Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к микроорганизму с повышенной продукцией полезных аминокислот вследствие улучшенной активности клеток и сокращенного времени культивирования клеток из-за повышения восстанавливающей способности и к способу получения L-аминокислот с его применением.
Уровень техники настоящего изобретения
Для микроорганизмов, которые продуцируют полезные продукты посредством ферментации, как известно, требуется большое количество энергии, как например, АТФ (аденозин-5′-трифосфат), или восстанавливающая способность, как например, НАДФ-Н (никотинамидадениндинуклеотидфосфат) для повышения продуктивности этого биосинтетического пути.
В ходе метаболизма микроорганизмов очень важным является внутриклеточный баланс НАДН (никотинамидадениндинуклеотид), используемого в катаболических реакциях, и НАДФ·Н (никотинамидадениндинуклеотидфосфат), используемого в анаболических реакциях. Баланс контролируется фосфорилированием НАД или дефосфорилированием НАДФ, как показано в следующей формуле:
НАД++АТФ→НАДФ++АДФ
НАДФ+→НАД++фосфат
У Е.coli фосфорилирование НАД, как известно, катализирует фермент, называемый НАД-киназой (КФ 2.7.1.23), кодируемый геном nadK (или yfjB). НАД-киназа использует Mg2+ в качестве кофактора ферментативной реакции и аллостерически ингибируется НАДФ·Н и НАДН. Известно, что значение Km для НАД+ составляет 2000 мкМ, а таковое для АТФ составляет 2500 мкМ (Eur. J. Biochem., (2001) 268:4359-4365).
Дефосфорилирование НАДФ редко исследовали, несмотря на его первоочередную важность в метаболическом пути. Хотя, как было показано, гомолог НАД-киназы у археи Methanococcus jannaschii обладает НАДФ-фосфатазной активностью, гены, кодирующие фермент с такой активностью, все еще не идентифицированы у источников из эукариот и эубактерий. У Е.coli продукт гена cysQ проявлял высокие уровни НАДФ- и НАДФ·Н-фосфатазных активностей, но исследования кинетики у очищенного фермента привели к предположению, что он не является истинной НАДФ-фосфатазой у этого организма (Biochem J., (2007) 402:205-218, Biosci. Biotechnol. Biochem., (2008) 72:919-930).
НАД-киназные активности обнаружены у многих микроорганизмов, а НАД-связывающий участок и активный центр НАД-киназы, которые являются важными для каталитической активности, показывают высокую консервативность аминокислотных последовательностей среди видов. Например, для различных микроорганизмов, в том числе грамположительных бактерий, показан высокий уровень гомологии при прогнозировании третичной структуры спиралей 2, 4 и 5 (каждая из которых обозначается Н2, Н4 и Н5) (Appl Microbiol Biotechnol (2010) 87:583-593).
НАДФ, образованный НАД-киназой, в конечном итоге обеспечивает восстанавливающую способность и, в частности, НАДФ+/НАДФ·Н, необходимые для массового продуцирования полезных продуктов у Е.coli, является существенным элементом для анаболических реакций (Biochem J., (2007) 402:205-218). У Е.coli НАДФ·Н в основном образуется 1) в окислительном пентозофосфатном пути, 2) НАДФ-зависимой изоцитратдегидрогеназой в цикле ТСА (ген icd) и 3) трансгидрогеназой (ген pntAB) (J Biol Chem., (2004) 279:6613-6619).
В этих реакциях образуется НАДФ·Н с использованием НАДФ в качестве субстрата и, таким образом, уровень НАДФ·Н может повышаться при повышении внутриклеточного уровня НАДФ. Таким образом, было предпринято много попыток для повышения внутриклеточного уровня НАДФ для промышленного производства различных метаболитов, например, 1) за счет сверхэкспрессии nadK у Е.coli повышалось продуцирование НАДФ·Н и тимидина (Biotechnol Lett., (2009) 31:19291936), 2) за счет сверхэкспрессии nadK у Е.coli повышалось продуцирование НАДФ·Н и РНВ (полигидроксибутирата) (Appl Microbiol Biotechnol., (2009) 83:939947), и 3) за счет сверхэкспрессии ppnK у Corynebacterium подобно сверхэкспрессии nadK у Е.coli повышалось продуцирование лизина. Ключевым моментом во всех вышеописанных случаях является повышение экспрессии гена nadK. Однако в каждом из этих случаев количество источника фосфата, такого как АТФ, также должно возрастать для повышения восстанавливающей способности посредством повышенного уровня НАДФ·Н, получаемого в результате повышения уровня НАДФ, обусловленного высоким уровнем экспрессии НАД-киназы.
У микроорганизмов АТФ в основном продуцируется системой переноса электронов или фосфорилированием на уровне субстрата. Образованная АТФ распадается с обеспечением клеток энергией и ее уровень восстанавливается через гликолиз или окислительное фосфорилирование. Исходя из этого факта, провели исследование применения бактериальной системы регенерации АТФ к процессу продуцирования с целью обеспечения энергией в ходе массового продуцирования полезных продуктов (Biosci Biotechnol Biochem., (1997) 61:840-845).
Однако, как описано выше, существует немного исследований, касающихся способа увеличения количества источника фосфата, который требуется для повышения восстанавливающей способности за счет высокого уровня экспрессии НАД-киназы и последующего увеличения количества продуктов биосинтеза. Кроме того, повышение обеспечения энергией посредством высокого уровня продукции АТФ было исследовано только в отношении обеспечения клеток энергией, и в уровне техники не было исследовано использование АТФ в качестве источника фосфата.
Раскрытие настоящего изобретения
Техническая проблема
Соответственно, для получения микроорганизмов, продуцирующих L-аминокислоты в высокой концентрации, авторами настоящего изобретения были проведены исследования в отношении генов, которые вовлечены в различные пути метаболизма, имеющие отношение к энергии и восстанавливающей способности. В результате этого они обнаружили, что микроорганизм с повышенной экспрессией НАД-киназы, кодируемой nadK, и инактивацией фермента с аминокислотной последовательностью SEQ ID NO 2, кодируемой геном tehB, способен эффективно продуцировать L-аминокислоты в высоких концентрациях, и, исходя из этого, количество АТФ в качестве источника фосфата можно эффективно повысить, таким образом осуществив настоящее изобретение.
Иными словами, настоящее изобретение относится к способу повышения продукции необходимой аминокислоты с помощью эффективного повышения восстанавливающей способности микроорганизма, при котором для повышения восстанавливающей способности рода Escherichia, продуцирующего L-аминокислоты, дополнительно обеспечивается АТФ, который нужно восстановить в ходе процесса биосинтеза НАДФ.
Таким образом, целью настоящего изобретения является получение микроорганизма рода Escherichia с повышенной продукцией L-аминокислот, при этом микроорганизм трансформирован так, чтобы он имел повышенную НАД-киназную активность и инактивированный фермент с аминокислотной последовательностью SEQ ID NO 2, кодируемой геном tehB, таким образом обладая повышенной восстанавливающей способностью.
Другой целью настоящего изобретения является обеспечение способа получения L-аминокислот с использованием микроорганизма рода Escherichia.
Решение проблемы
Для достижения поставленных выше целей настоящее изобретение предлагает микроорганизм рода Escherichia с повышенной продукцией L-аминокислот, при этом микроорганизм трансформирован так, чтобы он имел повышенную НАД-киназную активность и инактивированный фермент с аминокислотной последовательностью SEQ ID NO 2, кодируемой геном tehB.
Настоящее изобретение также обеспечивает способ получения L-аминокислот с применением микроорганизма рода Escherichia.
Преимущественные эффекты настоящего изобретения
В соответствии с настоящим изобретением дополнение восстанавливающим средством НАДФ·Н внутриклеточного энергетического метаболизма микроорганизма с продукцией L-аминокислот создается за счет повышения уровня НАДФ, а последующий недостаток АТФ дополняется за счет инактивации фермента с аминокислотной последовательностью SEQ ID NO 2, кодируемой геном tehB, и, следовательно, продукцию L-аминокислот можно улучшить за счет восстановления баланса энергетического метаболизма и повышения активности клеток и сокращения времени культивирования.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
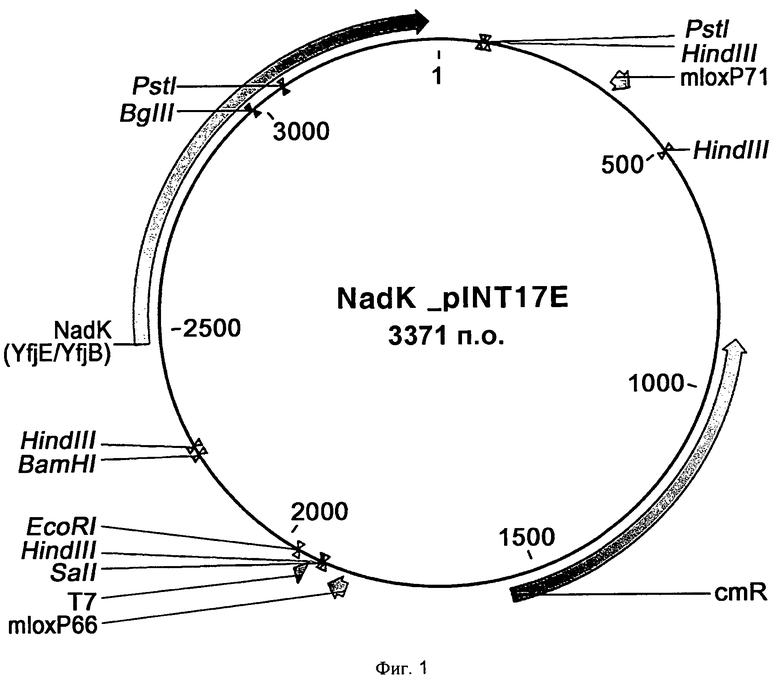
Фиг.1 представляет собой схему, на которой представлен вектор nadK-pINT17E для увеличения числа копий гена nadK в Е.coli.
Наилучший способ осуществления настоящего изобретения
Настоящее изобретение предполагает микроорганизм с повышенной продукцией L-аминокислот и способ получения L-аминокислот с его применением.
Продуцирующий L-аминокислоты микроорганизм согласно настоящему изобретению включает любой прокариотический или эукариотический микроорганизм, и его примеры включают штаммы микроорганизмов, принадлежащие к роду Escherichia, Erwinia, Serratia, Providencia, Corynebacterium и Brevibacterium. Микроорганизм согласно настоящему изобретению предпочтительно представляет собой микроорганизм, принадлежащий к роду Escherichia, и более предпочтительно Е.coli.
В настоящем изобретении L-аминокислота предпочтительно представляет собой L-треонин или L-триптофан.
В предпочтительном варианте осуществления настоящего изобретения предполагается микроорганизм рода Escherichia с повышенной продукцией L-аминокислот, при этом микроорганизм трансформирован так, чтобы он имел повышенную НАД-киназную активность и инактивированный фермент с аминокислотной последовательностью SEQ ID NO 2, кодируемой геном tehB, и таким образом обладая повышенной восстанавливающей способностью.
В настоящем изобретении НАД-киназа относится к ферменту с активностью для превращения НАД (никотинамидадениндинуклеотида) в НАДО (никотинамидадениндинуклеотидфосфат) с использованием фосфатной группы, полученной от АТФ или других соединений.
Последовательность белка с НАД-киназной активностью конкретно раскрыта в виде аминокислотной последовательности SEQ ID NO 4, а ген nadK, кодирующий НАД-киназу, предпочтительно представляет собой полинуклеотид с последовательностью оснований SEQ ID NO 3.
В настоящем изобретении повышение НАД-киназной активности рода Escherichia с продукцией L-аминокислот можно осуществить различными способами, хорошо известными в уровне техники. Например, способ может включать способ внедрения в хромосому собственно последовательности оснований, кодирующей НАД-киназу, или полинуклеотида, содержащего чужеродный участок, регулирующий экспрессию, способ увеличения числа копий при его введении в векторную систему или способ повышения ферментативной активности путем замены участка, регулирующего экспрессию гена, другой регуляторной последовательностью, модификацию всей или части последовательности, регулирующей экспрессию, или мутации гена самого по себе, но не ограничивается этим.
Более предпочтительно в настоящем изобретении можно использовать способ увеличения числа копий путем введения в хромосомную ДНК штамма последовательности оснований, кодирующей НАД-киназу, для повышения НАД-киназной активности микроорганизма, принадлежащего к роду Escherichia, который продуцирует L-аминокислоты.
Специалисту в данной области техники будет понятно, что увеличенное число копий НАД-киназы в пределах хромосомной ДНК проявляет такой же эффект, как и увеличенное число копий НАД-киназы с помощью внехромосомного вектора или как при повышенном уровне экспрессии с помощью модификации участка, регулирующего экспрессию кодирующего НАД-киназу гена nadK во внутри- или внехромосомном сайте или мутации гена самого по себе. Если используется вектор, род Escherichia с продукцией L-аминокислот трансформируют рекомбинантным вектором со встроенной последовательностью оснований, таким образом получая микроорганизм рода Escherichia с повышенной НАД-киназной активностью.
Вектор, который нужно использовать в настоящем изобретении, особым образом не ограничивается, и можно применять любой известный вектор экспрессии. Предпочтительно, можно применять вектор pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322 или pMW118.
Согласно одному варианту осуществления настоящего изобретения повышение НАД-киназной активности при трансформации повышает внутриклеточные уровни НАДФ и НАДФ·Н у штамма.
С целью повышения уровня продукции АТФ авторами настоящего изобретения также применялся и способ инактивации фермента, кодируемого геном tehB.
В настоящем изобретении ген tehB (идентификатор гена в базе NCBI: 945979) известен как ген, кодирующий белок устойчивости к теллуриту или предполагаемую S-аденозил-L-метионин-зависимую метилтрансферазу, но его функции все еще остаются невыясненными.
Однако в недавних исследованиях сообщалось, что при удалении гена tehB у Е.coli проявлялось 150% повышение продуцирования АТФ по сравнению с родительским штаммом, и предполагается, что этот результат связан со снижением количества АТФ, необходимого для биосинтеза S-аденозилметионина из метионина (FEMS Microbiol Lett., (2009) 297:217-224).
Более конкретно последовательность предполагаемой S-аденозил-L-метионин-зависимой метилтрансферазы может быть раскрыта с помощью аминокислотной последовательности SEQ ID NO 2. Кроме того, ген tehB, кодирующий фермент, происходит из Е.coli, и предпочтительно полинуклеотид имеет последовательность оснований SEQ ID NO 1.
Способ инактивации фермента, кодируемого геном tehB, охватывает все способы модификации соответствующего гена для предотвращения продуцирования фермента, кодируемого геном с последовательностью оснований SEQ ID NO 1. Способы могут быть подкреплены примерами удаление части или всего гена посредством гомологичной рекомбинации, подавления экспрессии фермента посредством вставки транспозона в пределах соответствующего гена, подавления экспрессии фермента посредством вставки генов устойчивости к антибиотикам или подобного, но не ограничиваясь этим.
Выражение "трансформация", как используется в данном документе, означает способ, с помощью которого в клетку-хозяина внедряют ген, который должен экспрессироваться в клетке-хозяине. Трансформированные гены, если они находятся в экспрессирующемся состоянии в клетке-хозяине, содержат любой из генов, внедренных в хромосому клетки-хозяина или расположенных в других частях хромосомы. Кроме того, ген содержит ДНК и РНК в качестве полинуклеотида, способного кодировать полипептид. При условии, что ген можно внедрить в клетку-хозяин и экспрессировать там, ген вводят в любом виде. Например, ген можно ввести в клетку-хозяин в виде кассеты экспрессии, которая представляет собой экспрессом полинуклеотида, который сама по себе содержит все элементы для экспрессии гена. Кассета экспрессии содержит промотор, который функционально соединен с геном, сигнал терминации транскрипции, сайт связывания рибосом и сигнал терминации трансляции. Кассета экспрессии может находиться в виде вектора экспрессии, способного к самоклонированию. Ген также можно ввести в клетку-хозяина сам по себе или в виде экспрессома полинуклеотида, который должен функционально соединяться с последовательностью, необходимой для экспрессии в клетке-хозяине.
В предпочтительном варианте осуществления настоящего изобретения микроорганизмом, трансформируемым с помощью способа, может быть Е.coli и предпочтительно Е.coli СА03-448(KCCM11167P), СА03-449(KCCM11168P) или СА04-2001(KCCM11166P).
Настоящее изобретение также предполагает способ получения L-аминокислот с применением микроорганизма рода Escherichia.
В предпочтительном варианте осуществления настоящее изобретение предполагает способ получения L-аминокислот путем культивирования рекомбинантного микроорганизма рода Escherichia с повышенной продукцией L-треонина или L-триптофана в среде, содержащей сахарозу или глюкозу в качестве основного источник углерода.
Конкретно, настоящее изобретение предполагает способ получения L-аминокислот, включающий этапы инокуляции и культивирования рекомбинантного микроорганизма рода Escherichia в культуральной среде, которая полностью или частично содержит сахарозу или глюкозу в качестве источника углерода, и отделения L-аминокислот от культуральной среды.
Процедуры культивирования согласно настоящему изобретению можно проводить в подходящих средах и в условиях культивирования, известных в уровне техники. В соответствии с применяемыми штаммами процедуры культивирования могут легко корректироваться, специалистом в данной области техники. Примеры процедур культивирования включают способы культивирования периодического типа, непрерывного типа и подпитываемого типа, но не ограничиваются ими. Среда, используемая в данном способе культивирования, предпочтительно должна удовлетворять требованиям конкретного штамма.
Среда, используемая в настоящем изобретении, содержит сахарозу или глюкозу в качестве основного источника углерода. Также в качестве источника углерода можно использовать мелассу, содержащую высокую концентрацию сахарозы, и среда может содержать подходящее количество различных источников углерода без ограничения. Примеры источников азота, которые можно использовать, включают органический источник азота, как например, пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт и соевую муку, и неорганический источник азота, как например, мочевину, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония, и их можно использовать либо по отдельности, либо в любой комбинации. В среду можно добавлять источники фосфора, как например, дигидрофосфат калия, гидроортофосфат калия или соответствующие натрийсодержащие соли. Помимо этого среда может содержать соли металлов, как например, сульфат магния и сульфат двухвалентного железа. Кроме того, среду можно дополнить аминокислотами, витаминами и подходящими предшественниками. Эти среды или предшественники можно добавить в культуры с использованием метода культивирования периодического типа или культуры непрерывного типа.
В ходе культивирования можно подходящим образом добавлять соединения, как например, гидроксид аммония, гидроксид калия, аммиак, фосфорную кислоту и серную кислоту, с тем, чтобы регулировать рН культур. Кроме того, в ходе культивирования можно подходящим образом добавлять пеногасители, как например, сложный полигликолевый эфир жирной кислоты, с тем, чтобы снизить ценообразование в культурах. Для поддержания культур в аэробных условиях в культуру можно подавать кислород или кислородосодержащий газ. Для поддержания культур в анаэробных и микроаэробных условиях можно не подавать газ в культуру или можно подавать азот, водород или углекислый газ.
Культуры поддерживают при 27-37°С и предпочтительно при 30-35°С. Культивирование можно продолжать до получения желаемого количества необходимого материала и предпочтительно в течение 10-100 часов.
Сбор и извлечение аминокислот, полученных на этапе культивирования согласно настоящему изобретению, можно осуществлять подходящим способом, известным в уровне техники, в зависимости от типа процедур культивирования, например, периодического типа, непрерывного типа или подпитываемого типа, с тем, чтобы собрать необходимые аминокислоты из культуральной среды
Способ осуществления настоящего изобретения
Настоящее изобретение будет описано ниже более подробно со ссылкой на примеры. Однако эти примеры приводятся только в целях иллюстрации, и не предполагается, что настоящее изобретение ограничивается этими примерами.
Пример 1. Получение продуцирующего L-треонин штамма с инактивированным ферментом, кодируемым происходящим из Е.coli геном tehB
Ген tehB продуцирующего L-треонин штамма Е.coli KCCM10541 (патент Кореи №10-0576342) удаляли посредством гомологичной рекомбинации.
Е.coli KCCM10541 представляет собой штамм, происходящий от продуцирующего L-треонин штамма Е.coli KFCC10718 (публикация патента Кореи №10-1992-0008365), а его родительский штамм Е.coli KFCC 10718 обладает устойчивостью к аналогу L-метионина, фенотипом ауксотрофа по метионину, устойчивостью к аналогу L-треонина, фенотипом неполного ауксотрофа по изолейцину, устойчивостью к аналогу L-лизина и устойчивостью к α-аминомасляной кислоте и способен продуцировать L-треонин.
Ген tehB (идентификатор гена в базе NCBI: 945979), который нужно удалить, как известно, кодирует предполагаемую S-аденозил-L-метионин-зависимую метилтрансферазу. Если ген удаляют, отсутствует потребность в АТФ, используемой при продуцировании S-аденозилметионина, и следовательно, ген tehB был выбран в качестве гена-мишени для сокращения потребления энергии, и он имеет последовательность оснований SEQ ID NO 1.
Для инактивации применяли одноэтапную инактивацию, которая представляет собой методику конструирования мутанта с использованием рекомбиназы Red фага лямбда, разработанную Datsenko KA с соавт. (Proc Natl Acad Sci USA., (2000) 97:6640 6645).
Для подтверждения внедрения в ген в качестве маркера использовали ген устойчивости к хлорамфениколу. Для удаления гена устойчивости к хлорамфениколу использовали систему Cre/loxP для сайт-специфической рекомбинации (ВМС Biotechnology(2001) 1:7).
Полимеразную цепную реакцию (называемую далее в данном документе ′ПЦР′) осуществляли с использованием вектора pMloxCm в качестве матрицы и праймера 1 и праймера 2, содержащих часть гена tehB и часть последовательности гена устойчивости к хлорамфениколу, в следующих условиях: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и элонгация при 72°С в течение 1 минуты, что приводило к амплификации фрагмента гена длиной приблизительно 1200 п.о.
Помимо этого, фрагмент ДНК, полученный посредством ПЦР-амплификации, подвергали электрофорезу в 0,8% агарозном геле, а затем элюировали и использовали в качестве матрицы для второй ПЦР. Вторую ПЦР осуществляли с использованием элюированного продукта первой ПЦР в качестве матрицы и праймера 3 и праймера 4, содержащих последовательность длиной 20 п.о., комплементарную 5′- и 3′-участкам первичного фрагмента ДНК, и дополнительно содержащих 5′- и 3′-участки гена tehB, в следующих условиях: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и элонгация при 72°С в течение 1 минуты, что приводило к амплификации фрагмента гена длиной приблизительно 1300 п.о. Фрагмент ДНК, полученный посредством вышеуказанной процедуры, подвергали электрофорезу на 0,8% агарозном геле, а затем элюировали и использовали в рекомбинации.
Штамм E.coli KCCM 10541, продуцирующий треонин, который трансформировали плазмидой pKD46 в соответствии со способом, разработанным Datsenko КА с соавт. (Proc. Natl. Acad. Sci. (2000) 97:6640-6645, GenBank №AY048746), подготавливали в качестве компетентного штамма, а трансформацию осуществляли путем введения фрагмента гена длиной 1300 п.о., который получали посредством ПЦР. Проводили селекцию полученных штаммов на среде LB, дополненной хлорамфениколом. Удаление гена tehB подтверждалось ПЦР-продуктом длиной приблизительно 2000 п.о., полученным посредством ПЦР с использованием следующих праймера 5 и праймера 6.
После удаления плазмиды pKD46 в первичный рекомбинантный штамм Е.coli с устойчивостью к хлорамфениколу внедряли плазмиду pJW168 с тем, чтобы удалить из штамма маркерный ген устойчивости к хлорамфениколу (Gene, (2000) 247, 255-264). Осуществляли ПЦР с использованием праймера 5 и праймера 6 с получением ПЦР-продукта длиной 832 п.о., это указывает на то, что полученный в конечном итоге штамм имел необходимое удаление.
Пример 2. Конструирование вектора для увеличения числа копий гена nadK в хромосоме Е.coli
Ген nadK амплифицировали посредством ПЦР с использованием в качестве матрицы хромосомы штамма Е.coli W3110 (номер доступа в GeneBank: AC000091), приобретенного у Американской коллекции типовых культур (АТСС).
В частности, ПЦР осуществляли с использованием праймера 7 и праймера 8 в следующих условиях: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и элонгация при 72°С в течение 1 минуты, что приводило к амплификации фрагмента гена длиной 1407 п.о. (SEQ ID NO 5).
Амплифицированная последовательность содержит кодирующую последовательность nadK, а также предполагаемый участок собственного промотора длиной 501 п.о. Помимо этого, праймер 7 содержит сайт рестрикции для EcoR I, a праймер 8 имеет сайт рестрикции для Xba I.
Полученный полинуклеотид обрабатывали рестрикционными ферментами Xba I и EcoR I и клонировали в сайты Xba I и EcoR I вектора pINT17E с последующей трансформацией в Е.coli BW25113. Затем клетки распределяли по поверхности плотной питательной среды LB Cm (LB + пластинка агаровой среды с хлорамфениколом). Клонированные векторы получали из колоний с использованием стандартных процедур mini-prep и обозначали nadK_pINT17E. Схема вектора показана на фиг.1.
Пример 3. Получение продуцирующего L-треонин штамма с инактивированным ферментом, кодируемым происходящим из Е.coli геном tehB и повышенной НАД-киназной активностью путем увеличения числа ее копий в хромосоме
Для увеличения числа копий НАД-киназы использовали штамм с удаленным геном tehB, полученный в соответствии со способом, описанным в примере 1, и вектор nadK_pINT17E, полученный в соответствии со способом, описанным в примере 2.
На первом этапе внедряли плазмиду pKD46 в штамм, имеющий удаленный ген tehB, полученный в соответствии со способом, описанным в примере 1, и подготовленный в качестве компетентного штамма, и штамм трансформировали вектором nadK_pINT17E. Для получения колоний осуществляли культивирование при 37°С в течение 1~2 дней. Чтобы подтвердить, внедрился ли ген в хромосому у полученных колоний, осуществляли ПЦР с использованием праймеров 8 и 9. ПЦР осуществляли в следующих условиях: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 45 секунд и элонгация при 72°С в течение 2 минут, что приводило к амплификации фрагмента гена длиной приблизительно 2000 п.о.
После удаления плазмиды pKD46 из первичного рекомбинантного штамма с устойчивостью к хлорамфениколу плазмиду pJW168 внедряли для удаления маркерного гена устойчивости к хлорамфениколу из штамма (Gene, (2000) 247, 255-264). Осуществляли ПЦР с использованием праймеров 10 и 11 с получением ПЦР-продукта длиной приблизительно 1500 п.о., что указывало на присутствие в хромосоме двух последовательных копий гена nadK в соответствии с ожиданиями.
Трансформированную Е.coli обозначили KCCM10541ΔtehB nadK 2 сору (СА03-448).
Пример 4. Получение продуцирующего L-триптофан штамма с инактивированным ферментом, кодируемым происходящим из Е.coli геном tehB, и повышенной активностью НАД-киназы путем увеличения числа ее копий в хромосоме
Трансформированную Е.coli получали таким же образом, как и в примерах 1-3, за исключением использования продуцирующего L-триптофан штамма Е.coli KCCM 10812.
Родительский штамм, используемый в этом примере, Е.coli KCCM 10812P, является штаммом, происходящим от мутанта Е.coli, продуцирующего L-фенилаланин (KFCC 10066), и отличается тем, что разблокирована ауксотрофия по триптофану, гены pheA, trpR, mtr и tnaAB инактивированы, а гены aroG и trpE мутированы в хромосоме (патент Кореи №10-0792095).
Трансформированную Е.coli обозначили KCCM10812ΔtehB nadK 2copy (СА04-2001).
Сравнительный пример 1. Получение продуцирующего L-треонин или L-триптофан штамма с инактивированным ферментом, кодируемым геном tehB
С целью удаления гена tehB использовали продуцирующий треонин штамм Е.coli KCCM10541 и продуцирующий триптофан штамм Е.coli KCCM10812, как описано в примере 1. Использовали одноэтапную инактивацию, которая представляет собой методику конструирования мутанта с использованием рекомбиназы Red фага лямбда, разработанную Datsenko KA с соавт. (Proc Natl Acad Sci USA., (2000) 97:6640-6645), и систему Cre/loxP для сайт-специфической рекомбинации (ВМС Biotechnology. (2001) 1:7).
Осуществляли ПЦР с использованием праймеров 5 и 6 с получением ПЦР-продуктов длиной 832 п.о., это указывает на то, что полученные в конечном итоге штаммы имеют нужное удаление, а штаммы обозначили соответственно KCCM 10541ΔtehB и KCCM10812PΔtehB.
Сравнительный пример 2. Получение продуцирующего L-треонин или L-триптофан штамма с повышенной НАД-киназной активностью
В соответствии со способом, описанным в примере 3, число копий гена nadK было увеличено до двух копий в хромосоме продуцирующих треонин и триптофан штаммов, так чтобы получить штаммы с повышенной НАД-киназной активностью.
В продуцирующий треонин штамм KCCM10541, а также продуцирующий триптофан штамм KCCM10812 вводили плазмиду pKD46 и подготавливали в качестве компетентных клеток и штаммы трансформировали вектором nadK_pINT17E. Затем осуществляли культивирование при 37°С в течение 1-2 дней для получения колоний. Чтобы подтвердить, внедрился ли ген в хромосому полученных колоний, осуществляли ПЦР с использованием праймеров 8 и 9.
После удаления плазмиды pKD46 из первичного рекомбинантного штамма с устойчивостью к хлорамфениколу вводили плазмиду pJW168 для удаления из штамма маркерного гена устойчивости к хлорамфениколу (Gene, (2000) 247, 255-264). ПЦР осуществляли с использованием праймеров 10 и 11 с получением ПЦР-продуктов длиной приблизительно 1500 п.о., что указывало на присутствие двух последовательных копий гена nadK в хромосоме в соответствии с ожиданиями. Полученные штаммы обозначили соответственно KCCM10541 nadK 2 сору и KCCM10812 nadK 2 сору.
Экспериментальный пример 1. Титрование продуцирующего L-треонин штамма с повышенной НАД-киназной активностью и инактивированным ферментом, кодируемым геном tehB
Прежде всего, для получения продуцирующего L-треонин штамма со способностью к ассимиляции сахарозы вектор pAcscBAR′-mak (публикация патента Кореи №10-2010-0092765) (SEQ ID NO 21), конструировали следующим образом.
После конструирования pAcscBAR, ген mak клонировали в pAcscBAR. Для конструирования pAcscBAR использовали праймеры 12 и 13, чтобы амплифицировать полинуклеотид участка cscB, где cscK был удален.
ПЦР осуществляли в следующих условиях: после денатурации при 94°С в течение 3 минут следовали 25 циклов денатурации при 94°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд и элонгация при 72°С в течение 1 минуты и 30 секунд, а затем элонгация при 72°С в течение 7 минут, что приводило к амплификации полинуклеотида длиной 1521 п.о. Полученный полинуклеотид и pAcscBAR обрабатывали рестрикционными ферментами EcoRV и EagI, соответственно, и клонировали, и трансформировали в Е.coli DH5α. Колонии, содержащие pAcscBAR, отбирали с помощью ПЦР с использованием колоний, выращенных на средах LB, а плазмиды получали с использованием стандартных процедур mini-prep для плазмид. Анализ последовательности cscBAR, связанной с сайтами XbaI и EagI полученной плазмиды pAcscBAR, не подтвердил ни одну мутацию.
Для амплификации полинуклеотида, содержащего ген mak, использовали праймеры 14 и 15 и хромосому Е.coli W3110 в качестве матрицы и клонировали в сайты рестрикции PstI и EagI в pAcscBAR с тем, чтобы сконструировать вектор pAcscBAR′-mak.
ПЦР осуществляли в следующих условиях: после денатурации при 94°С в течение 3 минут следовали 25 циклов денатурации при 94°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд и элонгация при 72°С в течение 1 минуты и 30 секунд, а затем элонгация при 72°С в течение 7 минут, что приводило к амплификации полинуклеотида длиной 1388 п.о. Полученный полинуклеотид и pAcscBAR обрабатывали рестрикционными ферментами PstI и EagI, соответственно, и клонировали, и трансформировали в Е.coli DH5α. Колонии, содержащие pAcscBAR′-mak отбирали с помощью ПЦР с использованием колоний, выращенных на средах LB, а плазмиды получали с использованием стандартных процедур mini-prep для плазмид. Анализ последовательности cscBAR-mak, связанной с сайтами XbaI и EagI полученной плазмиды pAcscBAR′-mak, не подтвердил ни одну мутацию.
Сконструированную pAcscBAR′-mak внедряли в рекомбинантный штамм Е.coli KCCM10541ΔtehB nadK 2 сору из примера 3, родительский штамм Е.coli KCCM10541 и штамм KCCM10541 с удаленным геном tehB (обозначенный KCCM10541ΔtehB), a также штамм KCCM10541 с увеличенным числом копий гена nadK (обозначенный KCCM10541 nadK 2 сору), полученные в сравнительных примерах 1 и 2, соответственно, а затем осуществляли титрование.
Каждый из штаммов с разными генетическими признаками культивировали на плотных питательных средах LB в термостате при 33°С в течение ночи. Затем 1 платиновую петлю инокулировали в 25 мл среды для титрования, содержащей сахарозу, как показано в следующей таблице 9, и культивировали в инкубаторе при 33°С и 200 об./мин. в течение 48 часов. Результаты показаны в таблице 10. Все результаты представлены в виде среднего значения, полученного от трех колб.
Как показано в таблице 10, если был удален только ген tehB, способность к ассимиляции сахарозы была подобной таковой у родительского штамма. Однако если повышалась НАД-киназная активность, способность ассимилировать сахарозу повышалась до приблизительно 2 г по сравнению с родительским штаммом.
Кроме того, если был удален ген tehB, плотность клеток уменьшалась приблизительно на 6% по сравнению с родительским штаммом, но продукция треонина была подобной таковой у родительского штамма. Однако если две мутации вводили одновременно, плотность клеток уменьшалась приблизительно на 8%, способность ассимилировать сахарозу повышалась приблизительно на 8%, а продукция треонина также повышалась на 9% по сравнению с родительским штаммом.
Более того, рекомбинантный штамм Е.coli KCCM10541ΔtehB nadK 2copy из примера 3, родительский штамм Е.coli KCCM10541, а также штамм KCCM10541ΔtehB и штамм KCCM10541 nadK 2copy, полученные в сравнительных примерах 1 и 2, анализировали посредством титрования с использованием глюкозы в качестве источника углерода. Каждый из штаммов с разными генетическими признаками культивировали на плотных питательных средах LB в инкубаторе при 33°С в течение ночи. Затем 1 платиновую петлю инокулировали в 25 мл среды для титрования, содержащей глюкозу, как показано в следующей таблице 11, и культивировали в термостате при 33°С и 200 об./мин. в течение 48 часов. Результаты показаны в таблице 12. Все результаты представлены в виде среднего значения, полученного от трех колб.
Штамм KCCM10541 ΔtehB nadK 2copy продуцирующей L-треонин E.coli, имеющий удаленный ген tehB и повышенную НАД-киназную активность со способностью ассимилировать глюкозу, обозначили СА03-448, а штамм KCCM10541 ΔtehB nadK 2copy/pAcscBAR′-mak, штамм KCCM10541 ΔtehB nadK 2copy с предоставленной способностью ассимилировать сахарозу, обозначили СА03-449. И их депонировали в международном органе по депонированию, Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms), который является филиалом корейской федерации коллекций культур (Korean Federation of Culture Collections) (расположен по адресу 361-221, Hongje-1-dong, Seodaemon-gu, Сеул, Корея), 10 января 2011 года, и им были присвоены депозитарные номера (доступа) KCCM11167P и KCCM11168P, соответственно.
Экспериментальный пример 2. Титрование продуцирующего L-триптофан штамма с повышенной НАД-киназной активностью и инактивированным ферментом, кодируемым геном tehB
Рекомбинантный штамм E.coli KCCM10812ΔtehB nadK 2copy из примера 4, родительский штамм E.coli KCCM10812, а также штамм KCCM10812ΔtehB и штамм KCCM10812 nadK 2copy, полученные в сравнительных примерах 1 и 2, анализировали посредством титрования с использованием глюкозы в качестве источника углерода.
Для титрования 1 платиновую петлю штамма инокулировали и культивировали на плотных питательных средах LB в течение ночи. Затем 1 платиновую петлю инокулировали в колбу с 25 мл среды для титрования с составом, представленным в следующей таблице 13, а затем культивировали при 37°С и 200 об./мин. в течение 48 часов. Результаты показаны в таблице 14. Все результаты представлены в виде среднего значения, полученного от трех колб.
Как показано в таблице 14, если был удален ген tehB, плотность клеток увеличивалась приблизительно на 10% по сравнению с родительским штаммом. Если повышалась НАД-киназная активность, способность к ассимиляции глюкозы улучшалась, но не было отличий в продукции триптофана по сравнению с родительским штаммом.
Однако если две мутации вводили одновременно, плотность клеток увеличивалась, способность к ассимиляции глюкозы также улучшалась, а продукция триптофана повышалась приблизительно на 14%.
Штамм KCCM10812P ΔtehB nadK 2copy продуцирующей L-триптофан Е.coli с удаленным геном tehB и повышенной НАД-киназной активностью обозначили СА04-2001, и его депонировали в международном органе по депонированию, корейском Центре культур микроорганизмов (Korean Culture Center of Microorganisms), который является филиалом Корейской федерации коллекций культур (Korean Federation of Culture Collections) (расположен по адресу 361-221, Hongje-1-dong, Seodaemon-gu, Сеул, Корея), 10 января 2011 года, и присвоили номер доступа KCCM11166P.
Специалисту в данной области техники будет понятно, что различные модификации и изменения можно осуществлять без отступления от объема и сущности настоящего изобретения. Таким образом, следует понимать, что вышеописанный вариант осуществления является не ограничивающим, а иллюстративным во всех аспектах. Объем настоящего изобретения определяется приложенной формулой изобретения, а не предшествующим ей описанием, и следовательно, все изменения и модификации, которые попадают в пределы и ограничения формулы изобретения или эквиваленты таких пределов или ограничений, таким образом, как предполагается, охватываются формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| МИКРООРГАНИЗМ, СПОСОБНЫЙ ПРОДУЦИРОВАТЬ L- АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ПРИМЕНЕНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2013 |
|
RU2600875C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| Вариант белка рецептора цАМФ и способ получения L-аминокислоты с его применением | 2019 |
|
RU2743745C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |
| Вариант белка-рецептора цАМФ и способ получения L-аминокислоты с его применением | 2019 |
|
RU2746860C1 |
| ВАРИАНТ БЕЛКА-РЕЦЕПТОРА цАМФ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ВАРИАНТА | 2019 |
|
RU2760536C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2014 |
|
RU2614258C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2014 |
|
RU2651461C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2250266C2 |












Изобретение относится к биотехнологии и представляет собой микроорганизм рода Escherichia, продукцирующий L-аминокислоты, причем микроорганизм трансформирован так, чтобы он имел повышенную НАД-киназную активность и инактивированный фермент с аминокислотной последовательностью SEQ ID NO 2, кодируемой геном tehB, и способ получения L-аминокислот с использованием такого микроорганизма рода Escherichia. Изобретение позволяет получать аминокислоты с высокой степенью эффективности. 2 н. и 9 з.п. ф-лы, 1 ил., 14 табл., 4 пр.
1. Микроорганизм рода Escherichia с повышенной продукцией L-аминокислот, где микроорганизм трансформирован так, чтобы он имел повышенную НАД-киназную активность и инактивированный фермент с аминокислотной последовательностью SEQ ID NO 2, кодируемой геном tehB.
2. Микроорганизм по п.1, где НАД-киназа представляет собой белок с аминокислотной последовательностью SEQ ID NO 4.
3. Микроорганизм по п.1, где НАД-киназная активность повышена с помощью одного или нескольких способов увеличения числа копий посредством хромосомной вставки или внедрения вектора, замены или модификации участка, регулирующего экспрессию, и генной мутации.
4. Микроорганизм по п.1, где инактивация осуществлена с помощью одного или нескольких способов удаления части или всего гена посредством гомологичной рекомбинации, подавления экспрессии фермента посредством вставки транспозона в пределах соответствующего гена и подавления экспрессии фермента посредством вставки генов устойчивости к антибиотикам.
5. Микроорганизм по п.1, где микроорганизм рода Escherichia представляет собой Е.coli.
6. Микроорганизм по п.1, где L-аминокислота представляет собой L-треонин или L-триптофан.
7. Микроорганизм по п.6, где микроорганизм рода Escherichia обеспечен способностью к ассимиляции сахарозы.
8. Микроорганизм по п.1, где микроорганизм рода Escherichia представляет собой продуцирующий L-треонин штамм Е.coli СА03-448 с № депонирования KCCM11167P или СА03-449 с № депонирования KCCM11168P.
9. Микроорганизм по п.1, где микроорганизм рода Escherichia представляет собой продуцирующий L-триптофан штамм Е.coli CA04-2001 с № депонирования KCCM11166P.
10. Способ получения L-аминокислот, включающий этапы инокуляции и культивирования микроорганизма рода Escherichia по любому из пп.1-9 в культуральной среде, которая полностью или частично содержит сахарозу или глюкозу в качестве источника углерода, и отделения L-аминокислоты от культуральной среды.
11. Способ по п.10, где L-аминокислота представляет собой L-треонин или L-триптофан.
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН rcsA | 2006 |
|
RU2359029C2 |
| WO 2010149574 A1, 29.12.2010 | |||
| WO 2010101359 A2, 10.09.2010 | |||

