Область техники
Настоящее изобретение относится к антигенсвязывающему фрагменту (Fab) и гибридному белку Fab-эффектор, содержащему указанный фрагмент.
Уровень техники
Препарат антигенсвязывающего фрагмента (Fab) является одним из наиболее эффективных терапевтических средств на основе моноклонального антитела. Например, абциксимаб (ReoPro®), ранибизумаб (люцентис®) и цетролизумаб пегол (Кимзиа ®) и другие уже были одобрены в качестве лекарственных препаратов во многих странах. Кроме того, препараты поликлональных Fab, включая абциксимаб (ReoPro®), ранибизумаб (люцентис®) и цетролизумаб пегол (Кимзиа®) существуют в продаже.
Конъюгация экзогенного эффекторного домена может обеспечивать терапевтическую эффективность фрагментов Fab, в случае когда они образуют форму гибрида Fab-эффектор. Таким образом, фактически, большое количество фрагментов антител, находящихся на стадии клинической разработки, конъюгированы с экзогенным функциональным фрагментом. В такой конструкции гибридного белка с Fab (или конструкции Fab-эффекторные компоненты), антигенсвязывающий фрагмент может обеспечивать направленную доставку к мишеням, и гибридный белок или (поли)пептид (эффекторный домен) может обладать терапевтическим действием. Домены для слияния, происходящие из прокариотических клеток, могут включать цитотоксины, например, деБоугаин (деиммунизированный токсин растений) (см. Entwistle et al., (2012) Cancer Biother Radiopharm. 27, 582-92), стафилококковый энтеротоксин (СЭ) (см. Ilack et al., (2003) Toxicology. 185, 161-174) или мутантную форму эндотоксина Pseudomonas (см. Choe et al., (1994) Cancer Res. 54, 3460-3467; см. Kreitman et al., (1994) Int. J. Cancer 57, 856-864). Кроме того, домены для слияния,содержащие полипептиды из эукариотических клеток, такие как scFv (см. Lu et al., (2002) J Immunolog Meth. 267, 213-226) или цитокин (см. Holzer et al., (1996) Cytokine. 8, 214-221; см. Sjogaard et al., (1999) Int J Oncol. 15, 873-882), могут функционировать как лекарственные препараты. Хотя как правило с фрагментом Fab или (Fab')2 обычно химически конъюгирован радиоактивный изотоп, цитотоксин, цитокин или фермент к Fab или (Fab')2 присоединяют генетическими способами. Известно, что молекулы Fab, в отличие от scFv, Fv или dsFv, можно легко получить в концентрациях до 1-2 г/л в растворимой форме в периплазме Е. coli (см. Humphreys et al., J. Immunol. Methods. 209, 193-202; Carter et al., Biotechnology (N Y). 10, 163167; Venturi et al., J Mol Biol. 315, 1-8; Donzeau et al., Methods Mol Biol. 378, 14-31), или даже в Pseudomonas fluorescens (см. Retallack et al., Prot Exp Purif. 81, 157-165). В настоящее время многие коммерчески доступные биологические агенты, такие как rhGH (рекомбинантный гормон роста человека), инсулин или разные типы цитокинов получают в клетках Е. coli (см. Graumann and Premstaller, (2006) Biotechnol J. 1, 164-186; Chadd and Chamow, (2001) Curr Opin Biotechnol. 12, 188-194). В данном отношении генетическое слияние терапевтического компонента с фрагментом Fab и другими терапевтическими агентами обеспечивает большое преимущество при разработке нового биологического лекарственного препарата, а также позволяет повысить эффективность существующих биологических препаратов. Кроме того, молекула Fab может быть гибридизована с другими фрагментами антител, такими как scFv, Fv, dsFv или dAb для получения молекулы биспецифического или триспецифического антитела (см. Lu et al., (2002) J Immunolog Meth. 267, 213-226). Однако экспрессия гибридных белков Fab-эффектор, в которых эффектор происходит из эукариотической клетки, в E.coli была затруднена, поскольку домен эффектора не может быть биологически функциональным вследствие неадекватной укладки (фолдинга) или отсутствия у E.coli процесса гликозилирования. Кроме того, оптимальная форма гибрида для получения гибридных белков Fab-эффектор в периплазме E.coli, еще не была тщательно изучена. Большинство белков сыворотки с молекулярной массой менее 50 кДа - 60 кДа, таких как цитокины и факторы роста, характеризуются малым периодом полувыведения in vivo, например, от нескольких минут до нескольких часов, в связи с выведением их почками. Так, увеличение периода полувыведения из сыворотки терапевтических полипептидов или белков является одной из самых интенсивно изучаемых областей биофармацевтических исследований (см. Kontermann, (2012) Wiley, ISBN: 978-3-527-32849-9). По этой причине были разработаны разные способы, включая пегилирование, полисиалирование, ХЕЗилирование, гликозилирование или слияние рекомбинантного аналога ПЭГ (полиэтиленгликоля) с гибкой и гидрофильной аминокислотной цепью (500-600 аминокислот) (см. Chapman, 2002; Adv Drug Deliv Rev. 54. 531-545; Schlapschy et al., (2007) Prot Eng Des Sel. 20, 273-283; Contermann (2011) Curr Op Biotechnol. 22, 868~876; Jevsevar et al., (2012) Methods Mol Biol. 901, 233-246). Кроме того, прямо или косвенно применяли FcRn-опосредуемый механизм рециклирования, для того чтобы увеличить период полувыведения терапевтических белков in vivo. Известно, что среди белков сыворотки сывороточный альбумин человека (САЧ) и иммуноглобулин (в частности, IgG) обладают исключительно длительным периодом полувыведения за счет FcRn-опосредуемого механизма рециклирования. В организме человека период полувыведения альбумина из сыворотки составляет 19 дней, а период полувыведения молекулы IgG составляет от одной недели до почти 4 недель, в зависимости от подкласса IgG. Таким образом, указанные молекулы применялись в качестве партнеров по слиянию для увеличения периода полувыведения терапевтических белков и/или (поли)пептидов.
Рекомбинантный hGH (~19кДа), полученный в цитоплазме или периплазме E.coli, применялся в клинической практике для лечения заболеваний, вызванных отсутствием гормонов роста у младенцев и взрослых, после процесса укладки in vitro (см. Blethen et al., (1997) J. Clin. Endocrinol. Metab. 82, 418-420). Одним из основных недостатков введения rhGH является необходимость ежедневных инъекций из-за короткого периода полувыведения (<30 минут). Чтобы увеличить период полувыведения hGH из сыворотки, были предприняты попытки химически конъюгации с полиэтиленгликолем (см. Clark et al., (1996) J. Biol. Chem. 271, 21969-21977; Pradhananga et al., 2002 J Mol Endocrinol. 29, 1114; Cho et al., 2011; Sondergaard et al., (2011) J Clin Endocrinol Metabol. 96, 681-688) и химической конъюгации модифицированного hGH с цепью Fab гуманизированного CovX-Body IgG (см. Palanki et al., (2013) Bioorg. Med. Chem. Lett. 23, 402-406). Кроме того, увеличения периода полувыведения hGH в сыворотке удалось достичь посредством генетического слияния сывороточного альбумина человека (САЧ) (Альбутропин®) или полипептидных последовательностей, содержащих сотни остатков Pro-Ala-Ser (PAS) (ПАСилирование) (см. Osborn et al., 2002 Eur J Pharmacol. 456, 149-158; Anderson et al., (2011) J Biol Chem. 286, 5234-5241; Sleep et al, (2013) Biochimica et Biophysica Acta. 1830, 5526-5534; Schlapschy et al., (2013) Protein Eng Des Sel 26, 489-501). Наиболее хорошо изученным в данной категории является VRS-317, который представляет собой rGH, генетически присоединенный к аминокислотным последовательностям XTEN на N-конце и С-конце, что позволяет применять режим лечения с введением раз в месяц (см. Schellenberger et al., (2007) Nat Biotech. 27, 1186-1190; Cleland et al., (2012) J Pharm Sci. 101, 2744-2754; Yuen et al., (2013) J Clin Endocrinol Metab. 98, 2595-2603). Также hGH ассоциирован с сосудистыми заболеваниями (см. Thomas J Merimee et. al., (1973), Diabetes, 22, 813-819) и болезнью Крейтцфельдта-Якоба (см. John Powell-Jackson et al., 1985, Lancet, 2, 244-246). Кроме того, IFN-γ ускоряет течение заболевания «трансплантат против хозяина» (см. Bruce R.Blazar et. al., 2003, The Journal of Immunology, 171, 1272-1277), a IFN-α связан с аутоиммунными заболеваниями (см. А Imagawa et al, 1995, The Journal of clinical endocrinology & metabolism, 80, 922-926). Также GSCF связан с аутоиммунными заболеваниями (см. Anke Franzke et al., 2003, Blood, 102, 734-739), a HCV (вирус гепатита С) ассоциирован с заболеваниями печени (см. Van Thiel DH et al., 1995, Hepato-gastroenterology, 42, 907-912).
Гибридный белок (или полипептид) с Fab обладает большим потенциалом в качестве терапевтического агента для лечения хронических заболеваний, которые требуют больших доз лекарственных препаратов в течение длительного периода времени, в частности, особенно, когда гибридный белок с Fab может быть получен с малыми затратами в системе экспрессии в микроорганизмах. Однако несмотря на такие возможные значимые преимущества применения Fab, не предпринималось попыток применить Fab антитела против сывороточного альбумина (СА) при разработке препарата на основе белка или (поли)пептида, обладающего увеличенным периодом полувыведения in vivo. Таким образом, авторы данного изобретения создали данное изобретение, сконструировав новые конструкции гибридных белков Fab против сывороточного альбумина (СА)-эффектор, и подтвердили продукцию с высоким выходом функциональных гибридных конструкций в периплазме E.coli.
Раскрытие изобретения
Техническая проблема
Техническая проблема, которую требовалось решить при помощи настоящего изобретения, состоит в получении нового антигенсвязывающего фрагмента (Fab), обладающего увеличенными периодом полувыведения in vivo.
Еще одна техническая проблема, которую требовалось решить при помощи настоящего изобретения, состоит в получении гибридных конструкций с фрагментами Fab-эффектор, которые обеспечивают оптимальную продукцию в периплазме клетки-хозяина.
Еще одна техническая проблема, которую требовалось решить при помощи настоящего изобретения, состоит в получении вектора экспрессии и клетки-хозяина для продукции конструкций Fab-эффектор в растворимой форме с высоким выходом.
Еще одна техническая проблема, которую требовалось решить при помощи настоящего изобретения, состоит в получении фармацевтической композиции, содержащей гибридные конструкции, описанные выше.
Техническое решение
Для решения перечисленных выше проблем в настоящем изобретении предложена оптимальная гибридная конструкция (или форма) Fab-эффектор для экспрессии в периплазме E.coli, причем указанный Fab содержит вариабельный домен тяжелой цепи, связывающийся с константным доменом 1 тяжелой цепи (CH1), и содержит вариабельный домен легкой цепи, связывающийся с константным доменом легкой цепи (CL).
Согласно одному варианту реализации настоящего изобретения в качестве фрагмента антитела был выбран Fab к СА человека, ввиду того, что было показано, что соединение разных терапевтических белков с альбумином или с альбуминсвязывающими фрагментами, такими как малые пептиды или доменные антитела (aAb), увеличивает период полувыведения терапевтических белков по FcRn-опосредуемому механизму рециклирования (см. Dennis et al., (2002) Biochimica et Biophysica Acta. 1830, 5526-5534; Sleep et al., (2013) Biochimica et Biophysica Acta. 1830, 5526-5534; Nguyen et al., (2006) Protein Eng Des Sel. 19, 291-297; Kontermann, (2011) Curr Op Biotechnol. 22, 868-876). Согласно данным предыдущих исследований фрагмент Fab обладает периодом полувыведения 16-20 ч у человека (см. Ujhelyi and Robert, (1995) Clin Pharmacokinet. 28, 483493) и ~3 ч у крыс после внутривенного введения (см. Nguyen et al., 2006 Protein Eng Des Sel. 19, 291-297). Неожиданно период полувыведения Fab (SL335) в настоящем изобретении составил 37 ч у крыс, что примерно в 12 раз превышает период полувыведения традиционных Fab человека, и, таким образом, логично предположить, что SL335 может иметь период полувыведения у человека по меньшей мере 160-200 часов (6-8 дней). В то же время было выяснено, что два домена Vk - dAbr3 и dAbr16 - со сродством 13 нМ и 1 мМ к САК (сывороточному альбумину кролика), соответственно, демонстрируют значения t1/2 53 ч (dAbr3) и 43 ч (dAbrl6) у крыс (см. Holt et al., (2008) Protein Eng Des Sel. 21, 283-288). Кроме того, t1/2b Ab Fab4D5-H со сродством к САК 92 нМ составило 26,9 ч (см. Nguyen et al., 2006). Следовательно, это значит, что функциональные свойства SL335 in vivo сопоставимы с таковыми у ранее описанных dAb и пептидов, специфичных в отношении СА. Примечательно, что VH и VL SL335 обладают гомологией аминокислотных последовательностей лишь 65-67% на уровне полной последовательности, и гомологией аминокислотных последовательностей ~50% на уровне гипервариабельных участков (CDR) с ранее описанными альбумин-специфичными dAb и пептидами (данные не показаны). Более конкретно, Fab, специфичный к сывороточному альбумину (СА), согласно одному варианту реализации настоящего изобретения содержит вариабельный домен тяжелой цепи, который обладает аминокислотной последовательностью, выбранной из группы, состоящей из 



вариабельный домен легкой цепи, который обладает аминокислотной последовательностью, выбранной из группы, состоящей из
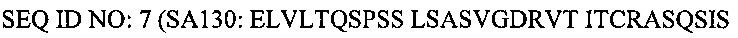

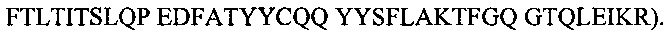
И домен VH Fab, описанного выше, связан с константным доменом 1 тяжелой цепи (домен CH1), а домен VL указанного Fab связан с константным доменом легкой цепи (домен CκL). Кроме того, Fab, специфичный в отношении сывороточного альбумина (СА) согласно настоящему изобретению содержит аминокислотные последовательности SEQ ID NO: 13 (CDR1)(AYSMN), 14 (CDR2) (SISSSGRYIHYADSVKG) и 15 (CDR3) (ETVMAGKALDY) в области VH SL335, и аминокислотные последовательности SEQ ID NO: 16 (CDR1)(RASQSVGSNLA), 17 (CDR2)(GASTGAT) и 18 (CDR3)(QQYYSFLAKT) в области VL SL335.
Согласно одному варианту реализации аминокислота цистеин в домене CH1 и домене CκL указанного Fab может быть удалена или заменена на остатки серина. В частности, как и для SK335, упоминаемого выше, аминокислота цистеин в домене CH1 является 233-ей аминокислотой, начиная с N-конца домена CH1, а цистеин в домене CκL является 214-ой аминокислотой, начиная с N-конца домена CκL, и они заменены на остатки серина. Чтобы избежать путаницы, Н-цепи и L-цепи, которые составляют Fab, были названы следующим образом: 1) Hcys: Н-цепь с цистеином в 233 положении, 2) Lcys: L-цепь с цистеином в 214 положении, 3) Hser: Н-цепь с серином в 233 положении, 4) Lser: L-цепь с серином в 214 положении.
Согласно другому варианту реализации настоящего изобретения гибрид Fab-эффектор конструируют путем соединения эффекторного домена с N- или С-концом либо Fd, либо легкой цепи молекулы Fab путем генетического слияния. Поскольку механизмы укладки и гетеродимеризации рекомбинантных белков в среде периплазмы E.coli значительно затруднены и в основном неизвестны, невозможно предсказать, какая форма гибрида Fab-эффектор оптимальна для функциональной экспрессии.
Также, согласно другому варианту реализации предложена гибридная конструкция из антигенсвязывающего фрагмента (Fab) и эффекторного домена (биологически активный эффекторный компонент), в которой аминокислота цистеин в домене CH1 и аминокислота цистеин в домене GκLFab удалены или заменены остатками серина; и в которой биологически активным эффекторным компонентом является белок или (поли)пептид; и в которой указанный Fab и биологически активный эффекторный компонент ковалентным образом связаны путем генетического слияния. Указанный Fab и биологически активный эффекторный компонент могут быть ковалентно связаны путем генетического слияния с применением пептидного линкера от 0 до 20 аминокислот. Среди шести форм (или конструкций) гибрида Fab-эффектор, содержащего hGH согласно настоящему изобретению, результаты явно демонстрируют, что HserG/Lser демонстрирует самый высокий выход экспрессии в E.coli. То есть, в соответствии с данным вариантом реализации удаление и Cys233 в домене CH1, и Cys214 в домене CLk путем либо делеции, либо замены на остаток другой аминокислоты повышает экспрессию в растворимой форме гибридных конструкций SL335 с эффектором в надосадочной жидкости питательной среды. Это позволяет решить три важных вопроса. Во-первых, соединение эффекторного компонента, например, hGH с С-концом CH1 более предпочтительно, чем соединение с С-концом CLk. Ранее Lu et al. сообщали, что генетическое соединение анти-Flt-1 scFv с С-концом CH1 анти-KDR Fab приводило к пятикратному увеличению выхода по сравнению с соединением с С-концом домена CL (см. Lu et al., (2002) J Immunolog Meth. 267, 213-226). Хотя указанные данные не были включены, вестерн-блоттинг, проводимый авторами заявки с применением полных лизатов Е. coli, выявил, что фрагменты Fd LcysG/Hcys и LserG/Hcys почти полностью разрушались, вследствие чего растворимая форма гибридных белков в надосадочной жидкости E.coli не определялась. Поскольку домены VH склонны к образованию агрегатов в E.coli (Dudgeon et al., (2009) Protein Eng Des Sel. 22, 217-220), можно предположить, что наличие эффекторного домена на С-конце CL может ограничивать взаимодействие домена VH с доменом VL, и домена CH1 с доменом CL, приводящее к быстрой агрегации и разрушению фрагментов Fd. Если сравнивать выход экспрессии в растворимой форме между LserG/Hcys и LserG/Hser, наличие Cys333 в домене CH1, по-видимому, ускоряет данный процесс, вероятно, из-за образования аномальных дисульфидных связей. После удаления Cys333 в домене CH1 присутствие эффекторного домена на конце CH1 могло бы оказывать положительное влияние на уменьшение агрегации домена VH за счет частичного блокирования гидрофобных поверхностей на домене VH перед спариванием VH-VL. Во-вторых, наличие Cys214 в домене CLk еще больше осложняет продукцию в растворимой форме гибридного белка SL335-hGH аддитивным образом. Более низкий выход HserG/Lcys, чем выход HserG/Lser может объясняться тенденцией L-Цепи к образованию гомодимеров, известных под названием белки Бен-Джонса (см. Kirsh et al., (2005) J Immunol Methods. 301, 173-185), в которых Cys214 CLk может оказывать действие на стабилизацию гомодимеров или участвовать в образовании аномальной дисульфидной связи(ей) с другими остатками цистеина в гибридном белке. Также было выяснено, что дисульфидные связи между С-концами CH1 и CL в Fab очень подвижны и обладают высокой степенью гибкости (см. Rothlisberger et al., (2005) J. Mol. Biol. 347, 773-789; Humphreys et al., (2007) Protein Eng Des Sel. 20, 227-234). В этом отношении согласно настоящему изобретению предложен антигенсвязывающий фрагмент (Fab) без Cys233 в константном домене тяжелой цепи 1 (CH1) и без Cys214 в константном домене легкой цепи (CLk). Аналогично, HerGF/Lser и HserIFNb/Lser проявляли самый высокий выход экспрессии у E.coli. В гибридной конструкции согласно настоящему изобретению молярное отношение биологически активного полипептида (или белка) к Fab составляет от 1:1 до 10:1, предпочтительно от 1:1 до 4:1. В-третьих, присутствие указанных двух С-концевых остатков цистеина в SL335 ограничивает не только экспрессию, но и в некоторой степени доступ анти-hGH антитела к домену hGH. Это может быть важно для терапевтической функции эффекторного домена гибрида Fab-эффектора, если создаются помехи для взаимодействия между эффекторным доменом и его лигандом. Авторы настоящего изобретения продемонстрировали, что использование FabΔds в качестве партнера по слиянию благоприятно не только для hGH, поскольку для других эффекторов, таких как G-SCF и INF-b были получены идентичные выводы.
Согласно еще одному аспекту настоящего изобретения для решения технических проблем предложены вектор экспрессии и мутантный штамм E.coli (КСТС 12657ВР) в качестве клетки-хозяина. Указанный штамм был создан путем случайного химического мутагенеза штамма E.coli MC1061, который был выбран, поскольку он происходит из штамма E.coli К12 - одного из основных штаммов-хозяев для получения коммерческих лекарственных биопрепаратов. Если сравнивать с исходным штаммом МС1061, применение мутантного штамма E.coli SUPEX5 в качестве хозяина для экспрессии благоприятно влияет на продукцию HserG/Lser. Не только для соединения SL335-hGH, но комбинации Fabds и штамма SUPEX5 Е. coli также дает преимущества при экспрессии в растворимой форме гибридного белка Fab-эффектора в целом, что явно продемонстрировали результаты, полученные для гибридов SL335-GCSF (SL335wt-GCSF и SL335Δds-GCSF), гибридов SL335-IFNβ (SL335wt-IFNb и SL335Δds-IFNβ) гибридов EGL4-hGH (EGL4wt-hGH и EGL4Δds-hGH), и гибридов 1β28-hGH (1β28wt-hGH и 1β28Δds-hGH). Следовательно, результаты подтверждают, что применение FabΔds - мутантной формы Fab без Cys233 в CH1 и без Cys214 в CLK, более выгодно, чем применение традиционного Fab при экспрессии в растворимой форме гибридных белков Fab-эффектор по меньшей мере в штамме E.coli - SUPEX5. Сопутствующая экспрессия белков-шаперонов или дисульфидизомеразы (FkpA, SurA, Skp, Sec A, Sec B, DsbA или Dsb С) может усиливать экспрессию в растворимой и функциональной форме SL335wt-GCSF или даже SL335Δds-GCSF, поскольку известно, что такие гибриды увеличивают выход продукта в периплазме для растворимых фрагментов Fab у E.coli (см. Schlapschy et al., (2006) Escherichia coli. Protein Eng Des Sel. 19, 385-390). Авторы изобретения считают, что применение FabΔds может быть выгодным, особенно когда шапероны и механизмы катализа для образования дисульфидных связей в эндоплазматическом ретикулуме перегружены из-за высокого уровня экспрессии гибридных белков Fab-эффектор в клетках-хозяевах.
Согласно одному варианту реализации настоящего изобретения SL335Δds-hGH получали приблизительно в концентрации 10 мг/л при применении колбы для культур, что представляет собой более высокий выход, чем сообщалось ранее, несмотря на 4-кратное увеличение размера молекулы согласно настоящем изобретению. Согласно предыдущим сообщениям исследования экспрессии в растворимой форме rhGH в периплазме E.coli показали, что выход составлял 0,64-2,57 мг/л для pelB-hGH и 0,32-2,29 мг/л для (см. Sockolosky and Szoka, (2013) Protein Exp Purif. 87, 129-135), хотя выход rhGH в значительной степени зависел от промоторов и штаммов E.coli-хозяина, которые применялись (см. Soares et al., (2003) Protein Engineering. 16, 1131-1138). Посредством простой оптимизации среды авторы настоящего изобретения в рабочем порядке получили выход ~50 мг/л в надосадочной жидкости при применении колбы для культур, которая позволяет создавать плотность клеток с OD600nm=~10-11 (рукопись на стадии подготовки), которая далее может быть увеличена достаточно для промышленных масштабов посредством тонкой корректировки состава среды и системы культуры с подпиткой.
Согласно еще одному аспекту настоящего изобретения белки SL335ds-эффектор демонстрируют повышенное сродство к САЧ. Согласно одному варианту реализации SL335ds-hGH демонстрирует пяти- - девятикратное увеличение отклика на САЧ (сывороточный альбумин человека) и 1,3 - 4-кратное увеличение отклика на САК (сывороточный альбумин крыс) в зависимости от рН, по сравнению с откликами исходного SL335. Генетическая связь фрагмента антитела и эффекторного домена может влиять на сродство связывания фрагмента антитела с антигеном, и изменения сродства могут в большой степени варьировать в зависимости от природы фрагмента антитела, эффекторного домена и того, как связаны эти два функциональных компонента. Также непонятно, возникают ли указанные различия из-за отсутствия дисульфидных связей между цепями или присутствия гибридного домена hGH. Тем не менее, влияние слияния с hGH на сродство связывания SL335Δds с антигенами, по-видимому, несущественно по сравнению с таким эффектом в IFN-a2b-DOM7 h-14, чье сродство к СА человека, мыши и крысы снижалось в 7,7, 22,3 и 15,8 раз по сравнению с исходным DOM7 h-14 (см. Walker et al., (2010) Protein Eng Des Sel. 23, 271-278). Следовательно, Fab мог превосходить домен Ab в сохранении сродства и укладки эффектора, поскольку домены CH1 и CL создают пространство для уменьшения стерического несоответствия между антигенсвязывающей областью и эффекторным доменом, который связывается с соответствующим лигандом.
Согласно другому варианту реализации настоящего изобретения SL335Δds-hGH обладает существенно увеличенным периодом полувыведения из сыворотки в том смысле, что его t1/2 (16,6 ч при внутривенном введении) было близко к t1/2 PEG5-hGH (250 кДа) (см. Clark et al., 1996). Интересно, что t1/2 SL335Δds-hGH было в 5,6 раз больше, чем t1/2 альбутропина® (t1/2=2,96 ч), и различие в t1/2 между SL335Δds-hGH и альбутропином® дополнительно увеличивалось при п/к (подкожном) введении до 16 раз (97,2 и 5,93 ч) (см. Osborn et al., 2002), хотя указанное сравнение является косвенным, если эксперименты не проводятся при одинаковых условиях. Аналогично t1/2 IFN-a2b-DOM7 h-14 также было приблизительно в 1,5 раз больше, чем у HSA-IFN-a2b (см. Walker et al., 2010). Следовательно, представляется вероятным, что гибрид альбуминсвязывающее вещество обеспечивает больший период полувыведения, чем гибрид с альбумином, и механизмы, лежащие в основе этого, еще предстоит выяснить. Примечательно, что t1/2 SL335Δds-hGH в сыворотке при в/в введении было близко к t1/2 VRS-317 (t1/2=15 ч) (Cleland et al., (2012) J Pharm Sci. 101, 27442754). Это может указывать на то, что возможно вводить SL335Δds-hGH реже, чем раз в неделю или даже раз в месяц (называется САФАтропин®).
Согласно другому варианту реализации настоящего изобретения фармакодинамические эффекты SL335Δds-hGH по-видимому сильно превосходят эффекты альбутропина®, и он является в 7 раз более мощным, чем гроутропин® на основании концентраций в молях с учетом режима введения раз в неделю. К сожалению, мы были вынуждены прекратить 2-недельно фармакодинамическое исследование на 11-ый день, поскольку некоторые крысы с удаленным гипофизом, особенно крысы, получавшие только вспомогательное вещество, рано погибли. Представляется вероятным, что животные перенесли тяжелый стресс из-за транспортировки на большие расстояния из Японии в Южную Корею после операции в августе, что проявилось в 5% снижении массы у крыс, получавших только вспомогательное вещество, и больших значениях стандартного отклонения, чем мы ожидали. Тем не менее, представляется очевидным, что SL335Δds-hGH имеет огромный потенциал в случае его разработки как hGH пролонгированного действия, и, следовательно, здесь и далее мы будем называть его САФАтропин®.
Согласно другому варианту реализации настоящего изобретения указанный биологически активный полипептид, соединенный с Fab, упоминаемым выше, выбран из группы, состоящей из гормона, цитокина, фермента, антитела, фактора роста, транскрипционного фактора, антигена крови, вакцины, структурного белка, белка-лиганда и рецептора.
Согласно еще одному варианту реализации настоящего изобретения биологически активный полипептид выбирают из группы, состоящей из фактора роста человека, соматотропин-релизинг фактора (GHRH), соматотропин-релизинг-пептида, интерферонов, рецепторов интерферонов, колониестимулирующих факторов (КСФ), глюкагоноподобных пептидов, рецепторов, сопряженных с G-белками, интерлейкинов, рецепторов интерлейкинов, ферментов, интерлейкинсвязывающих белков, цитокинсвязывающих белков, фактора активации макрофагов, пептида макрофагов, фактора В-лимфоцитов, фактора Т-лимфоцитов, белка А, ингибитора аллергии, гликопротеинов некроза клеток, иммунотоксина, лимфотоксина, фактора некроза опухоли, онкосупрессоров, фактора роста метастазов, антитрипсина альфа-1, альбумина, альфа-лактальбумина, аполипопротеина-Е, эритропоэтина, высоко гликозилированного эритропоэтина, ангиопоэтинов, гемоглобина, тромбина, пептида, активирующего рецептор тромбина, тромбомодулина, фактора VII, фактора VIIa, фактора VIII, фактора IX, фактора XIII, плазминогенактивирующего фактора, фибринсвязывающего пептида, урокиназы, стрептокиназы, гирудина, белка С, С-реактивного белка, ингибитора ренина, ингибитора коллагеназы, суперокисддисмутазы, лептина, тромбоцитарного фактора роста, фактора роста эпителия, фактора роста эпидермиса, ангиостатина, ангиотензина, фактора роста кости, белка, стимулирующего кости, кальцитонина, инсулина, атриопептина, хрящевого индуцирующего фактора импульсного ответа, элькатонина, фактора активации соединительной ткани, ингибитора пути тканевого фактора, фолликулостимулирующего гормона, лютеинизирующего гормона, релизинг-фактора лютеинизирующего гормона, факторов роста нервов, парат-гормона, релаксина, соматомедина, инсулиноподобного фактора роста, гормона коры надпочечников, глюкагона, холецистокинина, панкреатического полипептида, гастринвысвобождающего пептида, кортиколиберина, тиреотропного гормона, аутотаксина, лактоферрина, миостатина, рецепторов, антагонистов рецепторов, поверхностных антигенов клетки, вакцинных антигенов вирусного происхождения, моноклональных антител, поликлональных антител и фрагментов антител.
Согласно другому аспекту настоящего изобретения предложена фармацевтическая композиция, причем указанная композиция содержит гибридные конструкции Fab-эффектор согласно настоящему изобретению и фармацевтически приемлемое вспомогательное вещество, и обладает повышенной устойчивостью in vivo. Фармацевтическую композицию согласно настоящему изобретению можно вводить в организм разными путями, включая пероральный, чрескожный, подкожный, внутривенный или внутримышечный путь введения, и более предпочтительно можно вводить в виде препарата инъекционного типа. Также фармацевтическую композицию согласно настоящему изобретению можно преобразовать в лекарственную форму при помощи способа, хорошо известного специалистам в данной области техники, чтобы получить быстрое, длительное или отсроченное высвобождение активного ингредиента после ее введения. Указанные составы могут быть в виде таблеток, пилюль, порошка, саше, эликсира, суспензии, эмульсии, раствора, сиропа, аэрозоля, мягких и твердых желатиновых капсул, стерильных растворов для инъекций, стерильный порошок в упаковке и подобные. Примеры подходящих основ, вспомогательных веществ и растворителей включают лактозу, декстрозу, сахарозу, маннитол, ксилитол, эритритол, малтиол, крахмалы, аравийскую камедь, альгинаты, желатин, фосфат кальция, силикат кальция, целлюлозу, метилцеллюлозу, микрокристаллическую целлюлозу, поливинилпирролидон, воду, метилгидроксибензоаты, пропилгидроксибензоаты, тальк, стеарат магния и минеральный масла. Также указанные составы могут дополнительно включать наполнители, препятствующие агглютинации агенты, любриканты, смачивающие вещества, ароматизаторы, эмульгаторы, консерванты и подобные.
Следует понимать, что количество гибридного белка или полипептида, реального вводимого должно определяться исходя из разных значимых факторов, таких как заболевание, которое требуется лечить, избранный путь введения, возраст, пол и масса тела конкретного пациента, и тяжесть симптомов пациента: и типа биологически активного полипептида, применяемого в качестве активного ингредиента. Поскольку гибридный белок согласно настоящему изобретению обладает превосходной устойчивостью в крови, количество и частота введения препаратов пептидов, содержащих гибридный белок согласно настоящему изобретению, могут быть существенно снижены.
В настоящей заявке подразумевается, что единственная форма включает также множественную форму, если из контекста явно не следует другое. Кроме того, в случае, если в описании или формуле изобретения применяются термины «включающий», «включает», «обладающий», «обладает», «с», «такой как» или их варианты, такие термины не являются ограничивающими и должны быть всеобъемлющими, аналогично термину «содержащий».
Согласно настоящему изобретению под «биологически активным полипептидом или белком» понимают (поли)пептид или белок, проявляющий полезную биологическую активность при его введении млекопитающему, включая человека.
Согласно настоящему изобретению под «гибридной конструкцией (или формой) Fab-эффектор" понимают конструкцию, в которой биологически активный (поли)пептид или белок ковалентно связан с Fab. Также, подразумевается, что термин «гибридная конструкция (или форма) Fab-эффектор" включает гибридный белок Fab, гибридный (поли)пептид Fab, гибридные конструкции и гибридные формы.
В этой связи настоящее изобретение подробно описано в разделе «Примеры». Следует отметить, что описание примеров не ограничивает области изобретения, которое описано в предыдущем описании.
Полезные эффекты изобретения
Согласно настоящему изобретению предложена технология, ассоциированная с FabΔds к сывороточному альбумину, (SAFA) в качестве новой платформенной технологии для разработки биологических лекарственных препаратов пролонгированного действия. В этой связи настоящее изобретение имеет преимущество над другими традиционными технологиями, включая ПЭГилирование, Fc-присоединение, технологии «AlbudAb» и гибриды с альбумином в том, что касается пролонгированного действия in vivo, сохранения конформации эффекторного домена, сродства связывания и простоты производства и процедур с низкой стоимостью.
Описание чертежей
На фигуре 1 показаны результаты ELISA (твердофазный иммуноферментный анализ) с монолональными антителами и фагами для определения специфичности связывания Fan фаговых антител к СА.
На фигуре 2 показано определение антигенсвязывающей специфичности клонов Fab человека при помощи ELISA.
На фигуре 3 представлена фармакокинетика SL335 in vivo.
На фигуре 4 представлена схема, изображающая шесть гибридных форм SL335-hGH, сконструированных в данном исследовании.
На фигуре 5 показаны результаты ELISA по определению выхода и связывающей активности растворимых гибридов SL335-hGH в надосадочноой жидкости из питательной среды E.coli. Сигналы связывания визуализировались при помощи субстрата ТМВ (тетраметилбензидин), и поглощение на длине волны 450 нм измеряли при помощи ELISA-ридера. Данные представлены как среднее ±СО для трех экспериментов.
На фигуре 6 показан результат ELISA по определению экспрессии у хозяина E.coli и зависимой от температуры экспрессии вариантов SL335 и SL335-hGH (20°С, А; 25°С, В; или 30°С, С).
На фигуре 7 показан результат ELISA по определению выхода растворимых гибридных конструкций SL335-GCSF и SL335-IFNβ в надосадочной жидкости питательной среды E.coli.
На фигуре 8 показан результат ELISA по определению выхода растворимых гибридов EGL4-hGH (A), and 1β28-hGH (В) в надосадочной жидкости питательной среды E.coli.
На фигуре 9 показан результат анализа SL335wt-hGH и SL335ds-hGH при помощи SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) и вестерн-блоттинга.
На фигуре 10 показан результат анализа HcycG/Lcys и HserG/Lser при помощи капиллярного электрофореза, реализованного на чипе.
На фигуре 11 показан результат анализа HcycG/Lcys и HserG/Lser при помощи масс-спектрометрии.
На фигуре 12 показана очистка HserG/Lser посредством гель-фильтрации с применением FPLC (жидкостная экспресс-хроматография белков).
На фигуре 13 показано определение биологической активности hGH in vitro для SL335ds-hGH при помощи анализа пролиферации клеток Nb2-11.
На фигуре 14 показано определение стабильности SL335ds-hGH в сыворотке при помощи ELISA и анализа пролиферации клеток Nb2-11 in vitro.
На фигуре 15 показан анализ фармакокинетики гроутропина или SL335ds-hGH у крыс.
На фигуре 16 показан дозозависимый прирост массы у крыс с удаленным гипофизом, получавших гроутропин® или SL335Δds-hGH. N=3 крысы на группу, взвешивание крыс - один раз в день.
На фигуре 17 показано дозозависимое увеличение длины большеберцовой кости при лечении гроутропином® или SL335Δds-hGH. N=3-4 крысы на группу, одно измерение большеберцовой кости на крысу.
На фигуре 18 показан вектор рНЕКА согласно настоящему изобретению.
На фигуре 19 показана нуклеотидная последовательность вектора рНЕКА согласно настоящему изобретению.
На фигуре 20 показана предсказанная аминокислотная последовательность генов VH и VL, применяемых для клонов Fab к СА согласно настоящему изобретению.
На фигуре 21 показана последовательность ДНК генов VH (А) и VL (В), применяемых для клонов Fab к СА согласно настоящему изобретению.
На фигуре 22 показана информация о последовательности гибридных конструкций Fab-эффектор согласно настоящему изобретению. Линкер и эффекторные домены подчеркнуты, a CDR выделены жирным шрифтом.
Техническое выполнение изобретения
1. Материалы и анализ
1-(1) клонирование и штаммы
Все эксперименты по клонированию ДНК проводились в соответствии со стандартной процедурой (см. Sambrook et al., (1989) Molecular cloning: A laboratory manula, 2nd ed., (New Youk, USA: Cold Spring Harbor Laboratory Press)). Олигонуклеотиды степени чистоты для секвенирования и кодон-оптимизированные гены для конструирования гибридных конструкций SL335-эффектор синтезировались компанией «Bioneer», Тэджон, Южная Корея. Амплификацию путем ПЦР (полимеразной цепной реакции) проводили с применением ДНК-полимеразы Pyrobest или Ex-Taq («Takara», Цзу, Япония) при условиях - 25 циклов при 94°С в течение 1 мин, 58°С в течение 1 мин и 72°С в течение 1 мин, затем 72°С в течение 10 мин, если не указано другое. Рестриктазы, щелочная фосфатаза креветки (SIP) и ДНК-лигаза Т4 были приобретены у компании «Takara». Для клонирования применяли штамм Е. coli МС1061 [araD139 Del(araA-leu)7697 Del(lac)X74 galK16 galE15(GalS) лямбда- e14-mcrA0 relA1 rpsL150(strR) spoT1 mcrB1 hsdR2] (Американская коллекция типовых культур (АТСС), Манассас, США), а для экспрессии рекомбинантного белка применяли штамм Е. coli SUPEX5. Для препаратов рекомбинантного фага применяли штамм Е. coli TG1 {F' [traD36 proAB+lacIqlacZΔМ15]supE thi-1 Δ(lac-proAB) Δ(mcrB-hsdSM)5, (rK-mK-)}(«Agilent Technologies», Пало-Альто, США).
1-(2)-биопаннинг библиотеки антител HuDVFab-8L
Обогащение рекомбинантных фагов, связанных с целевыми антигенами, проводили, как было описано ранее (см. Joo et al., (2008) J. Immunol. Methods. 333, 24-37; Hur et al., (2010) Immunol Lett. 132, 24-30). Если кратко, тозилированные магнитные микроносители, конъюгированные с сывороточным альбумином человека, крысы или мыши (САЧ, САК или САМ, соответственно) («Sigma-Aldrich», Сент-Луис, Миссури, США), смешивали с 1010 фагов из библиотеки антител HuDVFab-8L («AprilBio», Чхунчхон, Южная Корея) в течение 4 ч при 4°С, и трижды отмывали фосфатно-солевым буфером, содержащим 0,02% Tween (ФСБТ). Антитела фага, которые связывались с микроносителями, десорбировали при помощи элюирующего буфера (0,1 М глицин, рН 2). Свежие клетки TG1, содержащие соответствующие легкие (L) (VL+CLk) цепи, инфицировали десорбированным фагом и выращивали на среде 2 YT, содержащей 25 мкг/мл ампициллина, 10 мкг/мл карбенициллина и 10 мкг/мл тетрациклина (2 × YT/ACT). Рекомбинантные фаги затем амплифицировали с применением фага-помощника Ех-12 («AprilBio») для последующего пэннинга. После финального пэннинга проводили ELISA с моноклональным фагом, чтобы идентифицировать положительные клоны. Гены Fd (VH+CH1) из положительных клонов субклонировали в вектор pHg3A-3 («AprilBio», Чхунчхон, Южная Корея), и была проведена оптимизация L цепи при помощи 1,4108 репертуара kL цепи хьюманнав в векторе-фагмиде pLf1T-3 («AprilBio»).
1-(3)- Анализ посредством секвенирования ДНК
Фагмиду pHf1g3A-2 («AprilBio») и плазмиду pLf1A-3 («AprilBio») выделяли из клеток Е. coli, продуцирующих молекулы Fab к СА при помощи набора Wizard Plasmid Miniprep Kit («Promega», Мадисон, Висконсин, США). Для считывания генов VH и VL применяли два разных секвенирующих праймера  (SEQ ID NO: 19) and
(SEQ ID NO: 19) and 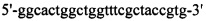 (SEQ ID NO: 20)), которые были комплементарны к pHf1g3A-2 или pLT-2, соответственно. Секвенирование ДНК проводили при помощи «SolGent», Тэджон, Южная Корея.
(SEQ ID NO: 20)), которые были комплементарны к pHf1g3A-2 или pLT-2, соответственно. Секвенирование ДНК проводили при помощи «SolGent», Тэджон, Южная Корея.
1-(4) Конструирование вектора экспрессии рНЕКА
Фрагмент ДНК №1, содержащий сайт рестрикции Bgl II + промотор trc + энхансер трансляции g10 - сайт связывания рибосомы (RBC) получали посредством ПЦР-амплификации с вектора pTrcHis-B («Invitrogen», Карлсбад, Калифорния, США) при помощи ДНК-полимеразы Pyrobest и набора ПЦР-праймеров №1  (SEQ ID NO: 21)) и №2
(SEQ ID NO: 21)) и №2 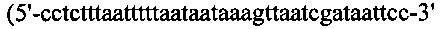 (SEQ ID NO: 22)). Фрагмент ДНК №2, содержащий энхансер трансляции g10 + RBC + BamH I + сайт множественного клонирования (MCS) + терминатор транскрипции получали посредством ПЦР-амплификации с той же матрицы, которая описана выше, при помощи набора праймеров №3
(SEQ ID NO: 22)). Фрагмент ДНК №2, содержащий энхансер трансляции g10 + RBC + BamH I + сайт множественного клонирования (MCS) + терминатор транскрипции получали посредством ПЦР-амплификации с той же матрицы, которая описана выше, при помощи набора праймеров №3  (SEQ ID NO: 23)) и №4
(SEQ ID NO: 23)) и №4  (SEQ ID NO: 24)). Для сборки указанных двух фрагментов ДНК применяли ПЦР-сборку с применением ДНК-полимеразы Ex-Taq и набора ПЦР №1 и праймеров №4. Образующийся в результате фрагмент ДНК ~520 по выделяли посредством электрофореза в агарозном геле. Затем продукт связывающей ПЦР и плазмиду рЕТ28а («Invitrogen») подвергали рестрикции с применением Bgl II и Dra III, и сшивали вместе при помощи ДНК-лигазы Т4 в течение 2 ч при КТ (комнатной температуре). После трансформации электрокомпетентных клеток МС1061 с 3 мл реакции сшивания, трансформанты E.coli подвергали селекции на планшетах с 2 YT, содержащих 50 мкг/мл канамицина («Sigma-Aldrich»). Для субклонирования генов Fab в вектор рНЕКА гены цепей Fd (VH+CH1) амплифицировали посредством ПЦР с вектора-фагмиды pHf1g3A-2 при помощи набора ПЦР-праймеров №5
(SEQ ID NO: 24)). Для сборки указанных двух фрагментов ДНК применяли ПЦР-сборку с применением ДНК-полимеразы Ex-Taq и набора ПЦР №1 и праймеров №4. Образующийся в результате фрагмент ДНК ~520 по выделяли посредством электрофореза в агарозном геле. Затем продукт связывающей ПЦР и плазмиду рЕТ28а («Invitrogen») подвергали рестрикции с применением Bgl II и Dra III, и сшивали вместе при помощи ДНК-лигазы Т4 в течение 2 ч при КТ (комнатной температуре). После трансформации электрокомпетентных клеток МС1061 с 3 мл реакции сшивания, трансформанты E.coli подвергали селекции на планшетах с 2 YT, содержащих 50 мкг/мл канамицина («Sigma-Aldrich»). Для субклонирования генов Fab в вектор рНЕКА гены цепей Fd (VH+CH1) амплифицировали посредством ПЦР с вектора-фагмиды pHf1g3A-2 при помощи набора ПЦР-праймеров №5  (SEQ ID NO: 25)) и №6
(SEQ ID NO: 25)) и №6  (SEQ ID NO: 26)), а гены L-цепи амплифицировали при помощи ПЦР с вектора-плазмиды pLT-2 при помощи набора ПЦР-праймеров №7
(SEQ ID NO: 26)), а гены L-цепи амплифицировали при помощи ПЦР с вектора-плазмиды pLT-2 при помощи набора ПЦР-праймеров №7  (SEQ ID NO: 27)) и №8
(SEQ ID NO: 27)) и №8 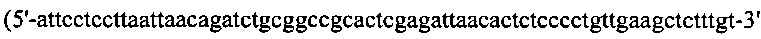 (SEQ ID NO: 28)). Образующиеся в результате фрагменты генов Fd И L-цепи собирали посредством ПЦР-сборки с применением набора ПЦР-праймеров №6 и №7, и образующийся продукт ПЦР размером ~1,4 кпо вырезали из агарозного геля. Затем указанный продукт ПЦР и плазмиду рНЕКА подвергали рестрикции с применением BamH I и Hind III, сшивали вместе при помощи ДНК-лигазы Т4 в течение 2 ч при КТ, и путем электропорации вводили в электрокомпетентные клетки МС1061 или SUPEX5. ПЦР-праймеры, применяемые при получении вектора экспрессии рНЕКА, перечислены в Таблице 1 ниже. На Фиг. 18 показана схема вектора экспрессии рНЕКА.
(SEQ ID NO: 28)). Образующиеся в результате фрагменты генов Fd И L-цепи собирали посредством ПЦР-сборки с применением набора ПЦР-праймеров №6 и №7, и образующийся продукт ПЦР размером ~1,4 кпо вырезали из агарозного геля. Затем указанный продукт ПЦР и плазмиду рНЕКА подвергали рестрикции с применением BamH I и Hind III, сшивали вместе при помощи ДНК-лигазы Т4 в течение 2 ч при КТ, и путем электропорации вводили в электрокомпетентные клетки МС1061 или SUPEX5. ПЦР-праймеры, применяемые при получении вектора экспрессии рНЕКА, перечислены в Таблице 1 ниже. На Фиг. 18 показана схема вектора экспрессии рНЕКА.

1-(5)- Создание мутантного штамма Е. Coli SUPEX5
Химический мутагенез проводили по существу, как было описано в предыдущей работе. Если кратко, клетки Е. coli MC1061, экспрессирующие scFv на комплекс дегидрогеназы альфа-кетокислот с разветвленными цепями человека-E2(BCKD-E2), соединенный со щелочной фосфатазой (ЩФ), выращивали на среде Лурия-Бертани (LB), содержащей 50 мкг/мл ампициллина, до плотности OD600 ~ 0,3. Клетки, содержащиеся в 5 мл культуры, отбирали посредством центрифугирования при 3000 g в течение 10 мин, дважды отмывали холодным 0,1 М цитратным буфером (рН 5,5). Затем клетки повторно суспендировали в 1,9 мл того же буфера и обрабатывали 50 мкг/мл N-метил-N'-нитро-N-нитрогуаниндина (MNNG) («Sigma-Adrich», Сент-Луис, Миссури, США) при 37°С в течение 15, 30 и 45 мин. После обработки MNNG клетки смешивали, дважды отмывали и повторно суспендировали в 2 мл среды LB. Затем проводили анализ высоты колоний с применением двухмембранной системы, как описано. Если кратко, чашки Петри с агаром, содержащим 50 мкг/мл ампициллина и 10 мкг/мл карбенициллина накрывали первыми нейлоновыми мембранами (0,45 м мембраной для блоттинга из нитрана - N нейлона) («GE Healthcare Life Science», Вауватоза, Висконсин, США) с низкой способностью связывать белки. Мутированные бактерии распределяли на мембране с плотностью 106 клеток/чашку Петри и выращивали в течение 8 ч при 37°С. При этом вторые нитроцеллюлозные мембраны (нитроцеллюлозная гибридизационная подложка Bio-Trace™ NT) («PALL», Порт Вашингтон, Нью-Йорк, США) загружали на чашки Петри со свежим агаром LB, содержащим 50 мкг/мл ампициллина, 10 мкг/мл карбенициллина и 1 мМ изопропил-D-1-тиогалактопиранозида (IPTG) («Sigma-Aldrich»). Первые нейлоновые мембраны снимали с агара LB в чашках Петри и помещали поверх вторых мембран, а затем инкубировали при 37°С в течение 5 ч. После инкубации первую мембрану (с колониями) удаляли, наносили на чашки Петри со свежим агаром LB, содержащим 50 мкг/мл ампициллина, 10 мкг/мл карбенициллина, и хранили при 4°С в течение позднего восстановления бактерий. Вторые мембраны промывали три раза в течение 10 мин в свежем фосфатно-солевом буфере, содержащем 0,1% TWEEN (объемные проценты) (ФСБ/Tween), и погружали в субстрат нитросиний тетразолий хлорид (NTB)/5-бром-4-хлор-3-индлолил-фосфат (BCIP) («Duchefa», Харлеем, Нидерланды) для визуализации ЩФ в колониях E.coli. Колонии E.coli, демонстрирующие различную активность ЩФ, отбирали с соответствующих первых фильтров, объединяли и проводили второй цикл мутагенеза и анализ высоты колоний. После второго цикла анализа высоты колоний гипотетически положительные клоны E.coli отбирали и выращивали в 10 мл среды 2 YT, содержащей 50 мкг/мл ампициллина и 10 мкг/мл карбенициллина, пока OD не достигала 0,5. IPTG добавляли в культуру в конечной концентрации 0,1 мМ, и клетки выращивали в течение ночи при 27°С. Затем надосадочную жидкость питательной среды отбирали посредством центрифугирования при 3300g в течение 20 мин. Для получения экстрактов периплазмы конгломерат клеток повторно суспендировали в буфере для экстракции периплазмы (матричный раствор: 200 мМ Tris-HCl, 20 мМ EDTA, 2 М NaCl, рН 7,4), замораживали и размораживали три раза и центрифугировали при 10000 g в течение 20 мин при 4°С. В итоге путем сбора надосадочной жидкости получали экстракт периплазмы, содержащий растворимый гибрид анти-BCKD-AP. Готовили последовательные разведения надосадочной жидкости питательной среды и экстракта периплазмы с применением ФСБ, содержащего 1% бычьего сывороточного альбумина (БСА) («Sigma-Aldrich»), и 50 мл пробы надосадочной жидкости питательной среды или экстракта периплазмы смешивали с 100 мл субстрата р-нитрофенила фосфата (pNPP) («Roche», Южный Сан-Франциско, Калифорния, США) в микротитрационном планшете на 96 лунок («SPL», Южная Корея). Через 5-10 мин в каждую лунку добавляли 25 мкл 3М NaOH для остановки реакции, и поглощение на длине волны 415 нм измеряли с применением ридера для ELISA («Bio-Rad», Геркулес, Калифорния, США). Четыре мутанта штаммов E.coli (М№5, М№7, М№54 и М№69), демонстрирующие повышенную экспрессию гибрида анти- BCKD-AP, выращивали в среде 2 YT без антибиотиков при 37°С в течение ночи. Затем клетки распределяли на чашках Петри с агаром LB при плотности ~103 клеток/лунку и выращивали при 37°С в течение ночи. Образующиеся колонии воспроизводили на чашках Петри с агаром LB, с добавлением или без добавления 50 мкг/мл ампициллина. Отбирали колонии E.coli, растущие на чашках Петри с агаром LB без антибиотика, но не способные расти на чашках Петри с агаром LB с антибиотиком, и выращивали их в среде 2 YT без антибиотиков, пока OD600 не достигала ~1,0. Запасы клеток готовили путем добавления глицерина (20%, объемные проценты) и хранили при 80°С. Для применения для клонирования электрокомпетентные клетки получали из мутантных штаммов согласно стандартному протоколу и хранили при 80°С. М№5, один из мутантных штаммов E.coli, был назван SUPEX5 (КСТС 12657ВР), и его применяли для экспрессии Fab и гибридных белков Fab-эффектор.
1-(6)-Твердофазный иммуноферментный анализ (ELISA)
Для ELISA с моноклональным фагом рекомбинантный фаг получали из положительных клонов E.coli путем «спасения» фага, и ~108 КОЕ/лунку добавляли в планшеты MaxiSorb ELISA («Nunc», Роскилле, Дания), в которые были нанесены 5 мкг/мл САЧ, САК, САМ и БСА. Фагу давали связаться с антигенами, либо при рН 6, либо при рН 7,4 в течение 1 ч при 37°С. В качестве вторичного антитела применяли антитело козы к каппа-L человека с HRPO («Sigma-Aldrich»). Сигналы связывания визуализировали при помощи субстрата ТМВ («BD Science», Сан-Хосе, Калифорния, США), поглощение на длине волны 450 нм измеряли с применением ридера для ELISA («Bio-Rad», Геркулес, Калифорния, США). Данные представляют собой среднее для трех экспериментов и стандартное отклонение. Для традиционного ELISA разные антигены [СА человека, СА крысы, СА мыши, СА обезьяны («Alpha diagnostic Intl», Сан-Антонио, Техас, США), СА собаки («CUSABIO», Ханьян, Хубей, Китай), СА кролика («Sigma-Aldrich»), рецептор фактора роста эпидермиса (EGFR) («R&D systems», Миннеаполис, Миннесота, США), молекулу адгезии клеток эпителия (ЕрСАМ) («R&D systems»), рецептор IL-15 α (IL-15Rα) («R&D systems»), IL-1β («eBioscience», Сан-Диего, Калифорния, США), CD 16а («R&D systems»), с-МЕТ («Sinobiological», Пекин, Китай)] в концентрациях 5 мкг/мл иммобилизовывали на микротирационных планшетах, и молекулам Fab давали связаться с антигенами, и определяли, как описано выше. Для определения концентрации растворимых Fab или гибридных белков Fab-hGH, проводили сэндвич-ELISA с применением mAb мыши на Fd IgG человека («AprilBio») в качестве захватывающего антитела и антитела козы pAb к каппа-L-цепьи человека, конъюгированного с HRPO («Sigma-Aldrich»), в качестве детектирующего антитела. Для построения стандартной кривой применяли фрагмент Fab человека («Bethyl», Монтгомери, Техас, США) с известной концентрацией. Для определения домена hGH, применяли Т-20 - pAb козы, специфичное к С-концу hGH («Santacruz Biotechnology», Даллас, Техас, США) и NYThGH - mAb мыши, специфичное в отношении полноразмерного hGH («Prospec», Ист-Брауншвейг, Нью-Джерси, США), затем применяли конъюгированный антикозий IgG pAb-HRPO кролика («Sigma-Aldrich») или конъюгированный антимышиный IgG pAb-HRPO козы («Sigma-Aldrich»), соответственно, в качестве вторичного антитела. Для определения домена G-CSF применяли pAb козы к GCSF человека («R&D Systems»), а для детекции домена IFN-β применяли pAb козы к IFN-β человека («PEPROTECH», Рокки Хилл, США).
1-(7)-Получение растворимых Fab и гибридных белков Fab-эффектор
Растворимые Fab и гибридные белки Fab-hGH получали путем выращивания клеток E.coli SUPEX5 в 10 мл или 1 л среды 2YT, содержащей 50 мкг/кг канамицина при 37°С, пока OD600нм не достигала 0,5, и последующего добавления 0,05 мМ IPTG. После 20-часовой инкубации при 20°С при интенсивном встряхивании надосадочную жидкость питательной среды и конгломерат клеток разделяли путем центрифугирования при 3 300 g в течение 20 мин. Экстракт периплазмы получали, как было описано выше. Для очистки надосадочную жидкость питательной среды и/или экстракты периплазмы затем пропускали через смолы Сеферозы 4В, на которых был иммобилизован САЧ («AprilBio»). После активного промывания молекулы Fab, связавшиеся со смолой, десорбировали при помощи элюирующего буфера (0,1 М глицина, 10% глицерин, рН 3), а затем сразу проводили нейтрализацию Tris - буфером (0,5 М Tris HCl, 2 М NaCl, рН 9,0). Также после аффинной очистки при помощи ЖХБР АКТА («GE Healthcare», Вауватоза, Висконсин, США) проводили гель-фильтрацию Hser/Lser. Если кратко, перезаполненную колонку Hiprep™ 16/60 Sephacryl™ S-200HRP уравновешивали при помощи уравновешивающего буфера (20 мМ HEPES, 150 мМ NaCl, рН 7,4), и загружали в нее 5 мкл HserG/Lser (гибрид SL335Δds-hGH). Десорбцию проводили при помощи уравновешивающего буфера при предельно-допустимом давлении 0,35 МПа и скорости потока 0,5 мкл/мин. Фракции под номерами 13, 16, 19 и 23 анализировали посредством SDS-PAGE, как было описано выше.
1-(8) измерение сродства посредством интерферометрии в биослое
Анализ связывания в режиме реального времени между очищенным SL335 и антигенами (СА человека, СА крысы и СА мыши) проводили при помощи интерферометрии в биослое с применением системы Octet RED («ForteBio», Менло-парк, Калифорния, США), как было описано ранее, за исключением применения сенсоров AR2G (реактивный амин второго поколения) (Costin et al., (2013) J Virol. 87, 52-66). Если кратко, заранее определенную концентрацию SL335 соединяли с биосенсорами AR2G кинетической степени очистки, и не связавшиеся фрагменты Fab удаляли с поверхностей сенсоров путем инкубации в кинетическом буфере (1М этаноламина, рН 8,5). Затем зондам давали связаться с СА человека, СА крысы или СА мыши в заранее определенных концентрациях при рН 6,0 или рН 7,4 (концентрация СА человека при рН 6 и рН 7,4: 200 нМ, 100 нМ, 50 нМ, 25 нМ и 12.5 нМ; концентрация СА крысы при рН 6: 4 мМ, 1 мМ, 500 нМ, 250 нМ и 125 нМ; концентрация СА крысы при рН 7.4: 4 мМ, 2 мМ, 1 мМ, 500 нМ и 125 нМ; концентрация СА мыши рН 6 и рН 7,4: 20 мМ, 10 мМ, 5 мМ, 2.5 мМ и 12.5 мМ), а затем проводили диссоциацию в ФСБ, содержащем 0,1% БСА,, рН 6 или рН 7.4. Кинетику связывания и диссоциации рассчитывали при помощи пакета программ «Octet QK», которая позволяет аппроксимировать наблюдаемые кривые связывания моделью связывания 1:1 для расчета констант скорости ассоциации. Константы скорости ассоциации или диссоциации рассчитывали по меньшей мере с применением трех разных концентраций СА человека, СА крысы или СА кролика. Равновесные константы диссоциации рассчитывали как кинетическую константу скорости диссоциации, деленную на кинетическую константу скорости ассоциации.
1-(9) Генерирование гибридных конструкций SL335-hGH
Для создания SL335ds мутантный Fd (замена Cys233 Ser233), называемый Hser, получали посредством ПЦР-амплификации с кодон-оптимизированного гена цепи Fd SK335 с применением набора ПЦР-праймеров №9 
 (SEQ ID NO: 29)) и №10
(SEQ ID NO: 29)) и №10 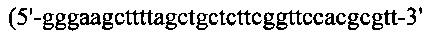 SEQ ID NO: 30)). Продукт ПЦР ~750 по обрабатывали EcoR I/ Hind III и сшивали при помощи рНЕКА. Мутантную L-Цепь (замена Cys214 → Ser214), называемый Lser, также получали посредством ПЦР-амплификации с кодон-оптимизированного гена L цепи SL335 с применением набора ПЦР-праймеров №11
SEQ ID NO: 30)). Продукт ПЦР ~750 по обрабатывали EcoR I/ Hind III и сшивали при помощи рНЕКА. Мутантную L-Цепь (замена Cys214 → Ser214), называемый Lser, также получали посредством ПЦР-амплификации с кодон-оптимизированного гена L цепи SL335 с применением набора ПЦР-праймеров №11 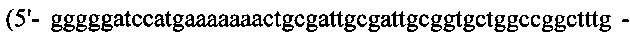
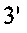 (SEQ ID NO: 31)) и №12
(SEQ ID NO: 31)) и №12  (SEQ ID NO: 32)), вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Hser. Процедуры клонирования для генерирования конструкции HcysG/Lcys, были следующими: Fd дикого типа с Cys233, называемых Hcys, подвергали ПЦР-амплификации с кодон-оптимизированного Fd или SL335 с применением набора ПЦР-праймеров №9 и №13
(SEQ ID NO: 32)), вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Hser. Процедуры клонирования для генерирования конструкции HcysG/Lcys, были следующими: Fd дикого типа с Cys233, называемых Hcys, подвергали ПЦР-амплификации с кодон-оптимизированного Fd или SL335 с применением набора ПЦР-праймеров №9 и №13  (SEQ ID NO: 33)), а также hGH-содержащую линкерную последовательность подвергали ПЦР-амплификации с кодон-оптимизированного гена hGH с применением набора ПЦР-праймеров №14
(SEQ ID NO: 33)), а также hGH-содержащую линкерную последовательность подвергали ПЦР-амплификации с кодон-оптимизированного гена hGH с применением набора ПЦР-праймеров №14  (SEQ ID NO: 34)) и №15
(SEQ ID NO: 34)) и №15 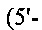
 (SEQ ID NO: 35)). Гены Hcys и hGH сшивали вместе и генерировали HcysG посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №15, вырезали при помощи EcoR I/Hind III и клонировали в рНЕКА, содержащий L-цепь дикого типа с Cys214 SL335, называемую Lcys. Для генерирования конструкции LcysG/Hcys, Leys подвергали ПЦР-амплификации с кодон-оптимизированной L-цепи SL335 с применением набора ПЦР-праймеров №11 и №16
(SEQ ID NO: 35)). Гены Hcys и hGH сшивали вместе и генерировали HcysG посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №15, вырезали при помощи EcoR I/Hind III и клонировали в рНЕКА, содержащий L-цепь дикого типа с Cys214 SL335, называемую Lcys. Для генерирования конструкции LcysG/Hcys, Leys подвергали ПЦР-амплификации с кодон-оптимизированной L-цепи SL335 с применением набора ПЦР-праймеров №11 и №16  (SEQ ID NO: 36)), и hGH, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена hGH с применением набора ПЦР-праймеров №14 и №17
(SEQ ID NO: 36)), и hGH, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена hGH с применением набора ПЦР-праймеров №14 и №17 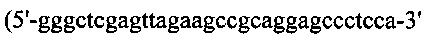 (SEQ ID NO: 37)). Leys и ген hGH сшивали вместе с образованием LcysG посредством ПЦР-сборки с применением набора ПЦР-праймеров №11 и №17, вырезали при помощи BamH/Xho I и клонировали в рНЕКА, содержащий Fd дикого типа. Для получения конструкции HserG/Lcys, Hser подвергали ПЦР-оптимизации с кодон-оптимизированного гена Fd-цепи дикого типа с применением набора ПЦР-праймеров №9 и №18
(SEQ ID NO: 37)). Leys и ген hGH сшивали вместе с образованием LcysG посредством ПЦР-сборки с применением набора ПЦР-праймеров №11 и №17, вырезали при помощи BamH/Xho I и клонировали в рНЕКА, содержащий Fd дикого типа. Для получения конструкции HserG/Lcys, Hser подвергали ПЦР-оптимизации с кодон-оптимизированного гена Fd-цепи дикого типа с применением набора ПЦР-праймеров №9 и №18  (SEQ ID NO: 38)). ПЦР-амплификацию hGH, содержащего линкерную последовательность, ПЦР-сборку и клонирование HserG, проводили как при получении конструкции HcysG/Lcys. Для получения конструкции Lser/Hcys подвергали ПЦР-оптимизации с кодон-оптимизированной L-цепью SL335 с применением набора ПЦР-праймеров №11 и №19
(SEQ ID NO: 38)). ПЦР-амплификацию hGH, содержащего линкерную последовательность, ПЦР-сборку и клонирование HserG, проводили как при получении конструкции HcysG/Lcys. Для получения конструкции Lser/Hcys подвергали ПЦР-оптимизации с кодон-оптимизированной L-цепью SL335 с применением набора ПЦР-праймеров №11 и №19 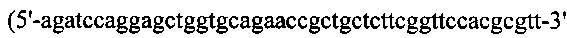 (SEQ ID NO: 39)). ПЦР-амплификацию hGH, содержащего линкерную последовательность, ПЦР-сборку и клонирование HserG, проводили как при получении конструкции HcysG/Lcys. Для получения конструкции HerG/Lser, ПЦР-амплификацию HserG и hGH, и ПЦР-сборку проводили, как при получении конструкции HserG/Lcys, за исключением того, что для клонирования применяли рНЕКА, содержащий Lser. LserG/Hser конструировали таким же образом, что получение конструкции LserG/Hcys, за исключением того, что для клонирования применяли рНЕКА, содержащий Hser. ПЦР-праймеры для получения гибридных конструкций SL335-hGH и гибридных конструкций SL335Δds-hGH перечислены в Таблице 2 ниже.
(SEQ ID NO: 39)). ПЦР-амплификацию hGH, содержащего линкерную последовательность, ПЦР-сборку и клонирование HserG, проводили как при получении конструкции HcysG/Lcys. Для получения конструкции HerG/Lser, ПЦР-амплификацию HserG и hGH, и ПЦР-сборку проводили, как при получении конструкции HserG/Lcys, за исключением того, что для клонирования применяли рНЕКА, содержащий Lser. LserG/Hser конструировали таким же образом, что получение конструкции LserG/Hcys, за исключением того, что для клонирования применяли рНЕКА, содержащий Hser. ПЦР-праймеры для получения гибридных конструкций SL335-hGH и гибридных конструкций SL335Δds-hGH перечислены в Таблице 2 ниже.

1-(10)- Генерирование гибридных конструкций SL335-GCSF
Процедуры клонирования для генерирования конструкции HcysGF/Lcys, были следующими: Hcys подвергали ПЦР-оптимизации с кодон-оптимизированного гена Н-цепи SL335 с применением набора ПЦР-праймеров №9 и №20  (SEQ ID NO: 40)), a G-CSF, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена G-CSF с применением набора ПЦР-праймеров №21
(SEQ ID NO: 40)), a G-CSF, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена G-CSF с применением набора ПЦР-праймеров №21 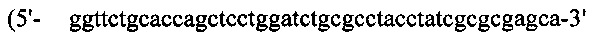 (SEQ ID NO: 41)) и №22
(SEQ ID NO: 41)) и №22 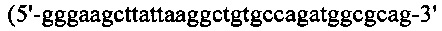 (SEQ ID NO: 42)). Гены Hcys и G-CSF сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №22, вырезали при помощи EcoR I/Hind III и клонировали в рНЕКА, содержащий L-цепь SL335. Для получения конструкции LcysGF/Hcys, Leys подвергали ПЦР-оптимизации с кодон-оптимизированного гена L-цепи SL335 с применением набора ПЦР-праймеров №11 и №23
(SEQ ID NO: 42)). Гены Hcys и G-CSF сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №22, вырезали при помощи EcoR I/Hind III и клонировали в рНЕКА, содержащий L-цепь SL335. Для получения конструкции LcysGF/Hcys, Leys подвергали ПЦР-оптимизации с кодон-оптимизированного гена L-цепи SL335 с применением набора ПЦР-праймеров №11 и №23  (SEQ ID NO: 43)), и G-CSF, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена G-CSF с применением набора ПЦР-праймеров №21 и №24
(SEQ ID NO: 43)), и G-CSF, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена G-CSF с применением набора ПЦР-праймеров №21 и №24  (SEQ ID NO: 44)). Гены Leys и G-CSF сшивали посредством ПЦР-сборки с применением набора ПЦР-праймеров №11 и №25
(SEQ ID NO: 44)). Гены Leys и G-CSF сшивали посредством ПЦР-сборки с применением набора ПЦР-праймеров №11 и №25  (SEQ ID NO: 45)), вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Fd SL335. Для получения конструкции HserGF/Lser, Hser подвергали ПЦР-оптимизации с кодон-оптимизированного Fd SL335 с применением набора ПЦР-праймеров №9 и №25. Гены Hser и G-CSF сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №22, вырезали при помощи EcoR I/Hind III, и клонировали в рНЕКА, содержащий Lser. Для получения конструкции HserGF/Hser, Lser подвергали ПЦР-оптимизации с кодон-оптимизированного гена L-цепи SL335 с применением набора ПЦР-праймеров №11 и №26
(SEQ ID NO: 45)), вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Fd SL335. Для получения конструкции HserGF/Lser, Hser подвергали ПЦР-оптимизации с кодон-оптимизированного Fd SL335 с применением набора ПЦР-праймеров №9 и №25. Гены Hser и G-CSF сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №22, вырезали при помощи EcoR I/Hind III, и клонировали в рНЕКА, содержащий Lser. Для получения конструкции HserGF/Hser, Lser подвергали ПЦР-оптимизации с кодон-оптимизированного гена L-цепи SL335 с применением набора ПЦР-праймеров №11 и №26  (SEQ ID NO: 46)), и G-CSF, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена G-CSF с применением набора ПЦР-праймеров №21 и №24. Гены Leys и G-CSF сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №11 и №25, вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Hser. ПЦР-праймеры для получения гибридных конструкций SL335-GCSH и гибридных конструкций SL335Δds-GCSF перечислены в Таблице 3 ниже.
(SEQ ID NO: 46)), и G-CSF, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена G-CSF с применением набора ПЦР-праймеров №21 и №24. Гены Leys и G-CSF сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №11 и №25, вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Hser. ПЦР-праймеры для получения гибридных конструкций SL335-GCSH и гибридных конструкций SL335Δds-GCSF перечислены в Таблице 3 ниже.

1-(11) Генерирование гибридных конструкций SL335-IFN-b
Процедуры клонирования для генерирования конструкции HcysIFNb/Lcys, были следующими. Hcys подвергали ПЦР-оптимизации с кодон-оптимизированного гена Н-цепи SL335 с применением набора ПЦР-праймеров №9 и №27  (SEQ ID NO: 47)), и IFN-b, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена IFN-b1a с применением набора ПЦР-праймеров №28
(SEQ ID NO: 47)), и IFN-b, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена IFN-b1a с применением набора ПЦР-праймеров №28 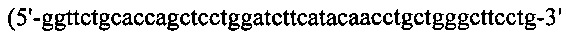 (SEQ ID NO: 48)) и №29
(SEQ ID NO: 48)) и №29 
 (SEQ ID NO: 49)). Гены Hcys и IFN-b1a сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №29, вырезали при помощи EcoR I/Hind III, и клонировали в рНЕКА, содержащий Lcys. Для получения конструкции HserIFN-b/Lser, Hser подвергали ПЦР-оптимизации с кодон-оптимизированного гена Н-цепи SL335 с применением набора ПЦР-праймеров №9 и №30
(SEQ ID NO: 49)). Гены Hcys и IFN-b1a сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №29, вырезали при помощи EcoR I/Hind III, и клонировали в рНЕКА, содержащий Lcys. Для получения конструкции HserIFN-b/Lser, Hser подвергали ПЦР-оптимизации с кодон-оптимизированного гена Н-цепи SL335 с применением набора ПЦР-праймеров №9 и №30  (SEQ ID NO: 50)). Гены Hser и IFN-b 1a сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №29, вырезали при помощи EcoR I/Hind III, и клонировали в рНЕКА, содержащий Lser. ПЦР-праймеры для получения гибридных конструкций SL334-IFNb и гибридных конструкций SL335Δds-IFNb перечислены в Таблице 4 ниже.
(SEQ ID NO: 50)). Гены Hser и IFN-b 1a сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №9 и №29, вырезали при помощи EcoR I/Hind III, и клонировали в рНЕКА, содержащий Lser. ПЦР-праймеры для получения гибридных конструкций SL334-IFNb и гибридных конструкций SL335Δds-IFNb перечислены в Таблице 4 ниже.

1-(12) Генерирование гибридных конструкций EGL4-hGH and the 1b28-hGH
EGL4 - Fab к EGFR человека - и 1b28 - Fab к IL-1b человека - выделяли из библиотеки антител (не опубликовано, «AprilBio Со.»). Для получения EGL4wt и EGL4Δds, Hcys и Hser подвергали ПЦР-амплификации с гена Н-цепи кДНК EGL4 с применением набора ПЦР-праймеров №5 и №6, и №5 и №31 (5'-gggaagcttattaactagatttgggctcaactctcttg-3' (SEQ ID NO: 51)), соответственно. Продукты ПЦР длиной ~750 по обрабатывали EcoR I/Hind III и сшивали при помощи рНЕКА, а затем трансформировали ими компетентные клетки МС1061. Lcys и Lser также подвергали ПЦР-амплификации с гена L-цепи кДНК EGL4 с применением набора ПЦР-праймеров №11 и №32  (SEQ ID NO: 52)), и №11 и №33
(SEQ ID NO: 52)), и №11 и №33 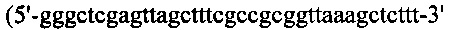 (SEQ ID NO: 53)), соответственно. Их вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Hcys или Hser of EGL4, соответственно. Для получения гибридной конструкции EGL4wt-hGH процедуры клонирования для генерирования конструкции HcysG/Lcys, были следующими. Hcys подвергали ПЦР-амплификации с гена Н-цепи кДНК EGL4 с применением набора ПЦР-праймеров №5 и №34
(SEQ ID NO: 53)), соответственно. Их вырезали при помощи BamH I/Xho I и клонировали в рНЕКА, содержащий Hcys или Hser of EGL4, соответственно. Для получения гибридной конструкции EGL4wt-hGH процедуры клонирования для генерирования конструкции HcysG/Lcys, были следующими. Hcys подвергали ПЦР-амплификации с гена Н-цепи кДНК EGL4 с применением набора ПЦР-праймеров №5 и №34  (SEQ ID NO: 54)), и hGH, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена hGH с применением набора ПЦР-праймеров №14 и №15. Гены Hcys и hGH сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №5 и №15, вырезали при помощи EcoR I/Hind III и клонировали в рНЕКА, содержащий Lcys EGL4. Для получения гибридной конструкции EGL4Δds-hGH Hser подвергали ПЦР-амплификации с гена Н-цепи кДНК EGL4 с применением набора ПЦР-праймеров №5 и №35
(SEQ ID NO: 54)), и hGH, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена hGH с применением набора ПЦР-праймеров №14 и №15. Гены Hcys и hGH сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №5 и №15, вырезали при помощи EcoR I/Hind III и клонировали в рНЕКА, содержащий Lcys EGL4. Для получения гибридной конструкции EGL4Δds-hGH Hser подвергали ПЦР-амплификации с гена Н-цепи кДНК EGL4 с применением набора ПЦР-праймеров №5 и №35 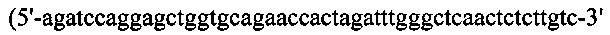 (SEQ ID NO: 55)), и hGH, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена HGH с применением набора ПЦР-праймеров №14 и №15. Гены Hser и hGH сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №5 и №15, вырезали при помощи EcoR I/ Hind III, и клонировали в рНЕКА, содержащий Lser EGL4Δds. 1b28wt, 1b28Δds, 1b28wt-hGH и 1b28Δds-hGH получали так же, как гибриды EGL4-hGH, с применением набора ПЦР-праймеров, за исключением того, что в качестве матрицы для ПЦР служила кДНК 1b28. ПЦР-праймеры для получения гибридных конструкций EGL4-hGH и 1b28-hGH перечислены в Таблице 5 ниже.
(SEQ ID NO: 55)), и hGH, содержащий линкерную последовательность, также подвергали ПЦР-амплификации с кодон-оптимизированного гена HGH с применением набора ПЦР-праймеров №14 и №15. Гены Hser и hGH сшивали вместе посредством ПЦР-сборки с применением набора ПЦР-праймеров №5 и №15, вырезали при помощи EcoR I/ Hind III, и клонировали в рНЕКА, содержащий Lser EGL4Δds. 1b28wt, 1b28Δds, 1b28wt-hGH и 1b28Δds-hGH получали так же, как гибриды EGL4-hGH, с применением набора ПЦР-праймеров, за исключением того, что в качестве матрицы для ПЦР служила кДНК 1b28. ПЦР-праймеры для получения гибридных конструкций EGL4-hGH и 1b28-hGH перечислены в Таблице 5 ниже.

1-(13) SDS-PAGE и вестерн-блоттинг
Для анализа посредством SDS-PAGE очищенные белки SL335wt-hGH и SL335Δds-hGH повторно суспендировали в буфере для проб NuPAGE® LDS («Invitrogen») с восстановителем для проб NuPAGE® («Invitrogen») или без него, и загружали на гель в концентрации 7 мкг/лунку. Полосы белка визуализировали путем окрашивания кумасси синим («Bio-Rad»). Для вестерн-блоттинга 500 нг афинно очищенного SL335wt-hGH и SL335Δds-hGH вносили в каждую лунку, как было описано выше, и переносили на нитроцеллюлозную мембрану. После блокирования 3% обезжиренным молоком («Bio-Rad») в ФСБ, содержащим 0,01 Tween («Sigma-Aldrich»), белки определяли путем инкубации с pAb антитела козы на каппа-L цепи человека, конъюгированным с ЩФ («Bethyl»). На мембрану добавляли субстрат нитросиний тетразолий хлорид (NBT)/5-бром-4-хлор-3-индлолил-фосфат (BCIP) («Duchefa», Харлеем, Нидерланды) для визуализации сигналов связывания.
1-(14) Капиллярный электрофорез, реализованный на чипе
Капиллярный электрофорез, реализованный на чипе, проводили с применением системы Agilent 2100 Bioanalyzer («Agilent Technologies», Санта-Клара, Калифорния, США). Пробы белка готовили в соответствии с прописью производителя и анализировали при помощи набора Protein 80, который рекомендован для анализа белков массой от 5 до 80 кДа. Если кратко, пробы смешивали с буфером для проб в присутствии или отсутствии ДТТ (дитиотреитола) для восстановительного или не восстановительного электрофореза, соответственно. Пробы подвергали денатурации при 95°С и загружали на чип, который был заполнен соответствующими реагентами, включая флуоресцентный краситель и раствор геля. Затем в систему вводили чип и запускали работу системы при помощи программы 2100. Результаты наносили на график зависимости интенсивности флуоресценции от размера белка.
1-(15) Масс-спектрометрия MALDI-TOF
Масс-спектрометрию MALDI-TOF (времяпролетная ионизация лазерной десорбцией с использованием матрицы) проводили на приборе Autoflex III Smartbeam («Bruker Daltonics», Биллерика, Массачусетс, США). Пробы смешивали с таким же объемом матрицы MALDI (10 мг/мл а-циано-4-гидроксикоричной кислоты) и наносили на отражательный экран MALDI. Внешнюю калибровку проводили при помощи набора для калибровки пептидов и белков MALDI-MS («Sigma-Aldrich»). Масс-спектры в диапазоне m/z 15000160000 и 1000070000 захватывали для гибрида SL335wt-hGH и гибрида SL335Δds-hGH, соответственно, в положительном режиме.
1-(16) Анализ биологической активности hGH in vitro
Клетки лимфомы крысы Nb2-ll («Sigma-Aldrich») выращивали в полной DMEM (минимальная эссенциальная среда Игла, модифицированная по способу Дульбекко) с добавлением 5% сыворотки лошади («Sigma-Aldrich») и 1% пенициллина-стрептомицина («Invitrogen») в инкубаторе с увлажненной атмосферой и 5% CO2 при 37°С (Tanakaef al., 1980). Клетки отмывали два раза средой DMEM, центрифугировали при 1000 g в течение 5 мин и повторно суспендировали в DMEM, содержащей 5% (по объему) сыворотки лошади при плотности 8×104 клеток/мл. Аликвоту суспензии клеток 50 мкг добавляли в каждую лунку планшетов на 96 лунок и инкубировали в течение ночи. Затем клетки инкубировали с возрастающими концентрациями (0-20 нМ) Гроутропина® (немодифицированного rhGH; «Dong-А Pharmaceuticals», Сеул, Южная Корея) или SL335Δds-hGH в 50 мл DMEM, содержащей 5% сыворотки лошади в течение 48 ч при 37°С. После инкубации в каждую лунку добавляли 10 мкл ССК-8 («Dojindo», Машики-Мачи Япония) и инкубировали в течение 4 ч. Поглощение регистрировали на микропланшетном ридере («Bio-Rad») на длине волны 450 нм.
1-(17) Стабильность SL335Δds-hGH в сыворотке
SL335wt и SL335Δds-hGH (в конечной концентрации 10 мкг/мл) повторно суспендировали в фетальной бычьей сыворотке (ФБС) («Thermo Scientific», Уолтем, Массачусетс, США), содержащей 0,03% азида натрия и инкубировали в течение 16 дней при 37°С. Каждый день отбирали небольшие аликвоты (50 мл) и хранили при -20°С до применения. Активность связывания с САЧ определяли посредством ELISA, а биологическую активность in vitro измеряли с применением клеток Nb2-11 cells («Sigma-Aldrich»), как описано выше.
1-(18) Анализ фармакокинетики in vivo
ФК исследования проводила сертифицированная компания - контрактная исследовательская организация («ChemOn», Сувон, Южная Корея). Животных содержали на стандартной диете для грызунов, и они получали пищу и воду в неограниченном количестве, в помещении поддерживали постоянную температуру и влажность при контролируемом цикле освещения (12 ч свет, затем 12 ч темнота). Если кратко, SL335 и отрицательный Fab (незначимый Fab человека) вводили внутривенно (в/в) или подкожно (п/к), по отдельности в группах из трех крыс Спраг-Доули в дозе 1 мг/кг, и пробы сыворотки отбирали в несколько моментов времени (5 мин, 15 мин, 30 мин, 1 ч, 2 ч, 4 ч, 8 ч, 24 ч, 48 ч, 96 ч и 144 ч для в/в введения, и 5 мин, 15 мин, 30 мин, 1 ч, 2 ч, 4 ч, 8 ч, 24 ч, 48 ч и 96 ч для п/к введения). Концентрацию SL335 и отрицательного Fab в пробах сыворотки измеряли посредством сэндвич-ELISA с применением mAb мыши к IgG Fd Человека и pAb козы к каппа-L-цепи человека, конъюгированного с HPRO, в качестве антител захвата и детекции, соответственно. Фрагменты Fab человека в известной концентрации также были включены в анализ для получения стандартной кривой. Кривые зависимости концентрации в сыворотке от времени строили для бескомпартментной модели с применением программы WinNonlin (SL335 и Neg Fab) и строили график при помощи программы Sigma Plot. Аналогично, гроутропин® и SL335Δds-hGH вводили внутривенно или подкожно по отдельности в группе из трех или четырех крыс. Дозы гроутропина® и SL335Δds-hGH для внутривенного введения составляли 0,3 мг/кг и для п/к введения составляли 0,6 мг/кг, соответственно. Пробы сыворотки отбирали в несколько моментов времени (5 мин, 15 мин, 30 мин, 1 ч, 2 ч, 3 ч, 4 ч, 6 ч и 8 ч для гроутропина® и 5 мин, 30 мин, 1 ч, 2 ч, 4 ч, 8 ч, 24 ч, 48 ч, 96 ч и 144 ч для SL335Δds-hGH. Количество гроутропина® в пробах сыворотки измеряли при помощи набора для детекции hGH посредством ELISA («Genway», Сан-Диего, Калифорния, США), а количество SL335Δds-hGH измеряли посредством сэндвич-ELISA, как было описано выше. Кривую зависимости концентрации в сыворотке от времени аппроксимировали однокомпартментной моделью с применением программы Phoenix™ WinNonlin (версия 6.2).
1-(19) Анализ фармакодинамики in vivo
Способность ежедневного введения гроутропина® и еженедельного введения SL335Δds-hGH обеспечивать увеличение массы анализировали у крыс с удаленным гипофизом, при введении препаратов п/к в ChenOn, как было описано ранее (см. Clark et al., (1996) J. Biol. Chem. 271, 21969-21977). Если кратко, приобретали молодых крыс Спраг-Доули с удаленным гипофизом («Harlan», Токио, Япония), и любое животное, прибавившее в весе более 7 г в первые 15 дней после операции исключали из исследования. Животных случайным образом распределяли в пять лечебных групп (только вспомогательное вещество, инъекция 1 р/д гроутропина® 0,3 мг/кг и инъекция 1 р/нед 0,6 мг/кг, 1,2 мг/кг или 2,4 мг/кг SL335Δds-hGH). Массу тела регистрировали раз в день после начала режима дозирования. Рост большеберцовой кости тщательно измеряли штангенциркулем для костей. Статистическое сравнение проводили посредством дисперсионного анализа, а затем применяли критерий множественного сравнения Даннета; значимыми считали p-значения менее 0,05.
2. Экспериментальные результаты
2-(1) Выделение клонов Fab к СА
В библиотеке антител HuDVFab-8L проводили селекцию в отношении магнитных микроносителей, конъюгированных с СА человека, СА крысы или СА мыши при рН 6 или рН 7,4. После трех циклов биопэннинга проводили ELISA С моноклональным фагом с целью идентификации клонов фагов-антител, которые были специфичны в отношении указанных антигенов. Посредством ELISA было идентифицировано более 60 позитивных клонов (данные не показаны), и анализ на основе секвенирования ДНК для генов VH и VL позволил идентифицировать восемь дискретных фагов-антител, которые были названы SA138, SA139, SA140, SA141, SL18, SL301, SL310 и SL335, соответственно. Активность связывания указанных клонов к СА человека, СА крысы или бычьему СА подтверждали посредством ELISA С моноклональным фагом при рН 6 или рН 7,4 (Фиг. 1А и 1В). Три клона фагов-антител - SA138, SA139 и SA141 - были активны только в отношении СА человека независимо от рН. SA140 также распознавал только СА человека при рН 7,4, но его активность связывания исчезала при рН 6. С другой стороны, SL18, SL310 и SL335 связывались с СА человека, СА крысы и СА мыши при обоих рН с несколько различными интенсивностями. SL301 обладал значительной активностью в отношении СА человека и СА крысы при обоих рН, и слабо связывался с СА мыши только при рН 7,4. Ни один из восьми клонов Fab не был активен в отношении бычьего СА. SL18, SL301, SL310 и SL335 также были охарактеризованы в связи с их перекрестной реактивностью в отношении СА, начиная по меньшей мере с двух разных видов. Гены Fd и L-цепи для четырех фагов-антител субклонировали в вектор рНЕКА для экспрессии в периплазме E.coli, и растворимые фрагменты Fab получали из надосадочной жидкости питательной среды или экстрактов периплазмы. После аффинной очистки проводили ELISA, чтобы сравнить активность связывания указанных фрагментов в отношении СА человека, СА крысы или СА мыши при рН 6 (Фиг. 2А) и рН 7,4 (Фиг. 2В). САЧ, САК, САМ и БСА в концентрации 50 мкг/мл иммобилизовывали в каждой лунке микротитрационных планшетов, и четырем очищенным молекулам Fab (SL18, SL301, SL310 и SL335) давали связаться с антигенами при рН 6,0 (Фиг. 2А) и при рН 7,4 (Фиг. 2В). В качестве вторичного антитела применяли конъюгат HRPO с pAb козы к каппа-L-цепи человека. Сигналы связывания визуализировали при помощи субстрата ТМВ, и поглощение измеряли на ридере для ELISA («Bio-Rad»). Данные представляют собой среднее ± стандартное отклонение для трех экспериментов. При связывании с СА человека порядок сигналов связывания был следующим SL335>SL310>SL301>SL18 при рН 6 и рН 7.4. При связывании с СА крысы порядок сигналов связывания был следующим SL335>SL310>SL301>SL18 при рН 6, и SL335=SL310>SL301=SL18 при рН 7,4. При связывании с СА мыши порядок сигналов связывания был следующим SL18>SL335>SL310 при рН 6 и SL335>SL310>SL18 при рН 7,4. В соответствии с Фиг. 2, SL301 не связывался с СА мыши при рН 6, и очень слабо связывался при рН 7,4. Было показано, что SL335 был наиболее эффективным связывающим веществом среди четырех клонов Fab, в отношении и СА человека, и СА крысы, независимо от рН. SL335 связывался с СА человека при рН 6,0 в два раза сильнее, чем при рН 7,4 (50% сигнал связывания при 20 нг/мл и 40 нг/мл), в 20 раз сильнее, чем с СА крысы при том же рН (50% сигнал связывания при 20 нг/мл и 400 нг/мл), и в четыре раза сильнее, чем с СА крысы при рН 7,4 (50% сигнал связывания при 40 нг/мл и 160 нг/мл).
2-(2) Перекрестная реактивность и сродство связывания SL335
Поскольку SL335 был наилучшим связывающим веществом среди четырех клонов Fab к СА человека, его перекрестную реактивность также анализировали посредством ELISA. Была воспроизведена активность связывания с СА человека, СА крысы и СА мыши, как показано на Фиг. 2. Также было показано, что SL335 интенсивно распознавал СА макаки-резус и слабо связывался с СА собаки. Однако SL335 не распознавал СА кролика, а также другие незначимые антигены, включая EGFR, ЕрСАМ, IL-15Ra, IL-1b, CD16a или с-МЕТ. Сродство связывания SL335 с СА человека, СА крысы и СА мыши при рН 6 или рН 7,4 также измеряли посредством интерферометрии в биослое путем пропускания через разную концентрацию антигенов на биосенсорах, которые были покрыты SL335 (см. Таблицу 6 ниже). Результаты хорошо коррелировали с данными ELISA на Фиг. 2 в том смысле, что константы диссоциации SL335 с САЧ составляли 9 нМ при рН 6 и 13 нМ при рН 7,4, соответственно, а константы диссоциации SL335 с САК составляли 122 нМ и 65 нМ при рН 6 и рН 7,4, соответственно. Сродство связывания SL335 с САМ составило приблизительно 10 мМ при рН 6 и 1,6 мМ при рН 7,4, но указанные данные не были включены в Таблицу 6 из-за недостаточной надежности.

Кинетика связывания и константы диссоциации рассчитывались при помощи пакета программ «Octet QK».
2-(3) Фармакокинетика SL335 in vivo
Из всех белков плазмы САЧ обладает исключительно большим периодом полувыведения благодаря FcRn-опосредуемому механизму рециклирования, и его обычно применяют как партнер по слиянию для увеличения периода полувыведения из сыворотки. Кроме того, было выяснено, что фрагменты антитела, которые ассоциированы с сывороточным альбумином, обладают увеличенным периодом полувыведения из сыворотки. Следовательно, проводили анализ фармакокинетики, чтобы проверить, обладает ли SL335 также большим периодом полувыведения. В качестве отрицательного контроля был включен Fab человека с неизвестной специфичностью связывания (отрицательный Fab). SL335 и отрицательный Fab вводили внутривенно или подкожно по отдельности в группе из трех крыс в дозе 1 мг/кг, и пробы сыворотки отбирали в несколько моментов времени (5 мин, 15 мин, 30 мин, 1 ч, 2 ч, 4 ч, 8 ч, 24 ч, 48 ч, 96 ч и 144 ч для в/в введения, и 5 мин, 15 мин, 30 мин, 1 ч, 2 ч, 4 ч, 8 ч, 24 ч, 48 ч, и 96 ч для подкожного введения). Концентрацию SL335 и Neg Fab в пробах сыворотки определяли посредством сэндвич-ELISA с применением mAb мыши к IgG Fd человека и pAb козы к каппа-L-цепи человека, конъюгированного с HRPO, в качестве антител захвата и детекции, соответственно. Фрагменты Fab человека известной концентрации также были включены в анализ для построения стандартной кривой. Кривые зависимости концентрации в сыворотке от времени аппроксимировали однокомпартментной моделью с применением программы WinNonlin (SL335 и Neg Fab) и двухкомпартментной моделью с применением программы Sigma Plot. При внутривенном введении терминальный период полувыведения (t1/2) SL335 составил 37 ч, а площадь под кривой зависимости концентрации от времени (AUC0→∞) составила 187 ч мг/мл, что соответствует десятикратному увеличению t1/2 и 26-кратному увеличению AUC0→∞ по сравнению с Neg Fab (3,8 ч и 7 ч, соответственно) (Фиг. 3А). Подкожная инъекция SL335 приводила к аналогичным показателям, включая девятикратное увеличение t1/2 (120 ч и 13 ч) и 44-кратное увеличение AUC0→∞ по сравнению с Neg Fab (87 и 2 ч мг/мл1) (Фиг. 3В). Указанные результаты явно показали увеличенный период полувыведения SL335 и подразумевают, что SSWL335 не должен мешать взаимодействию между САЧ и FcRn у крыс.
2-(4) Получение гибридов SL335-hGH
SL335 применяли для получения двух гибридов SL335-hGH и четырех дополнительных гибридов SL335-hGH путем генетического слияния рекомбинантного hGH (27 - 191 ак) с N- или С-концом Fd или L-цепи коротким пептидным линкером. кДНК рекомбинантного hGH (27 - 191 ак) соединяли с С-концом С- или L-цепи SL335wt в классической форме Fab при помощи короткого пептидного линкера, и получали конструкцию двух гибридных форм (HcysG/Lcys и LcysG/Hcys). Четыре дополнительных гибридных формы (HserG/Lcys, LserG/Hcys, HserG/Lser и LserG/Hser) конструировали так же, как описано выше, за исключением применения SL335 в нулевой форме (SL335null) или в форме ds Fab (SL335Δds), в которых Cys233 на С-конце CH1 и/или Cys214 на С-конце CLk были заменены на Ser. Для экспрессии гибридных белков в периплазме лидерную последовательность ompA (MKKTAIAIAVLAGFATVAQA (SEQ ID NO: 56)) располагали выше гибридов L-цепи или L-hGH и лидерную последовательность pelB (MKYLLPTAAAGLLLLAAQPAMA (SEQ IN NO: 57)) располагали выше гибридов Н-цепи и H-hGH. В данных предварительных экспериментах генетическое сшивание hGH с N-концом Fd или L-цепи приводило к низкой экспрессии или отсутствию экспрессии растворимых гибридных белков. Гибрид hGH с С-концом Fd также демонстрировал низкий выход экспрессии и, по-видимому, у него нарушалась укладка домена hGH, вероятно, из-за аномального образования дисульфидных связей в гибриде SL335-hGH (данные не показаны). Ранее сообщалось, что удаление дисульфидной связи между цепями Fab путем мутирования остатка Cys на С-конце в CH1 и CLk (Cys233 и Cys214, соответственно) не влияло на уровень периплазматической продукции, стабильность при экстракции и очистке, стабильность в сыворотке или период полувыведения из сыворотки (см. Kabat et al., (1991) Sequences of Proteins of Immunological Interest; Humphreys et al., (1997) J. Immunol. Methods. 209, 193202; Humphreys et al., (2007) Protein Eng Des Sel 20, 227234.). Заменив и Cys233 в CH1, и Cys214 в CLk на серин (замены Cys233 Ser233 и Cys214 Ser214), мы проверяли, модулируют ли указанные остатки Cys в SL335 экспрессию в растворимой форме и адекватную укладку гибридов SL335-hGH. На Фиг. 4 проиллюстрированы шесть гибридных конструкций SL335-hGH. Еще один вариант SL335, отличный от SL335wt и SL335Δds, названный SL335null, также был создан путем замены либо Cys233 CH1, либо Cys214 CLk, на Ser, чтобы выяснить влияние каждого из остатков цистеина (Cys233 или Cys214) по отдельности. Два производных гибрида SL335w включали HcysG/Lcys (гибрид HCys233-hGH, соединенный с LCys214) и LcysG/Hcys (гибрид LCys214-hGH, соединенный с HCys233), два производных гибрида SL335null включали HserG/Lcys (гибрид HSer233-hGH, соединенный с LCys214) и LserG/Hcys (гибрид LSer214-hGH, соединенный с HCys233). И наконец, два производных гибрида SL335Δds включали HserG/Lser (гибрид HSer233-hGH, соединенный с LSer214) и LserG/Hser (гибрид LSer214-hGH, соединенный с HSer233). Указанные шесть гибридных конструкций SL335-hGH экспрессировались в клетках-хозяевах Е. coli SUPEX5, выход и активность связывания с САЧ указанных шести гибридных белков SL335-hGH в надосадочной жидкости питательной среды анализировали посредством ELISA. Клоны E. coli, экспрессирующие гибридные белки SL335-hGH, выращивали при идентичных условиях в присутствии IPTG, и надосадочную жидкость питательной среды отбирали путем кратковременного центрифугирования. Концентрацию растворимых гибридов SL335-hGH измеряли посредством сэндвич-ELISA с применением mAb мыши к Fd человека в качестве захватывающего антитела и pAb козы к каппа-Ь-цепи человека, конъюгированного с HRPO, в качестве детектирующего антитела (Фиг. 5А). Никаких растворимых форм Fab ни из LcysG/Hcys, ни из LserG/Hcys не определялось. Хотя данные не были представлены, вестерн-блоттинг с применением лизата клеток E.coli выявил, что Cys233 Fd обусловливал сильную деградацию, и секреции фрагментов Fd не было, вероятно из-за агрегации белков. Выход HcysG/Lcys составил 0,5 мкг/мл, а выход для HserG/Lcys и LserG/Hser составил приблизительно 1,8 мг/мл и 1,4 мг/мл, соответственно (Фиг. 5А). Интересно, что выход HserG/Lser составил приблизительно 4 мкг/мл, что в восемь раз превышало выход для HcysG/Lcys. Экстракты периплазмы демонстрировали идентичный характер экспрессии, хотя суммарный выход составлял всего лишь ~30% от выхода, полученного для надосадочной жидкости питательной среды (данные не показаны). В повторных экспериментах было подтверждено, что различие выхода между HcysG/Lcys и HserG/Lser не зависело от вариации клонов или скорости роста клонов E. coli. Активность связывания гибридов SL335-hGH с САЧ была сопоставимой при применении микротитрационных планшетов, в которые было нанесено 5 мкг/мл САЧ, и инкубируемых с последовательными разведениями надосадочной жидкости, содержащей гибриды SL335-hGH. Гибриды SL335-hGH, связавшиеся с САЧ, затем детектировали при помощи pAb козы к каппа-L-цепи человека, конъюгированного с HRPO. Как и ожидалось, детекция HserG/Lser, который связался с САЧ, при помощи pAb к κL человека, выявила в восемь раз более высокий сигнал связывания, чем сигнал связывания для HcysG/Lcys, и приблизительно в четыре раза более высокий сигнал по сравнению с сигналом для HserG/Lcys и LserG/Hser (Фиг. 5 В). Сходный характер сигналов связывания также наблюдался, когда для детекции гибридов SL335-hGH применяли Т20 - pAb козы, специфичное в отношении С-конца hGH (Фиг. 5С). Однако при детекции при помощи NYThGH - mAb мыши, специфичного в отношении полноразмерного hGH, HserG/Lser создавал сигнал связывания, который в 30 раз превышал сигнал связывания HserG/Lcys и LserG/Hser, и в 60 раз превышал сигнал связывания HcysG/Lcys (Фиг. 5D), что свидетельствует о том, что связывание NYThGH с доменом hGH Hcys/Lcys нарушалось из-за присутствия дисульфидной связи между цепями в SL335. Поскольку Hcys/Lcys и Hser/Lser соответствуют применению SL335wtH SL335Δds для создания гибридов SL335-hGH, они были названы гибридом SL335wt-hGH и гибридом SL335Δds-hGH, соответственно, здесь и далее (Фиг. 5).
Чтобы определить, зависит ли высокий выход растворимого гибрида SL335Δds-hGH от удаления дисульфидной связи между цепями в SL335, штаммов E.coli или температуры индукции, гибрид SL335wt, SL335Δds, SL335wt-hGH и гибрид SL335Δds-hGH экспрессировали в исходном штамме МС1061, а также в мутантных клетках SUPEX5 при 20°С (Фиг. 6А), 25°С (Фиг. 6В) или 30°С (Фиг. 6С), и количество молекул Fab в надосадочной жидкости питательной среды измеряли посредством ELISA. Выход SL335wt, экспрессируемого в штамме МС1061, составил 1 мкг/мл при 20°С, что было приблизительно в три раза выше, чем при 25°С и 30°С. Это указывает на то, что индукция SL335wt при температуре ниже 25°С дает преимущество, особенно когда в качестве штамма-хозяина применяют МС1061. Аналогичные результаты были получены также для штамма SUPEX5. В случае SL335Δds выход составил приблизительно 1,3 мг/мл при 20°С, независимо от штаммов-хозяев E.coli и температуры индукции. Указанные результаты показали, что присутствие или отсутствие дисульфидной связи между цепями в Fab значимо не влияет на выход растворимого Fab при 20°С, независимо от штаммов-хозяев E.coli. Выход гибрида SL335wt-hGH составил приблизительно 0,3-0,5 мкг/мл, независимо от штаммов-хозяев E.coli и температуры индукции. С другой стороны выход гибрида SL335Δds-hGH, экспрессируемого в штамме МС1061, составил 1,8 мкг/мл при температурах 20°С и 25°С, и 1,5 мкг/мл при 30°С, что указывает на минимальную температурную зависимость, тогда как выход гибрида SL335Δds-hGH, экспрессируемого в штамме SUPEX5, составил 4,0 мкг/мл и при 20°С, и при 25°С, и 3,5 мкг/мл при 30°С. Указанные результаты означают, что применение формы SL335Δds и штамма E.coli SUPEX5 позволяет в 12 раз увеличить выход гибридного белка SL335-hGH по сравнению с сочетанием формы SL335wt и штамма E.coli МС1061.
2-(5) Генерирование гибридных конструкций SL335-GCSF, SL335-IFNb, EGL4-hGHu 1b28-hGH
Чтобы продемонстрировать благоприятное действие формы FabΔds и штамма SUPEX5 на улучшение экспрессии в растворимой форме гибридного белка Fab-эффектора, генерировали разные гибридные конструкции Fab-эффектора. Во-первых, чтобы определить влияние эффекторного домена, были созданы два гибридных варианта SL335-GCSF (HcysGCSF/Lcys, который назвали SL335wt-GCSF, HserGF/Lser, который назвали SL335Δds-GCSF) и два гибридных варианта SL335-IFNb (HcysIFNb/Lcys, который назвали SL335wt-IFNb, HserIFNb/Lser, который назвали SL335Δds-IFNb), тем же способом, которым генерировали гибриды SL335wt-hGH и SL335Δds-hGH. Температуру индукции устанавливали на оптимальном уровне 20°С, и выход экспрессии указанных двух гибридных белков в надосадочной жидкости культуры E.coli, сравнивали посредством ELISA. Выход SL335wt-GCSF составил 0,3 и 0,6 мг/мл у МС1061 и SUPEX5, соответственно, а для SL335Δds-GCSF составил 0,6 и 1,5 мг/мл у МС1061 и SUPEX5, соответственно (Фиг. 7А). В то же время выход SL335wt-IFNb составил приблизительно 0,16 мг/мл и у МС1061, и у SUPEX5, а выход SL335Δds-IFNb составил 0,2 мг/мл и 0,5 мг/мл у МС1061 и SUPEX5, соответственно (Фиг. 7В). Следовательно, сочетание гибрида SL335Δds-GCSF и штамма SUPEX5 приводит к приблизительно 5-кратному увеличению выхода формы гибрида SL335-GCSF по сравнению с сочетанием гибрида SL335wt-GCSF и штамма МС1061, а сочетание гибрида SL335Δds-IFNb и штамма SUPEX5 приводило к приблизительно 3-кратному увеличению количества формы гибрида SL335-IFNb по сравнению с сочетанием гибрида SL335wt-IFNb и штамма МС1061. Во-вторых, мы также создали две гибридных конструкции Fab-hGH с применением EGL4 - Fab человека к EFGR - и 1b28 - Fab человека к IL-1b, чтобы определить влияние Fab. Поскольку применялся тот же способ генерирования, что и для гибридов SL335wt-hGH и SL335Δds-hGH, двумя гибридными конструкциями EGL4-hGH были гибрид EGL4wt-hGH в форме HcysG/Lcys и гибрид EGL4Δds-hGH в форме HserG/Lser. Аналогично, гибридными конструкциями 1b284-hGH были гибрид 1b28wt-hGH в форме HcysG/Lcys и гибрид 1b28Δds-hGH в форме HserG/Lser.
Выход гибрида EGL4wt-hGH составил 8090 нг/мл у штаммов МС1061 и SUPEX5, а выход гибрида EGL4Δds-hGH составил 140 нг/мл у штамма МС1061 и 220 нг/мл у штамма SUPEX5 (Фиг. 8А), что указывает на то, что сочетание гибрида EGL4Δds-hGH и штамма клетки-хозяина SUPEX5 приводит к 2,4-кратному увеличению количества гибридного белка EGL4-hGH в надосадочной жидкости питательной среды по сравнению с сочетанием гибрида EGL4wt-hGH и клеток-хозяев МС1061. В случае гибридных конструкций 1b28-hGH выход гибрида 1b284wt-hGH составил 50 нг/мл у штамма МС1061 и 100 нг/мл у штамма SUPEX5, соответственно, а выход гибрида 1b28Δds-hGH составил 900 нг/мл у штамма МС1061 и 4 нг/мл у штамма SUPEX5 (Фиг. 8В), что указывает на то, что сочетание гибрида 1b28Δds-hGH и штамма клеток-хозяев SUPEX5 приводит к 800-кратному увеличению количества гибридной формы 1b28-hGH в надосадочной жидкости питательной среды по сравнению с сочетанием гибрида 1b28wt-hGH и клеток-хозяев МС1061.
2-(6) Описание молекул SL335wt-hGH и SL335Δds-hGH
Гибриды SL335wt-hGH и SL335Δds-hGH были дополнительно охарактеризованы на молекулярном уровне. Гибридные белки в надосадочной жидкости питательной среды подвергали аффинной очистки путем пропускания через смолы с нанесенным САЧ, и анализировали посредством SPS-PAGE и вестерн-блоттинга при восстановительных и невосстановительных условиях. HcysG/Lcys (дорожка 1) и HserG/Lser (дорожка 2) подвергали аффинной очистке из надосадочной жидкости питательной среды при помощи гранул сефарозы с иммобилизованным САЧ, и проводили SPS-PAGE с применением 4-12% Bis-Tris-геля при восстановительных и невосстановительных условиях. Полосы белков визуализировали при помощи окрашивания кумасси синим (Фиг. 9А). Белки с отдельных SPS-PAGE переносили на нитроцеллюлозную мембрану, и для детекции Leys и Lser применяли антитело овцы к каппа-L-цепи человека, конъюгированное с ЩФ (Фиг. 9В). Сигналы связывания визуализировали при помощи субстрата NTB/BCIP. При анализе SDS-PAGE и SL335wt-hGH, и SL335Δds-hGH давали крупные полосы белков по размеру 46 и 23 кДа, которые соответствовали гибридам Fd-hGH и L-цепям, соответственно, при восстановительных условиях. При невосстановительных условиях SL335Δds-hGH ожидаемо давал две идентичных полосы белков из-за отсутствия дисульфидной связи между цепями. В случае SL335wt-hGH была видна крупная полоса на 70 кДа, которая соответствует правильной гетеродимерной форме SL335wt-hGH. Еще также обнаруживались много производных SL335wt-hGH разного размера, включая четыре явных полосы белков в диапазоне от 24 кДА до 45 кДА неизвестной природы, а также пара слабых полос белков, соответствующих 100 кДа и 135 кДА по размеру. Также во всех пробах были видны белки с размерами 15 кДа и 12,5 кДа. Затем проводили вестерн-блоттинг с применением mAb к Fd человека, pAb к каппа-L-цепи и pAb к hGH - Т20. Блоттинг с mAb к Fd человека выявил только HhysG и HserG с размером 46 кДа как при невосстановительных, так и при восстановительных условиях (данные не показаны). С другой стороны четыре полосы белков с размером от 24 до 45 кДа, а также с более крупным размером 70 кДа в пробе SL335wt-hGH выявлялись при применении pAb к каппа-L-цепи при невосстановительных условиях (Фиг. 9В). Данный результат показывает, что Cys214 L-цепи обусловливает образование разных мультимерных L-цепей, по меньшей мере за счет образования аномальных дисульфидных связей. Блоттинг с pAb к hGH человека Т-20 выявлял гетеродимерную форму SL335wt-hGH размером 70 кДа и мономерный HerG SL335Δds-hGH ~ 45 кДа при невосстановительных условиях (Фиг. 9С). Белки с размерами 15 кДа и 12,5 кДа не определялись ни одним из антител, что указывает на то, что они представляли собой либо продукты распада гибридов, либо примеси из собственных белков Е. coli.
Капиллярный электрофорез, реализованный на чипе, подтвердил результаты анализа SPS-PAGE. HcysG/Lcys (Фиг. 10A) и HserG/Lser (Фиг. 10В) готовили в буфере для проб в присутствии или отсутствии ДТТ для электрофореза в восстановительных или невосстановительных условиях, и проводили капиллярный электрофорез, реализованный на чипе, в системе Agilent 2100 Bioanalyzer согласно протоколу производителя, с применением набора Protein 80. Результаты наносили на график, отражающий зависимость интенсивности флуоресценции от размера белка. SL335wt-hGH образовывал несколько производных SL335wt-hGH в диапазоне от 27,1 кДа до 52,4 кДа при невосстановительных условиях, и многие из них исчезали при восстановительных условиях в присутствии ДТТ (Фиг. 10A). SL335Δds-hGH давал почти идентичные пики белков между невосстановительными и восстановительными условиями, за исключением незначительных изменений молекулярных масс (Фиг. 10В).
SL335wt-hGH и SL335Δds-hGH также анализировали посредством масс-спектрометрии MALDI-TOF. Масс-спектрометрию MALDI-TOF проводили на приборе Autoflex III Smartbeam («Bruker Daltonics», Биллерика, Массачусетс, США). Подвергнутые афиинной очистке HcysG/Lcys (Фиг. 11А) и HserG/Lser (Фиг. 11В) смешивали с матрицей MALDI, и регистрировали спектры в диапазоне значений 10000-150000 Да в режимы положительно заряженных ионов. Для SL335Δds-hGH регистрировали масс-спектры в диапазоне значений 10000-70000. Для SL335wt-hGH, были получали спектры в диапазоне 15000-16000, поскольку проба SL335wt-hGH демонстрировала полосы белков с размером более 70 кДа, как показано на Фиг. 8А. Молекулярные массы Lcys, HcysG и SL335wt-hGH были идентифицированы как 23 226 Да, 46226 Да и 69 837 Да, соответственно (Фиг. 11А). Был показано, что размер трех раздельных белков, которые были больше нормального SL335wt-hGH, составил 92 824 Да, 117 455 Да и 139 Да. В случае SL335Δds-hGH, молекулярные массы Lser и HserG составили были идентифицированы как 23 334 Да и 46 667 Да, соответственно (Фиг. 11В). Низкий пик HserG по сравнению с Lser мог отражать более низкую эффективность ионизации у более крупных молекул, или наличие меньшего мольного отношения HserG и Lser в пробе.
Подвергнутый аффинной очистке SL335Δds-hGH дополнительно очищали, пропуская его через колонку SephacrylS-200HR посредством FPLC. После аффинной очистки проводили гель-филтрацию HserG/Lser с применением колонки Sephacryl™ S-200HR Prepacked и ACTA FPLC («GE Healthcare», Вауватоза, Висконсин, США). Колонку уравновешивали буфером для уравновешивания (20 мМ HEPES рН 7,4, содержащий 150 мМ NaCl), и загружали в нее подвергнутый аффинной очистке HserG/Lser. Десорбцию проводили при помощи буфера для уравновешивания при скорости потока 0,5 мл/мин. Стрелками показаны фракции, выбранные для анализа посредством SDS-PAGE (Фиг. 12А). Фракции №13, №16. №19 и №23, которые восстанавливали из двух различных пиков, анализировали при помощи геля 4-12% Bis-Tris при восстановительных условиях (Фиг. 12В). Полосы белков визуализировали посредством окрашивания кумасси синим. Два пика, которые соответствовали приблизительно 66 кДа и 25 кДа, выявлялись при анализе фракций №12-№27 (Фиг. 9А). Поэтому четыре фракции (фракции №13, №16, №19 и №23) анализировали посредством SDS-PAGE при восстановительных условиях, чтобы определить содержание белка в указанных фракциях (Фиг. 9В). Результаты показали, что фракции из пика 66 кДа (фракции №13, №16, и №19), содержащие гетеродимерный SL335Δds-hGH, и фракция из пика 25 кДа (фракция №23) содержали в основном мономерный Lser.
2-(7) описание функциональных свойств SL335Δds-hGH in vitro
Чтобы определить, влияет ли удаление дисульфидной связи между цепями в SL335wt и гибрида hGH на сродство связывания с САЧ и САК, проводили анализ посредством интерферометрии с применением SL335Δds-hGH при рН 6 и рН 7,4 (см. Таблицу 6 ниже). Константы диссоциации SL335Δds-hGH с САЧ составили 1,7 нМ при рН 6 и 1,5 нМ при рН 7,4, соответственно. Константы диссоциации с САК составили 499 нМ и 83,6 нМ при рН 6 и рН 7,4, т.е. было продемонстрировано 4,2-кратное и 1,3-кратное уменьшение сродства по сравнению со сродством SL335, соответственно.
Активность hGH in vitro у SL335ds-hGH также измеряли с применением клеток лимфомы крысы Nb2-11, которые пролиферируют при обработке hGH с зависимостью от концентрации. Клетки лимфомы крысы Nb2-11 повторно суспендировали в DMEM, содержащей 5% (по объему) сыворотки лошади с плотностью 8×104 клеток/лунку, и добавляли аликвоты суспензии клеток 50 мкл в каждую лунку планшетов на 96 лунок, а затем инкубировали в течение ночи. Затем клетки обрабатывали возрастающими концентрациями гроутропина® или HserG/Lser (0-20 нМ) в 50 мл DMEM, содержащей 5% сыворотки лошади, в течение 48 ч при 37°С. После инкубации в каждую лунку добавляли 10 мкл раствора ССК-8, и клетки инкубировали в течение 4 ч. Регистрировали поглощение на ридере для микропланшетов на длине волны 450 нм. Данные представлены как среднее ±СО для трех экспериментов. В отсутствии САЧ SL335Δds-hGH был способен стимулировать рост клеток Nb2-ll с кажущейся EC50 50 пМ (3,5 нг/мл (Фиг. 13А). Данное значение было в 6,7 раз больше, чем показатель для гроутропина ® - стандарта rhGH (7,5 пМ). В присутствии 10 мМ САЧ соответствующие активности гроутропина® и SL335Δds-hGH в основном оставались неизменными, хотя SL335Δds-hGH демонстрировал приблизительно пятикратное уменьшение активности по сравнению с активностью гроутропина® (Фиг. 13В). SL335, который применялся в качестве отрицательного контроля, не демонстрировал никакого пролиферативного действия. Указанные результаты явно демонстрируют, что SL335Δds-hGH обладает функциональной биологической активностью, характерной для hGH.
Затем определяли активность в сыворотке путем инкубации SL335Δds-hGH при 37°с в течение 16 дней. Вместо сыворотки человека для повторного суспендирования проб применяли ФБС, поскольку способность SL335Δds-hGH и SL335 связываться с САЧ в сыворотке человека может затруднять последующие эксперименты. Пробы отбирали раз в день, и активность связывания с САЧ и биологическую активность in vitro измеряли посредством ELISA (Фиг. 14А) и пробы на пролиферацию клеток Nb2-11 (Фиг. 14В), соответственно. Также инкубировали SL335 в качестве контроля. Аналогично SL335 активность связывания с САЧ и Nb2-11, пролиферативная активность SL335Δds-hGH не изменялась даже после 16 дней инкубации при 37°С, что демонстрирует, что SL335Δds-hGH так же стабилен, как и SL335. несмотря на отсутствие дисульфидной связи между цепями.
2-(8) Исследования фармакокинетики и фармакодинамики у крыс
Поскольку было показано, что SL335Δds-hGH является многообещающим кандидатом в качестве hGH пролонгированного действия, были проведены исследования эффективности in vivo. Во-первых, фармакокинетику гроутропина® и SL335Δds-hGH сравнивали у крыс путем измерения уровня каждого аналога в сыворотке как функцию от времени после однократной внутривенной или подкожной инъекции. Каждой группе крыс (четыре в группе) делали подкожную инъекцию (Фиг. 15А) однократной болюсной дозы 0,6 мг/кг гроутропина или САФАтропина, или внутривенную инъекцию (Фиг. 15В) однократной болюсной дозы 0,3 мг/кг гроутропина или САФАтропина. Пробы сыворотки отбирали с интервалами до 144 ч, в зависимости от белка. Пробы сыворотки анализировали в заданное время для гроутропина® или САФАтропина® посредством ELISA, как указано выше. Показатели фармакокинетики представлены в Таблице 7.
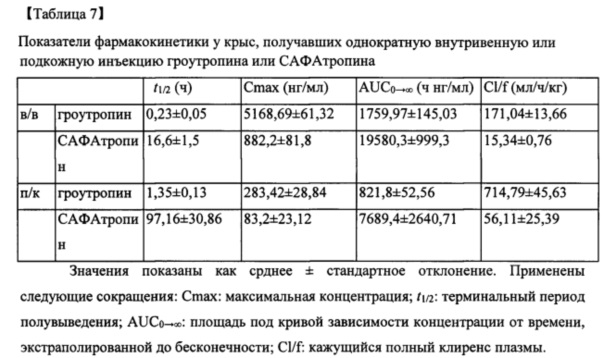
SL335Δds-hGH демонстрировал существенное увеличение t1/2 независимо от пути введения. При внутривенном введении SL335Δds-hGH проявлял 83-кратное увеличение t1/2 по сравнению с гроутропином (16,6 ч и 0,2 ч) и 69-кратное увеличение при подкожном введении (97,2 ч и 1,4 ч).
SL335Δds-hGH также проявлял ~10-краткное увеличение AUC0→∞ и более чем в 10 раз более медленный клиренс, чем гроутропин® независимо от пути введения. Каждая группа крыс (четыре в группе) получала подкожную инъекцию однократной болюсной дозы 0,6 мг/кг гроутропина или САФАтропина, или внутривенную инъекцию однократной болюсной дозы 0,3 мг/кг гроутропина или САФАтропина. Пробы сыворотки отбирали с интервалами до 144 ч в зависимости от белка. Пробы сыворотки анализировали в заданные моменты времени для гроутропина® или САФАтропина® посредством ELISA, как указано выше. Интересно, что значения Cmax для SL335Δds-hGH были в 6 и 3 раза меньше, чем значения для гроутропина® независимо от пути введения.
Затем скорость роста у крыс с удаленным гипофизом сравнивали в течение десяти дней после ежедневного п/к введения гроутропина® или вспомогательного буфера в качестве контроля (только вспомогательное вещество), или еженедельного п/к введения SL335ds-hGH. Крысы с удаленным гипофизом получали только вспомогательное вещество или 0,3 мг/кг гроутропина® раз в день или возрастающие дозы САФАтропина® в дни 0 и 7 (Фиг. 16). Сплошные линии показывают среднее процентное изменение массы тела. Планки погрешностей соответствуют стандартному отклонению. Крысы, получавшие вспомогательное вещество, демонстрировали приблизительно 5% снижение массы. В то же время, крысы, которым вводили гроутропин® 1 раз в день (0,3 мг/кг), демонстрировали 5% увеличение массы, что привело к суммарному увеличению массы на 10% по сравнению с группой только вспомогательного вещества. Инъекции SL335Δds-hGH один раз в неделю привели к дозозависимому увеличению массы: в группе 2,4 мг/кг - к 15% увеличению массы, а в группе 0,6 мг/кг - к 3,5% увеличению массы. Режим введения эквимолярной дозы SL335Δds-hGH (1,2 мг/мл) привел к 5% увеличению массы, что было сопоставимо с увеличением, достигнутом при ежедневных инъекциях гроутропина®.
На Фиг. 17 показано, что еженедельное введение 0,6 мг/кг SL335Δds-hGH приводило к увеличению длины большеберцовой кости, которое было эквивалентно увеличению, достигаемому при ежедневном введении гроутропина®. Заштрихованные столбики показывают среднее для измеренной длины большеберцовой кости. Планки погрешностей соответствуют стандартному отклонению.
Промышленная применимость
Настоящее изобретение можно применять для разработки лекарственных препаратов на основе биологически активных белков или полипептидов, поскольку можно получить гибридные конструкции согласно настоящему изобретению, которые будут содержать разные типы эффекторных компонентов, включая гормон роста человека, интерферон, эритропоэтин, колониестимулирующие факторы или их производные, и производные антител, и т.д.
Сведения, содержащиеся во всех патентах, опубликованных заявках и справочниках, цитируемых здесь, включены в настоящую заявку посредством ссылки на их полную версию.


























































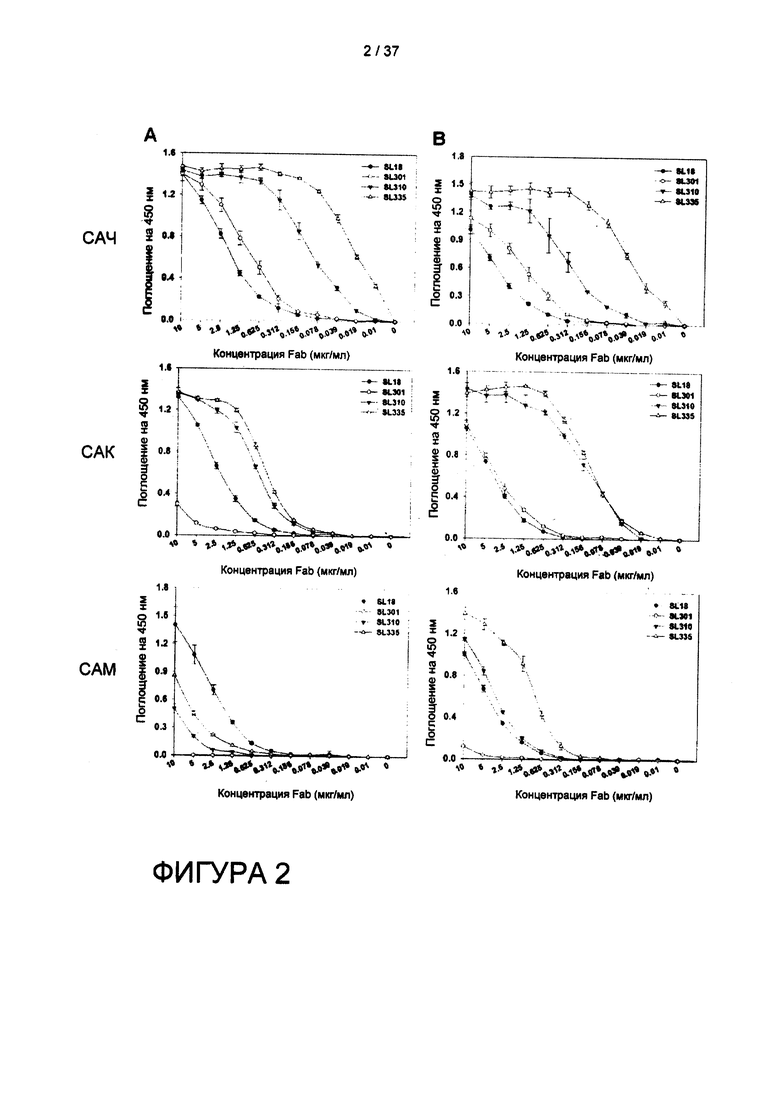
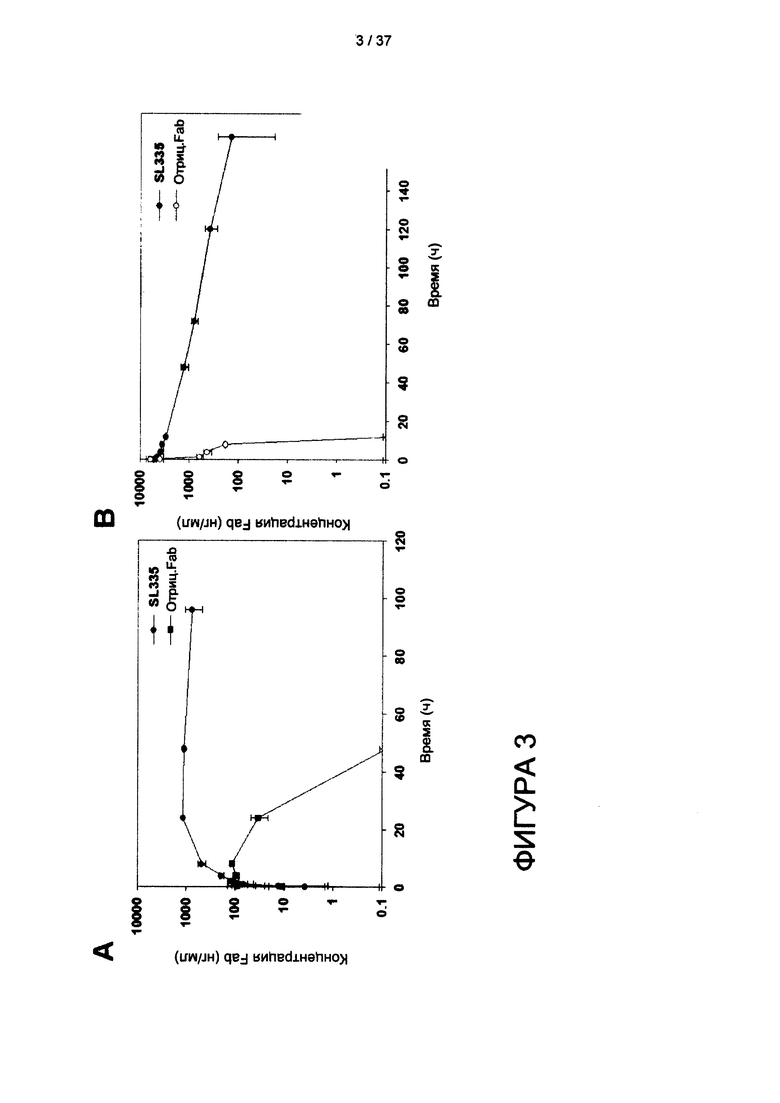
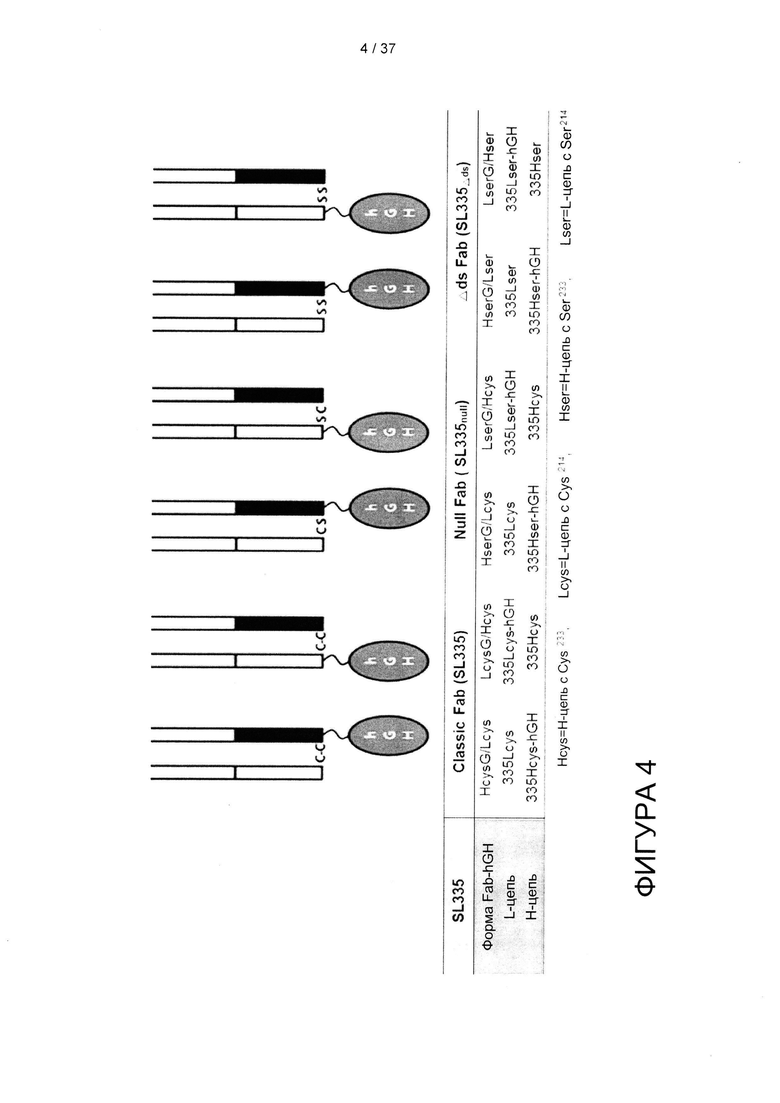
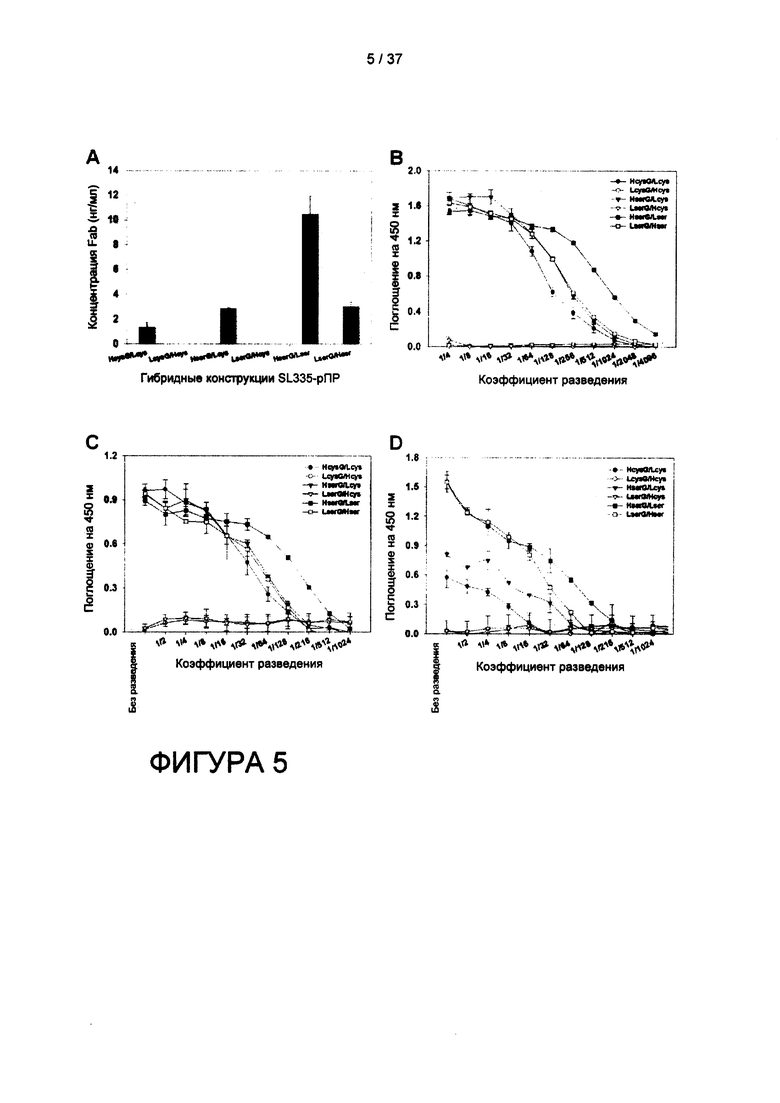

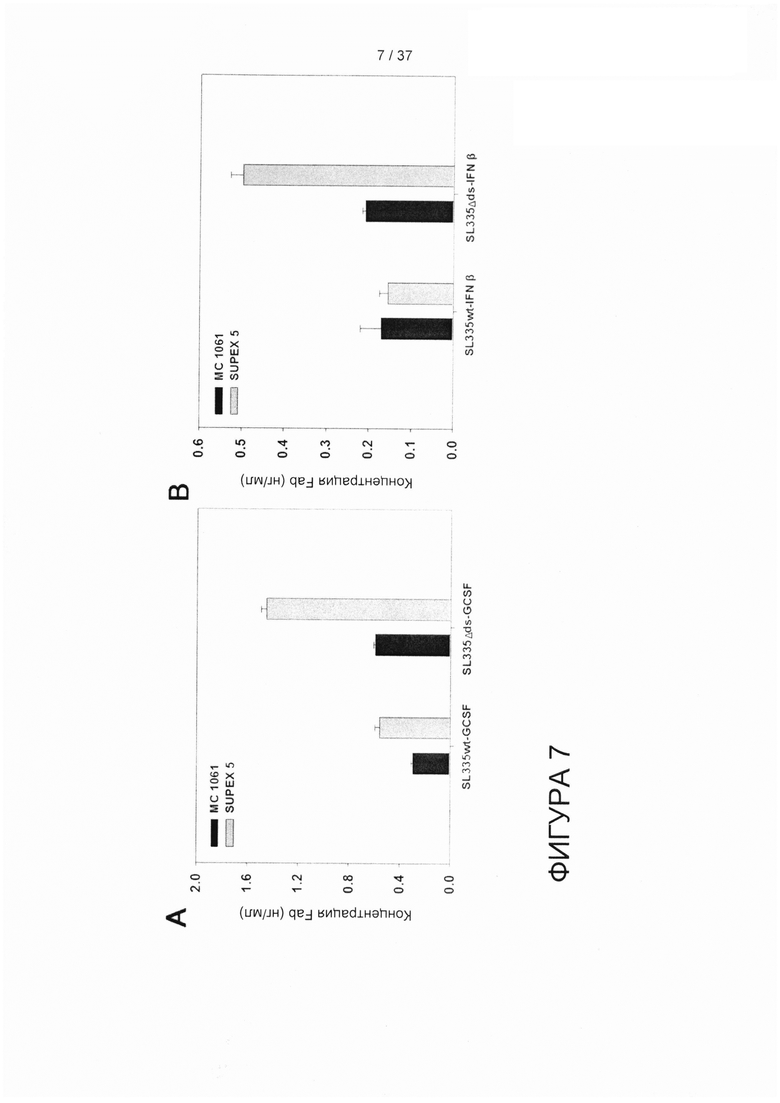


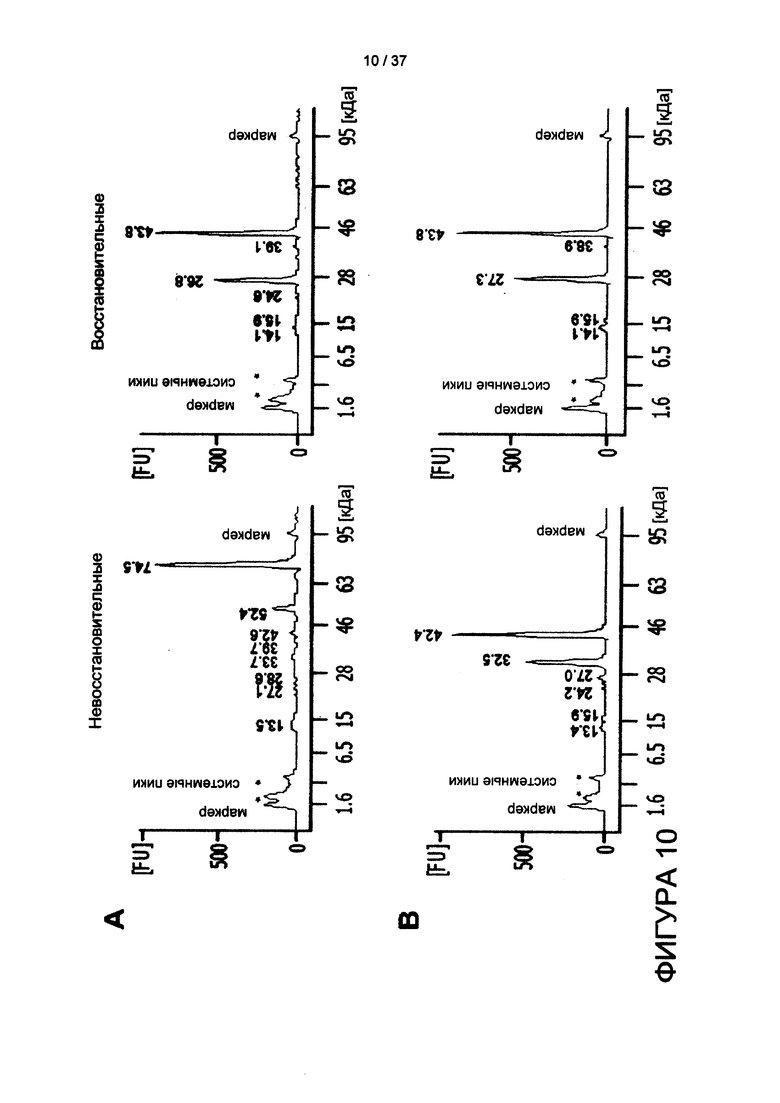


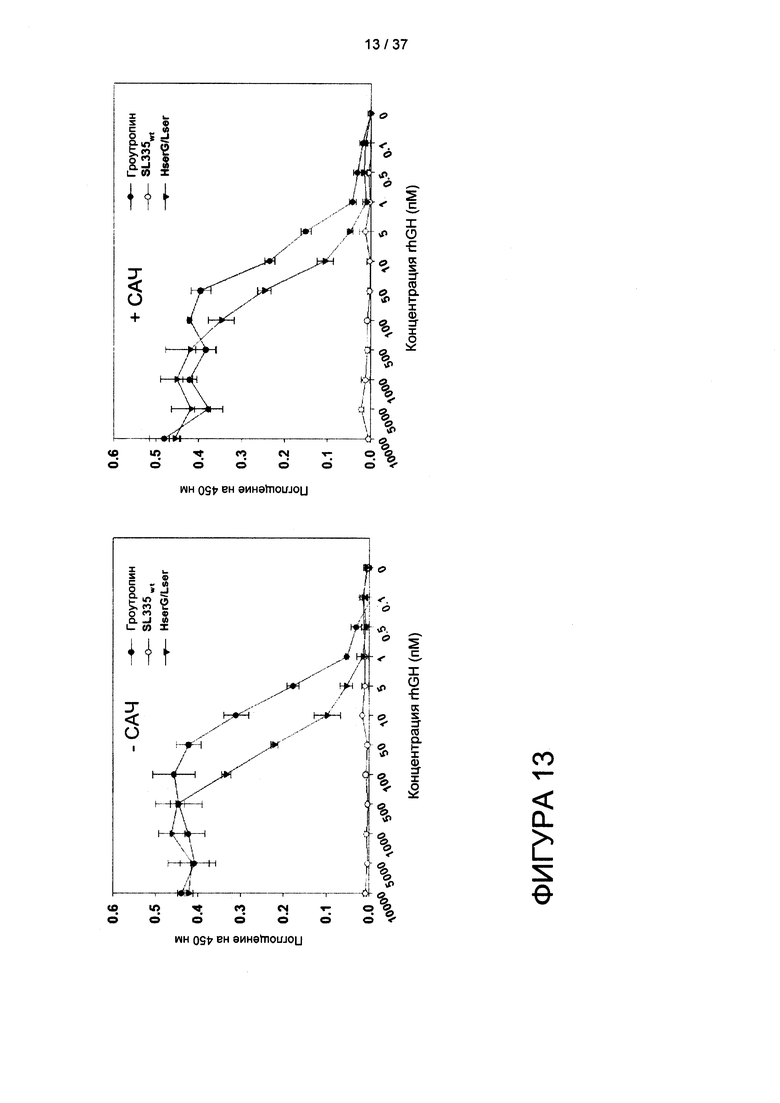


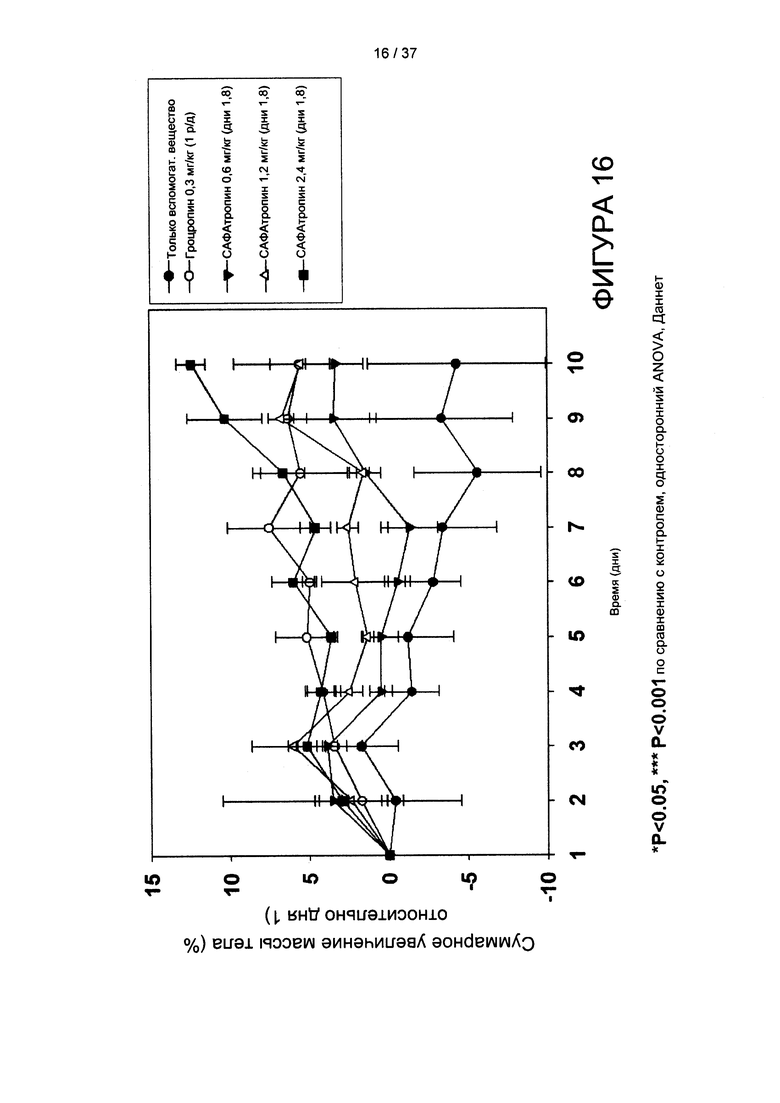
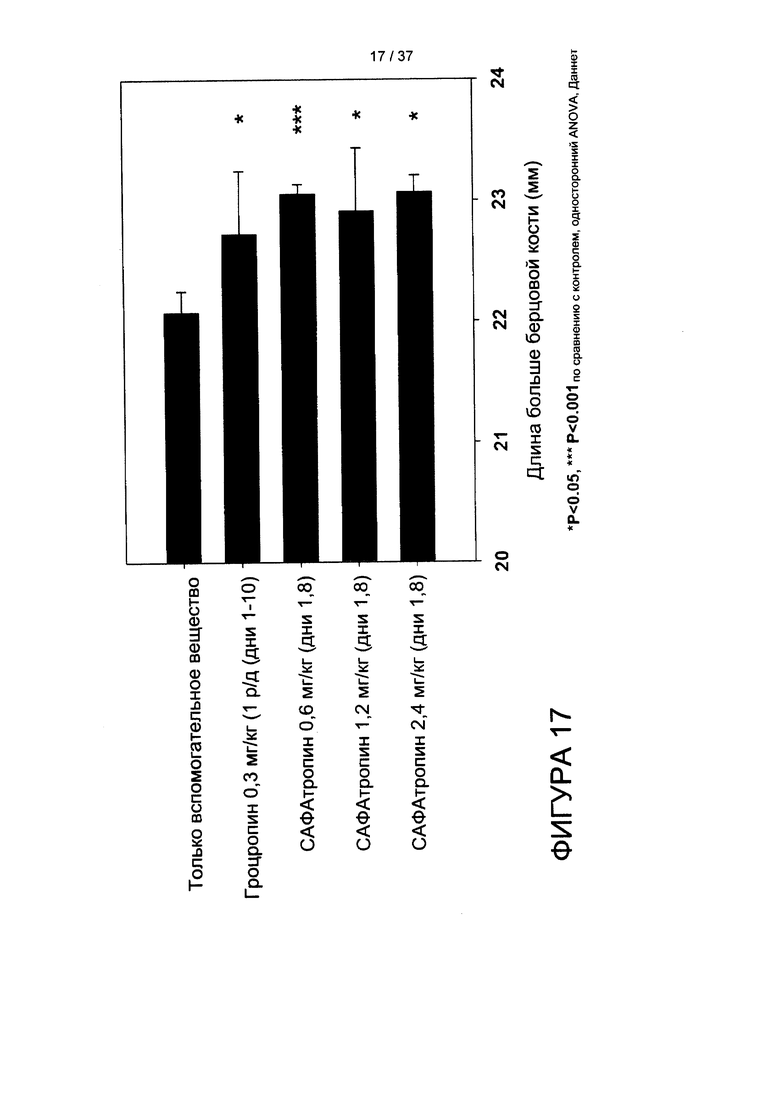




















Изобретение относится к области биотехнологии, конкретно к антигенсвязывающему фрагменту (Fab) для сывороточного альбумина (СА), что может быть использовано в медицине. Получают антигенсвязывающий фрагмент (Fab) и гибридную конструкцию из антигенсвязывающего фрагмента, которые используют в способе увеличения экспрессии в растворимой форме биологически активного белка, а также в способе увеличения периода полувыведения биологически активного белка. Изобретение позволяет увеличивать период полувыведения из сыворотки и экспрессию в растворимой форме эффекторного компонента. 7 н. и 17 з.п. ф-лы, 22 ил., 7 табл.
1. Антигенсвязывающий фрагмент (Fab) для сывороточного альбумина (СА) для увеличения периода полувыведения из сыворотки и увеличения экспрессии в растворимой форме эффекторного компонента, при этом указанный Fab содержит:
(a) вариабельный домен тяжелой цепи (домен VH) с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6, и
(b) вариабельный домен легкой цепи (домен VL) с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11 и SEQ ID NO: 12,
при этом аминокислота(ы), представляющая(ие) собой цистеин, удалена(ы) или заменена(ы) на любой(ые) другой(ие) аминокислотный(е) остаток(ки), за исключением цистеина;
при этом указанный(ые) удаленный(ые) или замещенный(ые) цистеин(ы) участвует(ют) в образовании S-S связи между тяжелой цепью и легкой цепью домена CH1, или участвует(ют) в образовании S-S связи между тяжелой цепью и легкой цепью домена CкL, или участвует(ют) в образовании S-S связи(ей) между тяжелой цепью и легкой цепью домена CH1 и между тяжелой цепью и легкой цепью домена CкL;
при этом указанный Fab специфически связывается с сывороточным альбумином.
2. Fab по п. 1, отличающийся тем, что указанный домен VH связан с константным доменом 1 тяжелой цепи (домен CH1), а указанный домен VL связан с константным доменом легкой цепи (домен CкL).
3. Fab по п. 1, отличающийся тем, что указанный домен VH имеет аминокислотную последовательность SEQ ID NO: 6, а указанный домен VL имеет аминокислотную последовательность SEQ ID NO: 12.
4. Антигенсвязывающий фрагмент (Fab) для сывороточного альбумина (СА) для увеличения периода полувыведения из сыворотки и увеличения экспрессии в растворимой форме эффекторного компонента, при этом указанный Fab содержит:
(a) аминокислотные последовательности SEQ ID NO: 13 (CDR1), 14 (CDR2) и 15 (CDR3), определяющие CDR домена VH, и
(b) аминокислотные последовательности SEQ ID NO: 16 (CDR1), 17 (CDR2) и 18 (CDR3), определяющие CDR домена VL,
и при этом аминокислота(ы), представляющая(ие) собой цистеин, удалена(ы) или заменена(ы) на любой(ые) другой(ие) аминокислотный(е) остаток(ки), за исключением цистеина;
при этом указанный(ые) удаленный(ые) или замещенный(ые) цистеин(ы) участвует(ют) в образовании S-S связи между тяжелой цепью и легкой цепью домена CH1, или участвует(ют) в образовании S-S связи между тяжелой цепью и легкой цепью домена CкL, или участвует(ют) в образовании S-S связи(ей) между тяжелой цепью и легкой цепью домена CH1 и между тяжелой цепью и легкой цепью домена CкL;
при этом указанный Fab специфически связывается с сывороточным альбумином.
5. Fab по п. 4, отличающийся тем, что указанный домен VH связан с константным доменом 1 тяжелой цепи (домен CH1), а указанный домен VL связан с константным доменом легкой цепи (домен CкL).
6. Fab по п. 4, отличающийся тем, что указанный домен VH имеет аминокислотную последовательность SEQ ID NO: 6, а указанный домен VL имеет аминокислотную последовательность SEQ ID NO: 12.
7. Гибридная конструкция из антигенсвязывающего фрагмента (Fab) по п. 1 или 4 и эффекторного компонента для увеличения периода полувыведения из сыворотки и увеличения экспрессии в растворимой форме указанного эффекторного компонента, при этом указанный эффекторный компонент представляет собой биологически активный эффекторный компонент, который представляет собой белок или (поли)пептид; и при этом Fab и указанный биологически активный эффекторный компонент ковалентным образом соединены посредством генетического слияния.
8. Гибридная конструкция по п. 7, отличающаяся тем, что аминокислота(ы), представляющая(ие) собой цистеин(ы) антигенсвязывающего фрагмента (Fab) по п. 1 или 4, заменена(ы) на серин.
9. Гибридная конструкция по любому из пп. 7, 8, отличающаяся тем, что указанный биологически активный эффекторный компонент представляет собой компонент, выбранный из группы, состоящей из гормона, цитокина, фермента, антитела, фактора роста, транскрипционного фактора, антигена крови, вакцины, структурного белка, белка-лиганда и рецептора.
10. Гибридная конструкция по пп. 7, 8, отличающаяся тем, что указанный биологически активный эффекторный компонент представляет собой компонент, выбранный из группы, состоящей из гормона роста человека (hGH), соматотропин-релизинг гормона (GHRH), соматотропин-релизинг-пептида, интерферонов (IFN), рецепторов интерферонов, колониестимулирующих факторов (КСФ), гранулоцитарных колониестимулирующих факторов (GCSF), глюкагоноподобных пептидов, рецептора, сопряженного с G-белком, интерлейкинов, рецепторов интерлейкинов, ферментов, интерлейкинсвязывающих белков, цитокинсвязывающих белков, фактора активации макрофагов, пептида макрофагов, фактора В-лимфоцитов, фактора Т-лимфоцитов, белка А, ингибитора аллергии, гликопротеинов некроза клеток, иммунотоксина, лимфотоксина, фактора некроза опухоли, онкосупрессоров, фактора роста метастазов, антитрипсина альфа-1, альбумина, альфа-лактальбумина, аполипопротеина-Е, эритропоэтина, высокогликозилированного эритропоэтина, ангиопоэтинов, гемоглобина, тромбина, пептида, активирующего рецептор тромбина, тромбомодулина, фактора VII, фактора VIIa, фактора VIII, фактора IX, фактора XIII, плазминогенактивирующего фактора, фибринсвязывающего пептида, урокиназы, стрептокиназы, гирудина, белка С, С-реактивного белка, ингибитора ренина, ингибитора коллагеназы, суперокcиддисмутазы, лептина, тромбоцитарного фактора роста, фактора роста эпителия, фактора роста эпидермиса, ангиостатина, ангиотензина, фактора роста кости, белка, стимулирующего кости, кальцитонина, инсулина, атриопептина, хрящевого индуцирующего фактора, элькатонина, фактора активации соединительной ткани, ингибитора пути тканевого фактора, фолликулостимулирующего гормона, лютеинизирующего гормона, релизинг-фактора лютеинизирующего гормона, факторов роста нервов, парат-гормона, релаксина, секретина, соматомедина, инсулиноподобного фактора роста, гормона коры надпочечников, глюкагона, холецистокинина, панкреатического (поли)пептида, гастринвысвобождающего пептида, кортиколиберина, тиреотропного гормона, аутотаксина, лактоферрина, миостатина, рецепторов, антагонистов рецепторов, поверхностных антигенов клетки, вакцинных антигенов вирусного происхождения, моноклональных антител, поликлональных антител и фрагментов антител.
11. Гибридная конструкция по п. 10, отличающаяся тем, что указанный биологически активный эффекторный компонент представляет собой hGH, GCSF или IFN.
12. Гибридная конструкция по п. 7, отличающаяся тем, что молярное отношение указанного биологически активного эффекторного компонента и Fab составляет от 1:1 до 10:1.
13. Гибридная конструкция по п. 12, отличающаяся тем, что молярное отношение указанного биологически активного эффекторного компонента и Fab составляет от 1:1 до 4:1.
14. Вектор экспрессии, содержащий: (а) промотор; (b) первую нуклеотидную последовательность, кодирующую Fab по любому из пп. 1 и 4; и (с) вторую нуклеотидную последовательность, кодирующую биологически активный (поли)пептид или белок и, необязательно, линкер, при этом указанные промотор, первая нуклеотидная последовательность и вторая нуклеотидная последовательность связаны функционально.
15. Клетка-хозяин для экспрессии Fab и гибридных конструкций, содержащая вектор экспрессии по п. 14.
16. Клетка-хозяин по п. 15, отличающаяся тем, что указанная клетка-хозяин представляет собой Е. coli.
17. Клетка-хозяин по п. 16, отличающаяся тем, что указанная клетка-хозяин представляет собой SUPEX5 (КСТС 12657 ВР).
18. Способ увеличения экспрессии в растворимой форме биологически активного белка или (поли)пептида в периплазме Е. coli, который включает введение вектора экспрессии по п. 14 в Е. coli.
19. Способ по п. 18, отличающийся тем, что указанной Е. coli является SUPEX5 (КСТС 12657 ВР).
20. Способ по п. 18, отличающийся тем, что указанный биологически активный белок или (поли)пептид представляет собой белок или (поли)пептид, выбранный из группы, состоящей из гормона, цитокина, фермента, антитела, фактора роста, транскрипционного фактора, антигена крови, вакцины, структурного белка, белка-лиганда и рецептора.
21. Способ по п. 18, отличающийся тем, что указанный биологически активный белок или (поли)пептид представляет собой белок или (поли)пептид, выбранный из группы, состоящей из гормона роста человека (hGH), соматотропин-релизинг гормона (GHRH), соматотропин-релизинг-пептида, интерферонов, рецепторов интерферонов, колониестимулирующих факторов (КСФ), гранулоцитарных колониестимулирующих факторов (GCSF), глюкагоноподобных пептидов, рецептора, сопряженного с G-белком, интерлейкинов, рецепторов интерлейкинов, ферментов, интерлейкинсвязывающих белков, цитокинсвязывающих белков, фактора активации макрофагов, пептида макрофагов, фактора В-лимфоцитов, фактора Т-лимфоцитов, белка А, ингибитора аллергии, гликопротеинов некроза клеток, иммунотоксина, лимфотоксина, фактора некроза опухоли, онкосупрессоров, фактора роста метастазов, антитрипсина альфа-1, альбумина, альфа-лактальбумина, аполипопротеина-Е, эритропоэтина, высокогликозилированного эритропоэтина, ангиопоэтинов, гемоглобина, тромбина, пептида, активирующего рецептор тромбина, тромбомодулина, фактора VII, фактора VIIa, фактора VIII, фактора IX, фактора XIII, плазминогенактивирующего фактора, фибринсвязывающего пептида, урокиназы, стрептокиназы, гирудина, белка С, С-реактивного белка, ингибитора ренина, ингибитора коллагеназы, суперокcиддисмутазы, лептина, тромбоцитарного фактора роста, фактора роста эпителия, фактора роста эпидермиса, ангиостатина, ангиотензина, фактора роста кости, белка, стимулирующего кости, кальцитонина, инсулина, атриопептина, хрящевого индуцирующего фактора, элькатонина, фактора активации соединительной ткани, ингибитора пути тканевого фактора, фолликулостимулирующего гормона, лютеинизирующего гормона, релизинг-фактора лютеинизирующего гормона, факторов роста нервов, парат-гормона, релаксина, секретина, соматомедина, инсулиноподобного фактора роста, гормона коры надпочечников, глюкагона, холецистокинина, панкреатического (поли)пептида, гастринвысвобождающего пептида, кортиколиберина, тиреотропного гормона, аутотаксина, лактоферрина, миостатина, рецепторов, антагонистов рецепторов, поверхностных антигенов клетки, вакцинных антигенов вирусного происхождения, моноклональных антител, поликлональных антител и фрагментов антител.
22. Способ увеличения периода полувыведения биологически активного белка или (поли)пептида in vivo, указанный способ включает соединение биологически активного белка или (поли)пептида с Fab по любому из пп. 1 или 4 посредством генетического слияния.
23. Способ по п. 22, отличающийся тем, что указанный биологически активный белок или (поли)пептид соединен с Fab пептидным линкером из 0 - 20 аминокислот.
24. Способ по п. 22 или 23, отличающийся тем, что указанный биологически активный белок или (поли)пептид представляет собой белок или (поли)пептид, выбранный из группы, состоящей из гормона роста человека (hGH), соматотропин-релизинг гормона (GHRH), соматотропин-релизинг-пептида, интерферонов (IFN), рецепторов интерферонов, колониестимулирующих факторов (КСФ), гранулоцитарных колониестимулирующих факторов (GCSF), глюкагоноподобных пептидов, рецепторов, сопряженных с G-белками, интерлейкинов, рецепторов интерлейкинов, ферментов, интерлейкинсвязывающих белков, цитокинсвязывающих белков, фактора активации макрофагов, пептида макрофагов, фактора В-лимфоцитов, фактора Т-лимфоцитов, белка А, ингибитора аллергии, гликопротеинов некроза клеток, иммунотоксина, лимфотоксина, фактора некроза опухоли, онкосупрессоров, фактора роста метастазов, антитрипсина альфа-1, альбумина, альфа-лактальбумина, аполипопротеина-Е, эритропоэтина, высокогликозилированного эритропоэтина, ангиопоэтинов, гемоглобина, тромбина, пептида, активирующего рецептор тромбина, тромбомодулина, фактора VII, фактора VIIa, фактора VIII, фактора IX, фактора XIII, плазминогенактивирующего фактора, фибринсвязывающего пептида, урокиназы, стрептокиназы, гирудина, белка С, С-реактивного белка, ингибитора ренина, ингибитора коллагеназы, суперокcиддисмутазы, лептина, тромбоцитарного фактора роста, фактора роста эпителия, фактора роста эпидермиса, ангиостатина, ангиотензина, фактора роста кости, белка, стимулирующего кости, кальцитонина, инсулина, атриопептина, хрящевого индуцирующего фактора, элькатонина, фактора активации соединительной ткани, ингибитора пути тканевого фактора, фолликулостимулирующего гормона, лютеинизирующего гормона, релизинг-фактора лютеинизирующего гормона, факторов роста нервов, парат-гормона, релаксина, секретина, соматомедина, инсулиноподобного фактора роста, гормона коры надпочечников, глюкагона, холецистокинина, панкреатического (поли)пептида, гастринвысвобождающего пептида, кортиколиберина, тиреотропного гормона, аутотаксина, лактоферрина, миостатина, рецепторов, антагонистов рецепторов, поверхностных антигенов клетки, вакцинных антигенов вирусного происхождения, моноклональных антител, поликлональных антител и фрагментов антител.
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| SMITH B.J | |||
| et al., Prolonged in Vivo Residence Times of Antibody Fragments Associated with Albumin, Bioconjugate Chem., 2001, v.12, p:750-756 | |||
| СПОСОБЫ ЛЕЧЕНИЯ РЕСПИРАТОРНОГО ЗАБОЛЕВАНИЯ С ПРИМЕНЕНИЕМ АНТАГОНИСТОВ РЕЦЕПТОРА ИНТЕРЛЕЙКИНА-1 ТИПА 1 | 2005 |
|
RU2411957C2 |
| Способ работы регенераторного устройства в паросиловых установках | 1926 |
|
SU12622A1 |

