ПЕРЕКРЕСТНАЯ ССЫЛКА НА СМЕЖНЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет по предварительным заявкам на патент США №№62/143,687 (подана 6 апреля 2015 г.), 62/158,804 (подана 8 мая 2015 г.) и 62/186,935 (подана 30 июня 2015 г.), причем каждая из заявок полностью включена в настоящий документ путем ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Официальная копия перечня последовательностей представлена в электронном виде через EFS-Web в виде файла с перечнем последовательностей в формате ASCII с наименованием 2016-04-06-10145WO01-SEQ-LIST_ST25.txt, созданного 6 апреля 2016 г., имеющего размер 56,7 килобайт и поданного одновременно с данным описанием. Перечень последовательностей, содержащийся в данном документе в формате ASCII, является частью спецификации и полностью включен в настоящий документ путем ссылки.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к не относящимся к человеку животным (например, грызунам, например мышам или крысам), обладающим способностью формировать по существу опосредованные человеческими (гуманизированными) Т-клетками иммунные ответы и экспрессировать: (i) один или более человеческих (гуманизированных) Т-клеточных корецепторов (например, CD4 и/или CD8 (например, CD8α и/или CD8β)); (ii) один или более человеческих (гуманизированных) главных комплексов гистосовместимости, которые ассоциируются с одним или более человеческими (гуманизированными) Т-клеточными корецепторами (например, МНС II (например, МНС α и/или МНС II β), и/или МНС I (например, МНС I α и/или β2-микроглобулин)); и/или (iii) человеческий (гуманизированный) Т-клеточный рецептор (TCR) (например, TCRα и/или TCRβ); эмбрионам, тканям, клеткам и/или нуклеиновым кислотам, выделенным из не относящихся к человеку животных; способам получения не относящихся к человеку животных и способам использования не относящихся к человеку животных для разработки терапевтических средств для человека.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
При адаптивном иммунном ответе инородные антигены распознаются молекулами рецепторов на В-лимфоцитах (например, иммуноглобулинами) и Т-лимфоцитах (например, Т-клеточным рецептором, который также обозначают TCR). Эти инородные антигены представляются на поверхности клеток в виде пептидных фрагментов специализированными белками, которые в общем называют молекулами главного комплекса гистосовместимости (МНС), а конкретно у человека называют человеческим лейкоцитарным антигеном (HLA). В ходе опосредованного Т-клетками ответа антигены, представляемые молекулами МНС, распознаются Т-клеточным рецептором. Однако для эффективного иммунного ответа, помимо распознавания Т-клеточным рецептором комплекса МНС-антиген, требуются дополнительные механизмы. Также необходимо связывание молекулы Т-клеточного корецептора (например, CD4 или CD8) с инвариантным участком МНС.
Т-клетки имеют несколько разновидностей, включая Т-хелперные клетки и цитотоксические Т-клетки. Т-хелперные клетки экспрессируют корецептор CD4 и распознают антигены, связанные с молекулами МНС II. Т-клетки CD4+ активируют другие эффекторные клетки в иммунной системе, например экспрессирующие МНС II В-клетки для продукции антител, экспрессирующие МНС II макрофаги для уничтожения патогенов и т.д. Связывание CD4 и Т-клеточного рецептора с одним и тем же чужеродным антигеном, представленным МНС II, делает Т-клетку гораздо более восприимчивой к этому антигену.
В противоположность этому цитотоксические Т-клетки (CTL) экспрессируют корецептор CD8 и распознают чужеродные антигены, связанные с молекулами МНС I. CTL специализируются на уничтожении любой клетки, несущей связанный МНС I пептид, распознаваемый их собственным связанным с мембраной TCR. При представлении клеткой пептидов, полученных из отсутствующих в норме клеточных белков (например, имеющих вирусное, опухолевое или другое чужеродное происхождение), такие пептиды распознаются CTL, которые активируются и уничтожают клетку, представляющую пептид. Аналогично CD4 задействование CD8 делает CTL более восприимчивыми к представленному МНС I антигену.
Не все антигены провоцируют Т-клеточную активацию вследствие механизмов толерантности. Однако при некоторых заболеваниях (например, раке, аутоиммунных заболеваниях) пептиды, полученные из собственных белков, становятся мишенью клеточного компонента иммунной системы, что приводит к разрушению клеток, представляющих такие пептиды. Имели место большие достижения в отношении распознавания клинически значимых антигенов (например, антигенов, ассоциированных с различными типами рака) и/или последовательностей TCR, которые связываются с клинически значимыми антигенами. Однако с целью улучшения идентификации и отбора клинически значимых пептидов, которые будут провоцировать приемлемый ответ в человеческой Т-клетке, и/или TCR, способных связываться с клинически значимыми антигенами (например, для адоптивной иммунотерапии рака, Т-клеточной вакцинации для лечения аутоиммунных заболеваний и т.д.), сохраняется потребность в in vivo и in vitro системах, которые имитируют аспекты человеческой иммунной системы. Таким образом, существует потребность в биологических системах (например, генетически модифицированных не относящихся к человеку животных и клетках), которые способны представлять компоненты человеческой иммунной системы, в частности компоненты Т-клеточного иммунного ответа.
ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Как описано в настоящем документе, тимус генетически модифицированных не относящихся к человеку животных, содержащих по существу гуманизированную Т-клеточную иммунную систему, имеет аналогичные абсолютные количества тимоцитов и Т-клеток CD3+, как у контрольных животных. Кроме того, эти клетки демонстрируют сопоставимое с контрольными животными развитие в одинарные положительные Т-клетки и способны формировать устойчивый человеческий клеточный иммунный ответ на антиген, например вирусный антиген. Человеческий клеточный ответ не относящихся к человеку животных, как правило, включает активированные нечеловеческие Т-клетки, экспрессирующие человеческие или гуманизированные вариабельные домены Т-клеточного рецептора (TCR), которые распознают антиген, представленный в связывающем пептид кармане, образованном внеклеточными доменами человеческого лейкоцитарного антигена (HLA), которые могут быть экспрессированы на поверхности клеток, представляющих нечеловеческий антиген. В некоторых вариантах осуществления по существу гуманизированная Т-клеточная иммунная система содержит:
(А) нечеловеческую Т-клетку, которая экспрессирует:
(i) Т-клеточный корецепторный полипептид, содержащий часть внеклеточного участка или весь внеклеточный участок человеческого Т-клеточного корецептора, например Т-клеточный корецепторный полипептид, содержащий один или более внеклеточных доменов человеческого Т-клеточного корецептора, в результате чего Т-клеточный корецепторный полипептид способен ассоциироваться и/или ассоциируется с:
(a) одним или более внеклеточными доменами молекулы человеческого или гуманизированного HLA (например, первым внеклеточным доменом человеческого HLA, который представляет собой сайт связывания для Т-клеточного корецепторного полипептида, и/или вторым внеклеточным доменом человеческого HLA, который образует связывающий пептид карман, например с третьим внеклеточным доменом человеческого HLA);
(b) внеклеточным доменом вариабельного домена человеческого или гуманизированного TCR (например, вариабельным доменом человеческого или гуманизированного TCRα и/или вариабельным доменом человеческого или гуманизированного TCRβ, который соответственно кодируется по меньшей мере одним сегментом гена вариабельного участка человеческого TCRα и/или TCRβ); и/или
(с) внеклеточным доменом константного домена человеческого TCR; и
(ii) Т-клеточный рецептор (TCR), содержащий по меньшей мере вариабельный домен человеческого TCR; и необязательно
(В) представляющую нечеловеческий антиген клетку, которая представляет антиген в связи с человеческим HLA, например представляющую нечеловеческий антиген клетку, которая экспрессирует на своей клеточной поверхности по меньшей мере одну молекулу МНС, которая содержит связывающий пептид карман, образованный двумя внеклеточными доменами человеческого HLA, и способна активировать и/или активирует нечеловеческую Т-клетку.
В одном аспекте нечеловеческая Т-клетка и представляющая нечеловеческий антиген клетка находятся в одном и том же не относящемся к человеку животном или выделены из него.
Соответственно, в настоящем документе предложены не относящиеся к человеку животные (например, грызуны, например мыши или крысы), созданные методами генной инженерии и экспрессирующие:
(A) человеческий или гуманизированный Т-клеточный корецептор (например, человеческий или гуманизированный CD4 и/или человеческий или гуманизированный CD8 (например, человеческий или гуманизированный CD8α и/или человеческий или гуманизированный CD8β));
(B) человеческий или гуманизированный главный комплекс гистосовместимости, который ассоциируется с человеческим или гуманизированным Т-клеточным корецептором (например, человеческим или гуманизированным МНС II (например, человеческим или гуманизированным МНС II α и/или человеческим или гуманизированным МНС II β), который связывается с человеческим или гуманизированным CD4 и/или человеческим или гуманизированным МНС I (например, человеческим или гуманизированным МНС Iα и необязательно человеческим или гуманизированным β2-микроглобулином), который связывается с человеческим или гуманизированным CD8); и/или
(C) человеческий или гуманизированный Т-клеточный рецептор (TCR);
а также эмбрионы, ткани и клетки, экспрессирующие их, и нуклеиновые кислоты, кодирующие вышеперечисленное. Также предложены способы получения и использования описанных не относящихся к человеку животных.
В одном аспекте предложено генетически модифицированное не относящееся к человеку животное, содержащее:
(A) гуманизированный корецептор CD4 и/или гуманизированный корецептор CD8, содержащий гуманизированный полипептид CD8α и гуманизированный полипептид CD8β (например, не относящееся к человеку животное содержит, например, в своем геноме зародышевой линии, первую нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD4, и/или вторую нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8α, и третью нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8β),
причем каждый гуманизированный Т-клеточный корецепторный полипептид содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого Т-клеточного корецептора, например, в котором гуманизированный корецептор CD4 содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого корецептора CD4 и/или гуманизированный корецептор CD8 содержит по меньшей мере трансмембранный и цитоплазматический домены полипептидов нечеловеческого CD8α и нечеловеческого CD8β,
при этом каждый химерный Т-клеточный корецепторный полипептид содержит часть внеклеточного участка или весь внеклеточный участок человеческого Т-клеточного корецептора, например один или более внеклеточных доменов человеческого Т-клеточного корецептора, например по меньшей мере внеклеточный домен человеческого Т-клеточного корецептора, который ассоциируется с молекулой HLA, например, в котором гуманизированный корецептор CD4 содержит внеклеточный участок (или его части, например внеклеточный (-ые) домен (-ы)) человеческого CD4, который ответственен за взаимодействие с МНС II, вариабельные домены Т-клеточного рецептора, константные домены Т-клеточного рецептора или их комбинацию, и/или, например, в котором гуманизированный корецептор CD8 содержит внеклеточные участки (или их части, например внеклеточные домены) человеческого CD8α и человеческого CD8β, которые ответственны за взаимодействие с МНС I, вариабельные домены Т-клеточного рецептора, константные домены Т-клеточного рецептора или их комбинацию;
(B) человеческий (гуманизированный) TCR (например, не относящееся к человеку животное содержит, например, в своем геноме зародышевой линии, неперестроенный вариабельный локус гена Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с нечеловеческой константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с нечеловеческой константной последовательностью гена TCRβ); и необязательно
(С) человеческий (гуманизированный) комплекс МНС II, который ассоциируется с гуманизированным корецептором CD4, и/или человеческий (гуманизированный) комплекс МНС I, который ассоциируется с гуманизированным корецептором CD8 (например, не относящееся к человеку животное содержит, например, в своем геноме зародышевой линии, первую последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС IIα, и вторую последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС IIβ, и/или третью последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС I),
при этом каждый химерный полипептид МНС содержит по меньшей мере внеклеточный участок (или его часть) человеческого полипептида МНС (например, полипептида HLA), который либо сам по себе (например, МНС I), либо в комплексе с другим химерным полипептидом МНС (например, МНС II α и МНС II β) соответственно способен ассоциироваться с человеческим (гуманизированным) корецептором CD8 или человеческим (гуманизированным) корецептором CD4 и представлять пептид в связи с HLA, например, в котором гуманизированный комплекс МНС II содержит: (i) химерный человеческий/нечеловеческий полипептид МНС II α, содержащий α1 и α2 домены человеческого полипептида HLA класса II α и трансмембранный и цитоплазматический домены нечеловеческого полипептида HLA класса II α; и (ii) химерный человеческий/нечеловеческий полипептид МНС II β, содержащий β1 и β2 домены человеческого полипептида HLA класса II β и трансмембранный и цитоплазматический домены нечеловеческого полипептида HLA класса II β, и/или в котором гуманизированный комплекс МНС I содержит α1, α2 и α3 домены человеческого полипептида МНС I, и необязательно человеческий (гуманизированный) β2 микроглобулин.
В некоторых вариантах осуществления не относящееся к человеку животное содержит:
(А) гуманизированный корецептор CD4 и гуманизированный корецептор CD8, содержащие гуманизированный полипептид CD8α или гуманизированный полипептид CD8β (например, не относящееся к человеку животное содержит, например, в своем геноме зародышевой линии, первую нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD4, вторую нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8α, и третью нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8β),
причем каждый гуманизированный Т-клеточный корецепторный полипептид содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого Т-клеточного корецептора, например, в котором гуманизированный корецептор CD4 содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого корецептора CD4, а гуманизированный корецептор CD8 содержит по меньшей мере трансмембранный и цитоплазматический домены полипептидов нечеловеческого CD8α и нечеловеческого CD8β,
при этом каждый химерный Т-клеточный корецепторный полипептид содержит часть внеклеточного участка или весь внеклеточный участок человеческого Т-клеточного корецептора, например один или более внеклеточных доменов человеческого Т-клеточного корецептора, например по меньшей мере внеклеточный домен человеческого Т-клеточного корецептора, который ассоциируется с молекулой HLA, например, в котором гуманизированный корецептор CD4 содержит внеклеточный участок (или его части, например внеклеточный (-ые) домен (-ы)) человеческого CD4, который ответственен за взаимодействие с МНС II, вариабельные домены Т-клеточного рецептора, константные домены Т-клеточного рецептора или их комбинацию, и/или, например, в котором гуманизированный корецептор CD8 содержит внеклеточные участки (или их части, например внеклеточные домены) человеческого CD8α и человеческого CD8β, которые ответственны за взаимодействие с МНС I, вариабельные домены Т-клеточного рецептора, константные домены Т-клеточного рецептора или их комбинацию;
(B) гуманизированный TCR (например, не относящееся к человеку животное содержит, например, в своем геноме зародышевой линии, неперестроенный вариабельный локус гена Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с нечеловеческой константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с нечеловеческой константной последовательностью гена TCRβ); и
(C) гуманизированный комплекс МНС II, который ассоциируется с гуманизированным корецептором CD4, и гуманизированный комплекс МНС I, который ассоциируется с гуманизированным корецептором CD8 (например, не относящееся к человеку животное содержит, например, в своем геноме зародышевой линии, первую последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС IIα, вторую последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС IIβ, и третью последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС I),
при этом каждый химерный полипептид МНС содержит по меньшей мере внеклеточный участок (или его часть) человеческого полипептида МНС (например, полипептида HLA), который либо сам по себе (например, МНС I), либо в комплексе с другим химерным полипептидом МНС (например, МНС II α и МНС II β) соответственно способен ассоциироваться с гуманизированным корецептором CD8 или гуманизированным корецептором CD4 и представлять пептид в случае HLA, например, в котором гуманизированный комплекс МНС II содержит: (i) химерный человеческий/нечеловеческий полипептид МНС II α, содержащий α1 и α2 домены человеческого полипептида HLA класса II α и трансмембранный и цитоплазматический домены нечеловеческого полипептида HLA класса II α; и (ii) химерный человеческий/нечеловеческий полипептид МНС II β, содержащий β1 и β2 домены человеческого полипептида HLA класса II β и трансмембранный и цитоплазматический домены нечеловеческого полипептида HLA класса II β; и (iii) гуманизированный комплекс МНС I содержит α1, α2 и α3 домены человеческого полипептида МНС I и необязательно человеческий (гуманизированный) β2 микроглобулин (например, не относящееся к человеку животное дополнительно содержит локус β2 микроглобулина, кодирующий полипептид, содержащий аминокислотную последовательность человеческого β2 микроглобулина или ее часть).
В некоторых вариантах осуществления первая нуклеотидная последовательность, кодирующая химерный полипептид Т-клеточного корецептора CD4, находится в эндогенном локусе Т-клеточного корецептора CD4 и/или вторая нуклеотидная последовательность, кодирующая химерный полипептид Т-клеточного корецептора CD8α, находится в эндогенном локусе Т-клеточного корецептора CD8α, а третья нуклеотидная последовательность, кодирующая химерный полипептид Т-клеточного корецептора CD8β, находится в эндогенном локусе Т-клеточного корецептора CD8β. Дополнительные варианты осуществления включают химерный человеческий/нечеловеческий полипептид CD4, кодируемый геном, изображенным на ФИГ. 5А (например, в котором человеческий участок полученного химерного человеческого/нечеловеческого полипептида Т-клеточного корецептора CD4 содержит по меньшей мере домены человеческого Ig1, человеческого Ig2 и человеческого Ig3, также соответственно называемые доменами D1, D2 и D3), и/или химерный корецептор CD8, кодируемый генами, изображенными на ФИГ. 5В (например, в котором человеческий участок химерного корецептора CD8 содержит весь или по существу весь внеклеточный участок человеческого полипептида CD8 (например, CD8α и/или CD8β), включая подобные человеческому иммуноглобулину V (IgV) домены α и β. В некоторых вариантах осуществления человеческий участок химерного полипептида Т-клеточного корецептора CD4 содержит один или более внеклеточных доменов человеческого полипептида CD4 (например, D1, D2, D3, D4 или любую их комбинацию), а нечеловеческий участок химерного полипептида Т-клеточного корецептора CD4 содержит трансмембранный и цитоплазматический домены нечеловеческого Т-клеточного корецептора CD4, человеческий участок химерного полипептида CD8α содержит внеклеточный домен (например, IgV-подобный домен) человеческого полипептида CD8α, а нечеловеческий участок химерного полипептида CD8α содержит трансмембранный и цитоплазматический домены нечеловеческого полипептида CD8α, и/или человеческий участок химерного полипептида CD8β содержит внеклеточный домен (например, IgV-подобный домен) человеческого полипептида CD8β, а нечеловеческий участок химерного полипептида Т-клеточного корецептора CD8β содержит трансмембранный и цитоплазматический домены нечеловеческого полипептида CD8β.
В некоторых вариантах осуществления первая последовательность нуклеиновых кислот, кодирующая человеческий (гуманизированный) МНС II α, находится в эндогенном нечеловеческом локусе МНС II α, вторая последовательность нуклеиновых кислот, кодирующая человеческий (гуманизированный) МНС II β, находится в эндогенном нечеловеческом локусе МНС II β и/или третья последовательность нуклеиновых кислот, кодирующая человеческий (гуманизированный) МНС I, находится в эндогенном нечеловеческом локусе МНС I. В одном аспекте человеческий (гуманизированный) полипептид МНС IIα содержит внеклеточный участок (или его часть) человеческого полипептида МНС IIα (например, полипептида HLA класса IIα), человеческий (гуманизированный) полипептид МНС IIβ содержит внеклеточный участок (или его часть) человеческого полипептида МНС IIβ (например, полипептида HLA класса Iβ) и/или человеческий (гуманизированный) полипептид МНС I содержит внеклеточный участок (или его часть) человеческого полипептида МНС I (например, полипептида HLA класса I). В некоторых вариантах осуществления гуманизированный полипептид МНС II α содержит человеческие домены МНС II α1 и α2, гуманизированный полипептид МНС II β содержит человеческие домены МНС II β1 и β2 и/или гуманизированный полипептид МНС I содержит человеческие домены МНС I α1, α2 и α3. В некоторых вариантах осуществления первая последовательность нуклеиновых кислот, кодирующая химерный человеческий/нечеловеческий полипептид МНС II α, экспрессируется под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов МНС II α, вторая последовательность нуклеиновых кислот, кодирующая химерный человеческий/нечеловеческий полипептид МНС II β, экспрессируется под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов МНС II β и/или третья последовательность нуклеиновых кислот, кодирующая химерный человеческий/нечеловеческий полипептид МНС I, экспресcируется под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов МНС I. В дополнительных вариантах осуществления нечеловеческий участок химерного человеческого/нечеловеческого полипептида MHC II α содержит трансмембранный и цитоплазматический домены эндогенного нечеловеческого полипептида МНС II α, нечеловеческий участок химерного человеческого/нечеловеческого полипептида МНС II β содержит трансмембранный и цитоплазматический домены эндогенного нечеловеческого полипептида МНС II β и/или нечеловеческий участок химерного человеческого/нечеловеческого полипептида МНС I содержит трансмембранный и цитоплазматический домены эндогенного нечеловеческого полипептида МНС I. Варианты осуществления включают не относящихся к человеку животных, у которых человеческий участок белков химерного человеческого/нечеловеческого комплекса МНС II получен из соответствующих человеческих белков HLA класса II, выбранных из группы, состоящей из HLA-DR, HLA-DQ и HLA-DP, и/или у которых человеческий участок третьего химерного человеческого/нечеловеческого полипептида МНС I получен из человеческого HLA-A, человеческого HLA-B или человеческого HLA-C. В качестве не имеющих ограничительного характера примеров в некоторых вариантах осуществления химерный полипептид МНС II α содержит внеклеточный участок (или его часть) белка HLA-DRα, белка HLA-DQ α или белка HLA-DP α, химерный полипептид МНС II β содержит внеклеточный участок (или его часть) белка HLA-DRβ, белка HLA-DQ β или белка HLA-DP β, и/или химерный полипептид МНС I содержит внеклеточный участок (или его часть) человеческого белка HLA-A, человеческого белка HLA-B или человеческого белка HLA-C. Также предложены не относящиеся к человеку животные, у которых человеческий участок белков химерного человеческого/нечеловеческого комплекса МНС II, полученный из соответствующих человеческих белков HLA-DR, например человеческий участок человеческого/нечеловеческого полипептида МНС II α, содержит домены α1 и α2 цепи α HLA-DR2, а человеческий участок человеческого/нечеловеческого полипептида МНС II β содержит домены β1 и β2 цепи β HLA-DR2, и/или у которых человеческий участок полипептида МНС I получен из человеческого полипептида HLA-A, например, человеческий участок человеческого/нечеловеческого полипептида МНС I содержит домены α1, α2 и α3 человеческого полипептида HLA-A2, например домены α1, α2 и α3 человеческого полипептида HLA-A2.1. Также предложены не относящиеся к человеку животные, у которых нечеловеческие участки комплекса МНС II получены из мышиной кодирующей последовательности Н-2Е и/или у которых нечеловеческие участки полипептида МНС I получены из мышиной кодирующей последовательности Н-2K. Например, химерный полипептид МНС II α содержит трансмембранный и цитоплазматический домены мышиного полипептида Н-2Е α, химерный полипептид МНС II β содержит трансмембранный и цитоплазматический домены мышиного полипептида Н-2Е β, а химерный полипептид МНС I содержит трансмембранный и цитоплазматический домены мышиного полипептида Н-2K.
В некоторых вариантах осуществления неперестроенный вариабельный локус гена TCRα находится в эндогенном вариабельном локусе гена TCRα, а неперестроенный вариабельный локус гена TCRβ находится в эндогенном вариабельном локусе гена TCRβ. В некоторых аспектах неперестроенный вариабельный локус гена TCRα содержит полный набор человеческих неперестроенных сегментов гена Vα и полный набор человеческих неперестроенных сегментов гена Jα и/или неперестроенный вариабельный локус гена TCRβ содержит полный набор человеческих неперестроенных сегментов гена Vβ, полный набор человеческих неперестроенных сегментов гена Dβ и полный набор человеческих неперестроенных сегментов гена Jβ. В некоторых вариантах осуществления человеческие неперестроенные сегменты гена Vα и Jα перестраиваются с образованием перестроенной человеческой последовательности Vα/Jα и/или человеческие неперестроенные сегменты гена Vβ, Dβ и Jβ перестраиваются с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ. В некоторых вариантах осуществления не относящееся к человеку животное, описанное в настоящем документе, экспрессирует Т-клеточный рецептор, содержащий человеческий вариабельный участок TCRα и/или человеческий вариабельный участок TCRβ на поверхности Т-клетки. В некоторых вариантах осуществления эндогенные нечеловеческие сегменты Vα и Jα неспособны перестраиваться с образованием перестроенной последовательности Vα/Jα и/или эндогенные нечеловеческие сегменты Vβ, Dβ и Jβ неспособны перестраиваться с образованием перестроенной последовательности Vβ/Dβ/Jβ, например, у животного может отсутствовать функциональный эндогенный нечеловеческий вариабельный локус TCRα и/или у животного может отсутствовать функциональный эндогенный нечеловеческий вариабельный локус TCRβ, например, животное содержит: (а) делецию всех или по существу всех функциональных эндогенных сегментов гена Vα, (b) делецию всех или по существу всех функциональных эндогенных сегментов гена Jα, (с) делецию всех или по существу всех функциональных эндогенных сегментов гена Vβ, (d) делецию всех или по существу всех функциональных эндогенных сегментов гена Dβ, (e) делецию всех или по существу всех функциональных эндогенных сегментов гена Jβ, и/или (f) их комбинацию. В некоторых вариантах осуществления в эндогенном нечеловеческом вариабельном локусе TCRα отсутствуют все или по существу все функциональные эндогенные сегменты гена Vα и/или отсутствуют все или по существу все функциональные эндогенные сегменты гена Jα, и/или в эндогенном нечеловеческом вариабельном локусе TCRβ: (а) отсутствуют все или по существу все функциональные эндогенные сегменты гена Vβ, (b) отсутствуют все или по существу все функциональные эндогенные сегменты гена Dβ, (с) отсутствуют все или по существу все функциональные эндогенные сегменты гена Jβ, или (d) любая комбинация (а), (b) и (с).
В некоторых вариантах осуществления первая, вторая и/или третья нуклеотидная (-ые) последовательность (-и), соответственно кодирующая (-ие) полипептид (-ы) химерного Т-клеточного корецептора CD4, CD8α и/или CD8β, находятся в эндогенных локусах Т-клеточного корецептора, например в эндогенных локусах корецептора CD4, CD8α и/или CD8β соответственно; неперестроенный вариабельный локус гена TCRα находится в эндогенном вариабельном локусе гена TCRα; неперестроенный вариабельный локус гена TCRβ находится в эндогенном вариабельном локусе гена TCRβ; и/или первая, вторая и/или третья последовательность (-и) нуклеиновых кислот, соответственно кодирующая (-ие) химерный (-ые) полипептид (-ы) МНС II α, МНС II β и/или МНС I, находится (находятся) в эндогенных локусах МНС; например в локусах МНС II α, МНС II β и/или МНС I соответственно. В некоторых вариантах осуществления нуклеотидная (-ые) последовательность (-и), кодирующая (-ие) химерный (-ые) Т-клеточный (-ые) корецептор (-ы), неперестроенный вариабельный локус гена TCRα, неперестроенный вариабельный локус гена TCRβ и/или последовательность (-и) нуклеиновых кислот, кодирующая (-ие) химерную (-ые) молекулу (-ы) МНС, может (могут) быть функционально связана (-ы) с нечеловеческими промоторными и регуляторными последовательностями. Например, первая нуклеотидная последовательность может быть экспрессирована под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов CD4, вторая нуклеотидная последовательность может быть экспрессирована под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов CD8α и/или третья нуклеотидная последовательность может быть экспрессирована под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов CD8β; неперестроенный вариабельный локус гена TCRα может быть экспрессирован под регуляторным контролем эндогенных регуляторных и промоторных элементов TCRα (вариабельный участок), а неперестроенный вариабельный локус гена TCRβ может быть экспрессирован под регуляторным контролем эндогенных регуляторных и промоторных элементов TCRβ (вариабельный участок); первая последовательность нуклеиновых кислот может быть экспрессирована под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов МНС II α, вторая последовательность нуклеиновых кислот может быть экспрессирована под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов МНС II β, а третья последовательность нуклеиновых кислот может быть экспрессирована под регуляторным контролем эндогенных нечеловеческих промоторных и регуляторных элементов МНС I.
В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая внеклеточный участок (или его части, например D1, D2, D3 и/или D4) человеческого полипептида CD4, замещает последовательность, кодирующую внеклеточный участок (или его части, например D1, D2, D3 и/или D4) эндогенного нечеловеческого (мышиного) полипептида корецептора CD4, и может быть функционально связана с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого (мышиного) CD4, в эндогенном нечеловеческом (мышином) локусе корецептора CD4; нуклеотидная последовательность, кодирующая весь внеклеточный участок или часть внеклеточного участка человеческого полипептида CD8α, замещает последовательность, кодирующую весь внеклеточный участок или часть внеклеточного участка эндогенного нечеловеческого (мышиного) полипептида Т-клеточного CD8α, и может быть функционально связана с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого (мышиного) CD8α, в эндогенном нечеловеческом (мышином) локусе CD8α; нуклеотидная последовательность, кодирующая весь внеклеточный домен или часть внеклеточного домена человеческого полипептида CD8β, замещает последовательность, кодирующую весь внеклеточный домен или часть внеклеточного домена эндогенного нечеловеческого (мышиного) полипептида Т-клеточного CD8β, и может быть функционально связана с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого CD8β, в эндогенном локусе CD8β; неперестроенный вариабельный локус гена TCRα замещает один или более эндогенных сегментов гена Vα и/или Jα в эндогенном нечеловеческом (мышином) вариабельном локусе гена TCRα; неперестроенный вариабельный локус гена TCRβ замещает один или более эндогенных сегментов гена Vβ, Dβ и/или Jα в эндогенном нечеловеческом (мышином) вариабельном локусе гена TCRβ; последовательность нуклеиновых кислот, кодирующая внеклеточный участок (или его части, например домены α1 и α2) человеческого полипептида МНС II α, замещает последовательность, кодирующую внеклеточный участок (или его части, например домены, α1 и α2) эндогенного нечеловеческого (мышиного) полипептида МНС II α, и может быть функционально связана с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого (мышиного) МНС II α, в эндогенном нечеловеческом (мышином) локусе МНС II α; последовательность нуклеиновых кислот, кодирующая внеклеточный участок (или его части, например домены β1 и β2) человеческого полипептида МНС II β, замещает последовательность, кодирующую внеклеточный участок (или его части, например домены β1 и β2) эндогенного нечеловеческого (мышиного) полипептида МНС II β, и может быть функционально связана с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого (мышиного) МНС IIβ, в эндогенном нечеловеческом (мышином) локусе МНС II β; и/или последовательность нуклеиновых кислот, кодирующая внеклеточный участок (или его части, например домены α1, α2 и/или α3) человеческого полипептида МНС I, замещает последовательность, кодирующую внеклеточный участок (или его части, например домены, α1, α2 и/или α3) эндогенного нечеловеческого (мышиного) полипептида МНС I, и может быть функционально связана с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого (мышиного) МНС I, в эндогенном нечеловеческом (мышином) локусе МНС I.
В некоторых вариантах осуществления у генетически модифицированного не относящегося к человеку животного, описанного в настоящем документе, не экспрессируется функциональный эндогенный нечеловеческий Т-клеточный корецептор CD4 из его эндогенного локуса, не экспрессируется функциональный эндогенный нечеловеческий Т-клеточный корецептор CD8 из эндогенного локуса CD8, не экспрессируется функциональный вариабельный домен TCRα из эндогенного вариабельного локуса TCRα, не экспрессируется функциональный вариабельный домен TCRβ из эндогенного вариабельного локуса TCRβ, не экспрессируется внеклеточный домен эндогенного комплекса МНС II из эндогенного локуса МНС II (например, на клеточной поверхности) и/или не экспрессируется внеклеточный домен эндогенного полипептида МНС I из эндогенного локуса МНС I (например, на клеточной поверхности).
Любое не относящееся к человеку животное, описанное в настоящем документе, может дополнительно содержать локус β2-микроглобулина, кодирующий полипептид, содержащий человеческую или гуманизированную аминокислотную последовательность β2-микроглобулина, причем у не относящегося к человеку животного экспрессируется человеческий или гуманизированный полипептид β2-микроглобулин. В некоторых вариантах осуществления не относящееся к человеку животное не экспрессирует функциональный эндогенный полипептид β2-микроглобулин не относящегося к человеку животного из эндогенного нечеловеческого локуса β2-микроглобулина. В некоторых вариантах осуществления локус β2-микроглобулина функционально связан с эндогенными нечеловеческими регуляторными элементами β2-микроглобулина. В одном варианте осуществления локус β2-микроглобулина содержит нуклеотидную последовательность, представленную в экзоне 2, экзоне 3 и экзоне 4 (например, экзонах 2-4) человеческого гена β2-микроглобулина, и необязательно локус β2-микроглобулина дополнительно содержит нуклеотидную последовательность, представленную в экзоне 1 нечеловеческого (например, грызуна) гена β2-микроглобулина.
Не относящиеся к человеку животные, описанные в настоящем документе, могут представлять собой грызуна, например мышь или крысу.
Также в настоящем документе предложена мышь, которая экспрессирует химерные человеческие/мышиные полипептиды Т-клеточных корецепторов CD4, CD8α и CD8β, каждый из которых соответственно содержит трансмембранный и цитоплазматический домены мышиного CD4, CD8α и CD8β; Т-клеточный рецептор, содержащий человеческий вариабельный участок TCRα и человеческий вариабельный участок TCRβ на поверхности Т-клетки; химерные человеческие/мышиные полипептиды МНС IIα, МНС IIβ и МНС I, каждый из которых соответственно содержит внеклеточные домены человеческого полипептида МНС II α (например, домены α1 и α2 человеческого HLA класса II), МНС II β (домены β1 и β2 человеческого HLA класса II) и МНС I (например, домены α1, α2 и α3 человеческого HLA класса I); и необязательно человеческий или гуманизированный полипептид β2-микроглобулин. В одном варианте осуществления в настоящем документе предложены не относящиеся к человеку животные, например мыши, у которых первая последовательность нуклеиновых кислот кодирует цепь α химерного человеческого/мышиного полипептида HLA-DR/H-2E, вторая последовательность нуклеиновых кислот кодирует цепь β химерного полипептида HLA-DR/H-2E, а третья последовательность нуклеиновых кислот кодирует химерный человеческий/мышиный полипептид HLA-A/H-2K, и причем у мыши экспрессируются белки HLA-A/H-2K и HLA-DR/H-2E.
Также в настоящем документе предложено не относящееся к человеку животное, содержащее по существу гуманизированную Т-клеточную иммунную систему, например, у которого по существу гуманизированная Т-клеточная иммунная система формирует по существу гуманизированный Т-клеточный иммунный ответ на антиген. В некоторых вариантах осуществления по существу гуманизированный Т-клеточный иммунный ответ включает активированные Т-клетки, экспрессирующие человеческие вариабельные домены Т-клеточного рецептора (TCR), которые распознают антиген, представленный в связи с внеклеточными доменами человеческого лейкоцитарного антигена (HLA), и/или антигенпредставляющие клетки, которые представляют антиген в связи с внеклеточными доменами HLA. В некоторых вариантах осуществления по существу гуманизированная Т-клеточная иммунная система содержит: (а) нечеловеческую Т-клетку, которая экспрессирует полипептид Т-клеточного корецептора, содержащий человеческий домен Т-клеточного корецептора, который связывается с человеческой молекулой HLA, и/или Т-клеточный рецептор (TCR), содержащий вариабельный домен TCR, который кодируется по меньшей мере одним человеческим сегментом гена вариабельного участка TCR; и (b) нечеловеческую антигенпредставляющую клетку, которая представляет антиген в связи с человеческим HLA и активирует нечеловеческую Т-клетку.
Также предложены способы получения и использования не относящегося к человеку животного, описанного в настоящем документе. Как правило, способы получения генетически модифицированного не относящегося к человеку животного, описанного в настоящем документе, включают: (а) внедрение в геном не относящегося к человеку животного первой нуклеотидной последовательности, кодирующей химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора (например, химерный полипептид CD4), и/или второй нуклеотидной последовательности, кодирующей второй химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора (например, химерный полипептид CD8α), и третьей нуклеотидной последовательности, кодирующей третий химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора (например, химерный полипептид CD8β), причем нечеловеческий участок каждого химерного полипептида Т-клеточного корецептора содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого Т-клеточного корецептора, и при этом человеческий участок каждого химерного полипептида содержит внеклеточный участок (или его часть, например один или более доменов) человеческого Т-клеточного рецептора; (b) вставку в геном не относящегося к человеку животного неперестроенного вариабельного локуса гена Т-клеточного рецептора (TCR) α, содержащего по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с нечеловеческой константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с нечеловеческой константной последовательностью гена TCRβ; и необязательно (с) включение в геном первой последовательности нуклеиновых кислот, кодирующей первый химерный человеческий/нечеловеческий полипептид МНС (например, химерный полипептид МНС IIα), второй последовательности нуклеиновых кислот, кодирующей второй химерный человеческий/нечеловеческий полипептид МНС (например, химерный полипептид МНС IIβ), и/или третьей последовательности нуклеиновых кислот, кодирующей третий химерный человеческий/нечеловеческий полипептид МНС (например, химерный полипептид МНС I); и/или (d) добавление в геном не относящегося к человеку животного локуса β2-микроглобулина, кодирующего человеческий или гуманизированный полипептид β2-микроглобулин. В некоторых вариантах осуществления первая нуклеотидная последовательность кодирует внеклеточный участок (или его часть) человеческого CD4, функционально связанный с по меньшей мере трансмембранным и цитоплазматическим доменами нечеловеческого корецептора CD4, вторая нуклеотидная последовательность кодирует внеклеточный участок (или его часть) человеческого CD8α и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого CD8α, третья нуклеотидная последовательность кодирует внеклеточный участок (или его часть) человеческого CD8β и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого CD8β, первая последовательность нуклеиновых кислот кодирует внеклеточный участок (или его часть) человеческого полипептида HLA класса II α и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого полипептида МНС II α, вторая последовательность нуклеиновых кислот кодирует внеклеточный участок (или его часть) человеческого полипептида HLA класса II β и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого полипептида МНС II β, третья последовательность нуклеиновых кислот кодирует внеклеточный участок (или его часть) человеческого полипептида HLA класса I и трансмембранный и цитоплазматический домены нечеловеческого полипептида МНС I, и локус β2-микроглобулина содержит нуклеотидную последовательность, представленную в экзонах 2-4 человеческого гена β2-микроглобулина, например нуклеотидную последовательность, представленную в экзонах 2, 3 и 4 человеческого гена β2-микроглобулина.
Способы получения не относящихся к человеку животных включают варианты осуществления, в которых: (а) внедрение первой, второй и/или третьей нуклеотидной (-ых) последовательности (-ей), кодирующей (-их) химерный (-ые) полипептид (-ы) Т-клеточного корецептора в геном не относящегося к человеку животного, включает замещение в эндогенном локусе CD4 нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD4, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD4, и/или замещение в эндогенном локусе CD8α нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD8α, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD8α, и замещение в эндогенном локусе CD8β нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD8β, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD8β; (b) вставка неперестроенного локуса TCRα и/или неперестроенного локуса TCRβ в геном животного включает замещение эндогенного нечеловеческого вариабельного локуса гена TCRα неперестроенным гуманизированным вариабельным локусом гена TCRα, содержащим по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, для создания гуманизированного вариабельного локуса гена TCRα, причем гуманизированный вариабельный локус гена TCRα функционально связан с эндогенным нечеловеческим константным участком TCRα, и/или замещение эндогенного нечеловеческого вариабельного локуса гена TCRβ неперестроенным гуманизированным вариабельным локусом гена TCRβ, содержащим по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, для создания гуманизированного вариабельного локуса гена TCRβ, при этом гуманизированный вариабельный локус гена TCRβ функционально связан с эндогенным нечеловеческим константным участком TCRβ; (с) включение первой, второй и/или третьей последовательности (-ей) нуклеиновых кислот, кодирующей (-их) химерный (-ые) полипептид (-ы) МНС в геном не относящегося к человеку животного, включает замещение в эндогенном нечеловеческом локусе МНС II нуклеотидной последовательности, кодирующей нечеловеческий комплекс МНС II, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий комплекс МНС II, и замещение в эндогенном нечеловеческом локусе МНС I нуклеотидной последовательности, кодирующей нечеловеческий полипептид МНС I, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид МНС I; и/или (d) добавление локуса β2-микроглобулина, кодирующего человеческий или гуманизированный полипептид β2-микроглобулин, в геном не относящегося к человеку животного включает замещение в эндогенном нечеловеческом локусе β2-микроглобулина нуклеотидной последовательности, кодирующей нечеловеческий полипептид β2-микроглобулин, нуклеотидной последовательностью, кодирующей человеческий или гуманизированный полипептид β2-микроглобулин.
В некоторых вариантах осуществления: (а) внедрение первой, второй и/или третьей нуклеотидной последовательности в геном не относящегося к человеку животного соответственно включает: (i) замещение в эндогенном локусе CD4 нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) эндогенного нечеловеческого полипептида CD4, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида CD4, в функциональной взаимосвязи с последовательностями, кодирующими транcмембранный и цитоплазматический домены эндогенного нечеловеческого CD4; (ii) замещение в эндогенном локусе CD8α нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) эндогенного нечеловеческого полипептида CD8α, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида CD8α, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого CD8α; и/или (iii) замещение в эндогенном локусе CD8β нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) эндогенного нечеловеческого полипептида CD8β, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида CD8β, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого CD8β; (b) вставка неперестроенного локуса TCRα и/или неперестроенного локуса TCRβ в геном животного соответственно включает: (i) замещение эндогенного нечеловеческого вариабельного локуса гена TCRα неперестроенным гуманизированным вариабельным локусом гена TCRα, содержащим по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, для создания гуманизированного вариабельного локуса гена TCRα, причем гуманизированный вариабельный локус гена TCRα функционально связан с эндогенным нечеловеческим константным участком TCRα; и/или (ii) замещение эндогенного нечеловеческого вариабельного локуса гена TCRβ неперестроенным гуманизированным вариабельным локусом гена TCRβ, содержащим по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, для создания гуманизированного вариабельного локуса гена TCRβ, при этом гуманизированный вариабельный локус гена TCRβ функционально связан с эндогенным нечеловеческим константным участком TCRβ; (с) включение первой, второй и/или третьей последовательности нуклеиновых кислот в геном не относящегося к человеку животного соответственно включает: (i) замещение в эндогенном нечеловеческом локусе МНС II α нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) нечеловеческого полипептида МНС II α, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида HLA класса II α, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого МНС II α; (ii) замещение в эндогенном нечеловеческом локусе МНС II β нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) нечеловеческого полипептида МНС II β, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида HLA класса II β, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого МНС II β; и/или (iii) замещение в эндогенном нечеловеческом локусе МНС I нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) нечеловеческого полипептида МНС I, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида HLA класса I, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого МНС I; и/или замещение в эндогенном локусе β2-микроглобулина нуклеотидной последовательности, представленной в экзонах 2-4, нуклеотидной последовательностью, содержащей экзоны 2, 3 и 4 человеческого гена β2-микроглобулина.
В одном варианте осуществления этап внедрения включает замещение у первого не относящегося к человеку животного в эндогенном локусе CD4 нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD4, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD4, замещение у второго не относящегося к человеку животного в эндогенном локусе CD8α нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD8α, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD8α, и замещение в эндогенном локусе CD8β нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD8β, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD8β. В некоторых вариантах осуществления этап внедрения включает замещение у первого не относящегося к человеку животного в эндогенном локусе CD4 нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) эндогенного нечеловеческого полипептида CD4, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида CD4, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого CD4, замещение у второго не относящегося к человеку животного в эндогенном локусе CD8α нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) эндогенного нечеловеческого полипептида CD8α, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида CD8α, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого CD8α, и замещение в эндогенном локусе CD8β нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) эндогенного нечеловеческого полипептида CD8β, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида CD8β, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого CD8β. В некоторых вариантах осуществления этапы замещения выполняют одновременно или в любом порядке.
В некоторых вариантах осуществления этап вставки включает замещение у третьего не относящегося к человеку животного эндогенного нечеловеческого вариабельного локуса гена TCRα неперестроенным гуманизированным вариабельным локусом гена TCRα, содержащим по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, для создания гуманизированного вариабельного локуса гена TCRα, причем гуманизированный вариабельный локус гена TCRα функционально связан с эндогенным нечеловеческим константным участком TCRα; замещение у четвертого не относящегося к человеку животного эндогенного нечеловеческого вариабельного локуса гена TCRβ неперестроенным гуманизированным вариабельным локусом гена TCRβ, содержащим по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, для создания гуманизированного вариабельного локуса гена TCRβ, при этом гуманизированный вариабельный локус гена TCRβ функционально связан с эндогенным нечеловеческим константным участком TCRβ. В некоторых вариантах осуществления этапы замещения выполняют одновременно или в любом порядке.
В некоторых вариантах осуществления этап размещения включает, в любом порядке, замещение у пятого не относящегося к человеку животного в эндогенном нечеловеческом локусе МНС II одной или более нуклеотидных последовательностей, кодирующих нечеловеческий комплекс МНС II, одной или более нуклеотидными последовательностями, кодирующими химерный человеческий/нечеловеческий комплекс МНС II; и замещение у пятого не относящегося к человеку животного в эндогенном нечеловеческом локусе МНС I нуклеотидной последовательности, кодирующей нечеловеческий полипептид МНС I, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид МНС I. В некоторых вариантах осуществления этап включения включает замещение у пятого не относящегося к человеку животного в эндогенном нечеловеческом локусе МНС II α нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) нечеловеческого полипептида МНС II α, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида МНС II α в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого МНС II α, и замещение в эндогенном нечеловеческом локусе МНС II β нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) нечеловеческого полипептида МНС II β, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида МНС II β, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого МНС II β; и замещение в эндогенном нечеловеческом локусе МНС I нуклеотидной последовательности, кодирующей внеклеточный участок (или его часть) нечеловеческого полипептида МНС I, нуклеотидной последовательностью, кодирующей внеклеточный участок (или его часть) человеческого полипептида МНС I, в функциональной взаимосвязи с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного нечеловеческого МНС I у пятого не относящегося к человеку животного. В некоторых вариантах осуществления этапы замещения выполняют одновременно или в любом порядке.
В некоторых вариантах осуществления этап добавления включает замещение у шестого не относящегося к человеку животного в эндогенном нечеловеческом локусе β2-микроглобулина нуклеотидной последовательности, кодирующей нечеловеческий полипептид β2-микроглобулин, нуклеотидной последовательностью, кодирующей человеческий или гуманизированный полипептид β2-микроглобулин. В некоторых вариантах осуществления человеческий или гуманизированный полипептид β2-микроглобулин кодируется нуклеотидной последовательностью, представленной в экзоне 2, экзоне 3 и экзоне 4 человеческого гена β2-микроглобулина.
Способы, описанные в настоящем документе, включают варианты осуществления, в которых: внедряют первую, вторую и/или третью нуклеотидную (-ые) последовательность (-и), кодирующую (-ие) химерный (-е) полипептид (-ы) Т-клеточного корецептора; вставляют локус TCRα и/или неперестроенный локус TCRβ; размещают первую, вторую и/или третью последовательность (-и) нуклеиновых кислот, кодирующую (-ие) химерный (-е) полипептид (-ы) МНС; и/или добавляют локус β2-микроглобулина посредством скрещивания не относящегося к человеку животного, содержащего одну или более генетических модификаций, описанных в настоящем документе, с другим (или несколькими) не относящимся к человеку животным того же вида, содержащим оставшиеся генетические модификации. Не имеющий ограничительного характера вариант осуществления включает скрещивание в любом порядке первого, второго, третьего, четвертого, пятого и шестого не относящихся к человеку животных, описанных выше.
Способы, описанные в настоящем документе, могут включать гомологичную рекомбинацию в нечеловеческих эмбриональных стволовых (ЭС) клетках. Способы, описанные в настоящем документе, можно использовать для создания мышей, описанных в настоящем документе. Не относящиеся к человеку животные, экспрессирующие химерные человеческие/нечеловеческие полипептиды Т-клеточных рецепторов CD4, CD8α и/или CD8β, человеческие (гуманизированные) белки TCR α/β и химерный комплекс МНС II и МНС I (с человеческим или гуманизированным β2-микроглобулином) могут быть созданы посредством: (а) сначала внедрения соответствующим образом каждого отдельного человеческого (гуманизированного) гена путем гомологичной рекомбинации в отдельные ЭС-клетки и создания каждого отдельного не относящегося к человеку животного из таких ЭС-клеток, и последующего скрещивания каждого полученного не относящегося к человеку животного в любом порядке; (b) внедрения всех человеческих (гуманизированных) генов путем последовательной гомологичной рекомбинации в одиночную ЭС-клетку, а затем создания не относящегося к человеку животного из такой ЭС-клетки; или (с) комбинирования последовательной гомологичной рекомбинации в некоторых локусах в ЭС-клетках и скрещивания. Животные, описанные в настоящем документе, могут также при необходимости быть созданы путем скрещивания потомства от начального скрещивания с другими животными. Скрещивание и/или гомологичная рекомбинация могут быть выполнены в любом предпочтительном порядке.
Также предложены способы выделения человеческих вариабельных доменов TCR, специфичных к антигену, из не относящегося к человеку животного, включающие выделение из не относящегося к человеку животного, предложенного в настоящем документе или полученного по способу, описанному в настоящем документе, Т-клетки или белка TCR, который связывается с антигеном. В некоторых вариантах осуществления способы могут дополнительно включать идентификацию первой и/или второй нуклеиновой кислоты, кодирующей вариабельные домены TCRα и/или TCRβ, которые связываются с антигеном, и/или культивирование клетки, содержащей один или более векторов в условиях, достаточных для экспрессии вектора (-ов), причем вектор (-ы) содержит (-ат) третью и/или четвертую нуклеиновую кислоту, соответственно идентичную или по существу идентичную первой и/или второй нуклеиновым кислотам, и при этом третью и/или четвертую нуклеиновую кислоту клонируют в пределах рамки считывания, например, с человеческим геном константного участка TCR, например геном константного участка TCRα и/или геном константного участка TCRβ соответственно. Также предложены ткани и клетки, содержащие генетические модификации, описанные в настоящем документе (которые могут включать перестроенные человеческие гены вариабельного участка TCRα и/или TCRβ), и нуклеиновые кислоты, кодирующие такие человеческие вариабельные домены TCR, экспрессируемые такими тканями или клетками, выделенными из не относящегося к человеку животного, модифицированного так, как описано в настоящем документе. Также включены: (1) рекомбинантные нуклеиновые кислоты, например экспрессионные векторы, содержащие последовательности нуклеиновых кислот, кодирующие человеческий вариабельный домен TCR, описанный в настоящем документе, например человеческий перестроенный ген вариабельного участка TCRα или TCRβ, клонированный в пределах рамки считывания с соответствующим человеческим геном константного участка TCR, например геном константного участка TCRα или геном константного участка TCRβ соответственно; (2) клетки-хозяева, содержащие такие нуклеиновые кислоты (например, экспрессионные векторы); и (3) TCR, экспрессированные клетками-хозяевами. В некоторых вариантах осуществления рекомбинантные нуклеиновые кислоты, предложенные в настоящем документе, содержат человеческий перестроенный ген вариабельного участка TCRδ или ген вариабельного участка TCRγ, например, полученный из не относящегося к человеку животного, генетически модифицированного так, как описано в настоящем документе, или ткани, выделенной из него, клонированный в пределах рамки считывания с человеческим геном константного участка TCRδ или геном константного участка TCRγ соответственно.
Также предложен способ создания гуманизированного Т-клеточного ответа у не относящегося к человеку животного, по существу включающий иммунизацию не относящегося к человеку животного, не относящегося к человеку животного, генетически модифицированного или имеющего по существу гуманизированную Т-клеточную иммунную систему, как описано в настоящем документе, антигеном, например человеческим антигеном, например человеческим опухолевым антигеном, человеческим бактериальным патогеном, человеческим вирусным патогеном и т.д. В некоторых вариантах осуществления иммунизированное не относящееся к человеку животное экспрессирует по меньшей мере 50% всех функциональных человеческих сегментов гена TCRVα и/или по меньшей мере 50% всех функциональных человеческих сегментов гена TCRVβ и/или содержит все или по существу все функциональные человеческие сегменты гена TCRVα и/или все или по существу все функциональные человеческие сегменты гена TCRVβ.
Также предложены способы in vitro выделения человеческого TCR, специфичного к антигену, которые, как правило, включают обнаружение активации первой клетки не относящегося к человеку животного после: (а) контакта со второй клеткой не относящегося к человеку животного; и (b) инкубации с антигеном; причем первая клетка экспрессирует химерный человеческий/нечеловеческий Т-клеточный корецептор и одно или оба из: (i) химерной человеческой/нечеловеческой цепи α TCR; и (ii) химерной человеческой/нечеловеческой цепи β TCR, и при этом вторая клетка экспрессирует химерный человеческий/нечеловеческий полипептид МНС. Способы могут дополнительно включать выделение TCR из первой клетки или кодирующих его нуклеиновых кислот.
В способах in vitro, описанных в настоящем документе, антиген может представлять собой опухолевый антиген, вирусный антиген, аутоантиген или бактериальный антиген. В некоторых вариантах осуществления не относящееся к человеку животное представляет собой грызуна, например крысу или мышь. Кроме того, в настоящем документе предложена ткань, Т-клетка, TCR (например, растворимый TCR) или нуклеиновая кислота, кодирующая весь TCR или его часть, которая выделена из не относящегося к человеку животного, генетически модифицированного или имеющего по существу гуманизированную Т-клеточную иммунную систему, как описано в настоящем документе, гибридома или квадрома, полученная из такой Т-клетки.
Также предложены композиции, например, содержащие первую и вторую клетки не относящегося к человеку животного; причем первая клетка экспрессирует химерный человеческий/нечеловеческий Т-клеточный корецептор и необязательно одну или обе из (i) химерной человеческой/нечеловеческой цепи α TCR и (ii) химерной человеческой/нечеловеческой цепи β TCR, и при этом вторая клетка экспрессирует химерный человеческий/нечеловеческий полипептид МНС, который ассоциируется с химерным человеческим/нечеловеческим Т-клеточным корецептором. В некоторых вариантах осуществления первая клетка представляет собой не относящуюся к человеку Т-клетку. В других вариантах осуществления вторая клетка представляет собой антигенпредставляющую клетку.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На ФИГ. 1 схематически изображен (без соблюдения масштаба) гуманизированный Т-клеточный рецепторный комплекс, содержащий гуманизированные белки TCR альфа и бета, гуманизированный МНС класса I в комплексе с гуманизированным β2-микроглобулином и гуманизированный гетеродимер CD8 (левая таблица); а также Т-клеточный рецепторный комплекс, содержащий гуманизированные белки TCR альфа и бета, гуманизированный гетеродимер МНС класса II и гуманизированный CD4 (правая таблица). Антиген, представляемый гуманизированным МНС, показан в виде окружности. Мышиные участки показаны в виде закрашенных фигур, а человеческие участки показаны в виде заштрихованных фигур.
На ФИГ. 2А-С схематически представлены (без соблюдения масштаба) примеры химерных локусов МНС I и МНС II, например химерный локус HLA-A2/H-2K (ФИГ. 2А), химерный локус HLA-DR2/H-2E (ФИГ. 2В) и гуманизированный локус β2М (ФИГ. 2С). При отсутствии иных указаний человеческие последовательности показаны в виде полых фигур, а мышиные последовательности показаны в виде закрашенных фигур. Заштрихованная фигура представляет собой экзон 1 Н-2Е, полученный из другой линии мышей по сравнению с эндогенным локусом (см. пример 1.3 и ФИГ. 3В). Фланкированная (-ые) loxP неомицинфосфотрансферазная (-ые) кассета (-ы) показана (-ы) со стрелками, отмеченными соответствующим образом.
На ФИГ. 3А-С показана стратегия создания гуманизированного локуса МНС, содержащего гуманизированные гены МНС I и МНС II. В конкретном варианте осуществления, показанном на ФИГ. 3А, локус МНС созданной мыши содержит химерные последовательности HLA-A2/H-2K и HLA-DR2/H-2E (H2-K+/1666 MHC-II+/6112), но в нем отсутствуют последовательность H2-D (H2-D+/delete) и последовательность Н-2А (метод генной инженерии также приводит к делеции Н-2А, см. пример 1.2). Большие нацеливающие векторы (LTVEC) или конструкт с рекомбиназой Cre, внедренные в ЭС-клетки на каждой стадии гуманизации, показаны справа от стрелок. MAID или 4-значные номера обозначают идентификационный номер модифицированной аллели. На ФИГ. 3В представлена принципиальная схема (без соблюдения масштаба) примера большого нацеливающего вектора HLA-DR2/H-2E. При отсутствии иных указаний человеческие последовательности показаны в виде полых фигур, а мышиные последовательности показаны в виде закрашенных фигур. Заштрихованная фигура представляет собой экзон 1 Н-2Е, полученный из другой линии мышей по сравнению с эндогенным локусом (см. пример 1.3). Фланкированная loxP гигромициновая кассета показана в виде стрелки, отмеченной соответствующим образом. На ФИГ. 3С схематически представлены (без соблюдения масштаба) примеры генотипов химерных человеческих/мышиных локусов МНС (** обозначает ген H-2L, который не присутствует во всех линиях мышей, например не присутствует в линиях мышей C57B L/6 или 129), где эндогенные мышиные локусы Н-2K и Н-2Е соответственно замещены химерными человеческими/мышиными локусами HLA-A2/H-2K и HLA-DR2/H-2E (заштрихованные фигуры), локусы Н-2А и H-2D были удалены (полые фигуры, обведенные пунктирными линиями), а остальные локусы представляют собой эндогенные мышиные гены (сплошные фигуры, обведенные сплошными линиями).
На ФИГ. 4А показана (без соблюдения масштаба) последовательная стратегия гуманизации мышиного локуса TCRα, в которой сегменты гена вариабельного участка TCRα последовательно добавляют ближе к 5'-концу от начальной гуманизации удаленного мышиного локуса (MAID 1540). Мышиная последовательность указана в виде закрашенных фигур, человеческая последовательность указана в виде полых фигур. MAID обозначает идентификационный номер модифицированной аллели. TRAV = сегмент Vα TCR, TRAJ = сегмент Jα TCR (hTRAJ = человеческий TRAJ), TRAC = домен Cα TCR, TCRD = TCRδ. На ФИГ. 4В показана (без соблюдения масштаба) последовательная стратегия гуманизации мышиного локуса TCRβ, в которой сегменты гена вариабельного участка TCRβ последовательно добавляют ближе к удаленному мышиному локусу TCRβ. Мышиная последовательность указана в виде закрашенных фигур, человеческая последовательность указана в виде полых фигур. MAID обозначает идентификационный номер модифицированной аллели. TRBV или TCRBV = сегмент V TCRβ.
На ФИГ. 5А представлено схематическое изображение (без соблюдения масштаба) химерного локуса CD4. Человеческие кодирующие экзоны представлены в виде заштрихованных фигур, мышиные кодирующие экзоны представлены в виде закрашенных фигур, а некодирующие экзоны представлены в виде полых фигур. Указаны экзоны, кодирующие иммуноглобулиноподобные домены (Ig), трансмембранный домен (ТМ), цитоплазматический домен (CYT) и сигнальный пептид (Signal), а также нетранслируемые области со стороны 3'-конца (UTR). Фланкированная loxP неомицинфосфотрансферазная (Pgk-neo) кассета показана со стрелками, отмеченными соответствующим образом. На ФИГ. 5В показано схематическое изображение (без соблюдения масштаба) химерных локусов CD8a и CD8b. Человеческие кодирующие экзоны представлены в виде заштрихованных фигур, мышиные кодирующие экзоны представлены в виде закрашенных фигур, а некодирующие экзоны представлены в виде полых фигур. Указаны экзоны, кодирующие иммуноглобулиноподобные домены (IgV), трансмембранный домен (ТМ), цитоплазматический домен (CYT) и сигнальный пептид (Signal), а также нетранслируемые области со стороны 3'-конца (UTR). Фланкированные loxP гигромициновая (Hyg) и неомицинфосфотранcферазная (Pgk-neo) кассеты изображены со стрелками, показанными соответствующим образом.
На ФИГ. 6А-С представлены контурные графики FACS клеток тимуса, выделенных из контрольной мыши или мыши, содержащей гуманизированные локусы МНС I, MHC II α и β, TCRα и β, CD4, CD8α и β, а также β2M (ТМ I/II В С4/8), сортированных по синглетам и меченных (ФИГ. 6А) антителами против мышиного CD19 и против мышиного CD3, (ФИГ. 6В) антителами против мышиного CD19 и против мышиного F4/80, или (ФИГ. 6С) антителами против мышиного CD8α и против мышиного CD4 (левая таблица), или антителами против человеческого CD8α и против человеческого CD4 (правая таблица).
На ФИГ. 7A-G представлены контурные графики FACS клеток тимуса, выделенных из контрольной мыши или мыши, содержащей гуманизированные локусы МНС I, MHC II α и β, TCRα и β, CD4, CD8α и β, а также β2М (ТМ I/II В С4/8), сортированных по клеткам CD19+, клеткам F4/80+ или клеткам CD3+ и меченных (ФИГ. 7А, 7В) антителами против человеческого В2М и против мышиного H-2D; (ФИГ. 7С, 7D) антителами против HLA-A2 или против HLA-DR; (ФИГ. 7Е, 7F) антителами против H-2D и против IAIE; или (ФИГ. 7G) антителами против мышиного CD4 и против человеческого CD4 (сверху), антителами против мышиного CD8α и против человеческого CD8α (посредине) и антителами против мышиного CD8β и против человеческого CD8β (снизу).
На ФИГ. 8 изображены контурные графики FACS клеток тимуса, выделенных из контрольной мыши или мыши, содержащей гуманизированные локусы MHC I, MHC II α и β, TCRα и β, CD4, CD8α и β, а также β2М (ТМ I/II В С4/8), сортированных по клеткам CD3+CD4+ и меченных антителами против мышиного FoxP3 и против мышиного CD25.
На ФИГ. 9А-Е изображены контурные графики FACS клеток селезенки, выделенных из контрольной мыши или мыши, содержащей гуманизированные локусы MHC I, MHC II α и β, TCRα и β, CD4, CD8α и β, а также β2М (ТМ I/II В С4/8), сортированных по синглетам, клеткам CD3+, Т-клеткам CD4+ или Т-клеткам CD8+ и меченных (ФИГ. 9А) антителами против мышиного CD19 и против мышиного CD3, (ФИГ. 9В) антителами против мышиного CD19 и против мышиного F4/80, или (ФИГ. 9С) антителами против мышиного CD4 и против мышиного CD8α (слева), или антителами против человеческого CD4 и против человеческого CD8α (справа), или (ФИГ. 9D, 9Е) антителами против мышиного CD44 и против мышиного CD62L.
На ФИГ. 10A-G изображены контурные графики FACS клеток селезенки, выделенных из контрольной мыши или мыши, содержащей гуманизированные локусы MHC I, MHC II α и β, TCRα и β, CD4, CD8α и β, а также β2М (ТМ I/II В С4/8), сортированных по клеткам CD19+, клеткам F4/80+ или клеткам CD3 и окрашенных (ФИГ. 10А, 10В) антителами против человеческого В2М или против мышиного H-2D, (ФИГ. 10С, 10D) антителами против HLA-A2 или против HLA-DR, (ФИГ. 10Е, 10F) антителами против H-2D и против IAIE или (ФИГ. 10G) антителами против мышиного CD4 и против человеческого CD4 (сверху), антителами против мышиного CD8α и против человеческого CD8α (посередине) и антителами против мышиного CD8β и против человеческого CD8β (снизу).
На ФИГ. 11 изображены контурные графики FACS клеток селезенки, выделенных из контрольной мыши или мыши, содержащей гуманизированные локусы MHC I, MHC II α и β, TCRα и β, CD4, CD8α и β, а также β2M (ТМ I/II В С4/8), сортированных по клеткам CD3+CD4+ и меченных антителами против мышиного FoxP3 и против мышиного CD25.
На ФИГ. 12 показано число клеток селезенки (пятен на лунку (среднее + СО); ось у), которые продуцируют ИФН-γ в анализе иммуноферментных пятен после выделения из контрольной мыши или мыши, содержащей гуманизированные локусы МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2M (ТМ I/II В С4/8), и инкубации при отсутствии пептида (только 200 тыс. клеток; ось х) или в присутствии 10 мкг/мл или 1 мкг/мл пептида MAGE-A3 (ось х).
На ФИГ. 13А показано развитие острой вирусной инфекции, вызванной штаммом Армстронга, либо у контрольной мыши, либо у мыши, содержащей гуманизированные локусы МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2M (ТМ I/II В С4/8); временная шкала эксперимента изображена в верхней части фигуры, а результаты измерения вирусных титров в разные дни после инфицирования для обоих штаммов мышей изображены на графике внизу. На ФИГ. 13В показано развитие хронической вирусной инфекции, вызванной штаммом Clone 13, либо у контрольных мышей, либо у мышей, содержащих гуманизированные локусы МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2M (ТМ I/II В С4/8); временная шкала эксперимента изображена в верхней части фигуры, а результаты измерения вирусных титров на 21-й день после инфицирования для обоих штаммов мышей изображены на графике внизу. Т-клетки из неинфицированных или хронически инфицированных мышей ТМ I/II В С4/8 или контрольных мышей В6 метили антителами против PD1, против Lag3 и против Tim3 (ФИГ. 13С; ось X); на фигуре показаны результаты количественного анализа меченых клеток (% положительных клеток; ось у).
На ФИГ. 14 показано развитие хронической вирусной инфекции, вызванной штаммом Clone 13, либо у контрольных мышей, либо у мышей ТМ I/II В С4/8 после предшествующей острой инфекции, вызванной штаммом Армстронга; временная шкала эксперимента изображена в верхней части фигуры, а результаты измерения вирусных титров на 31-й день после инфицирования изображены на графике внизу. Ложноинфицированных мышей включали в эксперимент в качестве дополнительного контроля.
На ФИГ. 15А-В изображено количество клеток CD8+ (ось y; ИФН-γ-положительные клетки), которые продуцировали ИФН-γ в ответ на введение пептидов LCMV, рестриктированных по HLA-A2 (GPC10-18; N69-77; Z49-58), рестриктированных по H2Db (GP33-41), овальбумина или только инкубацию, и были выделены либо из контрольных животных (ФИГ. 15А), либо из мышей, содержащих гуманизированные локусы МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2M (ТМ I/II В С4/8) (ФИГ. 15В), каждая из которых была ложноинфицирована (ложная инфекция; n=1 в каждой группе) или инфицирована острой инфекцией штамма Армстронга (Арм.; n=3 в каждой группе). % лимфоцитов CD8+ ИФН-γ+ (ось y) после стимуляции указанными пептидами (OVA, GP33, NP69, GPC10, GPC447 или Z49) в процессе развития инфекции (дни после инфицирования; ось х) у мышей, содержащих гуманизированные локусы МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2М (ТМ I/II В С4/8), или у контрольных животных В6 показан на ФИГ. 15С и 15D соответственно.
ПОДРОБНОЕ ОПИСАНИЕ
В настоящем документе описаны не относящиеся к человеку животные (например, грызуны, например мыши или крысы), созданные методами генной инженерии и экспрессирующие гуманизированный Т-клеточный корецептор (например, гуманизированный CD4 и/или CD8 (например, CD8α и/или CD8β)), человеческий или гуманизированный главный комплекс гистосовместимости (МНС), который связывается с гуманизированным Т-клеточным корецептором (например, человеческий или гуманизированный МНС II (например, цепи МНС II α и/или МНС II β) и/или МНС I (например, МНС Iα), и необязательно человеческий или гуманизированный β2-микроглобулин), и/или человеческий или гуманизированный Т-клеточный рецептор (TCR), а также эмбрионы, ткани и клетки, экспрессирующие вышеперечисленное. Развитие клеточного звена иммунной системы не относящихся к человеку животных, описанных в настоящем документе, и контрольных животных сопоставимо, например, тимус и селезенка содержат аналогичные абсолютные количества тимоцитов и клеток CD3+. Это существенно отличается от других не относящихся к человеку животных, модифицированных таким образом, чтобы они содержали человеческий TCR (α и β) и химерную человеческую/мышиную молекулу МНС I, см., например, Li (2010) Nature Medicine 16:1029-1035 и вспомогательные материалы. Такие животные демонстрировали снижение популяций Т-клеток по сравнению не только с контрольными животными дикого типа, но и с животными, модифицированными только человеческим TCR, и животными, модифицированными только химерной человеческой/мышиной молекулой МНС I, там же. Соответственно, в настоящем документе предложены не относящиеся к человеку животные, созданные таким образом, чтобы совместно экспрессировать гуманизированный корецептор CD4 и гуманизированный МНС II и/или гуманизированный корецептор CD8 и гуманизированный МНС I и необязательно гуманизированный TCR. Также предложены способы получения созданного методами генной инженерии животного, у которого экспрессируется по меньшей мере один гуманизированный Т-клеточный корецептор (например, гуманизированный CD4 и/или CD8), по меньшей мере один гуманизированный МНС, который ассоциируется с гуманизированным Т-клеточным корецептором (например, гуманизированный МНС II и/или МНС I, который ассоциируется с гуманизированным CD4 и/или CD8 соответственно), и/или гуманизированный TCR. Также предложены способы использования созданных методами генной инженерии животных, у которых формируется по существу гуманизированный Т-клеточный иммунный ответ, для разработки терапевтических средств для человека.
По существу гуманизированные Т-клеточные иммунные ответы
В настоящем документе описаны не относящиеся к человеку животные, которые были генетически модифицированы с возможностью формирования по существу гуманизированных Т-клеточных иммунных ответов. Мыши, описанные в настоящем документе, экспрессируют по меньшей мере один человеческий или гуманизированный Т-клеточный корецептор, по меньшей мере один человеческий или гуманизированный главный комплекс гистосовместимости (МНС), способный ассоциироваться с по меньшей мере одним человеческим или гуманизированным Т-клеточным корецептором, и/или человеческий или гуманизированный Т-клеточный рецептор (TCR), который предпочтительно способен распознавать антиген, представленный в связи с человеческим или гуманизированным МНС в комплексе с человеческим или гуманизированным Т-клеточным корецептором, и передавать сигналы активации на нечеловеческую клетку, например нечеловеческую Т-клетку, экспрессирующую человеческий или гуманизированный TCR. Человеческий или гуманизированный Т-клеточный корецептор, человеческий или гуманизированный TCR и/или человеческий или гуманизированный МНС может кодироваться геномом не относящегося к человеку животного. В предпочтительных вариантах осуществления при иммунизации антигеном не относящиеся к человеку животные представляют рестриктированные по HLA эпитопы антигена TCR, полученные из человеческих сегментов гена TCR, например человеческого сегмента V TCRα, человеческого сегмента J TCRα, человеческого сегмента V TCRβ, человеческого сегмента D TCRβ и/или человеческого сегмента J TCRβ.
Соответственно, в объем изобретения входит генетически модифицированное не относящееся к человеку животное, геном которого содержит (например, в эндогенном локусе) нуклеотидную последовательность, кодирующую гуманизированный полипептид Т-клеточного рецептора (например, полипептид CD4 или CD8), причем химерный полипептид Т-клеточного корецептора содержит консервативные аминокислотные замены в аминокислотной (-ых) последовательности (-ях), описанной (-ых) в настоящем документе, и/или последовательность нуклеиновых кислот, кодирующую гуманизированный полипептид МНС, который ассоциируется с гуманизированным полипептидом Т-клеточного корецептора, при этом гуманизированный полипептид МНС содержит консервативные аминокислотные замены в аминокислотой (-ых) последовательности (-ях), описанной (-ых) в настоящем документе.
Консервативная аминокислотная замена включает замену аминокислотного остатка другим аминокислотным остатком, имеющим R-группу боковой цепи с аналогичными химическими свойствами (например, зарядом или гидрофобностью). Консервативные аминокислотные замены могут быть выполнены посредством модификации нуклеотидной последовательности, направленной на внедрение изменения нуклеотидов, которые будут кодировать консервативную замену. В целом консервативная аминокислотная замена по существу не изменит интересующие функциональные свойства белка, например способность CD4 или CD8 ассоциироваться, например связываться с МНС II или МНС I соответственно, и может, например, повышать восприимчивость TCR к представленному МНС антигену. Примеры групп аминокислот, имеющих боковые цепи с аналогичными химическими свойствами, включают алифатические боковые цепи, такие как глицин, аланин, валин, лейцин и изолейцин; алифатические гидроксильные боковые цепи, такие как серин и треонин; амидосодержащие боковые цепи, такие как аспарагин и глутамин; ароматические боковые цепи, такие как фенилаланин, тирозин и триптофан; основные боковые цепи, такие как лизин, аргинин и гистидин; кислые боковые цепи, такие как аспарагиновая кислота и глутаминовая кислота; и серосодержащие боковые цепи, такие как цистеин и метионин. Группы консервативной аминокислотной замены включают, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат и аспарагин/глутамин. В некоторых вариантах осуществления консервативная аминокислотная замена может быть заменой любого нативного остатка в белке аланином, как, например, используется при аланин-сканирующем мутагенезе. В некоторых вариантах осуществления выполняется консервативная замена, которая имеет положительную величину в логарифмической матрице правдоподобия РАМ250, описанной в публикации Gonnet et al. ((1992) Exhaustive Matching of the Entire Protein Sequence Database, Science 256:1443-45), включенной в настоящий документ путем ссылки. В некоторых вариантах осуществления замена является умеренно консервативной и имеет неотрицательную величину в логарифмической матрице правдоподобия РАМ250.
Специалисту в данной области будет понятно, что в связи с вырождением генетического кода в дополнение к описанным в настоящем документе нуклеотидным остаткам, кодирующим гуманизированные полипептиды Т-клеточного корецептора, гуманизированные полипептиды МНС и/или вариабельные участки TCR, другие нуклеиновые кислоты могут кодировать полипептиды изобретения. Таким образом, в дополнение к генетически модифицированному не относящемуся к человеку животному, которое содержит в своем геноме нуклеотидную последовательность, кодирующую гуманизированный полипептид Т-клеточного корецептора (например, полипептид CD4 или CD8), неперестроенный вариабельный локус гена Т-клеточного рецептора (например, TCRα и/или TCRβ), содержащий человеческие неперестроенные сегменты гена, и/или последовательность нуклеиновых кислот, кодирующую гуманизированный полипептид МНС, способный ассоциироваться с гуманизированным полипептидом Т-клеточного рецептора с консервативными аминокислотными заменами, также предложено не относящееся к человеку животное, в геноме которого содержится нуклеотидная последовательность, кодирующая гуманизированный полипептид Т-клеточного рецептора (например, полипептид CD4 или CD8), неперестроенный вариабельный локус гена Т-клеточного рецептора (например, TCRα и/или TCRβ), содержащий человеческие неперестроенные сегменты гена, и/или последовательность нуклеиновых кислот, кодирующая гуманизированный полипептид MHC, способный ассоциироваться с гуманизированным полипептидом Т-клеточного рецептора, которая отличается от описанной в настоящем документе в связи с вырождением генетического кода.
Идентичность последовательности можно определять по ряду различных алгоритмов, известных в данной области, которые можно использовать для определения идентичности нуклеотидной и/или аминокислотной последовательности. В некоторых вариантах осуществления, описанных в настоящем документе, идентичности определяются с помощью программы выравнивания ClustalW, вер. 1.83 (медленное выравнивание) со штрафом за открытие гэпа 10,0 и штрафом за продолжение гэпа 0,1 и с помощью матрицы правдоподобия Гоннета (MacVector™ 10.0.2, MacVector Inc., 2008). Длина последовательностей, сравниваемых на идентичность последовательностей, будет зависеть от конкретных последовательностей. В различных вариантах осуществления идентичность определяется посредством сравнения последовательности зрелого белка от ее N-конца к ее С-концу. В различных вариантах осуществления при сравнении химерной человеческой/нечеловеческой последовательности с человеческой последовательностью человеческий участок химерной человеческой/нечеловеческой последовательности (но не нечеловеческий участок) используется для выполнения сравнения с целью подтверждения степени идентичности между человеческой последовательностью и человеческим участком химерной человеческой/нечеловеческой последовательности (например, сравнения человеческого эктодомена химерного человеческого/мышиного белка с человеческим эктодоменом человеческого белка).
Термины «гомология» или «гомологичный» по отношению к последовательностям, например нуклеотидным или аминокислотным последовательностям, означают две последовательности, у которых, например, по меньшей мере приблизительно 75% нуклеотидов или аминокислот, например по меньшей мере приблизительно 80% нуклеотидов или аминокислот, например по меньшей мере приблизительно 90-95% нуклеотидов или аминокислот, например более 97% нуклеотидов или аминокислот идентичны при оптимальном выравнивании и сравнении. Специалисту в данной области будет понятно, что для оптимального нацеливания на ген нацеливающий конструкт должен содержать плечи, гомологичные эндогенным последовательностям ДНК (т.е. «гомологичные плечи»). Следовательно, между нацеливающим конструктом и нацеленной эндогенной последовательностью может происходить гомологичная рекомбинация.
Термин «функционально связанный» относится к смежному положению, в котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать надлежащим образом. Так, последовательность нуклеиновых кислот, кодирующая белок, может быть функционально связана с регуляторными последовательностями (например, последовательностью промотора, энхансера, сайленсера и т.д), чтобы сохранить соответствующее регулирование транскрипции. Кроме того, различные участки химерного или гуманизированного белка изобретения могут быть функционально связаны между собой для сохранения приемлемых фолдинга, процессинга, нацеливания, экспрессии и других функциональных свойств белка в клетке. Если не указано иное, различные домены химерных или гуманизированных белков изобретения функционально связаны друг с другом.
Термин «замещение» в отношении замещения гена обозначает размещение экзогенного генетического материала в эндогенном генетическом локусе, что приводит к замещению всего или части эндогенного гена ортологичной или гомологичной последовательностью нуклеиновых кислот. Как продемонстрировано в примерах ниже, в одном варианте осуществления последовательности нуклеиновых кислот эндогенных локусов, кодирующие участки мышиных полипептидов CD4 или CD8 (CD8α и/или CD8β), были замещены нуклеотидными последовательностями, кодирующими участки человеческих полипептидов CD4 или CD8 (CD8α и/или CD8β) соответственно.
Термин «функциональный» в контексте настоящего документа, например по отношению к функциональному полипептиду, относится к полипептиду, который сохраняет по меньшей мере один вид биологического действия, в норме ассоциированный с нативным белком. Например, в некоторых вариантах осуществления изобретения замещение в эндогенном локусе (например, замещение в эндогенном нечеловеческом локусе CD4 или CD8) приводит к потере локусом способности экспрессировать функциональный эндогенный полипептид.
Гуманизированный (-ые) Т-клеточный (-ые) корецептор (-ы)
В настоящем документы описаны не относящиеся к человеку животные, которые экспреесируют по меньшей мере один человеческий или гуманизированный Т-клеточный корецептор, например, CD4, CD8α и/или CD8β. Соответственно, не относящееся к человеку животное, описанное в настоящем документе, содержит по меньшей мере одну из первой, второй и/или третьей нуклеотидной последовательности, каждая из которых кодирует разный человеческий или химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора, выбранный из человеческого или гуманизированного полипептида CD4, человеческого или гуманизированного полипептида CD8α и человеческого или гуманизированного полипептида CD8β. Использование обозначений «первая», «вторая», «третья» в настоящем документе не следует толковать как ограничивающее требование по наличию у не относящихся к человеку животных, описанных в настоящем документе, всех трех нуклеотидных последовательностей или наличию какой-либо из корецепторных нуклеотидных последовательностей в каком-либо порядке. Соответственно, не относящееся к человеку животное, описанное в настоящем документе, может содержать последовательность нуклеиновых кислот или последовательности нуклеиновых кислот, кодирующие полипептид (-ы) человеческого или гуманизированного CD4 и/или человеческого или гуманизированного CD8 (например, человеческого или гуманизированного CD8α и/или CD8β).
В одном варианте осуществления не относящееся к человеку животное, описанное в настоящем документе, содержит первую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD4. В другом варианте осуществления не относящееся к человеку животное, описанное в настоящем документе, содержит первую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD8α, и вторую нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD8β. В другом варианте осуществления не относящееся к человеку животное, описанное в настоящем документе, содержит первую и вторую нуклеотидные последовательности, кодирующие человеческие или гуманизированные полипептиды CD8α и CD8β, и дополнительно содержит третью нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD4.
Человеческий или гуманизированный CD4
В различных вариантах осуществления в изобретении по существу предложены генетически модифицированные не относящиеся к человеку животные, которые содержат в своем геноме, например в эндогенном локусе CD4, нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD4; следовательно, животные экспрессируют человеческий или гуманизированный полипептид CD4.
Человеческий ген CD4 находится в хромосоме 12, и считается, что он содержит 10 экзонов. Ген CD4 кодирует белок с аминоконцевой гидрофобной сигнальной последовательностью, кодируемой экзонами 2 и 3 гена. Белок содержит четыре внеклеточных иммуноглобулиноподобных домена Ig1-Ig4, также часто и соответственно обозначаемых доменами D1-D4. Maddon et al. (1987) Structure and expression of the human and mouse T4 genes, Proc. Natl. Acad. Sci. USA 84:9155-59. Считается, что домен D1 кодируется экзоном 3 (последовательность ближе к 3'-концу от сигнального пептида) и экзоном 4, а каждый из D2, D3 и D4 кодируется отдельным экзоном: экзонами 5, 6 и 7 соответственно (см. ФИГ. 5А: домены D1, D2, D3 и D4 кодируются последовательностями, обозначенными Ig1, Ig2, Ig3 и Ig4 соответственно). Littman (1987) The Structure of the CD4 and CD8 Genes, Ann. Rev. Immunol. 5:561-84; Hanna et al. (1994) Specific Expression of the Human CD4 Gene in Mature CD4+CD8- and Immature CD4+CD8+ Т cells and in Macrophages of Transgenic Mice, Mol. Cell. Biol. 14(2): 1084-94; Maddon et al., выше. В областях с высокой концентрацией белка, таких как область контакта Т-клетки с антигенпредставляющей клеткой, молекула стремится к гомодимеризации посредством взаимодействий между противоположными доменами D4. Zamoyska (1998) CD4 and CD8: modulators of Т cell receptor recognition of antigen and of immune responses? Curr. Opin. Immunol. 10:82-87; Wu et al. (1997) Dimeric association and segmental variability in the structure of human CD4, Nature 387:527; Moldovan et al. (2002) CD4 Dimers Constitute the Functional Component Required for Т Cell Activation, J. Immunol. 169:6261-68.
Домен D1 CD4 схож с вариабельным (V) доменом иммуноглобулина, и считается, что он вместе с частью домена D2 связывается (ассоциируется) с МНС II, например, в сайге связывания корецептора МНС II. Huang et al. (1997) Analysis of the contact sites on the CD4 Molecule with Class II МНС Molecule, J. Immunol. 158:216-25. В свою очередь, МНС II взаимодействует с Т-клеточным корецептором CD4 в гидрофобном кармане в месте соединения доменов МНС II α 2 и β2. Wang and Reinherz (2002) Structural Basis of Т Cell Recognition of Peptides Bound to МНС Molecules, Molecular Immunology, 38:1039-49.
Предполагается, что домены D3 и D4 корецептора CD4 взаимодействуют с комплексом TCR-CD3, так как замена этих двух доменов лишала CD4 способности связываться с TCR. Vignali et al. (1996) The Two Membrane Proximal Domains of CD4 Interact with the Т Cell Receptor, J. Exp. Med. 183:2097-2107. Молекула CD4 существует в виде димера, и считается, что остатки домена D4 молекулы ответственны за димеризацию CD4. Moldovan et al. (2002) CD4 Dimers Constitute the Functional Components Required for Т Cell Activation, J. Immunol. 169:6261-68.
Экзон 8 гена CD4 кодирует трансмембранный домен, а остальная часть гена кодирует цитоплазматический домен. Цитоплазматический домен CD4 выполняет множество разных функций. Например, цитоплазматический домен CD4 мобилизует тирозинкиназу Lck. Lck представляет собой киназу семейства Src, которая ассоциируется с цитоплазматическими доменами CD4 и CD8, и одновременное связывание корецепторов и TCR с тем же МНС приводит к увеличению фосфорилирования тирозина CD3 и цепи ζ комплекса TCR, что, в свою очередь, приводит к мобилизации других факторов, которые играют роль в Т-клеточной активации. Itano и коллеги после создания и тестирования экспрессии гибридного белка, содержащего внеклеточный домен CD8 и цитоплазматический хвост CD4 у трансгенных мышей, предположили, что цитоплазматический хвост CD4 также способствует дифференцировке Т-клеток CD4+CD8+ по линии CD4+. Itano et al. (1996) The Cytoplasmic Domain of CD4 Promotes the Development of CD4 Lineage Т Cells, J. Exp. Med. 183:731-41. Экспрессия гибридного белка приводила к развитию специфичных к МНС I Т-клеток линии CD4. Там же.
Корецептор CD4 является основным рецептором для вируса иммунодефицита человека (ВИЧ), а истощение численности Т-клеток CD4+ является индикатором прогрессирования заболевания. Цитоплазматический хвост CD4 является необходимым для передачи апоптотического сигнала на Т-клетки CD4+ при ВИЧ-индуцированном апоптозе. В частности, показано, что взаимодействие CD4 и Lck усиливает ВИЧ-индуцированный апоптоз в этих клетках. Corbeil et al. (1996) HIV-induced Apoptosis Requires the CD4 Receptor Cytoplasmic Tail and Is Accelerated by Interaction of CD4 with p561ck, J. Exp. Med. 183:39-48.
Т-клетки развиваются в тимусе, превращаясь из незрелых тимоцитов CD4-/CD8- (двойные отрицательные, или DN) в тимоциты CD4+/CD8+ (двойные положительные, или DP), которые в итоге подвергаются положительной селекции и становятся Т-клетками CD4+ или CD8+ (одинарные положительные, или SP). Тимоциты DP, которые принимают сигналы через рестриктированный по МНС I TCR, дифференцируются в Т-клетки CD8+, а тимоциты DP, которые принимают сигналы через рестриктированный по МНС II TCR, дифференцируются в Т-клетки CD4+. Стимулы, получаемые DP-клеткой, которые приводят к ее дифференцировке в Т-клетку CD4+ или CD8+, изучались в большом количестве исследований. Предложены различные модели выбора линии CD4/CD8, которые рассматриваются в публикации Singer et al. (2008) Lineage fate and intense debate: myths, models and mechanisms of CD4- versus CD8- lineage choice, Nat. Rev. Immunol. 8:788-801.
Инактивация специфического Т-клеточного рецептора в результате положительной селекции является следствием транскрипционной регуляции. В отношении CD4 было показано, что энхансер, расположенный на 13 т.п.н. ближе к 5'-концу от экзона 1 CD4, повышает экспрессию CD4 в Т-клетках CD4+ и CD8+. Killeen et al. (1993) Regulated expression of human CD4 rescues helper Т cell development in mice lacking expression of endogenous CD4, EMBO J. 12:1547-53. Действующий в цис-положении транскрипционный сайленсер, расположенный в пределах первого интрона мышиного гена CD4, вызывает отключение экспрессии CD4 в клетках, отличных от Т-клеток CD4+. Siu et al. (1994) A transcriptional silencer control the developmental expression of the CD4 gene, EMBO J. 13:3570-3579.
В связи с отсутствием в нескольких линиях ранее созданных трансгенных мышей, экспрессирующих человеческий CD4, важных транскрипционных регуляторов (например, промоторов, энхансеров, сайленсеров и т.д.), которые управляют выбором линии дифференцировки CD4, эти мыши были неспособны воспроизвести нормальное развитие по Т-клеточной линии дифференцировки и продуцировали иммунные клетки, отличные от Т-клеток CD4+, которые экспресcируют CD4. См., например, публикации Law et al. (1994) Human CD4 Restores Normal Т Cell Development and Function in Mice Deficient in CD4, J. Exp. Med. 179:1233-42 (экспрессия CD4 в Т-клетках CD8+ и В-клетках); Fugger et al. (1994) Expression of HLA-DR4 and human CD4 transgenes in mice determines the variable region p-chain T-cell repertoire and mediates an HLA-D-restricted immune response, Proc. Natl. Acad. Sci. USA, 91:6151-55 (CD4 экспрессировались на всех тимоцитах CD3+ и В-клетках). Таким образом, в одном варианте осуществления может иметь преимущество создание генетически модифицированного животного, которое сохраняет эндогенный мышиный промотор и другие регуляторные элементы, чтобы животное продуцировало Т-клетки, способные развиваться в Т-клетки и выбирать линию дифференцировки.
Таким образом, в различных вариантах осуществления в изобретении предложено генетически модифицированное не относящееся к человеку животное, содержащее, например, в своем эндогенном локусе Т-клеточного корецептора (например, локусе CD4) нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора. В одном варианте осуществления человеческий участок химерного полипептида содержит весь или по существу весь внеклеточный участок (или его часть, например один или более внеклеточных доменов, например по меньшей мере два последовательных внеклеточных домена) человеческого Т-клеточного корецептора. В одном варианте осуществления нечеловеческий участок химерного полипептида содержит трансмембранный и цитоплазматический домены нечеловеческого Т-клеточного корецептора. В одном варианте осуществления у не относящегося к человеку животного экспрессируется функциональный химерный полипептид Т-клеточного корецептора. Таким образом, в одном аспекте в изобретении предложено генетически модифицированное не относящееся к человеку животное, содержащее в своем эндогенном локусе CD4 нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD4, причем человеческий участок химерного полипептида содержит весь или по существу весь внеклеточный участок человеческого CD4, при этом нечеловеческий участок содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого CD4 и при этом животное экспрессирует функциональный полипептид CD4. В одном аспекте у не относящегося к человеку животного экспрессируется только гуманизированный полипептид CD4, т.е. химерный человеческий/нечеловеческий полипептид CD4, и не экспрессируется функциональный эндогенный нечеловеческий белок CD4 из его эндогенного локуса CD4.
В одном варианте осуществления человеческий участок химерного человеческого/нечеловеческого полипептида CD4 содержит весь или по существу весь внеклеточный участок человеческого полипептида CD4. В другом варианте осуществления человеческий участок химерного человеческого/нечеловеческого полипептида CD4 содержит по меньшей мере весь или по существу весь домен связывания с МНС II человеческого полипептида CD4 (например, значительный участок человеческих доменов D1 и D2); в одном варианте осуществления человеческий участок химерного человеческого/нечеловеческого полипептида CD4 содержит все или по существу все из доменов D1, D2 и D3 человеческого полипептида CD4; в еще одном варианте осуществления человеческий участок химерного человеческого/нечеловеческого полипептида CD4 содержит все или по существу все из иммуноглобулиноподобных доменов CD4, например доменов, обозначаемых D1, D2, D3 и D4. В еще одном варианте осуществления человеческий участок химерного человеческого/нечеловеческого полипептида CD4 содержит в своем человеческом участке всю или по существу всю человеческую последовательность CD4, которая несет ответственность за взаимодействие с МНС II и/или внеклеточным участком Т-клеточного рецептора. В еще одном варианте осуществления человеческий участок химерного человеческого/нечеловеческого полипептида CD4 содержит весь или по существу весь внеклеточный участок человеческого CD4, который несет ответственность за взаимодействие с МНС II и/или вариабельным доменом Т-клеточного рецептора. Таким образом, в одном варианте осуществления нуклеотидная последовательность, кодирующая человеческий участок химерного полипептида CD4, содержит всю или по существу всю кодирующую последовательность доменов D1-D2 человеческого CD4 (например, участок экзона 3 и экзоны 4-5 человеческого гена CD4); в другом варианте осуществления она содержит всю или по существу всю кодирующую последовательность D1-D3 человеческого CD4 (например, участок экзона 3 и экзоны 4-6 человеческого CD4). Таким образом, в одном варианте осуществления нуклеотидная последовательность, кодирующая химерный человеческий/нечеловеческий CD4, содержит нуклеотидные последовательности, кодирующие все или по существу все из доменов D1-D3 человеческого CD4. В другом варианте осуществления нуклеотидная последовательность, кодирующая человеческий участок химерного полипептида CD4, содержит кодирующую последовательность доменов D1-D4 человеческого гена CD4. В другом варианте осуществления нуклеотидная последовательность может содержать нуклеотидную последовательность, кодирующую мышиный сигнальный пептид CD4, например область, кодируемую участками экзонов 2-3 мышиного гена. В другом варианте осуществления нуклеотидная последовательность может содержать нуклеотидную последовательность, кодирующую человеческий сигнальный пептид CD4. В одном варианте осуществления химерный человеческий/нечеловеческий полипептид CD4 содержит аминокислотную последовательность, представленную в SEQ ID NO: 78, а человеческий участок химерного полипептида охватывает приблизительно аминокислоты 27-319 SEQ ID NO: 78 (представленные отдельно в SEQ ID NO: 79).
В одном варианте осуществления у не относящегося к человеку животного экспрессируется химерная человеческая/нечеловеческая последовательность полипептида CD4. В одном варианте осуществления человеческий участок химерной последовательности CD4 содержит одну или более консервативных или неконсервативных модификаций.
В одном аспекте предложено не относящееся к человеку животное, которое экспрессирует человеческую последовательность CD4, причем человеческая последовательность CD4 по меньшей мере приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентична человеческой последовательности CD4. В конкретном варианте осуществления человеческая последовательность CD4 по меньшей мере приблизительно на 90%, 95%, 96%, 97%, 98% или 99% идентична человеческой последовательности CD4, описанной в примерах. В одном варианте осуществления человеческая последовательность CD4 содержит одну или более консервативных замен. В одном варианте осуществления человеческая последовательность CD4 содержит одну или более неконсервативных замен.
В некоторых вариантах осуществления участок, например человеческий участок химерного CD4, может содержать по существу всю последовательность, указанную в настоящем документе (например, по существу весь белковый домен, указанный в настоящем документе). По существу вся последовательность включает 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислот, которые предположительно составляют конкретный участок белка (например, конкретный функциональный домен и т.д.). Специалисту в данной области будет понятно, что границы функционального домена могут немного варьировать в зависимости от используемых способов выравнивания и прогнозирования домена.
В одном аспекте нечеловеческий участок химерного человеческого/нечеловеческого полипептида CD4 содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого полипептида CD4. В связи с важными функциями, выполняемыми цитоплазматическим доменом CD4, сохранение эндогенной нечеловеческой (например, мышиной) последовательности у созданных методами генной инженерии животных обеспечивает сохранение правильной внутриклеточной сигнализации и других функций корецептора. В одном варианте осуществления не относящееся к человеку животное представляет собой мышь, а нечеловеческий полипептид CD4 представляет собой мышиный полипептид CD4. Хотя в примерах описана конкретная мышиная последовательность CD4, в область настоящего изобретения входит любая приемлемая последовательность, полученная из нее, например последовательность, содержащая консервативные/неконсервативные аминокислотные замены. В одном варианте осуществления нечеловеческий участок химерного корецептора CD4 содержит любую последовательность эндогенного CD4, которая не была гуманизирована.
Не относящееся к человеку животное, описанное в настоящем документе, может содержать в своем эндогенном локусе нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD4. В одном аспекте это приводит к замещению участка эндогенного гена CD4 нуклеотидной последовательностью, кодирующей участок человеческого полипептида CD4. В одном варианте осуществления такое замещение представляет собой замещение эндогенной нуклеотидной последовательности, кодирующей, например, весь или по существу весь внеклеточный домен нечеловеческого CD4, например последовательности, кодирующей по меньшей мере весь или по существу весь первый иммуноглобулиноподобный домен (т.е. D1) нечеловеческого CD4 (например, последовательности, кодирующей все или по существу все из доменов D1-D2 нечеловеческого CD4, например последовательности, кодирующей все или по существу все из доменов D1-D3 нечеловеческого CD4, например последовательности, кодирующей все или по существу все из доменов D1-D4 нечеловеческого CD4), человеческой нуклеотидной последовательностью, кодирующей вышеперечисленное. В одном варианте осуществления замещение приводит к образованию химерного белка, содержащего человеческую последовательность CD4, которая отвечает за взаимодействие с МНС II и/или внеклеточным участком Т-клеточного рецептора. В еще одном варианте осуществления замещение приводит к образованию химерного белка, содержащего человеческую последовательность CD4, которая отвечает за взаимодействие с МНС II и/или вариабельным доменом Т-клеточного рецептора. В одном варианте осуществления замещение не содержит замещения последовательности CD4, кодирующей по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого полипептида CD4. Таким образом, в одном аспекте у не относящегося к человеку животного экспрессируется химерный человеческий/нечеловеческий полипептид CD4 из эндогенного нечеловеческого локуса CD4. В еще одном варианте осуществления замещение приводит к образованию белка, содержащего последовательность полипептида, представленную в SEQ ID NO: 78.
В одном варианте осуществления предложена нуклеотидная последовательность химерного человеческого/нечеловеческого локуса CD4 (например, химерного локуса CD4 человека/грызуна, например химерного человеческого/мышиного локуса CD4), описанного в настоящем документе. В одном аспекте в связи с тем, что химерная человеческая/нечеловеческая (например, человека/грызуна, например человеческая/мышиная) последовательность CD4 размещена в эндогенном нечеловеческом (например, грызуна, например мышином) локусе CD4, она сохраняет энхансерный элемент CD4, расположенный ближе к 5'-концу от первого экзона CD4. В одном варианте осуществления замещение в эндогенном нечеловеческом (например, грызуна, например мышином) локусе CD4 содержит замещение, например, участка экзона 3, кодирующего D1, и экзонов 4-6, кодирующих остальную часть D1 и D2-D3 полипептида CD4; таким образом, в одном аспекте химерный локус CD4 сохраняет действующий в цис-положении сайленсер, расположенный в интроне 1 нечеловеческого (например, мышиного) гена CD4. Таким образом, в одном варианте осуществления химерный локус сохраняет эндогенные нечеловеческие (например, грызуна, например мышиные) промоторные и регуляторные элементы CD4. В другом варианте осуществления химерный локус может содержать человеческие промоторные и регуляторные элементы в количестве, достаточном для надлежащей экспрессии CD4, развития Т-клеток CD4+, выбора линии дифференцировки CD4 и функционирования корецептора. Таким образом, в некоторых аспектах животные изобретения содержат генетическую модификацию, которая не нарушает правильный выбор линии дифференцировки и развитие Т-клеток. В одном аспекте животные (например, грызуны, например мыши) изобретения не экспрессируют химерный полипептид CD4 на иммунных клетках, отличных от клеток, которые в норме экспрессируют CD4. В одном аспекте животные не экспрессируют CD4 на В-клетках или зрелых Т-клетках CD8+. В одном варианте осуществления замещение приводит к сохранению элементов, которые позволяют выполнять надлежащую пространственно-временную регуляцию экспрессии CD4.
В различных вариантах осуществления не относящееся к человеку животное (например, грызун, например мышь или крыса), которое экспрессирует функциональный химерный белок CD4 из химерного локуса CD4, как описано в настоящем документе, представляет химерный белок на клеточной поверхности, например Т-клеточной поверхности. В одном варианте осуществления у не относящееся к человеку животного экспрессируется химерный белок CD4 на клеточной поверхности с таким же клеточным распределением, которое наблюдается у человека. В одном аспекте белок CD4 изобретения способен взаимодействовать с белком МНС II, экспрессированным на поверхности второй клетки, например антигенпредставляющей клетки (АПК).
Человеческий или гуманизированный CD8
В различных вариантах осуществления в изобретении по существу предложены генетически модифицированные не относящиеся к человеку животные, которые содержат в своем геноме, например в эндогенном локусе CD8, нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD8; следовательно, животные экспрессируют человеческий или гуманизированный полипептид CD8. В различных вариантах осуществления в изобретении предложены не относящиеся к человеку животные, которые содержат в своем геноме, например в эндогенном локусе CD8, нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD8α, и/или нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид CD8β. Таким образом, генетически модифицированное не относящееся к человеку животное изобретения экспрессирует человеческий или гуманизированный полипептид CD8α и/или человеческий или гуманизированный полипептид CD8β.
Человеческий белок CD8, как правило, экспрессируется на клеточной поверхности в виде гетеродимера, состоящего из двух полипептидов (CD8α и CD8β), хотя также были обнаружены связанные дисульфидными мостиками гомодимеры и гомомультимеры (например, в NK-клетках и кишечных γδ Т-клетках, которые экспрессируют CD8αα). Гены, кодирующие человеческий CD8α и CD8β, расположены в непосредственной близости друг от друга на хромосоме 2. Nakayama et al. (1992) Recent Duplication of the Two Human CD8 β-chain genes, J. Immunol. 148:1919-27. Белок CD8α содержит лидерный пептид, область, подобную иммуноглобулину V, шарнирную область, трансмембранный домен и цитоплазматический хвост. Norment et а1. (1989) Alternatively Spliced mRNA Encodes a Secreted Form of Human CD8α. Characterization of the Human CD8α gene, J. Immunol. 142:3312-19. Экзоны/интроны гена CD8α схематически показаны на ФИГ. 5В.
Человеческий ген CD8β находится ближе к 5'-концу от гена CD8α на хромосоме 2. Описано множество изоформ, полученных путем альтернативного сплайсинга гена CD8β, причем предположительно одна изоформа не содержит трансмембранный домен и продуцирует секретируемый белок. Norment et а1. (1988) A second subunit of CD8 is expressed in human Т cells, EMBO J. 7:3433-39. Экзоны/интроны гена CD8β также схематически показаны на ФИГ. 5В.
Мембраносвязанный белок CD8β содержит N-концевую сигнальную последовательность, за которой расположены домен, подобный иммуноглобулину V, короткая внеклеточная шарнирная область и цитоплазматический хвост. См. публикацию Littman (1987) The structure of the CD4 and CD8 genes, Ann Rev. Immunol. 5:561-84. Шарнирная область представляет собой сайт обширного гликозилирования, который, как считается, поддерживает конформацию белка и защищает его от расщепления протеазами. Leahy (1995) A structural view of CD4 and CD8, FASEB J. 9:17-25.
Белок CD8, как правило, экспрессируется на цитотоксических Т-клетках и взаимодействует с молекулами МНС I. Взаимодействие опосредуется через связывание CD8 с доменом α3 МНС I. Хотя связывание МНС класса I с CD8 приблизительно в 100 раз слабее, чем связывание TCR с МНС класса I, связывание CD8 усиливает аффинность связывания TCR. Wooldridge et а1. (2010) МНС Class I Molecules with Superenhanced CD8 Binding Properties Bypass the Requirement for Cognate TCR Recognition and Nonspecifically Activate CTLs, J. Immunol. 184:3357-3366.
Связывание CD8 с молекулами МНС класса I является видоспецифичным. Показано, что мышиный гомолог CD8, Lyt-2, связывается с молекулами H-2Dd в домене α3, но не связывается с молекулами HLA-A. Connolly et al. (1988) The Lyt-2 Molecule Recognizes Residues in the Class I α3 Domain in Allogeneic Cytotoxic Т Cell Responses, J. Exp. Med. 168:325-341. Дифференцированное связывание происходило предположительно за счет CDR-подобных детерминант (CDR1- и CDR2-подобных) на CD8, которые не были сохранены у людей и мышей. Sanders et а1. (1991) Mutations in CD8 that Affect Interactions with HLA Class I and Monoclonal Anti-CD8 Antibodies, J. Exp. Med. 174:371-379; Vitiello et al. (1991) Analysis of the HLA-restricted Influenza-specific Cytotoxic Т Lymphocyte Response in Transgenie Mice Carrying a Chimeric Human-Mouse Class I Major Histocompatibility Complex, J. Exp. Med. 173:1007-1015 и Gao et а1. (1997) Crystal structure of the complex between human CD8αα and HLA-A2, Nature 387:630-634. Было описано, что CD8 связывается с HLA-A2 в консервативной области домена α3 (в положении 223-229). Одиночная замена (V245A) в HLA-A ослабляла связывание CD8 с HLA-A с одновременным значительным уменьшением опосредованного Т-клетками лизиса. Salter et al. (1989), Polymorphism in the α3 domain of HLA-A molecules affects binding to CDS, Nature 338:345-348. В целом полиморфизм в домене α3 молекул HLA-A также влиял на связывание с CD8. Там же. У мышей аминокислотная замена в остатке 227 в H-2Dd влияла на связывание мышиного Lyt-2 с H-2Dd, а клетки, трансфицированные мутантным H-2Dd, не лизировались клетками CD8+. Potter et al. (1989) Substitution at residue 227 of H-2 class I molecules abrogates recognition by CD8-dependent, but not CD8-independent, cytotoxic Т lymphocytes. Nature 337:73-75. Таким образом, экспрессия человеческого или гуманизированного CD8 может быть полезна для исследования Т-клеточных ответов на антиген, представляемый человеческим или гуманизированным МНС I.
Аналогично CD4 цитоплазматический домен CD8 взаимодействует с тирозинкиназой Lck, что, в свою очередь, приводит к Т-клеточной активации. Хотя Lck предположительно взаимодействует с цитоплазматичееким доменом CD8α, по-видимому, это взаимодействие регулируется присутствием цитоплазматического домена CD8β, так как мутации или делеции цитоплазматического домена CD8β приводили к снижению CD8α-ассоциированной активности Lck. Irie et al. (1998) The cytoplasmic domain of CD8β Regulates Lck Kinase Activation and CD8 Т cell Development, J. Immunol. 161:183-91. Снижение активности Lck было ассоциировано с нарушением развития Т-клеток. Там же.
Экспрессия CD8 на соответствующих клетках, например цитотоксических Т-клетках, жестко регулируется множеством энхансерных элементов, расположенных по всему локусу CD8. Например, в локусе CD8 было обнаружено по меньшей мере 4 области гиперчувствительности к ДНКазе I, которые часто ассоциируются со связыванием регулятора. Hosert et al. (1997) А CD8 genomic fragment that directs subset-specific expression of CD8 in transgenic mice, J. Immunol. 158:4270-81. С момента открытия этих областей гиперчувствительности к ДНКазе I в локусе CD8 было обнаружено по меньшей мере 5 энхансерных элементов, распределенных по всему локусу CD8, которые регулируют экспрессию CD8α и/или β в Т-клетках различных линий дифференцировки, включая Т-клетки DP, CD8 SP или клетки, экспрессирующие γδTCR. См., например, публикации Kioussis et а1. (2002) Chromatin and CD4, CD8A, and CD8B gene expression during thymic differentiation. Nature Rev. 2:909-919 и Online Erratum; Ellmeier et al. (1998) Multiple Development Stage-Specific Enhancers Regulate CD8 Expression in Developing Thymocytes and in Thymus-Independent Т cells. Immunity 9:485-96.
Таким образом, аналогично пользе, полученной за счет сохранения эндогенных промоторных и регуляторных элементов CD4 у генетически модифицированных животных с человеческим или гуманизированным CD4, в некоторых вариантах осуществления может быть полезно создание генетически модифицированного не относящегося к человеку животного, у которого сохранены эндогенные мышиные промоторные и регуляторные элементы, которые будут контролировать экспрессию человеческого или гуманизированного CD8. Особенно полезным может быть создание генетически модифицированных животных, содержащих замещение эндогенных нечеловеческих последовательностей, кодирующих белки CD8α и/или β, последовательностями, кодирующими человеческие или гуманизированные белки CD8α и/или β, как описано в настоящем документе.
В различных вариантах осуществления в изобретении предложено генетически модифицированное не относящееся к человеку животное, содержащее в своем геноме, например в своем эндогенном локусе CD8, по меньшей мере одну нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8 (например, полипептид CD8α и/или β), причем человеческий участок полипептида содержит весь или по существу весь внеклеточный участок (или его часть, например, внеклеточный домен) человеческого полипептида CD8 (например, CD8α и/или β), при этом нечеловеческий участок содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого CD8 (например, CD8α и/или β), и при этом животное экспрессирует химерный полипептид CD8 (например, полипептид CD8α и/или β). Таким образом, в одном варианте осуществления в изобретении предложено генетически модифицированное не относящееся к человеку животное, содержащее в своем эндогенном нечеловеческом локусе CD8 первую нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8α, и вторую нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8β, причем первая нуклеотидная последовательность содержит последовательность, которая кодирует весь или по существу весь внеклеточный участок человеческого полипептида CD8α и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого полипептида CD8α, и при этом вторая нуклеотидная последовательность содержит последовательность, которая кодирует весь или по существу весь внеклеточный участок человеческого полипептида CD8β и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого полипептида CDβ, при этом животное экспрессирует функциональный химерный человеческий/нечеловеческий белок CD8. В одном аспекте у не относящегося к человеку животного экспрессируется только гуманизированный полипептид CD8 (например, химерный человеческий/нечеловеческий полипептид CD8α и/или β) и не экспрессируется (-ются) соответствующий (-ие) функциональный (-ые) нечеловеческий (-ие) полипептид (-ы) CD8 из эндогенного локуса CD8.
В одном варианте осуществления химерный человеческий/нечеловеческий полипептид CD8α содержит в своем человеческом участке весь или по существу весь внеклеточный участок человеческого полипептида CD8α. В одном варианте осуществления человеческий участок химерного полипептида CD8α содержит по меньшей мере домен связывания с МНС I человеческого полипептида CD8α. В одном варианте осуществления человеческий участок химерного полипептида CD8α содержит последовательность по меньшей мере всего или по существу всего подобного иммуноглобулину V домена человеческого CD8α. В одном варианте осуществления нуклеотидная последовательность, кодирующая человеческий участок химерного полипептида CD8α, содержит по меньшей мере экзоны, которые кодируют внеклеточный участок человеческого полипептида CD8α. В одном варианте осуществления нуклеотидная последовательность содержит по меньшей мере экзоны, которые кодируют IgV-подобные домены. В одном варианте осуществления внеклеточный участок человеческого полипептида CD8α представляет собой область, охватывающую участок полипептида, который не является трансмембранным или цитоплазматическим доменом. В одном варианте осуществления нуклеотидная последовательность, кодирующая химерный человеческий/нечеловеческий полипептид CD8α, содержит последовательность, кодирующую нечеловеческий (например, грызуна, например мышиный) сигнальный пептид CD8α. Альтернативно нуклеотидная последовательность может содержать последовательность, кодирующую человеческую сигнальную последовательность CD8α. В одном варианте осуществления химерный человеческий/нечеловеческий полипептид CD8α содержит аминокислотную последовательность, представленную в SEQ ID NO: 88, а человеческий участок химерного полипептида включает аминокислоты 28-179 SEQ ID NO: 88 (представленные отдельно в SEQ ID NO: 89).
Аналогично в одном варианте осуществления химерный человеческий/нечеловеческий полипептид CD8β содержит в своем человеческом участке весь или по существу весь внеклеточный участок человеческого полипептида CD8β. В одном варианте осуществления человеческий участок химерного полипептида CD8β содержит последовательность всего или по существу всего подобного иммуноглобулину V домена человеческого CD8β. В одном варианте осуществления нуклеотидная последовательность, кодирующая человеческий участок химерного полипептида CD8β, содержит по меньшей мере экзоны, которые кодируют внеклеточный участок человеческого полипептида CD8β. В одном варианте осуществления нуклеотидная последовательность, кодирующая человеческий участок химерного человеческого/нечеловеческого полипептида CD8β, содержит по меньшей мере экзоны, которые кодируют IgV-подобный домен человеческого CD8β. В одном варианте осуществления нуклеотидная последовательность, кодирующая химерный человеческий/нечеловеческий полипептид CD8β, содержит последовательность, кодирующую нечеловеческий (например, грызуна, например мышиный) сигнальный пептид CD8β. Альтернативно нуклеотидная последовательность может содержать последовательность, кодирующую человеческую сигнальную последовательность CD8β. В одном варианте осуществления химерный человеческий/нечеловеческий полипептид CD8β содержит аминокислотную последовательность, представленную в SEQ ID NO: 83, а человеческий участок химерного полипептида включает аминокислоты 15-165 в SEQ ID NO: 83 (представленные отдельно в SEQ ID NO: 84).
В одном варианте осуществления у не относящегося к человеку животного экспрессируются химерные человеческие/нечеловеческие полипептиды CD8α и/или CD8β. В некоторых вариантах осуществления человеческий участок химерного человеческого/нечеловеческого полипептида CD8α и/или β содержит одну или более консервативных или неконсервативных модификаций.
В одном аспекте предложено не относящееся к человеку животное, которое экспрессирует человеческую последовательность полипептида CD8α и/или β, причем человеческая последовательность полипептида CD8α и/или β по меньшей мере приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентична человеческой последовательности полипептида CD8α и/или β соответственно. В конкретном варианте осуществления человеческая последовательность полипептида CD8α и/или β по меньшей мере приблизительно на 90%, 95%, 96%, 97%, 98% или 99% идентична соответствующей человеческой последовательности полипептида CD8α и/или β, описанной в примерах. В одном варианте осуществления человеческая последовательность полипептида CD8α и/или β содержит одну или более консервативных замен. В одном варианте осуществления человеческая последовательность полипептида CD8α и/или β содержит одну или более неконсервативных замен.
В некоторых вариантах осуществления участок, например человеческий участок химерного CD8, может содержать по существу всю последовательность, указанную в настоящем документе (например, по существу весь белковый домен, указанный в настоящем документе). По существу вся последовательность по существу включает 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислот, которые предположительно составляют конкретный участок белка (например, конкретный функциональный домен и т.д.). Специалисту в данной области будет понятно, что границы функционального домена могут немного варьировать в зависимости от используемых способов выравнивания и прогнозирования домена.
В одном аспекте нечеловеческий участок химерного человеческого/нечеловеческого полипептида CD8α и/или β содержит по меньшей мере трансмембранный и/или цитоплазматический домены нечеловеческого полипептида CD8α и/или β соответственно. В связи с важными функциями, выполняемыми цитоплазматическим доменом CD8, сохранение эндогенной нечеловеческой (например, мышиной) последовательности у созданных методами генной инженерии животных обеспечивает сохранение правильной внутриклеточной сигнализации и других функций корецептора. В одном варианте осуществления не относящееся к человеку животное представляет собой мышь, а нечеловеческий полипептид CD8α и/или β представляет собой мышиный полипептид CD8α и/или β соответственно. Хотя в примерах описаны конкретные мышиные последовательности CD8α и β, в объем настоящего изобретения входит любая приемлемая последовательность, полученная из нее, например последовательность, содержащая консервативные/неконсервативные аминокислотные замены. В одном варианте осуществления у не относящегося к человеку животного (например, грызуна, например мыши) сохранена эндогенная последовательность, которая не была гуманизирована.
Не относящееся к человеку животное, описанное в настоящем документе, может содержать в своем эндогенном локусе нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD8α и/или β. В одном аспекте это приводит к замещению участка эндогенного гена CD8α нуклеотидной последовательностью, кодирующей участок человеческого полипептида CD8α, и/или замещению участка эндогенного гена CD8β нуклеотидной последовательностью, кодирующей участок человеческого полипептида CD8β. В одном варианте осуществления такое замещение представляет собой замещение эндогенной нуклеотидной последовательности, кодирующей весь или по существу весь внеклеточный участок нечеловеческого CD8α и/или β, человеческой нуклеотидной последовательностью, кодирующей вышеперечисленное. В одном варианте осуществления такое замещение представляет собой замещение последовательности, кодирующей по меньшей мере весь или по существу весь подобный иммуноглобулину V домен нечеловеческого CD8α и/или β, человеческой нуклеотидной последовательностью, кодирующей вышеперечисленное. В одном варианте осуществления замещение не содержит замещения последовательности CD8α и/или β, кодирующей трансмембранный и цитоплазматический домены нечеловеческого полипептида CD8α и/или β. Таким образом, у не относящегося к человеку животного экспрессируется химерный человеческий/нечеловеческий полипептид CD8α и/или β из эндогенного нечеловеческого локуса CD8. В еще одном варианте осуществления замещение приводит к образованию белка CD8α и/или β, содержащего последовательность полипептида, представленную в SEQ ID NO: 88 и/или 84 соответственно.
В одном варианте осуществления предложена нуклеотидная последовательность химерного человеческого/нечеловеческого локуса CD8 (например, химерного локуса CD8 грызуна, например химерного мышиного локуса CD8). В одном аспекте в связи с тем, что химерная человеческая/нечеловеческая (например, человека/грызуна, например человека/мыши) последовательность CD8α и/или β находится в соответствующем эндогенном нечеловеческом (например, грызуна, например мышином) локусе CD8α и/или β, в ней сохранены эндогенные промоторные и регуляторные элементы CD8α и/или β. В другом варианте осуществления химерный локус может содержать человеческие промоторные и регуляторные элементы CD8α и/или β в количестве, достаточном для надлежащей экспрессии CD8α и/или β (надлежащей пространственно-временной экспрессии), развития Т-клеток CD8+, выбора линии дифференцировки CD8 и функционирования корецептора. Таким образом, в одном аспекте животные изобретения содержат генетическую модификацию, которая не нарушает правильный выбор линии дифференцировки и развитие Т-клеток. В одном аспекте животные (например, грызуны, например мыши) изобретения не экспрессируют химерный белок CD8 на иммунных клетках, отличных от клеток, которые в норме экспрессируют CD8, например, животные не экспрессируют CD8 на В-клетках или зрелых Т-клетках CD4+. В одном варианте осуществления замещение приводит к сохранению элементов, которые позволяют выполнять надлежащую пространственно-временную регуляцию экспрессии CD8α и/или β.
В различных вариантах осуществления не относящееся к человеку животное (например, грызун, например мышь или крыса), которое экспрессирует функциональный химерный белок CD8 (например, CD8αβ или CD8αα) из химерного локуса CD8, как описано в настоящем документе, представляет химерный белок на клеточной поверхности. В одном варианте осуществления у не относящегося к человеку животного экспрессируется химерный белок CD8 на клеточной поверхности с таким же клеточным распределением, которое наблюдается у человека. В одном аспекте белок CD8 изобретения способен взаимодействовать с белком МНС I, экспрессируемым на поверхности второй клетки.
Человеческий или гуманизированный Т-клеточный рецептор
В настоящем документе описаны генетически модифицированные не относящиеся к человеку животные, содержащие по существу гуманизированную Т-клеточную иммунную систему. В некотором варианте осуществления не относящееся к человеку животное, описанное в настоящем документе, содержит, например, в своем геноме: (а) нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий Т-клеточный корецептор, причем человеческий участок химерного полипептида Т-клеточного корецептора кодируется последовательностью, кодирующей внеклеточный домен человеческого Т-клеточного корецептора, и при этом последовательность, кодирующая внеклеточный домен человеческого Т-клеточного корецептора, функционально связана с нуклеотидом, содержащим последовательность, кодирующую трансмембранный и/или цитоплазматический домены нечеловеческого Т-клеточного корецептора; (b) неперестроенный ген вариабельного участка Т-клеточного рецептора, содержащий по меньшей мере один человеческий сегмент V, необязательно по меньшей мере один человеческий сегмент D и по меньшей мере один человеческий сегмент J, при этом неперестроенные сегменты V, необязательно D и J гена вариабельного участка TCR способны к рекомбинации с образованием перестроенного гена, функционально связанного с нечеловеческой константной последовательностью гена TCR; и (с) последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС, при этом человеческий участок химерного полипептида МНС содержит внеклеточный домен человеческого полипептида МНС, который ассоциируется с человеческим участком химерного полипептида Т-клеточного корецептора. Необязательно не относящееся к человеку животное также содержит человеческий или гуманизированный полипептид β2-микроглобулин.
Соответственно, в различных вариантах осуществления в изобретении по существу предложены генетически модифицированные не относящиеся к человеку животные, содержащие в геноме неперестроенные гуманизированные вариабельные локусы гена TCR, например неперестроенный человеческий вариабельный участок гена TCR, содержащий человеческие вариабельные сегменты TCR, способные к рекомбинации с образованием перестроенной вариабельной последовательности гена TCR. Локус TCR или локус гена TCR (например, локус TCRα или локус TCRβ) в настоящем документе означает геномную ДНК, содержащую кодирующий регион TCR, включающий весь кодирующий регион TCR, включая неперестроенные последовательности V(D)J, последовательность энхансера, константную (-ые) последовательность (-и) и любую расположенную ближе к 5'-концу или ближе к 3'-концу (нетранслируемые области, регуляторные области и т.д.) или вставочную последовательность ДНК (интроны и т.д.). Вариабельный локус TCR, вариабельный участок TCR или вариабельный локус гена TCR (например, вариабельный локус гена TCRα или вариабельный локус гена TCRβ) означает геномную ДНК, которая включает сегменты вариабельного участка TCR (область V(D)J), но не включает константные последовательности TCR и в различных вариантах осуществления последовательности энхансера. Другие последовательности могут быть включены в вариабельный локус гена TCR с целью выполнения генетических манипуляций (например, кассеты селекции, сайты рестрикции и т.д.), и эти последовательности входят в объем настоящего изобретения.
Т-клетки связываются с эпитопами на небольших антигенных детерминантах на поверхности антигенпредставляющих клеток, которые ассоциированы с главным комплексом гистосовместимости (МНС; у мышей) или человеческим лейкоцитарным антигеном (HLA; у людей). Т-клетки связываются с этими эпитопами посредством Т-клеточного рецепторного (TCR) комплекса на поверхности Т-клетки. Т-клеточные рецепторы представляют собой гетеродимерные структуры, состоящие из двух типов цепей: цепи α (альфа) и β (бета) или цепи γ (гамма) и δ (дельта). Цепь α кодируется последовательностью нуклеиновых кислот, расположенной в пределах локуса α (на человеческой или мышиной хромосоме 14), которая также охватывает весь локус δ, а цепь β кодируется последовательностью нуклеиновых кислот, расположенной в пределах локуса β (на мышиной хромосоме 6 или человеческой хромосоме 7). Большинство Т-клеток имеют αβ-TCR; а меньшинство Т-клеток несут γδ-TCR. Взаимодействия TCR с молекулами МНС класса I (представляющими Т-клеткам CD8+) и МНС класса II
(представляющими Т-клеткам CD4+) показаны на ФИГ. 1 (закрашенные символы обозначают нечеловеческие последовательности; заштрихованные обозначают человеческие последовательности, показывающие один конкретный вариант осуществления белка TCR настоящего изобретения).
α- и β-полипептиды Т-клеточного рецептора (и аналогично γ- и δ-полипептиды) связаны друг с другом дисульфидным мостиком. Каждый из двух полипептидов, составляющих TCR, включает внеклеточный домен, содержащий константный и вариабельный участки, трансмембранный домен и цитоплазматический хвост (трансмембранный домен и цитоплазматический хвост также являются частью константного участка). Вариабельный участок TCR определяет антигенную специфичность рецептора и по аналогии с иммуноглобулинами содержит три области, определяющие комплементарность (CDR). Также по аналогии с генами иммуноглобулина вариабельные локусы гена Т-клеточного рецептора (например, локусы TCRα и TCRβ) содержат ряд неперестроенных сегментов V(D)J (вариабельный (V), соединительный (J) и, в TCRβ и δ, дополнительный (D) сегменты). В процессе развития Т-клетки в тимусе вариабельный локус гена TCRα подвергается перестройке таким образом, что конечная цепь α TCR кодируется специфической комбинацией сегментов VJ (последовательность Vα/Jα); а вариабельный локус гена TCRβ подвергается перестройке таким образом, что конечная цепь β TCR кодируется специфической комбинацией сегментов VDJ (последовательность Vβ/Dβ/Jβ).
Взаимодействия со стромой тимуса активируют тимоциты, которые проходят несколько стадий развития, характеризуемых экспрессией различных маркеров на клеточной поверхности. Обзор характеристик маркеров клеточной поверхности на различных этапах развития в тимусе представлен в таблице 1. Перестройка в вариабельном локусе гена TCRβ начинается на стадии DN2 и завершается во время стадии DN4, а перестройка вариабельного локуса гена TCRα происходит на стадии DP. После завершения перестройки локуса TCRβ клетки экспрессируют на клеточной поверхности цепь β TCR, а также суррогатную цепь α, pТα. См. публикацию Janeway's Immunobiology, Chapter 7, 7th Ed., Murphy et al. eds., Garland Science, 2008.

Интактные Т-клетки CD4+ и CD8+ находятся в тимусе и попадают в периферические лимфоидные органы (например, селезенку), где при воздействии на них антигенов активируется клональное размножение и дифференцировка в ряд эффекторных Т-клеток (Teff), например цитотоксические Т-клетки, клетки TREG, клетки ТН17, клетки ТН1, клетки ТН2 и т.д. После инфицирования ряд Т-клеток сохраняется в виде Т-клеток памяти, которые могут быть двух типов: центральные Т-клетки памяти (Tcm) и эффекторные Т-клетки памяти (Tem). Sallusto et al. (1999) Two subsets of memory T lymphocytes with distinct homing potentials and effector functions, Nature 401:708-12 и Commentary by Mackay (1999) Dual personality of memory T cells, Nature 401:659-60. Sallusto и коллеги предположили, что после первичного инфицирования клетки Tem представляют собой легкодоступный пул примированных антигеном Т-клеток в периферических тканях с эффекторными функциями, а клетки Tcm представляют собой примированные антигеном Т-клетки в периферических лимфоидных органах, которые при вторичном введении антигена могут стать новыми эффекторными Т-клетками. Хотя все Т-клетки памяти экспрессируют изоформу CD45RO рецептора CD45 (интактные Т-клетки экспрессируют изоформу CD45RA), Tcm характеризуются экспрессией L-селектина (также известного как CD62L) и CCR7+, которые играют важную роль в связывании и сигнализации в периферических лимфоидных органах и лимфатических узлах. Там же. Таким образом, все Т-клетки, обнаруживаемые в периферических лимфоидных органах (например, интактные Т-клетки, клетки Tcm и т.д.), экспрессируют CD62L. Известно, что, кроме CD45RO, все Т-клетки памяти экспрессируют ряд различных маркеров клеточной поверхности, например CD44. Обзор различных маркеров клеточной поверхности на Т-клетках см. в Janeway's Immunobiology, Chapter 10, выше.
Хотя за распознавание антигена главным образом отвечает вариабельный домен TCR, внеклеточный участок константного домена, а также трансмембранный и цитоплазматический домены TCR также выполняют важные функции. Полный рецепторный комплекс TCR включает не только полипептиды α и β или γ и δ; необходимы дополнительные молекулы, такие как CD3γ, CD3δ и CD3ε, а также гомодимер цепи ζ, (ζζ). После завершения перестройки TCRβ (когда клетки экспрессируют TCRβ/pTα) данный неполный комплекс TCR находится на клеточной поверхности вместе с CD3. TCRα (или рТα) на клеточной поверхности имеет в своем трансмембранном домене два щелочных остатка, один из которых мобилизует гетеродимер CD3γε, а другой мобилизует ζζ с помощью соответствующих им кислых остатков. TCRβ имеет в своем трансмембранном домене дополнительный щелочной остаток, который, как считается, мобилизует гетеродимер CD3δε. См., например, публикации Kuhns et al. (2006) Deconstructing the Form and Function of the TCR/CD3 Complex, Immunity 24:133-39; Wucherpfennig et al. (2009) Structural Biology of the T-cell Receptor: Insights into Receptor Assembly, Ligand Recognition, and Initiation of Signaling, Cold Spring Harb. Perspect. Biol. 2:a005140. Собранный комплекс, содержащий гетеродимер TCRαβ, CD3γε, CD3δε и ζζ экспрессируется на поверхности Т-клетки. Было выдвинуто предположение, что полярные остатки в трансмембранном домене служат для контроля качества на выходе из эндоплазматического ретикулума. Было продемонстрировано, что при отсутствии субъединиц CD3 TCR-цепи остаются в эндоплазматическом ретикулуме и нацеливаются для разрушения. См., например, публикацию Call and Wucherpfennig (2005) The T Cell Receptor: Critical Role of the Membrane Environment in Receptor Assembly and Function, Annu. Rev. Immunol. 23:101-25.
Цепи CD3 и ζ собранного комплекса предоставляют компоненты для TCR-сигнализации, так как сам по себе гетеродимер TCRαβ (или гетеродимер TCRγδ) не обладает способностью к передаче сигнала. Каждая цепь CD3 имеет один иммунорецепторный тирозиновый активирующий мотив (ITAM), а цепь ζ содержит три последовательных ITAM. ITAM содержат тирозиновые остатки, которые могут подвергаться фосфорилированию соответствующими киназами. Таким образом, собранный комплекс TCR-CD3 содержит 10 мотивов ITAM. См., например, публикацию Love and Hayes (2010) ITAM-Mediated Signaling by the T-Cell Antigen Receptor, Cold Spring Harb. Perspect. Biol. 2:e002485. После задействования TCR мотивы ITAM подвергаются фосфорилированию тирозинкиназами семейства Scr (Lck и Fyn), которые инициируют сигнальный каскад, приводящий к активации Ras, мобилизации кальция, перестройкам актинового цитоскелета и активации факторов транскрипции, и все это в конечном счете приводит к Т-клеточной дифференцировке, пролиферации и эффекторным действиям. Там же, см. также Janeway's Immunobiology, выше; обе публикации включены в настоящий документ путем ссылки.
Кроме того, предполагается, что трансмембранный и цитоплазматический домены TCRβ играют роль в митохондриальном нацеливании и индукции апоптоза. Фактически в тимоцитах находятся усеченные со стороны N-конца молекулы TCRβ природного происхождения. Shani et al. (2009) Incomplete T-cell receptor--β peptides target the mitochondrion and induce apoptosis, Blood 113:3530-41. Таким образом, константный участок TCR (который в различных вариантах осуществления содержит участок внеклеточного, а также трансмембранного и цитоплазматического доменов) выполняет несколько важных функций, и в различных вариантах осуществления структуру этой области следует учитывать при создании гуманизированных TCR или экспрессирующих их генетически модифицированных не относящихся к человеку животных.
В данной области известны мыши, трансгенные по перестроенным последовательностям Т-клеточного рецептора. Настоящее изобретение относится к генетически модифицированным не относящимся к человеку животным (например, грызунам, например крысам, мышам), которые содержат неперестроенные человеческие или гуманизированные Т-клеточные вариабельные локусы гена, способные перестраиваться с образованием последовательностей нуклеиновых кислот, которые кодируют человеческие вариабельные домены Т-клеточного рецептора, включая животных, которые содержат Т-клетки, содержащие перестроенные человеческие вариабельные домены и нечеловеческие (например, мышиные или крысиные) константные участки. В настоящем изобретении также предложены не относящиеся к человеку животные (например, грызуны, например крысы, мыши), которые способны продуцировать разнородный набор человеческих последовательностей вариабельного участка Т-клеточного рецептора. Таким образом, в настоящем изобретении предложены не относящиеся к человеку животные, которые экспрессируют TCR с полностью человеческими вариабельными доменами в ответ на введение интересующего антигена, которые связываются с эпитопом интересующего антигена. В некоторых вариантах осуществления предложены не относящиеся к человеку животные, которые продуцируют разнородный набор Т-клеточных рецепторов, способных реагировать с различными антигенами, включая, без ограничений, антигены, представляемые АПК.
В одном варианте осуществления в изобретении предложены генетически модифицированные не относящиеся к человеку животные (например, грызуны, например крысы или мыши), которые содержат в своем геноме неперестроенные человеческие сегменты вариабельного участка TCR (сегменты V(D)J), при этом неперестроенные человеческие сегменты вариабельного участка TCR замещают в эндогенном нечеловеческом (например, грызуна) вариабельном локусе гена TCR (например, вариабельном локусе гена TCRα, β, δ и/или γ) эндогенные нечеловеческие сегменты вариабельного участка TCR. В одном варианте осуществления неперестроенный человеческий вариабельный локус гена TCR замещает эндогенный нечеловеческий вариабельный локус гена TCR.
В другом варианте осуществления в изобретении предложены генетически модифицированные не относящиеся к человеку животные (например, грызуны, например крысы, мыши), которые содержат в своем геноме неперестроенные человеческие сегменты вариабельного участка TCR (сегменты V(D)J), при этом неперестроенные человеческие сегменты вариабельного участка TCR функционально связаны с нечеловеческой последовательностью гена константного участка TCR, что приводит к образованию гуманизированного локуса TCR, при этом гуманизированный локус TCR находится в таком сайте в геноме, который отличен от эндогенного нечеловеческого локуса TCR. Таким образом, в одном варианте осуществления также предложено не относящееся к человеку животное (например, грызун, например мышь, крыса), содержащее трансген, который содержит неперестроенные человеческие сегменты вариабельного участка TCR, функционально связанные с нечеловеческой последовательностью гена константного участка TCR.
В одном аспекте генетически модифицированные не относящиеся к человеку животные изобретения содержат в своем геноме человеческие сегменты вариабельного участка TCR, при этом сохраняя нечеловеческую(-ие) (например, грызуна, например мыши, крысы) константную(-ые) последовательность(-и) гена TCR, которая(-ые) кодирует(-ют) константные домены TCR. В различных вариантах осуществления константный домен TCR включает трансмембранный домен и цитоплазматический хвост TCR. Таким образом, в различных вариантах осуществления настоящего изобретения генетически модифицированные не относящиеся к человеку животные сохраняют эндогенные нечеловеческие трансмембранный домен и цитоплазматический хвост TCR. В других вариантах осуществления не относящиеся к человеку животные содержат нечеловеческие неэндогенные константные последовательности гена TCR, например, кодирующие нечеловеческие неэндогенные трансмембранный домен и цитоплазматический хвост TCR. Как указано выше, константный домен TCR участвует в сигнальном каскаде, запущенном в процессе активации примированной антигеном Т-клетки. Таким образом, эндогенный константный домен TCR взаимодействует с разнообразными нечеловеческими якорными и сигнальными белками в Т-клетке. Таким образом, в одном аспекте генетически модифицированные не относящиеся к человеку животные изобретения экспрессируют гуманизированные Т-клеточные рецепторы, которые сохраняют способность мобилизовать разнообразные эндогенные нечеловеческие якорные или сигнальные молекулы, например молекулы CD3 (например, CD3γ, CD3δ, CD3ε), цепь ζ, Lck, Fyn, ZAP-70 и т.д. Не имеющий ограничительного характера список молекул, которые мобилизуются к комплексу TCR, описан в публикации Janeway's Immunobiology, выше. Считается, что возможность протекания процессов развития и дифференцировки Т-клеток у не относящихся к человеку животных и формирования устойчивого иммунного ответа может быть по меньшей мере частично связана с включением вариабельных участков в эндогенные мышиные локусы и сохранением мышиных константных доменов.
В некоторых вариантах осуществления предложено не относящееся к человеку животное, которое содержит в своем геноме неперестроенные человеческие сегменты вариабельного участка TCRα, при этом неперестроенные человеческие сегменты вариабельного участка TCRα функционально связаны с нечеловеческой последовательностью гена константного участка TCRα, что приводит к образованию гуманизированного локуса TCRα. В одном варианте осуществления гуманизированный локус TCRα находится в таком сайте в геноме, который не является эндогенным нечеловеческим локусом TCRα. В другом варианте осуществления неперестроенные человеческие сегменты вариабельного участка TCRα замещают эндогенные нечеловеческие сегменты вариабельного участка TCRα, при этом сохраняется(-ются) эндогенная(-ые) нечеловеческая(-ие) последовательность(-и) гена константного участка TCRα. В одном варианте осуществления неперестроенный человеческий вариабельный локус гена TCRα замещает эндогенный нечеловеческий вариабельный локус гена TCRα. В некоторых вариантах осуществления замещение эндогенного нечеловеческого локуса гена вариабельного участка TCRα неперестроенным человеческим вариабельным локусом гена TCRα включает делению или инактивацию вариабельного локуса гена TCRδ. В других вариантах осуществления замещение эндогенного нечеловеческого локуса гена вариабельного участка TCRα неперестроенным человеческим локусом гена TCRα включает замещение эндогенного вариабельного локуса гена TCRδ неперестроенными человеческими сегментами вариабельного участка TCRδ. В некоторых вариантах осуществления животное сохраняет эндогенные последовательности гена вариабельного участка и константного участка TCRβ. Таким образом, у животного экспрессируется TCR, который содержит химерную человеческую/нечеловеческую (т.е. гуманизированную) цепь α TCR и нечеловеческую цепь β TCR.
В некоторых вариантах осуществления предложено не относящееся к человеку животное, которое содержит в своем геноме неперестроенные человеческие сегменты вариабельного участка TCRδ, при этом неперестроенные человеческие сегменты вариабельного участка TCRδ функционально связаны с нечеловеческой последовательностью гена константного участка TCRδ, что приводит к образованию гуманизированного локуса TCRδ. В одном варианте осуществления гуманизированный локус TCRδ находится в таком сайте в геноме, который не является эндогенным нечеловеческим локусом TCRδ. В другом варианте осуществления неперестроенные человеческие сегменты вариабельного участка TCRδ замещают эндогенные нечеловеческие сегменты вариабельного участка TCRδ, при этом сохраняется(-ются) эндогенная(-ые) нечеловеческая(-ие) последовательность(-и) гена константного участка TCRδ. В одном варианте осуществления неперестроенный человеческий вариабельный локус гена TCRδ замещает эндогенный нечеловеческий вариабельный локус гена TCRδ.
В других вариантах осуществления предложено не относящееся к человеку животное, которое содержит в своем геноме неперестроенные человеческие сегменты вариабельного участка TCRβ, при этом неперестроенные человеческие сегменты вариабельного участка TCRβ функционально связаны с нечеловеческой последовательностью гена константного участка TCRβ, что приводит к образованию гуманизированного локуса TCRβ. В одном варианте осуществления гуманизированный локус TCRβ находится в таком сайте в геноме, который не является эндогенным нечеловеческим локусом TCRβ. В другом варианте осуществления неперестроенные человеческие сегменты вариабельного участка TCRβ замещают эндогенные нечеловеческие сегменты вариабельного участка TCRβ, при этом сохраняется(-ются) эндогенная(-ые) нечеловеческая(-ие) последовательность(-и) гена константного участка TCRβ В одном варианте осуществления неперестроенный человеческий вариабельный локус гена TCRβ замещает эндогенный нечеловеческий вариабельный локус гена TCRβ. В некоторых вариантах осуществления животное сохраняет эндогенные последовательности гена вариабельного участка и константного участка TCRα. Таким образом, у животного экспрессируется TCR, который содержит химерную человеческую/нечеловеческую (т.е. гуманизированную) цепь β TCR и нечеловеческую цепь α TCR.
В некоторых конкретных вариантах осуществления в изобретении предложено генетически модифицированное не относящееся к человеку животное (например, грызун, например мышь или крыса), которое содержит в своем геноме: (а) неперестроенный вариабельный локус гена Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанные с эндогенной(-ыми) нечеловеческой(-ими) (например, грызуна, например мыши или крысы) константной(-ыми) последовательностью(-ями) гена TCRα; (b) неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанные с эндогенной(-ыми) нечеловеческой(-ими) (например, грызуна, например мыши или крысы) последовательностью(-ями) гена константного участка TCRβ; и/или (с) неперестроенный вариабельный локус гена TCRδ, содержащий по меньшей мере один человеческий сегмент Vδ, по меньшей мере один человеческий сегмент Dδ и по меньшей мере один человеческий сегмент Jδ, функционально связанные с эндогенной нечеловеческой (например, грызуна, например мыши или крысы) последовательностью гена константного участка TCRδ. Другое не относящееся к человеку животное, предложенное в настоящем документе, содержит в своем геноме: (а) неперестроенный вариабельный локус гена Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанные с эндогенной(-ыми) нечеловеческой(-ими) (например, грызуна, например мыши или крысы) константной(-ыми) последовательностью(-ями) гена TCRα; (b) неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанные с эндогенной(-ыми) нечеловеческой(-ими) (например, грызуна, например мыши или крысы) последовательностью(-ями) гена константного участка TCRβ; (с) неперестроенный вариабельный локус гена TCRδ, содержащий по меньшей мере один человеческий сегмент Vδ, по меньшей мере один человеческий сегмент Dδ и по меньшей мере один человеческий сегмент Jδ, функционально связанные с эндогенной(-ыми) нечеловеческой(-ими) (например, грызуна, например мыши или крысы) последовательностью(-ями) гена константного участка TCRδ; и/или (d) неперестроенный вариабельный локус гена TCRγ, содержащий по меньшей мере один человеческий сегмент Vγ и по меньшей мере один человеческий сегмент Jγ, функционально связанные с эндогенной нечеловеческой (например, грызуна, например мыши или крысы) последовательностью гена константного участка TCRγ.
В различных вариантах осуществления изобретения неперестроенный человеческий или гуманизированный вариабельный локус гена TCR (например, вариабельный локус гена TCRα, TCRβ и/или TCRδ) содержится в зародышевой линии не относящегося к человеку животного (например, грызуна, например мыши или крысы). В различных вариантах осуществления замещения сегментов TCR V(D)J неперестроенными человеческими сегментами TCR V(D)J (например, сегментами Vα и Jα; Vβ и Dβ и Jβ; Vδ и Dδ и Jδ; Vγ и Jγ) выполняют в эндогенном нечеловеческом вариабельном локусе (или локусах) TCR, при этом неперестроенные человеческие сегменты V и J и/или V и D и J функционально связаны с нечеловеческими последовательностями гена константного участка TCR.
В некоторых вариантах осуществления изобретения не относящееся к человеку животное содержит две копии неперестроенного человеческого или гуманизированного вариабельного локуса гена TCRα, две копии неперестроенного человеческого или гуманизированного вариабельного локуса гена TCRβ и/или две копии неперестроенного человеческого или гуманизированного вариабельного локуса гена TCRδ. Таким образом, не относящееся к человеку животное является гомозиготным по одному или более неперестроенным человеческим или гуманизированным вариабельным локусам гена TCRα, TCRβ и/или TCRδ. В некоторых вариантах осуществления изобретения не относящееся к человеку животное содержит одну копию неперестроенного человеческого или гуманизированного вариабельного локуса гена TCRα, одну копию неперестроенного человеческого или гуманизированного вариабельного локуса гена TCRβ и/или одну копию неперестроенного человеческого или гуманизированного вариабельного локуса гена TCRδ. Таким образом, не относящееся к человеку животное является гетерозиготным по неперестроенному человеческому или гуманизированному вариабельному локусу гена TCRα, TCRβ и/или TCRδ. В другом варианте осуществления не относящееся к человеку животное является гетерозиготным или гомозиготным по неперестроенному человеческому или гуманизированному вариабельному локусу гена TCRγ.
В одном варианте осуществления неперестроенный вариабельный локус гена TCRα, содержащий человеческие сегменты вариабельного участка (например, человеческие сегменты Vα и Jα) расположен в нечеловеческом геноме таким образом, что человеческие сегменты вариабельного участка замещают соответствующие нечеловеческие сегменты вариабельного участка. В одном варианте осуществления неперестроенный вариабельный локус гена TCRα, содержащий человеческие сегменты вариабельного участка, замещает эндогенный вариабельный локус гена TCRα. В одном аспекте эндогенные нечеловеческие сегменты Vα и Jα неспособны к перестройке с образованием перестроенной последовательности Vα/Jα. Таким образом, в одном аспекте человеческие сегменты Vα и Jα в неперестроенном вариабельном локусе гена TCRα способны к перестройке с образованием перестроенной человеческой последовательности Vα/Jα.
Аналогичным образом, в одном варианте осуществления неперестроенный вариабельный локус гена TCRβ, содержащий человеческие сегменты вариабельного участка (например, человеческие сегменты Vβ, Dβ и Jβ), расположен в нечеловеческом геноме таким образом, что человеческие сегменты вариабельного участка замещают соответствующие нечеловеческие сегменты вариабельного участка. В одном варианте осуществления неперестроенный вариабельный локус гена TCRβ, содержащий человеческие сегменты вариабельного участка, замещает эндогенный вариабельный локус гена TCRβ. В одном аспекте эндогенные нечеловеческие сегменты Vβ, Dβ и Jβ способны к перестройке с образованием перестроенной последовательности Vβ/Dβ/Jβ. Таким образом, в одном аспекте человеческие сегменты Vβ, Dβ и Jβ в неперестроенном вариабельном локусе гена TCRβ способны к перестройке с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ.
В одном варианте осуществления неперестроенный вариабельный локус гена TCRδ, содержащий человеческие сегменты вариабельного участка (например, человеческие сегменты Vδ, Dδ и Jδ), расположен в нечеловеческом геноме таким образом, что человеческие сегменты вариабельного участка замещают соответствующие нечеловеческие сегменты вариабельного участка. В одном варианте осуществления неперестроенный вариабельный локус гена TCRδ, содержащий человеческие сегменты вариабельного участка, замещает эндогенный вариабельный локус гена TCRδ. В одном аспекте эндогенные нечеловеческие сегменты Vδ, Dδ и Jδ неспособны к перестройке с образованием перестроенной последовательности Vδ/Dδ/Jδ. Таким образом, в одном аспекте человеческие сегменты Vδ, Dδ и Jδ в неперестроенном вариабельном локусе гена TCRδ способны к перестройке с образованием перестроенной человеческой последовательности Vδ/Dδ/Jδ.
В одном варианте осуществления неперестроенный вариабельный локус гена TCRγ, содержащий человеческие сегменты вариабельного участка (например, человеческие сегменты Vγ и Jγ), расположен в нечеловеческом геноме таким образом, что человеческие сегменты вариабельного участка замещают соответствующие нечеловеческие сегменты вариабельного участка. В одном варианте осуществления неперестроенный вариабельный локус гена TCRγ, содержащий человеческие сегменты вариабельного участка, замещает эндогенный вариабельный локус гена TCRγ. В одном аспекте эндогенные нечеловеческие сегменты Vα и Jα неспособны к перестройке с образованием перестроенной последовательности Vγ/Jγ. Таким образом, в одном аспекте человеческие сегменты Vγ и Jγ в неперестроенном вариабельном локусе гена TCRγ способны к перестройке с образованием перестроенной человеческой последовательности Vγ/Jγ.
В еще одном варианте осуществления неперестроенные вариабельные локусы гена TCRα, β, δ и/или γ, содержащие человеческие сегменты вариабельного участка, замещают соответствующие эндогенные вариабельные локусы гена TCRα, β, δ и γ. В одном аспекте эндогенные нечеловеческие сегменты Vα и Jα неспособны к перестройке с образованием перестроенной последовательности Vα/Jα, эндогенные нечеловеческие сегменты Vβ, Dβ и Jβ неспособны к перестройке с образованием перестроенной последовательности Vβ/Dβ/Jβ, эндогенные сегменты Vδ, Dδ и Jδ неспособны к перестройке с образованием перестроенной последовательности Vδ/Dδ/Jδ и/или эндогенные сегменты Vγ и Jγ неспособны к перестройке с образованием перестроенной последовательности Vγ/Jγ. Таким образом, в одном аспекте человеческие сегменты Vα и Jα в неперестроенном вариабельном локусе гена TCRα способны к перестройке с образованием перестроенной человеческой последовательности Vα/Jα, человеческие сегменты Vβ, Dβ и Jβ в неперестроенном вариабельном локусе гена TCRβ способны к перестройке с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ человеческие сегменты Vδ, Dδ и Jδ в неперестроенном вариабельном локусе гена TCRδ способны к перестройке с образованием перестроенной человеческой последовательности Vδ/Dδ/Jδ и/или человеческие сегменты Vγ и Jγ в неперестроенном вариабельном локусе гена TCRα способны к перестройке с образованием перестроенной человеческой последовательности Vγ/Jγ.
В некоторых аспектах изобретения не относящееся к человеку животное, содержащее гуманизированный локус гена TCRα, TCRβ и/или TCRδ (содержащий неперестроенный человеческий вариабельный локус гена TCRα, TCRβ и/или TCRδ), сохраняет эндогенный нечеловеческий вариабельный локус гена TCRα, TCRβ и/или TCRδ. В одном варианте осуществления эндогенный нечеловеческий вариабельный локус гена TCRα, TCRβ и/или TCRδ представляет собой нефункциональный локус. В одном варианте осуществления нефункциональный локус представляет собой инактивированный локус, например инвертированный локус (например, кодирующая последовательность нуклеиновых кислот вариабельного локуса гена находится в инвертированной ориентации по отношению к последовательности константной области так, что успешные перестройки не могут быть выполнены с использованием сегментов вариабельного участка из инвертированного локуса). В одном варианте осуществления гуманизированный вариабельный локус гена TCRα, TCRβ и/или TCRδ расположен между эндогенным нечеловеческим вариабельным локусом гена TCRα, TCRβ и/или TCRδ и эндогенным нечеловеческим константным локусом гена TCRα, TCRβ и/или TCRδ соответственно. Аналогичные хромосомные перестройки могут быть выполнены для размещения человеческого или гуманизированного TCRγ в геном не относящегося к человеку животного, например, в локусе TCRγ.
Количество, номенклатура, положение, а также другие аспекты сегментов V и J и/или V, D и J человеческих и мышиных локусов TCR могут быть установлены с использованием базы данных IMGT, доступной на веб-сайте Международной иммуногенетической информационной системы (IMGT). Мышиный вариабельный локус TCRα имеет размер приблизительно 1,5 м. п. н. и содержит всего 110 сегментов Vα и 60 сегментов Jα. Человеческий вариабельный локус TCRα имеет размер приблизительно 1 м. п. н. и содержит всего 54 сегмента Vα и 61 сегмент Jα, при этом 45 Vα и 50 Jα считаются функциональными. Если не указано иное, количества человеческих сегментов V(D)J, упоминаемые в тексте описания, относятся к общему количеству сегментов V(D)J. В одном варианте осуществления изобретения генетически модифицированное не относящееся к человеку животное (например, грызун, например мышь или крыса) содержит по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα. В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 23, 25, 30, 35, 40, 45, 48, 50 или до 54 человеческих сегментов Vα. В некоторых вариантах осуществления гуманизированный локус TCRα содержит 2, 8, 23, 35, 48 или 54 человеческих сегмента Vα. Таким образом, в некоторых вариантах осуществления гуманизированный локус TCRα у не относящегося к человеку животного может содержать 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% 99% или 100% человеческих Vα; в некоторых вариантах осуществления он может содержать приблизительно 2%, приблизительно 3%, приблизительно 15%, приблизительно 65%, приблизительно 90% или 100% человеческих Vα.
В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Vα40-Vα41 (сегмент Vα также обозначается как TRAV или TCRAV), и фрагмент ДНК, содержащий непрерывную человеческую последовательность из 61 человеческого сегмента Jα (сегмент Jα также обозначается как TRAJ или TCRAJ). В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRAV35-TRAV41, и фрагмент ДНК, содержащий непрерывную человеческую последовательность из 61 человеческого TRAJ. В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRAV22-TRAV41, и фрагмент ДНК, содержащий непрерывную человеческую последовательность из 61 человеческого TRAJ. В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRAV13-2-TRAV41, и фрагмент ДНК, содержащий непрерывную человеческую последовательность из 61 человеческого TRAJ. В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRAV6-TRAV41 и 61 человеческого TRAJ. В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRAV1-1-TRAV41 и 61 человеческого TRAJ. В различных вариантах осуществления фрагменты ДНК, содержащие непрерывные человеческие последовательности из человеческих сегментов вариабельного участка TCRα, также содержат сайты рестрикционных ферментов, кассеты селекции, сайты эндонуклеаз и другие сайты, вставленные для содействия клонированию и селекции в процессе гуманизации локуса. В различных вариантах осуществления эти дополнительные сайты не нарушают надлежащее функционирование (например, перестройку, сплайсинг и т.д.) различных генов в локусе TCRα.
В одном варианте осуществления гуманизированный локус TCRα содержит 61 человеческий сегмент Jα или 100% человеческих сегментов Jα. В конкретном варианте осуществления гуманизированный локус TCRα содержит 8 человеческих сегментов Vα или 61 человеческий сегмент Jα; в другом конкретном варианте осуществления гуманизированный локус TCRα содержит 23 человеческих сегмента Vα или 61 человеческий сегмент Jα. В другом конкретном варианте осуществления гуманизированный локус TCRα содержит полный набор человеческих сегментов Vα и Jα, т.е. все человеческие сегменты гена вариабельного α-участка, кодируемые локусом α, или 54 человеческих сегмента Vα, или 61 человеческий сегмент Jα. В различных вариантах осуществления не относящееся к человеку животное не содержит каких-либо нечеловеческих сегментов Vα или Jα в локусе TCRα.
Мышиный вариабельный локус TCRβ имеет размер приблизительно 0,6 м. п. н. и содержит всего 33 сегмента Vβ, 2 сегмента Dβ и 14 сегментов Jβ. Человеческий вариабельный локус TCRβ имеет размер приблизительно 0,6 м. п. н. и содержит всего 67 сегментов Vβ, 2 сегмента Dβ и 14 сегментов Jβ. В одном варианте осуществления изобретения генетически модифицированное не относящееся к человеку животное (например, грызун, например мышь или крыса) содержит по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jα. В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20,23, 25, 30, 35, 40, 45, 48, 50, 55, 60 или до 67 человеческих сегментов Vβ. В некоторых вариантах осуществления гуманизированный локус TCRβ содержит 8, 14, 40, 66 или 67 человеческих сегментов Vβ. Таким образом, в некоторых вариантах осуществления гуманизированный локус TCRβ у не относящегося к человеку животного может содержать 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% 99% или 100% человеческих Vβ; в некоторых вариантах осуществления он может содержать приблизительно 20%, приблизительно 60%, приблизительно 15%, приблизительно 98% или 100% человеческих Vβ.
В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Vβ18-Vβ29-1 (сегмент Vβ также обозначается как TRBV или TCRBV). В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRBV18-TRBV29-1, отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ1-Jβ1 (т.е. человеческих сегментов Dβ1-Jβ1-Jβ1-6), и отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ2-Jβ2 (т.е. человеческих сегментов Dβ2-Jβ2-1-Jβ2-7). В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRBV6-5-TRBV29-1, отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ1-Jβ1 (т.е. человеческих сегментов Dβ1-Jβ1-1-Jβ1-6), и отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ2-Jβ2 (т.е. человеческих сегментов Dβ2-Hβ2-1-Jβ2-7). В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRBV1-TRBV29-1, отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ1-Jβ1, и отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ2-Jβ2. В одном варианте осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих TRBV1-TRBV29-1, отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ1-Jβ1, отдельный фрагмент ДНК, содержащий непрерывную человеческую последовательность из человеческих Dβ2-Jβ2 и отдельный фрагмент ДНК, содержащий последовательность человеческого TRBV30. В различных вариантах осуществления фрагменты ДНК, содержащие непрерывные человеческие последовательности из человеческих сегментов вариабельного участка TCRβ, также содержат сайты рестрикционных ферментов, кассеты селекции, сайты эндонуклеаз и другие сайты, вставленные для содействия клонированию и селекции в процессе гуманизации локуса. В различных вариантах осуществления эти дополнительные сайты не нарушают надлежащее функционирование (например, перестройку, сплайсинг и т.д.) различных генов в локусе TCRβ.
В одном варианте осуществления гуманизированный локус TCRβ содержит 14 человеческих сегментов Jβ или 100% человеческих сегментов Jβ и 2 человеческих сегмента Dβ или 100% человеческих сегментов Jβ. В другом варианте осуществления гуманизированный локус TCRβ содержит по меньшей мере один человеческий сегмент Vβ, например 14 человеческих сегментов Vβ, и все мышиные сегменты Dβ и Jβ. В конкретном варианте осуществления гуманизированный локус TCRβ содержит 14 человеческих сегментов Vβ, 2 человеческих сегмента Dβ или 14 человеческих сегментов Jβ. В другом конкретном варианте осуществления гуманизированный локус TCRβ содержит полный набор человеческих сегментов Vβ, Dβ и Jβ, т.е. все человеческие сегменты гена вариабельного участка β, кодируемые локусом β, или 67 человеческих сегментов Vβ, 2 человеческих Dβ или 14 человеческих сегментов Jβ. В одном варианте осуществления не относящееся к человеку животное содержит один (например, 5') нечеловеческий сегмент Vβ в гуманизированном локусе TCRβ. В различных вариантах осуществления не относящееся к человеку животное не содержит каких-либо нечеловеческих сегментов Vβ, Dβ или Jβ в локусе TCRβ.
В различных вариантах осуществления, в которых не относящееся к человеку животное (например, грызун) содержит набор сегментов вариабельного участка человеческих TCRα и TCRβ (и необязательно человеческих TCRδ и TCRγ) (например, полный набор сегментов вариабельного участка), набор различных сегментов (например, полный набор различных сегментов) используется животным для продукции разнородного набора молекул TCR для различных антигенов.
В различных аспектах не относящиеся к человеку животные содержат непрерывные участки человеческих геномных вариабельных локусов TCR, которые содержат сегменты V, D и J, или D и J, или V и J, или V, сгруппированные так же, как в неперестроенном человеческом геномном вариабельном локусе, например содержащие промоторные последовательности, лидерные последовательности, межгенные последовательности, регуляторные последовательности и т.д., сгруппированные так же, как в человеческом геномном вариабельном локусе TCR. В других аспектах различные сегменты сгруппированы так же, как в неперестроенном нечеловеческом геномном вариабельном локусе TCR. В различных вариантах осуществления гуманизированного локуса TCR α, β, δ и/или γ гуманизированный локус может содержать два или более геномных сегмента, которые не располагаются в человеческом геноме рядом друг с другом, например, фрагмент сегментов V человеческого вариабельного локуса, расположенный в человеческом геноме проксимально по отношению к константному участку, может располагаться рядом с фрагментом сегментов V человеческого вариабельного локуса, расположенного в человеческом геноме на 5'-конце человеческого вариабельного локуса.
Как у мыши, так и у человека сегменты гена TCRδ расположены с локусом TCRα (см. ФИГ. 4А сверху, область TCRD заключена в рамку). Сегменты J и D TCRδ расположены между сегментами Vα и Jα, а сегменты V TCRδ разбросаны по всему локусу TCRα, причем большинство расположено среди различных сегментов Vα. Количество и местоположения различных сегментов TCRδ можно определить по базе данных IMGT. В связи с расположением в геноме сегментов гена TCRδ в пределах локуса TCRα успешная перестройка в локусе TCRα может привести к делеции или инактивации сегментов гена TCRδ.
В некоторых вариантах осуществления изобретения не относящееся к человеку животное, содержащее неперестроенный человеческий вариабельный локус гена TCRα, также содержит по меньшей мере один человеческий сегмент Vδ, например вплоть до полного набора человеческих сегментов Vδ. Таким образом, в некоторых вариантах осуществления замещение эндогенного вариабельного локуса гена TCRα приводит к замещению по меньшей мере одного нечеловеческого сегмента Vδ человеческим сегментом Vδ. В других вариантах осуществления не относящееся к человеку животное изобретения содержит полный набор человеческих сегментов Vδ, Dδ и Jδ в неперестроенном гуманизированном локусе TCRα; в других вариантах осуществления не относящееся к человеку животное содержит полный неперестроенный человеческий локус TCRδ в неперестроенном гуманизированном локусе TCRα (т.е. локус TCRδ, включая человеческие сегменты вариабельного участка, а также человеческие энхансер и константный участок). Пример осуществления для конструирования неперестроенного гуманизированного локуса TCRα, содержащего полный неперестроенный локус TCRδ, показан в патенте США №9,113,616, включенном в настоящий документ путем ссылки.
В еще одном варианте осуществления не относящееся к человеку животное изобретения дополнительно содержит неперестроенный гуманизированный локус TCRγ, например локус TCRγ, содержащий по меньшей мере один человеческий сегмент Vγ и по меньшей мере один человеческий сегмент Jγ (например, полный набор человеческих сегментов вариабельного участка Vγ и Jγ). Человеческий локус TCRγ находится на человеческой хромосоме 7, а мышиный локус TCRγ находится на мышиной хромосоме 13. Более подробная информация о локусе TCRγ представлена в базе данных IMGT.
В одном аспекте не относящееся к человеку животное (например, грызун, например мышь или крыса), содержащее гуманизированные вариабельные локусы гена TCRα и β (и необязательно гуманизированные вариабельные локусы гена TCRδ/γ), описанные в настоящем документе, экспрессирует на поверхности Т-клетки гуманизированный Т-клеточный рецептор, содержащий человеческий вариабельный участок и нечеловеческий (например, грызуна, например мыши или крысы) константный участок. В некоторых аспектах не относящееся к человеку животное может экспрессировать или экспрессирует разнородный набор гуманизированных Т-клеточных рецепторов, которые распознают множество представленных антигенов.
В различных вариантах осуществления изобретения гуманизированные полипептиды Т-клеточного рецептора, описанные в настоящем документе, содержат человеческие лидерные последовательности. В альтернативных вариантах осуществления гуманизированные последовательности нуклеиновых кислот рецептора TCR созданы таким образом, что гуманизированные полипептиды TCR содержат нечеловеческие лидерные последовательности.
Гуманизированные полипептиды TCR, описанные в настоящем документе, могут экспрессироваться под контролем эндогенных нечеловеческих регуляторных элементов (например, регуляторных элементов грызуна), например промотора, сайленсера, энхансера и т.д. Гуманизированные полипептиды TCR, описанные в настоящем документе, могут альтернативно экспрессироваться под контролем человеческих регуляторных элементов. В различных вариантах осуществления не относящиеся к человеку животные, описанные в настоящем документе, дополнительно содержат все регуляторные и другие последовательности, в норме обнаруживаемые in situ в человеческом геноме.
В различных вариантах осуществления человеческий вариабельный участок гуманизированного белка TCR способен взаимодействовать с различными белками на поверхности той же клетки или другой клетки. В одном варианте осуществления человеческий вариабельный участок гуманизированного TCR взаимодействует с белками МНС (например, белками МНС класса I или II), представляющими антигены на поверхности второй клетки, например антигенпредставляющей клетки (АПК). В некоторых вариантах осуществления белок МНС I или II представляет собой нечеловеческий белок (например, грызуна, например мыши или крысы). В других вариантах осуществления белок МНС I или II представляет собой человеческий (гуманизированный) белок. В одном аспекте вторая клетка, например АПК, представляет собой эндогенную нечеловеческую клетку, экспрессирующую человеческую или гуманизированную молекулу МНС. В другом варианте осуществления вторая клетка представляет собой человеческую клетку, экспрессирующую человеческую молекулу МНС.
В одном аспекте у не относящегося к человеку животного на поверхности Т-клетки экспрессируется гуманизированный Т-клеточный рецептор с нечеловеческим константным участком, причем рецептор способен взаимодействовать с нечеловеческими молекулами, например якорными или сигнальными молекулами, экспрессирующимися в Т-клетке (например, молекулами CD3, цепью ζ, или другими белками, присоединенными к TCR посредством молекул CD3 или цепи ζ). Таким образом, в одном аспекте предложен клеточный комплекс, содержащий: (а) нечеловеческую Т-клетку, которая экспрессирует: (i) TCR, который содержит гуманизированную цепь α TCR, описанную в настоящем документе, и гуманизированную цепь β TCR, описанную в настоящем документе; и (ii) химерный корецептор, описанный в настоящем документе; и (b) нечеловеческую антигенпредставляющую клетку, содержащую антиген, связанный с химерным МНС I и/или химерным МНС II, описанным в настоящем документе. В одном варианте осуществления нечеловеческие константные цепи TCRα и TCRβ находятся в комплексе с нечеловеческими гомодимером зета (ζ) цепи и гетеродимерами CD3. В одном варианте осуществления клеточный комплекс представляет собой клеточный комплекс in vivo. В одном варианте осуществления клеточный комплекс представляет собой клеточный комплекс in vitro.
В различных вариантах осуществления не относящиеся к человеку животные (например, грызуны, например мыши или крысы), описанные в настоящем документе, продуцируют Т-клетки, которые способны подвергаться дифференцировке в тимусе, превращаясь из Т-клеток SP DN1 в DN2, в DN3, в DN4, в DP и в CD4 или CD8. Такие Т-клетки не относящегося к человеку животного изобретения экспрессируют молекулы клеточной поверхности, в норме продуцируемые Т-клеткой во время конкретной стадии дифференцировки в тимусе (например, CD25, CD44, Kit, CD3, pTα и т.д.). Таким образом, в одном варианте осуществления не относящиеся к человеку животные, описанные в настоящем документе, могут экспрессировать pTα в комплексе с TCRβ на стадии DN3 дифференцировки в тимусе. Не относящиеся к человеку животные, описанные в настоящем документе, экспрессируют Т-клетки, способные подвергаться дифференцировке в тимусе с образованием Т-клеток CD4+ и CD8+.
В различных вариантах осуществления не относящиеся к человеку животные, описанные в настоящем документе, продуцируют Т-клетки, которые способны подвергаться Т-клеточной дифференцировке в периферических органах. В некоторых вариантах осуществления не относящиеся к человеку животные, описанные в настоящем документе, способны продуцировать набор эффекторных Т-клеток, например CTL (цитотоксические Т-лимфоциты), ТН1, ТН2, TREG, ТН17 и т.д. Таким образом, в этих вариантах осуществления не относящиеся к человеку животные, описанные в настоящем документе, продуцируют Т-клетки, которые выполняют различные функции, типичные для конкретного типа Т-клеток, например распознают, связывают или формируют ответ на инородные антигены. В различных вариантах осуществления не относящиеся к человеку животные, описанные в настоящем документе, продуцируют эффекторные Т-клетки, которые уничтожают клетки, представляющие пептидные фрагменты цитоплазматических патогенов, экспрессированных в связи с молекулами МНС I; распознают пептиды, полученные из антигенов, разрушенных во внутриклеточных везикулах и представленных молекулами МНС II на поверхности макрофагов, и индуцируют уничтожение макрофагами микроорганизмов; продуцируют цитокины, которые запускают В-клеточную дифференцировку; активируют продукцию В-клетками опсонизирующих антител; индуцируют продукцию эпителиальными клетками хемокинов, которые мобилизуют нейтрофилы к очагам инфекции; и т.д.
В дополнительных вариантах осуществления не относящиеся к человеку животные, описанные в настоящем документе, содержат Т-клетки CD3+ в периферических органах, например в селезенке. В других аспектах не относящиеся к человеку животные, описанные в настоящем документе, способны продуцировать популяцию Т-клеток памяти в ответ на введение интересующего антигена. Например, не относящиеся к человеку животные продуцируют как центральные Т-клетки памяти (Tcm), так и эффекторные Т-клетки памяти (Tcm) в ответ на введение антигена, например интересующего антигена (например, антигена, тестируемого с целью разработки вакцины, и т.д.).
Клетки DN1 и DN2, которые не получают достаточно сигналов (например, сигналов Notch), могут развиваться в В-клетки, миелоидные клетки (например, дендритные клетки), тучные клетки и NK-клетки. См., например, публикацию Yashiro-Ohtani et al. (2010) Notch regulation of early thymocyte development, Seminars in Immunology 22:261-69. В некоторых вариантах осуществления у не относящихся к человеку животных, описанных в настоящем документе, развиваются В-клетки, миелоидные клетки (например, дендритные клетки), тучные клетки и NK-клетки. В некоторых вариантах осуществления у не относящихся к человеку животных, описанных в настоящем документе, развивается популяция дендритных клеток в тимусе.
Преимущественным типом Т-клеточных рецепторов, экспрессированных на поверхности Т-клеток, является TCRα/β, и лишь небольшая часть клеток экспрессирует TCRδ/γ. В некоторых вариантах осуществления изобретения Т-клетки не относящихся к человеку животных, содержащие гуманизированные локусы TCRα и/или β, демонстрируют задействование локусов TCRα/β и TCRδ/γ, например задействование локусов TCRα/β и TCRδ/γ, аналогичное таковому у животных дикого типа (например, Т-клетки не относящихся к человеку животных, описанных в настоящем документе, экспрессируют белки TCRα/β и TCRδ/γ в пропорциях, сопоставимых с таковыми, экспрессируемыми животными дикого типа). Таким образом, в некоторых вариантах осуществления не относящиеся к человеку животные, содержащие гуманизированные локусы TCRα/β и эндогенные нечеловеческие локусы TCRδ/γ, демонстрируют задействование всех локусов.
Человеческие или гуманизированные молекулы МНС
В различных вариантах осуществления в настоящем документе предложены генетически модифицированные не относящиеся к человеку животные, которые совместно экспрессируют по меньшей мере один гуманизированный Т-клеточный корецептор, по меньшей мере один гуманизированный МНС, который ассоциируется с гуманизированным Т-клеточным корецептором, и необязательно гуманизированный TCR, который при распознавании и связывании с пептидом, представленным гуманизированным МНС, и в сочетании с гуманизированным корецептором передает сигналы активации в клетку, экспрессирующую полипептиды гуманизированного TCR и химерного Т-клеточного корецептора. Соответственно, не относящееся к человеку животное, описанное в настоящем документе, содержит по меньшей мере одну из первой, второй и/или третьей последовательности нуклеиновых кислот, каждая из которых кодирует разные человеческие или гуманизированные полипептиды МНС, выбранные из группы, состоящей из человеческого или гуманизированного полипептида МНС II α, человеческого или гуманизированного полипептида МНС II β и человеческого или гуманизированного полипептида МНС I α; не относящееся к человеку животное также необязательно содержит человеческий или гуманизированный β2-микроглобулин. Использование обозначений «первая», «вторая» и «третья» в настоящем документе не следует толковать как требование обязательного наличия у не относящихся к человеку животных, описанных в настоящем документе, всех трех последовательностей нуклеиновых кислот или наличия какого-либо из человеческих или гуманизированных полипептидов МНС в каком-либо конкретном порядке.
Соответственно, в некоторых вариантах осуществления не относящееся к человеку животное, описанное в настоящем документе, может содержать, например, первую и вторую нуклеотидные последовательности, кодирующие, например, человеческий или химерный полипептид CD8α и человеческий или химерный полипептид CD8β, неперестроенный вариабельный локус гена Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с нечеловеческой константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с нечеловеческой константной последовательностью гена TCRβ, и необязательно первую и ворую последовательности нуклеиновых кислот, кодирующие, например, человеческий или гуманизированный полипептид МНС I α и человеческий или гуманизированный полипептид β2-микроглобулин. В других вариантах осуществления не относящееся к человеку животное, описанное в настоящем документе, может содержать первую нуклеотидную последовательность, кодирующую, например, химерный полипептид CD4; вариабельный локус гена неперестроенного Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с нечеловеческой константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с нечеловеческой константной последовательностью гена TCRβ; и необязательно первую и вторую последовательности нуклеиновых кислот, кодирующие, например, человеческий или гуманизированный полипептид МНС II α и человеческий или гуманизированный полипептид МНС II β. В некоторых вариантах осуществления не относящееся к человеку животное, описанное в настоящем документе, может содержать, например, первую, вторую и третью нуклеотидные последовательности, кодирующие, например, химерный полипептид CD4, химерный полипептид CD8α и химерный полипептид CD8β; вариабельный локус гена неперестроенного Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с нечеловеческой константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с нечеловеческой константной последовательностью гена TCRβ; и необязательно первую, вторую, третью и четвертую последовательности нуклеиновых кислот, кодирующие, например, человеческий или гуманизированный полипептпид МНС α, человеческий или гуманизированный полипептид МНС II β, человеческий или гуманизированный полипептид МНС I α и человеческий или гуманизированный полипептид β2-микроглобулин.
В различных вариантах осуществления в настоящем документе предложено генетически модифицированное не относящееся к человеку животное, например грызун (например, мышь или крыса), содержащее в своем геноме последовательность нуклеиновых кислот, кодирующую человеческий или гуманизированный полипептид МНС I, и/или последовательность нуклеиновых кислот, кодирующую человеческий или гуманизированный белок МНС II. Последовательность нуклеиновых кислот МНС I может кодировать полипептид МНС I, который является частично человеческим и частично нечеловеческим, например химерный человеческий/нечеловеческий полипептид МНС I, а последовательность нуклеиновых кислот МНС II может кодировать полипептид МНС II, который является частично человеческим и частично нечеловеческим, например химерный человеческий/нечеловеческий белок МНС II (например, содержащий химерные человеческие/нечеловеческие полипептиды МНС II α и β). В некоторых аспектах животное не экспрессирует эндогенные полипептиды МНС I и/или МНС II, например функциональные эндогенные полипептиды МНС I и/или МНС II, на клеточной поверхности. В некоторых вариантах осуществления только молекулы МНС I и/или МНС II, экспрессированные на клеточной поверхности животного, представляют собой химерные молекулы МНС I и/или МНС II.
Генетически модифицированное не относящееся к человеку животное, содержащее в своем геноме, например в эндогенном локусе, последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС I, описано в публикациях патентов США №20130111617 и №20130185819, полностью включенных в настоящий документ путем ссылки. Генетически модифицированное не относящееся к человеку животное, содержащее в своем геноме, например в эндогенном локусе, последовательность нуклеиновых кислот, кодирующую гуманизированные, например химерные человеческие/нечеловеческие полипептиды МНС II, описано в патенте США №8,847,005 и в публикации патента США №20130185820, каждый из которых полностью включен в настоящий документ путем ссылки. Генетически модифицированное не относящееся к человеку животное, содержащее в своем геноме, например в эндогенном локусе, последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС I, и содержащее в своем геноме, например в эндогенном локусе, последовательность нуклеиновых кислот, кодирующую гуманизированные, например химерные человеческие/нечеловеческие полипептиды МНС II, описано в публикации патента США №20140245467, полностью включенной в настоящий документ путем ссылки.
В различных вариантах осуществления в настоящем документе предложено генетически модифицированное не относящееся к человеку животное, содержащее в своем геноме, например в одном или более эндогенных локусах МНС, первую последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС I, причем человеческий участок химерного полипептида МНС I содержит внеклеточный участок (или его часть, например один или более внеклеточных доменов) человеческого полипептида МНС I; вторую последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС II α, при этом человеческий участок химерного полипептида МНС II α содержит внеклеточный участок (или его часть, например один или более внеклеточных доменов) человеческого полипептида МНС II α; и/или третью последовательность нуклеиновых кислот, кодирующую химерный человеческий/нечеловеческий полипептид МНС II β, при этом человеческий участок химерного полипептида МНС II β содержит внеклеточный участок (или его часть, например один или более внеклеточных доменов) человеческого полипептида МНС II β; при этом у не относящегося к человеку животного экспрессируются функциональные химерные человеческие/нечеловеческие белки МНС I и МНС II из его эндогенного нечеловеческого локуса МНС. В одном варианте осуществления первая, вторая и/или третья последовательности нуклеиновых кислот соответственно расположены в эндогенных нечеловеческих локусах МНС I, МНС II α и МНС II β. В одном варианте осуществления, в котором не относящееся к человеку животное представляет собой мышь, первая, вторая и/или третья последовательности нуклеиновых кислот расположены в эндогенном мышином локусе МНС на мышиной хромосоме 17. В одном варианте осуществления первая последовательность нуклеиновых кислот расположена в эндогенном нечеловеческом локусе МНС I. В одном варианте осуществления вторая последовательность нуклеиновых кислот расположена в эндогенном нечеловеческом локусе МНС II α. В одном варианте осуществления третья последовательность нуклеиновых кислот расположена в эндогенном нечеловеческом локусе МНС II β.
В одном варианте осуществления у не относящегося к человеку животного экспрессируются только химерные человеческие/нечеловеческие полипептиды МНС I, МНС II α и/или МНС β и не экспрессируются эндогенные нечеловеческие полипептиды МНС (например, функциональные эндогенные полипептиды МНС I, II α и/или II β) из эндогенного нечеловеческого локуса МНС. В одном варианте осуществления у животного, описанного в настоящем документе, экспрессируется функциональный химерный МНС I или функциональный химерный МНС II на поверхности его клеток, например антигенпредставляющих клеток и т.д. В одном варианте осуществления только МНС I и МНС II, экспрессируемые животным на клеточной поверхности, представляют собой химерный МНС I и химерный МНС II, а животное не экспрессирует какой-либо эндогенный МНС I и МНС II на клеточной поверхности.
В одном варианте осуществления химерный человеческий/нечеловеческий полипептид МНС I содержит в своем человеческом участке карман связывания с полипептидом, например человеческим полипептидом МНС I. В одном аспекте человеческий участок химерного полипептида содержит внеклеточный участок человеческого МНС I. В этом варианте осуществления человеческий участок химерного полипептида содержит внеклеточный домен цепи а человеческого МНС I. В одном варианте осуществления человеческий участок химерного полипептида содержит домены α1 и α2 человеческого МНС I. В другом варианте осуществления человеческий участок химерного полипептида содержит домены α1, α2 и α3 человеческого МНС I.
В одном аспекте человеческий участок химерного полипептида МНС II α и/или человеческий участок химерного полипептида МНС II β содержит пептид-связывающий домен человеческого полипептида МНС II α и/или человеческого полипептида МНС II β соответственно. В одном аспекте человеческий участок химерного полипептида МНС II α и/или β содержит внеклеточный участок человеческого полипептида МНС II α и/или β соответственно. В одном варианте осуществления человеческий участок химерного полипептида МНС II а содержит домен α1 человеческого полипептида МНС II α; в другом варианте осуществления человеческий участок химерного полипептида МНС II α содержит домены α1 и α2 человеческого полипептида МНС II α. В дополнительном варианте осуществления человеческий участок химерного полипептида МНС II β содержит домен β1 человеческого полипептида МНС II β; в другом варианте осуществления человеческий участок химерного полипептида МНС II β содержит домены β1 и β2 человеческого полипептида МНС II β.
В некоторых вариантах осуществления человеческий или гуманизированный полипептид МНС I может быть получен из функциональной человеческой молекулы HLA, кодируемой любым из локусов HLA-A, HLA-B, HLA-C, HLA-E, HLA-F или HLA-G. Человеческий или гуманизированный полипептид МНС II может быть получен из функциональной человеческой молекулы HLA, кодируемой локусами HLA-DP, -DQ и -DR. Список наиболее часто используемых антигенов и аллелей HLA приведен в публикации Shankarkumar et al. ((2004) The Human Leukocyte Antigen (HLA) System, Int. J. Hum. Genet. 4(2):91-103), включенной в настоящий документ путем ссылки. Shankarkumar et al. также представили краткое пояснение номенклатуры HLA, используемой в данной области. Дополнительную информацию о номенклатуре HLA и различных аллелях HLA можно найти в публикации Holdsworth et al. (2009) The HLA dictionary 2008: a summary of HLA-A, -В, -C, -DRB1/3/4/5, and DQB1 alleles and their association with serologically defined HLA-A, -В, -C, -DR, and -DQ antigens, Tissue Antigens 73:95-170 и недавней публикации Marsh et al. (2010) Nomenclature for factors of the HLA system, 2010, Tissue Antigens 75:291-455, обе из которых включены в настоящий документ путем ссылки. В некоторых вариантах осуществления полипептиды МНС I или МНС II могут быть получены из любых функциональных человеческих молекул HLA-A, В, С, DR или DQ. Таким образом, человеческие или гуманизированные полипептиды МНС I и/или II могут быть получены из любых функциональных человеческих молекул HLA, описанных в настоящем документе. В некоторых вариантах осуществления все полипептиды МНС I и МНС II, экспрессированные на клеточной поверхности, содержат участок, полученный из человеческих молекул HLA.
Особый интерес представляют человеческие молекулы HLA, специфические полиморфные аллели HLA, которые, как известно, ассоциированы с рядом человеческих заболеваний, например человеческими аутоиммунными заболеваниями. Фактически был обнаружен специфический полиморфизм в локусах HLA, который коррелирует с развитием ревматоидного артрита, сахарного диабета I типа, тиреоидита Хасимото, рассеянного склероза, тяжелой миастении, базедовой болезни, системной красной волчанки, целиакии, болезни Крона, язвенного колита и других аутоиммунных расстройств. См., например, публикации Wong and Wen (2004) What can the HLA transgenic mouse tell us about autoimmune diabetes?, Diabetologia 47:1476-87; Taneja and David (1998) HLA Transgenic Mice as Humanized Mouse Models of Disease and Immunity, J. Clin. Invest. 101:921-26; Bakker et al. (2006), A high-resolution HLA and SNP haplotype map for disease association studies in the extended human MHC, Nature Genetics 38:1166- 72 и Supplementary Information; и International MHC and Autoimmunity Genetics Network (2009) Mapping of multiple susceptibility variants within the MHC region for 7 immune-mediated diseases, Proc. Natl. Acad. Sci. USA 106:18680-85. Таким образом, человеческие или гуманизированные полипептиды МНС I и/или МНС II могут быть получены из человеческой молекулы HLA, которая ассоциирована с конкретным заболеванием, например аутоиммунным заболеванием.
В одном конкретном аспекте человеческий или гуманизированный полипептид МНС I получен из человеческого HLA-A. В конкретном варианте осуществления полипептид HLA-A представляет собой полипептид HLA-A2 (например, полипептид HLA-A2.1). В одном варианте осуществления полипептид HLA-A представляет собой полипептид, кодируемый аллелью HLA-A*0201, например аллелью HLA-A*02:01:01:01. Аллель HLA-A*0201 широко распространена среди североамериканской популяции. Хотя в настоящих примерах описана эта конкретная последовательность HLA, в объем настоящего изобретения входит любая приемлемая последовательность HLA-A, например полиморфные варианты HLA-A2, встречающиеся в человеческой популяции, последовательности с одной или более консервативными или неконсервативными аминокислотными модификациями, последовательности нуклеиновых кислот, отличающиеся от последовательности, описанной в настоящем документе, в связи с вырождением генетического кода, и т.д.
В другом конкретном аспекте человеческий участок химерного полипептида МНС I получен из человеческого МНС I, выбранного из HLA-B и HLA-C. В одном аспекте он получен из HLA-B, например HLA-B27. В другом аспекте он получен из HLA-A3, -В7, -Cw6 и т.д.
В одном конкретном аспекте человеческие участки гуманизированных полипептидов МНС IIα и β, описанные в настоящем документе, получены из человеческого HLA-DR, например HLA-DR2. Как правило, цепи α HLA-DR являются мономорфными, например, цепь α комплекса HLA-DR кодируется геном HLA-DRA (например, геном HLA-DRα*01). С другой стороны, цепь β HLA-DR является полиморфной. Таким образом, HLA-DR2 содержит цепь α, кодируемую геном HLA-DRA, а цепь β кодируется геном HLA-DR1β*1501. Хотя в настоящих примерах описаны эти конкретные последовательности HLA, в объем настоящего изобретения входят любые приемлемые последовательности HLA-DR, например полиморфные варианты, встречающиеся в человеческой популяции, последовательности с одной или более консервативными или неконсервативными аминокислотными модификациями, последовательности нуклеиновых кислот, отличающиеся от последовательностей, описанных в настоящем документе, в связи с вырождением генетического кода и т.д.
Человеческие участки химерного полипептида МНС II α и/или β могут кодироваться последовательностями нуклеиновых кислот аллелей HLA, которые ассоциированы с распространенными человеческими заболеваниями. Такие аллели HLA включают, без ограничений, HLA-DRB 1*0401, -DRB1*0301, -DQA1*0501, -DQB1*0201, DRB1*1501, -DRB1*1502, -DQB1*0602, -DQA1*0102, -DQA1*0201, -DQB1*0202, -DQA1*0501 и их комбинации. Обзор ассоциации аллелей HLA с заболеваниями см. в публикации Bakker et al. (2006), выше, включенной в настоящий документ путем ссылки.
В одном аспекте нечеловеческий участок полипептида(-ов) химерных человеческих МНС I, МНС II α и/или МНС II αβ содержит трансмембранный и/или цитоплазматический домены полипептида(-ов) эндогенных нечеловеческих (например, грызуна, например мыши, крысы и т.д.) МНС I, МНС II α и/или МНС II β соответственно. Таким образом, нечеловеческий участок химерного человеческого/нечеловеческого полипептида МНС I может содержать трансмембранный и/или цитоплазматический домены эндогенного нечеловеческого полипептида МНС I. Нечеловеческий участок химерного полипептида МНС II α может содержать трансмембранный и/или цитоплазматический домены эндогенного нечеловеческого полипептида МНС II α. Нечеловеческий участок химерного человеческого/нечеловеческого полипептида МНС IIβ может содержать трансмембранный и/или цитоплазматический домены эндогенного нечеловеческого полипептида МНС II β. В одном аспекте не относящееся к человеку животное представляет собой мышь, а нечеловеческий участок химерного полипептида МНС I получен из мышиного белка Н-2K. В одном аспекте животное представляет собой мышь, а нечеловеческие участки химерных полипептидов МНС II α и β получены из мышиного белка Н-2Е. Таким образом, нечеловеческий участок химерного полипептида МНС I может содержать трансмембранный и цитоплазматический домены, полученные из мышиного Н-2K, а нечеловеческие участки химерных полипептидов МНС II α и β могут содержать трансмембранный и цитоплазматический домены, полученные из мышиного белка Н-2Е. Хотя в примерах рассматриваются конкретные последовательности Н-2K и Н-2Е, область настоящего изобретения охватывает любые приемлемые последовательности, например полиморфные варианты, консервативные/неконсервативные аминокислотные замены и т.д. В одном аспекте не относящееся к человеку животное представляет собой мышь, и у мыши не экспрессируются функциональные эндогенные полипептиды МНС из ее локуса H-2D. В некоторых вариантах осуществления мышь конструируют таким образом, что у нее отсутствует весь эндогенный локус H-2D или его участок. В других аспектах у мыши не экспрессируются какие-либо функциональные мышиные МНС I и МНС II на клеточной поверхности.
Химерный человеческий/нечеловеческий полипептид может содержать человеческую или нечеловеческую лидерную (сигнальную) последовательность. В одном варианте осуществления химерный полипептид МНС I содержит нечеловеческую лидерную последовательность эндогенного полипептида МНС I. В одном варианте осуществления химерный полипептид МНС II α содержит нечеловеческую лидерную последовательность эндогенного полипептида МНС II α. В одном варианте осуществления химерный полипептид МНС II β содержит нечеловеческую лидерную последовательность эндогенного полипептида МНС II β. В альтернативном варианте осуществления химерный(-ые) полипептид(-ы) МНС I, МНС II α и/или МНС II β содержит(-ат) нечеловеческую лидерную последовательность полипептида(-ов) МНС I, МНС II α и/или МНС II β соответственно от другого не относящегося к человеку животного, например от другой линии грызуна или другой линии мыши. Таким образом, последовательность нуклеиновых кислот, кодирующая химерный полипептид МНС I, МНС II α и/или МНС II β, может быть функционально связана с последовательностью нуклеиновых кислот, кодирующей нечеловеческую лидерную последовательность МНС I, МНС II α и/или МНС II β соответственно. В еще одном варианте осуществления химерный(-ые) полипептид(-ы) МНС I, МНС II α и/или МНС II β содержит(-ат) человеческую лидерную последовательность полипептида человеческого МНС I, человеческого МНС II α и/или человеческого МНС II β соответственно (например, лидерную последовательность человеческого HLA-A2, человеческого HLA-DRα и/или человеческого HLA-DRβ1*1501 соответственно).
Химерный человеческий/нечеловеческий полипептид МНС I, МНС II α и/или МНС IIβ может содержать в своем человеческом участке полный или по существу полный внеклеточный домен полипептида человеческого МНС I, человеческого МНС II α и/или человеческого МНС II β соответственно. Таким образом, человеческий участок может содержать по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, например 95% или более, аминокислот, кодирующих внеклеточный домен полипептида человеческого МНС I, человеческого МНС II α и/или человеческого МНС IIβ (например, человеческого HLA-A2, человеческого HLA-DRα и/или человеческого HLA-DRβ1*1501). В одном примере в по существу полном внеклеточном домене полипептида человеческого МНС I, человеческого МНС II α и/или человеческого МНС II β отсутствует человеческая лидерная последовательность. В другом примере полипептид химерного человеческого/нечеловеческого МНС I, химерного человеческого/нечеловеческого МНС II α и/или химерного человеческого/нечеловеческого МНС II β содержит человеческую лидерную последовательность.
Более того, химерный полипептид МНС I, МНС II α и/или МНС II β может быть функционально связан (например, может экспрессироваться под регуляторным контролем) с эндогенными нечеловеческими промоторными и регуляторными элементами, например мышиными регуляторными элементами МНС I, МНС IIα и/или МНС II β соответственно. Такое сочетание будет способствовать правильной экспрессии химерных полипептидов МНС I и/или МНС II γ не относящегося к человеку животного, например, в ходе иммунного ответа у не относящегося к человеку животного.
В дополнительном варианте осуществления не относящееся к человеку животное изобретения, например грызун, например мышь, содержит (например, в эндогенном локусе β2-микроглобулина) последовательность нуклеиновых кислот, кодирующую человеческий или гуманизированный β2 микроглобулин. β2-микроглобулин или легкая цепь комплекса МНС класса I (также обозначаемая β2М) представляет собой небольшой (12 кДа) негликозилированный белок, основная функция которого заключается в стабилизации цепи α МНС I. Создание животных с человеческим или гуманизированным β2-микроглобулином подробно описано в публикации патента США №20130111617, которая включена в настоящий документ путем ссылки.
Нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2-микроглобулин, может содержать остатки нуклеиновых кислот, соответствующие полному человеческому гену β2-микроглобулина. Альтернативно нуклеотидная последовательность может содержать остатки нуклеиновых кислот, кодирующие аминокислотную последовательность, идентичную аминокислотам 21-119 человеческого белка β2-микроглобулина (т.е. аминокислотные остатки, соответствующие зрелому человеческому β2-микроглобулину). В альтернативном варианте осуществления нуклеотидная последовательность может содержать остатки нуклеиновых кислот, кодирующие аминокислотную последовательность, идентичную аминокислотам 23-115 человеческого белка β2-микроглобулина, например, аминокислотную последовательность, идентичную аминокислотам 23-119 человеческого белка β2-микроглобулина. Нуклеотидные и аминокислотные последовательности человеческого β2-микроглобулина описаны в публикации Gussow et al., выше, включенной в настоящий документ путем ссылки.
Таким образом, человеческий или гуманизированный полипептид β2-микроглобулин может содержать аминокислотную последовательность, аналогичную аминокислотам 23-115 человеческого полипептида β2-микроглобулина, например аминокислотную последовательность, аналогичную аминокислотам 23-119 человеческого полипептида β2-микроглобулина, например аминокислотную последовательность, аналогичную аминокислотам 21-119 человеческого полипептида β2-микроглобулина. Альтернативно человеческий β2-микроглобулин может содержать аминокислоты 1-119 человеческого полипептида β2-микроглобулина.
В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный β2-микроглобулин, содержит нуклеотидную последовательность, представленную в экзонах 2-4 человеческого гена β2-микроглобулина. Альтернативно нуклеотидная последовательность содержит нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 человеческого гена β2-микроглобулина. В этом варианте осуществления нуклеотидные последовательности, представленные в экзонах 2, 3 и 4, функционально связаны для обеспечения нормальной транскрипции и трансляции гена. Таким образом, в одном варианте осуществления человеческая последовательность содержит нуклеотидную последовательность, соответствующую экзонам 2-4 человеческого гена β2-микроглобулина. В конкретном варианте осуществления человеческая последовательность содержит нуклеотидную последовательность, соответствующую последовательности, начинающейся от экзона 2 и до приблизительно 267 п. н. после экзона 4 человеческого гена β2-микроглобулина. В конкретном варианте осуществления человеческая последовательность содержит приблизительно 2,8 т. п. н. человеческого гена β2-микроглобулина.
Таким образом, человеческий или гуманизированный полипептид β2-микроглобулин может кодироваться нуклеотидной последовательностью, представленной в экзонах 2-4 человеческого β2-микроглобулина, например нуклеотидной последовательностью, соответствующей экзонам 2-4 человеческого гена β2-микроглобулина. Альтернативно полипептид может кодироваться нуклеотидной последовательностью, содержащей нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 человеческого гена β2-микроглобулина. В конкретном варианте осуществления человеческий или гуманизированный полипептид β2-микроглобулин кодируется нуклеотидной последовательностью, соответствующей последовательности, начинающейся от экзона 2 и до приблизительно 267 п. н. после экзона 4 человеческого гена β2-микроглобулина. В другом конкретном варианте осуществления человеческий или гуманизированный полипептид кодируется нуклеотидной последовательностью, содержащей приблизительно 2,8 т. п. н. человеческого гена β2-микроглобулина. Так как экзон 4 гена β2-микроглобулина содержит нетранслируемую область на 5'-конце, человеческий или гуманизированный полипептид может кодироваться нуклеотидной последовательностью, содержащей экзоны 2 и 3 гена β2-микроглобулина.
Обычному специалисту в данной области будет понятно, что хотя в настоящем документе описаны конкретные нуклеотидные и аминокислотные последовательности для получения созданных методами генной инженерии животных, также предусмотрены последовательности с одной или более консервативными или неконсервативными аминокислотными заменами или последовательности, отличающиеся от описанных в настоящем документе в связи с вырождением генетического кода.
Таким образом, предложено не относящееся к человеку животное, которое экспрессирует человеческую последовательность β2-микроглобулина, причем последовательность β2-микроглобулина по меньшей мере приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентична человеческой последовательности β2-микроглобулина. В конкретном варианте осуществления последовательность β2-микроглобулина по меньшей мере приблизительно на 90%, 95%, 96%, 97%, 98% или 99% идентична человеческой последовательности β2-микроглобулина, описанной в настоящем документе. В одном варианте осуществления человеческая последовательность β2-микроглобулина содержит одну или более консервативных замен. В одном варианте осуществления человеческая последовательность β2-микроглобулина содержит одну или более неконсервативных замен.
Кроме того, предложены не относящиеся к человеку животные, у которых нуклеотидная последовательность, кодирующая человеческий или гуманизированный β2-микроглобулин, также содержит нуклеотидную последовательность, представленную в экзоне 1 нечеловеческого гена β2-микроглобулина. Таким образом, в конкретном варианте осуществления не относящееся к человеку животное содержит в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный β2-микроглобулин, причем нуклеотидная последовательность содержит экзон 1 нечеловеческого гена β2-микроглобулина и экзоны 2, 3 и 4 человеческого гена β2-микроглобулина. Таким образом, человеческий или гуманизированный полипептид β2-микроглобулин кодируется экзоном 1 нечеловеческого гена β2-микроглобулина и экзонами 2, 3 и 4 человеческого гена β2-микроглобулина (например, экзонами 2 и 3 человеческого гена β2-микроглобулина).
В одном варианте осуществления не относящееся к человеку животное (например, грызун, например мышь) изобретения в дополнение к нуклеотидной последовательности, кодирующей химерный белок CD8, дополнительно содержит последовательность нуклеиновых кислот, кодирующую человеческий или гуманизированный белок МНС I, таким образом, что химерный белок CD8, экспрессированный на поверхности Т-клетки, способен ассоциироваться, связываться и/или взаимодействовать с человеческим или гуманизированным МНС I, экспрессирующимся на поверхности второй клетки, например антигенпредставляющей клетки. В одном варианте осуществления белок МНС I содержит внеклеточный домен человеческого полипептида МНС I. В одном варианте осуществления животное дополнительно содержит человеческий или гуманизированный полипептид β2-микроглобулин. Примеры генетически модифицированных животных, экспрессирующих человеческий или гуманизированный полипептид МНС I и/или полипептид β2-микроглобулин, описаны в публикациях патентов США №№20130111617 и 20130185819, полностью включенных в настоящий документ путем ссылки. Таким образом, в одном варианте осуществления животное, содержащее химерный белок CD8, описанный в настоящем документе, может дополнительно содержать гуманизированный комплекс МНС I, причем гуманизированный комплекс МНС I содержит: (1) гуманизированный полипептид МНС I, например, при этом гуманизированный полипептид МНС I содержит внеклеточный домен человеческого МНС I и трансмембранный и цитоплазматический домены эндогенного (например, мышиного) МНС I, например, при этом гуманизированный МНС I содержит домены α1, α2 и α3 человеческого полипептида МНС I; и (2) человеческий или гуманизированный β2-микроглобулин (например, животное содержит в своем геноме нуклеотидную последовательность, представленную в экзонах 2, 3 и 4 человеческого β2-микроглобулина). В одном аспекте полипептиды как гуманизированного МНС I, так и человеческого или гуманизированного β2-микроглобулина кодируются нуклеотидными последовательностями, расположенными в эндогенных локусах МНС I и β2-микроглобулина соответственно; в одном аспекте животное не экспрессирует функциональные эндогенные полипептиды МНС I и β2-микроглобулин. Таким образом, МНС I, экспрессируемый животными, может представлять собой химерный человеческий/нечеловеческий полипептид МНС I (например, человека/грызуна, например человека/мыши). Человеческий участок химерного полипептида МНС I может быть получен из человеческого белка HLA класса I, выбранного из группы, состоящей из HLA-A, HLA-B и HLA-C, например HLA-A2, HLA-B27, HLA-B7, HLA-Cw6, или любой другой молекулы HLA класса I, присутствующей в человеческой популяции. В варианте осуществления, в котором животное представляет собой мышь, нечеловеческий (т.е. мышиный) участок химерного полипептида МНС I может быть получен из мышиного белка МНС I, выбранного из H-2D, Н-2K и H-2L.
В одном варианте осуществления не относящееся к человеку животное (например, грызун, например мышь) изобретения дополнительно содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный белок МНС II таким образом, что химерный белок CD4, экспрессирующийся на поверхности Т-клетки, способен взаимодействовать с человеческим или гуманизированным МНС II, экспрессирующимся на поверхности второй клетки, например антигенпредставляющей клетки. В одном варианте осуществления белок МНС II содержит внеклеточный домен человеческого полипептида МНС II α и внеклеточный домен человеческого полипептида МНС II β. Примеры генетически модифицированных животных, экспрессирующих человеческий или гуманизированный полипептид МНС II, описаны в патенте США №8,847,005, выданном 30 сентября 2014 г., и публикации патента США №20130185820, которые полностью включены в настоящий документ путем ссылки. Таким образом, в одном варианте осуществления животное, содержащее химерный белок CD4, описанный в настоящем документе, может дополнительно содержать гуманизированный комплекс МНС II, причем гуманизированный белок МНС II содержит: (1) гуманизированный полипептид МНС II α, содержащий внеклеточный домен человеческого МНС II α и трансмембранный и цитоплазматический домены эндогенного (например, мышиного) МНС II, при этом человеческий внеклеточный домен МНС II α содержит домены α1 и α2 человеческого МНС II α; и (2) гуманизированный полипептид МНС II β, содержащий внеклеточный домен человеческого МНС II β и трансмембранный и цитоплазматический домены эндогенного (например, мышиного) МНС II, при этом человеческий внеклеточный домен МНС II β содержит домены β1 и β2 человеческого МНС II β. В одном аспекте оба гуманизированных полипептида МНС II α и β кодируются последовательностями нуклеиновых кислот, расположенными в эндогенных локусах МНС II α и β соответственно; в одном аспекте у животного не экспрессируются функциональные эндогенные полипептиды МНС II α и β. Таким образом, МНС II, экспрессируемый у животных, может представлять собой химерный человеческий/нечеловеческий белок МНС II (например, человека/грызуна, например человека/мыши). Человеческий участок химерного белка МНС II может быть получен из человеческого белка HLA класса II, выбранного из группы, состоящей из HLA-DR, HLA-DQ и HLA-DP, например HLA-DR4, HLA-DR2, HLA-DQ2.5, HLA-DQ8, или любой другой молекулы HLA класса II, присутствующей в человеческой популяции. В варианте осуществления, в котором животное представляет собой мышь, нечеловеческий (т.е. мышиный) участок химерного полипептида МНС II может быть получен из мышиного белка МНС II, выбранного из Н-2Е и Н-2А.
Специалисту в данной области будут очевидны различные другие варианты осуществления генетически модифицированного не относящегося к человеку животного, например грызуна, например мыши или крысы, на основании настоящего описания и на основании описания публикаций патентов США №№20130111617, 20130185819 и 20130185820 и патента США №8,847,005, включенных в настоящий документ путем ссылки.
В различных вариантах осуществления генетически модифицированные не относящиеся к человеку животные, описанные в настоящем документе, продуцируют клетки, например АПК, на поверхности которых находятся человеческий или гуманизированный МНС I или II, и в результате представляют пептиды в качестве эпитопов для Т-клеток так, как это происходит у человека, так как по существу все из компонентов комплекса являются человеческими или гуманизированными. Генетически модифицированные не относящиеся к человеку животные изобретения могут использоваться для изучения функции человеческой иммунной системы у гуманизированного животного; для выявления антигенов и эпитопов антигенов, которые вызывают иммунный ответ (например, эпитопов Т-клеток, например уникальных человеческих раковых эпитопов), например, с целью использования для разработки вакцины; для оценки потенциальных вакцин и других стратегий вакцинации; для изучения человеческих аутоиммунных реакций; для изучения человеческих инфекционных заболеваний и иным образом для разработки улучшенных терапевтических стратегий на основании экспрессии человеческого МНС.
Не относящиеся к человеку животные, ткани и клетки
Не относящееся к человеку генетически модифицированное животное изобретения можно выбрать из группы, состоящей из мыши, крысы, кролика, свиньи, крупного рогатого скота (например, коровы, быка, буйвола), оленя, овцы, козы, курицы, кота, собаки, хорька, примата (например, мартышки, макаки-резус). В случае не относящихся к человеку животных, для которых отсутствуют легкодоступные приемлемые генетически модифицируемые ЭС-клетки, применяют другие способы создания не относящегося к человеку животного, содержащего генетическую модификацию. Такие способы включают, например, модификацию генома другой клетки, кроме ЭС-клетки (например, фибробласта или индуцированной плюрипотентной клетки), и использование ядерного переноса для переноса модифицированного генома в приемлемую клетку, например ооцит, и постепенное созревание модифицированной клетки (например, модифицированного ооцита) в не относящемся к человеку животном в приемлемых условиях для формирования эмбриона.
В одном аспекте не относящееся к человеку животное представляет собой млекопитающее. В одном аспекте не относящееся к человеку животное представляет собой мелкое млекопитающее, например, из надсемейства Dipodoidea или Muroidea. В одном варианте осуществления генетически модифицированное животное представляет собой грызуна. В одном варианте осуществления грызуна выбирают из мыши, крысы и хомяка. В одном варианте осуществления грызуна выбирают из надсемейства Muroidea. В одном варианте осуществления генетически модифицированное животное относится к семейству, которое выбирают из Calomyscidae (например, мышевидные хомяки), Cricetidae (например, хомяк, мыши и крысы Нового света, полевка), Muridae (например, мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (например, рипидомисы, скальные крысы, белохвостые крысы, мадагаскарские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В конкретном варианте осуществления генетически модифицированного грызуна выбирают из мыши или крысы (семейство Muridae), песчанки, иглистой мыши и косматого хомяка. В одном варианте осуществления генетически модифицированная мышь является представителем семейства Muridae. В одном варианте осуществления животное представляет собой грызуна. В конкретном варианте осуществления грызуна выбирают из мыши и крысы. В одном варианте осуществления не относящееся к человеку животное представляет собой мышь.
В конкретном варианте осуществления не относящееся к человеку животное представляет собой грызуна, который является мышью линии C57BL, выбранной из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. В другом варианте осуществления мышь представляет собой мышь линии 129, которую выбирают из группы, состоящей из линий 129Р1, 129Р2, 129РЗ, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (см., например, публикацию Festing et al. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10:836, см. также публикацию Auerbach et al (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). В одном варианте осуществления генетически модифицированная мышь представляет собой смесь вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. В другом конкретном варианте осуществления мышь представляет собой смесь вышеупомянутых линий 129 или смесь вышеупомянутых линий BL/6. В конкретном варианте осуществления линия 129 смеси представляет собой линию 129S6 (129/SvEvTac). В другом варианте осуществления мышь представляет собой мышь линии BALB, например, линии BALB/c. В другом варианте осуществления мышь представляет собой смесь линии BALB с другой вышеупомянутой линией. Не относящиеся к человеку животные, предложенные в настоящем документе, могут представлять собой мышь, полученную от любой комбинации вышеупомянутых линий.
В одном варианте осуществления не относящееся к человеку животное представляет собой крысу. В конкретном варианте осуществления крысу выбирают из крысы линии Вистар, линии LEA, линии Спрег-Доули, линии Фишер, F344, F6 и темной агути. В одном варианте осуществления линия крыс представляет собой смесь из двух или более линий, которые выбирают из группы, состоящей из линий Вистар, LEA, Спрег-Доули, Фишер, F344, F6 и темной агути.
Таким образом, в одном варианте осуществления изобретения предложена генетически модифицированная мышь, причем мышь содержит, например, в своем геноме, например в своем геноме зародышевой линии: (а) первую нуклеотидную последовательность, кодирующую первый химерный человеческий/мышиный полипептид Т-клеточного корецептора (например, CD4), вторую нуклеотидную последовательность, кодирующую второй химерный человеческий/мышиный полипептид Т-клеточного корецептора (например, CD8α), и/или третью нуклеотидную последовательность, кодирующую третий химерный человеческий/мышиный полипептид Т-клеточного корецептора (например, CD8β), при этом мышиный участок каждого химерного полипептида Т-клеточного корецептора содержит по меньшей мере трансмембранный и цитоплазматический домены мышиного Т-клеточного корецептора, при этом человеческий участок каждого химерного полипептида содержит внеклеточный участок (или его часть, например один или более внеклеточных доменов) человеческого Т-клеточного корецептора, и при этом у мыши экспрессируются первый, второй и/или третий химерный полипептид Т-клеточного корецептора; (b) вариабельный локус гена неперестроенного Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с мышиной константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с мышиной константной последовательностью гена TCRβ; и необязательно (с) первую последовательность нуклеиновых кислот, кодирующую первый химерный человеческий/мышиный полипептид МНС (например, МНС II α), вторую последовательность нуклеиновых кислот, кодирующую второй химерный человеческий/мышиный полипептид МНС (например, МНС II β), и/или третью последовательность нуклеиновых кислот, кодирующую третий химерный человеческий/мышиный полипептид МНС (например, МНС I), и локус β2-микроглобулина, кодирующий человеческий или гуманизированный β2-микроглобулин, при этом человеческий участок каждого химерного полипептида МНС содержит внеклеточный домен человеческого полипептида МНС, который ассоциируется с первым, вторым и/или третьим химерным полипептидом Т-клеточного корецептора (например, при этом человеческий участок химерного комплекса МНС II (например, гуманизированных полипептидов МНС II α и β) ассоциируется с химерным полипептидом CD4, и/или человеческий участок химерного полипептида МНС I (или комплекса МНС I, например гуманизированного МНС Iα и человеческого (гуманизированного) β2-микроглобулина) ассоциируется с химерным корецептором CD8 (например, гуманизированными полипептидами CD8 α и β).
В настоящем документе предложена генетически модифицированная мышь, содержащая в своем геноме, например в своем эндогенном локусе CD4, нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид CD4, причем мышиный участок химерного полипептида содержит по меньшей мере трансмембранный и цитоплазматический домены мышиного полипептида CD4, и при этом у мыши экспрессируется химерный человеческий/мышиный CD4. В одном варианте осуществления человеческий участок химерного полипептида содержит по меньшей мере весь или по существу весь внеклеточный домен человеческого полипептида CD4. В одном варианте осуществления человеческий участок химерного полипептида содержит по меньшей мере весь или по существу весь домен D1 человеческого белка CD4. В одном варианте осуществления человеческий участок химерного полипептида содержит по меньшей мере все или по существу все из доменов D1-D2 человеческого белка CD4, например по меньшей мере все или по существу все из доменов D1-D3 человеческого белка CD4, например все или по существу все из доменов D1-D4 человеческого белка CD4. Таким образом, в одном варианте осуществления мышь содержит в эндогенном локусе CD4 нуклеотидную последовательность, содержащую по меньшей мере все или по существу все из экзонов 4, 5 и 6 человеческого гена CD4, например последовательность экзона 3 человеческого гена CD4, кодирующую участок домена D1 человеческого CD4, и экзоны 4-6 человеческого гена CD4. В одном варианте осуществления мышь содержит в эндогенном локусе CD4 химерный человеческий/мышиный CD4, который содержит человеческую последовательность CD4, которая отвечает за взаимодействие с МНС II и/или внеклеточным участком Т-клеточного рецептора. В другом варианте осуществления мышь содержит в эндогенном локусе CD4 химерный человеческий/мышиный CD4, который содержит человеческую последовательность CD4, отвечающую за взаимодействие с МНС II и/или вариабельным доменом Т-клеточного рецептора. В одном варианте осуществления нуклеотидная последовательность содержит последовательность, кодирующую мышиный сигнальный пептид CD4. В одном варианте осуществления мышь содержит замещение нуклеотидной последовательности, кодирующее мышиный внеклеточный домен CD4, нуклеотидной последовательностью, кодирующей человеческий внеклеточный домен CD4. В другом варианте осуществления мышь содержит замещение нуклеотидной последовательности, кодирующей по меньшей мере весь или по существу весь мышиный домен D1 CD4, например нуклеотидной последовательности, кодирующей по меньшей мере все или по существу все из мышиных доменов D1-D2 CD4, например нуклеотидной последовательности, кодирующей по меньшей мере все или по существу все из мышиных доменов D1-D3 CD4, человеческой нуклеотидной последовательностью, кодирующей вышеперечисленное. В одном варианте осуществления домены химерного полипептида CD4 кодируются нуклеотидной последовательностью, которая схематически представлена на ФИГ. 5А.
В одном варианте осуществления у мыши не экспрессируется функциональный эндогенный CD4 из ее эндогенного мышиного локуса CD4. В одном варианте осуществления мышь, описанная в настоящем документе, содержит химерную человеческую/мышиную нуклеотидную последовательность CD4 в своей зародышевой линии.
В одном варианте осуществления у мыши сохранены любые эндогенные последовательности, которые не были гуманизированы, например, в варианте осуществления, в котором мышь содержит замещение нуклеотидной последовательности, кодирующей все или по существу все из мышиных доменов D1-D3, у мыши сохранена эндогенная нуклеотидная последовательность, кодирующая мышиный домен D4 CD4, а также нуклеотидная последовательность, кодирующая трансмембранный и цитоплазматический домены мышиного CD4.
В одном аспекте у мыши, экспрессирующей химерный человеческий/мышиный белок CD4, сохранены мышиные промоторные и регуляторные последовательности CD4, например нуклеотидная последовательность у мыши, кодирующая химерный человеческий/мышиный CD4, функционально связана с эндогенными мышиными промоторными и регуляторными последовательностями CD4. В одном аспекте эти мышиные регуляторные последовательности, сохраненные у созданного методами генной инженерии животного по изобретению, включают последовательности, которые регулируют экспрессию химерного белка на соответствующих стадиях в процессе развития Т-клетки. Таким образом, в одном аспекте у мыши не экспрессируется CD4 на В-клетках или зрелых Т-клетках CD8+. В одном аспекте у мыши также не экспрессируется химерный CD4 на клетках любого типа, например иммунных клетках любого типа, которые в норме не экспрессируют эндогенный CD4.
Генетически модифицированная мышь, описанная в настоящем документе, может содержать в своем геноме, например в своем эндогенном локусе CD8, первую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид CD8α, и вторую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид CD8β. В одном варианте осуществления первая нуклеотидная последовательность содержит последовательность, которая кодирует весь или по существу весь внеклеточный участок человеческого полипептида CD8α и по меньшей мере трансмембранный и цитоплазматический домены мышиного полипептида CD8α, а вторая нуклеотидная последовательность содержит последовательность, которая кодирует весь или по существу весь внеклеточный участок человеческого полипептида CD8β и по меньшей мере трансмембранный и цитоплазматический домены мышиного полипептида CD8β, и при этом у мыши экспрессируется функциональный химерный человеческий/мышиный белок CD8. В одном варианте осуществления первая нуклеотидная последовательность содержит последовательность, которая кодирует по меньшей мере подобный иммуноглобулину V домен человеческого полипептида CD8α и остальные последовательности мышиного полипептида CD8α, а вторая нуклеотидная последовательность содержит последовательность, которая кодирует по меньшей мере подобный иммуноглобулину V домен человеческого полипептида CD8β и остальные последовательности мышиного полипептида CD8β. В одном варианте осуществления первая нуклеотидная последовательность содержит по меньшей мере домен связывания с МНС I человеческого полипептида CD8α. В одном варианте осуществления первая и вторая нуклеотидные последовательности содержат по меньшей мере экзоны, которые кодируют внеклеточный участок человеческого полипептида CD8α и/или полипептида CD8β соответственно. В одном варианте осуществления внеклеточный участок человеческого полипептида CD8α и/или полипептида CD8β представляет собой область, охватывающую участок человеческого полипептида CD8α и/или полипептида CD8β, который не является трансмембранным или цитоплазматическим доменом. В одном варианте осуществления домены химерного полипептида CD8α кодируются нуклеотидной последовательностью, которая схематически представлена на ФИГ. 5В. В одном варианте осуществления домены химерного полипептида CD8β кодируются нуклеотидной последовательностью, которая схематически представлена на ФИГ. 5В. В одном варианте осуществления нуклеотидная последовательность, кодирующая химерный человеческий/мышиный полипептид CD8α и/или полипептид CD8β, содержит последовательность, кодирующую мышиный сигнальный пептид CD8α и/или CD8β соответственно. Альтернативно нуклеотидная последовательность может содержать последовательность, кодирующую человеческую сигнальную последовательность CD8α и/или CD8β. В одном варианте осуществления мышь содержит замещение нуклеотидной последовательности, кодирующей весь или по существу весь мышиный внеклеточный домен CD8α и/или CD8β, нуклеотидной последовательностью, кодирующей весь или по существу весь человеческий внеклеточный домен CD8a и/или CD8P соответственно.
В одном варианте осуществления у мыши не экспрессируется функциональный эндогенный мышиный полипептид CD8α и/или CD8β из ее эндогенного локуса CD8. В одном варианте осуществления мышь, описанная в настоящем документе, содержит химерную человеческую/мышиную последовательность CD8 в своей зародышевой линии.
В одном аспекте у мыши, экспрессирующей химерный человеческий/мышиный полипептид CD8α и/или CD8β, сохранены мышиные промоторные и регуляторные последовательности CD8α и/или CD8β, например, нуклеотидная последовательность у мыши, кодирующая химерный человеческий/мышиный CD8, функционально связана с эндогенными мышиными промоторными и регуляторными последовательностями CD8. В одном аспекте регуляторные последовательности, сохраненные у мыши, включают последовательности, регулирующие экспрессию белка CD8 на соответствующих стадиях развития Т-клетки. В одном аспекте у генетически модифицированной мыши не экспрессируется химерный CD8 на В-клетках, или зрелых Т-клетках CD4+, или любых клетках, например иммунных клетках, которые в норме не экспрессируют эндогенный CD8.
В изобретении также предложена генетически модифицированная мышь, содержащая в своем геноме неперестроенный человеческий или гуманизированный вариабельный локус гена TCR, например вариабельный локус гена TCRα, TCRβ, TCRδ и/или TCRγ. В некоторых вариантах осуществления неперестроенный человеческий или гуманизированный вариабельный локус гена TCR замещает эндогенный мышиный вариабельный локус гена TCR. В других вариантах осуществления неперестроенный человеческий или гуманизированный вариабельный локус гена TCR находится в таком сайте в геноме, который отличен от соответствующего эндогенного мышиного локуса TCR. В некоторых вариантах осуществления человеческий или гуманизированный неперестроенный вариабельный локус гена TCR функционально связан с мышиным константным участком TCR.
В одном варианте осуществления предложена генетически модифицированная мышь, причем мышь содержит в своем геноме неперестроенный вариабельный локус гена Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с мышиной константной последовательностью гена TCRα, и неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с мышиной константной последовательностью гена TCRβ. В одном конкретном варианте осуществления мышь содержит в своем геноме неперестроенный вариабельный локус гена TCR α, содержащий полный набор человеческих сегментов Vα и полный набор человеческих сегментов Jα, функционально связанных с мышиной константной последовательностью гена TCRα, и неперестроенный вариабельный локус гена TCRβ, содержащий полный набор человеческих сегментов Vβ, полный набор человеческих сегментов Dβ и полный набор человеческих сегментов Jβ, функционально связанных с мышиной константной последовательностью гена TCRβ.
В некоторых вариантах осуществления неперестроенный вариабельный локус гена TCRα, содержащий человеческие сегменты вариабельного участка TCRα, замещает эндогенный мышиный вариабельный локус гена TCRα, а неперестроенный вариабельный локус гена TCRβ, содержащий человеческие сегменты вариабельного участка TCRβ, замещает эндогенный мышиный вариабельный локус гена TCRβ. В некоторых вариантах осуществления эндогенные мышиные сегменты Vα и Jα неспособны перестраиваться с образованием перестроенной последовательности Vα/Jα, а эндогенные мышиные сегменты Vβ, Dβ и Jβ неспособны перестраиваться с образованием перестроенной последовательности Vβ/Dβ/Jβ. В некоторых вариантах осуществления человеческие сегменты Vα и Jα перестраиваются с образованием перестроенной человеческой последовательности Vα/Jα, а человеческие сегменты Vβ, Dβ и Jβ перестраиваются с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ.
Изобретение также относится к генетически модифицированной мыши, которая содержит в своем геноме последовательность нуклеиновых кислот, кодирующую химерный полипептид МНС, причем человеческий участок химерного полипептида МНС ассоциируется с человеческим внеклеточным доменом химерного Т-клеточного корецептора, как описано в настоящем документе. Генетически модифицированные мыши, описанные в настоящем документе, могут содержать первую последовательность нуклеиновых кислот, кодирующую химерный человеческий/мышиный полипептид МНС I, вторую последовательность нуклеиновых кислот, кодирующую химерный человеческий/мышиный полипептид МНС II α, и/или третью последовательность нуклеиновых кислот, кодирующую химерный человеческий/мышиный полипептид МНС II β. Человеческий участок химерного МНС I, МНС II α и/или МНС II β может содержать внеклеточный домен человеческого МНС I, МНС II α и/или МНС II β соответственно. В одном варианте осуществления у мыши экспрессируются функциональные химерные человеческие/мышиные полипептиды МНС I, МНС II α и/или МНС II β из ее эндогенного мышиного локуса МНС. В одном варианте осуществления у мыши не экспрессируются функциональные мышиные полипептиды МНС, например функциональные мышиные полипептиды МНС I, МНС II α и/или МНС II β, из ее эндогенного мышиного локуса МНС. В других вариантах осуществления единственные МНС I и МНС II, экспрессируемые мышью на клеточной поверхности, представляют собой химерные МНС I и МНС II.
В одном варианте осуществления человеческий участок химерного человеческого/мышиного полипептида МНС I содержит домен связывания с пептидом или внеклеточный домен человеческого МНС I (например, человеческого HLA-A, например человеческого HLA-A2, например человеческого HLA-A2.1). В некоторых вариантах осуществления у мыши не экспрессируется домен связывания с пептидом или внеклеточный домен эндогенного мышиного полипептида МНС I из ее эндогенного мышиного локуса МНС I. Домен связывания с пептидом человеческого МНС I может содержать домены α1 и α2. Альтернативно домен связывания с пептидом человеческого МНС I может содержать домены α1, α2 и α3. В одном аспекте внеклеточный домен человеческого МНС I содержит внеклеточный домен цепи α человеческого МНС I. В одном варианте осуществления эндогенный мышиный локус МНС I представляет собой локус Н-2K (например, Н-2Kb), а мышиный участок химерного полипептида МНС I содержит трансмембранный и цитоплазматический домены мышиного полипептида Н-2K (например, Н-2Kb). Таким образом, в одном варианте осуществления мышь изобретения содержит в своем эндогенном мышином локусе МНС I последовательность нуклеиновых кислот, кодирующую химерный человеческий/мышиный МНС I, причем человеческий участок химерного полипептида содержит внеклеточный домен человеческого полипептида HLA-A2 (например, HLA-A2.1), а мышиный участок содержит трансмембранный и цитоплазматический домены мышиного полипептида Н-2K (например, Н-2Kb), и у мыши экспрессируется химерный человеческий/мышиный белок HLA-A2/H-2K. В другом варианте осуществления мышиный участок химерного полипептида МНС I может быть получен из другого мышиного МНС I, например H-2D, H-2L и т.д.; а человеческий участок химерного полипептида МНС I может быть получен из другого человеческого МНС I, например HLA-B, HLA-C и т.д. В одном аспекте у мыши не экспрессируется функциональный эндогенный полипептид Н-2K из ее эндогенного мышиного локуса Н-2K. В одном варианте осуществления у мыши не экспрессируются функциональные эндогенные полипептиды МНС из ее эндогенного локуса H-2D. В некоторых вариантах осуществления мышь конструируют таким образом, что у нее отсутствует весь эндогенный локус H-2D или его участок. В других вариантах осуществления единственные полипептиды МНС I, экспрессируемые у мыши на клеточной поверхности, представляют собой химерные человеческие/мышиные полипептиды МНС I.
В одном варианте осуществления человеческий участок химерного человеческого/мышиного полипептида МНС II α содержит человеческий домен связывания с пептидом или внеклеточный домен МНС II α, а человеческий участок химерного человеческого/мышиного полипептида МНС II β содержит человеческий домен связывания с пептидом или внеклеточный домен МНС II β. В некоторых вариантах осуществления у мыши не экспрессируется домен связывания с пептидом или внеклеточный домен эндогенного мышиного полипептида а и/или р из эндогенного мышиного локуса (например, локуса Н-2А и/или Н-2Е). В некоторых вариантах осуществления мышь содержит геном, в котором отсутствует ген, который кодирует функциональную молекулу МНС класса II, содержащую Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2, Н-2Еа и их комбинацию. В некоторых вариантах осуществления единственные полипептиды МНС II, экспрессируемые у мыши на клеточной поверхности, представляют собой химерные человеческие/мышиные полипептиды МНС II. Домен связывания с пептидом человеческого полипептида МНС II α может содержать домен α1, а домен связывания с пептидом человеческого полипептида МНС II β может содержать домен β1; таким образом, домен связывания с пептидом химерного комплекса МНС II может содержать человеческие домены α1 и β1. Внеклеточный домен человеческого полипептида МНС II α может содержать домены α1 и α2, а внеклеточный домен человеческого полипептида МНС II β может содержать домены β1 и β2; таким образом, внеклеточный домен химерного комплекса МНС II может содержать человеческие домены α1, α2, β1 и β2. В одном варианте осуществления мышиный участок химерного комплекса МНС II содержит трансмембранный и цитоплазматический домены мышиного МНС II, например мышиного Н-2Е (например, трансмембранный и цитоплазматический домен мышиных цепей α и β Н-2Е). Таким образом, в одном варианте осуществления мышь изобретения содержит в своем эндогенном мышином локусе МНС II последовательность нуклеиновых кислот, кодирующую химерный человеческий/мышиный МНС II α, причем человеческий участок химерного полипептида МНС II α содержит внеклеточный домен, полученный из цепи α человеческого МНС II (например, цепи α HLA-DR2), а мышиный участок содержит трансмембранный и цитоплазматический домены, полученные из цепи а мышиного МНС II (например, Н-2Е); и мышь содержит в своем эндогенном мышином локусе МНС II последовательность нуклеиновых кислот, кодирующую химерный человеческий/мышиный МНС II β, при этом человеческий участок химерного полипептида МНС II β содержит внеклеточный домен, полученный из цепи β человеческого МНС II (например, цепи β HLA-DR2), а мышиный участок содержит трансмембранный и цитоплазматический домены, полученные из цепи β мышиного МНС II (например, Н-2Е); например, при этом у мыши экспрессируется химерный человеческий/мышиный белок HLA-DR2/H-2E. В другом варианте осуществления мышиный участок химерного белка МНС II может быть получен из другого мышиного МНС II, например Н-2А и т.д.; а человеческий участок химерного белка МНС II может быть получен из другого человеческого МНС II, например, HLA-DQ и т.д. В одном аспекте у мыши не экспрессируются функциональные эндогенные полипептиды Н-2А и Н-2Е из ее эндогенных мышиных локусов (например, у мыши не экспрессируются полипептиды Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа). В некоторых вариантах осуществления у мыши не экспрессируется эндогенная молекула МНС I или МНС II на клеточной поверхности.
В различных аспектах человеческий или гуманизированный β2-микроглобулин, экспрессируемый генетически модифицированным не относящимся к человеку животным или клетками, эмбрионами или тканями, полученными от не относящегося к человеку животного, сохраняет все функциональные аспекты эндогенного и/или человеческого β2-микроглобулина. Например, предпочтительно, чтобы человеческий или гуманизированный β2-микроглобулин связывался с цепью а полипептида МНС I (например, эндогенным нечеловеческим или человеческим полипептидом МНС I). Человеческий или гуманизированный полипептид β2-микроглобулин может связываться или иным образом ассоциироваться с любыми другими молекулами, например, рецепторными, якорными или сигнальными молекулами, которые ассоциируются с эндогенным нечеловеческим и/или человеческим β2-микроглобулином (например, HFE и т.д.), или мобилизовать их.
Кроме генетически модифицированных животных (например, грызунов, например мышей или крыс) также предложена ткань или клетка, причем ткань или клетка получена из не относящегося к человеку животного, описанного в настоящем документе, и содержит гетерологичный ген β2-микроглобулина или последовательность β2-микроглобулина, т.е. нуклеотидную и/или аминокислотную последовательность. В одном варианте осуществления гетерологичный ген β2-микроглобулина или последовательность β2-микроглобулина представляет собой человеческий или гуманизированный ген β2-микроглобулина или человеческую или гуманизированную последовательность β2-микроглобулина. Предпочтительно клетка представляет собой клетку с ядром. Клетка может являться любой клеткой, экспрессирующей комплекс МНС, например антигенпредставляющей клеткой. Человеческий или гуманизированный полипептид β2-микроглобулин, экспрессируемый указанной клеткой, может взаимодействовать с эндогенным нечеловеческим МНС I (например, МНС I грызуна) с образованием функционального комплекса МНС I. Полученный комплекс МНС I может быть способен взаимодействовать с Т-клеткой, например цитотоксической Т-клеткой. Таким образом, также предложен комплекс in vitro, состоящий из клетки из не относящегося к человеку животного, описанного в настоящем документе, и Т-клетки.
Также предложены нечеловеческие клетки, которые содержат человеческий или гуманизированный ген или последовательность β2-микроглобулина и дополнительную человеческую или гуманизированную последовательность, например описанный в настоящем документе химерный полипептид МНС I. В таком случае человеческий или гуманизированный полипептид β2-микроглобулин может взаимодействовать, например, с химерным человеческим/нечеловеческим полипептидом МНС I, и может образовываться функциональный комплекс МНС I. В некоторых аспектах такой комплекс способен взаимодействовать с TCR на Т-клетке, например человеческой или нечеловеческой Т-клетке. Таким образом, также предложен комплекс in vitro из клетки из не относящегося к человеку животного, описанного в настоящем документе, и человеческой или нечеловеческой Т-клетки.
В другом аспекте описания предложен эмбрион грызуна (например, эмбрион мыши или крысы), содержащий гетерологичный ген β2-микроглобулина или последовательность β2-микроглобулина, как описано в настоящем документе. В одном варианте осуществления эмбрион содержит донорскую ЭС-клетку, которая содержит гетерологичный ген β2-микроглобулина или последовательность β2-микроглобулина, и клетки эмбриона-хозяина. Гетерологичный ген β2-микроглобулина или последовательность β2-микроглобулина представляет собой человеческий или гуманизированный ген β2-микроглобулина или последовательность β2-микроглобулина.
В объем настоящего изобретения также входит нечеловеческая клетка, содержащая хромосому или фрагмент хромосомы не относящегося к человеку животного, описанного в настоящем документе (например, причем хромосома или ее фрагмент содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид β2-микроглобулин). Нечеловеческая клетка может содержать ядро не относящегося к человеку животного, описанного в настоящем документе. В одном варианте осуществления в результате переноса ядра нечеловеческая клетка содержит хромосому или ее фрагмент.
В одном аспекте предложена нечеловеческая индуцированная плюрипотентная клетка, содержащая гетерологичный ген β2-микроглобулина или последовательность β2-микроглобулина. В одном варианте осуществления индуцированная плюрипотентная клетка получена из не относящегося к человеку животного, описанного в настоящем документе. В одном варианте осуществления гетерологичный ген β2-микроглобулина или последовательность β2-микроглобулина представляет собой человеческий или гуманизированный ген или последовательность.
В некоторых вариантах осуществления изобретения у мыши, описанной в настоящем документе, экспрессируется химерный человеческий/мышиный МНС II только на профессиональных антигенпредставляющих клетках, например В-клетках, моноцитах/макрофагах и/или дендритных клетках мыши. В некоторых вариантах осуществления у мыши, описанной в настоящем документе, формируется иммунный ответ, например клеточный иммунный ответ, на один или более человеческих антигенов. В некоторых вариантах осуществления у мыши, описанной в настоящем документе, формируется гуманизированный Т-клеточный ответ на один или более человеческих антигенов.
Кроме созданного методами генной инженерии не относящегося к человеку животного также предложен нечеловеческий эмбрион (например, эмбрион грызуна, например мышиный или крысиный эмбрион), содержащий донорскую ЭС-клетку, которая получена от не относящегося к человеку животного (например, грызуна, например мыши или крысы), как описано в настоящем документе. В одном аспекте эмбрион содержит донорскую ЭС-клетку, которая содержит химерный ген CD4, химерный ген CD8 (например, CD8α и/или CD8β), гуманизированную последовательность нуклеиновых кислот МНС I (например, МНС I α), гуманизированную последовательность нуклеиновых кислот МНС II (например, МНС II α и/или МНС II β), неперестроенный гуманизированный локус TCR (например, TCRα и/или TCRβ или TCRδ и/или TCRγ) и/или человеческую или гуманизированную последовательность гена β2-микроглобулина, и клетки эмбриона-хозяина.
Также предложена ткань, которая получена от не относящегося к человеку животного (например, грызуна, например мыши или крысы), описанного в настоящем документе, и экспрессирует химерный белок CD4, химерный белок CD8 (например, белок CD8α и/или CD8β), гуманизированный полипептид TCR (например, полипептид TCRα и/или TCRβ или TCRδ и/или TCRγ), гуманизированный полипептид МНС I (например, МНС I α), гуманизированный полипептид МНС II (например, полипептид МНС II α и/или МНС II β) и/или человеческий или гуманизированный β2-микроглобулин.
В одном аспекте предложен способ получения химерной человеческой/нечеловеческой молекулы CD4, включающий экспрессию в одной клетке химерного белка CD4 из нуклеотидного конструкта, как описано в настоящем документе. В одном варианте осуществления нуклеотидный конструкт представляет собой вирусный вектор; в определенном варианте осуществления вирусный вектор представляет собой лентивирусный вектор. В одном варианте осуществления клетку выбирают из СНО, COS, 293, HeLa и клетки сетчатки, экспрессирующей вирусную последовательность нуклеиновых кислот (например, клетки PERC.6™).
В одном аспекте предложена клетка, которая экспрессирует химерный белок CD4. В одном варианте осуществления клетка содержит экспрессионный вектор, содержащий химерную последовательность CD4, как описано в настоящем документе. В одном варианте осуществления клетку выбирают из СНО, COS, 293, HeLa и клетки сетчатки, экспрессирующей вирусную последовательность нуклеиновых кислот (например, клетки PERC.6™).
Также предложена химерная молекула CD4, продуцируемая не относящимся к человеку животным, как описано в настоящем документе, причем в одном варианте осуществления химерная молекула CD4 содержит аминокислотную последовательность всего или по существу всего внеклеточного домена человеческого белка CD4 и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого белка CD4, например мышиного белка CD4. В другом варианте осуществления предложена химерная молекула CD4, продуцируемая не относящимся к человеку животным, описанным в настоящем документе, причем химерная молекула CD4 содержит аминокислотную последовательность по меньшей мере всего или по существу всего домена D1 человеческого CD4, например по меньшей мере всех или по существу всех доменов D1-D2 человеческого CD4, например по меньшей мере всех или по существу всех доменов D1-D3 человеческого CD4, например аминокислотную последовательность человеческого CD4, которая отвечает за связывание с МНС II и/или внеклеточным доменом TCR, например аминокислотную последовательность человеческого CD4, которая отвечает за связывание с МНС II и/или вариабельным доменом TCR; и при этом остальная часть белка (например, трансмембранный домен, цитоплазматический домен, любой участок внеклеточного домена, который не был гуманизирован) получена из эндогенной нечеловеческой последовательность белка. Пример химерного человеческого/нечеловеческого полипептида CD4 содержит аминокислотную последовательность, представленную в SEQ ID NO: 78, а человеческий участок химерного полипептида охватывает приблизительно аминокислоты 27-319 SEQ ID NO: 78 (представленные отдельно в SEQ ID NO: 79).
В одном аспекте предложен способ получения химерной человеческой/нечеловеческой молекулы CD8 (например, CD8α и/или CD8β), включающий экспрессию в одной клетке химерного(-ых) полипептида(-ов) CD8 из нуклеотидного(-ых) конструкта(-ов), как описано в настоящем документе. В одном варианте осуществления нуклеотидный конструкт представляет собой вирусный вектор; в определенном варианте осуществления вирусный вектор представляет собой лентивирусный вектор. В одном варианте осуществления клетку выбирают из СНО, COS, 293, HeLa и клетки сетчатки, экспрессирующей вирусную последовательность нуклеиновых кислот (например, клетки PERC.6™).
В одном аспекте предложена клетка, которая экспрессирует химерный белок CD8. В одном варианте осуществления клетка содержит экспрессионный вектор, содержащий химерную(-ые) последовательность(-и) CD8, как описано в настоящем документе. В одном варианте осуществления клетку выбирают из СНО, COS, 293, HeLa и клетки сетчатки, экспрессирующей вирусную последовательность нуклеиновых кислот (например, клетки PERC.6™).
Также предложена химерная молекула CD8, продуцируемая не относящимся к человеку животным, описанным в настоящем документе, причем химерная молекула CD8 содержит весь или по существу весь внеклеточный домен человеческого белка CD8 (например, CD8α и/или CD8β) и по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого белка CD8, например мышиного белка CD8. Пример химерного полипептида CD8α представлен в SEQ ID NO: 88, а пример химерного белка CD8β представлен в SEQ ID NO: 83.
Также предложен гуманизированный белок TCR, продуцируемый не относящимся к человеку животным (например, грызуном, например мышью или крысой), описанным настоящем документе, содержащий человеческий вариабельный участок и нечеловеческий константный участок. Таким образом, гуманизированный белок TCR содержит человеческие участки, определяющие комплементарность (т.е. человеческие CDR1, 2 и 3), в своем вариабельном домене и нечеловеческий константный участок. Также предложены нуклеиновые кислоты, которые кодируют человеческие вариабельные домены TCR, продуцируемые не относящимся к человеку животным, описанным в настоящем документе.
Кроме того, предложена нечеловеческая клетка, выделенная из не относящегося к человеку животного, как описано в настоящем документе. В одном варианте осуществления клетка представляет собой ЭС-клетку. В одном варианте осуществления клетка представляет собой Т-клетку, например Т-клетку CD4+. В одном варианте осуществления клетка представляет собой хелперную Т-клетку (клетку ТН). В одном варианте осуществления клетка ТН представляет собой эффекторную клетку ТН, например клетку ТН1 или клетку ТН2. В одном варианте осуществления клетка представляет собой Т-клетку CD8+. В одном варианте осуществления клетка представляет собой цитотоксическую Т-клетку. Также предложена нечеловеческая клетка, которая экспрессирует белок TCR, содержащий человеческий вариабельный участок и нечеловеческий константный участок. Белок TCR может содержать TCRα, TCRβ или их комбинацию. В одном варианте осуществления клетка представляет собой Т-клетку, например Т-клетку CD4+ или CD8+. Кроме того, нечеловеческие Т-клетки, предложенные в настоящем документе, могут экспрессировать на своей клеточной поверхности: (а) химерный человеческий/нечеловеческий Т-клеточный корецептор, например химерный полипептид CD4 или химерный полипептид CD8, содержащий внеклеточный домен человеческого Т-клеточного корецептора, функционально связанный с нечеловеческим трансмембранным и/или внутриклеточным доменом Т-клеточного корецептора; и (b) белок TCR, содержащий человеческий вариабельный участок и нечеловеческий константный участок.
В другом варианте осуществления клетка представляет собой антигенпредставляющую клетку. В одном варианте осуществления антигенпредставляющая клетка представляет антиген на гуманизированных молекулах МНС I. В другом варианте осуществления антигенпредставляющая клетка является профессиональной антигенпредставляющей клеткой, например В-клеткой, дендритной клеткой и макрофагом. В другом варианте осуществления антигенпредставляющая клетка представляет антиген на гуманизированных молекулах МНС I и/или МНС II.
В одном аспекте предложена клетка, которая экспрессирует химерные человеческие/нечеловеческие белки МНС I и МНС II (например, белки HLA-A2/H-2K и HLA-DR2/H-2E). В одном аспекте клетка представляет собой мышиную клетку, которая не экспрессирует функциональные эндогенные полипептиды МНС из своего локуса H-2D. В некоторых вариантах осуществления клетка представляет собой мышиную клетку, сконструированную таким образом, что в ней отсутствует весь эндогенный локус H-2D или его участок. В некоторых вариантах осуществления клетка представляет собой мышиную клетку, которая не экспрессирует на своей поверхности какой-либо функциональный эндогенный полипептид МНС I и МНС II. В одном варианте осуществления клетка содержит экспрессионный вектор, содержащий химерную последовательность МНС класса I и химерную последовательность МНС класса II, как описано в настоящем документе. В одном варианте осуществления клетку выбирают из СНО, COS, 293, HeLa и клетки сетчатки, экспрессирующей вирусную последовательность нуклеиновых кислот (например, клетки PERC.6™).
Химерный комплекс МНС II, содержащий внеклеточный домен HLA-DR2, описанный в настоящем документе, может быть обнаружен антителами против HLA-DR. Таким образом, клетка, представляющая химерный человеческий/нечеловеческий полипептид МНС II, может быть обнаружена и/или отобрана с использованием антитела против HLA-DR. Химерный комплекс МНС I, содержащий внеклеточный домен HLA-A2, описанный в настоящем документе, может быть обнаружен с использованием антител против HLA-A, например против HLA-A2. Таким образом, клетка, представляющая химерный человеческий/нечеловеческий полипептид МНС I, может быть обнаружена и/или отобрана с использованием антитела против HLA-A. Антитела, которые распознают другие аллели HLA, могут быть приобретены на рынке или могут быть получены и использованы для обнаружения/отбора.
Хотя в примерах ниже описано созданное методами генной инженерии животное, в геноме которого содержится замещение последовательности нуклеиновых кислот, кодирующей мышиные белки Н-2K, Н-2А и Н-2Е, последовательностью нуклеиновых кислот, кодирующей химерные человеческие/мышиные белки HLA-A2/H-2K и HLA-DR2/H-2E соответственно, специалисту в данной области будет понятно, что аналогичную стратегию можно использовать для внедрения химер, содержащих другие человеческие гены МНС I и II (другие гены HLA-A, HLA-B и HLA-C; а также другие гены HLA-DR, HLA-DP и HLA-DQ). Также предложены такие животные, содержащие множество химерных человеческих/нечеловеческих генов МНС I и МНС II (например, человека/грызуна, например человека/мыши) в эндогенных локусах МНС. Примеры таких химерных белков МНС I и МНС II описаны в публикациях патентов США №№20130111617, 20130185819, 20130185820 и 20140245467 и патенте США №8,847,005, которые включены в настоящий документ путем ссылки.
Также предложена нечеловеческая клетка, содержащая хромосому или фрагмент хромосомы не относящегося к человеку животного, описанного в настоящем документе. В одном варианте осуществления нечеловеческая клетка содержит ядро не относящегося к человеку животного, описанного в настоящем документе. В одном варианте осуществления в результате переноса ядра нечеловеческая клетка содержит хромосому или ее фрагмент.
В одном аспекте предложена нечеловеческая индуцированная плюрипотентная клетка, содержащая ген, кодирующий химерный полипептид CD4, ген, кодирующий химерный полипептид CD8 (например, полипептид CD8α и/или CD8β), ген, кодирующий гуманизированный полипептид МНС I (например, МНС I α и/или β2-микроглобулина), ген, кодирующий гуманизированный полипептид МНС II (например, МНС II α и/или МНС II β), и/или неперестроенный гуманизированный локус TCR, кодирующий гуманизированный полипептид TCRα и/или TCRβ, описанные в настоящем документе. В одном варианте осуществления индуцированная плюрипотентная клетка получена из не относящегося к человеку животного, описанного в настоящем документе.
В одном аспекте предложена гибридома или квадрома, полученная из не относящегося к человеку животного, описанного в настоящем документе. В одном варианте осуществления не относящееся к человеку животное представляет собой мышь или крысу.
Получение генетически модифицированных не относящихся к человеку животных, которые формируют по существу гуманизированные Т-клеточные иммунные ответы
Также предложен способ получения созданного методами генной инженерии не относящегося к человеку животного (например, сконструированного методами генной инженерии грызуна, например мыши или крысы), описанного в настоящем документе. Как правило, способы включают: (а) внедрение в геном не относящегося к человеку животного первой нуклеотидной последовательности, кодирующей химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора, второй нуклеотидной последовательности, кодирующей второй химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора, и/или третьей нуклеотидной последовательности, кодирующей третий химерный человеческий/нечеловеческий полипептид Т-клеточного корецептора, причем нечеловеческий участок каждого химерного полипептида Т-клеточного корецептора содержит по меньшей мере трансмембранный и цитоплазматический домены нечеловеческого Т-клеточного корецептора, и при этом человеческий участок каждого химерного полипептида содержит внеклеточный участок (или его часть) человеческого Т-клеточного рецептора; (b) вставку в геном не относящегося к человеку животного неперестроенного вариабельного локуса гена Т-клеточного рецептора (TCR) α, содержащего по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, функционально связанный с нечеловеческой константной последовательностью гена TCRα, и/или неперестроенный вариабельный локус гена TCRβ, содержащий по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, функционально связанный с нечеловеческой константной последовательностью гена TCRβ; и необязательно (с) включение в геном первой последовательности нуклеиновых кислот, кодирующей первый химерный человеческий/нечеловеческий полипептид МНС, второй последовательности нуклеиновых кислот, кодирующей второй химерный человеческий/нечеловеческий полипептид МНС, и/или третьей последовательности нуклеиновых кислот, кодирующей третий химерный человеческий/нечеловеческий полипептид МНС; и/или (d) добавление в геном не относящегося к человеку животного локуса β2-микроглобулина, кодирующего человеческий или гуманизированный полипептид β2-микроглобулин. В некоторых вариантах осуществления этапы внедрения, вставки и/или включения включают нацеливание на последовательности, кодирующие внеклеточный(-ые) домен(-ы) Т-клеточного корецептора, вариабельный(-ые) домен(-ы) TCR, внеклеточный(-ые) домен(-ы) полипептида МНС или участок β2-микроглобулина, и замещение их последовательностями, кодирующими человеческий(-ие) внеклеточный(-ые) домен(-ы) Т-клеточного корецептора, человеческие вариабельные домены TCR, человеческий(-ие) внеклеточный(-ые) домен(-ы) МНС и/или человеческий участок β2-микроглобулина соответственно.
В других вариантах осуществления внедрение, вставка, включение и/или добавление могут включать разведение, например скрещивание, животных одного вида. В других вариантах осуществления внедрение, вставка, размещение и/или добавление включают последовательную гомологичную рекомбинацию ЭС-клеток. В некоторых вариантах осуществления ЭС-клетки получены из не относящихся к человеку животных, генетически модифицированных так, что они содержат одну или более, но не все нужные генетические модификации, и гомологичная рекомбинация в таких ЭС-клетках завершает генетическую модификацию. В других вариантах осуществления внедрение, вставка, размещение и/или добавление могут включать комбинацию скрещивания и гомологичной рекомбинации в ЭС-клетках, например скрещивание животного с другим (или несколькими) животным одного вида, причем некоторые или все из животных могут быть получены из ЭС-клеток, генетически модифицированных посредством одиночной гомологичной рекомбинации или последовательных этапов гомологичной рекомбинации, и при этом ЭС-клетка может быть выделена из не относящегося к человеку животного, содержащего одну или более генетических модификаций, описанных в настоящем документе.
В некоторых вариантах осуществления в способе применяется нацеливающий конструкт, полученный по технологии VELOCIGENE®, причем конструкт внедряют в ЭС-клетки, а нацеленные ЭС-клеточные клоны внедряют в мышиный эмбрион по технологии VELOCIMOUSE®, как описано в примерах. Нацеливающий конструкт может содержать гомологичные плечи на 5'- и/или 3'-конце, которые нацеливают на подлежащую замещению эндогенную последовательность, вставляемые последовательности (которые замещают эндогенную последовательность) и одну или более кассет селекции. Кассета селекции представляет собой нуклеотидную последовательность, вставленную в нацеливающий конструкт для упрощения отбора клеток (например, ЭС-клеток), в которые был интегрирован интересующий конструкт. В данной области известно множество приемлемых кассет селекции. Как правило, кассета селекции позволяет осуществить положительный отбор в присутствии конкретного антибиотика (например, Neo, Hyg, Pur, CM, SPEC и т.д.). Кроме того, кассета селекции может быть фланкирована сайтами рекомбинации, которые позволяют осуществлять делецию кассеты селекции путем обработки рекомбиназными ферментами. Наиболее распространенными сайтами рекомбинации являются loxP и Frt, распознаваемые ферментами Cre и Flp соответственно, но в данной области также известны другие сайты рекомбинации. Кассета селекции может быть расположена в любом месте конструкта за пределами кодирующего региона. В одном варианте осуществления кассета селекции расположена на 5'-конце человеческого фрагмента ДНК. В другом варианте осуществления кассета селекции расположена на 3'-конце человеческого фрагмента ДНК. В другом варианте осуществления кассета селекции расположена в пределах человеческого фрагмента ДНК. В другом варианте осуществления кассета селекции расположена в пределах интрона человеческого фрагмента ДНК. В другом варианте осуществления кассета селекции расположена в участке соединения человеческого и мышиного фрагментов ДНК.
В одном варианте осуществления способ получения созданного методами генной инженерии не относящегося к человеку животного приводит к получению животного, которое содержит в эндогенном локусе CD4 нуклеотидную последовательность, кодирующую химерный человеческий/нечеловеческий полипептид CD4. В одном варианте осуществления в изобретении предложен способ модификации локуса CD4 не относящегося к человеку животного так, чтобы обеспечить экспрессию химерного человеческого/нечеловеческого полипептида CD4, описанного в настоящем документе. В одном варианте осуществления в изобретении предложен способ модификации локуса CD4 мыши таким образом, чтобы обеспечить экспрессию химерного человеческого/мышиного полипептида CD4, включающий внедрение, например замещение в эндогенном локусе CD4 не относящегося к человеку животного, например мыши, нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD4, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD4. В одном аспекте способа химерный человеческий/мышиный полипептид CD4 содержит весь или по существу весь внеклеточный домен человеческого полипептида CD4 и по меньшей мере трансмембранный и цитоплазматический домены эндогенного мышиного полипептида CD4. В другом аспекте способа химерный человеческий/нечеловеческий полипептид CD4 содержит все или по существу все из доменов D1-D2 человеческого полипептида CD4. В еще одном варианте осуществления химерный человеческий/мышиный полипептид CD4 содержит все или по существу все из доменов D1-D3 человеческого полипептида CD4. В еще одном варианте осуществления химерный человеческий/мышиный полипептид CD4 содержит всю или по существу всю аминокислотную последовательность человеческого CD4, которая отвечает за взаимодействие с МНС II и/или внеклеточным доменом Т-клеточного рецептора. В еще одном варианте осуществления химерный человеческий/мышиный полипептид CD4 содержит всю или по существу всю аминокислотную последовательность человеческого CD4, которая отвечает за взаимодействие с МНС II и/или вариабельным доменом Т-клеточного рецептора.
Таким образом, предложен нуклеотидный конструкт для создания генетически модифицированных животных, содержащих химерный человеческий/нечеловеческий CD4. В одном аспекте нуклеотидная последовательность содержит гомологичные плечи на 5'- и 3'-концах, фрагмент ДНК, содержащий человеческую последовательность гена CD4 (например, человеческую последовательность гена внеклеточного домена CD4, например последовательность гена всех или по существу всех из доменов D1-D2 человеческого CD4, например последовательность гена всех или по существу всех из доменов D1-D3 и/или D2-D3 человеческого CD4, например последовательность гена всех или по существу всех из доменов D1-D4 человеческого CD4), и кассету селекции, фланкированную сайтами рекомбинации. В одном варианте осуществления человеческая последовательность гена CD4 представляет собой геномную последовательность, которая содержит интроны и экзоны человеческого CD4. В одном варианте осуществления гомологичные плечи гомологичны нечеловеческой (например, мышиной) геномной последовательности CD4. Пример конструкта изобретения показан на ФИГ. 5А.
В некоторых вариантах осуществления способ приводит к получению животного, которое содержит в эндогенном локусе CD8 нуклеотидную(-ые) последовательность(-и), кодирующую(-ие) химерный человеческий/нечеловеческий полипептид CD8α и/или CD8β. В одном варианте осуществления в изобретении предложен способ модификации локуса CD8 не относящегося к человеку животного таким образом, чтобы обеспечить экспрессию химерного человеческого/нечеловеческого полипептида CD8, описанного в настоящем документе. В одном аспекте предложен способ модификации локуса CD8 мыши таким образом, чтобы обеспечить экспрессию химерного человеческого/мышиного полипептида CD8, включающий внедрение, например замещение в эндогенном локусе CD8 не относящегося к человеку животного, например мыши, нуклеотидной последовательности, кодирующей эндогенный нечеловеческий полипептид CD8, нуклеотидной последовательностью, кодирующей химерный человеческий/нечеловеческий полипептид CD8. Полипептид CD8 может быть выбран из CD8α, CD8β и их комбинации. В одном аспекте химерный полипептид содержит весь или по существу весь внеклеточный домен человеческого полипептида CD8 и по меньшей мере трансмембранный и цитоплазматический домены эндогенного мышиного полипептида CD8.
Таким образом, также предложен нуклеотидный конструкт для создания генетически модифицированных животных, содержащих человеческий/нечеловеческий CD8. В одном аспекте последовательность нуклеотидного конструкта содержит гомологичные плечи на 5'- и 3'-концах, фрагмент ДНК, содержащий человеческую последовательность CD8α или CD8β, и кассету селекции, фланкированную сайтами рекомбинации. В некоторых вариантах осуществления человеческая последовательность содержит интроны и экзоны человеческого CD8α или CD8β, например экзоны, кодирующие внеклеточный домен человеческого CD8α или CD8β соответственно. В одном варианте осуществления гомологичные плечи гомологичны нечеловеческой последовательности CD8α или CD8β. Примеры конструктов для CD8α и CD8β показаны на ФИГ. 5В.
В связи с близким хромосомным расположением генов, кодирующих CD8α и CD8β, последовательное нацеливание двух генов повышает шансы на успешную гуманизацию. В одном варианте осуществления стратегия нацеливания включает внедрение химерного конструкта CD8β, описанного в настоящем документе, в ЭС-клетки, создание мыши из нацеленных ЭС-клеток, получение генетически модифицированных ЭС-клеток из указанной мыши и внедрение химерного конструкта CD8α, описанного в настоящем документе, в указанные генетически модифицированные ЭС-клетки. В другом варианте осуществления стратегия нацеливания включает внедрение химерного конструкта CD8β, описанного в настоящем документе, в ЭС-клетки, отбор клеток, в которые был внедрен химерный конструкт CD8β, внедрение химерного конструкта CD8α, описанного в настоящем документе, в ЭС-клетки, в которые был внедрен химерный конструкт CD8β и которые его содержат, и отбор клеток, в которые были внедрены оба химерных рецептора: CD8β и CD8α. В одном аспекте данного варианта осуществления этапы отбора выполняют с использованием разных селективных маркеров. В альтернативных вариантах осуществления сначала может быть выполнена гуманизация CD8α. После завершения нацеливания гена ЭС-клетки генетически модифицированных не относящихся к человеку животных могут быть подвергнуты скринингу для подтверждения успешного внедрения интересующей экзогенной нуклеотидной последовательности или экспрессии экзогенного полипептида с использованием разнообразных способов, известных в данной области (например, модифицированного анализа аллелей, описанного в публикации Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21(6):652-659).
В некоторых вариантах осуществления способ создания генетически модифицированного не относящегося к человеку животного приводит к получению животного, в геноме которого содержится гуманизированный неперестроенный локус TCR (например, гуманизированный неперестроенный локус TCRα, TCRβ, TCRδ и/или TCRγ). В одном варианте осуществления предложен способ создания генетически модифицированного не относящегося к человеку животного (например, грызуна, например мыши или крысы), которое экспрессирует Т-клеточный рецептор, содержащий человеческий вариабельный участок и нечеловеческий (например, грызуна, например мыши или крысы) константный участок на поверхности Т-клетки, причем способ включает вставку, например замещение, у первого не относящегося к человеку животного эндогенного нечеловеческого вариабельного локуса гена TCRα неперестроенным гуманизированным вариабельном локусом гена TCRα, содержащим по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, при этом гуманизированный вариабельный локус гена TCRα функционально связан с эндогенным константным участком TCRα; вставку, например замещение, у второго не относящегося к человеку животного эндогенного нечеловеческого вариабельного локуса гена TCRβ неперестроенным гуманизированным вариабельным локусом гена TCRβ, содержащим по меньшей мере один человеческий сегмент Vβ, один человеческий сегмент Dβ и один человеческий сегмент Jβ, при этом гуманизированный вариабельный локус гена TCRβ функционально связан с эндогенным константным участком TCRβ; и скрещивание первого и второго не относящихся к человеку животных с получением не относящегося к человеку животного, у которого экспрессируется Т-клеточный рецептор, содержащий человеческий вариабельный участок и нечеловеческий константный участок. В других вариантах осуществления в изобретении предложены способы получения генетически модифицированного не относящегося к человеку животного, в геноме которого содержится гуманизированный неперестроенный локус TCRα, или не относящегося к человеку животного, в геноме которого содержится гуманизированный неперестроенный локус TCRβ. В различных вариантах осуществления замещения выполнены в эндогенных локусах. В различных вариантах осуществления способ включает последовательную стратегию гуманизации, при которой конструкт, содержащий дополнительные сегменты вариабельного участка, внедряют в ЭС-клетки на каждом последующем этапе гуманизации, что в конечном итоге приводит к получению мыши, содержащей полный набор человеческих сегментов вариабельного участка (см., например, ФИГ. 4А и 4В).
В описании также предложен способ модификации вариабельного локуса гена TCR (например, локуса гена TCRα, TCRβ, TCRδ и/или TCRγ) не относящегося к человеку животного так, чтобы обеспечить экспрессию гуманизированного белка TCR, описанного в настоящем документе. В одном варианте осуществления в изобретении предложен способ модификации вариабельного локуса гена TCR таким образом, чтобы обеспечить экспрессию гуманизированного белка TCR на поверхности Т-клетки, который включает вставку, например замещение, у не относящегося к человеку животного эндогенного нечеловеческого вариабельного локуса гена TCR неперестроенным гуманизированным вариабельным локусом гена TCR. В одном варианте осуществления, в котором вариабельный локус гена TCR представляет собой вариабельный локус гена TCRα, неперестроенный гуманизированный вариабельный локус гена TCR содержит по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмет Jα. В одном варианте осуществления, в котором вариабельный локус гена TCR представляет собой вариабельный локус гена TCRβ, неперестроенный гуманизированный вариабельный локус гена TCR содержит по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ. В различных аспектах неперестроенный гуманизированный вариабельный локус гена TCR функционально связан с соответствующим эндогенным нечеловеческим константным участком TCR.
Таким образом, также предложены нуклеотидные конструкты для создания генетически модифицированных животных, содержащих гуманизированные гены вариабельного участка TCR. В одном аспекте нуклеотидный конструкт содержит: гомологичные плечи на 5'- и 3'-концах, фрагмент ДНК, содержащий человеческий (-ие) сегмент (-ы) гена вариабельного участка TCR, и кассету селекции, фланкированную сайтами рекомбинации. В одном варианте осуществления человеческий фрагмент ДНК представляет собой фрагмент гена TCRα, и он содержит по меньшей мере один человеческий сегмент вариабельного участка TCRα. В другом варианте осуществления человеческий фрагмент ДНК представляет собой фрагмент TCRβ, и он содержит по меньшей мере один человеческий сегмент гена вариабельного участка TCRβ. В одном аспекте по меньшей мере одно гомологичное плечо представляет собой нечеловеческое гомологичное плечо, и оно гомологично нечеловеческому локусу TCR (например, нечеловеческому локусу TCRα или TCRβ).
В различных аспектах изобретения последовательность (-и), кодирующая (-ие) химерные человеческие/нечеловеческие полипептиды МНС I и МНС II, расположена (-ы) в эндогенном нечеловеческом локусе МНС (например, мышином локусе Н-2K и/или Н-2Е). В одном варианте осуществления это приводит к размещению, например замещению, эндогенного (-ых) гена (-ов) МНС или его (их) участка (-ов) последовательностью (-ями) нуклеиновых кислот, кодирующей (-ими) человеческие или гуманизированные полипептиды МНС I. Так как последовательности нуклеиновых кислот, кодирующие полипептиды МНС I, МНС II α и МНС β, расположены вблизи друг друга на хромосоме, для достижения наилучшего результата в отношении гуманизации обоих МНС I и МНС II у одного животного локусы нацеливания МНС I и МНС II следует проводить последовательно. Таким образом, также в настоящем документе предложены способы создания генетически модифицированного не относящегося к человеку животного, содержащего последовательности нуклеиновых кислот, кодирующие химерные человеческие/нечеловеческие полипептиды МНС I, МНС II α и МНС II β, как описано в настоящем документе.
Таким образом, предложен нуклеотидный конструкт для создания генетически модифицированных животных, содержащих химерный человеческий/нечеловеческий МНС. В одном аспекте нуклеотидный конструкт содержит: гомологичные плечи на 5'- и 3'-концах, человеческий фрагмент ДНК, содержащий человеческие последовательности гена МНС (например, человеческие последовательности гена HLA-A2 или HLA-DR), и кассету селекции, фланкированную сайтами рекомбинации. В одном варианте осуществления человеческий фрагмент ДНК представляет собой геномный фрагмент, который содержит как интроны, так и экзоны человеческого гена МНС (например, человеческого гена HLA-A2 или HLA-DR2). В одном варианте осуществления нечеловеческие гомологичные плечи гомологичны нечеловеческому локусу МНС (например, локусу МНС I или МНС II).
В одном варианте осуществления нечеловеческие гомологичные плечи на 5'- и 3'-концах содержат геномную последовательность на 5'- и 3'-концах соответственно эндогенного нечеловеческого (например, мышиного) локуса гена МНС класса I или МНС класса II (например, в направлении 5' от первой лидерной последовательности и 3' от экзона α3 мышиного гена МНС I или ближе к 5'-концу от мышиного гена Н-2Ab1 и ближе к 3' концу от мышиного гена Н-2Еа). В одном варианте осуществления эндогенный локус МНС класса I выбирают из мышиного Н-2K, H-2D и H-2L. В конкретном варианте осуществления эндогенный локус МНС класса I представляет собой мышиный Н-2K. В одном варианте осуществления эндогенный локус МНС II выбирают из мышиного Н-2Е и Н-2А. В одном варианте осуществления созданный конструкт МНС II позволяет осуществлять замещение обоих мышиных генов Н-2Е и Н-2А. В одном варианте осуществления у мыши не экспрессируются функциональные эндогенные полипептиды МНС из ее эндогенного локуса Н-2D. В некоторых вариантах осуществления мышь конструируют таким образом, что у нее отсутствует весь эндогенный локус H-2D или его участок. В другом варианте осуществления у мыши не экспрессируются какие-либо функциональные эндогенные полипептиды МНС I и МНС II на клеточной поверхности. В одном варианте осуществления единственные МНС I и МНС II, экспрессируемые у мыши на клеточной поверхности, представляют собой химерные человеческие/мышиные МНС I и МНС II.
В описании также предложены способы получения созданного методами генной инженерии не относящегося к человеку животного (например, созданного методами генной инженерии грызуна, например мыши или крысы), в геноме которого содержится локус β2-микроглобулина, кодирующий человеческий или гуманизированный полипептид β2-микроглобулин. В одном аспекте способы приводят к получению созданного методами генной инженерии грызуна, например мыши, в геноме которого содержится в эндогенном локусе β2-микроглобулина нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2-микроглобулин. В некоторых примерах у мыши не экспрессируется функциональный мышиный β2-микроглобулин из эндогенного мышиного локуса β2-микроглобулина. В некоторых аспектах в способах применяется нацеливающий конструкт, например, полученный по технологии VELOCIGENE®, причем конструкт внедряют в ЭС-клетки, а нацеленные ЭС-клеточные клоны внедряют в мышиный эмбрион, например, по технологии VELOCIMOUSE®, как описано в настоящем документе.
Также предложен нуклеотидный конструкт, используемый для получения созданных методами генной инженерии не относящихся к человеку животных. Нуклеотидный конструкт может содержать: гомологичные плечи на 5'- и 3'-концах, фрагмент ДНК, содержащий человеческие последовательности β2-микроглобулина, и кассету селекции, фланкированную сайтами рекомбинации. В одном варианте осуществления человеческий фрагмент ДНК представляет собой геномный фрагмент, который содержит как интроны, так и экзоны человеческого гена β2-микроглобулина. В одном варианте осуществления нечеловеческие гомологичные плечи гомологичны нечеловеческому локусу β2-микроглобулина. Геномный фрагмент может содержать экзоны 2, 3 и 4 человеческого гена β2-микроглобулина. В одном примере геномный фрагмент содержит (от 5' к 3'): экзон 2, интрон, экзон 3, интрон и экзон 4 человеческой последовательности β2-микроглобулина. Кассета селекции может быть расположена в любом месте в конструкте за пределами кодирующего региона β2-микроглобулина, например, она может быть расположена в направлении 3' от экзона 4 человеческого β2-микроглобулина. Нечеловеческие гомологичные плечи на 5'- и 3'-концах могут содержать геномную последовательность на 5'- и 3'-концах эндогенного нечеловеческого гена β2-микроглобулина соответственно. В другом варианте осуществления нечеловеческие гомологичные плечи на 5'- и 3'-концах содержат геномную последовательность на 5'-конце экзона 2 и на 3'-конце экзона 4 эндогенного нечеловеческого гена соответственно.
Другой аспект изобретения относится к способу модификации локуса β2-микроглобулина не относящегося к человеку животного (например, грызуна, например мыши или крысы) так, чтобы обеспечить экспрессию человеческого или гуманизированного полипептида β2-микроглобулина, описанного в настоящем документе. Один способ модификации локуса β2-микроглобулина не относящегося к человеку животного, например мыши, так, чтобы обеспечить экспрессию человеческого или гуманизированного полипептида β2-микроглобулина, включает замещение в эндогенном локусе β2-микроглобулина нуклеотидной последовательности, кодирующей мышиный β2-микроглобулин, нуклеотидной последовательностью, кодирующей человеческий или гуманизированный полипептид β2-микроглобулин. В одном варианте осуществления такого способа не относящееся к человеку животное, например мышь, не экспрессирует функциональный полипептид β2-микроглобулин из эндогенного нечеловеческого, например мышиного, локуса β2-микроглобулина. В некоторых конкретных вариантах осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2-микроглобулин, содержит нуклеотидную последовательность, представленную в экзонах 2-4 человеческого гена β2-микроглобулина. В других вариантах осуществления нуклеотидная последовательность, кодирующая человеческий или гуманизированный полипептид β2-микроглобулин, содержит нуклеотидные последовательности, представленные в экзонах 2, 3 и 4 человеческого гена β2-микроглобулина.
Различные примеры осуществления гуманизированных локусов, описанных в настоящем документе, представлены на ФИГ. 2-5.
После завершения нацеливания на ген ЭС-клетки или генетически модифицированных не относящихся к человеку животных подвергают скринингу для подтверждения успешного введения интересующей экзогенной нуклеотидной последовательности или экспрессии экзогенного полипептида. Специалистам в данной области известно множество способов, включая (без ограничений) саузерн-блоттинг, ПЦР длинных фрагментов, количественную ПЦР (например, ПНР в реальном времени с использованием TAQMAN®), флуоресцентную гибридизацию in situ, нозерн-блоттинг, проточную цитометрию, вестерн-блоттинг, иммуноцитохимические способы, иммуногистохимические способы и т.д. В одном примере не относящихся к человеку животных (например, мышей), несущих интересующую генетическую модификацию, идентифицируют путем скрининга на потерю мышиного аллеля и/или приобретение человеческого аллеля с помощью модифицированного анализа аллелей, описанного в публикации Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21(6):652-659. Специалистам в данной области известны другие анализы, выявляющие специфическую нуклеотидную или аминокислотную последовательность у генетически модифицированных животных.
В некоторых вариантах осуществления животных получают путем скрещивания.
В одном не имеющем ограничительного характера аспекте, например, не относящееся к человеку животное, содержащее химерный человеческий/нечеловеческий CD8, описанный в настоящем документе, и гуманизированный МНС I и/или β2-микроглобулин, может быть получено путем скрещивания животного, содержащего химерный локус CD8 (например, химерный локус CD8α и/или β), как описано в настоящем документе, с животным, содержащим гуманизированный локус МНС I и/или β2-микроглобулина. Животное также может быть получено путем внедрения в ЭС-клетки животного, содержащего гуманизированный локус МНС I и/или β2-микроглобулина, нуклеотидной последовательности, кодирующей химерный CD8 (например, химерный CD8α и/или β), например, для замещения в эндогенном локусе CD8 (например, химерном локусе CD8α и/или β); или внедрения в ЭС-клетки животного, содержащего химерный локус CD8 (например, химерный локус CD8α и/или β), нуклеотидной (-ых) последовательности (-ей), кодирующей (-их) гуманизированный МНС I и/или β2-микроглобулин.
В некоторых вариантах осуществления животное, содержащее химерный локус CD8, может быть сначала скрещено с животным, содержащим гуманизированный вариабельный локус гена TCR, для получения животного, содержащего гуманизированные локусы CD8 и вариабельного участка TCR, которое после этого может быть скрещено с животным, содержащим гуманизированные локусы МНС I и/или β2 микроглобулина, для получения животного, содержащего гуманизированные локусы МНС I, вариабельного гена TCR и/или β2-микроглобулина. Альтернативно животное, содержащее гуманизированные локусы МНС I и/или β2-микроглобулина, может быть сначала скрещено с животным, содержащим гуманизированный вариабельный локус гена TCR, для получения животного, содержащего гуманизированные локусы МНС I и вариабельного участка TCR, которое после этого может быть скрещено с животным, содержащим химерный локус CD8, для получения животного, содержащего гуманизированные локусы МНС I, вариабельного гена TCR и/или β2-микроглобулина соответственно.
В одном аспекте не относящееся к человеку животное, содержащее химерный человеческий/нечеловеческий CD4 и гуманизированный МНС II, может быть получено путем скрещивания животного, содержащего химерный локус CD4, так, как описано в настоящем документе, с животным, содержащим гуманизированный локус МНС II. Животное также может быть получено путем внедрения в ЭС-клетки животного, содержащего гуманизированный локус МНС II, нуклеотидной последовательности, кодирующей химерный CD4, например, для замещения в эндогенном локусе CD4; или внедрения в ЭС-клетки животного, содержащего химерный локус CD4, нуклеотидной последовательности, кодирующей гуманизированный МНС II.
В некоторых вариантах осуществления животное, содержащее химерный локус CD4, может быть сначала скрещено с животным, содержащим гуманизированный вариабельный локус гена TCR, для получения животного, содержащего гуманизированные локусы CD4 и вариабельного участка TCR, которое после этого может быть скрещено с животным, содержащим гуманизированный локус МНС II, для получения животного, содержащего гуманизированные локусы CD4, МНС II и вариабельного гена TCR. Альтернативно животное, содержащее гуманизированный локус МНС II, может быть сначала скрещено с животным, содержащим гуманизированный вариабельный локус гена TCR, для получения животного, содержащего гуманизированные локусы МНС II и вариабельного участка TCR, которое после этого может быть скрещено с животным, содержащим химерный локус CD4, для получения животного, содержащего гуманизированные локусы МНС II, вариабельного гена TCR и/или β2-микроглобулина соответственно.
В некоторых вариантах осуществления не относящееся к человеку животное, содержащее химерный человеческий/нечеловеческий CD8, описанный в настоящем документе, и гуманизированный МНС I и/или β2-микроглобулин, скрещивают с животным, содержащим химерный локус CD4, описанный в настоящем документе, и животным, содержащим гуманизированный локус МНС II, для получения не относящегося к человеку животного, содержащего химерные полипептиды CD4 и CD8 и гуманизированные молекулы МНС I (и/или β2-микроглобулина) и МНС II. В некоторых вариантах осуществления животное, содержащее химерные человеческие/нечеловеческие полипептиды CD4 и CD8 и гуманизированные молекулы МНС I и МНС II, скрещивают с животным, содержащим гуманизированный вариабельный домен TCR, для получения животного, содержащего по существу гуманизированную Т-клеточную иммунную систему, например химерные человеческие/нечеловеческие полипептиды CD4 и CD8, гуманизированные молекулы МНС I (и/или β2-микроглобулина) и МНС II и гуманизированные вариабельные домены TCR.
Любое генетически модифицированное не относящееся к человеку животное (например, мышь), описанное в настоящем документе, может содержать одну или две копии генов, кодирующих химерный человеческий/нечеловеческий CD8 (например, CD8α и/или CD8β); химерный человеческий/нечеловеческий CD4; человеческий или гуманизированный МНС I; человеческий или гуманизированный β2-микроглобулин; человеческий или гуманизированный МНС II (например, МНС IIα и/или МНС IIβ) и человеческий или гуманизированный TCR (например, TCRα и/или TCRβ). Соответственно, животное может быть гетерозиготным или гомозиготным по любому или всем из этих генов.
Использование генетически модифицированных не относящихся к человеку животных, которые Формируют по существу гуманизированные Т-клеточные иммунные ответы
Генетически модифицированные не относящиеся к человеку животные, например грызуны, например мыши или крысы, содержащие гуманизированные CD4 и МНС II и/или гуманизированные CD8 и МНС I (и β2-микроглобулин), представляют пептиды Т-клеткам (Т-клеткам CD4+ или CD8+ соответственно) так, как это происходит у человека, поскольку по существу все из компонентов комплекса являются человеческими или гуманизированными. Генетически модифицированные не относящиеся к человеку животные изобретения могут использоваться для изучения функции человеческой иммунной системы у гуманизированного животного; для выявления антигенов и эпитопов антигенов, которые вызывают иммунный ответ (например, эпитопов Т-клеток, например уникальных человеческих раковых эпитопов), например, с целью использования для разработки вакцины; для выявления Т-клеток с высокой аффинностью к человеческим патогенам или раковым антигенам (т.е. Т-клеток, которые связываются с антигеном в связи с человеческим комплексом МНС I с высокой авидностью), например, для использования в адаптивной Т-клеточной терапии; для оценки потенциальных вакцин и других стратегий вакцинации; для изучения человеческих аутоиммунных реакций; для изучения человеческих инфекционных заболеваний и иным образом для разработки улучшенных терапевтических стратегий на основании экспрессии человеческих МНС и CD4/CD8.
Таким образом, в различных вариантах осуществления созданные методами генной инженерии животные настоящего изобретения используются, среди прочего, для оценки способности антигена инициировать иммунный ответ у человека и для создания разнообразия антигенов и выявления конкретного антигена, который можно использовать для разработки вакцины для человека.
В одном аспекте предложен способ определения того, будет ли пептид провоцировать клеточный иммунный ответ у человека, включающий воздействие на генетически модифицированное не относящееся к человеку животное, описанное в настоящем документе, пептида, что позволяет сформировать у не относящегося к человеку животного иммунный ответ, и обнаружение у не относящегося к человеку животного клетки (например, Т-клетки CD8+ или CD4+, содержащей человеческий CD8 или CD4 соответственно), которая связывается с последовательностью пептида, представленной химерной человеческой/нечеловеческой молекулой МНС I или II, как описано в настоящем документе. В одном варианте осуществления не относящееся к человеку животное после воздействия содержит рестриктированный по МНС класса I цитотоксический Т-лимфоцит (CTL) CD8+, который связывается с пептидом. В другом варианте осуществления не относящееся к человеку животное после воздействия содержит рестриктированную по МНС класса II цитотоксическую Т-клетку CD4+, которая связывается с пептидом.
В одном аспекте предложен способ выявления человеческого Т-клеточного эпитопа, включающий воздействие на не относящееся к человеку животное, описанное в настоящем документе, антигена, содержащего гипотетический Т-клеточный эпитоп, что позволяет не относящемуся к человеку животному формировать иммунный ответ, выделение из не относящегося к человеку животного рестриктированной по МНС класса I или МНС класса II T-клетки, которая связывается с эпитопом, и выявление эпитопа, связанного с указанной Т-клеткой.
В одном аспекте предложен способ выявления антигена, который формирует Т-клеточный ответ у человека, включающий воздействие гипотетического антигена на мышь, описанную в настоящем документе, что позволяет формировать у мыши иммунный ответ, и выявление антигена, связанного с рестриктированной по HLA класса I или класса II молекулой.
В одном аспекте предложен способ определения того, содержит ли гипотетический антиген эпитоп, который при воздействии на человеческую иммунную систему будет формировать рестриктированный по HLA класса I или класса II иммунный ответ, включающий воздействие на мышь, описанную в настоящем документе, гипотетического антигена и измерение антиген-специфичного рестриктированного по HLA класса I или HLA класса II иммунного ответа у мыши.
Кроме того, созданные методами генной инженерии не относящиеся к человеку животные, описанные в настоящем документе, могут использоваться для выявления Т-клеточных рецепторов, например Т-клеточных рецепторов с высокой авидностью, которые распознают интересующий антиген, например опухолевый антиген или антиген другого заболевания. Способ может включать: воздействие на не относящееся к человеку животное, описанное в настоящем документе, антигена, что позволяет формировать у не относящегося к человеку животного иммунный ответ на антиген, выделение из не относящегося к человеку животного Т-клетки, содержащей Т-клеточный рецептор, который связывается с антигеном, представленным человеческим или гуманизированным МНС I или МНС II, и определение последовательности указанного Т-клеточного рецептора.
Не относящиеся к человеку животные, экспрессирующие разнообразный набор функциональных человеческих сегментов гена TCR V(D)J, могут использоваться для изучения заболеваний у человека. Соответственно, в одном варианте осуществления созданные методами генной инженерии не относящиеся к человеку животные, описанные в настоящем документе, могут экспрессировать набор TCR, по существу аналогичный набору TCR, экспрессируемому у человека, например, набор TCR не относящегося к человеку животного, описанного в настоящем документе, может быть получен из по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 97% или по меньшей мере приблизительно 99% всех функциональных человеческих сегментов гена TCRα, TCRβ, TCRγ и/или TCRδ. В некоторых вариантах осуществления у не относящегося к человеку животного, описанного в настоящем документе, экспрессируется набор TCR, полученный из:
(i) по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 97% или по меньшей мере приблизительно 99% всех функциональных человеческих сегментов гена TCR Vα;
(ii) по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 97% или по меньшей мере приблизительно 99% всех функциональных человеческих сегментов гена TCR Jα;
(iii) по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 97% или по меньшей мере приблизительно 99% всех функциональных человеческих сегментов гена TCR Vβ;
(iv) по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 97% или по меньшей мере приблизительно 99% всех функциональных человеческих сегментов гена TCR Dβ; и/или
(v) по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 97% или по меньшей мере приблизительно 99% всех функциональных человеческих сегментов гена TCR Jβ.
В одном варианте осуществления у мыши продуцируется набор Т-клеток, содержащий все или по существу все функциональные человеческие сегменты гена TCR Vα и содержащий все или по существу все функциональные человеческие сегменты гена TCR Vβ. В одном варианте осуществления у мыши, предложенной в настоящем документе, человеческие гены TCR Vα и/или Vβ используются с частотой, аналогичной частоте использования человеческих генов TCR Vα и/или Vβ соответственно человеческими Т-клетками у человека. Способы обнаружения сегментов гена, экспрессируемых в наборе TCR не относящегося к человеку животного,включают, например, способы проточной цитометрии и/или секвенирования (например, ПЦР в реальном времени, секвенирование следующего поколения и т.д.).
В одном варианте осуществления предложен способ определения Т-клеточной активации гипотетическим терапевтическим средством для человека, включающий воздействие на генетически модифицированное животное, описанное в настоящем документе, гипотетического терапевтического средства для человека (или, например, воздействие на клетку, экспрессирующую человеческий или гуманизированный МНС II или МНС I, такого животного пептидной последовательности гипотетического терапевтического средства), воздействие на клетку генетически модифицированного животного, у которого экспрессируется человеческий или гуманизированный комплекс МНС/пептид Т-клетки, содержащей химерный человеческий/нечеловеческий (например, человеческий/мышиный) CD4 или CD8, способный связываться с клеткой генетически модифицированного животного, и измерение активации Т-клетки, индуцированной пептидпредставляющей клеткой генетически модифицированного животного.
Помимо возможности выявления антигенов и эпитопов антигена из человеческих патогенов или новообразований, генетически модифицированные животные изобретения могут использоваться для выявления аутоантигенов, связанных с аутоиммунными заболеваниями у человека, например сахарным диабетом I типа, рассеянным склерозом и т.д. Также генетически модифицированные животные изобретения могут использоваться для изучения различных аспектов аутоиммунных заболеваний у человека и могут применяться в качестве моделей аутоиммунных заболеваний.
В различных вариантах осуществления генетически модифицированные не относящиеся к человеку животные изобретения продуцируют Т-клетки с гуманизированными молекулами TCR на их поверхности и в результате смогут распознавать пептиды, представленные им комплексами МНС, так же, как это происходит у человека. Генетически модифицированные не относящиеся к человеку животные, описанные в настоящем документе, могут использоваться для изучения развития и функционирования человеческих Т-клеток и процессов иммунологической толерантности; для тестирования потенциальных вакцин для человека; для создания TCR с определенной специфичностью для генной терапии TCR; для создания библиотек TCR к антигенам, ассоциированным с заболеваниями (например, опухолеассоциированным антигенам (ТАА)); и т.д.
В данной области наблюдается растущий интерес к Т-клеточной терапии, так как Т-клетки (например, цитотоксические Т-клетки) могут быть направлены на атаку и уничтожение интересующего антигена, например вирусного антигена, бактериального антигена, опухолевого антигена и т.д., или представляющей его клетки. Начальные исследования в области Т-клеточной терапии рака были направлены на выделение инфильтрирующих опухоль лимфоцитов (TIL; популяции лимфоцитов в опухолевой массе, которые предположительно содержат Т-клетки, реагирующие на опухолевые антигены) из массы опухолевых клеток, выращивание их in vitro с использованием Т-клеточных факторов роста и перенос их обратно пациенту в рамках процесса, который называется адоптивным Т-клеточным переносом. См., например, публикации Restifo et al. (2012) Adoptive immunotherapy for cancer: harnessing the T cell response, Nature Reviews 12:269-81; Linnermann et al. (2011) T-Cell Receptor Gene Therapy: Critical Parameters for Clinical Success, J. Invest. Dermatol. 131:1806-16. Однако успех этих видов терапии до настоящего времени был достигнут только при меланоме и почечноклеточной карциноме; и адоптивный перенос TIL специфически не направлен на конкретные опухолеассоциированные антигены (TAA). Linnermann et al., выше.
Были предприняты попытки по проведению генной терапии TCR, при которой Т-клетки либо отбирали, либо программировали на нацеливание на интересующий антиген, например ТАА. Текущая генная терапия TCR основана на выявлении последовательностей TCR, которые направлены на конкретные антигены, например опухолеассоциированные антигены. Например, Rosenberg и коллеги опубликовали несколько исследований, в которых они трансдуцировали лимфоциты периферической крови, полученные от пациента с меланомой, генами, кодирующими цепи α и β TCR, специфичного к эпитопам ассоциированного с меланомой антигена MART-1, и использовали полученные выращенные лимфоциты для адоптивной Т-клеточной терапии. Johnson et al. (2009) Gene therapy with human and mouse T-cell receptors mediates cancer regression and targets normal tissues expressing cognate antigen, Blood 114:535-46; Morgan et al. (2006) Cancer Regression in Patients After Transfer of Genetically Engineered Lymphocytes, Science 314:126-29. TCR, специфичные к MART-1, выделяли у пациентов, у которых наблюдалась регрессия опухоли после терапии TIL. Однако выявление таких TCR, в особенности TCR с высокой авидностью (которые с наибольшей вероятностью можно использовать в терапевтических целях), осложняется тем, что большинство опухолевых антигенов являются аутоантигенами, и TCR, нацеливающиеся на эти антигены, часто либо удаляются, либо обладают субоптимальной аффинностью, главным образом в связи с иммунологической толерантностью.
В различных вариантах осуществления в настоящем изобретении эта проблема решается путем обеспечения созданных методами генной инженерии не относящихся к человеку животных, содержащих в своем геноме неперестроенный человеческий вариабельный локус гена TCR. Не относящееся к человеку животное, описанное в настоящем документе, способно продуцировать Т-клетки с разнообразным набором гуманизированных Т-клеточных рецепторов. Таким образом, не относящиеся к человеку животные, описанные в настоящем документе, могут являться источником разнородного набора гуманизированных Т-клеточных рецепторов, например гуманизированных Т-клеточных рецепторов с высокой аффинностью, предназначенных для использования в адоптивном Т-клеточном переносе.
Таким образом, в одном варианте осуществления в настоящем изобретении предложен способ создания Т-клеточного рецептора к человеческому антигену, включающий иммунизацию не относящегося к человеку животного (например, грызуна, например мыши или крысы), описанного в настоящем документе, интересующим антигеном, что позволяет формировать у животного иммунный ответ, выделение из животного активированной Т-клетки со специфичностью к интересующему антигену и определение последовательности нуклеиновых кислот Т-клеточного рецептора, экспрессированного антиген-специфичной Т-клеткой.
В одном варианте осуществления в изобретении предложен способ продуцирования человеческого Т-клеточного рецептора, специфичного к интересующему антигену (например, ассоциированному с заболеванием антигену), включающий иммунизацию не относящегося к человеку животного, описанного в настоящем документе, интересующим антигеном; что позволяет формировать у животного иммунный ответ; выделение из животного Т-клетки, реагирующей на интересующий антиген; определение последовательности нуклеиновых кислот человеческого вариабельного участка TCR, экспрессированного Т-клеткой; клонирование человеческого вариабельного участка TCR в нуклеотидный конструкт, содержащий последовательность нуклеиновых кислот человеческого константного участка TCR, так, что человеческий вариабельный участок TCR функционально связан с человеческим константным участком TCR; и экспрессирование из конструкта человеческого Т-клеточного рецептора, специфичного к интересующему антигену. В одном варианте осуществления этапы выделения Т-клетки, определения последовательности нуклеиновых кислот человеческого вариабельного участка TCR, экспрессированного Т-клеткой, клонирования человеческого вариабельного участка TCR в нуклеотидный конструкт, содержащий последовательность нуклеиновых кислот человеческого константного участка TCR, и экспрессии человеческого Т-клеточного рецептора выполняют с использованием стандартных техник, известных специалистам в данной области.
В одном варианте осуществления нуклеотидная последовательность, кодирующая Т-клеточный рецептор, специфичный к интересующему антигену, экспрессируется в клетке. В одном варианте осуществления клетку, экспрессирующую TCR, выбирают из клетки СНО, COS, 293, HeLa, PERC.6™ и т.д.
Интересующий антиген может представлять собой антиген, который вызывает заболевание или состояние или ассоциирован с ним, например опухолеассоциированный антиген; антиген вирусного, бактериального или другого патогенного происхождения и т.д. В данной области известно множество опухолеассоциированных антигенов. Подборка опухолеассоциированных антигенов представлена в базе данных пептидов (archive.cancerimmunity.org/peptidedatabase/Tcellepitopes.htm) журнала Cancer Immunity (журнал Института исследований рака). В некоторых вариантах осуществления изобретения интересующий антиген представляет собой человеческий антиген, например человеческий опухолеассоциированный антиген. В некоторых вариантах осуществления антиген представляет собой внутриклеточный антиген, специфичный к определенному типу клетки, а Т-клеточный рецептор используется для уничтожения клетки, экспрессирующей антиген.
В одном варианте осуществления в настоящем документе предложен способ выявления Т-клетки со специфичностью к интересующему антигену, например опухолеассоциированному антигену, включающий иммунизацию не относящегося к человеку животного, описанного в настоящем документе, интересующим антигеном, что позволяет формировать у животного иммунный ответ, и выделение из не относящегося к человеку животного Т-клетки со специфичностью к указанному антигену.
В настоящем изобретении предложены новые способы адоптивной Т-клеточной терапии. Таким образом, в настоящем документе предложен способ лечения или облегчения тяжести заболевания или состояния (например, рака) у субъекта (например, млекопитающего субъекта, например субъекта-человека), включающий иммунизацию не относящегося к человеку животного, описанного в настоящем документе, антигеном, ассоциированным с заболеванием или состоянием, что позволяет формировать у животного иммунный ответ, выделение из животного популяции антиген-специфичных Т-клеток и инфузию выделенных антиген-специфичных Т-клеток субъекту. В одном варианте осуществления в изобретении предложен способ лечения или облегчения тяжести заболевания или состояния у субъекта-человека, включающий иммунизацию не относящегося к человеку животного, описанного в настоящем документе, интересующим антигеном (например, антигеном, ассоциированным с заболеванием или состоянием, например опухолеассоциированным антигеном), что позволяет формировать у животного иммунный ответ, выделение из животного популяции антиген-специфичных Т-клеток, определение последовательности нуклеиновых кислот Т-клеточного рецептора (например, первой и/или второй последовательности нуклеиновых кислот, кодирующей человеческий перестроенный ген вариабельного участка TCRα и/или TCRβ); третьей и/или четвертой последовательности нуклеиновых кислот, кодирующей человеческий перестроенный ген вариабельного участка TCRδ или ген вариабельного участка TCRγ, экспрессируемый антиген-специфичными Т-клетками, клонирование последовательности нуклеиновых кислот Т-клеточного рецептора, например первой, второй, третьей и/или четвертой последовательности нуклеиновых кислот, соответственно кодирующей человеческий перестроенный ген вариабельного участка TCRα человеческий перестроенный ген вариабельного участка TCRβ, ген вариабельного участка, TCRδ или ген вариабельного участка TCRγ, в экспрессионный вектор (например, ретровирусный вектор), например, необязательно при этом первую, вторую, третью и/или четвертую последовательность нуклеиновых кислот, соответственно кодирующую человеческий перестроенный ген вариабельного участка TCRα, человеческий перестроенный ген вариабельного участка TCRβ, ген вариабельного участка TCRδ или ген вариабельного участка TCRγ, соответственно клонируют в пределах рамки считывания с человеческим константным геном TCRα, человеческим константным геном TCRβ, константным геном TCRδ или константным геном TCRγ, внедряя вектор в Т-клетки, полученные от субъекта таким образом, что Т-клетки экспрессируют антиген-специфичный Т-клеточный рецептор, и инфузию Т-клеток субъекту. В одном варианте осуществления последовательность нуклеиновых кислот Т-клеточного рецептора дополнительно гуманизируют перед внедрением в Т-клетки, полученные от субъекта, например последовательность, кодирующую человеческий константный участок, модифицируют таким образом, чтобы она имела большее сходство с человеческим константным участком TCR (например, нечеловеческий константный участок замещают человеческим константным участком). В некоторых вариантах осуществления заболевание или состояние представляет собой рак. В некоторых вариантах осуществления антиген-специфичную Т-клеточную популяцию выращивают перед инфузией субъекту. В некоторых вариантах осуществления популяцию иммунных клеток субъекта подвергают иммуноистощению перед инфузией антиген-специфичных Т-клеток. В некоторых вариантах осуществления антиген-специфичный TCR представляет собой TCR с высокой авидностью, например TCR с высокой авидностью к опухолеассоциированному антигену. В некоторых вариантах осуществления Т-клетка представляет собой цитотоксическую Т-клетку. В других вариантах осуществления заболевание или состояние вызвано вирусом или бактерией.
В другом варианте осуществления заболевание или состояние представляет собой аутоиммунное заболевание. Клетки TREG представляют собой субпопуляцию Т-клеток, которая сохраняет толерантность к аутоантигенам и предотвращает патологическую аутореактивность. Таким образом, в настоящем документе также предложены способы лечения аутоиммунного заболевания, которые основаны на создании антиген-специфичных клеток TREG у не относящегося к человеку животного изобретения, описанного в настоящем документе.
Также в настоящем документе предложен способ лечения или облегчения тяжести заболевания или состояния (например, рака) у субъекта, включающий внедрение клеток, пораженных заболеванием или состоянием (например, раковых клеток), от субъекта не относящемуся к человеку животному, что позволяет формировать у животного иммунный ответ на клетки, выделение из животного популяции Т-клеток, реагирующих на клетки, определение последовательности нуклеиновых кислот вариабельного домена Т-клеточного рецептора, экспрессируемого Т-клетками, клонирование вариабельного домена Т-клеточного рецептора, кодирующего последовательность, в вектор (например, находящийся в пределах рамки считывания и функционально связанный с человеческим константным геном TCR), внедрение вектора в Т-клетки, полученные от субъекта, и инфузию Т-клеток субъекта, несущих Т-клеточный рецептор, субъекту.
Кроме того, в настоящем документе предложено использование не относящегося к человеку животного, описанного в настоящем документе, для получения последовательностей нуклеиновых кислот, кодирующих человеческие вариабельные домены TCR (например, вариабельные домены TCR α и/или β). В одном варианте осуществления предложен способ получения последовательности нуклеиновых кислот, кодирующей человеческий вариабельный домен TCR, включающий иммунизацию не относящегося к человеку животного, описанного в настоящем документе, интересующим антигеном, что позволяет формировать у животного иммунный ответ на интересующий антиген, и получение в результате последовательности нуклеиновых кислот, кодирующей человеческий вариабельный домен TCR, который связывается с интересующим антигеном. В одном варианте осуществления способ дополнительно включает получение последовательности нуклеиновых кислот, кодирующей человеческий вариабельный домен TCR, который функционально связан с нечеловеческой константной областью TCR, выделение Т-клетки из не относящегося к человеку животного, описанного в настоящем документе, и получение в результате последовательности нуклеиновых кислот, кодирующей вариабельный домен TCR, связанный с нечеловеческим константным участком TCR, а также клонирование последовательности (-ей) нуклеиновых кислот, кодирующей (-их) вариабельный домен TCR (например, первой, второй, третьей или четвертой последовательности нуклеиновых кислот соответственно, кодирующей человеческий перестроенный ген вариабельного участка TCRα, человеческий перестроенный ген вариабельного участка TCRβ, ген вариабельного участка TCRδ или ген вариабельного участка TCRγ) в пределах рамки считывания с соответствующим человеческим геном константного участка (например, человеческим геном константного участка TCRα, человеческим геном константного участка TCRβ, человеческим геном константного участка TCRδ или геном вариабельного участка TCRγ соответственно).
Таким образом, в настоящем документе предложены последовательности нуклеиновых кислот вариабельного участка TCR, такие как перестроенные вариабельные последовательности нуклеиновых кислот TCR, например перестроенные последовательности нуклеиновых кислот вариабельного участка TCRα и/или TCRβ, продуцируемые не относящимися к человеку животными, описанными в настоящем документе, и кодируемые соответственно, например, человеческой перестроенной последовательностью гена Vα/Jα и человеческой перестроенной последовательностью гена VβDβJβ. Также предложены аминокислотные последовательности вариабельного участка TCR, кодируемые такими перестроенными последовательностями нуклеиновых кислот вариабельного участка TCR. Такие перестроенные последовательности нуклеиновых кислот вариабельного участка TCR (последовательности нуклеиновых кислот вариабельного участка TCRα и/или TCRβ), полученные от не относящихся к человеку животных, описанных в настоящем документе, могут быть клонированы в функциональной взаимосвязи с человеческим константным участком TCR (константный участок TCRα и/или TCRβ) и использованы в различных областях применения, описанных в настоящем документе, например, в качестве терапевтических средств для человека, у людей.
Кроме того, в настоящем документе предложено использование не относящегося к человеку животного, описанного в настоящем документе, для получения терапевтического средства для человека, включающее иммунизацию не относящегося к человеку животного интересующим антигеном (например, опухолеассоциированным антигеном), что позволяет формировать иммунный ответ у не относящегося к человеку животного, получение от животного Т-клеток, реагирующих на интересующий антиген, получение из Т-клеток последовательности (-ей) нуклеиновых кислот, кодирующей (-их) гуманизированный белок TCR или человеческий вариабельный домен TCR, который связывается с интересующим антигеном, и использование последовательности (-ей) нуклеиновых кислот, кодирующей (-их) гуманизированный белок TCR или человеческий вариабельный домен TCR, для создания терапевтического средства для человека.
Таким образом, также предложен способ получения терапевтического средства для человека, включающий иммунизацию не относящегося к человеку животного, описанного в настоящем документе, интересующим антигеном, что позволяет формировать иммунный ответ у не относящегося к человеку животного, получение от животного Т-клеток, реагирующих на интересующий антиген, получение из Т-клеток последовательности (-ей) нуклеиновых кислот, кодирующей (-их) гуманизированный Т-клеточный рецептор, который связывается с интересующим антигеном, и использование гуманизированного (или полностью человеческого) Т-клеточного рецептора в терапевтическом средстве для человека.
В одном варианте осуществления терапевтическое средство для человека представляет собой Т-клетку (например, человеческую Т-клетку, например Т-клетку, полученную от субъекта-человека), включающую интересующую последовательность нуклеиновых кислот (например, в которую трансфицирована, трансдуцирована или иным образом внедрена интересующая нуклеотидная последовательность) таким образом, что Т-клетка экспрессирует гуманизированный белок TCR с аффинностью к интересующему антигену. В одном аспекте субъект, у которого применяется терапевтическое средство, нуждается в терапии конкретного заболевания или состояния, а антиген ассоциирован с заболеванием или состоянием. В одном аспекте Т-клетка представляет собой цитотоксическую Т-клетку, антиген представляет собой опухолеассоциированный антиген, а заболевание или состояние представляет собой рак. В одном аспекте Т-клетка получена от субъекта.
В другом варианте осуществления терапевтическое средство для человека представляет собой Т-клеточный рецептор. В одном варианте осуществления терапевтический рецептор представляет собой растворимый Т-клеточный рецептор. Для создания растворимых Т-клеточных рецепторов или вариабельных участков TCR для использования в качестве терапевтических агентов были приложены значительные усилия. Создание растворимых Т-клеточных рецепторов зависит от получения перестроенных вариабельных участков TCR. Одним из подходов является создание одноцепочечных TCR, содержащих TCRα и TCRβ, и аналогично структуре иммуноглобулина scFv их сливание между собой посредством линкера (см., например, международную заявку № WO 2011/044186). Полученный scTv, если он аналогичен scFv, будет представлять собой термостабильную и растворимую форму связывающего белка TCRα/β. Альтернативные подходы включали создание растворимого TCR, имеющего константные домены TCRβ (см., например, публикацию Chung et al., (1994) Functional three-domain single-chain T-cell receptors, Proc. Natl. Acad. Sci. USA. 91:12654-58); a также обеспечение ненативной дисульфидной связи между константными доменами TCR (рассмотрено в публикации Boulter and Jakobsen (2005) Stable, soluble, high-affinity, engineered T cell receptors: novel antibody-like proteins for specific targeting of peptide antigens, Clinical and Experimental Immunology 142:454-60; см. также патент США №7,569,664). Описаны другие структуры растворимых Т-клеточных рецепторов. Не относящиеся к человеку животные, описанные в настоящем документе, могут использоваться для определения последовательности Т-клеточного рецептора, который связывается с высокой аффинностью с интересующим антигеном, с последующим созданием растворимого Т-клеточного рецептора на основе последовательности.
Растворимый Т-клеточный рецептор, полученный из последовательности Т-клеточного рецептора, экспрессируемого не относящимся к человеку животным, может использоваться для блокировки функции интересующего белка, например вирусного, бактериального или опухолеассоциированного белка. Альтернативно растворимый Т-клеточный рецептор может быть слит с функциональной группой, которая уничтожает инфицированную или раковую клетку, например, цитотоксическими молекулами (например, химиотерапевтическим препаратом), токсином, радионуклидом, пролекарством, антителом и т.д. Растворимый Т-клеточный рецептор также может быть слит с иммуномодулирующей молекулой, например цитокином и т.д. Растворимый Т-клеточный рецептор также может быть слит с иммуноингибирующей молекулой, например, молекулой, которая предотвращает уничтожение Т-клеткой других клеток, несущих антиген, распознаваемый Т-клеткой. Такие растворимые Т-клеточные рецепторы, слитые с иммуноингибирующими молекулами, могут использоваться, например, при блокировке аутоиммунного процесса. Различные примеры иммуноингибирующих молекул, которые могут быть слиты с растворимым Т-клеточным рецептором, рассматриваются в публикации Ravetch and Lanier (2000) Immune Inhibitory Receptors, Science 290:84-89, включенной в настоящий документ путем ссылки.
В настоящем изобретении также предложены способы изучения иммунологического ответа в связи с человеческим TCR, включая перестройку человеческого TCR, развитие Т-клетки, активацию Т-клетки, иммунологическую толерантность и т.д.
Также предложены способы тестирования потенциальных вакцин. В одном варианте осуществления в настоящем документе предложен способ определения того, будет ли вакцина активировать иммунологический ответ (например, пролиферацию Т-клеток, высвобождение цитокинов и т.д.) и приводить к образованию эффектора, а также Т-клеток памяти (например, центральных и эффекторных Т-клеток памяти).
В одном аспекте предложен препарат in vitro, который содержит Т-клетку, которая несет на своей поверхности химерный белок CD8, и вторую клетку, которая связывается с химерным CD8. В одном варианте осуществления вторая клетка представляет собой клетку, экспрессирующую полипептид МНС I, например химерный человеческий/нечеловеческий белок МНС I. В одном варианте осуществления химерный CD8 на поверхности первой клетки взаимодействует с химерным МНС I на поверхности второй клетки. В одном варианте осуществления химерный белок CD8 сохраняет способность к взаимодействию с эндогенными цитоплазматическими молекулами, например эндогенными цитоплазматическими сигнальными молекулами (например, эндогенным Lck и т.д.).
В одном аспекте предложен препарат in vitro, который содержит Т-клетку, которая несет на своей поверхности химерный белок CD4, и вторую клетку, которая связывается с химерным CD4. В одном варианте осуществления вторая клетка представляет собой клетку, например АПК, экспрессирующую полипептид МНС I, например химерный
человеческий/нечеловеческий белок МНС II. В одном варианте осуществления химерный CD4 на поверхности первой клетки взаимодействует с химерным МНС II на поверхности второй клетки. В одном варианте осуществления химерный белок CD4 сохраняет способность к взаимодействию с эндогенными цитоплазматическими молекулами, например эндогенными цитоплазматическими сигнальными молекулами (например, эндогенным Lck и т.д.).
Другие области применения генетически модифицированных животных, описанных в настоящем документе, т.е. животных, содержащих человеческий или гуманизированный Т-клеточный корецептор (например, химерный человеческий/нечеловеческий CD4 или CD8), необязательно дополнительно содержащих человеческий или гуманизированный белок МНС II или I, будут очевидны из настоящего описания.
ПРИМЕРЫ
Следующие примеры приведены для того, чтобы предоставить обычным специалистам в данной области описание того, как получать и использовать способы и композиции изобретения. Примеры не предназначены для ограничения объема того, что авторы рассматривают в качестве их изобретения. Были приложены усилия для обеспечения точности использованных чисел (например, количеств, температуры и т.д.), но следует учитывать некоторые погрешности и отклонения в экспериментах. Примеры не включают подробные описания стандартных способов, которые хорошо известны обычным специалистам в данной области (методики молекулярного клонирования и т.д.). При отсутствии особых указаний части представляют собой массовые части, молекулярная масса представляет собой средневесовую молекулярную массу, температура указана в градусах по шкале Цельсия, а давление является атмосферным или близким к атмосферному.
Пример 1. Создание мышей с гуманизированным МНС
Различные этапы конструирования мыши, содержащей гуманизированные локусы МНС I и МНС II с соответствующими и дополнительными делециями эндогенных локусов МНС I и МНС II (HLA-A2/H-2K, HLA-DR2/H-2E, H-2A-del, H-2D-del), показаны на ФИГ. 3А. Подробное описание этапов приведено ниже.
Пример 1.1. Создание и определение характеристик мышей с гуманизированным МНС I
Создание мышей с гуманизированным МНС I было описано ранее в публикации патента США №20130111617, включенной в настоящий документ путем ссылки. Вкратце, мышиный ген Н-2K был гуманизирован в одну стадию посредством конструирования уникального нацеливающего вектора из ДНК человеческой и мышиной бактериальной искусственной хромосомы (ВАС) по технологии VELOCIGENE® (см., например, патент США №6,586,251 и публикацию Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nat. Biotech. Nat. Biotech. 21(6): 652-659). ДНК из мышиного клона ВАС RP23-173k21 (Invitrogen) модифицировали путем гомологичной рекомбинации с замещением геномной ДНК, кодирующей домены αl, α2 и α3 мышиного гена Н-2K, человеческой геномной ДНК, кодирующей субъединицы α1, α2 и α3 человеческого гена HLA-A (ФИГ. 2А).
В частности, геномную последовательность, кодирующую мышиные субъединицы α1, α2 и α3 гена Н-2K, замещали человеческой геномной ДНК, кодирующей домены α1, α2 и α3 человеческого гена HLA-A*0201, путем одиночного нацеливания с использованием нацеливающего вектора, содержащего гигромициновую кассету, фланкированную сайтами loxP, гомологичное плечо на 5'конце, содержащее последовательность 5' мышиного локуса Н-2K, включающую нетранслируемую область (UTR) 5', и гомологичное плечо на 3'-конце, содержащее геномную последовательность 3' мышиной кодирующей последовательности Н-2K α3.
Конечный конструкт для нацеливания на эндогенный локус гена Н-2K включал (от 5' к 3'): (1) гомологичное плечо на 5'-конце, содержащее ~ 200 п.н. мышиной геномной последовательности 5' эндогенного гена Н-2K, включая 5'UTR; (2) ~ 1339 п.н. человеческой геномной последовательности, включая лидерную последовательность HLA-A*0201, лидерную последовательность/α1 интрон HLA-A*0201, экзон α1 HLA-A*0201, интрон α1-α2 HLA-А*0201, экзон α2 HLA-A*0201, ~ 316 п.н. 5'-конца интрона α2-α3; (3) сайт 5' loxP; (4) гигромициновую кассету; (5) сайт 3' loxP; (6) ~ 580 п.н. человеческой геномной последовательности, включающей ~ 304 п.н. 3'-конца интрона α2-α3, экзон α3 HLA-A*0201; и (7) гомологичное плечо на 3'-конце, содержащее ~ 200 п.н. мышиной геномной последовательности, включая интрон между последовательностями, кодирующими мышиный Н-2K α3 и трансмембранный домен. Последовательность из 149 нуклеотидов в участке соединения мышиной/человеческой последовательностей на 5'-конце нацеливающего вектора представлена в SEQ ID NO: 90, а последовательность из 159 нуклеотидов в участке соединения человеческой/мышиной последовательностей на 3'-конце нацеливающего вектора представлена в SEQ ID NO: 91. Гомологичная рекомбинация с этим нацеливающим вектором позволила создать модифицированный мышиный локус Н-2K, содержащий человеческую геномную ДНК, кодирующую домены α1, α2 и α3 гена HLA-A*0201, функционально связанные с последовательностями, кодирующими трансмембранный и цитоплазматический домены эндогенного мышиного Н-2K, что при трансляции приводит к образованию химерного человеческого/мышиного белка МНС класса I. Кассета селекции, присутствующая в нацеливающем конструкте, может быть впоследствии удалена различными способами, известными в данной области.
Нацеленную ДНК ВАС использовали для электропорации мышиных ЭС-клеток F1H4 с созданием модифицированных ЭС-клеток для получения мышей, у которых экспрессируется химерный белок МНС класса I на поверхности ядросодержащих клеток (например, Т- и В-лимфоцитов, макрофагов, нейтрофилов) (см., например, этап 1 в схеме, показанной на ФИГ. 3А). ЭС-клетки, содержащие вставку человеческих последовательностей HLA, выявляли путем количественного анализа TAQMAN™ (Valenzuela et al. (2003), выше).
Для создания мышей, экспрессирующих химерный МНС I, нацеленные ЭС-клетки, описанные в настоящем документе, использовали в качестве донорских ЭС-клеток и внедряли в мышиный эмбрион на стадии 8 клеток по способу VELOCIMOUSE® (см., например, патент США №7,294,754 и публикацию Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1):91-99). Мышей VELOCIMICE® (мыши поколения F0, полностью происходящие из донорской ЭС-клетки), независимо несущих химерный ген МНС класса I, определяют путем генотипирования с помощью анализа на модификацию аллеля (Valenzuela et al., выше), в котором обнаруживают присутствие уникальных последовательностей человеческого гена HLA-A*0201. Гетерозиготных мышей, полученных этим способом, разводили до достижения гомозиготности. Экспрессию химерного HLA-A2/H-2K подтверждали с помощью проточной цитометрии с использованием антител, специфичных к HLA-A и Н-2K.
Описанные выше нацеленные ЭС-клетки, содержащие химерный HLA-A2/H-2K, использовали на дополнительных этапах конструирования, описанных в примерах 1.2-1.3, для создания мышей, содержащих гуманизированные локусы МНС I и МНС II и не содержащих эндогенные локусы МНС I и МНС II (см. ФИГ. 3А).
Пример 1.2. Создание мышиных ЭС-клеток, содержащих делеции локусов МНС I и МНС II
Делеция эндогенных локусов МНС II описана в публикации заявки на патент США №20130111616, включенной в настоящий документ путем ссылки. Вкратце, нацеливающий вектор для внедрения делеции эндогенных генов МНС класса II Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа был получен с использованием генно-инженерной технологии VELOCIGENE® (см., например, патент США №6,586,251 и публикацию Valenzuela et al., выше). ДНК искусственной бактериальной хромосомы (ВАС) RP23-458i22 (Invitrogen) была модифицирована с удалением эндогенных генов МНС класса II Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа.
В частности, нижележащее и вышележащее гомологичные плечи были получены с помощью ПЦР мышиной ДНК ВАС из местоположений 5' от гена Н-2Ab1 и 3' от гена Н-2Еа соответственно. Эти гомологичные плечи использовали для создания кассеты, которая удалила ~ 79 т.п.н. RP23-458i22, содержащих гены Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа локуса МНС класса II путем бактериальной гомологичной рекомбинации (BHR). Данную область заместили неомициновой кассетой, фланкированной сайтами lox2372. Конечный нацеливающий вектор включал (от 5' к 3') гомологичное плечо 26 т.п.н., содержащее мышиную геномную последовательность ближе к 5'-концу от гена Н-2Ab1 эндогенного локуса МНС класса II, сайт lox2372 5', неомициновую кассету, сайт lox2372 3' и гомологичное плечо 63 т.п.н., содержащее мышиную геномную последовательность ближе к 3'-концу от гена Н-2Еа эндогенного локуса МНС класса II.
Нацеливающий вектор ДНК ВАС (описанный выше) использовали для электропорации мышиных ЭС-клеток, содержащих гуманизированный локус МНС I (из примера 1.1 выше; см., например, этап 2 на ФИГ. 3А), для создания модифицированных ЭС-клеток, содержащих делецию эндогенного локуса МНС класса II (удалены как Н-2А, так и Н-2Е). Положительные ЭС-клетки, содержащие удаленный эндогенный локус МНС класса II, идентифицировали посредством количественного анализа ПЦР с использованием зондов TAQMAN™ (Lie and Petropoulos (1998) Curr. Opin. Biotechnology 9:43-48). Область ближе к 5'-концу от удаленного локуса подтверждали посредством ПЦР с использованием праймеров 5111U F (CAGAACGCCAGGCTGTAAC; SEQ ID NO: 1) и 5111U R (GGAGAGCAGGGTCAGTCAAC; SEQ ID NO: 2) и зонда 5111U Р (CACCGCCACTCACAGCTCCTTACA; SEQ ID NO: 3), а область ближе к 3'-концу от удаленного локуса подтверждали с использованием праймеров 511ID F (GTGGGCACCATCTTCATCATTC; SEQ ID NO: 4) и 511ID R (CTTCCTTTCCAGGGTGTGACTC; SEQ ID NO: 5) и зонда 511ID P (AGGCCTGCGATCAGGTGGCACCT; SEQ ID NO: 6). Присутствие неомициновой кассеты из нацеливающего вектора подтверждали с использованием NEOF (GGTGGAGAGGCTATTCGGC; SEQ ID NO: 7) и NEOR (GAACACGGCGGCATCAG;SEQ ID NO: 8) и зонда NEOP (TGGGCACAACAGACAATCGGCTG; SEQ ID NO: 9). Нуклеотидная последовательность, проходящая через точку делеции ближе к 5'-концу (SEQ ID NO: 10), включала нижеследующее, что обозначает эндогенную мышиную последовательность ближе к 5'-концу от точки делеции (находящейся в скобках ниже), непрерывно связанную с последовательностью кассеты, находящейся в точке делеции: (TTTGTAAACA AAGTCTACCC AGAGACAGAT GACAGACTTC AGCTCCAATG CTGATTGGTT CCTCACTTGG GACCAACCCT) ACCGGTATAA CTTCGTATAA GGTATCCTAT ACGAAGTTAT ATGCATGGCC TCCGCGCCGG. Нуклеотидная последовательность, проходящая через точку делеции ближе к 3'-концу (SEQ ID NO: 11), включала нижеследующее, что обозначает последовательность кассеты, смежную с эндогенной мышиной последовательностью ближе к 3'-концу от точки делеции (находящейся в скобках ниже): CGACCTGCAG CCGGCGCGCC ATAACTTCGT ATAAGGTATC CTATACGAAG TTATCTCGAG (CACAGGCATT TGGGTGGGCA GGGATGGACG GTGACTGGGA CAATCGGGAT GGAAGAGCAT AGAATGGGAG TTAGGGAAGA).
После создания ЭС-клеток, содержащих как гуманизацию МНС I, так и делецию эндогенного МНС II, описанную выше, фланкированную сайтами loxP, неомициновую кассету удаляли с использованием CRE (см., например, этап 3 на ФИГ. 3А). В частности, плазмиду, кодирующую рекомбиназу Cre, электропорировали в ЭС-клетки для удаления неомициновой кассеты. Неомициновая кассета также может быть удалена с использованием других способов, известных в данной области.
При удалении мышиного локуса H-2D BHR использовали для модификации мышиного клона ВАС bMQ-218H21 (Sanger Institute), заместив 3756 п.н. гена H-2D (от старт-кодона ATG до 3 п.н. ближе к 3'-концу от стоп-кодона TGA, экзоны 1-8 мышиного H-2D) 6085 п.н. кассеты, содержащей (от 5' к 3'): ген LacZ в пределах рамки считывания с сайтом loxp 5', промотор UbC, ген неомицина и сайт loxp 3'.
Нацеливающий вектор ДНК ВАС (описанный выше) использовали для электропорации мышиных ЭС-клеток, содержащих гуманизированный локус МНС I и делецию мышиного МНС II, описанных выше (см., например, этап 4 на ФИГ. 3А). Положительные ЭС-клетки, содержащие удаленный эндогенный локус H-2D, выявляли посредством анализа количественной ПЦР, как описано выше. В таблице 2 приведены праймеры и зонды, используемые для анализа количественной ПЦР.
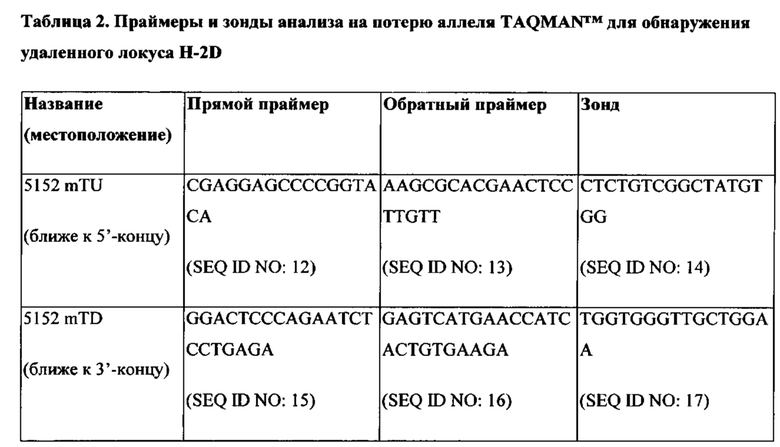
Пример 1.3. Внедрение химерного человеческого/мышиного локуса МНС II
Для создания вектора, содержащего гуманизированный HLA-DR2/H-2E, сначала мышиный ген Н-2Еа модифицировали в соответствии с описанием в патенте США №8,847,005, выданном 30 сентября 2014 г. и включенном в настоящий документ путем ссылки, для создания вектора, содержащего последовательность, кодирующую химерный белок H-2Ea/HLA-DRA1*01.
Что касается мышиного гена Н-2Eb, синтезированную человеческую цепь β HLA-DR2 (DRB1*1501) использовали для создания вектора, содержащего экзоны и интроны DRβ1*02(1501), и внедряли путем замены с помощью бактериальной гомологичной рекомбинации в вектор, содержащий химерный белок H-2Ea/HLA-DRA1*01. Ген Н-2Eb1 модифицировали по существу так, как описано в публикации патента США №20130185820 и патенте США №8,847,005, каждый из которых включен в настоящий документ путем ссылки. Использовали гигромициновую кассету селекции.
Полученный большой нацеливающий вектор (LTVEC) HLA-DR2/H-2E показан на ФИГ. 2В и 3В. Различные участки соединения нуклеотидных последовательностей полученных LTVEC (например, участки соединения мышиной/человеческой последовательностей, участки соединения человеческой/мышиной последовательностей или участки соединения мышиной или человеческой последовательности с кассетами селекции) приведены ниже в таблице 3 и перечислены в перечне последовательностей; их местоположения показаны на принципиальной схеме на ФИГ. 3В. В таблице 3 ниже, за исключением последовательностей, отмеченных звездочками (*, см. легенду таблицы), мышиные последовательности указаны обычным шрифтом; человеческие последовательности указаны в скобках; последовательности Lox указаны курсивом; а сайты рестрикции, внедренные в ходе этапов клонирования, и другие векторные последовательности (например, сайты множественного клонирования и т.д.) указаны жирным шрифтом.


Последовательности, отмеченные звездочкой, представляют собой последовательности участка соединения C57BL/6-BALB/c, где последовательности C57BL/6 приведены в скобках. В ходе клонирования химерного гена Н-2Еа экзон 1 и остальную часть интрона 1 аллеля C57BL/6 Н-2Еа замещали эквивалентной областью 2616 п.н., начиная от аллеля BALB/c Н-2Еа. Это выполнялось в связи с тем, что экзон 1 аллеля C57BL/6 Н-2Еа содержит делению, которая делает ген нефункциональным, тогда как экзон 1 аллеля BALB/c Н-2Еа является функциональным. Более подробное описание см. в патенте США №8,847,005, включенном в настоящий документ путем ссылки.
Нацеленную ДНК ВАС, описанную выше, использовали для электропорации мышиных ЭС-клеток, содержащих гуманизированный МНС I (HLA-A2), а также делецию МНС II и H-2D, для получения модифицированных ЭС-клеток с целью создания мышей, которые экспрессируют химерные гены МНС I и МНС II и у которых отсутствует функциональные эндогенные мышиные локусы Н-2Е, Н-2А, Н-2K и H-2D (см., например, этап 5 на ФИГ. 3А). ЭС-клетки, содержащие вставку человеческих последовательностей HLA, выявляли путем количественной ПЦР (TAQMAN™) с использованием праймеров и зондов из таблицы 4.

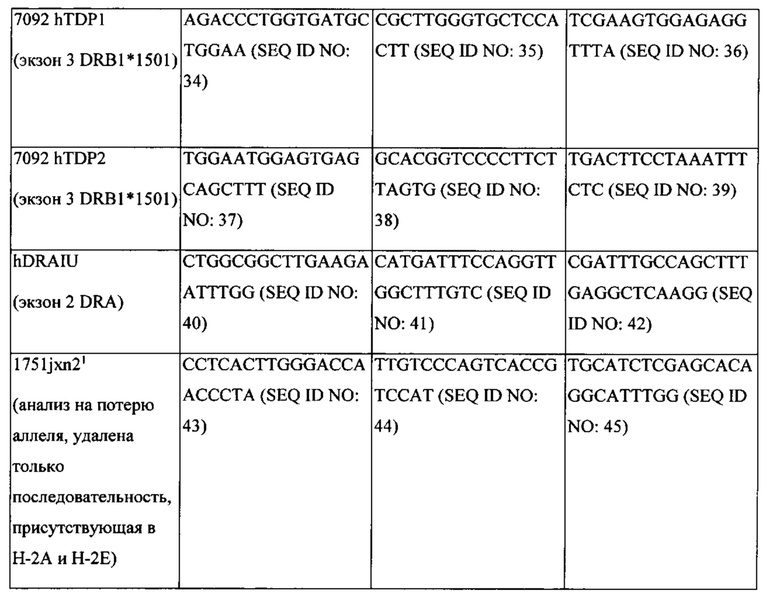
1Все последовательности, кроме этой, используются в анализе на приобретение аллеля.
Кассету селекции можно удалить способами, известными специалистам в данной области. Например, ЭС-клетки, несущие химерный человеческий/мышиный локус МНС класса I, могут быть трансфицированы конструктом, который экспрессирует Cre, для удаления фланкированной loxP кассеты селекции, внедренной путем вставки нацеливающего конструкта (см., например, этап 6 на ФИГ. 3А). Кассета селекции может быть необязательно удалена посредством выведения мышей, которые экспрессируют рекомбиназу Cre. Необязательно кассета селекции у мышей может быть сохранена.
Нацеленные ЭС-клетки, содержащие все из модификаций, описанных в настоящем документе (HLA-A2/H-2K, HLA-DR2/H-2E, H-2A-del, H-2D-del на ФИГ. 3А), проверяли с помощью количественного анализа TAQMAN®, описанного выше, с использованием наборов праймер/зонд, описанных в настоящем документе, для отдельных модификаций. Дополнительный набор праймер/зонд использовали для определения того, что в ходе этапа делеции кассеты не был создан инвертированный клон из-за наличия сайтов 1ох в противоположной ориентации.
Нацеленные ЭС-клетки, описанные выше, использовали в качестве донорских ЭС-клеток и внедряли в мышиный эмбрион на стадии 8 клеток по способу VELOCIMOUSE® (см., например, патент США №7,294,754 и публикацию Poueymirou et al. (2007), выше). Мышей VELOCIMICE® (мыши поколения F0, полностью происходящие из донорской ЭС-клетки), независимо несущих химерный ген МНС класса I и МНС класса II, определяли путем генотипирования с помощью анализа на модификацию аллеля (Valenzuela et al, выше), в котором обнаруживают присутствие уникальных последовательностей человеческого гена. Схематически генотип локусов МНС у полученных мышей представлен на ФИГ. 3С (** обозначает ген H-2L, который отсутствует во всех мышиных штаммах). Экспрессию обоих химерных человеческих/мышиных белков МНС I и МНС II подтверждают с использованием антител, специфичных к человеческим HLA-DR2 и HLA-A2. Гетерозиготных мышей разводили до достижения гомозиготности.
Пример 1.4. Создание мышей с гуманизированным β2-микроглобулином
Создание мышей с β2-микроглобулином описано в публикации заявки на патент США №20130111617, включенной в настоящий документ путем ссылки. Вкратце, мышиный ген β2-микроглобулина (β2m) гуманизировали за один этап посредством конструирования уникального нацеливающего вектора из ДНК человеческой и мышиной бактериальных искусственных хромосом (ВАС) по технологии VELOCIGENE® (см., например, патент США №6,586,251 и публикацию Valenzuela et al., выше).
В частности, нацеливающий вектор был создан путем бактериальной гомологичной рекомбинации, включающей вышележащее и нижележащее гомологичные плечи мышиного β2m из клона ВАС 89С24 из библиотеки RPCI-23 (Invitrogen). Мышиные гомологичные плечи были разработаны таким образом, чтобы ограничивать человеческий фрагмент ДНК β2m 2,8 т.п.н., проходящий от экзона 2 до приблизительно 267 нуклеотидов ближе к 3'-концу от некодирующего экзона 4 (ФИГ. 2С). Кассету селекции лекарственными средствами (неомицином), фланкированную сайтами распознавания рекомбиназы (например, сайтами loxP), внедряли в нацеливающий вектор, чтобы обеспечить возможность последующей селекции. Конечный нацеливающий вектор линеаризовывали и электропорировали в мышиную линию ЭС-клеток F1H4 (Valenzuela et al., выше).
Нацеленные ЭС-клеточные клоны с удаленной кассетой селекции лекарственными средствами (путем внедрения рекомбиназы Сге) внедряли в мышиный эмбрион на стадии 8 клеток по способу VELOCIMOUSE® (см., например, патент США №7,294,754 и публикацию Poueymirou et al., выше). Мышей VelociMice® (мыши F0, полученные полностью из донорской ЭС-клетки), несущих гуманизированный ген β2m, выявляли путем скрининга для определения утраты мышиного аллеля и приобретения человеческого аллеля с использованием анализа на модификацию аллеля (см. Valenzuela et al., выше). Гетерозиготных мышей разводили до достижения гомозиготности. Экспрессию человеческого β2-микроглобулина подтверждали посредством проточной цитометрии с использованием антител, специфичных к человеческому β2-микроглобулину.
Пример 2. Создание мышей с гуманизированным Т-клеточным рецептором
Мышей, содержащих делецию эндогенных вариабельных локусов TCR (α или β) и замещение эндогенных сегментов V и J или V, D и J, получают с использованием генно-инженерной технологии VELOCIGENE® (см., например, патент США №6,586,251 и публикацию Valenzuela, D.M., et al. (2003), выше), причем человеческие последовательности, полученные из библиотек ВАС с помощью бактериальной гомологичной рекомбинации, используют для создания больших нацеливающих векторов (LTVEC), содержащих геномные фрагменты человеческих вариабельных локусов TCR, фланкированных нацеливающими плечами, для нацеливания LTVEC на эндогенные мышиные вариабельные локусы TCR в мышиных ЭС-клетках. Подробное описание гуманизации альфа- и бета-локусов TCR приведено в патенте США №9,113,616, включенном в настоящий документ путем ссылки. LTVEC повторно линеаризовывали и электропорировали в линию мышиных ЭС-клеток в соответствии с публикацией Valenzuela et al. ЭС-клетки подвергали селекции на основании резистентности к гигромицину или неомицину и скринингу на потерю аллеля или приобретение аллеля.
Нацеленные ЭС-клеточные клоны внедряли в мышиные эмбрионы на стадии 8 клеток (или ранее) по способу VELOCIMOUSE® (Poueymirou, W.T. et al. (2007), выше). Мышей VELOCIMICE® (мыши F0, полученные полностью из донорской ЭС-клетки), несущих гуманизированные локусы TCR, выявляют путем скрининга на потерю эндогенного вариабельного аллеля TCR и приобретение человеческого аллеля с использованием анализа на модификацию аллеля (см. Valenzuela et al., выше). Детенышей F0 генотипировали и разводили до достижения гомозиготности. Мышей, гомозиготных по гуманизированным вариабельным локусам TCRα и/или TCRβ, получают так, как описано в настоящем документе.
Пример 2.1. Гуманизация алъфа-локуса TCR
1,5 м.п.н. ДНК в мышином локусе TCRα, соответствующих 110 V и 60 J мышиным сегментам, замещали 1 м.п.н. ДНК, соответствующих 54 V- и 61 J-сегментам человеческого TCRα, с использованием стратегии последовательной гуманизации, изображенной на ФИГ. 4А и описанной в патенте США №9,113,616. Соединительные последовательности нуклеиновых кислот различных нацеливающих векторов, используемых для стратегии последовательной гуманизации локуса TCRα, приведены в таблице 5 и включены в перечень последовательностей.

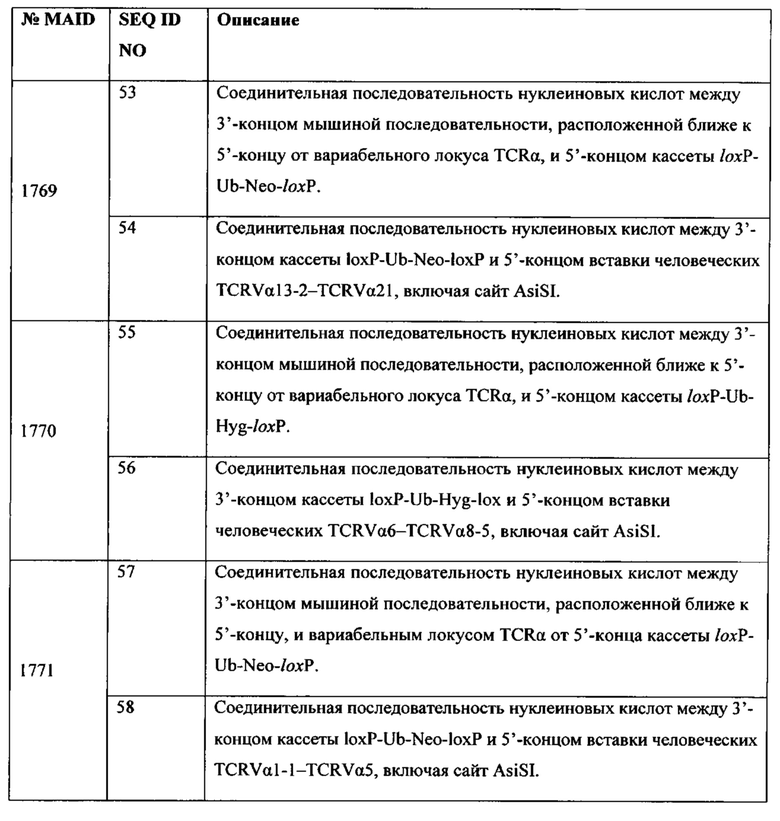
Человеческие сегменты вариабельного участка TCRα пронумерованы так же, как в базе данных IMGT. По меньшей мере 100 п.н. в каждом участке соединения (приблизительно 50 п.н. с каждого конца) включены в перечень последовательностей.
Сначала ДНК из мышиного клона ВАС RP23-6A14 (Invitrogen) модифицировали путем гомологичной рекомбинации и использовали в качестве нацеливающего вектора для замещения области TCRAJ1-TCRAJ28 эндогенного мышиного локуса TCRα кассетой Ub-гигромицин с последующим сайтом loxP. ДНК из мышиного клона ВАС RP23-117i19 (Invitrogen) модифицировали путем гомологичной рекомбинации и использовали в качестве нацеливающего вектора для замещения области ~ 15 т.п.н., окружающей (и включающей) TCRAV1 эндогенного мышиного локуса TCRα и δ, кассетой PGK-неомицин с последующим сайтом loxP. ЭС-клетки, несущие дважды нацеленную хромосому (т.е. один эндогенный мышиный локус TCRα нацеливали обоими из этих нацеливающих векторов), идентифицировали путем кариотипирования и способов скрининга (например, TAQMAN™), известных в данной области. Модифицированные ЭС-клетки обрабатывали рекомбиназой CRE, таким образом опосредуя делецию области между двумя сайтами loxP (т.е. области, состоящей из эндогенного мышиного локуса TCRα от TCRAV1 до TCRAJ1) и оставляя только один сайт loxP, неомициновую кассету и мышиные константный и энхансерный участки. Эта стратегия приводила к созданию удаленного мышиного локуса TCR α/δ (MAID 1540, ФИГ. 4А, вторая схема).
Первый человеческий нацеливающий вектор с TCRα содержал 191 660 п.н. человеческой ДНК из клонов ВАС CTD2216p1 и CTD2285m07 (Invitrogen), которые содержали первые два последовательных человеческих сегмента гена V TCRα (TRAV40 и 41) и 61 сегмент гена J (50 функциональных) TCRa. ВАС модифицировали путем гомологичной рекомбинации с включением 403 п.н. сайта Not1 ближе к 3'-концу от сегмента гена J1 TCRα для лигирования мышиного гомологичного плеча 3' и сайта AsiSI 5' для лигирования мышиного гомологичного плеча 5'. Два разных гомологичных плеча использовали для лигирования с этим человеческим фрагментом: гомологичное плечо 3' содержало эндогенные мышиные последовательности TCRα из клона ВАС RP23-6A14, а гомологичное плечо 5' содержало эндогенную последовательность TCRα ближе к 5'-концу от мышиного TCRαV из мышиного клона ВАС RP23-117i19. Эту мышиную-человеческую химерную ВАС использовали в качестве нацеливающего вектора (MAID 1626) для выполнения начальной вставки человеческих сегментов гена TCRα и кассеты loxp-ub-hygromycin-loxp ближе к 5'-концу в мышиных локусах TCRα. Соединительные последовательности нуклеиновых кислот (SEQ ID NO: 46-48) для нацеливающего вектора MAID 1626 описаны в таблице 5.
Впоследствии был получен ряд человеческих нацеливающих векторов, в которых использовалось то же мышиное плечо 5', которое включало эндогенную последовательность TCRα ближе к 5'-концу от мышиного TCRαV, из мышиного клона ВАС RP23-117i19 с чередующимися кассетами селекции loxpP-неомицин-loxP и loxP-гигромицин-loxP (или frt-гигромицин-frt в случае MAID 1979). Конкретные конструкты описаны в патенте США №9,113,616 и показаны на ФИГ. 4А, а соединительные последовательности для каждой вставки включены в таблицу 5 и перечень последовательностей. Конечный локус TCRα содержал кассету loxp-ub-неомицин-loxP 5' и всего 54 человеческих сегмента гена TCRaV (45 функциональных) и 61 человеческий сегмент гена TCRαJ, функционально связанных с мышиными константными генами и энхансерами TCRα. Соединительные последовательности нуклеиновых кислот (SEQ ID NO: 57 и 58) для нацеливающего вектора MAID 1771 описаны в таблице 5.
На любом из последовательных этапов гуманизации кассеты селекции удаляли путем делеции рекомбиназой CRe или Flp. Кроме того, человеческий локус TCRδ может быть повторно внедрен в последовательность TCR альфа.
Пример 2.2. Гуманизация вариабельного локуса TCRβ
0,6 м.п.н. ДНК в мышином локусе TCRβ, соответствующих мышиным сегментам 33 V, 2 D и 14 J, замещали 0,6 м.п.н. ДНК, соответствующих сегментам 67 V, 2 D и 14 J человеческого TCRβ, с использованием стратегии последовательной гуманизации, изображенной на ФИГ. 4В и подробно описанной в патенте США №9,113,616. Соединительные последовательности нуклеиновых кислот различных нацеливающих векторов, используемых для стратегии последовательной гуманизации локуса TCRβ, приведены в таблице 6 и включены в перечень последовательностей.
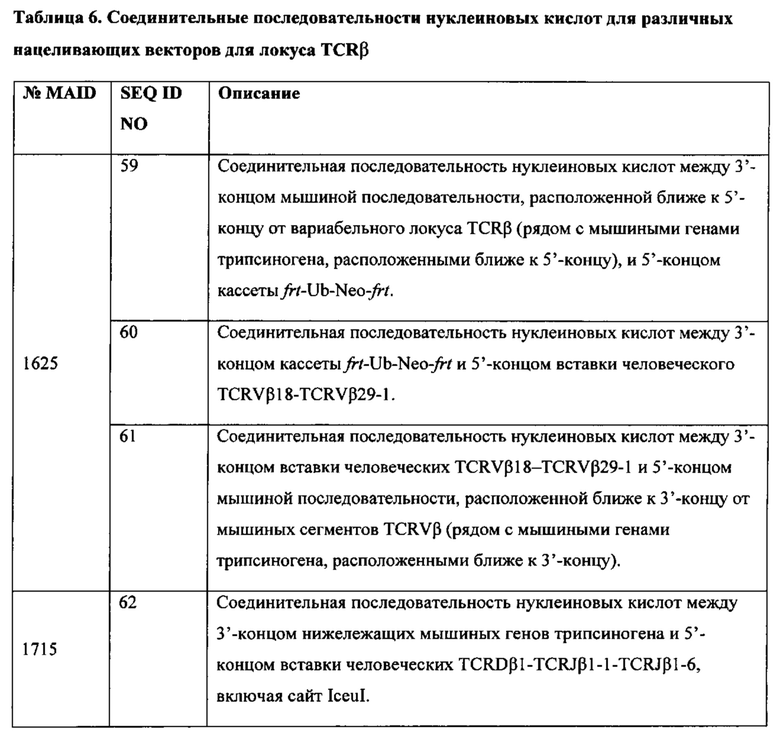


Человеческие сегменты вариабельного участка TCRβ пронумерованы так же, как в базе данных IMGT. По меньшей мере 100 п.н. в каждом участке соединения (приблизительно 50 п.н. с каждого конца) включены в перечень последовательностей.
В частности, ДНК из мышиного клона ВАС RP23-153p19 (Invitrogen) модифицировали путем гомологичной рекомбинации и использовали в качестве нацеливающего вектора для замещения области 17 т.п.н. (включая TCRBV30), прилегающей ближе к 5'-концу к кластеру гена трипсиногена (TRY) 3' в эндогенном мышином локусе TCRβ, кассетой PGK-неомицин с последующим сайтом loxP. ДНК из мышиного клона ВАС RP23-461h15 (Invitrogen) модифицировали путем гомологичной рекомбинации и использовали в качестве нацеливающего вектора для замещения области 8355 п.н. (включая TCRBV2 и TCRBV3) ближе к 3'-концу к кластеру гена трипсиногена (TRY) 5' в эндогенном мышином локусе TCRβ, кассетой Ub-гигромицин с последующим сайтом loxP. ЭС-клетки, несущие дважды нацеленную хромосому (т.е. один эндогенный мышиный локус TCRβ, нацеленный обоими нацеливающими векторами), идентифицировали с помощью кариотипирования и способов скрининга (например, TAQMAN™), известных в данной области. Модифицированные ЭС-клетки обрабатывали рекомбиназой CRE, опосредуя делецию области между сайтами loxP 5' и 3' (состоящими из эндогенного мышиного локуса TCRβ от TCRBV2 до TCRBV30) и оставляя только один сайт loxP, гигромициновую кассету и мышиные TCRBD, TCRBJ, константную и энхансерную последовательности. Один мышиный TCRVβ был оставлен ближе к 5'-концу от кластера 5' генов трипсиногена, и один мышиный TCRVβ был оставлен ближе к 3'-концу от мышиного Еβ, как показано на Фиг. 4В.
Первый человеческий нацеливающий вектор с TCRβ содержал 125 781 п.н. человеческой ДНК из клона ВАС CTD2559j2 (Invitrogen), который содержал первые 14 последовательных человеческих сегмента гена TCRβV (TRBV18-TRBV29-1); соединительные последовательности нуклеиновых кислот (SEQ ID NO: 59-61) для нацеливающего вектора MAID 1625 описаны в таблице 6.
Для замещения мышиных сегментов TCRβ D и J человеческими сегментами TCRβ D и J ДНК из мышиного клона ВАС RP23-302p18 (Invitrogen) и из человеческого клона ВАС RP11-701D14 (Invitrogen) модифицировали путем гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1715) в ЭС-клетках, которые содержали небольшой локус TCRβV, описанный выше (т.е. MAID 1625). В результате этой модификации были замещены ~ 18540 п.н. (от 100 п.н. ближе к 3'-концу от poly A генов трипсиногена 3' до 100 п.н. ближе к 3'-концу от сегментов J в кластере D2, который включал мышиный TCRBD1-J1, мышиный константный 1 и мышиный TCRBD2-J2) в эндогенном мышином локусе TCRβ, при этом ~ 25 425 п.н. последовательности содержали человеческий TCRBD1-J1, кассету loxP Ub-гигромицин-loxP, мышиный константный 1, человеческий TCRBD2-J2. ЭС-клетки, несущие дважды нацеленную хромосому (т.е. один эндогенный мышиный локус TCRβ, нацеленный обоими нацеливающими векторами), идентифицировали с помощью кариотипирования и способов скрининга (например, TAQMAN™), известных в данной области. Модифицированные ЭС-клетки обрабатывали рекомбиназой CRE, таким образом опосредуя делецию гигромициновой кассеты и оставляя только один сайт loxP ближе к 3'-концу от человеческих сегментов J в кластере D1J. Соединительные последовательности нуклеиновых кислот (SEQ ID NO: 62-66) для нацеливающего вектора MAID 1715 описаны в таблице 6.
Впоследствии получили ряд человеческих нацеливающих векторов, в которых использовалось то же мышиное плечо 5', которое содержало эндогенную последовательность TCRβ, окружающую мышиные гены трипсиногена, расположенные ближе к 5'-концу, из мышиного клона ВАС RP23-461h15 с чередующимися кассетами селекции. Конкретные конструкты описаны в патенте США №9,113,616 и показаны на ФИГ. 4В, а соединительные последовательности для каждой вставки включены в таблицу 6 и перечень последовательностей.
В конечном счете был создан человеческий небольшой локус TCRβ, содержащий всего 66 человеческих сегментов TCRPV (47 функциональных) и человеческих сегментов TCRβ D и J (MAID 1792), функционально связанных с мышиными константными генами и энхансерами TCRβ. Соединительные последовательности нуклеиновых кислот (SEQ ID NO: 69 и 70) для нацеливающего вектора MAID 1792 описаны в таблице 6.
Мышиный TCRBV31 расположен приблизительно на 9,4 т.п.н. ближе к 3'-концу от TCRBV2 (вторая последовательность константного участка TCRB) и находится в противоположной ориентации по отношению к другим сегментам TCRBV. Эквивалентный человеческий сегмент V представляет собой TCRBV30, который находится в аналогичном положении в человеческом локусе TCRB. Для гуманизации TCRBV31 мышиный клон ВАС, содержащий мышиный TCRBV31, модифицировали путем бактериальной гомологичной рекомбинации с получением LTVEC MAID 6192. Весь кодирующий регион, начинающийся со старт-кодона в экзоне 1, интрон, 3' UTR и рекомбинационные сигнальные последовательности (RSS) TCRBV31 замещали гомологичными человеческими последовательностями TCRBV30. На ФИГ. 4В показана кассета селекции, расположенная в интроне между экзоном 1 и экзоном 2 гена hTCRBV30.
Соединительные последовательности нуклеиновых кислот (SEQ ID NO: 71 и 72) для нацеливающего вектора MAID 6192 описаны в таблице 6. ДНК MAID 6192 электропорировали в ЭС-клетки MAID 1792, и клетки подвергали скринингу на потерю мышиного аллеля TCRB31 и приобретение человеческого аллеля TCRB30.
Аналогичная генно-инженерная стратегия используется для необязательной делеции оставшегося 5' мышиного сегмента V TCRβ.
На любом из вышеуказанных этапов кассеты селекции удаляли путем делеции рекомбиназой Cre или Flp.
Мышей, гомозиготных по гуманизированному вариабельному локусу TCRα, скрещивали с мышами, гомозиготными по гуманизированному вариабельному локусу TCRβ, с получением потомства, содержащего гуманизированные вариабельные локусы TCRα и TCRβ. Потомство разводили до достижения гомозиготности по гуманизированным локусам TCRα и TCRβ.
Подтверждали, что у мышей, содержащих гуманизированные вариабельные локусы TCRα и TCRβ, протекает нормальное развитие Т-клеток, и они содержат Т-клеточные рецепторы, которые экспрессируют вариабельные домены, полученные из множества вариабельных сегментов гена.
Пример 3. Гуманизация локусов Т-клеточного корецептора
Гуманизация локусов CD4 и CD8 (локусы как CD8 альфа, так и CD8 бета) подробно описана в публикации заявки на патент США №20140245466, полностью включенной в настоящий документ путем ссылки.
Пример 3.1. Гуманизация локуса CD4
В частности, мышиный локус CD4 гуманизировали за один этап путем конструирования уникального нацеливающего вектора из ДНК человеческой и мышиной бактериальных искусственных хромосом (ВАС) по технологии VELOCIGENE® (см., например, патент США №6,586,251 и публикацию Valenzuela et al. (2003), выше). Для получения нацеливающего вектора выполняли ряд бактериальных гомологичных рекомбинаций (BHR) с использованием ДНК искусственной бактериальной хромосомы (ВАС), а также другие этапы генной инженерии, как подробно описано в публикации заявки на патент США №20140245466.
Нацеливающий вектор, содержащий человеческий CD4, линеаризовывали с помощью NotI и электропорировали в мышиные ЭС-клетки F1H4. Нацеленные ЭС-клетки, несущие гуманизированный локус CD4, выявляли путем генотипирования с использованием анализа на модификацию аллеля (Valenzuela et al.), который обнаруживал присутствие неомициновой кассеты и человеческого гена CD4, а также одной копии мышиного гена CD4.
Конечный гуманизированный локус CD4, полученный в результате успешного встраивания гуманизированного нацеливающего вектора CD4 в ЭС-клетки, показан на ФИГ. 5А. Последовательность участка соединения человеческого интрона 3 с кассетой lox-neo (5'-конец кассеты) представлена в SEQ ID NO: 75, а последовательность участка соединения кассеты lox-neo (3' конец кассеты) с человеческим интроном 3 представлена в SEQ ID NO: 76; обе последовательности также перечислены в таблице 7. Полная последовательность нуклеиновых кислот гуманизированной части CD4, включая кассету pgk-neo, показанную на Фиг. 5А, представлена в SEQ ID NO: 77. Кассета pgk-neo охватывает остатки 307-2176 в SEQ ID NO: 77, два сайта 1ох расположены в пределах остатков 267-300 и 2182-2215, а человеческая последовательность охватывает остатки 1-234 и 2222-18263. Аминокислотная последовательность полностью гуманизированного белка CD4 представлена в SEQ ID NO: 78, причем человеческая последовательность охватывает аминокислоты 27-319 (представлены в SEQ ID NO: 79).


Человеческие последовательности приведены в скобках, а последовательность, содержащая сайт рестриктазы (PI-Sce I), указана жирным шрифтом. Последовательности кассеты селекции указаны курсивом.
Фланкированную сайтами loxP кассету резистентности к неомицину удаляли путем электропорации плазмиды, экспрессирующей рекомбиназу Cre, в ЭС-клетки, содержащие гуманизированный локус CD4.
Нацеленные ЭС-клетки, несущие гуманизированный локус CD4 без маркера резистентности, выявляли путем генотипирования, которое обнаруживало отсутствие неомициновой кассеты, присутствие одной копии человеческого гена CD4 и одной копии мышиного гена CD4.
Нацеленные ЭС-клетки, описанные выше, использовали в качестве донорских ЭС-клеток и внедряли в мышиный эмбрион на стадии 8 клеток по способу VELOCIMOUSE® (см., например, патент США №7,294,754 и публикацию Poueymirou et al. (2007), выше). Мышей VELOCDVIICE® (мыши поколения F0, полностью происходящие из донорской ЭС-клетки), независимо несущих химерный ген CD4, выявляли путем генотипирования с помощью анализа на модификацию аллеля (Valenzuela et al., выше), в котором обнаруживают присутствие уникальных последовательностей человеческого гена CD4. Экспрессию гуманизированных белков CD4 на поверхности Т-клеток обнаруживали с использованием антител против человеческого CD4. Мышей, гетерозиготных по гуманизированному белку CD4, описанных в настоящем документе, разводили до достижения гомозиготности.
Пример 3.2. Гуманизация локусов CD8
Гены CD8α и CD8β колокализуют в геноме, например, на мышиной хромосоме 6 они расположены на расстоянии приблизительно 37 т.п.н. друг от друга. В связи с плотным сцеплением выполняют последовательное нацеливание путем сначала внедрения одного гена, например, CD8β, с последующим внедрением второго гена, например, CD8α. Конкретные подробные этапы гуманизации описаны в публикации заявки на патент США №20140245466, включенной в настоящий документ путем ссылки.
Вкратце, мышиный локус CD8β гуманизировали за один этап путем конструирования уникального нацеливающего вектора из ДНК мышиной бактериальной искусственной хромосомы по технологии VELOCIGENE®. ДНК из ВАС RP23-431M6 модифицировали путем BHR с получением большого нацеливающего вектора (LTVEC) (MAID 1737), содержащего замещение мышиных экзонов 2-3, кодирующих эктодомен CD8 (от участка соединения 5' в интроне 1 до участка соединения 3' в интроне 3), гомологичными человеческими последовательностями (ФИГ. 5В). Кассету loxp-Ub-Hyg вставляли на участке соединения 3' в интроне 3. Нуклеотидные последовательности в различных участках соединения полученного вектора перечислены в таблице 8 и представлены в перечне последовательностей. Полная аминокислотная последовательность гуманизированного белка CD8β представлена в SEQ ID NO: 83; причем человеческие последовательности охватывают аминокислоты 15-165 (представлены в SEQ ID NO: 84).


Человеческие последовательности приведены в скобках, сайты lox указаны курсивом, а сайты рестриктаз, сайты множественного клонирования и полученные из вектора последовательности указаны жирным шрифтом.
Нацеливающий вектор электропорировали в мышиные ЭС-клетки F1H4. Нацеленные ЭС-клетки, несущие гуманизированный локус CD8β, выявляли путем генотипирования с использованием анализа на модификацию аллеля (Valenzuela et al.), который обнаруживал присутствие человеческого гена CD8β.
Мышиный локус CD8α гуманизировали за один этап путем конструирования уникального нацеливающего вектора из ДНК мышиной бактериальной искусственной хромосомы по технологии VELOCIGENE®. ДНК из ВАС RP23-431М6 модифицировали путем BHR с получением большого нацеливающего вектора (LTVEC) (MAID 1738), содержащего замещение мышиных экзонов 1-2, кодирующих эктодомен CD8α (от участка соединения 5' в кодоне 27 Ala в мышином экзоне 1 до участка соединения 3' в мышином интроне 2), гомологичными человеческими последовательностями (от участка соединения 5' в человеческом экзоне 2 до участка соединения 3' в интроне 3 (Фиг. 5А)). Это обеспечивает сохранение мышиной лидерной последовательности в начале экзона 1. Кассету lox2372-Ub-Neo вставляли на участке соединения 3' человеческой/мышиной последовательностей. Нуклеотидные последовательности в различных участках соединения полученного вектора перечислены в таблице 9 и представлены в перечне последовательностей. Полные аминокислотные последовательности гуманизированного полипептида CD8α представлены SEQ ID NO: 88, причем человеческая последовательность охватывает аминокислоты 28-179 (представлены в SEQ ID NO: 89).


Человеческие последовательности приведены в скобках, сайты lox указаны курсивом, а сайты рестриктаз, сайты множественного клонирования и полученные из вектора последовательности указаны жирным шрифтом.
Нацеливающий вектор, содержащий гуманизированный CD8α, описанный выше, электропорировали в мышиные ЭС-клетки, которые содержали гуманизированный локус CD8b, для получения модифицированных ЭС-клеток, которые содержат гуманизированные локусы CD8b и CD8a (Фиг. 5В). Нацеленные ЭС-клетки, несущие гуманизированные локусы CD8a и CD8b, выявляли путем генотипирования с использованием анализа на модификацию аллеля (Valenzuela et al.).
Нацеленные ЭС-клетки, описанные выше, использовали в качестве донорских ЭС-клеток и внедряли в мышиный эмбрион на стадии 8 клеток по способу VELOCIMOUSE® (см., например, патент США №7,294,754 и публикацию Poueymirou et al., выше). Мышей VELOCIMICE® (мыши поколения F0, полностью происходящие из донорской ЭС-клетки), независимо несущих химерный ген CD8b и химерный ген CD8a, выявляли путем генотипирования с помощью анализа на модификацию аллеля (Valenzuela etal, выше), в котором обнаруживают присутствие уникальных последовательностей человеческих генов CD8b и CD8a.
Кассеты селекции в локусах CD8α и CD8β можно удалить способами, известными специалистам в данной области. Мышей, гетерозиготных по гуманизированным локусам CD8α и CD8β, как описано в настоящем документе, разводили до достижения гомозиготности. Экспрессию гуманизированных белков CD8α и CD8β на поверхности Т-клеток обнаруживали с использованием антител против человеческого CD8.
Пример 4. Создание мышей, содержащих гуманизированные компоненты клеточной иммунной системы Для создания мышей, содержащих гуманизированные компоненты клеточной иммунной системы, мыши, гомозиготные по гуманизации различных компонентов, например МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также Р2М, могут быть скрещены между собой в любой комбинации с получением мышей, у которых различные компоненты Т-клеточного иммунного ответа являются гуманизированными. Например, мышь, содержащая гуманизированный МНС I, может быть скрещена с мышью, содержащей гуманизированный р2М, для получения мыши, экспрессирующей гуманизированный МНС 1/р2М. Мышей, гомозиготных по гуманизации различных компонентов, например МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также Р2М, скрещивают друг с другом способами, известными в данной области, для получения мыши, содержащей все девять гуманизации (мыши «ТМ I/II В С4/8»). Мышей разводят до достижения гомозиготности способами, известными в данной области. Альтернативно нацеливающие векторы, содержащие каждый гуманизированный ген, могут быть внедрены посредством последовательного нацеливания в ту же ЭС-клетку для получения ЭС-клетки, содержащей все девять гуманизации, и полученную ЭС-клетку внедряют в мышиный эмбрион на стадии 8 клеток по способу VELOCBVIOUSE®, описанному в примерах 1-3 выше.
Пример 5. Определение характеристик мышей, содержащих гуманизированные компоненты клеточной иммунной системы
Определяли характеристики мышей, гомозиготных по гуманизированным МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β и по гуманизированному Р2М. В частности, собирали селезенку и тимусы мышей и получали суспензии одиночных клеток. Суспензии центрифугировали при 1200 об./мин в течение 5 мин при 4°С для осаждения клеток, и клетки из каждой ткани лизировали 4 мл лизирующего буферного раствора ACK (GIBCO) для лизирования эритроцитов. Клетки фильтровали через клеточный фильтр, центрифугировали до осаждения, ресуспендировали в среде и подсчитывали.
Экспрессию на клеточной поверхности CD19, CD3, CD4 и CD8α, показанную на ФИГ. 6А-С и ФИГ. 9А-С, анализировали с помощью цитометрии посредством сортировки клеток с активацией флуоресценции (FACS) с использованием антител, конъюгированных с флуорохромом: против мышиного CD3 (17А2, BD), против мышиного CD19 (1D3, BD), против мышиного F4/80 (ВМ8, Biolegend), против мышиного CD8α (53-6.7, BD), против мышиного CD4 (RM4-5, eBioscience), против человеческого CD8α (SK1, BD) и против человеческого CD4 (RPA-Т4, BD). Экспрессию на клеточной поверхности мышиного H2Db, человеческих молекул HLA (HLA-A2, B2m и HLA-DR) и мышиных молекул МНС IAIE, показанных на ФИГ. 7A-F и 10A-F, анализировали с помощью FACS с использованием антител, конъюгированных с флуорохромом: против мышиного CD 19 (6D5, Biolegend), против мышиного F4/80 (ВМ8, Biolegend), против мышиного H2Db (KH95, Biolegend), против человеческого HLA-A2 (ВВ7.2, BD), против человеческого HLA-DR (G46-6, BD), против человеческого В2-микроглобулина (2М2, Biolegend) и против мышиного IAIE (М5/114.15.2, eBioscience). Экспрессию на клеточной поверхности мышиного и человеческого CD4 и CD8, изображенных на ФИГ. 7G и ФИГ. 10G, анализировали с помощью FACS с использованием антител, конъюгированных с флуорохромом: против мышиного CD3 (17А2, Biolegend), против мышиного CD4 (GK1.5, eBiosciences), против мышиного CD8α (53-6.7, BD 2), против мышиного CD80 (Н35-17.2, eBioscience), против человеческого CD4 (OKT4, eBioscience), против человеческого CD8α (RPA-T8, BD 6), против человеческого CD8β (2ST8.5H7, BD). Экспрессию на клеточной поверхности FoxP3 и CD25, показанных на ФИГ. 8 или ФИГ. 11, анализировали с помощью FACS с использованием антител против FoxP3 (FJK-16s, eBioscience) и против CD25 (РС61, Biolegend). Экспрессию на клеточной поверхности CD44 и CD62L, показанных на ФИГ. 9D-9E, анализировали с использованием антител против CD44 (IM7, BD) и против CD62L (MEL-14, Biolegend).
Всю проточную цитометрию выполняли с использованием BD Fortessa. Данные анализировали с использованием FlowJo.
Экспрессия в тимусе показана на ФИГ. 6А-С, 7A-G и 8. Абсолютные количества тимоцитов и клеток CD3+ и общее развитие Т-клеток тимуса были сопоставимы у контрольных мышей и гуманизированных мышей ТМ I/II В С4/8 (данные не показаны). На ФИГ. 6А показано, что пропорция В-клеток и Т-клеток в тимусах мышей, имеющих гуманизированную клеточную иммунную систему (ТМ I/II В С4/8), аналогична пропорции, обнаруженной у контрольных мышей. Частота появления и количество клеток F4/80 в тимусах мышей ТМ I/II В С4/8 были сопоставимы с таковыми у контрольных мышей (ФИГ. 6В, данные не показаны). Кроме того, гуманизированные CD4 и CD8 экспрессируются на клетках тимуса мыши, гуманизированной по всем девяти генам клеточного иммунитета (ТМ I/IIB С4/8), аналогично экспрессии мышиных CD4 и CD8 у негуманизированных контрольных мышей (ФИГ. 6С). Гуманизированный β2М экспрессируется на поверхности В-клеток и макрофагов у гуманизированных мышей ТМ I/II В С4/8, тогда как в В-клетках и макрофагах контрольных мышей его экспрессия отсутствует (ФИГ. 7А и 7В). Аналогичным образом, гуманизированные МНС I и II присутствуют на поверхности как В-клеток, так и макрофагов гуманизированных мышей ТМ I/II В С4/8 (ФИГ. 7С и 7D), а мышиные молекулы МНС класса I и II не были обнаружены (ФИГ. 7Е и 7F). Гуманизированные CD4, CD8α и CD8β экспрессируются на поверхности клеток тимуса CD3+, полученных из гуманизированных мышей ТМ I/II В С4/8, но отсутствуют в клетках тимуса CD3+ контрольных мышей (ФИГ. 7G). Гуманизированные мыши ТМ I/II В CD4/8 экспрессируют Т-клетки (Treg) (ФИГ. 8), NK-клетки (CD335+ CD3-) и моноциты (CD11b+) (данные не показаны).
Экспрессия в селезенке показана на ФИГ. 9A-D и 10A-10G. Селезенки мышей, гуманизированных по компонентам клеточной иммунной системы (ТМ I/II В CD4/8), содержали сопоставимые абсолютные количества клеток CD3+ и практически нормальную пропорцию В- и Т-клеток (ФИГ. 9А и данные не показаны). Частота появления и количество клеток F4/80 в селезенках мышей ТМ I/II В С4/8 были сопоставимы с таковыми у контрольных мышей (ФИГ. 9В и данные не показаны). Мыши, гуманизированные по компонентам клеточной иммунной системы (ТМ I/II В CD4/8), экспрессировали гуманизированные CD4 и CD8α на клетках CD3+ селезенки (ФИГ. 9С). Гуманизированные мыши ТМ I/II/ В CD4/8 содержали эффекторные Т-клетки памяти CD4+ и CD8+ (CD44+CD62L-) и центральные Т-клетки памяти CD8+ (CD44+ CD62L+) (ФИГ. 9D и 9Е).
Как показано на ФИГ. 10А и 10В, гуманизированный β2М экспрессируется на поверхности В-клеток и макрофагов в селезенке гуманизированных мышей ТМ I/II В С4/В, тогда как на В-клетках и макрофагах в селезенке контрольных мышей его экспрессия и экспрессия мышиных молекул МНС отсутствует. Аналогичным образом, гуманизированные МНС I и II присутствуют на поверхности как В-клеток, так и макрофагов в селезенке гуманизированных мышей ТМ I/II В С4/В (ФИГ. 10С и 10D), тогда как мышиные молекулы МНС класса I и II не были обнаружены (ФИГ. 10Е и 10F). Гуманизированные CD4, CD8α и CD8β экспрессируются на поверхности клеток селезенки CD3+, полученных из гуманизированных мышей ТМ I/II В С4/8, но отсутствуют в клетках селезенки CD3+ контрольных мышей (ФИГ. 10G). Мыши ТМ I/II В С4/8 имеют практически нормальную экспрессию регуляторных Т-клеток селезенки по сравнению с контрольными мышами (ФИГ. 11) и экспрессируют селезеночные NK-клетки (CD335+CD3-) и моноциты (CD11b+).
Пример 6. Оценка представления и активации Т-клеток человеческим пептидом
Для определения того, формируют ли мыши, содержащие гуманизированные компоненты клеточной иммунной системы, гуманизированные Т-клеточные иммунные ответы, протестировали способность спленоцитов от мышей, гуманизированных по компонентам клеточной иммунной системы (ТМ I/II В CD4/8), представлять MAGE-А3, пептид, представляемый исключительно человеческим HLA-A2, и реагировать на него.
MAGE-A3, пептид, представляемый исключительно человеческим HLA-A2, синтезируют (Celtek Biosciences), разводят в фосфатно-солевом буфере (PBS) и смешивают в равных объемах с полным адъювантом Фрейнда (CFA; Chondrex, Inc.) таким образом, что 200 мкг MAGE-A3 содержатся в 200 мкл эмульсии. 50 мкл эмульсии вводят в 4 зоны на каждом животном. Две зоны находятся в заднем паху и 2 зоны находятся рядом с каждым плечом мышей, гомозиготных по гуманизированным МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2М (ТМ I/II В CD4/8), или контрольных мышей, которые экспрессируют эндогенные МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2М.
Получают и разделяют суспензии селезенок от иммунизированных мышей. Эритроциты лизируют в лизирующем буферном растворе ACK (Life Technologies), а спленоциты суспендируют в полной среде RPMI. 2 × 105 выделенных спленоцитов при отсутствии или в присутствии 10 мкг/мл или 1 мкг/мл разведенного пептида MAGE-A3 тестируют в каждой лунке планшетов PVDF (Millipore), покрытых 5 мкг/мл мышиного захватного антитела ИФН-γ (BD Biosciences) в анализе ELISPOT. После 16-20 часов инкубации с пептидом планшеты промывали и инкубировали с биотинилированным детекторным антителом (BD Biosciences), промывали, обрабатывали конъюгатом стрептавидин-пероксидаза хрена (HRP) (MabTech), промывали, проявляли субстратом ТМВ (Mabtech) и подсчитывали с помощью считывателя AID Elispot.
Несмотря на то что для одного генотипа показана только одна мышь, исследовали несколько мышей каждого генотипа, и все образцы анализировали в трех повторностях с указанием стандартного отклонения в виде планок погрешности. Как показано на ФИГ. 12, только образцы от мышей, гомозиготных по каждому из гуманизированных МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2М (ТМ I/II В CD4/8), реагировали секрецией ИФН-γ на обработку HLA-А2-специфичным пептидом MAGE-A3, что указывает на активацию Т-клеток от этих мышей после представления MAGE-A3 гуманизированным HLA-A2.
Пример 7. Оценка Т-клеточной функции с использованием модели инфекции LCMV
Чтобы определить, могут ли мыши, содержащие гуманизированные компоненты клеточной иммунной системы, формировать нормальный ответ на инфекцию, исследовали способность гуманизированных мышей уничтожать вирус лимфоцитарного хориоменингита (LCMV). LCMV представляет собой вирус, воздействующий на мышей, причем развитие инфекции зависит от вирусного штамма. Воздействие штамма Армстронга приводит к острой инфекции, при которой мыши быстро формируют Т-клеточный ответ на вирус и уничтожают инфекцию приблизительно в течение недели. С другой стороны, вирус Clone 13 не может быть уничтожен, Т-клетки становятся «истощенными» (экспрессируя маркеры, ассоциированные с Т-клеточным истощением, например, PD1, Lag3, Tim3) и развивается хроническая инфекция. Показано, что инфицирование мышей, истощенных по CD8 или дефицитных по МНС класса I, штаммом Армстронга приводит к сохранению высоких титров вируса (J. Virol. 68:8056-63 (1994)). Поскольку вирусная инфекция зависит от активности Т-клеток, LCMV представляет собой идеальную модель для исследования Т-клеточной функции.
Для определения того, обладают ли мыши, содержащие гуманизированные компоненты клеточной иммунной системы, например МНС I, МНС II α и β, TCRα и β, CD4, CD8α и β, а также β2М, нормальной Т-клеточной функцией, контрольных и гуманизированных (ТМ I/II В С4/8) мышей инфицировали 2 × 105 БОЕ вируса штамма Армстронга интраперитонеально (и/п) в день 0. В дни 3, 6, 9 и 12 собирали органы и измеряли титры вируса. Как показано на ФИГ. 13А, как контрольные, так и гуманизированные мыши обладали способностью к уничтожению инфекции Армстронга.
Как контрольных, так и гуманизированных мышей также инфицировали 4,5 × 105 БОЕ вируса Clone 13 внутривенно (в/в) в день 0, а в день 21 собирали органы и измеряли титры вируса. Как показано на ФИГ. 13В, хроническая инфекция LCMV могла развиваться в обеих линиях мышей. Также измеряли способность гуманизированных мышей экспрессировать маркеры PD1, Lag3 и Tim3 Т-клеточного истощения. Кровь собирали у неинфицированных мышей и инфицированных гуманизированных мышей через 3 недели после инфицирования и метили, используя проточную цитометрию, антителами против PD1, конъюгированными с РЕ-Су7 (BIOLEGEND), антителами Lag3, конъюгированными с PerCpCy5.5 (BIOLEGEND), и антителами Tim3, конъюгированными с РЕ (R&D Systems). Данные на ФИГ. 13С представляют собой результаты количественного определения меченых клеток, положительных в отношении указанных рецепторов. Как гуманизированные (ТМ I/II В С4/8), так и контрольные мыши В6 экспрессировали все три маркера Т-клеточного истощения через 3 недели после инфицирования хроническим штаммом Clone 13 LCMV.
Для оценки появления Т-клеток памяти у мышей, гуманизированных по компонентам клеточной иммунной системы, 5 контрольных и 4 гуманизированных мыши инфицировали 2 × 105 БОЕ штамма Армстронга, а в день 17 суперинфицировали 4,5 × 105 БОЕ штамма Clone 13 (по 2 каждых из гуманизированных и контрольных мышей ложно инфицировали в качестве дополнительного контроля). В день 31 после первичного инфицирования собирали органы и анализировали титры вируса. Как показано на ФИГ. 14, у 5/5 контрольных мышей и 3/4 гуманизированных мышей, которые перенесли острую инфекцию LCMV, впоследствии не развивалась хроническая инфекция, что указывает на появление у этих животных интактных Т-клеток памяти.
Для анализа природы клеточных ответов контрольных и гуманизированных мышей инфицировали в день 0 2 × 105 БОЕ вируса штамма Армстронга. В день 10 (ФИГ. 15А-В) или в указанные временные отметки после инфицирования (ФИГ. 15C-D) специфичность клеточного ответа анализировали с использованием трех рестриктированных по HLA-A2 пептидов, которые, как известно, активируют человеческие Т-клетки CD8+ (GPC10-18, N69-77 или Z49-58), см. публикацию Botten et al. (2007) J. Virol. 81:2307-17, или gp33, представляющий собой иммунодоминантный пептид LCMV, распознаваемый мышами на основе H-2Db. В частности, Т-клетки CD8+ выделяли из собранных селезенок и активировали пептидами. Клетки CD8+, продуцирующие интерферон-у (ИФНγ), измеряли посредством ELISpot (ФИГ. 15А-В) или посредством мечения для определения внутриклеточного ИФНγ (ФИГ. 15C-D).
Т-клетки CD8+, выделенные из контрольных животных, специфически активируются пептидом gp33 (ФИГ. 15А), а Т-клетки CD8+, выделенные из гуманизированных животных, активируются рестриктированными по HLA-A2 пептидами (ФИГ. 15В). Временная динамика активации Т-клеток CD8+, отслеживаемая по их способности экспрессировать ИФНγ после стимуляции пептидами, показывает, что как у контрольных, так и у гуманизированных мышей Т-клетки CD8+ растут в течение первых двух недель после инфицирования и перестают обнаруживаться после уничтожения вируса (ФИГ. 15C-D). Хотя реакция пептида gp33 оказалась сильнее у контрольных животных, следует отметить, что gp33 является известным иммунодоминантный эпитопом LCMV, тогда как иммунодоминантный рестриктированный по HLA-A2 эпитоп LCMV не выявлен. В заключение: животные, содержащие гуманизированную или по существу гуманизированную Т-клеточную иммунную систему, способны обрабатывать экспрессируемый LCMV белок, представляя его на гуманизированных молекулах МНС и активируя Т-клетки посредством гуманизированного Т-клеточного рецептора.
Эквиваленты
Специалисты в данной области смогут определить или с помощью лишь стандартных экспериментов смогут установить множество эквивалентов конкретных вариантов осуществления изобретения, описанных в настоящем документе. Подразумевается, что такие эквиваленты охвачены следующей формулой изобретения.
Все содержание всех непатентных документов, патентных заявок и патентов, упоминаемых в настоящей заявке, полностью включено в настоящий документ путем ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Regeneron Pharmaceuticals, Inc.
<120> ОПОСРЕДОВАННЫЕ ГУМАНИЗИРОВАННЫМИ Т-КЛЕТКАМИ ИММУННЫЕ ОТВЕТЫ У НЕ ОТНОСЯЩИХСЯ К ЧЕЛОВЕКУ ЖИВОТНЫХ
<130> 10145WO01
<150> 62/143,687
<151> 2015-04-06
<150> 62/158,804
<151> 2015-05-08
<150> 62/186,935
<151> 2015-06-30
<160> 91
<170> PatentIn, версия 3.5
<210> 1
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 1
cagaacgcca ggctgtaac 19
<210> 2
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 2
ggagagcagg gtcagtcaac 20
<210> 3
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 3
caccgccact cacagctcct taca 24
<210> 4
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 4
gtgggcacca tcttcatcat tc 22
<210> 5
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 5
cttcctttcc agggtgtgac tc 22
<210> 6
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 6
aggcctgcga tcaggtggca cct 23
<210> 7
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 7
ggtggagagg ctattcggc 19
<210> 8
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 8
gaacacggcg gcatcag 17
<210> 9
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 9
tgggcacaac agacaatcgg ctg 23
<210> 10
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 10
tttgtaaaca aagtctaccc agagacagat gacagacttc agctccaatg ctgattggtt 60
cctcacttgg gaccaaccct accggtataa cttcgtataa ggtatcctat acgaagttat 120
atgcatggcc tccgcgccgg 140
<210> 11
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 11
cgacctgcag ccggcgcgcc ataacttcgt ataaggtatc ctatacgaag ttatctcgag 60
cacaggcatt tgggtgggca gggatggacg gtgactggga caatcgggat ggaagagcat 120
agaatgggag ttagggaaga 140
<210> 12
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 12
cgaggagccc cggtaca 17
<210> 13
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 13
aagcgcacga actccttgtt 20
<210> 14
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 14
ctctgtcggc tatgtgg 17
<210> 15
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 15
ggactcccag aatctcctga ga 22
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 16
gagtcatgaa ccatcactgt gaaga 25
<210> 17
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 17
tggtgggttg ctggaa 16
<210> 18
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 18
ctgtttcttc cctaactccc attctatgct cttccatccc gaccgcggcc caatctctct 60
ccactacttc ctgcctacat gtatgtaggt 90
<210> 19
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 19
caaggtttcc tcctatgatg cttgtgtgaa actcggggcc ggccagcatt taacagtaca 60
gggatgggag cacagctcac 80
<210> 20
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 20
gaaagcagtc ttcccagcct tcacactcag aggtacaaat ccccattttc atattagcga 60
ttttaattta ttctagcctc 80
<210> 21
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 21
tcttccctaa ctcccattct atgctcttcc atcccgaccg cggcccaatc tctctccact 60
acttcctgcc tacatgtatg 80
<210> 22
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 22
gagttcctcc atcacttcac tgggtagcac agctgtaact gtccagcctg tcctgggctg 60
caggtggtgg gcgttgcggg tggggccggt taaggttcca 100
<210> 23
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 23
tcccacatcc tattttaatt tgctccatgt tctcatctcc atcagcacag ctcgagataa 60
cttcgtataa tgtatgctat acgaagttat atgcatggcc 100
<210> 24
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 24
atacgaagtt atgctagtaa ctataacggt cctaaggtag cgagtggctt acaggtaggt 60
gcgtgaagct tctacaagca cagttgcccc ctgggaagca 100
<210> 25
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 25
tgcggccgat cttagcc 17
<210> 26
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 26
ttgaccgatt ccttgcgg 18
<210> 27
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 27
acgagcgggt tcggcccatt c 21
<210> 28
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 28
ccccacagca cgtttcct 18
<210> 29
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 29
cgtcccattg aagaaatgac act 23
<210> 30
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 30
tggcagccta agagg 15
<210> 31
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 31
ccccacagca cgtttcct 18
<210> 32
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 32
acccgctccg tcccatt 17
<210> 33
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 33
agcctaagag ggagtgtc 18
<210> 34
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 34
agaccctggt gatgctggaa 20
<210> 35
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 35
cgcttgggtg ctccactt 18
<210> 36
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 36
tcgaagtgga gaggttta 18
<210> 37
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 37
tggaatggag tgagcagctt t 21
<210> 38
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 38
gcacggtccc cttcttagtg 20
<210> 39
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 39
tgacttccta aatttctc 18
<210> 40
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 40
ctggcggctt gaagaatttg g 21
<210> 41
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 41
catgatttcc aggttggctt tgtc 24
<210> 42
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 42
cgatttgcca gctttgaggc tcaagg 26
<210> 43
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 43
cctcacttgg gaccaaccct a 21
<210> 44
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 44
ttgtcccagt caccgtccat 20
<210> 45
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 45
tgcatctcga gcacaggcat ttgg 24
<210> 46
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 46
atggagtagt cagaacacac tcttcagaag ggactcctga tttcaaaggg ggtaccgggc 60
cccccctcga ggtcgacata acttcgtata gcatacatta 100
<210> 47
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 47
ggccatgcat ataacttcgt atagcataca ttatacgaag ttataccggt gcgatcgcgc 60
gcttccctct tctaaccact aattcaaaaa ggattgtaag taatgttt 108
<210> 48
<211> 145
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 48
agacagaccc ctaaacacct ccaaattaaa agcggcaaag agataaggtt ggagctccac 60
cgcggtggcg gccgccaccg cggtggagct cgaggtttcc ggtacttaac aacagagcac 120
agatttagtg gtgagggact ctctc 145
<210> 49
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 49
atggagtagt cagaacacac tcttcagaag ggactcctga tttcaaaggg ggtaccgggc 60
cccccctcga ggtcgacata acttcgtata gcatacatta 100
<210> 50
<211> 109
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 50
ggccatgcat ataacttcgt atagcataca ttatacgaag ttataccggt gcgatcgctc 60
aagcatgcaa gggtaacata tgttatgaga ttatattttc tttatctca 109
<210> 51
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 51
atggagtagt cagaacacac tcttcagaag ggactcctga tttcaaaggg gggtaccggg 60
ccccccctcg agaagttcct attccgaagt tcctattctc 100
<210> 52
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 52
gttcctattc cgaagttcct attctctaga aagtatagga acttcctagg gcgatcgctc 60
ctctccaggc tcgaattagt attacagttg aggcacgttg tcctcccg 108
<210> 53
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 53
atggagtagt cagaacacac tcttcagaag ggactcctga tttcaaaggg ggtaccgggc 60
cccccctcga ggtcgacata acttcgtata gcatacatta 100
<210> 54
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 54
ggccatgcat ataacttcgt atagcataca ttatacgaag ttataccggt gcgatcgccg 60
cctccatttc cttcatagga aacatgaagt gaatggggct gtgtgtgt 108
<210> 55
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 55
atggagtagt cagaacacac tcttcagaag ggactcctga tttcaaaggg ggtaccgggc 60
cccccctcga ggtcgacata acttcgtata gcatacatta 100
<210> 56
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 56
ggccatgcat ataacttcgt atagcataca ttatacgaag ttataccggt gcgatcgctg 60
ggagcacgtt ccattattat aacaactttc tgaacacaag agggcagt 108
<210> 57
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 57
atggagtagt cagaacacac tcttcagaag ggactcctga tttcaaaggg ggtaccgggc 60
cccccctcga ggtcgacata acttcgtata gcatacatta 100
<210> 58
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 58
ggccatgcat ataacttcgt atagcataca ttatacgaag ttataccggt gcgatcgctt 60
taaggtgagg aggcaggcaa taccccctct ccaccgcatt ctcaatcc 108
<210> 59
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 59
gggggggtgg ggtggaggag gagggtacag catctcctct ccttcctctc tggtaccgaa 60
gttcctattc cgaagttcct attctctaga aagtatagga 100
<210> 60
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 60
gaagttccta ttctctagaa agtataggaa cttcctaggg tttcaccggt gcgatcgcgt 60
gaatatacta aaaaccactt aattatatat ttgaaagggt ggatgtta 108
<210> 61
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 61
ctctctccta cccagctcct ctcacacgag cctgaaggcc ctgccaaggt ggcgcgcctt 60
tcaaattgtt gttgagttca aagtgggcaa cagaaaaggg ggtgtgag 108
<210> 62
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 62
aataaatagt aaatttctgt agaatcataa tgaggtctag acccccgggc tcgataacta 60
taacggtcct aaggtagcga aatggcgcgt aatcaagccc agctcttcat gctgcatttt 120
tatcttcttt 130
<210> 63
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 63
ttgactcggg ggtgcctggg tttgactgca atgatcagtt gctgggaagg accggtataa 60
cttcgtataa tgtatgctat acgaagttat atgcatggcc 100
<210> 64
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 64
ccggcgcgcc ataacttcgt ataatgtatg ctatacgaag ttatgtcgac ataaggtaag 60
acagagtcgt cccttcccat ctggaaccct ctacctttct 100
<210> 65
<211> 107
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 65
gttgatgaat cataaaagaa gagatattca agaaaaggat ggccacactg cggccgcaga 60
ggtattcaag gaaaatgcag actcttcacg taagagggat gaggggc 107
<210> 66
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 66
tccccggagt cggagggtgg accggagctg gaggagctgc cgcggtggcg gccgatgcca 60
tttcattacc tctttctccg cacccgacat agataaagct 100
<210> 67
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 67
gggggggtgg ggtggaggag gagggtacag catctcctct ccttcctctc tggtaccgaa 60
gttcctattc cgaagttcct attctctaga aagtatagga 100
<210> 68
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 68
gaagttccta ttctctagaa agtataggaa cttcctaggg tttcaccggt gcgatcgcga 60
agcaattaac tgcccctggt ccagttgcct cctctgataa tgcattgt 108
<210> 69
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 69
gggggggtgg ggtggaggag gagggtacag catctcctct ccttcctctc tggtaccgaa 60
gttcctattc cgaagttcct attctctaga aagtatagga 100
<210> 70
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 70
gaagttccta ttctctagaa agtataggaa cttcctaggg tttcaccggt gcgatcgcgt 60
tatctagtag acttaattaa ggatcgatcc ggcgcgccaa tagtcatg 108
<210> 71
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 71
gttttccaga cttcaacttg actatcagcc agaaattcag tggcaaaccc ccacccagtc 60
cctaagtgaa ggcccctggg gagtatggtt agggctcagg 100
<210> 72
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 72
cacccaccaa agaaagtgcc caggagaagg gcaaggagag agcagagcat agttcaagat 60
ggtctttgtc taggcttgtc tactctgcac ttgtacttcc 100
<210> 73
<211> 202
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 73
aggggaaacc cgcaaaggat gggacatagg gagacagctg ttaacatctg aaacatgacc 60
ttcttttctg tgcagcacaa ctcctagctg tcactcaagg gaagaaagtg gtgctgggca 120
aaaaagggga tacagtggaa ctgacctgta cagcttccca gaagaagagc atacaattcc 180
actggaaaaa ctccaaccag at 202
<210> 74
<211> 240
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 74
ctggtcacct ggatgaagtg agggagggcc ctctgggttt ggggctggtt ttgaactgag 60
acatccatga gccagcctgg ggctggcttc actgaagatc atctatgtcg ggtgcggaga 120
aagaggtaat gaaatggcac atgctatgta caaactctat tgctgagcag cacccagtcc 180
tgagctggct ctgaattgag ggtgaaattc acacattctc ccccaacatc tataatctgg 240
<210> 75
<211> 151
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 75
tatggagtga aagcctttgg tgtctgagat ctggtcttag ttaaactctg ggatcggcgc 60
gccgaattcc tgcagcccgg gctcgagata acttcgtata atgtatgcta tacgaagtta 120
tatgcatccg ggtaggggag gcgcttttcc c 151
<210> 76
<211> 151
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 76
agtattgttt tgccaagttc taattccatc agacctcgac ctgcagccct agataacttc 60
gtataatgta tgctatacga agttatccta ggccagaggg cttgggttga cagaaactca 120
gtggcattct tatccagagt ttctctacac c 151
<210> 77
<211> 18263
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<220>
<221> прочие приложения
<222> (2957)..(2957)
<223> n = A,T,C или G
<220>
<221> прочие приложения
<222> (3193)..(3193)
<223> n = A,T,C или G
<400> 77
aagaaagtgg tgctgggcaa aaaaggggat acagtggaac tgacctgtac agcttcccag 60
aagaagagca tacaattcca ctggaaaaac tccaaccaga taaagattct gggaaatcag 120
ggctccttct taactaaagg tagggttgcc tggctcccca tccagggagg aaaacacact 180
atggagtgaa agcctttggt gtctgagatc tggtcttagt taaactctgg gatcggcgcg 240
ccgaattcct gcagcccggg ctcgagataa cttcgtataa tgtatgctat acgaagttat 300
atgcatccgg gtaggggagg cgcttttccc aaggcagtct ggagcatgcg ctttagcagc 360
cccgctgggc acttggcgct acacaagtgg cctctggcct cgcacacatt ccacatccac 420
cggtaggcgc caaccggctc cgttctttgg tggccccttc gcgccacctt ctactcctcc 480
cctagtcagg aagttccccc ccgccccgca gctcgcgtcg tgcaggacgt gacaaatgga 540
agtagcacgt ctcactagtc tcgtgcagat ggacagcacc gctgagcaat ggaagcgggt 600
aggcctttgg ggcagcggcc aatagcagct ttgctccttc gctttctggg ctcagaggct 660
gggaaggggt gggtccgggg gcgggctcag gggcgggctc aggggcgggg cgggcgcccg 720
aaggtcctcc ggaggcccgg cattctgcac gcttcaaaag cgcacgtctg ccgcgctgtt 780
ctcctcttcc tcatctccgg gcctttcgac ctgcagccaa ttgttgacaa ttaatcatcg 840
gcatagtata tcggcatagt ataatacgac aaggtgagga actaaaccat gggatcggcc 900
attgaacaag atggattgca cgcaggttct ccggccgctt gggtggagag gctattcggc 960
tatgactggg cacaacagac aatcggctgc tctgatgccg ccgtgttccg gctgtcagcg 1020
caggggcgcc cggttctttt tgtcaagacc gacctgtccg gtgccctgaa tgaactgcag 1080
gacgaggcag cgcggctatc gtggctggcc acgacgggcg ttccttgcgc agctgtgctc 1140
gacgttgtca ctgaagcggg aagggactgg ctgctattgg gcgaagtgcc ggggcaggat 1200
ctcctgtcat ctcaccttgc tcctgccgag aaagtatcca tcatggctga tgcaatgcgg 1260
cggctgcata cgcttgatcc ggctacctgc ccattcgacc accaagcgaa acatcgcatc 1320
gagcgagcac gtactcggat ggaagccggt cttgtcgatc aggatgatct ggacgaagag 1380
catcaggggc tcgcgccagc cgaactgttc gccaggctca aggcgcgcat gcccgacggc 1440
gatgatctcg tcgtgaccca tggcgatgcc tgcttgccga atatcatggt ggaaaatggc 1500
cgcttttctg gattcatcga ctgtggccgg ctgggtgtgg cggaccgcta tcaggacata 1560
gcgttggcta cccgtgatat tgctgaagag cttggcggcg aatgggctga ccgcttcctc 1620
gtgctttacg gtatcgccgc tcccgattcg cagcgcatcg ccttctatcg ccttcttgac 1680
gagttcttct gaggggatcc gctgtaagtc tgcagaaatt gatgatctat taaacaataa 1740
agatgtccac taaaatggaa gtttttcctg tcatactttg ttaagaaggg tgagaacaga 1800
gtacctacat tttgaatgga aggattggag ctacgggggt gggggtgggg tgggattaga 1860
taaatgcctg ctctttactg aaggctcttt actattgctt tatgataatg tttcatagtt 1920
ggatatcata atttaaacaa gcaaaaccaa attaagggcc agctcattcc tcccactcat 1980
gatctataga tctatagatc tctcgtggga tcattgtttt tctcttgatt cccactttgt 2040
ggttctaagt actgtggttt ccaaatgtgt cagtttcata gcctgaagaa cgagatcagc 2100
agcctctgtt ccacatacac ttcattctca gtattgtttt gccaagttct aattccatca 2160
gacctcgacc tgcagcccta gataacttcg tataatgtat gctatacgaa gttatcctag 2220
gccagagggc ttgggttgac agaaactcag tggcattctt atccagagtt tctctacacc 2280
aactgctggt ggcccaggga aaggtggtat gtgaatttca atattttaat atttaatatt 2340
catgaactta ttttagtgag ttttagaaca atcactatca cttaaaaccc gtgatttctt 2400
gagtattgtt gctacagacc tatgtagata atactttgca cagtgactca tatgtataat 2460
cctagcactg tgggaggctg aggccggagg attgcttgag tccaggagtt caagaccagc 2520
ctgaacaaca tagtgagact ctgtctctat gaaaaaaaat atatatatat tttttttgga 2580
gacaaggtct agttctatca cccaggctcc agtgcagtgg tgtgatctcg gctcactgca 2640
atctccacct cccaggctca agtcatcatc ccacctcagc ctcccaagta gctgggacta 2700
caggcatgca ccaccatgcc aggctaattt ttgtattttt tatagagaca gggtttcacc 2760
atgttggcca ggctggtctc gaactcatga gctcaagtga tccactcacc ttggcctctc 2820
agagtgctgg aattacaggt gtgtgtcact atgcctagcc aaaaaaaatt tttttaatta 2880
aaaaaaaaaa ggccggctgt agtggctcac acctgtaatc cagaactttg ggagtttgag 2940
gtgggcagat caccggnggt caggagttca agaccagtct ggccaacatg gtgaaacccg 3000
gtctctacta aaaatacaaa aattagccag gtgtgggggt gcagtcctgt acttccagct 3060
actcaggagg ctgaggcagg agactcgctt gaacctggga ggcaaaggct gcagtgagct 3120
gagattgcac cactgcactc cagcctgggt gacagagcaa gacttcatct caaaaaaaaa 3180
aaaaaagctg canatttatt attattatta ttagtttatt tatttatttt tttgagacag 3240
agtctcgttc tgtcgcccag gctggagtgc ggtggcgtga tcttggctca ttgcaacctc 3300
cacctcccgg gttcaagtga ttctcctgcc tcagcctccc gagtagctgg gactacaggc 3360
gtatgccacc atgcctggct aattttttgt acttttagta gagacagagt ttcacggtgt 3420
tagccaggct ggtcttgatc tcctgacctc gtgatttacc ctccttggcc tcccaaagtg 3480
ctgggattac aggcgtgagt cactgtgccc ggcccagaat catttttttc tacttttttt 3540
tttttgaggc aaactctcga tctgttgccc aggctggagt gcagtgggca tgatcttggc 3600
tcactgcaag ctctgcctcc caggttcaag caattctcct gcctcagcct cctgagtagc 3660
tgggactaca ggcgtgtgcc accatgcccg gctaatttgc gtatttttag tagagaccgg 3720
ttttcatcat attggccagg ctggtcttga actcctgacc tcaagtgatt ctcccacctt 3780
agcctcccaa agtgctggga ttacaggcat gagctactgc acttggcctt ttctcctggt 3840
tttaaaacta ttatatgctc attacaaaat atttggtcaa tgaagaaaag aatatggaag 3900
aaaatcaaat gcatgcatac ttctatcact cagagatatc ctctgctaac attttgattg 3960
attttcttcc aatctttttt tttttttttc tttttgagac agggtctcac tctgctgccc 4020
aggctggagt acagtggcat gaccacaaca catcacagcc tcaagtgatc ttcccacttc 4080
agccttccca gtagctggga ctacaggtgc acgccaccat gttcacctaa ttttttactt 4140
tttgtagaga tgagacttca ccatgttgct caggctggtc ttgaattcct aggctcaagt 4200
gatcttcccg ctttggcctc ccaaagtgct gggattatag gtatgagcca ctgcatgtgg 4260
cctattttct tccactgttg ttcggcgtgg agaatattat atacataatt acgtaaatga 4320
tatcatactg tatatacctt ttttcctact ccttccttaa gttatatcat aatgagacta 4380
ccaattatta gacttttttt cttttttttg agacggagtc tcggtctgtc acctaggctg 4440
gagtgcaatg gcgcgatctc agctcgctgc aacctctgcc tcccaggttc aagcaattct 4500
gcctcagcct cccgagtagc tgggactaca gacacgtgcc accatgccca gctaactttt 4560
ttattttttt attagagaca gggttccacc atgctagcag gatggtctca atctctcgac 4620
ttcgtgatca gcccggcttg gcctcccaaa gtgctgggag tacaggtgtg agccaccgca 4680
ctcggcctag actaactatt taaagtaatc tggcaatgtt taacgaatac aaaactctaa 4740
aacccttgga cctaataata gctattttgg aaagtctact tgacagaaat aaaattgtga 4800
atattctttt ttgttgtttt tttgagacag agtctcattt ggacgcctag gctggagtgc 4860
agtggcatga tctcggctaa ctgcaacctc cacctcctgg gttcaagtga ttctcctgcc 4920
tcagcctcct gagcagctgg gattacaggt gtgcaccacc atgtctggct aatttttgca 4980
tttttagtag atggggtttc accatgttga ccagggtggt ctggaacttc taccctcaag 5040
tgatctaccc accttggcct cccaaagtgc tgggattaca ggtgtgagcc accacgcctg 5100
accagtgaac acttaataat atctatggaa aggtgttatt ataagaattg cttgtggggc 5160
cgggcgtggt ggctcacgcc tgtaatccca gcactttggg aggctgtggc aggcggatca 5220
cgaggtcagg agatcaagat catcctggct aacacggtga aaccccgtct ctactaaaaa 5280
taccaaaaaa ttagccaggc gtggtggcgg gcacttgtaa tcccagctat ccaggaggct 5340
gaggcaggag aattgcgtga acccaggagg cggaggtcgc agtgagctga gaccgtgcca 5400
ttgcactcca gcctgagtga cagagtgaga ctccatcaca aaaaataaat aaataaataa 5460
ataaaatata aataagtaaa taaaggtcag gagtggtggc tcacgcctgt aatcccagca 5520
ctttgggagg ccgaggtgga cagatcatga ggtcatgaga tcaagaccat cctggctaac 5580
acagtgaaac cctgcctcta ctaaaaatac aaaaagtcat ccaggtgtgg tggcacacac 5640
ctatagtccc agctacttgg gaggctgagg caggagaatc acttgaaccc aggaggcaga 5700
ggttgcagtg agctgagatc gcgccactac actccagcct aggcgacaga gcaagactct 5760
gtctcaaaat aaataaataa ataaataaat aaataaataa ataaataaaa taaaaagcac 5820
acacacacac acacacacac acacacaatg caaaagaccc accctactac aactaacatt 5880
atatttaatg gtgaaaaact gaattctttc tccctaagtg caggaataag acaaagatgt 5940
ctgctcttac tactcttatt caacataata ctgcaatccc ttgccagtgc aataaggcaa 6000
gaaaaatgaa ataaaaggaa aactgatcag aaagaaagaa ataaaactgt tcctatttgt 6060
ggatgacatg attacataga aaatctcaaa gaatctgtaa gaaacttctt agaattaata 6120
aatgaattca tcaaggttgc agaatataag ataaacataa aaaatctatt gtatttctat 6180
atattagcaa ggaacatgtg tacacagaaa ttaaaactac aataccattt ataattgctc 6240
aaaaaggcca ggcatggtgg ctcacacctg taattcctgc actttgggag gccaaggtgg 6300
gaagattgct taagcccagg agttcaagac cagcccgggc aacatagtga gaccttgtct 6360
ctacaaaaag taaaaaatta gctgagcatg gccgggtgca gtggctcact cctgtaaccc 6420
caacactttg ggaggctgag gcgggcggat catgaggtca ggagatcgag accatcctgg 6480
ctaacacggt gaaaccctgt ctctactaaa aacacaaaaa attagctgga tgtggtggca 6540
ggcgcctgta gacccagcta ctcgggaagc tgaggcagga gaatggcgtg aacctgggag 6600
gcggagcttg cagtgagctg agattgtgcc actgcactcc agcctgggtg acacagtgag 6660
actacgtctc aaaaaaaaaa aaaaaaaaat tagctgagca ttatggtgta tgcctgtagt 6720
cccagctact ggggaggctg aggtgggagg attgcttgag ccctaggagg gcaaggctgc 6780
agtgagccat gatcacacca ctgctttcca gcctcggtag gagagcaaga ccctatctca 6840
aaaaaaaaaa aaaaaaaaaa agaaaagaaa agaaaagaaa agaaaaagaa agagagaaag 6900
aaatacttag gtgtaaatct aaaaaacatg cgtagggcca ggtgcagtgg ctcatgcctg 6960
taatcccagc actttgggaa gttgaggctg gcggatcact tgaagtcggg agtttgagac 7020
cagcctggcc aacatggtga aaccccgtct ctactaaaaa tgcaaaaatt aggcaggtgt 7080
tgtggcgcat gcctgatccc agctactttg gaggctgagg caggagaatt gcttcaaccc 7140
gggaggcaga ggttgcagtg agccaagact gttccactgc actccagcct gggcaacaga 7200
gtaagagtct gtctcccgaa aaaaaaaaaa agaaaaaaga aagcattgaa ttgtatgcta 7260
aaaactacac gatgctgatt aaagaagtca aagaagatct aaatatatgg agagacatgc 7320
tgtactcatg gattgatgga ttggaagact caacataaga cagatatcaa ttttccccaa 7380
attaatatac aagtttaatc caattcctat aaaaatacca gcaagatttt ttgtagatat 7440
aaacaagttg gccaggtgta gtggcttaca cctgtaatcc tagcactttg ggaggctgag 7500
gtgggaagat cgcttgagcc caggtgttca cgactgcagt gagctatgat tgtgtcactg 7560
cattccagct ggcactccag cctaagtgac aaagggagac cctgtctcaa aaacaaaaac 7620
aaaaccaaaa taattttgct ctgcaaaatc cctattaaga agaagaaaag aggctgggca 7680
cagtggctca ccgctgtaat cccagcacgt tgggaggctg aggcaggctg atcacttcag 7740
cccagaagtt tgagatcagc ctgggcaaca tgaggaaacc ccgtctctac caaaaaaaaa 7800
aaaaggtaca tacacacaca cacacacaca cacacacata cacaagtata tacacatata 7860
tatacacata caggtgaata gatgtatata catctattta ttgtgaatat acatctatac 7920
acacacgtgt gtgtacacat atatttaaaa tttattttta tttatttatt tatttttgag 7980
acagagtctt gctctgtcac ccaggctggg tgcacctgta ttcccaacga cacaggaggc 8040
tgaggtggga gaatcactga gccagggagg cagaggttgc agtgagccaa gatgttgcct 8100
ggttgcctgg gcaacagagc gagaccctat atcaaaaaag aagaataata agaaaagaca 8160
gtttacagaa tataagaaaa tatattcaca atccacatac ttagcaaagg actggtatct 8220
agaatatgat aaacaactct caaaactcaa aaccaaaaaa atgaacaatt caattagaaa 8280
acaggccgaa aaggacatac agttggcaaa taagcacatg aaaagttgtt caacatcatt 8340
aatcattagg gatatgtaca ttaaaaccac aataggctat cactaaacct atcagaatgg 8400
ctaaatacaa aattggaaca ccaccaaatg ctgatgagga tgtggagaaa ctgggtcatt 8460
cttccaatat tggtgggagg ctaaaatggc aaagccactc tggaaaacag tttgatagtt 8520
tcttataaaa caaaacatgc ggccgggcgc ggtagctcac gcctgtaatc ccagcacttt 8580
gggaggccga ggcgggtgga tcacgaggtc aggagatcga gaccatcctg gctaacacgg 8640
tgaaaccctg tctctactaa aaatacaaaa aattagccgg gcgtggtggc gggcgcctgt 8700
agtcccagct actcgggagg ctgaggcagg agaatggtgt gaacccggga ggcggagctt 8760
gcagtgagcc gagatcgcgc cattgcactc caacctggga gacggaggga gactccgtct 8820
caaaaaaaca aaaacaaaca aacaaaaaac atgcaacaat ccagcaatat tgcaccccta 8880
ggcatttatc ctagagcaat gaagacttat gcccacacaa aaagctgcac acaaatgttc 8940
atagcagctt tattcatggt agccaacaat tagaaacaat ctagatgtcc ttcaactggt 9000
gaatgattac atccatacca cgaaatactt ttcagcaata aaaaggatga atcatagtac 9060
acaccacaac ctggatgaat ctccagggaa ttatgctgag tgaaaaaaag ccaatctcaa 9120
aaggtaatat actgtattaa tccatttata taacattctt aaaataacta attatagaaa 9180
tggagaacag atgagtgatt gccaggggtt aaggggctca gggatgggga ggggaagggg 9240
tatggctaca aaaagcaaca accttatggc gccggaaatg ttctgtattc tgattgtgtc 9300
aatgtgagca tactggttga gatatagtgc tacagttttg caagttatta ccatcagagt 9360
aaactggata gagggcacat aggatttctc tgtattactt cttacaactg caagtgaatc 9420
tacaattatc tcaaaataat aagtttagtt taatgctagg cgtggtggct cacatctgta 9480
atctcagctc tttgggaggc tgagacgggt ggatggcttg agtccaggag ttcgagacca 9540
gcctggccaa catggcaaaa ccggtctcta ctaaaaatac aaaaattagc tgggcgtggt 9600
ggcaagtgcc tgtagtccca gctactcggg aggctgaggc aggagaattg cttgaacccg 9660
ggaggtggag gttgcagtga gccgagatca cgccactaca ctgtagcttg ggcgacagag 9720
tgaggctctt tctcaaaaaa aaaaaaaaaa aaaaaaaagc aggcaggcag ggccaggaaa 9780
gcgtataatt tttgtagttc aaatgactaa cctaaaaagt gaagattggc caggcgcagt 9840
ggctcacgcc tgtaatccca gcactttggg aggccaaggc gggtggatca cgaggtcagg 9900
agattgagcc actctggcta acacagtgaa accccgtctc tactaaaata caaaaaatta 9960
gctgggcgtg gtggcacccg cctgtagttg cagctacttg ggaggctgag gcaggagaat 10020
cacttgaacc caggaggcga agttgcagcg agccgagatc acactactgc actccagcct 10080
gggtgacaaa gtgagattct gtctcaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaagtg 10140
aagtttacct ttttttttaa atttttcttc ttttccttcc ctactttgtg agataatttt 10200
cttcttttta aaaagccaag agcttacttc tgtaagtaaa gattatctta agacaactta 10260
gaaatgtata ttattagtat tttctatttc attgtaagtt atttgtaaat attggttttg 10320
gtgctaacct agaattccat caaattaatt gtcccctaat atatggccat tatcattttg 10380
tctaacattg tatcctatta acaatgctgt aagtattatt tttgtagcta aattatggtt 10440
tgcattttaa aattattgtt ttaaggataa agttccagaa atgaaattaa ggatatgaac 10500
tttttgagca catcttgtca gcactgagta gtattattta aaacttttgg ggggggcaat 10560
tttataattg aaaaatatat cattgtttta atttgcattt ctttcactgc ctatgagatt 10620
aaaacaatgc actactttcc aaaaattctt aagtcttttg tgttgatgct tttgttctgt 10680
ttctatggat ctcatcttcc ttcagaacag ctccccttcc caacttcctg atttctaaca 10740
ataacagtat caccctcctt gttctcccaa tttctgaaac acagagtcat gtttttttct 10800
ctgcttcaat ccctggtttc ctatcgtcat caattatgac ctttccttgc tttgaaagtg 10860
ttttgggccg ggcatgatgt ctgccaccta ttgtaatcct agcactttgg gaggctgagg 10920
cggctggatg acttgacctg aggatttcga gaccagcctg ggcaacaggg cgaaacctcg 10980
tctctacaaa aaatacaaaa gttagtcggg agtggtggca catgcttgta gtcccagtta 11040
cttggggggc tgaggtggca ggatctcttg agcccacgag gtagatgttg cagtgagccg 11100
tgattgcgcc actgcacccc agcctaggtg acagagtgag accctgtctc aaaaaaaaaa 11160
aaatgttcta gtttcttcct cttctttgtt cccatgggaa tgccaccatc accagccaag 11220
gctcacatac ctcccacctg gattacagtg agcttccagg taatttggtc tgctactagt 11280
ctcgcctact tggatttccc ttccccctgc tgcagcattg ccttccaaag ccatgctttg 11340
cacatgccac atcctagccc attagactaa gcctagaagc ctctgcagga cgttcaccct 11400
ctcagcgcca ctgctcagtt tcccagtgga aacctctgca cccaggaggt ttccccacag 11460
cttgcctgtg ctgcctctct ggagcttttc tcccttcctg taatgtcctt gctgctcccc 11520
gtctctagtc cattgcctat acctcttttt tttttttttt ttgagatgga gtctctctct 11580
ctcatccagg ctggagtgca gtggcgcgat ctcggctcac tgcaaccttt gtctcctggg 11640
ttcaagggat tctcctgcct cagcctcccg agtaactggg attacaggcg tgcaccacca 11700
ttcctggcta atttttgtat ttttagtaaa gactgggttt caccatgttg gccaggctgg 11760
tcttgaactc ctgccctcag gtgatccacc tgcctcggcc tcccagagtg ctgggattac 11820
aggcgtgagc caccgcacct gccacaggcc catacctctt ttaagtcttc attcaatacc 11880
agttgtccca tgaatttgtc ccagactcac tcatatgctt agacctttca tattatcttg 11940
ccatagcttt ttcaaagtat gggacagcat ggacaagcag gccatggttt tcttttgaag 12000
agaagcaagg aggcagagtt attttaggag gagggttata catttcattt tgaaccaatt 12060
gcgtttgggg tgatggcagg atattaacat aaacttattt cttggaccat tggaaatgtg 12120
tgcctagaac tgaggagaga ggtcagggct ggcagtaaca acttggccac aatctgcaga 12180
gctgactggg gatgaggtgg aatttagaat gtctgtagaa acggggaaga gaaccaaaga 12240
cagagtctgg gacaacacct aaatgtagat gtcagagcaa gagttcaaga cgaagaaaaa 12300
cgaatcatac ttagaaatgg aggggaggaa caaaagaggc ggagcaaagt ggggcagaac 12360
cagagtaggc cacgctttta agaagtttgg taaaggaact gtgaaaggaa tgtagttgaa 12420
tttcagggta agctggggaa ttaaagcagt gtgtagatcc agggcaaaca gcaagtaggg 12480
caggaaccac tgaaggaaca aataaagggg gaggttgggt ccaggttgtc ttgagtaggg 12540
aagttttttt aaaaagtgtg aaactgaagg tgtggggtgg attgggtgcc tgccgtgctc 12600
tgaggaagct tggggcaact gtgtgctgag gctgtgaggt tgtctggaag gggctcctgg 12660
acagtaagag ctgagcagtg gggaagagga ctgtgtggtc tggaagagga gagaaaggag 12720
agtgagtgac tgaactggta tccaggctcc cacaccaagg cagaaagagg gagaggacct 12780
gggcatctca gggaggcaga ggcagtacca agcagggtga gaggctttag tcttagccac 12840
ctttgcccca ttcctccaaa tatacattct aagtaaaaac aaaacaaaac agaactgttt 12900
gctatgtaaa tttagcttct aaagccctgt tctacagaga ttttggagct tccactgcac 12960
ccagaaaatg cacagctaaa gagaaaactt cccttggtga tggttattag attttacaag 13020
aagaggccaa aggagacaca tacttatgcc agaagaactt tccagagata gcattgcata 13080
gcgaaatagc ctgaattatt tttatttttt aaaacatttt ttcttttctt ttttcttttc 13140
tttttctttt tttttttttt tttttgagac agagtctcac tctgtcaccc aggctggagt 13200
gcagtggcgt gatcttggct cactgcaatc tccacctccc gggttcaagc cattctcctc 13260
cctcagcctc ccaagtagct gggattacag gcatgcgtca ctatgctctg gctaattttt 13320
tttttctttt tttttttggt atttttagta gagatggggt ttcaccatgt tggccaggct 13380
ggtcttgaac tcctgacctc aagtgatcca ccgccttggc ctcccaaagt gctgggattt 13440
caggcgtgag ccaccgcacc cggccaaaaa tttcttttct ttaagatgag gcctcactct 13500
gttgcccagg ctggagtgca gtgttacaat catagctcac tgtaactttg aactcctggg 13560
ctcaagtgat cctcctgctt cagcctctca agtagctggg attacaggca tgtgccacca 13620
cacccagcta atttttttta aaataatttt ttttagagac gagggtctcg attggctgcc 13680
taggttggtc ccagactcct gacgggctgc attttaatcc tagctccacc acttacggga 13740
gtcaaaattc aaaagataga aaagggcata taggctgggt gcagtggctc acacctgcaa 13800
tcccagcaat ttgggaggct gaggtgggcg ggttgcttga ggtcaggagt tcgagatcag 13860
cctgggcaac atggcaaaac ttgtatctac taaaaataca aaaattagcc agatgtggtg 13920
gtgtacacct gtaatcccag ctactccgaa ggctgaggca agagaatccc ttgaactcag 13980
gaggcagagg ttacaatgag cagagatcga acactcgact ccataaaaac aaacaaacaa 14040
aaaaagaaag caggctgggt gtggtggctc acgcctgtaa ccccagcact tcgggaggcc 14100
aaggcgagcg gatcacctga ggttgggcat tcgagaccag cctgaccaac aaggagaaac 14160
cctgtctcta ctgaaaatac aaaattagcc gggcttggtt gcgcatgcct gtaatctcag 14220
ctactcggga ggcagaggca agataattgc ttgaacccgg gaggcggagg ttgcggtgag 14280
ccaagatcat gccattgcac tccaacctgg gcaacaatag cgaaactcca tctcaaaaaa 14340
aaaaaagcaa agggcatata gtgaaaagct ttcttcctac acatgagtat tcacttcctc 14400
ttcctagagg caaccaaggt tatttttgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 14460
tgttttggga cagtctcact ctctcaccaa ggctggaatg cagtggtgcg atctcactgc 14520
aaactctgcc tcccagtctc aagcgatctt gtgcctcagc ctcccagttt ttttttcttt 14580
taaatggggt ctcattctgt cgcccagggt ggagtgcagt ggcatgatca tagctcactg 14640
cagcctcgac ctcctgggtc aggttatcct cccacctcag cctccggcat agctggggct 14700
actggcatgc accaccacac tcagttaatt ttttttcttt tttgagacag agtctcactc 14760
tgtcacctag actggagtgc agtggtgcca tctcatttgt ttcactgcaa cctttgactt 14820
ctgggctcaa gtgattctcc cacctcagcc tcccaaggcg gctaattaaa aaaaattttt 14880
tttttttttt ttttagagat ggggtttcgc catgttgccc aggctgatct cgaactcctg 14940
ggcacaaaca atctttccac ctcgatcttt caaagagctg ggatgagaga tttccaccat 15000
gcctggcctc attttctttt ttaatttttt tttagacatt atagctcttt ttaatggcct 15060
cattttctta tgtttaattc gagaattatt cttttcatat acaaagaata tattttctcc 15120
acctttaaaa acaaatagta gactgtttaa catctcgctt tattcagtta gtgatgtttc 15180
ttagatacgg gtccaaatta gtacacaaag cacttcctca ttcctctctt acggctgcat 15240
agcagtccac tgaatgggtg agctatgatc tatttaacct attctttatt gatggacatt 15300
tggttttgta tatacatttg taattctgta tagattacaa atcaccatcc aaagaaattg 15360
tactggttta ttctcctaca atgtgtgaga gttgggtaat tacttaatct caatatgtga 15420
gagtttaggc agttacctaa tctctctgag tctcagtttc tctatctgca aaataaacaa 15480
aacagtgttg acagtatcta tttctcggaa ttattgtgga gattactgag atgatgcctg 15540
taaagtattt ggcatgtagg agttggtgct ctccaaataa ggatatgatt ttatttgtat 15600
ttgtgagcta ctgtcccagc caggtaaatg gatatgatga gacctccttg ccagaccggg 15660
tttctctgat tagaacgagg agcagatgtt gcaggaaatt agcaactgat atcagaagag 15720
ccgtgggcat tctcttgcca gaggtgccct gtctccaggg cgcctcagtc cccccccata 15780
tgtcttctgc tcccaggtcc atccaagctg aatgatcgcg ctgactcaag aagaagcctt 15840
tgggaccaag gaaactttcc cctgatcatc aagaatctta agatagaaga ctcagatact 15900
tacatctgtg aagtggagga ccagaaggag gaggtgcaat tgctagtgtt cggatgtgag 15960
tggggcaggt ggggatgagg atacctcctg cctggttccc ttccccacta ctcccacccc 16020
tgcaccaaat ccagcctgag ctggtgatac cgcagcagcc ccaagaggac caggctgtca 16080
aactggcctc caaatgtctt aaaacccttc ttgatcaggt gagggatgct ggtgggcgga 16140
ggagggaaga ggccttggga aaaggaaaga aaagggaagg aggcaaggga aggagggaga 16200
gagactgggg aagagaggat gaggggagag gaggaaagaa gagagagagg aggggagagg 16260
gaaaccctat cttggctggg ggtgcgcagc tgggtgctgg gaggaaggag atgttgggac 16320
ggcgataatg gagagatgtt gttggtttcc tgttgtctgc ccttctcctt ggggatggta 16380
tgtgtgtgac acagctggcc tttccctcca cagtgactgc caactctgac acccacctgc 16440
ttcaggggca gagcctgacc ctgaccttgg agagcccccc tggtagtagc ccctcagtgc 16500
aatgtaggag tccaaggggt aaaaacatac agggggggaa gaccctctcc gtgtctcagc 16560
tggagctcca ggatagtggc acctggacat gcactgtctt gcagaaccag aagaaggtgg 16620
agttcaaaat agacatcgtg gtgctaggta agggaagccc ctcttcgcgc agtctcctcc 16680
ctgccccagg ggctgacagc ccctccctct gctctgactg ccctgtttct ggttctggtg 16740
ctgggaggtc aggagtggag aagactaggt cccctagagc tgaggcctgt cttgaaggac 16800
tcactggggc cctcatcctc agggggctga ttggcagcca cccctcagtg tggtggacat 16860
ggagaaagga aaggctgggg aaggtaagga tgctagaggc ccgagtctcc tttggaggcc 16920
ccaaaggagg aatgtcaggg agcttacttt ctttgttgcc tcagctccac acccctacca 16980
agttggcaaa tccacttact cagggacact aacaccagta agccaaccct gatgatgttc 17040
tatgttgtac ctctggacct ctaagccagg ccactgtggg gagaccaagg tcctacccca 17100
gatcctgtcc cctgggtgct tatgtgactt aaggtagaca taaggtagtg tgccagttta 17160
gtgcatgtac gctgattgaa atcctggttc tgccacaacc atgtgacctt gggtgagtta 17220
ctaaacctct ctgcaccttg gtttcagcct ctgtgaaatg gggatgatgt taactgccat 17280
agtgactacc tcgtattaag ttgaggactg atatacgtaa ggcactgaaa atggtgcctg 17340
gcacagagta agccctagtt aagtgttcgc tgttattttg tgaagggtga tgaatacgcc 17400
tctaaggagt ggaggccaaa tggcttctgt ggtccaggaa tcctaaggac agcaaggatc 17460
ccctgtggct gggctgctct gtgatggctt ccgggaggag ggaggtggcc tgctgtagga 17520
aaatgctggg tggaagaagg gagagaaggc tggagaggta ggaaggaact gaagtatctg 17580
aagtgacaag gtgggtgtct ggactcgtcg ggtccccttc catctccctg ctgcctccac 17640
atgccaaccc cactcgtgca ccctcatctt cctatctcct cacccagggt ctctcccttc 17700
ccacctccag ctttccagaa ggcctccagc atagtctata agaaagaggg ggaacaggtg 17760
gagttctcct tcccactcgc ctttacagtt gaaaagctga cgggcagtgg cgagctgtgg 17820
tggcaggcgg agagggcttc ctcctccaag tcttggatca cctttgacct gaagaacaag 17880
gaagtgtctg taaaacgggt tacccaggac cctaagctcc agatgggcaa gaagctcccg 17940
ctccacctca ccctgcccca ggccttgcct cagtatgctg gctctggaaa cctcaccctg 18000
gcccttgaag cgaaaacagg aaagttgcat caggaagtga acctggtggt gatgagaggt 18060
gaggggccag gccagggagg ggtgggcagg ggaaggagtt ggaggggcct ggcccagggc 18120
tccctctgag gcaagccagg ccccaagagg ggatgcctag gccctggtca cctggatgaa 18180
gtgagggagg gccctctggg tttggggctg gttttgaact gagacatcca tgagccagcc 18240
tggggctggc ttcactgaag atc 18263
<210> 78
<211> 458
<212> PRT
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 78
Met Cys Arg Ala Ile Ser Leu Arg Arg Leu Leu Leu Leu Leu Leu Gln
1 5 10 15
Leu Ser Gln Leu Leu Ala Val Thr Gln Gly Lys Lys Val Val Leu Gly
20 25 30
Lys Lys Gly Asp Thr Val Glu Leu Thr Cys Thr Ala Ser Gln Lys Lys
35 40 45
Ser Ile Gln Phe His Trp Lys Asn Ser Asn Gln Ile Lys Ile Leu Gly
50 55 60
Asn Gln Gly Ser Phe Leu Thr Lys Gly Pro Ser Lys Leu Asn Asp Arg
65 70 75 80
Ala Asp Ser Arg Arg Ser Leu Trp Asp Gln Gly Asn Phe Pro Leu Ile
85 90 95
Ile Lys Asn Leu Lys Ile Glu Asp Ser Asp Thr Tyr Ile Cys Glu Val
100 105 110
Glu Asp Gln Lys Glu Glu Val Gln Leu Leu Val Phe Gly Leu Thr Ala
115 120 125
Asn Ser Asp Thr His Leu Leu Gln Gly Gln Ser Leu Thr Leu Thr Leu
130 135 140
Glu Ser Pro Pro Gly Ser Ser Pro Ser Val Gln Cys Arg Ser Pro Arg
145 150 155 160
Gly Lys Asn Ile Gln Gly Gly Lys Thr Leu Ser Val Ser Gln Leu Glu
165 170 175
Leu Gln Asp Ser Gly Thr Trp Thr Cys Thr Val Leu Gln Asn Gln Lys
180 185 190
Lys Val Glu Phe Lys Ile Asp Ile Val Val Leu Ala Phe Gln Lys Ala
195 200 205
Ser Ser Ile Val Tyr Lys Lys Glu Gly Glu Gln Val Glu Phe Ser Phe
210 215 220
Pro Leu Ala Phe Thr Val Glu Lys Leu Thr Gly Ser Gly Glu Leu Trp
225 230 235 240
Trp Gln Ala Glu Arg Ala Ser Ser Ser Lys Ser Trp Ile Thr Phe Asp
245 250 255
Leu Lys Asn Lys Glu Val Ser Val Lys Arg Val Thr Gln Asp Pro Lys
260 265 270
Leu Gln Met Gly Lys Lys Leu Pro Leu His Leu Thr Leu Pro Gln Ala
275 280 285
Leu Pro Gln Tyr Ala Gly Ser Gly Asn Leu Thr Leu Ala Leu Glu Ala
290 295 300
Lys Thr Gly Lys Leu His Gln Glu Val Asn Leu Val Val Met Arg Val
305 310 315 320
Ala Gln Leu Asn Asn Thr Leu Thr Cys Glu Val Met Gly Pro Thr Ser
325 330 335
Pro Lys Met Arg Leu Thr Leu Lys Gln Glu Asn Gln Glu Ala Arg Val
340 345 350
Ser Glu Glu Gln Lys Val Val Gln Val Val Ala Pro Glu Thr Gly Leu
355 360 365
Trp Gln Cys Leu Leu Ser Glu Gly Asp Lys Val Lys Met Asp Ser Arg
370 375 380
Ile Gln Val Leu Ser Arg Gly Val Asn Gln Thr Val Phe Leu Ala Cys
385 390 395 400
Val Leu Gly Gly Ser Phe Gly Phe Leu Gly Phe Leu Gly Leu Cys Ile
405 410 415
Leu Cys Cys Val Arg Cys Arg His Gln Gln Arg Gln Ala Ala Arg Met
420 425 430
Ser Gln Ile Lys Arg Leu Leu Ser Glu Lys Lys Thr Cys Gln Cys Pro
435 440 445
His Arg Met Gln Lys Ser His Asn Leu Ile
450 455
<210> 79
<211> 293
<212> PRT
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 79
Lys Lys Val Val Leu Gly Lys Lys Gly Asp Thr Val Glu Leu Thr Cys
1 5 10 15
Thr Ala Ser Gln Lys Lys Ser Ile Gln Phe His Trp Lys Asn Ser Asn
20 25 30
Gln Ile Lys Ile Leu Gly Asn Gln Gly Ser Phe Leu Thr Lys Gly Pro
35 40 45
Ser Lys Leu Asn Asp Arg Ala Asp Ser Arg Arg Ser Leu Trp Asp Gln
50 55 60
Gly Asn Phe Pro Leu Ile Ile Lys Asn Leu Lys Ile Glu Asp Ser Asp
65 70 75 80
Thr Tyr Ile Cys Glu Val Glu Asp Gln Lys Glu Glu Val Gln Leu Leu
85 90 95
Val Phe Gly Leu Thr Ala Asn Ser Asp Thr His Leu Leu Gln Gly Gln
100 105 110
Ser Leu Thr Leu Thr Leu Glu Ser Pro Pro Gly Ser Ser Pro Ser Val
115 120 125
Gln Cys Arg Ser Pro Arg Gly Lys Asn Ile Gln Gly Gly Lys Thr Leu
130 135 140
Ser Val Ser Gln Leu Glu Leu Gln Asp Ser Gly Thr Trp Thr Cys Thr
145 150 155 160
Val Leu Gln Asn Gln Lys Lys Val Glu Phe Lys Ile Asp Ile Val Val
165 170 175
Leu Ala Phe Gln Lys Ala Ser Ser Ile Val Tyr Lys Lys Glu Gly Glu
180 185 190
Gln Val Glu Phe Ser Phe Pro Leu Ala Phe Thr Val Glu Lys Leu Thr
195 200 205
Gly Ser Gly Glu Leu Trp Trp Gln Ala Glu Arg Ala Ser Ser Ser Lys
210 215 220
Ser Trp Ile Thr Phe Asp Leu Lys Asn Lys Glu Val Ser Val Lys Arg
225 230 235 240
Val Thr Gln Asp Pro Lys Leu Gln Met Gly Lys Lys Leu Pro Leu His
245 250 255
Leu Thr Leu Pro Gln Ala Leu Pro Gln Tyr Ala Gly Ser Gly Asn Leu
260 265 270
Thr Leu Ala Leu Glu Ala Lys Thr Gly Lys Leu His Gln Glu Val Asn
275 280 285
Leu Val Val Met Arg
290
<210> 80
<211> 142
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 80
tgtttgcctg tgacatgaac tcattgtgac acaaaccact gtgctagggg ggatccacta 60
gtaacggccg ccagtgtgct ggaattcgcc ctcgcaaggg ccaggcatat aagtacacaa 120
taaacaaatg gcagctctct cc 142
<210> 81
<211> 99
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 81
cccctccttc cttccccagg cactttccaa gtgtcaactc tagagcctat cgcggccgca 60
ccggtataac ttcgtataat gtatgctata cgaagttat 99
<210> 82
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 82
ataacttcgt ataatgtatg ctatacgaag ttatgtcgac gtagcctatt tctctagatc 60
caaaatgatg acaacaaaag gtaccttgtg 90
<210> 83
<211> 210
<212> PRT
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 83
Met Gln Pro Trp Leu Trp Leu Val Phe Ser Met Lys Leu Ala Val Leu
1 5 10 15
His Gly Asn Ser Val Leu Gln Gln Thr Pro Ala Tyr Ile Lys Val Gln
20 25 30
Thr Asn Lys Met Val Met Leu Ser Cys Glu Ala Lys Ile Ser Leu Ser
35 40 45
Asn Met Arg Ile Tyr Trp Leu Arg Gln Arg Gln Ala Pro Ser Ser Asp
50 55 60
Ser His His Glu Phe Leu Ala Leu Trp Asp Ser Ala Lys Gly Thr Ile
65 70 75 80
His Gly Glu Glu Val Glu Gln Glu Lys Ile Ala Val Phe Arg Asp Ala
85 90 95
Ser Arg Phe Ile Leu Asn Leu Thr Ser Val Lys Pro Glu Asp Ser Gly
100 105 110
Ile Tyr Phe Cys Met Ile Val Gly Ser Pro Glu Leu Thr Phe Gly Lys
115 120 125
Gly Thr Gln Leu Ser Val Val Asp Phe Leu Pro Thr Thr Ala Gln Pro
130 135 140
Thr Lys Lys Ser Thr Leu Lys Lys Arg Val Cys Arg Leu Pro Arg Pro
145 150 155 160
Glu Thr Gln Lys Gly Leu Thr Cys Ser Leu Thr Thr Leu Ser Leu Leu
165 170 175
Val Val Cys Ile Leu Leu Leu Leu Ala Phe Leu Gly Val Ala Val Tyr
180 185 190
Phe Tyr Cys Val Arg Arg Arg Ala Arg Ile His Phe Met Lys Gln Phe
195 200 205
His Lys
210
<210> 84
<211> 151
<212> PRT
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 84
Val Leu His Gly Asn Ser Val Leu Gln Gln Thr Pro Ala Tyr Ile Lys
1 5 10 15
Val Gln Thr Asn Lys Met Val Met Leu Ser Cys Glu Ala Lys Ile Ser
20 25 30
Leu Ser Asn Met Arg Ile Tyr Trp Leu Arg Gln Arg Gln Ala Pro Ser
35 40 45
Ser Asp Ser His His Glu Phe Leu Ala Leu Trp Asp Ser Ala Lys Gly
50 55 60
Thr Ile His Gly Glu Glu Val Glu Gln Glu Lys Ile Ala Val Phe Arg
65 70 75 80
Asp Ala Ser Arg Phe Ile Leu Asn Leu Thr Ser Val Lys Pro Glu Asp
85 90 95
Ser Gly Ile Tyr Phe Cys Met Ile Val Gly Ser Pro Glu Leu Thr Phe
100 105 110
Gly Lys Gly Thr Gln Leu Ser Val Val Asp Phe Leu Pro Thr Thr Ala
115 120 125
Gln Pro Thr Lys Lys Ser Thr Leu Lys Lys Arg Val Cys Arg Leu Pro
130 135 140
Arg Pro Glu Thr Gln Lys Gly
145 150
<210> 85
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 85
tgaacctgct gctgctgggt gagtcgatta tcctggggag tggagaagct aggccgagcc 60
agttccgggt gtcgccgctg gatcggacct ggaacctggg 100
<210> 86
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 86
atgccaggga cagccctgat actgtaggta gagtcaaggg ctgtccaagt accggtataa 60
cttcgtataa ggtatcctat acgaagttat 90
<210> 87
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 87
ataacttcgt ataaggtatc ctatacgaag ttatctcgac ctgatcttgg agggagacct 60
ggaccgggag acgtgctggg ggcagggtt 89
<210> 88
<211> 243
<212> PRT
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 88
Met Ala Ser Pro Leu Thr Arg Phe Leu Ser Leu Asn Leu Leu Leu Leu
1 5 10 15
Gly Glu Ser Ile Ile Leu Gly Ser Gly Glu Ala Arg Pro Ser Gln Phe
20 25 30
Arg Val Ser Pro Leu Asp Arg Thr Trp Asn Leu Gly Glu Thr Val Glu
35 40 45
Leu Lys Cys Gln Val Leu Leu Ser Asn Pro Thr Ser Gly Cys Ser Trp
50 55 60
Leu Phe Gln Pro Arg Gly Ala Ala Ala Ser Pro Thr Phe Leu Leu Tyr
65 70 75 80
Leu Ser Gln Asn Lys Pro Lys Ala Ala Glu Gly Leu Asp Thr Gln Arg
85 90 95
Phe Ser Gly Lys Arg Leu Gly Asp Thr Phe Val Leu Thr Leu Ser Asp
100 105 110
Phe Arg Arg Glu Asn Glu Gly Tyr Tyr Phe Cys Ser Ala Leu Ser Asn
115 120 125
Ser Ile Met Tyr Phe Ser His Phe Val Pro Val Phe Leu Pro Ala Lys
130 135 140
Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile
145 150 155 160
Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala
165 170 175
Gly Gly Ala Val Lys Gly Thr Gly Leu Asp Phe Ala Cys Asp Ile Tyr
180 185 190
Ile Trp Ala Pro Leu Ala Gly Ile Cys Val Ala Leu Leu Leu Ser Leu
195 200 205
Ile Ile Thr Leu Ile Cys Tyr His Arg Ser Arg Lys Arg Val Cys Lys
210 215 220
Cys Pro Arg Pro Leu Val Arg Gln Glu Gly Lys Pro Arg Pro Ser Glu
225 230 235 240
Lys Ile Val
<210> 89
<211> 152
<212> PRT
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 89
Arg Pro Ser Gln Phe Arg Val Ser Pro Leu Asp Arg Thr Trp Asn Leu
1 5 10 15
Gly Glu Thr Val Glu Leu Lys Cys Gln Val Leu Leu Ser Asn Pro Thr
20 25 30
Ser Gly Cys Ser Trp Leu Phe Gln Pro Arg Gly Ala Ala Ala Ser Pro
35 40 45
Thr Phe Leu Leu Tyr Leu Ser Gln Asn Lys Pro Lys Ala Ala Glu Gly
50 55 60
Leu Asp Thr Gln Arg Phe Ser Gly Lys Arg Leu Gly Asp Thr Phe Val
65 70 75 80
Leu Thr Leu Ser Asp Phe Arg Arg Glu Asn Glu Gly Tyr Tyr Phe Cys
85 90 95
Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His Phe Val Pro Val
100 105 110
Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr
115 120 125
Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala
130 135 140
Cys Arg Pro Ala Ala Gly Gly Ala
145 150
<210> 90
<211> 149
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность химерного человеческого/мышиного локуса MHC I в участке
соединения 5’ мышиной/человеческой последовательностей
<400> 90
agtgtcgccg cggacgctgg atataaagtc cacgcagccc gcagaactca gaagtcgcga 60
atcgccgaca ggtgcgatgg ccgtcatggc gccccgaacc ctcgtcctgc tactctcggg 120
ggctctggcc ctgacccaga cctgggcgg 149
<210> 91
<211> 159
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность химерного человеческого/мышиного локуса MHC I в участке
соединения 3’ человеческой/мышиной последовательностей
<400> 91
ggtggtgcct tctggacagg agcagagata cacctgccat gtgcagcatg agggtttgcc 60
caagcccctc accctgagat ggggtaagga gagtgtgggt gcagagctgg ggtcagggaa 120
agctggagct ttctgcagac cctgagctgc tcagggctg 159
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ Т-КЛЕТОЧНОГО РЕЦЕПТОРА МЫШИ | 2012 |
|
RU2661106C2 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ЭКСПРЕССИРУЮЩИЕ ЭКЗОГЕННУЮ ТЕРМИНАЛЬНУЮ ДЕЗОКСИНУКЛЕОТИДИЛТРАНСФЕРАЗУ | 2017 |
|
RU2753585C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ МЫШИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ МОЛЕКУЛЫ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ | 2012 |
|
RU2660564C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ДЛЯ ВЫРАБОТКИ ТЕРАПЕВТИЧЕСКИХ АНТИТЕЛ ПРОТИВ КОМПЛЕКСОВ ПЕПТИД-MHC, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ВАРИАНТЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2819525C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ МЫШИ | 2012 |
|
RU2653433C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ МЫШИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ МОЛЕКУЛЫ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ | 2012 |
|
RU2797549C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР | 2013 |
|
RU2665548C2 |
| НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ ГУМАНИЗИРОВАННЫЙ КОМПЛЕКС CD3 | 2015 |
|
RU2726446C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ МЫШИ | 2012 |
|
RU2783984C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ T-КЛЕТОЧНОГО РЕЦЕПТОРА МЫШИ | 2012 |
|
RU2797272C1 |
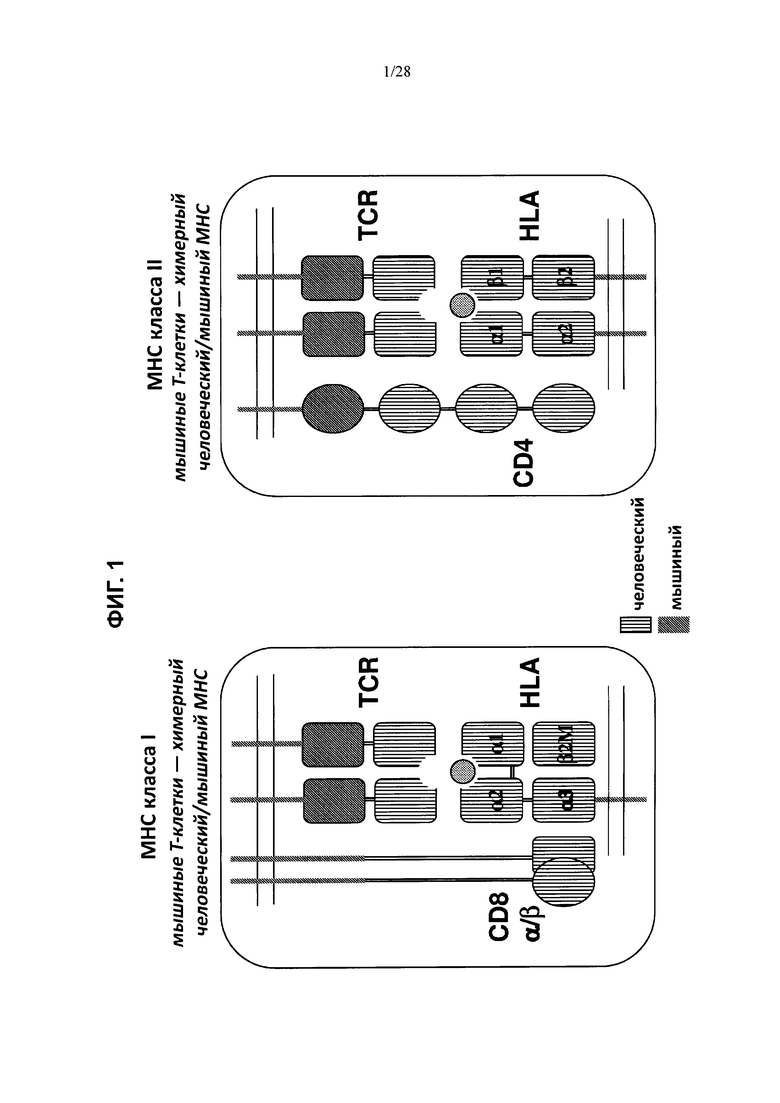


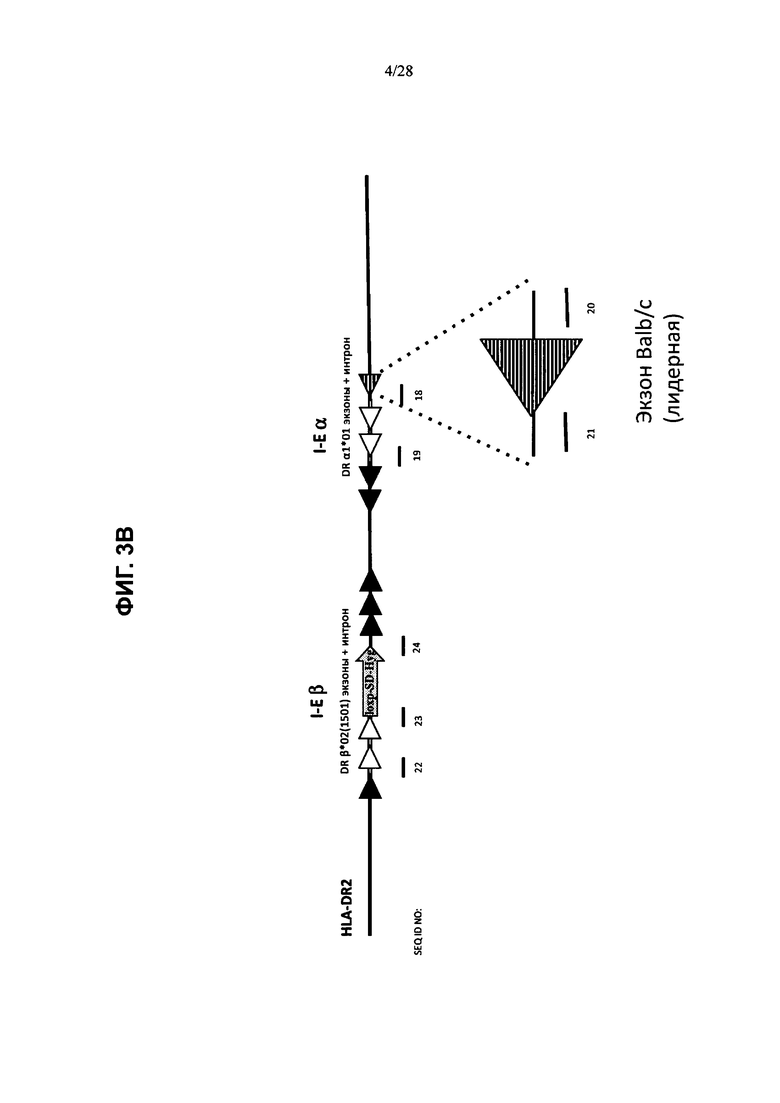
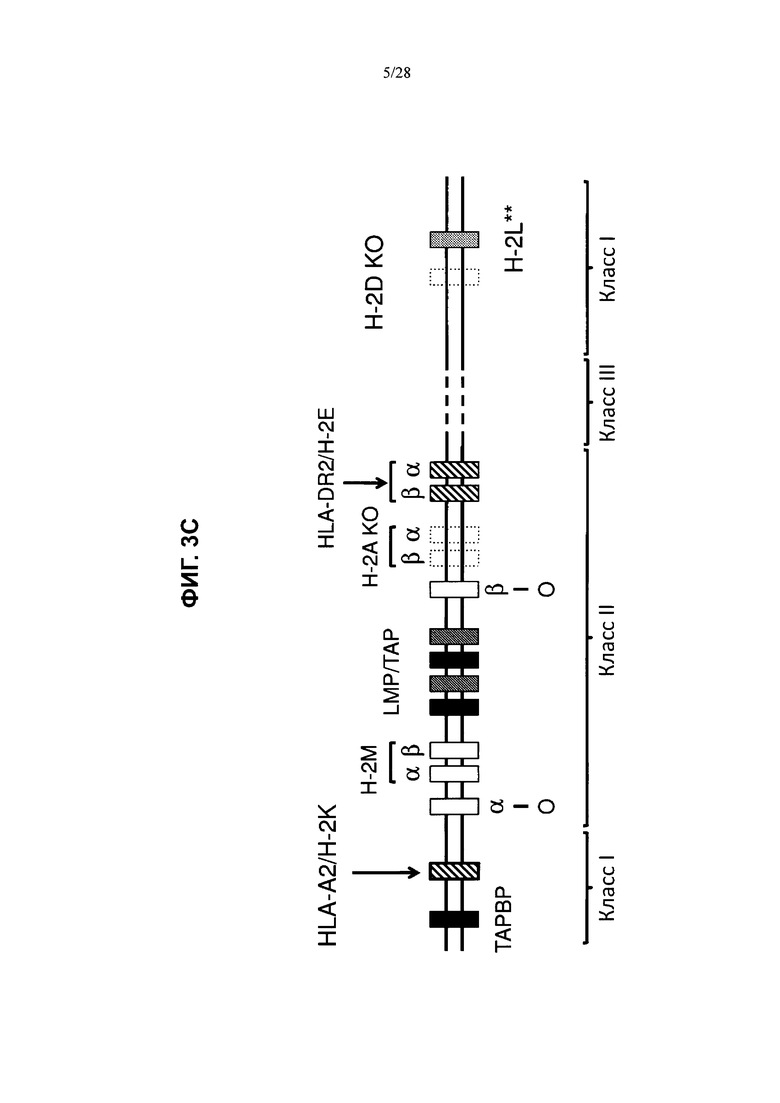

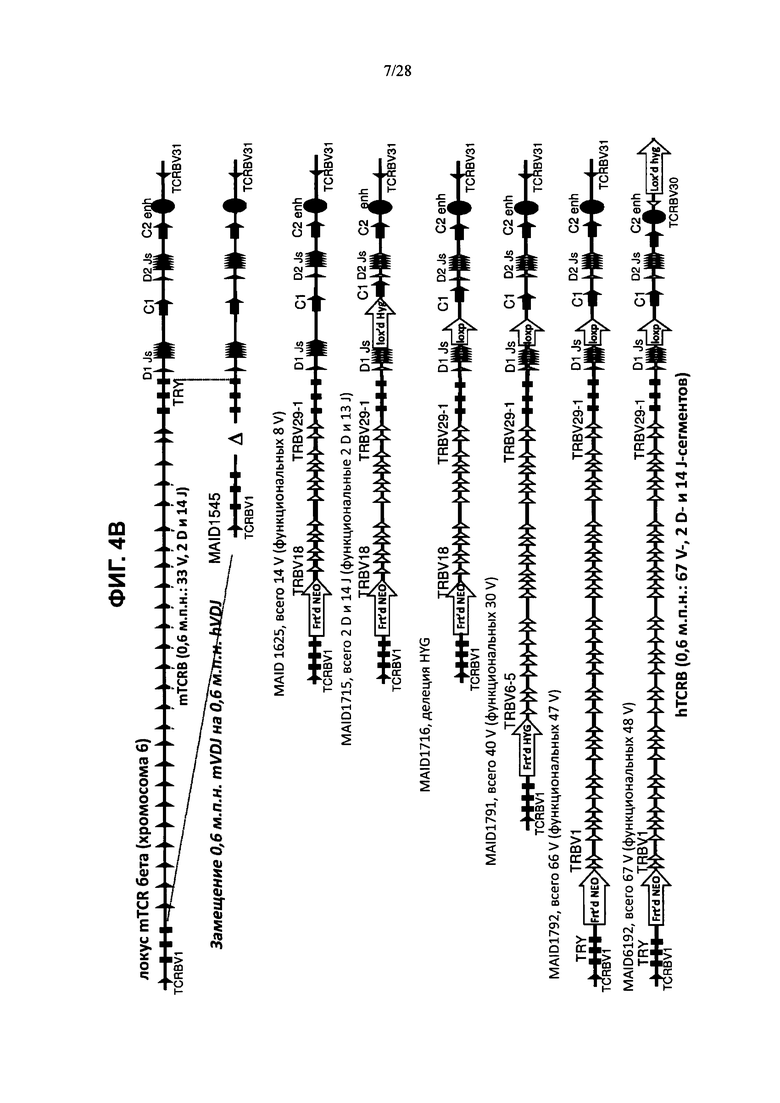

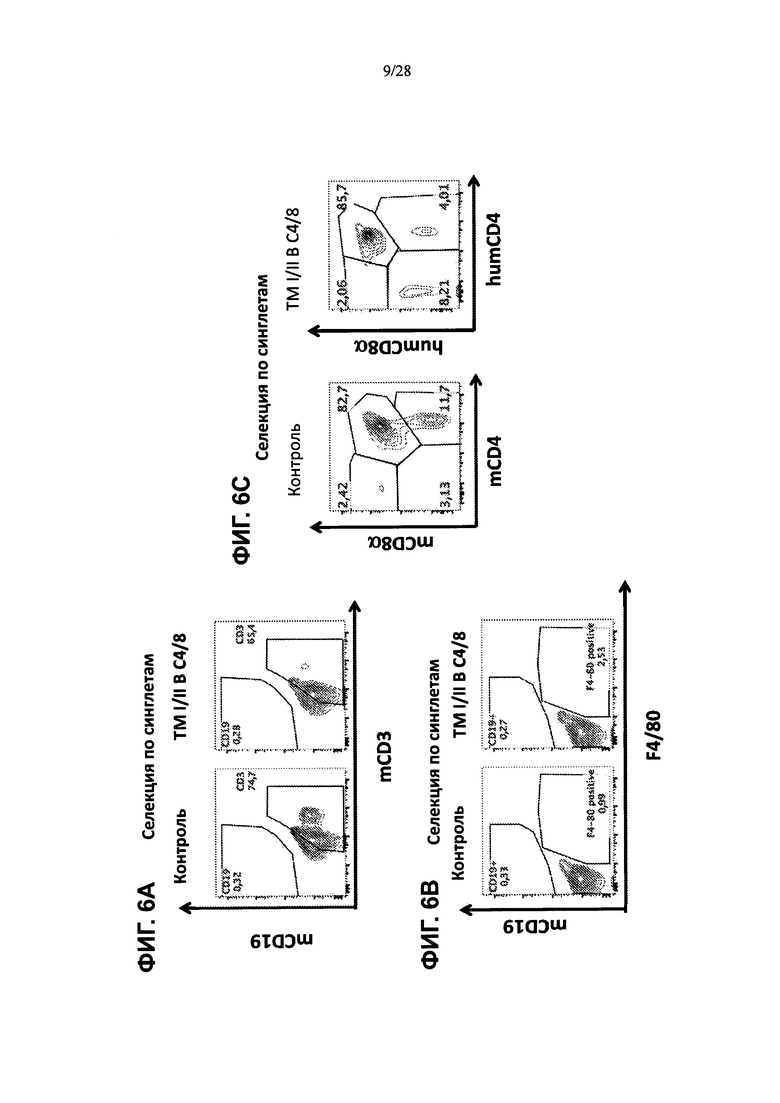

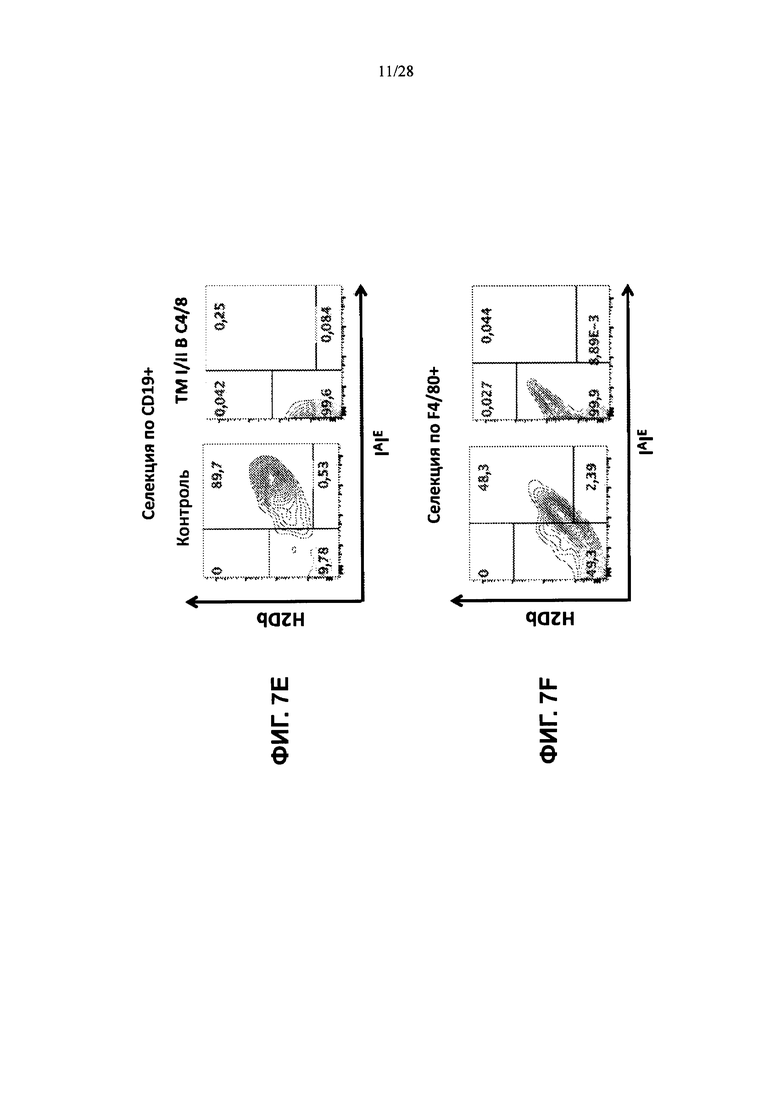
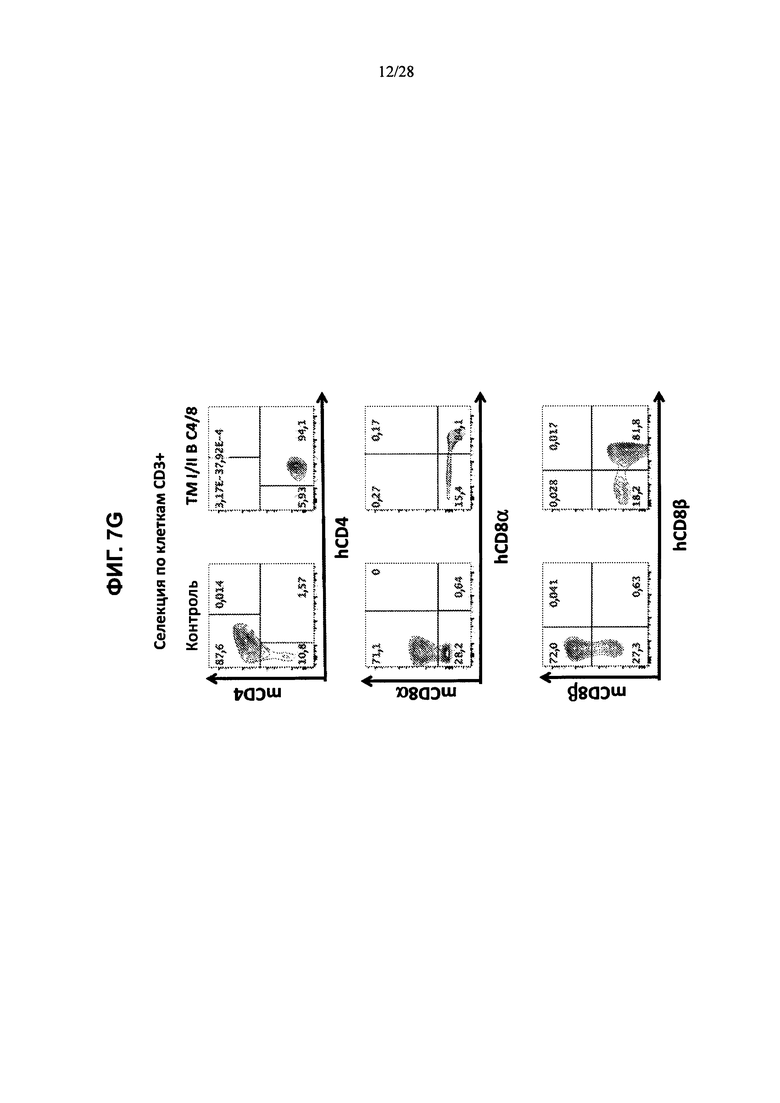
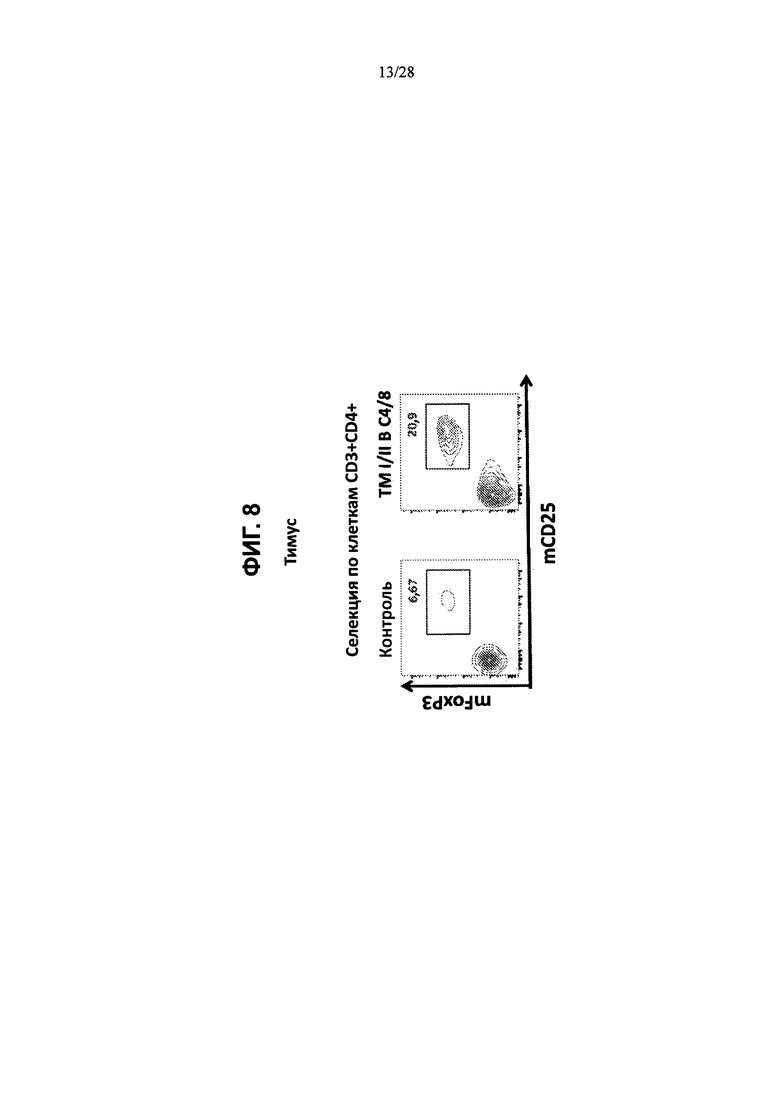

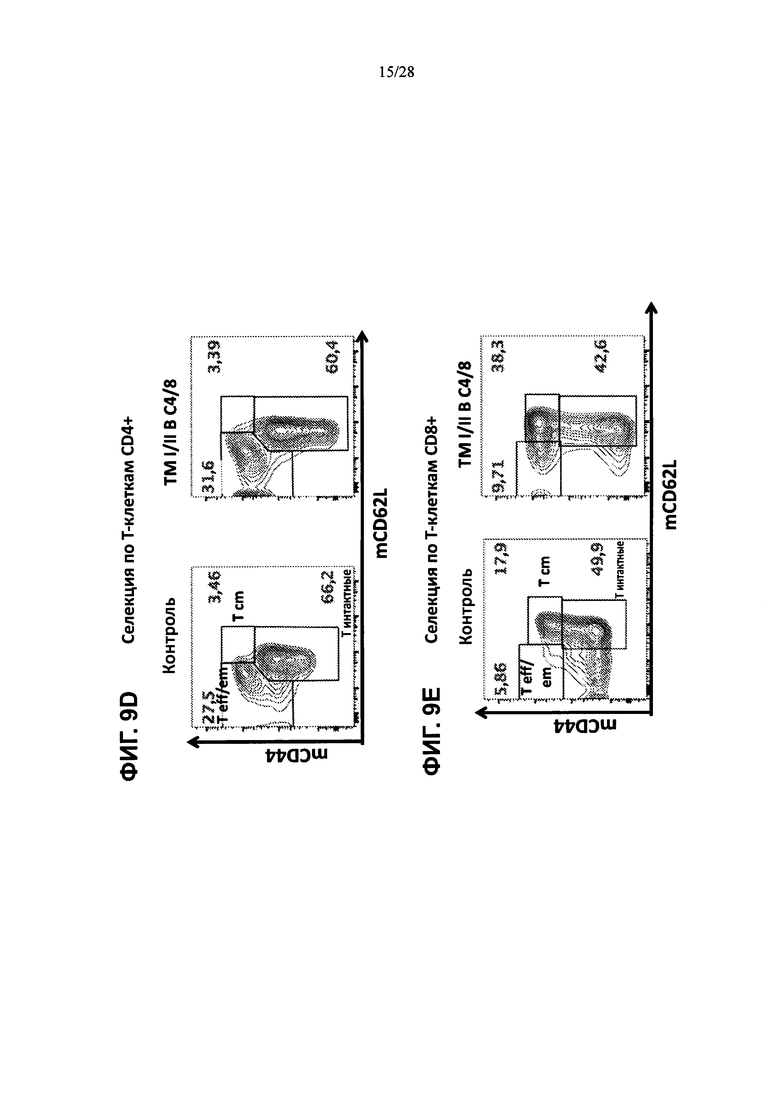
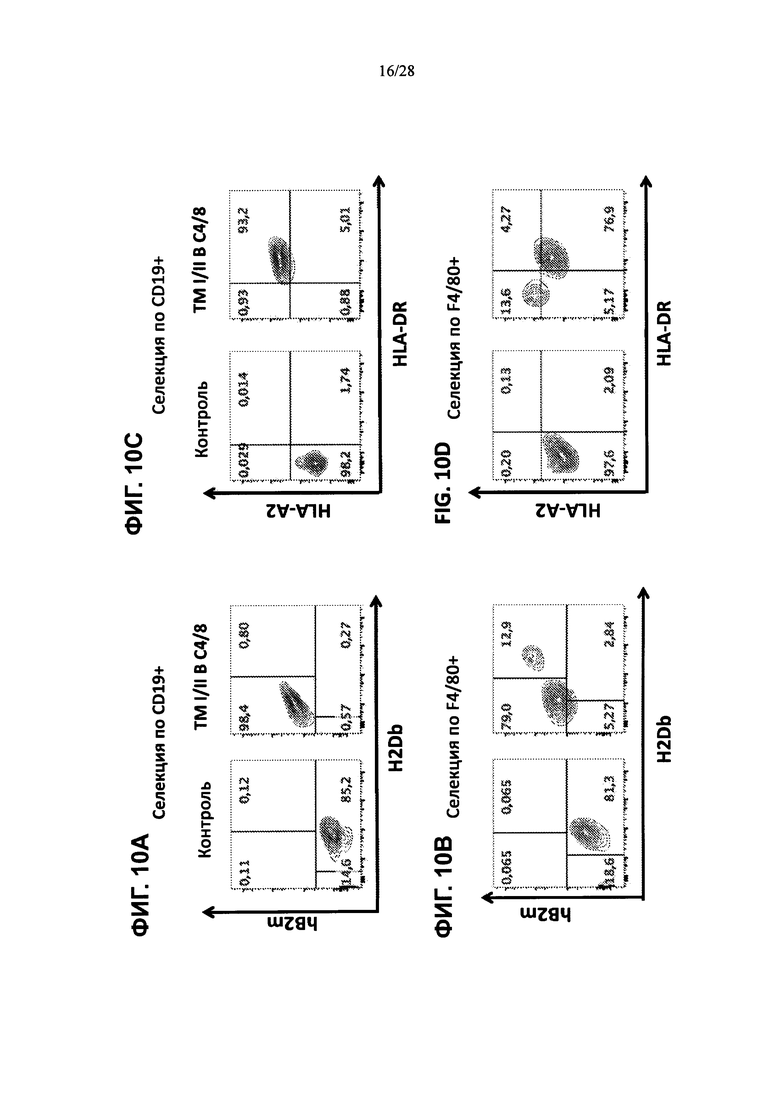

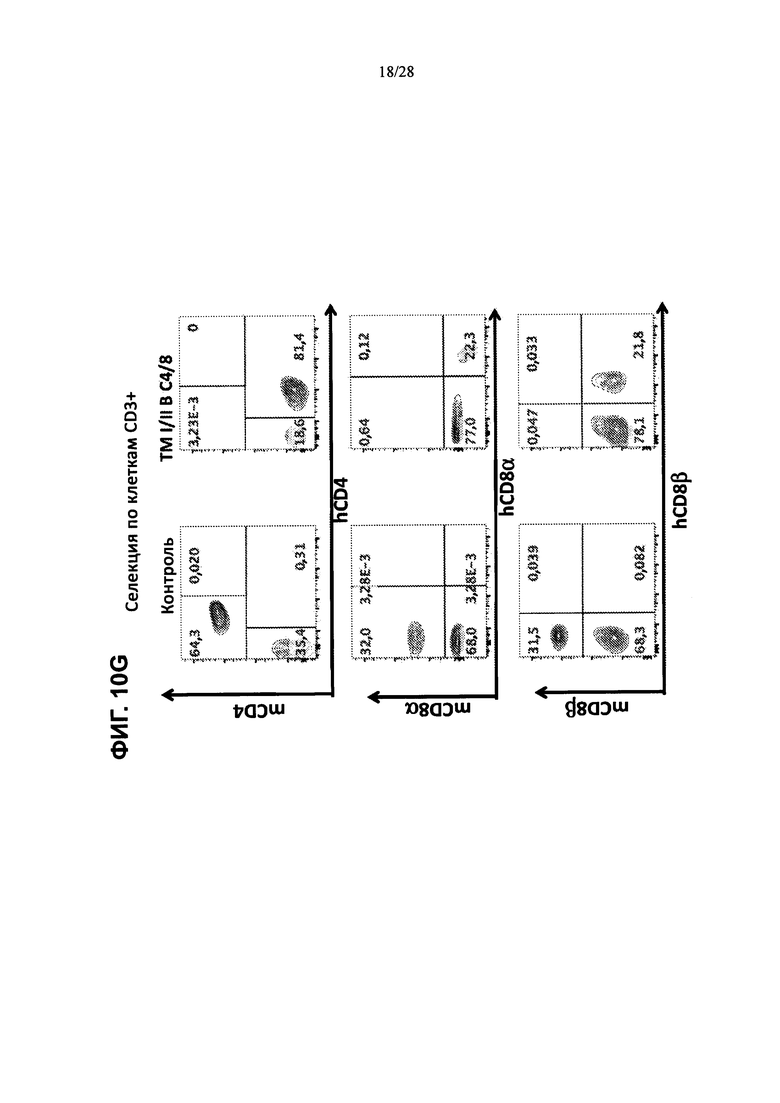
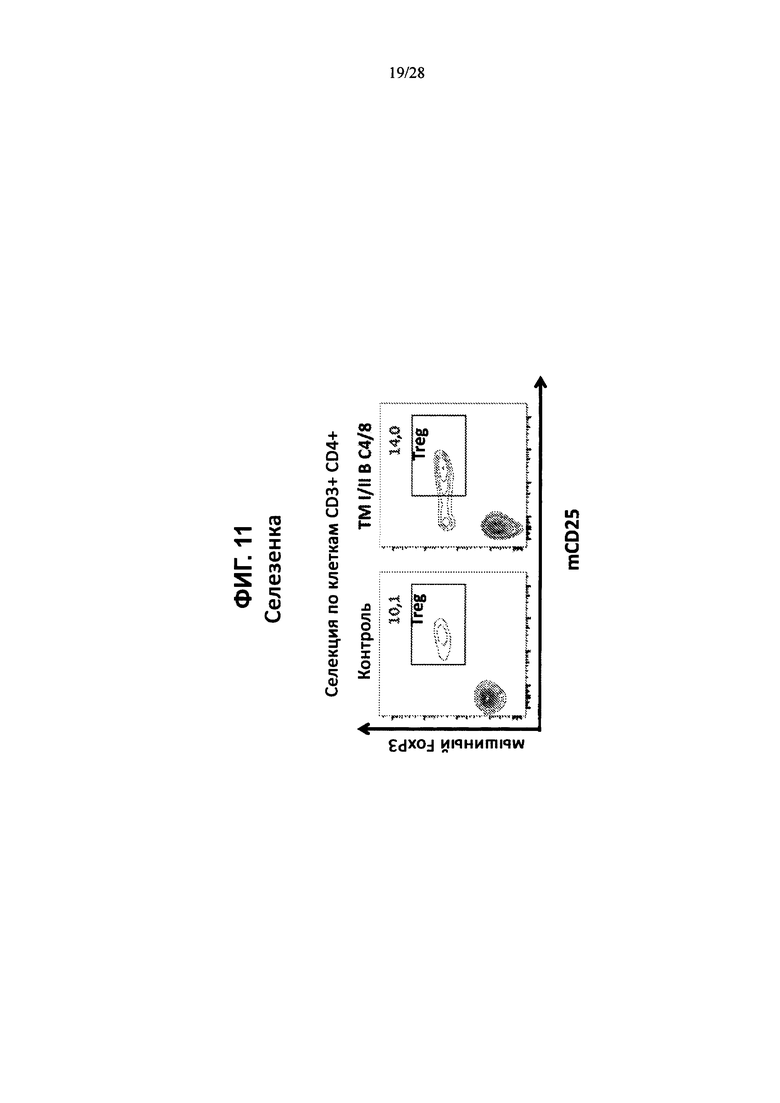
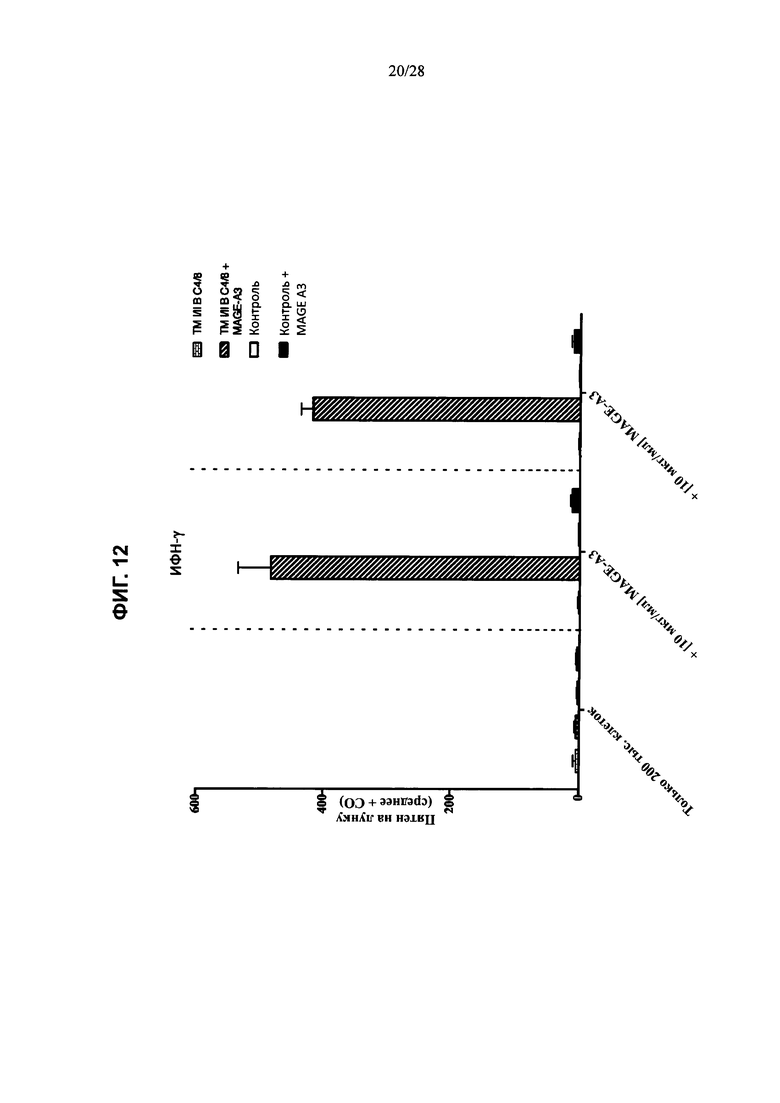
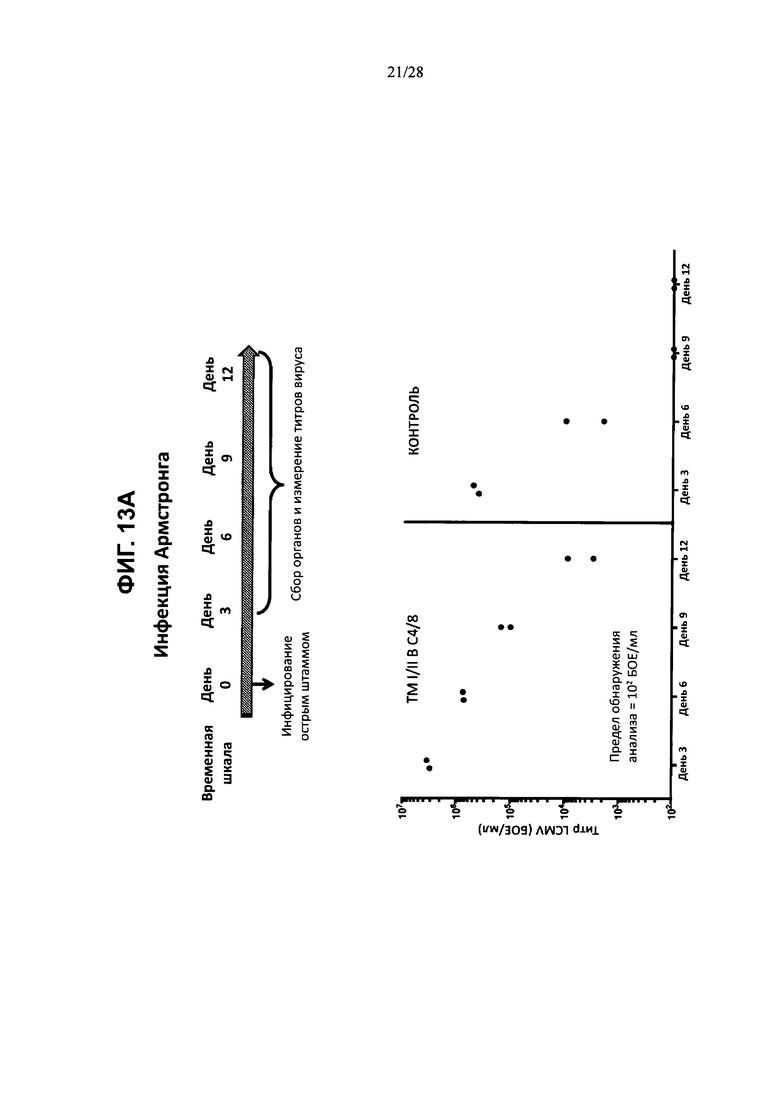
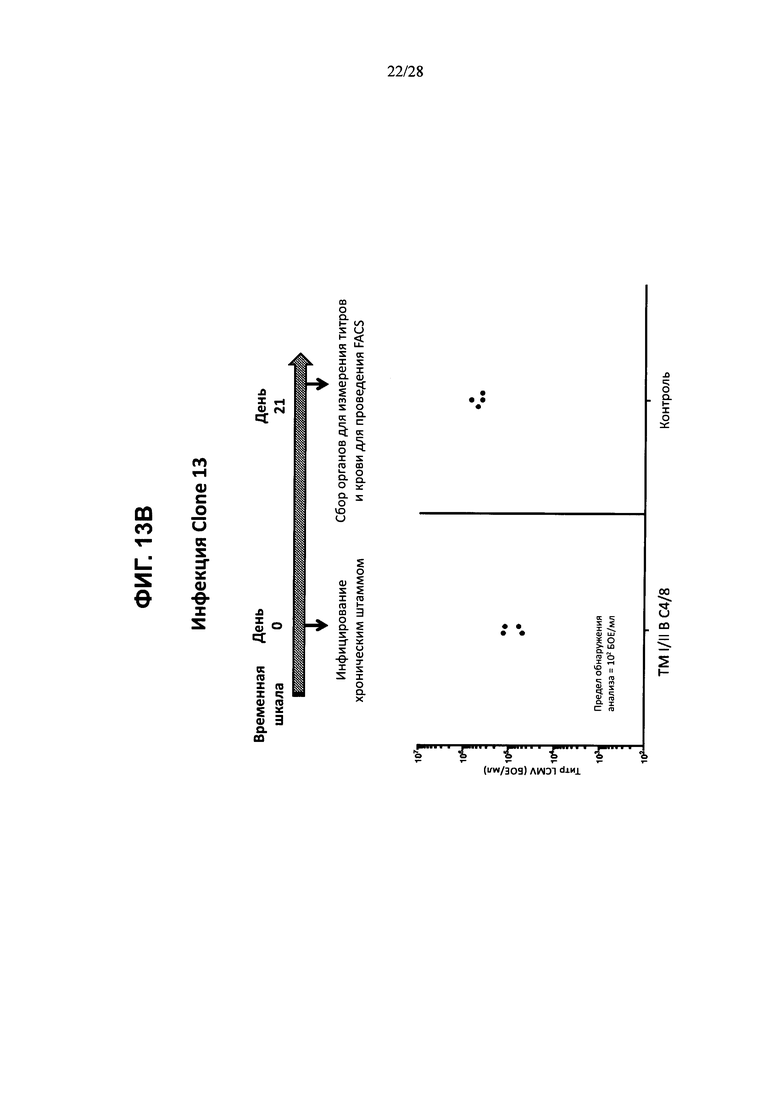
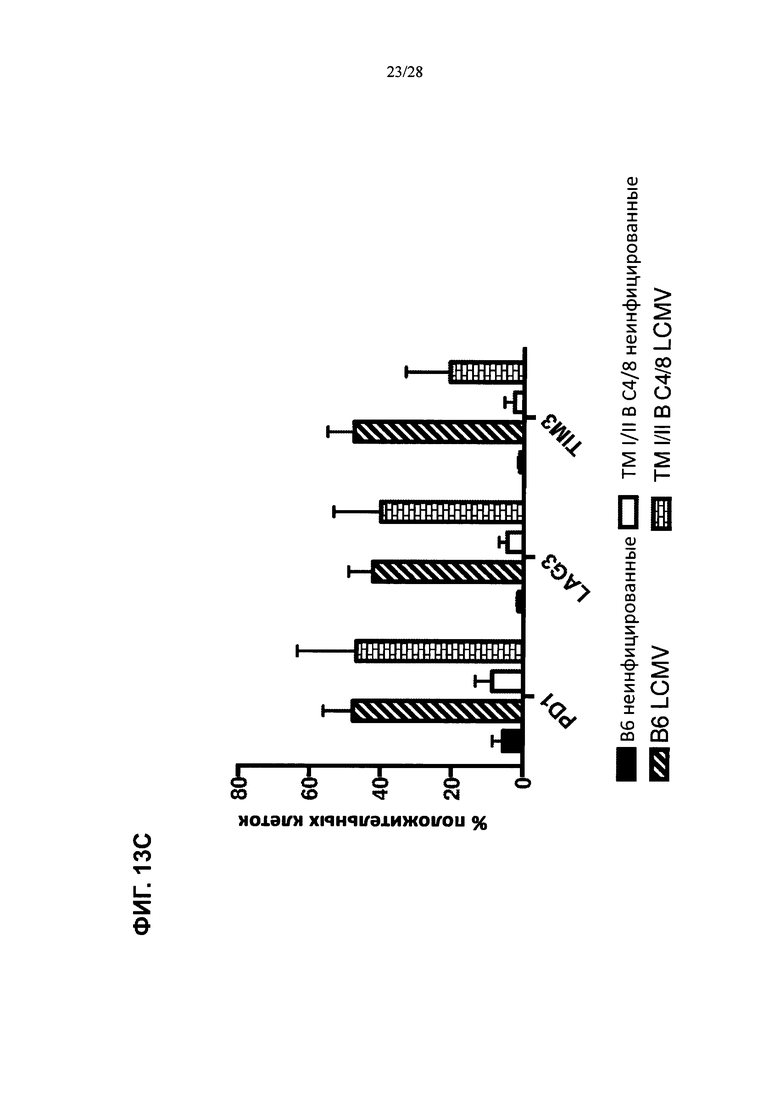
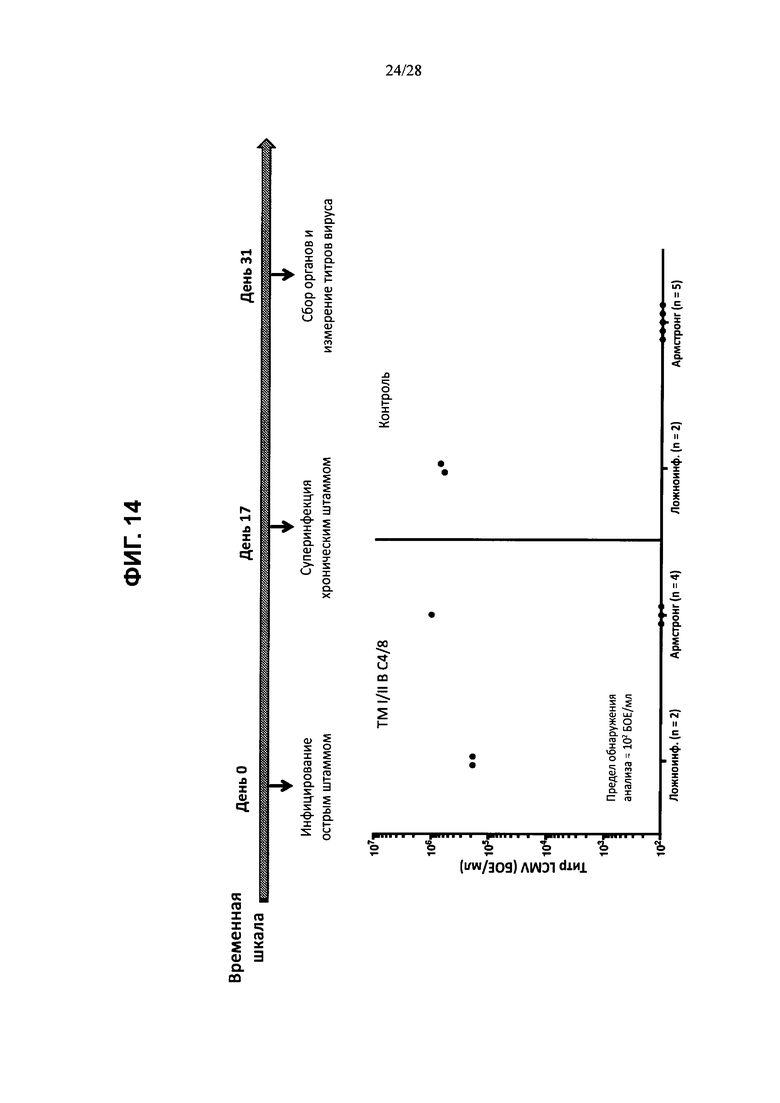
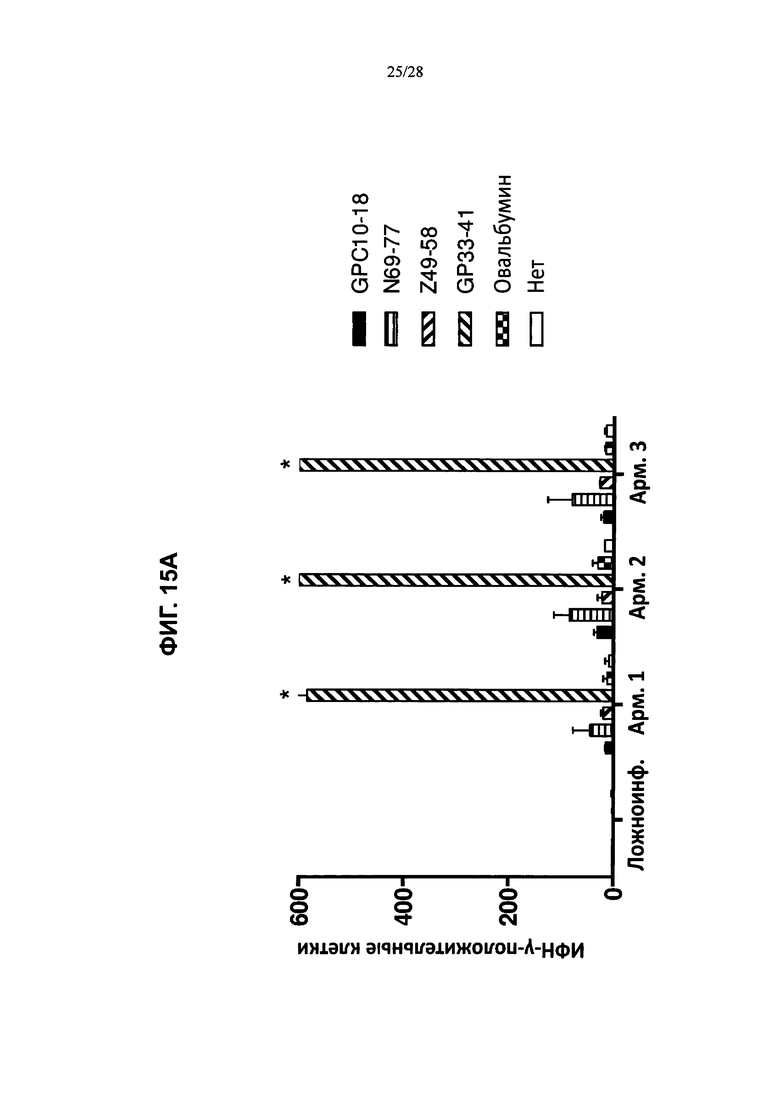
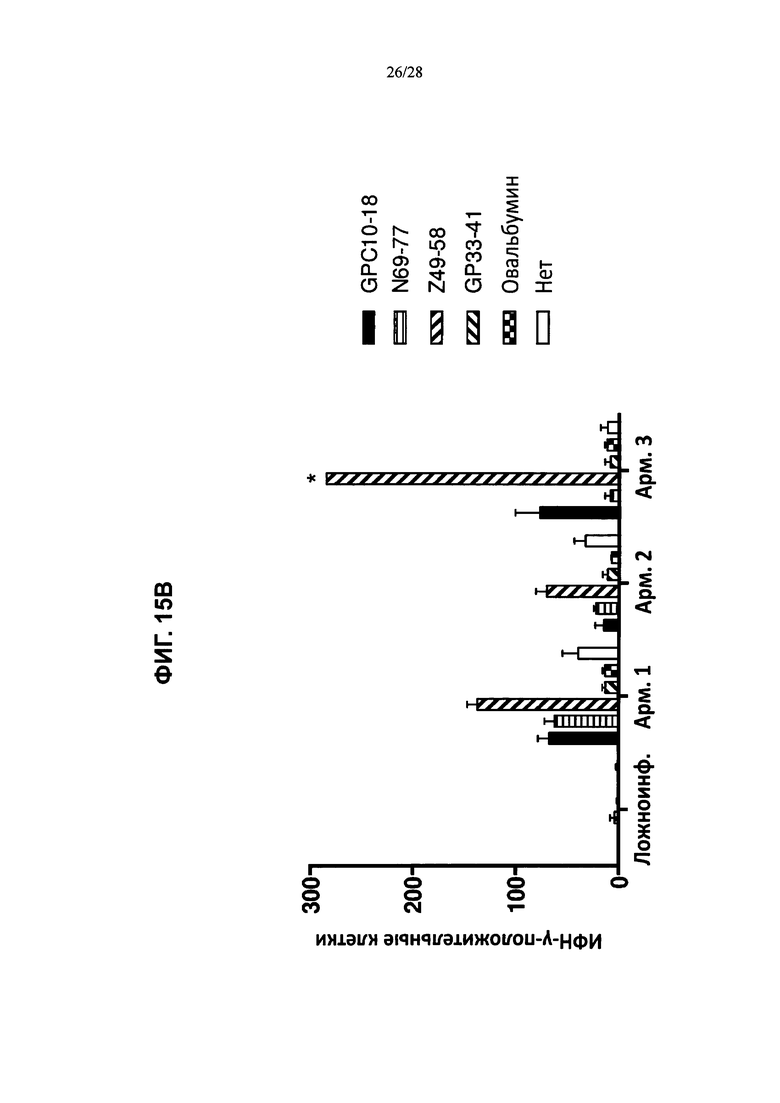
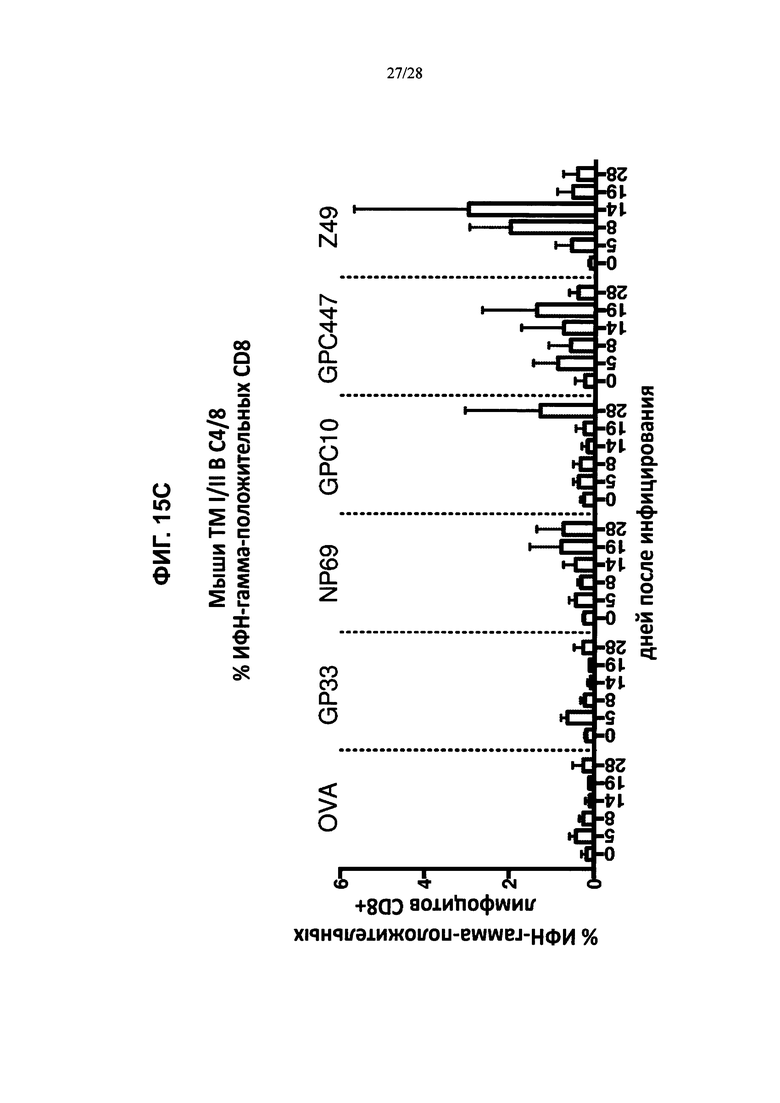
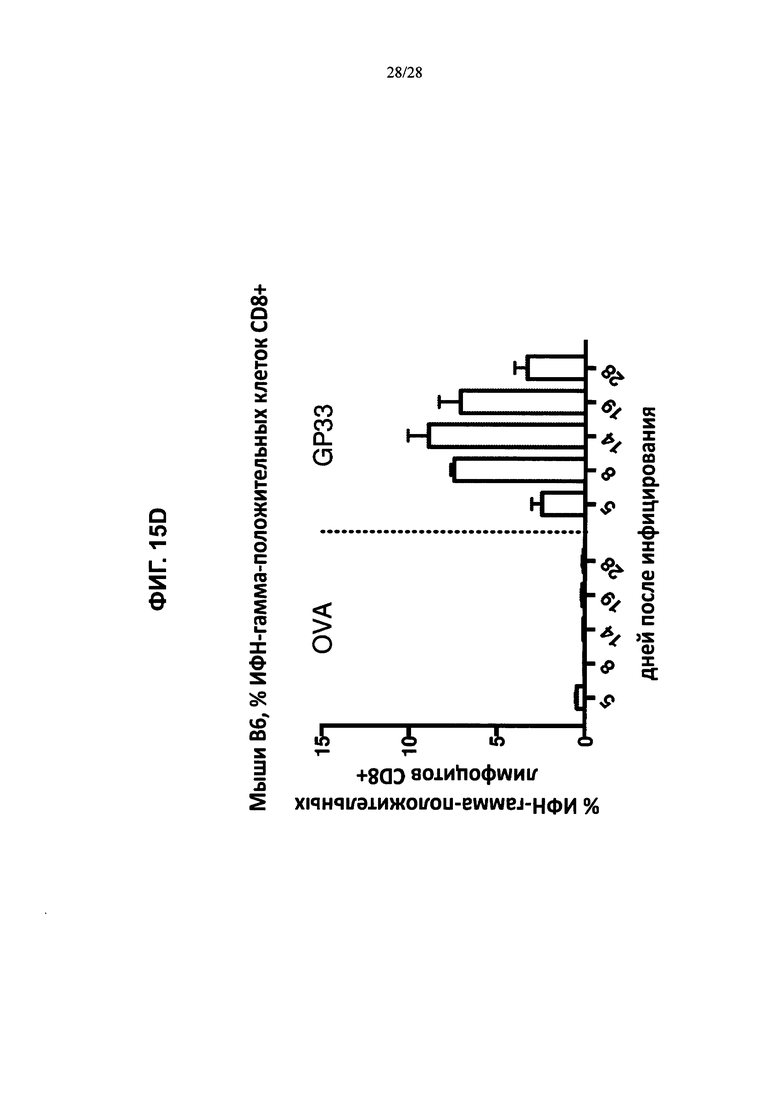
Изобретение относится к области биохимии, в частности к генетически модифицированной мыши для формирования опосредованных Т-клетками иммунных ответов, а также к способу ее получения. Также раскрыты Т-клетка, гибридома и эмбриональная стволовая клетка, полученные из вышеуказанной мыши, а также способ получения белка TCR, который специфичен к антигену и содержит человеческий вариабельный домен TCR, способ получения человеческого вариабельного домена TCR белка TCR, который специфичен к антигену, а также способ получения последовательности нуклеиновой кислоты, кодирующей человеческий вариабельный домен TCR белка TCR. Изобретение также относится к композиции для оценки гуманизированного Т-клеточного иммунного ответа среди мышиных клеток, причем композиция содержит первую и вторую клетку вышеуказанной мыши. Изобретение возможно эффективно использовать для разработки терапевтических средств для человека. 10 н. и 31 з.п. ф-лы, 15 ил., 9 табл., 7 пр.
1. Генетически модифицированная мышь для формирования опосредованных Т-клетками иммунных ответов, причем мышь содержит в своем геноме:
(a) первую нуклеотидную последовательность, кодирующую химерный корецептор CD4, который содержит домены D1, D2 и D3 человеческого полипептида CD4 и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD4 мыши,
при этом мышь экспрессирует химерный корецептор CD4;
(b) вторую нуклеотидную последовательность и третью нуклеотидную последовательность, соответственно кодирующие химерный полипептид CD8α и химерный полипептид CD8β,
при этом химерный полипептид CD8α содержит внеклеточный участок человеческого полипептида CD8α и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8α мыши,
при этом химерный полипептид CD8β содержит внеклеточный участок человеческого полипептида CD8β и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8β мыши, и
при этом мышь экспрессирует химерный корецептор CD8, содержащий химерный полипептид CD8α и химерный полипептид CD8β;
(c) первую последовательность нуклеиновой кислоты и вторую последовательность нуклеиновой кислоты, соответственно кодирующие химерный полипептид МНС II α и химерный полипептид МНС II β,
при этом химерный полипептид МНС II α содержит внеклеточный участок человеческого полипептида HLA класса II α и по меньшей мере трансмембранный и цитоплазматический домены полипептида МНС II α мыши,
при этом химерный полипептид МНС II β содержит внеклеточный участок человеческого полипептида HLA класса II β и по меньшей мере трансмембранный и цитоплазматический домены полипептида МНС II β мыши,
при этом у мыши экспрессируется химерный комплекс МНС II, содержащий химерный полипептид МНС II α и химерный полипептид МНС II β; и
при этом химерный комплекс МНС II ассоциируется с химерным корецептором CD4;
(d) третью последовательность нуклеиновой кислоты, кодирующую химерный полипептид МНС I,
при этом химерный полипептид МНС I содержит внеклеточный участок человеческого полипептида HLA класса I и трансмембранный и цитоплазматический домены полипептида МНС I мыши,
при этом у мыши экспрессируется химерный полипептид МНС I, и
при этом химерный полипептид МНС I ассоциируется с химерным корецептором CD8;
(e) неперестроенную последовательность вариабельного участка Т-клеточного рецептора (TCR) α, содержащую по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vα/Jα, функционально связанной с последовательностью константного участка TCRα мыши; и неперестроенную последовательность вариабельного участка TCRβ, содержащую по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ, функционально связанной с последовательностью константного участка TCRβ мыши,
при этом перестроенная человеческая последовательность Vα/Jα, функционально связанная с последовательностью константного участка TCRα мыши, кодирует гуманизированную цепь TCRα, содержащую человеческий вариабельный домен TCRα, функционально связанный с константным доменом TCRα мыши,
при этом перестроенная человеческая последовательность Vβ/Dβ/Jβ, функционально связанная с последовательностью константного участка TCRβ мыши, кодирует гуманизированную цепь TCRβ, содержащую человеческий вариабельный домен TCRβ, функционально связанный с константным доменом TCRβ мыши, и
при этом у мыши экспрессируется Т-клеточный рецептор на поверхности Т-клетки, причем Т-клеточный рецептор содержит гуманизированную цепь TCRα и гуманизированную цепь TCRβ, и
(f) необязательно, локус β2-микроглобулина, кодирующий человеческий или гуманизированный полипептид β2-микроглобулин.
2. Генетически модифицированная мышь по п. 1, содержащая в своем геноме зародышевой линии:
(a) первую нуклеотидную последовательность, кодирующую химерный корецептор CD4;
(b) вторую и третью нуклеотидные последовательности, соответственно кодирующие химерный полипептид CD8α и химерный полипептид CD8β;
(c) первую и вторую последовательности нуклеиновых кислот, соответственно кодирующие химерный полипептид МНС II α и химерный полипептид МНС II β;
(d) третью последовательность нуклеиновой кислоты, кодирующую химерный полипептид МНС I; и
(e) неперестроенную последовательность вариабельного участка TCRα, содержащую по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vα/Jα, функционально связанной с последовательностью константного участка TCRα мыши, и неперестроенную последовательность вариабельного участка TCRβ, содержащую по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ, функционально связанной с последовательностью константного участка TCRβ мыши.
3. Генетически модифицированная мышь по п. 1, у которой:
(а) первая нуклеотидная последовательность находится в эндогенном локусе Т-клеточного корецептора CD4;
(b) вторая нуклеотидная последовательность находится в эндогенном локусе Т-клеточного корецептора CD8α и третья нуклеотидная последовательность находится в эндогенном локусе Т-клеточного корецептора CD8β;
(c) первая последовательность нуклеиновой кислоты находится в эндогенном локусе МНС II α мыши и вторая последовательность нуклеиновой кислоты находится в эндогенном локусе МНС II β мыши;
(d) третья последовательность нуклеиновой кислоты находится в эндогенном локусе МНС I мыши; и/или
(е) неперестроенная последовательность вариабельного участка TCRα находится в эндогенном локусе вариабельного участка TCRα и неперестроенная последовательность вариабельного участка TCRβ находится в эндогенном локусе вариабельного участка TCRβ.
4. Генетически модифицированная мышь по п. 3, у которой:
(a) первая нуклеотидная последовательность экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов корецептора CD4 мыши;
(b) вторая нуклеотидная последовательность экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов полипептида CD8α мыши и третья нуклеотидная последовательность экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов полипептида CD8β мыши;
(c) первая последовательность нуклеиновой кислоты экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов МНС II α мыши и вторая последовательность нуклеиновой кислоты экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов МНС II β мыши;
(d) третья последовательность нуклеиновой кислоты экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов МНС I мыши; и/или
(e) неперестроенная последовательность вариабельного участка TCRα экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов TCRα и неперестроенная последовательность вариабельного участка TCRβ экспрессируется под регуляторным контролем эндогенных промоторных и регуляторных элементов TCRβ.
5. Генетически модифицированная мышь по п. 1, у которой:
(a) химерный корецептор CD4 содержит домены D1, D2 и D3 человеческого полипептида CD4, функционально связанные с D4, трансмембранным и цитоплазматическим доменами эндогенного полипептида CD4 мыши,
(b) химерный полипептид CD8α содержит IgV-подобный домен человеческого полипептида CD8α, функционально связанный с трансмембранным и цитоплазматическим доменами эндогенного полипептида CD8α мыши, и химерный полипептид CD8β содержит IgV-подобный домен человеческого полипептида CD8β, функционально связанный с трансмембранным и цитоплазматическим доменами эндогенного полипептида CD8β мыши;
(c) химерный полипептид МНС II α содержит домены α1 и α2 человеческого HLA класса II, функционально связанные с трансмембранным и цитоплазматическим доменами мыши эндогенного полипептида МНС II α мыши, и химерный полипептид МНС II β содержит домены β1 и β2 человеческого HLA класса II, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС II β мыши; и/или
(d) химерный полипептид МНС I содержит домены α1, α2 и α3 человеческого HLA класса I, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС I мыши.
6. Генетически модифицированная мышь по п. 1, у которой человеческий внеклеточный участок химерного полипептида МНС II α кодируется геном человеческого лейкоцитарного антигена (HLA) класса II, выбранным из группы, состоящей из любого гена цепи α HLA-DR, HLA-DQ и HLA-DP;
при этом человеческий внеклеточный участок химерного полипептида МНС II β кодируется геном HLA класса II, выбранным из группы, состоящей из любого гена цепи β HLA-DR, HLA-DQ и HLA-DP; и/или
при этом внеклеточный участок химерного полипептида МНС I кодируется человеческим геном HLA-A, человеческим геном HLA-B или человеческим геном HLA-C.
7. Генетически модифицированная мышь по п. 6, у которой человеческие внеклеточные участки химерного полипептида МНС II α и химерного полипептида МНС II β соответственно кодируются геном цепи α и геном цепи β HLA-DR, и
при этом внеклеточный участок химерного полипептида МНС I кодируется человеческим геном HLA-A.
8. Генетически модифицированная мышь по п. 7, у которой химерный комплекс МНС II содержит домены α1, α2, β1 и β2 человеческого белка HLA-DR2.
9. Генетически модифицированная мышь по п. 7, у которой химерный полипептид МНС I содержит домены α1, α2 и α3 человеческого белка HLA-A2.
10. Генетически модифицированная мышь по п. 1, у которой неперестроенная последовательность вариабельного участка TCRα содержит набор человеческих сегментов гена Vα и набор человеческих сегментов гена Jα и/или неперестроенная последовательность вариабельного участка TCRβ содержит набор человеческих сегментов гена Vβ, набор человеческих сегментов гена Dβ и набор человеческих сегментов гена Jβ.
11. Генетически модифицированная мышь по п. 1, причем в эндогенном вариабельном локусе TCRα мыши отсутствуют все или по существу все функциональные эндогенные сегменты гена Vα и/или отсутствуют все или по существу все функциональные эндогенные сегменты гена Jα; и/или
причем в эндогенном вариабельном локусе TCRβ мыши (а) отсутствуют все или по существу все функциональные эндогенные сегменты гена Vβ, (b) отсутствуют все или по существу все функциональные эндогенные сегменты гена Dβ, (с) отсутствуют все или по существу все функциональные эндогенные сегменты гена Jβ или (d) любая комбинация (а), (b) и (с).
12. Генетически модифицированная мышь по п. 1, у которой:
(a) первая нуклеотидная последовательность содержит последовательность, кодирующую домены D1, D2 и D3 человеческого полипептида CD4, которая (i) замещает последовательность, кодирующую домены D1, D2 и D3 эндогенного полипептида корецептора CD4 мыши, и (ii) функционально связана с эндогенными последовательностями, кодирующими D4, трансмембранный и цитоплазматический домены CD4 мыши, в эндогенном локусе корецептора CD4 мыши;
(b) вторая нуклеотидная последовательность содержит последовательность, кодирующую внеклеточный участок человеческого полипептида CD8α, которая (i) замещает последовательность, кодирующую внеклеточный участок эндогенного полипептида CD8α мыши, и (ii) функционально связана с эндогенными последовательностями, кодирующими трансмембранный и цитоплазматический домены CD8α мыши, в эндогенном локусе полипептида CD8α мыши, и
третья нуклеотидная последовательность содержит последовательность, кодирующую внеклеточный участок человеческого полипептида CD8β, которая (i) замещает последовательность, кодирующую внеклеточный участок эндогенного полипептида CD8β мыши, и (ii) функционально связана с эндогенными последовательностями, кодирующими трансмембранный и цитоплазматический домены CD8β мыши, в эндогенном локусе полипептида CD8β;
(c) первая последовательность нуклеиновой кислоты содержит последовательность, кодирующую внеклеточный участок человеческого полипептида HLA класса II α, которая (i) замещает последовательность, кодирующую внеклеточный участок эндогенного полипептида МНС II α мыши, и (ii) функционально связана с эндогенными последовательностями, кодирующими трансмембранный и цитоплазматический домены полипептида МНС II α, в эндогенном локусе МНС II α мыши, и
вторая нуклеиновая кислота содержит последовательность, кодирующую внеклеточный участок человеческого полипептида HLA класса II β, которая (i) замещает последовательность, кодирующую внеклеточный участок эндогенного полипептида МНС II β мыши, и (ii) функционально связана с эндогенными последовательностями, кодирующими трансмембранный и цитоплазматический домены полипептида МНС II β, в эндогенном локусе МНС II β мыши;
(d) третья последовательность нуклеиновой кислоты содержит последовательность, кодирующую внеклеточный участок человеческого полипептида HLA класса I, которая (i) замещает последовательность, кодирующую внеклеточный участок эндогенного полипептида МНС I мыши, и (ii) функционально связана с эндогенными последовательностями, кодирующими трансмембранный и цитоплазматический домены полипептида МНС I, в эндогенном локусе МНС I мыши; и/или
(е) неперестроенная последовательность вариабельного участка TCRα замещает один или более эндогенных сегментов гена Vα и/или Jα в эндогенном локусе вариабельного участка TCRα и неперестроенная последовательность вариабельного участка TCRβ замещает один или более эндогенных сегментов гена Vβ, Dβ и/или Jβ в эндогенном локусе вариабельного участка TCRβ.
13. Генетически модифицированная мышь по п. 1, причем у мыши не экспрессируется (а) функциональный эндогенный корецептор CD4 и/или CD8 мыши из эндогенных локусов корецептора CD4 и/или CD8 соответственно, (b) эндогенный вариабельный домен TCRα из эндогенного локуса TCRα, (с) эндогенный вариабельный домен TCRβ из эндогенного локуса TCRβ и/или (d) внеклеточный домен эндогенного полипептида классического МНС класса I и/или класса II из своего эндогенного локуса МНС на клеточной поверхности.
14. Генетически модифицированная мышь по п. 1, дополнительно содержащая (f) локус β2-микроглобулина, кодирующий полипептид, содержащий человеческую аминокислотную последовательность β2-микроглобулина, причем у мыши экспрессируется человеческий или гуманизированный полипептид β2-микроглобулин.
15. Генетически модифицированная мышь по п. 14, причем у мыши не экспрессируется функциональный эндогенный полипептид β2-микроглобулин мыши из эндогенного локуса β2-микроглобулина мыши.
16. Генетически модифицированная мышь по п. 14, у которой локус β2-микроглобулина функционально связан с эндогенными регуляторными элементами β2-микроглобулина мыши.
17. Генетически модифицированная мышь по п. 14, у которой локус β2-микроглобулина содержит нуклеотидную последовательность, представленную в экзоне 2, экзоне 3 и экзоне 4 человеческого гена β2-микроглобулина.
18. Генетически модифицированная мышь по п. 17, у которой локус β2-микроглобулина дополнительно содержит нуклеотидную последовательность, представленную в экзоне 1 гена β2-микроглобулина мыши.
19. Генетически модифицированная мышь по п. 1,
при этом мышь экспрессирует:
химерный человеческий/мышиный корецептор CD4, содержащий домены D1, D2 и D3 человеческого полипептида CD4, функционально связанные с по меньшей мере трансмембранным и цитоплазматическим доменами мышиного полипептида CD4,
полипептиды корецепторов CD8α и CD8β, каждый из которых соответственно содержит внеклеточный участок человеческого полипептида корецептора CD8α и CD8β, функционально связанный с мышиными трансмембранным и цитоплазматическим доменами CD8α и CD8β;
химерный человеческий/мышиный Т-клеточный рецептор, содержащий человеческий вариабельный участок TCRα и человеческий вариабельный участок TCRβ, каждый из которых соответственно функционально связан с мышиным константным участком TCRα и мышиным константным участком TCRβ на поверхности Т-клетки;
химерные человеческие/мышиные полипептиды МНС II α, МНС II β и МНС I, каждый из которых соответственно содержит внеклеточный участок человеческого полипептида HLA класса II α, HLA класса II β и HLA класса I, функционально связанный с мышиными трансмембранным и цитоплазматическим доменами мышиного полипептида МНС класса II α, МНС класса II β и МНС класса I; и
гуманизированный полипептид β2-микроглобулин.
20. Генетически модифицированная мышь по п. 1, причем у мыши экспрессируются по меньшей мере 50% всех функциональных человеческих сегментов гена TCRVα и/или по меньшей мере 50% всех функциональных человеческих сегментов гена TCRVβ.
21. Генетически модифицированная мышь по п. 19, у которой:
(a) химерный человеческий/мышиный корецептор CD4 содержит домены D1, D2 и D3 человеческого полипептида CD4, функционально связанные с D4, трансмембранным и цитоплазматическим доменами эндогенного мышиного полипептида CD4,
(b) химерный человеческий/мышиный полипептид CD8α содержит IgV-подобный домен человеческого полипептида CD8α, функционально связанный с трансмембранным и цитоплазматическим доменами эндогенного мышиного полипептида CD8α, и химерный человеческий/мышиный полипептид корецептора CD8β содержит IgV-подобный домен человеческого полипептида CD8β, функционально связанный с трансмембранным и цитоплазматическим доменами эндогенного мышиного полипептида CD8β;
(c) химерный человеческий/мышиный полипептид МНС II α содержит домены α1 и α2 человеческого HLA класса II, функционально связанные с мышиными трансмембранным и цитоплазматическим доменами эндогенного мышиного полипептида МНС II α, и химерный человеческий/мышиный полипептид МНС II β содержит домены β1 и β2 человеческого HLA класса II, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного мышиного полипептида МНС II β,
(d) химерный полипептид МНС I содержит домены α1, α2 и α3 человеческого HLA класса I, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного мышиного полипептида МНС I; и/или
(e) гуманизированный полипептид β2-микроглобулин, кодируемый нуклеотидной последовательностью, содержащей нуклеотидную последовательность, представленную в экзоне 1 эндогенного мышиного гена β2-микроглобулина, функционально связанную с нуклеотидной последовательностью, представленной в экзоне 2, экзоне 3 и экзоне 4 человеческого гена β2-микроглобулина.
22. Генетически модифицированная мышь по п. 21, у которой химерный человеческий/мышиный полипептид МНС II α представляет собой химерный полипептид HLA-DR/H-2E α, химерный полипептид МНС II β представляет собой химерный человеческий/мышиный полипептид HLA-DR/H-2E β и химерный полипептид МНС I представляет собой химерный человеческий/мышиный полипептид HLA-A/H-2K, и причем у мыши экспрессируются белки HLA-A/H-2K и HLA-DR/H-2E.
23. Способ создания генетически модифицированной мыши по п. 1, включающий модификацию генома мыши для содержания:
(a) первой нуклеотидной последовательности, кодирующей химерный корецептор CD4;
(b) второй и третьей нуклеотидных последовательностей, соответственно кодирующих химерный полипептид CD8α и химерный полипептид CD8β;
(c) первой и второй последовательностей нуклеиновых кислот, соответственно кодирующих химерный полипептид МНС II α и химерный полипептид МНС II β;
(d) третьей последовательности нуклеиновой кислоты, кодирующей химерный полипептид МНС I;
(e) неперестроенной последовательности вариабельного участка TCRα, содержащей по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vα/Jα, функционально связанной с последовательностью константного участка TCRα мыши, и неперестроенной последовательности вариабельного участка TCRβ, содержащей по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ, функционально связанной с последовательностью константного участка TCRβ мыши; и
(f) необязательно локуса β2-микроглобулина, кодирующего человеческий или гуманизированный полипептид β2-микроглобулин.
24. Способ по п. 23, в котором модификация генома включает гомологичную рекомбинацию в одной или более эмбриональных стволовых клетках мыши с тем, чтобы в геном эмбриональной стволовой клетки мыши добавить в любом порядке первую, вторую и третью нуклеотидные последовательности, неперестроенную последовательность вариабельного участка TCRα и неперестроенную последовательность вариабельного участка TCRβ, первую, вторую и третью последовательность нуклеиновой кислоты и необязательно локус β2-микроглобулина.
25. Способ по п. 24, дополнительно включающий создание мыши из эмбриональной стволовой клетки мыши, содержащей
первую, вторую и третью нуклеотидные последовательности, неперестроенную последовательность вариабельного участка TCRα и неперестроенную последовательность вариабельного участка TCRβ, первую, вторую и третью последовательность нуклеиновой кислоты и необязательно локус β2-микроглобулина.
26. Способ получения клетки, которая экспрессирует белок TCR, который специфичен к антигену и содержит человеческий вариабельный домен TCR,
причем способ включает
выделение из мыши по п. 1 Т-клетки, экспрессирующей белок TCR, который специфичен к антигену и содержит как человеческий вариабельный домен TCR α, так и человеческий вариабельный домен TCR β.
27. Способ получения белка TCR, который специфичен к антигену и содержит человеческий вариабельный домен TCR,
причем способ включает
получение из мыши по п. 1 Т-клетки, экспрессирующей белок TCR, который специфичен к антигену и содержит как человеческий вариабельный домен TCR α, так и человеческий вариабельный домен TCR β, и выделение белка TCR.
28. Способ получения человеческого вариабельного домена TCR белка TCR, который специфичен к антигену,
причем способ включает
получение из мыши по п. 1 белка TCR, который специфичен к антигену и содержит как человеческий вариабельный домен TCR α, так и человеческий вариабельный домен TCR β, и выделение человеческого вариабельного домена TCR α и/или человеческого вариабельного домена TCR β.
29. Способ получения последовательности нуклеиновой кислоты, кодирующей человеческий вариабельный домен TCR белка TCR, который специфичен к антигену,
причем способ включает получение из мыши по п. 1 белка TCR, который специфичен к антигену и содержит как человеческий вариабельный домен TCR α, так и человеческий вариабельный домен TCR β, и выделение последовательности нуклеиновой кислоты, кодирующей человеческий вариабельный домен TCR α, и/или последовательности нуклеиновой кислоты, кодирующей человеческий вариабельный домен TCR β.
30. Способ по любому из пп. 26-28, дополнительно включающий:
внедрение в клетку-хозяина одного или более экспрессионных векторов, содержащих (i) последовательность нуклеиновой кислоты, кодирующую человеческий вариабельный домен TCR α в функциональной взаимосвязи с человеческим константным участком TCR α, и/или (ii) последовательность нуклеиновой кислоты, кодирующую человеческий вариабельный домен TCR β в функциональной взаимосвязи с человеческим константным участком TCR β,
культивирование клетки-хозяина в условиях, достаточных для экспрессии (i) и/или (ii),
причем последовательности нуклеиновой кислоты, кодирующие человеческий вариабельный домен TCR α и человеческий вариабельный домен TCR β находятся на одном или разных экспрессионных векторах.
31. Способ по п. 30, в котором один или более экспрессионных векторов содержат последовательность нуклеиновой кислоты, кодирующую человеческий вариабельный домен TCRα, функционально связанную с последовательностью, кодирующей человеческий константный участок TCRα, и последовательность нуклеиновой кислоты, кодирующую человеческий вариабельный домен TCRβ, функционально связанную с последовательностью, кодирующей человеческий константный участок TCRβ.
32. Способ по любому из пп. 26-31, в котором антиген представляет собой опухолевый антиген, вирусный антиген или бактериальный антиген.
33. Т-клетка, экспрессирующая белок TCR, содержащий человеческий вариабельный домен TCR, специфичный к антигену, содержащая в своем геноме
(а) первую нуклеотидную последовательность, кодирующую химерный корецептор CD4, который содержит домены D1, D2 и D3 человеческого полипептида CD4 и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD4 мыши,
(b) вторую нуклеотидную последовательность и третью нуклеотидную последовательность, соответственно кодирующие химерный полипептид CD8α и химерный полипептид CD8β,
при этом химерный полипептид CD8α содержит внеклеточный участок человеческого полипептида CD8α и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8α мыши,
при этом химерный полипептид CD8β содержит внеклеточный участок человеческого полипептида CD8β и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8β мыши,
(c) первую последовательность нуклеиновой кислоты и вторую последовательность нуклеиновой кислоты, соответственно кодирующие химерный полипептид МНС II α и химерный полипептид МНС II β,
при этом химерный полипептид МНС II α содержит внеклеточный участок человеческого полипептида HLA класса II α и по меньшей мере трансмембранный и цитоплазматический домены полипептида МНС II α мыши,
при этом химерный полипептид МНС II β содержит внеклеточный участок человеческого полипептида HLA класса II β и по меньшей мере трансмембранный и цитоплазматический домены полипептида МНС II β мыши,
(d) третью последовательность нуклеиновой кислоты, кодирующую химерный полипептид МНС I,
при этом химерный полипептид МНС I содержит внеклеточный участок человеческого полипептида HLA класса I и трансмембранный и цитоплазматический домены полипептида МНС I мыши, и
(e) перестроенную человеческую последовательность Vα/Jα, функционально связанную с последовательностью константного участка TCRα мыши, которая кодирует гуманизированную цепь TCRα, содержащую человеческий вариабельный домен TCRα, функционально связанный с константным доменом TCRα мыши; и перестроенную человеческую последовательность Vβ/Dβ/Jβ, функционально связанную с последовательностью константного участка TCRβ мыши, которая кодирует гуманизированную цепь TCRβ, содержащую человеческий вариабельный домен TCRβ, функционально связанный с константным доменом TCRβ мыши, и где белок TCR содержит гуманизированную цепь TCRα и гуманизированную цепь TCRβ.
34. Гибридома, экспрессирующая белок TCR, содержащий человеческий вариабельный домен TCR, специфичный к антигену, причем гибридома получена из Т-клетки, экспрессирующей белок TCR, содержащий человеческий вариабельный домен TCR, специфичный к антигену, и
при этом гибридома содержит:
(a) первую нуклеотидную последовательность, кодирующую химерный корецептор CD4, которая содержит домены D1, D2 и D3 человеческого полипептида CD4 и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD4 мыши;
(b) вторую и третью нуклеотидные последовательности, соответственно кодирующие химерный полипептид CD8α и химерный полипептид CD8β;
при этом химерный полипептид CD8α содержит внеклеточный участок человеческого полипептида CD8α и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8α мыши,
при этом химерный полипептид CD8β содержит внеклеточный участок человеческого полипептида CD8β и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8β мыши, и
(c) перестроенную человеческую последовательность Vα/Jα, функционально связанную с последовательностью константного участка TCRα мыши, которая кодирует гуманизированную цепь TCRα, содержащую человеческий вариабельный домен TCRα, функционально связанный с константным доменом TCRα мыши, и перестроенную человеческую последовательность Vβ/Dβ/Jβ, функционально связанную с последовательностью константного участка TCRβ мыши, которая кодирует гуманизированную цепь TCRβ, содержащую человеческий вариабельный домен TCRβ, функционально связанный с константным доменом TCRβ мыши, и
при этом гибридома экспрессирует Т-клеточный рецептор на своей поверхности, причем Т-клеточный рецептор содержит гуманизированную цепь TCRα и гуманизированную цепь TCRβ.
35. Композиция для оценки гуманизированного Т-клеточного иммунного ответа среди мышиных клеток, причем композиция содержит первую и вторую клетку мыши по п. 1, причем первая и вторая клетки, каждая, содержат:
(a) первую нуклеотидную последовательность, кодирующую химерный корецептор CD4;
(b) вторую и третью нуклеотидные последовательности, соответственно кодирующие химерный полипептид CD8α и химерный полипептид CD8β;
(c) первую и вторую последовательности нуклеиновых кислот, соответственно кодирующие химерный полипептид МНС II α и химерный полипептид МНС II β;
(d) третью последовательность нуклеиновой кислоты, кодирующую химерный полипептид МНС I;
(e) неперестроенную последовательность вариабельного участка TCRα, содержащую по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vα/Jα, функционально связанной с последовательностью константного участка TCRα мыши, и неперестроенную последовательность вариабельного участка TCRβ, содержащую по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, которые перестраиваются в Т-клетке с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ, функционально связанной с последовательностью константного участка TCRβ мыши, и
f) необязательно, локус β2-микроглобулина, кодирующий человеческий или гуманизированный полипептид β2-микроглобулин,
причем первая клетка экспрессирует
(a) химерный корецептор CD4 и/или химерный корецептор CD8, содержащий химерный полипептид CD8α и химерный полипептид CD8β, и
(b) химерный TCR человека/мыши, содержащий одну или обе из (i) химерной цепи TCRα человека/мыши, кодируемой перестроенной человеческой последовательностью Vα/Jα, функционально связанной с последовательностью константного участка TCRα мыши, и (ii) химерной цепи TCRβ человека/мыши, кодируемой перестроенной человеческой последовательностью Vβ/Dβ/Jβ, функционально связанной с последовательностью константного участка TCRβ мыши, и
причем у второй клетки экспрессируется химерный полипептид МНС I и, необязательно, человеческий или гуманизированный β2-микроглобулин.
36. Композиция по п. 35, в которой первая клетка представляет собой Т-клетку мыши.
37. Композиция по п. 36, в которой вторая клетка представляет собой антиген представляющую клетку мыши, и причем вторая клетка дополнительно экспрессирует химерный комплекс МНС II, содержащий химерный полипептид МНС II α и химерный полипептид МНС II β.
38. Эмбриональная стволовая клетка генетически модифицированной мыши для создания мыши, которая формирует опосредованные Т-клетками иммунные ответы, причем эмбриональная стволовая клетка мыши содержит в своем геноме:
(a) первую нуклеотидную последовательность, кодирующую химерный корецептор CD4, который содержит домены D1, D2 и D3 человеческого полипептида CD4 и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD4 мыши,
(b) вторую нуклеотидную последовательность и третью нуклеотидную последовательность, соответственно кодирующие химерный полипептид CD8α и химерный полипептид CD8β,
при этом химерный полипептид CD8α содержит внеклеточный участок человеческого полипептида CD8α и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8α мыши,
при этом химерный полипептид CD8β содержит внеклеточный участок человеческого полипептида CD8β и по меньшей мере трансмембранный и цитоплазматический домены полипептида CD8β мыши,
(c) первую последовательность нуклеиновой кислоты и вторую последовательность нуклеиновой кислоты, соответственно кодирующие химерный полипептид МНС II α и химерный полипептид МНС II β,
при этом химерный полипептид МНС II α содержит внеклеточный участок человеческого полипептида HLA класса II α и по меньшей мере трансмембранный и цитоплазматический домены полипептида МНС II α мыши,
при этом химерный полипептид МНС II β содержит внеклеточный участок человеческого полипептида HLA класса II β и по меньшей мере трансмембранный и цитоплазматический домены полипептида МНС II β мыши,
(d) третью последовательность нуклеиновой кислоты, кодирующую химерный полипептид МНС I,
при этом химерный полипептид МНС I содержит внеклеточный участок человеческого полипептида HLA класса I и трансмембранный и цитоплазматический домены полипептида МНС I мыши, и
(e) неперестроенную последовательность вариабельного участка Т-клеточного рецептора (TCR) α, содержащую по меньшей мере один человеческий сегмент Vα и по меньшей мере один человеческий сегмент Jα, которые способны перестраиваться в Т-клетке с образованием перестроенной человеческой последовательности Vα/Jα, функционально связанной с последовательностью константного участка TCRα мыши; и неперестроенную последовательность вариабельного участка TCRβ, содержащую по меньшей мере один человеческий сегмент Vβ, по меньшей мере один человеческий сегмент Dβ и по меньшей мере один человеческий сегмент Jβ, которые способны перестраиваться в Т-клетке с образованием перестроенной человеческой последовательности Vβ/Dβ/Jβ, функционально связанной с последовательностью константного участка TCRβ мыши.
39. Эмбриональная стволовая клетка генетически модифицированной мыши по п. 38, у которой:
(а) первая нуклеотидная последовательность находится в эндогенном локусе Т-клеточного корецептора CD4;
(b) вторая нуклеотидная последовательность находится в эндогенном локусе Т-клеточного корецептора CD8α и третья нуклеотидная последовательность находится в эндогенном локусе Т-клеточного корецептора CD8β;
(c) первая последовательность нуклеиновой кислоты находится в эндогенном локусе МНС II α мыши и вторая последовательность нуклеиновой кислоты находится в эндогенном локусе МНС II β мыши;
(d) третья последовательность нуклеиновой кислоты находится в эндогенном локусе МНС I мыши; и/или
(е) неперестроенная последовательность вариабельного участка TCRα находится в эндогенном локусе вариабельного участка TCRα и неперестроенная последовательность вариабельного участка TCRβ находится в эндогенном локусе вариабельного участка TCRβ.
40. Эмбриональная стволовая клетка генетически модифицированной мыши по п. 39, у которой:
(a) первая нуклеотидная последовательность функционально связана с эндогенными промоторными и регуляторными элементами корецептора CD4 мыши;
(b) вторая нуклеотидная последовательность функционально связана с эндогенными промоторными и регуляторными элементами полипептида CD8α мыши и третья нуклеотидная последовательность функционально связана с эндогенными промоторными и регуляторными элементами полипептида CD8β мыши;
(c) первая последовательность нуклеиновой кислоты функционально связана с эндогенными промоторными и регуляторными элементами МНС II α мыши и вторая последовательность нуклеиновой кислоты функционально связана с эндогенными промоторными и регуляторными элементами МНС II β мыши;
(d) третья последовательность нуклеиновой кислоты функционально связана с эндогенными промоторными и регуляторными элементами МНС I мыши; и/или
(e) неперестроенная последовательность вариабельного участка TCRα функционально связана с эндогенными промоторными и регуляторными элементами TCRα и неперестроенная последовательность вариабельного участка TCRβ функционально связана с эндогенными промоторными и регуляторными элементами TCRβ.
41. Эмбриональная стволовая клетка генетически модифицированной мыши по п. 38, у которой:
(a) первая нуклеотидная последовательность кодирует химерный корецептор CD4, содержащий домены D1, D2 и D3 человеческого полипептида CD4, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного полипептида CD4 мыши,
(b) вторая нуклеотидная последовательность кодирует химерный полипептид CD8α, содержащий IgV-подобный домен человеческого полипептида CD8α, функционально связанный с трансмембранным и цитоплазматическим доменами эндогенного полипептида CD8α мыши, и третья нуклеотидная последовательность кодируют химерный полипептид CD8β, содержащий IgV-подобный домен человеческого полипептида CD8β, функционально связанный с трансмембранным и цитоплазматическим доменами эндогенного полипептида CD8β мыши;
(c) первая последовательность нуклеиновой кислоты кодирует химерный полипептид МНС II α, содержащий домены α1 и α2 человеческого HLA класса II, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС II α мыши, и вторая последовательность нуклеиновой кислоты кодирует химерный полипептид МНС II β, содержащий домены β1 и β2 человеческого HLA класса II, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС II β мыши; и/или
(d) третья последовательность нуклеиновой кислоты кодирует химерный полипептид МНС I, содержащий домены α1, α2 и α3 человеческого HLA класса I, функционально связанные с трансмембранным и цитоплазматическим доменами эндогенного полипептида МНС I мыши.
| WO 02059263 A2, 01.08.2002 | |||
| WO 2013063361 A1, 02.05.2013 | |||
| WO 2014164640 A1, 09.10.2014 | |||
| WO 2014130667 A1, 28.08.2014 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЫШЕЙ | 2009 |
|
RU2425880C2 |

