Область, к которой относится изобретение
Настоящее изобретение относится к иммуноглобулинам, специфически связывающимся с интерлейкином-13 (IL-13), в частности с человеческим IL-13 (hIL-13). В одном из своих вариантов настоящее изобретение относится к антителам, специфически связывающимся с hIL-13. Настоящее изобретение также относится к способам лечения заболеваний или расстройств с использованием указанных иммуноглобулинов, к фармацевтическим композициям, содержащим указанные иммуноглобулины, и к способам их получения. Другие аспекты настоящего изобретения будут очевидны из нижеследующего описания.
Предшествующий уровень техники
Интерлейкин-13 (IL-13)
IL-13 представляет собой секретируемый цитокин размером 12 кДа, который первоначально был описан как Т-клеточный цитокин и который ингибирует продуцирование воспалительных цитокинов. Исследования его структуры показали, что он представляет собой четырехспиральный пучок, связанный двумя дисульфидными связями. Хотя IL-13 имеет четыре потенциальных сайта гликозилирования, однако анализ нативного IL-13, выделенного из легких крыс, показал, что он продуцируется в виде негликозилированной молекулы. Экспрессия человеческого IL-13 в клетках NSO и СОS-7 подтверждает это наблюдение (Eisenmesser et al., J. Mol. Biol. 2001, 310(1):231-241; Moy et al., J. Mol. Biol. 2001, 310(1):219-230; Cannon-Carlson et al., Protein Expression and Purification, 1998, 12(2):239-248).
IL-13 представляет собой плейотропный цитокин, продуцируемый клетками различных типов, включая активированные Th2-клетки, тучные клетки, базофилы, дендритные клетки, кератиноциты и NKT-клетки. Он может также продуцироваться Th0-клетками, Th1-клетками, CD8-клетками и “необученными” CD45RA+-T-клетками. IL-13 обладает иммунорегуляторной активностью, которая частично перекрывается с активностью IL-4, и такое дублирование может быть объяснено наличием общих компонентов в рецепторах IL-4 и IL-13. IL-13 передает сигналы посредством рецептора IL-4 типа II, который представляет собой гетеродимер, состоящий из IL4Rα- и IL-13Rα1-цепей. IL-13Rα1 связывается с IL-13 с низкой аффинностью (Kd = 2-10 нМ), но при спаривании с IL4Rα он связывается с IL-13 с высокой аффинностью (Kd = 400 пМ) и образует функциональный рецептор IL-13 (человеческий рецептор, далее обозначаемый “hIL-13R”), который передает сигналы, приводящие к активации путей JAK/STAT- и IRS-1/IRS-2. Была также охарактеризована и другая цепь рецептора IL-13 (IL-13Rα2), которая связывается с IL-13 c высокой аффинностью (Kd = 250 пМ), но не передает сигнал, а вместо этого она, вероятно, действует как рецептор-ловушка. Функциональные рецепторы IL-13 экспрессируются на клетках широкого ряда, включая клетки эпителия дыхательных путей, клетки гладких мышц, тучные клетки, эозинофилы, базофилы, В-клетки, фибробласты, моноциты и макрофаги. Т-клетки не имеют функциональных рецепторов для IL-13 (Hilton et al., PNAS, 1996, 93(1):497-501; Caput et al., J. Biol. Chem. 1996, 271(28):16921-16926; Hershey G.K., J. Allergy Clin. Immunol. 2003 111(4):677-690).
IL-13 и IL-4 модифицируют иммунные и воспалительные ответы путем стимуляции ассоциированного с аллергией воспаления и подавления воспаления, вызванного бактериями, вирусами и внутриклеточными патогенами. Основными биологическими эффектами IL-13 являются индуцирование пролиферации В-клеток и регуляции переключения изотипа на IgE; индуцирование экспрессии МНС II и CD23 на В-клетках и моноцитах; позитивная регуляция VCAM-1 на эндотелиальных клетках; регуляция продуцирования хемокинов; активация функции тучных клеток, эозинофилов и нейтрофилов, а также ингибирование экспрессии про-воспалительных генов в популяциях моноцитов и макрофагов. IL-13 не оказывает какого-либо пролиферативного действия на Т-клетки. Таким образом, IL-13 в отличие от IL-4, вероятно, не играет важной роли в начальной дифференцировке CD4-T-клеток в клетки типа Th2, но тем не менее очевидно, что он играет важную роль в эффекторной фазе аллергического воспаления (McKenzie et al., PNAS 1993 90(8):3735-3739; Wynn T.A., Annu. Rev. Immunol. 2003 21:425-456).
IL-13 и астма
Астма представляет собой хроническое заболевание легких, вызываемое воспалением нижних дыхательных путей, и характеризуется рецидивирующими затруднениями дыхания. Дыхательные пути пациента становятся сенсибилизированными и опухают, либо в них появляются некоторые очаги воспаления в течение всего периода заболевания, даже если оно протекает бессимптомно. Воспаление приводит к сужению дыхательных путей и к снижению поступления потока воздуха в легкие и из легких, что приводит к затруднению дыхания и к возникновению свистящих хрипов, стеснения в груди и кашля. Астма вызывается повышенной чувствительностью к аллергенам (например, к клещам домашней пыли, пыльце и плесени), к раздражителям (например, к курению, вдыханию дыма и к резким запахам), к респираторным инфекциям, а также к физическим упражнениям и к сухой погоде. Такие провоцирующие факторы приводят к раздражению дыхательных путей, после чего выстилка дыхательных путей опухает и становится еще более воспаленной, слизь в дыхательных путях образует сгустки, мышцы, окружающие дыхательные пути, сжимаются и дыхание становится затрудненным и более напряженным, что указывает на появление симптомов астмы.
Данные исследований, проводимых на животных-моделях, и результаты обследования пациентов со всей очевидностью показали, что астматическое воспаление и другие патологии вызываются нарушениями регуляции Th2-ответов на аэроаллергены и другие раздражители (Busse et al., Am. J. Resp. Crit. Care Med. 1995, 152(1):388-393). В частности, очевидно, что IL-13 представляет собой главный эффекторный цитокин, инициирующий различные клеточные ответы в легких, включая гиперреактивность дыхательных путей, эозинофилию, метаплазию бокаловидных клеток и повышенную секрецию слизистой.
Клинические данные о роли IL-13 в развитии астмы
Ген, кодирующий IL-13, находится на хромосоме 5q31. Эта область также содержит гены, кодирующие IL-3, IL-4, IL-5, IL-9 и GM-CSF и ассоциирующиеся с развитием астмы. Генетические варианты IL-13, ассоциированного с развитием астмы и атопии, были обнаружены как в промоторной, так и в кодирующей областях (Vercelli D., Curr. Opin. Allergy Clin. Immunol. 2002 2(5):389-393). Были получены данные функциональных исследований для кодирующего варианта Q130 IL-13 (далее обозначаемого “Q130 IL-13”). Полиморфизм по одному нуклеотиду +2044 G → А (SNP), обнаруженный в четвертом экзоне, приводит к замене аргинина на глутамин в положении 130 (Q130 IL-13). Кроме того, в последовательности SEQ ID NO:9 такая замена является эквивалентной замене в положении 110, где первый аминокислотный остаток “G” в начале зрелой аминокислотной последовательности IL-13 находится в положении 1. По данным обследования людей, проживающих в Японии и в Европе, было обнаружено, что такой вариант ассоциируется с развитием астмы, с увеличением уровней IgE и с развитием атопического дерматита. Очевидно, что Q130 IL-13 обладает более высокой стабильностью по сравнению с IL-13 дикого типа. Он также обладает несколько более низкой аффинностью по отношению к рецептору-ловушке IL-13Rα2, и согласно этим наблюдениям более высокие средние уровни IL-13 в сыворотке были обнаружены у пациентов, которые являются гомозиготными по вариантам Q130 IL-13, по сравнению с не-гомозиготными пациентами. Эти результаты показали, что Q130 IL-13 может оказывать влияние на локальную и системную концентрации IL-13 (Kazuhiko et al., J. Allergy Clin. Immunol. 2002 109(6):980-987).
Повышенные уровни IL-13 были обнаружены у пациентов с атопической и не-атопической бронхиальной астмой. В одном исследовании, у пациентов, страдающих астмой, были измерены средние уровни IL-13 в сыворотке, составляющие 50 пг/мл, тогда как у здоровых контрольных индивидуумов такие уровни составляли 8 пг/мл (Lee et al., J. Asthma 2001 38(8):665-671). Повышенные уровни IL-13 были также выявлены в плазме, в жидкости бронхоальвеолярного лаважа, в образцах биоптата легких и в мокроте (Berry et al., J. Allergy Clin. Immunol. 2004 114(5):1106-1109; Kroegel et al., Eur. Respir. J. 1996 9(5):899-904; Huang et al., J. Immunol. 1995 155(5):2668-2694; Humbert et al., J. Allergy Clin. Immunol. 1997 99(5):657-665).
In vivo данные, подтверждающие участие IL-13 в развитии астмы
В ряде исследований была определена важная эффекторная роль IL-13 в развитии патологии у мышей, которые были использованы в качестве моделей острой и хронической аллергической астмы. У этих моделей для нейтрализации биологической активности мышиного IL-13 был использован высокоаффинный рецептор IL-13 (IL-13Rα2) или поликлональные антитела против IL-13. Блокада IL-13 во время заражения аллергеном полностью ингибировала OVA-индуцированную гиперреактивность дыхательных путей, эозинофилию и метаплазию бокаловидных клеток. В противоположность этому введение антитела против IL-4 после сенсибилизации и во время заражения аллергеном лишь частично ослабляло астматический фенотип. Таким образом, хотя экзогенные IL-4 и IL-13 обладают способностью индуцировать фенотип, подобный фенотипу астмы, однако очевидно, что эффекторная активность IL-13 превосходит активность IL-4. Эти данные дают основание предположить, что IL-4 играет главную роль в индуцировании иммунного ответа (в частности, образования Th2-клеток и их рекрутинга в дыхательных путях, и продуцирования IgE), тогда как IL-13, вероятно, главным образом ответственен за индуцирование различных эффекторных функций, включая гиперреактивность дыхательных путей, повышенное выделение слизи и воспалительные процессы в клетках (Wills-Karp et al., Science 1998, 282:2258-2261; Grunig et al., Science 1998, 282:2261-2263; Taube et al., J. Immunol. 2002 169:6482-6489; Blease et al., J. Immunol. 2001 166(8):5219-5224).
В дополнительных экспериментах уровни IL-13 в легких были увеличены в результате сверхэкспрессии у трансгенных мышей или в результате введения в трахею белка IL-13 мышей дикого типа. В обоих случаях были индуцированы признаки, подобные признакам, наблюдаемым при астме, а именно неспецифическая гиперреактивность дыхательных путей в ответ на стимуляцию холинергическим рецептором, легочная эозинофилия, гиперплазия эпителиальных клеток, метаплазия клеток слизи, субэпителиальный фиброз, стеноз дыхательных путей и образование кристаллов, подобных кристаллам Шарко-Ляйдена. Кроме того, было обнаружено, что IL-13 является сильным стимулятором действия металлопротеиназ матрикса и катепсиновых протеаз в легких, приводящего к эмфизематозным изменениям и к метаплазии слизистой. Следовательно, IL-13 может представлять собой эффекторную молекулу, играющую важную роль в развитии фенотипов астмы и фенотипов заболевания ХОБЛ (Zhu et al., J. Clin. Invest. 1999 103(6):779-788; Zheng et al., J. Clin. Invest. 2000 106(9):1081-1093).
Эти данные показали, что активность IL-13 является необходимой и достаточной для развития некоторых основных клинических и патологических признаков аллергической астмы у животных-моделей, на что указывают данные, подтвержденные путем клинических исследований.
Хроническая обструктивная болезнь легких (ХОБЛ)
ХОБЛ является общим термином, который охватывает несколько клинических синдромов, включая эмфизему и хронический бронхит. Симптомы ХОБЛ сходны с симптомами астмы, а поэтому ХОБЛ может быть подвергнута лечению такими же лекарственными средствами. ХОБЛ характеризуется хроническим, прогрессирующим и в основном необратимым стенозом дыхательных путей. Причина возникновения такого заболевания у человека пока не известна, однако считается, что в 90% случаев оно вызывается курением сигарет. Симптомами такого заболевания являются кашель, хронический бронхит, одышка и респираторные инфекции. В конечном счете такое заболевание может приводить почти к полной нетрудоспособности и к летальному исходу. Хронический бронхит диагностируется у пациентов, у которых в течение длительного периода, например по меньшей мере от 3 месяцев до 2 лет и более, наблюдается ничем необъяснимый кашель или образование мокроты. Эмфизема легких характеризуется аномальным перманентным увеличением объема дыхательных путей и деструкцией стенок альвеол.
В развитии ХОБЛ определенную роль может играть IL-13. У курильщиков, у которых развивается ХОБЛ, в паренхиме легких присутствуют воспалительные клетки многих типов (нейтрофилы, макрофаги, эозинофилы). IL-13 представляет собой провоспалительный Th2-цитокин, а поэтому он используется для моделирования прогрессирующей эмфиземы; так, например, в работе Zheng et al. рассматривается сверхэкспрессия IL-13 в эпителии дыхательных путей у IL-13-трансгенной мыши. У этих животных возникает воспаление паренхимы дыхательных путей и легких и развивается эмфизема. У этих мышей развивается также метаплазия слизистой, напоминающая хронический бронхит (J. Clin. Invest. 2000 106(9):1081-1093).
Также сообщалось, что полиморфизм промотора IL-13 (-1055 С → Т), который ассоциируется с аллергической астмой, более часто встречается у пациентов с ХОБЛ, чем у здоровых контрольных индивидуумов. Это указывает на функциональную роль полиморфизма промотора IL-13 в повышении риска развития ХОБЛ (Kraan et al., Genes and Immunity 2002 3:436-439). Кроме того, увеличение числа IL-13- и IL-4-позитивных клеток наблюдалось у курильщиков с хроническим бронхитом, но не наблюдалось у курильщиков, у которых отсутствовали симптомы такого заболевания (Miotto et al., Eur. Resp. J. 2003, 22:602-608). Однако в недавно проводимых исследованиях по оценке уровня экспрессии IL-13 в легких пациентов с тяжелой эмфиземой не было выявлено какой-либо взаимосвязи между уровнями IL-13 и развитием этого заболевания (Boutten et al., Thorax 2004, 59:850-854).
Аллергическое заболевание, включая атопический дерматит и аллергический ринит
IL-13 также участвует в развитии атопических расстройств, таких как атопический ринит и атопический дерматит. В США аллергический ринит является наиболее распространенным атопическим заболеванием, и по оценкам специалистов таким заболеванием страдают до 25% взрослых и более чем 40% детей. Между аллергическим ринитом и астмой существует тесная взаимосвязь. Оба эти состояния имеют общую иммунную патологию и патофизиологию, и при этих заболеваниях наблюдаются сходные иммунологические процессы, в которых определенную роль играют эозинофилы и Th2-лимфоциты, присутствующие в тканях носа и бронхов. Считается, что избыточное продуцирование Th2-цитокинов, в частности IL-4 и IL-5, играет главную роль в патогенезе аллергического заболевания. IL-13 и IL-4 имеют некоторые общие свойства и эффекторные функции, и этот факт, в сочетании с перекрыванием их функций при их взаимодействии с рецепторами IL-4 и IL-13 и с компонентами, передающими внутриклеточные сигналы, а также генетическая организация IL-13 дают неоспоримые доказательства (хотя и косвенные) того, что этот цитокин играет определенную роль в стимуляции или в обеспечении in vivo гиперчувствительности немедленного типа в организме человека. Эти данные были подтверждены Li и др. (Li et al., J. Immunol. 1998; 161:7007), которые продемонстрировали, что у индивидуумов с атопическим заболеванием, таким как сезонный аллергический ринит, наблюдаются значительно более сильные IL-13-ответы на Ag-зависимую, но не поликлональную активацию.
Атопический дерматит представляет собой широко распространенное хроническое, рецидивирующее, воспалительное заболевание кожи с сильным зудом. Поражение кожи у пациентов с атопическим дерматитом гистологически характеризуются воспалительным Т-клеточным инфильтратом, и это поражение в своей острой фазе ассоциируется с преобладающей экспрессией IL-4, IL-5 и IL-13 (Simon et al., J. Allergy Clin. Immunol. 2004; 114:887; Hamid et al., J. Allergy Clin. Immunol. 1996; 98:225). Кроме того, Tazawa и др. продемонстрировали, что мРНК IL-13 (но не IL-4) в значительной степени активируются у пациентов, страдающих атопическим дерматитом с подострыми и хроническими поражениями кожи (Tazawa et al., Arch. Derm. Res. 2004; 296:459). У этих пациентов также значительно увеличивается уровень IL-13-экспрессирующих циркулирующих CD4+- и CD8+-Т-клеток (Aleksza et al., British J. Dermatol. 2002; 147:1135). Такая повышенная активность IL-13, очевидно, приводит к продуцированию повышенных уровней сывороточного IgЕ, что также способствует патогенезу атопического дерматита. Кроме того, повышенные уровни продуцирования IL-13 неонатальными CD4+-Т-клетками являются ценным маркером для идентификации новорожденных с высоким риском последующего развития у них аллергических заболеваний, в частности атопического дерматита (Ohshima et al., Pediatr. Res. 2002; 51:195). Дополнительное свидетельство важной роли IL-13 в этиологии атопического дерматита приводится в работе Simon et al. (Simon et al., J. Allergy Clin. Immunol. 2004; 114:887); причем местное лечение мазью такролимус (иммунодепрессантом, ингибирующим внутриклеточные пути передачи сигнала продуцирования цитокинов), приводит к значительному клиническому и гистологическому ослаблению атопических поражений кожи, сопровождающемуся значительным снижением уровней локальной экспрессии Th2-цитокинов, включая IL-13. Кроме того, было обнаружено, что IL-13Rα1 (белок клеточной поверхности, который вместе с IL-4Rα образует функциональный рецептор IL-13) сверхэкспрессируется на супрабазальных кератиноцитах в коже пациентов с атопическим дерматитом, а IL-13 способен стимулировать мРНК IL-13Rα1 in vitro (Wongpiyabovorn et al., J. Dermatol. Science 2003;33:31).
В целом эти данные указывают на то, что использование IL-13 в качестве мишени, включая моноклональное антитело против IL-13, может быть эффективным способом лечения аллергического заболевания у человека.
Эзофагальная эозинофилия
Аккумуляция эозинофилов в пищеводе является широко распространенным явлением, встречающимся у пациентов с различными заболеваниями, включая гастроэзофагальный рефлюкс, эозинофильный эзофагит, эозинофильный гастроэнтерит и паразитарные инфекции. Эзофагальная эозинофилия ассоциируется с аллергическими реакциями, а повторное заражение мышей аэроаллергенами позволяет установить связь между аллергическим воспалением дыхательных путей и эзофагальной эозинофилией. Считается, что Th2-клетки индуцируют ассоциированное с эозинофилами воспаление посредством секреции ряда цитокинов, включая IL-4 и IL-13, которые непосредственно и опосредованно активируют воспалительные и эффекторные пути. Очевидно, что в такой активации IL-13 играет особенно важную роль, поскольку он продуцируется в больших количествах Th2-клетками и регулирует множество признаков аллергического заболевания (например, продуцирование IgЕ, сверхпродуцирование слизи, рекрутинг и выживание эозинофилов и гиперреактивность дыхательных путей). Эозинофилы могут генерировать функционально активный IL-13 после их обработки цитокинами GM-CSF и/или IL-5 в условиях in vitro, ex vivo и in vivo при эозинофильных воспалительных реакциях (Schmid-Grendelmeier J. Immunology, 2002, 169:1021-1027). Было установлено, что при доставке IL-13 путем его введения через трахею в легкие мышей дикого типа, мышей, дефицитных по STAT-6, эотаксину-1 или IL-5, воспаление легких, запускаемое IL-13, ассоциируется с развитием эзофагальной эозинофилии (Mishra et al., Gastroenterol. 2003;125:1419). В целом эти данные подтверждают роль IL-13 в развитии эзофагальной эозинофилии.
Показания в онкологии
Другой важной сферой интересов исследователей является доставка IL-13 или рецепторов IL-13 в опухоли некоторых типов для ингибирования их роста. Защита хозяина, опосредуемая Т-клетками типа 1, очевидно, обеспечивает оптимальное отторжение опухоли in vivo, а переключение на ответ Th2-типа может способствовать блокированию отторжения опухоли и/или стимуляции рецидивов опухоли (Kobayashi M. et al., J. Immunol. 1998;160:5869). Исследования, проводимые на нескольких животных с использованием трансплантируемых опухолевых клеточных линий, подтвердили это мнение путем установления того факта, что Stat-6, IL-4 и IL-13 (продуцируемые частично NКТ-клетками) обладают способностью ингибировать отторжение опухоли (Terabe et al., Nat. Immunol. 2000; 1:515; Kacha et al., J. Immunol. 2000; 165:6024-28; Ostrand-Rosenberg et al., J. Immunol. 2000;165:6015). Очевидно, что сильная противоопухолевая активность в отсутствие Stat-6 обусловлена усилением продуцирования опухолеспецифического IFNg и СТL-активности. Кроме того, было показано, что отсутствие клеток NKТ приводит к снижению уровня продуцирования IL-13 и одновременно к увеличению риска рецидива опухоли, что указывает на то, что IL-13, продуцируемый частично клетками NKТ, имеет важное значение для иммунного надзора (Terabe et al., Nat. Immunol. 2000; 1:515). По существу, эти данные подтверждают тот факт, что ингибиторы IL-13 или новые антагонисты IL-13, включая mAb против IL-13, могут служить эффективными противораковыми иммунотерапевтическими средствами, которые действуют путем ингибирования негативной регуляции цитокина IL-13, играющего определенную роль в ингибировании иммунных ответов на опухолевые клетки.
Помимо усиления противоопухолевой защиты, ассоциированной с клетками типа Th-1, ингибиторы IL-13 также могут непосредственно блокировать рост опухолевых клеток. Так, например, при В-клеточном хроническом лимфоцитарном лейкозе (В-ХЛЛ) и болезни Ходжкина IL-13 либо блокирует апоптоз, либо стимулирует пролиферацию опухолевых клеток (Chaouchi et al., Blood 1996; 87:1022; Kapp et al., J. Exp. Med. 1999; 189:1939). В-ХЛЛ представляет собой клинически гетерогенное заболевание, вызываемое В-лимфоцитами, которые участвуют в нарушении апоптоза лейкозных клеток. Очевидно, что IL-13 не действует непосредственно как фактор роста, а лишь защищает опухолевые клетки от in vitro спонтанного апоптоза (Chaouchi et al., Blood 1996; 87:1022; Lai et al., J. Immunol. 1999; 162:78) и может способствовать развитию В-ХЛЛ посредством предотвращения гибели неопластических клеток.
Болезнь Ходжкина представляет собой один из типов лимфомы, которая поражает главным образом молодых людей, и, по оценкам специалистов, в США это заболевание диагностируется примерно у 7500 человек в год. Этот вид рака характеризуется присутствием крупных многоядерных клеток Ходжкина/Рида-Штернберга (Н/RS). В подавляющем большинстве случаев эта популяция злокачественных клеток происходит от В-клеток. Некоторые клеточные линии, ассоциированные с болезнью Ходжкина, а также ткани лимфоузлов, взятые у пациентов с ходжкинской лимфомой, сверхэкспрессируют IL-13 и/или рецепторы IL-13. (Kapp et al., J. Exp. Med. 1999;189:1939; Billard et al., Eur. Cytokine Netw. 1997; 8:19; Skinnider et al., Blood 2001; 97:250; Oshima et al., Cell Immunol. 2001; 211:37). Было показано, что нейтрализующие анти-IL-13-mAb или антагонисты IL-13 ингибируют дозозависимую пролиферацию клеток Н/RS (Kapp et al., J. Exp. Med. 1999; 189:1939; Oshima et al., Cell Immunol. 2001; 211:37). Аналогичным образом, введение растворимого рецептора-ловушки IL-13Rα2 мышам NОD/SCID с имплантированной клеточной линией, ассоциированной с болезнью Ходжкина, приводит к замедлению начала развития и роста опухоли и повышает выживаемость этих мышей, что свидетельствует о том, что нейтрализация IL-13 может подавлять рост ходжкинской лимфомы in vitro и in vivo (Trieu et al., Cancer Research 2004;64:3271). В целом эти исследования показали, что IL-13 стимулирует аутокринную пролиферацию клеток Н/RS (Kapp et al., J. Exp. Med. 1999;189:1939; Ohshima et al., Histopathology 2001;38:368).
Поэтому нейтрализация IL-13 представляет собой привлекательный и эффективный способ лечения болезни Ходжкина и других ассоциированных с В-клетками раковых заболеваний путем подавления роста опухолевых клеток при одновременном усилении противоопухолевой защиты.
Воспалительные заболевания кишечника
Возможно, что IL-13 играет определенную роль в патогенезе воспалительного заболевания кишечника (ВЗК). Воспалительное заболевание кишечника включает в себя ряд заболеваний, клинически классифицируемых как язвенный колит, болезнь Крона и колит неясной этиологии. Главными манифестациями таких заболеваний является хроническое воспаление тонкой кишки, вызванное усилением иммунного ответа с одновременным дисбалансом активации Th1- и Th2-лимфоцитов в слизистой тонкой кишки. Такие манифестации были продемонстрированы на животных моделях болезни Крона (Bamias et al., Gastroenterol 2005; 128:657) и язвенного колита (Heller et al., Immunity 2002; 17:629). Нейтрализация IL-13 путем введения IL-13Rα2-Fc приводит к предотвращению развития колита у мышей с Th2-моделью человеческого язвенного колита (Heller et al., Immunity 2002; 17:629). Кроме того, в этой модели продуцирование IL-13 происходит быстрее, чем продуцирование IL-4, и такое продуцирование IL-13 может быть индуцировано путем стимуляции NKТ-клеток, что позволяет предположить, что наблюдаемое разрушение ткани может быть результатом токсического воздействия IL-13 на эпителиальные клетки. При обследовании некоторых пациентов были получены некоторые данные, подтверждающие эти выводы, а именно частота встречаемости IL-13 в IL-13-позитивных биоптатах, взятых из прямой кишки пациентов с язвенным колитом, значительно превышала частоту встречаемости IL-13 у индивидуумов с воспалением и у контрольных индивидуумов, при этом более высокий уровень экспрессии IL-4 и IL-13 наблюдался при остром язвенном колите, а не при обычном язвенном колите (Inoue et al., Am. J. Gastroenterol. 1999;94:2441). Кроме того, в работе Akido et al. была охарактеризована иммунная активность во внешней мышечной пластинке (muscularis externa), выделенной из сегментов тонкой кишки пациентов с болезнью Крона, и было обнаружено, что IL-4 и IL-13 опосредуют увеличение сокращения клеток гладких мышц тонкой кишки по STAT-6-пути. Авторами был сделан вывод, что этот путь может способствовать гиперактивному сокращению мышц тонкой кишки у пациентов с болезнью Крона (Akiho et al., Am. J. Physiol. Gastrointest. Liver. Physiol. 2005;288:619).
Таким образом, использование mAb против IL-13, возможно, в комбинации с молекулами, направленными на другие цитокины, может представлять собой способ прекращения или замедления прогрессирования ВЗК.
Псориаз и псориатический артрит
Псориаз представляет собой хроническое кожное заболевание, характеризующееся гиперпролиферацией кератиноцитов и иммунологической клеточной инфильтрацией, включая активацию Т-клеток, продуцирующих различные цитокины, которые могут влиять на фенотип эпидермальных кератиноцитов. CDw60 представляет собой углевод-несущую молекулу, которая активируется на поверхности псориатических базальных и супрабазальных кератиноцитов псориатической кожи. Было показано, что IL-4 и IL-13, секретированные Т-клетками, происходящими из участков, пораженных псориазом, в высокой степени стимулируют экспрессию CDw60 на кератиноцитах (Skov et al., Am. J. Pathol. 1997;15:675), тогда как интерферон-гамма блокирует IL-4/IL-13-опосредуемую индукцию CDw60 на культивируемых кератиноцитах (Huang et al., J. Invest. Dermatol. 2001;116:305). Таким образом, считается, что экспрессия CDw60 на псориатических эпидермальных кератиноцитах индуцируется, по меньшей мере частично, цитокином IL-13, секретируемым активированными Т-клетками в области поражения. Кроме того, IL-13Rα1 и IL-4Rα, то есть белки клеточной поверхности, которые вместе образуют рецепторный комплекс IL-13, в различной степени экспрессируются в кожных биоптатах, взятых у пациентов, страдающих и не страдающих псориазом (Cancino-Diaz et al., J. Invest. Dermatol. 2002;119:1114; Wongpiyabovorn et al., J. Dermatol. Science 2003;33:31), при этом эксперименты in vitro продемонстрировали, что IL-13 (но не IL-4) может активировать экспрессию IL-13Rα1 (Wongpiyabovorn et al., J. Dermatol. Science 2003;33:31). Поскольку IL-13 влияет на клетки различных типов, то данные этих исследований позволяют предположить, что рецептор IL-13 может играть определенную роль в ранней стадии воспалительного процесса при псориазе.
Псориатический артрит характеризуется синовитом, который опосредован провоспалительными и противовоспалительными цитокинами. В настоящее время роль IL-13 в возникновении различных форм артрита представляет все возрастающий интерес. Spadaro и сотрудники продемонстрировали, что уровни IL-13 в синовиальной жидкости пациентов с псориатическим артритом значительно выше, чем уровни IL-13 у пациентов с остеоартритом. Кроме того, уровни IL-13 в синовиальной жидкости значительно превышали уровни IL-13 в сыворотке пациентов с псориатическим артритом, а отношение IL-13 в синовиальной жидкости к IL-13 в сыворотке значительно выше у пациентов с псориатическим артритом, чем у пациентов с ревматоидным артритом, что дает основание предположить, что в развитии псориатического артрита, возможно, определенную роль играет IL-13, локально продуцируемый в синовиальных тканях пациентов (Spadaro et al., Ann. Rheum. Dis. 2002;61:174).
Возможная роль IL-13 в развитии других состояний
Острая реакция “трансплантат против хозяина” является основной причиной заболеваемости и смертности после трансплантации стволовых клеток и непосредственно ассоциируется со степенью несовместимости человеческого лейкоцитарного антигена (HLA) у донора и реципиента. Jordan и др. впервые идентифицировали IL-13 как типичный Th2-цитокин, который обильно продуцируется в процессе реакций смешанной культуры неродственных несовместимых лимфоцитов (МLR) (реакция смешанной культуры лимфоцитов; in vitro-анализ для тонкого отбора донора после предварительного НLA-типирования)(Jordan et al., J. Immunol. Methods; 2002;260:1). Затем этой группой исследователей было обнаружено, что продуцирование IL-13 донорными Т-клетками после трансплантации донорных стволовых клеток является показателем острой реакции “трансплантат против хозяина” (aGVHD) (Jordan et al., Blood 2004;103:717). Все пациенты, у которых после трансплантации стволовых клеток наблюдалась тяжелая острая aGVHD третьей степени, были донорами, у которых продуцировались очень сильные ответы на IL-13 перед трансплантацией, что указывало на тесную взаимосвязь между уровнями IL-13 и aGVHD и на увеличение вероятности того, что IL-13 может быть непосредственно ответственен за некоторые из aGVHD-ассоциированных патологий. Следовательно, терапия, основанная на специфическом блокировании IL-13, может быть использована для лечения aGVHD после трансплантации стволовых клеток.
В западных странах диабетическая нефропатия является одной из наиболее главных причин почечного заболевания конечной стадии. Хотя число случаев заболеваний нефропатией, ассоциированных с диабетом типа 1, снижается, однако в настоящее время сахарный диабет типа 2 является одной из наиболее распространенных причин почечной недостаточности в США, Японии и в Европе. Кроме того, эта группа пациентов имеет очень плохой прогноз относительно результатов поддерживающего диализа вследствие исключительно высокой смертности от приступов сердечно-сосудистых заболеваний. В настоящее время становится все более очевидно, что гемодинамические, метаболические и структурные изменения взаимосвязаны друг с другом, и было установлено, что различные ферменты, факторы транскрипции и факторы роста играют определенную роль в патогенезе этих заболеваний. В частности, ТGF-β играет важную роль в развитии гипертрофии почек и накоплении внеклеточных компонентов матрикса, при этом считается, что этот цитокин играет решающую роль в опосредовании образования коллагена в почках (Cooper. Diabetologia 2001; 44:1957; Wolf. Eur. J. Clin. Invest. 2004; 34(12):785). При диабетической нефропатии у человека и у экспериментальных животных биологическая активность TGF-1 возрастает, и введение антител против ТGF-β1 мышам с диабетом приводит к улучшению почечной функции и к снижению накопления внеклеточного матрикса. Недавно было показано, что в трансгенной мышиной модели фиброза легких действие IL-13 направлено, по крайней мере частично, на регуляцию продуцирования и активации ТGF-β1 и отложение коллагена (Lee et al., J. Exp. Med. 2001; 194:809; Zhu et al., J. Clin. Invest. 1999;103:779), что указывает на прямую функциональную взаимосвязь между IL-13 и ТGF-β. Следовательно, можно сделать вывод об аналогичной роли IL-13 в регуляции активности ТGF-β1 в почках у больных диабетом, а поэтому подавление действия IL-13 может, вероятно, иметь определенный эффект при лечении диабетической нефропатии.
Фиброзные состояния
Фиброз легких представляет собой состояние, характеризующееся нежелательным и опасным образованием рубцов в легких, приводящим к инвалидности, а часто и к летальному исходу. Этот термин охватывает ряд состояний различной этиологии, патологии и восприимчивости к лечению. В некоторых случаях была выявлена причина фиброза. Такими причинами являются (1) вдыхание профибротического вещества, такого как асбестовая или кремниевая пыль, или порошок тяжелых металлов, (2) вдыхание органического вещества, которое вызывает у пациента идиосинкратический иммунный ответ, приводящий к фиброзу (например, к состоянию, называемому “легкие фермера”), (3) употребление лекарственных средств, таких как нитрофурантоин, амиодарон и метотрексат, (4) наличие системного воспалительного заболевания, такого как системный склероз или ревматоидный артрит.
Однако во многих случаях причина этого заболевания или лежащее в его основе бессимптомное состояние остаются неизвестными. У многих таких пациентов диагностируют идиопатический фиброз легких (ИФЛ). Это заболевание встречается довольно редко (преимущественно 20 случаев на 100000 человек). Этот диагноз обычно устанавливают по отсутствию явной причины заболевания в комбинации с некоторыми радиологическими и патологическими признаками, в частности по сотовидной структуре легких, определяемой с помощью компьютерной томографии, (КТ) или биопсии легких. Это заболевание обычно встречается у пожилых людей (>50) и часто сопровождается стойким прогрессирующим поражением легких, приводящим к смерти, причем, по оценкам специалистов, такое состояние может длиться от 2 до 5 лет. Более того, пациенты испытывают крайне неприятные ощущения в виде одышки, прогрессирующей в течение нескольких месяцев или лет. Такое состояние сначала приводит к ограничению физической активности, но на конечной стадии заболевания, которая может продолжаться несколько месяцев, пациент начинает задыхаться даже в состоянии покоя, и его самочувствие все больше зависит от поступления кислорода.
В настоящее время не существует удовлетворительного лечения этого заболевания. Современные способы лечения обычно предусматривают введение кортикостероидов и иммунодепрессантов, таких как азатиоприн. Однако для многих пациентов кортикостероиды могут оказаться неэффективными, а их побочные эффекты могут даже ухудшать состояние пациента. Существует множество возможных средств лечения, находящихся на стадии исследования, включая интерферон-гамма, который, как показали последние крупномасштабные исследования, может увеличивать продолжительность жизни, и перфенидон.
Имеются данные, свидетельствующие о том, что IL-13 и цитокины, ассоциированные с Th2-фенотипом, участвуют в репарации ткани, пораженной фиброзом (Wynn T.A., Nat. Rev. Immunol. 2004 4:583-594; Jakubzick et al., Am. J. Pathol. 2004 164(6):1989-2001; Jakubzick et al., Immunol. Res. 2004 30(3):339-349; Jakubzick et al., J. Clin. Pathol. 2004, 57:477-486). IL-13 и IL-4 участвуют в развитии различных фиброзных состояний. Фиброз печени, индуцированный Schistosoma, вероятно, является IL-13-зависимым, и имеются немногочисленные данные, указывающие на то, что IL-13 участвует в патогенезе склеродермии (Hasegawa et al., J. Rheumatol. 1997 24:328-332; Riccieri et al., Clin. Rheumatol. 2003:22:102-106).
Что касается фиброза легких, то in vitro исследования показали, что IL-13 стимулирует фиброгенный фенотип. Исследования, проводимые на животных, обнаруживали повышенные уровни экспрессии IL-13 у моделей с искусственно индуцированным фиброзом, а также показали, что степень фиброза может быть уменьшена путем элиминации IL-13.
IL-13 стимулирует профибротический фенотип. На клеточном уровне существует несколько механизмов, посредством которых IL-13 может стимулировать фиброз. Пути передачи сигнала и важная роль этих различных механизмов пока еще точно не определены.
Имеются данные, свидетельствующие о том, что IL-13 действует на фибробласты и тем самым стимулирует продуцирование коллагена и ингибирует его разрушение, что благоприятствует развитию фибротического фенотипа. Фибробласты кожи имеют рецепторы IL-13, и обработка культивируемых фибробластов кожи цитокином IL-13 приводит к активации образования коллагена (Oriente et al., J. Pharmacol. Exp. Ther. 2000 292:988-994). IL-4 обладает таким же, но более кратковременным действием. Клеточная линия фибробластов легких человека (ICIG7) экспрессирует рецептор IL-4 типа II (Jinnin et al., J. Biol. Chem. 2004, 279:41783-41791). Обработка этих клеток цитокином IL-13 стимулирует секрецию различных воспалительных и профибротических медиаторов: GM-CSF, G-CSF и интегрина VСАМ-бета 1 (Doucet et al., Int. Immunol. 1998 10(10):1421-1433).
IL-13 ингибирует IL-1а-индуцируемую продукцию белков 1 и 3 матриксных металлопротеиназ фибробластами кожи, которые способны снижать уровень разрушения внеклеточного (ВК) матрикса (Oriente et al., J. Pharmacol. Exp. Ther. 2000 292:988-994). IL-13 вместе с TGF-β действуют синергически на человеческие фибробласты, полученные путем биопсии дыхательных путей при астме, стимулируя тем самым экспрессию тканевого ингибитора металлопротеиназы 1 (ТIMP-1). Разрушение внеклеточного матрикса осуществляется под действием матриксных металлопротеиназ, которые ингибируются TIMP-1. Таким образом, это действие IL-13 должно приводить к снижению разрушения матрикса (Zhou et al., Am. J. Physiol. Cell. Physiol. 2005:288:С435-С442).
Сверхэкспрессия IL-13 у трансгенных мышей приводит к субэпителиальному фиброзу, гипертрофии эпителиальных клеток, гиперплазии бокаловидных клеток, отложению кристаллов (кислотной хитиназы млекопитающих), геперреактивности дыхательных путей, интерстициальному фиброзу, гипертрофии клеток типа 2 и накоплению поверхностно-активных веществ (Zhu et al., J. Clin. Invest. 1999 103(6):779-788).
Различные виды мышей имеют различную восприимчивость к индуцированному блеомицином фиброзу легких. У восприимчивых мышей С57BL/6J наблюдается быстрая активация IL-13, IL-13Rα и IL-4 (а также ТGFβ, TNFRα и IL-1Rs) в ответ на блеомицин. У мышей BALB/c, которые не являются восприимчивыми, активации IL-13 не наблюдается.
Belpero и др. (Am. J. Respir. Cell. Mol. Biol. 2002:27:419-427) проводили исследования по экспрессии и роли IL-13, IL-4 и CC-хемокина С10 в развитии фиброза у мышей с моделью фиброза, индуцированного блеомицином. Уровни IL-13 и IL-4 в тканях легких увеличивались в ответ на блеомицин. Предварительная нейтрализация IL-13 с использованием поликлональных антител против IL-13 приводила к значительному снижению степени фиброза легких, индуцированного блеомицином, как было оценено по уровням гидроксипролина в легких. Несмотря на повышенный уровень экспрессии IL-4 в той же самой модели, нейтрализация IL-4 не оказывала какого-либо влияния на фиброз легких.
В другой модели острого фиброза легких, индуцированного ФИТЦ у мышей BALB/c, отсутствие IL-13 (у дефицитных мышей), но не IL-4, предотвращало фиброз легких. При этом дефицит IL-4 у IL-13-дефицитных мышей не давал какой-либо дополнительной защиты (Kolodsick et al., J. Immunol. 2004 172:4068-4076). Протективный эффект отсутствия IL-13 не связан с различиями в рекрутинге клеток в легкие, то есть у всех дефицитных мышей и мышей BALB/c общее число обновленных (рекрутированных) клеток было одинаковым, а поэтому очевидно, что исходный воспалительный компонент оставался неизменным. Рекрутинг эозинофилов у IL-4- и IL-13-дефицитных мышей был ниже, чем у мышей BALB/c, но поскольку IL-4-/--мыши не были защищены от фиброза, это не может служить объяснением различия в степени их фиброза. Вероятно, может показаться неожиданным, что у IL-13+/+- и IL-13-/--мышей отсутствует какая-либо разница в уровнях цитокинов, включая IL-10, МСР-1, гамма-интерферон, TGF-i. Кроме того, из легких различных животных после их обработки ФИТЦ было выделено одинаковое число фибробластов, однако у IL-13-/--мышей уровень продуцирования коллагена I снижался. Это указывает на то, что отсутствие IL-13 не просто предупреждает воспалительный ответ, но даже играет более специфическую антифибротическую роль. Было высказано предположение, что IL-13 может оказывать фиброгенное действие посредством TGF-i (Lee et al., J. Exp. Med. 2001 194:809-821). Однако в этой ФИТЦ-модели экспрессия TGF-i не снижалась у IL-13-дефицитных мышей.
Было высказано предположение, что интерлейкин-4 обладает таким же действием, как и IL-13, и оба они действуют посредством одного и того же рецептора. IL-4 в значительной степени активируется в легких мышей с блеомицин-индуцированным фиброзом легких (Gharaee-Kermani et al., Cytokine 2001 15:138-147). Однако по сравнению с блеомицин-индуцированным фиброзом легких у мышей С57BL/6J, у которых наблюдается сверхэкспрессия IL-4, у IL-4-дефицитных мышей и у мышей дикого типа (Izbicki et al. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002 283(5):L1110-L1116) не было обнаружено каких-либо признаков, свидетельствующих о том, что IL-4 участвует в развитии фиброза легких. У IL-4-дефицитных мышей не наблюдалось снижения степени фиброза, тогда как у мышей со сверхэкспрессией IL-4 наблюдалось увеличение степени фиброза.
Уровни цитокина IL-13 в бронхоальвеолярном лаваже (БАЛ) от пациентов с различными формами фиброза легких были значительно увеличены, хотя и в различной степени. Экспрессия IL-13 значительно повышалась в альвеолярных макрофагах, полученных от пациентов с фиброзом легких.
Наиболее убедительные клинические данные были получены в результате исследований, проведенных в Мичиганском университете. Jakubzick и коллеги исследовали экспрессию гена IL-13 и IL-4 и их рецепторов в биоптатах легких, полученных после хирургической операции у пациентов с фиброзом легких. Экспрессия гена IL-13 в образцах, взятых из легких, пораженных ИФЛ, была заметно выше, чем в нормальных легких или в легких, пораженных другими фиброзными состояниями. Культивируемые фибробласты, взятые у пациентов с ИФЛ/ОИП, обнаруживали значительно более высокий уровень экспрессии рецептора IL-13 и IL-4, чем ткани и фибробласты, полученные путем биопсии ткани пациентов с нормальными легкими или с другими формами фиброза легких. В частности, фибробласты, сконцентрированные на участках, которые предположительно были эпицентрами активности заболевания, давали наиболее яркую окраску, указывающую на присутствие этих рецепторов (Jakubzick et al., J. Immunol. 2003 171:2684-2693; Jakubzick et al., Am. J. Pathol. 2003 162:1475-1486; Jakubzick et al., Am. J. Pathol. 2004 164(6):1989-2001; Jakubzick et al., Immunol. Res. 2004 30(3):339-349; Jakubzick et al., J. Clin. Pathol. 2004 57:477-486).
Имеются убедительные данные in vitro, указывающие на то, что в основном Th2-цитокины, в частности IL-13, стимулируют профибротический фенотип. Было показано, что по меньшей мере у 2 животных-моделей химически индуцированный фиброз может быть ослаблен путем элиминации IL-13 (осуществляемой либо путем удаления гена, либо с использованием антител против IL-13). Некоторые данные указывают на то, что IL-13 играет более важную роль в стимуляции фиброза легких, чем IL-4. Клинические данные относительно роли IL-13 в развитии фиброза легких дают основание предполагать, что в легких пациентов с ИФЛ отсутствует регуляция IL-13 и его рецепторов.
Все возрастающий объем данных позволяет предположить, что лекарственные средства, полученные на основе IL-13, являются достаточно эффективными для лечения ряда фиброзных состояний, включая индуцированный шистосомозом фиброз печени, и различных форм фиброза легких (например, ИФЛ [обсуждаемый в настоящем описании] и склеродермия).
Эксперименты, в которых независимо ингибировались IL-4 и IL-13, показали, что IL-13 является доминантным эффекторным цитокином фиброза в некоторых моделях (Chiaramonte et al., J. Clin. Invest. 1999; 104:777-785; Blease et al. J. Immunol. 2001; 166:5219; Kumar et al., Clin. Exp. Allergy 2002; 32:1104). В случае шистосомоза, индуцированный в яйцеклетке воспалительный ответ не изменялся при блокаде IL-13, однако, несмотря на продолжающееся и неснижающееся продуцирование IL-4, отложение коллагена у животных с хронической инфекцией снижалось более чем на 85% (Chiaramonte et al. J. Clin. Invest. 1999; 104:777; Chiaramonte et al., Hepatology 2001; 34:273).
Аминокислотная последовательность hIL-13 представлена в SEQ ID NO:9 (Эта последовательность представляет собой последовательность зрелого белка, то есть последовательность, в которой отсутствует сигнальная последовательность).
кДНК, кодирующая hIL-13, представлена в SEQ ID NO:10 (Эта последовательность ДНК кодирует последовательность зрелого белка, то есть последовательность, в которой отсутствует сигнальная последовательность).
Все ссылки на патенты и литературу, приведенные в настоящей заявке, точно и во всей своей полноте вводятся в настоящее описание посредством ссылки (включая любую патентную заявку, приоритет которой испрашивается в настоящей заявке).
Полученные недавно вакцины, вырабатывающие иммунные ответы против IL-13 и предназначенные для лечения астмы, были описаны в литературе (WO 02/070711). Недавно была также описана роль IL-13 в восприимчивости кожи к внешним аллергенам (Herrick et al., The Journal of Immunology, 2003, 170:2488-2495).
Настоящее изобретение, помимо прочего, относится к антителу, обозначенному 6А1. Как продемонстрировано ниже, связывание 6А1 с hIL-13, очевидно, зависит от присутствия аргинина в положении 107 SEQ ID NO:9. Сообщалось, что аргинин в положении 107 SEQ ID NO:9 представляет собой остаток, играющий важную роль во взаимодействии hIL-13 с hIL-13R. См. публикацию Thompson J.P. & Debinski W. (1999) J. Biol. Chem. vol.24, № 42, pp.29944-29950, озаглавленную “Glutamic Acids at position 13 and 16 in hIL13α-helix A, arginine and serine at positions 66 and 69 in helix C, and arginine at position 109 in helix D were found to be important in inducing biological signalling since their specific mutation resulted in loss and/or gain of function phenomena” (см. реферат и все описание). В этой работе аргинин в положении 109 эквивалентен аргинину в положении 107 SEQ ID NO:9 настоящей заявки, что обусловлено различием в системах нумерации, используемых авторами настоящего изобретения и авторами указанной работы. Таким образом, антитело 6А1, связывающееся с hIL-13, включает один из указанных остатков hIL-13, который был ранее идентифицирован как остаток, который играет важную роль во взаимодействии hIL-13 с hIL-13R, а следовательно, участвует в биологической передаче сигнала по IL-13-пути.
Описание сущности изобретения
Следовательно, настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и нейтрализует активность hIL-13. См., например, таблицу А, представленную ниже.
Термин “специфически связывается”, используемый в настоящем описании по отношению к антителам и к их антигенсвязывающим фрагментам согласно изобретению, означает, что указанное антитело связывается с hIL-13, но при этом не связывается или связывается в незначительной степени с другими человеческими белками, в частности с человеческим IL-4. Однако этот термин фактически не включает антитела согласно изобретению, которые могут перекрестно реагировать с IL-13 обезьян cynomolgus.
В другом своем аспекте, настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, которое специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R. Такое ингибирование включает в себя, но не ограничиваются им, конкурентное ингибирование. В некоторых вариантах изобретения антитела согласно изобретению по меньшей мере ингибируют взаимодействие hIL-13 с hIL-13R, но могут также блокировать взаимодействие hIL-13 с hIL-13R и тем самым блокировать путь передачи сигнала hIL-13/hIL-13R.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и содержит область CDRН3, имеющую последовательность, представленную в SEQ ID NO:3.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и содержит область CDRН3, представляющую собой вариант последовательности, представленной в SEQ ID NO:3, где один или два остатка указанной области CDRН3 данного варианта отличаются от остатков в соответствующем положении SEQ ID NO:3.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и содержит следующие области CDR:
CDRН1: SEQ ID NO:1,
CDRН2: SEQ ID NO:2,
CDRН3: SEQ ID NO:3,
CDRL1: SEQ ID NO:4,
CDRL2: SEQ ID NO:5,
CDRL3: SEQ ID NO:6.
В настоящем описании аминокислотные остатки в последовательностях антител пронумерованы в соответствии с нумерацией Кабата. Аналогичным образом, термины “CDR”, “CDRL1”, “CDRL2”, “CDRL3”, “CDRН1”, “CDRН2”, “CDRН3” соответствуют системе нумерации Кабата и др., описанной в публикации “Sequences of proteins of Immunological Interest NIH, 1987”. Термин “CDRН1” означает фрагмент последовательности, который включает фрагмент CDRН1, определенный по Кабату (остатки 31-35В), а также фрагмент CDRН1, определенный по Chothia (Chothia et al.(1989); Conformations of immunoglobulins hypervariable regions; Nature 342, р877-883), который включает фрагмент 26-32 по Кабату. Поэтому в соответствии с настоящим изобретением CDR были определены следующим образом:
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, включающему домен VH, имеющий последовательность, представленную в SEQ ID NO:7, и домен VL, имеющий последовательность, представленную в SEQ ID NO:8.
В другом своем аспекте настоящее изобретение относится к выделенному домену VH антитела, содержащего (или, по существу, состоящего, или состоящего из) SEQ ID NO:7 или 11, 12, 13, 14.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, включающему домен VH, выбранный из группы, состоящей из SEQ ID NO:7 или SEQ ID NO: 11, 12, 13, 14.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, который конкурентно ингибирует связывание терапевтического антитела, содержащего CDRН3 SEQ ID NO:3 с hIL-13.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, который конкурентно ингибирует связывание терапевтического антитела, содержащего CDR SEQ ID NO:1, 2, 3, 4, 5 и 6 с hIL-13.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу или к его антигенсвязывающему фрагменту, который конкурентно ингибирует связывание терапевтического антитела, содержащего тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22, с hIL-13.
В соответствии с этим настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему домен VH, выбранный из группы, состоящей из SEQ ID NO: 11, 12, 13, 14, и домен VL, выбранный из группы, состоящей из SEQ ID NO: 15, 16.
В другом своем аспекте настоящее изобретение относится к способу лечения человека, страдающего заболеванием или расстройством, восприимчивым к модуляции взаимодействия hIL-13 с hIL-13R (таким как астма, ХОБЛ, аллергический ринит, атопический дерматит), где указанный способ включает в себя стадию введения указанному пациенту терапевтически эффективного количества описанного здесь терапевтического антитела или его антигенсвязывающего фрагмента.
Настоящее изобретение также относится к применению антитела согласно изобретению в целях изготовления лекарственного средства для лечения заболевания или расстройства, восприимчивого к модуляции взаимодействия hIL-13 с hIL-13R.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу, которое специфически связывается с человеческим IL-13, где указанное антитело специфически связывается с человеческим IL-13 в положении остатков 97-108 SEQ ID NO:9. Исходя из результатов, приведенных ниже, очевидно, что термин “в положении остатков 97-108 SEQ ID NO:9” подразумевает, включая и остатки в положениях 97 и 108.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу, которое конкурентно ингибирует связывание терапевтического антитела, имеющего область CDRН3 SEQ ID NO:3, с человеческим IL-13 (такому как терапевтическое антитело, содержащее тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22), где указанное конкурентное антитело специфически связывается с человеческим IL-13 в положениях остатков 97-108 SEQ ID NO:9.
В другом своем аспекте настоящее изобретение относится к терапевтическому антителу, которое специфически связывается с человеческим IL-13 в положениях остатков 103-107 SEQ ID NO:9 включительно и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R.
В одном из своих вариантов настоящее изобретение относится к фармацевтической композиции, содержащей множество моноклональных терапевтических антител (обычно человеческих или гуманизованных), которые специфически связываются с человеческим IL-13 в положениях остатков 103-107 SEQ ID NO:9 и модулируют (например, ингибируют или блокируют) взаимодействие hIL-13 с hIL-13R, и фармацевтически приемлемый носитель.
В другом своем аспекте настоящее изобретение относится к способу продуцирования терапевтического антитела, которое специфически связывается с hIL-13 в положениях остатков 103-107 SEQ ID NO:9 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 и hIL-13R, где указанный способ включает в себя стадию культивирования в бессывороточной культуральной среде рекомбинантной клетки-хозяина, содержащей первый и второй векторы, где указанный первый вектор содержит полинуклеотид, кодирующий тяжелую цепь указанного антитела, а указанный второй вектор содержит полинуклеотид, кодирующий легкую цепь указанного антитела. Исходя из результатов, приведенных ниже, очевидно, что термин “в положении остатков 103-107 SEQ ID NO:9” включает остатки в положениях 103 и 107.
В другом своем варианте настоящее изобретение относится к способу продуцирования терапевтического антитела, которое специфически связывается с hIL-13 в положениях остатков 97-108 SEQ ID NO:9 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 и hIL-13R, где указанный способ включает в себя стадию культивирования в бессывороточной культуральной среде рекомбинантной клетки-хозяина, содержащей первый и второй векторы, где указанный первый вектор содержит полинуклеотид, кодирующий тяжелую цепь указанного антитела, а указанный второй вектор содержит полинуклеотид, кодирующий легкую цепь указанного антитела.
В другом своем варианте настоящее изобретение относится к интактному терапевтическому антителу, которое связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R, где указанное антитело взаимодействует с остатком 107 SEQ ID NO:9.
В другом своем варианте настоящее изобретение относится к интактному терапевтическому антителу, которое связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R, где указанное связывание терапевтического антитела и hIL-13 зависит от присутствия аргининового остатка (или позитивно коррелирует) в положении 107 SEQ ID NO:9.
В другом своем варианте настоящее изобретение относится к терапевтическому антителу, которое специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R, и имеет константу диссоциации k off в пределах от 1,4 × 10-4 до 8,22 × 10-5 с-1 (например, как было измерено в анализе BiacoreTM). Такое антитело может содержать CDRH3 SEQ ID NO:3 или его вариант и, помимо SEQ ID NO:3 или его варианта, оно может также содержать SEQ ID NO:1, 2, 4, 5 и 6.
В другом своем варианте настоящее изобретение относится к антителу, которое специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R, где указанное антитело содержит CDRH3 SEQ ID NO:3 и, необязательно, содержит каждую из областей CDRH1 SEQ ID NO:1, CDRH2 SEQ ID NO:2, CDRL1 SEQ ID NO:4, CDRL2 SEQ ID NO:5 и CDRL3 SEQ ID NO:6, и где указанное антитело также перекрестно реагирует с IL-13 обезьян cynomolgus (cIL-13).
Краткое описание чертежей
Фигура 1:
Сэндвич-ELISA, иллюстрирующий способность моноклонального антитела 6А1 связываться с рекомбинантным IL-13, экспрессируемым в E.coli при возрастающих концентрациях.
Фигура 2a:
ELISA, иллюстрирующий способность моноклонального антитела 6А1, при возрастающих концентрациях, ингибировать связывание рекомбинантного человеческого IL-13, экспрессируемого в E.coli, с цепью α1 человеческого рецептора IL-13.
Фигура 2b:
ELISA, иллюстрирующий способность моноклонального антитела 6А1, при возрастающих концентрациях, ингибировать связывание рекомбинантного человеческого IL-13, экспрессируемого в E.coli, с цепью α2 человеческого рецептора IL-13.
Фигура 3:
Анализ на нейтрализацию, иллюстрирующий способность моноклонального антитела 6А1, при возрастающих концентрациях, ингибировать биологическую активность рекомбинантного человеческого IL-13 и IL-13 обезьян cynomolgus, экспрессируемых в E.coli, в анализе на пролиферацию клеток ТF-1.
Фигура 4:
Анализ на нейтрализацию, иллюстрирующий способность моноклонального антитела 6А1, при возрастающих концентрациях, ингибировать биологическую активность человеческого IL-13, экспрессируемого в клетках млекопитающих (в клетках СНО), в анализе на пролиферацию клеток ТF-1.
Фигура 5:
Анализ на нейтрализацию, иллюстрирующий способность моноклонального антитела 6А1, при возрастающих концентрациях, ингибировать биологическую активность рекомбинантного человеческого IL-13, экспрессируемого в E.coli, в положении Q130, в анализе на пролиферацию клеток ТF-1.
Фигура 6:
Сэндвич-ELISA, который продемонстрировал, что 6А1 не связывается с рекомбинантным человеческим IL-4, экспрессируемым в E.coli.
Фигура 7:
Анализ на нейтрализацию IL5, который продемонстрировал, что 6А1 не ингибируют биологическую активность рекомбинантного человеческого IL-5, экспрессируемого в E.coli, в анализе на пролиферацию клеток ТF-1.
Фигура 8:
Сэндвич-ELISA, иллюстрирующий способность химерного mAb 6А1 связываться с рекомбинантным человеческим IL-13 и IL-13 обезьян cynomolgus, экспрессируемыми в E.coli, при возрастающих концентрациях.
Фигура 9:
Сэндвич-ELISA, иллюстрирующий способность 8 гуманизованных mAb против человеческого IL-13 связываться с рекомбинантным человеческим IL-13, экспрессируемым в E.coli, при возрастающих концентрациях.
Фигура 10а:
Сэндвич-ELISA, иллюстрирующий способность химерного 6А1, L1 + А1 и L2 + А1 связываться с рекомбинантным человеческим IL-13, экспрессируемым в E.coli, при возрастающих концентрациях.
Фигура 10b:
Сэндвич-ELISA, иллюстрирующий способность химерного 6А1, L1 + А1 и L2 + А1 связываться с рекомбинантным IL-13 обезьян cynomolgus, экспрессируемым в E.coli, при возрастающих концентрациях.
Фигура 11:
Сэндвич-ELISA, иллюстрирующий способность химерного 6А1, L1 + А1 и L2 + А1 связываться с нативным человеческим IL-13 при возрастающих концентрациях.
Фигура 12а:
ELISA, иллюстрирующий способность моноклонального антитела 6А1, химерного антитела 6А1, L1 + А1 и L2 + А1 при возрастающих концентрациях ингибировать связывание рекомбинантного человеческого IL-13, экспрессируемого в E.coli, с цепью α1 человеческого рецептора IL-13.
Фигура 12b:
ELISA, иллюстрирующий способность моноклонального антитела 6А1, химерного антитела 6А1, L1 + А1 и L2 + А1 при возрастающих концентрациях ингибировать связывание рекомбинантного человеческого IL-13, экспрессируемого в E.coli, с цепью α2 человеческого рецептора IL-13.
Фигура 13а
Анализ на нейтрализацию, иллюстрирующий способность антитела 6А1, химерного антитела 6А1, L1 + А1 и L2 + А1 при возрастающих концентрациях ингибировать биологическую активность рекомбинантного человеческого IL-13, экспрессируемого в E.coli, в анализе на пролиферацию клеток ТF-1.
Фигура 13b
Анализ на нейтрализацию, иллюстрирующий способность антитела 6А1, химерного антитела 6А1, L1 + А1 и L2 + А1 при возрастающих концентрациях ингибировать биологическую активность рекомбинантного IL-13 обезьян cynomolgus, экспрессируемого в E.coli, в анализе на пролиферацию клеток ТF-1.
Фигура 13с
Анализ на нейтрализацию, иллюстрирующий способность антитела 6А1, химерного антитела 6А1, L1 + А1 и L2 + А1 при возрастающих концентрациях ингибировать биологическую активность рекомбинантного человеческого IL-13, экспрессируемого в E.coli в положении Q130, в анализе на пролиферацию клеток ТF-1.
Фигура 13d:
Анализ на нейтрализацию, иллюстрирующий способность антитела 6А1, химерного антитела 6А1, L1 + А1 и L2 + А1 при возрастающих концентрациях ингибировать биологическую активность человеческого IL-13, экспрессируемого у млекопитающих (в клетках СНО), в анализе на пролиферацию клеток ТF-1.
Фигура 14а
Сэндвич-ELISA, который продемонстрировал, что антитело 6А1, химерное антитело 6А1, L1 + А1 и L2 + А1 не связываются с рекомбинантным человеческим IL-4, экспрессируемым в E.coli.
Фигура 14b
Сэндвич-ELISA, который продемонстрировал, что антитело 6А1, химерное антитело 6А1, L1 + А1 и L2 + А1 не связываются с рекомбинантным человеческим GM-CSF, экспрессируемым в E.coli.
Фигура 14с:
Анализ на нейтрализацию IL5, который продемонстрировал, что антитело 6А1, химерное антитело 6А1, L1 + А1 и L2 + А1 не ингибируют биологическую активность рекомбинантного человеческого IL-5, экспрессируемого в E.coli, в анализе на пролиферацию клеток ТF-1.
Фигура 15
Картирование эпитопа с помощью ELISA для определения эпитопа, с которым связывается антитело 6А1, на человеческом IL-13 или на IL-13 обезьян cynomolgus.
Фигура 16а
Картирование эпитопа с помощью ELISA для идентификации высокой специфичности связывания антитела 6А1 с человеческим IL-13.
Фигура 16b
Картирование эпитопа с помощью ELISA для идентификации высокой специфичности связывания антитела 6А1 с IL-13 обезьян cynomolgus.
Фигура 17а
Картирование эпитопа с помощью ELISA для определения ключевых аминокислотных остатков, необходимых для связывания 6А1 с человеческим IL-13.
Фигура 17b
Картирование эпитопа с помощью ELISA для определения ключевых аминокислотных остатков, необходимых для связывания L1+A1 с человеческим IL-13.
На фигурах 17с и 17d представлены графики, иллюстрирующие аланин-сканирующий анализ родительского (мышиного) антигена 6А1 (фиг. 17с) и гуманизованного антитела L1-А1.
Подробное описание изобретения
1. Структура антител
1.1. Интактные антитела
Интактными антителами являются гетеромультимерные гликопротеины, содержащие по меньшей мере две тяжелые и две легкие цепи. Интактные антитела, кроме IgМ, обычно представляют собой гетеротетрамерные гликопротеины размером приблизительно в 150 кДа, состоящие из двух идентичных легких цепей (L) и двух идентичных тяжелых (Н) цепей. Обычно каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, и число дисульфидных связей между тяжелыми цепями иммуноглобулинов различных изотипов варьирует. Каждая тяжелая и легкая цепи также имеют внутирицепьевые дисульфидные мостики. Каждая тяжелая цепь на одном своем конце имеет вариабельный домен (VH), за которым следует ряд константных областей. Каждая легкая цепь на другом своем конце имеет вариабельный домен (VL) и константную область, где указанная константная область легкой цепи соответствует первой константной области тяжелой цепи, а вариабельный домен легкой цепи соответствует вариабельному домену тяжелой цепи. Легкие цепи антител большинства позвоночных, исходя из их аминокислотной последовательности константной области, могут быть отнесены к одному их двух типов легких цепей, называемых каппа и лямбда. В зависимости от аминокислотной последовательности константной области тяжелых цепей человеческие антитела могут относиться к пяти различным классам, IgА, IgD, IgЕ, IgG и IgМ. Антитела IgG и IgА могут быть затем подразделены на подклассы IgG1, IgG2, IgG3 и IgG4, а также IgА1 и IgА2. У мышей и крыс имеются другие разновидности антител, по меньшей мере, IgG2а и IgG2b. Вариабельный домен антитела сообщает антителу способность специфически связываться с некоторыми областями, обнаруживающими особенно высокую вариабельность и называемыми гипервариабельными областями (комплементарность-определяющими областями, CDR). Более консервативные части вариабельной области называются каркасными областями (FR). Вариабельные домены каждой из тяжелых и легких цепей интактных антител содержат четыре FR, соединенные тремя CDR. CDR в каждой цепи находятся в непосредственной близости с FR-областями и с CDR другой цепи и участвуют в образовании антигенсвязывающего сайта антител. Константные области не принимают непосредственного участия в связывании антитела с антигеном, но обладают различными эффекторными функциями, такими как участие в антитело-зависимой клеточно-опосредуемой цитотоксичности (АDCC), фагоцитоз посредством связывания с рецептором Fcγ, изменение времени полужизни/скорости выведения посредством неонатального рецептора Fc (FcRn) и участие в комплемент-зависимой цитотоксичности посредством компонента С1q каскада комплемента.
Поэтому в одном из своих вариантов настоящее изобретение относится к интактному терапевтическому антителу, которое специфически связывается с hIL-13 и которое модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R. Интактное терапевтическое антитело может содержать константную область любого изотипа или любого подкласса, описанных выше. В одном из вариантов изобретения указанное антитело имеет изотип IgG, в частности IgG1. Указанным антителом может быть крысиное антитело, мышиное антитело, кроличье антитело, антитело приматов или человеческое антитело. В одном из репрезентативных вариантов изобретения указанным антителом является антитело приматов (таких как обезьяны cynomolgus, мартышки или человекообразные обезьяны, см., например, WO 99/55369, WO93/02108) или человека.
В другом своем варианте настоящее изобретение относится к выделенному интактному терапевтическому антителу, содержащему CDRН3 SEQ ID NO:3. В другом своем варианте настоящее изобретение относится к выделенному интактному терапевтическому антителу, содержащему вариабельную область, имеющему CDR SEQ ID NO:1, 2, 3, 4, 5 и 6.
В другом своем варианте настоящее изобретение относится к выделенному мышиному интактному терапевтическому антителу или к его антигенсвязывающему фрагменту, содержащему домен VH, имеющий последовательность SEQ ID NO:7, и домен VL, имеющий последовательность SEQ ID NO:8.
1.1.2. Человеческие антитела
Человеческие антитела могут быть получены различными методами, известными специалистам. Человеческие антитела могут быть получены гибридомным методом с использованием человеческих миеломных клеточных линий или гетеромиеломных клеточных линий “мышь-человек”, см. Kozbor J. Immunol. 133, 3001 (1984) и Brodeur, Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker Inc., 1987). Альтернативные методы предусматривают применение фаговых библиотек или трансгенных мышей, имеющих набор человеческих V-областей (см., Winter G. (1994) Annu. Rev. Immunol. 12, 433-455, Green L.L. (1999), J. Immunol. Methods 231, 11-23).
В настоящее время существует несколько штаммов трансгенных мышей, у которых мышиные иммуноглобулиновые локусы были заменены сегментами гена человеческого иммуноглобулина (см. Tomizuka K. (2000) PNAS 97, 722-727; Fishwild D.M. (1996) Nature Biotechnol. 14, 845-851, Mendez M.J. 1997, Nature Genetics, 15, 146-156). После сенсибилизации антигеном такие мыши способны продуцировать спектр человеческих антител, из которых могут быть выбраны представляющие интерес антитела. Особого внимания заслуживает система TrimeraTM (см. Eren R. et al. (1998) Immunology 93:154-161), в которой человеческие лимфоциты были трансплантированы облученным мышам, то есть система антител отобранных лимфоцитов (SLAM, см., Babcook et al. (1996) 93:7843-7848), где человеческие лимфоциты (или лимфоциты других видов) были подвергнуты эффективной процедуре пропускания через массив генерации антител, собранных in vitro, процедуре лимитирующего разведения и процедуре отбора и Xenomouse IIТМ (Abgenix Inc). Описание альтернативного способа с использованием технологии MorphdomaТМ можно найти в Morphotek Inc.
Для продуцирования человеческих антител (и их фрагментов) может быть использована технология фагового представления, см. McCafferty; Nature, 348, 552-553 (1990) и Griffiths A.D. et al. (1994) EMBO 13:3245-3260. В соответствии с этой технологией гены V-домена антитела клонируют с сохранением рамки считывания в ген большого или малого белка оболочки нитчатого бактериофага, такого как М13 или fd, и представляют (обычно с помощью хелперного фага) на поверхности фаговой частицы в виде функциональных фрагментов антител. Отбор, проводимый на основе функциональных свойств антитела, позволяет проводить отбор гена, кодирующего антитело, обладающего этими свойствами. Технология фагового представления может быть использована для отбора антигенспецифических антител из библиотеки, полученной из человеческих В-клеток, взятых от индивидуумов, страдающих заболеванием или расстройством, описанным выше, или, альтернативно, от неиммунизованных доноров (см, Marks, J. Mol.Bio. 222, 581-597, 1991). Если желательно, чтобы интактное человеческое антитело содержало Fc-домен, то фаг, представляющий полученный фрагмент, необходимо субклонировать в экспрессирующие векторы млекопитающего, содержащие нужные константные области, и в клеточные линии со стабильной экспрессией.
Для повышения аффинности связывания может быть использована техника созревания аффинности (Marks, Bio/Technol. 10, 779-783, 1992)), где аффинность исходного человеческого антитела может быть увеличена путем последующей замены V-областей Н и L природными вариантами и путем отбора исходя из повышенной аффинности связывания. Также описаны и другие варианты этого метода, такие как методы, основанные на “эпитопном импринтинге”, см., WO 93/06213. См. также Waterhouse; Nucl. Acids. Res. 21, 2265-2266 (1993).
Таким образом, в другом своем варианте настоящее изобретение относится к выделенному человеческому интактному терапевтическому антителу или его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R.
В другом своем аспекте настоящее изобретение относится к выделенному человеческому интактному терапевтическому антителу или его антигенсвязывающему фрагменту, содержащему CDRН3 SEQ ID NO:3, который специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R. В другом своем аспекте настоящее изобретение относится к выделенному человеческому интактному терапевтическому антителу или его антигенсвязывающему фрагменту, содержащему вариабельную область, имеющую CDR SEQ ID NO:1, 2, 3, 4, 5 и 6, определенные выше.
1.2. Химерные и гуманизованные антитела
Применение интактных не-человеческих антител для лечения человеческих заболеваний или расстройств может быть связано с хорошо известными в настоящее время проблемами иммуногенности, которые заключаются в том, что иммунная система пациента может распознавать не-человеческое интактное антитело как “чужое” и вырабатывать нейтрализующий ответ. Это особенно очевидно после многократного введения человеку не-человеческого антитела. Для решения этих проблем в последние годы были разработаны различные методы, которые в основном направлены на снижение уровня не-человеческих аминокислотных последовательностей в интактном антителе при сохранении относительной легкости получения не-человеческих антител от иммунизованного животного, например мыши, крысы или кролика. В широком смысле для достижения этой цели могут быть применены два метода. В первом методе получают химерные антитела, которые в основном содержат не-человеческий вариабельный домен (например, домен, происходящий от грызунов, таких как мышь), присоединенный к человеческой константной области. Поскольку антигенсвязывающий сайт антитела локализован в вариабельных областях, то химерное антитело сохраняет свою аффинность связывания с антигеном, но приобретает эффекторные функции человеческой константной области, а поэтому оно способно осуществлять эффекторные функции, описанные выше. Химерные антитела обычно получают методами рекомбинантных ДНК. ДНК, кодирующую антитела (например, кДНК), выделяют и секвенируют в соответствии со стандартными процедурами (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими Н- и L-цепи антитела согласно изобретению, например с ДНК, кодирующей SEQ ID NO: 1, 2, 3, 4, 5 и 6, описанные выше). Типичным источником такой ДНК служат гибридомные клетки. После выделения ДНК вводят в экспрессирующие векторы, которые затем переносят в клетки-хозяева, такие как клетки E.coli, клетки СОS, клетки СНО или миеломные клетки, которые в естественных условиях не продуцируют белок иммуноглобулина, необходимый для синтеза антитела. Такая ДНК может быть модифицирована путем замены последовательности, кодирующей константные области Н- и L-цепи не-человеческого (например, мышиного) антитела последовательностью, кодирующей соответствующие константные области человеческих Н- и L-цепей, см., например, Morrison; PNAS 81, 6851 (1984).
Второй метод предусматривает генерирование гуманизованных антител, в которых содержание не-человеческих антител снижено в результате гуманизации вариабельных областей. Эти два метода гуманизации приобретают особую популярность. Первым методом является гуманизация посредством CDR-прививки. CDR образуют петли в непосредственной близости к N-концу антитела, где они формируют поверхность, расположенную на носителе, образуемом каркасными областями. Специфичность связывания антигена с антителом определяется главным образом топографическими и химическими свойствами его CDR-поверхности. Эти признаки, в свою очередь, определяются конформацией отдельных CDR, относительным расположением CDR, а также природой и расположением боковых цепей остатков, составляющих CDR. Значительное снижение иммуногенности может быть достигнуто путем присоединения только CDR не-человеческих (например, мышиных) антител (“донорных” антител) к человеческим каркасным (“акцепторным каркасным”) и константным областям (см. Jones et al. (1986) Nature 321, 522-525 и Verhoeyen M. et al. (1988) Science 239, 1534-1536). Однако присоединение CDR per se не позволяет полностью сохранить антигенсвязывающие свойства антитела, и часто оказывается, что для сохранения значительного уровня аффинности связывания гуманизованного антитела с антигеном необходимо сохранить некоторые каркасные остатки (иногда называемые “остатками с обратными мутациями”) донорного антитела (см. Queen C. et al. (1989) PNAS 86, 10029-10033, Co, M. et al. (1991) Nature 351, 501-502). В этом случае, для получения человеческой каркасной области (FR), из базы данных отбирают человеческие V-области, обнаруживающие наибольшую гомологию своей последовательности с последовательностью не-человеческого донорного антитела. Человеческие FR могут быть выбраны либо из FR человеческих консенсусных антител, либо из отдельных человеческих антител. Если это необходимо для сохранения конформаций CDR, то ключевые остатки донорного антитела заменяют остатками человеческой акцепторной каркасной области. Для облегчения идентификации таких структурно важных остатков может быть применено компьютерное моделирование антител, см., WO 99/48523.
Альтернативно, гуманизация может быть осуществлена методом “маскировки”. Статистический анализ уникальных вариабельных областей тяжелой и легкой цепей человеческого и мышиного иммуноглобулина выявил, что точные профили поверхностных остатков у человеческих антител и у мышиных антител отличаются и большинство отдельных положений поверхностных остатков имеют строгую предпочтительность по отношению к небольшому числу других остатков (см. Padlan E.A. et al. (1991) Mol. Immunol. 28, 489-498 и Pedersen J.T. et al. (1994) J. Mol. Biol. 235;959-973). Поэтому иммуногенность не-человеческого Fv может быть снижена путем замены поверхностных остатков в каркасных областях, которые отличаются от областей, обычно присутствующих в человеческих антителах. Поскольку антигенность белков может коррелировать с доступностью поверхности, то замена поверхностных остатков может оказаться достаточной для того, чтобы “скрыть” мышиную вариабельную область от надзора человеческой иммунной системы (см. также Mark G.E. et al. (1994), Handbook of Experimental Pharmacology vol.113: The pharmacology of monoclonal Antibodies, Springer-Verlag, pp.105-134). Эта процедура гуманизации называется “маскировкой”, поскольку она позволяет изменить только поверхность антитела, но при этом основные остатки остаются неизменными.
Таким образом, в другом своем варианте настоящее изобретение относится к химерному терапевтическому антителу, содержащему не-человеческий вариабельный домен (например, грызунов), присоединенный к человеческой константной области (которая может принадлежать к изотипу IgG, например IgG1), которая специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R.
В другом своем аспекте настоящее изобретение относится к химерному терапевтическому антителу, содержащему не-человеческую вариабельную область (например, грызунов) и человеческую константную область (которая может принадлежать к изотипу IgG, например IgG1), которая специфически связывается с hIL-13, где указанное антитело также содержит CDRН3 SEQ ID NO:3. Такие антитела могут также содержать человеческую константную область изотипа IgG, например IgG1.
В другом своем варианте настоящее изобретение относится к химерному терапевтическому антителу, содержащему не-человеческую вариабельную область (например, грызунов) и человеческую константную область (которая может принадлежать к изотипу IgG, например IgG1), которая специфически связывается с hIL-13, имеющим CDR SEQ ID NO: 1, 2, 3, 4, 5 и 6.
В другом своем варианте настоящее изобретение относится к химерному терапевтическому антителу, содержащему VH-домен SEQ ID NO:7 и VL-домен SEQ ID NO:8 и человеческую константную область изотипа IgG, например IgG1, которая специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу или его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу или его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и содержит CDRН3 SEQ ID NO:3. Такие антитела могут также содержать человеческую константную область изотипа IgG, например IgG1.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу или его антигенсвязывающему фрагменту, который специфически связывается с hIL-13 и содержит CDR SEQ ID NO: 1, 2, 3, 4, 5 и 6. Такие антитела могут также содержать человеческую константную область изотипа IgG, например IgG1.
В соответствии с этим настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен, выбранный из группы, состоящей из SEQ ID NO:11, 12, 13, 14, и VL-домен, выбранный из группы, состоящей из SEQ ID NO:15, 16. Такие антитела могут также содержать человеческую константную область изотипа IgG, например IgG1.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:11 и VL-домен SEQ ID NO:15.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:12 и VL-домен SEQ ID NO:15.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:13 и VL-домен SEQ ID NO:15.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:14 и VL-домен SEQ ID NO:15.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:11 и VL-домен SEQ ID NO:16.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:12 и VL-домен SEQ ID NO:16.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:13 и VL-домен SEQ ID NO:16.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему VH-домен SEQ ID NO:14 и VL-домен SEQ ID NO:16.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу или его антигенсвязывающему фрагменту, который специфически связывается с hIL-13, где указанное антитело или его фрагмент содержит CDRН3 (SEQ ID NO:3), а также необязательно содержит CDR SEQ ID NO: 1, 2, 4, 5 и 6, где остатки выбраны из группы, состоящей из остатков 19, 38, 73 и 81 акцепторной каркасной области человеческой тяжелой цепи, а остаток в положении 85 акцепторной каркасной области человеческой легкой цепи заменен соответствующими остатками, имеющимися в каркасной области донорного антитела, от которой происходит CDRН3.
Для специалиста в данной области очевидно, что термин “происходящий” относится не только к источнику, в смысле, к физическому источнику данного материала, но также и к материалу, который структурно идентичен (по первичной аминокислотной последовательности) указанному материалу, но который не происходит от указанного источника. Таким образом, “остатки, присутствующие в донорном антителе, от которого происходит CDRН3”, не обязательно должны быть выделены из донорного антитела.
В другом своем варианте настоящее изобретение относится к гуманизованному терапевтическому антителу или к его антигенсвязывающему фрагменту, который специфически связывается с hIL-13, где указанное антитело или его фрагмент содержит CDRН3 SEQ ID NO:3, а также необязательно содержит CDR SEQ ID NO: 1, 2, 4, 5 и 6, где указанная каркасная область человеческой тяжелой цепи содержит один или несколько (например, все) из нижеследующих остатков (или их консервативных замен):
а человеческая легкая цепь содержит:
Специалисту в данной области хорошо известно, что некоторые аминокислотные замены рассматриваются как “консервативные”. Аминокислоты подразделяются на группы исходя из общих свойств их боковых цепей и замен внутри групп, которые полностью или почти полностью сохраняют аффинность связывания антитела согласно изобретению или его антигенсвязывающего фрагмента и которые рассматриваются как консервативные замены, например, указанные в нижеследующей таблице:
В соответствии с этим настоящее изобретение относится к гуманизованному терапевтическому антителу, содержащему тяжелую цепь, выбранную из группы, состоящей из SEQ ID NO:18, 19, 20, 21, и легкую цепь, выбранную из группы, состоящей из SEQ ID NO:22, 23.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:19 и легкую цепь SEQ ID NO:22.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:20 и легкую цепь SEQ ID NO:22.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:21 и легкую цепь SEQ ID NO:22.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:23.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:19 и легкую цепь SEQ ID NO:23.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:20 и легкую цепь SEQ ID NO:23.
В одном из своих вариантов настоящее изобретение относится к гуманизованному терапевтическому антителу, которое специфически связывается с hIL-13 и содержит тяжелую цепь SEQ ID NO:21 и легкую цепь SEQ ID NO:23.
1.3. Биспецифические антитела
Биспецифическое антитело представляет собой антитело, специфически связывающееся по меньшей мере с двумя различными эпитопами. Методы получения таких антител известны специалистам. Обычно рекомбинантное продуцирование биспецифических антител основано на коэкспрессии двух пар “Н-цепь - L-цепь” иммуноглобулина, где две Н-цепи обладают различными специфичностями связывания, см., Millstein et al., Nature 305:537-539 (1983), WO 93/08829 и Traunecker et al., EMBO, 10, 1991, 3655-3659. Из-за случайного ассортимента Н- и L-цепей продуцируется потенциальная смесь из десяти антител с различной структурой, из которых только одно антитело обладает нужной специфичностью связывания. Альтернативный метод включает в себя присоединение вариабельных доменов с нужными специфичностями связывания к константной области тяжелой цепи, содержащей, по меньшей мере, часть шарнирной области, области СН2 и области СН3. При этом предпочтительно, чтобы присутствовала область СН1, содержащая сайт, необходимый для связывания с легкой цепью и имеющийся, по меньшей мере, в одном из гибридов. ДНК, кодирующую эти гибриды, и, если это необходимо, L-цепь, встраивают в отдельные экспрессирующие векторы, а затем вместе вводят в организм подходящего хозяина. В один экспрессирующий вектор могут быть введены последовательности, кодирующие две или все три такие цепи. В одном из предпочтительных методов биспецифическое антитело состоит из Н-цепи, имеющей первую специфичность связывания на одной ветви, и пару Н-L-цепей, имеющих вторую специфичность связывания на другой ветви, см. WO 94/04690. Также см. публикацию Suresh et al., Methods in Enzymology, 121, 210, 1986.
В одном из своих вариантов настоящее изобретение относится к биспецифическому терапевтическому антителу, которое имеет по меньшей мере одну специфичность связывания с hIL-13, где указанное антитело модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R. Такие антитела могут также содержать человеческую константную область изотипа IgG, например IgG1. В некоторых вариантах изобретения биспецифическое терапевтическое антитело имеет первую специфичность связывания с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R и вторую специфичность связывания с hIL-4 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-4 с рецептором hIL-4.
В одном из своих вариантов настоящее изобретение относится к биспецифическому терапевтическому антителу, которое имеет по меньшей мере одну специфичность связывания с hIL-13, где указанное антитело содержит CDRН3 SEQ ID NO:3. Такие антитела могут также содержать человеческую константную область изотипа IgG, например IgG1.
В одном из своих вариантов настоящее изобретение относится к биспецифическому терапевтическому антителу, которое имеет по меньшей мере одну специфичность связывания с hIL-13, где указанное антитело содержит по меньшей мере CDR SEQ ID NO:1, 2, 3, 4, 5 и 6. Такие антитела могут также содержать человеческую константную область изотипа IgG, например IgG1.
1.4. Фрагменты антител
В некоторых своих вариантах настоящее изобретение относится к фрагментам терапевтических антител, которые модулируют взаимодействие hIL-13 с hIL-13R. Такими фрагментами могут быть функциональные антигенсвязывающие фрагменты интактных и/или гуманизованных и/или химерных антител, такие как Fab-, Fab'-, F(ab')2-, Fv-, scFv-фрагменты антител, описанных выше. Обычно такие фрагменты получают путем протеолитического гидролиза интактных антител, например путем гидролиза папаином (см., например, WO 94/29348), но они могут быть также получены непосредственно из рекомбинантно трансформированных клеток-хозяев. Описание способа продуцирования scFv можно найти в работе Bird et al. (1988) Science, 242, 423-426. Кроме того, фрагменты антител могут быть получены различными методами генной инженерии, описанными ниже.
Fv-фрагменты, очевидно, обладают меньшей энергией взаимодействия своих двух цепей, чем Fab-фрагменты. Для стабилизации связи между доменами VH и VL они присоединены посредством пептидных связей (Bird et al. (1988) Science, 242, 423-426; Huston et al., PNAS, 85, 5879-5883), дисульфидных мостиков (Glockshuber et al. (1990) Biochemistry 29, 1362-1367) и посредством введения мутаций “knob in hole” (Zhu et al. (1997) Protein Sci., 6, 781-788). scFv-фрагменты могут быть получены методами, хорошо известными специалистам, cм., Whitlow et al. (1991) Methods companion Methods Enzymol, 2, 97-105 & Huston et al. (1993) Int. Rev. Immunol. 10, 195-217. scFv-фрагменты могут быть продуцированы в бактериальных клетках, таких как E.coli, но более предпочтительно в эукариотических клетках. Одним из недостатков scFv является их одновалентность, которая не позволяет повышать авидность, что может быть достигнуто за счет поливалентного связывания, и их короткое время полужизни. В попытке решить эти проблемы из scFv', содержащего дополнительный С-концевой цистеин, был продуцирован двухвалентный (scFv')2 путем химического присоединения (Adams et al. (1993) Can. Res. 53, 4026-4034 & McCartney et al. (1995) Protein Eng. 8, 301-314) или путем спонтанной сайт-специфической димеризации scFv, содержащего неспаренный С-концевой цистеиновый остаток (см. Kipriyanov et al. (1995) Cell. Biophys. 26, 187-204). Альтернативно, scFv может быть искусственно получен в форме мультимеров путем укорочения пептидного линкера до 3-12 остатков с образованием “ди-антител”, см. Holliger et al. PNAS (1993), 90, 6444-6448. Еще большее укорочение этого линкера может также приводить к образованию scFv-тримеров (“три-антител”, см. Kortt et al., (1997) Protein Eng. 10, 423-433) и тетрамеров (“тетра-антител”, см. Le Gall et al. (1999) FEBS Lett, 453, 164-168). Конструирование двухвалентных scFv-молекул может быть осуществлено также путем генетического сцепления с белок-димеризующими мотивами, с образованием “мини-антител” (см., Pack et al. (1992) Biochemistry 31, 1579-1584) и “мини-телец” (см., Hu et al. (1996) Cancer Res. 56, 3055-3061). scFv-scFv-тандемы ((scFv)2) могут быть также получены путем присоединения двух scFv-фрагментов посредством третьего пептидного линкера, см. Kurucz et al. (1995) J. Immunol. 154, 4576-4582. Биспецифические ди-антитела могут быть получены посредством нековалентного связывания двух одноцепочечных гибридных продуктов, состоящих из VH-домена одного антитела, соединенного коротким линкером с VL-доменом другого антитела, см., Kipriyanov et al. (1998) Int. J. Can. 77, 763-772. Стабильность таких биспецифических ди-антител может быть повышена путем введения дисульфидных мостиков или мутаций в конфигурации типа “knob in hole”, описанных выше, или путем образования одноцепочечных ди-антител (scDb), где два гибридных scFv-фрагмента соединены пептидным линкером, см. работу Kontermann et al., (1999) J. Immunol. Methods 226:179-188. Четырехвалентные биспецифические молекулы могут быть получены, например, путем присоединения scFv-фрагмента к СН3-домену молекулы IgG или к Fab-фрагменту посредством шарнирной области, см. Coloma et al. (1997) Nature Biotechnol. 15, 159-163. Альтернативно, четырехвалентные биспецифические молекулы могут быть получены путем присоединения биспецифических одноцепочечных ди-антител (см. Alt et al., (1999) FEBS Lett 454, 90-94). Менее крупные четырехвалентные биспецифические молекулы могут быть также получены путем димеризации либо scFv-scFv-тандемов с линкером, содержащим мотив “спираль-петля-спираль” (мини-антитела DiBi, см., Muller et al. (1998) FEBS Lett. 432, 45-49), либо с одноцепочечной молекулой, содержащей четыре вариабельных домена антитела (VH и VL), в ориентации, предотвращающей внутримолекулярное спаривание (тандемное ди-антитело, см., Kipriyanov et al. (1999) J. Mol. Biol. 293, 41-56). Биспецифические F(ab')2-фрагменты могут быть созданы путем химического присоединения Fab'-фрагментов или путем гетеродимеризации посредством “лейциновых молний” (см., Shalaby et al. (1992) J. Exp. Med. 175, 217-225 и Kostelny et al. (1992), J. Immunol. 148, 1547-1553). Могут быть также получены отдельные VH- и VL-домены (Domantis plc), см. патенты США 6248516, 6291158, 6172197.
В одном из своих вариантов настоящее изобретение относится к фрагменту терапевтического антитела (например, scFv, Fab, Fab', F(ab')2) или к сконструированному фрагменту антитела, описанному выше, который специфически связывается с hIL-13 и модулирует (например, ингибирует или блокирует) взаимодействие hIL-13 с hIL-13R. Фрагмент терапевтического антитела обычно содержит CDRН3, имеющий последовательность SEQ ID NO:3, необязательно вместе с CDR, имеющими последовательности, представленные в SEQ ID NO:1, 2, 4, 5 и 6.
1.5. Гетероконъюгированные антитела
Одним из вариантов настоящего изобретения также являются гетероконъюгированные антитела. Гетероконъюгированные антитела состоят из двух ковалентно связанных антител, образованных любыми стандартными методами перекрестного связывания. См., например, патент США 4676980.
1.6. Другие модификации
Взаимодействие между Fc-областью антитела и различными Fc-рецепторами (FcγR), очевидно, опосредует эффекторные функции антитела, включая антитело-зависимую клеточную цитотоксичность (АDCC), фиксацию комплемента, фагоцитоз и изменение времени полужизни/скорости выведения антитела. Различные модификации в Fc-области антител согласно изобретению могут быть осуществлены в зависимости от нужного свойства. Так, например, в Fc-область могут быть введены специфические мутации, превращающие литическое антитело в нелитическое антитело, как подробно описано в ЕР 0629240В1 и ЕР 0307434В2, для увеличения его времени полужизни в сыворотке, либо может быть введен эпитоп, связывающийся с “рецептором спасения” см. патент США 57399277. В настоящее время известно пять человеческих Fcγ-рецепторов, а именно FcγR(I), FcγRIIа, FcγRIIb, FcγRIIIа и неонатальный FcRn. В работе Shields et al. (2001) J. Biol. Chem. 276, 6591-6604 продемонстрировано, что в связывании всех FcγR участвует общая группа остатков IgG1, а в связывании FcγRII и FcγRIII участвуют другие остатки, отличающиеся от остатков этой группы. Одна группа остатков IgG1, при замене остатков Pro-238, Asp-265, Asp-270, Asn-297 и Pro-239 аланином, снижает уровень связывания со всеми FcγR. Все они присутствуют в СН2-домене IgG и образуют кластер возле шарнирной области, соединяющей СН1 и СН2. В связывании с FcγRI участвует только общая группа остатков IgG1, а FcγRII и FcγRIII, помимо указанной общей группы остатков, взаимодействуют и с другими остатками. Модификации в некоторых остатках снижают уровень связывания только с FcγRII (например, Arg-292) или FcγRIII (например, Glu-293). Некоторые варианты обнаруживают повышенный уровень связывания с FcγRII или FcγRIII, но не влияют на связывание с другим рецептором (например, замена Ser-267Ala приводит к повышению уровня связывания с FcγRII, но не влияет на связывание с FcγRIII). Другие варианты обнаруживают повышенный уровень связывания с FcγRII или с FcγRIII, но пониженный уровень связывания с другим рецептором (например, замена Ser-298Ala приводит к повышению уровня связывания с FcγRIII и к снижению уровня связывания с FcγRII). Что касается рецептора FcγRIIIа, то наилучшее связывание вариантов IgG1 происходит при объединении аланиновых замен Ser-298, Glu-333 и Lys-334. Неонатальный рецептор FcRn, очевидно, участвует в клиренсе антител и в трансцитозе через ткани (см. Junghans R.P. (1997) Immunol. Res. 16. 29-57 и Ghetie et al. (2000) Annu. Rev. Immunol. 18, 739-766). Остатками человеческих IgG1, которые, как было определено, непосредственно взаимодействуют с человеческим FcRn, являются Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435. Замены в любом из этих положений, описанных в данном разделе, могут увеличивать время полужизни в сыворотке и/или изменять эффекторные свойства антител согласно изобретению.
Другими модификациями являются варианты гликозилирования антител согласно изобретению. Известно, что гликозилирование антител в консервативных положениях в их константных областях оказывает значительное влияние на функцию антитела, в частности на эффекторную функцию, такую как функция, описанная выше, см. например, Boyd et al., (1996) Mol. Immunol. 32, 1311-1318. Рассматриваются также варианты гликозилирования терапевтических антител или их антигенсвязывающих фрагментов согласно изобретению, где были добавлены, заменены, делетированы или модифицированы одна или несколько углеводных частей. Введение мотива “аспарагин-Х-серин” или “аспарагин-Х-треонин” приводит к образованию потенциального сайта ферментативного присоединения углеводных частей, а поэтому такое введение может применяться для изменения характера гликозилирования антитела. В работе Raju et al. (2001) Biochemistry 40, 8868-8876 продемонстрировано, что уровень концевого сиалилирования иммуноадгезина TNFR-IgG повышается в процессе регалактозилирования и/или реасиалилирования под действием бета-1,4-галактозилтрансферазы и/или альфа-2,3-сиалилтрансферазы. Повышение уровня концевого сиалилирования, вероятно, приводит к увеличению времени полужизни иммуноглобулина. Антитела, наряду с большинством гликопротеинов, обычно продуцируются в виде смеси гликоформ. Эта смесь является особенно явной, если антитела продуцируются в эукариотических клетках, в частности в клетках млекопитающих. Для получения определенных гликоформ был разработан ряд методов, см. Zhang et al. Science (2004), 303, 371, Sears et al., Science (2001) 291, 2344, Wacker et al. (2002) Science 298, 1790, Davis et al., (2002) Chem. Rev. 102, 579, Hang et al. (2001) Acc. Chem. Res. 34, 727. Таким образом, в настоящем изобретении рассматривается множество терапевтических (моноклональных) антител (которые могут принадлежать к изотипу IgG, например, IgG1), описанных в настоящей заявке и содержащих определенное число (например, 7 или менее, например 5 или менее, а именно две или одну) гликоформ указанных антител или их антигенсвязывающих фрагментов.
Другие варианты настоящего изобретения включают терапевтические антитела согласно изобретению или их антигенсвязывающие фрагменты, связанные с не-белковым полимером, таким как полиэтиленгликоль (ПЭГ), полипропиленгликоль или полиоксиалкилен. Конъюгирование белков с ПЭГ представляет собой известный метод увеличения времени полужизни белков, а также снижения антигенности и иммуногенности белков. Применение ПЭГилирования с использованием молекул с различными молекулярными массами и структурами (линейными или разветвленными) было исследовано на интактных антителах, а также на Fab'-фрагментах, см. Koumenis I.L. et al. (2000) Int. J. Pharmaceut. 198:83-95.
2. Конкурентные антитела
Настоящее изобретение также относится к антителам и к их антигенсвязывающим фрагментам, которые специфически связываются с hIL-13 и конкурентно ингибируют связывание c IL-13 терапевтического антитела согласно изобретению или его антигенсвязывающего фрагмента, содержащего CDRН3 SEQ ID NO:3, и/или терапевтического антитела согласно изобретению или его антигенсвязывающего фрагмента, содержащего CDR SEQ ID NO:1, 2, 3, 4, 5 и 6, с hIL-13. В некоторых вариантах изобретения указанным терапевтическим антителом является мышиное антитело, содержащее VH-домен SEQ ID NO:7 и VL-домен SEQ ID NO:8. Такие конкурентные антитела связываются с тем же эпитопом, или с перекрывающимся эпитопом hIL-13, или с пространственно смежным эпитопом hIL-13, который связывается с терапевтическим антителом, содержащим CDR SEQ ID NO:1, 2, 3, 4, 5 и 6. Конкурентное антитело или фрагмент такого антитела, в эквимолярных концентрациях, обеспечивает, по меньшей мере, 25%-ное, обычно 35%-ное или более, а в основном, по меньшей мере, 50%-ное ингибирование.
Таким образом, в одном из своих вариантов настоящее изобретение относится к способу скрининга антитела-кандидата или его фрагмента, проводимому для того, чтобы определить, является ли антитело-кандидат или его фрагмент конкурентным антителом, описанным в настоящей заявке, где указанный способ включает стадии: (а) инкубирования антитела-кандидата или его фрагмента с терапевтическим антителом, содержащим CDRН3 SEQ ID NO:3, а также необязательно содержащим CDR SEQ ID NO:1, 2, 4, 5 и 6 (таким как мышиное терапевтическое антитело, имеющее VH-домен SEQ ID NO:7 и VL-домен SEQ ID NO:8, или гуманизованное терапевтическое антитело, имеющее тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22, или гуманизованное терапевтическое антитело, имеющее тяжелую цепь SEQ ID NO:19 и легкую цепь SEQ ID NO:23), или с его антигенсвязывающим фрагментом; (b) определения того факта, может ли указанное антитело-кандидат или его фрагмент стадии (а) конкурентно ингибировать связывание указанного терапевтического антитела или его антигенсвязывающего фрагмента с hIL-13.
Настоящее изобретение также относится к конкурентному антителу или к его антигенсвязывающему фрагменту, который конкурентно ингибирует связывание терапевтического антитела или его антигенсвязывающего фрагмента, содержащего область CDR, имеющую последовательности, представленные в SEQ ID NO: 1, 2, 3, 4, 5 и 6.
В другом своем варианте настоящее изобретение относится к конкурентному антителу или к его антигенсвязывающему фрагменту, который конкурентно ингибирует связывание терапевтического антитела согласно изобретению с hIL-13, где указанное терапевтическое антитело содержит тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22.
Конкурентным антителом или его антигенсвязывающим фрагментом может быть любое антитело, имеющее вышеуказанные структуры. Так, например, конкурентным антителом может быть интактное антитело приматов или человека или гуманизованное антитело, предпочтительно имеющее изотип IgG, например, IgG1 или IgG4. Фрагментами конкурентного антитела могут быть Fab, Fab', F(ab')2, scFv и т.п. Конкурентное антитело может быть получено методами, описанными в настоящей заявке.
3. Методы продуцирования антител
Антитела согласно изобретению могут быть продуцированы в виде поликлональной популяции, но более предпочтительно в виде моноклональной популяции (то есть в виде в основном гомогенной популяции идентичных антител, направленных против специфического антигенсвязывающего сайта). Для специалиста в данной области очевидно, что такая популяция включает более чем одну молекулу антитела. Антитела согласно изобретению могут быть продуцированы в трансгенных организмах, таких как козы (см. Pollock et al. (1999) J. Immunol. Methods 231:147-157), куры (см. Morrow K.J.J. (2000) Genet. Eng. News 20:1-55), мыши (см. Pollock et al.) или растения (см. Doran P.M. (2000) Curr. Opinion Biotechnol. 11, 199-204, Ma JK-C (1998), Nat. Med. 4; 601-606, Baez J. et al., BioPharm (2000) 13:50-54, Stoger E. et al. (2000) Plant. Mol. Biol. 42:583-590). Антитела могут быть также продуцированы методом химического синтеза. Однако антитела согласно изобретению обычно получают с применением рекомбинантной технологии культивирования клеток, хорошо известной специалистам. Полинуклеотид, кодирующий антитело, выделяют и встраивают в реплицируемый вектор, такой как плазмида, для последующего клонирования (амплификации) или экспрессии. Одной из обычно используемых систем экспрессии является система глутамат-синтетазы (такая как система, поставляемая фирмой Lonza Biologics), где клеткой-хозяином, в частности, является клетка СНО или NSO (см. ниже). Полинуклеотид, кодирующий антитело, может быть легко выделен и секвенирован в соответствии со стандартными процедурами (например, с использованием олигонуклеотидных зондов). Используемыми векторами могут быть плазмида, вирус, фаг, транспозоны, минихромосомы, из которых типичным вариантом являются плазмиды. Обычно, такие векторы дополнительно включают сигнальную последовательность, ориджин репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательности терминации транскрипции, функционально присоединенные к полинуклеотиду легкой цепи и/или тяжелой цепи для облегчения экспрессии. Полинуклеотид, кодирующий легкие и тяжелые цепи, может быть встроен в отдельные векторы и трансфицирован в одну и ту же клетку-хозяина, либо, если это необходимо, тяжелая и легкая цепи могут быть встроены в один и тот же вектор для трансфекции в клетку-хозяина. Таким образом, в соответствии с одним из своих аспектов настоящее изобретение относится к способу конструирования вектора, кодирующего легкую и/или тяжелую цепи терапевтического антитела и/или его антигенсвязывающего фрагмента согласно изобретению, где указанный способ включает встраивание в вектор полинуклеотида, кодирующего легкую и/или тяжелую цепь терапевтического антитела согласно изобретению.
В другом своем аспекте настоящее изобретение относится к полинуклеотиду, кодирующему мышиный VH-домен, имеющий последовательность, представленную в SEQ ID NO:24.
В другом своем аспекте настоящее изобретение относится к полинуклеотиду, кодирующему мышиный VL-домен, имеющий последовательность, представленную в SEQ ID NO:25.
В другом своем варианте настоящее изобретение относится к полинуклеотиду, кодирующему VH-домен, имеющий последовательность, выбранную из группы, состоящей из SEQ ID NO:26, 27, 28, 29.
В другом своем варианте настоящее изобретение относится к полинуклеотиду, кодирующему VL-домен, имеющий последовательность, выбранную из группы, состоящей из SEQ ID NO:30, 31.
В соответствии с этим настоящее изобретение относится к полинуклеотиду, кодирующему тяжелую цепь согласно изобретению, где указанный полинуклеотид выбран из группы, состоящей из SEQ ID NO:32, 33, 34, 35.
В соответствии с этим настоящее изобретение относится к полинуклеотиду, кодирующему легкую цепь согласно изобретению, где указанный полинуклеотид выбран из группы, состоящей из SEQ ID NO:36, 37.
Как совершенно очевидно для специалиста в данной области, вследствие вырожденности генетического кода могут быть также получены полинуклеотиды, которые являются альтернативой описанным здесь полинуклеотидам, кодирующим полипептиды согласно изобретению.
3.1. Сигнальные последовательности
Антитела согласно изобретению могут быть продуцированы в виде гибридного белка с гетерологичной сигнальной последовательностью, имеющей специфический сайт отщепления у N-конца зрелого белка. Сигнальная последовательность должна распознаваться и процессироваться клеткой-хозяином. Для прокариотических клеток-хозяев сигнальными последовательностями могут быть лидерные последовательности щелочной фосфатазы, пенициллиназы или термостабильного энтеротоксина II. Для секреции в дрожжах сигнальными последовательностями могут быть лидерная последовательность инвертазы дрожжей, лидерная последовательность α-фактора или лидерные последовательности кислой фосфатазы, см., например, WO 90/13646. В клеточных системах млекопитающих могут присутствовать вирусные секреторные лидерные последовательности, такие как сигнал gD вируса простого герпеса и сигнальная последовательность нативного иммуноглобулина. Обычно, сигнальную последовательность лигируют в той же самой рамке считывания с ДНК, кодирующей антитело согласно изобретению.
3.2. Ориджин репликации
Ориджины репликации хорошо известны специалистам, при этом для большинства грамотрицательных бактерий, подходящим ориджином репликации является рВR322; для большинства дрожжей подходящим ориджином репликации является плазмида 2µ, а для большинства клеток млекопитающих подходящими являются различные ориджины репликации вирусов, таких как SV40, полиомавирус, аденовирус, VSV или BPV. Обычно, для экспрессирующих векторов млекопитающих ориджин репликации не требуется, однако в этих векторах может быть использован SV40, поскольку он содержит ранний промотор.
3.3. Селективный маркер
Типичные селективные гены кодируют белки, которые (а) сообщают резистентность к антибиотикам или к другим токсинам, например к ампициллину, неомицину, метотрексату или тетрациклину, или (b) ауксотрофную недостаточность комплемента или поступление питательных веществ, отсутствующих в комплексных средах. Схема отбора может включать прекращение роста клетки-хозяина. Клетки, которые были успешно трансформированы генами, кодирующими терапевтическое антитело согласно изобретению, выживают, например, благодаря своей резистентности к лекарственному средству, сообщаемой селективным маркером. Другим примером является так называемый маркер DНFR-отбора, при котором трансформанты культивируют в присутствии метотрексата. В типичных вариантах, клетки культивируют в присутствии возрастающих количеств метотрексата для амплификации числа копий представляющего интерес экзогенного гена. Для DНFR-отбора особенно подходящими клетками-мишенями являются клетки СНО. Другим примером является экспрессионная система глутамат-синтетазы (Lonza Biologics). Для использования в дрожжах подходящим селективным геном является ген trp1, см., Stinchcomb et al., Nature 282, 38, 1979.
3.4.Промоторы
Промоторы, подходящие для экспрессии антител согласно изобретению, функционально присоединяют к ДНК/полинуклеотиду, кодирующему данное антитело. Подходящими промоторами для прокариотических хозяев являются промотор phoА, промоторные системы бета-лактамазы и лактозы, промоторы щелочной фосфатазы, триптофановый промотор и гибридные промоторы, такие как Тас. Промоторами, подходящими для экспрессии в дрожжевых клетках, являются промоторы 3-фосфоглицерат-киназы или других гликолитических ферментов, например энолазы, глицеральдегид-3-фосфат-дегидрогеназы, гексокиназы, пируват-декарбоксилазы, фосфофруктокиназы, глюкозо-6-фосфат-изомеразы, 3-фосфоглицерат-мутазы и глюкокиназы. Индуцибельными дрожжевыми промоторами являются промоторы алкоголь-дегидрогеназы 2, изоцитохрома С, кислой фосфатазы, металлотионеина и ферментов, ответственных за метаболизм азота или утилизацию мальтозы/галактозы.
Промоторами для экспрессии в клеточных системах млекопитающих являются вирусные промоторы, такие как промоторы полиомавируса, вируса оспы домашних птиц и аденовируса (например, аденовируса 2), вируса коровьей папилломы, вируса саркомы птиц, цитомегаловируса (в частности, промотор предраннего гена), ретровируса, вируса гепатита В, актина, вируса саркомы Рауса (RSV) и раннего или позднего обезьяньего вируса 40. Очевидно, что выбор промотора зависит от совместимости с клеткой-хозяином, используемой для экспрессии. Поэтому в одном из своих вариантов настоящее изобретение относится к первой плазмиде, содержащей промотор вируса RSV и/или SV40 и/или CMV, ДНК, кодирующую V-область легкой цепи (VL) согласно изобретению, область κС вместе с маркерами для отбора на резистентность к неомицину и ампициллину, и ко второй плазмиде, содержащей промотор вируса RSV или SV40, ДНК, кодирующую V-область тяжелой цепи (VН) согласно изобретению, ДНК, кодирующую константную область γ1, DHFR-маркер и маркер резистентности к ампициллину.
3.5. Энхансерный элемент
Если это необходимо, например, для экспрессии в высших эукариотах, то может быть использован энхансерный элемент, функционально присоединенный к промоторному элементу в векторе. Подходящими энхансерными последовательностями млекопитающих являются энхансерные элементы, происходящие от генов глобина, эластазы, альбумина, фетопротеина и инсулина. Альтернативно, может быть использован энхансерный элемент, происходящий от вируса эукариотических клеток, такой как энхансер SV40 (п.н. 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомавируса, энхансер бакуловируса или энхансер локуса мышиного IgG2а (см. WO 04/009823). Энхансер предпочтительно локализован на векторе в сайте, расположенном выше промотора.
3.6. Клетки-хозяева
Для клонирования или экспрессии векторов, кодирующих антитела согласно изобретению, подходящими клетками-хозяевами являются прокариотические, дрожжевые или высшие эукариотические клетки. Подходящими прокариотическими клетками являются эубактерии, например энтеробактерии, такие как Esсherichia, например E.coli (например, АТСС 31446; 31537; 27325), Enterobacter, Erwinia, Klebsiella proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans и Shigella, а также бациллы, такие как B. subtilis и Bacillus licheniformis (см. DD 266710), Pseudomonas, такие как Р.aeruginosa и Streptomyces. Из дрожжевых клеток-хозяев также рассматриваются Saccharomyces cerevisiae, schizosaccharomyces pombe, Kluyveromyces (например, АТСС 16045; 12424; 24178; 56500), yarrowia (EP 402226), Pichia pastoris (ЕР 183070, см. также Peng et al., J. Biotechnol. 108 (2004) 185-192), Candida, Trichoderma reesia (EP244, 234), Penicillin, Tolypocladium и Aspergillus, такие как A. nidulans и A. niger.
Хотя в настоящем изобретении конкретно рассматриваются прокариотические и дрожжевые клетки-хозяева, однако предпочтительными клетками-хозяевами согласно изобретению являются высшие эукариотические клетки. Подходящими высшими эукариотическими клетками-хозяевами являются клетки млекопитающих, такие как СОS-1 (АТСС № СRL 1650), СОS-7 (АТСС № СRL 1651), клеточная линия почек человеческого эмбриона 293, клетки почек детеныша хомячка (ВНК)(АТСС № СRL 1632), ВНК570 (АТСС № СRL 10314), 293 (АТСС № СRL 1573), клетки яичника китайского хомячка СНО (например, СНО-К1, АТСС № ССL 61, клеточная линия DHFR-СНО, такая как DG44 (см. Urlaub et al., (1986) Somatic Cell Mol. Genet. 12, 555-556)), в частности клеточные линии СНО, адаптированные для выращивания в суспензионной культуре, мышиные клетки Сертоли, клетки почек обезьяны, клетки почек африканской зеленой мартышки (АТСС № СRL 1587), клетки НЕLА, клетки почек собак (АТСС ССL 34), клетки почек человека (АТСС ССL 75), клетки Нер G2 и клетки миеломы или лимфомы, например NS0 (см. патент США № 5807715), Sр2/0 и Y0.
Таким образом, в одном из своих вариантов настоящее изобретение относится к стабильно трансформированной клеточной линии, содержащей вектор, кодирующий тяжелую и/или легкую цепь терапевтического антитела или его антигенсвязывающего фрагмента, описанных в настоящей заявке. Предпочтительно такие клетки-хозяева содержат первый вектор, кодирующий легкую цепь, и второй вектор, кодирующий указанную тяжелую цепь.
Бактериальная ферментация
Для экспрессии фрагментов антител особенно подходящими являются бактериальные системы. Такие фрагменты локализуются внутри клеток или в периплазме. Нерастворимые периплазматические белки могут быть экстрагированы и подвергнуты рефолдингу с образованием активных белков методами, известными специалистам, см. Sanchez et al. (1999) J. Biotechnol. 72, 13-20 и Cupit P.M. et al. (1999) Lett. Appl. Microbiol. 29, 273-277.
3.7. Методы культивирования клеток
Клетки-хозяева, трансформированные векторами, кодирующими терапевтические антитела согласно изобретению или их антигенсвязывающие фрагменты, могут быть культивированы любым методом, известным специалистам. Клетки-хозяева могут быть культивированы в центрифужных колбах, в роллер-флаконах или в системах из полого волокна, но для крупномасштабного продуцирования предпочтительно использовать чаны-реакторы для смешения, особенно для суспензионных культур. При этом предпочтительно, чтобы чаны для смешения были адаптированы для аэрации с использованием, например, барботеров-распылителей, перегородок или лопастных мешалок с малым сдвиговым усилием. В колонках-барботерах и в аэролифтных реакторах можно проводить аэрацию непосредственно путем барботирования воздуха или кислорода. Если клетки-хозяева культивируются в бессывороточной культуральной среде, то предпочтительно, чтобы в эту среду был добавлен агент для защиты клеток, такой как плюроник F-68, в целях предупреждения повреждения клеток в процессе аэрации. В зависимости от свойств клеток-хозяев могут быть использованы микроносители в качестве субстратов для роста субстрат-зависимых клеточных линий, либо эти клетки могут быть адаптированы для суспензионной культуры (которая обычно применяется для этих целей). Для культивирования клеток-хозяев, в частности клеток-хозяев безпозвоночных, могут быть применены различные операционные методы, такие как культивирование с подпиткой, периодическое культивирование (см., Drapeau et al., (1994) Cytotechnology 15:103-109), непрерывное культивирование или перфузионное культивирование. Хотя рекомбинантно трансформированные клетки-хозяева млекопитающих могут быть культивированы в среде, содержащей сыворотку, такую как фетальная телячья сыворотка (FCS), однако предпочтительно, чтобы такие клетки-хозяева были культивированы в синтетической бессывороточной среде, такой как среда, описанная Keen et al. (1995) Cytotechnology 17:153-163, или в коммерчески доступной среде, такой как ProCНО-СDM или UltraCHOТМ (Cambrex, NJ, USA), в которую, при необходимости, добавляют источник энергии, такой как глюкоза, и синтетические факторы роста, такие как рекомбинантный инсулин. Для культивирования клеток-хозяев в бессывороточной среде необходимо, чтобы эти клетки были адаптированы для роста в бессывороточных условиях. Одним из способов адаптации является культивирование таких клеток-хозяев в среде, содержащей сыворотку, с последующей периодической заменой 80% этой культуральной среды бессывороточной средой, так, чтобы клетки-хозяева адаптировались к условиям культивирования в бессывороточной среде (см. например, Scharfenberg K. et al. (1995) Animal Cell Technology: Developments towards the 21st century (Beuvery E.C. et al., eds.), pp.619-623, Kluwer Academic publishers).
Антитела согласно изобретению, секретированные в указанную среду, могут быть выделены и очищены различными методами, позволяющими проводить очистку до нужной степени. Так, например, для лечения человека в основном используются терапевтические антитела согласно изобретению, имеющие, по меньшей мере, 95% чистоту, а обычно 98% или 99% чистоту или более (по сравнению с неочищенной культуральной средой). В первом случае клеточный дебрис обычно удаляют из культуральной среды путем центрифугирования с последующим проведением стадии очистки супернатанта, например путем микрофильтрации, ультрафильтрации и/или глубокой фильтрации. Существуют также различные другие методы очистки, такие как диализ и электрофорез в геле, а также хроматографические методы, такие как хроматография на гидроксиаппатитах (ГА), аффинная хроматография (необязательно, включающая систему аффинного мечения, такую как полигистидин) и/или гидрофобная хроматография (ГФХ, см. патент США № 5429746). В одном из вариантов настоящего изобретения антитела согласно изобретению после проведения различных стадий очистки иммобилизуют с помощью аффинной хроматографии на белке А или G-белке, а затем проводят дополнительные хроматографические стадии, такие как ионообменная хроматография и/или хроматография на гидроксиаппатитах (ГА), анионо- или катионообменная хроматография, эксклюзионная хроматография и преципитация сульфатом аммония. Кроме того, обычно проводят различные стадии элиминации вирусов (например, посредством нанофильтрации с использованием, например, фильтра DV-20). После проведения указанных различных стадий получают очищенный (предпочтительно моноклональный) препарат, содержащий по меньшей мере, 75 мг/мл или более, например 100 мг/мл или более, указанного антитела согласно изобретению или его антигенсвязывающего фрагмента, и такой препарат является одним из вариантов настоящего изобретения. Предпочтительно, чтобы такие препараты, по существу, не содержали агрегированных форм антител согласно изобретению.
4. Фармацевтические композиции
Очищенные препараты антител согласно изобретению (в частности, препараты моноклональных антител), описанные выше, могут быть введены в фармацевтические композиции, предназначенные для лечения заболеваний и расстройств у человека, таких как атопические заболевания, например астма, аллергический ринит, ХОБЛ. Обычно такие композиции содержат известный фармацевтически приемлемый носитель, применяемый в фармацевтической практике, см., например, Remington's Pharmaceutical Sciences, 16th edition (1980), Mack Publishing. Co. Примерами таких носителей являются стерилизованные носители, такие как физиологический раствор, раствор Рингера или раствор декстрозы, забуференные подходящими буферами до рН 5-8. Фармацевтические композиции для инъекций (например, внутривенных, внутрибрюшинных, внутрикожных, подкожных, внутримышечных или внутрипортальных инъекций) или для непрерывной инфузии обычно не содержат видимых крупных частиц и могут содержать от 0,1 нг до 100 мг антитела, предпочтительно 5 мг - 25 мг антитела. Методы получения таких фармацевтических композиций хорошо известны специалистам. В одном из вариантов изобретения фармацевтические композиции содержат от 0,1 нг до 100 мг терапевтических антител согласно изобретению в унифицированной лекарственной форме, необязательно вместе с прилагаемыми инструкциями по их применению. Фармацевтические композиции согласно изобретению могут быть лиофилизованы (осушены вымораживанием), а затем перед введением разведены методами, хорошо известными или очевидными для специалиста в данной области. Если варианты настоящего изобретения включают антитела изотипа IgG1, то в фармацевтическую композицию может быть добавлен агент, образующий хелатный комплекс с медью, а именно такой агент, как цитрат (например, цитрат натрия) или EDTA или гистидин для уменьшения степени опосредуемой медью деградации антител этого изотипа, см. ЕР 0612251. Введение антитела против hIL-13 может быть осуществлено перорально путем ингаляции и местно (например, путем внутриглазного введения, интраназального введения, ректального введения и введения в раны на коже).
Эффективные дозы и схемы введения для лечения антителом согласно изобретению обычно определяют эмпирически, и эти дозы и схемы зависят от таких факторов, как возраст, масса и состояние здоровья пациента, а также от заболевания или расстройства, подвергаемого лечению. Такие факторы должны учитываться лечащим врачом. Руководство по определению подходящих доз можно найти в публикации Smith et al. (1977) Antibodies in human diagnosis and therapy, Raven Press, New York, и обычно такие дозы составляют от 1 мг до 1000 мг.
В зависимости от заболевания или расстройства, подвергаемого лечению (в частности, астмы), фармацевтические композиции, содержащие терапевтически эффективное количество антитела согласно изобретению, могут быть введены одновременно, отдельно или последовательно в сочетании с эффективным количеством другого лекарственного средства, такого как противовоспалительные средства (например, кортикостероид или НСПВС), антихолинергические средства (в частности, антагонисты рецептора М1/М2/М3), агонисты β2-адренорецептора, противоинфекционные средства (например, антибиотики, противовирусные средства), антигистамины и ингибитор PDE4. Примерами агонистов β2-адренорецептора являются салметерол, салбутамол, формотерол, салмефамол, фенотерол, тербуталин. Предпочтительными агонистами β2-адренорецептора пролонгированного действия являются агонисты, описанные в WO 02/66422A, WO 02/270490, WO 02/076933, WO 03/024439 и WO 03/072539. Подходящими кортикостероидами являются метилпреднизолон, преднизолон, дексаметазон, пропионат флутиказона, S-фторметиловый эфир 6α,9α-дифтор-17α-[(2-фуранилкарбонил)окси]-11β-гидрокси-16α-метил-3-оксо-андроста-1,4-диен-17β-карботионовой кислоты, S-2-оксо-тетрагидрофуран-3S-иловый эфир 6α,9α-дифтор-11β-гидрокси-16α-метил-3-оксо-17α-пропионилокси-андроста-1,4-диен-17β-карботионовой кислоты, сложные эфиры беклометазона (например, эфир 17-пропионовой кислоты или эфир 17,21-дипропионовой кислоты), будезонид, флунизолид, сложные эфиры мометазона (например, сложный эфир фуроата), ацетонид триамцинолона, рофлепонид, циклезонид (16α, 17-[[(R)-циклогексилметилен]бис(окси)]-11β,21-дигидроксипрегна-1,4-диен-3,20-дион), пропионат бутиксокорта, RPR-106541 и ST-126. Предпочтительными кортикостероидами являются пропионат флутиказона, S-фторметиловый эфир 6α,9α-дифтор-11β-гидрокси-16α-метил-17α-[(4-метил-1,3-тиазол-5-карбонил)окси]-3-оксо-андроста-1,4-диен-17β-карботионовой кислоты и S-фторметиловый эфир 6α,9α-дифтор-17α-[(2-фуранилкарбонил)окси]-11β-гидрокси-16α-метил-3-оксо-андроста-1,4-диен-17β-карботионовой кислоты, а более предпочтительно S-фторметиловый эфир 6α,9α-дифтор-17α-[(2-фуранилкарбонил)окси]-11β-гидрокси-16α-метил-3-оксо-андроста-1,4-диен-17β-карботионовой кислоты.
Нестероидные соединения, которые являются агонистами глюкокортикоидов и обладают селективностью по отношению к трансрепрессии посредством трансактивации и которые могут быть использованы в комбинированной терапии, были описаны в следующих патентах: WO 03/082827, WO 01/10143, WO 98/54159, WO 04/005229, WO 04/009016, WO 04/009017, WO 04/018429, WO 03/104195, WO 03/082787, WO 03/082280, WO 03/059899, WO 03/101932, WO 02/02565, WO 01/16128, WO 00/66590, WO 03/086294, WO 04/026248, WO 03/061651, WO 03/08277.
Подходящими противовоспалительными средствами являются нестероидные противовоспалительные средства (НСПВС).
Подходящими НСПВС являются хромогликат натрия, недокромил-натрий, ингибиторы фосфодиэстеразы (PDE)(например, теофиллин, ингибиторы PDE4 или смешанные ингибиторы PDE3/PDE4), антагонисты лейкотриена, ингибиторы синтеза лейкотриена (например, монтелукаст), ингибиторы iNOS, ингибиторы триптазы и эластазы, антагонисты интегрина бета-2 и агонисты или антагонисты аденозинового рецептора (например, агонисты аденозина 2а), антагонисты цитокина (например, антагонисты хемокина, такие как антагонист ССR3) или ингибиторы синтеза цитокинов или ингибиторы 5-липоксигеназы. Другими подходящими агонистами β2-адренорецепторов являются салметерол (например, в виде ксинафоата), салбутамол (например, в виде сульфата или свободного основания), формотерол (например, в виде фумарата), фенотерол или тербуталин и их соли. Для перорального введения предпочтительным является iNOS (индуцибельный ингибитор (окись азота)-синтазы). Подходящими ингибиторами iNOS являются ингибиторы, описанные в WO 93/13055, WO 98/30537, WO 02/50021, WO 95/34534 и WO 99/62875. Подходящими ингибиторами ССR3 являются ингибиторы, описанные в WO 02/26722.
Особый интерес представляет использование антител согласно изобретению в комбинации с ингибитором фосфодиэстеразы 4 (PDE4). PDE-4-специфическим ингибитором, используемым в данном аспекте настоящего изобретения, могут быть любые соединения, которые, как известно, ингибируют фермент PDE4 или, как было установлено, действуют как ингибитор PDE4 и которые представляют собой только ингибиторы PDE4, но не соединения, ингибирующие другие члены семейства PDE, такие как PDE3 и PDE5, а также PDE4.
Представляющими интерес соединениями являются цис-4-циано-4-(3-циклопентилокси-4-метоксифенил)циклогексан-1-карбоновая кислота, 2-карбометокси-4-циано-4-(3-циклопропилметокси-4-дифторметоксифенил)циклогексан-1-он и цис-[4-циано-4-(3-циклопропилметокси-4-дифторметоксифенил)циклогексан-1-ол]. Кроме того, цис-4-циано-4-[3-(циклопентилокси)-4-метоксифенил]циклогексан-1-карбоновая кислота (также известная как циломиласт) и ее соли, сложные эфиры, пролекарства и физические формы описаны в патенте США 5552438, выданном 3 сентября 1996; этот патент и описанные в нем соединения во всей своей полноте вводятся в настоящее описание посредством ссылки.
AWD-12-281 поставляется фирмой Elbion (Hofgen, N. et al. 15th EFMC Int. Symp. Med. Chem (Sept. 6-10, Edinburgh) 1998, Abst.P.98; реферат CAS № 247584020-9); производное 9-бензиладенина обозначено NCS-613 (INSERM); D-4418 поставляется фирмами Chiroscience и Schering-Plough; ингибитор бензодиазепина PDE4 обозначается CI-1018 (PD-168787) и поставляется фирмой Pfizer; производное бензодиоксола описано Kyowa Hakko в WO 99/16766; К-34 описано у Kyowa Hakko; V-11294А поставляется фирмой Napp (Landells, L.J. et al., Eur. Resp. J.[Annu. Cong. Eur. Resp. Soc (Sept 19-23, Geneva), 1998], 12 (Suppl. 28): Abst. P2393); рофлумиласт (реферат CAS № 162401-32-3) и фталазинон (WO 99/47505, описание которой вводится в настоящее изобретение посредством ссылки) поставляются фирмой Byk-Gulden; пумафентрин, (-)-п-[(4aR*,10bS*)-9-этокси-1,2,3,4,4а,10b-гексагидро-8-метокси-2-метилбензо[c][1,6]нафтиридин-6-ил]-N,N-диизопропилбензамид, который представляет собой смешанный ингибитор PDE3/PDE4, был получен, как описано в руководстве Byk-Gulden, now Altanta; арофиллин находится на стадии разработки у Almirall-Prodesfarma; VM554/UM565 поставляются фирмой Vernalis; Т-440 описан Tanabe Seiyaku; Fuji K. et al., J. Pharmacol. Exp. Ther, 1998, 284(1):162, а также описан Т2585.
Другие представляющие интерес соединения описаны в опубликованной Международной патентной заявке WO 04/024728 (Glaxo Group Ltd), PCT/EP2003/014867 (Glaxo Group Ltd) и РСТ/ЕР2004/005494 (Glaxo Group Ltd).
Подходящими антихолинергическими средствами являются соединения, которые действуют как антагонисты мускариновых рецепторов, в частности соединения, которые являются антагонистами рецепторов М1 или М3, двойными антагонистами рецепторов М1/М3 или М2/М3 или пан-антагонистами рецепторов М1/М2/М3. Репрезентативными соединениями для введения путем ингаляции являются ипратропий (например, в виде бромида, CAS 22254-24-6, имеющегося в продаже под торговым знаком Atrovent), окситропий (например, в виде бромида, CAS 30286-75-0) и тиотропий (например, в виде бромида, CAS 136310-93-5, имеющегося в продаже под торговым знаком Spiriva). Представляющими интерес соединениями также являются реватропат (например, в виде гидробромида, CAS 262586-79-8) и LAS-34273, описанные в WO 01/04118. Репрезентативными соединениями для перорального введения являются пирензепин (CAS 28797-61-7), дарифенацин (CAS 133099-04-4 или CAS 133099-07-7 в виде гидробромида, имеющегося в продаже под торговым знаком Enablex), оксибутинин (CAS 5633-20-5, имеющийся в продаже под торговым знаком Ditropan), теродилин (CAS 15793-40-5), толтеродин (CAS 124937-51-5 или CAS 124937-52-6 в виде тартрата, имеющегося в продаже под торговым знаком Detrol), отилоний (например, в виде бромида, CAS 26095-59-0, имеющегося в продаже под торговым знаком Spasmomen), хлорид троспия (CAS 10405-02-4) и солифенацин (CAS 242478-37-1 или CAS 242478-38-2 в виде сукцината, также известного как YM-905 и имеющегося в продаже под торговым знаком Vesicare).
Другими подходящими антихолинергическими средствами являются соединения формулы (ХХI), описанные в заявке на патент США 60/487981:
где предпочтительной ориентацией алкильной цепи, присоединенной к тропановому кольцу, является эндо-ориентация;
R31 и R32 независимо выбраны из группы, состоящей из прямых или разветвленных низших алкильных групп, имеющих предпочтительно от 1 до 6 атомов углерода, циклоалкильных групп, имеющих от 5 до 6 атомов углерода, циклоалкилалкила, имеющего от 6 до 10 атомов углерода, 2-тиенила, 2-пиридила, фенила, фенила, замещенного алкильной группой, имеющей не более 4 атомов углерода, и фенила, замещенного алкоксигруппой, имеющей не более 4 атомов углерода;
Х- представляет собой анион, ассоциированный с положительно заряженным атомом N;
Х- может представлять собой, но не ограничивается ими, хлорид, бромид, иодид, сульфат, бензолсульфонат и толуолсульфонат, включая, например:
(3-эндо)-3-(2,2-ди-2-тиенилэтенил)-8,8-диметил-8-азониабицикло[3.2.1]октанбромид;
(3-эндо)-3-(2,2-дифенилэтенил)-8,8-диметил-8-азониабицикло[3.2.1]октанбромид;
(3-эндо)-3-(2,2-дифенилэтенил)-8,8-диметил-8-азониабицикло[3.2.1]октан-4-метилбензолсульфонат;
(3-эндо)-8,8-диметил-3-[2-фенил-2-(2-тиенил)этенил]-8-азониабицикло[3.2.1]октанбромид; и/или
(3-эндо)-8,8-диметил-3-[2-фенил-2-(2-пиридинил)этенил]-8-азониабицикло[3.2.1]октанбромид.
Другими подходящими антихолинергическими средствами являются соединения формул (ХХII) или (XXIII), описанные в заявке на патент США 60/511009:
где
атом Н находится в экзо-положении;
R41- представляет собой анион, ассоциированный с положительно заряженным атомом N. R41- может представлять собой, но не ограничивается ими, хлорид, бромид, иодид, сульфат, бензолсульфонат и толуолсульфонат;
R42 и R43 независимо выбраны из группы, состоящей из прямых или разветвленных низших алкильных групп (имеющих предпочтительно от 1 до 6 атомов углерода), циклоалкильных групп (имеющих от 5 до 6 атомов углерода), циклоалкилалкила (имеющего от 6 до 10 атомов углерода), гетероциклоалкила (имеющего от 5 до 6 атомов углерода) и N или О в качестве гетероатома, гетероциклоалкилалкила (имеющего от 6 до 10 атомов углерода) и N или О в качестве гетероатома, арила, необязательно замещенного арила, гетероарила и необязательно замещенного гетероарила;
R44 выбран из группы, состоящей из (С1-С6)алкила, (С3-С12)циклоалкила, (С3-С7)гетероциклоалкила, (С1-С6)алкил(С3-С12)циклоалкила, (С1-С6)алкил(С3-С7)гетероциклоалкила, арила, гетероарила, (С1-С6)алкиларила, (С1-С6)алкилгетероарила, -ОR45, -СН2ОR45, -СН2ОН, -СN, -СF3, -СН2О(СО)R46, -СО2R47, -СН2NH2, -СН2N(R47)SО2R45, -SО2N(R47)(R48), -СОN(R47)R48, -СН2N(R48)СО(R46), -СН2N(R48)SО2(R46), -СН2N(R48)СО2(R45), -СН2N(R48)СОNН(R47);
R45 выбран из группы, состоящей из (С1-С6)алкила, (С1-С6)алкил(С3-С12)циклоалкила, (С1-С6)алкил(С3-С7)гетероциклоалкила, (С1-С6)алкиларила, (С1-С6)алкилгетероарила;
R46 выбран из группы, состоящей из (С1-С6)алкила, (С3-С12)циклоалкила, (С3-С7)гетероциклоалкила, (С1-С6)алкил(С3-С12)циклоалкила, (С1-С6)алкила(С3-С7)гетероциклоалкила, арила, гетероарила, (С1-С6)алкиларила, (С1-С6)алкилгетероарила;
R47 и R48 независимо выбраны из группы, состоящей из Н, (С1-С6)алкила, (С3-С12)циклоалкила, (С3-С7)гетероциклоалкила, (С1-С6)алкил(С3-С12)циклоалкила, (С1-С6)алкил(С3-С7)гетероциклоалкила, (С1-С6)алкиларила и (С1-С6)алкилгетероарила, включая, например,
(эндо)-3-(2-метокси-2,2-дитиофен-2-ил-этил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид;
3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенил-пропионитрил;
(эндо)-8-метил-3-(2,2,2-трифенилэтил)-8-аза-бицикло[3.2.1]октан;
3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенил-пропионамид;
3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенил-пропионовую кислоту;
(эндо)-3-(2-циано-2,2-дифенилэтил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид;
(эндо)-3-(2-циано-2,2-дифенилэтил)-8,8-диметил-8-азониабицикло[3.2.1]октанбромид;
3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенил-пропан-1-ол;
N-бензил-3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенил-пропионамид;
(эндо)-3-(2-карбамоил-2,2-дифенилэтил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид;
1-бензил-3-[3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенилпропил]мочевину;
1-этил-3-[3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенилпропил]мочевину;
N-[3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенилпропил]ацетамид;
N-[3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенилпропил]бензамид;
3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-ди-тиофен-2-ил-пропионитрил;
(эндо)-3-(2-циано-2,2-дитиофен-2-ил-этил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид;
N-[3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенилпропил]бензолсульфонамид;
[3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенилпропил]мочевину;
N-[3-((эндо)-8-метил-8-аза-бицикло[3.2.1]окт-3-ил)-2,2-дифенилпропил]метансульфонамид; и/или
(эндо)-3-{2,2-дифенил-3-[(1-фенилметаноил)амино]пропил}-8,8-диметил-8-азониабицикло[3.2.1]октанбромид.
Более предпочтительными соединениями, которые могут быть использованы в настоящем изобретении, являются:
(эндо)-3-(2-метокси-2,2-дитиофен-2-ил-этил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид;
(эндо)-3-(2-циано-2,2-дифенилэтил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид;
(эндо)-3-(2-циано-2,2-дифенилэтил)-8,8-диметил-8-азониабицикло[3.2.1]октанбромид;
(эндо)-3-(2-карбамоил-2,2-дифенилэтил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид;
(эндо)-3-(2-циано-2,2-дитиофен-2-ил-этил)-8,8-диметил-8-азониабицикло[3.2.1]октаниодид; и/или
(эндо)-3-{2,2-дифенил-3-[(1-фенилметаноил)амино]пропил}-8,8-диметил-8-азониабицикло[3.2.1]октанбромид.
Подходящими антигистаминами (также называемыми антагонистами Н1-рецептора) являются любые один или несколько различных известных антагонистов, которые ингибируют Н1-рецепторы и являются безопасными для человека. Антагонистами первого ряда являются производные этаноламинов, этилендиаминов и алкиламинов, например дифенилгидрамин, пириламин, клемастин, хлорфенирамин. Антагонисты второго ряда, которые не являются седативными средствами, включают лоратидин, деслоратидин, терфенадин, астемизол, акривастин, азеластин, левоцетиризин, фексофенадин и цетиризин.
Предпочтительными примерами антигистаминов являются лоратидин, деслоратидин, фексофенадин и цетиризин.
Другими рассматриваемыми комбинациями являются комбинации антител согласно изобретению со средством против IL-4 (например, антителом против IL-4, таким как пасколизумаб) и/или средством против IL-5 (например, антителом против IL-5, таким как меполизумаб) и/или со средством против IgЕ (например, антителом против IgЕ, таким как омализумаб (XolairТМ) или тализумаб).
В настоящем изобретении также рассматривается фармацевтическая композиция, содержащая набор, включающий такие компоненты, как антитело согласно изобретению или его антигенсвязывающий фрагмент вместе с вышеуказанными другими лекарственными средствами, и, но необязательно, инструкции по его применению.
Кроме того, в настоящем изобретении рассматривается фармацевтическая композиция, содержащая терапевтически эффективное количество описанного здесь моноклонального терапевтического антитела или его антигенсвязывающего фрагмента и используемая для лечения заболеваний, восприимчивых к модуляции взаимодействия hIL-13 с hIL-13R.
В соответствии с этим настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество моноклонального гуманизованного терапевтического антитела, где указанное антитело содержит VH-домен, выбранный из группы, состоящей из SEQ ID NO:11, 12, 13, 14, и VL-домен, выбранный из группы, состоящей из SEQ ID NO:15, 16.
В соответствии с этим настоящее изобретение относится к фармацевтической композиции, включающей моноклональное терапевтическое антитело, содержащее тяжелую цепь, выбранную из группы, состоящей из SEQ ID NO:18, 19, 20, 21, и легкую цепь, выбранную из группы, состоящей из SEQ ID NO:22, 23.
В соответствии с этим настоящее изобретение относится к фармацевтической композиции, включающей моноклональное терапевтическое антитело, содержащее тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22, и фармацевтически приемлемый носитель.
В соответствии с этим настоящее изобретение относится к фармацевтической композиции, включающей моноклональное антитело, содержащее тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22 (или, по существу, состоящее из них), и фармацевтически приемлемый носитель.
В соответствии с этим настоящее изобретение относится к фармацевтической композиции, содержащей фармацевтически приемлемый носитель и терапевтически эффективное количество популяции моноклональных терапевтических антител, где указанное терапевтическое антитело содержит тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22.
5. Клиническое применение
Антитела согласно изобретению могут быть использованы для лечения атопических заболевания/расстройств и хронических воспалительных заболеваний/расстройств. Особый интерес представляет их использование для лечения астмы, такой как аллергическая астма, в частности тяжелой астмы (то есть астмы, которая является не восприимчивой к лечению современными методами, включая системное введение кортикостероидов; см., Busse W.W. et al., J. Allergy Clin. immunol. 2000, 106:1033-1042), “трудно поддающейся” лечению астмы (определяемой как астматический фенотип, характеризующийся отсутствием эффекта, несмотря на введение максимальных рекомендованных доз прописываемых стероидов, вводимых путем ингаляции, см. Barnes P.J. (1998), Eur. Respir. J. 12:1208-1218), “нестабильной” астмы (определяемой как тяжелая нестабильная астма у подгруппы пациентов, у которых наблюдается широкая вариабельность максимальной скорости выдоха, несмотря на высокие дозы стероидов, вводимых путем ингаляции, см. Ayres J.G. et al. (1998) Thorax 58:315-321), “ночной” астмы, предменструальной астмы, стероид-резистентной астмы (см. Woodcock A.J. (1993) Eur. Respir. J. 6:743-747), стероид-зависимой астмы (определяемой как астма, которая может поддаваться лечению только высокими дозами перорально вводимых стероидов), индуцированной аспирином астмы, астмы взрослых и детской астмы. Антитела согласно изобретению могут быть использованы для предотвращения, снижения частоты или остроты приступов астмы (астматического статуса). Антитела согласно изобретению могут быть также использованы для снижения дозы других лекарственных средств (определяемой как количество или частота введения дозы), необходимых для лечения астмы. Так, например, антитела согласно изобретению могут быть использованы для снижения дозы стероидов, таких как кортикостероидов, требуемой для лечения астмы (“для воздержания от применения стероидов”). Другими заболеваниями или расстройствами, которые могут быть подвергнуты лечению антителами согласно изобретению, являются атопический дерматит, аллергический ринит, болезнь Крона, хроническая обструктивная болезнь легких (ХОБЛ), эозинофильный эзофагит, фиброзные заболевания или расстройства, такие как идиопатический фиброз легких, прогрессирующий системный склероз (склеродермия), фиброз печени, гранулемы печени, шистосомоз, лейшманиоз, и заболевания, ассоциированные с нарушением регуляции клеточного цикла, например болезнь Ходжкина и В-клеточный хронический лимфоцитарный лейкоз. Другие заболевания или расстройства, которые могут быть подвергнуты лечению антителами согласно изобретению, подробно описаны в вышеуказанном разделе “Предшествующий уровень техники”.
В одном из своих вариантов настоящее изобретение относится к способу лечения пациента (человека), страдающего астматическим состоянием, которое не поддается лечению кортикостероидами, где указанный способ включает стадию введения указанному пациенту терапевтически эффективного количества антитела согласно изобретению.
В другом своем варианте настоящее изобретение относится к способу предупреждения острых приступов астмы у пациента (человека), где указанный способ включает стадию введения указанному пациенту терапевтически эффективного количества антитела согласно изобретению.
В другом своем варианте настоящее изобретение относится к способу снижения частоты и/или остроты приступов астмы у пациента (человека), где указанный способ включает стадию введения указанному пациенту терапевтически эффективного количества антитела согласно изобретению.
В другом своем варианте настоящее изобретение относится к способу смещения Т-хелперного клеточного ответа в сторону ответа Th1-типа после воспалительного и/или аллергического приступа у человека, где указанный способ включает стадию введения указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента согласно изобретению.
В другом своем варианте настоящее изобретение относится к способу лечения пациента (человека), имеющего вариант Q130hIL-13 и страдающего астмой, такой как тяжелая астма, где указанный способ включает стадию введения указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента согласно изобретению.
Хотя настоящее изобретение описано в основном применительно к лечению заболеваний или расстройств у человека, однако настоящее изобретение может быть также использовано для лечения аналогичных заболеваний или расстройств у млекопитающих, не являющихся человеком.
Представленное ниже описание настоящего изобретения приводится лишь в качестве примера.
Примеры
1. Продуцирование моноклональных антител и характеризация мышиного моноклонального антитела 6А1
Моноклональные антитела (mAb) получали из гибридомных клеток в основном в соответствии с методикой, описанной в руководстве E. Harlow и D. Lane, Antibodies a Laboratory Manual, Cold Spring Harbor Laboratory, 1988. В соответствии с этой методикой моноклональные антитела получали в результате слияния мышиных миеломных клеток с В-лимфоцитами, взятыми у мышей, иммунизованных антигеном-мишенью. Гибридомная клетка становится иммортализованной благодаря миеломному партнеру по слиянию, а способность к вырабатыванию антител сообщается В-лимфоцитами.
Пять мышей SJL иммунизировали путем внутрибрюшинной инъекции каждой мыши 2 мкг рекомбинантного человеческого IL-13, происходящего от E.coli (Cambridge Bioscience, Cat. № СН-013). Для вырабатывания иммунного ответа у мышей с высоким титром антител против человеческого IL-13 была разработана соответствующая схема иммунизации. После 5 иммунизаций в течение 64 дней мышей умерщвляли и собирали клетки селезенки. Для продуцирования гибридом у 3 из этих мышей брали клетки селезенки и В-лимфоциты подвергали слиянию с мышиными миеломными клетками, полученными от клеток Р3Х, с использованием ПЭГ1500 (Boehringer). Отдельные гибридомные клеточные линии клонировали путем лимитирующего разведения (E. Harlow & D. Lane, см.выше). Лунки, содержащие моноколонии, идентифицировали под микроскопом и супернатант тестировали на активность. Клетки от большинства активных клонов размножали для криоконсервации для продуцирования антител и т.п.
Сначала супернатанты гибридомной культуры скринировали на активность связывания с det-1-меченным рекомбинантным человеческим белком IL-13, экспрессированным в E.coli (полученным в лабораторных условиях) в формате “сэндвич”-анализа. Второй скрининг позитивных по связыванию культур проводили методом BIAcoreTM для детекции на связывание с det-1-меченным человеческим белком IL-13. Затем образцы этих гибридом тестировали на способность нейтрализовать биологическую активность рекомбинантного человеческого IL-13, экспрессированного в E.coli (Cambridge Bioscience, cat. № СН-013) в биоанализе клеток ТF-1.
Шесть позитивных клонов, идентифицированных в биоанализе на нейтрализацию человеческого IL-13, субклонировали путем лимитирующего разведения с получением стабильных моноклональных клеточных линий. Иммуноглобулины, выделенные из этих гибридом, культивированных в клеточных “фабриках” в бессывороточной среде, очищали с использованием колонок с иммобилизованным белком А. Затем эти очищенные mAb повторно скринировали в следующих аналитических системах анализов:
- Анализа на связывание с рекомбинантным человеческим IL-13, экспрессируемым в E.coli (в формате сэндвич-ELISA)
- Анализа на ингибирование связывания det-1-меченного рекомбинантного человеческого IL-13, экспрессируемого в E.coli (в формате сэндвич-ELISA)
- Анализа на нейтрализацию рекомбинантного IL-13 человека или IL-13 обезьян cynomolgus, экспрессируемого в E.coli (в биоанализе на клетки ТF-1)
- Анализа на нейтрализацию Q130-варианта рекомбинантного человеческого IL-13, экспрессируемого в E.coli (в биоанализе на клетки ТF-1).
- Анализа на нейтрализацию рекомбинантного IL-13 человека или IL-13 обезьян cynomolgus, экспрессируемого в E.coli (в биоанализе на клетки ТF-1)
- Анализа на специфичность связывания с человеческим IL-13, определяемую путем оценки перекрестной реактивности mAb с человеческим IL-4 в ELISA и путем оценки перекрестной реактивности mAb с человеческим IL-5 в биоанализе на нейтрализацию IL-5.
- Анализа BIAcoreТМ для определения уровня аффинного связывания с человеческим IL-13.
Моноклональное антитело 6А1 было идентифицировано как антитело, нейтрализующее биологическую активность IL-13 человека и обезьян cynomolgus. В нижеследующих анализах описан профиль моноклонального антитела 6А1.
1.1. Связывание с рекомбинантным человеческим IL-13, экспрессируемым в E.coli
Связывание 6А1 с рекомбинантным человеческим IL-13, экспрессируемым в E.coli, оценивали в “сэндвич”-ELISA методом, по существу, описанным в разделе 7. См. фигуру 1.
1.2. Ингибирование связывания det-1-меченного рекомбинантного человеческого IL-13, экспрессируемого в E.coli, с IL-13Rα1 и IL-13Rα2 в формате ELISA
Антитело 6А1 ингибировало связывание det-1-меченного рекомбинантного человеческого IL-13, экспрессируемого в E.coli, с обеими цепями рецептора человеческого IL-13. Кроме того, это антитело ингибировало связывание более эффективно, чем коммерчески доступное поликлональное антитело против человеческого IL-13 и моноклональное антитело против человеческого IL-13 (R&D Systems). Величина IC50, которая составляла 0,165 мкг/мл, была вычислена для оценки ингибирования моноклональным антителом 6А1 связывания человеческого IL-13 с человеческим IL-13Rα1. Величина IC50, которая составляла 0,056 мкг/мл, была вычислена для оценки ингибирования моноклональным антителом 6А1 связывания человеческого IL-13 с человеческим IL-13Rα2. См. фигуры 2a и 2b. Контрольный IgG с иррелевантной специфичностью не обнаруживал детектируемой активности.
1.3. Нейтрализация рекомбинантного IL-13 человека и IL-13 обезьян cynomolgus, экспрессируемых в E.coli в биоанализе на пролиферацию клеток ТF-1
Клетки ТF-1 могут пролиферироваться в ответ на человеческий IL-13 и IL-13 обезьян cynomolgus. Был разработан биоанализ для оценки способности mAb против IL-13 нейтрализовать пролиферацию клеток ТF-1, индуцированную IL-13 человека и обезьян cynomolgus. В биоанализе на пролиферацию клеток ТF-1 антитело 6А1 нейтрализовало биологическую активность рекомбинантного IL-13 человека и обезьян cynomolgus. Кроме того, это антитело нейтрализовало IL-13 человека и обезьян cynomolgus более эффективно, чем коммерчески доступное поликлональное антитело против человеческого IL-13 и моноклональное антитело против человеческого IL-13 (полученного от R&D Systems). См. фигуру 3.
Средняя величина ND50, которая составляла 0,0783 мкг/мл, была вычислена для оценки нейтрализации моноклональным антителом 6А1 биологической активности 5 нг/мл рекомбинантного человеческого IL-13, экспрессируемого в E.coli, в биоанализе на пролиферацию клеток ТF-1. Величина ND50, которая составляла 0,04 мкг/мл, была вычислена для оценки нейтрализации моноклональным антителом 6А1 биологической активности 5 нг/мл рекомбинантного IL-13 обезьян cynomolgus, экспрессируемого в E.coli, в биоанализе на пролиферацию клеток ТF-1. [Величина ND50 (нейтрализующая доза) представляет собой концентрацию моноклонального антитела, необходимую для снижения пролиферации клеток ТF-1 на 50%, индуцируемой определенной концентрацией IL-13].
1.4. Нейтрализация человеческого IL-13, экспрессируемого в клетках млекопитающих (в клетках СНО), в биоанализе на пролиферацию клеток ТF-1
В анализе на пролиферацию клеток ТF-1 оценивали способность моноклонального антитела 6А1 нейтрализовать человеческий IL-13, экспрессируемый в клетках СНО. Антитело 6А1 нейтрализовало человеческий IL-13, экспрессируемый в клетках млекопитающих, более эффективно, чем коммерчески доступное поликлональное антитело против человеческого IL-13, как было определено по величинам ND50. Величина ND50, которая составляла 0,037 мкг/мл, была вычислена для оценки нейтрализации моноклональным антителом 6А1 приблизительно 50 нг/мл человеческого IL-13, экспрессируемого у млекопитающих, в биоанализе на пролиферацию клеток ТF-1. См. фигуру 4.
1.5. Нейтрализация Q130-варианта рекомбинантного человеческого IL-13 в биоанализе на пролиферацию клеток ТF-1
В анализе на пролиферацию клеток ТF-1 оценивали способность моноклонального антитела 6А1 нейтрализовать Q130-вариант рекомбинантного человеческого IL-13, экспрессируемого в E.coli (Peprotech, Cat № 200-13А). Антитело 6А1 нейтрализовало Q130-вариант человеческого IL-13 более эффективно, чем коммерчески доступное поликлональное антитело против человеческого IL-13. Величина ND50, которая составляла 0,11 мкг/мл, была вычислена для оценки нейтрализации моноклональным антителом 6А1 биологической активности 60 нг/мл Q130-варианта человеческого IL-13. См. фигуру 5.
1.6. Специфичность связывания с человеческим IL-13
Поскольку человеческий IL-4 по своим структурным и функциональным свойствам является наиболее сходным с человеческим IL-13, то специфичность связывания моноклонального антитела 6А1 с человеческим IL-13 оценивали в ELISA на связывание с человеческим IL-4. Антитело 6А1 не обнаруживало связывания с рекомбинантным человеческим IL-4, экспрессируемым в E.coli, что указывало на высокий уровень специфичности связывания указанного моноклонального антитела с человеческим IL-13. Кроме того, антитело 6А1 не обнаруживало способности перекрестно нейтрализовывать биологическую активность рекомбинантного человеческого IL-5, экспрессируемого в E.coli, в биоанализе на пролиферацию клеток ТF-1. См. фигуры 6 и 7.
1.7. Анализ BIAcore ТМ
Аффинность связывания 6А1 с рекомбинантным IL-13 человека и обезьян cynomolgus оценивали в анализе BIAcoreТМ. См. таблицу 1.
Эти данные показали, что антитело 6А1 не обладает аффинностью связывания с IL-13 человека и IL-13 обезьян cynomolgus. [В этом анализе использовали два различных образца человеческого IL-13 (которые оба были продуцированы в E.coli). IL-13, по существу, является нерастворимым, если он продуцируется в E.coli, однако он может быть солюбилизирован, а затем подвергнут рефорлингу in vitro. Различия в качестве двух образцов IL-13, подвергнутых рефолдингу, могут служить объяснением различий в аффинностях связывания каждого из этих образцов человеческого IL-13].
2. Клонирование вариабельных областей клона 6А1
Полноразмерную РНК экстрагировали из гибридомных клеток клона 6А1, а кДНК вариабельных доменов тяжелой и легкой цепей продуцировали путем проведения реакции обратной транскрипции с использованием праймеров, специфичных к мышиным лидерным последовательностям и к константным областям антител, соответствующим предварительно определенному изотипу (IgG1/κ). Затем кДНК вариабельных доменов тяжелой и легкой цепей клонировали в вектор рCR2.1 для секвенирования.
2.1. Экстракция РНК
Полноразмерную РНК экстрагировали из осадка приблизительно 106 клеток гибридомного клона 6А1 с использованием системы выделения полноразмерной РНК SV (Promega) в соответствии с инструкциями производителя.
2.2. Обратная транскрипция
РНК подвергали обратной транскрипции для продуцирования кДНК вариабельных доменов тяжелой и легкой цепей с использованием праймеров, специфичных к мышиным лидерным последовательностям и к мышиным константным областям IgG1/κ. Используемая смесь праймеров описана в публикации Jones ST и Bendig MM Bio/technology 9:88-89 (1991).
Пулы прямых праймеров лидерной последовательности мышиных VН и VL получали в концентрации 50 мкМ. Растворы обратных праймеров мышиных константных областей IgG1 и κ также получали в концентрации 50 мкМ.
2.3. ПЦР с обратной транскриптазой (ОТ-ПЦР)
Обратную транскрипцию РНК, кодирующей вариабельные области тяжелой и легкой цепей, проводили в дубликатах с использованием системы Access RT-PCR (Promega) в соответствии с инструкциями производителя. Прямой и обратный праймеры VH и VL описаны выше.
2.4. Гель-очистка продукта ОТ-ПЦР
Продукты ОТ-ПЦР (2 × VН и 2 × VL) загружали в раствор для загрузки препаративного 1% агарозного геля, содержащего 0,01% этидийбромида, и проводили электрофорез в буфере ТАЕ при 100 В в течение 1 часа, а затем полосы V-области вырезали. Для идентификации VН- и VL-полос в этом геле проводили электрофорез 100 п.н. ДНК-лэддера.
Фрагменты ДНК экстрагировали и очищали из геля с использованием набора для экстракции из геля QIAquickTM (Qiagen) в соответствии с инструкциями производителя.
2.5. Лигирование
Очищенные ОТ-ПЦР-фрагменты (2 × VН и 2 × VL) клонировали в вектор pCR2.1 с использованием набора для клонирования ТА (от Invitrogen) в соответствии с инструкциями производителей.
2.6. Трансформация
Лигированные плазмиды переносили в клетки TOP10F' в соответствии с инструкциями, прилагаемыми к набору для клонирования ТА. 50 мкл и 200 мкл трансформированных клеток высевали на планшеты с L-агаром, содержащие 100 мкг/мл ампициллина и сенсибилизированные 8 мкл 500 мМ раствора IPTG и 16 мкл 50 мг/мл раствора Х-Gal в ДМФ. Планшеты инкубировали в течение ночи при 37°С.
2.7. Секвенирование
Колонии высевали и культивировали в течение ночи при 37°С в 5 мл среды LВ, в которую было добавлено 100 мкг/мл ампициллина. Плазмиды рСR2.1, содержащие VН- и VL-домены 6А1, экстрагировали и очищали с использованием набора для приготовления мини-препаратов Qiagen QIAprep Spin Miniprep в соответствии с инструкциями производителя. VН- и VL-домены секвенировали с использованием праймеров Т7, прямого праймера М13 и обратного праймера М13.
Аминокислотная последовательность VН-области антитела 6А1 (консенсусная последовательность 10 клонов от 2 ОТ-ПЦР-реакций) представляет собой SEQ ID NO:7.
Аминокислотная последовательность VL-области антитела 6А1 (консенсусная последовательность 10 клонов от 2 ОТ-ПЦР-реакций) представляет собой SEQ ID NO:8.
3. Химерное антитело
Химерное антитело, состоящее из родительских мышиных V-областей (описанных в разделе 2.7), присоединенных к С-областям человеческого IgG1/κ дикого типа, было сконструировано для подтверждения клонирования соответствующих мышиных V-областей, а также для его использования в качестве стандарта при тестировании гуманизованных конструкций. Химерное антитело было экспрессировано в клетках СНО, очищено и протестировано на связывание с человеческим IL-13 с помощью ELISA.
3.1. ПЦР-амплификация
Клонированные мышиные V-области амплифицировали посредством ПЦР для введения рестрикционных сайтов, необходимых для клонирования в экспрессирующие векторы млекопитающих Rld и Rln. В VН-домене создавали HindIII- и SрeI-сайты в той же рамке считывания и осуществляли клонирование в модифицированный Rld-вектор, содержащий С-область цепи γ1 человеческого антитела дикого типа. В VL-домене создавали HindIII- и BsiwI-сайты в той же рамке считывания и осуществляли клонирование в модифицированный Rln-вектор, содержащий С-область цепи κ человеческого антитела дикого типа.
Прямой праймер для VН:
5'-GAT GAA GCT T GC CAC CAT GAA ATG CAG CTG GGT CAT C-3'
(SEQ ID NO:86)
Рестрикционный HindIII-сайт подчеркнут, а последовательность Козака выделена жирным шрифтом.
Обратный праймер для VН:
5'-GAT GGA CTA GTG TTC CTT GAC CCC AGT A-3' (SEQ ID NO:87)
Рестрикционный SрeI-сайт подчеркнут.
Прямой праймер для VL:
5'-GAT GAA GCT T GC CAC CAT GAA GTT GCC TGT TAG GCT G-3'
(SEQ ID NO:88)
Рестрикционный HindIII-сайт подчеркнут, а последовательность Козака выделена жирным шрифтом.
Обратный праймер для VL:
5'-GAT GCG TAC GTT TGA TTT CCA GCT TGG TGC C-3' (SEQ ID NO:89)
Рестрикционный BsiWI-сайт подчеркнут.
Праймер 1: прямой VН- или VL-праймер
Праймер 2: обратный VН- или VL-праймер
Очищенная плазмида: VН- или VL-содержащая плазмида рСR2.1, очищенная с использованием набора Qiagen Minipreps (с разведением 200 х)
3.2. Клонирование экспрессирующих векторов в млекопитающих
ПЦР-продукты очищали с использованием набора для ПЦР-очистки MinElute от Qiagen в соответствии с инструкциями производителя.
VН-продукт ПЦР и экспрессирующий вектор млекопитающих RldhCγ1wt гидролизовали ферментами HindIII и SрeI:
ДНК: очищенный VН-продукт ПЦР или экспрессирующий вектор млекопитающих RldhCγ1wt (при 0,25 мг/мл) инкубировали в течение 2 ч при 37оС.
VL-продукт ПЦР и экспрессирующий вектор млекопитающих RlnhCκ гидролизовали ферментами HindIII и BsiWI:
ДНК: очищенный VL-продукт ПЦР или вектор Rln hCκ (при 0,25 мг/мл) инкубировали в течение 2 ч при 37оС. Затем добавляли 2 мкл BsiWI (NЕВ) и инкубировали 2 часа при 55°С.
Продукты гидролиза рестриктирующими ферментами загружали в раствор для загрузки на препаративный 1% агарозный гель, содержащий 0,01% этидийбромида, и проводили электрофорез в буфере ТАЕ при 100 В в течение 1 часа, а затем полосы векторов Rld и Rln, а также полосы VН- и VL-ПЦР-фрагментов вырезали. Для идентификации полос VН, VL и вектора в этом геле проводили электрофорез 100 п.н. ДНК-лэддера. ДНК экстрагировали и очищали из геля с использованием набора для экстракции из геля QIAquickTM (Qiagen) в соответствии с инструкциями производителя. VН-ПЦР-фрагмент, гидролизованный ферментами HindIII и SрeI, лигировали в HindIII-SрeI-гидролизованный вектор Rld hCγ1wt. VL-ПЦР-фрагмент, гидролизованный ферментами HindIII и BsiWI, лигировали в HindIII-BsiWI-гидролизованный вектор RlnhCκ. Лигирование проводили с использованием системы быстрого лигирования ДНК LigaFast (Promega) в соответствии с инструкциями производителей, где:
VН: вектор: RldhCγ1wt, гидролизованный ферментами HindIII и SрeI,
вставка: VН-ПЦР-фрагмент, гидролизованный ферментами HindIII и SрeI
VL: вектор: RlnhCκ, гидролизованный ферментами HindIII и BsiWI,
вставка: VL-ПЦР-фрагмент, гидролизованный ферментами HindIII и BsiWI
Лигированные продукты переносили в компетентные клетки DH5α. 200 мкл-сосуды с DH5α оттаивали на льду. 50 мкл-аликвоты получали в пробирках для трансформации. Затем добавляли 2 мкл-смеси для лигирования и смесь слегка перемешивали кончиком пипетки, а затем инкубировали в течение 30 минут на льду. Смесь инкубировали при встряхивании в течение 45 секунд при 42°С. Затем эту смесь переносили на лед на 2 минуты. После этого добавляли 450 мкл среды SОС и пробирки инкубировали в течение 1 часа при 37°С на шейкерном инкубаторе. 100 мкл-культуры высевали на планшеты с L-агаром, в которые было добавлено 100 мкг/мл ампициллина, и инкубировали в течение ночи при 37оС.
3.3. Секвенирование
VН- и VL-клоны культивировали в течение ночи при 37°С в 5 мл среды LВ, в которую было добавлено 100 мкг/мл ампициллина. Плазмиды Rld и Rln, содержащие VН- и VL-домены соответственно, экстрагировали и очищали с использованием набора для приготовления мини-препаратов Qiagen QIAprep Spin Miniprep в соответствии с инструкциями производителя. VН-область секвенировали с использованием прямых праймеров в векторе Rld и с использованием сигнальной последовательности и обратного праймера в человеческой области Сγ1.
VL-область секвенировали с использованием прямых праймеров в векторе Rln и с использованием сигнальной последовательности и обратного праймера в человеческой области Сκ. Клоны, имеющие соответствующие последовательности VН и VL, идентифицировали и получали плазмиды для экспрессии в клетках СНО.
3.4. Экспрессия химерного антитела в клетках СНО
Плазмиды Rld и Rln, содержащие VН- и VL-домены антитела 6А1 соответственно, подвергали временной котрансфекции в клетки СНО и экспрессировали. Полученное химерное антитело выделяли из супернатанта клеточной культуры с помощью аффинной хроматографии на белке А - сефарозе.
3.4.1. Очистка плазмиды
Клетки DH5α, содержащие Rld-6A1VН и Rln-6A1VL, культивировали в 5 мл среды LB, в которую было добавлено 100 мкг/мл ампициллина, в течение 8 часов при 37°С в шейкерном инкубаторе. 200 мл среды LB, в которую было добавлено 100 мкг/мл ампициллина, инокулировали 1 миллилитром дневной культуры и инкубировали в течение ночи при 37°С в шейкерном инкубаторе. Плазмиды экстрагировали и очищали с использованием набора для приготовления мини-препаратов Qiagen QIAfilter Plasmid Miniprep в соответствии с инструкциями производителей. Этаноловый осадок ресуспендировали в 200 мкл буфера ТЕ и концентрацию плазмиды определяли путем измерения оптической плотности при 260 нм после 100-кратного разведения маточного раствора.
3.4.2. Трансфекция
Клетки СНО культивировали до конфлюэнтности в модифицированной по способу Дульбекко среде MEM c глутамаксом-1 (DMEM), в которую были добавлены фетальная бычья сыворотка сверхнизкой концентрации (Ultra low) 1% пенициллин-стрептомицин, в 4 × 175 см2-колбах для культивирования ткани BD Falcon при 37°С.
В каждую колбу, а именно в 50 мл-пробирку Falcon, добавляли, интенсивно перемешивая, следующие реагенты:
8 мл Optimem 1 с глутамаксом-1
20 мкг очищенной плазмиды Rld-6A1VН
20 мкг очищенной плазмиды Rln-6A1VL
240 мкл реагента для трансфекции TransFast.
Полученную смесь инкубировали в течение 10-15 минут при комнатной температуре. Затем среду DMEM удаляли из колбы, смесь интенсивно перемешивали и добавляли в колбу. После этого смесь инкубировали при 37°С в течение 1 часа. В колбу добавляли 32 мл Optimem и смесь инкубировали при 37°С в течение 48-72 часов.
3.4.3. Очистка химерного антитела
Среды, взятые из колбы 175 см2, объединяли и центрифугировали при 1500 об/мин в течение 3 мин на приборе МSE Mistral 2000, и супернатант пропускали через фильтровальную систему 0,22 мкм СА объемом 500 мл. Антитело выделяли из осветленного супернатанта на устройстве Amersham Biosciences Akta Explorer с использованием компьютерной программы Unicorn. На используемую колонку наносили 1 мл рекомбинантного белка А HiTrap с сефарозой FF. Скорость потока составляла 1 мл/мин.
Колонку уравновешивали 10 колоночными объемами PBS Дульбекко, а затем в колонку загружали осветленный супернатант с помощью насоса А. Колонку промывали 20 колоночными объемами PBS Дульбекко, после чего насос А промывали для удаления отходов, а затем через колонку пропускали 10 колоночных объемов PBS Дульбекко для гарантии полного клиренса супернатанта. Антитело элюировали 10 колоночными объемами буфера для элюирования IgG ImmunoPure (Pierce) и собирали в 1 мл-фракции, содержащие 100 мкл нейтрализующего буфера 1М Trizma-HCl, pH 8,0. Затем колонку снова уравновешивали 5 колоночными объемами PBS Дульбекко.
Антитело во фракциях элюата количественно оценивали путем считывания оптической плотности при 280 нм по сравнению с контрольным раствором, содержащим 10 объемов буфера для элюирования IgG ImmunoPure + 1 объем 1М Trizma-HCl, pH 8,0, и фракции, содержащие достаточное количество чистого антитела, объединяли и хранили в 100 мкл-аликвотах при -20°С.
3.4.4. Анализ химерного антитела
Супернатант и очищенное химерное антитело 6А1 (6А1с) анализировали на связывание с IL-13 человека и обезьян cynomolgus с помощью ELISA. Супернатант от клеток СНО, временно трансфицированный химерным моноклональным антителом 6А1, связывался с рекомбинантным IL-13 человека и IL-13 обезьян cynomolgus, экспрессируемыми в E.coli в “сэндвич”-ELISA. Очищенное антитело также связывалось с рекомбинантным человеческим IL-13 и IL-13 обезьян cynomolgus, экспрессируемыми в E.coli, в “сэндвич”-ELISA (данные не приводятся). См. фигуру 8.
Аминокислотная последовательность и последовательность кДНК IL-13 обезьян cynomolgus (включая сигнальную последовательность) представлены в SEQ ID NO:90 и 91 соответственно.
Эти результаты подтвердили, что нужные вариабельные области 6А1 были успешно клонированы и продуцировали антигенсвязывающее химерное антитело, способное связываться с IL-13 человека и обезьян cynomolgus.
4. Гуманизация клона 6А1
4.1. Стратегия гуманизации
4.1.1. Поиск в базе данных мышиных последовательностей
12 мышиных последовательностей, имеющих наибольшую гомологию с аминокислотной последовательностью VН 6А1 и 11 мышиных последовательностей, имеющих наибольшую гомологию с аминокислотной последовательностью VL, идентифицировали путем поиска в базе данных пептидных последовательностей (Genbank).
Аминокислотную последовательность VН 6А1 сравнивали со всеми 12 мышиными последовательностями путем поиска в базе данных и идентифицировали нижеследующие каркасные остатки:
Положения даны в соответствии с системой нумерации по Кабату и др.
Аминокислотную последовательность Vl 6А1 сравнивали с 11 мышиными последовательностями путем поиска в базе данных и идентифицировали каркасные остатки.
4.1.2. Поиск в базе данных человеческих последовательностей
Человеческие каркасные последовательности, имеющие наибольшую гомологию с каркасными последовательностями VН и VL 6А1, идентифицировали путем поиска в базе данных пептидных последовательностей с использованием программы EasyBlast.
Была идентифицирована одна серия человеческих последовательностей для VН 6А1, из которых для гуманизации была выбрана каркасная последовательность SEQ ID NO:92.
Нижеследующие каркасные остатки были идентифицированы как остатки, имеющие потенциально важное значение для восстановления аффинности, и эти остатки могут потребовать обратной мутации:
Были сконструированы 4 гуманизованные VН-конструкции с различными обратными мутациями, одна из которых представляла собой прямую мутацию, а другие представляли собой различные обратные мутации (А2, А3, А4). А именно:
А2 представляла собой А1 плюс R381
А3 представляла собой А2 плюс Е73Т
А4 представляла собой А3 плюс К19R плюс Е81R
Была идентифицирована одна серия человеческих последовательностей для VL 6А1, из которых для гуманизации была выбрана каркасная последовательность SEQ ID NO:93.
Нижеследующие остатки были идентифицированы как остатки, имеющие потенциально важное значение для восстановления аффинности, и эти остатки могут потребовать обратной мутации:
Были получены две конструкции, одна из которых имела прямую мутацию (L1), а другая имела обратную мутацию (L2)(то есть L1 плюс 185V).
Гуманизованная V Н -конструкция А1:
SEQ ID NO:11
Гуманизованная V Н -конструкция А2:
SEQ ID NO:12
Гуманизованная V Н -конструкция А3:
SEQ ID NO:13
Гуманизованная V Н -конструкция А4:
SEQ ID NO:14
Гуманизованная V L -конструкция L1:
SEQ ID NO:15
Гуманизованная V L -конструкция L2:
SEQ ID NO:16
4.2. Гуманизация 6А1:
Гуманизованные VН- и VL-конструкции были получены de novo путем конструирования перекрывающихся олигонуклеотидов, включая рестрикционные сайты для клонирования в экспрессионные сайты Rld и Rln млекопитающих, а также человеческую сигнальную последовательность. Рестрикционные HindIII- и SрeI-сайты были введены с сохранением рамки считывания в VН-домен, содержащий человеческую сигнальную последовательность, для клонирования в вектор Rld, содержащий человеческую константную область γ1 дикого типа. Рестрикционные HindIII- и BsiWI-сайты были введены с сохранением рамки считывания в VL-домен, содержащий человеческую сигнальную последовательность, для клонирования в вектор Rln, содержащий человеческую константную область каппа.
Человеческая сигнальная последовательность представляет собой последовательность SEQ ID NO:17.
Были сконструированы 4 гуманизованные VН-конструкции и две гуманизованные VL-конструкции. В результате такого конструирования получали 8 различных комбинаций тяжелых и легких цепей.
Для создания конструкций было получено приблизительно 10 олигонуклеотидов длиной примерно в 60 оснований, с перекрываниями примерно в 18 оснований.
4.2.1. Конструирование олигонуклеотидов
Растворы олигонуклеотидных пулов получали из 5 мкл каждого из олигонуклеотидных маточных растворов при концентрации 100 мкМ. Синтез гуманизованных VН- и VL-генов путем конструирования перекрывающихся олигонуклеотидов проводили в основном, как описано в публикации Stemmer W.P. et al. (1995) Gene 164(1):49-53, с помощью компьютерной программы, описанной Ertl P.F. et al. (2003) Methods 31:199-206.
4.2.1.1. Репрезентативная реакция ПЦР-сборки:
10х ПЦР-буфер ProofStart
dNTP (10 мМ)
Олигонуклеотидный пул
ДНК-полимераза ProofStart
Общий объем
5 мкл
1,5 мкл
1 мкл
1 мкл
50 мкл
Цикл ПЦР-сборки:
1-94°С, 2 минуты
2-94°С, 30 секунд
3-40°С, 2 минуты
4-72°С, 10 секунд
5-94°С, 15 секунд
6-40°С, 30 секунд
7-72°С, 20 секунд + 3 секунды/цикл; стадии 4-7 повторяли 25 раз.
4.2.1.2. Репрезентативная реакция ПЦР-восстановления:
Праймеры 1 и 2 представляют собой первый верхний и нижний олигонуклеотиды, используемые в ПЦР-сборке. Реакция ПЦР-восстановления позволяет амплифицировать полноразмерный ген V.
10х ПЦР-буфер ProofStart
dNTP (10 мМ)
Праймер 1 (100 мкМ)
Праймер 2 (100 мкМ)
Реагент для ПЦР-сборки
ДНК-полимераза ProofStart
Общий объем
4 мкл
1,5 мкл
0,5 мкл
0,5 мкл
1 мкл
0,5 мкл
50 мкл
Цикл ПЦР-восстановления:
1-94°С, 2 минуты
2-94°С, 45 секунд
3-60°С, 30 секунд
4-72°С, 2 минуты
5-72°С, 4 минуты
стадии 2-4 повторяли 25 раз.
Продукты ПЦР-восстановления очищали с использованием набора для ПЦР-очистки MinElute (Qiagen) в соответствии с инструкциями производителей.
4.2.2. Гидролиз рестриктирующими ферментами
Гуманизованные VН-конструкции А1, А2, А3 и А4 антитела 6А1 гидролизовали ферментами HindIII-SpeI, а гуманизованные VL-конструкции L1, L2 антитела 6А1 гидролизовали ферментами HindIII-BsiWI, как описано в разделе 3.
4.2.3. Гель-очистка
Продукты гидролиза рестриктирующими ферментами очищали способом, аналогичным способу, описанному в разделе 3.
4.2.4. Лигирование
Гуманизованные VН-фрагменты антитела 6А1, гидролизованные ферментами HindIII-SpeI, лигировали в HindIII-SpeI-гидролизованный вектор Rld hCγ1wt.
Гуманизованные VL-фрагменты антитела 6А1, гидролизованные ферментами HindIII-BsiWI, лигировали в HindIII-BsiWI-гидролизованный вектор Rln hCκ.
Лигирование осуществляли с использованием системы быстрого лигирования ДНК LigaFast (Promega) в соответствии с инструкциями производителей.
4.2.5. Трансформация
Трансформацию проводили способом, аналогичным способу, описанному в разделе 3.
4.2.6. Репрезентативный метод секвенирования
Колонии из каждого реакционного планшета культивировали в течение ночи при 37°С в 5 мл среды LB, в которую было добавлено 100 мкг/мл ампициллина. Затем плазмиды экстрагировали, очищали с использованием набора для получения минипрепаратов QIAprep Spin Miniprep (Qiagen) в соответствии с инструкциями производителей и секвенировали с использованием праймеров, описанных в разделе 3. Клоны, содержащие соответствующие гуманизованные последовательности VН и VL, идентифицировали и получали плазмиды для экспрессии в клетках СНО.
5. Экспрессия и характеризация гуманизованных антител
Четыре гуманизованные VН-конструкции (А1, А2, А3, А4) и две гуманизованные VL-конструкции (L1 и L2) получали в экспрессирующих векторах Rld hCγ1wt и Rln hCκ млекопитающих. Восемь плазмидных комбинаций тяжелой цепи-легкой цепи (А1L1, A1L2, A2L1, A2L2, A3L1, A3L2, A4L1, A4L2) подвергали временной ко-трансфекции в клетки СНО и экспрессировали в лабораторном масштабе с получением 8 различных гуманизованных антител. Антитела, продуцированные в супернатанте клеток СНО, анализировали в ELISA на связывание с человеческим IL-13.
5.1. Репрезентативный метод очистки плазмид
Клетки DH5α, содержащие одну из плазмид, описанных выше, культивировали в 5 мл среды LB, в которую было добавлено 100 мкг/мл ампициллина, в течение 8 часов при 37°С в шейкерном инкубаторе. 200 мл среды LB, в которую было добавлено 100 мкг/мл ампициллина, инокулировали 1 миллилитром дневной культуры и инкубировали в течение ночи при 37°С в шейкерном инкубаторе. Плазмиды экстрагировали и очищали с использованием набора QIAfilter Plasmid Maxi (Qiagen) в соответствии с инструкциями производителей. Этаноловый осадок ресуспендировали в 200 мкл буфера ТЕ и концентрацию плазмиды измеряли по оптической плотности при 260 нм после 100-кратного разведения маточного раствора.
5.2. Репрезентативный метод трансфекции
В 9 лунок 96-луночного планшета Corning Costar 3506 высевали 106 клеток СНО и культивировали в течение ночи в модифицированной по способу Дульбекко среде МЕМ с глутамаксом-1 (DMEM), в которую были добавлены фетальная бычья сыворотка сверхнизкой концентрации (Ultra low) и 1% пенициллин-стрептомицин при 37°С.
Затем в каждую лунку в 5 мл cреды Bijou добавляли, при встряхивании, нижеперечисленные реагенты так, чтобы каждая смесь для трансфекции содержала различные комбинации легких и тяжелых цепей, а именно:
1 мл Optimem 1 с глутамаксом-1;
5 мкг плазмиды, содержащей гуманизованную VН;
5 мкг плазмиды, содержащей гуманизованную VL;
30 мкг реагента для трансфекции TransFast.
Инкубирование проводили в течение 10-15 минут при комнатной температуре. Затем из лунок удаляли среду DMEM, смесь встряхивали и добавляли в соответствующую лунку. После этого проводили инкубирование при 37°С в течение 1 ч. Затем добавляли 2 мл Optimem на лунку и смесь инкубировали при 37°С в течение 48-72 часов.
5.3. Анализ гуманизованных антител
Среды из каждой лунки восстанавливали и центрифугировали при 13000 об/мин в течение 1 минуты на лабораторной настольной центрифуге Эппендорфа 5415R и супернатант пропускали через 0,2 мкм-фильтр-шприц Pall Acrodisc (25 мм). Клеточный супернатант анализировали в ELISA на связывание с человеческим IL-13. ELISA-анализ показал, что все 8 гуманизованных антител связывались с человеческим IL-13 с профилем, аналогичным профилю связывания химерного антитела 6А1 с человеческим IL-13. См. фигуру 9.
Гуманизованные антитела L1+A1 и L2+A1 были отобраны для крупномасштабной экспрессии, очистки и для проведения дополнительных анализов.
6. Анализ гуманизованных антител L1+A1 и L2+A1 против человеческого IL-13
6.1. Активность связывания IL-13 человека и обезьян cynomolgus в ELISA
Антитела L1+A1 и L2+A1 были успешно продуцированы в результате крупномасштабной экспрессии и оценены в ELISA-анализе на связывание IL-13 человека и обезьян cynomolgus, экспрессируемым в E.coli. См. фигуры 10а и 10b и таблицу В.
Профили связывания антител L1+A1 и L2+A1, экспрессируемыми в E.coli, с человеческим IL-13 и с IL-13 обезьян cynomolgus, экспрессируемым в E.coli, были аналогичными. Величины ЕС50 (полученные путем построения кривой зависимости с использованием компьютерной программы Excel “Robosage”) показали, что уровень активности связывания таких антител почти аналогичен уровню активности связывания химерного стандартного mAb 6A1.
6.2. Анализ на связывание антител L1+A1 и L2+A1 с нативным человеческим IL-13 (выделенным из МКПК)
Для анализа на связывание химерного антитела mAb 6A1 антитело L1+A1 и антитело L2+A1 с нативным человеческим IL-13 (выделенным из МКПК) использовали супернатанты CD4+-Th2-клеток (генерированные из культур человеческих МКПК), стимулированных антителами против CD23 и CD28. ELISA-анализ показал, что все 3 антитела связывались с нативным человеческим IL-13 в супернатанте Th2-клеток с эффективностью, почти аналогичной эффективности связывания родительского mAb 6A1. См. фигуру 11.
Кроме того, для определения уровня нативного человеческого IL-13, присутствующего в супернатанте Th2-клеток, строили стандартную кривую с использованием коммерчески доступных реагентов. Все 3 антитела и коммерчески доступное mAb против человеческого IL-13 обнаруживали эквивалентное количество IL-13 в образце супернатанта Th2-клеток. См. ниже таблицу 2.
6.3. Активность ингибирования связывания L1+A1 и L2+A1 с человеческим IL-13 в ELISA-анализах на связывание с рецептором IL-13
Родительское мышиное mAb 6A1, химерное 6А1, антитело L1+A1 и антитело L2+A1 анализировали на их способность ингибировать связывание человеческого IL-13 c цепями IL-13Rα1 и с IL-13Rα2 в ELISA-анализе на конкурентное связывание. См. фигуры 12а и 12b, а также таблицу 3, представленную ниже.
Все антитела ингибировали связывание det1-меченного человеческого IL-13, экспрессируемого в E.coli, с человеческим IL-13Rα1 с аналогичным профилем. Аналогичным образом, все антитела ингибировали связывание det1-меченного человеческого IL-13, экспрессируемого в E.coli, с человеческим IL-13Rα2, хотя, как показал данный анализ, антитела L1+A1 и L2+A1 обладали несколько пониженной активностью (величины ЕС50 были получены по кривой зависимости функциональной зависимости с использованием компьютерной программы Excel “Robosage”).
6.4. Оценка аффинности связывания антител L1+A1 и L2+A1 с человеческим IL-13
Кинетику связывания L1+A1 и L2+A1 с человеческим IL-13 оценивали с использованием системы BIAcoreTM. Описание применяемых методов приводится ниже в разделе 7.
Анализ 1:
Проводили анализ на связывание с IL-13 человека и с IL-13 обезьян cynomolgus (белком, экспрессируемым в E.coli). Приведенные в таблице величины KD означают среднее из 5 кривых для различных концентраций IL-13 (данные получены с тремя повторностями). Следует отметить, что в этом анализе наблюдались продукты переноса массы и что модифицированный протокол эксперимента (с поправкой на этот продукт) был осуществлен в анализе 4 (где не присутствовали продукты переноса массы). См. таблицу 4.
Анализ 2:
Проводили анализ на связывание человеческого IL-13 (белка, экспрессируемого в E.coli) с антителом L1+A1. См. таблицу 5.
Анализ 3:
Проводили анализ на связывание с 16-мерным биотинилированным человеческим пептидом IL-13 № 24 (идентифицированным как линейный эпитоп, связывающийся с родительским mAb 6A1, см. раздел 6.7). Следует отметить, что абсолютные величины KD, полученные для связывания с пептидными лигандами, часто совершенно отличаются от величин, полученных для связывания с полноразмерными белками-мишенями. Однако, очевидно, что эти данные совпадают с данным, полученными для полноразмерного белка, и с данными по нейтрализации IL-13 (в биоанализе TF-1), которые указывают примерно на 3-кратное снижение уровня аффинности связывания химерного антитела 6А1 и аффинности связывания антитела L1+A1. См. таблицу 6.
Анализ 4:
Проводили анализ на связывание с IL-13 человека и с IL-13 обезьян cynomolgus (с белком, экспрессируемым в E.coli). Приведенные в таблице величины KD означают среднее из 5 кривых для различных концентраций IL-13 (данные получены с тремя повторностями). Следует отметить, что в этом наборе данных отсутствовали какие-либо продукты переноса массы.
Результаты показали, что какое-либо значимое различие между гуманизованными конструкциями L1+A1 и L2+A1 отсутствует.
Аффинность связывания L1+A1 с человеческим IL-13 составляет приблизительно 168 пМ. Такая кинетика реакции была доминирующей из-за исключительно низкой скорости диссоциации, как было предсказано исходя из значительной нейтрализующей активности антитела. Значения констант ассоциации k on были постоянными и составляли примерно 6 × 105 М-1с-1. Значения констант диссоциации k off были более вариабельными и составляли в пределах от 1,4 × 10-4 до 8,22 × 10-5 с-1, что обусловлено некоторыми техническими проблемами, не позволяющими точно вычислить значения для низкой скорости диссоциации.
6.5. Активность L1+A1 и L2+A1 в биологических анализах на нейтрализацию IL-13
Родительское мышиное mAb 6A1, химерное антитело 6А1, антитело L1+A1 и антитело L2+A1 оценивали на активность нейтрализации IL-13 в биологическом анализе клеток TF-1 in vitro (этот биологический анализ является промышленным стандартным анализом для оценки биологической активности IL-13 и для оценки нейтрализующей активности коммерчески доступных анти-IL-13 антител). В этом анализе оценивали ряд вариантов IL-13, включая человеческий IL-13, экспрессируемый в E.coli, IL-13 человекоподобных обезьян, экспрессируемый в E.coli, Q130-вариант человеческого IL-13, экспрессируемый в E.coli (вариант, ассоциированный с развитием астмы) и человеческий IL-13, экспрессируемый в клетках СНО млекопитающих (примечание: нативный человеческий IL-13 в образце супернатанта Th2-клеток не может быть использован в этом биоанализе, поскольку этот супернатант также содержит другие цитокины, способные вызывать пролиферацию клеток TF-1). См. фигуры 13а, 13b, 13c и 13d.
Все антитела, протестированные в этой биоаналитической системе, нейтрализовали биологическую активность всех вариантов IL-13; при этом определяли способность каждого антитела нейтрализовать каждый из вариантов IL-13 и эту способность выражали как величину ND50. См. таблицу 8.
Примечание: поскольку в этом биоанализе для пролиферации клеток TF-1 в одинаковой степени требуются различные количества каждого из вариантов IL-13, то нежелательно проводить сравнение величин ND50, полученных для одного конкретного антитела, направленного против каждого из используемых вариантов IL-13. Однако необходимо проводить сравнение величин ND50, полученных для каждого из антител, направленного против одного варианта IL-13.
По существу, уровень нейтрализации, достигаемый родительским mAb 6A1 и химерным 6А1, был одинаковым, что указывает на отсутствие какой-либо детектируемой потери активности родительского mAb и химерного антитела. Однако активность L1+A1 и L2+A1 в отношении каждого отдельного протестированного варианта IL-13 была значительно ниже в среднем примерно в 3-4 раза по сравнению с активностью родительского mAb 6A1 и химерного 6A1. Эти данные очень тесно коррелируют с данными, полученными в анализе BIAcoreTM.
6.6. Специфичность связывания антител L1+A1 и L2+A1 с человеческим IL-13
Специфичность связывания антител L1+A1 и L2+A1 с человеческим IL-13 оценивали с помощью анализа на потенциальную перекрестную реактивность с человеческим IL-4 и человеческим GM-CSF в ELISA-анализе на связывание. См. фигуры 14а и 14b.
Было обнаружено, что mAb обладают специфичностью связывания с IL-13 и не обладают перекрестной реактивностью с человеческим IL-4 или с человеческим GM-CSF при концентрациях mAb до 30 мкг/мл. Кроме того, эти mAb не обладают способностью перекрестно нейтрализовать биологическую активность человеческого IL-5 в биоанализе IL-5. См. фигуру 14с.
6.7. Картирование эпитопов для 6А1 с использованием биотинилированных пептидов
Белки, а именно человеческий IL-13 и IL-13 обезьян cynomolgus, подвергали электрофорезу в денатурирующем ПААГ с ДСН. Вестерн-блот-анализ с использованием мышиного mAb 6A1 выявил полосы ожидаемого размера как для человеческого белка IL-13 (экспрессируемого в E.coli, в лабораторном масштабе), так и для белка IL-13 обезьян cynomolgus (экспрессируемого в E.coli, в лабораторном масштабе). Антитело 6А1 не детектировало hIL-13 (экспрессируемый в E.coli, Cambridge Bioscience), что, вероятно, связано с техническими проблемами. Этот анализ дал основание предположить, что mAb 6A1 распознает линейный пептидный эпитоп в последовательностях IL-13 человека и обезьян cynomolgus (данные не приводятся).
Биотинилированные 16-мерные пептиды со сдвигом на 4 остатка были синтезированы для создания карты локализации В-клеточного эпитопа, распознаваемого mAb 6А1, на IL-13 человека и IL-13 обезьян cynomolgus. Для детекции связывания иммобилизованного биотинилированного пептида с родительским mAb 6A1 был применен метод ELISA.
16-мерные специально сконструированные пептиды имеют следующие параметры: 88 × 16-меры со сдвигом на 4 остатка (такие пептиды поставляются фирмой Mimotopes, Australia).
Пример: типичный 96-луночный планшет, применяемый для проведения этого анализа.
NB: Числа обозначают пептид в каждой лунке; числа в скобках обозначают степень разведения контрольного антитела.
Оптические плотности при 490 нм в 96 лунках
Этот результат (один из ряда попыток) коррелирует с положительным результатом для пептидов 24, 25, 43 и 44, указанных ниже (а также позитивные контрольные пептиды). См. фигуру 15. Все попытки продемонстрировали, что пептиды 24, 25, 43 и 44 были позитивными.
Пептиды 24 и 25 происходили от hIL-13. Пептиды 43 и 44 происходили от IL-13 обезьян cynomolgus.
Кроме того, все антитела, а именно химерное антитело 6А1, антитело L1+A1 и антитело L2+A1, связывались с одним и тем же линейным эпитопом в С-концевой области IL-13 человека и обезьян cynomolgus (данные для mAb, таких как химерное 6А1, L1+A1 и L2+A1 не приводятся).
В итоге, ELISA-результаты показали, что все антитела, а именно родительское мышиное mAb 6A1, химерное 6А1, L1+A1 и L2+A1, связываются в нижеследующей последовательности, происходящей от человеческого белка IL-13, такой как:
DLLLHLKKLFRE (SEQ ID NO:84)
и в нижеследующей последовательности, происходящей от белка IL-13 обезьян cynomolgus, такой как:
DLLVHLKKLFRE (SEQ ID NO:85).
NB: Выделение жирным шрифтом означает, что в этом положении остаток человеческого IL-13 отличается от остатка ортолога IL-13 обезьян cynomolgus.
В соответствии с этим было установлено, что родительское мышиное mAb 6A1, химерное 6А1, L1+A1 и L2+A1 иммуноспецифически связываются с человеческим IL-13 в положениях остатков 97-108 SEQ ID NO:9.
6.8. Тонкое картирование эпитопов для 6А1 с использованием биотинилированных пептидов
Эпитоп, связывающийся с mAb 6A1, определяли путем проведения анализа на связывание пептидов, имеющих приблизительную последовательность KDLLLHLKKLFREG, с человеческим IL-13, и пептидов, имеющих приблизительную последовательность KDLLVHLKKLFREG, с IL-13 обезьян cynomolgus. Для точного определения линейного эпитопа, связывающегося с mAb 6A1, к пептидам присоединяли 1 аминокислоту, которую затем удаляли либо с N-, либо с С-конца этих родительских пептидных последовательностей (т.е. KDLLLHLKKLFREG или KDLLVHLKKLFREG).
Для детекции связывания иммобилизованного биотинилированного пептида с родительским mAb 6A1 применяли метод ELISA.
Идентификационные номера пептидов (413-447) и их соответствующие последовательности приводятся ниже.
Пептидные последовательности:
Пример: 96-луночный планшет, применяемый для проведения этого анализа.
NB: Числа означают пептид в каждой лунке.
Оптические плотности при 490 нм в 96 лунках
См. фигуры 16а и 16b. Эти результаты показали, что родительское mab 6A1 связывается с линейным эпитопом, имеющим аминокислотную последовательность KKLFR, в С-концевой области человеческого IL-13 и ортолога IL-13 обезьян cynomolgus.
Кроме того, все антитела, а именно химерное антитело mAb 6A1, антитело L1+A1 и антитело L2+A1, связывались с одним и тем же линейным эпитопом (т.е., KKLFR) в С-концевой области человеческого IL-13 (данные для химерного mAb 6A1, L1+A1 и L2+A1 не приводятся). Кроме того, было обнаружено, что родительское mAb 6A1 связывается с тем же эпитопом IL-13 обезьян cynomolgus.
В итоге ELISA-результаты показали, что все антитела, а именно родительское мышиное mAb 6A1, химерное 6А1, L1+A1 и L2+A1, связываются в последовательности, происходящей от человеческого белка IL-13, а именно в KKLFR.
6.9. Аланиновое сканирование эпитопа, связывающегося с 6А1, с использованием биотинилированных пептидов
Для идентификации некоторых ключевых остатков, участвующих во взаимодействии IL-13 с mAb 6A1, был разработан метод аланинового сканирования, проводимый с использованием родительской пептидной последовательности, содержащей связывающийся эпитоп KKLFR (т.е. QFVKDLLLHLKKLFREGRFN). Для осуществления этого анализа получали пептиды (поставляемые фирмой AnaSpec Inc.), где одну аминокислоту последовательно заменяли аланиновым остатком в каждом положении аминокислоты в эпитопе KKLFR (а также каждую из аминокислот, непосредственно ограничивающих этот эпитоп).
Для детекции связывания иммобилизованного биотинилированного пептида с родительским mAb 6A1 и L1+A1 применяли метод ELISA.
Пептиды, полученные для этого анализа, и их соответствующие идентификационные номера приводятся ниже:
Результаты: оптические плотности при 490 нм.
Усредненные результаты теста (n=2).
Данные для родительского (мышиного) mAb 6A1:
Данные для L1+A1:
См. фигуры 17а и 17b.
Эти данные дают основание предположить, что ключевыми аминокислотными остатками, участвующими во взаимодействии mAb 6A1 или L1+A1 с человеческим IL-13, являются аргинин (R) в положении 107 и лизин (К) в положении 103.
Для подтверждения этого эффекта в данном интервале разведения mAb этот анализ повторяли, но с использованием mAb 6A1 и L1+A1 при различных концентрациях.
Родительское мышиное mAb 6A1 (фигура 17с) и гуманизованный кандидат L1+A1 (фигура 17d) анализировали на связывание с пептидами, используемыми для аланинового сканирования (SEQ ID NO:129, 131-135) при различных концентрациях. Поскольку пептиды были распределены по двум 96-луночным планшетам, то родительскую пептидную последовательность, не содержащую аланиновых замен (SEQ ID NO:129), анализировали на обоих планшетах, а поэтому получали два результата на график. Это было сделано для того, чтобы определить, имеются ли какие-либо основные отличия между планшетами, и в обоих случаях не наблюдалось каких-либо видимых отличий.
Было обнаружено, что пептиды, содержащие замены К103А, L105A и F106A (SEQ ID NO:131, 133 и 134 соответственно; нумерация остатков представлена в SEQ ID NO:9), имели почти такой же профиль связывания с mAb, как и родительский пептид (SEQ ID NO:129), а следовательно, эти остатки не играют решающей роли в связывании 6А1/L1+A1 с IL-13. Однако было обнаружено, что пептиды, содержащие замены К104А и R107A (SEQ ID NO:132 и 135 соответственно; нумерация остатков представлена в SEQ ID NO:9), имели пониженную способность связывания с 6А1/L1+A1, чем родительский пептид (SEQ ID NO:129), в частности, при более низких концентрациях, что указывает на то, что эти остатки не играют решающей роли в оптимальном связывании 6А1/L1+A1 с IL-13.
См. фигуры 17с и 17d.
Эти данные показали, что ключевыми аминокислотными остатками, участвующими во взаимодействии родительского (т.е. мышиного) 6A1 или L1+A1 с человеческим IL-13, являются аргинин (R) в положении 107 и лизин (К) в положении 103 SEQ ID NO:9.
Раздел 7. Материалы и методы
В данном разделе, где это необходимо, приводятся нижеследующие используемые материалы и методы. Эти материалы и методы являются репрезентативными. При повторном проведении экспериментов в указанные материалы и методы могут быть внесены небольшие изменения.
Материалы:
Система выделения полноразмерной РНК SV: Promega Z3100;
Система ОТ-ПЦР с удлинением цепи: Promega A1250;
Набор для экстракции из геля QIAquick: Qiagen 28704;
Раствор для загрузки геля: Sigma G7654;
Агароза: Invitrogen 15510-019;
Этидийбромид: Sigma E1510;
Буфер ТАЕ: получаемый в лаборатории;
ДНК-лэддер размером 100 п.н.: New England BioLabs N3231S;
Набор для клонирования ТА: Invitrogen 45-0046;
Клетки TOP10F': Invitrogen 44-0300;
L-агар + 100 мкг/мл ампициллина: получаемый в лаборатории;
X-Gal, 50 мг/мл в ДМФ: Promega V394A;
ДНК-полимераза AmpliTaq: Applied Biosystems;
10х ПЦР-буфер: Applied Biosystems;
Е-гель, 1,2% агароза: Invitrogen G501801;
Среда LB + 100 мкг/мл ампициллина: получаемая в лаборатории;
Набор для получения мини-препаратов QIAprep Spin Miniprep: Qiagen 27106;
Набор для ПЦР-очистки MinElute: Qiagen 28004;
NE-буфер 2, 10х конц.: New England Biolabs B7002S;
Очищенный BSA, 100x конц.: New England Biolabs В9001S;
BsiWI: New England Biolabs R0553L;
Hind III: Promega R604A;
Spe I: New England Biolabs R0133S;
Система быстрого лигирования ДНК LigaFast: Promega M8225;
Химически компетентные клетки DH5α с максимальной эффективностью: Invitrogen 18258-012;
Среда SOC: получаемая в лаборатории;
Набор QIAfilter Plasmid Maxi: Qiagen 12263;
Среда МЕМ Дульбекко с глутамаксом-1: Invitrogen 31966-021;
Среда Optimem 1 с глутамаксом-1: Invitrogen 51985-026;
Реагент для трансфекции TransFast: Promega E2431;
1 мл рекомбинантного белка А - Сефарозы FF HiTrap: Amersham Biosciences 17-5079-01;
PBS Дульбекко: Sigma D8537;
Буфер для элюирования IgG ImmunoPure: Pierce 21009;
1М Trizma-HCl, pH 8,0: Sigma T2694;
ДНК-полимераза ProofStart: Qiagen 1016816;
ПЦР-буфер ProofStart: Qiagen 1016961.
7.1. ELISA-анализ на связывание с IL-13 человека или IL-13 обезьян cynomolgus
Этот анализ представляет собой ELISA, позволяющий детектировать связывание антитела с IL-13 человека или обезьян cynomolgus. Данный анализ проводили в формате “сэндвич”-ELISA.
7.1.1. Материалы:
1. Иммунопланшет Nunc Immunoplate 1 F96 Maxisorp (Life Technologies, 4-39454A);
2. Человеческий IL-13 (Cambridge Biosciences, cat.№ СН1-013);
3. IL-13 обезьян cynomolgus (изготавливаемый фирмой GlaxoSmithkline);
4. Козье поликлональное антитело против человеческого IL-13 (R+D Systems, cat. № AF-213-NA);
5. ПХ-конъюгированное антитело против человеческого IgG (Sigma, Cat. № А-6029);
6. ПХ-конъюгированное антитело против мышиного IgG (Sigma, Cat. № А-9309);
7. Карбонатный/бикарбонатный буфер (Sigma; cat. № С-3041);
8. TBST [забуференный трисом физиологический раствор (6,06 г триса + 8,06 г NаСl + 0,2 г КСl + H2O до объема 1 литр) + 0,05% твина-20];
9. BSA (Sigma A-7030);
10. OPD (Sigma, Cat.№ Р-9187);
11. Серная кислота.
7.1.2. Метод
1. Блокирующим раствором является 3% BSA+TBST.
2. Промывочным раствором является TBST.
3. ELISA-планшеты “Nunc Maxisorp” сенсибилизировали 50 мкл 5 мкг/мл козьего поликлонального антитела против человеческого IL-13 (R+D Systems, cat.№ AF-213-NA. Раствор приготавливали при исходной концентрации 500 мкг/мл в соответствии с инструкциями производителей и хранили в аликвотах при -20°С) в карбонатном/бикарбонатном буфере (Sigma; cat.№ С-3041, приготовленном в соответствии с инструкциями производителей), а затем планшеты закрывали с помощью приспособления для заклеивания планшетов и инкубировали в течение ночи при 4°С.
4. Блокировали 100 мкл 3% BSA/TBST и инкубировали при комнатной температуре в течение 1 часа.
5. Промывали 3 раза в TBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
6. Добавляли 20 нг на лунку (в объеме 50 мкл) человеческого IL-13 (Cambridge Biosciences, cat. № СН1-013. Раствор приготавливали при исходной концентрации 100 нг/мл в соответствии с инструкциями производителей, и хранили в аликвотах при -20°С) или 20 нг на лунку IL-13 обезьян cynomolgus в блокирующем растворе, и инкубировали при комнатной температуре в течение 1 ч.
7. Промывали 3 раза в TBST.
8. Добавляли 50 мкл образца антитела (если это необходимо, проводили титрование до получения данных титра в конечной точке) в блокирующем растворе и инкубировали при комнатной температуре в течение 1 часа.
9. Промывали 3 раза в TBST.
10. Для химерного антитела или гуманизованного антитела 6А1 связывание детектировали с использованием 50 мкл на лунку ПХ-конъюгированного антитела против человеческого IgG (Sigma, Cat. № А-6029) при разведении 1/2000 в блокирующем растворе в течение 1 часа при комнатной температуре. Для мышиного моноклонального антитела 6А1 связывание детектировали с использованием 50 мкл на лунку ПХ-конъюгированного антитела против мышиного IgG (Sigma, Cat. № А-9309) при разведении 1/1000 в блокирующем растворе в течение 1 часа при комнатной температуре.
11. Промывали 3 раза в TBST.
12. Проявляли 100 микролитрами OPD (Sigma, Cat. № Р-9187, приготовленного в соответствии с инструкциями производителей), реакцию прекращали добавлением 50 мкл 3М Н2SO4 и планшеты считывали при оптической плотности 490 нм. Время проявления составляло примерно 12 минут.
7.2. ELISA-анализ на связывание человеческого IL-13 с человеческой IL-13Rα1-цепью
Этот ELISA-анализ позволяет определить, может ли антитело ингибировать связывание человеческого IL-13 с человеческой IL-13Rα1-цепью.
7.2.1. Материалы:
1. Иммунопланшет Nunc Immunoplate 1 F96 Maxisorp (Life Technologies, 4-39454A);
2. Человеческий IL-13Rα1-Fc (R&D Systems, cat.№ 146-IR);
3. Det1-меченный человеческий IL-13 (приготовленный в лабораторных условиях);
4. Биотинилированное антитело против человеческого IL-13 (R&D Systems, cat.№ BAF213);
5. Стрептавидин-ПХ;
6. Карбонатный/бикарбонатный буфер (Sigma; cat. № С-3041);
7. TBST [забуференный трисом физиологический раствор (6,06 г триса + 8,06 г NаСl + 0,2 г КСl + H2O до объема 1 литр) + 0,05% твина-20];
8. BSA (Sigma A-7030);
9. OPD (Sigma, Cat.№ Р-9187);
10. Серная кислота.
7.2.2. Метод
1. Блокирующим раствором является 3% BSA+TBST.
2. Промывочным раствором является TBST.
3. ELISA-планшеты “Nunc Maxisorp” сенсибилизировали 50 мкл 5 нг/мл человеческого IL-13Rα1-Fc в карбонатном/бикарбонатном буфере. Планшеты закрывали с помощью приспособления для заклеивания планшетов и инкубировали в течение ночи при 4°С.
4. Блокировали 100 микролитрами 3% BSA/TBST и инкубировали при комнатной температуре в течение 1 часа.
5. Промывали 3 раза в TBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
6. В полном объеме 50 мкл предварительно инкубировали 0,04 нг/мкл det-1-меченного человеческого IL-13 с образцом антитела (титрованного) в блокирующем растворе в течение 30 минут. Затем предварительно инкубированный образец добавляли в сенсибилизированный рецептором ELISA-планшет и инкубировали при комнатной температуре в течение 1 ч.
7. Промывали 3 раза в TBST.
8. Детектировали связывание человеческого IL-13 с использованием 50 мкл на лунку биотинилированного антитела против человеческого IL-13, разведенного при 1 мкг/мл. Затем инкубировали в течение 1 часа при комнатной температуре.
9. Промывали 3 раза в TBST.
10. Добавляли 50 мкл на лунку конъюгата “стрептавидин-ПХ” при разведении 1/1000. Затем инкубировали в течение 1 часа при комнатной температуре.
11. Промывали 3 раза в TBST.
12. Проявляли 100 микролитрами/лунку OPD (Sigma, Cat. № Р-9187, приготовленного в соответствии с инструкциями производителей), реакцию прекращали добавлением 50 мкл на лунку 3М Н2SO4 и планшеты считывали при оптической плотности 490 нм. Время проявления составляло примерно 2 минуты.
7.3. ELISA-анализ на связывание человеческого IL-13 с человеческой IL-13Rα2-цепью
Этот ELISA-анализ позволяет определить, может ли антитело ингибировать связывание человеческого IL-13 с человеческой IL-13Rα2-цепью.
7.3.1. Материалы:
1. Иммунопланшет Nunc Immunoplate 1 F96 Maxisorp (Life Technologies, 4-39454A);
2. Антитело против человеческого IgG (Sigma, cat.№ I-3382);
3. Человеческий IL-13Rα2-Fc (R&D Systems, cat.№ 614-IR);
4. Det1-меченный человеческий IL-13 (приготовленный в лабораторных условиях);
5. Биотинилированное антитело против человеческого IL-13 (R&D Systems, cat.№ BAF213);
6. Стрептавидин-ПХ
7. Карбонатный/бикарбонатный буфер (Sigma; cat. № С-3041);
8. TBST [забуференный трисом физиологический раствор (6,06 г триса + 8,06 г NаСl + 0,2 г КСl + H2O до объема 1 литр) + 0,05% твина-20];
9. BSA (Sigma A-7030);
10. OPD (Sigma, Cat.№ Р-9187);
11. Серная кислота.
7.3.2. Метод
1. Блокирующим раствором является 3% BSA+TBST.
2. Промывочным раствором является TBST.
3. ELISA-планшеты “Nunc Maxisorp” сенсибилизировали 50 мкл антитела против человеческого IgG, разведенного до 1/1000 в карбонатном/бикарбонатном буфере. Планшеты закрывали с помощью приспособления для заклеивания планшетов и инкубировали в течение ночи при 4°С.
4. Блокировали 100 микролитрами 3% BSA/TBST и инкубировали при комнатной температуре в течение 1 часа.
5. Промывали 3 раза в TBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
6. Добавляли 50 мкл на лунку 1 мкг/мл человеческого IL-13Rα2-Fc в блокирующем растворе. Планшеты закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре в течение 1 ч.
7. Промывали 3 раза в TBST.
8. В полном объеме 50 мкл предварительно инкубировали 0,004 нг/мкл det1-меченного человеческого IL-13 с образцом антитела (титрованного) в блокирующем растворе в течение 30 минут. Затем предварительно инкубированный образец добавляли в сенсибилизированный рецептором ELISA-планшет и инкубировали при комнатной температуре в течение 1 ч.
9. Промывали 3 раза в TBST.
10. Детектировали связывание человеческого IL-13 с использованием 50 мкл на лунку биотинилированного антитела против человеческого IL-13, разведенного при 1 мкг/мл. Затем инкубировали в течение 1 часа при комнатной температуре.
11. Промывали 3 раза в TBST.
12. Добавляли 50 мкл на лунку конъюгата “стрептавидин-ПХ” при разведении 1/1000. Затем инкубировали в течение 1 часа при комнатной температуре.
13. Промывали 3 раза в TBST.
14. Проявляли 100 микролитрами/лунку OPD (Sigma, Cat. № Р-9187, приготовленного в соответствии с инструкциями производителей), реакцию прекращали добавлением 50 мкл на лунку 3М Н2SO4 и планшеты считывали при оптической плотности 490 нм. Время проявления составляло примерно 2 минуты.
7.4. Биоанализ на нейтрализацию IL-13 (анализ на пролиферацию клеток TF-1)
Этот анализ представляет собой биоанализ IL-13, который может быть применен для определения нейтрализующей способности антитела против IL-13. В описанном ниже методе используется рекомбинантный человеческий IL-13 или IL-13 обезьян cynomolgus. В этом анализе может быть также использован экспрессируемый у млекопитающего человеческий IL-13 или Q130-вариант человеческого IL-13. (Клетки TF-1 также пролиферируются в ответ на человеческий IL-5. Этот анализ может быть также применен для оценки способности антитела 6А1 нейтрализовать биологическую активность человеческого IL-5).
7.4.1. Материалы:
1. Клеточная линия TF-1 (полученная в лаборатории).
2. 96-луночные планшеты для культивирования ткани (Invitrogen).
3. Человеческий IL-13 (Cambridge Bioscience, cat. № СН1-013).
4. Анализ на пролиферацию нерадиоактивных клеток CellTiter 96 (Promega, Cat. № G4000).
7.4.2. Метод
1. Осуществляли метод определения способности mAb против человеческого IL-13 нейтрализовать биологическую активность рекомбинантного IL-13 человека или обезьян cynomolgus в биоанализе клеток TF-1 (клеточная линия TF-1 была получена в лаборатории и не имеет регистрационного номера АТСС).
2. Этот анализ проводили в стерильных 96-луночных планшетах для культивирования тканей (Invitrogen) в стерильных условиях. Все тесты проводили с тремя повторностями.
3. Предварительно инкубировали 10 нг/мл человеческого IL-13 (Cambridge Bioscience, cat.№ СН1-013. Маточный раствор приготавливали при концентрации 100 нг/мкл в соответствии с инструкциями производителей в стерильных условиях в шкафу для культивирования тканей класса 2 и хранили в небольших аликвотах при -20°С) или 10 нг/мл IL-13 обезьян cynomolgus (получаемого в лабораторных условиях из СА) с различными разведениями mAb против человеческого IL-13 (в 3-кратных разведениях от 6 мкг/мл до 0,025 мкг/мл) в полном объеме 50 мкл в течение 1 часа при 37°С. В эксперимент также были включены позитивные контрольные лунки, содержащие IL-13, но не mAb против человеческого IL-13. Кроме того, в эксперимент были также включены негативные контрольные лунки, не содержащие IL-13 и mAb против человеческого IL-13. Такое предварительное инкубирование проводили в стерильном круглодонном 96-луночном планшете с низким уровнем связывания белка. (Следует отметить, что при добавлении клеток на более поздней стадии IL-13 и mAb против человеческого IL-13 использовали в половинной концентрации).
4. 50 мкл клеток TF-1 высевали при плотности 2 × 105/мл в стерильный 96-луночный планшет для культивирования тканей. После предварительного инкубирования в течение 1 часа к клеткам добавляли образец IL-13 и mAb против человеческого IL-13. Полученный продукт в конечном аналитическом объеме 100 мкл, содержащий различные разведения mAb против человеческого IL-13, рекомбинантный IL-13 и клетки TF-1, инкубировали при 37°С в течение примерно 70 часов в СО2-инкубаторе с повышенной влажностью.
5. Примерно через 66 часов лунки сканировали для подтверждения их стерильности и отсутствия в них бактериальной инфекции.
6. В течение последних 4 часов инкубирования добавляли 15 мкл/лунку стерилизованного на фильтре субстрата МТТ (Сat.№ G4000, Promega, приготовленного в соответствии с инструкциями производителя).
7. Реакцию прекращали добавлением 100 мкл раствора для прекращения реакции (поставляемого в наборе с МТТ) для солюбилизации метаболизированного продукта формазанового синего. Затем продукт оставляли, по меньшей мере, на 2 часа, после чего пипетку с продуктом встряхивали для облегчения растворения кристаллов. Альтернативно, планшет закрывали с помощью приспособления для заклеивания планшетов и оставляли на ночь при 4°С, а затем, на следующий день, пипетку встряхивали (такая процедура облегчает пипетирование).
8. Считывали оптическую плотность раствора в каждой лунке 96-луночного планшет-ридера на длине волны 570 нм.
9. Способность mAb против человеческого IL-13 нейтрализовать биоактивность IL-13 человека или обезьян cynomolgus выражали как концентрацию mAb против человеческого IL-13, необходимую для нейтрализации биологической активности определенного количества IL-13 человека или обезьян cynomolgus (5 нг/мл)/50% (=ND50). Чем меньше требуемая концентрация, тем эффективнее способность нейтрализации.
Пример: 96-луночный планшет, используемый в этом анализе
TF-1
↓
↓
7.5. ELISA-анализ на связывание с человеческим IL-4
Этот анализ представляет собой ELISA, позволяющий детектировать связывание антитела с человеческим IL-4. Данный анализ проводили в формате “сэндвич”-ELISA.
7.5.1. Материалы:
1. Иммунопланшет Nunc Immunoplate 1 F96 Maxisorp (Life Technologies, 4-39454A).
2. Человеческий IL-4 (R+D Systems, cat.№).
3. Козье поликлональное антитело против человеческого IL-4 (R+D Systems, Cat.№ AF-204-NA).
4. Биотинилированное крысиное моноклональное антитело против человеческого IL-4 (BD/Pharmingen, Cat.№).
5. ПХ-конъюгированное антитело против мышиного IgG (Dako, Cat. № P0260).
6. ПХ-конъюгированное антитело против мышиного IgG (Sigma, Cat. № А-9309).
7. Карбонатный/бикарбонатный буфер (Sigma; cat. № С-3041);
8. PBST (PBS + 0,05% твина-20).
9. BSA (Sigma A-7030).
10. OPD (Sigma, Cat.№ Р-9187).
11. Серная кислота.
7.5.2. Метод
1. Блокирующим раствором является 3% BSA+РBST.
2. Промывочным раствором является РBST.
3. ELISA-планшеты “Nunc Maxisorp” сенсибилизировали 50 мкл 5 мкг/мл козьего поликлонального антитела против человеческого IL-4 (R+D Systems, cat.№ AF-204-NA. Маточный раствор приготавливали при концентрации 500 мкг/мл в соответствии с инструкциями производителей и хранили в аликвотах при -20°С) в карбонатном/бикарбонатном буфере (Sigma; cat.№ С-3041, приготовленном в соответствии с инструкциями производителей), а затем планшеты закрывали с помощью приспособления для заклеивания планшетов и инкубировали в течение ночи при 4°С.
4. Блокировали 100 микролитрами 3% BSA/РBST и инкубировали при комнатной температуре и при нормальном давлении в течение 1 часа.
5. Промывали 3 раза в РBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
6. Добавляли 1 нг/мл (в объеме 50 мкл) человеческого IL-4 в блокирующем растворе и инкубировали при комнатной температуре в течение 1 ч.
7. Промывали 3 раза в РBST.
8. Добавляли 50 мкл образца антитела (если это необходимо, проводили титрование для получения титра в конечной точке) в блокирующем растворе и инкубировали при комнатной температуре и при нормальном давлении в течение 1 часа. В анализе на связывание с человеческим IL-4 в качестве позитивного контроля использовали биотинилированное моноклональное антитело против человеческого IL-4 (титрованное).
9. Промывали 3 раза в РBST.
10. Для мышиного моноклонального антитела 6А1 связывание детектировали с использованием 50 мкл на лунку ПХ-конъюгированного антитела против мышиного IgG (Sigma, Cat. № А-9309) при разведении 1/1000 в блокирующем растворе в течение 1 часа при комнатной температуре и при нормальном давлении. Для химерного антитела или гуманизованного антитела 6А1 связывание детектировали с использованием 50 мкл на лунку ПХ-конъюгированного антитела против человеческого IgG (Sigma, Cat. № А-6029) при разведении 1/2000 в блокирующем растворе в течение 1 часа при комнатной температуре и при нормальном давлении. Для биотинилированного крысиного моноклонального антитела против человеческого IL-4, используемого в качестве позитивного контроля, детекцию проводили с использованием антитела, конъюгированного со стрептавидином-ПХ. (Альтернативно, с использованием ПХ-конъюгированного антитела против мышиного IgG, P0260 могут быть детектированы антитело 6А1 и биотинилированное крысиное моноклональное антитело против человеческого IL-4).
11. Промывали 3 раза в РBST.
12. Проявляли 100 микролитрами OPD (Sigma, Cat. № Р-9187, приготовленного в соответствии с инструкциями производителей), реакцию прекращали добавлением 50 мкл 3М Н2SO4 и планшеты считывали при оптической плотности 490 нм.
7.6. ELISA-анализ на картирование эпитопов
Этот анализ представляет собой ELISA, позволяющий детектировать связывание мышиного mAb 6A1 с пептидами IL-13 человека или обезьян cynomolgus.
7.6.1. Материалы:
1. Иммунопланшет Nunc Immunoplate 1 F96 Maxisorp (Life Technologies, 4-39454A).
2. Стрептавидин ImmunoPure® (Pierce, cat. № 21125).
3. PBST (забуференный фосфатом физиологический раствор + 0,05% твина-20).
4. BSA (Sigma A-7030).
5. 16-мерные пептиды IL-13 человека и обезьян cynomolgus, сдвиг = 4 (специально изготовленные фирмой Mimotopes)
6. 20-мерные пептиды, используемые в качестве позитивного и негативного контроля (специально изготовленные фирмой Mimotopes).
7. Mab 6A1.
8. Контрольное Ab (специально изготовленное фирмой Mimotopes).
9. ПХ-конъюгированное кроличье антитело против мышиных Ig (DAKO, кодовый номер Р0260).
10. OPD (Sigma, Cat.№ Р-9187).
11. 3М серная кислота.
7.6.2. Метод
1. Блокирующим раствором является 3% BSA+РBST.
2. Промывочным раствором является РBST.
3. ELISA-планшеты “Nunc Maxisorp” сенсибилизировали 100 микролитрами 5 мкг/мл стрептавидина ImmunoPure® (Pierce, cat. № 21125; препарат приготавливали при исходной концентрации 1 мг/мл в соответствии с инструкциями производителей и хранили в аликвотах при +4°С) с использованием PBST в качестве разбавителя. Затем планшеты инкубировали в течение ночи при 37°С для сушки раствора.
4. Блокировали 200 микролитрами 3% BSA/РBST. После этого планшет закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре и при нормальном давлении в течение 1 ч.
5. Промывали 3 раза в PBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
6. В лунки, взятые в дубликате, в которых в качестве разбавителя использовали PBST, добавляли 100 мкл на лунку (за исключением контрольных лунок) 1000-кратных разведений каждого пептида (которые растворяли в соответствии с инструкциями производителей в 200 мкл 40% ацетонитрила/60% воды, а затем разделяли на аликвоты в 10-кратных разведениях в том же самом растворителе и хранили при -20°С).
7. В контрольные лунки, взятые в дубликате, в которых в качестве разбавителя использовали PBST, добавляли 100 мкл на лунку 10-кратных разведений контрольных пептидов (которые растворяли в соответствии с инструкциями производителей в 1 мл 40% ацетонитрила/60% воды, а затем хранили при -20°С). Планшеты закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре и при нормальном давлении в течение 1 часа на шюттль-аппарате с горизонтальной платформой.
8. Промывали 3 раза в РBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
9. Добавляли 100 мкл на лунку (за исключением контрольных лунок) 1,506 мкг/мл мышиного mAb в PBST.
10. Добавляли 100 мкл на лунку (только в контрольные лунки) 4-, 16- и 32-кратных разведений контрольного антитела (которое использовали в соответствии с инструкциями производителей и хранили при -20°С) с использованием PBST в качестве разбавителя. Планшет закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре и при нормальном давлении в течение 1 часа на шюттль-аппарате с горизонтальной платформой.
11. Промывали 3 раза в PBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
12. Добавляли 100 мкл на лунку 2000-кратных разведений ПХ-конъюгированного кроличьего антитела против мышиного Ig (DAKO, кодовый номер Р0260, которое использовали в соответствии с инструкциями производителей и хранили при +4°С) с использованием PBST в качестве разбавителя. Планшет закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре и при нормальном давлении в течение 1 часа на шюттль-аппарате с горизонтальной платформой.
13. Промывали 3 раза в PBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
14. Проявляли 100 микролитрами OPD (Sigma, Cat. № Р-9187, приготовленного в соответствии с инструкциями производителей), реакцию прекращали добавлением 50 мкл 3М Н2SO4 и планшеты считывали при оптической плотности 490 нм. Время проявления составляло примерно 10 минут.
7.7. ELISA-анализ на тонкое картирование эпитопов
Этот анализ представляет собой ELISA, позволяющий детектировать связывание mAb 6A1 с пептидами IL-13 человека или обезьян cynomolgus.
7.7.1. Материалы:
1. Иммунопланшет Nunc Immunoplate 1 F96 Maxisorp (Life Technologies, 4-39454A).
2. Стрептавидин ImmunoPure® (Pierce, cat. № 21125).
3. PBST (забуференный фосфатом физиологический раствор + 0,05% твина-20).
4. BSA (Sigma A-7030).
5. Пептиды человеческого IL-13 и IL-13 обезьян cynomolgus, синтезированные методом “сети парциальных окон” (“partial window net”) (14-мерный пептид, усеченный по одной аминокислоте у N- и у С-конца одновременно; этот пептид был специально изготовлен фирмой Mimotopes).
6. 16-мерный пептид, используемый в качестве позитивного контроля (предварительно изготовленный на заказ фирмой Mimotopes).
7. Mab 6A1 (изготовленное в лаборатории).
8. ПХ-конъюгированное козье антитело против мышиного IgG (Fc-специфичное)(Sigma A-9309).
9. OPD (Sigma, Cat.№ Р-9187).
10. 3М серная кислота.
7.7.2. Метод
1. Блокирующим раствором является 3% BSA+РBST.
2. Промывочным раствором является РBST.
3. ELISA-планшеты “Nunc Maxisorp” сенсибилизировали 100 мкл 5 мкг/мл стрептавидина ImmunoPure® в сверхчистой воде (Pierce, cat. № 21125; препарат приготавливали при исходной концентрации 1 мг/мл в соответствии с инструкциями производителей и хранили при +4°С). Затем планшеты инкубировали в течение ночи при 37°С.
4. Блокировали 200 микролитрами 3% BSA/РBST. После этого планшет закрывали с помощью приспособления для заклеивания планшетов и инкубировали в течение ночи при +4°С.
5. Промывали 3 раза в РBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
6. В лунки, взятые в дубликате, в которых в качестве разбавителя использовали PBST, добавляли 100 мкл на лунку 1000-кратных разведений каждого пептида (которые растворяли в соответствии с инструкциями производителей в 200 мкл 40% ацетонитрила/60% воды, а затем хранили при -20°С). Планшет закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре в течение 1 часа на шюттль-аппарате с горизонтальной платформой.
7. Промывали 3 раза в РBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
8. Добавляли 100 мкл на лунку 3 мкг/мл 6А1, разведенного в 3% BSA в PBST. Планшет закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре в течение 1 часа на шюттль-аппарате с горизонтальной платформой.
9. Промывали 3 раза в РBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
10. Добавляли 100 мкл/лунку 1000-кратного разведения ПХ-конъюгированного козьего антитела против мышиных IgG (Sigma A-9309, которое использовали в соответствии с инструкциями производителей и хранили при +4°С) с использованием 3% BSA в PBST в качестве разбавителя. Планшет закрывали с помощью приспособления для заклеивания планшетов и инкубировали при комнатной температуре в течение 1 часа на шюттль-аппарате с горизонтальной платформой.
11. Промывали 3 раза в PBST (по меньшей мере, 200 мкл промывочного раствора/лунку/промывку).
12. Проявляли 100 микролитрами OPD (Sigma, Cat. № Р-9187, приготовленного в соответствии с инструкциями производителей), реакцию прекращали добавлением 50 мкл 3М Н2SO4 и планшеты считывали при оптической плотности 490 нм. Время проявления составляло примерно 10 минут.
7.8. Метод Biacore TM для анализа гуманизованных конструкций на связывание анти-IL-13 антитела с полноразмерным IL-13
Кинетический анализ проводили на аппарате Biacore 3000 с применением метода иммобилизации антител. Вкратце, для конструкций химерного 6А1 и гуманизованного антитела применяли метод захвата на белке А, а для родительского мышиного антитела 6А1 применяли метод захвата антителом против Fc мышиного IgG, поставляемым фирмой Biacore. Вкратце, этот метод заключался в следующем: Лиганд для захвата иммобилизовали на биосенсорном чипе СМ5 путем присоединения первичного амина в соответствии со стандартными протоколами Biacore и с использованием реагентов, поставляемых в наборе для присоединения первичного амина Biacore. Этот метод включает активацию сенсорной поверхности СМ5 путем пропускания через такую поверхность раствора 50 мМ N-гидроксисукцинимида (NHS) и 200 мМ N-этил-N'-диметиламинопропилкарбонида (EDC). Затем лиганд для захвата (растворенный в ацетатном буфере, рН 5 или рН 4,5) был присоединен к активированной сенсорной поверхности, после чего любые все еще активированные сложные эфиры были блокированы путем введения 1М гидрохлорида этаноламина, рН 8,5. Затем антитело-кандидат пропускали через поверхность, на которой были иммобилизованы либо белок А, либо антитело против Fc-фрагмента мышиного иммуноглобулина, в зависимости от того, является ли оно человеческим или мышиным, а затем его иммобилизовали. После регистрации стабильного сигнала связывания IL-13 пропускали через поверхность, на которой было иммобилизовано антитело, при различных определенных концентрациях. Затем кривые связывания анализировали с помощью аналитической компьютерной программы Biacore, BIAeval v.4.1, для определения кинетики реакций. Эксперименты проводили с использованием HBS-EP-буфера Biacore.
7.8.1. Метод Biacore TM для анализа связывания анти-IL-13 антитела с пептидом
Кинетический анализ проводили на аппарате Biacore 3000 с применением метода прямого связывания антитела с иммобилизованным пептидом IL-13. Вкратце, биотинилированный пептид IL-13 иммобилизовали с использованием биосенсорного чипа Biacore SA (стрептавидина). После этого антитела пропускали через сенсорную поверхность при различных концентрациях. Затем кривые связывания анализировали с помощью аналитической компьютерной программы Biacore, BIAeval v.4.1, для определения кинетики реакций. Эксперименты проводили с использованием HBS-EP-буфера Biacore.
8. Эффективность гуманизованного анти-IL-13-mAb L1+A1 у обезьян cynomolgus с моделью астмы
В этом разделе описан прогностический метод.
Модель легочного бронхоспазма у обезьян cynomolgus (Macaca fascicularis), индуцированного Ascaris suum (A.suum), известна как не-клиническая модель астмы или модель заболевания, ассоциированного с астмой у человека (Patterson R. et al., Trans. Assoc. Am. Physicians 1980 93:317-325; Patterson R. et al., J. Lab. Clin. Med. 1983, 101:864-872).
В этой модели животных с врожденной восприимчивостью легких к инфекции, вызываемой A.suum, заражали антигеном A.suum путем ингаляции для индуцирования у этих животных астматического ответа. Этот астматический ответ может быть охарактеризован путем измерения уровня гиперчувствительности дыхательных путей (AHR), инфильтрации клеток, определяемой в жидкости после бронхоальвеолярного лаважа (БАЛ) и уровней IgE в сыворотке. Экспериментальные методы аналогичны методам, описанным ранее в публикациях Mauser P. et al., Am. J. Resp. Crit. Care Med. 1995 204:467-472 и Evanoff H. et al., Immunologic Investigation 1992:21:39.
В этом исследовании участвовали 30 животных, предварительно отобранных и включенных в группу исследований, у которых наблюдался позитивный бронхоспазм в ответ на введение специфической дозы антигена A.suum. Антиген A.suum вводили каждому животному в дозе, вызывающей оптимальную реакцию доза-ответ (ORD). Такая доза представляет собой предварительно определенную дозу A.suum, которая индуцирует увеличение RL (легочной резистентности), по меньшей мере, на 40% и уменьшение CDYN (динамического изменения объема легких при колебаниях давления), по меньшей мере, на 35%, при ее введении путем аэрозольной ингаляции (для введения одной дозы с помощью ингалятора потребовалось свыше 15 вдохов).
Это исследование проводили в 2 фазы. В фазе 1 оценивали AHR в ответ на внутривенную (i.v.) инъекцию гистамина (дозы гистамина, достаточной для индуцирования увеличения RL, по меньшей мере, на 30% по сравнению с фоновым уровнем (PC30)) до введения (фоновая оценка функции легких на день 1) и после введения (на день 11) антигена A.suum (на дни 9 и 10, где A.suum вводили каждому животному в оптимальной предварительно определенной дозе путем аэрозольной ингаляции).
Фаза 2 идентична фазе 1, за исключением того, что животным вводили антитело (см. ниже), где каждое антитело вводили путем внутривенного вливания (i.v.) в виде 3 доз приблизительно по 30 мг/кг на дни 1, 5 и 9.
Группа 1 (n=12): L1+A1 (гуманизованное mAb против IL-13, SEQ ID NO:18 и SEQ ID NO:22).
Группа 2 (n=12): L1+A1 (гуманизованное mAb против IL-13, 30 мг/кг) и пасколизумаб (гуманизованное mAb против IL-4, 30 мг/кг).
Группа 3 (n=6): вводили только носитель, используемый в качестве негативного контроля.
AHR-данные фаз 1 и 2 вычисляли путем измерения давления и регистрации параметров состояния дыхательных путей, а именно легочной резистентности (RL) и динамического изменения объема легких при колебаниях давления (CDYN) в ответ на гистамин, с использованием механической системы дыхания Buxco. Максимальное процентное изменение фонового значения после введения антигена A.suum [для легочной резистентности (RL) и динамического изменения объема легких при колебаниях давления (CDYN)] сравнивали в фазах 1 и 2, т.е. с обработкой или без обработки антителом, и эти данные использовали для оценки фенотипа AHR.
Кроме того, образцы BAL брали на дни 1 и 11 в фазах 1 и 2 для определения уровня инфильтрации клеток, в частности эозинофилии. Для мониторинга уровней IgE также брали образцы сыворотки.
полинуклеотидов (PN)
(SEQ ID NO:)
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2605327C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2016 |
|
RU2650767C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2007 |
|
RU2472807C2 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА ЧЕЛОВЕКА К РЕЦЕПТОРУ IL-4 ЧЕЛОВЕКА | 2007 |
|
RU2445318C2 |
| АНТИ-IL-22R-АНТИТЕЛА | 2017 |
|
RU2758721C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА С ВЫСОКОЙ АФФИННОСТЬЮ К РЕЦЕПТОРАМ IL-4 ЧЕЛОВЕКА | 2009 |
|
RU2539774C2 |
| ИММУНОГЛОБУЛИН С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ДОМЕНАМИ И ЕГО ПРИМЕНЕНИЯ | 2006 |
|
RU2515108C2 |
| IL-17A-СВЯЗЫВАЮЩИЙ АГЕНТ И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2014 |
|
RU2682046C1 |
| Антитело против IL-4R и его применение | 2018 |
|
RU2758091C1 |
| ПОЛИПЕПТИДНЫЕ ВАРИАНТЫ С ИЗМЕНЕННОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2005 |
|
RU2367667C2 |


































































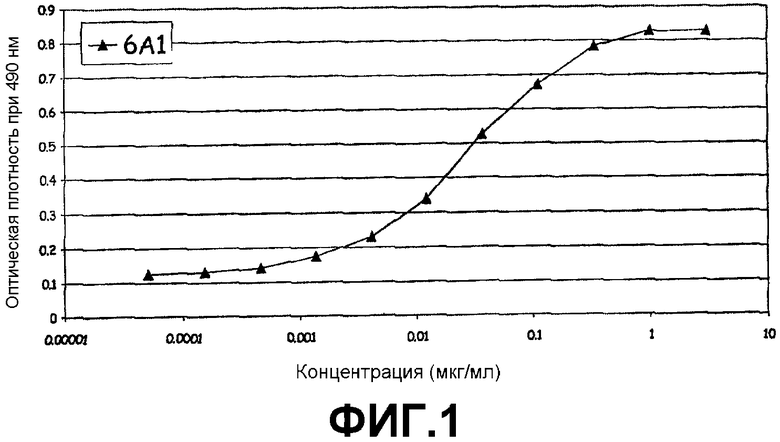
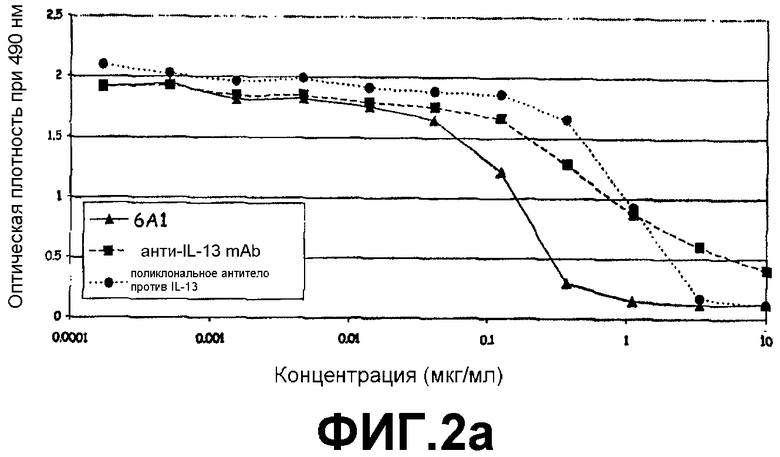
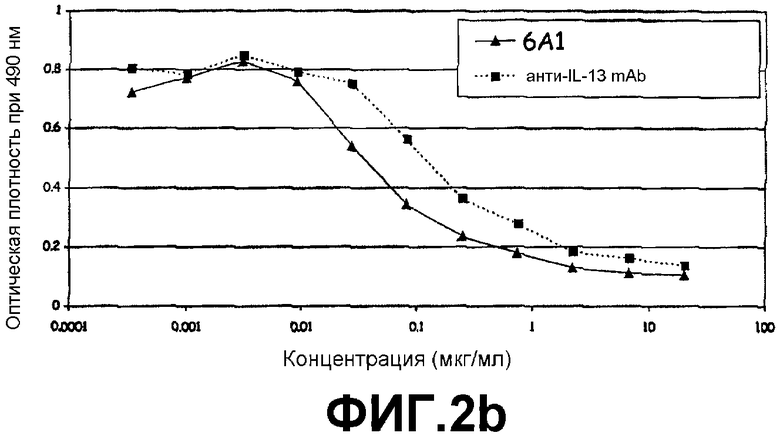
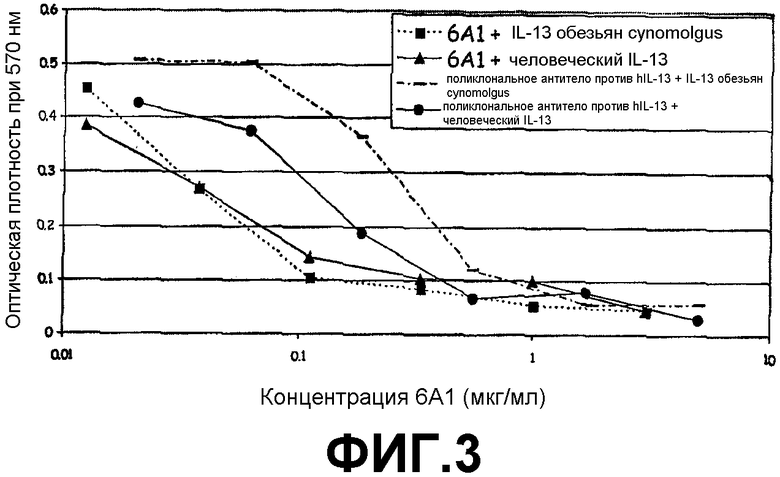
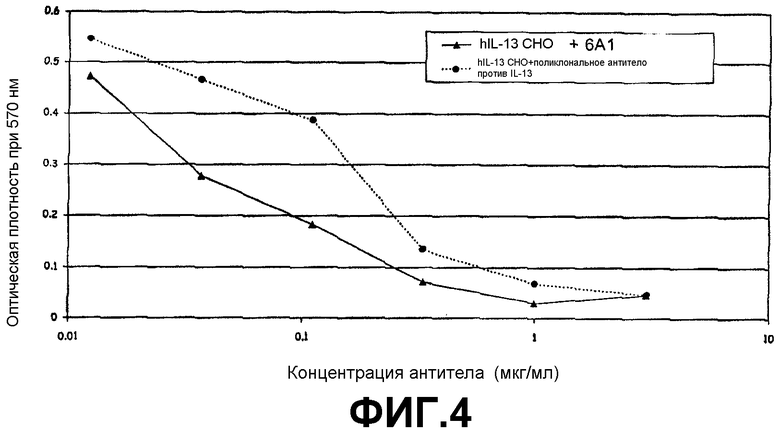
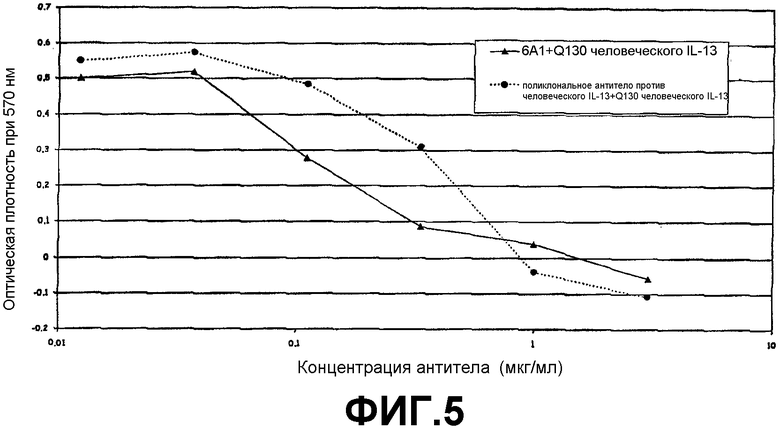
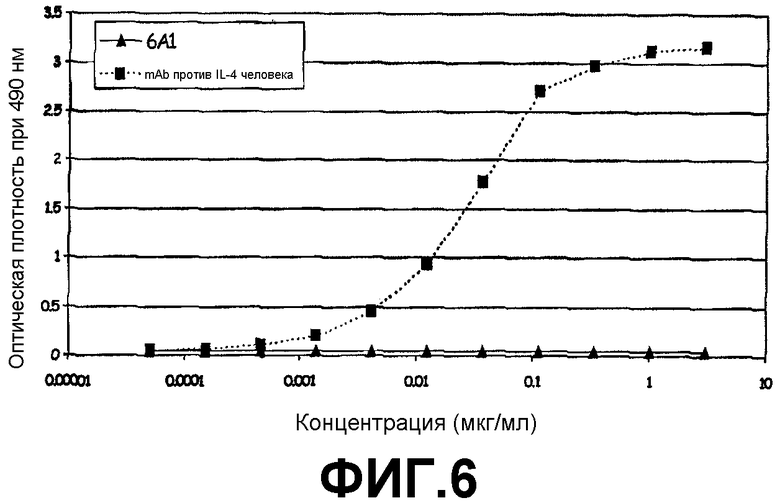
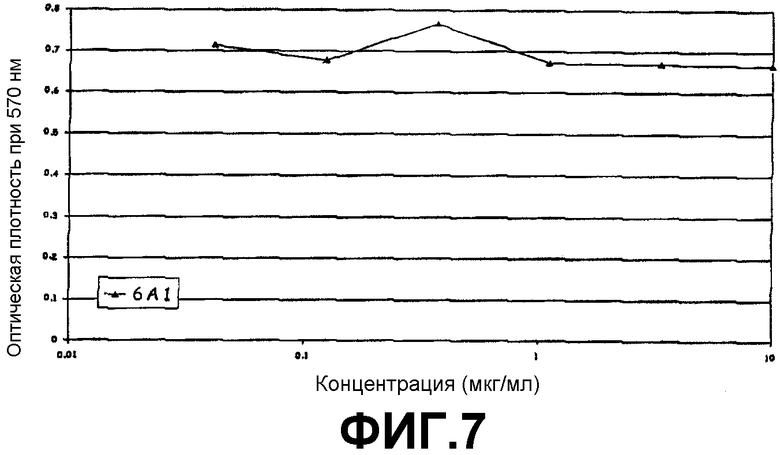
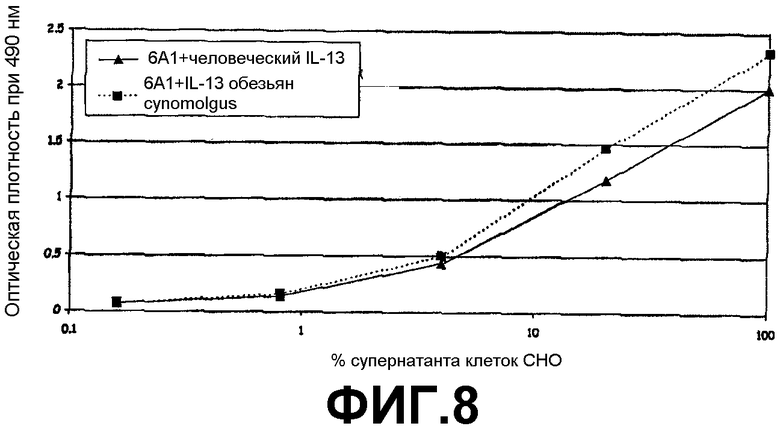
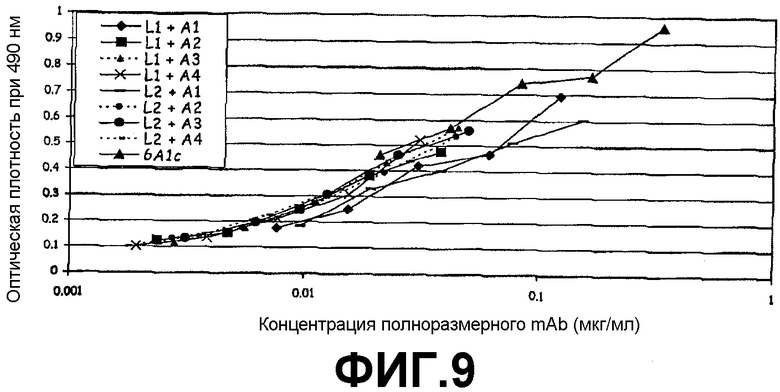
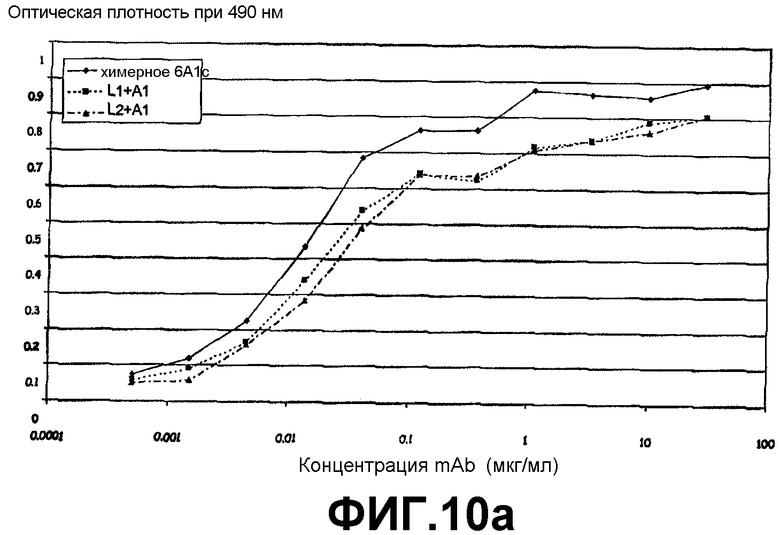
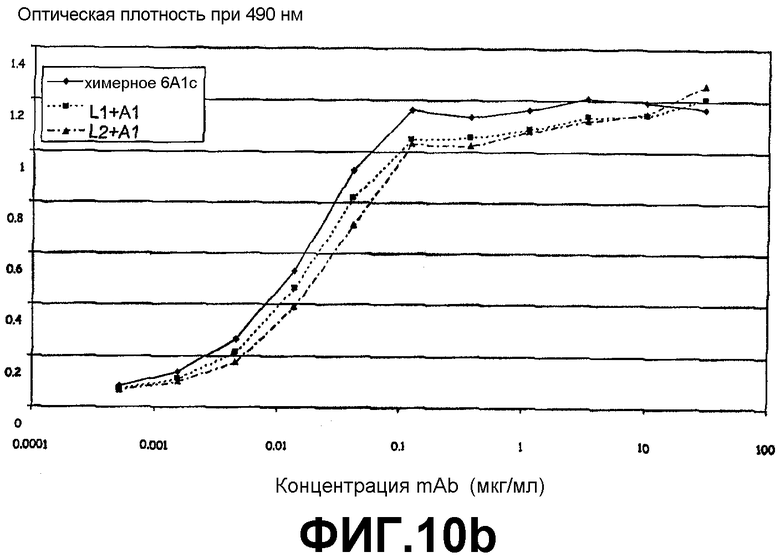
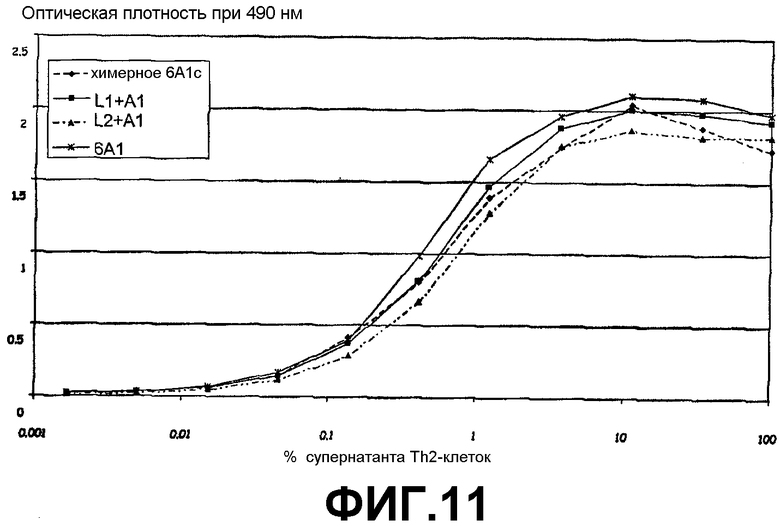
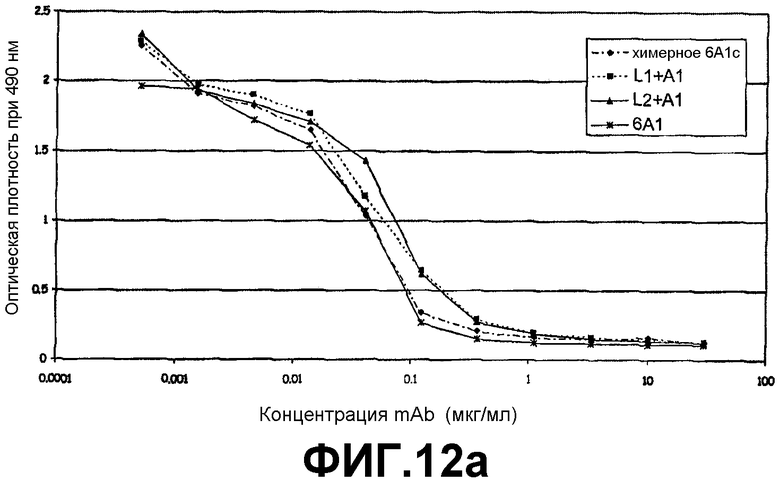
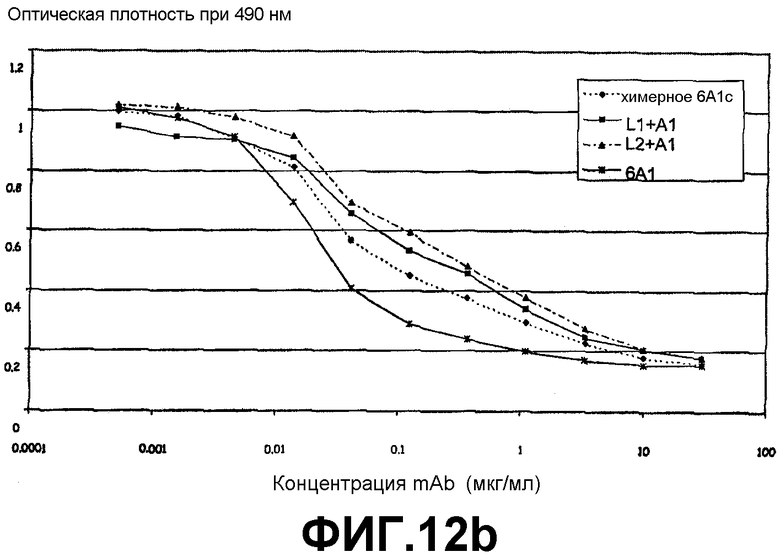
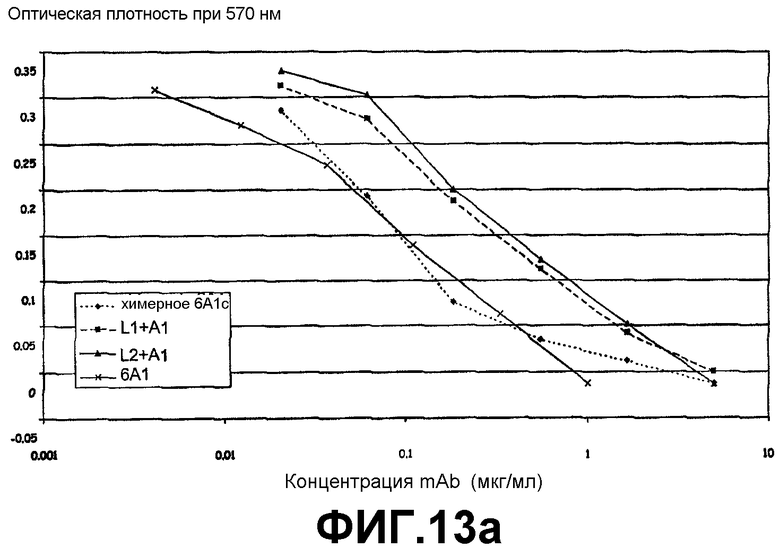
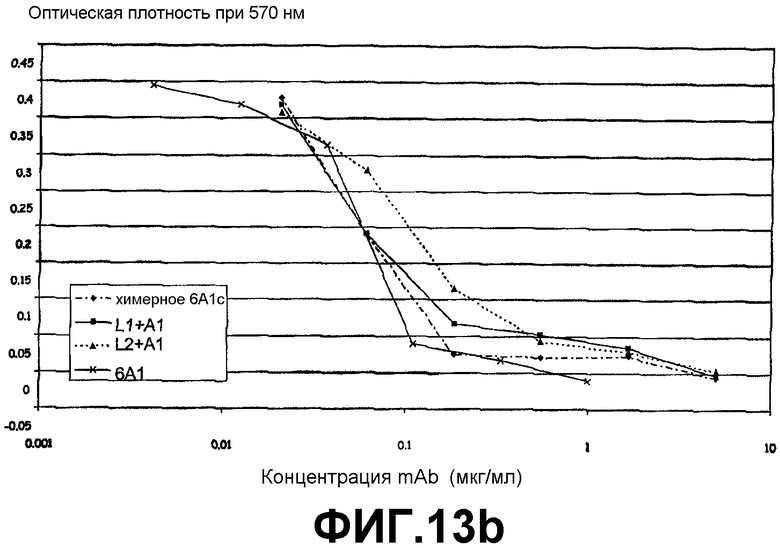
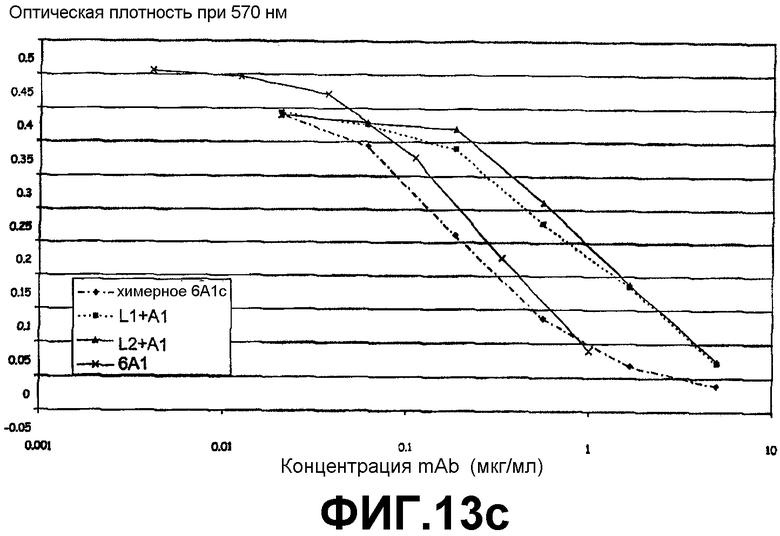
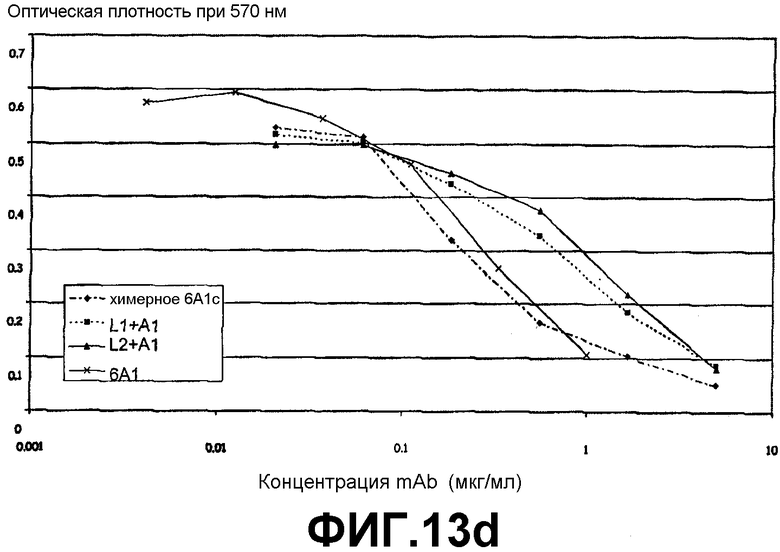
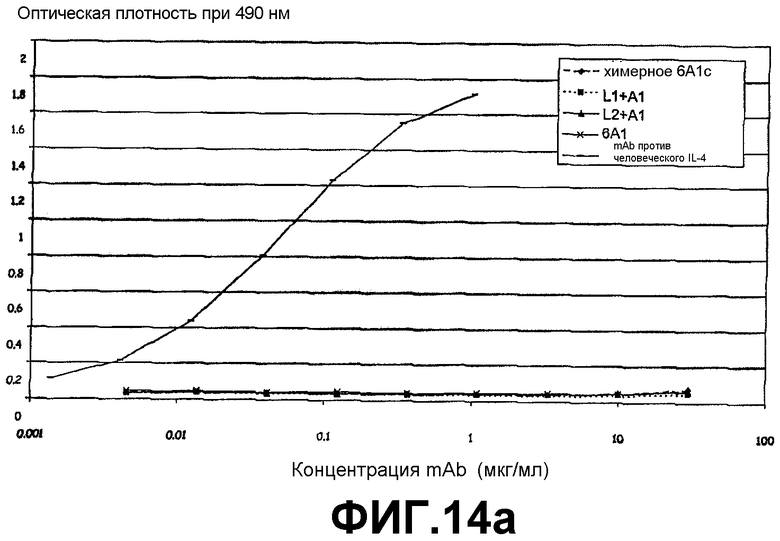
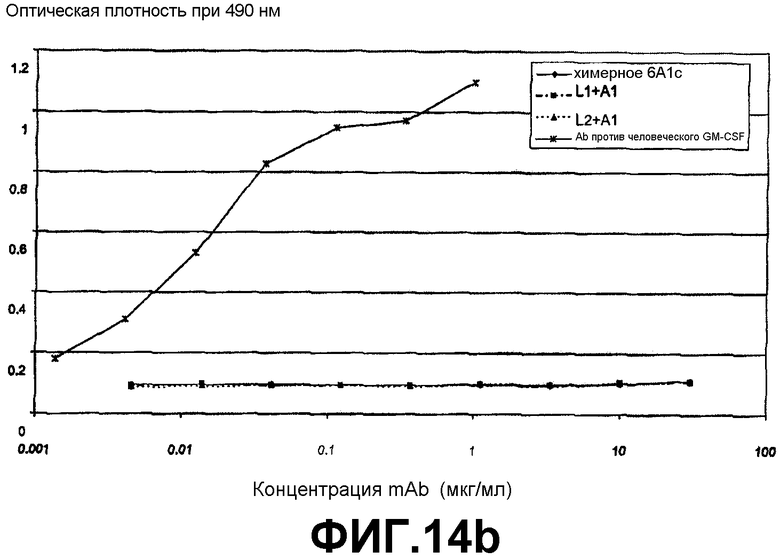
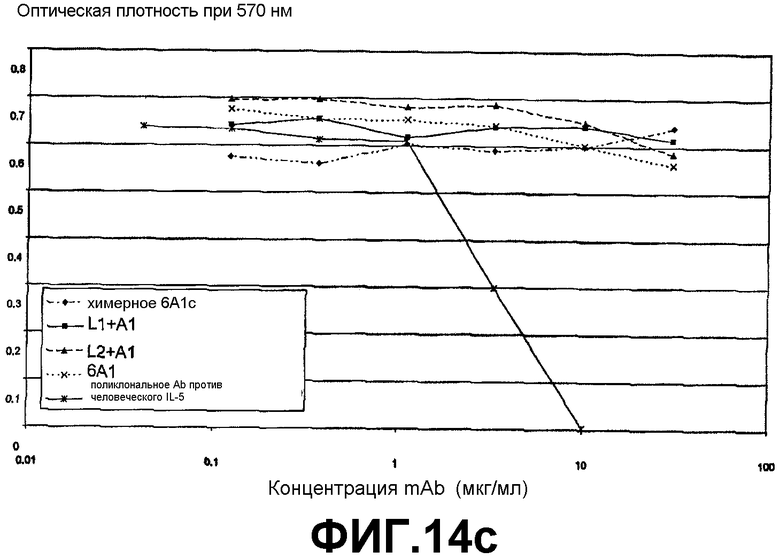
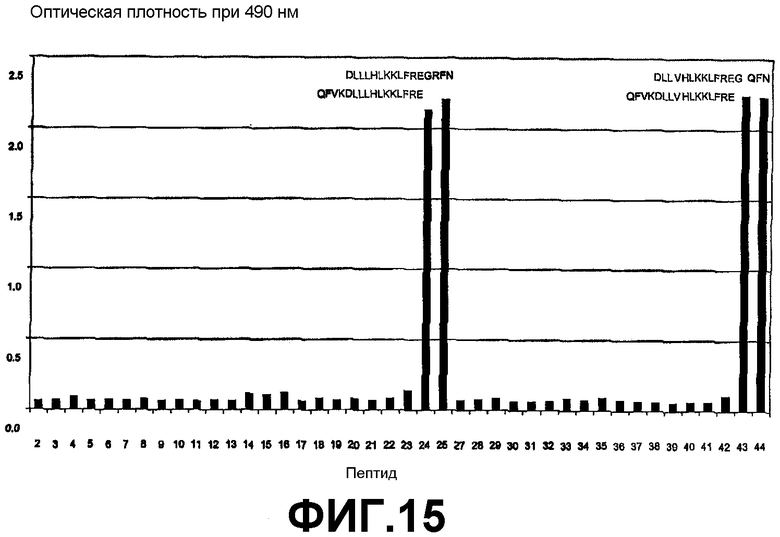
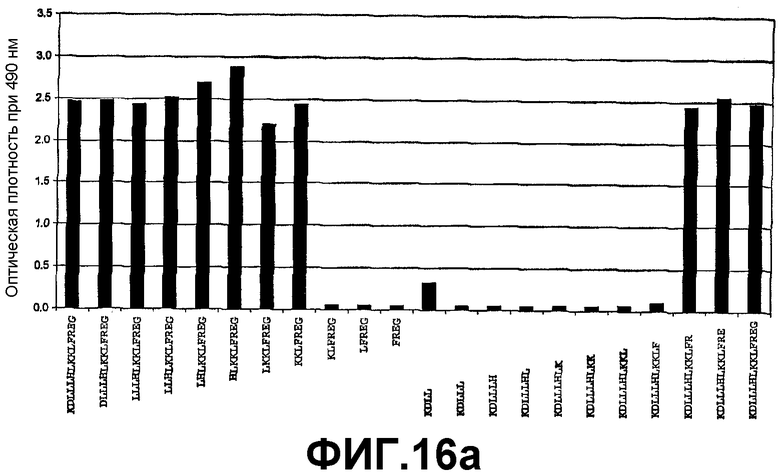
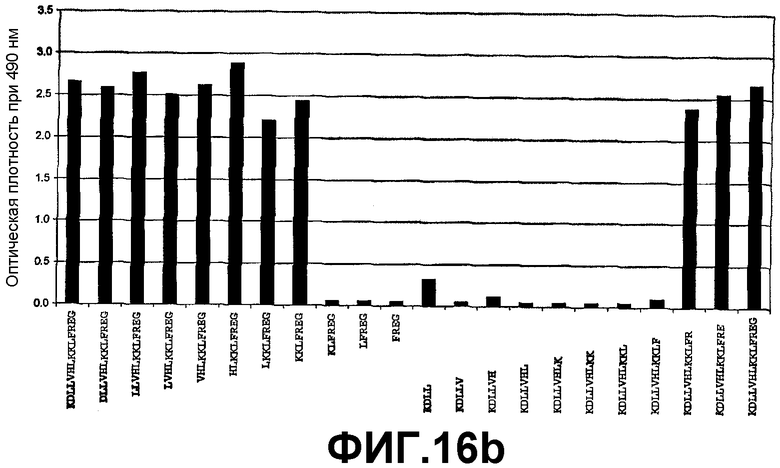
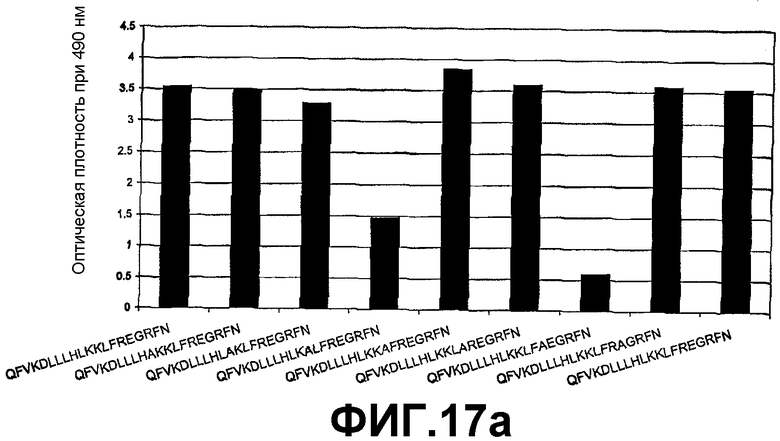
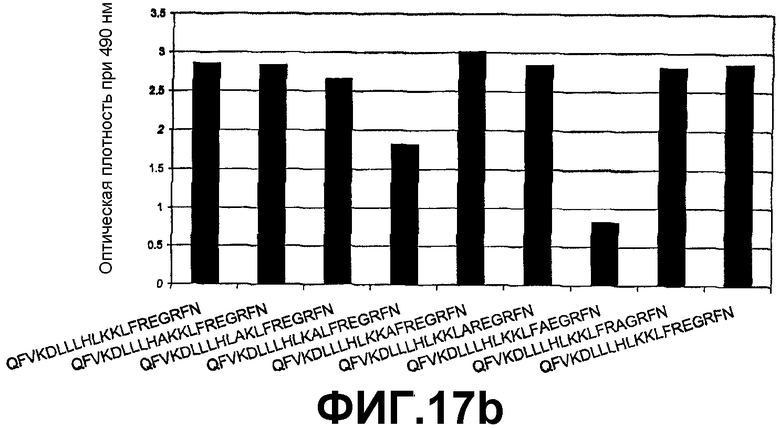
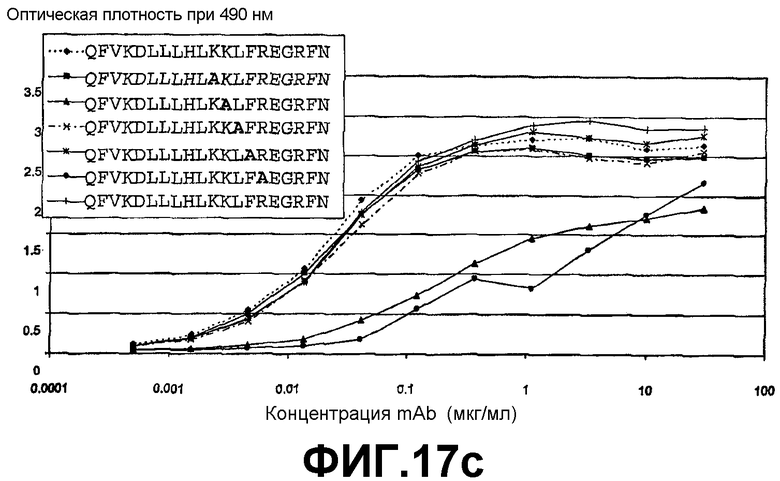

Изобретение относится к биотехнологии и представляет собой иммуноглобулины, в частности антитела, которые специфически связываются с человеческим интерлейкином 13 (hIL-13). Предложены рекомбинантная или трансфицированная клетка-хозяин млекопитающего для клонирования, а также для экспрессии векторов, кодирующих антитела. Предложены фармацевтическая композиция, содержащая антитело, набор, содержащий такую фармацевтическую композицию, а также набор, содержащий такую фармацевтическую композицию и фармацевтическую композицию, содержащую моноклональное антитело против IL-4, такое как пасколизумаб. Предложены способ получения антитела, а также применения таких антител. Антитела могут быть использованы для лечения различных заболеваний или расстройств, отвечающих на модуляцию взаимодействия hIL-13 с рецептором человеческого IL-13. Такими заболеваниями являются тяжелая астма, атопический дерматит, ХОБЛ и различные фиброзные заболевания. 11 н. и 40 з.п. ф-лы, 29 ил., 22 табл.
1. Терапевтическое антитело или его антигенсвязывающий фрагмент, который специфически связывается с hIL-13 и ингибирует взаимодействие hIL-13 с hIL-13R и содержит нижеследующие CDR:
CDRH1: SEQ ID NO:1
CDRH2: SEQ ID NO:2
CDRH3: SEQ ID NO:3
CDRL1: SEQ ID NO:4
CDRL2: SEQ ID NO:5
CDRL3: SEQ ID NO:6.
2. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.1, который специфически связывается с эпитопом, представленным в последовательности SEQ ID NO:84, содержащейся в SEQ ID NO:9, и ингибирует взаимодействие hIL-13 с hIL-13R.
3. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.2, где связывание указанного антитела имеет место между остатками в положениях 103-107 включительно в последовательности SEQ ID NO:9.
4. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.3, где связывание антитела с hIL-13 зависит от присутствия аргининового остатка в положении 107 последовательности SEQ ID NO:9.
5. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.4, где замена аргининового остатка в положении 107 последовательности SEQ ID NO:9 аланиновым остатком приводит к потере связывания указанного антитела с hIL-13 по сравнению со связыванием указанного антитела и его антигенсвязывающего фрагмента с последовательностью SEQ ID NO:9 в отсутствие указанной замены в положении 107 последовательности SEQ ID NO:9.
6. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.1, которое является интактным антителом.
7. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.6, которое является крысиным антителом, мышиным антителом, антителом приматов (например, обезьян cynomolgus, мартышек или человекообразных обезьян) или человеческим антителом.
8. Терапевтическое антитело по п.1, которое является гуманизированным или химерным антителом.
9. Антитело по п.8, содержащее человеческую константную область.
10. Антитело по п.9, содержащее константную область изотипа IgG.
11. Антитело по п.10, которое представляет собой IgG1 или IgG4.
12. Терапевтическое антитело по п.7, которое является мышиным антителом, содержащим VH-домен последовательности SEQ ID NO:7 и VL-домен последовательности SEQ ID NO:8.
13. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:11 и VL-домен последовательности SEQ ID NO:15.
14. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:12 и VL-домен последовательности SEQ ID NO:15.
15. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:13 и VL-домен последовательности SEQ ID NO:15.
16. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:14 и VL-домен последовательности SRQ ID NO:15.
17. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:11 и VL-домен последовательности SEQ ID NO:16.
18. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:12 и VL-домен последовательности SEQ ID NO:16.
19. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:13 и VL-домен последовательности SEQ ID NO:16.
20. Терапевтическое антитело по п.8, которое является гуманизированным антителом, содержащим VH-домен последовательности SEQ ID NO:14 и VL-домен последовательности SEQ ID NO:16.
21. Терапевтическое антитело по любому из п.п.13-20, дополнительно содержащее человеческую константную область изотипа IgG (например, IgG1 или IgG4).
22. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:22.
23. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:19 и легкую цепь SEQ ID NO:22.
24. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:20 и легкую цепь SEQ ID NO:22.
25. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:21 и легкую цепь SEQ ID NO:22.
26. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:18 и легкую цепь SEQ ID NO:23.
27. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:19 и легкую цепь SEQ ID NO:23.
28. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:20 и легкую цепь SEQ ID NO:23.
29. Терапевтическое антитело по п.1, которое является гуманизированным антителом, содержащим тяжелую цепь SEQ ID NO:21 и легкую цепь SEQ ID NO:23.
30. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.1, где указанное антитело является гуманизированным и где остатки, выбранные из группы, состоящей из остатков 19, 38, 73 и 81 акцепторной каркасной области человеческой тяжелой цепи и остатка в положении 85 акцепторной каркасной области человеческой легкой цепи, заменены соответствующими остатками, имеющимися в каркасной области донорного антитела, от которой происходит CDRH3.
31. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.1, где указанное антитело является гуманизированным и где каркасная область человеческой тяжелой цепи содержит один или несколько (например, все) из нижеследующих остатков (или их консервативных замен):
а человеческая легкая цепь содержит
32. Антигенсвязывающий фрагмент по любому из пп.1-7, где указанным фрагментом является Fab, Fab', F(ab')2, Fv, ди-антитело, три-антитело, тетра-антитело, мини-антитело, мини-тельце, выделенный VH или выделенный VL.
33. Антитело по п.9, содержащее мутированную Fc-область, такую, что указанное антитело имеет пониженную АDСС и/или активацию комплемента.
34. Терапевтическое антитело или его антигенсвязывающий фрагмент по любому из пп.1-20, 22-31 или 33, которое конкурентно ингибирует связывание hIL-13 с hIL-13R.
35. Терапевтическое антитело или его антигенсвязывающий фрагмент по п.34, где антитело блокирует связывание hIL-13 с hIL-13R.
36. Терапевтическое антитело по п.1, которое специфически связывается с hIL-13 и ингибирует взаимодействие между hIL-13 и hIL-13R и имеет константу диссоциации koff в пределах 1,4 · 10-4 - 8,22 · 10-5 с-1 (например, как было измерено в анализе Biacore™).
37. Терапевтическое антитело по п.36, где указанное антитело является гуманизованным.
38. Рекомбинантная трансформированная или трансфицированная клетка-хозяин млекопитающего для клонирования векторов, кодирующих антитела по изобретению, содержащая первый и второй векторы, где указанный первый вектор содержит полинуклеотид, кодирующий тяжелую цепь антитела по любому из предшествующих пунктов, а указанный второй вектор содержит полинуклеотид, кодирующий легкую цепь по любому из предшествующих пунктов.
39. Рекомбинантная трансформированная или трансфицированная клетка-хозяин млекопитающего для экспрессии векторов, кодирующих антитела по изобретению, содержащая первый и второй векторы, где указанный первый вектор содержит полинуклеотид, кодирующий тяжелую цепь антитела по любому из предшествующих пунктов, а указанный второй вектор содержит полинуклеотид, кодирующий легкую цепь по любому из предшествующих пунктов.
40. Клетка-хозяин по п.38 или 39, где указанный первый вектор содержит полинуклеотид, выбранный из группы, состоящей из SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:28, SEQ ID NO:29, SEQ ID NO:32, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:35, а указанный второй вектор содержит полинуклеотид, выбранный из группы, состоящей из SEQ ID NO:30, SEQ ID NO:31, SEQ ID NO:36 и SEQ ID NO:37.
41. Клетка-хозяин по п.40, где указанной клеткой является СНО или NSO.
42. Способ получения терапевтического антитела по любому из пп.1-20, 22-31, 33 или 36 и 37, где указанный способ включает в себя стадию культивирования клетки-хозяина по любому из пп.38-41 в бессывороточной культуральной среде.
43. Способ по п.42, где указанное антитело секретируется указанной клеткой-хозяином в указанную культуральную среду.
44. Способ по п.43, где указанное антитело дополнительно очищено до степени чистоты, составляющей по меньшей мере 95% или более (например, 98% или более) по отношению к указанной содержащей антитело культуральной среде.
45. Фармацевтическая композиция для применения при лечении заболеваний, восприимчивых к модуляции взаимодействия hIL-13 с hIL-13R, содержащая терапевтически эффективное количество терапевтического антитела или его антигенсвязывающего фрагмента по любому из пп.1-20, 22-31, 33 или 36 и 37 и фармацевтически приемлемый носитель.
46. Набор для применения при лечении заболеваний, восприимчивых к модуляции взаимодействия hIL-13 с hIL-13R, частью которого является композиция по п.45, вместе с инструкциями по применению.
47. Применение терапевтического антитела или его антигенсвязывающего фрагмента по любому из пп.1-20, 22-31, 33 или 36 и 37 в целях изготовления лекарственного средства для лечения заболевания или расстройства, выбранного из группы, состоящей из аллергической астмы, тяжелой формы астмы, астмы, трудно поддающейся лечению, нестабильной астмы, "ночной" астмы, предменструальной астмы, стероид-резистентной астмы, стероид-зависимой астмы, индуцированной аспирином астмы, астмы взрослых, детской астмы, атонического дерматита, аллергического ринита, болезни Крона, ХОБЛ, фиброзных заболеваний или расстройств, таких, как идиопатический фиброз легких, прогрессирующий системный склероз, фиброз печени, гранулема печени, шистосомоз и лейшманиоз, и заболеваний, ассоциированных с нарушением регуляции клеточного цикла, таких, как болезнь Ходжкина, В-клеточный хронический лимфоцитарный лейкоз.
48. Применение антитела по любому из пп.1-20, 22-31, 33 или 36 и 37 и моноклонального антитела против IL-4, такого как пасколизумаб, в целях изготовления лекарственного средства для лечения заболевания или расстройства, выбранного из группы, состоящей из аллергической астмы, тяжелой формы астмы, астмы, трудно поддающейся лечению, нестабильной астмы, "ночной" астмы, предменструальной астмы, стероид-резистентной астмы, стероид-зависимой астмы, индуцированной аспирином астмы, астмы взрослых, детской астмы, атонического дерматита, аллергического ринита, болезни Крона, ХОБЛ, фиброзных заболеваний или расстройств, таких, как идиопатический фиброз легких, прогрессирующий системный склероз, фиброз печени, гранулема печени, шистосомоз и лейшманиоз, и заболеваний, ассоциированных с нарушением регуляции клеточного цикла, таких, как болезнь Ходжкина и В-клеточный хронический лимфоцитарный лейкоз.
49. Применение антитела по любому из пп.1-20, 22-31, 33 или 36 и 37 и моноклонального антитела против IL-4, такого как пасколизумаб, в целях приготовления набора из нескольких частей для лечения заболевания или расстройства, выбранного из группы, состоящей из аллергической астмы, тяжелой формы астмы, астмы, трудно поддающейся лечению, нестабильной астмы, "ночной" астмы, предменструальной астмы, стероид-резистентной астмы, стероид-зависимой астмы, индуцированной аспирином астмы, астмы взрослых, детской астмы, атопического дерматита, аллергического ринита, болезни Крона, ХОБЛ, фиброзных заболеваний или расстройств, таких, как идиопатический фиброз легких, прогрессирующий системный склероз, фиброз печени, гранулема печени, шистосомоз и лейшманиоз, и заболеваний, ассоциированных с нарушением регуляции клеточного цикла, таких, как болезнь Ходжкина и В-клеточный хронический лимфоцитарный лейкоз.
50. Набор из нескольких частей для применения при лечении заболеваний, восприимчивых к модуляции взаимодействия hIL-13 с hIL-13R, включающий в себя первую фармацевтическую композицию, содержащую терапевтически эффективное количество антитела по любому из пп.1-20, 22-31, 33 или 36 и 37 и фармацевтически приемлемый носитель, и вторую фармацевтическую композицию, содержащую моноклональное антитело против IL-4, такое, как пасколизумаб, и фармацевтически приемлемый носитель, необязательно вместе с прилагаемыми инструкциями по применению.
51. Фармацевтическая композиция для применения при лечении заболеваний, восприимчивых к модуляции взаимодействия hIL-13 с hIL-13R, содержащая терапевтически эффективное количество первого антитела по любому из пп.1-20, 22-31, 33 или 36 и 37, и терапевтически эффективное количество второго антитела, где указанным вторым антителом является анти-IL-4-антитело, такое, как пасколизумаб, и фармацевтически приемлемый носитель.
| PUNNONEN J | |||
| ET AL | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| J Allergy Clin Immunol | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| РОЙТ А | |||
| и др | |||
| Иммунология | |||
| - М.: Мир, 2000, с.168-193. | |||

