Ссылки на родственные заявки
Согласно настоящей заявке испрашивается приоритет в соответствии с предварительными заявками на выдачу патента США №№61/552582 и 61/552587, поданными 28 октября 2011 г., и предварительной заявкой на выдачу патента США №61/700908, поданной 14 сентября 2012 г., полное содержание которых включено в настоящий документ посредством ссылки.
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к генетически модифицированному не относящемуся к человеку животному, например, грызуну (например, мыши или крысе), которое экспрессирует человеческую или гуманизированную молекулу главного комплекса гистосовместимости (MHC) I класса. Настоящее изобретение также относится к генетически модифицированному не относящемуся к человеку животному, например, мыши или крысе, которое экспрессирует человеческий или гуманизированный белок МНС I (например, α цепь МНС I) и/или человеческий или гуманизированный β2 микроглобулин; а также зародышам, тканям и клеткам, их экспрессирующим. Настоящее изобретение дополнительно относится к способам получения генетически модифицированного не относящегося к человеку животного, которое экспрессирует человеческий или гуманизированный белок МНС I класса (например, α цепь МНС I) и/или β2 микроглобулин. Также предусмотрены способы идентификации и оценки пептидов в контексте гуманизированной клеточной иммунной системы in vitro или у генетически модифицированного не относящегося к человеку животного, и способы модификации локуса МНС I и/или локуса β2 микроглобулина не относящегося к человеку животного, например, мыши или крысы, для экспрессии человеческого или гуманизированного МНС I и/или β2 микроглобулина.
Предшествующий уровень техники настоящего изобретения
В приобретенном иммунном ответе чужеродные антигены распознаются рецепторными молекулами на В-лимфоцитах (например, иммуноглобулинами) и Т-лимфоцитах (например, Т-клеточным рецептором, или TCR). Эти чужеродные антигены презентируются на поверхности клеток в виде пептидных фрагментов специализированными белками, имеющими общее название молекулы главного комплекса гистосовместимости (МНС). Молекулы МНС кодируются множественными локусами, которые встречаются в виде соединенного кластера генов, который охватывает приблизительно 4 м.п.н. У мышей гены MHC находятся на 17 хромосоме, и исторически они называются гены гистосовместимости 2 (Н-2). У людей гены находятся на 6 хромосоме и имеют название гены антигена лейкоцита человека (HLA). Локусы у мышей и людей являются полигенными; они включают в себя три высоко полиморфных класса генов MHC (I, II и III класс), которые проявляют сходную организацию в геноме человека и мыши (смотрите фиг. 2 и фиг. 3, соответственно).
Локусы МНС проявляют самый высокий полиморфизм в геноме; некоторые гены представлены >300 аллелями (например, HLA-DRβ человека и HLA-B человека). Все гены МНС I и II класса могут презентировать пептидные фрагменты, но каждый ген экспрессирует белок с различными характеристиками связывания, отражая полиморфизмы и аллельные варианты. Любой рассматриваемый индивидуум содержит уникальный спектр пептидных фрагментов, которые могут быть презентированы на клеточной поверхности В- и Т-клеткам в ходе иммунного ответа.
Как люди, так и мыши содержат гены МНС I класса (смотрите фигуры 2 и 3). У людей классические гены МНС I называются HLA-A, HLA-B и HLA-C, тогда как у мышей они представляют собой Н-2K, H-2D и H-2L. Молекулы I класса состоят из двух полипептидных цепей: полиморфной α цепи (в некоторых случаях имеющей название тяжелая цепь) и меньшей цепи, которая называется β2 микроглобулин (также известная как легкая цепь), которая в основном не является полиморфной (фиг. 1). Эти две цепи образуют нековалентный гетеродимер на клеточной поверхности. α цепь содержит три домена (α1, α2 и α3). Экзон 1 гена α цепи кодирует лидерную последовательность, экзоны 2 и 3 кодируют α1 и α2 домены, экзон 4 кодирует α3 домен, экзон 5 кодирует трансмембранный домен, и экзоны 6 и 7 кодируют цитоплазматический хвост. α цепь образует пептидсвязывающую бороздку, включающую в себя α1 и α2 домены (которые напоминают Ig-подобные домены), за которыми следует α3 домен, который аналогичен β2 микроглобулину.
β2 микроглобулин представляет собой негликозилированный 12 кДа белок; одной из его функций является стабилизация α цепи МНС I класса. В отличие от α цепи β2 микроглобулин не пересекает мембрану. Локус β2 микроглобулина человека находится на 15 хромосоме, тогда как локус мыши находится на 2 хромосоме. Ген β2 микроглобулина состоит из 4 экзонов и 3 интронов. Циркулирующие формы β2 микроглобулина присутствуют в сыворотке, моче и других биологических жидкостях; таким образом, нековалентно связанный с МНС I β2 микроглобулин может обмениваться на циркулирующий β2 микроглобулин при физиологических условиях.
Молекулы МНС I класса экспрессируются на всех ядросодержащих клетках, включающих в себя опухолевые клетки. Они экспрессируются специфически на Т- и В-лимфоцитах, макрофагах, дендритных клетках и нейтрофилах, среди прочих клеток, и функционируют для представления пептидных фрагментов (как правило 8-10 аминокислот в длину) на поверхности CD8+ цитотоксическим Т-лимфоцитам (CTL). CTL специализируются на цитолизе любой клетки, которая несет связанный с МНС I пептид, распознанный ее собственным мембраносвязанным TCR. Когда клетка представляет пептиды, происходящие из клеточных белков, не присутствующих в норме (например, вирусного, опухолевого или другого не являющегося собственным происхождения), такие пептиды распознаются CTL, которые активируются и лизируют представляющую пептид клетку.
Как правило, презентация нормальных (т.е., собственных) белков в контексте молекул МНС I не вызывает активации CTL вследствие механизмов толерантности. Тем не менее, при некоторых заболеваниях (например, злокачественной опухоли, аутоиммунных заболеваниях), происходящие из собственных белков пептиды становятся мишенью клеточного компонента иммунной системы, что приводит к разрушению клеток, презентирующих такие пептиды. Несмотря на достигнутый прогресс в распознавании собственных антигенов, которые вызывают клеточный иммунный ответ (например, антигенов, связанных с различными злокачественными опухолями), для улучшения идентификации пептидов, распознаваемых CTL человека посредством молекул МНС I класса, сохраняется потребность в системах как in vivo, так и in vitro, которые имитируют аспекты системы клеточного иммунитета человека. Системы, которые имитируют систему клеточного иммунитета человека, могут использоваться в идентификации связанных с заболеваниями антигенов для разработки терапевтических средств для людей, например, вакцин и других биологических средств. Системы оценки распознавания антигенов в контексте иммунной системы человека могут содействовать в идентификации терапевтически применимых популяций CTL (например, применимых для исследования и борьбы с заболеванием человека). Такие системы также могут содействовать в усилении активности популяций CTL человека для более эффективной борьбы с инфекциями и чужеродными несущими антигены соединениями. Таким образом, существует необходимость в биологических системах (например, генетически сконструированных животных), которые могут создавать иммунную систему, которая проявляет компоненты, имитирующие функцию иммунной системы человека.
Краткое раскрытие настоящего изобретения
Предусмотрена биологическая система для получения или идентификации пептидов, которые ассоциируются с белками МНС I класса человека и их химерами и связываются с CD8+ Т-клетками. Предусмотрены не относящиеся к человеку животные, содержащее не относящиеся к человеку клетки, которые экспрессируют человеческие или гуманизированные молекулы, функционирующие в клеточном иммунном ответе. Также предусмотрены гуманизированные локусы грызуна, которые кодируют человеческие или гуманизированные белки МНС I и β2 микроглобулина. Также предусмотрены гуманизированные клетки грызуна, которые экспрессируют человеческие или гуманизированные молекулы МНС и β2 микроглобулина. Предусмотрены системы in vivo и in vitro, которые содержат гуманизированные клетки грызуна, причем клетки грызуна экспрессируют одну или несколько человеческих или гуманизированных молекул иммунной системы.
В настоящем документе предусмотрено не относящееся к человеку животное, например, грызун (например, мышь или крыса), содержащее в своем геноме нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку (например, человеческий/относящийся к грызуну, например, человеческий/мышиный или человеческий/крысиный) полипептид МНС I, причем человеческая часть химерного полипептид содержит внеклеточный домен полипептид МНС I человека. В частности, в настоящем документе предусмотрено не относящееся к человеку животное, содержащее на эндогенном локусе МНС I нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, причем человеческая часть химерного полипептид содержит внеклеточный домен полипептида МНС I человека, и причем животное экспрессирует химерный человеческий/не относящийся к человеку полипептид МНС I. Согласно одному аспекту животное не экспрессирует внеклеточный домен эндогенного не относящегося к человеку полипептида МНС I из эндогенного не относящегося к человеку локуса МНС I. Согласно одному аспекту настоящего изобретения не относящееся к человеку животное (например, грызун, например, мышь или крыса) содержит две копии локуса МНС I, содержащего нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку (например, человеческий/относящийся к грызуну, например, человеческий/мышиный или человеческий/крысиный) полипептид МНС I. Согласно другому аспекту настоящего изобретения животное содержит одну копию локуса МНС I, содержащего нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I. Таким образом, животное может являться гомозиготным или гетерозиготным в отношении локуса МНС I, содержащего нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I. Согласно различным вариантам осуществления нуклеотидная последовательность, кодирующая химерный человеческий/не относящийся к человеку полипептид МНС I, содержится в зародышевой линии не относящегося к человеку животного (например, грызуна, например, крысы или мыши).
Согласно одному аспекту нуклеотидная последовательность, кодирующая химерный человеческий/не относящийся к человеку МНС I, функционально связана с эндогенными не относящимися к человеку регуляторными элементами, например, промотором, энхансером, сайленсером и т.д. Согласно одному варианту осуществления человеческая часть химерного полипептид содержит лидерную последовательность человека. Согласно дополнительному варианту осуществления человеческая часть химерного полипептид содержит α1, α2 и α3 домены полипептида МНС I человека. Полипептид МНС I человека может быть выбран из группы, состоящей из HLA-A, HLA-B и HLA-C. Согласно одному варианту осуществления полипептид МНС I человека представляет собой полипептид HLA-A2, например, полипептид HLA-A2.1.
Согласно одному аспекту генетически сконструированное не относящееся к человеку животное представляет собой грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Таким образом, согласно одному варианту осуществления эндогенный не относящийся к человеку локус представляет собой локус мыши, например, локус Н-2K, H-2D или H-2L мыши. Согласно одному варианту осуществления не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида МНС I содержит трансмембранный и цитоплазматический домены эндогенного не относящегося к человеку полипептида МНС I. Таким образом, согласно варианту осуществления, в котором не относящееся к человеку животное представляет собой мышь, эндогенный не относящийся к человеку локус МНС I может представлять собой локус Н-2K (например, локус Н-2Kb), и эндогенный не относящийся к человеку полипептид МНС I может представлять собой полипептид Н-2K; следовательно, химерный человеческий/не относящийся к человеку полипептид МНС I может содержать трансмембранный и цитоплазматический домены полипептида Н-2K. Согласно другому варианту осуществления, в котором не относящееся к человеку животное представляет собой мышь, эндогенный не относящийся к человеку локус МНС I может представлять собой локус H-2D, и эндогенный не относящийся к человеку полипептид МНС I может представлять собой полипептид H-2D; следовательно, химерный человеческий/не относящийся к человеку полипептид МНС I может содержать трансмембранный и цитоплазматический домены полипептида H-2D. Аналогично, согласно другому варианту осуществления эндогенный не относящийся к человеку МНС I локус может представлять собой локус H-2L, и эндогенный не относящийся к человеку полипептид МНС I может представлять собой полипептид H-2L; следовательно, химерный человеческий/не относящийся к человеку полипептид МНС I может содержать трансмембранный и цитоплазматический домены полипептида H-2L.
Также в настоящем документе предусмотрена мышь, содержащая на эндогенном локусе Н-2K нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид МНС I, причем человеческая часть химерного полипептида содержит внеклеточный домен HLA-A полипептида человека (например, HLA-A2), и относящаяся к мыши часть содержит трансмембранный и цитоплазматический домены полипептида Н-2K мыши, и причем мышь экспрессирует химерный человеческий/мышиный полипептид МНС I. Согласно некоторым вариантам осуществления мышь не экспрессирует внеклеточный домен полипептида Н-2K мыши из эндогенного локуса Н-2K. Согласно одному аспекту нуклеотидная последовательность, кодирующая химерный человеческий/мышиный полипептид МНС I, функционально связана с эндогенными регуляторными элементами мыши. Человеческая часть химерного полипептида может содержать лидерную последовательность человека. Она также может содержать α1, α2 и α3 домены полипептида МНС I человека. Полипептид МНС I человека может представлять собой полипептид HLA-A, например, полипептид HLA-A2.1. Согласно одному аспекту локус Н-2K мыши представляет собой локус Н-2Kb.
Другой аспект настоящего изобретения относится к не относящемуся к человеку животному, например, грызуну (например, мыши или крысе), содержащему в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Таким образом, в настоящем документе предусмотрено не относящееся к человеку животное, содержащее на эндогенном не относящемся к человеку локусе β2 микроглобулина нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина, причем животное экспрессирует человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному аспекту животное не экспрессирует функциональный эндогенный не относящийся к человеку полипептид β2 микроглобулина из эндогенного не относящегося к человеку локуса β2 микроглобулина. Согласно одному аспекту животное содержит две копии локуса β2 микроглобулин, кодирующего человеческий или гуманизированный полипептид β2 микроглобулина; согласно другому варианту осуществления животное содержит одну копию локуса β2 микроглобулина, кодирующего человеческий или гуманизированный полипептид β2 микроглобулина. Таким образом, животное может являться гомозиготным или гетерозиготным в отношении локуса β2 микроглобулина, кодирующего человеческий или гуманизированный полипептид β2 микроглобулина. Согласно различным вариантам осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, содержится в зародышевой линии не относящегося к человеку животного (например, грызуна, например, крысы или мыши). Согласно одному варианту осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина содержит нуклеотидную последовательность, кодирующую полипептид, содержащий аминокислотную последовательность β2 микроглобулина человека. Согласно одному варианту осуществления полипептид способен связываться с белком МНС I.
Согласно некоторым вариантам осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, функционально связана с эндогенными не относящимися к человеку регуляторными элементами β2 микроглобулина. Согласно одному аспекту нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, содержит нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека. Согласно другому аспекту нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, содержит нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 гена β2 микроглобулина человека. Согласно дополнительному аспекту нуклеотидная последовательность также содержит нуклеотидную последовательность, представленную в экзоне 1 не относящегося к человеку гена β2 микроглобулина. Согласно некоторым вариантам осуществления не относящееся к человеку животное представляет собой грызуна (например, мышь или крысу); таким образом, не относящийся к человеку локус β2 микроглобулина представляет собой локус β2 микроглобулина грызуна (например, мыши или крысы).
Также предусмотрена мышь, содержащая на эндогенном локусе β2 микроглобулина нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина, причем мышь экспрессирует человеческий или гуманизированный полипептид β2 микроглобулина. Согласно некоторым вариантам осуществления мышь не экспрессирует функциональный эндогенный β2 микроглобулин мыши из эндогенного локуса β2 микроглобулина. Нуклеотидная последовательность может быть связана с эндогенными регуляторными элементами мыши. Согласно одному аспекту нуклеотидная последовательность содержит нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека. Альтернативно, нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, может содержать нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 гена β2 микроглобулина человека. Нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, может дополнительно содержать нуклеотидную последовательность экзона 1 гена β2 микроглобулина мыши. Согласно одному варианту осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, содержит нуклеотидную последовательность, кодирующую полипептид, содержащий аминокислотную последовательность β2 микроглобулина человека. Согласно одному варианту осуществления полипептид способен связываться с белком МНС I.
Настоящее изобретение дополнительно относится к не относящемуся к человеку животному (например, грызуну, например, мыши или крысе), содержащему в своем геноме нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, и нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному варианту осуществления настоящее изобретение относится к не относящемуся к человеку животному, содержащему в своем геноме первую нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, причем человеческая часть химерного полипептида содержит внеклеточный домен полипептида МНС I человека; и вторую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина, причем первая нуклеотидная последовательность расположена на эндогенном не относящемся к человеку локусе МНС I, и вторая нуклеотидная последовательность расположена на эндогенном не относящемся к человеку локусе β2 микроглобулина, и причем животное экспрессирует химерный человеческий/не относящийся к человеку полипептид МНС I и человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному аспекту животное представляет собой мышь. Таким образом, эндогенный локус МНС I может быть выбран из группы, состоящей из локуса Н-2K, H-2D и H-2L. Согласно одному варианту осуществления эндогенный локус мыши представляет собой локус Н-2K (например, локус Н-2Kb). Согласно одному варианту осуществления полипептид МНС I человека выбран из группы, состоящей из полипептида HLA-A, HLA-B и HLA-C. Согласно одному аспекту полипептид МНС I человека представляет собой HLA-A, например, HLA-A2 (например, HLA-A2.1). Согласно различным вариантам осуществления первая и вторая нуклеотидные последовательности содержатся в зародышевой линии не относящегося к человеку животного (например, грызуна, например, мыши или крысы).
Следовательно, настоящее изобретение относится к мыши, содержащей в своем геноме первую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный МНС I полипептид, причем человеческая часть химерного полипептида содержит внеклеточный домен HLA-A человека (например, HLA-A2), и мышиная часть содержит трансмембранный и цитоплазматический домены Н-2K мыши; и вторую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина, причем первая нуклеотидная последовательность расположена на эндогенном локусе Н-2K, и вторая нуклеотидная последовательность расположена на эндогенном локусе β2 микроглобулина мыши, и причем мышь экспрессирует химерный человеческий/мышиный полипептид МНС I и человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному варианту осуществления не относящееся к человеку животное (например, мышь), содержащее как химерный полипептид МНС I, так и человеческий или гуманизированный полипептид β2 микроглобулина, не экспрессирует внеклеточный домен эндогенного не относящегося к человеку полипептида МНС I (например, полипептид Н-2K мыши) и/или функциональные эндогенные не относящиеся к человеку (например, мышиные) полипептиды β2 микроглобулина из их соответствующих эндогенных локусов. Согласно одному аспекту животное (например, мышь) содержит две копии каждой из первой и второй нуклеотидной последовательности. Согласно другому аспекту животное (например, мышь) содержит одну копию первой и одну копию второй нуклеотидных последовательностей. Таким образом, животное может являться гомозиготным или гетерозиготным в отношении как первой, так и второй нуклеотидных последовательностей.
Согласно одному аспекту первая нуклеотидная последовательность функционально связана с эндогенными не относящимися к человеку (например, мышиными) регуляторными элементами МНС I, и вторая нуклеотидная последовательность функционально связана с эндогенными не относящимися к человеку (например, мышиными) элементами β2 микроглобулина. Человеческая часть химерного полипептида может содержать α1, α2 и α3 домены полипептида MHC I человека. Вторая нуклеотидная последовательность может содержать нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека. Альтернативно, вторая нуклеотидная последовательность может содержать нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 гена β2 микроглобулина человека. Согласно одному аспекту мышь, содержащая как химерный полипептид МНС I, так и человеческий или гуманизированный полипептид β2 микроглобулина, может быть такой, чтобы экспрессия человеческого или гуманизированного β2 микроглобулина увеличивала экспрессию химерного человеческого/мышиного полипептида МНС I по сравнению с экспрессией химерного человеческого/мышиного полипептида МНС I при отсутствии экспрессии человеческого или гуманизированного полипептида β2 микроглобулина.
Также предусмотрены способы получения описанных в настоящем документе генетически сконструированных не относящихся к человеку животных (например, грызунов, например, мышей или крыс). Таким образом, согласно одному варианту осуществления предусмотрен способ модификации локуса МНС I грызуна (например, мыши или крысы) для экспрессии химерного человеческого/относящегося к грызуну (например, человеческого/мышиного или человеческого/крысиного) полипептида МНС I, причем способ предусматривает замещение на эндогенном локусе МНС I нуклеотидной последовательности, кодирующей внеклеточный домен полипептида МНС I грызуна, нуклеотидной последовательностью, кодирующей внеклеточный домен полипептида МНС I человека. Согласно другому варианту осуществления предусмотрен способ модификации локуса β2 микроглобулина грызуна (например, мыши или крысы) для экспрессии человеческого или гуманизированного полипептида β2 микроглобулина, причем способ предусматривает замещение на эндогенном локусе β2 микроглобулина грызуна (например, мыши или крысы) нуклеотидной последовательности, кодирующей полипептид β2 микроглобулина грызуна (например, мыши или крысы), нуклеотидной последовательностью, кодирующей человеческий или гуманизированный полипептид β2 микроглобулина. Согласно таким способам замещение может быть проведено в одной ES (эмбриональной стволовой) клетке, и одна ES клетка может быть введена грызуну (например, мыши или крысе) для получения зародыша. Полученный грызун (например, мышь или крыса) может подвергаться скрещиванию для получения дважды гуманизированного животного.
Таким образом, настоящее изобретение также относится к способам получения дважды гуманизированных животных, например, грызунов (например, мышей или крыс).
Согласно одному варианту осуществления предусмотрен способ получения генетически модифицированной мыши, предусматривающий (а) модификацию локуса МНС I первой мыши для экспрессии химерного человеческого/мышиного полипептида МНС I, предусматривающую замещение на эндогенном локусе МНС I мыши нуклеотидной последовательности, кодирующей внеклеточный домен полипептида МНС I мыши, нуклеотидной последовательностью, кодирующей внеклеточный домен полипептида МНС I человека, (b) модификацию локуса β2 микроглобулина второй мыши для экспрессии человеческого или гуманизированного полипептида β2 микроглобулина, предусматривающую замещение на эндогенном локусе β2 микроглобулина мыши нуклеотидной последовательности, кодирующей полипептид β2 микроглобулина мыши, нуклеотидной последовательностью, кодирующей человеческий или гуманизированный полипептид β2 микроглобулина; и (с) скрещивание первой и второй мыши для получения генетически модифицированной мыши, содержащей в своем геноме первую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид МНС I, и вторую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина, причем генетически модифицированная мышь экспрессирует химерный человеческий/мышиный полипептид МНС I и человеческий или гуманизированный полипептид β2 микроглобулина. Согласно некоторым вариантам осуществления локус МНС I выбран из Н-2K, H-2D и H-2L; согласно некоторым вариантам осуществления полипептид МНС I человека выбран из HLA-A, HLA-B и HLA-C. Согласно одному варианту осуществления локус МНС I представляет собой локус Н-2K, полипептид МНС I человека представляет собой HLA-A (например, HLA-A2), и мышь экспрессирует химерный полипептид HLA-А/Н-2K (например, полипептид HLA-A2/H-2K). Согласно одному аспекту химерный полипептид HLA-A2/H-2K содержит внеклеточный домен полипептида HLA-A2 и цитоплазматический и трансмембранный домены полипептида Н-2K. Согласно одному аспекту вторая нуклеотидная последовательность содержит нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 (например, экзоне 2 - экзоне 4) гена β2 микроглобулина человека, и нуклеотидную последовательность, представленную в экзоне 1 гена β2 микроглобулина мыши.
Также в настоящем документе предусмотрены клетки, например, выделенные антигенпрезентирующие клетки, полученные от описанных в настоящем документе не относящихся к человеку животных (например, грызунов, например, мышей или крыс). Также предусмотрены ткани и зародыши, полученные от описанных в настоящем документе не относящихся к человеку животных.
Согласно другому варианту осуществления настоящее изобретение относится к способам идентификации антигенов или антигенных эпитопов, которые вызывают иммунный ответ, способам оценки кандидатной вакцины, способам идентификации высокоаффинных Т-клеток к патогенам человека или антигенам злокачественных опухолей.
Любой из описанных в настоящем документе вариантов осуществления и аспектов может использоваться совместно друг с другом, если иное не указано или не очевидно из контекста. Другие варианты осуществления станут очевидными специалистам в настоящей области техники из обзора последующего подробного раскрытия. Последующее подробное раскрытие включает в себя иллюстративные представления различных вариантов осуществления настоящего изобретения, которые не ограничивают заявленное настоящее изобретение. Прилагаемые фигуры составляют часть настоящего описания изобретения и вместе с описанием служат исключительно для иллюстрации вариантов осуществления, а не для ограничения настоящего изобретения.
Краткое описание графических материалов
На фиг. 1 представлено схематическое изображение четырех доменов молекулы MHC I класса: α цепь, содержащая α1, α2 и α3 домены и нековалентно связанный четвертый домен, β2 микроглобулин (β2m). Серый кружок представляет пептид, связанный в пептидсвязывающей бороздке.
На фиг. 2 представлено схематическое представление (без соблюдения масштаба) относительной геномной структуры HLA человека, показывающее гены I, II и III класса.
На фиг. 3 представлено схематическое представление (без соблюдения масштаба) относительной геномной структуры МНС мыши, показывающее гены I, II и III класса.
На фиг. 4 проиллюстрирована вирусная векторная конструкция, содержащая кДНК, кодирующую химерный полипептид HLA-A/H-2K с репортером IRES-GFP (А); и гистограммы, на которых сравнивают экспрессию HLA-A2 человека в MG87 клетках, трансдуцированных с HLA-A2 (пунктирная линия), HLA-A2/H-2K (точечная линия) или без трансдукции (сплошная линия) или отдельно (слева), или котрансдуцированных с гуманизированным β2 микроглобулином (справа) (В). Данные из горизонтальных интервалов, представленных графически в (В), показаны как процент клеток, экспрессирующих конструкцию в таблице в (С).
На фиг. 5 представляет собой схематическую диаграмму (без соблюдения масштаба) стратегии нацеливания, используемой для получения химерного локуса Н-2K, который экспрессирует внеклеточную область белка HLA-A2 человека.
Последовательности мыши представлены черным цветом, а последовательности человека представлены белым цветом. L = лидерная последовательность, UTR = нетранслируемая область, ТМ = трансмембранный домен, CYT = цитоплазматический домен, HYG = гигромицин.
На фиг. 6А показана экспрессия (% от числа всех клеток) HLA-A2 (слева) и Н-2K (справа) в клетках, выделенных либо из мыши дикого типа (WT), либо из гетерозиготной (НЕТ) мыши, несущей химерный локус HLA-A2/H-2K (HLA-A/H-2K НЕТ).
На фиг. 6 В представляет собой точечную диаграмму экспрессии in vivo химерного белка HLA-A2/H-2K у гетерозиготной мыши, несущей химерный белок HLA-A2/H-2K.
На фиг. 7 показана стратегия нацеленного воздействия (без соблюдения масштаба) для гуманизации гена β2 микроглобулина на локусе β2 микроглобулина мыши. Последовательности мыши представлены черным цветом, а последовательности человека представлены белым цветом. NEO = неомицин.
На фиг. 8 показана репрезентативная точечная диаграмма экспрессии HLA I класса и β2 микроглобулина человека в клетках, выделенных из крови мышей дикого типа (WT), мышей, гетерозиготных в отношении химерного HLA-A2/H-2K, и мышей, гетерозиготных в отношении химерного HLA-A2/H-2K и гетерозиготных в отношении гуманизированного β2 микроглобулина (двойные гетерозиготы; НЕТ в отношении I класса /β2m).
На фиг. 9 показана репрезентативная гистограмма экспрессии HLA I класса человека (Х-ось) в клетках, выделенных из крови мышей дикого типа (WT), мышей, гетерозиготных в отношении химерного HLA-A2/H-2K (НЕТ I класса) и двойных гетерозиготных в отношении химерного HLA-А2/H2K/гуманизированного β2 микроглобулина (НЕТ I класса/β2m).
На фиг. 10 показаны результаты анализа методом иммуноферментных пятен IFNγ Т-клеток человека, подвергнутых воздействию антигенпрезентирующих клеток (АПК) от мышей дикого типа (АПК WT) или мышей, гетерозиготных в отношении как химерного HLA-A2/H-2K, так и гуманизированного β2 микроглобулина (двойные НЕТ АПК) в присутствии пептидов гриппа (слева) или ЭБВ (вируса Эпштейна-Барра) (справа). Статистический анализ проводили с использованием однофакторного дисперсионного анализа с апостериорным критерием множественного сравнения Тьюки.
Подробное раскрытие настоящего изобретения
Определения
Настоящее изобретение относится к генетически модифицированным не относящимся к человеку животным (например, мышам, крысам, кроликам и т.д.), которые экспрессируют человеческие или гуманизированные полипептиды МНС I и/или β2 микроглобулина; зародышам, клеткам и тканям, их содержащим; способам их получения; а также способам их применения. Если не указано иное, все используемые в настоящем документе термины и фразы включают в себя значения, которые подразумеваются под терминами и фразами в настоящей области техники, если противоположное ясно не указано или ясно не следует из контекста, в котором используется термин или фраза.
Термин "консервативная", используемый для описания консервативной аминокислотной замены, включает в себя замену аминокислотного остатка другим аминокислотным остатком с R-группой боковой цепи со сходными химическими свойствами (например, зарядом или гидрофобностью). Консервативные аминокислотные замены могут быть достигнуты путем модификации нуклеотидной последовательности так, чтобы ввести изменение нуклеотида, которое будет кодировать консервативную замену. Как правило, консервативная аминокислотная замена не будет существенно изменять представляющие интерес функциональные свойства белка, например, способность МНС I к презентации представляющего интерес пептида. Примеры групп аминокислот, которые содержат боковые цепи со сходными химическими свойствами, включают в себя такие алифатические боковые цепи глицин, аланин, валин, лейцин и изолейцин; такие алифатические гидроксильные боковые цепи серин и треонин; такие амидсодержащие боковые цепи, как аспарагин и глутамин; такие ароматические боковые цепи, как фенилаланин, тирозин и триптофан; такие основные боковые цепи, как лизин, аргинин и гистидин; такие кислотные боковые цепи, как аспарагиновая кислота и глутаминовая кислота; и такие серосодержащие боковые цепи, как цистеин и метионин. Группы консервативных аминокислотных замен включают в себя, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат, и аспарагин/глутамин. Согласно некоторым вариантам осуществления консервативная аминокислотная замена может представлять собой замену любого нативного остатка в белке на аланин, что используется, например, в сканирующем аланином мутагенезе. Согласно некоторым вариантам осуществления проводят консервативную замену, которая характеризуется положительным значением в матрице логарифмического правдоподобия РАМ250, раскрытой в Gonnet et al. ((1992) Exhaustive Matching of the Entire Protein Sequence Database, Science 256: 1443-45), включенной в настоящий документ посредством ссылки. Согласно некоторым вариантам осуществления представляет собой умеренно консервативную замену, причем замена характеризуется неотрицательным значением в матрице логарифмического правдоподобия РАМ250.
Таким образом, также настоящим изобретением предусмотрено генетически модифицированное не относящееся к человеку животное, чей геном содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид МНС I и/или полипептид β2 микроглобулина, причем полипептид(ы) содержит(ат) консервативные аминокислотные замены описанной(ых) в настоящем документе аминокислотной(ых) последовательности(ей).
Специалисту в настоящей области техники понятно, что в дополнение к остаткам нуклеиновой кислоты, кодирующим описанный в настоящем документе человеческий или гуманизированный полипептид МНС I и/или β2 микроглобулина, вследствие вырожденности генетического кода, другие нуклеиновые кислоты могут кодировать полипептид(ы) по настоящему изобретению. Следовательно, в дополнение к генетически модифицированному не относящемуся к человеку животному, которое содержит в своем геноме нуклеотидную последовательность, кодирующую полипептид(ы) МНС I и/или β2 микроглобулина с консервативными аминокислотными заменами, также предусмотрено не относящееся к человеку животное, чей геном содержит нуклеотидную(ые) последовательность(и), которая(ые) отличает(ют)ся от описанной(ых) в настоящем документе вследствие вырожденности генетического кода.
Термин "идентичность" при использовании в связи с последовательностью включает в себя идентичность, определяемую с помощью набора различных известных в настоящей области техники алгоритмов, которые могут использоваться для измерения идентичности нуклеотидной и/или аминокислотной последовательности. Согласно некоторым описанным в настоящем документе вариантам осуществления идентичности определяют с использованием ClustalW v. 1.83 (медленного) выравнивания с использованием штрафа за открытие делеции, составляющего 10,0, штрафа за продление делеции, составляющего 0,1, и с использованием матрицы сравнения согласно Gonnet (Mac Vector™ 10.0.2, Mac Vector Inc., 2008). Длина последовательностей, сравниваемых в отношении идентичности последовательностей, будет зависеть от конкретных последовательностей. Согласно различным вариантам осуществления идентичность определяют путем сравнения последовательности зрелого белка в направлении от его N-конца к его С-концу. Согласно различным вариантам осуществления при сравнении химерной человеческой/не относящейся к человеку последовательности с последовательностью человека, человеческая часть химерной человеческой/не относящейся к человеку последовательности (но не часть, не относящаяся к человеку) используется в осуществлении сравнения с целью выяснения уровня идентичности между последовательностью человека и человеческой части химерной человеческой/не относящейся к человеку последовательности (например, сравнивая эктодомен человека химерного человеческого/мышиного белка с эктодоменом человека из белка человека).
Термины "гомология" или "гомологичный" в отношении последовательностей, например, нуклеотидных или аминокислотных последовательностей, означает две последовательности, которые при оптимальном выравнивании и сравнении являются идентичными по меньшей мере приблизительно в 75% нуклеотидов или аминокислот, по меньшей мере приблизительно в 80% нуклеотидов или аминокислот, по меньшей мере приблизительно в 90-95% нуклеотидов или аминокислот, например, больше чем в 97% нуклеотидов или аминокислот. Специалисту в настоящей области техники будет понятно, что для оптимального нацеленного воздействия на ген нацеливающая конструкция должна содержать плечи, гомологичные эндогенным последовательностям ДНК (т.е., "плечи гомологии"); таким образом, может происходить гомологичная рекомбинация между нацеливающей конструкцией и нацеленной эндогенной последовательностью.
Термин "функционально связанный" относится к смежному положению, причем описанные таким образом компоненты находятся во взаимодействии, позволяющем им функционировать предусмотренным для них образом. В связи с этим, кодирующая белок последовательность нуклеиновой кислоты может быть функционально связана с регуляторными последовательностями (например, последовательностью промотора, энхансера, сайленсера и т.д.) так, чтобы сохранять надлежащую транскрипционную регуляцию. Кроме того, различные части химерного или гуманизированного белка по настоящему изобретению могут быть функционально связаны для сохранения надлежащей укладки, процессинга, нацеленного воздействия, экспрессии и других функциональных свойств белка в клетке. Если не указано иное, различные домены химерного или гуманизированного белка по настоящему изобретению функционально связаны друг с другом.
Используемый в настоящем документе термин "комплекс МНС I" или подобное включает в себя комплекс между полипептидом α цепи МНС I и полипептидом β2 микроглобулина. Используемый в настоящем документе термин "полипептид МНС I" или подобное включает в себя полипептид α цепи МНС I отдельно. Как правило, термины "МНС человека" и "HLA" могут использоваться взаимозаменяемо.
Термин "замещение" в отношении замещения гена относится к размещению экзогенного генетического материала на эндогенном генетическом локусе, тем самым замещая весь эндогенный ген или его часть ортологичной или гомологичной последовательностью нуклеиновой кислоты. Как показано в примерах ниже, последовательности нуклеиновой кислоты эндогенных локусов, кодирующих части полипептидов MHC I и β2 микроглобулина мыши замещали нуклеотидными последовательностями, кодирующими части полипептиды МНС I и β2 микроглобулина человека, соответственно.
Используемый в настоящем документе термин "функциональный", например, по отношению к функциональному полипептиду, относится к полипептиду, который сохраняет по меньшей мере одну биологическую активность, в норме связанную с нативным белком. Например, согласно некоторым вариантам осуществления настоящего изобретения замещение на эндогенном локусе (например, замещение на эндогенном не относящемся к человеку локусе МНС I и/или β2 микроглобулин) дает в результате локус, который не способен экспрессировать функциональный эндогенный полипептид.
Некоторые аспекты, описанные в настоящем документе ниже для генетически модифицированных в отношении МНС I не относящихся к человеку животных, например, тип животного; линии животного; типы клеток; способы скрининга, обнаружения и другие способы; способы применения и т.д., будут применимыми к генетически сконструированным в отношении β2 микроглобулина и в отношении МНС I/β2 микроглобулина животным.
Генетически модифицированные в отношении МНС I животные
Согласно различным вариантам осуществления настоящее изобретение в основном относится к генетически модифицированным не относящимся к человеку животным, которые содержат в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид МНС I; таким образом, животные экспрессируют человеческий или гуманизированный полипептид МНС I.
Гены МНС относятся к трем классам: I классу, II классу и III классу, причем все указанные классы кодируются либо на 6 хромосоме человека, либо на 17 хромосоме мыши. Схематическая иллюстрация относительной организации классов МНС человека и мыши представлена на фигурах 2 и 3, соответственно. Гены МНС находятся среди наиболее полиморфных генов геномов мыши и человека. Предполагают, что полиморфизмы МНС являются важными в обеспечении эволюционного преимущества; изменения в последовательности могут привести к различиям в связывании пептида, что обеспечивает возможность лучшей презентации патогенов цитотоксическим Т-клеткам.
Белок МНС I класса содержит внеклеточный домен (который содержит три домена: α1, α2 и α3), трансмембранный домен и цитоплазматический хвост, α1 и α2 домены образуют пептидсвязывающую бороздку, тогда как аз взаимодействует с β2 микроглобулином.
Кроме своего взаимодействия с β2 микроглобулином α3 домен взаимодействует с корецептором TCR CD8, облегчая антигенспецифическую активацию. Несмотря на то, что связывание МНС I класса с CD8 приблизительно в 100 раз слабее, чем связывание TCR с МНС I класса, связывание CD8 усиливает аффинность связывания TCR. Wooldridge et al. (2010) МНС Class I Molecules with Superenhanced CD8 Binding Properties Bypass the Requirement for Cognate TCR Recognition and Nonspecifically Activate CTLs, J. Immunol. 184: 3357-3366. Интересно, что увеличение связывания МНС I класса с CD8 отменяет антигенную специфичность в активации CTL (там же).
Связывание CD8 с молекулами МНС I класса является видоспецифическим; показали, что мышиный гомолог CD8, Lyt-2, связывает молекулы H-2Dd на α3 домене, но не связывает молекулы HLA-A. Connolly et al. (1988) The Lyt-2 Molecule Recognizes Residues in the Class I α3 Domain in Allogeneic Cytotoxic Т Cell Responses, J. Exp. Med. 168: 325-341. Предполагают, что различное связывание обусловлено CDR-подобными детерминантами (CDR1- и CDR2-подобными) на CD8, которые не являются консервативными и для людей и мышей. Sanders et al. (1991) Mutations in CD8 that Affect Interactions with HLA Class I and Monoclonal Anti-CD8 Antibodies, J. Exp. Med. 174: 371-379; Vitiello et al. (1991) Analysis of the HLA-restricted Influenza-specific Cytotoxic Т Lymphocyte Response in Transgenic Mice Carrying a Chimeric Human-Mouse Class I Major Histocompatibility Complex, J. Exp. Med. 173: 1007-1015; and, Gao et al. (1997) Crystal structure of the complex between human CD8αα and HLA-A2, Nature 387: 630-634. Сообщалось, что CD8 связывает HLA-A2 в консервативной области α3 домена (в положении 223-229). Одна замена (V245A) в HLA-A снижала связывание CD8 с HLA-A с сопутствующим сильным снижением опосредованного Т-клетками лизиса. Salter et al. (1989), Polymorphism in the α3 domain of HLA-A molecules affects binding to CD8, Nature 338: 345-348. В общем, полиморфизм в α3 домене молекул HLA-A также оказывал влияние на связывание с CD8 (там же). У мышей аминокислотная замена на остатке 227 в H-2Dd оказывала воздействие на связывание Lyt-2 мыши с H-2Dd, и клетки, трансфектированные с мутантным H-2D4, не лизировались CD8+ Т-клетками. Potter et al. (1989) Substitution at residue 227 of H-2 class I molecules abrogates recognition by CD8-dependent, but not CD8-independent, cytotoxic Т lymphocytes. Nature 337: 73-75.
Следовательно, вследствие видоспецифичности взаимодействия между α3 доменом МНС I класса и CD8, комплекс МНС I, содержащий замещение α3 домена Н-2K на α3 домене HLA-A2 человека, был нефункциональным у мыши (т.е., in vivo) при отсутствии CD8 человека. У животных, трансгенных в отношении HLA-A2, замена α3 домена человека на α3 домен мыши приводила к восстановлению Т-клеточного ответа. Irwin et al. (1989) Species-restricted interactions between CDS and the α3 domain of class I influence the magnitude of the xenogeneic response, J. Exp. Med. 170: 1091-1101; Vitiello et al. (1991), ранее.
Трансмембранный домен и цитоплазматический хвост белков МНС I класса мыши также характеризуются важными функциями. Одной функцией трансмембранного домена МНС I является облегчение модуляции с помощью HLA-A2 гомотипической клеточной адгезии (для усиления или ингибирования адгезии), предположительно, в результате перекрестного сшивания (или лигирования) поверхностных молекул МНС. Wagner et al. (1994) Ligation of МНС Class I and Class II Molecules Can Lead to Heterologous Desensitization of Signal Transduction Pathways That Regulate Homotypic Adhesion in Human Lymphocytes, J. Immunol. 152: 5275-5287. На клеточную адгезию могут оказывать воздействие моноклональные антитела, которые связываются с различными эпитопами молекулы HLA-A2, что позволяет предположить, что существует множественные сайты на HLA-A2, вовлеченные в модуляцию гомотипической клеточной адгезии; в зависимости от связанного эпитопа воздействие может усиливать или ингибировать зависимую от HLA-A2 адгезию (там же).
Сообщалось, что цитоплазматический хвост, кодируемый экзонами 6 и 7 гена МНС I, необходим для надлежащей экспрессии на клеточной поверхности и для опосредованного LIR1 ингибирования NK-клеточной цитотоксичности. Gruda et al. (2007) Intracellular Cysteine Residues in the Tail of МНС Class I Proteins Are Crucial for Extracellular Recognition by Leukocyte Ig-Like Receptor 1, J. Immunol. 179: 3655-3661. Цитоплазматический хвост необходим для мультимеризации по меньшей мере некоторых молекул МНС I посредством образования дисульфидных связей на его цистеиновых остатках и, таким образом, может играть роль в кластеризации и в распознавании NK-клетками. Lynch et al. (2009) Novel МНС Class I Structures on Exosomes, J. Immunol. 183: 1884-1891.
Цитоплазматический домен HLA-A2 содержит конститутивно фосфорилированный остаток серина и фосфорилируемый тирозин, хотя - в Т-клетках линии Jurkat - мутантные молекулы HLA-A2, не содержащие цитоплазматический домен, оказались нормальными в отношении экспрессии, ассоциации цитоскелета, агрегации и эндоцитозной интернализации. Gur et al. (1997) Structural Analysis of Class I МНС Molecules: The Cytoplasmic Domain Is Not Required for Cytoskeletal Association, Aggregation, and Intemalization, Mol. Immunol. 34(2): 125-132. По-видимому, процессированные молекулы HLA-A2, не содержащие цитоплазматический домен, в большинстве случаев экспрессировались и ассоциировались с β2 микроглобулином (там же).
Тем не менее, некоторые исследования показали, что цитоплазматический хвост является критически важным для внутриклеточного транспорта, опосредованной дендритными клетками (DC) презентации антигена и примирования CTL. Показали, что остаток тирозина, кодируемый экзоном 6, необходим для транспорта МНС I через эндосомальные компартменты, презентации экзогенных антигенов и примирования CTL;
тогда как делеция экзона 7 вызывала усиление антивирусных ответов CTL. Lizee et al. (2003) Control of Dendritic Cross-Presentation by the Major Histocompatibility Complex Class I Cytoplasmic Domain, Nature Immunol. 4: 1065-73; Basha et al. (2008) МНС Class I Endosomal and Lysosomal Trafficking Coincides with Exogenous Antigen Loading in Dendritic Cells, PLoS ONE 3: e3247; and Rodriguez-Cruz et al. (2011) Natural Splice Variant of МНС Class I Cytoplasmic Tail Enhances Dendritic Cell-Induced CD8+ T-Cell Responses and Boosts Anti-Tumor Immunity, PLoS ONE 6:e22939.
Согласно различным вариантам осуществления настоящее изобретение относится к генетически модифицированному не относящемуся к человеку животному (например, мыши, крысе, кролику и т.д.), которое содержит в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид МНС I класса. Не относящееся к человеку животное может содержать в своем геноме нуклеотидную последовательность, которая кодирует полипептид МНС I, который является частично человеческим и частично не относящимся к человеку, например, не относящееся к человеку животное, которое экспрессирует химерный человеческий/не относящийся к человеку полипептид МНС I. Согласно одному аспекту не относящееся к человеку животное экспрессирует только человеческий или гуманизированный полипептид МНС I, например, химерный человеческий/не относящийся к человеку полипептид МНС I, и не экспрессирует эндогенный не относящийся к человеку белок МНС I из эндогенного локуса МНС I.
Согласно одному варианту осуществления химерный человеческий/не относящийся к человеку полипептид МНС I содержит в своей человеческой части пептидсвязывающий домен полипептида МНС I человека. Согласно одному аспекту человеческая часть химерного полипептида содержит внеклеточный домен МНС I человека. Согласно настоящему варианту осуществления человеческая часть химерного полипептида содержит внеклеточный домен α цепи МНС I человека. Согласно одному варианту осуществления человеческая часть химерного полипептида содержит α1 и α2 домены МНС I человека. Согласно другому варианту осуществления человеческая часть химерного полипептида содержит α1, α2 и α3 домены МНС I человека.
Человеческий или гуманизированный полипептид МНС I может происходить из функциональной молекулы HLA человека, кодируемой любым из локусов HLA-A, HLA-В, HLA-C, HLA-E, HLA-F или HLA-G. Список широко распространенных антигенов HLA описан в Shankarkumar et al. ((2004) The Human Leukocyte Antigen (HLA) System, Int. J. Hum. Genet. 4(2): 91-103), включенной в настоящий документ посредством ссылки. Shankarkumar с соавт. также представляют краткое объяснение используемой в настоящей области техники номенклатуры HLA. Дополнительную информацию относительно номенклатуры HLA и различных аллелей HLA можно найти в Holdsworth et al. (2009) The HLA dictionary 2008: a summary of HLA-A, -B, -C, -DRB 1/3/4/5, and DQB1 alleles and their association with serologically defined HLA-A, -B, -C, -DR, and -DQ antigens. Tissue Antigens 73: 95-170, и в недавно обновленной редакции Marsh et al. (2010) Nomenclature for factors of the HLA system, 2010, Tissue Antigens 75: 291-455, включенных в настоящий документ посредством ссылки. Таким образом, человеческий или гуманизированный полипептид МНС I может происходить из любых описанных в настоящем документе функциональных молекул HLA I класса человека.
Согласно одному конкретному аспекту человеческий или гуманизированный полипептид МНС I происходит из HLA-A человека. Согласно конкретному варианту осуществления полипептид HLA-A представляет собой полипептид HLA-A2 (например, полипептид HLA-A2.1). Согласно одному варианту осуществления полипептид HLA-A представляет собой полипептид, кодируемый аллелем HLA-A*0201, например, аллелем HLA-A*02:01:01:01. Аллель HLA-A*0201 является широко распространенным среди североамериканской популяции. Несмотря на то, что настоящие примеры описывают эту конкретную последовательность HLA, в настоящем документе предусмотрена любая подходящая последовательность HLA-A, например, полиморфные варианты HLA-A2, встречающиеся в человеческой популяции, последовательности с одной или несколькими консервативными или неконсервативными аминокислотными модификациями, последовательности нуклеиновой кислоты, отличающиеся от описанной в настоящем документе последовательности вследствие вырожденности генетического кода и т.д.
Согласно одному аспекту предусмотрено не относящееся к человеку животное, которое экспрессирует последовательность HLA-A2 человека, причем последовательность HLA-A2 человека содержит одну или несколько консервативных или неконсервативных модификаций.
Согласно одному аспекту предусмотрено не относящееся к человеку животное, которое экспрессирует последовательность HLA-A2 человека, причем последовательность HLA-A2 человека по меньшей мере приблизительно на 85%, 90%, 95%, 96%, 97%, 98%, или 99% идентична последовательности HLA-A2 человека. Согласно конкретному варианту осуществления последовательность HLA-A2 человека по меньшей мере приблизительно на 90%, 95%, 96%, 97%, 98% или 99% идентична описанной в примерах последовательности HLA-A2 человека. Согласно одному варианту осуществления последовательность HLA-A2 человека содержит одну или несколько консервативных замен. Согласно одному варианту осуществления последовательность HLA-A2 человека содержит одну или несколько неконсервативных замен.
Согласно другому конкретному аспекту человеческий или гуманизированный полипептид МНС I происходит из МНС I человека, выбранного из HLA-B и HLA-C. Согласно одному аспекту человеческий или гуманизированный МНС I происходит из HLA-B, например, HLA-B27.
Согласно одному аспекту не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида МНС I содержит трансмембранный и/или цитоплазматический домены не относящегося к человеку полипептида МНС I. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь, и не относящийся к человеку полипептид МНС I выбран из Н-2K, H-2D и H-2L. Согласно одному варианту осуществления не относящийся к человеку полипептид МНС I представляет собой Н-2K, например, Н-2Kb. Несмотря на то, что в примерах описаны конкретные последовательности Н-2K, в настоящем документе предусмотрены любые подходящие последовательности Н-2K, например, полиморфные варианты, консервативные/неконсервативные аминокислотные замены и т.д.
Описанное в настоящем документе не относящееся к человеку животное может содержать в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид МНС I, например, химерный человеческий/не относящийся к человеку полипептид МНС I, причем кодирующая такой полипептид нуклеотидная последовательность расположена на эндогенном не относящемся к человеку локусе МНС I (например, локусе Н-2K). Согласно одному аспекту это приводит к замещению эндогенного гена МНС I или его части нуклеотидной последовательностью, кодирующей человеческий или гуманизированный полипептид МНС I, например, химерным геном, кодирующим химерный человеческий/не относящийся к человеку полипептид МНС I, описанный в настоящем документе. Согласно одному варианту осуществления замещение предусматривает замещение эндогенной нуклеотидной последовательности, кодирующей не относящийся к человеку пептидсвязывающий домен МНС I или не относящийся к человеку внеклеточный домен МНС I, нуклеотидной последовательностью человека (например, нуклеотидной последовательностью HLA-A2), кодирующей указанный домен. Согласно настоящему варианту осуществления замещение не предусматривает замещение последовательности МНС I, кодирующей трансмембранный и/или цитоплазматический домены не относящегося к человеку полипептида МНС I (например, полипептида Н-2K). Таким образом, не относящееся к человеку животное содержит химерную человеческую/не относящуюся к человеку нуклеотидную последовательность на эндогенном не относящемся к человеку локусе МНС I, и экспрессирует химерный человеческий/не относящийся к человеку полипептид МНС из эндогенного не относящегося к человеку локуса МНС I.
Химерный человеческий/не относящийся к человеку полипептид может быть таким, что он содержит человеческую или не относящуюся к человеку лидерную (сигнальную) последовательность. Согласно одному варианту осуществления химерный полипептид содержит не относящуюся к человеку лидерную последовательность эндогенного белка МНС I. Согласно другому варианту осуществления химерный полипептид содержит лидерную последовательность белка МНС I человека, например, белка HLA-A2 (например, лидерную последовательность HLA-A2.1). Таким образом, нуклеотидная последовательность, кодирующая химерный полипептид МНС I, может быть функционально связана с нуклеотидной последовательностью, кодирующей лидерную последовательность МНС I человека.
Химерный человеческий/не относящийся к человеку полипептид МНС I может содержать в своей человеческой части полный или по существу полный внеклеточный домен полипептида МНС I человека. Таким образом, человеческая часть может содержать по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, например, 95% или больше аминокислот, кодирующих внеклеточный домен полипептида МНС I человека (например, полипептида HLA-A2). Согласно одному примеру по существу полный внеклеточный домен полипептида МНС I человека не содержит лидерную последовательность МНС I человека. Согласно другому примеру химерный человеческий/не относящийся к человеку полипептид МНС I содержит лидерную последовательность МНС I человека.
Более того, химерный полипептид МНС I может экспрессироваться под контролем эндогенных не относящихся к человеку регуляторных элементов, например, регуляторных элементов МНС I грызуна. Такое расположение будет облегчать надлежащую экспрессию химерного полипептида МНС I у не относящегося к человеку животного, например, в ходе иммунного ответа у не относящегося к человеку животного.
Генетически модифицированное не относящееся к человеку животное может быть выбрано из группы, состоящей из мыши, крысы, кролика, свиньи, крупного рогатого скота (например, коровы, быка, буйвола), оленя, овцы, козы, курицы, кошки, собаки, хорька, примата (например, игрунки, макака-резуса). Для не относящихся к человеку животных, у которых подходящие генетически модифицируемые ES клетки не являются общедоступными, используются другие способы для получения не относящегося к человеку животного, содержащего генетическую модификацию. Такие способы включают в себя, например, модификацию генома не относящихся к ES клеток (например, фибробласта или индуцированной плюрипотентной клетки) и использование ядерного транспорта для переноса модифицированного генома в подходящую клетку, например, ооцит, и гестацию модифицированной клетки (например, модифицированного ооцита) в не относящемся к человеку животном при подходящих условиях для образования зародыша.
Согласно одному аспекту не относящееся к человеку животное представляет собой млекопитающее. Согласно одному аспекту не относящееся к человеку животное представляет собой небольшое млекопитающее, например, из надсемейства Dipodoidea или Muroidea. Согласно одному варианту осуществления генетически модифицированное животное представляет собой грызуна. Согласно одному варианту осуществления грызуна выбирают из мыши, крысы и хомяка. Согласно одному варианту осуществления грызуна выбирают из надсемейства Muroidea. Согласно одному варианту осуществления генетически модифицированное животное происходит из семейства, выбранного из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомячок, крысы и мыши Нового Света, полевки), Muridae (настоящие мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (рипидомисы, скалистые хомячки, белохвостые крысы, магадаскарские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). Согласно конкретному варианту осуществления генетически модифицированного грызуна выбирают из настоящей мыши или крысы (семейство Muridae), карликовой песчанки, иглистой мыши и косматого хомяка. Согласно одному варианту осуществления генетически модифицированную мышь получают из представителя семейства Muridae. Согласно одному варианту осуществления животное представляет собой грызуна. Согласно конкретному варианту осуществления грызуна выбирают из мыши и крысы. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь.
Согласно конкретному варианту осуществления не относящееся к человеку животное представляет собой грызуна, который представляет собой мышь линии C57BL, выбранной из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. Согласно другому варианту осуществления мышь представляет собой мышь линии 129, выбранной из группы, состоящей из линии 129Р1, 129Р2, 129РЗ, 129Х1, 129S1 (например, 129S1/SV, 129Sl/Svhn), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (смотрите, например, Festing et al. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10:836, смотрите также, Auerbach et al (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). Согласно конкретному варианту осуществления генетически модифицированная мышь представляет собой сочетание вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. Согласно другому конкретному варианту осуществления мышь представляет собой сочетание вышеупомянутых линий 129 или сочетание вышеупомянутых линий BL/6. Согласно конкретному варианту осуществления линия 129 сочетания представляет собой линию 129S6 (129/SvEvTac). Согласно другому варианту осуществления мышь представляет собой мышь линии BALB, например, линии BALB/c. Согласно другому варианту осуществления мышь представляет собой сочетание линии BALB и другой вышеупомянутой линии.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой крысу. Согласно одному варианту осуществления крысу выбирают из крысы линии Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. Согласно одному варианту осуществления линия крысы представляет собой сочетание двух или больше линий, выбранных из группы, состоящей из Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
Таким образом, согласно одному варианту осуществления настоящее изобретение относится к генетически модифицированной мыши, которая содержит в своем геноме нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид МНС I, причем человеческая часть химерного полипептида содержит пептидсвязывающий домен или внеклеточный домен MHC I человека (например, HLA-A человека, например, HLA-A2 человека, например, HLA-A2.1 человека). Согласно некоторым вариантам осуществления мышь не экспрессирует пептидсвязывающий или внеклеточный домен эндогенного полипептида мыши из его эндогенного локуса мыши. Пептидсвязывающий домен МНС I человека может содержать α1 и α2 домены. Альтернативно, пептидсвязывающий домен МНС I человека может содержать α1, α2 и α3 домены. Согласно одному аспекту внеклеточный домен МНС I человека содержит внеклеточный домен α цепи МНС I человека. Согласно одному варианту осуществления эндогенный локус мыши представляет собой локус Н-2K (например, Н-2Kb), и мышиная часть химерного полипептида содержит трансмембранный и цитоплазматический домены полипептида Н-2K (например, Н-2Kb) мыши.
Таким образом, согласно одному варианту осуществления предусмотрена генетически модифицированная мышь, причем мышь содержит на эндогенном локусе Н-2K (например, Н-2Kb) нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид МНС I, причем человеческая часть химерного полипептида содержит внеклеточный домен полипептида HLA-A2 человека (например, HLA-A2.1), и мышиная часть содержит трансмембранный и цитоплазматический домены полипептида Н-2K (например, Н-2Kb) мыши. Согласно одному аспекту мышь не экспрессирует внеклеточный домен полипептида Н-2K (например, Н-2Kb) мыши из эндогенного локуса МНС I. Согласно одному варианту осуществления мышь экспрессирует химерный полипептид HLA-A2/H-2K (например, химерный HLA-A2.1/Н-2Kb) из эндогенного локуса Н-2K (например, Н-2Kb). Согласно различным вариантам осуществления экспрессия химерного гена находится под контролем эндогенных регуляторных элементов МНС I класса мыши. Согласно некоторым аспектам мышь содержит две копии химерного локуса МНС I, содержащего нуклеотидную последовательность, кодирующую химерный полипептид HLA-A2/H-2K; тогда как согласно другим аспектам мышь содержит одну копию химерного локуса МНС I, содержащего нуклеотидную последовательность, кодирующую химерный полипептид HLA-A2/H-2K. Таким образом, мышь может являться гомозиготной или гетерозиготной в отношении нуклеотидной последовательности, кодирующей химерный полипептид HLA-A2/H-2K.
Согласно некоторым описанным в настоящем документе вариантам осуществления предусмотрена мышь, которая содержит химерный локус МНС I, расположенный на эндогенном локусе Н-2K мыши. Химерный локус содержит нуклеотидную последовательность, которая кодирует внеклеточный домен белка HLA-A2 человека, например, α1, α2 и α3 домены гена HLA-A2 человека. Химерный локус не содержит нуклеотидную последовательность, которая кодирует внеклеточный домен белка Н-2K мышь (например, α1, α2 и α3 домены Н-2K мыши). Согласно одному аспекту химерный локус не содержит нуклеотидную последовательность, которая кодирует лидерный пептид, α1, α2 и α3 домены Н-2K мыши; и содержит лидерный пептид, α1, α2 и α3 домены HLA-A2 человека, и трансмембранный и цитоплазматический домены Н-2K мыши.
Различные домены химерного локуса функционально связаны друг с другом так, чтобы химерный локус экспрессировал функциональный химерный человеческий/мышиный белок МНС I.
Согласно различным вариантам осуществления не относящееся к человеку животное, например, грызун (например, мышь или крыса), которое экспрессирует функциональный химерный белок МНС I из химерного локуса МНС I, описанного в настоящем документе, представляет химерный белок на клеточной поверхности. Согласно одному варианту осуществления не относящееся к человеку животное экспрессировало химерный белок МНС I на клеточной поверхности в клеточной локализации, которая аналогична наблюдаемой у человека. Согласно одному аспекту клетка представляет пептидный фрагмент (фрагмент антигена), связанный с внеклеточной частью (например, внеклеточной частью HLA-A2 человека) химерного белка МНС I. Согласно варианту осуществления внеклеточная часть такого химерного белка взаимодействует с другими белками на поверхности указанной клетки, например, β2 микроглобулина.
Согласно различным вариантам осуществления клетка, представляющая химерный белок МНС I, например, белок HLA-A2/H-2K, представляет собой ядросодержащую клетку. Согласно различным аспектам клетка представляет собой антигенпрезентирующую клетку (АПК). Хотя большинство клеток в организме могут презентировать антиген в контексте МНС I, некоторые не ограничивающие примеры антигенпрезентирующих клеток включают в себя макрофаги, дендритные клетки и В-клетки. Другие антигенпрезентирующие клетки, включающие в себя профессиональные и непрофессиональные АПК, известны в настоящей области техники и предусмотрены в настоящем документе. Согласно некоторым вариантам осуществления представляющая химерный белок МНС I клетка представляет собой опухолевую клетку, и пептидный фрагмент, презентируемый химерным белком, происходит из опухоли. Согласно другим вариантам осуществления пептидный фрагмент, презентируемый химерным белком МНС I, происходит из патогена, например, бактерии или вируса.
Описанный в настоящем документе химерный белок МНС I может взаимодействовать с другими белками на поверхности той же клетки или второй клетки. Согласно некоторым вариантам осуществления химерный белок МНС I взаимодействует с эндогенными не относящимися к человеку белками на поверхности указанной клетки. Химерный белок МНС I также может взаимодействовать с человеческими или гуманизированными белками на поверхности той же клетки или второй клетки.
На одной и той же клетке молекулы HLA класса I могут функционально взаимодействовать как с β2 микроглобулином, не относящимся к человеку (например, грызуна, например, мыши или крысы), так и с β2 микроглобулином человека. Таким образом, согласно одному варианту осуществления химерный белок MHC I, например, белок HLA-A2/H-2K, взаимодействует с β2 микроглобулином мыши. Хотя взаимодействие между некоторыми молекулами HLA класса I человека и β2 микроглобулином мыши является возможным, тем не менее, оно может сильно снижаться по сравнению с взаимодействием между HLA класса I человека и β2 микроглобулином человека. Таким образом, при отсутствии β2 микроглобулина человека может снижаться экспрессия MHC I человека на клеточной поверхности. Peramau et al. (1988) Human β2-microglobulin Specifically Enhances Cell-Surface Expression of HLA Class I Molecules in Transfected Murine Cells, J. Immunol. 141: 1383-89. Другие молекулы HLA, например, HLA-B27, не взаимодействуют с β2 микроглобулином мыши; смотрите, например, Tishon et al. (2000) Transgenic Mice Expressing Human HLA and CD8 Molecules Generate HLA-Restricted Measles Virus Cytotoxic Т Lymphocytes of the Same Specificity as Humans with Natural Measles Virus Infection, Virology 275: 286-293, в которой сообщается, что функция HLA-В27 у трансгенных мышей нуждается как в β2 микроглобулине человека, так и CD8 человека. Следовательно, согласно другому варианту осуществления химерный белок МНС I взаимодействует с человеческим или гуманизированным β2 микроглобулином. Согласно таким вариантам осуществления, которые описаны в настоящем документе ниже, не относящееся к человеку животное, например, грызун (например, мышь или крыса), содержит в своем геноме человеческий или гуманизированный ген β2 микроглобулина, и животное экспрессирует функциональный человеческий или гуманизированный полипептид β2 микроглобулина; следовательно, химерный белок МНС I взаимодействует с человеческим или гуманизированным полипептидом β2 микроглобулина.
Согласно различным аспектам химерный белок (например, белок HLA-A2/H-2K) также взаимодействует с белками на поверхности второй клетки (посредством своей внеклеточной части). Вторая клетка может представлять собой клетку, происходящую от не относящегося к человеку животного, например, мыши, или от человека. Вторая клетка может быть получена от того же не относящегося к человеку животного или того же вида не относящегося к человеку животного, что и экспрессирующая химерный полипептид МНС I клетка. Не ограничивающие примеры белков, с которыми может взаимодействовать внеклеточный часть химерного белка (например, HLA-A2/H-2K), включают в себя Т-клеточный рецептор (TCR) и его корецептор CD8. Таким образом, вторая клетка может представлять собой Т-клетку. Кроме того, внеклеточная часть химерного белка МНС I может связываться с белком на поверхности клеток естественных киллеров (NK), например, с иммуноглобулиноподобные рецепторы клеток-киллеров (KIR) на поверхности NK-клеток.
Т-клетка или NK-клетка может связывать комплекс, образованный между химерным полипептидом МНС I и его представленным пептидным фрагментом. Такое связывание может приводить к активации Т-клеток или ингибированию опосредованного NK-клетками цитолиза, соответственно. Согласно одной гипотезе NK-клетки вовлечены в цитолиз либо инфицированных, либо опухолевых клеток, которые избежали опосредованной Т-клетками цитотоксичности путем отрицательной регуляции их комплекса MHC I. Тем не менее, когда комплекс МНС I экспрессируется на клеточной поверхности, рецепторы NK-клетки распознают его, и опосредованный NK- клетками цитолиз ингибируется. Таким образом, согласно некоторым аспектам, когда NK-клетка связывается с комплексом, образованным между химерным полипептидом МНС I (например, полипептидом HLA-A2/H-2K) и представленным пептидным фрагментом на поверхности инфицированной или опухолевой клетки, опосредованный NK- клетками цитолиз ингибируется.
Согласно одному примеру описанный в настоящем документе химерный полипептид МНС I, например, химерный полипептид HLA-A2/H-2K, взаимодействует с белком CD8 на поверхности второй клетки. Согласно одному варианту осуществления химерный полипептид HLA-A2/H-2K взаимодействует с эндогенным белком CD8 грызуна (например, мыши или крысы) на поверхности второй клетки. Согласно одному варианту осуществления вторая клетка представляет собой Т-клетку. Согласно другому варианту осуществления вторая клетка сконструирована для экспрессии CD8. Согласно определенным аспектам химерный полипептид HLA-A2/H-2K взаимодействует с CD8 человека на поверхности второй клетки (например, клетки человека или клетки грызуна). Согласно некоторым таким вариантам осуществления не относящееся к человеку животное, например, мышь или крыса, содержит трансген CD8 человека, и мышь или крыса экспрессирует функциональный белок CD8 человека.
Описанный в настоящем документе химерный полипептид МНС I также может взаимодействовать с не относящимся к человеку (например, мышиным или крысиным) TCR, TCR человека или гуманизированным TCR на второй клетке. Химерный полипептид МНС I может взаимодействовать с эндогенным TCR (например, TCR мыши или крысы) на поверхности второй клетки. Химерный полипептид МНС I также может взаимодействовать с человеческим или гуманизированным TCR, экспрессированным на поверхности второй клетки, причем клетка получена от того же животного или того же вида животного (например, мыши или крысы), что и экспрессирующая химерный полипептид МНС I клетка. Химерный полипептид МНС I может взаимодействовать с TCR человек, экспрессированным на поверхности клетки человека.
В дополнение к генетически сконструированным не относящимся к человеку животным, также предусмотрен не относящийся к человеку зародыш (например, зародыш грызуна, например, зародыш мыши или крысы), причем зародыш содержит донорную ES клетку, которая получена от не относящегося к человеку животного (например, грызуна, например, мыши или крысы), описанного в настоящем документе. Согласно одному аспекту зародыш содержит донорную ES клетку, содержащую химерный ген МНС I и клетки зародыша-хозяина.
Также предусмотрена ткань, причем ткань получена от не относящегося к человеку животного (например, мыши или крысы), описанного в настоящем документе, и экспрессирует химерный полипептид МНС I (например, полипептид HLA-A2/H-2K).
Кроме того предусмотрена не относящаяся к человеку клетка, выделенная из описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления клетка представляет собой ES клетку. Согласно одному варианту осуществления клетка представляет собой антигенпрезентирующую клетку, например, дендритную клетку, макрофаг, В-клетку. Согласно одному варианту осуществления клетка представляет собой иммунную клетку. Согласно одному варианту осуществления иммунная клетка представляет собой лимфоцит.
Также предусмотрена не относящаяся к человеку клетка, содержащая хромосому или ее фрагмент описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит ядро описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит хромосому или ее фрагмент как результат ядерного транспорта.
Согласно одному аспекту предусмотрена не относящаяся к человеку индуцированная плюрипотентная клетка, содержащая ген, кодирующий химерный полипептид МНС I (например, полипептид HLA-A2/H-2K), описанный в настоящем документе. Согласно одному варианту осуществления индуцированная плюрипотентная клетка получена от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусмотрена гибридома или квадрома, происходящая из клетки описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь или крыса.
Также предусмотрен способ получения описанного в настоящем документе генетически сконструированного не относящегося к человеку животного (например, генетически сконструированного грызуна, например, мыши или крысы). Способ получения генетически сконструированного не относящегося к человеку животного дает в результате животного, геном которого содержит нуклеотидную последовательность, кодирующую химерный полипептид МНС I. Согласно одному варианту осуществления способ дает в результате генетически сконструированную мышь, геном которой содержит на эндогенном локусе МНС I, например, локусе Н-2K, нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид МНС I, причем человеческая часть химерного полипептида МНС I содержит внеклеточный домен HLA-А2 человека, и мышиная часть содержит трансмембранный и цитоплазматический домены Н-2K мыши. Согласно некоторым вариантам осуществления в способе используется нацеливающая конструкция, полученная с использованием технологии VELOCIGENE®, введение конструкции в ES клетки и введение нацеленных ES клеточный клонов к зародыш мыши с использованием описанной в примерах технологии VELOCIMOUSE®. Согласно одному варианту осуществления ES клетки представляют собой сочетание линий мышей 129 и C57BL/6; согласно другому варианту осуществления ES клетки представляют собой сочетание линий мышей BALB/c и 129.
Таким образом, также предусмотрена нуклеотидная конструкция, используемая для создания описанных в настоящем документе генетически сконструированных не относящихся к человеку животных. Согласно одному аспекту нуклеотидная конструкция содержит: 5' и 3' не относящиеся к человеку плечи гомологии, фрагмент ДНК человека, содержащий генные последовательности HLA-A человека, и кассету селекции, фланкированную сайтами рекомбинации. Согласно одному варианту осуществления фрагмент ДНК человека представляет собой геномный фрагмент, который содержит как интроны, так и экзоны гена HLA-A человека. Согласно одному варианту осуществления не относящиеся к человеку плечи гомологии гомологичны не относящемуся к человеку локусу МНС I класса (например, локусу Н- 2K мыши).
Согласно одному варианту осуществления геномный фрагмент содержит лидерную последовательность HLA-A человека, кодирующую α1 домен, α2 домен и α3 домен последовательность. Согласно одному варианту осуществления фрагмент ДНК человека содержит, в направлении 5'-3': лидерную последовательность HLA-A, интрон, лидерную последовательность HLA-A/α1, экзон HLA-A α1, интрон HLA-Aα1-α2, экзон HLA-A α2, интрон HLA-A α2-α3 и экзон HLA-A α3.
Кассета селекции представляет собой нуклеотидную последовательность, вставленную в нацеливающую конструкцию для облегчения селекции клеток (например, ES клеток), которые интегрировали представляющую интерес конструкцию. В настоящей области техники известен ряд подходящих кассет селекции. Как правило, кассета селекции обеспечивает положительную селекцию в присутствии конкретного антибиотика (например, Neo, Hyg, Pur, CM, Spec и т.д.). Кроме того, кассета селекции может быть фланкирована сайтами рекомбинации, которые обеспечивают делецию кассеты селекции при обработке ферментами рекомбиназами. Наиболее широко используемые сайты рекомбинации представляют собой loxP и Frt, распознаваемые ферментами Cre и Flp, соответственно, но в настоящей области техники известны и другие.
Согласно одному варианту осуществления кассета селекции расположена на 5'-конце фрагмента ДНК человека. Согласно другому варианту осуществления кассета селекции расположена на 3'-конце фрагмента ДНК человека. Согласно другому варианту осуществления кассета селекции расположена в пределах фрагмента ДНК человека. Согласно другому варианту осуществления кассета селекции расположена в пределах интрона фрагмента ДНК человека. Согласно другому варианту осуществления кассета селекции расположена в пределах интрона α2-α3.
Согласно одному варианту осуществления 5' и 3' не относящиеся к человеку плечи гомологии содержат геномную последовательность на 5' и 3' положениях эндогенного не относящегося к человеку (например, мышиного) генного локуса МНС I класса, соответственно (например, 5' по отношению к первой лидерной последовательности и 3' по отношению к экзону α3 не относящегося к человеку гена МНС I). Согласно одному варианту осуществления эндогенный локус МНС I класса выбран из Н-2K, H-2D и H-2L мыши. Согласно конкретному варианту осуществления эндогенный локус МНС I класса представляет собой Н-2K мыши.
Согласно одному аспекту предусмотрена нуклеотидная конструкция, содержащая, в направлении 5'-3': 5' плечо гомологии, содержащее геномную последовательность мыши 5' по отношению к эндогенному локусу Н-2K мыши, первый фрагмент ДНК человека, содержащий первую геномную последовательность гена HLA-A, 5' сайт последовательности рекомбинации (например, loxP), кассету селекции, 3' сайт последовательности рекомбинации (например, loxP), второй фрагмент ДНК человека, содержащий вторую геномную последовательность гена HLA-A, и 3' плечо гомологии, содержащее геномную последовательность мыши 3' по отношению к эндогенному экзону Н-2K α3. Согласно одному варианту осуществления нуклеотидная конструкция содержит, в направлении 5'-3': 5' плечо гомологии, содержащее геномную последовательность мыши 5' по отношению к эндогенному локусу Н-2K. мыши, геномную последовательность человека, включающую лидерную последовательность HLA-A, интрон, лидерную последовательность HLA-A/α1, экзон HLA-A α1, интрон HLA-A α1-α2, экзон HLA-A α2, первую 5' часть интрона α2-α3, кассету селекции, фланкированную сайтами рекомбинации, вторую 3' часть интрона α2-α3, экзон HLA-A α3 и 3' плечо гомологии, содержащее не относящуюся к мыши геномную последовательность 3' по отношению к эндогенному экзону Н-2K α3 мыши. Согласно одному варианту осуществления последовательность 5' плеча гомологии представлена в SEQ ID NO: 1, и последовательность 3' плеча гомологии представлена в SEQ ID NO: 2.
После завершения нацеленного воздействия на ген ES клетки или генетически модифицированных не относящихся к человеку животных подвергают скринингу для подтверждения успешного встраивания представляющей интерес экзогенной нуклеотидной последовательности или экспрессии экзогенного полипептида. Специалистам в настоящей области техники известны различные техники, и они включают в себя (без ограничения) саузерн-блоттинг, ПЦР длинных фрагментов, количественную ПЦР (например, ПЦР в реальном времени с использованием TAQMAN®), флуоресцентную гибридизацию in situ, нозерн-блоттинг, проточную цитометрию, вестерн-блоттинг, иммуноцитохимию, иммуногистохимию и т.д. Согласно одному примеру не относящиеся к человеку животные (например, мыши), несущие представляющую интерес генетическую модификацию, могут быть идентифицированы путем скрининга в отношении потери аллеля мыши и/или приобретения аллеля человека с использованием модификации аллельного анализа, описанного в Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nature Biotech. 21(6): 652-659. Специалистам в настоящей области техники известны другие анализы, которые идентифицируют конкретную нуклеотидную или аминокислотную последовательность у генетически модифицированных животных.
Настоящее раскрытие также относится к способу модификации локуса МНС I не относящегося к человеку животного для экспрессии описанного в настоящем документе химерного человеческого/не относящегося к человеку полипептида МНС I. Согласно одному варианту осуществления настоящее изобретение относится к способу модификации локуса МНС I мыши для экспрессии химерного человеческого/мышиного полипептида МНС I, причем способ предусматривает замещение на эндогенном локусе МНС I нуклеотидной последовательности, кодирующей пептидсвязывающий домен полипептида МНС мыши, нуклеотидной последовательностью, кодирующей пептидсвязывающий домен полипептида МНС I человека. Согласно некоторым аспектам нуклеотидную последовательность внеклеточного домена МНС I мыши замещают нуклеотидной последовательностью внеклеточного домена МНС I человека. Мышь может быть не способна экспрессировать пептидсвязывающий или внеклеточный домен МНС I мыши из эндогенного локуса МНС I. Согласно некоторым вариантам осуществления нуклеотидную последовательность внеклеточного домена Н-2K мыши замещают нуклеотидной последовательностью внеклеточного домена HLA-A2 человека так, чтобы модифицированный локус МНС I мыши экспрессировал химерный полипептид HLA-А2/Н-2K.
Согласно одному аспекту предусмотрен способ получения химерной человеческой HLA I класса/не относящейся к человеку молекулы МНС I класса, предусматривающий экспрессию в одной клетке химерного белка HLA-A/H-2K из нуклеотидной конструкции, причем нуклеотидная конструкция содержит последовательность кДНК, которая кодирует α1, α2 и α3 домен белка HLA-A и трансмембранный и цитоплазматический домен не относящегося к человеку белка Н-2K, например, белка Н-2K мыши. Согласно одному варианту осуществления нуклеотидная конструкция представляет собой вирусный вектор;
согласно конкретному варианту осуществления вирусный вектор представляет собой лентивирусный вектор. Согласно одному варианту осуществления клетку выбирают из СНО, COS, 293, HeLa и ретинальной клетки, экспрессирующей вирусную последовательность нуклеиновой кислоты (например, клетки PERC.6™).
Согласно одному аспекту предусмотрена клетка, которая экспрессирует химерный человеческий HLA I класса/не относящийся к человеку белок МНС I (например, белок HLA-A/H-2K). Согласно одному варианту осуществления клетка содержит вектор экспрессии, содержащий химерный ген МНС I класса, причем химерный ген МНС I класса содержит последовательность гена HLA-A человека, слитую в функциональной связи с последовательностью не относящегося к человеку гена Н-2K, например, гена Н-2K мыши. Согласно одному варианту осуществления последовательность гена HLA-A человека содержит экзоны, которые кодируют α1, α2 и α3 домены белка HLA-A. Согласно одному варианту осуществления последовательность не относящегося к человеку гена Н-2K содержит экзоны, которые кодируют трансмембранный и цитоплазматический домены белка Н-2K. Согласно одному варианту осуществления клетку выбирают из СНО, COS, 293, HeLa и ретинальной клетки, экспрессирующей вирусную последовательность нуклеиновой кислоты (например, клетки PERC.6™).
Также предусмотрена химерная молекула МНС I класса, произведенная описанным в настоящем документе не относящимся к человеку животным, причем химерная молекула МНС I класса содержит α1, α2, и α3 домены из белка HLA-A человека и трансмембранный и цитоплазматический домены из не относящегося к человеку, например, мышиного белка Н-2K. Описанный в настоящем документе химерный полипептид MHC I можно обнаружить с помощью антител к HLA-A. Таким образом, клетку, представляющую химерный человеческий/не относящийся к человеку полипептид MHC I, можно обнаружить и/или выбрать с использованием антитела к HLA-A. В некоторых случаях описанный в настоящем документе химерный полипептид МНС I можно обнаружить с помощью антитела к HLA-A2.
Несмотря на то, что представленные ниже примеры описывают генетически сконструированное животное, геном которого содержит замещение нуклеотидной последовательности, кодирующей внеклеточный домен полипептида Н-2K мыши, последовательностью, кодирующей внеклеточный домен HLA-A человека на эндогенном локусе Н-2K мыши, специалисту в настоящей области техники понятно, что аналогичная стратегия может использоваться для замещения других локусов МНС I мыши (H-2D, H-2L и т.д.) соответствующими им локусами HLA человека (HLA-B, HLA-C и т.д.). Таким образом, также предусмотрено не относящееся к человеку животное, содержащее в своем геноме нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, причем человеческая часть полипептида происходит из другого белка HLA класса I. Также предусмотрено замещение множественных локусов МНС I.
Генетически модифицированные в отношении β2 микроглобулина животные
Настоящее изобретение в основном относится к генетически модифицированным не относящимся к человеку животным, которые содержат в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина; таким образом, животные экспрессируют человеческий или гуманизированный полипептид β2 микроглобулина.
β2 микроглобулин или легкая цепь комплекса МНС I класса (также имеющая сокращение "β2М") представляет собой небольшой (12 кДа) негликозилированный белок, который функционирует преимущественно для стабилизации α цепи МНС I. Ген β2 микроглобулина человека кодирует белок из 119 аминокислот, с 20 N-концевыми аминокислотами, кодирующими лидерную последовательность. Зрелый белок содержит 99 аминокислот. Ген содержит 4 экзона, причем первый экзон содержит 5' нетранслируемую область, полную лидерную последовательность и первые две аминокислоты зрелого полипептида; второй экзон кодирует основную часть зрелого белка; третий экзон кодирует последние четыре аминокислоты зрелого белка и стоп-кодон; и четвертый экзон содержит 3' нетранслируемую область. Gussow et al. (1987) The β2-Microglobulin Gene. Primary Structure and Definition of the Transcriptional Unit, J. Immunol. 139: 3131-38. β2 микроглобулин нековалентно ассоциирован с МНС I. Несвязанный β2 микроглобулин обнаруживается в таких биологических жидкостях, как плазма, и переносится к почкам для выведения из организма. Нарушение функции почек вызывает накопление β2 микроглобулина, что может быть патогенным (например, вызывать диализный амилоидоз); накопленный белок образует нитевидные фибриллы, напоминающие амилоидные бляшки в суставах и соединительных тканях.
Кроме диализного амилоидоза β2 микроглобулин был вовлечен в ряд других нарушений. Повышенное содержание β2 микроглобулина обнаружили в лимфоцитарных злокачественных опухолях, например, неходжкинской лимфоме и множественной миеломе. Смотрите, например, Shi et al. (2009) β2 Microglobulin: Emerging as a Promising Cancer Therapeutic Target, Drug Discovery Today 14: 25-30. Некоторые другие злокачественные опухоли с повышенным содержанием β2 микроглобулина включают в себя рак молочной железы, рак предстательной железы, рак легких, рак почки, рак желудочно-кишечного тракта и рак носоглотки. Предположили, что избыточная экспрессия β2 микроглобулина характеризуется стимулирующими эффектами на рост опухоли (там же). Также недавно было показано, что β2 микроглобулин управляет превращением эпителиальных клеток в мезенхимальные, стимулируя образование метастазов в костях и мягких тканях при раке молочной железы, раке предстательной железы, раке легких и раке почки. Josson et al. (2011) β2 microglobulin Induces Epitelial to Mesenchymal Transition and Confers Cancer Lethality and Bone Metastasis in Human Cancer Cells. Cancer Res. 71(7): 1-11. β2 микроглобулин взаимодействует с неклассическим представителем МНС I, белком гемохроматоза (HFE), и с рецептором трансферрина, и модулирует гомеостаз железа (там же). Вовлечение β2 микроглобулина в другие признаки злокачественности (самообновление, усиление ангиогенеза, устойчивость к лечению) обширно подтверждено документально в настоящей области техники.
Сообщалось о мышах с недостаточностью в отношении β2 микроглобулина. Смотрите, Koller et al. (1990) Normal development of mice deficient in β2m, МНС class I proteins, and CD8+ Т cells. Science 248: 1227-1230. Как сообщают Koller с соавт., эти мыши оказались здоровыми, тем не менее, не была обнаружена экспрессия МНС I класса. Кроме того, большинство Т-клеточных популяций в некоторых тканях оказались нормальными, тогда как в других тканях наблюдалось значительной снижение количества CD8+ Т-клеток. Эта предполагаемое отсутствие экспрессии МНС I противоречит предыдущим результатам, полученным Alien et al. ((1986) β2 microglobulin Is Not Required for Cell Surface Expression of the Marine Class I Histocompatibility Antigen H-2Db or of a Truncated H-2D1', Proc. Natl. Acad. Sci. USA 83: 7447-7451). Alien с соавт. показали, что β2 микроглобулин не являлся безусловно необходимым для экспрессии всех комплексов МНС I на клеточной поверхности, поскольку клетки, не содержащие β2 микроглобулин, были способны экспрессировать H-2Db. Тем не менее, функция H-2Db в этих клетках была предположительно дефектной, и конформация H-2Db отличалась от нативного белка, что объясняет тот факт, что Koller и коллеги не смогли обнаружить этот белок с использованием антител к нативному H-2Db. Тем не менее сообщалось, что клетки, не содержащие β2 микроглобулин, могут презентировать эндогенный антиген CD8+ Т-клеткам (включающим в себя экзогенные CD8+ Т-клетки от нормальных мышей), и сообщалось, что β2 микроглобулин не является необходимым для выработки высокого содержания рестриктированных по Н-2d МНС I класса CD8+ CTL в ответ на антигенную стимуляцию у мышей, хотя он необходим для обеспечения эффективного иммунного ответа. Quinn et al. (1997) Virus-Specific, CD8+ Major Histocompatibility Complex Class I-Restricted Cytotoxic Т Lymphocytes in Lymphocytic Choriomeningitis Virus-Infected β2-Microglobulin-Deficient Mice, J. Virol. 71: 8392-8396. Следует отметить, что способность производить высокое содержание таких Т-клеток при отсутствии β2 микроглобулина, по имеющимся данным, ограничена рестриктированным по Н-2d МНС I класса ответом. Сообщалось, что мыши с недостаточностью в отношении β2 микроглобулина обладали такими выраженными характеристиками, как, например, увеличенная предрасположенность к некоторым паразитарным заболеваниям, увеличенная предрасположенность к гепатитным инфекциями, недостаточность метаболизма железа и нарушенный фенотип в отношении скрещивания. Cooper et al. (2007) An impaired breeding phenotype in mice with a genetic deletion of Beta-2 microglobulin and diminished МНС class I expression: Role in reproductive fitness, Biol. Reprod. 77: 274-279.
Сообщалось о мышах, которые экспрессируют β2 микроглобулин человека, а также молекулы HLA класс I человека (т.е., HLA-B7) в случайном образом вставленном трансгене. Chamberlain et al. (1988) Tissue-specific and cell surface expression of human major histocompatibility complex class I heavy (HLA-B7) and light (β2-microglobulin) chain genes in transgenic mice, Proc. Natl. Acad. Sci. USA 85: 7690-7694. Экспрессия HLA класса I человека была сопоставима с экспрессией эндогенного класса I со значительным снижением в печени (там же). Экспрессия β2 микроглобулина человека также была сопоставима с экспрессией эндогенного β2 микроглобулином, тогда как экспрессия молекулы HLA класса I человека увеличивалась в 10-17 раз у двойных трансгенных мышей (там же). Тем не менее, авторы не предприняли попыток заместить эндогенный локус β2 микроглобулина мыши локусом β2 микроглобулина человек.
Следовательно, в настоящем документе раскрыто генетически сконструированное не относящееся к человеку животное (например, грызун, например, мышь или крыса), геном которого содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному аспекту животное не экспрессирует эндогенный не относящийся к человеку β2 микроглобулин из эндогенного не относящегося к человеку локуса β2 микроглобулина. Согласно некоторым вариантам осуществления нуклеотидная последовательность кодирует полипептид β2 микроглобулина, который частично является человеческим и частично не относящимся к человеку, например, он содержит некоторые аминокислоты, которые соответствуют человеческому β2 микроглобулину, и некоторые аминокислоты, которые соответствуют не относящемуся к человеку β2 микроглобулину. Согласно одному аспекту не относящееся к человеку животное не экспрессирует эндогенный не относящийся к человеку полипептид β2 микроглобулина из эндогенного не относящегося к человеку локуса, и экспрессирует только человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному примеру не относящееся к человеку животное не экспрессирует полный эндогенный не относящийся к человеку полипептид β2 микроглобулина, но экспрессирует только часть не относящегося к человеку эндогенного полипептида β2 микроглобулина из эндогенного локуса β2 микроглобулина. Таким образом, согласно различным вариантам осуществления животное не экспрессирует функциональный не относящийся к человеку полипептид β2 микроглобулина из эндогенного не относящегося к человеку локуса β2 микроглобулина. Согласно конкретному аспекту нуклеотидная последовательность, кодирующая человеческий или гуманизированный β2 микроглобулин, расположена на эндогенном не относящемся к человеку локусе β2 микроглобулина. Согласно одному аспекту животное содержит две копии локуса β2 микроглобулина, содержащего нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Согласно другому аспекту животное содержит одну копию локуса β2 микроглобулина, содержащего нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Таким образом, животное может являться гомозиготным или гетерозиготным в отношении локуса β2 микроглобулина, содержащего нуклеотидную последовательность, которая кодирует человеческий или гуманизированный полипептид β2 микроглобулина. Нуклеотидная последовательность человеческого или гуманизированного β2 микроглобулина может происходить из совокупности последовательностей β2 микроглобулина, которые встречаются в природе в человеческих популяциях. Согласно различным вариантам осуществления генетически сконструированное не относящееся к человеку животное по настоящему изобретению содержит в своей зародышевой линии нуклеотидную последовательность, кодирующую человеческий или гуманизированный β2 микроглобулин. Согласно одному варианту осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, содержит нуклеотидную последовательность, кодирующую полипептид, содержащий аминокислотную последовательность β2 микроглобулина человека. Согласно одному варианту осуществления полипептид способен связываться с белком МНС I.
Нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, может содержать остатки нуклеиновой кислоты, соответствующие полному гену β2 микроглобулина человека. Альтернативно, нуклеотидная последовательность может содержать остатки нуклеиновой кислоты, кодирующие аминокислотную последовательность, представленную в аминокислотах 21-119 белка β2 микроглобулина человека (т.е., аминокислотные остатки, соответствующие зрелому β2 микроглобулину человека). Согласно альтернативному варианту осуществления нуклеотидная последовательность может содержать остатки нуклеиновой кислоты, кодирующие аминокислотную последовательность, представленную в аминокислотах 23-115 белка β2 микроглобулина человека, например, аминокислотную последовательность, представленную в аминокислотах 23-119 белка β2 микроглобулина человека. Последовательности нуклеиновой кислоты и аминокислотные последовательности β2 микроглобулина человека описаны в Gussow et al., ранее, включенной в настоящий документ посредством ссылки.
Таким образом, человеческий или гуманизированный полипептид β2 микроглобулина может содержать аминокислотную последовательность, представленную в аминокислотах 23-115 полипептида β2 микроглобулина человека, например, аминокислотную последовательность, представленную в аминокислотах 23-119 полипептида β2 микроглобулина человека, например, аминокислотную последовательность, представленную в аминокислотах 21-119 полипептида β2 микроглобулина человека. Альтернативно, β2 микроглобулин человека может содержать аминокислоты 1-119 полипептида β2 микроглобулина человека.
Согласно некоторым вариантам осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный β2 микроглобулин, содержит нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека. Альтернативно, нуклеотидная последовательность содержит нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 гена β2 микроглобулина человека. Согласно настоящему варианту осуществления нуклеотидные последовательности, представленные в экзонах 2, 3 и 4, функционально связаны для обеспечения нормальной транскрипции и трансляции гена. Таким образом, согласно одному варианту осуществления последовательность человека содержит нуклеотидную последовательность, соответствующую экзону 2 - экзону 4 гена β2 микроглобулина человека. Согласно конкретному варианту осуществления последовательность человека содержит нуклеотидную последовательность, соответствующую экзону 2 до приблизительно 267 п.н. после экзона 4 гена β2 микроглобулина человека. Согласно конкретному варианту осуществления последовательность человека содержит приблизительно 2,8 т.п.н. гена β2 микроглобулина человека.
Таким образом, человеческий или гуманизированный полипептид β2 микроглобулина может кодироваться нуклеотидной последовательностью, содержащей нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 человека β2 микроглобулин, например, нуклеотидную последовательность, соответствующую экзону 2 - экзону 4 гена β2 микроглобулина человека. Альтернативно, полипептид может кодироваться нуклеотидной последовательностью, содержащей нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 гена β2 микроглобулина человека. Согласно конкретному варианту осуществления человеческий или гуманизированный полипептид β2 микроглобулина кодируется нуклеотидной последовательностью, соответствующей экзону 2 до приблизительно 267 п.н. после экзона 4 гена β2 микроглобулина человека. Согласно другому конкретному варианту осуществления человеческий или гуманизированный полипептид кодируется нуклеотидной последовательностью, содержащей приблизительно 2,8 т.п.н. гена β2 микроглобулина человека. Поскольку экзон 4 гена β2 микроглобулина содержит 5' нетранслируемую область, человеческий или гуманизированный полипептид может кодироваться нуклеотидной последовательностью, содержащей экзоны 2 и 3 гена β2 микроглобулина.
Специалистам в настоящей области техники будет понятно, что хотя в настоящих примерах описаны конкретные последовательности нуклеиновой кислоты и аминокислотные последовательности для создания генетически сконструированных животных, также предусмотрены последовательности одной или нескольких консервативных или неконсервативных аминокислотных замен, или последовательности, отличающиеся от описанных в настоящем документе вследствие вырожденности генетического кода.
Следовательно, предусмотрено не относящееся к человеку животное, которое экспрессирует последовательность β2 микроглобулина человека, причем последовательность β2 микроглобулина по меньшей мере приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентична последовательности β2 микроглобулина человека. Согласно конкретному варианту осуществления последовательность β2 микроглобулина по меньшей мере приблизительно на 90%, 95%, 96%, 97%, 98% или 99% идентична описанной в примерах последовательности β2 микроглобулина человека. Согласно одному варианту осуществления последовательность β2 микроглобулина человека содержит одну или несколько консервативных замен. Согласно одному варианту осуществления последовательность β2 микроглобулина человека содержит одну или несколько неконсервативных замен.
Кроме того, предусмотрены не относящиеся к человеку животные, причем нуклеотидная последовательность, кодирующая человеческий или гуманизированный белок β2 микроглобулина, также содержит нуклеотидную последовательность, представленную в экзоне 1 не относящегося к человеку гена β2 микроглобулина. Таким образом, согласно конкретному варианту осуществления не относящееся к человеку животное содержит в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный β2 микроглобулин, причем нуклеотидная последовательность содержит экзон 1 не относящегося к человеку гена β2 микроглобулина и экзоны 2, 3 и 4 гена β2 микроглобулина человека. Таким образом, человеческий или гуманизированный полипептид β2 микроглобулина кодируется экзоном 1 не относящегося к человеку гена β2 микроглобулина и экзонами 2, 3 и 4 гена β2 микроглобулина человека (например, экзонами 2 и 3 гена β2 микроглобулина человека).
Аналогично не относящемуся к человеку животному, содержащему нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, не относящееся к человеку животное, содержащее нуклеотидную последовательность, кодирующую человеческий или гуманизированный β2 микроглобулин, может быть выбрано из группы, состоящей из мыши, крысы, кролика, свиньи, крупного рогатого скота (например, коровы, быка, буйвола), оленя, овцы, козы, курицы, кошки, собаки, хорька, примата (например, игрунки, макака-резуса). Согласно некоторым вариантам осуществления настоящего изобретения, не относящееся к человеку животное представляет собой млекопитающее. Согласно конкретному варианту осуществления не относящееся к человеку животное представляет собой представитель семейства мышиных, например, грызуна (например, мышь или крысу). Согласно одному варианту осуществления животное представляет собой мышь.
Таким образом, согласно некоторым аспектам предусмотрена генетически сконструированная мышь, причем мышь содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина, описанный в настоящем документе. Предусмотрена генетически сконструированная мышь, причем мышь содержит на своем эндогенном локусе β2 микроглобулина нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина (например, человеческий или по существу человеческий полипептид β2 микроглобулина). Согласно некоторым вариантам осуществления мышь не экспрессирует эндогенный полипептид β2 микроглобулина (например, функциональный эндогенный полипептид β2 микроглобулина) из эндогенного локуса β2 микроглобулина. Согласно некоторым вариантам осуществления генетически сконструированная мышь содержит нуклеотидную последовательность, содержащую экзон 1 гена β2 микроглобулина мыши и экзоны 2, 3 и 4 гена β2 микроглобулина человека. Согласно некоторым вариантам осуществления мышь экспрессирует человеческий или гуманизированный полипептид β2 микроглобулина.
Согласно одному аспекту предусмотрен модифицированный не относящийся к человеку локус β2 микроглобулина, которые содержит гетерологичную последовательность β2 микроглобулина. Согласно одному варианту осуществления гетерологичная последовательность β2 микроглобулина представляет собой человеческую или гуманизированную последовательность.
Согласно одному варианту осуществления модифицированный локус представляет собой локус грызуна. Согласно конкретному варианту осуществления локус грызуна выбран из локуса мыши или крысы. Согласно одному варианту осуществления не относящийся к человеку локус модифицирован с помощью по меньшей мере одной кодирующей β2 микроглобулин человека последовательности.
Согласно одному варианту осуществления гетерологичная последовательность β2 микроглобулина функционально связана с эндогенными регуляторными элементами, например, эндогенным промотором и/или последовательностью контроля экспрессии. Согласно конкретному варианту осуществления гетерологичная последовательность β2 микроглобулина представляет собой последовательность человека и последовательность человека функционально связана с эндогенным промотором и/или последовательностью контроля экспрессии.
Согласно одному аспекту предусмотрен модифицированный не относящийся к человеку локус β2 микроглобулина, который содержит последовательностью человека, функционально связанную с эндогенным промотором и/или последовательностью контроля экспресии.
Согласно различным аспектам человеческий или гуманизированный β2 микроглобулин, экспрессируемый генетически модифицированным не относящимся к человеку животным, или клетками, зародышами или тканями, полученными от не относящегося к человеку животного, сохраняет все функциональные аспекты эндогенного и/или относящегося к человеку β2 микроглобулина. Например, предпочтительно, что человеческий или гуманизированный β2 микроглобулин связывается сα цепью полипептида МНС I (например, эндогенного не относящегося к человеку или человеческого полипептида МНС I). Человеческий или гуманизированный полипептид β2 микроглобулина может связываться, проводить рекрутинг или иным образом ассоциироваться с любыми другими молекулами, например, рецептором, якорной молекулой или молекулой передачи сигнала, которые ассоциируются с эндогенным не относящимся к человеку и/или относящимся к человеку β2 микроглобулином (например, HFE и т.д.).
В дополнение к генетически модифицированным животным (например, грызунам, например, мышам или крысам), также предусмотрена ткань или клетка, причем ткань или клетка получена от не относящегося к человеку животного, описанного в настоящем документе, и содержит гетерологичный ген β2 микроглобулина или последовательность β2 микроглобулина, т.е., нуклеотидную и/или аминокислотную последовательность. Согласно одному варианту осуществления гетерологичный ген β2 микроглобулина или последовательность β2 микроглобулина представляет собой человеческий или гуманизированный ген β2 микроглобулина или человеческую или гуманизированную последовательность β2 микроглобулина. Предпочтительно, клетка представляет собой ядросодержащую клетку. Клетка может представлять собой любую клетку, которая, как известно, экспрессирует комплекс МНС I, например, антигенпрезентирующую клетку. Человеческий или гуманизированный полипептид β2 микроглобулина, экспрессируемый указанной клеткой может взаимодействовать с эндогенным не относящимся к человеку МНС I (например, МНС I грызуна), для образования функционального комплекса МНС I. Полученный комплекс МНС I может быть способен к взаимодействию с Т-клеткой, например, цитотоксической Т-клеткой. Таким образом, также предусмотрен in vitro комплекс клетки от описанного в настоящем документе не относящегося к человеку животного и Т-клетки.
Также предусмотрены не относящиеся к человеку клетки, которые содержат человеческий или гуманизированный ген β2 микроглобулина или последовательность, и дополнительную человеческую или гуманизированную последовательность, например, раскрытый в настоящем документе химерный полипептид МНС I. В таком случае человеческий или гуманизированный полипептид β2 микроглобулина может взаимодействовать, например, с химерным человеческим/не относящимся к человеку полипептидом MHC I, и может быть образован функциональный комплекс МНС I. Согласно некоторым аспектам такой комплекс способен к взаимодействию с TCR на Т-клетке, например, человеческой или не относящейся к человеку Т-клетке. Таким образом, также предусмотрен in vitro комплекс клетки от описанного в настоящем документе не относящегося к человеку животного и человеческой или не относящейся к человеку Т-клетки.
Другой аспект настоящего раскрытия относится к зародышу грызуна (например, зародышу мыши или крысы), содержащему гетерологичный ген β2 микроглобулина или последовательность β2 микроглобулина, описанные в настоящем документе. Согласно одному варианту осуществления зародыш содержит ES донорную клетку, которая содержит гетерологичный ген β2 микроглобулина или последовательность β2 микроглобулина и клетки зародыша-хозяина. Гетерологичный ген β2 микроглобулина или последовательность β2 микроглобулина представляет собой человеческий или гуманизированный ген β2 микроглобулина или последовательность β2 микроглобулина.
Настоящее изобретение также предусматривает не относящуюся к человеку клетку, содержащую хромосому или ее фрагмент описанного в настоящем документе не относящегося к человеку животного (например, причем хромосома или ее фрагмент содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина). Не относящаяся к человеку клетка может содержать ядро описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит хромосому или ее фрагмент как результат ядерного транспорта.
Согласно одному аспекту предусмотрена не относящаяся к человеку индуцированная плюрипотентная клетка, содержащая гетерологичный ген β2 микроглобулина или последовательность β2 микроглобулина. Согласно одному варианту осуществления индуцированная плюрипотентная клетка происходит от описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления гетерологичный ген β2 микроглобулина или последовательность β2 микроглобулина представляет собой человеческий или гуманизированный ген или последовательность.
Также предусмотрена гибридома или квадрома, происходящая из клетки описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь или крысу.
Настоящее раскрытие также относится к способам получения описанного в настоящем документе генетически сконструированного не относящегося к человеку животного (например, генетически сконструированного грызуна, например, мыши или крысы) Способы дают в результате животного, геном которого содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному аспекту способы дают в результате генетически сконструированную мышь, геном которой содержит на эндогенном локусе β2 микроглобулина нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. В некоторых случаях мышь не экспрессирует функциональный β2 микроглобулин мыши из эндогенного локуса β2 микроглобулина мышь. Согласно некоторым вариантам осуществления в способе используется нацеливающая конструкция, полученная с использованием технологии VELOCIGENE®, введение конструкции в ES клетки и введение нацеленных ES клеточный клонов в мышиный зародыш с использованием описанной в примерах технологии VELOCIMOUSE®. Согласно одному варианту осуществления ES клетки представляют собой сочетание линий мышей 129 и C57BL/6; согласно другому варианту осуществления ES клетки представляют собой сочетание линий мышей BALB/c и 129.
Также предусмотрена нуклеотидная конструкция, используемая для создания генетически сконструированных не относящихся к человеку животных. Нуклеотидная конструкция может содержать: 5' и 3' не относящиеся к человеку плечи гомологии, фрагмент ДНК человека, содержащий последовательности β2 микроглобулина человека, и кассету селекции, фланкированную сайтами рекомбинации. Согласно одному варианту осуществления фрагмент ДНК человека представляет собой геномный фрагмент, который содержит как интроны, так и экзоны гена β2 микроглобулина человека. Согласно одному варианту осуществления не относящиеся к человеку плечи гомологии гомологичны не относящемуся к человеку локусу β2 микроглобулина. Геномный фрагмент может содержать экзоны 2, 3 и 4 гена β2 микроглобулина человека. В одном случае геномный фрагмент содержит, в направлении 5'-3': экзон 2, интрон, экзон 3, интрон и экзон 4, все из последовательностей β2 микроглобулина человека. Кассета селекции может быть расположена где-либо в конструкции за пределами кодирующей области β2 микроглобулина, например, она может быть расположена 3' по отношению к экзону 4 гена β2 микроглобулина человека. 5' и 3' не относящиеся к человеку плечи гомологии могут содержать геномную последовательность 5' и 3' от эндогенного не относящегося к человеку гена β2 микроглобулин, соответственно. Согласно другому варианту осуществления 5' и 3' не относящиеся к человеку плечи гомологии содержат геномную последовательность 5' по отношению к экзону 2 и 3' по отношению к экзону 4 эндогенного не относящегося к человеку гена, соответственно.
Другой аспект настоящего изобретения относится к способу модификации локуса β2 микроглобулина не относящегося к человеку животного (например, грызуна, например, мыши или крысы) для экспрессии описанного в настоящем документе относящегося к человеку или гуманизированного полипептида β2 микроглобулина. Один способ модификации локуса β2 микроглобулина мыши для экспрессии относящегося к человеку или гуманизированного полипептида β2 микроглобулина предусматривает замещение на эндогенном локусе β2 микроглобулина нуклеотидной последовательности, кодирующей β2 микроглобулин мыши, нуклеотидной последовательностью, кодирующей относящийся к человеку или гуманизированный полипептид β2 микроглобулина. Согласно одному варианту осуществления такого способа мышь не экспрессирует функциональный полипептид β2 микроглобулина из эндогенного локуса β2 микроглобулина мыши. Согласно некоторым конкретным вариантам осуществления нуклеотидная последовательность, кодирующая относящийся к человеку или гуманизированный полипептид β2 микроглобулина, содержит нуклеотидную последовательность, представленную в экзонах 2-4 гена β2 микроглобулина человека. Согласно другим вариантам осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2 микроглобулина, содержит нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 гена β2 микроглобулина человека.
Генетически модифицированные в отношении МНС I/β2 микроглобулина животные
Согласно различным вариантам осуществления настоящее изобретение в основном относится к генетически модифицированным не относящимся к человеку животным, которые содержат в своем геноме нуклеотидные последовательности, кодирующие человеческие или гуманизированные полипептиды как МНС I, так и β2 микроглобулина; таким образом, животные экспрессируют человеческие или гуманизированные полипептиды как МНС I, так и β2 микроглобулина.
При применении смешанных человеческих/не относящихся к человеку системных компонентов возникают функциональные различия. HLA I класса связывается с β2 микроглобулином сильнее, чем класс I мыши. Bemabeu (1984) β2-microgobulin from serum associates with MHC class I antigens on the surface of cultured cells. Nature 308: 642-645. Попытки устранить функциональные различия отражаются в конструировании конкретных гуманизированных в отношении MHC мышей. Были разработаны нокаутные в отношении Н-2 I класса и II класса мыши (в нокаутном в отношении мышиного β2 микроглобулина генетического фона), которые экспрессируют случайным образом интегрированный человеческий HLA-A2.1/HLA-DR1 химерный трансген, содержащий α1 и α2 HLA-A2.1 человека, и α3 H-2Db мыши, прикрепленный на своем N-конце посредством линкера к С-концу β2 микроглобулина человека. Смотрите, например, Pajot et al. (2004) A mouse model of human adaptive immune functions: HLA-A2.1-/HLA-DR1-transgenic Н-2 class I-/class II-knockout mice, Eur. J. Immunol. 34: 3060-3069. Как сообщалось, эти мыши производили антигенспецифическое антитело и CTL ответы против вируса гепатита В, что не наблюдалось для мышей, трансгенных только в отношении HLA-A2.1 или нокаутных в отношении Н-2 класса I/класса II мышей. Недостаток мышей, которые являются трансгенными исключительно в отношении указанных генов, предположительно обусловлена способностью таких мышей использовать эндогенные гены I класса и/или II класса, чтобы избежать возникновения какого-либо трансгена, причем такая возможность недоступна для нокаутных в отношении MHC мышей. Тем не менее, мыши могут экспрессировать по меньшей мере Н-2Db, предположительно вследствие скрещиваний до нокаутного в отношении мышиного β2 микроглобулина мышиного генетического фона (смотрите, Pajot et al., ранее; которые вероятно содержали интактные эндогенный локус I класса и II класса).
Как сообщалось, экспрессия на клеточной поверхности химерного слияния с β2 микроглобулином человека ниже, чем экспрессия эндогенного MHC, но не сообщалось о выживаемости/скорости NK-цитотоксичности, а также о скорости NK-аутоцитотоксичности. Pajot et al., ранее. Некоторое повышение количества CD8+ Т-клеток наблюдалось для дефицитных по MHC I класса нокаутных в отношении β2 микроглобулина мышей (2-3% от общего числа спленоцитов по сравнению с 0,6-1% у нокаутных в отношении β2 мышей). Тем не менее, частота использования вариабельной области Т-клетки проявляла измененные профили для генных сегментов BV 5.1, BV 5.2 и BV 11. Сообщалось, что CD8+ и CD4+ Т-клеточные ответы были рестриктированы по соответствующему антигену гепатита В, используемому для иммунизации мышей, хотя по меньшей мере две мыши, у которых наблюдался лизис клеток, несущих любой из антигенов, представляли собой мышей, иммунизированных только одним антигеном, что может быть следствием отсутствия ингибирования NK-клеток или отсутствием NK-клеточной селективности.
Как упоминалось выше, мыши, трансгенные в отношении как МНС I человека, как и β2 микроглобулина человека, содержат нуклеотидную последовательность, кодирующую химерный белок MHC I/β2 микроглобулина, причем части МНС I и β2 микроглобулина содержаться в пределах одной полипептидной цепи, давая в результатеα цепь МНС I и β2 микроглобулин, ковалентно связанные друг с другом и тем самым прикрепленные к клеточной поверхности. Предусмотрена мышь, которая содержит в своем геноме две независимые нуклеотидные последовательности, одну, кодирующую человеческий или гуманизированный полипептид МНС I, и другую, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Предусмотренная в настоящем документе мышь будет экспрессировать комплекс МНС I, который характеризуется более полным сходством с присутствующим в природе комплексом МНС I, причемα цепь МНС I и β2 микроглобулин представлены на двух отдельных полипептидных цепях, причем β2 микроглобулин нековалентно связан сα цепью МНС I.
Таким образом, настоящее раскрытие относится к не относящемуся к человеку животному, содержащему в своем геноме: первую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид МНС I, и вторую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина. Согласно одному аспекту предусмотрено не относящееся к человеку животное, содержащее в своем геноме: (а) первую нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, причем человеческая часть химерного полипептида содержит пептидсвязывающий домен или внеклеточный домен МНС I человека (например, HLA-A, HLA-B или HLA-C; например, HLA-A2), и (b) вторую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина.
Первая нуклеотидная последовательность может быть расположена на эндогенном не относящемся к человеку локусе МНС I так, чтобы животное содержало в своем геноме замещение на локусе МНС I всего эндогенного гена МНС I или его части (например, части, кодирующей пептидсвязывающий домен или внеклеточный домен) соответствующей последовательностью МНС I человека. Таким образом, животное может содержать на эндогенном локусе МНС I нуклеотидную последовательность, кодирующую внеклеточный домен МНС I человека (например, HLA-A, HLA-B или HLA-C; например, HLA-A2) и трансмембранный и цитоплазматический домены эндогенного не относящегося к человеку МНС I (например, Н-2K, H-2D и т.д., например, Н-2Kb). Согласно одному аспекту животное представляет собой мышь, и первая нуклеотидная последовательность содержит нуклеотидную последовательность, кодирующую внеклеточный домен HLA-A2 человека (например, HLA-A2.1) и трансмембранный и цитоплазматический домены Н-2K мыши (например, Н-2Kb).
Вторая нуклеотидная последовательность может быть расположена на эндогенном не относящемся к человеку локусе β2 микроглобулина так, чтобы животное содержало в своем геноме замещение на локусе β2 микроглобулина всего эндогенного гена β2 микроглобулина или его части соответствующей последовательностью β2 микроглобулина человека. Вторая нуклеотидная последовательность может содержать нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека. Альтернативно, вторая нуклеотидная последовательность может содержать нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 гена β2 микроглобулина человека. Согласно настоящему варианту осуществления нуклеотидные последовательности функционально связаны друг с другом. Вторая нуклеотидная последовательность может дополнительно содержать последовательность экзона 1 не относящегося к человеку гена β2 микроглобулина.
Согласно одному аспекту животное не экспрессирует функциональный МНС I из эндогенного не относящегося к человеку локуса МНС I (например, не экспрессирует или пептидсвязывающий домен, или внеклеточный домен эндогенного МНС I), и животное не экспрессирует функциональный полипептид β2 микроглобулина из эндогенного не относящегося к человеку локуса β2 микроглобулина. Согласно некоторым аспектам животное является гомозиготным в отношении как локуса МНС I, содержащего нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, так и локуса β2 микроглобулина, содержащего нуклеотидную последовательность, кодирующую человеческий или гуманизированный β2 микроглобулин. Согласно другим аспектам животное является гетерозиготным в отношении как локуса МНС I, содержащего нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид МНС I, так и локуса β2 микроглобулина, содержащего нуклеотидную последовательность, кодирующую человеческий или гуманизированный β2 микроглобулин.
Предпочтительно, первая и вторая нуклеотидные последовательности функционально связаны с эндогенными элементами контроля экспрессии (например, промоторами, энхансерами, сайленсерами и т.д.).
Различные другие варианты осуществления первой и второй нуклеотидных последовательностей (и полипептидов, которые они кодируют), предусмотренных в настоящем документе, могут легко стать понятными из описанных в настоящем описании изобретения вариантов осуществления, например, вариантов осуществления, описанных в разделах, относящихся к генетически сконструированным в отношении МНС I животным и генетически сконструированным в отношении β2 микроглобулина животным.
Согласно одному аспекту раскрытие относится к мыши, содержащей в своем геноме (а) первую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид МНС I (в частности, полипептид HLA-A2/H-2K), причем человеческая часть химерного полипептида содержит внеклеточный домен HLA-А2 человека, и мышиная часть содержит трансмембранный и цитоплазматический домены H-2K мыши, и (b) вторую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2 микроглобулина (например, причем нуклеотидная последовательность содержит нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека, или нуклеотидные последовательности, представленные в экзоне 2, 3 и 4 гена β2 микроглобулина человека), причем первая нуклеотидная последовательность расположена на эндогенном локусе Н-2K, и вторая последовательность расположена на эндогенном локусе β2 микроглобулина. Согласно одному варианту осуществления мышь не экспрессирует функциональные полипептиды Н-2K и β2 микроглобулина мыши из их соответствующих эндогенных локусов. Согласно одному варианту осуществления мышь экспрессирует как химерный человеческий/мышиный полипептид МНС I, так и человеческий или гуманизированный полипептид β2 микроглобулина.
Как показано в последующих примерах, животные, генетически сконструированные для коэкспрессии как человеческого или гуманизированного МНС I, так и β2 микроглобулина, проявляют увеличенную экспрессию химерного МНС I класса на клеточной поверхности по сравнению с животными, гуманизированными в отношении отдельно МНС I. Согласно некоторым вариантам осуществления коэкспрессия человеческого или гуманизированного МНС I и β2 микроглобулина увеличивает экспрессию на клеточной поверхности человеческого или гуманизированного МНС I больше чем приблизительно на 10%, например, больше чем приблизительно на 20%, например, приблизительно на 50% или больше, например, приблизительно на 70%, по сравнению с экспрессией человеческого или гуманизированного МНС I при отсутствии человеческого или гуманизированного β2 микроглобулина.
Настоящее раскрытие также относится к способу получения генетически сконструированных не относящихся к человеку животных (например, грызунов, например, крыс или мышей), геном которых содержит описанную в настоящем документе первую и вторую нуклеотидную последовательность. В основном способ предусматривает получение первого генетически сконструированного не относящегося к человеку животного, геном которого содержит описанную в настоящем документе первую нуклеотидную последовательность (т.е., человеческую или гуманизированную последовательность МНС I), получение второго генетически сконструированного не относящегося к человеку животного, геном которого содержит описанную в настоящем документе вторую нуклеотидную последовательность (т.е., человеческую или гуманизированную последовательность β2 микроглобулина), и скрещивание первого и второго животного для получения потомства, геном которого содержит обе нуклеотидных последовательности. Согласно одному варианту осуществления первое и второе животное являются гетерозиготными в отношении первой и второй нуклеотидной последовательности, соответственно. Согласно одному варианту осуществления первое и второе животное являются гомозиготными в отношении первой и второй нуклеотидной последовательности, соответственно. Согласно одному варианту осуществления первого и второго животных получают посредством замещения эндогенных не относящихся к человеку локусов первой и второй нуклеотидными последовательностями, соответственно. Согласно одному аспекту первое и второе животное получают посредством использования конструкций, созданных с помощью технологии VELOCIGENE®, и путем введения нацеленных ES клеточных клонов, несущих такие конструкции, в зародыш (например, зародыш грызуна, например, зародыш мыши или крысы) способом VELOCIMOUSE®.
Применение генетически модифицированных животных
Согласно различным вариантам осуществления описанные в настоящем документе генетически модифицированные не относящиеся к человеку животные производят АПК с человеческим или гуманизированным МНС I и/или β2 микроглобулином на клеточной поверхности и, как результат, презентируют пептиды, происходящие из цитозольных белков в качестве эпитопов для CTL характерным для человека образом, поскольку по существу все компоненты комплекса являются человеческими или гуманизированными. Генетически модифицированные не относящиеся к человеку животные по настоящему изобретению могут использоваться для исследования функции иммунной системы человека у гуманизированного животного; для идентификации антигенов и антигенных эпитопов, которые вызывают иммунный ответ (например, Т-клеточных эпитопов, например, уникальных эпитопов злокачественных опухолей человека), например, для применения в разработке вакцин; для идентификации высокоаффинных Т-клеток по отношению к патогенам человека или антигенам злокачественных опухолей (т.е., Т-клеток, которые связываются с антигеном в контексте комплекса MHC I человека с высокой авидностью), например, для применения в терапии на основе адаптивных Т-клеток; для оценки кандидатных вакцин и других стратегий разработки вакцин; для изучения аутоиммунитета человека; для исследования инфекционных заболеваний человека; и в других случаях для разработки лучших терапевтических стратегий на основании экспрессии MHC человека.
Комплекс MHC I связывает пептиды и презентирует их на клеточной поверхности. Будучи презентированными на поверхности в контексте такого комплекса, пептиды распознаются Т-клетками. Например, когда пептид происходит из патогена или другого представляющего интерес антигена (например, опухолевого антигена), распознавание Т-клетками может привести к активации Т-клеток, опосредованному макрофагами цитолизу клеток, несущих презентированную пептидную последовательность, и В-клеточной активации антител, которые связывают презентированную последовательность.
Т-клетки взаимодействуют с клетками, экспрессирующими комплекс МНС I, посредством связанного с пептидом эктодомена МНС I класса и Т-клеточного эктодомена CD8. CD8+ Т-клетки, которые встретились с АПК, содержащими подходящие антигены, связанные с молекулой МНС I класса, станут цитотоксическими Т-клетками. Таким образом, антигены, которые в контексте МНС I класса связываются с высокой авидностью с Т-клеточным рецептором, являются потенциально важными в разработке способов лечения для патологий человека. Тем не менее, презентация антигенов в контексте МНС I мыши лишь в некоторой степени имеет отношение к заболеванию человека, поскольку комплексы МНС человека и мыши распознают антигены различным образом, например, МНС I мыши может не распознавать одинаковые антигены или может презентировать другие эпитопы по сравнению с МНС I человека. Таким образом, наиболее релевантные данные для патологий человека получают посредством исследования презентации антигенных эпитопов МНС I человека.
Таким образом, согласно различным вариантам осуществления генетически сконструированные животные по настоящему изобретению являются применимыми, среди прочего, для оценки способности антигена инициировать иммунный ответ у человека, и для получения разнообразных антигенов и идентификации специфического антигена, который может использоваться для разработки вакцины для человека.
Согласно одному аспекту предусмотрен способ определения у человека антигенности пептидной последовательности, предусматривающий воздействие на описанное в настоящем документе генетически модифицированное не относящееся к человеку животное молекулы, содержащей пептидную последовательность, предоставление не относящемуся к человеку животному возможности развить иммунный ответ и обнаружение у не относящегося к человеку животного клетки, которая связывает последовательность пептида, презентируемого описанным в настоящем документе описанным в настоящем документе химерным человеческим/не относящимся к человеку МНС I, или гуманизированным комплексом МНС I (содержащим химерный человеческий/не относящийся к человеку МНС I и человеческий или гуманизированный β2 микроглобулин).
Согласно одному аспекту предусмотрен способ определения того, будет ли пептид вызывать иммунный ответ у человека, предусматривающий воздействие пептида на описанное в настоящем документе генетически модифицированное не относящееся к человеку животное, предоставление не относящемуся к человеку животному возможности развить иммунный ответ и обнаружение у не относящегося к человеку животного клетки, которая связывает последовательность пептида описанной в настоящем документе химерной человеческой/не относящейся к человеку молекулой МНС I класса. Согласно одному варианту осуществления не относящееся к человеку животное после воздействия содержит рестриктированный по МНС I класса CD8+цитотоксический Т-лимфоцит (CTL), который связывает пептид. Согласно одному варианту осуществления CTL лизируют несущую пептид клетку.
Согласно одному аспекту предусмотрен способ идентификации эпитопа CTL человека, предусматривающий воздействие на описанное в настоящем документе не относящееся к человеку животное антигена, содержащего предполагаемый CTL эпитоп, предоставление не относящемуся к человеку животному возможности развить иммунный ответ, выделение из не относящегося к человеку животного рестриктированного по МНС I класса CD8+ CTL, который связывает эпитоп, и идентификацию эпитопа, связанного рестриктированным по МНС I класса CD8+ CTL.
Согласно одному аспекту предусмотрен способ идентификации рестриктированного по HLA класса I пептида, презентация которого клеткой человека и связывание лимфоцитом человека (например, Т-клеткой человека) будет приводить к цитотоксичности несущей пептид клетки, предусматривающий воздействие на описанное в настоящем документе не относящееся к человеку животное (или его экспрессирующую МНС I класса клетку) молекулы, содержащей представляющий интерес пептид, выделение клетки не относящегося к человеку животного, которая экспрессирует химерную человеческую/не относящуюся к человеку молекулу I класса, которая связывает представляющий интерес пептид, воздействие на клетку лимфоцита человека, который способен осуществить рестриктированную по HLA I класса цитотоксичность, и измерение индуцированной пептидом цитотоксичности.
Согласно одному аспекту предусмотрен способ идентификации антигена, который вызывает цитотоксический Т-клеточный ответ у человека, предусматривающий воздействие предполагаемого антигена на описанную в настоящем документе мышь, предоставление мыши возможности развить иммунный ответ и идентификацию антигена, связанного с рестриктированной по HLA-A молекулой.
Согласно одному варианту осуществления антиген содержит бактериальный или вирусный поверхностный или оболочечный белок. Согласно одному варианту осуществления антиген предусматривает антиген на поверхности опухолевой клетки человека. Согласно одному варианту осуществления антиген предусматривает предполагаемую вакцину для применения у человека. Согласно одному варианту осуществления антиген содержит эпитоп человека, который вызывает образование антител у человека. Согласно другому варианту осуществления антиген содержит эпитоп человека, который вызывает образование высокоаффинных CTL, нацеленных на комплекс эпитоп/МНС I.
Согласно одному аспекту предусмотрен способ определения того, содержит ли предполагаемый антиген эпитоп, который при воздействии на иммунную систему человека будет вызывать рестриктированный по HLA-A иммунный ответ (например, рестриктированный по HLA-A2 ответ), предусматривающий воздействие на описанную в настоящем документе мышь предполагаемого антигена и измерение антигенспецифического рестриктированного по HLA-A (например, рестриктированного по HLA-A2) иммунного ответа у мыши.
Согласно одному варианту осуществления предполагаемый антиген выбран из биофармацевтического средства или его фрагмента, «чужого» белка, поверхностного антигена «чужой» клетки, поверхностного антигена опухолевой клетки, поверхностного антигена бактериальной или дрожжевой или грибковой клетки, поверхностного антигена или оболочечного белка вируса.
Кроме того, описанные в настоящем документе генетически сконструированные не относящиеся к человеку животные могут быть применимы для идентификации Т-клеточных рецепторов, например, Т-клеточных рецепторов с высокой авидностью, которые распознают представляющий интерес антиген, например, опухолевый антиген или антиген другого заболевания. Способ может предусматривать: воздействие антигена на описанное в настоящем документе не относящееся к человеку животное, предоставление не относящемуся к человеку животному возможности развить иммунный ответ на антиген, выделение из не относящегося к человеку животного Т-клетки, содержащей Т-клеточный рецептор, который связывает антиген, презентируемый человеческим или гуманизированным МНС I, и определение последовательности указанного Т-клеточного рецептора.
Согласно одному аспекту предусмотрен способ идентификации вариабельного домена Т-клеточного рецептора с высокой аффинностью к опухолевому антигену человека, предусматривающий воздействие опухолевого антигена человека на мышь, содержащую гуманизированные α1, α2, и α3 домены (например, HLA-A2 α1, α2, и α3 домены) МНС I; предоставление мыши возможности развить иммунный ответ; и выделение из мыши последовательности нуклеиновой кислоты, кодирующей вариабельный домен Т-клеточного рецептора, причем вариабельный домен Т-клеточного рецептора связывает опухолевый антиген человека с KD не выше чем приблизительно 1 нМ.
Согласно одному варианту осуществления мышь дополнительно содержит замещение на эндогенном генном локусе вариабельной области Т-клеточного рецептора мыши множеством переаранжированных генных сегментов вариабельной области Т-клеточного рецептора человека, причем переаранжированные генные сегменты вариабельной области Т-клеточного рецептора человека рекомбинируются для кодирования химерного человеческого/мышиного гена Т-клеточного рецептора, содержащего вариабельную область человека и константную область мыши. Согласно одному варианту осуществления мышь содержит трансгенный CD8 человека, и мышь экспрессирует функциональный белок CD8 человека.
Т-клеточные рецепторы с высокой авидностью к опухолевым антигенам применимы в терапевтических средствах на основе клеток. Популяции Т-клеток с высокой авидностью к опухолевым антигенам человека были получены путем воздействия на Т-клетки человека HLA-A2, который был мутирован для минимизации CD8 связывания с α3 субъединицей для выбора только тех Т-клеток, которые обладают наибольшей авидностью к опухолевому антигену (т.е., Т-клеточных клонов, которые распознают антиген несмотря на неспособность CD8 связываться с α3). Смотрите, Pittet et al. (2003) α3 Domain Mutants of Peptide/MHC Class I Multimers Allow the Selective Isolation of High Avidity Tumor-Reactive CD8 Т Cells, J. Immunol. 171: 1844-1849. He относящиеся к человеку животные и клетки не относящихся к человеку животных применимы для идентификации пептидов, которые будут образовывать комплекс с HLA класса I человека, который будет связываться с высокой авидностью с Т-клеточным рецептором, или активировать лимфоцит, несущий Т-клеточный рецептор.
Связывание комплекса антиген/HLA I класса с Т-клеткой, или активацию Т-клетки, можно измерить любым подходящим известным в настоящей области техники способом. Можно измерить пептидспецифическое связывание АПК и Т-клеток и их активацию. Например, сообщалось, что вовлечение Т-клеток антигенпрезентирующими клетками, которые экспрессируют HLA-A2, вызывает накопление PIP2 на иммунном синапсе, тогда как поперечное сшивание молекул МНС I класса этого не вызывает. Смотрите, Fooksman et al. (2009) Cutting Edge: Phosphatidylinositol 4,5-Bisphosphate Concentration at the APC Side of the Immunological Synapse Is Required for Effector Т Cell Function, J. Immunol. 182: 5179-5182.
Также можно измерить функциональные последствия взаимодействия лимфоцита, несущего TCR, и экспрессирующей молекулу I класса АПК, и они включают в себя цитолиз с помощью лимфоцита. Например, сообщалось, что точки контакта на α2 субъединице HLA-A2 с помощью CD8+ CTL создают сигнал для Fas-независимого цитолиза. Экспрессирующие HLA-A2 Т-клетки линии Jurkat подвергаются апоптозу при контакте (с помощью антител) эпитопов на молекуле HLA-A2, которая, как известно (из кристаллографических исследований) контактирует с CD8, вне какой-либо видимой зависимости от цитоплазматического домена. Смотрите, Pettersen et al. (1998) The TCR-Binding Region of the HLA Class I α2 Domain Signals Rapid Fas-Independent Cell Death: A Direct Pathway for Т Cell-Mediated Killing of Target Cells? J. Immunol. 160: 4343-4352. Предположили, что быстрый цитолиз, индуцированный контактом HLA-A2 α2 с CD8 CD8+ CTL, преимущественно может быть обусловлен этим Fas-независимым опосредованным HLA-A2 путем (там же), что отличается от TCR-независимого опосредованного α3 доменом цитолиза, который сам по себе может индуцировать апоптоз (смотрите, Woodle et al. (1997) Anti-human class I МНС antibodies induce apoptosis by a pathway that is distinct from the Fas antigen-mediated pathway, J. Immunol. 158: 2156-2164).
Последствие взаимодействия между Т-клеткой и АПК, представляющей пептид в контексте МНС I, также можно измерить с помощью анализа Т-клеточной пролиферации. Альтернативно, его можно определить путем измерения высвобождения цитокинов, зачастую связанного с активацией иммунного ответа. Согласно одному варианту осуществления IFNγ ELISPOT (способ иммуноферментных пятен) может использоваться для мониторинга и количественного определения активации CD8+ Т-клеток.
Как описано в настоящем документе, активация CD8+ Т-клеток может быть затруднена у описанных в настоящем документе генетически модифицированных не относящихся к человеку животных вследствие видоспецифического связывания CD8 с MHC I. Для вариантов осуществления, где необходимо видоспецифическое взаимодействие CD8, клетку описанного в настоящем документе генетически модифицированного животного (например, грызуна, например, мыши или крысы) подвергают воздействию (например, in vitro) с помощью клетки человека, например, несущей CD8 клетки человека, например, Т-клетки человека. Согласно одному варианту осуществления экспрессирующую МНС I класса клетку описанной в настоящем документе мыши подвергают воздействию in vitro с помощью Т-клетки, которая содержит CD8 человека и Т-клеточный рецептор. Согласно конкретному варианту осуществления Т-клетка представляет собой Т-клетку человека. Согласно одному варианту осуществления экспрессирующая МНС I класса клетка мыши содержит пептид, связанный с химерным человеческим/мышиным комплексом МНС I или гуманизированным комплексом МНС I (который включает в себя β2 микроглобулин человека), Т-клетка представляет собой Т-клетку человека, и определяют способность Т-клетки связывать представляющую пептид клетку мыши. Согласно одному варианту осуществления определяют активацию Т-клетки человека представляющей пептид клеткой мыши. Согласно одному варианту осуществления предусмотрен способ in vitro измерения активации Т-клетки человека представляющей пептид клеткой, предусматривающий воздействие на описанную в настоящем документе мышь или клетку мыши представляющего интерес антигена, воздействие Т-клетки человека на клетку от указанной мыши или указанную клетку мыши (предположительно несущую пептид, происходящий из антигена в комплексе с человеческим или гуманизированным МНС I) и измерение активации Т-клетки человека. Согласно одному варианту осуществления способ используется для идентификации Т-клеточного эпитопа патогена человека или опухоли человека. Согласно одному варианту осуществления способ используется для идентификации эпитопа для вакцины.
Согласно одному варианту осуществления предусмотрен способ определения активации Т-клетки предполагаемым терапевтическим средством для человека, предусматривающий воздействие на описанное в настоящем документе генетически модифицированное животное с помощью предполагаемого терапевтического средства для человека (или например, воздействие на экспрессирующую человеческий или гуманизированный МНС I клетку такого животного с помощью пептидной последовательности предполагаемого терапевтического средства), воздействие на клетку генетически модифицированного животного, которая представляет комплекс человеческого или гуманизированного МНС I/пептида с помощью Т-клетки, содержащей Т-клеточный рецептор человека и CD8, способный связываться с клеткой генетически модифицированного животного, и измерение активации Т-клетки человека, которая индуцирована представляющей пептид клеткой генетически модифицированного животного.
Согласно различным вариантам осуществления комплекс, образованный между экспрессирующей человеческий или гуманизированный МНС I класса клеткой от описанного в настоящем документе животного образуется с Т-клеткой, которая содержит последовательность СD8 человека, например, Т-клеткой человека, или Т-клеткой не относящегося к человеку животного, которая содержит трансген, который кодирует CD8 человека. В настоящей области техники известны мыши, трансгенные в отношении CD8 человека. Tishon et al. (2000) Trangenic Mice Expressing Human HLA and CDS Molecules Generate HLA-Restricted Measles Virus Cytotoxic Т Lymphocytes of the Same Specificity as Humans with Natural Measles Virus Infection, Virology 275(2): 286-293; а также, LaFace et al. (1995) Human CDS Transgene Regulation of HLA Recognition by Murine Т Cells, J. Exp. Med. 182: 1315-1325.
В дополнение к способности идентифицировать антигены и антигенные эпитопы из патогенов или опухолей человека, генетически модифицированные животные по настоящему изобретению могут использоваться для идентификации аутоантигенов имеющих отношение к аутоиммунным заболеваниям человека, например, диабету I типа, рассеянному склерозу и т.д. Например, в Takaki et al. ((2006) HLA-A*0201-Restricted T Cells from Humanized NOD Mice Recognize Autoantigens of Potential Clinical Relevance to Type 1 Diabetes, J. Immunol. 176: 3257-65) описано применение NOD мышей, несущих моноцепь HLA/β2 микроглобулина в идентификации аутоантигенов диабета 1 типа. Кроме того, генетически модифицированные животные по настоящему изобретению могут использоваться для исследования различных аспектов аутоиммунного заболевания человека. Поскольку известно, что некоторые полиморфные аллели МНС I человека связаны с развитием определенных заболеваний, например, аутоиммунных заболеваний (например, диффузного токсического зоба, тяжелой миастении, псориаза и т.д.; смотрите Bakker et al. (2006) A high-resolution HLA and SNP haplotype map for disease association studies in the extended human МНС, Nature Genetics 38: 1166-72 и Supplementary Information and International МНС and Autoimmunity Genetics Network (2009) Mapping of multiple susceptibility variants within the МНС region for 7 immune-mediated diseases, Proc. Natl. Acad. Sci. USA 106: 18680-85Supplementary Information and International МНС and Autoimmunity Genetics Network (2009) Mapping of multiple susceptibility variants within the MHC region for 7 immune-mediated diseases, Proc. Natl. Acad. Sci. USA 106: 18680-85, включенные в настоящий документ посредством ссылки), генетически модифицированное животное по настоящему изобретению, содержащее гуманизированный локус MHC I, включающий в себя такой аллель, может быть применимо в качестве модели аутоиммунного заболевания. Согласно одному варианту осуществления аллель заболевания представляет собой HLA-B27, и заболевание представляет собой анкилозирующий спондилит или реактивный артрит; таким образом, согласно одному варианту осуществления животное, используемое для исследования указанных заболеваний, содержит человеческий или гуманизированный HLA-B27.
В настоящей области техники известны другие аспекты клеточного иммунитета, которые включают в себя комплексы MHC I; следовательно, описанные в настоящем документе генетически сконструированные не относящиеся к человеку животные могут использоваться для исследования этих аспектов биологии иммунитета. Например, связывание TCR с MHC I класса модулируется in vivo с помощью дополнительных факторов. Представитель подсемейства В лейкоцитарных иммуноглобулинподобных рецепторов (LILRB1, или LIR-1) экспрессируется на рестриктированных по MHC I класса CTL и отрицательно регулирует стимуляцию Т-клеток путем связывания специфической детерминанты на α3 субъединице молекулы MHC I класса на АПК. Структурные исследования показывают, что сайт связывания для LIR-1 и CD8 перекрываются, указывая на то, что ингибирующий LIR-1 конкурирует со стимулирующим CD8 за связывание с молекулами MHC I класса. Willcox et al. (2003) Crystal structure of HLA-A2 bound to LIR-1, a host and viral major histocompatibility complex receptor, Nature Immunology 4(9): 913-919; a также, Shirioshi et al. (2003) Human inhibitory receptors Ig-like transcript 2 (ILT2) and ILT4 compete with CD8 for MHC class I binding and bind preferentially to HLA-G, Proc. Natl. Acad. Sci. USA 100(15): 8856-886L LIR-1 передает ингибирующие сигналы через свой (внутриклеточный) иммунорецепторный тирозиновый ингибирующий мотив (ITIM). Исследования показали, что в NK-клетках KIR (ингибирующие Ig-подобные рецепторы клеток-киллеров), не содержащие ITIM (в норме неспособные ингибировать) могут ингибировать в присутствии LIR-1 (предположительно посредством LIR-1 ITIM), связанного с α3 доменом молекулой MHC I класса (смотрите, Kirwin et al. (2005) Killer Cell Ig-Like Receptor-Dependent Signaling by Ig-Like Transcript 2 (ILT2/CD85J/LILRB1/LIR-1) J. Immunol. 175: 5006-5015), указывая на взаимодействие между LIR-1, связанным с MHC I класса, и KIR и, таким образом, указывая на роль связывания α3 домена HLA в модуляции ингибирования NK-клеток.
Как описано выше, молекулы МНС взаимодействуют с клетками, которые не экспрессируют TCR. В числе таких клеток находятся NK-клетки. NK-клетки представляют собой цитотоксические лимфоциты (которые отличаются от CTL, или цитотоксических Т-лимфоцитов), которые играют центральную роль в клеточном иммунном ответе и, в частности, врожденном иммунитете. NK-клетки представляют собой первую линию защиты от инвазивных микроорганизмов, вирусов и других «чужих» (например, опухолевых) соединений. NK-клетки активируются или ингибируются через поверхностные рецепторы, и они экспрессируют CD8, но не экспрессируют TCR. NK-клетки могут взаимодействовать с клетками, которые экспрессируют МНС I класса, но взаимодействие осуществляется через CD8-связывающий α3 домен, а не через TCR-связывающие, несущие пептид α1 и α3 домены. Главная функция NK-клеток заключается в разрушении клеток, которые не содержат достаточное количество поверхностного белка МНС I класса.
Поперечное сшивание молекул МНС I класса на поверхности клеток естественных киллеров (NK) человека приводит к внутриклеточному фосфорилированию тирозина, миграции молекулы МНС I класса из иммунного синапса и отрицательной регуляции цитолиза опухолевой клетки. Rubio et al. (2004) Cross-linking of МНС class I molecules on human NK cells inhibits NK cell function, segregates МНС I from the NK cell synapse, and induces intracellular phosphotyrosines, J. Leukocyte Biol. 76: 116-124.
Другая функция МНС I класса в NK-клетках, вероятно, заключается в предотвращении аутоцитотоксичности. NK-клетки несут как активирующий рецептор 2В4, так и 2В4 лиганд CD48; по-видимому, МНС I класса связывает 2В4 и предотвращает его активацию с помощью CD48. Betser-Cohen (2010) The Association of МНС Class I Proteins with the 2В4 Receptor Inhibits Self-Killing of Human NK Cells, J. Immunol. 184: 2761-2768.
Таким образом, описанные в настоящем документе генетически сконструированные не относящиеся к человеку животные могут использоваться для исследования этих не опосредованных TCR или CTL процессов и для разработки подходов для их модуляции.
ПРИМЕРЫ
Настоящее изобретение будет дополнительно проиллюстрированы представленными ниже не ограничивающими примерами. Эти примеры представлены для содействия в понимании настоящего изобретения, но не предусмотрены для ограничения и не должны рассматриваться как ограничивающие его объем каким-либо образом.
Примеры не включают в себя подробные описания общепринятых способов, которые будут хорошо известны специалистам в настоящей области техники (техники молекулярного клонирования и т.д.). Если не указано иное, части представляют собой части по весу, молекулярный вес представляет собой средний молекулярный вес, температура указана в градусах Цельсия и давление равно или близко атмосферному.
Пример 1. Конструирование и определение характеристик генетически модифицированных в отношении HLA-A2 мышей
Пример 1.1: Экспрессия HLA-A2/H-2K в MG87 клетках.
Вирусную конструкцию, содержащую химерную генную последовательность HLA-A2/H-2K (фиг. 4А), получали с использованием стандартных методов молекулярного клонирования, известных специалисту в настоящей области техники для анализа экспрессии химерного человеческого/мышиного МНС I в трансфицированных клетках.
Кратко, химерную вирусную конструкцию HLA-A человека/Н-2K мыши получали с использованием последовательностей экзона, кодирующих α1, α2 и α3 домены α цепи, и клонируя их в рамку считывания с кодирующими последовательностями мыши для трансмембранного и цитоплазматического доменов из гена Н-2K (фиг. 4А, pMIG-HLA-А2/Н2К). Как показано на фиг. 4, конструкция содержала репортерную последовательность IRES-GFP, которая обеспечивала возможность определения того, сохраняла ли конструкция способность к экспрессии в клетках после трансфекции.
Получали содержащие описанную выше химерную конструкцию вирусы и размножали их в клетках эмбриональной почки человека 293 (293Т). 293Т-клетки помещали на 10 см чашки и выращивали до 95% конфлюентности. Смесь для трансфекции ДНК получали с помощью 25 мкг pMIG-HLA-A2/H2K, pMIG- HLA человека -А2, или pMIG-гуманизированный β2 микроглобулин, и 5 мкг pMDG (оболочечная плазмида), 15 мкг pCL-Eco (упаковывающая конструкция без сигнала упаковки Т), 1 мл Opti-MEM (Invitrogen). К этой 1 мл смеси ДНК добавляли 80 мкл липофектамина-2000 (Invitrogen) в 1 мл Opti-MEM, которые предварительно перемешивали и инкубировали при комнатной температуре в течение 5 минут. Смесь липофектамина/ДНК инкубировали в течение дополнительных 20 минут при комнатной температуре и затем добавляли в 10 см чашки и планшеты инкубировали при 37°С. Среду из клеток собирали через 24 часа и к клеткам добавляли 10 мл R10 (RPMI 1640 + 10% FBS) свежей среды. Эту смену среды повторяли дважды. Через полных четыре дня собранные среды объединяли, центрифугировали и пропускали через стерильный фильтр для удаления клеточного дебриса.
Полученные выше размноженные вирусы использовали для трансдукции клеток MG87 (фибробласт мыши). Клетки MG87 из одной колбы Т-75 отмывали один раз PBS. 3 мл 0,25% трипсина + EDTA добавляли к клеткам и инкубировали при комнатной температуре в течение 3 минут. 7 мл D 10 (DMEM с высоким содержанием глюкозы; 10% фетальной бычьей сыворотки) добавляли к смеси клеток и трипсина и переносили в 15 мл пробирку для центрифугирования при 1300 об/мин в течение пяти минут. После центрифугирования клеток среду аспирировали и клетки ресуспендировали в 5 мл D10. Клетки подсчитывали и ~3,0×105 клеток на лунку помещали в 6-луночный планшет. pMIG-HLA-A2 человека или pMIG-HLA-A2/H-2K либо отдельно, либо с вирусом pMIG-гуманизированный β2 микроглобулин добавляли в лункам, с нетрансдуцированными клетками в качестве контроля. Клетки инкубировали при 37°С с 5% CO2 в течение 2 дней. Получали клетки для анализа FACS (с использованием антитела к HLA-A2, клон ВВ7.2) для экспрессия HLA-A2 с β2 микроглобулином или без него.
На графиках (фиг. 4В), а также таблице, обобщающей данные, полученные из графиков (фиг. 4С), показано, что котрансдукция с гуманизированным β2 микроглобулином увеличивает экспрессию HLA-A2 человека или химерного человеческого/не относящегося к человеку HLA-A2/H-2K, что показано сдвигом кривых вправо.
Пример 1.2. Конструирование химерного локуса HLA-A2/H-2K. Ген Н-2K мыши гуманизировали за одну стадию путем конструирования уникального нацеливающего вектора из ДНК бактериальной искусственной хромосомы (ВАС) человека и мыши с использованием технологии VELOCIGENE® (смотрите, например, патент США №6586251 и Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nat. Biotech. 21(6): 652-659). ДНК из ВАС клона RP23-173k21 мыши (Invitrogen) модифицировали с помощью гомологичной рекомбинации для замещения геномной ДНК, кодирующей α1, α2 и α3 домены гена Н-2K мыши, геномной ДНК человека, кодирующей α1, α2 и α3 субъединицы гена HLA-A человека (фиг. 5).
Кратко, геномную последовательность, кодирующую α1, α2 и α3 субъединицы мыши гена Н-2K, замещали геномной ДНК человека, кодирующей α1, α2 и α3 домены гена HLA-A*0201 человека за одно событие нацеленного воздействия с использованием нацеливающего вектора, содержащего гигромициновую кассету, фланкированную сайтами loxP с 5' плечом гомологии мыши, содержащим последовательность 5' от локуса Н-2K мыши, включающую в себя 5' нетранслируемую область (UTR; 5' плечо гомологии представлено в SEQ ID NO: 1) и 3' плечом гомологии мыши, содержащим геномную последовательность 3' от кодирующей последовательности Н-2K α3 мыши (3' плечо гомологии представлено в SEQ ID NO: 2).
Конечная конструкция для нацеливания на эндогенный генный локус Н-2K в направлении 5' - 3' включала в себя (1) 5' плечо гомологии, содержащее -200 п.н. геномной последовательности мыши 5' от эндогенного гена Н-2K, включающей в себя 5'UTR, (2) ~1339 п.н. геномной последовательности человека, включающей в себя лидерную последовательность HLA-A*0201, 1 интрон лидерная последовательность HLA-А*0201/α, экзон HLA-A*0201 α1, интрон HLA-A*0201 α1-α2, экзон HLA-A*0201 α2, -316 п.н. 5' конца интрона α2-α3, (3) сайт 5' loxP, (4) гигромициновую кассету, (5) сайт 3' loxP, (6) -580 п.н. геномной последовательности человека, включающей в себя -304 п.н. 3' конца интрона α2-α3, экзон HLA-A*0201 α3 и (7) 3' плечо гомологии, содержащее -200 п.н. геномной последовательности мыши, включающей в себя интрон между Н-2K α3 мыши и трансмембранными кодирующими последовательностями (смотрите фиг. 5 для схематического представления Н-2K нацеливающего вектора). Последовательность из 149 нуклеотидов на стыке последовательностей мыши и человека в направлении 5' от нацеливающего вектора представлена в SEQ ID NO: 3, и последовательность из 159 нуклеотидов на стыке последовательностей мыши и человека в направлении 3' от нацеливающего вектора представлена в SEQ ID NO: 4. Гомологичная рекомбинация с указанным нацеливающим вектором создавала модифицированный локус Н-2K мыши, содержащий геномную ДНК человека, кодирующую α1, α2 и α3 домены гена HLA-А*0201, функционально связанную с эндогенными последовательностями, кодирующими Н-2K трансмембранный и цитоплазматический домены мыши, которая при трансляции приводит к образованию химерного человеческого/мышиного белка МНС I класса.
Нацеленную ВАС ДНК использовали для электропорации клеток F1H4 ES мыши для получения модифицированных ES клеток для создания мышей, которые экспрессируют химерный белок МНС I класса на поверхности ядросодержащих клеток (например. Т- и В-лимфоцитов, макрофагов, нейтрофилов). ES клетки, содержащие вставку последовательностей HLA человека идентифицировали с помощью количественного анализа TAQMAN™. Наборы специфических праймеров и зонды разрабатывали для обнаружения вставки последовательностей HLA человек и ассоциированных кассет селекции (приобретение аллеля, GOA) и потери эндогенных последовательностей мыши (потеря аллеля, LOA). В таблице 1 указаны названия и положения, обнаруженные для каждого из зондов, используемых в анализах количественной ПЦР.

Кассету селекции могут удалить способами, известными специалисту в настоящей области техники. Например, ES клетки, несущие химерный человеческий/мышиный локус MHC I класса могут трансфектировать конструкцией, которая экспрессирует Cre для удаления фланкированной сайтом lox гигромициновой кассеты, введенной путем вставки нацеливающей конструкции, содержащей генные последовательности HLA-A*0201 человека (смотрите фиг. 5). Гигромициновую кассету необязательно могут удалить путем скрещивания мышей, которые экспрессируют Cre рекомбиназу. Необязательно у мыши сохраняют гигромициновую кассету.
Описанные выше нацеленные ES использовали в качестве донорных ES клеток и вводили в зародыш мыши на стадии 8 клеток способом VELOCIMOUSE® (смотрите, например, патент США №7294754 и Poueymirou et al. (2007) FO generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1): 91-99). VELOCIMICE® (FO мышей, полученные полностью из донорной ES клетки), независимо несущих химерный ген МНС I класса, идентифицировали путем генотипирования с использованием модификации аллельного анализа (Valenzuela et al., ранее), который определяет присутствие уникальных генных последовательностей HLA-A*0201 человека.
Пример 1.3. Экспрессия in vivo химерного HLA-A/H-2K у генетически модифицированных мышей.
Гетерозиготную мышь, несущую генетически модифицированный локус Н-2K, описанный в примере 1.2, анализировали в отношении экспрессии химерного белка HLA-A/H-2K в клетках животного.
Кровь получали отдельно от мыши дикого типа и химерной в отношении HLA-A/H-2K гетерозиготной (А2/H2K) мыши. Клетки окрашивали в отношении HLA-A2 человека с помощью конъюгированного с фикоэритрином (РЕ) антитела к HLA-A и подвергали воздействию конъюгированного с аллофикоцианином антитела к Н-2Kb в течение одного часа при 4°С. Клетки анализировали в отношении экспрессии с помощью проточной цитометрии с использованием антител, специфических для HLA-A и Н-2Kb. На фиг. 6А показана экспрессия Н-2Kb и HLA-A2 у мыши дикого типа и химерной гетерозиготной мыши, причем химерная гетерозиготная мышь экспрессировала оба белка. На фиг. 6В показана экспрессия как Н-2Kb, так и химерного HLA-A2/H2K у гетерозиготной мыши.
Пример 2: Конструирование и определение характеристик генетически модифицированных в отношении β2 микроглобулина мышей
Пример 2.1: Конструирование гуманизированного локуса β2 микроглобулина Ген β2 микроглобулина (β2m) мыши гуманизировали за одну стадию путем конструирования уникального нацеливающего вектора из ДНК бактериальной искусственной хромосомы (ВАС) человека и мыши с использованием технологии VELOCIGENE® (смотрите, например, патент США №6586251 и Valenzuela et al., ранее).
Кратко, путем бактериальной гомологичной рекомбинации получали нацеливающий вектор, содержащий выше- и нижележащие плечи гомологии β2m мыши из ВАС клона 89С24 из библиотеки RPCI-23 (Invitrogen). Плечи гомологии мыши сконструировали для фланкирования 2,8 т.п.н. фрагмента ДНК β2m человека, расположенного от экзона 2 до приблизительно 267 нуклеотидов по ходу транскрипции от некодирующего экзона 4 (фиг. 7). Кассету селекции по лекарственному средству (неомицину), фланкированную сайтами распознавания рекомбиназы (например, сайтами loxP) вводили в нацеливающий вектор для обеспечения последующей селекции. Конечный нацеливающий вектор линеаризировали и вводили электропорацией в ES клеточную линию F1H4 мышей (Valenzuela et al., ранее).
Нацеленные ES клеточные клоны с удаленной лекарственной кассетой (путем введения рекомбиназы Cre) вводили в зародыш мыши на стадии 8 клеток способом VELOCIMOUSE® (смотрите, например, патент США №7294754 и Poueymirou et al., ранее). VELOCIMICE® (FO мышей, полученные полностью из донорной ES клетки), несущих гуманизированный ген β2m, идентифицировали путем скрининга в отношении потери аллеля мыши и приобретения аллеля человека с использованием модификации аллельного анализа (Valenzuela et al., ранее).
Пример 2.2: Определение характеристик гуманизированных в отношении β2 микроглобулина мышей
Мышей, гетерозиготных в отношении гуманизированного гена β2 микроглобулина (β2m), оценивали в отношении экспрессии с использованием проточной цитометрии (фигуры 8. и 9).
Кратко, кровь получали от группы (n=4 на группу) мышей дикого типа, гуманизированных в отношении β2m, гуманизированных в отношении МНС (т.е., HLA человека) класс I и двойных гуманизированных в отношении β2m и МНС I класса мышей с использованием методов, известных в настоящей области техники. Кровь от каждой группы мышей обрабатывали лизирующим буфером ACK (Lonza Walkersville) для удаления эритроцитов. Оставшиеся клетки окрашивали с использованием конъюгированных с флуорохромом антител к CD3 (17А2), к CD 19 (1D3), к CD11b (M1/70), к HLA I класса человека и к β2 микроглобулину человека (2М2). Проточную цитометрию проводили с использованием BD-FACSCANTO™ (BD Biosciences).
Экспрессию HLA I класса человека обнаруживали на клетках от однократно гуманизированных и двойных гуманизированных животных, тогда как экспрессию β2 микроглобулина обнаруживали только на клетках от дважды гуманизированных мышей (фиг. 8). Коэкспрессия β2m человека и HLA I класса человека приводила к увеличению обнаруживаемого количества HLA I класса человека на клеточной поверхности по сравнению с экспрессией HLA I класса человека при отсутствии β2m человека (фиг. 9; средняя интенсивность флуоресценции 2370 по сравнению с 1387).
Пример 3. Иммунный ответ на пептиды вируса гриппа (Flu) и вируса Эпштейна-Барра (EBV), презентируемый АПК от генетически модифицированных мышей, экспрессирующих HLA-A2/H-2K и гуманизированный β2 микроглобулин.
РВМС от нескольких доноров-людей подвергали скринингу в отношении как экспрессии HLA-A2, так и их способности развивать ответ на пептиды вируса гриппа и EBV. Одного донора выбрали для последующих экспериментов.
Т-клетки человека выделяли из РВМС выбранного донора с использованием отрицательной селекции. Не относящиеся к Т-клеткам клетки селезенки выделяли из мыши, гетерозиготной в отношении химерного HLA-A2/H-2K и гетерозиготной в отношении гуманизированного гена β2 микроглобулина, и мыши дикого типа. Приблизительно 50000 не относящихся к Т-клеткам клеток селезенки от мышей добавляли на планшет для анализа иммуноферментных пятен, покрытый антителом к IFNγ человека. Добавляли пептид Flu (10 мкМ) или пул пептидов EBV (каждый по 5 мкМ). Poly IC добавляли в концентрации 25 мкг/лунка и лунки инкубировали в течение трех часов при 37°С в 5% CO2. Т-клетки человека (50000) и анти-CD28 человека добавляли к не относящимся к Т-клеткам клеткам селезенки и пептиды, и лунки инкубировали в течение 40 часов при 37°С в 5% CO2, после чего проводили анализ иммуноферментных пятен IFNγ.
Как показано на фиг. 10, Т-клетки человека были способны развивать ответ на пептиды flu и EBV, презентированные АПК мыши, которые экспрессировали химерный HLA-A2/H-2K и гуманизированный β2 микроглобулин на своей поверхности.
Эквиваленты
Специалистам в настоящей области техники будут понятны, или с использованием не более чем рутинного экспериментирования они смогут установить многие эквиваленты описанных в настоящем документе конкретных вариантов осуществления настоящего изобретения. Предусматривается, что такие эквиваленты включены в представленную ниже формулу изобретения.
Полное содержание всех процитированных в настоящей заявке непатентных документов, патентных заявок и патентов полностью включено посредством ссылки в настоящем документе.



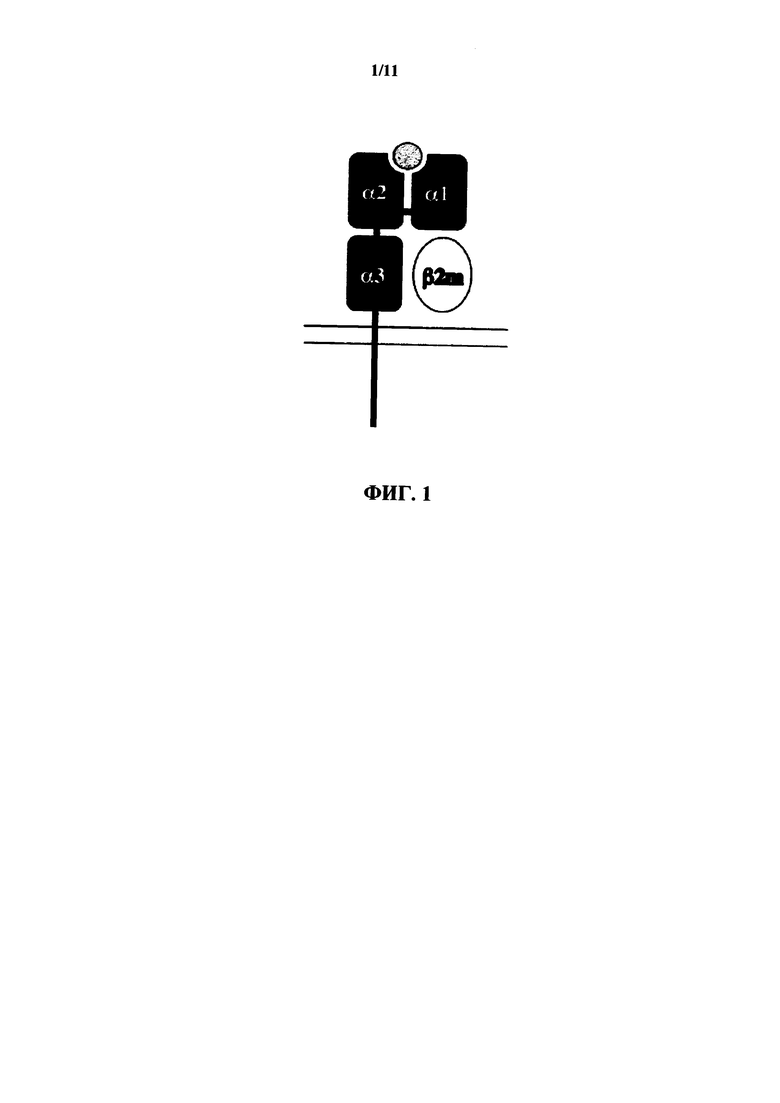
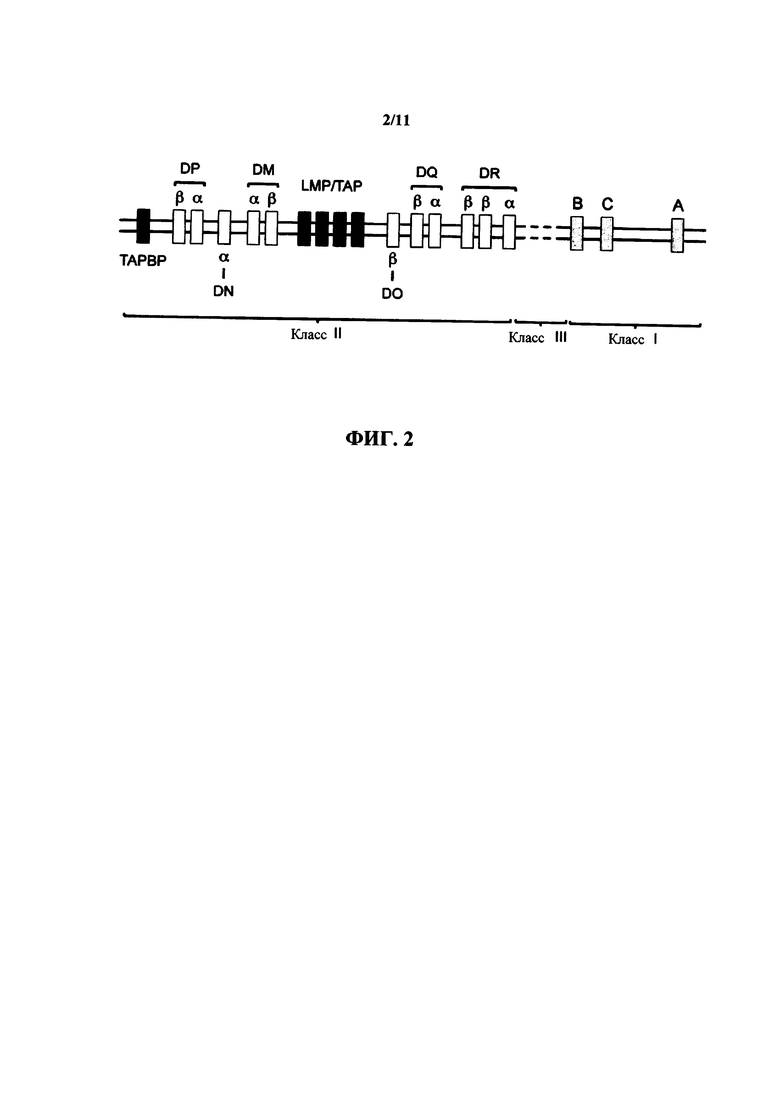

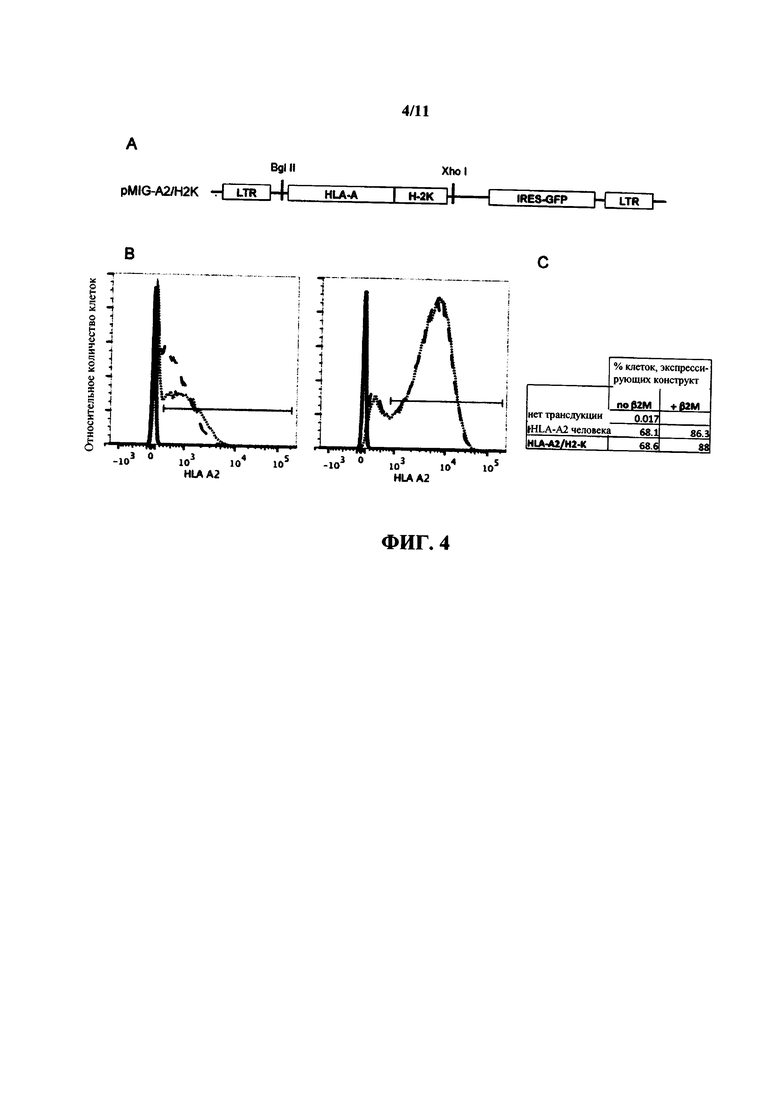
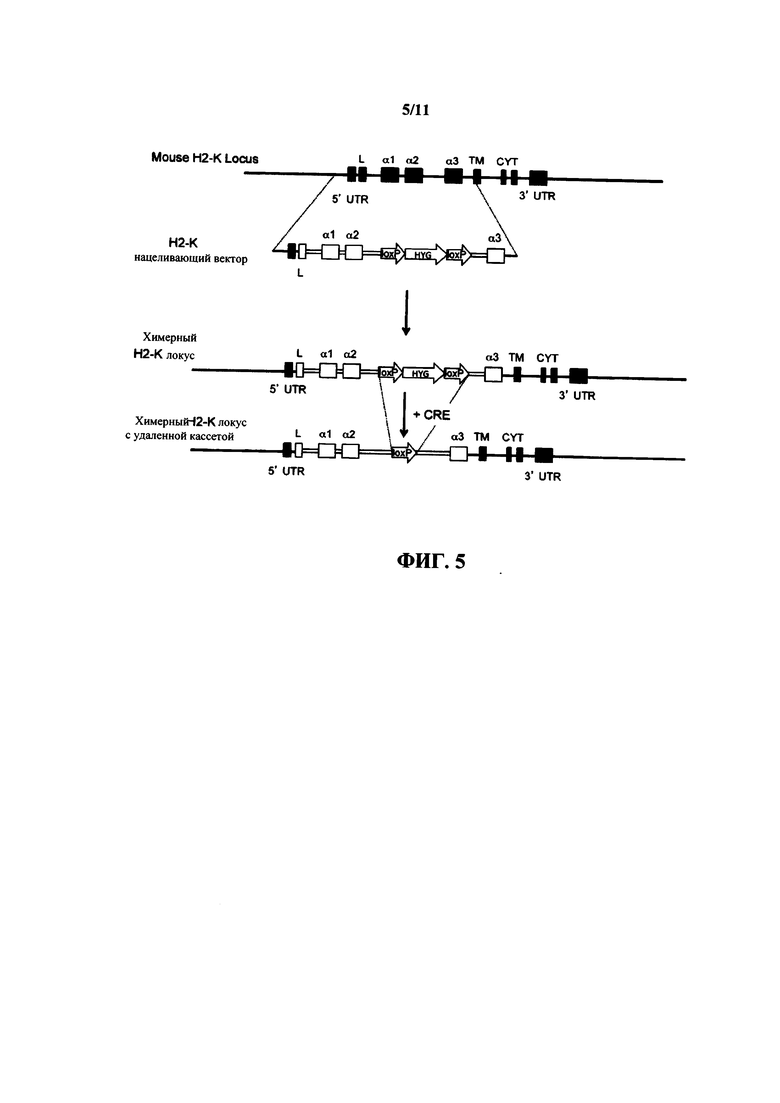
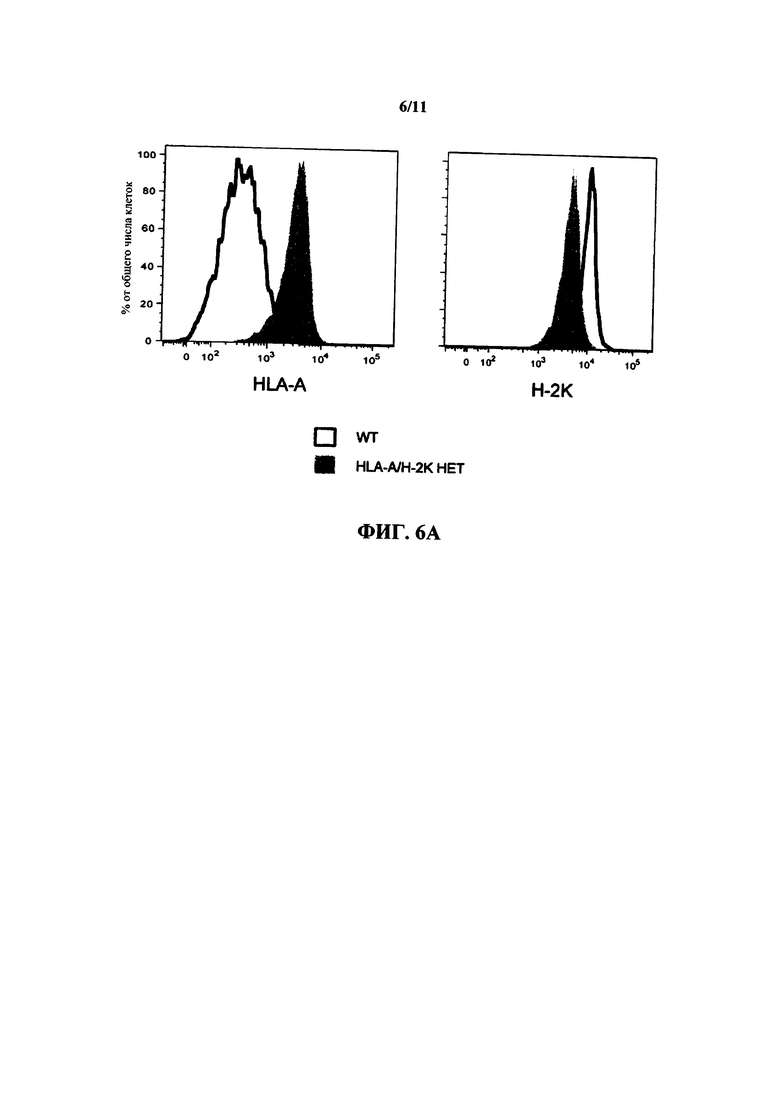
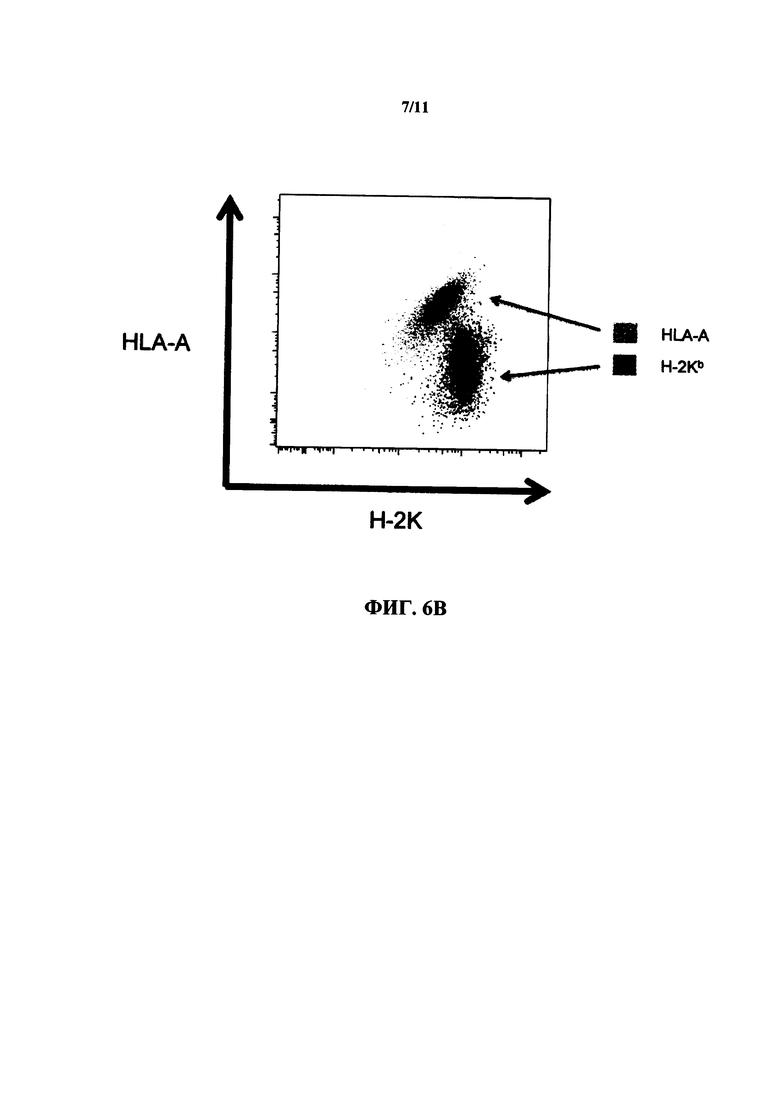
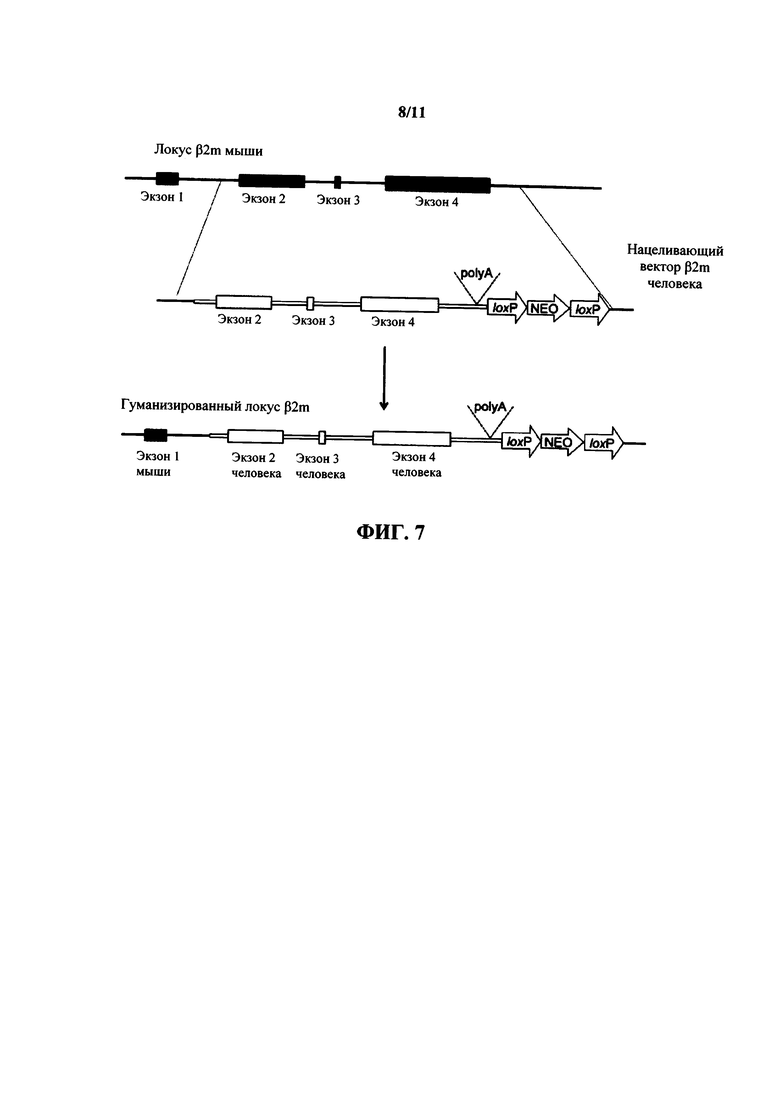
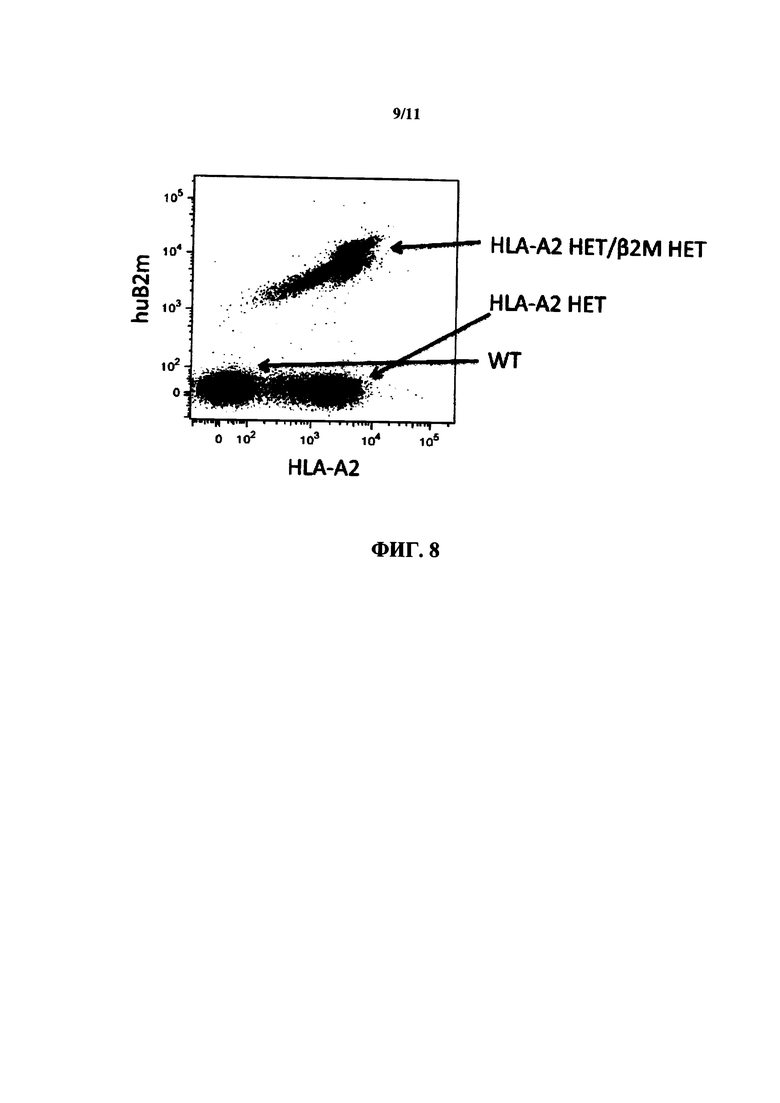
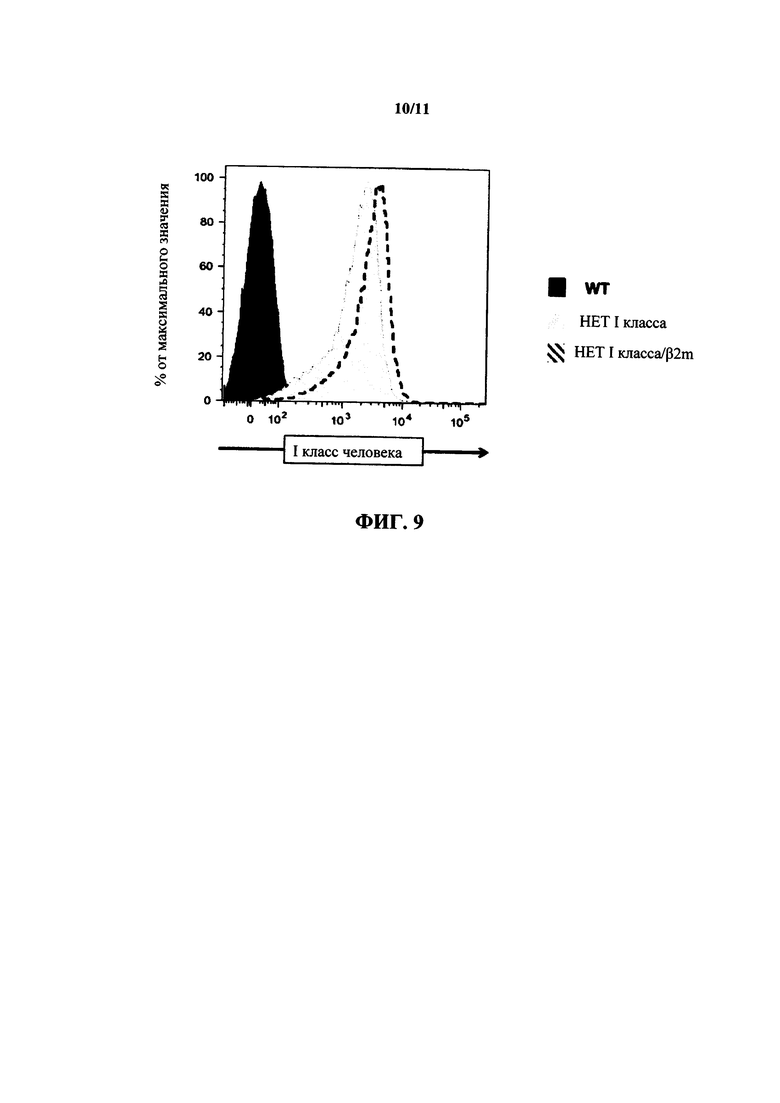
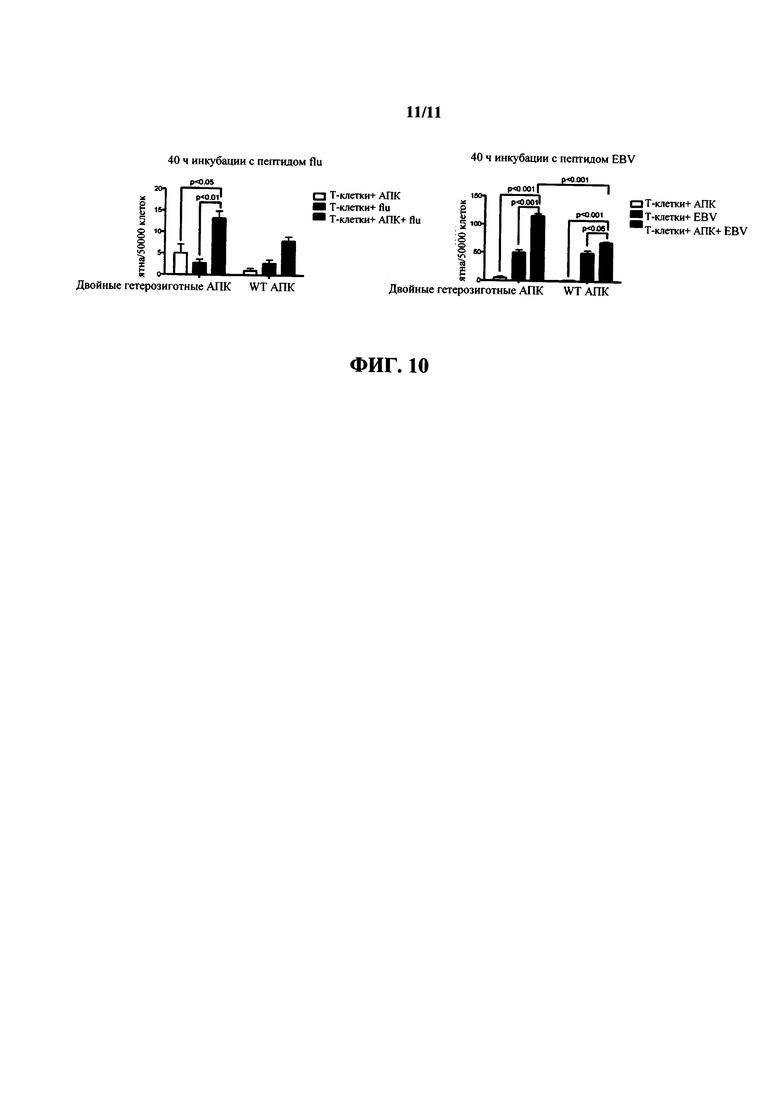
Настоящее изобретение относится к биотехнологии и генетической инженерии, в частности к генетически модифицированным грызунам, таким как мышь и крыса, которые экспрессируют химерный человеческий/не относящийся к человеку полипептид МНС I и/или человеческий полипептид β2 микроглобулина, а также к клеткам таких грызунов. Настоящее изобретение раскрывает способы получения указанных животных, а также способ идентификации эпитопа CTL и способ идентификации рестриктированного по HLA класса I пептида с их использованием. Настоящее изобретение позволяет получать биологические системы, которые будут создавать иммунную систему, которая проявляет компоненты, имитирующие функцию иммунной системы человека. 10 н. и 52 з.п. ф-лы, 10 ил., 1 табл., 3 пр.
1. Грызун, который экспрессирует химерный человеческий/относящийся к грызуну полипептид МНС I, содержащий в эндогенном локусе главного комплекса гистосовместимости I (МНС I) нуклеотидную последовательность, кодирующую химерный человеческий/относящийся к грызуну полипептид МНС I,
причем нуклеотидная последовательность функционально связана с эндогенными регуляторными элементами МНС класса I грызуна,
причем химерный полипептид содержит α1, α2 и α3 домены полипептида МНС I человека,
причем грызун экспрессирует химерный человеческий/относящийся к грызуну полипептид МНС I на поверхности клетки, и
при этом грызун представляет собой крысу или мышь.
2. Грызун по п. 1, причем грызун не экспрессирует внеклеточный домен эндогенного полипептида МНС I грызуна из эндогенного локуса МНС I грызуна.
3. Грызун по п. 2, у которого химерный полипептид содержит лидерную последовательность человека.
4. Грызун по п. 2, причем грызун представляет собой крысу.
5. Грызун по п. 4, у которого α1, α2 и α3 домены полипептида МНС I человека функционально связаны с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС I крысы.
6. Грызун по п. 2, причем грызун представляет собой мышь.
7. Грызун по п. 6, у которого α1, α2 и α3 домены полипептида МНС I человека функционально связаны с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС I мыши.
8. Грызун по п. 7, причем эндогенный локус МНС I грызуна представляет собой локус Н-2K мыши, и эндогенный полипептид МНС I мыши представляет собой Н-2K.
9. Грызун по п. 2, у которого полипептид МНС I человека выбран из группы, состоящей из HLA-A, HLA-B и HLA-C.
10. Грызун по п. 9, у которого полипептид МНС I человека представляет собой полипептид HLA-A.
11. Грызун по п. 10, у которого полипептид МНС I человека представляет собой полипептид HLA-A2.
12. Грызун по п. 10, причем полипептид HLA-A2 человека представляет собой полипептид HLA-A2.1.
13. Грызун по п. 12, причем грызун представляет собой мышь, эндогенный локус МНС I грызуна представляет собой локус Н-2K мыши, и эндогенный полипептид МНС I мыши представляет собой Н-2K.
14. Грызун по п. 13, причем локус Н-2K представляет собой локус Н-2Kb и эндогенный полипептид МНС I мыши представляет собой Н-2Kb.
15. Способ модификации локуса МНС I мыши для экспрессии химерного человеческого/мышиного полипептида МНС I, при этом способ предусматривает замещение в эндогенном локусе МНС I нуклеотидной последовательности, кодирующей α1, α2 и α3 домены полипептида МНС I мыши, нуклеотидной последовательностью, кодирующей α1, α2 и α3 домены полипептида МНС I человека.
16. Способ по п. 15, в котором мышь не экспрессирует внеклеточный домен полипептида МНС I мыши из эндогенного локуса МНС I мыши.
17. Способ по п. 15, в котором локус МНС I мыши представляет собой локус Н-2K, и полипептид МНС I мыши представляет собой полипептид Н-2K.
18. Способ по п. 15, в котором полипептид МНС I человека представляет собой полипептид HLA-A.
19. Способ по п. 15, в котором мышь экспрессирует трансмембранный и цитоплазматический домены полипептида МНС I мыши.
20. Способ по п. 15, в котором замещение проводят в одной ES клетке и одну ES клетку вводят в зародыш мыши для получения мыши.
21. Грызун, который экспрессирует гуманизированный β2 микроглобулин и химерный человеческий/относящийся к грызуну полипептид МНС I, при этом грызун дополнительно содержит в эндогенном локусе β2 микроглобулина грызуна первую нуклеотидную последовательность, кодирующую полипептид, содержащий аминокислотную последовательность β2 микроглобулина человека,
причем первая нуклеотидная последовательность содержит нуклеотидную последовательность, представленную в экзонах 2, 3 и 4 гена β2 микроглобулина человека, и в эндогенном локусе главного комплекса гистосовместимости I (МНС I) вторую нуклеотидную последовательность, кодирующую химерный человеческий/относящийся к грызуну полипептид МНС I, который содержит α1, α2 и α3 домены полипептида МНС I человека,
причем первая нуклеотидная последовательность функционально связана с эндогенными регуляторными элементами β2 микроглобулина грызуна,
причем вторая нуклеотидная последовательность функционально связана с эндогенными регуляторными элементами МНС класса I грызуна,
причем грызун экспрессирует гуманизированный полипептид β2 микроглобулина и химерный человеческий/относящийся к грызуну полипептид МНС I на поверхности клетки,
и при этом грызун представляет собой крысу или мышь.
22. Грызун по п. 21, причем грызун представляет собой крысу.
23. Грызун по п. 21, причем грызун не экспрессирует функциональный эндогенный полипептид β2 микроглобулина грызуна из эндогенного локуса β2 микроглобулина грызуна.
24. Грызун по п. 21, у которого первая нуклеотидная последовательность содержит нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека.
25. Грызун по п. 21, у которого первая нуклеотидная последовательность кодирует полипептид, содержащий аминокислоты 23-119 белка β2 микроглобулина человека.
26. Грызун по п. 24, у которого первая нуклеотидная последовательность дополнительно содержит нуклеотидную последовательность, представленную в экзоне 1 гена β2 микроглобулина грызуна.
27. Грызун по п. 25, у которого первая нуклеотидная последовательность дополнительно содержит нуклеотидную последовательность, представленную в экзоне 1 гена β2 микроглобулина грызуна.
28. Грызун по п. 21, причем грызун представляет собой мышь.
29. Грызун по п. 28, причем мышь не экспрессирует функциональный эндогенный β2 микроглобулин мыши из эндогенного локуса β2 микроглобулина мыши.
30. Грызун по п. 28, у которого первая нуклеотидная последовательность содержит нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека.
31. Грызун по п. 28, у которого первая нуклеотидная последовательность кодирует полипептид, содержащий аминокислоты 23-119 белка β2 микроглобулина человека.
32. Грызун по п. 30, у которого первая нуклеотидная последовательность дополнительно содержит нуклеотидную последовательность, представленную в экзоне 1 гена β2 микроглобулина мыши.
33. Грызун по любому из пп. 21-32, причем грызун не экспрессирует внеклеточный домен эндогенного полипептида МНС I грызуна из эндогенного локуса МНС I грызуна.
34. Грызун по любому из пп. 21-32, у которого вторая нуклеотидная последовательность дополнительно содержит последовательность, кодирующую лидерную последовательность человека.
35. Грызун по любому из пп. 21-32, у которого α1, α2 и α3 домены полипептида МНС I человека функционально связаны с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС I грызуна.
36. Способ получения мыши, которая экспрессирует гуманизированный полипептид β2 микроглобулина и химерный человеческий/мышиный полипептид МНС I, предусматривающий замещение в эндогенном локусе β2 микроглобулина мыши нуклеотидной последовательности, кодирующей полипептид β2 микроглобулина мыши, первой нуклеотидной последовательностью, кодирующей гуманизированный полипептид β2 микроглобулина, причем первая нуклеотидная последовательность содержит нуклеотидную последовательность, представленную в экзонах 2, 3 и 4 гена β2 микроглобулина человека, при этом первая нуклеотидная последовательность функционально связана с эндогенными регуляторными элементами β2 микроглобулина мыши, и
замещение в эндогенном МНС I локусе нуклеотидной последовательности, кодирующей α1, α2 и α3 домены полипептида МНС I мыши, второй нуклеотидной последовательностью, кодирующей α1, α2 и α3 домены полипептида МНС I человека, при этом вторая нуклеотидная последовательность функционально связана с эндогенными регуляторными элементами МНС класса I мыши.
37. Способ по п. 36, в котором мышь не экспрессирует функциональный полипептид β2 микроглобулина мыши из эндогенного локуса β2 микроглобулина.
38. Способ по п. 36, в котором первая нуклеотидная последовательность, кодирующая гуманизированный полипептид β2 микроглобулина, содержит нуклеотидную последовательность, представленную в экзоне 2 - экзоне 4 гена β2 микроглобулина человека.
39. Способ по п. 36, в котором нуклеотидная последовательность, кодирующая гуманизированный полипептид β2 микроглобулина, кодирует полипептид, содержащий аминокислоты 23-119 белка β2 микроглобулина человека.
40. Способ по п. 36, в котором модифицированный локус β2 микроглобулина сохраняет нуклеотидную последовательность экзона 1 гена β2 микроглобулина мыши.
41. Способ по п. 36, в котором замещение проводят в одной ES клетке и одну ES клетку вводят в зародыш мыши для получения мыши.
42. Способ по любому из пп. 36-41, в котором мышь не экспрессирует внеклеточный домен эндогенного полипептида МНС I мыши из эндогенного локуса МНС I мыши.
43. Способ по любому из пп. 36-41, в котором α1, α2 и α3 домены полипептида МНС I человека функционально связаны с трансмембранным и цитоплазматическим доменами полипептида МНС I мыши.
44. Способ получения генетически модифицированной мыши, которая экспрессирует химерный человеческий/мышиный полипептид МНС I и гуманизированный полипептид β2 микроглобулина, предусматривающий:
модификацию локуса МНС I первой мыши для экспрессии химерного человеческого/мышиного полипептида МНС I, предусматривающую замещение в эндогенном локусе МНС I мыши нуклеотидной последовательности, кодирующей α1, α2 и α3 домены полипептида МНС I мыши, нуклеотидной последовательностью, кодирующей α1, α2 и α3 домены полипептида МНС I человека;
модификацию локуса β2 микроглобулина второй мыши для экспрессии гуманизированного полипептида β2 микроглобулина, предусматривающую замещение в эндогенном локусе β2 микроглобулина мыши нуклеотидной последовательности, кодирующей полипептид β2 микроглобулина мыши, нуклеотидной последовательностью, кодирующей гуманизированный полипептид β2 микроглобулина, причем нуклеотидная последовательность, кодирующая гуманизированный β2 микроглобулин, содержит нуклеотидную последовательность, представленную в экзонах 2, 3 и 4 гена β2 микроглобулина человека; и
скрещивание первой и второй мыши для получения генетически модифицированной мыши, содержащей в своем геноме первую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид МНС I, и вторую нуклеотидную последовательность, кодирующую гуманизированный полипептид β2 микроглобулина, причем генетически модифицированная мышь экспрессирует химерный человеческий/мышиный полипептид МНС I и гуманизированный полипептид β2 микроглобулина.
45. Способ по п. 44, в котором локус МНС I представляет собой локус Н-2K, полипептид МНС I человека представляет собой HLA-A2, и мышь экспрессирует химерный полипептид HLA-A2/H-2K.
46. Способ по п. 45, в котором химерный полипептид HLA-A2/H-2K содержит внеклеточный домен полипептида HLA-A2 и цитоплазматический и трансмембранный домены полипептида Н-2K.
47. Способ по п. 44, в котором вторая нуклеотидная последовательность содержит нуклеотидные последовательности, представленные в экзоне 2 - экзоне 4 гена β2 микроглобулина человека, и нуклеотидную последовательность, представленную в экзоне 1 гена β2 микроглобулина мыши.
48. Нуклеиновая кислота, кодирующая химерный человеческий/относящийся к грызуну полипептид МНС I для оценки способности антигена инициировать иммунный ответ у человека, при этом нуклеиновая кислота содержит последовательность, кодирующую α1, α2 и α3 домены полипептида МНС I человека, функционально связанную с последовательностью, кодирующей трансмембранный и цитоплазматический домены полипептида МНС I грызуна.
49. Нуклеиновая кислота по п. 48, дополнительно характеризующаяся наличием регуляторных элементов МНС I грызуна, функционально связанных с последовательностью, кодирующей химерный человеческий/относящийся к грызуну полипептид МНС I.
50. Нуклеиновая кислота по п. 48, при этом полипептид МНС I грызуна представляет собой полипептид МНС I мыши.
51. Химерный человеческий/относящийся к грызуну полипептид МНС I для оценки способности антигена инициировать иммунный ответ у человека, содержащий α1, α2 и α3 домены полипептида МНС I человека, функционально связанные с трансмембранным и цитоплазматическим доменами полипептида МНС I грызуна.
52. Химерный человеческий/относящийся к грызуну полипептид МНС I по п. 51, причем грызун представляет собой крысу.
53. Химерный человеческий/относящийся к грызуну полипептид МНС I по п. 52, причем грызун представляет собой мышь.
54. Химерный человеческий/относящийся к грызуну полипептид МНС I по п. 53, причем человеческий полипептид МНС I представляет собой полипептид HLA-A2 и полипептид МНС I мыши представляет собой полипептид Н-2K.
55. Химерный человеческий/относящийся к грызуну полипептид МНС I по любому из пп. 51-54, причем полипептид МНС I нековалентно связан с человеческим или гуманизированным β2 микроглобулином.
56. Способ идентификации эпитопа CTL человека, предусматривающий:
воздействие на грызуна по любому из пп. 1-14 антигена, содержащего предполагаемый эпитоп для цитотоксических Т-лимфоцитов (CTL), предоставление грызуну возможности развить иммунный ответ,
выделение из грызуна рестриктированного по МНС класса I CD8+ CTL, который связывает эпитоп, и
идентификацию эпитопа, связанного рестриктированным по МНС класса I CD8+ CTL.
57. Способ идентификации рестриктированного по HLA класса I пептида, презентация которого клеткой человека и связывание лимфоцитом человека будет приводить к цитотоксичности несущей пептид клетки, предусматривающий:
воздействие на грызуна по любому из пп. 1-14 молекулой, содержащей представляющий интерес пептид,
выделение клетки грызуна, которая экспрессирует химерную человеческую/относящуюся к грызуну молекулу класса I, которая связывает представляющий интерес пептид,
воздействие на клетку лимфоцита человека, который способен осуществить рестриктированную по HLA класса I цитотоксичность, и
измерение индуцированной пептидом цитотоксичности для идентификации рестриктированного по HLA класса I пептида.
58. Выделенная клетка грызуна для экспрессии химерного человеческого/относящегося к грызуну полипептида МНС I, содержащая в эндогенном локусе МНС I нуклеотидную последовательность, кодирующую химерный человеческий/относящийся к грызуну полипептид МНС I, который содержит α1, α2 и α3 домены полипептида МНС I человека, причем нуклеотидная последовательность функционально связана с эндогенными регуляторными элементами МНС класса I грызуна, при этом грызун представляет собой крысу или мышь.
59. Выделенная клетка по п. 58, причем α1, α2 и α3 домены полипептида МНС I человека функционально связаны с трансмембранным и цитоплазматическим доменами полипептида МНС I грызуна.
60. Выделенная клетка по п. 59, причем полипептид МНС I грызуна представляет собой полипептид МНС I мыши.
61. Выделенная клетка по п. 58, причем полипептид МНС I человека представляет собой полипептид HLA-A2 и полипептид МНС I мыши представляет собой полипептид Н-2K.
62. Выделенная клетка по любому из пп. 58-61, причем клетка представляет собой антигенпрезентирующую клетку или эмбриональную стволовую клетку.
| WO 03006639 A1, 23.01.2003 | |||
| KALINKE U | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| VITIELLO A | |||
| et al | |||
| Analysis of the hla-restricted influenza-specific cytotoxic t lymphocyte response in transgenic mice carrying a chimeric human-mouse class i major histocompatibility complex, J Exp Med., 1991, 173(4), p.1007-1015 | |||
| РЫБЧИН В.Н | |||
| Основы генетической инженерии, Учебник для ВУЗов, Санкт-Петербург, Издательство СПбГТУ, 2002, 522 с., с.410-417. | |||

