть любая из SEQ ID NO: 5-12 или гомологичный ей ген. Указанный гомологичный ген обладает по меньшей мере 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательностей в отношении любой из SEQ ID NO: 5-12.
Нуклеиновая кислота, кодирующая m6A-деметилазу, определяемую в настоящем документе, может не быть нуклеиновой кислотой полной длины. Часть нуклеиновой кислоты, кодирующей m6A-деметилазу, определяемую в настоящем документе, может быть получена путем осуществления одной или нескольких делеций из нуклеиновой кислоты полной длины.
Другим вариантом нуклеиновой кислоты, используемой в соответствии с настоящим изобретением, является нуклеиновая кислота, которая гибридизируется с нуклеиновой кислотой, кодирующей m6A-деметилазу, определяемую в настоящем документе, или с частью нуклеиновой кислоты, кодирующей m6A-деметилазу, определяемую в настоящем документе, в жестких условиях.
Соответствующими жесткими условиями, которые обеспечивают гибридизацию ДНК, например, 6,0× хлорид натрия/цитрат натрия (SSC) при приблизительно 45°С, с последующим промыванием 2,0×SSC при 50°С, известны специалистам в данной области или могут быть найдены в Current Protocols in Molecular Biology (1989). Например, концентрация соли на стадии промывания может быть выбрана от низкой жесткости приблизительно 2,0×SSC при 50°С до высокой жесткости приблизительно 0,2×SSC при 50°С. Кроме того, температура на стадии промывания может быть повышенной от условий низкой жесткости при комнатной температуре, приблизительно 22°С, до условий высокой жесткости при приблизительно 65°С. И температуру, и соль можно варьировать, или либо температуру, либо концентрацию соли можно держать постоянной при изменении другой переменной. Нуклеиновая кислота, используемая в соответствии с настоящим изобретением, может специфически гибридизироваться с нуклеиновой кислотой, кодирующей белок FTO, например, с любой из SEQ ID NO: 5-12 или их частью, при указанных условиях.
Все или часть нуклеиновых кислот в соответствии с настоящим изобретением могут быть синтезированы с использованием кодонов, предпочтительных для выбранного хозяина. Предпочтительные в отношении вида кодоны могут быть определены, например, из кодонов, используемых наиболее часто в белках, экспрессированных у конкретного вида-хозяина. Поэтому, нуклеиновые кислоты в соответствии с настоящим изобретением включают в себя полученные путем создания кодон-оптимизации для натуральных белков FTO с целью экспрессии в растениях. Другие модификации нуклеотидных последовательностей могут приводить к мутантам, обладающим слегка измененной активностью.
Настоящее изобретение относится к трансгенному растению, в которого вводят ген, кодирующий указанную выше m6A-деметилазу FTO, или его гомолог, или к его растению-потомку. Настоящее изобретение также относится к клетке, ткани, органу, пыльце, семени, зерну или плоду указанного растения.
Термин «введенный» в контексте вставки фрагмента нуклеиновой кислоты (например, рекомбинантной ДНК-конструкции) в клетку означает «трансфекцию», или «трансформацию», или «трансдукцию» и включает в себя упоминание включения фрагмента нуклеиновой кислоты в эукариотическую или прокариотическую клетку, при этом фрагмент нуклеиновой кислоты может быть включен в геном клетки (например, хромосому, плазмиду, пластидную или митохондриальную ДНК), превращен в автономный репликон или транзиентно экспрессирован (например, трансфицированная mRNA).
Термин «растение» включает в себя упоминание целых растений, растительных органов, растительных тканей, семян и растительных клеток, а также их потомства. Растительные клетки включают в себя без ограничения клетки из семян, суспензионные культуры, зародыши, меристемные участки, каллюсную ткань, листья, корни, побеги, гаметофиты, спорофиты, пыльцу и микроспоры. Термин «потомство» предусматривает любое последующее поколение растения.
Согласно применению растения в соответствии с настоящим изобретением могут быть пищевыми сельскохозяйственными культурами, промышленными сельскохозяйственными культурами, овощными сельскохозяйственными культурами, плодовыми, цветочными, травами, деревьями, промышленными сырьевыми сельскохозяйственными культурами, кормовыми сельскохозяйственными культурами или медицинскими сельскохозяйственными культурами. В частности, указанные пищевые сельскохозяйственные культуры включают в себя рис, кукурузу, сою, бобы, ямс, картофель, голозерный ячмень, кормовые бобы, пшеницу, ячмень, просо, рожь, овес, сорго и т.д.; Указанные промышленные сельскохозяйственные культуры включают в себя масло чайного дерева, рапс, семя рапса, лен, ложный лен (Camelina sativa), арахис, масло льна (Linum usitatissimum), коноплю посевную (Cannabis sativa), подсолнечник, табак, хлопчатник, свеклу, сахарный тростник и т.д.; указанные овощные сельскохозяйственные культуры включают в себя редис, капусту китайскую, томат, огурец, острый перец, морковь и т.д.; указанные плодовые включают в себя грушу, яблоню, грецкий орех, вишню, землянику, китайский финик или персик; указанные цветочные включают в себя декоративные цветочные, например, орхидею, хризантему, гвоздику, розу, растения для озеленения и т.д., указанные травы и деревья включают в себя тополь, гевею бразильскую, Taxus chinensis, а также растения для городского озеленения или растения, живущие в пустынях и суровых условиях, таких как засуха; указанные сырьевые сельскохозяйственные культуры включают в себя кок-сагыз, гваюлу, Jatropha curcas и т.д., указанные кормовые сельскохозяйственные культуры включают в себя кормовые культуры для сельскохозяйственных животных, например, люцерну и т.д.; указанные лекарственные сельскохозяйственные культуры включают в себя женьшень, дудник, ганодерму и т.д.
Термин «трансгенное растение» предусматривает упоминание растения, которое содержит в своем геноме гетерологичный полинуклеотид. Предпочтительно гетерологичный полинуклеотид стабильно вставляется в геном так, что полинуклеотид передается следующим поколениям. Гетерологичный полинуклеотид может быть вставлен в геном самостоятельно или как часть рекомбинантной ДНК-конструкции.
Термин «гетерологичная» в отношении последовательности означает последовательность, которая походит от чужого вида или, если из того же вида, существенно модифицирована от своей нативной формы в составе и/или в геномном локусе путем преднамеренного вмешательства человека.
В соответствии с настоящим изобретением получают ДНК-конструкцию, которая содержит молекулу нуклеиновой кислоты, кодирующую белок FTO, описываемый в настоящем документе. Конструкция может содержать молекулу нуклеиновой кислоты, кодирующую белок FTO, описываемый в настоящем документе, необязательно функционально связанный с промоторной последовательностью, которая функционирует в клетке-хозяине. Другие компоненты конструкции могут включать в себя дополнительные регуляторные элементы, такие как 5'-лидерные последовательности и интроны для усиления транскрипции, 3'-нетранслируемые области (такие как сигналы и сайты полиаденилирования), ДНК для транзита, сигнальные пептиды или один или несколько селективных маркерных генов.
Используемый в настоящем документе термин «промотор» означает регуляторную ДНК для интернализации транскрипции. Растительным промотором является промотор, способный интернализировать транскрипцию в клетках растения, независимо от того, походит ли он из растительной клетки, например, хорошо известно, что промоторы Agrobacterium являются функциональными в растительных клетках. Таким образом, растительные промоторы включают в себя промоторную ДНК, полученную из растений, растительных вирусов и бактерий, таких как бактерии Agrobacterium и Bradyrhizobium. Примеры промоторов под контролем развития включают в себя промоторы, которые предпочтительно инициируют транскрипцию в определенных тканях, таких как листья, корни или семена. Такие промоторы упоминаются как «предпочтительные по отношению к ткани». Промоторы, которые инициируют транскрипцию только в определенных тканях, называются «специфические по отношению к ткани». Специфический по отношению к «типу клеток» промотор в первую очередь управляет экспрессией в определенных типах клеток в одном или нескольких органах, например, в сосудистых клетках в корнях или листьях. «Индуцибельным» или «репрессируемым» промотором является промотор, который находится под контролем окружающей среды. Примеры условий окружающей среды, которые могут влиять на транскрипцию с помощью индуцибельных промоторов, включают в себя анаэробные условия, или определенные химические вещества, или наличие света. Специфические по отношению к ткани, предпочтительные по отношению к ткани, специфические по отношению к типу клеток и индуцибельные промоторы составляют класс «неконститутивных» промоторов. «Конститутивным» промотором является промотор, который активен при большинстве условий. Промоторы, применимые в соответствии с настоящим изобретением, специально не ограничены. Специалисты в данной области смогут выбрать подходящие промоторы в соответствии с их опытом.
Используемый в настоящем документе термин «функционально связанный» означает ассоциацию двух или более фрагментов ДНК в ДНК-конструкции таким образом, что функция одного, например, кодирующей белок ДНК, контролируется другим, например, промотором.
ДНК-конструкция, как правило, содержит селективный маркерный ген. Селективные маркерные гены используют для обеспечения эффективной системы для идентификации тех клеток, которые стабильно трансформированы путем получения и встраивания трансгенной ДНК-конструкции в их геномы. Предпочтительные маркерные гены обеспечивают селективные маркеры, которые придают устойчивость к селективному средству, такому как антибиотик или гербицид. Потенциально трансформированные клетки подвергают воздействию селективного средства. В популяции выживших клеток останутся те клетки, в которых, как правило, придающий устойчивость ген встраивается и экспрессируется при достаточных уровнях для обеспечения выживаемости клеток. Клетки могут быть далее тестированы для подтверждения стабильного встраивания экзогенной ДНК. Широко используемые селективные маркерные гены включают в себя гены, обеспечивающие устойчивость к антибиотикам, таким как канамицин и паромомицин (nptll), гигромицин В (aph IV) и гентамицин (аас3 и аасС4), или устойчивость к гербицидам, таким как глифосинат (bar или pat) и глифосат (aroA или EPSPS). Примеры таких селективных маркеров представлены в патентах США №№5550318, 5633435, 5780708 и 6118047, все из которых включены в настоящий документ посредством ссылки. Также могут быть использованы селективные маркеры, которые обеспечивают возможность визуально идентифицировать трансформантов, например, ген, экспрессирующий цветной или флуоресцентный белок, такой как люцифераза или зеленый флуоресцентный белок (GFP), или ген, экспрессирующий бета-глюкуронидазу, или ген uidA (GUS), для которого известны различные хромогенные субстраты.
Введение рекомбинантной ДНК-конструкции в растения может быть выполнено с помощью любой подходящей методики, в том числе без ограничения прямое поглощение ДНК, химическая обработка, электропорация, микроинъекция, клеточное слияние, инфицирование, опосредованный вектором перенос ДНК, бомбардировка или опосредованная Agrobacterium трансформация. Для основанной на Agrobacterium tumefaciens системы трансформации растения дополнительные элементы, присутствующие в трансформационных конструкциях, будут включать в себя левые и правые пограничные последовательности Т-ДНК для облегчения включения рекомбинантного полинуклеотида в растительный геном.
В целом, полезно вводить рекомбинантную ДНК рандомно, т.е. в неспецифическую локализацию, в геном целевой линии растения. В особых случаях может быть полезным нацеливание вставки рекомбинантной ДНК для достижения сайт-специфического встраивания, например, для замены существующего гена в геноме, для применения существующего промотора в растительном геноме или для вставки рекомбинантного полинуклеотида в предварительно определенный сайт, который, как известно, активен для генной экспрессии. Имеется несколько сайт-специфических рекомбинационных систем, которые, как известно, функционируют в растениях и включают в себя cre-lox, раскрываемую в патенте США №4959317, а также FLP-FRT, раскрываемую в патенте США №5527695, оба из которых включены в настоящий документ посредством ссылки.
Способы трансформации в соответствии с настоящим изобретением предпочтительно осуществляют на тканевой культуре в средах и в контролируемой окружающей среде. Термин «среды» относится к многочисленным питательным смесям, которые используют для выращивания клеток in vitro, то есть вне интактного живого организма. Цели реципиентных клеток включают в себя без ограничения меристемные клетки, каллюс, незрелые зародыши и гаметные клетки, такие как микроспоры, пыльца, спермин и яйцеклетки. Следует иметь в виду, что любая клетка, из которой может регенерироваться фертильное растение, применима в качестве реципиентной клетки. Каллюс может быть инициирован из тканевых источников, в том числе без ограничения из незрелых зародышей, апикальных меристем проростков, микроспор и т.п. Клетки, способные пролиферировать как каллюс, также являются реципиентными клетками для генетической трансформации. Используемые на практике способы трансформации и материалы для получения трансгенных растений в соответствии с настоящим изобретением, например, различные среды и реципиентные целевые клетки, трансформация незрелых зародышевых клеток и последующая регенерация фертильных трансгенных растений, раскрываются в патентах США №№6194636 и 6232526, которые включены в настоящий документ посредством ссылки.
Развитие или регенерация растений, содержащих чужеродный, экзогенный выделенный фрагмент нуклеиновой кислоты, который кодирует представляющий интерес белок, хорошо известны в уровне техники. Регенерированные растения самоопыляются с обеспечением гомозиготных трансгенных растений. В противоположном случае, пыльцу, полученную из регенерированных растений, скрещивают с выращенными из семени растениями агрономически важных линий. Напротив, пыльцу из растений этих важных линий используют для опыления регенерированных растений. Трансгенное растение в соответствии с настоящим изобретением, содержащее молекулу нуклеиновой кислоты, кодирующую белок FTO, культивируют с использованием способов, хорошо известных специалисту в данной области.
Семена трансгенных растений могут быть собраны из фертильных трансгенных растений и использованы для выращивания поколений потомков трансформированных растений в соответствии с настоящим изобретением, в том числе линии гибридных растений, для селекции растений, имеющих усиленный признак. Кроме прямой трансформации растения молекулой нуклеиновой кислоты, кодирующей белок FTO, трансгенные растения могут быть получены путем скрещивания первого растения, имеющего молекулу нуклеиновой кислоты, кодирующую белок FTO, со вторым растением, не имеющим молекулы нуклеиновой кислоты. Например, молекула нуклеиновой кислоты, кодирующая белок FTO, может быть введена в первую линию растений, которая поддается трансформации, для получения трансгенного растения, которое может быть скрещено со второй линией растений с интрогрессией молекулы нуклеиновой кислоты, кодирующей белок FTO, во вторую линию растений.
Трансгенное растение, полученное из растительной клетки в соответствии с настоящим изобретением, культивируют для получения повышенной урожайности и/или биомассы по сравнению с контрольным растением. Используемый в настоящем документе термин «контрольное растение» означает растение, которое не содержит молекулу нуклеиновой кислоты, кодирующую белок FTO. Контрольное растение служит для идентификации и отбора трансгенного растения, которое характеризуется повышенной урожайностью и/или биомассой. Подходящим контрольным растением может быть нетрансгенное растение родительской линии, используемой для создания трансгенного растения, т.е. лишенное молекулы нуклеиновой кислоты, кодирующей белок FTO. Подходящим контрольным растением в некоторых случаях может быть потомок гемизиготной линии трансгенных растений, который не содержит молекулу нуклеиновой кислоты, кодирующую белок FTO, известный как негативный сегрегант.
Термин «урожайность» трансгенного растения, описываемого в настоящем документе, означает собранное количество желаемого продукта путем культивирования. Стандарты оценки урожайностей различных растений различны. Например, субъектом оценивания урожайностей зерновых сельскохозяйственных культур (риса, пшеницы, кукурузы и т.д.), бобов и масличных сельскохозяйственных культур (сои, арахиса, рапса и т.д.) является семя (зерно); для хлопчатника - это хлопковое семя или хлопок-волокно; для ямсовых сельскохозяйственных культур (сладкого картофеля, картофеля, маниока и т.д.) - это клубневидный корень или клубень; для лубяных сельскохозяйственных культур - это волокно стеблей или волокно листьев; для сахарного тростника - это стебель; для свеклы - это корень; для табака - это лист; для сидерационных сельскохозяйственных культур (люцерны, клевера и т.д.) - это стебель и лист, и т.д. Значение урожайности одного и того же растения отличается, когда его культивируют для разных целей. Например, если кукурузу культивируют в качестве пищевой и кормовой сельскохозяйственной культуры, то урожайность представляет собой собранное количество зерен, а если ее культивируют в качестве силоса, то урожайность включает в себя общее собранное количество стеблей, листьев и початков.
Повышенная урожайность трансгенного растения в соответствии с настоящим изобретением может быть измерена многими средствами, включающими в себя измерение массы, числа семян на растение, массы семени, массы клубня, числа семян на единицу площади (т.е. семян или массы семян на акр), бушелей на акр, метрических тонн на акр, тонн на акр, килограмм на гектар.
Специалисты в данной области смогут определить значение урожайности для каждого растения и стандарт для ее оценивания согласно известному уровню техники.
Биомасса означает суммарную массу имеющихся органических материалов организма. Ее выражают как сухую массу, сырую массу, число корневых отростков и т.д. в соответствии с настоящим изобретением. Увеличенная биомасса трансгенного растения в соответствии с настоящим изобретением может быть измерена многими средствами, включающими в себя измерение массы, сухой массы или сырой массы надземных частей на растение, числа корневых отростков, сухой массы надземных частей на единицу площади (т.е. сухой массы или сырой массы на акр), бушелей на акр, метрических тонн на акр, тонн на акр, килограмм на гектар.
Примеры
Далее настоящее изобретение иллюстрируется следующими примерами. Следует учитывать, что данные примеры наряду с тем, что описывают варианты осуществления настоящего изобретения, приводятся исключительно с иллюстративной целью. По приведенному выше обсуждению и данным примерам специалист в данной области сможет установить основные характеристики настоящего изобретения и без отступления от его признаков и объема сможет выполнить различные изменения и модификации настоящего изобретения для адаптации его к различным применениям и условиям. Кроме того, различные модификации настоящего изобретения в дополнение к показанным и описываемым в настоящем документе будут очевидны специалистам в данной области из приведенного выше описания. Такие модификации также предусматриваются объемом прилагаемой формулы изобретения.
В следующих примерах использовали следующие материалы.
1. Состав бульона YEP для роста Agrobacterium (на литр): 10 г/л дрожжевого экстракта+10 г/л пептона+5 г/л NaCl, рН 7,2. Для твердой среды добавляли 15 г/л агара.
2. Бульон ААМ для повторного суспендирования Agrobacterium: 50 мл 20× макроэлемента АА, 10 мл 100×FeEDTA, 10 мл 100× макроэлемента В5, 10 мл 100× витамина В5, 100 мл 10× аминокислоты АА, 1 мл 100 мМ ацетосирингона, 68,5 г сахарозы, 36 г глюкозы, 0,5 г гидролизированного казеина. Объем доводили до 1000 мл. рН доводили до 5,2. Стерилизовали с помощью 0,2-мм мембраны из ацетата целлюлозы.
3. 20× макроэлемент АА: 59 г KCl, 3 г CaCl2⋅2Н2О, 10 г MgSО4⋅7H2О и 3 г NaH2PO4⋅H2О. Объем доводили дистиллированной водой до 1 л. Хранили при 4°С.
4. 10× АА аминокислота: 8,76 г Gin, 2,66 г Asp, 1,74 г Arg и 75 мг Gly. Объем доводили дистиллированной водой до 1 л. Стерилизовали с помощью 0,2-мм мембраны из ацетата целлюлозы. Хранили при 4°C.
5. 10 мл 100× витамина В5: 10 г миоинозита, 1 г тиамина гидрохлорида, 100 мг пиридоксина гидрохлорида и 100 мг никотиновой кислоты. Объем доводили дистиллированной водой до 1 л. Хранили при 4°С
6. 100× макроэлемент В5: 1,320 мг MnSO4⋅4H2О, 200 мг ZnSO4⋅7H2О, 2,5 мг CuSО4⋅5H2О, 25 мг Na2MoО4⋅2H2О, 2,5 мг CoCl2⋅6Н2О, 300 мг Н3ВО3 и 75 мг KI. Объем доводили дистиллированной водой до 1 л. Хранили при 4°C.
7. Среда NB: макроэлементы и микроэлементы из среды N6, органические элементы из среды В5, 300 мг/л гидролизированного казеина, 500 мг/л глутамина, 30 г/л сахарозы и 8 г/л агара.
Пример 1. Получение cDNA из гена m6A-деметилазы
Гены FTO искали в базе данных Национального центра биотехнологической информации (NCBI). Получали аминокислотные последовательности (SEQ ID NO: 1-4) и последовательности нуклеиновой кислоты (SEQ ID NO: 5, 7, 9 и 11) FTO человека (Homo sapiens), FTO свиньи (Sus scrofa), FTO быка (Bos Taurus) и FTO водоросли (Ostreococcus lucimarinus CCE9901). Соответствующие cDNA приобретали или синтезировали с помощью GenScript, если cDNA не было в продаже. Аминокислотные последовательности из SEQ ID NO: 1-4 оптимизировали к кодонам растения для синтеза кодон-оптимизированных последовательностей нуклеиновой кислоты SEQ ID NO: 6, 8, 10 и 12.
В следующих примерах использовали cDNA из гена FTO SEQ ID NO: 5, 7, 9 и 11.
Пример 2. Клонирование гена m6A-деметилазы
cDNA из гена FTO (SEQ ID NO: 5, 7, 9, 11) клонировали в растительный бинарный вектор pCAMBIA1307. Средний размер вставленных последовательностей гена составлял 1,5 т.п.н. Полученная плазмида показана на фиг. 1.
Пример 3. Трансформация растений
3.1 Введение гена FTO из примера 2 в рис с помощью трансгенных технологий
I. Индуцирование каллюсов со зрелыми зародышами риса в качестве тестируемых материалов
1. Стерилизация
Зрелые семена риса (Japonica nipponbare) не были искусственно облущены. Семена, которые были полными, блестящими, чистыми и без бактериальных бляшек, отбирали и помещали в 100-мл стерилизованную колбу. Добавляли 70% спирт в колбу для стерилизации на 2 минут. Затем спирт декантировали и добавляли 20% NaClO для замачивания на 30 минут. Затем NaClO декантировали и семена ополаскивали 4-5 раз стерилизованной дистиллированной водой. Наконец семена замачивали в стерилизованной дистиллированной воде на 30 минут.
2. Введение в культуру (в асептических условиях)
Стерилизованные семена помещали на стерилизованную фильтровальную бумагу. После абсорбирования воды на поверхности фильтровальной бумагой семена помещали в среду NB (рН 5,8), содержащую 2,0 мг/л 2,4-D, с плотностью 12-14 семян на чашку Петри. Для гарантии хорошей степени индуцирования прорастание ориентировали параллельно среде или слегка вниз, а не вверх или вертикально вверх. Затем чашку Петри запечатывали мембраной, индуцировали и инкубировали на протяжении 20-30 суток в инкубаторе со светом при 30%, 50% влажности до очевидного появления рыхлых каллюсов. Затем выполняли пересев.
3. Пересев (в асептических условиях)
Чашку Петри открывали на сверхчистом операционном столе. Каллюсы, которые естественно делились, интенсивно росли, которые были твердыми и ярко-желтыми и которые имели диаметр приблизительно 3 мм, помещали в среду NB (рН 5,8), содержащую 2,0 мг/л 2,4-D и 0,5 мг/л 6-ВА, с плотностью 10 каллюсов на чашку Петри и инкубировали в темноте при 30°С. Если каллюсы становились сильно мягкими, их перемещали на свет для пересева. Пересев выполняли дважды, каждые 10-15 суток (время следующего пересева определяли в зависимости от того, насколько каллюсы вырастали).
II. Культивирование Agrobacterium
Вектор pCAMBIA1307, несущий ген FTO и ген устойчивости к гигромицину, показанный на фиг. 1, трансфицировали в Agrobacterium LBA4404. Затем Agrobacterium LBA4404 высевали на твердую среду YEP, содержащую 20 мг/л рифампина (Rif) и 50 мг/л канамицина (Kan). После культивирования при 28°C в течение двух суток моноклоны Agrobacterium пикировали, чтобы подвергнуть ПЦР для отбора колоний с целью тестирования переноса FTO в Agrobacterium. Положительные моноклоны отбирали, помещали в 4 мл бульона YEP (содержащего 50 мг/л Kan и 20 мг/л Rif) и культивировали со взбалтыванием при 28°C, 220 оборотов в минуту, в течение 20-36 часов до OD600 бактериального раствора 0,8-1,0.
III. Инфицирование и совместное культивирование
1. Раствор культивируемой Agrobacterium центрифугировали при 4°С 4000 оборотов в минуту, в течение 10 минут и готовили суспензию с бульоном ААМ, содержащим 100 мкмоль/л ацетосирингона. Конечная OD600 бактериального раствора составляла приблизительно 0,2.
2. Каллюсы определенного размера пикировали и помещали в суспензию Agrobacterium для инфицирования на 20-30 минут.
3. Каллюсы вынимали и помещали на стерилизованную фильтровальную бумагу для сушки на 20-30 минут с тем, чтобы предупредить чрезмерное повреждение каллюсов, вызываемое чрезмерным ростом Agrobacterium в процессе совместного культивирования.
4. Каллюсы помещали в среду NB (рН 5,2), содержащую 2,0 мг/л 2,4-D и 100 мкмоль/л ацетосирингона, и культивировали при 25°С в течение 48-72 часов в темноте.
IV. Скрининг каллюсов с устойчивостью
Каллюсы вынимали и ополаскивали при встряхивании со стерилизованной водой 5-6 раз. Затем каллюсы помещали на стерилизованную фильтровальную бумагу для сушки, а затем помещали равномерно в среду NB (рН 5,8), содержащую 50 мг/л гигромицина, для первого скрининга. Каллюсы культивировали в темноте при 28°С и, если наблюдался рост плесени или Agrobacterium, каллюсы сразу же переносили в новую чашку для скрининга.
После скрининга на протяжении приблизительно 30 суток получали новые каллюсы. Их переносили в новую среду NB (рН 5,8), содержащую 50 мг/л гигромицина, для еще одного скрининга на протяжении 7-10 суток. Если наблюдали рост, то их считали положительными каллюсами. Если рост отсутствовал (даже если не было коричневого цвета или отмирания), вероятно они были ложноположительными.
V. Интродукция дифференцировки каллюсов с устойчивостью и укоренение
Энергично растущие желтые каллюсы после второго скрининга (для гарантии того, что используемые для дифференцировки каллюсы не были дефективными) помещали в среду NB (рН 5,8), содержащую 2,0 мг/л 6-ВА, 0,5 мг/л кинетина и 50 мг/л гигромицина, с плотностью 2-3 положительных клона на бутылку. Было достаточно поместить немного каллюсов на клон. Помещенные каллюсы должны были характеризоваться высоким качеством, а не большим количеством. После культивирования при 21% в темноте на протяжении 10 суток обеспечивали их дифференцировку на свету на протяжении 10-20 суток. После появления зеленых листьев обеспечивали их укоренение при плотности излучения 4000 люкс, 14 часов/сутки.
Сильные проростки, дифференцированные из каждого клона, помещали в среду 1/2N6 (рН 5,8), содержащую 0,5 мг/л нафтилуксусной кислоты и 50 мг/л гигромицина, для индуцирования укоренения с плотностью 2-3 проростка в каждой бутылке. Проростки культивировали при 27-30°С на свету при плотности излучения 4000 люкс, 14 часов/сутки. Через 7-10 суток закупоривающие мембраны открывали для добавления соответствующего количества воды. Через 2-3 суток проростки пересаживали.
VI. Закалка и пересаживание трансгенных проростков
Проростки риса с хорошо дифференцированными корнями, стеблями и листьями пикировали (когда проростки дорастали до верхней части тестовых пробирок, крышки должны были быть вовремя открыты). Закупоривающие мембраны открывали и добавляли соответствующее количество дистиллированной воды или стерилизованной воды (для предупреждения роста бактерий в среде). Проростки закаляли на протяжении от приблизительно 3 суток до одной недели. После вымывания агара проростки пересаживали в емкости с почвой в теплице для выращивания и тестирования.
Пример 4. Тестирование трансформированных проростков
1. Выявление с помощью ПЦР: схема праймеров гена FTO Праймеры для выявления FTO человека (hFTO):
hFTO-F: 5'-ATGAAGCGCACCCCGACTG-3' (SEQ ID NO: 13);
hFTO-R: 5'-GGGTTTTGCTTCCAGAAGCTGA-3' (SEQ ID NO: 14).
Праймеры для выявления FTO быка (cFTO):
cFTO-F: 5'-ATGAAGCGGACCCCGACG-3' (SEQ ID NO: 15);
cFTO-R: 5'-GGGCCTGGTTTCCAGAAGCAG-3' (SEQ ID NO: 16).
Праймеры для выявления FTO свиньи (pFTO):
pFTO-F: 5'-ATGAAGCGAACCCCAACCGC-3' (SEQ ID NO: 17);
pFTO-R: 5'-GGGTTTGGCTTCCAGAAGCAGAC-3' (SEQ ID NO: 18).
Праймеры для выявления FTO Ostreococcus lucimarinus (olFTO):
olFTO-F: 5'-ATGTCGCCGTCATCCTCCG-3' (SEQ ID NO: 19);
olFTO-R: 5'-CACTTTGTTTTGCTCCTCCTCGAGAAA-3' (SEQ ID NO: 20).
Программа ПЦР-реакции была следующая: 35 циклов 95°С 5 минут, 95°С 15 секунд, 58°С 15 секунд, 72°С 30 секунд. Удлиняли при 72°С в течение 10 минут, хранили при 4°С. После реакции ПЦР-продукты подвергали анализу электрофореза в 1% агарозном геле.
2. Способ быстрого выявления трансгенных проростков. Свежие зеленые листья приблизительно 1 см отрезали и собирали у проростков, подлежащих тестированию (обе стороны имели разрезы), помещали плоской стороной на среду для тестирования (0,7% агара, 1 мл/л 6-ВА, 50 мг/л гигромицина) и культивировали при 28"С в течение 48 часов (16 часов света/8 часов темноты каждые сутки). Растения, у которых были свежие зеленые листья, были положительными, а листья отрицательных проростков имели бляшки некроза.
Пример 5. Оценка эффектов генов FTO из некоторых видов в отношении риса
Результаты тестирования риса из примера 3 представлены в таблице 1 (сорт Japonica nipponbare, в который не вводили ген FTO, служил контролем). Данные получали из 50 трансгенных и контрольных растений риса на стадии созревания. Измеряли сухую массу после помещения в печь 105°С на 20 минут и в печь 80°С на 20 часов.
В таблице 1 показано, что после того, как нуклеиновые кислоты, кодирующие FTO некоторых видов, были введены в рис, биомасса и урожайность семян повышались.
Автор настоящего изобретения тем же способом также тестировал кодон-оптимизированные нуклеиновые кислоты, кодирующие FTO (SEQ ID NO: 6, 8, 10 и 12). Данные подобны указанным выше и, поэтому, не показаны в настоящем документе.
Пример 6. Оценка эффекта FTO в отношении ряда растений
Ряд растений трансформировали нуклеиновой кислотой, кодирующей FTO человека (SEQ ID NO: 5), для получения ряда трансгенных растительных клеток и растений, в которых была введена нуклеиновая кислота, кодирующая FTO. Способы являются следующими.
Способ генетической трансформации табака
1. Экспериментальные материалы
Материалы: сорт табака: K326; Agrobacterium: LBA4404; ген HF (FTO человека) клонировали в управляемый 35S промотором вектор pCAMBIA2300. СкринируеМой устойчивостью в эукариотических растениях была устойчивость к канамицину.
Химические реагенты: макроэлементы MS, микроэлементы MS, соли железа MS, индолуксусная кислота (IAA), 6-бензиламиноаденин (6-ВА), инозитола никотинат (В1 В6), сахароза, агар, цефалоспорин (Cef), карбенициллин (Carb), канамицин (Кп), гентамицин, рифампицин.
Среда MS (1 л): макроэлементов (20×) 50 мл, микроэлементов (100×) 10 мл, Fe2+ (100×) 10 мл, сахарозы 30 г, агары 8 г. рН составлял приблизительно 6,0.
Среда предварительного культивирования (1 л): макроэлементов (20×) 50 мл, микроэлементов (100×) 10 мл, Fe2+ (100×) 10 мл, 6-ВА (1000×) 2 мл, B1B6 (200×) 5 мл, глицина (1000×) 1 мл, агара 8 г.рН составлял приблизительно 6,0. После стерилизации высокой температурой добавляли 1 мл IAA (0,2 мг/л).
Культуральная среда для дифференцировки (1 л): на основе среды предварительного культивирования, добавляли цефалоспорина 2 мл, карбенициллина 1 мл и канамицина 1 мл.
Культуральная среда для укоренения (1 л): 1/2MS, IAA 2 мг/л, сахарозы 30 г/л, агара 5,8 г/л, рН=5,8.
Бульон LB (1 л): триптона 10 г, дрожжевого экстракта 5 г, NaCl 10 г.
Среда MS0: культуральная среда MS, без добавления агара и содержащая только макроэлементы.
2. Трансформация табака (способ с использованием дисков из листьев) Табак трансформировали способом с использованием дисков из листьев.
Вырезанные диски из листьев табака помещали в среду предварительного культивирования на 1-2 суток, а затем замачивали в суспензии Agrobacterium (MS0 суспендировали, разбавляли в 50-100 раз) в течение 3-5 минут. Затем диски вынимали и использовали стерильную фильтровальную бумагу для абсорбирования жидкости на их поверхностях. Инфицированные диски из листьев соответственно инокулировали на среде для предварительного культивирования, покрытой двумя слоями фильтровальной бумаги, и культивировали при 26°С в течение 20-24 часов в темноте. Диски ополаскивали стерилизованной водой с добавлением цефалоспорина и карбенициллина (разбавленного в 1000 раз маточного раствора) в течение 10 минут. Наконец их ополаскивали стерилизованной водой в течение 10 минут. Потом использовали стерильную фильтровальную бумагу для абсорбирования жидкости на их поверхностях, а затем диски помещали в культуральную среду для дифференцировки с целью дифференцировки культуры. На ранних стадиях пересев выполняли каждые 2-3 суток, и каждый пересев должен был осуществляться в асептических условиях; после повторения пересева три раза его выполняли каждые две недели. Совместное культивирование осуществляли до появления первой почки. Почки переносили в сосуды для культуры ткани с целью культивирования. Когда почки вырастали до 2 см, все каллюсы на базальных частях почек и базальные листья разрезали на суперчистом операционном столе, а затем почки помещали в культуральную среду для укоренения. Когда корни вырастали до 3 см, стерильные проростки вынимали; твердую культуральную среду осторожно удаляли, а оставшуюся культуральную среду вымывали. После этого стерильные проростки помещали в почву, покрытую чистыми пластиковыми пакетами (проколотыми), и культивировали на протяжении приблизительно одной недели, а затем переносили на открытый воздух (выращивая в темноте первые 3 суток).
Способ трансформации картофеля
1. Материалы
Сорт картофеля: Dongnong 303; Agrobacterium: LBA4404; ген HF (FTO человека) клонировали в управляемый 35S промотором вектор pCAMBIA2300. Скринируемой устойчивостью в эукариотических растениях была устойчивость к канамицину.
2. Трансформация картофеля
Картофелины очищали путем ополаскивания дистиллированной водой, замачивали в 75% спирте на 30 секунд, замачивали в 0,1% хлориде ртути на 10 минут, промывали стерилизованной водой 5 раз, а затем очищали с нарезанием срезов толщиной около 1 мм. Срезы смешивали с раствором Agrobacterium при осторожном встряхивании для обеспечения контакта раствора Agrobacterium с эксплантатами в достаточной степени. Стерильную фильтровальную бумагу использовали для абсорбирования лишнего раствора Agrobacterium. Затем кусочки картофеля помещали в середу для совместного культивирования для культивирования в темноте. После завершения совместного культивирования кусочки картофеля очищали 3 раза стерилизованной водой и жидкой культуральной средой MS, соответственно. Лишний раствор Agrobacterium смывали. Кусочки картофеля переносили в культуральную среду для регенерации. Затем кусочки картофеля переносили в культуральную среду для укоренения на срок, пока почки не вырастали до 1,5-2,0 мм, и разрезали для скринингового размножения путем укоренения.
Генетическая трансформация сои
1. Материалы
Штаммом Agrobacterium был LBA4404; массы зародышей соматических клеток сои Dongnong L13 на стадии глобулы; трансгенная плазмида представляла собой управляемый 35S промотором вектор pCAMBIA2300, в который клонировали ген HF. Скринируемой устойчивостью в эукариотических растениях была устойчивость к канамицину.
Среда для культивирования растительных тканей и условия культивирования
Среда для пересева: MS+15 мг/л+20 мг/л 2,4-D+0,8% агара+3% сахарозы, рН=5,8, естественное освещение.
Культуральная среда для повторного суспендирования: среда для пересева (жидкая)+AS (0-100 мкмоль/л), рН=5,8, естественное освещение.
Среда для совместного культивирования: культуральная среда для инфицирования+AS (0-100 мкмоль/л), рН=5,6.
Культуральная среда для скрининга со стерилизацией: среда для пересева+(50-300 мг/л) цефалоспорин+25-50 мг/л канамицина.
Культуральная среда для прорастания 1: MS+1% активированного угля+0,8% агара+10% сахарозы, освещение 16 часов/сутки.
Культуральная среда для прорастания 2: MS+0,8% агара+10% сахарозы, освещение 16 часов/сутки.
Культуральная среда для укрепления проростков: MSB+0,8% агара+3% глюкозы, рН=7, освещение 16 часов/сутки.
(В последних трех культуральных средах канамицин, 0-50 мг/л, и цефалоспорин, 0-300 мг/л, добавляли одновременно.)
2. Трансформация инфицированием Agrobacterium и регенерация растений Массу зародышей соматических клеток сои диаметром около 3 мм помещали в
приготовленную жидкость для инфицирования с Agrobacterium с целью инфицирования. Затем бактериальный раствор сливали и использовали стерильную фильтровальную бумагу для абсорбирования лишней жидкости при высушивании зародышевых масс. Потом зародышевые массы инокулировали в среду для совместного культивирования, затем в культуральную среду для скрининга со стерилизацией, содержащую 50 мг/л канамицина и 300 мг/л цефалоспорина (концентрации снижали последовательно в зависимости от ситуации, при условии, что бактерии не росли). Время инфицирования составляло 10 минут. Время совместного культивирования составляло 2-3 суток. Ацетосирингон составлял 100 мкмоль/л. OD600 концентрации Agrobacterium составляла 0,5-0,7. Пересев выполняли каждые 15-20 суток. Число устойчивых масс зародышей соматических клеток (с диаметрами около 3 мм) по мере образования проверяли в течение 3 месяцев. Вычисляли степень трансформации (число устойчивых масс зародышей соматических клеток/число инокулированных масс зародышей соматических клеток ×100). Затем устойчивые массы зародышей соматических клеток переносили в культуральную среду для прорастания 1, а затем в культуральную среду для прорастания 2 после прорастания на протяжении 20 суток. До вырастания регенерированных устойчивых небольших растений указанные растения переносили в культуральную среду для укрепления проростков с целью получения трансформированных растений.
Генетическая трансформация люцерны
1. Материалы
Сорт люцерны: Gongnon-1; Agrobacterium: LBA4404; растительный бинарный вектор экспрессии pCAMBIA2300, в который клонировали ген HF, и устойчивый к канамицину.
Культуральной средой была культуральная среда MS с добавлением различных регулирующих рост веществ, со значением рН приблизительно 5,8. Температура культивирования составляла 23-25°C. Интенсивность освещения составляла 2000 люкс; время освещения составляло 18 часов/сутки. Пересев выполняли приблизительно каждые 20 суток.
2. Получение каллюсов
Гипокотили использовали в качестве эксплантатов для индуцирования каллюсов и дифференцировки соматических зародышей. После созревания соматические зародыши переносили в MS0 для тестирования прорастания.
3. Суспензионная культура эмбриогенных каллюсов
3 г каллюсов инокулировали в 150-мл колбу Эрленмейера, содержащую 40 мл раствора суспензионной культуры для выполнения суспензионного культивирования. Свежую культуральную среду использовали для замены каждые 7 суток. Скорость вращения шейкера составляла 150 оборотов/минута. Для освещения использовали естественный рассеянный свет. Через 15 суток определяли скорость роста клеток. Подходящие каллюсы инокулировали в бульон для индуцирования наилучших эмбриогенных каллюсов, которые были отобраны. Скорость вращения шейкера устанавливали на 120 оборотов/минута. Свежую культуральную среду использовали для замены каждые 7 суток. Освещение естественным рассеянным светом осуществляли приблизительно 30 суток.
4. Трансформация с помощью Agrobacterium
Эмбриогенные каллюсы помещали в 2,0-мл небольшую центрифужную пробирку, содержащую 600 мкл жидкой среды для совместного культивирования, для обработки ультразвуковой волной 8 секунд с параметром 100 мГц, а затем совместно культивировали 4 суток. Инфицированные суспензионные эмбриогенные каллюсы помещали перпендикулярно к плоской поверхности на полутвердую среду для совместного культивирования, содержащую 100 мкмоль/л ацетосирингона для культивирования в темноте на протяжении 4 суток.
5. Скрининг и регенерация трансформированных растений
Совместно культивируемые эмбриогенные каллюсы инокулировали в культуральную среду для дифференцировки с целью культивирования на протяжении 50-60 суток. После 4 зародышевых стадий эмбриогенные каллюсы вырастали в зрелые соматические зародыши. Затем их переносили в культуральную среду для прорастания соматических зародышей. Через приблизительно 20-30 суток, когда растения вырастали до приблизительно 10 см, проростки пересаживали, при этом применяемая скрининговая концентрация канамицина составляла 30 мг/л.
Генетическая трансформация рапса
1. Материалы
Сорт рапса капустного типа CY2 для генетической трансформации; штамм Agrobacterium ЕНА105; растительный бинарный вектор экспрессии pCAMBIA1301, в который клонировали ген HF.
2. Трансформация с помощью Agrobacterium
Свежие бактериальные колонии отбирали с помощью зубочистки для культивирования в бульоне YEB, содержащем 50 мг/л канамицина и 50 мг/л рифампицина при встряхивании при 28°C, до средней фазы логарифмического деления (OD600=0,3). Бактериальный раствор отбирали для центрифугирования при 12000 оборотов/минута в течение 1 минуты, промывали один раз культуральной средой MS при центрифугировании, а затем разбавляли в 10 раз. Реципиентные родительские семена CY2 стерилизовали и инокулировали в культуральную среду 1/2MS с целью культивирования стерильных проростков. Через 5-7 суток гипокотили разрезали на небольшие срезы приблизительно 1 см длиной, которые служили эксплантатами для опосредованной Agrobacterium tumefaciens трансформации с плазмидой pCAMBIA1301, несущей ген HF. Трансформацию осуществляли согласно методу Wang Fulin et al. (Journal of Nuclear Agricultural Sciences, 5(26): 1129-1134(2011)). Трансформанты отбирали с помощью гигромицина (Hyg), 10 мг/л. Отобранные устойчивые к Hyg проростки укореняли и акклиматизировали в тепличных горшках, а затем пересаживали на поля.
Генетическая трансформация хлопчатника
1. Материалы и реагенты
Сорт хлопчатника: Zhong 521; штамм Agrobacterium GV3101; растительный бинарный вектор экспрессии pCAMBIA2300, в который клонировали ген HF, и устойчивый к канамицину.
Культуральная среда MBS: неорганический ингредиент MS+органический ингредиент В5 в качестве основной культуральной среды, с добавлением различных гормональных ингредиентов.
Жидкость для инфицирования: неорганические соли MS+органические ингредиенты В5+0,1 мг/л 2,4-D+0,1 мг/л КТ+100 мкмоль/л AS (ацетосирингона)+30 г/л глюкозы.
Совместная культура: неорганические соли MS+органические ингредиенты В5+0,1 мг/л 2,4-D+0,1 мг/л КТ+100 мкмоль/л AS (ацетосирингона)+30 г/л глюкозы+2,5 г/л растительного геля.
Культуральная среда для скрининга: неорганические соли MS+органические ингредиенты В5+0,1 мг/л 2,4-D+0,1 мг/л КТ+30 г/л глюкозы+2,5 г/л растительного геля+50 мг/л канамицина+150 мг/л карбенициллина.
2. Культивирование эксплантатов
Семена хлопчатника, обеспушенные концентрированной серной кислотой, очищали водопроводной водой; ядра семян вынимали. Затем обеспушенные семена помещали в стерилизованную колбу Эрленмейера на сверхчистом операционном столе. Семена замачивали в 70% - 75% спирте на 1 минуту, ополаскивали стерилизованной водой, а затем замачивали в 2% гипохлорите натрия на 60 минут. После этого гипохлорит натрия сливали, семена ополаскивали стерилизованной водой несколько раз и замачивали в стерилизованной воде на 24 часа. После того, как семена вытаскивали, оболочки семян отделяли. Семена инокулировали в культуральную среду для прорастания 1/2MS с целью культивирования в темноте при 28°C. Проростки возрастом 5 суток или около того вынимали и разрезали на небольшие кусочки 0,5-0,6 см, которые служили эксплантатами.
3. Совместное культивирование эксплантатов и Agrobacterium
Гипокотили стерильных проростков разрезали на небольшие срезы по 0,5 см или около того и замачивали в растворе Agrobacterium на 30-40 минут. Использовали стерильную фильтровальную бумагу для абсорбирования бактериального раствора. Указанные небольшие срезы помещали на твердую культуральную среду MSB для совместного культивирования на протяжении 48 часов.
4. Индуцирование и отбор каллюсов, а также регенерация растений
Совместно культивируемые отрезки гипокотилей переносили на твердую культуральную среду для скрининга MSB, содержащую 50 мг/л канамицина и 500 мг/л карбенициллина. На протяжении культивирования в течение 60 суток отбирали массы положительных каллюсов, устойчивых к канамицину, и переносили в упомянутую выше культуральную среду без антибиотика для пересева. После этого согласно состоянию каллюсов получали эмбриогенные каллюсы в форме частиц путем регулирования концентрации гормонов в культуральной среде, а затем эмбриоиды дифференцировали в регенерированные в культуре растения.
Генетическая трансформация пшеницы (эмбриогенные каллюсы)
1. Тестируемые материалы
Для культивирование использовали ткани незрелых зародышей. Тестируемым сортом был Henong 827. Штамм Agrobacterium С58. Растительный бинарный вектор экспрессии pCAMBIA2300, в который клонировали ген HF, и устойчивый к канамицину.
Культуральные среды, используемые в процессе генетической трансформации пшеницы, включали в себя:
культуральную среду для индуцирования каллюсов MSW0: основа MS+2 мг/л 2,4-D+4 мг/л пиклорида+0,5 г/л глутамина+0,75 г/л MgCl2+0,1 г/л гидролизированного казеина+1,95 г/л MES+100 мг/л аскорбиновой кислоты+40 г/л мальтозы+4,5 г/л агара, рН 5,8;
инфекционную жидкость РСМ: основа MS+2 мг/л 2,4-D+4 мг/л пиклорида+0,5 г/л глутамина+0,75 г/л MgCl2+0,1 г/л гидролизированного казеина+1,95 г/л MES+100 мг/л аскорбиновой кислоты+200 мкмоль/л ацетосирингона+40 г/л мальтозы+4,5 г/л агара, рН 5,8;
культуральную среду для скрининга каллюса SM: основа MS+2 мг/л 2,4-D+4 мг/л пиклорида+0,5 г/л глутамина+0,75 г/л MgCl2+0,1 г/л гидролизированного казеина+1,95 г/л MES+100 мг/л аскорбиновой кислоты+250 мг/л карбенициллина+25 мг/л G418+40 г/л мальтозы+4,5 г/л агара, рН 5,8;
культуральную среду для дифференцировки устойчивых каллюсов RSM: основа MS+2 мг/л 2,4-D+4 мг/л пиклорида+0,5 г/л глутамина+0,75 г/л MgCl2+0,1 г/л гидролизированного казеина+1,95 г/л MES+100 мг/л аскорбиновой кислоты+250 мг/л карбенициллина+25 мг/л G418+0,5 мг/л кинетина+0,2 мг/л нафталинуксусной кислоты+40 г/л мальтозы+4,5 г/л агара, рН 5,8.
2. Стерилизация незрелых зерен и инокуляция незрелых зародышей Незрелые зерна брали через 12-15 суток после опыления пшеницы, поверхность
стерилизовали 70% спиртом в течение 30 секунд, стерилизовали 0,1% хлоридом ртути в течение 8 минут и очищали стерилизованной водой 4-5 раз. Незрелые зародыши извлекали препаровальными иглами и соответственно инокулировали на культуральную среду для индуцирования каллюсов MSW0 щитками вверх с целью культивирования в темноте на 2 недели при 25°C, а затем переносили в культуральную среду для дифференцировки с целью культивирования с освещением 16 часов при 25°C и в темноте 8 часов на протяжении 4-6 недель.
3. Опосредованные Agrobacterium процедуры генетической трансформации
Agrobacterium С58, содержащую HF, инокулировали равномерно аппликатором на твердую культуральную среду LB (рН 7,0, содержащую 50 мг/л канамицина и 50 мг/л рифампицина) для культивирования на протяжение 3 суток при 28°С с последующим культивированием 1 сутки при 23°С. После этого незначительное количество Agrobacterium соскабливали с культуральной среды для переноса и инокулирования в бульоне YEP, содержащем указанный выше антибиотик, с целью культивирования на протяжении ночи при 28°C при скорости вращения шейкера 250 оборотов/минута. Agrobacterium собирали по достижении OD600 1,0 и повторно суспендировали в инфекционной жидкости РСМ. Повторную суспензию использовали для замачивания каллюсов в течение 3 часов. Затем бактериальный раствор сливали. Каллюсы переносили в культуральную чашку, застеленную стерильной фильтровальной бумагой, для совместного культивирования на протяжении 3 суток, потом переносили в культуральную среду для скрининга с целью культивирования в темноте на протяжении 2 недель при 25°C, а затем переносили в культуральную среду для дифференцировки с целью культивирования с освещением при 25°C.
Генетическая трансформация проса
Использовали тот же способ, что и для пшеницы.
Генетическая трансформация льна
1. Материалы
Лен: Heiya 7; штамм Agrobacterium tumefaciens ЕНА105; растительный бинарный вектор экспрессии pCAMBIA1301, в который клонировали ген HF, и устойчивый к канамицину.
2. Получение эксплантатов
Отбирали выполненные и блестящие семена Heiya 7, замачивали в 75% спирте на 5 минут, замачивали в 20% супернатанте хлорной извести на 20 минут, ополаскивали стерилизованной водой три раза, а затем инокулировали в культуральную среду MS для культивирования в темноте на протяжении 5-7 суток при 25°C. За 2 суток до применения семена помещали под освещение на 16 часов каждые сутки при 22°C для применения.
3. Получение бактериального раствора Agrobacterium
Размножение штамма ЕНА105, содержащего представляющий интерес ген, выполняли с использованием культуральной среды YEP, содержащей 10 г/л пептона, 10 г/л дрожжевого экстракта, 5 г/л NaCl и 50 мг/л канамицина. После инокуляции выполняли культивирование со встряхиванием при 28°C на протяжении 2 суток. Верхнюю фазу удаляли центрифугированием в течение 10 минут при 3000 оборотов/минута. Бактерии суспендировали в бульоне 1/2MS (OD600=0,5) для трансформации.
4. Тест давления отбора
Стерильные гипокотили льна разрезали на небольшие кусочки по 0,3-0,5 мм. Указанные небольшие кусочки замачивали в стерилизованной воде на 10 минут, влагу абсорбировали досуха стерильной фильтровальной бумагой и соответственно инокулировали в культуральную среду MS с 50 мг/л канамицина для культивирования при 24-26°C. Период освещения составлял 16 часов каждые сутки.
5. Совместное культивирование
Гипокотили льна разрезали на небольшие кусочки по 0,3-0,5 мм. Указанные небольшие кусочки замачивали в суспензии Agrobacterium ЕНА105 на 10-20 минут, влагу абсорбировали досуха стерильной фильтровальной бумагой и соответственно инокулировали в культуральную среду MS, В5 или N6 с 2 мг/л КТ, 3,5 мг/л IAA или 150 мг/л HL. Условия культивирования были таким же, как и указанные выше.
6. Скрининговое культивирование и укоренение
Культуральная среда для скрининга была такой же, что и для совместного культивирования, с единственным отличием, заключающимся в добавлении 50 мг/л канамицина и 1000 мг/л цефалоспорина. Эксплантаты, совместно культивируемые на протяжении 3 суток с Agrobacterium, замачивали с раствором 2000 мг/л цефалоспорина на 10-20 минут, влагу абсорбировали досуха стерильной фильтровальной бумагой, а затем инокулировали в культуральную среду для скрининга с целью культивирования при тех же условиях, что показаны выше. Стерилизацию и образование каллюсов проверяли через одну неделю и две недели после инокуляции, соответственно. Отобранные устойчивые почки переносили в культуральную среду для укоренения с целью индуцирования укоренения.
Генетическая трансформация подсолнечника
1. Материалы
Helianthusannuus Xinkuiza 6; штамм Agrobacterium tumefaciens ЕНА105; растительный вектор экспрессии pCAMBIA1301, в который клонировали ген HF.
2. Культивирование штамма
Свежие одиночные колонии Agrobacterium tumefaciens отбирали для инокуляции в бульон YEP (1% дрожжевого экстракта+1% триптона+0,5% говяжьего экстракта) со встряхиванием при 28°C на протяжение ночи. На следующие сутки их переносили и инокулировали в 20 мл бульона YEP, содержащего антибиотик, при 1% величине инокуляции для дальнейшей вибрации и культивирования до фазы логарифмического роста; бактерии собирали центрифугированием и разбавляли жидкостью MES до тех пор, пока OD600 не достигала 0,8, в качестве рабочей концентрации для применения.
3. Культуральные среды
Среда MS0: основные ингредиенты MS+2% сахарозы+0,8% агара, рН 5,8.
Основная GBA: MS0+0,5 мг/л ВАР+0,25 мг/л IAA+0,1 мг/л GA3+30 г/л сахарозы+0,8% агара, рН 5,8.
Среда для совместного культивирования (МС): GBA+ацетосирингон (ACS) (100 моль/л)+30 г/л сахарозы+0,8% агара, рН 5,8.
Культуральная среда для скрининга (MB): GBA+400 мг/л Carb+10 мг/л гигромицина+30 г/л сахарозы+0,8% агара, рН 5,8.
Для укоренения (MR): l/2MS0+0,2 мг/л NAA+250 мг/л Carb+5 мг/л гигромицина+30 г/л сахарозы+0,8% агара, рН 5,8.
4. Получение эксплантатов
Отбирали семена, которые были выполненными, однородными по размеру и не имели вредителей и болезней, отделяли оболочку, замачивали в 70% этаноле на 1 минуту, дважды ополаскивали стерилизованной водой, стерилизовали 1% AgNO3 в течение 3 минут и ополаскивали стерилизованной водой три раза. Семена высевали на твердую культуральную среду MS0 и проращивали в темноте при 28°C получали стерильные проростки после культивирования в течение 36-48 часов. Корни, семядоли и филлоподии стерильных проростков разрезали для обнажения кончиков стеблей, а затем разрезали продольно. Полученные эксплантаты содержали половину меристем кончика стебля и две половины пазушных почек семядоли.
5. Инфицирование
Полученные из кончиков стеблей эксплантаты замачивали в бактериальном растворе достаточным образом в течение 10 минут, а затем вынимали. Использовали стерильную фильтровальную бумагу для абсорбирования достаточным образом лишнего бактериального раствора с поверхностей эксплантатов. Эксплантаты помещали в среду для совместного культивирования МС с целью совместного культивирования на протяжении 3 суток в темноте при и обеспечивали контроль.
6. Скрининг трансформантов и регенерация растений
Эксплантаты, совместно культивируемые на протяжении 3 суток, переносили в культуральную среду для скрининга MB, содержащую 10 мг/мл гигромицина (Hyg), с целью культивирования на протяжении 2 недель, а затем отбирали в 2-3 цикла (2 недели на каждый цикл). Отобранные устойчивые почки переносили на культуральную среду для укоренения MR с целью индуцирования укоренения.
Генетическая трансформация Taraxacum kok-saghyz Rodin (так называемого кок-сагыза)
1. Материалы
Taraxacum kok-saghyz Rodin; штамм Agrobacterium: GV3101; плазмида (pCAMBIA2300-35S-HF), которая представляла собой бинарный вектор pCAMBIA2300, несущий ген HF и устойчивый к канамицину.
2. Генетическая трансформация и регенерация Taraxacum kok-saghyz Rodin
(1) Отбирали хорошо растущие из тканевой культуры проростки. Края листьев удаляли. Отрезали стебель Taraxacum kok-saghyz Rodin длиной 2 см. Листья нарезали на кусочки размером 1 см2. Их помещали в культуральную среду MS с добавлением растительных гормонов 6-ВА и NAA для культивирования в темноте на протяжении 2 суток.
(2) Из пробирки с глицерином брали 200 мкл Agrobacterium GV3101, хранившейся при -70°C и содержащей плазмиду pCAMBIA2300-35S-HF, для инокулирования в 50 мл жидкости LB (50 мг/л Gen+100 мг/л Rif+50 мг/л Kan) с целью культивирования на протяжении ночи.
(3) Бактериальный раствор, культивируемый на протяжении ночи, сеяли штрихом на твердую культуральную среду LB (50 мг/л Gen+100 мг/л Rif+50 мг/л Kan), помещали вверх дном для культивирования на протяжении 2 суток при 28°C.
(4) Отбирали моноклональные колонии для инокуляции в бульон LB (50 мг/л Gen+100 мг/л Rif+50 мг/л Kan) с целью культивирования со встряхиванием на протяжении 2 суток при 28°C.
(5) Культивируемые бактериальные растворы соответственно инокулировали в 100 мл жидкости LB (50 мг/л Gen+100 мг/л Rif+50 мг/л Kan) при отношении 1:100 с целью наращивания культуры и активировали до OD260 приблизительно 0,6 для инфицирования.
(6) Два указанных выше бактериальных раствора соответственно помещали в 250-мл стерильные большие центрифужные пробирки и центрифугировали в течение 10 минут при 5000 оборотах в минуту.
(7) Супернатант сливали. Бактерии суспендировали в 100 мл бульона MS и культивировали в течение 5 минут при 28°C.
(8) Указанные выше эксплантаты культивировали в темноте на протяжении 2 суток, помещали в отобранный бактериальный раствор для культивирования со встряхиванием в течение 20 минут при 28°С.
(9) Инфицированные эксплантаты распределяли на стерильной сухой фильтровальной бумаге, лишний бактериальный раствор абсорбировали для культивирования в темноте на протяжении 2 суток.
(10) Через 2 суток эксплантаты забирали и помещали на твердую культуральную среду MS (1 мг/л 6-ВА+0.1 мг/л NAA+400 мг/л Сb (карбенициллина)+50 мг/л Kan (канамицина)); чашку заливали каждые пол месяца.
(11) Когда адвентивные побеги дорастали до 2 см, отдельные побеги отламывали и вставляли в культуральную среду для укоренения 1/2MS (0,2 мг/л NAA+50 мг/л Kan+400 мг/л Сb) для выращивания.
(12) Через один месяц сильные проростки из тканевой культуры акклиматизировали на протяжении 1 суток, пересаживали в питательную почву (торфяная почва:вермикулит=3:1), покрытую пленкой, на 1 неделю и помещали в культивационное помещение для дальнейшего культивирования.
Генетическая трансформация кукурузы
1. Материалы и реагенты
Сорт кукурузы: Qi 319; штамм Agrobacterium: GV3101; плазмида (pCAMBIA2300-35S-HF), которая представляла собой бинарный вектор pCAMBIA2300, несущий ген HF и устойчивый к канамицину.
Культуральная среда D: 10 мл/л NaFeEDTA+50 мл/л макроэлементов N6+10 мл/л микроэлементов В5+1 мл/л дикамбы (2,4-D) (5 мл/л)+10 мл/л RTV+0,5 г/л касаминовой кислоты (казеингидролазы)+700 мг/л L-Prine+100 мг/л инозита+20 г/л сахарозы, рН 5,8.
Культуральная среда D-Inf: 10 мл/л NaFeEDTA+50 мл/л макроэлементов N6+10 мл/л микроэлементов В5+1 мл/л дикамбы (2,4-D)+10 мл/л RTV+0,5 г/л касаминовой кислоты (казеингидролазы)+700 мг/л L-Prine+100 мг/л инозита+68,5 г/л сахарозы+36 г/л глюкозы, рН 5,2.
Культуральная среда D-AS: культуральная среда D+10 г/л глюкозы+8 г/л порошка агара+200 мкл AS (0,5 М)+1 мл/л AgNO3 (и AS, и AgNO3 характеризовались конечной концентрацией 100 мкМ и были добавлены после стерилизации культуральной среды при легком охлаждении для однородного смешивания).
Культуральная среда D-Cef: культуральная среда D+1 мл/л AgNO3+1 мл/л cef.
2. Генетическая трансформация и регенерация кукурузы
(1) Получение незрелого зародыша кукурузы: листья кукурузы, столбики кукурузы и некоторые ненужные части сдирали; нож вставляли в верхнюю часть для добавления 70% этанола; кукурузу помещали суперчистый операционный стол; через 30 секунд ее брали и сушили потоком воздуха на суперчистом операционном столе в течение приблизительно 15-20 минут, 2/3 поверхности зерна сдирали, очищали незрелый зародыш.
(2) Бактериальный раствор, добавленный в D-Inf (содержащую AS), разбавляли до OD600 0,3-0,5 и выдерживали в течение более чем 1 часа. Незрелый зародыш промывали один раз D-Inf (без AS), а затем замачивали в бактериальном растворе поворотом руки в течение 30 секунд и выдерживали в течение 5 минут. Осмотренный на предмет отсутствия повреждений незрелый зародыш вынимали, влагу абсорбировали стерильной фильтровальной бумагой и помещали на твердую культуральную среду D-AS для совместного культивирования на протяжении 3 суток в темноте при 25°С,
(3) Стадия восстановительного культивирования: Незрелый зародыш, совместно культивируемый на протяжении 3 суток, ополаскивали стерилизованной водой (с добавлением 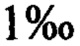 cef) три раза, каждый раз по 20 минут, а затем влагу абсорбировали фильтровальной бумагой, переносили на твердую культуральную среду D-Cef и выполняли восстановительное культивирование в темноте при 25°С на протяжении 7 суток. После этого выполняли стадию скрининга давлением: 4 цикла с интервалом 2 недели между любыми двумя.
cef) три раза, каждый раз по 20 минут, а затем влагу абсорбировали фильтровальной бумагой, переносили на твердую культуральную среду D-Cef и выполняли восстановительное культивирование в темноте при 25°С на протяжении 7 суток. После этого выполняли стадию скрининга давлением: 4 цикла с интервалом 2 недели между любыми двумя.
1ый цикл: культуральная среда D+cef ()+РРТ (5 мг/мл).
2ой цикл: культуральная среда D+cef ()+РРТ (10 мг/мл).
3ий цикл: культуральная среда D+cef ()+РРТ (10 мг/мл).
4ый цикл: культуральная среда D+cef ()+РРТ (10 мг/мл).
(4) Стадия восстановления после скрининга: культуральная среда D+6-ВА (5 мг/л), при этом концентрацию 2,4-D или дикамбы уменьшали в 5 раз, а сахароза составляла 30 г/л (или 20 г/л сахарозы, 10 г/л глюкозы); период составлял 2 недели; культивировали в темноте.
(5) Стадия индуцирования после скрининга: культуральная среда D+6-ВА (5 мг/л), при этом концентрацию 2,4-D или дикамбы уменьшали в 5 раз, а сахароза составляла 50 г/л, без добавления глюкозы,+1 мл/л cef; культивировали в темноте.
(6) Стадия дифференцировки: культуральная среда D, при этом без добавления какого-либо гормона; концентрация сахарозы составляла 30 мг/л, без добавления глюкозы,+1 мл/л cef; культивировали при освещении.
(7) Стадия укоренения: культуральная среда 1/2MS.
Генетическая трансформация ложного льна (Camelina sativa) (с помощью способа капания на цветки)
1. Материалы и реагенты
Camelina sativa; штамм Agrobacterium: GV3101; плазмида (pCAMBIA2300-35S-HF), которая представляла собой бинарный вектор pCAMBIA2300, несущий ген HF и устойчивый к канамицину.
2. Генетическая трансформация Camelina sativa с помощью способа капания на цветки
(1) Camelina sativa дикого типа высевали на питательную почву (гумус : вермикулит : перлит=4:2:1), 5 зерен/горшок (диаметр 9 см) и помещали в культивационное помещение для культивирования. Температура в культивационном помещении составляла 16-20°C, а относительная влажность 60%. Интенсивность освещения составляла 4000 люкс. Период освещения составлял 16 часов света/8 часов темноты. Когда главный стебель растения Camelina sativa дорастал до 5 см, а боковые ветви были похожи на почку, верхние соцветия удаляли для обеспечения роста и развития вторичных ветвей. Когда растения достигали фазы полного цветения, они были готовы к трансформации. За 2 суток до трансформации сформированные стручки срезали.
(2) Когда Agrobacterium GV3101, содержащую плазмиду pCAMBIA2300-35S-HF, культивировали до достижения значения OD600 0,8, она изометрически инфильтрировалась в культуральную среду (1/2MS, 5% сахарозы, 200 мкл/л Silwet L-77) для повторного суспендирования бактерий и трансформации Camelina sativa. При трансформации использовали пипеточный дозатор для отсасывания повторной суспензии Agrobacterium для инфильтрации в культуральную среду и капания в цветочные почки Camelina sativa, с гарантией того, что каждая цветочная почка будет орошена. После этого использовали защитные пленки для покрытия трансформированных растений с целью поддержания влажности. Растения оставляли в темноте в культивационном помещении для культивирования на протяжении 1 суток, а затем культивировали при нормальных условиях.
(3) После созревания растений семена собирали и обозначали как семена поколения ТО. Использовали 50 мг/л канамицина для скрининга полученных семян Camelina sativa поколения ТО с получением положительных трансгенных растений.
Измеряли урожайность и биомассу трансгенных растений, содержащих нуклеиновую кислоту, кодирующую FTO, полученных указанными выше способами, и контрольных растений, которые не содержали FTO. Результаты показаны в таблице 2.
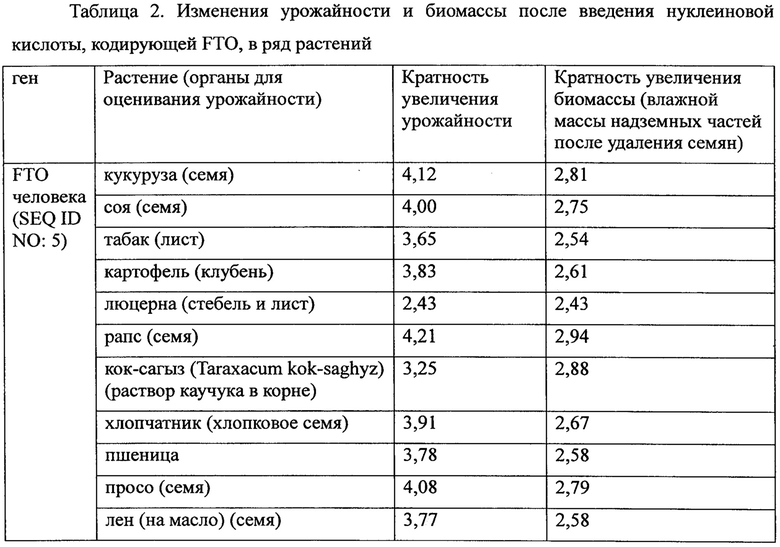

Из таблицы 2 можно видеть, что после введения FTO в ряд растений урожайность и биомасса повысились.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> EpiPlanta Biotech Ltd.
<120> Трансгенное растение и способ его получения
<130> FPCH17160008US
<160> 20
<170> Патентная версия 3.5
<210> 1
<211> 505
<212> Белок
<213> Homo sapiens
<400> 1
Met Lys Arg Thr Pro Thr Ala Glu Glu Arg Glu Arg Glu Ala Lys Lys
1 5 10 15
Leu Arg Leu Leu Glu Glu Leu Glu Asp Thr Trp Leu Pro Tyr Leu Thr
20 25 30
Pro Lys Asp Asp Glu Phe Tyr Gln Gln Trp Gln Leu Lys Tyr Pro Lys
35 40 45
Leu Ile Leu Arg Glu Ala Ser Ser Val Ser Glu Glu Leu His Lys Glu
50 55 60
Val Gln Glu Ala Phe Leu Thr Leu His Lys His Gly Cys Leu Phe Arg
65 70 75 80
Asp Leu Val Arg Ile Gln Gly Lys Asp Leu Leu Thr Pro Val Ser Arg
85 90 95
Ile Leu Ile Gly Asn Pro Gly Cys Thr Tyr Lys Tyr Leu Asn Thr Arg
100 105 110
Leu Phe Thr Val Pro Trp Pro Val Lys Gly Ser Asn Ile Lys His Thr
115 120 125
Glu Ala Glu Ile Ala Ala Ala Cys Glu Thr Phe Leu Lys Leu Asn Asp
130 135 140
Tyr Leu Gln Ile Glu Thr Ile Gln Ala Leu Glu Glu Leu Ala Ala Lys
145 150 155 160
Glu Lys Ala Asn Glu Asp Ala Val Pro Leu Cys Met Ser Ala Asp Phe
165 170 175
Pro Arg Val Gly Met Gly Ser Ser Tyr Asn Gly Gln Asp Glu Val Asp
180 185 190
Ile Lys Ser Arg Ala Ala Tyr Asn Val Thr Leu Leu Asn Phe Met Asp
195 200 205
Pro Gln Lys Met Pro Tyr Leu Lys Glu Glu Pro Tyr Phe Gly Met Gly
210 215 220
Lys Met Ala Val Ser Trp His His Asp Glu Asn Leu Val Asp Arg Ser
225 230 235 240
Ala Val Ala Val Tyr Ser Tyr Ser Cys Glu Gly Pro Glu Glu Glu Ser
245 250 255
Glu Asp Asp Ser His Leu Glu Gly Arg Asp Pro Asp Ile Trp His Val
260 265 270
Gly Phe Lys Ile Ser Trp Asp Ile Glu Thr Pro Gly Leu Ala Ile Pro
275 280 285
Leu His Gln Gly Asp Cys Tyr Phe Met Leu Asp Asp Leu Asn Ala Thr
290 295 300
His Gln His Cys Val Leu Ala Gly Ser Gln Pro Arg Phe Ser Ser Thr
305 310 315 320
His Arg Val Ala Glu Cys Ser Thr Gly Thr Leu Asp Tyr Ile Leu Gln
325 330 335
Arg Cys Gln Leu Ala Leu Gln Asn Val Cys Asp Asp Val Asp Asn Asp
340 345 350
Asp Val Ser Leu Lys Ser Phe Glu Pro Ala Val Leu Lys Gln Gly Glu
355 360 365
Glu Ile His Asn Glu Val Glu Phe Glu Trp Leu Arg Gln Phe Trp Phe
370 375 380
Gln Gly Asn Arg Tyr Arg Lys Cys Thr Asp Trp Trp Cys Gln Pro Met
385 390 395 400
Ala Gln Leu Glu Ala Leu Trp Lys Lys Met Glu Gly Val Thr Asn Ala
405 410 415
Val Leu His Glu Val Lys Arg Glu Gly Leu Pro Val Glu Gln Arg Asn
420 425 430
Glu Ile Leu Thr Ala Ile Leu Ala Ser Leu Thr Ala Arg Gln Asn Leu
435 440 445
Arg Arg Glu Trp His Ala Arg Cys Gln Ser Arg Ile Ala Arg Thr Leu
450 455 460
Pro Ala Asp Gln Lys Pro Glu Cys Arg Pro Tyr Trp Glu Lys Asp Asp
465 470 475 480
Ala Ser Met Pro Leu Pro Phe Asp Leu Thr Asp Ile Val Ser Glu Leu
485 490 495
Arg Gly Gln Leu Leu Glu Ala Lys Pro
500 505
<210> 2
<211> 505
<212> Белок
<213> Sus scrofa
<400> 2
Met Lys Arg Thr Pro Thr Ala Glu Glu Arg Glu Arg Gly Ala Lys Lys
1 5 10 15
Leu Arg Leu Leu Glu Glu Leu Glu Asp Thr Trp Leu Pro Tyr Leu Thr
20 25 30
Pro Lys Asp Asp Glu Phe Tyr Gln Gln Trp Gln Leu Lys Tyr Pro Lys
35 40 45
Leu Ile Leu Arg Glu Ala Gly Ser Val Pro Glu Gly Leu His Lys Glu
50 55 60
Val Gln Glu Ala Phe Leu Ala Leu His Lys His Gly Cys Leu Phe Arg
65 70 75 80
Asp Leu Val Arg Ile Gln Gly Lys Asp Leu Leu Thr Pro Val Ser Arg
85 90 95
Leu Leu Ile Gly Asn Pro Gly Cys Thr Tyr Lys Tyr Leu Asn Thr Arg
100 105 110
Leu Phe Thr Val Pro Trp Pro Val Lys Gly Ser Asp Ala Lys Tyr Asn
115 120 125
Glu Ala Glu Ile Gly Ala Ala Cys Gln Thr Phe Leu Lys Leu Asn Asp
130 135 140
Tyr Leu Gln Ile Glu Thr Ile Gln Ala Leu Glu Glu Leu Ala Ala Lys
145 150 155 160
Glu Lys Ala Asn Ile Asp Thr Val Pro Ala Cys Ile Gly Pro Asp Phe
165 170 175
Pro Arg Val Gly Met Gly Ser Ser Phe Asp Gly His Asp Glu Val Asp
180 185 190
Arg Lys Ser Arg Ala Ala Tyr Asn Leu Thr Leu Leu Ser Phe Met Asp
195 200 205
Pro Gln Lys Met Pro Tyr Leu Lys Glu Glu Pro Tyr Phe Gly Met Gly
210 215 220
Lys Met Ala Val Ser Trp His His Asp Glu Asn Leu Val Asp Arg Ser
225 230 235 240
Ala Val Ala Val Tyr Asn Tyr Ser Cys Glu Gly Pro Glu Glu Glu Ser
245 250 255
Glu Asp Asp Pro Gln Leu Glu Gly Arg Asn Pro Asp Val Trp His Val
260 265 270
Gly Phe Lys Ile Ser Trp Asp Ile Glu Thr Pro Gly Leu Ala Ile Pro
275 280 285
Leu His Gln Gly Asp Cys Tyr Phe Met Leu Asp Asp Leu Asn Ala Thr
290 295 300
His Gln His Cys Val Leu Ala Gly Leu Pro Pro Arg Phe Ser Ser Thr
305 310 315 320
His Arg Val Ala Glu Cys Ser Thr Gly Ala Leu Asp Tyr Ile Leu Gln
325 330 335
Arg Cys Gln Leu Ala Leu Gln Asn Val Arg Asp Glu Ala Asp Ser Gly
340 345 350
Glu Val Ser Leu Lys Ser Leu Glu Pro Ala Val Leu Lys Gln Gly Glu
355 360 365
Glu Ile His Asn Glu Val Glu Phe Glu Trp Leu Arg Gln Phe Trp Phe
370 375 380
Gln Gly Asn Arg Tyr Lys Lys Cys Thr Asp Trp Trp Cys Gln Pro Met
385 390 395 400
Thr Gln Leu Glu Glu Leu Trp Lys Lys Met Glu Gly Ala Thr His Ala
405 410 415
Val Leu Arg Glu Val Arg Arg Glu Gly Ala Pro Val Glu Gln Ser Ser
420 425 430
Asp Ile Leu Thr Ala Ile Leu Ala Val Leu Thr Thr Arg Gln Asn Leu
435 440 445
Arg Arg Glu Trp His Ala Arg Cys Gln Ser Arg Ile Ala Arg Thr Leu
450 455 460
Pro Val Asp Gln Lys Pro Glu Cys Arg Pro Tyr Trp Glu Lys Asp Asp
465 470 475 480
Pro Ser Met Pro Leu Pro Phe Asp Leu Thr Asp Thr Val Ala Glu Leu
485 490 495
Arg Gly Leu Leu Leu Glu Ala Lys Pro
500 505
<210> 3
<211> 505
<212> Белок
<213> Bos taurus
<400> 3
Met Lys Arg Thr Pro Thr Ala Glu Glu Arg Glu Arg Glu Ala Lys Lys
1 5 10 15
Leu Arg Leu Leu Glu Glu Leu Glu Asp Thr Trp Leu Pro Tyr Leu Thr
20 25 30
Pro Lys Asp Asp Glu Phe Tyr Gln Gln Trp Gln Leu Lys Tyr Pro Lys
35 40 45
Leu Ile Leu Arg Glu Ala Ala Ser Val Pro Glu Leu Leu His Lys Glu
50 55 60
Val Gln Gln Ala Phe Leu Thr Leu His Lys His Gly Cys Leu Phe Arg
65 70 75 80
Asp Leu Val Arg Ile Gln Gly Lys Asp Leu Leu Thr Pro Val Ser Arg
85 90 95
Ile Leu Ile Gly Asn Pro Gly Cys Thr Tyr Lys Tyr Leu Asn Thr Arg
100 105 110
Leu Phe Thr Val Pro Trp Pro Val Lys Gly Ser Asp Ala Lys Tyr Asn
115 120 125
Glu Ala Glu Ile Ala Ala Ala Cys Gln Thr Phe Leu Lys Leu Asn Ser
130 135 140
Tyr Leu Gln Val Glu Thr Ile Gln Ala Leu Glu Glu Leu Ala Ala Lys
145 150 155 160
Glu Lys Ala Asn Ile Asp Ala Val Pro Val Cys Ile Gly Pro Asp Phe
165 170 175
Pro Arg Val Gly Met Gly Ser Ser Phe Asp Gly His Asp Glu Ile Asp
180 185 190
Met Lys Asn Arg Ala Ala Tyr Asn Val Thr Leu Leu Asn Phe Met Asp
195 200 205
Pro Gln Lys Met Pro Tyr Leu Lys Glu Glu Pro Tyr Phe Gly Met Gly
210 215 220
Lys Met Ala Val Ser Trp His His Asp Glu Asn Leu Val Asp Arg Ser
225 230 235 240
Ala Val Ala Val Tyr Ser Tyr Ser Cys Glu Gly Pro Glu Glu Glu Ser
245 250 255
Glu Asp Asp Pro Gln Leu Glu Gly Arg Asp Pro Asp Ile Trp His Val
260 265 270
Gly Phe Lys Ile Ser Trp Asp Ile Glu Thr Pro Gly Leu Ala Ile Pro
275 280 285
Leu His Gln Gly Asp Cys Tyr Phe Met Leu Asp Asp Leu Asn Ala Thr
290 295 300
His Gln His Cys Val Leu Ala Gly Leu Pro Pro Arg Phe Ser Ser Thr
305 310 315 320
His Arg Val Ala Glu Cys Ser Thr Gly Thr Leu Glu Tyr Ile Leu Gln
325 330 335
Arg Cys Gln Val Ala Leu Gln Asn Val Arg Glu Glu Ala Asp Asn Gly
340 345 350
Glu Ile Ser Leu Lys Ser Leu Glu Ser Val Val Leu Lys Gln Gly Glu
355 360 365
Glu Ile His Asn Glu Val Glu Phe Glu Trp Leu Arg Gln Phe Trp Phe
370 375 380
Gln Gly Ser Arg Tyr Lys Lys Cys Thr Asp Trp Trp Cys Gln Pro Met
385 390 395 400
Ser Gln Leu Glu Glu Met Trp Arg Lys Met Glu Trp Leu Thr Ser Ala
405 410 415
Val Leu Arg Glu Val Arg Arg Glu Gly Val Pro Met Glu Gln Lys Asn
420 425 430
Glu Met Leu Thr Ser Ile Leu Ala Ser Ile Thr Thr Arg Gln Asn Leu
435 440 445
Arg Arg Glu Trp His Ala Arg Cys Gln Ser Arg Ile Ala Arg Thr Leu
450 455 460
Pro Ala Asp Gln Lys Pro Glu Cys Arg Pro Tyr Trp Glu Lys Gly Asp
465 470 475 480
Pro Ser Met Pro Leu Pro Phe Asp Leu Thr Glu Ile Val Ser Glu Leu
485 490 495
Arg Gly Leu Leu Leu Glu Thr Arg Pro
500 505
<210> 4
<211> 419
<212> Белок
<213> Ostreococcus lucimarinus
<400> 4
Met Ser Pro Ser Ser Ser Val Leu Glu Pro Glu Asp Gly Glu Pro Phe
1 5 10 15
Ala Arg Val His Arg Ala His Tyr Arg Gly Phe Val Val Asp Ala Pro
20 25 30
Ser Val Leu Pro Ala Ser Leu His Asp Asp Val Glu Arg Ala Phe Asp
35 40 45
Asp Met Arg Ser Arg Gly Glu Phe Thr His Asp Val Val Ser Ala Gly
50 55 60
Asn Lys Val Ser Thr Thr Tyr Val Arg Arg Cys Leu Leu Gly Glu Asp
65 70 75 80
Gly Met Thr Tyr His Tyr Gln Lys Leu Arg Leu Phe Ala Gln Pro Trp
85 90 95
Arg Gly Arg Glu Ala Tyr Glu Val Val Arg Arg Leu Asn Glu Thr Leu
100 105 110
Thr Arg Ser Ala Arg Glu Arg Cys Glu Lys Leu Gly Gly Ala Phe Ala
115 120 125
Glu Ser Glu Cys Glu Tyr Asn Val Thr Leu Ile Asn Tyr Met Glu Thr
130 135 140
Glu Gly Glu Ser Glu Ile Glu Leu Arg Asn Glu Glu Lys Phe Asp Leu
145 150 155 160
Gly Thr Thr Ser Val Ser Trp His Ser Asp Ser Ser Leu Arg Glu Asn
165 170 175
Ser Thr Val Ala Val Tyr His Thr Tyr Glu Ala Pro Glu Arg Lys Asp
180 185 190
Trp Arg Val Ala Leu Arg Ala Leu Asn Ala Glu Cys Glu Val Leu Cys
195 200 205
Val Pro Leu Glu Asp Lys Ala Thr Tyr Tyr Met Cys Gly Glu Phe Asn
210 215 220
Ala Thr His His His Ala Val Leu Thr Gly Ser Ser Ala Arg Tyr Ser
225 230 235 240
Ser Thr His Arg Val Ala Val Val Ala Lys Asp Thr Phe Gln Tyr Ile
245 250 255
Lys Arg Arg Cys Ile Asp Ala Leu Ala Ile Val Pro Asp Leu Glu Arg
260 265 270
Glu Asn Lys Pro Leu Asp Ala Lys Gln Ile Gln Phe Leu Ala Asp Val
275 280 285
His Arg Glu Val Glu Phe Gln Trp Ile Arg Met Phe His Leu Gln Gly
290 295 300
Glu Ala His Ala Ala Trp His Asp Thr Tyr Trp Thr Arg Lys Ile Ala
305 310 315 320
Glu Leu Thr Glu Ala Trp Asp Arg Met Glu Ala Cys Phe Arg Val Ile
325 330 335
Leu Ser Lys Leu Lys Arg Ser Ser Arg Ser Pro Glu Ser Pro Pro Arg
340 345 350
Ala Tyr Ala Met Leu Leu Tyr Ala Leu Lys Thr Val Lys Glu Leu Arg
355 360 365
Asp Glu Tyr Thr Lys Arg Thr Lys Ala Ser Ala Tyr Ala Ser Leu Pro
370 375 380
Pro Ser Gln Arg Pro Val Asp Leu Pro Ala Tyr Asp Asn Thr Ser Pro
385 390 395 400
Leu Pro Phe Glu Leu Lys Pro Val Ile Tyr Phe Leu Glu Glu Glu Gln
405 410 415
Asn Lys Val
<210> 5
<211> 1518
<212> ДНК
<213> Homo sapiens
<400> 5
atgaagcgca ccccgactgc cgaggaacga gagcgcgaag ctaagaaact gaggcttctt 60
gaagagcttg aagacacttg gctcccttat ctgaccccca aagatgatga attctatcag 120
cagtggcagc tgaaatatcc taaactaatt ctccgagaag ccagcagtgt atctgaggag 180
ctccataaag aggttcaaga agcctttctc acactgcaca agcatggctg cttatttcgg 240
gacctggtta ggatccaagg caaagatctg ctcactccgg tatctcgcat cctcattggt 300
aatccaggct gcacctacaa gtacctgaac accaggctct ttacggtccc ctggccagtg 360
aaagggtcta atataaaaca caccgaggct gaaatagccg ctgcttgtga gaccttcctc 420
aagctcaatg actacctgca gatagaaacc atccaggctt tggaagaact tgctgccaaa 480
gagaaggcta atgaggatgc tgtgccattg tgtatgtctg cagatttccc cagggttggg 540
atgggttcat cctacaacgg acaagatgaa gtggacatta agagcagagc agcatacaac 600
gtaactttgc tgaatttcat ggatcctcag aaaatgccat acctgaaaga ggaaccttat 660
tttggcatgg ggaaaatggc agtgagctgg catcatgatg aaaatctggt ggacaggtca 720
gcggtggcag tgtacagtta tagctgtgaa ggccctgaag aggaaagtga ggatgactct 780
catctcgaag gcagggatcc tgatatttgg catgttggtt ttaagatctc atgggacata 840
gagacacctg gtttggcgat accccttcac caaggagact gctatttcat gcttgatgat 900
ctcaatgcca cccaccaaca ctgtgttttg gccggttcac aacctcggtt tagttccacc 960
caccgagtgg cagagtgctc aacaggaacc ttggattata ttttacaacg ctgtcagttg 1020
gctctgcaga atgtctgtga cgatgtggac aatgatgatg tctctttgaa atcctttgag 1080
cctgcagttt tgaaacaagg agaagaaatt cataatgagg tcgagtttga gtggctgagg 1140
cagttttggt ttcaaggcaa tcgatacaga aagtgcactg actggtggtg tcaacccatg 1200
gctcaactgg aagcactgtg gaagaagatg gagggtgtga caaatgctgt gcttcatgaa 1260
gttaaaagag aggggctccc cgtggaacaa aggaatgaaa tcttgactgc catccttgcc 1320
tcgctcactg cacgccagaa cctgaggaga gaatggcatg ccaggtgcca gtcacgaatt 1380
gcccgaacat tacctgctga tcagaagcca gaatgtcggc catactggga aaaggatgat 1440
gcttcgatgc ctctgccgtt tgacctcaca gacatcgttt cagaactcag aggtcagctt 1500
ctggaagcaa aaccctag 1518
<210> 6
<211> 1515
<212> ДНК
<213> Homo sapiens
<400> 6
atgaagcgca cccctactgc tgaggagaga gagagggagg ccaagaagct cagactcctg 60
gaggagctgg aggatacctg gctcccttac ctgactccca aggatgacga gttctaccag 120
caatggcagc tcaagtaccc caagctcatt ctgagggagg cttcctctgt ttccgaggag 180
ctgcacaagg aggtgcaaga ggccttcctt actttgcaca agcatggatg ccttttcaga 240
gatttggtta ggatccaggg caaggacctt ttgacccccg tgagcaggat tctcatcgga 300
aacccaggtt gcacttacaa gtacctcaat actcgcctgt tcacagtgcc atggcctgtt 360
aagggctcaa acattaagca tactgaggct gagatcgccg ctgcctgcga gacattcctc 420
aagctgaatg attacctcca gattgagaca atccaagctc ttgaggagtt ggctgccaag 480
gagaaggcta acgaggatgc cgttccactg tgcatgtccg ccgacttccc tagagtgggc 540
atgggaagct catacaatgg ccaagatgag gttgacatta agtctagggc tgcctacaac 600
gtgaccctcc tgaatttcat ggacccacag aagatgcctt accttaagga ggagccatac 660
ttcggtatgg gcaagatggc tgtttcctgg caccatgatg agaacttggt ggacaggtct 720
gctgtggccg tttacagcta ctcatgcgag ggacctgagg aggagagcga ggatgactca 780
cacctggagg gtcgcgatcc cgacatttgg catgttggct tcaagatttc ttgggatatc 840
gagacccctg gccttgccat ccccttgcac cagggagact gctacttcat gctcgatgac 900
ctgaatgcta cacaccagca ttgcgttctt gccggttccc aaccacgctt ctcctctacc 960
catagagtgg ctgagtgctc tacaggcacc ctcgattaca ttctgcagag atgccaactt 1020
gccttgcaaa acgtgtgcga tgacgttgac aatgatgacg tttcccttaa gtctttcgag 1080
cctgctgtgt tgaagcaggg agaggagatc cacaacgagg ttgagttcga gtggctgcgc 1140
cagttctggt tccaaggtaa tcgctacaga aagtgcactg attggtggtg ccagccaatg 1200
gctcaacttg aggccttgtg gaagaagatg gagggcgtga caaacgccgt tctccacgag 1260
gtgaagaggg agggactgcc tgtggagcaa cgcaacgaga ttcttacagc tatcctcgcc 1320
agcctgaccg ctagacagaa tttgaggcgc gagtggcatg ccaggtgcca atcaaggatt 1380
gctcgcaccc ttccagctga ccagaagcca gagtgcaggc catactggga gaaggatgac 1440
gctagcatgc cccttccatt cgatttgact gacatcgtgt cagagttgag aggccagctt 1500
ttggaggcca agcca 1515
<210> 7
<211> 1518
<212> ДНК
<213> Sus scrofa
<400> 7
atgaagcgaa ccccaaccgc cgaggaacga gagcgcggag ctaagaaact gaggcttctt 60
gaagagctgg aagacacttg gcttccttat ctgaccccca aagatgatga attctatcag 120
cagtggcagc tgaaataccc taagctaatt ctccgagaag caggcagcgt ccctgaggga 180
ctccacaaag aggttcaaga agccttcctc gcactgcaca agcatggctg cttatttcgg 240
gacctggtca ggatccaagg caaagatttg ctcacgccag tatctcgcct cctcattggt 300
aaccccggct gcacctacaa gtacctgaac accaggctct tcacggtccc ctggccagtg 360
aagggctctg atgcaaagta caatgaggcc gagataggcg ccgcctgcca gaccttcctc 420
aagctcaacg actacctgca gattgagacc atccaggcgc tggaggaact cgctgccaag 480
gagaaagcca atatcgacac cgtgccggcg tgtataggtc cagatttccc cagggtcggc 540
atggggtcat cctttgacgg ccatgacgag gtggacagga agagcagagc cgcctacaac 600
ctaactttgt tgagcttcat ggatccccag aaaatgccgt acctgaaaga ggagccctac 660
tttggcatgg ggaagatggc tgtgagctgg catcacgatg aaaatctggt ggacaggtca 720
gcggtggcag tgtacaatta tagctgtgaa ggccctgaag aggaaagcga ggatgatccc 780
cagctcgaag gcagaaatcc cgatgtgtgg catgttggct ttaagatctc atgggacata 840
gagacccctg gtttggcgat accccttcac caaggagact gctactttat gctggatgat 900
ctcaatgcca cccaccaaca ctgtgttttg gctggtttac caccccggtt tagttccacc 960
caccgagtgg ccgagtgctc gacgggagcc ttggattaca tcttacagcg ctgccagttg 1020
gccctgcaga atgtccgtga tgaggcggac agtggtgaag tctctttgaa atccttggag 1080
cctgcggttt tgaaacaagg agaagaaatc cacaacgagg tcgagtttga gtggctgaga 1140
cagttttggt ttcaaggcaa tcgatacaaa aagtgcaccg attggtggtg tcaacccatg 1200
actcagctgg aagagctttg gaagaagatg gaaggtgcga cccatgctgt gcttcgtgaa 1260
gttagaagag agggggcccc tgtggaacag agcagtgaca tcctgactgc catcctagcc 1320
gtgctcacca ctcgccagaa cctgaggagg gagtggcatg ccaggtgcca gtcccgaatt 1380
gcccgaactc tgcctgtgga ccagaagcca gaatgccggc cgtattggga aaaggatgat 1440
ccctccatgc ctctgccgtt tgatctcaca gacactgtgg ctgaactcag aggtctgctt 1500
ctggaagcca aaccctag 1518
<210> 8
<211> 1515
<212> ДНК
<213> Sus scrofa
<400> 8
atgaagagaa caccaaccgc tgaggagagg gagagaggag ctaagaagtt gaggctcctg 60
gaggagctgg aggacacatg gttgccctac ctcaccccaa aggatgacga gttctaccag 120
caatggcaat tgaagtaccc caagttgatt ctcagggagg ctggatctgt gccagaggga 180
ctgcacaagg aggttcaaga ggccttcctg gctcttcaca agcatggctg cctgttccgc 240
gaccttgtta gaattcaggg aaaggatctt ttgactccag tgtctcgcct cctgatcgga 300
aaccctggtt gcacatacaa gtacttgaat accagactct tcactgtgcc ctggccagtt 360
aagggtagcg acgccaagta caacgaggct gagatcggcg ccgcttgcca gacattcttg 420
aagctcaatg attacctgca gattgagacc atccaagctc tggaggagct tgccgctaag 480
gagaaggcca acattgacac tgttcctgct tgcatcggcc cagatttccc tagggtgggc 540
atgggatcct ctttcgatgg acatgacgag gttgatagga agtcacgcgc cgcttacaat 600
ttgacacttt tgtccttcat ggatcctcaa aagatgccct acctcaagga ggagccctac 660
ttcggtatgg gcaagatggc cgtgtcatgg caccatgacg agaaccttgt tgatagatcc 720
gccgtggctg tttacaatta ctcttgcgag ggccctgagg aggagagcga ggatgaccct 780
cagctggagg gaaggaaccc cgacgtgtgg cacgttggtt tcaagattag ctgggatatc 840
gagacccccg gtctggctat tccacttcat caaggcgact gctacttcat gttggatgac 900
ctcaatgcca ctcaccagca ttgcgttctg gctggacttc cacctaggtt cagctcaact 960
caccgcgtgg ccgagtgctc tacaggtgct ttggattaca ttctccagcg ctgccaactg 1020
gcccttcaaa acgtgagaga cgaggctgat agcggcgagg ttagcttgaa gtccctggag 1080
ccagctgtgc tgaagcaggg agaggagatc cataacgagg tggagttcga gtggcttaga 1140
cagttctggt tccaaggcaa taggtacaag aagtgcaccg actggtggtg ccagccaatg 1200
actcaactgg aggagctttg gaagaagatg gagggagcca cacacgctgt gctcagggag 1260
gttaggcgcg agggtgctcc tgttgagcag tcctctgaca ttctgaccgc catcttggct 1320
gtgctcacca ctaggcagaa tcttagaagg gagtggcatg cccgctgcca atcaagaatc 1380
gctaggaccc tgcctgttga tcagaagcca gagtgcaggc cttactggga gaaggatgac 1440
ccctccatgc ctctgccctt cgaccttact gatacagtgg ccgagcttcg cggactcctg 1500
cttgaggcta agcct 1515
<210> 9
<211> 1518
<212> ДНК
<213> Bos taurus
<400> 9
atgaagcgga ccccgacggc cgaggaacga gagcgcgaag ctaagaaact gaggcttctt 60
gaagagcttg aagacacgtg gcttccctac ctgaccccca aagacgatga attctatcag 120
cagtggcaac tgaaatatcc taaactaatt ctccgagaag ctgccagcgt gcctgagttg 180
ctccataaag aggttcaaca agcctttctc acgctgcaca agcacggctg tttatttcgg 240
gatctggtga ggatccaggg caaagacttg ctcactccag tctctcgcat cctcatcggt 300
aaccccggct gcacctacaa gtacctgaac accaggctct tcacggtacc ctggcccgtg 360
aaaggctctg atgcaaagta caatgaggcc gaaatagctg ccgcctgtca aacgttcctc 420
aagctcaaca gctacctgca ggtagagacc atccaggctt tggaagagct tgctgccaag 480
gagaaagcca acatcgatgc cgtgccagtg tgcataggtc cagatttccc cagggttggc 540
atggggtcct cctttgacgg gcacgatgag attgacatga agaaccgagc agcgtacaac 600
gtcactttgt tgaatttcat ggatccccag aagatgccat acctgaaaga ggaaccgtat 660
tttggcatgg ggaaaatggc cgtgagctgg catcatgatg aaaatctggt ggacaggtca 720
gcggtggcag tgtacagtta tagctgtgaa ggccctgaag aggaaagtga ggacgaccct 780
cagcttgaag gcagagatcc tgatatttgg cacgttggtt ttaagatctc gtgggacata 840
gagacacctg gtttggcgat accccttcac caaggagact gctattttat gcttgatgat 900
ctcaatgcca cccaccaaca ctgtgttttg gctggtttac cacctcggtt tagttccacc 960
caccgagtgg cagagtgctc aacagggacc ttggagtaca tcttacagcg ctgccaggtg 1020
gccctgcaga atgtccgcga ggaggcagac aacggtgaaa tctctttgaa atccttggag 1080
tcagtggttt tgaaacaagg agaagaaatc cacaacgagg tcgagtttga atggctgaga 1140
cagttttggt ttcaaggcag tcgatacaaa aagtgcactg actggtggtg tcagcccatg 1200
agtcagctgg aagagatgtg gagaaagatg gagtggttga caagcgctgt gcttcgcgaa 1260
gttagaagag agggggtccc catggaacaa aagaatgaga tgttgacatc catcctcgcc 1320
tcgatcacca ctcgccagaa cctgaggaga gaatggcatg ccaggtgcca gtctcgaatt 1380
gcccgaactt tacccgctga tcagaagcca gaatgccggc cgtactggga aaagggtgat 1440
ccttcgatgc ctctgccatt tgatctcaca gagatcgttt ctgaactcag aggtctgctt 1500
ctggaaacca ggccctag 1518
<210> 10
<211> 1515
<212> ДНК
<213> Bos taurus
<400> 10
atgaagcgca caccaaccgc tgaggagcgc gagagagagg ccaagaagct gagactcctg 60
gaggagcttg aggacacatg gctgccctac cttaccccaa aggatgacga gttctaccag 120
caatggcaac ttaagtaccc caagttgatt ctcagggagg ccgctagcgt gccagagctt 180
ttgcacaagg aggttcagca ggccttcctc acccttcaca agcatggatg cttgttcaga 240
gacctcgtga ggatccaggg caaggatctc ctgaccccag tttcaaggat tctgatcgga 300
aatcctggtt gcacttacaa gtacctgaac acccgccttt tcactgtgcc ctggccagtt 360
aagggaagcg acgctaagta caatgaggcc gagattgccg ctgcctgcca gactttcttg 420
aagctcaact cataccttca ggtggagaca atccaggccc tggaggagct cgctgccaag 480
gagaaggcta atattgacgc cgtgcctgtt tgcatcggcc cagatttccc tagagtgggc 540
atgggatcct ctttcgatgg acatgacgag attgatatga agaacagggc tgcctacaat 600
gttactcttt tgaacttcat ggatcctcaa aagatgccct acttgaagga ggagccctac 660
ttcggtatgg gcaagatggc cgtgtcatgg caccatgacg agaatctcgt tgatcgctcc 720
gctgtggccg tttactccta ctcttgcgag ggccctgagg aggagtccga ggatgaccct 780
cagctggagg gtagagatcc cgacatttgg cacgttggct tcaagatttc ttgggacatc 840
gagacacccg gcttggctat cccactccat caaggagatt gctacttcat gctggatgac 900
cttaacgcta ctcaccagca ttgcgtgttg gccggactcc cacctcgctt cagctcaaca 960
cacagagttg ccgagtgctc cactggtaca ctggagtaca tccttcagcg ctgccaagtg 1020
gctttgcaaa atgttagaga ggaggccgac aacggtgaaa ttagcctgaa gtcacttgag 1080
tccgtggttt tgaagcaggg cgaggagatc cataacgagg tggagttcga gtggctcaga 1140
cagttctggt tccaaggttc aaggtacaag aagtgcaccg attggtggtg ccagccaatg 1200
tcccaactgg aggagatgtg gagaaagatg gagtggttga cttctgctgt gctcagggag 1260
gttaggaggg agggcgttcc tatggagcag aagaatgaga tgttgacatc tattctcgct 1320
agcatcacca ctaggcagaa cctgagaagg gagtggcacg cccgctgcca atctaggatt 1380
gctcgcaccc ttcctgccga tcagaagcca gagtgccgcc cttactggga gaagggcgac 1440
ccctctatgc ctctgccctt cgatcttaca gagatcgtga gcgagctgag gggactcctg 1500
cttgagaccc gccca 1515
<210> 11
<211> 1260
<212> ДНК
<213> Ostreococcus lucimarinus
<400> 11
atgtcgccgt catcctccgt gctcgaaccc gaggacggcg aaccgttcgc gcgcgtccat 60
cgagcgcact accgcggttt cgtcgtggac gcgccctcgg tgcttcccgc gtcgcttcac 120
gacgacgtcg aacgcgcgtt cgacgacatg cgcagccgag gcgaattcac gcacgacgtc 180
gtttccgccg gaaataaagt gtccacgacg tacgtgcggc gctgcctgct cggtgaggac 240
ggaatgacgt atcattatca gaagttacgc ctgtttgcgc aaccgtggcg cggacgggag 300
gcgtacgaag tggtgagacg gttgaacgag acgctgacga ggagcgcgcg agagcgatgc 360
gaaaagctcg gcggagcgtt cgcggagagt gagtgcgaat acaacgtgac gctgattaat 420
tacatggaaa cagagggtga gagcgaaata gagctacgga atgaggagaa gttcgatctc 480
gggacgacgt cggtgagttg gcacagcgat tcgtcgttgc gagaaaactc caccgtggcg 540
gtgtatcaca catacgaggc gccagagaga aaagactggc gagtggcgct gagggcgttg 600
aacgcggagt gcgaggtgct ttgcgtgcct ttggaggata aagcgacgta ttacatgtgt 660
ggtgagttca acgccactca ccatcacgcc gtgctcaccg gctcgagtgc gagatattct 720
tcgacacatc gagtcgccgt ggtggccaag gatacgttcc agtacatcaa aaggcggtgt 780
atcgatgcgc tcgcaattgt accagactta gagcgcgaga ataaaccttt ggatgcgaaa 840
cagatacagt tcttagccga cgtccatcgc gaggttgaat ttcaatggat tagaatgttt 900
cacctgcaag gcgaagcaca cgcggcgtgg cacgatacgt attggacgag aaaaatcgcc 960
gagctcaccg aggcgtggga tcgcatggag gcttgctttc gagtgatttt gtccaagctc 1020
aagcgttcat ctcgctcgcc cgagtcgccg cctcgagcgt acgcgatgct gctttacgcg 1080
ctgaaaaccg tcaaggagtt gcgagacgag tacacgaagc gaaccaaggc gtcggcgtac 1140
gcgtcgctac cgccgtcgca acgtccagtt gatctgcccg cgtacgacaa tacttctccc 1200
cttccgttcg agctcaaacc cgtgatttac tttctcgagg aggagcaaaa caaagtgtga 1260
<210> 12
<211> 1257
<212> ДНК
<213> Ostreococcus lucimarinus
<400> 12
atgagcccat cctctagcgt gctggagcct gaggatggcg agcccttcgc cagggttcat 60
cgcgctcact acaggggatt cgtggttgac gccccaagcg tgcttcctgc ttcattgcat 120
gatgacgttg agcgcgcctt cgatgacatg aggtcacgcg gagagttcac ccacgatgtg 180
gtttccgctg gtaacaaggt gtctaccact tacgttaggc gctgcctcct gggcgaggac 240
ggcatgactt accattacca gaagctccgc ctgttcgccc aaccttggag aggcagggag 300
gcttacgagg tggttagaag gctcaatgag actctgacaa gatccgcccg cgagagatgc 360
gagaagctcg gaggagcctt cgctgagtct gagtgcgagt acaacgtgac cctgatcaat 420
tacatggaga ctgagggaga gtccgagatt gagcttagaa acgaggagaa gttcgatttg 480
ggtacaacct cagtttcctg gcatagcgac tcatccctta gggagaattc aactgtggcc 540
gtttaccaca catacgaggc tcccgagcgc aaggattgga gagtggctct tagggccttg 600
aacgctgagt gcgaggtgct ttgcgttcca ttggaggaca aggccactta ctacatgtgc 660
ggcgagttca atgccacaca ccatcacgct gtgctgaccg gctctagcgc tcgctactca 720
tccactcaca gagttgccgt ggttgctaag gatacattcc agtacattaa gcgcagatgc 780
atcgatgccc ttgctattgt gccagacttg gagagggaga acaagcctct cgatgccaag 840
cagatccaat tcctggctga cgtgcatcgc gaggttgagt tccagtggat tagaatgttc 900
caccttcaag gagaggccca tgccgcttgg cacgatacct actggactag gaagatcgcc 960
gagttgaccg aggcttggga caggatggag gcttgcttcc gcgttattct ctccaagctg 1020
aagagatcta gcaggtctcc tgagagccca cctcgcgcct acgctatgct tttgtacgcc 1080
ctcaagacag tgaaggagct gagagacgag tacacaaaga ggaccaaggc ctcagcttac 1140
gcctcccttc ccccatctca gaggcccgtt gatttgccag cttacgacaa cacctctcct 1200
ctccccttcg agctgaagcc cgtgatctac ttcctggagg aggagcaaaa taaggtt 1257
<210> 13
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 13
atgaagcgca ccccgactg 19
<210> 14
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 14
gggttttgct tccagaagct ga 22
<210> 15
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 15
atgaagcgga ccccgacg 18
<210> 16
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 16
gggcctggtt tccagaagca g 21
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 17
atgaagcgaa ccccaaccgc 20
<210> 18
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 18
gggtttggct tccagaagca gac 23
<210> 19
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 19
atgtcgccgt catcctccg 19
<210> 20
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 20
cactttgttt tgctcctcct cgagaaa 27
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СОЗДАНИЯ НОВЫХ МУТАЦИЙ В ОРГАНИЗМАХ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2833881C1 |
| МУТАНТНЫЙ ПОЛИПЕПТИД ГИДРОКСИФЕНИЛПИРУВАТДИОКСИГЕНАЗА, КОДИРУЮЩИЙ ЕГО ГЕН И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2822892C1 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| ГЕНЫ ТОКСИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2825305C2 |
| ГЕНЫ ТОКСИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2723717C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2741833C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2745322C2 |
| СПОСОБЫ УСОВЕРШЕНСТВОВАНИЯ РАСТЕНИЙ | 2015 |
|
RU2727424C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2816526C2 |

Изобретение относится к области биохимии, в частности к трансгенному растению, которое содержит белок FTO и демонстрирует увеличенную биомассу. Также раскрыты клетка для экспрессии белка FTO и способ получения указанного трансгенного растения. Изобретение позволяет эффективно увеличивать биомассу, получаемую из указанного растения. 3 н. и 6 з.п. ф-лы, 1 ил., 2 табл., 6 пр.
1. Трансгенное растение, которое содержит молекулу нуклеиновой кислоты, кодирующую белок FTO, как показано в любой из последовательностей SEQ ID NO: 1-4, где указанное растение демонстрирует увеличенную биомассу, повышенную урожайность или их комбинацию по сравнению с контрольным растением, которое не содержит молекулу нуклеиновой кислоты, кодирующую белок FTO.
2. Трансгенное растение по п. 1, в котором указанная молекула нуклеиновой кислоты представлена в любой из SEQ ID NO: 5-12.
3. Трансгенное растение по п. 1 или 2, при этом указанное растение выбрано из группы, состоящей из риса, кукурузы (Zea mays), сои, табака, картофеля, люцерны (Medicago sativa), рапса (Brassica), кок-сагыза (Taraxacum Kok-saghyz), хлопчатника, пшеницы, проса (Panicum miliaceum), льна, подсолнечника и ложного льна (Camelina sativa).
4. Растительная клетка для экспрессии белка FTO, причем указанная растительная клетка содержит молекулу нуклеиновой кислоты, кодирующую белок FTO, как показано в любой из последовательностей SEQ ID NO: 1-4.
5. Растительная клетка по п. 4, в которой указанная молекула нуклеиновой кислоты представлена в любой из SEQ ID NO: 5-12.
6. Способ получения трансгенного растения, демонстрирующего увеличенную биомассу, повышенную урожайность или их комбинацию, при этом указанный способ предусматривает:
a) введение молекулы нуклеиновой кислоты, кодирующей белок FTO, как показано в любой из последовательностей SEQ ID NO: 1-4, в способную к регенерации клетку растения; и
b) регенерацию трансгенного растения из способной к регенерации клетки растения, при этом трансгенное растение содержит в своем геноме указанную молекулу нуклеиновой кислоты, кодирующую m6A-деметилазу, и демонстрирует увеличенную биомассу, повышенную урожайность или их комбинацию по сравнению с контрольным растением, которое не содержит молекулу нуклеиновой кислоты, кодирующую белок FTO.
7. Способ по п. 6, при этом указанный способ дополнительно предусматривает:
c) получение растения-потомка, получаемого из трансгенного растения стадии b), при этом указанное растение-потомок содержит в своем геноме указанную молекулу нуклеиновой кислоты, кодирующую белок FTO, и демонстрирует увеличенную биомассу, повышенную урожайность или их комбинацию по сравнению с контрольным растением, которое не содержит молекулу нуклеиновой кислоты, кодирующую белок FTO.
8. Способ по любому из пп. 5-7, при котором указанная молекула нуклеиновой кислоты представлена в любой из SEQ ID NO: 5-12.
9. Способ по любому из пп. 5-8, при котором указанное растение выбрано из группы, состоящей из риса, кукурузы (Zea mays), сои, табака, картофеля, люцерны (Medicago sativa), рапса (Brassica), кок-сагыза (Taraxacum Kok-saghyz), хлопчатника, пшеницы, проса (Panicum miliaceum), льна, подсолнечника и ложного льна (Camelina sativa).
| RU 99123057 A, 27.08.2001 | |||
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ В | 2011 |
|
RU2608643C2 |
| Подъемник для кирпичей | 1930 |
|
SU22869A1 |

