Перекрестные ссылки на родственные заявки
Отсутствует.
Федеральная финансовая поддержка исследования или разработки
Отсутствует
Предпосылки создания изобретения
1. Область техники изобретения
Настоящее изобретение относится к глобальным регуляторам бактериальных патогенных генов и к их применению для придания устойчивости к болезням.
2. Описание прототипов
Список литературы приведен после раздела “Подробное описание изобретения”. Перечисленные документы полностью включены в настоящее описание в качестве ссылок.
Взаимодействие клетка-клетка посредством малых сигнальных молекул является жизненно важным не только для живых многоклеточных организмов, таких как животные и растения, но оно также играет важную роль в координации функций одноклеточных организмов, таких как бактерии. Интенсивные исследования, проведенные в последние годы, позволили точно установить, что N-ацилгомосеринлактоны, называемые аутоиндукторами (АИ), представляют собой высококонсервативиые сигнальные молекулы в грамотрицательных бактериях. Сначала было установлено, что АИ участвуют в регуляции биолюминисценции морских бактерий вида Vibrio (Eberhard и др., 1981; Сао и Meighen, 1989). В последние годы АИ были обнаружены во многих видах грамотрицательных бактерий. Было установлено, что АИ участвуют в регуляции разнообразных биологических функций, включая опосредуемый Ti-плазмидой конъюгативный перенос в Agrobacterium tumefaciens (Zhang и др., 1993), индукцию вирулентных генов в Erwinia carotovora, Pseudomonas aeruginosa, Erwinia stewartii, Xenorhabdus nematophilus, Erwinia chrysanthemi, Pseudomonas solanacerum и Xanthomonas campestris (Jones и др., 1993; Passador и др., 1993; Pirhonen и др., 1993; Pearson и др., 1994; Beck von Bodman и Farrand, 1995; Barber и др., 1997; dough и др., 1997; Costa и Loper, 1997; Dunphy и др., 1997; Nasser и др., 1998), регуляцию производства антибиотиков в Pseudomonas aureofaciens и Erwinia carotovora (Pierson и др., 1994; Costa и Loper, 1997), регуляцию движения расселительной стадии у Serratia liquifaciens (Eberi и др., 1996) и образование биологических пленок у Pseudomonas fluorescens и Р. aeruginosa (Allison и др., 1998; Davies и др., 1998). Известно, что многие другие виды бактерий продуцируют АИ, но опосредуемые ими биологические функции до сих пор не выяснены (Bassler и др., 1997; Dumenyo и др., 1998; Cha и др., 1998).
Различные виды бактерий могут продуцировать различные АИ. Все производные АИ содержат одинаковые гомосеринлактоновые фрагменты, но могут иметь различия в длине и структуре содержащихся в них ацильных групп. Основными компонентами в регуляторных системах опосредуемых АИ генов, являются протеины типа Luxl и LuxR. При создании настоящего изобретения было установлено, что протеин типа Luxl служит в качестве аутоиндукторной синтазы, которая использует в качестве субстратов ацил-АСР и AdoMet (S-аденозилметионин) (More и др., 1996; Schaefer и др., 1996). Предполагается, что протеин типа LuxR является как рецептором для АИ, так и зависящим от АИ регулятором транскрипции, который связывается с ДНК непосредственно против хода транскрипции от промотора lux (Meighen, 1994; Sitnikov и др., 1995). Был идентифицирован состоящий из 20 нуклеотидов инвертированный повтор, который расположен внутри состоящей из 44 нуклеотидов области, расположенной против хода транскрипции от сайта инициации транскрипции люминисцентного оперона. Эта последовательность, называемая lux-боксом, необходима для активации транскрипции с помощью LuxR и, по-видимому, представляет собой сайт связывания LuxR (Fugua и др., 1994). Аналогичные tra-боксы, состоящие из 18 пар оснований, были обнаружены против хода транскрипции относительно по крайней мере трех TraR-регулируемых промоторов, и было установлено, что разрушение этих элементов приводит к ликвидации активации транскрипции с помощью TraR (Fugus и Winans, 1996a).
Протеины LuxR-типа, по-видимому, состоят из двух модулей (Choi и Greenberg, 1991; Hanzelka и Greenberg, 1995). Их С-концевые области содержат консервативную короткую последовательность, состоящую из 19 аминокислот, предположительно представляющую собой мотив спираль-виток-спираль типа зонда, которая предположительно участвует в связывании с промоторами-мишенями. Был предложен механизм активации, согласно которому N-концевой домен протеина LuxR-типа обладает отрицательной активностью, препятствуя взаимодействию его С-концевого домена с сайтами связывания ДНК-мишени. Это ингибирование может уменьшаться под действием лиганда аутоиндуктора. Убедительное доказательство этого заключается в том, что делеция N-концевого домена LuxR приводит к образованию обладающих конститутивной активностью аллелей luxR, в то время как более крупные делеции, в результате которых удаляется часть предсказанного ДНК-связывающего домена, приводят к ликвидации активации транскрипции (Choi и Greenberg, 1991). Однако другие представители этих протеинов могут обладать иными механизмами действия. Результаты генетических исследований, выполненных в последние годы, свидетельствуют о том, что EsaR и EspR, по-видимому, являются репрессорами, а не активаторами соответствующих генов-мишеней. Экспрессия генов, репрессорами которых являются EsaR и EspR, увеличивается благодаря действию аутоиндукторов (Beckvon Bodman и Farrand, 1995; Throup и др., 1995). По-видимому, связывание этих протеинов с их сайтами-мишенями в области промотора приводит к репрессорному действию, следовательно лиганды аутоиндуктора могут вызывать уменьшение аффинности связывания.
Было установлено, что сайт связывания аутоиндуктора располагается в аминоконцевом домене протеина LuxR (Hanzelka и Greenberg, 1995). Аллели LuxR, имеющие мутацию в аминоконцевой области, требуют более высокого уровня этого сигнала, чем протеин дикого типа, что свидетельствует о том, что эта область необходима для взаимодействия с лигандом (Slock и др., 1990; Shadel и др., 1990). Эта область (аминокислоты 79-127) и область, располагающаяся внутри ДНК-связывающего домена (аминокислоты 180-230), обладают более высоким уровнем консервативности среди LuxR и его гомологов (обладающих приблизительно 50%-ной идентичностью), чем другие участки этих полипептидов. Однако до настоящего времени не выявлено предполагаемое взаимодействие типа протеин-лиганд между LuxR и аутоиндуктором. Анализ меродиплоидных штаммов E.coli, содержащих аллели LuxR дикого типа и мутантные аллели, позволил предположить, что LuxR обладает функциями гомомультимера и что область, необходимая для мультимеризации, располагается между аминокислотными остатками 116 и 161 (Choi и Greenberg, 1992).
Краткое изложение сущности изобретения
Одним из объектов настоящего изобретения является выделенная молекула нуклеиновой кислоты, кодирующая протеин инактивации бактериальных аутоиндукторов.
Следующим объектом настоящего изобретения является экспрессионный вектор, содержащий молекулу нуклеиновой кислоты, кодирующую протеин инактивации бактериальных аутоиндукторов, где экспрессионный вектор обладает способностью размножаться в прокариотических или эукариотических клетках.
Еще одним объектом настоящего изобретения является прокариотическая или эукариотическая клетка, трансформированная или трансфектированная экспрессионным вектором по настоящему изобретению.
Еще одним объектом настоящего изобретения является выделенный протеин, обладающий способностью инактивировать бактериальную аутоиндукцию, где протеин включает аминокислотную последовательность SEQ ID NO:2.
Еще одним объектом настоящего изобретения является способ повышения устойчивости к болезням растения или животного, заключающийся в том, что в клетку такого растения или животного вводят нуклеотидную последовательность, которая кодирует протеин инактивации бактериальных аутоиндукторов, таким образом, что указанная клетка обладает способностью экспрессировать указанную нуклеотидную последовательность.
Еще одним объектом настоящего изобретения является способ предупреждения или уменьшения вызванного бактериями повреждения растения или животного, заключающийся в том, что в растение или животное, нуждающееся в таком предупреждении или уменьшении, вводят эффективное количество протеина инактивации бактериальных аутоиндукторов.
Еще одним объектом настоящего изобретения является композиция, предназначенная для уменьшения повреждения бактериями растения или животного, включающая
а) эффективное количество протеина инактивации бактериальных аутоиндукторов; и
б) пригодный носитель.
Краткое описание чертежей
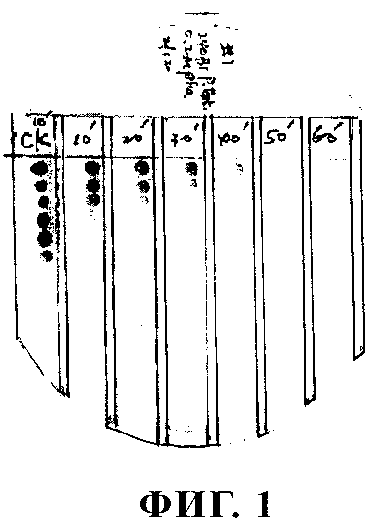
На фиг.1 представлена зависимость от времени инактивации АИ клеточными экстрактами штамма Bacillus sp. 240BI. Клеточные экстракты в 0,2М фосфатном буфере (рН 7,0), содержащем 100 мкг общего протеина, добавляли к такому же буферу, содержащему OHHL (N-β -оксогексаноил-L-гомосеринлактон) в конечной концентрации 20 мкМ. Реакцию осуществляли в центрифужной пробирке Эппендорфа объемом 1,5 мл в конечном объеме 200 мкл и смесь инкубировали при 28°С. В качестве контроля использовали такую же концентрацию OHHL в фосфатном буфере. Образцы отбирали с 10-минутными интервалами в течение 60 мин и реакцию прекращали путем кипячения в течение 3 мин. Образцы центрифугировали в течение 5 мин на лабораторной центрифуге при максимальной скорости и затем анализировали в отношении АИ-активности согласно описанному ранее методу (Zhang, 1993). Голубой цвет колоний свидетельствовал о наличии АИ, активирующего репортерный ген lacZ, в то время как белый цвет колоний свидетельствовал об отсутствии АИ. Ряды слева направо: 1 - OHHL-контроль, не содержащий протеинового экстракта; 2 - 7 - образцы после проведения ферментативной реакции в течение 10, 20, 30, 40, 50, 60 мин.
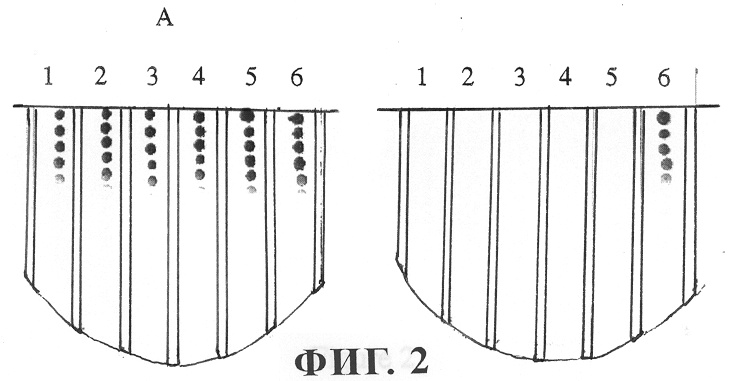
На фиг.2 представлены результаты оценки молекулярной массы фермента инактивации АИ. Аликвоты клеточных экстрактов по 600 мкл вносили в устройство типа Centricon 30 (фирма Amicon) и центрифугировали при скорости 5000 xg в течение 30 мин при 4°С. Фракцию, прошедшую через фильтр (прошедшую фракцию) (550 мкл), и непрошедшую фракцию (50 мкл) по отдельности доводили до конечного объема 600 мкл путем добавления 0,2М фосфатного буфера (рН 7,0). Для осуществления биологического анализа в пробирки, содержащие OHHL в конечной концентрации 20 мкМ добавляли различные количества образцов протеина. Для рядов с 1 по 6 добавляли 2, 4, 6, 8, 10 и 0 мкл образцов протеина, при этом конечный объем для каждой реакционной смеси составлял 20 мкл. Панель А: прошедшая фракция, панель Б: непрошедшая фракция.
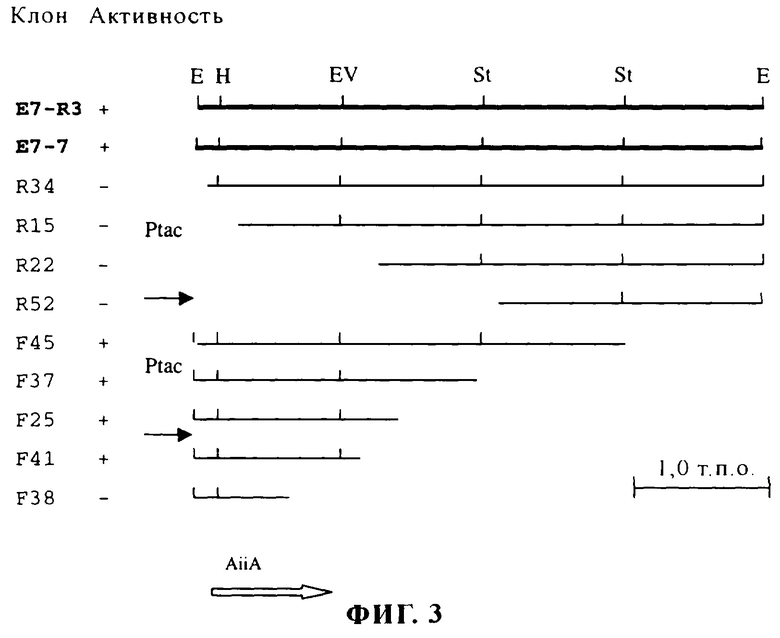
На фиг.3 представлены результаты клонирования и делеционного анализа области инактивации АИ штамма Bacillus sp. 240B 1. Космидный клон E7-R3 содержит EcoRI-фрагмент длиной 4,3 т.п.н., идентифицированный с помощью рестрикционного анализа перекрывающихся космидных клонов. Для осуществления делеционного анализа этот фрагмент клонировали в клонирующем векторе pGEM-7Zf(+) с целью получения клона Е7-7. Субклоны, содержащие делеции, получали из клона Е7-7 путем ферментативного расщепления и обработки ДНказой I. Положение и ориентация Ptac-промоторов в космиде и в клоне pGEM-7Zf(+) указаны стрелками. Данные о способности клонов к инактивации АИ показаны во второй колонке: "+" обозначает наличие инактивирующей активности в отношении АИ, "-" обозначает отсутствие инактивирующей активности в отношении АИ. Рестриктазы: Е - EcoRl; Н - Hin-dlll; Ev - EcoRV, St - Styl. Местоположение и направление транскрипции OPC aiiA показаны незакрашенной стрелкой.
На фиг.4А представлена нуклеотидная последовательность гена aiiA (SEQ ID NO: 1). Возможная последовательность сайта связывания рибосомы и элемент промотора-10 подчеркнуты одной и двумя линиями соответственно. Кодирующая область начинается от основания 1. Предполагаемый независящий от фактора сайт терминации подчеркнут жирной линией. На фиг.4Б представлена предполагаемая аминокислотная последовательность продукта гена aiiA (SEQ ID NО:2). Подчеркнута короткая последовательность пептида, сходная с консенсусным мотивом активного сайта аспартилпротеазы.
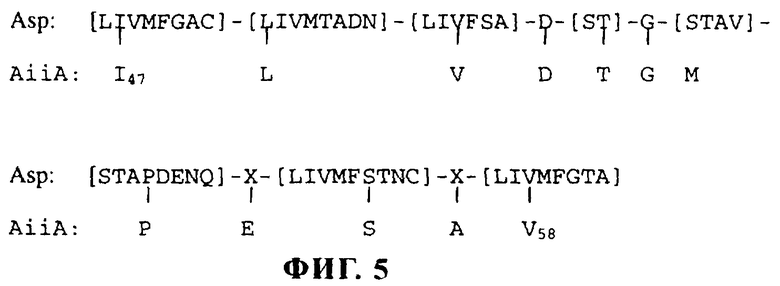
На фиг.5 показано наилучшее совпадение аминокислотной последовательности продукта гена aiiA (AiiA) с консенсусным мотивом активного сайта аспартилпротеазы (Asp). Символ Х обозначает любую аминокислоту. Вертикальная линия обозначает идеальное совпадение.
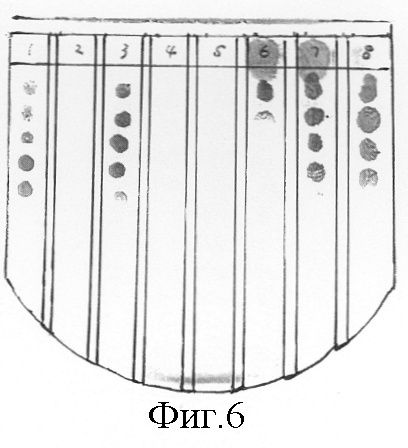
На фиг.6 представлены результаты биологического анализа активностей в отношении инактивации АИ в клоне Bacillus sp. 240B 11, клонах Ecoli и активности производства АИ в штаммах Erwinia carotovora. Ряд 1: OHHL-контроль, ряд 2: Bacillus sp. 240B11; ряд 3: штамм E.coli DH5α ; ряд 4: штамм E.coli DH5α (pE7-R3); ряд 5: штамм E.coli DH5α (pE41); ряд 6: штамм Erw. carotovora SCG1 (pE7R3); ряд 7: штамм Erw. carotovora SCG1 (pLAFR3); ряд 8: штамм Е. carotovora SCG1. При осуществлении биологического анализа OHHL добавляли к образцам, соответствующим рядам 1-5 в конечной концентрации 20 мкМ. К образцам, соответствующим рядам 6-8, экзогенный АИ не добавляли.
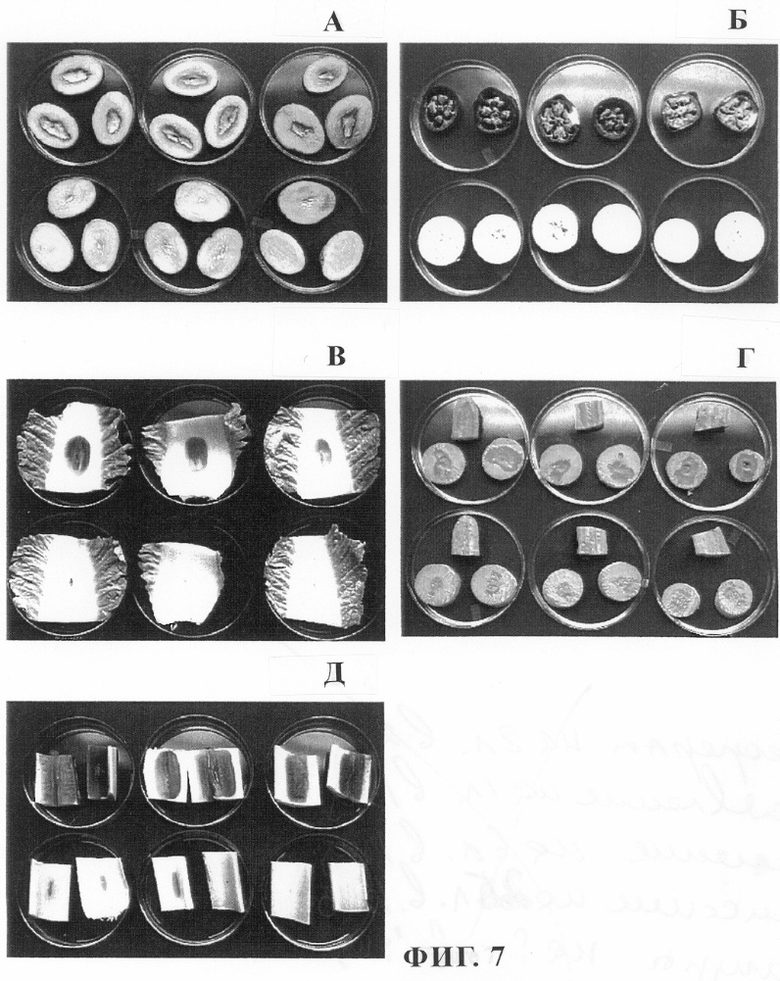
На фиг.7 проиллюстрировано влияние экспрессии гена aiiA в Erw. carotovora на патогенность в отношении (А) картофеля; (Б) баклажана; (В) капусты китайской; (Г) моркови и (Д) сельдерея. Вверху: растительные ткани, инокулированные штаммом Erw. carotovora SCG1. Внизу: растительные ткани, инокулированные штаммом Erw. carotovora SCG1 (pE7-R3). Активно растущие бактерии центрифугировали в течение 1 мин при 3000 xg, ресуспендировали с помощью жидкой среды YEB до достижения оптической плотности ОП600=1,3 (2× 109 КОЕ/мл), и продукт обозначали как инокулят 10° . Инокулят 100 разбавляли в 5 и 10 раз соответственно, получая разведения 10-1/2- и 10-1 соответственно. Ткани растений инокулировали путем добавления 4 мкл инокулята бактерий на поверхность свежего среза или в область повреждения, образованного путем протыкания ткани кончиком пипетки. Концентрации инокулята составляли (слева направо): 100, 10-1/2 и 10-1. Инокулированные ткани растений помещали в пластиковые чашки и инкубировали при 28°С. Фотографирование осуществляли через 48 ч после инокуляции.
Подробное описание изобретения
Настоящее изобретение основано на открытии того, что протеин, имеющий последовательность, представленную в SEQ ID NO: 2, обладает способностью уменьшать или устранять активность бактериальных аутоиндукторов (АИ). Благодаря этому протеин и любая нуклеотидная последовательность, кодирующая протеин, могут использоваться в различных ситуациях, когда требуется уменьшить или устранить воздействие таких бактерий.
Одним из предпочтительных объектов настоящего изобретения является молекула нуклеиновой кислоты, выбранная из группы, включающей:
а) нуклеиновую кислоту, имеющую последовательность SEQ ID NO:1;
б) нуклеиновую кислоту, кодирующую аминокислотную последовательность SEQ ID NO:2; и
в) нуклеиновую кислоту, которая гибридизуется с нуклеиновыми кислотами, указанными в подпунктах а) или б), при этом позитивный сигнал гибридизации наблюдается после однократной промывки SSC (IxSSC) и 0,1%-ным ДСН при 55°С в течение 1 ч. Нуклеиновая кислота необязательно дополнительно содержит кодирующую область сигнального пептида любой из указанных последовательностей.
Нуклеотидную последовательность можно использовать для придания устойчивости растений или животных к бактериям. Нуклеиновую кислоту, кодирующую протеин инактивации бактериальных аутоиндукторов, можно встраивать в клетку таким образом, чтобы осуществлялась экспрессия инактивирующего протеина в клетках растения или животного.
Нуклеотидную последовательность можно использовать для придания устойчивости к болезням, при которых экспрессия патогенных генов регулируется аутоиндукторами, таким как болезни, вызываемые Pseudomonas aeruginosa, Erwinia stewartii, Xenorhabdus nematophilus, Erwinia chrysanthemi, Pseudomonas solanacerum и Xanthomonas campestris (Passador и др., 1994; Pirhonen и др., 1993; Pearson и др., 1994; Beck von Bodman и Farrand, 1995; Barber и др., 1997; Clough и др., 1997, Costa и Loper, 1997; Dunphy и др., 1997; Nasser и др., 1998). Предпочтительно в сельскохозяйственной области указанные последовательности можно применять для придания устойчивости к заболеванию мокрой гилью чувствительным растениям, таким как картофель, баклажаны, китайская капуста, морковь и сельдерей.
Последовательность можно встраивать в клетки растений или животных хорошо известными методами. Методы трансформации или трансфекции эукариотических клеток экзогенными нуклеотидными последовательностями включают трансфекцию, бомбардировку микроснарядами, электропорацию или заражение с использованием Agrobacterium tumefaciens. Эти методы известны специалистам в области молекулярной биологии и биотехнологии и не требуют подробных пояснений. Поскольку клетки патогенных бактерий располагаются во внутриклеточной области растительных тканей, то целесообразно осуществлять направленный перенос протеина AiiA во внутриклеточное пространство. Это можно осуществлять путем слияния сигнального пептида секреции с протеином AiiA (Sato и др., 1995; Firek и др., 1993; Conrad и Fiedler, 1998; Borisjuk и др., 1999). В альтернативном варианте мотив, обеспечивающий прикрепление к мембране клетки растения, можно встраивать в пептидную последовательность AiiA для связывания AiiA-фермента с внешней поверхностью мембраны растительной клетки.
В настоящем изобретении предложена новая стратегия создания устойчивости к болезням. В частности, мишенью этой стратегии являются N-ацилгомосеринлактоновые аутоиндукторы, которые индуцируют экспрессию патогенных генов многих бактериальных патогенов в пороговой концентрации. Эта стратегия может применяться для всех болезней растений, животных или млекопитающих, при которых экспрессия патогенных генов бактериальных патогенов индуцируется N-ацилгомосеринлактоновыми аутоиндукторами.
В настоящем изобретении также предложено использовать протеин инактивации бактериальных аутоиндукторов непосредственно для лечения или предупреждения бактериального повреждения. Например, протеин можно вводить непосредственно в растения, если это необходимо для такого лечения или предупреждения. В предпочтительном варианте протеин применяют в форме композиции, которая содержит эффективное количество протеина и пригодный носитель. Композиции могут иметь разнообразные формы, включая растворы, порошки, эмульсии, дисперсии, пасты, аэрозоли и т.д.
Протеин инактивации бактериальных аутоиндукторов может также применяться для лечения бактериальных инфекций у животных, включая людей. При таком применении животному, нуждающемуся в таком лечении, вводят эффективное количество действующего вещества.
Для терапевтического лечения на основе действующего вещества можно приготавливать фармацевтическую композицию, которая может включать в дополнение к эффективному количеству действующего вещества фармацевтически приемлемые носители, разбавители, буферы, консерванты, поверхностно-активные вещества и т.п. Композиции могут также включать одно или несколько других действующих веществ, если это является необходимым или целесообразным.
Фармацевтические композиции по настоящему изобретению можно вводить различными путями, что является очевидным для специалиста в данной области. Например, можно использовать местное, пероральное введение, введение путем ингаляции или парентеральное введение.
Композиции для местного применения могут включать мази, лосьоны, кремы, гели, капли, суппозитории, спрей, жидкости и порошки. Композиции для перорального введения включают, например, порошки, гранулы, суспензии или растворы в воде или неводных средах, капсулы или таблетки. При необходимости можно использовать загустители, корригенты, разбавители, эмульгаторы, диспергирующие или связующие вещества.
Композиции для парентерального введения могут включать стерильные водные растворы, которые могут также содержать буферы, разбавители и другие пригодные добавки.
Режим доз должен зависеть от многих факторов, которые легко могут быть установлены, таких как серьезность и реактивность состояния, подлежащего лечению.
Ниже изобретение проиллюстрировано на примерах, не ограничивающих его объем.
Пример 1
Бактериальный изолят 240В 1 выделяли из суспензии почвы на основе его способности инактивировать N-β -оксогексаноил-L-гомосеринлактон (OHHL) и N-β -оксооктаноил-L-гомосеринлактон (OOHL) и N-β -оксодеканоил-L-гомосеринлактон (ODHL) (Zhang и др., 1993). Если не указано иное, для стандартного биологического анализа использовали OHHL. Штамм Erwinia carotovora SCG1 выделяли из листа китайской капусты, имеющего симптомы мокрой гнили. Было установлено, что штамм SCG1 продуцирует АИ и вызывает заболевание мокрой гнилью картофеля и китайской капусты. В качестве хозяина для клонирования и субклонирования ДНК использовали штамм Escherichia coli DH5α . В качестве индикаторного штамма при биологическом анализе АИ-активности использовали штамм Agrobacterium tumefaciens NT1 (traR; tra::lacZ749) (Piper и др., 1993). Штамм E.coli выращивали в среде Луриа-Бертани (LB) при 37°С, а другие штаммы культивировали в LB-среде (Miller, 1972) или в среде YEB (которая содержала на 1 л: 10 г гидролизата казеина, 5 г дрожжевого экстракта, 10 г NaCl, 5 г сахарозы, 0,5 г MgSO4·7H20, 15 г агара, рН 7,2) при 28°С. Для биологического анализа OHHL (Petit и Тетре, 1978) использовали минимальноую солевую среду, дополненную маннитом и (NH4)2SО4 в качестве источника углерода и азота. Добавляли соответствующие антибиотики в следующих концентрациях: ампициллин - 100 мкг/мл; тетрациклин - 20 мкг/мл и канамицин - 50 мкг/мл.
Биологический анализ активности АИ
Ранее были описаны качественные и количественные методы биологического анализа, которые могут применяться для определения активности АИ (Zhang, 1993). Для определения способности штамма дикого типа и генетически модифицированных штаммов Erwinia продуцировать АИ применяли такой же метод биологического анализа, за исключением того, что в бактериальные культуры не добавляли OHHL.
Клонирование и секвенирование гена AiiA
Геномную ДНК из штамма 240В 1 частично расщепляли с помощью EcoRI. Фрагменты ДНК встраивали путем лигирования в дефосфорилированный сайт EcoRI космидного вектора pLAFR3 (Staskawicz и др., 1987). Встроенную путем лигирования ДНК упаковывали с помощью набора Gigapack III XL Packaging Extract (фирма Stratagene) и продуктом трансфектировали штамм E.coli DH5α . Космидные клоны, обладающие инактивирующей активностью в отношении OHHL, идентифицировали согласно описанному выше методу биологического анализа. Субклонирование в векторе для секвенирования pGEM-7Zf(+) осуществляли с помощью стандартных методов (Sambrook и др., 1989). Делеционный анализ осуществляли с использованием ДНК-азы I с помощью метода, описанного у Lin и др. (1985). Секвенирование обеих цепей осуществляли с помощью набора АВ1 PRISM™ dRhodamine Terminator Cycle Sequencing Ready Reaction Kit (фирма РЕ Applied Biosystems). Данные, касающиеся нуклеотидной последовательности и выведенных на ее основе аминокислотных последовательностей, анализировали с использованием пакета программ для анализа последовательностей DNASTAR™ (фирма DNASTAR Inc.), а поиск в базе данных осуществляли с помощью алгоритма поиска BLASTA (Altschul и др., 1990).
Генетическая модификация штамма Erwinia SCG1
Плазмиду E7-R3, несущую ген aiiA в космидном векторе pLAER3, переносили в штамм Erwinia SCG1 путем трехродительского спаривания с использованием штамма-хелпера RK2013 (Ditta и др., 1980). Трансконъюганты отбирали на планшетах с минимальной средой, дополненной тетрациклином, и проверяли с помощью ПЦР с использованием праймеров, специфичных для гена aiiA.
Исследование вирулентности
Вирулентность штамма Erw. carolovora SCG1 дикого типа и трансформанта, несущего ген aiiA SCG1 (E7-R3), оценивали с помощью инокуляции. На разрезанные поверхности или поврежденные места тканей растений наносили 4 мкл суспензии бактерий, находящихся на ранней стационарной фазе (содержащей ~2× 109 клеток/мл) или разбавленной суспензии бактерий. Инокулированные ткани растения инкубировали в течение ночи в чашке Петри при 28°С. Серьезность поражения мягкой гнилью оценивали через 48 ч после инкубации.
Результаты
Скрининг бактерий, обладающих способностью к инактивации АИ
Бактериальные изоляты из образцов растений и почвы подвергали скринингу в отношении их способности к ферментативной инактивации АИ. Бактериальный изолят 240В 1, у которого была выявлена выраженная способность устранять активность АИ, был отобран для дальнейших исследований. Экстракты общего протеина из изолята 240В 1 полностью устраняли активность АИ после инкубации в течение 1 ч (фиг.1), при этом экстракт протеина терял способность инактивировать АИ после обработки в течение 1 ч протеиназой или кипячения в течение 5 мин. Эти результаты свидетельствуют о том, что бактериальный изолят 240В 1 инактивировал АИ в результате ферментативного действия. Изолят был таксономически охарактеризован как Bacillus sp. на основе следующих его характеристик: грамположительный, палочкообразный, позитивный в отношении каталазы, факультативно анаэробный и имеет последовательность 16рРНК, гомологичную соответствующим последовательностям других бактерий Bacillus (данные не приведены).
Молекулярная масса фермента, обладающего способностью инактивировать АИ, по-видимому, превышает 30 кДа. Его активность терялась после пропускания экстракта протеина через фильтрующее устройство типа Centriсоn 30 (фирма Amicon), однако активность восстанавливалась у подвергнутой ресуспендированию фракции, которая задерживалась при прохождении через Centricon 30 (фиг.2).
Клонирование и локализация области инактивации АИ
Для идентификации гена, кодирующего инактивацию АИ, конструировали космидную библиотеку с использованием геномной ДНК штамма Listera sp. 240B 1. 1200 клонов подвергали скринингу в отношении способности инактивировать АИ. Были выявлены 3 клона, обладавшие активностью в отношении инактивации АИ. Рестрикционный анализ позволил установить, что 3 клона обладали одним общим фрагментом длиной 4,3 т.п.н., получаемым в результате расщепления с помощью EcoRI. Биологический анализ субклона Е7-7, содержащего этот EcoRI-фрагмент длиной 4,3 т.п.н., подтвердил, что данный фрагмент кодирует функцию инактивации АИ (фиг.3). Для идентификации минимального размера и местоположения гена инактивации АИ (aiiA) с помощью метода с использованием ДНКазыI (Lin и др., 1985) создавали серию делеционных клонов, имеющих делеции на обоих концах рассматриваемого фрагмента длиной 4,3 т.п.н. Результаты этого анализа позволили установить, что ген aiiA расположен в фрагменте длиной 1,2 т.п.н. клона F41 (фиг.3).
Ген aiiA кодирует новый протеин
Были полностью секвенированы обе цепочки ДНК-вставки длиной 1,2 т.п.н. клона F41. Нуклеотидная последовательность aiiA и предсказанная аминокислотная последовательность представлены на фиг.4. Полная последовательность ДНК-вставки состоит из 1222 пары оснований и содержит внутри себя 4 возможные открытые рамки считывания (ОРС), начинающихся с нуклеотидов в положениях 1, 42, 156 и 228 соответственно (фиг.4). Делеционный анализ позволил установить, что только наиболее длинная ОРС кодирует функцию инактивации АИ, поскольку клон R34, в котором были исключены промоторная область длиной 48 пар оснований и нуклеотиды с 1 по 13 из наиболее длинной ОРС, полностью терял функцию инактивации АИ, хотя оставшаяся часть ДНК-вставки находилась под контролем функционального Ptac-промотора (фиг.3). Это подтверждали путем слияния наиболее длинной ОРС с геном глутатион-S-трансферазы в этой же ОРС и тестирования инактивирующей активности очищенного слитого протеина в отношении АИ (данные не приведены). Эта ОРС содержит 750 пар нуклеотидов и кодирует протеин, состоящий из 250 аминокислот, который характеризуется предсказанной величиной молекулярной массы 28036 Да и изоэлектрической точкой 4,7 вследствие наличия 19 выраженно основных и 39 выраженно кислотных аминокислотных остатков. Перед предполагаемым инициирующим кодоном на расстоянии 7 пар оснований от него располагается возможная связывающая рибосому последовательность (AAGGTGG), комплементарная 3'-концу последовательности 16S-рРНК E.coli. Наилучшее совпадение последовательности (TATTGT) с консенсусным - 10-промоторным элементом (ТАТААТ) оказалось в области, расположенной против хода транскрипции на расстоянии 35 пар оснований от инициирующего кодона. По ходу транскрипции относительно терминирующего кодона (Brendel, 1986) был обнаружен ТСТТ-бокс, располагающийся за областью с высоким содержанием Т-остатков, которая напоминает потенциальный независящий от фактора сайт терминации. Общее содержание GC-остатков в гене аiiА составляет 37%, а содержание GC в третьем положении кодона составляет 27%.
Поиск в базе данных, проведенный путем анализа с помощью алгоритмов FASTA и BLAST как на уровне нуклеотидной последовательности, так и на уровне пептидной последовательности, не выявил существенного сходства с известными последовательностями из основных баз данных (GenBank, European Molecular Biology Laboratory, Protein Information Resource и Swiss-Prot), что свидетельствует о том, что протеин AiiA является новым. Поиск консенсусного мотива протеина с использованием программы MOTIF фирмы Genetics Computer Group (Мэдисон, штат Висконсин) показал, что короткая пептидная последовательность “1LVDTGMPESAV”, располагающаяся в AiiA от положения 47 до положения 58, была сходной, но не идентичной последовательности активного сайта аспартилпротеазы (Rawlings и Barrett, 1995) (фиг.5).
Экспрессия гена aiiA в Erwinia carotovora уменьшает высвобождение АИ и снижает вирулентность
Космидный клон E7-R3 переносили в штамм Erwinia carotovora SCG1 путем трехродительского спаривания. Вектор pLAFR3 стабильно встраивали в Erwinia carotovora без использования давления отбора. Биологический анализ позволил установить, что уровни АИ, высвобождаемые штаммом Erwinia carotovora (E7-R3), были существенно пониженными (фиг.6, полоса 6), в то время как присутствие только одного космидного вектора pLAFR3 в Erwinia carotovora не оказывало влияния на производство АИ (фиг.6, полоса 7). Эти данные свидетельствуют о том, что большая часть АИ, продуцируемых штаммом Erwinia carotovora SCG1, инактивировалась продуктом гена aiiA.
Штамм Erwinia carotovora SCG1 (E7-R3), экспрессирующий протеин AiiA, не вызывал совсем или вызывал лишь незначительные симптомы заболевания мокрой гнилью у картофеля, баклажанов, китайской капусты, моркови и сельдерея, тогда как его родительский штамм вызывал появление серьезных симптомов (фиг.7А, Б, В, Г, Д). С целью исключения экспериментальных ошибок, обусловленных генетическими изменениями, случайным образом отбирали четыре колонии штамма Erwinia carotovora SCG1 и его трансформанты, содержащие ген aiiA, для оценки уровня производства АИ и вирулентности в отношении картофеля. В обоих экспериментах были получены аналогичные результаты. Штамм Erwinia carotovora SCG1 (pLAFR3), который содержал только космидный вектор, приводил к такому же уровню серьезности симптомов заболевания, что и его родительский штамм Erwinia carotovora SCG1 (фиг.7).
Обсуждение
Бактериальный изолят 240В 1, который был идентифицирован как Bacillus sp., продуцирует фермент, который обладает способностью эффективно инактивировать три подвергнутых тестированию АИ, а именно N-β -оксогексаноил-L-гомосеринлактон, N-β -оксооктаноил-L-гомосеринлактон и N-β -оксодеканоил-L-гомосеринлактон. Клонировали и полностью секвенировали ген (aiiA), кодирующий фермент инактивации АИ. Наличие экспрессии гена aiiA в трансформированной E.coli и патогенной бактерии Erwinia carotovora обусловливает способность к инактивации АИ и существенно уменьшает высвобождение АИ клетками Erwinia carotovora. Согласно имеющимся сведениям рассматриваемый протеин представляет собой первый идентифицированный протеин, обладающий способностью ферментативным путем инактивировать N-ацилгомосеринлактоны, являющиеся аутоиндукторами глобальной регуляции генов в различных видах бактерий.
AiiA представляет собой новый протеин. Не обнаружено существенного гомологического сходства с известными протеинами в основных базах данных. Он обладает сходством с консенсусной последовательностью активного сайта аспартилпротеаз (Rawlings и Barret, 1995). Аспартилпротеазы, также известные как кислые протеазы, широко распространены в клетках позвоночных, грибов, растений, ретровирусах и некоторых растительных вирусах. Аспартилпротеазы из большинства ретровирусов и некоторых растительных вирусов являются гомодимерами. Молекулярная масса протеина AiiA составляет приблизительно 28 кДа, однако он не проходит через молекулярное сито с предельной величиной пропускаемой молекулярной массы 30 кДа, что свидетельствует о возможности того, что протеин AiiA в естественных условиях существует в виде гомодимера или гомомультимера. Однако также существует возможность того, что мономер AiiA имеет нерегулярную трехмерную структуру, которая препятствует его прохождению через молекулярное сито. Аспартилпротеазы представляют собой эндопептидазы и они обладают способностью гидролизовать амидные связи протеинов. Кристаллографическое исследование позволило установить, что ферменты, относящиеся к семейству аспартилпротеаз, представляют собой двудольные молекулы с находящейся между долями “расщелиной”, которая содержит активный сайт, при этом каждая доля включает пару остатков аспарагиновой кислоты, обусловливающих каталитическую активность (Sielecki и др., 1991).
Erwinia carotovora представляет собой растительный патоген, который продуцирует и секретирует экзоферменты, действующие в качестве вирулентных детерминант для заболеваний мокрой гнилью различных растений, включая картофель, баклажан, томат, перец стручковый, морковь, сельдерей, лук и салат (Kotoujansky, 1987).
У мутантов, у которых нарушена способность продуцировать N-β -оксогексаноил-L-гомосеринлактон, также нарушена спомобность синтезировать такие экзоферменты как пектиназа, целлюлаза и протеаза. Эти мутанты не могут индуцировать заболевание мокрой гнилью клубней картофеля (Jones и др., 1993). Установлено, что ген expl, который гомологичен гену luxl Vibrio fischeri, кодирует производство аутоиндукторов в Erwinia carotovora. Мутант expl оказался авирулентным при его инокуляции в лист табака, однако вирулентность восстанавливалась после добавления внешнего аутоиндуктора (Pirhonen и др., 1993). Очевидно, что аутоиндукторы представляют собой возможную мишень при создании генетической устойчивости растений к заболевания мягкой гнилью. В качестве подхода, позволяющего осуществить предварительное тестирование и проверку концепции, в штамм Erwinia carotovora SCG1 встраивали космидный клон, содержащий ген aiiA. Экспрессия фермента AiiA в Erwinia carotovora приводила к существенному уменьшению высвобождения аутоиндукторов и генетически модифицировала Erwinia carotovora таким образом, что экспрессируемый AiiA совсем не обладал способностью индуцировать или индуцировал лишь в незначительной степени симптомы заболевания мокрой гнилью у всех тестированных растений, включая картофель, баклажан, китайскую капусту, морковь и сельдерей. Результаты, полученные при создании изобретения, подтверждают важную роль аутоиндукторов в регуляции экспрессии вирулентных генов в Erwinia carotovora и потенциальную способность гена aiiA обусловливать устойчивость к заболеванию мокрой гнилью и другими болезнями, при которых аутоиндукторы принимают участие в регуляции экспрессии патогенных генов.
В настоящем изобретении предложена новая стратегия создания устойчивости к болезням. В частности, мишенью этой стратегии являются N-ацилгомосеринлактоновые аутоиндукторы, которые индуцируют экспрессию патогенных генов многих бактериальных патогенов в предельных концентрациях. При использовании подхода, основанного на вышеизложенной концепции, при создании настоящего изобретения было продемонстрировано, что уменьшение или устранение аутоиндукторов, продуцируемых патогенными бактериями, с помощью фермента инактивации аутоиндукторов существенно уменьшает патогенность бактериальных патогенов, которые в иных условиях являются вирулентными. Поскольку экспрессия патогенных генов в патогенных бактериях требует наличия предельной концентрации, то рассматриваемая стратегия, основанная на инактивации АИ, применима ко всем заболеваниям растений, животных или млекопитающих, при которых экспрессия патогенных генов бактериальных патогенов индуцируется N-ацилгомосеринлактоновыми аутоиндукторами.
Ген aiiA можно также применять в качестве “инструмента” для исследования роли АИ для тех бактерий, в которых еще не выявлены биологические функции, регулируемые АИ. В последние годы было установлено, что намного большее количество видов бактерий продуцируют АИ (Bassler и др., 1997; Dumenyo и др., 1998; Cha и др., 1998; Surette и др., 1999). Некоторые из них представляют собой имеющие важное значение патогены растений, такие как различные виды Pseudomonas и Xanthomonas. Подход к “выключению” гена, основанный на гомологии последовательностей, может быть трудноосуществимым. Общие уровни сходства последовательностей синтазы АИ и родственного регуляторного протеина из различных родов являются сравнительно низкими, часто идентичность между протеинами Luxl-типа не превышает 28-35%, а для протеинов LuxR-типа она составляет 18-25% (Fugua и др., 1996). Однако можно, и это оказывается легко осуществить, встраивать ген aiiA в эти бактерии для выявления биологических функций, регулируемых АИ.
Литература
1. Allison D. G. R., B. Sanjose, C. Jaspe, A. Gilbert P. (1998). Extracellular products as mediators of the formation and detachment of Pseudomonas fluorescens biofilms. FEMS Microbiology Letters 167, 179-184.
2. Altschul S. F., Gish W., Miller W., Myers E. W. и Lipman D. J. (1990). Basic local alignment search tool. Journal Molecular Biology 215, 403-410.
3. Barber С. E., Tang J. L., Feng J. X., Pan M. Q., Wilson T. J., Slater H., Dow J. M., Williams P. и Daniels M. J. (1997). A novel regulatory system required for pathogenicity of Xantbomonas campestris is mediated by a small diffusible signal molecule. Molecular Microbiology 24, 555-566.
4. Bassler В. L., Greenberg E. P. и Stevens A. M. (1997). Cross-species induction of luminescence in the quorum-sensing bacterium Vibrio harveyi. Journal of Bacteriology 179, 4043-4045.
5. Beck von Bodman S. и Farrand S. K. (1995). Capsular polysaccharide biosynthesis and pathogenicity in Erwinia slewarfii require induction by an Nacylho-moserine lactone autoinducer. Journal of Bacteriology 177, 5000-5008.
6. Borisjuk N. V., Borisjuk L. G., Logendra S., Logendra S., Petersen F., Gleba Y., Raskin 1. 1999. Production of recombinant proteins in plant root exudates. Nature Biotechnology 17, 466-469.
7. Brendel V. и Trifonov E. N. (1984). A computer algorithm for testing potential prokaryotic terminators. Nucleic Acids Research 12, 4411-4427.
8. Cao J. G. и Meighen E. A. (1989). Purification and structural identification of an autoinducer for the luminescence system of Vibrio harveyi. Journal of Biological Chemistry 264, 21670-21676.
9. Cha С., Gao Р., Chen Y. С., Shaw P. D. и Farrand S. К. (1998). Production ofacyl-homoserine lactone quorum-sensing signals by gram-negative plantassociated bacteria. Molecular and Plant Microbe Interactions 11, 1119-1129.
10. Choi S. H., Greenberg E. P. (1991). The C-terminal region of the Vibrio fischeri LuxR protein contains an inducer-independent lux gene activating domain. Proc. Natl. Acad. Sci. USA 88, 11115-11119.
11. Choi S. H., Greenberg E. P. (1992). Genetic evidence for multimerization of LuxR, the transcriptional activator of Vibrio fischeri luminescence. Molec. Mar. Вiol. Biotech. 1: 408-413.
12. Clough S. J., Lee K. E., Schell M. А. и Denny T. P. (1997). A two-component system in Ralstonia (Pseudomonas) solanacearum modulates production of PhcAregulated virulence factors in response to 3-hydroxypalmitic acid methyl ester..7. Bacterial. 179, 3639-3648.
13. Collmer А. К., N. Т. (1986). The role ofpectic enzymes in plant pathogenesis. Annual Review of Phytopathology 24, 383-409.
14. Conrad U. и Fiedler U. 1998. Compartment-specific accumulation of recombinant imnmnoglobulins in plant cells: an essential tool for antibody production and immunomodulation of physiological functions and pathogen activity. Plant Molecular Biology 38, 101-109.
15. Costa J. M. и Loper J. E. (1997). Ecbl and EcbR: homologs of Luxl and LuxR affecting antibiotic and exoenzyme production by Erwinia carotovora subsp. betavasculorum. Can../. Microhiol. 43, 1164-71.
16. Davies D. G., Parsek M. R., Pearson J. P., Iglewski B. H., Costerton J. W и Greenberg E. P. 1998. The involvement of cell-to-cell signals in the develoment of a bacterial biofilm. Science 280, 295-298.
17. Ditta G., StanFleld S., Corbin D. и Helinski D. R. (1980). Broad host range DNA cloning system for gram-negative bacteria: construction of a gene bank of Rhizohium meliloti. Proc. Natl. Acad. Sci. USA 77, 7347-7351.
18. Dumenyo С. К. М., A. Chun W., Chatterjee А. К. (1998). Genetic and physiological evidence for the production of N-acyi homoserine lactones by Pseudomonas syringae pv. syringae and other fluorescent plant pathogenic Pseudomonas species. European Journal of Plant Pathology 104(6). 1998. 569-582. 104, 569-582.
19. Dunphy G., Miyamoto С. и Meighen E. (1997). A homoserine lactone autoinducer regulates virulence of an insect-pathogenic bacterium, Xenorhabdus nematophilus (Enterobacteriaceae). J. Bacterial. 179, 5288-5291.
20. Eberhard A., Burlingame A. L., Eberhard С., Kenyon G. L., Nealson K. H. и Oppenheimer, N. J. (1981). Structural identification of autoinducer of Phopfohacterlum fischeri luciferase. Biochemistry 20, 2444-2449.
21. Eberi L., Winson M. K., Sternberg C., Steward G. S. А. В., Christiansen G., Chhabra S. R., Bycroft В., Williams P., Molin S. и Givskov M. (1996). Involvement of N-acyl-L-homoserine lactone autoinducers in controlling the multicellular behaviour of Serratia liquefaciens. Molecular Microbiology 20, 127- 136.
22. Firek S., Draper J., Owen M. R. L. Gandecha A., Cockburn В. и Whitelam G. C. 1993. Secretion of a functional single-chain Fv protein in transgenic tobacco plants and cell suspension cultures. Plant Molecular Biology 23, 861-870.
23. Fuqua C., Winans S. C. (1996). Conserved cis-acting promoter elements are required for density-dependent transcription of Agrobacterium tumefaciens conjugal transfer genes.J. Bacterial. 178, 435-440.
24. Fuqua W. C., Winans S. C., Greenberg E. P. (1994). Quorum sensing in bacteria: the LuxR/LuxI family of cell density-responsive transcriptional regulators.J. Bacterial. 176, 269-75.
25. Fuqua С., Winans S. С. и Greenberg E. P. (1996). Census and consensus in bacterial ecosystems: The LuxR-LuxI family of quorum-sensing transcriptional regulators. Annu. Rev. Microbiol. 50, 727-751.
26. Hanzelka В. L., Greenberg E. P. (1995). Evidence that the N-terminal region of the Vibrio fiscneri LuxR protein constitutes an autoinducer-binding domain. J. Bacterial. 177, 815-817.
27. Jones S. M., Yu В., Bainton N. J., Birdsall M., Bycroft B. W., Chhabra S. R., Cox A. J. R., Golby P., Reeves P. J., Stephens S., Winson M. K., Salmond G. P C., Stewart G. S. А. В. и Williams P. (1993). The Lux autoinducer regulates the production of exoenzyme virulence determination in Erwinia carotovora and Pseudomonas aeruginosa. EMBO J., 12, 2477-2482.
28. Kotoujansky A. (1987). Molecular genetics of pathogenesis by soft-rot Erwinias. Annual Review of Phylopathology 25, 405-430.
29. Lin H. С., Lei S. P. и Wilcox G. (1985). An improved DNA sequencing strategy. A nal. Biochem. 147, 114-119.
30. Meighen E. A. (1994). Genetics of bacterial luminescence. Annu. Rev. Genet. 28, 117-139.
31. More M. 1., Finger L. D., Stryker J. L., Fuqua C., Eberhard А. и Winans S. C. (1996). Enzymatic synthesis of a quorum-sensing autoinducer through use of defined substrates. Science 272, 1655-1658.
32. Nasser W., Bouillant M. L., Salmond G. и Reverchon S. (1998). Characterization of the Erwinia chrysanthemi expl-expR locus directing the synthesis of two N-acyl-homoserine lactone signal molecules. Molecular Microbiology 29,1391-1405.
33. Passador L., Cook J. M., Gambello M. J.. Rust L., Iglewski В. Н. (1993). Expression of Pseudomonas aeruginosa virulence genes requires cell-to-cell communication. Science 260, 1127-1130.
34. Pearson J. P., Gray К. M., Passador L., Tucker K. D., Eberhard A., Iglewski В. Н. и Greenberg E. P. (1994). Structure of the autoinducer required for expression of Pseudomonas aeruginosa virulence genes. Proc. Natl. Acad. Sci. USA 91, 197-201.
35. Petit А. и Tempe J. (1978). Isolation of Agrobacterium Ti plasmid regulatory mutants. Mol. Gen. Genet. 167, 147-155.
36. Pierson L. S., 3-й, Keppenne V. D. и Wood D. W. (1994). Phenazine antibiotic biosynthesis in Pseudomonas aureofaciens 30-84 is regulated by PhzR in response to cell density. J. Bacterial. 176, 3966-74.
37. Piper К. R., Beck von Bodman S. и Farrand S. K. (1993). Conjugation factor of Agrobacterium tumefaciens regulates Ti plasmid transfer by autoinduc-tion. Nature 362, 448-450.
38. Pirhonen M., Flego D., Heikinheimo R. и Palva E. (1993). A small diffusible signal molecule is responsible for the global control of virulence and exoen-zyme production in the plant pathogen Erwinia carotovora. EMBO J. 12, 2467-2476.
39. Rawlings N. D. В., А, J. (1995). Families ofaspartic peptidases, and those of unknown catalytic mechanism. In Methods in Enzymology, стр. 105-180. Edited by A. J. Barrett. New York: Academic Press.
40. Sambrook J. F., E. F. Maniatis T. (1989). Molecular Cloning, 2-изд., New York: Cold Spring Harbor Laboratory Press.
41 Sato F., Koiwa H., Sakai Y., Yamada Y. (1995). Synthesis and secretion of tobacco neutral PR-5 protein by transgenic tobacco and yeast. Biochemical & Biophysical Research Communications. 211, 909-913.
42. Schaefer A. L. V., D L. Hanzelka B. L. Cronan J. E. Jr. Greenberg E. P. (1996). Generation of cell-to-cell signals in quorum sensing: Acyi homoserine lac-tone synthase activity of a purified Vibrio fischeri Luxl protein. Proceedings of the National Academy of Sciences of the United States of America 93, 9505-9509.
43. Shadel G. S., Young R., Baldwin T. 0. (1990). Use of regulated cell lysis in a lethal genetic selection in Escherichia coli: identification of the autoinducer-binding region of the LuxR protein from Vibrio fischeri ATCC 7744. J. Bacterial. 172,39803987.
44. Sielecki A. R., Fujinaga M., Read R. J. и James M. N. (1991). Refined structure of porcine pepsinogen at 1.8 A resolution..7. Mol. Blol. 219, 671-692.
45. Sitnikov D., Schineller J. В., Baldwin T. 0. (1995). Transcriptional regulation ofbioluminescence genes from Vibrio fischeri. Mol. Microhlol. 17, 801-12.
46. Slock J., Kolibachuk D., Greenberg E.P. (1990). Critical regions of the Vibrio fischeri LuxR protein defined by mutational analysis. J. Bacterial. 172: 3974-3979.
47. Staskawicz В. D., D. Keen N. Т. и Napoli С. (1987). Molecular characterization of cloned avirulence genes from race 0 and race I of Pseudonionas syringae pv. glycinea. Journal of Bacteriology 169, 5789-5794.
48. Surette M. G. В., В. L. (1998). Quorum sensing in Escherichia coli and Salmonella typhimurium. Proceedings of the National Academy of Sciences of the United Slates of America. 95, 7046-7050.
49. Throup J. P., Camara M., Briggs G. S., Winson M.K., Chhabra S. R. и др. (1995). Characterisation of the yenl/yenR locus from Yersenia enterocolitica mediating the synthesis of two N-acylhomoserine lactone signal molecules. Mol. Microhiol. 17, 345-356.
50. Zhang L.-H. (1993). Molecular biology and biochemistry of a novel conjugation factor in Agrobacterium. Докторская диссертация. The Adelaide University, Australia.
51. Zhang L.-H., Murphy P. J., Kerr А. и Tate M. E. (1993). Agrobacterium conjugation and gene regulation by N-acyl-L-homoserine lactones. Nature 362, 446-447.


Изобретение относится к биотехнологии, в частности к глобальным регуляторам бактериальных патогенных генов, применяемых для придания устойчивости растений к болезням. Выделяют нуклеиновую кислоту, кодирующую протеин инактивации бактериальных аутоиндукторов установленной структуры. Конструируют экспрессионный вектор, в состав которого включают вышеуказанную нуклеиновую кислоту, и трансформируют им растение. Выделяют белок, образующийся в результате экспрессии упомянутой нуклеиновой кислоты, который используют в составе композиции, предназначенной для предупреждения или уменьшения повреждения растения или животного, вызванного бактериями, путем его введения или нанесение на поврежденные органы. Проводят скрининг бактериальных изолятов на способность инактивации их аутоиндукторов путем выделения бактериальной культуры из образцов почвы и скрининга на способность культуры к инактивации аутоиндукторов при экспрессии указанного протеина. Выделение нуклеиновой кислоты проводят путем создания банка генов из организма-донора, содержащего данную нуклеиновую кислоту, скрининга клонов, содержащихся в банке генов и их выделения. Определяют влияние нуклеиновой кислоты на биологическую функцию клетки путем встраивания нуклеиновой кислоты в клетку и скрининга трансформированных клеток. Изобретение позволяет предупреждать или уменьшать повреждения растений или животных, тем самым снижает производственные затраты и повышает качество сельскохозяйственной продукции. 9 н. и 11 з.п. ф-лы, 7 ил.
1l. Способ предупреждения или уменьшения повреждения у растения или животного, вызванного бактериями, заключающийся в том, что в растение или животное, нуждающееся в таком предупреждении или уменьшении, вводят эффективное количество протеина инактивации бактериальных аутоиндукторов, где протеин включает последовательность SEQ ID NO:2.
| HWANG I et al | |||
| J Bacteriol | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Кулисный парораспределительный механизм | 1920 |
|
SU177A1 |
| Abstracts of the general meeting of the American society for microbiology | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| Journal Bacteriology | |||
| Способ получения кодеина | 1922 |
|
SU178A1 |
| Archives of microbiology | |||
| vol | |||
| Приспособление, увеличивающее число оборотов движущихся колес паровоза | 1919 |
|
SU146A1 |
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| Journal of bacteriology | |||
| vol | |||
| Переносный кухонный очаг | 1919 |
|
SU180A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Приспособление для отбивки кос | 1926 |
|
SU5398A1 |

