В данном документе приведены новые комбинации элементов экспрессионного вектора, таких как промотор, поли(А)-сигнальная последовательность, терминатор транскрипции, структуры экспрессионного вектора, их комбинации, а также новые способы получения линий клеток-продуцентов, такие как новые, способы трансфекции и селекции, а также применение данных экспрессионных векторов и линий клеток-продуцентов для рекомбинантного получения интересующих полипептидов.
Предшествующий уровень техники
Уровень транскрипции гена может оказывать сильное влияние на его уровень экспрессии и таким образом определять продуктивность клетки. В основном он обусловлен тремя элементами вектора: промотором, поли(А)-сигнальной последовательностью и (если присутствует) терминатором транскрипции.
Нуклеиновая кислота, кодирующая тяжелую цепь антитела, обычно содержит лидерную последовательность (сигнальную
последовательность) (приблизительно 57 п. н./19 а.к.), удаляемую при созревании белка, вариабельный участок, VH (приблизительно 350 п. н./115 а.к.), и константный участок, СН (приблизительно 990 п. н./330 а.к.). Нуклеиновая кислота, кодирующая легкую цепь антитела, обычно состоит из лидерной последовательности (приблизительно 66 п. н./22 а.к.), которая удаляется при созревании белка, вариабельного участка, VK или VL (приблизительно 350 п. н./115 а.к.) и константного участка, CK или CL (приблизительно 321 п. н./107 а.к.).
Рекомбинантное получение антител в эукариотических клетках обычно включает создание экспрессионных систем (см., McCafferty, J., et al., (eds.), Antibody Engineering, A Practical Approach., IRL Press (1997)). При разработке систем экспрессии антител создают экспрессионную кассету, содержащую нуклеиновую кислоту, кодирующую легкую цепь, фланкированную промотором и участком полиаденилирования (поли(А)). Также создают экспрессионную кассету тяжелой цепи, содержащую нуклеиновую кислоту, кодирующую тяжелую цепь, фланкированную промотором и участком поли(А). Экспрессионная кассета тяжелой цепи может быть скомбинирована с экспрессионной кассетой легкой цепи в единый вектор, содержащий экспрессионный кассеты как тяжелой, так и легкой цепи, или кассеты могут быть встроены в два различных вектора.
Молекулы ДНК-кассет иммуноглобулинов, конструкты монотел, способы их получения и применения описаны в US 7,053,202. В US 5,168,062 описаны трансфекционные вектора и микроорганизмы, содержащие регуляторную последовательность ДНК предраннего промотора цитомегаловируса. Фрагмент ДНК, содержащий промоторный участок фактора элонгации-1α полипептидной цепи человека, его основная последовательность и экспрессионные плазмиды, содержащие фрагмент ДНК, имеющий высокую совместимость с широким рядом хозяйских клеток с высокой экспрессионной способностью, описан в US 5,225,348. В US 5,266,491 описаны экспрессионные плазмиды, содержащие ориджин репликации SV40 и фрагмент ДНК, содержащий промоторный участок для фактора элонгации-1α полипептидной цепи человека. Рекомбинантные соединения ДНК и экспрессия полипептидов, таких как tPA описаны в US 5,122,458. В US 7,422,874 описан экспрессионный вектор для животных клеток.
Sanna Pietro, Р., описывает экспрессию Fab-фрагментов антитела и целый иммуноглобулин в клетках млекопитающих (Meth. Mol. Biol. 178 (2002) 389-395). Библиотека клеточного дисплея для генного клонирования вариабельных участков антител человека к поверхностному антигену вируса гепатита описана в Higuchi, K., et al. (J. Immunol. Meth. 202 (1997) 193-204). Kim, D., et al., описывают улучшенные экспрессионные системы млекопитающих, полученные за счет манипулирования участками терминации транскрипции (Biotechnol. Progress 19 (2003) 1620-1622). Руководство по клеточному инженированию для получения моноклональных антител описано Costa, R.A., et al. (Eur. J. Pharmaceut. Biopharmaceut. 74 (2010) 127-138). Kim, D.W., et al., описывают применение промотора фактора элонгации 1 альфа человека в качестве универсальной и эффективной экспрессионной системы (Gene 91 (1990) 217-223). Сравнение интрон-зависимой и интрон-независимой генной экспрессии описано Buchman, A.R., et al., (Mol. Cell. Biol. 8 (1988) 4395-4405). Wang, F., et al., описывают экспрессию антител в клетках млекопитающих (в Therapeutic monoclonal antibodies - From bench to clinic, Wiley (2009) pages 557-572). Сравнительное исследование различных дизайнов векторов для экспрессии в клетках млекопитающих рекомбинантных антител IgG описано Li et al. (J. Immunol. Meth. 318 (2007) 113-124). Ho, S.C.L., et al. описывают "IRES-опосредованные трицистронные вектора для увеличения получения клеточных линий СНО с высокой экспрессией моноклональных антител (J. Biotechnol. 157 (2011) 130-139). Продукция рекомбинантными клетками животных химерных антител к CD2 описана Hotta, Α., et al. (J. Biosci. Bioeng. 98 (2004) 298-303). Lee, J-C, et al. описывают высокоэффективную экспрессию белка, опосредованную внутренней посадкой рибосомы энтеровируса 71 (Biotechnol. Bioeng. 90 (2005) 656-662). В WO 2008/142124 описано получение рекомбинантного белка в клетках Avian ЕВХ®. Краткое описание изобретения
Обнаружено, что создание экспрессионного вектора сильно зависит от его предназначения и что лучшие вектора для кратковременных трансфекций, стабильных пулов и селекции единичных клонов различаются.
Для ясности основные открытия: для кратковременной трансфекций выигрышными являются разнонаправленная экспрессия легкой и тяжелой цепи антитела и применение полноразмерного промотора hCMV, включая интрон А. Однако для стабильной трансфекций была выигрышна последовательность 1) легкая цепь антитела, 2) тяжелая цепь антитела и 3) селекционный маркер.
Промотор hEF1α явно превосходит промотор hCMV в стабильных пулах, однако был обнаружено явно противоположное влияние на уровне единичных клонов. В данном документе были получены клоны цитомегаловируса человека с предранним промотором/энхансером (hCMV) с наиболее высокой продуктивностью.
Более того, получение промотора hCMV может быть в дальнейшем улучшено за счет комбинации его с bGH поли(А)-сигнальной и терминальной последовательностью гена гастрина человека (hGT), который повышает как продуктивность, так и стабильность экспрессии.
Обнаружено, что применение экспрессионного вектора, содержащего экспрессионные кассеты для тяжелой цепи антитела и экспрессионную кассету для легкой цепи антитела, каждую содержащую промотор, структурный ген и поли(А)-сигнальную последовательность и возможно последовательность терминатора, приводит к более высокому числу клеточных клонов продуцирующих/секретирующих антитело после трансфекции если i) промотор представляет собой промотор цитомегаловируса человека (hCMV), поли(А)-сигнальная последовательность представляет собой поли(А)-сигнальную последовательность коровьего гормона роста (bGH поли(А)), и последовательность терминатора представляет собой
последовательность терминатора транскрипции гена гастрина человека (hGT), или 2) промотор представляет собой промотор фактора элонгации 1 альфа человека (hEF1alpha), поли(А)-сигнальная последовательность представляет собой поли(А)-сигнальную последовательность коровьего гормона роста (bGH поли(А)), и последовательность терминатора отсутствует.
Посредством применения экспрессионного вектора, как описано выше, может быть получено более высокое число клеток, продуцирующих/секретирующих антитело, после трансфекции, и, таким образом, снижены необходимые усилия для идентификации высокопродуцирующей клетки, приемлемой для крупномасштабного получения рекомбиантных антител.
Таким образом, один аспект, как описано в данном документе, представляет собой способ селекции рекомбинантных клеток млекопитающих, включающий следующий этап:
а) трансфекция клеток млекопитающих экспрессионным вектором, содержащим
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последователь и hGT последовательность терминатора и
таким образом, получают множество рекомбинантных клеток млекопитающих,
b) селекция множества рекомбинантных клеток млекопитающих (единичных).
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении для селекции стабильно трансфецированной клетки.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в разных направлениях для селекции кратковременно трансфецированных клеток.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер. В одном воплощении экспрессионные кассеты и селекционный маркер расположены в одном направлении. В одном воплощении экспрессионные кассеты расположены в последовательности LC-HC-SM.
В одном воплощении клетки млекопитающих выбраны из клеток CHO, HEK, BHK, NS0 и клеток SP2/0. В одном воплощении клетки млекопитающих представляют собой клетки СНО для селекции стабильно трансфецированных клеток. В одном воплощении клетки млекопитающих представляют собой клетки HEK для селекции кратковременно трансфецированных клеток.
Один аспект, как описано в данном документе, представляет собой способ получения антител, включающий следующий этап:
а) культивирование клеток млекопитающих, содержащих
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, bGH поли(А)-сигнальную последовательность, и hGT последовательность терминатора,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора, и
b) получение антитела из клетки или культуральной среды.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении для стабильной продукции антитела.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в разных направлениях для кратковременной продукции антитела.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер. В одном воплощении экспрессионные кассеты и селекционный маркер расположены в одном направлении. В одном воплощении экспрессионные кассеты расположены в последовательности LC-HC-SM.
В одном воплощении клетки млекопитающих выбраны из клеток CHO, HEK, BHK, NS0 и клеток SP2/0. В одном воплощении клетки млекопитающих представляют собой клетки СНО для стабильной продукции антитела. В одном воплощении клетки млекопитающих представляют собой клетки HEK для кратковременной продукции антитела.
Один аспект, как описано в данном документе, представляет собой экспрессионный вектор, содержащий
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении для селекции стабильно трансфецированных клеток.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в разных направлениях для селекции кратковременно трансфецированных клеток.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер. В одном воплощении экспрессионные кассеты и селекционный маркер расположены в одном направлении. В одном воплощении экспрессионные кассеты расположены в последовательности LC-HC-SM.
Обнаружено, что для получения клеточных линий, экспрессирующих/секретирующих стабильное рекомбинантное антитело, при применении промотора фактора элонгации человека 1 альфа (hEF1 alpha) в сочетании с bGH поли(А)-сигнальной последовательностью, присутствие hGT последовательности терминатора уменьшает получаемый экспрессионный выход.
Один аспект, как описано в данном документе, представляет собой способ селекции рекомбинантных клеток млекопитающих, включающий следующий этап:
a) трансфекция клеток млекопитающих экспрессионным вектором, содержащим
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hEF1alpha, нуклеиновую кислоту, кодирующую легкую цепь антитела, и bGH поли(А)-сигнальную последовательность.
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hEF1alpha, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последовательность, и
таким образом, получая множество рекомбинантных клеток млекопитающих,
b) селекция множества рекомбинантных клеток млекопитающих (единичных).
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении для селекции стабильно трансфецированных клеток.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в разных направлениях для селекции кратковременно трансфецированных клеток.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер. В одном воплощении экспрессионные кассеты и селекционный маркер расположены в одном направлении. В одном воплощении экспрессионные кассеты расположены в последовательности LC-HC-SM.
В одном воплощении промотор фактора элонгации человека 1 альфа содержит интрон А.
В одном воплощении экспрессионный вектор является свободным от каких-либо последовательностей терминатора транскрипции. В одном воплощении последовательность терминатора представляет собой hGT последовательность.
В одном воплощении клетки млекопитающих выбраны из клеток CHO, HEK, BHK, NS0 и клеток SP2/0. В одном воплощении клетки млекопитающих представляют собой клетки СНО для селекции стабильно трансфецированных клеток. В одном воплощении клетки млекопитающих представляют собой клетки HEK для селекции кратковременно трансфецированных клеток.
Один аспект, как описано в данном документе, представляет собой способ получения антитела, содержащий следующий этап:
a) культивирование клеток млекопитающих, содержащих
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hEF1alpha, нуклеиновую кислоту, кодирующую легкую цепь антитела, и bGH поли(А)-сигнальную последовательность,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hEF1alpha, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, и bGH поли(А)-сигнальную последовательность, и
b) получение антитела из клеток или культуральной среды.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении для селекции стабильно трансфецированных клеток.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в разных направлениях для селекции кратковременно трансфецированных клеток.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер. В одном воплощении экспрессионные кассеты и селекционный маркер расположены в одном направлении. В одном воплощении экспрессионные кассеты расположены в последовательности LC-HC-SM.
В одном воплощении промотор фактора элонгации человека 1 альфа содержит интрон А.
В одном воплощении экспрессионный вектор свободен от каких-либо последовательностей терминатора транскрипции. В одном воплощении последовательность терминатора представляет собой hGT последовательность.
В одном воплощении клетки млекопитающих выбраны из клеток СНО, HEK, BHK, NS0 и клеток SP2/0. В одном воплощении клетки млекопитающих представляют собой клетки СНО для стабильной продукции антитела. В одном воплощении клетки млекопитающих представляют собой клетки HEK для кратковременной продукции антитела.
Один аспект, как описано в данном документе, представляет собой экспрессионный вектор, содержащий
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ альфа промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, и bGH поли(А)-сигнальную последовательность, и
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор hEF1alpha, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, и bGH поли(А)-сигнальную последовательность.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении для селекции стабильно трансфецированных клеток.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в разных направлениях для селекции кратковременно трансфецированных клеток.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер. В одном воплощении экспрессионные кассеты и селекционный маркер расположены в одном направлении. В одном воплощении экспрессионные кассеты расположены в последовательности LC-HC-SM.
В одном воплощении промотора фактора элонгации человека 1 альфа содержит интрон А.
В одном воплощении экспрессионный вектор свободен от каких-либо последовательностей терминатора транскрипции. В одном воплощении последовательность терминатора представляет собой hGT последовательность.
Обнаружено, что для стабильной рекомбинантной продукции антитела применение экспрессионного вектора, содержащего
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность и возможно первую последовательность терминатора транскрипции,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность и возможно вторую последовательность терминатора транскрипции, и
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно третью последовательность терминатора транскрипции,
в соответствии с чем три экспрессионные кассеты расположены противоположно и в последовательности первая экспрессионная кассета-вторая экспрессионная кассета-третья экспрессионная кассета, приводит к улучшенному экспрессионному выходу в стабильно трансфецированных рекомбинантных клетках млекопитающих.
В противоположность вышеуказанному обнаружено, что для кратковременной рекомбинантной продукции антитела, применение экспрессионного вектора, содержащего
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность и возможно первую последовательность терминатора транскрипции,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность и возможно вторую последовательность терминатора транскрипции, и
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно третью последовательность терминатора транскрипции,
В соответствии с этим экспрессионные кассеты расположены в двух направлениях, в соответствии с чем первая экспрессионная кассета и вторая экспрессионная кассета расположены в противоположном направлении, что приводит к улучшению экспрессионного выхода в кратковременно трансфецированных рекомбинантных клетках млекопитающих.
Термин в противоположном направлении означает, что одна экспрессионная кассета транскрибируется в направлении 5ʹ->3ʹ и одна экспрессионная кассета транскрибируется в направлении 3ʹ->5ʹ.
Таким образом, один аспект, как описано в данном документе, представляет собой применение экспрессионного вектора, содержащего
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность и возможно первую последовательность терминатора транскрипции,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность и возможно вторую последовательность терминатора транскрипции, и
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно третью последовательность терминатора транскрипции,
В соответствии с чем три экспрессионные кассеты расположены в противоположном направлении в последовательности первая экспрессионная кассета-вторая экспрессионная кассета-третья экспрессионная кассета, для стабильной рекомбинантной продукции антитела в клетках млекопитающих.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первый и второй промотор представляют собой промотор hCMV, первая и вторая поли(А)-сигнальная последовательность представляют собой bGH поли(А)-сигнальную последовательность, и последовательность терминатора транскрипции присутствует и представляет собой hGT последовательность терминатора.
В одном воплощении первый и второй промотор представляют собой промотор hEF1alpha, первая и вторая поли(А)-сигнальная последовательность представляют собой bGH поли(А)-сигнальную последовательность и экспрессионные кассеты являются свободными от какой-либо последовательности терминатора транскрипции.
В одном воплощении клетки млекопитающих выбраны из клеток CHO, HEK, BHK, NS0 и клеток SP2/0. В одном воплощении клетки млекопитающих представляют собой клетки СНО.
Один аспект, как описано в данном документе, представляет собой экспрессионный вектор, содержащий
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность и возможно первую последовательность терминатора транскрипции,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность, и возможно последовательность терминатора транскрипции, и
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно третью последовательность терминатора транскрипции,
В соответствии с чем экспрессионные кассеты расположены в противоположном направлении и в последовательности первая экспрессионная кассета-вторая экспрессионная кассета-третья экспрессионная кассета.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первый и второй промотор представляют собой промотор hCMV, первая и вторая поли(А)-сигнальная последовательность представляют собой bGH поли(А)-сигнальную последовательность и последовательность терминатора транскрипции присутствует и представляет собой hGT последовательность терминатора.
В одном воплощении первый и второй промотор представляют собой промотор hEF1alpha, первая и вторая поли(А)-сигнальная последовательность представляет собой bGH поли(А)-сигнальную последовательность и экспрессионные кассеты являются свободными от последовательности терминатора транскрипции.
Один аспект, как описано в данном документе, представляет собой применение экспрессионного вектора, содержащего
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность и возможно первую последовательность терминатора транскрипции,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность и возможно вторую последовательность терминатора транскрипции,
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно третью последовательность терминатора транскрипции,
в соответствии с чем экспрессионные кассеты расположены в двух направлениях, в соответствии с чем первая экспрессионная кассета и вторая экспрессионная кассета расположены в противоположном направлении для кратковременной рекомбинантной продукции антитела в клетках млекопитающих.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первый и второй промотор представляют собой промотор hCMV, первая и вторая поли(А)-сигнальная последовательность представляют собой bGH поли(А)-сигнальную последовательность и последовательность терминатора транскрипции присутствует и представляет собой hGT последовательность терминатора.
В одном воплощении первый и второй промотор представляют собой промотор hEF1alpha, первая и вторая поли(А)-сигнальная последовательность представляет собой bGH поли(А)-сигнальную последовательность, и экспрессионные кассеты свободны от последовательности терминатора транскрипции.
В одном воплощении клетки млекопитающих выбраны из клеток CHO, HEK, BHK, NS0 и клеток SP2/0. В одном воплощении клетки млекопитающих представляют собой клетки HEK.
Один аспект, как описано в данном документе, представляет собой экспрессионный вектор, содержащий
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность и возможно последовательность терминатора транскрипции,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность и возможно вторую последовательность терминатора транскрипции, и
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно третью последовательность терминатора транскрипции,
в соответствии с чем экспрессионные кассеты расположены в двух направлениях, в соответствии с чем первая экспрессионная кассета и вторая экспрессионная кассета расположены в противоположном направлении.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
В одном воплощении первый и второй промотор представляют собой промотор hCMV, первая и вторая поли(А)-сигнальная последовательность представляют собой bGH поли(А)-сигнальную последовательность и последовательность терминатора транскрипции присутствует и представляет собой hGT последовательность терминатора.
В одном воплощении первый и второй промотор представляют собой промотор hEF1alpha, первая и вторая поли(А)-сигнальная последовательность представляют собой bGH поли(А)-сигнальную последовательность и экспрессионные кассеты свободны от последовательности терминатора транскрипции.
Кроме того, обнаружено среди прочего, что экспрессионные вектора, содержащие промотор hCMV и промотор Ef1α с интроном А вместо короткого промотора CMV человека без интрона А, усиливают кратковременную экспрессию и экспрессию пула генов.
Один аспект, как описано в данном документе, представляет собой экспрессионную плазмиду, содержащую
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, и первую поли(А)-сигнальную последовательность,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела и вторую поли(А)-сигнальную последовательность,
где одна или обе экспрессионные кассеты после поли(А)-сигнальной последовательности дополнительно содержат последовательность терминатора гастрина человека.
В одном воплощении первая и вторая поли(А)-сигнальная последовательность выбраны независимо друг от друга из SV40 поли(А)-сигнальной последовательности и поли(А)-сигнальной
последовательности коровьего гормона роста.
В одном воплощении первый и второй промотор выбраны независимо друг от друга из промотора CMV человека, промотора SV40 и промотора фактора элонгации человека 1 альфа.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержат по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляют собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер. В одном воплощении экспрессионные кассеты и селекционный маркер расположены в разных направлениях.
Один аспект, как описано в данном документе, представляет собой применение экспрессионной плазмиды, как описано в данном документе, для кратковременной экспрессии антитела или стабильной экспрессии антитела.
Один аспект, как описано в данном документе, представляет собой эукариотические клетки, содержащие экспрессионную плазмиду, как описано в данном документе.
Один аспект, как описано в данном документе, представляет собой способ продукции антитела, включающий этапы
- культивирование эукариотических клеток, содержащих экспрессионную плазмиду, как описано в данном документе,
- получение антитела из эукариотических клеток или культуральной среды.
В одном воплощении эукариотические клетки представляют собой клетки млекопитающих. В одном воплощении клетки млекопитающих выбраны из клеток CHO, HEK, BHK, NS0 и клеток SP2/0.
Один аспект, как описано в данном документе, представляет собой экспрессионную плазмиду, содержащую
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ первый промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, и первую поли(А)-сигнальную последовательность,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ второй промотор, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, и вторую поли(А)-сигнальную последовательность,
где первый и/или второй промотор представляют собой промотор фактора элонгации человека 1 альфа.
В одном воплощении одна или обе экспрессионные кассеты не содержат дополнительно после поли(А)-сигнальной последовательности последовательность терминатора гастрина человека.
В одном воплощении одна или обе экспрессионные кассеты свободны от последовательности терминатора гастрина человека.
В одном воплощении промотор фактора элонгации человека 1 альфа содержит интрон А.
В одном воплощении первая и вторая поли(А)-сигнальная последовательность выбраны независимо друг от друга из SV40 поли(А)-сигнальной последовательности и поли(А)-сигнальной последовательности коровьего гормона роста.
В одном воплощении первый и второй промотор выбраны независимо друг от друга из промотора CMV человека, промотора SV40 и промотора фактора элонгации человека 1 альфа.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
В одном воплощении нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляют собой кДНК.
В одном воплощении первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении.
В одном воплощении экспрессионная плазмида дополнительно содержит селекционный маркер.
В одном воплощении экспрессионные кассеты и селекционный маркер расположены в разных направлениях.
Один аспект, как описано в данном документе, представляет собой применение экспрессионной плазмиды, как описано в данном документе, для кратковременной экспрессии антитела или стабильной экспрессии антитела.
Один аспект, как описано в данном документе, представляет собой эукариотические клетки, содержащие экспрессионную плазмиду, как описано в данном документе.
Один аспект, как описано в данном документе, представляет собой способ получения антитела, включающий этапы культивирование эукариотических клеток, содержащих экспрессионную плазмиду, как описано в данном документе, или клеток, как описано в данном документе,
получение антитела из эукариотических клеток или культуральной среды.
В одном воплощении эукариотические клетки представляют собой клетки млекопитающих. В одном воплощении клетки млекопитающих выбраны из клеток CHO, HEK, BHK, NS0 и клеток SP2/0.
Один аспект, как описано в данном документе, представляет собой экспрессионную плазмиду, содержащую в направлении 5ʹ-3ʹ последовательность промотора, нуклеиновую кислоту, кодирующую тяжелую цепь антитела или легкую цепь антитела, IRES элемент, нуклеиновую кислоту, кодирующую селекционный маркер и поли(А)-сигнальную последовательность, в соответствии с чем IRES элемент представляет собой EMCV-IRES элемент.
В одном воплощении нуклеиновая кислота кодирует тяжелую цепь антитела.
В одном воплощении селекционный маркер представляет собой слитый белок формулы A-C-S, при этом А является детектируемым полипептидом, С представляет собой протеолитическую сигнальную последовательность и S представляет собой селекционный маркер.
В одном воплощении протеолитическая сигнальная последовательность представляет собой последовательность PEST орнитин декарбоксилазы.
В одном воплощении детектируемый полипептид представляет собой зеленый флуоресцентный белок.
В одном воплощении селекционный маркер представляет собой неомицин.
Один аспект, как описано в данном документе, представляет собой нуклеиновую кислоту, кодирующую полипептид, содержащий в направлении от N-конца к С-концу зеленый флуоресцентный белок, последовательность PEST орнитин декарбоксилазы и неомицин.
Один аспект, как описано в данном документе, представляет собой применение нуклеиновой кислоты, кодирующей полипептид, содержащий в направлении от N-конца к С-концу зеленый флуоресцентный белок, последовательность PEST орнитин декарбоксиллазы и неомицин для селекции клеток, секретирующих антитело.
Один аспект, как описано в данном документе, представляет собой применение экспрессионной кассеты, содержащей в направлении 5ʹ-3ʹ последовательность промотора, нуклеиновую кислоту, кодирующую тяжелую цепь антитела или легкую цепь антитела, IRES элемент, нуклеиновую кислоту, кодирующую селекционный маркер и поли(А)-сигнальную последовательность для селекции клеток, продуцирующих антитело, при этом IRES элемент представляет собой EMCV-IRES элемент.
В одном воплощении нуклеиновая кислота кодирует тяжелую цепь антитела.
В одном воплощении селекционный маркер представляет собой слитый белок формулы A-C-S, при этом А представляет собой детектируемый полипептид, С представляет собой протеолитическую сигнальную последовательность и S представляет собой селекционный маркер.
В одном воплощении протеолитическая сигнальная последовательность представляет собой последовательность PEST орнитин декарбоксилазы.
В одном воплощении детектируемый полипептид представляет собой зеленый флуоресцентный белок.
В одном воплощении селекционный маркер представляет собой неомицин.
Один аспект, как описано в данном документе, представляет собой способ селекции эукариотических клеток, экспрессирующих антитело, включающий следующие этапы:
- культивирование эукариотических клеток, содержащих i) экспрессионную плазмиду, как описано в данном аспекте и ii) нуклеиновую кислоту, кодирующую соответствующую другую цепь антитела, не кодируемую экспрессионной плазмидой, как описано в данном аспекте,
- селекция клеток, экспрессирующих детектируемый полипептид.
Один аспект, как описано в данном документе, представляет собой экспрессионную плазмиду, содержащую в направлении 5ʹ-3ʹ последовательность промотора, нуклеиновую кислоту, кодирующую легкую цепь антитела, IRES элемент, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, и поли(А)-сигнальную последовательность, при этом IRES элемент представляет собой EV71-IRES элемент.
В одном воплощении последовательность промотора выбрана из последовательности промотора CMV человека с или без интрона А, последовательности промотора SV40, и последовательности промотора фактора элонгации человека 1 альфа с или без интрона А.
В одном воплощении поли(А)-сигнальная последовательность выбрана из поли(А)-сигнальной последовательности коровьего гормона роста и SV40 поли(А)-сигнальной последовательности.
В одном воплощении плазмида содержит от 3ʹ к поли(А)-сигнальной последовательности последовательность терминатора гастрина человека.
Один аспект, как описано в данном документе, представляет собой применение экспрессионной плазмиды, содержащей в направлении 5ʹ-3ʹ последовательность промотора, нуклеиновую кислоту, кодирующую легкую цепь антитела, IRES элемент, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, и поли(А)-сигнальную последовательность, для экспрессии антитела, при этом IRES элемент представляет собой EV71-IRES элемент.
В одном воплощении последовательность промотора выбрана из последовательности промотора CMV человека с или без интрона А, последовательности промотора SV40, и последовательности промотора фактора элонгации человека 1 альфа с или без интрона А.
В одном воплощении поли(А)-сигнальная последовательность выбрана из поли(А)-сигнальной последовательности коровьего гормона роста и SV40 поли(А)-сигнальной последовательности.
В одном воплощении плазмида содержит от 3ʹ к поли(А)-сигнальной последовательности последовательность терминатора гастрина человека.
Один аспект, как описано в данном документе, представляет собой способ получения антитела, содержащий следующие этапы:
- культивирование эукариотических клеток, содержащих экспрессионную плазмиду, как описано в данном документе,
- получение антитела из клеток или культуральной среды и таким образом получение антитела.
В одном воплощении промотор hCMV обладает последовательностью SEQ ID NO: 01. Она представляет собой промотор hCMV без интрона А и без 5ʹUTR.
В одном воплощении промотор hCMV обладает последовательностью SEQ ID NO: 02. Она представляет собой промотор hCMV без интрона А и с 5ʹUTR.
В одном воплощении промотор hCMV обладает последовательностью SEQ ID NO: 03. Она представляет собой полноразмерный промотор hCMV без интрона А.
В одном воплощении промотор фактора элонгации человека 1 альфа обладает последовательностью SEQ ID NO: 04. Она представляет собой промотор hEF1 без интрона А.
В одном воплощении промотор фактора элонгации человека 1 альфа обладает последовательностью SEQ ID NO: 05. Она представляет собой промотор hEF1alpha с интроном А.
В одном воплощении промотор фактора элонгации человека 1 альфа обладает последовательностью SEQ ID NO: 06. Она представляет собой короткий промотор hEF1alpha без интрона А и с 5ʹUTR.
В одном воплощении промотор CMV крысы обладает последовательностью SEQ ID NO: 07.
В одном воплощении SV40 поли(А)-сигнальная последовательность обладает последовательностью SEQ ID NO: 08.
В одном воплощении поли(А)-сигнальная последовательность коровьего гормона роста обладает последовательностью SEQ ID NO: 09.
В одном воплощении терминатор гастрина человека обладает последовательностью SEQ ID NO: 10.
В одном воплощении промотор SV40 обладает последовательностью SEQ ID NO: 11.
В одном воплощении последовательность PEST орнитин декарбоксилазы кодируется последовательностью SEQ ID NO: 12.
В одном воплощении последовательность GFP кодируется последовательностью SEQ ID NO: 13.
В одном воплощении селекционный маркер неомицин обладает последовательностью SEQ ID NO: 14.
В одном воплощении GFP-PEST-NEO слитый полипептид кодируется последовательностью SEQ ID NO: 15.
В одном воплощении EMCV-IRES обладает последовательностью SEQ ID NO: 16.
В одном воплощении EV71-IRES обладает последовательностью SEQ ID NO: 17.
В одном воплощении всех аспектов, как описано в данном документе, антитело представляет собой биспецифичное антитело.
В одном воплощении биспецифичное антитело обладает первой специфичностью связывания или сайтом связывания, который специфично связывается с первым антигеном или первым эпитопом на антигене, и биспецифичное антитело обладает второй специфичностью связывания или вторым сайтом связывания, который специфично связывается со вторым антигеном или со вторым эпитопом на антигене.
Если в одном воплощении экспрессионный вектор содержит или
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор, нуклеиновую кислоту, кодирующую первую легкую цепь антитела, поли(А)-сигнальную последовательность и возможно последовательность терминатора,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор, нуклеиновую кислоту, кодирующую вторую легкую цепь антитела, поли(А)-сигнальную последовательность и возможно последовательность терминатора,
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор, нуклеиновую кислоту, кодирующую первую тяжелую цепь антитела, поли(А)-сигнальную последовательность и возможно последовательность терминатора,
- четвертую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор, нуклеиновую кислоту, кодирующую вторую тяжелую цепь антитела, поли(А)-сигнальную последовательность и возможно последовательность терминатора,
или
- первую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор, нуклеиновую кислоту, кодирующую легкую цепь антитела, поли(А)-сигнальную последовательность и возможно последовательность терминатора,
- вторую экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор, нуклеиновую кислоту, кодирующую первую тяжелую цепь антитела, поли(А)-сигнальную последовательность и возможно последовательность терминатора, и
- третью экспрессионную кассету, содержащую в направлении 5ʹ-3ʹ промотор, нуклеиновую кислоту, кодирующую вторую тяжелую цепь антитела, поли(А)-сигнальную последовательность и возможно последовательность терминатора,
при этом легкая цепь антитела является общей легкой цепью для обеих тяжелых цепей антитела.
В одном воплощении всех аспектов, как описано в данном документе, экспрессионный вектор содержит
- экспрессионную кассету с легкой цепью антитела,
- экспрессионную кассету с первой тяжелой цепью антитела,
- экспрессионную кассету со второй тяжелой цепью антитела, и
- экспрессионную кассету с селекционным маркером,
где по меньшей мере одна из экспрессионных кассет с тяжелой цепью антитела и экспрессионная кассета с селекционным маркером расположены в одном направлении, и
где расположенные в одном направлении экспрессионные кассеты расположены в направлении 5ʹ-3ʹ в последовательности экспрессионная кассета с тяжелой цепь антитела, экспрессионная кассета с легкой цепью антитела и экспрессионная кассета с селекционным маркером или расположенные в одном направлении экспрессионные кассеты расположены в направлении 5ʹ-3ʹ в последовательности экспрессионная кассета с легкой цепью антитела, экспрессионная кассета с тяжелой цепью антитела и экспрессионная кассета с селекционным маркером.
В одном воплощении всех аспектов, как описано в данном документе, экспрессионный вектор содержит
- экспрессионную кассету с легкой цепью антитела,
- экспрессионную кассету со второй легкой цепью антитела,
- экспрессионную кассету с первой тяжелой цепью антитела
- экспрессионную кассету со второй тяжелой цепью антитела, и
- экспрессионную кассету с селекционным маркером,
где одна из экспрессионных кассет с тяжелой цепью антитела, одна из экспрессионных кассет с легкой цепью антитела и экспрессионная кассета с селекционным маркером расположены в одном направлении, и
где расположенные в разных направлениях экспрессионные кассеты расположены в направлении 5ʹ-3ʹ в последовательности экспрессионная кассета с тяжелой цепью антитела, экспрессионная кассета с легкой цепью антитела и экспрессионная кассета с селекционным маркером или расположенные в разном направлении экспрессионные кассеты расположены в направлении 5ʹ-3ʹ в последовательности экспрессионная кассета с легкой цепью антитела, экспрессионная кассета с тяжелой цепью антитела и экспрессионная кассета с селекционным маркером.
В одном воплощении кодирует одну из экспрессионных кассет с тяжелой цепью антитела, тяжелой цепью антитела, содержащей hole-мутацию.
В одном воплощении кодирует экспрессионные кассеты с тяжелой цепью антитела, тяжелой цепью, содержащей knob-мутацию.
В одном воплощении кодирует одну из экспрессионных кассет с легкой цепью антитела, содержащей вариабельный домен легкой цепи антитела, и домен СН1 тяжелой цепи антитела в качестве константного домена, и/или одну из экспрессионных кассет с легкой цепью антитела, содержащей варибельный домен легкой цепи антитела и домен CL легкой цепи антитела в качестве константного домена.
В одном воплощении одна из экспрессионных кассет с тяжелой цепью антитела кодирует тяжелую цепь антитела, содержащую в качестве первого константного домена константный домен легкой цепи антитела (CL), и/или одна из экспрессионных кассет с тяжелой цепью антитела кодирует тяжелую цепь антитела, содержащую в качестве первого константного домена домен СН1 тяжелой цепи антитела.
Подробное описание изобретение
I. Общие аспекты
Как известно специалистам в области техники применение технологии рекомбинантной ДНК делает возможным получение множества производных нуклеиновой кислоты и/или полипептида. Такие производные, например, могут быть модифицированы по одной или нескольким позициям при помощи замещения, изменения, обмена, делеции или инсерции. Модификация или получение производных, например, могут быть выполнены при помощи сайт-направленного мутагенеза. Такие модификации могут быть легко выполнены специалистами в области техники (см, например. Sambrook, J., et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York, USA (1999)). Применение рекомбинантной технологии позволяет специалистам в области техники трансформировать различные хозяйские клетки при помощи гетерологичной нуклеиновой кислоты(от). Хотя транскрипционный и тансляционный, т.е. экспрессионный, аппарат различных клеток применяют сходные элементы, клетки, относящиеся к различным видам, могут обладать среди прочего, различной так называемой частотой использования кодона. Таким образом идентичные полипептиды (в соответствии с аминокислотной последовательностью) могут кодироваться различными нуклеиновыми кислотами. Также, из-за вырожденности генетического кода различные нуклеиновые кислоты могут кодировать одинаковый полипептид.
Применение технологии рекомбинантной ДНК делает возможным получение множества производных нуклеиновой кислоты и/или полипептида. Такие производные могут, например, быть модифицированы по одной или нескольким позициям при помощи замещения, изменения, обмена, делеции или инсерции. Модификация или получение производных могут, например, быть выполнены при помощи сайт-направленного мутагенеза. Такие модификации могут быть легко выполнены специалистами в области техники (см., например, Sambrook, J., et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York, USA (1999); Hames, B.D. and Higgins, S.J., Nucleic acid hybridization - a practical approach, IRL Press, Oxford, England (1985)).
Применение рекомбинантной технологии делает возможной трансформацию различных хозяйских клеток гетерологичной нуклеиновой кислотой(ми). Хотя транскрипционный и трансляционный, т.е. экспрессионный, аппарат различных клеток применяет сходные элементы, клетки, относящиеся к разным видам, могут среди прочего обладать различной так называемой частотой использования кодона. Таким образом идентичные полипептиды (в соответствии с аминокислотной последовательностью) могут кодироваться различной нуклеиновой кислотой(ми). Также, из-за вырожденности генетического кода различные нуклеиновые кислоты могут кодировать один и тот же полипептид.
Определения
«Аффинность зрелого» антитела относится к антителу с одним или более изменениями в одном или более гипервариабельном участке (HVRs), по сравнению с родительским антителом, которое не имеет таких изменений, приводящих к улучшению аффинности антитела к антигену.
Термин «антитело» в данном документе применяют в самом широком смысле, и он охватывает различные структуры антител, включая, но не ограничено, моноклональные антитела, поликлональные антитела, мультиспецифичные антитела (например, биспецифичные антитела), и такие фрагменты антител, которые проявляют желаемую антиген связывающую активность.
«Фрагмент антитела» относится к молекуле другой, нежели интактное антитело, которая содержит часть интактного антитела, связывающую антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничены, Fv, Fab, Fabʹ, Fabʹ-SH, F(abʹ)2; диантитела; линейные антитела; одноцепочечные молекулы антител (например, scFv); и мультиспецифичные антитела, формируемые из фрагментов антител.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получают из определенного источника или видов, в то время как тяжелую и/или легкую цепь получают из различных источников или видов.
«Класс» антитела относится к типу константного домена или константного участка, охватываемого тяжелой цепью. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM и несколько из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, соответствующих различным классам иммуноглобулинов называют α, δ, ε, γ и μ, соответственно.
Термин «экспрессия», как употреблено в данном документе, относится к процессам транскрипции и/или трансляции, протекающим в клетке. Уровень транскрипции интересующей последовательности нуклеиновой кислоты в клетке может быть определен на основании количества соответствующей мРНК, которая присутствует в клетке. Например, мРНК, транскрибируемая с интересующей последовательности, может быть подсчитана при помощи ОТ-ПЦР или при помощи Нозерн-гибридизации (см. Sambrook et al., 1999, supra). Полипептиды, кодируемые интересующей нуклеиновой кислотой, могут быть подсчитаны различными способами, например, при помощи ELISA, при помощи исследования биологической активности полипептида или при помощи применения исследований, не зависящих от такой активности, таких как Вестерн-блот или радиоиммуноанализ, при помощи иммуноглобуллинов, которые распознают и связывают полипептид (см. Sambrook et al., 1999, supra).
«Экспрессионная кассета» относится к конструкции, содержащей необходимые регуляторные элементы, такие как промотор и сайт полиаденилирования, для экспрессии по меньшей мере нуклеиновой кислоты, содержащейся в клетке.
«Экспрессионный вектор» представляет собой нуклеиновую кислоту, обеспечивающую все необходимые элементы для экспрессии содержащегося структурного гена(ов) в хозяйской клетке. Обычно экспрессионная плазмида содержит прокариотическую плазмидную единицу воспроизводства, например, для Е. coli, содержащей ориджин репликации, и селекционный маркер, эукариотический селекционный маркер и одну или более экспрессионные кассеты для экспрессии интересующего структурного гена(ов), каждая содержащая промотор, структурный ген и сигнал полиаденилирования (поли(А)-сигнальную последовательность). Экспрессию гена обычно помещают под контроль промотора, и такой структурный ген называют «операционно связанным» с промотором. Аналогично, регуляторный элемент и коровый промотор операционно связаны, если регуляторный элемент модулирует активность корового промотора.
Термин «Fc-участок» в данном документе применяют для определения С-терминального участка тяжелой цепи иммуноглобулина, содержащей часть константного участка. Термин включает нативную последовательность Fc-участков и вариантные Fc-участки. В одном воплощении Fc-участок тяжелой цепи IgG человека простирается от Cys226 или от Pro230, до карбоксильного конца тяжелой цепи. Однако, С-терминальный лизин (Lys447) Fc-участка может присутствовать или нет. Если не указано иное, в данном документе, нумерация аминокислотных остатков в Fc-участке или константном участке соответствует системе нумерации EU, также называемой «EU index», как описано в Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242.
«Fc-участок» представляет собой термин хорошо известный и определяемый на основании расщепления папаином тяжелой цепи антитела. Комплексы, как описано в данном документе, могут содержать в одном воплощении в качестве полипептида петлевой части тяжелой цепи антитела человека Fc-участок человека или Fc-участок человеческого происхождения. В дополнительном воплощении Fc-участок представляет собой или Fc-участок антитела человека подкласса IgG4 или Fc-участок антитела человека подкласса IgG1, IgG2 или IgG3, который модифицируют таким образом, что ни связывание Fcγ рецептора (например, FcγRIIIa) и/или ни связывание C1q не могут быть детектированы. В одном воплощении Fc-участок представляет собой Fc-участок человека и в частности или подкласса IgG4 человека или от мутированного Fc-участка подкласса IgG1 человека. В одном воплощении Fc-участок представляет собой подкласс IgG1 человека с мутациями L234A и L235A. В тоже время IgG4 проявляет пониженное связывание с Fcγ рецептором (FcγRIIIa), антитела других подклассов IgG проявляют сильное связывание. Однако Pro238, Asp265, Asp270, Asn297 (утрата Fc карбогидрата), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 или/и His435 представляют собой остатки, которые, при замене, обеспечивают также пониженное связывание с Fcγ-рецептором (Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604; Lund, J., et al., FASEB J. 9 (1995) 115-119; Morgan, Α., et al., Immunology 86 (1995) 319-324; ЕР 0307434). В одном воплощении антитело, экспрессируемое в аспекте в отношении связывания с Fcγ-рецептором подкласс IgG4 или подкласс IgG1 или подкласс IgG2, с мутацией в L234, L235 и/или D265, и/или содержит мутацию PVA236. В одном воплощении мутации представляют собой S228P, L234A, L235A, L235E и/или PVA236 (PVA236 обозначает, что аминокислотная последовательность ELLG (заданная в однобуквенном аминокислотном коде) от аминокислотной позиции 233 до 236 IgG1 или EFLG IgG4 замещена PVA). В одном воплощении мутации представляют собой S228P IgG4 и L234A и L235A IgG1. Fc-участок антитела непосредственно вовлечены в ADCC (антитело-зависимую клеточно-опосредованную цитотоксичность) и CDC (комплемент-зависимую цитотоксичность). Комплекс, который не связывается с Fcγ-рецептором и/или фактор комплемента C1q, не вызывает антитело-зависимую клеточную цитотоксичность (ADCC) и/или комплемент-зависимую цитотоксичность (CDC). Knob-модификация обозначает мутацию T366W в домене СН3 антитела (нумерация согласно Kabat). Hole-модификация обозначает мутации T366S, L368A и Y407V в домене СН3 антитела. В дополнение к knob- и hole-модификации может присутствовать мутация S354C в одном домене СН3, мутация Y349C в другом домене СН3.
"Каркас" или "FR" относится к остаткам вариабельного домена другим нежели остатки гипервариабельного участка (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, HVR и FR последовательности обычно расположены в следующей последовательности VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термины «полноразмерное антитело», «интактое антитело» и «целое антитело» применяют в данном документе взаимозаменяемо в отношении антитела, обладающего структурой по существу сходной со структурой нативного антитела или имеющему тяжелые цепи, содержащие Fc-участок, как определено в данном документе.
«Ген» означает нуклеиновую кислоту, которая представляет собой сегмент, например, на хромосоме или на плазмиде, которая может влиять на экспрессию пептида, полипептида или белка. Помимо кодирующего участка, т.е. структурного гена, ген содержит другие функциональные элементы, например, сигнальную последовательность, промотор(ы), интроны и/или терминаторы.
Термины «хозяйская клетка», «хозяйская клеточная линия» и «хозяйская клеточная культура» применяют взаимозаменяемо и в отношении клеток, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Хозяйские клетки включают «трансформантов» и «трансформированные клетки», включая первично трансформированные клетки и потомство, полученное от них вне зависимости от числа пассажей. Потомство может быть полностью идентичным по содержанию нуклеиновой кислоты родительской клетке, но может содержать мутации. Мутантное потомство, обладающее такой же функцией или биологической активностью, в качестве скринируемого или селекционируемого в исходно трансформированной клетке включено в данный документ.
«Антитело человека» представляет собой антитело, обладающее аминокислотной последовательностью, соответствующей таковой антитела, получаемого от человека или клетки человека или получаемого из источника, не являющегося человеком, в котором использован набор антител человека или другие последовательности, кодирующие антитела. Данное определение антитела человека определенно исключает гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
«Гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из FR человека. В конкретных воплощениях, гуманизированное антитело будет содержать по существу все из по меньшей мере одного и обычно двух вариабельных доменов, в которых все или по существу все HVRs (например, CDRs) соответствуют таковым нечеловеческого антитела, и все или по существу все FRs соответствуют таковым антитела человека. Гуманизированное антитело может дополнительно содержать по меньшей мере часть константного участка антитела, полученную из антитела человека. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подвергали гуманизации.
Термин «гипервариабельный участок» или «HVR», как употреблено в данном документе, относится к каждому из участков вариабельного домена антитела, который является гипервариабельным в последовательности и/или формирует структурно определенные петли («гипервариабельные петли»). Обычно, нативные четерех-цепочечные антитела содержат шесть HVRs; три в VH (H1, H2, H3) и три в VL (L1, L2, L3). HVRs обычно содержат аминокислотные остатки из гипервариабельных петель и/или из «участков, определяющих комплементарность» (CDRs), последние имеют наиболее вариабельную последовательность и/или вовлечены в распознавание антигена. Типичные гипервариабельные петли возникают в аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (Н1), 53-55 (Н2), and 96-101 (Н3) (Chothia, С. and Lesk, A.M., J. Mol. Biol. 196 (1987) 901-917). Типичные CDRs (CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2, and CDR-H3) возникают в аминокислотных остатках 24-34 L1, 50-56 L2, 89-97 L3, 31-35 В Н1, 50-65 Н2, и 95-102 H3 (Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242). За исключением CDR1 в VH, CDRs обычно содержат аминокислотные остатки, формирующие гипервариабельные петли. CDRs также содержат «остатки определяющие специфичность» или «SDRs», которые представляют собой остатки, контактирующие с антигеном. SDRs заключены внутри участков CDRs, называемых сокращенных-CDRs или a-CDRs (abbreviated-CDRs). Типичные a-CDRs (a-CDR-L1, a-CDR-L2, a-CDR-L3, a-CDR-H1, a-CDR-H2 и a-CDR-H3) возникают в аминокислотных остатках 31-34 L1, 50-55 L2, 89-96 L3, 31-35 В Н1, 50-58 Н2 и 95-102 Н3 (Almagro, J.C. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633). Если не указано иное HVR остатки и другие остатки в вариабельном домене (например, FR-остатки) нумеруют в данном документе согласно Kabat et al., supra.
«Сайт внутренней посадки рибосомы» или «IRES» описывают последовательность, которая функционально обеспечивает инициацию трансляции независимо от гена 5ʹ IRES и позволяет двум цистронам (открытым рамкам считывания) быть транслированными с одного транскрипта в животной клетке. IRES обеспечивает сайт внутренней посадки рибосомы для трансляции открытой рамки считывания непосредственно ниже (ниже применяют взаимозаменяемо в данном документе с 3ʹ) участка. В отличие от бактериальной мРНК, которая может быть полицистронной, т.е. кодировать несколько различных полипептидов, которые танслируются последовательно с мРНК, большинство мРНК животных клеток являются моноцистронными и кодируют для синтеза только один белок. С полицистронного транскрипта в эукариотической клетке трансляция будет иницироваться с 5ʹ большинства сайтов инициации трансляции, терминироваться первым стоп-кодоном, и транскрипт будет высвобождаться из рибосомы, приводя к трансляции первого кодируемого полипептида в мРНК. В эукариотической клетке, полицистронный транскрипт, обладающий IRES, операционно связанным со второй или последующей открытой рамкой считывания в транскрипте делает возможным последовательную трансляцию открытой рамки считывания расположенной ниже для получения двух или более полипептидов, кодируемых тем же транскриптом. Применение IRES элементов в векторной конструкции описано ранее, см.например, Pelletier, J., et al., Nature 334 (1988) 320-325; Jang, S.K., et al., J. Virol. 63 (1989) 1651-1660; Davies, M.V., et al., J. Virol. 66 (1992) 1924-1932; Adam, M.A., et al., J. Virol. 65 (1991) 4985-4990; Morgan, R.A., et al. Nucl. Acids Res. 20 (1992) 1293-1299; Sugimoto, Y, et al., Biotechnology 12 (1994) 694-698; Ramesh, Ν., et al., Nucl. Acids Res. 24 (1996) 2697-2700; и Mosser, D.D., et al., BioTechniques 22 (1997) 150-152).
Термин «моноклональное антитело» как употреблено в данном документе относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, содержащие популяцию являются идентичными и/или связывают один и тот же эпитоп, исключая возможные вариантные антитела, например, содержащие мутации, возникшие естественным путем или в течение получения препарата моноклональных антител, такие варианты обычно присутствуют в минорных количествах. По сравнению с препаратами поликлональных антител, которые обычно включают различные антитела к различным детерминантнам (эпитопы), каждое моноклональное антитело препарата моноклональных антител направлено против единственной детерминанты на антигене. Таким образом, модификатор «моноклональное» означает характер антитела в качестве полученного из по существу гомогенной популяции антител, и не должен толковаться в качестве необходимости получения антитела каким-либо конкретным способом. Например, моноклональные антитела, подлежащие применению в соответствии с настоящим изобретением, могут быть получены при помощи различных техник, включая, но не ограничено гибридомным способом, способами рекомбинантной ДНК, способами фагофого отображения и способами, при которых используют трансгенных животных, содержащих весь или часть локуса иммуноглобулина человека, такие способы и другие примерны способвы получения моноклональных антител описаны в данном документе.
«Нативные антитела» относится к молекулам иммуноглобулина естественного происхождения с различными структурами. Например, нативные антитела к IgG представляют собой гетеротетрамерные гликопротеины приблизительно 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидными связями. От N- к С-концу каждая тяжелая цепь имеет вариабельный участок (VH), также называемый вариабельный тяжелый домен или вариабельный домен тяжелой цепи, с последующими тремя константными доменами (СН1, СН2 и СН3). Аналогично от N- до С-конца, каждая легкая цепь имеет вариабельный участок (VL), также называемый вариабельный легкий домен или вариабельный домен легкой цепи, с последующим константным доменом легкой цепи (CL). Легкая цепь антитела может быть обозначена одним из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности ее константного домена.
«Нуклеиновая кислота» как употреблено в данном документе относится к полимерной молекуле, состоящей из отдельных нуклеотидов (также называемых основаниями) а, с, g и t (или и в РНК), например, к ДНК, РНК или их модификациям. Данная полинуклеотидная молекула может представлять собой полинуклеотидную молекулу естественного происхождения или синтетическую полинуклеотидную молекулу или комбинацию одной или более полинуклеотидных молекул естественного происхождения с одной или более синтетическими полинуклеотидными молекулами. Также данное определение охватывает полинуклеотидные молекулы естественного происхождения, в которых один или более нуклеотидов заменены (например, в результате мутагенеза), делетированы или добавлены. Нуклеиновая кислота может быть или изолирована или интегрирована в другую нуклеиновую кислоту, например, в экспрессионную кассету, плазмиду или хромосому хозяйской клетки. Также нуклеиновую кислоту характеризуют при помощи последовательности нуклеиновой кислоты, состоящей из отдельных нуклеотидов.
Специалистам в области техники хорошо известны процедуры и способы перевода аминокислотной последовательности, например, полипептида в соответствующую последовательность нуклеиновой кислоты, кодирующую данную аминокислотную последовательность. Таким образом, нуклеиновую кислоту характеризуют при помощи ее нуклеотидной последовательности, состоящей из отдельных нуклеотидов, а также при помощи аминокислотной последовательности таким образом кодируемого полипептида.
«Нуклеиновая кислота», как употреблено в данном документе, относится к нуклеиновой кислоте естественного происхождения или частично или полностью неестественного происхождения, кодирующей полипептид, который может быть получен рекомбинантно. Нуклеиновая кислота может быть построена из фрагментов ДНК, которые или изолируют или синтезируют химическими способами. Нуклеиновая кислота может быть интегрирована в другую нуклеиновую кислоту, например, в экспрессионную плазмиду или геном/хромосому эукариотической хозяйской клетки. Плазмида включает шаттл и экспрессионные плазмиды.
Обычно плазмида также будет содержать прокариотическую единицу воспроизводства, содержаую ориджин репликации (например, ориджин репликации ColE1) и селекционный маркер (например, ген устойчивости к ампицилину или тетрациклину), для репликации и селекции соответственно, плазмиды в прокариотах.
«Операционно связанный» относится к смежному положению двух или более компонентов, где таким образом описанные компоненты находятся во взаимодействии, позволяющем им функционировать предназначенным им образом. Например, промотор и/или энхансер операционно связаны с кодирующей последовательностью, при его действие в цис-ориентации, для контроля и модуляции транскрипции связанной последовательности. Обычно, но не необходимо, ДНК последовательности, которые «операционно связаны», являются соседними и при необходимости соединяют два участка, кодирующих белок, таких как секреторный лидер и полипептид, соседними и внутри рамки (считывания). Однако хотя операционно связанный промотор обычно локализован выше кодирующей последовательности, он не обязательно прилегает к ней. Энхансеры не обязательно должны быть смежно расположены. Энхансер операционно связан с кодирующей последовательностью, в случае если энхансер усиливает транскрипцию кодирующей последовательности. Операционно связанные энхансеры могут быть локализованы выше, внутри или ниже кодирующих последовательностей и на значительном расстоянии от промотора. Сайт полиаденилирования операционно связан с кодирующей последовательностью, если он локализован ниже кодирующей последовательности, так что транскрипция продолжается через кодирующей последовательности в последовательность полиаденилирования. Трансляционный стоп-кодон операционно связан с экзонной последовательностью нуклеиновой кислоты, в случае если она локализована ниже (3ʹ конец) кодирующей последовательности, так что трансляция продолжается через кодирующую последовательность до стоп-кодона и терминируется здесь. Связи достигают при помощи способов рекомбинации, известных в области техники, например, при помощи ПЦР и/или лигирования по подходящим рестрикционным сайтам.
Если приемлемые рестрикционные сайты отсутствуют, применяют синтетические олигонуклеотидные адапторы или линкеры в соответствии с обычной практикой.
«Полицистронная транскрипционная единица» представляет собой транскрипционную единицу, в которой более чем один структурный ген находится под контролем одного и того же промотера.
Термин «сигнал полиаденилирования» как употребелно в данной заявке обозначает последовательность нуклеиновой кислоты, применяемую для индукции расщепления и полиаденилирования первичных транскриптов определенного сегмента последовательности нуклеиновой кислоты. 3ʹ нетранслируемый участок, содержащий сигнал полиаденилирования может быть выбран из группы, состоящей из 3ʹ нетранслируемого участка, содержащего сигналы полиаденилирования, полученные из SV40, гена коровьего гормона роста (bGH), генов иммуноглобулинов и гена тимидин киназы (tk, например, сигнал полиаденилирования тимидин киназа Herpes Simplex).
«Промотор» относится к полинуклеотидной последовательности, контролирующей транскрипцию гена/структурного гена или последовательности нуклеиновой кислоты, с которой он операционно связан. Промотор включает сигналы для связывания РНК-полимеразы и инициации транскрипции. Применяемые промоторы будут функциональны в хозяйских клетках, в которых предполагается экспрессия выбранной последовательности. Большое число промоторов, включая конститутивные, индуцибельные и репрессируемые промоторы из многообразия различных источников, хорошо известно в области техники (и идентифицированы в базах данных, таких как GenBank) и являются доступными в качестве или внутри клонированных полинуклеотидов (из, например, хранилищ, таких как АТСС, а также других коммерческих или индивидуальных источников).
«Промотор» содержит нуклеотидную последовательность, которая направляет транскрипцию структурного гена. Обычно, промотор локализован в 5ʹ некодирующем или нетранслируемом участке гена, проксимальном по отношению к сайту начала транскрипции структурного гена. Элементы последовательности внутри промотора, функционирующие при инициации транскрипции, часто характеризуют при помощи консенсусных нуклеотидных последовательнстей. Данные промоторные элементы включают сайты связывания РНК-полимеразы, TATA последовательности, СААТ последовательности, элементы, специфичные для дифференцировки (differentiation-specific elements, DSEs; McGehee, R.E., et al., Mol. Endocrinol. 7 (1993) 551), цАМФ-чувствительные элементы (cyclic AMP response elements, CREs), элементы, чувствительные к сыворотке (serum response elements, SREs; Treisman, R., Seminars in Cancer Biol. 1 (1990) 47), элементы, чувствительные к глюкокортикоидам (glucocorticoid response elements, GREs) и сайты связывания для других транскрипционных факторов, таких как CRE/ATF (OʹReilly, М.А., et al., J. Biol. Chem. 267 (1992) 19938), AP2 (Ye, J., et al., J. Biol. Chem. 269 (1994) 25728), SP1, белок, связывающийся с цАМФ-чувствительным элементом (cAMP response element binding protein, CREB; Loeken, M.R., Gene Expr. 3 (1993) 253) и октамерные факторы (см. Watson, et al., (eds.), Molecular Biology of the Gene, 4th ed. (The Benjamin/Cummings Publishing Company, Inc. (1987)), и Lemaigre, F.P. and Rousseau, G.G., Biochem. J. 303 (1994) 1-14). Если промотор представляет собой индуцибельный промотор, то скорость транскрипции увеличивается в ответ на индуцирующий агент. Для сравнения, скорость транскрипции не регулируется индуцирующим агентом, если промотор является конститутивным промотором. Также известны репрессируемые промоторы. Например, c-fos промотор специфично активируется при связывании гормона роста с его рецептором на клеточной поверхности. Экспрессия, регулируемая тетрациклином (tet) может быть достигнута за счет искусственного гибридного промотора, который состоит, например, из промотора CMV с последующими двумя Tet-операторными сайтами. Tet-репрессор связывается с двумя Tet-операторными сайтами и блокирует транскрипцию. При добавлении индуцирующего тетрациклина, Tet-репрессор высвобождается из Tet-операторных сайтов и транскрипция продолжается (Gossen, M. and Bujard, H., PNAS 89 (1992) 5547-5551). В отношении других индуцибельных промоторов, включая промоторы металлотионеинов и белков теплового шока, см, например, Sambrook et al. (supra) and Gossen et al., Curr. Opin. Biotech. 5 (1994) 516-520. Среди эукариотических промоторов, в качестве сильных промоторов с высоким уровнем экспрессии идентифицированы ранний промотор SV40, основной поздний промотор аденовируса, промотор мышиного металлотионеина-1, длинный концевой повтор вируса саркомы Рауса, альфа промотор фактора элонгации 1 китайских хомячков (CHEF-1, см, например, US 5,888,809), альфа промотор EF-1 человека, промотор убиквитина и предранний промотор цитомегаловируса человека (CMV IE).
«Промотор» может быть конститутивным или индуцибельным. Энхансер (т.е. цис-действующий элемент ДНК, который воздействует на промотор повышая транскрипцию) может быть необходим для функционирования вместе с промотором для повышения уровня экспрессии, достигаемого при помощи одного промотора, и может быть включен в качестве транскрипционного регуляторного элемента. Часто, полинуклеотидный сегмент, содержащий промотор, будет также включать энхансерные последовательности (например, CMV или SV40).
Термины «стабильно трансформированный», «стабильно трансфецированный» или стабильный, как употреблено внутри данной заявки означают наследуемую и стабильную интеграцию экзогенной нуклеиновой кислоты в геном/хромосому хозяйской клетки. Стабильно трансфецированные клетки получают после процесса клеточной селекции в селективных условиях роста, т.е. в присутствии одного или более селекционных маркеров.
«Структурный ген» означает участок гена без сигнальной последовательности, т.е. кодирующий участок.
Термин «терминатор транскрипции» означает последовательность ДНК из 50-750 пар оснований в длину, которая дает РНК-полимеразе сигнал для терминации синтеза мРНК. Очень эффективные (сильные) терминаторы на 3ʹ-конце экспрессионной кассеты являются подходящими для предотвращения работы РНК-полимеразы, в частности при применении сильных промоторов. Неэффективные терминаторы транскрипции могут приводить к образованию оперон-подобной мРНК, которая может быть причиной нежелательной, например, плазмид-кодируемой, генной экспрессии.
Внутри объема настоящего изобретения трансфецированные клетки могут быть получены по существу при помощи любого типа способа трансфекций, известного в области техники. Например, нуклеиновая кислота может быть введена в клетки при помощи средств электропорации или микроинъекции. Альтернативно, могут быть применены реагенты для липофекции, такие как FuGENE 6 (Roche Diagnostics GmbH, Germany), X-tremeGENE (Roche Diagnostics GmbH, Germany) и LipofectAmine (Invitrogen Corp., USA). Кроме того, альтернативно, нуклеиновая кислота может быть введена в клетку при помощи соответствующих вирусных векторных систем, основанных на ретровирусах, лентивирусах, аденовирусах или адено-ассоциированных вирусах (Singer, О., Proc. Natl. Acad. Sci. USA 101 (2004) 5313-5314).
Термин «кратковременная трансфекция», как употреблено в данной заявке, означает способ, при котором нуклеиновая кислота, введенная в хозяйскую клетку, не интегрирует в геномную или хромосомную ДНК этой клетки. Она фактически поддерживается в качестве внехромосомного элемента, например в виде эписомы, в клетке. Это не влияет на транскрипционные процессы нуклеиновой кислоты эписомы и, например, кодируемый нуклеиновой кислотой эписомы белок продуцируют. Кратковременная трансфекция приводит к «кратковременно трансфецированной» клетке.
Термин «вариабельный участок» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который вовлечен в связывание антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно обладают сходными структурами, с каждым доменом, содержащим четыре консервативных каркасных участка (FRs) и три гипервариабельных участка (HVRs) (см., например, Kindt, T.J., et al., Kuby Immunology, 6th ed., W.H. Freeman and Co., N.Y. (2007), page 91). Единственного домена VH или VL может быть достаточно для придания антиген-связывающей специфичности. Кроме того, антитела, которые связывают конкретный антиген могут быть изолированы при помощи домена VH или VL от антитела, которое связывает антиген для скрининга библиотеки комплементарности доменов VL или VH, соответственно (см, например, Portolano, S., et al., J. Immunol. 150 (1993) 880-887; Clackson, T., et al., Nature 352 (1991) 624-628).
Термин «вектор», как употреблено в данном документе, относится к молекуле нуклеиновой кислоты, способной к воспроизводству другой нуклеиновой кислоты, с которой она связана. Термин включает вектор в качестве самореплицирующейся нуклеокислотной структуры, а также вектора, инкорпорированный в геном хозяйской клетки, в которую он введен. Конкретные вектора способны к направлению экспрессии нуклеиновых кислот, с которыми они операционно связаны. Такие вектора рассматривают в данном документе в качестве «экспрессионных векторов».
Антитела
Способы и композиции, предложенные в данном документе, являются таковыми для продукции рекомбинантных моноклональных антител. Антитела могут представлять собой различные структуры, такие как, но не ограничено, моноспецифичные антитела (например, биспецифичные антитела), фрагменты антител, моновалентные антитела, мультивалентные антитела (например, бивалентные антитела).
В конкретных воплощениях антитело представляет собой фрагмент антитела. Фрагменты антитела включают, но не ограничены Fab, Fabʹ, Fabʹ-SH, F(abʹ)2, Fv и scFv фрагменты, и другие фрагменты, описанные ниже. Для обзора определенных фрагментов антител, см. Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134. Для обзора scFv фрагментов, см., например, Plueckthun, Α., In: The Pharmacology of Monoclonal Antibodies, Vol.113, Rosenburg and Moore (eds.), Springer-Verlag, New York (1994), pp.269-315; см., также WO 1993/16185; и US 5,571,894 и US 5,587,458. Для обсуждения Fab и F(abʹ)2 фрагментов, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих повышенный in vivo срок жизни, см. US 5,869,046.
Диантитела представляют собой фрагменты с двумя антиген-связывающими сайтами, которые могут быть бивалентными или биспецифичными (см. например, ЕР 0404097; WO 1993/01161; Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134; and Holliger, P., et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448). Триантитела и тетраантитела также описаны в Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134).
Однодоменные антитела представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В конкретных воплощениях однодоменное антитело представляет собой однодоменное антитело человека (Domantis, Inc., Waltham, MA; see, e.g., US 6,248,516).
Фрагменты антител могут быть получены при помощи различных техник, включая, но не ограничено, протеолитическое расщепление интактного антитела, а также продукцию рекомбинантных хозяйских клеток (например, Е. coli или фаг), как описано в данном документе.
В конкретных воплощениях антитело представляет собой химерное антитело. Конкретные химерные антитела описаны, например, в US 4,816,567; и Morrison, S.L., et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855. В одном примере химерное антитело содержит нечеловеческий вариабельный участок (например, вариабельный участок, полученный из мыши, крысы, хомяка, кролика или примата, не являющегося человеком, такого как обезьяна) и константный участок человека. В дополнительном примере химерное антитело представляет собой «класс-свич» антитело, в котором класс или подкласс заменен от такового родительского антитела. Химерные антитела включают их антиген-связывающие фрагменты.
В конкретных воплощениях химерное антитело представляет собой гуманизированное антитело. Обычно, нечеловеческое антитело является гуманизированным для уменьшения иммуногенности к людям, в тоже время сохраняя специфичность и аффинность парентерального нечеловеческого антитела. Обычно, гуманизированное антитело содержит один или более вариабельный домен, в котором HVRs, например, CDRs, (или их части) получают из нечеловеческого антитела и FRs (или их части) получают из последовательностей антитела человека. Гуманизированное антитело возможно также будет содержать по меньшей мере часть константного участка человека. В некоторых воплощениях, некоторые остатки FR в гуманизированном антителе замещены соответствующими остатками из нечеловеческого антитела (например, антитело, из которого получают остатки HVR), например, для восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и их способы получения рассмотрены, например, в Almagro, J.С. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633, и кроме того описаны, например, в Riechmann, I., et al., Nature 332 (1988) 323-329; Queen, С, et al., Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033; US 5,821,337, US 7,527,791, US 6,982,321, and US 7,087,409; Kashmiri, S.V., et al., Methods 36 (2005) 25-34 (describing SDR (a-CDR) grafting); Padlan, E.A., Mol. Immunol. 28 (1991) 489-498 (describing "resurfacing"); DallʹAcqua, W.F., et al., Methods 36 (2005) 43-60 (describing "FR shuffling"); and Osbourn, J., et al., Methods 36 (2005) 61-68 and Klimka, Α., et al., Br. J. Cancer 83 (2000) 252-260 (описание подхода «направленной селекции» для FR-перетасовки).
Каркасные участки человека, которые могут быть применены для гуманизации, включают, но не ограничены: каркасные участки, выбранные при помощи способа "наилучшего соответствия" (см., например, Sims, M.J., et al., J. Immunol. 151 (1993) 2296-2308); каркасные участки, полученные из консенсусной последовательности антител человека в частности подгруппы вариабельных участков легкой или тяжелой цепи (см., например, Carter, P., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289; and Presta, L.G., et al., J. Immunol. 151 (1993) 2623-2632); зрелые (соматически мутированные) каркасные участки человека или гаметические каркасные участки человека (см., например, Almagro, J.С. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633); и каркасные участки, полученные в результате скрининга FR-библиотек (см., например, Baca, M., et al., J. Biol. Chem. 272 (1997) 10678-10684 and Rosok, M.J., et al., J. Biol. Chem. 271 (19969 22611-22618).
В конкретных воплощениях антитело представляет собой гуманизированное антитело. Человеческие антитела могут быть получены при помощи различных техник, известных в области техники. Человеческие антитела описаны в целом van Dijk, М.А. and van de Winkel, J.G., Curr. Opin. Pharmacol. 5 (2001) 368-374 and Lonberg, N., Curr. Opin. Immunol. 20 (2008) 450-459.
Человеческие антитела могут быть получены при помощи введения иммуногена трансгенному животному, которое модифицировано для получения интактных человеческих антител или интактных антител с вариабельными участками человека в ответ на антигенный сигнал. Такие животные обычно содержат весь или часть локуса иммуноглобулина человека, который замещает локус эндогенного иммуноглобулина, или который присутствует экстрахромосомно или интегрирован случайным образом в хромосомы животного. В такой трансгенной мыши, локус эндогенного иммуноглобулина обычно инактивирован. Для обзора способов получения человеческих антител из трансгенных животных, см. Lonberg, N., Nat. Biotech. 23 (2005) 1117-1125, а также, например, US 6,075,181 и US 6,150,584, описывающий технологию XENOMOUSE™ US 5,770,429, описывающий технологию HuMab®; US 7,041,870, описывающий технологию K-М MOUSE®, и US 2007/0061900, описывающий технологию VelociMouse®. Человеческие вариабельные участки из интактных антител, полученных посредством таких животных могут быть дополнительно модифицированы, например, за счет комбинирования с различными константными участками человека.
Антитела человека также могут быть получены при помощи способов, основанных на гибридомах. Описаны клеточные линии миеломы человека и гетеромиеломы мыши-человека для получения моноклональных антител человека (см., например, Kozbor, D., J. Immunol. 133 (1984) 3001-3005; Brodeur, B.R., et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York (1987), pp.51-63; and Boerner, P., et al., J. Immunol. 147 (1991) 86-95). Человеческие антитела, получаемые при помощи гибридомной технологии с применением В-клеток человека, также описаны в Li, J., et al., Proc. Natl. Acad. Sci. USA 103 (2006) 3557-3562. Дополнительные способы включают таковые, описанные, например, в US 7,189,826 (описывающем получение моноклональных человеческих антител к IgM из гибридомных клеточных линий) и Ni, J., Xiandai Mianyixue 26 (2006) 265-268 (описывающем гибридомы человеκ-человек). Технология гибридома человека (триомная технология) также описана в Vollmers, H.P. and Brandlein, S., Histology and Histopathology 20 (2005) 927-937 and Vollmers, H.P. and Brandlein, S., Methods and Findings in Experimental and Clinical Pharmacology 27 (2005) 185-191.
Человеческие антитела могут быть получены при помощи выделения Fv клонированной последовательности вариабельного домена и из библиотек фагового отображения человека. Такие последовательности вариабельных доменов затем могут быть скомбинированы с желаемым константным доменом человека. Техники селекции антител человека из библиотек антител описаны ниже.
Антитела могут быть выделены при помощи скрининга комбинированных библиотек антител с желаемой активностью или активностями. Например, разнообразие способов описано в области техники для получения библиотек фагового отображения и скрининга таких библиотек для антител, обладающих желаемыми характеристиками связывания. Такие способы описаны, например, в Hoogenboom, H.R., et al., Methods in Molecular Biology 178 (2001) 1-37 и дополнительно описаны, например, в McCafferty, J., et al., Nature 348 (1990) 552-554; Clackson, T., et al., Nature 352 (1991) 624-628; Marks, J.D., et al., J. Mol. Biol. 222 (1992) 581-597; Marks, J.D. and Bradbury, Α., Methods in Molecular Biology 248 (2003) 161-175; Sidhu, S.S., et al., J. Mol. Biol. 338 (2004) 299-310; Lee, C.V., et al., J. Mol. Biol. 340 (2004) 1073-1093; Fellouse, F.A., Proc. Natl. Acad. Sci. USA 101 (2004) 12467-12472; и Lee, C.V., et al., J. Immunol. Methods 284 (2004) 119-132.
В конкретных способах фагового отображения, наборы генов VH и VL отдельно клонированы при помощи полимеразной цепной реакции (ПЦР) и рекомбинированы случайно в фаговых библиотеках, которые могут быть затем скринированы в отношении антиген-связывающего фага, как описано в Winter, G., et al., Ann. Rev. Immunol. 12 (1994) 433-455. Фаги обычно проявляют фрагменты антитела, или в качестве одноцепочечных Fv (scFv) фрагментов или в качестве Fab-фрагментов. Библиотеки из иммунизированных источников обеспечивают высокоафинные антитела к иммуногену без необходимости построения гибридомов. Альтернативно интактный антигенный репертуар может быть клонирован (например, из человека) для обеспечения единственного источника антител с широким диапазоном неаутоантигенов, а также аутоантигенов без какой-либо иммунизации, как описано Griffiths, A.D., et al., EMBO J. 12 (1993) 725-734. И наконец, интактные библиотеки также могут быть получены синтетически при помощи клонирования неаранжированнных сегментов V-гена из стволовых клеток, и при помощи праймеров ПЦР, содержащих случайную последовательность, для кодирования вариабельных участков CDR3 и выполнения реаранжировки in vitro, как описано Hoogenboom, H.R. and Winter, G., J. Mol. Biol. 227 (1992) 381-388. Патенты, описывающие фаговые библиотеки антител человека, включают, например, US 5,750,373, и US 2005/0079574, US 2005/0119455, US 2005/0266000, US 2007/0117126, US 2007/0160598, US 2007/0237764, US 2007/0292936, и US 2009/0002360.
Антитела и фрагменты антител, изолированные из библиотек антител человека, представляют собой антитела человека или фрагменты антител человека, рассмотренные в данном докмуенте.
В конкретных воплощениях, антитело представляет собой мультиспецифичное антитело, например, биспецифичное антитело. Мультиспецифичные антитела представляют собой моноклональные антитела, обладающие специфичностями связывания с по меньшей мере двумя различными сайтами. В конкретных воплощениях одна из специфичностей связывания представляет собой связывание с первым антигеном, а другая с отличным вторым антигеном. В конкретных воплощениях биспецифичные антитела могут связываться с двумя различными эпитопами одного и того же антигена. Биспецифичные антитела также могут быть применены для локализации цитотоксических агентов в клетках, которые экспрессируют антиген. Биспецифичные антитела могут быть получены в качестве полноразмерных антител или фрагментов антител.
Техники получения мультиспецифичных антител включают, но не ограничены, рекомбинантную соэкспрессию пар тяжелых и легких цепей иммуноглобулина (см. Milstein, С. and Cuello, А.С., Nature 305 (1983) 537-540, WO 93/08829, and Traunecker, Α., et al., EMBO J. 10 (1991) 3655-3659), и "knob-in-hole" инжинирование (см. например, US 5,731,168). Мультиспецифичные антитела также могут быть получены при помощи электростатических взаимодействий для получения Fc-гетеродимерных молекул антител (WO 2009/089004); перекрестного связывания двух или более антител или фрагментов (см., например, US 4,676,980 и Brennan, M., et al., Science 229 (1985) 81-83); при помощи лейциновой «молнии» для получения биспецифичных антител (см., например, Kostelny, S.A., et al., J. Immunol. 148 (1992) 1547-1553; при помощи технологии «диатело» для получения фрагментов биспецифичных антител (см, например, Holliger, Р., et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448); и при помощи одноцепочечных Fv (sFv) димеров (см., например, Gruber, M., et al., J. Immunol. 152 (1994) 5368-5374); и получения триспецифических антител, как описано в Tutt, Α., et al., J. Immunol. 147 (1991) 60-69).
Инжинированные антитела с тремя или более функциональными антигенсвязывающими сайтами, включая «антитела-осьминоги», также включены в данный документ (см., например, US 2006/0025576).
Антитела могут представлять собой «Dual Acting Fab» или «DAF», содержащий антиген связывающий сайт, который связывается с первым антигеном, а также другим отличным антигеном (см., например, US 2008/0069820).
Антитело или фрагмент также может быть мультиспецифичным антителом, как описано в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, WO 2010/112193, WO 2010/115589, WO 2010/136172, WO 2010/145792 или WO 2010/145793.
Способы
В конкретных воплощениях, способы, предложенные в данном документе применяют для изменения, т.е. повышения или понижения, степени, с которой антитело гликозилируют.
В случае, когда антитело содержит Fc-участок, прикрепленный к нему углеводород может быть изменен. Нативные антитела, получаемые в клетках млекопитающих, обычно содержат разветвленный биантенарный олигосахарид, который обычно прикреплен N-связью к Asn297 домена СН2 Fc-участка (см., например, Wright, A. and Morrison, S.L., TIBTECH 15 (1997) 26-32). Олигосахарид может включать различные углеводороды, например, манозу, N-ацетилглюкозамин (GlcNAc), галактозу, и салициловую кислоту, а также фукозу, прикрепленную к GlcNAc в «стебле» биантенарной олигосахаридной структуры. В некоторых воплощениях, модификации олигосахарида в антителе по изобретению могут быть получены с целью создания вариантов антитела с определенными конкретными свойствами.
В одном воплощении предложенный способ приводит к получению антител, обладающих углеводородной структурой, утратившей фукозу, прикрепленную (прямо или непрямо) к Fc-участку. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяют при помощи вычисления среднего количества фукозы внутри сахарной цепи при Asn297, относительно суммы гликоструктур, прикрепленных к Asn 297 (например, комплексные, гибридные и высокоманозные структуры), как измерено при помощи MALDI-TOF масс-спектрометрии, как описано, например, в WO 2008/077546. Asn297 относится к остатку аспарагина, локализованному приблизительно в положении 297 в Fc-участке (EU нумерация согласно Kabat остатков Fc-участка); однако, Asn297 может также быть локализован приблизительно ±3 аминокислоты выше или ниже позиции 297, т.е. между позициями 294 и 300, из-за минорных вариаций последовательности в антителах. Такие варианты фукозилирования могут улучшать ADCC функцию (см., например, US 2003/0157108; US 2004/0093621). Примеры публикаций, относящихся к «дефукозилированным» или «фукозо-дефицитным» вариантам антитела включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; WO2002/031140; Okazaki, Α., et al., J. Mol. Biol. 336 (2004) 1239-1249; Yamane-Ohnuki, N., et al., Biotech. Bioeng. 87 (2004) 614-622. Примеры клеточных линий, способных к продукции дефукозилированных антител, включают Lec13 CHO клетки, дефицитные по фукозилированию белка (Ripka, J., et al., Arch. Biochem. Biophys. 249 (1986) 533-545; US 2003/0157108; и WO 2004/056312, в частности в примере 11), и нокаутные клеточные линии, такие как ген альфа-1,6-фукозилтрансферазы, FUT8, нокаутные клетки СНО (см., например, Yamane-Ohnuki, N., et al., Biotech. Bioeng. 87 (2004) 614-622; Kanda, Y., et al., Biotechnol. Bioeng. 94 (2006) 680-688; и WO 2003/085107).
В конкретных воплощениях предложенные способы могут быть применены для получения антител с бисекционными олигосахаридами, например, в которых биантенарный олигосахарид, прикрепленный к Fc-участку антитела является бисекционным за счет GlcNAc. Такие варианты антител могут обладать пониженным фукозилированием и/или функцией ADCC. Примеры таких вариантов антител описаны, например, в WO 2003/011878; US 6,602,684 и US 2005/0123546. Также могут быть получены варианты антител с по меньшей мере одним остатком галактозы в олигосахариде, прикрепленном к Fc-участку. Такие варианты антител могут обладать улучшенной CDC-функцией. Такие варианты антител описаны, например, в WO 1997/30087; WO 1998/58964 и WO 1999/22764.
Антитела могут быть получены при помощи рекомбинантных способов и композиций, например как описано в US 4,816,567. Нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В дополнительном воплощении предложены один или более векторов (например, экспрессионных векторов), содержащих такую нуклеиновую кислоту. В дополнительном воплощении предложена хозяйская клетка, содержащая такую нуклеиновую кислоту. В одном таком воплощении хозяйская клетка содержит (например, трансформированная при помощи): (1) вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL антитела и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL антитела и второй вектор, содержащий нуклеиновую кислоту, кодируюущю аминокислотную последовательность, содержащую VH антитела. В одном воплощении хозяйская клетка является эукариотической, например, клеткой яичника китайского хомячка (СНО) или лимфоидной клеткой (например, YO, NS0, Sp2/0). В одном воплощении предложен способ получения антитела, где способ содержит культивирование хозяйской клетки, содержащей нуклеиновую кислоту, кодирующую антитело, как предложено выше, при условиях приемлемых для экспрессии антитела и возможно выделения антитела из хозяйской клетки (или среды для культивирования хозяйской клетки).
Для рекомбинантной продукции антитела нуклеиновую кислоту, кодирующую антитело, выделяют и встраивают в один или более векторов для дополнительного клонирования и/или экспрессии в хозяйской клетке. Такая нуклеиновая кислота может быть быстро выделена и секвенирована при помощи обычных способов (например, при помощи олигонуклеотидных проб, способных к специфичному связыванию с генами, кодирующими тяжелую и легкую цепи антитела).
Приемлемые хозяйские клетки для клонирования или экспрессии векторов, кодирующих антитело, включают прокариотические и эукариотические клетки, описанные в данном документе. Например, антитела могут быть получены в бактериях, в частности при необходимости гликозилирования и эффекторной функции Fc-участка. Для экспрессии фрагментов антител и полипептидов в бактериях, см., например, US 5,648,237, US 5,789,199 и US 5,840,523; также см. Charlton, K.A., In: Methods in Molecular Biology, Vol.248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2003), pp.245-254, описывающий экспрессию фрагментов антител в Ε. coli. После экспрессии, антитело может быть выделено из массы бактериальных клеток в растворимой фракции и может быть дополнительно очищено.
Приемлемыми хозяевами для клонирования или экспрессии помимо прокариот являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, включая штаммы грибов и дрожжей, в которых пути гликозилирования были «гуманизированы», что привело к получению антител с частично или полностью гуманизированным паттерном гликозилирования (см. Gerngross, T.U., Nat. Biotech. 22 (2004) 1409-1414; and Li, H., et al., Nat. Biotech. 24 (2006) 210-215).
Приемлемые хозяйские клетки для экспрессии гликозилированных антител также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Идентифицировано множество штаммов бакуловируса, которые могут быть применены совместно с клетками насекомых, в частности для трансфекции клеток Spodoptera frugiperda.
Культуры растительных клеток также могут быть применены в качестве хозяйских (см., например, US 5,959,177, US 6,040,498, US 6,420,548, US 7,125,978 и US 6,417,429 (описывающий технологию PLANTIBODIES™ получения антител в трансгенных растениях)).
Клетки позвоночных также могут быть применены в качестве хозяев. Например, могут быть применены клеточные линии млекопитающих, адаптированные для роста в суспензии. Другие примеры полезных хозяйских клеточных линий млекопитающих представляют собой линию почки макаки CV1, трансформированую SV40 (COS-7); линию почки эмбриона человека (293 или 293 клетки, как описано, например, в Graham, F.L., et al., J. Gen Virol. 36 (1977) 59-74); клетки почки новорожденного хомяка (BHK); клетки Сертоли мыши (ТМ4 клетки, как описано, например, в Mather, J.P., Biol. Reprod. 23 (1980) 243-252); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK; клетки печени серой крысы (BRL 3А); клетки легких человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); TRI клетки, как описано, например, в Mather, J.P., et al., Annals N.Y. Acad. Sci. 383 (1982) 44-68; MRC 5 клетки; и FS4 клетки. Другие полезные клеточные линии млекопитающих включают клетки яичника китайского хомячка (СНО), включая DHFR негативные (DHFR-) СНО клетки (Uriaub, G., et al., Proc. Natl. Acad. Sci. USA 77 (1980) 4216-4220); и клеточные линии миеломы, такие как YO, NS0 и Sp2/0. Для обзора конкретных хозяйских клеточных линий млекопитающих, приемлемых для получения антитела, см., например, Yazaki, P. and Wu, A.M., Methods in Molecular Biology, Vol.248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2004), pp. 255-268.
II. Конкретные аспекты изобретения
Обнаружено, что в зависимости от организации вектора построение экспрессионного вектора отличается для (1) дизайна вектора и (2) идентичных векторов между кратковременной и стабильной трансфекциями.
Обнаружено, что оптимальная организация вектора для кратковременной и стабильной трансфекций может сильно отличаться. Без связи с какой-либо теорией этим различиям может способствовать: 1) феномен транскрипционной интерференции между копиями интегрированного вектора в хозяйском геноме, который зависит от и является специфичным для соответствующего дизайна вектора и который не существует в кратковременной системе, 2) влияние процесса селекции и строгости селекции, которая зависит от соответствующей организации вектора и играет важную роль в стабильности, но не при кратковременной системе, и 3) оптимальное LC-HC полипептидное соотношение, которое явно отличается при кратковременной и стабильной экспрессии моноклональных антител и которая является более низкой при кратковременной, чем при стабильной трансфекций.
Обнаружено, что для кратковременных трансфекций двунаправленная экспрессия LC и НС давала самый высокий титр продукта из всех тестируемых организаций вектора. Без связи с какой-либо теорией предполагается, что дизайн вектора удовлетворяет обоим критериям для оптимальной экспрессии IgG А) высокие уровни экспрессии LC- и НС-полипептидов и В) оптимальное соотношение «кратковременных» LC - НС полипептидов, и С) двунаправленная экспрессия LC и НС так же может минимизировать механизмы транскрипционной интерференции из-за эффектов прочтения РНК-полимеразы.
Обнаружено в отношении стабильных трансфекций, однако, двунаправленная экспрессия LC и НС была хуже, чем при организации в ряд LC-HC-SM. Без связи с данной теорией 1) конвергентная организация экспрессионных кассет для LC, НС и SM может снижать феномен транскрипционной интерференции между интегрированными копиями вектора, 2) LC выше НС заметно усиливает соотношение LC к НС полипептида, которое является (более) оптимальным для стабильных трансфекций, и 3) позиция ниже селекционного маркера заметно увеличивает строгость селекции. Однако обнаружено, что процент клеток, продуцирующих IgG, и продуктивность клеточных линий повышен. Повышение концентрации селекционного давления для векторов, содержащих селекционный маркер в разных направлениях выше экспрессионных кассет с антителом не повышает продуктивность стабильных пулов или клонов хотя строгость селекции заметно повышена (данные не приведены).
Обнаружено, что конкоминантная замена промотора hCMV и поли(А)-сигнала SV40 промотором hEF1α и поли(А)-сигналом bGH существенно повышала титры кратковременных продуктов, в случае, когда кассеты для экспрессии антитела расположены в одном направлении, но понижала титры продуктов, в случае, когда LC- и НС-экспрессионные кассеты расположены в разных направлениях.
Обнаружено, что промотор hEF1α генерирует большое количество хорошо продуцирующих и очень малое число не- или слабопродуцирующих клонов. Однако титры продуктов для лучших отдельных клонов промотора hEF1α при периодическом культивировании с подпиткой были более низкими, чем для клонов промотора hCMV. Но общее число лучших клонов для промотора hCMV является относительно низким и их идентификация обычно требует значительных усилий при скрининге.
Обнаружено, что применение hGT существенно повышало продуктивность векторов, содержащих промотор hCMV при комбинировании с SV40 или bGH поли(А)-сигналом при кратковременной, а также при стабильной трансфекций. Для векторов, содержащих промотер hEf1α, однако, его влияние на титр продукта был незначителен при комбинировании с bGH поли(А)-сигналом. Таким образом, обнаружено, что влияние hGT на свойства вектора зависит от примененного промотора.
Выбор соответствующих клонов для конечной оценки при полностью контролируемых промышленных ферментациях обычно основан на периодическом культивировании и периодическом культивировании с подпиткой в перемешиваемых сосудах. Обнаружено, что присутствуют различия в свойствах и порядке некоторых экспрессионных векторов или элементов между периодическим культивированием и периодическим культивированием с подпиткой. Замещение SV40 поли(А) bGH поли(А)-сигналом повышало продуктивность векторов, содержащих промотор hCMV при периодическом культивировании, но не при периодическом культивировании с подпиткой. hGT не оказывает существенного влияния на титр продукта клонов при периодическом культивировании, но не при периодическом культивировании с подпиткой. Различия между векторами, отличающимися позицией селекционного маркера или промоторами (промотор hEf1α или hCMV) были умеренными при периодическом культивировании, но явно сильными при периодическом культивировании с подпиткой. Экспрессионные уровни и конкретные скорости продукции были выше при периодическом культивировании с подпиткой, чем при периодическом культивировании.
Для большинства клонов обнаружена хорошая корреляция в свойствах при периодическом культивировании с подпиткой и 2L ферментации, не только на уровне абсолютных титров продуктов, но также между различными векторами и клонами.
Обнаружено, что расположение ниже селекционного маркера несколько снижало потерю продуктивности по сравнению с двунаправленным расположением селекционного маркера выше кассет для экспрессии антитела. Без связи с теорией, это может быть обусловлено повышенной строгостью селекции и таким образом более высоким уровнем мРНК или улучшенным соотношением LC-НС мРНК или полипептида. Оба фактора могут приводить к более высокой толерантности в отношении изменений в продуктивности.
bGH поли(А)-сигнал существенно понижал стабильность экспрессии антитела в клонах по сравнению с SV40 поли(А)-сигналом. Однако инсерция hGT ниже bGH поли(А)-сигнала заметно увеличивала стабильность экспрессии. Положительный эффект hGT на стабильность был наиболее заметен при отсутствии селекционного давления.
Анализ стабильности стабильных пулов выявил, что клетки быстро теряли продуктивность в случае промотора hCMV, но не в случае с промотором hEf1α. Неожиданно, хотя hGT понижал продуктивность клонов для векторов, содержащих промотор hEf1α, он незначительно повышал их стабильность.
Только небольшая часть клонов существенно продуцирует антитела после процесса селекции. Однако модификации в организации вектора и/или элементах существенно повышала соотношение IgG продуцирующих и не продуцирующих клонов. Различные организации вектора и таким образом различные уровни экспресии селекционного маркера, определяющего строгость селекции, также явно влияют на процент клеток, продуцирующих IgG. Статистические моделирования, основанные на данных процесса скрининга, демонстрировали, что некоторые экспрессионные вектора также несут потенциал для снижения рабочей нагрузки в течение процесса скрининга значительно. Данный факт оказывает важное влияние на стоимость для биофармацевтических компаний.
Улучшение транскрипционных процессов
Инициация транскрипции при помощи промотора
Промотор определяет уровень транскрипции гена и таким образом оказывает сильное влияние на продуктивность и стабильность клеточных линий:
- инициация транскрипции, опосредованная промотором, определяет количество мРНК, которая может быть транслирована в рекомбинантный белок;
- несмотря на стабильную интеграцию длительная продуктивность клонов может быстро снижаться (как результат сайленсинга генов (например, при помощи метилирования промотора));
- активность промотора зависит от типа клеток.
Описаны промоторы, проявившие себя сильными промоторами в различных клеточных линиях. Известно, что интроны могут содержать энхансеро-подобные элементы. Часто первый интрон (интрон А) содержит большинство из регуляторных элементов. Данный интрон А представляет собой первый интрон внутри последовательности/организации полноразмерного естественного промотора.
Полиаденилирование мРНК, опосредованное поли(А)-сигналом
Полиаденилирование мРНК имеет различные функции. Оно сильно влияет на экспорт мРНК из ядра, инициацию трансляции и стабильность мРНК. Эффективность процесса полиаденилирования таким образом оказывает сильное влияние на уровень экспрессии гена и таким образом на продуктивность.
Некоторые публикации показали, что сигнал полиаденилирования может оказывать сильное влияние на экспрессию белка. Описано, что поли(А)-сигнал коровьего гормона роста (bGH поли(А)-сигнал), может приводить к повышенной экспрессии белка замещения SV40 поли(А)-сигнала.
Обнаружено, что поли(А)-сигнал коровьего гормона роста может обладать улучшенными свойствами при рекомбинантном получении цепей антител.
Терминация транскрипции, опосредованная терминатором транскрипции.
Элементы, содержащиеся в экспрессионном векторе, могут сильно интерферировать друг с другом (транскрипционная интерференция). Это обусловлено конкуренцией при процессе транскрипции. Например, субоптимальным присоединением к промотору транскрипционных факторов и/или РНК-полимеразы из-за стерических ограничений и пониженной доступности транскрипционного аппарата. Интерференция также может возникать между различными близкорасположенными экспрессионными кассетами. Например, при прочтении РНК-полимеразой с первой по вторую экспрессионную единицу может возникать интерференция в результате неэффективной терминации транскрипции.
Описано, что неэффективная терминация транскрипции может приводить к транскрипционной интерференции, которая может приводить к неэффективной экспрессии гена. Применение терминатора транскрипции гена гастрина человека (hGT) может улучшить терминацию транскрипции и таким образом способствовать усилению экспрессии рекомбинантных белков. Этот эффект зависит от промотора, с которым скомбинирован hGT. Известны дополнительные эффективные терминаторы транскрипции.
Обнаружено, что посредством инсерции последовательности терминатора транскрипции гена гастрина человека после раннего SV40 поли(А)-сигнала кассеты, экспрессирующей антитело, может быть достигнута повышенная терминация транскрипции и предотвращение транскрипционной интерференции между экспрессионными кассетами, например, легкой цепи антитела и тяжелой цепи антитела.
Результаты
В данном документе описаны экспрессионные вектора для усиления экспрессии одной или более кодирующих нуклеиновых кислот, например, структурных генов, кодирующих цепи антитела.
Обнаружено, что продуктивность рекомбинантной клетки, экспрессирующей структурный ген, улучшена в случае, когда генетические элементы, необходимые для транскрипции, выбраны и расположены соответствующим образом.
- поли(А)-сигнальная последовательность и последовательность терминатора транскрипции
В векторе р5068 экспрессионные кассеты терминированы при помощи SV40 поли(А)-сигнала.
В векторе рх6001 экспрессионные кассеты содержали терминатор транскрипции гастрина человека (hGT), помещенный ниже SV40 поли(А)-сигнала.
В векторе рх6007 экспрессионные кассеты терминированы при помощи bGH поли(А)-сигнала и терминатора транскрипции hGT.
В векторе рх6008 экспрессионные кассеты терминированы при помощи bGH поли(А)-сигнала без дополнительного терминатора транскрипции.
Данные вектора кратковременно трансфецировали в CHO-K1 клетки и спустя шесть дней после трансфекции супернатанты клеточной культуры собирали и количество продуцированного антитела в культуральном супернатанте определяли при помощи ELISA.
Замещение SV40 поли(А)-сигнала bGH поли(А)-сигналом и добавление/инсерция терминатора гастрина человека (hGT) после SV40 поли(А)-сигнала (вектор рх6001) приводило к повышению продуктивности в кратковременной экспрессии приблизительно 45%-30% по сравнению с контрольным вектором р5068. Комбинация bGH поли(А)-сигнала и терминатора гастрина человека (hGT) приводила к самому большому увеличению титра (+58%) по сравнению с контрольным вектором р5068.
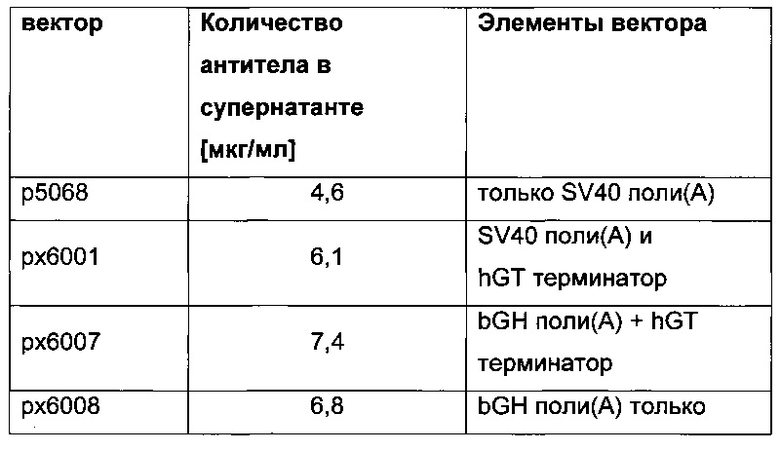
Таким образом, обнаружено, что в кратковременных экспрессионных системах полиаденилирование при помощи bGH поли(А)-сигнала и улучшение терминации транскрипции при помощи добавления терминатора транскрипции гастрина человека увеличивает секрецию антитела.
Вектора р5068, px6001, px6007 и px6008 применяли для получения клеточных линий, стабильно экспрессирующих антитела.
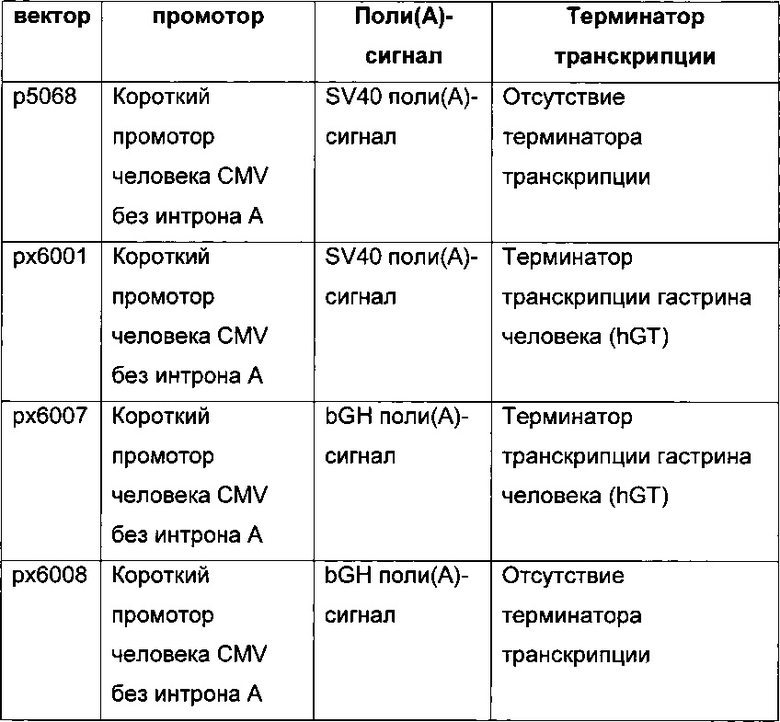
Средняя продуктивность лучших 15 единичных клонов, полученных при помощи вектора р5068 в серии продукции антител составляла 624 мкг/мл. Клоны, полученные при помощи вектора px6001 (содержащего терминатор транскрипции (hGT)) или вектора px6007 (содержащего комбинацию bGH поли(А)-сигнала и hGT) имели продуктивность, повышенную на 23% и 40%, соответственно (770 мкг/мл для вектора px6001 и 872 мкг/мл для вектора рх6007) по сравнению с вектором p5068 (см. Фигура 1). Данное увеличение продуктивности также отражено на уровне лучших клонов (+23% для вектора рх6001 (939 мкг/мл) и +31% для вектора px6007 (1001 мкг/мл); значения вычисляли для трех лучших клонов, полученных при помощи каждого вектора).
Клоны, полученные при помощи вектора px6008, имели среднюю продуктивность 15 лучших клонов 576 мкг/мл.
Средняя продуктивность трех лучших клонов для вектора рх6008 составляет 760 мкг/мл.
- последовательность промотора
Экспрессионные кассеты содержали px6051 полноразмерный промотор человека CMV с интроном А и SV40 поли(А)-сигналом.
В векторе рх6052 экспрессионные кассеты содержали Ef1α и SV40 поли(А)-сигнал.
В векторе px6062 экспрессионные кассеты содержали полноразмерный промотор человека CMV с интроном А и bGH поли(А)-сигнал и терминатор гастрина человека.
В векторе px6063 экспрессионные кассеты содержали промотор человека Ef1α и bGH поли(А)-сигнал и терминатор гастрина человека.
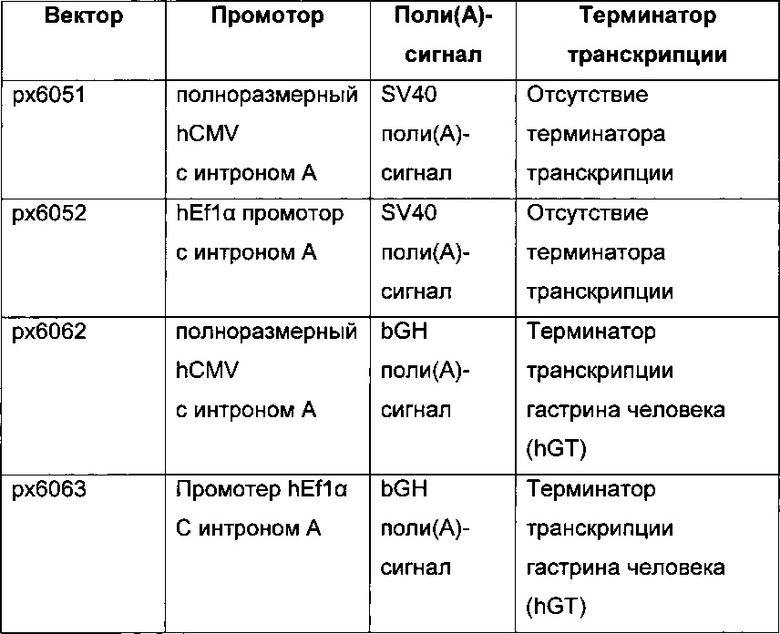
Идентифицировали лучшие 18 стабильных клонов, полученных для каждого вектора, и тестировали продуктивность клонов при периодическом культивировании.
Замена SV40 поли(А)-сигнала на комбинацию bGH поли(А)-сигнала и hGT повышает среднюю продуктивность: в случае промотора hEF1α приблизительно на 19% (505 мкг/мл для вектора px6063 по сравнению с 426 мкг/мл для вектора 6052) и в случае полноразмерного промотора hCMV на приблизительно 71% (333 мкг/мл для вектора рх6062 по сравнению с 195 мкг/мл вектора рх6051).
Повышение продуктивности может также наблюдаться на уровне лучших клонов (+36% для вектора px6063 (693 мкг/мл) по сравнению с вектором рх6052 (511 мкг/мл) и +102% для вектора рх6062 (529 мкг/мл) по сравнению с вектором рх6051 (262 мкг/мл), значения вычисляли для трех лучших клонов каждого вектора) (см. фигура 2).
Таким образом, обнаружено, что комбинация промотора hEF1alpha с bGH поли(А)-сигналом и hGT увеличивает продуктивность при стабильной трансфекции при помощи двунаправленной организации вектора с экспрессионной кассетой для селекционного маркера в одном направлении транскрипции и экспрессионные кассеты для легкой цепи выше экспрессионной кассеты для тяжелой цепи соответственно в другом направлении транскрипции (см. рх6052).
Кроме того, обнаружено, что комбинация bGH поли(А)-сигнала и hGT повышает продуктивность при стабильной трансфекции для промотора hCMV при помощи или двунаправленной векторной организации с экспрессионной кассетой для легкой цепи выше экспрессионной кассеты для тяжелой цепи, которая в свою очередь локализована выше экспрессионной кассеты с селекционным маркером (5ʹ->3ʹ ориентация LC-HC-SM экспрессионных кассет) (см. рх9005), или двунаправленной организации вектора с экспрессионной кассетой для селекционного маркера в одном направлении транскрипции и экспрессионной кассеты для легкой цепи выше экспрессионной кассеты для тяжелой цепи в соответственно другом направлении транскрипции (см. рх6007).
Комбинацию bGH поли(А)-сигнала и hGT применяли при способе кратковременной трансфекции. Вектора рх6062 и px6063 (содержащие полноразмерный промотор hCMV или hEF1α с комбинацией bGH поли(А)-сигнала и hGT) непосредственно сравнивали с векторами рх6051 и рх6052 (содержащими полноразмерный промотор hCMV или hEF1α с SV40 поли(А)-сигналом).
По сравнению со стабильными трансфекциями комбинация bGH поли(А)-сигнала и терминатора гастрина человека (hGT) не увеличивает продуктивность при кратковременных трансфекциях, ни в случае полноразмерного промотора CMV с интроном А ни в случае с промотором человека EF1α с интроном А (2,38 мкг/мл для вектора px6052 по сравнению с 2,32 мкг/мл для вектора px6063). В случае полноразмерного промотора человека CMV с интроном А комбинация bGH поли(А)-сигнала и терминатора гастрина человека (hGT) приводила к продуктивности 2,74 мкг/мл для вектора рх6062А и 3,64 мкг/мл для вектора px6051.
В клетках, полученных при помощи кратковременной трансфекции, обнаружено, что комбинация короткого промотора hCMV с последовательностью поли(А)-сигнала коровьего гормона роста и терминатором гастрина человека приводит к улучшенному экспрессионному выходу антитела.
В клеточных линиях, полученных при помощи трансфекции и селекции стабильных клеточных клонов обнаружено, что комбинация последовательности поли(А)-сигнала коровьего гормона роста и терминатора гастрина человека независимо примененного промотора приводит к улучшенному экспрессионному выходу антитела.
В векторе р5068 экспрессия генов, кодирующих как легкую (LC), так и тяжелую (НС) цепи антитела, осуществляется при помощи короткого промотора человека CMV (hCMV). Данный промотор активен во множестве типов клеток и обычно его применяют в плазмидах, экспрессирующихся в млекопитающих. Учитывая, что активность промотора сильно зависит от типа клеток и поскольку известно, что промотор hCMV чувствителен к эффектам сайленсинга генов, в клетках СНО-K1 присутствует более сильное метилирование промотора и/или более устойчивые промоторы.
Были сконструированы экспрессионные вектора, содержащие различные промоторы: i) содержащие короткий hCMV без интрона A, ii) полноразмерный промотор hCMV с интроном А, iii) полноразмерный промотор ratCMV с интроном А, и iv) промотор hEF1alpha с интроном А.
Вектора, содержащие полноразмерный промотор hCMV (рх6051), промотор hEf1α (px6052) и промотор крысы CMV (px6053) сконструированы и их применяют для кратковременных и стабильных трансфекций.
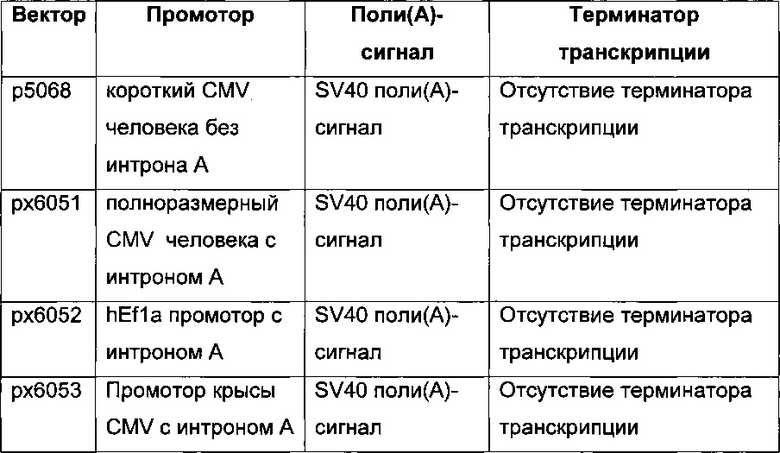
Экспрессионные вектора р5068, рх6051, рх6052 и рх6053 кратковременно трансфецировали в клетки СНО-K1 при помощи нуклеофекции, и спустя четыре дня после трансфекций супернатант культур клеток собирали, и продуктивность определяли при помощи ELISA.
Вектора, содержащие промотор hEF1α с интроном А (рх6052) или полноразмерный промотор hCMV с интроном А (рх6051), имели продуктивность повышенную на 53% и 134%, соответственно, по сравнению с экспрессионным вектором p5068 (3,64 мкг/мл для вектора px6051, 2,38 мкг/мл для вектора px6052, 1,56 мкг/мл для вектора р5068). Вектор px6053, содержащий промотор ratCMV, однако показал продуктивность приблизительно на 50% ниже (0,8 мкг/мл для р5068) (см. фигуру 3).
Вектора, содержащие полноразмерный промотор человека CMV (вектор рх6051 и рх6062) или промотор hEF1alpha с интроном А (вектора рх6052 и px6063) также применяют для стабильных трансфекций.
Экспрессионные вектора р5068, рх6051, рх6052 и рх6053 трансфецировали в клетки СНО-K1 при помощи нуклеофекции и отбирали стабильные пулы. Продуктивность пулов анализировали при периодическом культивировании.
Анализ при периодическом культивировании стабильных пулов показал, что титры антител из клеток, трансфецированных вектором, содержащим hEF1alpha промотор с интроном А (вектор рх6052), были более чем в 4 раза выше, чем таковые клеток, трансфецированных вектором, содержащим короткий промотор hCMV (вектор р5068) или вектором, содержащим промотор hCMV с интроном А (вектор рх6051) (78 мкг/мл для вектора рх6052, 14 мкг/мл для вектора р5068, 17 мкг/мл для вектора рх6051, 8,46 мкг/мл для вектора рх6053) (фигура 4).
При применении экспрессионного вектора р5068, рх6051 и рх6052 получали стабильные клоны. Кроме того применяли вектора рх6062 и рх6063 (оба содержащие комбинацию bGH поли(А)-сигнала и терминатор гастрина человека вместо SV40 поли(А)-сигнала).
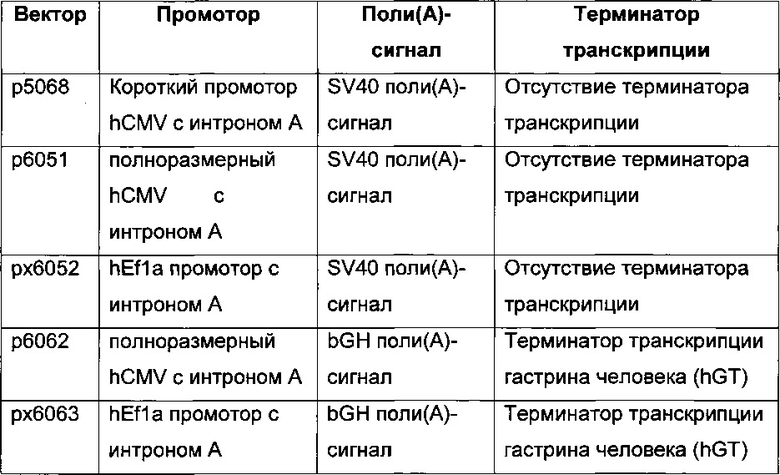
Анализ периодического культивирования лучших 54 клонов, полученных для каждого вектора, показал, что средняя продуктивность клеток, трансфецированных вектором рх6052 составляла 316 мкг/мл, и для вектора р5068 341 мкг/мл. Клоны, полученные при помощи векторов рх6051 и рх6062, содержащих полноразмерный промотор hCMV, имели среднюю продуктивность 141 мкг/мл для вектора рх6051 и 220 мкг/мл для вектора рх6062.
Средняя продуктивность 54 тестированных клонов, трансфецированных вектором px6063 (содержащим промотор hEF1alpha в сочетании с bGH поли(А)-сигналом и hGT) при периодическом культивировании составила 367 мкг/мл. Лучшие клоны с наиболее высоким титром получают из вектора px6063 (748 и 731 мкг/мл, соответственно).
Обнаружено, что клоны, полученные при помощи вектора, содержащего промотор EF1α человека (рх6052 и px6063), проявляют пониженное число низкопродуцирующих клонов.
Клоны тестировали на стабильность продукции антитела. Двенадцать клонов, полученных при помощи векторов р5068 и рх6052, культивировали в присутствии и отсутствии Гигромицина B в течение трех пассажей.
В отсутствии селекционного давления (гигромицин В) в течение трех пассажей, наблюдали 35% снижение средней продуктивности 12 тестированных клонов для короткого промотора hCMV, утратившего интрон А (р5068; 264 мкг/мл по сравнению с 410 мкг/мл). Для сравнения только 18% снижение средней продуктивности наблюдали для промотора hEF1-alpha с интроном А (рх6052; 256 мкг/мл по сравнению с 312 мкг/мл).
В присутствии селекционного давления клоны, полученные при помощи вектора р5068, показали снижение средней продуктивности на 27% (298 мкг/мл по сравнению с 410 мкг/мл) и титры клонов, полученных при помощи вектора рх6052, содержащего промотор hEF1-alpha с интроном А, снижались на 15% (266 мкг/мл по сравнению с 312 мкг/мл).
Определяли число стабильных клонов, трансфецированных при помощи вектора р5068, содержащего короткий промотор hCMV, утративший интрон А, или при помощи вектора рх6052, содержащего промотор hEF1-alpha с интроном А, после трех пассажей с или без селекционного давления. Для определения того, будет ли клон «стабильным» после трех пассажей, титры IgG из пассажа ноль считали за 100% и 80% считали пороговым значением. Клоны, проявляющие относительные титры IgG после трех пассажей с или без селекционного давления ниже 80% титра IgG при пассаже ноль (пассаж 0) определяли как нестабильные.
Относительные титры антител после трех пассажей с или без селекционного давления всех клонов, трансфецированных при помощи р5068, были ниже 80% по сравнению с титром антител пассажа ноль. Стабильный клон клеток представляет собой клональную клеточную популяцию, продуцирующую антитело с титром 80% или более по сравнению с титром антител пассажа ноль в присутствии селекционного давления, а также в отсутствии селекционного давления.
Относительные титры после трех пассажей с или без селекционного давления клонов, трансфецированных рх6052, были ниже 80% по сравнению с титром IgG пассажа ноль в 8 из 12 случаев, но четыре клона были определены как «стабильные». По сравнению с клонами, полученными при помощи вектора р5068, 8 из 12 клонов, полученных при помощи вектора рх6052, были «стабильны» в присутствии селекционного давления. Для сравнения только один клон, полученный при помощи вектора р5068, стабилен в присутствии селекционного давления.
Вектора рх6014, рх6014А и рх6014 В содержат промотор hEF1α впереди от легкой цепи антитела и короткий промотор hCMV впереди от тяжелой цепи. Данные вектора отличаются в 5ʹ UTR легкой цепи (рх6014: 5ʹ UTR промотора hEF1α, содержащего сайт рестрикции Pmel; рх6014А: 5ʹ UTR промотора hEF1α без сайта рестрикции Pmel; рх6014 В: 5ʹ UTR промотора hEF1α плюс 5ʹ UTR вектора р5068). Вектора кратковременно трансфецировали в клетки СНО-K1 при помощи нуклеофекции.
Вектора рх6014А и рх6014 В проявляют повышенную продуктивность на 20% и 40%, соответственно, по сравнению с вектором рх6014 (2,01 мкг/мл для вектора рх6014 В, 1,71 мкг/мл для вектора рх6014А, 1,41 мкг/мл для вектора рх6014).
Обнаружено, что при кратковременных трансфекциях вектора, содержащие промотор EF1α человека или полноразмерный промотор CMV (оба с интроном А), проявляет улучшенную продуктивность по сравнению с вектором р5068.
Обнаружено, что стабильные пулы, полученные при помощи векторов, содержащих промотор EF1α человека, проявляет повышенную продуктивность при периодическом культивировании по сравнению с вектором р5068.
Обнаружено, что клеточные клоны, полученные посредством стабильной трансфекции при помощи векторов, содержащих промотор EF1α человека с интроном А, проявляют пониженное число низкопродуцирующих клонов.
Обнаружено, что клеточные клоны, полученные посредством стабильной трансфекции, при помощи векторов, содержащих полноразмерный промотор CMV человека с интроном А, проявляют сильно сниженную продуктивность как на уровне средних, так и лучших клонов.
Таким образом, обнаружено, что комбинация bGH поли(А)- сигнала и hGT повышает продуктивность по сравнению с SV40 поли(А) при стабильной трансфекции независимо от примененного промотора при помощи двунаправленной организации вектора с экспрессионной кассетой селекционного маркера в одном направлении транскрипции и экспрессионной кассетой легкой цепи выше экспрессионной кассеты тяжелой цепи соответственно в другом направлении транскрипции (см. рх6052).
Кроме того, обнаружено, что комбинация bGH поли(А)-сигнала и hGT увеличивает продуктивность при стабильной трансфекции для промотора hCMV при применении двунаправленной организации вектора с экспрессинной кассетой легкой цепи выше экспрессионной кассеты тяжелой цепи, которая наоборот, локализована выше экспрессионной кассеты селекционного маркера (5ʹ->3ʹ ориентация LC-HC-SM экспрессионных кассет).
Улучшение процесса селекции
Из-за высокого числа непродуцирующих и низкопродуцирующих клонов и низкой стабильности клеточной линии, экспрессирующей ген, разработка в настоящее время является затратной по времени и стоимости. В тысяче клонов обычно вы можете обнаружить только несколько стабильных «высоко продуцирующих».
Без связи с теорией, возможно низкая селективность и строгость селекционной стратегии обусловлена раздельной экспрессией антитела и селекционного маркера в примененной векторной системе, что не оказывает селекционного давления на экспрессию антитела (см., например, вектор р5069).
Обнаружено, что данная проблема может быть преодолена при применении IRES элемента.
IRES элементы представляют собой элементы ДНК, функционирующие на уровне мРНК в качестве сайта внутренней посадки рибосомы, и таким образом делают возможной экспрессию двух генов с одной мРНК.
IRES-связанная экспрессия селекционного маркера и цепи антитела оказывает селекционное давление на общую экспрессию антитела (т.е. одна мРНК, кодирующая как селекционный маркер, так и цепь антитела), и таким образом, обуславливает селективность селекции. Стабильность экспрессии гена может быть повышена.
Применение IRES элемента со слабой активностью IRES делает возможным слабую экспрессию селекционного маркера. Данная слабая экспрессия селекционного маркера может повышать строгость селекции.
Посредством IRES-связанной соэкспрессии селекционного маркера и цепи антитела селекционный процесс может быть улучшен за счет
- повышения числа продуцирующих клеток,
- понижения стабильности экспрессии гена, и
- повышения строгости селекционного процесса.
Для IRES-связанной соэкспрессии антитела и селекционного маркера примененный IRES элемент должен удовлетворять двум требованиям:
- IRES элемент может не оказывать или оказывать только минимальное влияние на стабильность мРНК/экспрессию антитела, и
- IRES элемент должен проявлять слабую активность IRES так, что селекционный маркер экспрессируется очень незначительно.
Легкая и тяжелая цепь, кодирующие нуклеиновую кислоту, связаны несколькими IRES элементами, EV71-IRES, ELF4G-IRES и EMCV-IRES элементами (вектора рх6015А, px6015B и рх6015С). Экспрессионная кассета содержит в направлении 5ʹ-3ʹ промотор CMV человека, легкую цепь, кодирующую нуклеиновую кислоту, IRES, тяжелую цепь, кодирующую нуклеиновую кислоту, и поли(А)-сайт.
Вектор р5068 без IRES элемента в качестве сравнения и вектора рх6015А, рх6015 В и рх6015С, кратковременно трансфецировали в клетки CHO-K1 и продуктивность определяли при помощи ELISA.
Вектора рх6015 В и рх6015С (содержащие ELF4G и EMCV-IRES соответственно) проявляют экспрессию IgG 0,1 мкг/мл - 0,15 мкг/мл. Вектор р5068 проявляет экспрессию IgG 2 мкг/мл. Вектор рх6015А, содержащий EV71-IRES, проявляет продуктивность 1,7 мкг/мл.
Альтернативно селекционный маркер неомицин может быть связан при помощи IRES элемента непосредственно с тяжелой цепью антитела. Вектор содержит в направлении 5ʹ-3ʹ элементы промотора CMV человека, легкую цепь, кодирующую нуклеиновую кислоту, поли(А)-сайт, промотор CMV человека, тяжелую цепь, кодирующую нуклеиновую кислоту, IRES элемент, нуклеиновую кислоту, кодирующую селекционный маркер неомицин и поли(А)- сайт.
Вектора px6010A, px6010B и px6010C кратковременно трансфецировали в клетки СНО-K1, как описано, и продуктивность определяли при помощи ELISA.
Вектор px6010C, содержащий EMCV-IRES, проявляет продуктивность 13 мкг/мл. Вектор р5069 проявляет продуктивность 14 мкг/мл. Вектора, содержащие EV71-IRES (вектор px6010A) или ELF4G-IRES (вектор px6010B) проявляют продуктивность 4,3 мкг/мл и 2,4 мкг/мл соответственно.
Таким образом, обнаружено, что EMCV-IRES элемент удовлетворяет требованиям, необходимым для IRES-связанной экспрессии селекционного маркера:
- слабая активность IRES (так, что селекционный маркер только незначительно экспрессирован)
- при наименьшем влиянии IRES элемента - соответственно IRES-связанной экспрессии селекционного маркера - на экспрессию антитела.
Таким образом, селекционный маркер неомицин (опосредует устойчивость к G418) может быть связан с тяжелой цепью, кодирующей нуклеиновую кислоту при помощи EMCV-IRES.
Вектора р5069 и px6010C тестировали на продуктивность и стабильность при стабильной трансфекций, как на уровне пула, так и на уровне единичных клонов.
Вектора трансфецировали в клетки СНО-K1 при помощи технологии нуклеофекции. Приемлемые пулы отбирали, и продуктивность пулов анализировали при периодическом культивировании.
Пулы, полученные при помощи вектора px6010C, проявляют продуктивность при периодическом культивировании 14 мкг/мл. Пулы, полученные при помощи вектора р5069, проявляют продуктивность при периодическом культивировании 5,4 мкг/мл (фигура 5).
Обнаружено, что пулы, полученные при помощи вектора px6010C, состоят из более продуцирующих клеток (или лучше продуцирующих клеток) и/или стабильность экспрессии IgG повышена (по сравнению с вектором р5069).
Обнаружено, что IRES-связанная экспрессия селекционного маркера неомицина в векторе px6010C также приводит к стабильности пула. Стабильные пулы культивировали в присутствии и в отсутствии селекционного давления в течение 30 поколений. В начале (= поколение 0) и в конце (= поколение 30) теста на стабильность определяли продуктивность пулов при периодическом культивировании.
Пулы, полученные при помощи вектора px6010C, проявляют повышенную стабильность экспрессии IgG при периодическом культивировании по сравнению с пулами, полученными при помощи вектора р5069. Продуктивность пулов, полученных при помощи вектора р5069, при периодическом культивировании сильно снижена (>80%; значения ниже детектируемого предела) после 30 поколений, как в присутствии, так и в отсутствии селекционного давления. Продуктивность пулов, полученных при помощи вектора px6010C, также снижена приблизительно на 70% в отсутствии селекционного давления. Однако в присутствии селекционного давления продуктивность пулов, полученных при помощи вектора px6010C, снижена только на 10% (фигура 6).
Вектора р5069 и px6010C трансфецировали в клетки СНО-K1 при помощи нуклеофекции, и стабильные клоны получали как описано выше. Клоны скринировали, и продуктивность лучших 15 клонов каждой трансфекции анализировали при периодическом культивировании. Проводили две независимых трансфекции каждого вектора.
Клоны, полученные при помощи вектора px6010C, проявляют среднюю продуктивность 348 мкг/мл. Клоны, полученные при помощи вектора р5069, проявляют среднюю продуктивность 239 мкг/мл. Повышенная средняя продуктивность клонов, полученных при помощи вектора px6010C, обусловлена не лучшими клонами, а явно сниженным числом низкопродуцирующих клонов. По сравнению с вектором р5069, клоны, полученные при помощи вектора px6010C, не проявляют продуктивности ниже, чем 200 мкг/мл при периодическом культивировании (фигура 7).
Стабильность экспрессии антитела тестировали для 17 клонов (р5069) и 14 клонов (px6010C), соответственно, при помощи культивирования в присутствии и в отсутствии селекционного давления в течение 45 поколений. Продуктивность клонов измеряли в начале теста на стабильность (поколение 0) и в поколении 45 при периодическом культивировании. Титр IgG поколения 45 сравнивали с продуктивностью клонов в начале теста на стабильность.
Средняя продуктивность 17 клонов, полученных при помощи вектора р5069, снижалась после 45 поколений, как в присутствии, так и в отсутствии селекционного давления (приблизительно 43% потеря продуктивности). Средняя продуктивность 14 клонов, полученных при помощи вектора px6010C, снижалась в отсутствии селекционного давления приблизительно на 45%. Однако в присутствии селекционного маркера снижение продуктивности для данных клонов составляет 4%.
В присутствии селекционного давления только 6 из 17 тестированных клонов, полученных при помощи вектора р5069, проявляют продуктивность, которая все еще составляет свыше 80% продуктивности поколения 0. Для сравнения 10 из 14 тестированных клонов, полученных при помощи вектора px6010C, проявляют продуктивность свыше 80% от продуктивности поколения 0.
В отсутствии селекционного давления нет значительной разницы в стабильности между клонами, полученными при помощи векторов р5069 и px6010C, соответственно.
Улучшение процесса селекции и скрининга
Из-за значительных усилий при скрининге развитие клеточной линии в настоящее время является затратным по времени и стоимости. В тысяче клонах вы обычно найдете только несколько стабильно высоких продуцентов.
В настоящее время существуют несколько стратегий скрининга для идентификации высокопродуцирующих клонов:
- стратегии скрининга, основанные на ELISA, для прямой детекции продуцируемого белка (время и высокая стоимость);
- стратегии скрининга, основанные на флуоресценции для прямой детекции продуцируемого белка (время и стоимость);
- сортинг клеток, основанный на FACS, клеток соэкспрессирующих количественный скрининговый маркер (поверхностный белок или флуоресцентный маркер как GFP) для непрямой детекции/вычисления продуцируемого белка.
В большинстве случаев экспрессия флуоресцентного маркера не связана с экспрессией антитела. Это ограничивает зависимость интенсивности флуоресценции и продуктивности, так как часто отсутствует хорошая корреляция между интенсивностью флуоресценции и продуктивностью.
GFP-белок (как пример флуоресцентного маркера) является стабильным и имеет тенденцию накапливаться. Наблюдается (в большинстве случаев) отсутствие хорошей корреляции между уровнем экспрессии флуоресцентного маркера в клетках (интенсивность флуоресценции) и их продуктивностью.
Обнаружено, что за счет связи экспрессии антитела и экспрессии флуоресцентного маркера, комбинированная стратегия селекции и скрининга для идентификации клеточных клонов, имеющих повышенную стабильность и продуктивность, является простой и делает возможной быструю и простую идентификацию сильных продуцентов.
Обнаружено, что IRES-связанная экспрессия слитого белка действует и как селекционный и как количественный скрининговый маркер.
Сконструирован слитый белок зеленого флуоресцирующего белка (GFP) и селекционный маркер неомицин, который непосредственно связан с тяжелой цепью антитела за счет IRES элемента. Зеленый флуоресцентный белок и селекционный маркер неомицин в слитом белке разделены последовательностью PEST (последовательность, соответствующая кодонам 423-449 гена орнитиндекарбоксилазы мыши (mODC)). Данная последовательность PEST работает на уровне белка в качестве последовательности сильного протеолитического сигнала, таким образом, явно снижая время полужизни белка.
GFP-PEST-Neo слитый белок функционирует не только в качестве селекционного, но также и в качестве количественного скринингового маркера.
Применение последовательности PEST обеспечивает улучшенную селекцию клонов из-за PEST-опосредованного сниженного времени полужизни слитого белка (повышенная строгость селекции).
Применение PEST последовательности обеспечивает хорошую корреляцию между GFP интенсивностью флуоресценции слитого белка и уровнем экспрессии антитела из-за IRES-связанной соэкспрессии антитела и селекционного маркера.
Применение PEST последовательности снижает аккумуляцию слитого белка из-за сниженного времени полужизни белка.
Уровень экспрессии GFP стабильных единичных клонов анализировали при помощи FACS и коррелировали с продуктивностью при периодическом культивировании. Популяции с различными уровнями экспрессии GFP из стабильных пулов сортировали при помощи FACS, и анализировали продуктивность отсортированных популяций.
IRES элементы, которые полностью удовлетворяют требованиям для данного подхода:
- должны обеспечить слабую IRES активность, так чтобы слитый белок был незначительно экспрессирован;
- не должен влиять на уровень экспрессии антитела.
IRES активность и силу определяли за счет связи экспрессии легкой и тяжелой цепи несколькими IRES элементами, EV71-IRES, ELF4G-IRES и EMCV-IRES элементами (см. вектора рх6015А, рх6015В, и рх6015С). Вектор р5068 (без IRES элемента) в качестве сравнения и вектора рх6015А, рх6015В и рх6015С, кратковременно трансфецировали в клетки СНО-K1, и продуктивность измеряли при помощи ELISA.
Вектора рх6015 В и рх6015С (содержащие ELF4G и EMCV-IRES, соответственно) проявляют экспрессию IgG 0,1-0,15 мкг/мл. Экспрессия IgG 2 мкг/мл была показана для вектора р5068. Вектор рх6015А, содержащий EV71-IRES, проявлял продуктивность 1,7 мкг/мл, свидетельствуя, что EV71-IRES элемент обладает сильной IRES активностью в клетках СНО-K1.
GFP-PEST-Neo слитый белок связан при помощи IRES элемента непосредственно с тяжелой цепью, кодирующей нуклеиновую кислоту антитела (см. фигуру 8).
Конструкцию, в которой IRES элемент связывал селекционный маркер с тяжелой цепью антитела, содержащуюся в векторах рх6011А, рх6011 В и рх6011С кратковременно трансфецировали в клетки СНО-K1, как описано, и продуктивность определяли при помощи ELISA.
Вектор рх6011С, содержащий EMCV-IRES, проявляет IgG продуктивность 8,7 мкг/мл и вектор р5059 проявлял IgG продуктивность 10,8 мкг/мл. Вектор рх6011А, содержащий EV71-IRES, проявлял IgG продуктивность 3,9 мкг/мл. Вектор рх6011 В, содержащий ELF4G-IRES, не проявлял значимой продуктивности.
Вектор рх6011С с IRES-связанным GFP-PEST-Neo слитым белком, вектора р5069 и px6010C тестировали при периодическом культивировании стабильных пулов и стабильных клонов.
Вектора р5069, px6010C и рх6011С трансфецировали в клетки СНО-K1 при помощи нуклеофекции. Стабильные пулы выбирали, как описано выше, и продуктивность стабильных пулов анализировали при периодическом культивировании.
Стабильные пулы, полученные при помощи вектора px6010C, содержащего IRES-связанный селекционный маркер, проявляли IgG-продуктивность 9,5 мкг/мл. Стабильные пулы, полученные при помощи вектора р5069, проявляли IgG-продуктивность 5,4 мкг/мл. Пулы, полученные при помощи вектора рх6011С, содержащего IRES-связанный слитый белок проявляли продуктивность 36,3 мкг/мл (фигура 9).
Эти данные подтверждают, что пулы, полученные при помощи векторов px6010C и рх6011С, видимо состоят из намного лучше продуцирующих или даже хорошо продуцирующих клеток, чем пулы (полученные при помощи вектора р5069). Повышенная стабильность экспрессии IgG в пулах px6010C и рх6011С также способствует повышенной продуктивности.
Для сравнения продуктивности экспрессионного вектора р5068 с векторами px6010C и рх6011С в стабильных клонах вектора трансфецировали в клетки СНО-K1 при помощи нуклеофекции. Стабильные клоны выбирали, как описано выше, и скринировали. Продуктивность лучших 15 клонов каждого вектора анализировали при периодическом культивировании.
Для вектора р5068 95 из всех 3072 лунок в 384-луночных планшетах показана продукция IgG свыше 2 мкг/мл (предшествующий уровень техники). IRES-связанная экспрессия селекционного маркера (px6010C) удваивала число продуцирующих клонов/лунок до 195 и в случае IRES-связанного слитого белка (рх6011С) число продуцирующих клеток/лунок в дальнейшем повышалось до свыше 280.
Более того, не только число продуцирующих клонов, но также средняя продуктивность IRES содержащих клонов (полученных при помощи векторов px6010C и рх6011С) составляет выше 3,4 мкг/мл для вектора р5069, 4,2 мкг/мл для вектора px6010C, 8,1 мкг/мл для вектора рх6011С для лучших 100 клонов каждого вектора; 2,2 мг/мл для вектора р5069, 3,0 мкг/мл для вектора px6010C, 5,1 мкг/мл для вектора рх6011С для лучших 250 клонов каждого вектора)).
Клоны, полученные при помощи IRES-векторов px6010C и рх6011С и р5059, проявляют среднюю продуктивность при 24-луночном скрининге 212 мкг/мл для вектора рх6011С, 178 мкг/мл для вектора px6010C и 118 мкг/мл для вектора р5069 (определенную в 24-луночных культурах с неопределенным числом клеток на 4 день после деления клеток). Для векторов px6010C и рх6011С можно было видеть пониженное число непродуцирующих и низкопродуцирующих клонов и повышенное число хорошо продуцирующих клеток (фигура 10).
Анализ при периодическом культивировании единичных клонов показал, что средняя продуктивность 15 лучших клонов, полученных при помощи вектора px6010C, составляет 348 мкг/мл и 239 мкг/мл для вектора р5069. Средняя продуктивность 15 лучших клонов, полученных при помощи вектора px6010C, составляет 404 мкг/мл. Клоны с наивысшим общим титром были получены из трансфектантов при помощи вектора рх6011С. Клоны, полученные при помощи вектора px6010C и вектора рх6011С, не проявляют продуктивности ниже, чем 200 мкг/мл при периодическом культивировании.
Для тестирования клонов на стабильность экспрессии антитела, 14-19 клонов, полученных при помощи каждого вектора, культивировали в присутствии и в отсутствии селекционного давления (G418) для 45 поколений. Продуктивность клонов измеряли в начале теста на стабильность (поколение 0) и в поколении 45 при периодическом культивировании. Титр IgG поколения 45 сравнивали с продуктивностью клонов в начале теста на стабильность в поколении 0 (значения принимали за 100%).
Средняя продуктивность 17 клонов, полученных при помощи вектора р5069, снижалась после 45 поколений, как в присутствии, так и в отсутствии селекционного давления (приблизительно 43% потеря продуктивности). Средняя продуктивность 14-19 клонов, полученных при помощи векторов px6010C и рх6011С, соответственно, также снижалась в отсутствии селекционного давления (приблизительно 45-35%» потеря продуктивности), но в присутствии селекционного маркера G418 снижение средней продуктивности для данных клонов составляет только 0-4% после 45 поколений.
В присутствии селекционного давления G418 6 из 17 тестируемых клонов, полученных при помощи вектора р5069, проявляли продуктивность свыше 80% от продуктивности поколения 0. Для сравнения 10 из 14 тестируемых клонов, полученных при помощи вектора px6010C, и 17 из 19 тестируемых клонов, полученных при помощи вектора рх6011С, проявляли продуктивность свыше 80% от продуктивности поколения 0.
В отсутствии селекционного давления клоны, полученные при помощи векторов р5069, px6010C и рх6011С, проявляют сравнимые характеристики.
Обнаружено, что IRES-содержащие вектора и таким образом, клоны, полученные с их помощью, обладают повышенной стабильностью в присутствии селекционного маркера.
Однако в отсутствии селекционного маркера не существует разницы в стабильности между клонами, полученными при помощи IRES-содержащего вектора или IRES-свободного вектора.
GFP-PEST-Neo слитый белок полезен в качестве количественного скринингового маркера. Уровень флуоресценции GFP клонов, полученных при помощи вектора рх6011С, является прогностическим в отношении их продуктивности.
Уровень экспрессии/интенсивности флуоресценции GFP единичных клонов определяли и результаты коррелировали с продуктивностью клонов при периодическом культивировании, популяции с различными уровнями экспрессии/интенсивности флуоресценции GFP отсортировывали от стабильных пулов при помощи FACS (1,000 клеток каждый вектор) и затем продуктивность данных различных популяций анализировали при периодическом культивировании. Данные различные популяции отсортировывали:
Популяция 1: отсутствие геометрического среднего экспрессии GFP (GM) FL1-H (=GFP)0-4.
Популяция 2: низкий уровень экспрессии GFP, GM 4-5,5.
Популяция 3: высокая экспрессия GFP, GM 5,5-7.
Анализ при периодическом культивировании и FACS-анализ показывают, что как на уровне единичного клона, так и на уровне пула существует значительная корреляция между флуоресценцией GFP клонов/пулов и их продуктивностью. Клоны и пулы с высокой флуоресценцией GFP обычно проявляют более высокую продуктивность, чем клетки, проявляющие низкую флуоресценцию GFP. Вообщем, продуктивность клона/пула возрастает с повышением их интенсивности флуоресценции (фигура 11 и 12).
Высоко продуцирующие клоны могут быть прямо идентифицированы при помощи FACS-сортинга единичных клонов с высокой флуоресценцией GFP из стабильных пулов. Единичные клоны, не проявляющие или проявляющие высокую экспрессию GFP, отсортировывали при помощи FACS. Клоны помещали в покачиваемые 6-луночные планшеты, и продуктивность клонов определяли при периодическом культивировании.
Обнаружено, что EMCV-IRES-связанная экспрессия селекционного маркера или GFP-PEST-Neo слитого белка явно повышает селективность селекции (приводя к более продуцирующим клонам), и таким образом, повышает строгость селекции (средняя продуктивность продуцирующих клонов является более высокой). Таким образом, усилия при скрининге явно снижены.
Более сильный эффект IRES-связанного слитого белка (рх6011С) по сравнению с IRES-связанным селекционным маркером (px6010C) возможно обусловлена последовательностью PEST в слитом белке, которая опосредует сниженное время полужизни слитого белка и/или меньшую аффинность слитого белка для селекционного агента - оба фактора, которые по-видимому повышают строгость селекции. Элементы вектора в комбинации с организацией вектора Следующие вектора тестировали в хозяйских клеточных линиях СНО-K1 при кратковременных трансфекциях, в стабильных пулах и некоторые на уровне единичных клонов.
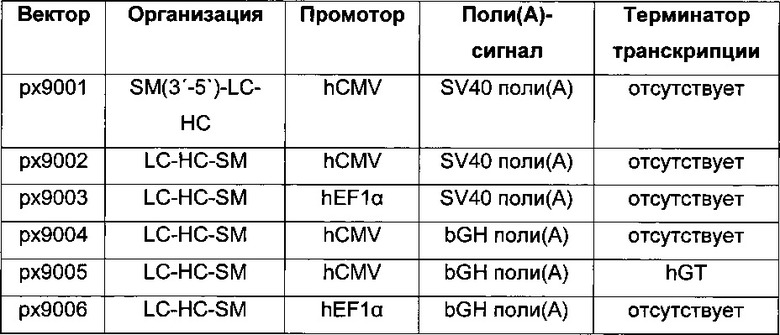

Свойства различных векторов при кратковременных трансфекциях тестировали после нуклеофекции в клетки СНО-K1.
Вектора, содержащие промотор фактора элонгации человека 1 альфа (hEF1α) (на основании организации вектора LC-HC-SM), обладают повышенной продуктивностью приблизительно +34% (рх9003 по сравнению с рх9002; SV40 поли(А)-сигнальная последовательность) и +30% (рх9006 по сравнению с рх9004; bGH поли(А)-сигнальной последовательностью), в зависимости от примененной поли(А)-сигнальной последовательности, соответственно, по сравнению с применением промотора hCMV.
Добавление терминатора гастрина человека (hGT) к bGH поли(А)-сигнальной последовательности оказывает положительное влияние на продуктивность для векторов, содержащих hCMV- (рх9005 по сравнению с рх9004; +13%).
Экспрессионные вектора, основанные на двунаправленной экспрессии легкой и тяжелой цепи антитела, проявляют улучшенные свойства. Титры продукта составляют приблизительно 2,7-3,4-кратное повышение по сравнению с контрольным вектором рх9001 в зависимости от примененного промотора (hEF1α или hCMV) и примененной поли(А)-сигнальной последовательности (SV40 или bGH поли(А)-сигнальная последовательность).
Обнаружено, что применение промотора фактора элонгации человека 1 альфа и bGH поли(А)-сигнальной последовательности оказывает положительное влияние на продуктивность при организации вектора LC-HC-SM (+50%; сравнение рх9006 и рх9002), но не при организации вектора LC(3ʹ-5ʹ)-HC-SM (- 28%; сравнение рх9010 и рх9011).
Для сравнения продуктивности экспрессионного вектора рх9002 с векторами рх9003-9007 вектора трансфецировали в клетки СНО-K1 при помощи нуклеофекции, и стабильные пулы отбирали. Продуктивность пулов анализировали при периодическом культивировании (см. Фигуру 10).
Анализ периодического культивирования стабильных пулов показал, что титры антител пулов, трансфецированных при помощи вектора, содержащего промотор фактора элонгации человека 1 альфа (рх9003, рх9006, рх9007), были приблизительно в 7-8 кратно выше, чем таковые клеток, трансфецированных при помощи вектора сравнения, содержащего короткий промотор hCMV (вектор рх9002) (для сравнения 97,5 мкг/мл, 112,5 мкг/мл и 95,6 мкг/мл и для векторов px9003, рх9006 и рх9007 по сравнению с 14,0 мкг/мл для вектора рх9002.
Вектора рх9001, рх9002 и рх9004-рх9007 трансфецировали в клетки СНО-K1 при помощи нуклеофекции и лучшие единичные клоны идентифицировали при помощи классических способов скрининга. Продуктивность лучших 36 клонов каждого вектора анализировали при периодическом культивировании и лучшие (при периодическом культивировании) 15 клонов каждого вектора тестировали при периодическом культивировании с подпиткой.
Средняя продуктивность лучших 36 единичных клонов, полученных при помощи вектора рх9001, при периодическом культивировании составляет 356 мкг/мл. Клоны, полученные при помощи вектора рх9002 или векторов рх9004 и 9005 (дополнительно содержащих bGH поли(А)-сигнал (один или в сочетании с HGT) вместо SV40 поли(А)-сигнала) проявляют 37% (рх9002) соответственно 61% (рх9004) - 53% (рх9005) повышение продуктивности по сравнению с контрольным вектором рх9001. Клоны вектора рх9006 и рх9007 проявляют повышение в продуктивности приблизительно 19% и 7% соответственно.
В экспериментах по периодическому культивированию с подпиткой лучшие 15 клонов, полученных при помощи каждого вектора при периодическом культивировании, тестировали при периодическом культивировании с подпиткой свыше 14 дней.
Средняя продуктивность лучших единичных 15 клонов (полученных при помощи вектора рх9001), при периодическом культивировании с подпиткой составляет 1345 мкг/мл. Клоны, полученные при помощи вектора рх9002 или при помощи векторов рх9004 и рх9005 (дополнительно, содержащих bGH поли(А)-сигнальную
последовательность (одну или в сочетании с hGT) вместо SV40 поли(А)-сигнальной последовательности), проявляют 80% (рх9002) 58% (рх9004) -92% (рх9005), соответственно, повышение продуктивности по сравнению с контрольным вектором рх9001.
Также помимо средней продуктивности (см. выше) свойства лучших клонов сильно улучшены. Вектора рх9002, рх9004 и рх9005 проявляют - в отношении лучших 5 клонов повышение продуктивности приблизительно 64% (рх9002), 50% (рх9004) и 88% (рх9005) по сравнению с контрольным вектором рх9001.
Непродуцирующие клетки для каждых векторов прямо сравнивали в следующих процентах продуктивности, соответственно. 14,2% клонов, полученных при помощи вектора рх9001, продуцируют антитело, в остальных устойчивых клонах экспрессия антитела или отсутствовала или клоны имели другие дефекты.
Организация вектора рх9002 всегда удваивала процент продуцирующих клеток (26,0%). Вектор, дополнительно содержащий bGH поли(А)-сигнальную последовательность одну вместо SV40 поли(А)-сигнальной последовательности или один (вектор рх9004) или в комбинации с hGT (вектор рх9005) проявляет приблизительно 3-кратное повышение в проценте продуцирующих клеток (39% и 43%, соответственно).
Применение промотора фактора элонгации человека 1 альфа вместо промотора hCMV повышало число продуцирующих клеток 5-кратно (более чем 70% клонов, полученных после способа селекции реально продуцировали антитело).
15 лучших клонов, полученных посредством трансфекции при помощи векторов рх9001-9007 (на основании результатов периодического культивирования с подпиткой), культивировали в присутствии и в отсутствии гигромицина в течение 15 пассажей (= приблизительно 60 поколений). Титры продуктов клонов при периодическом культивировании после 15 пассажей сравнивали с титром продукта клонов при периодическом культивировании в начале теста на стабильность.
В присутствии селекционного давления изменение в титре продукта между 15 клонами каждого вектора варьирует от - 14,7% для вектора рх9007 и +0,2% для вектора рх9002 после 15 пассажей.
В отсутствии селекционного давления снижение титра продукта варьирует от 25,5% для вектора рх9004 до 5,9% для вектора рх9005.
Число клонов, удовлетворяющих определенным критериям стабильности, например >80% титра продукта при периодическом культивировании по сравнению со значениями в начальной точке (G0) как в присутствии, так и в отсутствии селекционного маркера, варьирует от 4 до 10. Вектора рх9005, рх9007 и рх9002 обеспечивают наивысшее число стабильных клонов также в присутствии и в отсутствии селективного давления/селекционного маркера (рх9005: 10; рх9007: 7; рх9002:6)).
Таким образом, обнаружено, что организация векторов рх9005 оказывает положительные эффекты на стабильность и повышение числа стабильных клонов, в частности в отсутствии селекционного маркера.
Обнаружено, что комбинация bGH поли(А) и hGT по сравнению с SV40 поли(А) без терминатора транскрипции (hGT) явно повышает продуктивность стабильных клонов независимо от примененного промотора.
Кратковременные трансфекций:
- применение промотора фактора элонгации человека 1 альфа (с интроном А) обеспечивает повышенную продуктивность (при организации LC-HC-SM)
- применение поли(А)-сигнальной последовательности коровьего гормона роста обеспечивает повышенную продуктивность по сравнению с применением SV40 поли(А)-сигнальной последовательности;
- добавление HGT к bGH поли(А)-сигнальной последовательности приводит к повышенной продуктивности в векторах, содержащих промотор hCMV;
- организация вектора LC(3ʹ-5ʹ)-HC-SM приводит к улучшенной экспрессии.
Стабильные пулы
- пулы, полученные при помощи векторов, содержащих промотор hEF1α, проявляют повышенную продуктивность при периодическом культивировании;
- клоны, полученные при помощи векторов, содержащих промотор hEF1α, проявляют пониженное число низко продуцирующих клонов;
- клоны, полученные при помощи векторов, содержащих промотор hEF1α, проявляют более высокую стабильность экспрессии IgG.
Единичные клоны
- организация вектора с нижним (downstream) расположением селекционного маркера (LC-HC-SM) оказывает положительное влияние на продуктивность стабильных единичных клонов;
- клоны, полученные при помощи векторов, содержащих bGH поли(А)-сигнальную последовательность и hGT, обладают более высокими продуктивностью и стабильностью.
Краткое описание изобретения
Некоторые различные транскрипционно значимые генетические элементы и их комбинации сравнивали с комбинацией генетического элемента сравнения. На основании сравнительных кратковременных экспериментов следующие результаты получали для двунаправленной организации вектора с экспрессионной кассетой селекционного маркера в противоположном направлении по отношению к экспрессионным кассетам легкой и тяжелой цепи (экспрессионная касета легкой цепи выше экспрессионной кассеты тяжелой цепи) (см. таблицу ниже).
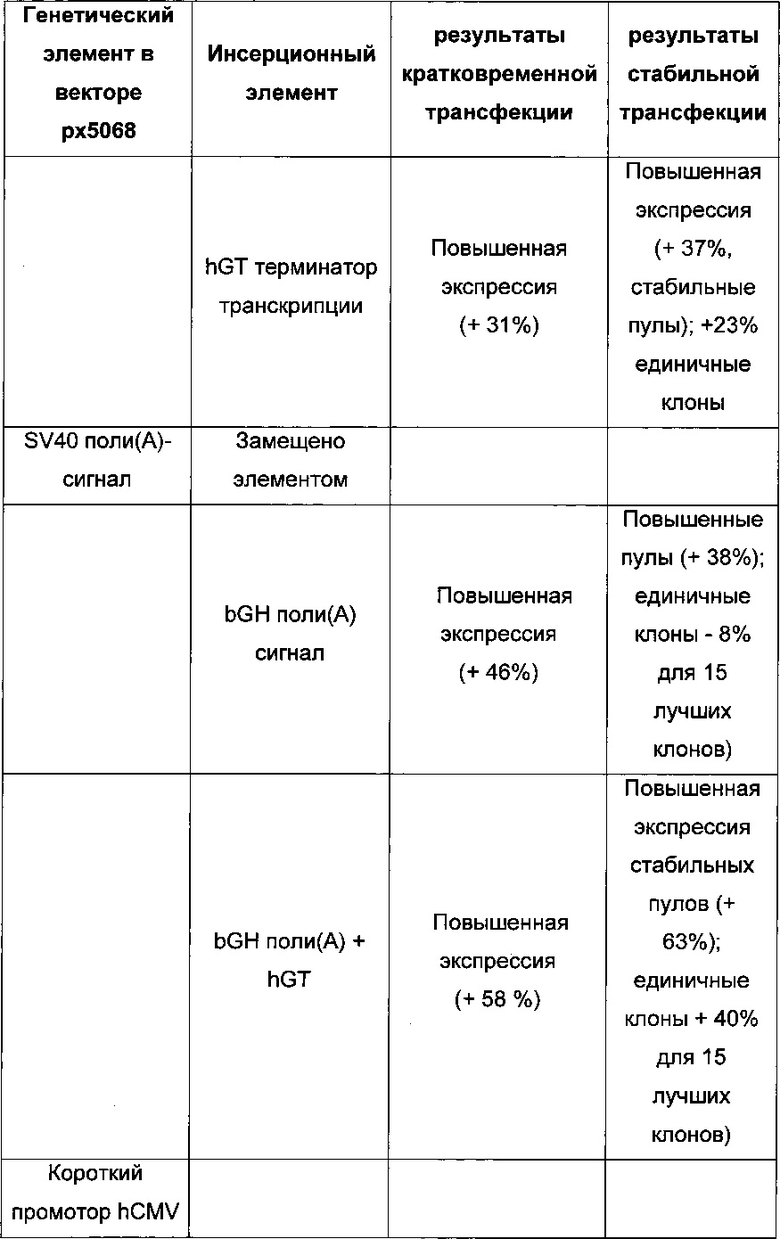
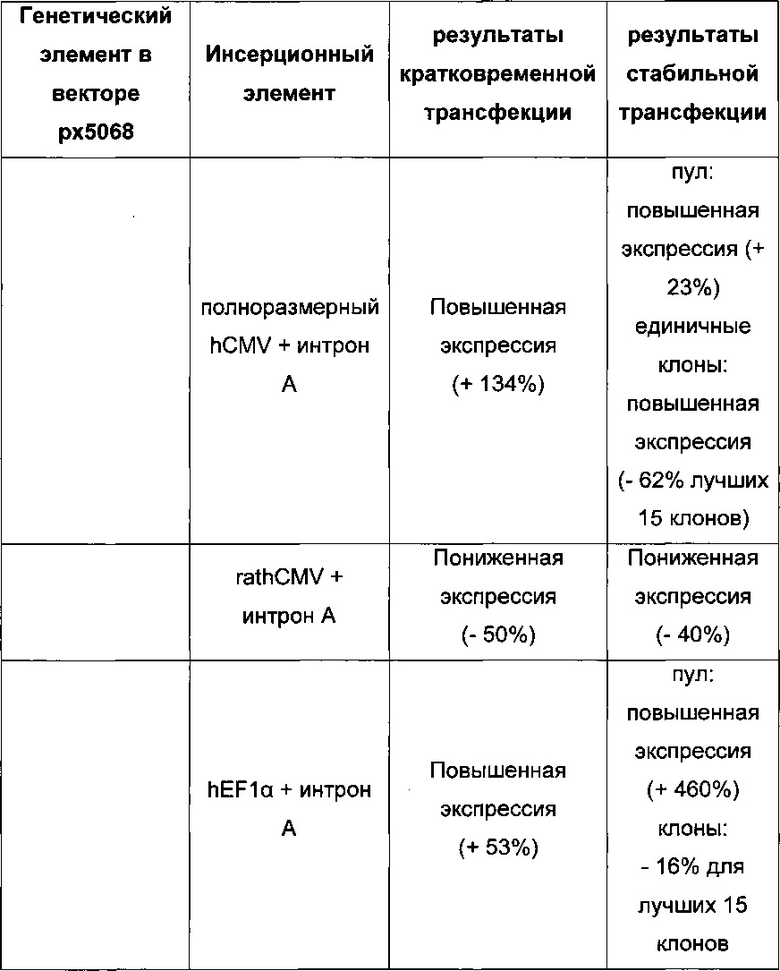
Различные промоторы комбинировали с bGH поли(А)-сигналом и hGT терминатор транскрипции (см. таблицу ниже).

Некоторые транскрипционно связанные генетические элементы и их комбинации сравнивали с вектором сравнения (рх9001, организация вектора SM (3ʹ-5ʹ ориентация)-LC-НС (5ʹ-3ʹ ориентация)). На основании сравнительных экспериментов были получены следующие результаты для однонаправленной организации вектора с экспрессионной кассетой легкой и тяжелой цепи (экспрессионная кассета легкой цепи выше экспрессионной кассеты тяжелой цепи) и селекционного маркера в том же направлении (см. таблицу ниже) по сравнению с вектором сравнения (рх9001, двунаправленная организация вектора SM (3ʹ-5ʹ)-LC-HC (5ʹ-3ʹ)).
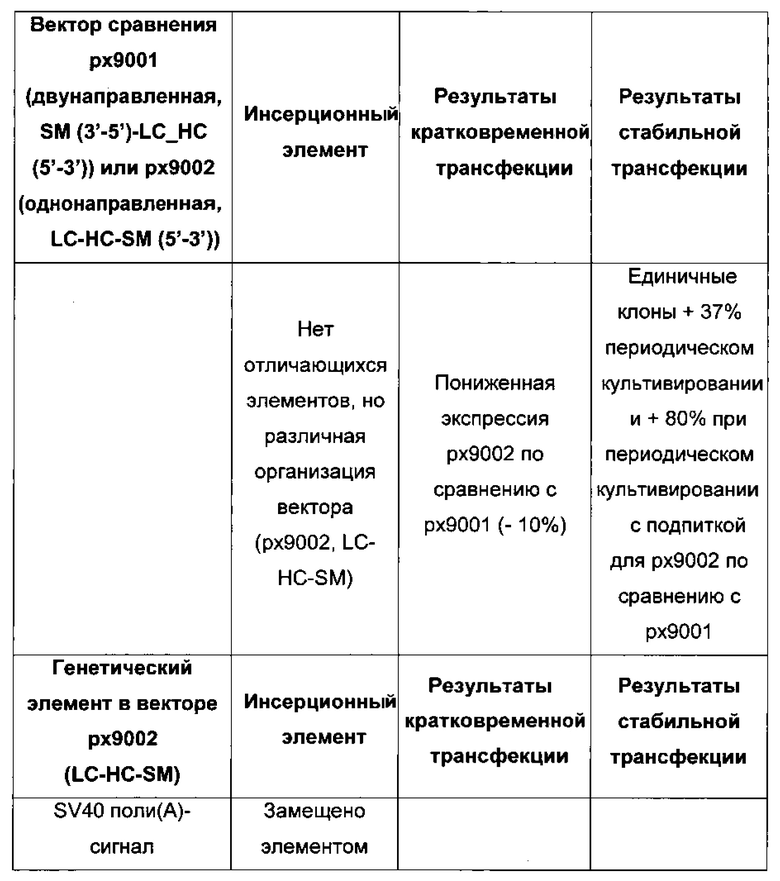
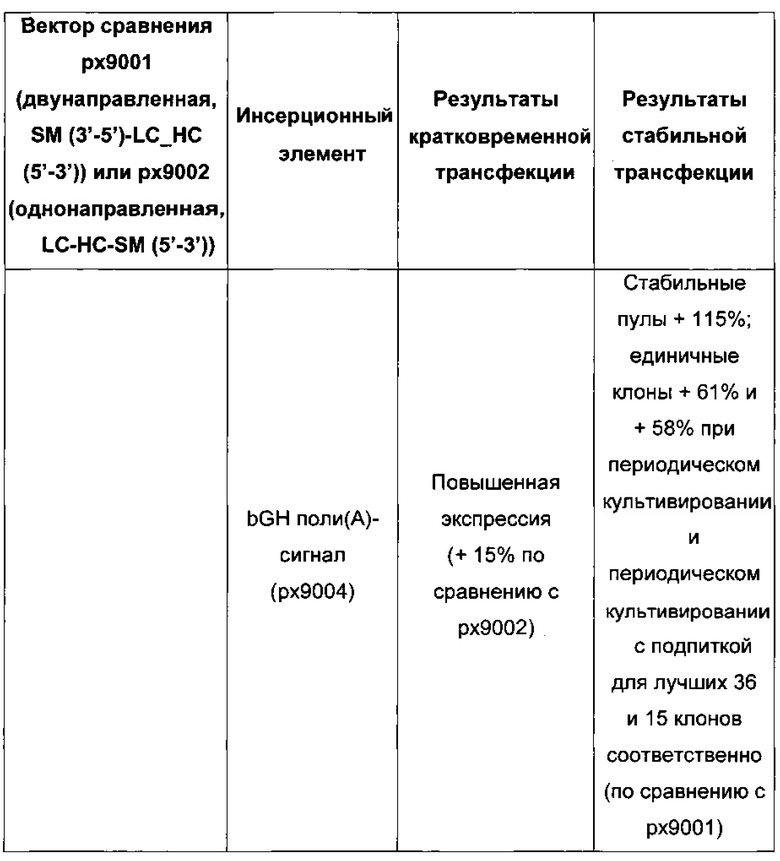
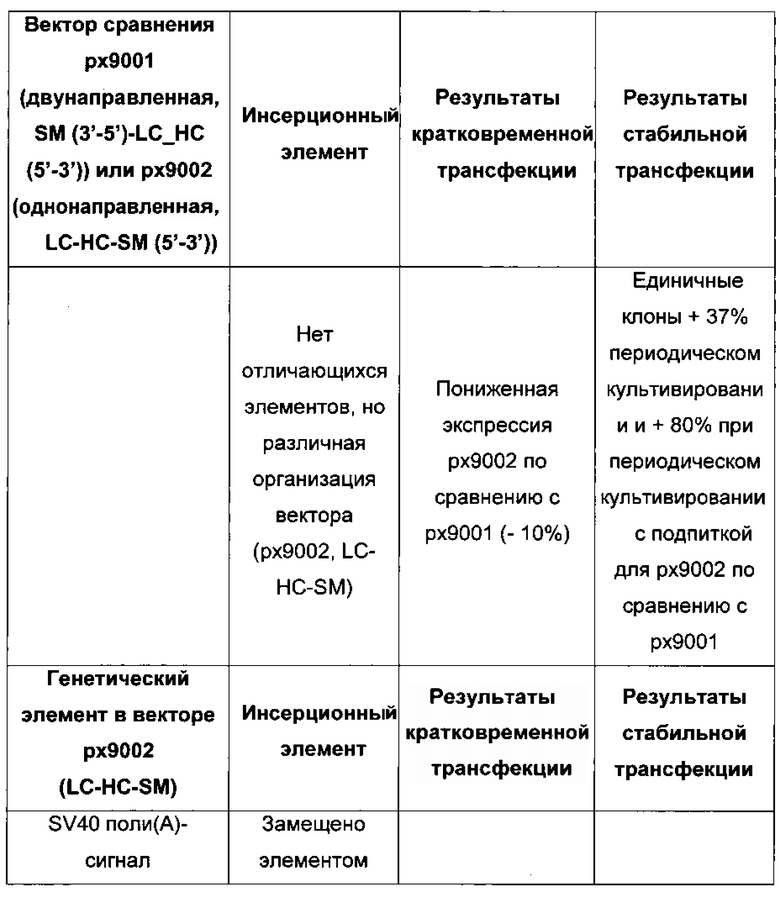
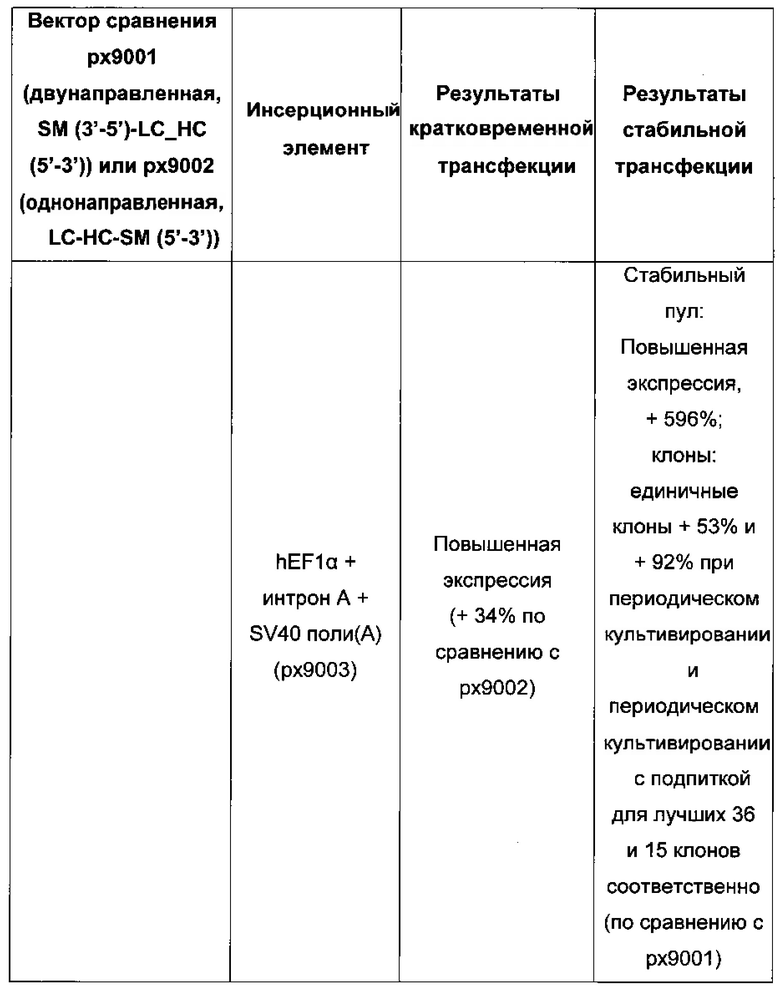
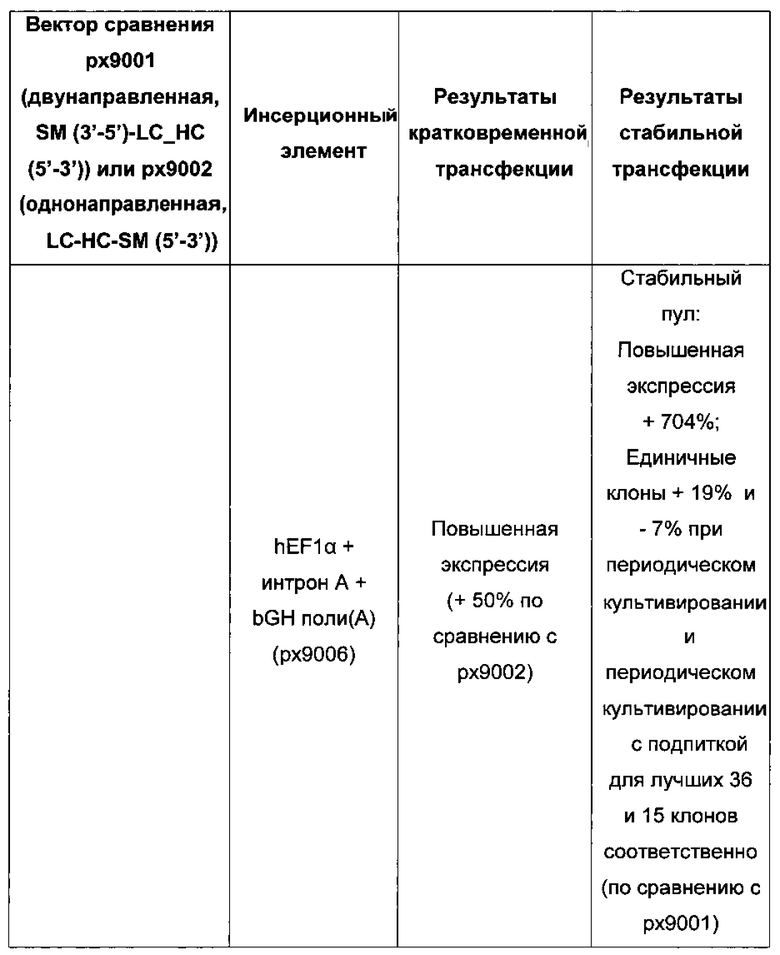
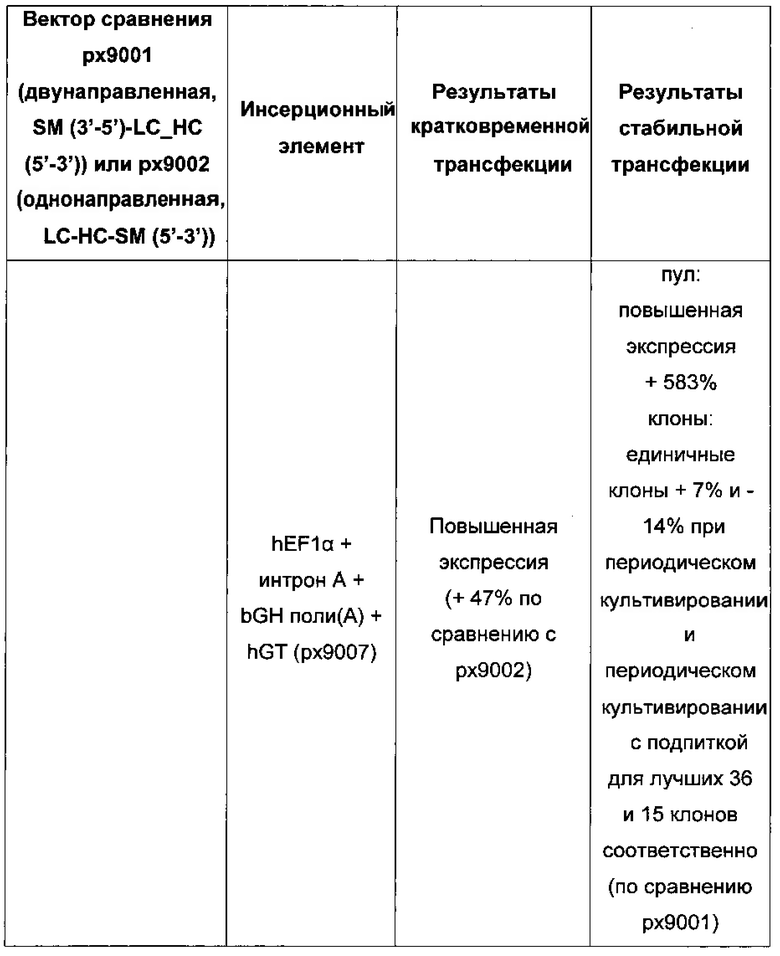
Примененные вектора:
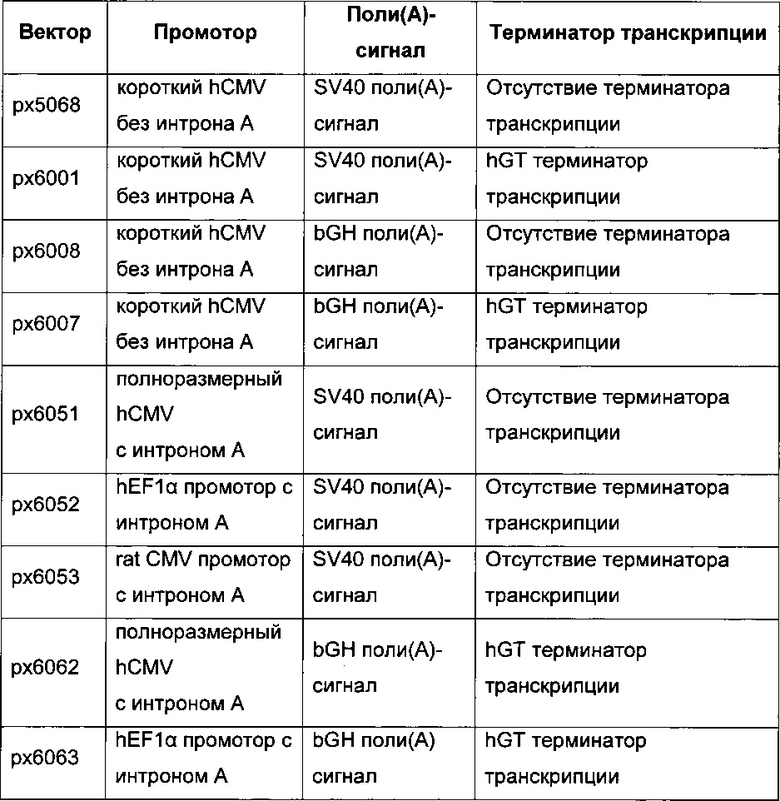
Обнаружено, что повышенная экспрессия (продуктивность) может быть достигнута за счет применения элементов вектора/комбинации элементов, как описано в данном документе:
Промотор человека CMV:
Xu et al., J. Control. Release, 81 (2002) 155-163.
Xia et al., Prot. Expr. Purif. 45 (2006) 115-124.
Промотер крысы CMV:
Xia et al., Prot. Expr. Purif. 45 (2006) 115-124.
Промотер человека EF1α:
Teschendorf et al., Anticancer Res. 22 (2002) 3325-3330.
Li et al., J. Immunol. Methods 318 (2007) 113-124.
MPSV промотор:
Xia et al., Prot. Expr. Purif. 45 (2006) 115-124.
Artelt et al., Gene 68 (1988) 213-219.
Stocking et al., Proc. Natl. Acad. Sci. USA 82 (1985) 5746-5750. Lin et al., Gene 147 (1994) 287-292.
MPSV-CMV гибридный промотор:
Liu et al., Anal. Biochem. 246 (1996) 150-152.
Высокая селективность и высокая строгость способа селекции может быть обеспечена при помощи IRES-связанной экспрессионной кассеты селекционного маркера:
- селекционное давление на экспрессию антитела приводит к высокой селективности;
- связанная экспрессия антитела и селекционного маркера приводит к высокой селективности;
- обнаружено, что применение IRES элемента со слабой активностью приводит к высокой строгости, т.е. высокой продукции антитела и низкой продукции селекционного маркера;
- связывание экспрессии антитела и селекционного маркера посредством IRES элементов;
- идентификация IRES элементов (EMCV/Gtx) со слабой активностью, которая очень мало изменяет экспрессию IgG;
- применение слитого белка, действующего в качестве селекционного и скринингового маркера;
- бифункциональный GFP-неомицин слитый белок;
- PEST последовательность орнитин декарбоксилазы является сильной протеолитической сигнальной последовательностью и способствует сниженному времени полужизни белка;
- IRES-связанная экспрессия слитого белка приводит к высокой селективности;
- короткое время полужизни за счет протеолитической сигнальной последовательности приводит к высокой строгости;
- из-за слабой экспрессии и короткого времени полужизни слитого белка необходима сильная экспрессия;
- быстрая идентификация высоких продуцентов посредстовм FACS (сортинг высоко экспрессирующих GFP клонов обеспечивают селекцию сиьльных продуцентов)
Селекционный маркер, связанный с тяжелой цепью антитела посредством различных IRES элементов
- рх5068 (без IRES): 100% экспрессия антитела (сравнение)
GFP-Neo слитый белок, связанный с тяжелой цепью антитела посредстовм различных элементов IRES
Gtx-IRES:
Komuro et al., EMBO J. 12 (1993) 1387-1401.
EMCV-IRES:
Mountford et al., Proc. Natl. Acad. Sci. USA 91 (1991) 4303-4307.
EV71-IRES:
Lee et al., Biotechnol. Bioeng. 90 (2005) 656-662.
ELF4G-IRES:
Wong et al., Gene Ther. 9 (2002) 337-344.
Gtx (синтетический)-IRES:
Chappell et al., Proc. Natl. Acad. Sci. USA97 (2000) 1536-1541.
Обнаружено, что повышенная экспрессия (продуктивность) с применением связи экспрессионной кассеты легкой цепи с экспрессионной кассетой тяжелой цепи может быть достигнута при помощи EV71-IRES элемента:
Один аспект, как описано в данном документе, представляет собой оптимизированный промотор фактора элонгации человека 1 альфа, содержащий оптимизированный 5ʹUTR, который обладает последовательностью SEQ ID NO: 06.
Следующие примеры, фигуры и последовательности предложены с целью понимания настоящего изобретения, реальный объем которого приведен в прилагаемой формуле изобретения. Следует понимать, что в приведенных способах могут быть сделаны модификации без отклонения от сущности изобретения.
Последовательности
SEQ ID NO: 01 короткий промотор человека CMV без интрона А
SEQ ID NO: 02 короткий промотор человека CMV без интрона А с 5ʹUTR
SEQ ID NO: 03 полноразмерный промотор человека CMV с интроном А
SEQ ID NO: 04 полноразмерный промотор EF1alpha человека без интрона А
SEQ ID NO: 05 полноразмерный промотор EF1alpha с интроном А
SEQ ID NO: 06 короткий промотор EF1alpha человека с интроном А с 5ʹUTR
SEQ ID NO: 07 полноразмерный промотор CMV крысы с интроном А
SEQ ID NO: 08 SV40 поли(А)-сигнальная последовательность
SEQ ID NO: 09 bGH поли(А)-сигнальная последовательность
SEQ ID NO: 10 hGT терминатор транскрипции
SEQ ID NO: 11 SV40 промотор
SEQ ID NO: 12 PEST последовательность орнитин декарбоксилазы
SEQ ID NO: 13 последовательность нуклеиновой кислоты, кодирующая GFP
SEQ ID NO: 14 селекционный маркер неомицин
SEQ ID NO: 15 нуклеиновая кислота, кодирующая слитый полипептид GFP-PEST-NEO
SEQ ID NO: 16 EMCV-IRES
SEQ ID NO: 17 EV71-IRES
Фигуры
Фиг. 1: продуктивность стабильных клонов, полученных при помощи векторов р5068, рх6001, рх6008 и рх6007. Показанное представляет собой среднюю продуктивность лучших 15 клонов каждого вектора при периодическом культивировании всех трех независимых трансфекций.
Фиг. 2: продуктивность стабильных клонов, полученных при помощи векторов рх6051, рх6062, рх6052 и px6063. Показанное представляет собой среднюю продуктивость лучших 18 клонов каждого вектора при периодическом культивировании всех трех независимых тррансфекций.
Фиг. 3: продуктивность векторов р5068, рх6051, рх6052 и рх6053 при кратковременных трансфекциях с применением 96-луночной шатл-системы от Amaxa. Показанное представляет собой среднюю продуктивность восьми независимых трансфекций каждого вектора на 4 день после трансфекций, измеренной при помощи ELISA.
Фиг. 4: продуктивность различных стабильных пулов, полученных при помощи векторов р5068, рх6051, рх6052 и рх6053 при периодическом культивировании. Показанное представляет собой среднюю продуктивность трех пулов каждого вектора на день 7.
Фигура 5: продуктивность стабильных пулов, полученных при помощи векторов р5069 и px6010C. Показанное представляет собой среднюю продуктивность двух (рх5069) относительно трех (px6010C) различных пулов каждого вектора при периодическом культивировании на день 7.
Фиг. 6: стабильность генной экспрессии стабильных пулов, полученных при помощи векторов р5069 и px6010C.Показанное представляет собой среднюю продуктивность двух различных пулов каждого вектора при периодическом культивировании на день 7 пассажа 0 (принято за 100%, черный столбик) и поколении 30 (белый столбик) с и без селекционного (узорчатый столбик) давления (G418).
Фиг. 7: продуктивность лучших 15 клонов, полученных при помощи векторов р5069 и px6010C. Средняя продуктивность лучших 15 клонов каждого вектора при периодическом культивировании всех двух независимых трансфекций.
Фиг. 8: схематическое изображение дизайна вектора рх6011С, опосредующего IRES-опосредованную экспрессию GFP-PEST-NEO слитого белка. GFP-PEST-Neo слитый белок связан посредством EMCV-IRES с тяжелой цепью антитела. Кодирующие последовательности тяжелой цепи антитела и слитый белок транскрибируются с короткого CMV промотора человека с одной мРНК. В результате трансляции данной мРНК образуется тяжелая цепь антитела и GFP-PEST-NEO слитый белок.
Фиг. 9: продуктивность различных стабильных пулов, полученных при помощи векторов р5069, рх6011С и px6010C при периодическом культивировании. Показанное представляет собой среднюю продуктивность двух раличных пулов каждого вектора на день 7.
Фиг. 10: продуктивность лучших 15 клонов, полученных при помощи векторов р5069, px6010C (вектор, экспрессирующий селекционный маркер неомицин за счет EMCV-IRES элемента, связанного с тяжелой цепью антитела) и рх6011С (вектор, экспрессирующий слитый белок GFP-PEST-неомицин за счет EMCV-IRES элемента, связанного с тяжелой цепью антитела). (А) Распределение продуктивности лучших 15 клонов каждого вектора при периодическом культивировании из всех двух независимых трансфекций. (В) Средняя продуктивность лучших 15 клонов каждого вектора при периодическом культивировании всего двух независимых трансфекций.
Фиг. 11: показанное представляет собой зависимость уровня экспрессии/интенсивности флуоресценции GFP и продуктивности при периодическом культивировании для 11 клонов, полученных при помощи вектора рх6011С. Клоны отбирали случайно в 24-луночном формате скрининга, затем наращивали и окончательно анализировали при периодическом культивировании. Геометрическое среднее (ГС) интенсивности флуоресценции GFP и процент GFP-положительных клеток каждого клона определяли при помощи FACS. (А) Зависимость интенсивности флуоресценции GFP и продуктивности при периодическом культивировании для 11 единичных клонов (В). Зависимость процента GFP-позитивных клеток каждого клона и продуктивности при периодическом культивировании для 11 единичных клонов.
Фиг. 12: продуктивность стабильных пулов с различными интенсивностями флуоресценции GFP при периодическом культивировании. Клетки с различными уровнями экспрессии/интенсивностями флуоресценции GFP (низкий (1), средний (2) и высокий (3)) сортировали при помощи FACS. Пулы наращивали и продуктивность пулов определяли при периодическом культивировании на день 7.
Фиг. 13: карта плазмиды рх6007.
Фиг. 14: карта плазмиды рх6053.
Фиг. 15: карта плазмиды рх6062.
Примеры
Экспрессионный вектор р5068 и р5069
Экспрессионные плазмиды р5068 и р5069 содержат экспрессионные кассеты для экспрессии антител к P-селектину (геномно организованная экспрессионная кассета с фиксированной экзон-интронной организацией), как описано в WO 2005/100402.
Гены, кодирующие легкую и тяжелую цепь HuMab антитела к P-селектину, отдельно собирают в экспрессионных векторах клеток млекопитающих.
Таким образом, сегменты гена, кодирующего вариабельный (VL) участок легкой цепи HuMab антитела к P-селектину и константный участок κ-легкой цепи человека (CL) соединяли в качестве сегментов гена для вариабельного участка тяжелой цепи (VH) HuMab антитела к Р-селектину и константного участка γI-тяжелой цепи человека или константного участка γ4-тяжелой цепи человека (CH1-Hinge-CH2-CH3).
Общая информация, касающаяся нуклеотидных последовательностей легкой и тяжелой цепей человека, из которой применение кодона может быть выведено, приведена в: Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication No 91-3242.
Транскрипционная единица κ-легкой цепи HuMab антитела к P-селектину состоит из следующих элементов:
- предранний промотор и промотор цитомегаловируса человека (hCMV),
- синтетическая 5ʹ-UT, включая последовательность Козака,
- сигнальная последовательность тяжелой цепи иммуноглобулина мыши, включая интрон сигнальной последовательности,
- клонированная кДНК вариабельной легкой цепи HuMab антитела к P-селектину, расположенная с уникальным сайтом рестрикции Bsml на 5ʹ-конце и донорный сайт сплайсинга и уникальный сайт рестрикции Notl на 3ʹ-конце,
- константный участок геномного κ-гена человека, включая интрон 2 энхансера Ig-κ мыши (Picard, D., and Schaffner, W. Nature 307 (1984) 80-82), и
- сигнальная последовательность κ-полиаденилирования ("поли(А)") иммуноглобулина человека.
Транскрипционная единица γΙ-тяжелой цепи HuMab антитела к Р-селектину состоит из следующих элементов:
- предранний энхансер и промотор цитомегаловируса человека (hCMV),
- синтетическая 5ʹ-UT, включая последовательность Козака,
- модифицированная сигнальная последовательность тяжелой цепи иммуноглобулина мыши, включая интрон сигнальной последовательности,
- клонированная кДНК вариабельной тяжелой цепи HuMab антитела к Р-селектину, расположенная с уникальным сайтом рестрикции Bsml на 5ʹ-конце и донорный сайт сплайсинга и уникальный сайт рестрикции Notl на 3ʹ-конце,
- константный участок геномного гена γI-тяжелой цепи человека, включая μ-энхансер иммуноглобулина мыши (Neuberger, M.S., EMBO J. 2 (1983) 1373-1378),
- сигнальная последовательность для полиаденилирования ("поли(А)") γI-иммуноглобулина человека.
Помимо экспрессионной кассеты κ-легкой или γI-тяжелой цепи HuMab антитела к Р-селектину данные плазмиды содержат
- ген устойчивости к гигромицину,
- ориджин репликации, oriP, вируса Эбштейн-Барр (EBV),
- ориджин репликации вектора pUC18, который делает возможной репликацию данной плазмиды в Е. coli, и
- ген β-лактамазы, способствующий устойчивости к ампициллину в
Е. coli.
Техники рекомбинации ДНК
Клонирование проводили при помощи стандартных техник клонирования, как описано в Sambrook et al., 1999 (supra). Все молекулярно-биологические реагенты были коммерчески доступны (если не указано иное) и применены согласно инструкциям производителя.
Синтез нуклеиновой кислоты
ДНК различных генетических элементов синтезировали Geneart AG, Regensburg.
Определение последовательности нуклеиновой кислоты
Последовательности ДНК определяли при помощи секвенирования обеих цепей ДНК, выполненного на SequiServe (SequiServe GmbH, Germany).
Анализ последовательности ДНК и белка и обработка данных секвенирования
Vector NTI Advance suite version 9.0 применяли для создания последовательности, картирования, анализа, аннотации и иллюстрации.
Техники культивирования клеток
Клетки СНО-K1 выращивали в среде CD-CHO (Invitrogen Corp., Gibco®, Cat. No. 10743-011), дополненной 1x HT supplement (Invitrogen Corp., Gibco®, Cat. No. 11067-030).
Для селекции стабильно трансфецированных пулов/клеточных линий СНО-K1 добавляли 400-800 мкг/мл G418 или 200-400 мкг/мл гигромицина (Roche Diagnostics GmbH, Roche Applied Sciences, Germany, Cat. No.: 843555).
Все клеточные линии поддерживали во влажной камере при 37°C с 5% CO2 при постоянном покачивании при 120-140 об/мин. Каждый 3-4 день клетки помещали в свежую среду. Плотность и жизнеспособность культур определяли при помощи цитометра Casey ТТ или Cedex Hires (Roche innovates AG, Bielefeld). Трансфекцию клеток проводили при помощи технологии нуклеофекции Amaxa (Lonza GmbH, Germany).
Более того, стандартные техники клеточного культивирования применяли, как описано в Bonifacino, J.S., et al., (eds.), Current Protocols in Cell Biology, John Wiley and Sons, Inc. (2000).
Подсчет клеток и определение жизнеспособности клеток
a) Система подсчета клеток в электрическом поле (CASY)
CASY® Technology Cell Counter, Model TT (Roche Innovatis AG, Bielefeld) использует электрическое напряжение для подсчета клеток. Анализ площади импульса применяли для получения информации исходя из сигналов, созданных при прохождении клеток через измеряющую пору в поле низкого напряжения. Структурная целостность клеточной мембраны является степенью жизнеспособности клетки. Таким образом, для определения жизнеспособности не требуются красители, такие как трипановый синий.
b) Автоматизированный способ вытеснения трипанового синего (Cedex)
Cedex HiRes system (Roche Innovatis AG, Bielefeld) применяли для определения жизнеспособности клеток в течение селекции пулов и для автоматизированного подсчета клеток.
Трипановый синий представляет собой краситель, не способный проникать в клетки с интактной клеточной мембраной. Только клетки с поврежденной клеточной мембраной окрашиваются и учитываются как мертвые. Процесс окраски, подсчет клеток и графический анализ результатов выполняли автоматически при помощи системы Cedex посредством распознования цифрового изображения. Другие измеряемые параметры представляли собой размер клетки, морфологию и скорость аггрегации. При помощи мультисэмплера до 20 образцов измеряли последовательно.
Получение плазмиды и проверка качества для точного сравнения плазмиды при трансфекциях
Эффективность трансфекции и таким образом продуктивность сильно зависит от нескольких факторов, таких как количество и качество ДНК. Для обеспечения эквивалентных начальных условий для каждого вектора количество и качество ДНК всех векторов тщательно проверяли перед трасфекцией.
- одновременное получение экспрессионных векторов
Все вектора одновременно получали при помощи High Speed Maxi plasmid isolation Kit (Qiagen GMBH, Hilden) согласно инструкциям производителя.
- Фенол-хлороформенная очистка и осаждение этанолом
Все вектора одновременно очищали при помощи фенол-хлороформа. 500 мкг каждой линеаризованной плазмидной ДНК смешивали с 200 мкл 50% (о/о) забуференного Трисом фенола, 48% (о/о) хлороформа, 2% (о/о) раствора изоамилового спирта и центрифугировали в течение 1 минуты при 13000 об/мин. Верхнюю водную фазу затем переносили в новые пробирки и смешивали с 200 мкл 96% (о/о) хлороформа, 4% (о/о) изоамилового спирта и центрифугировали в течение 1 минуты при 13000 об/мин. Верхнюю фазу снова переносили в новую пробирку и смешивали с 1/10 (общий объем) ЗМ ацетата натрия (рН 5,2) и 2,5-кратным объемом (общий объем) 100% этанола. Для осаждения ДНК после перемешивания и инкубирования реакции в течение 5 мин при комнатной температуре смеси центрифугировали в течение 5 мин при 13000 об/мин. Супернатант удаляли и осадок промывали 900 мкл 70% (о/о) этанола и инкубировали в течение 5 мин при комнатной температуре. После этапа заключительного центрифугирования на максимальной скорости в течение 5 минут супернатанты удаляли и осадки высушивали и ресуспендировали в стерильной H2O.
- определение ДНК.
Количество ДНК каждого вектора определяли при помощи BioPhotometer (Eppendorf; Hamburg). Измерение ДНК всегда выполняли в трех повторностях при помощи разведения 1:20 в Трисе (Tris рН 8,0).
- Агарозный гель
Качество ДНК каждой плазмиды проверяли в 0,8% агарозном геле. Определяли деградацию ДНК, конформацию векторов и концентрации ДНК. Вектора с сопоставимыми количеством и качеством (отсутствие деградации ДНК, сходная форма суперспирализации (ссс), сходные количества ДНК на геле), применяли для кратковременных и стабильных трансфекций.
Кратковременные трансфекций
Все вектора трансфецировали в клетки CHO-K1 при помощи 96-луночной шатл-системы Amaxa (Lonza GmbH, Germany) согласно инструкции производителя. Каждый вектор трансфецировали в 8 повторностях. Количества ДНК трансфецируемого вектора нормализовали до эквивалентных молярных количеств/числа копий в соответствии с 1 мкг экспрессионной плазмиды сравнения (р5068 или р5069). Для определения продуктивности супернатант клеточной культуры, свободный от клеток, анализировали в отношении титра IgG на день 4-7 после трансфекций при помощи одноэтапоного универсального ELISA (Dianova). 96-луночная шатл-система Amaxa:
Клетки CHO-K1, выращиваемые в роллерных колбах, осаждали центрифугированием при 850 об/мин в течение 5 мин и ресуспендировали в среде для культивирования. Кольцевые плазмиды помещали в 96-луночные планшеты для нуклеофекции в эквимолярных концентрациях в соответствии с 1 мкг экспрессионного вектора сравнения р5068 или р5069. Затем клетки добавляли в планшеты при концентрации 4×105 клеток на лунку. Трансфекцию проводили при помощи Amaxa program DN-137. Клетки инкубировали в течение 10 минут после трансфекций и затем переносили в 96-луночные плоскодонные планшеты для инкубирования, содержащие 200 мкл среды для культивирования. Затем клетки статически культивировали. На день 4-6 после трансфекций уровни IgG определяли при помощи одноэтапного универсального ELISA.
Стабильные трансфекций и получение рекомбинантных клеточных линий СНО
Стабильные трансфекций проводили при помощи технологии нуклеофекции (Amaxa Biosystems, Lonza cologne AG) согласно инструкции производителя. Перед трансфекцией плазмиды линеаризовали при помощи рестриктазы SgrA I. Каждую плазмиду трансфецировали в двух или трех повторностях. 5×106 клеток и 1,2 пкмоль линеаризованной плазмиды применяли для одной трансфекций (Nucleofector Kit Τ, Amaxa program A33).
Для трансфекций клетки ресуспендировали в Nucleofector solution Τ и разделяли на аликвоты по 2 мл пробиркам. После добавления плазмиды трансфекцию проводили при помощи импульса. Затем клетки переносили во флаконы для культивирования Т25, содержащие 4 мл предварительно нагретой свежей среды и 4 мл кондиционированной среды. Селективное давление оказывали спустя 24 часа после трансфекций за счет добавления 250 мкг/мл гигромицина В.
Получение стабильных пулов
Вектора трансфецировали в клетки CHO-K1 при помощи технологии нуклеофекции Amaxa и стабильные пулы отбирали применяя гигромицин В или G418 в качестве селекционного агента. Каждую трансфекцию выполняли в трех повторностях. Для получения стабильных пулов все плазмиды одинаково линеаризовали при помощи рестриктазы SgrA I. Nucleofector Kit Τ от Amaxa применяли для выполнения стабильных трансфекций и каждую плазмиду трансфецировали в трех повторностях.
Стабильные пулы получали следующим образом: 5×106 клеток и 1,2 пкмоль линеаризованной плазмиды применяли для каждой трансфекции. Клетки ресуспендировлаи в растворе Τ и разделяли на аликвоты в 2 мл пробирки. После добавления плазмиды трансфекцию проводили при помощи импульса (Amaxa program А33). Трансфецированные клеточные пулы статически культвировали во флаконах для культивирования тканей Т25, содержащих 4 мл предварительно нагретой свежей среды и 4 мл кондиционированной среды.
Спустя 24 часа после трансфекции оказывали селекционное давление: клетки центрифугировали в течение 5 минут при 800 об/мин, ресуспендировали в 3 мл среды для культивирования, содержащей 300 мкг/мл гигромицина В. Клетки переносили в плоскодонные 6-луночные планшеты спустя 3 дня после трансфекции. Затем клетки культивировали в течение двух недель пока жизнеспособность клеток не снизится до минимума и не поднимится снова свыше 99%. Число клеток и жизнеспособность постоянно определяли при помощи системы Cedex HiRes (Innovatis, Bielefeld). В течение культивирования, клеточный детрит удаляли центрифугированием, и клетки ресуспендировали в 3 мл свежей среды.
Получение стабильных клонов при помощи роботизированной системы Caliper
Вектора трансфецировали в клетки CHO-K1, как описано выше. Спустя 48 часов после трансфекции оказывали селекционное давление (гигромицин В или G418) и клетки засевали в 384-луночные плоскодонные планшеты в концентрации 350-700 клеток на лунку при помощи автоматизированной системы локализации клонов с высокой пропускной способностью (Sciclone ALH 3000 workstation, Caliper Life Sciences GmbH, Mainz).
Спустя 10-14 дней 384-луночные планшеты скринировали в отношении уровней IgG при помощи высокопроизводительного скрининга, основанного на ELISA (ELISA based ultrahigh-throughput screening, ELSIA uHTS). На основании первичного скрининга отбирали клоны с лучшей продуктивностью и переносили в плоскодонные 96-луночные планшеты. Спустя 3-6 дней клетки скринировали в отношении уровней IgG во второй раз. Клоны с лучшей продуктивностью снова отбирали и вручную переносили в плоскодонные 24-луночные планшеты. После дополнительного этапа скрининга, основанного на ELISA, лучшие клоны отбирали и переносили в плоскодонные 6-луночные планшеты. Уровни IgG в 6-луночных планшетах определяли при помощи ProtA измерения идентичности конечных лучших клонов для периодической культуры в 6-луночных планшетах с перемешиванием.
Периодическое культивирование пулов/единичных клонов
С целью детекции различий в продуктивности и стабильности, число клеточных клонов/пулов подсчитывали при помощи счетчика клеток Casey и одновременно засевали в плоскодонные 6-луночные планшеты с концентрацией 3×105 клеток/мл и общим объемом 3,0 мл. Все периодические культуры культивировали в течение 12 дней и супернатанты клеточных культур скринировали в отношении уровней IgG человека на день 4, 7, 9, 11 или 12.
Колличественная оценка IgG
Титр IgG в кратковременных экспериментах и в форматах скрининга (384-24-луночный) определяли при помощи одноэтапного универсального ELISA. Продуктивность стабильных пулов и стабильных единичных клонов при периодическом культивировании определяли при помощи ВЭЖХ белка А.
Одноэтапный универсальный ELISA
Одноэтапный универсальный ELISA (Dianova) применяли для определения уровней IgG человека супернатантов клеточных культур. Стандартную кривую получали при помощи серийных разведений антител к P-селектину (F. Hoffmann-La Roche AG, Basle, Switzerland) с диапазоном 0,3125-20 нг/мл при помощи буфера для разведения (PBS + 5% (м/о) RPLA1). 95 мкл смеси антител, содержащей 0,5 мкг/мл биотинилированных антител к F(abʹ)2 Fc-фрагмента человека (Jackson laboratories) и 0,1 мкг/мл конъюгированных с пероксидазой антител к F(abʹ)2 Fcγ (Jackson laboratories; Suffolk) добавляли к 96-луночному микропланшету, покрытому стрептавидином (StreptaWell, Roche Diagnostics GmbH). 5 мкл супернатанта клеточной культуры, раведенного 1:20000, добавляли в планшеты и инкубировали в течение 1 часа. Планшеты, покрытые антителами, промывали три раза 200 мл промывочного буфера (PBS + 0,05% (о/о) Tween20). 100 мкл ABTS (Roche Diagnostics GmbH, Mannheim, Germany) добавляли к планшетам и абсорбцию измеряли при 405 нм с длиной волны сравнения 492 нм. Измерение белка А
Титр IgG при периодическом культивировании определяли при помощи белка А с применением ВЭЖХ в сочетании с одноэтапным универсальным ELISA.
FACS
Сортировку флуорисцентно-активированных клеток применяли для определения эффективности трансфекций (на основании клеток, экспрессирующих GFP) или уровней экспрессии GFP стабильно или кратковременно трансфецированных клеток. Обычно 5×106 клеток каждого клона или пула измеряли при помощи FACS Calibur Flow Cytometer (BD Biosciences, San Diego, CA). Размер клеток, жизнеспособность и морфологию клеток определяли на основании данных прямого и бокового светорассеяния.
















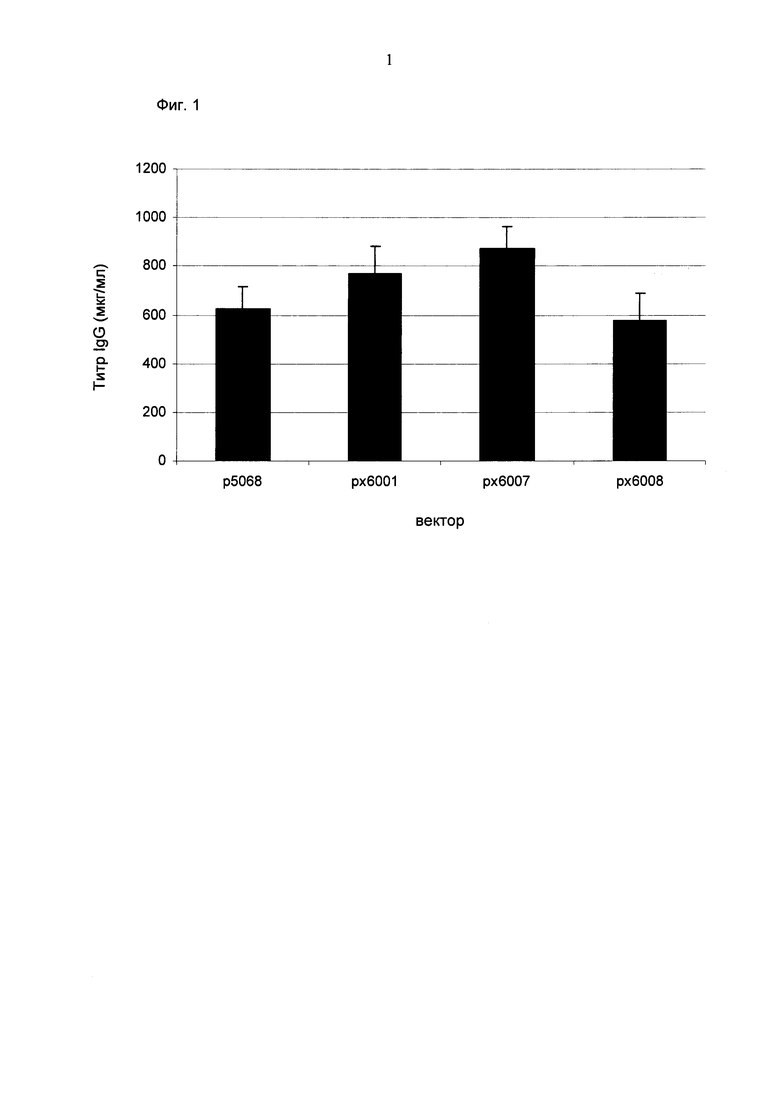


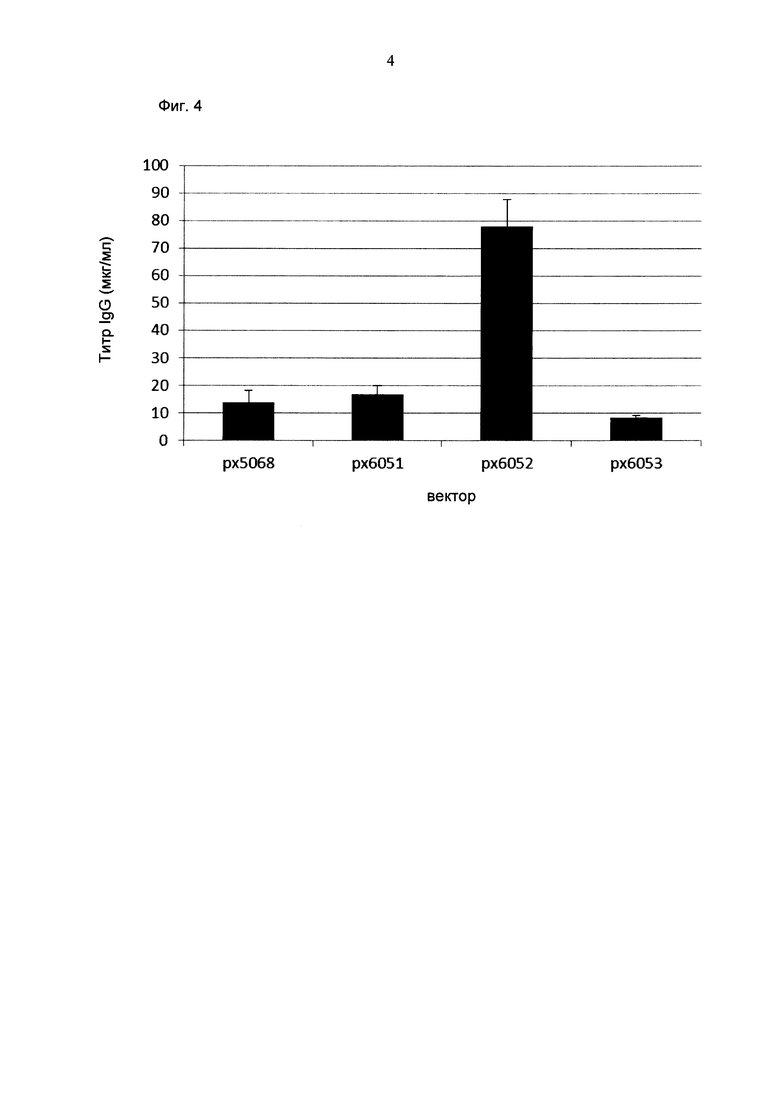
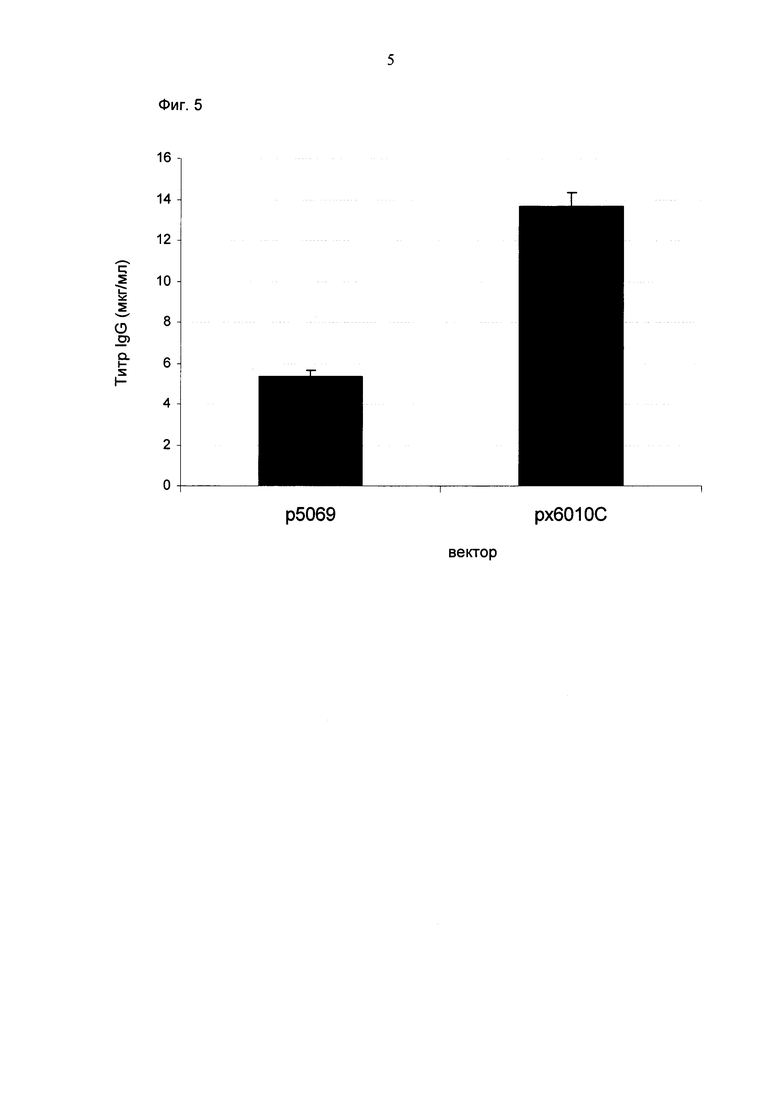
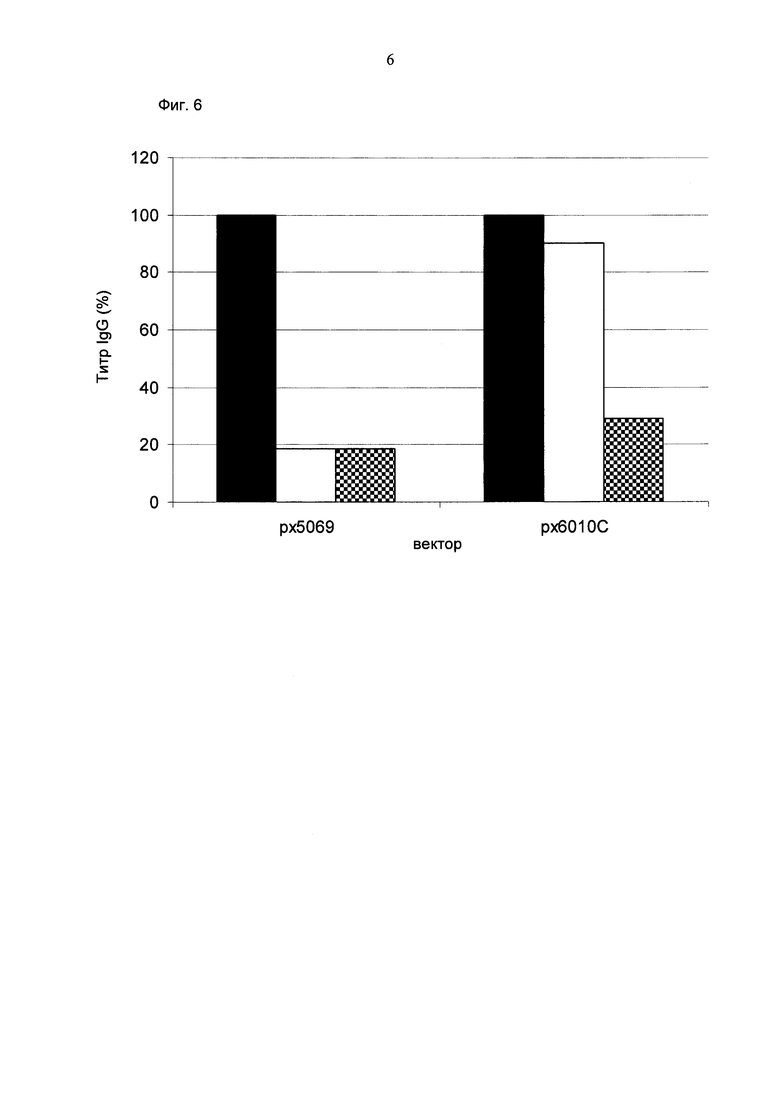
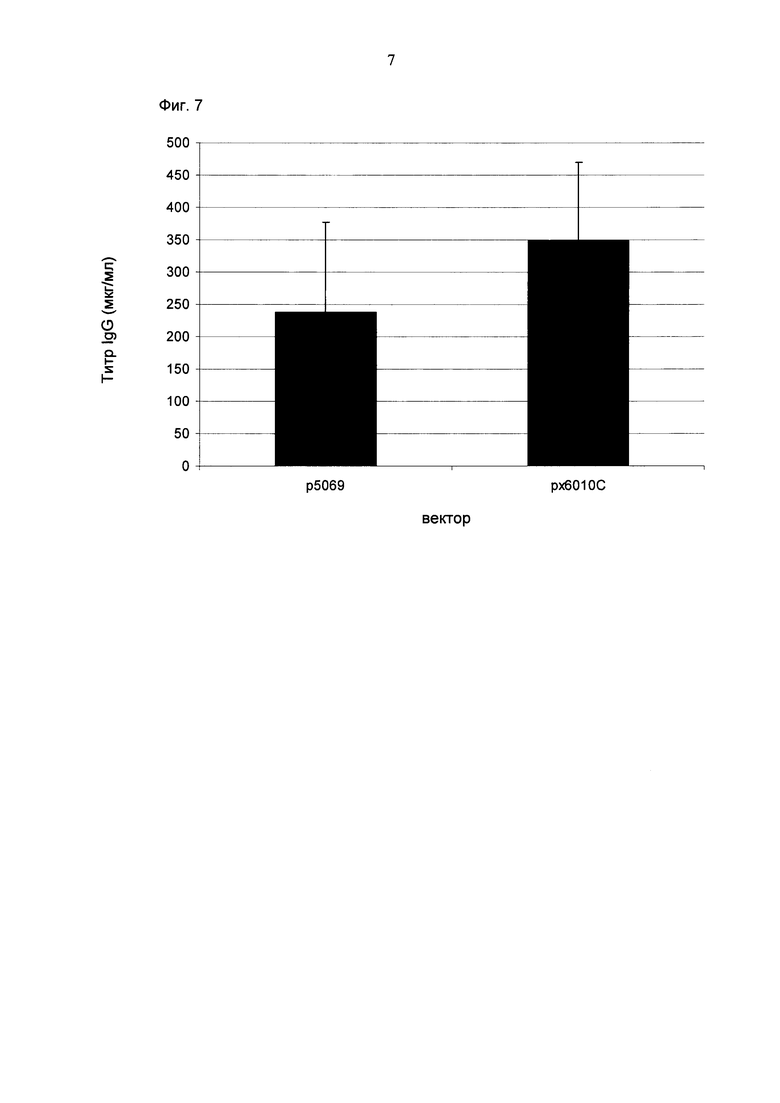
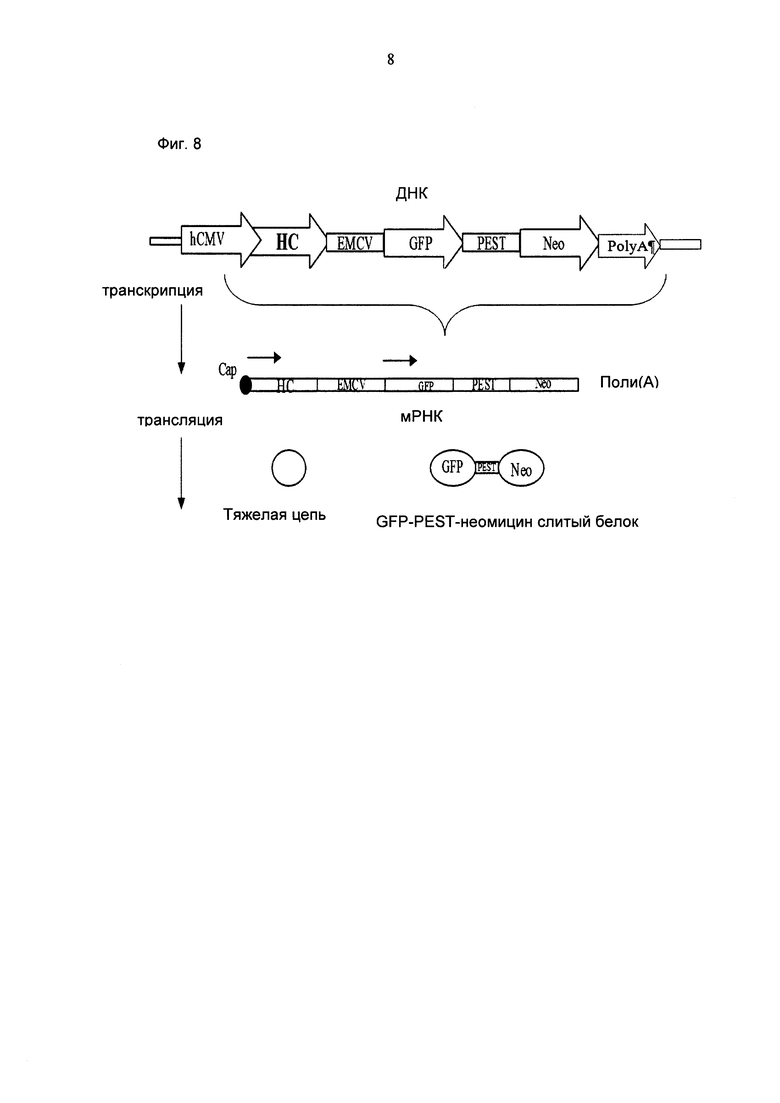
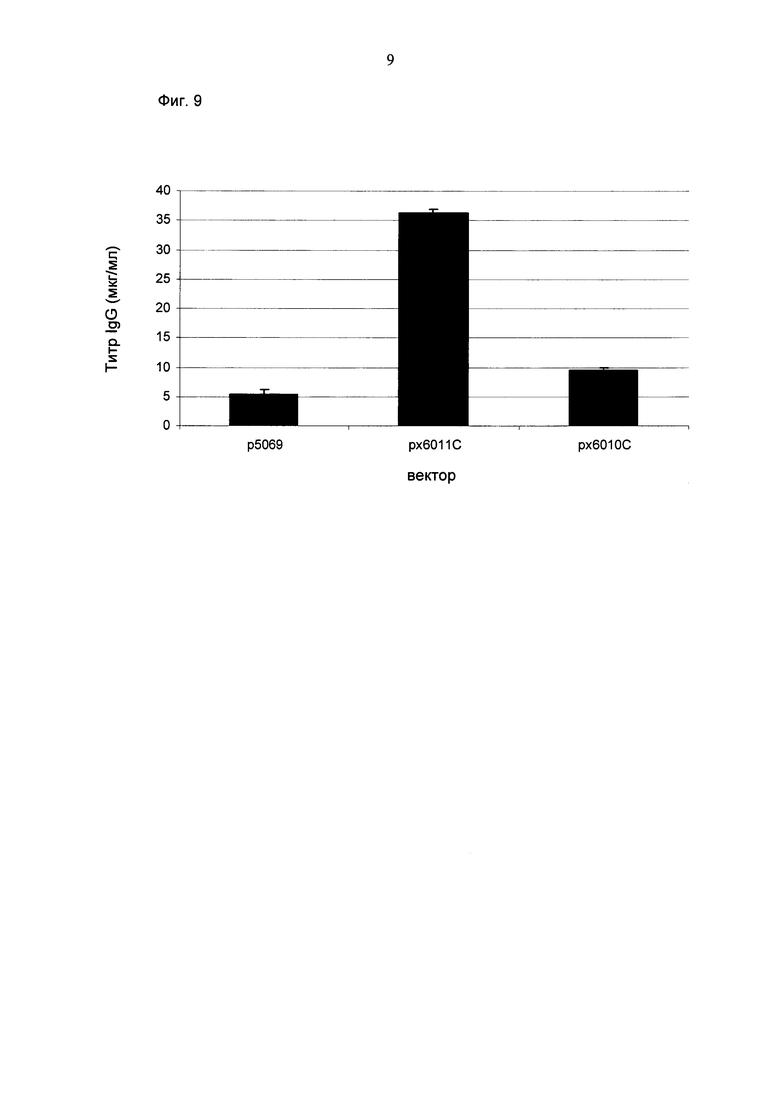
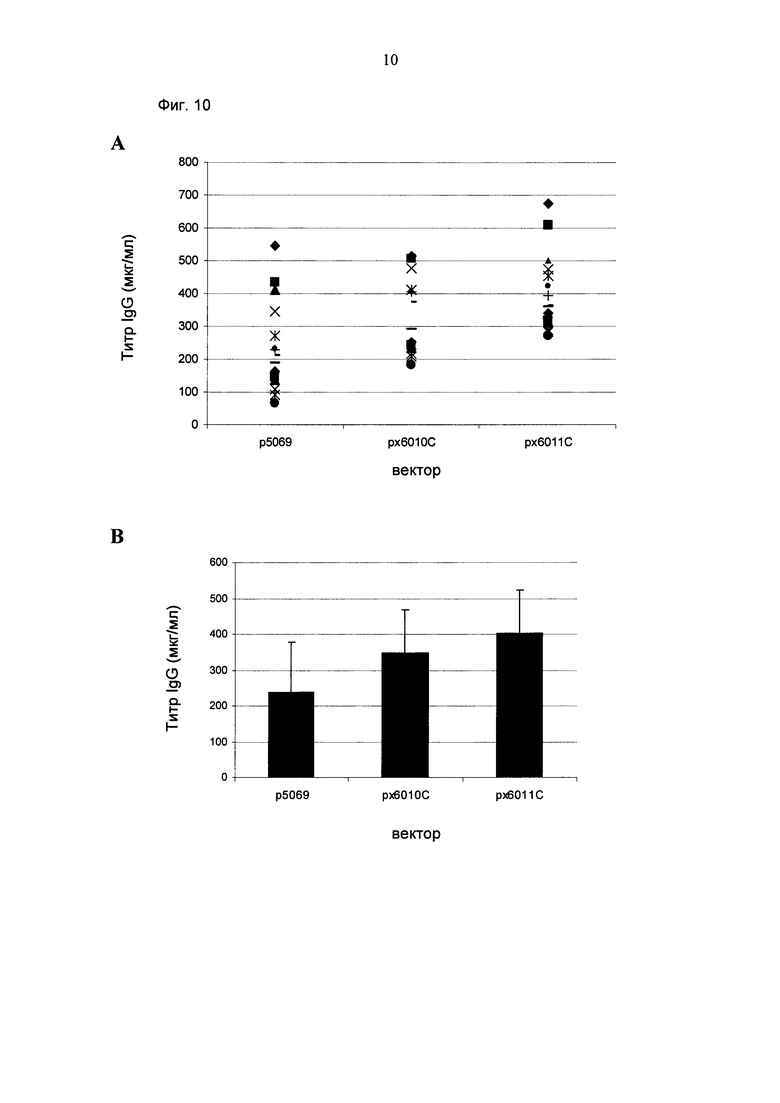

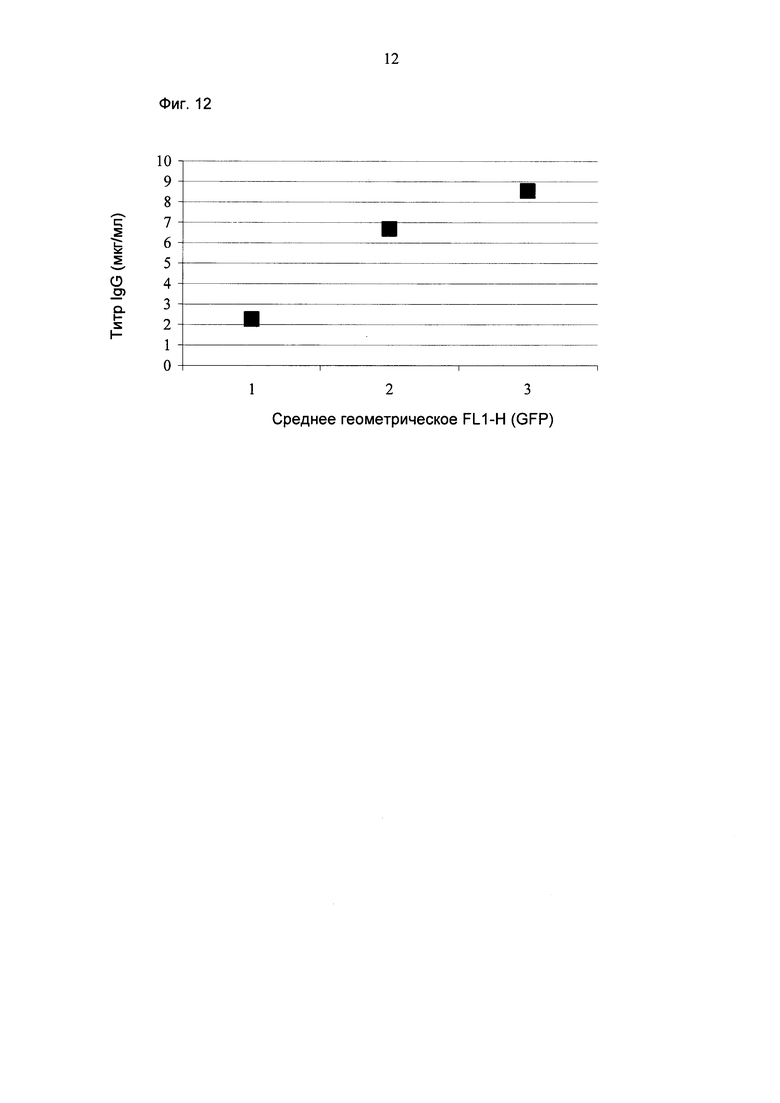
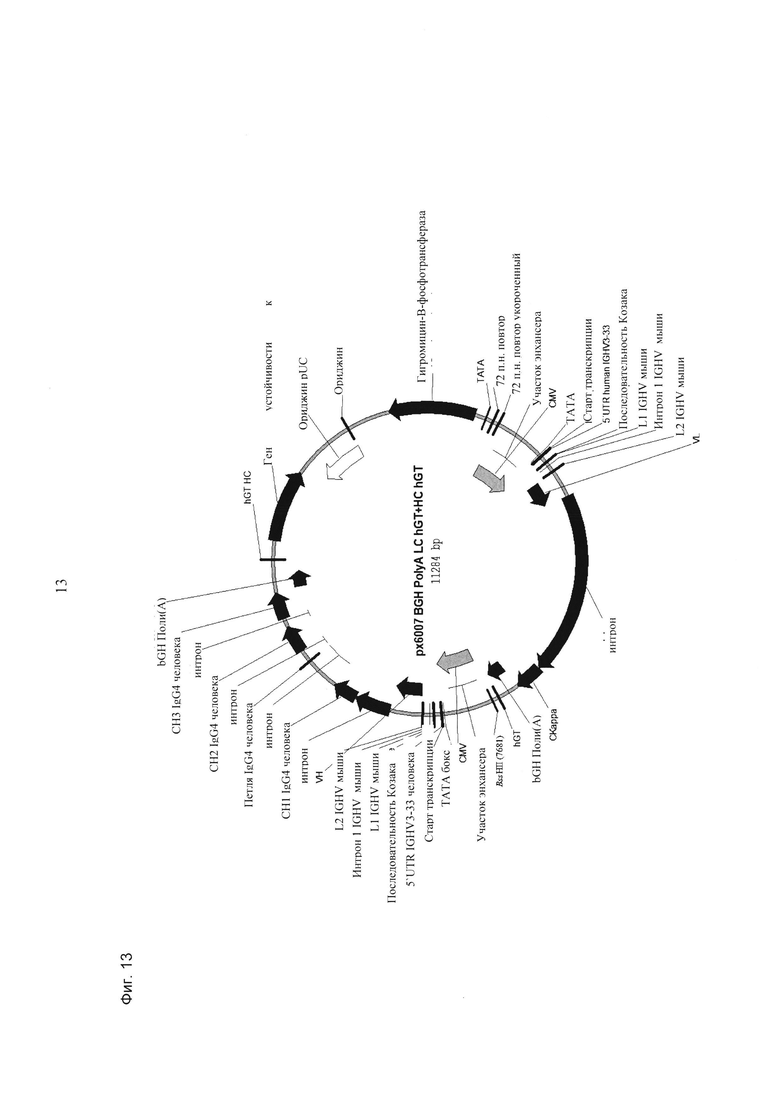
Представлены способ селекции рекомбинантных клеток млекопитающих, способ получения антитела, экспрессионный вектор и его применение. Способы включают трансфекцию клеток млекопитающего экспрессионным вектором, содержащим
- первую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, bGH поли(А)-сигнальную последовательность и последовательность терминатора hGT, и
- вторую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последовательность и последовательность терминатора hGT,
где первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении, и где указанный экспрессионный вектор дополнительно содержит селекционный маркер, и получение множества рекомбинантных клеток млекопитающего с последующей селекцией из этого множества единичных рекомбинантных клеток млекопитающего. Изобретения позволяют получать стабильные антитела. 5 н и22 з.п.ф-лы, 15 ил.
1. Способ селекции рекомбинантных клеток млекопитающих, включающий следующие этапы:
a) трансфекция клеток млекопитающего экспрессионным вектором, содержащим
- первую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, bGH поли(А)-сигнальную последовательность и последовательность терминатора hGT,
- вторую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последовательность и последовательность терминатора hGT,
где первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении и где указанный экспрессионный вектор дополнительно содержит селекционный маркер,
и, таким образом, получение множества рекомбинантных клеток млекопитающего,
b) селекция из множества рекомбинантных клеток млекопитающего (единичных) рекомбинантных клеток млекопитающего.
2. Способ по п. 1, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
3. Способ по любому из пп. 1, 2, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
4. Способ по п. 1, отличающийся тем, что экспрессионные кассеты и селекционный маркер расположены в одном направлении.
5. Способ по п. 1, отличающийся тем, что экспрессионные кассеты расположены в последовательности LC-HC-SM.
6. Способ по п. 1, отличающийся тем, что клетки млекопитающих выбраны из клеток СНО, HEK, BHK, NS0 и клеток SP2/0.
7. Способ по п. 1, отличающийся тем, что клетки млекопитающих представляют собой клетки СНО для селекции стабильно трансфецированных клеток.
8. Способ получения антитела, включающий следующие этапы:
a) культивирование клеток млекопитающих, содержащих
- первую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора,
- вторую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора,
и где экспрессионный вектор дополнительно содержит селекционный маркер, и
b) получение антитела из клетки или культуральной среды,
где первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении для селекции стабильно трансфецированных клеток.
9. Способ по п. 8, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
10. Способ по любому из пп. 8, 9, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
11. Способ по п. 8, отличающийся тем, что экспрессионные кассеты и селекционный маркер расположены в одном направлении.
12. Способ по п. 8, отличающийся тем, что экспрессионные кассеты расположены в последовательности LC-HC-SM.
13. Способ по п. 8, отличающийся тем, что клетки млекопитающих выбраны из клеток СНО, HEK, BHK, NS0 и клеток SP2/0.
14. Способ по п. 8, отличающийся тем, что клетки млекопитающих представляют собой клетки СНО для стабильной продукции антитела.
15. Экспрессионный вектор, содержащий
- первую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора,
- вторую экспрессионную кассету, содержащую в направлении 5'-3' промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, bGH поли(А)-сигнальную последовательность и hGT последовательность терминатора,
- селекционный маркер,
где первая экспрессионная кассета и вторая экспрессионная кассета расположены в одном направлении.
16. Экспрессионный вектор по п. 15, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
17. Экспрессионный вектор по любому из пп. 15, 16, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
18. Экспрессионный вектор по п. 15, отличающийся тем, что экспрессионные кассеты и селекционный маркер расположены в одном направлении.
19. Экспрессионный вектор по п. 15, отличающийся тем, что экспрессионные кассеты расположены в последовательности LC-HC-SM.
20. Применение экспрессионного вектора, содержащего
- первую экспрессионную кассету, содержащую в направлении 5'-3' первый промотор, представляющий собой промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность, представляющую собой bGH поли(А)-сигнальную последовательность, и последовательность первого терминатора транскрипции, представляющего собой терминатор hGT,
- вторую экспрессионную кассету, содержащую в направлении 5'-3' второй промотор, представляющий собой промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность, представляющую собой bGH поли(А)-сигнальную последовательность, и последовательность второго терминатора транскрипции, представляющего собой терминатор hGT, и
- третью экспрессионную кассету, содержащую в направлении 5'-3' третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно последовательность третьего терминатора транскрипции, где три экспрессионные кассеты расположены в одном направлении и в последовательности: первая экспрессионная кассета-вторая экспрессионная кассета-третья экспрессионная кассета, для стабильной рекомбинантной продукции антитела в клетках млекопитающих.
21. Применение по п. 20, характеризующееся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
22. Применение по любому из пп. 20, 21, характеризующееся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляет собой кДНК.
23. Применение по п. 20, характеризующееся тем, что клетки млекопитающих выбраны из клеток СНО, HEK, BHK, NS0 и клеток SP2/0.
24. Применение по п. 20, характеризующееся тем, что клетки млекопитающих представляют собой клетки СНО.
25. Экспрессионный вектор, содержащий
- первую экспрессионную кассету, содержащую в направлении 5'-3' первый промотор, представляющий собой промотор hCMV, нуклеиновую кислоту, кодирующую легкую цепь антитела, первую поли(А)-сигнальную последовательность, представляющую собой bGH поли(А)-сигнальную последовательность, и последовательность первого терминатора транскрипции, представляющего собой терминатор hGT,
- вторую экспрессионную кассету, содержащую в направлении 5'-3' второй промотор, представляющий собой промотор hCMV, нуклеиновую кислоту, кодирующую тяжелую цепь антитела, вторую поли(А)-сигнальную последовательность, представляющую собой bGH поли(А)-сигнальную последовательность, и последовательность второго терминатора транскрипции, представляющего собой терминатор hGT, и
- третью экспрессионную кассету, содержащую в направлении 5'-3' третий промотор, нуклеиновую кислоту, придающую устойчивость к селекционному агенту, третью поли(А)-сигнальную последовательность и возможно последовательность третьего терминатора транскрипции,
где три экспрессионные кассеты расположены в одном направлении и в последовательности: первая экспрессионная кассета-вторая экспрессионная кассета-третья экспресионная кассета.
26. Экспрессионный вектор по п. 25, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержит по меньшей мере один интрон.
27. Экспрессионный вектор по любому из пп. 25, 26, отличающийся тем, что нуклеиновая кислота, кодирующая легкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжелую цепь антитела, представляют собой кДНК.
| PIETRO P | |||
| SANNA et al | |||
| Expression of Antibody Fab Fragments and Whole Immunoglobulin in Mammalian Cells, Methods in Molecular Biology 2002, vol | |||
| Способ получения кодеина | 1922 |
|
SU178A1 |
| Гидравлический подъемник | 1922 |
|
SU389A1 |
| PAUL WINNARD et al | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Пишущая машина | 1922 |
|
SU37A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Разборное колесо | 1921 |
|
SU370A1 |
| STEVEN C.L | |||
| HO et al | |||
| IRES-mediated Tricistronic vectors for enhancing generation of high monoclonal antibody expressing CHO cell lines, Journal of Biotechnology 2012, vol.157, p | |||
| Реверсивный дисковый культиватор для тросовой тяги | 1923 |
|
SU130A1 |
| JIANDONG LI et al | |||
| A comparative study of different vector designs for the mammalian;expression of recombinant IgG antibodies, Journal of Immunological Methods 2007, vol.318, p | |||
| Способ обработки грубых шерстей на различных аппаратах для мериносовой шерсти | 1920 |
|
SU113A1 |
| Приспособление к прессу для периодической подачи материала посредством вальцев | 1926 |
|
SU7905A1 |

