Описанный в данном документе способ относится к области отбора (селекции) клеток и экспрессии/продукции полипептидов. Более подробно, в данном документе описан способ отбора клетки, долгосрочно экспрессирующей или секретирующей полипептид, на основе определения ацилирования гистона.
Уровень техники
ДНК представляет собой макромолекулу, которая кодирует инструкции всех известных живых организмов (Avery, О.Т., et al., J. Exp. Med. 79 (1944) 137-158). В клетках человека ДНК имеет длину приблизительно два метра и, в основном, хранится в ядре, которое имеет диаметр 10 мкм (Turner, В.М., Cell 111 (2002) 285-291). Для организации этого количества информации ДНК должна быть очень компактно организована. Группа консервативных небольших базовых белков, называемых гистонами, формирует комплексы с ДНК, создавая упорядоченную и компактную структуру, называемую хроматином. Эти положительно заряженные белки взаимодействуют с отрицательно заряженным фосфодиэфирным остовом двойной спирали ДНК (Alberts, B.J.A., et al., Molecular Biology of the Cell; Meyers, R.A., Epigenetic Regulation and Epigenomics, WILEY-BLACKWELL, 1; Olins, E.D., Nat. Rev. Mol. Cell Biol. (2003)). Четыре "коровых" (осевых) гистона - Н2А, Н2В, Н3 и Н4 - объединяются с ДНК, формируя базовое повторяющееся звено хроматина, называемое нуклеосомой (Thomas, J.О. and Kornberg, R.D., Proc. Natl. Acad. Sci. USA 72 (1975) 2626-2630). Базовая структура нуклеосомы весом 225 кДа состоит приблизительно из 147 пар оснований ДНК, обернутых вокруг гистонового октамера, состоящего из двух димеров Н2А/Н2В и тетрамера Н3/Н4, вокрут которого сверхспиральная ДНК делает 1,67 оборота в левую сторону (Arents, G., et al., Proc. Natl. Acad. Sci. USA 88 (1991) 10148-10152; Arents, G. and Moudrianakis, E.N., Proc. Natl. Acad. Sci. USA 90 (1993), 10489-10493; Richmond, T.J., Scientist 13 (1999) 15-15). В зависимости от доступности ДНК хроматин подразделяют на два типа: высоко компактный гетерохроматин, который является менее доступным для транскрипции, и свободно упакованный транскрипционно активный эухроматин. Факультативный гетерохроматин может образовываться в других местах в ядре, часто локализуясь в промоторах, и устанавливается либо обусловленным эволюцией образом, либо в ответ на экологические триггеры (Chen, Т. and Dent, S.Y., Nat. Rev. Genet. 15 (2014), 93-106).
Постепенная потеря продуктивности в долгоживущей культуре является общей проблемой в разработке продуцирующих клеточных линий. Снижение экспрессии рекомбинантного белка может быть связано с потерей копий трансгена и/или молчанием (сайленсингом) промотора трансгена. Молчание вызвано эпигенетическими модификациями хроматина, такими как прямое метилирование промотора ДНК на CpG-сайтах и посттрансляционные модификации гистонов, которые являются основными белковыми компонентами хроматина. Инактивирующие модификации гистонов противодействуют другим модификациям, которые являются активирующими.
Barnes, L.M., et al. сообщают о молекулярном определении прогнозирующих показателей стабильной экспрессии белка в рекомбинантных клетках миеломы NS0 (Biotechnol. Bioeng. 85 (2004) 115-121). Корреляция, о которой сообщают Barnes et al., является слабой и недостаточной для предсказания стабильности.
В WO 2004/056986 описаны средства и способы получения белка с помощью веществ, "открывающих" хроматин, которые способны делать хроматин более доступным для факторов транскрипции.
В WO 2011/128377 сообщалось, что прямое метилирование главного немедленно-раннего промотора цитомегаловируса человека (human cytomegalovirus major-immediately-early promoter, hCMV-MIE) можно использовать в качестве раннего маркера для предсказания нестабильности продукции рекомбинантных клеточных линий клеток яичника китайского хомячка (СНО, от англ. - chinese hamster ovary).
Osterlehner, A., et al. сообщают о том, что метилирование промотора и число копий трансгена предсказывают нестабильную продукцию белка в рекомбинантных клеточных линиях яичников китайского хомяка (Biotechnol. Bioeng. 108 (2011) 2670-2681).
Сущность изобретения
Было установлено, что определение степени метилирования конкретного CpG-сайта в промоторной нуклеиновой кислоте, функционально связанной со структурным геном, кодирующим полипептид, в клетке или клеточной линии, используемой для продукции соответствующего полипептида, в комбинации с определением ацилирования гистона, расположенного вблизи промотора, т.е. в хроматине промотора, могут быть использованы для предсказания снижения продуктивности при долгосрочном культивировании. Кроме того, может быть определено число копий экспрессионной кассеты легкой цепи, интегрированной в геном.
Одним из аспектов, описанных в данном документе, является способ отбора клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, при этом указанный способ включает следующие этапы:
a) определение (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты для первого множества клеточных клонов/клеточных линий, и
b) отбор клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, которая имеет (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3, определенный на этапе а), равный 0,1 или более.
Одним из аспектов, описанных в данном документе, является способ отбора клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, при этом указанный способ включает следующие этапы:
a) определение для каждого клеточного клона/клеточной линии первого множества клеточных клонов/клеточных линий, где каждый клон/клеточная линия содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, среднего (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты на основании (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3), определенного по меньшей мере в 10 клетках, полученных при культивировании каждого клеточного клона/клеточной линии, и
b) отбор клеточного клона/клеточной линии, которая имеет средний (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3, равный 0,1 или более.
Одним из аспектов, описанных в данном документе, является способ отбора клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, которая имеет нуклеиновокислотную последовательность SEQ ID NO 01, при этом указанный способ включает следующие этапы:
a) определение (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO 01, для каждого клона первого множества клеточных клонов/клеточных линий, и
b) определение частоты метилирования CpG-сайта в позиции 425 в SEQ ID NO 01 для каждого клона второго множества клеточных клонов/клеточных линий, и
c) отбор клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеиновокислотную последовательность SEQ ID NO 01, которая имеет (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3, определенный на этапе а), равный 0,1 или более, и которая имеет частоту метилирования CpG-сайта в позиции 425, определенную на этапе b), менее 5%.
Одним из аспектов, описанных в данном документе, является способ отбора клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, которая имеет нуклеиновокислотную последовательность SEQ ID NO 01, при этом указанный способ включает следующие этапы:
a) определение для каждого клеточного клона/клеточной линии первого множества клеточных клонов/клеточных линий, где каждый клон/клеточная линия содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеиновокислотную последовательность SEQ ID NO 01, среднего (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO 01, на основании (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3), определенного по меньшей мере в 10 клетках, полученных при культивировании каждого клеточного клона/клеточной линии, и
b) определение для каждого клеточного клона/клеточной линии второго множества клеточных клонов/клеточных линий, где каждый клон/клеточная линия содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO 01, средней частоты метилирования CpG-сайта в позиции 425 в SEQ ID NO 01, на основании метилирования, определенного по меньшей мере в 10 клетках, полученных при культивировании каждого клеточного клона/клеточной линии, и
с) отбор клеточного клона/клеточной линии, которая имеет средний (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3, равный 0,1 или более, и которая имеет частоту метилирования в позиции 425 менее 5%.
В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,2 или более. В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,5 или более. В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,75 или более. В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 1,0 или более.
В одном предпочтительном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,5 или более.
В одном воплощении промоторная нуклеиновая кислота имеет последовательность SEQ ID NO 01 или содержит ее функциональный фрагмент или функциональный вариант. В одном воплощении CpG-сайт находится в позиции 425 в SEQ ID NO 01 или в соответствующей позиции в ее фрагменте или варианте.
В одном из воплощений всех аспектов изобретения указанный способ дополнительно содержит следующий этап:
ab) определение числа (копий) стабильно интегрированных экспрессионных кассет легкой цепи,
где этап с) представляет собой:
отбор клеточного клона/клеточной линии, которая имеет уровень ацетилирования гистона 3 относительно уровня гистона 3, определенный на этапе а), более 0,5, которая имеет частоту метилирования CpG-сайта в позиции 425, определенную на этапе b), менее 5%, и которая имеет число (копий) стабильно интегрированных экспрессионных кассет легкой цепи, определенное на этапе ab), равное 10 или менее.
В одном предпочтительном воплощении число копий стабильно интегрированных экспрессионных кассет легкой цепи, определенное на этапе ab), составляет 6 или меньше.
В одном воплощении средний (относительный) уровень ацетилирования гистона 3 представляет собой средний (относительный) уровень ацетилирования гистона 3 на остатках лизина в позиции 4, и/или 9, и/или 14, и/или 18, и/или 27. В одном воплощении остатки указанные лизина находятся в позиции 9 и/или 14 и/или 27.
В одном из воплощений всех аспектов указанный способ включает в качестве первого этапа:
- трансфекцию популяции клеток нуклеиновой кислотой, содержащей структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, которая имеет нуклеиновокислотную последовательность SEQ ID NO 01, и получение из нее первого и второго и, возможно, третьего множество клеточных клонов/клеточных линий.
В одном воплощении (отобранная) клеточная линия после 30-60 поколений в ходе культивирования имеет скорость продукции, составляющую более 60% от скорости продукции в начале культивирования.
Одним из аспектов, описанных в данном документе, является способ отбора клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий по меньшей мере легкую цепь антитела, функционально связанный с промоторной нуклеиновой кислотой, которая имеет нуклеиновокислотную последовательность SEQ ID NO 01, при этом указанный способ включает следующие этапы:
a) определение (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO 01, для каждого клона первого множества клеточных клонов/клеточных линий, и
b) определение числа (копий) стабильно интегрированных экспрессионных кассет легкой цепи для каждого клона второго множества клеточных клонов/клеточных линий, и
c) определение частоты метилирования CpG-сайта в позиции 425 в SEQ ID NO 01 для каждого клона третьего множества клеточных клонов/клеточных линий, и
d) отбор клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеиновокислотную последовательность SEQ ID NO 01, i) которая имеет (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3, определенный на этапе а), равный 0,1 или более, ii) которая имеет частоту метилирования CpG-сайта в позиции 425, определенную на этапе b), менее 5%, и iii) которая имеет число (копий) стабильно интегрированных экспрессионных кассет легкой цепи, определенное на этапе ab), равное 10 или менее.
Одним из аспектов, описанных в данном документе, является способ отбора клеточного клона/клеточной линии, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий по меньшей мере легкую цепь антитела, функционально связанный с промоторной нуклеиновой кислотой, которая имеет нуклеиновокислотную последовательность SEQ ID NO 01, при этом указанный способ включает следующие этапы:
a) определение для каждого клеточного клона/клеточной линии множества клеточных клонов/клеточных линий, где каждый клон/клеточная линия содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO 01, среднего (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO 01, на основании (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3), определенного по меньшей мере в 10 клетках, полученных при культивировании каждого клеточного клона/клеточной линии, и
b) определение для каждого клеточного клона/клеточной линии множества клеточных клонов/клеточных линий числа (копий) стабильно интегрированных экспрессионных кассет легкой цепи по меньшей мере в 10 клетках, полученных при культивировании каждого клеточного клона/клеточной линии, и
c) определение для каждого клеточного клона/клеточной линии множества клеточных клонов/клеточных линий, где каждый клон/клеточная линия содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO 01, средней частоты метилирования CpG-сайта в позиции 425 в SEQ ID NO 01 на основании метилирования, определенного по меньшей мере в 10 клетках, полученных при культивировании каждого клеточного клона/клеточной линии, и
d) отбор клеточного клона/клеточной линии, i) которая имеет (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3), равный 0,1 или более, ii) которая имеет частоту метилирования в позиции 425 менее 5%, и iii) которая имеет число (копий) стабильно интегрированных экспрессионных кассет легкой цепи, определенное на этапе ab), равное 10 или менее.
В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,2 или более. В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,5 или более. В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,75 или более. В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 1,0 или более.
В одном предпочтительном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 составляет 0,5 или более.
В одном воплощении средний (относительный) уровень ацетилирования гистона 3 представляет собой средний (относительный) уровень ацетилирования гистона 3 на остатках лизина в позиции 4, и/или 9, и/или 14, и/или 18, и/или 27. В одном воплощении указанные остатки лизина находятся в позиции 9 и/или 14 и/или 27.
В одном воплощении промоторная нуклеиновая кислота имеет последовательность SEQ ID NO 01 или содержит ее функциональный фрагмент или функциональный вариант. В одном воплощении CpG-сайт находится в позиции 425 в SEQ ID NO 01 или в соответствующей позиции в ее фрагменте или варианте.
В одном предпочтительном воплощении число (копий) стабильно интегрированных экспрессионных кассет легкой цепи, определенное на этапе b), составляет 6 или меньше.
В одном воплощении определение (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) включает следующие этапы:
1) выделение хроматина из каждого клеточного клона/клеточной линии,
2) обработка первой аликвоты хроматина антителом, которое подходит для определения (относительного) уровня ацетилирования гистона 3, таким как, например, антитело, специфическое к ацетилированию гистона 3, и формирование преципитата "антитело - хроматин", и обработка второй аликвоты хроматина антителом, которое подходит для определения уровня гистона 3, таким как, например, антитело, специфическое к гистону 3, и формирование преципитата "антитело - хроматин",
3) амплификация геномной ДНК из третьей необработанной аликвоты хроматина и из первой и второй обработанных аликвот путем количественной ПЦР в режиме реального времени,
4) определение с помощью результата, полученного на этапе 3), (относительного) уровня ацетилирования гистона 3 относительно уровня гистона 3.
В одном воплощении определение частоты метилирования включает следующие этапы:
1) выделение ДНК из каждого клеточного клона/клеточной линии,
2) проведение для каждой выделенной ДНК отдельной полимеразной цепной реакции, специфической в отношении метилирования,
3) определение с помощью результата, полученного на этапе 2), частоты метилирования.
В одном воплощении этап 2) представляет собой
2) проведение для каждой выделенной ДНК отдельной полимеразной цепной реакции с использованием праймера, специфического в отношении метилирования, и универсального праймера.
Также воплощением этапа 2) является
2) расщепление по отдельности выделенной ДНК с помощью рестрикционного фермента и проведение полимеразной цепной реакции для каждой из расщепленных ДНК с использованием праймера, специфического в отношении метилирования, и универсального праймера.
В одном воплощении праймеры независимо друг от друга выбраны из группы, включающей SEQ ID NO 06-20.
В одном воплощении праймеры выбраны из группы, состоящей из SEQ ID NO 11, 14 и 15, универсальный праймер имеет последовательность SEQ ID NO 09, а праймер, специфический в отношении метилирования, выбран из группы, состоящей из SEQ ID NO 17, 18 и 19.
В одном воплощении универсальные праймеры имеют последовательность SEQ ID NO 09 и 11, а праймеры, специфические в отношении метилирования, имеют последовательность SEQ ID NO 11 и 18.
В одном воплощении (относительный) уровень ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) нормализован по референсному гену. В одном воплощении референсный ген представляет собой Gusb.
В одном воплощении второе множество клеточных клонов/клеточных линий содержит по меньшей мере один клеточный клон/клеточную линию, также включенную в первое множество клеточных клонов/клеточных линий. В одном воплощении второе множество клеточных клонов/клеточных линий идентично первому множеству клеточных клонов/клеточных линий.
В одном воплощении клеточный клон/клеточная линия является эукариотическим клеточным клоном/клеточной линией. В одном воплощении клеточный клон/клеточная линия представляет собой клеточный клон/клеточную линию млекопитающего. В одном воплощении клеточный клон/клеточная линия выбраны среди СНО, BHK (от англ. baby hamster kidney - клетки почки новорожденного хомяка), НЕК (от англ. human embryonic kidney - клетки эмбриональных почек человека) и Sp2/0. В одном воплощении клеточный клон/клеточная линия представляет собой клеточный клон/клеточную линию СНО. В одном воплощении клеточный клон/клеточная линия представляет собой клеточный клон/клеточную линию СНО K1.
В одном предпочтительном воплощении клеточный клон/клеточная линия представляет собой клеточный клон/клеточную линию СНО.
В одном воплощении полипептид представляет собой i) антитело, или ii) фрагмент антитела, или iii) конъюгат антитела, или iv) легкую цепь антитела и тяжелую цепь антитела. В одном воплощении изобретения антитело представляет собой биспецифическое антитело.
Одним из аспектов, описанных в данном документе, является способ получения полипептида, который включает следующие этапы:
a) отбор клеточного клона/клеточной линии с помощью способа, описанного в данном документе,
b) культивирование отобранного клеточного клона/клеточной линии, и
c) выделение полипептида из среды культивирования и/или клеточного клона/клеточной линии и, таким образом, получение полипептида.
В одном воплощении указанный способ включает перед этапом а) следующие этапы:
а-3) получение клетки млекопитающего, не являющейся клеткой человека,
а-2) трансфекция полученной клетки нуклеиновой кислотой, которая содержит нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой, которая имеет нуклеиновокислотную последовательность SEQ ID NO 01,
а-1) i) возможно, культивирование трансфицированного клеточного клона/клеточной линии в присутствии селективного агента, ii) одиночное депонирование трансфицированных клеток, и iii) культивирование одиночных депонированных трансфицированных клеток в присутствии селективного агента.
Одним из аспектов, описанных в данном документе, является способ получения полипептида, который включает следующие этапы:
a) выбор клеточного клона/клеточной линии с помощью способа, описанного в данном документе,
b) культивирование отобранного клеточного клона/клеточной линии, и
c) выделение полипептида из среды культивирования и/или клеток и, таким образом, получение полипептида.
В одном воплощении указанный способ также включает этап
d) очистки извлеченного полипептида.
В одном воплощении указанный способ включает перед этапом а) следующие этапы:
а-3) получение клетки,
а-2) трансфекция полученной клетки нуклеиновой кислотой, которая содержит структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой,
а-1) i) возможно, культивирование и размножение трансфицированной клетки в присутствии селективного агента, ii) одиночное депонирование клеток, и iii) культивирование одиночных депонированных трансфицированных клеток в присутствии селективного агента.
В одном воплощении этап а) включает:
i) получение по меньшей мере одной клетки, содержащей нуклеиновую кислоту, содержащую структурный ген, кодирующий полипептид, функционально связанный с промоторной нуклеиновой кислотой,
ii) определение метилирования CpG-сайта в позиции 425 в пределах промоторной нуклеиновой кислоты SEQ ID NO 01, и
iii) отбор клетки, продуцирующей полипептид, где метилирование, определенное на этапе b), находится ниже порогового значения.
Подробное описание изобретения
Экономные клеточные линии необходимы, чтобы обеспечить высокую продуктивность и стабильный уровень продукции при переходе от малых масштабов к большим. Снижение продуктивности в ходе увеличения масштабов представляет собой серьезную опасность во время развития клеточной линии (Barnes, L. М., et al., Biotechnol. Bioeng. 81 (2003) 631-639). Одной из главных причин нестабильности продукции является снижение числа активных копий в течение клеточного цикла, что может быть отнесено к хромосомному нарушению/перегруппировке как неотъемлемой характеристики клеток СНО (Kim, М., et al., Biotechnol. Bioeng. 108 (2011) 2434-2446) и/или индукции за счет процесса амплификации гена (Kaufman, R. J., et al., Mol. Cell. Biol. (1983) 699-711). Снижение количества мРНК при постоянном количестве копий рекомбинантных генов является еще одной важной причиной падения продуктивности (Barnes, L.М., et al., Biotechnol. Bioeng. 85 (2004) 115-121; Chusainow, J., et al., Biotechnol. Bioeng. 102 (2009) 1182-1196; Strutzenberger, K., et al., J. Biotechnol. 69 (1999) 215-226). Разумное объяснение этого явления заключается в эпигенетическом молчании за счет метилирования промотора (Osterlehner, A., et al., Biotechnol. Bioeng. 108 (2011) 2670-2681; Yang, Y., et al., J. Biotechnol. 147 (2010) 180-185) и модификации гистонов как типичных примеров деацетилирования и специфического метилирования (Mutskov, V. and Felsenfeld, G., EMBO J. 23 (2004) 138-149; Paredes, V., et al., Biotechnol. Lett. 35 (2013) 987-993). Кроме того, сама рекомбинантная, последовательность, формирование тандемных повторов и геномное расположение трансгенов предполагаются как инициаторы молчания генов (Kaufman, W. L, et al., Nuc. Acids Res. 36 (2008) e111). Поэтому влияние соседнего хроматина на интеграционные сайты называют "позиционным влиянием" (Lattenmayer, С, et al., Cytotechnol. 51 (2006) 171-182; Yin, Z., et al., Genet. Mol. Res. 11 (2012) 355-369).
Промотор, находящийся выше по последовательности относительно рекомбинантного гена, инициирует транскрипцию гена и способен влиять на уровень экспрессии и стабильность гена (Kaufman, W.L, et al., Nuc. Acids Res. 36 (2008) e111). Основной немедленно-ранний промотор гена цитомегаловируса человека (hCMV-MIE) обычно используется для управления рекомбинантной экспрессией в клетках млекопитающих в исследованиях и производстве для получения высоких уровней экспрессии при временных и стабильных трансфекциях (Boshart, М., et al., Cell 41 (1985) 521-530; Chapman, В.S., et al., Nuc. Acids Res. 19 (1991) 3979-3986; Foecking, M.K. and Hofstetter, H. Gene 45 (1986) 101-105; Wright, A., et al., Hum. Gene Ther. 16 (2005) 881-892). Хотя промотор hCMV-MIE обеспечивает высокий уровень экспрессии генов, многие исследования сообщали о снижении продуктивности при долгосрочном культивировании (Bailey, L.A., et al., Biotechnol. Bioeng. 109 (2012) 2093-2103; Barnes, L.M., et al., Biotechnol. Bioeng. 73 (2001) 261-270; He, L, et al., Biotechnol. Bioeng. 109 (2012) 1713-1722.). Молчание промотора hCMV-MIE (в дополнение к потере числа копий) в значительной степени объясняется эпигенетическими событиями метилирования промотора ДНК и модификации гистонов (Brooks, A.R., et al., J. Gene Med. 6 (2004) 395-404.; Kim, M., et al., Biotechnol. Bioeng. 108 (2011) 2434-2446; Osterlehner, A., et al., Biotechnol. Bioeng. 108 (2011) 2670-2681; Paredes, V., et al., Biotechnol. Lett. 35 (2013) 987-993; Yang, Y., et al., J. Biotechnol. 147 (2010) 180-185).
Хроматин, как известно, участвует в регуляции экспрессии генов. Модификация ДНК или хроматина и/или ассоциированных с хроматином белков, таких как гистоны, оказывает влияние на структуру хроматина и, следовательно, на экспрессию генов. Модификация ДНК в клетках млекопитающих может представлять собой метилирование остатков цитозина в динуклеотидах CpG. Гистоны, в особенности их N-концевые части, могут быть модифицированы путем ацетилирования, метилирования, фосфорилирования или убиквитинилирования.
Нуклеиновокислотное содержимое клетки, т.е. ее ДНК, находится в ядре клетки в уплотненной форме вместе с гистонами. Гистоны упаковывают и упорядочивают ДНК в нуклеосомы. У человека есть пять основных гистоновых белков. Гистоны являются высоко щелочными белками. Гистон Н3 (histone 3) имеет главный глобулярный домен и N-концевой хвост. Он имеет размер 137 аминокислотных остатков. В целом, функция гистонов заключается в обеспечении осевой структурой, вокруг которой располагается ДНК, и в регуляции экспрессии генов.
Гистоны могут быть модифицированы многими различными способами, в основном вдоль остатков N-концевой аминокислотной последовательности, которая имеет от 13 до 40 аминокислот в длину в зависимости от конкретного гистона. Такое большое число модификаций даже возрастает из-за того, что некоторые модификации, такие как метилирование лизина, содержат вплоть до трех различных состояний (Kouzarides, Т., Cell 128 (2007) 693-705). Было обнаружено более 100 модификаций гистонов (Zentner, G.Е. and Henikoff, S., Nat. Struct. Mol. Biol. 20 (2013) 259-266). Ацетилирование и метилирование хвостовых остатков гистона Н3 и гистона Н4 являются наиболее изученными модификациями. В целом, было найдено 14 различных типов модификаций (Dawson, М.A. and Kouzarides, Т., Cell 150 (2012) 12-27), и они перечислены в следующей таблице.
Таблица: K, лизин; R, аргинин; S, серии; Т, треонин; Е, глутаминовая кислота; Р, пролин; ас, ацетилирование; me, метилирование; cit, цитруллин; cr, кротонилирование; pr, пропионилирование; bu, бутирилирование; fo, формилирование; oh, гидроксилирование; а, асимметричный; s, симметричный; >, конверсия. Адаптировано из (Dawson & Kouzarides, 2012).
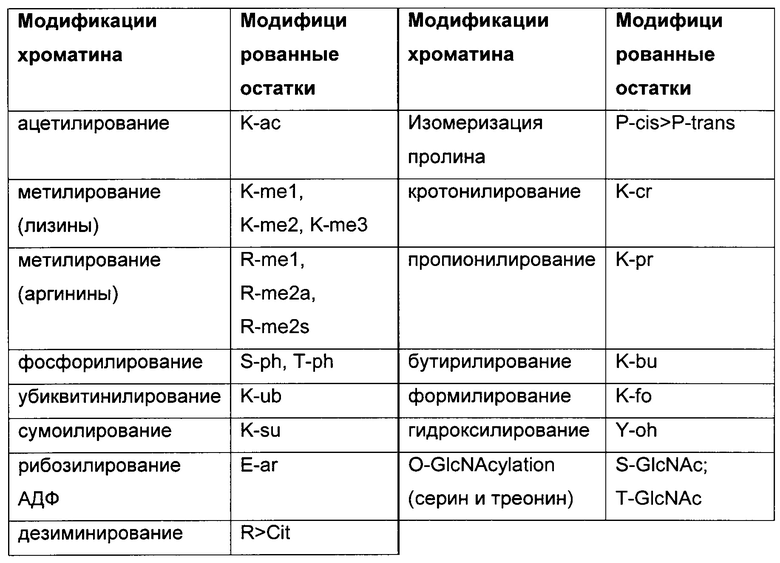
Комбинации этих модификаций, называемые мотивами РТМ (посттрансляционная модификация), больше связаны с функциями, чем отдельные маркеры. Исследование комбинаций модификаций предлагает более 200 различных РТМ-мотивов (Feller, С, et al., Mol. Cell 57 (2015) 559-571). Модификации носят обратимый характер.
Известно, что метилирование гистона Н3 на лизине 9 (H3K9) и 27 (H3K27) является модификациями, приводящими к репрессии гена. Модификации гистона, по-видимому, обеспечивают лабильную транскрипционную репрессию, тогда как метилирование ДНК является высокостабильным маркером молчания, который не так легко восстановим (Cedar, Н. and Bergman, Y., Nat. Rev. Gen. 10 (2009) 295-304).
Ацетилирование гистонов было обнаружено в 1961 году как первая модификация гистонов (Phillips, D.М., Biochem. J. 87 (1963) 258-263). Ранние исследования связывают активные транскрибируемые гены с гиперацетилированием гистонов, что указывает на функцию ацетилирования в процессе транскрипции (Allfrey, V.G., et al., Proc. Natl. Acad. Sci. USA 51 (1964) 786-794). Во время S-фазы наблюдали глобальное увеличение числа сайтов ацетилирования, таких как H3K56ac, с последующим его снижением во время G2-фазы. Это привело к выводу, что ацетилирование гистонов может способствовать встраиванию вновь синтезированных гистонов (Miller, K.М., et al., Cell Cycle 5 (2006) 2561-2565). В дополнение к своей глобальной роли в процессе репликации ДНК, ацетилирование гистонов образует специфические геномные паттерны, которые коррелируют с активной транскрипцией. Таким образом, области гетерохроматина являются гипоацетилированными, а транскрипционно активные гены эухроматина являются высоко ацетилированными (Kouzarides, Т., Cell 128 (2007) 693-705). Пики ацетилирования находятся на определенных сайтах в промоторе вблизи сайтов начала транскрипции (transcription start sites, TSS) (Wang, Z., et al., Nat. Genet. 40 (2008) 897-903). Остатки лизина N-концевого хвоста гистона 3 в позициях 4, 9, 14, 18 и остатки лизина гистона 4 в позициях 5, 8 и 12 преимущественно склонны к ацетилированию. Все вместе - регуляция экспрессии генов, репликации ДНК, репарация и рекомбинация - находятся под влиянием различных состояний ацетилирования гистонов (Dawson, М.A. and Kouzarides, Т., Cell 150 (2012) 12-27).
Гистонацетилтрансферазы (histone acetyltransferase, HAT), как правило, опосредуют экспрессию генов и активацию транскрипции (Cheung, P., et al., Cell 103 (2000) 263-271). Используя иммунопреципитацию хроматина, ученые обнаружили, что неметилированная ДНК в основном собрана в нуклеосомы, которые содержат ацетилированные гистоны, которые связаны с открытым хроматином, тогда как присутствие метильных групп на идентичных последовательностях ДНК коррелирует со сборкой нуклеосом, содержащих неацетилированные гистоны Н3 и Н4, что приводит к образованию более компактного хроматина (Cedar, Н. and Bergman, Y., Nat. Rev. Gen. 10 (2009) 295-304; Eden, S., et al., Nature 394 (1998) 842-843; Hashimshony, Т., et al., Nat. Gen. 34 (2003) 187-192). Гистон Н3 является наиболее чрезмерно модифицированным гистоном из пяти природных гистонов.
В областях высокой транскрипционной активности можно обнаружить высокую степень ацетилирования гистонов. Ацетилирование гистонов катализируется ферментом гистонацетилтрансферазой (HAT), которая переносит ацетильную часть ацетил-КоА к ε-аминогруппе конкретных остатков лизина в N-концевой области гистона. Ацетилирование гистонов имеет место исключительно место на остатках лизина, таких как, например, H3K9 (лизин в позиции 9 гистона 3), H3K14 и H3K27 (Koch, СМ., et al., Gen. Res. 17 (2007) 691-707; Creyghton, M.P., et al., Proc. Natl. Acad. Sci. USA 107 (2010) 21931-21936).
В ходе разработки клеточной линии и при больших масштабах производства стандартным наблюдением является молчание экспрессии рекомбинантного гена при длительном (крупномасштабном) культивировании клетки-хозяина. В клетках млекопитающих это может быть связано, например, с формированием производных хроматина, которые предотвращают транскрипцию рекомбинантного гена.
Это приводит к образованию гетерогенной популяции клеток после длительного времени культивирования, например, при крупномасштабном производстве (включая систему посевных ферментации и основную ферментацию). В этой гетерогенной популяции клеток некоторые клетки продолжают экспрессировать на высоком уровне рекомбинантный белок, представляющий интерес, в то время как другие клетки экспрессируют на низком уровне или вовсе не экспрессируют белок, представляющий интерес (см., например, Martin, D.I. and Whitelaw, E., Bioessays 18, (1996) 919-923; McBurney, M.W., et al., Exp. Cell. Res. 274 (2002) 1-8).
Продуцирующие клеточные линии, как правило, являются потомками одной родительской клетки. Число этих клеток часто увеличивают в процессе культивирования, и культивируют их в течение длительного времени в крупномасштабных ферментациях (система посевных ферментации и основная ферментация), получая объемы культивирования до 25000 литров и клеточные плотности часто более одного миллиона клеток на миллилитр. Такие крупномасштабные ферментации могут показать резкое сокращение продуктивности (Migliaccio, A.R., et al., Gene 256 (2000) 197-214; Strutzenberger, K., et al., J. Biotechnol. 69 (1999) 215-226).
При выборе продуцирующих клеточных линий/клеточных клонов, т.е. клеточных клонов/клеточных линий, которые предназначены для использования в крупномасштабной продукции рекомбинантного полипептида, такого как, например антитело, важное значение имеет стабильность продукции/экспрессии, т.е. потеря клеточным клоном/клеточном линией продуктивности из поколения в поколение. Таким образом, клеточные линии млекопитающих для продукции рекомбинантных белков должны поддерживать свою продуктивность в течение длительного времени культивирования. Как правило, стабильность продукции клеточного клона/клеточной линии определяют путем культивирования клеточного клона/клеточной линии в течение длительного периода времени, т.е. в течение множества поколений. Через регулярные промежутки времени среду разводят свежей средой, на основании титра продукта и плотности жизнеспособных клеток определяют удельную продуктивность на одну клетку. Изменение (как правило, уменьшение) удельной продуктивности свидетельствует о долгосрочной стабильности продукции клеточного клона/клеточной линии.
Исследования долгосрочной стабильности требуют времени и ресурсов, но широко проводятся для выявления и устранения нестабильных кандидатов в ходе разработки клеточной линии. В зависимости от целевых критериев исследование охватывает от 30 до 60 делений клеток, что обычно соответствует периоду от 30 до 70 дней. Таким образом, для этого требуются необходимые материалы и рабочее время. Кроме того, до 75% анализируемых клеточных линий/клеточных клонов не являются стабильными и, таким образом, нужно оценить большое число клеточных линий/клеточных клонов.
Помимо потери продуктивности трансгена нестабильность производства клеточных линий может быть связана с метилированием и молчанием гетерологичного промотора, используемого для экспрессии трансгена (см., например, Escher, G., et al. J. Lipid Res. 46 (2005) 356-365; Krishnan, M., et al., FASEB J. 20 (2006) 106-108; Yang, Y., et al., J. Biotechnol. 147 (2010) 180-185). Молчание промотора может быть результатом эпигенетической модификации хроматина (комбинации или комплекса ДНК и белков, которые составляют содержимое ядра клетки). Это может быть прямое метилирование CpG-динуклеотидов в пределах промотора, такого как, например, главный немедленно-ранний промотор/энхансер цитомегаловируса человека (hCMV-MIE), и/или посттрансляционная модификация гистонов. Кроме того, известны инактивирующие модификации гистонов, такие как метилирование лизина 9 или 27 гистона 3, активирующие модификации, такие как метилирование лизина 4 гистона 3, или глобальное ацетилирование гистона 3 и 4 (см., например, Cedar, Н. and Bergman, Y., Nat. Rev. Genet. 10 (2009) 295-304).
Эпигенетическая модификация CpG-метилирования промотора hCMV-MIE была использована в качестве индикатора долгосрочной стабильности продукции (см. Osterlehner et al.).
В данном изобретении было выявлено местное ацетилирование лизина (H3ac) в качестве потенциального маркера для предсказания долгосрочного молчания трансгена.
Было установлено, что динуклеотид CpG в позиции 425 главного немедленно-раннего промотора/энхансера цитомегаловируса человека (hCMV-MIE) (SEQ ID NO 01) часто является метилированным в клеточных линиях яичника китайского хомяка (СНО), нестабильно продуцирующих антитела. Для обеспечения быстрого и чувствительного измерения метилирования hCMV-MIE была разработана кПЦР в режиме реального времени, специфическая в отношении метилирования.
Кроме того, было обнаружено, что посттрансляционная модификация гистона, примыкающего к промотору, может быть использована в качестве дополнительного маркера для идентификации стабильно экспрессирующих/продуцирующих клеточных линий. Наличие инактивирующей модификации является маркером того, что соответствующий клеточный клон/клеточная линия является очень нестабильной и покажет снижение продуктивности при дальнейшем культивировании выше среднего (в долгосрочной перспективе нестабильно продуцирующий клеточный клон/клеточная линия). С другой стороны, наличие активирующей модификации является маркером того, что соответствующий клеточный клон/клеточная линия является очень стабильной и покажет снижение продуктивности при дальнейшем культивировании ниже среднего (в долгосрочной перспективе стабильно продуцирующий клеточный клон/клеточная линия).
Было обнаружено, что при использовании комбинации обоих указанных выше маркеров, т.е. метилирования в позиции 425 в SEQ ID NO 01 и относительного уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты, можно достичь дальнейшего улучшения в выявлении долгосрочных стабильно продуцирующих рекомбинантных клеточных линий/клеточных клонов. Дальнейшее улучшение возможно, если в случае антитела также включить число (копий) стабильно интегрированных экспрессионных кассет легкой цепи.
Таким образом, в данном документе описаны способы отбора клеточного клона/клеточной линии, а также способ получения полипептида с использованием такого клеточного клона/клеточной линии. Отобранный клеточный клон/клеточная линия представляет собой долгосрочно продуцирующую клетку. Такой клеточный клон/клеточная линия может быть отобрана, как описано в данном документе, на основании i) относительного уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты, или ii) метилирования главного немедленно-раннего промотора/энхансера CMV человека (hCMV-MIE) в позиции 425, функционально связанного со структурным геном, кодирующий полипептид, или iii) комбинации i) и ii).
А. Определения
Термин "почти" означает, что значение после этого выражения является центральным значением с определенной вариабельностью. В одном воплощении изменчивость находится в пределах плюс/минус 40% от данного значения, в другом воплощении вариабельность находится в пределах плюс/минус 30%, а в еще одном воплощении изменчивость находится в пределах плюс/минус 20%. Таким образом, термин "почти" постоянно означает, что указанное значение находится в одном из воплощений в диапазоне от 60% до 140%, в другом воплощении в диапазоне от 70% до 130%, а в еще одном воплощении в диапазоне от 80% до 120%.
Термин "антитело" обозначает молекулу, содержащую по меньшей мере два так называемых полипептида легкой цепи (легкая цепь) и два так называемых полипептида тяжелых цепей (тяжелая цепь). Каждый из полипептидов тяжелой и легкой цепи содержит вариабельный домен (вариабельную область) (как правило, амино-концевую часть полипептидной цепи), который включает связывающие области, которые способны взаимодействовать с антигеном. Каждый из полипептидов тяжелой и легкой цепи также содержит константную область (как правило, карбокси-концевую часть). Константная область тяжелой цепи опосредует связывание антитела i) с клетками, несущими Fc-гамма-рецептор (FcγR), такими как фагоциты, или ii) с клетками, несущими неонатальный Fc-рецептор (FcRn), также известный как рецептор Брамбелла. Она также опосредует связывание с некоторыми факторами, включая факторы классической системы комплемента, такими как компонент (C1q).
Термин "антитело" в данном описании используется в самом широком смысле и включает различные структуры антитела, в том числе, но не ограничиваясь ими, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител до тех пор, пока они проявляют нужную антигенсвязывающую активность.
В зависимости от аминокислотной последовательности константной области тяжелых цепей антитела разделяют на различные классы: класс IgA, класс IgD, класс IgE, класс IgG и класс IgM. Некоторые из этих классов далее разделяют на подклассы (изотипы), т.е. IgG на IgG1, IgG2, IgG3 и IgG4, или IgA на IgA1 и IgA2. Согласно классу, к которому принадлежит антитело, константные области тяжелой цепи называются α (IgA), δ (IgD), ε (IgE), γ (IgG) и μ (IgM), соответственно. В одном воплощении антитело представляет собой антитело класса IgG. В другом воплощении изобретения антитело имеет человеческую константную область или константную область человеческого происхождения. В другом воплощении антитело относится к подклассу IgG4 или подклассу IgG1, IgG2 или IgG3, который модифицирован таким образом, что ни связывание Fcγ-рецептора (например, FcγRIIIa), ни связывание C1q не может быть обнаружено. В одном воплощении антитело является человеческим антителом подкласса IgG4 или мутантным человеческим антителом подкласса IgG1. В одном воплощении изобретения антитело является человеческим антителом подкласса IgG1 с мутациями L234A и L235A. В другом воплощении антитело относится к Fcγ-рецепторному связыванию подкласса IgG4 или подкласса IgG1 или IgG2 с мутацией в L234, L235 и/или D265, и/или содержит мутацию PVA236. В другом воплощении антитело имеет мутацию, выбранную среди S228P, L234A, L235A, L235E, SPLE (S228P и L235E) и/или PVA236 (PVA236 означает, что аминокислотная последовательность ELLG (дана в однобуквенном коде аминокислот) в позиции аминокислот с 233 по 236 в IgG1 или EFLG в IgG4 заменяется на PVA). В одном воплощении антитело относится к подклассу IgG4 и имеет мутации S228P в IgG4, или антитело относится к подклассу IgG1 и имеет мутации L234A и L235A.
Вариабельный домен легкой или тяжелой цепи иммуноглобулина, в свою очередь, включает различные сегменты, т.е. четыре каркасные области (framework region, FR) и три гипервариабельные области (от англ. - complementarity determining regions (CDR) - участок, определяющий комплементарность).
Термин "обработка бисульфитом" обозначает реакцию превращения (конверсии) оснований цитозина в нуклеиновой кислоте в основания урацила в присутствии бисульфит-ионов, в результате чего 5-метилцитозиновые основания незначительно преобразуются. Эта реакция для обнаружения метилированного цитозина подробно описана в Frommer et al. (Frommer, M., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 1827-1831) и в Grigg and Clark (Grigg, G.W. and Clark, S., Bioessays 16 (1994) 431-436; Grigg, G.W., DNA Seq. 6 (1996) 189-198). Бисульфитная реакция включает этап дезаминирования и этап десульфонирования, которые могут быть проведены по отдельности или одновременно. Утверждение, что 5-метилцитозиновые основания незначительно преобразуется, только принимает во внимание тот факт, что нельзя исключить, что небольшой процент 5-метилцитозиновых оснований превращается в урацил, хотя они предназначены для конверсии только и исключительно в (неметилированные) основания цитозина.
Термин "клетка" обозначает клетку, в которую введена или может быть введена нуклеиновая кислота (которой клетка трансфицирована), например, кодирующая полипептид, возможно, гетерологичный. Термин "клетка" включает как прокариотические клетки, которые используются для размножения плазмид, так и эукариотические клетки, которые используются для экспрессии нуклеиновой кислоты. В одном воплощении клетка является эукариотической клеткой, а в другом воплощении эукариотическая клетка является клеткой млекопитающего. В другом воплощении клетка млекопитающего выбрана из группы клеток млекопитающих, включающей клетки СНО (например, СНО К1, СНО DG44), клетки BHK, клетки NS0, клетки SP2/0, клетки НЕК 293, клетки НЕК 293 EBNA, клетки PER.C6® и клетки COS. Используемое в данном документе выражение "клетка" включает рассматриваемую клетку и ее потомство. Таким образом, термин "клетка" обозначает первичную рассматриваемую клетку и культуры, полученные из нее, независимо от числа переносов. Также нужно понимать, что все потомство может не быть абсолютно идентичным по содержанию ДНК из-за случайных или неслучайных мутаций. В данный документ включено вариантное потомство, которое имеет такую же функцию или биологическую активность, которая показана для исходно трансформированной клетки.
Термин "CpG-сайт" обозначает динуклеотид CG в пределах нуклеиновой кислоты, который может быть распознан метилирующими ферментами клетки, и в котором цитозин может быть преобразован в 5-метилцитозин. В одном воплощении CpG-сайт находится в пределах промоторной нуклеиновой кислоты.
Термин "экспрессионная кассета" обозначает конструкцию, содержащую необходимые регуляторные элементы, такие как промотор и сайт полиаденилирования, для экспрессии в клетке по меньшей мере содержащейся нуклеиновой кислоты.
Термин "экспрессионная плазмида" обозначает нуклеиновую кислоту, которая предоставляет все необходимые элементы для экспрессии в клетке содержащегося структурного гена (генов). Как правило, экспрессионная плазмида содержит блок для прокариотического размножения плазмиды, например для Е. coli, содержащий точку начала репликации (ориджин) и маркер для селекции, маркер для эукариотической селекции и одну или более чем одну экспрессионную кассету для экспрессии представляющего интерес структурного гена (генов), каждая из которых содержит промоторную нуклеиновую кислоту, структурный ген и терминатор транскрипции, включая сигнал полиаденилирования. Экспрессия гена, как правило, находится под контролем промоторной нуклеиновой кислоты, и такой структурный ген называется "функционально связанным" с промоторной нуклеиновой кислотой. Аналогичным образом, регуляторный элемент и осевая промоторная нуклеиновая кислота функционально связаны, если регуляторный элемент модулирует активность осевой промоторной нуклеиновой кислоты.
Термин "время одного поколения" означает время, необходимое клетке для деления и производства дочерней клетки. Таким образом, клетка, которая один раз поделилась, имеет возраст одного поколения. Термин "поколение" обозначает номер клеточного деления клетки.
Термин "высокая частота" означает, что на этом сайте метилирования цитозин метилируется чаще, чем на других сайтах метилирования, что определяется на основании анализа метилирования статистически значимого количества отдельных клеток или ДНК-клонов, соответственно. Это статистически значимое число в одном воплощении составляет по меньшей мере 10 отдельных клеток или ДНК-клонов, соответственно, в другом воплощении по меньшей мере 15 отдельных клеток или ДНК-клонов, соответственно, и в другом воплощении по меньшей мере 20 отдельных клеток или ДНК-клонов, соответственно. В одном воплощении проанализированы максимум 400 клеток или ДНК-клонов, соответственно.
Термин "долгосрочно продуцирующая клетка" обозначает клетку, которая продуцирует полипептид, в одном воплощении гетерологичный полипептид, где удельная скорость продукции клеткой является почти постоянной в течение 30 поколений. В одном воплощении долгосрочно продуцирующая клетка имеет удельную скорость продукции, которая является почти постоянной в течение 30 поколений, в другом воплощении по меньшей мере в течение 45 поколений, и в другом воплощении по меньшей мере в течение 60 поколений. В одном воплощении долгосрочно продуцирующая клетка имеет удельную скорость продукции, которая является почти постоянной в течение 60 поколений, в другом воплощении в течение 75 поколений, и в другом воплощении в течение 90 поколений.
Термин "метилирование" обозначает процесс в клетке, которая была трансфицирована нуклеиновой кислотой, содержащей структурный ген, кодирующий полипептид, функционально связанный с промотором, во время которого цитозин промоторной нуклеиновой кислоты преобразуется в 5-метилцитозин. Промоторная нуклеиновая кислота, в которой по меньшей мере один цитозин преобразован в 5-метилцитозин, обозначается как "метилированная" нуклеиновая кислота.
Термин "моноклональное антитело", используемый в данном документе, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантных антител, например, содержащих природные мутации или возникающих в процессе производства препарата моноклональных антител, при этом такие варианты, как правило, присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело препарата моноклонального антитела направлено против одной и той же детерминанты на антигене. Таким образом, понятие "моноклональное" указывает на характер антитела, полученного из по существу гомогенной популяции антител, и не должно быть истолковано как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела, которые будут использоваться в соответствии с данным изобретением, могут быть получены с помощью различных методик, включая, но не ограничиваясь ими, гибридомную методику, методики рекомбинантной ДНК, методики фагового дисплея, а также методики, использующие трансгенных животных, содержащих все или часть локусов человеческого иммуноглобулина, при этом такие методики и другие иллюстративные методики получения моноклональных антител описаны в данном документе. В одном воплощении антитело представляет собой моноклональное антитело.
Термин "функционально связанный" обозначает сопоставление двух или более чем двух компонентов, где компоненты, описанные таким образом, находятся в отношениях, которые позволяют им функционировать предназначенным им образом. Например, промотор и/или энхансер функционально связан с кодирующей последовательностью, если он контролирует или модулирует транскрипцию связанной последовательности. Как правило, но не обязательно, "функционально связанные" последовательности ДНК являются смежными и, когда необходимо соединить две области, кодирующие белок, например, секреторный лидер и полипептид, они являются смежными и находятся в одной рамке (считывания). Тем не менее, несмотря на то, что функционально связанный промотор, как правило, расположен выше кодирующей последовательности, он не обязательно должен быть смежным с ней. Энхансеры не обязательно должны быть смежными. Энхансер функционально связан с кодирующей последовательностью, если он увеличивает транскрипцию кодирующей последовательности. Функционально связанные энхансеры могут быть расположены выше, в пределах или ниже кодирующих последовательностей, а также на значительном расстоянии от промотора. Сайт полиаденилирования функционально связан с кодирующей последовательностью, если он расположен на нижнем конце кодирующей последовательности, так что транскрипция проходит через кодирующую последовательность в последовательность полиаденилирования. Стоп-кодон трансляции функционально связан с экзонной нуклеиновокислотной последовательностью, если он находится на нижнем конце (3'-конце) кодирующей последовательности, так что трансляция проходит через кодирующую последовательность до стоп-кодона и заканчивается на нем. Связывание осуществляется с помощью рекомбинантных способов, известных в данной области, например, с помощью методики ПЦР или путем лигирования на подходящих сайтах рестрикции. Если подходящих сайтов рестрикции нет, то в соответствии с обычной практикой используются синтетические олигонуклеотидные адаптеры или линкеры.
Термин "полипептид" обозначает полимер, состоящий из аминокислот, соединенных пептидными связями, полученный природным или синтетическим путем. Полипептиды длиной менее примерно 20 аминокислотных остатков могут быть названы "пептидами", в то время как молекулы, содержащие два или более двух полипептидов или содержащие один полипептид длиной более 100 аминокислотных остатков, могут быть названы "белками". Полипептид может также содержать неаминокислотные компоненты, такие как углеводные группы, ионы металлов или сложные эфиры карбоновых кислот. Неаминокислотные компоненты могут быть добавлены клеткой, в которой экспрессируется полипептид, и они могут варьировать в зависимости от типа клетки. Полипептиды в данном документе определены в плане структуры их аминокислотных остовов или нуклеиновых кислот, их кодирующих. Добавки, такие как углеводные группы, как правило, не уточняются, но могут, тем не менее, присутствовать.
Термин "вариант" промоторной нуклеиновой кислоты означает, что в промоторной нуклеиновой кислоте один или более чем один нуклеотид изменен без влияния на функцию промоторной нуклеиновой кислоты. Такое изменение может быть проведено для удаления или введения рестрикционного сайта.
Термин "продукция" обозначает экспрессию в клетке структурного гена, вставленного в экспрессионную кассету. Этот термин включает процессы транскрипции и трансляции нуклеиновой кислоты. Продукция осуществляется в соответствующих прокариотических или эукариотических клетках, и экспрессированный, т.е. продуцированный, полипептид может быть выделен из клеток после их лизиса или из культурального супернатанта.
Термин "промоторная нуклеиновая кислота" обозначает полинуклеотидную последовательность, которая контролирует транскрипцию гена/структурного гена или нуклеиновокислотной последовательности, с которой она функционально связана. Промоторная нуклеиновая кислота включает сигналы для связывания РНК-полимеразы и для инициации транскрипции. Используемая промоторная нуклеиновая кислота будет функционировать в клетке, в которой предполагается экспрессия выбранного структурного гена. Большое количество промоторных нуклеиновых кислот, включая конститутивные, индуцируемые и репрессируемые промоторы из различных источников, хорошо известно в данной области (и определено в базах данных, таких как GenBank) и доступно в виде клонированных полинуклеотидов или в их пределах (например, из таких депозитариев как АТСС, а также других коммерческих или индивидуальных источников).
Как правило, промоторная нуклеиновая кислота находится в 5'-некодирующей или нетранслируемой области гена, проксимальнее сайта начала транскрипции структурного гена. Элементы последовательности в пределах промоторных нуклеиновых кислот, которые функционируют в инициации транскрипции, часто характеризуются консенсусными нуклеотидными последовательностями. Эти элементы включают сайты связывания РНК-полимеразы, ТАТА-последовательности, СААТ-последовательности, специфические для дифференцировки элементы (differentiation-specific element, DSE), цАМФ-отвечающие элементы (cyclic AMP response element, CRE), элементы ответа на сыворотки (serum response element, SRE), элементы ответа на глюкокортикоиды (glucocorticoid response element, GRE), а также сайты связывания других транскрипционных факторов, таких как CRE/ATF, АР2, SP1, белок, связывающий цАМФ-отвечающие элементы (cAMP response element binding protein, CREB) и октамерные факторы. Если промоторная нуклеиновая кислота является индуцируемой промоторной нуклеиновой кислотой, то скорость транскрипции увеличивается в ответ на индуцирующий агент, например, нуклеиновая кислота промотора CMV, за которой следуют два тет-операторных сайта, нуклеиновые кислоты промоторов металлотионеина и гена теплового шока. Скорость транскрипции не регулируется индуцирующим агентом, если промоторная нуклеиновая кислота является конститутивно активной промоторной нуклеиновой кислотой. Среди эукариотических промоторных нуклеиновых кислот, которые были определены как сильные промоторные нуклеиновые кислоты для экспрессии, имеется нуклеиновая кислота раннего промотора SV40, нуклеиновая кислота главного позднего промотора аденовируса, нуклеиновая кислота промотора мышиного металлотионеина-I, длинный концевой повтор вируса саркомы Рауса, фактор элонгации 1 альфа китайского хомяка (CHEF-1), EF-1 альфа человека, убиквитин и нуклеиновая кислота главного немедленно-раннего промотора цитомегаловируса человека (hCMV-MIE).
Термин "селективный маркер" означает нуклеиновую кислоту, которая позволяет клеткам, несущим ее, быть специфически отобранными, положительно или отрицательно, в присутствии соответствующего селективного агента. Как правило, селективный маркер придает устойчивость к лекарственному препарату или компенсирует метаболический или катаболический дефект в клетке, в которую он введен. Селективный маркер может быть положительным, отрицательным или бифункциональным. Используемым положительным селективным маркером является ген устойчивости к антибиотику, который позволяет отбирать клетки, трансформированные им, в присутствии соответствующего селективного агента, например, антибиотика. Нетрансформированная клетка не способна расти и выживать в селективных условиях, т.е. в присутствии селективного агента. Отрицательные селективные маркеры позволяют селективно устранять клетки, несущие маркер. Селективные маркеры, используемые с эукариотическими клетками, включают, например, структурные гены, кодирующие фосфотрансферазу аминогликозидов (aminoglycoside phosphotransferase, АРН), гигромицин (hyg), неомицин (neo) и селективный маркер G418, дигидрофолатредуктазу (dihydrofolate reductase, DHFR), тимидинкиназу (thymidine kinase, tk), глутаминсинтетазу (glutamine synthetase, GS), аспарагинсинтетазу, триптофансинтетазу (селективный агент индол), гистидинолдегидрогеназу (селективный агент гистидинол D), и нуклеиновые кислоты, придающие устойчивость к пуромицину, блеомицину, флеомицину, хлорамфениколу, зеоцину и микофенольной кислоте.
Термин "скорость краткосрочной продукции" обозначает количество полипептида, произведенного одной клеткой в течение одного дня, определенное из количества полипептида, произведенного в течение определенного периода времени, и плотности жизнеспособных клеток, где период времени является коротким. В одном воплощении краткосрочным является культивирование длительностью от 2 до 20 дней, в другом воплощении от 4 до 15 дней, в еще одном воплощении от 10 до 14 дней.
Термин "удельная скорость продукции" или "скорость продукции" обозначает количество полипептида, произведенного одной клеткой в течение одного дня, определенное из количества полипептида, произведенного в течение определенного периода времени, и плотности жизнеспособных клеток. Удельная скорость продукции (SPR) может быть рассчитана по следующей формуле:

где
SPR [пг/клетка/день]: удельная скорость продукции,
P1 [мкг/мл]: концентрация полипептида в начале периода времени,
Р2 [мкг/мл]: концентрация полипептида в конце периода времени,
D1 [клеток/мл]: плотность жизнеспособных клеток в начале периода времени,
D2 [клеток/мл]: плотность жизнеспособных клеток в конце периода времени,
Δt [дни]: продолжительность периода времени.
Термин "структурный ген" обозначает область гена без сигнальной последовательности, т.е. кодирующую область.
В. Способы, описанные в данном документе
Клеточные линии/клеточные клоны, продуцирующие полипептид, т.е. клетки, трансфицированные экспрессионной кассетой, содержащей нуклеиновую кислоту, содержащую структурный ген, кодирующий гетерологичный полипептид, могут быть сгруппированы в разные классы: в первом классе клеток удельная скорость продукции является почти постоянной в течение множества поколений. Напротив, во втором классе клеток удельная скорость продукции снижается, особенно монотонно убывает, в течение множества поколений. Не привязываясь к этой теории, убывающая продуктивность продуцирующих полипептид клеток и клеточных линий, соответственно, обусловлена, по меньшей мере частично, неуклонным увеличением метилирования и связанной с ним инактивацией/молчанием промоторной нуклеиновой кислоты, функционально связанной со структурным геном, кодирующим полипептид, представляющий интерес. Также уменьшение может быть связано с потерей копий структурного гена, кодирующего белок, представляющий интерес.
Данное изобретение основано, по меньшей мере частично, на обнаружении того факта, что CpG-динуклеотид в позиции 425 главного немедленно-раннего промотора/энхансера CMV человека (hCMV-MIE) (SEQ ID NO 01) часто является метилированным в клеточных линиях яичника китайского хомяка (СНО), нестабильно продуцирующих антитела. Это можно использовать в качестве маркера для предсказания долгосрочной стабильности рекомбинантной продукции рекомбинантным клеточным клоном/клеточной линией, экспрессирующей представляющий интерес белок, кодируемый структурным геном, функционально связанным с промотором hCMV-MIE (например, SEQ ID NO 01).
Данное изобретение основано, по меньшей мере частично, на обнаружении того факта, что посттрансляционная модификация гистона, примыкающего к промотору, может быть использована в качестве другого маркера для выявления стабильно экспрессирующих/продуцирующих клеточных линий. Наличие инактивирующей модификации является маркером того, что соответствующий клеточный клон/клеточная линия является очень нестабильной и покажет снижение продуктивности при дальнейшем культивировании выше среднего (в долгосрочной перспективе нестабильно продуцирующий клеточный клон/клеточная линия). С другой стороны, наличие активирующей модификации является маркером того, что соответствующий клеточный клон/клеточная линия является очень стабильной и покажет снижение продуктивности при дальнейшем культивировании ниже среднего (в долгосрочной перспективе стабильно продуцирующий клеточный клон/клеточная линия).
Данное изобретение основано, по меньшей мере частично, на обнаружении того факта, что, используя комбинацию метилирования в позиции 425 в SEQ ID NO 01 и относительного уровня ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) вблизи промоторной нуклеиновой кислоты, можно достичь дальнейшего улучшения в выявлении долгосрочных стабильно продуцирующих рекомбинантных клеточных линий/клеточных клонов.
В.1. Метилирование промотора
Было обнаружено, что присутствие обнаруживаемого метилирования в позиции 425 в нуклеиновой кислоте MIE-промотора CMV человека, которая функционально связана со структурным геном, кодирующим полипептид, предоставляет информацию о долгосрочной продуктивности клеточной линии или клеточного клона, соответственно.
Каждая промоторная нуклеиновая кислота, используемая для экспрессии структурного гена, включает сайты, склонные к метилированию ферментами клетки, в которую она была введена, если промоторная нуклеиновая кислота не экранируется защитными элементами. Сайт, поддающийся метилированию, называется CpG-сайтом и включает/состоит из динуклеотида CG. Но не все CpG-сайты метилируются с одной и той же относительной частотой - некоторые из CpG-сайтов метилируются чаще, чем другие. Было обнаружено, что некоторые сайты, например, в MIE-промоторе CMV человека, метилированы с разной частотой и имеют различное влияние на молчание промотора.
Для определения CpG-сайта в нуклеиновокислотной последовательности может быть использован следующий способ:
1) получение клетки с такой скоростью продукции полипептида, которая после культивирования клетки в течение 30 поколений в отсутствие селективного агента составляет менее 90% от скорости продукции клеткой после первого поколения культивирования,
2) отдельное выделение ДНК по меньшей мере из 10 клеток, полученных путем культивирования клетки из этапа 1),
3) модификация цитозина выделенной ДНК путем обработки бисульфитом,
4) определение CpG-сайта в промоторной нуклеиновой кислоте, функционально связанной со структурным геном, кодирующим полипептид, с частотой метилирования по меньшей мере 0,2 на основе ДНК, полученной в этапе 3), и тем самым выявление CpG-сайта.
Одним из критериев (т.е. в одном воплощении) нестабильного клеточного клона/клеточной линии является скорость продукции клеточного клона/клеточной линии после культивирования в течение 30-60 поколений, которая составляет 60% или меньше от скорости продукции клеточного клона/клеточной линии после первого поколения культивирования.
В одном воплощении клеточная линия имеет скорость продукции после культивирования в течение 30-60 поколений более 60% от скорости продукции в начале культивирования.
Этап модификации цитозина выделенной ДНК путем обработки бисульфитом может включать следующие этапы:
3-а) инкубация выделенной ДНК в присутствии ионов сульфита, в результате чего ДНК дезаминируется, и
3-b) инкубация дезаминированной ДНК в щелочных условиях, в результате чего дезаминированная ДНК десульфонируется.
Способ получения клеточного клона/клеточной линии, продуцирующей полипептид, представляет собой процесс, включающий по меньшей мере один этап трансфекции и по меньшей мере один этап отбора, в том числе депонирование отдельных успешно трансфицированных клеток либо непосредственно после трансфекции, либо после выращивания в присутствии селективного агента. На этапе отбора клетки выявляли на основании их краткосрочной удельной скорости продукции, т.е. на основании концентрации полипептида в супернатанте после краткосрочного культивирования. Среди отобранных клеток некоторые обладают удельной скоростью продукции, которая является почти постоянной в течение множества поколений, а другие обладают удельной скоростью продукции, которая монотонно убывает в течение множества поколений. Таким образом, с помощью обычно применяемых (краткосрочных) критериев отбора нельзя провести специфический отбор клетки или клеток со способностью к стабильной долгосрочной продукции.
В способе, о котором сообщается в данном документе, можно проанализировать любой клеточный клон/клеточную линию, полученную путем трансфекции экспрессионной плазмидой, содержащей экспрессионную кассету, которая содержит промоторную нуклеиновую кислоту, функционально связанную со структурным геном, кодирующим полипептид, представляющий интерес, который будет продуцироваться трансфицированной клеткой. Экспрессионная плазмида, как правило, также включает селективный маркер. Таким образом, в одном воплощении клетки культивируют в присутствии селективного агента после этапа трансфекции и до этапа отбора. Альтернативно, способ включает культивирование клеток без предварительного депонирования отдельных клеток или ограниченного разведения, т.е. в виде пула, в присутствии селективного агента. Другой альтернативный способ может включать культивирование клеток после депонирования отдельных клеток или ограниченного разведения.
После культивирования/отбора пула должно быть выполнено депонированиеотдельной клетки. Если депонирование отдельной клетки выполняется после этапа культивирования пула, то клетки также культивируют после депонирования отдельной клетки.
Для того чтобы выявить клеточные линии или клеточные клоны с удельной скоростью продукции, которая является почти постоянной в течение множества поколений, нужно определить концентрацию полипептида в супернатанте и плотность жизнеспособных клеток в определенные моменты культивирования при длительном культивировании в течение множества поколений. В данной области известны способы, подходящие для этого, такие как иммуноферментный анализ (ELISA, от англ. - enzyme-linked immunosorbent assay) и сортировка флуоресцентно-активированных клеток (FACS, от англ. - fluorescence-activated cell sorting), соответственно.
CpG-сайт с высокой частотой метилирования можно выявить путем обработки одноцепочечной ДНК бисульфитом, например, при рН 5, с последующим щелочным десульфонированием. При этом можно различить метилированные и неметилированные CpG-сайты. В соответствии с конкретными условиями обработки цитозин, в отличие от 5-метилцитозина, дезаминируется в позиции 4 N-гетероцикла и превращается в урацил. Комплементарные нити ДНК превращаются в две нити, нить А и нить В, которые больше не являются комплементарными. Определение можно провести на основании любой из этих нитей.
Это длительное культивирование можно выполнить только один раз для промоторной нуклеиновой кислоты или комбинации промоторной нуклеиновой кислоты и клеточной линии. Если та же промоторная нуклеиновая кислота или комбинация используется второй раз, то отбор может быть основан на уже собранных данных.
Чтобы показать различия в последовательностях метилированных и неметилированных аллелей после обработки бисульфитом, можно использовать ряд методик. Например, последовательность, представляющая интерес (нить А или нить В), может быть амплифицирована с помощью ПЦР в неспецифических в отношении метилирования условиях, т.е. с праймером, не чувствительным к сайтам метилирования, а затем проанализирована такими способами как секвенирование ДНК (с клонированием или без него), высокочувствительный анализ плавления или микрочиповый анализ. В другом воплощении может быть выполнена количественная ПЦР (кПЦР) с чувствительным в отношении метилирования или специфическим в отношении метилирования праймером или зондом. В альтернативном подходе метилированную ДНК осаждают 5-метилцитозин-специфическими антителами, после чего проводят количественную полимеразную цепную реакцию.
Специфическая в отношении метилирования ПЦР (methylation specific PCR, MSP) также может быть использована для работы с последовательностью ДНК, обработанной бисульфитом, непосредственно без предварительной ПЦР-амплификации интересующей области. Праймеры, используемые в MSP, будут содержать один или более чем один CpG-сайт. Они будут либо комплементарны неизмененному 5-метилцитозину для обнаружения метилированной ДНК, либо комплементарны урацилу, образованному из цитозина, для обнаружения неметилированной ДНК.
Метилирование CpG-сайтов определяют для некоторого числа клеток. В одном воплощении число клеток составляет по меньшей мере 10, в другом воплощении по меньшей мере 15, а в другом воплощении по меньшей мере 20. Затем для каждого CpG-сайта рассчитывают частоту метилирования, т.е. для каждого CpG-сайта рассчитывают число клеток, метилированных на этом CpG-сайте, и делят на общее число проанализированных клеток. CpG-сайт с высокой частотой метилирования представляет собой CpG-сайт, который имеет частоту метилирования по меньшей мере 0,2, в одном воплощении по меньшей мере 0,25, в одном воплощении по меньшей мере 0,4, и в одном предпочтительном воплощении по меньшей мере 0,5.
Было обнаружено, что даже низкие уровни метилированной промоторной нуклеиновой кислоты, т.е. выше заданного порогового значения, в клетке или клеточной линии, используемой для продукции полипептида, могут быть использованы для предсказания снижения продуктивности при долгосрочном культивировании.
На фиг. 1 показан ряд метилированных CpG-сайтов одной и той же промоторной нуклеиновой кислоты, полученной из различных клеток.
Промотор цитомегаловируса человека CMV-MIE в виде нуклеиновокислотной последовательности, показанной далее (CpG-сайты подчеркнуты):

(SEQ ID NO 01).
В SEQ ID NO 01 присутствует тридцать три CpG-сайта, которые являются потенциальными сайтами для метилирования нуклеиновой кислоты. С помощью способа, описанного выше, в промоторе цитомегаловируса человека CMV-MIE можно выявить остатки цитозина, которые преимущественно метилированы.
Нить А со всеми сохраненными CpG-сайтами имеет нуклеотидную последовательность

(SEQ ID NO 02), а полностью дезаминированная нить А имеет нуклеотидную последовательность

(SEQ ID NO 03). Нить В со всеми сохраненными CpG-сайтами имеет нуклеотидную последовательность

(SEQ ID NO 04), а нить В в полностью дезаминированной форме имеет нуклеотидную последовательность


(SEQ ID NO 05).
На фиг. 4 показана частота метилирования отдельных CpG-сайтов в различных клеточных линиях. Количества были определены на основе анализа от 19 до 22 различных клонов, полученных из разных родительских клеточных линий СНО после трансфекции плазмидой, содержащей экспрессионную кассету для экспрессии полипептида, представляющего интерес. Показаны характер (паттерн) метилирования отдельных ДНК (внизу) и частота метилирования отдельных CpG-сайтов (наверху) для каждой клеточной линии. Клеточная линия K18.1, например, является высоко метилированной (фиг. 4А). Частота метилирования не одинаковая на различных CpG-сайтах, но, по-видимому, есть центры в 3 кластерах, т.е. на 5'-конце, 3'-конце и в районе позиции (или нуклеотида, соответственно) 400. 14 из 22 секвенированных вставок имели цитозин в позиции 425. На фиг. 4Е показано метилирование промоторной нуклеиновой кислоты в клеточной линии 43-16 А10. Распределение метилирования похоже на распределение, наблюдаемое для клеточной линии K18.1. Как и у K18.1, позиция 425 является наиболее часто метилированной: пять из двадцати секвенированных вставок содержали цитозин в этой позиции.
В трех других анализируемых клеточных линиях цитозин обнаруживался спорадически, т.е. как отдельные события, в разных CpG-сайтах (фиг. 4В, 4С и 4D). Для получения статистического значения для клеточных линий с низким общим метилированием потребовалось бы секвенирование сотен вставок. Кроме того, эти отдельные события могут также представлять ложноположительные события в связи с неполным дезаминированием цитозина, а не фактическим метилированием промотора.
Для надежного определения специфического метилирования позиции CpG был разработан способ ПЦР, специфический в отношении метилирования. Для специфической в отношении метилирования ПЦР могут быть использованы праймеры, указанные в следующей таблице 1. Эти праймеры либо самостоятельно, либо в комбинации являются аспектом, о котором сообщается в данном документе


В ходе оценки праймеров было установлено, что пары праймеров, специфические в отношении метилирования, которые являются высоко селективными для дезаминированной ДНК промотора CMV с цитозином в позиции 425, отличаются по своим свойствам (см. фиг. 6). В одном воплощении праймеры для специфической в отношении метилирования ПЦР имеют нуклеотидную последовательность SEQ ID NO 14 и SEQ ID NO 18.
Таким образом, в одном воплощении способов, о которых сообщается в данном документе, промоторная нуклеиновая кислота является нуклеиновой кислотой промотора CMV человека SEQ ID NO 01. В одном воплощении CpG-сайт с высокой частотой метилирования представляет собой CpG-сайт в позиции (bp) 425 в SEQ ID NO 01.


Универсальная пара праймеров должна амплифицировать все четыре матрицы, в то время как пары праймеров MSP должны селективно амплифицировать матрицу №62 (SEQ ID NO 22) и матрицу №01 (SEQ ID NO 23). Значение ΔCp между парой праймеров MSP и универсальной парой праймеров на матрице №62 и на матрице №01 должно быть как можно меньше. Напротив, значения Ср, полученные с парой праймеров MSP на матрицах №11 (SEQ ID NO 21) и №04 (SEQ ID NO 24), должны быть как можно выше, т.е. ΔСр по сравнению с амплификацией с универсальной парой праймеров должно быть максимальным.
Специфический в отношении метилирования праймер должен быть способен обнаруживать 5-метилцитозин селективно в позиции 425, хотя в позициях 416 и 437 присутствуют еще две другие позиции метилирования.
Определение метилирования является возможным при частоте метилирования от 1% до 100%.
При специфической в отношении метилирования ПЦР и при клонировании и секвенировании были обнаружены сопоставимые результаты по степени выявленного метилирования, но специфическая в отношении метилирования ПЦР является гораздо более чувствительной. Клеточные линии со сниженной продуктивностью при долгосрочной продукции имеют метилирование в позиции 425 с частотой метилирования выше порогового значения. Пороговое значение в одном воплощении в два раза превышает фоновый шум способа определения. Клеточная линия с долгосрочной стабильной продуктивностью имеет частоту метилирования на CpG-сайте ниже порогового значения.
Для высокочувствительного анализа плавления промоторную нуклеиновую кислоту амплифицируют, начиная с позиции 334 и до позиции 487, т.е. 154 пар оснований. Иллюстративный анализ плавления показан на фиг. 10А, а его первая производная на фиг. 10 В. Видно, что при высокочувствительном анализе плавления метилированную промоторную нуклеиновую кислоту (матрица №16, SEQ ID NO 25) можно отличить от неметилированной промоторной нуклеиновой кислоты (матрица №11). Метилированный промоторный нуклеиновокислотный фрагмент можно обнаружить с относительной частотой 50% или более. Неметилированный промоторный фрагмент можно обнаружить с относительной частотой 10% и более.
Эти данные показывают, что стабильность продукции рекомбинантных клеточных линий СНО, которые содержат рекомбинантно введенные гены, экспрессия которых управляется главным немедленно-ранним промотором/энхансером CMV человека, может быть предсказана путем измерения статуса метилирования цитозина в позиции 425.
Таким образом, было установлено, что определение метилирования С425 может использоваться в качестве прогностического маркера для определения стабильности экспрессии полипептида в созданных рекомбинантных клеточных клонах. Это позволяет отбирать при разработке клеточной линии стабильные клоны со стабильной продуктивностью. Кроме того, было обнаружено, что метилирование С425 на уровне 5% или менее является подходящим критерием для отбора стабильных клеточных клонов (нужно учитывать пороговые значения используемых способов обнаружения). Также было установлено, что фракцию клеточных клонов, которые ложно предсказаны как стабильные (ложноотрицательные клеточные клоны), можно уменьшить путем их культивирования в течение некоторого времени в отсутствие метотрексата (МТХ, от англ. - methotrexate) перед анализом.
Таким образом, в данном документе описан способ обогащения клеточными линиями, долгосрочно (стабильно) экспрессирующими рекомбинантный полипептид, популяции трансфицированных клеток путем отбора клеток, имеющих относительное метилирование промотора в позиции 425 в SEQ ID NO 01, составляющее 5% или меньше. Эту относительную частоту метилирования можно определить с использованием специфического в отношении метилирования способа кПЦР, описанного в данном документе, с помощью праймеров, описанных в данном документе.
Имея принятый чувствительный и точный способ ПЦР для количественной оценки метилирования нуклеотида hCMV-MIE на сайте CpG 425, оценивали метилирование в рекомбинантных клеточных линиях СНО K18.1, 43 16 А10 и G45-2. Обработанную бисульфитом ДНК, которая были проанализирована путем секвенирования, использовали в качестве матрицы в специфической в отношении метилирования кПЦР в режиме реального времени либо непосредственно, либо после ПЦР-амплификации полной области hCMV-MIE (фиг. 8А и 8В). Эти две схемы анализа обеспечивали сопоставимые результаты. Более того, они хорошо коррелировали с результатами бисульфитного секвенирования. Это показывает, что специфический в отношении метилирования анализ кПЦР CpG-сайта 425 можно использовать для измерения метилирования hCMV-MIE на сайте CpG-425 в рекомбинантных клеточных линиях СНО и можно использовать без предварительной ПЦР-амплификации ДНК-мишени.
В принципе, чтобы предсказать нестабильность продукции, могут быть изучены другие CpG-сайты в ДНК главного немедленно-раннего промотора/энхансера CMV. Тем не менее, метилирование С425 оказалось примерно в 5 раз выше, чем среднее метилирование на всех сайтах CpG. Кроме того, некоторые сайты не были метилированы вообще даже в высоко метилированных клеточных клонах, например, С280 и С289 (фиг. 4F, 4J и 13). При выборе для анализа метилирования промотора правильного, т.е. часто модифицированного, CpG-сайта анализ становится более чувствительным. Значительное метилирование клонов G25-17, G25-10 и А10 43-16, которое составляет примерно 10%, скорее всего, будет пропущено, если CpG-сайт для анализа будет выбран случайно.
Прогноз стабильности в отношении статуса метилирования релевантных CpG-сайтов в промоторной нуклеиновой кислоте может быть также использован для других гетерогенных промоторных нуклеиновых кислот.
Чтобы оценить потенциальную корреляцию метилирования раннего промотора и нестабильности продукции, в начале исследования стабильности метилирование CpG-сайта 425 откладывали на графике в виде зависимости от относительного изменения qP в присутствии или в отсутствие селективного агента (МТХ). Графики корреляции показаны на фиг. 11А (с МТХ) и 11В (без МТХ). Для оценки площадь графика разделяли на четыре компартмента с границами на 5%-ном метилировании (равном двух-трехкратному фону измеренного метилирования, т.е. пределу обнаружения) и на 40%-ном снижении qP, представляющем допустимый предел стабильности продукции (это также является воплощением способа, описанного в данном документе). Было определено число клонов в разных компартментах. Анализ сопряженности статуса стабильности и статуса метилирования с помощью теста хи-квадрат по Пирсону демонстрировал значимую связь с р-значением 0,05 для культивирования с МТХ и тенденцию с р-значением 0,13 для культивирования без МТХ. Оказалось, что большинство клонов с менее чем 5% метилированием CpG-сайта 425 входят во фракцию стабильных клонов (менее чем 40% снижение qP с МТХ или без него, показанное в верхних левых компартментах).
Таким образом, резюмируя сказанное выше, потеря продуктивности при долгосрочном культивировании и масштабном производстве является основным риском в разработке продуцирующих клеточных линий. Таким образом, существует необходимость молекулярных маркеров нестабильности продукции, которые могут быть легко и быстро определены и проверены. Было установлено, что метилирование промотора может быть использовано для предсказания потери продуктивности в рекомбинантных клеточных линиях СНО. Для оценки этого анализировали метилирование ДНК на 33 CpG-сайтах в области длиной 603 пар оснований широко используемого промотора/энхансера hCMV-MIE. Общий уровень метилирования исследуемой области колебался примерно от 1% до 18% от всех CpG-сайтов. Кажущееся метилирование 1% представляет собой технический фон, который возникает при неполном дезаминировании неметилированных цитозинов. Кроме того, в метилированных промоторах уровень метилирования сильно различается между отдельными CpG-сайтами и накапливается в трех кластерах с пшксимумом на CpG-сайте 425. Метилирование на CpG-сайте 425 оказалось примерно в 5 раз выше, чем средняя степень метилирования всех других CpG-сайтов. С другой стороны, некоторые CpG-сайты, по-видимому, полностью неметилированы даже в высоко метилированных клеточных линиях. Общее метилирование hCMV-MIE, а также распределение метилирования между отдельными CpG-сайтами, могут значительно различаться между клеточными типами и тканями (см., например, Kong, Q., et al., PLoS One 4 (2009) e6679; Krishnan, M., et al., FASEB J. 20 (2006) 106-108; Mehta, A.K., et al., Gene 428 (2009) 20-24).
Было установлено, что доминантное метилирование CpG-сайта 425 является подходящим маркером метилирования hCMV-MIE. Специфическая в отношении метилирования кПЦР на CpG-сайте 425 была принята в качестве быстрого и чувствительного способа со средней продуктивностью. При анализе большого числа клеточных линий посредством кПЦР, специфической в отношении метилирования CpG-сайта 425, было обнаружено, что большинство нестабильных продуцентов демонстрируют более чем 5% метилирование на CpG-сайте 425 даже до длительного культивирования, тогда как большинство стабильных продуцентов демонстрируют менее чем 5% метилирование на этом сайте.
Раннее метилирование CpG-сайта 425 было обнаружено только у клонов, несущих более 10 копий гетерологичной плазмиды. Предыдущие доклады представили некоторые доказательства того, что тандемные повторы множественных копий трансгенов более чувствительны к метилированию и молчанию в клетках млекопитающих (Garrick, D., et al., Nat. Genet. 18 (1998) 56-59, McBurney, M.W., et al., Exp Cell Res 274 (2002) 1-8).
B.2. Ацилирование гистонов
Определяли относительные количества модификаций гистона, расположенного вблизи промотора hCMV-MIE. Рассматривали маркеры Н3ас и H3K4me3 (активирующие маркеры) и H3K9me3, а также маркеры H3K27me3 (репрессирующие) маркеры. Эти модификации гистонов проверяли в позициях от 404 до 507 в SEQ ID NO 01 (последовательность MIE-промотора CMV человека; tacatca atgggcgtgg atagcggttt gaetcacggg gatttccaag tctccacccc attgacgtca atgggagttt gttttggcac caaaatcaac gggactt), которая показывает последовательность от -97 до -200 п.о. до сайта инициации транскрипции и содержит сайт CpG в позиции С-425 (т.е. -179 п.н. выше сайта инициации транскрипции), и которая может быть амплифицирована с помощью пары праймеров 396/397.
Уровни относительного ацетилирования гистона 3 относительно уровня гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека исследовали в качестве маркеров для предсказания стабильности продукции с селективным агентом и без него.
Две иллюстративные замороженные клеточные линии из исходной точки культивирования (первичный посевной банк) размораживали и оценивали на относительные количества указанных модификаций гистонов вблизи промотора hCMV-MIE. Проект Т и проект Н выполняли независимо, и каждую клеточную линию культивировали в течение по меньшей мере 60 поколений в условиях с селективным агентом (+) (250 нМ метотрексат) и без него (-).
Получали данные о скорости метилирования промотора hCMV-MIE (mC-425), о числе копий на клетку (PSB, phosphate-buffered saline - фосфатно-солевой буфер), а также о процентном изменении удельной продуктивности (ΔqP) в течение 60 поколений. Запись данных проводили так, как было описано ранее в Osterlehner et al.
Таблица: Данные для 12 клеточных линий проекта Т. mC-425 представляет собой среднее процентное метилирования цитозина в позиции 425 выше сайта инициации транскрипции (transcription start site, TSS; -179 по отношению к TSS) в hCMV-MIE в начале фазы культивирования (PSB); ΔqP представляет собой процентное изменение удельной продуктивности (qP) в течение 60 поколений клеток; среднее число копий легкой цепи (LC) IgG-трансгена измеряли в начале культивирования (PSB).
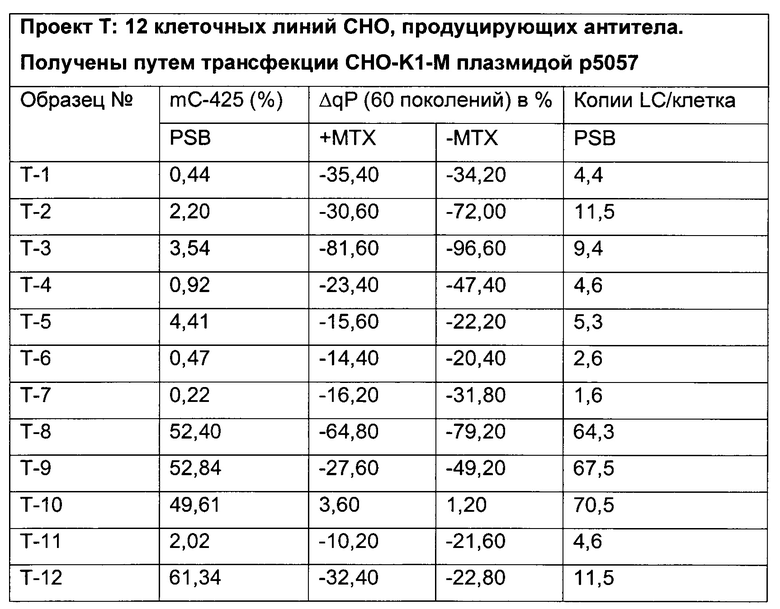
Таблица: Данные для 20 клеточных линий проекта Н. mC-425 представляет собой среднее процентное метилирования цитозина в позиции 425 выше TSS в hCMV-MIE в начале фазы культивирования (PSB); ΔqP представляет собой процентное изменение удельной продуктивности (qP) в течение 60 поколений клеток; среднее число копий легкой цепи (LC) IgG-трансгена измеряли в начале культивирования (PSB).
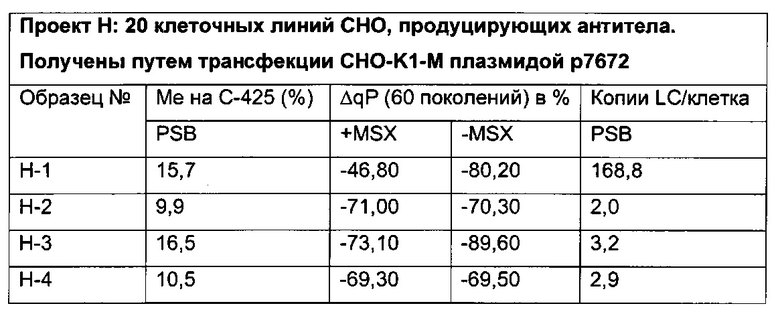
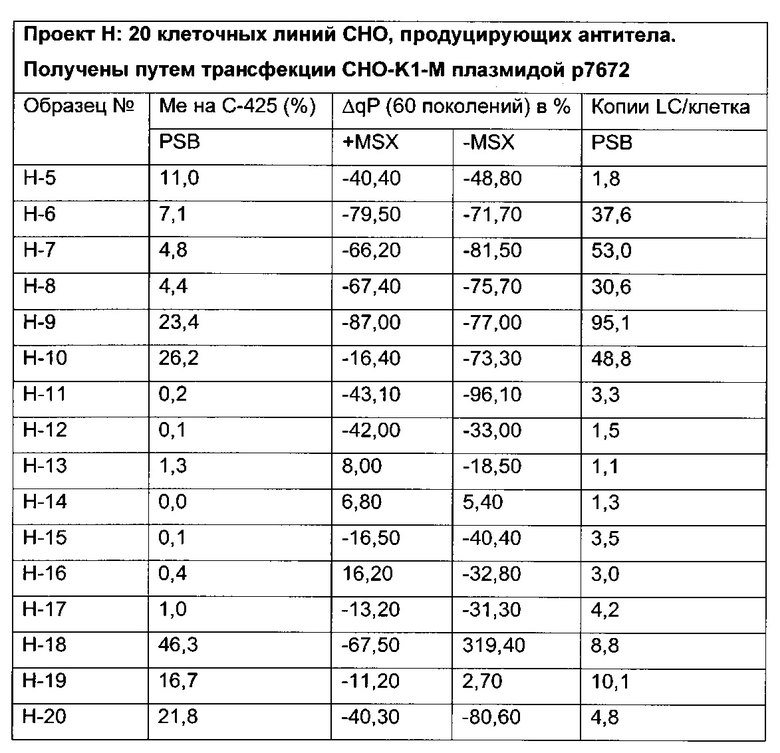
Чтобы исследовать относительное количество модификаций гистона, собирали жизнеспособные клетки СНО, которые несли рекомбинантный ген под контролем промотора hCMV-MIE. После фиксации в 3,7% растворе формальдегида лизированный хроматин подвергали воздействию ультразвука, и сшитые комплексы "ДНК - гистон" очищали согласно соответствующей модификации гистона. Затем накопленные комплексы "ДНК - гистон" подвергали деградации с помощью протеиназы К и элюировали фрагменты ДНК. Количества различных фрагментов ДНК сравнивали с помощью кПЦР, используя специфические пары праймеров.
Для проверки надлежащего связывания антитела и гистона в связи с соответствующей модификацией в референсной области различные гены анализировали на накопление конкретной модификации гистона. Вначале количество отфильтрованных с помощью антитела модификаций гистона вблизи референсных областей сравнивали с очищенным образцом неспецифического контроля без антитела (имитационный контроль) в проекте Т и проекта Н, каждый в одном образце. ДНК-фрагменты очищенных с помощью антитела образцов, имитационного и необработанного входящего образца использовали в качестве матриц для кПЦР. К каждому образцу добавляли пары праймеров для потенциальных референсных областей и проводили кПЦР в трех повторах. Полученные значения Cq сравнивали в процентах от значений для входящего образца. В обоих проектах очищенные с помощью антитела образцы давали более высокие значения, чем имитационный контроль, в соответствующей области. Этот факт подтверждал надлежащее связывание. Кроме того, маркеры активации Н3ас и H3K4me3 накапливались в активных областях Eif3i и Gusb. Напротив, молчащая область Fox2a имела более высокую концентрацию H3K9me3, тогда как H3K27me3 поразительно располагался в области Gata5. Различные модификации гистона совпадали с соответствующими контрольными областями в обоих проектах.
Оценка относительного количества модификаций гистона в целевой области строго зависит от стабильно модифицированной референсной области.
Нормализация конкретной модификации гистона вблизи промотора hCMV-MIE по референсной последовательности позволяет сравнивать различные клеточные линии. Для этого референсная последовательность должна иметь надежные и воспроизводимые уровни соответствующей модификации гистона.
В следующей таблице приведены средние значения Cq и коэффициент вариации (coefficient of variation, CV) для всех клеточных линий проекта Т и проекта Н, включая три биологических повтора.
Таблица: Референсные области проекта Т и Н. Средние значения и соответствующий коэффициент вариации (CV) значений Cq для всех ChIP-образцов, включая биологические трехкратные повторы, рассчитывали для обоих проектов; строки отображают различные референсные области в геноме; столбцы содержат конкретные модификации гистонов; выбранные контрольные области для соответствующих эпигенетических модификаций выделены жирным шрифтом; поэтому расстояние Cq до имитационного контроля и CV было решающим; в обоих проектах маркеры активации Н3ас, H3K4me3, а также общий гистон 3 стабильно накапливаются в областях Gusb и Eif3i (CV 2%); нижние значения Cq определяют Gusb как референс для маркеров активации; все потенциальные референсные области имели стабильные значения общего гистона 3.

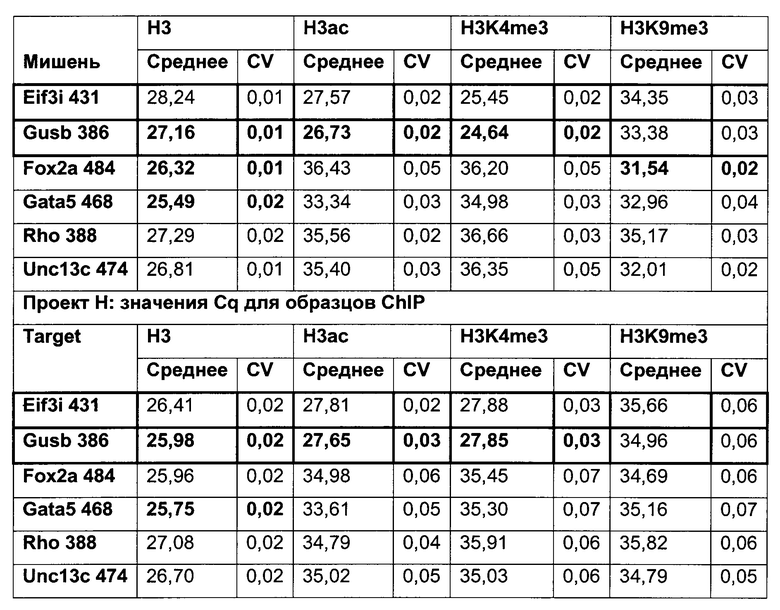
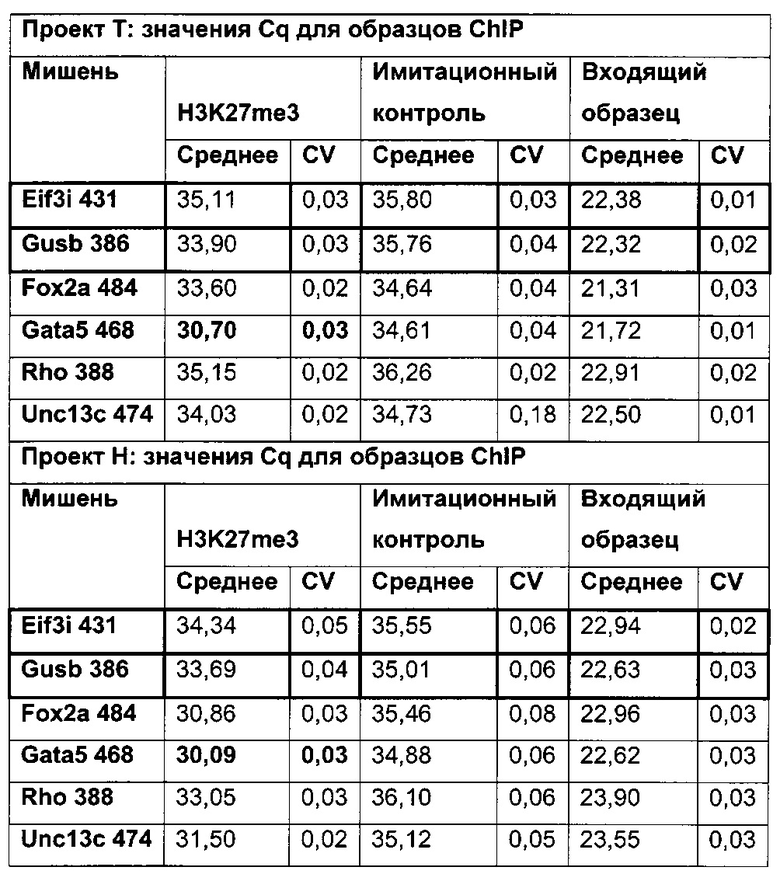
Gusb в качестве референсной области для активной модификации гистона Н3ас и H3K4me3 и Gata5 в качестве стабильной области для H3K27me3 отличались от соответствующего имитационного контроля и имели согласованные значения Cq. Активные контрольные области Eif3i и Gusb обладали высокой стабильностью. Для всех контрольных областей наблюдалось стабильное накопление общего гистона 3 (Н3).
Были проанализированы биологические трехкратные повторы клеточных линий СНО, рекомбинантно экспрессирующих проектное антитело, на предмет модификации гистона вблизи локализации промотора hCMV-MIE. Для этого фрагменты хроматина очищали с использованием антител против конкретных модификаций гистонов, расщепляли для получения геномной ДНК, свободной от белка, и количественно оценивали с помощью ПЦР в режиме реального времени (см. фиг. 26). Нормализацию выполняли для каждой используемой референсной области. Для определения эффективности амплификации каждой пары праймеров использовали программное обеспечение LinReg PCR, версии 2014.5 (Ruijter, 2009, LinRegPCR). Эффективность амплификации всех пар "праймер - матрица" была очень похожей. Таким образом, для нормализации использовали способ Livak, который предполагает аналогичные эффективности амплификации.
Таблица: Эффективность праймеров. Эффективности амплификации определяли для всех клеточных линий, в том числе для биологических трехкратных повторов; использовали пару праймеров (396/397); для этой цели необработанные данные кПЦР обрабатывали с помощью программного обеспечения LinRegPCR, версия 2014,5, и рассчитывали эффективности; строки отличаются различными парами праймеров; столбец "среднее" содержит среднюю эффективность амплификации каждой пары "праймер - матрица", а столбец "ст. откл." содержит соответствующее стандартное отклонение; различия между всеми эффективностями амплификации составляли менее 2%; таким образом, для нормализации использовали способ Livak.
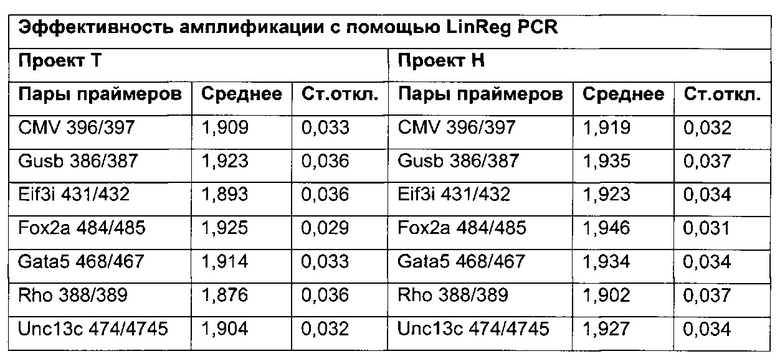
Нормализация по Livak включает два этапа нормализации:
- на первом этапе количество (Cq) модификации гистона 3 вблизи промотора hCMV-MIE нормализовали по количеству (Cq) модификации гистона 3 в референсной области →ΔCq
- на втором этапе нормализованную модификацию гистона 3 применяли для нормализованного общего гистона 3 вблизи промотора hCMV-MIE→ΔΔCq
Это значение, наконец, отображает относительное количество модификации в расчете на гистон 3 вблизи от промотора hCMV-MIE.
Общий уровень гистона 3 является стабильным во всех референсных областях, позволяя нормализовать модификацию гистона и общий уровень гистона 3 по одной и той же референсной области. Это сводит к минимуму вариации в процессе нормализации. Например, относительное ацетилирование гистона в расчете на гистон 3 нормализовали по контрольной области Gusb, так как область Gusb содержит стабильное количество модифицированного гистона и общего гистона 3. Для расчета использовали следующую формулу:
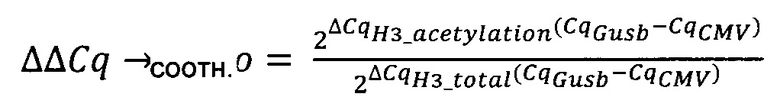
Относительные количества модификации гистона в расчете на гистон 3 (ΔΔCq) и общий гистон 3 (ΔCq) вблизи промотора hCMV-MIE откладывали на графике в виде зависимости от значений ΔqP. Общий гистон 3 вблизи промотора hCMV-MIE нормализовали по Gusb и откладывали на графике в виде зависимости от значений ΔqP. Кроме того, исследовали гистон 3 в расчете на число копий (Н3/rCN) и имитационный контроль в расчете на гистон3 (Мосk/Н3). Н3/rCN вычитает в идеале число копий и является мерой относительной плотности гистона 3 (H3D) вблизи промотора hCMV-MIE. Mock/Н3 представляет неспецифическое влияние фонового шума в расчете на гистон 3.
Корреляции изучали с помощью стандартной модели наименьших квадратов. С этой целью использовали программу JMP10 (JMP®10.0.1 Release: 2, 64-битная версия; SAS Institute Inc.).
Таблица: Измерение влияния эпигенетических модификаций на долгосрочную стабильности продукции. Р-значения влияний на ΔqP с селекционным давлением (+) (250 нМ МТХ) и без него (-) приведены в столбцах; все влияния измерены вблизи промотора hCMV-MIE и показаны в строках; влияния нормализованного гистона 3 (H3), плотность гистона 3 в расчете на число копий (H3/rCN), фоновый шум в расчете на гистон 3 (Mock/H3) и модификации в расчете на гистон 3 (Н3ас/Н3, H3K4me3/H3, H3K27me3/H3 и H3K9me3/H3) вычисляли с помощью стандартной модели наименьших квадратов с использованием программного обеспечения JMP10; в проекте Н ацетилирование в расчете на гистон 3 (Н3ас/Н3) имеет весьма существенное влияние на ΔqP, если клетки культивируют под селекционным давлением (+), также события Н3/rCN, имитация/H3, H3K4me3, H3K27me3 и Н3 показали влияния от умеренного до высокого в условиях ΔqP+.
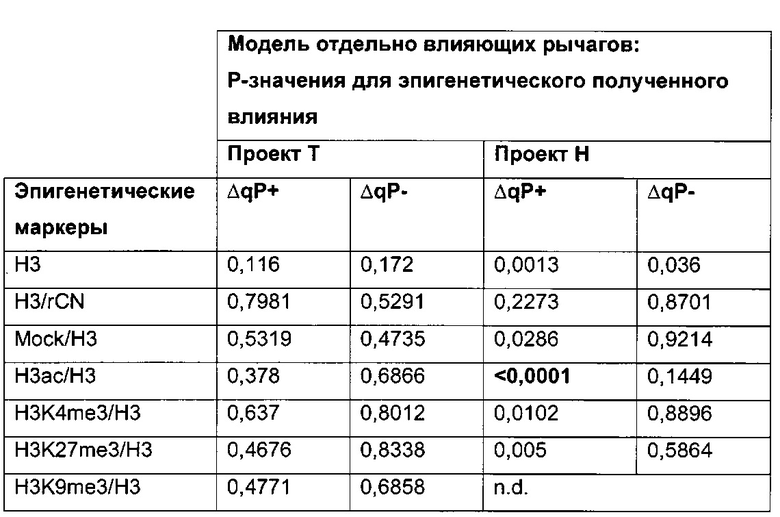
n.d. - не определено
Степень ацетилирования в расчете на гистон 3 (Н3ас/Н3) на промоторе hCMV-MIE имеет весьма существенное влияние на потерю продуктивности, полученную в течение 60 поколений в присутствии селективного агента МТХ.
Далее наблюдались значительные влияния (перечисленные от самых слабых к самым сильным влияниям) для Н3/rCN, имитация/H3, H3K4me3/H3, H3K27me3/H3 и Н3. При рассмотрении уровней влияний для Mock/H3 и Н3 в одиночку, Н3 в одиночку имеет доминирующее влияние на комбинированное влияние Х/H3. Было обнаружено, что эпигенетические события с большими эффектами, чем Н3 в одиночку, являются хорошим маркером предсказания: →Н3ас/Н3.
Проводили анализ выбросов с помощью методики "складного ножа". При этом находили один выброс в трех повторах путем сравнения ΔqP в обоих условиях касательно селективного агента. Что касается анализа, клеточную линию Н-18 исключали из измерений влияния. После этого образцы из проекта Н снова анализировали на модели скрининга влияния после исключения клеточной линии Н-18. Было обнаружено, что падение показателя ацетилирования в расчете на гистон 3 (Н3ас/Н3) на промоторе hCMV-MIE имеет наиболее существенное влияние на падение продуктивности, полученной в течение 60 поколений, в обоих условиях селекции.
Таблица Измерения влияния модификаций на долгосрочную стабильность в проекте Н после исключения клеточной линии-выброса Н-18. Р-значения влияний на ΔqP с селекционным давлением (+) (250 нМ МТХ) и без него (-) показаны в столбцах; все влияния измерены вблизи промотора hCMV-MIE и показаны в строках; влияния нормализованного гистона 3 (H3), плотности гистона 3 в расчете на относительное число копий (H3/rCN), фонового шума в расчете на гистон 3 (имитация/H3) и модификаций в расчете на гистон 3" (Н3ас/Н3, H3K4me3/H3, H3K27me3/H3 и H3K9me3/H3) вычисляли с помощью стандартной модели наименьших квадратов с использованием программного обеспечения JMP10; наиболее существенное влияние на ΔqP можно было наблюдать для Н3ас/Н3 в обоих условиях селекции.
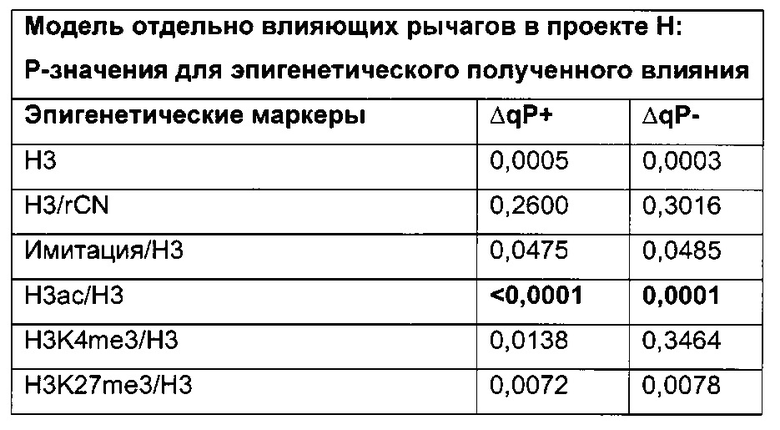
В проекте Т график Н3ас/Н3 против ΔqP показывает одновременное повышение долгосрочной стабильности и ацетилирование в расчете на гистон 3, особенно под селекционным давлением.
Чтобы учесть различные уровни гистона 3 на промоторе hCMV-MIE, ацетилирование гистона 3 дополнительно нормализовали по гистону 3 (Н3ас/Н3).
Относительные уровни ацетилирования гистона 3 (Н3ас/Н3) сравнивали с изменениями удельной продуктивности (ΔqP) в течение 60 поколений в присутствии (+) и в отсутствие (-) селективного агента MSX. После анализа выбросов использовали стандартную модель наименьших квадратов со значениями Н3ас/Н3 биологических повторов и значениями ΔqP от 19 клеточных линий СНО. Была обнаружена значительная корреляция Н3ас/Н3 и ΔqP.
Для значений Н3ас/Н3 с помощью программного обеспечения JMP рассчитывали дерево решений. Лучший узел расщепления и, следовательно, лучший фильтр рассчитывали с помощью статистического анализа LogWorth. Лучшее расщепление в условиях (+) MSX было рассчитано как 0,58 ацетилирования гистона 3 относительно уровня гистона 3 (Н3ас/Н3) для образцов Н. Для того чтобы уменьшить число ложноотрицательных образцов, фильтр был установлен на более низкое значение А>0,5 Н3ас/Н3.
Средние значения дельта-SPR (ΔqP) в условиях с селективным агентом МТХ или MSX или без него, соответственно, вычисляли для каждого положительного гейта и сравнивали со средним значением без фильтра; полученные значения представлены ниже:
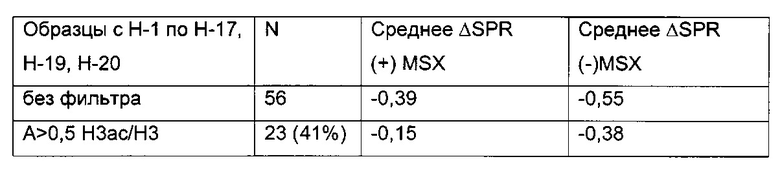
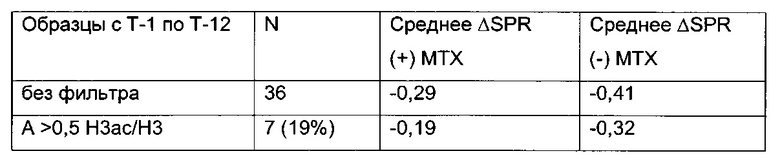
Фильтр А>0,5 Н3ас/Н3 увеличивает среднее значение дельта-SPR по сравнению со значением без фильтра.
В.3. Сочетание метилирования промотора и ацетилирования гистона
Относительные уровни ацетилирования гистона 3 относительно уровня гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека и процент метилирования С-425 в главном немедленно-раннем промоторе/энхансере CMV человека исследовали в качестве маркеров предсказания стабильности продукции с использованием селективного агента и без него.
Средние значения дельта-SPR (ΔqP) в условиях с селективным агентом или без него вычисляли для каждого положительного гейта и сравнивали со значением без фильтра; полученные значения представлены ниже:
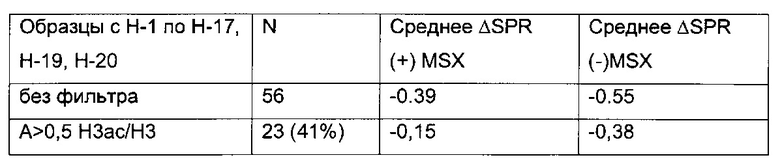
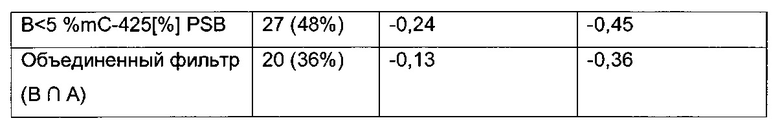
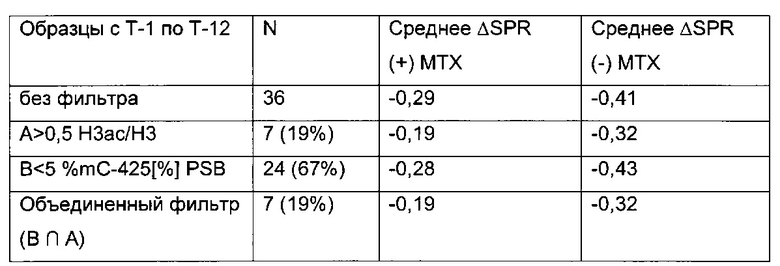
Объединение фильтра А>0,5 H3ac/H3 с фильтром В менее 5%mC-425[%] PSB увеличивает среднее значение дельта-SPR по сравнению со значением без фильтра.
Более подробно, для определения порогового значения соотношения H3ac/H3, чтобы исключить плохих продуцентов, с помощью программного обеспечения Jmp 10 рассчитывали дерево решений. Для определения лучшего узла расщепления использовали статистический анализ LogWorth. Logworth рассчитывали как: -log10 (р-значение), где скорректированное значение p рассчитывали сложным образом, учитывая огромное число различных способов расщеплений, которые могут произойти (Sall, 2002, Monte Carlo Calibration of Distributions of Partition Statistics; информационный документ SAS). Лучшее расщепление для условия ΔqP+ составляло 0,47 ацетилирования в расчете на гистон 3. Для того чтобы минимизировать ложноположительные значения, пороговое значение устанавливали на 0,5 H3ac/H3 и переносили на оба условия в проектах Н и Т.
Кроме того, определяли порог 5% для метилирования промотора hCMV-MIE (%mC-425) из записанных данных о метилировании (см. таблицы выше), чтобы использовать (синергетическую) комбинацию обоих маркеров предсказания. Таким образом, были исключены образцы с более чем 5% метилированием ДНК или менее чем 0,5 ацетилированием гистона 3 в промоторе hCMV-MIE.
Среднее значение ΔqP рассчитывали для положительных фильтрованных образцов и сравнивали со средним значением ΔqP образцов без фильтра. Это делали с учетом того, был ли добавлен МТХ к культуральной среде или не был.
Использование маркера предсказания H3ac/H3 приводит к увеличению долговременной стабильности включенных образцов.
Средние значения ΔqP после фильтрации были почти идентичны в обоих проектах.
Таблица: Исключение плохих продуцентов. Образцы со значениями выше 0,5 H3ac/H3 и ниже 5%mC-425 использовали для расчета среднего ΔqP; интересно, что фильтрованные значения ΔqP были почти идентичны для обоих проектов.
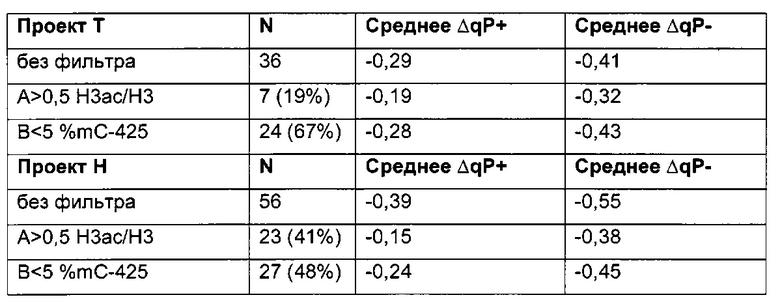
Степень ацетилирования гистона 3 на промотора hCMV-MIE оказывает существенное влияние на стабильность. Кроме того, количество общего H3 и степень метилирования в позиции С-425 (%mC-425) имели влияние на долгосрочную стабильность. Кроме того, обнаруженная отрицательная корреляция H3ac/H3 и %mC-425 подтверждает достоверность этих маркеров.
Установка порогового значения приводит к исключению всех плохих продуцентов. Клетки выше этого значения являются преимущественно стабильными.
В.4. Сочетание метилирования промотора, ацетилирования гистона и числа копий трансгена
Корреляция ацетилирования гистона 3 с метилированием ДНК и числом копий была значительной, что доказывает взаимосвязь числа копий, метилирования ДНК и ацетилирования гистона 3. Было установлено, что степень метилирования ДНК является обратной ацетилированию гистона 3. Кроме того, было обнаружено, что число копий в отличие от ацетилирования гистона 3 и метилирования ДНК обеспечивает тот факт, что клеточные линии с более высоким числом копий, в основном, являются неацетилированными и склонны к метилированию ДНК. Была обнаружена корреляция числа копий и изменения долгосрочной продуктивности в присутствии и в отсутствие селективного агента. Таким образом, число копий взаимодействует со степенью маркеров, которые, в свою очередь, оказывают влияние на долгосрочную стабильность продукции.
Кроме того, непреднамеренное накопление нестабильных клеточных линий усиливается при разработке стандартной клеточной линии за счет изначального отбора клеточных линий в соответствии с титром их антител. Предполагается, что клеточные линии с низким числом копий должны быть преимущественно активными, тогда как продуценты с высоким числом копий могут иметь множество различных эпигенетических состояний от отсутствия активности через умеренную активность вплоть до высокой активности у каждого отдельного трансгена до тех пор, пока сумма транскриптов сравнима. Учитывая это, при первой селекции в соответствии с титром антител предпочтение отдается либо клеточным линиям с высоким числом копий, которые по случайности могут иметь стабильные сайты интеграции, либо клеточным линиям с низким числом копий, которые слегка вынуждены иметь их. Число копий в начале фазы культивирования откладывали на графике в виде зависимости от стабильности, и было обнаружено, что клеточные линии с высоким числом трансгенов склонны к нестабильности.
Таким образом, в данном документе в качестве одного из аспектов описан трехэтапный способ/процесс селекции клеточной линии. На первом этапе экспрессирующие антитело клетки с низким числом копий легкой цепи размножают, на втором этапе проводят селекцию в соответствии со степенью их ацетилирования гистона 3. Затем выполняют быстрое снижение уровня селективного агента. Таким образом получают клеточные линии со стабильными сайтами интеграции, сниженным риском молчания генов и снижения числа копий, а также проводят идентификацию субпопуляций с предварительно напряженными дестабилизирующими механизмами.
В одном воплощении число копий стабильно интегрированной экспрессионной кассеты легкой цепи составляет менее 50. В другом воплощении число копий стабильно интегрированной экспрессионной кассеты легкой цепи составляет менее 25. В другом воплощении число копий стабильно интегрированной экспрессионной кассеты легкой цепи составляет менее 10. В другом воплощении число копий стабильно интегрированной экспрессионной кассеты легкой цепи составляет менее 6.
Следующие примеры, список последовательностей и графические материалы приведены для облегчения понимания данного изобретения, истинные масштабы которого изложены в приведенной формуле изобретения. Нужно понимать, что в изложенных процедурах могут быть сделаны изменения без отхода от сущности изобретения.
Описание графических материалов
Фиг. 1 Число метилированных CpG-сайтов в промоторе/энхансере hCMV-MIE, полученном из различных клеточных линий.
Фиг. 2 Карта плазмиды р5057.
Фиг. 3 А: Удельная скорость продукции клеточной линии K18.1 в отсутствие селективного агента в течение многократных поколений при долгосрочной продукции.
В: Удельная скорость продукции клеточной линии G25-10 в отсутствие селективного агента в течение многократных поколений при долгосрочной продукции.
С: Удельная скорость продукции клеточной линии G25-17 в отсутствие селективного агента в течение многократных поколений при долгосрочной продукции.
D: Удельная скорость продукции клеточной линии G42-5 в отсутствие селективного агента в течение многократных поколений при долгосрочной продукции.
Е: Удельная скорость продукции в отсутствие селективного агента клеточной линии 43-16 А10 в течение многократных поколений при долгосрочной продукции.
Фиг. 4 Верхняя фигура: частота метилирования ДНК в промоторе/энхансере hCMV-MIE из рекомбинантных клеточных линий СНО на разных сайтах метилирования, определенная путем анализа 19-22 отдельных промоторных нуклеиновых кислот; нижняя фигура: схематическое представление метилированных CpG-сайтов в ДНК промотора/энхансера hCMV-MIE из рекомбинантных клеточных линий СНО - сайты метилирования показаны черным - позиция 425 выделена стрелкой.
А: клеточная линия K18.1 - метилирование всех CpG-сайтов: 12%, метилирование сайта С425: 64%, метилирование сайта С591: 27%, метилирование сайта С96: 32%;
В: клеточная линия G25-10 - метилирование всех CpG-сайтов: 0,5%, метилирование сайта С425: 0%, метилирование сайта С591: 5%, метилирование сайта С96: 0%;
С: клеточная линия G25-17 - метилирование всех CpG-сайтов: 0,3%, метилирование сайта С425: 0%, метилирование сайта С591: 0%, метилирование сайта С96: 0%;
D: клеточная линия G42-5 - метилирование всех CpG-сайтов: 0,6%, метилирование сайта С425: 0%, метилирование сайта С591: 0%, метилирование сайта С96: 0%;
Е: клеточная линия 43-16 А10 - метилирование всех CpG-сайтов: 4,4%, метилирование сайта С425: 25%, метилирование сайта С591: 15%, метилирование сайта С96: 10%.
Фиг. 5 Отделение промотора/энхансера hCMV-MIE, который метилирован на сайте 425, от hCMV-MIE, который не метилирован на сайте 425, с помощью специфической в отношении метилирования кПЦР в режиме реального времени; кривые ПЦР-амплификации для матриц №11 (А), №62 (В), №01 (С) и №04 (D).
Фиг. 6 Кривые ПЦР-амплификации для различных праймеров, специфических в отношении метилирования, и пар праймеров.
Фиг. 7 Выделение промотора/энхансера hCMV-MIE, метилированного на сайте 425, на фоне неметилированного промотора/энхансера hCMV-MIE с помощью специфической в отношении метилирования кПЦР в режиме реального времени.
Фиг. 8 Метилирование промоторной/энхансерной нуклеиновой кислоты hCMV-MIE на сайте метилирования 425, полученное с праймерами №239 и №267, с предварительной амплификацией (А) и непосредственно из обработанной бисульфитом геномной ДНК (В).
Фиг. 9 Метилирование промоторной/энхансерной нуклеиновой кислоты hCMV-MIE на сайте метилирования 425, полученное с праймерами (черный) 239 плюс 237 и 239 плюс 267, (белый) 263 плюс 237 и 263 плюс 267, (горизонтальные линии) 264 плюс 237 и 264 плюс 267, и (вертикальные линии) 239 плюс 237 и 239 плюс 266.
Фиг. 10 Пример высокоточного анализа нормализованных кривых плавления (А) и их первых производных, т.е. пиков плавления (В).
Фиг.11 Корреляция степени метилирования С425 перед долгосрочным культивированием и относительного изменения SPR после 60 поколений культивирования с 250 нМ МТХ (А) или без МТХ (В).
Фиг. 12 Корреляция степени метилирования на С425 перед долгосрочным культивированием и степени метилирования после 60 поколений культивирования с 250 нМ МТХ.
Фиг. 13 Схематическое изображение метилированных CpG-сайтов в ДНК промотора/энхансера hCMV-MIE из клона 44-28. Метилированные сайты показаны черным цветом. Нуклеотидная позиция 425 выделена стрелкой. Метилирование всех CpG-сайтов: 18%; метилирование на С425: 80%; метилирование на С591: 70%; метилирование на С96: 60%.
Фиг. 14 Число копий гена легкой цепи до и после анализа стабильности без МТХ.
Фиг. 15 Относительные уровни ацетилирования гистона 3 относительно уровня гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека, нормализованные по референсному гену Gusb. Отклонения в трех биологических повторах отображаются стандартным отклонением.
Фиг. 16 Относительные уровни трехкратного метилирования лизина 4 гистона 3 относительно уровня гистона 3 вблизи фрагмента главного немедленнораннего промотора/энхансера CMV человека, нормализованные по референсному гену Gusb. Отклонения в трех биологических повторах отображаются стандартным отклонением.
Фиг. 17 График сравнения прогнозируемых значений с фактическими значениями влияния H3K4me3/H3 для образцов с Н-1 по Н-20 в трех биологических повторах. Кривые доверительного интервала для линии соответствия показаны пунктирными линиями, чтобы визуально показать, является ли анализ, представляющий интерес, значимым на уровне 5%. Для значимости кривые должны пересекать горизонтальную линию.
Фиг. 18 График сравнения прогнозируемых значений с фактическими значениями влияния H3ac/H3 для образцов с Н-1 по Н-20 в трех биологических повторах. Кривые доверительного интервала для линии соответствия показаны пунктирными линиями, чтобы визуально показать, является ли анализ, представляющий интерес, значимым на уровне 5%. Для значимости кривые должны пересекать горизонтальную линию.
Фиг. 19 График сравнения прогнозируемых значений с фактическими значениями влияния mC-425[%] для образцов с Н-1 по Н-20. Кривые доверительного интервала для линии соответствия показаны пунктирными линиями, чтобы визуально показать, является ли анализ, представляющий интерес, значимым на уровне 5%. Для значимости кривые должны пересекать горизонтальную линию.
Фиг. 20 Анализ выбросов проекта Н с расстояниями по методу "складного ножа" для дельта-SPR с MSX и без него в качестве условия. Образец Н-18 находится далеко за верхним контрольным пределом (Upper Control Limit, UCL), что явно указывает на образец как на выброс.
Фиг. 21 График сравнения прогнозируемых значений с фактическими значениями влияния H3K4me3/H3 для образцов с Н-1 по Н-17, Н-19 и Н-20 в трех биологических повторах. Кривые доверительного интервала для линии соответствия показаны пунктирными линиями, чтобы визуально показать, является ли анализ, представляющий интерес, значимым на уровне 5%. Для значимости кривые должны пересекать горизонтальную линию.
Фиг. 22 График сравнения прогнозируемых значений с фактическими значениями влияния H3ac/H3 для образцов с Н-1 по Н-17, Н-19 и Н-20 в трех биологических повторах. Кривые доверительного интервала для линии соответствия показаны пунктирными линиями, чтобы визуально показать, является ли анализ, представляющий интерес, значимым на уровне 5%. Для значимости кривые должны пересекать горизонтальную линию.
Фиг. 23 График сравнения прогнозируемых значений с фактическими значениями влияния mC-425[%] для образцов с Н-1 по Н-17, Н-19 и Н-20 в трех биологических повторах. Кривые доверительного интервала для линии соответствия показаны пунктирными линиями, чтобы визуально показать, является ли анализ, представляющий интерес, значимым на уровне 5%. Для значимости кривые должны пересекать горизонтальную линию.
Фиг. 24 Значения дельта-SPR для образцов с Н-1 по Н-17, Н-19 и Н-20, отложенные на графике в виде гистограмм. Отображены различные условия селекции и фильтрации. Горизонтальная линия внутри диаграммы ("ящика с усами") представляет собой медиану. Ромб достоверности содержит среднее значение, верхний и нижний 95% пределы от среднего значения. Середина ромба представляет собой среднее значение. Верхние и нижние точки ромба представляют собой 1-й и 3-й квартили. Ящик имеет линии, которые тянутся от каждого конца и называются "усами". Усы тянутся от концов ящика до крайней точки данных, которая попадает в расстояние, вычисленное как расстояние между значением "1-й квартиль - 1,5*(межквартильный диапазон)" и значением "3-й квартиль плюс 1,5*(межквартильный диапазон)". Если точки данных не достигают вычисленных диапазонов, то усы определяются верхними и нижними значениями точек данных (не включая выбросы). Скоба за пределами ящика показывает самую короткую половину, которую составляют наиболее плотные 50% наблюдений (Rousseuw and Leroy 1987).
Фиг. 25 Значения дельта-SPR проекта Т отложены на графике в виде гистограмм. Отображены различные условия селекции и фильтрации.
Фиг. 26 Схема иммунопреципитации хроматина (chromatin immunoprecipitation, ChIP) ДНК клеточной линии СНО.
Фиг. 27 Корреляционные исследования эпигенетических маркеров, числа копий и стабильности. А: График метилирования С-425 в виде зависимости от ацетилирования гистона 3 на промоторе hCMV-MIE. В: график изменения удельной продуктивности в течение 60 поколений клеток СНО в отсутствие (-) и в присутствии (+) селективного агента MSX после исключения клеточной линии Н-18. Была определена высокая корреляция стабильности клеточных линий с селективным агентом и без него. С: График показывает корреляцию числа копий и ацетилирования гистона 3, указывая, что только клеточные линии с низким числом копий были ацетилированы в гистоне 3. D: График показывает корреляцию числа копий и метилирования ДНК, тогда как только клеточные линии с высоким числом копий преимущественно метилированы на промоторе hCMV-MIE. Е и F: графики показывают соотношение числа копий и изменения продуктивности в течение 60 поколений в присутствии (+) и в отсутствие (-) селективного агента. Поэтому клеточные линии с высоким числом копий склонны к молчанию.
Фиг. 28 Экспрессионная плазмида для человеческого антитела класса IgG. Экспрессионные кассеты легкой и тяжелой цепи человеческого иммуноглобулина находятся под контролем главного немедленно-раннего промотора и энхансера CMV человека (SEQ ID NO 01) (hCMV-MIE).
Пример 1
Общие методики
Растворы



Методики рекомбинантной ДНК
Для манипуляций с ДНК использовали стандартные способы, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989). Молекулярно-биологические реагенты использовали в соответствии с инструкцией изготовителя.
Определение последовательности ДНК
Секвенирование ДНК проводили на SequiServe GmbH (Ватерстеттен, Германия).
Анализ последовательности ДНК и белков и управление данными о последовательностях
Пакет программного обеспечения EMBOSS (European Molecular Biology Open Software Suite) и Vector NTI версии 9.1 от Invitrogen использовали для создания, картирования, анализа, аннотирования и иллюстрирования последовательностей.
Определение белков
Для количественной оценки антитела, присутствующего в образце, использовали хроматографический способ. Использовали колонку PorosA, которая связывает Fc-область антитела. Антитело связывали с колонкой и затем элюировали в условиях низкого рН. Концентрацию белка определяли путем определения оптической плотности (OD) при 280 нм с референсной длиной волны 320 нм, используя коэффициент молярной экстинкции, рассчитанный на основе аминокислотной последовательности.
Электрофорез в агарозном геле
Электрофорез в агарозном геле проводили для анализа качества, размера и количества линейных фрагментов ДНК (Chong, 2001). В соответствии с размером фрагментов ДНК 1% (вес/объем) раствор агарозы растворяли в 1× растворе ТАЕ путем кипячения. Гели содержали этидия бромид в конечной концентрации 0,5 мкг/мл Образцы готовили путем добавления 1/6 (объем/объем) 6× красителя для загрузки ДНК. ДНК-маркер использовали в качестве стандарта размеров. Электрофорез проводили в 1× ТАЕ, прикладывая ток 10 В на 1 см длины геля (не более 250 В). После разделения ДНК исследовали под действием УФ-излучения (254-366 нм) в системе гель-документации (Intas Science Imaging Instruments GmbH, Геттинген, Германия).
Выделение из геля
После электрофореза в агарозном геле кусок геля с нужной полосой вырезали скальпелем, и далее из этого куска геля экстрагировали ДНК с помощью набора QIAquick® Gel Extraction (Qiagen, Хильден, Германия). При этом к 1 объему геля (100 мг ~ 100 мкл) добавляли 3 объема буфера QG и инкубировали в течение 10 минут при 50°С. К растворенному гелю добавляли один объем изопропанола и перемешивали. Спин-колонку QIAquick с фиксированными на ней фрагментами ДНК в течение 1 минуты центрифугировали со скоростью 13000 об/мин, промывали 0,75 мл буфера РЕ и снова центрифугировали (13000 оборотов об/мин, 1 мин). Промывали второй раз, собирая прошедшую фракцию в другую пробирку, удаляя таким образом оставшийся промывочный буфер. Наконец, ДНК элюировали в 30 мкл безРНКазной H2O, а затем колонку центрифугировали (13000 об/мин, 1 мин), собирая ДНК в пробирку для образца. ДНК может быть проверена на системе электрофореза FlashGel™ (Lonza, Кельн, Германия).
Количественная оценка ДНК
Для оценки концентрации ДНК использовали NanoDrop 2000 (Peqlab Biotechnologie GmbH, Эрланген, Германия) путем измерения оптической плотности (OD) при длине волны 260 нм. О чистоте ДНК можно судить по отношению OD 260/280 и 260/230. Чистые препараты ДНК должны обладать соотношением более или равно 1,8 и 2,0.
Трансформация компетентных бактерий
45 мкл химически компетентных клеток Е. coli NEB 5 alpha (NEB, Германия) размораживали на льду в течение 10 минут, а затем инкубировали с 1 мкг-100 нг плазмидной ДНК (1-5 мкл) в течение 30 минут на льду. Суспензию клеток подвергали тепловому шоку при 42°С в течение 45 секунд и немедленно охлаждали на льду в течение 5 минут. Добавляли 950 мкл среды SOC или LB комнатной температуры и суспензию инкубировали при 37°С в течение 60 минут в условиях встряхивания (250 об/мин). Трансформированные бактерии высевали на предварительно согретые чашки с агаром, содержащим ампициллин. Чашки инкубировали в течение ночи при 37°С.
Получение плазмид
Плазмиды получали с помощью наборов для выделения плазмид Qiagen Plasmid Mini и Qiagen Plasmid Maxi (Qiagen, Хильден, Германия), следуя инструкциям производителя.
Очистка ДНК
Геномную ДНК выделяли и очищали с помощью наборов Allprep DNA/RNA Mini Kit (50) (кат. №80204, Qiagen, Хильден, Германия) и DNAse Blood & Tissue Kit (Qiagen, Хильден, Германия) в соответствии с рекомендациями производителя. Очистку фрагментов проводили с помощью набора для очистки ПЦР-продуктов High Pure PCR Product Purification Kit (Roche Diagnostics GmbH, Мангейм, Германия) в соответствии с инструкциями производителя.
Экстракция фенолом-хлороформом
Концентрирование и очистку плазмид проводили путем экстракции смесью фенола/хлороформа. Все этапы выполняли в химическом вытяжном шкафу. 500 мкл линеаризованной плазмиды смешивали с одним объемом смеси фенол/хлороформ/изоамиловый спирт (25:24:1) Roti® (кат. № А156, Рот, Германия) и перемешивали на вортексе в течение 30 секунд. Затем эмульсию переносили в предварительно центрифугированные пробирки Phase Lock Gel™ Light (кат. №0032005.101, Eppendorf, Гамбург, Германия) и центрифугировали (13000 об/мин, 1 мин). Верхнюю водную фазу переносили в новую пробирку и повторяли экстракцию. После этого верхнюю водную фазу смешивали с 500 мкл смеси хлороформа/изоамилового спирта (24:1) и этапы экстракции повторяли еще один раз. Верхнюю фазу переносили в новую пробирку и смешивали с 0,1 исходного объема 3 моль/л буферного раствора ацетата натрия (рН 4,8-5,2) и 0,7 исходного объема 100% изопропанола (20-30°С) путем переворачивания пробирки 4-6 раз. После 20-минутной инкубации при -80°С с последующим центрифугированием (13000 об/мин, 30 минут при 4°С) супернатант отбрасывали. Осадок промывали 1 мл охлажденного 70% этанола, центрифугировали (13000 об/мин, 5 минут при 4°С), и супернатант отбрасывали. Этап промывания повторяли в стерильных условиях, и остаточный супернатант полностью удаляли. Осадок высушивали в течение 5-10 минут в стерильном воздухе, а затем ресуспендировали в 0,5 исходного объема дважды дистиллированной воды.
Определение числа копий трансгена
Определение числа копий трансгена, а также скорости метилирования С-425 в промоторе hCMV-MIE выполняли в соответствии с рекомендациями Osterlehner et al. (см. выше).
Относительное число копий (relative copy number, rCN) рассчитывали как два в степени, представляющей разность значения Cq для целевого входящего образца промотора hCMV-MIE и значения Cq для референсного входящего образца (например, Gusb).

Иммунопреципитация хроматина
Собирали клеточные линии СНО с жизнеспособностью выше 90%, предпочтительно выше 97%. Перед фиксацией готовили гранулы для предварительной очистки и преципитации. Гранулы для предварительной очистки получали путем инкубации (1 ч, RT, в условиях вращения) 15 мкл агарозной суспензии с белком A (Roche Diagnostics GmbH, Мангейм, Германия) с 2 мкл очищенного кроличьего IgG (кат. № РР64 В, Millipore, Германия) в 120 мкл буфера для разведения ChIP на каждый образец. Для очистки проводили два этапа промывания одним объемом буфера для разведения ChIP, а затем гранулы ресуспендировали в 85 мкл буфера для разведения ChIP. 15 мкл агарозных гранул для преципитации блокировали в 1 мл буфера для разведения ChIP с 5 мг/мл BSA (Roche Diagnostics GmbH, Мангейм, Germany) и 100 мкг/мл предварительно нагретой ДНК спермы лосося (10 минут при 95°С и 5 минут на льду; кат. №15632-011, Invitrogen, Германия). Ресуспендированный осадок инкубировали в течение 4-5 часов при 4°С в условиях вращения, затем промывали три раза и помещали в 2-кратный объем буфера для разведения ChIP.
Примерно 1×107 клеток на образец фиксировали путем добавления в среду формальдегида до конечной концентрации 3,7% и инкубации в течение 10 минут при комнатной температуре, как описано ранее (Beneke, S., et al. PLoS One 7 (2012) e32914). Фиксацию останавливали добавлением глицина до 1-кратного раствора. После двух этапов промывания ледяным PBS и центрифугирования при 2000 g в течение 3 минут осадок ресуспендировали в 1 мл PBS, содержащего ингибитор протеазы Roche Complete (Roche Diagnostics GmbH, Мангейм, Германия). Суспензию осаждали центрифугированием (3000 g, 5 мин, 4°С). Супернатант отбрасывали.
Лизис осадка проводили путем добавления 1 мл буфера для лизиса клеток с добавлением Roche Complete (Roche Diagnostics GmbH, Мангейм, Германия) и инкубации на льду в течение 10 минут. После центрифугирования (2300 g, 4 мин, 4°С) осадок ядер ресуспендировали в 300 мкл раствора для лизиса ядер и обрабатывали ультразвуком (выход 5, рабочий цикл 90%, 15 секунд обработки ультразвуком с последующей 2-3-минутной инкубацией на льду, 6 циклов; Branson Sonifier В15, Дитценбах, Германия). К обработанным ультразвуком ядрам добавляли 700 мкл раствора для лизиса ядер и затем центрифугировали (13000 об/мин, 15 мин, 4°С). Супернатант переносили в новую пробирку для хранения при -80°С до использования. После расщепления белков степень обработки ультразвуком оценивали в агарозном геле.
Хроматин предварительно очищали путем инкубации с 80 мкл приготовленной агарозной суспензии с белком А (2 ч при 4°С в условиях вращения) и центрифугирования (13000 об/мин, 6 мин, 4°С, Eppendorf, Гамбург, Германия). Супернатант переносили в новую пробирку и определяли концентрацию белка (набор Pierce® ВСА Protein Assay, Thermo Scientific, Рокфорд, США). 25-100 мкг хроматина на один анализ иммунопреципитации (IP) вводили в 200 мкл буфера для лизиса ядер и добавляли к 300 мкл буфера для разведения ChIP, содержащего 3 мкг соответствующего антитела. Иммунопреципитацию проводили в течение ночи при 4°С в условиях вращения. Неразведенные входящие образцы хранили при -20°С. После инкубации в течение ночи 40 мкл блокированных агарозных гранул с белком А добавляли к раствору IP и инкубировали (1 ч при 4°С в условиях вращения). Преципитат промывали два раза низкосолевым, один раз высокосолевым промывочным буфером, один раз промывочным буфером с LiCl и один раз промывочным ТЕ-буфером. Каждое промывание проводили с помощью 1 мл буфера в течение 5 минут на вращающейся платформе с последующим центрифугированием (500 g, 30 сек, 4°С). После последнего этапа промывания гранулы центрифугировали со скоростью 500 g в течение 1 минуты. Осадки гранул объединяли с 200 мкл элюирующего буфера IP, при этом одновременно 25-100 мкг входящего образца хроматина заполняли элюирующим буфером IP в объеме до 200 мкл, и инкубировали в течение 30 минут при 65°С в условиях встряхивания. Затем в каждую пробирку добавляли 0,5 мкл безДНКазной РНКазы (Roche Diagnostics GmbH, Мангейм, Германия) и инкубировали в течение 30 минут при 37°С. Для окончательного расщепления белка к реакционным образцам добавляли 10 мкл 4 М раствора NaCl (конечная конц. 0,2 М) и 2 мкл протеиназы К (конечная конц. 100-200 мкг/мл, Roche Diagnostics GmbH, Мангейм, Германия) и инкубировали в течение 1,5 ч при 65°С. ДНК очищали с помощью набора Roche PCR purification kit (Roche Diagnostics GmbH, Мангейм, Германия).
Бисульфитная конверсия
Для конверсии неметилированных цитозинов в урацил использовали набор EZ-96 DNA methylation-lightning™ (глубоколуночный формат) (ZymoResarch, Фрайбург, Германия) в соответствии с инструкциями производителя. Вкратце, гДНК выделяли с помощью набора DNAse blood & tissue kit (Qiagen, Хильден, Германия) и измеряли концентрацию с помощью Nanodrop 2000 (Peqlab Biotechnologie GmbH, Эрланген, Германия), чтобы получить 350 нг гДНК в 20 мкл дважды дистиллированной воды. 20 мкл гДНК смешивали с 130 мкл реагента для конверсии Lightning Conversion Reagent в планшете для конверсии. Планшеты инкубировали при 98°С в течение 8 минут, а затем при 54°С в течение 60 минут и, наконец, временно хранили при температуре 4°С. Планшет для связывания Zymo-Spin™ I-96, содержащий 600 мкл М-связывающего буфера в каждой лунке, устанавливали на планшете для сбора материала. Образцы из планшета для конверсии добавляли в планшет для связывания Zymo-Spin™ I-96, перемешивали и центрифугировали (3000 g в течение 5 минут). Планшеты промывали с помощью 400 мкл М-промывочного буфера и центрифугировали (3000 g, 5 минут). В каждую лунку добавляли 200 мкл L-десульфонирующего буфера, инкубировали (20-30°С, 20 минут), а затем центрифугировали (3000 g в течение 5 минут). Дополнительный этап промывания завершали 10-минутным центрифугированием при 3000 g, и преобразованную ДНК элюировали в 30 мкл М-элюирующего буфера. Преобразованные ДНК хранили при -20°С до использования.
Амплификация преобразованного и интегрированного промотора hCMV-MIE
Для исследования метилирования CpG в промоторе hCMV-MIE были разработаны следующие пары праймеров к дистальной (F1 и R1) и проксимальной (F2 и R2) промоторной области:

10 пмоль каждого праймера (0,5 мкл) добавляли к 12,5 мкл смеси ZymoTaq™ DNA polymerase (ZymoResarch, Германия) и 3 мкл матрицы гДНК, и объем доводили до 25 мкл дважды дистиллированной водой. Полимеразную цепную реакцию (ПЦР) проводили с помощью Mastercyler nexus Х1 (Eppendorf, Гамбург, Германия); условия ПЦР были следующими:
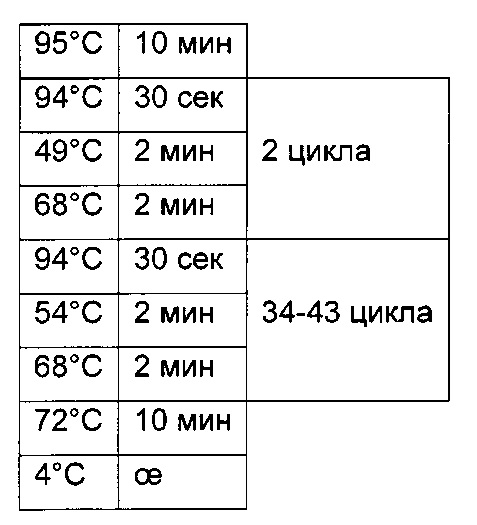
Ампликоны очищали с помощью набора для очистки ПЦР-смеси Roche PCR purification (Roche Diagnostics GmbH, Мангейм, Германия), элюируя их в 30-40 мкл элюирующего буфера. Концентрацию определяли с помощью Nanodrop 2000 (Peqlab Biotechnologie GmbH, Эрланген, Германия), а размер ампликонов определяли с помощью электрофореза в агарозном геле.
Обратная ПЦР
Обратная (инвертированная) полимеразная цепная реакция (inverse polymerase chain reaction, iPCR, оПЦР) представляет собой способ определения сайтов интеграции случайно интегрированного вектора. Благодаря ей известная последовательность вектора вблизи участка разрезания будет использоваться для создания праймеров длиной по меньшей мере 20 п.о. Эти праймеры ориентированы в противоположных направлениях.
Геном расщепляли часто режущими ферментами (которые распознают участок длиной 4 п.о.), которые не режут его между сайтами связывания праймеров и на линеаризованном участке вектора. Полученные фрагменты лигировали в очень разведенных условиях, чтобы усилить внутримолекулярное лигирование. Теперь циклическую последовательность можно амплифицировать с помощью инвертированных праймеров.
Геномную ДНК выделяли с помощью набора Qiagen blood & cell culture DNA prep, midi (Qiagen, Хильден, Германия) из 2×107 клеток (примерно 90 мкг ДНК) на каждое условие (геномную ДНК можно хранить при -20°С в течение нескольких месяцев) и растворяли в 90 мкл дважды дистиллированной воды.
Выделенную геномную ДНК расщепляли различными рестрикционными эндонуклеазами при соответствующей рабочей температуре в течение минимум 16 часов. Для расщепления гДНК использовали ферменты CviQI, MseI и MspI для оПЦР выше сайта интеграции PvuI и ферменты MseI, BfaI и MluCI для оПЦР ниже сайта интеграции PvuI. В каждом расщеплении использовали 10 мкг геномной ДНК (3,8×106 копий).
Расщепление:
Расщепленную ДНК очищали с помощью набора для очистки Roche PCR-purification и разводили в × мкл (200 мкл) элюирующего буфера. Высокая степень разведения ДНК способствует внутримолекулярному связыванию.
Лигирование:


Для того чтобы создать кольцевые фрагменты ДНК, расщепленную ДНК лигировали в течение ночи при 16°С с использованием ДНК-лигазы Т4 (NEB, Франкфурт-на-Майне, Германия). Лигированную ДНК элюировали в 50 мкл буфера для элюирования из набора для очистки High Pure PCR Purification (Roche Diagnostics GmbH, Мангейм, Германия) и добавляли к 10 нг ДНК-матрицы. В качестве контрольной группы исследовали клеточную линию СНО, не трансфицированную гДНК.
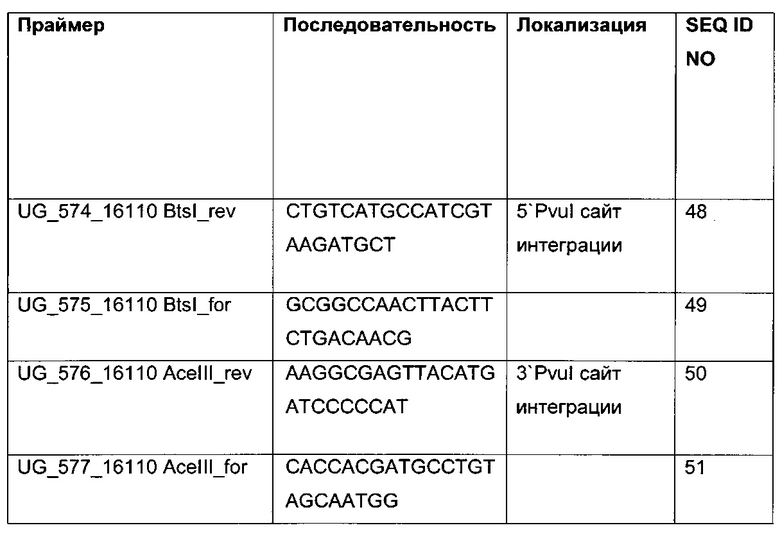
Условия оПЦР для 5'-оПЦР были следующими.
5 мкл ПЦР-фрагмента смешивали с буфером Orange G (5 мкл) и дважды дистиллированной водой (10 мкл) и наносили на агарозный гель для проверки распределения фрагментов. Для 5'-оПЦР фрагментов, порезанных MseI и MluCI, были получены хорошие результаты, а в контрольных дорожках не было обнаружено неспецифических полос.
6'-ампликоны каждого образца (порезанные MseI и MluCI) объединяли и очищали с помощью набора для очистки High Pure PCR Purification (Roche Diagnostics GmbH, Мангейм, Германия), элюируя в 40 мкл буфера для элюирования.
Количественная ПЦР в режиме реального времени
Для каждой кПЦР использовали неспецифическую флуоресцентную метку LightCycler®480 SYBR Green I Master в системе LightCycler®480 Instrument II (Roche Diagnostics GmbH (Roche Diagnostics GmbH, Мангейм, Германия). SYBR Green возбуждается при использовании синего света (λmax=488 нм) и излучает зеленый свет λmax=522 нм). Метка SYBR Green связывается со всей двухцепочечной ДНК в ПЦР. В результате интенсивность флуоресценции и количество ДНК-продукта увеличиваются одновременно, что можно регистрировать с помощью системы LightCycler® после каждого цикла. Для количественной оценки флуоресценцию откладывали на графике в виде зависимости от числа циклов на логарифмической шкале. Для обнаружения флуоресценции на основе ДНК в системе LightCycler® задавали порог немного выше излучаемого фона. Число циклов, при котором флуоресценция проходит порог, называется пороговым циклом (quantification cycle, Cq) (Bustin, S. A., et al., Clin. Chem. 55 (2009) 611-22). Во время экспоненциальной фазы амплификации ожидается удвоение ДНК-мишени в каждом новом цикле. Тем не менее, эффективность амплификации часто меняется в зависимости от праймеров и матриц. Анализ температуры плавления амплифицированных фрагментов ДНК дает первые намеки на специфичность используемых пар праймеров и количество димеров праймеров.
Эффективность комбинации праймера и матрицы можно оценить в эксперименте с титрованием, создавая стандартную кривую, или с помощью программы для расчета эффективности, такой как LinRegPCR. Программа использует данные из системы LightCycler® без коррекции по исходным условиям, выполняет базовую коррекцию по отдельности для каждого образца, определяет окно линейности, а затем использует анализ линейной регрессии, чтобы подогнать прямую линию к набору ПЦР-данных. Из наклона этой линии вычисляют эффективность ПЦР для каждого отдельного образца.
Для относительной количественной оценки количество целевой последовательности сравнивали с контрольной последовательностью путем вычитания Cq (мишени) из Cq (референса). Эта нормализация называется ΔCq-способом (Schefe, J. Н., et al., J. Mol. Med. (Berlin) 84 (2006) 901-910). Референсная последовательность должна иметь очень стабильные значения, значительно выше фоновых значений (имитационного контроля). Поэтому для каждой референсной последовательности рассчитывали среднее значение и коэффициент вариации всех образцов в рамках одного проекта и сравнивали с фоновыми уровнями. Значения Cq для разных условий показывают в % входящего образца, визуализируя расстояние до имитационного контроля.
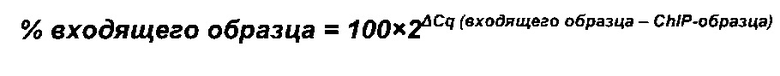
Что касается большого количества образцов, все кПЦР выполняли в 384-луночных планшетах. В каждую лунку переносили следующие ингредиенты.
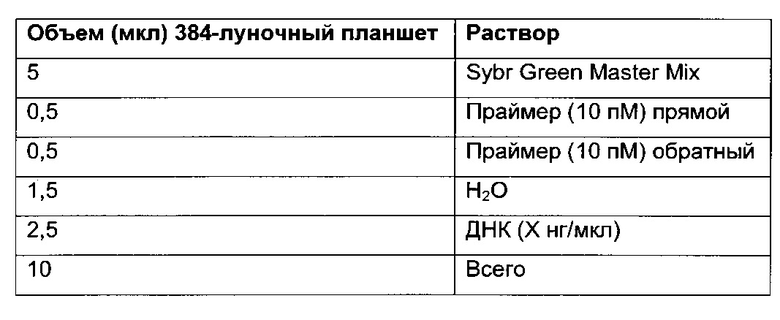
Программа кПЦР была разработана следующим образом:
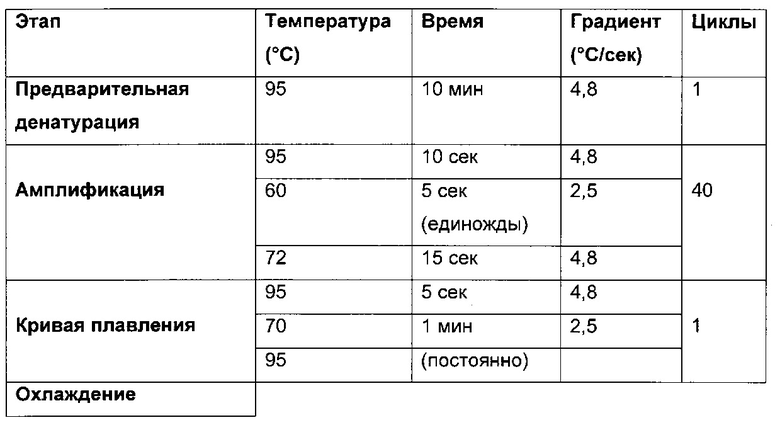
В случае относительной количественной оценки модификаций гистонов нормализация образца (обработанного) по входящему образцу (необработанному) непосредственно заменяли нормализациейм обработанной целевой последовательности по обработанной референсной последовательности. При дальнейшем вычислении модификации гистона проводили нормализацию по избыточному входящему образцу.
Относительное количество конкретной модификации гистона вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека оценивали с помощью способа Livak, также известного как способ "дельта дельта Cq" (ΔΔCq) (Livak, K.J. and Schmittgen, Т.D., Methods 25 (2001) 402-408), до тех пор, пока эффективности "праймер - матрица" были близки к значению 2 и близки друг к другу (менее 2% разница со средним Cq).
Данный способ может быть использован для относительной количественной оценки мишени относительно референса. Относительная количественная оценка ацетилирования гистона 3 и уровней гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека нормализовали по ацетилированию гистона 3 и уровням гистона 3 вблизи референсного гена в два этапа. Первое значение ΔCq рассчитывали следующим образом:
ΔCq = контроль - образец
ΔCq представляет собой расстояние между пороговыми циклами (Cq) для контроля (например, значение ChIP-образца для Gusb) и мишени (например, значение ChIP-образца для hCMV-MIE) в одном и том же образце, амплифицированном с разными парами праймеров. Для того чтобы определить уровень модификации гистона 3 в расчете на гистон 3, использовали способ ΔΔCq.
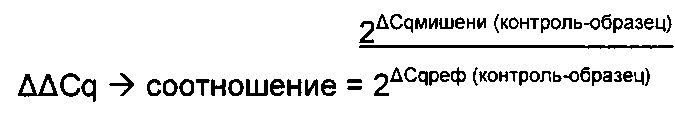
Способы секвенирования
Секвенирование по Сэнгеру проводили в компании SequiServe (SequiServe GmbH, Фатерштеттен, Германия).
Секвенирование нового поколения проводили в GATC (GATC-biotech, Констанц, Германия) и Active Motif (Ла-Юльп, Бельгия) с использованием методики Illumina Sequencing.
Количественная оценка белка
Концентрации белка оценивали в сравнении со стандартом белка с помощью набора для анализа Pierce® ВСА protein assay kit (Thermo Scientific, Рокфорд, США).
Фракционирования белка в компартментах
Белки фракционировали на мембранные/цитоплазматические и ядерные белки (Misawa, Y, 2006). Для этого клетки СНО (1×107 клеток на каждый образец) осаждали центрифугированием (300 g, 3 мин, 4°С) и осадок дважды промывали ледяным PBS с добавлением Roche Complete (Roche Diagnostics GmbH, Мангейм, Германия). Промытый осадок лизировали с помощью 1 мл буфера для лизиса с 0,5% Triton Х-100 и инкубировали на льду в течение 15 минут. Нерастворимые ядра отделяли центрифугированием (13000 об/мин, 15 минут, 4°С). Супернатант, содержащий мембрану/цитоплазму, переносили в новую пробирку и охлаждали на льду. Осадок ядер один раз промывали буфером для лизиса и ресуспендировали в 300 мкл буфера для лизиса, содержащего 0,5% SDS, а затем обрабатывали ультразвуком (5 секунд, выход 2, рабочий цикл 90%, Branson Sonifier 450, Дитценбах, Германия). Добавляли 700 мкл буфера для лизиса и образец центрифугировали 15 минут со скоростью 13000 об/мин при 4°С. Супернатант, содержащий ядра, переносили в новые пробирки и белковые фракции хранили при -80°С.
SDS-электрофорез в полиакриламидном геле (SDS-PAGE)
Для денатурирующего SDS-электрофореза в полиакриламидном геле использовали NuPAGE-гели (4-12%) (10% Bis-Tris гель 1,0 мм × 12 лунок, NuPAGE® Novex®, кат. № NP0302BOX, Invitrogen, Германия) в комбинации с системой электрофореза NuPAGE®. В соответствии с Quick Reference Card (NuPAGE ® Bis-Tris Mini Gels) гель фиксировали в камере и внутреннюю кассету заполняли рабочим 1× буфером MOPS (20× рабочий буфер NuPAGE MOPS SDS, кат. № NP0001, Invitrogen, Германия). Внешнюю камеру также заполняли на две трети ее объема. Гребенку удаляли и дорожки промывали с помощью маленькой пипетки. 30 мкл образца добавляли к 10 мкл 4× LDS-буфера для образцов NuPAGE (кат. № NP0007, Invitrogen, Германия), 2 мкл 1 М DTT (конечная концентрация 50 мМ), и инкубировали при 95°С в течение 5 минут. Образцы (20 мкл/дорожка) и маркеры ((10 мкл/дорожка) предварительно окрашенного SeeBlue Marker, кат. № LC 5625, Invitrogen; (5 мкл/дорожка) MagicMark ХР anti IgG, кат. № LC5602, Invitrogen, Германия) вносили в дорожки и к гелю прикладывали напряжение 30 вольт в течение 50 минут с помощью электрофорезной системы NuPAGE®.
SDS-PAGE и Вестерн-блот (WB)
SDS-PAGE-индуцированные разделенные белки переносили на метанол-активированную нитроцеллюлозную мембрану в соответствии с протоколом NuPAGE для денатурирующего электрофореза (техническое руководство NuPAGE® Technical Guide, часть IM1001, Invitrogen, Германия) в блот-модуле XCell II Blot-Module (Invitrogen, Германия) с соответствующими реагентами. Блоттинг проводили в SDS-PAGE-камере при напряжении 30 вольт в течение 50 минут. Мембрану с перенесенными белками переносили в новую камеру и промывали 3 раза в течение 10 минут в 1× промывочном TBS-буфере (10× TBS: 0,5 М Tris-Base, 1,5 М NaCl, рН 7,5), а затем 5 минут инкубировали в растворе Ponceau (0,1%) для обнаружения общего белка. Раствор Ponceau удаляли и остатки смывали двумя этапами промывки TBS в течение 1 минуты.
Мембрану блокировали в течение 1 часа при температуре 20-30°С в 1% блокирующем растворе (блокирующий раствор: 1/10 казеин (Roche Diagnostics GmbH, Мангейм, Германия), 1× TBS). Затем мембрану инкубировали в растворе первичного антитела (1% блокирующий раствор плюс антитело) при 4°С в течение ночи в условиях встряхивания. Мембрану промывали три раза в 1× TBST в течение 10 минут при температуре 20-30°С, а затем промывали в течение 1 минуты в 1× TBS. Мембрану инкубировали в растворе вторичного антитела (1% блокирующий раствор плюс антитело) в течение 45 минут при температуре 20-30°С или при температуре 4°С в течение ночи в условиях перемешивания. Мембрану промывали три раза в 1× TBST в течение 10 минут при температуре 20-30°С, а затем промывали два раза в течение 1 минуты в 1× TBS, чтобы удалить Tween.
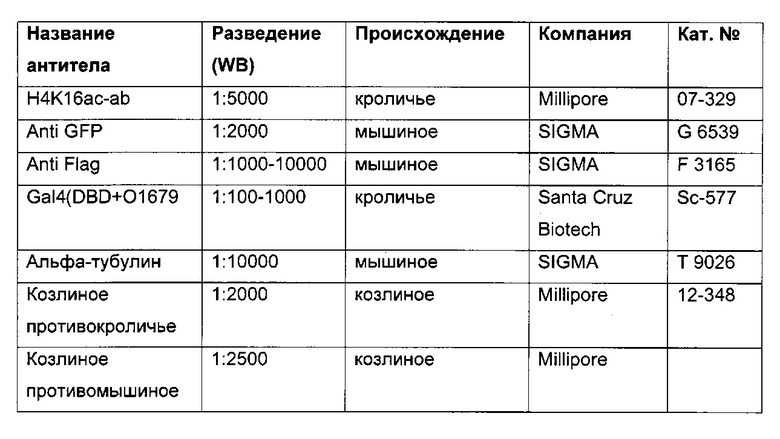
Обнаружение связывания антител проводили с помощью системы для вестерн-блоттинга Lumi-LightPLUS (Roche, Пенцберг, Германия).
Размораживание клеточных линий СНО
Готовили пробирки Falcon с 5 мл соответствующей среды. Криопробирки хранили на сухом льду до размораживания в водяной бане (37°С). Затем клетки СНО переносили в подготовленные пробирки Falcon и центрифугировали в течение 3 минут со скоростью 500 g для удаления остаточного DMSO. Осадок ресуспендировали в 5 мл среды и культивировали в одноразовых 125 мл колбах (содержащих 20 мл соответствующей среды) в стандартных увлажненных условиях (95% относительной влажности, 37°С, от 5% до 8% CO2) при постоянной скорости встряхивания от 120 об/мин до 150 об/мин. Жизнеспособность (более 95%) и плотность клеток можно было проверить с помощью Cedex HiRes Analyzer (Roche Diagnostics GmbH, Мангейм, Германия).
Культивирование клеточных линий СНО
Нетрансфицированные клетки СНО рассеивали каждые 3-4 дня и высевали с плотностью 2-3×105 клеток/мл в среде для культивирования. Трансфицированные клетки высевали с различными концентрациями метотрексата (МТХ) или метионина сульфоксимина (MSX) в качестве селективного агента. После восстановления жизнеспособности клеток (3-4 недели) стабильно трансфицированные клетки отбирали в среде без тимидина и белка, содержащей от 20 нМ до 1200 нМ метотрексата (МТХ) или 140-160 мкМ метионина сульфоксимина (MSX) в качестве селективного агента. Клетки размножали в 125 мл вентилируемых колбах в стандартных увлажненных условиях (95% относительной влажности, 37°С и 5% CO2) с постоянной скоростью встряхивания от 120 об/мин до 150 об/мин. Через каждые 3-4 дня клетки рассеивали на свежих средах с плотностью клеток 2-3×105 клеток/мл. Плотность и жизнеспособность культур определяли с помощью счетчика клеток CASY ТТ или Cedex HiRes (Roche Innovates AG, Билефельд, Германия).
Временная и стабильная трансфекция
Трансфекцию проводили с помощью набора Amaxa® Cell line Nucleofector® Kit V (Lonza, Кельн, Германия) и трансфекционных платформ Nucleofector™2b Device и 96-луночной системы Shuttle™ (Lonza, Кельн, Германия).
5×106 клеток СНО на каждую трансфекцию центрифугировали (200 g, 5 минут) и супернатант отбрасывали. Осадок ресуспендировали в 100 мкл дополненного раствора Nucleofector Solution V и добавляли 1,2 пмоль стерильной плазмиды, пипетируя вверх и вниз. Плазмиду линеаризовали для стабильной трансфекции и сохраняли кольцевой для временной трансфекции. Суспензию помещали в кювету без пузырьков и активировали программу U-24 для трансфекции клеточной линии СНО. После импульса в кювету помещали 500 мкл предварительно согретой среды и всю суспензию переносили в 8 мл предварительно согретой среды. Клетки переносили в инкубатор, и через два дня после этого клетки были готовы к первым оценкам.
Оценка числа клеток с помощью Cellavista
60 мкл суспензии клеток СНО смешивали с 60 мкл трипанового синего (0,1 мкм) в 96-луночном круглодонном планшете и инкубировали при 20-30°C в течение 5 минут. Затем обработанную суспензию клеток СНО разводили средой 1:50, и 200 мкл переносили в 96-луночный плоскодонный планшет (Greiner, Германия). Медленное центрифугирование (500 g, 5 мин) приводило к быстрому осаждению клеток.
Для точной оценки числа клеток получали изображения середины лунки, чтобы избежать ошибок изображения вблизи краев каждой лунки. Для подсчета клеток использовали систему Imager Cellavista Imager (SynenTec, Мюнхен, Германия). Каждый образец анализировали в повторах, и всего для подсчета числа клеток использовали 10 изображений.
Оценка числа клеток с помощью анализатора Cedex HiRes Analyzer
Для подсчета числа клеток в образце числом до 40 использовали анализатор Cedex HiRes Analyzer (Roche, Германия). Для этого клетки СНО разводили в соотношении 1:5 в среде HiRes, и 300 мкл переносили в пробирки. Подсчет клеток проводили в соответствии с рекомендациями производителя. Для оценки числа клеток образцы анализировали в двух повторах.
Подготовка образцов для анализа с антителами
Рассчитывали плотность клеток и центрифугировали по 2 мл на каждый образец (500 g, 5 мин, 20-30°С). Супернатант переносили в новые 96-луночные глубоколуночные планшеты и хранили при -20°С до использования. Замороженный супернатант размораживали в течение ночи при температуре 4°С, 6 раз переворачивали и центрифугировали (4000 об/мин, 30 минут, 20-30°С). 310 мкл фильтровали путем центрифугирования (1200 об/мин, 3 мин, 20-30°С) с помощью планшета Multiscreen Millipore, установленного на 96-луночный круглодонный планшет со штрих-кодом.
Количественная оценка продукции антител с помощью HPLC
Для количественной оценки антитела, присутствующего в образце, использовали хроматографический способ. Использовали колонку Poros А, которая связывает Fc-область антитела. Антитело фиксируют на колонке и затем элюируют при низких значениях рН. Концентрацию белка оценивают путем определения оптической плотности (OD) при 280 нм с референсной длиной волны 320 нм, используя коэффициент молярной экстинкции, рассчитанный на основании аминокислотной последовательности.
Количественная оценка продукции антител путем ELISA
Методика ELISA (Enzyme Linked Immunosorbent Assay, иммуноферментный анализ, ИФА) основана на принципе "сэндвича из антител". Антитело захвата, специфическое для анализируемого соединения, представляющего интерес, например, для Fc-части IgG, фиксировали на планшете для микротитрования, создавая твердую фазу. Затем, после этапов блокировки и промывания, образцы, стандарты (серию разведений референсного антитела) и контроли инкубировали с антителом на твердой фазе, которое захватывает анализируемое соединение. После отмывания несвязанного анализируемого соединения добавляли конъюгированное антитело для обнаружения (например, конъюгированное с POD). Это антитело для обнаружения связывалось с другим эпитопом на измеряемой молекуле, завершая образование "сэндвича". Субстрат POD - ВМ Chemiluminescence ELISA Substrate (Roche Diagnostics GmbH, Мангейм, Германия) - представлял собой раствора субстрата системы вторичного обнаружения на основе пероксидазы (POD, HRP). Скорость образования сигнала в иммунологическом анализе была прямо пропорциональна количеству фермента-маркера, связанного с твердой фазой. Концентрацию антитела рассчитывали по наклону стандартного разведения.
Создание рекомбинантных клеточных линий СНО, содержащих промотор hCMV-MIE
Рекомбинантные клеточные линии, экспрессирующие конструкции человеческого антитела класса IgG, получали путем временной или стабильной трансфекции клеток СНО-K1, растущих в суспензии. Экспрессионные кассеты легкой и тяжелой цепей человеческого иммуноглобулина находились под контролем главного немедленно-раннего промотора и энхансера CMV человека.
Вектор также содержал нуклеиновокислотную последовательность, кодирующую мышиную дигидрофолатредуктазу (dihydrofolate reductase, DHFR). Трансфекцию клеток проводили с помощью системы Amaxa nucleofection (Lonza Cologne GmbH, Кельн, Германия).
Суспензию СНО-К1 М трансфицировали кольцевой плазмидной ДНК для временной экспрессии антитела, используя устройство Nucleofector в сочетании с набором Nucleofector Kit V (Lonza Cologne GmbH, Кельн, Германия) в соответствии с протоколами производителей. Суспензии временно трансфицированных клеток высевали в 96-луночные планшеты и инкубировали в течение 5 дней.
Суспензии стабильно трансфицированных клеток высевали в 384- или 96-луночные планшеты, содержащие бестимидиновую среду с метотрексатом (МТХ) в качестве селективного агента в концентрации от 250 до 1600 нМ. Через 3-4 недели пулы клеток, экспрессирующих антитела, оценивали на долгосрочную стабильность в течение периода времени от одного до трех месяцев. Отдельные клеточные клоны, экспрессирующие антитела, высевали в 384- и 96-луночные планшеты. Через 3 недели клеточные линии, экспрессирующие антитела, определяли путем измерения титров антител в культуральной среде путем ELISA. Растущие лунки отбирали случайным образом, и для анализа долгосрочной стабильности клеточные клоны размножали в больших объемах (3 мл на лунку в 6-луночных планшетах) и в конце каждого пассажа определяли концентрацию антител с помощью HPLC на белке А и ELISA.
Клетки размножали в одноразовых 125 мл вентилируемых встряхиваемых колбах или в 6-луночных планшетах в стандартных условиях влажности (95% относительной влажности, 37°С, от 5% до 8% CO2) при постоянной скорости встряхивания от 120 об/мин до 150 об/мин. Каждые 3-4 дня клетки разделяли и переносили в свежую среду. Плотность и жизнеспособность культур определяли с помощью счетчика клеток Cedex HiRes (Roche Innovates AG, Билефельд, Германия) или Cellavista CV3.1 (SynenTec Bio Services GmbH, Мюнхен, Германия).
Долгосрочное культивирование и продукция на клеточных линиях СНО, содержащих промотор hCMV-MIE
Клетки проверяли на фенотипическую стабильность (т.е. продуктивность) в течение 2-3 месяцев после трансфекции в присутствии селективного агента МТХ. Клетки непрерывно культивировали в вентилируемых 125 мл колбах, содержащих 20-40 мл среды с селективным агентом, или в 6-луночных планшетах, содержащих 2-4 мл среды с селективным агентом, и разводили два раза в неделю свежей средой. Плотность посева составляла 2-3×105 клеток/мл. Перед пассажем определяли жизнеспособность и плотность жизнеспособных клеток.
Концентрацию антител в супернатанте (титр антител) определяли в конце каждого пассажа с помощью HPLC на белке А и ELISA. Исходя из этих данных, удельную продуктивность клетки (qP) для каждого пассажа рассчитывали по следующей формуле:
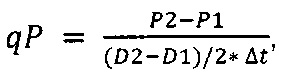 где:
где:
qP [пг/клетка/день]: удельная продуктивность клетки,
P1 [мкг/мл]: титр антитела в начале пассажа,
Р2 [мкг/мл]: титр антитела в конце пассажа,
D1 [клеток/мл]: плотность жизнеспособных клеток в начале пассажа,
D2 [клеток/мл]: плотность жизнеспособных клеток в конце пассажа,
Δt [d]: продолжительность пассажа.
Значения qP откладывали на графике в виде зависимости от возраста культуры в конце соответствующего пассажа в поколениях. Линейную линию тренда рассчитывали по всем точкам данных qP и вычисляли относительное изменение qP (в процентах) в течение периода времени по следующему уравнению:
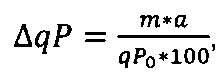 где
где
ΔqP [%]: процентное изменение qP,
m [пг/клетка/день/поколение]: наклон линейной линии тренда,
а [число поколений]: возраст культуры,
qP0: пересечение Y-оси и линейной линии тренда.
В связи с меньшим числом точек данных, полученных для каждого образца, среднее значение трех последних значений qP делили на среднее значение первых двух значений qP и отображали в процентах, чтобы получить ΔqP.
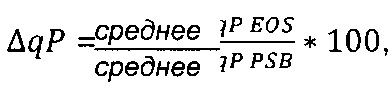 где
где
Среднее qP EOS: среднее значение от последних трех значений qP
Среднее qP PSB: среднее значение от первых двух значений qP
Обработка рекомбинантных клеточных линий СНО, используемых для определения эпигенетического маркера
32 клеточные линии СНО из двух проектов использовали для определения эпигенетических маркеров, прогнозирующих экспрессию генов-мишеней на два месяца вперед. Для обоих проектов в течение 60 поколений (примерно 2 месяца) собирали данные, включая концентрацию антител, статус метилирования С-425 в промоторе hCMV-MIE и число копий. Проект Н состоял из 20 клеточных линий СНО-К1, экспрессирующих человеческое антитело класса IgG под контролем промотора hCMV-MIE. Также вектор содержал нуклеиновокислотную последовательность, кодирующую глутаминсинтетазу, что делало клетки восприимчивыми к селекции на сульфоксимине метионина (MSX). Проект Т состоял из 12 клеточных линий СНО-К1, экспрессирующих человеческое антитело класса IgG под контролем промотора hCMV-MIE. Эти клеточные линии подвержены селекции на метотрексате (МТХ) из-за трансгена мышиной дигидрофолатредуктазы. Замороженные исходные культуры размораживали и культивировали в соответствующих средах, содержащих 250 нМ МТХ или 140 мкМ MSX. Через две недели культивирования (временная точка PSB) клетки собирали и проводили иммунопреципитацию хроматина со следующими антителами.
Список для проекта Т
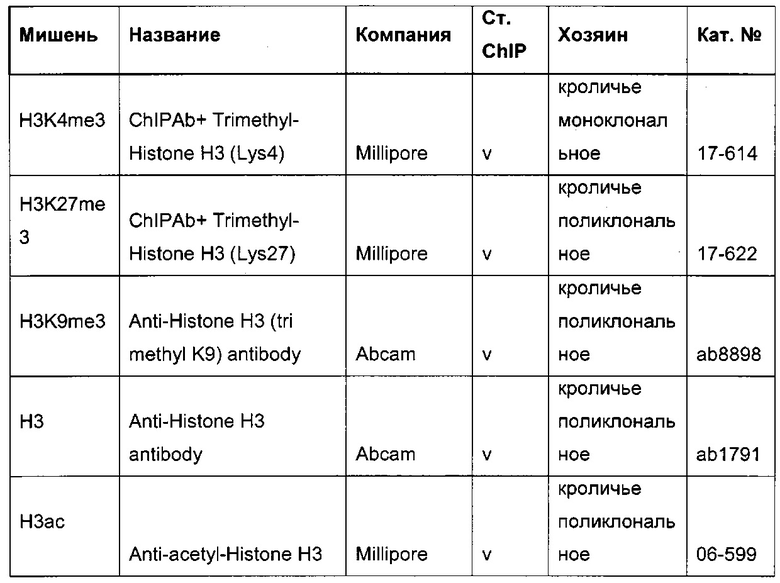
Список для проекта Н
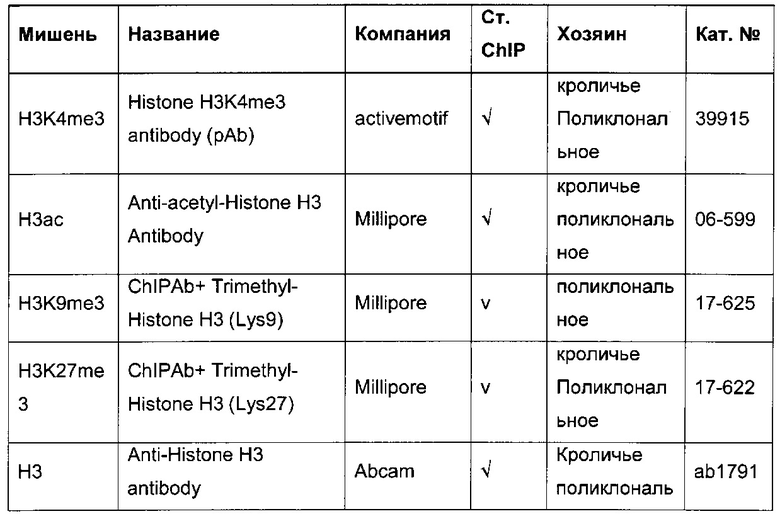

Количественную ПЦР проводили со следующими праймерами:
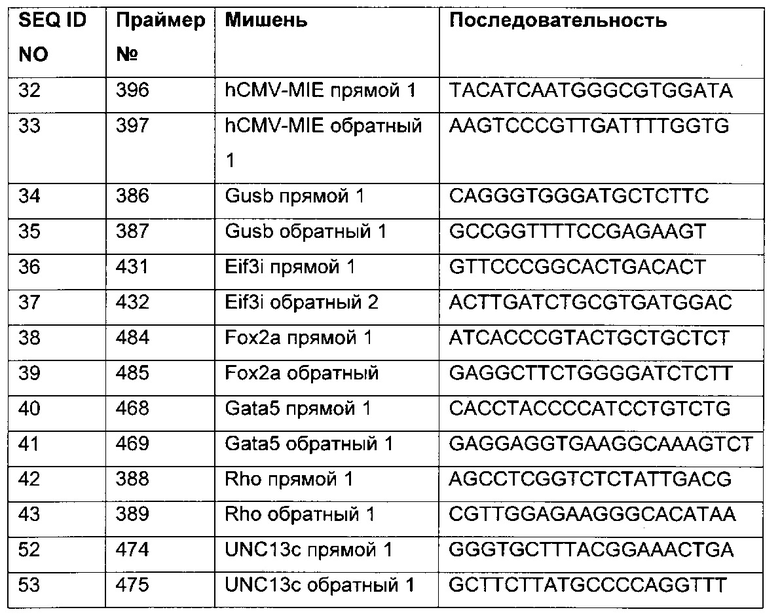
Определение числа копий трансгена
Определение числа копий трансгена проводили так, как описано ранее (Osterlehner, et al. 2011). Число копий на образец (LCS или HCS) экстраполировали из стандартной кривой серии разбавлений. Если предположить, что каждая клетка содержит примерно 6 пг ДНК, то количество трансгена на клетку (LCcell) рассчитывали следующим образом.
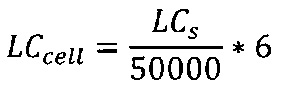
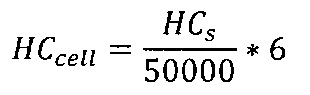
Компьютерный анализ с JMP
Процент метилирования С-179 и ацетилирования в расчете на гистон 3 вблизи hCMV-MIE в начале фазы культивирования откладывали на графике в виде зависимости от изменений удельной продуктивности в течение 60 поколений (ΔqP) на стандартной модели наименьших квадратов. Для этого использовали программное обеспечение JMP10 (JMP®10.0.1 Release: 2, 64-битная версия; SAS Institute Inc.) и рассчитывали Р-значения на графике влияния рычагов. Также выполняли анализы выбросов с использованием методики складного ножа. Для определения лучшего узла расщепления использовали статистический анализ LogWorth, разделяя клеточные популяции по стабильности и степени эпигенетической модификации (Sail 2002).
Пример 2
Создание рекомбинантных клеточных линий СНО
Рекомбинантные клеточные линии, экспрессирующие человеческое антитело класса IgG, получали путем стабильной трансфекции клеток СНО-К1 или CHO-DG44, растущих в суспензии, вектором, кодирующим легкую и тяжелую цепь антитела, содержащим легкую каппа-цепь иммуноглобулина человека и тяжелую гамма-1- или гамма-4-цепь иммуноглобулина человека. Вектор также содержал нуклеиновую кислоту, кодирующую мышиную дигидрофолатредуктазу (DHFR) или глутаминсинтетазу (GS). Экспрессионные кассеты легкой и тяжелой цепей находились под контролем главного немедленно-раннего промотора и энхансера CMV человека (SEQ ID NO 01). Трансфекцию клеток выполняли либо путем нуклеофекции (Lonza Cologne GmbH или Amaxa Biosystems), либо путем электропорации с использованием Gene Pulser XCell (BIO-RAD).
Например, суспензию СНО-К1 или CHO-DG44 трансфицировали линеаризованной плазмидной ДНК, используя устройство Nucleofector в сочетании с набором Nucleofector Kit V (Lonza Кельн GmbH, Кельн, Германия), или путем электропорации с использованием Gene Pulser XCell (Bio-Rad, Hercules, CA) в соответствии с протоколами производителей. Трансфицированные клетки высевали в 96-или 384-луночные планшеты, содержащие бестимидиновую среду с различными концентрациями метотрексата (МТХ) в качестве селективного агента или метионина сульфоксимина (MSX) в качестве селективного агента. Через три недели клеточные линии, экспрессирующие антитела, определяли путем измерения титров антител в культуральной среде с помощью ELISA. Самые мощные продуценты размножали до больших объемов, субклонировали путем ограниченного разведения и криоконсервировали.
Стабильно трансфицированные клетки отбирали на среде без тимидина и белка, содержащей 20-1200 нМ метотрексата (МТХ) или 140-160 мкМ метионина сульфоксимина (MSX) в качестве селективного агента. Клетки или клеточные линии, экспрессирующие антитело, определяли путем измерения титров антител в культуральной среде и субклонировали путем ограниченного разведения и/или FACS отдельно депонированных клеток.
Клетки размножали в одноразовых 50 мл или 125 мл вентилируемых встряхиваемых колбах в стандартных условиях влажности (95% относительной влажности, 37°С, от 5% до 8% CO2) с постоянной скоростью встряхивания от 120 об/мин до 150 об/мин. Каждые 3-4 дня клетки разделяли и переносили в свежую среду. Плотность и жизнеспособность культур определяли с помощью счетчика клеток Casy ТТ или Cedex HiRes (Roche Innovates AG, Билефельд, Германия).
Кроме того, применяли стандартные методики культивирования клеток, описанные, например, в Current Protocols in Cell Biology, Bonifacino, J.S. et al. (eds), John Wiley & Sons, Inc., New York (2000).
Пример 3
Долгосрочное культивирование и продукция
Клеточные линии СНО, полученные в соответствии с примером 2, исследовали на предмет долгосрочной продуктивности.
Клетки анализировали на фенотипическую стабильность, т.е. продуктивность, в течение 35-70 поколений в отсутствие селективного агента. Клетки непрерывно культивировали в вентилируемых встряхиваемых колбах объемом 125 мл, содержащих 50 мл среды без селективного агента, и разводили два раза в неделю свежей средой. Плотность посева составляла 2-3×105 клеток/мл. Перед пассажем определяли жизнеспособность и плотность жизнеспособных клеток. Возраст культуры в поколениях в конце каждого пассажа рассчитывали по следующему уравнению:

где
а2 [число поколений]: возраст культуры в конце пассажа,
a1 [число поколений]: возраст культуры в начале пассажа, т.е. возраст культуры в конце предыдущего пассажа,
D1 [клеток/мл]: плотность жизнеспособных клеток в начале пассажа,
D2 [клеток/мл]: плотность жизнеспособных клеток в конце пассажа.
Концентрацию антитела в супернатанте (титр антитела) определяли путем HPLC на белке А в конце каждого пассажа. Исходя из этих данных, удельную скорость продукции (SPR) для каждого пассажа рассчитывали по следующей формуле:
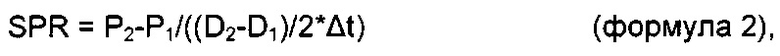
где
SPR [пг/клетка/д]: удельная скорость продукции,
P1 [мкг/мл]: титр антитела в начале пассажа,
Р2 [мкг/мл]: титр антитела в конце пассажа,
D1 [клеток/мл]: плотность жизнеспособных клеток в начале пассажа,
D2 [клеток/мл]: плотность жизнеспособных клеток в конце пассажа,
Δt [дни]: продолжительность пассажа.
Значения SPR откладывали на графике в виде зависимости от возраста культуры в конце соответствующего пассажа в поколениях. Линейную линию тренда рассчитывали по всем точкам данных SPR, и рассчитывали относительное изменение SPR (в процентах) за исследуемый период по следующему уравнению:

где
ΔSPR [%]: процентное изменение SPR,
m [пг/клетка/дни/поколение]: наклон линейной линии тренда,
а [число поколений]: возраст культуры,
SPR0: пересечение Y-оси с линейной линией тренда.
Почти все клеточные линии показали уменьшение продуктивности, при этом потеря продуктивности имела тенденцию становиться сильнее в условиях без селективного агента
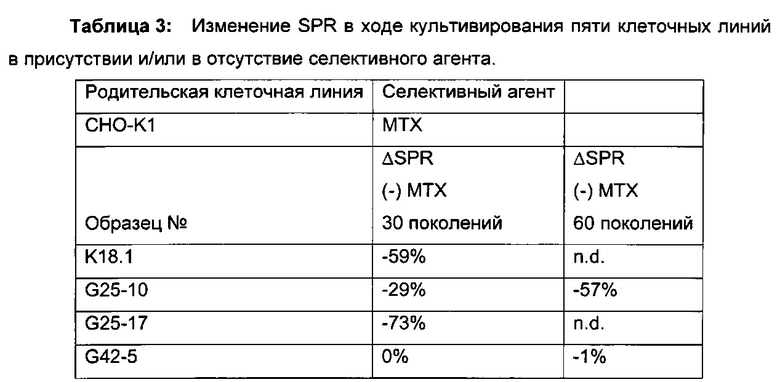
n.d. - не определено

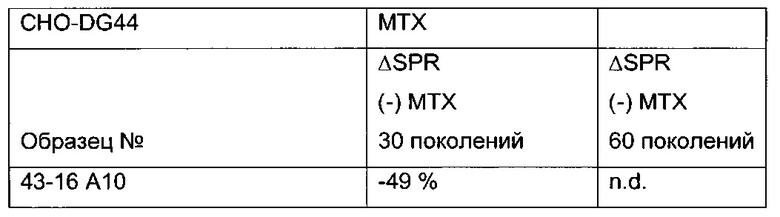
n.d. - не определено
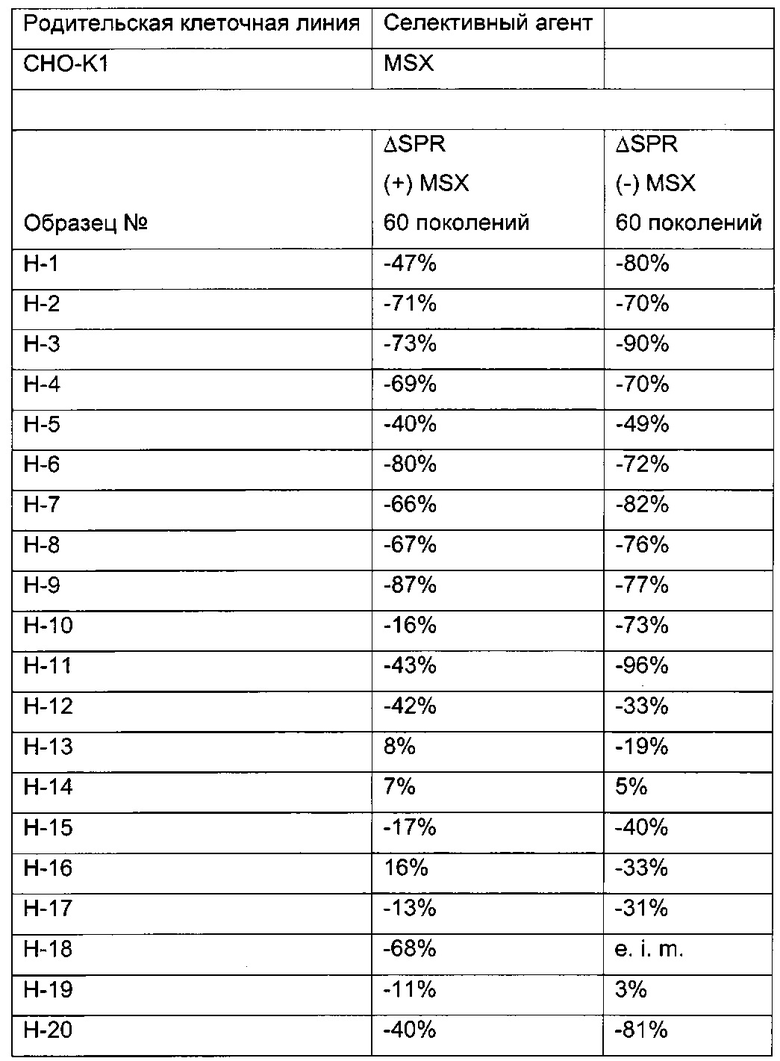
е.i.m. - ошибка в измерении
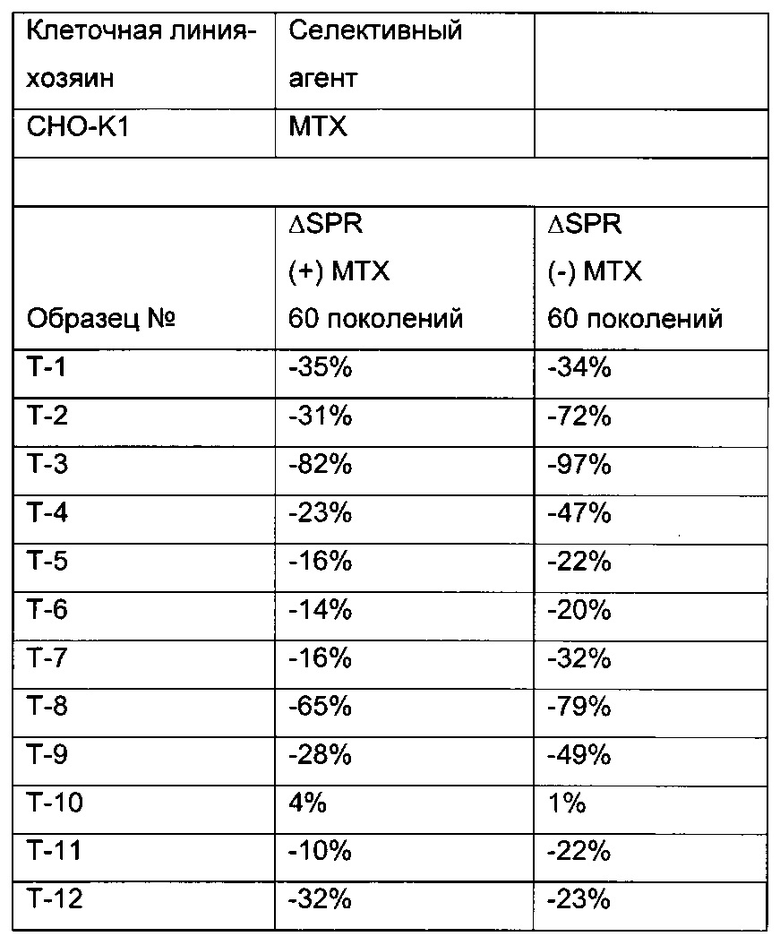
Пример 4
Определение метилированных CpG-сайтов в ДНК главного немедленно-раннего промотора/энхансера CMV человека путем обработки бисульфитом и секвенирования ДНК
Фрагмент главного немедленно-раннего промотора/энхансера CMV человека (SEQ ID NO 01), используемый для экспрессии генов легких и тяжелых цепей антитела, содержит 33 CpG-сайта.
Геномную ДНК выделяли из клеточных линий СНО K18.1, G25-10, G25-17, G42-5 и 43-16 А10 с помощью набора Allprep DNA/RNA Mini Kit от Qiagen (Хильден, Германия). 5 мкг ДНК расщепляли ферментом Dral и количественно оценивали путем измерения экстинкции при 260 нм. 100 нг ДНК подвергали обработке бисульфитом и очищали, используя набор EpiTect Bisulfite Kit (Qiagen, Хильден, Германия). Обработанную бисульфитом ДНК растворяли в 20 мкл свободной от РНКаз воды (Qiagen, Хильден, Германия).
Для амплификации нити А (прямой) фрагмента главного немедленно-раннего промотора/энхансера CMV человека (SEQ ID NO 02) 1 мкл обработанной бисульфитом ДНК объединяли с 24 мкл ПЦР-смеси, и проводили ПЦР с помощью GeneAmp® PCR System 9700 (Applied Biosystems Inc., США).
24 мкл ПЦР-смеси включали:
1 мкл прямого праймера 227 с SEQ ID NO 06 (10 пмоль/мкл), 1 мкл обратного праймера 229 с SEQ ID NO 08 (10 пмоль/мкл), 22 мкл смеси Platinum® PCR SuperMix HighFidelty (Invitrogen Corp., США). Прямой праймер 227 является комплементарным 5'-концу фрагмента главного немедленно-раннего промотора/энхансера CMV человека. Обратный праймер 229 связывается ниже в пределах 5'-UTR генов иммуноглобулина. Условия ПЦР были следующими:

ПЦР-продукт проверяли на размер и чистоту с помощью электрофореза в агарозном геле и клонировали в вектор pCR4 (Invitrogen Corp., США) с использованием набора ТОРО ТА Cloning Kit (Invitrogen Corp., США). Плазмидные клоны выделяли и анализировали путем расщепления рестриктазами и электрофореза в агарозном геле. Для каждой клеточной линии секвенировали от 19 до 22 плазмид, содержащих вставку. Для того чтобы оценить эффективность дезаминирования He-CpG-сайтов при обработке бисульфитом, определяли количество остаточного цитозина на нe-CpG-сайтах. Процентную эффективность дезаминирования рассчитывали следующим образом:

где
Emod [%]: эффективность дезаминирования,
Cres: количество остаточного цитозина на нe-CpG-сайтах во всех проанализированных вставках, исключая сайты ПЦР-праймеров,
Ctotal: количество цитозина в не обработанном бисульфитом фрагменте промотора/энхансера CMV, исключая сайты ПЦР-праймеров, умноженное на количество проанализированных вставок, т.е. 107*20.
Было установлено, что эффективность дезаминирования цитозина на не-CpG была больше 99% во всех образцах (таб. 4).
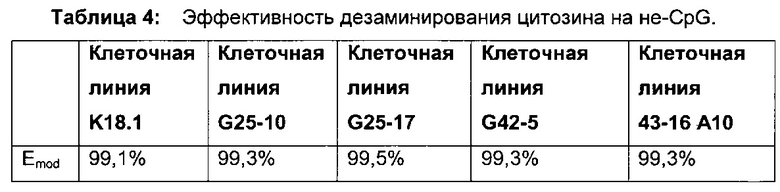
Защита 5-метилцитозина от дезаминирования подтверждали путем обработки бисульфитом и последующего клонирования и секвенирования плазмидной ДНК, выделенной из dcm+ Е. coli. Dcm+ Е. coli метилируют внутренние остатки цитозина в последовательностях CCAGG или CCTGG (dcm-сайты). Было установлено, что эффективность дезаминирования составляла 99% на не-dcm-сайтах и менее 5% на внутреннем цитозине на dcm-сайтах.
Чтобы оценить степень метилирования ДНК на CpG-сайтах в обработанном бисульфитом фрагменте промотора/энхансера CMV, определяли количество цитозина на каждом CpG-сайте и строили график для каждой анализируемой клетки.
Клеточная линия K18.1 является высоко метилированной (фиг. 3А). Частота метилирования накапливается в 3 кластерах, один на 5'-конце, один на 3'-конце и один в районе позиции 400. Самая высокая степень метилирования была обнаружена в позиции 425. Четырнадцать из двадцати двух секвенированных вставок имели цитозин в этой позиции.
Заметным было метилирование промотора CMV из клеточной линии 43-16 А10 (фиг. 3Е). Распределение метилирования было подобно распределению, наблюдаемому для клеточной линии K18.1. Позиция 425 была метилирована наиболее часто. Пять из двадцати секвенированных вставок содержали цитозин в этой позиции.
В трех других исследованных клеточных линиях цитозин обнаруживался лишь эпизодически в CpG-сайтах (фиг. 3В, 3С и 3D).
Пример 5
Количественная специфическая в отношении метилирования ПЦР с обработанной бисульфитом ДНК главного немедленно-раннего промотора/энхансера CMV человека
В этом примере описана специфическая в отношении метилирования кПЦР в режиме реального времени в качестве способа обнаружения метилирования в позиции CpG, более точно в позиции 425, в нуклеиновой кислоте промотора hCMV.
Были разработаны два набора праймеров:
- пара специфических в отношении метилирования праймеров (пара праймеров MSP), которые селективно аплифицируют дезаминированную ДНК промотора CMV с цитозином в позиции 425, представляя ДНК, которая метилирована в позиции 425, и
- пара универсальных праймеров, которые аплифицируют дезаминированную ДНК промотора CMV независимо от статуса метилирования.
Универсальную пару праймеров использовали для нормализации. Чтобы использовать их в одной и той же ПЦР, обе пары праймеров должны иметь одинаковые температуры плавления.
Дизайн праймеров, чувствительных в отношении метилирования в позиции 425, был осложнен наличием двух дополнительных CpG-сайтов в непосредственной близости (в позиции 416 и позиции 437). Чувствительные в отношении метилирования праймеры должны обнаруживать 5mC425 независимо от статуса метилирования в позиции 416 и позиции 437.
Четыре дезаминированных фрагмента главного немедленно-раннего промотора/энхансера CMV человека, выделенные в примере 4, представляющие возможные вариации последовательности в позициях 425 и 437: №11, №62, №01 и №04 (таблица 5, SEQ ID NO 19, SEQ ID NO 20, SEQ ID NO 21, SEQ ID NO 22), использовали в качестве кПЦР-матриц в специфической в отношении метилирования ПЦР и в ПЦР с универсальной парой праймеров.
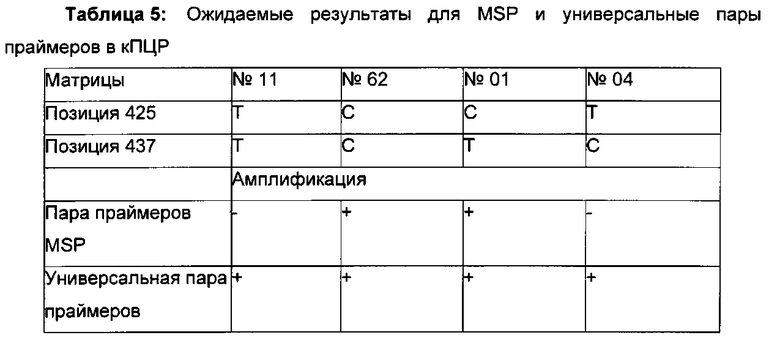
С универсальным праймером все четыре матрицы могут быть сравнимо амплифицированы, тогда как пара специфических в отношении метилирования праймеров может селективно амплифицировать матрицу №62 и матрицу №01. Значение ΔСр должно быть как можно меньше между парой специфических в отношении метилирования праймеров и парой универсальных праймеров на матрицах №62 и №01. Значения Ср, полученные с парой специфических в отношении метилирования праймеров на матрицах №11 и №04, должны быть как можно выше, т.е. значение ΔСр по сравнению с амплификацией с универсальной парой праймеров должно быть максимальным.
Для кПЦР использовали систему LightCycler® 480 II (Roche Diagnostics GmbH, Мангейм, Германия), а образцы получали с помощью LightCycler® 480 SYBR Green I Master (Roche Diagnostics GmbH, Мангейм, Германия). 5 мкл раствора матрицы, содержащего 0,05 нг ДНК, объединяли с 15 мкл ПЦР-смеси в лунке 96-луночного планшета.
15 мкл ПЦР смеси включали: 4,2 мкл воды,
- 0,4 мкл прямого праймера 239, 263 или 264 (также возможны праймеры 227, 228, 240, 238) (10 пмоль/мкл),
- 0,4 мкл обратного праймера 237, 254, 262, 265, 266, 267 или 268 (также возможен праймер 229) (10 пмоль/мкл),
- 10 мкл SYBR Green I Master.
Многолуночный планшет запечатывали герметизирующей фольгой LightCycler® 480 (Roche Diagnostics GmbH, Мангейм, Германия) и центрифугировали со скоростью 1500 g в течение 2 минут. После этого планшет устанавливали в систему LightCycler® 480 и подвергали кПЦР. Каждый образец анализировали в двух, трех или четырех повторах. Чтобы определить абсолютное число копий, для LC- и НС-трансгенов создавали стандартные кривые, используя линеаризованные экспрессионные плазмиды в качестве стандарта. Стандартные разведения содержали 2,5×107, 2,5×106, 2,5×105, 2,5×104 или 2,5×103 копий плазмиды. Геномную ДНК анализировали в трех повторах; стандарты вводили в четырех повторах.
Условия ПЦР были следующими:
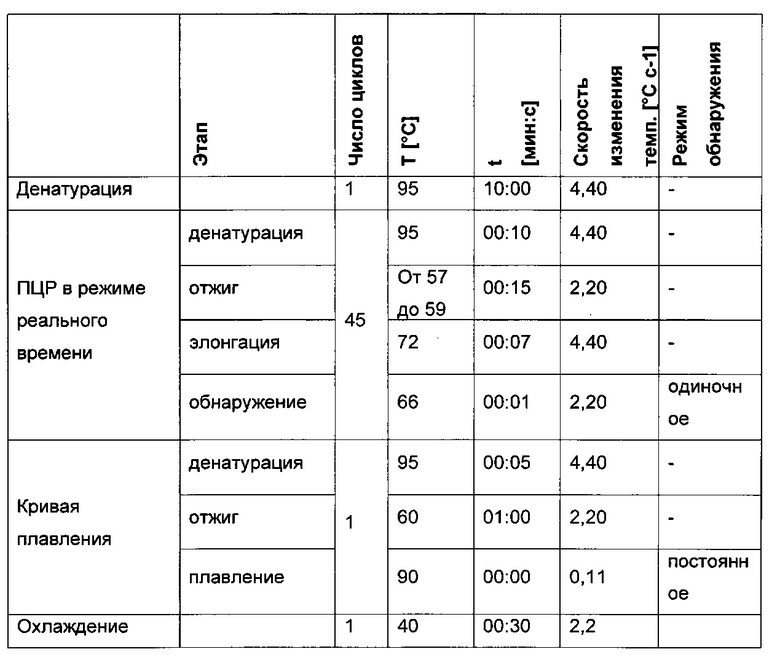
Сбор и анализ данных проводили с помощью программного обеспечения LightCycler® 480 версии 1.5. Кажущуюся степень метилирования рассчитывали по следующей формуле, где предполагается идеальная эффективность амплификации (Е=2).

где
mCapp [%]: кажущаяся степень метилирования,
Cp(t): значение Ср, полученное с универсальными праймерами,
Ср(m): значение Ср, полученное с праймерами, специфическими в отношении метилирования.
При оценке праймеров было установлено, что разработку праймеров, специфических в отношении метилирования, которые высоко селективны для дезаминированной ДНК промотора CMV с цитозином в позиции 425 ("метилированной"), нужно проводить с осторожностью. Праймеры 266 и 267 продемонстрировали максимальную разницу между Ср (№11) и Ср (№62), т.е. наибольшую селективность для "метилированной" ДНК. Праймеры 265, 268 и 254 продемонстрировали незначительную селективность. Универсальная пара праймеров 263/237 анализировали в качестве контроля для минимального ΔСр (фиг. 6 и таблица 6).
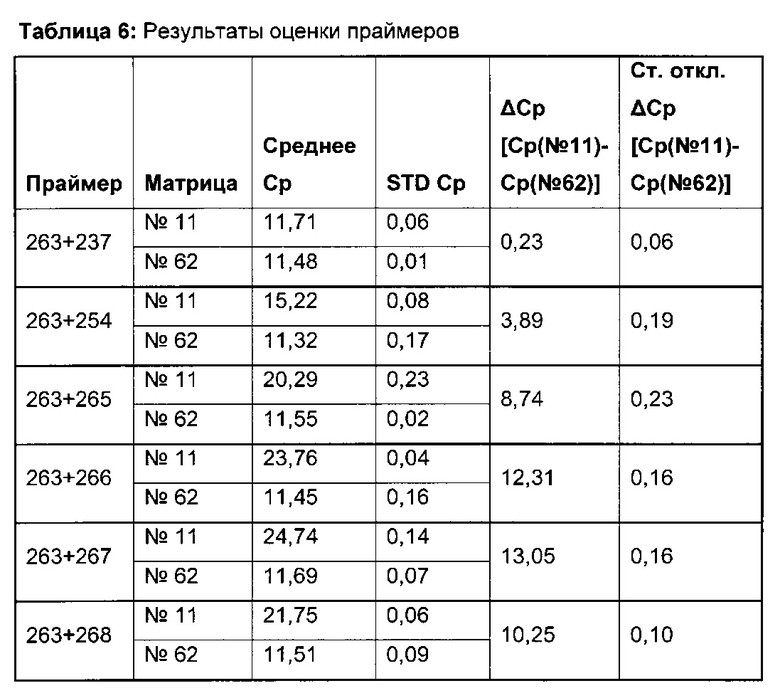
На фиг. 5 показаны результаты, полученные с парой специфических в отношении метилирования праймеров 239/267 (SEQ ID NO 11, SEQ ID NO 18) в сочетании с универсальной парой праймеров 239/237 (SEQ ID NO 11, SEQ ID NO 09). Универсальная пара праймеров 239/237 амплифицировала все четыре матрицы примерно одинаково хорошо, в то время как пара специфических в отношении метилирования праймеров 239/267 амплифицировала матрицы №62 и №01. Матрицы №11 и №4 плохо амплифицировались только парой праймеров 239/267.
Частота метилирования, рассчитанная из значений Ср, составляла почти 100% для матрицы №62 и матрицы №01 и почти 0% для матрицы №11 и матрицы №04 (Таблица 7А). Это показывает, что пара специфических в отношении метилирования праймеров 239/267 в сочетании с универсальной парой праймеров 239/237 может быть использована для различения ДНК промотора CMV, которая метилирована в позиции 425, и ДНК промотора CMV, которая не метилирована в позиции 425, с помощью кПЦР в режиме реального времени.
Были обнаружены дополнительные пары универсальных и специфических в отношении метилирования праймеров, они охарактеризованы в таб. 7А и 7В.
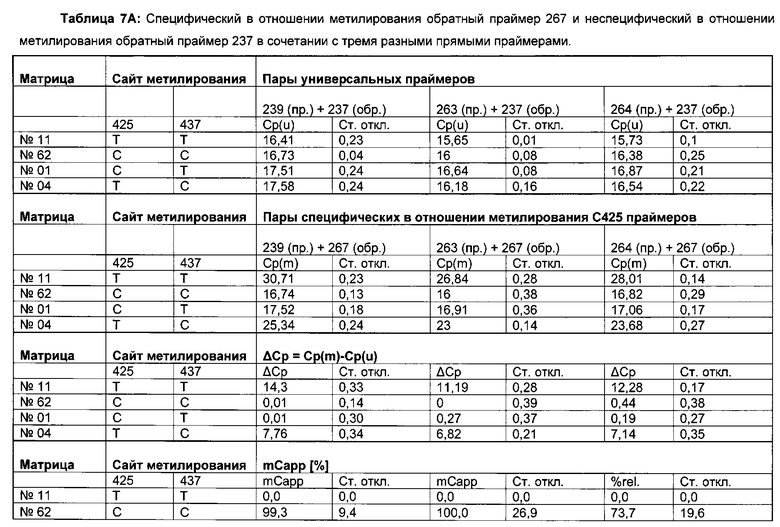

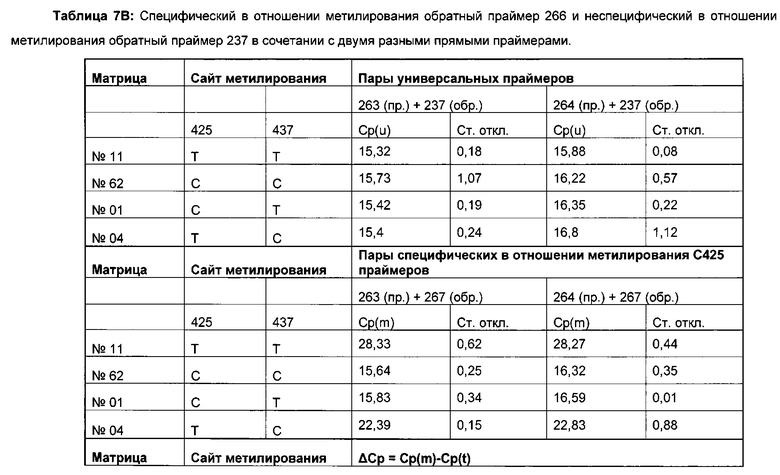
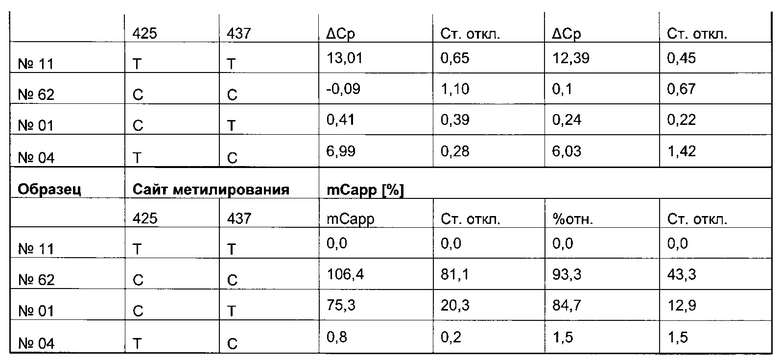
Для количественной оценки степени метилирования в широком диапазоне матрицу №62 смешивали в различных соотношениях с матрицей №11. кПЦР проводили, как описано выше, с использованием пар праймеров 239/237 и 239/267 и рассчитывали матрицу №62 на фоне матрицы №11.
Чтобы рассчитать фракцию ДНК матрицы №62, определяли эффективность амплификации пар праймеров в используемых условиях. Серийные разведения матриц №62 и №11 от п равного 0,005 нг до 0,5 нг ДНК подвергали кПЦР, и полученные значения Ср откладывали на графике в виде зависимости от log (n). Линейную линию регрессии рассчитывали с использованием XLfit (Microsoft). Эффективность амплификации рассчитывали по следующей формуле:

где
Е: эффективность амплификации,
m: наклон линейной линии тренда.
Рассчитанная эффективность амплификации обеих пар праймеров составляла примерно 1,7. Для расчета фракции ДНК матрицы №62 использовали следующую формулу:

где
mC [%]: доля ДНК, метилированной в позиции 425,
Cp(t): значение Ср, полученное с универсальной парой праймеров,
Ср(m): значение Ср, полученное с парой специфических в отношении метилирования праймеров.
Полученные фракции ДНК матрицы №62 из двух независимых экспериментов откладывали на графике в виде зависимости от ожидаемых значений (фиг. 7). Количественная оценка метилирования может составлять от 1% до 100%.
Пример 6
Определения статуса метилирования С-425 в главном немедленно-раннем промоторе/энхансере CMV человека путем обработки бисульфитом и количественной ПЦР в режиме реального времени Обработка бисульфитом геномной ДНК клеточных линий СНО
Анализ проводили, как описано в Osterlehner et al. (см. выше). Геномную ДНК выделяли из клеточных линий СНО с помощью набора Allprep DNA/RNA Mini Kit от Qiagen (Хильден, Германия). 5 мкг ДНК расщепляли ферментом Dral и количественно оценивали путем измерения экстинкции при 260 нм. 100 нг ДНК подвергали обработке бисульфитом и очищали, используя набор EpiTect Bisulfite Kit (Qiagen, Хильден, Германия). Обработанную бисульфитом ДНК растворяли в 20 мкл свободной от РНКаз воды (Qiagen, Хильден, Германия).
Чтобы амплифицировать нить А (прямой) фрагмента главного немедленно-раннего промотора/энхансера CMV человека (SEQ ID NO 02), 1 мкл обработанной бисульфитом ДНК объединяли с 24 мкл ПЦР-смеси, и проводили ПЦР с помощью GeneAmp® PCR System 9700 (Applied Biosystems Inc., США).
24 мкл ПЦР-смеси включали:
- 1 мкл прямого праймера 227 с SEQ ID NO 06 (10 пмоль/мкл),
- 1 мкл обратного праймера 229 с SEQ ID NO 08 (10 пмоль/мкл),
- 22 мкл смеси Platinum® PCR SuperMix HighFidelty (Invitrogen Corp., США).
Прямой праймер 227 является компелементарным 5'-концу фрагмента главного немедленно-раннего промотора/энхансера CMV человека. Обратный праймер 229 связывается ниже в пределах 5'-UTR генов иммуноглобулинов.
Условия ПЦР были следующими:

ПЦР-продукт проверяли на размер и чистоту с помощью электрофореза в агарозном геле.
Количественная ПЦР в режиме реального времени, специфическая в отношении метилирования
Для проведения кПЦР использовали систему LightCycler® 480 II (Roche Diagnostics GmbH, Мангейм, Германия) в сочетании с LightCycler® 480 SYBR Green I Master (Roche Diagnostics GmbH, Мангейм, Германия). В качестве матриц использовали 50 пг плазмидной ДНК, 3 мкл обработанной бисульфитом геномной ДНК или 5 мкл разведенного 1:50000 ПЦР-продукта, полученного с использованием праймеров 227 и 229 (SEQ ID NO 06/08) и обработанной бисульфитом геномной ДНК. Для кПЦР матрицы смешивали с 4 или 12 пмоль прямого и обратного праймера, 10 мкл LightCycler 480 SYBR Green I Master и безнуклеазной водой до получения общего объема 20 мкл. Чтобы количественно оценить общее количество ДНК фрагмента главного немедленно-раннего промотора/энхансера CMV человека, использовали пару праймеров 239/237 (SEQ ID NO 11/09), а метилированную в позиции С-425 ДНК (SEQ ID NO 01) обнаруживали с помощью пары праймеров 239/267 (SEQ ID NO 11/18). ПЦР-смесь вносили в многолуночный планшет.
Многолуночный планшет запечатывали герметизирующей фольгой LightCycler® 480 (Roche Diagnostics GmbH, Мангейм, Германия) и центрифугировали со скоростью 1500 g в течение 2 минут. После этого планшет устанавливали в систему LightCycler® 480 и подвергали кПЦР. Каждый образец анализировали в двух, трех или четырех повторах.
Условия ПЦР были следующими:
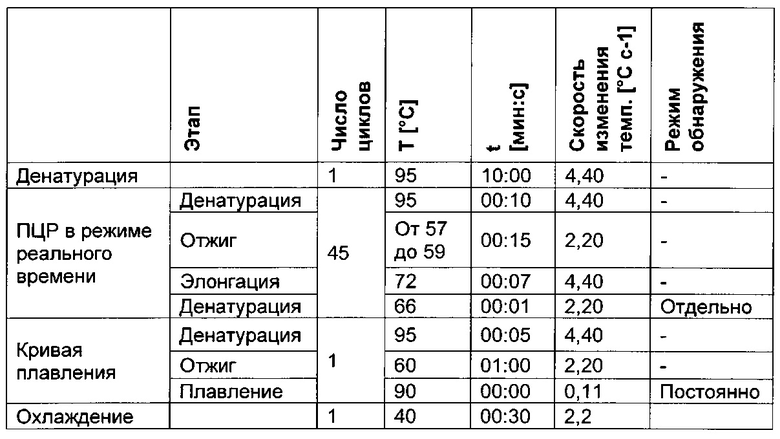
Сбор и анализ данных проводили с помощью программного обеспечения LightCycler® 480 версии 1.5. (Roche Diagnostics GmbH, Мангейм, Германия). Метилирование mC выражали в процентах по следующей формуле:
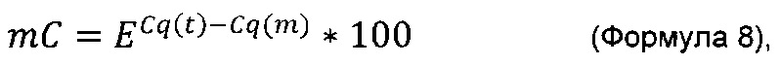
где Cq(t) является пороговым циклом, полученным с универсальной парой праймеров 239/237, Cq(m) представляет собой пороговый цикл, полученный с парой праймеров 239/267, специфических в отношении метилирования, а Е является эффективностью амплификации (MIQE Guidelines). Е составляла примерно 1,7 при концентрации праймеров 0,2 пмоль/мкл и 2,0 при концентрации праймеров 0,6 пмоль/мкл.
Пример 7
Корреляция метилирования главного немедленно-раннего промотора/энхансера CMV человека и долгосрочной продуктивности
Специфическая в отношении метилирования кПЦР с предварительной амплификацией нити А главного немедленно-раннего промотора/энхансера CMV человека
Как сообщалось в примере 4, геномную ДНК выделяли из клеточных линий СНО K18.1, G25-10, G25-17, G42-5 и 43-16 А10, расщепляли ферментом Dral и дезаминировали путем обработки бисульфитом. Нить А главного немедленно-раннего промотора/энхансера CMV человека амплифицировали с помощью праймеров 227 и 229.
ПЦР-продукт разводили 1:50000. 5 мкл каждого разведения использовали для кПЦР в режиме реального времени. Праймеры 239 и 237 использовали для количественной оценки общей ДНК промотора CMV, праймеры 239 и 267 использовали для количественной оценки ДНК промотора/энхансера CMV, метилированной в позиции 425. Образцы анализировали в трех повторах. Матрицы №11, №62 и №01 использовали в качестве контролей.
кПЦР проводили так же, как описано в примере 5, объединяя 5 мкл матрицы с 15 мкл ПЦР-смеси. Условия ПЦР были такими же, как описано в примере 5. Температура отжига праймеров составляла 58°С.
Специфическая в отношении метилирования кПЦР с обработанной бисульфитом геномной ДНК
Геномную ДНК выделяли из клеточных линий СНО K18.1, G25-10, G25-17, G42-5 и 43-16 А10, расщепляли ферментом Dral и дезаминировали путем обработки бисульфитом. 2 мкл дезаминированной ДНК, разведенные в 3 мкл воды, использовали в качестве матрицы в кПЦР в режиме реального времени с применением пары праймеров 239/237 для амплификации общей нити А промотора/энхансера CMV, либо пары праймеров 239/267 для амплификации нити А промотора/энхансера CMV, метилированной в позиции 425. Образцы анализировали в трех повторах. Матрицы №11 и №62 использовали в качестве контролей.
кПЦР проводили так же, как описано в примере 5. Температура отжига праймеров составляла 58°С.
Для а) и b) долю промоторной ДНК, метилированной в позиции 425, рассчитывали следующим образом:

где
mC [%]: доля ДНК, метилированной в позиции 425,
Cp(t): значение Ср, полученное с универсальной парой праймеров 239/237,
Ср(m): значение Ср, полученное с парой специфических в отношении метилирования праймеров 239/267.
Оба проведенных анализа давали сопоставимые результаты. Стандартное отклонение в трех повторах было выше без предварительной амплификации нити А промотора CMV (фиг. 8А и 8 В). Метилирование в позиции 425 нуклеиновой кислоты промотора CMV в клеточных линиях K18.1, G25-10, G25-17 и 43-16 А10 было выше фона неполного дезаминирования, который составлял примерно 1%. Метилирование в клеточной линии G42-5 находилось ниже или максимум на уровне фона. В клеточной линии K18.1 был выявлен самый высокий уровень метилирования (более 60%).
Анализ с настройками а) проводили с другими универсальными и специфическими в отношении метилирования парами праймеров. Для расчета доли ДНК, метилированной в позиции 425, эффективность амплификации для всех пар праймеров предполагалась как 2:

где
mC [%]: доля ДНК, метилированной в позиции 425,
Cp(t): значение Ср, полученное с универсальной парой праймеров,
Ср(m): значение Ср, полученное с парой специфических в отношении метилирования праймеров.
Таблица 8 показывает сводку комбинаций пар праймеров, которые анализировали либо на клонированных ДНК-матрицах, либо на геномной ДНК с предварительной амплификацией ДНК промотора CMV или без нее.

Пример 8
Метилирование главного немедленно-раннего промотора/энхансера CMV человека и прогноз нестабильности продукции рекомбинантной клеточной линии СНО
Клетки СНО-К1 трансфицировали плазмидой, кодирующей человеческое антитело IgG4, и отбирали стабильные клоны с помощью системы DHFR/MTX. Высокопродуцирующие родительские клоны субклонировали путем ограниченного разведения. МТХ сохраняли в ростовой среде в течение всего процесса создания клеточной линии. От 10 родительских клонов было отобрано 16 субклонов.
Выбранные клетки вновь культивировали с 250 нМ МТХ. Как только они демонстрировали стабильный рост, их проверяли на долгосрочную стабильность продукции в течение 60-80 поколений в присутствии и в отсутствие МТХ. Рассчитывали относительное изменение SPR в течение 60 поколений. Метилирование С435 определяли в начале исследования в клетках, выращенных с МТХ, и в конце исследования в клетках, культивируемых без МТХ.
Метилирование С425 в начале исследования откладывали на графике в виде зависимости от относительного изменения SPR в присутствии (фиг. 11А) и в отсутствие МТХ (фиг. 11В). Большинство клонов с менее чем 5% метилированием на С425 могли быть найдены во фракции стабильных клонов (менее чем 40% снижение SPR с МТХ или без него), в то время как большинство клонов с более чем 5% метилированием на С425 были сосредоточены во фракции нестабильных клонов. Независимо от условий метилирование коррелировало со стабильностью в присутствии или в отсутствие МТХ (см. также таблицу 9). Большинство стабильных клонов, которые теряли менее 20% продуктивности с МТХ и менее 30% продуктивности без МТХ, демонстрировали метилирование на С425 менее 5%.
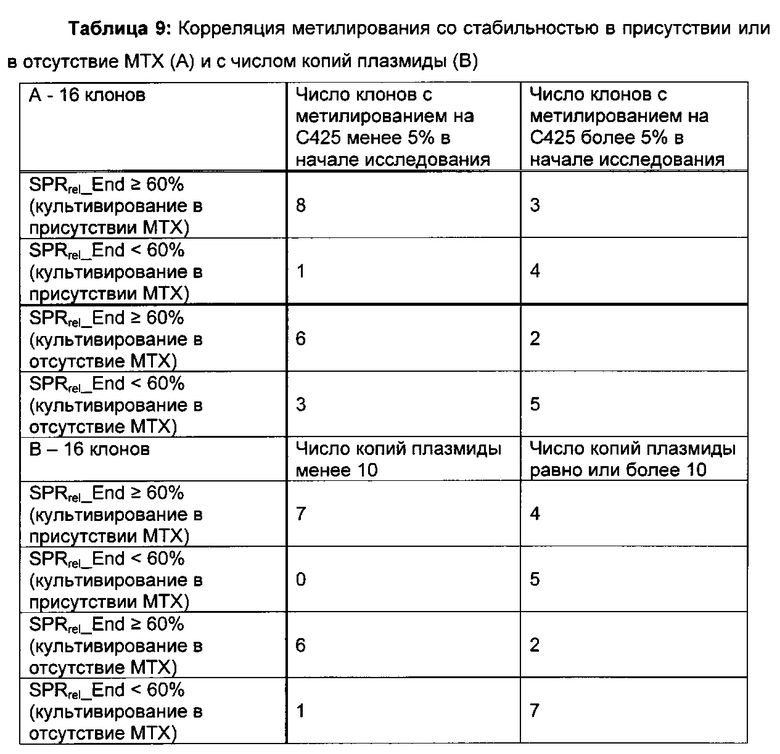
Это открытие показывает, что определение метилирования С425 может использоваться в качестве прогностического маркера для определения стабильности экспрессии полипептида в создаваемых клеточных клонах и тем самым позволяет отбирать стабильные клоны со стабильной продуктивностью при разработке клеточной линии. Было установлено, что метилирование С425 на уровне 5% или менее является подходящим критерием для отбора стабильных клеточных клонов.
Кроме того, было обнаружено, что два умеренно стабильных клеточных клона, а также два очень нестабильных клона, которые были неметилированными в начале исследования, поднимали уровень метилирования выше 5% во время анализа стабильности без МТХ (см. также фиг. 12). Это показывает, что доля клеточных клонов, которые ложно предсказаны как стабильные (ложноотрицательные клеточные клоны), может быть уменьшена путем культивирования их в течение некоторого времени в отсутствие МТХ перед анализом.
Для того чтобы подтвердить метилирование С425 вторым способом, мы проводили бисульфитное секвенирование высокометилированного клона 44-28 (фиг. 13). Степень метилирования С425 составляла 80%. Это согласовалось с результатами специфической в отношении метилирования ПЦР с учетом вариабельности обоих анализов. Как и в клонах K18.1 и 43-16 А10 (фиг. 4), С425 был наиболее часто метилирован среди всех CpG-сайтов в ДНК главного немедленно-раннего промотора/энхансера CMV человека. Другие события метилирования группировались на 5'-конце и на 3'-конце. Средняя степень метилирования на всех сайтах составляла 18%.
Пример 9
Раннее метилирование совпадает с большим числом копий трансгенов
Мы определяли интегрированные копии генов легкой и тяжелой цепи иммуноглобулина в начале и в конце анализа стабильности, используя многокомпонентную кПЦР, основанную на принципе TaqMan. Использовали два набора праймеров, состоящие из прямого праймера, обратного праймера и гидролизируемого зонда: один из них был специфическим для гена человеческой каппа-цепи, другой был специфическим для генов человеческой тяжелой гамма-цепи. Для определения абсолютного числа копий в качестве стандарта мы использовали линеаризованную экспрессионную плазмиду, которую использовали для трансфекции. Равная эффективность амплификации образцов и стандартов была достоверной.
Для кПЦР использовали систему LightCycler® 480 II (Roche Diagnostics GmbH, Мангейм, Германия), а образцы готовили с использованием LightCycler® 480 Probes Master (Roche Diagnostics GmbH, Мангейм, Германия).
5 мкл раствора матрицы, содержащего 50 нг геномной ДНК, объединяли с 15 мкл ПЦР-смеси в лунке 96-луночного планшета для микротитрования. В случае стандартов раствор матрицы содержал 2,5×107, 2,5×106, 2,5×105, 2,5×104 или 2,5×103 копий соответствующей линеаризованной плазмидной ДНК.
15 мкл ПЦР-смеси включали:
- 10 мкл LightCycler® 480 Probes Master
- 1 мкл прямого праймера №133 (10 пмоль/мкл)
- 1 мкл обратного праймера №132 (10 пмоль/мкл)
- 0,5 мкл зонда №166 (10 пмоль/мкл)
- 1 мкл прямого праймера №178 (10 пмоль/мкл)
- 1 мкл обратного праймера №180 (10 пмоль/мкл)
- 0,5 мкл зонда №185 (10 пмоль/мкл)
Планшет запечатывали герметизирующей фольгой LightCycler® 480 (Roche Diagnostics GmbH, Мангейм, Германия) и центрифугировали со скоростью 1500 g в течение 2 минут. После этого планшет устанавливали в систему LightCycler® 480 и подвергали кПЦР. Каждый образец анализировали в трех повторах, стандарты вводили в четырех повторах.
Условия ПЦР были следующими:
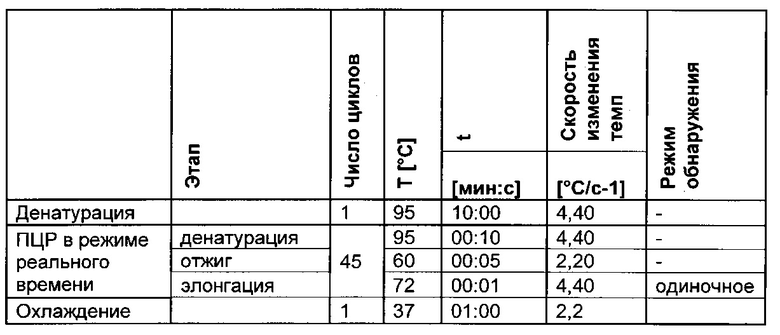
Сбор и анализ данных осуществляли с помощью программного обеспечения LightCycler® 480 версии 1.5. По существу, средние значения Ср стандартных разведений плазмиды откладывали на графике в виде зависимости от соответствующего числа копий гена, получая стандартную кривую, из которой экстраполировали число трансгенов в образце.
Рассчитывали число трансгенов на одну клетку, предполагая, что среднее содержание ДНК в клетке составляет:
Nc=Ns/50000*6, где
Nc: число копий трансгенов на клетку
Ns: число копий трансгенов в образце


На фиг. 14 показано число копий гена легкой цепи в метилированных и неметилированных клетках до и после анализа стабильности. Неудивительно, что были обнаружены идентичные количества копий гена тяжелой цепи, так как клетки были трансфицированы вектором с двойным геном, несущим оба гена (данные не показаны). Раннее метилирование, т.е. метилирование до анализа стабильности, было обнаружено исключительно в клетках, несущих более 10 копий трансгена, в то время как некоторые клоны с менее чем 10 копиями трансгена приобретали метилирование в ходе анализа стабильности. Как следствие, отобранные клоны с низким числом копий трансгена перед анализом стабильности в равной степени обогащали клоны со стабильной продуктивностью (таб. 11).

Мы также отметили, что нестабильность продукции, как правило, не связана с потерей копий трансгена. Клоны 1А5-05, 1А5-21, 1А5-24 и 2 В1-02 теряли копии трансгена, 2В-13 была стабильной, а 14-13 и 14-23 увеличивали число копий гена (см. также фиг. 11В).
Пример 10
Выявление присутствующих модификаций гистона вблизи ДНК главного немедленно-раннего промотора/энхансера CMV человека путем иммунопреципитации хроматина и ПЦР в режиме реального времени
Иммунопреципитации хроматина клеточных линий СНО
Фрагмент главного немедленно-раннего промотора/энхансера CMV человека (SEQ ID NO 01), используемый для экспрессии легкой и тяжелой цепи антитела, находится рядом с ДНК, уплотненной гистонами (хроматином), в стабильно продуцирующих клеточных линиях. Накопление модифицированных гистонов вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека будет измеряться с помощью анализа иммунопреципитации хроматина (ChIP).
Тридцать две клеточные линии СНО (образцы с Н-1 по Н-20 и с Т-1 по Т-12, см. таблицу 3) фиксировали в 3,7% формальдегиде в течение 10 минут при комнатной температуре. Фиксацию останавливали в 10% глицине в течение 5 минут. Клетки подвергали лизису в течение 10 минут на льду в буфере для лизиса клеток, центрифугировали со скоростью 5000 об/мин, 4 мин, 4°С. Осадки ресуспендировали в буфере для лизиса ядер и хроматин обрабатывали ультразвуком с помощью Branson Sonifier В15 до получения среднего размера фрагментов 200-500 п.о. Концентрацию фрагментов хроматина определяли с помощью набора Pierce® ВСА Protein Assay Kit (Thermo Scientific, Рокфорд, США). 2,5-100 мкг фрагментов хроматина добавляли к 1,5 объемам буфера для разведения IP, содержащего 3-12 мкл антител, и инкубировали в течение ночи при 4°С со скоростью вращения 25 об/мин.
Агарозные гранулы с белком A (Roche Diagnostics GmbH, Мангейм, Германия) блокировали в буфере для разведения IP со 100 мкг/мл ДНК спермы лосося (Invitrogen, Карлсбад, США) и 5 мг/мл BSA (бычий сывороточный альбумин, Roche Diagnostics GmbH, Мангейм, Германия) в течение ночи при температуре 4°С. Преципитаты "антитело - хроматин" очищали путем инкубации в блокированных агарозных гранулах в течение 1 часа, затем инкубировали в течение 5 минут при 4°С в 2× низкосолевом буфере, 1× высокосолевом буфере, 1× LiCl и 1× промывочном ТЕ-буфере. Антитела элюировали из агарозных гранул с помощью 200 мкл буфера для элюирования IP в течение 30 минут при 65°С. Добавляли 0,5 мкл безДНКазной РНКазы (Roche Diagnostics GmbH, Мангейм, Германия) для расщепления РНК при 37°С в течение 30 минут. Для расщепления белков добавляли NaCl (конечная концентрация 0,2 М) и протеиназу К (конечная концентрация 100-200 мкг/мл) (Roche Diagnostics GmbH, Мангейм, Германия) и раствор инкубировали в течение 1,5 ч при температуре 65°С. ДНК восстанавливали с помощью набора для очистки ДНК Roche PCR Purification Kit в 150 мкл H2O класса ПЦР (Roche Diagnostics GmbH, Мангейм, Германия).


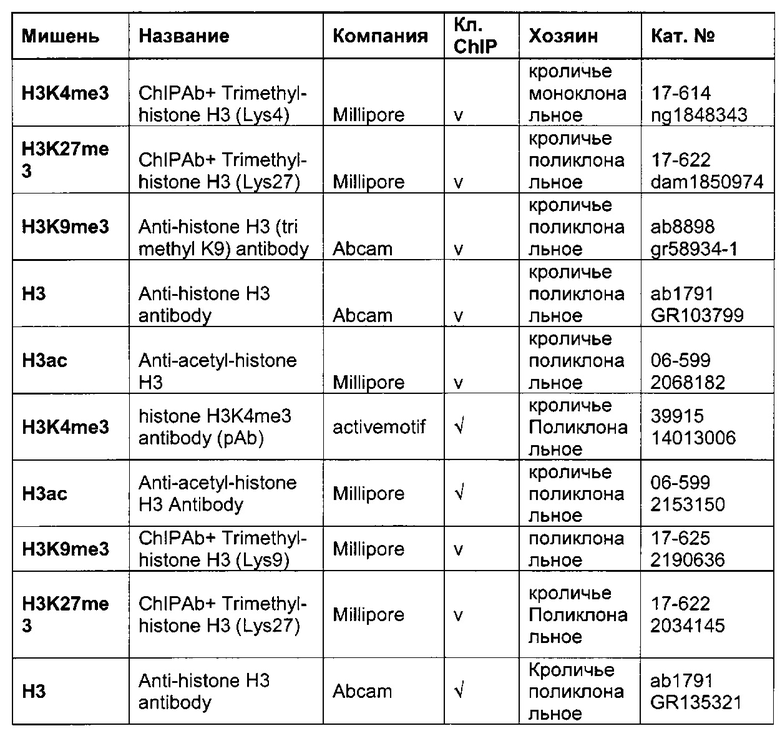
Количественная ПЦР в режиме реального времени ChIP-ДНК
Количественную ПЦР в режиме реального времени использовали для выявления накопления специфических модификаций гистона во фрагменте главного немедленно-раннего промотора/энхансера CMV человека (SEQ ID NO 01) в клеточных линиях СНО путем измерения относительного количества фрагмента главного немедленно-раннего промотора/энхансера CMV человека в расчете на иммунопреципитацию хроматина.
Геномную ДНК из необработанного входящего образца (input sample, IS) и фрагментов хроматина, очищенных с помощью антитела, амплифицировали путем количественной ПЦР в режиме реального времени с помощью LightCycler 480 SYBR Green I Master (Roche Diagnostics GmbH, Мангейм, Германия) с использованием системы LightCycler 480 II (Roche Diagnostics GmbH, Мангейм, Германия). Для того чтобы определить количество фрагмента главного немедленно-раннего промотора/энхансера CMV человека (SEQ ID NO 01), использовали специфический праймер на возможные референсные гены и на фрагмент главного немедленно-раннего промотора/энхансера CMV человека.
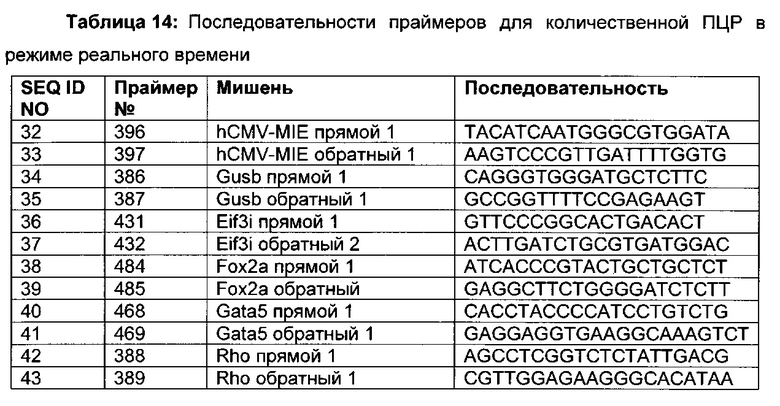
2,5 мкл элюированных фрагментов ДНК, 1 мкл (10 пмоль/мкл) прямого праймера 396 (SEQ ID NO 32) и 1 мкл (10 пмоль/мкл) обратного праймера 397 (SEQ ID NO 33) к фрагменту главного немедленно-раннего промотора/энхансера CMV человека (SEQ ID NO 01), 5 мкл LightCycler 480 SYBR Green I Master и безнуклеазную воду смешивали в общем объеме 10 мкл. В этом же планшете количественно оценивали референсные гены. Смесь вносили в 384-луночные планшеты (Roche Diagnostics GmbH, Мангейм, Германия).
Планшет запечатывали герметизирующей фольгой LightCycler® 480 (Roche Diagnostics GmbH, Мангейм, Германия) и центрифугировали со скоростью 1200 g в течение 3 минут. После этого планшет устанавливали в систему LightCycler® 480 и подвергали кПЦР. Каждый образец анализировали в трех повторах.
Условия ПЦР были следующими:
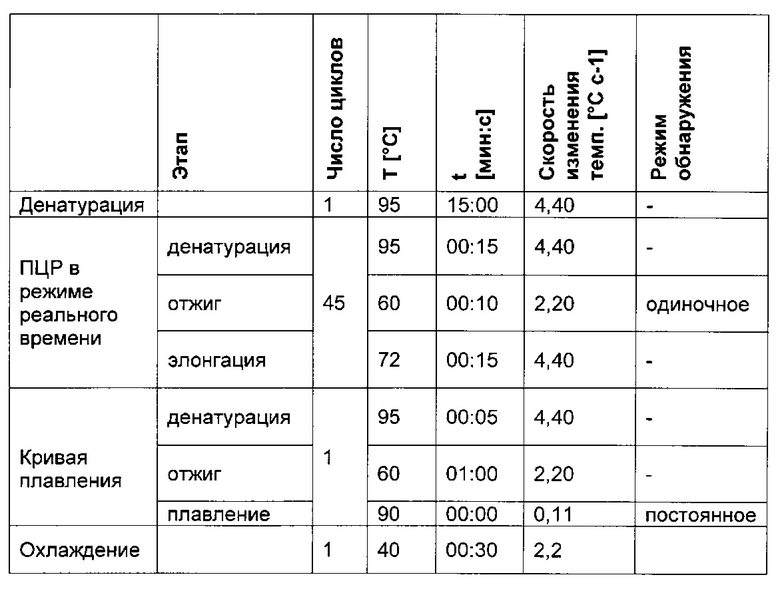
Сбор и анализ данных проводили с помощью программного обеспечения LightCycler® 480 версии 1.5.
Пример 11
Относительная количественная оценка фрагмента главного немедленно-раннего промотора/энхансера CMV человека на модификации гистона в ПЦР в режиме реального времени
Стабильность референсных генов
В этом примере стабильность потенциальных референсных генов для специфических модификаций гистона рассчитывали с помощью программы "Normfinder". Модель и статистическая база, лежащие в основе "NormFinder", описаны в Andersen, C.L., et al. Cancer Res.64 (2004) 5245-5250.
Инструмент "NormFinder add on" для Excel дает значение стабильности для каждого гена, которое является прямой мерой предполагаемой вариации экспрессии. Ген с наименьшим значением стабильности является наиболее стабильным геном. Предполагали, что входные данные находятся на линейной шкале, поэтому использовали подход Percent Input Method, чтобы линеаризовать значения шкалы экспрессии с помощью следующей формулы:.
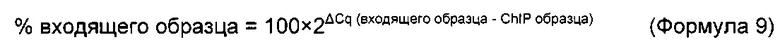
Пять генов (см. таблицу 15) анализировали с помощью "Normfinder". Двадцать образцов (с Н-1 по H20) в трех биологических повторах исследовали на стабильность ацетилирования гистона 3, трехкратное метилирование лизина 4 гистона 3 и уровни гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека и референсных генов.
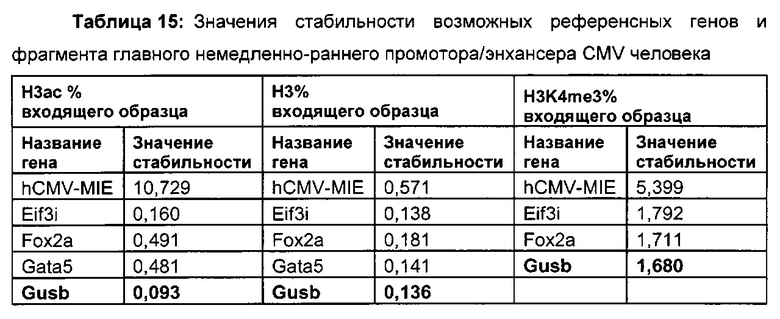
Референсный ген Gusb является более стабильным в выбранных ChIP-условиях, чем другие контроли. Gusb использовали в качестве референсного гена для нормализации ацетилирования гистона 3, трехкратного метилирования лизина 4 гистона 3 и уровней гистона 3.
Оценка эффективности амплификации пар праймеров с помощью программного обеспечения LinRegPCR
В данном примере эффективность амплификации пары праймеров 396/397 для фрагмента главного немедленно-раннего промотора/энхансера CMV человека и пары праймеров 386/387 для референсного гена Gusb оценивали с помощью программного обеспечения LinRegPCR (http://www.hartfaalcentrum.nl) для образцов с Н1 по H20. Программа определяет базальный уровень флуоресценции и выполняет вычитание базального уровня. Затем устанавливает окно линейности и рассчитывает эффективность ПЦР для каждого образца (см. Ramakers, et al., NeuroSci Lett. 2003; Ruijter et al., Nucleic Acids Research 2009).

Индивидуальную эффективность ПЦР (Eff) выводят из наклона линейной линии регрессии и определяют кратность увеличения в каждом цикле. Она может находиться в диапазоне от 1 до 2. Эффективность 2 представляет собой идеальное удвоение ампликона в каждом цикле.

Пары праймеров 396/397 и 386/387 давали стабильные и сопоставимые эффективности амплификации с 20 образцами и 3 биологическими повторами независимо от того, измерялись ли входящие ChIP-образцы или ДНК, преципитированные антителами против гистона 3, ацетилированного гистона 3 или трехкратного метилирования лизина 4 гистона 3. Пары праймеров 396/397 и 386/387 использовали для относительной количественной оценки ацетилирования гистона 3, трехкратного метилирования лизина 4 гистона 3 и уровней гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека относительно уровней гистона 3 и модификаций гистона 3 вблизи референсного гена Gusb.
Способ дельта-дельта-Cq для относительной количественной оценки модификации гистона относительно уровня гистона
Относительную количественную оценку ацетилирования гистона 3, трехкратного метилирования лизина 4 гистона 3 и уровней гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека оценивали с помощью способа Livak, также известного как способ дельта-дельта-Cq (ΔΔCq).
Данный способ может быть использован для относительной количественной оценки мишени относительно референса. В этом примере относительную количественную оценку ацетилирования гистона 3, трехкратного метилирования лизина 4 гистона 3 и уровней гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека нормализовали по модификации гистона 3 и уровням гистона 3 вблизи референсного гена Gusb в два этапа.
Первое значение дельта-Cq рассчитывали следующим образом:

Значение дельта-Cq (ΔCq) представляет собой разность между пороговым циклом (Cq) необработанного (входящего образца) и обработанного (ChIP-образца) состояния одного и того же образца, которые амплифицированы с помощью одной и той же пары праймеров. Значение дельта-Cq используется для отображения значения относительно необработанного состояния в пределах одного и того же образца. Для сравнения различных образцов использовали способ дельта-дельта-Cq.
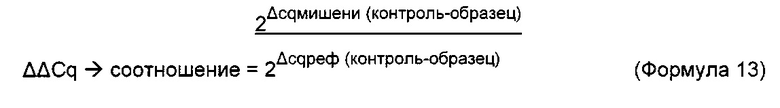
В этом примере дельта-дельта-Cq представляет собой разность между значением дельта-Cq мишени hCMV-MIE и значением дельта-Cq референсного Gusb для всех образцов с Н-1 по Н-20. Относительное количество ДНК, определенное с помощью значений дельта-дельта-Cq, можно сравнивать между образцами.
Наконец, уровни ацетилирования гистона 3 и трехкратного метилирования лизина 4 гистона 3 нормализовали по уровню гистона 3 для 20 клеточных линий СНО с Н-1 по Н-20 в трех биологических повторах. Результаты со стандартным отклонением приведены на фиг. 15 и 16.
Пример 12
Ацетилирование гистона 3 и трехкратное метилирование лизина 4 гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека, а также метилирование С-425 фрагмента главного немедленно-раннего промотора/энхансера CMV человека коррелирует с долгосрочной стабильностью
Уровни относительного ацетилирования и трехкратного метилирования лизина 4 относительно уровня гистона 3 (H3ac/H3 и H3K4me3/H3) вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека, а также процент метилирования С-425 фрагмента главного немедленно-раннего промотора/энхансера CMV человека (mC-425[%]) исследовали на подходящей модели "стандартных наименьших квадратов" с акцентом на "модель влияния рычагов", чтобы обнаружить влияния на долгосрочную стабильность продуцирующих клеточных линий с Н-1 по Н-20. Для расчета использовали программное обеспечение JMP версии 10 (SAS, Беблинген, Германия).
Значения модификаций гистона и процентное метилирование С-425 для образцов с Н-1 по Н-20 подавали в виде влияния на модель по отдельности. Процентное изменение удельной скорости продукции (дельта-SPR) после 60 поколений подавали в качестве ответа. Полученные в результате графики рычагов показаны на фиг. 17-19. Число параметров, связанных с влиянием (Nparm), и степеней свободы составляет 1.
RSquare (RSq) оценивает долю вариации в ответе, которую можно отнести к модели, а не к случайной ошибке. R2 ближе к 1 указывает на лучшее совпадение. R2 ближе к 0 указывает на то, что совпадение предсказывает ответ не лучше, чем средний общий ответ. Среднеквадратичная ошибка (root mean square error, RMSE) оценивает стандартное отклонение случайной ошибки. Она представляет собой квадратный корень из среднего квадрата ошибки в отчете анализа вариативности. Prob>F перечисляет р-значения для анализа влияния, которые информируют о значимости влияния.

Таблица 17 показывает значимость mC-425[%] в условиях (-) МТХ. Влияния H3ac/H3 и Н3К4-те3/H3 являются значимыми в условиях (+) MSX.
Фактические значения дельта-SPR откладывали в виде зависимости от прогнозируемых значений на графиках рычагов, касающихся влияния и условий селекции в течение более 60 поколений (фиг. 17-19). Этот график зависимости фактических значений от прогнозируемых показывает, насколько хорошо модель соответствует данным. Самые низкие р-значения были обнаружены для влияния H3ac/H3 с селективным агентом (фиг. 18), а затем следовало влияние mC 425[%]без селективного агента (фиг. 19).
Анализ выбросов, сравнивающий условия с селективным агентом MSX и без него, для значений дельта-SPR образцов с Н-1 по Н-20
Графики рычагов условия без селективного агента ((-) MSX) имеют заметно отклоняющиеся значения для дельта-SPR в образце Н-18. Поэтому был проведен анализ выбросов, показывающий расстояния по методу "складного ножа" (фиг. 20).
Он подтвердил, что увеличение дельта-SPR в образце Н-18 в условиях (-) MSX является аномальным по сравнению со всеми другими образцами. Поэтому анализ рычагов повторяли без образца Н-18. Вычисленные р-значения для анализа влияния на 19 образцов с Н-1 по Н-17 и с Н-19 по Н-20 представлены в таблице 18.

Таблица 18 показывает высокую значимость H3ac/H3 при обоих условиях селекции. Влияние H3K4-me3/H3 является значимым в условиях (+) MSX.
Фактические значения дельта-SPR откладывали в виде зависимости от прогнозируемых значений на графиках рычагов, касающихся влияния и условий селекции в течение более 60 поколений (фиг. 20-22). Этот график зависимости фактических значений от прогнозируемых показывает, насколько хорошо модель соответствует данным. Самые низкие р-значения были обнаружены для влияния H3ac/H3 (фиг. 21).
Пример 13
Предсказание нестабильности продукции в рекомбинантных клеточных линиях СНО с помощью модификаций гистона вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека, а также метилирования С-425 фрагмента главного немедленно-раннего промотора/энхансера CMV человека в качестве маркера
Увеличение среднего значения дельта-SPR образцов проекта Н путем отклонения плохо продуцирующих клеточных линий с настройками фильтрации
Как сообщалось в примере 12, значения образца-выброса Н-18 были исключены из дальнейших исследований по установлению маркера предсказания.
Уровни ацетилирования гистона 3 относительно уровня гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека и процент метилирования С-425 фрагмента главного немедленно-раннего промотора/энхансера CMV человека исследовали в качестве маркеров предсказания стабильности продукции с селективным агентом и без него.
Как описано в Osterlehner et al., фильтр для значений mC-425[%] был установлен на 5%. Значения выше 5% отклоняли, чтобы увеличить среднее значение дельта-SPR оставшихся образцов.
Для значений H3ac/H3 образцов с Н-1 по Н-17, Н-19 и Н-20 дерево решений рассчитывали с помощью программного обеспечения JMP. Лучший узел расщепления и поэтому лучший фильтр рассчитывали с помощью статистического анализа LogWorth. Logworth рассчитывали как:

где скорректированное значение р рассчитывали сложным образом, учитывая большое число различных способов расщеплений, которые могут произойти. Этот расчет является очень справедливым по сравнению с нерегулируемым р-значением, которое благоприятствует X с большим числом уровней, и р-значением Бонферрони, которое благоприятствует X с малым числом уровней (информационный документ: "Monte Carlo Calibration of Distributions of Partition Statistics").
Лучшее расщепление в условиях (+) MSX для образцов с Н-1 по Н-17, Н-19 и Н-20 составляло 0,58 для ацетилирования гистона 3 относительно уровня гистона 3 (H3ac/H3). Чтобы уменьшить число ложноотрицательных образцов, фильтр устанавливали на нижнее значение А более 0,5 для H3ac/H3.
Оба фильтра использовали по отдельности и в комбинации. Средние значения дельта-SPR в условиях с селективным агентом MSX или без него вычисляли для каждого положительного гейта и сравнивали с нефильтрованным средним значением (таблица 19).
Таблица 19: Рассчитанные средние значения дельта-SPR с селективным агентом и без него и в условиях фильтрации А более 0,5 для H3ac/H3 и В менее 5% для mC-425[%] PSP. Значения рассчитывали для образцов с Н-1 по Н-17, Н-19 и Н-20 относительно настроек фильтра.
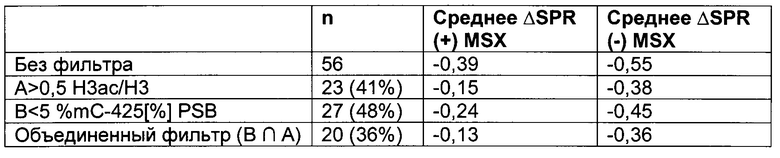
Каждый фильтр может увеличить среднее значение дельта-SPR по сравнению с условиями без фильтра. Пересечение фильтров А и В увеличивает среднее значение дельта-SPR фильтра А более 0,5 для H3ac/H3. Для визуализации измененных значений дельта-SPR гистограммы каждого условия и фильтра показаны на фиг. 23.
Принятые настройки фильтра для проекта Н были адоптированы к проекту Т. чтобы подтвердить положительное влияние
Двенадцать клеточных линий (с Т-1 по Т-12) также анализировали с использованием настроек фильтра, определенных выше (см. таблицу 20). Для визуализации измененных значений дельта-SPR гистограммы каждого условия и фильтра показаны на фиг. 25.
Таблица 20: Рассчитанные средние значения дельта-SPR с селективным агентом и без него. Значения рассчитывали для 12 образцов с Т-1 по Т-12 относительно настроек фильтра
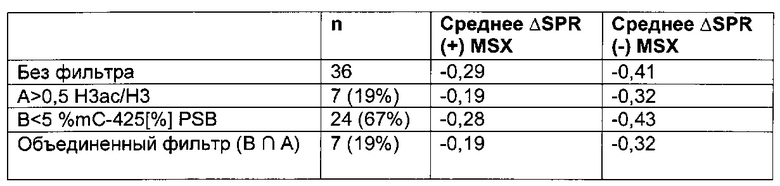
Наблюдаемое увеличение средних значений дельта-SPR сравнивали для фильтрованных и нефильтрованных образцов в условиях (-) МТХ.
Использование настроек фильтра А более 0,5 для H3ac/H3 приводит к увеличению среднего значения дельта-SPR по сравнению с нефильтрованными данными.
Это показывает, что ацетилирование гистона 3 относительно уровня гистона 3 вблизи фрагмента главного немедленно-раннего промотора/энхансера CMV человека является ценным маркером предсказания.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНАЦИИ ЭЛЕМЕНТОВ ЭКСПРЕССИОННОГО ВЕКТОРА, НОВЫЕ СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК-ПРОДУЦЕНТОВ И ИХ ПРИМЕНЕНИЕ ДЛЯ РЕКОМБИНАНТНОГО ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ | 2012 |
|
RU2639519C2 |
| КОМБИНАЦИИ ЭЛЕМЕНТОВ ЭКСПРЕССИОННОГО ВЕКТОРА, НОВЫЕ СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК-ПРОДУЦЕНТОВ И ИХ ПРИМЕНЕНИЕ ДЛЯ РЕКОМБИНАНТНОГО ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ | 2012 |
|
RU2756910C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ, КОДИРУЮЩАЯ БЕЛОК GAG ВИЧ-1, СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ, ВЕКТОР, СОДЕРЖАЩИЙ ЕЕ, БЕЛОК, КОДИРУЕМЫЙ ЕЮ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ И/ИЛИ ЛЕЧЕНИЯ ВИЧ-ИНФЕКЦИИ И СПИДА | 2002 |
|
RU2312896C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ | 2015 |
|
RU2699715C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA1 | 2016 |
|
RU2656142C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA2m1F16-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA2m1 | 2016 |
|
RU2671477C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-Intht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IgA1 | 2016 |
|
RU2664184C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА IGA2m1-ИЗОТИПА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2022 |
|
RU2801178C1 |
| Способ получения димерной формы иммуноглобулина IgA1-изотипа в клетках млекопитающих | 2023 |
|
RU2822890C1 |
| Способ получения димерной формы мутантного иммуноглобулина IgA2m1-изотипа в клетках млекопитающих | 2023 |
|
RU2822889C1 |

















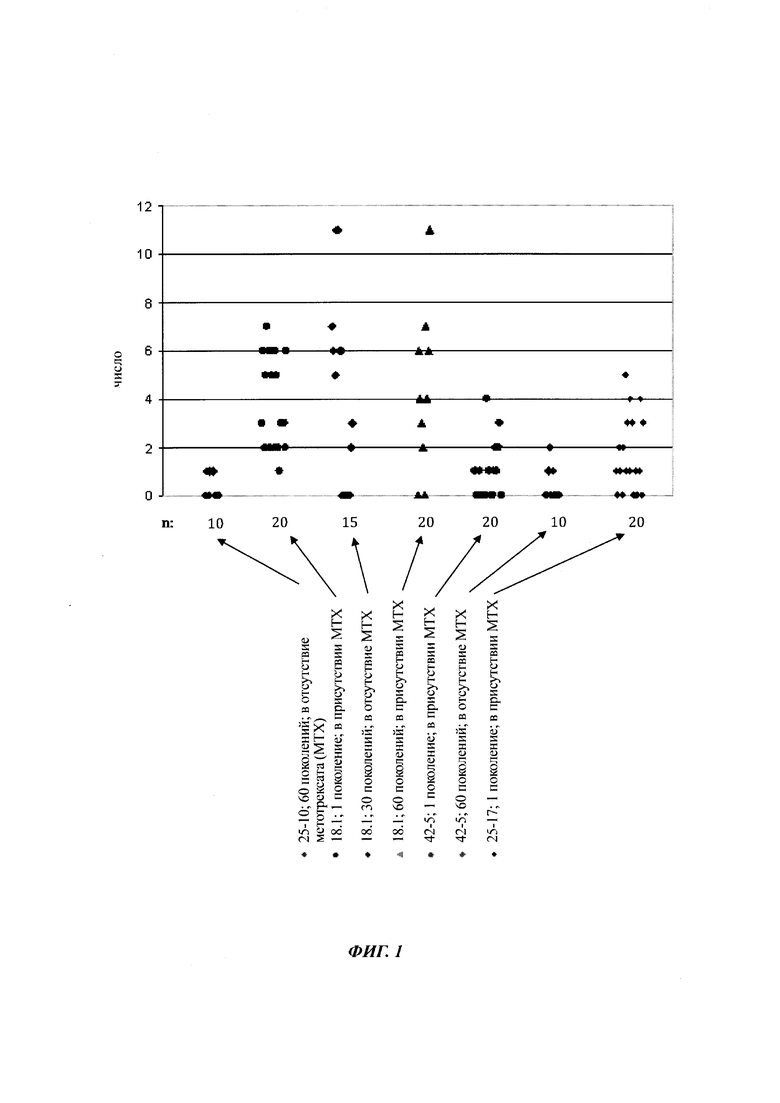
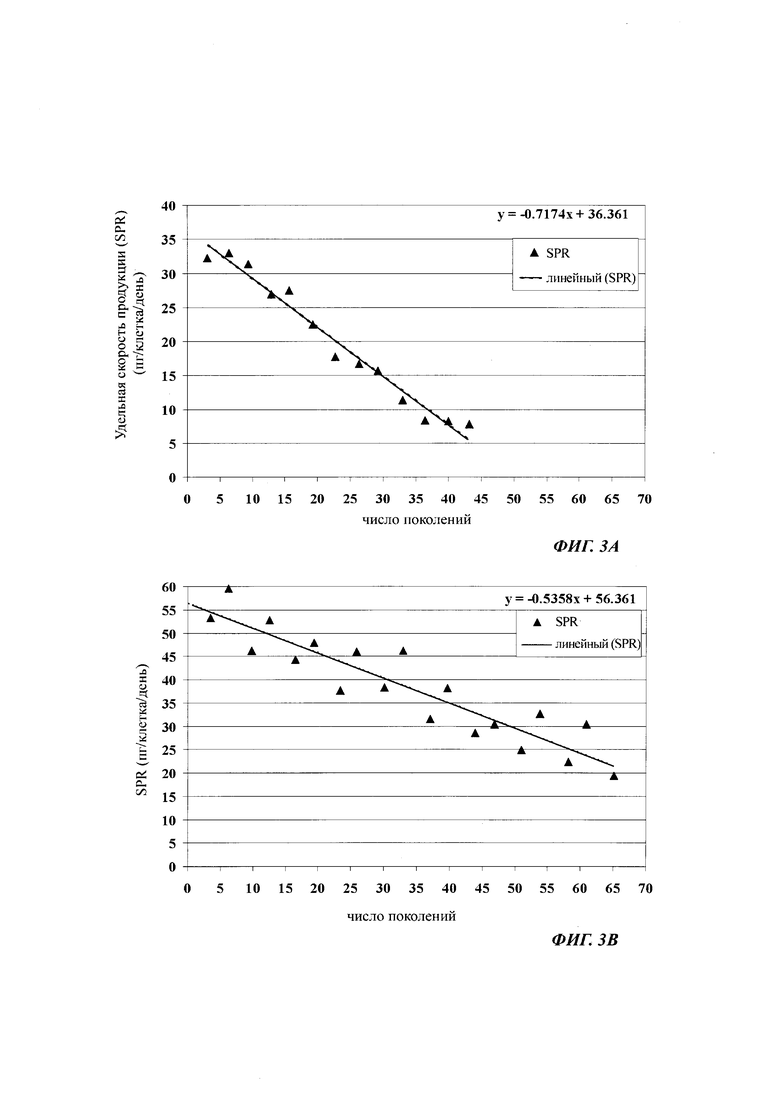
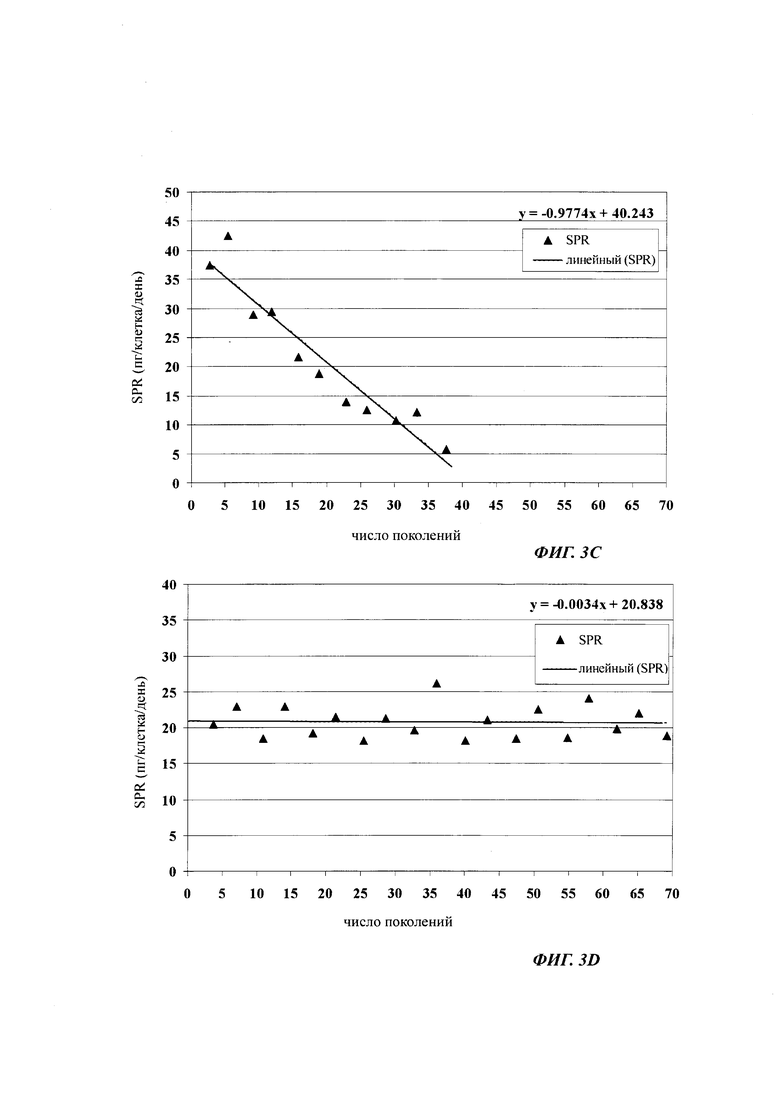
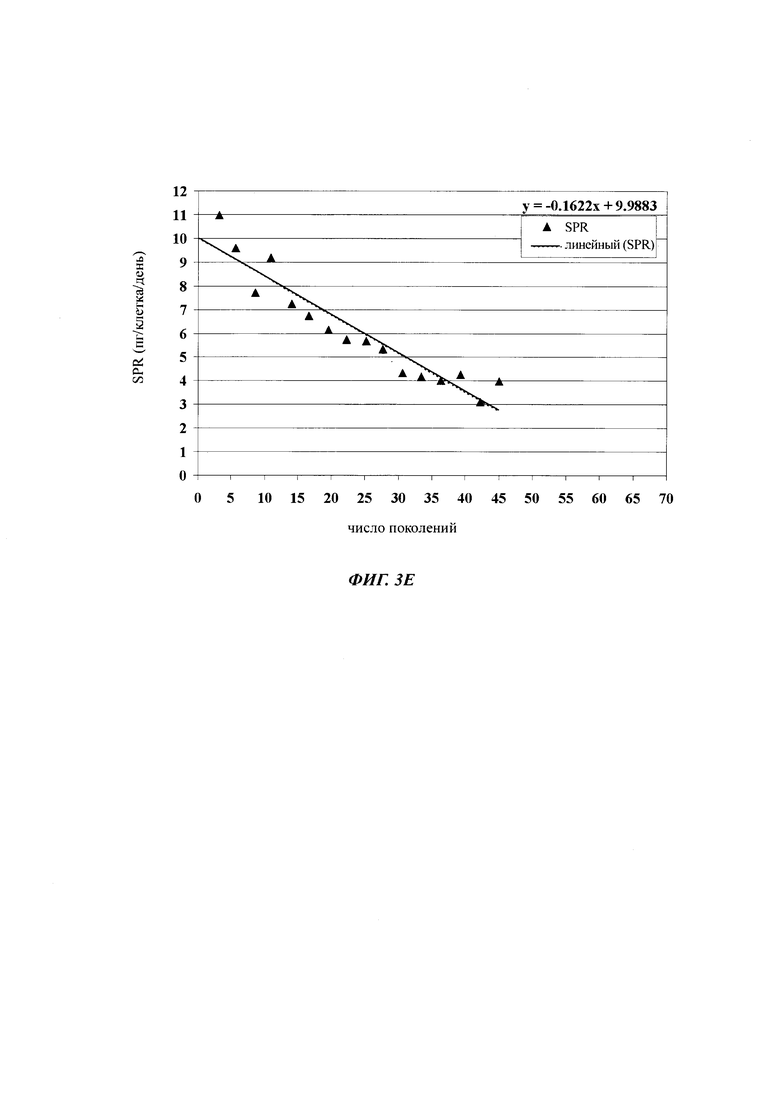
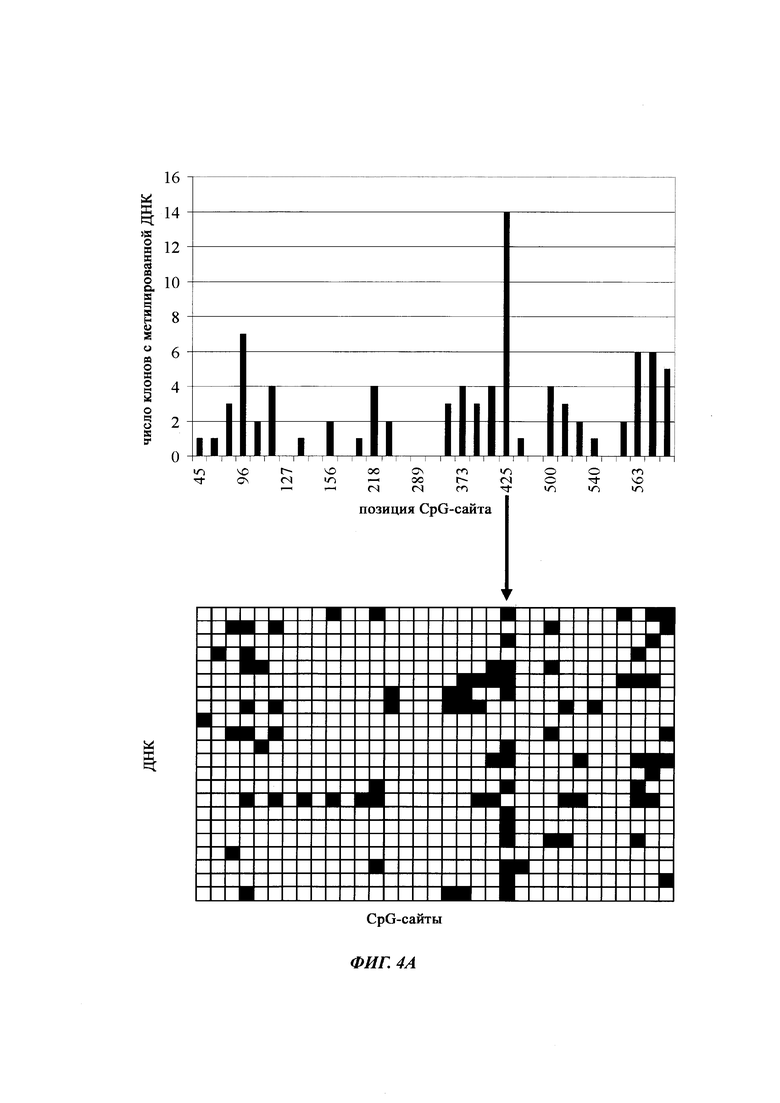
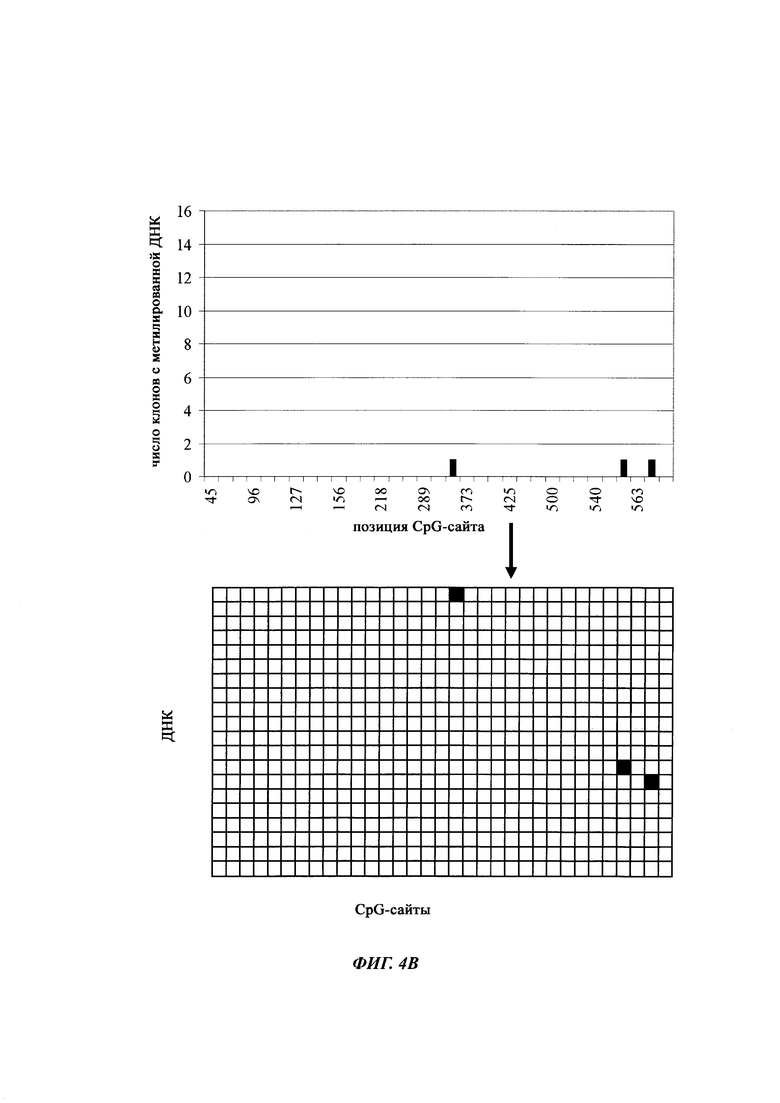
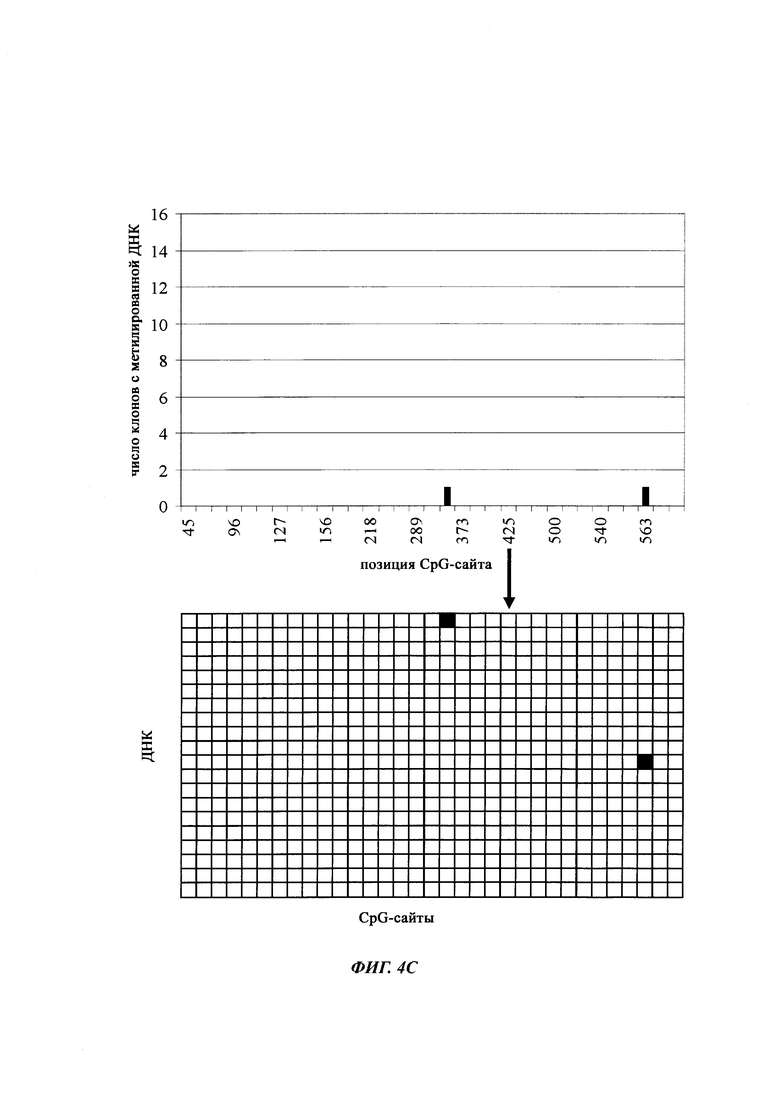
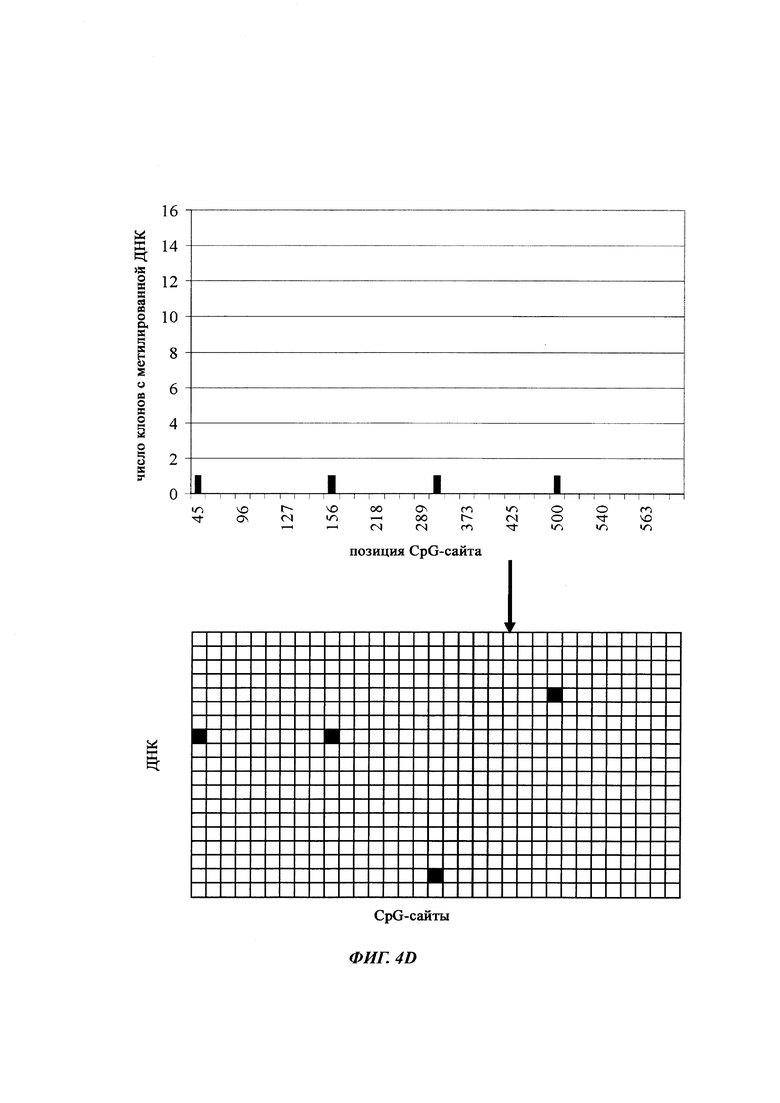
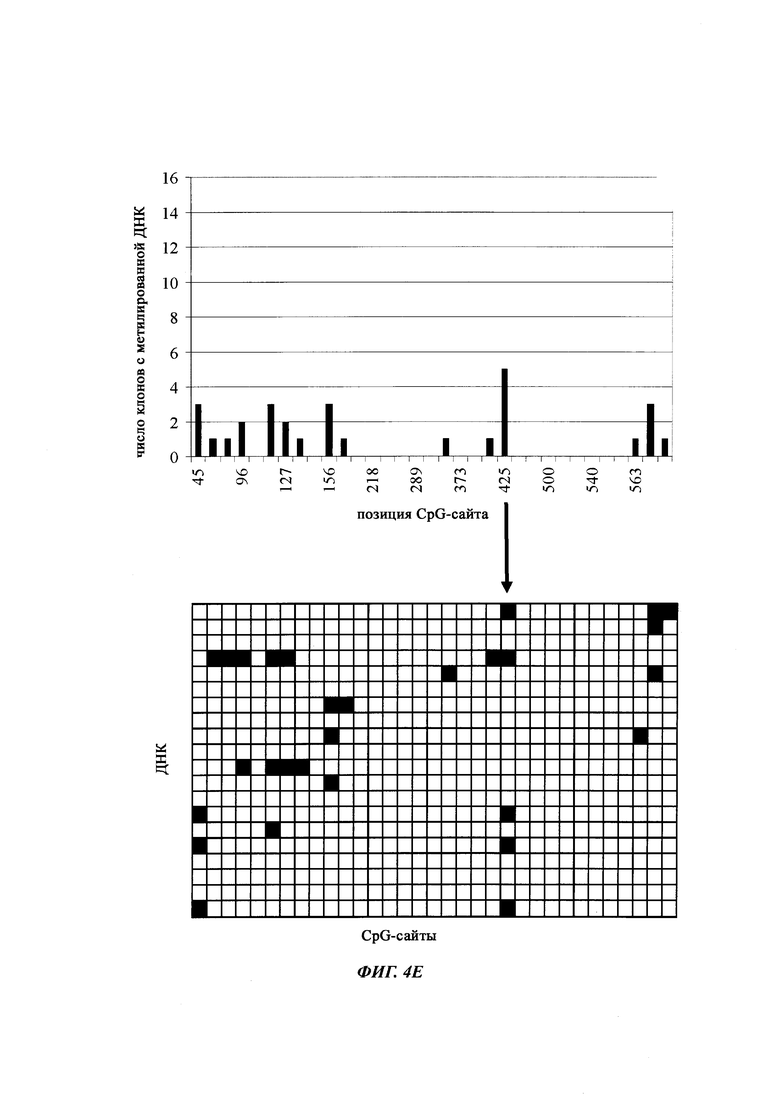
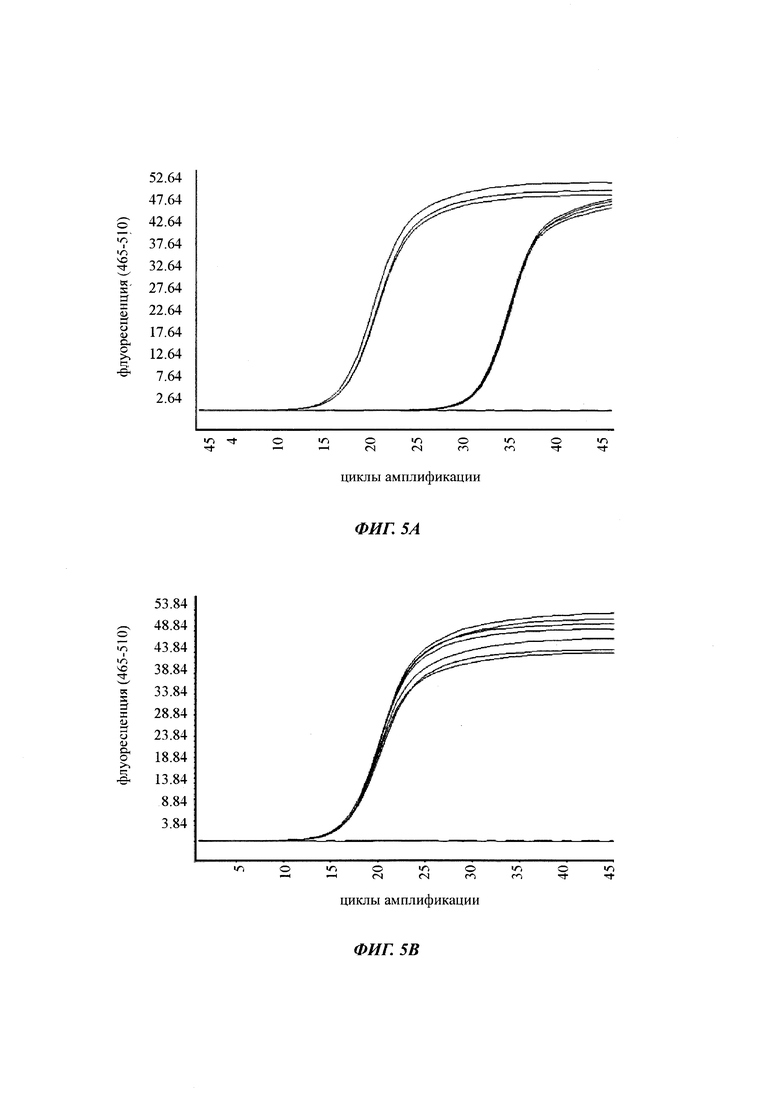
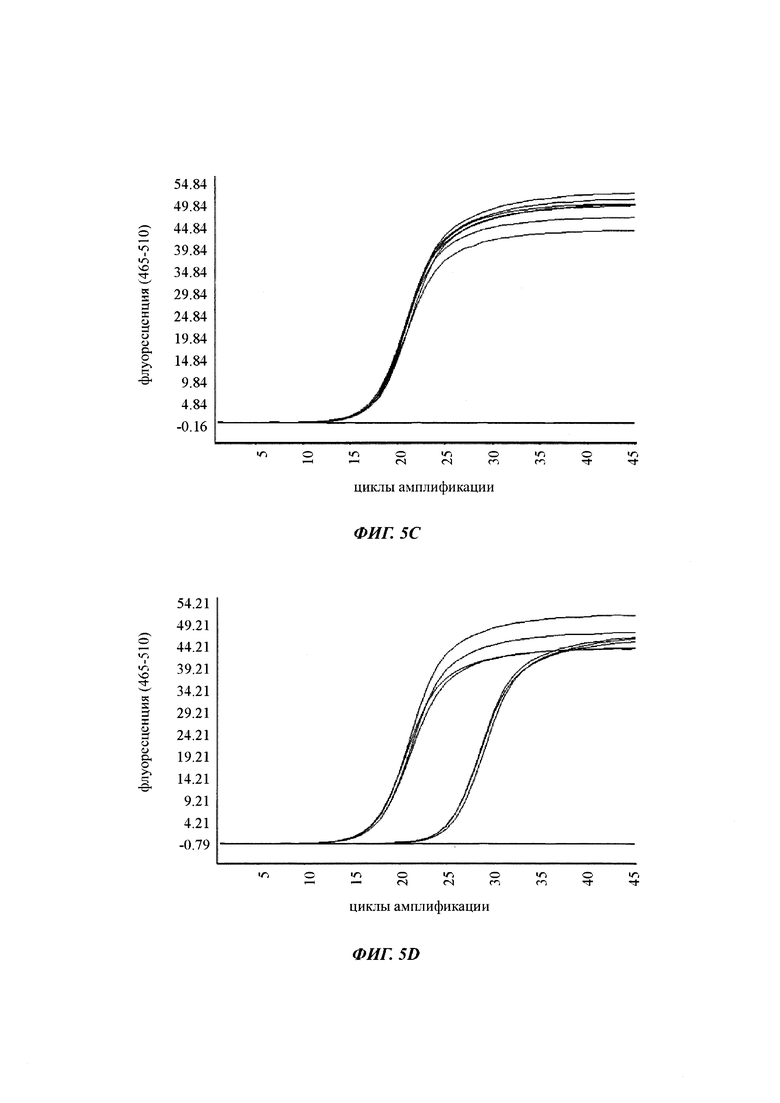
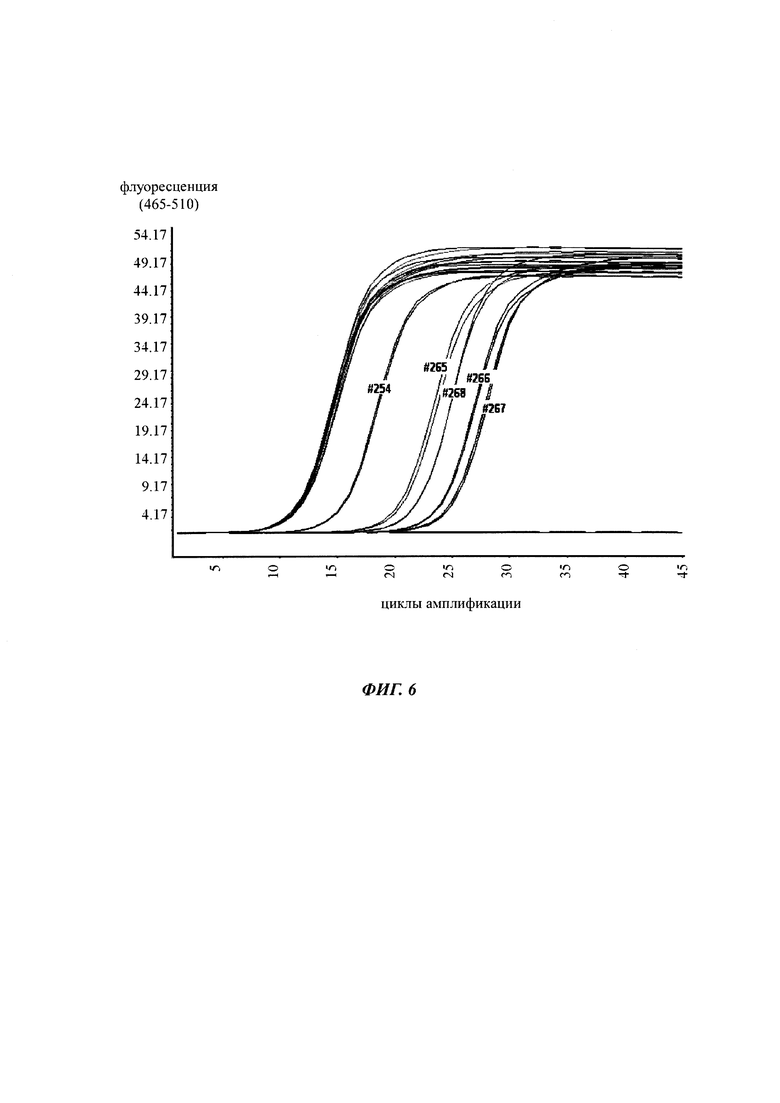
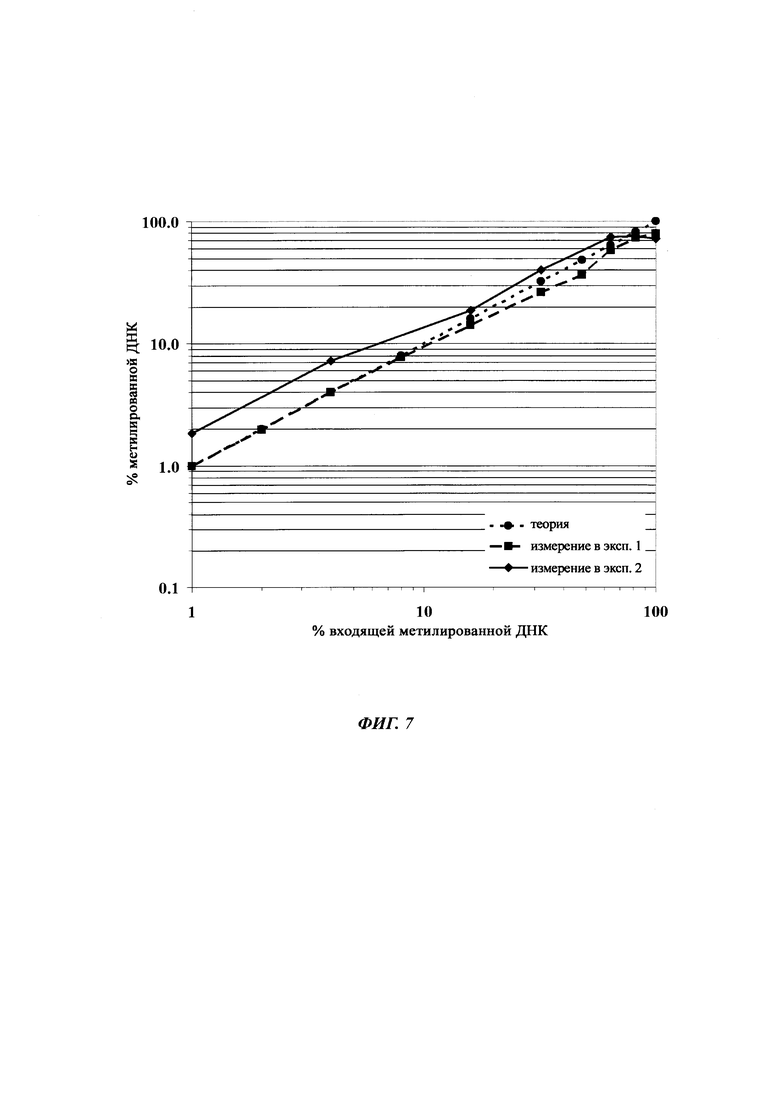
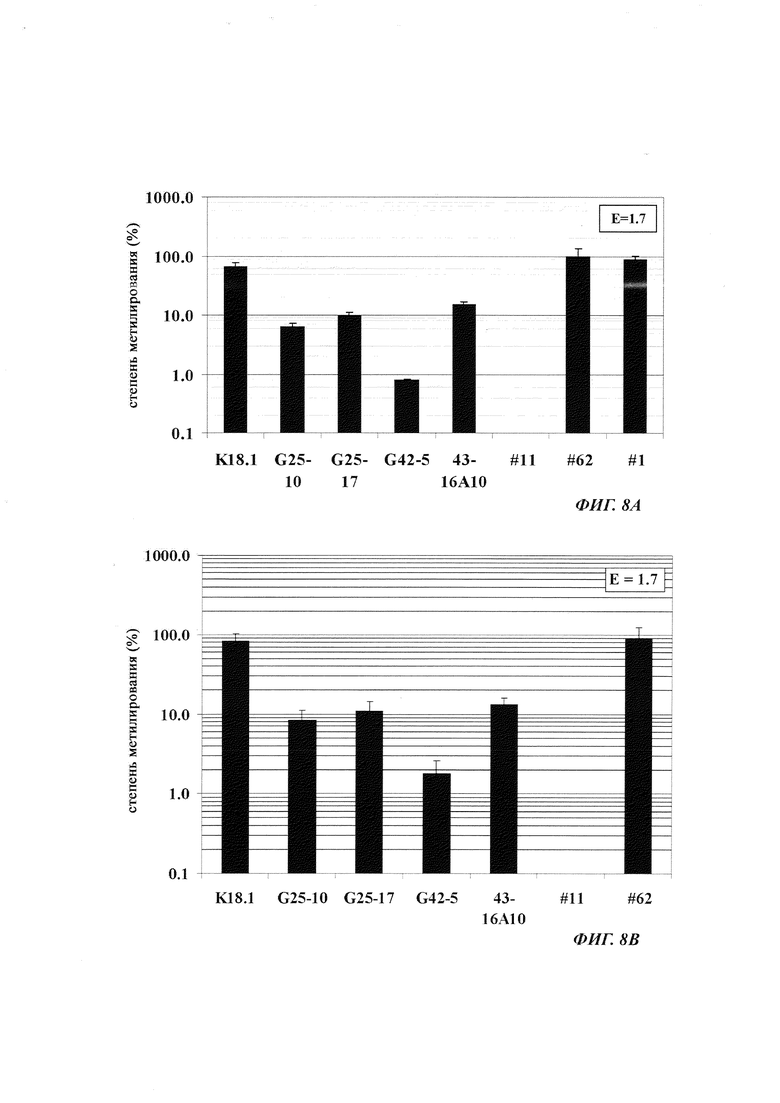
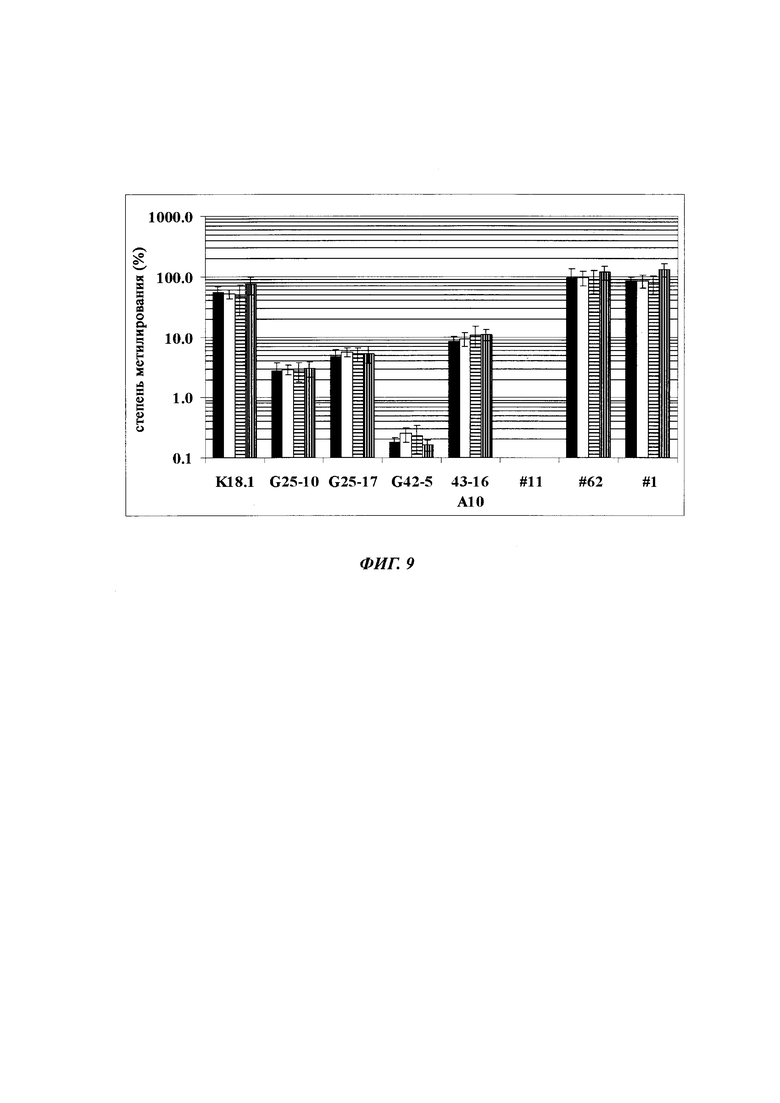
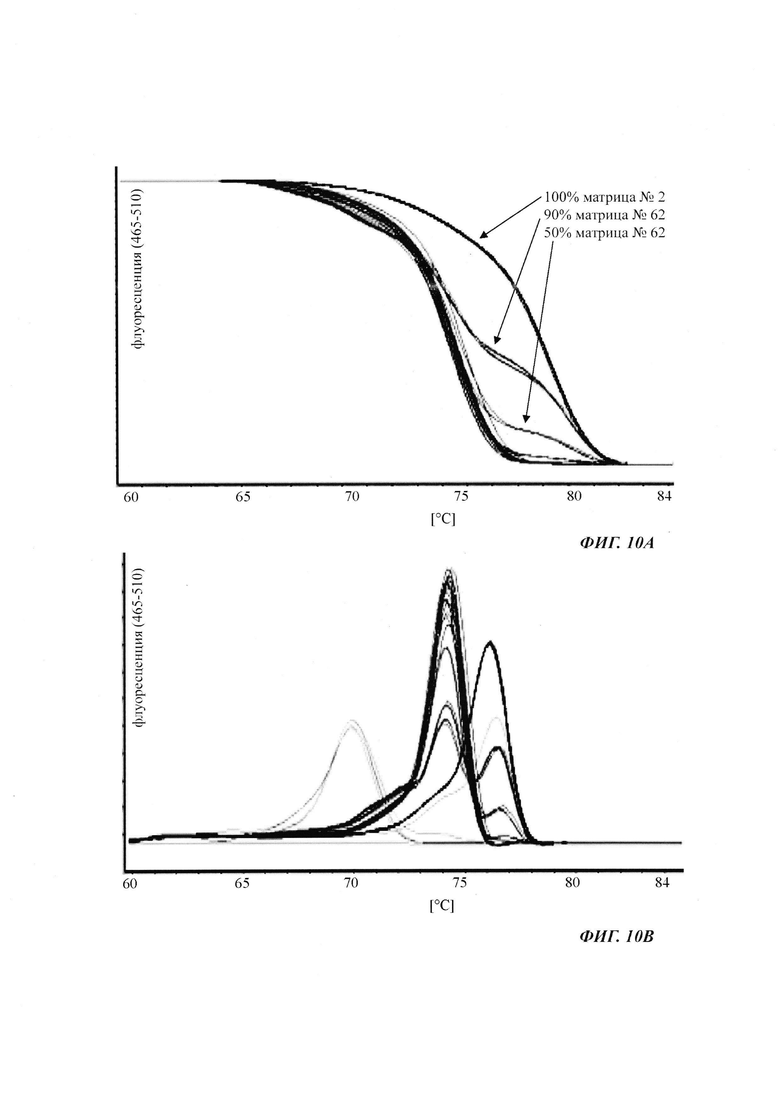
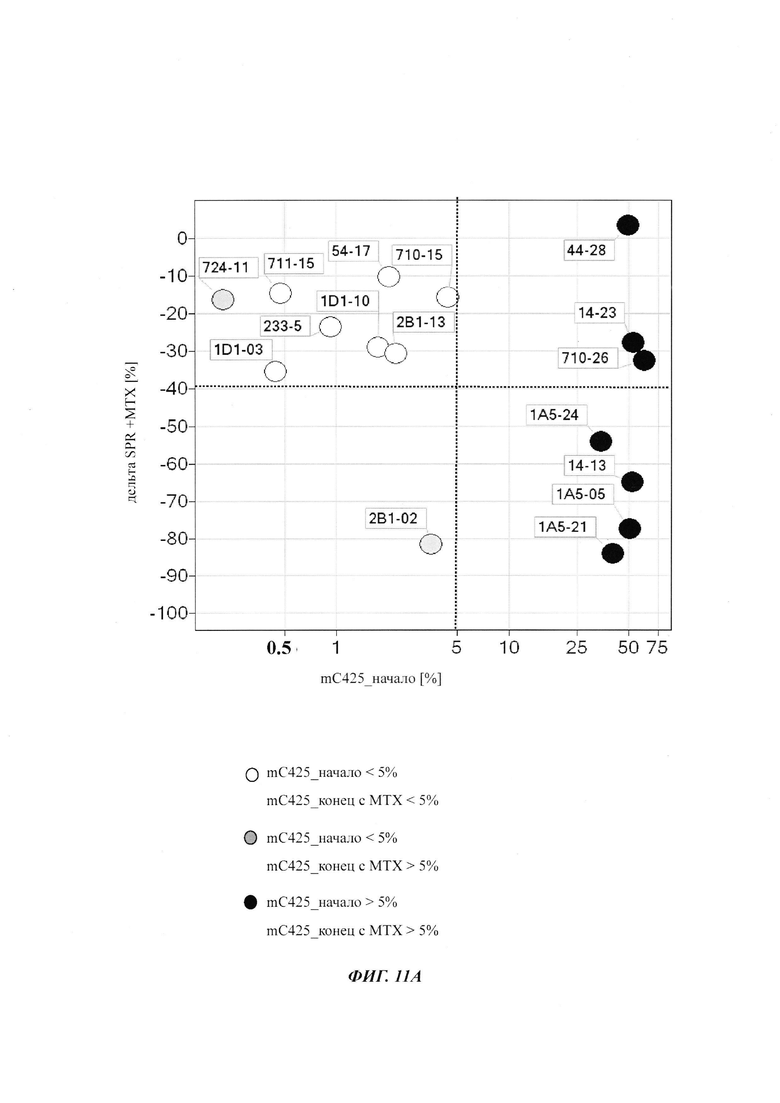
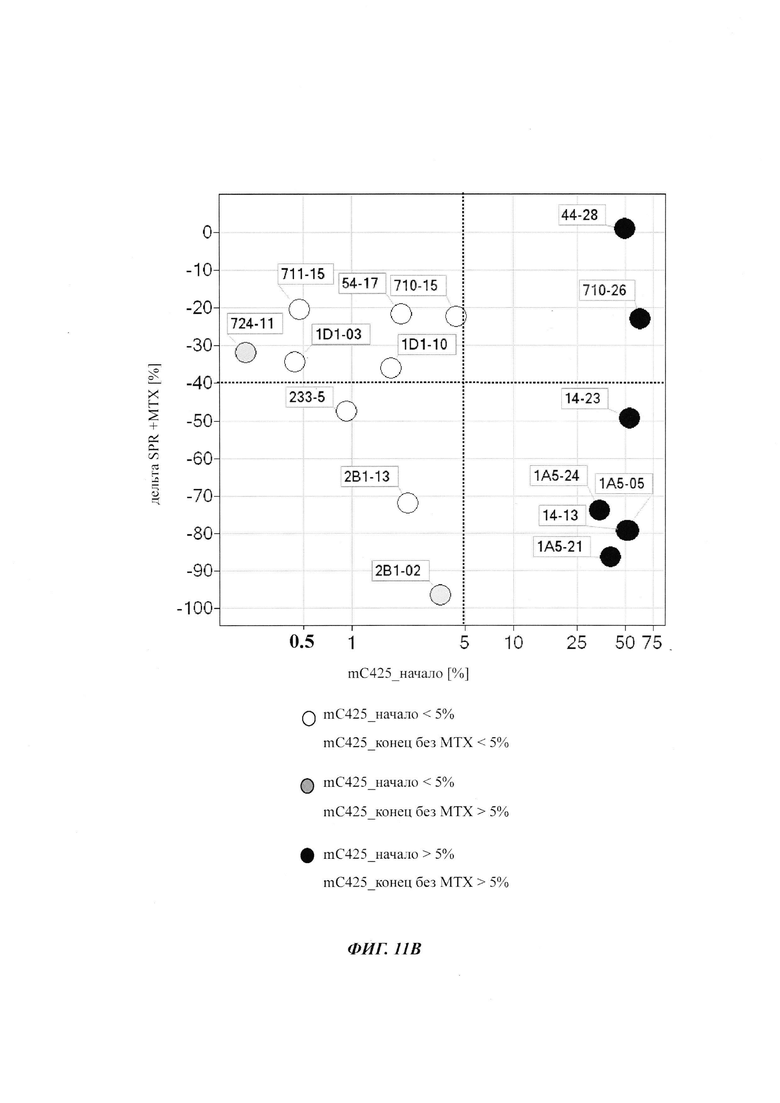
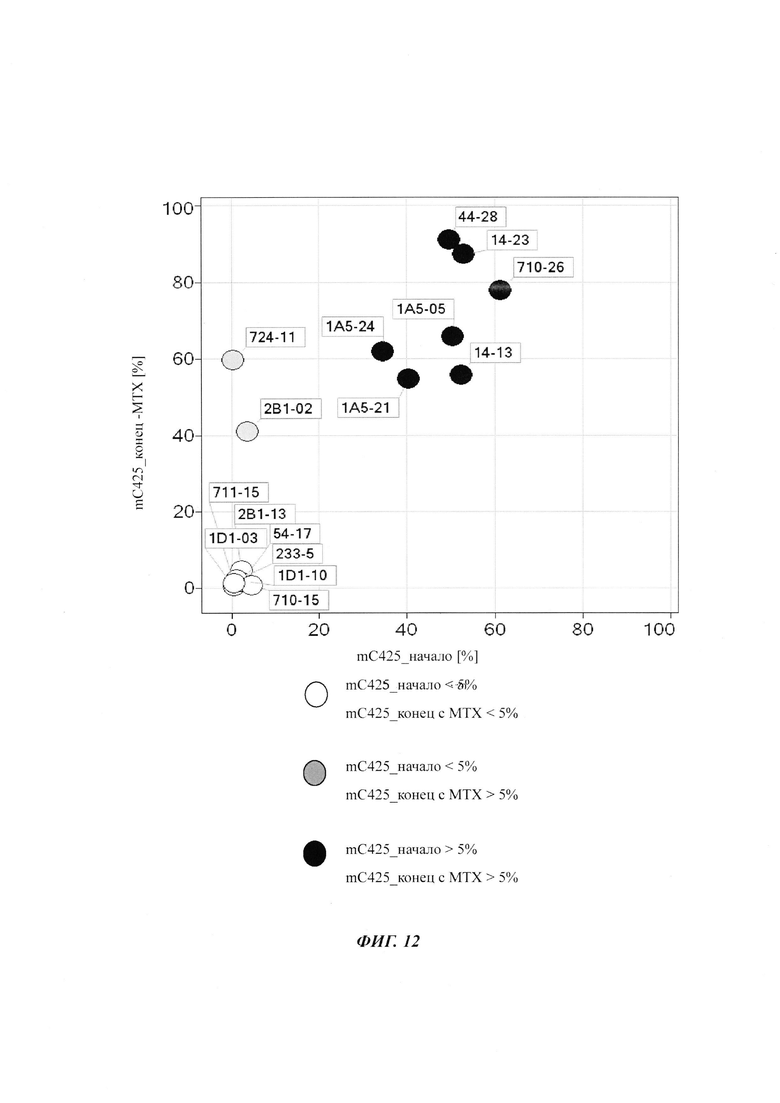
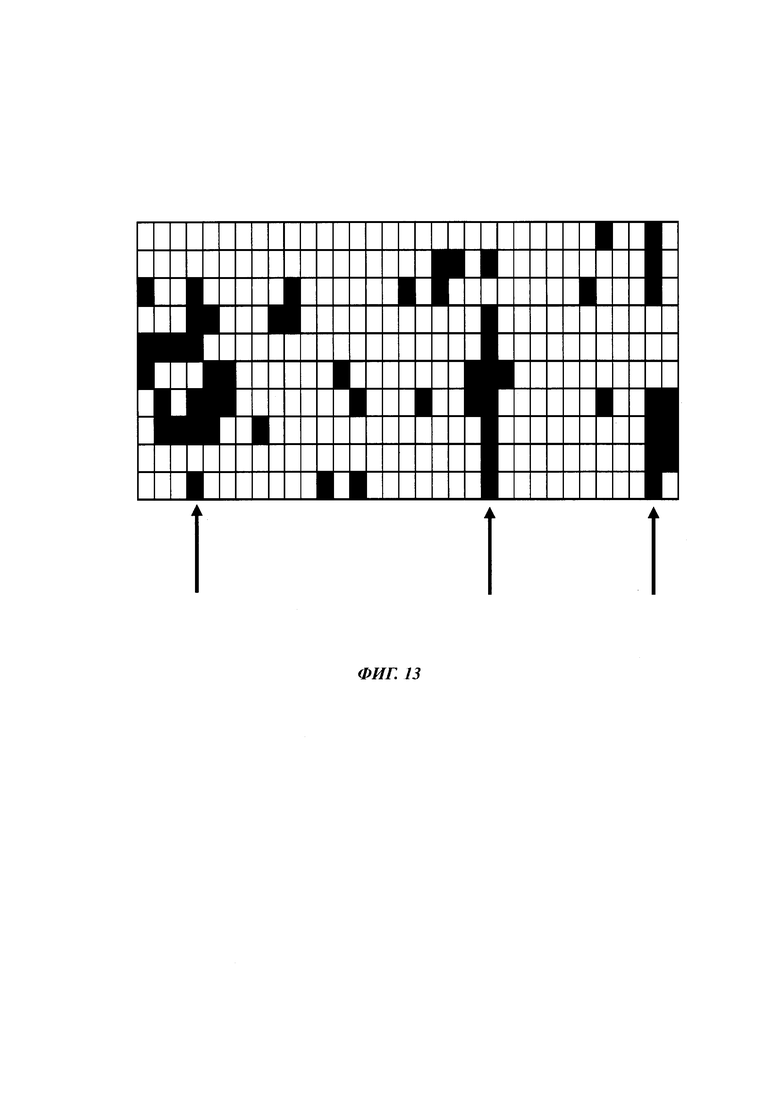
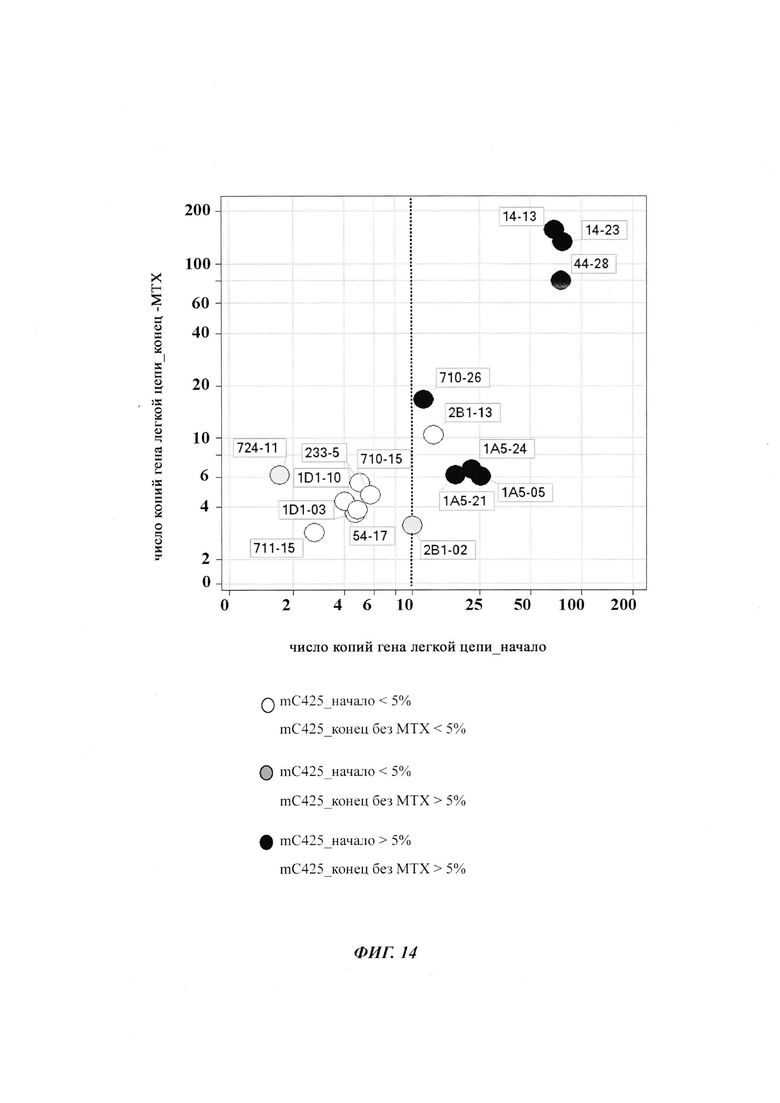
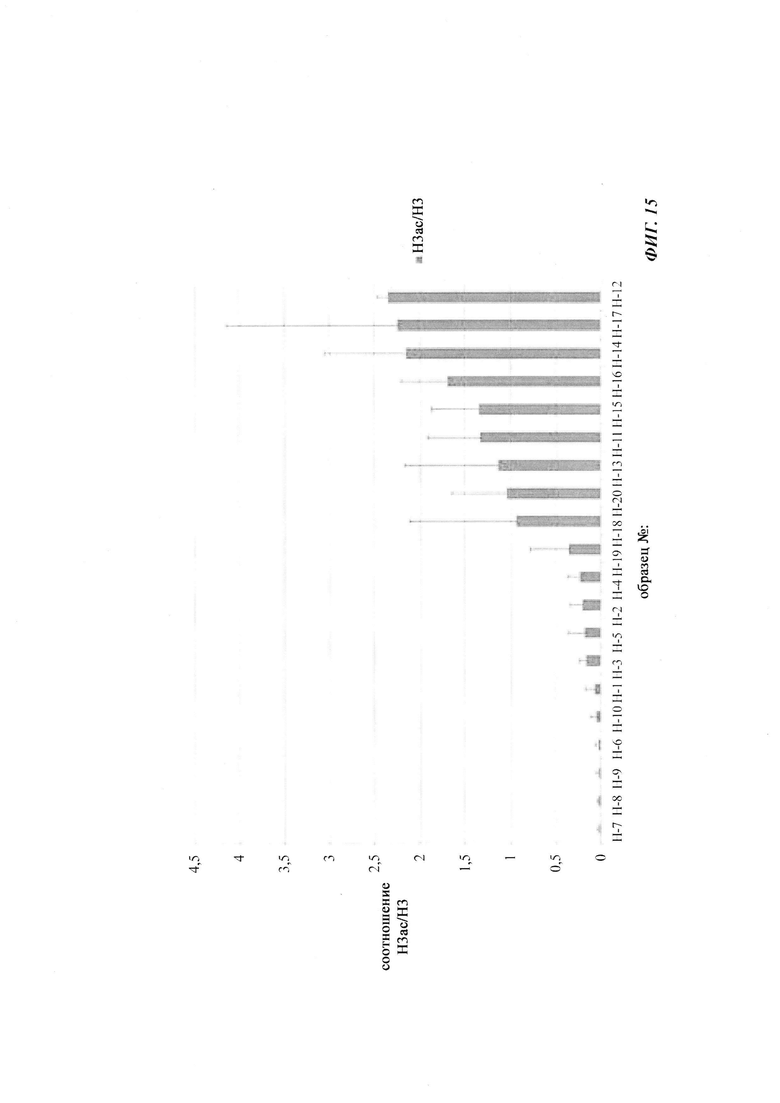
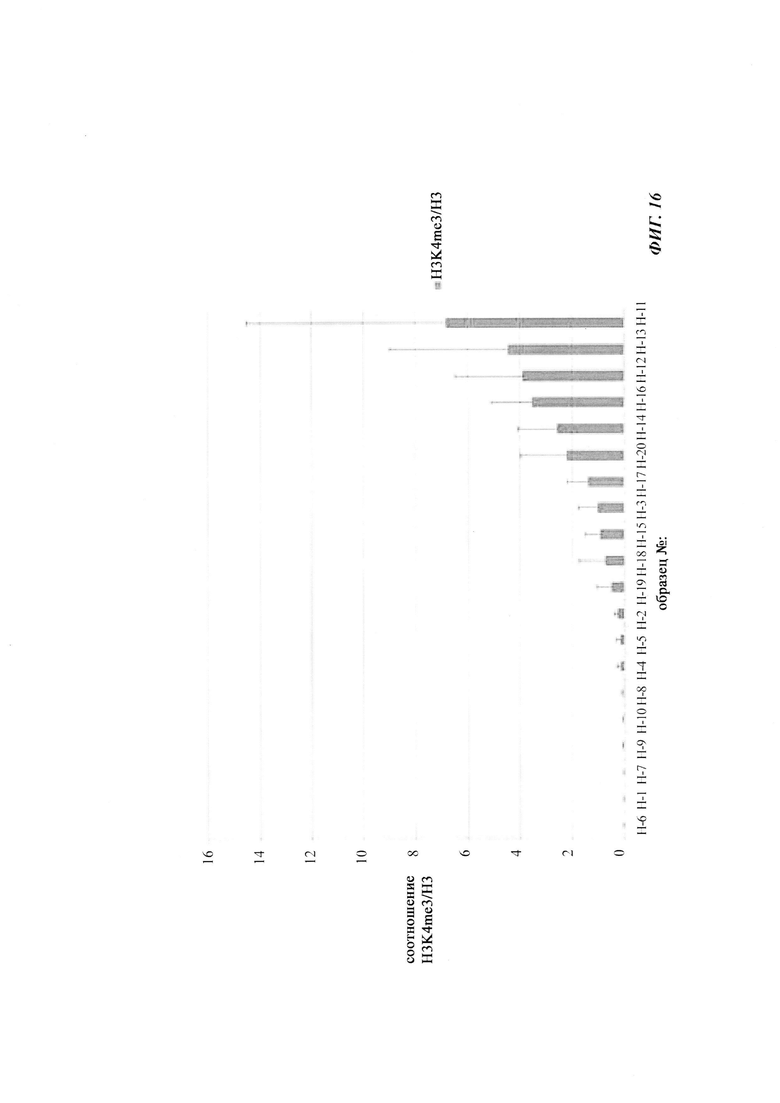
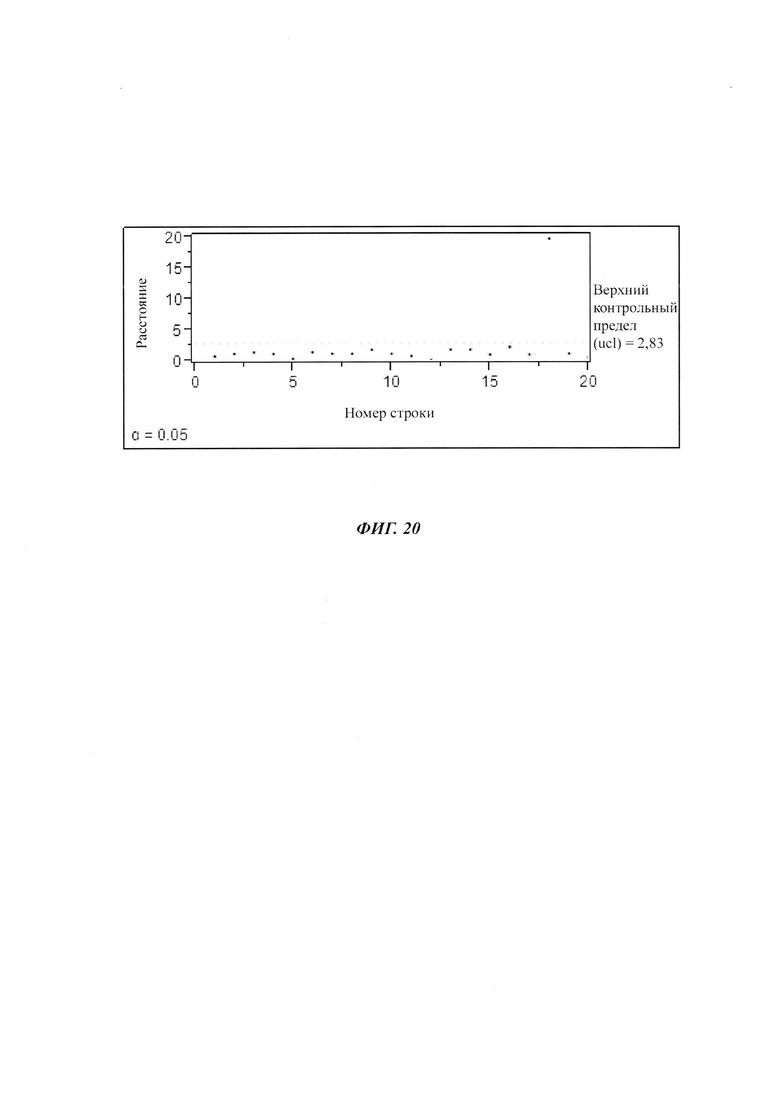
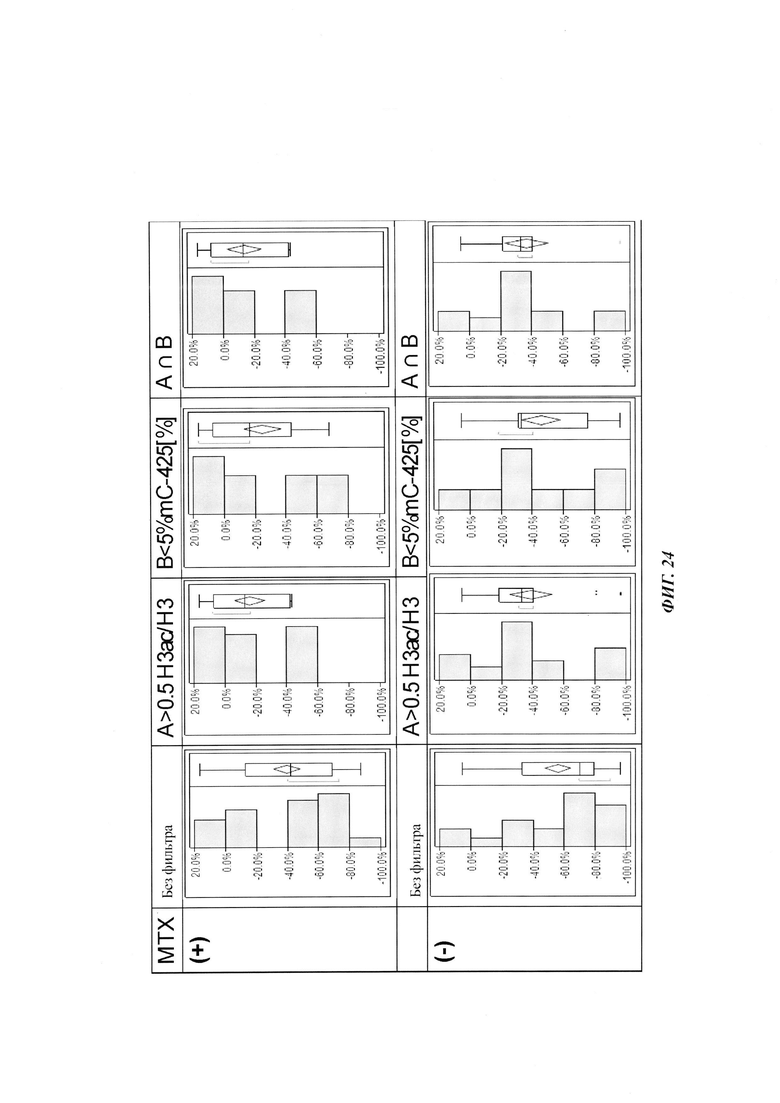
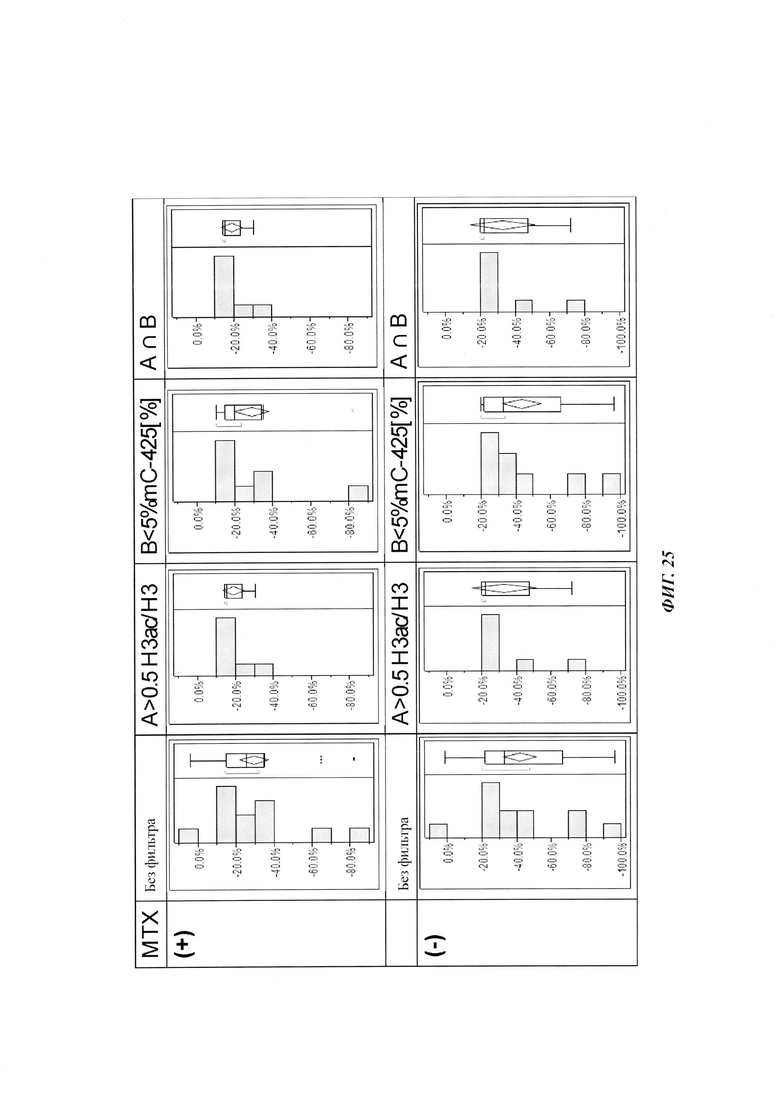
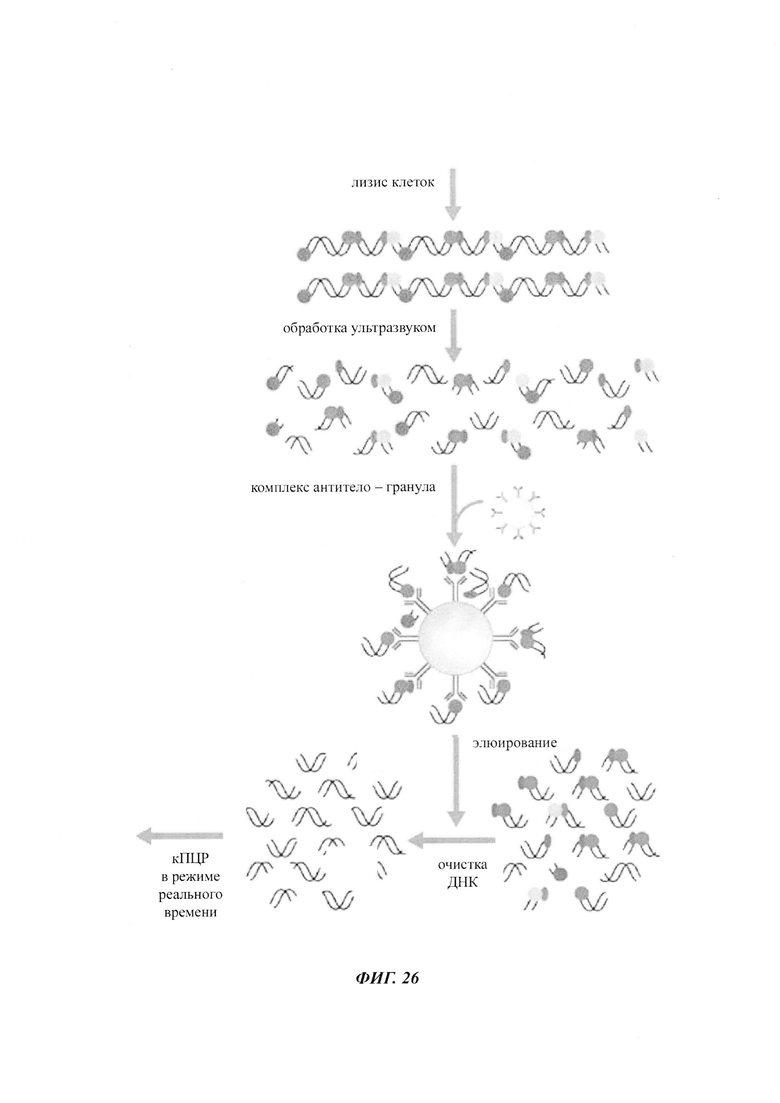
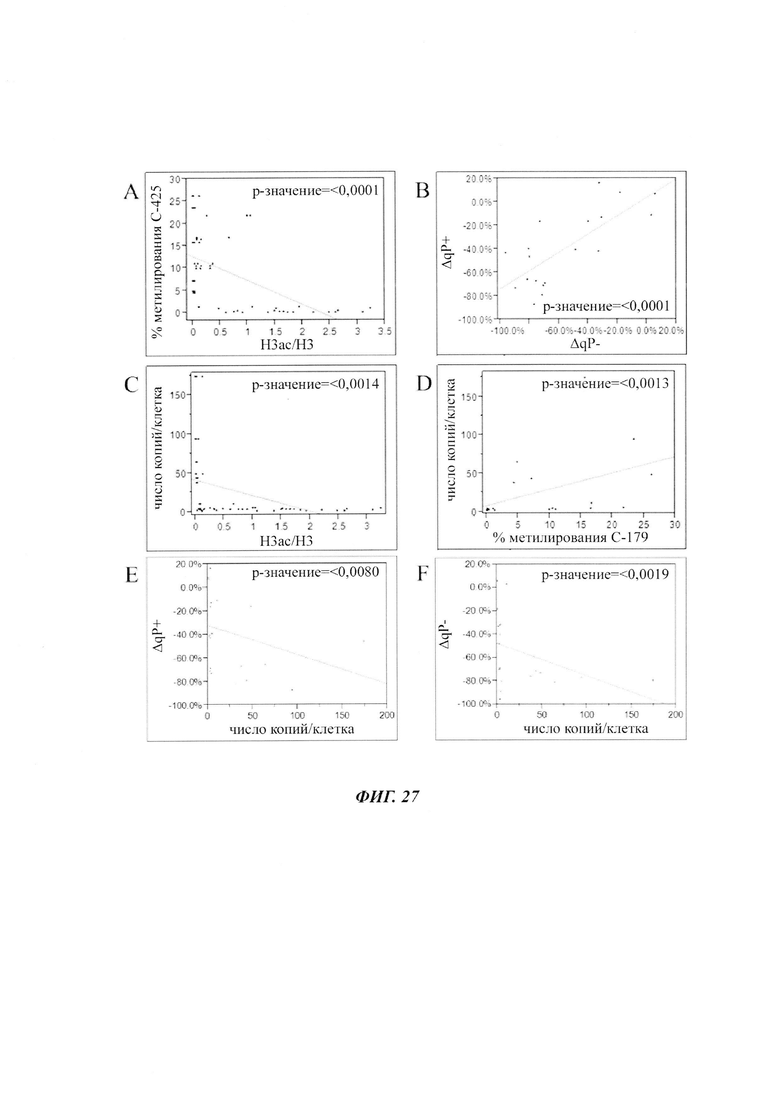
Изобретение относится к области биотехнологии, конкретно к способу отбора стабильной продуцирующей антитело под контролем промотора CMV человека с SEQ ID NO: 1 клетки яичника китайского хомячка СНО-К1. Заявленный способ отбора клеток-продуцентов антитела на основе определения уровня ацетилирования гистона 3 и частоты метилирования CpG-сайта в позиции 425 в SEQ ID NO: 1 позволяет уменьшить количество клонов, подлежащих тестированию при культивировании нескольких поколений. 2 н. и 5 з.п. ф-лы, 28 ил., 20 табл., 13 пр.
1. Способ отбора стабильной продуцирующей антитело клетки яичника китайского хомячка СНО-К1, которая содержит интегрированную в геном нуклеиновую кислоту, содержащую структурный ген, кодирующий антитело, функционально связанный с промоторной нуклеиновой кислотой, которая имеет нуклеотидную последовательность SEQ ID NO: 1, при этом указанный способ включает следующие этапы:
а) определение для клеток СНО-К1 первого множества клеток, где клетки СНО-К1 трансфицированы нуклеиновой кислотой, содержащей экспрессионную кассету, содержащую структурный ген, кодирующий указанное антитело, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 1, среднего уровня ацетилирования гистона 3 относительно уровня гистона 3 (H3ac/H3) в хроматине промоторной нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO: 1, на основании уровня ацетилирования гистона 3 относительно уровня гистона 3 (H3ac/Н3), определенного по меньшей мере в 10 клетках СНО-К1, полученных при культивировании клеток СНО-К1, и
b) определение для клеток СНО-К1 второго множества клеток, где клетки СНО-К1 трансфицированы нуклеиновой кислотой, содержащей экспрессионную кассету, содержащую структурный ген, кодирующий указанное антитело, функционально связанный с промоторной нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 1, средней частоты метилирования CpG-сайта в позиции 425 в SEQ ID NO: 1, на основании метилирования, определенного по меньшей мере в 10 клетках СНО-К1, полученных при культивировании клеток СНО-К1, и
c) отбор клетки СНО-К1, которая имеет средний уровень ацетилирования гистона 3 относительно уровня гистона 3, равный 0,5 или более, и которая имеет частоту метилирования в позиции 425 менее 5%,
при этом первое множество клеток и второе множество клеток являются одним и тем же множеством клеток СНО-К1,
и где определение уровня ацетилирования гистона 3 относительно уровня гистона 3 (H3ac/Н3) включает следующие этапы:
1) выделение хроматина из клетки СНО-К1,
2) обработка первой аликвоты хроматина антителом, специфическим в отношении ацетилирования гистона 3 или гистона 3, и формирование преципитата "антитело – хроматин", и обработка второй аликвоты хроматина антителом, специфическим в отношении гистона 3, и формирование преципитата "антитело – хроматин",
3) амплификация геномной ДНК из третьей необработанной аликвоты хроматина и из первой и второй обработанных аликвот путем количественной ПЦР в режиме реального времени,
4) определение с помощью результата, полученного на этапе 3), уровня ацетилирования гистона 3 относительно уровня гистона 3,
и где определение частоты метилирования включает следующие этапы:
(1) выделение ДНК из клеток СНО-К1,
(2) проведение для выделенной ДНК отдельной полимеразной цепной реакции, специфической в отношении метилирования,
(3) определение с помощью результата, полученного на этапе (2), частоты метилирования.
2. Способ по п. 1, в котором этап (2) представляет собой:
(2) проведение для выделенной ДНК отдельной полимеразной цепной реакции с использованием праймера, специфического в отношении метилирования, и универсального праймера.
3. Способ по п. 2, в котором универсальные праймеры имеют последовательности SEQ ID NO: 9 и 11, а праймеры, специфические в отношении метилирования, имеют последовательности SEQ ID NO: 11 и 18.
4. Способ по п. 1, в котором уровень ацетилирования гистона 3 относительно уровня гистона 3 (H3ac/Н3) нормализован по референсному гену.
5. Способ по п. 4, характеризующийся тем, что референсный ген представляет собой Gusb.
6. Способ по п. 1, в котором антитело представляет собой биспецифическое антитело.
7. Способ получения антитела, который включает следующие этапы:
а) отбор стабильной продуцирующей антитело клетки яичника китайского хомячка СНО-К1 с помощью способа по любому из пп. 1-6,
b) культивирование отобранной клетки СНО-К1, и
c) выделение антитела из среды культивирования и/или клетки СНО-К1 и, таким образом, получение антитела.
| WO 2011128377, 20.10.2011 | |||
| PAREDES V | |||
| et al., Unstable expression of recombinant antibody during long-term culture of CHO cells is accompanied by histone H3 hypoacetylation, Biotechnology letters, 2013, V | |||
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| MEILINGER D | |||
| et al., Np95 interacts with de novo DNA methyltransferases, Dnmt3a and Dnmt3b, and mediates epigenetic | |||

