ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение раскрывает ген изопропилмалат синтазы из Nicotiana tabacum и его варианты, гомологи и фрагменты. В частности, здесь описана модификация экспрессии этого гена или активности кодируемого им белка с целью изменения состава сложных эфиров сахарозы растения - такого как растение табака.
УРОВЕНЬ ТЕХНИКИ
Сложные эфиры сахарозы общепризнанны в качестве обладающих инсектицидными свойствами в растениях, для них показана токсичность в отношении мягкотелых насекомых, в том числе тлей, клещей, медяницы грушевой и белокрылок, а также антибиотическая активность. Сложные эфиры сахарозы также рассматриваются как предшественники ароматических веществ, они накапливаются на поверхности листа в течение жизни растения. Они представляют собой устойчивые соединения, которые могут быть идентифицированы в сушеных и курительных изделиях. Этерифицированные низшие карбоновые кислоты высвобождаются из сложных эфиров сахарозы при нагревании. Эти низшие карбоновые кислоты представляют собой очень сильные ароматические молекулы и, как полагают, частично ответственны за восточный аромат табака. Сложные эфиры сахарозы образуются в клетках железистых трихом растения. Клетки железистых трихом также являются местом синтеза других экссудатов из листьев - таких как белки (филлопланины) и дитерпеноиды (цембреноиды и лабденоиды). Сахароза и небольшие карбоновые кислоты, которые этерифицируют сахарозу с образованием сложных эфиров сахарозы, образуются в двух различных метаболических путях.
Сорта табака различаются по количеству и качеству сложных эфиров сахарозы, присутствующих на поверхности листа. Как правило, сорта дымовой сушки Burley и Maryland накапливают небольшие количества сложных эфиров сахарозы с ацильными группами до пяти атомов углерода в длину, тогда как большинство восточных сортов и многие типы сигарного табака накапливают большие количества сложных эфиров сахарозы, имеющих ацильные группы до шести атомов углерода в длину и, в меньшей степени, в них обнаруживаются эфиры, имеющие ацильные группы до семи атомов углерода в длину. Дихотомия, наблюдающаяся в сложных эфирах сахарозы, образуемых сортами табака, связана с одним доминантным локусом, названным BMVSE (бета-метилвалерил содержащие сложные эфиры сахарозы) для его функционального аллеля и bmvse для его нефункционального аллеля. Геномный локус BMVSE расположен на хромосоме А табака.
Kroumova and Wagner (2009) General and Applied Plant Physiology 35, 3-4, p95-110 описывают попытки изменить содержание ацильных групп сложного эфира сахарозы в различных растениях с использованием обратного генетического подхода подавления (нокдауна) экспрессии изопропилмалат синтазы с помощью двухцепочечной интерферирующей РНК. В этом исследовании ген изопропилмалат синтазы из Solanum pennellii использовали для амплификации части гена из кДНК. Затем они были введены в смысловой и антисмысловой ориентации в конструкцию двухцепочечной интерферирующей РНК и трансформированы посредством Agrobacterium в Nicotiana tabacum T.I. 1068, Nicotiana glutinosa cv. 24a и S. pennellii. Результаты, полученные с использованием растений N. tabacum, обнаружили некоторые изменения в количестве ацильных групп сложных эфиров сахарозы вследствие пониженного бета-метилвалерил- и повышенного 2-метилбутирил-ацилирования по сравнению с нетрансформированными контролями. Тем не менее, полученные растения N. tabacum были фенотипически ослаблены, так как были хлоротичными и некоторые из них имели закручивающиеся листья. Аналогичным образом, трансформированные растения N. glutinosa и S. pennellii были фенотипически ослабленными. Авторы приходят к выводу, что изопропилмалат синтаза является ключевым ферментом и что ее ослабление, вероятно, будет приводить к больному растению.
Существует потребность в области техники в растениях, в которых композиция сложных эфиров сахарозы модулируется при минимизации нежелательных эффектов для растения. Задачей настоящего изобретения является удовлетворить этой потребности.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано, по меньшей мере, частично, на неожиданном открытии того, что модулирование активности или экспрессии гена изопропилмалат синтазы из Nicotiana tabacum приводит к изменению состава сложных эфиров сахарозы с меньшим числом нежелательных эффектов на общий метаболизм растений. Большинство этих эффектов осуществляется в трихомах растения и относятся к вторичному метаболизму. Таким образом, модулирование активности или экспрессии гена изопропилмалат синтазы Nicotiana tabacum, no существу, не приводит к изменениям во внешнем виде растения по сравнению с контрольным растением. Это выгодно, потому что растения могут быть использованы для коммерческого производства различных продуктов, при этом изменения во внешнем виде будут либо не приемлемы для промышленности, либо могут привести к неприемлемому снижению урожайности. Предпочтительно, чтобы профиль аромата аэрозоля, полученного из табака, мог быть модифицирован таким образом, чтобы можно было создать новые вкусовые профили. Кроме того, у растений, профиль сложных эфиров сахарозы которых был модифицирован, может быть изменена устойчивость к вредителям. Композиции сложных эфиров сахарозы могут быть выделены из растений для различных применений - например, в лекарствах, пищевых добавках, курительных ароматизаторах и в качестве компонентов органических пестицидов и тому подобном.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Аспекты и варианты воплощения настоящего изобретения приведены в прилагаемой формуле изобретения.
В первом аспекте здесь предусмотрен выделенный полинуклеотид, содержащий или состоящий или состоящий в основном из последовательности, кодирующей изопропилмалат синтазу и, по меньшей мере, на 60% идентичной последовательности SEQ ID NO:1 или SEQ ID NO:10 или SEQ ID NO:12 или SEQ ID NO:14.
В другом аспекте здесь предусмотрен выделенный полипептид, кодируемый этим полинуклеотидом.
В другом аспекте здесь предусмотрен выделенный полипептид, последовательность которого, по меньшей мере, на 60% идентична SEQ ID NO:2 или SEQ ID NO:11 или SEQ ID NO:13 или SEQ ID NO:15.
Конструкция, вектор или вектор экспрессии, содержащий выделенную полинуклеотидную последовательность, где указанная конструкция, вектор или вектор экспрессии необязательно дополнительно содержат промотор, содержащий, состоящий или состоящий в основном из последовательности, приведенной в SEQ ID NO:8 или его вариант, по меньшей мере, на 60% идентичный ему, или промотор трихомы или природный промотор изопропилмалат синтазы.
В другом аспекте здесь предусмотрен выделенный полинуклеотид, содержащий, состоящий или состоящий в основном из SEQ ID NO:8, или его вариант, который, по меньшей мере, на 60% ему идентичен.
В другом аспекте здесь предусмотрен мутант, не встречающаяся в природе или трансгенная растительная клетка, содержащая, по меньшей мере, один из полинулеотидов, по меньшей мере, один из полипептидов, или конструкцию, вектор или вектор экспрессии из настоящего изобретения.
В другом аспекте здесь предусмотрен мутант, не встречающееся в природе или трансгенное растение, содержащие растительную клетку из настоящего изобретения.
В другом аспекте здесь предусмотрен способ модулирования количества сложных эфиров сахарозы в части растения, включающий стадии: (i) модулирования экспрессии или активности изопропилмалат синтазы в растении, где изопропилмалат синтаза предпочтительно содержит полинуклеотидную последовательность или полипептидную последовательность, описанные здесь; (ii) измерения количества сложных эфиров сахарозы, по меньшей мере, в части мутанта, не встречающегося в природе или трансгенного растения, полученных на стадии (i); и (iii) определение мутанта, не встречающегося в природе или трансгенного растения, в котором количество сложных эфиров сахарозы было изменено по сравнению с контрольным растением, в котором экспрессия или активность изопропилмалат синтазы не модулировалась, и, предпочтительно, где внешний вид указанного мутанта, не встречающегося в природе или трансгенного растения, по существу, такой же, как у контрольного растения.
В другом аспекте здесь предусмотрен мутант, не встречающееся в природе или трансгенное растение, полученное или получаемое этим способом.
В другом аспекте здесь предусмотрен мутант, не встречающееся в природе или трансгенное растение, в котором экспрессия гена изопропилмалат синтазы или активность белка, кодируемого им, модулирована и, по меньшей мере, у части растения изменен состав сложных эфиров сахарозы по сравнению с контрольным растением, в котором экспрессия или активность изопропилмалат синтазы не была модулирована, и где внешний вид указанного растения, по существу, такой же, как у контрольного растения.
В другом аспекте здесь предусмотрен растительный материал, включающий биомассу, семена или листья, содержащие клетки или ткани из этого растения.
В другом аспекте здесь предусмотрено табачное изделие, содержащее часть растения или растительного материала в соответствии в настоящим изобретением.
В другом аспекте здесь предусмотрен способ производства композиции сложных эфиров сахарозы, включающий стадии: (i) получения, по меньшей мере, части мутанта, не встречающегося в природе или трансгенного растения, растительного материала или табачного изделия в соответствии с настоящим изобретением; (ii) выделения из них (например, одного или нескольких) сложных эфиров сахарозы; и (iii) необязательно, очистки выделенных сложных эфиров сахарозы.
Также предусмотрена композиция сложных эфиров сахарозы, полученная или получаемая указанным способом.
Соответственно, один или несколько сложных эфиров сахарозы имеют структуру, такую как показано на Фигуре 5, и где R3 представляет собой ацетил или водород, предпочтительно ацетил; один или несколько из R1, R2 и R4 содержат, по меньшей мере, одну ацильную цепь с 6 атомами углерода, предпочтительно бета-метилвалерил; и R5 представляет собой ацетил или водород, предпочтительно водород.
Соответственно, один или несколько сложных эфиров сахарозы выбирают из группы, состоящей из сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гесканоил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил (С2С14:0); сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил (С2С15:0); сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой валерил или его изомер, R4 представляет собой гексаноил или его изомер и R5 представляет собой атом водорода или ацетильную группу (С2С16:0); или сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой пропионил или его изомер, R2 представляет собой гексаноил или его изомер, R4 представляет собой гексаноил или его изомер, и R5 представляет собой атом водорода или ацетильную группу (С2С16:0); сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу (С2С17:0); или сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, где R3 представляет собой ацетил, R1 представляет собой гексаноил, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу (С2С18:0), или их комбинацию.
Соответственно, один или несколько сложных эфиров сахарозы выбирают из группы, состоящей из сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил (С2С15:0); сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой валерил или его изомер, R4 представляет собой гексаноил или его изомер и R5 представляет собой атом водорода или ацетильную группу (С2С16:0); или сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой пропионил или его изомер, R2 представляет собой гексаноил или его изомер, R4 представляет собой гексаноил или его изомер, и R5 представляет собой атом водорода или ацетильную группу (С2С16:0); сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу (С2С17:0); или сложного эфира сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой гексаноил, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу (С2С18:0), или их комбинацию.
В еще одном аспекте здесь предусмотрен способ модулирования аромата табака или табачного изделия, включающий: (i) добавление к табаку или табачному изделию части растения, предпочтительно листьев, из мутанта, не встречающегося в природе или трансгенного растения, или растительного материала, как описано здесь, предпочтительно, где указанный мутант, не встречающееся в природе или трансгенное растение или растительный материал принадлежат к роду Nicotinia; или (ii) добавление к табаку или табачному изделию композиции, включающей содержащие бета-метилвалерил сложные эфиры сахарозы, полученной или получаемой способом, описанным здесь.
В еще одном аспекте здесь предусмотрен способ получения бета-метилвалериановой кислоты, включающий стадии: (i) получения, по меньшей мере, части мутанта, не встречающегося в природе или трансгенного растения, растительного материала или табачного изделия; (ii) гидролиза материала, полученного на стадии (i), или его экстракта; и (iii) необязательного выделения или очистки бета-метилвалериановой кислоты. Соответственно, растение представляет собой растение табака - например, из рода Nicotiana или вида Nicotiana tabacum.
Соответственно, внешний вид растения, по существу, такой же, как у контрольного растения. Соответственно, внешний вид растения, по существу, такой же, как у контрольного растения спустя три месяца после высадки в поле или спустя 36 дней после укорачивания верхушки, преимущественно, такой, что высота стебля мутантов, не встречающихся в природе или трансгенных растений, по существу, такая же, как высота стебля контрольного растения спустя три месяца после высадки в поле или спустя 36 дней после укорачивания верхушки, и/или содержание хлорофилла в мутанте, не встречающимся в природе или в трансгенном растении, по существу, такое же, как содержание хлорофилла в контрольных растениях спустя три месяца после высадки в поле или спустя 36 дней после укорачивания верхушки. В других вариантах воплощения любая одна или несколько из таких характеристик как степень зрелости, число листьев на растение, высота стебля, угол вставки листа, размер листа (ширина и длина), расстояние между междоузлиями, отношение пластинка/жилки и окраска листьев у мутанта, не встречающегося в природе или трансгенного растения, по существу, такая же, как у контрольных растений спустя три месяца после высадки в поле или спустя 36 дней после укорачивании верхушки.
Еще один аспект относится к биомассе, семенам или листьям, содержащим клетки и ткани из растения (растений), описанных здесь, представляющим собой табачное изделие, содержащее часть растений или биомассы, семян или листьев.
Также в еще одном аспекте данного описания предусмотрен табачный материал, определенный или определяемый описанным здесь способом.
Устойчивость растения к вредителям может быть изменена.
Дополнительные аспекты настоящего изобретения приведены ниже.
Предусмотрен химерный ген, содержащий выделенный полинуклеотид, который функционально связан с одним или несколькими регуляторными последовательностями.
Предусмотрена полинуклеотидная конструкция изопропилмалат синтазы, содержащая, состоящая или состоящая преимущественно из, по меньшей мере, 15-30 нуклеотидов, 30-50 нуклеотидов 50-100 нуклеотидов, 100-150 нуклеотидов, 150-200 нуклеотидов, 200-300 нуклеотидов, 300-400 нуклеотидов, 400-500 нуклеотидов, 500-600 нуклеотидов, 600-700 нуклеотидов, 700-1000 нуклеотидов, 1000-1300 нуклеотидов, 1300-1500 нуклеотидов или 1500-1900 нуклеотидов.
Предусмотрен продукт, который включает или в котором использованы растительный материал, биомасса, семена или листья в соответствии с настоящим изобретением.
Предусмотрена клетка или клеточная линия, содержащая выделенный полинуклеотид, химерный ген, полинуклеотидную конструкцию, двухцепочечную РНК, конъюгат или вектор экспрессии и тому подобное в соответствии с настоящим изобретением.
Предусмотрен способ модулирования экспрессии изопропилмалат синтазы или активности белка, кодируемого таким образом в клетке, где указанный способ включает введение химерного гена, полинуклеотидной конструкции, двухцепочечной РНК, конъюгата или вектора экспрессии, описанных здесь.
Способ обнаружения, выделения, амплификации или анализа полинуклеотида изопропилмалат синтазы, где способ включает стадии получения образца, содержащего полинуклеотид, и гибридизации указанного полинуклеотида с молекулой полинуклеотида, содержащей нуклеотидную последовательность, по меньшей мере, из 10 последовательных нуклеотидов из выделенной нуклеотидной последовательности в соответствии с настоящим изобретением.
Применение агента, который модулирует экспрессию изопропилмалат синтазы или активность белка, кодируемого таким образом, для модулирования содержания сложных эфиров сахарозы, по меньшей мере, в части растения - например, в листьях.
В одном варианте воплощения агент является нуклеиновой кислотой изопропилмалат синтазы, химерным геном изопропилмалат синтазы, полинуклеотидной конструкцией, содержащей полинуклеотид изопропилмалат синтазы, антисмысловой РНК, двухцепочечной РНК, кДНК, конъюгатом, содержащим полинуклеотид изопропилмалат синтазы и, по меньшей мере, один не нуклеотидный фрагмент или не полинуклеотидный фрагмент, ковалентно присоединенный к нему, рибозимом, мутагеном, белком с цинковым пальцем, низкомолекулярным соединением или мегануклеазой, или является их производными.
В другом варианте воплощения полинуклеотидный фрагмент (фрагменты) кодируют антисмысловую нуклеиновую кислоту, рибозим, РНК, которая влияет на сплайсосома-опосредованный транс-сплайсинг, интерферирующую РНК (РНКи), гидовую РНК (guide RNA) или другую не транслируемую РНК и тому подобное. В другом варианте воплощения полинуклеотидный фрагмент (фрагменты) кодирует РНКи.
Еще один аспект относится к способу получения табачного продукта, включающего стадии: (а) получения семян от трансгенного, мутантного или не встречающегося в природе растения, описанного здесь; (b) посадки и выращивания семян; (с) сбора урожая растения; и (d) производства табачного продукта из собранного урожая растения.
Вышеупомянутые варианты воплощения раскрыты в качестве вариантов воплощения каждого из аспектов, описанных выше.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 иллюстрирует подсчитанное количество изомеров сложных эфиров сахарозы, измеренное в экссудатах из листьев. Измерение проводили в ацетон/метанольных смывах с зеленых дисков листа сорта табака bmvse Hicks Broadleaf (HBL) и сорта табака BMVSE Red Russian (RR). Количество оценивали с добавлением октаацетата сахарозы в качестве внешнего стандарта. n=4 для каждого сорта. С2С12:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = пропионил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = ацетил, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил. С2С13:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = бутирил, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = пропионил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = пропионил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил. С2С14:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложного эфира сахарозы, такого, как показано на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил. С2С15:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахароз, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; С2С16:0 включает (насыщенный) сложный эфир сахарозы, который имею общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой валерил или его изомер, R4 представляет собой гексаноил или его изомер и R5 представляет собой атом водорода или ацетильную группу; или где сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой пропионил или его изомер, R2 представляет собой гексаноил или его изомер, R4 представляет собой гексаноил или его изомер и R5 представляет собой атом водорода или ацетильную группу. С2С17:0 включает (насыщенный) сложный эфир сахарозы такой, как показан на Фигуре 5, где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу. С2С18:0 включает (насыщенный) сложный эфир сахарозы такой, как показан на Фигуре 5, где R3 представляет собой ацетил, R1 представляет собой гексаноил, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу.
На Фигуре 2 показаны основные изомеры сложных эфиров сахарозы, упорядоченные по молекулярной массе и их встречаемости в сортах табака, продуцирующих или не продуцирующих сложные эфиры сахарозы, содержащие бета-метилвалериановые эфиры. Не делается различия между 2- и 3-метилбутириловыми или 3- и 4 метилвалериановыми эфирами, перечисленными как С5 и С6, соответственно. Следующая номенклатура используется для определения состава ацильных групп. Например, обозначение С2С4С4С5 относится к числу атомов углерода в ацильных группах в R3, R1, R2 и R4, соответственно. С2 может быть ацетилом; С3 может быть пропионилом или его изомером; С4 может быть бутирилом или изобутирилом; С5 может быть валерилом (пентаноилом), или его изомером, или 2-метил-бутирилом, или изовалерилом, или изопентеноилом, или пентеноилом; С6 может быть гексаноилом, или его изомером, или 2-метилвалерилом, или бета-метилвалерилом, или 4-метилвалерилом. В R5 (не показано) может присутствовать ацетил или водород, в данном случае соответственно водород.
Фигура 3 иллюстрирует путь синтеза аминокислот с разветвленной цепью и образование ацильных цепей, этерифицирующих сахарозу. Заключенные в рамку стадии представляют основные стадии пути синтеза разветвленной аминокислотной цепи, где углеродные цепи удлиняются на один атом углерода в цикле конденсации изомеризации и декарбоксилирования. 1: ацетолактат синтаза [2.2.1.6]; 2: изомероредуктаза ацетогидроксикислот [1.1.1.86]; 3: дегидратаза 3,4-дигидроксикислоты [4.2.1.9]; 4: аминотрансфераза аминокислот с разветвленными цепями [2.6.1.42]; 5: дегидрогеназный комплекс кетокислот с разветвленными цепями (BCKD) ([1.2.4.4] [2.3.1.168] [1.8.1.4]); 6: синтаза изопропилмалата [2.3.3.13]; 7: изомераза изопропилмалата [4.2.1.33]; 8: дегидрогеназа изопропилмалата [1.1.1.85]; 9: деаминаза треонина [4.3.1.19]. С4: эфир изобутановой кислоты; iC5: эфир 3-метил-бутановой кислоты (изовалериановый эфир); aiC5: эфир 2-метил-бутановой кислоты (антеизо-валериановый эфир); iC6: 4-метил-валериановый эфир (эфир изокаприловой кислоты); aiC6: 3-метил-валериановый эфир (эфир антеизо-каприловой кислоты), β-метилвалериановый эфир, BMV). Карбоновые кислоты представлены в нейтральной форме, хотя названия отражают также растворимые формы. Стадии, предположительно катализируемые IPMS2, обведены кружком.
На Фигуре 4 приведена оценка содержания тетраэфиров сахарозы в экссудате с поверхности листа трансформированного растения. Листья собирали с одномесячных регенерирующих растений, полученных в результате независимых трансформаций. Внутренний стандарт (октаацетат сахарозы) добавляли в количестве 2,5 нг/мг сырой массы. Площадь сигнала, записанного для иона, специфичного для каждого сложного эфира сахарозы, измеряли и отображали как отношение к площади сигнала внутреннего стандарта. Отношение к площади сигнала от внутреннего стандарта позволяет нормализовать сигналы на сырую массу листа. Представленная здесь номенклатура сложных эфиров сахарозы описана на Фигуре 2. А: состав экссудатов из листьев исходного сорта K326 (сорт дымовой сушки, n=3) и трансформантов, экспрессирующих SEQ ID NO:1 под контролем специфичного промотора трихомы; В: состав экссудатов из листьев исходного сорта K326 и трансформантов, экспрессирующих SEQ ID NO:1 под контролем вирусного промотора; С: состав экссудатов из листьев исходного сорта Basma (восточный, n=4) и трансформантов, экспрессирующих конструкцию РНКи SEQ ID NO:9 под контролем специфичного промотора трихомы; и D: состав экссудатов из листьев исходного сорта Basma и трансформантов, экспрессирующих конструкцию РНКи SEQ ID NO:9 под контролем вирусного промотора. С2С14:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил. С2С15:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 представляет собой водород или ацетил; или сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил; С2С16:0 включает (насыщенный) сложный эфир сахарозы, который имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой валерил или его изомер, R4 представляет собой гексаноил или его изомер и R5 представляет собой атом водорода или ацетильную группу; или где сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, и где R3 представляет собой ацетил, R1 представляет собой пропионил или его изомер, R2 представляет собой гексаноил или его изомер, R4 представляет собой гексаноил или его изомер и R5 представляет собой атом водорода или ацетильную группу. С2С17:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 представляет собой ацетил, R1 представляет собой валерил или его изомер, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу. С2С18:0 включает (насыщенный) сложный эфир сахарозы, такой, как показан на Фигуре 5, где R3 представляет собой ацетил, R1 представляет собой гексаноил, R2 представляет собой гексаноил, R4 представляет собой гексаноил и R5 представляет собой атом водорода или ацетильную группу.
Фигура 5 иллюстрирует структуру остова сложного эфира сахарозы. R1 - R5 представляют собой либо атом водорода, либо ацильную группу, присоединенную эфирной связью.
Фигура 6 иллюстрирует ацильные цепи, этерифицированные в сложные эфиры сахарозы. R' представляет молекулу сахарозы. 1. ацетил, 2. бутирил, 3. пропионил, 4. изобутирил, 5. валерил (пентаноил), 6. 2-метил-бутирил, 7. изовалерил, 8. изопентеноил, 9. пентеноил, 10. гексаноил или его изомер 11. 2-метилвалерил, 12. бета-метилвалерил, 13. 4-метилвалерил. Также рассматриваются их изомеры и производные. Таким образом, в качестве примера, изомером может быть изомер пропионила - например, изобутирил. В качестве еще одного примера, изомером может изомер валерила (пентаноила) - например, 2-метил-бутирил или изовалерил; или изомер валерила (пентаноила) с двойной связью - например, изопентеноил или пентеноил. В качестве еще одного примера, изомером может быть изомер гексаноила - например, 2-метилвалерил, бета-метилвалерил или 4-метилвалерил. Изомер может содержать или может не содержать одну или несколько двойных связей.
ОПРЕДЕЛЕНИЯ
Технические термины и выражения, использованные в рамках данной заявки, как правило, имеют значение, обычно применяемое для них в данной области биологии растений и молекулярной биологии. Все следующие определения терминов приводятся для полноты содержания заявки. Слово "содержащий" не исключает другие элементы или стадии, и формы единственного числа не исключают множественного. Одна стадия может соответствовать функциям нескольких признаков, перечисленных в формуле изобретения. Термины "примерно", "в основном" и "приблизительно", в данном контексте, обозначающие величину или диапазон, относятся к величине или диапазону в пределах 20%, в пределах 10% или в пределах 5%, 4%, 3%, 2% или 1% от заданного количества или диапазона.
Термин "выделенный" относится к любому объекту полученному из его природной среды, при этом термин не означает какую-либо степень очистки.
"Вектор" относится к переносчику нуклеиновой кислоты, который содержит комбинацию компонентов нуклеиновой кислоты, обеспечивающих транспорт нуклеиновой кислоты, конструкции нуклеиновой кислоты и конъюгатов нуклеиновой кислоты и тому подобное. Подходящие векторы включают эписомальные векторы, способные к экстра-хромосомальной репликации, например, кольцевые, двухцепочечные плазмиды нуклеиновых кислот; линеаризованные двухцепочечные плазмиды нуклеиновых кислот; и другие векторы любого происхождения.
"Вектор экспрессии" представляет собой переносчик нуклеиновой кислоты, который содержит комбинацию компонентов нуклеиновой кислоты, обеспечивающих экспрессию нуклеиновой кислоты, конструкции нуклеиновой кислоты и конъюгатов нуклеиновой кислоты и тому подобное. Подходящие векторы экспрессии включают эписомы, способные к экстра-хромосомной репликации, например, кольцевые, двухцепочечные плазмиды нуклеиновых кислот; линеаризованные двухцепочечные плазмиды нуклеиновых кислот; и другие функционально эквивалентные векторы экспрессии любого происхождения. Вектор экспрессии содержит, по меньшей мере, промотор, расположенный до нуклеиновой кислоты, конструкции нуклеиновой кислоты или конъюгата нуклеиновой кислоты и функционально связанный с ними, как определено ниже.
Термин "конструкция" относится к фрагменту двухцепочечной рекомбинантной нуклеиновой кислоты, содержащему один или несколько полинуклеотидов. Конструкция содержит "матричную цепь", спаренную с комплементарной "смысловой или кодирующей цепью". Данная конструкция может быть вставлена в вектор в двух возможных ориентациях, либо в той же (или смысловой) ориентации или в обратной (или анти-смысловой) ориентации по отношению к ориентации промотора, расположенного в векторе, таком как вектор экспрессии. Пример конструкции приведен в SEQ ID NO:9.
"Промотор" относится к элементу/последовательности нуклеиновой кислоты, обычно расположенному до фрагмента двухцепочечной нуклеиновой кислоты и функционально связанному с ним. Промоторы могут быть целиком получены из областей, прилегающих к представляющему интерес нативному гену, или могут быть составлены из различных элементов, полученных из различных нативных промоторов или синтетических сегментов нуклеиновой кислоты. Пример промотора приведен в SEQ ID NO:8.
Термин "гомология, идентичность или сходство" относится к степени сходства последовательностей двух полипептидов или двух молекул нуклеиновых кислот, сравниваемых по выравниванию последовательностей. Степень гомологии двух отдельных сравниваемых последовательностей нуклеиновых кислот представляет собой функцию числа идентичных, или совпадающих, нукпеотидов в сравниваемых позициях. Процент идентичности можно определить путем визуального просмотра и математического расчета. Альтернативно, процент идентичности двух последовательностей нуклеиновых кислот можно определить путем сравнения информации о последовательностях с использованием компьютерной программы, такой как ClustalW, ClustalX, BLAST, FASTA или Smith-Waterman.
Термин "растение" относится к любому растению на любой стадии его жизненного цикла или развития, и к его потомству. В одном варианте воплощения растение представляет собой растение табака, которое относится к растению, принадлежащему к роду Nicotiana. Предпочтительные виды, сорта, гибриды и разновидности растения табака описаны здесь.
"Растительная клетка" относится к структурной и физиологической единице растения. Растительная клетка может представлять собой протопласт без клеточной стенки, выделенную одиночную клетку или культивируемую клетку, или может быть частью более организованной структуры, такой как, например, растительная ткань, орган растения или целое растение, но не ограничиваясь ими.
Термин "растительный материал" относится к любой твердой, жидкой или газообразной композиции, или их комбинации, полученной из растения, в том числе из биомассы, листьев, листовой пластинки, средней жилки, стеблей, корней, цветков или частей цветка, плодов, пыльцы, яйцеклеток, зигот, семян, отходов (обрезков), секретов, экстрактов, культур клеток или тканей, или любых других частей или продуктов растения. В одном варианте воплощения растительный материал содержит или состоит из биомассы, семян и листьев. В другом варианте воплощения растительный материал содержит или состоит из листьев.
Термин "сорт" относится к популяции растений, имеющих постоянные характеристики, которые отделяют их от других растений того же вида. Обладая одним или несколькими отличительными признаками, сорт кроме того характеризуется очень небольшими общими вариациями между индивидуумами в пределах сорта. Сорт часто реализуется коммерчески.
Использованный здесь термин "линия" или "линия разведения" обозначает группу растений, которую используют в процессе разведения растений. Линия отличается от сорта, поскольку она проявляет небольшие вариации между индивидуумами в одном или нескольких представляющих интерес признаках, хотя могут быть некоторые вариации между индивидуумами и по другим признакам.
Термин "модулирование" может относиться к снижению, ингибированию, повышению или другому влиянию на активность полипептида. Термин может также относиться к снижению, ингибированию, повышению или другому влиянию на активность гена, кодирующего полипептид, что может включать модулирование транскрипционной активности, но не ограничивается этим.
Использованные здесь термины "снижать" или "пониженный" относятся к снижению от примерно 10% до примерно 99%, или снижению, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98%, по меньшей мере, на 99% или, по меньшей мере, на 100% или более от количества или активности, например, активности полипептида, транскрипционной активности и/или белковой экспрессии, но не ограничиваясь ими.
Использованные здесь термины "ингибировать" или "ингибированный" относятся к снижению от примерно 98% до примерно 100%, или снижению, по меньшей мере, на 98%, по меньшей мере, на 99%, но частности до 100% от количества или активности, например, активности полипептида, транскрипционной активности и/или белковой экспрессии, но не ограничиваясь ими.
Использованные здесь термины "повышение" или "повышенный" относятся к повышению от примерно 10% до примерно 99%, или повышению, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98%, по меньшей мере, на 99%, по меньшей мере, на 100%, по меньшей мере, на 150% или, по меньшей мере, на 200% или больше от количества или активности, например, активности полипептида, транскрипционной активности и/или белковой экспрессии, но не ограничиваясь ими.
Термин "контроль" в контексте контрольного растения или контрольных растительных клеток означает растение или растительные клетки, в которых экспрессия или активность изопропилмалат синтазы не была модифицирована (например, повышена или понижена), так что это может обеспечить сравнение с растением, в котором экспрессия или активность изопропилмалат синтазы была модифицирована. Контрольное растение может содержать пустой вектор. Контрольное растение может представлять собой растение дикого типа.
В одном аспекте обеспечивается выделенный полинуклеотид, содержащий, состоящий или состоящий по существу из полинуклеотидной последовательности, и идентичный, по меньшей мере, на 60% любой из описанных здесь последовательностей, и, в том числе любому из полинукпеотидов, представленных в перечне последовательностей. Соответственно, выделенные полинуклеотиды включают, состоят или по существу состоят из последовательности, имеющей, по меньшей мере, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с указанными последовательностями.
В одном аспекте обеспечивается выделенный полинуклеотид, содержащий, состоящий или состоящий по существу из полинукпеотидной последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, 60% идентичность последовательности с SEQ ID NO:1. Соответственно, выделенные полинуклеотиды включают, состоят или по существу состоят из последовательности, имеющей, по меньшей мере, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:1.
В другом аспекте предложен выделенный полинуклеотид, содержащий, состоящий или состоящий по существу из полинуклеотидной последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:10.
В другом аспекте предложен выделенный полинуклеотид, содержащий, состоящий или состоящий по существу из полинуклеотидной последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, 60%, 61%, 62%, 63%, 64%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:12.
В другом аспекте предложен выделенный полинуклеотид, содержащий, состоящий или состоящий по существу из полинуклеотидной последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:14.
В другом аспекте предложен выделенный полинуклеотид, содержащий, состоящий или состоящий по существу из полинуклеотидной последовательности, имеющей, по меньшей мере, 60%, 61%, 62%, 63%, 64%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:8. Последовательность SEQ ID NO:8 кодирует предпочтительный промотор.
Термин "полинуклеотид " относится к полимеру из нуклеотидов, который может быть не модифицированной или модифицированной дезоксирибонуклеиновой кислотой (ДНК), либо рибонуклеиновой кислотой (РНК). Соответственно, полинуклеотид может быть, без ограничения, геномной ДНК, комплементарной ДНК (кДНК), мРНК или антисмысловой РНК или их фрагментом. Кроме того, полинуклеотид может быть одноцепочечной или двухцепочечной нуклеиновой кислотой, которая представляет собой смесь одноцепочечных и двухцепочечных участков, гибридной молекулой, содержащей ДНК и РНК, или гибридной молекулой со смесью одноцепочечных и двухцепочечных участков или их фрагментом (фрагментами). Кроме того, полинуклеотид может состоять из трехцепочечных участков, включающих ДНК, РНК или обе, или их фрагмент (фрагменты). Полинуклеотид может содержать одно или несколько модифицированных оснований, таких как фосфотиоаты, и может представлять собой пептидную нуклеиновую кислоту (PNA). Как правило, полинуклеотиды могут быть собраны из выделенных или клонированных фрагментов кДНК, геномной ДНК, олигонуклеотидов или отдельных нуклеотидов, или комбинаций вышеперечисленного. Хотя полинуклеотидные последовательности, описанные здесь, представлены в виде последовательностей ДНК, последовательности включают и соответствующие последовательности РНК, а также комплементарные им (например, полностью комплементарные) последовательности ДНК или РНК, в том числе и комплементарные им противоположно направленные последовательности.
Термин "полинуклеотид изопропилмалат синтазы" включает в себя полинуклеотиды, кодирующие изопропилмалат синтазу (IMPS) из Nicotiana tabacum, и включает полинуклеотиды, содержащие, состоящие или состоящие по существу из полинуклеотидов с существенной гомологией (то есть, сходством последовательности) или значительной идентичностью с SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14; полинуклеотидные варианты, имеющие, по меньшей мере, приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность последовательности с SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14; фрагменты полинуклеотидов, в том числе фрагменты SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14, и фрагменты SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14 с существенной гомологией (то есть, сходством последовательности) или значительной идентичностью с ними, которые имеют, по меньшей мере, около 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с соответствующими фрагментами SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14. Полинуклеотид изопропилмалат синтазы включает также последовательности, имеющие достаточную или значительную степень идентичности или сходства с SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14, чтобы кодировать полипептид, который функционирует как изопропилмалат синтаза. В одном варианте осуществления термин "полинуклеотид изопропилмалат синтазы" относится к полимеру из нуклеотидов, который включает, состоит или по существу состоит из полинуклеотида, обозначенного здесь как SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14.
Полинуклеотид, как описано здесь, обычно содержит фосфодиэфирные связи, хотя в некоторых случаях, включены аналоги полинуклеотидов, которые могут иметь альтернативный остов, включая, например, фосфорамидатные, фосфоротиоатные, фосфородитиоатные, или О-метилфофороамидитные связи, и пептид-полинукпеотидные остовы и связи. Другие аналоги полинуклеотидов включают аналоги с положительно зараженным остовом; неионным остовом и не рибозным остовом. Модификация рибозо-фосфатного остова может быть выполнена по различным причинам, например, для увеличения стабильности и периода полураспада таких молекул в физиологических средах или при использовании в качестве зондов на биочипе. Могут быть сделаны смеси встречающихся в природе полинуклеотидов и аналогов; альтернативно, могут быть сделаны смеси различных аналогов полинуклеотидов, и смеси встречающихся в природе полинуклеотидов.
Известны разнообразные аналоги полинуклеотидов, в том числе, например, фосфоамидатные, фосфоротиоатные, фосфородитиоатные, и/или O-метилфофороамидитные связи, и пептид-полинуклеотидные остовы и связи. Другие аналоги полинуклеотидов включают аналоги с положительными остовами; неионными остовами и на рибозными остовами. Также включены полинуклеотиды, содержащие один или несколько карбоциклических углеводов.
Другие аналоги включают пептидные нуклеиновые кислоты, являющиеся пептидными аналогами полинуклеотидов. Эти остовы являются по существу незаряженными в нейтральных условиях, в отличие от высоко заряженных фосфодиэфирных остовов встречающихся в природе полинуклеотидов. Это может привести к преимуществам. Во-первых, пептидный полинуклеотидный остов может обладать улучшенной кинетикой гибридизации. Пептидные нуклеиновые кислоты имеют большие изменения в температуре плавления для несоответствий по сравнению с совершенно комплементарными парами оснований. ДНК и РНК обычно имеют понижение температуры плавления в 2-4°С для несоответствия внутри фрагмента. С незаряженным пептидным полинуклеотидным основанием понижение близко к 7-9°С. Аналогичным образом из-за их незаряженности гибридизация оснований, присоединенных к этим основаниями, относительно нечувствительна к концентрации соли. Кроме того, пептидные нуклеиновые кислоты не могут быть деградированы или подвергаются деградации в меньшей степени клеточными ферментами, и, таким образом, могут быть более стабильными.
Среди различных применений описанных полинуклеотидов и комбинаций их фрагментов выделяют использование фрагментов в качестве зондов в анализах гибридизации нуклеиновой кислоты или в качестве праймеров в анализах амплификации нуклеиновой кислоты. Такие фрагменты обычно включают, по меньшей мере, примерно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или более последовательных нуклеотидов последовательности ДНК. В других вариантах осуществления фрагмент ДНК содержит, по меньшей мере, примерно 10, 15, 20, 30, 40, 50 или 60 или более последовательных нуклеотидов последовательности ДНК. Эти методы также могут быть использованы для идентификации растений BMVSE и bmvse, так как функциональный ген изопропилмалат синтазы, как показано здесь, необходим для формирования сложных эфиров сахарозы. Близость генетических маркеров и аллелей гена изопропилмалат синтазы можно использовать для предсказания выработки растением эфиров сахарозы. Такой тест может быть использован для отслеживания сортов растений или предсказания свойств табака или однородности партии продукта. Таким образом, в одном аспекте, также предложен способ для обнаружения полинуклеотида изопропилмалат синтазы, как описано здесь, включающий применение зондов или праймеров, способных специфически обнаруживать или специфически амплифицировать указанный полинуклеотид или специфически обнаруживать и специфически амплифицировать указанный полинуклеотид. В еще одном аспекте предложен способ идентификации растения, которое способно производить сложные эфиры сахарозы, включающий следующие стадии: (а) получение образца, содержащего нуклеиновую кислоту, из интересующего растения, и (б) определение присутствия полинуклеотида изопропилмалат синтазы, как описано здесь, где наличие указанного полинуклеотида в указанном растении демонстрирует, что указанное растение способно производить сложные эфиры сахарозы. Также предусмотрены наборы для обнаружения, по меньшей мере, части полинуклеотида изопропилмалат синтазы, которые содержат один или несколько праймеров или зондов для специфичного обнаружения, по меньшей мере, части полинуклеотида изопропилмалат синтазы. Набор может содержать реагенты для амплификации полинуклеотида, например, полимеразной цепной реакции (ПЦР), или реагенты для техники обнаружения гибридизации зонда нуклеиновой кислоты, например, Саузерн блоттинга, Нозерн блоттинга, гибридизации in situ или микрочипов. Набор может содержать реагенты для технологий обнаружения связывания антител, таких как Вестерн блоттинг, ELISA, SELDI масс-спектрометрия или тест-полоски. Набор может содержать реагенты для секвенирования ДНК. Набор может содержать реагенты и/или инструкции для определения содержания эфиров сахарозы. В некоторых вариантах осуществления набор может включать инструкции для одного или более из описанных способов. Описанные наборы могут быть полезны для определения генетической идентичности, филогенетического исследования, генотипирования, гаплотипирования, анализа родословной или селекции растений, особенно с кодоминантной оценкой. Методы и наборы могут быть использованы в подходе селекции, основывающемся на скрещивании BMVSE и bmvse сортов и селекции растений в потомстве на основе последовательностей. Как правило, будут выбраны растения, демонстрирующие наличие функциональной генетической области изопропилмалат синтазы, которая присутствовала в производящем BMVSE сорте, или растения, демонстрирующие наличие нефункциональной генетической области изопропилмалат синтазы, присутствующей в не производящих bmvse растениях.
Основные параметры, влияющие на выбор условий гибридизации, и рекомендации для подбора подходящих условий описаны в Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.). С применением знаний о генетическом коде в сочетании с аминокислотными последовательностями, описанными здесь, могут быть получены наборы вырожденных олигонуклеотидов. Такие олигонуклеотиды могут быть использованы в качестве праймеров, например, в ПЦР, где ДНК-фрагменты выделяют и амплифицируют. В некоторых вариантах осуществления вырожденные праймеры могут быть использованы в качестве зондов для генетических библиотек. Такие библиотеки будут включать, но не ограничиваться ими, библиотеки кДНК, геномные библиотеки, и даже электронные библиотеки маркерных экспрессирующихся последовательностей или ДНК. Гомологичные последовательности, идентифицированные этим способом, будут затем использованы в качестве зондов для определения гомологов последовательностей, определенных в данном документе.
Также потенциально могут быть использованы полинуклеотиды и олигонуклеотиды (например, праймеры или зонды), которые гибридизуются в условиях пониженной жесткости, как правило, в умеренно жестких условиях, и обычно высоко жестких условиях с полинуклеотидом (полинуклеотидами), как описано здесь. Основные параметры, влияющие на выбор условий гибридизации, и рекомендации для подбора подходящих условий изложены в Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.), и могут быть легко определены обычным специалистом в данной области техники на основе, например, длины или состава оснований полинуклеотида.
Один из путей достижения умеренно строгих условий включает в себя применение предпромывочного раствора, содержащего 5 × Стандартный раствор цитрата натрия, 0,5% додецилсульфата натрия, 1,0 мМ этилендиаминтетрауксусную кислоту (рН 8,0), буфера гибридизации, содержащего около 50% формамида, 6 × Стандартный раствор цитрата натрия, и температуру гибридизации около 55°С (или других подобных растворов гибридизации, таких как содержащий около 50% формамида раствор, с температурой гибридизации около 42°С) и условия промывания при около 60°С в 0,5 × стандартном цитрате натрия, 0,1% додецилсульфате натрия. Как правило, очень строгие условия определены как условия гибридизации, как описано выше, но с промывкой при примерно 68°С, 0,2 × Стандартным цитратом натрия, 0,1% додецилсульфатом натрия. В буферах гибридизации и промывания SSPE (1×SSPE является раствором 0,15 М хлорида натрия, 10 мМ фосфата натрия и 1,25 мМ этилендиаминтетрауксусной кислоты, рН 7,4) может быть заменен на стандартный цитрат натрия (1 × Стандартный цитрат натрия является раствором 0,15 М хлорида натрия и 15 мМ цитрата натрия); промывка осуществляется в течение 15 минут после завершения гибридизации. Следует понимать, что при необходимости температуру промывания и концентрацию солей для промывания можно регулировать, чтобы достичь желаемой степени жесткости с применением основных принципов, которые регулируют реакцию гибридизации и стабильность дуплекса, как известно специалистам в данной области и описано ниже (см., например, Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.). При гибридизации полинуклеотида к целевому полинуклеотиду неизвестной последовательности длина гибрида предполагается такой же, что и у гибридизующегося полинуклеотида. Когда гибридизуются полинуклеотиды известной последовательности, длина гибрида может быть определена путем выравнивания последовательностей полинуклеотидов и идентификации области или областей оптимальной комплементарное™ последовательности. Температура гибридизации гибридов, длина которых ожидается меньше, чем 50 пар оснований в длину, должна быть на 5-10°С меньше, чем температура плавления гибрида, где температура плавления определяется в соответствии со следующими уравнениями. Для гибридов менее 18 пар оснований в длину, температура плавления (°С) = 2 (число оснований А+Т) + 4 (число оснований G+С). Для гибридов больше 18 пар оснований в длину, температура плавления (°С) = 81,5+16,6 (log 10 [Na+]) 0,41 (% G+С) - (600/N), где N это количество оснований в гибриде, и [Na+] обозначает концентрацию ионов натрия в буфере гибридизации ([Na+] для 1 × Стандартного цитрата натрия = 0,165 М). Как правило, каждый такой гибридизующийся полинуклеотид имеет длину, которая, по меньшей мере, составляет 25% (как правило, по меньшей мере, 50%, 60% или 70% и наиболее обычно, по меньшей мере, 80%) от длины полинуклеотида, к которому он гибрид изуется, и имеет идентичность последовательности с полинуклеотидом, к которому он гибридизуется, по меньшей мере, 60% (например, по меньшей мере, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100%).
Как будет понятно специалисту в данной области, линейная ДНК имеет две возможные ориентации: направление 3'- к - 5' и направление 5'- к - 3'. Например, если последовательность сравнения расположена в направлении 5'- к 3' -, и если вторая последовательность расположена в направлении 5'- к 3' в пределах одной молекулы/цепи полинуклеотида, то последовательность сравнения и вторая последовательность ориентированы в том же направлении, или имеют одинаковую ориентацию. Как правило, последовательность промотора интересующего гена, регулируемого данным промотором, расположена в той же ориентации. Тем не менее, по отношению к последовательности сравнения, расположенной в направлении 5'- к 3', если вторая последовательность расположена в направлении 3'- к - 5' в пределах одной молекулы/цепи полинуклеотида, то последовательность сравнения и вторая последовательность ориентированы в антисмысловом направлении, или антисмысловой ориентации. Две последовательности, имеющие антисмысловые ориентации по отношению друг к другу, могут быть альтернативно описаны как имеющие ту же ориентацию, если последовательность сравнения (направление 5'- к - 3') и обратная комплементарная последовательность последовательности сравнения (последовательность сравнения расположена в 5'- к -3') расположены в пределах одной молекулы полинуклеотида/цепи. Последовательности, описанные в настоящем документе, приведены в направлении 5'- к - 3'.
Рекомбинантные конструкции, представленные здесь, могут быть использованы для трансформации растений или растительных клеток, чтобы модулировать уровень экспрессии белка изопропилмалат синтазы. Рекомбинантная полинуклеотидная конструкция может включать в себя полинуклеотид, кодирующий полинуклеотид изопропилмалат синтазы, как описано здесь, функционально связанный с регуляторной областью, подходящей для экспрессии полипептида изопропилмалат синтазы в растении или клетке. Таким образом, полинуклеотид может содержать кодирующую последовательность, которая кодирует полипептид изопропилмалат синтазы, как описано здесь.
Полипептид изопропилмалат синтазы, кодируемый рекомбинантным полинуклеотидом, может быть нативным полипептидом изопропилмалат синтазы, или может быть гетерологичным по отношению к клетке. В некоторых случаях, рекомбинантная конструкция содержит полинуклеотид, который снижает или ингибирует экспрессию полипептида, модулирующего изопропилмалат синтазу, функционально связанный с регуляторной областью. Примеры подходящих регуляторных областей описаны в данном документе.
Также предусмотрены векторы, содержащие рекомбинантные полинуклеотидные конструкции, такие как те, что описаны здесь. Подходящие векторные остовы, включают, например, те, что обычно используются в данной области, например, плазмиды, вирусы, искусственные хромосомы, ВАС, YAC, или РАС. Подходящие векторы экспрессии включают, без ограничения, плазмиды и вирусные векторы, полученные, например, из бактериофагов, бакуловирусов и ретровирусов. Многочисленные векторы и системы экспрессии являются коммерчески доступными.
Векторы могут также включать, например, сайты инициации репликации, районы прикрепления скэффолда или маркеры. Маркерный ген может придать селектируемый фенотип растительной клетке. Например, маркер может придавать устойчивость к биоциду, например, устойчивость к антибиотику (например, канамицину, G418, блеомицину или гигромицину), или гербициду (например, глифосату, хлорсульфурону или фосфинотрицину). Кроме того, вектор экспрессии может включать последовательность метки, предназначенную для облегчения манипуляции или обнаружения (например, для очистки или локализации) экспрессированного полипептида. Последовательности меток, такие как последовательности люциферазы, бета-глюкуронидазы (GUS), зеленого флуоресцентного белка (GFP), глутатион-S-трансферазы (GST), полигистидина, последовательности с-Мус или гемагглютинина, как правило, экспрессируются в виде белка слияния с кодируемым полипептидом. Такие метки могут быть вставлены в любом месте в пределах полипептида, в том числе на карбоксильном или амино-конце.
Растение или растительная клетка может быть трансформирована путем интегрирования рекомбинантного полинуклеотида в геном, чтобы стать стабильно трансформированным. Стабильно трансформированные клетки обычно сохраняют введенный полинуклеотид с каждым клеточным делением. Растение или растительная клетка также может быть временно трансформировано таким образом, что рекомбинантный полинуклеотид не интегрирован в геном. Временно трансформированные клетки обычно теряют весь или некоторую часть введенного рекомбинантного полинуклеотида с каждым клеточным делением таким образом, что введенный рекомбинантный полинуклеотид не может быть обнаружен в дочерних клетках после достаточного количества клеточных делений.
Существует ряд доступных в данной области методов для трансформации растительной клетки, которые все охватываются настоящим изобретением, в том числе биолистика, методы генной пушки, Agrobacterium-опосредованная трансформация, трансформация, опосредованная вирусным вектором, и электропорация. Система Agrobacterium для интеграции чужеродной нуклеиновой кислоты в растительные хромосомы была тщательно изучена, модифицирована и введена в эксплуатацию в генной инженерии растений. Голые (депротеинизированные) рекомбинантные молекулы нуклеиновых кислот, содержащие последовательности нуклеиновых кислот, соответствующие конкретному очищенному белку табака, функционально связывают в смысловой или антисмысловой ориентации с регуляторными последовательностями, соединенными с соответствующими последовательностями Т-ДНК, обычными способами. Их вводят в протопласты табака с помощью полиэтиленгликолевого метода или с помощью метода электропорации, где оба метода являются стандартными. Альтернативно, такие векторы, содержащие рекомбинантные молекулы нуклеиновых кислот, кодирующие конкретный очищенный белок, вводят в живые клетки Agrobacterium, которые затем передают нуклеиновые кислоты в клетки растений. Трансформация голой ДНК без сопутствующих векторных последовательностей Т-ДНК может быть осуществлена с помощью слияния протопластов табака с липосомами, содержащими нуклеиновую кислоту, или посредством электропорации. Голая ДНК без сопровождения векторных последовательностей Т-ДНК также может быть использована для трансформации клеток табака с помощью инертных высокоскоростных микроснарядов.
Если клетка или культивируемая ткань используется в качестве реципиентной ткани для трансформации, растения при желании могут быть регенерированы из трансформированных культур с помощью методик, известных специалистам в данной области.
Выбор регуляторных областей, которые будут включены в рекомбинантную конструкцию, зависит от нескольких факторов, включая, но не ограничиваясь этим, от эффективности, селективности, индуцибельности, желаемого уровня экспрессии и предпочтительной экспресии в клетках или тканях. Модулирование экспрессии кодирующей последовательности путем соответствующего выбора и размещения регуляторной области по отношению к кодирующей последовательности является обычной процедурой для специалиста в данной области техники. Транскрипцию полинуклеотида можно модулировать аналогичным образом. Некоторые подходящие регуляторные области инициируют транскрипцию исключительно или преимущественно в определенных типах клеток. Методы идентификации и характеристики регуляторных областей в растительной геномной ДНК хорошо известны в данной области техники.
Подходящие промоторы включают тканеспецифичные промоторы, узнаваемые тканеспецифичными факторами, присутствующими в различных тканях или типах клеток (например, специфичные для корней промоторы, специфичные для побегов промоторы, специфичные для ксилемы промоторы), или присутствующими на разных стадиях развития, или появляющимися в ответ на различные условия окружающей среды. Подходящие промоторы включают конститутивные промоторы, которые можно активировать в большинстве типов клеток, не требующие конкретных индукторов. Примеры подходящих промоторов для управления продуцированном РНКи полипептида изопропилмалат синтазы включают промоторы вируса мозаики цветной капусты 35S (CaMV/35S), SSU, OCS, lib4, usp, STLS1, В33, nos, или промоторы убиквитина или фазеолина. Специалисты в этой области техники способны создавать множество вариантов рекомбинантных промоторов.
Тканеспецифичные промоторы - элементы контроля транскрипции, которые являются активными только в особых клетках или тканях в определенное время развития растений, таких как вегетативные ткани или репродуктивные ткани. Тканеспецифичная экспрессия может быть выгодна, например, когда экспрессия полинуклеотидов в определенных тканях является предпочтительной. Примеры тканеспецифических промоторов под контролем развития включают промоторы, которые могут инициировать транскрипцию только (или преимущественно только) в определенных тканях, таких как вегетативные ткани, например, корни или листья, или в репродуктивных тканях, таких как плоды, яйцеклетки, семена, пыльца, пестики, цветы, или любая эмбриональная ткань. Репродуктивные тканеспецифичные промоторы могут быть, например, специфичными для пыльника, специфичными для яйцеклетки, специфичными для зародыша, специфичными для эндосперма, специфичными для интегумента, специфичными для семени и семенной кожуры, специфичными для пыльцы, специфичными для лепестков, специфичными для чашелистиков или их комбинациями.
Подходящие специфичные для листа промоторы включают промотор пируват ортофосфат дикиназы (ПФДК) от С4 растения (кукуруза), cab-m1Ca+2 промотор из кукурузы, промотор myb-ассоциированного гена Arabidopsis thaliana (Atmyb5), промотор рибулозобифосфаткарбоксилазы (RBCS) (например, генов томата RBCS1, RBCS2 и RBCS3A, экспрессируемых в листьях и выращенных на свету проростках, RBCS1 и RBCS2, экспрессируемых в развивающихся плодах томата, или промотор рибулозобисфосфаткарбоксилазы, экспрессируемый почти исключительно в клетках мезофилла в листовых пластинках и листовых влагалищах на высоких уровнях).
Подходящие специфичные для стадии старения промоторы включают промотор томата, активный во время созревания плодов, старения и опадения листьев, промотор гена кукурузы, кодирующего цистеиновую протеазу. Могут быть использованы специфичные для пыльника промоторы. Могут быть выбраны предпочтительные для корней промоторы, известные специалистам в данной области. Подходящие предпочтительные для семян промоторы включают в себя как специфичные для семян промоторы (т.е. промоторы, активные во время развития семян, такие как промоторы запасных белков семян) и промоторы прорастающего семени (т.е. промоторы, активные во время прорастания семян). Такие предпочтительные для семян промоторы включают, но не ограничиваются ими, Cim1 (цитокинин-индуцированный сигналинг); CZ19B1 (19 кДа зеин кукурузы); milps (миоинозит-1-фосфат-синтаза); mZE40-2, также известный как ZM-40; nude и celA (целлюлозосинтаза). Гамма-зеин является эндосперм-специфичным промотором. Glob-1 является специфичным для зародыша промотором. Для двудольных специфичные для семян промоторы включают, но не ограничиваются ими, бета-фазеолин фасоли, напин, β-конглицинин, лектин сои, круциферин, и тому подобное. Для однодольных, специфичные для семян промоторы включают, но не ограничиваются ими, промотор 15 кДа зеина кукурузы, промотор 22 кДа зеина, промотор 27 кДа зеина, промотор гамма-зеина, промотор 27 кДа γ-зеина (например, промотор gzw64A, см. Genbank, номер S78780), промотор waxy, промотор shrunken 1, промотор shrunken 2, промотор глобулина 1 (см. GenBank, номер L22344), промотор ltp2, промотор cim1, промоторы end1 и end2 кукурузы, промотор NUC1, промотор ZM40, еер1 и еер2; lec1, промотор тиоредоксина Н; промотор mlip15, промотор PCNA2 и промотор shrunken-2.
Примеры индуцируемых промоторов включают промоторы, реагирующие на атаки патогена, анаэробные условия, повышенную температуру, свет, засуху, низкую температуру или высокую концентрацию соли. Индуцируемые патогенном промоторы включают промоторы патогенеззависимых белков (PR-белков), которые индуцируются после инфицирования патогеном (например, PR-белки, белки SAR, бета-1,3-глюканазы, хитиназы).
В дополнение к растительным промоторам могут быть использованы подходящие промоторы бактериального происхождения, например, промотор октопинсинтазы, промотор нопалинсинтазы и другие промоторы, полученные из плазмиды Ti, или вирусные промоторы (например, промоторы 35S и 19S РНК вируса мозаики цветной капусты (CaMV), конститутивные промоторы вируса табачной мозаики, 19S и 35S промоторы вируса мозаики цветной капусты (CaMV), или промотор 35S вируса мозаики норичника).
Термин "полипептид изопропилмалат синтазы" относится к полипептиду, кодирующему изопропилмалат синтазу (IPMS) из Nicotiana tabacum, и включает в себя варианты полипептидов, содержащие, состоящие или в основном состоящие из аминокислотной последовательности, кодируемой полинуклеотидным вариантом, по меньшей мере, приблизительно на 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичной последовательности SEQ ID NO:2, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15; фрагментов полипептида (полипептидов) изопропилмалат синтазы, и фрагментов SEQ ID NO:2, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15, которые, по меньшей мере, приблизительно на 60%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичны последовательности соответствующих фрагментов SEQ ID NO:2, SEQ ID NO:11, SEQ ID NO:13 и SEQ ID NO:15. Полипептид изопропилмалат синтазы также включает последовательности, имеющие достаточную или значительную степень идентичности или сходства с SEQ ID NO:2, SEQ ID NO:11, SEQ ID NO:13 и SEQ ID NO:15, чтобы функционировать в качестве изопропилмалат синтазы. Фрагменты полипептида изопропилмалат синтазы обычно сохраняют активность изопропилмалат синтазы. Полипептиды изопропилмалат синтазы также включают мутанты, полученные путем введения любого типа изменений (например, вставки, удаления или замены аминокислот, изменений в состояний гликозилирования; изменений, которые влияют на рефолдинг или изомеризацию, трехмерную структуру или состояния самоассоциации), которые могут быть получены преднамеренно или выделены при естественном происхождении при условии, что они все еще функционируют в качестве изопропилмалат синтазы. Полипептиды изопропилмалат синтазы могут быть в линейной форме или могут быть циклизованы с применением известных методов. Термин "полипептид изопропилмалат синтазы" также может относиться к полипептиду, кодируемому SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:12 или SEQ ID NO:14, который имеет 60%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с ними, или полипептиду, содержащему, состоящему или состоящему в основном из последовательности, представленной в SEQ ID NO:2, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15, который имеет 100% идентичность последовательности с ними.
В другом аспекте предложен выделенный полипептид, содержащий, состоящий или состоящий по существу из полипептидной последовательности, имеющей идентичность последовательности, по меньшей мере, 60% с любой из описанных здесь последовательностей, в том числе с любым из полипептидов, представленных в перечне последовательностей. Соответственно, выделенные полипептиды включают, состоят или по существу состоят из последовательности, имеющей, по меньшей мере, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с ними.
Полипептиды включают в себя варианты, полученные путем введения любого типа изменений (например, вставки, удаления или замены аминокислот, изменений в состояниях гликозилирования; изменений, которые влияют на рефолдинг или изомеризацию, трехмерную структуру, или состояния самоассоциации), которые могут быть получены преднамеренно или выделены при естественном происхождении. Вариант может иметь изменения, которые ведут к молчащей замене и приводят к функционально эквивалентному белку. Преднамеренные аминокислотные замены могут быть сделаны на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и амфипатической природы остатков при условии, что сохраняется вторичная активность связывания вещества. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают лизин и аргинин, и аминокислоты с незаряженными полярными головными группами, имеющими аналогичные величины гидрофильности, включают лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин. Консервативные замены могут быть сделаны, например, в соответствии с приведенной ниже таблицей. Аминокислоты в том же блоке во втором столбце и предпочтительно в той же строке в третьем столбце могут быть заменены друг на друга:

Полипептид может быть зрелым белком или незрелым белком, или белком, полученным из незрелого белка. Полипептиды могут быть в линейной форме или циклизованы с применением известных методов. Полипептиды обычно содержат, по меньшей мере, 10, по меньшей мере, 20, по меньшей мере, 30, или, по меньшей мере, 40 последовательных аминокислот.
В другом аспекте предложен выделенный полипептид изопропилмалат синтазы, содержащий, состоящий или состоящий по существу из последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, примерно 60%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:2.
В другом аспекте предложен выделенный полипептид изопропилмалат синтазы, содержащий, состоящий или состоящий по существу из последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, примерно 60%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:11.
В другом аспекте предложен выделенный полипептид изопропилмалат синтазы, содержащий, состоящий или состоящий по существу из последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, примерно 60%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с SEQ ID NO:13.
В другом аспекте предложен выделенный полипептид изопропилмалат синтазы, содержащий, состоящий или состоящий по существу из последовательности, кодирующей изопропилмалат синтазу и имеющей, по меньшей мере, примерно 60%, 65%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% % или 100% идентичность последовательности c SEQ ID NO:15.
Фрагменты полипептидных последовательностей также раскрыты в данном описании, соответственно, такие фрагменты сохраняют активность полноразмерной последовательности.
Мутантные полипептиды могут быть использованы для создания мутантных растений, не встречающихся в природе растений и трансгенных растений, содержащих мутантный полипептид. Соответственно, мутантный полипептид сохраняет активность не мутантного полипептида. Активность мутантного полипептида может быть выше, ниже или примерно такой же, как не мутантного полипептида.
Полипептид может быть получен путем культивирования трансформированных или рекомбинантных клеток-хозяев в условиях культивирования, подходящих для экспрессии полипептида. Полученный экспрессированный полипептид может быть затем очищен из такой культуры с применением известных способов очистки. Очистка полипептида может включать аффинную колонку, содержащую вещества, которые связываются с полипептидом, один или более этапов очистки на колонке с такими аффинными смолами; одну или несколько стадий с участием гидрофобной хроматографии, или иммуноаффинную хроматографию. Альтернативно, полипептид может быть также экспрессирован в форме, которая облегчает очистку. Например, он может быть экспрессирован в виде слитного полипептида, например, со связывающим мальтозу полипептидом, глутатион-5-трансферазой или тиоредоксином. Наборы для экспрессии и очистки слитных полипептидов являются коммерчески доступными. Полипептид может быть помечен эпитопом, а затем очищен с применением специфичного антитела к такому эпитопу. Одну или более стадий жидкостной хроматографии такой, как обращенно-фазовая высокоэффективная жидкостная хроматография, можно применять для дальнейшей очистки полипептида. Чтобы обеспечить получение практически гомогенного рекомбинантного полипептида могут быть использованы некоторые или все из вышеуказанных стадий очистки в различных комбинациях. Очищенный таким образом полипептид может быть по существу свободным от других полипептидов и определяется здесь как «по существу очищенный полипептид»; такие очищенные полипептиды включают полипептиды, фрагменты, варианты и тому подобное. Экспрессия, выделение и очистка полипептидов и их фрагментов могут быть достигнуты любым подходящим способом, включая методы, описанные в данном документе, но не ограничиваясь ими.
Кроме того, для аффинной очистки экспрессированных полипептидов можно использовать аффинную колонку, например, с монокпональным антителом, созданным против полипептидов. Эти полипептиды могут быть удалены с аффинной колонки с применением обычных способов, например, буфером для элюции с высокой концентрацией соли с последующим диализом против буфера с низкой концентрацией соли для дальнейшего использования, или изменением рН или других компонентов в зависимости от используемой аффинной матрицы, или могут быть конкурентно удалены с применением природного субстрата аффинной части.
Полипептид может быть также получен известным обычным химическим синтезом. Способы конструирования полипептидов или их фрагментов синтетическим путем известны специалистам в данной области. Синтетически полученные полипептидные последовательности в силу общей первичной, вторичной или третичной структуры, или конформационных характеристик с нативным полипептидом, могут обладать общими с ним биологическими свойствами, в том числе биологической активностью.
Термин "неприродные", используемый здесь, описывает сущность (например, полинуклеотид, генетические мутации, полипептид, растение, клетки растений и растительные материалы), которая не формируется в природе или не существует в природе. Такие не встречающиеся в природе сущности или искусственные объекты могут быть сделаны, синтезированы, инициированы, изменены, получены или модифицированы с помощью методов, описанных здесь, или методов, которые известны в данной области. Таким образом, в качестве примера не встречающееся в природе растение, не встречающаяся в природе растительная клетка или не встречающийся в природе растительный материал могут быть получены с применением традиционных методов селекции растений, таких как обратное скрещивание, или технологии генетических манипуляций, такиой как антисмысловая РНК, интерферирующие РНК, мегануклеаза и тому подобное. В качестве еще одного примера, не встречающееся в природе растение, не встречающаяся в природе растительная клетка или не встречающийся в природе растительный материал могут быть получены интрогрессией или путем передачи одной или нескольких генетических мутаций (например одного или более полиморфизмов) от первого растения или растительной клетки во второй растение или растительную клетку (которое само может быть встречающимся в природе), так что полученное растение, клетка растения или растительный материал, или их потомство включает в себя генетические характеристики (например, геном, хромосомы или их сегмент), которые не сформированы в природе или не существует в природе. Полученное растение, растительная клетка или растительный материал, таким образом, являются искусственными или не встречающимися в природе. Соответственно, искусственные или не встречающиеся в природе растение или растительная клетка могут быть получены путем модификации генетической последовательности в первом искусственном или природном растении или растительной клетке, что приводит к измененной генетической последовательности, (i) которая является искусственной или (ii) которая встречается в природе во втором растений или растительной клетке, при условии, что второе растение или растительная клетка содержит генетический фон, отличный от первого растения или растительной клетки. Различия в генетическом фоне растения или растительной клетки могут быть обнаружены фенотипическими различиями или методами молекулярной биологии, известными в данной области, такими как секвенирование нуклеиновой кислоты, наличие или отсутствие генетических маркеров (например, микросателлитных маркеров РНК).
Ряд методов на основе полинуклеотидов может быть использован для увеличения экспрессии гена в растениях. В качестве примера, могут быть созданы конструкция, вектор или вектор экспрессии, совместимые с растением, которое будет трансформировано, содержащие интеерсующий ген вместе с промотором, расположенным до гена, который способен гиперэкспрессировать ген в растении. Примеры промоторов описаны здесь. После трансформации и при выращивании в подходящих условиях, промотор может стимулировать экспрессию для модулирования (например, увеличения) уровня этого фермента в растении или в его специфической ткани. В одном примере варианта осуществления, вектор, несущий изопропилмалат синтазу, создается для сверхэкспрессии гена в растении. Вектор несет пригодный промотор до гена изопропилмалат синтазы, направляющий ее экспрессию во всех тканях растения. Вектор может также нести ген устойчивости к антибиотику для проведения селекции трансформированных каллусов и клеточных линий. В одном варианте осуществления промотор способствует транскрипционной активности в трихомах растения. В одном варианте осуществления промотор имеет транскрипционную активность в трихомах растения. В другом варианте осуществления промотор имеет специфичную транскрипционную активность в трихомах растения без активности или по существу без активности в любых других частях растения. Соответственно, промотор содержит, состоит или по существу состоит из последовательности, представленной в SEQ ID NO:8; или последовательности промотора трихомы или нативного промотора изопропилмалат синтазы. Нативный промотор изопропилмалат синтазы расположен на 5' конце гена изопропилмалат синтазы. В SEQ ID NO:6, например, эндогенный промотор изопропилмалат синтазы будет расположен до нуклеотида 2826. Соответственно, ген изопропилмалат синтазы может быть сверхэкспрессирован в трихомах растения. Удивительно, но авторы настоящего изобретения обнаружили, что конститутивная экспрессия не приводит к высокому уровню накопления сложных эфиров сахарозы по сравнению с уровнем, достигнутым при помощи специфичной для трихом экспрессии (см. Фиг.4 и их описание).
В соответствии с одним из вариантов осуществления изобретения, экспрессия или активность изопропилмалат синтазы увеличивается в растении, уже способном продуцировать содержащие бета-метилвалерил сложные эфиры сахарозы, в результате чего наблюдается повышенное накопление изопропилмалат синтазы в нем и, следовательно, увеличение продукции содержащих бета-метилвалерил сложных эфиров сахарозы. Согласно другому варианту осуществления изобретения экспрессия или активность изопропилмалат синтазы увеличивается в растении, которое не способно производить содержащие бета-метилвалерил сложные эфиры сахарозы, в результате чего в нем наблюдается накопление изопропилмалат синтазы и, таким образом, происходит продукция содержащих бета-метилвалерил сложных эфиров сахарозы.
Существует ряд методов, основанных на полинуклеотидах, в том числе применение антисмысловой РНК, направленное рибозимом расщепление РНК, посттранскрипционный сайленсинг генов, например, РНК-интерференция, и транскрипционный сайленсинг генов, которые могут быть использованы для модулирования экспрессии гена в растениях. В то время как такие подходы классически используются для снижения или ингибирования экспрессии гена, они также могут быть использованы для увеличения экспрессии гена за счет увеличения клеточной производительности и качества белков, которые экспрессируются. Такие подходы могут включать сайленсинг экспрессии генов, связанных с апоптозом, экспрессию генов белков, связанных с гликозилированием, лактатдегидрогеназы, участвующей в клеточном метаболизме, и дигидрофолатредуктазы, используемой для амплификации генов. Для снижения или ингибирования множества мишеней, участвующих в различных клеточных путях, могут быть применены другие подходы для изменения регуляции генов в клетках растений.
В некоторых вариантах осуществления может быть использована комплементарная последовательность полноразмерного полинуклеотида или его фрагмента. Как правило, фрагмент включает, по меньшей мере, 10 последовательных нуклеотидов, например, по меньшей мере, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 30, 35, 40, 50, 80, 100, 200, 500 последовательных нуклеотидов или более. В одном варианте осуществления изобретения полинуклеотидный фрагмент (фрагменты) может снижать экспрессию или активность и может кодировать, например, антисмысловую нуклеиновую кислоту, рибозим, РНК, оказывающую эффект на опосредованный сплайсосомой транс-сплайсинг, интерферирующие РНК, гидовые РНК, или другие нетранслируемые РНК и т.п., как описано в данном документе. В другом варианте осуществления полинуклеотидный фрагмент (фрагменты) кодирует интерферирующие РНК.
Соединения, которые могут снизить экспрессию или активность изопропилмалат синтазы, включают, но не ограничиваются ими, последовательности конкретных полинуклеотидов, которые могут препятствовать транскрипции одного или более эндогенного гена изопропилмалат синтазы (S), последовательности конкретных полинуклеотидов, которые могут препятствовать трансляции РНК-транскриптов изопропилмалат синтазы (например, двухцепочечные РНК, миРНК, рибозимы); специфичные к последовательности полипептиды, которые могут препятствовать стабильности белков изопропилмалат синтазы; специфичные к последовательности полинуклеотиды, которые могут препятствовать ферментативной активности белка изопропилмалат синтазы или активности связывания белка изопропилмалат синтазы по отношению к субстратам или регуляторным белкам; антитела, которые обладают специфичностью к белку изопропилмалат синтазы; низкомолекулярные соединения, которые могут препятствовать стабильности белка изопропилмалат синтазы или ферментативной активности белка изопропилмалат синтазы или связывающей активности белка изопропилмалат синтазы; белки с цинковыми пальцами, которые связывают полинуклеотид изопропилмалат синтазы, и мегануклеазы, которые имеют активность по отношению к полинуклеотиду изопропилмалат синтазы.
Антисмысловая технология является одним из известных методов, который можно использовать для модуляции (например, снижения или ингибирования) экспрессии полипептида. Подавляемый полинуклеотид клонируют и функционально связывают с регуляторной областью и последовательностью терминации транскрипции, так что транскрибируется антисмысловая цепь РНК. Рекомбинантную конструкцию затем трансформируют в растения, и синтезируется антисмысловая цепь РНК. Полинукпеотид может не быть всей последовательностью гена, который будет репрессирован, но обычно будет существенно комплементарен, по меньшей мере, части смысловой цепи гена, который будет подавляться.
Полинуклеотид может быть транскрибирован в рибозим, или каталитическую РНК, которая влияет на экспрессию мРНК. Рибозимы могут быть разработаны для специфического спаривания с практически любой целевой РНК и расщепления фосфодиэфирной связи в определенном месте, что приводит к функциональной инактивации РНК-мишени. Гетерологичные полинуклеотиды могут кодировать рибозимы, направленные на расщепление специфических транскриптов мРНК, тем самым предотвращающие экспрессию полипептида. Для уничтожения конкретных мРНК могут быть использованы рибозимы Hammerhead, хотя могут быть использованы различные рибозимы, которые расщепляют мРНК в сайт-специфических последовательностях узнавания,. Рибозимы Hammerhead расщепляют мРНК в местах, определяемых фланкирующими участками, которые формируют комплементарные пары оснований с мРНК-мишенью. Единственным требованием является то, что РНК-мишень должна содержать нуклеотидную последовательность 5'- UG- 3'. Конструирование и изготовление рибозимов Hammerhead известно в данной области. Последовательности рибозимов Hammerhead могут быть встроены в стабильную РНК, такую как транспортная РНК, для повышения эффективности расщепления in vivo.
В одном варианте осуществления изобретения, специфичные к последовательности нуклеотиды, которые могут препятствовать трансляции РНК-транскриптов, являются интерферирующими РНК. РНК-интерференция или РНК-сайленсинг является эволюционно консервативным процессом, при котором специфичные мРНК могут быть направлены для ферментативного расщепления. Двухцепочечная РНК должна быть введена или синтезирована в клетке (например, вирусная двухцепочечная РНК, или полинуклеотиды интерферирующей РНК), чтобы инициировать пути интерференции РНК. Двухцепочечная РНК может быть преобразована в нескольких небольших интерферирующих РНК-дуплексов длиной 21-23 п.о. РНКазой III, которая является эндонуклеазой, специфичной к двухцепочечной РНК. Короткие интерферирующие РНК могут быть впоследствии узнаны РНК-индуцируемыми комплексами сайленсинга, которые способствуют раскручиванию коротких интерферирующих РНК через АТФ-зависимый процесс. Раскрученная антисмысловая цепь коротких интерферирующих РНК направляет РНК-индуцируемые комплексы сайленсинга к целевой мРНК (мишени), содержащей последовательность, комплементарную антисмысловой цепи данных коротких интерферирующих РНК. Целевая мРНК и антисмысловая цепь могут образовать А-спиральную форму, и главная борозда А-спиральной формы может быть узнана активированными РНК-индуцируемыми комплексами сайленсинга. мишени мРНК могут быть расщеплены активированными РНК-индуцируемыми комплексами сайленсинга в одном месте, определяемом как сайт связывания 5'-конца цепи коротких интерферирующих РНК. Активированные РНК-индуцируемые комплексы сайленсинга могут быть рецикпизованы, чтобы катализировать следующий акт расщепления.
Векторы экспрессии интерферирующих РНК могут содержать конструкции интерферирующих РНК, кодирующие полинуклеотиды интерферирующих РНК, которые демонстрируют активность РНК-интерференции, снижая уровень экспрессии мРНК, пре-мРНК, или связанных с ними вариантов РНК. Векторы экспрессии могут включать промотор, расположенный перед конструкциями интерферирующих РНК и функционально связанный с ними, как дополнительно описано в данном документе. Векторы экспрессии интерферирующих РНК могут содержать подходящий минимальный коровый промотор, конструкцию интересующей интерферирующей РНК, расположенную выше по цепи (5') регуляторную область, расположенную ниже по цепи (3') регуляторную область, включая сигналы терминации транскрипции и сигналы полиаденилирования, и другие последовательности, известные специалистам в данной области, например, различные маркеры селекции.
Полинуклеотиды могут быть получены в различных формах, в том числе как двухцепочечные структуры (то есть, двухцепочечная молекула РНК, содержащая антисмысловую цепь и дополнительную смысловую цепь), двухцепочечные шпилько-подобные структуры, или одноцепочечные структуры (то есть, одноцепочечные молекулы РНК, состоящие только из антисмысловой цепи). Структуры могут содержать дуплекс, асимметричный дуплекс, шпильку или асимметричную шпилечную вторичную структуру, имеющие самокомплементарные смысловые и антисмысловые цепи. Двухцепочечные интерферирующие РНК могут быть ферментативно превращены в двухцепочечные короткие интерферирующие РНК. Одна из нитей малой интерферирующей РНК дуплекса может отжигаться на комплементарной последовательности в мишени мРНК и родственных вариантах РНК. Дуплексы коротких интерферирующих РНК/мРНК узнаются РНК-индуцируемыми комплексами сайленсинга, которые могут расщеплять РНК в нескольких местах зависимым от последовательности образом, в результате чего деградируют мишени мРНК и родственные варианты РНК.
Молекулы двухцепочечных РНК могут включать молекулы коротких интерферирующих РНК, собранные из одного олигонуклеотида в структуру стебель-петля, в которой самокомплементарные смысловые и антисмысловые районы молекулы коротких интерферирующих РНК связаны посредством основанного на полинуклеотиде или не основанного на полинуклеотиде линкера (линкеров), а также кольцевую одноцепочечную РНК, имеющую две или более структуры петли, и стебель, содержащий самокомплементарные смысловые и антисмысловые цепи, где кольцевая РНК может быть процессирована in vivo или in vitro, давая молекулы активных коротких интерферирующих РНК, способных опосредовать РНК интерференцию.
Также в настоящем документе предусмотрено применение коротких шпилечных молекул РНК, которые содержат специфическую антисмысловую последовательность в дополнение к обратной комплементарной (смысловой) последовательности, как правило, разделенные спейсером или последовательностью петли. Расщепление спейсера или петли дает одноцепочечную молекулу РНК и ее обратную комплементарную последовательность, так что они могут отжигаться с образованием двухцепочечной молекулы РНК (необязательно с дополнительными стадиями обработки, которые могут привести к добавлению или удалению одного, двух, трех или более нуклеотидов с 3'-конца или 5'-конца одной или обеих цепей). Спейсер может иметь достаточную длину, чтобы позволить антисмысловой и смысловой последовательностям отжечься и образовать двухцепочечную структуру (или стебель) до расщепления спейсера (и, возможно, последующих стадий обработки, которые могут привести к добавлению или удалению одного, двух, трех, четырех или более нуклеотидов на 5'-конце или 3'-конце одной или обеих цепей). Последовательность спейсера, как правило, представляет собой не родственную нуклеотидную последовательность, которая находится между двумя комплементарными районами нуклеотидной последовательности, которые при отжиге в двухцепочечный полинуклеотид, дают малую шпилечную РНК. Последовательность спейсера обычно содержит от примерно 3 до примерно 100 нуклеотидов.
Любой интересующий полинуклеотид РНК может быть получен путем выбора подходящего состава последовательности, размера петли и длины стебля для получения дуплекса шпильки. Подходящий диапазон для подбора длины стебля дуплекса шпильки включает в себя стебель длиной, по меньшей мере, приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидов, например, около 14-30 нуклеотидов, около 30-50 нуклеотидов, около 50-100 нуклеотидов, около 100-150 нуклеотидов, около 150-200 нуклеотидов, около 200-300 нуклеотидов, около 300-400 нукпеотидов, около 400-500 нуклеотидов, около 500-600 нуклеотидов и около 600-700 нуклеотидов. Подходящий диапазон для подбора длины петли шпильки дуплекса включает в себя длину петли примерно 4-25 нуклеотидов, 25-50 нуклеотидов, или больше, если длина стебля дуплекса является существенной. Двухцепочечная или одноцепочечная молекула РНК составляет от приблизительно 15 до приблизительно 40 нуклеотидов в длину. Молекула короткой интерферирующей РНК может быть двухцепочечной или одноцепочечной молекулой РНК и имеет длину между около 15 и около 35 нуклеотидов. Молекула короткой интерферирующей РНК может быть двухцепочечной или одноцепочечной молекулой РНК длиной между около 17 и около 30 нуклеотидов. Молекула короткой интерферирующей РНК может быть двухцепочечной или одноцепочечной молекулой РНК длиной между около 19 и около 25 нуклеотидов. Молекула короткой интерферирующей РНК представляет собой молекулу двухцепочечной или одноцепочечной РНК длиной от около 21 до около 23 нукпеотидов. Шпилечные структуры с двухцепочечными областями длиной более чем 21 нуклеотид могут обеспечивать эффективный направляемый короткими интерферирующими РНК сайленсингу, независимо от последовательности петли и длины.
Последовательность мишени мРНК может быть между примерно от 14 до примерно 50 нуклеотидов в длину. Мишени мРНК, следовательно, могут быть исследованы на предмет участков между около 14 и около 50 нуклеотидов в длину, которые предпочтительно соответствуют одному или нескольким из следующих критериев последовательности мишени: соотношение A+T/G+C в интервале от примерно 2:1 до примерно 1:2; присутствие динуклеотида АА или динуклеотида СА на 5'-конце последовательности-мишени; последовательность, по меньшей мере, из 10 последовательных нуклеотидов, уникальных для мишени мРНК (т.е. последовательность не присутствует в других последовательностей мРНК из того же растения), отсутствуют "участки" из более, чем трех последовательных гуанинов (G) подряд или более трех цитозинов (С) подряд. Эти критерии могут быть оценены с помощью различных способов, известных в данной области, например, компьютерных программ, таких как BLAST, которая может быть использована для поиска в общедоступных базах данных, чтобы определить, является ли выбранная последовательность-мишень уникальной для мишени мРНК. Альтернативно, последовательность-мишень может быть выбрана (и создана последовательность короткой интерферирующей РНК) с применением коммерчески доступного компьютерного программного обеспечения (например, OligoEngine, Target Finder и Small interfering RNA Design Tool, которые коммерчески доступны).
Могут быть выбраны последовательности мишени мРНК, которые имеют длину между около 14 и около 30 нуклеотидов, которые удовлетворяют одному или более из вышеперечисленных критериев, или могут быть выбраны последовательности мишени, которые имеют длину между около 16 и около 30 нуклеотидов в длину, которые отвечают одному или более вышеуказанным критериям. Могут быть выбраны последовательности мишени, которые имеют длину между около 19 и около 30 нукпеотидов, которые удовлетворяют одному или более из вышеперечисленных критериев. Выбирали последовательности мишени между около 19 и около 25 нуклеотидов в длину, которые удовлетворяли одному или более из вышеперечисленных критериев.
Молекулы коротких интерферирующих РНК могут содержать специфичную антисмысловую последовательность, комплементарную, по меньшей мере, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более смежным нуклеотидам любой одной из полинуклеотидных последовательностей, описанных в настоящем документе.
Специфичная антисмысловая последовательность, включенная в молекулу короткой интерферирующей РНК, может быть идентична или по существу идентична с комплементарной последовательностью последовательности мишени. Специфичная антисмысловая последовательность, включенная в молекулу короткой интерферирующей РНК, может быть, по меньшей мере, приблизительно на 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична комплементарной последовательности мишени мРНК. Методы определения идентичности последовательности известны в данной области, и идентичность может быть определена, например, с помощью программы BLASTN от University of Wisconsin Computer Group (GCG), представленной на веб-сайте NCBI.
Конкретная антисмысловая последовательность молекул короткой интерферирующей РНК может демонстрировать изменчивость, отличаясь (например, путем замены нуклеотидов, в том числе транзиции или трансверсии) на один, два, три, четыре или более нуклеотидов из последовательности мишени мРНК. Когда такие нукпеотидные замены присутствуют в антисмысловой цепи двухцепочечной молекулы РНК, комплементарный нуклеотид в смысловой цепи, с которым замещенный нуклеотид, как правило, должен образовывать спаривание оснований через водородные связи, может быть или может не быть соответственно замещен. Также предусмотрена двухцепочечная молекула РНК, в которой одна или более нуклеотидная замена входит в смысловую последовательность, а не в антисмысловую цепь. Когда антисмысловая последовательность молекулы короткой интерферирующей РНК содержит одно или несколько несовпадений между нуклеотидной последовательностью короткой интерферирующей РНК и нуклеотидной последовательностью-мишенью, как описано выше, несоответствия могут присутствовать в 3'-конце, 5'-конце или в центральной части антисмысловой последовательности.
Молекулы короткой интерферирующей РНК могут содержать специфичную антисмысловую последовательность, которая способна селективно гибридизоваться в жестких условиях с частью природного гена-мишени или мРНК-мишенью. Как известно специалистам в данной области, вариации жесткости условий гибридизации могут быть достигнуты путем изменения времени, температуры или концентрации растворов, используемых для этапов гибридизации и отмывки. Подходящие условия могут также частично зависеть от конкретных используемых нуклеотидных последовательностей, например, последовательностей мишени мРНК или гена.
Одним из методов для стимулирования сайленсинга двухцепочечной РНК в растениях является трансформация генетической конструкцией для создания шпильки РНК (см. Smith et al. (2000) Nature, 407, 319-320). Такие конструкции содержат инвертированные районы последовательности гена-мишени, разделенные с помощью соответствующего спейсера. Введение функциональной области интрона растения как спейсерного фрагмента дополнительно повышает эффективность индукции сайленсинга гена за счет образования шпилечной РНК со сплайсированным интроном (Wesley et al. (2001) Plant J., 27, 581-590). Соответственно, длина стебля составляет от около 50 нуклеотидов до примерно 1 тысячи пар оснований в длину. Способы получения шпилечной РНК со сплайсированным интроном хорошо описаны в данной области (см., например, Bioscience, Biotechnology, and Biochemistry (2008) 72, 2, 615-617).
Молекулы интерферирующих РНК, имеющие дуплексные или двухцепочечные структуры, например, двухцепочечные РНК или короткие шпилечные РНК, могут иметь тупые концы, или могут иметь 3' или 5' выступы. Внастоящем документе "выступ" относится к неспаренному нуклеотиду или нуклеотидам, которые выступают из дуплексной структуры, когда 3'-конец одной цепи РНК выходит за пределы 5'-конца другой цепи (3'-выступ), или наоборот (5'-выступ). Нуклеотиды, составляющие выступ, могут быть рибонуклеотидами, дезоксирибонуклеотидами или их модифицированными версиями. В одном варианте осуществления изобретения, по меньшей мере, одна цепь молекулы интерферирующей РНК имеет 3'-выступ от около 1 до около 6 нуклеотидов в длину. В других вариантах осуществления изобретения 3'-выступ составляет от примерно 1 до примерно 5 нуклеотидов, от около 1 до около 3 нуклеотидов и от около 2 до около 4 нуклеотидов в длину.
Когда молекула интерферирующей РНК включает в себя 3'-выступ на одном конце молекулы, другой конец может быть тупым концом или так же иметь выступ (5' или 3'). Когда молекула интерферирующей РНК включает в себя выступ на обоих концах молекулы, длина выступающих элементов может быть одинаковой или разной. В одном варианте осуществления изобретения, молекула интерферирующей РНК включает в себя 3'-выступ от около 1 до около 3 нуклеотидов на обоих концах молекулы. В дополнительном варианте осуществления молекула интерферирующей РНК является двухцепочечной РНК, имеющей 3'-выступ из 2 нуклеотидов на обоих концах молекулы. В еще одном варианте осуществления изобретения, нуклеотиды, содержащие выступ интерферирующей РНК, представляют собой динуклеотид ТТ или динуклеотид UU.
При определении процента идентичности молекул интерферирующих РНК, содержащих один или более выступов, с последовательностью мишени мРНК, выступ (выступы) могут быть или может не быть принят во внимание. Например, нуклеотиды из 3'-выступа и до 2 нуклеотидов из 5'- или 3'-конца двойной цепи могут быть изменены без значительной потери активности молекул короткой интерферирующей РНК.
Молекулы интерферирующих РНК могут содержать одну или более 5'- или 3'-кэп структур. Молекула интерферирующей РНК может содержать структуру кэпа на 3'-конце смысловой цепи, антисмысловой цепи, или как смысловой, так и антисмысловой цепей, или на 5'-конце смысловой цепи, антисмысловой цепи, или обеих смысловой и антисмысловой цепей молекулы интерферирующей РНК. Кроме того, молекула интерферирующей РНК может содержать структуру кэпа как на 3'-конце, так и на 5'-конце молекулы интерферирующей РНК. Термин "кэп-структура" относится к химической модификации, включенной в любой конец олигонуклеотида, которая защищает молекулу от экзонуклеазной деградации, и может также облегчить доставку или локализацию в клетке.
Другой модификацией, применимой к молекуле интерферирующей РНК, является химическая связь с молекулой интерферирующей РНК одного или нескольких фрагментов или конъюгатов, которые повышают активность, клеточное распределение, поглощение клетками, биодоступность и стабильность молекулы интерферирующей РНК. Полинуклеотиды могут быть синтезированы или модифицированы способами, хорошо известными в данной области. Химические модификации могут включать, но не ограничиваются ими, 2'-модификации, вставки неприродных оснований, ковалентное присоединение к лиганду и замену фосфатных связей на тиофосфатные связи. В этом варианте целостность дуплексной структуры усилена, по меньшей мере, одной, а обычно двумя химическими связями. Химическое связывание может быть достигнуто с помощью любого из различных хорошо известных методов, например, путем введения ковалентных, ионных или водородных связей; гидрофобных взаимодействий, Ван-дер-Ваальсовых или стэкинг-взаимодействий; посредством координации ионами металла, или путем использования аналогов пурина.
В еще одном варианте осуществления нуклеотиды в одной или обеих из двух отдельных цепей могут быть модифицированы, чтобы снизить или ингибировать активацию клеточных ферментов, таких как, например, определенные нуклеазы, но не ограничиваясь ими. Методы снижения или ингибирования активации клеточных ферментов, как известно в данной области, включают, но не ограничиваются ими, 2'-амино модификации, 2'-фтор модификации, 2'-алкильные модификации, модификации незаряженного остова, морфолиновые модификации, 2'-O-метил модификации и введение фосфорамидата. Таким образом, по меньшей мере, одна 2'-гидроксильная группа из нуклеотидов двухцепочечной РНК заменена химической группой. Кроме того, по меньшей мере, один нуклеотид может быть изменен, чтобы сформировать заблокированный нуклеотид. Такой заблокированный нуклеотид содержит метиленовый или этиленовый мост, соединяющий 2'-кислород рибозы с 4'-углеродом рибозы. Введение заблокированного нуклеотида в олигонуклеотид улучшает сродство к комплементарным последовательностям и повышает температуру плавления на несколько градусов.
Лиганды могут быть конъюгированы с молекулой интерферирующей РНК, например, чтобы повысить ее поглощение клетками. В некоторых вариантах с молекулой конъюгируют гидрофобный лиганд, чтобы облегчить прямое проникновение через клеточную мембрану. Эти подходы были использованы, чтобы облегчить проникновение в клетки антисмысловых олигонуклеотидов. В некоторых случаях сопряжение катионного лиганда с олигонуклеотидами часто приводит к улучшенной устойчивости к действию нуклеаз. Типичные примеры катионных лигандов включают пропиламмоний и диметилпропиламмоний. Антисмысловые олигонуклеотиды могут сохранять свою высокую аффинность связывания с мРНК, когда катионный лиганд рассредоточен по всей длине олигонуклеотида.
В соответствии с одним из вариантов осуществления экспрессия или активность изопропилмалат синтазы уменьшается в растении, уже способном продуцировать содержащие бета-метилвалерил сложные эфиры сахарозы, в результате чего снижается накопление в нем изопропилмалат синтазы и, таким образом, снижается уровень продукции содержащих бета-метилвалерил сложных эфиров сахарозы.
Молекулы и нукпеотиды, описанные здесь, могут быть получены с применением хорошо известных методов твердофазного синтеза. Дополнительно или альтернативно могут быть использованы любые другие средства для такого синтеза, известные в технике.
Различные варианты осуществления направлены на экспрессионные вектора, содержащие один или более полинуклеотидов, описанных здесь, или конструкции интерферирующих РНК, которые содержат один или более полинукпеотидов.
Различные варианты осуществления направлены на экспрессионные вектора, содержащие один или более полинуклеотидов или одну или более конструкций интерферирующих РНК.
Различные варианты осуществления направлены на экспрессионные вектора, содержащие один или более полинуклеотидов или одну или более конструкций интерферирующих РНК, кодирующих один или более полинуклеотидов интерферирующих РНК, способных самоотжигаться, чтобы сформировать структуру шпильки, где эта конструкция включает: (а) один или более полинуклеотидов, описанных здесь, (б) вторую последовательность, кодирующую спейсерный элемент, который включает интрон, который будучи сплайсированным, образует петлю структуры шпильки, и (в) третью последовательность, содержащую обратную комплементарную последовательность по отношению к первой последовательности, расположенную в той же ориентации, что и первая последовательность, где вторая последовательность расположена между первой последовательностью и третьей последовательностью и вторая последовательность функционально связана с первой последовательностью и с третьей последовательностью.
Описанные последовательности могут быть использованы для построения различных полинуклеотидов, которые не образуют структуру шпильки. Например, двухцепочечная РНК может быть сформирована путем (1) транскрибирования первой цепи ДНК путем функционального связывания с первым промотором, и (2) транскрибирования обратной комплементарной последовательности для первой цепи ДНК-фрагмента путем функционального связывания со вторым промотором. Каждая цепь полинуклеотида могут быть транскрибирована с того же вектора экспрессии, или с различных векторов экспрессии. РНК дуплекс, имеющий активность интерферирующей РНК, может быть ферментативно преобразован в короткие интерферирующие РНК для снижения уровня РНК.
Таким образом, различные варианты осуществления направлены на вектора экспрессии, содержащие полинуклеотид или конструкции интерферирующих РНК, кодирующие полинукпеотиды интерферирующих РНК, способные самоотжигаться, где конструкция включает (а) один или более из описанных здесь полинуклеотидов, и (б) вторую последовательность, содержащую комплементарную (например, обратную комплементарную) последовательность для первой последовательности, расположенную в той же ориентации, что и первая последовательность.
Предложены различные композиции и способы для уменьшения уровня эндогенной экспрессии изопропилмалат синтазы путем косупрессии экспрессии гена. Явление косупрессии происходит в результате введения нескольких копий трансгена в растительную клетку-хозяина. Интеграция нескольких копий трансгена может привести к снижению экспрессии трансгена и эндогенного гена-мишени. Степень косупрессии зависит от степени идентичности последовательностей трансгена и эндогенного гена-мишени. Сайленсинг как эндогенного гена, так и трансгена, может происходить путем обширного метилирования локуса, подвергаемого сайленсингу (то есть эндогенного промотора и эндогенного интересующего гена), которое может препятствовать транскрипции. Кроме того, в некоторых случаях, косупрессия эндогенного гена и трансгена может происходить путем пост-транскрипционного сайленсинга гена, при котором транскрипты могут образовываться, но повышенная скорость деградации исключает накопление транскриптов. Механизм косупрессии путем пост-транскрипционного сайленсинга генов, как полагают, похож на РНК-интерференцию, в отношении того, что РНК, по-видимому, является важным инициатором и мишенью в этих процессах, и механизм может быть опосредован, по меньшей мере, частично, одними и теми же молекулярными механизмами, возможно, через направляемую РНК деградацию мРНК.
Косупрессия нуклеиновых кислот может быть достигнута путем интеграции в геном интрересующего растения нескольких копий нуклеиновой кислоты или ее фрагментов, таких как трансгены. Растение-хозяин может быть трансформировано с помощью экспрессионного вектора, содержащего промотор, функционально связанный с нуклеиновой кислотой или ее фрагментами. Различные варианты осуществления направлены на экспрессионные векторы для обеспечения косупрессии эндогенных генов, включающие промотор, функционально связанный с полинуклеотидом.
Также описаны способы получения консервативных мутантов полинуклеотидов и полипептидов. Любое интрересующее растение, в том числе клетки растений или растительный материал, могут быть генетически модифицированы различными способами, индуцирующими мутагенез, в том числе сайт-направленным мутагенезом, олигонуклеотид-направленным мутагенезом, химически индуцированным мутагенезом, мутагенезом, индуцированным облучением, мутагенезом с использованием модифицированных оснований, мутагенезом с использованием дуплекса ДНК с разрывами, мутагенезом с двуцепочечными разрывами, мутагенезом с использованием штаммов-хозяев с дефектами в системе репарации, мутагенезом полным синтезом генов, перетасовкой ДНК и другими эквивалентными методами.
С другой стороны, гены могут быть инактивированы путем введения транспозонов (например, IS элементов) в геномы растений, представляющих интерес. Эти мобильные генетические элементы могут быть введены при перекрестном опылении, и инсерционные мутанты могут быть исследованы на потерю активности белка. Поврежденный ген из родительского растения может быть введен в другие растения путем скрещивания родительского растения с растением, не подвергнутым мутагенезу, индуцированному транспозонами, например, путем перекрестного опыления. Могут быть использованы любые стандартные методы селекции, известные специалистам в данной области. В одном варианте осуществления один или несколько генов могут быть инактивированы путем введения одного или более транспозонов. Мутации могут привести к гомозиготному повреждению одного или нескольких генов, к гетерозиготному повреждению одного или нескольких генов или комбинации гомозиготных и гетерозиготных повреждений, если повреждают более чем один ген. Подходящие мобильные элементы включают ретротранспозоны, ретропозоны и элементы семейства SINE. Такие способы известны специалистам в данной области.
С другой стороны, гены могут быть инактивированы путем введения рибозимов, полученных из ряда небольших кольцевых РНК, которые способны к саморасщеплению, и репликациии в растениях. Эти РНК могут реплицироваться либо самостоятельно (вироидные РНК), либо со вспомогательным вирусом (сателлитные РНК). Примеры подходящих РНК включают производные вироида солнечных пятен авокадо и сателлитные РНК, полученные из вируса кольцевой пятнистости табака, вируса временной полосатости люцерны, вируса бархатистой крапчатости табака, вируса мозаики черного паслена и вируса мозаики подземного клевера. Различные направленные РНК-специфичные рибозимы известны специалистам в данной области.
В некоторых вариантах осуществления экспрессию полипептида изопропилмалат синтазы можно модулировать не трансгенными способами, такими как создание мутации в гене. Методы, которые вводят случайную мутацию в последовательность гена, могут включать химический мутагенез, EMS мутагенез и радиационный мутагенез. Методы, которые вводят одну или более направленных мутаций в клетку, включают, но не ограничиваются ими, технологии редактирования генома, в частности мутагенез с помощью нуклеазы с цинковым пальцем, TILLING (targeting induced local lesions in genomes, нацеливание индуцированных локальных повреждений в геноме), гомологичную рекомбинацию, олигонуклеотид-направленный мутагенез и опосредованный мегануклеазой мутагенез.
Некоторые не ограничивающие примеры мутаций - это делеции, вставки и миссенс-мутации, по меньшей мере, одного нуклеотида, однонуклеотидные полиморфизмы и простые повторяющиеся последовательности. После мутации может быть выполнен скрининг, чтобы определить мутации, которые создают преждевременные стоп-кодоны или иным образом приводят к нефункциональным генам. После мутации может быть выполнен скрининг для идентификации мутаций, которые создают функциональные гены, которые способны экспрессироваться на повышенном уровне. Скрининг мутантов может быть осуществлен путем секвенирования или с использованием зондов или праймеров, специфичных для гена или белка. Также в полинуклеотиде могут быть созданы конкретные мутации, что может привести к снижению или повышению экспрессии генов, снижению или повышению стабильности мРНК или снижению или повышению стабильности белка. Такие растения упоминаются здесь как мутантные растения.
Мутантные растения могут иметь любое сочетание одной или нескольких мутаций, которые приводят к модуляции уровня полипептида. Например, мутантные растения могут иметь одну мутацию в одном гене; множественные мутации в одном гене; одну мутацию в двух или более или трех или более генах, или множественные мутации в двух или более или трех или более генах. Соответственно, описаны мутантные растения, содержащие варианты мутантных полипептидов.
В одном варианте семена растений подвергали мутагенезу, а затем выращивали мутантные растения первого поколения. Растениям первого поколения позволяли самоопыляться, и из семян от растений первого поколения выращивали растения второго поколения, которые затем проверяли на мутации в их локусах. Хотя скринингу на наличие мутаций можно подвергать подвергнутый мутагенезу растительный материал, преимуществом скрининга растений второго поколения является то, что все соматические мутации соответствуют мутациям зародышевой линии. Специалисту в области техники понятно, что для создания мутантных растений можно подвергать мутагенезу различные растительные материалы, включая, но не ограничиваясь ими, семена, пыльцу, растительные ткани или растительные клетки. Однако тип подвергнутого мутагенезу растительного материала может быть важен, когда нуклеиновые кислоты растения подвергают скринингу на наличие мутаций. Например, когда мутагенезу подвергают пыльцу перед опылением не мутантного растения, семена, являющиеся результатом этого опыления, выращивают в растения первого поколения. Каждая клетка растений первого поколения будет содержать мутации, возникшие в пыльце, поэтому эти растения первого поколения могут затем быть подвергнуты скринингу на мутации вместо того, чтобы ждать второго поколения.
Для создания мутации могут быть использованы мутагены, которые создают точечные мутации, короткие делеции, вставки, трансверсии или транзиции, в том числе химические мутагены или радиация. Мутагены включают, но не ограничиваются ими, этилметансульфонат, метилметансульфонат, N-этил-N-нитрозомочевину, триэтилмеламин, N-метил-N-нитрозомочевину, прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, мономерный акриламид, мелфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил -N'-нитро-нитрозогуанидин, нитрозогуанидин, 2-аминопурин, 7,12-диметил-бенз(а)антрацен, этиленоксид, гексаметилфосфорамид, бусульфан, диэпоксиалканы (диэпоксиоктан, диэпоксибутан и тому подобное), 2-метокси-6-хлор-9[3-(этил-2-хлор-этил)аминопропиламин]акридина дигидрохлорид и формальдегид. Также предусмотрены спонтанные мутации в локусе, которые не могут быть непосредственно вызваны мутагеном, при условии, что они приводят к желательному фенотипу. Подходящие мутагенные агенты также включают, например, ионизирующее излучение, например, рентгеновское излучение, гамма-лучи, облучение быстрыми нейтронами и УФ-излучение.
Любой способ получения нуклеиновой кислоты из растения, известный в данной области техники, может быть использован для получения нуклеиновой кислоты растения для скрининга мутации.
Полученные нуклеиновые кислоты из отдельных растений, растительных клеток или растительного материала могут быть необязательно объединены в целях ускорения скрининга на наличие мутации в гене изопропилмалат синтазы популяции растений, происходящей из подвергнутой мутагенезу растительной ткани, клетки или материала. Одно или несколько последующих поколений растений, растительных клеток или растительного материала могут быть подвергнуты скринингу. Размер необязательно объединенной группы зависит от чувствительности используемого метода скрининга.
После того, как образцы нуклеиновых кислот, необязательно, объединены, они могут быть подвергнуты специфичным для последовательности полинукпеотидов методам амплификации, например, полимеразной цепной реакции (ПЦР). Любой один или более праймеров или зондов, специфичных для гена изопропилмалат синтазы или последовательностей в непосредственной близости от него, могут быть использованы для амплификации последовательностей в необязательно объединенных образцах нуклеиновой кислоты. Предпочтительно, чтобы один или несколько праймеров были предназначены для амплификации участка локуса изопропилмалат синтазы, где полезные мутации возникнут вероятнее всего. Наиболее предпочтительно, чтобы праймер был предназначен для обнаружения мутаций в участках полинуклеотида изопропилмалат синтазы. Кроме того, предпочтительно, чтобы праймер (праймеры) не соответствовали известным участкам полиморфизма, чтобы облегчить скрининг точечных мутаций. Для облегчения обнаружения продуктов амплификации, один или более праймеров или зондов могут быть помечены с помощью любого обычного способа мечения. Праймер (праймеры) или зонды могут быть сконструированы на основе последовательностей, описанных здесь, с использованием способов, которые хорошо известны специалистам в данной области техники. Полиморфизмы могут быть идентифицированы с помощью известных в данной области способов. В одном аспекте предложен способ получения мутантного растения. Способ включает обеспечение, по меньшей мере, одной клетки растения, содержащей ген, кодирующий функциональный полипептид изопропилмалат синтазы. Далее, по меньшей мере, одну клетку растения обрабатывают в условиях, эффективных для модулирования активности гена изопропилмалат синтазы, по меньшей мере, одна мутантная растительная клетка затем выращивается в мутантное растение, где мутантное растение имеет измененный уровень полипептида изопропилмалат синтазы по сравнению с контрольным растением. В одном варианте осуществления этого способа получения мутантного растения стадия обработки включает воздействие, по меньшей мере, на одну клетку химическим мутирующим агентом, описанным выше, в условиях, эффективных для получения, по меньшей мере, одной мутантной растительной клетки. В другом варианте осуществления этого способа стадия обработки включает этап воздействия, по меньшей мере, на одну клетку источника излучения в условиях, эффективных для получения, по меньшей мере, одной мутантной растительной клетки. Термин «мутантное растение» включает мутированные растения, у которых генотип модифицирован по сравнению с контрольным растением, соответственно, способами отличными от генной инженерии или генетической модификации.
В некоторых вариантах осуществления мутантные растения, мутантные растительные клетки или мутантный растительный материал могут содержать одну или несколько мутаций, которые возникли естественно в другом растении, растительной клетке или растительном материале и придают желаемый признак. Эта мутация может быть введена (например, интрогрессией) в другое растение, растительную клетку или растительный материал (например, растение, растительную клетку или растительный материал с генетическим фоном, отличным от растения, из которого была получена мутация), чтобы придать ему данный признак. Таким образом, в качестве примера, мутация, которая возникла естественно в первом растении, может быть введена во второе растение, такое как второе растение с генетическим фоном, отличным от первого растения. Специалист, следовательно, способен найти и определить растение, несущее естественно в своем геноме один или несколько мутантных аллелей гена изопропилмалат синтазы, которые придают желаемый признак. Мутантный аллель (аллели), который возник естественно, может быть передан второму растению с помощью различных методов, включая селекцию, возвратное скрещивание и интрогрессию, для получения линий, сортов или гибридов, которые имеют одну или несколько мутаций в гене изопропилмалат синтазы. Растения, демонстрирующие желаемый признак, могут быть выбраны из пула мутантных растений. Соответственно, выбор осуществляется с использованием знаний о нуклеотидных последовательностях изопропилмалат синтазы, описанных здесь. Следовательно, можно отбирать по генетическому признаку, являющемуся показателем изменений состава сложных эфиров сахарозы по сравнению с контролем. Такой способ скрининга может предусматривать применение обычной амплификации нуклеиновой кислоты и/или методов гибридизации, описанных здесь. Таким образом, еще один аспект настоящего изобретения относится к способу идентификации мутантного растения, включающему следующие стадии: (а) получение образца, содержащего полинуклеотид изопропилмалат синтазы, из растения, и (б) определение последовательности нуклеиновой кислоты полинуклеотида изопропилмалат синтазы, где разница в последовательности полинуклеотида изопропилмалат синтазы по сравнению с полинуклеотидом изопропилмалат синтазы контрольного растения показывает, что указанное растение является мутантным растением по гену изопропилмалат синтазы. В другом аспекте предлагается способ идентификации мутантного растения, которое имеет изменения состава сложных эфиров сахарозы по сравнению с контрольным растением, включающий стадии: (а) получение образца из растения, которое скринируют, (б) определение, содержит ли указанный образец одну или несколько мутаций в полинуклеотиде изопропилмалат синтазы, и (с) определение состава сложных эфиров сахарозы в указанном растении; где, если образец содержит одну или несколько мутаций в полинуклеотиде изопропилмалат синтазы, которые модулируют экспрессию или активность кодируемого белка по сравнению с контрольным растением, и часть растения имеет изменения состава сложных эфиров сахарозы по сравнению с контрольным растением, в котором не снижена экспрессия или активность изопропилмалат синтазы, то это указывает на мутантное растение, которое имеет изменения состава сложных эфиров сахарозы. В другом аспекте предлагается способ получения мутантного растения, которое имеет изменения состава сложных эфиров сахарозы по сравнению с контрольным растением, включающий следующие стадии: (а) получение образца из первого растения, (б) определение, содержит ли указанный образец одну или несколько мутаций в полинуклеотиде изопропилмалат синтазы, которые приводят к изменению состава сложных эфиров сахарозы в нем, и (с) перенесение одной или нескольких мутаций во второе растение. Мутация (мутации) могут быть переданы во второе растение с использованием различных методов, известных в данной области, таких как использование генной инженерии, генетических манипуляций, интрогрессии, селекции растений, возвратного скрещивания и тому подобного. В одном варианте осуществления первое растение является природным растением. В одном варианте осуществления второе растение имеет другой генетический фон по сравнению с первым растением. В другом аспекте предлагается способ получения мутантного растения, которое имеет изменения состава сложных эфиров сахарозы по сравнению с контрольным растением, включающий следующие стадии: (а) получение образца из первого растения, (б) определение, содержит ли указанный образец одну или несколько мутаций в полинуклеотиде изопропилмалат синтазы, которые приводят к изменению состава сложных эфиров сахарозы в нем, и (с) интрогрессию одной или нескольких мутаций из первого растения во второе растение. В одном варианте осуществления стадия интрогрессии включает селекцию растений, в том числе, необязательно, возвратное скрещивание и тому подобное. В одном варианте осуществления первое растение является природным растением. В одном варианте осуществления второе растение имеет другой генетический фон по сравнению с первым растением. В одном варианте осуществления первое растение не принадлежит к культурному или элитному сорту. В одном варианте осуществления второе растение не принадлежит к культурному или элитному сорту. Еще один аспект относится к мутантным растениям (в том числе к мутантам растений культурных сортов или элитных сортов), полученным или получаемым способами, описанными в данном документе. В некоторых вариантах осуществления мутантные растения могут иметь одну или несколько мутаций, локализованных только в конкретном участке растения, таком как последовательность полинуклеотида изопропилмалат синтазы. Согласно этому варианту осуществления оставшаяся геномная последовательность мутантного растения будет такой же или по существу такой же, как у растения до мутагенеза.
В некоторых вариантах осуществления мутантное растение может иметь одну или несколько мутаций, локализованных в более чем одной области растения, такой как последовательность полинуклеотида изопропилмалат синтазы, и в одной или нескольких других областях генома. Согласно этому варианту осуществления, оставшаяся последовательность генома мутантного растения не будет такой же, или не будет по существу такой же, как у растения до мутагенеза. В некоторых вариантах осуществления мутантное растение не имеет одну или несколько мутаций в одном или более, двух или более, трех или более, четырех или более или пяти или более экзонах полинуклеотида изопропилмалат синтазы, или в одном или более, двух или более, трех или более, четырех или более или пяти или более интронах полинуклеотида изопропилмалат синтазы; в промотере полинуклеотида изопропилмалат синтазы, в 3'-нетранслируемой области полинуклеотида изопропилмалат синтазы, в 5'-нетранслируемой области полинуклеотида изопропилмалат синтазы; в кодирующей области полинуклеотида изопропилмалат синтазы, или в некодирующей области полинуклеотида изопропилмалат синтазы или любой комбинации двух или более, трех или более, четырех или более, пяти или более или шести или более из них и их частей. В еще одном аспекте предложен способ идентификации растения, растительной клетки или растительного материала, содержащего мутацию в гене, кодирующем изопропилмалат синтазу, включающий: (а) мутагенное воздействие на растение, растительные клетки или растительный материал (б) получение образца нуклеиновой кислоты из указанного растения, растительной клетки или растительного материала или их потомства, и (с) определение нуклеотидной последовательности гена, кодирующего изопропилмалат синтазу, или его варианта, или его фрагмента, причем разница в указанной последовательности свидетельствует об одной или нескольких мутациях в гене.
Чтобы модулировать экспрессию или активность изопропилмалат синтазы, могут быть использованы белки с цинковыми пальцами. В различных вариантах осуществления последовательность геномной ДНК, содержащую часть или всю кодирующую последовательность полинуклеотида, модифицируют с помощью мутагенеза нуклеазой с цинковым пальцем. В геномной последовательности ДНК ищут уникальный сайт для связывания с белком с цинковым пальцем. Альтернативно, в последовательности геномной ДНК ищут два уникальных сайта для связывания белком с цинковым пальцем, где оба сайта находятся на противоположных нитях и близко друг к другу, например, в 1, 2, 3, 4, 5, 6 или более парах оснований друг от друга. Соответственно, предусмотрены белки с цинковыми пальцами, которые связываются с полинуклеотидами, описанными здесь.
Белок с цинковым пальцем может быть сконструирован таким образом, чтобы узнавать выбранный сайт-мишень в гене изопропилмалат синтазы. Белок с цинковым пальцем может содержать любую комбинацию мотивов, полученных из природных ДНК-связывающих доменов белков с цинковыми пальцами и искусственных ДНК-связывающих доменов белков с цинковыми пальцами, полученную путем вырезания или удлинения, или с использованием сайт-направленного мутагенеза в сочетании со способами отбора, включая, но не ограничиваясь ими, фаговый дисплей, бактериальная двухгибридная или бактериальная одногибридная селекция. Термин «искусственный ДНК-связывающий домен цинковый палец» относится к ДНК-связывающим доменам цинковый палец, которые связывают последовательность из трех пар оснований в ДНК-мишени и которые не образуются в клетке или организме, включающем ДНК, которую нужно изменить. Методы конструкции белков с цинковым пальцами, которые связывают конкретные нуклеотидные последовательности, которые являются уникальными для гена-мишени, известны в данной области.
Нуклеазы с цинковым пальцем могут быть получены путем соединения первого полинуклеотида, кодирующего белок с цинковым пальцем, который связывается с полинуклеотидом, и второго полинуклеотида, кодирующего неспецифическую эндонукпеазу, такую как, эндонуклеаза типа IIS, но не ограничиваясь ими. Слитый белок, в котором соединены белок с цинковым пальцем и нуклеаза, может содержать спейсер, состоящий из двух пар оснований или, альтернативно, спейсер может содержать три, четыре, пять, шесть или семь или более пар оснований. В различных вариантах осуществления нуклеазы с цинковым пальцем вводят двухцепочечный разрыв в регуляторной области, кодирующей области или некодирующей области последовательности полинуклеотида в геномной ДНК и приводят к снижению уровня экспрессии полинуклеотида или снижению активности белка, кодируемого им. Расщепление с помощью нуклеазы с цинковым пальцем часто приводит к удалению ДНК в сайте расщепления при последующей репарации ДНК путем негомологичного соединения концов.
В других вариантах осуществления белок с цинковым пальцем может быть выбран для связывания с регуляторной последовательностью полинуклеотида изопропилмалат синтазы. В частности, регуляторная последовательность может содержать сайт инициации транскрипции, стартовый кодон, область экзона, границу экзон-интрон, терминатор или стоп-кодон. Соответственно, изобретение обеспечивает мутантные, не встречающиеся в природе или трансгенные растения или растительные клетки, полученные с использованием мутагенеза с помощью нуклеазы с цинковым пальцем в непосредственной близости или внутри гена изопропилмалат синтазы, и способы получения таких растений или растительных клеток с использованием мутагенеза с помощью нуклеазы с цинковым пальцем. Способы доставки в растение белка с цинковым пальцем и нуклеазы с цинковым пальцем, аналогичны тем, которые описаны ниже для доставки мегануклеазы.
В другом аспекте предложены способы получения рекомбинантных, мутантных, не встречающихся в природе или трансгенных или иным образом генетически модифицированных растений с использованием мегануклеазы, такой как мегануклеаза I-Crel. Для специфичного создания разрыва в одном месте или в относительно небольшом количестве мест в геномной ДНК растения, чтобы в полинуклеотиде изопропилмалат синтазы образовался двухцепочечный разрыв, могут быть использованы природные меганукпеазы, а также рекомбинантные мегануклеазы. Может быть сконструирована меганукпеаза с измененными свойствами распознавания ДНК. Белки-мегануклеазы могут быть доставлены в клетки растений с помощью множества различных механизмов, известных в данной области.
Раскрываемая информация охватывает применение мегануклеазы для инактивации в клетках растений или растениях одного или более из описанных здесь полинуклеотидов. Аспекты также относятся к способу инактивации полинуклеотида в растении с использованием мегануклеазы, включающему: (а) получение растительной клетки, содержащей один или более из описанных здесь полинуклеотидов, (б) введение мегануклеазы или конструкции, кодирующей мегануклеазу, в указанную растительную клетку; и (в) предоставление меганукпеазе возможность существенно инактивировать полинуклеотид.
Мегануклеазы могут быть использованы для расщепления сайтов узнавания мегануклеазы внутри кодирующих областей полинуклеотида. Такое расщепление часто приводит к удалению ДНК в сайте узнавания мегануклеазой с последующей мутагенной репарацией ДНК путем негомологичного соединения концов. Таких мутаций в кодирующей последовательности гена, как правило, достаточно, чтобы инактивировать ген. Этот способ изменения растительной клетки включает, во-первых, доставку экспрессионной кассеты мегануклеазы в растительную клетку с использованием подходящего метода трансформации. Для достижения максимальной эффективности желательно связать экспрессионную кассету мегануклеазы с маркером селекции и отбирать успешно трансформированные клетки в присутствии селективного агента. Такой подход приводит к интеграции экспрессионной кассеты мегануклеазы в геном, что, однако, может быть нежелательно, если для такого растения, что вероятно, потребуется одобрение регулирующих органов. В таких случаях экспрессионная кассета мегануклеазы (и связанный ген маркера селекции) может быть удалена в последующих поколениях растений с использованием традиционных методов селекции. Кроме того, растительные клетки могут быть первоначально трансформированы экспрессионной кассетой мегануклеазы без маркера селекции и могут быть выращены на средах без селективного агента. В таких условиях доля обработанных клеток получит экспрессионную кассету мегануклеазы и будет экспрессировать сконструированную мегануклеазу временно, без интеграции экспрессионной кассеты мегануклеазы в геном. Поскольку это не учитывает эффективности трансформации, эта последняя процедура трансформации требует скрининга большего количества обработанных клеток для получения желаемой модификации генома. Данный подход может быть применен для модификации растительной клетки при использовании белка с цинковым пальцем или нуклеазы с цинковым пальцем.
После доставки экспрессионной кассеты мегануклеазы на начальном этапе растительные клетки выращивают в условиях, характерных для конкретной процедуры трансформации, которая была использована. Это может означать выращивание трансформированных клеток в среде при температурах ниже 26°С, часто в темноте. Такие стандартные условия могут быть использованы в течение некоторого периода времени, предпочтительно 1-4 дня, чтобы позволить растительным клеткам восстановиться после процесса трансформации. В любой момент после этого начального периода восстановления температура роста может быть увеличена до температуры, стимулирующей активность сконструированной мегануклеазы расщеплять и мутировать сайт узнавания мегануклеазы.
Для некоторых применений может быть желательным точно удалить полинуклеотид из генома растений. Такие применения осуществимы с помощью пары модифицированных мегануклеаз, каждая из которых расщепляет сайт узнавания мегануклеазы с одной из сторон от предполагаемой делеции.
Мегануклеазы и другие стратегии репарации ДНК также могут быть использованы, чтобы вызвать гетерологичную рекомбинацию между ДНК, содержащейся в растительной клетке, и целевой ДНК. Целевая ДНК обычно содержит, по меньшей мере, один, а предпочтительно два участка, которые гомологичны ДНК, содержащейся в растительной клетке. После репарации ДНК клеткой, гомологичные концы соединяются, и в результате целевая ДНК вводится в клетку.
В дополнительном варианте осуществления уровень экспрессии изопропилмалат синтазы в растении может быть увеличен путем вставки гена непосредственно в геном хлоропласта, так как изопропилмалат синтаза является хлоропластным белком, кодируемым в ядре растительной клетки. Кроме того, увеличение транспорта или импорта белка изопропилмалат синтазы в хлоропласты также может модулировать паттерн активности.
Одной целью является получение мутантных, трансгенных или не встречающихся в природе растений, которые демонстрирует измененный уровень сложных эфиров сахарозы при сохранении по существу такого же внешнего вида по сравнению с контрольным растением. Соответственно, здесь описаны мутантные или трансгенные, не встречающиеся в природе растения или клетки, которые имеют измененный уровень сложных эфиров сахарозы по сравнению с контрольными клетками или контрольными растениями. Мутантные, трансгенные или не встречающиеся в природе растения или клетки были изменены, чтобы модулировать синтез или активность изопропилмалат синтазы, описанной здесь, путем модуляции экспрессии одного или более полипептидов, кодируемых полинуклеотидными последовательностями, описанными здесь.
Еще один аспект относится к мутантным, не встречающимся в природе или трансгенным растениям или клеткам, у которых экспрессия или активность изопропилмалат синтазы изменена, и часть растения имеет, по меньшей мере, на 5% увеличенный или уменьшенный уровень сложных эфиров сахарозы по сравнению с контрольным растением, в котором экспрессия или активность указанного фермента не изменена. Еще один аспект относится к мутантным, не встречающимся в природе или трансгенным растениям или клеткам, где экспрессия изопропилмалат синтазы или активность кодируемого белка модулирована, и в которых уровень сложных эфиров сахарозы в аэрозоле увеличен или уменьшен, по меньшей мере, на 5% по сравнению с аэрозолем из контрольного растения.
Изменение содержания сложных эфиров сахарозы по сравнению с контрольным растением может быть, по меньшей мере, приблизительно на 5%, по меньшей мере, приблизительно на 10%, по меньшей мере, приблизительно на 20%, по меньшей мере, приблизительно на 25%, по меньшей мере, приблизительно на 30%, по меньшей мере, приблизительно на 40%, по меньшей мере, приблизительно на 50%, по меньшей мере, приблизительно на 60%, по меньшей мере, приблизительно на 70%, по меньшей мере, приблизительно на 75%, по меньшей мере, приблизительно на 80%, по меньшей мере, приблизительно на 90%, по меньшей мере, приблизительно на 95%, по меньшей мере, приблизительно на 96%, по меньшей мере, приблизительно на 97%, по меньшей мере, приблизительно на 98%, по меньшей мере, приблизительно на 99% или приблизительно на 100% или более.
Растение может быть нагрето до 100°С или выше, например, по меньшей мере, до 125°С, по меньшей мере, до 150°С, по меньшей мере, до 175°С или, по меньшей мере, до 200°С для выделения аэрозоля.
Растения, подходящие для использования для генной модификации, включают одно- и двудольные растения и системы клеток растений, в том числе виды из одного из следующих семейств: Acanthaceae, Alliaceae, Alstroemeriaceae, Amaryllidaceae, Apocynaceae, Arecaceae, Asteraceae, Berberidaceae, Bixaceae, Brassicaceae, Bromeliaceae, Cannabaceae, Caryophyllaceae, Cephalotaxaceae, Chenopodiaceae, Colchicaceae, Cucurbitaceae, Dioscoreaceae, Ephedraceae, Erythroxylaceae, Euphorbiaceae, Fabaceae, Lamiaceae, Linaceae, Lycopodiaceae, Malvaceae, Melanthiaceae, Musaceae, Myrtaceae, Nyssaceae, Papaveraceae, Pinaceae, Plantaginaceae, Poaceae, Rosaceae, Rubiaceae, Salicaceae, Sapindaceae, Solanaceae, Taxaceae, Theaceae или Vitaceae.
Подходящие виды включают представителей родов Abelmoschus, Abies, Acer, Agrostis, Allium, Alstroemeria, Ananas, Andrographis, Andropogon, Artemisia, Arundo, Atropa, Berberis, Beta, Bixa, Brassica, Calendula, Camellia, Camptotheca, Cannabis, Capsicum, Carthamus, Catharanthus, Cephalotaxus, Chrysanthemum, Cinchona, Citrullus, Coffea, Colchicum, Coleus, Cucumis, Cucurbita, Cynodon, Datura, Dianthus, Digitalis, Dioscorea, Elaeis, Ephedra, Erianthus, Erythroxylum, Eucalyptus, Festuca, Fragaria, Galanthus, Glycine, Gossypium, Helianthus, Hevea, Hordeum, Hyoscyamus, Jatropha, Lactuca, Linum, Lolium, Lupinus, Lycopersicon, Lycopodium, Manihot, Medicago, Mentha, Miscanthus, Musa, Nicotiana, Oryza, Panicum, Papaver, Parthenium, Pennisetum, Petunia, Phalaris, Phleum, Pinus, Poa, Poinsettia, Populus, Rauwolfia, Ricinus, Rosa, Saccharum, Salix, Sanguinaria, Scopolia, Secale, Solanum, Sorghum, Spartina, Spinacea, Tanacetum, Taxus, Theobroma, Triticosecale, Triticum, Uniola, Veratrum, Vinca, Vitis и Zea.
Подходящие виды могут включать Panicum spp., Sorghum spp., Miscanthus spp., Saccharum spp., Erianthus spp., Populus spp., Andropogon gerardii (бородач), Pennisetum purpureum (слоновая трава), Phalaris arundinacea (канареечник трубковидный), Cynodon dactylon (бермудская трава), Festuca arundinacea (овсяница тростниковая), Spartina pectinata (спартина перистая), Medicago sativa (люцерна), Arundo donax (арундо тростниковый), Secale cereale (рожь), Salix spp. (ива), Eucalyptus spp. (эвкалипт), Triticosecale, бамбук, Helianthus annuus (подсолнечник), Carthamus tinctorius (сафлор), Jatropha curcas (ятрофа), Ricinus communis (клещевина), Elaeis guineensis (масличная пальма), Linum usitatissimum (лен), Brassica juncea, Beta vulgaris (сахарная свекла), Manihot esculenta (маниок съедобный), Lycopersicon esculentum (томат), Lactuca sativa (салат-латук), Musa paradisiaca (банан), Solanum tuberosum (картофель), Brassica oleracea (брокколи, цветная капуста, брюссельская капуста), Camellia sinensis (чай), Fragaria ananassa (клубника), Theobroma cacao (какао), Coffea arabica (кофе), Vitis vinifera (виноград), Ananas comosus (ананас), Capsicum annum (острый и сладкий перец), Allium сера (лук), Cucumis melo (дыня), Cucumis sativus (огурец), Cucurbita maxima (тыква гигантская), Cucurbita moschata (тыква мускатная), Spinacea oleracea (шпинат), Citrullus lanatus (арбуз), Abelmoschus esculentus (окра), Solanum melongena (баклажан), Rosa spp. (роза), Dianthus caryophyllus (гвоздика), Petunia spp. (петунья), Poinsettia pulcherrima (молочай), Lupinus albus (люпин), Uniola paniculata (овес), полевица (Agrostis spp.), Populus tremuloides (тополь), Pinus spp. (сосна), Abies spp. (ель), Acer spp. (клен), Hordeum vulgare (ячмень), Poa pratensis (мятлик), Lolium spp. (плевел) и Phleum pratense (тимофеевка), Panicum virgatum (просо), Sorghum bicolor (сорго, суданка), Miscanthus giganteus (мискантус), Saccharum sp. (тростник), Populus balsamifera (тополь), Zea mays (кукуруза), Glycine max (соя), Brassica napus (канола), Triticum aestivum (пшеница), Gossypium hirsutum (хлопок), Oryza sativa (рис), Helianthus annuus (подсолнечник), Medicago sativa (люцерна), Beta vulgaris (сахарная свекла) или Pennisetum glaucum (просо африканское).
Различные варианты осуществления направлены на мутантные растения, не встречающиеся в природе растения или трансгенные растения, модифицированные для уменьшения уровня экспрессии гена изопропилмалат синтазы, тем самым производя растения, такие, как растения табака, в которых уровень экспрессии изопропилмалат синтазы в тканях интрересующего растения снижен по сравнению с контрольным растением. Описанные композиции и способы могут быть применены к любым видам рода Nicotiana, в том числе N. rustica и N. tabacum (например, LA B21, LN KY171, TI 1406, Basma, Galpao, Perique, Beinhart 1000-1, K326, Hicks Broadleaf и Petico). Другие виды включают N. acaulis, N. acuminata, N. acuminata van multiflora, N. africana, N. alata, N. amplexicaulis, N. arentsii, N. attenuata, N. benavidesii, N. benthamiana, N. bigelovii, N. bonariensis, N. cavicola, N. clevelandii, N. cordifolia, N. corymbosa, N. debneyi, N. excelsior, N. forgetiana, N. fragrans, N. glauca, N. glutinosa, N. goodspeedii, N. gossei, N. hybrid, N. ingulba, N. kawakamii, N. knightiana, N. langsdorffii, N. linearis, N. longiflora, N. maritima, N. megalosiphon, N. miersii, N. noctiflora, N. nudicaulis, N. obtusifolia, N. occidentalis, N. occidentalis subsp. hesperis, N. otophora, N. paniculata, N. pauciflora, N. petunioides, N. plumbaginifolia, N. quadrivalvis, N. raimondii, N. repanda, N. rosulata, N. rosulata subsp. ingulba, N. rotundifolia, N. setchellii, N. simulans, N. solanifolia, N. spegazzinii, N. stocktonii, N. suaveolens, N. sylvestris, N. thyrsiflora, N. tomentosa, N. tomentosiformis, N. trigonophylla, N. umbratica, N. undulata, N. velutina, N. Wigandioides и N. x sanderae. В наиболее предпочтительном варианте осуществления изобретения растение является растением табака, таким как растения из рода Nicotiana или вида Nicotiana tabacum.
Здесь также предусмотрено использование культурных сортов табака и элитных сортов табака. Трансгенными, не встречающимися в природе или мутантными растениями могут быть растения культурного сорта табака или элитного сорта табака, которые содержат один или более трансгенов, или одну или несколько генетических мутаций, или их комбинацию. Генетическая мутация (мутации) (например, один или более полиморфизмов) может быть мутацией, которая не существует в природе в индивидуальной разновидности табака или культурном сорте табака (например, элитном сорте табака), или может быть генетической мутацией (мутациями), которая встречается в природе, при условии, что мутация не встречается в природе в индивидуальной разновидности табака или культурном сорте табака (например, элитном сорте табака). Особенно полезные сорта Nicotiana tabacum включают сорт Burley (Берли), темный табак, табак дымовой сушки и Oriental (Ориенталь). Не ограничивающими примерами культурных сортов или элитных сортов, являются: BD 64, СС 101, СС 200, СС 27, СС 301, СС 400, СС 500, СС 600, СС 700, СС 800, СС 900, Coker 176, Coker 319, Coker 371 Gold, CokeR48, CD 263, DF911, DT 538 LC Galpao tobacco, GL 26H, GL 350, GL 600, GL 737, GL 939, GL 973, HB 04P, HB 04P LC, HB3307PLC, Гибрид 403LC, Гибрид 404LC, Гибрид 501 LC, K 149, K 326, K 346, K 358, K 394, K 399, K 730, KDH 959, KT 200, KT204LC, KYW, KY14, KY 160, KY 17, KY 171, KY 907, KY907LC, KTY14xL8 LC, Little Crittenden, McNair 373, McNair 944, msKY 14xL8, Narrow Leaf Madole, Narrow Leaf Madole LC, NBH 98, N-126, N-777LC, N-7371 LC, NC 100, NC 102, NC 2000, NC 291, NC 297, NC 299, NC 3, NC 4, NC 5, NC 6, NC 7, NC 606, NC 71, NC 72, NC 810, NC BH 129, NC 2002, Neal Smith Madole, OXFORD 207, PD 7302 LC, PD 7309 LC, PD 7312 LC, 'Perique', PVH03, PVH09, PVH19, PVH50, PVH51, R 610, R 630, R 7-11, R 7-12, RG 17, RG 81, RG H51, RGH 4, RGH 51, RS 1410, Speight 168, Speight 172, Speight 179, Speight 210, Speight 220, Speight 225, Speight 227, Speight 234, Speight G-28, Speight G-70, Speight H-6, Speight H20, Speight NF3, TI 1406, TI 1269, TN 86, TN86LC, TN 90, TN 97, TN97LC, TN D94, TN D950, TR (Tom Rosson) Madole, VA 309, VA359, AA 37-1, В 13Р, Xanthi (Ксанти) (Mitchell-Mor), Bel-W3, 79-615, Samsun Holmes NN, KTRDC номер 2 Гибрид 49, Burley 21 (Берли), KY 8959, KY 9, MD 609, PG 01, PG 04, PO1, PO2, РО3, RG 11, RG 8, VA 509, AS44, Banket A1, Basma Drama B84/31, Basma I Zichna ZP4/B, Basma Xanthi BX 2A, Batek, Besuki Jember, C104, Coker 347, Criollo Misionero, Delcrest, Djebel 81, DVH 405, Galpao Comum, HB04P, Hicks Broadleaf, Kabakulak Elassona, Kutsage E1, LA BU 21, NC 2326, NC 297, PVH 2110, Red Russian, Samsun, Saplak, Simmaba, Talgar 28, Wislica, Yayaldag, Prilep HC-72, Prilep P23, Prilep PB 156/1, Prilep P12-2/1, Yaka JK-48, Yaka JB 125/3, TI-1068, KDH-960, TI-1070, TW136, Basma, TKF 4028, L8, TKF 2002, GR141, Basma xanthi, GR149, GR153, Petit Havana. Незначительно измененные подтипы вышеуказанных сортов, даже если не были конкретно приведены здесь, также предусмотрены. В некоторых вариантах осуществления предпочтительными являются культурные сорта и элитные сорта. В некоторых других вариантах осуществления могут быть использованы виды Nicotiana tabacum. Может быть использован сорт Берли Nicotiana tabacum.
Варианты осуществления также относятся к композициям и способам получения мутантных, не встречающихся в природе или трансгенных растений, которые были модифицированы, чтобы модулировать экспрессию или активность изопропилмалат синтазы, в результате чего могут быть получены растения или растительные компоненты с модулированным уровнем сложных эфиров сахарозы по сравнению с контролем. Модуляция уровня сложных эфиров сахарозы в растениях может быть использована для создания растений, в которых профиль аромата аэрозоля, полученного из растения, изменен. Следовательно, настоящее изобретение может обеспечить новые возможности смешивания для получения желаемого аромата табака. Таким образом, в качестве примера, содержание сложных эфиров сахарозы в растении может быть увеличено за счет увеличения экспрессии или активности изопропилмалат синтазы, что может привести к получению растения, способного высвобождать повышенный уровень ароматических молекул, что делает его более похожими на табак сорта Ориенталь. Таким образом, в качестве дополнительного примера, содержание сложных эфиров сахарозы в растении может быть уменьшено путем уменьшения экспрессии или активности изопропилмалат синтазы, что может привести к получению растения, способного высвобождать меньше ароматических молекул, что делает его более похожим на табак дымовой сушки.
В еще одном аспекте предложен способ смешивания табака, включающий стадию замены или добавления к одному или более типам табака в табачной смеси табака (например, табачных листьев или композиции, полученной или получаемой из листьев табака), полученного из трансгенного, мутантного или не встречающегося в природе растения, в котором содержание сложных эфиров сахарозы было изменено (например, увеличено или уменьшено) путем модуляции (предпочтительно, увеличения) экспрессии или активности изопропилмалат синтазы, как описано здесь. В другом аспекте предлагается способ смешивания табака, включающий стадии замены, уменьшения или исключения табака сорта Ориенталь из табачной смеси и добавления табака (например, табачных листьев или композиции, полученной или получаемой из листьев табака), полученного из трансгенного, мутантного или не встречающегося в природе растения, в котором содержание сложных эфиров сахарозы было модулировано (предпочтительно, увеличено) путем изменения (предпочтительно, увеличения) экспрессии или активности изопропилмалат синтазы, как описано здесь. Замена, уменьшение или добавление табака, такого, как табак сорта Ориенталь, может соответствовать примерно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% от табака, количество которого будет изменено, снижено или добавлено.
Еще один аспект относится к способу модуляции вкуса табака или табачных изделий, включающему добавление к табаку (например, табачным листьям или композициям, полученным или получаемым из листьев табака) или табачным изделиям табака, полученного или получаемого из трансгенных, мутантных или не встречающиеся в природе растений, в которых содержание сложных эфиров сахарозы было модулировано, как описано здесь.
Дополнительный аспект относится к применению агента, который модулирует экспрессию или активность изопропилмалат синтазы, для модуляции вкуса табака или табачных изделий.
Модуляция уровня сложных эфиров сахарозы в растениях также может быть использована для создания растений, у которых изменена устойчивость к вредителям. Устойчивость растения к вредителям может быть увеличена путем увеличения уровня образования сложных эфиров сахарозы, описанных здесь. Устойчивость растений к вредителям может быть уменьшена за счет снижения уровня образования сложных эфиров сахарозы, описанных здесь.
Композиции сложных эфиров сахарозы, которые образуются в растениях, описанные здесь, могут быть извлечены и, необязательно, очищены для различного дальнейшего использования, например, в лекарственных средствах, пищевых добавках, курительных ароматизаторах, в качестве компонентов органических пестицидов и тому подобное. В одном варианте осуществления сложные эфиры сахарозы извлекают из листьев или с поверхности листьев. Соответственно, продукты, такие как лекарственные средства, пищевые добавки, курительные ароматизаторы и органические пестициды, содержащие сложные эфиры сахарозы, являются еще одним аспектом настоящего изобретения.
Сложные эфиры сахарозы в соответствии с настоящим изобретением имеют общую структуру, показанную на Фигуре 5, где R1, R2, R3, R4 и R5 представляют собой либо атомы водорода, либо ацильные цепи. Ацильные цепи, которые могут быть этерифицированы сахарозой, приведены на Фигуре 6. Некоторые примеры конкретных сложных эфиров сахарозы приведены в Таблице 1 и на Фигуре 2.
Общее число атомов углерода в ацильных цепях R1, R2 и R4 в каждом одном или нескольких сложных эфирах сахарозы может быть, например, 12, 13, 14, 15, 16, 17 или 18; например, 14, 15, 16, 17 или 18; например, 16, 17 или 18; или, например, 14 или 15. При условии, что это общее число атомов углерода присутствует в каждом из сложных эфиров сахарозы, ацильные цепи могут быть выбраны из любых, которые приведены на Фигуре 6. Подходящий эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий эфир сахарозы имеет ацетил в R3 и водород в R5. Подходящие одна или более, две или более или три или более ацильных цепей представляют собой бета-метилвалерил, особенно когда присутствует 6-углеродная ацильная цепь.
В одном варианте осуществления сложным эфиром сахарозы является сложный эфир сахарозы, имеющий ацетил или водород в R3 и R5 и ацильную цепь с 3, 4, 5 или 6 атомами углерода в любом из положений R1, R2 и R4. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5. В другом варианте осуществления сложный эфир сахарозы имеет ацетил или водород в R3 и R5 и ацильную цепь с 4, 5 или 6 атомами углерода в любом из положений R1, R2 и R4. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий эфир сахарозы имеет ацетил в R3 и водород в R5. В другом варианте осуществления сложный эфир сахарозы имеет ацетил или водород в R3 и R5 и ацильную цепь с 5 или 6 атомами углерода в любом из положений R1, R2 и R4. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5. В другом варианте осуществления сложный эфир сахарозы имеет ацетил или водород в R3 и R5 и ацильную цепь с 6 атомами углерода в положениях R1, R2 и R4. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5.
В одном варианте осуществления сложный эфир сахарозы имеет ацетил или водород в R3 и R5 и ацильную цепь с 6 атомами углерода, по меньшей мере, в одной из позиций из R1, R2 и R4. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5.
В одном варианте осуществления сложный эфир сахарозы имеет ацетил или водород в R3 и R5 и ацильную цепь с 6 атомами углерода в положении R1, ацильную цепь с 6 атомами углерода в положении R2 и ацильную цепь с 6 атомами углерода в положении R4. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5.
В одном варианте осуществления сложный эфир сахарозы имеет ацетил или водород в R3 и R5 и ацильную цепь с 6 атомами углерода в положении R1, ацильную цепь с 6 атомами углерода в положении R2 и ацильную цепь с 6 атомами углерода в положении R4. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5.
В одном варианте сложным эфиром сахарозы является сложный эфир сахарозы, содержащий бета-метилвалерил (с разветвленной цепью).
В одном варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где каждый из R1-R5 представляет собой любую из ацильных цепей, выбранных из группы, состоящей из ацетила, бутирила, пропионила или его изомера, изобутирила, валерила (пентаноила), 2-метил-бутирила, изовалерила, изопентеноила, пентеноила, гексаноила или его изомера 2-метилвалерила, бета-метилвалерила и 4-метилвалерила. В другом варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 означает ацетил, и каждый из R1, R2, R4 и R5 представляет собой любую из ацильных цепей, выбранных из группы, состоящей из ацетила, бутирила, пропионила или его изомера, изобутирила, валерила (пентаноила), 2-метил-бутирила, изовалерила, изопентеноила, пентеноила, гексаноила или его изомера 2-метилвалерила, бета-метилвалерила и 4-метилвалерила. В одном варианте осуществления предпочтительно, чтобы ацильные цепи с 6 атомами углерода были разветвленными цепями. В другом варианте осуществления предпочтительно, чтобы ацильные цепи с 6 атомами углерода представляли собой бета-метилвалерил.
В другом варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 обозначает водород или ацетил, R5 обозначает водород или ацетил, и каждый из R1, R2 и R4 представляет собой любую из ацильных цепей, выбранных из группы, состоящей из ацетила, бутирила, пропионила или его изомера, изобутирила, валерила (пентаноила) или его изомера, 2-метил-бутирила, изовалерила, изопентеноила, пентеноила, гексаноила или его изомера 2-метилвалерила, бета-метилвалерила и 4-метилвалерила. Отдельные сложные эфиры сахарозы раскрыты в Таблице 1 и на Фигуре 2.
В другом варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 обозначает ацетил, R5 обозначает водород или ацетил и каждый из R1, R2 и R4 представляет собой любую из ацильных цепей, выбранных из группы, состоящей из ацетила, бутирила, пропионила или его изомера, изобутирила, валерила (пентаноила) или его изомера, 2-метил-бутирила, изовалерила, изопентеноила, пентеноила, гексаноила или его изомера 2-метилвалерила, бета-метилвалерила и 4-метилвалерила.
В другом варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 означает ацетил, R5 означает водород или ацетил; R1 представляет собой бутирил, пропионил или его изомер, валерил (пентаноил) или его изомер, или гексаноил или его изомер, R2 обозначает пропионил или его изомер, валерил (пентаноил) или его изомер, или гексаноил или его изомер, и R4 представляет собой валерил (пентаноил) или его изомер или гексаноил или его изомер.
В другом варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 обозначает ацетил, R5 обозначает водород или ацетил, R1 обозначает пропионил или его изомер, валерил (пентаноил) или его изомер, или гексаноил или его изомер, R2 обозначает валерил (пентаноил) или его изомер, или гексаноил или его изомер, и R4 представляет собой гексаноил или его изомер.
В одном варианте осуществления модулируемый сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил, или сложный эфир сахарозы имеет структуру, показанную на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил, или сложный эфир сахарозы имеет структуру, показанную на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С14:0).
В одном варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил; или сложный эфир сахарозы имеет структуру, показанную на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил, или сложный эфир сахарозы имеет структуру, показанную на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С12С15:0).
В одном варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 обозначает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает валерил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0)
В одном варианте осуществления модулируемый сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 обозначает ацетил, R1 обозначает пропионил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0).
В одном варианте осуществления сложный эфир сахарозы имеет общую структуру, показанную на Фигуре 5, где R3 обозначает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С17:0).
В одном варианте осуществления сложный эфир сахарозы, содержание которого модулируют, имеет общую структуру, показанную на Фигуре 5, где R3 обозначает ацетил, R1 обозначает гексаноил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С18:0).
В другом варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 594 дальтон до 692 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 608 дальтон до 692 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 622 дальтон до 692 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 636 дальтон до 692 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу от около 650 дальтон до 692 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу от около 650 дальтон до 678 дальтон. В другом варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 594 дальтон до 678 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 608 дальтон до 678 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 622 дальтон до 678 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 636 дальтон до 678 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу от около 650 дальтон до 678 дальтон. В одном варианте осуществления сложный эфир сахарозы имеет молекулярную массу примерно от 664 дальтон до 678 дальтон.
Еще один аспект относится к способу модуляции вкуса табака или табачного изделия, включающему добавление к ним композиции сложных эфиров сахарозы, описанных здесь.
В другом варианте осуществления концентрация, уровень или образование сложных эфиров сахарозы в растении увеличены. В другом варианте осуществления концентрация, уровень или образование сложных эфиров сахарозы в растении уменьшены.
В одном конкретном варианте осуществления раскрыты композиции и способы получения мутантного, не встречающегося в природе или трансгенного растения, которое было модифицировано, чтобы увеличить экспрессию или активность изопропилмалат синтазы, которые могут привести к получению растения или растительного материала, в котором концентрация, уровень или образование сложных эфиров сахарозы увеличены. В частности, уровень или образование сложных эфиров сахарозы, описанных здесь, увеличиваются или индуцируются.
В другом конкретном варианте осуществления раскрыты композиции и способы получения мутантного, не встречающегося в природе или трансгенного растения, которое было модифицировано, чтобы уменьшить или ингибировать экспрессию или активность изопропилмалат синтазы, которые могут привести к получению растения или растительного материала, в котором концентрация, уровень или образование сложных эфиров сахарозы уменьшены. В частности, уровень сложных эфиров сахарозы, описанных здесь, уменьшается или исчезает.
Преимущественно, мутантные, не встречающиеся в природе или трансгенные растения, которые получают в соответствии со способами, описанными здесь, аналогичны или имеют по существу такой же внешний вид, как и контрольные растения. В одном варианте осуществления высота стебля мутантных, не встречающихся в природе или трансгенных растений, по существу такая же, как у контрольного растения после, например, одного, двух или трех или более месяцев после высадки или 10, 20, 30 или 36 или более дней после прищипывания верхушек. Например, высота стебля мутантных, не встречающихся в природе или трансгенных растений не меньше, чем высота стебля контрольного растения. В другом варианте осуществления содержание хлорофилла в мутантных, не встречающихся в природе или трансгенных растениях по существу такое же, как у контрольного растения. В другом варианте осуществления высота стебля мутантных, не встречающихся в природе или трансгенных растений по существу такая же, как у контрольного растения, и содержание хлорофилла в мутантных, не встречающихся в природе или трансгенных растениях по существу такое же, как у контрольного растения. В других вариантах осуществления, размер, или форма, или число или окраска листьев мутантных, не встречающихся в природе или трансгенных растений по существу такие же, как у контрольного растения. Подходящее растение - это растение табака.
В другом аспекте предложен способ модуляции содержания сложных эфиров сахарозы, такого как бета-метилвалериловый сложный эфир сахарозы, по меньшей мере, в части растения, включающий следующие стадии: (I) модулирование экспрессии или активности изопропилмалат синтазы в растении, предпочтительно, где изопропилмалат синтаза представлена полинуклеотидной последовательностью, описанной в настоящем документе, или полипептидной последовательностью, описанной в данном документе; (II) необязательно, измерение содержания сложного эфира сахарозы, по меньшей мере, в части мутантного, не встречающегося в природе или полученного трансгенно растения на стадии (I); и (III) идентификация мутантных, не встречающихся в природе или трансгенных растений, в которых содержание сложных эфиров сахарозы было изменено в сравнении с контрольным растением. Соответственно, внешний вид указанного мутантного, не встречающегося в природе или трансгенного растения является по существу таким же, как у контрольного растения. Подходящее растение - это растение табака.
В другом аспекте предложен способ повышения содержания сложных эфиров сахарозы, таких как бета-метилвалериловый сложный эфир сахарозы, по меньшей мере, в части растения (например, листьях), включающий следующие стадии; (I) увеличение экспрессии или активности изопропилмалат синтазы в растении, предпочтительно, где изопропилмалат синтаза представлена полинуклеотидной последовательностью, описанной в настоящем документе, или полипептидной последовательностью, описанной в данном документе; (II) необязательно, измерение содержания сложных эфиров сахарозы, по меньшей мере, в части мутантного, не встречающегося в природе или трансгенного растения, полученного на стадии (I); и (III) идентификация мутантных, не встречающихся в природе или трансгенных растений, в которых содержание сложных эфиров сахарозы было увеличено по сравнению с контрольным растением.
В другом аспекте предложен способ уменьшения содержания сложных эфиров сахарозы, таких как бета-метилвалериловый сложный эфир сахарозы, по меньшей мере, в части растения, включающий следующие стадии: (I) уменьшение экспрессии или активности изопропилмалат синтазы в растении, предпочтительно, где изопропилмалат синтаза представлена полинуклеотидной последовательностью, описанной в настоящем документе, или полипептидной последовательностью, описанной в данном документе; (II) необязательно, измерение содержания сложных эфиров сахарозы, по меньшей мере, в части мутантного, не встречающегося в природе или полученного трансгенно растения на стадии (I); и (III) идентификация мутантных, не встречающихся в природе или трансгенных растений, в которых содержание сложных эфиров сахарозы было уменьшено по сравнению с контрольным растением.
Предпочтительно, способы, описанные здесь, могут также быть использованы для создания растений, имеющих сходство с сортом Ориенталь, путем увеличения экспрессии или активности изопропилмалат синтазы. Таким образом, согласно этому варианту осуществления, растения не сорта Ориенталь, такие как сорта дымовой сушки или Берли, могут быть изменены так, чтобы экспрессия или активность изопропилмалат синтазы в них была увеличена. Это может придавать растению признаки сорта Ориенталь, что может быть выгодно, так как это может изменить (например, увеличить или улучшить) в них оттенок вкуса, свойственный сорту Ориенталь, тем самым давая новые ароматические линии растений, сорта или гибриды.
Способы, описанные здесь, могут также быть использованы для создания растений, в которых черты сходства с сортом Ориенталь уменьшены, снижены или устранены путем уменьшения экспрессии или активности изопропилмалат синтазы. Таким образом, согласно этому варианту осуществления, растения сорта Ориенталь могут быть изменены таким образом, что экспрессия или активность изопропилмалат синтазы была в них уменьшена. Уменьшение или устранение у растения черт сходства с растениями сорта Ориенталь может быть выгодно, так как это может изменить или уменьшить в них оттенок вкуса, свойственный сорту Ориенталь, тем самым давая новые ароматические линии растений, сорта или гибриды.
Увеличение экспрессии изопропилмалат синтазы по сравнению с контрольным растением может составлять от приблизительно 5% до приблизительно 100% или более, или увеличение может составлять, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98%, на 100%, на 150%, или на 200% или более, что включает увеличение транскрипционной активности или экспрессии белка, или и то, и другое.
Увеличение активности изопропилмалат синтазы по сравнению с контрольным растением может составлять от приблизительно 5% до приблизительно 100% или более, или увеличение может составлять, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98%, на 100%, на 150%, или на 200% или больше и более.
Снижение экспрессии изопропилмалат синтазы по сравнению с контрольным растением может составлять от приблизительно 5% до приблизительно 100%, или уменьшение может составлять, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98% или на 100%, что включает снижение транскрипционной активности или экспрессии белка, или и то, и другое.
Снижение активности изопропилмалат синтазы по сравнению с контрольным растением может составлять от приблизительно 5% до приблизительно 100%, или уменьшение может составлять, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98% или на 100% или больше.
Увеличение содержания сложных эфиров сахарозы по сравнению с контрольным растением может составлять от примерно 5% до примерно 100%, или увеличение может составлять, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98% или до 100%.
Снижение содержания сложных эфиров сахарозы по сравнению с контрольным растением может составлять от приблизительно 5% до приблизительно 100%, или уменьшение может составлять, по меньшей мере, на 10%, по меньшей мере, на 20%, по меньшей мере, на 25%, по меньшей мере, на 30%, по меньшей мере, на 40%, по меньшей мере, 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 98% или до 100%.
Полинуклеотиды и рекомбинантные конструкции, описанные здесь, могут использоваться для модуляции экспрессии изопропилмалат синтазы, описанной здесь, в видах растений, представляющих интерес, например, в табаке.
Одним из аспектов является семя раскрытого мутантного растения, не встречающегося в природе растения, гибридного растения, трансгенного растения. Предпочтительно, чтобы семя являлось семенем табака. Еще одним аспектом являются пыльца или яйцеклетка раскрытого мутантного растения, не встречающегося в природе растения, гибридного растения, трансгенного растения. Кроме того, настоящее изобретение обеспечивает описанное мутантное растение, не встречающееся в природе растение, гибридное растение, трансгенное растение, которое дополнительно включает нуклеиновую кислоту, придающую мужскую стерильность.
В описании также предложены культуры тканей регенерируемых клеток мутантных растений, не встречающихся в природе растений, гибридных растений или трансгенных растений или их частей, являющихся объектами настоящего изобретения, где регенерированные из клеток растения способны показывать все морфологические и физиологические характеристики родителя Регенерируемые клетки включают, но не ограничиваются, клетки листьев, пыльцы, зародышей, семядолей, гипокотиля, корней, кончиков корней, пыльников, цветов и их частей, семязачатков, побегов, стеблей, цветоножки, сердцевины и семенных коробочек или каллуса, или протопласты, полученные из них.
Согласно описанию, растение, несущее мутантный аллель изопропилмалат синтазы, может использоваться в программе разведения растений для создания полезных линий, сортов и гибридов. В частности, мутантный аллель изопропилмалат синтазы может быть интрогрессирован в коммерчески важные разновидности, описанные выше. Таким образом, предложены способы размножения растений, которые включают скрещивание, селекцию или интрогрессию мутантного растения, не встречающегося в природе растения или трансгенного растения, описанного здесь, с другим растением, например, растением с другой генетической идентичностью, другим генетическим фоном, другого сорта, другой линии или другого гибрида. Один способ включает скрещивание, селекцию или интрогрессию мутантного растения, не встречающегося в природе растения или трансгенного растения, описанного здесь, с растением культурного сорта или элитного сорта растения рода Nicotiana, таким как Nicotiana tabacum (например, сорт Берли). Способ может дополнительно включать скрещивание дочернего растения с другим растением, и, возможно, повторение скрещивания до получения потомства с желаемыми генетическими признаками или генетическим фоном. Способ может дополнительно включать стадию идентификации растения, в котором модулирована экспрессия или активность изопропилмалат синтазы. Способ может дополнительно включать стадию селекции растения, в котором экспрессия или активность изопропилмалат синтазы модулирована по сравнению с контрольным растением. Одной из целей таких методов селекции является введение желаемого генетического признака в растения других разновидностей, линий, гибридов или сортов, особенно тех, которые имеют коммерческий интерес. Другой целью является сочетание генетических модификаций различных генов в растении одной разновидности, линии, гибрида или сорта. Рассматриваются как внутривидовое, так и межвидовое скрещивание. Потомство растений, которые возникают из таких скрещиваний, также называемые селекционными линиями, являются примерами раскрытых не встречающихся в природе растений.
В одном варианте осуществления обеспечивается способ получения не встречающихся в природе растений, включающий: (а) скрещивания мутантного или трансгенного растения со вторым растением с получением дочерних семян табака, (б) выращивание дочерних семян табака, в условиях роста растений, с получением не встречающихся в природе растений. Способ может дополнительно включать дополнительный этап идентификации растения, в котором изменена экспрессия или активность изопропилмалат синтазы. Способ может дополнительно включать дополнительный этап отбора растения, в котором экспрессия или активность изопропилмалат синтазы модулирована по сравнению с контрольным растением. Способ может дополнительно включать: (в) скрещивание предыдущего поколения не встречающегося в природе растения с ним самим или с еще одним растением с получением дочерних семян табака, (г) выращивание дочерних семян табака из стадии (в) в условиях роста растений, с получением дополнительных не встречающихся в природе растений, и (д) повторение этапов скрещивания и выращивания (б) и (г) несколько раз, чтобы получить следующие поколения не встречающихся в природе растений. Способ может дополнительно включать перед стадией (а) этап обеспечения родительского растения, которое представлено генетической идентичностью, которая охарактеризована и которая не идентична мутантному или трансгенному растению. В некоторых вариантах осуществления, в зависимости от программы разведения, этапы скрещивания и выращивания повторяются от 0 до 2 раза, от 0 до 3 раз, от 0 до 4 раза, от 0 до 5 раз, от 0 до 6 раз, от 0 до 7 раз, от 0 до 8 раз, от 0 до 9 раз или от 0 до 10 раз, в целях получения поколений не встречающихся в природе растений. Возвратное скрещивание является примером такого способа, в котором потомство скрещивается с одним из своих родителей или другим растением, генетически аналогичным его родителю, чтобы получить дочернее растение следующего поколения, которое имеет генетическую идентичность, которая ближе к одному из родителей. Методы селекции растений, в частности селекции растений табака, хорошо известны и могут быть использованы в описанных способах. Описание дополнительно обеспечивает не встречающиеся в природе растения, произведенные, полученные или получаемые с помощью этих методов.
В некоторых вариантах осуществления описанных здесь способов, линии, полученные в результате селекции и скрининга вариантов гена изопропилмалат синтазы, оцениваются на посадках с использованием стандартных методик полевых работ. Контрольные генотипы, включая исходный родительский не подвергнутый мутагенезу генотип, также включают в исследование, и претендентов располагают на посадке по рандомизированному полному блочному плану или другому подходящему плану посадки. Для табака используются стандартные агрономические методы, например, табак собирают, взвешивают и отбирают образцы для химических и других общих испытаний до и во время сушки. Для подтверждения сходства выбранных линий и родительской линии проводят статистический анализ данных. Необязательно, проводят цитогенетические анализы отобранных растений, чтобы подтвердить родство по хромосомному набору и конъюгации хромосом.
В одном варианте осуществления растения, которые получены или получаемы методами скрещивания, разведения или интрогрессии, описанными выше, не будут производить цис-абиенол, преимущественно не будут производить цис-абиенол или не будет производить цис-абиенол в детектируемых количествах. В другом варианте осуществления композиции сложных эфиров сахарозы и композиции сложных эфиров сахарозы, полученные или получаемые с помощью способов, описанных здесь, не будут включать или содержать цис-абиенол, преимущественно не будут содержать цис-абиенол или не будет содержать цис-абиенол в детектируемых количествах. Таким образом, свойство содержать цис-абиенол может присутствовать или может отсутствовать.
Для программы скрещивания с селекцией с помощью маркеров (MAS) для переноса или скрещивания мутантных аллелей гена изопропилмалат синтазы в другие растения табака, как описано здесь, могут быть использованы ДНК-фингерпринтинг, однонуклеотидный полиморфизм, микросателлитные маркеры или аналогичные технологии. Например, производитель может создать отдельные популяции при гибридизации генотипа, содержащего мутантный аллель, с агрономически желаемым генотипом. Растения поколения F2 или потомки возвратного скрещивания могут быть подвергнуты скринингу с помощью маркера, сконструированного из геномной последовательности изопропилмалат синтазы или ее фрагмента, с использованием одного из методов, перечисленных в данном документе. Растения, идентифицированные как обладающие мутантным аллелем, могут быть подвергнуты возвратному скрещиванию или самоопылению, чтобы получить вторую популяцию для скрининга. В зависимости от предполагаемой схемы наследования или используемой технологии MAS, может быть необходимо самоопыление отобранных растений перед каждым циклом возвратного скрещивания для облегчения идентификации отдельных желаемых растений. Возвратное скрещивание или другая процедура разведения может быть повторена, пока не будет восстановлен желаемый фенотип рекуррентного родителя.
Согласно описанию, в программе скрещивания успешные скрещивания приводят к появлению фертильных растений F1. Отобранные растения F1 могут быть скрещены с одним из родителей, и растения первого поколения возвратного скрещивания самоопыляют, чтобы получить популяцию, которую снова скринируют на варианты экспрессии гена изопропилмалат синтазы (например, нулевую экспрессию гена изопропилмалат синтазы). Процесс возвратного скрещивания, самоопыления и отбора повторяют, например, по меньшей мере, 4 раза до тех пор, пока финальный скрининг не обнаружит растение, которое является фертильным и достаточно похожим на рекуррентного родителя. Это растение, при желании, самоопыляют, и потомство впоследствии скринируют еще раз для подтверждения того, что растения демонстрируют вариант экспрессии гена изопропилмалат синтазы. В некоторых вариантах осуществления популяции растений поколения F2 скринируют на варианты экспрессии гена изопропилмалат синтазы, например, растение определяют, как не способное экспрессировать изопропилмалат синтазу из-за отсутствия гена изопропилмалат синтазы, в соответствии со стандартными способами, например, с помощью метода ПЦР с праймерами, основанными на информации о нуклеотидной последовательности изопропилмалат синтазы, описанной здесь.
Гибридные сорта табака могут быть получены путем предотвращения самоопыления женских родительских растений (то есть, материнских растений) первого сорта, что позволяет оплодотворить женские родительские растения пыльцой мужских родительских растений второго сорта, и позволяет сформироваться гибридным семенам F1 на женских растениях. Самоопыление женских растений может быть предотвращено путем удаления тычинок на ранней стадии развития цветка. Кроме того, формирование пыльцы можно предотвратить на женских родительских растениях, используя форму мужского бесплодия. Например, мужское бесплодие может быть получено путем цитоплазматической мужской стерильности (CMS) или трансгенной мужской стерильности, где трансген ингибирует микроспорогенез и/или образование пыльцы, или самонесовместимости. Женские родительские растения, содержащие CMS, особенно полезны. В вариантах, в которых женские родительские растения обладают CMS, пыльца собирается с мужского фертильного растения и наносится вручную на рыльца CMS-женского родительского растения, и в результате получают семена F1.
Сорта и линии, описанные здесь, могут быть использованы для получения гибридов табака F1 после однократного скрещивания. В таких вариантах осуществления растения родительских сортов можно выращивать в виде преимущественно однородных соседних популяций, чтобы облегчить естественное перекрестное опыление от мужских родительских растений к женским родительским растениям. Семена F1, сформировавшиеся на женских родительских растениях, избирательно собирают с помощью обычных средств. Можно также выращивать два сорта родительского растения вместе и собирать урожай смеси гибридных семян F1, сформированных на женских растениях, и семян, образующихся на отцовских растениях в результате самоопыления. Кроме того, можно проводить тройственное скрещивание, в котором гибрид F1 после одиночного скрещивания используется в качестве женского родителя и скрещивается с другим отцовским родителем. В качестве другой альтернативы, могут быть получены гибриды от двойного скрещивания, в котором потомство F1 двух различных одиночных скрещиваний скрещиваются между собой.
Популяция мутантных, не встречающихся в природе или трансгенных растений может быть подвергнута скринингу или отбору тех членов популяции, которые имеют желаемый признак или фенотип. Например, в популяции потомства одного события трансформации могут быть подвергнуты скринингу те растения, которые имеют желаемый уровень экспрессии полипептида или полинукпеотида. Для идентификации уровня экспрессии могут быть использованы физические и биохимические методы. Они включают Саузерн-анализ или ПЦР-амплификацию для обнаружения полинуклеотида; Нозерн блоттинг, защиту от РНКазы S1, достройку праймера или ОТ-ПЦР-амплификацию для обнаружения РНК-транскриптов; ферментативные анализы для обнаружения ферментной или рибозимной активности полипептида и полинуклеотида, а также белковый гель-электрофорез, Вестерн-блот, иммунопреципитацию и ИФА анализы для обнаружения полипептидов. Другие методы, такие как in situ гибридизация, энзиматическое окрашивание и иммунное окрашивание могут также использоваться для обнаружения присутствия или экспрессии полипептидов или полинуклеотидов.
Мутантные, не встречающиеся в природе или трансгенные клетки растений и растения, описанные здесь, содержат один или несколько рекомбинантных полинуклеотидов, таких как один или более выделенных полинуклеотидов, описанных здесь, одну или несколько полинуклеотидных конструкций, одну или несколько двухцепочечных РНК, один или более конъюгатов или один или более векторов/экспрессионных векторов.
Экспрессия гена может быть оценена с использованием методов, включающих, например, ОТ-ПЦР, Нозерн-блот, защиту от РНКазы, достройку праймеров, Вестерн-блот, гель-электрофорез белков, иммунопреципитацию, ИФА, чипы и масс-спектрометрию. Следует отметить, что если полипептид экспрессируется под контролем тканеспецифичного или широко экспрессирующегося промотора, экспрессия может быть оценена во всем растении или в выбранной ткани. Аналогичным образом, если полипептид экспрессируется в определенное время, например, в определенный момент времени в развитии или после индукции, экспрессия может быть оценена выборочно в желаемый период времени.
Без ограничения, растения, описанные здесь, могут быть изменены для других целей до или после того, как была модулирована экспрессия или активность. Одна или несколько из следующих генетических модификаций может присутствовать в мутантном, не встречающемся в природе или трансгенном растении настоящего описания. В одном варианте осуществления модифицированы один или несколько генов, которые вовлечены в поглощение тяжелых металлов или транспорт тяжелых металлов, в результате чего растение или части растений (например, листья), имеют более низкое содержание тяжелых металлов, чем контрольное растение или его части без модификации (модификаций). Не ограничивающие примеры включают гены семейства посредников диффузии катионов (CDF), семейства Zrt-, Irt-подобных белков (ZIP), семейство белков катионообменников (САХ), семейство транспортеров меди (СОРТ), семейство транспортирующих тяжелые металлы АТФаз Р-типа (HMAS, как описано в WO2009074325), семейство гомологов белков, связанных с естественной устойчивостью макрофагов (NRAMP), и семейство транспортеров с АТФ-связывающей кассетой (АВС), которые участвуют в транспорте тяжелых металлов, таких как кадмий. Термин тяжелый металл, используемый здесь, включает переходные металлы. В другом варианте осуществления модифицирован один или несколько генов, которые участвуют в конверсии промежуточных метаболитов азота, в результате чего растение или части растений (например, листья) при нагревании производят более низкие уровни, по меньшей мере, одного из специфичных для табака нитрозаминов (например, 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанон, N-нитрозонорникотин, N-нитрозоанатабин, и N-нитрозоанабазин), чем контрольное растение или его части. Неограничивающие примеры генов, которые могут быть изменены, включают гены, кодирующие деметилазы никотина, такие как CYP82E4, CYP82E5 и CYP82E10, которые участвуют в конверсии никотина в норникотин и описаны в WO2006091194, WO2008070274, WO2009064771 и PCT/US2011/021088.
Примеры других модификаций включают устойчивость к гербицидам, например, глифосат является активным ингредиентом многих гербицидов широкого спектра действия. Устойчивые к глифосату трансгенные растения были разработаны путем переноса гена AroA (глифосат EPSP синтетаза из Salmonella Typhimurium и E. coli). Устойчивые к сульфонилмочевине растения были произведены путем трансформирования мутантного гена ALS (ацетолактатсинтетаза) из Arabidopsis. Белок OB фотосистемы II из мутанта Amaranthus hybridus был перенесен растениям для получения трансгенных растений, устойчивых к атразину, и трансгенные растения устойчивые к бромоксинилу были произведены путем вставки гена Ьхп из бактерии Klebsiella pneumoniae. Другой пример модификации приводит к получению растений, которые устойчивы к насекомым. Токсины Bacillus thuringiensis (ВТ) могут обеспечить эффективный способ задержки появления Bt-резистентных насекомых-вредителей, как недавно показано для брокколи, где введенные совместно Bt-гены cry1Ac и cry1C контролировали капустную моль, устойчивую к любому одному белку, и значительно задерживали развитие устойчивых насекомых. Другой пример модификации приводит к получению растений, которые устойчивы к заболеваниям, вызванным патогенными микроорганизмами (например, вирусами, бактериями, грибками). Были созданы растения, экспрессирующие ген Ха21 (устойчивость к бактериальным некрозам), и растения, экспрессирующие и слитый ген Bt, и ген хитиназы (устойчивость к желтому стеблевому точильщику и устойчивость к заболеваниям эпидермиса. Другой пример модификации приводит к измененному репродуктивному потенциалу, такому как мужская стерильность. Другой пример модификации приводит к получению растений, которые устойчивы к абиотическим стрессам (например, засухе, температуре, солености), устойчивые трансгенные растения были произведены путем передачи гена фермента ацилглицерол фосфат ацилтрансферазы от Arabidopsis; генов, кодирующих маннитдегидрогеназу и сорбитолдегидрогеназу, которые участвуют в синтезе маннита и сорбитола, улучшают засухоустойчивость. Другой пример модификации приводит к получению растений, которые производят белки, которые имеют благоприятные иммуногенные свойства для использования в организме человека. Например, могут быть использованы растения, способные продуцировать белки, в N-гликанах которых по существу отсутствуют остатки фукозы с альфа-1,3 связью, остатки ксилозы с бета-1,2 связью, или оба. Другие примеры модификаций могут привести к созданию растений с улучшенным запасанием белков и масел, растениям с повышенной эффективностью фотосинтеза, растениям с длительным сроком хранения, растениям с повышенным содержанием углеводов и растениям, устойчивых к грибкам; растениям, кодирующим фермент, участвующий в биосинтезе алкалоидов. Также рассматриваются трансгенные растения, в которых модулирована экспрессия S-аденозил-L-метионин (SAM) и/или цистатионин-гамма-синтазы (CGS).
Один или несколько таких признаков могут быть интрогрессированы в мутантные, не встречающиеся в природе или трансгенные растения из другого растения или сорта табака, или могут быть непосредственно трансформированы в него. Интрогрессия признака (признаков) в мутантные, не встречающиеся в природе или трансгенные растения может быть достигнута с помощью любого способа селекции растений, известного в данной области, например, чистопородного скрещивания, возвратного скрещивания, скрещивания с удвоением гаплоидного набора хромосом и т.п. (см., Wernsman, E. A, and Rufty, R. С. 1987. Chapter Seventeen. Tobacco. Pages 669-698 In: Cultivar Development. Crop Species. W. H. Fehr (ed.), MacMillan Publishing Co, Inc., New York, N.Y 761 pp.). Для того чтобы идентифицировать потомков таких возвратных скрещиваний, наиболее генетически идентичных реккурентному родителю, могут быть использованы методы на основе молекулярной биологии, описанные выше, в частности RFLP и микросателлитные маркеры. Это позволяет ускорить получение табачных сортов, генетически идентичных, по меньшей мере, на 90%, предпочтительно, по меньшей мере, на 95%, более предпочтительно, по меньшей мере, на 99% рекуррентному родителю, еще более предпочтительно генетически идентичных рекуррентному родителю, и дополнительно включающих признак (признаки), интрогрессированные из донорского родителя. Такое определение генетической идентичности может быть основано на молекулярных маркерах, известных в данной области техники.
Последнее поколение возвратного скрещивания может быть самоопылено с получением потомства чистопородного разведения для передаваемой нуклеиновой кислоты (кислот). Полученные в результате растения, как правило, имеют практически все морфологические и физиологические характеристики мутантного, не встречающегося в природе или трансгенного растения настоящего описания, в дополнение к передаваемому признаку (признакам) (например, один или более отдельный генетический признак). Точный протокол возвратного скрещивания будет зависеть от признака, который нужно изменить, чтобы подобрать соответствующий протокол тестирования. Хотя методы возвратного скрещивание упрощаются, когда передаваемый признак является доминантным аллелем, рецессивный аллель также может быть передан. В данном случае, возможно, необходимо ввести тестирование потомства, чтобы определить, был ли успешно передан желаемый признак.
Растение или растительная клетка могут быть трансформированы путем интеграции рекомбинантного полинуклеотида в геном, что приводит к стабильной трансформации. Стабильно трансформированные клетки обычно сохраняют введенный полинуклеотид с каждым клеточным делением. Растение или растительная клетка также могут быть временно трансформированы так, что рекомбинантный полинуклеотид не интегрирован в геном. Временно трансформированные клетки обычно теряют все или некоторые части введенного рекомбинантного полинуклеотида при каждом клеточном делении, таким образом, введенный рекомбинантный полинуклеотид не может быть обнаружен в дочерних клетках после достаточного количества клеточных делений.
Раскрыты конъюгированные фрагменты, в том числе высокомолекулярные соединения, такие как белки (например, антитела), цепи жирных кислот, остатки сахаров, гликопротеины, полимеры (например, полиэтиленгликоль) или их комбинации. Олигонуклеотиды могут быть конъюгированы с фрагментом, который увеличивает поглощение олигонуклеотида клеткой. Неограничивающие примеры фрагментов включают, но не ограничиваются ими, антитела, полипептиды, липидные группы, такие как фрагмент холестерина, холевая кислота, тиоэфиры, например, гексил-S-тритилтиол, тиохолестерол, алифатические цепи, например, додекандиоловый или ундециловый остатки, фосфолипиды, полиамины или полиэтиленгликоли, остаток адамантан уксусной кислоты, пальмитиловый остаток, октадециламино- или гексиламино-карбонил-оксихолестероловый фрагменты. Фрагмент может быть положительно заряженным полимером, таким как положительно заряженный пептид, который, например, имеет длину примерно от 1 до 50 аминокислотных остатков, или полиалкиленоксидом, таким как полиэтиленгликоль (ПЭГ) или полипропиленгликоль. Подходящий положительно заряженный полимер, такой как полиалкиленоксид, может быть прикреплен к олигомеру через линкер, такой как расщепляемый линкер.
Когда измеряют экспрессию полипептида, определение количества мРНК, кодирующей полипептид, в клетке может быть проведено количественно с помощью, например, ПЦР или нозерн-блоттинга. В случае, когда измеряют изменение количества полипептида в образце, может быть использовано его обнаружение с помощью антител для определения количества полипептида в клетке с использованием известных методик. Альтернативно, биологическая активность фермента может быть измерена с использованием методов, которые хорошо известны в данной области.
В другом варианте осуществления предложены антитела, которые иммунореактивны с полипептидами, описанными здесь. Полипептиды, фрагменты, варианты, слитые полипептиды и тому подобное, описанные в настоящем документе, могут быть использованы в качестве «иммуногена», чтобы продуцировать антитела, иммунореактивные к ним. Такие антитела могут специфично связываться с полипептидом через антигенсвязывающие сайты антитела. В частности, связывающими антителами являются те, которые специфически распознают и связывают полипептид, его гомологи и варианты, но не другие молекулы. В одном варианте осуществления антитела специфичны к полипептиду, имеющему аминокислотную последовательность, указанную в данном документе, и не вступают в перекрестную реакцию с другими полипептидами.
Более конкретно, полипептиды, фрагменты, варианты, слитые полипептиды и тому подобное, содержат антигенные детерминанты или эпитопы, которые вызывают образование антител. Эти антигенные детерминанты или эпитопы могут быть либо линейными, либо конформационными (прерывистыми). Линейные эпитопы состоят из аминокислот из одного участка полипептидной цепи, а конформационные, или прерывистые, эпитопы состоят из аминокислотных участков из различных участков полипептидной цепи, которые оказываются в непосредственной близости при сворачивании полипептида. Эпитопы могут быть определены любым из способов, известных в данной области. Кроме того, эпитопы полипептидов могут быть использованы в качестве научно-исследовательских реагентов, в анализах, для очистки специфических связывающих антител из таких веществ как поликлональная сыворотка или супернатант от культивируемых гибридом. Такие эпитопы или их варианты могут быть получены с использованием способов, известных в данной области, таких как твердофазный синтез, химическое или ферментативное расщепление полипептида или с помощью технологии рекомбинантной ДНК.
Как поликлональные, так и моноклональные антитела к полипептидам могут быть получены обычными методами. Клеточные линии гибридомы, которые продуцируют моноклональные антитела, специфичные к полипептидам, также предусмотрены в данном описании. Такие гибридомы могут быть получены и идентифицированы с помощью обычных методов. Для получения антител различные животные-хозяева могут быть иммунизированы инъекцией полипептида, фрагмента, варианта или его мутанта. Такие животные-хозяева в качестве примера, включают кроликов, мышей и крыс, но не ограничиваются ими. Для увеличения иммунного ответа могут быть использованы различные адъюванты. В зависимости от вида хозяина, такие адъютанты включают, но не ограничиваются ими, адъювант Фрейнда (полный и неполный), минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, плюроновые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианин лимфы улитки, динитрофенол и потенциально полезные адъюванты для человека, такие как BCG (бацилла Кальметта-Герена) и Corynebacterium parvum. Моноклональные антитела могут быть выделены с помощью обычных методов. Такие моноклональные антитела могут принадлежать любому классу иммуноглобулинов, в том числе IgG, IgM, IgE, IgA, IgD и любому их подклассу.
Антитела могут быть также использованы в анализах для обнаружения присутствия полипептидов или их фрагментов, in vitro или in vivo. Антитела могут быть также использованы для очистки полипептидов или фрагментов с помощью иммуноаффинной хроматографии.
Различные варианты осуществления обеспечивают мутантные растения, не встречающиеся в природе растения или трансгенные растения, а также биомассу, в которой модулирован уровень экспрессии полинуклеотида изопропилмалат синтазы.
Части таких растений, в частности растений табака, а более конкретно листовые пластинки и жилки табачных растений, могут быть включены или использованы при изготовлении различных расходных материалов, включая, но не ограничиваясь ими, вещества, образующие аэрозоль, образующие аэрозоль устройства, курительные изделия, изделия, которые можно курить, бездымные продукты и табачные изделия. Примеры веществ, образующих аэрозоль, включают, но не ограничиваются ими, композиции табака, табак, экстракт табака, резаный табак, скрошенный высушенный табак, взорванный табак, гомогенизированный табак, восстановленный табак и трубочный табак. Курительные изделия и изделия, которые можно курить, являются типами формирующих аэрозоль устройств. Примеры курительных изделий и изделий, которые можно курить, включают, но не ограничиваются ими, сигареты, сигариллы и сигары. Примеры бездымных продуктов включают жевательные табаки и нюхательные табаки. В некоторых образующих аэрозоль устройствах, композиции табака или другого вещества, образующих аэрозоль, нагревают одним или несколькими электрическими нагревательными элементами для получения аэрозоля, а не сжигают.В другом типе устройств, образующих аэрозоль подогревом, аэрозоль получают путем передачи тепла от горючего топливного элемента или источника тепла к физически отделенному образующему аэрозоль веществу, которое может быть расположено внутри, рядом или ниже по потоку от источника тепла. Изделия из бездымного табака и различные образующие аэрозоль вещества, содержащие табак, могут содержать табак в любой форме, в том числе в виде высушенных частиц, кусков, гранул, порошков или суспензии, нанесенных, смешанных, окруженных или иным образом совмещенных с другими ингредиентами в любом формате, например, хлопьев, пленок, смесей, пены или пузырей. Используемый здесь термин «дым» используется для описания типа аэрозоля, который произведен курительными изделиями, такими как сигареты, или при сжигании образующего аэрозоль вещества.
В одном варианте осуществления предлагается также высушенный материал мутантных, трансгенных и не встречающихся в природе растений, описанных здесь. Процессы высушивания зеленых листьев табака известны специалистам в данной области техники, и включают, но не ограничиваются, воздушную сушку, дымовую сушку и солнечную сушку. Процесс высушивания зеленых листьев табака зависит от типа собираемого табака. Например, табак Virginia flue (bright), как правило, подвергают дымовой сушке, табак Берли и некоторые темные сорта обычно подвергают теневой сушке, а трубочный табак, жевательный табак и нюхательный табак, как правило, подвергают огневой сушке.
В другом варианте осуществления описаны табачные изделия, включая содержащие табак образующие аэрозоль вещества, включая листья, предпочтительно высушенные листья мутантных растений табака, трансгенных растений табака или не встречающихся в природе растений табака, описанных здесь. Табачные изделия, описанные здесь, могут быть смешанными табачными изделиями, которые могут дополнительно содержать не модифицированный табак.
Процент содержания сложных эфиров сахарозы в этих образующих аэрозоль веществах или композициях табака может быть больше, по меньшей мере, на приблизительно на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% и на 100%, на 150% или на 200% или больше по сравнению с образующими аэрозоль веществами или композициями, полученными из не мутантных, встречающихся в природе или не трансгенных аналогичных растений табака.
Процент содержания сложных эфиров сахарозы в этих образующих аэрозоль веществах или композициях табака может быть, по меньшей мере, на приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% и 100% ниже по сравнению с образующими аэрозоль веществами или композициями, полученными из не мутантных, встречающихся в природе или не трансгенных аналогичных растений табака.
Мутантные, не встречающиеся в природе или трансгенные растения могут иметь другие применения, например, в сельском хозяйстве. Например, мутантные, не встречающиеся в природе или трансгенные растения, описанные здесь, могут быть использованы для изготовления корма и продуктов питания человека.
Изобретение также относится к способам получения семян, охватывающим выращивание мутантных растений, не встречающихся в природе растений или трансгенных растений, описанных здесь, и сбор семян выращенных растений. Семена растений, описанных здесь, могут быть кондиционированы и упакованы в упаковочный материал с помощью известных в данной области технологий, чтобы сформировать изделие. Упаковочный материал может быть таким, как бумага и ткани, хорошо известные в данной области техники. Пакет семян может иметь метку, например, ярлык или этикетку, прикрепленную к упаковочному материалу, этикетку, напечатанную на упаковочном материале, или этикетку, вложенную в пакет, которая описывает природу семян в упаковке.
Еще один аспект относится к способу получения композиции, содержащей сложные эфиры сахарозы, включающему следующие стадии: (а) получение частей мутантного, не встречающегося в природе или трансгенного растения; биомассы, семян или листьев, или табачного продукта, описанных здесь; и (б) извлечение оттуда сложных эфиры сахарозы и (с) необязательно, очистку сложных эфиров сахарозы.
Еще один аспект относится к способу получения бета-метилвалериановой кислоты, включающему следующие стадии: (а) получение частей мутантного, не встречающегося в природе или трансгенного растения; биомассы, или листьев, или веществ, образующих аэрозоль, как описано здесь; (б) гидролиз материала, полученного на стадии (а), или его экстракта, и (с) необязательно, сбора бета-метилвалериановой кислоты. Бета-метилвалериановая кислота может быть использована в производстве полигидроксиалканоатов и тому подобное, что может привести к получению материалов, включая пластмассы, с полезными свойствами. Пластмассы, которые могут быть получены, могут быть биоразлагаемыми, и поэтому могут быть использованы в производстве биопластика. Могут быть созданы бактерии, такие E. coli или Pseudomonas, которые экспрессируют изопропилмалат синтазу, описанную в настоящем документе. Соответственно, используемая бактерия может быть изменена, чтобы также производить полигидроксиалканоаты. Методы производства полигидроксиалканоатов с использованием микробных систем описаны, например, в US5750848 и US6492134. Таким образом, один аспект относится к бактерии, содержащей изопропилмалат синтазу и, необязательно, один или несколько генов для производства полигидроксиалканоатов. Еще один аспект относится к способу получения полигидроксиалканоатов, включающему использование бактерии.
В одном варианте осуществления предлагается также высушенный материал из мутантных, трансгенных и не встречающихся в природе растений, описанных здесь. Например, процессы высушивания зеленых листьев табака известны специалистам в этой области, и включают воздушную сушку, огневую сушку, дымовую сушку и солнечную сушку, но не ограничиваются ими. Процесс высушивания зеленых листьев табака зависит от типа собранного табака. Например, сорт табака Virginia flue (bright), как правило, подвергают дымовой сушке, сорт Берли и некоторые темные сорта обычно подвергают теневой сушке, а трубочный табак, жевательный табак, и нюхательный табак, как правило, подвергают огневой сушке.
В другом варианте осуществления описаны табачные изделия, содержащие листья, например, высушенные листья мутантных, трансгенных и не встречающихся в природе растений, описанных здесь, или которые получают с помощью способов, описанных здесь. Табачные изделия, описанные здесь, могут дополнительно содержать не модифицированный табак.
В другом варианте осуществления описаны табачные изделия, содержащие растительный материал, предпочтительно листья, такие как высушенные листья, мутантных, трансгенных и не встречающихся в природе растений, описанных здесь. Например, растительный материал может быть добавлен к внутренней или наружной части табачного изделия и, таким образом, при сжигании высвобождается желаемый аромат. Табачное изделие согласно этому варианту осуществления может быть даже не модифицированным табаком или модифицированным табаком. Табачные изделия согласно этому варианту осуществления даже могут быть получены из мутантных трансгенных или не встречающихся в природе растений, которые имеют модификации в одном или нескольких других генах, в сравнении с раскрытыми в данном документе генами.
Еще один аспект относится к (выделенным) композициям сложных эфиров сахарозы, включающим, состоящим или по существу состоящим из одного или нескольких сложных эфиров сахарозы, описанных здесь. Сложный эфир сахарозы может быть сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 равно 14; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 равно 15; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 равно 16; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 17, и сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 18. При условии, что это общее число атомов углерода выбрано, ацильные цепи могут быть выбраны из любых, представленных на Фигуре 6. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5. Соответственно, одна или более, две или более или три или более ацильных цепей представляют собой бета-метилвалерил. Таким образом, в качестве примера, сложный эфир сахарозы может быть сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гексаноил или его изомер, и R5 представляет собой водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил; или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С14:0); сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С15:0); сложным эфиром сахарозы, имеющим общую структуру, показанную на Фигуре 5, где R3 означает ацетил, R1 представляет собой валерил или его изомер его, R2 обозначает валерил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0), или сложным эфиром сахарозы, имеющим общую структуру, показанную на Фигуре 5, где R3 представляет собой ацетил, R1 обозначает пропионил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0); сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 обозначает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С17:0), и сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 обозначает ацетил, R1 обозначает гексаноил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С18:0).
Альтернативно, сложный эфир сахарозы может быть сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 14; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 15; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 16; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 17; и сложным эфиром сахарозы, в котором отсутствует, по существу не присутствует или не присутствует в обнаруживаемом количестве сложный эфир сахарозы, в котором число атомов углерода в ацильных цепях R1, R2 и R4 составляет 18 (С2С18:0). При условии, что общее число атомов углерода определено, ацильные цепи могут быть выбраны из любых, приведенных на Фигуре 6. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Соответственно, эфир сахарозы имеет ацетил в R3 и водород в R5. Соответственно, одна или более, две или более или три или более ацильных цепей представляют собой бета-метилвалерил. Таким образом, в качестве примера, сложный эфир сахарозы может быть сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил; или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С14:0); сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С15:0); сложным эфиром сахарозы, который имеет общую структуру, показанную на Фигуре 5, где R3 означает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает валерил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0), или сложным эфиром сахарозы, который имеет общую структуру, показанную на Фигуре 5, где R3 представляет собой ацетил, R1 обозначает пропионил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0); сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 обозначает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С17:0), и где сложный эфир сахарозы, показанный на Фигуре 5, где R3 обозначает ацетил, R1 обозначает гексаноил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную часть, отсутствует, по существу не присутствует или не присутствует в детектируемых количествах (С2С18:0).
Альтернативно, сложный эфир сахарозы может быть сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 14; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 15; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 16; сложным эфиром сахарозы, в котором отсутствует, по существу не присутствует или не присутствует в обнаруживаемом количестве сложный эфир сахарозы, в котором число атомов углерода в ацильных цепях R1, R2 и R4 составляет 17; и сложным эфиром сахарозы, в котором отсутствует, по существу не присутствует или не присутствует в обнаруживаемом количестве сложный эфир сахарозы, в котором число атомов углерода в ацильных цепях R1, R2 и R4 составляет 18. При условии, что общее число атомов углерода определено, ацильные цепи могут быть выбраны из любых, приведенных на Фигуре 6. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5. Соответственно, одна или более, две или более, или три или более ацильных цепей представляют собой бета-метилвалерил. Таким образом, в качестве примера, сложный эфир сахарозы может быть сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4=валерил или его изомер и R5 обозначает водород или ацетил; или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С14:0); сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С15:0); сложным эфиром сахарозы, который имеет общую структуру, показанную на Фигуре 5, где R3 означает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает валерил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0), или сложным эфиром сахарозы, который имеет общую структуру, показанную на Фигуре 5, где R3 представляет собой ацетил, R1 обозначает пропионил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу (С2С16:0); и в котором сложный эфир сахарозы, показанный на Фигуре 5, где R3 обозначает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу, отсутствует, по существу не присутствует или не присутствует в детектируемых количествах (С2С17:0), и в котором сложный эфир сахарозы, показанный на Фигуре 5, где R3 обозначает ацетил, R1 обозначает гексаноил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу, отсутствует, по существу не присутствует или не присутствует в детектируемых количествах (С2С18:0).
Альтернативно, сложный эфир сахарозы может быть сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 14; сложным эфиром сахарозы, в котором общее число атомов углерода в ацильных цепях R1, R2 и R4 составляет 15; сложным эфиром сахарозы, в котором отсутствует, по существу не присутствует или не присутствует в обнаруживаемом количестве сложный эфир сахарозы, в котором число атомов углерода в ацильных цепях R1, R2 и R4 составляет 16; сложным эфиром сахарозы, в котором отсутствует, по существу не присутствует или не присутствует в обнаруживаемом количестве сложный эфир сахарозы, в котором число атомов углерода в ацильных цепях R1, R2 и R4 составляет 17, и сложным эфиром сахарозы, в котором отсутствует, по существу не присутствует или не присутствует в обнаруживаемом количестве сложный эфир сахарозы, в котором число атомов углерода в ацильных цепях R1, R2 и R4 составляет 18. При условии, что общее число атомов углерода определено, ацильные цепи могут быть выбраны из любых, приведенных на Фигуре 6. Подходящий сложный эфир сахарозы имеет ацетил в R3 и ацетил или водород в R5. Подходящий сложный эфир сахарозы имеет ацетил в R3 и водород в R5. Соответственно, одна или более, две или более, или три или более ацильных цепей представляют собой бета-метилвалерил. Таким образом, в качестве примера, сложный эфир сахарозы может быть сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = пропионил или его изомер, R4 = гексаноил или его изомер и R5 представляет собой водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил; или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С14:0); сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = пропионил или его изомер, R2 = валерил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = валерил или его изомер, R2 = валерил или его изомер, R4 = валерил или его изомер и R5 обозначает водород или ацетил, или сложным эфиром сахарозы, таким как показанный на Фигуре 5, где R3 = ацетил, R1 = бутирил, R2 = гексаноил или его изомер, R4 = гексаноил или его изомер и R5 обозначает водород или ацетил (С2С15:0); и в котором сложный эфир сахарозы, который имеет общую структуру, показанную на Фигуре 5, где R3 означает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает валерил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу, отсутствует, по существу не присутствует или не присутствует в детектируемых количествах (С2С16:0); или в котором сложный эфир сахарозы, который имеет общую структуру, показанную на Фигуре 5, где R3 означает ацетил, R1 обозначает пропионил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу, отсутствует, по существу не присутствует или не присутствует в детектируемых количествах (С2С16:0); или и в котором сложный эфир сахарозы, показанный на Фигуре 5, где R3 обозначает ацетил, R1 представляет собой валерил или его изомер, R2 обозначает гексаноил или его изомер его, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу, отсутствует, по существу не присутствует или не присутствует в детектируемых количествах (С2С17:0), и в котором сложный эфир сахарозы, показанный на Фигуре 5, где R3 обозначает ацетил, R1 обозначает гексаноил или его изомер, R2 обозначает гексаноил или его изомер, R4 обозначает гексаноил или его изомер и R5 обозначает либо атом водорода, либо ацетильную группу, отсутствует, по существу не присутствует или не присутствует в детектируемых количествах (С2С18:0).
Еще один аспект относится к мутантным, не встречающимся в природе или трансгенным растениям (соответственно, растениям табака), у которых экспрессия гена изопропилмалат синтазы, описанного здесь, или активность белка, кодируемого им, увеличена, по сравнению с контрольным растением, у которого экспрессия гена изопропилмалат синтазы или активность белка, кодируемого им, не была увеличена, и где указанное растение образует содержащие бета-метилвалерил сложные эфиры сахарозы в больших количествах, чем указанное контрольное растение. Еще один аспект относится к мутантным, не встречающимся в природе или трансгенным растениям (соответственно, растениям табака), у которых экспрессия гена изопропилмалат синтазы, описанного здесь, или активность белка, кодируемого им, уменьшена, по сравнению с контрольным растением, у которого экспрессия гена изопропилмалат синтазы или активность белка, кодируемого им, не была уменьшена, и где указанное растение образует содержащие бета-метилвалерил сложные эфиры сахарозы в меньших количествах, чем указанное контрольное растение.
Еще один аспект относится к мутантным, не встречающимся в природе или трансгенным растениям табака дымовой сушки у которых экспрессия гена изопропилмалат синтазы, описанного здесь, или активность белка, кодируемого им, увеличена, по сравнению с контрольным растением, у которого экспрессия гена изопропилмалат синтазы или активность белка, кодируемого им, не была увеличена, и где указанное растение образует содержащие бета-метилвалерил сложные эфиры сахарозы в больших количествах, чем указанное контрольное растение.
Еще один аспект относится к мутантным, не встречающимся в природе или трансгенным растениям табака сорта Ориенталь, у которых экспрессия гена изопропилмалат синтазы, описанного здесь, или активность белка, кодируемого им, уменьшена, по сравнению с контрольным растением, у которого экспрессия гена изопропилмалат синтазы или активность белка, кодируемого им, не была уменьшена, и где указанное растение образует содержащие бета-метилвалерил сложные эфиры сахарозы в меньших количествах, чем указанное контрольное растение.
Мутантные, не встречающиеся в природе или трансгенные растения табака, описанные здесь, могут быть собраны, и полученный из них табак может быть обработан или высушен с использованием методов, известных для выбранного типа табака. Поверхностные соединения листьев могут быть собраны с использованием, например, промывки водой или органическими растворителями, а затем высушены. Полученный порошок затем может быть использован для усиления вкусовых свойств курительного материала. Табачный материал, остающийся после промывок, затем может использоваться в качестве материала, например, для взорванного табака.
В еще одном аспекте предложен способ изменения устойчивости к вредителями, по меньшей мере, частей растения, включающий следующие стадии: (I) модулирование экспрессии или активности изопропилмалат синтазы в растении, предпочтительно, где изопропилмалат синтаза представлена нуклеотидной последовательностью или аминокислотной последовательностью, описанными в данном документе; (II) необязательно, измерение количества бета-метилвалериловых сложных эфиров сахарозы, по меньшей мере, в части мутантного, не встречающегося в природе или трансгенного растения, полученного на стадии (I), (III) идентификация мутантного, не встречающегося в природе или трансгенного растения, в котором количество содержащих бета-метилвалерил сложных эфиров сахарозы изменилось по сравнению с контрольным растением, в котором экспрессия или активность изопропилмалат синтазы не модулировали и, предпочтительно, чтобы внешний вид упомянутого мутантного, не встречающегося в природе или трансгенного растения являлся по существу таким же, как у контрольного растения, и (IV) получение растения, у которого устойчивость к вредителям модулирована.
Соответственно, предложен способ повышения устойчивости к вредителям, по меньшей мере, частей растения, включающий следующие стадии: (I) увеличение экспрессии или активности изопропилмалат синтазы в растении, предпочтительно, где изопропилмалат синтаза представлена нуклеотидной последовательность или аминокислотной последовательностью, описанными здесь; (II) необязательно, измерение количества содержащих бета-метилвалерил сложных эфиров сахарозы, по меньшей мере, в части мутантного, не встречающегося в природе или трансгенного растения, полученного на стадии (I), (III) идентификация мутантного, не встречающегося в природе или трансгенного растения, в котором количество содержащих бета-метилвалерил сложных эфиров сахарозы увеличено, по сравнению с контрольным растением, в котором экспрессия или активность изопропилмалат синтазы не увеличивали, и, предпочтительно, чтобы внешний вид упомянутого мутантного, не встречающегося в природе или трансгенного растения являлся по существу таким же, как у контрольного растения, и (IV) получение растения, у которого устойчивость к его вредителям увеличена.
Также предусмотрены растение или растительный материал, полученные или получаемые данным способом.
Еще один аспект относится к мутантным, не встречающимся в природе или трансгенным растениям, в которых экспрессия гена изопропилмалат синтазы или активность белка, кодируемого этим геном, модулированы, и, по меньшей мере, часть растения имеет изменения состава содержащих бета-метилвалерил сложных эфиров сахарозы по сравнению с контрольным растением, у которого экспрессия или активность изопропилмалат синтазы не была изменена, и внешний вид указанных растений является по существу таким же, как у контрольного растения, и у которых устойчивость к вредителям также изменена.
Соответственно, раскрыты мутантные, не встречающиеся в природе или трансгенные растения, в которых экспрессия гена изопропилмалат синтазы или активность белка, кодируемого этим геном, увеличена, и, по меньшей мере, часть растения имеет изменения состава содержащего бета-метилвалерил сложного эфира (эфиров) сахарозы по сравнению с контрольным растением, у которого экспрессия или активность изопропилмалат синтазы не была увеличена, и внешний вид указанных растений является по существу таким же, как у контрольного растения, и у которых устойчивость к вредителям также увеличена.
Охвачены композиции, способы и наборы для генотипирования растений для идентификации, отбора, или разведения, которые могут содержать средство обнаружения присутствия полинуклеотида изопропилмалат синтазы в образце. Соответственно, описана композиция, содержащая один или несколько праймеров для специфической амплификации, по меньшей мере, части полинуклеотида изопропилмалат синтазы и, необязательно, один или более зондов и, возможно, один или несколько реагентов для проведения амплификации или обнаружения. Раскрыты специфичные для гена олигонуклеотидные праймеры или зонды, содержащие около 10 или более последовательных полинуклеотидов, соответствующих полинуклеотиду изопропилмалат синтазы. Указанные праймеры или зонды могут включать или состоять из около 15, 20, 25, 30, 40, 45 или более 50 последовательных полинуклеотидов, которые гибридизуются (например, специфично гибрид изуются, например, при жестких условиях гибридизации) с полинуклеотидом изопропилмалат синтазы. В некоторых вариантах осуществления праймеры или зонды могут включать или состоять из приблизительно от 10 до 50 последовательных нуклеотидов, примерно от 10 до 40 последовательных нуклеотидов, примерно от 10 до 30 последовательных нуклеотидов или примерно от 15 до 30 последовательных нуклеотидов, которые могут быть использованы в зависимых от последовательности методах идентификации гена (например, гибридизации по Саузерну) или выделения (например, при in situ гибридизации бактериальных колоний или бляшек бактериофага) или детекции гена (например, как один или более праймеров для амплификации при амплификации или обнаружении нуклеиновой кислоты). Один или более специфических праймеров или зондов могут быть разработаны и использованы для амплификации или детекции части или всего полинуклеотида изопропилмалат синтазы. В качестве конкретного примера, два праймера могут быть использованы для осуществления полимеразной цепной реакции для амплификации фрагмента нуклеиновой кислоты, кодирующей изопропилмалат синтазу, такой как ДНК или РНК. Полимеразная цепная реакция также может быть выполнена с использованием одного праймера, который является производным от последовательности нуклеиновой кислоты изопропилмалат синтазы, и второго праймера, который гибридизуется с последовательностью перед или после нуклеотидной последовательности изопропилмалат синтазы, например, с последовательностью промотора изопропилмалат синтазы, 3'-концом мРНК-предшественника или с последовательностью, находящейся в векторе и тому подобное. Примеры изотермических способов и способов с изменением температуры, используемых для амплификации полинуклеотидов in vitro, хорошо известны в данной области. Образец может представлять собой растение, растительные клетки или растительный материал, или табачное изделие, изготовленное или полученное из растений, растительные клетки или растительный материал, описанные здесь, или может быть получен из них.
Таким образом, в дополнительном аспекте предлагается также способ обнаружения полинуклеотида изопропилмалат синтазы в образце, включающий стадии: (а) получение образца, содержащего, или предположительно содержащего полинуклеотид, (б) контактирование указанного образца с одним или более праймерами или одним или более зондами для специфичного обнаружения, по меньшей мере, части полинукпеотида изопропилмалат синтазы, и (с) обнаружение присутствия продукта амплификации, причем присутствие продукта амплификации свидетельствует о присутствии полинуклеотида изопропилмалат синтазы в образце. В дополнительном аспекте также предложено применение одного или более праймеров или зондов для специфичного обнаружения, по меньшей мере, части полинуклеотида изопропилмалат синтазы. Также предложены наборы для обнаружения, по меньшей мере, части полинуклеотида изопропилмалат синтазы, которые содержат один или несколько праймеров или зондов для специфичного обнаружения, по меньшей мере, части полинуклеотида изопропилмалат синтазы. Набор может содержать реагенты для амплификации полинуклеотида, например, полимеразной цепной реакции (ПЦР), или реагенты для технологий обнаружения нуклеиновых кислот путем гибридизации с зондом, например, Саузерн-блоттинга, Нозерн-блоттинга, in situ гибридизации или микрочипов. Набор может содержать реагенты для технологий обнаружения путем связывания антителами, таких как Вестерн-блоттинг, ИФА, масс-спектрометрия SELDI или тест-полоски. Набор может включать реагенты для секвенирования нуклеиновой кислоты. Набор может содержать реагенты и/или инструкции для определения содержания сложных эфиров сахарозы. В некоторых вариантах осуществления набор может содержать инструкции для одного или более описанных способов. Описанные наборы могут быть полезны для определения генетической идентичности, филогенетических исследований, генотипирования, гаплотипирования, анализа родословной или селекции растений, особенно с кодоминантным анализом.
Настоящее изобретение также обеспечивает способ генотипирования растения, клеток растения или растительного материала, содержащих полинуклеотид изопропилмалат синтазы. Генотипирование предоставляет средства различения гомологов в паре хромосом и может быть использовано для дифференциации сегрегантов в популяции растений. Методы молекулярных маркеров можно использовать для филогенетических исследований, характеризующих генетические отношения между сортами сельскохозяйственных культур, выявления кросс-гибридов или соматических гибридов, локализации хромосомных сегментов, влияющих на моногенные признаки, клонирования, основанного на карте, и изучения. Конкретный метод генотипирования может использовать любое количество технологий, анализирующих молекулярные маркеры, включая полиморфизм длины амплифицированных фрагментов (AFLP). AFLP является результатом аллельных различий между амплифицируемыми фрагментами, вызванных изменчивостью нуклеотидной последовательности. Таким образом, настоящее изобретение дополнительно предоставляет средства для последующей сегрегации гена изопропилмалат синтазы или нуклеиновой кислоты, а также хромосомных последовательностей, генетически связанных с этими генами или нуклеиновыми кислотами, с использованием таких методов, как анализ AFLP.
Описание дополнительно раскрыто в приведенных ниже примерах, которые приведены для более подробного раскрытия описания. Эти примеры, в которых изложены предпочтительные варианты осуществления, предусмотренные для реализации изобретения, предназначены для иллюстрации и не ограничивают настоящее изобретение.
ПРИМЕРЫ
Пример 1: Анализ сложных эфиров сахарозы в сортах Nicotinia tabacum, сорте дымовой сушки Hicks Broadleaf (HBL) и первичном Red Russian (RR)
Для изучения синтеза сложных эфиров сахарозы в табаке выбрали две сортовые модели: сорт дымовой сушки Hicks Broadleaf (HBL) и первичный сорт Red Russian (RR). В соответствии с исходными данными, доступными из программы национальных генетических ресурсов National Genetic Resources Program (Germplasm Resources Information Network - (GRIN), National Germplasm Resources Laboratory, Beltsville, Maryland) предполагается различное содержание содержащих бета-метилвалерил сложных эфиров сахарозы в этих двух линиях.
Анализировали состав сложных эфиров сахарозы в экссудатах из листьев HBL и RR, результаты представлены на Фигуре 1. Состав сложных эфиров сахарозы RR отличается от состава сложных эфиров сахарозы HBL. В HBL сложные эфиры сахарозы в основном состоят из С2С15:0, тогда как в RR обнаружено присутствие более высокомолекулярных вариантов, например, С2С16:0, С2С17:0 и С2С18:0. Как показано на Фигуре 2, различия между RR и HBL объясняются присутствием или отсутствием метилвалериановых эфиров в молекулах. Таким образом, можно сделать вывод, что RR представляет собой BMVSE сорт, а HBL представляет собой bmvse сорт.
Пример 2: Анализ локуса количественного признака
Состав сложных эфиров сахарозы в экссудатах из листьев анализировали в 136 популяциях восьми F3 растений, происходящих, соответственно, от 136 самоопыленных генотипированных растений F2, которые были получены от скрещивания HBL и RR. Изучение локуса количественного признака проводят для того, чтобы соотнести микросателлитные маркеры с фенотипом содержания содержащих бета-метилвалерил сложных эфиров сахарозы. Данные анализировали с использованием программы оценки локусов количественного признака Quantitative trait locus cartographer. Такой признак как содержание содержащих бета-метилвалерил сложных эфиров сахарозы связан с единственным локусом в геноме табака. Этотлокус находится в группе сцепления 15 (хромосома А) табака.
Пример 3: Идентификация гена, связанного с содержащими бета-метилвалерил сложными эфирами сахарозы
Транскрипционный уровень экзонов табака в трихоме изучали с целью выбора предпочтительной генной мишени среди всех потенциальных генов, определяющих функции, описанные на Фигуре 3. Экспрессию изучали в нескольких сортах, выбранных по их профилю продукции содержащих бета-метилвалерил сложных эфиров сахарозы, с использованием микрочипов.
По результатам были идентифицированы образцы экзонов, связанных с изопропилмалат синтазой, поскольку их экспрессия коррелирует с подсчитанным количеством содержащих бета-метилвалерил сложных эфиров сахарозы, измеренным в каждом экстракте каждого сорта (R>0,95).
Были идентифицированы четыре последовательности табака, кодирующие гомологи изпропилмалат синтаз томата IPMSA и IPMSB. Они были названы NtIPMS1A (SEQ ID NO:10), NtIPMS1Bv1 (SEQ ID NO:12), NtIPMS1Bv2 (SEQ ID NO:14) и NtIPMS2 (SEQ ID NO:1). Согласно базам данным EST, в качестве кандидата была выбрана SEQ ID NO:1, поскольку в трихомах можно наблюдать наибольшую степень ее экспрессии.
Библиотеку ВАС, полученную из N. tabacum сорта HBL, скринировали с использованием кДНК NtIPMS2 в качестве затравки для ПЦР, что позволило идентифицировать ВАС, названную PISOGE. Указанную ВАС секвенировали методом shotgun. В результате сборки получили несколько непрерывных последовательностей (контигов), среди которых pisoge1 и pisoge2, которые содержали последовательности, связанные с NtIPMS2.
Интрон-экзонная структура является консервативной для изопропилмалат синтаз из табака и томата. Для NtIPMS1Bv1, NtIPMS1Bv2, SIIPMSB и SIIPMSA было обнаружено, что гены разделены на двенадцать экзонов. Ожидается также, что NtIPMS2 состоит из двенадцати экзонов, но поскольку первый экзон SIIPMSA соответствует двум экзонам на pisoge1, постулируется, что NtIPMS2 содержит тринадцать экзонов.
Надежность предсказания геномной последовательности проверяли с помощью ПЦР. В целом, предсказанную последовательность из сборки pisoge верифицировали с использованием очищенной PISOGE ВАС или геномной ДНК, выделенной либо из растений Hicks Broadleaf, либо из Red Russian. Была идентифицирована одна полноразмерная геномная структура, соответствующая NtIPMS2. Кроме того, были обнаружены две другие геноподобные структуры, и они, вероятно, представляют собой псевдогены, образованные дупликацией исходного гена. Структуры, предложенные с использованием PISOGE ВАС, были частично верифицированы, но были обнаружены некоторые расхождения между ВАС структурой и HBL из геномной ДНК.
Для того, чтобы различить аллели NtIPMS2 е RR и HBL, в качестве генетического маркера использовали ПЦР. Маркер IPMS2 использовали в ПЦР амплификации геномной ДНК всех растений из F2, использованных в картировании локуса количественного признака. Локус количественного признака сложных эфиров сахарозы пересчитали с этим дополнительным маркером. Пик количественного признака локуса идентифицирует позицию, для которой вероятность ассоциации между признаком и маркером является максимальной. Маркер IPMS2, совместно с маркером простых повторяющихся последовательностей РТ30172, являются маркерами, для которых вероятность влияния на признак содержания содержащих бета-метилвалерил сложных эфиров сахарозы является самой высокой.
Пример 4: Демонстрация активности NtIPMS2 как ключевого фактора в синтезе содержащих бета-метилвалерил сложных эфиров сахарозы
Метки экспрессируемой последовательности, соответствующие NtIPMS2, выделили из библиотек EST трихомы в форме GT09L09 (происхождение: трихомы Galpao [TI1068]) и ОТ05С20 (происхождение: трихомы Orinocco [TI81]). Обе молекулы имеют идентичные нуклеотидные последовательности и, поэтому только кДНК ОТ05С20 использовали в экспериментах по определению экспрессии. Конструкции РНКи создавали на основе консенсусной последовательности первого экзона и первого интрона NtIPMS2. Обе конструкции переносили в бинарный вектор, содержащий либо промотор CPS2p (CPS2 относится к копалил синтазе 2, вовлеченной в синтез цис-абиенола, который осуществляется только в трихомах), специфичный для трихом, либо вирусный промотор MMVp (MMV относится к вирусу мозаики ночной красавицы (mirabilis mosaic virus). Стабильные трансформации с использованием Agrobactehum tumefaciens AGL1, содержащей соответствующие бинарные векторы, проводили на сорте восточного табака Basma xanthi и сорте табака дымовой сушки K326.
Листья собирали с одномесячных регенерирующих растений, полученных в результате независимых трансформаций. Внутренний стандарт (октаацетат сахарозы) добавляли в количестве 2.5 нг/мг сырой массы. Площадь сигнала, записанного для иона, специфичного для каждого сложного эфира сахарозы, измеряли и отображали как отношение к площади пика внутреннего стандарта. Сигнал варьирует для разных типов молекул, так что сравнение является достоверным только в пределах каждого типа молекул. Отношение к площади сигнала внутреннего стандарта позволяет нормализовать сигналы на сырую массу листа. Представленная здесь номенклатура сложных эфиров сахарозы описана на Фигуре 2. В отношении Фигуры 4: (А) состав экссудатов из листьев исходного сорта K326 (дымовой сушки, n=3) и трансформантов, экспрессирующих NtIPMS2 под контролем специфичного промотора трихомы; (В) состав экссудатов из листьев исходного сорта K326 и трансформантов, экспрессирующих NtIPMS2 под контролем вирусного промотора; (С) состав экссудатов из листьев исходного сорта Basma (восточный, n=4) и трансформантов, экспрессирующих конструкцию NtIPMS2-RNAi под контролем специфичного промотора трихомы; (D): состав экссудатов из листьев исходного сорта Basma и трансформантов, экспрессирующих конструкцию NtIPMS2-RNAi под контролем вирусного промотора.
Представленные данные достаточны для демонстрации того, что NtIPMS2 является ключевым геном в продукции сложных эфиров сахарозы, содержащих бета-метилвалериановую кислоту. Супрессия IPMS2 в сорте (восточный), содержащим сложный эфир сахарозы, содержащий бета-метилвалериановую кислоту, понижает или прекращает продукцию сложного эфира сахарозы, содержащего бета-метилвалериаовую кислоту (см. С и D), что убеждает в том, что IPMS2 является ключевым компонентом биосинтетического пути. Конститутивная и специфичная для трихомы экспрессия конструкции для РНК-интерференции приводит к прекращению накопления сложных эфиров сахарозы С2С16:0, С2С17:0 и С2С18:0, являющихся эфирами, которые могут содержать бета-метилвалериановую кислоту.
Введение NtIPMS2 в сорт bmvse (дымовой сушки) может индуцировать продукцию сложных эфиров сахарозы (см. А и В). Это доказывает то, что экспрессия кДНК NtIPMS2 как таковой достаточна для восстановления продукции сложных эфиров сахарозы. Здесь конститутивная и специфичная для трихомы экспрессия кДНК NtIPMS2 приводит к накоплению de novo сложных эфиров сахарозы С2С16:0, С2С17:0 и С2С18:0, являющихся эфирами, которые содержат бета-метилвалериановую кислоту. Интересно, что конститутивная экспрессия кДНК (В) не привела к высокому накоплению сложных эфиров сахарозы, тогда как специфичная для трихомы экспрессия привела к накоплению сложных эфиров сахарозы на уровне, сравнимом с тем, который мог бы наблюдаться, например, для сорта, содержащего сложный эфир сахарозы, содержащий бета-метилвалериановую кислоту.
Пример 5: Протокол поиска, проводимого для выбора сайтов-мишеней нуклеаз с «цинковыми пальцами»
Этот пример иллюстрирует то, как надо осуществлять поиск по гену изопропилмалат синтазы для скрининга уникальных сайтов-мишеней в пределах последовательности данного гена по сравнению с заданной геномной базой данных для разработки средств модификации экспрессии гена. Сайты-мишени, идентифицированные способами из описания, фрагменты последовательностей и применение любых сайтов или фрагментов для модификации соответствующей последовательности гена в растении, таком как табак, включены в описание.
Алгоритм поиска. Разработана компьютерная программа, позволяющая проводить скрининг нуклеотидной последовательности запроса (мишени) на встречаемость двух подпоследовательностей фрагментов ДНК фиксированной длины, разделенных спейсером заданного размера, с использованием массива суфиксов в пределах базы данных ДНК. Для конструирования массива суффиксов и поиска используется общедоступная программа libdivsufsort library-2.0.0 (http://code.google.com/p/libdivsufsort/), которая преобразует любую вводимую строку напрямую в преобразованную строку Барроуза-Уилера. Программа сканирует полную вводимую нуклеотидную последовательность (мишень) и выдает все комбинации подпоследовательностей, встречающиеся меньше выбранного числа раз в выбранной базе данных ДНК.
Выбор в интересующей последовательности сайта-мишени для мутагенеза, опосредованного нуклеазой с «цинковыми пальцами».
ДНК-связывающий домен с цинковыми пальцами узнает нуклеотидную последовательность трех пар оснований. Нуклеазы с «цинковыми пальцами» включают белок с цинковыми пальцами, содержащий один, два, три, четыре, пять, шесть или больше ДНК-связывающих доменов, и не специфичную нуклеазу фермент рестрикции типа IIS. Нуклеазы с «цинковыми пальцами» могут быть использованы для внесения двухцепочечного разрыва в целевую последовательность. Для внесения двухцепочечного разрыва необходима пара нукпеаз с «цинковыми пальцами», одна из которых связывается с позитивной (верхней) цепью целевой последовательности, а другая с минус (нижней) цепью той же самой целевой последвательности, разделенными 0, 1, 2, 3, 4, 5, 6 или большим числом нуклеотидов. С использованием различных чисел входных данных 3 для каждой из двух подпоследовательностей ДНК-фрагментов фиксированной длины, программу можно использовать для идентификации двух сайтов-мишеней белка с цинковыми пальцами, разделенных спейсером заданной длины.
Входные данные программы:
1. Целевая вводимая последовательность ДНК
2. База данных ДНК для осуществления поиска
3. Фиксированный размер первой подпоследовательности фрагмента ДНК
4. Фиксированный размер спейсера
5. Фиксированный размер второй подпоследовательности фрагмента ДНК
6. Пороговое число встречаемости комбинации входных данных программы 3 и 5, разделенных входным данным программы 4 в выбранной базе данных ДНК согласно входным данным программы 2
Результат выполнения программы:
список нуклеотидных последовательностей, где для каждой последовательности указано число раз, которое данная последовательность встречается в базе данных ДНК с максимальным пороговым значением (вводимые данные программы 6).
Любая публикация, цитированная или описанная здесь, предоставляет релевантную информацию, раскрытую до даты регистрации настоящей заявки. Утверждения здесь не должны истолковываться как допущения того, что изобретатели не имеют права датировать задним числом такие раскрытия. Все публикации, упомянутые в вышеприведенном описании, включены здесь посредством ссылки. Различные модификации и вариации раскрытия будут очевидны специалистам в данной области техники без отступления от объема и сущности раскрытия. Хотя раскрытие было описано в связи с конкретными предпочтительными воплощениями, следует понимать, что изобретение, как заявлено, не должно быть неправильно ограничено такими конкретными вариантами воплощениями. Действительно, различные модификации описанных способов воплощения изобретения, которые очевидны специалистам в области клеточной, молекулярной и растительной биологии или смежных областей, как предполагается, охватываются следующей формулой изобретения.
ПОСЛЕДОВАТЕЛЬНОСТИ
SEQ ID NO:1 (нуклеотидная последовательность NtIPMS2)


SEQ ID NO:2 (аминокислотная последовательность NtIPMS2)

SEQ ID NO:3 (последовательность кДНК NtIPMS2, использованная для комплементации)

SEQ ID NO:4 (Последовательность района геномной ДНК NtIPMS2, охватывающего экзоны XI, XII и XIII, из Red Russian. Совпадающие районы кДНК подчеркнуты. Жирным шрифтом выделены консервативные районы ДНК между последовательностями ДНК сортов Red Russian и Hicks Broadleaf)


SEQ ID NO:5 (Последовательность района геномной ДНК NtIPMS2, охватывающего экзоны XI и XIII из Hicks Broadleaf. Совпадающие районы кДНК подчеркнуты. Жирным шрифтом выделены консервативные районы ДНК между последовательностями ДНК сортов Red Russian и Hicks Broadleaf.)


SEQ ID NO:6 (Соответствующая NtIPMS2 последовательность ВАС (общая длина ВАС составляет 76832 bp. Приведено только 14232bp)







SEQ ID NO:7 (Соответствующая NtIPMS2 последовательность ВАС (общая длина ВАС составляет 10082bp)






SEQ ID NO:8 (Последовательность промотора, используемого для специфичной для трихом экспрессии. Промотор был выделен из Nicotinia tabacum, где он был расположен на расстоянии 1,5 kb до гена, кодирующего копалил-синтазу, вовлеченную в синтез цис-абиенола.)


SEQ ID NO:9 (Искусственная конструкция, используемая для запуска супрессии экспрессии гена NtIPMS2 через РНКи механизм. Состоя из фрагмента первого интрона NtIPMS2 с первым экзоном, фрагмент первого интрона повторно присутствует после экзона в инвертированной форме. Трансляция и сплайсинг конструкции теоретически приводит к двухцепочечной РНК в шпилечной структур)

выделено жирным шрифтом и подчеркнуто: экзоны 1,21-229
жирный шрифт: граница сплайсинга
курсив: сайт сплайсинга
курсив и подчеркнуто: интрон 1
SEQ ID NO:10 (нуклеотидная последовательность NtIPMS1A)

SEQ ID NO:11 (аминокислотная последовательность NtIPMS1A)

SEQ ID NO:12 (нуклеотидная последовательность NtIPMS1Bv1)


SEQ ID NO:13 (аминокислотная последовательность NtIPMS1Bv1)

SEQ ID NO:14 (нуклеотидная последовательность NtIPMS1Bv2)


SEQ ID NO:15 (аминокислотная последовательность NtIPMS1Bv2)
















С2 обозначает ацетил; С3 обозначает бутирил; С4 обозначает пропионил; C4i обозначает изобутирил; С5 обозначает валерил (пентаноил); C5ai обозначает 2-метил-бутирил; C5i обозначает изовалерил; C5:1i обозначает изопентеноил; С5:1 обозначает пентеноил; С6 обозначает гексаноил; С6(2) обозначает 2-метилвалерил; C6ai обозначает бета-метилвалерил; C6i обозначает 4-метилвалерил; - обычно представляет собой водород.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРЕОНИНСИНТАЗА ИЗ NICOTIANA TABACUM И ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ ПРИМЕНЕНИЯ | 2012 |
|
RU2689719C2 |
| СНИЖЕНИЕ ПРЕВРАЩЕНИЯ НИКОТИНА В НОРНИКОТИН В РАСТЕНИЯХ | 2015 |
|
RU2733837C2 |
| МОДУЛИРОВАНИЕ БЕТА-ДАМАСЦЕНОНА У РАСТЕНИЙ | 2012 |
|
RU2681497C2 |
| СПОСОБ УВЕЛИЧЕНИЯ СОДЕРЖАНИЯ СЛОЖНОГО ЭФИРА САХАРОЗЫ В РАСТЕНИЯХ ТАБАКА | 2018 |
|
RU2758117C2 |
| ГЕНЫ И БЕЛКИ ДЛЯ СИНТЕЗА АЛКАНОИЛ-СОА | 2012 |
|
RU2650766C2 |
| СПОСОБ И КОНСТРУКТ ДЛЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА SCBV | 2012 |
|
RU2627595C2 |
| КОНСТРУКЦИЯ И СПОСОБ КОНСТРУИРОВАНИЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА UBI1 | 2012 |
|
RU2639538C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ АМИНОКИСЛОТ В РАСТЕНИИ | 2019 |
|
RU2799785C2 |
| РАСТЕНИЯ СО СНИЖЕННЫМ СОДЕРЖАНИЕМ АСПАРАГИНА | 2016 |
|
RU2742725C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ РЕДУЦИРУЮЩИХ САХАРОВ В РАСТЕНИИ | 2019 |
|
RU2801948C2 |


















































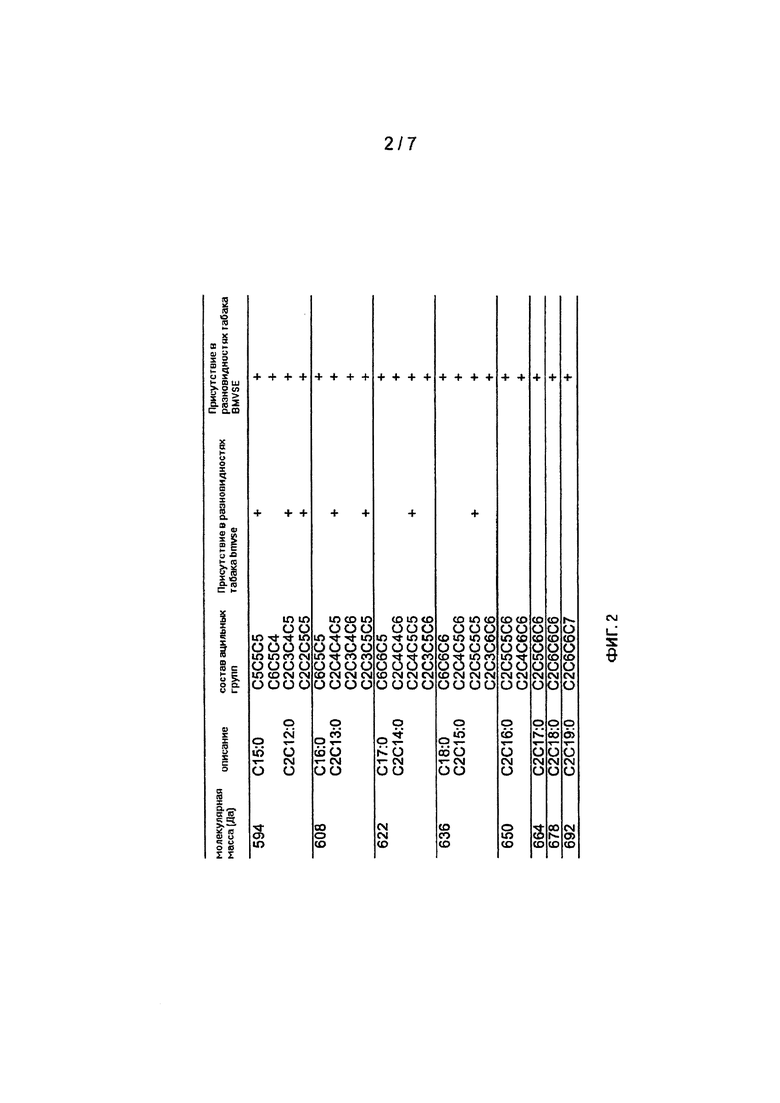





Изобретение относится к области биохимии, в частности к трансгенной растительной клетке табака, трансгенному растению табака, растительному материалу табака для получения повышенного количества С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы, а также к курительному изделию, содержащему вышеуказанный растительный материал или часть вышеуказанного растения. Также раскрыты способ повышения количества С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы, по меньшей мере, в части растения табака, а также способ получения композиции, содержащей один или несколько сложных эфиров сахарозы. Изобретение также относится к способу получения бета-метилвалериановой кислоты. Изобретение позволяет эффективно увеличивать количество сложных эфиров сахарозы в растении табака. 10 н. и 6 з.п. ф-лы, 6 ил., 1 табл., 5 пр.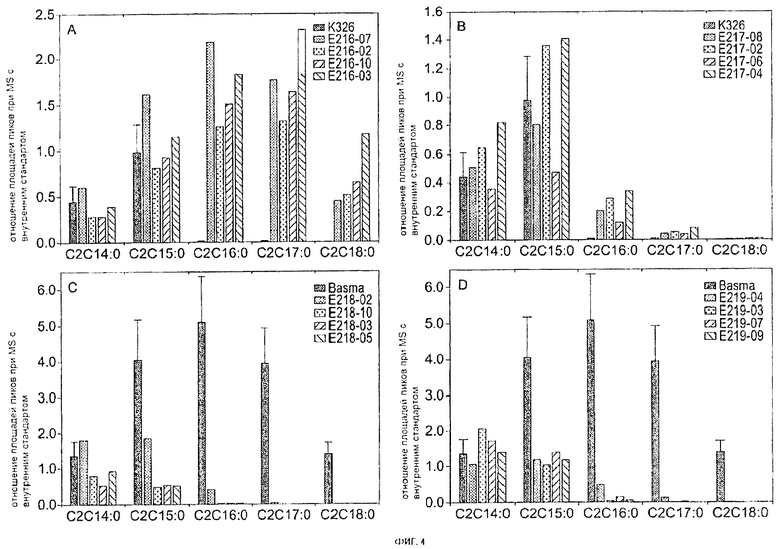
1. Трансгенная растительная клетка табака для получения повышенного количества С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы по сравнению с клеткой растения табака дикого типа, содержащая:
(i) по меньшей мере, один полинуклеотид, состоящий из последовательности, кодирующей изопропилмалат синтазу, и, по меньшей мере, на 90% идентичный SEQ ID NO: 1, или SEQ ID NO: 10, или SEQ ID NO: 12, или SEQ ID NO: 14; или
(ii) полипептид, кодируемый указанным полинуклеотидом (полинуклеотидами); или
(iii) полипептид, по меньшей мере, на 90% идентичный последовательности SEQ ID NO: 2, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 15, и имеющий активность изопропилмалат синтазы; или
(iv) конструкцию, вектор или вектор экспрессии, содержащие указанную последовательность (последовательности) полинуклеотида, где указанная конструкция, вектор или вектор экспрессии дополнительно содержат промотор, содержащий, состоящий или состоящий в основном из последовательности, приведенной в SEQ ID NO: 8, или его вариант, который, по меньшей мере, на 90% идентичен ему, или промотор трихомы, или встречающийся в природе промотор изопропилмалат синтазы.
2. Трансгенное растение табака для получения повышенного количества С2С16:0, С2С17:0 и С2С18:0 эфиров сахарозы по сравнению с растением табака дикого типа, содержащее растительную клетку по п. 1.
3. Способ повышения количества С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы, по меньшей мере, в части растения табака, включающий стадии:
(i) повышения экспрессии или активности изопропилмалат синтазы в растении, предпочтительно, где изопропилмалат синтаза характеризуется полинуклеотидом, состоящим из последовательности, кодирующей указанную изопропилмалат синтазу, и имеющим, по меньшей мере, 90% идентичность последовательности SEQ ID NO: 1, или SEQ ID NO: 10, или SEQ ID NO: 12, или SEQ ID NO: 14, или полипептидом, кодируемым указанным (указанными) полинуклеотидом (полинуклеотидами); или где изопропилмалат синтаза характеризуется полипептидом, последовательность которого, по меньшей мере, на 90% идентична последовательности SEQ ID NO: 2, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 15;
(ii) необязательно, измерения количества или определения типа одного или нескольких сложных эфиров сахарозы, по меньшей мере, в части мутантного, не встречающегося в природе или трансгенного растения, полученного на стадии (i); и
(iii) идентификации трансгенного растения, в котором количество С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы было повышено по сравнению с растением дикого типа, в котором экспрессия или активность изопропилмалат синтазы не модулировалась и, предпочтительно, где внешний вид указанного мутантного, не встречающегося в природе или трансгенного растения такой же, как у растения дикого типа.
4. Способ повышения состава С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы, по меньшей мере, в части растения табака, включающий стадии:
(i) повышения экспрессии или активности изопропилмалат синтазы в растении, предпочтительно, где изопропилмалат синтаза характеризуется полинуклеотидом, состоящим из последовательности, кодирующей указанную изопропилмалат синтазу, и имеющим, по меньшей мере, 90% идентичность последовательности SEQ ID NO: 1, или SEQ ID NO: 10, или SEQ ID NO: 12, или SEQ ID NO: 14, или полипептидом, кодируемым указанным (указанными) полинуклеотидом (полинуклеотидами); или где изопропилмалат синтаза характеризуется полипептидом, последовательность которого, по меньшей мере, на 90% идентична последовательности SEQ ID NO: 2, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 15;
(ii) необязательно, измерения количества или определения типа одного или нескольких сложных эфиров сахарозы, по меньшей мере, в части мутантного, не встречающегося в природе или трансгенного растения, полученного на стадии (i); и
(iii) идентификации трансгенного растения, в котором количество С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы было повышено по сравнению с растением дикого типа, в котором экспрессия или активность изопропилмалат синтазы не модулировалась и, предпочтительно, где внешний вид указанного мутантного, не встречающегося в природе или трансгенного растения такой же, как у растения дикого типа.
5. Способ по п.3, где один или несколько сложных эфиров сахарозы имеют структуру:
где R3 представляет собой ацетил или водород, предпочтительно ацетил; один или несколько из R1, R2 и R4 содержат ацильную цепь с 6 атомами углерода, предпочтительно бета-метилвалерил; и R5 представляет собой ацетил или водород, предпочтительно, водород.
6. Трансгенное растение табака для получения повышенного количества С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы, полученное или получаемое способом по п. 4, где экспрессия или активность изопропилмалат синтазы повышена в растении по сравнению с растением дикого типа, предпочтительно, где изопропилмалат синтаза характеризуется полинуклеотидом, состоящим из последовательности, кодирующей указанную изопропилмалат синтазу, и имеющим, по меньшей мере, 90% идентичность последовательности SEQ ID NO: 1, или SEQ ID NO: 10, или SEQ ID NO: 12, или SEQ ID NO: 14, или полипептидом, кодируемым указанным (указанными) полинуклеотидом (полинуклеотидами); или где изопропилмалат синтаза характеризуется полипептидом, последовательность которого, по меньшей мере, на 90% идентична последовательности SEQ ID NO: 2, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 15.
7. Трансгенное растение табака по п. 6, в котором один или несколько сложных эфиров сахарозы имеют структуру:

где R3 представляет собой ацетил или водород, предпочтительно ацетил; один или несколько из R1, R2 и R4 содержат ацильную цепь с 6 атомами углерода, предпочтительно бета-метилвалерил; и R5 представляет собой ацетил или водород, предпочтительно, водород.
8. Трансгенное растение табака по п. 2, в котором модулируется экспрессия изопропилмалат синтазы или активность кодируемого таким образом белка и, по меньшей мере, в части растения изменен состав одного или нескольких сложных эфиров сахарозы по сравнению с контрольным растением, в котором экспрессия или активность изопропилмалат синтазы не модулировалась, и где внешний вид указанного растения по существу такой же, как у контрольного растения.
9. Трансгенное растение табака по п. 2, в котором модулируется экспрессия изопропилмалат синтазы или активность кодируемого таким образом белка и, по меньшей мере, в части растения изменено количество одного или нескольких сложных эфиров сахарозы по сравнению с контрольным растением, в котором экспрессия или активность изопропилмалат синтазы не модулировалась, и где внешний вид указанного растения по существу такой же, как у контрольного растения.
10. Трансгенное растение табака по п. 8, в котором один или несколько сложных эфиров сахарозы имеют структуру:

где R3 представляет собой ацетил или водород, предпочтительно ацетил; один или несколько из R1, R2 и R4 содержат ацильную цепь с 6 атомами углерода, предпочтительно бета-метилвалерил; и R5 представляет собой ацетил или водород, предпочтительно, водород.
11. Растительный материал, включающий биомассу, семена или листья, для получения повышенного количества С2С16:0, С2С17:0 и С2С18:0 сложных эфиров сахарозы по сравнению с растительным материалом растения дикого типа, содержащий клетки или ткани из растения по п.6 или 8, где экспрессия или активность изопропилмалат синтазы повышена в клетках или тканях из растения по сравнению с растением дикого типа, предпочтительно, где изопропилмалат синтаза характеризуется полинуклеотидом, состоящим из последовательности, кодирующей указанную изопропилмалат синтазу, и имеющим, по меньшей мере, 90% идентичность последовательности SEQ ID NO: 1, или SEQ ID NO: 10, или SEQ ID NO: 12, или SEQ ID NO: 14, или полипептидом, кодируемым указанным (указанными) полинуклеотидом (полинуклеотидами); или где изопропилмалат синтаза характеризуется полипептидом, последовательность которого, по меньшей мере, на 90% идентична последовательности SEQ ID NO: 2, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 15.
12. Курительное изделие, которым является сигарета, сигарилла, или сигара, содержащее часть растения по п. 6 или 8 или растительный материал по п. 11.
13. Способ получения композиции, содержащей один или несколько сложных эфиров сахарозы, включающий стадии:
(i) получения, по меньшей мере, трансгенного растения табака по п. 6 или 8, растительного материала по п. 11 или курительного изделия по п. 12;
(ii) экстракции из них сложного эфира (эфиров) сахарозы;
(iii) необязательно, выделения или очистки экстрагированного сложного эфира (эфиров) сахарозы; и
(iv) получения композиции, содержащей один или несколько сложных эфиров сахарозы.
14. Способ по п. 13, где один или несколько сложных эфиров сахарозы имеют структуру:

где R3 представляет собой ацетил или водород, предпочтительно ацетил; один или несколько из R1, R2 и R4 содержат ацильную цепь с 6 атомами углерода, предпочтительно бета-метилвалерил; и R5 представляет собой ацетил или водород, предпочтительно, водород.
15. Способ повышения количества С2С16:0, С2С17:0 и С2С18:0 сложных эфиров в табаке или табачном изделии, включающий: (i) добавление к табаку или табачному изделию части растения, предпочтительно листьев, из трансгенного растения табака по пп. 2, 6 или 8 или растительного материала по п. 11.
16. Способ получения бета-метилвалериановой кислоты, включающий стадии:
(i) получения, по меньшей мере, трансгенного растения табака по любому из пп. 2, 6 или 8, растительного материала по п. 11 или курительного изделия по п. 12;
(ii) гидролиза материала или его экстракта, полученных на стадии (i); и
(iii) необязательно, выделения или очистки бета-метилвалериановой кислоты.
| KROUMOVA A | |||
| et al., PATHWAYS FOR SYNTHESIS, AND POSSIBILITIES FOR GENETIC MODIFICATION OF SUGAR ESTER ACYL GROUPS PRODUCED BY TRICHOMES OF SOLANACEOUS SPECIES, General and Applied Plant Physiology, 2009, Vol | |||
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| US 2007137663 A1, 21.06.2007 | |||
| MAZOUREK M | |||
| et al., A Dynamic Interface for Capsaicinoid Systems Biology, Plant Physiology, 2009, vol.150, no | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Способ подготовки глины для глинобитных, саманных и т.п. построек из необожженной глины | 1924 |
|
SU1806A1 |
| FIELD B.et al., Expression of a Brassica Isopropylmalate Synthase Gene in Arabidopsis Perturbs Both Glucosinolate and Amino Acid Metabolism, Plant Molecular Biology, 2006, Vol | |||
| Способ получения молочной кислоты | 1922 |
|
SU60A1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324737C1 |

