Область техники
Изобретение относится к медицине и фармакологии, а именно к биологически активным пептидам, которые модифицируют пуринергическую сигнализацию и могут применяться для профилактики и лечения заболеваний, фармакологической мишенью при которых являются пуринергические рецепторы.
Уровень техники
Изучение и лечение боли являются существенными аспектами улучшения качества жизни пациентов. Лечение боли основывается главным образом на назначении противовоспалительных препаратов, которые могут быть нестероидными противовоспалительными препаратами (НПВП) или стероидными (кортикостероидными) препаратами, антидепрессантов и антиконвульсантов, а также сильных или слабых опиатов. НПВП образуют наиболее назначаемый в мире класс терапевтических препаратов в связи с их высокой эффективностью как в отношении воспалительного процесса, так и в отношении самой боли. Их применяют при всех типах воспалительных болей, как острых, так и хронических. Хотя НПВП являются очень эффективными средствами, тем не менее, они остаются серьезным источником нежелательных побочных эффектов, что ограничивает применение НПВП в многочисленных клинических ситуациях. Несмотря на большое количество существующих терапевтических средств, многие виды боли остаются малочувствительными к известным анальгетикам, поэтому разработка новых анальгетических средств с новым механизмом действия является важной и актуальной задачей.
Ионотропные пуринергические рецепторы (Р2Х) являются лиганд-управляемыми ионными каналами, природным агонистом которых выступает аденозинтрифосфат (АТФ), прикладываемый с внеклеточной стороны. Эти рецепторы встречаются в различных органах и тканях, особое физиологическое значение имеет их функция в нервной системе [Surprenant A., North R.A. Signaling at purinergic P2X receptors // Annu. Rev. Physiol., 2009, Vol. 71, p. 333-359]. Они экспрессируются в чувствительных нейронах, и АТФ, освобожденный из поврежденных или воспаленных тканей, активирует Р2Х рецепторы и инициирует болевые сигналы. Полагают, что пуринергическая сигнализация играет важную роль в болевых состояниях, связанных с травмами, опухолями, воспалением, мигренью и респираторными заболеваниями. У человека известно семь изоформ Р2Х рецепторов, среди которых изоформы Р2Х2, Р2Х3, Р2Х4 и Р2Х7, как полагают, участвуют в системе восприятия боли [Donnelly-Roberts D. et al. Painful purinergic receptors // J. Pharmacol. Exp. Ther., 2008, Vol. 324, p. 409-415]. Среди семи изоформ пуринергических рецепторов Р2Х3 представляется важным и хорошо изученным «участником» болевых состояний [Wirkner К. et al. Р2Х3 receptor involvement in pain states // Mol. Neurobiol., 2007, Vol. 36, p. 165-183].
Селективные модуляторы функции пуринергических рецепторов, которые способны модулировать работу только определенных изоформ, являются основой для создания лекарственных препаратов для лечения заболеваний, в развитии которых принимают участие данные рецепторы, а также необходимы для изучения функций пуринергических рецепторов. В случае Р2Х3 рецепторов наибольшим фармакологическим потенциалом обладают антагонисты (ингибиторы) рецепторов, поскольку именно снижение их активности требуется для терапии целого ряда болевых синдромов.
В настоящее время известны многие низкомолекулярные антагонисты Р2ХЗ рецепторов, например 2',3'-O-(2,4,6-тринитрофенил)-АТФ [Virginio С. et al. Trinitrophenyl-substituted nucleotides are potent antagonists selective for P2X1, P2X3, and heteromeric P2X2/3 receptors // Mol. Pharmacol., 1998, Vol.53, p.969-973], другие производные моно- и динуклеотидов [Ford K.K. et al. The P2X3 antagonist P1, P5-di[inosine-5'] pentaphosphate binds to the desensitized state of the receptor in rat dorsal root ganglion neurons // J. Pharmacol. Exp. Ther., 2005, Vol. 315, p. 405-413; US 6,881,725], аналоги сурамина [Hausmann R. et al. The suramin analog 4,4',4'',4'''-(carbonylbis(imino-5,1,3-benzenetriylbis (carbonylimino)))tetra-kis-benzenesulfonic acid (NF110) potently blocks P2X3 receptors: subtype selectivity is determined by location of sulfonic acid groups // Mol. Pharmacol., 2006, Vol. 69, p. 2058-2067], антагонист A-317491 [Jarvis M.F. et al. A-317491, a novel potent and selective non-nucleotide antagonist of P2X3 and P2X2/3 receptors, reduces chronic inflammatory and neuropathic pain in the rat // Proc. Natl. Acad. Sci. U.S.A., 2002, Vol. 99, p. 17179-17184], производные диаминопиримидина [Carter D.S. et al. Identification and SAR of novel diaminopyrimidines. Part 1: The discovery of RO-4, a dual P2X3/P2X2/3 antagonist for the treatment of pain // Bioorg. Med. Chem. Lett. 2009, Vol 19, p. 1628-1631; Jahangir A. et al. Identification and SAR of novel diaminopyrimidines. Part 2: The discovery of RO-51, a potent and selective, dual P2X3/P2X2/3 antagonist for the treatment of pain // Bioorg. Med. Chem. Lett., 2009, Vol. 19, p. 1632-1635; US 7,589,090; US 7,531,547], тетразола и ариламидов [US 7,595,405], а также пиперазина и фенилтиенопиразола [Brotherton-Pleiss СЕ. Discovery and optimization of RO-85, a novel drug-like, potent, and selective P2X3 receptor antagonist // Bioorg. Med. Chem. Lett., 2010, Vol. 20, p. 1031-1036; US 7,491,821].
Среди ингибиторов P2X3 рецепторов известны также вещества белковой природы: спинорфин, фрагмент В-цепи гемоглобина, воздействующий также на другие мишени [Jung K.Y. et al. Structure-activity relationship studies of spinorphin as a potent and selective human P2X3 receptor antagonist // J. Med. Chem., 2007, Vol. 50, p. 4543-4547], и два компонента яда паука Alopecosa marikovskyi - пуротоксин-1 (РТ1) [Grishin Е.V. et al. Novel peptide from spider venom inhibits P2X3 receptors and inflammatory pain // Ann. Neurol., 2010, Vol. 67, p. 680-683; RU 2422459] и пуротоксин-2 (PT2) [Kabanova N.V. et al. Modulation of P2X3 receptors by spider toxins // Biochim. Biophys. Acta, 2012, Vol. 1818, p. 2868-2875]. PT1 является наиболее близким аналогом заявляемых соединений. Этот пептид состоит из 35 аминокислотных остатков и селективно угнетает активность Р2Х3 рецепторов за счет стабилизации их десенситизированного состояния. Рекомбинантный РТ1 может быть получен в бактериальной системе экспрессии [Grishin E.V. et al. Novel peptide from spider venom inhibits P2X3 receptors and inflammatory pain // Ann. Neurol., 2010, Vol. 67, p. 680-683; RU 2422459; RU 257194].
Несмотря на достаточно большое количество известных соединений, способных ингибировать Р2Х3 рецепторы, в настоящее время, тем не менее, нет ни одного препарата, действующего как модулятор Р2ХЗ, разрешенного для применения в клинической практике. В то же время, поиск новых лекарственных средств, эффективно и безопасно купирующих боль различного происхождения, является актуальной задачей современной медицины. Одним из подходов к решению данной задачи может быть создание и внедрение в клинику новых эффективных и селективных модуляторов Р2Х3 рецепторов.
Раскрытие изобретения
Задачей данного изобретения является разработка и получение новых эффективных антагонистов пуринергических рецепторов Р2Х3, перспективных для применения в клинической практике.
Техническим результатом изобретения является получение новых эффективных пептидов, модулирующих активность пуринергических рецепторов Р2Х3, а также обладающих селективностью по отношению к Р2Х3, высокой стабильностью и являющихся перспективными для применения в терапии заболеваний или состояний млекопитающего, фармакологической мишенью при которых являются пуринергические рецепторы Р2Х3, в частности, болей различной этиологии.
Кроме того, дополнительным техническим результатом является то, что пептиды по изобретению отличаются простотой структуры по сравнению с прототипом и могут быть легко получены в бактериальной системе экспрессии, поэтому пептиды по изобретению являются перспективными для создания на их основе лекарственных препаратов.
Указанный технический результат достигается посредством разработки и получения пептида, модулирующего активность пуринергических рецепторов Р2Х3, имеющего следующую аминокислотную последовательность:
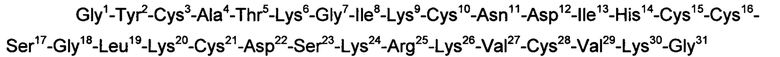
или последовательность, гомологичную ей, по меньшей мере, на 90%.
В некоторых вариантах воплощения изобретения пептид, модулирующий активность пуринергических рецепторов Р2Х3, имеет следующую аминокислотную последовательность:
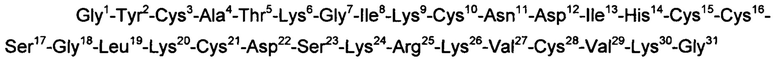
или последовательность, гомологичную ей, по меньшей мере, на 95%.
Еще в одном варианте воплощения изобретения пептид, модулирующий активность пуринергических рецепторов Р2ХЗ, имеет следующую аминокислотную последовательность:
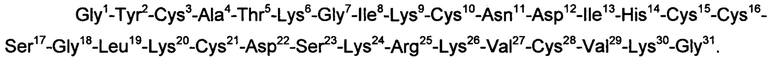
Настоящее изобретение также относится к применению пептидов, являющихся предметом изобретения, в качестве модуляторов пуринергических рецепторов Р2Х3.
Настоящее изобретение также относится к применению пептидов, являющихся предметом изобретения, в качестве антагонистов пуринергических рецепторов Р2Х3.
Настоящее изобретение также относится к применению пептидов, являющихся предметом изобретения, для получения фармацевтической композиции для лечения и/или предотвращения состояний, ассоциированных с пуринергическими рецепторами Р2Х3, например, в терапии болей различной этиологии и/или симптомов боли, в частности боли при воспалении, постоперационной боли, висцеральной боли, зубной боли, предменструальный боли, болей, вызванных ожогами, мигрени или кластерной головной боли, боли при повреждении нерва, неврите, невралгии, боли при онкологических заболеваниях или боли, связанной с синдромом раздраженного кишечника.
Изобретение также включает получение пептидов, являющихся предметом изобретения. Указанные пептиды могут быть получены генно-инженерным способом или химическим синтезом.
Определение и термины
Как используется в настоящем документе, термин «пептид РТ6» относится к пептиду, имеющему следующую аминокислотную последовательность:

Термин «модулировать» в настоящем документе означает изменять функциональные характеристики пуринергических рецепторов Р2Х (в частности ингибировать, блокировать, снижать или предотвращать вызываемые связыванием агониста (в том числе эндогенного агониста) с рецептором физиологические эффекты), в частности Р2Х3 рецепторов.
Термин «модулятор» в настоящем документе означает вещество, способное модулировать, то есть изменять функциональные характеристики пуринергических рецепторов Р2Х (в частности, ингибировать, блокировать, снижать или предотвращать вызываемые связыванием агониста (в том числе эндогенного агониста) с рецептором физиологические эффекты), в частности Р2Х3 рецепторов.
Под «антагонистом» (антагонистом рецепторов) в биохимии и фармакологии подразумевается подтип лигандов к клеточным рецепторам. Лиганд, обладающий свойствами антагониста рецепторов, - это такой лиганд, который ингибирует, блокирует, снижает или предотвращает вызываемые связыванием агониста (в том числе эндогенного агониста) с рецептором физиологические эффекты.
«Селективным» антагонист называют в том случае, если он ингибирует конкретный рецептор либо подтип рецепторов. Степень селективности может различаться.
Термин «гомологичный» (или «гомологичность») в настоящем изобретении является синонимом термину «идентичность» и означает сходство последовательностей пептидных молекул. В настоящем изобретении степень идентичности последовательностей означает сравнение между аминокислотными последовательностями и определяется путем сравнения двух оптимально выровненных последовательностей в окне сравнения, при этом часть аминокислотной последовательности в окне сравнения может содержать вставки или делеции (т.е. пробелы) в сравнении с контрольной последовательностью (не содержащей вставок или делеции) для оптимально выравнивания двух последовательностей. Степень идентичности можно рассчитать путем определения числа положений, по которым в обеих последовательностях находятся идентичные аминокислотные остатки, получая число совпадающих положений, деления числа совпадающих положений на общее число положений в окне сравнения и умножения результата на 100, получая степень идентичности последовательностей в процентах. Специалистам должно быть известно, что существует много установленных алгоритмов для выравнивания двух последовательностей. Примерами алгоритмов, подходящих для определения степени идентичности последовательностей, являются алгоритмы BLAST и BLAST 2.0 [Altschul S.F. et al. Basic local alignment search tool // J. Mol. Biol., 1990, Vol. 215, p. 403-410; Altschul S.F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs // Nucleic Acids Res., 1997, Vol. 25, p. 3389-3402].
Гомологичные пептиды имеют идентичные или сходные аминокислотные последовательности. В контексте настоящего изобретения сходные остатки представляют собой «консервативные замены» или «разрешенные точечные мутации» соответствующих аминокислотных остатков в контрольной последовательности. Консервативные замены остатков в контрольной последовательности являются такими заменами, которые физически или функционально сходны с соответствующим контрольным остатком, например, имеют сходный размер, форму, электрический заряд, химические свойства, включая способность к образованию ковалентных или водородных связей, и т.п. Для специалиста в данной области техники очевидно, что все гомологичные пептидные последовательности, являющиеся предметом изобретения, обладают способностью эффективно модулировать активность Р2Х3 рецепторов.
В используемом здесь значении термин «лечение» означает способ получения благоприятных или желаемых клинических результатов. В соответствии с целями настоящего изобретения благоприятные или желаемые клинические результаты включают, не ограничиваясь ими, один или несколько нижеследующих аспектов: уменьшение любого проявления, в частности, болей различной этиологии, в том числе интенсивности боли, ослабление одного или нескольких симптомов боли, включая любой признак боли, такой как сокращение продолжительности боли и/или уменьшение восприятия или ощущения боли, временное облегчение и/или замедление развития боли.
Термин «уменьшение проявления» боли означает любые способы ослабления боли (которые могут включать уменьшение потребности в лекарственных средствах и/или сокращение количества других лекарственных средств, обычно применяемых в данных условиях), уменьшения продолжительности, интенсивности и/или частоты возникновения боли (включая, например, увеличение времени до появления у субъекта боли различной этиологии).
Термин «уменьшение интенсивности» боли различной этиологии означает ослабление или улучшение одного или нескольких симптомов боли по сравнению с болью, имеющей место без введения модулятора рецепторов Р2Х3. Термин «уменьшение интенсивности» означает также уменьшение продолжительности симптома.
Термин «временное облегчение» боли различной этиологии или одного или нескольких симптомов боли означает уменьшение степени проявления одного или нескольких нежелательных клинических признаков боли у одного или нескольких субъектов, подвергаемых лечению модулятором рецепторов Р2Х3 в соответствии с настоящим изобретением.
В используемом здесь значении термин «замедление» развития боли различной этиологии, означает торможение, препятствование, задержку, стабилизацию и/или отсрочку прогрессирования боли. Подобное замедление может быть различным по времени в зависимости от истории болезни и/или субъекта, подлежащего лечению. Как должно быть очевидно специалисту в данной области, существенное или значительное замедление может в действительности включать предотвращение боли, выражающееся в том, что у субъекта не возникает боль. Способ «замедления» развития симптома является способом, который уменьшает вероятность возникновения симптома в данный период времени и/или уменьшает степень проявления симптома в данный период времени по сравнению с ситуацией, когда данный способ не применяется. Такие сравнения обычно основаны на клинических исследованиях с участием числа субъектов, достаточного для получения статистически значимого результата.
Описание чертежей
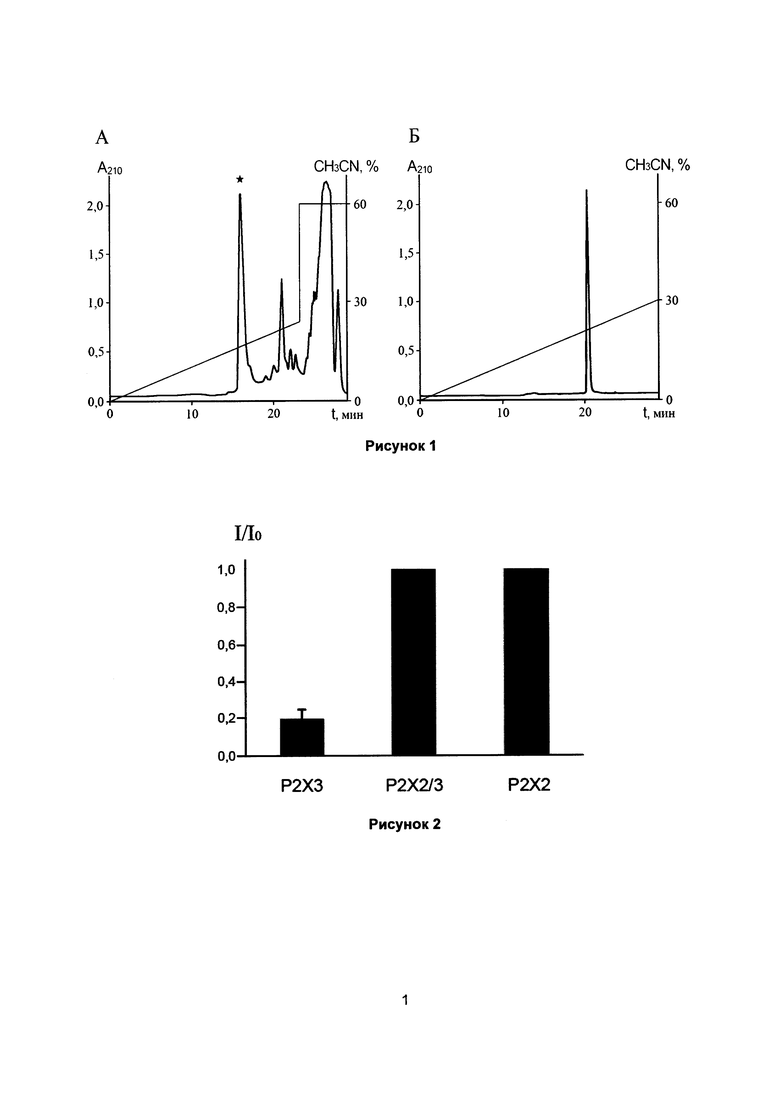
Рисунок 1. Получение рекомбинантного пептида РТ6:
(А) Разделение продуктов гидролиза гибридного белка энтеропептидазой на колонке Jupiter С4 (10×250 мм). Отмечена фракция, соответствующая РТ6 (*).
(Б) Повторная хроматография выделенного РТ6 на колонке Vydac С18 (4,6*250 мм).
Рисунок 2. Влияние пептида РТ6 на токи, опосредованные Р2Х рецепторами в культуре нейронов из ганглиев дорсальных корешков спинного мозга крыс. Гистограмма показывает относительную величину остаточных токов (I), опосредованных гомомерными Р2Х3, гетеромерными Р2Х2/3 и гомомерными Р2Х2 рецепторами, при приложении РТ6 в концентрации 50 нМ по сравнению с контрольными токами в отсутствии РТ6 (10).
Подробное раскрытие изобретения
Поставленная задача данного изобретения решается за счет получения пептидов по изобретению, модулирующих активность пуринергических рецепторов Р2Х3. Так, в частности, последовательность пептида РТ6 определена посредством анализа мРНК из ядовитых желез аранеоморфных пауков, а именно, указанная последовательность пептида РТ6 определена посредством анализа библиотеки кДНК желез паука Thomisus onustus. Пептиды по изобретению, и в частности пептид РТ6, являются веществами полипептидной (белковой) природы и могут быть получены химическим синтезом или генно-инженерным способом.
Неожиданно было обнаружено, что заявляемые пептиды способны ингибировать пуринергические рецепторы Р2Х3. Так, в частности, пептид РТ6 в концентрации 50 нМ вызывает 80%-е ингибирование токов, опосредованных Р2Х3 рецепторами в сенсорных нейронах крыс. РТ6 является селективным антагонистом пуринергических рецепторов Р2Х3.
Препараты на основе пептидов по изобретению являются перспективными для применения в терапии болей различной этиологии, в частности, боли при воспалении, постоперационной боли, висцеральной боли, зубной боли, предменструальный боли, болей, вызванных ожогами, мигрени или кластерной головной боли, боли при повреждении нерва, неврите, невралгии, боли при онкологических заболеваниях или боли, связанной с синдромом раздраженного кишечника. Пептиды по изобретению могут применяться, в частности, для уменьшения проявления боли, для уменьшения интенсивности боли, для временного облегчения боли или для замедления развития боли различной этиологии.
Кроме того, пептид РТ6 характеризуется чрезвычайно стабильной структурой (содержит мотив так называемого «цистинового узла»), легко может быть получен в бактериальной системе экспрессии и отличается простотой структуры по сравнению с прототипом (пептид РТ1, RU 2422459): он короче на четыре остатка и содержит на одну дисульфидную связь меньше, нежели пептид РТ1, РТ6 содержит шесть остатков цистеина, образующих три внутримолекулярные дисульфидные связи. Это служит основанием для предположения высокого потенциала РТ6 с точки зрения создания лекарственных препаратов.
Осуществление изобретения
Возможность объективного достижения технического результата при осуществлении изобретения подтверждена достоверными данными, приведенными в примерах, содержащих сведения экспериментального характера. Следует понимать, что эти и все приведенные в материалах заявки примеры не являются ограничивающими и приведены только для иллюстрации настоящего изобретения.
Получение рекомбинантного пептида РТ6
Аминокислотную последовательность пептида РТ6 преобразуют в соответствующую ей нуклеотидную последовательность с учетом частоты использования кодонов у Escherichia coli. Синтезируют несколько олигонуклеотидных фрагментов длиной до 50 нуклеотидов, перекрывающие полную последовательность гена РТ6, при этом концевые из них содержат сайты рестрикции для клонирования в плазмиду pET-32b (Novagen, США):

Синтез полноразмерной последовательности гена, кодирующего пептид РТ6, осуществляют с помощью методов лигирования олигонуклеотидов друг с другом и полимеразной цепной реакции (ПЦР). Для этого проводят фосфорилирование 3'-концов фрагментов с помощью полинуклеотидкиназы (Promega, США) при 37°C в течение 30 мин, лигирование осуществляют с использованием ДНК-лигазы фага Т4 (Promega, США) в течение 18 ч при 16°C. ПЦР проводят по стандартной процедуре на ПЦР-амплификаторе РТС-200 (MJ Research, США). Реакционная смесь содержит буферный раствор для ПЦР (50 мМ KCl, 1,5 мМ MgCl2, 0,1% Твин 20, 50 мМ Трис-HCl, pH 8,6), смесь четырех дезоксинуклеотидов (Promega, США), праймерные (концевые) олигонуклеотиды (5'-GGCTATTGCGCGACCA-3' и 5'-TTAGCCTTTCACGCACAC-S'), матричную ДНК (лигазную смесь) и Taq-полимеразу. Условия ПЦР: денатурация (96°C) - 20 сек.; отжиг (50°C) - 20 сек.; элонгация (72°C) - 30 сек., 25 циклов. Затем проводят рестрикцию ПЦР-продукта и плазмиды рЕТ-32b в буфере MultiCore, оптимальном для работы ферментов BamHI и KpnI (Promega, США). В качестве контроля проводят рестрикцию плазмиды каждым из ферментов по отдельности, результат оценивают по изменению подвижности вектора при электрофорезе в агарозном геле. Очистку рестрицированных фрагментов ДНК и вектора проводят с помощью электрофореза в агарозном геле с последующим выделением ДНК из геля с использованием набора реактивов Wizard SV Gel and PCR Clean-Up System (Promega, США) в соответствии с методикой фирмы-производителя. Синтетический ген РТ6 и вектор рЕТ-32b лигируют друг с другом с помощью ДНК-лигазы фага Т4. В результате получают плазмиду, кодирующую химеру белка тиоредоксина с РТ6 под контролем Т7 промотора.
Полученные плазмиды используют для трансформации клеток Е. coli штамма XL1 Blue с помощью электропоратора Cellject (Eurogentec, Бельгия). Параметры электроимпульса выставляют согласно рекомендациям фирмы-производителя электропоратора. Суспензию клеток переносят на чашку Петри с твердой средой LB (1% бактотриптона, 0,5% дрожжевого экстракта, 1% NaCl, 1,5% агара, 10 мМ Трис-HCl, pH 7,6) с добавлением ампициллина (70 мкг/мл) в качестве селективного фактора. Чашки инкубируют при 37°C в течение 18 ч. Колонии E. coli XL1 Blue, полученные на селективной среде после проведения трансформации, рассеивают штрихами на новой чашке Петри с твердой средой LB и ампициллином (70 мкг/мл). Чашки инкубируют 18 ч при 37°C. Проводят ПЦР со специфичными к интересующей последовательности олигонуклеотидными праймерами (5'-GGCTATTGCGCGACCA-3' и 5'-TTAGCCTTTCACGCACAC-3'), в качестве матрицы используют тотальную нуклеиновую кислоту из полученной клеточной массы. Отобранные трансформанты используют для получения ночной жидкой культуры в 5 мл жидкой среды LB с добавлением ампициллина (70 мкг/мл), суспензию инкубируют 18 ч при 37°C. Процедуру препаративного выделения плазмидной ДНК проводят с использованием набора реактивов Wizard Plus SV Minipreps DNA Purification System (Promega, США) в соответствии с методикой фирмы-производителя.
Правильность сборки и лигирования проверяют секвенированием по методу Сэнгера на приборе Model 373 DNA Sequencer (Applied Biosystems, США) с использованием реактивов ABI PRISM BigDye Terminator Cycle Sequencing Ready reaction kit, ДНК-полимеразы AmpliTaq DNA polymerase, FS (Perkin Elmer, США) и праймера pET-HindSeq (5'-CTTCCTTTCGGGCTTTG-3') в соответствии с рекомендациями указанных производителей.
Наработку химерного белка проводят с помощью контролируемой экспрессии соответствующего гена в клетках Е. coli штамма BL21(DE3). Ночную культуру клеток, трансформированных экспрессионным вектором рЕТ-32b, содержащим вставку гена РТ6, разбавляют в 200 раз жидкой питательной средой LB с ампициллином (100 мкг/мл). Клетки инкубируют при 37°C и перемешивании (200 об/мин) до достижения оптической плотности OD620=0,6, после чего к культуре добавляют индуктор (изопропил-β-D-1-тиогалактопиранозид, ИПТГ; 0,2 мМ) и инкубируют при комнатной температуре в течение 18 ч.
Клетки осаждают центрифугированием (10 мин, 4500 об/мин) и ресуспендируют в 25 мл стартового буфера для аффинной хроматографии (см. ниже) с добавлением 1% Тритона Х-100. Разрушение клеточных структур проводят с использованием ультразвукового дезинтегратора СРХ 750 (Cole-Parmer Instruments, США). Выделение гибридного белка проводят с помощью аффинной хроматографии. Клеточный лизат наносят в стартовом буфере (50 мМ Трис-HCl, pH 8, 300 мМ NaCl) на колонку объемом 3 мл со смолой TALON Superflow Metal Affinity Resin (Clontech, США) со скоростью потока 1 мл/мин. Для элюции целевого белка используют буфер, содержащий 50 мМ Трис-HCl, pH 8, 300 мМ NaCl, 150 мМ имидазол. Детекцию осуществляют по оптическому поглощению элюата при 280 нм. Контроль за выделением и очисткой гибридного белка ведут с помощью электрофореза аликвот получаемых фракций в полиакриламидном геле.
Полученный белок подвергают обессоливанию на колонке Jupiter С4 (10×250 мм,  , 10 мкм; Phenomenex, США). Элюцию осуществляют ступенчатым изменением концентрации ацетонитрила (0-70%) в 0,1%-ной трифторуксусной кислоте (ТФУ) со скоростью потока 2 мл/мин. Белок высушивают на вакуумном концентраторе, растворяют в 50 мМ Трис-HCl, pH 8, конечная концентрация белка составляет ~1 мг/мл. К раствору белка добавляют раствор каталитической субъединицы энтеропептидазы человека из расчета 1 ед. на 1 мг белка, гидролиз проводят при комнатной температуре в течение 18 ч. Разделение продуктов гидролиза гибридного белка проводят на колонке Jupiter С4 (250×10 мм, , 10 мкм; Phenomenex, США) в линейном градиенте концентрации ацетонитрила (0-24% за 24 мин, 23-60% за 1 мин) в 0,1%-ной ТФУ при скорости потока 2 мл/мин. Детекцию осуществляют по оптическому поглощению элюата при 210 нм (рисунок 1А). Индивидуальность полученного препарата РТ6 подтверждают повторной хроматографией на колонке Vydac 218ТР54 С18 column (4,6×250 мм, , 5 мкм; Separations Group, США) в линейном градиенте концентрации ацетонитрила (0-30% за 30 мин) в 0,1%-ной ТФУ при скорости потока 1 мл/мин (рисунок 1Б). В результате получают выход рекомбинантного РТ6 5 мг с 1 л бактериальной культуры. Структуру рекомбинантного пептида подтверждают при помощи масс-спектрометрии и N-концевого секвенирования.
, 10 мкм; Phenomenex, США). Элюцию осуществляют ступенчатым изменением концентрации ацетонитрила (0-70%) в 0,1%-ной трифторуксусной кислоте (ТФУ) со скоростью потока 2 мл/мин. Белок высушивают на вакуумном концентраторе, растворяют в 50 мМ Трис-HCl, pH 8, конечная концентрация белка составляет ~1 мг/мл. К раствору белка добавляют раствор каталитической субъединицы энтеропептидазы человека из расчета 1 ед. на 1 мг белка, гидролиз проводят при комнатной температуре в течение 18 ч. Разделение продуктов гидролиза гибридного белка проводят на колонке Jupiter С4 (250×10 мм, , 10 мкм; Phenomenex, США) в линейном градиенте концентрации ацетонитрила (0-24% за 24 мин, 23-60% за 1 мин) в 0,1%-ной ТФУ при скорости потока 2 мл/мин. Детекцию осуществляют по оптическому поглощению элюата при 210 нм (рисунок 1А). Индивидуальность полученного препарата РТ6 подтверждают повторной хроматографией на колонке Vydac 218ТР54 С18 column (4,6×250 мм, , 5 мкм; Separations Group, США) в линейном градиенте концентрации ацетонитрила (0-30% за 30 мин) в 0,1%-ной ТФУ при скорости потока 1 мл/мин (рисунок 1Б). В результате получают выход рекомбинантного РТ6 5 мг с 1 л бактериальной культуры. Структуру рекомбинантного пептида подтверждают при помощи масс-спектрометрии и N-концевого секвенирования.
Для специалиста в данной области техники понятно, что все пептиды, являющиеся предметом изобретения и имеющие последовательность, гомологичную последовательности пептида РТ6, по меньшей мере, на 90%, в частности, на 95%, могут быть получены аналогичным способом.
Исследование активности РТ6 в отношении Р2Х рецепторов
Пептиды по изобретению исследовались на способность модулировать активность Р2Х рецепторов. Нейроны для экспериментов выделяют из ганглиев дорсальных корешков спинного мозга (заднекорешковых ганглиев) 9-12-дневных крыс линии Wistar. Ганглии помещают в чашку Петри, наполненную внеклеточным раствором (130 мМ NaCl, 5 мМ KCl, 2 мМ CaCl2, 2 мМ MgCl2, 20 мМ HEPES, pH 7,4) комнатной температуры, а затем - в раствор ферментов (1 мг трипсина и 2 мг коллагеназы в 2 мл внеклеточного раствора), нагретый до 35°C, инкубируют в течение 10 мин. Затем ганглии тщательно промывают чистым (без ферментов) внеклеточным раствором в другой чашке Петри. Из промытых ганглиев выделяют одиночные клетки с помощью тонких игл. Чашка с одиночными клетками переносится в электрофизиологическую установку. Эксперименты начинают после того, как все клетки осядут на дно чашки через 30 мин. после выделения.
Электрофизиологическое тестирование проводится с использованием классического метода локальной фиксации потенциала (patch clamp) в конфигурации отведения напряжения от целой клетки (whole cell) с применением подхода быстрого приложения вещества (агониста), которое связывается с Р2Х3 рецепторами и вызывает открытие ионного канала. Электрофизиологическая установка состоит из трех частей: микроскопа, системы приложения веществ на клетку («прыгающий столик»; Pharma Robot, Украина) и системы регистрации токов, протекающих через каналы в плазматической мембране клетки. Выделенные нейроны в чашке Петри располагают под микроскопом с фазовым контрастом на установке. Для эксперимента выбирают клетки диаметром 8-12 мкм. К выбранной клетке подводят стеклянную пипетку-электрод, заполненную внутриклеточным раствором (120 мМ CsF, 10 мМ Трис-HCl, pH 7,2), в результате формируется плотный контакт между пипеткой и клеточной мембраной с сопротивлением порядка ГОм. Затем клетку на пипетке поднимают со дна чашки и помещают в аппликационную трубку. Растворы, используемые в эксперименте, находятся в камерах прыгающего столика. В опыте используются четыре типа растворов: раствор 1 - базовый внеклеточный раствор, в котором хранятся полученные клетки, раствор 2 - базовый раствор с добавлением агониста в необходимой концентрации, раствор 3 - базовый раствор с пептидом РТ6 в необходимой концентрации, раствор 4 - базовый раствор с агонистом и пептидом РТ6 в необходимых концентрациях. Процедуру отмывки проводят не менее 15 раз на протяжении 3 мин. Через 3 мин после приложения агониста процедуру повторяют. После получения контрольных токов к клетке прикладывают пептид РТ6. Далее повторяют процедуру прикладывания агониста вместе с РТ6. После прикладывания агониста вместе с РТ6 отмывку клетки от агониста также проводят на фоне РТ6. Последовательное приложение агониста с РТ6 прекращают, когда амплитуда Р2Х-опосредованого тока остается неизменной. Отношение амплитуды контрольного тока к установившейся амплитуде тока на фоне определенной концентрации РТ6 является показателем эффективности действия данной концентрации РТ6. Токи регистрируют при поддерживаемом потенциале -60 мВ и комнатной температуре с помощью усилителя модели 2400 (А-М Systems, США). Электроды для электрофизиологических исследований изготовляют из боросиликатного стекла на установке Р-97 Flamming/Brown (Sutter Instrument, США), их сопротивление составляет 3-5 МОм. В качестве агониста используют цитидинтрифосфат (ЦТФ) в насыщающей концентрации (100 мкМ). Ток с быстрой фазой нарастания (до 12 мсек) и быстрой фазой спада (до 500 мсек) соответствует току, опосредованному Р2ХЗ рецепторами. Время активации рецепторов составляет 2-4 мсек, время десенситизации - 20-100 мсек, а время выхода рецепторов из десенситизации - порядка 2 мин для ЦТФ.
В результате проведенного эксперимента неожиданно обнаружено, что связывание пептида РТ6 с Р2Х3 рецепторами в десенситизированном состоянии приводит к значительному уменьшению амплитуды тока. В концентрации 50 нМ РТ6 приводит к уменьшению амплитуды тока, опосредованного Р2Х3 рецепторами, на 80% (рисунок 2). Таким образом, пептид РТ6 является модулятором, а именно антагонистом, пуринергических рецепторов Р2Х3.
В мембранах нейронов ганглиев дорсальных корешков спинного мозга кроме Р2Х3 рецепторов присутствуют также Р2Х2 и Р2Х2/3 рецепторы [Burgard Е.С. et al. Р2Х receptor-mediated ionic currents in dorsal root ganglion neurons // J. Neurophysiol., 1999, V. 82, P. 1590-1598]. Время выхода из десенситизации Р2Х3 рецепторов очень велико по сравнению с Р2Х2 и Р2Х2/3 рецепторами. При приложении агониста с периодом в 1 мин все наблюдаемые ответы, кроме первого, опосредованы Р2Х2 и Р2Х2/3 рецепторами. Чувствительность Р2Х2/3 рецептора к АТФ и к α,β-метилен-АТФ приблизительно одинакова (Kd ~30 мкМ), а минимальная концентрация α,β-метилен-АТФ, необходимая для активации Р2Х2 рецепторов, составляет не менее 100 мкМ. Эквивалентность ионных токов, наблюдаемых в ответ на поочередное приложение АТФ и α,β-метилен-АТФ, свидетельствует о значительном преобладании Р2Х2/3 рецепторов в мембране клетки. В клетках, которые генерируют ионные токи разной амплитуды, большей - в ответ на приложение АТФ и очень малой - в ответ на приложение α,β-метилен-АТФ, мембрана содержит в основном Р2Х2 рецепторы. В результате эксперимента было обнаружено, что РТ6 не влияет на ионные токи, опосредованные Р2Х2/3 и Р2Х2 рецепторами (рисунок 2). Таким образом, неожиданно было установлено, что пептид РТ6 действует селективно на рецепторы изоформы Р2Х3 и является селективным антагонистом пуринергических рецепторов Р2Х3.
Несмотря на то, что изобретение описано со ссылкой на раскрываемые варианты воплощения, для специалистов в данной области должно быть очевидно, что конкретные подробно описанные эксперименты приведены лишь в целях иллюстрирования настоящего изобретения и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения. Должно быть понятно, что возможно осуществление различных модификаций без отступления от сути настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПЕПТИДНЫЙ МОДУЛЯТОР ПУРИНЕРГИЧЕСКИХ РЕЦЕПТОРОВ | 2010 |
|
RU2422459C1 |
| Способ получения пептида, модулирующего активность пуринергических рецепторов | 2019 |
|
RU2714114C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНАЛЬГЕТИЧЕСКОГО ПЕПТИДА | 2013 |
|
RU2571942C2 |
| АНАЛЬГЕТИЧЕСКИЙ ПЕПТИД ИЗ МОРСКОЙ АНЕМОНЫ | 2016 |
|
RU2614759C1 |
| ПРОИЗВОДНЫЕ БЕНЗИМИДАЗОЛА, ПОЛЕЗНЫЕ ПРИ ЛЕЧЕНИИ СОСТОЯНИЙ, АССОЦИИРОВАННЫХ С АКТИВНОСТЬЮ Р2Х3 ИЛИ Р2Х2/3 | 2010 |
|
RU2542582C2 |
| ИНГИБИРУЮЩИЙ РАСТВОР И СПОСОБ ПОДАВЛЕНИЯ БОЛИ, ВОСПАЛЕНИЯ ИЛИ СПАЗМА | 1995 |
|
RU2180852C2 |
| СПОСОБ ЛЕЧЕНИЯ ОСТРОГО, ХРОНИЧЕСКОГО И ПОДОСТРОГО КАШЛЯ И НЕПРЕОДОЛИМОГО ЖЕЛАНИЯ ОТКАШЛЯТЬСЯ | 2014 |
|
RU2650118C2 |
| Средство пролонгированного анальгетического действия и лекарственный препарат на его основе | 2020 |
|
RU2755206C1 |
| РАСТВОРЫ И СПОСОБЫ ИНГИБИРОВАНИЯ БОЛИ, ВОСПАЛЕНИЯ И РАЗРУШЕНИЯ ХРЯЩА | 2000 |
|
RU2271825C2 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ И ВОСПАЛЕНИЯ В НЕЙРОННОЙ ТКАНИ С ПРИМЕНЕНИЕМ АНТАГОНИСТОВ IL-31 | 2007 |
|
RU2440130C2 |
Изобретение относится к области биотехнологии, конкретно к биологически активным пептидам, модулирующим пуринергическую сигнализацию, и может быть использовано для разработки на их основе новых лекарственных средств, например анальгетиков, для изучения механизмов возникновения боли, для обнаружения и тестирования новых модуляторов рецептора Р2ХЗ. Предложен пептид, имеющий следующую аминокислотную последовательность: Gly1-Tyr2-Cys3-Ala4-Thr5-Lys6-Gly7-Ile8-Lys9-Cys10-Asn11-Asp12-Ile13-His14-Cys15-Cys16-Ser17-Gly18-Leu19-Lys20-Cys21-Asp22-Ser23-Lys24-Arg25-Lys26-Val27-Cys28-Val29-Lys30-Gly31 - SEQ ID NO: 1. Изобретение позволяет получить эффективный антагонист пуринергических рецепторов Р2Х3. 2 н.п. ф-лы, 2 ил.
1. Пептид, являющийся антагонистом пуринергических рецепторов Р2Х3 и имеющий следующую аминокислотную последовательность:
Gly1-Tyr2-Cys3-Ala4-Thr5-Lys6-Gly7-Ile8-Lys9-Cys10-Asn11-Asp12-Ile13-His14-Cys15-Cys16-Ser17-Gly18-Leu19-Lys20-Cys21-Asp22-Ser23-Lys24-Arg25-Lys26-Val27-Cys28-Val29-Lys30-Gly31 - SEQ ID NO: 1.
2. Применение пептида по п. 1 в качестве антагониста пуринергических рецепторов Р2Х3.
| GRISHIN E.V | |||
| et al., Novel Peptide from Spider Venom Inhibits P2X3 Receptors and Inflammatory Pain, ANNALS of Neurology, 2010, V.67, N.5, p.680-683 | |||
| ПЕПТИДНЫЙ МОДУЛЯТОР ПУРИНЕРГИЧЕСКИХ РЕЦЕПТОРОВ | 2010 |
|
RU2422459C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНАЛЬГЕТИЧЕСКОГО ПЕПТИДА | 2013 |
|
RU2571942C2 |
| TOKURIKI N | |||
| ET AL., Stability effects of mutations and protein evolvability, Curr | |||
| Opin | |||
| Struct | |||
| Biol., 2009, v.19, n.5, p.596-604. | |||

