Перекрестная ссылка на родственную заявку
Настоящая заявка относится к патентным притязаниям по Предварительной Заявке США с номером USSN 61/507,331, поданной 13 июля 2011 г., содержание которой полностью включено в настоящий документ.
Область изобретения
Настоящее изобретение относится к молекулам нуклеиновой кислоты и белков, участвующих в синтезе тиоэфиров алканоил-кофермента-А, и к использованию молекул и белков нуклеиновой кислоты для инженерного биосинтеза каннабиоидов в растениях, микроорганизмах или бесклеточных системах и для получения конопли с повышенным или пониженным содержанием каннабиоидов.
Уровень техники
Конопля посевная L. (каннабис, конопля, марихуана) - это одно из самых распространенных древнейших одомашненных растений, которое в настоящее время используется в медицинских, пищевых, косметических и промышленных целях. Конопля также известна тем, что она применяется в качестве нелегального наркотического средства по причине содержания в ней психоактивных каннабиоидов (например, Δ9-тетрагидроканнабинол, Δ9-ТНС). Проводится исследование терапевтического применения каннабиоидов и прочих наркотических веществ, действующих через каннабиоидные рецепторы млекопитающих, для лечения различных состояний, например, хронических болей, рассеянного склероза и эпилепсии.
Биосинтез каннабиоидов происходит как в поликетидном, так и в терпеноидном метаболизме. Канабиоиды называются терпенофенолами или пренилированными поликетидами (Page J., Nagel J. (2006) Биосинтез терпенофенолов в хмеле и конопле. JT Romeo, ред. Интегративная растительная биохимия. Том 40. Elsevier, Оксфорд, стр. 179-210.). Биосинтез каннабиоидов в основном происходит в грандулярных трихомах, которые плотно покрывают женские цветы. Каннабиоиды образуются в биосинтетическом процессе, состоящем из трех этапов: образование поликетида, ароматическое пренилирование и циклизация (см. Фигуру 1 (Figure 1)).
Первый энзимный этап в биосинтезе каннабиоидов - это образование оливетоловой кислоты с помощью фермента поликетид-синтазы, которая катализирует конденсацию гексаноил-кофермента-А (СоА) тремя молекулами малонил-кофермента-А. Основные каннабиоиды, в т.ч. Δ9-тетрагидроканнабиноловая и каннабидиоловая кислоты, образуются из прекурсора гексаноил-кофермент-А, который является жирным акил-коферментом А средней цепи (см. Фигуру 1). Прочие каннабиоиды с разными боковыми цепями образуются из альфатических коферментов А разной длины (например, Δ9-тетрагидроканнабивариновая кислота образуется из затравки n-бутирил-кофермент-А).
Гексаноил-кофермент-А и прочие акил-кофермент-А-теоэфиры в растениях синтезируются акил-активирующими энзимами (ААЭ, также называемыми акил-кофермент-А-синтетазами), которые катализируют активность субстратов карбоновой кислоты с помощью АТР. Эти энзимы активизируются на разных карбоксилатных кислотах короткой, средней, длинной и очень длинной цепи, прекурсорах жасмоната, кислотах производных фенилпропаноида (например, коричной кислоты) и прочих органических кислотах, например, малоновой, ацетатной и лимонной. В природе найдено очень мало акил-кофермент-А-синтетаз средней цепи. Было выявлено, что три растительных энзима A. thaliana, ААЕ7, At4g05160 и At5g63380 образуют гексаноил-кофермент-А из гексаноата (Шнейдер К и др. (2005). Новый тип пероксимальной акил-кофермент-А-синтетазы из Arabidopsis thaliana обладает каталитической способностью активировать биосинтетические прекурсоры жасмоновой кислоты. Журнал биологической химии 280: 13962-72; Shockey JM, Fulda MS, Browse J (2003). Arabidopsis содержит крупное суперсемейство акил-активирующих ферментов. Филогенетический и биохимический анализ выявил новый класс акил-коэнзим-синтетаз. Физиология растений 132: 1065-76.) Было выявлено, что акил-кофермент-А-синтетазы в Pseudomonas spp. активируются на жирных кислотах средней цепи, например, на гексаноате (Fernandez-Valverde M, Reglero A, Martinez-Bianco H, Luengo JM (1993) Очищение акил-коэнзим-А-лигазы Pseudomonas putida активируется целым рядом алифатических и ароматических субстрат. Прикладная микробиология окружающей среды 59: 1149-1154.)
Каннабиоиды - это ценные природные продукты. Гено-кодирующие энзимы, которые участвуют в биосинтезе каннабиоидов, пригодны для метаболической инженерии конопли с целью получения растений, которые содержат очень низкий уровень или нулевой уровень ТНСА (tetrahydrocannabinolic acid) и прочих каннабиоидов, в результате реализации целевого мутагенеза (например, через «TILLING») или прочих методов нокаута генов. Такие гены могут также пригодиться для создания, посредством маркерного отбора, специфичных разновидностей конопли, используемых для каннабиоидо-содержащих лекарственных средств, или для биосинтеза каннабиоидов в таких гетерологичных организмах, как бактерии или дрожжи, или для получения каннабиоидов в бесклеточных системах, с применением рекомбинантных белков.
В биосинтезе каннабиоидов, энзимы, участвующие в кодировании генов, также могут применяться для синтеза аналогов и прекурсоров каннабиоидов. В прошлом уже были синтезированы аналоги каннабиоидов, используемые в фармацевтике.
В науке до сих пор существует потребность идентифицировать энзимы и нуклеотидные последовательности, кодирующие эти энзимы, которые участвуют в синтезе ароматических поликетидов.
Сущность изобретения
В конопле были обнаружены два новых гена, которые кодируют ранее неизвестные алканоил-кофермент-А-синтетазы. Эти две новые алканоил-кофермент-А-синтетазы в настоящем документе обозначены как «Конопля посевная гексаноил-кофермент-А-синтетаза 1» (или «CsHCS1») и «Конопля посевная гексаноил-кофермент-А-синтетаза 2» (или «CsHCS2»).
Таким образом, в первом аспекте изобретения представлена изолированная или очищенная молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, имеющую идентичность с последовательностью SEQ ID NO: 1 по меньшей мере на 75%, или кодоновую вырожденную последовательность.
В втором аспекте изобретения представлена изолированная или очищенная молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, имеющую идентичность с последовательностью SEQ ID NO: 3 по меньшей мере на 75%, или кодоновую вырожденную последовательность.
В третьем аспекте изобретения представлен изолированный или очищенный полипептид, содержащий аминокислотную последовательность, имеющую идентичность с последовательностью SEQ ID NO: 2 по меньшей мере на 85%, или консервативно замещенную аминокислотную последовательность.
В четвертом аспекте изобретения представлен изолированный или очищенный полипептид, содержащий аминокислотную последовательность, имеющую идентичность с последовательностью SEQ ID NO: 4 по меньшей мере на 85%, или консервативно замещенную аминокислотную последовательность.
В пятом аспекте изобретения представлен вектор, конструкция или экспрессирующая система, содержащие молекулу нуклеиновой кислоты изобретения.
В шестом аспекте изобретения представлена клетка-акцептор, трансформированная молекулой нуклеиновой кислоты изобретения.
В седьмом аспекте изобретения представлен процесс синтезирования алканоил-кофермента-А в присутствии энзима изобретения.
В восьмом аспекте изобретения представлен процесс изменения уровня каннабиоидных соединений в организме, клетке или ткани с помощью молекулы нуклеиновой кислоты настоящего изобретения, или ее части, для подавления в организме, клетке или ткани гена, который кодирует энзим, катализирующий синтез алканоил-кофермента-А.
В девятом аспекте изобретения представлен процесс изменения уровня каннабиоидных соединений в организме, клетке или ткани, мутированием генов в организме, клетке или ткани, и с помощью молекулы нуклеиновой кислоты настоящего изобретения, с целью отбора организмов, клеток или тканей, которые содержат мутанты или варианты генов, кодирующих энзим, которые катализируют синтез алканоил-кофермента-А.
В десятом аспекте изобретения представлен процесс изменения уровня каннабиоидных соединений в организме, клетке или ткани, с помощью экспрессии или сверхэкспресии молекулы нуклеиновой кислоты изобретения в организме, клетке или ткани, относительно такого же вида организмов, клеток или тканей, выращиваемых в таких же условиях, но без экспрессии или сверхэкспрессии молекулы нуклеиновой кислоты.
В одиннадцатом аспекте изобретения представлен процесс изменения уровня каннабиоидных соединений в организме, клетке или ткани, с помощью экспрессии или сверхэкспрессии молекула нуклеиновой кислоты, кодирующей полипептид изобретения в организме, клетке или ткани, относительно такого же вида организмов, клеток или тканей, выращиваемых в таких же условиях, но без экспрессии или сверхэкспрессии молекулы нуклеиновой кислоты.
В двенадцатом аспекте изобретения представлен процесс синтезирования а естественных каннабиоидного соединения или искусственного аналога каннабиоидного соединения в организме, клетке или ткани, с помощью экспрессии молекулы нуклеиновой кислоты изобретения в организме, клетке или ткани в присутствии карбоновой кислоты и кофермента-А.
В тринадцатом аспекте настоящего изобретения представлен процесс синтезирования алканоил-кофермента-А с помощью бесклеточной реакции in vitro. В этот процесс входит реакция карбоновой кислоты с в присутствии энзима изобретения: коэнзима А.
К настоящему моменту были идентифицированы и описаны полипептиды, являющиеся энзимами, которые катализируют синтез алканоил-кофермента-А, и нуклеотидные последовательности, которые кодируют эти энзимы. Нуклеотидные последовательности могут применяться для получения, через выведение, селекцию или генетическую инженерию, конопли, которая имеет большее или меньшее содержание каннабиоидных соединений, их аналогов или комбинаций. Эти нуклеотидные последовательности могут также применяться, отдельно или в сочетании с генами, которые кодируют прочие этапы синтеза каннабиоидов, для инженерного биосинтеза каннабиоидов в прочих растениях или микроорганизмах (например, дрожжи, бактерии, грибки) или прочих прокариотических или эукариотических организмах или бесклеточных системах. Кроме того, блокирование или снижение экспрессии этих генов в конопле может применяться для блокирования биосинтеза каннабиоидов, тем самым снижая содержание каннабиоидов.
Дополнительные характеристики изобретения будут описаны или станут очевидны по ходу изложения нижеследующего детального описания.
Краткое описание чертежей
Чтобы изобретение было более понятным, далее детально описаны его воплощения в виде примеров, со ссылкой на сопроводительные чертежи, а именно:
На фигуре 1 изображен предлагаемый каскад реакций для получения основных каннабиоидов типа Конопля посевная. Аббревиатуры: ТНСА-синтаза - это синтаза Δ9-тетрагидроканнабиноловой кислоты; CBDA-синтаза - это синтаза каннабидиоловой кислоты; СВСА-синтаза - это синтаза каннабихроменовой кислоты.
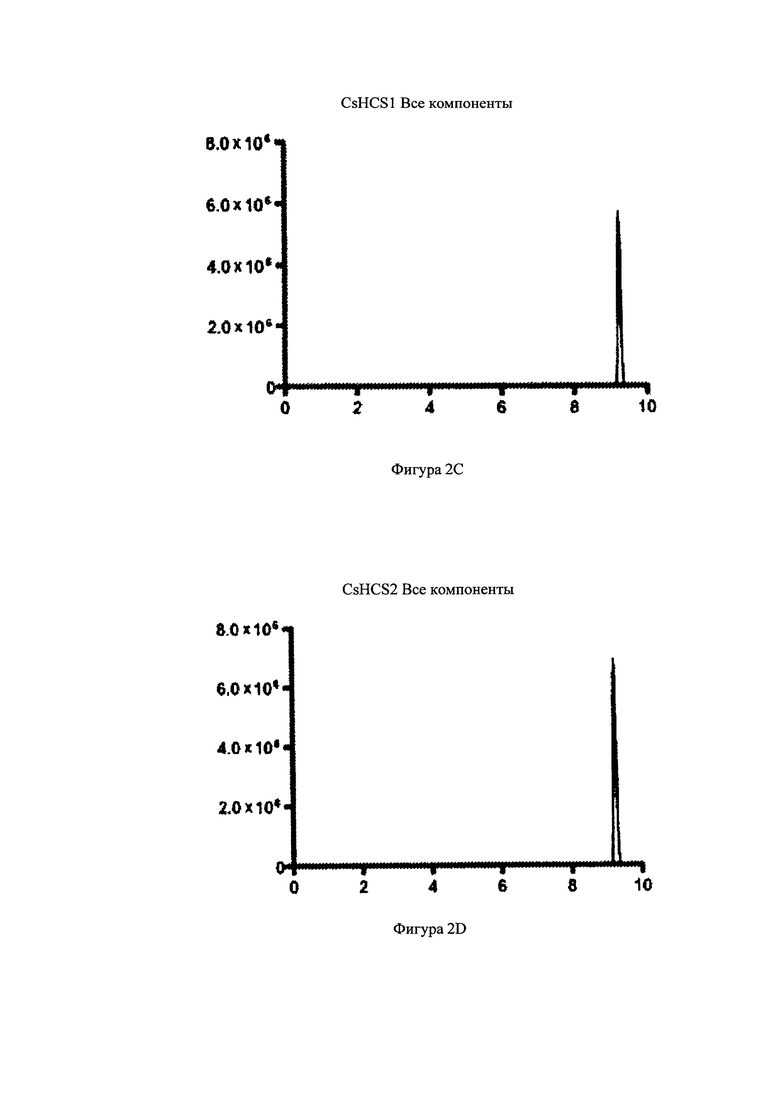
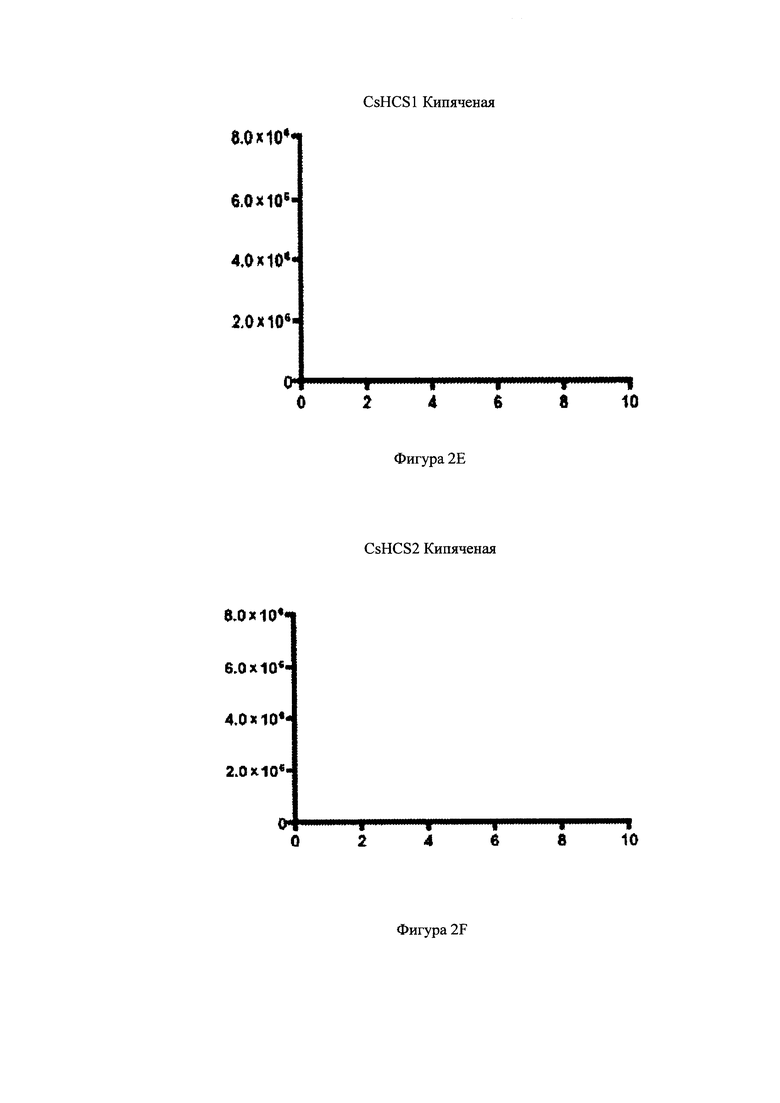
На фигурах 2A-2F изображен анализ с помощью метода «хроматография жидкости-спектрометрия массы/спектрометрия массы», или LC-MS/MS-анализ, энзимной активности Конопли посевной гексаноил-кофермент-А-синтаз. На фигурах 2A-2F показаны распространенность ионов (m/z 866>359) по вертикальной оси и время (минуты) по горизонтальной оси. На Фигурах 2А и 2B изображено время удержания аутентичного стандарта гексаноил-кофермента-А. На фигуре 2С изображены анализ содержания белка CsHCS1, кофермента-А, MgCl2, натрия гексаноата, АТР, и буфера HEPES, где был получен и выявлен гексаноил-кофермент-А. На Фигуре 2D изображен анализ содержания, в котором находится белок CsHCS2, кофермент-А, MgCl2, натрий гексаноат, АТР, и буфер HEPES, где был получен гексаноил-кофермент-А. На фигуре 2Е изображен анализ, в котором находится белок CsHCS1 дезактивированный кипячением при температуре °C в течение 15 минут, кофермент-А, натрий гексаноат, АТР, и буфер HEPES, где не был получен гексаноил-кофермент-А. На фигуре 2F изображен анализ содержания, в котором находится белок CsHCS1 дезактивированный кипячением при температуре 95°С в течение 15 минут, кофермент-А, натрий гексаноат, АТР, и буфер HEPES, где не был получен гексаноил-кофермент-А.
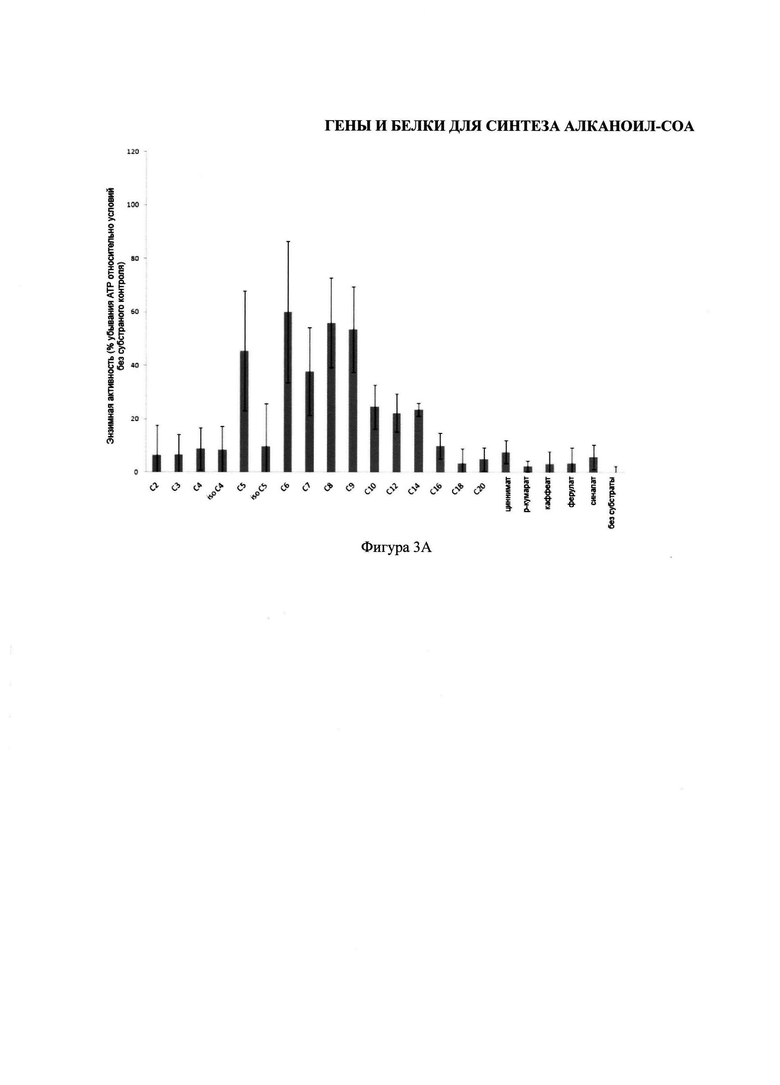
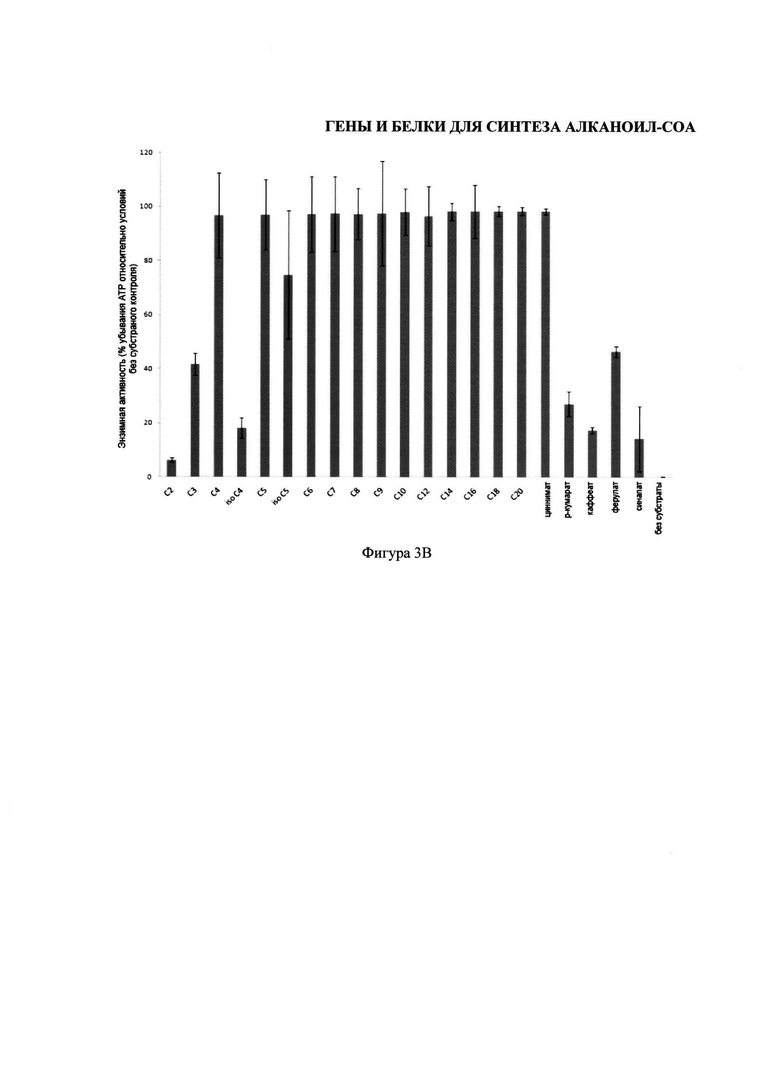
На фигуре 3 изображены два графика, иллюстрирующих субстраты карбоновой кислоты, используемых энзимами изобретения. На Фигуре 3А изображены субстраты карбоновой кислоты, используемые CsHCS1. На Фигуре 3В изображены субстраты карбоновой кислоты, используемые CsHCS2.
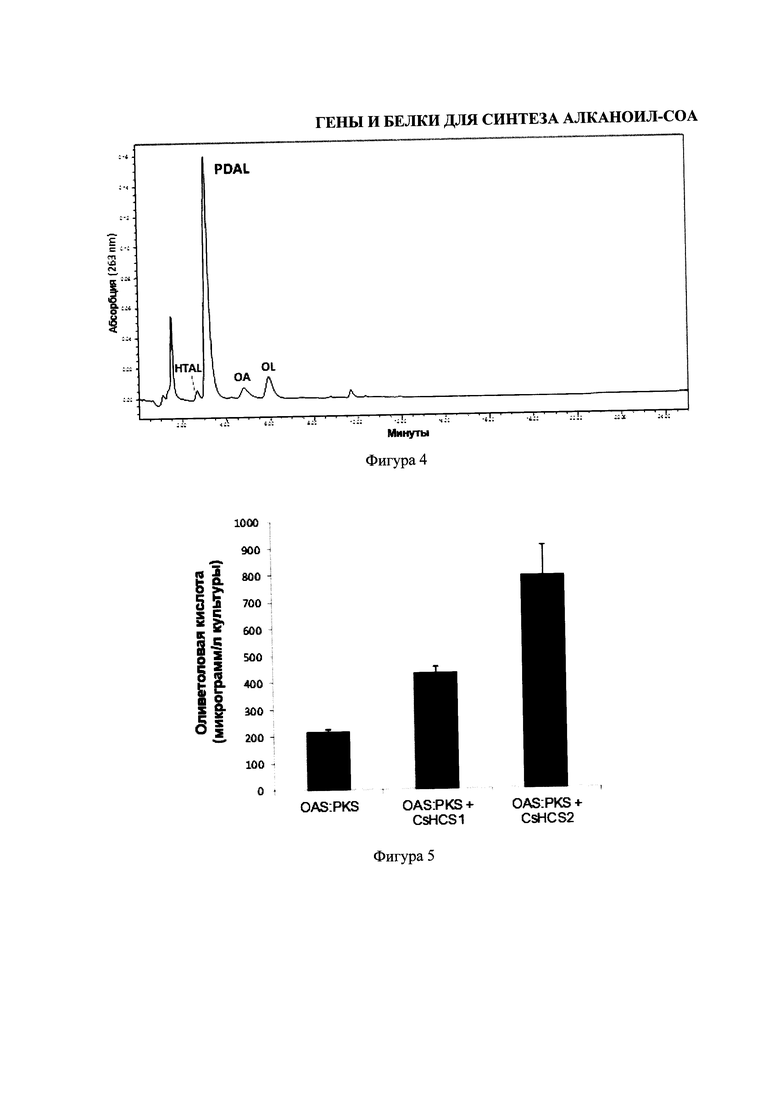
На фигуре 4 изображен высокоэффективный хроматографический анализ жидкости продуктов, полученных сдвоенным анализом энзимного содержания, состоящего из Конопли посевной гексаноил-кофермент-А-синтетазы CsHCS2, малонил-кофермент-А-синтетазы (MCS), Конопли посевной оливетол-синтаза/поликетид-синтазы, и Конопли посевной синтазы оливетоловой кислоты. Элютированные соединения были выявлены с помощью адсорбции на уровне 263 нм и идентифицированы по одинаковому времени удержания в качестве изолированных стандартов, и по их массе, с помощью одиночного квадрупольного масс-детектора. Выявление оливетола и оливетоловой кислоты означает, что CsHCS2 способно обеспечить достаточно субстраты гексаноил-кофермента-А для синтеза оливетоловой кислоты. При проведении анализа без CsHCS2, кофермента-А или гексаноата, поликетидные продукты не были получены. HTAL = гексаноил-ацетил-ацето-уксусный лактон, PDAL = рентил-ацето-уксусный лактон, OA = оливетоловая кислота, OL = оливетол.
На Фигуре 5 изображен график получения оливетоловой кислоты в клетках дрожжей, модифицированных для выработки оливетоловой кислоты, используя CsHCS1 и CsHCS2 для синтеза гексаноил-кофермента-А, а также соединения коноплянной «оливетол-синтазы»/поликетид-синтазы (PKS) и синтазы оливетоловой кислоты (OAS) для формирования оливетоловой кислоты.
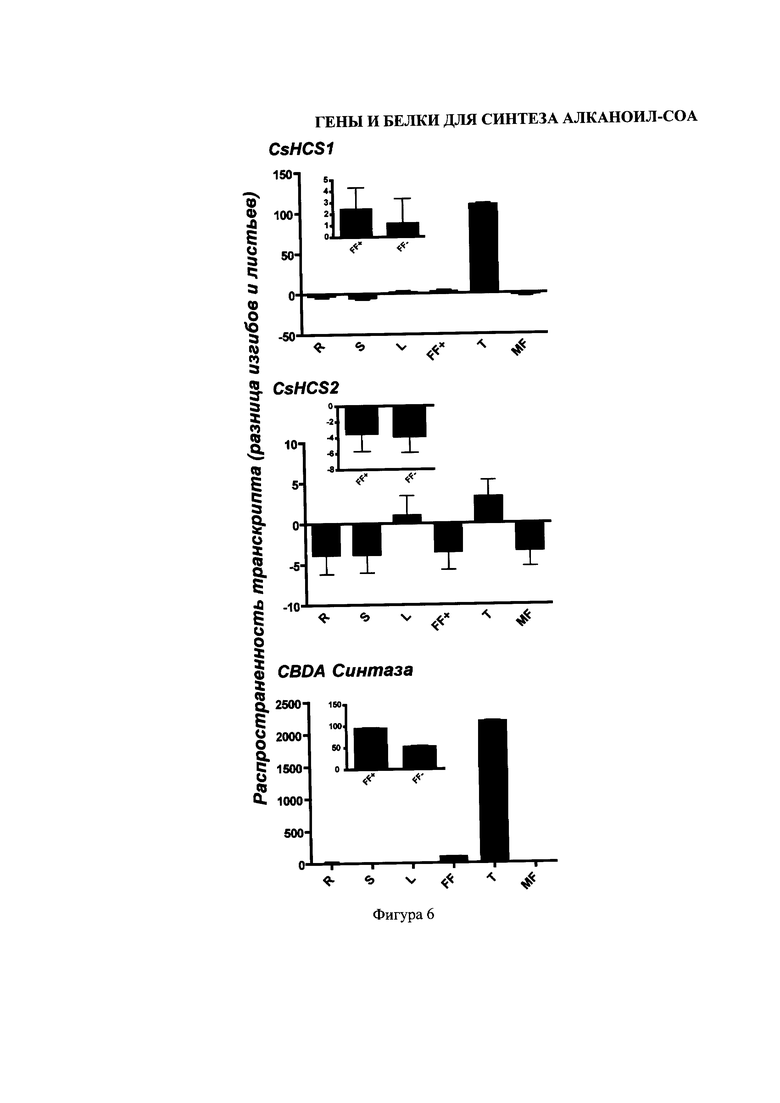
На Фигуре 6 изображен qRT-PCR-анализ экспрессии синтаз CsHCS1, CsHCS2 и CBDA в разных тканях культурного сорта конопли «Finola». Значения экспрессии гена относительно актина указаны как разница в складках и листьях, с принятым значением экспрессией в листе = 1. Во вставках изображена генная экспрессия в женских цветках с трихомами и без них, со значениями, также указанными, как разница в складках и листьях, R, корнях; S, стеблях; L, листьях; FF+, женских цветках с трихомами; FF-, женских цветках, где трихомы удалены по методу Бидбитера; Т, трихом; MF, мужских цветках. Средния значения: ±SD, n=3.
Описание предпочтительных воплощений
Была установлена последовательность генотеки трихом-специфичных сДНК конопли, с получением 9157 маркерных экспрессирующих последовательностей («EST»), которые были собраны в 4113 уникальные последовательности (1227 контигов, 2886 синглтонов). Унигены были аннотированы через сравнение с базой данных белков UniProt™ с помощью поиска в интернете и инструмента сравнения, который называется «blastx». Конопляные акил-активирующие энзимные белки были идентифицированы с помощью Arabidopsis акил-активирующих энзимных последовательностей, для осуществления запроса по собранным конопляным EST в интернет-поисковике и инструменте сравнения, который называется «tblastn». Были определены одиннадцать акил-активирующих энзима, и названы в соответствии с их транскриптной распространенностью в генотеке сДНК. Синтаза CsHCS1 была самым распространенным акил-активирующим энзимом по уровню транскрипции (42 EST); CsHCS2 имела меньшую распространенность (5 EST). В виду более высокого уровня транскрипции в трихомах и локализации CsHCS1 в цитоплазме, вероятно, этот энзим является акил-активирующим энзимом, участвующим в поставке гексаноил-кофермента-А в каннабиоидный каскад реакций. CsHCS2, которая локализуется в пероксисомах, возможно не участвует в образовании каннабиоидов. Однако, благодаря своим кинетическим свойствам, это -энзим, пригодный для синтезирования гексаноил-кофермента-А в гетерологичных акцепторах или бесклеточных системах.
Далее показана последовательность гена CsHCS1:
Конопля посевная CsHCS1-2163 bp (SEQ ID NO: 1)


Далее показана последовательность гена CsHCS2:
Конопля посевная CsHCS2-1547 bp (SEQ ID NO: 3)

CsHCS1 и CsHCS2 были усилены с помощью PCR, как описано в Примере 1 и активность алканоил-кофермента-А-синтетазы или кофермент-А-лигазы была измерена, как описано в Примере 2. Как показано на Фигуре 2, CsHCS1 и CsHCS2 катализируют получение алканоил-кофермента-А из карбоновой кислоты и кофермента-А.
Некоторые воплощения настоящего изобретения относятся к изолированной или очищенной молекуле нуклеиновой кислоты, молекуле, в которой содержится последовательность SEQ ID NO: 1 или имеется идентичность с SEQ ID NO: 1 по меньшей мере на 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%.
Некоторые воплощения настоящего изобретения относятся к изолированной или очищенной молекуле нуклеиновой кислоты, в которой содержится последовательность SEQ ID NO: 3 или имеется идентичность с SEQ ID NO: 3 по меньшей мере на 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%.
Кроме того, сюда включены молекулы нуклеиновой кислоты, которые гибридизируются с вышеописанными последовательностями нуклеиновой кислоты. Условия гибридизации могут быть ужесточены тем, что гибридизация будет происходить, только если будет идентичность, по меньшей мере на 90%, 95% или 97%, с последовательностью в молекуле нуклеиновой кислоты, которая кодирует энзим настоящего изобретения. Ужесточенные условия могут означать также условия, применяемые в известных «южных» гибридизациях, таких как, например, инкубация в течение ночи при температуре 42°С в растворе 50% формамида, 5× SSC (150 mM NaCl, 15 mМ тринатрий-цитрата), 50 mM натрий-фосфат (pH 7.6), 5× раствор Денгардта, 10% декстран-сульфат и 20 микрограмм/миллиметр денатурированный ДНК из молок лососевых, с последующим вымыванием опоры гибридизации в 0.1× SSC при температуре около 65°С. Прочие условия гибридизации хорошо известны и описаны в литературе Sambrook и др., Молекулярное клонирование: Лабораторный справочник, Третье издание, Cold Spring Harbor, N.Y. (2001).
Специалист данной сферы заметит, что незначительные изменения последовательности нуклеиновой кислоты не обязательно изменят аминокислотную последовательность закодированного полипептида. Специалисты данной сферы поймут, что изменения в идентичности нуклеотидов в специфичной генной последовательности, которые изменяют аминокислотную последовательность закодированного полипептида, могут привести к снижению или улучшению эффективности генов, и что, в некоторых применениях (например, анти-смысловая цепь, косупрессия, или PHKi), частичные последовательности зачастую работают так же эффективно, как и при полной последовательности. Способы, которыми нуклеотидная последовательность может быть изменена или укорочена, хорошо известны специалистам данной области, так же, как и способы определения эффективности измененных генов. В некоторых воплощениях, эффективность может быть легко протестирована, например, с помощью традиционной газовой хроматографии. Таким образом, в описание настоящего патентного раскрытия входят все такие вариации генов.
Как будет подтверждено специалистом данной сферы, длина молекулы нуклеиновой кислоты, описанная выше, будет зависеть от планируемого применения. Например, если планируемое применение - это использование в качестве затравки или пробы, например, для усиления с помощью PCR или для скрининга генотеки, длина молекулы нуклеиновой кислоты будет меньше, чем длина всей последовательности, например, 15-50 нуклеотидов. В этих воплощениях, затравки или пробы могут быть значительно идентичны высоко-сохранившейся части последовательности нуклеиновой кислоты или могут быть значительно идентичны цепи на 5' или 3' от конца последовательности ДНК. В некоторых случаях, эти затравки или пробы могут использовать универсальные базы в некоторых позициях, становясь «значительно идентичными», но все же обеспечивающими гибкость при распознании последовательности. Необходимо отметить, что в этой сфере хорошо известны подходящие состояния гибридизации затравок и проб.
В настоящее изобретение также включен энзим CsHCS1. Этот энзим обладает следующей аминокислотной последовательностью (SEQ ID NO: 2):

Некоторые воплощения относятся к изолированному или очищенному полипептиду, в котором присутствует SEQ ID NO: 2, или имеется идентичность с аминокислотной последовательностью SEQ ID NO: 2 по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%.
В настоящее изобретение также входит энзим CsHCS2. Этот энзим имеет следующую аминокислотную последовательность (SEQ ID NO: 4)

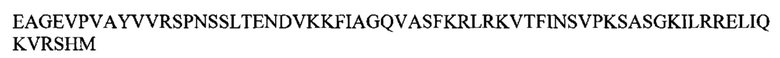
Некоторые воплощения относятся к изолированному или очищенному полипептиду, в котором присутствует SEQ ID NO: 4, или имеется идентичность аминокислотной последовательности SEQ ID NO: 4 по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%.
Некоторые воплощения относятся к вектору, конструкции или экспрессирующей системе, содержащих изолированный или очищенный полинуклеотид, в котором присутствует последовательность SEQ ID NO: 1 или SEQ ID NO: 3, или имеется идентичность последовательности SEQ ID NO: 1 или SEQ ID NO: 3 по меньшей мере на 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Также, представлен метод подготовки вектора, конструкции или экспрессирующей системы, включающих такую последовательность, или ее часть, для введения в клетку последовательности или части последовательности в смысловой или анти-смысловой ориентации, или в дополнении.
В некоторых воплощениях, изолированная и/или очищенная молекула нуклеиновых кислот, или векторы, конструкции или экспрессирующие системы, в которые входят эти изолированные и/или очищенные молекулы нуклеиновых кислот, могут применяться для получения трансгенных организмов или клеток организмов, которые образуют полипептиды, катализирующие синтез ароматических поликетидов. Таким образом, одно из воплощений относится к трансгенным организмам, клеткам или микробным тканям организма, в которые входит изолированная и/или очищенная молекула нуклеиновой кислоты, с последовательностью SEQ ID NO: 1 или SEQ ID NO: 3, или с последовательностью, имеющей идентичность с SEQ ID NO: 1 или SEQ ID NO: 3 по меньшей мере на 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%.
В качестве организма рекомендуется выбрать растение, микроорганизм или насекомое. Из растений рекомендуется выбирать Cannabis, например, Конопля посевная L., Cannabis indica Lam. и Cannabis ruderalis Janisch. Особенно рекомендуется растение Конопля посевная. Из микроорганизмов рекомендуются бактерии (например, Escherichia coli) или дрожжи (например, Saccharomyces cerevisiae). Из насекомых рекомендуется Spodoptera frugiperda.
Организмы, клетки и микробные ткани этого воплощения могут изменить уровень содержания каннабиоидных соединений. По Фигуре 1 специалист данной области определит, что экспрессия или сверхэкспрессия молекулы нуклеиновой кислоты настоящего изобретения приводит к экспрессии или сверхэкспрессии энзима, который катализирует синтез гексаноил-кофермента-А, который, в сочетании с прочими энзимами, может привести к образованию или повышенному образованию каннабиоидных соединений, таких как каннабигероловая кислота (CBGA), Δ9-тетрагидроканнабиноловая кислота (ТНСА), каннабидиоловая кислота (CBDA), каннабихроменовая кислота (СВСА), Δ9-тетрагидроканнабинол (ТНС), каннабидиол (CBD), каннабихромен (CBC) и т.д. Подобным же образом, в зависимости от используемой субстраты, экспрессия или сверхэкспрессия молекулы нуклеиновых кислот настоящего изобретения, приводящая к экспрессии или сверхэкспрессии энзима, которая катализирует синтез гексаноил-кофермента-А, может привести к образованию или повышенному образованию аналогов каннабиоидных соединений, или аналогов прекурсоров таких соединений.
Подавление гена в организме, клетке или ткани приведет к недостаточной экспрессии энзима, что может привести к аккумуляции таких прекурсоров, как гексановая кислота (шесть углеродов), октановая кислота (восемь углеродов), нонановая кислота (девять углеродов), валериановая кислота (паять углеродов), гептановая кислота (семь углеродов) или прочие карбоновая кислоты, и/или снижение таких каннабиоидов, как ТНСА (прекурсор ТНС) или CBDA (прекурсор CBD).
Настоящее изобретение включает себя процесс изменения уровня каннабиоидных соединений в организме, клетке или ткани через экспрессию или сверхэкспрессию экзогенного энзима изобретения в организме, клетке или ткани, относительно такого же вида организма, клетки или ткани, выращиваемых в тех же условиях, но без экспрессии или сверхэкспрессии экзогенного энзима изобретения.
Экспрессия или сверхэкспрессия молекул нуклеиновых кислот изобретения может быть осуществлена в сочетании с экспрессией или сверхэкспрессией одной и нескольких прочих нуклеиновых кислот, которые кодируют один и более энзимов в каннабиоидном биосинтетическом каскаде реакций. Некоторые примеры прочих нуклеиновых кислот включают в себя кислоты, которые кодируют: поликетид-синтазу III типа, поликетид-циклазу, ароматическую пренилтрансферазу и каннабиоид-формирующую оксидоциклазу. Конкретные примеры этих энзимов включают в себя «оливетол-синтазу»/поликетид-синтазу, синтазу оливетоловой кислоты, геранилпирофосфатюливетолат геранилтрансферазу, синтазу Δ9-тетрагидроканнабиноловой кислоты, синтазу каннабидиоловой кислоты или синтазу каннабихроменовой кислоты. Синтез алканоил-кофермента-А в присутствии энзимного полипептида настоящего изобретения может быть осуществлено in vivo или in vitro. Как упомянуто выше, такие синтезы in vivo могут осуществляться через экспрессию или сверхэкспрессию молекул нуклеиновой кислоты изобретения в организме, клетке или ткани.
Синтез алканоил-кофермента-А in vitro может осуществляться в бесклеточной системе. В бесклеточной системе in vitro, карбоновая кислота и энзим настоящего изобретения могут быть смешаны вместе в посуде, подходящей для реакций.
In vitro, полипептиды настоящего изобретения могут применяться в сочетании с прочими энзимами для осуществления полного синтеза каннабиоидных соединений из прекурсоров. Например, такие прочие энзимы могут быть использованы в каннабиоидном биосинтезе, как описано на Фигуре 1, (например, «оливетол-синтаза»/PKS, синтаза оливетоловой кислоты, ароматическая пренилтрансфераза, ТНСА-синтаза, CBDA-синтаза, СВСА-синтаза).
Полипептиды настоящего изобретения могут применяться, in vivo или in vitro, для синтеза аналогов каннабиоидных соединений, который не возникают в акцепторах в естественной среде. Такие аналоги могут быть получены с помощью соединений карбоновой кислоты, за исключением тех, которые применяются для получения природных каннабиоидных соединений в растениях. Например, уксусная кислота, маслянная кислота, октановая кислота, декановая кислота, лауровая кислота, миристиновая кислота, пальмитиновая кислота; кислоты с ответленными цепями, например, изовалерьяновая кислота; и гидрокси-коричные кислоты, например, коричная кислота.
Термины:
Для упрощения описания патентуемых воплощений, далее приведены разъяснения специфических терминов:
Алканоил-кофермент-А (СоА): Алканоил-кофермент-А - это алифатическое карбониловое соединение, в котором половина коэнзима-А связана с атомом углерода карбониловой группы сульфидным мостом. Предпочтительные соединения алканоил-кофермента-А включают в себя от 2 до 10 атомов углерода в алифатической карбониловой части соединения. Более предпочтительно, когда алканоил-кофермент-А - это кофермент-А-S-С(O)-(СН2)n-CH3, где n - это целое число от 0 до 8. Несколько примеров соединений алканоил-коферментов-А: ацетил-кофермент-А, бутирил-кофермент-А, гексаноил-кофермент-А и октаноил-кофермент-А. Использование ацетил-кофермента-А дает метиловую боковую цепь в полученном ароматическом поликетиде; использование бутирил-кофермента-А дает пропиловую боковую цепь; и использование гексаноил-кофермента-А дает пентиловую боковую цепь. Гексаноил-кофермент-А высоко предпочтителен. В конопле также присутствуют каннабиоиды с более короткими боковыми цепями (например, тетрагидроканнабивариновая кислота, в которой вместо пентиловой боковой цепи ТНСА находится пропиловая боковая цепь).
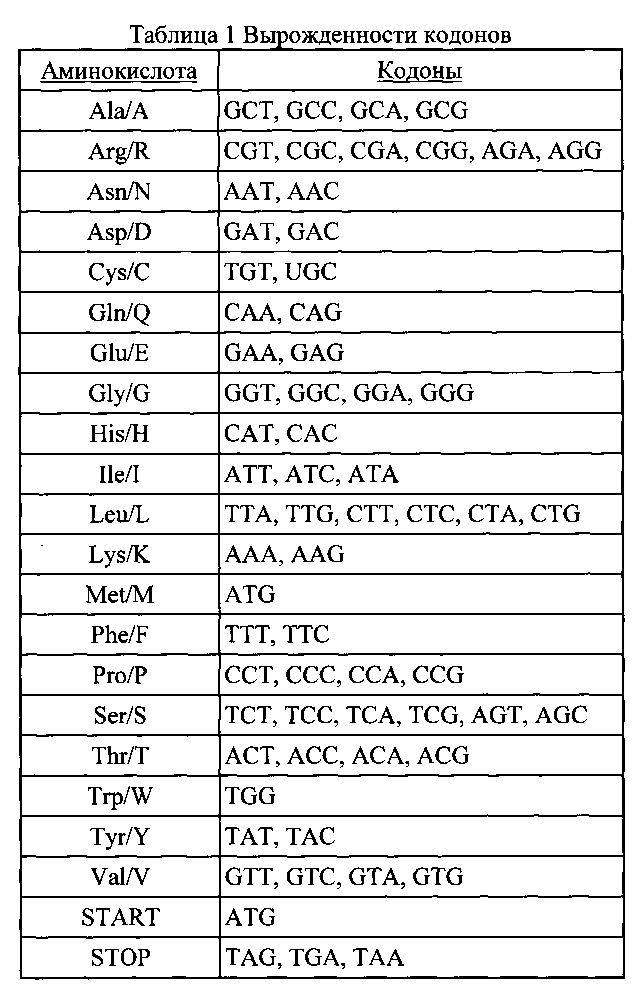
Вырожденность кодона: В данное патентное раскрытие включено применение вырожденности кодона, как станет ясно для специалиста данной области и как показано в Таблице 1.
Комплементарная нуклеотидная последовательность: «Комплементарная нуклеотидная последовательность» в последовательности - это любая молекула нуклеиновой кислоты, нуклеотиды которой комплементарны к молекулам последовательности, раскрытым в настоящем документе, и ориентация которых обратная (антипараллельная последовательность).
Консервативные замещения: Специалист данной области понимает под термином «консервативные замещения» аминокислотную последовательность полипептида без нарушения трехмерной структуры и функции полипептида. Соответственно, настоящее изобретение включает в себя полипептиды, содержащие консервативно замещенные CsHCS1 и CsHCS2. Консервативные замещения осуществляются специалистом данной области путем замещения одной аминокислоты на другую с одинаковой гидрофобностью, полярностью, и длиной R-цепи. Кроме того, при сравнении выровненных последовательностей гомологичных белков, взятых от разных видов, консервативные замещения могут быть идентифицированы с помощью локализации остатков аминокислот, которые мутировали между видами без изменения базовых функций закодированных белков. В таблице 2 приведен пример консервативных замещений.

Степень или доля гомологии в последовательности: Термин «степень или дола гомологии в последовательности» относится к степени или доли идентичных частей в двух последовательностях после оптимального выравнивания.
Гомологичная изолированная и/или очищенная последовательность: «Гомологичная изолированная и/или очищенная последовательность» - это изолированная и/или очищенная последовательность, в которой имеется частичная идентичность с основными нуклеотидными последовательностями, или последовательностями аминокислотных полипептидов, по меньшей на 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99.5%, 99.6%, или 99.7%. Эта доля исключительно статистическая, и разницу можно распределить между двумя нуклеотидными или аминокислотными последовательностями случайным образом или по всей их длине. Идентичность последовательности может быть определена, например, с помощью компьютерных программ, предназначенных для единичного или множественного выравнивания последовательностей.
Увеличение, уменьшение, модулирование, изменение или т.п.: Специалист данной сферы знает, что эти термины относятся к сравнению схожих вариаций или штаммов, выращиваемых в одинаковых условиях, но без модификации, которая приводит к увеличению, уменьшению, модуляции или изменению. В отдельных случаях, это может нетрансформированный контроль, симулируемый трансформированный контроль, или векторный трансформированный контроль.
Изолированные: Специалисты рассматриваемой сферы используют термин «изолированные» в отношении полипептидов или нуклеиновых кислот, которые были «изолированы» из их родной среды.
Последовательность нуклеотидов, полинуклеотидов, или нуклеиновой кислоты: «Последовательность нуклеотидов, полинуклеотидов, или нуклеиновой кислоты» - это двутяжевые или однотяжевые цепи в мономерной или димерной (так называемой, «тандемной») форме и транскрипция их продуктов.
Идентичность последовательностей: Две аминокислоты или нуклеотидные последовательности называются «идентичными», если последовательности аминокислот или нуклеотидов в двух последовательностях одинаковы, когда они выравнены для установления максимального соответствия, как описано ниже. Доля идентичности последовательности (или степень идентичности) определяется с помощью сравнения двух оптимально выравненных последовательностей в сравнительном окне, где часть пептидной или полинуклеотидной последовательности в сравнительном окне может содержать дополнительные или отсутствующие элементы (т.е. пробелы) в сравнении с образцовой последовательностью (т.е. той, которая не содержит дополнительные или отсутствующие элементы) для оптимального выравнивания двух последовательностей. Эта доля рассчитывается путем определения количества позиций, в которых находятся идентичные остатки аминокислот или основ нуклеиновой кислоты в оба/обеих последовательностях, для получения количества совпадающих позиций, с делением количества совпадающих позиций на общее количество позиций в окне сравнения и умножением результата на 100, для получения процентного выражения доли идентичной последовательности.
Оптимальное выравнивание последовательностей для проведения сравнения может быть сделано с помощью алгоритма локальной гомологии (Смит и Уотерман, Продвинутая прикладная математика 2: 482 (1981), Алгоритм выравнивания гомологии Нидлмана и Вунша, J. Мол. Биол. 48: 443 (1970), Метод поиска схожестей Пирсона и Липмэна, Proc. Natl. Acad. Sci. (США) 85: 2444 (1988), компьютерная реализация этих алгоритмов (GAP, BESTFIT, FASTA, и TFASTA, Пакет ПО Висконсина по генетике, Генетическая компьютерная группа (GCG), 575 Доктор наук, Madison, Wis.) или визуальной проверки.
Определение идентичности последовательности, описанное выше, осуществляется специалистом данной области. Сам процесс определения не нуждается в каком-либо особом алгоритме и перечисленные алгоритмы используются только для достижения оптимальных выравниваний последовательностей, а не для расчета идентичности последовательностей.
Из вышеприведенного определения следует, что существует четко установленное и единственное значение идентичности последовательностей в двух сравниваемых последовательностях, значение которой соответствует значению, полученному при наилучшем или оптимальном выравнивании.
Гибридизация в строгих условиях: Гибридизация в строгих условиях с нуклеотидной последовательностью - это гибридизация в таких условиях выбранной температуры и ионной силы, которые позволяют проводить гибридизацию двух фрагментов комплементарных молекул нуклеиновой кислоты.
Гомологи новых генов, описанных в настоящем документе, полученные из других организмов, например, растений, могут быть получены скринингом соответствующих генотек, содержащих гомологи, при этом проводится скрининг нуклеотидной последовательности специфических генов изобретения, или их частей/проб, или определяются поиском гомологии последовательностей с помощью программ выравнивания последовательностей, например, «BLAST» или «FASTA».
Изоляция и клонирование нуклеиновой кислоты - это хорошо изученная процедура. Подобным образом, изолированный ген может быть вставлен в вектор и трансформирован в клетку с помощью традиционных методов, известных специалистам рассматриваемой области. Молекулы нуклеиновой кислоты могут быть трансформированы в организм. Как известно специалистам, существует ряд способов, которыми гены, векторы, конструкции и системы экспрессии могут быть внедрены в организмы, и сочетание методов трансформация и культивации тканей были успешно интегрированы в эффективные стратегии создания трансгенных организмов. Эти методы, который могут применяться в изобретении, были описаны в другой литературе (Potrykus I (1991) Перевод генов в растения: Оценка опубликованных подходов и результатов. Ежегодный обзор физиологии растений, молекулярной биологии растений 42: 205-225; Василь И.К. (1994) Молекулярное улучшение зерновых. Растительная мол. биол. 25: 925-937. Walden R, Wingender R (1995) Генный трансферт и методы регенерации растений. Тенденции в биотехнологии 13: 324-331; Songstad DD, Somers DA, Griesbach RJ (1995) Прогресс в альтернативных методах получения ДНК. Культ, клетки растительной ткани. 40: 1-15), и они хорошо известны специалистам данной области.
Подходящие векторы известны специалистам данной области и описаны в общих технических справочниках, например, Pouwels и др., Векторы клонирования. Лабораторный справочник, Elsevier, Амстердам (1986). Подходящие векторы включают в себя Ti-плазмид-векторы. Например, специалист данной области знает, что кроме опосредованной агробактериями трансформации Arabidopsis с помощью вакуумной инфильтрации (Bechtold Ν, Ellis J, Pelletier G (1993), опосредованного растительными агробактериями генного трансферта, с помощью инфильтрации взрослых растений Arabidopsis thaliana. С R Acad Sci Paris, Sciences de la vie/Life sciences 316: 1194-1199.), прививки надрезом (Katavic V, Haughn GW, Reed D, Martin M, Kunst L (1994), растительной трансформации Arabidopsis thaliana. Мол. ген. генет. 245: 363-370.), также возможно трансформировать прочие виды растений, с помощью Agrobacterium Ti-плазмид (например, гипокотил (DeBlock M, DeBrouwer D, Terming Ρ (1989) Трансформация Brassica napus и Brassica olerácea с помощью Agrobacterium tumefaciens и экспрессии ген-бара и нео-генов в трансгенных растениях. Физиол. Растений 91: 694-701) или семядольном черешке (Moloney MM, Walker JM, Sharma KK (1989), высокоэффективной трансформации Brassica napus, с помощью векторов Agrobacterium. Растительная клетка Rep. 8: 238-242.), с помощью заражения надреза, бомбардировки частицами/биолистических методов (Sanford JC, Klein ТМ, Wolf ED, Allen Ν (1987), Введение вещество в клетки и ткани, с помощью процесса бомбардировки частицами. J. Part. Sci. Технол. 5: 27-37.) или опосредованных полиэтилен-гликолем методов трансформации протопласта (Rhodes СА, Pierce DA, Mettler IJ, Mascarenhas D, Detmer JJ (1988) Генетически-трансформированные растения кукурузы из протопласт. Science 240: 204-207).
Также, как очевидно для специалистов данной области, и описано в прочей литературе (Meyer Ρ (1995) Понимание и контролирование трансгенной экспрессии. Тенденции в биотехнологии 13: 332-337; Datla R, Anderson JW, Selvaraj G (1997) Растительные промоторы в трансгенной экспрессии. Биотехнологический ежегодный обзор 3: 269-296.), возможно использование промоторов, оперативно связанных с молекулами нуклеиновой кислоты, для направления любого преднамеренного регулирования вверх или вниз трансгенной экспрессии с помощью нерегулируемых (т.е., конститутивных) промоторов (например, основанных на CaMV35S), или с помощью промоторов, которые могут направить генную экспрессию в конкретные клетки, ткани (например, напиновый промотор для экспрессии трансгенов в развивающихся семенных котиледонах), органы (например, корни), в конкретную стадию развития, или в ответ на конкретный внешний стимул (например, тепловой шок).
Промоторы, используемые в изобретении, могут быть индуцибельные, конститутивные, или тканеспецифичные или иметь разные сочетания этих характеристик. Пригодные промоторы включают в себя, среди прочего, конститутивные промоторы, например, промотор гвоздичного вируса кольцевой гравировки (CERV), промотор капустного мозаичного вируса (CaMV) 35S, или, более конкретно, двойной улучшенный промотор капустного мозаичного вируса, содержащий два промотора CaMV 35S в тандеме (называется «двойной промотор 35S»). Возможно будет целесообразным использование тканеспецифичных или регулируемых развитием промоторов вместо конститутивных промоторов в некоторых случаях. Тканеспецифичные промоторы позволяют добиться сверхэкспрессии в некоторых тканях, без изменения экспрессии в прочих тканях.
Промоторы и терминальные регуляторные участки будут функционировать в клетке-акцепторе и могут быть гетерологичны (т.е, не встречаться в природе) или гомологичны (производные из акцепторов) по отношению к клетке и гену.
Терминальный регуляторный участок может получен из 3' участка гена, в котором был получен промотор или из другого гена. Пригодные терминальные участки хорошо известны специалистам и включают в себя терминатор Agrobacterium tumefaciens нопалин-синтазы (Tnos), A. tumefaciens маннопин-синтазы (Tmas) и терминатор CaMV 35S (T35S). Предпочтительные терминальные участки, используемые в настоящем изобретении, включают в себя малую подъедницу терминального участка гороховой рибулоза-бисфорсфат-карбоксилазы (TrbcS) или терминальный участок Tnos. Генная конструкция, используемая в изобретении, может быть проверена в отношении активности, например, трансформацией в растение-акцептор через Agrobacterium и скринингом изменений в уровне каннабиоидов.
Молекулы нуклеиновой кислоты изобретения, или ее фрагменты, могут применяться для блокировки биосинтеза каннабиоидов в организмах, которые образуют каннабиоидные соединения в естественной среде. Подавление с помощью молекулы нуклеиновой кислоты изобретения может осуществляться целым рядом способов, известных специалистам, например, методом вмешательства в РНК (PHKi), методами искусственного микроРНК, вирусного генного подавления (VIGS), антисмысловыми методами, смысловыми методами косупрессии и целевыми мутагенными методами.
Методы PHKi означает стабильную трансформацию с помощью РНК-интерференции (PHKi) в конструкцию плазмида (Helliwell СА, Waterhouse РМ (2005) Конструкции и методы подавления генов, опосредованного шпилькообразным РНК, в растениях. Методы Этимологии 392: 24-35). Такие плазмиды составлены из фрагмента целевого гена, подлежащего подавлению, в инвертированной повторяющейся структуре. Инвертированные повторы разделены спейсером, в основном интроном. Конструкция PHKi, управляемая соответствующим промотором, например, промотором мозаичного вируса цветной капусты (CaMV) 35S, интегрируется в геном растения и последующая транскрипция трансгена приводит к созданию молекулы РНК, которая сворачивается, формируя двутяжевую шпилькообразную РНК. Эта двутяжевая структура РНК признается растением и разрезается на малые РНК (длиной около 21 нуклеотидов), которые называются малыми интерферирующими РНК (миРНК). Эти миРНК связываются с белковым комплексом, (RISC), и происходит переход к прямой деградации мРНК в целевом гене.
Методы искусственного микроРНК (имиРНК) используют каскад реакций микроРНК (миРНК), который подавляет эндогенные гены в растениях и прочих эукариотах (Schwab R, Ossowski S, Riester M, Warthmann N, Weigel D (2006). Высокоспецифичное подавление гена искусственными микроРНК в Arabidopsis. Растительная клетка 18: 1121-33; Альварес Х.П., Пеккер И., Голдшмидт Α., Блум Е., Амзеллем Ц., Эшед Й. (2006 г.) Эндогенные и синтетические микроРНК стимулируют одновременное, эффективное, и локализованное регулирование множества направлений в разных растениях. Растительная клетка 18: 1134-51). В этом методе, 21 нуклеотидных длинных фрагмента подавляемого гена вводятся в предварительный ген миРНК для формирования предварительной конструкции имиРНК. Предварительная конструкция имиРНК переходит в геном организма с помощью методов трансформации, известных специалисту. После транскрипции предварительных имиРНК, появляются имиРНК, направленные на гены, которые имеют нуклеотидную идентичность с последовательностью имиРНК с 21 нуклеотидами.
В методах подавления PHKi, на выбор длины фрагмента могут повлиять два фактора. Чем короче фрагмент, чем реже будет достигаться эффективное подавление, но слишком длинные РНК-шпильки увеличивают вероятность рекомбинации в бактериальных штаммах-акцепторах. Эффективность подавления также зависит от гена и может отражать доступность целевого мРНК или относительную распространненость целевого мРНК и шпилькообразного РНК в клетке с активным геном. Фрагмент длиной от 100 до 800 bp, а лучше от 300 до 600 bp, как правило, подходит для максимальной эффективности подавления. Другое соображение касается целевой части гена. С одинаково хорошими результатами могут применяться фрагменты 5° UTR, участок кодирования, и 3' UTR. Так как механизм подавления зависит от гомологии последовательности, существует потенциал перекрестного подавления связанных последовательностей мРНК. Там, где это нежелательно, должен быть выбран участок с последовательностью мало схожей с прочими последовательностями, например, 5' или 3' UTR. Чтобы избежать подавления перекрестной гомологии, необходимо использовать последовательности, которые не содержат блоки идентичных последовательностей в больше 20 баз конструкций и нецелевых генных последовательностей. Многие их этих принципов применимы к выбору целевых участков при составлении имиРНК.
Методы вирусного подавления генов (VIGS) - это вариация метода PHKi, в которой используется эндогенная антивирусная защита растений. Заражение растений рекомбинантными вирусами VIGS, содержащими фрагменты ДНК-акцептора, приводят к пост-транскрипционногому генному подавлению в целевом гене. В одном воплощении, может применяться табачный вирус погремковости (TRV) на базе системы VIGS с нуклеотидными последовательностями настоящего изобретения.
Антисмысловые методы включают в себя введение в растение антисмыслового олигонуклеотида, который связывается с информационным РНК (мРНК), созданным интересующим нас геном. «Антисмысловой» олигонуклеотид имеет базовую последовательность комплементарную информационному РНК гена (мРНК), который называется «смысловой» последовательностью. Активность смыслового сегмента мРНК блокирутеся анти-смысловым сегментом мРНК, тем самым эффективно дезактивирую экспрессию гена. Применения антисмыслового приема для подавления гена в растениях описано более детально в литературе: Stam M, de Bruin R, van Blokland R, van der Hoorn RA, Mol JN, Kooter JM (2000), Отличительные черты пост-транскрипционного подавления гена антисмысловыми трансгенами в единой копии и повторяющихся местах инвертированного Т-ДНК. Plant J. 21: 27-42
В смысловые методы косупрессии входит введение высоко-экспрессирующего смыслового трансгена в растение, что приводит к снижению экспрессии в трансгене и в эндогеном гене (Depicker A, Montagu MV (1997) Пост-транскрипционное подавление гена в растениях. Curr Opin Cell Biol. 9: 373-82). Эффект зависит от идентичности последовательностей трансгенного и эндогенного генов.
Целевые мутагенные методы, например, «TILLING» (Targeting Induced Local Lesions in Genomes - целевые вызванные локальные поражения в геномах) и бомбардировка быстрым нейтроном для «уничтожения гена», могут применяться для нокаутирования генной функции в организме (Henikoff S, Till BJ, Comai L (2004) TILLING. Традиционный мутаген встречает функциональный геном. Физиология растений 135: 630-6; Li X, Lassner M, Zhang Y. (2002). Уничтожение гена: система нокаутирования гена на основе мутагенного уничтожения быстрым нейтроном в растениях. Comp Funct Genomics. 3: 158-60). В «TILLING» входит обработка микроб-плазмы или отдельных клеток мутагеном, чтобы вызвать точечные мутации, которые затем обнаруживаются в интересующих генах, с помощью чувствительного метода выявления мутации одиночного нуклеотида. Выявление желаемых мутаций (например, мутации, приводящий к дезактивации интересующего генного продукта) может проводиться, например, с помощью методов PCR. Например, могут быть подготовлены олигонуклеотидные затравки, полученные из интересующего гена, и PCR может быть использовано для амплификации участков интересующего гена организмов в мутагенизированной популяции. Можно комплементарно присоединить усиленные мутантные гены к генам дикого растения для нахождения несоответствий между мутантными генами и генами дикого растения. Разница может быть отслежена по организму, содержащему мутантный ген, тем самым показывая, в котором мутагенизированном организме будет желаемая экспрессия (например, подавление интересующего гена). Затем эти организмы могут быть селективно культивированы для получения популяции с желаемой экспрессией. «TILLING» может привести к аллельным сериям, которые содержат несмысловые и нокаутирующие мутации со сниженной экспрессией целевого гена. «TILLING» предлагается в качестве возможного подхода к нокауту гена, в котором не применяется введение трансгенов, и, следовательно, может быть более приемлемым методом. Бомбардировка быстрыми нейтронами вызывает мутации, т.е., уничтожения, в геномах организма, которые могут также быть выявлены с помощью PCR так же, как и в методе «TILLING».
Специалисты поймут, что процессы изобретения могут также осуществляться в бесклеточной среде в присутствии одной или более карбоновых кислот.
Воплощения изобретения склонны к разным модификациям и альтернативным формам, кроме конкретных примеров, описанных в настоящем документе. Таким образом, воплощения изобретения не ограничиваются только конкретными раскрытыми формами.
Примеры:
Пример 1: Амплификация и клонирование CsHCS1 и CsHCS2
CsHCS1 и CsHCS2 были PCR-амплифицированы из клонов сДНК плазмид с помощью затравок, перечисленных в Таблице 3 и полимеразы Phusion™ (финнзимы). РСК продукты были очищены и клонированы во входной вектор pCR8/GW/ТОРО (Инвитроген). После трансформации в клетки Е.coli TOP10 (Инвитроген), отдельные клоны были проверены с помощью определения последовательности. Гены CsHCS1 и CsHCS2 были рекомбинированы в вектор назначения pHIS8/GW с помощью LR-рекомбиназы (Инвитроген). Продукты реакции LR были трансформированы в клетки TOP10 и проверены с помощью определения последовательности.

pHIS8/GW-CsHCS1 и pHIS8/GW-CsHCS2 были трансформированы в клетки E.coli Rosetta 2 (Merck). Были использованы отдельные колонии для инокуляции мелкомасшабных культур в жидкой LB-среде, содержащей хлорамфениколь и канамицин, которые использовались инокуляции 500 мл жидкой LB-среды без антибиотиков. По достижению роста OD600 до 0.6, экспрессия была вызвана добавлением IPTG в 0.2 μМ. Культуры, экспрессирующие CsHCS1, затем выращивались при температуре 12°C с тряской в течение 24 часов, а культуры с CsHCS2 выращивались при температуре 37°С в течение 16 часов. Использовались разные температуры, т.к. было замечено, что CsHCS1 не формировала растворимый белок при более высоких температурах.
Клетки были собраны центрифугой и ресуспезированы в 10 мл His-tag лизисном буфере (50 mM Tris-HCl pH 7, 500 mM NaCl, 2.5 mM имидазол, 10% v/v глицерол, 10 mM β-меркаптоэтенол, 1% v/v Tween™ 20, и 750 мг/мл лизоцим). Ресуспензированные клетки инкубировались на льду в течение 1 часа, затем были разрушены ультразвуком. После центрифугирования в течение 20 мин. (12000 g, 4°С), была добавлена фракция растворимого белка в смолу 160 μл Talon™ (Clontech), которая до этого была промыта буфером для промывания His-tag (HWB; 50 mM Tris-HCl pH 7, 500 mM NaCl, 2.5 mM идидазол, 10% глицерол, 10 mM β-меркаптоэтанол). Образцы инкубировались при мягком покачивании и температуре 4°С, после чего смола была изолирована центрифугированием (700 g в течение 5 мин.). Смола была ресуспензирована в буфере HWB и промыта при мягком покачивании и температуре 4°С, затем центрифугирована. Этап промывания затем был повторен дважды и ресуспензированная смола была загружена в колону хроматографии и осущена. После промывании смолы в 10 мл буфера HWB, His-tag белки были элютированы с помощью добавления 2.5 мл His-tag буфера элюции (50 mM Tris-HCl рН 7, 500 mM NaCl, 250 mM имдазол, 10% v/v глицерол, 10 mМ β-меркаптоэтанол). Элюаты были помещены через буферный обмен в буфер хранения (50 mM HEPES pH 9, 10% v/v глицерол, 2 mM MgCl2, и 2 mM дитиотреитол), с помощью колонн PD10 (Amersham Biosciences). Чистота изолированных белков была проверена с помощью SDS-PAGE, и концентрация белка была определена с помощью метода Бредфорда.
Пример 2: Анализ активности гексаноил-кофермент-А-синтетазы
Активность гексаноил-кофермент-А-синтетазы была измерена инкубацией 0.1 мг энзима в 20 μл реактивной смеси, содержащей 50 mM HEPES pH 9, 8 mM АТР, 10 mM MgCl2, 0.5 mM кофермент-А, и 4 mM натрий гексаноат. Реакции проводились в инкубации в течение 10 мин. при температуре 40°С, закончились с с 2 μл 1 N HCl и хранились на льду до проведения анализа.
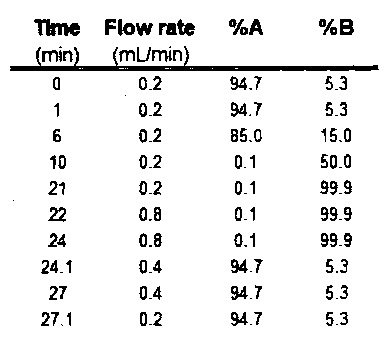
Реакционные смеси были разведены водой в пропорции 1:100 и затем отделены с помощью системы «Waters Acquity UPLC», снабженной колонной «Acquity UPLC ВЕН C18» (размер частиц 1.7 μm, колонна 2.1×50 mm), и проанализированы с помощью метода MS/MS, с использованием тройного квадрупольного спектрометра массы «Micromass Quattro Ultima». Растворителем был буфер А: 5 mM TEA и 3 mM уксусная кислота в воде, и буфер В: 5 mM TEA и 3 mM уксусной кислоты в 95:5 метанол : вода. Программа течения показана в Таблице 4. Установки спектрометра массы были следующими: ESI-положительный режим, энергия столкновения 27 V, конус 135 V, сканирование переходов 866>359.
Таблица 4: Программа потока жидкости хроматографии
Как показано на Фигуре 2, CsHCS1 и CsHCS2 катализировали образование гексаноил-кофермента-А из гексаноата и кофермента-А. На фигурах 2А и 2В показана элюция аутентичного стандарта гексаноил-кофермента-А. На Фигуре 2С показан полный анализ, состоящий из CsHCS1, 50 mM HEPES pH 9, 8 mM ATP, 10 mM MgCl2, 0.5 mM кофермент-А, и 4 mM натрий гексаноат. Сложные соединения с такими же массовыми переходами и временем элюции, как и аутентичный стандарт гексаноил-кофермента-А, могут быть увидены через 9.25 минут. На Фигуре 2B показан полный анализ, содержащий CsHCS2, 50 mM HEPES pH 9, 8 mM АТР, 10 mM MgCl2, 0.5 mM кофермент-А, и 4 mM натрий гексаноат. Сложные соединения с такими же массовыми переходами и временем элюции, как и аутентичный стандарт гексаноил-кофермента-А, могут быть увидены через 9.25 минут. На фигурах 2Е и 2F показаны негативные контроли с дезактивизированными (после кипячения) энзимами CsHCS1 и CsHCS2. На фигурах 2Е и 2F видно, что эти в этих тестах синтез гексаноил-кофермента-А не наблюдался.
Как CsHCS1, так и CsHCS2 показали оптимальную температуру и pH: 40°С и pH 9, соответственно. При тестировании ряда дивалентных катионов, CsHCS1 оптимально использовала Mg2+ и в меньшей степени Mn2+ и Со2+. Активность CsHCS2 была самой высокой при использовании Co2+, но также наблюдалась высокая активность с Mg2+, Mn2+, и в меньшей степени с Ca2+. Биологическая релевантность высокой активности с Со2+ еще пока не ясна, и Mg2+ использовалась во всех остальных анализах.
С гексаноатом, у CsHCS1 было Km=6.1±1.0 mM, Vmax=15.6±1.7 pKat и kcat=f 4.5 сек-1. У CsHCS2 было Km=320 нм, Vmax=1.7 pKat, и kcat=57.6 сек-1.
Пример 3: Тестирование с разными карбоновыми кислотами
Для тестирования ряда карбоновых кислот, которые могут активизировать CsHCS1 и CsHCS2, были осуществлены энзимные анализы целого ряда карбоновых кислот и ограничивающих АТР. Условия анализа были такими же как описано у Шнейдера К. и др. (2005). Новый тип пероксимальной акил-коэнзим-А синтетазы из Arabidopsis thaliana обладает каталитической способностью активировать биосинтетические прекурсоры жасмоновой кислоты. Журнал биологической химии, 280: 13962-72. Описывая процесс кратко, очищенный HCS-энзим (1 мг) инкубировался в 500 μM субстраты карбоновой кислоты и 100 μM кофермента-А при проведении анализа, содержащем 100 mM HEPES pH 9, 250 μM MgCl2, 50 μM ATP и 1 mM DTT. Все субстраты карбоновой кислоты были растворены в 2% v/v Triton™ Х-100, с получением конечной концентрации 0.05% Triton Х-100 при проведении анализа. После реакции, проводившейся в течение 3 часов при температуре 29°С, 10 μл аликвоты реакций были перемещены в 96-луночные плашки для измерения неиспользованного АТР с помощью люциферина/люциферазы. Плашки были проанализированы с помощью счетчика 1420 Multilabel (PerkinElmer). В каждую лугку были введены 90 μл раствора 100 mM Tris pH 7.8, 1 mM MgCl2, 2.3 мг люциферина, и 0.5 мг люциферазы, и после встряхивания в течение 2 секунд, была измерена люминесценция в течение 15 секунд без фильтра. Нижние значения, относительно реакций без субстраты карбоновой кислоты, показали высокий уровень энзимной активности, и, следовательно, использование субстраты. Результаты показаны на Фигуре 3, в котором столбики показывают процентную ошибку соотношения, n=3.
Как показано на Фигуре 3A, CsHCS1 использует гексаноат (шесть углеродов), октаноат (восемь углеродов), и нонаноат (девять углеродов), и в меньшей степени валериат (пять углеродов) и гептаноат (семь углеродов), в качестве субстратов. И напротив, как показано на Фигуре 3B, CsHCS2 демонстрирует большую неразборчивость, и способна использовать целый ряд субстратов, от пропаноата (С3) до арахидоата (С20), и ряд фенилпропаноидов (циннамат, ферулят, и в меньшей степени p-кумарат).
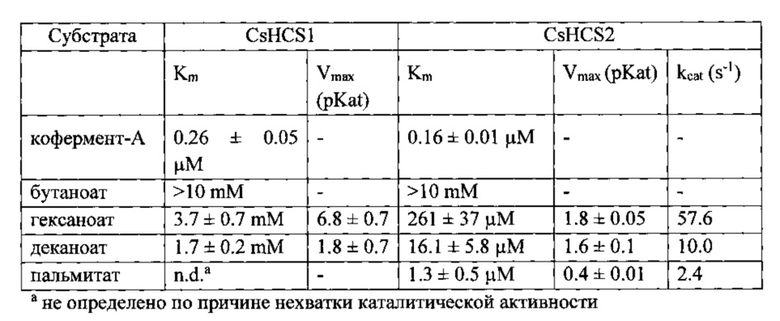
В отдельном эксперименте, кинетические свойства CsHCS1 и CsHCS3 были более точно измерены в отношении кофермента-А и репрезентативных жирных кислот короткой (бутаноат), средней (гексаноат и деканоат), и длинной (пальмитат) цепи (Таблица 5). Высокие концентрации кофермента-А ингибировали CsHCS1. При помощи нелинейной регрессирующей модели ингибирования субстраты, было оценено, что Ki в CsHCS1 равно 5.101±1.8 mM. Кофермент-А не ингибировал CsHCS2 в тестируемых концентрациях. Декановая кислота ингибировала CsHCS2 (Ki=120.8±47.9 μM) и незначительно ингибировала CsHCS1 в концентрациях выше 4 mM (Ki не измерялась). Эти данные свидетельствуют о таких же тенденциях, что и кинетические данные, представленные выше.
Таблица 5: Кинетические свойства CsHCS1 и CsHCS2.
Пример 4: Синтез оливетоловой кислоты, с помощью CsHCS2
CsHCS2 использовалась для хемоэнзимного синтеза ароматической поликетидной оливетоловой кислоты. Оливетоловая кислота - это первый специализированный прекурсор биосинтеза каннабиоидов. Именно в in vitro синтезе были применены четыре рекомбинантных энзима: CsHCS2, малонил-кофермент-А синтетаза (MCS) из Rhizobium leguminosarum, «оливетол-синтаза»/поликетид-синтаза из конопли, и синтаза оливетоловой кислоты из конопли.
Малонил-кофермент-А-синтетаза (MCS) была усилена из геномного ДНК Rhizobium leguminosarum затравкой прямой MCS и обратной MCS (см. Таблица 3). Продукт PCR был клонирован в вектор pCR8/GW/TOPO (Инвитроген), рекомбинированный в вектор pHIS8/GW. После верификации с помощью определения последовательности, плазмид был трансформирован в E.coli Rosetta II (DE3). Рекомбинантный MCS был экспрессирован и очищен, как описано в отношении CsHCS-энзимов.
Клонирование, экспрессия и очищение «оливетол-синтазы»/поликетид-синтазы и синтазы оливетоловой кислоты были реализованы следующим образом. В отношении экспрессии в клетках E.coli, открытые рамки считывания поликетид-синтазы/оливетол-синтазы и синтазы оливетоловой кислоты были усилены с помощью PCR, клонированы в pHIS8/GW при поликетид-синтазе/оливетол-синтазе, или pET100 (Инвитроген) при синтазе оливетоловой кислоты и трансформированы в E.coli BL21 (DE3) (Инвитроген). Клонирование было проверено с помощью определения последовательности.
Синтаза оливетоловой кислоты была экспрессирована в 200 мл бульонной культуре «terrifie broth», в то время как поликетид-синтаза/оливетол-синтаза выращивались в 1 L культуры. Обе культуры были инкубированы при температуре 30°С/150 rpm со встряхиванием, индуцированы с помощью 0.5 μM IPTG и выращены за ночь. Культуры прошли центрифугирование - 16,000 g в течение 20 мин., и пеллеты были лизированы лизозимом и ультразвуком. Очищенные лизаты были смешаны со смолой Talon (200 μL для синтазы оливетоловой кислоты, 1 мл для поликетид-синтазы/оливетол-синтазы; Clontech), промыты в 5 мл буфера для промывания His-tag (50 mM Tris-HCl (pH 7), 150 mM NaCl, 20 mM имдазол, 10 mM β-меркаптоэтанол) и рекомбинантные белки были элютированы с помощью буфера для элюции His-tag (20 mM Tris HCl (pH 7), 150 mM NaCl, 100 mM имдазол, 10 mM β-меркаптоэтанол). Элюат был сконцентрирован с помощью концентратора YM10 и через буферный обмен помещен в буфер хранения (20 mM HEPES (pH 7.5), 25 mМ NaCl, 10% глицерол, 5 mM DTT). Количество конечных белковых растворов были подсчитаны с помощью инструмента для анализа белков RC/DC (Bio-Rad), который установил концентрации белков на уровне 0.5 mg/мл (синтаза оливетоловой кислоты) и 5.6 mg/мл (поликетид синтаза/оливетол-синтаза). Гелевый анализ SDS-PAGE подтвердил чистоту обоих белков.
Возможность объединения синтеза гексаноил-кофермента-А с синтезом ароматического поликетида с помощью недорогих реагентов была проверена проведением энзимных анализов, в который входили 4 mM гексаноат, 8 mM малонат, 0.4 mM кофермент-А, 0.4 mM АТР, 5 mM MgCl2, 2 mM DTT, 20 mM HEPES pH 7.5, 0.3 мг CsHCS2, 12 мг MCS, 8 мг «оливетол-синтаза»/PKS и 10 мг OAS. Реакция была инкубирована при комнатной температуре в течение 16 часов, окислена и экстрагирована в этил-ацетат. Полярная фракция была восстановлена, выпарена до сухости и ресуспензирована в 50 μл метанола. Аликвота (5 μл) была проанализирована с помощью LCMS, и продукты были идентифицированы по их времени удержания и массам (см. Фигура 4).
Пример 5: Синтез оливетоловой кислоты в дрожжах с помощью CsHCS1 и CsHCS2
Дрожжи (Saccharomyces cerevisiae) были модифицированы для получения оливетоловой кислоты с помощью CsHCS1 и CsHCS2, для синтеза гексаноил-кофермента-А, и фузии «оливетол-синтазы»/поликетид-синтазы конопли (PKS) и синтазы оливетоловой кислоты (OAS) с целью получения оливетоловой кислоты.
OAS была клонирована в рамку с «оливетол-синтазой»/PKS, с помощью синтетической соединяющей последовательности, кодирующей аминокислоты AATSGSTGSTGSTGSGRSTGSTGSTGSGRSHMV (SEQ ID NO: 11) в дрожжевом экспрессирующем векторе pESC-Trp (Стратаген) под контролем промотора GAL10. Открытая рамка считывания CsHCS1 была клонирована в дрожжевой экспрессирующий вектор pYESDEST52-Ura (Инвитроген) с помощью технологии Шлюза. Открытая рамка считывания CsHCS2 была клонирована в pESC-Trp дрожжевой экспрессирующий вектор (Stratagene) под контролем промотора GAL1.
Дрожжевые клетки (InVSc I, Инвитроген) трансформировались вышеуказанной конструкцией (только фузия OAS:«оливетол-синтаза»/PKS, фузия OAS:«оливетол-синтаза»/PKS и CsHCS1; фузия OAS:«оливетол-синтаза»/PKS и CsHCS2) и трансформанты выращивались в плашке SD-Trp в течение 3 дней при температуре 28°С. В каждом случае, единственная колония была инокулирована в 3 мл SD-Trp глюкозную среду и инкубирована со встряхиванием при температуре 28°С в течение 2 дней. Аликвота 0.5 мл стартовой культуры использовалась для инокуляции 10 мл SD-Trp галактозной среды, содержащей 1 mM натрия гексаноата и инкубирована при температуре 20°С в течение 4 дней. Вся культура была экстрагирована этил-ацетатом, высушена и остаток был ресуспензирован в 100 μл раствора состоящего из: 30% ацетонитрил/70% вода/0.05% муравьиная кислота. Продукты были проанализированы с помощью LCMS (см. Фигура 5).
Пример 6: Роль CsHSC1 и CsHSC2 в биосинтезе каннабиоидов в растениях
В ходе анализа последовательности трихом массива данных EST и биохимического анализа пяти ААЕ, две проявили активность гексаноил-кофермент-А-синтетазы (CsHSC1 и CsHSC2). Для определения, которая из них с наибольшей вероятностью участвует в каскаде реакций биосинтеза каннабиоидов, были проведены qRT-PCR и эксперименты суб-клеточной локализации.
qRT-PCR экспрессии CsHSC1 и CsHSC2
Растения «Finola» были выращены со стадии семени до стадии среднего цветения. Были отобраны корни, стебли, листья, женские цветки (с трихомами и после изоляции трихом с помощью «Beadbeater»), клетки трихом и мужские цветки трех растений. Целая РНК была изолирована с помощью вышеописанного процесса. В этом РНК было: Abs260:Abs280 более 1.9, с проявлением четких рибосомных полос на денатурированном геле. сДНК первой нити были синтезированы с помощью 0.5 мг РНК, используя инструментарий «QuanitTect DNA Synthesis» (Qiagen). Каждый образец 20 μл сДНК был разведен водой 1:4, и 1 μл использовался как шаблон PCR. Были приготовлены ген-специфичные затравки для получения ампликонов 90-200 bp. Реакции PCR (20 μл) были проведны в 96-луночных плашках с помощью анализа на основе «SYBR Green» (набор «QuantiFast SYBR Green», Qiagen) вместе с инструментов «StepOne Plus» (Applied Biosystems). Использовались циклические параметры: 95°С в течение 5 мин., затем 40 циклов при 95°С в течение 10 с, 60°С в течение 30 с, и стандартный протокол диссоциации (95°С в течение 15 с, 60°С в течение 1 мин., 60-95°C с инкрементами по 0.3°С в течение 15 с). Эксперименты были осуществлены, используя сДНК трех растений с двумя техническими репликатами. Actin, который проявил стабильную экспрессию в всех тестируемых тканях, использовался как референс-ген. Эффективность во всех парах затравки была 90-110%, согласно расчету по стандартному методу кривой. Значения G были рассчитаны с помощью программного обеспечения «StepOne» (Applied Biosystems). Метод 2-ΔΔCt использовался для анализа относительной экспрессии гена.
Субклеточная локализация CsHSC1 и CsHSC2
Фузии YFP:CsHSC1 и YFP:CsHSC2 были сонструированы рекомбинацией в pEARLYGATE104 (Earley, K.W., Haag, J.R., Pontes, О., Opper, K., Juehne, Т., Song, K. и Pikaard, С.S. (2006). Векторы совместимые со шлюзами для растительных функциональных геномик и протеомик. Plant J. 45, 616-629.) с помощью LR-рекомбиназы (Инвитроген). Для генерации конструкции OLS:CFP, стоп-кодон с недостающим OLS был клонирован в pCR8/GW/TOPO до рекомбинации в pEARLYGATE102 с помощью LR-рекомбиназы. Пероксисомный маркер РХ-СK (Nelson, B.K., Cai, X. и Nebenführ, Α. (2007). Разноцветный набор in vivo органоидных маркеров для изучения колокализации в Arabidopsis и прочих растениях. Plant J. 51, 1126-1136.) был взят у ABRC (www.arabidopsis.org). Плазмиды были трансформированы в Agrobacterium tumefaciens GV3101 электропорацией и отобраны на LB плашках с 10 мг/мл рифампацина и 50 мг/мл канамицина. Листья двухнедельного растения Nicotiana benthamiana были инфильтрированы раствором Agrobacterium при OD600=0.02 (Sparkes, LA., Runions, J., Kearns, А. и Hawes, С. (2006). Скорая, проходящая экспрессия белков флуоресцентной фузии в растении табака и генерация стабильно трансформированных растений. Nat. Protocol. 1, 2019-2025.). Через два дня инфильтрации, эпидермальные клетки листьев были визуализированы с помощью конфокального микроскопа Zeiss LSM510. CFP была визуализирована возбуждением 458 нм и сбором изображения с полосовым фильтром 475-525 нм; YFP=514 нм с полосовым фильтром 530-600 нм. Изображения были собраны и проанализированы с помощью программного обеспечения Zeiss LSM.
Данные свидетельствуют о том, что CsHSC1 - это энзим, участвующий в биосинтезе каннабиоидов. CsHSC1 была самым распространенными транскриптом в каталоге EST, и данные qRT-PCR показывают, что ее экспрессия более, чем в 100 в трихомных клетках, чем в прочих тканях (Фигура 6). Кроме того, CsHSC1 локализуется в цитозоле, как было показано в эксперименте суб-клеточной локализации, т.е. в том же месте, где гипотетически локализуется каннабиоидный энзим OLS. Предпочтение субстраты CsHSC1 обеспечивает дополнительные сведения о ее роли в биосинтезе каннабиоидов, так как она демонстрирует большую специфичность для гексаноата и прочих короткоцепных жирных акил-кофакторов-А, чем CsHSC2 (Фигура 3).
Хотя CsHSC1 - это энзим, который с большей вероятностью будет участвовать в биосинтезе каннабиоидов в растениях, CsHSC2 более эффективна, чем CsHSC1 при синтезировании гексаноил-кофермента-А. Однако, CsHSC2 локализуется в пероксисоме, и не ясно, каким образом гексаноил-кофермент-А, сформированный в этом отсеке, может быть перенесен цитоплазму, где происходит фаза синтеза поликетидов в каннабиоидных реакциях. CsHSC2 принимает очень широкий ряд субстрат, и это означает, что это более обобщеная акил-кофермент-А-синтетаза, которая может функционировать в пероксимальном β-окислении.
CsHSC1 и CsHSC2 являются ценными промышленными инструментами. Нокаутирование CsHSC1 в конопле может привести к значительному снижению каннабиоидов в растении, что очень желательно для разведения конопли. Сверхэкспрессия CsHSC1 в конопле может привести к повышенному уровню каннабиоидов, что пригодно для фармацевтических целей. С другой стороны, CsHSC2 особенно пригодна при образовании каннабиоидов в микроорганизмах или в системе in vitro.
Пример 7: Генерация мутаций в генах CsHSC1 и CsHSC2, с помощью метода Целевых вызванных локальных поражений в геномах (TILLING)
Идентификация растения конопли с мутациями в генах CsHSC1 или CsHCS2 может быть осуществлена с помощью метода «TILLING». Мутагенизированная популяция конопли проходит скрининг с помощью олигонуклеотидных затравок и PCR с целью усиления интересующих генов. Усиленные мутантные гены комплементарно присоединяются к природным генам с целью нахождения несоответствий между мутантными генами и природными гены. Выявленные разницы используются для определения растений, которые содержат мутации в одном из генов - CsHSC1 или CsHCS2. Растения, содержащие мутации, которые ведут к изменению аминокислот в позициях, которые важны для стабильности или активность алканоил-кофермент-А-синтетазы белков CsHSC1 или CsHCS2, неспособны вырабатывать прекурсоры алканоил-кофермента-А для биосинтеза каннабиоидов. Получившиеся растения содержат сниженный или изменненный уровень каннабиоидов.
В настоящем изобретении описываются гены, который кодируют два энзима алканоил-кофермент-А-синтетазы конопли. Эти гены могут применяться для создания, через культивацию, целенаправленную мутагенную или генетическую инженерию, растений конопли с улучшенным образованием каннабиоидов. Кроме того, дезактивация или подавление гена изобретения в конопле может применяться для блокирования биосинтеза каннабиоидов, тем самым снижая формирование таких каннабиоидов, как ТНСА, прекурсора ТНС, в растениях конопли (например, в промышленно выращиваемой конопле). Гены настоящего изобретения могут применяться, в сочетании с генами, кодирующими прочие энзимы в каннабиоидном каскаде реакций, для осуществления биосинтеза каннабиоидов в прочих растениях, микроорганизмах или бесклеточных системах, или для получения аналогов каннабиоидных соединений или каннабиоидных прекурсоров.
В настоящем патентном раскрытии даются ссылки на публикации, содержание которых полностью включено в настоящий документ по упоминанию.
Прочие преимущества, которые характеризуют изобретение, очевидны специалисту. Воплощения, описанные в настоящем документе, иллюстративны и не должны ограничивать сферу изобретения только пунктами формулы изобретения. Вариации нижеописанных воплощений очевидны для обыкновенного специалиста, и включены изобретателями в следующие пункты формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СИНТАЗА КАННАБИХРОМЕНОВОЙ КИСЛОТЫ ИЗ КОНОПЛИ ПОСЕВНОЙ | 2015 |
|
RU2708144C2 |
| СПОСОБ ПОЛУЧЕНИЯ 3-ГИДРОКСИ-3-МЕТИЛМАСЛЯНОЙ КИСЛОТЫ ИЗ АЦЕТОНА И АЦЕТИЛ-СОА | 2010 |
|
RU2573389C2 |
| ПОЛУЧЕНИЕ ДГК И ДРУГИХ ДЦ-ПНЖК В РАСТЕНИЯХ | 2012 |
|
RU2644241C2 |
| ИЗОПРОПИЛМАЛАТ СИНТАЗА ИЗ NICOTIANA TABACUM И СПОСОБЫ И ЕЕ ПРИМЕНЕНИЕ | 2012 |
|
RU2644238C2 |
| ТРАНСГЕННЫЕ РАСТЕНИЯ, ЭКСПРЕССИРУЮЩИЕ ГЕН АЦЕТИЛ-СОА-КАРБОКСИЛАЗЫ | 1996 |
|
RU2187555C2 |
| ПРОМОТОР ЦЕЛОГО СЕМЕНИ | 2010 |
|
RU2561463C2 |
| ГЛОБАЛЬНЫЕ РЕГУЛЯТОРЫ БАКТЕРИАЛЬНЫХ ПАТОГЕННЫХ ГЕНОВ, ПРОТЕИН ИНАКТИВАЦИИ БАКТЕРИАЛЬНЫХ АУТОИНДУКТОРОВ, ЯВЛЯЮЩИХСЯ МИШЕНЯМИ ДЛЯ СОЗДАНИЯ УСТОЙЧИВОСТИ К БОЛЕЗНЯМ | 1999 |
|
RU2236462C2 |
| РАСТЕНИЯ ПШЕНИЦЫ С ПОВЫШЕННОЙ УСТОЙЧИВОСТЬЮ К ИМИДАЗОЛИНОНОВЫМ ГЕРБИЦИДАМ | 2002 |
|
RU2337532C2 |
| РАСТЕНИЯ ПШЕНИЦЫ С ПОВЫШЕННОЙ УСТОЙЧИВОСТЬЮ К ИМИДАЗОЛИНОНОВЫМ ГЕРБИЦИДАМ | 2002 |
|
RU2337531C2 |
| СПОСОБ УВЕЛИЧЕНИЯ СОДЕРЖАНИЯ СЛОЖНОГО ЭФИРА САХАРОЗЫ В РАСТЕНИЯХ ТАБАКА | 2018 |
|
RU2758117C2 |












Изобретение относится к области биотехнологии и биохимии, а именно к молекулам нуклеиновой кислоты и белков, участвующих в синтезе тиоэфиров алканоил-кофермента-А, и к использованию молекул и белков нуклеиновой кислоты для инженерного биосинтеза каннабиоидов в растениях, микроорганизмах или бесклеточных системах. Изобретение позволяет контролировать синтез каннабиноидов в организме, клетке или ткани, и получить, в зависимости от цели, организм клетку или ткань с повышенным или пониженным содержанием каннабиоидов. 11 н. и 9 з.п. ф-лы, 6 ил., 5 табл., 7 пр.
1. Изолированная или очищенная молекула нуклеиновой кислоты, характеризующаяся нуклеотидной последовательностью SEQ ID NO: 1 или ее кодоновой вырожденной нуклеотидной последовательностью, при этом молекула нуклеиновой кислоты кодирует полипептид, катализирующий синтез алканоил-кофермента-А (алканоил-СоА) из кофермента-А (СоА) и алканоата, выбранного из гексаноата, октаноата, нонаноата, валериата и гептаноата.
2. Молекула нуклеиновой кислоты по п. 1, характеризующаяся тем, что полипептид имеет активность гексаноил-кофермент-А-синтетазы.
3. Изолированный или очищенный полипептид, характеризующийся аминокислотной последовательностью SEQ ID NO: 2, или консервативно замещенной аминокислотной последовательностью, по меньшей мере на 85% идентичной последовательности SEQ ID NO: 2, при этом полипептид катализирует синтез алканоил-кофермента-А (алканоил-СоА) из кофермента-А (СоА) и алканоата, выбранного из гексаноата, октаноата, нонаноата, валериата и гептаноата.
4. Полипептид по п. 3, характеризующийся тем, что он имеет активность гексаноил-кофермент-А-синтетазы.
5. Экспрессирующий вектор, включающий молекулу нуклеиновой кислоты по любому из пп. 1 и 2.
6. Экспрессирующая система, включающая молекулу нуклеиновой кислоты по любому из пп. 1 и 2.
7. Клетка-акцептор, трансформированная молекулой нуклеиновой кислоты, по любому из пп. 1 и 2 для синтезирования каннабиоидного соединения.
8. Способ синтезирования алканоил-СоА, включающий реакцию карбоновой кислоты с СоА в присутствии полипептида по любому из пп. 3 и 4, при этом в качестве карбоновой кислоты используют гексаноат, октаноат, нонаноат, валериат или гептаноат.
9. Способ по п. 10, характеризующийся тем, что алканоил-СоА содержит гексаноил-СоА.
10. Способ снижения уровня каннабиоидных соединений в организме, клетке или ткани, включающий использование RNAi молекулы нуклеиновой кислоты комплементарно участку SEQ ID N0:1 молекулы нуклеиновой кислоты по любому из пп. 1 и 2, или ее части, при этом RNAi молекула нуклеиновой кислоты содержит фрагмент, по меньшей мере, в 21 нуклеотид, функционально способный подавлять экспрессию генов в организме, клетке или ткани, который кодирует энзим, катализирующий синтез алканоил-СоА, относительно подобного вида организма, клетки или ткани, выращенных в одинаковых условиях без использования RNAi молекулы нуклеиновой кислоты комплементарно участку SEQ ID NO: 1.
11. Способ повышения уровня каннабиоидных соединений в организме, клетке или ткани, путем экспрессии или сверхэкспрессии экзогенной молекулы нуклеиновой кислоты по любому из п. 1, 2, в организме, клетке или ткани, относительно подобного вида организма, клетки или ткани, выращенных в одинаковых условиях, но без экспрессии или сверхэкспрессии экзогенной молекулы нуклеиновой кислоты, при этом организм, клетка или ткань способны к биосинтезу каннабиоидных соединений.
12. Способ повышения уровня каннабиоидных соединений в организме, клетке или ткани, путем экспрессии или сверхэкспрессии экзогенной молекулы нуклеиновой кислоты, кодирующей полипептид, по любому из пп. 3 и 4, в организме, клетке или ткани, относительно подобного вида организма, клетки или ткани, выращенных в одинаковых условиях, но без экспрессии или сверхэкспрессии экзогенной молекулы нуклеиновой кислоты, при этом организм, клетка или ткань способны к биосинтезу каннабиоидных соединений.
13. Способ синтезирования каннабиоидного соединения или аналога каннабиоидного соединения в организме, клетке или ткани, с помощью экспрессии молекулы нуклеиновой кислоты по любому из пп. 1 и 2 в организме, клетке или ткани в присутствии карбоновой кислоты и СоА, при этом в качестве карбоновой кислоты использован гексаноат, октаноат, нонаноат, валериат или гептаноат.
14. Способ по п. 11-13, характеризующийся тем, что организм или клетка представляют собой микроорганизм.
15. Способ по п. 14, характеризующийся тем, что в качестве микроорганизма используют дрожжи Saccharomyces cerevisiae или Е. coli.
16. Способ по п. 11-13, 15, характеризующийся тем, что молекула нуклеиновой кислоты экспрессируется или сверхэкспрессируется, в сочетании с экспрессией или сверхэкспрессией одной или нескольких других нуклеиновых кислот, кодирующих один или несколько энзимов в каннабиоидном биосинтетическом каскаде реакций.
17. Способ по п. 16, характеризующийся тем, что один или несколько энзимов в каннабиоидном биосинтетическом каскаде реакций представлены одной или несколькими синтазами оливетоловой кислоты, поликетид-синтазой III типа, поликетид-циклазой, ароматической пренилтрансферазой или каннабиоид-формирующей оксидоциклазоой.
18. Способ по п. 17, характеризующийся тем, что один или несколько энзимов в каннабиоидном биосинтетическом каскаде реакций - это одна или несколько поликетид-синтаз III типа/оливетол-синтаза, геранилпирофосфат: оливетолат-геранилтрансфераза, синтаза Δ9-тетрагидроканнабиноловой кислоты, синтаза каннабидиоловой кислоты или синтаза каннабихроменовой кислоты.
19. Способ по пп. 10 и 13, 15, 17, 18, характеризующийся тем, что каннабиоидное соединение - это одна или несколько каннабигероловых кислот, Δ9-тетрагидроканнабиноловая кислота, каннабидиоловая кислота, каннабихроменовая кислота, Δ9-тетрагидроканнабинол, каннабидиол или каннабихромен или его аналог, содержащий боковую цепь длиною в 1-9 атомов углерода.
20. Способ синтезирования алканоил-СоА в бесклеточной реакции in vitro, включающий реагирование карбоновой кислоты с коэнзимом-А с помощью использования полипептида по любому из пп. 3 и 4, при этом в качестве карбоновой кислоты использован гексаноат, октаноат, нонаноат, валериат или гептаноат.
| PAGE, J.E., NAGEL J., Biosynthesis of terpenophenolic metabolites in hop and cannabis, Recent Advances in Phytochemistry, 2006 | |||
| FELLERMEIER M | |||
| et al., Biosynthesis of cannabinoids, Eur | |||
| J | |||
| Biochem., 2001 | |||
| RU 94002209 A1, 10.04.1996. |

