Притязание на приоритет: В настоящей заявке испрашивается приоритет предварительной заявки на патент США 61/663700, поданной 25 июня, 2012. Содержание этой заявки во всей своей полноте вводится в настоящее описание посредством ссылки.
Предшествующий уровень техники
Гены, кодирующие Cry1F и Cry1Ac synpro (Cry1Ac), способны сообщать трансгенным растениям резистентность к насекомым, например, резистентность к чешуекрылым насекомым, а ген, кодирующий PAT (фосфинотрицин-ацетилтрансферазу), способен сообщать трансгенным растениям устойчивость к гербициду фосфинотрицину (глюфозинату). Ген PAT был успешно экспрессирован в сое и использован в качестве селективного маркера для промышленного продуцирования резистентных к насекомым трансгенных культур и для сообщения этим трансгенным растениям устойчивости к гербициду глюфозинату.
Известно, что экспрессия трансгенов в растениях зависит от локализации трансгенов в геноме растения, что, вероятно, обусловлено хроматиновой структурой (например, гетерохроматином) или близостью элементов регуляции транскрипции (например, энхансеров) к сайту интеграции (Weising et al., Ann. Rev. Genet 22:421-477, 1988). В то же время, присутствие трансгена в различных участках генома так или иначе зависит от общего фенотипа растения. По этой причине часто бывает необходимым прибегать к скринингу большого числа трансген-содержащих трансформантов для идентификации трансген-специфического трансформанта, характеризующегося оптимальной экспрессией представляющего интерес встроенного гена. Так, например, в этих растениях и в других организмах может наблюдаться широкая вариабельность в уровнях экспрессии гена, встроенного в эти трансформанты. Могут также наблюдаться различия в пространственных или временных профилях экспрессии, например, различия в относительных уровнях экспрессии трансгена в различных тканях растений, где указанные различия могут не соответствовать предполагаемым профилям элементов регуляции транскрипции, присутствующих во встроенной генной конструкции. По этой причине, обычно продуцируют от ста до тысячи различных трансформантов и скринируют их в поиске одного трансформанта, который имеет нужные уровни и профили экспрессии трансгена, и который может быть использован в промышленных целях. Трансформант, который имеет нужные уровни или профили экспрессии трансгена, может быть использован для интрогрессии этого трансгена в генетическое окружение другого вида путем полового ауткроссинга с применением стандартных методов скрещивания. Потомство таких кроссов сохраняет уровни экспрессии трансгена, характерные для исходного трансформанта. Эта стратегия применяется для обеспечения надежной экспрессии гена в различных сортах, хорошо адаптированных к условиям культивирования в определенной местности.
Для того, чтобы определить, содержит ли потомство полового кросса представляющий интерес трансген или группу таких трансгенов, желательно иметь возможность детектировать наличие конкретной генетической трансформации. Кроме того, способ детектирования конкретного трансформанта должен удовлетворять нормам, установленным Регуляторными органами для его апробации до выхода на рынок, а пищевые продукты, полученные из рекомбинантных сельскохозяйственных культур, должны, например, иметь соответствующую маркировку, либо такой способ может быть применен для мониторинга качества окружающей среды, мониторинга качества сельскохозяйственных культур, выращенных в полевых условиях, или для мониторинга продуктов, полученных после сбора урожая, а также его проведение должно удовлетворять нормам, утвержденным Регуляторными органами или сторонами договора.
При этом, присутствие трансформанта может быть детектировано любым методом детектирования нуклеиновых кислот, известным специалистам, включая, но не ограничиваясь ими, полимеразную цепную реакцию (ПЦР) или гибридизацию ДНК с использованием нуклеиновокислотных зондов. Такие методы детектирования обычно направлены на часто используемые генетические элементы, такие как промоторы, терминаторы, маркерные гены и т.п., поскольку для многих ДНК-конструкций, кодирующие области являются одинаковыми. Поэтому, такие методы не могут быть использованы для дифференциации различных трансформантов, особенно тех трансформантов, которые продуцируются с использованием одних и тех же ДНК-конструкций или почти аналогичных конструкций, за исключением лишь тех случаев, когда известна последовательность фланкирующей ДНК, смежной со встроенной гетерологичной ДНК. Так, например, в заявке на патент США 2006/0070139, относящийся к трансформанту кукурузы DAS-59122-7, описан трансформант-специфический ПЦР-анализ. Исходя из вышесказанного, очевидно, что необходимо разработать простой и дифференциальный метод идентификации трансформанта сои PDAB9582.816.15.1.
Краткое описание сущности изобретения
В некоторых своих вариантах, настоящее изобретение относится к новым трансгенным растениям трансформированной сои, резистентным к насекомым и устойчивым к гербицидам, а в частности, настоящее изобретение относится к трансформанту сои, обозначенному pDAB9582.816.15.1 и содержащему описанные здесь гены crylF v3 (cryIF), crylAc synpro (cry1Ac) и pat v6 (pat), встроенные в специфический сайт в геноме клеток сои. Репрезентативные семена сои были депонированы в Американской коллекции типовых культур (АТСС) под депозитарным номером No. РТА-12588 23 февраля 2012 г. компанией Dow AgroSciences LLC.
ДНК растений сои, содержащих такой трансформант, включает последовательности стыка/фланкирующие последовательности, описанные в настоящей заявке, которые указывают на локализацию встроенной ДНК в геноме сои. SEQ ID NO: 1 и SEQ ID NO: 2 являются диагностическим признаком присутствия трансформанта сои pDAB9582.816.15.1. Более конкретно, последовательности, окружающие область стыка в положениях п.о. 1273/1274 SEQ ID NO: 1, и в положениях п.о. 175/176 и п.о. 316/317 SEQ ID NO: 2, являются диагностическим признаком присутствия трансформанта сои pDAB9582.816.15.1. Ниже описаны примеры последовательностей, содержащих такие стыки, которые являются характерными для ДНК сои, содержащей трансформант сои pDAB9582.816.15.1. В одном из своих вариантов, настоящее изобретение относится к растению сои или к его части, которые являются резистентными к Pseudoplusia includens (соевой пяденице), и которые имеют геном, содержащий одну или более последовательностей, выбранных из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов. В другом своем варианте, настоящее изобретение относится к семенам таких растений.
В другом своем варианте, настоящее изобретение относится к способу борьбы с насекомыми, который включает заражение насекомыми растений сои, резистентных к насекомым, где указанные растения сои имеют геном, содержащий одну или более последовательностей, выбранных из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов, где указанные последовательности является характерными признаками присутствия трансформанта сои pDAB9582.816.15.1, резистентного к насекомым. Присутствие генов cryIF v3 {cryIF) и cry1Ac synpro (crylAc) в трансформанте сои pDAB9582.816.15.1 сообщает такому растению резистентность, например, к Pseudoplusia includens (соевой пяденице), Anticarsia gemmatalis (листовертке фасоли), Epinotia aporema, Omoides indicatus, Rachiplusia nu, Spodoptera frugiperda, Spodoptera cosmoides, Spodoptera eridania, Heliothis virescens, Heliocoverpa zea, Spilosoma virginica и Elasmopalpus lignosellus.
В другом своем варианте, настоящее изобретение относится к способу борьбы с сорняками сои, который включает обработку сельскохозяйственной культуры сои гербицидом глюфозинатом, где указанная сельскохозяйственная культура сои включает растения сои, имеющие геном, содержащий одну или более последовательностей, выбранных из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NО: 1; п.о. в положениях 1173-1373 SEQ ID NО: 1; п.о. в положениях 1073-1473 SEQ ID NО: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов, где указанные последовательности являются диагностическими признаками присутствия трансформанта сои pDAB9582.816.15.1. Присутствие гена pat в трансформанте сои pDAB9582.816.15.1 сообщает устойчивость к гербициду глюфозинату.
В другом своем варианте, настоящее изобретение относится к способу детектирования трансформанта сои pDAB9582.816.15.1 в образце, содержащем соевую ДНК, где указанный способ включает:
(a) контактирование указанного образца с первым праймером длиной по меньшей мере в 10 п.о., который селективно связывается с фланкирующей последовательностью в положениях п.о. 1-1273 SEQ ID NO: 1 или с ее комплементом, и со вторым праймером длиной по меньшей мере в 10 п.о., который селективно связывается с последовательностью вставки в положениях п.о. 1274-1577 SEQ ID NO: 1 или с ее комплементом; и проведение анализа на ампликон, образующийся между указанными праймерами; или
(b) контактирование указанного образца с первым праймером длиной по меньшей мере в 10 п.о., который селективно связывается с последовательностью вставки в положениях п.о. 1-175 SEQ ID NO: 2 или с ее комплементом, и со вторым праймером длиной по меньшей мере в 10 п.о., который селективно связывается с фланкирующей последовательностью в положениях п.о. 176-1687 SEQ ID NO: 2 или с ее комплементом; и
(c) проведение анализа на ампликон, образующийся между указанными праймерами.
В другом своем варианте, настоящее изобретение относится к способу детектирования трансформанта сои pDAB9582.816.15.1, где указанный способ включает:
(a) контактирование указанного образца с первым праймером, который селективно связывается с фланкирующей последовательностью, выбранной из группы, состоящей из п.о. в положениях 1-1273 SEQ ID NO: 1 и п.о. в положениях 176-1687 SEQ ID NO: 2, или с их комплементами, и со вторым праймером, который селективно связывается с SEQ ID NО: 3 или с ее комплементом;
b) осуществление полимеразной цепной реакции указанного образца; и
c) проведение анализа на ампликон, образующийся между указанными праймерами.
В другом своем варианте, настоящее изобретение относится к способу скрещивания растения сои, где указанный способ включает скрещивание первого растения сои со вторым растением сои с получением третьего растения сои, где указанное первое растение содержит ДНК, включающую одну или более последовательностей, выбранных из группы, состоящей из в п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов; и проведение анализа указанного третьего растения сои на присутствие ДНК, включающей одну или более последовательностей, выбранных из группы, состоящей из в п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов.
В другом своем варианте, настоящее изобретение относится к выделенной молекуле ДНК, которая является диагностическим признаком присутствия трансформанта сои pDAB9582.816.15.1. Такими молекулами, помимо SEQ ID NOS: 1 и 2, являются молекулы длиной по меньшей мере 25 п.о., содержащие п.о. в положениях 1273-1274 SEQ ID NO: 1 и по меньшей мере 10 п.о. SEQ ID NO: 1 в каждом направлении от п.о. 1273/1274 в области стыка; ампликоны длиной по меньшей мере 25 п.о., содержащие п.о. в положениях 175 - 176 SEQ ID NO: 2 и по меньшей мере 10 п.о. SEQ ID NO: 2 в каждом направлении от п.о. 175/176 в области стыка. Примерами являются п.о. в положениях l258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементы.
В другом своем варианте, настоящее изобретение относится к способу борьбы с насекомыми-вредителями соевых бобов, семян или муки из семян, в которых содержится трансформант сои pDAB9582.816.15.1, где указанные бобы, семена или мука из семян содержат ДНК, включающую одну или более последовательностей, выбранных из группы, состоящей из п.о. в положениях l258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов.
Варианты осуществления изобретения также относятся к клеткам растений сои и к частям растений сои, включая, но не ограничиваясь ими, пыльцу, овули, цветки, побеги, корни и листья, ядра вегетативных клеток, клетки пыльцы, семена и муку из семян, а также яйцеклетку, содержащие трансформант сои pDAB 9582.816.15.1.
В некоторых вариантах осуществления изобретения, трансформанту сои pDAB 9582.816.15.1 могут быть сообщены и другие признаки, включая, например, другой(ие) ген(ы) устойчивости к гербицидам и/или белки-ингибиторы насекомых и последовательности регуляции транскрипции (то есть, РНК-интерферирующий агент, дцРНК, факторы транскрипции и т.п.). Дополнительные признаки могут быть внесены в геном растения путем скрещивания растений, ретрансформации трансгенного растения, содержащего трансформант сои pDAB 9582.816.15.1, либо путем сообщения новых признаков благодаря нацеленной интеграции посредством гомологичной рекомбинации.
Другие варианты осуществления изобретения включают вырезание полинуклеотидных последовательностей, которые содержат трансформант сои pDAB9582.816.15.1, включая, например, кластер для экспрессии гена pat. После вырезания полинуклеотидной последовательности, модифицированный трансформант может быть перенацелен в специфический сайт хромосомы, где дополнительные полинуклеотидные последовательности присутствуют вместе с трансформантом сои pDAB9582.816.15.1.
В одном из своих вариантов, настоящее изобретение охватывает сайт-мишень хромосомы сои, локализованный на хромасоме 03 между фланкирующими последовательностями, представленными в SEQ ID NOS: 1 и 2.
В одном из своих вариантов, настоящее изобретение относится к способу получения трансгенного растения сои, где указанный способ включает инсерцию гетерологичной нуклеиновой кислоты в положении на хромосоме 03 между геномными последовательностями, представленными в SEQ ID NOS: 1 и 2, то есть, между п.о. в положениях 1-1273 SEQ ID NO: 1 и п.о. в положениях 176-1687 SEQ ID NO: 2.
Кроме того, варианты осуществления изобретения также относятся к анализам, включающим детектирование присутствия рассматриваемого трансформанта в образце (например, сои). Эти анализы могут быть осуществлены на основе последовательности ДНК рекомбинантной конструкции, встроенной в геном сои, и на основе геномных последовательностей, фланкирующих сайт инсерции. Настоящее изобретение также относится к наборам для проведения анализов в соответствующих условиях.
В частности, варианты осуществления изобретения также относятся к клонированию и анализу последовательностей ДНК граничных областей, полученных после инсерции Т-ДНК от pDAB9582 в линиях трансгенной сои. Эти последовательности являются уникальными. Исходя из последовательностей вставки и стыка могут быть идентифицированы и сконструированы трансформант-специфические праймеры. ПЦР-анализ продемонстрировал, что эти трансформанты могут быть идентифицированы с помощью анализа ПЦР-ампликонов, полученных с использованием наборов трансформант-специфических праймеров. Таким образом, эти и другие родственные методы могут быть применены для уникальной идентификации линий сои, содержащих трансформант согласно изобретению.
Один из вариантов осуществления изобретения относится к способу борьбы с насекомыми, который включает заражение насекомыми растений сои, резистентных к насекомым, где указанные растения сои включают ДНК, содержащую последовательность, выбранную из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, где указанные последовательности является диагностическими признаками присутствия трансформанта сои pDAB9582.816.15.1, резистентного к насекомым.
Один из вариантов осуществления изобретения относится к способу борьбы с насекомыми Pseudoplusia includens, Anticarsia gemmatalis или Spodoptera frugiperda, где указанный способ включает заражение резистентных к насекомым растений сои насекомыми Pseudoplusia includens, Anticarsia gemmatalis, Heliothis virescens или Spodoptera frugiperda, где указанные растения сои включают ДНК, содержащую последовательность, выбранную из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, где указанная последовательность является диагностическим признаком присутствия трансформанта сои pDAB9582.816.15.1, резистентного к насекомым.
Один из вариантов осуществления изобретения относится к способу борьбы с сорняками сои, который включает обработку сельскохозяйственной культуры сои гербицидом глюфозинатом, где указанная сельскохозяйственная культура сои включает растения сои, содержащие ДНК, включающую последовательность, выбранную из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NО: 1; п.о. в положениях 1173-1373 SEQ ID NО: 1; п.о. в положениях 1073-1473 SEQ ID NО: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, где указанная последовательность является диагностическим признаком присутствия трансформанта сои pDAB9582.816.15.1.
Один из вариантов осуществления изобретения относится к выделенной последовательности ДНК, содержащей одну или более последовательностей, выбранных из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NО: 1; п.о. в положениях 1173-1373 SEQ ID NО: 1; п.о. в положениях 1073-1473 SEQ ID NО: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2.
Один из вариантов осуществления изобретения относится к способу скрещивания растения сои, где указанный способ включает скрещивание первого растения сои со вторым растением сои с получением третьего растения сои, где указанное первое растение содержит ДНК, включающую одну или более последовательностей, выбранных из группы, состоящей из в п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов; и проведение анализа указанного третьего растения сои на присутствие ДНК, включающей одну или более последовательностей, выбранных из группы, состоящей из в п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов.
Один из вариантов осуществления изобретения относится к выделенной молекуле ДНК, содержащей последовательность стыка, включающую по меньшей мере одну последовательность, выбранную из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NО: 1; п.о. в положениях 1173-1373 SEQ ID NО: 1; п.о. в положениях 1073-1473 SEQ ID NО: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов. Один из вариантов осуществления изобретения относится к растению сои или его части, которые являются резистентными к Pseudoplusia includens (соевой пяденице) и содержат ДНК, имеющую по меньшей мере одну нуклеотидную последовательность, выбранную из группы, состоящей из п.о. в положениях 1258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NО: 1; п.о. в положениях 1173-1373 SEQ ID NО: 1; п.о. в положениях 1073-1473 SEQ ID NО: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов.
Один из вариантов осуществления изобретения относится к композиции, полученной из растения сои или его частей, где указанной композицией является пищевой продукт, выбранный из группы, состоящей из кормовой соевой муки, пищевой муки, белкового концентрата и масла.
Один из вариантов осуществления изобретения относится к способу борьбы с насекомыми-вредителями соевых бобов, семян, кормовой муки или пищевой муки, в которых содержится трансформант сои pDAB9582.816.15.1, где указанные бобы, семена, кормовая мука или пищевая мука содержат ДНК, включающую одну или более последовательностей, выбранных из группы, состоящей из п.о. в положениях l258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и их комплементов.
Один из вариантов осуществления изобретения относится к семенам сои, содержащим в своем геноме последовательность ДНК, выбранную из группы, состоящей из п.о. в положениях l258-1288 SEQ ID NO: 1; п.о. в положениях 1223-1323 SEQ ID NO: 1; п.о. в положениях 1173-1373 SEQ ID NO: 1; п.о. в положениях 1073-1473 SEQ ID NO: 1; п.о. в положениях 160-190 SEQ ID NO: 2; п.о. в положениях 125-225 SEQ ID NO: 2; и п.о. в положениях 75-275 SEQ ID NO: 2, и ее комплементы. Другой вариант осуществления изобретения относится к семенам сои, содержащим в своем геноме гены crylF, crylAc и pat трансформанта сои pDAB9582.816.15.1, и к репрезентативным семенам сои, депонированным в Американской коллекции типовых культур под регистрационным номером PTA-12588. Другой вариант осуществления изобретения относится к растению сои, выращенному путем культивирования семян сои согласно любому из этих двух вариантов. Другой вариант осуществления изобретения относится к семенам сои, продуцированным этим растением сои, где указанные семена содержат в своем геноме гены crylF, crylAc и pat трансформанта сои pDAB9582.816.15.1, которые присутствуют в семенах сои, депонированных в Американской коллекции типовых культур под регистрационным номером PTA-12588. Другой вариант осуществления изобретения относится к части этого растения сои, где указанная часть выбрана из группы, состоящей из пыльцы, овулей, цветков, побегов, корней и листьев, и где указанная часть содержит указанный трансформант. Другой вариант осуществления изобретения относится к композиции, полученной из растения сои или его части, где указанной композицией является пищевой продукт, выбранный из группы, состоящей из кормовой соевой муки, пищевой муки и масла.
В другом варианте осуществления изобретения, растение сои содержит последовательность ДНК, которая по меньшей мере на 95% идентична последовательности SEQ ID NO: 14. Один из вариантов осуществления изобретения относится к потомству растения сои согласно вышеуказанному варианту, где указанное растение обладает устойчивостью к гербициду глюфозинату, где указанная устойчивость обусловлена экспрессией белка, кодируемого в указанном трансформанте или в указанном геноме.
Другой вариант осуществления изобретения относится к семенам сои, содержащим геном, включающий последовательность ДНК, которая по меньшей мере на 95% идентична последовательности SEQ ID NO: 14. Другой вариант осуществления изобретения относится к растению, выращенному путем культивирования этих семян сои.
Один из вариантов осуществления изобретения относится к трансгенному растению сои или к его части, включающим трансформант сои pDAB9582.816.15.1, где репрезентативные семена сои, содержащие трансформант сои pDAB9582.816.15.1, были депонированы в Американской коллекции типовых культур под регистрационным номером PTA-12588.
Депонирование семян
За время разработки настоящего изобретения, по меньшей мере 2500 семян линии сои, содержащей трансформант pDAB9582.816.15.1, были депонированы в Американской коллекции типовых культур (ATCC), 10801 University Boulevard, Manassas, VA, 20110, и являются общедоступными без каких-либо ограничений (но в соответствии с патентным правом). Этот депозит был зарегистрирован в ATCC под депозитарным номером No. PTA-12588 23 февраля 2012 г. компанией Dow AgroSciences LLC. Указанный депозит был положен на хранение согласно Будапештскому договору о депонировании семян в целях проведения патентной процедуры.
Краткое описание последовательностей
SEQ ID NO: 1 представляет собой 5'-фланкирующую граничную последовательность ДНК трансформанта сои pDAB9582.816.15.1. Нуклеотиды 1-1273 составляют геномную последовательность. Нуклеотиды 1274-1577 составляют последовательность вставки.
SEQ ID NO: 2 представляет собой 3'-фланкирующую граничную последовательность ДНК трансформанта сои pDAB9582.816.15.1. Нуклеотиды 1-175 составляют последовательность вставки. Нуклеотиды 176-316 составляют реаранжированную последовательность pDAB9582. Нуклеотиды 317-1687 составляют геномную последовательность.
SEQ ID NO: 3 представляет собой последовательность T-цепи ДНК pDAB9582, которая указана ниже в таблице 1.
SEQ ID NO: 4 представляет собой олигонуклеотидный праймер 81615_FW2 для подтверждения 5'-граничной геномной ДНК.
SEQ ID NO: 5 представляет собой олигонуклеотидный праймер 81516_RV1 для подтверждения 3'-граничной геномной ДНК.
SEQ ID NO: 6 представляет собой олигонуклеотидный праймер 81516_RV2 для подтверждения 3'-граничной геномной ДНК.
SEQ ID NO: 7 представляет собой олигонуклеотид 81516_RV3 для подтверждения 3'-граничной геномной ДНК.
SEQ ID NO: 8 представляет собой олигонуклеотидный праймер 5'IREnd-01 для подтверждения 5'-граничной геномной ДНК.
SEQ ID NO: 9 представляет собой олигонуклеотидный праймер 5'IREnd-02 для подтверждения 5'-граничной геномной ДНК.
SEQ ID NO: 10 представляет собой олигонуклеотидный праймер AtUbi10RV1 для подтверждения 5'-граничной геномной ДНК.
SEQ ID NO: 11 представляет собой олигонуклеотидный праймер AtUbi10RV2 для подтверждения 5'-граничной геномной ДНК.
SEQ ID NO: 12 представляет собой олигонуклеотидный праймер 3'ATEnd05 для подтверждения 3'-граничной геномной ДНК.
SEQ ID NO: 13 представляет собой олигонуклеотидный праймер 3'TATEnd06 для подтверждения 3'-граничной геномной ДНК.
SEQ ID NO: 14 представляет собой предполагаемую последовательность трансформанта сои pDAB9582.816.15.1. Эта последовательность включает геномную 5'-фланкирующую последовательность, последовательность-вставку T-цепи pDAB9582 и 3'-фланкирующую геномную последовательность.
Краткое описание графического материала
На фигуре 1 представлена карта плазмиды pDAB9582, содержащей кластеры экспрессии генов cry1F v3, cry1Ac synpro и pat v6.
На фигуре 2 указана локализация праймеров для подтверждения 5'- и 3'-граничной последовательности трансформанта сои pDAB9582.816.15.1.
На фигуре 3 представлена геномная последовательность, расположенная в определенном порядке в трансформанте сои pDAB9582.816.15.1.
Подробное описание настоящего изобретения
Оба конца последовательности-вставки трансформанта сои pDAB9582.816.15.1 были секвенированы и охарактеризованы. Были разработаны трансформант-специфические анализы. Этот трансформант был также картирован по геному сои (хромосоме сои 03). Данный трансформант может быть подвергнут интрогрессии с получением более элитных линий.
Как упоминалось выше в разделе «Предшествующий уровень техники», встраивание и интеграция трансгена в геном растения включает некоторые случайные события (и следовательно, слово «событие» включает такое встраивание). То есть, многие методы трансформации, такие как трансформация агробактерией, биобаллистическая трансформация (то есть, посредством «выстреливания» генов) и трансформация, опосредуемая карбидом кремния (то есть, WHISKERS™), не позволяют предсказать, в какой участок генома встраивается данный трансген. Таким образом, идентификация фланкирующей геномной ДНК растения на обеих сторонах вставки может иметь важное значение для идентификации растения, имеющего данную вставку. Так, например, могут быть сконструированы ПЦР-праймеры, которые генерируют ПЦР-ампликон по всей области стыка вставки и генома хозяина. Этот ПЦР-ампликон может быть использован для идентификации уникальной вставки или вставки другого типа.
Для лучшего понимания вариантов изобретения и для облегчения практического осуществления этих вариантов специалистом в данной области, в описании настоящего изобретения приводятся определения терминов и примеры. Если это не оговорено особо, то такие термины имеют общепринятые значения, известные специалисту в области, к которой относится настоящее изобретение. Номенклатура оснований ДНК приводится в соответствии со ст. 37 Кодекса законов США (CFR) в §1.822.
Используемый здесь термин «потомство» означает потомство любого поколения родительского растения, которое содержит трансформант сои pDAB9582.816.15.1.
«Трансгенное растение с генной модификацией» [или «трансформант»] («event») получают путем трансформации клеток растений гетерологичной ДНК, то есть, конструкцией нуклеиновой кислоты, включающей представляющие интерес трансгены; регенерации популяции растений, полученных после встраивания трансгена в геном растения; и отбора конкретного растения, отличающегося тем, что оно имеет инсерцию в конкретном положении генома. Термин «трансгенное растение с генной модификацией» («event») означает исходный трансформант и потомство трансформанта, которые включают гетерологичную ДНК. Термин «трансгенное растение с генной модификацией» («event») также означает потомство растения, продуцированного посредством полового ауткроссинга трансформанта и растения другого сорта, включающего геномную/трансгенную ДНК. Даже после повторного возвратного скрещивания с рекуррентным родителем, встроенная трансгенная ДНК и фланкирующая геномная ДНК (геномная/трансгенная ДНК), происходящая от трансформированного родителя, присутствует в потомстве этого кросса в том же самом хромосомном участке. Термин «трансформант» («event») также означает ДНК исходного трансформанта и его потомства, содержащих встроенную ДНК и фланкирующую геномную последовательность, расположенную в непосредственной близости от встроенной ДНК, которая, как предполагается, должна передаваться потомству, приобретающему эту встроенную ДНК, включая представляющий интерес трансген, в результате полового скрещивания одной родительской линии, которая содержит встроенную ДНК (например, исходного трансформанта и его потомства, продуцируемого после самоопыления), и родительской линии, которая не содержит встроенную ДНК.
«Последовательность стыка» или «граничная последовательность» охватывает положение, в котором ДНК, встроенная в геном, присоединена к ДНК генома нативного растения сои, фланкирующего положение вставки, причем, идентификация или детектирование одной или другой последовательности стыка в растительном генетическом материале будут достаточными для выявления такого трансформанта. Настоящее изобретение включает последовательности ДНК, которые охватывают инсерции в описанных здесь трансформантах сои и фланкирующую ДНК аналогичной длины. Конкретные примеры таких диагностических последовательностей описаны в настоящей заявке, однако, в настоящем изобретении могут быть использованы и другие диагностические последовательности, которые перекрываются с последовательностями стыка указанных вставок или с последовательностями стыка указанных вставок и геномной последовательности, и такие последовательности могут быть использованы в соответствии с вариантами осуществления изобретения.
Варианты осуществления изобретения, в частности, относятся к идентификации событий трансформации с использованием таких фланкирующих последовательностей, последовательностей стыка и последовательностей вставки. Варианты осуществления изобретения включают использование родственных ПЦР-праймеров и ампликонов. В соответствии с вариантами осуществления изобретения, для детектирования или идентификации коммерчески доступных сортов или линий трансгенной сои, выведенных из запатентованных линий трансгенной сои, могут быть применены методы ПЦР-анализа с использованием ампликонов, которые охватывают встроенную ДНК и ее граничные области.
Фланкирующие последовательности/последовательности стыка используются в качестве диагностических признаков присутствия трансформантов сои pDAB9582.816.15.1. На основе этих последовательностей были получены трансформант-специфические праймеры. ПЦР-анализ показал, что эти линии растения сои могут быть идентифицированы по различным генотипам сои путем анализа ПЦР-ампликонов, полученных с использованием наборов этих трансформант-специфических праймеров. Таким образом, эти и другие аналогичные процедуры могут быть применены для идентификации уникальности этих линий растения сои. Идентифицированные здесь последовательности являются уникальными.
Способы детектирования согласно вариантам изобретения являются особенно ценными, если их применять в комбинации со скрещиванием растений, где после определения какое именно потомство растения содержит данный трансформант, родительское растение, содержащее представляющий интерес трансформант, может быть скрещено с линией другого растения для сообщения потомству одного или более дополнительных представляющих интерес признаков. Эти методы с применением ПЦР-анализов являются предпочтительными для реализации программ по скрещиванию растений сои, а также по контролю качества, в частности, товарного семенного материала трансгенной сои. В настоящее время существуют и используются наборы для ПЦР-детектирования этих линий трансгеной сои. Это позволяет также ускорить регистрацию продукта и его поступление на реализацию.
Кроме того, для идентификации конкретной геномной локализции каждой вставки могут быть использованы фланкирующие/геномные последовательности сои. Эта информация может быть использована для получения молекулярных маркерных систем, специфичных для каждого трансформанта. Такие системы могут быть использованы для разработки ускоренной стратегии скрещивания и получения данных по сцеплению.
Кроме того, информация о фланкирующей последовательности может быть использована для исследования и характеризации процессов интеграции трансгенов, свойств сайта геномной интеграции, отбора трансформантов, стабильности трансгенов и их фланкирующих последовательностей, и экспрессии генов, (а в частности, процессов, относящихся к сайленсингу генов, профилю метилирования трансгенов, эффектам положений и к потенциальным элементам, ассоциированным с экспрессией, таким как MARS [области присоединения к матрице] и т.п.).
Исходя из всего вышесказанного, совершенно очевидно, что варианты осуществления изобретения включают семена, депонированные в АТСС под депозитарным номером No. РТА-12588 23 февраля 2012 г.компанией Dow AgroSciences LLC. Варианты осуществления изобретения также включают устойчивое к гербицидам растение сои, выращенное из семян, депонированных в АТСС под депозитарным номером No. РТА-12588 23 февраля 2012 г. компанией Dow AgroSciences LLC. Варианты осуществления изобретения также включают части указанного растения, такие как листья, образцы ткани, семена, полученные от указанного растения, пыльца и т.п. (где указанные части растения содержат cryIF, cry1Ac, pat и SEQ ID NO: 1 и 2).
Кроме того, варианты осуществления изобретения также включают потомство растений и/или потомство растений, выращенных из депонированных семян, а предпочтительно, семян растения сои, резистентного к гербицидам, где указанное растение имеет геном, содержащий описанную здесь детектируемую последовательность «стыка»/фланкирующую последовательность дикого типа. Используемый здесь термин «соя» означает растение Glycine max и включает все его сорта, которые могут быть выведены путем скрещивания одного растения сои с другим растением сои.
Настоящее изобретение также включает способы получения кроссов с использованием растения согласно изобретению в качестве по меньшей мере одного родителя. Так, например, настоящее изобретение включает F1-гибрид растения, имеющий, в качестве одного или двух родителей, любые описанные здесь растения. Настоящее изобретение также относится к семенам, продуцированным такими F1-гибридами согласно изобретению. Настоящее изобретение включает способ продуцирования семян F1-гибрида путем скрещивания описанного здесь растения с другим растением (например, инбредным родителем) и сбора полученных гибридных семян. Настоящее изобретение включает описанное здесь растение, которое является либо женским родительским растением, либо мужским родительским растением. Характеризация полученных растений может быть проведена более эффективно после тщательного исследования растений-родителей.
Растение сои согласно изобретению, резистентное к насекомым/устойчивое к глюфозинату, может быть выведено путем первого полового скрещивания первого родительского растения сои, выращенного из семян любой из указанных здесь линий, со вторым родительским растением сои, с получением множества растений-потомков первого поколения, после чего проводят отбор потомства первого поколения, которое является резистентным к глюфозинату; самоопыление потомства первого поколения с продуцированием множества растений-потомков второго поколения, а затем отбор из потомства второго поколения растения, которое является резистентным к глюфозинату. Эти стадии могут также включать возвратное скрещивание растения-потомка первого поколения или растения-потомка второго поколения со вторым родительским растением сои или с третьим родительским растением сои. Затем может быть засеяна культура сои, дающая семена сои или ее потомства согласно изобретению.
Следует также отметить, что два различных трансгенных растения могут быть также скрещены с получением потомства, которое содержит два независимо сегрегирующихся добавленных экзогенных гена. После самоопыления соответствующего потомства могут быть получены растения, которые являются гомозиготными по обоим добавленным экзогенным генам. Возвратное скрещивание с родительским растением и ауткроссинг с нетрансгенным растением также рассматривается как вегетативное размножение. Специалистам известны и другие методы скрещивания, обычно применяемые для сообщения различных признаков и выращивания различных культур. Возвратное скрещивание было использовано для простой передачи хорошо наследуемого признака путем переноса генов в нужные гомозиготные сорта или в инбредную линию, которая является рекуррентным родителем. Источник передаваемого признака называют родителем-донором. Как и ожидалось, полученное растение имеет признаки рекуррентного родителя (например, сорта), и этот желательный признак передается от родителя-донора. После первого скрещивания отбирают растения, имеющие фенотип родителя-донора, и эти растения повторно скрещивают (подвергают возвратному скрещиванию) с рекуррентным родителем. Как и ожидалось, полученное растение имеет признаки рекуррентного родителя (например, сорта), и этот желательный признак передается от родителя-донора.
Аналогичным образом, резистентное к насекомым/устойчивое к глюфозинату растение сои согласно варианту осуществления изобретения может быть трансформировано дополнительными трансгенами с применением известных методов. Методы трансформации, такие как трансформация агробактерией, биобаллистическая трансформация (то есть, посредством «выстреливания» генов) и трансформация, опосредуемая карбидом кремния (то есть, WHISKERS™), могут быть применены для введения дополнительного(ых) трансгена(ов) в геном трансформанта сои pDAB9582.816.15.1. Отбор и характеризация трансгенных растений, содержащих новые встроенные трансгены, могут быть осуществлены для идентификации растений, которые содержат, помимо генов cry1F, cry1Ac, pat согласно вариантам осуществления изобретения, стабильный интегрант нового трансгена.
Молекулы ДНК согласно вариантам осуществления изобретения могут быть использованы в качестве молекулярных маркеров в методе скрещивания с применением маркеров (MAB). Молекулы ДНК согласно вариантам осуществления изобретения могут быть использованы в методах с применением маркеров (таких как AFLP-, RFLP-, RAPD-, SNP- и SSR-маркеры), которые позволяют идентифицировать агрономически ценные и генетически сцепленные признаки, известные специалистам. Признаки резистентности к насекомым и устойчивости к гербицидам могут быть прослежены у потомства кросса, полученного путем скрещивания с растением сои согласно вариантам осуществления изобретения (или с их потомством и любым другим сортом или другой разновидностью растения сои), с применением MAB-методов. Маркерами для указанных признаков являются молекулы ДНК, и MAB-методы, хорошо известные специалистам, могут быть применены для прослеживания признака(ов) устойчивости к гербицидам у растений сои, где по меньшей мере одна линия растения сои согласно вариантам осуществления изобретения или его потомства является родителем или предком. Способы согласно вариантам осуществления изобретения могут быть применены для идентификации растения сои любого сорта, имеющего рассматриваемый трансформант.
Варианты осуществления изобретения включают способ получения растения сои, резистентного к насекомым и устойчивого к гербицидам, где указанный способ включает скрещивание с растением согласно изобретению. Более конкретно, указанные способы могут включать скрещивание двух растений согласно изобретению или одного растения согласно изобретению с любым другим растением. Предпочтительные способы также включают отбор потомства указанного кросса путем анализа указанного потомства на трангенную модификацию, детектируемую в соответствии с вариантом осуществления изобретения, и на желательную продуктивность данного сорта (например, урожайность). Так, например, варианты осуществления изобретения могут быть применены для прослеживания рассматриваемого события трансформации во всех циклах скрещивания с растениями, имеющими другие желательные признаки, такие как агрономические показатели, устойчивость или резистентность к болезням, устойчивость или резистентность к нематодам и время созревания семян. Растения, содержащие рассматриваемый трансформант и имеющие нужный признак, могут быть детектированы, идентифицированы, отобраны и сразу использованы, например, в последующих раундах скрещивания. Рассматриваемый трансформант/признак может быть также сцеплен с другим(ими) признаком(ами) резистентности к насекомым и/или с другими признаками устойчивости к гербицидами посредством скрещивания, а затем может быть проведен мониторинг такого трансформанта/признака в соответствии с вариантами осуществления изобретения. Одним из вариантов последнего аспекта являются растения, содержащие рассматриваемый трансген, объединенный с генами cryIF и cry1Ac, сообщающими резистентность к чешуекрылым, которыми являются Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Epinotia aporema, Omoides indicatus, Rachiplusia nu, Spodoptera frugiperda, Spodoptera cosmoides, Spodoptera eridania, Heliothis virescens, Heliocoverpa zea, Spilosoma virginica и Elasmopalpus lignosellus.
Таким образом, варианты осуществления изобретения могут быть объединены, например, с дополнительными признаками, кодирующими резистентность к глифосату (например, резистентность к растительным или бактериальным EPSPS, GOX, GAT), резистентность к глюфозинату (например, резистентность к dsm-2, bar), резистентность к гербициду, ингибирующему ацетолактат-синтазу (ALS) (например, к имидазолинонам [таким как имазетапир], к сульфонилмочевинам, к сульфоанилиду триазолпиримидина, к пиримидинилтиобензоатам и к другим химическим веществам [Csr1, SurA и т.п.]); резистентность к бромоксинилу (например, Bxn); резистентность к ингибиторам фермента HPPD (4-гидроксифенил-пируват-диоксигеназы); резистентность к ингибиторам фитоендезатуразы (PDS); резистентность к гербицидам, ингибирующим фотосистему II (например, psbA); резистентность к гербицидам, ингибирующим фотосистему I; резистентность к гербицидам, ингибирующим протопорфириногеноксидазу IX (PPO) (например, РРО-1), резистентность к гербицидам на основе фенилмочевины (например, CYP76B1), резистентность к ферментам, разлагающим дикамбу (см., например, US20030135879), и с другими признаками, которые могут быть сцеплены с одним признаком или во множестве их комбинаций в целях эффективного уничтожения или предупреждения распространения заносных сорняков и/или сообщения резистентности к любым гербицидам вышеупомянутых классов.
Кроме того, трансформант сои pDAB9582.816.15.1 может быть сцеплен с одним или более дополнительными признаками (например, с резистентностью к насекомым, резистентностью к заражению патогенами или устойчивостью к стрессам и т.п.) или с продуктивными признаками (например, с повышенной урожайностью, с повышенной маслянистостью, улучшенным качеством волокна и т.п.). Таким образом, варианты осуществления изобретения могут быть применены для выращивания сельскохозяйственных культур с полным набором улучшенных агрономических признаков, таких как качество культуры, ее приспособляемость и экономический показатель «затраты - эффективность борьбы с любыми вредителями сельскохозяйственных культур».
Методы интеграции полинуклеотидной последовательности в специфический сайт хромосомы растительной клетки посредством гомологичной рекомбинации описаны в литературе. Так, например, сайт-специфическая интеграция, описанная в публикации патентной заявки на патент США No. 2009/0111188 A1, которая вводится в настоящее описание посредством ссылки, включает использование рекомбиназ или интеграз, опосредующих введение донорной полинуклеотидной последовательности в хромосому мишени. Кроме того, в международной патентной заявке No. WO 2008/021207, которая вводится в настоящее описание посредством ссылки, описана гомологичная рекомбинация, опосредуемая белками «цинковые пальцы» и осуществляемая для интеграции одной или более донорных полинуклеотидных последовательностей в специфические положения генома. Рекомбиназы, такие как FLP/FRT, описанные в патенте США № 6720475, который вводится в настоящее описание посредством ссылки, или CRE/LOX, описанные в патенте США № 5658772, который вводится в настоящее описание посредством ссылки, могут быть использованы для интеграции полинуклеотидной последовательности в специфический сайт хромосомы. И наконец, использование мегануклеаз для встраивания донорных полинуклеотидов в специфический сайт хромосомы было описано в публикации Puchta et al., PNAS USA 93 (1996) pp. 5055-5060).
Также существуют и, в основном, применяются другие, по существу, известные методы сайт-специфической интеграции в клетки растений (Kumar et al, Trends in Plant Set 6(4) (2001) pp. 155-159). Кроме того, системы сайт-специфической рекомбинации, которые были идентифицированы у некоторых прокариотических и низших эукариотических организмов, могут быть использованы и в растениях. Примерами таких систем являются, но не ограничиваются ими, система рекомбиназ R/RS плазмиды pSR1 дрожжей Zygosaccharomyces rouxii (Araki et al. (1985) J. Mol. Biol. 182: 191-203) и система Gin/Gix фага Mu (Maeser and Kahlmann (1991) Mol. Gen. Genet. 230: 170-176).
В некоторых вариантах осуществления изобретения могут оказаться желательными интеграция или сцепление с новым(ыми) трансгеном(ами) в непосредственной близости от существующей трансгенной вставки. Трансгенная вставка может рассматриваться как предпочтительный геномный локус, который был выбран исходя из уникальных признаков, таких как единственный инсерционный сайт, нормальная менделевская сегрегация и стабильная экспрессия, и преимущественная комбинация свойств, включая устойчивость к гербицидам и агрономическая продуктивность в отдельно взятом и в различных других географических регионах. Потомство растений, содержащее интегрированные трансгены, должно сохранять экспрессионные свойства трансгена в уже существующих трансформантах. Кроме того, поскольку уже были идентифицированы геномные фланкирующие последовательности и их локализация в хромосоме потомства растений, содержащих такой трансген, то могут быть проведены разработанные анализы для детектирования и подтверждения этого трансгена в потомстве растений, содержащих интегрированный трансген. И наконец, интеграция нового трансгена в специфическое положение хромосомы, которое тесно сцеплено с существующим трансгеном, будет облегчать интрогрессию трансгенов в другой генетический фон посредством полового ауткроссинга с применением стандартных методов скрещивания.
В некоторых вариантах осуществления изобретения, может оказаться желательным вырезание полинуклеотидных последовательностей из трансгенной вставки. Так, например, вырезание трансгена, описанное в публикации заявки на патент США No. 2011/191877, которая вводится в настоящее описание посредством ссылки, осуществляют с использованием нуклеаз «цинковые пальцы» для удаления полинуклеотидной последовательности, состоящей из генного экспрессионного кластера, из интегрированной в хромосому трансгеной вставки. Удаляемой полинуклеотидной последовательностью может быть селективный маркер. После вырезания и удаления полинуклеотидной последовательности, модифицированная трансгенная вставка может быть снова встроена путем инсерции полинуклеотидной последовательности. Вырезание полинуклеотидной последовательности и последующее повторное встраивание модифицированной трансгенноой вставки дает определенные преимущества, такие как возможность повторного использования селективного маркера или предупреждение спонтанных замен в транскриптоме растения, которые могут происходить в результате экспрессии специфических генов.
В настоящем изобретении описан специфический сайт на хромосоме 03 в геноме сои, который может быть использован для встраивания в него гетерологичных нуклеиновых кислот. Таким образом, варианты осуществления изобретения относятся к способам введения представляющих интерес гетерологичных нуклеиновых кислот в этот предварительно определенный сайт-мишень или в область, расположенную поблизости от этого сайта-мишени. Варианты осуществления настоящего изобретения также охватывают семена сои и/или растение сои, содержащие любую гетерологичную нуклеотидную последовательность, встроенную в описанный здесь сайт-мишень или в обычное окружение этого сайта. Одним из средств достижения такой нацеленной интеграции является вырезание и/или введение другой вставки вместо описанного здесь pat-экспрессирующего кластера. Вообще говоря, в вариантах осуществления изобретения может быть применен любой метод, которым является, например, но не ограничивается им, направленная гомологичная рекомбинация.
Используемый здесь термин «сцепление» гена, трансгена или признака означает объединение желательных признаков в одной трансгенной линии. Сцепление трансгенных признаков осуществляются селекционерами посредством скрещивания родителей, каждый из которых имеет один из нужных признаков, и последующей идентификации потомства, которое имеет оба этих нужных признака. Другим способом сцепления генов является перенос двух или более генов в ядро клетки растения во время трансформации. Другим способом сцепления генов является повторная трансформация трансгенного растения другим представляющим интерес геном. Так, например, сцепление генов может быть применено для объединения двух или более различных признаков, включая, например, два или более различных признаков, таких как, признак(и) резистентности к насекомым и признак(и) резистентности к болезням, а также два или более признаков резистентности к гербицидам, и/или признак(и) резистентности к насекомым и признак(и) резистентности к гербицидам. При «сцеплении генов», помимо использования представляющего интерес гена, можно также использовать селективный маркер.
Термин «гомологичная рекомбинация» означает реакцию между любой парой нуклеотидных последовательностей, имеющих соответствующие сайты, содержащие аналогичные нуклеотидные последовательности, посредством которых две нуклеотидных последовательности могут взаимодействовать (подвергаться рекомбинации) с образованием новой последовательности рекомбинантной ДНК. Сайты каждой из аналогичных нуклеотидных последовательностей называются здесь «гомологичными последовательностями». Вообще говоря, частота гомологичной рекомбинации возрастает по мере увеличения длины гомологичной последовательности. Таким образом, если гомологичная рекомбинация происходит между двумя нуклеотидными последовательностями, имеющими меньшую степень идентичности, то частота (или эффективность) рекомбинации снижается по мере увеличения дивергентности этих двух последовательностей. Рекомбинация может быть осуществлена с использованием одной гомологичной последовательности на каждого донора и молекулу-мишень, в результате чего может быть получен продукт рекомбинации «простого кроссинговера». Альтернативно, две гомологичные последовательности могут находиться на каждой из нуклеотидных последовательностей мишени и донора. Рекомбинация между двумя гомологичными последовательностями на доноре и двумя гомологичными последовательностями на мишени приводит к получению продукта рекомбинации «двойного кроссинговера». Если гомологичные последовательности на донорной молекуле фланкируют последовательность, подвергаемую модификации (например, представляющую интерес последовательность), то рекомбинация посредством двойного кроссинговера с молекулой-мишенью будет приводить к образованию продукта рекомбинации, в котором представляющая интерес последовательность будет заменять последовательность ДНК, которая первоначально находилась между гомологичными последовательностями на молекуле-мишени. Обмен последовательностями ДНК между мишенью и донором, который происходит в результате события рекомбинации с двойным кроссинговером, называется «заменой последовательностей».
Предпочтительные растения или семена согласно вариантам осуществления изобретения содержат в своем геноме идентифицированные здесь операторные нуклеотидные последовательности cry1F, cry1Ac synpro и pat вместе по меньшей мере с 20-500 или более смежными фланкирующими нуклеотидами, находящимися на обеих сторонах вставки, как описано в настоящей заявке. Если это не оговорено особо, то термин «фланкирующие последовательности» означает последовательности, идентифицированные в SEQ ID NO: 1 и SEQ ID NO: 2. Как и ожидалось, все эти фланкирующие последовательности или их части могут передаваться потомству, которое будет иметь встроенную ДНК благодаря половому скрещиванию с родительской линией, включающей указанный трансген.
Варианты осуществления изобретения включают тканевые культуры регенерируемых клеток растения согласно вариантам осуществления изобретения. Варианты осуществления изобретения также включают растение, регенерируемое из указанной тканевой культуры, а в частности, растение, которое обладает способностью экспрессировать все морфологические и физиологические признаки репрезентативного сорта. Предпочтительные растения согласно вариантам осуществления изобретения обладают всеми физиологическими и морфологическими признаками растения, выращенного из депонированных семян. Варианты осуществления изобретения также включают потомство таких семян и семян, обладающих представляющими интерес качественными признаками.
Используемый здесь термин «линия» означает группу растений, которые имеют незначительные различия между собой по меньшей мере по одному признаку или вообще не имеют родовых различий. Такие линии могут быть созданы в результате самоопыления и отбора или вегетативного размножения нескольких поколений из одного родителя с применением методов культивирования тканей или клеток.
Используемые здесь термины «сорт» и «разновидность» являются синонимами и означают линию, используемую в промышленном производстве.
Термины «стабильность» или «стабильный», если они относятся к данному компоненту, означают, что данный компонент сохраняется от поколения к поколению, а предпочтительно, по меньшей мере по трем поколениям.
Термин «коммерческая ценность» растения означает, что данное растение имеет хорошую всхожесть и высокую фертильность, что позволяет фермерам выращивать эту культуру на фермах со стандартным сельскохозяйственным оборудованием, а масло с описанными здесь компонентами может быть экстрагировано из семян на стандартном оборудовании для помола и экстракции.
Термин «агрономически элитный» относится к линии, которая имеет желательные агрономические свойства, такие как высокая урожайность, зрелость, резистентность к болезням и т.п., а также резистентность к насекомым и устойчивость к гербицидам, сообщаемые рассматриваемому(ым) трансформанту(ам). Любые и все описанные агрономические признаки и контрольные точки могут быть использованы для идентификации таких растений либо в виде оценочного балла, либо любой граничной точки или обеих граничных точек интервала значений, используемых для определения свойств таких растений.
Как будет очевидно из данного описания, предпочтительные варианты наборов для детектирования могут, например, включать зонды и/или праймеры, связанные с «последовательностями стыка» или «последовательностями транзиции» и/или содержащие «последовательности стыка» или «последовательности транзиции» (где геномная фланкирующая последовательность сои присоединена к последовательности вставки). Так, например, эти наборы включают полинуклеотидные зонды, праймеры и/или ампликоны, сконструированные для идентификации одной или обеих последовательностей стыка (где вставка присоединена к фланкирующей последовательности). Одной из стандартных конструкций является конструкция, имеющая один праймер, который гибридизуется во фланкирующей области, и один праймер, который гибридизуется во вставке. Такие праймеры обычно имеют длину по меньшей мере приблизительно 15 остатков. При таком расположении, праймеры могут быть использованы для генерирования/амплификации детектируемого ампликона, который указывает на присутствие трансгена согласно варианту осуществления изобретения. Эти праймеры могут быть использованы для генерирования ампликона, который охватывает (и включает) последовательность стыка, указанную выше.
Праймер(ы), «присоединяющийся(еся)» к фланкирующей последовательности, обычно не гибридизуется(ются) в области, находящейся за пределами приблизительно 1200 оснований или за пределами области стыка. Таким образом, типичные фланкирующие праймеры должны быть сконструированы так, чтобы они содержали по меньшей мере 15 остатков любой цепи в пределах 1200 оснований во фланкирующих последовательностях от начала вставки. То есть, праймеры, содержащие последовательность соответствующего размера, состоящую из остатков (или гибридизующуюся с ними) в области, составляющей в пределах от пар оснований 1 до 1273 SEQ ID NО: 1 и/или пар оснований 176-1687 SEQ ID NО: 2, входят в объем вариантов осуществления изобретения. Аналогичным образом, праймеры вставки могут быть сконструированы в любом участке вставки, однако, пары оснований 1273-1873 и 13058-13658 SEQ ID NО: 14 могут быть использованы, например, для конструирования каждого праймера без исключения.
Для специалиста в данной области также очевидно, что праймеры и зонды могут быть сконструированы для гибридизации в стандартных условиях гибридизация и/или ПЦР-условиях, где указанный праймер или зонд не является полностью комплементарным репрезентативной последовательности. То есть, в данном случае может быть допустима определенная степень несоответствия или вырожденности. Что касается праймера, состоящего приблизительно из 20 нуклеотидов, то, обычно, например, один, два или т.п. нуклеотидов необязательно будут связываться с противоположной цепью, если несоответствующее основание находится внутри или на конце праймера, противоположного ампликону. Различные подходящие условия гибридизации описаны ниже. В зондах могут быть также использованы синтетические нуклеотидные аналоги, такие как инозин. Могут быть также использованы зонды на основе пептид-содержащей нуклеиновой кислоты (PNA), а также ДНК- и РНК-зонды. Важно отметить, что такие зонды и праймеры являются диагностическими показателями (то есть, способны идентфицировать и дифференцировать уникальные признаки) присутствия трансформанта согласно варианту осуществления изобретения.
Следует отметить, что при ПЦР-амплификации могут возникать ошибки, которые могут приводить, например, к незначительным ошибкам при секвенировании. То есть, если это не оговорено особо, то описанные здесь последовательности были определены путем конструирования длинных ампликонов, происходящих от геномных ДНК сои, с последующим клонированием и секвенированием этих ампликонов. Поэтому неудивительно, что в последовательностях, полученных и определенных таким способом, могут быть обнаружены незначительные различия и некоторые несоответствия, если принять во внимание необходимость проведения множества раундов амплификации для получения ампликона, достаточного для секвенирования геномных ДНК. Следует заметить и принять во внимание, что любые уточнения, необходимые для исправления ошибок или несоответствий такого типа, которые обычно возникают при секвенировании, входят в объем вариантов осуществления изобретения.
Следует также отметить, что некоторые геномные последовательности нередко являются делетированными, например, при встраивании последовательности в процессе создания трансформанта. Таким образом, между рассматриваемыми фланкирующими последовательностями и геномными последовательностями, имеющимися, например, в базе данных GENBANK, могут наблюдаться некоторые различия.
Компоненты последовательности ДНК-«вставки» представлены на фигурах и более подробно обсуждаются ниже в примерах. Полинуклеотидные последовательности ДНК этих компонентов или их фрагменты могут быть использованы в качестве ДНК-праймеров или зондов в способах согласно вариантам осуществления изобретения.
В некоторых вариантах осуществления изобретения, композиции и способы применяются для детектирования присутствия трансгенной/геномной области вставки в растениях сои и в ее семенах и т.п. Были получены последовательности ДНК, содержащие описанную здесь 5'-трансгенную/геномную последовательность стыка в области вставки (между парами оснований 1-1273 SEQ ID NО: 1 и 1-1273 SEQ ID NО: 14); их сегменты и комплементы любых репрезентативных последовательностей и любых их сегментов. Были получены последовательности ДНК, содержащие описанную здесь 3'-трансгенную/геномную последовательность стыка в области вставки (между парами оснований 176-1687 SEQ ID NО: 2 и 13659-15170 SEQ ID NО: 14); их сегменты и комплементы любых репрезентативных последовательностей и любых их сегментов. Последовательность стыка области вставки охватывает последовательность стыка, расположенную между гетерологичной ДНК, встроенной в геном, и ДНК, присутствующей в клетках растения сои и фланкирующей инсерционный сайт. Такие последовательности позволяют диагностировать данный трансформант.
На основе этих последовательностей вставки и граничных последовательностей могут быть сконструированы праймеры, специфичные для данного трансформанта. ПЦР-анализ показал, что линии растения сои согласно вариантам осуществления изобретения могут быть идентифицированы на различные генотипы сои путем проведения анализа ПЦР-ампликонов, полученных с использованием наборов этих праймеров, специфичных для данного трансформанта. Эти и другие родственные методы могут быть применены для идентификации уникальных свойств этих линий растений сои, содержащих трансформант сои pDAB9582.816.15.1. Таким образом, ПЦР-ампликоны, происходящие от таких пар праймеров, являются уникальными и могут быть использованы для идентификации этих линий растения сои.
В некоторых вариантах осуществления изобретения, последовательности ДНК, которые содержат смежный фрагмент новой трансгенной/геномной области вставки, также являются аспектом настоящего изобретения. Настоящее изобретение включает последовательности ДНК, которые содержат полинуклеотиды трансгенной последовательности вставки достаточной длины, и полинуклеотиды геномной последовательности сои достаточной длины, происходящие от одного или более из вышеупомянутых растений сои, и/или последовательности, которые могут служить в качестве праймеров для продуцирования продукта-ампликона, используемого для идентификации одного или более из этих растений сои.
Родственные варианты осуществления изобретения относятся к последовательностям ДНК, которые содержат по меньшей мере 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более смежных нуклеотидов трансгенной части идентифицированной здесь последовательности ДНК (такой как SEQ ID NО: 1 и ее сегмент) или ее комплементов, и фланкирующую последовательность ДНК сои аналогичной длины, происходящую от указанных последовательностей или их комплементов. Такие последовательности могут быть использованы в качестве ДНК-праймеров в методах амплификации ДНК. Ампликоны, продуцированные с использованием этих праймеров, позволяют идентифицировать любые описанные здесь трансгенные растения сои. Поэтому, варианты осуществления изобретения также включают ампликоны, продуцированные с использованием таких ДНК-праймеров.
Варианты осуществления изобретения также включают способы детектирования присутствия ДНК в образце, который соответствует описанному здесь трансформанту сои. Такие способы могут включать: (a) контактирование образца, содержащего ДНК, с набором праймеров, которые, при их использовании в реакции амплификации нуклеиновой кислоты с ДНК, происходящей по меньшей мере от одного из этих трансформантов сои, продуцируют ампликон, позволяющий идентифицировать указанный(е) трансформант(ы); (b) осуществление реакции амплификации нуклеиновой кислоты с получением ампликона, и (c) детектирование ампликона.
Другие способы детектирования согласно вариантам осуществления изобретения включают способ детектирования присутствия ДНК в образце, соответствующем указанному трансформанту, где указанный способ включает: (a) контактирование образца, содержащего ДНК, с зондом, который гибридизуется в жестких условиях гибридизации с ДНК, происходящей по меньшей мере от одного из указанных трансформантов сои, и который не гибридизуется в жестких условиях гибридизации с ДНК контрольного растения сои (не содержащего представляющую интерес ДНК-вставку); (b) гибридизацию образца и зонда в жестких условиях; и (c) детектирование гибридизации зонда с ДНК.
В других своих вариантах, настоящее изобретение включает способы получения растения сои, содержащего трансформант pDAB9582.816.15.1 согласно варианту осуществления изобретения, где указанный способ включает стадии: (a) полового скрещивания первой родительской линии сои (содержащей экспрессионный кластер согласно варианту осуществления изобретения, сообщающий растениям указанной линии признак устойчивости к глюфозинату) со второй родительской линией сои (не имеющей такого признака устойчивости к гербицидам) с получением множества потомств растения; и (b) отбора потомства растения с использованием молекулярных маркеров. Такие способы могут включать, но необязательно, дополнительную стадию возвратного скрещивания потомства растения со второй родительской линией сои с получением чистосортного растения сои, которое имеет указанный признак резистентности к насекомым и устойчивости к глюфозинату.
В соответствии с другим своим аспектом, настоящее изобретение относится к способам определения зиготности потомства кроссов с указанным трансформантом. Такие способы могут включать контактирование образца, содержащего ДНК сои, с набором праймеров согласно варианту осуществления изобретения. Указанные праймеры, при их использовании в реакции амплификация нуклеиновой кислоты с геномной ДНК, происходящей от указанных трансформантов сои, продуцируют первый ампликон, позволяющий идентифицировать указанные трансформанты сои. Такие способы также включают осуществление реакции амплификации нуклеиновой кислоты с продуцированием первого ампликона; детектирование первого ампликона и контактирование образца, содержащего ДНК сои, со втором набором праймеров (указанный второй набор праймеров, если он используется в реакции амплификации нуклеиновой кислоты с геномной ДНК растений сои, продуцирует второй ампликон, включающий эндогенную последовательность геномной ДНК природной сои, которая не содержит полинуклеотидной последовательности указанного трансформанта); и осуществление реакции амплификации нуклеиновой кислоты с продуцированием второго ампликона. Указанные способы также включают детектирование второго ампликона и сравнение первого и второго ампликонов в образце, где присутствие обоих ампликонов указывает на зиготность трансгенной вставки.
Наборы для детектирования ДНК могут быть разработаны с применением описанных здесь композиций и методов, хорошо известных специалистам в области детектирования ДНК. Такие наборы являются подходящими для идентификации рассматриваемой ДНК трансгенной сои в образце, и могут быть применены в методах скрещивания растений сои, содержащих эту ДНК. Указанные наборы содержат последовательности ДНК, комплементарные ампликонам, например, описанным здесь ампликонам, или последовательностям ДНК, которые являются комплементарными ДНК, содержащейся в трансгенных генетических элементах рассматриваемых трансформантов. Эти последовательности ДНК могут быть использованы в реакциях амплификации ДНК или в качестве зондов в методе гибридизации ДНК. Указанные наборы могут также содержать реагенты и материалы, необходимые для осуществления метода детектирования.
«Зонд» представляет собой выделенную молекулу нуклеиновой кислоты, к которой присоединена стандартная детектируемая метка или репортерная молекула (такая как радиоактивный изотоп, лиганд, хемилюминесцентный агент или фермент). Такой зонд может гибридизоваться с цепью нуклеиновой кислоты-мишени, а в случае вариантов осуществления изобретения, с цепью геномной ДНК одного из указанных трансформантов сои, независимо от того, происходят ли они от самого растения сои или образца, включающего ДНК трансформанта. Зондами согласно вариантам осуществления изобретения являются не только дезоксирибонуклеиновые или рибонуклеиновые кислоты, но также и полиамиды и другие материалы, которые специфически связываются с последовательностью ДНК-мишени и могут быть использованы для детектирования присутствия этой последовательности ДНК-мишени.
«Праймеры» представляют собой выделенные/синтезированные нуклеиновые кислоты, которые были подвергнуты отжигу с цепью ДНК-мишени посредством гибридизации нуклеиновых кислот с образованием гибрида «праймер - цепь ДНК-мишени» и с последующим удлинением цепи ДНК-мишени под действием полимеразы, например, ДНК-полимеразы. Пары праймеров согласно вариантам осуществления изобретения могут быть использованы для амплификации последовательности нуклеиновой кислоты-мишени, например, с помощью полимеразной цепной реакции (ПЦР) или с применением других стандартных методов амплификации нуклеиновых кислот.
Зонды и праймеры обычно имеют длину в 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189,190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225,226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500 или 1000 или 2000 или 5000 или более полинуклеотидов. Такие зонды и праймеры специфически гибридизуются с последовательностью-мишенью посредством гибридизации в условиях высокой жесткости. Предпочтительно, зонды и праймеры согласно вариантам осуществления изобретения имеют последовательности, полностью идентичные последовательности-мишени, хотя могут быть сконструированы зонды, которые отличаются от последовательности-мишени и сохраняют способность гибридизоваться с последовательностями-мишенями, и такие зонды могут быть сконструированы стандартными методами.
Методы получения и использования зондов и праймеров описаны, например, в руководстве Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989. Пары ПЦР-праймеров могут быть получены из известной последовательности, например, с использованием компьютерных программ, разработанных для этой цели.
Праймеры и зонды, полученные на основе описанных здесь фланкирующих последовательностей ДНК и последовательностей вставок, могут быть использованы для подтверждения (и если это необходимо, для коррекции) описанных последовательностей стандартными методами, например, путем повторного клонирования и секвенирования таких последовательностей.
Нуклеиновокислотные зонды и праймеры согласно вариантам осуществления изобретения гибридизуются с последовательностью ДНК-мишени в жестких условиях. Для детектирования присутствия ДНК трансгенной вставки в образце может быть применен стандартный метод гибридизации нуклеиновых кислот или метод амплификации нуклеиновых кислот. Молекулы нуклеиновой кислоты или их фрагменты способны специфически гибридизоваться с другими молекулами нуклеиновой кислоты в определенных условиях. Считается, что две молекулы нуклеиновой кислоты способны специфически гибридизоваться друг с другом, если эти две молекулы способны образовывать антипараллельную структуру двухцепочечной нуклеиновой кислоты. Считается, что молекула нуклеиновой кислоты «комплементарна» другой молекуле нуклеиновой кислоты, если эти молекулы обладают полной комплементарностью. Считается, что молекулы являются «полностью комплементарными», если каждый нуклеотид одной из этих молекул комплементарен нуклеотиду другой молекулы. Молекулы, которые являются «полностью комплементарными», по существу гибридизуются друг с другом, но при этом, сохраняют стабильность, достаточную для их гибридизации друг с другом по меньшей мере в стандартных условиях «высокой жесткости». Стандартные условия высокой жесткости описаны в руководстве Sambrook et al., 1989.
Две молекулы считаются «минимально комплементарными», если они могут гибридизоваться друг с другом, но при этом, сохранять стабильность, достаточную для их гибридизации друг с другом по меньшей мере в стандартных условиях «низкой жесткости». Стандартные условия низкой жесткости описаны в руководстве Sambrook et al., 1989. Для того чтобы молекула нуклеиновой кислоты могла служить в качестве праймера или зонда, необходимо, чтобы она была лишь минимально комплементарна последовательности, способной образовывать стабильную двухцепочечную структуру в присутствии конкретного растворителя и в присутствии соли в определенной концентрации.
Термин «жесткие условия» или «условия жесткости» функционально определен с точки зрения гибридизации нуклеиновокислотного зонда с нуклеиновой кислотой-мишенью (то есть, с конкретной представляющей интерес последовательностью нуклеиновой кислоты), проводимой в соответствии с процедурой специфической гибридизации, обсуждаемой в руководстве Sambrook et al., 1989, 9.52-9.55. См. также, Sambrook et al., 1989, 9.47-9.52 и 9.56-9.58.
В зависимости от цели применения, условия гибридизации могут варьироваться для достижения различных степеней селективности зонда или праймера с вырожденной полинуклеотидной последовательностью по отношению к последовательности-мишени. В тех случаях, когда требуется высокая селективность, обычно используются относительно жесткие условия образования гибридов одной полинуклеотидной последовательности с другой полинуклеотидной последовательностью, например, в данном случае следует использовать относительно низкие концентрации соли и/или высокую температуру, например, концентрация NaCl может составлять приблизительно от 0,02 M до 0,15 M, а температура может составлять приблизительно от 50°C до 70°C. Жесткие условия, например, могут включать по меньшей мере 2-кратную промывку гибридизационного фильтра промывочным буфером высокой жесткости (0,2×SSC, 0,1% ДСН, 65°C). Соответствующие условия жесткости, стимулирующие гибридизацию ДНК, например, обработка 6,0× хлоридом натрия/цитратом натрия (SSC) приблизительно при 45°C, с последующей промывкой 2,0×SSC при 50°C, известны специалистам в данной области. Так, например, концентрация соли в стадии промывки может быть выбрана из концентраций соли, обычно используемых в условиях низкой жесткости, а именно, приблизительно 2,0×SSC при 50°C, и концентраций соли, обычно используемых в условиях высокой жесткости, а именно, приблизительно 0,2×SSC при 50°C. Кроме того, температура в стадии промывки, которая в условиях низкой жесткости составляет приблизительно 22°C, то есть, комнатную температуру, может быть повышена до температуры гибридизации в условиях высокой жесткости, то есть, приблизительно до 65°C. Температура и концентрация соли могут варьироваться, либо один из этих параметров может оставаться постоянным, а другой может изменяться. При таких селективных условиях могут наблюдаться, если они вообще имеются, незначительные несоответствия, между зондом и матрицей или цепью-мишенью. Детектирование последовательностей ДНК посредством гибридизации хорошо известно специалистам, и репрезентативные методы анализов посредством гибридизации описаны в патентах США №№ 4965188 и 5176995.
В особенно предпочтительном варианте изобретения, нуклеиновая кислота согласно варианту осуществления изобретения может специфически гибридизоваться с одним или более проиллюстрированными или описанными здесь праймерами (или ампликонами или другими последовательностями), включая комплементы и их фрагменты, в условиях высокой жесткости. В одном из аспектов настоящего изобретения, маркерная молекула нуклеиновой кислоты согласно варианту осуществления изобретения имеет последовательность нуклеиновой кислоты, представленную здесь в одной из репрезентативных последовательностей, или комплементы и/или фрагменты этой последовательности.
В другом аспекте настоящего изобретения, маркерная молекула нуклеиновой кислоты согласно варианту осуществления изобретения имеет последовательность, которая на 80%-100% или 90%-100% идентична указанным последовательностям нуклеиновой кислоты. В другом аспекте настоящего изобретения, маркерная молекула нуклеиновой кислоты согласно варианту осуществления изобретения имеет последовательность, которая на 95%-100% идентична такой последовательности нуклеиновой кислоты. Эти последовательности могут быть использованы в качестве маркеров в методах скрещивания растений для идентификации потомства генетических кроссов. Гибридизация зонда с молекулой ДНК-мишени может быть детектирована любыми методами, известными специалистам, и такими методами могут быть, но не ограничиваются ими, методы, осуществляемые с использованием флуоресцентных меток, радиоактивных меток, меток на основе антител и хемилюминесцентных меток.
Что касается амплификации последовательности нуклеиновой кислоты-мишени (например, с помощью ПЦР), проводимой с использованием конкретной пары праймеров для амплификации, то термин «жесткие условия» означает условия, которые позволяют паре праймеров гибридизоваться только с последовательностью нуклеиновой кислоты-мишени, с которой может связываться праймер, имеющий соответствующую последовательность дикого типа (или ее комплемент), и предпочтительно, продуцировать уникальный продукт амплификации, то есть, ампликон.
Термин «специфичный к (последовательности-мишени)» означает, что зонд или праймер гибридизуются в жестких условиях гибридизации только с последовательностью-мишенью в образце, содержащем такую последовательность-мишень.
Используемые здесь термины «амплифицированная ДНК» или «ампликон» означают продукт амплификации последовательности нуклеиновой кислоты-мишени, которая является частью нуклеиновокислотной матрицы. Так, например, для того, чтобы определить, может ли растение сои, полученное в результате полового скрещивания, содержать трансгенную геномную ДНК растения-трансформанты сои согласно варианту осуществления изобретения, ДНК, экстрагированная из образца ткани растения сои, может быть подвергнута амплификации нуклеиновой кислоты с использованием пары праймеров, которая включает праймер, происходящий от фланкирующей последовательности, присутствующей в геноме растения и находящейся поблизости от инсерционного сайта встроенной гетерологичной ДНК, и второй праймер, происходящий от встроенной гетерологичной ДНК, в результате чего может быть получен ампликон, который может быть использован для детектирования присутствия трансгенной ДНК. Ампликон имеет длину и последовательность, которые также являются подходящими для детектирования такого трансгена. Длина ампликона может варьироваться в пределах от общей длины пар праймеров плюс одна пара нуклеотидных оснований, и/или общей длины пар праймеров плюс приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500, 750, 1000, 1250, 1500, 1750, 2000 или более пар нуклеотидных оснований (± любые из перечисленных выше приращений). Альтернативно, пара праймеров может происходить от фланкирующей последовательности, расположенной на обеих сторонах встроенной ДНК, в результате чего может продуцироваться ампликон, который включает полноразмерную нуклеотидную последовательность вставки. Один из членов пары праймеров, происходящий от геномной последовательности растения, может находиться на определенном расстоянии от встроенной последовательности ДНК. Такое расстояние может включать интервал от одной пары нуклеотидных оснований и приблизительно до двадцати тысяч пар нуклеотидных оснований. В используемое здесь понятие «ампликон», в частности, не входят димеры праймеров, которые могут образовываться в реакции термоамплификации ДНК.
Амплификация нуклеиновой кислоты может быть осуществлена любым из различных методов амплификации нуклеиновой кислоты, известных специалистам, включая полимеразную цепную реакцию (ПЦР). Специалистам известны различные методы амплификации, и эти методы описаны, inter alia, в патенте США No. 4683195 и в патенте США № 4683202. Методы ПЦР-амплификации были разработаны для амплификации геномной ДНК длиной до 22 т.п.о. Эти методы, а также другие методы амплификации ДНК, известные специалистам, могут быть применены для практического осуществления настоящего изобретения. Последовательность гетерологичной трансгенной ДНК-вставки или фланкирующая геномная последовательность рассматриваемого трансформанта сои могут быть подтверждены (и скорректированы, если это необходимо) путем амплификации таких последовательностей, происходящих от указанного трансформанта, с использованием праймеров, происходящих от описанных здесь последовательностей, с последующим проведением стандартного секвенирования ПЦР-ампликона или клонированной ДНК.
Ампликон, полученный такими методами, может быть детектирован различными способами. Наиболее хорошо известными методами детектирования ДНК-ампликонов являются электрофорез в агарозном геле и окрашивание этидийбромидом. Другим таким методом является генетический элементный анализ, где сконструированный ДНК-олигонуклеотид перекрывается со смежной фланкирующей последовательностью геномной ДНК и со встроенной последовательностью ДНК. Этот олигонуклеотид иммобилизуют на лунках микротитрационного луночного планшета. После ПЦР представляющей интерес области (проводимой с использованием одного праймера во встроенной последовательности и одного праймера в смежной фланкирующей геномной последовательности), одноцепочечный ПЦР-продукт может быть гибридизован с иммобилизованным олигонуклеотидом и может служить в качестве матрицы для проведения реакции удлинения на одно основание с использованием ДНК-полимераз и меченных ddNTP, специфичных к предполагаемому следующему основанию. Анализ связанного продукта может быть осуществлен путем количественной оценки флуоресцентного сигнала. Флуоресцентный сигнал указывает на присутствие вставки/фланкирующей последовательности, что свидетельствует об успешном осуществлении амплификации, гибридизации и удлинении на одно основание.
Другим методом является метод пиросеквенирования, описанный Winge (Innov. Pharma. Tech. 00: 18-24, 2000). В этом методе, сконструированный олигонуклеотид перекрывается со смежной геномной ДНК и с последовательностью стыка ДНК-вставки. Олигонуклеотид подвергают гибридизации с одноцепочечным ПЦР-продуктом представляющей интерес области (проводимой с использованием одного праймера во встроенной последовательности и одного праймера во фланкирующей геномной последовательности), и инкубируют в присутствии ДНК-полимеразы, ATP, сульфурилазы, люциферазы, апиразы, аденозин-5'-фосфосульфата и люциферина. Затем отдельно добавляют DNTP, и такое добавление приводит к продуцированию детектируемого светового сигнала. Световой сигнал указывает на присутствие трансгенной вставки/фланкирующей последовательности, что свидетельствует об успешном осуществлении амплификации, гибридизации и удлинении на одно или несколько оснований.
Другим методом, который может быть применен для детектирования ампликона согласно варианту осуществления изобретения, является поляризация флуоресценции. В этом методе конструируют олигонуклеотид, который перекрывается с геномной фланкирующей последовательностью и с последовательностью стыка ДНК-вставки. Олигонуклеотид подвергают гибридизации с одноцепочечным ПЦР-продуктом представляющей интерес области (проводимой с использованием одного праймера во встроенной ДНК и одного праймера во фланкирующей геномной последовательности ДНК), и инкубируют в присутствии ДНК-полимеразы и флуоресцентно меченного ddNTP. Удлинение на одно основание приводит к включению ddNTP. Такое включение флуоресцентно меченного ddNTP может быть определено по изменению поляризации на флуориметре. Изменение поляризации указывает на присутствие трансгенной вставки/фланкирующей последовательности, что свидетельствует об успешном осуществлении амплификации, гибридизации и удлинении на одно основание.
TAQMAN® (PE Applied Biosystems, Foster City, Calif.) представляет собой метод детектирования присутствия ДНК и количественной оценки последовательности ДНК. Вкратце, был сконструирован олигонуклеотидный зонд FRET, который перекрывается с геномной фланкирующей последовательностью и с последовательностью стыка ДНК-вставки. Зонд FRET и ПЦР-праймеры (один праймер во встроенной последовательности ДНК и один во фланкирующей геномной последовательности) подвергают реакции циклизации в присутствии термостабильной полимеразы и dNTP. В процессе специфической амплификации, ДНК-полимераза Taq гидролизуется по механизму «коррекции», в результате чего, из гасящей молекулы на зонде FRET высвобождается флуоресцентная молекула. Флуоресцентный сигнал указывает на присутствие фланкирующей последовательности/трансгенной последовательности-вставки, что свидетельствует об успешном осуществлении амплификации и гибридизации.
Для детектирования полинуклеотидной последовательности могут быть применены описанные методы «молекулярных маяков» (Beacons). Вкратце, был сконструирован олигонуклеотидный зонд FRET, который перекрывается с фланкирующей геномной последовательностью и с последовательностью стыка ДНК-вставки. Уникальность структуры зонда FRET состоит в том, что этот зонд имеет вторичную структуру, поддерживающую флуоресцентную и гасящую молекулы в непосредственной близости друг от друга. Зонд FRET и ПЦР-праймеры (один праймер во встроенной последовательности ДНК и один во фланкирующей геномной последовательности) подвергают реакции циклизации в присутствии термостабильной полимеразы и dNTP. После успешной ПЦР-амплификации, гибридизация зонда FRET с последовательностью-мишенью приводит к удалению вторичной структуры зонда и к пространственному разделению флуоресцентной и гасящей молекул. В результате возникает флуоресцентный сигнал. Флуоресцентный сигнал указывает на присутствие фланкирующей геномной последовательности/трансгенной последовательности-вставки, что свидетельствует об успешном осуществлении амплификации и гибридизации.
Принимая во внимание тот факт, что определенная локализация в геноме сои является особенно подходящей для инсерции, можно сказать, что варианты осуществления изобретения также включают семена сои и/или растение сои, содержащие по меньшей мере одну не-соевую pDAB9582.816.15.1-вставку-трансформант, находящуюся, в основном, поблизости от этого положения генома. Одним из вариантов является замена одного из описанных здесь трансформантов сои pDAB9582.816.15.1 на другую вставку. Вообще говоря, в конкретных вариантах осуществления изобретения, может быть, например, осуществлена направленная гомологичная рекомбинация. Технология такого типа рассматривается, например, в WO 03/080809 A2 и в соответствующей опубликованной заявке на патент США (US 20030232410). Таким образом, варианты осуществления изобретения включают растения и клетки растений, содержащие гетерологичную вставку (введенную вместо множества копий генов cry1F, cry1Ac или pat или вместе с множеством копий этих генов), фланкированную полноразмерными идентифицированными здесь фланкирующими последовательностями (п.о. в положениях 1-1273 SEQ ID NO: 1 и п.о. в положениях 176-1687 SEQ ID NO: 2) или распознаваемой частью этих последовательностей. С применением такого(их) способа(ов) может (могут) быть также встроена(ы) дополнительная копия (или дополнительные копии) cry1F, cry1Ac или pat.
Все цитируемые здесь патенты, патентные заявки, предварительные заявки и публикации во всей своей полноте вводятся в настоящее описание посредством ссылки в той области, которая конкретно соответствует описанию настоящего изобретения. Нижеследующие примеры приводятся для иллюстрации методов практического осуществления вариантов изобретения и описания некоторых предпочтительных вариантов изобретения. Эти примеры не должны рассматриваться как ограничение настоящего изобретения. При этом, для специалиста в данной области очевидно, что методы, описанные в нижеследующих примерах, представляют собой конкретно применяемые подходы, представленные для иллюстрации предпочтительных вариантов практического осуществления изобретения. Однако, специалисту в данной области, исходя из описания изобретения, будет очевидно, что в эти конкретные варианты осуществления изобретения может быть внесено множество изменений, позволяющих получить такой же или аналогичный результат и не выходящих за рамки существа и объема изобретения. Если это не оговорено особо, то все проценты даны по массе, а все соотношения растворителей в смеси даны по объему, если это не указано конкретно.
Если это не оговорено особо, то нижеследующие сокращения означают:
Примеры
Пример 1: Трансформация и отбор растений-трансформантов сои pDAB9582.816.15J, содержащих гены CrylF, CrylAc и PAT
Трансгенную сою (Glycine max), содержащую генную модификацию pDAB9582.816.15.1, получали посредством Agrobacterium-опосредуемой трансформации эксплантатов в утолщениях в семядоле сои. Для инициации трансформации использовали инактивированный штамм EHA101 Agrobacterium (Hood et al., 1993), несущий бинарный вектор pDAB9582 (фигура 1), содержащий селективный маркер, pat v6 и представляющие интерес гены cry1F v3 и cryl Ac synpro, в области T-цепи ДНК. Последовательность T-цепи ДНК для pDAB9582 представлена в SEQ ID NO:3 и описана ниже в таблице 1.
Генные элементы, локализованные на pDAB9582
Agrobacterium-опосредуемую трансформацию осуществляли модифицированным методом, описанным Zeng et al. (2004). Вкратце, семена сои (cорта Maverick) проращивали на базальной среде, а затем выделяли утолщения семядоли, и эти утолщения инфицировали агробактерией. В среды для стимуляции образования, удлинения и укоренения побегов добавляли цефотаксим, тиментин и ванкомицин в целях удаления агробактерий. Для ингибирования роста нетрансформированных побегов проводили отбор с использованием глюфозината. Отобранные побеги переносили в среду для укоренения в целях инициации образования корней, а затем переносили в почвенную смесь для акклиматизации проростков.
Верхушечные листья отобранных проростков окрашивали глюфозинатом для скрининга на предполагаемые трансформанты. Скринированные проростки переносили в теплицу, оставляли для акклиматизации, а затем листья окрашивали глюфозинатом для повторного подтверждения устойчивости к гербицидам и детектирования присутствия предполагаемых трансформантов. После этого брали образцы скринированных растений и проводили их молекулярный анализ для подтверждения присутствия селективного маркерного гена и/или представляющего интерес гена. Растения T0 оставляли в теплице для самоопыления и получали семена Тi.
Этот трансформант, то есть, трансформант сои pDAB9582.816.15.1, получали из независимо трансформированного изолята. Растения Т1 подвергали возвратному скрещиванию и интрогрессии данного трансформанта в элитные сорта последующих поколений. Данный трансформант отбирали исходя из его уникальных свойств, таких как единственный инсерционный сайт, нормальная менделевская сегрегация, стабильная экспрессия и превосходная комбинация свойств, включая резистентность к насекомым, устойчивость к гербицидам и агрономические показатели. В нижеследующих примерах представлены данные, которые были использованы для характеризации трансформанта сои pDAB9582.816.15.1.
Пример 2: Характеризация экспрессии белка в трансформанте сои pDAB9582.816.15.J
Были охарактеризованы биохимические свойства рекомбинантных белков CrylF, CrylAc и PAT в трансформанте сои pDAB9582.816.15.1. Количественный твердофазный иммуноферментный анализ (ELISA) представляет собой известный биохимический анализ, который может быть осуществлен в целях характеризации биохимических свойств белков и подтверждения экспрессии этих белков в трансформанте сои pDAB9582.816.15.1.
Пример 2.1: Экспрессия белка PAT, CrylF и CrylAc в тканях растения
Образцы ткани сои выделяли из тестируемых растений и подготавливали для анализа на экспрессию. Белок PAT экстрагировали из тканей растений сои с использованием забуференного фосфатом физиологического раствора, содержащего детергент твин-20 (PBST), включающий 0,5% альбумин бычьей сыворотки (BSA). Ткань растения центрифугировали, затем водный супернатант собирали, разводили соответствующим буфером, если это необходимо, и анализировали с использованием набора для «сэндвич»-ELISA PAT. Этот набор использовали в соответствии с протоколом, рекомендованным производителями (Envirologix, Portland, ME). С помощью этого анализа измеряли концентрации экспрессированного белка PAT.
Белок Cry1F экстрагировали из тканей растений сои с использованием забуференного фосфатом физиологического раствора, содержащего детергент твин-20 (PBST). Ткань растения центрифугировали, затем водный супернатант собирали, разводили соответствующим буфером, если это необходимо, и анализировали с использованием набора для «сэндвич»-ELISA Cry1F. Этот набор использовали в соответствии с протоколом, рекомендованным производителями (Strategic Diagnostics Inc., Newark, DE). С помощью этого анализа измеряли концентрации экспрессированного белка CrylF.
Белок Cry1Ac экстрагировали из тканей растений сои с использованием забуференного фосфатом физиологического раствора, содержащего детергент твин-20 (PBST), включающий 0,5% альбумин бычьей сыворотки (BSA). Ткань растения центрифугировали, затем водный супернатант, собирали, разводили соответствующим буфером, если это необходимо, и анализировали с использованием набора для «сэндвич»-ELISA CrylAc. Этот набор использовали в соответствии с протоколом, рекомендованным производителями (Strategic Diagnostics Inc., Newark, DE). С помощью этого анализа измеряли концентрации экспрессированного белка Cry1Ac.
Анализ по детектированию осуществляли для оценки стабильности экспрессии, а также вертикального наследования (между поколениями) и горизонтального наследования (между линиями внутри поколения) трансформанта pDAB9582.816.15.1 в растениях сои.
Пример 2.2: Экспрессия белков CrylF, CrylAc и PAT в тканях растения
Уровни белков CrylF, CrylAc и PAT определяли в трансформанте сои pDAB9582.816.15.1 в соответствии с протоколами, описанными выше. Растворимые белки, экстрагированные из ткани листьев сои, анализировали с помощью количественного твердофазного иммуноферментного анализа (ELISA). Поколения трансформантов сои с генной модификацией pDAB9582.816.15.1 от T2 до T6 имели стабильную экспрессию (не сегрегировались), и такая экспрессия наблюдалась во всех линиях. В таблице 2 представлен средний уровень экспрессии трансгенных белков в трансформанте сои pDAB9582.816.15.1.
Средний уровень экспрессии различных трансгенных белков в трансформанте сои pDAB9582.816.15.1.
Пример 3: Клонирование и характеризация последовательности ДНК во вставке и во фланкирующих граничных областях трансформанта сои pDAB9582.816.15.1
Для характеризации и описания геномного инсерционного сайта определяли последовательность граничных областей фланкирующей геномной Т-ДНК трансформанта сои pDAB9582.816.15.1. Была подтверждена геномная последовательность трансформанта сои pDAB9582.816.15.1, включающая 1273 п.о. 5'-фланкирующей граничной последовательности (SEQ ID NO: 1) и 1371 п.о. 3'-фланкирующей граничной последовательности (SEQ ID NO: 2). ПЦР-амплификация граничных последовательностей трансформанта сои pDAB9582.816.15.1 подтвердила, что граничные области происходят от исходного растения сои, а области стыка представляют собой уникальные последовательности трансформанта сои pDAB9582.816.15.1. Области стыка могут быть использованы для трансген-специфической идентификации трансформанта сои pDAB9582.816.15.1. Кроме того, инсерционный сайт Т-цепи был охарактеризован путем амплификации геномного фрагмента, соответствующего области идентифицированных фланкирующих граничных последовательностей, происходящих от генома нетрансформированной сои. Сравнение трансформанта сои pDAB9582.816.15.1 с растением, имеющим нетрансформированную геномную последовательность, выявило делецию оригинального локуса приблизительно в 21 п.о., образованную в процессе интеграции Т-цепи. В целом, характеризация вставки и граничной последовательности трансформанта сои pDAB9582.816.15.1 показала, что в геноме сои присутствует интактная копия Т-цепи pDAB9582.
Список праймеров и их последовательностей, используемых для подтверждения геномной ДНК трансформанта сои pDAB9582.816.15.1
Условия проведения стандартной ПЦР-амплификации граничных областей и трансген-специфических последовательностей в трансформанте сои pDAB9582.816.15.1
ПЦР-смесь для стандартной ПЦР-амплификации граничных областей и трансген-специфических последовательностей в трансформанте сои pDAB9582.816.15.1
(10 мкМ)
Пример 3.1: Подтверждение геномных последовательностей сои
Выравнивание 5'- и 3'-фланкирующих граничных областей с полноразмерной геномной «шотган»-последовательностью хромосомы 03 Glycine max показало, что трансген трансформанта сои pDAB9582.816.15.1 был встроен в хромосому 03 генома сои. Для подтверждения инсерционного сайта в геноме трансформанта сои pDAB9582.816.15.1 осуществляли ПЦР с использованием различных пар праймеров (фигура 2, таблица 3, таблица 4 и таблица 5). Геномную ДНК трансформанта сои pDAB9582.816.15.1 и другие трансгенные или нетрансгенные линии сои использовали в качестве матрицы. Для подтверждения того, что 5'-граничные последовательности являются скорректированными, был выбран праймер, сконструированный так, чтобы он связывался с элементом промотора гена AtUbi10, например, AtUbi10RV1, и праймер, сконструированный так, чтобы он связывался с клонированной 5'-концевой граничной последовательностью на хромосоме 03 генома сои, где указанный праймер был обозначен 81615_FW2, и эти праймеры были использованы в целях амплификации ДНК-сегмента, который охватывает элемент промотора гена AtUbi10 до 5'-концевой граничной последовательности. Аналогогичным образом, для подтверждения клонированной 3'-концевой граничной последовательности, pat-специфический праймер, например, 3'PATEnd05, и праймеры, сконструированные для клонированной 3'-концевой граничной последовательности и обозначенные 81615_RV1 и 81615_RV2, были использованы в целях амплификации ДНК-сегментов, которые охватывают ген pat до 3'-концевой граничной последовательности. ДНК-фрагменты предсказанного размера были амплифицированы только из геномной ДНК трансформанта сои pDAB9582.816.15.1. с использованием каждой пары праймеров, но не из образцов ДНК, происходящих от других трансгенных линий сои или нетрансгенного контроля. Полученные результаты показали, что клонированные 5'- и 3'-граничные последовательности представляют собой фланкирующие граничные последовательности вставки T-цепи для трансформанта сои pDAB9582.816.15.1.
Для дополнительного подтверждения присутствия в геноме сои ДНК-вставки, геномную ДНК, не содержащую вставки Т-цепи в трансформанте сои pDAB9582.816.15.1, подвергали ПЦР-амплификации по всем граничным последовательностям растения сои. Праймер 81615_FW2, сконструированный для 5'-концевой граничной последовательности, и один праймер 81615_RV3, сконструированный для 3'-концевой граничной последовательности, использовали в целях амплификации ДНК-сегментов, содержащих локус, в который была интегрирована Т-цепь pDAB9582. Как и ожидалось, ПЦР-реакции, проводимые с использованием пары праймеров 81615_FW2 и 81615_RV3, позволяли получить приблизительно 1,8 т.п.о. ДНК-фрагмента, происходящего от всех контрольных линий сои, но не из pDAB9582.816.15.1. Выравнивание идентифицированных 5'- и 3'-граничных последовательностей трансформанта сои pDAB9582.816.15.1 с полноразмерной геномной «шотган»-последовательностью хромосомы 03 Glycine max показало, что в локусе исходного генома присутствовала делеция приблизительно в 21 п.о. (фигура 3). Полученные результаты показали, что трансген трансформанта сои pDAB9582.816.15.1 был встроен в сайт хромосомы 03 генома сои.
Пример 4: Характеризация трансформанта сои pDAB9582.816.15.1 с помощью саузерн-блот-анализа
Саузерн-блот-анализ проводили в целях определения профиля интеграции в трансформанте сои pDAB9582.816.15.1. В результате этих экспериментов были получены данные, которые продемонстрировали интеграцию и целостность трансгенов crylAc и crylF в геноме сои. Трансформант сои pDAB9582.816.15.1 был охарактеризован как полноразмерный трансформант, образованный посредством простой интеграции и содержащий одну копию PTU crylAc и crylF, происходящую от плазмиды pDAB9582.
Данные Саузерн-блот-анализа позволяют предположить, что фрагмент Т-цепи был встроен в геном трансформанта сои pDAB9582.816.15.1. Более тщательный Саузерн-блот-анализ был проведен с использованием зондов, специфичных к генам crylAc и crylF, содержащимся в области интеграции Т-цепи pDAB9582.816.15.1, и с использованием описанных здесь рестриктирующих ферментов, которые имеют сайты расщепления, локализованные в плазмиде, и продуцируют гибридизующиеся фрагменты, которые находятся внутри плазмиды, или фрагменты, которые охватывают область стыка плазмиды и геномной ДНК сои (граничные фрагменты). Молекулярные массы, полученные исходя из Саузерн-гибридизации для комбинации рестриктирующего фермента и зонда, были уникальными для данного трансформанта, что обеспечивало его точную идентификаицю. Эти анализы также показали, что данный плазмидный фрагмент был встроен в геномную ДНК сои без реаранжировки PTU crylAc и crylF.
Пример 4.1: Сбор образцов листьев сои и выделение геномной ДНК (гДНК)
Геномную ДНК экстрагировали из ткани листьев, собранных от отдельных растений сои, содержащих трансформант сои pDAB9582.816.15.1. Кроме того, гДНК выделяли из обычных растений сои сорта Maverick, имеющих генетический фон, который является репрезентативным для линии данного растения, где отсутствуют гены cry1Ac и cry1F. Отдельную геномную ДНК экстрагировали из лиофилизованной ткани листьев в соответствии со стандартным методом CTAB. После экстракции, ДНК количественно оценивали на спектрофлуориметре с использованием реагента PICO GREEN (Invitrogen, Carlsbad, CA).
Пример 4.2: Гидролиз и разделение ДНК
Для молекулярной характеризации трансформанта сои pDAB9582.816.15.1 с помощью Саузерн-блот-анализа, десять микрограммов (10 мкг) геномной ДНК подвергали гидролизу. Геномную ДНК, происходящую от трансформанта сои pDAB9582.816.15.1 и от нетрансгенной линии сои сорта Maverick, гидролизовали путем добавления приблизительно пяти единиц выбранного рестриктирующего фермента на мкг ДНК и соответствующего реакционного буфера к каждому образцу ДНК. Каждый образец инкубировали приблизительно при 37°C в течение ночи. Рестриктирующие ферменты Asel, HindIII, Nsil и Ndel использовали отдельно для каждого гидролизата (New England Biolabs, Ipswich, MA). Рестриктирующие ферменты Notl и ApaLI использовали вместе для двойного гидролиза (New England Biolabs, Ipswih, MA). Кроме того, позитивный по гибридизации контрольный образец получали путем объединения плазмидной ДНК, pDAB9582, с геномной ДНК нетрансгенной сои сорта Maverick. «Коктейль» из плазмидной ДНК/геномной ДНК гидролизовали с применением тех же самых методов и с использованием того же самого рестриктирующего фермента, которые были выбраны для тест-образцов.
После инкубирования гидролизатов в течение ночи добавляли 25 мкл раствора Quiсk-Precip Plus (EdgeBiosystems, Gaithersburg, MD), и гидролизованные образцы ДНК осаждали изопропанолом. Осажденную ДНК ресуспендировали в 15 мкл 1× буфера для загрузки (0,01% бромфенолового синего, 10,0 мМ EDTA, 10,0% глицерина, 1,0 мМ триса, pH 7,5). Образцы ДНК и маркеры молекулярной массы подвергали электрофорезу в 0,85%> агарозных гелях с 0,4× буфера TAE (Fisher Scientific, Pittsburgh, PA) при 35 вольт приблизительно в течение 18-22 часов для разделения фрагментов. Гели окрашивали этидийбромидом (Invitrogen, Carlsbad, CA), и ДНК визуализировали путем облучения ультрафиолетом (УФ).
Пример 4.3: Перенос с помощью Саузерн-блот-анализа и обработка мембраны
Саузерн-блот-анализ проводили, в основном, как описано в публикации Memelink, et al. (1994). Вкратце, после электрофоретического разделения и визуализации ДНК-фрагментов, гели подвергали депуринизации путем добавления 0,25M HCl приблизительно в течение 20 минут, а затем обрабатывали денатурирующим раствором (0,4 M NaOH, 1,5 M NaCl) приблизительно в течение 30 минут и нейтрализующим раствором (1,5 M NaCl, 0,5 M триса, pH 7,5) в течение по меньшей мере 30 минут. Перенос с помощью Саузерн-блот-анализа проводили в течение ночи на найлоновых мембранах с использованием капиллярной системы, содержащей 10×SSC. После переноса, ДНК связывали с мембраной с использованием реагентов для перекрестного сшивания при УФ-излучении, а затем мембрану быстро промывали 2×SSC-раствором. С применением такого способа получали Саузерн-блот-мембраны, готовые для гибридизации.
Пример 4.4: Мечение ДНК-зонда и гибридизация
ДНК-фрагменты, связанные с найлоновой мембраной, детектировали с использованием меченого зонда (таблица 6). Зонды получали путем ПЦР-включения нуклеотида, меченного дигоксигенином (DIG), [DIG-11]-dUTP, в ДНК-фрагмент, амплифицированный из плазмиды pDAB9582 с использованием праймеров, специфичных к генным элементам. Генерирование ДНК-зондов путем ПЦР-синтеза осуществляли с использованием набора для синтеза ПЦР-зонда, меченного DIG (Roche Diagnostics, Indianapolis, IN), в соответствии с процедурами, рекомендованными производителями.
Меченые зонды анализировали с помощью электрофореза в агарозном геле для определения их качества и количества. Затем нужное количество меченого зонда использовали для гибридизации с ДНК-мишенью на найлоновых мембранах в целях детектирования специфических фрагментов методами, в основном, описанными для получения раствора DIG EASY HYB (Roche Diagnostics, Indianapolis, IN). Вкратце, блоты на найлоновой мембране, содержащие иммобилизованную ДНК, быстро промывали 2×SSC и предварительно гибридизовали с 20-25 мл предварительно нагретого раствора DIG EASY HYB в сосудах для гибридизации приблизительно при 45-55°C в течение приблизительно 2 часов в печи для гибридизации. Затем раствор для предварительной гибридизации декантировали и заменяли на ~15 мл предварительно нагретого раствора DIG EASY HYB, содержащего нужное количество специфических зондов, денатурированных путем нагревания в термоячейке в течение приблизительно пяти минут. Затем проводили стадию гибридизации приблизительно при 45-55°C в течение ночи в печи для гибридизации.
По окончании гибридизации зонда, растворы DIG EASY HYB, содержащие зонды, декантировали в очищенные пробирки и хранили приблизительно при -20°C. Эти зонды могут быть повторно использованы два раза в соответствующей процедуре, рекомендованной производителями. Блоты на мембранах быстро ополаскивали и два раза промывали в очищенных пластиковых контейнерах промывочным буфером в условиях низкой жесткости (2×SSC, 0,1% ДСН) приблизительно в течение пяти минут при комнатной температуре, а затем два раза промывали промывочным буфером в условиях высокой жесткости (0,1×SSC, 0,1% ДСН), каждый раз в течение 15 минут приблизительно при 65°С. Блоты на мембранах быстро промывали 1×малеиновокислотным буфером, взятым из набора промывочных и блокирующих буферов DIG (Roche Diagnostics, Indianapolis, IN), приблизительно в течение 5 минут. Затем проводили блокирование в 1×блокирующем буфере в течение 2 часов и инкубирование с антителом против DIG-AP (щелочной фосфатазы) (Roche Diagnostics, Indianapolis, ΓΝ) в 1× блокирующем буфере в течение минимум 30 минут. После 2-3 промывок 1× промывочным буфером, специфические ДНК-зонды оставались связанными с блотами на мембране, и DIG-меченные ДНК-стандарты визуализировали с использованием хемилюминесцентной системы детектирования нуклеиновых кислот (CDP-STAR) (Roche Diagnostics, Indianapolis, IN)) в соответствии с рекомендациями производителей. Блоты экспонировали с хемилюминесцентной пленкой один или несколько раз для детектирования гибридизующихся фрагментов и для визуализации стандартов молекулярных масс. Пленки проявляли в проявителе пленок ALL-PRO 100 PLUS (Konica Minolta, Osaka, Japan), и изображения сканировали. Число и размер детектированных полос были задокументированы для каждого зонда. DIG-меченный маркер молекулярной массы ДНК II (DIG MWM II) и DIG-меченный маркер молекулярной массы ДНК VII (DIG MWM VII) визуально наблюдались после детектирования DIG, как описано в настоящей заявке, а поэтому они были использованы для определения размера гибридизующегося фрагмента на Саузерн-блотах.
Локализация и длина зондов, используемых в Саузерн-блот-анализе
Пример 4.5: Результаты Саузерн-блот-анализа
Размеры предполагаемых и наблюдаемых фрагментов, а также конкретные гидролизаты и зонд, где указанные гидролизаты и зонд были получены на основе известных рестрикционных сайтов ферментов для PTU сrylAc и сrylF представлены в таблице 7. Исходя из этих гидролизатов и гибридизации были идентифицированы два типа фрагментов: внутренние фрагменты, в которых известные сайты ферментов фланкируют область зонда и полностью входят в состав инсерционной области PTU crylAc и crylF; и граничные фрагменты, в которых один известный сайт фермента локализован на одном конце области зонда, а другой сайт, как и ожидалось, локализован в геноме сои. Размеры граничных фрагментов варьируются в зависимости от типа трансформанта, поскольку в большинстве случаев, сайты интеграции ДНК-фрагментов являются уникальными для каждого трансформанта. Граничные фрагменты представляет собой средство для определения локализации рестрикционных сайтов ферментов по отношению к интегрированной ДНК и для оценки числа ДНК-вставок. Саузерн-блот-анализы, проводимые на многих поколениях сои, содержащей трансформант pDAB9582.816.15.1, давали результаты, которые позволяют предположить, что низкокопийные интактные PTU crylAc и crylF, происходящие от плазмиды pDAB9582, были встроены в геном трансформанта сои pDAB9582.816.15.1.
Предсказанные и наблюдаемые гибридизующиеся фрагменты в Саузерн-блот-анализе
1. Размеры предполагаемых фрагментов получены на основе карты плазмиды pDAB9582. 2. Размеры наблюдаемых фрагментов были вычислены приблизительно по данным, полученным в этих анализах и по указанным размерам фрагментов маркера II и маркера VII молекулярной массы DIG-меченной ДНК.
Рестриктирующие ферменты Asel и Nsil связываются с уникальными рестрикционными сайтами в плазмиде pDAB9582 и расщепляют их. Затем, эти ферменты отбирали для характеризации вставки гена сrylAc в трансформанте сои pDAB9582.816.15.1. Было предсказано, что граничные фрагменты размером >7286 п.о. или >9479 п.о. гибридизуются с зондом после гидролиза ферментами Asel и Nsil, соответственно (таблица 7). Одиночные полосы гибридизации сrylAc размером приблизительно 8500 п.о. и >10000 п.о. наблюдались в случае использования ферментов Asel и Nsil, соответственно. Гибридизация зонда с полосами такого размера, предположительно, указывает на присутствие одного сайта инсерции гена сrylAc в геноме трансформанта сои pDAB9582.816.15.1. Рестриктирующие ферменты Notl и ApaLl были выбраны для двойного гидролиза и выделения фрагмента, содержащего транскрипционную единицу сrylAc растения (PTU; промотор/ген/терминатор)(таблица 7). Предсказанные 4550 п.о. - фрагменты наблюдались в случае использования зонда после двойного гидролиза ферментами Notl и ApaLl. Результаты, полученные при ферментативном гидролизе образцов pDAB9582.816.15.1 с последующей гибридизацией зонда показали, что интактная PTU сrylAc, происходящая от плазмиды pDAB9582, была встроена в геном трансформанта сои pDAB9582.816.15.1.
Рестриктирующие ферменты Ndel и Nsil связываются с рестрикционными сайтами в плазмиде pDAB9582 и расщепляют их. Затем, эти ферменты отбирали для характеризации вставки гена сrylF в трансформанте сои pDAB9582.816.15.1. Было предсказано, что граничные фрагменты размером >5569 п.о. и >9479 п.о. гибридизуются с зондом после гидролиза ферментами Ndel и Nsil, соответственно (таблица 7). Одиночные полосы гибридизации сrylF размером ~7500 п.о. и >10000 п.о. наблюдались в случае использования ферментов Ndel и Nsil, соответственно. Гибридизация зонда с полосами такого размера, предположительно, указывает на присутствие одного сайта инсерции гена сrylF в геноме трансформанта сои pDAB9582.816.15.1. Рестриктирующий фермент HindIII был выбран для выделения фрагмента, содержащего транскрипционную единицу сrylF растения (PTU; промотор/ген/терминатор)(таблица 7). Предсказанный 7732 п.о. - фрагмент наблюдался в случае использования зонда после гидролиза ферментом HindIII. Результаты, полученные при ферментативном гидролизе образцов pDAB9582.816.15.1 с последующей гибридизацией зонда показали, что интактная PTU сrylF, происходящая от плазмиды pDAB9582, была встроена в геном трансформанта сои pDAB9582.816.15.1.
Пример 4.6: Отсутствие последовательностей остова
Саузерн-блот-анализ также проводили в целях подтверждения отсутствия гена резистентности к спектиномицину (specK), элемента Ori Rep и белка инициации репликации trfA (элемента trf A) в трансформанте сои pDAB9582.816.15.1. Как и ожидалось, в образцах трансформанта сои pDAB9582.816.15.1, какой-либо специфической гибридизации с геном резистентности к спектиномицину, с элементом Ori Rep или с элементом trfA не наблюдалось в том случае, если в Саузерн-блот-анализ были включены соответствующие позитивный контроль (с pDAB9582, добавленной к геномной ДНК Maverick) и негативный контроль (геномная ДНК Maverick). После гидролиза ферментом Nsil и гибридизации со specR-специфическим зондом, в образце позитивного контроля (с pDAB9582, добавленной к геномной ДНК Maverick) наблюдалась одна ожидаемая полоса размером 15320 п.о.). SpecR-зонд не гибридизовался с образцами негативного контроля и с трансформантом сои pDAB9582.816.15.1. Аналогичным образом, в образце позитивного контроля (с pDAB9582 плюс Maverick) детектировалась одна ожидаемая полоса размером 15320 п.о., но эта полоса отсутствовала в образцах негативного контроля и в трансформанте сои pDAB9582.816.15.1 после Nsil-гидролиза и гибридизации с trfA-зондом. Другая ожидаемая полоса размером 5329 п.о. детектировалась в образце позитивного контроля (с pDAB9582, добавленной к геномной ДНК Maverick), но отсутствовала в образцах негативного контроля и в трансформанте сои pDAB9582.816.15.1 после Ndel-гидролиза и гибридизации с OriRep-специфическим зондом. Эти данные указывают на отсутствие гена резистентности к спектиномицину, элемента Ori Rep и белка инициации репликации trfA в трансформанте сои pDAB9582.816.15.1.
Пример 5: Испытания для оценки агрономических показателей и испытания в полевых условиях на урожайность и устойчивость к гербицидам
Было проведено несколько агрономических испытаний для сравнения агрономических показателей трансформанта сои pDAB9582.816.15.1 и нулевой изолинии - Maverick. Большинство испытаний в полевых условиях проводили в различных географических регионах в Соединенных Штатах, где культивировали сорт сои, содержащей трансген pDAB9582.816.15.1. Были проведены дополнительные испытания в полевых условиях за пределами этих регионов, и отобранные сорта сои, содержащие трансформант pDAB9582.816.15.1, помещали в потенциально стрессовые условия, которые наблюдаются при выращивании культур в регионах, менее благоприятных для выращивания сои. Из-за изменения окружающей среды при проведении небольшого числа испытаний в полевых условиях, на некоторых участках испытания были прерваны.
Эксперимент проводили по схеме полных рандомизированных блоков с двумя повторностями для каждого расположения делянки. Было использовано восемь делянок, включающих трансформант сои pDAB9582.816.15.1. Каждая делянка состояла из двух рядов длиной 12,5 футов, а всего было засеяно 30 дюймов. В течение всего сезона, полевые делянки возделывали в соответствии с обычной агрономической практикой и с уничтожением сорняков.
Семена для испытаний получали в зимнем питомнике в Пуэрто-Рико. Смена трансформанта сои pDAB9582.816.15.1 и Maverick выращивали в том же самом питомнике и обрабатывали аналогичным образом для минимизации какого-либо различия в источнике происхождения семян. Затем семена возвращали в Северную Америку, где их упаковывали и распределяли по различным регионам для посева. В течение всего сезона оценивали различные агрономические свойства. Свойства и стадии роста, если для них были получены данные, указаны в таблице 8.
Список агрономических показателей, определенных в испытаниях в полевых условиях, проводимых для сравнения трансформантов сои pDAB9582.816.15.1 с растениями Maverick
По окончании вегетационного периода, данные, полученные со всех участков, объединяли и проводили анализ по всем участкам. Анализ данных проводили с использованием JMP® Pro 9.0.3 (SAS, Cary, NC). Для анализа использовали смешанную модель, где элемент (участок) рассматривался как фиксированный эффект, а расположение, то есть, расположение блока, и повторность рассматривались как случайные эффекты. Исходя из этого анализа вычисляли величины для различных показателей культур с применением метода наименьших квадратов как показано в таблице 9. В случае переменных, для которых измеряли значимый эффект элемента, осуществляли разделение средних с применением t-критерия Стъюдента в целях проведения сравнения между растениями Maverick и трансформантами сои pDAB9582.816.15.1 сои. Величина вероятности для определения значимости составляла p=0,05.
Величины, определенные с применением метода наименьших квадратов по результатам анализа для всех участков в целях сравнения трансформантов сои pDAB9582.816.15.1 и растений Maverick. Для каждого признака, величины, за которыми не стоит та же самая буква, значимо отличаются по t-критерию Стьюдента (р=0,05).
Все оцененные признаки, за исключением таких параметров, как дни до цветения, созревание и масса 100 семян, были одинаковыми для трансформанта сои pDAB9582.816.15.1 и для растения сорта Maverick. Трансформант сои pDAB.816.15.1 зацветал приблизительно на 2 дня позже, чем растение сорта Maverick. Такое различие в 2 дня не является слишком существенным для фермеров и не влияет на продуктивность данной культуры. Делянки рассматривались как цветущие, если приблизительно 50% растений на делянке имело распустившиеся цветки. Трансформант сои pDAB9582.816.15.1 также созревал на один день позже, чем растение сорта Maverick в конце сезона, но с агрономической точки зрения, такое замедление созревания не дает значимого эффекта, который мог бы влиять на продуктивность данной культуры. Аналогичнным образом, масса 100 семя трансформанта сои pDAB9582.816.15.1 статистически отличалась от массы 100 семян растения сорта Maverick, но такое отличие не приводило к значительному снижению урожайности. Полученные результаты показали, что трансформант сои pDAB9582.816.15.1 может развиваться не так, как растение сорта Maverick с точки зрения некоторых агрономических показателей, однако, такие различия являются минимальными и не выходят за пределы обычных показателей соевых культур при их возделывании на стандартных фермах.
Для проведения теста на устойчивость трансформанта сои pDAB9582.816.15.1 к гербициду, этот трансформант высевали на опытные поля штата Санта-Исабель, Пуэрто Рико. Растение сорта Maverick, которое было впервые трансформировано для продуцирования трансформанта сои pDAB9582.816.15.1, высевали в каждый питомник и использовали в экспериментах в качестве контроля. Семена для T3-питомника получали после отбора семян растения поколения T2 из одной клетки, а семена для T4-питомника получали после отбора семян поколения T3 из одной клетки. Четыре линии трансформанта тестировали для каждого поколения. Каждую линию высевали на делянку шириной в 4 ряда и длиной 7,5 футов. Расстояние между рядами составляло 30 дюймов. Растения на делянках выращивали при освещении приблизительно в течение 2,5 недель для компенсации короткого светового дня в Пуэрто Рико. Каждый питомник опрыскивали глюфозинатом в норме 411 г э.к./га. Одну делянку с контрольными растениями Maverick опрыскивали глюфозинатом в той же норме, а другую делянку не опрыскивали и использовали в качестве контроля для сравнения с трансформантом. Трансформант сои pDAB9582.816.15.1 обнаруживал устойчивость к гербициду глюфозинату. В противоположность этому, ни одно из растений Maverick не было устойчивыми к обработке гербицидами.
Пример 6: Характеризация инсектицидной активности трансформанта pDAB9582.816.15.1
Испытания в полевых условиях и в теплице проводили для характеризации уровня защиты растения, обеспечиваемой белками Cry1Ac и Cry1F в трансформанте сои pDAB9582.816.15.1, от вредителей сои, включая чешуекрылых насекомых (Lepidopteran), включая Anticarsia gemmatalis (листовертку фасоли), Pseudoplusia includens (соевую пяденицу), Spodopterafrugiperda (капустную совку) и Heliothis virescens (табачную совку).
Испытания в теплице проводили приблизительно на 4-недельных растениях. Пятнадцать растений использовали для оценки трансформантов сои pDAB9582.816.15.1 и контрольных растений Maverick. Для испытания на поражение насекомыми каждого тестируемого вида (новорожденными личинками A. gemmatalis, P.includes и S. frugiperda), из каждого растения вырезали три листовых диска, и всего было использовано 45 листовых дисков на растение и на вид насекомого. Вырезанный перфоратором 1,4 см-лист, зараженный одной новорожденной личинкой, помещали на тест-площадку на поверхности 2% водного агара и закрывали перфорированной пластиковой крышкой.
Смертность насекомых и степень повреждения листьев оценивали после заражения личинками. Личинки, которые не реагировали на легкое прикасание, считались погибшими. Смертность насекомых, которые находились на растительном материале, содержащем трансформант сои pDAB9582.816.15.1, была значительно выше (смертность для Spodopterafrugiperda составляла 86%, смертность для Anticarsia gemmatalis составляла 100% и смертность для Pseudoplusia includens составляла 100%), чем смертность насекомых, присутствующих на контрольных растениях Maverick. Таблица 10. Повреждение листьев определяли визуально путем оценки процента поедания листового диска насекомыми. Результаты, полученные путем проведения экспериментов в теплице, показали, что на трансформанте сои pDAB9582.816.15.1 наблюдалось значительно меньшее повреждение листьев и значительно большая смертность насекомых, чем на контрольных растениях Maverick, зараженных личинками Anticarsia gemmatalis, Pseudoplusia includens и Spodoptera frugiperda.
Оценку эффективности растений-трансформантов сои pDAB9582.816.15.1, выращенных в полевых условиях, проводили путем сбора образцов листьев с делянок питомника растений со зрелыми семенами в Санта-Исабель, Пуэрто Рико, с последующей отсылкой этих листьев в Индианаполис, IN, для биоанализа. Делянка питомника для растений-трансформантов сои pDAB9582.816.15.1 поколения T3 была засеяна приблизительно 180 растениями в четыре ряда. Каждый ряд имел длину 2,3 м, а расстояние между рядами составляло 76,2 см, причем, в каждом ряду, расстояние между отдельными растениями составляло 5,1 см. Биоанализы проводили на одном полностью сформировавшемся тройчатом листе главного стебля, имеющего приблизительно четыре узла, расположенных ниже меристемы. Ткань тройчатого листа вырезали у 10 отдельных растений сои, несущих pDAB9582.816.15.1, и у 10 отдельных растений Maverick. Затем листья упаковывали и отдавали в лабораторию. В этой лаборатории, из каждого тройчатого листа перфоратором вырезали один или два диска диаметром 3,33 см, и всего было получено 16 листовых дисков. Каждый листовой диск помещали на тест-площадку на поверхности 2% агара, заражали одной новорожденной личинкой S. frugiperda, и закрывали перфорированной пластиковой крышкой. Листовые диски хранили в камере с регулируемыми условиями окружающей среды в течение 7 дней, а затем оценивали количество погибших насекомых и степень повреждения листа. Личинки, которые не реагировали на легкое прикасание, считались погибшими. Повреждение листьев определяли визуально путем оценки процента поедания листового диска насекомыми.
Смертность насекомых и степень повреждения листьев оценивали после заражения личинками. Личинки, которые не реагировали на легкое прикасание, считались погибшими. Смертность насекомых, которые находились на растительном материале, содержащем трансформант сои pDAB9582.816.15.1, была значительно выше (смертность для Spodopterafrugiperda составляла 68%), чем смертность насекомых, присутствующих на контрольных растениях Maverick (смертность для Spodopterafrugiperda составляла 0%). Таблица 10. Повреждение листьев определяли визуально путем оценки процента поедания листового диска насекомыми. Результаты, полученные путем проведения биоанализа листьев, показали, что личинки Spodoptera frugiperda, находящиеся на трансформанте сои pDAB9582.816.15.1, в значительно меньшей степени повреждали листья и имели более низкую выживаемость (также определяемую как более высокая смертность насекомых), чем личинки Spodoptera frugiperda, находящиеся на контрольных растениях Maverick.
Эффективность трансформантов сои pDAB9582.816.15.1 оценивали в первом испытании в полевых условиях (первое испытание в полевых условиях описано в таблице 10). Семена сои поколения T4, содержащие трансформант сои pDAB9582.816.15.1, и семена нетрансформированной сои сорта Maverick высевали на произвольно расположенных делянках с 2 повторностями. Каждая делянка-дубликат имела 2 ряда длиной 2,3 м, а расстояние между рядами составляло 0,76 м. Было высажено 40 семян на ряд с расстоянием между растениями этого ряда 5,7 см. В соответствии с планом испытаний, одну делянку из двух делянок-дубликатов опрыскивали гербицидом глюфозинатом в норме 411 г э.к./га, а другую делянку из двух дубликатов не опрыскивали, и в результате чего, только неопрысканные растения Maverick оставались живыми и подвергались биоанализу.
Листья для биоанализа собирали с растений сои на стадии роста R2. За несколько дней до сбора листьев для биоанализа, из одного нижнего листового узла (самого старого листа) на одних и тех же растениях вырезали перфоратором листовые диски, и эти диски анализировали на экспрессию белков Cry1Ac и CryIF ELISA-методом, аналогичным методу, описанному в примере 2 (таблица 12). Полностью сформировавшиеся тройчатые листья основного стебля, не обнаруживающие каких-либо признаков поражения или обесцвечивания и находящиеся в четырех узлах ниже меристемы, отрезали и использовали для биоанализа. От каждого из 15 растений отрезали один тройчатый лист в качестве дубликата. Эти листья хранили при 15°C и подвергали биоанализу. Листочки каждого тройчатого листа отрезали, после чего из центра каждого листочка трансформанта сои pDAB9582.816.15.1 вырезали один диск диаметром 3,33 см, а из каждого листочка растения Maverick вырезали два диска диаметром 3,33 см. Эти листовые диски помещали в отдельные маркированные лунки 32-луночных пластиковых планшетов для биоанализа, где каждая лунка содержала тонкий слой агара. На каждый листовой диск сажали одну новорожденную личинку P. includens, новорожденную личинку A. gemmatalis или новорожденную личинку S. frugiperda. Планшеты для биоанализа закрывали клейкими пластиковыми листами, которые были перфорированы для обеспечения вентиляции. Ткани листьев трансформанта сои pDAB9582.816.15.1 заражали 30 личинками каждого вида, и такими же 30 личинками заражали ткани листьев растения Maverick. Пластиковые планшеты, содержащие зараженные листовые диски, выдерживали при 25°C и при относительной влажности 40% (RH). Через 7 дней, личинки были идентифицированы как погибшие личинки (то есть, они не двигались при их стимуляции острым зондом), как личинки, которые не развивались (они имели меньший размер, чем личинки, присутствующие на листьях растения Maverick), или как живые личинки (то есть, они имели нормальный размер и были чувствительны к раздражителю).
Смертность насекомых оценивали после заражения личинками. Смертность насекомых, которые находились на растительном материале, содержащем трансформант сои pDAB9582.816.15.1, была значительно выше (смертность для Spodopterafrugiperda составляла 97%, смертность для Anticarsia gemmatalis составляла 100%, и смертность для Pseudoplusia includens составляла 100%), чем смертность насекомых, присутствующих на контрольных растениях Maverick. Таблица 10. Результаты, полученные путем проведения биоанализа листьев, показали, что личинки Spodoptera frugiperda, Anticarsia gemmatalis и Pseudoplusia includens, находящиеся на трансформанте сои pDAB9582.816.15.1, значительно меньше повреждали листья и имели более низкую выживаемость (также определяемую как более высокая смертность насекомых), чем личинки Spodoptera frugiperda, Anticarsia gemmatalis и Pseudoplusia includens, находящиеся на контрольных растениях Maverick.
Эффективность трансформанта сои pDAB9582.816.15.1 оценивали во втором отдельном испытании в полевых условиях (второе испытание в полевых условиях описано в таблице 10). Семена сои поколения T4, содержащие трансформант сои pDAB9582.816.15.1, и семена нетрансформированной сои сорта Maverick высевали на произвольно расположенных делянках с 4 повторностями. Каждая дублированная делянка имела 4 ряда длиной 6,1 м, а расстояние между рядами составляло 1,02 м. Было высажено 160 семян на ряд с расстоянием между растениями этого ряда 3,8 см. Между испытуемыми делянками и вокруг них высаживали дополнительные ряды растений Maverick для приманки природных популяций насекомых-вредителей.
Листья для биоанализа собирали с растений сои на стадии роста R2. Листья для дополнительного биоанализа собирали с тех же растений сои на стадии роста R5. За несколько дней до сбора листьев для биоанализа, из одного нижнего листового узла на одних и тех же растениях вырезали перфоратором листовые диски, и эти диски анализировали на экспрессию белков Cry1Ac и CryIF ELISA-методом, аналогичным методу, описанному в примере 2 (таблица 12). Полностью сформировавшиеся тройчатые листья основного стебля, не обнаруживающие каких-либо признаков поражения или обесцвечивания и находящиеся в четырех узлах ниже меристемы, отрезали и использовали для биоанализа. От каждого из 15 растений отрезали один тройчатый лист в качестве дубликата, где 4 тройчатых листа каждого дубликата использовали для биоанализа на P. includens, 4 тройчатых листа каждого дубликата использовали для биоанализа на S. frugiperda, 4 тройчатых листа на дубликат использовали для биоанализа на H. virescens и 3 тройчатых листа каждого дубликата использовали для биоанализа на A. gemmatalis. Затем, от каждого тройчатого листа отрезали два боковых листочка, и эти листочки помещали в отдельные маркированные чашки Петри, содержащие тонкий слой агара. На каждый листочек сажали личинки в двух других возрастных стадиях, а именно, P. includens, A. gemmatalis, S. frugiperda или H. virescens. Ткани листьев трансформанта сои pDAB9582.816.15.1 и ткани листьев растения Maverick заражали 64 личинками P. includens, S. frugiperda и H. virescens. Ткани листьев трансформанта сои pDAB9582.816.15.1 и контрольных растений Maverick заражали 48 личинками A. gemmatalis. Чашки Петри, содержащие зараженные листочки, закрывали крышками и выдерживали при 25°C и при относительной влажности 40% (RH). Через 4 дня, личинки были идентифицированы как погибшие личинки (то есть, они не двигались при их стимуляции острым зондом), как нежизнеспособные личинки (они реагировали на раздражитель, но были неспособны самостоятельно принимать прежнее положение, если их переворачивали), как личинки, которые не развивались (они имели меньший размер, чем личинки, присутствующие на листьях растения Maverick), или как живые личинки (то есть, они имели нормальный размер и были чувствительны к раздражителю).
Процедура биоанализа растений на стадии роста R5 была аналогична процедуре биоанализа растений на стадии роста R2, за исключением того, что для биоанализа S. frugiperda и H. virescens, все три листочка отрезали от каждого тройчатого листа, и на каждый листочек сажали одну личинку во второй возрастной стадии. В результате этого, на листочках трансформанта сои pDAB9582.816.15.1 и растения Maverick находилось 48 личинок S. frugiperda и H. virescens.
Смертность насекомых оценивали после заражения личинками для биоанализов листьев растения на стадиях роста R2 и R5. Смертность насекомых, которые находились на растительном материале, содержащем трансформант сои pDAB9582.816.15.1, была значительно выше (смертность для Spodopterafrugiperda в биоанализе листьев на стадии R2 составляла 69%, а в биоанализе на стадии R5 составляла 54%; смертность для Anticarsia gemmatalis в биоанализе листьев на стадиях R2 и R5 составляла 100%; смертность для Heliothis virescens в биоанализе листьев на стадии R2 составляла 95%, а на стадии R5-70%; а смертность для Pseudoplusia includens в биоанализе листьев на стадии R2 составляла 100%, а на стадии R5-98%), чем смертность насекомых, присутствующих на контрольных растениях Maverick. Таблица 10. Результаты, полученные в биоанализе листьев, показали, что личинки Spodoptera frugiperda, Anticarsia gemmatalis, Pseudoplusia includens и Heliothis virescens, находящиеся на трансформанте сои pDAB9582.816.15.1, значительно меньше повреждали листья и имели более низкую выживаемость (также определяемую как более высокая смертность насекомых), чем личинки Spodoptera frugiperda, Anticarsia gemmatalis, Pseudoplusia includens и Heliothis virescens, находящиеся на контрольных растениях Maverick.
Бобы сои собирали с растений трансформанта сои pDAB9582.816.15.1 и с растений Maverick, выращенных во втором испытании в полевых условиях, и подвергали биоанализу на выживаемость личинок H. virescens. Два самых верхних боба на главном стебле отрезали у шести растений, выбранных произвольно в каждой дублированной делянке. Каждую партию бобов помещали в пластиковые чашки Петри и заражали одной личинкой H. virescens во второй возрастной стадии. Этот эксперимент проводили так, чтобы на партии бобов, собранных с трансформанта сои pDAB9582.816.15.1 и с контрольных растений Maverick, находилось 24 личинки. Чашки Петри выдерживали в тех же условиях, которые были описаны ранее для биоанализа, проводимого на отрезанных листьях. Через 2 дня оценивали выживаемость личинок H. virescens в соответствии с процедурой, описанной для биоанализов листьев.
Смертность насекомых оценивали после заражения соевых бобов личинками. Смертность насекомых, которые находились на соевых бобах, содержащих трансформант сои pDAB9582.816.15.1, была значительно выше (смертность для Heliothis virescens в биоанализе соевых бобов составляла 50%), чем смертность насекомых, присутствующих на соевых бобах контрольных растений Maverick. Таблица 10. Результаты, полученные в этом анализе на соевых бобах, выращенных в полевых условиях, показали, что личинки Heliothis virescens, находящиеся на трансформанте сои pDAB9582.816.15.1, имели значительно более низкую выживаемость (также определяемую как более высокая смертность насекомых), чем личинки Heliothis virescens, находящиеся на контрольных растениях Maverick.
Верхушки (самую верхнюю часть главного стебля, несущую два или три сформировавшихся тройчатых листа и гроздь незрелых бобов) трансформанта сои pDAB9582.816.15.1 и контрольных растений сои Maverick заражали в полевых условиях яйцами H. virescens. Части сырной салфетки, несущей приблизительно двадцать яиц H. virescens, помещали на верхушки пяти произвольно выбранных растений в каждой дублированной делянке (всего было протестировано 20 растений), и закрепляли в определенном месте пластиковым зажимом для бумаг. На верхушки растений надевали мешки из ситовой ткани, а свободные концы сетчатого мешка завязывали узлом вокруг основного стебля. Мониторинг вылупления из яиц проводили ежедневно в репрезентативной партии сетчатых мешков. После вылупления личинок из всех яиц, в каждом сетчатом мешке, надетом на каждое из пяти растений, подсчитывали число живых личинок H. virescens.
Среднее число живых личинок насекомых, помещенных на верхушки растений-трансформантов сои pDAB9582.816.15.1, было значительно ниже (0,00 насекомых Heliothis virescens в биоанализе верхушек растения сои), чем число личинок насекомых, помещенных на верхушки контрольных растений Maverick. Таблица 11. Результаты, полученные в этом анализе показали, что личинки Heliothis virescens, находящиеся на трансформанте сои pDAB9582.816.15.1, имели значительно более низкую выживаемость, чем личинки Heliothis virescens, находящиеся на контрольных растениях Maverick.
Число природных личинок P. includens в испытаниях на делянках подсчитывали один раз в неделю в течение четырех недель. Образцы брали с двух центральных рядов каждой делянки. В произвольно выбранных участках между центральными двумя рядами помещали белый лоскут ткани размером 91 см × 91 см. Растения, находящиеся на участке ряда, расположенного поблизости от одного края лоскута, перегибали через этот лоскут, а затем 15 раз трясли для стряхивания всех имеющихся там насекомых. Эту процедуру повторяли для ряда с противоположного края лоскута. Личинки оценивали по виду и размеру: личинки длиной <6 мм были определены как небольшие личинки, а личинки длиной ≥6 мм были определены как крупные личинки. Все насекомые были удалены с лоскута для последующего анализа. Затем лоскут переносили во второе, произвольно выбранное положение между двумя центральными рядами, и процесс взятия образцов повторяли, в результате чего получали два субобразца на делянку в каждый момент взятия образца.
Среднее число насекомых, подсчитанных в ряду длиной 1,82 м, засеянном трансформантом pDAB9582.816.15.1, было значительно ниже (0,00 насекомых Pseudoplusia includens), чем число насекомых, подсчитанных в ряду длиной 1,82 м, засеянном контрольными растениями Maverick. Таблица 11. Результаты, полученные в этом анализе, показали, что заражение трансформанта сои pDAB9582.816.15.1 личинками Pseudoplusia includens было значительно ниже, чем заражение личинками Pseudoplusia includens, находящимися на контрольных растениях Maverick.
Результаты, полученные в этих экспериментах с повторностями, показали, что личинки чешуекрылых (Lepidoptera), находящиеся на трансформанте сои pDAB9582.816.15.1, имели значительно более низкую выживаемость, чем личинки насекомых всех тестируемых видов, присутствующие на контрольных растениях Maverick. Таким образом, трансформант сои pDAB9582.816.15.1 обладал инсектицидной активностью по отношению к насекомым-вредителям данного широкого ряда.
Оценка числа погибших чешуекрылых насекомых, проводимая путем биоанализа листьев и бобов сои, полученных от трансформанта сои pDAB9582.816.15.1, по сравнению с контрольными растениями Maverick
Среднее число живых чешуекрылых насекомых, присутствующих на трансформанте сои pDAB9582.816.15.1 по сравнению с контрольными растениями Maverick.
Средний уровень экспрессии различных трансгенных белков, выделенных из растительных материалов трансформанта сои pDAB9582.816.15.1, используемых для биоанализа на инсектицидность
ткань
Пример 7: Предполагаемая последовательность трансформанта сои pDAB9582.816.15.1
SEQ ID NO: 14 представляет собой предполагаемую последовательность трансформанта сои pDAB9582.816.15.1. Эта последовательность содержит геномную 5'-фланкирующую последовательность, предполагаемую вставку T-цепи pDAB9582 и геномную 3'-фланкирующую последовательность. Что касается SEQ ID NO: 14, то остатки 1-1273 составляют геномную 5'-фланкирующую последовательность, остатки 1274-13658 представляют собой остатки вставки T-цепи pDAB9582, остатки 13659-13821 представляют собой остатки, образующиеся после реаранжировки плазмиды pDAB9582, а остатки 13822-15170 составляют 3'-фланкирующую последовательность. Таким образом, последовательность стыка или транзиция у 5'-конца вставки находится в положениях остатков 1273-1274 SEQ ID NO: 14. Последовательность стыка или транзиция у 3'-конца вставки находится в положениях остатков 13658-13659 SEQ ID NO: 14.
Следует отметить, что SEQ ID NO: 14 представляет собой предполагаемую последовательность трансформанта сои pDAB9582.816.15.1, и такая последовательность была собрана после выравнивания последовательностей SEQ ID NO: 1, SEQ ID NO: 2 и t-цепи pDAB9582. Фактическая последовательность вставки T-цепи трансформанта сои pDAB9582.816.15.1 может несколько отличаться от SEQ ID NO: 14. В процессе трансформации, а именно, введения вставки T-цепи в геном клетки растения, нередко образуются некоторые делеции или другие альтерации вставки. Кроме того, при ПЦР-амплификации могут возникать ошибки, которые могут приводить к незначительным ошибкам секвенирования. Так, например, перечисленные здесь фланкирующие последовательности были определены путем генерирования ампликонов из геномных ДНК сои, с последующим клонированием и секвенированием этих ампликонов. При этом, нередко, для выявления незначительных различий и небольших расхождений в последовательностях, полученных и определенных таким способом, может потребоваться проведение множества раундов амплификации, необходимых для продуцирования ампликона, длина которого является достаточной для его секвенирования из геномных ДНК. Специалисту в данной области очевидно и следует обратить внимание, что любые коррекции, которые необходимо проводить из-за ошибок или расхождений, часто встречающихся при секвенировании такого типа, входят в объем настоящего изобретения. Таким образом, рассматриваемый здесь релевантный сегмент плазмидной последовательности может содержать некоторые незначительные изменения. Так, например, растение, содержащее полинуклеотид, имеющий последовательность, которая в некоторой степени идентична рассматриваемой здесь последовательности вставки, входит в объем настоящего изобретения. Что касается идентичности последовательности SEQ ID NO: 14, то такая полинуклеотидная последовательность должна быть по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентична рассматриваемой или описанной здесь последовательности. Фланкирующие последовательности плюс последовательность вставки могут быть подтверждены как описано в разделе «Депонированные семена». Таким образом, могут быть идентифицированы некоторые различия между последовательностью SEQ ID NO: 14 и фактической вставкой T-цепи трансформанта сои pDAB9582.816.15.1.
При ознакомлении с проиллюстрированными и описанными здесь идеями изобретения, специалисту в данной области должно быть очевидно, что в структуру и детали описания настоящего изобретения могут быть внесены изменения, не выходящие за рамки основной концепции изобретения. Авторами заявлены все модификации, не выходящие за рамки существа и объема, определенного в прилагаемой формуле изобретения.
Все публикации и опубликованные патентные документы, цитируемые в настоящей заявке, вводятся в настоящее описание посредством ссылки так, как если бы каждая конкретная публикация или патентная заявка была конкретно и отдельно введена посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОБЪЕКТ СОИ 9582.814.19.1, ПРИДАЮЩИЙ УСТОЙЧИВОСТЬ К НАСЕКОМЫМ И УСТОЙЧИВОСТЬ К ГЕРБИЦИДАМ | 2012 |
|
RU2628099C2 |
| УСТОЙЧИВЫЙ К НАСЕКОМЫМ-ВРЕДИТЕЛЯМ И ТОЛЕРАНТНЫЙ К ГЕРБИЦИДАМ СЕЛЕКЦИОННЫЙ ГИБРИД ТРАНСФОРМАНТА СОИ pDAB9582.814.19.1 и pDAB4468.04.16.1 | 2012 |
|
RU2627161C2 |
| СПОСОБ ОБНАРУЖЕНИЯ ТРАНСФОРМАНТА СОИ рDAB9582.814.19.1 | 2012 |
|
RU2622079C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ ВЫЯВЛЕНИЯ РАСТЕНИЯ СОИ DBN8002 И СПОСОБ ЕЕ ВЫЯВЛЕНИЯ | 2019 |
|
RU2818368C2 |
| ДЕТЕКТИРОВАНИЕ AAD-12-СОБЫТИЯ 416 У СОИ | 2010 |
|
RU2573898C2 |
| РАСТЕНИЕ МАИСА DBN9936 И СПОСОБ ПРИМЕНЕНИЯ В ДЕТЕКТИРОВАНИИ ЕГО ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ | 2016 |
|
RU2707527C2 |
| СОБЫТИЕ 416 AAD-12, РОДСТВЕННЫЕ ЛИНИИ ТРАНСГЕННОЙ СОИ И ИХ СОБЫТИЕСПЕЦИФИЧНАЯ ИДЕНТИФИКАЦИЯ | 2010 |
|
RU2603252C2 |
| РАСТЕНИЯ-ТРАНСФОРМАНТЫ КУКУРУЗЫ PV-ZMIR13 (MON863) И КОМПОЗИЦИИ И СПОСОБЫ ИХ ОБНАРУЖЕНИЯ | 2003 |
|
RU2352638C2 |




















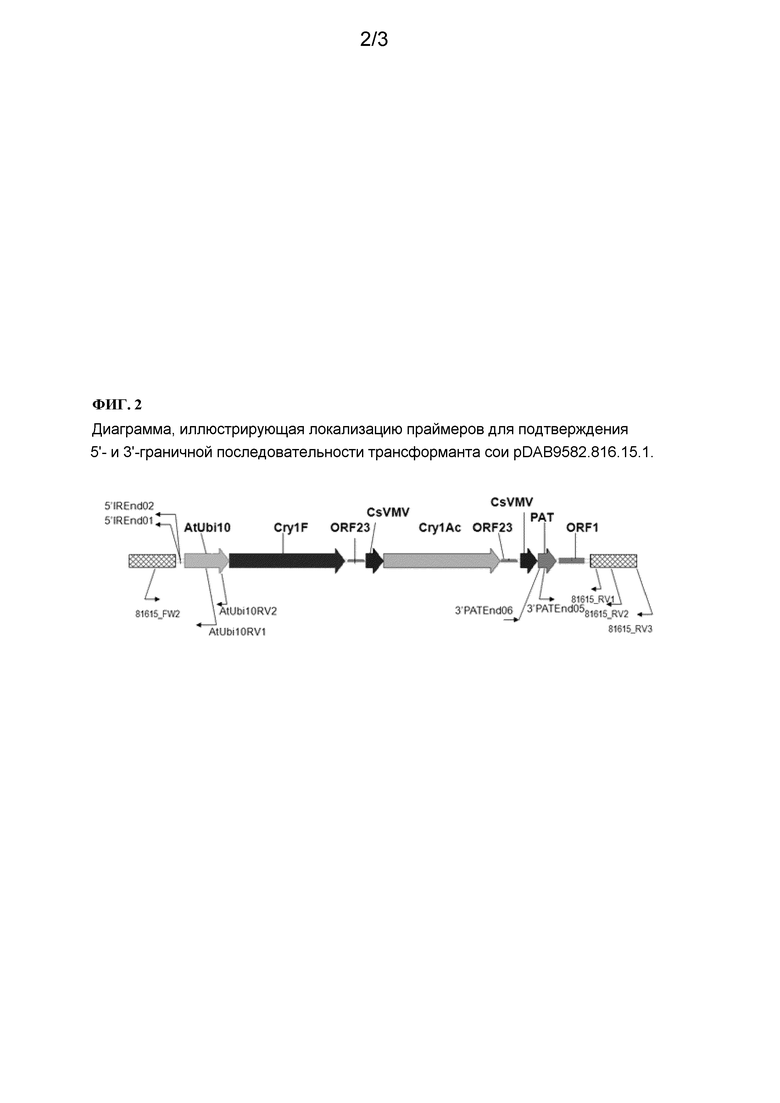

Изобретение относится к области биохимии, в частности к способу борьбы с насекомыми, выбранными из группы, состоящей из Pseudoplusia includens, Anticarsia gemmatalis, Spodoptera frugiperda и Heliothis virescens, а также к способу борьбы с сорняками сельскохозяйственной культуры сои, который включает обработку сельскохозяйственной культуры сои гербицидом глюфосинатом. Также раскрыты выделенная молекула ДНК для борьбы с вышеуказанными насекомыми, растение, семя, часть растения, товарный продукт, ее содержащие, а также способ получения вышеуказанного растения. Изобретение позволяет эффективно получать растение, устойчивое к насекомым и гербициду. 12 н. и 4 з.п. ф-лы, 3 ил., 12 табл., 7 пр.
1. Способ борьбы с насекомыми, который включает заражение насекомыми растений сои, резистентных к насекомым, где указанные растения сои содержат последовательность ДНК, состоящую из SEQ ID NO:14, таким образом осуществляя борьбу с насекомыми, где указанные насекомые выбраны из группы, состоящей из Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка).
2. Способ по п.1, где указанными насекомыми являются Pseudoplusia includens (соевая пяденица).
3. Способ по п.1, где указанными насекомыми являются Anticarsia gemmatalis (листовертка фасоли).
4. Способ по п.1, где указанными насекомыми являются Spodoptera frugiperda (капустная совка).
5. Способ по п.1, где указанными насекомыми являются Heliothis virescens (табачная совка).
6. Способ борьбы с сорняками сельскохозяйственной культуры сои, который включает обработку сельскохозяйственной культуры сои гербицидом глюфосинатом, где указанная сельскохозяйственная культура сои включает растения сои, содержащие последовательность ДНК, состоящую из SEQ ID NO:14.
7. Способ получения трансгенного растения сои, имеющего устойчивость к насекомым и устойчивость к гербицидам, где указанный способ включает скрещивание первого растения со вторым растением сои с получением третьего растения сои, где указанное первое растение сои содержит последовательность ДНК, состоящую из SEQ ID NO:14, и проведение анализа указанного третьего растения сои на присутствие последовательности ДНК, состоящей из SEQ ID NO:14, где указанные насекомые выбраны из группы, состоящей из Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка), и где указанным гербицидом является глюфосинат.
8. Выделенная молекула ДНК, состоящая из SEQ ID NO:14, для борьбы с насекомыми, где указанные насекомые выбраны из группы, состоящей из Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка).
9. Растение сои, которое являются резистентным к Pseudoplusia includens (соевой пяденице) и содержит последовательность ДНК, состоящую из SEQ ID NO:14.
10. Семя растения по п. 9 для борьбы с Pseudoplusia includens (соевая пяденица), где указанное семя содержит последовательность ДНК, состоящую из SEQ ID NO:14.
11. Товарный продукт, полученный из растения сои по п. 9, где указанный товарный продукт выбран из группы, состоящей из кормовой соевой муки, пищевой соевой муки и белкового концентрата сои, где указанный товарный продукт содержит последовательность ДНК, состоящую из SEQ ID NO:14, и где указанный товарный продукт является резистентным к Pseudoplusia includens (соевой пяденице).
12. Способ борьбы с насекомыми-вредителями соевых бобов, семян, кормовой муки или пищевой муки, который включает введение SEQ ID NO:14 в указанные бобы, семена, кормовую муку или пищевую муку, где указанные насекомые-вредители выбраны из группы, состоящей из Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка).
13. Семя сои для борьбы с насекомыми, содержащее в своем геноме последовательность ДНК, состоящую из SEQ ID NO:14, где указанные насекомые выбраны из группы, состоящей из Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка).
14. Растение сои, выращенное из семени сои по п.13, для борьбы с насекомыми, где указанное растение содержит последовательность ДНК, состоящую из SEQ ID NO:14, где указанные насекомые выбраны из группы, состоящей из Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка).
15. Часть растения сои по п.14 для борьбы с насекомыми, где указанная часть выбрана из группы, состоящей из пыльцы, овулей, цветков, побегов, корней и листьев, и где указанная часть содержит SEQ ID NO:14, где указанные насекомые выбраны из группы, состоящей из Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка).
16. Товарный продукт, полученный из растения сои по п.14, где указанный товарный продукт выбран из группы, состоящей из кормовой соевой муки и пищевой соевой муки, где указанный товарный продукт содержит последовательность ДНК, состоящую из SEQ ID NO:14, и где указанный товарный продукт является резистентным к Pseudoplusia includens (соевая пяденица), Anticarsia gemmatalis (листовертка фасоли), Spodoptera frugiperda (капустная совка) и Heliothis virescens (табачная совка).
| WO 2012075426 A1, 07.06.2012 | |||
| US 2009041887 A1, 12.02.2009 | |||
| WO 2012075429 A1, 07.06.2012 | |||
| СПОСОБЫ ТРАНСФОРМАЦИИ РАСТЕНИЙ ДЛЯ ЭКСПРЕССИИ ДЕЛЬТА-ЭНДОТОКСИНОВ BACILLUS THURINGIENSIS | 1999 |
|
RU2234531C2 |

