Область техники
Настоящее изобретение относится к способам получения генетически модифицированных птиц и/или птиц дикого типа, например, кур, из половых клеток, и к птицам, полученным с применением таких способов.
Уровень техники
Получение пород птиц из выделенных половых клеток в настоящее время является в значительной степени неэффективным и сложным процессом. Особой проблемой, связанной с получением генетически модифицированных птиц, является стабильная передача половых клеток, содержащих генетическую модификацию, потомству птиц и последующим поколениям таких птиц. В частности, при использовании обычных способов передача зародышевых клеток является особенно неэффективной из-за конкуренции в формировании функциональных гамет между донорскими зародышевыми клетками, которые могут быть или не быть генетически модифицированными, и эндогенными зародышевыми клетками.
Хотя диплоидные зародышевые клетки могут быть трансплантированы от одной птицы птице-хозяину, доля потомства, полученного из образовавшихся впоследствии гамет, варьирует, при этом все потомство или его часть образуется из гамет, полученных из эндогенных (принадлежащих птице-хозяину) зародышевых клеток (Nakamura et al (2010) Reprod Fert Dev 22(8): 1237-1246; Song et al (2014) Biol Reprod 90(1): 15). Эндогенные (хозяйские) зародышевые клетки могут быть уничтожены с использованием облучения или химиотерапевтических реагентов, таких как бусульфан, но эти токсичные реагенты также могут убить животное вместе с эндогенными зародышевыми клетками. Tagami использовал химическую обработку для получения стерильных суррогатных кур-хозяев (Nakamura et al (2010) Biol Reprod 83(1):130-7). Авторы изобретения и Nakamura использовали гамма-облучение для уничтожения эндогенных зародышевых клеток (MacDonald et al (2010) Plos One 5(11): e15518; Nakamura et al, (2012) J Reprod Dev 58(4): 432-437). Однако, хотя количество потомства, полученного из донорских зародышевых клеток, увеличилось после обработки, не все потомство было получено из донорских зародышевых клеток, и обработка убила многих кур-хозяев.
Созданы трансгенные линии стерильных млекопитающих и рыб, которые экспрессируют продукт гена (нитроредуктазу, Ntr) в зародышевых клетках, который будет уничтожать зародышевые клетки в присутствии пролекарства. Ген iC9 (индуцибельная каспаза 9) использовали для уничтожения стволовых клеток у людей и мышей и для уничтожения эндотелиальных клеток у трансгенных мышей. Нельзя ожидать, что такие методы можно будет напрямую перенести на птиц, учитывая разные результаты, полученные с использованием методов модификации зародышевых клеток при применении у млекопитающих и птиц. Например, в предыдущей работе авторов изобретения была получена курица-самка без зародышевых клеток в результате генетической мутации в гене DDX4 с помощью технологии редактирования генома. (Taylor et al., Development 2017). Авторы изобретения не ожидали, что курица-самка будет стерильной, поскольку, в то время как самцы млекопитающих с мутантным геном Ddx4 являются стерильными, самки млекопитающих с мутантным Ddx4 имеют нормальную репродуктивную способность. Таким образом, методы модификации зародышевых клеток могут оказывать совершенно разные эффекты у млекопитающих и птиц. Стерильность также зависит от того, на какой стадии развития погибают эндогенные половые клетки. У птиц стерильные самки DDX4 содержат половые клетки до вылупления, которые могут конкурировать с донорскими зародышевыми клетками, инъецированными в генетически модифицированный хозяйский эмбрион.
Краткое описание изобретения
Настоящее изобретение решает многие проблемы предшествующего уровня техники. Авторы изобретения неожиданно показали, что при применении генной инженерии для экспрессии рекомбинантного белка в зародышевых клетках, который будет селективно уничтожать хозяйские зародышевые клетки по требованию, например, при воздействии определенного пролекарства или индуктора, не уничтожая другие клетки курицы-хозяина, может быть достигнута высокоэффективная интеграция донорских половых клеток. Этой птице-хозяину можно переносить зародышевую плазму (половые клетки) от разных видов птиц, и генетика потомства будет формироваться из перенесенного материала (фиг. 1).
Хотя были созданы трансгенные линии стерильных млекопитающих и рыб, экспрессирующие этот продукт гена, различия в активности и функции белка у представителей птиц и млекопитающих и различия в функции и активности генов между представителями птиц и млекопитающих означают, что нельзя ожидать, что методы, используемые у млекопитающих и рыб, могут быть перенесены на птиц, и, таким образом, методы модификации клеток могут оказывать совершенно разные эффекты у птиц и млекопитающих. Стерильность также зависит от того, когда эндогенные половые клетки погибают во время развития (временный характер активности гена или белка). Необходимо проанализировать много различных локусов у птиц, чтобы определить, будут ли они экспрессировать трансген на подходящей стадии развития, то есть, могут ли зародышевые клетки быть удалены на 100% во время эмбрионального развития.
Соответственно, первый аспект настоящего изобретения относится к трансгенной птице, содержащей трансген в зародышевых клетках указанной птицы, где активность белка, кодируемого указанным трансгеном, индуцируется посредством экзогенного индуцирующего агента, и при индукции активность указанного белка вызывает гибель указанных зародышевых клеток.
Второй аспект изобретения относится к трансгенной конструкции, содержащей (i) первую нуклеотидную последовательность, где активность белка, кодируемого указанной первой нуклеотидной последовательностью, вызывает гибель зародышевых клеток в присутствии экзогенного индуцирующего агента, и (ii) вторую нуклеотидную последовательность, которая нацеливает указанную конструкцию на птичьи зародышевые клетки.
Третий аспект относится к способу модификации зародышевой плазмы птицы, включающему введение трансгенной конструкции в оплодотворенное яйцо указанной птицы и инкубацию указанного яйца, где указанная трансгенная конструкция представляет собой трансгенную конструкцию согласно второму аспекту изобретения, и где указанная трансгенная конструкция встроена в зародышевые клетки указанного эмбриона.
Четвертый аспект изобретения относится к трансгенной птице, содержащей трансгенную конструкцию согласно второму аспекту изобретения или полученной способом согласно третьему аспекту изобретения.
Согласно изобретению птица может представлять собой любую подходящую птицу. Например, птица может принадлежать к порядку galliformes (курообразные), aseriformes (гусеобразные), passeriformes (воробьинообразные), gruiformes (журавлеобразные), Struthioniformes (страусообразные), rheiformes (нандуобразные), casuariformes (казуарообразные), apyerygiformes (кивиобразные), otidiformes (дрофообразные), columbiformes (голубеобразные), sphenisciformes (пингвинообразные), cathartiformes (катартиды), accipitriformes (ястребообразные), strigiformes (совообразные), psittaciformes (попугаеобразные), charadriiformes (ржанкообразные) или falconiformes (соколообразные). В подходящем варианте птица представляет собой курицу, индейку, утку, гуся, перепела, фазана, тетерева, цесарку, голубя, страуса, эму, певчую птицу, попугая, зяблика, воробья, пингвина или сокола. В одном из вариантов осуществления птица представляет собой курицу.
В изобретении трансгенная конструкция согласно изобретению и для применения в нем нацелена на зародышевые клетки. В одном из вариантов осуществления зародышевые клетки представляют собой примордиальные зародышевые клетки. В еще одном варианте осуществления зародышевые клетки представляют собой зародышевые клетки взрослого. Предпочтительно указанная конструкция нацелена на локус птичьего генома, который избирательно экспрессируется в примордиальных зародышевых клетках или зародышевых клетках в гонаде эмбриона, или в семенниках и яичниках взрослой птицы. Предпочтительно локус экспрессируется только в примордиальных зародышевых клетках или зародышевых клетках в гонаде эмбриона или взрослой птицы. В одном из вариантов осуществления трансгенная конструкция нацелена на один из следующих локусов, которые у птицы экспрессируются только в половых клетках: DAZL, DDX4, DMRT1, MIR383, TDRD15, TDRD5, FKB6, GASZ, DMRTB1, TDRD9, GTSF1, MOV10L1, STK31, RNF17, FDFT1, GNG10, DDX43, KCNH7, SOX21 TUBA1B или PNLDC1. В еще одном варианте осуществления указанная конструкция нацелена на один или более из следующих генов, кодирующих РНК: MSTRG.9846 (2:40789480-40848190), MSTRG.10457(2:71880785-71991485), MSTRG.17017 (3: 85453009-85462029). В подходящем варианте трансгенная конструкция нацелена на один из следующих локусов: DAZL, RNF17, TUBA1B, TUBA1C, STK31, FDFT1, gga-mir-6611. Предпочтительно трансген нацелен на локусы, которые на наиболее высоком уровне экспрессируются в куриных ПЗК. Предпочтительно трансген нацелен на локусы, которые на наиболее высоком уровне экспрессируются в ПЗК определенных птиц, например, гуся, утки и т. п.
Сравнение транскриптома РНК примордиальных зародышевых клеток различных видов птиц позволило определить гены, которые экспрессируются на высоких уровнях в зародышевых клетках большинства видов птиц. Этот анализ показал, что DAZL имеет самый высокий уровень у большинства видов птиц, GTSF1 - второй по величине для гусиных зародышевых клеток, а TDRD9 - второй по величине для утиных зародышевых клеток. Таким образом, разные участки генома могут лучше функционировать у отдельных видов птиц. На фиг. 21 показаны уровни экспрессии у курицы, утки и гуся, соответственно.
В подходящем варианте трансген нацелен на локусы, которые экспрессируются в правильное время в процессе роста и репродуктивном цикле птиц, чтобы обеспечить развитие птицы и репродуктивной системы птицы, но минимизировать конкуренцию донорских и хозяйских половых клеток. Например, в вариантах осуществления изобретения авторы изобретения определили, что из-за временного характера обеспечиваемой экспрессии стерильные самки DDX4 содержали половые клетки до вылупления, которые могут конкурировать с донорскими зародышевыми клетками. Зародышевые клетки погибали только после вылупления. Понятно, что возможность обеспечения возможности экспрессии до тех пор, пока не будет применен экзогенный агент, модулирования экспрессии посредством предоставления экзогенного агента и возможность учитывать временной характер экспрессии может обеспечить более выгодную систему, чем простой нокаут полового гена в организме хозяина.
В подходящем варианте абляционный ген гибели клеток и локус, на который он нацелен, могут быть выбраны для абляции зародышевых клеток во время эмбрионального развития в зависимости от требований.
Абляционный трансген гибели клеток должен быть достаточно активным, чтобы обеспечивать абляцию большей части зародышевых клеток во время эмбрионального развития. DDX4 может быть выбран в качестве второй по предпочтительности альтернативы dazl в качестве локуса у кур. Понятно, что обеспечение трансгена, вызывающего абляцию клеток, обладающего повышенной апоптотической активностью могло бы преодолеть недостаточность абляции, о которой сообщалось при использовании нитроредуктазы. Например, повышенная активность iCaspase9 позволила бы использовать локус DDX4. Как будет понятно специалистам в данной области техники, для идентификации изменений аминокислот в белках, которые обеспечивают абляцию или потенциальную абляцию (Caspase9 или нитроредуктаза, или любой другой ген, индуцирующий гибель клеток), может быть использован скрининговый анализ, и чтобы сделать такие белки достаточно активными или более активными для обеспечения абляции зародышевых клеток на функционально полезных уровнях/с повышенной эффективностью может быть использован скрининг. Затем такие белки могут быть введены в любой из перечисленных локусов, специфичных для зародышевых клеток.
В вариантах осуществления изобретения было обнаружено, что ген каспазы 9 (версия аминокислот млекопитающего или модифицированной последовательностями курицы («куринизированная») версия (aviCaspase9), т.е. версия, приближенная по последовательности аминокислот к куриному белку) является особенно подходящим для обеспечения абляции хозяйских клеток. Как отмечено в примерах, было определено, что aviCaspase9 не вызывает абляцию всех зародышевых клеток, когда он присутствует в культуре клеток при введении в локус DDX4 куриной клетки.
В подходящем варианте трансген каспазы 9 с аминокислотами млекопитающего может быть предложен для абляции зародышевых клеток при нацеливании на локус DAZL. В подходящем варианте абляция обеспечивает получение по существу чистого донорского потомства, например, более 75%, более 80%, более 85%, более 90%, более 95%, более 97%, более 99%, 100%. В подходящем варианте iCaspase9 может быть использован у кур для стерилизующей абляции. Было установлено, что ген iCaspase9 обеспечивает полную стерилизующую абляцию у кур (фиг. 23).
В частном варианте осуществления трансген может представлять собой кДНК, нацеленную на птичий локус DAZL, в частности, на с-концевую область птичьего локуса DAZL. Авторы изобретения определили, что DAZL не только экспрессируется на высоких уровнях в примордиальных зародышевых клетках и, таким образом, обеспечивает селективную абляцию клеток, но также обеспечивает достаточную экспрессию токсичного белка для обеспечения абляции. Более того, авторы изобретения определили, что помимо уровня экспрессии, обеспечиваемого локусами, важно время осуществления экспрессии (временной характер экспрессии). DAZL экспрессируется на ранней стадии в примордиальных зародышевых клетках птичьего эмбриона. Следовательно, временная экспрессия DAZL также выгодна для его использования при экспрессии токсичных белков или белков, индуцирующих апоптоз. В подходящем варианте iCaspase9 под управлением DAZL является особенно предпочтительным. Например, авторы изобретения определили, что трансген iCaspase9 хорошо функционирует в локусе DAZL курицы.
Уровень экспрессии, обеспечиваемый конструкцией, и время осуществления экспрессии считаются важными факторами в контексте экспрессии токсичных белков или белков, индуцирующих апоптоз (например, каспазы), в отличие от использования нокаутов, когда может быть достаточным разрушение локуса. Степень абляции, производимой трансгеном, может регулироваться генетическим расположением трансгена, например, когда он встроен на 5'-конце гена или встроен на 3'-конце гена, или как независимый трансген, экспрессия которого управляется регуляторными областями генов, экспрессируемых только в половых клетках птиц.
Как описано в настоящем документе, трансгенная конструкция согласно изобретению и для применения в нем содержит первую нуклеотидную последовательность, экспрессируемый с которой белок вызывает гибель зародышевых клеток в присутствии экзогенного индуцирующего агента. В некоторых вариантах осуществления трансген кодирует белок, активность или экспрессия которого индуцируются посредством экзогенного индуцирующего агента, где экспрессированный белок вызывает гибель указанных зародышевых клеток в присутствии экзогенного индуцирующего агента.
В подходящем варианте трансген может содержать часть, которая кодирует индуцируемый домен димеризации и домен, индуцирующий апоптоз. В подходящем варианте индуцируемый домен димеризации может представлять собой химически индуцируемый домен димеризации. В присутствии агента, индуцирующего димеризацию, например, химического соединения, индуцирующего димеризацию, экспрессированный белок димеризуется, вызывая апоптоз эндогенных зародышевых клеток.
В изобретении может быть использован любой подходящий домен, индуцирующий апоптоз. В одном из вариантов осуществления домен, индуцирующий апоптоз, содержит ген каспазы, кодирующий белок каспазы, или состоит из него. Такими белками каспазы являются каспаза 2, 3, 4, 6, 7, 8, 9 или 10. Такой белок каспазы может содержать аминокислотные последовательности млекопитающих или птиц, или других позвоночных. В одном из вариантов осуществления каспаза представляет собой каспазу 9.
Соответственно, в подходящем варианте ген индуцибельная каспаза 9 (iC9) может экспрессироваться в зародышевых клетках птицы, например, курицы. При воздействии препарата для химически индуцированной димеризации (CID) каспаза 9 будет димеризоваться и активироваться, а затем вызовет апоптоз зародышевых клеток, содержащих димеризованную каспазу 9.
В подходящем варианте экспрессия указанного гена каспазы может быть индуцирована применением агента-индуктора димеризации посредством домена димеризации, индуцирующего димеризацию. Подходящие агенты-индукторы димеризации, которые могут быть использованы в изобретении, включают лиганд AP20187 (молекула B/B, Takara) или химические варианты этого продукта FK1012, AP1501, AP1903.
В одном таком варианте осуществления трансген представляет собой кДНК, кодирующую домен димеризации FKBP12 и ген каспазы 9, нацеленный на генетический локус, выбранный из DAZL или DDX4, в частности, DAZL или DDX4 у курицы.
В еще одном варианте осуществления указанный трансген может кодировать домен димеризации, слитый с доменом, индуцирующим апоптоз, например, геном каспазы, который может приводить к димеризации кодируемого белка после доставки препарата-индуктора стабильной димеризации. Например, трансген может кодировать стабилизируемый полипептидный линкер (SPL), присоединенный к молекуле каспазы. Добавление соединения, такого как асунапревир и телапревир, может стабилизировать домен димеризации молекул каспазы, что приведет к активации молекулы каспазы и активации гибели клеток (Jacobs et al (2018) Nature Methods 15: 523-526).
В альтернативном варианте осуществления трансген может кодировать фермент, который превращает пролекарства в цитотоксические метаболиты. В таком примере в эндогенные зародышевые клетки может быть введено пролекарство (которое действует как экзогенный индуцирующий агент), и экспрессия трансгена, например кДНК, кодирующей ген бактериальной нитроредуктазы, может обеспечить фермент, который превращает пролекарство в цитотоксический метаболит.
В такой системе могут быть использованы любой подходящий фермент и пролекарство, активируемое указанным ферментом. Например, когда фермент представляет собой нитроредуктазу, пролекарство может представлять собой CB1954 или метронидазол.
В одном таком варианте осуществления трансген представляет собой кДНК, кодирующую ген нитроредуктазы, нацеленный на генетический локус, выбранный из DAZL или DDX4, в частности, DAZL или DDX4 у курицы.
В одном из вариантов осуществления трансгенная конструкция согласно изобретению и для применения в нем содержит кДНК и последовательность 2A или IRES (участок внутренней посадки рибосомы), так что рекомбинантный белок экспрессируется на одинаковых уровнях с эндогенным геном. Например, пептидная последовательность 2А может быть связана с кДНК, так что рекомбинантный белок экспрессируется на одинаковых уровнях с эндогенным геном.
В одном из вариантов осуществления нуклеотидная последовательность из локуса, который у птицы экспрессируется только в зародышевых клетках или половых клетках (примеры таких локусов, включая DAZL и DDX4, приведены выше), где указанная нуклеотидная последовательность содержит регуляторные области и первый экзон до первого кодирующего метионина, связана с кДНК. Эта область ДНК может быть введена в организм птицы любым подходящим способом, который будет обеспечивать специфическую экспрессию рекомбинантного белка в зародышевых клетках, например, в транспозоне.
Примеры матриц репарации DAZL и DDX4 для применения в изобретении показаны на фиг. 15, 16, 17 и 18. Такие матрицы репарации составляют дополнительные независимые аспекты изобретения.
В способах согласно изобретению для нацеливания трансгенной конструкции на зародышевые клетки могут быть использованы любые подходящие средства. В подходящем варианте для нацеливания трансгена на зародышевые клетки может быть использована система на основе CRISPR, такая как система на основе CRISPR/cas или CRISPR/cfp. В такой системе гидовая РНК может нацеливать конструкцию на зародышевые клетки.
В соответствии с дополнительным аспектом настоящего изобретения предложен набор, содержащий трансгенную конструкцию согласно второму аспекту изобретения и сайт-специфическую нуклеазу, такую как Cas9, для нацеливания указанной конструкции на генетический локус, специфически транскрибируемый в зародышевых клетках.
В альтернативном варианте осуществления системы на основе CRISPR для нацеливания трансгенной конструкции на зародышевые клетки может быть использована система на основе транспозона для нацеливания трансгенной конструкции на зародышевые клетки. В такой системе нацеливание может быть достигнуто путем включения в транспозон (i) регуляторных областей из гена, избирательно экспрессируемого, предпочтительно исключительно экспрессируемого, в зародышевых клетках, вместе с (ii) указанной первой нуклеотидной последовательностью. Например, такой транспозон может содержать регуляторные области из DDX4 или DAZL, ген каспазы 9 и домен димеризации (в подходящем варианте - DAZL и icaspase9). Ожидается, что первая нуклеотидная последовательность будет экспрессироваться только в зародышевых клетках. Таким образом, при применении экзогенного индуцирующего агента можно ожидать гибели только зародышевых клеток.
Трансгенная конструкция согласно второму аспекту изобретения может быть использована для модификации зародышевой плазмы птицы. Путем нацеливания трансгенной конструкции на зародышевые клетки и введения индуцирующего агента трансген активируется таким образом, что он может селективно уничтожать эндогенные зародышевые клетки. В подходящем варианте активация трансгена зародышевых клеток вызывает снижение количества зародышевых клеток по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 50%, по меньшей мере на 70%, по меньшей мере на 90%, до 100% от нормальных значений. В подходящем варианте процесс обеспечивает птицу, у которой отсутствуют эндогенные половые клетки.
В подходящем варианте при индукции трансген может экспрессироваться в присутствии экзогенного индуцирующего агента на уровне, достаточном для того, чтобы вызвать гибель зародышевых клеток при культивировании in vitro.
После или во время уничтожения эндогенных зародышевых клеток посредством, например, активации белка, кодируемого доменом, индуцирующим апоптоз, или превращения пролекарства в цитотоксический метаболит ферментом, кодируемым трансгеном, птице, лишенной зародышевых клеток (птице-хозяину), могут быть введены клетки (трансплантированные клетки), например, зародышевые клетки от другой птицы того же или другого вида птиц, так что птица-хозяин производит потомство с генетикой трансплантированных клеток. В подходящем варианте способ может включать этап трансплантации зародышевых клеток от птицы-донора суррогатной птице.
В подходящем варианте индукция активности белка, кодируемого трансгеном, может быть остановлена до трансплантации донорских клеток.
В подходящем варианте трансплантированные клетки от птицы-донора могут быть получены из замороженных клеток.
Понятно, что зародышевые клетки, трансплантированные суррогатному хозяину от птицы-донора, будут иметь повышенный шанс конкурировать с эндогенными зародышевыми клетками, которые присутствовали в организме суррогатного хозяина, из-за действия трансгена.
В подходящем варианте трансплантированные клетки могут представлять собой половые клетки с отредактированным геномом. В подходящем варианте половые клетки с отредактированным геномом могут происходить от того же или другого вида птиц, что и птица-хозяин.
В подходящем варианте способ может включать этап обеспечения птицы с геномом трансплантированных клеток. В подходящем варианте суррогатная птица-хозяин может применяться для получения множества, например, стаи, птиц с отредактированным геномом из половых клеток с отредактированным геномом этого же вида птиц или другого вида птиц.
В одном из аспектов изобретения предложен способ получения суррогатной птицы-хозяина, включающий встраивание трансгенной конструкции в оплодотворенные яйца птицы и инкубацию указанных яиц до вылупления, где указанную трансгенную конструкцию встраивают в зародышевые клетки указанного эмбриона и белок, экспрессируемый с этой трансгенной конструкции, вызывает гибель указанных зародышевых клеток в присутствии экзогенного индуцирующего агента. Указанный способ позволяет встраивать указанную трансгенную конструкцию не только в зародышевые клетки указанного эмбриона, но также и в зародышевые клетки всего потомства, полученного впоследствии от птицы, полученной из указанного эмбриона.
В подходящем варианте способ может дополнительно включать обработку полученного суррогатного хозяина экзогенным индуцирующим агентом, чтобы вызвать гибель указанных эндогенных зародышевых клеток. В подходящем варианте способ дополнительно включает трансплантацию экзогенных половых клеток указанному суррогатному хозяину.
В подходящем варианте способ дополнительно включает скрещивание самцов и самок-потомков от одной или более указанных суррогатных птиц-хозяев с получением птиц-потомков с зародышевыми клетками, имеющими генетические характеристики трансплантированных зародышевых клеток. Обнаружение потомства от трансплантированных зародышевых клеток может быть осуществлено стандартными методами геномного секвенирования.
Суррогатная птица-хозяин может быть использована для трансплантации клеток, в частности, зародышевых клеток, от других видов птиц. Зародышевые клетки могут представлять собой примордиальные зародышевые клетки, эмбриональные зародышевые клетки, гоноциты. Зародышевые клетки могут быть трансплантированы путем трансплантации семенников или яичников взрослой особи. Суррогатная птица-хозяин производит потомство с генетикой трансплантированных клеток.
Суррогатная птица-хозяин может быть использована для воссоздания видов птиц из замороженного генетического материала, хранящегося в форме половых клеток. Воссозданные виды птиц будут иметь геном замороженных половых клеток.
Варианты осуществления настоящего изобретения далее будут описаны только в качестве примера со ссылкой на прилагаемые графические материалы, где:
Фиг. 1. Схема трансплантации зародышевых клеток стерильному хозяину.
Фиг. 2. Схема, показывающая нацеливание NTR и iCaspase9 (iC9) на локус DDX4. Матрица репарации NTR, нацеленная на локус DDX4, и матрица репарации ic9, нацеленная на локус DDX4.
Фиг. 3. Схема, показывающая нацеливание NTR и iCaspase9 (iC9) на локус DAZL. Матрица репарации NTR, нацеленная на локус DAZL, и матрица репарации ic9, нацеленная на локус DAZL.
Фиг. 4. ПЗК, нацеленные на локус DAZL, экспрессируют примерно в 4 раза больше GFP, чем клетки, нацеленные на локус DDX4. ПЗК, содержащие GFP, нацеленные на локус Dazl, экспрессируют в 3,8 раза большую флуоресценцию GFP, чем ПЗК, содержащие GFP, нацеленные на локус Ddx4. A. Анализ флуоресценции GFP методом проточной цитометрии B. Микрофотография нацеленных ПЗК.
Фиг. 5. 500 ПЗК (контроль, CAG-NTR или Dazl-NTR) культивировали в присутствии или в отсутствие пролекарства нитроредуктазы CB1954 (n=2 для каждой концентрации пролекарства). Через 10 дней подсчитывали общее количество клеток в каждой лунке. N=6 для каждого значения из трех независимых экспериментов.
Фиг. 6. 500 ПЗК (dazl-GFP, dazl-каспаза-человека или dazl-каспаза-курицы) культивировали в присутствии или в отсутствие молекулы-индуктора димеризации B/B. Через 10 дней подсчитывали общее количество клеток в каждой лунке. N=6 для каждого значения из трех независимых экспериментов.
Фиг. 7. Обработка B/B вызывает абляцию инъецированных ПЗК, нацеленных посредством DAZL-iCasp9. Эмбрионам на стадии 16 HH инъецировали 3500 ПЗК, трансфицированных реагентами CRISPR для вставки гена индуцибельной каспазы и GFP в локус dazl, и 3500 ПЗК, трансфицированных транспозоном для случайной вставки кассеты для экспрессии TdTomato в геном. После инъекции эмбрионам вводили 50 мкл 1x пенициллина/стрептомицина с или без 25 нМ препарата-индуктора димеризации B/B. На 8 день развития гонады иссекали и просматривали при флуоресценции. Изображения представляют собой три независимых инъекции для каждого варианта обработки.
Фиг. 8. Семь вариантов потомков G1, нацеленных посредством iCaspase9 и aviCaspase9
DAZL icaspase9, Dazl aviCaspase9 (курицы) инъецировали в оплодотворенные яйца от гетерозиготных по DDX4 самцов кур, скрещенных с самками дикого типа. 3000 ПЗК инъецировали в эмбрионы стадии 16 HH через сделанные в яйцах отверстия, яйца запечатывали и инкубировали до вылупления. Разведение этих кур-основателей дало семь трансгенных потомков G1, содержащих нацеленный трансген. Показанные на фигуре положительные потомки имеют номера 42, 4, 18. A. ПЦР с праймерами, специфическими к Caspase9; MW, маркеры молекулярной массы, A, Нацеленные ПЗК, содержащие aviCaspase9, B, Нацеленные ПЗК, содержащие iCaspase9. B. ПЦР с праймерами, специфическими к GFP.
Фиг. 9. Экспрессия GFP на 6 и 10 день эмбрионов G2 iCaspas9 и aviCaspase9.
Эмбрионы G2, положительные по ПЦР на гены iCaspase9 и aviCaspase9, визуализировали на предмет флуоресценции GFP.
Фиг. 10. Экспрессия GFP является специфичной для зародышевых клеток у эмбрионов G2 iCaspas9 и aviCaspase9 на 10 день. Левые панели; Флуоресценция GFP в эмбрионах iCaspase9 и aviCaspase9, которым инъецировали B/B. Правые панели: иммунофлуоресценция на белок DDX4 (красный).
Фиг. 11. Эмбрионы iCaspase9 и aviCaspase9 G2, получившие B/B, не имеют зародышевых клеток.
B/B инъецировали в спинную аорту куриных эмбрионов на стадии 16 (день 2,5). Эмбрионы инкубировали и исследовали на 10 день на предмет флуоресценции GFP и экспрессии DDX4. Левые панели; Флуоресценция GFP в эмбрионах iCaspase9 и aviCaspase9, которым инъецировали B/B. Правые панели: иммунофлуоресценция для белка DDX4 (красный).
Фиг. 12. Обработка трансгенных эмбрионов aviCaspase9 препаратом B/B вызывает абляцию хозяйских зародышевых клеток и позволяет трансплантированным донорским клеткам заселять гонады хозяина. Эмбрионам на стадии 16 HH, содержащим трансген aviCaspase9, нацеленный на локус DAZL, инъецировали 200 донорских ПЗК, трансфицированных транспозоном для случайной вставки кассеты для экспрессии TdTomato в геном. Инъецированные клетки находились в 1 мкл раствора, содержащего 0,5 мМ (конечная концентрация) препарата-индуктора димеризации B/B. После инъецирования в яйца добавляли вводили 50 мкл 1x пенициллина/стретпомицина, содержащего 15 мкМ (конечная концентрация) препарата-индуктора димеризации B/B, и инкубировали в течение 8 дней. На 10 день развития гонады иссекали и просматривали при флуоресценции. Все зародышевые клетки в эмбрионе aviCaspase9, получившем препарат-индуктор димеризации, были из донорских клеток с TdTomato, эндогенные (хозяйские) зародышевые клетки не были обнаружены.
Фиг. 13. ДНК, синтезированная для трансгена iCaspase9 человека.
Фиг. 14. ДНК, синтезированная для трансгена iCaspase9 курицы.
Фиг. 15. Последовательность ДНК для матрицы репарации DDX, содержащей оптимизированный для кур ген нитроредуктазы.
Фиг. 16. Последовательность ДНК для матрицы репарации DAZL, содержащей aviCaspase9 курицы.
Фиг. 17. Последовательность ДНК для матрицы репарации DAZL, содержащей iCaspase9 человека.
Фиг. 18. Последовательность ДНК для матрицы репарации DDX4, содержащей iCaspase9 человека.
Фиг. 19. ДНК для оптимизированного для кур гена нитроредуктазы. В верхней строке этой таблицы указан однобуквенный аминокислотный код для каждой аминокислоты, присутствующей в последовательности Ntr E.coli и оптимизированных по кодонам последовательностях Ntr. Во второй строке указана последовательность оснований в последовательности Ntr E.coli, а в третьей строке таблицы выделены изменения, которые были выполнены при оптимизации гена Ntr для курицы. Выделенные столбцы показывают, где были выполнены три мутации для создания конструкции 3AAS Ntr: треонин в 41 кодоне был мутирован в глутамин (CAG); аспарагин в положении 71 кодона был мутирован в серин (AGC), а фенилаланин в положении 124 кодона был мутирован в треонин (ACC).
На фиг. 20 представлена таблица транскриптома РНК куриных примордиальных зародышевых клеток по сравнению с другими тканями куриного эмбриона и плюрипотентными клетками для идентификации генов, которые на этих эмбриональных стадиях экспрессируются только в зародышевых клетках и на высоких уровнях. ESC - куриные эмбриональные стволовые клетки, EGKX - клетки куриных эмбрионов на стадии отложенного яйца, Не плюри - совокупность 66 неплюрипотентных тканей и клеточных линий взрослых кур.
Фиг. 21. Экспрессия генов, специфичных для зародышевых клеток, в птичьих ПЗК. На графике показана относительная экспрессия генов, специфичных для зародышевых клеток, в куриных, гусиных и утиных ПЗК. Среднее значение нормированных значений экспрессии получено из пакета DESeq2. Эти значения экспрессии нормированы к общему количеству операций чтения для всех образцов. Зарегистрированная экспрессия TDRD9 и TUBA1B является более высокой в утиных ПЗК, GASZ и RNF17 экспрессируются на высоком уровне в гусиных ПЗК, а остальные гены экспрессируются на более высоком уровне в куриных ПЗК.
Фиг. 22. 500 ПЗК (dazl-GFP, dazl-iCaspase, dazl-aviCaspase, ddx4-iCaspase9) культивировали в присутствии или в отсутствие молекулы-индуктора димеризации B/B. Через 10 дней подсчитывали общее количество клеток в каждой лунке. N=2 для каждого значения.
На фиг. 23 проиллюстрировано применение абляции клеток хозяина посредством caspase9 с использованием донорских ПЗК от китайских шелковых кур с черной кожей, инъецированных в суррогатные хозяйские эмбрионы iCaspase9, которые вылупляются и подвергаются разведению для получения чистого потомства. A) Потомство (эмбрионы) от суррогата Dazl-iCaspase9 не содержало трансгена GFP, что указывает на то, что большая часть хозяйских зародышевых клеток не дала потомства (50% потомства от эндогенных зародышевых клеток должны быть GFP+, если они не были подвергнуты абляции). Два потомка от суррогата Dazl-aviCaspase9 содержали трансген GFP, что указывает на то, что некоторые из потомков произошли от эндогенных зародышевых клеток. (A-C) Потомство (эмбрионы и куры) имеет черную кожу, что указывает на то, что они произошли от донорских зародышевых клеток.
Подробное описание изобретения
FK-связывающий белок (FKBP) FKBP12 принадлежит к рецепторному семейству иммунофилинов, и его аминокислотные последовательности являются высоко консервативными у млекопитающих и кур (Yazawa et al (2003) Comparative Biochem. Physiol: Mol. Integ. Physiol 136(2):391-399). Он представляет собой цитозольный рецептор иммуносупрессивного препарата FK506 и является мишенью для селективного контроля клеточной сигнализации посредством димеризации белка.
Димерные варианты FKBP12, FK1012, были синтезированы Spencer и коллегами для опосредования контроля клеточной сигнализации посредством димеризации или олигомеризации внутриклеточных белков (Spencer et al (1993) Science 262:1019-1024). Позже был создан специфический связывающий карман в FKBP12 путем замены объемного фенилаланина меньшим остатком валина (FKBP12F36V). Модернизированные лиганды FK1012, в том числе AP1903 и близкородственный AP20187, были разработаны с высокой аффинностью и селективностью по отношению к FKBP12F36V и минимальным взаимодействием с эндогенными FKBP (Clackson et al (1998) PNAS 95(18):10437-10442; Nör et al (2002) Gene Ther 9(7):444-51).
Белки каспазы, 2, 3, 4, 7, 8, 9 или 10, представляют собой встречающиеся в природе белки, которые, как известно, индуцируют программируемую гибель клеток. Каспаза 9 (Casp9) активируется при димеризации и приводит к апоптозу клеток. Casp9 может быть усечена для удаления ее домена димеризации (CARD). Путем слияния домена димеризации FKBP12F36V с Casp9 может быть селективно индуцирована абляция клеток при введении лиганда. Эта система называется системой индуцибельной каспазы 9 (iCas9 (iC9)). AP20187 представлен на рынке как препарат-индуктор гомодимеризации B/B. Он использовался для создания системы, индуцирующей абляцию клеток. Слитый белок использовался для устранения клеток in vitro (Carlotti et al (2005) Cancer Gene Ther 12(7):627-39). Он также использовался для устранения клеток in vivo у мышей, шпорцевых лягушек и данио-рерио (Mallet et al (2002) Nat Biotechnol 20(12):1234-1239; Pajvani et al (2005) Nat Med 11(7):797-803; Hamm et al (2009) Invest Ophthalmol Vis Sci 50(2):885-92; Weber et al (2016) Development 143(22):4279-4287; Shimokawa (2017) Nature 11; 545(7653):187-192).
Пример 1
Селективная абляция куриных примордиальных зародышевых клеток (ПЗК) посредством модификации куриного генома трансгеном индуцибельная каспаза 9 (iC9).
Получение конструкций ДНК, использованных для получения ПЗК, трансфицированных iCasp9
Для специфической экспрессии индуцибельной Casp9 в ПЗК было использовано редактирование генома с помощью технологии CRISPR/Cas9 для опосредования вставки последовательности в следующие локусы, которые специфически экспрессируются в ПЗК:
DDX4: Куриный гомолог vasa гена, РНК-хеликаза, специфически экспрессируется в зародышевых клетках из локуса DDX4. Направленная вставка экзогенных генов в стартовый кодон DDX4 (ATG) была предпринята для достижения специфичной для зародышевых клеток экспрессии соответствующих белков.
DAZL: Локус DAZL управляет экспрессией РНК-связывающего белка DAZL. Экспрессия DAZL также является специфичной для зародышевых клеток и согласно измерениям является по меньшей мере в 10 раз более высокой, чем экспрессия DDX4 (Jean et al. 2014). Направленная вставка экзогенных кДНК в стоп-кодон (TGA) локуса DAZL была использована для достижения специфической для зародышевых клеток экспрессии соответствующих белков.
Схема стратегии нацеливания представлена на фиг. 2 и фиг. 3.
Индуцибельная каспаза 9
Плазмида pMSCV-F-del Casp9.IRES.GFP (https://www.addgene.org/15567/), депонированная в Spencer lab (Straathof et al (2005) Blood 105(11):4247-54), содержит последовательность кДНК для FKBP12F36V, слитую с усеченной Casp9 человека (усеченной таким образом, что ее домен димеризации был удален для снижения базальной активности), с HA-меткой, слитой с C-концом Casp9. Эта последовательность ДНК называется в настоящем документе iCasp9.
Последовательность iCasp9 была химически синтезирована с предшествующей последовательностью P2A, с фланкирующими сайтами рестрикции BamH1 и с удаленным путем замены кодонов сайтом BamH1 внутри последовательности. Последовательность iCasp9, фланкированная сайтом BamH1 (iCasp9_человека), была синтезирована в векторе pMA.
Кроме того, домен Casp9 iCasp9 человека (аминокислоты 135-417 в NP_001220.2) был заменен на гомологичную аминокислотную область последовательности белка Casp9 курицы (аминокислоты 169-450 в XP_424580.6) с получением последовательности iCasp9_курицы. Последовательность iCasp9 курицы была химически синтезирована с предшествующей последовательностью P2A, с фланкирующими сайтами рестрикции BamH1 и с удаленным путем замены кодонов сайтом BamH1 внутри последовательности. Последовательность iCasp9, фланкированная сайтом BamH1 (iCasp9_курицы), была синтезирована в векторе pMA. Эта последовательность ДНК называется в настоящем документе aviCasp9.
Трансген нитроредуктазы
Ген нитроредуктазы присутствует у некоторых видов бактерий и восстанавливает нитрогруппу определенных химических соединений до цитотоксических метаболитов. Ген нитроредуктазы из E. coli был оптимизирован по кодонам для экспрессии у кур. Были произведены три аминокислотные замены, которые, как было показано, обеспечивают более высокую удельную активность (см. фиг. 19) в присутствии пролекарства-субстрата CB1954.
Матрица репарации DDX4-GFP
Матрица репарации DDX4 изначально была сконструирована с использованием клонирования Гибсона, которое позволяет лигировать несколько перекрывающихся двухцепочечных фрагментов ДНК. Фрагменты для этой плазмиды получали с помощью ПЦР (праймеры Invitrogen) или рестрикционного гидролиза (ферменты NEB). Между генами были включены последовательности 2A с рибосомным проскоком, чтобы избежать трансляции слитых белков. Эти пептиды 2A были сконструированы с линкерами GSG на их аминоконцах, последовательность которых включает сайт вырезания рестриктазой BamH1, позволяющий встраивать дополнительные 2A-связанные гены.
Основной фрагмент был получен с использованием последовательности из вектора pGEM-T (Promega), который содержит кассеты устойчивости к ампициллину и сайтов множественного клонирования (MCS). Последовательность pGEM-Т размером 3 тыс. п. н., наряду с 3 тыс. п. н. гомологии до стартового кодона DDX4 (левое нацеливающее плечо), была получена из ранее сконструированного вектора, нацеливающего на DDX4 (HOMOL pGEM-T левое плечо и правое плечо ddx4+GFPpuropolyA), с использованием Xcm1 и Nco1 для вырезания фрагмента размером 6 тыс. п. н., который был очищен с помощью очистки из геля.
Правое нацеливающее плечо, состоящее из 1,5 тыс. п. н. гомологии от стартового кодона DDX4, было синтезировано с помощью ПЦР с использованием геномной ДНК, полученной из куриных ПЗК (клетки Y2, полученные из яиц, полученных от NARF (National Avian Research Facility, Национальный центр исследования птиц)). Прямой праймер для этой реакции (CGGTGACGTCGAGGAGAATCCTGGACCTATGGAGGAGGATTGGGATACCGAACTCGAGCAGGAGGCGGCAGCGGC, 75 п. н.), SEQ ID NO: 7, содержал частичную последовательность для T2A с рибосомным проскоком и замены кодонов в сайте нацеливания CRISPR/Cas9, чтобы матрица репарации не могла быть разрезана белком Cas9. Обратный праймер для этой реакции (GAAATCCAGCTTCCAGTTCCCACCTGGCCAGACAAGGGGCTGCTTGG, 47 п. н.), SEQ ID NO: 8, содержал выступ размером 20 п. н. от последовательности вектора pGEM-T вместе с нуклеотидами, которые повторно вставляли сайт разрезания для Xcm1 после выступа.
Последний фрагмент (800 п. н.) для матрицы репарации DDX4 содержал последовательность для eGFP, которая была снова синтезирована с помощью ПЦР из ранее сконструированного вектора, нацеливающего на DDX4, HOMOL pGEM-T левое плечо и правое плечо ddx4+GFPpuropolyA. Прямой праймер (GGTGGGCTGCTGGCATTCGCCATGGTGAGCAAGGGCGAGGA, 41 п. н.), SEQ ID NO: 9, для этой реакции содержал выступ размером 20 п. н. от левого нацеливающего плеча вместе с нуклеотидами, которые повторно вставляли сайт разрезания для Nco1 после выступа. Обратный праймер для этой реакции (GATTCTCCTCGACGTCACCGCATGTTAGCAGACTTCCTCTGCCCTCTCCGGATCCCTTGTACAGCTCGTCCATGCC, 76 п. н.), SEQ ID NO: 10, содержал оставшуюся последовательность T2A плюс 20 п. н. выступа для частичной последовательности T2A во фрагменте правого плеча.
Для лигирования фрагментов смесь 100 нг основного фрагмента вместе с эквимолярными количествами других фрагментов инкубировали с ферментом Gibson HiFi DNA Assembly Master Mix enzyme (NEB) в течение 1 часа при 50 °C. Компетентные клетки XL-10 Gold трансформировали 2 мкл лигированной плазмиды. Мини-препараты, полученные из трансформированных клеток, проверяли рестрикционным гидролизом, и получали макси-препараты.
Матрица репарации DAZL была первоначально сконструирована с использованием клонирования Гибсона с фрагментами плазмиды, полученными с помощью ПЦР (праймеры IDT) или рестрикционного гидролиза (ферменты NEB). IDT позволяет синтезировать праймеры длиной > 100 п. н., которые были необходимы для этой работы. Также была сконструирована матрица репарации, включающая оптимизированный для кур ген NTR, продукт которого может быть использован для селективной абляции клеток при введении пролекарства.
Основным фрагментом для нацеливающей матрицы была последовательность pGEM-T размером 3 тыс. п. н., которая была получена из описанной выше матрицы репарации DDX4-GFP с использованием Xcm1 и Not1 для вырезания ДНК.
Левое нацеливающее плечо, состоящее из 1,5 тыс. п. н. гомологии до стоп-кодона DAZL (но не включая его), было синтезировано с помощью ПЦР с использованием геномной ДНК, полученной из куриных ПЗК (клетки Y2). Прямой праймер для этой реакции
(TCTCCCATATGGTCGACCTGCAGGCGGCCGCGAATTCACTAGTGATTCTTCGTGGTT, 67 п. н.), SEQ ID NO: 11, содержал выступ размером 25 п. н. от последовательности вектора pGEM-T вместе с нуклеотидами, которые повторно вставляли сайт разрезания для Not1 после выступа. Обратный праймер для этой реакции (AGGCTGAAGTTAGTAGCTCCGGATCCAACACTTTTGAGCACTGCTCTT, 48 п. н.), SEQ ID NO: 12, содержал выступ размером 25 п. н. от последовательности P2A с рибосомным проскоком.
Третий фрагмент (600 п. н.) содержал последовательность P2A, за которой следовал NTR, который был вырезан с использованием BamH1 из матрицы репарации DDX4-GFP-NTR, которая, в свою очередь, была сконструирована с использованием матрицы репарации DDX4-GFP (линеаризованной с помощью BamH1 и лигированной со вставкой, содержащей последовательность P2A-NTR).
Четвертый фрагмент (800 п. н.) содержал последовательность для eGFP, которая была синтезирована с помощью ПЦР из матрицы репарации DDX4-GFP. Прямой праймер
(CAGAACATCACCCTGACCGAGGTGGGATCCGGAGAGGGCAGAGGAAGTCTGCTAACATGCGGTGACGTCGAGGAGAATCCTGGACCTATGGTGAGCAAGGGCGAGGA, 107 п. н.), SEQ ID NO: 13, для этой реакции содержал выступ размером 25 п. н. от гена NTR и последовательность для T2A с рибосомным проскоком. Обратный праймер для этой реакции (CTTGTACAGCTCGTCCATGCCG, 22 п. н.), SEQ ID NO: 14, не содержал выступов.
Пятый и последний фрагмент для нацеливающей матрицы DAZL-GFP представлял собой правое нацеливающее плечо, состоящее из 1,5 тыс. п. н. гомологии от стоп-кодона DAZL (включительно), синтезированное с помощью ПЦР с использованием геномной ДНК, полученной из куриных ПЗК (клетки Y2). Прямой праймер для этой реакции (TCTCGGCATGGACGAGCTGTACAAGTGATGAACAAAGACTTTGAAGTACATAAATGTATTACTTTGATGTTAATACAGTTCAGTTTAGTAAGAT, 94 п. н.), SEQ ID NO: 15, содержал выступ размером 25 п. н. в ген eGFP и мутации в сайте PAM для соответствующей плазмиды CRISPR/Cas9. Обратный праймер для этой реакции (CTCTTCGAAATCCAGCTTCCAGTTCCCACCTGGCAATACTATTAAAGCAATAGGT, 55 п. н.), SEQ ID NO: 16, содержал выступ размером 25 п. н. в последовательности вектора pGEM-T вместе с нуклеотидами, которые повторно вставляли сайт разрезания для Xcm1 после выступа.
Для лигирования фрагментов смесь 100 нг основного фрагмента вместе с эквимолярными количествами других фрагментов инкубировали с ферментом Gibson HiFi DNA Assembly Master Mix enzyme (NEB) в течение 2 часов при 50 °C. Компетентные клетки XL-10 Gold трансформировали 2 мкл лигированной плазмиды. Мини-препараты, полученные из трансформированных клеток, проверяли рестрикционным гидролизом, и получали макси-препараты.
BamH1 использовали для вырезания оптимизированной для кур последовательности NTR из матрицы репарации DAZL-GFP. Последовательности 2A-iCasp9 человека и курицы (iCasp9 и aviCasp9) вырезали с использованием BamH1 из соответствующих векторов pMA и вставляли в открытую матрицу репарации DAZL-GFP путем лигирования Т7. Компетентные клетки XL-10 Gold трансформировали 2 мкл лигированной плазмиды. Мини-препараты, полученные из трансформированных клеток, проверяли рестрикционным гидролизом, и получали макси-препараты для матрицы репарации DAZL-aviCasp9-GFP (курицы) и матрицы репарации DAZL- iCasp9-GFP (человека).
Последовательности гидовой РНК (гРНК) в пределах 150 п. н. от стоп-кодона локуса DAZL были запрошены с использованием инструмента для конструирования CRISPR, доступного на сайте crispr.mit.edu. Прямые и обратные олигонуклеотиды (IDT) для 5 лучших гидовых РНК (с заменой двух первых оснований гидовой последовательности на GG) были синтезированы с липкими концами Bbs1. Олигонуклеотиды отжигали с помощью ПЦР и лигировали в pSpCas9(BB)-2A-Puro (PX459) V2.0 (https://www.addgene.org/62988/), используя смесь для гидролиза/лигирования в ПЦР. Компетентные клетки трансформировали 2 мкл лигированной плазмиды и получали макси-препараты из трансформированных колоний. Макси-препараты плазмид DAZL-PX459 (№ 1-5) были проверены с помощью ПЦР с использованием прямого олигонуклеотида и обратного праймера, комплементарного плазмиде PX459 на расстоянии 400 п. н. по ходу транскрипции от сайта разрезания для гидовой вставки (Bpi1).
Все последовательности ДНК показаны на фиг. 13-19.
Процесс трансфекции
Приблизительно 150 000 ПЗК трансфицировали 1 мкг матрицы репарации DAZL-iCasp9-GFP (курицы или человека) и 1 мкг DAZL-PX459 -4 или -5 с использованием Lipofectamine 3000. Через 5 часов в растворе Lipofectamine ПЗК осаждали и ресуспендировали в полной (FAOT) среде. Через 24 часа ПЗК получали свежую среду и в каждую лунку добавляли 2 мкл 0,1 мг/мл раствора пуромицина (конечная концентрация 0,04 мкг/мл). ПЗК инкубировали с пуромицином в течение 48 часов, один раз промывали и ресуспендировали в свежей среде. Клетки культивировали 1-2 недели для достижения популяции 200 000-400 000 клеток, а затем сортировали с использованием FACS для сбора успешно модифицированных (GFP-положительных) клеток. GFP-положительные клетки получали из ПЗК, трансфицированных DAZL-PX459 -4 или DAZL-PX459 -5, хотя только ПЗК, трансфицированные DAZL-PX459 -5, использовали для получения кур.
DAZL-PX459 -4 содержит следующую гидовую последовательность: GGTCCTATTCCAGGAGAGGA SEQ ID NO: 17. Сайт PAM для этой гидовой последовательности находится на прямой цепи генома, на расстоянии 44 п. н. против хода транскрипции от стоп-кодона локуса DAZL.
DAZL-PX459 -5 содержит в качестве своей гидовой последовательности: GGCTTACTAAACTGAACTGT SEQ ID NO: 18. Сайт PAM для этой гидовой последовательности находится на обратной цепи генома, на расстоянии 46 п. н. по ходу транскрипции от стоп-кодона локуса DAZL. В то время как последовательность сайта PAM для гидовой последовательности в DAZL-PX459 -5 была мутирована в плече гомологии конечной матрицы репарации DAZL-iCasp9-GFP, следует отметить, что последовательность сайта PAM для гидовой последовательности в DAZL-PX459 -4 не была мутирована.
ПЗК культивировали в течение трех недель для отбора клеток, которые были стабильно нацелены конструкциями, экспрессирующими GFP. Женские ПЗК, нацеленные посредством ddx4_GFP и dazl_GFP, очищали методом проточной цитометрии с использованием FACS-ARIA с гейтированием по флуоресценции GFP. Очищенные клетки наращивали в культуре и анализировали методом проточной цитометрии для количественной оценки уровня флуоресценции GFP. Клетки с GFP, нацеленные на локус DAZL, были в 3,75 раза более флуоресцентными, чем клетки с GFP, нацеленные на локус DDX4 (фиг. 4).
Женские ПЗК, нацеленные посредством DDX-Ntr, DDX-icaspase9, DAZL-Ntr, DAZL-icaspase9 (человека), Dazl-icaspase9 (курицы), очищали аналогичным образом.
ПЗК, содержащие нацеленный ген нитроредуктазы, обрабатывали пролекарством CB1954. Под воздействием препарата ПЗК погибали. Клетки, содержащие NTR, нацеленные на локус dazl, демонстрировали уменьшение количества клеток по сравнению с контрольными клетками (фиг. 5).
ПЗК, содержащие нацеленный ген icaspase9, обрабатывали соединением-индуктором димеризации B/B. Контрольные ненацеленные ПЗК не демонстрировали уменьшение количеств ПЗК при обработке препаратом. Клетки, содержащие гены caspase9 человека и курицы, нацеленные на локус dazl, демонстрировали значительно уменьшенные количества ПЗК (фиг. 6). Клетки, содержащие iCaspase 9, нацеленный на локус ddx4, демонстрировали немного меньшее количество ПЗК. Эти клетки смешивали с контрольными ПЗК с красной флуоресценцией и инъецировали куриным эмбрионам. Куриные эмбрионы обрабатывали соединением-индуктором димеризации B/B. У эмбрионов были визуально обнаружимы только красные ПЗК и контрольные ПЗК GFP. ПЗК с Dazl icaspase9 были уничтожены (фиг. 7).
Получение курицы, нацеленной посредством Dazl icaspase9
Dazl iCaspase9 (человека) или Dazl aviCaspase9 (курицы), или обе из них в смеси инъецировали в оплодотворенные яйца от гетерозиготных по DDX4 (Z-Z) самцов кур, скрещенных с самками дикого типа. 3000 ПЗК инъецировали в эмбрионы стадии 16 HH через сделанные в яйцах отверстия, яйца запечатывали и инкубировали до вылупления. Разведение вылупившихся самок кур-основательниц (Z-W) дало трансгенное потомство, содержащее нацеленный трансген (фиг. 8).
Анализ абляции нацеленных посредством Dazl-iCaspase9 и Dazl-aviCaspase9 куриных и зародышевых клеток с использованием реагента-индуктора димеризации B/B
Кур G1 Dazl-icaspase9 и Dazl-aviCaspase9 выращивали до половой зрелости и спаривали с курами дикого типа. Оплодотворенные яйца от спариваний (эмбрионы G2) инкубировали и исследовали на предмет экспрессии GFP в гонадах. Зародышевые клетки в гонадах эмбрионов G2 как Dazl-icaspase9, так и Dazl-avicaspase9 содержали GFP+ клетки в гонаде (фиг. 9). Криосрезы и иммуноокрашивание антителом к маркеру зародышевых клеток DDX4 показали, что клетки, экспрессирующие GFP, являются зародышевыми клетками (фиг. 10). Эмбрионы G2 были протестированы на предмет абляции зародышевых клеток. 1,0 мкл 0,1 мМ реагента-индуктора димеризации B/B (Takara Bio, Inc) инъецировали в кровоток куриных эмбрионов на 2,5 день (стадия 16 по Гамбургеру и Гамильтону (HH)). Эмбрионы инкубировали в течение 8 дней, подвергали скринингу с помощью ПЦР для идентификации эмбрионов iCaspase9 и исследовали на предмет экспрессии GFP в гонадах. Обработанные препаратом эмбрионы G2 Dazl-iCaspase9 и Dazl-aviCaspase9 не имеют видимой экспрессии GFP (фиг. 11). Криосрезы и иммуноокрашивание антителом к маркеру зародышевых клеток DDX4 демонстрируют, что в гонадах эмбрионов Dazl-icaspase9 и Dazl-aviCaspase9 почти нет идентифицируемых DDX4-положительных или GFP+ клеток (фиг. 11). Чтобы показать, что экзогенные (донорские) ПЗК могут колонизировать стерилизованные хозяйские эмбрионы G2 Dazl-aviCaspase9, в эмбрионы инъецировали донорские красные флуоресцентные зародышевые клетки и препарат B/B. Эмбрионы инкубировали в течение 8 дней и исследовали на предмет флуоресценции и зародышевых клеток. В гонаде эмбриона G2 aviCaspase9 присутствовали только донорские зародышевые клетки, эндогенные (хозяйские) зародышевые клетки не были обнаружены. (фиг. 12).
Абляция зародышевых клеток у трансгенных кур
Трасген iCaspase9 был нацелен на локус DDX4 или DAZL в ПЗК, а затем клетки подвергались действию препарата-индуктора димеризации. Было определено, что клетки, содержащие трансген, вставленный в локус DAZL, были ингибированы/уничтожены. Не ограничиваясь какой-либо конкретной теорией, авторы изобретения полагают, что уровни экспрессии на стадиях эмбрионального развития являются важными для абляции зародышевых клеток.
Это проиллюстрировано на фиг. 22.
Используя экспрессию каспазы 9, нацеленную на локус DAZL в ПЗК, тестировали абляцию хозяйских зародышевых клеток и получение потомства от донорской породы кур с использованием донорских куриных ПЗК от породы китайских шелковых кур с черной кожей. Были протестированы донорские ПЗК, инъецированные в хозяйские эмбрионы с каспазой, которые затем выращивали до половой зрелости и подвергали разведению. Как показано на фиг. 23, у потомства от хозяев Dazl-Caspase9, которым инъецировали ПЗК китайских шелковых кур и вводили препарат B/B, была обнаружена черная кожа, что указывает на то, что они произошли от донорских зародышевых клеток.
Более подробно, в этом варианте осуществления ПЗК китайских шелковых кур с черной кожей смешивали с препаратом-индуктором димеризации B/B и инъецировали хозяйским эмбрионам Dazl-Caspase, когда они находились в яйце. Яйца запечатывали, и эмбрионы вылуплялись, а затем были скрещены друг с другом по достижении половой зрелости.
50% потомства должны быть GFP-положительными, если они получены из эндогенных хозяйских GFP+ ПЗК с caspase9. Некоторые представители потомства от хозяина Dazl-aviCaspase9 были GFP+, и ни один из представителей потомства Dazl-iCaspase9 не был GFP+ (показано в таблице на фиг. 23 A).
Кроме того, эмбриональное потомство от хозяина Dazl-Caspase демонстрировало фенотип черной кожи донорских ПЗК китайских шелковых кур (показано на фиг. 23 B). Более того, вылупившееся потомство от хозяина Dazl-Caspase демонстрировало фенотип черной кожи донорских ПЗК китайских шелковых кур (показано на фиг. 23 C).
Хотя изобретение было конкретно проиллюстрировано и описано со ссылкой на конкретные примеры, специалистам в данной области техники будет ясно, что в него могут быть внесены различные изменения в отношении формы и деталей, не выходящие за пределы объема настоящего изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНСТЕЙ
<110> The University Court of the University of Edinburgh
<120> ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ СТЕРИЛЬНЫЕ ПТИЦЫ И СПОСОБ ИХ
ВОСПРОИЗВОДСТВА
<130> P191530.WO.01
<150> GB1816633.0
<151> 2018-10-12
<160> 18
<170> PatentIn version 3.5
<210> 1
<211> 1317
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутантный сайт 2a-iCaspase9-Bam H1
<400> 1
ggatccggag ctactaactt cagcctgctg aagcaggctg gagacgtgga ggagaaccct
60
ggacctatgc tcgagggagt gcaggtggag actatctccc caggagacgg gcgcaccttc
120
cccaagcgcg gccagacctg cgtggtgcac tacaccggga tgcttgaaga tggaaagaaa
180
gttgattcct cccgggacag aaacaagccc tttaagttta tgctaggcaa gcaggaggtg
240
atccgaggct gggaagaagg ggttgcccag atgagtgtgg gtcagagagc caaactgact
300
atatctccag attatgccta tggtgccact gggcacccag gcatcatccc accacatgcc
360
actctcgtct tcgatgtgga gcttctaaaa ctggaatctg gcggtggttc cggagtcgac
420
ggatttggtg atgtcggtgc tcttgagagt ttgaggggaa atgcagattt ggcttacatc
480
ctgagcatgg agccctgtgg ccactgcctc attatcaaca atgtgaactt ctgccgtgag
540
tccgggctcc gcacccgcac tggctccaac atcgactgtg agaagttgcg gcgtcgcttc
600
tcctcgctgc atttcatggt ggaggtgaag ggcgacctga ctgccaagaa aatggtgctg
660
gctttgctgg agctggcgcg gcaggaccac ggtgctctgg actgctgcgt ggtggtcatt
720
ctctctcacg gctgtcaggc cagccacctg cagttcccag gggctgtcta cggcacagat
780
ggatgccctg tgtcggtcga gaagattgtg aacatcttca atgggaccag ctgccccagc
840
ctgggaggga agcccaagct ctttttcatc caggcctgtg gtggggagca gaaagaccat
900
gggtttgagg tggcctccac ttcccctgaa gacgagtccc ctggcagtaa ccccgagcca
960
gatgccaccc cgttccagga aggtttgagg accttcgacc agctggacgc catatctagt
1020
ttgcccacac ccagtgacat ctttgtgtcc tactctactt tcccaggttt tgtttcctgg
1080
agggacccca agagtggctc ctggtacgtt gagaccctgg acgacatctt tgagcagtgg
1140
gctcactctg aagacctgca gtccctcctg cttagggtcg ctaatgctgt ttcggtgaaa
1200
gggatttata aacagatgcc tggttgcttt aatttcctcc ggaaaaaact tttctttaaa
1260
acatcagtcg actatccgta cgacgtacca gactacgcac tcgacctcga cggatcc
1317
<210> 2
<211> 1315
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 2a-aviCaspase9
<400> 2
agggatccgg agctactaac ttcagcctgc tgaagcaggc tggagacgtg gaggagaacc
60
ctggacctat gctcgaggga gtgcaggtgg agactatctc cccaggagac gggcgcacct
120
tccccaagcg cggccagacc tgcgtggtgc actacaccgg gatgcttgaa gatggaaaga
180
aagttgattc ctcccgggac agaaacaagc cctttaagtt tatgcttggc aagcaggagg
240
tgatccgagg ctgggaagaa ggggttgccc agatgagtgt gggtcagaga gccaaactga
300
ctatatctcc agattatgcc tatggtgcca ctgggcaccc aggcatcatc ccaccacatg
360
ccactctcgt cttcgatgtg gagcttctaa aactggaatc tggcggtggt tccggagtcg
420
acggtgtctc tgtgaattgc agaccagcta ggatgcatgc tagtgcatgc caggtgtacc
480
agctgcgagc agacccttgt gggcactgcc tgatcttcaa caatgtcagc ttcagcagag
540
actctgatct gtcgactcga gctggctctg acatagactg tgagaagctg gagaagcgtt
600
tcaggtccct gtgcttccac gtccggaccc tgcggaacct caaagctcag gaaattgatg
660
tggagctgcg gaagctggcg cggctcgacc acagtgccct ggactgctgc ctcgtggtca
720
tcctctccca tggttgccag acaagccata ttcagtttcc cggagggatt tatggaacag
780
atggcaaaat cattccaatc gaaaggattg tgaactattt caatgggtcc cagtgcccga
840
gtttgagagg aaaacccaaa ctcttcttca tccaggcctg tggaggagaa caaaaggacc
900
aaggatttga ggtggattgt gaatcacccc aagatgaaac ttgccgacgt tccatagagt
960
cggatgcgat tcctttccag gctccatcag ggaatgagga cgagccagac gccgtcgcca
1020
gtttgcccac tcctggtgac atcttggtgt cctattcaac ttttccaggt tttgtgtcct
1080
ggagggacaa ggtgagtggc tcgtggtacg tggaaacctt ggacagcgta ctggaacatt
1140
acgcccgttc tgaagacctg cttaccatgc tacttcgggt gtcagacatc gtatccacca
1200
aggggaggta caagcagatc ccgggctgtt tcaacttcct tcgtaaaaaa ttcttcttcc
1260
tgtgcaaggt cgactatccg tacgacgtac cagactacgc actcgacgga tccaa
1315
<210> 3
<211> 5985
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Leftarm-GFP-2a-Nitroreductase-T2a-Right arm
<400> 3
tttttttctt tgagatgagt atcttattgc agctctgtgc ataccccttg aaaatccgta
60
ggtgatgtat ttatgttgac tgaaaatgga gtcattcaaa aattgatgac ttcttccttt
120
tctcttttta atctcgaaac ctgttctcag attcctttat cagaacaaaa agcacggctg
180
catgtttttc accgttcaca cgactgtgtt tagccagcgt atctaaatgc ataacatgat
240
ttgccataac ttgagcctgc cataatcaga gcctcatttt gaaagctgtc atggtctgaa
300
acatagcatg ggtaagatga ttttcacctt gatgatgcat gtttaactca tttcaacgag
360
aggtcaaatc catccaaaaa tgctaaatct atcaaccagt tttcatgttc aagttctggc
420
acaaattttc gttctgactc catacgtgat tagatttcat tttgcaaatc atgaaaacta
480
ttgccaaaac atacaatttt aagtcagcag ttttactgcc caaatttgca caagcacatg
540
taacgtgaca gacaaaaacc cactggacag agacaaaata ctgggtttgg ccagtatggt
600
ttggctaagc cacgtttttc agggcaagaa ttgcctcacc tttttttata ttggaatcaa
660
attctcaagt gtaacagcct ggcgccatct cagctgccat taagtagcat gtgctgcaac
720
aagatgtcta ccacacactc tgctctgggt ggtgggggca caccagtgtg gacggcaacc
780
atgacacaag catagagtct tggcagaggt ccattgggtg catttatagt ggttcacact
840
ggggggttat cacagaatca tggaggttgg aaggtacctc cgggggtcat caaatccaac
900
cacttgacaa agcaggttcc ctaaagtaga ttgctcaaga acagtcacag cactgctgct
960
cacagaaaat ctctagttgg taaggttacc tttttttttg ttgtcgctcc cagttaaacc
1020
aaggtgggaa cacatccttt tcatagttgt attgtaggga tgtttatagc tattgctagg
1080
gaaaactgaa cagcgtgcaa ggaagtcagc actgacacag ctttcccacg tgagagagct
1140
atttcaaagc aagatcagca catatcccaa tctgtacttc ctcaaacaca gcccaaaacc
1200
gatagcaagg ccaagcagca gcactgctcc ttcaaggaaa ctgctgcata atctgaattt
1260
cagacggcag gaggaaacaa ggcagcagac tgatgaccta ccaccctgag tctcagctac
1320
atgcgatccg agccactcca actctgcttc ctgcagccct tccttcacgc tcatcttttg
1380
caccttaggc actcttataa ttcaagtact ttgtggcttt gggatatttg aagagcttgg
1440
tcagttagtc acaagtctgg ccacgtgcta tcatattagt ttgaaaagca atggagacac
1500
catctgctga tgctcaaagt ggttacaacc aaaacacaaa aaagcagagc tgtgggaaga
1560
attcaacatt ttgattatgc aagaagctag tcccagcctt gaaatccacc atctgcatca
1620
tgaaagacct aagtagttaa agccacagca gacatacagc ttctatttcc ttaccttctt
1680
catcattaac tacaggtctt ggaagatttt tgctgggaaa aagcttttat tgcaagaact
1740
gtaatttatt aacagggaaa catgaaataa atgtgtaaat tctcctgcac tcccactgtc
1800
attaaaaacg gctttaagaa agagtattca gtaactgcct gcattgtgtt gtgactttct
1860
actctgtgac acacagcctt ctgggcaaag cacatattct gccatgcatg tgggtcgtgc
1920
cttggaaaat gggaacccac attcactgag ggcctcttga tgagctttcc ccttgagaac
1980
agcgaggtct catggatatc ttctcttctc caagccaaat aagcccagat ccctcagctt
2040
tccttcatgg agaggctcca gccctctgat catgcctgca gcccttctct ggacctgctc
2100
cagcagcccc acacgctcct catgctggga ccccagatct gcacatggta ttgcacatgg
2160
ggcctcacaa cggcagagct gagagggaca attccctccc tcagccttta attctgcacg
2220
tacttaattt tgtctgtatt ttttttgcaa tagaatagcc tgcatgctag ttgctgtgtc
2280
gcacagctga attcactagg ttcctgtgaa acaaagggca atcccacagc cactgtagca
2340
cgtgaggagc cacacaggtg ctctaattcc tacaggacag gcctagggac agagcggccc
2400
taggactgcc ctcccgcacc tcctagcttc accttttgcc cgcgatcttt aagtaccctg
2460
aagcataaac agggaaagcg ccgcccgcag cctcaccctg cttcccgccc aacgcggcac
2520
cgcccaggcc gcagccgccc actaagagca ctagcgccac cttctcaccc cacccccacc
2580
acgcgttcct agcggccctg agagctgctg cgcatgcgcc gcctgacgat tcgccctccc
2640
attggctggc ggtcgaagca cggcgggggc acgcggtggc ggctatataa ggcgtctcgg
2700
aagacggcgc catgctattt ggagcggaga gtgaaagtta cagttcctgg tgctggtagg
2760
gagtgtggcg cggagcggag cgctgcggct catcggaacc acaatggagc catagcagag
2820
ccgggcgtgg gggcaagggc agtgcgtgct ggggagggct ccgtcgcgtg gccacgtcgc
2880
gagagccgtc gggatggtgg cgtcagggcg gggggtgctg ctaacgtgct cctggtcctg
2940
caggtgggct gctggcattc gccatggtga gcaagggcga ggagctgttc accggggtgg
3000
tgcccatcct ggtcgagctg gacggcgacg taaacggcca caagttcagc gtgtccggcg
3060
agggcgaggg cgatgccacc tacggcaagc tgaccctgaa gttcatctgc accaccggca
3120
agctgcccgt gccctggccc accctcgtga ccaccctgac ctacggcgtg cagtgcttca
3180
gccgctaccc cgaccacatg aagcagcacg acttcttcaa gtccgccatg cccgaaggct
3240
acgtccagga gcgcaccatc ttcttcaagg acgacggcaa ctacaagacc cgcgccgagg
3300
tgaagttcga gggcgacacc ctggtgaacc gcatcgagct gaagggcatc gacttcaagg
3360
aggacggcaa catcctgggg cacaagctgg agtacaacta caacagccac aacgtctata
3420
tcatggccga caagcagaag aacggcatca aggtgaactt caagatccgc cacaacatcg
3480
aggacggcag cgtgcagctc gccgaccact accagcagaa cacccccatc ggcgacggcc
3540
ccgtgctgct gcccgacaac cactacctga gcacccagtc cgccctgagc aaagacccca
3600
acgagaagcg cgatcacatg gtcctgctgg agttcgtgac cgccgccggg atcactctcg
3660
gcatggacga gctgtacaag ggatccggag ctactaactt cagcctgctg aagcaggctg
3720
gagacgtgga ggagaaccct ggacctatgg atatcatcag cgtggccctg aagagacaca
3780
gcaccaaggc cttcgatgcc agcaagaagc tgacccccga gcaggccgag cagatcaaga
3840
ccctgctgca gtacagcccc agcagccaga acagccagcc ctggcacttc atcgtggcca
3900
gcaccgagga aggcaaggcc agagtggcca agagcgccgc cggcaactac gtgttcagcg
3960
agagaaagat gctggatgcc agccacgtgg tggtgttctg cgccaagacc gctatggatg
4020
atgtgtggct gaagctggtg gtggatcagg aggatgccga tggcagattc gccacccccg
4080
aggccaaggc cgccaacgat aagggcagaa agttcaccgc cgatatgcac agaaaggatc
4140
tgcacgatga tgccgagtgg atggccaagc aggtgtacct gaacgtgggc aacttcctgc
4200
tcggcgtggc cgccctgggc ctggatgccg tgcccatcga gggcttcgat gccgccatcc
4260
tggatgccga gttcggcctg aaggagaagg gctacaccag cctggtggtg gtgcccgtgg
4320
gccaccacag cgtggaggat ttcaacgcca ccctccccaa gagcagactg ccccagaaca
4380
tcaccctgac cgaggtggga tccggagagg gcagaggaag tctgctaaca tgcggtgacg
4440
tcgaggagaa tcctggacct atggaggagg attgggatac cgaactcgag caggaggcgg
4500
cagcggcttc ccaggggcgt tctgaggagc aggcgtggat ggtgagctgt gtccagggga
4560
gggcggtgcg gcagggagcg gggcactggg atggcccggt ctgggaaggg gaggccgaga
4620
ggccttcgca gtgcttcctg cagctccccg agcagtgcga agaaggaggt gccgggctct
4680
gctgtgggag ccgaaggcac ggagctgcct ggggagggaa tggctgtgtg cctgaggtgc
4740
cagagaccgg caagggctgt acagagaggc aacggttttg tgtacaaatc atttgtatgg
4800
gaaaccacaa aatcttgaag ctttattaca tggcagcgaa atactttggt gtgaagtaaa
4860
ggataagata gaggagtgta atgagagaca gctaaataat attttactac ttgtgggatg
4920
agtgaatatc aggaggaact gctgtaaatt tcaggaggac ctgttgtgaa tcttcagtag
4980
ttggcgccgc tcttacacat cttacagatg ccccttgagc aaagggggat aaggagagat
5040
gaacgggtta tccaaacagg tgatgagcta aaagtacagt tgcctaaaga agtagtagca
5100
tgtgctatca gaatgataat tttgttagtt tggggttagt ttcctgtagt ggtagatagc
5160
cacaacaaga aaaccgctta agtttttgta aaacaaaaaa agcacgatcc agaagtaaaa
5220
aatatgggta gttttttttg atgttcctct ctcacctggt gctttggcat actaatatgt
5280
gtctaattgt attaaacagg agaaatttaa acctaggctt tgctggaaat aaaatgttac
5340
aatgctacaa tgtgaaaagt aggtgctatt ctgaactgtt ttgggtggag tatctgaatc
5400
tttgaataat ttaagaggga ctgacatatt taaaatactt aaggataatc tgtagccatg
5460
ctgtaaaaga acaacagaaa tgcagttggg aaggtgatgg aaatagtttt attcatgtta
5520
ctggtggtct gaaacctttc taagcttaaa ctgtagaaaa aaattgcttc aaaagattgc
5580
actattactt tgggcgacaa atgtttttaa ttggttttaa gtgtttgtta gcaaagtgaa
5640
gttgatgcca ccataagtct gacaggaggc aagataaact tgtcttcata ctgcttgtgt
5700
ataacttggt tttgatgaca tttgtgtgtg aacattatgc acttcagtgt agcgaagttt
5760
aagaaacttt gaacagaata acttgaaaga gtgtgcacat gggtgcagaa gtcactttat
5820
ttcagttggg agacttagca cctaaatgca ctgttagttc acatacactt tgcttggcct
5880
gaaggtaaca ttgtgatgtc gctttttttc cctgtaggct aactctggca gaccaaacag
5940
cccatccctc cgcttctcca gcagaccaag cagccccttg tctgg
5985
<210> 4
<211> 5155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Left arm-2a-aviCaspase9-T2a-GFP-Right arm
<400> 4
cttcgtggct tggtgaagag atcttctgac ctcttcatca ccccctagaa ttaaggctgt
60
aatcatcatt tcagccattg cttaatccac tataatttct gcatgatttc agaaggctat
120
cttgcttcct tgaaactaat aatacctagt ttgtagtgtt tagggatgtg taaaatgtgt
180
ttatgtcaag attttagtta gtctgataag aattgtggaa gaattgcagc tactaaaaag
240
gcagactggg tagcacttcc caccttactc agatttagga gtctcaagac atgtagatgg
300
gtgaaattta ttattctttt ctctgcttgc ttttctggaa agctacctct atttggccag
360
cagttggctc tgaattcctt tctttaaagg agacctgact gctctcctgt ctgcctctta
420
cacagctgta ctggtgtgtc actgatagaa acgtttacct tgaaagtgac aagtgggggc
480
gttgataccc attaacaaca ttaaagggaa aatttcagga aacaagaaaa ctgagaggag
540
ttcagaatga aatgtaagcc ctgtgacaga ctcacactga tggtagtagg gtaactgatg
600
gagatgcatc tataacaatt ctaactttta tttcagaagt ctgtggacag gagcatacaa
660
acagtagtat cttgtctgtt taaccctgaa aaccgtctga ggaacacctt tgtatcacaa
720
gaagactact tcagggtatg tagtgaaagc aaaactaaat ctctaaaacc agctgatctt
780
aactttgttc atatacagga tgccatgaag acaaaggttt atctgtactt gcttgagatt
840
ttactttttg tatgtgcatt gagatgaatg actggttagc tctcagttgg ttaatactca
900
atgaaaattg cagaaattga gttcagtgga tttaagtgca ttgatacaag gctgcataag
960
aactgctgag cagtcagcta ttggatattt agagctaatc atccctcagg agcaatctgc
1020
tgaaaggtaa gcagattgca tcttgtaaac gagagagcag agttaaacgt ttgtggtctt
1080
tgctccactt aagtacccac agtggatgtc tatctgtact tttggttaaa aaatagataa
1140
tttctacaac agactgtagt taggtcagta cctggtcatc aggtattaat ggaaagcttt
1200
ttgctgtgca ttcaatcttc atagaggtct gctttccaat tacattctct ttgttttaag
1260
tataactttg aaactcaaat ataaaagcct gctacttttt tttgttctaa aaagacagat
1320
gtggactagg cacctgtact gtgaaaccag aatagagaag ctgttgggca gatttgaaga
1380
ggattgagta atatagagaa tgtcttgaga tatttgtgaa gcttttattg ctttgttgca
1440
attgttgtct agaaacatgg gctttttctg ttttttctcc tattccagga gaggagggcg
1500
catcacttca gaaaaggaag agcagtgctc aaaagtgttg gatccggagc tactaacttc
1560
agcctgctga agcaggctgg agacgtggag gagaaccctg gacctatgct cgagggagtg
1620
caggtggaga ctatctcccc aggagacggg cgcaccttcc ccaagcgcgg ccagacctgc
1680
gtggtgcact acaccgggat gcttgaagat ggaaagaaag ttgattcctc ccgggacaga
1740
aacaagccct ttaagtttat gcttggcaag caggaggtga tccgaggctg ggaagaaggg
1800
gttgcccaga tgagtgtggg tcagagagcc aaactgacta tatctccaga ttatgcctat
1860
ggtgccactg ggcacccagg catcatccca ccacatgcca ctctcgtctt cgatgtggag
1920
cttctaaaac tggaatctgg cggtggttcc ggagtcgacg gtgtctctgt gaattgcaga
1980
ccagctagga tgcatgctag tgcatgccag gtgtaccagc tgcgagcaga cccttgtggg
2040
cactgcctga tcttcaacaa tgtcagcttc agcagagact ctgatctgtc gactcgagct
2100
ggctctgaca tagactgtga gaagctggag aagcgtttca ggtccctgtg cttccacgtc
2160
cggaccctgc ggaacctcaa agctcaggaa attgatgtgg agctgcggaa gctggcgcgg
2220
ctcgaccaca gtgccctgga ctgctgcctc gtggtcatcc tctcccatgg ttgccagaca
2280
agccatattc agtttcccgg agggatttat ggaacagatg gcaaaatcat tccaatcgaa
2340
aggattgtga actatttcaa tgggtcccag tgcccgagtt tgagaggaaa acccaaactc
2400
ttcttcatcc aggcctgtgg aggagaacaa aaggaccaag gatttgaggt ggattgtgaa
2460
tcaccccaag atgaaacttg ccgacgttcc atagagtcgg atgcgattcc tttccaggct
2520
ccatcaggga atgaggacga gccagacgcc gtcgccagtt tgcccactcc tggtgacatc
2580
ttggtgtcct attcaacttt tccaggtttt gtgtcctgga gggacaaggt gagtggctcg
2640
tggtacgtgg aaaccttgga cagcgtactg gaacattacg cccgttctga agacctgctt
2700
accatgctac ttcgggtgtc agacatcgta tccaccaagg ggaggtacaa gcagatcccg
2760
ggctgtttca acttccttcg taaaaaattc ttcttcctgt gcaaggtcga ctatccgtac
2820
gacgtaccag actacgcact cgacggatcc ggagagggca gaggaagtct gctaacatgc
2880
ggtgacgtcg aggagaatcc tggacctatg gtgagcaagg gcgaggagct gttcaccggg
2940
gtggtgccca tcctggtcga gctggacggc gacgtaaacg gccacaagtt cagcgtgtcc
3000
ggcgagggcg agggcgatgc cacctacggc aagctgaccc tgaagttcat ctgcaccacc
3060
ggcaagctgc ccgtgccctg gcccaccctc gtgaccaccc tgacctacgg cgtgcagtgc
3120
ttcagccgct accccgacca catgaagcag cacgacttct tcaagtccgc catgcccgaa
3180
ggctacgtcc aggagcgcac catcttcttc aaggacgacg gcaactacaa gacccgcgcc
3240
gaggtgaagt tcgagggcga caccctggtg aaccgcatcg agctgaaggg catcgacttc
3300
aaggaggacg gcaacatcct ggggcacaag ctggagtaca actacaacag ccacaacgtc
3360
tatatcatgg ccgacaagca gaagaacggc atcaaggtga acttcaagat ccgccacaac
3420
atcgaggacg gcagcgtgca gctcgccgac cactaccagc agaacacccc catcggcgac
3480
ggccccgtgc tgctgcccga caaccactac ctgagcaccc agtccgccct gagcaaagac
3540
cccaacgaga agcgcgatca catggtcctg ctggagttcg tgaccgccgc cgggatcact
3600
ctcggcatgg acgagctgta caagtgatga acaaagactt tgaagtacat aaatgtatta
3660
ctttgatgtt aatacagttc agtttagtaa gatgtgtagt aaaaagtgta accttgttca
3720
aaaagttgct tcaagttgat gtttgtgttc tgttttacct gttccagaat agctattttt
3780
gcttgagaag tttgaagttg taagagttga aatatttcca ggttttatta ctagcttgca
3840
tgcttttcct gctaactaac tgaaatgcta atcttaagga atttatatgg ggaaggggaa
3900
aaaagaaaaa cactttgttt ggtacgtgtg gattttcttc tgagctttaa ggtacagttt
3960
gttgcatgtt aaaatttagt tcttattaaa ccacaacttt aagttactaa cgtcaaccag
4020
ttacctcttg cagttcaaaa gttgaagcag ttccttgtcc aagatggagt attttaaaac
4080
tgagctctta atcagtggaa cagaagacgt cacggtgtaa ctcaactgaa gccctttaag
4140
tcccggttct ctttagacta cctaatcaat gtctttgttt gctaacgaca gtttatctat
4200
gtgaatccta aaattcctat atgtaactta agatgcaaga atgtaattag ttacattggc
4260
tgctcagtgg agtatgactt ttttttttac tggattaatt ttagcaatac ctgtatctta
4320
aaattgtgag aaaatactgc atttaaaata tgcctaactt tgtgatgcaa tatgttaatc
4380
aaagaataca tgtaagcata ttttaataat aattatgtag attttagtca tgtattttga
4440
aacaattaaa atttttaatt ttgacttacc ttcccagtgt gagtgacatc ctaatataat
4500
acttctaaat cttaagctgc tttgagaaag gcatgcagcg tatttattga aggaattgaa
4560
gatttcttac cctacataag aattccagtt aggacaagtt tatagcaaca aactttcaca
4620
tttgctgtta gttctacctg cgattttgaa ggagtacaac tgagaacagc actcagtctt
4680
gtatgtgtgt tgggtcctag tctgattcat ttttcttatt actaccctta tacctcagtc
4740
tccaagtaaa aaaggaaata actcctcctt tgtagacgtg tatatgtgaa tgaatagaat
4800
ggcatgtccc acttcaaatg tctagaagta gatgttggtg aaacatgcaa taagagctga
4860
gttgctctgt acctggatag tgggctgtaa gatgcagcac aggagtgtcc caggactgtt
4920
gttcaggagt aggagtcagg ggagggaggc agggcttggg ggatagcaat aatagttggt
4980
gccgttctcc ataaaattac tcagaagcaa tgttctggca gcataaatcg acctaaagtt
5040
gttaactttt tctctgctca gaccgttccc tagcactaaa gatgctgtgt ccttaagtgt
5100
gtttccatac tatccatcac catgtttttc ccaacctatt gctttaatag tattg
5155
<210> 5
<211> 5161
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Left arm-2a-iCaspase9-T2a-GFP-Right arm
<400> 5
cttcgtggct tggtgaagag atcttctgac ctcttcatca ccccctagaa ttaaggctgt
60
aatcatcatt tcagccattg cttaatccac tataatttct gcatgatttc agaaggctat
120
cttgcttcct tgaaactaat aatacctagt ttgtagtgtt tagggatgtg taaaatgtgt
180
ttatgtcaag attttagtta gtctgataag aattgtggaa gaattgcagc tactaaaaag
240
gcagactggg tagcacttcc caccttactc agatttagga gtctcaagac atgtagatgg
300
gtgaaattta ttattctttt ctctgcttgc ttttctggaa agctacctct atttggccag
360
cagttggctc tgaattcctt tctttaaagg agacctgact gctctcctgt ctgcctctta
420
cacagctgta ctggtgtgtc actgatagaa acgtttacct tgaaagtgac aagtgggggc
480
gttgataccc attaacaaca ttaaagggaa aatttcagga aacaagaaaa ctgagaggag
540
ttcagaatga aatgtaagcc ctgtgacaga ctcacactga tggtagtagg gtaactgatg
600
gagatgcatc tataacaatt ctaactttta tttcagaagt ctgtggacag gagcatacaa
660
acagtagtat cttgtctgtt taaccctgaa aaccgtctga ggaacacctt tgtatcacaa
720
gaagactact tcagggtatg tagtgaaagc aaaactaaat ctctaaaacc agctgatctt
780
aactttgttc atatacagga tgccatgaag acaaaggttt atctgtactt gcttgagatt
840
ttactttttg tatgtgcatt gagatgaatg actggttagc tctcagttgg ttaatactca
900
atgaaaattg cagaaattga gttcagtgga tttaagtgca ttgatacaag gctgcataag
960
aactgctgag cagtcagcta ttggatattt agagctaatc atccctcagg agcaatctgc
1020
tgaaaggtaa gcagattgca tcttgtaaac gagagagcag agttaaacgt ttgtggtctt
1080
tgctccactt aagtacccac agtggatgtc tatctgtact tttggttaaa aaatagataa
1140
tttctacaac agactgtagt taggtcagta cctggtcatc aggtattaat ggaaagcttt
1200
ttgctgtgca ttcaatcttc atagaggtct gctttccaat tacattctct ttgttttaag
1260
tataactttg aaactcaaat ataaaagcct gctacttttt tttgttctaa aaagacagat
1320
gtggactagg cacctgtact gtgaaaccag aatagagaag ctgttgggca gatttgaaga
1380
ggattgagta atatagagaa tgtcttgaga tatttgtgaa gcttttattg ctttgttgca
1440
attgttgtct agaaacatgg gctttttctg ttttttctcc tattccagga gaggagggcg
1500
catcacttca gaaaaggaag agcagtgctc aaaagtgttg gatccggagc tactaacttc
1560
agcctgctga agcaggctgg agacgtggag gagaaccctg gacctatgct cgagggagtg
1620
caggtggaga ctatctcccc aggagacggg cgcaccttcc ccaagcgcgg ccagacctgc
1680
gtggtgcact acaccgggat gcttgaagat ggaaagaaag ttgattcctc ccgggacaga
1740
aacaagccct ttaagtttat gctaggcaag caggaggtga tccgaggctg ggaagaaggg
1800
gttgcccaga tgagtgtggg tcagagagcc aaactgacta tatctccaga ttatgcctat
1860
ggtgccactg ggcacccagg catcatccca ccacatgcca ctctcgtctt cgatgtggag
1920
cttctaaaac tggaatctgg cggtggttcc ggagtcgacg gatttggtga tgtcggtgct
1980
cttgagagtt tgaggggaaa tgcagatttg gcttacatcc tgagcatgga gccctgtggc
2040
cactgcctca ttatcaacaa tgtgaacttc tgccgtgagt ccgggctccg cacccgcact
2100
ggctccaaca tcgactgtga gaagttgcgg cgtcgcttct cctcgctgca tttcatggtg
2160
gaggtgaagg gcgacctgac tgccaagaaa atggtgctgg ctttgctgga gctggcgcgg
2220
caggaccacg gtgctctgga ctgctgcgtg gtggtcattc tctctcacgg ctgtcaggcc
2280
agccacctgc agttcccagg ggctgtctac ggcacagatg gatgccctgt gtcggtcgag
2340
aagattgtga acatcttcaa tgggaccagc tgccccagcc tgggagggaa gcccaagctc
2400
tttttcatcc aggcctgtgg tggggagcag aaagaccatg ggtttgaggt ggcctccact
2460
tcccctgaag acgagtcccc tggcagtaac cccgagccag atgccacccc gttccaggaa
2520
ggtttgagga ccttcgacca gctggacgcc atatctagtt tgcccacacc cagtgacatc
2580
tttgtgtcct actctacttt cccaggtttt gtttcctgga gggaccccaa gagtggctcc
2640
tggtacgttg agaccctgga cgacatcttt gagcagtggg ctcactctga agacctgcag
2700
tccctcctgc ttagggtcgc taatgctgtt tcggtgaaag ggatttataa acagatgcct
2760
ggttgcttta atttcctccg gaaaaaactt ttctttaaaa catcagtcga ctatccgtac
2820
gacgtaccag actacgcact cgacctcgac ggatccggag agggcagagg aagtctgcta
2880
acatgcggtg acgtcgagga gaatcctgga cctatggtga gcaagggcga ggagctgttc
2940
accggggtgg tgcccatcct ggtcgagctg gacggcgacg taaacggcca caagttcagc
3000
gtgtccggcg agggcgaggg cgatgccacc tacggcaagc tgaccctgaa gttcatctgc
3060
accaccggca agctgcccgt gccctggccc accctcgtga ccaccctgac ctacggcgtg
3120
cagtgcttca gccgctaccc cgaccacatg aagcagcacg acttcttcaa gtccgccatg
3180
cccgaaggct acgtccagga gcgcaccatc ttcttcaagg acgacggcaa ctacaagacc
3240
cgcgccgagg tgaagttcga gggcgacacc ctggtgaacc gcatcgagct gaagggcatc
3300
gacttcaagg aggacggcaa catcctgggg cacaagctgg agtacaacta caacagccac
3360
aacgtctata tcatggccga caagcagaag aacggcatca aggtgaactt caagatccgc
3420
cacaacatcg aggacggcag cgtgcagctc gccgaccact accagcagaa cacccccatc
3480
ggcgacggcc ccgtgctgct gcccgacaac cactacctga gcacccagtc cgccctgagc
3540
aaagacccca acgagaagcg cgatcacatg gtcctgctgg agttcgtgac cgccgccggg
3600
atcactctcg gcatggacga gctgtacaag tgatgaacaa agactttgaa gtacataaat
3660
gtattacttt gatgttaata cagttcagtt tagtaagatg tgtagtaaaa agtgtaacct
3720
tgttcaaaaa gttgcttcaa gttgatgttt gtgttctgtt ttacctgttc cagaatagct
3780
atttttgctt gagaagtttg aagttgtaag agttgaaata tttccaggtt ttattactag
3840
cttgcatgct tttcctgcta actaactgaa atgctaatct taaggaattt atatggggaa
3900
ggggaaaaaa gaaaaacact ttgtttggta cgtgtggatt ttcttctgag ctttaaggta
3960
cagtttgttg catgttaaaa tttagttctt attaaaccac aactttaagt tactaacgtc
4020
aaccagttac ctcttgcagt tcaaaagttg aagcagttcc ttgtccaaga tggagtattt
4080
taaaactgag ctcttaatca gtggaacaga agacgtcacg gtgtaactca actgaagccc
4140
tttaagtccc ggttctcttt agactaccta atcaatgtct ttgtttgcta acgacagttt
4200
atctatgtga atcctaaaat tcctatatgt aacttaagat gcaagaatgt aattagttac
4260
attggctgct cagtggagta tgactttttt ttttactgga ttaattttag caatacctgt
4320
atcttaaaat tgtgagaaaa tactgcattt aaaatatgcc taactttgtg atgcaatatg
4380
ttaatcaaag aatacatgta agcatatttt aataataatt atgtagattt tagtcatgta
4440
ttttgaaaca attaaaattt ttaattttga cttaccttcc cagtgtgagt gacatcctaa
4500
tataatactt ctaaatctta agctgctttg agaaaggcat gcagcgtatt tattgaagga
4560
attgaagatt tcttacccta cataagaatt ccagttagga caagtttata gcaacaaact
4620
ttcacatttg ctgttagttc tacctgcgat tttgaaggag tacaactgag aacagcactc
4680
agtcttgtat gtgtgttggg tcctagtctg attcattttt cttattacta cccttatacc
4740
tcagtctcca agtaaaaaag gaaataactc ctcctttgta gacgtgtata tgtgaatgaa
4800
tagaatggca tgtcccactt caaatgtcta gaagtagatg ttggtgaaac atgcaataag
4860
agctgagttg ctctgtacct ggatagtggg ctgtaagatg cagcacagga gtgtcccagg
4920
actgttgttc aggagtagga gtcaggggag ggaggcaggg cttgggggat agcaataata
4980
gttggtgccg ttctccataa aattactcag aagcaatgtt ctggcagcat aaatcgacct
5040
aaagttgtta actttttctc tgctcagacc gttccctagc actaaagatg ctgtgtcctt
5100
aagtgtgttt ccatactatc catcaccatg tttttcccaa cctattgctt taatagtatt
5160
g
5161
<210> 6
<211> 6579
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Left arm-GFP-2a-iCaspase9-T2a-Right arm
<400> 6
tttttttctt tgagatgagt atcttattgc agctctgtgc ataccccttg aaaatccgta
60
ggtgatgtat ttatgttgac tgaaaatgga gtcattcaaa aattgatgac ttcttccttt
120
tctcttttta atctcgaaac ctgttctcag attcctttat cagaacaaaa agcacggctg
180
catgtttttc accgttcaca cgactgtgtt tagccagcgt atctaaatgc ataacatgat
240
ttgccataac ttgagcctgc cataatcaga gcctcatttt gaaagctgtc atggtctgaa
300
acatagcatg ggtaagatga ttttcacctt gatgatgcat gtttaactca tttcaacgag
360
aggtcaaatc catccaaaaa tgctaaatct atcaaccagt tttcatgttc aagttctggc
420
acaaattttc gttctgactc catacgtgat tagatttcat tttgcaaatc atgaaaacta
480
ttgccaaaac atacaatttt aagtcagcag ttttactgcc caaatttgca caagcacatg
540
taacgtgaca gacaaaaacc cactggacag agacaaaata ctgggtttgg ccagtatggt
600
ttggctaagc cacgtttttc agggcaagaa ttgcctcacc tttttttata ttggaatcaa
660
attctcaagt gtaacagcct ggcgccatct cagctgccat taagtagcat gtgctgcaac
720
aagatgtcta ccacacactc tgctctgggt ggtgggggca caccagtgtg gacggcaacc
780
atgacacaag catagagtct tggcagaggt ccattgggtg catttatagt ggttcacact
840
ggggggttat cacagaatca tggaggttgg aaggtacctc cgggggtcat caaatccaac
900
cacttgacaa agcaggttcc ctaaagtaga ttgctcaaga acagtcacag cactgctgct
960
cacagaaaat ctctagttgg taaggttacc tttttttttg ttgtcgctcc cagttaaacc
1020
aaggtgggaa cacatccttt tcatagttgt attgtaggga tgtttatagc tattgctagg
1080
gaaaactgaa cagcgtgcaa ggaagtcagc actgacacag ctttcccacg tgagagagct
1140
atttcaaagc aagatcagca catatcccaa tctgtacttc ctcaaacaca gcccaaaacc
1200
gatagcaagg ccaagcagca gcactgctcc ttcaaggaaa ctgctgcata atctgaattt
1260
cagacggcag gaggaaacaa ggcagcagac tgatgaccta ccaccctgag tctcagctac
1320
atgcgatccg agccactcca actctgcttc ctgcagccct tccttcacgc tcatcttttg
1380
caccttaggc actcttataa ttcaagtact ttgtggcttt gggatatttg aagagcttgg
1440
tcagttagtc acaagtctgg ccacgtgcta tcatattagt ttgaaaagca atggagacac
1500
catctgctga tgctcaaagt ggttacaacc aaaacacaaa aaagcagagc tgtgggaaga
1560
attcaacatt ttgattatgc aagaagctag tcccagcctt gaaatccacc atctgcatca
1620
tgaaagacct aagtagttaa agccacagca gacatacagc ttctatttcc ttaccttctt
1680
catcattaac tacaggtctt ggaagatttt tgctgggaaa aagcttttat tgcaagaact
1740
gtaatttatt aacagggaaa catgaaataa atgtgtaaat tctcctgcac tcccactgtc
1800
attaaaaacg gctttaagaa agagtattca gtaactgcct gcattgtgtt gtgactttct
1860
actctgtgac acacagcctt ctgggcaaag cacatattct gccatgcatg tgggtcgtgc
1920
cttggaaaat gggaacccac attcactgag ggcctcttga tgagctttcc ccttgagaac
1980
agcgaggtct catggatatc ttctcttctc caagccaaat aagcccagat ccctcagctt
2040
tccttcatgg agaggctcca gccctctgat catgcctgca gcccttctct ggacctgctc
2100
cagcagcccc acacgctcct catgctggga ccccagatct gcacatggta ttgcacatgg
2160
ggcctcacaa cggcagagct gagagggaca attccctccc tcagccttta attctgcacg
2220
tacttaattt tgtctgtatt ttttttgcaa tagaatagcc tgcatgctag ttgctgtgtc
2280
gcacagctga attcactagg ttcctgtgaa acaaagggca atcccacagc cactgtagca
2340
cgtgaggagc cacacaggtg ctctaattcc tacaggacag gcctagggac agagcggccc
2400
taggactgcc ctcccgcacc tcctagcttc accttttgcc cgcgatcttt aagtaccctg
2460
aagcataaac agggaaagcg ccgcccgcag cctcaccctg cttcccgccc aacgcggcac
2520
cgcccaggcc gcagccgccc actaagagca ctagcgccac cttctcaccc cacccccacc
2580
acgcgttcct agcggccctg agagctgctg cgcatgcgcc gcctgacgat tcgccctccc
2640
attggctggc ggtcgaagca cggcgggggc acgcggtggc ggctatataa ggcgtctcgg
2700
aagacggcgc catgctattt ggagcggaga gtgaaagtta cagttcctgg tgctggtagg
2760
gagtgtggcg cggagcggag cgctgcggct catcggaacc acaatggagc catagcagag
2820
ccgggcgtgg gggcaagggc agtgcgtgct ggggagggct ccgtcgcgtg gccacgtcgc
2880
gagagccgtc gggatggtgg cgtcagggcg gggggtgctg ctaacgtgct cctggtcctg
2940
caggtgggct gctggcattc gccatggtga gcaagggcga ggagctgttc accggggtgg
3000
tgcccatcct ggtcgagctg gacggcgacg taaacggcca caagttcagc gtgtccggcg
3060
agggcgaggg cgatgccacc tacggcaagc tgaccctgaa gttcatctgc accaccggca
3120
agctgcccgt gccctggccc accctcgtga ccaccctgac ctacggcgtg cagtgcttca
3180
gccgctaccc cgaccacatg aagcagcacg acttcttcaa gtccgccatg cccgaaggct
3240
acgtccagga gcgcaccatc ttcttcaagg acgacggcaa ctacaagacc cgcgccgagg
3300
tgaagttcga gggcgacacc ctggtgaacc gcatcgagct gaagggcatc gacttcaagg
3360
aggacggcaa catcctgggg cacaagctgg agtacaacta caacagccac aacgtctata
3420
tcatggccga caagcagaag aacggcatca aggtgaactt caagatccgc cacaacatcg
3480
aggacggcag cgtgcagctc gccgaccact accagcagaa cacccccatc ggcgacggcc
3540
ccgtgctgct gcccgacaac cactacctga gcacccagtc cgccctgagc aaagacccca
3600
acgagaagcg cgatcacatg gtcctgctgg agttcgtgac cgccgccggg atcactctcg
3660
gcatggacga gctgtacaag ggatccggag ctactaactt cagcctgctg aagcaggctg
3720
gagacgtgga ggagaaccct ggacctatgc tcgagggagt gcaggtggag actatctccc
3780
caggagacgg gcgcaccttc cccaagcgcg gccagacctg cgtggtgcac tacaccggga
3840
tgcttgaaga tggaaagaaa gttgattcct cccgggacag aaacaagccc tttaagttta
3900
tgctaggcaa gcaggaggtg atccgaggct gggaagaagg ggttgcccag atgagtgtgg
3960
gtcagagagc caaactgact atatctccag attatgccta tggtgccact gggcacccag
4020
gcatcatccc accacatgcc actctcgtct tcgatgtgga gcttctaaaa ctggaatctg
4080
gcggtggatc cggagtcgac ggatttggtg atgtcggtgc tcttgagagt ttgaggggaa
4140
atgcagattt ggcttacatc ctgagcatgg agccctgtgg ccactgcctc attatcaaca
4200
atgtgaactt ctgccgtgag tccgggctcc gcacccgcac tggctccaac atcgactgtg
4260
agaagttgcg gcgtcgcttc tcctcgctgc atttcatggt ggaggtgaag ggcgacctga
4320
ctgccaagaa aatggtgctg gctttgctgg agctggcgcg gcaggaccac ggtgctctgg
4380
actgctgcgt ggtggtcatt ctctctcacg gctgtcaggc cagccacctg cagttcccag
4440
gggctgtcta cggcacagat ggatgccctg tgtcggtcga gaagattgtg aacatcttca
4500
atgggaccag ctgccccagc ctgggaggga agcccaagct ctttttcatc caggcctgtg
4560
gtggggagca gaaagaccat gggtttgagg tggcctccac ttcccctgaa gacgagtccc
4620
ctggcagtaa ccccgagcca gatgccaccc cgttccagga aggtttgagg accttcgacc
4680
agctggacgc catatctagt ttgcccacac ccagtgacat ctttgtgtcc tactctactt
4740
tcccaggttt tgtttcctgg agggacccca agagtggctc ctggtacgtt gagaccctgg
4800
acgacatctt tgagcagtgg gctcactctg aagacctgca gtccctcctg cttagggtcg
4860
ctaatgctgt ttcggtgaaa gggatttata aacagatgcc tggttgcttt aatttcctcc
4920
ggaaaaaact tttctttaaa acatcagtcg actatccgta cgacgtacca gactacgcac
4980
tcgacctcga cggatccgga gagggcagag gaagtctgct aacatgcggt gacgtcgagg
5040
agaatcctgg acctatggag gaggattggg ataccgaact cgagcaggag gcggcagcgg
5100
cttcccaggg gcgttctgag gagcaggcgt ggatggtgag ctgtgtccag gggagggcgg
5160
tgcggcaggg agcggggcac tgggatggcc cggtctggga aggggaggcc gagaggcctt
5220
cgcagtgctt cctgcagctc cccgagcagt gcgaagaagg aggtgccggg ctctgctgtg
5280
ggagccgaag gcacggagct gcctggggag ggaatggctg tgtgcctgag gtgccagaga
5340
ccggcaaggg ctgtacagag aggcaacggt tttgtgtaca aatcatttgt atgggaaacc
5400
acaaaatctt gaagctttat tacatggcag cgaaatactt tggtgtgaag taaaggataa
5460
gatagaggag tgtaatgaga gacagctaaa taatatttta ctacttgtgg gatgagtgaa
5520
tatcaggagg aactgctgta aatttcagga ggacctgttg tgaatcttca gtagttggcg
5580
ccgctcttac acatcttaca gatgcccctt gagcaaaggg ggataaggag agatgaacgg
5640
gttatccaaa caggtgatga gctaaaagta cagttgccta aagaagtagt agcatgtgct
5700
atcagaatga taattttgtt agtttggggt tagtttcctg tagtggtaga tagccacaac
5760
aagaaaaccg cttaagtttt tgtaaaacaa aaaaagcacg atccagaagt aaaaaatatg
5820
ggtagttttt tttgatgttc ctctctcacc tggtgctttg gcatactaat atgtgtctaa
5880
ttgtattaaa caggagaaat ttaaacctag gctttgctgg aaataaaatg ttacaatgct
5940
acaatgtgaa aagtaggtgc tattctgaac tgttttgggt ggagtatctg aatctttgaa
6000
taatttaaga gggactgaca tatttaaaat acttaaggat aatctgtagc catgctgtaa
6060
aagaacaaca gaaatgcagt tgggaaggtg atggaaatag ttttattcat gttactggtg
6120
gtctgaaacc tttctaagct taaactgtag aaaaaaattg cttcaaaaga ttgcactatt
6180
actttgggcg acaaatgttt ttaattggtt ttaagtgttt gttagcaaag tgaagttgat
6240
gccaccataa gtctgacagg aggcaagata aacttgtctt catactgctt gtgtataact
6300
tggttttgat gacatttgtg tgtgaacatt atgcacttca gtgtagcgaa gtttaagaaa
6360
ctttgaacag aataacttga aagagtgtgc acatgggtgc agaagtcact ttatttcagt
6420
tgggagactt agcacctaaa tgcactgtta gttcacatac actttgcttg gcctgaaggt
6480
aacattgtga tgtcgctttt tttccctgta ggctaactct ggcagaccaa acagcccatc
6540
cctccgcttc tccagcagac caagcagccc cttgtctgg
6579
<210> 7
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер
<400> 7
cggtgacgtc gaggagaatc ctggacctat ggaggaggat tgggataccg aactcgagca
60
ggaggcggca gcggc
75
<210> 8
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер
<400> 8
gaaatccagc ttccagttcc cacctggcca gacaaggggc tgcttgg
47
<210> 9
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер
<400> 9
ggtgggctgc tggcattcgc catggtgagc aagggcgagg a
41
<210> 10
<211> 76
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер
<400> 10
gattctcctc gacgtcaccg catgttagca gacttcctct gccctctccg gatcccttgt
60
acagctcgtc catgcc
76
<210> 11
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер
<400> 11
tctcccatat ggtcgacctg caggcggccg cgaattcact agtgattctt cgtggtt
57
<210> 12
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер
<400> 12
aggctgaagt tagtagctcc ggatccaaca cttttgagca ctgctctt
48
<210> 13
<211> 107
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер
<400> 13
cagaacatca ccctgaccga ggtgggatcc ggagagggca gaggaagtct gctaacatgc
60
ggtgacgtcg aggagaatcc tggacctatg gtgagcaagg gcgagga
107
<210> 14
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер
<400> 14
cttgtacagc tcgtccatgc cg
22
<210> 15
<211> 94
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер
<400> 15
tctcggcatg gacgagctgt acaagtgatg aacaaagact ttgaagtaca taaatgtatt
60
actttgatgt taatacagtt cagtttagta agat
94
<210> 16
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер
<400> 16
ctcttcgaaa tccagcttcc agttcccacc tggcaatact attaaagcaa taggt
55
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> направляющая последовательность
<400> 17
ggtcctattc caggagagga
20
<210> 18
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> направляющая последовательность
<400> 18
ggcttactaa actgaactgt
20
<---
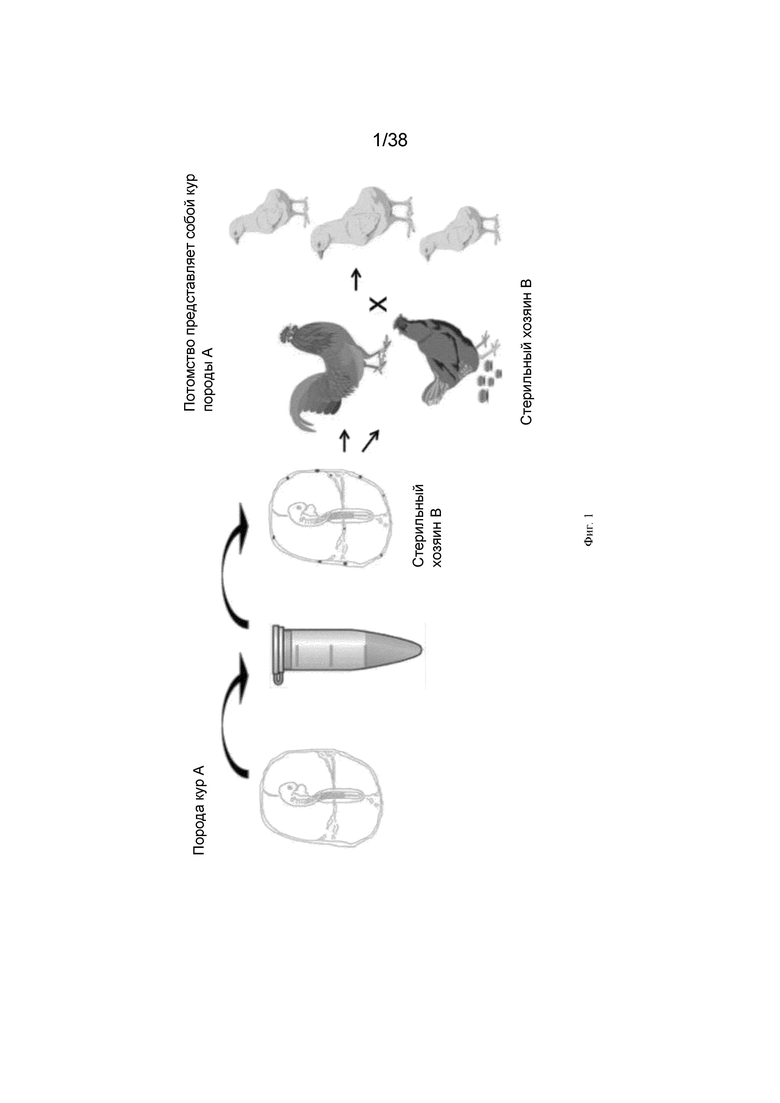
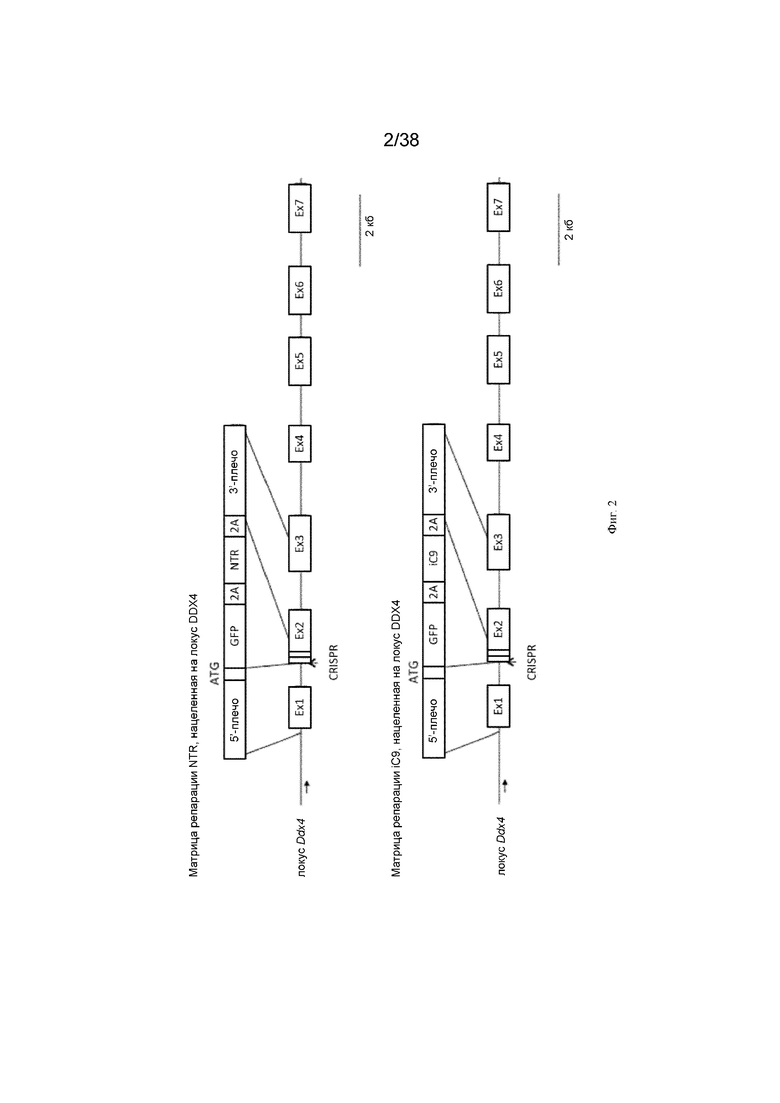
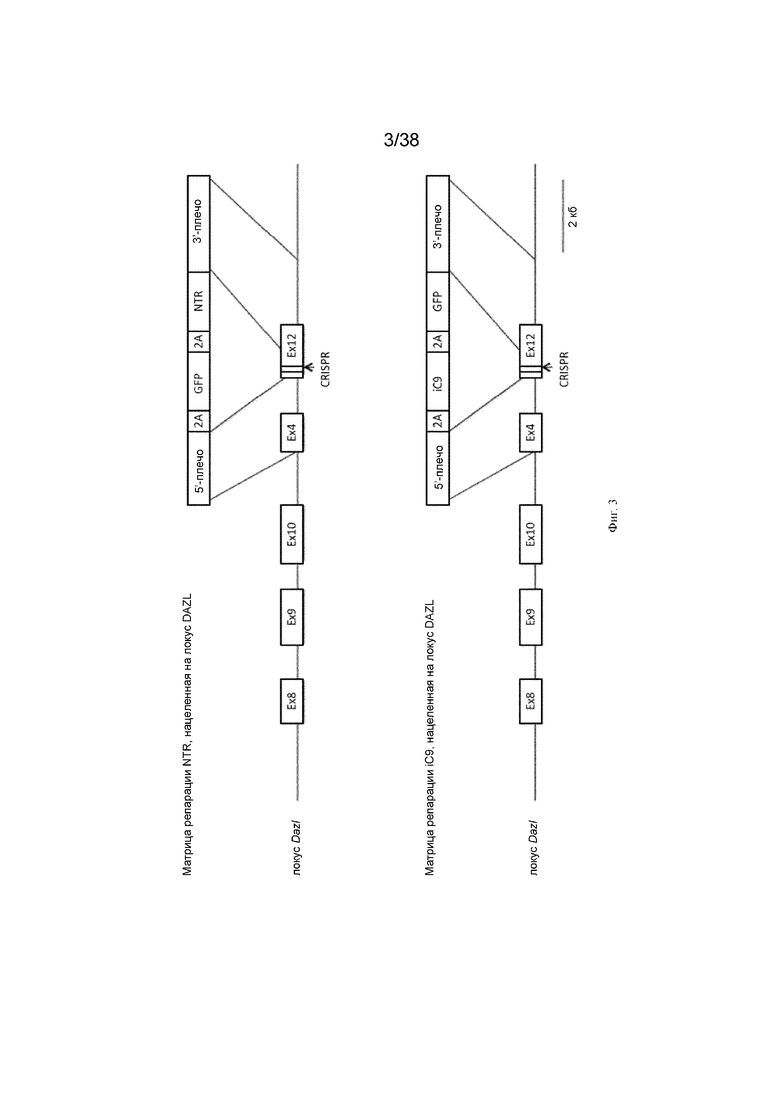
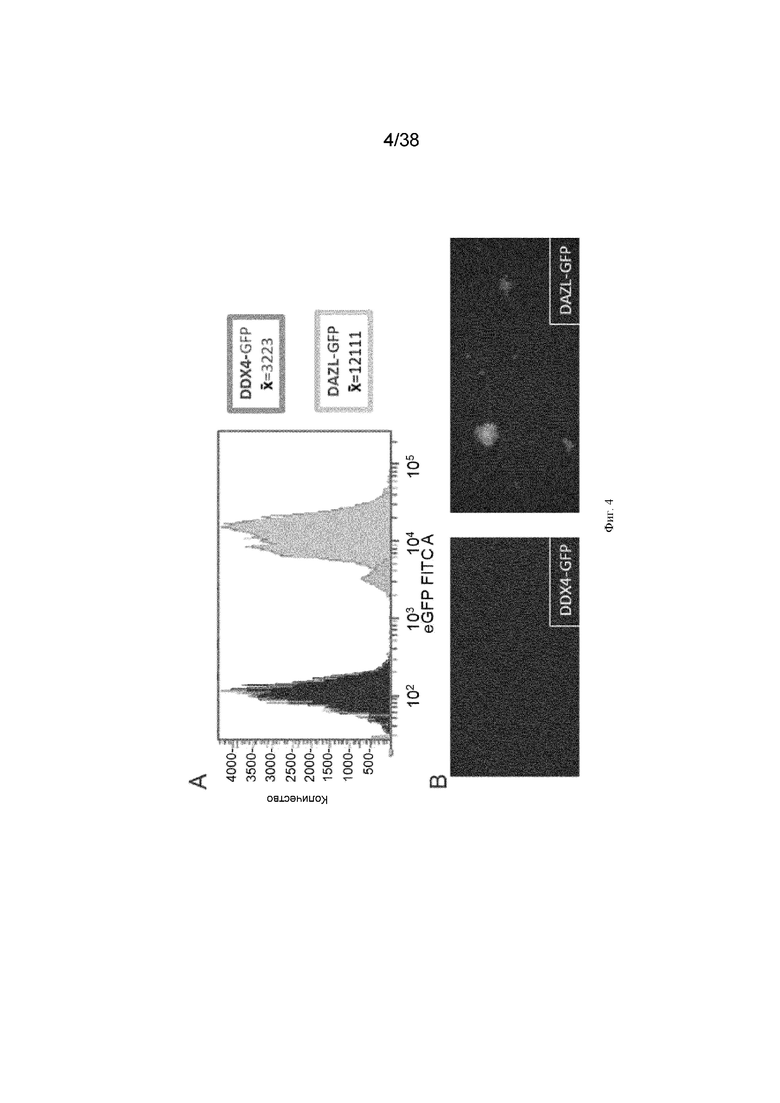
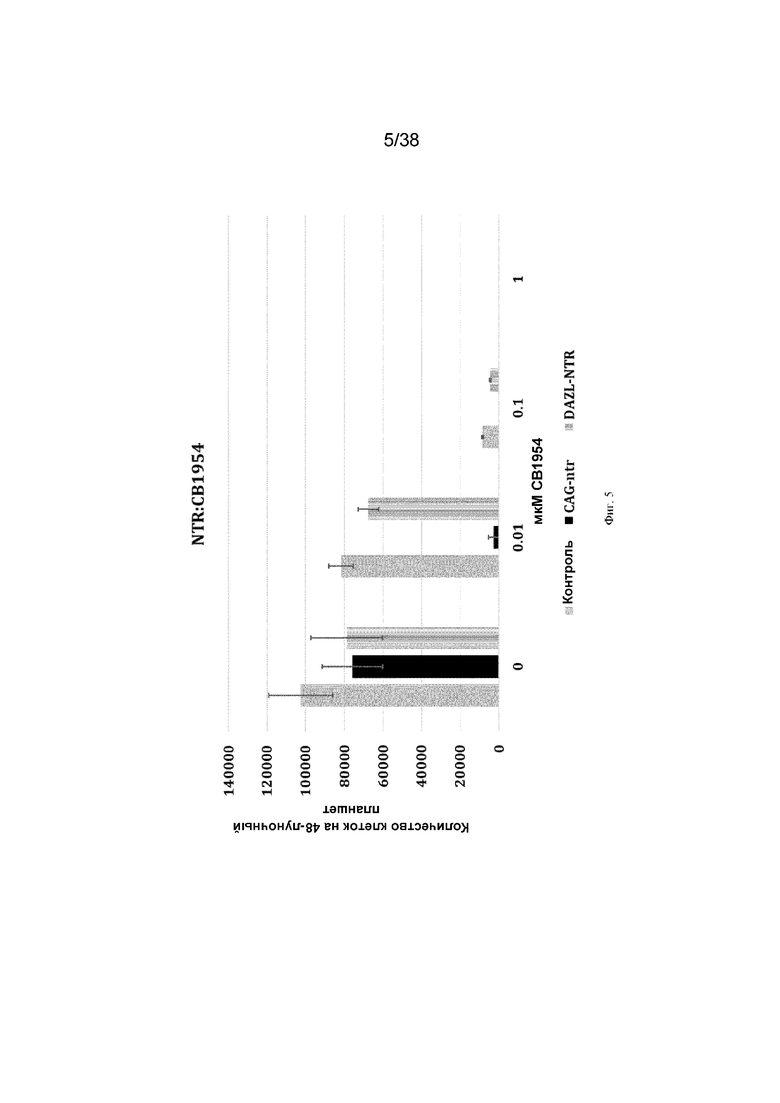
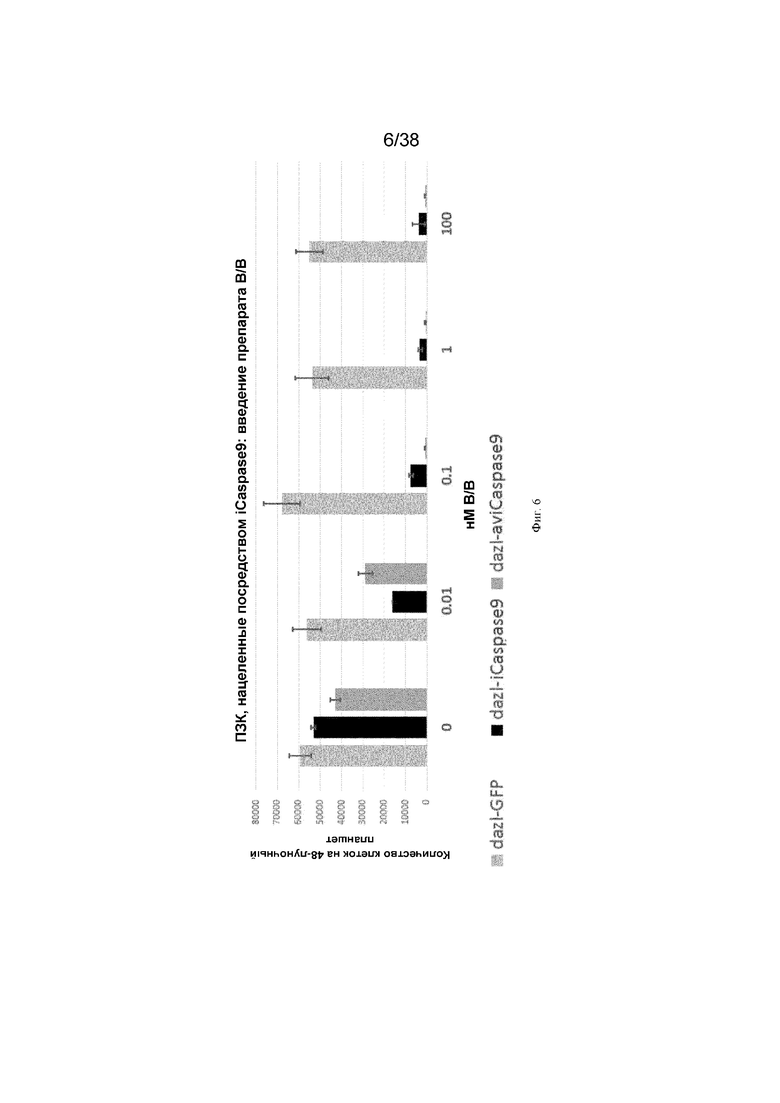
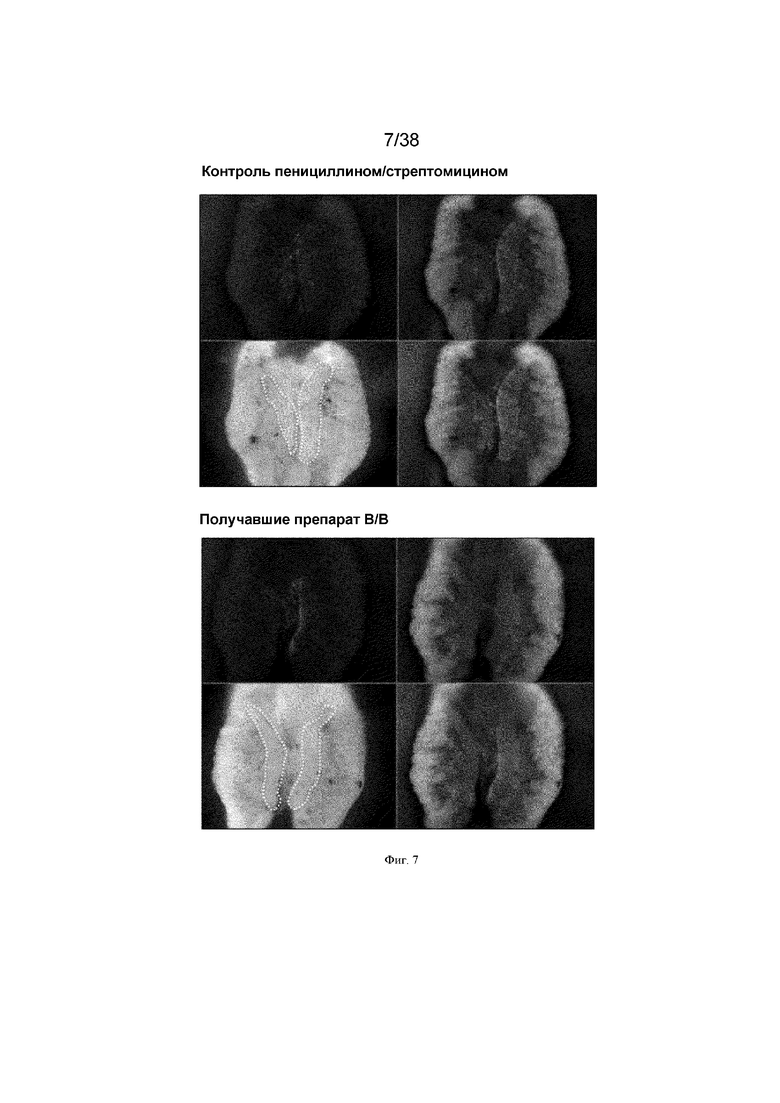
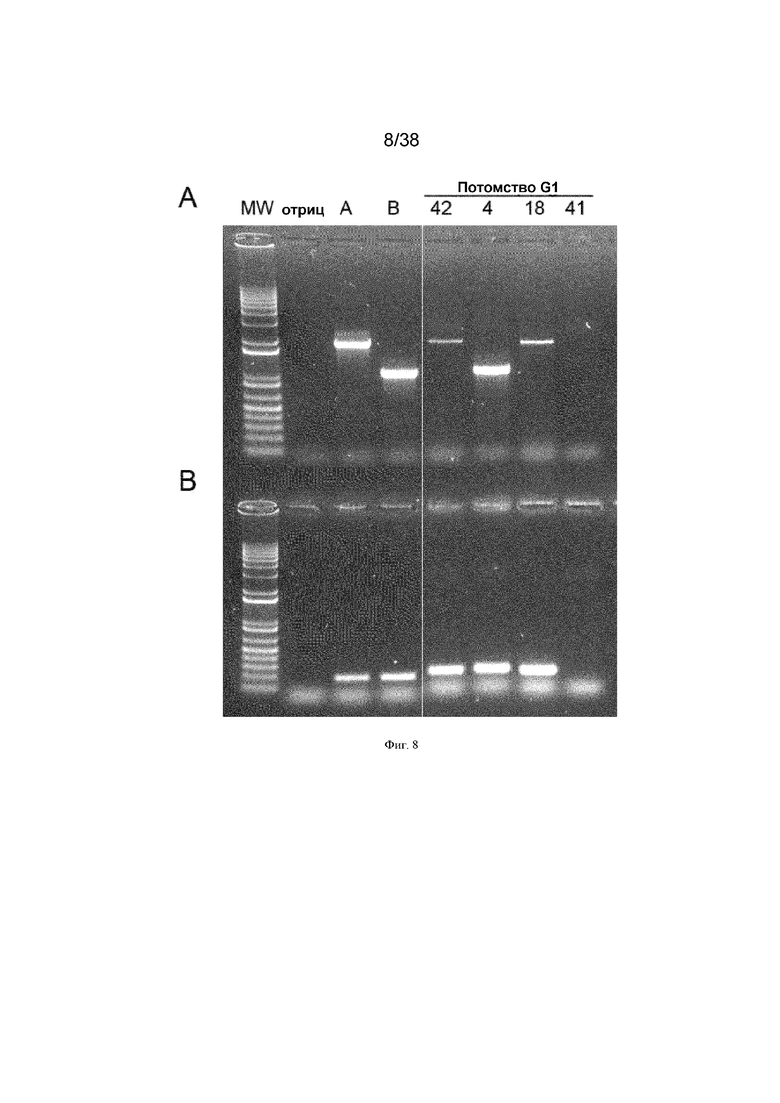
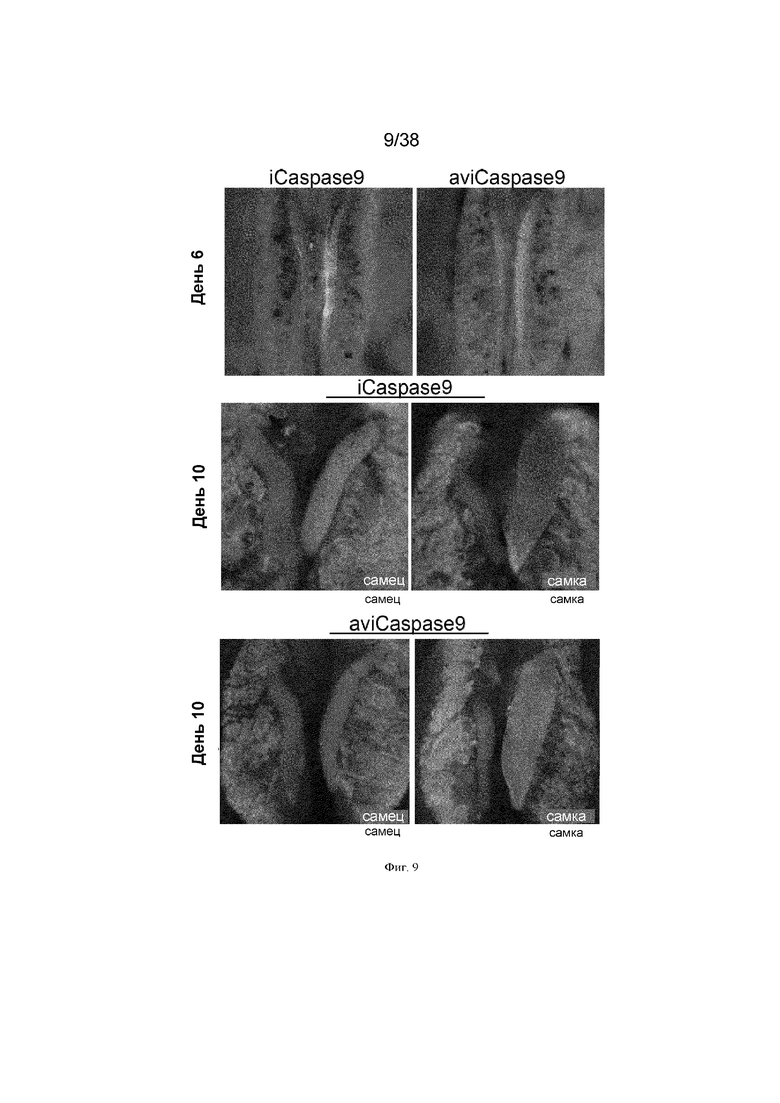
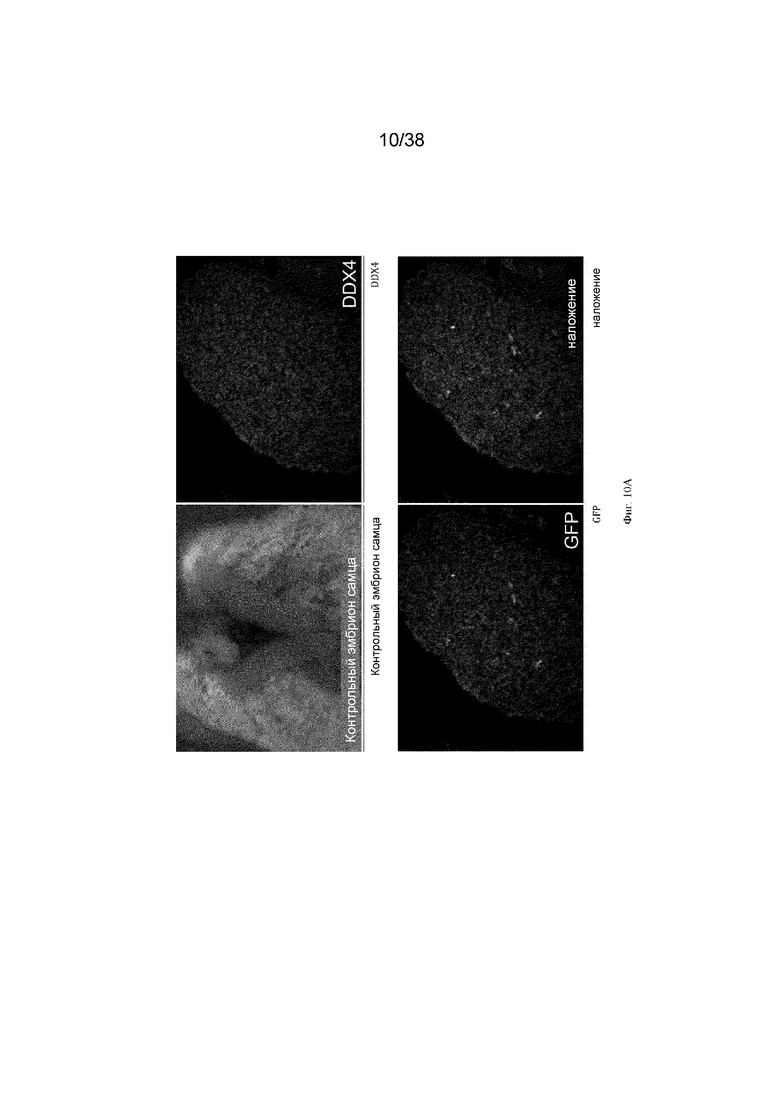
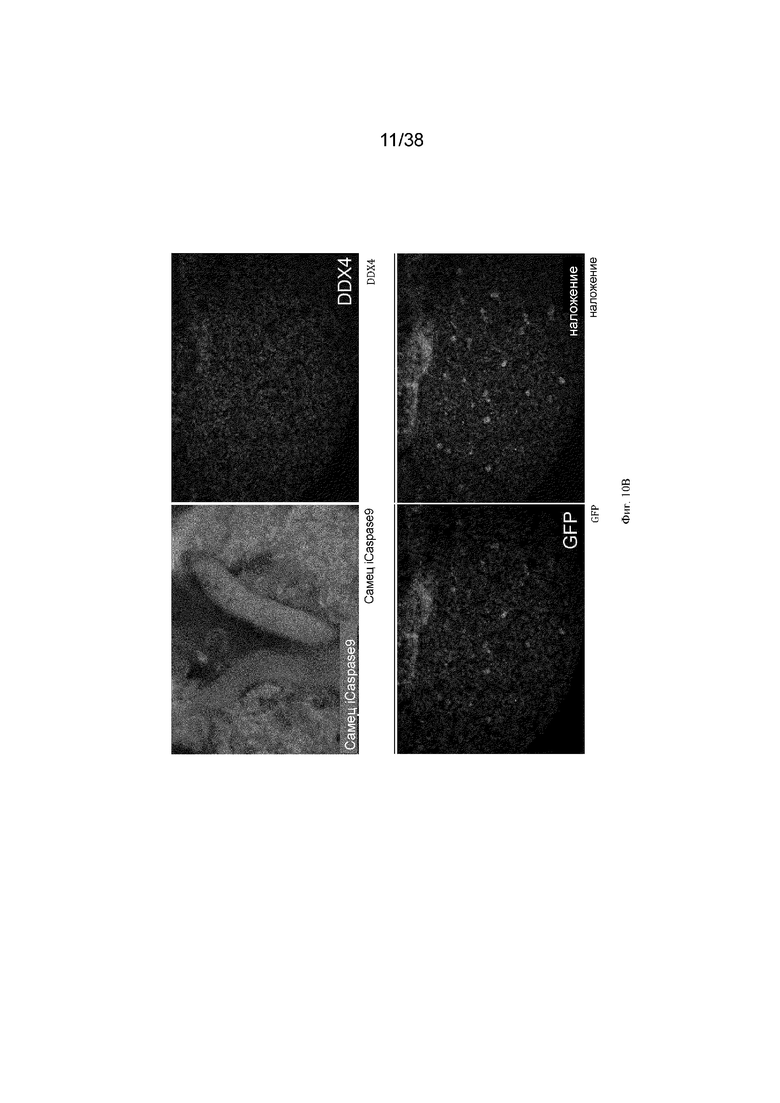
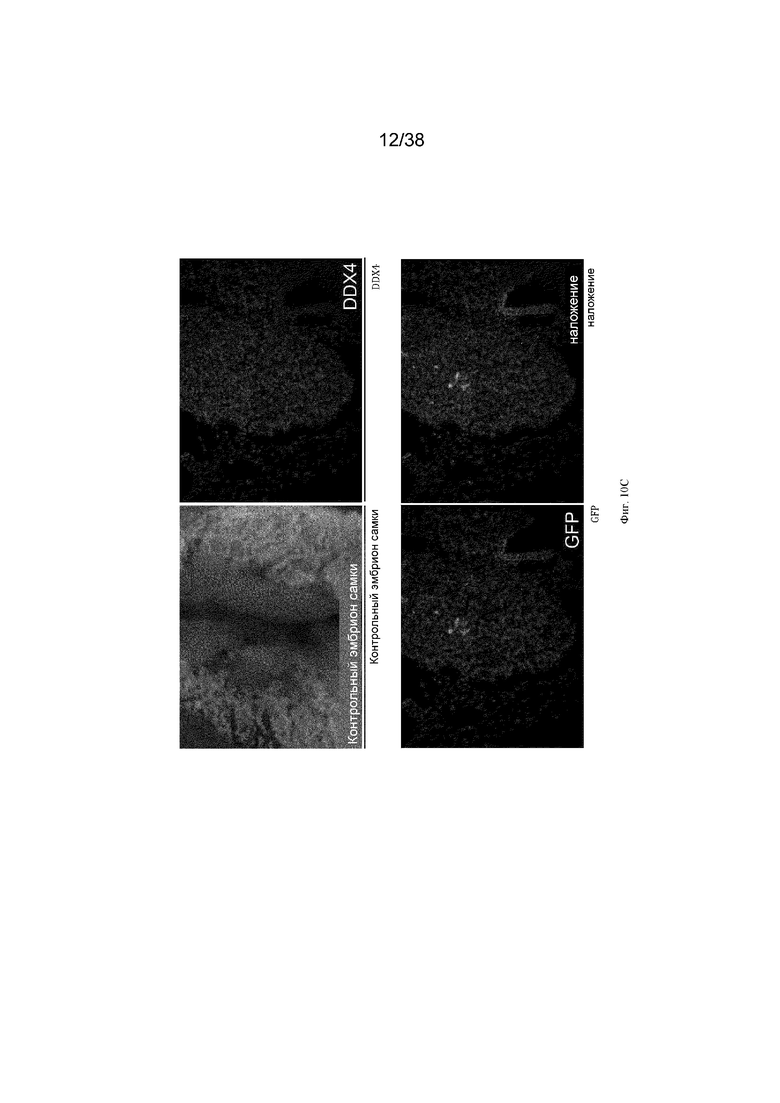
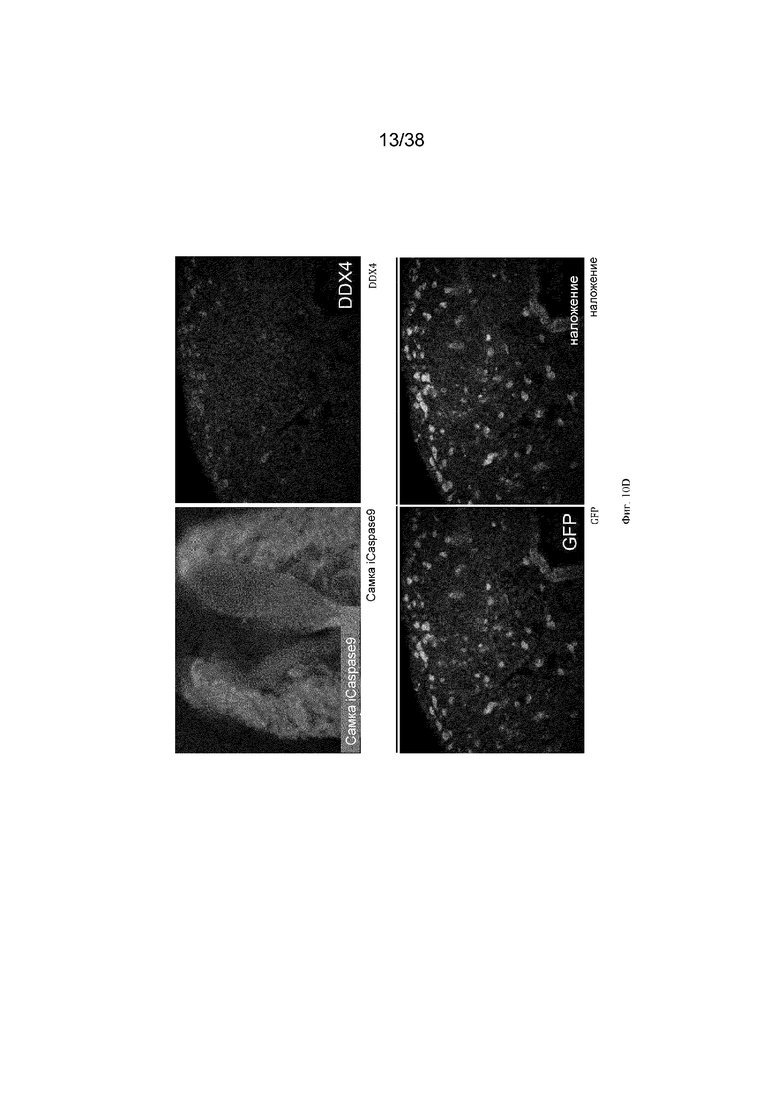


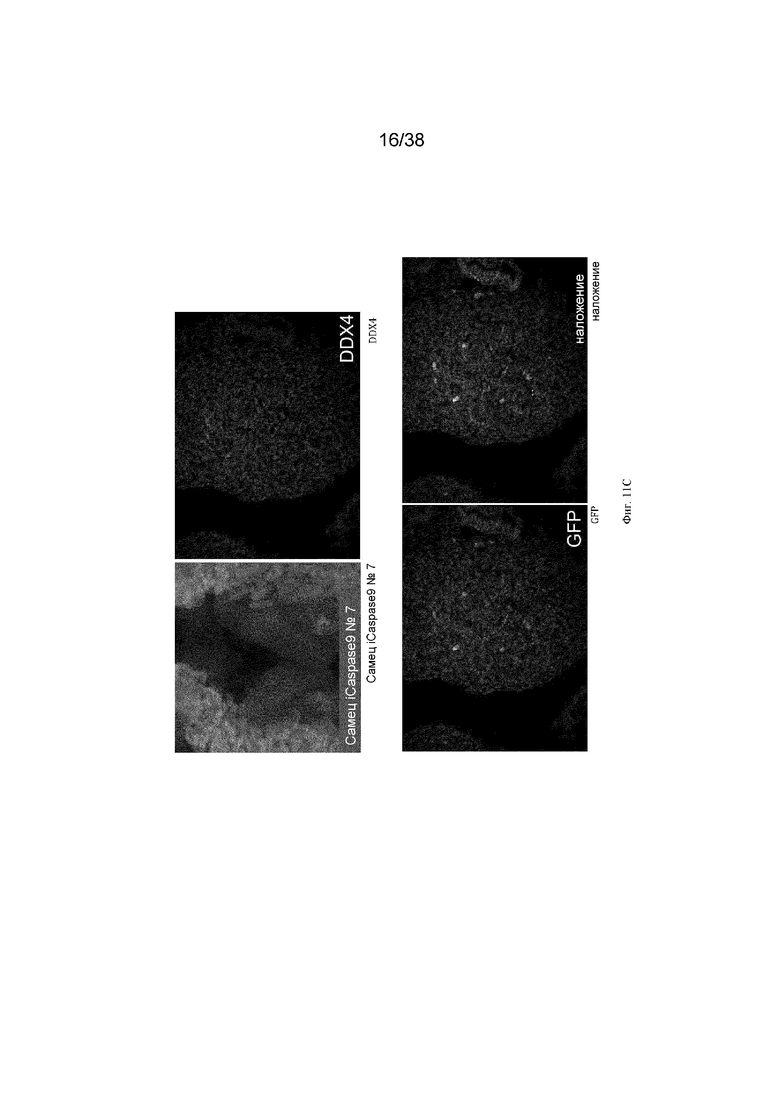
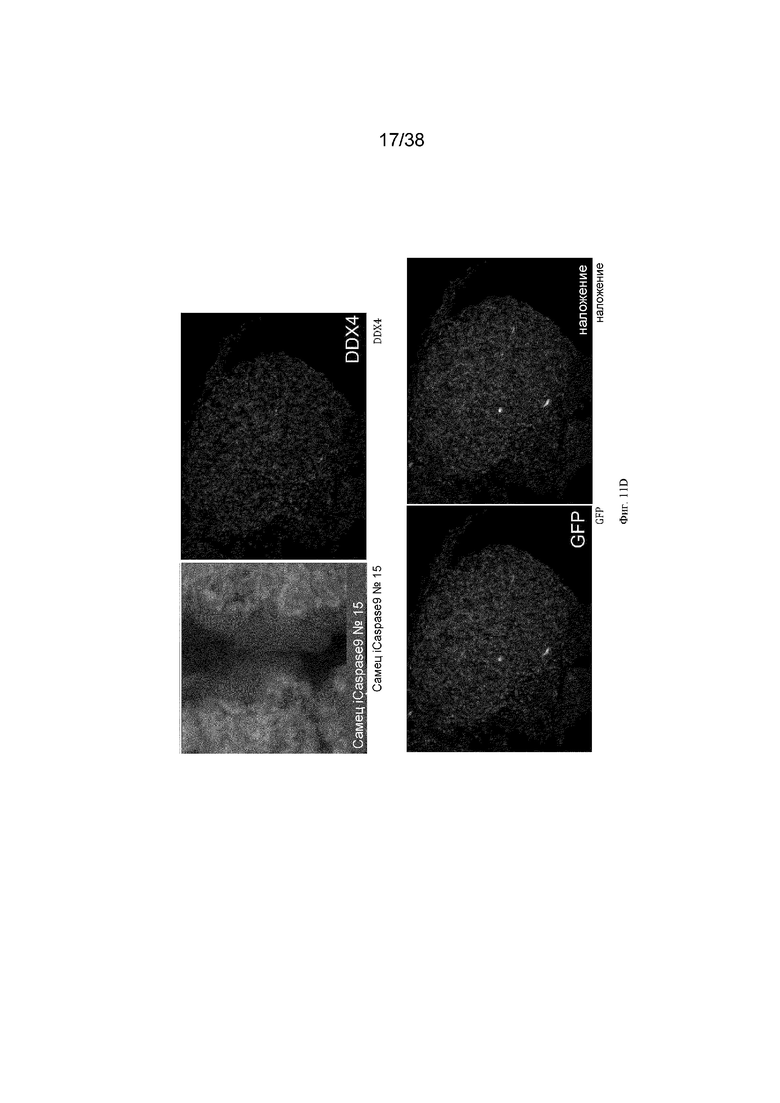
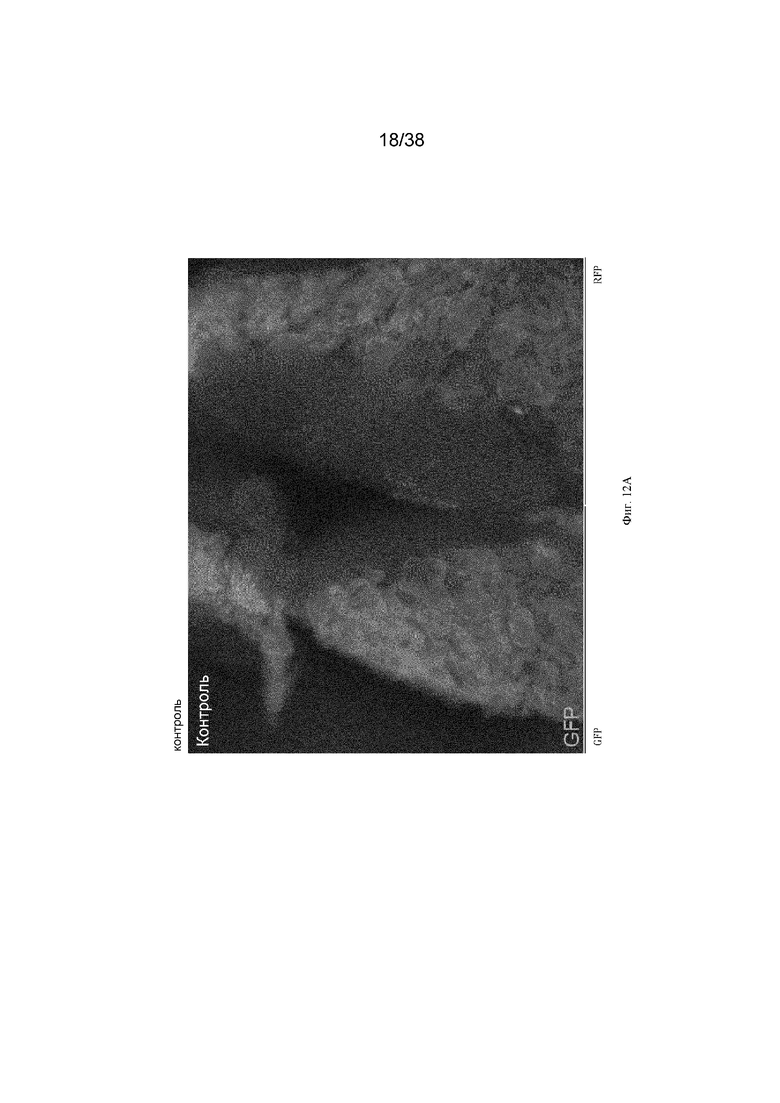
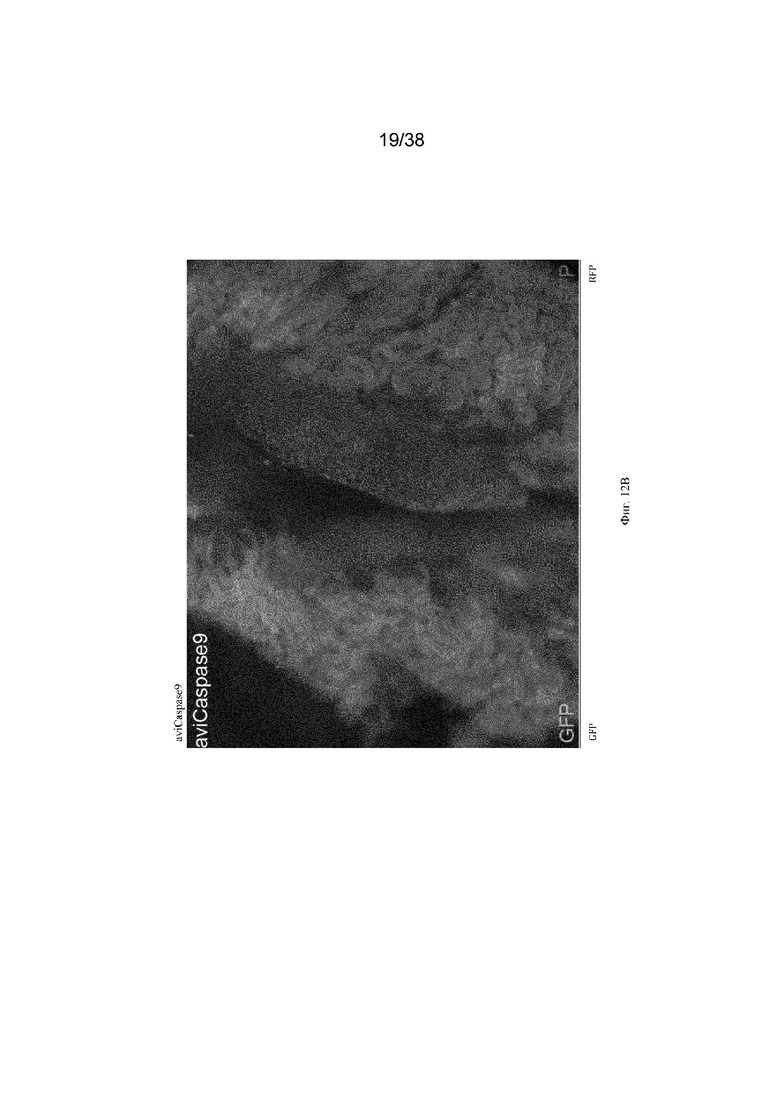
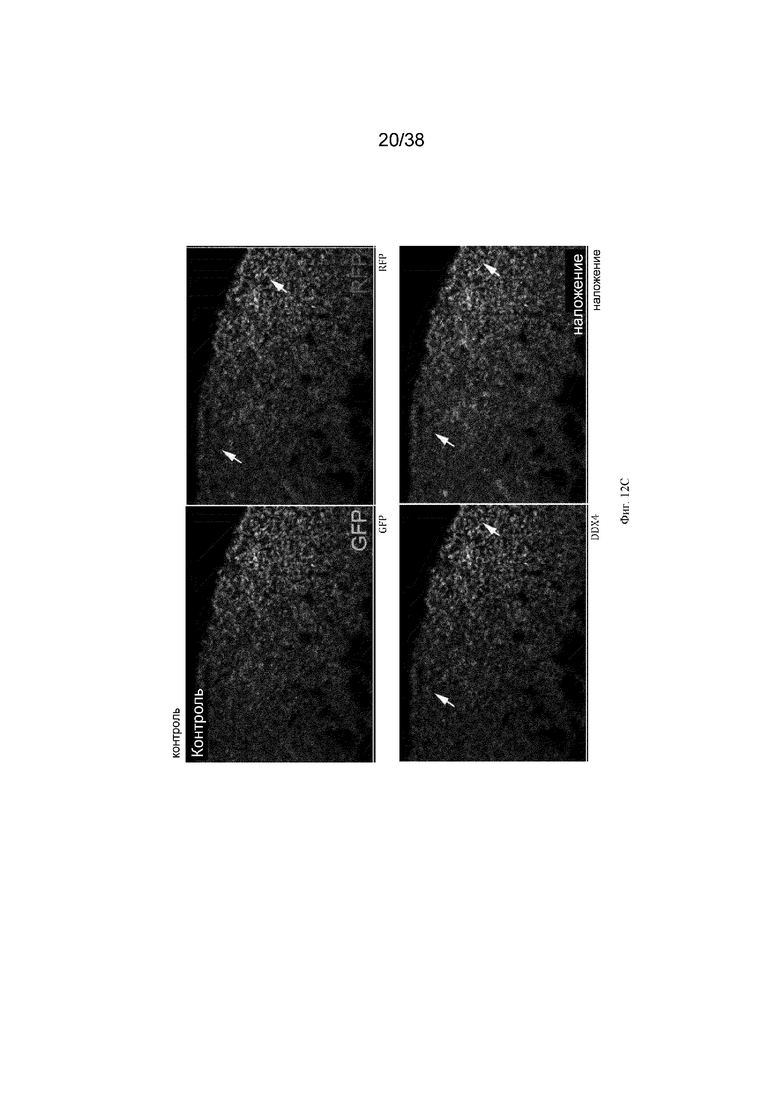
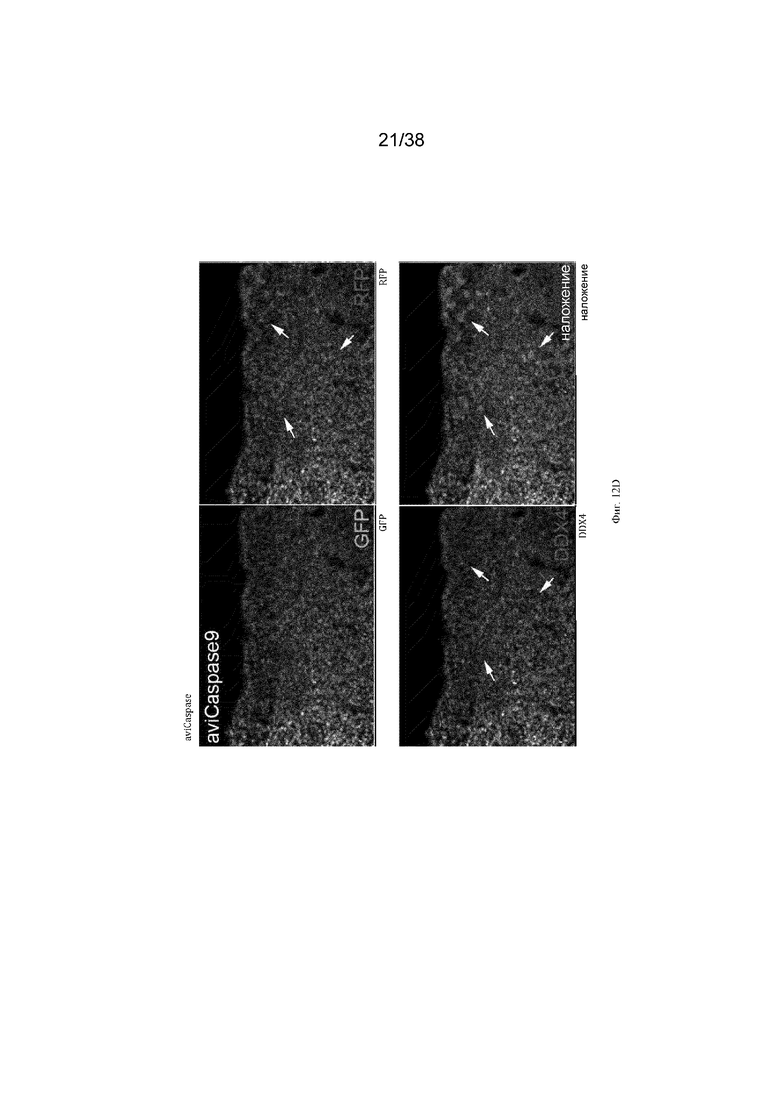










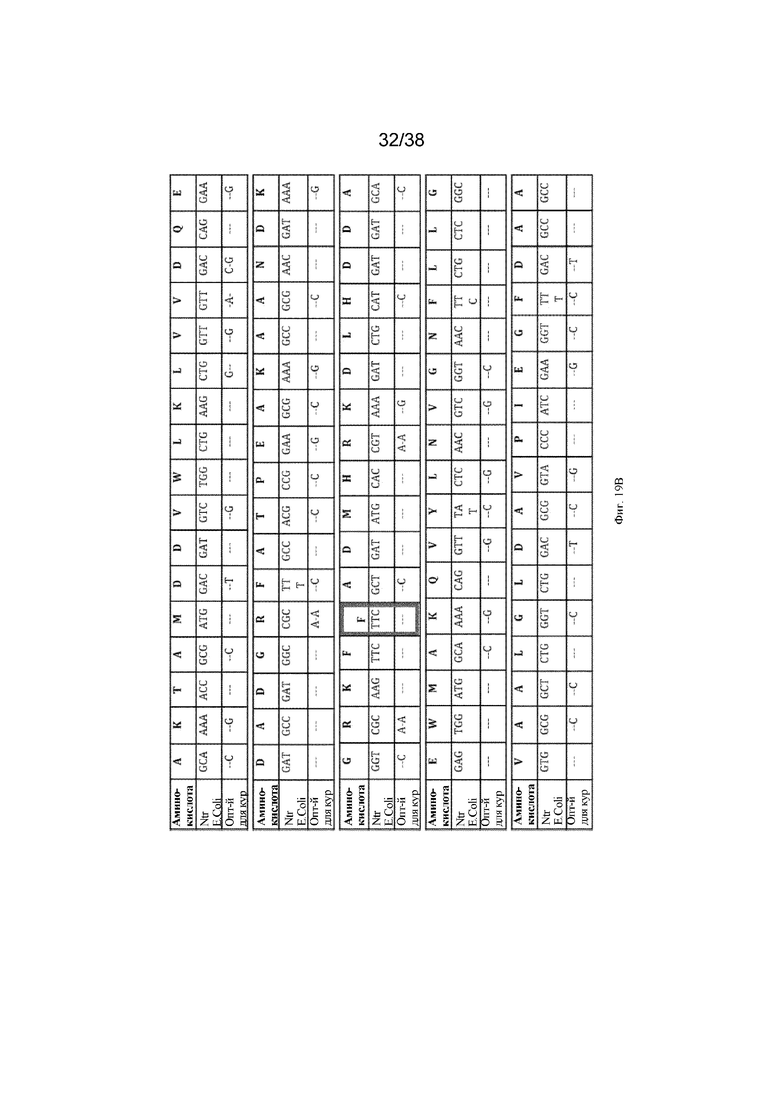

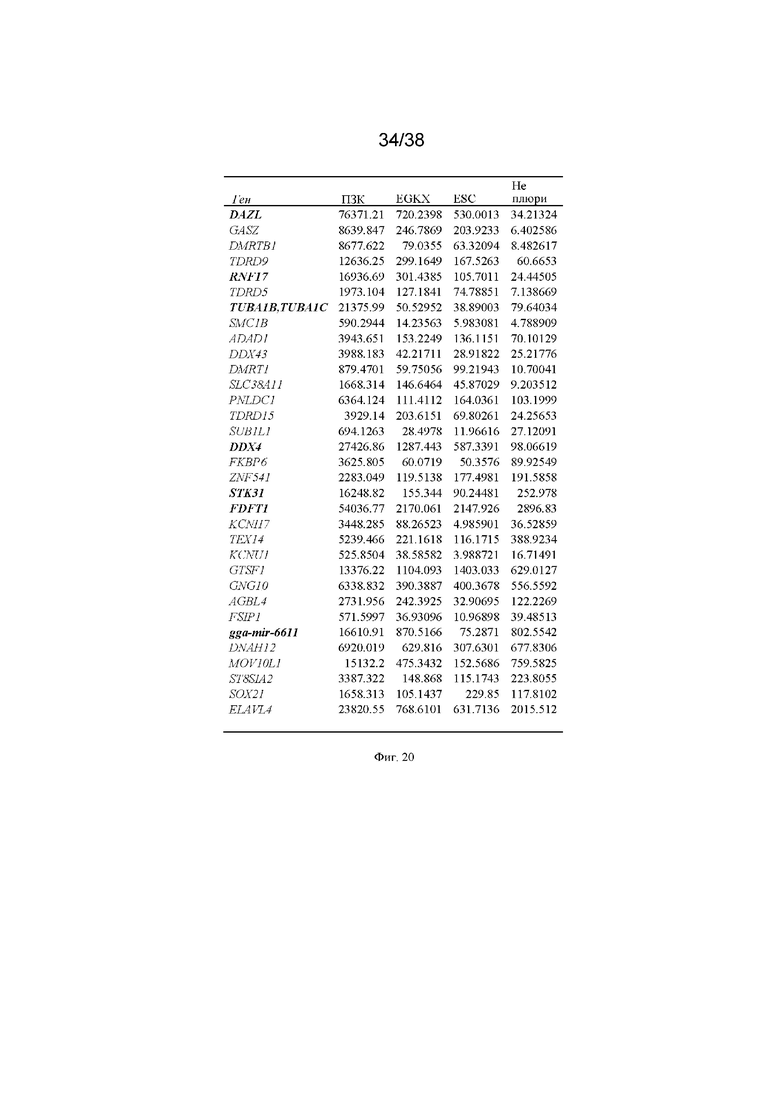
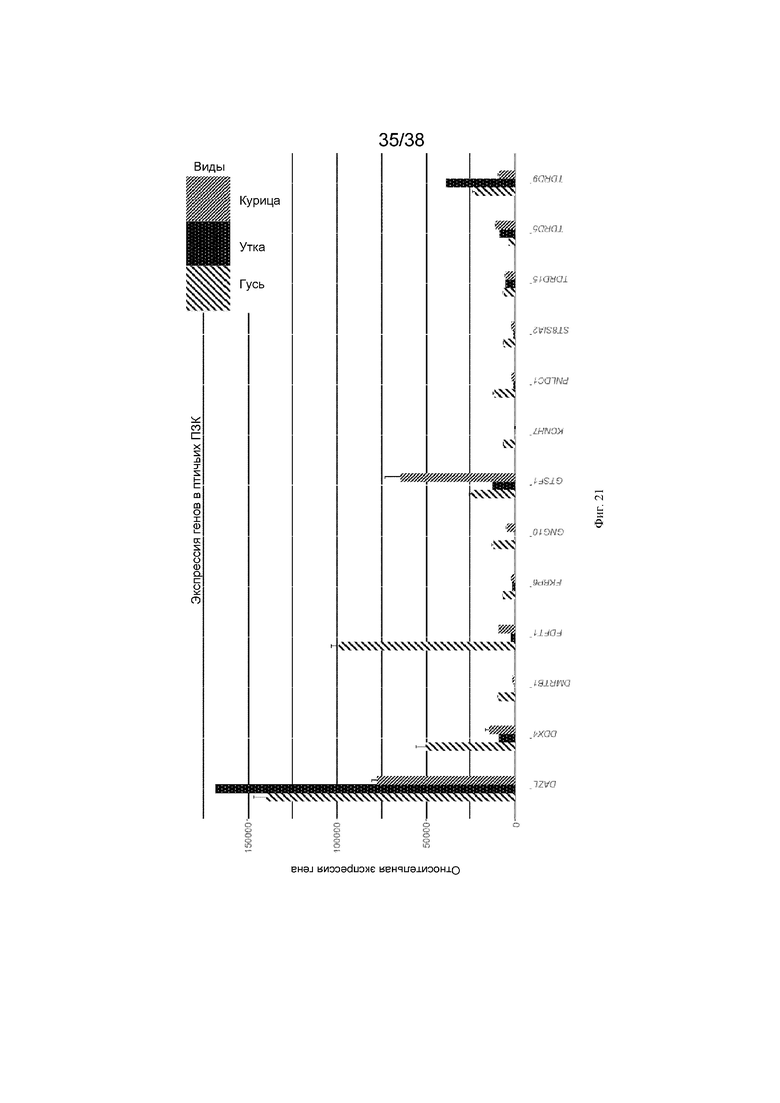
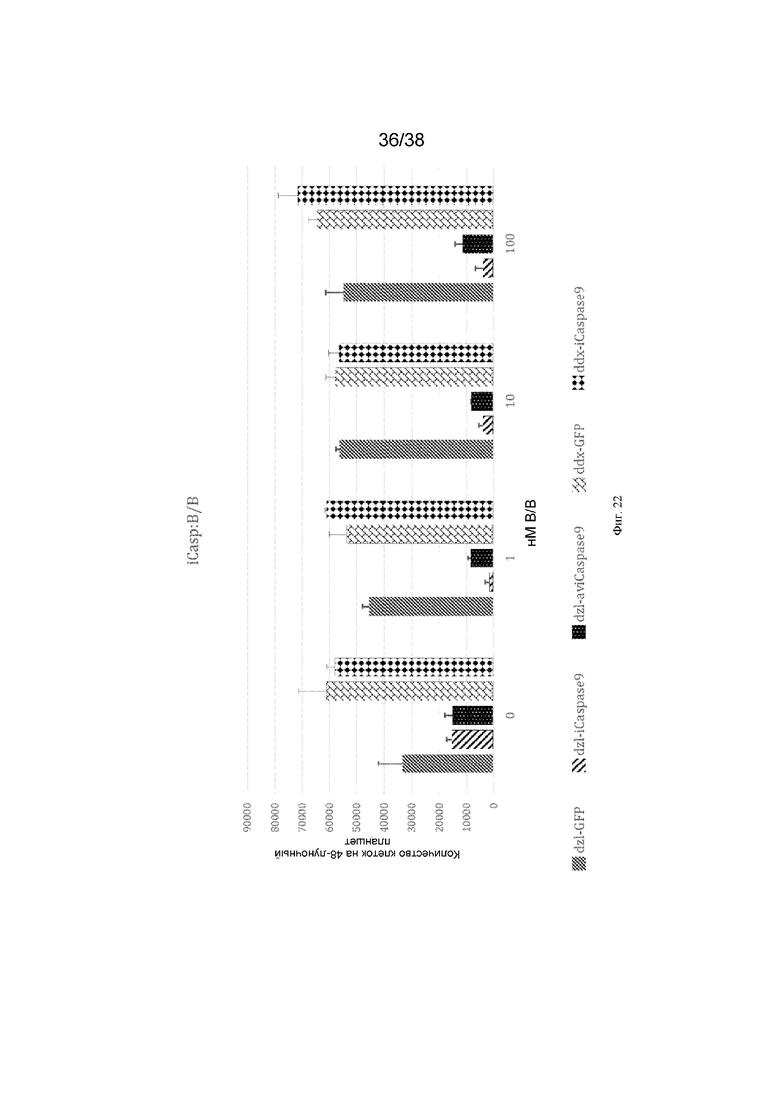
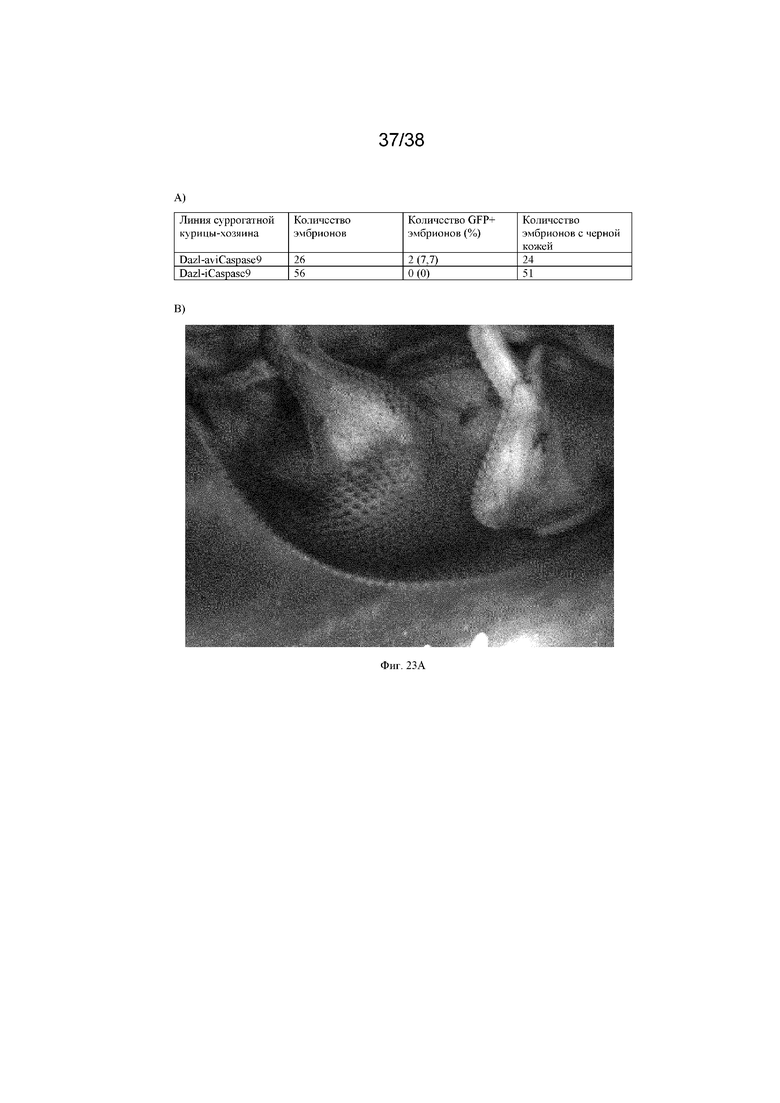

Изобретение относится к биотехнологии. Предложена трансгенная конструкция для экспрессии в зародышевой клетке птицы-хозяина. Указанная конструкция содержит (i) первую нуклеотидную последовательность, где указанная первая нуклеотидная последовательность кодирует фермент, индуцирующий апоптоз, причем указанный фермент, индуцирующий апоптоз, представляет собой каспазу, и (ii) вторую нуклеотидную последовательность, которая нацеливает указанную конструкцию на птичьи зародышевые клетки, причем указанная вторая нуклеотидная последовательность нацелена на локус DAZL. Также предложены способ получения суррогатной птицы-хозяина путем модификации зародышевой плазмы указанной птицы с использованием указанной конструкции и трансгенная птица, содержащая указанную конструкцию и предназначенная для применения в качестве суррогатной птицы-хозяина. Изобретение обеспечивает селективное уничтожение зародышевых клеток птицы-хозяина по требованию без уничтожения других клеток в зародышевой клетке птицы-хозяина и позволяет достигнуть высокоэффективной интеграции донорских половых клеток. 3 н. и 8 з.п. ф-лы, 23 ил., 1 пр.
1. Трансгенная конструкция для экспрессии в зародышевой клетке птицы-хозяина, причем трансгенная конструкция сконфигурирована для селективного уничтожения зародышевых клеток птицы-хозяина по требованию без уничтожения других клеток в зародышевой клетке птицы-хозяина, содержащая
(i) первую нуклеотидную последовательность, где указанная первая нуклеотидная последовательность кодирует фермент, индуцирующий апоптоз, причем указанный фермент, индуцирующий апоптоз, представляет собой каспазу, где активность белка, кодируемого указанной первой нуклеотидной последовательностью, вызывает гибель зародышевых клеток в присутствии экзогенного индуцирующего агента; и
(ii) вторую нуклеотидную последовательность, которая нацеливает указанную конструкцию на птичьи зародышевые клетки,
причем указанная вторая нуклеотидная последовательность нацеливает указанную трансгенную конструкцию на птичьи зародышевые клетки, причем указанная вторая нуклеотидная последовательность нацелена на локус DAZL.
2. Трансгенная конструкция по п.1, где указанная первая нуклеотидная последовательность содержит домен, индуцирующий апоптоз, и где указанный экзогенный индуцирующий агент представляет собой агент димеризации, где экзогенный индуцирующий агент индуцирует активность указанного домена апопоптоза, что вызывает апоптоз зародышевых клеток.
3. Трансгенная конструкция по п.1, где указанный экзогенный индуцирующий агент выбран из группы, состоящей из лиганда AP20187, FK1012 или AP1903.
4. Трансгенная конструкция по п.2, где указанный экзогенный индуцирующий агент представляет собой лекарственное средство для стабильной димеризации.
5. Трансгенная конструкция по п.1, где указанный фермент, индуцирующий апоптоз, представляет собой каспазу, при этом необязательно указанная каспаза выбрана из каспазы 9 млекопитающих, iCaspase9 или aviCaspase9.
6. Трансгенная конструкция по любому из пп. 1-5, где указанная вторая нуклеотидная последовательность нацеливает указанную трансгенную конструкцию на зародышевые клетки посредством гомологичной рекомбинации в генетическом локусе, экспрессируемом только в птичьих зародышевых клетках.
7. Трансгенная конструкция по любому из пп. 1-6, где указанная вторая нуклеотидная последовательность нацелена на зародышевые клетки посредством системы CRISPR/Cas, где указанная трансгенная конструкция содержит гидовую РНК, нацеленную на последовательность, специфичную для зародышевых клеток.
8. Способ получения суррогатной птицы-хозяина путем модификации зародышевой плазмы указанной птицы, включающий следующие стадии:
(i) введение трансгенной конструкции в оплодотворенное яйцо или культивируемые зародышевые клетки птицы и инкубацию указанного яйца, содержащего указанную трансгенную конструкцию или инъецированное зародышевыми клетками, содержащими указанную трансгенную конструкцию, где указанная трансгенная конструкция представляет собой трансгенную конструкцию по любому из пп. 1-7, и при этом указанная трансгенная конструкция интегрирована в зародышевые клетки указанного эмбриона; и
(ii) введение указанного экзогенного индуцирующего агента с индукцией гибели указанных зародышевых клеток.
9. Способ по п.8, дополнительно включающий:
(iii) трансплантацию зародышевых клеток от птицы-донора в указанное оплодотворенное яйцо и инкубацию до вылупления с получением птиц-потомков, имеющих генетическую идентичность трансплантированных зародышевых клеток.
10. Способ по п.9, дополнительно включающий:
(iv) скрещивание самцов и самок - потомков из указанных птиц-потомков с получением одного или более дополнительных поколений птиц-потомков с зародышевыми клетками, имеющими генетическую идентичность трансплантированных зародышевых клеток.
11. Трансгенная птица для применения в качестве суррогатной птицы-хозяина, содержащая трансгенную конструкцию по любому из пп. 1-7 и полученная способом по любому из пп. 8-10, причем трансгенная конструкция экспрессируется в зародышевой клетке-хозяине и сконфигурирована для селективного уничтожения зародышевых клеток-хозяев по требованию без уничтожения других клеток в птице-хозяине.
| TAYLOR L | |||
| ET AL | |||
| Efficient TALEN-mediated gene targeting of chicken primordial germ cells | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| LI ZHOU ET AL | |||
