Изобретение относится к биохимии, конкретно к биологически активным полипептидам, которые обладают модулирующим действием на участвующие в генерации болевого сигнала клеточные рецепторы и могут найти применение в медицине и научных исследованиях.
Изобретение решает задачу получения рекомбинантного пептида РТ1, модулирующего активность пуринергических рецепторов Р2Х3 и проявляющего анальгетическую активность в животных моделях.
РТ1 имеет следующую аминокислотную последовательность:
Gly1-Tyr2-Cys3-Ala4-Glu5-Lys6-Gly7-Ile8-Arg9-Cys10-Asp11-Asp12-Ile13-His14-Cys15-Cys16-Thr17-Gly18-Leu19-Lys20-Cus21-Cys23-Asn24-Ala25-Ser26-Gly27-Tyr28-Asn29-Cus30-Val31-Cys32-Arg33-Lys34-Lys35
Пептид РТ1 состоит из 35 аминокислотных остатков, может быть выделен хроматографическими методами из яда паука Alopecosa marikovskyi (ранее вид обозначался как Geolycosa sp.) или получен с помощью пептидного синтеза, а также генно-инженерными методами [Grishin E.V. et al. Novel peptide from spider venom inhibits P2X3 receptors and inflammatory pain // Ann. Neurol., 2010, Vol. 67, рp. 680-683; патент РФ №2422459]. Пептид PT1 способен селективно ингибировать токи, опосредованные пуринергическими рецепторами Р2Х3 в сенсорных нейронах крыс, а также в условиях экспрессии генов Р2Х3 рецепторов человека в клетках НЕК 293. Показано, что при действии наномолярных концентраций пептида значительно уменьшается активирующий эффект природного агониста АТФ, что связано, по-видимому, со стабилизацией десенситизированного состояния рецепторов. Концентрация РТ1, вызывающая 50% ингибирование токов, опосредованных Р2Х3 рецепторами в сенсорных нейронах крыс, (IC50) составляет 12±2 нМ при коэффициенте Хилла (nH) 1,1±0,2. Исследования активности РТ1 в отношении ряда ионных каналов и ионотропных рецепторов подтверждают специфичность действия пептида на Р2Х3 рецепторы.
Препараты на основе РТ1 могут оказаться эффективными в терапии болей различной этиологии. В настоящее время известно большое разнообразие низкомолекулярных соединений, модифицирующих активность рецепторов Р2Х3 [Andreev Y.A. et al. Molecules to selectively target receptors for treatment of pain and neurogenic inflammation // Recent Pat. Inflamm. Allergy Drag Discov., 2012, Vol. 6, pp. 35-45]. PT1 является первым соединением полипептидной природы, способным оказывать селективное модулирующие действие на Р2Х3 рецепторы. Пептид характеризуется чрезвычайно стабильной структурой (содержит мотив так называемого «цистинового узла»). Это служит основанием для предположения высокого потенциала РТ1 с точки зрения создания лекарственных препаратов. Обнаруженный недавно пептид РТ2 обладает сходными с РТ1 свойствами [Kabanova N.V. et al. Modulation of P2X3 receptors by spider toxins // Biochim. Biophys. Acta, 2012, Vol. 1818, pp. 2868-2875], однако существенно большими размерами, что затрудняет его получение в необходимых для медицинских целей количествах.
Большинство терапевтических полипептидов, присутствующих на рынке, являются рекомбинантными [Dimitrov D.S. Therapeutic proteins // Methods Mol. Biol., 2012, Vol. 899, pp. 1-26]. А порядка 30% из них получают в Escherichia coli [Huang C.J. et al. Industrial production of recombinant therapeutics in Escherichia coli and its recent advancements // J. Ind. Microbiol. BiotechnoL, 2012, Vol. 39, pp. 383-399]. Рекомбинантный PT1 также был получен в Е. coli с выходом 3 мг с 1 л бактериальной культуры [патент РФ №2422459]. Для этого ген, кодирующий РТ1, был клонирован в составе экспрессионного вектора рЕТ-32b (Novagen, США). Искомый пептид получали в виде химеры с белком-помощником (тиоредоксином). Отделение пептида РТ1 от тиоредоксина проводили с помощью гидролиза химеры каталитической субъединицей энтерокиназы человека. Такой способ-прототип заявляемого легко реализуем и востребован в масштабах исследовательской лаборатории, однако оказывается слишком дорогостоящим и неудобным для масштабирования.
Поставленная задача получения пептида РТ1 решается за счет того, что ген анальгетического пептида РТ1 сливают с последовательностью, кодирующей хитин -связывающий домен из Bacillus circulans [Chong S. et al. Single-column purification of free recombinant proteins using a self-cleavable affinity tag derived from a protein splicing element // Gene, 1997, Vol. 192, pp. 271-281] и интеина DnaB из Synechocystis sp.[Yan S.S. et al. Production of native protein by using Synechocystis sp. PCC6803 DnaB mini-intein in Escherichia coli II Protein Expr. Purif., 2005, Vol. 40, pp. 340-345]. Это позволяет проводить очистку химерного белка с помощью аффинной хроматографии на хитине, а также отделять искомый рекомбинантный РТ1 от вспомогательных элементов с помощью автокаталитического расщепления химерного белка. Указанный способ позволяет получать анальгетический пептид РТ1 с высоким выходом для использования в медицинских целях и устраняет недостатки прототипа.
Способ осуществляют следующим образом:
1) последовательность, кодирующую анальгетический пептид РТ1, клонируют в составе вектора pTWIN1 (New England Biolabs, США), в результате чего получают векторную конструкцию pER-PT1,
2) векторную конструкцию pER-PT1 используют для трансформации штамма Е. coli ER2566, в результате чего получают штамм-продуцент Е. coli ER2566/pER-PT1,
3) проводят контролируемую экспрессию химерного гена,
4) выделяют химерный белок DnaB-PT1, состоящий из хитин-связывающего домена, интеина DnaB и пептида РТ1, с помощью аффинной хроматографии на хитиновом сорбенте,
5) проводят автокаталитическое расщепление слитного белка DnaB-PT1,
6) очищают рекомбинантный РТ1 с помощью обращенно-фазовой хроматографии.
Технический результат изобретения состоит в:
1) получении химерного белка DnaB-PT1, содержащего хитин-связывающий домен, интеин DnaB и пептид РТ1, вследствие экспрессии химерного гена в составе векторной конструкции pER-PT1, использованной для создания штамма-продуцента Е. coli ER2566/pER-PT1;
2) получении рекомбинантного РТ1 вследствие автокаталитического расщепления слитного белка DnaB-PT1.
Изобретение иллюстрируют графические материалы:
Рисунок 1. А) Нуклеотидная последовательность, кодирующая пептид РТ1. Б) Праймеры, используемые при амплификации. Подчеркнуты сайты рестрикции KpnI, BamHI и SapI.
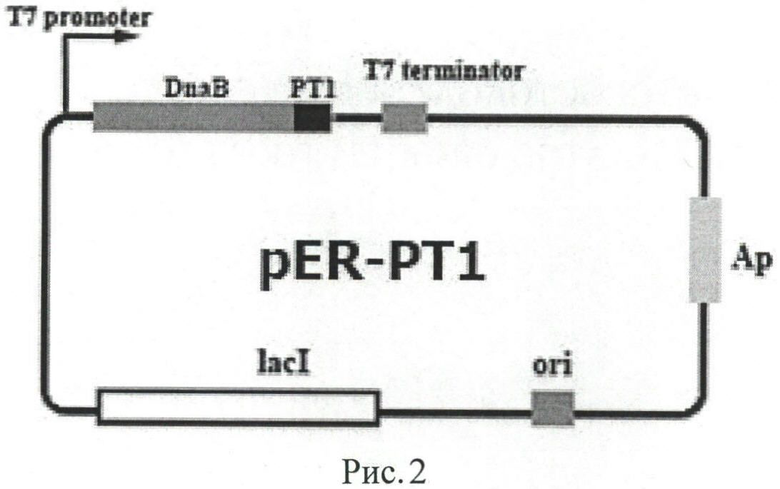
Рисунок 2. Карта экспрессионного вектора pER-PT1. DnaB- ген DnaB мини-интеина из Synechocystis sp.РТ1 - ген РТ1.
Рисунок 3. Аминокислотная последовательность гибридного белка DnaB-PT1.
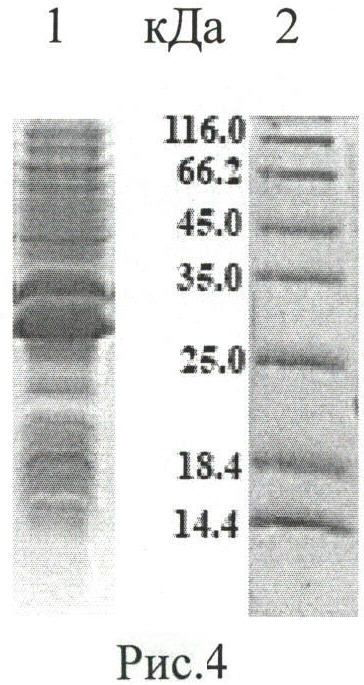
Рисунок 4. Анализ продуктивности штамма-продуцента РТ1. Электрофорез в 15% ПААГ в денатурирующих условиях. 1 - тотальный клеточный лизат Е. coli ER2566/pER-РТ1; 2 - стандарты молекулярных масс.
Рисунок 5. Подбор условий культивирования продуцента Е. coli ER2566/pER-PTl. Электрофорез в 15% ПААГ в денатурирующих условиях. 1, 4, 7- культивирование при 20°C в течение 2, 3 и 4 ч с момента добавления индуктора; 2, 5, 8 - культивирование при 30°C в течение 2, 3 и 4 ч с момента добавления индуктора; 3, 6, 9 - культивирование при 37° в течение 2, 3 и 4 ч с момента добавления индуктора; 10 - стандарты молекулярных масс.
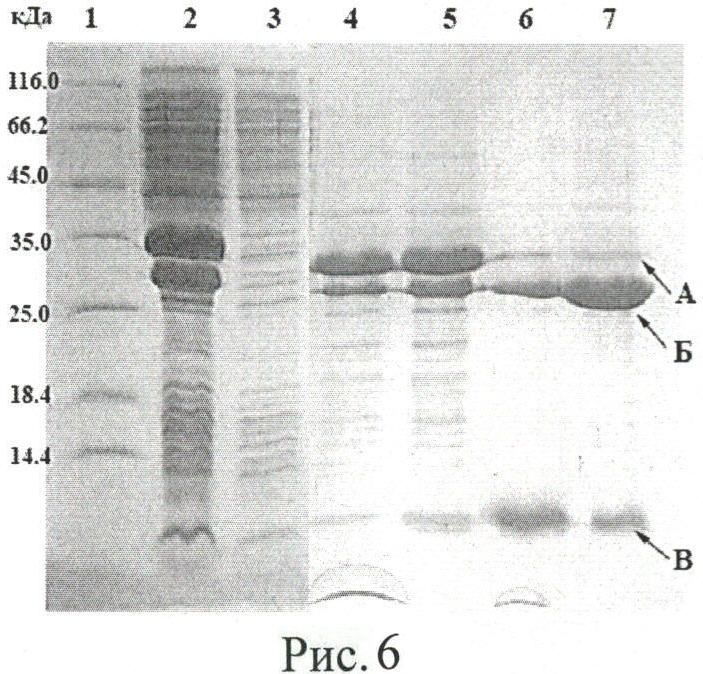
Рисунок 6. Анализ автокаталитического расщепления гибридного белка DnaB-PT1. Электрофорез в 15% ПААГ в денатурирующих условиях. 1 - стандарты молекулярных масс; 2 - тотальный клеточный лизат после 4 ч культивирования продуцента с момента добавления индуктора; 3 - фракция, не сорбировавшаяся на аффинном сорбенте; 4 - сорбированный на хитине гибридный белок; 5 - гибридный белок после 4 ч инкубации; 6 - растворимая фракция гибридного белка после 12 ч инкубации; 7 - сорбированный на хитине гибридный белок после 12 ч инкубации; А - DnaB-PT1; Б - DnaB, В - РТ1.
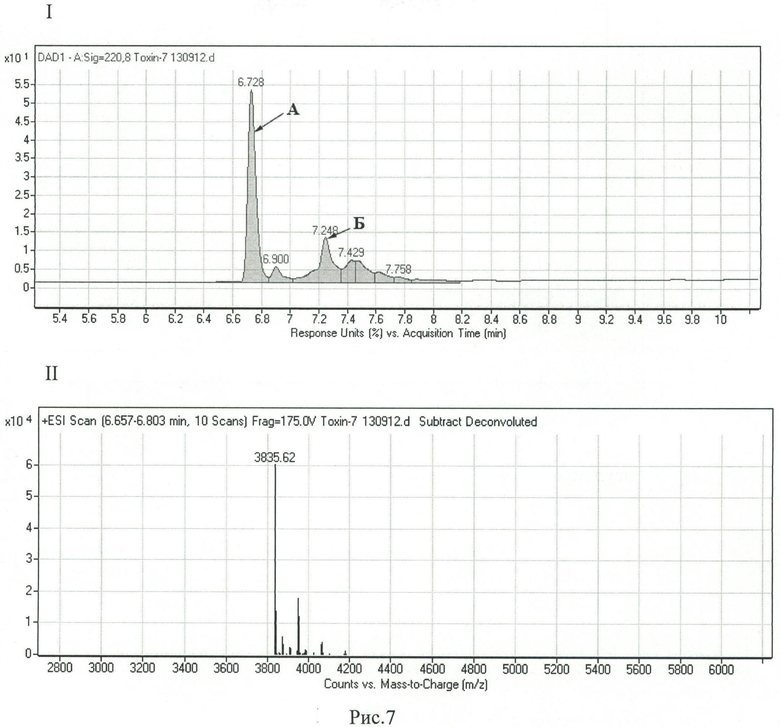
Рисунок 7. Хромато-масс-спектрометрический анализ объединенной фракции, полученной после аффинной хроматографии и содержащей РТ1. Использован прибор 6224 ESI-TOF (Agilent, США) с колонкой YMC-Pack С8 (150×2,1 мм; YMC Europe, Германия). I - хроматограмма; II - масс-спектр пика А.
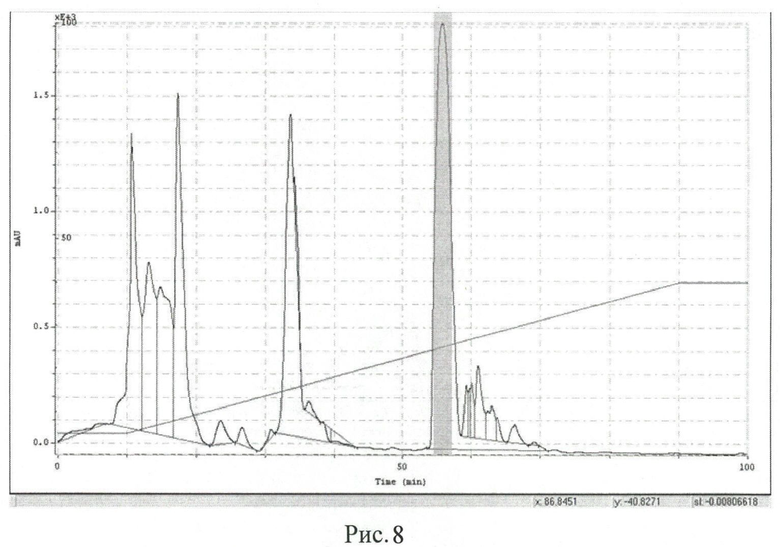
Рисунок 8. Очистка РТ1 с помощью обращенно-фазовой хроматографии на колонке Диасорб 130 С16Т. Серым выделены отобранные фракции, содержащие РТ1.
Изобретение иллюстрируют примеры.
Пример 1. Создание методами генетической инженерии экспрессионного вектора, содержащего гибридный ген со встроенной последовательностью, кодирующей РТ1
Ген пептида РТ1 (рисунок 1А), модулирующего активность пуринергических рецепторов Р2Х3, амплифицируют посредством ПЦР (по программе: 92°C - 1 мин, затем 30 циклов: 92°C - 30 с, 60°C - 30 с, 72°C - 30 с) с плазмидной ДНК рЕТ-32b-РТ1 [патент РФ №2422459] в качестве матрицы и олигонуклеотидами PTIN-1 и PTEV-2 (рисунок 1Б) в качестве праймеров.
Амплификаты обрабатывают эндонуклеазами рестрикции SapI, BamHI и лигируют с вектором pTWIN1, также предварительно обработанным вышеупомянутыми эндонуклеазами. Лигазной смесью трансформируют компетентные клетки Е. coli ER2566, которые затем выращивают на агаризованной среде YT, содержащей ампициллин в концентрации 100 мкг/мл, в течение ночи (16 ч) при 37°C. Отбор трансформантов осуществляют посредством анализа с помощью электрофореза в 15% полиакриламидном геле (ПААГ) в денатурирующих условиях. В результате получают рекомбинантный вектор pER-PT1 (рисунок 2), несущий последовательность гена гибридного белка DnaB-PT1, представленную на рисунке 3. При экспрессии гибридного гена, кодирующего вспомогательный мини-интеин DnaB, должен образовываться слитный белок, способный при определенных условиях к автокаталитическому расщеплению и высвобождению целевого пептида РТ1.
Полученным рекомбинантным вектором трансформируют клетки Е. coli ER2566, получают штамм-продуцент Е. coli ER2566/pER-PT1.
Пример 2. Оценка уровня экспрессии гибридного гена, кодирующего РТ1, в полученном штамме-продуценте
Среду YT, содержащую 100 мкг/мл ампициллина, засевают ночной культурой (1%) штамма Е. coli ER2566/pER-PT1. Культивацию осуществляют в колбах при 37°C до оптического поглощения культуральной среды OD600=0,6. Добавляют индуктор изопропил-β-D-1-тиогалактопиранозид (ИПТГ; Sigma, США) до конечной концентрации 0,4 мМ, культуру инкубируют 4 ч при 37°C. С 1 л культуры получают 5 г влажных клеток. Продуктивность оценивают посредством анализа с помощью электрофореза в 15% ПААГ в денатурирующих условиях (рисунок 4). Для штамма Е. coli ER2566/pER-PT1 наблюдается значительное автокаталитическое расщепление гибридного белка in vivo.
Пример 3. Условия экспрессии химерного гена и выделения целевого продукта
Важным этапом является проверка способности полученного гибридного белка, содержащего интеин, к автокаталитическому расщеплению и высвобождению целевого продукта.
На стадии оценки эффективности полученного штамма-продуцента было отмечено, что при стандартных условиях культивирования гибридный белок претерпевает автокаталитическое расщепление in vivo примерно на 50% (рисунок 4). Поэтому вначале проводят оптимизацию условий роста культуры, варьируют температуру и время культивирования (рисунок 5). Оптимальными условиями оказываются: добавление индуктора при OD600=2 и дальнейшее культивирование при 30°C в течение 2,5 часов. При этом гибридный белок претерпевает автокаталитическое расщепление in vivo на 15-20%.
Среду YT, содержащую 100 мкг/мл ампициллина, засевают ночной культурой (1%) штамма Е. coli ER2566/pER-PT1. Культивацию осуществляют в колбах при 37°C до оптического поглощения культуральной среды OD600=2. Добавляют индуктор ИПТГ до конечной концентрации 0,4 мМ, культуру инкубируют 2,5 ч при 30°C. С 1 л культуры получают 5 г влажных клеток.
Клетки (100 мг) ресуспендируют в 2 мл буфера А (50 мМ Трис-HCl, pH 8,5, 50 мМ ЭДТА, 1 мМ PMSF) и затем лизируют с помощью ультразвукового дезинтегратора. Клеточный лизат центрифугируют, супернатант отделяют от осажденных компонентов, разводят в 2 раза буфером Б (50 мМ Трис-HCl, pH 8,5, 1 мМ ЭДТА, 1 мМ PMSF). 4 мл полученного раствора целевого гибридного белка наносят на хроматографическую колонку, содержащую 1 мл хитинового сорбента и предварительно уравновешенную буфером Б. Промывку колонки осуществляют 10 мл буфера Б, после чего подают 3 мл буфера В (50 мМ Трис-HCl, pH 6,5, 1 мМ ЭДТА, 1 мМ PMSF). Колонку инкубируют 16 ч при 30°C. Получают, что степень расщепления гибридного белка достигает ~100% (рисунок 6).
Отщепленный от гибридного белка пептид РТ1 элюируют 1 мл буфера В и анализируют посредством обращенно-фазовой высокоэффективной жидкостной хроматографии (ОФ-ВЭЖХ) (рисунок 7). В ходе анализа устанавливают, что, наряду с целевым продуктом (А на рисунке 7), элюат содержит продукты неполного окисления и побочный продукт с молекулярной массой, идентичной РТ1 (Б на рисунке 7). Продукт А очищают посредством ОФ-ВЭЖХ на колонке Macrosphere RP 300 Cig (250×4,6 мм; Alltech, США). Масс-спектрометрический анализ подтверждает, что продукт А соответствует РТ1.
Фракции, получаемые после аффинной хроматографии и содержащие целевой пептид, объединяют, наносят на колонку Macrosphere RP 300 C18, предварительно уравновешенную раствором Г (5% ацетонитрил, 0,1% трифторуксусная кислота). Хроматографическое разделение осуществляют в линейном градиенте концентрации ацетонитрила (5-40%) в течение 25 мин со скоростью 0,7 мл/мин. Фракции, содержащие целевой пептид РТ1 с чистотой не менее 98%, объединяют и лиофильно высушивают. Выход РТ1 составляет 12 мг с 1 л бактериальной культуры.
Основной недостаток в протоколе - автокаталитическое расщепление гибридного белка DnaB-PT1 in vivo, однако оптимизация условий культивирования позволяет значительно снизить степень расщепления (до 10-15%). Таким образом, протокол выделения с использованием штамма-продуцента Е. coli ER2566/pER-PT1 оказывается эффективным.
Пример 4. Оптимизированный протокол получения рекомбинантного пептидного анальгетика РТ1 с использованием штамма-продуцента Е. coli ER2566/pER-PT1
20 г клеток продуцента ресуспендируют в 150 мл буфера А и затем лизируют с помощью ультразвукового дезинтегратора (20 с импульс / 20 с охлаждение, 20 циклов). Клеточный лизат центрифугируют при 12000 g в течение 30 мин. Супернатант отделяют от осажденных компонентов, разводят в 2 раза буфером Б. 300 мл полученного раствора гибридного белка наносят на хроматографическую колонку, содержащую 50 мл хитинового сорбента и предварительно уравновешенную буфером Б. Промывку колонки осуществляют 200 мл буфера Б, после чего подают 100 мл буфера В. Колонку инкубируют 16 ч при 30°C. Степень расщепления гибридного белка составляет ~100%. Отщепленный от гибридного белка пептид РТ1 элюируют 80 мл буфера В. Фракции, содержащие РТ1, объединяют.
Затем раствор РТ1 наносят на хроматографическую колонку Диасорб 130 С16Т (250×16 мм; Элсико, Россия), предварительно уравновешенную раствором Г, со скоростью 1,5 мл/мин. Разделение осуществляют в линейном градиенте концентрации ацетонитрила (5-40%) в течение 80 мин со скоростью 3 мл/мин (рисунок 8). Фракции, содержащие пептид РТ1 чистотой не менее 98%, объединяют и лиофильно высушивают. Выход составляет 57 мг с 20 г влажных клеток (14,3 мг с 1 л культуры).
Пример 5. Наработка и анализ опытных образцов РТ1 на содержание примесей
Наработку РТ1 проводят в соответствии с разработанным оптимизированным протоколом с использованием созданного штамма-продуцента Е. coli ER2566/pER-PT1.
Анализ опытных образцов, проведенный в контрольно-аналитической лаборатории ИБХ РАН, показывает, что полученный препарат по показателям «Бактериальные эндотоксины», «Остаточные ДНК штамма-продуцента» и «Уровень содержания бактериальных эндотоксинов» является безопасным и соответствует фармакопейным требованиям.
Аминокислотную последовательность рекомбинантного РТ1 устанавливают с помощью автоматического секвенирования по Эдману. Она соответствует природной: GYCAEKGIRCDDIHCCTGLKCKCNASGYNCVCRKK. Молекулярную массу рекомбинантного РТ1 измеряют с помощью масс-спектрометрии на приборе Ultraflex II (Bruker, Германия). Она составляет 3833,6 Да и равна расчетному значению.
Пример 6. Определение биологической активности рекомбинантного РТ1
Полученный препарат РТ1 тестируют на рекомбинантных Р2Х3 рецепторах человека. Для этого ген Р2Х3 человека, клонированный в вектор pIRES2-EGFP (Clonteth), используют для трансфекции клеток НЕК 293. Трансфекцию проводят по стандартной методике с использованием липофектамина (Invitrogen, США). Электрофизиологическое тестирование проводят с использованием классического метода локальной фиксации потенциала ("patch clamp") в конфигурации отведения напряжения от целой клетки ("whole cell") с применением подхода быстрого приложения агониста. Токи регистрируют при комнатной температуре с помощью усилителя модели 2400 (А-М Systems, США) при поддерживаемом потенциале -60 мВ. В качестве агонистов, которые вызывают Р2Х3-опосредованые токи, используют аденозинтрифосфат (АТФ) и цитидинтрифосфат (ЦТФ). Приложение РТ1 к рецепторам в нечувствительном (десенситизированном) состоянии приводит к значительному уменьшению амплитуды тока при последующей активации. Экспериментальные данные анализируют по модели Хилла. Концентрация РТ1, при которой наблюдается уменьшение амплитуды тока в два раза по сравнению с контрольным значением, (IC50) составляет 20 нМ.
Таким образом, заявляемый способ позволяет получить рекомбинантный пептидный анальгетик РТ1, полностью соответствующий нативному пептиду по структуре и биологической активности.



Изобретения касаются векторной конструкции, штамма Escherichia coli, включающего такую векторную конструкцию, и способа получения рекомбинантного анальгетического пептида РТ1. Векторная конструкция получена на основе вектора pTWIN1 и содержит ген химерного белка DnaB-PT1, который состоит из хитин-связывающего домена из Bacillus circulans, интеина DnaB из Synechocystis sp. и анальгетического пептида РТ1. Охарактеризованный способ получения включает проведение контролируемой экспрессии гена химерного белка DnaB-PT1 в составе охарактеризованной векторной конструкции в штамме-продуценте Е. coli ER2566 с использованием в качестве индуктора изопропил-β-D-1-тиогалактопиранозида. Далее проводят лизис клеток, отделение растворимых компонентов, очистку химерного белка DnaB-PT1 с помощью аффинной хроматографии на хитиновом сорбенте, автокаталитическое расщепление химерного белка DnaB-PT1 и очистку пептида РТ1 с помощью обращенно-фазовой хроматографии. Представленная группа изобретений позволяет получить анальгетический пептид с высоким выходом для использования в медицинских целях. 3 н.п. ф-лы, 8 ил., 6 пр.
1. Векторная конструкция, полученная на основе вектора pTWIN1 и содержащая ген химерного белка DnaB-PT1, который состоит из хитин-связывающего домена из Bacillus circulans, интеина DnaB из Synechocystis sp. и анальгетического пептида РТ1, и позволяющая производить контролируемую экспрессию слитного гена.
2. Штамм Escherichia coli - продуцент химерного белка DnaB-PT1, представляющий собой штамм Е. coli ER2566 с внедренной векторной конструкцией по п. 1.
3. Способ получения рекомбинантного анальгетического пептида РТ1, включающий проведение контролируемой экспрессии гена химерного белка DnaB-PT1 в составе векторной конструкции по п. 1 в штамме-продуценте Е. coli по п. 2 с использованием в качестве индуктора изопропил-β-D-1-тиогалактопиранозида, лизис клеток, отделение растворимых компонентов, очистку химерного белка DnaB-PT1 с помощью аффинной хроматографии на хитиновом сорбенте, автокаталитическое расщепление химерного белка DnaB-PT1, очистку пептида РТ1 с помощью обращенно-фазовой хроматографии.
| ПЕПТИДНЫЙ МОДУЛЯТОР ПУРИНЕРГИЧЕСКИХ РЕЦЕПТОРОВ | 2010 |
|
RU2422459C1 |
| EUGENE V | |||
| GRISHIN et al., Novel Peptide from Spider Venom Inhibits P2X3 Receptors and Inflammatory Pain, ANNALS of Neurology, 2010 May, Volume 67, No | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |

