В настоящем документе описаны новое строение векторов, новые способы получения линий клеток-продуцентов, такие как новые способы трансфекции или селекции, а также применение этих экспрессионных векторов и линий клеток-продуцентов для рекомбинантного получения интересующих полипептидов.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Уровень транскрипции гена может иметь сильное влияние на уровень его экспрессии и, следовательно, определять продуктивность клетки. Главным образом, на уровень транскрипции гена влияют три элемента вектора: промотор, поли(А)-сигнальная последовательность и (если имеется) терминатор транскрипции.
Нуклеиновая кислота, кодирующая тяжёлую цепь антитела, как правило, содержит лидерную последовательность (сигнальную последовательность) (приблизительно 57 п. н./19 а. о.), которая удаляется при созревании белка, вариабельный район, VH (приблизительно 350 п. н./115 а. о.) и константный район, CH (приблизительно 990 п. н./330 а. о.). Нуклеиновая кислота, кодирующая лёгкую цепь антитела, как правило, состоит из лидерной последовательности (приблизительно 66 п. н./22 а. о.), которая удаляется при созревании белка, вариабельного района, VK или VL (приблизительно 350 п. н./115 а. о.) и константного района, CK или CL (приблизительно 321 п. н./107 а. о.).
Рекомбинантное получение антител в эукариотических клетках включает получение экспрессионных систем (см. McCafferty, J., et al., (eds.), Antibody Engineering, A Practical Approach, IRL Press (1997)). Для разработки экспрессионных систем антитела создают экспрессионную кассету, содержащую нуклеиновую кислоту, кодирующую лёгкую цепь, фланкированную районами промотора и сигнала полиаденилирования (полиA). Также создают экспрессионную кассету тяжёлой цепи, содержащую нуклеиновую кислоту, кодирующую тяжёлую цепь, фланкированную районами промотора и полиA. Экспрессионная кассета тяжёлой цепи может быть скомбинирована с экспрессионной кассетой лёгкой цепи в одном векторе, который в итоге содержит обе указанные экспрессионные кассеты; или они могут быть интегрированы в два отдельных вектора.
Кассеты молекул ДНК иммуноглобулина, конструкции монотел, способы их получения и способы их применения описаны в патенте США номер 7,053,202. В патенте США номер 5,168,062 описаны вектора для переноса и микроорганизмы, содержащие немедленно-раннюю промоторную регуляторную последовательность ДНК цитомегаловируса. Фрагмент ДНК, содержащий промоторный район человеческого фактора элонгации полипептидной цепи 1α , его последовательность и экспрессионные плазмиды, содержащие фрагмент ДНК, имеющие высокую применимость к широкому спектру клеток-хозяев с высоким экспрессионным потенциалом, описаны в патенте США номер 5,225,348. В патенте США номер 5,266,491 описаны экспрессионные плазмиды, содержащие точку начала репликации SV40 и фрагмент ДНК, содержащий промоторный район гена человеческого фактора элонгации полипептидной цепи 1α . Соединения рекомбинантной ДНК и экспрессия полипептидов, таких как tPA, описаны в патенте США номер 5,122,458. В патенте США номер 7,422,874 описан экспрессионный вектор для животной клетки.
В Kim, D., et al. описаны улучшенные посредством манипулирования районами терминации транскрипции экспрессионные системы в клетках млекопитающих (Biotechnol. Prog. 19 (2003) 1620-1622). Описанный 9-нуклеотидный участок клеточной мРНК может функционировать как внутренний сайт посадки рибосомы (IRES), и, когда он представлен в виде множества соединённых копий, происходит значительное усиление IRES-активности, что описано в Chappell, S.A., et al. (Proc. Natl. Acad. Sci. USA 97 (2000) 1536-1541. В Corish, P. and Tyler-Smith, C. описано уменьшение времени полужизни зелёного флуоресцентного белка в клетках млекопитающих (Prot. Eng. 12 (1999) 1035-1040). Новый вектор GFPneo, разработанный для выделения и анализа энхансерных элементов в трансфецированных клетках млекопитающих, описан в Primig, M., et al. (Gene 215 (1998) 181-189). В Ng, S.K., et al. описано применение дестабилизирующих последовательностей на маркере селекции для улучения продуктивности рекомбинантного белка в CHO-DG44 (Metabol. Eng. 9 (2007) 304-316).
В Sanna Pietro, P. описана экспрессия фрагментов Fab антитела и полноразмерного иммуноглобулина в клетках млекопитающих (Meth. Mol. Biol. 178 (2002) 389-395). Библиотека клеточного дисплея для клонирования генов различных районов человеческих антител к поверхностному антигену гепатита B описана в Higuchi, K., et al. (J. Immunol. Meth. 202 (1997) 193-204). В Kim, D., et al. описаны улучшенные посредством манипулирования районами терминации транскрипции экспрессионные системы млекопитающих (Biotechnol. Progress 19 (2003) 1620-1622). Методические указания по конструированию клеток для получения моноклональных антител описаны в Costa, R.A., et al. (Eur. J. Pharmaceut. Biopharmaceut. 74 (2010) 127-138). В Kim, D.W., et al. описано применение промотора фактора элонгации 1α человека как универсальной и эффективной экспрессионной системы (Gene 91 (1990) 217-223). Сравнение интрон-зависимой и интрон-независимой экспрессии генов описано в Buchman, A.R., et al., (Mol. Cell. Biol. 8 (1988) 4395-4405). В Wang, F., et al. описана экспрессия антител в клетках млекопитающих (в Therapeutic monoclonal antibodies - From bench to clinic, Wiley (2009), страницы 557-572). Сравнительное исследование разных конструкций векторов для экспрессии в млекопитающих рекомбинантных антител IgG описано в Li et al. (J. Immunol. Meth. 318 (2007) 113-124). В Ho, S.C.L., et al. описаны IRES-опосредованные трицистронные вектора для улучшенного получения клеточных линий CHO c высоким уровнем экспрессии моноклональных антител (J. Biotechnol. 157 (2011) 130-139). Получение химерного анти-CD2 антитела рекомбинантными животными клетками описано в Hotta, A., et al. (J. Biosci. Bioeng. 98 (2004) 298-303). В Lee, J-C., et al. описана высокоэффективная экспрессия белка, опосредованная последовательностью внутренней посадки рибосомы энтеровируса 71 (Biotechnol. Bioeng. 90 (2005) 656-662). В WO 2008/142124 описана получение рекомбинантного белка в птичьих клетках EBX®.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Обнаружено, что при получении рекомбинантных антител расположение экспрессионной кассеты тяжёлой цепи перед экспрессионной кассетой лёгкой цепи (HC-LC (5'-3')) обеспечивает лучшие результаты экспрессии по сравнению с обратным порядком (LC-HC (5'-3')). Кроме того, обнаружено, что расположение маркера селекции после обеих экспрессионных кассет антитела обеспечивает лучшие результаты экспрессии (HC-LC-SM (5'-3')) по сравнению с двунаправленным расположением перед первой цепью антитела (SM (3'-5')-HC-LC (5'-3')).
Обнаружено, что для стабильных трансфекций является оптимальным расположение в порядке 1) тяжёлая цепь антитела, 2) лёгкая цепь антитела и 3) маркер селекции. Но хотя промотор hEF1α , несомненно, превосходит промотор hCMV в стабильных пулах, мы наблюдали прямо противоположный эффект на уровне отдельных клонов. А именно, немедленно-ранний промотор/энхансер цитомегаловируса человека (hCMV) давал клоны с наиболее высокой продуктивностью. Более того, его действие может быть также усилено с помощью его комбинирования с поли(А)-сигнальной последовательностью из bGH и с терминаторной последовательностью гена гастрина человека (hGT), который усиливает и продуктивность, и стабильность экспрессии.
Обнаружено, что применение экспрессионного вектора, включающего в себя экспрессионную кассету тяжёлой цепи антитела и экспрессионную кассету лёгкой цепи антитела, каждая из которых содержит промотор, структурный ген и поли(А)-сигнальную последовательность, и, необязательно, терминаторную последовательность, приводит к большему количеству клеточных клонов, продуцирующих/секретирующих антитело, после трансфекции в случае, если 1) промотором является промотор цитомегаловируса человека (hCMV), поли(A)-сигнальной последовательностью является сигнальная последовательность полиA бычьего гормона роста (полиA bGH) и терминаторной последовательностью является транскрипционная терминаторная последовательность гена гастрина человека (hGT) или 2) промотором является промотор фактора элонгации 1α человека (EF1α), сигнальной последовательностью полиA является поли(A)-сигнальная последовательность бычьего гормона роста (полиA bGH) и терминаторная последовательность отсутствует.
При использовании экспрессионного вектора, соответствующего описанию выше, после трансфекции может быть получено большее количество клеток, продуцирующих/секретирующих антитело, и, таким образом, снижены необходимые усилия для обнаружения высокопродуктивной клетки, подходящей для крупномасштабного получения рекомбинантных антител.
Одним аспектом настоящего изобретения является экспрессионный вектор, содержащий
экспрессионную кассету лёгкой цепи антитела
экспрессионную кассету тяжёлой цепи антитела и
экспрессионную кассету маркера селекции,
где экспрессионные кассеты расположены однонаправлено и
где экспрессионные кассеты расположены по направлению от 5'- к 3'-концу последовательности в следующем порядке: экспрессионная кассета тяжёлой цепи антитела, экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета маркера селекции.
В одном варианте экспрессионная кассета лёгкой цепи антитела и/или экспрессионная кассета тяжёлой цепи антитела, и/или экспрессионная кассета маркера селекции независимо друг от друга содержат промотор, выбранный из промотора фактора элонгации 1α , человека, промотора CMV человека и промотора SV40.
В одном варианте одна, две или все три экспрессионные кассеты содержат промотор фактора элонгации 1α человека. В одном варианте экспрессионная кассета лёгкой цепи антитела и/или экспрессионная кассета тяжёлой цепи антитела и/или кассета маркера селекции независимо друг от друга содержат промотор фактора элонгации 1α человека. В одном варианте экспрессионная кассета не содержит терминаторную последовательность, т.е. экспрессионная кассета свободна от терминаторной последовательности. В одном варианте терминаторной последовательностью является транскрипционная терминаторная последовательность гена гастрина человека (hGT).
В одном варианте одна, две или все три экспрессионные кассеты содержат промотор CMV человека. В одном варианте экспрессионная кассета лёгкой цепи антитела и/или экспрессионная кассета тяжёлой цепи антитела и/или кассета маркера селекции независимо друг от друга содержат промотор CMV человека.
В одном варианте одна, две или три экспрессионные кассеты содержат сигнальную последовательность полиA бычьего гормона роста. В одном варианте экспрессионная кассета лёгкой цепи антитела и/или экспрессионная кассета тяжёлой цепи антитела, и/или кассета маркера селекции независимо друг от друга содержат сигнальную последовательность полиA бычьего гормона роста.
В одном варианте экспрессионная кассета лёгкой цепи антитела и/или экспрессионная кассета тяжёлой цепи антитела и/или кассета маркера селекции независимо друг от друга содержат одну из следующих сигнальных последовательностей полиA: сигнальная последовательность полиA бычьего гормона роста и сигнальная последовательность SV40.
В одном варианте одна, две или все три экспрессионные кассеты содержат терминаторную последовательность гастрина человека после сигнальной последовательности полиA с условием, что экспрессионные кассеты не содержат промотор фактора элонгации 1α человека. В одном варианте экспрессионная кассета лёгкой цепи антитела и/или экспрессионная кассета тяжёлой цепи антитела, и/или кассета маркера селекции независимо друг от друга содержат терминаторную последовательность гастрина человека после сигнальной последовательности полиA.
В одном варианте экспрессионная кассета легкой цепи антитела и/или экспрессионная кассета тяжёлой цепи антитела и/или кассета маркера селекции независимо друг от друга содержат в направлении от 5'- к 3'-концу сигнальную последовательность полиA бычьего гормона роста и терминаторную последовательность гастрина человека с условием, что экспрессионные кассеты не содержат промотор фактора элонгации 1α человека.
В одном варианте промотор одной, двух или всех трёх экспрессионных кассет содержит интрон A.
В одном варианте одна, две или все три экспрессионные кассеты содержат сигнальную последовательность полиA SV40.
В одном варианте одна, две или все три экспрессионные кассеты содержат промотор SV40.
В одном варианте нуклеиновая кислота, кодирующая лёгкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжёлую цепь антитела, содержит, по меньшей мере, один интрон.
В одном варианте нуклеиновая кислота, кодирующая лёгкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжёлую цепь антитела, является кДНК.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, для рекомбинантного получения антитела.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, для получения стабильной клеточной линии.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, для получения линии клеток-продуцентов.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, включающего в себя, по меньшей мере, одну экспрессионную кассету, содержащую промотор фактора элонгации 1α человека, для получения стабильной клеточной линии.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, включающего в себя, по меньшей мере, одну экспрессионную кассету, содержащую промотор фактора элонгации 1α человека, для получения линии клеток-продуцентов.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, включающего в себя, по меньшей мере, одну экспрессионную кассету, содержащую промотор фактора элонгации 1α человека, для рекомбинантного получения антитела.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, включающего в себя, по меньшей мере, одну экспрессионную кассету, содержащую промотор CMV человека, для получения линии клеток-продуцентов.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, включающего в себя, по меньшей мере, одну экспрессионную кассету, содержащую промотор CMV человека, для получения стабильной клеточной линии.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, описанного в настоящем документе, включающего в себя, по меньшей мере, одну экспрессионную кассету, содержащую промотор CMV человека, для получения антитела.
Одним аспектом настоящего изобретения является способ трансфекции эукариотической клетки экспрессионным вектором, который характеризуется тем, что он линеаризован перед трансфекцией путём разрезания в точке начала репликации прокариот.
В одном варианте точка начала репликации прокариот находится между экспрессионной кассетой лёгкой цепи антитела и экспрессионной кассетой тяжёлой цепи антитела.
Одним аспектом настоящего изобретения является применение способа, описанного в настоящем документе, для получения эукариотической клетки для рекомбинантного получения антитела.
Одним аспектом настоящего изобретения является способ селекции эукариотических клеток, содержащих нуклеиновую кислоту, кодирующую антитело, который характеризуется тем, что агент для селекции в первый раз добавляют в культуральную среду примерно через 24 часа после трансфекции.
Одним аспектом настоящего изобретения является применение способа, описанного в настоящем документе, для получения эукариотической клетки для рекомбинантного получения антитела.
Одним аспектом настоящего изобретения является применение клетки, селектированной с помощью способа, описанного в настоящем документе, для рекомбинантного получения антитела.
Одним аспектом настоящего изобретения является способ получения антитела, включающий следующие этапы:
культивирование эукариотической клетки, содержащей экспрессионный вектор согласно настоящему документу, и
выделение антитела из клетки или из культуральной среды.
Одним аспектом настоящего изобретения является способ получения антитела, включающий следующий этап:
культивирование эукариотической клетки, полученной путём трансфекции экспрессионным вектором, который был линеаризован перед трансфекцией путём разрезания в точке начала репликации прокариот, и
выделение антитела из клетки или из культуральной среды.
Одним аспектом настоящего изобретения является способ получения антитела, включающий следующий этап:
культивирование эукариотической клетки, селектированной с помощью добавления в культуральную среду агента для селекции примерно через 24 часа после трансфекции, и
выделение антитела из клетки или из культуральной среды.
Одним аспектом настоящего изобретения является способ трансфекции эукариотической клетки экспрессионным вектором, содержащим прокариотические и эукариотические последовательности нуклеиновых кислот, который характеризуется тем, что прокариотические последовательности нуклеиновых кислот удаляют из экспрессионного вектора перед трансфекцией эукариотической клетки экспрессионным вектором.
Одним аспектом настоящего изобретения является применение линеаризованного экспрессионного вектора, не содержащего прокариотические последовательности нуклеиновых кислот, для трансфекции эукариотической клетки.
Одним аспектом настоящего изобретения является применение экспрессионного вектора, содержащего только эукариотические последовательности нуклеиновых кислот, для получения эукариотической клетки для рекомбинантного получения антитела.
В одном варианте всех аспектов, заявленных в настоящем документе, антитело является биспецифическим.
В одном варианте биспецифическое антитело имеет первую специфичность связывания или сайт связывания, который специфично связывается с первым антигеном или с первым эпитопом на антигене, и биспецифическое антитело имеет вторую специфичность связывания или сайт связывания, который специфично связывается со вторым антигеном или со вторым эпитопом на антигене.
В одном варианте экспрессионный вектор содержит или
первую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую первую лёгкую цепь антитела, сигнальную последовательность полиA, и, необязательно, терминаторную последовательность,
вторую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую вторую лёгкую цепь антитела, сигнальную последовательность полиA, и, необязательно, терминаторную последовательность,
третью экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую первую тяжёлую цепь антитела, сигнальную последовательность полиA, и, необязательно, терминаторную последовательность,
четвёртую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую вторую тяжёлую цепь антитела, сигнальную последовательность поли A, и, необязательно, терминаторную последовательность,
или
первую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую лёгкую цепь антитела, сигнальную последовательность поли A, и, необязательно, терминаторную последовательность,
вторую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую первую тяжёлую цепь антитела, сигнальную последовательность поли A, и, необязательно, терминаторную последовательность,
третью экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую вторую тяжёлую цепь антитела, сигнальную последовательность поли A, и, необязательно, терминаторную последовательность,
где лёгкая цепь антитела является общей для обеих тяжёлых цепей антитела.
В одном варианте всех аспектов, заявленных в настоящем документе, экспрессионный вектор содержит
экспрессионную кассету лёгкой цепи антитела,
экспрессионную кассету первой тяжёлой цепи антитела,
экспрессионную кассету второй тяжёлой цепи антитела, и
экспрессионную кассету маркера селекции,
где, по меньшей мере, одна из экспрессионной кассеты тяжёлой цепи антитела, экспрессионной кассеты лёгкой цепи антитела и экспрессионная кассета маркера селекции расположены однонаправленно, и
где однонапраленные экспрессионные кассеты расположены по направлению от 5'- к 3'-концу последовательности в следующем порядке: экспрессионная кассета тяжёлой цепи антитела, экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета маркера селекции.
В одном варианте всех аспектов, заявленных в настоящем изобретении, экспрессионный вектор содержит
экспрессионную кассету первой лёгкой цепи антитела,
экспрессионную кассету второй лёгкой цепи антитела,
экспрессионную кассету первой тяжёлой цепи антитела,
экспрессионную кассету второй тяжёлой цепи антитела, и
экспрессионную кассету маркера селекции,
где одна из экспрессионных кассет тяжёлой цепи антитела, одна из экспрессионных кассет лёгкой цепи антитела и экспрессионная кассета маркера селекции расположены однонаправленно, и
где однонапраленные экспрессионные кассеты расположены по направлению от 5'- к 3'-концу последовательности в следующем порядке: экспрессионная кассета тяжёлой цепи антитела, экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета маркера селекции.
В одном варианте одна из экспрессионных кассет тяжёлой цепи антитела кодирует тяжёлую цепь антитела, содержащую мутацию типа «впадина».
В одном варианте одна из экспрессионных кассет тяжёлой цепи антитела кодирует тяжёлую цепь антитела, содержащую мутацию типа «выступ».
В одном варианте одна из экспрессионных кассет лёгкой цепи антитела кодирует лёгкую цепь антитела, содержащую вариабельный домен лёгкой цепи антитела и домен CH1 тяжёлой цепи антитела в качестве константного домена, и/или одна из экспрессионных кассет лёгкой цепи антитела кодирует лёгкую цепь антитела, содержащую вариабельный домен лёгкой цепи антитела и домен CL лёгкой цепи антитела в качестве константного домена.
В одном варианте одна из экспрессионных кассет тяжёлой цепи антитела кодирует тяжёлую цепь антитела, содержащую в качестве первого константного домена константный домен лёгкой цепи (CL), и/или одна из экспрессионных кассет тяжёлой цепи антитела кодирует тяжёлую цепь антитела, содержащую в качестве первого константного домена константный домен CH1 тяжёлой цепи.
В одном варианте промотор hCMV имеет последовательность SEQ ID NO: 01. Это промотор hCMV без интрона A и без 5'-НТО.
В одном варианте промотор hCMV имеет последовательность SEQ ID NO: 02. Это промотор hCMV без интрона A и с 5'-НТО.
В одном варианте промотор hCMV имеет последовательность SEQ ID NO: 03. Это полноразмерный промотор hCMV с интроном A.
В одном варианте промотор фактора элонгации 1α человека имеет последовательность SEQ ID NO: 04. Это промотор hEFα без интрона A.
В одном варианте промотор фактора элонгации 1α человека имеет последовательность SEQ ID NO: 05. Это промотор hEFα c интроном A.
В одном варианте промотор фактора элонгации 1α человека имеет последовательность SEQ ID NO: 06. Это короткий промотор hEFα с интроном A и с 5'-НТО.
В одном варианте промотор CMV крысы имеет последовательность SEQ ID NO: 07.
В одном варианте сигнальная последовательность полиA SV40 имеет последовательность SEQ ID NO: 08.
В одном варианте сигнальная последовательность полиA бычьего гормона роста имеет последовательность SEQ ID NO: 09.
В одном варианте терминатор гастрина человека имеет последовательность SEQ ID NO: 10.
В одном варианте промотор SV40 имеет последовательность SEQ ID NO: 11.
В одном варианте последовательность PEST орнитиндекарбоксилазы кодируется последовательностью SEQ ID NO: 12.
В одном варианте последовательность GFP кодируется последовательностью SEQ ID NO: 13.
В одном варианте маркер селекции для неомицина имеет последовательность SEQ ID NO: 14.
В одном варианте слитый полипептид GFP-PEST-NEO кодируется последовательностью SEQ ID NO: 15.
В одном варианте EMCV-IRES имеет последовательность SEQ ID NO: 16.
В одном варианте EV71-IRES имеет последовательность SEQ ID NO: 17.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
I. Основные аспекты
Как известно специалисту в данной области техники, применение технологии рекомбинантной ДНК делает возможным получение множества производных нуклеиновой кислоты и/или полипептида. Такие производные могут быть, например, модифицированы в одной отдельной позиции или в разных позициях путём замещения, изменения, обмена, делеции или вставки. Данная модификация может быть, например, выполнена посредством сайт-направленного мутагенеза. Такие модификации могуть быть легко выполнены специалистом в данной области техники (см., например, Sambrook, J., et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Нью-Йорк, США (1999)). Применение рекомбинантной технологии позволяет специалисту в данной области техники трансформировать различные клетки-хозяева гетерологичными нуклеиновыми кислотами. Хотя механизм транскрипции и трансляции, т. е. экспрессии, в различных клетках использует одинаковые элементы, клетки, принадлежащие к разным видам, могут иметь, помимо прочего, отличаться так называемой частотой использования кодонов. В связи с этим, идентичные полипептиды (по аминокислотной последовательности) могут кодироваться различными нуклеиновыми кислотами. Также, в связи с вырожденностью генетического кода, различные нуклеиновые кислоты могут кодировать один и тот же полипептид.
Применение технологии рекомбинантной ДНК позволяет создавать множество производных нуклеиновой кислоты и/или полипептида. Такие производные могут быть, например, модифицированы в одной отдельной или в нескольких позициях путём замещения, изменения, обмена, делеции или вставки. Данная модификация или создание производных могут быть, например, выполнены посредством сайт-направленного мутагенеза. Такие модификации могуть быть легко выполнены специалистом в данной области техники (см., например, Sambrook, J., et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Нью-Йорк, США (1999); Hames, B.D., and Higgins, S.J., Nucleic acid hybridization - a practical approach, IRL Press, Оксфорд, Англия (1985)).
Применение рекомбинантной технологии позволяет трансформировать различные клетки-хозяева гетерологичными нуклеиновыми кислотами. Хотя механизм транскрипции и трансляции, т. е. экспрессии, в различных клетках использует одинаковые элементы, клетки, принадлежащие к разным видам, могут иметь, помимо прочего, отличаться так называемой частотой использования кодонов. В связи с этим, идентичные полипептиды (по аминокислотной последовательности) могут кодироваться различными нуклеиновыми кислотами. Также, в связи с вырожденностью генетического кода, различные нуклеиновые кислоты могут кодировать один и тот же полипептид.
Определения
Термин антитело «с созревшей аффинностью» подразумевает антитело с одним или более изменениями в одном или более гипервариабельных районах (HVR) по сравнению с исходным антителом, которое не обладает такими изменениями; такие изменения приводят к улучению аффинности антитела к антигену.
Термин «антитело» в настоящем документе используется в широком смысле и охватывает различные структуры антител, влючая моноклональные антитела, поликлональные антитела, мультиспецифичные антитела (например, биспецифические антитела) и фрагменты антител, но не ограничиваясь ими, при условии, что они демонстрируют желаемую антиген-связывающую активность.
Термин «фрагмент антитела» подразумевает молекулу, отличающуюся от интактного антитела, которая содержит часть интактного антитела, которая связывает антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничиваются ими, Fv, Fab, Fab', Fab'-SH,. F(ab')2; димерные антитела; линейные антитела; одноцепочечные молекулы антитела (например, scFv); и мультиспецифичные антитела, сформированные из фрагментов антител.
Термин «химерное» антитело подразумевает антитело, в котором часть тяжёлой и/или лёгкой цепи получена из одного источника или вида, тогда как оставшаяся часть тяжёлой и/или лёгкой цепи получена из другого источника или вида.
Термин «класс» антитела подразумевает тип константного домена или константного района, который имеет его тяжёлая цепь. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и несколько из них могут быть подразделены на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжёлой цепи, соответствующие различным классам иммуноглобулинов, называются α, δ, ε, γ, и μ, соответственно.
Термин «экспрессия» в настоящем документе подразумевает процессы транскрипции и/или трансляции, происходящие внутри клетки. Уровень транскрипции интересующей последовательности нуклеиновой кислоты в клетке может быть определён на основе количества соответствующей мРНК, присутствующей в клетке. Например, мРНК, транскрибированная с интересующей последовательности, может быть количественно определена с помощью ПЦР в реальном времени или путём Нозерн-гибридизации (см. Sambrook et al., 1999, выше). Полипептиды, кодируемые интересующей нуклеиновой кислотой, могут быть количественно определены различными способами, например, с помощью ИФА, с помощью анализа на биологическую активность данного полипептида или путём применения методов анализа, не зависящих от подобной активности, таких как Вестерн-блоттинг или радиоиммуноанализ, с применением иммуноглобулинов, которые распознают данный полипептид и связываются с ним (см. Sambrook et al., 1999, выше).
Термин «экспрессионная кассета» подразумевает конструкцию, содержащую необходимые регуляторные элементы, такие как промотор и сайт полиаденилирования, для экспрессии в клетке, по меньшей мере, содержащейся в ней нуклеиновой кислоты.
Термин «экспрессионный вектор» обозначает нуклеиновую кислоту, имеющую все необходимые элементы для экспрессии содержащихся в ней структурных генов в клетке-хозяине. Как правило, экспрессионная плазмида содержит элемент для размножения плазмиды в клетках прокариот, например, в E. coli, содержит точку начала репликации и маркер селекции, эукариотический маркер селекции и одну или более экспрессионных кассет для экспрессии интересующих структурных генов, каждая из которых содержит промотор, структурный ген и терминатор транскрипции, включая сигнал полиаденилирования. Экспрессия гена обычно находится под контролем промотора, и такой структурный ген называют «функционально связанным» с данным промотором. Аналогично, регуляторный элемент и коровый промотор являются функционально связанными, если регуляторный элемент модулирует активность корового промотора.
Термин «Fc-район» в настоящем документе используется для определения C-концевого района тяжёлой цепи иммуноглобулина, содержащего, по меньшей мере, часть константного района. Данный термин включает нативные последовательности Fc-районов и варианты Fc-районов. В одном варианте, Fc-район тяжёлой цепи IgG человека располагается от Cys226 или от Pro230 до карбоксильного конца тяжёлой цепи. Однако C-концевой лизин (Lys447) Fc-района может присутствовать или может отсутствовать. Если в настоящем документе не указано иное, нумерация аминокислотных остатков Fc-района или константного района происходит согласно европейской (EU) системе нумерации, также называемой EU-индексом, как описано в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Бетесда, Мэриленд 1991, NIH Publication 91-3242.
Термин «Fc-район» является хорошо известным и определяется на основе разрезания папаином тяжёлой цепи антитела. В одном варианте комплексы, зявленные в настоящем документе, могут содержать в качестве полипептида шарнирного района тяжёлой цепи антитела человеческий Fc-район или производное Fc-района человеческого происхождения. В другом варианте Fc-район представляет собой Fc-район человеческого антитела подкласса IgG4 или представляет собой Fc-район человеческого антитела подкласса IgG1, IgG2 или IgG3, который модифицирован таким образом, что не обнаруживает связывания с рецептором Fcγ (например, FcγRIIIa) и/или с C1q. В одном варианте Fc-район представляет собой Fc-район человека и, в частности, принадлежит к подклассу IgG4 человека или является мутантным Fc-районом из подкласса IgG1 человека. В одном варианте данный Fc-район принадлежит к подклассу IgG1 человека и содержит мутации L234A и L235A. Тогда как IgG4 демонстрирует сниженное связывание с рецептором Fcγ (FcγIIIa), антитела других подклассов IgG демонстрируют сильное связывание с данным рецептором. Однако аминокислотные остатки Pro238, Asp265, Asp270, Asn297 (отсутствие углевода Fc), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434, и/или His435, в случае, если они изменены, также обеспечивают сниженное связывание с рецептором Fcγ (Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604; Lund, J., et al., FASEB J. 9 (1995) 115-119; Morgan, A., et al., Immunology 86 (1995) 319-324; EP 0307434). В одном варианте антитело для экспрессии согласно аспекту настоящего документа в отношении связывания с рецептором Fcγ относится к подклассу IgG4 или к подклассам IgG1 или IgG2 с мутацией в позиции L234, L235 и/или D265, и/или с мутацией PVA236. В одном варианте данные мутации представляют собой S228P, L234A, L235A, L235E, и/или PVA236 (PVA236 обозначает, что аминокислотная последовательность ELLG (приведена в однобуквенном аминокислотном коде) в аминокислотных позициях от 233 до 236 IgG1 или EFLG в IgG4 замещена на PVA). В одном варианте данные мутации представляют собой S228P в IgG4 и L234A и L235A в IgG1. Fc-район антитела непосредственно вовлечён в ADCC (антителозависимую клеточноопосредованную цитотоксичность) и CDC (комплементзависимую цитотоксичность). Комплекс, который не связывает рецептор Fcγ и/или фактор комплемента C1q, не вызывает антителозависимую клеточноопосредованную цитотоксичность (ADCC) и/или комплементзависимую цитотоксичность (CDC). Модификация «выступа» обозначает мутацию T366W в домене CH3 антитела (нумерация в соответствии с Kabat). Модификация «впадины» обозначает мутации T366S, L368A и Y407V в домене CH3 антитела. В дополнение к модификациям «выступа» и «впадины» может присутствовать мутация S354C в одном домене CH3 и мутация Y349C в другом домене CH3.
Термин «каркасный район» или «FR» подразумевает аминокислотные остатки вариабельного домена, отличающиеся от остатков гипервариабельного района (HVR). FR вариабельного домена обычно состоит из четырёх FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR в составе VH (или VL) обычно расположены в следующем порядке: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термины «полноразмерное антитело», «интактное антитело» и «целое антитело» используются в настоящем документе взаимозаменяемо для обозначения антитела, имеющего по существу сходную структуру со структурой нативного антитела или имеющего тяжёлые цепи, которые содержат Fc-район, как он определён в настоящем документе.
Термин «ген» обозначает нуклеиновую кислоту, являющуюся сегментом, например, хромосомы или плазмиды, которая может вызывать экспрессию пептида, полипептида или белка. Помимо кодирующего района, т. е. структурного гена, ген содержит другие функциональные элементы, например, сигнальную последовательность, промотор (промоторы), интроны и/или терминаторы.
Термины «клетка-хозяин», линия «клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и подразумевают клетки, в которые введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Термин клетка-хозяин включает «трансформантов» и «трансформированные клетки», которые включают первично трансформированные клетки и полученное от них потомство независимо от количества пассажей. Потомство может быть не полностью идентично родительской клетке по составу нуклеиновых кислот, может содержать мутации. Мутантное потомство, обладающее теми же функциональными характеристиками или той же биологической активностью, которые отбирали или селектировали в изначально трансформированных клетках, также включено в данные термины.
Термин «человеческое антитело» означает антитело, имеющее аминокислотную последовательность, соответствующую последовательности антитела, производимого организмом человека или человеческой клеткой, или полученного из источника, который не является человеком, но который использует репертуары человеческих антител или другие последовательности, кодирующие человеческие антитела. Данное определение человеческого антитела специально исключает гуманизированное антитело, содержащее не человеческие аминокислотные остатки, связывающие антиген.
Термин «гуманизированное» антитело подразумевает химерное антитело, содержащее аминокислотные остатки из не человеческих HVR и аминокислотные остатки из человеческих FR. В конкретных вариантах гуманизированное антитело содержит по существу все из, по меньшей мере, одного, и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют HVR не человеческого антитела. Гуманизированное антитело, необязательно, может содержать, по меньшей мере, часть константного района антитела, полученного из человеческого антитела. Термин «гуманизированная форма» антитела, например, не человеческого антитела, подразумевает антитело, прошедшее гуманизацию.
Термин «гипервариабельный район» или «HVR» в настоящем документе подразумевает каждый из районов вариабельного домена антитела, который является гипервариабельным по последовательности и/или формирует петли с определённой структурой («гипервариабельные петли»). Как правило, нативные антитела, состоящие из четырёх цепей, содержат шесть HVR; три в VH (H1, H2, H3) и три в VL (L1, L2 и L3). Как правило, HVR содержат аминокислотные остатки из гипервариабельных петель и/или из «районов, определяющих комплементарность» (CDR); последние имеют наибольшее разнообразие последовательностей и/или участвуют в распознавании антигена. Примеры гипервариабельных петель расположены в области аминокислотных остатков 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (H2) и 96-101 (H3) (Chothia, С. and Lesk, A.M., J. Mol. Biol. 196 (1987) 901-917). Примеры CDR (CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3) расположены в области аминокислотных остатков 24-34 L1, 50-56 L2, 89-97 L3, 31-35B H1, 50-65 H2 и 95-102 H3 (Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed. Public Health Service, National Institutes of Health, Бетесда, Мэриленд (1991), NIH Publication 91-3242). За исключением CDR1 VH, CDR обычно содержат аминокислотные остатки, которые формируют гипервариабельные петли. CDR также содержат «остатки, определяющие специфичность» или «SDR», которые контактируют с антигеном. SDR находятся внутри районов CDR, называемых укороченными CDR или a-CDR. Примеры a-CDR (a-CDR-L1, a-CDR-L2, a-CDR-L3, a-CDR-H1, a-CDR-H2 и a-CDR-H3) расположены в области аминокислотных остатков 31-34 L1, 50-55 L2, 89-96 L3, 31-35B H1, 50-58 H2 и 95-102 H3 (Almagro, J.C. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633). Если не указано иное, аминокислотные остатки HVR и другие остатки вариабельного домена (например, FR-остатки) нумеруются в настоящем документе согласно Kabat et al., см. выше.
Термин «внутренний сайт посадки рибосомы» или «IRES» описывает последовательность, которая функционально стимулирует инициацию трансляции независимо от 5'-конца данного гена и обеспечивает трансляцию двух цистронов (открытых рамок считывания) с одного транскрипта в животной клетке. IRES обеспечивает независимый сайт посадки рибосомы для трансляции открытой рамки считывания непосредственно после IRES (в настоящем документе «после» используется взаимозаменяемо с «по направлению к 3'-концу»). В отличие от бактериальной мРНК, которая может быть полицистронной, т.е. кодировать несколько раличных полипептидов, которые последовательно транслируются с мРНК, большинство мРНК животных клеток являются моноцистронными и кодируют получение только одного белка. С полицистронного транскрипта в эукариотической клетке трансляция инициируется с сайта инициации трансляции, расположенного ближе всего к 5'-концу молекулы, терминируется на первом стоп-кодоне, и данный транскрипт освобождается от рибосомы, что приводит к трансляции только первого полипептида, кодируемого данной мРНК. В эукариотической клетке полицистронный транскрипт, имеющий IRES, функционально связанный со второй или последующей открытой рамкой считывания данного транскрипта, позволяет последовательно транслировать данную открытую рамку считывания с получением двух или более полипептидов, кодируемых одним и тем же транскриптом. Применение IRES-элементов в конструировании векторов было описано ранее; см., например, Pelletier, J., et al., Nature 334 (1988) 320-325; Jang, S.K., et al., J. Virol. 63 (1989) 1651-1660; Davies, M.V., et al., J. Virol. 66 (1992) 1924-1932; Adam, M.A., et al., J. Virol. 65 (1991) 4985-4990; Morgan, R.A., et al., Nucl. Acids Res. 20 (1992) 1293-1299; Sugimoto, Y., et al., Biotechnology 12 (1994) 694-698; Ramesh, N., et al., Nucl. Acids Res. 24 (1996) 2697-2700; и Mosser, D.D., et al., BioTechniques 22 (1997) 150-152).
Термин «моноклональное антитело» в настоящем документе подразумевает антитело, полученное из популяции практически гомогенных антител, т. е. отдельные антитела, образующие данную популяцию, являются идентичными и/или связывают тот же эпитоп; исключение составляют возможные варианты антитела, например, содержащие мутации, которые являются природными или появляются в процессе получения препарата моноклональных антител, такие варианты, как правило, представлены в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно содержат разные антитела против разных детерминант (эпитопов), каждое моноклональное антитело в составе препарата моноклональных антител распознаёт одну детерминанту антигена. Таким образом, понятие «моноклональное» указывает тип антитела, как полученного из практически гомогенной популяции антител, и оно не может быть истолковано как требующее получения антитела каким-либо определённым способом. Например, моноклональные антитела для применения согласно настоящему изобретению могут быть получены с помощью различных методик, включая, но не ограничиваясь ими, гибридомную технологию, способы с применением рекомбинантной ДНК, способы фагового дисплея и способы применения трансгенных животных, имеющих все локусы человеческого иммуноглобулина или их часть; данные способы и другие примеры способов для получения моноклональных антител описаны в настоящем документе.
Термин «нуклеиновая кислота» в настоящем документе подразумевает полимерную молекулу, состоящую из отдельных нуклеотидов (также называемых азотистыми основаниями) а, c, g и t (или u в РНК), например, ДНК, РНК или их модификации. Данная полинуклеотидная молекула может быть природной полинуклеотидной молекулой или синтетической полинуклеотидной молекулой, или комбинацией одной или более природных молекул с одной или более синтетическими полинуклеотидными молекулами. Данное определение также охватывает природные полинуклеотидные молекулы, в которых один или более нуклеотид замещён (например, путём мутагенеза), делетирован или вставлен. Нуклеиновая кислота может быть или отдельной или встроенной в другую нуклеиновую кислоту, например, в экспрессионную кассету, плазмиду или в хромосому клетки-хозяина. Нуклеиновая кислота, как правило, характеризуется по её последовательности, состоящей из отдельных нуклеотидов.
Специалисту в данной области техники хорошо известны процедуры и способы для преобразования аминокислотной последовательности, например, полипептида, в соответствующую последовательность нуклеиновой кислоты, кодирующую данную аминокислотную последовательность. Следовательно, нуклеиновая кислота характеризуется своей последовательностью, состоящей из отдельных нуклеотидов и, сходным образом, по аминокислотной последовательности кодируемого ей полипептида.
Термин «нуклеиновая кислота» в настоящем документе также подразумевает природную нуклеиновую кислоту или частично или полностью не природную нуклеиновую кислоту, кодирующую полипептид, который может быть получен рекомбинантными методами. Данная нуклеиновая кислота может состоять из фрагментов ДНК, которые или являются выделенными, или являются синтезированными химическими методами. Данная нуклеиновая кислота может быть интегрирована в другую нуклеиновую кислоту, например, в экспрессионную плазмиду или в геном/хромосому эукариотической клетки-хозяина. Термин «плазмида» включает челночную и экспрессионную плазмиды. Как правило, плазмида также содержит элемент для размножения в прокариотических клетках, содержащий точку начала репликации (например, точка начала репликации ColE1) и маркер селекции (например, ген устойчивости к ампициллину или тетрациклину), соответственно, для репликации и селекции плазмиды в прокариотах.
Термин «функционально связанный» подразумевает совмещение двух или более компонентов, где данные компоненты, описываемые таким образом, находятся во взамодействии, которое позволяет им функционировать предназначенным образом. Например, промотор и/или энхансер функционально связаны с кодирующей последовательностью, если они действуют на неё, контролируя или модулируя транскрипцию данной последовательности. Как правило, но не обязательно, последовательности ДНК, которые «функционально связаны», являются смежными и, где необходимо соединить два района, кодирующих белок, такие как секреторный лидер и полипептид, последовательности ДНК являются смежными и находятся в одной рамке (считывания). Однако, хотя функционально связанный промотор, как правило, расположен до кодирующей последовательности, он не обязательно является смежным с ней. Энхансеры не обязательно должны быть смежными. Энхансер является функционально связанным с кодирующей последовательностью, если он усиливает транскрипцию кодирующей последовательности. Функционально связанные энхансеры могут быть расположены до, внутри или после кодирующих последовательностей и на значительном расстоянии от промотора. Сайт полиаденилирования функционально связан с кодирующей последовательностью, если он расположен на конце после кодирующей последовательности, так чтобы процесс транскрипции проходил через кодирующую последовательность до последовательности полиаденилирования. Стоп-кодон трансляции функционально связан с экзонной последовательностью нуклеиновой кислоты, если он расположен на конце после кодирующей последовательности (3'-конце), чтобы процесс трансляции проходил через кодирующую последовательность до стоп-кодона и терминировался на нём. Связывание выполняют рекомбинантными способами, известными в уровне техники, например, с помощью методик ПЦР и/или путём лигирования с использованием подходящих сайтов рестрикции. Если подходящих сайтов рестрикции не существует, используют синтетические олигонуклеотидные адаптеры или линкеры, в соответствии с общепринятой практикой.
Термин «полицистронная транскрипционная единица» означает транскрипционную единицу, в которой более одного структурного гена находится под контролем одного промотора.
Термин «сигнал полиаденилирования» (сигнал полиA) в данной заявке обозначает последовательность нуклеиновой кислоты, обладающую способностью индуцировать разрезание и полиаденилирование первичных транскриптов определённого сегмента последовательности нуклеиновой кислоты. 3'-нетранслируемая область, содержащая сигнал полиаденилирования, может быть выбрана из группы 3'-нетранслируемых областей, содержащих сигналы полиаденилирования, полученные из SV40, гена бычьего гормона роста (bGH), генов иммуноглобулинов и гена тимидинкиназы (tk, например, сигнал полиаденилирования тимидинкиназы простого герпеса).
Термин «промотор» подразумевает полинуклеотидную последовательность, контролирующую транскрипцию гена/структурного гена или последовательности нуклеиновой кислоты, с которой он функционально связан. Термин «промотор» включает сигнальные последовательности для связывания РНК-полимеразы и инициации транскрипции. Используемые промоторы являются функциональными в таком типе клетки-хозяина, в которой предусмотрена экспрессия выбранной последовательности. В уровне техники хорошо известно (и приведено в базах данных, таких как GenBank) большое количество промоторов, включая конститутивные, индуцибельные и репрессируемые промоторы из множества различных источников, и они являются доступными в виде клонированных полинуклеотидов (например, из депозитариев, таких как ATCC, а также из других коммерческих или индивидуальных источников).
Термин «промотор» включает нуклеотидную последовательность, которая направляет транскрипцию структурного гена. Обычно промотор расположен в 5'-некодирующей или нетранслируемой области гена, недалеко от сайта инициации транскрипции структурного гена. Элементы последовательности внутри промоторов, которые участвуют в инициации транскрипции, часто характеризуются как консенсусные нуклеотидные последовательности. Эти элементы промотора включают сайты связывания РНК-полимеразы, последовательности TATA, последовательности CAAT, специфичные для дифференцировки элементы (DSE; McGehee, R.E., et al., Mol. Endocrinol. 7 (1993) 551), элементы ответа на циклоАМФ (CRE), элементы ответа на сыворотку (SRE; Treisman, R., Seminars in Cancer Biol. 1 (1990) 47), элементы ответа на глюкокортикоды (GRE) и сайты связывания других транскипционных факторов, таких как CRE/ATF (O'Reilly, M.A., et al., J. Biol. Chem. 267 (1992) 19938), AP2 (Ye, J., et al., J. Biol. Chem. 269 (1994) 25728), SP1, белок, связывающий элемент ответа на цАМФ (CREB; Loeken, M.R., Gene Expr. 3 (1993) 253), и факторов, связывающих октамерный бокс (см. основные положения в Watson et al., (eds.), Molecular Biology of the Gene, 4th ed. (The Benjamin/Cummings Publishing Company, Inc. (1987)), и Lemaigre, F.P. and Rousseau, G.G., Biochem. J. 303 (1994) 1-14). Если промотор является индуцибельным, то уровень транскрипции возрастает в ответ на добавление индуцирующего агента. Напротив, если промотор является конститутивным, уровень транскрипции не регулируется индуцирующим агентом. Также известны репрессируемые промоторы. Например, промотор c-fos специфично активируется при связывании гормона роста со своим рецептором на клеточной поверхности. Экспрессия, регулируемая тетрациклином (tet), может быть получена с применением искусственных гибридных промоторов, которые состоят, например, из промотора CMV, за которым следуют два сайта Tet-оператора. Tet-репрессор связывается с двумя данными Tet-операторами и блокирует транскрипцию. После добавления индукторного тетрациклина Tet-репрессор высвобождается с сайтов Tet-оператора и транскрипция становится возможной (Gossen, M. and Bujard, H. PNAS 89 (1992) 5547-5551). Для обзора других индуцибельных промоторов, включая промоторы металлотионеина и теплового шока, см., например, Sambrook et al. (выше) и Gossen et al., Curr. Opin. Biotech. 5 (1994) 516-520. Среди эукариотических промоторов, являющихся сильными промоторами для высокоуровневой экспрессии, имеются ранний промотор SV40, главный поздний промотор аденовируса, промотор металлотионеина-I мыши, длинный концевой повтор вируса саркомы Рауса, промотор фактора элонгации 1α китайского хомячка (CHEF-1, см., например, патент США номер 5,888,809), EF-1α человека, убиквитина и немедленно-ранний промотор цитомегаловируса человека (CMV IE).
«Промотор» может подразумевать конститутивный или индуцибельный промотор. Энхансер (т. е. цис-действующий элемент ДНК, действие которого на промотор усиливает транскрипцию) может быть необходим совместно с промотором для увеличения уровня экспрессии относительно одиночного промотора, и он может быть включён в качестве элемента регуляции транскрипции. Часто полинуклеотидный сегмент, содержащий промотор, также содержит энхансер (например, CMV или SV40).
Термины «стабильно трансформированный», «стабильно трансфецированный» или «стабильный» в настоящем документе обозначают наследуемое и устойчивое внедрение экзогенной нуклеиновой кислоты в геном/хромосому клетки-хозяина. Стабильно трансформированную клетку получают после процесса селекции клеток в селективных условиях роста, т. е. в присутствии одного или более маркера селекции.
Термин «структурный ген» обозначает район гена без сигнальной последовательности, т. е. кодирующий район.
Термин «терминатор транскрипции» обозначает последовательность ДНК из 50-750 пар нуклеотидов в длину, которая даёт РНК-полимеразе сигнал терминации синтеза мРНК. Рекомендуется использовать очень эффективные (сильные) терминаторы на 3'-конце экспрессионной кассеты для предотвращения пропуска их РНК-полимеразой, особенно в случае применения сильных промоторов. Неэффективные терминаторы транскрипции могут приводить к образованию мРНК, подобной оперонам, которая может стать причиной нежелаемой экспрессии генов, например, кодируемых плазмидой.
В рамках настоящего изобретения трансфецированные клетки могут быть получены практически любым способом трансфекции, известным в уровне техники. Например, нуклеиновая кислота может быть введена в клетки с помощью электропорации или микроинъекции. Альтернативно, могут применяться реагенты для липофекции, такие как FuGENE 6 (Roche Diagnostics GmbH, Германия), X-tremeGENE (Roche Diagnostics GmbH, Германия) и LipofectAmine (Invitrogen Corp., США). Также, альтернативно, нуклеиновая кислота может быть введена в клетку с помощью подходящих систем вирусных векторов, основанных на ретровирусах, лентивирусах, аденовирусах или аденоассоциированных вирусах (Singer О., Proc. Natl. Acad. Sci. USA 101 (2004) 5313-5314).
Термин «транзиентная трансфекция», как он использован в настоящем документе, обозначает процесс, в котором нуклеиновая кислота, введённая в клетку, не внедряется в геном или в хромосомную ДНК данной клетки. Данная нуклеиновая кислота, фактически, сохраняется в виде внехромосомного элемента в клетке, например, эписомы. Процессы транскрипции нуклеиновой кислоты эписомы не нарушаются, и, например, синтезируется белок, кодируемый нуклеиновой кислотой эписомы. В результате транзиентной трансфекции получается «транзиентно трансфецированная» клетка.
Термин «вариабельный район» или «вариабельный домен» подразумевает домен тяжёлой или лёгкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжёлой и лёгкой цепей (VH и VL соответственно) нативного антитела, как правило, имеют сходные структуры, где каждый домен содержит четыре консервативных каркасных района (FR) и три гипервариабельных района (HVR) (см., например, Kindt, T.J., et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., Нью-Йорк (2007), страница 91). Отдельный домен VH или VL может быть достаточным для специфичного связывания антигена. Более того, антитела, связывающиеся с определённым антигеном, могут быть выделены с применением домена VH или VL из антитела, связывающегося с данным антигеном, для скрининга библиотеки комплементарных VL или VH доменов, соответственно (см., например, Portolano, S., et al., J. Immunol. 150 (1993) 880-887; Clarkson, T., et al., Nature 352 (1991) 624-628).
Термин «вектор» в настоящем документе подразумевает молекулу нуклеиновой кислоты, способную размножать другую нуклеиновую кислоту, с которой она соединена. Данный термин включает вектор как самореплицирующуюся структуру нуклеиновой кислоты, а также вектор, встроенный в геном клетки-хозяина, в которую он введён. Определённые вектора способны направлять экспрессию нуклеиновых кислот, с которыми они функционально связаны. Такие вектора обозначаются в настоящем документе как «экспрессионные вектора».
Антитело
Способы и композиции, предусмотренные в настоящем документе, предназначены для получения рекомбинантных моноклональных антител. Антитело может иметь различную структуру, такую как моноспецифичные антитела, мультиспецифичные антитела (например, биспецифические антитела), фрагменты антител, моновалентные антитела, мультивалентные антитела (например, бивалентные антитела), но не ограничиваясь ими.
В конкретных вариантах антитело является фрагментом антитела. Фрагменты антитела включают фрагменты Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv, и другие фрагменты, описанные ниже, но не ограничиваются ими. Для обзора определённых фрагментов антитела, см. Hudson, P.J., et al. Nat. Med. 9 (2003) 129-134. Для обзора фрагментов scFv, см., например, Pluckthun, A., In The Pharmacology of Monoclonal Antibodies, Vol. 113, Rosenburg и Moore (eds.), Springer-Verlag, Нью-Йорк, стр. 269-315; см. также WO 1993/16185; и патенты США номер 5,571,894 и 5,587,458. Описание фрагментов Fab и F(ab')2, содержащих аминокислотные остатки эпитопа, связывающего рецептор реутилизации, и имеющих увеличенное время полужизни in vivo, см. патент США номер 5,869,046.
Димерные антитела являются фрагментами антитела с двумя антиген-связывающими участками, которые могут быть бивалентными или биспецифическими (см., например, EP 0404097; WO 1993/01161; Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134; и Holliger, P., et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448). Тримерные и тетрамерные антитела также описаны в Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134).
Однодоменные антитела являются фрагментами антитела, содержащими весь вариабельный домен тяжёлой цепи антитела или его часть, или весь вариабельный домен лёгкой цепи антитела или его часть. В конкртных вариантах однодоменное антитело является человеческим однодоменным антителом (Domantis, Inc., Уолтем, Массачусетс; см, например, патент США номер 6,248,516).
Фрагменты антител могут быть созданы с помощью различных методик, включая протеолитическое расщепление интактного антитела, а также получение с помощью рекомбинантных клеток-хозяев (например, E. coli или фага), как описано в настоящем документе, но не ограничиваясь этим.
В конкретных вариантах антитело является химерным антителом. Определённые химерные антитела описаны, например, в патенте США номер 4,816,567; и в Morrison, S.L., et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855. В одном примере химерное антитело содержит не человеческий вариабельный район (например, вариабельный район, полученный из мыши, крысы, хомячка, кролика или примата, не являющегося человеком, такого как мартышка) и человеческий константный район. В другом примере класс химерного антитела является «переключённым», класс или подкласс такого антитела отличается от класса или подкласса родительского антитела. Термин химерные антитела включает их антиген-связывающие фрагменты.
В конкретных вариантах химерное антитело является гуманизированным антителом. Обычно не человеческие антитела гуманизируют для снижения иммуногенности для людей, сохраняя при этом специфичность и аффинность исходного не гуманизированного антитела. Как правило, гуманизированное антитело содержит один или более вариабельный домен, в котором районы HVR, например, CDR (или их части) получены из не человеческого антитела, и районы FR (или их части) получены из последовательностей человеческих антител. Гуманизированное антитело, необязательно, также содержит, по меньшей мере, часть человеческого константного района. В некоторых вариантах некоторые аминокислотные остатки FR гуманизированного антитела замещены соответствующими остатками не человеческого антитела (например, антитела, из которого получены амнокислотные остатки HVR), например, для восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения рассмотрены, например, в Almagro, J.C. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633, и более подробно описаны, например, в Riechmann, I., et al., Nature 332 (1988) 323-329; Queen, C., et al., Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033; в патентах США номер 5,821,337, 7,527,791, 6,982,321 и 7,087,409; Kashmiri, S.V., et al., Methods 36 (2005) 25-34 (описание переноса SDR (a-CDR)); Padlan, E.A., Mol. Immunol. 28 (1991) 489-498 (описание «изменения поверхности»); Dall'Acqua, W.F., et al., Methods 36 (2005) 43-60 (описание «перестановки FR»); и Osbourn, J., et al., Methods 36 (2005) 61-68 и Klimka, A., et al., Br. J. Cancer 83 (2000) 252-260 (описание подхода «управляемой селекции» в отношении перестановки FR).
Человеческие каркасные районы, которые могут быть использованы для гуманизации, включают каркасные районы, выбранные с применением способа «наилучшей пригодности» (см., например, Sims, M.J., et al., J. Immunol. 151 (1993) 2296-2308); каркасные районы, полученные из консенсусных последовательностей человеческих антител, имеющих определённую подгруппу вариабельный районов тяжёлой или лёгкой цепи (см., например, see, e.g., Carter, P., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289; и Presta, L.G., et al., J. Immunol. 151 (1993) 2623-2632); созревшие (соматически изменённые) человеческие каркасные районы или человеческие каркасные районы зародышевой линии (см., например, Almagro, J.C. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633); и каркасные районы, полученные путём скрининга библиотек FR (см., например, Baca, M., et al., J. Biol. Chem. 272 (1997) 10678-10684 и Rosok, M.J., et al., J. Biol. Chem. 271 (19969 22611-22618), но не ограничиваются ими.
В конкретных вариантах антитело является человеческим. Человеческие анитела могут быть созданы с применением различных методик, известных в уровне техники. Человеческие антитела в общем описаны в van Dijk, M.A. and van de Winkel, J.G., Curr. Opin. Pharmacol. 5 (2001) 368-374 и Lonberg, N., Curr. Opin. Immunol. 20 (2008) 450-459.
Человеческие антитела могут быть получены путём введения иммуногена трансгенному животному, модифицированному для получения интактных человеческих антител или интактных антител с человеческими вариабельными районами в ответ на введение антигена. Такие животные обычно имеют все локусы человеческого иммуноглобулина или их часть, которые замещают локусы эндогенного иммуноглобулина или которые находятся вне хромосом, или встроены в хромосомы данного животного случайным образом. В таких трансгенных мышах локусы эндогенного иммуноглобулина, как правило, инактивированы. Для обзора способов получения человеческих антител из трансгенных животных, см. Lonberg, N., Nat. Biotech. 23 (2005) 1117-1125, а также, например, патенты США номер 6,075,181 и 6,150,584, где описана технология XENOMOUSE™; патент США номер 5,770,429, где описана технология HuMab®, патент США номер 7,041,870, где описана технология K-M MOUSE®, и US 2007/0061900, где описана технология VelociMouse® . Человеческие вариабельные районы инактных антител, произведённых такими животными, могут быть затем модифицированы, например, путём комбинирования с другим человеческим константным районом.
Человеческие антитела также могут быть созданы способами, основанными на гибридоме. Для получения человеческих моноклональных антител описаны человеческие миеломные клеточные линии и гетеромиеломные клеточные линии человека-мыши (см., например, Kozbor, D., J. Immunol. 133 (1984) 3001-3005; Brodeur, B.R., et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York (1987), pp. 51-63; и Boerner, P., et al., J. Immunol. 147 (1991) 86-95). Человеческие антитела, созданные с применением технологии гибридомы B-клеток человека, также описаны в et al., Proc. Natl. Li, J., Acad. Sci. USA 103 (2006) 3557-3562. Дополнительные способы влючают способы, описанные, например, в патенте США номер 7,189,826 (где описано получение моноклональных человеческих антител IgM из гибридомных клеточных линий) и в Ni, J., Xiandai Mianyixue 26 (2006) 265-268 (где описаны человек-человеческие гибридомы). Технология человеческой гибридомы (технология триомы) также описана в Vollmers, H.P. and Brandlein, S., Histology and Histopathology 20 (2005) 927-937 и Vollmers, H.P. and Brandlein, S., Methods and Findings in Experimental and Clinical Pharmacology 27 (2005) 185-191.
Человеческие антитела также могут быть получены путём выделения последовательностей клона вариабельного домена Fv, выбранных из библиотек фагового дисплея, полученных из человека. Такие последовательности вариабельного домена затем могут быть скомбинированы с желаемым человеческим константным доменом. Методики выбора человеческих антител из библиотек антител описаны ниже.
Антитела могут быть выделены с помощью скрининга комбинаторных библиотек для поиска антител с желаемой активностью или активностями. Например, в уровне техники известно множество способов получения библиотек фагового дисплея и скрининга таких библиотек на наличие антител, обладающих желаемыми характеристиками связывания. Такие способы рассмотрены, например, в Hoogenboom, H.R., et al., Methods in Molecular Biology 178 (2001) 1-37 и более подробно описаны, например, в McCafferty, J., et al., Nature 348 (1990) 552-554; Clackson, T., et al., Nature 352 (1991) 624-628; Marks, J.D., et al., J. Mol. Biol. 222 (1992) 581-597; Marks, J.D. and Bradbury, A., Methods in Molecular Biology 248 (2003) 161-175; Sidhu, S.S., et al., J. Mol. Biol. 338 (2004) 299-310; Lee, C.V., et al., J. Mol. Biol. 340 (2004) 1073-1093; Fellouse, F.A., Proc. Natl. Acad. Sci. USA 101 (2004) 12467-12472; и Lee, C.V., et al., J. Immunol. Methods 284 (2004) 119-132.
В некоторых способах фагового дисплея наборы генов VH и VL раздельно клонируют с помощью полимеразной цепной реакции (ПЦР) и случайным образом рекомбинируют в фаговые библиотеки, которые затем могут быть скринированы на наличие антиген-связывающего фага, как описано в Winter, G., et al., Ann. Rev. Immunol. 12 (1994) 433-455. Фаг, как правило, несет фрагменты антитела, такие как одноцепочечные фрагменты Fv (scFv) или фрагменты Fab. Библиотеки из иммунизированных источников обеспечивают высокоаффинные к иммуногену антитела без необходимости конструирования гибридом. Альтернативно, для получения единого источника антител к широкому спектру чужеродных антигенов, а также аутоантигенов может быть клонирован наивный репертуар (например, человеческий) без какой-либо иммунизации, как описано в Griffiths, A.D., et al., EMBO J. 12 (1993) 725-734. Наконец, наивные библиотеки также могут быть созданы искусственно путём клонирования неперестроенных сегментов V-гена стволовых клеток и применения праймеров для ПЦР, содержащих случайную последовательность для кодирования высоковариабельных районов CDR3, с последующей перестройкой in vitro, как описано в Hoogenboom, H.R. and Winter, G., J. Mol. Biol. 227 (1992) 381-388. Патентная литература, описывающая фаговые библиотеки человеческих антител, включает, например, патент США номер 5,750,373 и US 2005/0079574, US 2005/0119455, US 2005/0266000, US 2007/0117126, US 2007/0160598, US 2007/0237764, US 2007/0292936 и US 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, в настоящем документе рассматриваются как человеческие антитела или фрагменты человеческих антител.
В конкретных вариантах антитело является мультиспецифичным, например, биспецифическим антителом. Мультиспецифичные антитела являются моноклональными антителами, имеющими специфичность связывания, по меньшей мере, к двум различным сайтам. В конкретных вариантах одна специфичность связывания относится к первому антигену, а другая специфичность связывания относится к отличному от него другому антигену. В конкретных вариантах биспецифические антитела могут связываться с двумя различными эпитопами одного антигена. Биспецифические антитела также могут быть использованы для локализации цитотоксических агентов в клетках, экспрессирующих данный антиген. Биспецифические антитела могут быть получены как полноразмерные антитела или как фрагменты антител.
Методики получения мультиспецифичных антител включают, но не ограничиваются ими, рекомбинантную коэкспрессию двух пар лёгкой цепи и тяжёлой цепи иммуноглобулина, имеющих разные специфичности (см. Milstein, C. and Cuello, A.C., Nature 305 (1983) 537-540, WO 93/08829 и Traunecker, A., et al., EMBO J. 10 (1991) 3655-3659), и конструирование по типу «выступ-во-впадину» (см., например, патент США номер 5,731,168). Мультиспецифичные антитела также могут быть созданы путём получения Fc-гетеродимерных молекул антитела за счёт электростатических эффектов (WO 2009/089004); перекрёстного сшивания двух или более антител или их фрагментов (см., например, патент США номер 4,676,980 и Brennan, M., et al., Science 229 (1985) 81-83); с применением лейциновых застёжек для получения биспецифических антител (см., например, Kostelny, S.A., et al., J. Immunol. 148 (1992) 1547-1553; с применением технологии «диател» для получения фрагментов биспецифических антител (см., например, Holliger, P., et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448); с применением димеров одноцепочечных Fv (scFv) фрагментов (см., например, Gruber, M., et al., J. Immunol. 152 (1994) 5368-5374); и путём получения триспецифичных антител, как описано, например, в Tutt, A., et al., J. Immunol. 147 (1991) 60-69).
Сконструированные антитела с тремя или более функциональными антиген-связывающими участками, включая «Octopus-антитела», также включены в настоящий документ (см., например, US 2006/0025576).
Антитело может быть представлено в виде «Fab двойного действия» или «DAF», содержащего антиген-связывающий участок, который связывается с первым антигеном, а также с другим, отличным от него антигеном (см., например, US 2008/0069820).
Антитело или его фрагмент также могут быть мультиспецифичным антителом, как описано в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, WO 2010/112193, WO 2010/115589, WO 2010/136172, WO 2010/145792 или в WO 2010/145793.
Способы
В конкретных вариантах способы, предусмотренные в настоящем документе, использованы для изменения, т. е. для увеличения или уменьшения гликозилирования антитела.
В случае, если данное антитело содержит Fc-район, углевод, присоеднённый к нему, может быть изменён. Нативные антитела, синтезируемые клетками млекопитающих, обычно содержат разветвлённый биантенный олигосахарид, который, как правило, соединён посредством N-связи с Asn297 домена CH2 Fc-района. (см., например, Wright, A. and Morrison, S.L., TIBTECH 15 (1997) 26-32). Данный олигосахарид может включать различные углеводные компоненты, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, соединённую с GlcNAc в «стержне» биантенной олигосахаридной структуры. В некоторых вариантах могут быть произведены модификации олигосахарида в составе антитела настоящего изобретения для получения вариантов антитела с определёнными улучешенными свойствами.
В одном варианте предусмотренный способ позволяет получать антитела, имеющие углеводную структуру, которая не содержит фукозы, соединённой (напрямую или не напрямую) с Fc-районом. Например, количество фукозы в таком антителе может быть в диапазоне от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяют путём вычисления среднего количества фукозы в составе углеводной цепи Asn297 относительно суммы всех углеводных структур, соединённых с Asn297 (например, комплексных структур, гибридных структур и структур с высоким содержанием маннозы), измеренного с помощью масс-спектрометрии MALDI-TOF, как описано, например, в WO 2008/077546. Asn297 относится к остатку аспарагина, расположенному примерно в позиции 297 Fc-района (Европейская нумерация остатков Fc-района согласно Kabat); однако Asn297 также может быть локализован примерно в ± 3 аминокислотах до или после позиции 297, т. е. между позициями 294 и 300, вследствие незначительных отличий последовательностей антител. Такие варианты с измененным фукозилированием могут иметь улучшенную функцию ADCC. (cм., например, US 2003/0157108; US 2004/0093621). Примеры публикаций, имеющих отношение к «дефукозилированным» или «дефицитным по фукозе» вариантам антител, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; WO2002/031140; Okazaki, A., et al., J. Mol. Biol. 336 (2004) 1239-1249; Yamane-Ohnuki, N., et al., Biotech. Bioeng. 87 (2004) 614-622. Примеры клеточных линий, способных синтезировать дефукозилированные антитела, включают клетки Lec13 CHO, неспособные к фукозилированию белков (Ripka, J., et al., Arch. Biochem. Biophys. 249 (1986) 533-545; US 2003/0157108; и WO 2004/056312, главным образом, в примере 11), и нокаутные клеточные линии, такие как нокаутные по гену α -1,6-фукозилтрансферазы FUT8 клетки CHO (см, например, Yamane-Ohnuki, N., et al., Biotech. Bioeng. 87 (2004) 614-622; Kanda, Y., et al., Biotechnol. Bioeng. 94 (2006) 680-688; и WO 2003/085107).
В конкретных вариантах предусмотренные способы могут быть использованы для получения антител с олигосахаридами, разветвлёнными на две части, например, в которых биантенный олигосахарид, прикреплённый к Fc-району антитела, является разветвлённым на две части за счёт GlcNAc. Такие варианты антител могут иметь пониженное фукозилирование и/или улучшенную функцию ADCC. Примеры таких вариантов антитела описаны, например, в WO 2003/011878; в патенте США номер 6,602,684; и в US 2005/0123546. Также могут быть созданы варианты антител, по меньшей мере, с одним остатком галактозы в составе олигосахарида, соединённого с Fc-районом. Такие варианты антитела могут иметь улучшенную функцию CDC. Такие варианты антител описаны, например, в WO 1997/30087; WO 1998/58964; и WO 1999/22764.
Антитела могут быть созданы с применением рекомбинантных способов и композиций, например, как описано в патенте США номер 4,816,567. Нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, лёгкую и/или тяжёлую цепи антитела). В другом варианте предусмотрен один или более вектор (например, экспрессионный вектор), содержащий такую нуклеиновую кислоту. В другом варианте предусмотрена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном таком варианте клетка-хозяин включает в себя (1) вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL данного антитела, и аминокислотную последовательность, содержащую VH данного антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL данного антитела, и второй вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VH данного антитела (например, клетка-хозяин была трансформирована указанными векторами). В одном варианте клетка-хозяин является эукариотической клеткой, например, клеткой яичника китайского хомячка или лимфоидной клеткой (например, Y0, NS0, Sp2/0). В одном варианте предусмотрен способ получения антитела, включающий культивирование клеток-хозяев, содержащих нуклеиновую кислоту, кодирующую данное антитело, как предусмотрено выше, в подходящих условиях для экспрессии данного антитела, и, необязательно, выделение данного антитела из клетки-хозяина (или культуральной среды клеток-хозяев).
Для рекомбинантного получения антитела нуклеиновую кислоту, кодирующую антитело, выделяют и вставляют в один или более вектор для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такая нуклеиновая кислота может быть легко выделена и секвенирована с применением общепринятых процедур (например, путём применения олигонуклеотидных зондов, способных специфично связываться с генами, кодирующими тяжёлую и лёгкую цепи данного антитела).
Подходящие клетки-хозяева для клонирования и экспрессии векторов, кодирующих антитело, включают прокариотические или эукариотические клетки, описанные в настоящем документе. Например, антитела могут быть получены в бактериях, в частности, когда гликозилирование и эффекторная функция Fc-района не являются необходимыми. Для обзора способов экспрессии фрагментов антитела и полипептидов в бактериях, см., например, патент США номер 5,648,237, 5,789,199 и 5,840,523; см. также Charlton, K.A., In: Methods in Molecular Biology, Vol. 248, Lo, B.K.C. (ed.), Humana Press, Тотова, Нью-Джерси, 2003), стр. 245-254, где описана экспрессия фрагментов антитела в E. coli). После экспрессии антитело может быть выделено из массы бактериальных клеток в растворимую фракцию и может быть далее очищено.
В дополнение к прокариотам подходящими экспрессионными хозяевами для векторов, кодирующих антитело, являются эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, включая штаммы грибов и дрожжей с «гуманизированными» каскадами гликозилирования, синтезирующие антитело с частично или полностью человеческой структурой гликозилирования. (см., Gerngross, T.U., Nat. Biotech. 22 (2004) 1409-1414; и Li, H., et al., Nat. Biotech. 24 (2006) 210-215).
Подходящие клетки для экспрессии гликозилированного антитела также могут быть получены из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Описано множество штаммов бакуловируса, которые могут быть использованы совместно с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
Клеточные культуры растений также могут быть использованы в качестве хозяев (см., например, патент США номер 5,959,177, 6,040,498, 6,420,548, 7,125,978 и 6,417,429 (с описанием технологии PLANTIBODIESTM для получения антител в трансгенных растениях)).
Клетки позвоночных также могут быть использованы в качестве хозяев. Например, могут быть применены клеточные линии млекопитающих, адаптированные к росту в суспензии. Другими примерами применимых линий клеток-хозяев млекопитающих являются клеточная линия почки обезьяны CV1, трансформированная SV40 (COS-7); клеточная линия почки эмбриона человека (293 или клетки293, как описано, например, в Graham, F.L., et al., J. Gen Virol. 36 (1977) 59-74); клетки почки детёныша хомячка (BHK); клетки Сертоли мыши (клетки TM4, как описано, например, в Mather, J.P., Biol. Reprod. 23 (1980) 243-252); клетки почки обезьяны (CV1); клетки почки африканской зелёной мартышки (VERO-76); клетки карциномы шейки матки человека (HeLa); клетки почки собаки (MDCK); клетки печени крысы линии Буффало (BRL 3A); клетки лёгкого человека (W138); клетки печени человека (Hep G2); опухолевые клетки молочной железы мыши (MMT 060562); клетки TRI, как описано, например, в Mather, J.P., et al., Annals N.Y. Acad. Sci. 383 (1982) 44-68; клетки MRC 5; и клетки FS4. Другими применимыми линиями клеток-хозяев млекопитающих являются клетки яичника китайского хомячка (CHO), включая DHFR отрицательные клетки (DHFR-) CHO (Urlaub, G., et al., Proc. Natl. Acad. Sci. USA 77 (1980) 4216-4220); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Для обзора конкретных линий клеток-хозяев млекопитающих, применимых для получения антител, см., например, Yazaki, P. and Wu, A.M., Methods in Molecular Biology, Vol. 248, Lo, B.K.C. (ed.), Humana Press, Тотова, Нью-Джерси), стр. 255-268.
II. Специфичные аспекты настоящего изобретения
Обнаружено, что в зависимости от строения вектора производительность экспрессионного вектора различается в зависимости от конструкции вектора в стабильных трансфекциях.
Независимо от теоретических представлений было обнаружено, что на производительность вектора для стабильных трансфекций могут влиять несколько аспектов строения вектора: 1) феномен взаимного влияния на транскрипцию между копиями вектора, интегрированными в геном хозяина, который зависит от конструкции соответствующего вектора и является специфичным для неё, и где данный феномен не существует в транзиентных системах, 2) влияние процесса селекции и жёсткости селекции, которые зависят от строения соответствующего вектора и которые играют важную роль в стабильной системе, и 3) оптимальное соотношение полипептидов LC и HC.
Однако обнаружено, что для стабильных трансфекций двунаправленная экспрессия LC и HC давала худшие результаты по сравнению со строением в порядке HC-LC-SM. Не желая быть связанными теорией, авторы предполагют, что 1) конвергентное строение экспрессионных кассет для LC, HC и SM может уменьшать взаимное влияние на транскрипцию между интегрированными копиями вектора, 2) расположение HC до LC, очевидно, обеспечивает соотношение полипептидов LC и HC, которое является (более) оптимальным для стабильных трансфекций, и 3) расположение маркера селекции после других упомянутых элементов, очевидно, увеличивает жёсткость селекции. Более того, обнаружено увеличение процентного показателя клеток, продуцирующих IgG, и продуктивности клеточных линий. Для векторов, содержащих маркер селекции, расположенный до экспрессионных кассет антитела, увеличение селекционного давления не увеличивало продуктивность стабильных пулов или клонов, хотя жёсткость селекции, очевидно, возрастала (данные не представлены).
Обнаружено, что промотор hEF1α даёт большое количество высокопродуктивных клонов и очень малое количество низкопродуктивных или непродуктивных клонов. Однако в анализе на подпитываемых культурах титры продукта для лучших отдельных клонов с промотором hEF1α были ниже, чем для клонов с промотором hCMV. Но общее количество лучших клонов с промотором hCMV сравнительно ниже и их идентификация обычно требует больших усилий при скрининге.
Обнаружено, что применение hGT значительно увеличивает продуктивность для векторов, содержащих промотор hCMV, при комбинировании с сигнальной последовательностью полиA SV40 или bGH в стабильных трансфекциях. Однако для векторов, содержащих промотор hEF1α , эффект hGT на титр продукта был незначительным при комбинировании с сигнальной последовательностью полиA bGH. Таким образом, обнаружено, что влияние hGT на производительность вектора зависит от используемого промотора.
Выбор подходящих клонов для конечной оценки в полностью контролируемом широкомасштабном культивировании, как правило, основан на анализе периодической культуры или на анализе подпитываемой культуры в перемешиваемых колбах. Обнаружено, что существуют различия в продуктивности и ранжировании некоторых экспрессионных векторов или элементов при анализе периодической культуры и анализе подпитываемой культуры. Замещение сигнала полиA SV40 на сигнал полиA bGH увеличивает продуктивность векторов, содержащих промотор hCMV, в анализе периодической культуры, но не в анализе подпитываемой культуры. hGT не имееет существенного влияния на титр продукта клонов в анализе периодической культуры, но имеет влияние в анализе подпитываемой культуры. Абсолютные значения различий производительности векторов между периодической культурой и подпитываемой культурой варьировали. Различия между векторами, отличающимися по позиции маркера селекции или по позиции промотора (hEF1α или hCMV), были умеренно выражены в анализе периодических культур и были очевидны в анализе подпитываемой культуры. Уровни экспрессии и скорости получения продукта были выше при использовании подпитываемых культур, чем при использовании периодических культур.
Для большинства клонов была обнаружена хорошая корреляция при выполнении анализов на подпитываемых культурах и при анализе культивирования в объеме 2 л, причем не только для абсолютного титра продукта, но также для ранжирования различных векторов и клонов.
Обнаружено, что 3'-расположение маркера селекции немного снижает потерю продуктивности по сравнению с двунаправленным расположением маркера селекции до экспрессионных кассет антитела. Не желая быть связанными теорией, авторы предполагают, что это может быть связано с повышением жёсткости селекции и, таким образом, большим уровнем мРНК, или с улучшенным соотношением LC к HC на уровне мРНК или полипептида. Оба фактора могут приводить к более высокой устойчивости к изменениям продуктивности.
Сигнал полиA bGH существенно снижает стабильность экспрессии антитела в клонах по сравнению с сигналом полиA SV40. Однако вставка hGT после сигнала полиA bGH явно повышает стабильность экспрессии. Положительный эффект hGT на стабильность проявлялся наибольшим образом при отстутствии селекционного давления. Анализ стабильности в стабильных пулах выявил, что клетки быстро теряли продуктивность, когда их получали с промотором hCMV, но не с промотором hEF1α. К удивлению, хотя hGT снижал продуктивность клонов для вектора, содержащего промотор hEF1α , он немного увеличивал ее стабильность.
Только небольшая часть клонов в существенном количестве продуцирует антитело после процесса селекции. Однако модификации строения вектора и/или его элементов значительно увеличили отношение клонов, продуцирующих IgG, к непродуцирующим клонам. Различные строения вектора и, таким образом, различные уровни экспрессии маркера селекции, определяющего жёсткость селекции, также определённо влияют на процентный показатель клеток, продуцирующих IgG. Статистические моделирования, основанные на данных скрининга, продемонстрировали, что некоторые экспрессионные вектора также имеют потенциал для значительного снижения трудоёмкости процесса скрининга. Этот факт имеет главное влияние на стоимость для биофармацевтичечких компаний.
Строение экспрессионной кассеты
Четыре различных вектора с различным расположением лёгкой и тяжёлой цепи антитела и/или маркера селекции были проанализированы на продуктивность (детали см. на Фиг. 1).
Вектора p5137, p5156, p5158 и p5159 были проанализированы в транзиентных трансфекциях и в анализе периодических культур стабильных пулов. Вектора p5137 и p5156 также были проанализированы в анализе периодических культур отдельных стабильных клонов.
Вектора p5137, p5156, p5158 и p5159 транзиентно трансфецировали в клетки CHO-K1 и определяли продуктивность с помощью ИФА на 5 день после трансфекции (Фиг. 2).
Обнаружено, что для транзиентной трансфекции расположение тяжёлой цепи антитела перед лёгкой цепью антитела обеспечивает лучшие результаты экспрессии по сравнению с расположением в обратном порядке. Таким образом, может быть достигнута большая продуктивность: вектор p5137 - 11,6 мкг/мл; вектор p5156 - 7,1 мкг/мл; вектор p5159 - 8,0 мкг/мл; вектор p5158 - 4,2 мкг/мл.
Обнаружено, что расположение маркера селекции после обеих цепей антитела обеспечивает лучшие результаты экспрессии по сравнению с двунаправленным расположением перед первой цепью антитела. Таким образом, может быть достигнута большая продуктивность.
Вектора p5137, p5156, p5158 и p5159 трансфецировали в клетки CHO-K1 путём нуклеофекции и отбирали стабильные пулы. Продуктивность стабильных пулов определяли с помощью анализа периодических культур.
Обнаружено, что расположение маркера селекции после обеих цепей антитела (в 5'-3' направлении) обеспечивает лучшие результаты экспрессии по сравнению с его двунаправленным расположением (в 3'-5' направлении) перед первой цепью антитела. Таким образом, может быть достигнута большая продуктивность: вектор p5156 - 18,0 мкг/мл; вектор p5158 - 9,1 мкг/мл; вектор p5137 - 15,6 мкг/мл; вектор p5159 - 5,7 мкг/мл (Фиг. 3).
Вектора p5137 и p5156 трансфецировали в клетки CHO-K1 путём нуклеофекции. Отбирали стабильные трансфецированные клетки и анализировали продуктивность лучших 15 клонов с помощью анализа периодических культур.
Средняя продуктивность лучших 15 клонов, созданных с помощью вектора p5137 и вектора p5156, составила 159 мкг/мл и 141 мкг/мл соответственно. Разброс продуктивности среди лучших 15 клонов был сходным для каждого вектора в анализе периодических культур. Продуктивности клонов в анализе периодических культур варьируют примерно от 50 мкг/мл до 300 мкг/мл (одно исключение: самый лучший клон, созданный с помощью вектора p5137, достигал продуктивности >450 мкг/мл).
Расположение маркера селекции после обеих цепей антитела (в 5'-3' направлении) приводит к значительно более высоким продуктивностям, чем расположение в двунаправленное 3'-5'-расположение данного маркера селекции перед первой цепью антитела, как в транзиентных трансфекциях, так и в анализе периодических культур стабильных пулов.
Обнаружено, что в транзиентных трансфекциях расположение тяжёлой цепи антитела перед лёгкой цепью антитела (обе в 5'-3' направлении) обеспечивает лучшую экспрессию по сравнению с расположением в обратном порядке.
Обнаружено, что расположение/порядок лёгкой и тяжёлой цепей антитела не имеет значительного влияния на продуктивность стабильных пулов и отдельных клонов. Необходимо обеспечить небольшой избыток экспрессии лёгкой цепи. Таким образом, последовательности экспрессионных кассет цепей антитела расположены таким образом, чтобы данное условие было выполнено.
Обнаружено, что расположение экспрессионной кассеты цепи антитела (в любом порядке) перед экспрессионной кассетой маркера селекции (экспрессионная кассета маркера селекции следует за экспрессионной кассетой цепи антитела) (все в 5'-3' направлении) является особенно подходящим.
Если два гена экспрессируются непосредственно один за другим, второй ген, как правило, экспрессируется в меньшей степени. Прохождение РНК-полимеразы через вторую транскрипционную единицу отрицательно влияет на инициацию транскрипции с промотора второй транскрипционной единицы.
В векторе px6068 экспрессионные кассеты лёгкой и тяжёлой цепи анитела расположены двунаправленно (Фиг. 4).
Для предотвращения конкурирования или взаимного влияния промоторов (неэффективной инициации транскрипции вследствие тесной близости двух промоторов, т. е. стерических затруднений для доступа к промотору транскрипционных факторов и РНК-полимеразы, сниженного доступа ресурсов транскрипционной машинерии), промоторы, управляющие экспрессией тяжёлой и лёгкой цепей антитела, были пространственно разделены (точкой начала репликации puc).
Вектора p5068 и px6068 были транзиентно трансфецированы в клетки CHO-K1 путём нуклеофекции.
Вектор px6068 продемонстрировал повышенную продуктивность в транзиентных трансфекциях по сравнению с экспрессионным вектором p5068: 9,0 мкг/мл для вектора px6068, 4,5 мкг/мл для вектора p5068 (Фиг. 5).
Обаружено, что в транзиентной трансфекции двунаправленное и разобщённое расположение лёгкой и тяжёлой цепи антитела обеспечивает улучшенную экспрессию антитела по сравнению с расположением лёгкой и тяжёлой цепи типа «спина к спине».
Вектора p5068 и px6068 трансфецировали в клетки CHO-K1 путём нуклеофекции и отбирали стабильные пулы. Продуктивность стабильных пулов определяли с помощью анализа периодических культур.
Анализ периодических культур стабильных пулов показал, что продуктивность векторов p5068 и px6068 является сходной в стабильных пулах: 12,5 мкг/мл для вектора p5068; 12,5 мкг/мл для вектора px6068.
Протокол трансфекции
Для стабильных трансфекций вектора линеаризовали путём рестрикции ферментами, разрезающими остов экспрессионного вектора. В случае вектора px6068 приемлемыми являются два сайта рестрикции:
1. между транскрипционными единицами лёгкой цепи и маркера селекции (SgrAI);
2 в точке начала репликации puc между промоторами hCMV лёгкой и тяжёлой цепи (BssHII).
Вектор px6068, линеаризованный с помощью рестрикции SgrAI или BssHII, трансфецировали в клетки CHO-K1 путём нуклеофекции и отбирали стабильные пулы. Продуктивность стабильных пулов определяли посредством анализа периодических культур.
Обнаружено, что расположение сайта линеаризации в экспрессионном векторе определённо имеет влияние на продуктивность векторов в анализе периодических культур стабильных пулов.
Пулы, созданные с применением вектора px6068, линеаризованного рестриктазой BssHII, демонстрируют более высокую продуктивность по сравнению со стабильными пулами, созданными с применением вектора px6068, линеаризованного рестриктазой SgrAI: первый эксперимент: 6,4 мкг/мл для вектора px6068, линеаризованного рестриктазой BssHII, и 2,8 мкг/мл для вектора px6068, линеаризованного рестриктазой SgrAI; второй экспреимент: 12,5 мкг/мл для вектора px6068, линеаризованного рестриктазой BssHII, и 9,6 мкг/мл для вектора px6068, линеаризованного рестриктазой SgrAI (см. Фиг. 6).
Момент начала селекционного давления
Селекционное давление для получения стабильных клеточных клонов начинали в разные моменты времени после трансфекции, т. е. через 0 часов, 4 часа, 8 часов, 24 часа и 48 часов.
Обнаружено, что начало селекционного давления через 24 часа после трансфекции независимо от концентрации агента для селекции приводит к наиболее высокому титру антитела в супернатанте культуральной среды (см. Фиг. 7).
Влияние остова вектора на производительность клеточных линий
Как правило, экспрессионный вектор линеаризуют перед трансфекцией в эукариотическую клетку. Кроме того, экспрессионные вектора содержат прокариотические последовательности, необходимые для амплификации экспрессионного вектора в прокариотических клетках.
Обнаружено, что удаление прокариотических элементов из линеаризованного экспрессионного вектора перед трансфекцией в эукариотические клетки приводит к:
- сокращению необходимого времени селекции для получения стабильных клеточных клонов (Фиг. 8a),
- повышению продуктивности пулов (Фиг. 8b) и отдельных клонов (Фиг. 8c),
- ускорению восстановления числа клеток (Фиг. 8d), и
- повышению скорости роста клеток (Фиг. 8e).
Векторные элементы и ориентация экспрессионных кассет
Несколько различных значимых для транскрипции генетических элементов и их комбинаций сравнили с комбинацией генетических элементов сравнения. На основе сравнительных транзиентных экспериментов были получены следующие результаты (см. таблицу ниже, двунаправленное строение вектора, SM (3'-5')-LC-HC (5'-3')).
(+ 31 %)
+ 37 % в стабильных пулах; + 23 % в отдельных клонах
(+ 46 %)
(+ 58 %)
(+ 134 %)
усиление экспрессии (+ 23 %)
в отдельных клонах:
ослабление экспрессии
(- 62 % для лучших 15 клонов)
(- 50 %)
(- 40 %)
(+ 53 %)
усиление экспрессии
(+ 460 %)
в клонах: -16 % для лучших 15 клонов (в периодической культуре)
Различные промоторы были скомбинированы с сигналом полиA bGH и терминатором транскрипции hGT (см. таблицу ниже).
сигнал полиA SV40
с интроном A
сигнал полиA bGH
терминатор транскрипции hGT
(+ 75 %)
(- 34 % для лучших 15 клонов)
с интроном A
сигнал полиA bGH
терминатор транскрипции hGT
(+ 20 %)
Использованные вектора:
Обнаружено, что может быть достигнуто усиление экспрессии (продуктивности) с применением векторных элементов/комбинации элементов, раскрытых в настоящем документе:
промотор CMV человека:
Xu et al., J. Control. Release, 81 (2002) 155-163.
Xia et al., Prot. Expr. Purif. 45 (2006) 115-124.
промотор CMV крысы:
Xia et al., Prot. Expr. Purif. 45 (2006) 115-124.
промотор EF1α человека:
Teschendorf et al., Anticancer Res. 22 (2002) 3325-3330.
Li et al., J. Immunol. Methods 318 (2007) 113-124.
промотор MPSV:
Xia et al., Prot. Expr. Purif. 45 (2006) 115-124.
Artelt et al., Gene 68 (1988) 213-219.
Stocking et al., Proc. Natl. Acad. Sci. USA 82 (1985) 5746-5750.
Lin et al., Gene 147 (1994) 287-292.
гибридный промотор MPSV-CMV:
Liu et al., Anal. Biochem. 246 (1996) 150-152.
Высокая селективность и высокая жёсткость селекции могут быть обеспечены с применением экспрессионной кассеты, связанной с IRES, для экспрессии маркера селекции:
селекционное давление на экспрессию антитела приводит к высокой селективности
связанная экспрессия антитела и маркера селекции приводят к высокой селективности
обнаружено, что применение IRES-элемента со слабой активностью приводит к высокой жёсткости, т. е. высокому уровню получения антитела и низкому уровню получения маркера селекции
связывание экспрессии антитела и маркера селекции IRES-элементами
идентификация IRES-элементов (EMCV/Gtx) со слабой активностью, которые минимально изменяют уровень экспрессии IgG
применение слитого белка, работающего как маркер селекции и маркер для скрининга
бифункциональный слитый белок GFP-неомицин
последовательность PEST орнитиндекарбоксилазы является сильной протеолитической сигнальной последовательностью и снижает время полужизни данного белка
экспрессия слитого белка, связанная с IRES, приводит к высокой селективности
уменьшенное время полужизни за счёт протеолитической сигнальной последовательности приводит к высокой жёсткости селекции
вследствие слабой экспрессии и короткого времени полужизни слитого белка, необходима сильная экспрессия
быстрая идентификация высокопродуктивных клонов посредством FACS (сортинг клонов с высокой экспрессией GFP обеспечивает селекцию высокопродуктивных клонов)
Маркер селекции, связанный с тяжёлой цепью антитела посредством различных IRES-элементов
Слитый белок GFP-Neo, связанный с тяжёлой цепью антитела различными IRES-элементами
Gtx-IRES:
Komuro et al., EMBO J. 12 (1993) 1387-1401.
EMCV-IRES:
Mountford et al., Proc. Natl. Acad. Sci. USA 91 (1991) 4303-4307.
EV71-IRES:
Lee et al., Biotechnol. Bioeng. 90 (2005) 656-662.
ELF4G-IRES:
Wong et al., Gene Ther. 9 (2002) 337-344.
Gtx (синтетический)-IRES:
Chappell et al., Proc. Natl. Acad. Sci. USA97 (2000) 1536-1541.
Обнаружено, что усиленная экспрессия (продуктивность) с применением связывания экспрессионной кассеты лёгкой цепи с экспрессионной кассетой тяжёлой цепи может быть достигнута за счёт IRES-элемента EV71:
Векторные элементы в комбинации со строением вектора
В клеточной линии CHO-K1 в транзиентных трансфекциях, в стабильных пулах и на уровне отдельных клонов были проанализированы следующие вектора:
Несколько генетических элементов, связанных с транскрипцией, и их комбинаций сравнивали с вектором сравнения (px9001, строение вектора SM (3'-5' ориентация)-LC-HC (5'-3' ориентация)). На основе сравнительных экспериментов получены следующие результаты для однонаправленного строения вектора с экспрессионными кассетами лёгкой и тяжёлой цепи (экспрессионная кассета лёгкой цепи до экспрессионной кассеты тяжёлой цепи) и маркера селекции, ориентированными в одном направлении (см. таблицу ниже), относительно вектора сравнения (px9001, двунаправленное строение вектора SM (3'-5')-LC-HC (5'-3')).
(двунаправленное строение, SM (3'-5')-LC-HC (5'-3')) или px9002 (однонаправленное строение, LC-HC-SM (5'-3'))
(двунаправленное строение, SM (3'-5')-LC-HC (5'-3')) или px9002 (однонаправленное строение, LC-HC-SM (5'-3'))
(двунаправленное строение, SM (3'-5')-LC-HC (5'-3')) или px9002 (однонаправленное строение, LC-HC-SM (5'-3'))
(двунаправленное строение, SM (3'-5')-LC-HC (5'-3')) или px9002 (однонаправленное строение, LC-HC-SM (5'-3'))
(+ 30 % по сравнению с px9002)
(двунаправленное строение, SM (3'-5')-LC-HC (5'-3')) или px9002 (однонаправленное строение, LC-HC-SM (5'-3'))
(+ 34 % по сравнению с px9002)
в клонах:
в отдельных клонах + 53% и + 92% в анализе периодических культур и в анализе подпитываемых культур для лучших 36 и 15 клонов соответственно (по сравнению с px9001)
(двунаправленное строение, SM (3'-5')-LC-HC (5'-3')) или px9002 (однонаправленное строение, LC-HC-SM (5'-3'))
(+ 50% по сравнению с px9002)
в отдельных клонах + 19% и - 7% в анализе периодических культур и в анализе подпитываемых культур для лучших 36 и 15 клонов соответственно (по сравнению с px9001)
(двунаправленное строение, SM (3'-5')-LC-HC (5'-3')) или px9002 (однонаправленное строение, LC-HC-SM (5'-3'))
(+ 47% по сравнению с px9002)
в клонах:
в отдельных клонах + 7% и - 14% в анализе периодических культур и в анализе подпитываемых культур для лучших 36 и 15 клонов соответственно (по сравнению с px9001)
Производительность различных векторов в транзиентных трансфекциях анализировали после нуклеофекции в клетки CHO-K1 (см. Фиг. 9).
Вектора, содержащие промотор фактора элонгации 1α человека (hEF1α ) (на основе строения вектора LC-HC-SM), обладали повышенной продуктивностью примерно на 34% (px9003 относительно px9002; сигнальная последовательность полиA SV40) и + 30% (px9006 относительно px9004; сигнальная последовательность полиA bGH), в зависимости от используемого сигнала полиA, соответственно, по сравнению с применением промотора hCMV.
Присоединение терминатора гастрина человека (hGT) к сигнальной последовательности полиA bGH давало положительный эффект на продуктивность векторов, содержащих hCMV (px9005 относительно px9004; + 13%).
Экспрессионные вектора, основанные на двунаправленной экспрессии тяжёлой и лёгкой цепи антитела, проявляют увеличенную производительность. Титры продукта увеличены примерно от 2,7- до 3,4-раз по сравнению с контрольным вектором px9001, в зависимости от используемого промотора (hEF1α или hCMV) и используемой сигнальной последовательности полиA (SV40 или bGH).
Обнаружено, что применение промотора фактора элонгации 1α человека и сигнальной последовательности полиA bGH даёт положительный эффект на продуктивность при строения вектора LC-HC-SM (+ 50%; сравнение px9006 и px9002), но не при строения вектора LC(3'-5')-HC-SM (- 28%; сравнение px9010 и px9011).
Для сравнения продуктивности экспрессионного вектора px9002 с векторами px9003-9007, вектора трансфецировали в клетки CHO-K1 путём нуклеофекции и селектировали стабильные пулы. Продуктивность данных пулов анализировали с помощью анализа периодических культур (см. Фиг. 10).
Анализ периодических культур стабильных пулов показал, что титры антител из пулов, трансфецированных вектором, содержащим промотор фактора элонгации 1α человека (px9003, px9006, px9007), были приблизительно в 7-8 раз больше, чем титры для клеток, трансфецированных вектором сравнения, содержащим короткий промотор hCMV (вектор px9002) (сравнение 97,5 мкг/мл, 112,5 мкг/мл и 95,6 мкг/мл для векторов px9003, px9006 и px9007 соответственно, относительно 14,0 мкг/мл для вектора px9002).
Вектора px9001, px9002 и px9004-9007 трансфецировали в клетки CHO-K1 путём нуклеофекции и идентифицировали лучшие отдельные клоны с помощью обыкновенного процесса скрининга. Продуктивность лучших 36 клонов для каждого вектора анализировали посредством анализа периодических культур, и лучшие 15 клонов из анализа периодических культур анализировали посредством анализа подпитываемых культур.
Средняя продуктивность лучших 36 отдельных клонов, созданных с применением вектора px9001, в анализе периодических культур составляет 356 мкг/мл. Клоны, созданные с применением вектора px9002 или векторов px9004 и px9005 (дополнительно содержащих сигнал полиA bGH (отдельно или в комбинации c hGT) вместо сигнала полиA SV40), демонстрируют, соответственно, 37% (px9002), 61% (px9004) и 53% (px9005) увеличение продуктивности по сравнению с контрольным вектором px9001. Клоны векторов px9006 и px9007 демонстрируют увеличение продуктивности примерно на 19% и 7%, соответственно.
В экспериментах с подпитываемой культурой лучшие 15 клонов, полученные в анализе периодических культур с применением каждого вектора, анализировали с помощью анализа подпитываемых культур в течение 14 дней.
Средняя продуктивность лучших 15 отдельных клонов собственного получения (созданных с применением вектора px9001) в анализе подпитываемых культур составляет 1345 мкг/мл. Клоны, созданные с применением вектора px9002 или векторов px9004 и px9005 (дополнительно содержащих сигнальную последовательность полиA bGH (отдельно или в комбинации c hGT) вместо сигнальной последовательности полиA SV40), демонстрируют, соответственно, 80% (px9002), 58% (px9004) и 92% (px9005) увеличение продуктивности по сравнению с контрольным вектором px9001.
Помимо увеличенной средней продуктивности (см. выше), также намного улучшена производительность лучших клонов. Вектора px9002, px9004 и px9005 демонстрируют в отношении продуктивности лучших 5 клонов увеличение примерно на 64% (px9002), 50% (px9004) и 88% (px9005) по сравнению с контрольным вектором px9001.
Далее непосредственно сравнивали процентный показатель продуцирующих клеток относительно не продуцирующих клеток для каждого из различных векторов. 14,2% клонов, созданных с применением вектора px9001, продуцируют антитело, в остальных устойчивых клонах экспрессия антитела или сайленсирована, или клоны имеют другие дефекты.
Строение вектора px9002 практически удваивает процентный показатель продуцирующих клеток (до 26,0%). Вектор, дополнительно содержащий только сигнальную последовательность полиA bGH вместо сигнальной последовательности полиA SV40 (вектор px9004) или сигнальной последовательности полиA SV40 в комбинации с hGT (вектор px9005), демонстрирует приблизительно 3-кратное увеличение процентного показателя продуцирующих клеток (39% и 43% соответственно).
Применение промотора фактора элонгации 1α человека вместо промотора hCMV увеличивает количество продуцирующих клеток до 5 раз (более 70% клонов, полученных после процесса селекции, продуцируют антитело).
Лучшие 15 клонов, полученные в результате трансфекции векторами px9001-9007 (на основе анализа подпитываемых культур), культивировали в присутствии и в отсутствии гигромицина B в течение 15 пассажей (= приблизительно 60 поколений). Титры продукта клонов в анализе периодических культур после 15 пассажей сравнивали с титром продукта клонов в группе в момент начала теста на стабильность.
В присутствии селекционного давления изменения в титре продукта между 15 клонами каждого вектора после 15 пассажей варьируют от -14,7% для вектора px9007 до +0,2% для вектора px9002.
В отсутствии селекционного давления уменьшение титра продуктов варьирует от 25,5% для вектора px9004 до 5,9% для вектора px9005.
Количество клонов, соответствующих установленным критериям стабильности, таким как >80% титра продукта в анализе периодических культур по сравнению со значениями в исходной точке (G0), как в присутствии маркера селекции, так и в его отсутствии, варьирует в пределах от 4 до 10. Вектора px9005, px 9007 и px9002 обеспечивают наибольшее количество стабильных клонов как в присутствии, так и в отсутствии селекционного давления/маркера селекции (px9005: 10; px9007: 7; px9002: 6).'
Таким образом, обнаружено, что строение векторов px9005 демонстрирует положительные эффекты на стабильность и увеличивает количество стабильных клонов, особенно в отсутствии селекционного давления.
Обнаружено, что комбинация полиA bGH и hGT по сравнению с полиA SV40 без терминатора транскрипции (hGT) явно увеличивает продуктивность стабильных клонов независимо от используемого промотора.
Транзиентные трансфекции:
применение промотора фактора элонгации 1α человека (с интроном A) обеспечивает улучшенную продуктивность (при строении LC-HC-SM)
применение сигнальной последовательности полиA бычьего гормона роста обеспечивает улучшенную продуктивность по сравнению с применением сигнальной последовательности полиA SV40
добавление hGT к сигнальной последовательности полиA bGH приводит к улучшенной продуктивности для векторов, содержащих промотор hCMV
строение вектора LC(3'-5')-HC-SM приводит к улучшенной экспрессии
Стабильные пулы
пулы, созданные с применением векторов, содержащих промотор hEF1α , демонстрируют улучшенную продуктивность в анализе периодических культур
клоны, созданные с применением векторов, содержащих промотор hEF1α , демонстрируют уменьшенное количество низкопродуктивных клонов
клоны, созданные с применением векторов, содержащих промотор hEF1α , демонстрируют повышенную стабильность экспрессии IgG
Отдельные клоны
строение вектора с 3' положением маркера селекции (LC-HC-SM) даёт положительный эффект на продуктивность отдельных клонов
клоны, созданные с применением векторов, содержащих сигнальную последовательность полиA bGH и hGT, обладают более высокой продуктивностью и стабильностью.
Нижеследующие примеры, фигуры и последовательности приведены для лучшего понимания настоящего изобретения, объем которого определяется прилагаемой формулой изобретения. Следует понимать, что в описанные процедуры могут быть внесены модификации без отклонения от сути настоящего изобретения.
Последовательности
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1
Схематическое представление различных конструкций векторов, проанализированных в транзиентных трансфекциях, в стабильных пулах и на уровне отдельных клонов. Вектора p5158, p5137, p5156 и p5159 различаются по позиции лёгкой цепи (LC) и тяжёлой цепи (HC) соответственно и/или по позиции маркера селекции (SM).
Фиг. 2
Продуктивность векторов p5137, p5156, p5158 и p5159 в транзиентно трансфецированных клетках CHO-K1. Показана средняя продуктивность восьми независимых трансфекций каждым вектором на 5 день после трансфекции, измеренная с помощью ИФА.
Фиг. 3
Продуктивность стабильных пулов, созданных с применением векторов p5137, p5156, p5158 и p5159, в анализе на периодических культурах. Показана средняя продуктивность трёх пулов для каждого вектора на 7 день.
Фиг. 4
Схематическое представление конструкции экспрессионного вектора p5068 и вектора px6068. Указаны LC (лёгкая цепь), HC (тяжёлая цепь), SM (маркер селекции (под контролем промотора SV40)) и ori (точка начала репликации).
Фиг. 5
Продуктивность векторов p5068 и px6068 в транзиентно трансфецированных клетках CHO-K1. Показана средняя продуктивность восьми независимых трансфекций каждым вектором на 7 день после трансфекции, измеренная с помощью ИФА.
Фиг. 6
Продуктивность стабильных пулов, созданных с применением вектора px6068, линеаризованного рестриктазой SgrAI или рестриктазой BssHII. Показана средняя продуктивность двух пулов для каждого вектора в анализе периодических культур на 7 день.
Фиг. 7
Зависимость титра IgG в стабильных клеточных пулах от момента начала селекции после трансфекции.
Фиг. 8
(A) время селекции для получения стабильных клонов клеток; (B) продуктивность пулов; (C) продуктивность отдельных клонов; (D) процесс восстановления плотности жизнеспособных клеток; (E) плотность жизнеспособных клеток через 4, 7 и 10 дней культивирования; препарат вектора A - линеаризованный полноразмерный вектор, препарат вектора B - вырезаны прокариотические элементы вектора, препарат вектора C - вырезаны и удалены прокариотические элементы вектора.
Фиг. 9
Продуктивность векторов px9001-px9011 в транзиентных трансфекциях путём нуклеофекции: показана средняя продуктивность восьми независимых трансфекций каждым вектором на 6 день после трансфекции; значения нормированы на значения для вектора сравнения px9001 (заданного как 100%).
Фиг. 10
Обзор титров продукта стабильных пулов, созданных с применением векторов px9001-px9007, в анализе периодических культур на 10 день. Показано среднее значение для двух (px9001 и px9002) или трёх независимых трансфекций каждым вектором.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Экспрессионные вектора p5068 и p5069
Экспрессионные плазмиды p5068 и p5069 содержат экспрессионные кассеты для экспрессии анти-P-селектин антитела (геномное строение экспрессионной кассеты с сохранением экзонно-интронного строения), как описано в WO 2005/100402.
Гены, кодирующие лёгкую и тяжёлую цепи данного анти-P-селектин HuMab, были раздельно собраны в экспрессионных векторах для клеток млекопитающих.
Таким образом, были соединены сегменты гена, кодирующие вариабельный район лёгкой цепи анти-P-селектин HuMab (VL) и константный район κ-лёгкой цепи (CL) человека, а также сегменты гена, кодирующие вариабельный район тяжёлой цепи анти-P-селектин HuMab (VH) и константный район γ1-тяжёлой цепи человека или константный район γ4-тяжёлой цепи человека (CH1-шарнирная область-CH2-CH3).
Основная информация о нуклеотидных последовательностях человеческих лёгких и тяжёлых цепей, из которых могут быть выведены данные о частоте использования кодонов, приведена в: Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Бетесда, Мэриленд (1991), NIH Publication No 91-3242.
Транскрипционная единица κ-лёгкой цепи анти-P-селектин HuMab состоит из следующих элементов:
немедленно-раннего энхансера и промотора цитомегаловируса человека (hCMV),
синтетической 5'-НТО, включающей последовательность Козак,
сигнальной последовательности тяжёлой цепи мышиного иммуноглобулина, включающей интрон данной сигнальной последовательности,
клонированной кДНК вариабельного района лёгкой цепи анти-P-селектин HuMab с уникальным сайтом рестрикции BsmI на 5'-конце и с донорным сайтом сплайсинга и уникальным сайтом рестрикции NotI на 3'-конце,
гена κ-константного района человеческого генома, включающего энхансер мышиного Ig-κ из интрона 2 (Picard, D., and Schaffner, W. Nature 307 (1984) 80-82), и
сигнальной последовательности полиаденилирования («полиA») человеческого κ-иммуноглобулина
Транскрипционная единица γ1-тяжёлой цепи анти-P-селектин HuMab состоит из следующих элементов:
немедленно-раннего энхансера и промотора цитомегаловируса человека (hCMV),
синтетической 5'-НТО, включающей последовательность Козак,
модифицированной сигнальной последовательности тяжёлой цепи мышиного иммуноглобулина, включающей интрон данной сигнальной последовательности,
клонированной кДНК вариабельного района тяжёлой цепи анти-P-селектин HuMab с уникальным сайтом рестрикции BsmI на 5'-конце и с донорным сайтом сплайсинга и уникальным сайтом рестрикции NotI на 3'-конце,
гена константного района γ1 тяжёлой цепи человеческого генома, включающего энхансер мышиного Ig-µ (Neuberger, M.S., EMBO J. 2 (1983) 1373-1378), и
сигнальной последовательности полиаденилирования («полиA») человеческого γ1 -иммуноглобулина.
Помимо экспрессионных кассет κ-лёгкой цепи или γl-тяжёлой цепи анти-P-селектин HuMab, данные плазмиды содержат
ген устойчивости к гигромицину,
точка начала репликации oriP вируса Эпштейна-Барр (EBV),
точка начала репликации вектора pUC18, обеспечивающий репликацию данной плазмиды в E. сoli и
ген β-лактамазы, который придаёт E. сoli устойчивость к ампициллину.
Методы рекомбинантных ДНК
Клонирование выполняли с применением стандартных методик клонирования, как описано в Sambrook et al., 1999 (см. выше). Все реагенты для методов молекулярной биологии были коммерчески доступными (если не указано иное), и их использовали согласно инструкциям производителя.
Синтез нуклеиновых кислот
ДНК различных генетических элементов синтезировали в компании Geneart AG, Регенсбург.
Определение последовательности нуклеиновых кислот
Последовательности ДНК определяли посредством секвенирования двойной цепи, которое проводили в компании SequiServe (SequiServe GmbH, Германия).
Анализ последовательностей ДНК и белка и анализ данных о последовательностях
Для получения, картирования, анализа, аннотирования и иллюстрирования последовательностей использовали программный пакет Vector NTI Advance версии 9.0.
Методы клеточных культур
Клетки CHO-K1 выращивали на среде CD-CHO (Invitrogen Corp., Gibco®, Cat. No. 10743-011) с добавлением 1 x HT (Invitrogen Corp., Gibco® , Cat. No. 11067-030).
Для селекции стабильно трансфецированных пулов/клеточных линий CHO-K1 добавляли от 400 до 800 мкг/мл G418 или от 200 до 400 мкг/мл гигромицина (Roche Diagnostics GmbH, Roche Applied Sciences, Германия, Cat. No.: 843555).
Все клеточные линии содержали в увлажняемых инкубаторах при 37°C с 5% содержанием CO2 при постоянном перемешивании при от 120 до 140 об/мин. Каждые 3-4 дня клетки пересевали в свежую среду. Плотность и жизнеспособность культур определяли с применением счётчика клеток Casey TT или Cedex Hires (Roche innovates AG, Билефельд). Трансфекцию клеток выполняли посредством технологии нуклеофекции Amaxa (Lonza GmbH, Германия).
Кроме того, применяли стандартные методы культивирования клеток, как описано, например, в Bonifacino, J.S., et al., (eds.), Current Protocols in Cell Biology, John Wiley and Sons, Inc. (2000).
Подсчёт клеток и определение их жизнеспособности
a) Система подсчёта клеток, основанная на анализе электрического поля (CASY)
Счётчик клеток CASY® Technology Cell Counter, модель TT (Roche Innovatis AG, Билефельд) использует электрический ток для подсчёта клеток. Для получения информации из сигналов, получаемых при прохождении клеткой через измерительную пору в поле низкого напряжения, использовали анализ площади импульса. Структурная целостность клеточной мембраны является показателем жизнеспособности клетки. Таким образом для определения жизнеспособности клеток не требуются красители, такие как трипановый синий.
b) Автоматизированный способ окрашивания трипановым синим (Cedex)
Для определения жизнеспособности клеток в ходе селекции пулов, а также для автоматизированного подсчёта клеток использовали систему Cedex HiRes (Roche Innovatis AG, Билефельд).
Трипановый синий является красителем, который не может входить в клетки через интактные клеточные мембраны. Окрашиваются и маркируются как мёртвые только те клетки, которые имеют повреждённую клеточную мембрану. Процесс окрашивания, подсчёта клеток и графического анализа результата выполняли автоматически системой Cedex с помощью цифрового распознавания изображений. Другими измеряемыми параметрами являются размер клетки, морфология и степень агрегации. С применением множественного анализа образцов проводили последовательный анализ до 20 образцов.
Подготовка плазмид и проверка качества для достоверного сравнения плазмид в трансфекциях
На эффективность трансфекции и, следовательно, на продуктивность сильно влияют несколько факторов, таких как количество ДНК и её качество. Для гарантии одинаковых начальных условий для каждого вектора перед трансфекцией тщательно проверяли качество и количество ДНК всех векторов.
Одновременное получение экспрессионных векторов
Все вектора получали одновременно с помощью набора для выделения плазмид High Speed Maxi (Qiagen GMBH, Хильден) согласно инструкциям производителя.
Очистка фенол/хлороформом и осаждение этанолом
Все вектора одновременно очищали фенол/хлороформом. 500 мкг каждой линеаризованной плазмидной ДНК смешивали с 200 мкл раствора 50% (объем/объем) фенола, 48% (объем/объем) хлороформа, 2% (объем/объем) изоамилового спирта в трис-буфере и центрифугировали в течение 1 мин при 13000 об/мин. Затем верхнюю водную фазу переносили в чистую пробирку и смешивали с 200 мкл 96% (объем/объем) хлороформа, 4% (объем/объем) изоамилового спирта, и центрифугировали в течение 1 мин при 13000 об/мин. Верхнюю фазу снова переносили в чистую пробирку и смешивали с 1/10 объёма (от общего объёма) 3M ацетата натрия (pH 5,2) и с 2,5-кратным объёмом (от общего объёма) 100% этанола. После перемешивания и инкубирования данной смеси в течение 5 мин при комнатной температуре её центрифугировали в течение 5 мин при 13000 об/мин для осаждения ДНК. Супернатант удаляли, и осадок промывали 900 мкл 70% (объем/объем) этанола и инкубировали в течение 5 минут при комнатной температуре. После заключительного этапа центрифугирования при максимальных оборотах в течение 5 мин супернатанты удаляли и осадки высушивали и растворяли в стерильной H2O.
Определение количества ДНК
Количество ДНК каждого вектора определяли с помощью прибора BioPhotometer (Eppendorf; Гамбург). Измерения ДНК всегда выполняли в трех повторностях с применением разведения 1:20 в трис-буфере pH 8,0.
Агарозный гель
Качество ДНК каждой плазмиды проверяли в 0,8% агарозном геле. Определяли уровень деградации ДНК, конформацию вектора и концентрацию ДНК. Вектора, представленные в сравнимых количествах и качествах (отсутствие деградации ДНК, сходные суперспирализованные (ccc) формы, сходные количества ДНК в геле) использовали для транзиентных и стабильных трансфекций.
Транзиентные трансфекции
Все вектора трансфецировали в клетки CHO-K1 с помощью 96-луночной системы Amaxa (Lonza GmbH, Germany) согласно инструкциям производителя. Каждый вектор трансфецировали в 8 повторностях. Количества ДНК трансфецированного вектора нормировали для получения эквивалентного молярного количества/количества копий, соответствующего 1 мкг экспрессионной плазмиды сравнения (p5068 или p5069). Для определения продуктивности клеток культуральный супернатант, не содержащий клеток, анализировали на титр IgG через 4-7 дней после трансфекции с помощью одностадийного универсального ИФА (Dianova).
96-луночная система Amaxa
Клетки, растущие в перемешиваемых колбах, осаждали центрифугированием при 850 об/мин в течение 5 мин и ресуспендировали в культуральной среде. Кольцевые плазмиды помещали в 96-луночные планшеты для нуклеофекции в концентрациях, эквимолярных 1 мкг экспрессионного вектора сравнения p5068 или p5069. Затем в данные планшеты добавляли клетки в концентрации 4 x 105 клеток на лунку. Трансфекцию выполняли по программе Amaxa DN-137. Клетки инкубировали в течение 10 мин после трансфекции и затем переносили в 96-луночные планшеты с плоским дном для инкубирования, содержащие 200 мкл культуральной среды. Затем клетки культивировали в статичных условиях. На 4-6 день после трансфекции определяли уровни IgG c помощью одностадийного универсального ИФА.
Стабильные трансфекции и получение рекомбинантных клеточных линий CHO
Стабильные трансфекции выполняли с помощью технологии нуклеофекции (Amaxa Biosystems, Lonza cologne AG) согласно инструкциям производителя. Перед трансфекцией плазмиды линеаризовали рестриктазой SgrAI. Каждую плазмиду трансфецировали в двух или в трёх повторностях. Для каждой отдельной трансфекции использовали 5 x 106 клеток и 1,2 пмоль линеаризованной плазмиды (набор Nucleofector T, программа Amaxa A33).
Для трансфекции клетки ресуспендировали в растворе Nucleofector T и отбирали аликвоты в 2 мл пробирки. После добавления плазмиды трансфекцию выполняли путём применения импульса. Затем клетки переносили в колбы для клеточной культуры T25, содержащие 4 мл предварительно нагретой свежей среды и 4 мл кондиционированной среды. Селекционное давление применяли через 24 часа после трансфекции путём добавления 250 мкг/мл гигромицина B.
Получение стабильных пулов
Вектора трансфецировали в клетки CHO-K1 с помощью технологии нуклеофекции Amaxa и селектировали стабильные пулы с применением гигромицина B или G418 в качестве агента для селекции. Каждую трансфекцию выполняли в трёх повторностях. Для получения стабильных пулов все плазмидны единообразно линеаризовали рестриктазой SgrAI. Для выполнения стабильных трансфекций использовали набор для нуклеофекции Nucleofector T Amaxa, и каждую плазмиду трансфецировали в трёх повторностях.
Стабильные пулы получали следующим образом: для каждой трансфекции использовали 5×106 клеток и 1,2 пмоль линеаризованной плазмиды. Клетки ресуспендировали в растворе T и отбирали аликвоты в 2 мл пробирки. После добавления плазмиды трансфекцию выполняли путём применения импульса (программа Amaxa A33). Трансфецированные пулы клеток культивировали в статических условиях в колбах для культивирования клеток T25, содержащих 4 мл предварительно нагретой свежей среды и 4 мл кондиционированной среды.
Через 24 ч после трансфекции применяли селекционное давление: клетки центрифугировали в течение 5 минут при 800 об/мин, ресуспендировали в 3 мл культуральной среды, содержащей 300 мкг/мл гигромицина B. Через 3 дня после трансфекции клетки переносили в 6-луночные планшеты с плоским дном. Затем клетки инкубировали в течение двух недель, пока жизнеспособность клеток не снижалась до минимума и снова не возрастала до значения свыше 99%. Количество клеток и их жизнеспособность непрерывно определяли с помощью системы Cedex HiRes (Innovatis, Билефельд). В ходе культивирования остатки клеток удаляли центрифугированием, и клетки каждый раз ресуспендировали в 3 мл свежей среды.
Получение стабильных клонов с применением роботизированной системы Caliper
Вектора трансфецировали в клетки CHO-K1, как описано выше. Через 48 часов после трансфекции применяли селекционное давление (гигромицин B или G418), и клетки высевали на 384-луночные планшеты с плоским дном в концентрации от 350 до 700 клеток на лунку с применением автоматизированной высокопроизводительной системы выделения клонов (Sciclone ALH 3000 workstation, Caliper Life Sciences GmbH, Майнц).
Через 10-14 дней проводили скрининг 384-луночных планшетов на уровни IgG c применением сверхвысокопроизводительного скрининга, основанного на ИФА (ELISA uHTS). При первичном скрининге отбирали лучшие продуцирующие клоны и переносили их в 96-луночные планшеты с плоским дном. Через 3-6 дней проводили скрининг клеток на уровни IgG второй раз. Лучшие продуцирующие клоны снова отбирали и вручную переносили в 24-луночные планшеты с плоским дном. После еще одного этапа скрининга, основанного на ИФА, отбирали лучшие клоны и переносили их в 6-луночные планшеты с плоским дном. Уровни IgG в данных 6-луночных планшетах определяли с помощью измерения ProtA для идентификации лучших клонов для периодического культивирования в перемешиваемых 6-луночных планшетах.
Анализ периодических культур для пулов/отдельных клонов
Для обнаружения различий в продуктивности и стабильности, количества клеток для клонов/пулов подсчитывали с применением счётчика клеток Casey, и их единообразно высевали в 6-луночные планшеты с плоским дном в концентрации 3×105 клеток/мл в конечном объёме 3,0 мл. Все культуры культивировали в течение 12 дней и проводили скрининг супернатантов клеточных культур на уровни человеческого IgG в день 4, 7, 9, 11 или 12.
Определение количества IgG
Титр IgG в транзиентных экспериментах в различных форматах скрининга (от 384-луночного до 24-луночного) измеряли с помощью одностадийного универсального ИФА. Продуктивность стабильных пулов и стабильных отдельных клонов в серийных экспериментах определяли посредством ВЭЖХ с белком A.
Одностадийный универсальный ИФА
Одностадийный универсальный ИФА (Dianova) использовали для определения уровней человеческого IgG в супернатантах клеточных культур. Стандартную кривую получали с применением серии разведений анти-P-селектин антитела (F. Hoffmann-La Roche AG, Базель, Швейцария) в диапазоне 0,3125-20 нг/мл с применением буфера для разведения (PBS + 5% (вес/объем) RPLA1). 95 мкл смеси антител, содержащей 0,5 мкг/мл биотинилированного F(ab')2-анти-(человеческий Fc) антитела (Jackson laboratories) и 0,1 мкг/мл F(ab')2-анти-(человеческий Fcγ) антитела, конъюгированного с пероксидазой (Jackson laboratories; Саффолк), наносили на 96-луночный микротитрационный планшет, покрытый стрептавидином (StreptaWell, Roche Diagnostics GmbH). В данные планшеты добавляли 5 мкл супернатанта клеточной культуры, разведённого в 20000 раз, и инкубировали в течение часа. Планшеты, покрытые антителом, промывали три раза 200 мкл промывочного буфера (PBS + 0,05 % (объем/объем) твин 20). В планшеты добавляли 100 мкл ABTS (Roche Diagnostics GmbH, Мангейм, Германия) и измеряли оптическую плотность при 405 нм с длиной волны сравнения в 492 нм.
Измерение с белком A
Титр IgG в анализе периодических культур измеряли с помощью белка A с применением хроматографии, основанной на ВЭЖХ, в комбинации с одностадийным универсальным ИФА.
FACS
Для определения эффективностей трансфекции (на основе клеток, экспрессирующих GFP) или уровней экспрессии GFP в стабильно или транзиентно трансфецированных клетках использовали сортировку клеток с активированной флуоресценцией. В общем, анализировали 5 x 106 клеток каждого клона или пула с применением проточного цитометра FACSCalibur (BD Biosciences, Сан-Диего, Калифорния). Данные о прямом и боковом рассеянии использовали для определения размера клетки, её жизнеспособности и морфологии.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> F. Hoffmann-La Roche AG
<120> СТРОЕНИЕ ЭКСПРЕССИОННОГО ВЕКТОРА, НОВЫЕ СПОСОБЫ ПОЛУЧЕНИЯ
КЛЕТОК-ПРОДУЦЕНТОВ И ИХ ПРИМЕНЕНИЕ ДЛЯ РЕКОМБИНАНТНОГО ПОЛУЧЕНИЯ
ПОЛИПЕПТИДОВ
<130> 30790 WO
<150> EP11195363.4
<151> 2011-12-22
<160> 17
<170> PatentIn version 3.5
<210> 1
<211> 608
<212> DNA
<213> Human cytomegalovirus
<400> 1
gttgacattg attattgact agttattaat agtaatcaat tacggggtca ttagttcata 60
gcccatatat ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc 120
ccaacgaccc ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag 180
ggactttcca ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac 240
atcaagtgta tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg 300
cctggcatta tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg 360
tattagtcat cgctattagc atggtgatgc ggttttggca gtacatcaat gggcgtggat 420
agcggtttga ctcacgggga tttccaagtc tccaccccat tgacgtcaat gggagtttgt 480
tttggcacca aaatcaacgg gactttccaa aatgtcgtaa caactccgcc ccattgacgc 540
aaatgggcgg taggcgtgta cggtgggagg tctatataag cagagctccg tttagtgaac 600
gtcagatc 608
<210> 2
<211> 696
<212> DNA
<213> Human cytomegalovirus
<400> 2
gttgacattg attattgact agttattaat agtaatcaat tacggggtca ttagttcata 60
gcccatatat ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc 120
ccaacgaccc ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag 180
ggactttcca ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac 240
atcaagtgta tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg 300
cctggcatta tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg 360
tattagtcat cgctattagc atggtgatgc ggttttggca gtacatcaat gggcgtggat 420
agcggtttga ctcacgggga tttccaagtc tccaccccat tgacgtcaat gggagtttgt 480
tttggcacca aaatcaacgg gactttccaa aatgtcgtaa caactccgcc ccattgacgc 540
aaatgggcgg taggcgtgta cggtgggagg tctatataag cagagctccg tttagtgaac 600
gtcagatcta gctctgggag aggagcccag cactagaagt cggcggtgtt tccattcggt 660
gatcagcact gaacacagag gaagcttgcc gccacc 696
<210> 3
<211> 2125
<212> DNA
<213> Human cytomegalovirus
<400> 3
ctgcagtgaa taataaaatg tgtgtttgtc cgaaatacgc gttttgagat ttctgtcgcc 60
gactaaattc atgtcgcgcg atagtggtgt ttatcgccga tagagatggc gatattggaa 120
aaatcgatat ttgaaaatat ggcatattga aaatgtcgcc gatgtgagtt tctgtgtaac 180
tgatatcgcc atttttccaa aagtgatttt tgggcatacg cgatatctgg cgatagcgct 240
tatatcgttt acgggggatg gcgatagacg actttggtga cttgggcgat tctgtgtgtc 300
gcaaatatcg cagtttcgat ataggtgaca gacgatatga ggctatatcg ccgatagagg 360
cgacatcaag ctggcacatg gccaatgcat atcgatctat acattgaatc aatattggcc 420
attagccata ttattcattg gttatatagc ataaatcaat attggctatt ggccattgca 480
tacgttgtat ccatatcata atatgtacat ttatattggc tcatgtccaa cattaccgcc 540
atgttgacat tgattattga ctagttatta atagtaatca attacggggt cattagttca 600
tagcccatat atggagttcc gcgttacata acttacggta aatggcccgc ctggctgacc 660
gcccaacgac ccccgcccat tgacgtcaat aatgacgtat gttcccatag taacgccaat 720
agggactttc cattgacgtc aatgggtgga gtatttacgg taaactgccc acttggcagt 780
acatcaagtg tatcatatgc caagtacgcc ccctattgac gtcaatgacg gtaaatggcc 840
cgcctggcat tatgcccagt acatgacctt atgggacttt cctacttggc agtacatcta 900
cgtattagtc atcgctatta ccatggtgat gcggttttgg cagtacatca atgggcgtgg 960
atagcggttt gactcacggg gatttccaag tctccacccc attgacgtca atgggagttt 1020
gttttggcac caaaatcaac gggactttcc aaaatgtcgt aacaactccg ccccattgac 1080
gcaaatgggc ggtaggcgtg tacggtggga ggtctatata agcagagctc gtttagtgaa 1140
ccgtcagatc gcctggagac gccatccacg ctgttttgac ctccatagaa gacaccggga 1200
ccgatccagc ctccgcggcc gggaacggtg cattggaacg cggattcccc gtgccaagag 1260
tgacgtaagt accgcctata gagtctatag gcccaccccc ttggcttctt atgcatgcta 1320
tactgttttt ggcttggggt ctatacaccc ccgcttcctc atgttatagg tgatggtata 1380
gcttagccta taggtgtggg ttattgacca ttattgacca ctcccctatt ggtgacgata 1440
ctttccatta ctaatccata acatggctct ttgccacaac tctctttatt ggctatatgc 1500
caatacactg tccttcagag actgacacgg actctgtatt tttacaggat ggggtctcat 1560
ttattattta caaattcaca tatacaacac caccgtcccc agtgcccgca gtttttatta 1620
aacataacgt gggatctcca cgcgaatctc gggtacgtgt tccggacatg ggctcttctc 1680
cggtagcggc ggagcttcta catccgagcc ctgctcccat gcctccagcg actcatggtc 1740
gctcggcagc tccttgctcc taacagtgga ggccagactt aggcacagca cgatgcccac 1800
caccaccagt gtgccgcaca aggccgtggc ggtagggtat gtgtctgaaa atgagctcgg 1860
ggagcgggct tgcaccgctg acgcatttgg aagacttaag gcagcggcag aagaagatgc 1920
aggcagctga gttgttgtgt tctgataaga gtcagaggta actcccgttg cggtgctgtt 1980
aacggtggag ggcagtgtag tctgagcagt actcgttgct gccgcgcgcg ccaccagaca 2040
taatagctga cagactaaca gactgttcct ttccatgggt cttttctgca gtcaccgtcc 2100
ttgacacggt ttaaacgccg ccacc 2125
<210> 4
<211> 575
<212> DNA
<213> Homo sapiens
<400> 4
cccgggctgg gctgagaccc gcagaggaag acgctctagg gatttgtccc ggactagcga 60
gatggcaagg ctgaggacgg gaggctgatt gagaggcgaa ggtacaccct aatctcaata 120
caacctttgg agctaagcca gcaatggtag agggaagatt ctgcacgtcc cttccaggcg 180
gcctccccgt caccaccccc cccaacccgc cccgaccgga gctgagagta attcatacaa 240
aaggactcgc ccctgccttg gggaatccca gggaccgtcg ttaaactccc actaacgtag 300
aacccagaga tcgctgcgtt cccgccccct cacccgcccg ctctcgtcat cactgaggtg 360
gagaagagca tgcgtgaggc tccggtgccc gtcagtgggc agagcgcaca tcgcccacag 420
tccccgagaa gttgggggga ggggtcggca attgaaccgg tgcctagaga aggtggcgcg 480
gggtaaactg ggaaagtgat gtcgtgtact ggctccgcct ttttcccgag ggtgggggag 540
aaccgtatat aagtgcagta gtcgccgtga acgtt 575
<210> 5
<211> 1571
<212> DNA
<213> Homo sapiens
<400> 5
cccgggctgg gctgagaccc gcagaggaag acgctctagg gatttgtccc ggactagcga 60
gatggcaagg ctgaggacgg gaggctgatt gagaggcgaa ggtacaccct aatctcaata 120
caacctttgg agctaagcca gcaatggtag agggaagatt ctgcacgtcc cttccaggcg 180
gcctccccgt caccaccccc cccaacccgc cccgaccgga gctgagagta attcatacaa 240
aaggactcgc ccctgccttg gggaatccca gggaccgtcg ttaaactccc actaacgtag 300
aacccagaga tcgctgcgtt cccgccccct cacccgcccg ctctcgtcat cactgaggtg 360
gagaagagca tgcgtgaggc tccggtgccc gtcagtgggc agagcgcaca tcgcccacag 420
tccccgagaa gttgggggga ggggtcggca attgaaccgg tgcctagaga aggtggcgcg 480
gggtaaactg ggaaagtgat gtcgtgtact ggctccgcct ttttcccgag ggtgggggag 540
aaccgtatat aagtgcagta gtcgccgtga acgttctttt tcgcaacggg tttgccgcca 600
gaacacaggt aagtgccgtg tgtggttccc gcgggcctgg cctctttacg ggttatggcc 660
cttgcgtgcc ttgaattact tccacgcccc tggctgcagt acgtgattct tgatcccgag 720
cttcgggttg gaagtgggtg ggagagttcg aggccttgcg cttaaggagc cccttcgcct 780
cgtgcttgag ttgaggcctg gcctgggcgc tggggccgcc gcgtgcgaat ctggtggcac 840
cttcgcgcct gtctcgctgc tttcgataag tctctagcca tttaaaattt ttgatgacct 900
gctgcgacgc tttttttctg gcaagatagt cttgtaaatg cgggccaaga tctgcacact 960
ggtatttcgg tttttggggc cgcgggcggc gacggggccc gtgcgtccca gcgcacatgt 1020
tcggcgaggc ggggcctgcg agcgcggcca ccgagaatcg gacgggggta gtctcaagct 1080
ggccggcctg ctctggtgcc tggcctcgcg ccgccgtgta tcgccccgcc ctgggcggca 1140
aggctggccc ggtcggcacc agttgcgtga gcggaaagat ggccgcttcc cggccctgct 1200
gcagggagct caaaatggag gacgcggcgc tcgggagagc gggcgggtga gtcacccaca 1260
caaaggaaaa gggcctttcc gtcctcagcc gtcgcttcat gtgactccac ggagtaccgg 1320
gcgccgtcca ggcacctcga ttagttctcg atcttttgga gtacgtcgtc tttaggttgg 1380
ggggaggggt tttatgcgat ggagtttccc cacactgagt gggtggagac tgaagttagg 1440
ccagcttggc acttgatgta attctccttg gaatttgccc tttttgagtt tggatcttgg 1500
ttcattctca agcctcagac agtggttcaa agtttttttc ttccatttca ggtggtttaa 1560
acgccgccac c 1571
<210> 6
<211> 1653
<212> DNA
<213> Artificial Sequence
<220>
<223> human elongation factor 1 alpha promoter with Intron A and with
optimized 5'UTR
<400> 6
cccgggctgg gctgagaccc gcagaggaag acgctctagg gatttgtccc ggactagcga 60
gatggcaagg ctgaggacgg gaggctgatt gagaggcgaa ggtacaccct aatctcaata 120
caacctttgg agctaagcca gcaatggtag agggaagatt ctgcacgtcc cttccaggcg 180
gcctccccgt caccaccccc cccaacccgc cccgaccgga gctgagagta attcatacaa 240
aaggactcgc ccctgccttg gggaatccca gggaccgtcg ttaaactccc actaacgtag 300
aacccagaga tcgctgcgtt cccgccccct cacccgcccg ctctcgtcat cactgaggtg 360
gagaagagca tgcgtgaggc tccggtgccc gtcagtgggc agagcgcaca tcgcccacag 420
tccccgagaa gttgggggga ggggtcggca attgaaccgg tgcctagaga aggtggcgcg 480
gggtaaactg ggaaagtgat gtcgtgtact ggctccgcct ttttcccgag ggtgggggag 540
aaccgtatat aagtgcagta gtcgccgtga acgttctttt tcgcaacggg tttgccgcca 600
gaacacaggt aagtgccgtg tgtggttccc gcgggcctgg cctctttacg ggttatggcc 660
cttgcgtgcc ttgaattact tccacgcccc tggctgcagt acgtgattct tgatcccgag 720
cttcgggttg gaagtgggtg ggagagttcg aggccttgcg cttaaggagc cccttcgcct 780
cgtgcttgag ttgaggcctg gcctgggcgc tggggccgcc gcgtgcgaat ctggtggcac 840
cttcgcgcct gtctcgctgc tttcgataag tctctagcca tttaaaattt ttgatgacct 900
gctgcgacgc tttttttctg gcaagatagt cttgtaaatg cgggccaaga tctgcacact 960
ggtatttcgg tttttggggc cgcgggcggc gacggggccc gtgcgtccca gcgcacatgt 1020
tcggcgaggc ggggcctgcg agcgcggcca ccgagaatcg gacgggggta gtctcaagct 1080
ggccggcctg ctctggtgcc tggcctcgcg ccgccgtgta tcgccccgcc ctgggcggca 1140
aggctggccc ggtcggcacc agttgcgtga gcggaaagat ggccgcttcc cggccctgct 1200
gcagggagct caaaatggag gacgcggcgc tcgggagagc gggcgggtga gtcacccaca 1260
caaaggaaaa gggcctttcc gtcctcagcc gtcgcttcat gtgactccac ggagtaccgg 1320
gcgccgtcca ggcacctcga ttagttctcg atcttttgga gtacgtcgtc tttaggttgg 1380
ggggaggggt tttatgcgat ggagtttccc cacactgagt gggtggagac tgaagttagg 1440
ccagcttggc acttgatgta attctccttg gaatttgccc tttttgagtt tggatcttgg 1500
ttcattctca agcctcagac agtggttcaa agtttttttc ttccatttca ggtgtcgtga 1560
ggaattagct ctgggagagg agcccagcac tagaagtcgg cggtgtttcc attcggtgat 1620
cagcactgaa cacagaggaa gcttgccgcc acc 1653
<210> 7
<211> 2473
<212> DNA
<213> Rattus norvegicus
<400> 7
gatattttta tggaaatttt aaaaaattct ggtaagctat ttaaaaaaat gaactttatt 60
atgaaactat tgcccttttc tctaaaaaac aacacaattt cacggaatat cctatgatta 120
attatgacct tttagccagt tcccatatta agaatgagtt atagatgact ctctttaaaa 180
aattattcga tttaaaccat ctgttttaaa gcacagcatt tgtgaataat gtgaagaact 240
tagaagtata atctactcca aggtctgatg tatttttcaa ggccacgtta aagtgtatgc 300
ttgtaacaga gtgcttacat tcaagccaaa tgttaatata acaatcctga attcgtacat 360
aatgtgaata agacactcaa ctctatttaa atccagatct aaatagttac ttttatctaa 420
atgtcaccat ctgtttctac ttagaataat aaacttctta aaggtcacgt atcgggctga 480
ttataaatca ttataattat aacaaaacag atgatttgtt taaaggtcac atcccgttcc 540
gtggtctttt tagtcgaaat aactattaat cttcattatg tttctgagaa agtttaaata 600
tcacgatttc cacccataac agtcattatg agtcagtggg agtcatactg aatcagggta 660
ttttaactgg aaattttttg aaaaacatga gtttttctta aggtcaacat ctggtcttat 720
aaacagaact gagatttatg gccggtaatt accactggac gatttcccgg gaaatcgcta 780
tgggaacggc ccgttttgca acttctttga ccaaaatata tcgagttaag caacttttaa 840
ggccaagtca ctatgactat gccaaataaa gcaactatta aggtcatttc actatggaaa 900
cacccaattc agcaacattg taagccaaat ctccatagaa acctcataag tcagccaaaa 960
gtcaacgacc taccatctgt ttctgcttat ttctctaatt ttaattgcag actttgtcat 1020
tttatgttcc tcttattctg agaatacgtg acgcccgctc gttaaggaca ccgaaactgc 1080
ataagagtca cgttgactca gatgacctcg acatctggtc tggtttttct gccaattttt 1140
cgtctaaact gtggaaaatc cccacagatg acctacaaaa ctccgatttc tattggacga 1200
tgaccgtcag acgtaggtat aaatctccta acgccgttcg ggcagtcaca gtcttcggat 1260
cggacgccgt ggaacgcagt tctcagcgaa gaaggacacc gcccgactcc agaagacacc 1320
gctgcccgaa gaagagaaga cttcatcggt aagagaccca gcttctcctc cccggagctt 1380
cggccacgcc gctccacacc cgggaaccga ggcttcggag cccgataccc ggacagaagc 1440
ttctccccgg ccgctccaca tcagggagcc ttgaccggcg agcctgctat ccgggtagag 1500
actgtcctgc ggccgcttca gcagctccac gatcgacgac tgtgaccgtt gagcccgccg 1560
tttaggcaga ggctccgctt caactaccct accgacacat tcgcggttct tcctccagaa 1620
catcttaccc tctactcggc cactctacaa ggaccggtaa gcaattttta tatactagac 1680
ttaaatgttt ctatgatcat tatgtggtga tggttctgtg tatgaagaga gctaggtgga 1740
ggctatcttt cgcttcggtg atggaacact actcttacaa tggcggctct aatgacggtt 1800
ttctcaacat cggtggcggc tctaattacg gttctctcaa catcggtggt ggtcttcgca 1860
tgcgagctct agattttttt tatctgtaaa ataagattga agatggttga ctgtgtatca 1920
attctttttc ataggcatca gatcttgtca accgttatta atctttagga tcagatgaac 1980
ttgcgagctc gatatctaga atagaatccc cgtgactgct aagatcatct ccgttcatac 2040
accagatgtt acaggccacg gctaccatta tgaatccaaa catgaacaga attgccagaa 2100
tggtgctcaa tggttgtatc catctcgctg gtctattttc tctcaccgac gagaccccaa 2160
catcgagagt tccgtttatt tcatgagtcg accttttagt tcgtgattta ttttctgtgt 2220
taagaaaatc agtgagatca attattgtca gtctatacga ttacaataat gtctgaatta 2280
tcgacgtgca taagatcgtc tcacccggcg cagattccaa cagatctttg tcgccatgcc 2340
ttccgttaga aaggtagtat agtaatatga taccagcaat gcacagaatc gaacatttga 2400
taacaatttt gttgatgtcg tatatctgtt aaaaattaat aaatatatta cagtcagttt 2460
aaacgccgcc acc 2473
<210> 8
<211> 129
<212> DNA
<213> Simian virus 40
<400> 8
aacttgttta ttgcagctta taatggttac aaataaagca atagcatcac aaatttcaca 60
aataaagcat ttttttcacc attctagttg tggtttgtcc aaactcatca atgtatctta 120
tcatgtctg 129
<210> 9
<211> 225
<212> DNA
<213> Bos taurus
<400> 9
ctgtgccttc tagttgccag ccatctgttg tttgcccctc ccccgtgcct tccttgaccc 60
tggaaggtgc cactcccact gtcctttcct aataaaatga ggaaattgca tcgcattgtc 120
tgagtaggtg tcattctatt ctggggggtg gggtggggca ggacagcaag ggggaggatt 180
gggaagacaa tagcaggcat gctggggatg cggtgggctc tatgg 225
<210> 10
<211> 73
<212> DNA
<213> Homo sapiens
<400> 10
caggataata tatggtaggg ttcatagcca gagtaacctt tttttttaat ttttatttta 60
ttttattttt gag 73
<210> 11
<211> 288
<212> DNA
<213> Simian virus 40
<400> 11
agtcagcaac caggtgtgga aagtccccag gctccccagc aggcagaagt atgcaaagca 60
tgcatctcaa ttagtcagca accatagtcc cgcccctaac tccgcccatc ccgcccctaa 120
ctccgcccag ttccgcccat tctccgcccc atggctgact aatttttttt atttatgcag 180
aggccgaggc cgcctctgcc tctgagctat tccagaagta gtgaggaggc ttttttggag 240
gcctaggctt ttgcaaaaag ctcccgggag cttgtatatc cattttcg 288
<210> 12
<211> 81
<212> DNA
<213> Homo sapiens
<400> 12
catggcttcc cgccggaggt ggaggagcag gatgatggca cgctgcccat gtcttgtgcc 60
caggagagcg ggatggaccg t 81
<210> 13
<211> 798
<212> DNA
<213> Artificial Sequence
<220>
<223> green fluorescent protein encoding nucleic acid
<400> 13
atggtgagca agggcgagga gctgttcacc ggggtggtgc ccatcctggt cgagctggac 60
ggcgacgtaa acggccacaa gttcagcgtg tccggcgagg gcgagggcga tgccacctac 120
ggcaagctga ccctgaagtt catctgcacc accggcaagc tgcccgtgcc ctggcccacc 180
ctcgtgacca ccctgaccta cggcgtgcag tgcttcagcc gctaccccga ccacatgaag 240
cagcacgact tcttcaagtc cgccatgccc gaaggctacg tccaggagcg caccatcttc 300
ttcaaggacg acggcaacta caagacccgc gccgaggtga agttcgaggg cgacaccctg 360
gtgaaccgca tcgagctgaa gggcatcgac ttcaaggagg acggcaacat cctggggcac 420
aagctggagt acaactacaa cagccacaac gtctatatca tggccgacaa gcagaagaac 480
ggcatcaagg tgaacttcaa gatccgccac aacatcgagg acggcagcgt gcagctcgcc 540
gaccactacc agcagaacac ccccatcggc gacggccccg tgctgctgcc cgacaaccac 600
tacctgagca cccagtccgc cctgagcaaa gaccccaacg agaagcgcga tcacatggtc 660
ctgctggagt tcgtgaccgc cgccgggatc actctcggca tggacgagct gtacaagtcc 720
ggactcagat ctcgagctca agcttcgaat tctgcagtcg acggtaccgc gggcccggga 780
tccaccggat ctagatga 798
<210> 14
<211> 795
<212> DNA
<213> Artificial Sequence
<220>
<223> neomycin selection marker
<400> 14
atgattgaac aagatggatt gcacgcaggt tctccggccg cttgggtgga gaggctattc 60
ggctatgact gggcacaaca gacaatcggc tgctctgatg ccgccgtgtt ccggctgtca 120
gcgcaggggc gcccggttct ttttgtcaag accgacctgt ccggtgccct gaatgaactg 180
caggacgagg cagcgcggct atcgtggctg gccacgacgg gcgttccttg cgcagctgtg 240
ctcgacgttg tcactgaagc gggaagggac tggctgctat tgggcgaagt gccggggcag 300
gatctcctgt catctcacct tgctcctgcc gagaaagtat ccatcatggc tgatgcaatg 360
cggcggctgc atacgcttga tccggctacc tgcccattcg accaccaagc gaaacatcgc 420
atcgagcgag cacgtactcg gatggaagcc ggtcttgtcg atcaggatga tctggacgaa 480
gagcatcagg ggctcgcgcc agccgaactg ttcgccaggc tcaaggcgcg catgcccgac 540
ggcgaggatc tcgtcgtgac ccatggcgat gcctgcttgc cgaatatcat ggtggaaaat 600
ggccgctttt ctggattcat cgactgtggc cggctgggtg tggcggaccg ctatcaggac 660
atagcgttgg ctacccgtga tattgctgaa gagcttggcg gcgaatgggc tgaccgcttc 720
ctcgtgcttt acggtatcgc cgctcccgat tcgcagcgca tcgccttcta tcgccttctt 780
gacgagttct tctga 795
<210> 15
<211> 1677
<212> DNA
<213> Artificial Sequence
<220>
<223> GFP-PEST-NEO fusion polypeptide encoding nucleic acid
<400> 15
atggtgagca agggcgagga gctgttcacc ggggtggtgc ccatcctggt cgagctggac 60
ggcgacgtaa acggccacaa gttcagcgtg tccggcgagg gcgagggcga tgccacctac 120
ggcaagctga ccctgaagtt catctgcacc accggcaagc tgcccgtgcc ctggcccacc 180
ctcgtgacca ccctgaccta cggcgtgcag tgcttcagcc gctaccccga ccacatgaag 240
cagcacgact tcttcaagtc cgccatgccc gaaggctacg tccaggagcg caccatcttc 300
ttcaaggacg acggcaacta caagacccgc gccgaggtga agttcgaggg cgacaccctg 360
gtgaaccgca tcgagctgaa gggcatcgac ttcaaggagg acggcaacat cctggggcac 420
aagctggagt acaactacaa cagccacaac gtctatatca tggccgacaa gcagaagaac 480
ggcatcaagg tgaacttcaa gatccgccac aacatcgagg acggcagcgt gcagctcgcc 540
gaccactacc agcagaacac ccccatcggc gacggccccg tgctgctgcc cgacaaccac 600
tacctgagca cccagtccgc cctgagcaaa gaccccaacg agaagcgcga tcacatggtc 660
ctgctggagt tcgtgaccgc cgccgggatc actctcggca tggacgagct gtacaagtcc 720
ggactcagat ctcgagctca agcttcgaat tctgcagtcg acggtaccgc gggcccggga 780
tccaccggat ctagacatgg cttcccgccg gaggtggagg agcaggatga tggcacgctg 840
cccatgtctt gtgcccagga gagcgggatg gaccgtagtt taaacattga acaagatgga 900
ttgcacgcag gttctccggc cgcttgggtg gagaggctat tcggctatga ctgggcacaa 960
cagacaatcg gctgctctga tgccgccgtg ttccggctgt cagcgcaggg gcgcccggtt 1020
ctttttgtca agaccgacct gtccggtgcc ctgaatgaac tgcaggacga ggcagcgcgg 1080
ctatcgtggc tggccacgac gggcgttcct tgcgcagctg tgctcgacgt tgtcactgaa 1140
gcgggaaggg actggctgct attgggcgaa gtgccggggc aggatctcct gtcatctcac 1200
cttgctcctg ccgagaaagt atccatcatg gctgatgcaa tgcggcggct gcatacgctt 1260
gatccggcta cctgcccatt cgaccaccaa gcgaaacatc gcatcgagcg agcacgtact 1320
cggatggaag ccggtcttgt cgatcaggat gatctggacg aagagcatca ggggctcgcg 1380
ccagccgaac tgttcgccag gctcaaggcg cgcatgcccg acggcgagga tctcgtcgtg 1440
acccatggcg atgcctgctt gccgaatatc atggtggaaa atggccgctt ttctggattc 1500
atcgactgtg gccggctggg tgtggcggac cgctatcagg acatagcgtt ggctacccgt 1560
gatattgctg aagagcttgg cggcgaatgg gctgaccgct tcctcgtgct ttacggtatc 1620
gccgctcccg attcgcagcg catcgccttc tatcgccttc ttgacgagtt cttctga 1677
<210> 16
<211> 583
<212> DNA
<213> Encephalomyocarditis virus
<400> 16
ggcgcgcccc cctctccctc ccccccccct aacgttactg gccgaagccg cttggaataa 60
ggccggtgtg cgtttgtcta tatgtgattt tccaccatat tgccgtcttt tggcaatgtg 120
agggcccgga aacctggccc tgtcttcttg acgagcattc ctaggggtct ttcccctctc 180
gccaaaggaa tgcaaggtct gttgaatgtc gtgaaggaag cagttcctct ggaagcttct 240
tgaagacaaa caacgtctgt agcgaccctt tgcaggcagc ggaacccccc acctggcgac 300
aggtgcctct gcggccaaaa gccacgtgta taagatacac ctgcaaaggc ggcacaaccc 360
cagtgccacg ttgtgagttg gatagttgtg gaaagagtca aatggctctc ctcaagcgta 420
ttcaacaagg ggctgaagga tgcccagaag gtaccccatt gtatgggatc tgatctgggg 480
cctcggtgca catgctttac atgtgtttag tcgaggttaa aaaaacgtct aggccccccg 540
aaccacgggg acgtggtttt cctttgaaaa acacgatgga tcc 583
<210> 17
<211> 655
<212> DNA
<213> Enterovirus 71
<400> 17
ggcgcgcccc cgaagtaact tagaagctgt aaatcaacga tcaatagcag gtgtggcaca 60
ccagtcatac cttgatcaag cacttctgtt tccccggact gagtatcaat aggctgctcg 120
cgcggctgaa ggagaaaacg ttcgttaccc gaccaactac ttcgagaagc ttagtaccac 180
catgaacgag gcagggtgtt tcgctcagca caaccccagt gtagatcagg ctgatgagtc 240
actgcaaccc ccatgggcga ccatggcagt ggctgcgttg gcggcctgcc catggagaaa 300
tccatgggac gctctaattc tgacatggtg tgaagagcct attgagctag ctggtagtcc 360
tccggcccct gaatgcggct aatcctaact gcggagcaca tgctcacaaa ccagtgggtg 420
gtgtgtcgta acgggcaact ctgcagcgga accgactact ttgggtgtcc gtgtttcctt 480
ttattcctat attggctgct tatggtgaca atcaaaaagt tgttaccata tagctattgg 540
attggccatc cggtgtgcaa cagggcaatt gtttacctat ttattggttt tgtaccatta 600
tcactgaagt ctgtgatcac tctcaaattc attttgaccc tcaacacaat caaac 655
<---
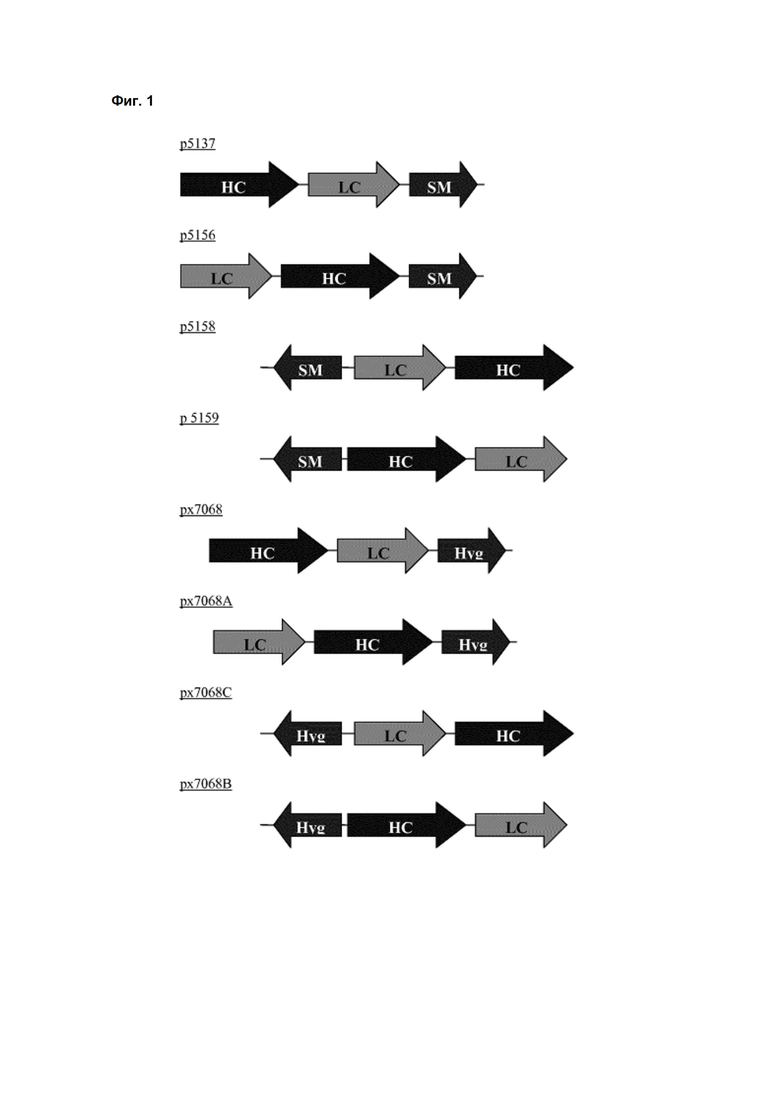
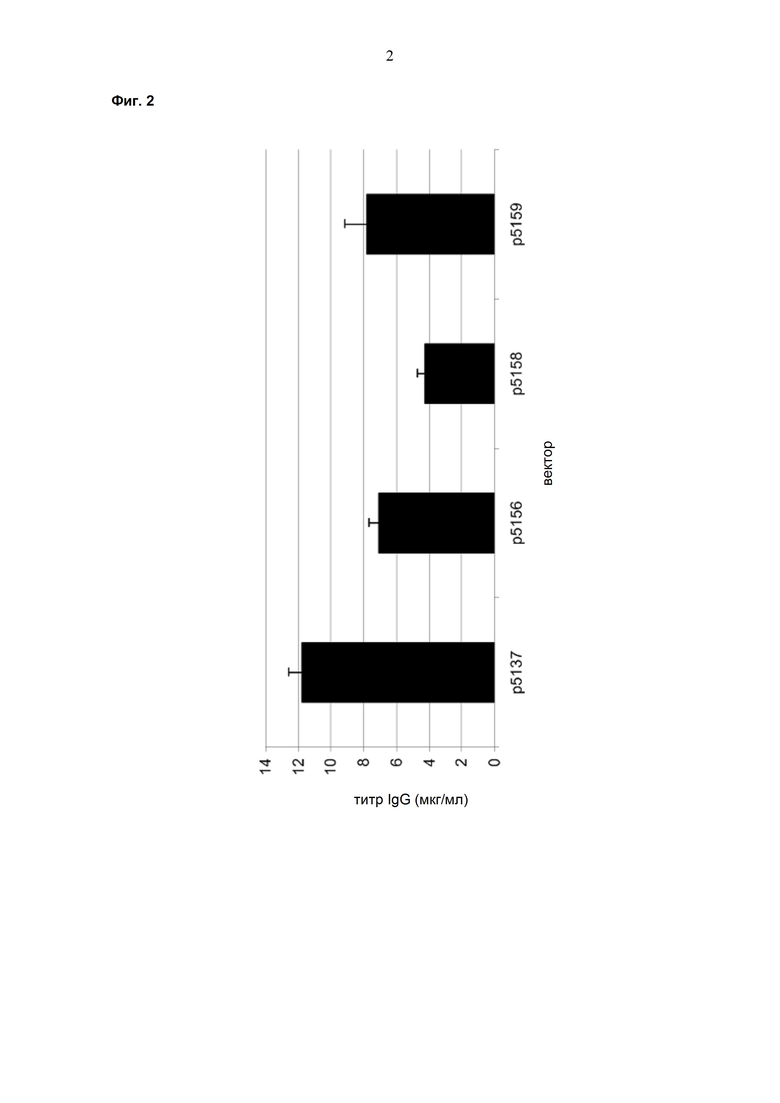
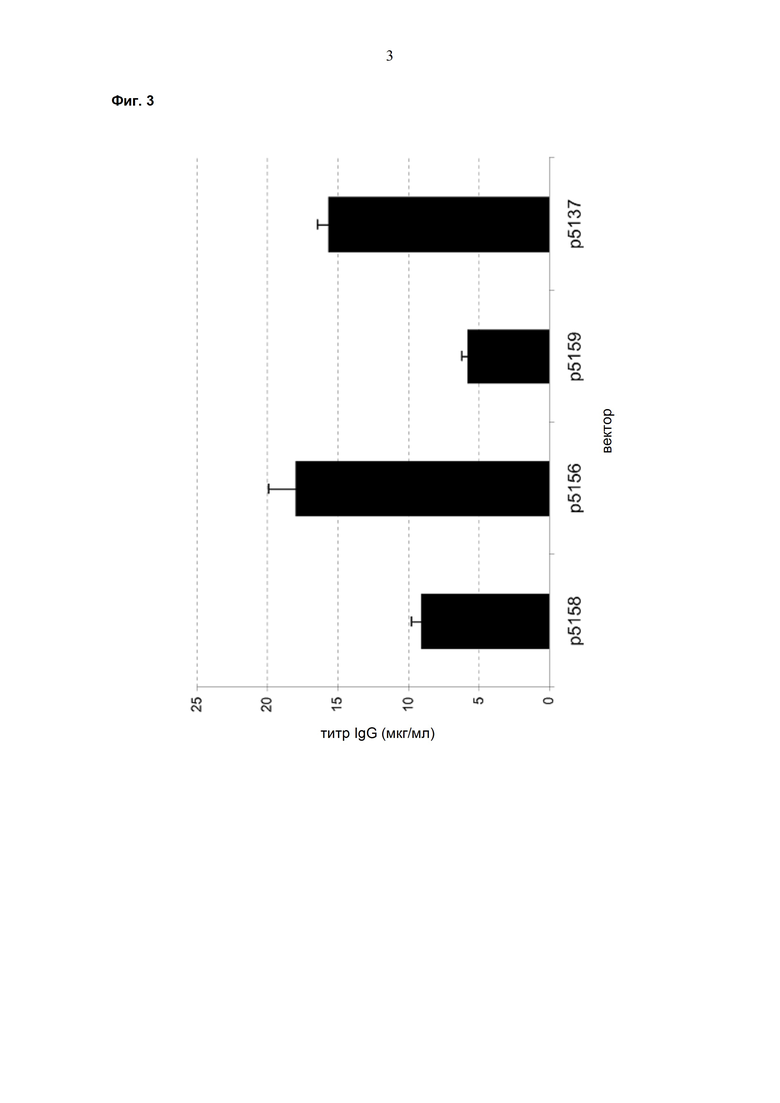
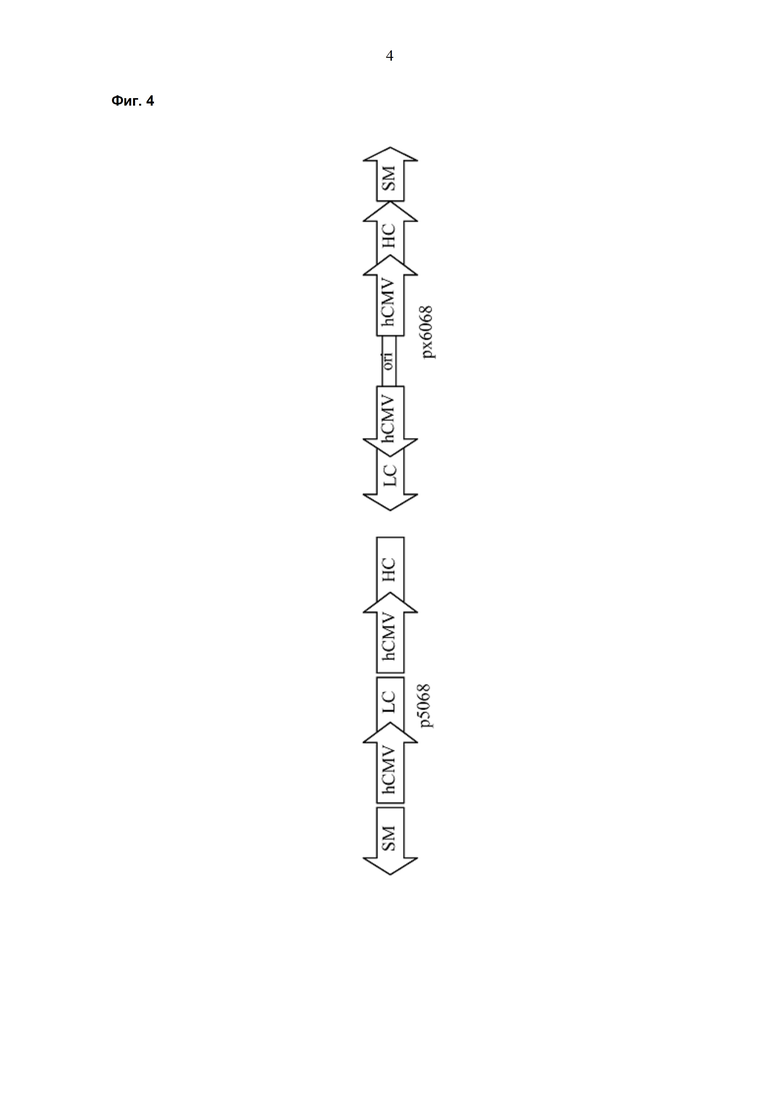
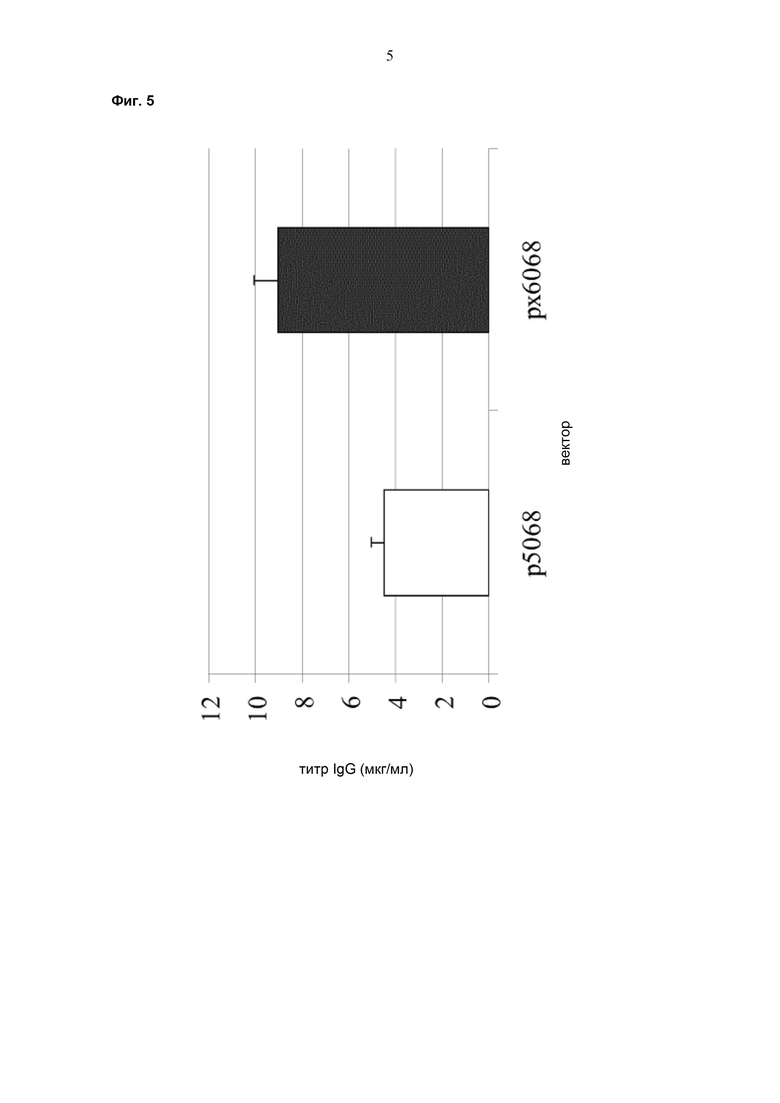
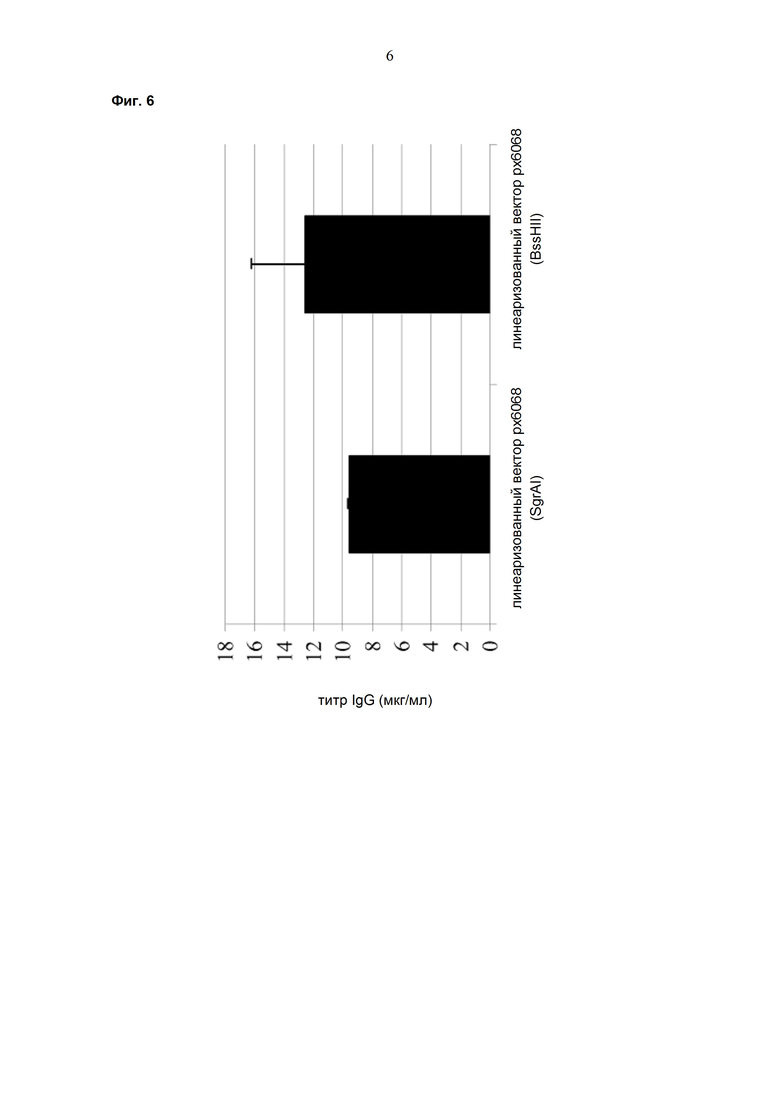
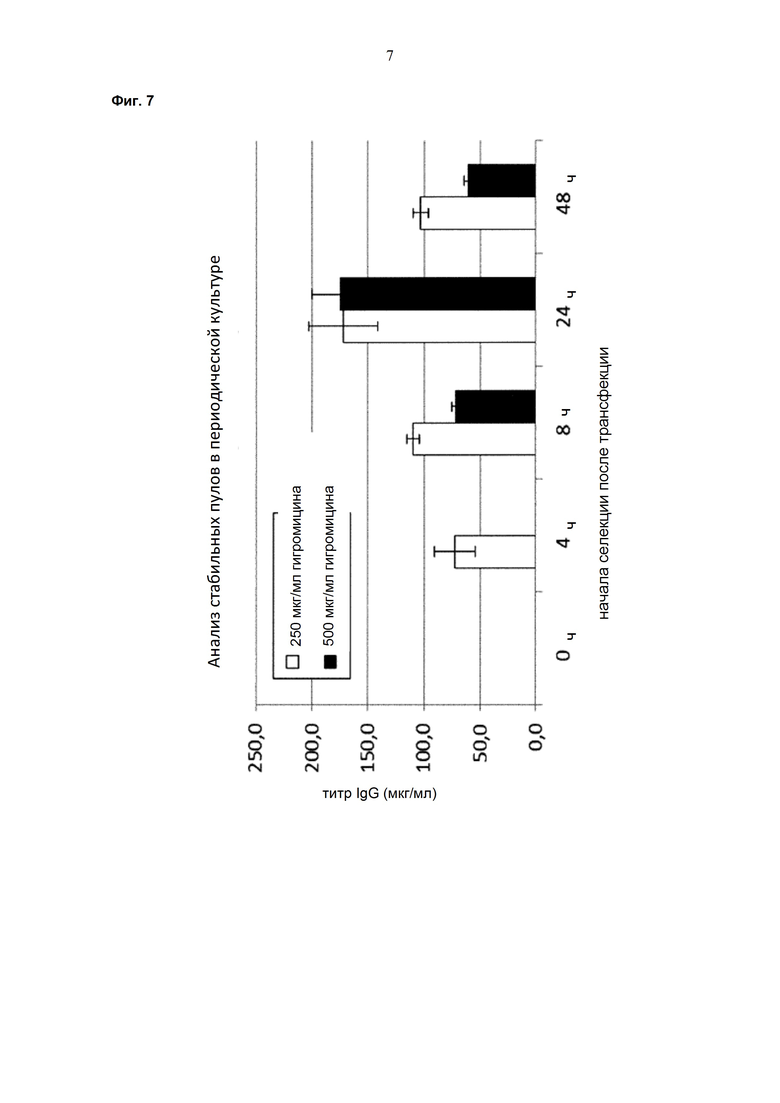
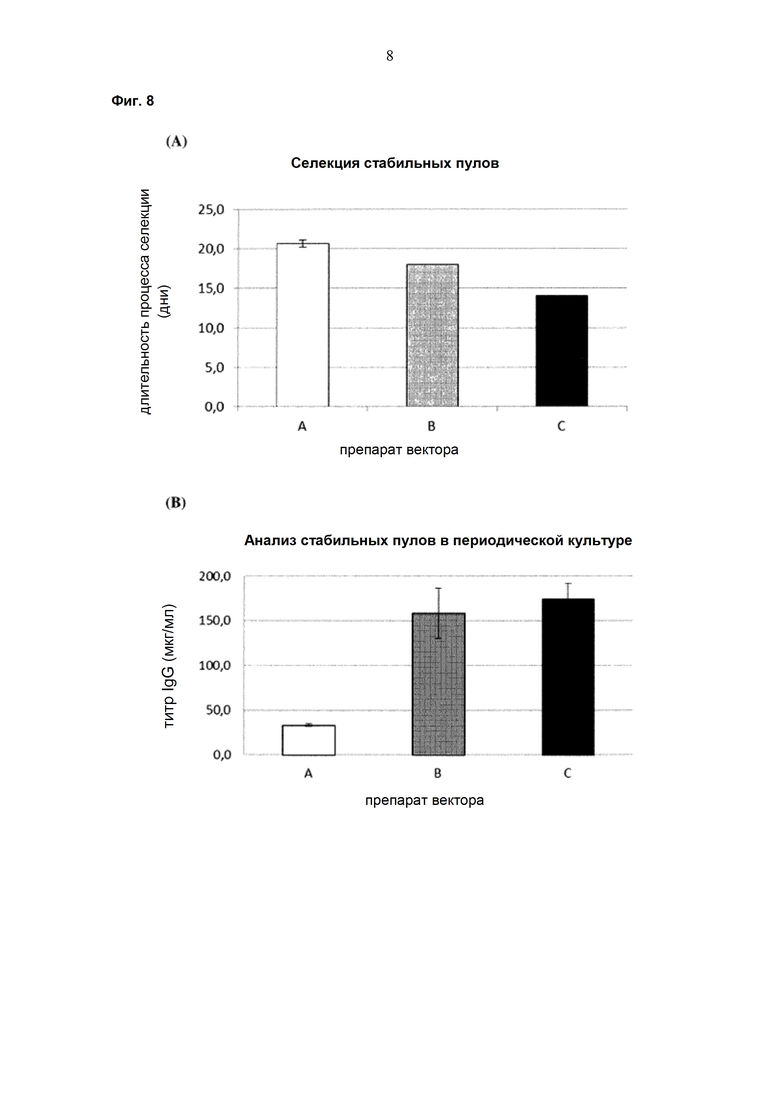
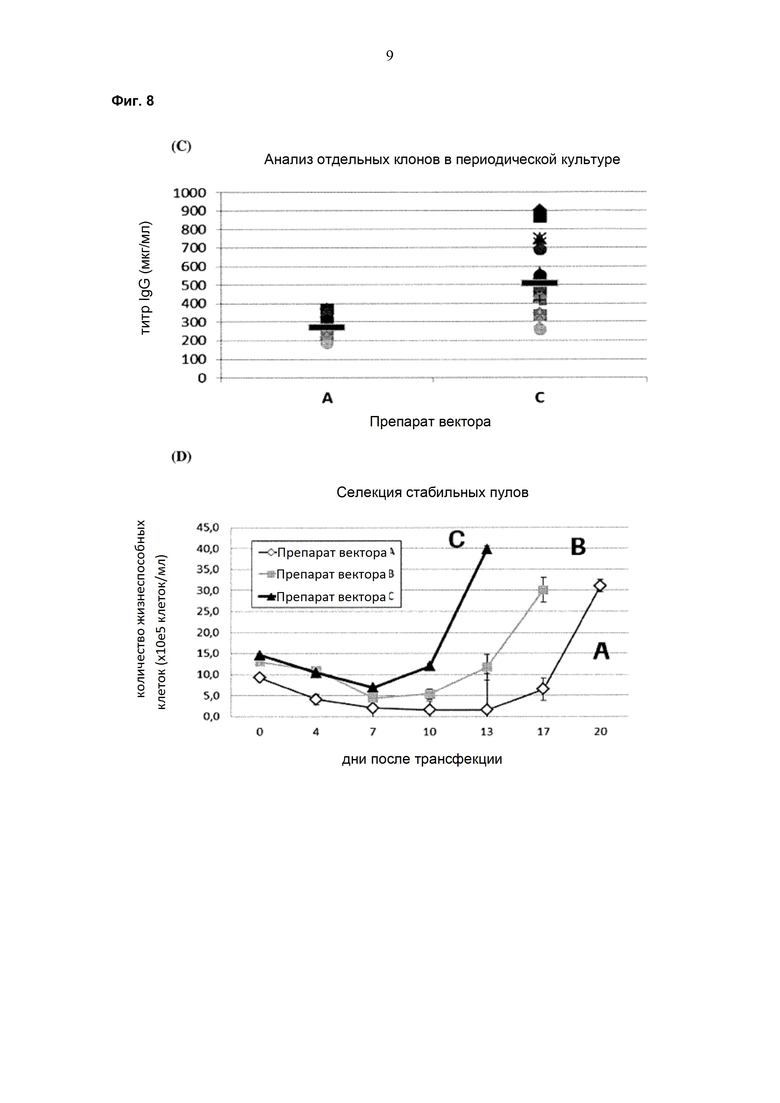
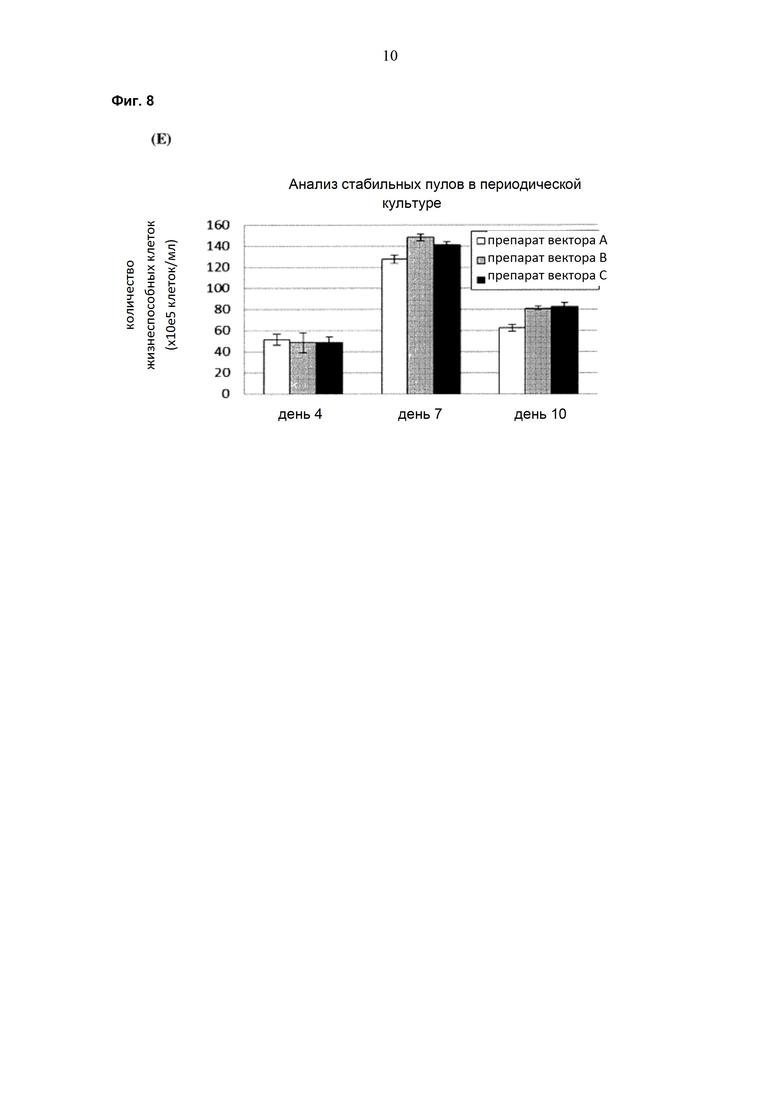
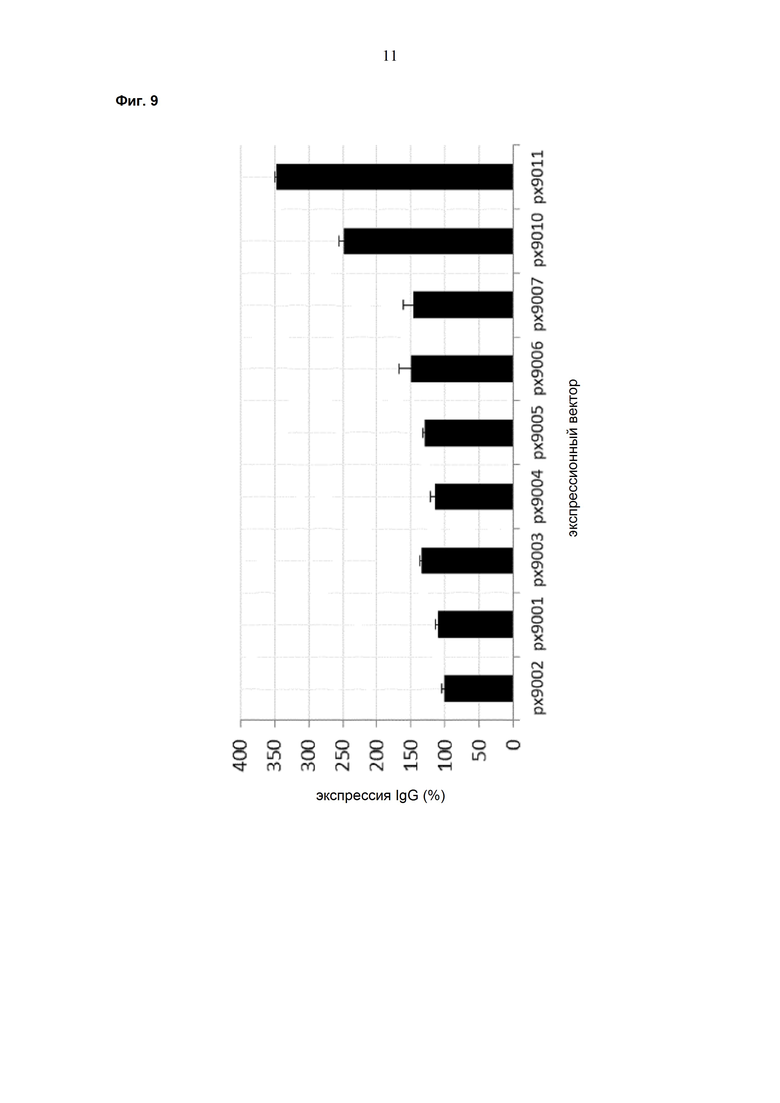
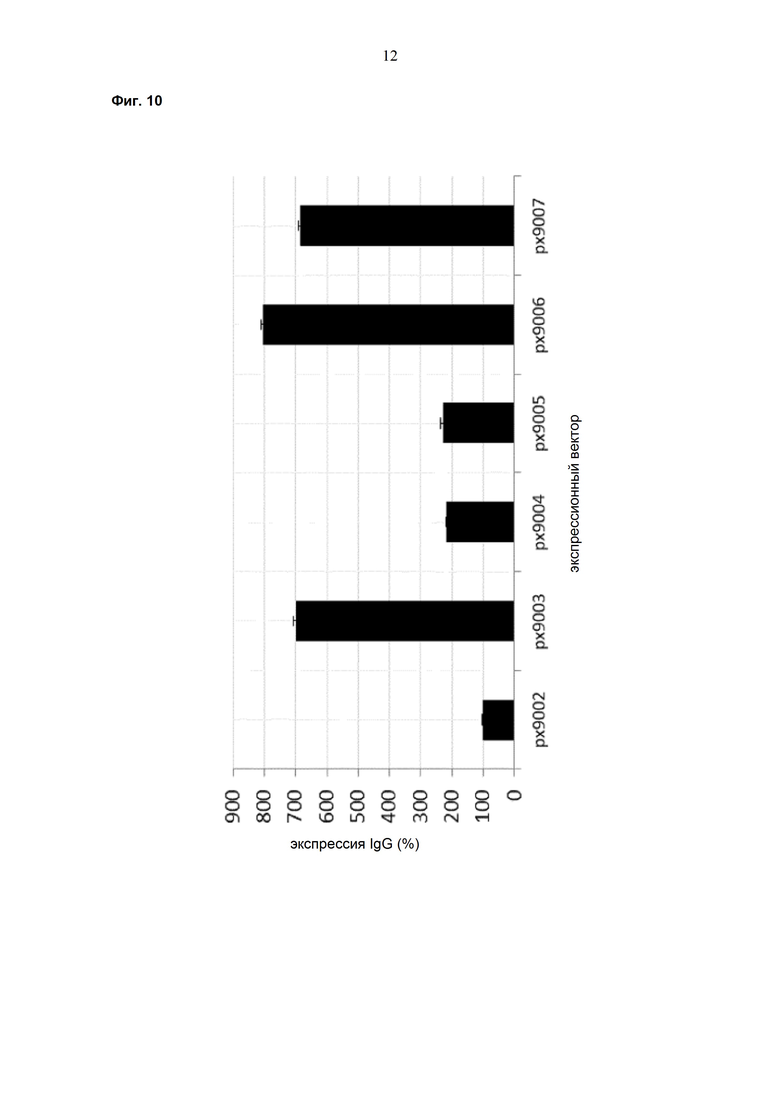
Изобретение относится к области биотехнологии. Предложен экспрессионный вектор, содержащий экспрессионные кассеты, расположенные однонаправленно по направлению от 5'- к 3'-концу последовательности в следующем порядке: экспрессионная кассета тяжёлой цепи антитела, экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета маркера селекции. Экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета тяжёлой цепи антитела содержат промотор CMV человека, поли(A)-сигнальную последовательность бычьего гормона роста и терминаторную последовательность гастрина человека. Экспрессионная кассета маркера селекции содержит промотор CMV человека или промотор SV40 и поли(A)-сигнальную последовательность SV40. Также раскрыты способы получения продуцирующих антитела клеток и применение этих клеток для рекомбинантного получения антител. Изобретение обеспечивает получение большого количества высокопродуктивных клонов для продуцирования рекомбинантных антител. 8 н. и 12 з.п. ф-лы, 10 ил.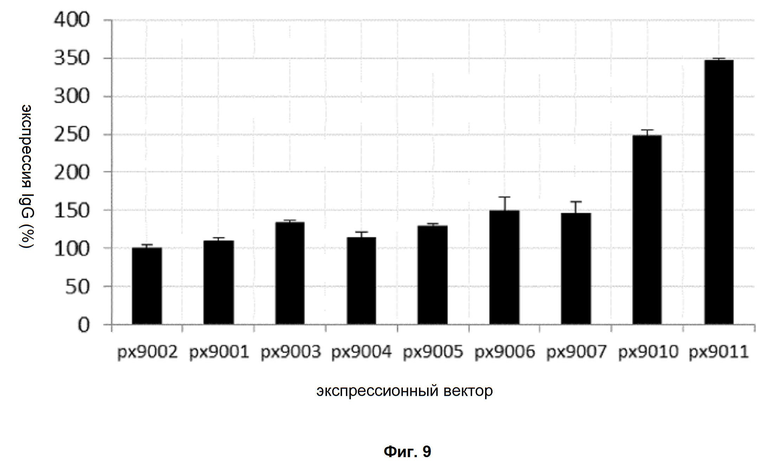
1. Экспрессионный вектор, содержащий:
экспрессионную кассету лёгкой цепи антитела,
экспрессионную кассету тяжёлой цепи антитела и
экспрессионную кассету маркера селекции,
где экспрессионные кассеты расположены однонаправленно и где экспрессионные кассеты расположены по направлению от 5'- к 3'-концу последовательности в следующем порядке: экспрессионная кассета тяжёлой цепи антитела, экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета маркера селекции, где экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета тяжёлой цепи антитела содержат промотор CMV человека, поли(A)-сигнальную последовательность бычьего гормона роста и терминаторную последовательность гастрина человека, а экспрессионная кассета маркера селекции содержит промотор CMV человека или промотор SV40 и поли(A)-сигнальную последовательность SV40.
2. Экспрессионный вектор по п. 1, в котором нуклеиновая кислота, кодирующая лёгкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжёлую цепь антитела, содержит по меньшей мере один интрон.
3. Экспрессионный вектор по п. 1, в котором нуклеиновая кислота, кодирующая лёгкую цепь антитела, и/или нуклеиновая кислота, кодирующая тяжёлую цепь антитела, является кДНК.
4. Применение экспрессионного вектора по любому из пп. 1-3 для рекомбинантного получения антитела.
5. Применение экспрессионного вектора по любому из пп. 1-3 для получения клеточной линии, экспрессирующей антитело.
6. Способ трансфекции эукариотической клетки экспрессионным вектором по любому из пп. 1-3, где экспрессионный вектор линеаризован перед трансфекцией путём разрезания в точке начала репликации прокариот.
7. Способ по п. 6, в котором точка начала репликации прокариот находится между экспрессионной кассетой лёгкой цепи антитела и экспрессионной кассетой тяжёлой цепи антитела.
8. Способ получения антитела, включающий следующие этапы: культивирование эукариотической клетки, которая была трансфецирована экспрессионным вектором по любому из пп. 1-3, и выделение антитела из клетки или из культуральной среды.
9. Способ получения антитела, включающий следующий этап: культивирование эукариотической клетки, полученной путём трансфекции экспрессионным вектором по любому из пп. 1-3, который был линеаризован перед трансфекцией путём разрезания в точке начала репликации прокариот, и выделение антитела из клетки или из культуральной среды.
10. Способ получения антитела, включающий следующий этап: культивирование эукариотической клетки, которая была трансфецирована экспрессионным вектором по любому из пп. 1-3 и которая была селектирована с помощью добавления в культуральную среду агента для селекции примерно через 24 часа после трансфекции, и выделение антитела из клетки или из культуральной среды.
11. Способ трансфекции эукариотической клетки экспрессионным вектором по любому из пп. 1-3, где экспрессионный вектор содержит прокариотические и эукариотические последовательности нуклеиновых кислот, в котором прокариотические последовательности нуклеиновых кислот удаляют из экспрессионного вектора перед трансфекцией эукариотической клетки экспрессионным вектором.
12. Экспрессионный вектор по п. 1, где экспрессионный вектор кодирует биспецифическое антитело.
13. Экспрессионный вектор по п. 12, в котором биспецифическое антитело имеет первую специфичность связывания или сайт связывания, который специфично связывается с первым антигеном или с первым эпитопом на антигене, и биспецифическое антитело имеет вторую специфичность связывания или сайт связывания, который специфично связывается со вторым антигеном или со вторым эпитопом на антигене.
14. Экспрессионный вектор по п. 1, где экспрессионный вектор содержит первую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую первую лёгкую цепь антитела, поли(A)-сигнальную последовательность и терминаторную последовательность, вторую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую вторую лёгкую цепь антитела, поли(A)-сигнальную последовательность,и терминаторную последовательность, третью экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую первую тяжёлую цепь антитела, поли(A)-сигнальную последовательность и терминаторную последовательность, четвёртую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую вторую тяжёлую цепь антитела, поли(A)-сигнальную последовательность и терминаторную последовательность, или первую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую лёгкую цепь антитела, поли(A)- сигнальную последовательность, и терминаторную последовательность, вторую экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую первую тяжёлую цепь антитела, поли(A)-сигнальную последовательность, и терминаторную последовательность, третью экспрессионную кассету, содержащую по направлению от 5'- к 3'-концу промотор, нуклеиновую кислоту, кодирующую вторую тяжёлую цепь антитела, поли(A)-сигнальную последовательность и терминаторную последовательность, где лёгкая цепь антитела является общей для обеих тяжёлых цепей антитела.
15. Экспрессионный вектор по п. 1, где экспрессионный вектор содержит экспрессионную кассету лёгкой цепи антитела, экспрессионную кассету первой тяжёлой цепи антитела, экспрессионную кассету второй тяжёлой цепи антитела и экспрессионную кассету маркера селекции, где по меньшей мере одна из экспрессионной кассеты тяжёлой цепи антитела и экспрессионной кассеты лёгкой цепи антитела и экспрессионная кассета маркера селекции расположены однонаправленно и где однонапраленные экспрессионные кассеты расположены по направлению от 5'- к 3'-концу последовательности в следующем порядке: экспрессионная кассета тяжёлой цепи антитела, экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета маркера селекции.
16. Экспрессионный вектор по п. 1, где экспрессионный вектор содержит экспрессионную кассету первой лёгкой цепи антитела, экспрессионную кассету второй лёгкой цепи антитела, экспрессионную кассету первой тяжёлой цепи антитела, экспрессионную кассету второй тяжёлой цепи антитела и экспрессионную кассету маркера селекции, где одна из экспрессионных кассет тяжёлой цепи антитела, одна из экспрессионных кассет лёгкой цепи антитела и экспрессионная кассета маркера селекции расположены однонаправленно и где однонаправленные экспрессионные кассеты расположены по направлению от 5'- к 3'-концу последовательности в следующем порядке: экспрессионная кассета тяжёлой цепи антитела, экспрессионная кассета лёгкой цепи антитела и экспрессионная кассета маркера селекции.
17. Экспрессионный вектор по п. 12, в котором одна из экспрессионных кассет тяжёлой цепи антитела кодирует тяжёлую цепь антитела, содержащую мутацию типа «впадина».
18. Экспрессионный вектор по п. 12, в котором одна из экспрессионных кассет тяжёлой цепи антитела кодирует тяжёлую цепь антитела, содержащую мутацию типа «выступ».
19. Экспрессионный вектор по п. 12, в котором одна из экспрессионных кассет лёгкой цепи антитела кодирует вариант лёгкой цепи антитела, содержащий вариабельный домен лёгкой цепи антитела и домен CH1 тяжёлой цепи антитела в качестве константного домена, и/или одна из экспрессионных кассет лёгкой цепи антитела кодирует лёгкую цепь антитела, содержащую вариабельный домен лёгкой цепи антитела и домен CL лёгкой цепи антитела в качестве константного домена.
20. Экспрессионный вектор по п. 12, в котором одна из экспрессионных кассет тяжёлой цепи антитела кодирует вариант тяжёлой цепи антитела, содержащий в качестве первого константного домена константный домен лёгкой цепи (CL), и/или одна из экспрессионных кассет тяжёлой цепи антитела кодирует тяжёлую цепь антитела, содержащую в качестве первого константного домена константный домен CH1 тяжёлой цепи.
| WO 2008142124 A1, 27.11.2008 | |||
| KENNARD M.L | |||
| et al., The Generation of Stable, High MAb Expressing CHO Cell Lines Based on the Artificial Chromosome Expression (ACE) Technology, BIOTECHNOLOGY AND BIOENGINEERING, 2009, vol | |||
| Счетная таблица | 1919 |
|
SU104A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| РЕКОМБИНАНТНЫЙ ЛЕНТИВИРУСНЫЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ТРАНСДУЦИРОВАННАЯ ЛЕНТИВИРУСНЫМ ВЕКТОРОМ, СПОСОБ ЕЕ ТРАНСДУКЦИИ И ПРИМЕНЕНИЕ | 2002 |
|
RU2305708C2 |
| US 6207455 B1, 27.03.2001 | |||
| SCHLATTER S | |||
| et al., On the Optimal Ratio of | |||

