Изобретение, относящееся к медицине, конкретно к фармакологии, результаты которого могут быть использованы для коррекции нарушений в иммунной системе при патологических состояниях, связанных с недостаточностью Th1-зависимого иммунного ответа (хронические, вялотекущие и рецидивирующие инфекционные, а также онкологические заболевания).
Гуминовые вещества - основная органическая составляющая почвы, воды, твердых полезных ископаемых, образующихся при разложении растительных и животных остатков под действием микроорганизмов и абиотических факторов окружающей среды. Запасы гуминовых веществ планеты можно считать неограниченными, поскольку гумификация - это «всеобщий процесс постмортального превращения органических остатков, он может протекать в почвах, природных водах, илах, торфах, при углеобразовании и образовании горючих сланцев, любых каустобиолитов… его нельзя ограничивать только почвенной средой» [13]. Гумификация является глобальным явлением, свойственным любой почве, начиная от старых почв арктических пустынь, черноземов и тропиков до довольно молодых на техногенных отвалах или вулканических пеплах. В процессе гумификации образуется особый класс высокомолекулярных органических азотсодержащих природных соединений, не существующих в живых организмах - гумусовых кислот (гидрокси-, оксокарбоновых кислот с ароматическим ядром), делящихся, в свою очередь, на темноокрашенные гуминовые кислоты (ГК), растворимые в щелочах и нерастворимые в кислотах, гиматомелановые кислоты, растворимые в этаноле и фульвовые кислоты, растворимые в водных растворах кислот [3].
Состав гумусовых веществ торфяников, содержание функциональных групп и молекулярных фрагментов (по данным 13С-ЯМР) зависит от ботанического вида исходных растительных остатков, степени гумификации (разложения) растительного опада, а также климатических условий периода формирования почвенного органического вещества [4, 6, 12, 15, 29].
Гуминовые кислоты торфа - высокомолекулярные азотсодержащие соединения циклического строения, представляющие собой смесь темноокрашенных органических, высокомолекулярных, в основном ароматических, метоксисодержащих, гидрокси- и оксокарбоновых кислот, объединенных общим типом строения, но имеющих некоторые различия, определяемые их происхождением [11].
Несмотря на недостаточную изученность, вещества гуминовой природы давно и широко применяются во многих отраслях деятельности человека: в промышленности при нефте- и газодобыче, для ремедиации территорий, загрязненных отходами производства, в сельском хозяйстве в качестве ветеринарных препаратов [24] и компонентов органоминеральных удобрений [16].
В последние годы и в экспериментальной медицине наблюдается повышение интереса к использованию ГК. Получены весомые результаты, как подтверждающие их фармакологические противоопухолевые, гепатопротекторные, ранозаживляющие свойства, так и предполагаемые механизмы действия [2, 9, 27, 28]. Показано, что ГК различной этиологии и разных физико-химических характеристик обладают широким спектром плейотропных иммунологических эффектов - оказывают влияние на поляризацию иммунокомпетентных клеток по классическому или альтернативному пути за счет селективной секреции про- и противовоспалительных цитокинов (ИЛ-1β, ИЛ-2, ИЛ-6, ФНО-α и ИНФ-γ) [18, 25], обладают противовирусной активностью против вирусов герпеса 1 и 2 типов (HSV1, HSV2), респираторного и цитомегаловируса человека (HCMV, RSV) in vitro [17] и иммунокоррегирующими свойствами на фоне антибактериальной терапии рядом антибиотиков (ампициллин, амикацин, доксициклин, рифампицин, рифамицин), а также способствует значительной локализации воспаления и усилению сосудообразования при ксенотрансплантации [22]. Также ГК способны действовать как мощные антиприонные агенты при лечении нейродегенеративных расстройств у животных [21] и проявлять синергический цитотоксический эффект на β-амилоидный белок на модели SK-N-MC нервных клеток человека [19]. Кроме того, ГК не проявляют токсических эффектов в широкой линейке доз у экспериментальных животных при пероральном или накожном применении [8, 26].
Известно, что эффективный иммунный ответ зависит от поляризации антигенпрезентирующих клеток, прежде всего макрофагов. Функциональное состояние МФ определяется способом утилизации L-аргинина и изменением цитокинового профиля: Th1-цитокины (ИЛ-1, ИЛ-2, ИЛ-6, ИЛ-12, ИНФ-γ и ФНО-α), стимулируя цитотоксические лимфоциты и натуральные киллеры, являются основными индукторами клеточно-опосредованного иммунного ответа и ассоциированных с ним воспалительных реакций; Тh2-цитокины (ИЛ-4, ИЛ-5, ИЛ-10, ИЛ-13, ТФР-β активируют В-лимфоциты и определяют таким образом гуморальный иммунный ответ. Классически активированные МФ преобразуют L-аргинин с помощью индуцибельной NO-синтазы в оксид азота и цитруллин, а альтернативно активированные - с помощью аргиназы в мочевину и орнитин, приводя, соответственно, к развитию Th1 или Th2 типа иммунного ответа [20, 23].
Однако применение гуминовых веществ в медицине на текущий момент является сложной задачей. Это связано с тем, что ГК не имеют привычного стехиометрического состава и регулярного строения, обладают полидисперсностью и широкой гетерогенностью структурных элементов, а также могут значительно отличаться между собой по свойствам в зависимости не только от вида исходного сырья, но и от способа их извлечения - выщелачивающего реагента [1, 7]. Так, извлечение ГК из торфа водными растворами гидроксидов щелочных металлов представляет собой процесс образования солей, имеющих большую степень ионизации в водных растворах, чем сами ГК. Гидроокись натрия среди всех известных экстрагентов ГК имеет самую наибольшую степень диссоциации в водных растворах, поэтому данный экстрагент обладает высокой извлекающей способностью, т.к. растворимость ГК усиливается повышением содержания гидроксильных ионов. При использовании другого щелочного экстрагента - децимолярного раствора пирофосфата натрия происходит внутрисферное замещение лигандов в металло-гуминовых комплексах, поэтому вся специфичность действия пирофосфата натрия сводится к образованию нерастворимых осадков с кальцием и другими многовалентными катионами. Пирофосфат натрия, за счет своих комплексоообразующих свойств, способен разрушать комплексы ковалентного и ионного типов. Пирофосфаты кальция, железа и алюминия труднорастворимы, поэтому параллельно протекает процесс декальцинирования, что способствует более полной экстракции именно свободных (истинных) ГК [10]. Полученные таким образом ГК торфа представляют собой водорастворимый аморфный порошок темно-коричневого цвета, без запаха.
В предыдущих исследованиях нами показано, что ГК, как базовый компонент гуминовых веществ, способны дозозависимо усиливать продукцию оксида азота, не влияя на активность аргиназы, перитонеальными макрофагами (МФ), что предполагает наличие у них потенциальной иммуномодулирующей активности [5].
Задачей данного изобретения является расширение арсенала иммуномодулирующих средств природного происхождения, обладающих способностью избирательной стимуляции продукции макрофагами интерлейкина-12, фактора некроза опухоли-альфа и лимфоцитами интерферона-гамма.
Поставленная задача решается путем применения водорастворимых гуминовых кислот, выделенных экстракцией пирофосфатом натрия из верхового сосново-сфагново-пушицевого торфа, отобранного с глубины 10-50 см на торфяном месторождении «Васюганское» (участок Высокий рям) Бакчарского района Томской области, обладающие способностью к избирательной стимуляции продукции ИЛ-12, ФНО-α и ИФН-γ, для коррекции нарушений в иммунной системе при патологических состояниях, связанных с недостаточностью Th1-зависимого типа иммунного ответа (хронические, вялотекущие и рецидивирующие инфекционные, а также онкологические заболевания).
Принципиально новым в предлагаемом изобретении является применение в качестве иммуномодулирующего средства водорастворимых ГК, выделенных из верхового сосново-сфагново-пушицевого торфа экстракцией пирофосфатом натрия. Новое свойство водорастворимых ГК, выделенных при экстракции пирофосфатом натрия из верхового сосново-сфагново-пушицевого торфа, было обнаружено в результате экспериментальных исследований и для специалиста явным образом не вытекает из уровня техники, и описание этого свойства не обнаружено авторами в патентной и научно-медицинской литературе.
Таким образом, предлагаемое техническое решение соответствует критериям изобретения, а именно - «новизна», «изобретательский уровень» и «промышленная применимость».
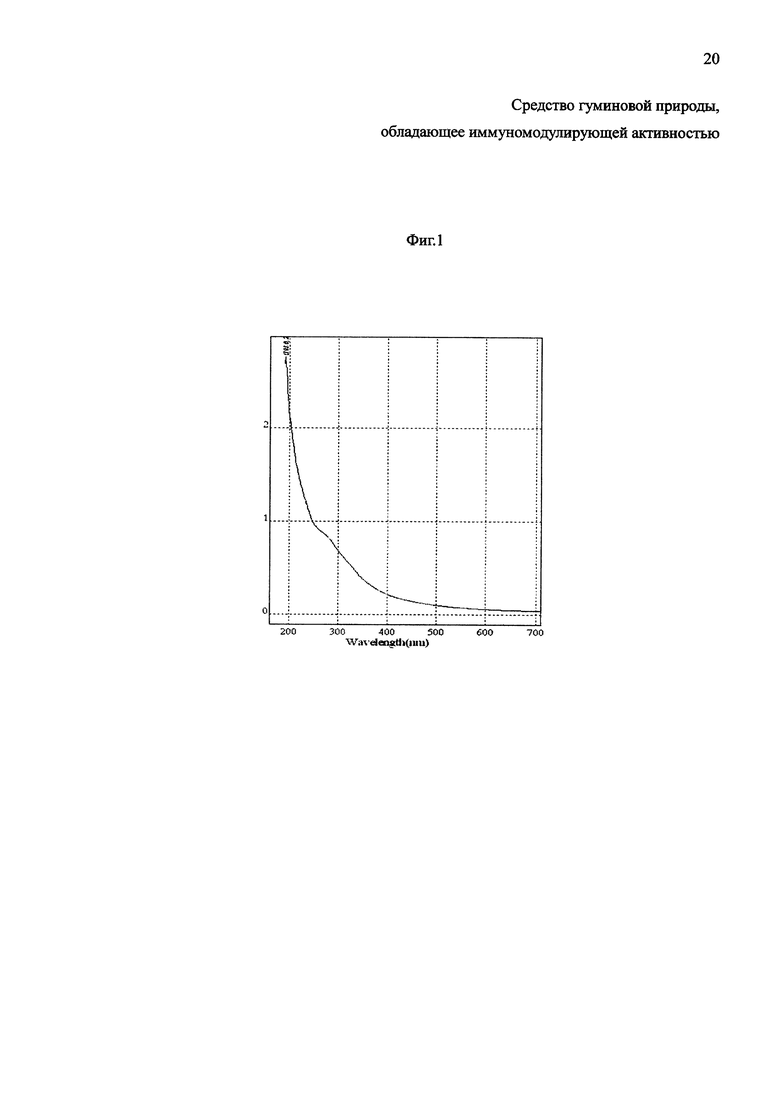
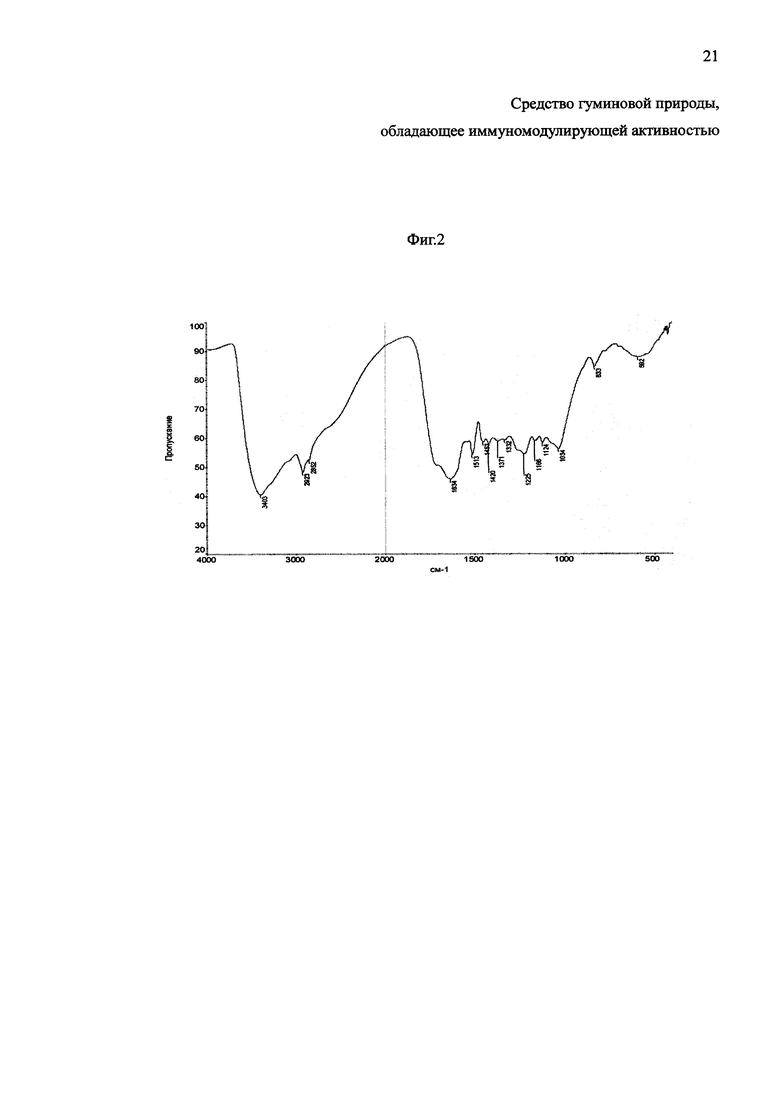
Изобретение будет понятно из следующего описания и приложенных фигур 1 и 2. На фиг. 1 представлен абсорбционный спектр гуминовых кислот, выделенных из верхового сосново-сфагново-пушицевого торфа. На фиг. 2 представлен молекулярный спектр гуминовых кислот верхового сосново-сфагново-пушицевого торфа.
С целью получения представительного среднего образца торфа, в котором наиболее полно отражена неоднородность химического состава всей партии анализируемого материала, отбор пробы торфа проводили буром ТБГ-1 в генетическом центре торфяного месторождения «Васюганское» (участок Высокий рям) Бакчарского района Томской области, из середины однородного по ботаническому составу горизонта с глубины 10-50 см, в летний период (июнь-август) согласно ГОСТ 17644-83 «Торф. Методы отбора проб из залежи и обработки их для лабораторных испытаний». В скважине с каждой глубины отбирали только один раз, масса одной пробы составляла не менее 600 г, количество скважин - не менее 50, средняя масса пробы торфа составляла более 30 кг. Согласно проведенному ботаническому и гравиметрическому анализу, а также общетехнической характеристики, отобранный торф охарактеризован как верховой тип, сосново-сфагново-пушицевый вид, степень разложения растительных остатков 30-35%, содержание минеральной примеси (общей золы) - не более 7,2%, содержание биологически активных гуминовых кислот 13,2 масс. %.
Партию торфа высушивали при комнатной температуре на воздухе в хорошо проветриваемом помещении, периодически перемешивая, до воздушно-сухого состояния, измельчали в роторно-ножевой мельнице и просеивали через сито с диаметром отверстий 3 мм, обрабатывали 0,1 моль/л раствором пирофосфата натрия (Na4P2O7) в массовом соотношении 1:50-1:100 в течение 8 часов при постоянном перемешивании в реакторе Р-100 при температуре 25-27°C, отделяли жидкую фазу (экстракт ГК) от осадка (остатка торфа) фильтрованием при помощи системы вакуумной фильтрации (нутч-фильтр), для осаждения ГК из экстракта жидкую фазу обрабатывали хлороводородной кислотой до pH 1-2, выделившиеся ГК разделяли в поле центробежных сил (центрифугированием), отмывали водой очищенной до pH 7 и высушивали при комнатной температуре.
Полученные вышеописанным способом гуминовые кислоты из верхового сосново-сфагново-пушицевого торфа были охарактеризованы по физико-химическим параметрам: элементному составу, электронным и молекулярным спектральным свойствам, молекулярному весу.
1. Элементный состав ГК определяли методом сожжения на C,H,N - анализаторе «Carlo Erba Strumentazione» модель 1106, содержание кислорода определяли по разности.
Элементный состав ГК верхового сосново-сфагново-пушицевого торфа, представленный в таблице 1, показывает распределение основных конституционных элементов молекулярной структуры вещества и является диагностическим показателем для характеристики субстанции. Атомное отношение элементов углерода и водорода (Н/С=1,11) указывает на ароматический характер молекулы ГК, т.к. содержание водорода всего на 11% выше углерода, следовательно, алифатическая часть молекулы выражена незначительно и/или является полизамещенной различными кислород и азотсодержащими функциональными группами. Соотношение содержания кислорода в ГК относительно углерода порядка 45% свидетельствуют о большом количестве кислородсодержащих функциональных групп, в основном фенольных, карбонильных и карбоксильных групп. Содержание азота в молекуле ГК относительно количества углерода незначительно (около 4%).
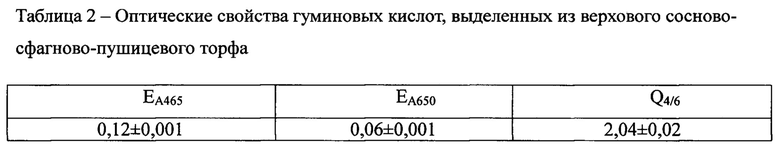
2. Регистрацию электронных спектров поглощения 0,001% водных растворов ГК проводили на спектрофотометре Unico 2800 (производство США) в диапазоне длин волн 190-800 нм в кварцевой кювете толщиной 1 см. Из полученных спектров определяли коэффициенты экстинкции, которые характеризуют оптические плотности раствора ГК для слоя 1 см, при концентрации ГК 0,001% при длине волны 465 нм (А465) и 650 нм (A650) и вычисляли коэффициент цветности Q4/6 по Е. Вельте [30] как отношение оптических плотностей при длинах волн 465 и 650 нм (А465/А665).

Электронный спектр ГК верхового сосново-сфагново-пушицевого торфа представлен на фиг. 1, согласно которой в видимой области (400-800 нм) спектр не имеет четко выраженных максимумов поглощения, и выглядит как пологая кривая, характеризующая сплошное поглощение с постепенным уменьшением оптической плотности по мере увеличения длины волны. В ультрафиолетовой области спектра (200-400 нм) поглощение света резко возрастает в коротковолновую сторону. При этом в области 275 нм выделяется плечо, характерное для фенольных, карбонильных, карбоксильных групп и полиеновых цепей, что, вероятнее всего, соответствуют n→π*-переходам, присущим для ароматических альдегидов, кетонов, карбоновых кислот и их функциональных и гетерофункциональных производных. Кроме того, в этой же области расположены: полосы поглощения 1) n→π*-переходов вызванные ауксохромными группами, такими как -O-R, -NHR, -NR2 и др., находящимися в p-π- и π-p-π-сопряжениях с бензольными кольцами; 2). π→π*- и n→π*-переходами в виду наличия в структуре ГК фрагментов с цепью π-π-сопряжений ароматического кольца и карбонильных групп с кратными C=C-связями (структурные фрагменты стильбенового типа, коричных кислот и т.п.), ката-конденсированных и периконденсированных ароматических систем типа «аценов», «фенов», пиренов, периленов и др. Характер n→π*-перехода в ГК подтверждается также смещением полосы (275 нм) в коротковолновую область спектра при протонировании молекулы ГК.
Важным диагностическим параметром ГК, как темноокрашенных соединений [13], является интенсивность поглощения света. Полученные гуминовые кислоты характеризовались невысокими коэффициентами цветности (Q4/6) и экстинкции (ЕA465, ЕA650), обуславливающими высокую поляризованность молекулы ГК (Таблица 2).
3. Исследование молекулярных параметров структуры ГК проводили методом инфракрасной (ИК) спектроскопии на ИК - Фурье - спектрометре ФСМ 1201 (ООО «Инфраспек») в таблетках с KBr (в соотношении 1:100 соответственно), в интервале значений частоты от 500 до 4000 см-1. Для количественной оценки интенсивности полос поглощения и относительных концентраций функциональных групп использован метод относительных оптических плотностей полос поглощения (ОППП).
Молекулярный спектр ГК верхового сосново-сфагново-пушицевого торфа (фиг. 2), выявил широкую интенсивную полосу поглощения с максимумом при 3500-3300 см-1, обусловленную переменными валентными колебаниями гидроксильных групп (ион) алифатического и ароматического характера, связанных преимущественно водородными связями, наличие которых проявляется максимумом поглощения в интервале 1270-1220 и 1170-1040 см-1. В длинноволновом крыле главной полосы при 3250-3200 см-1 обнаруживается поглощение средней интенсивности, имеющее вид уступа (перегиба) и отвечающее колебаниям N-H (υNH) в структуре амида, аминов, связанных водородными связями. Полосы средней интенсивности хорошо видны при 2928-2921 см-1 и 2855-2842 см-1 за счет переменных валентных колебаний -CH3 и -CH2 групп боковых цепей в молекулах ГК, в том числе связанных с ароматическими фрагментами, судя по наличию полосы поглощения в области 1385 см-1. Отмечается слабое поглощение при 2700-2400 см-1 присущее димерам карбоновых кислот. Отчетливый максимум сильной интенсивности обнаруживается в интервале 1725 см-1-1710 см-1, свойственном для валентных колебаний карбонильных групп (υС=О), которые могут быть представлены кетонами, альдегидами, карбоновыми кислотами и их функциональными производными. В ИК-спектре ГК обнаруживается сильная полоса в области 1650-1600 см-1, обусловленная плоскостными валентными колебаниями сопряженных углерод-углеродных (ароматические, υС=С) и углерод-кислородных связей (карбонилы, связанные водородными связями, карбоксилат-ионы, υC=O) в ароматическом скелете и хинонах. Полоса поглощения около 1513 см-1 (υС-С) указывает на наличие неконденсированных моноароматических структур. В данной области наблюдаются также колебания связей полипептидов в составе ГК, связанных с атомами азота и кислорода (N-H, N-C=O): первичной (1580-1632 см-1) и вторичной (1512-1560 см-1) аминогрупп. Колебания в области 1264-1225 см-1 определяются в основном валентными (υС-О) и деформационными колебаниями (δО-Н) связей недиссоциированных карбоновых кислот и их функциональных производных (в основном сложных эфиров как арильного, так и алкильного типов). За поглощение излучения в коротковолновой части спектра в области 1175-1000 см-1 ответственны валентные колебания гидроксильных групп (ион) спиртов и углеводов. В интервале около 1075-1013 см-1 (υС-О) поглощают излучение первичные спирты, при 1125-1100 см-1 (υС-О) - вторичные спирты и при 1175-1150 см-1 (υС-О) - третичные спирты. Поглощение в данной области (1175-1000 см-1) может быть также обусловлено валентными колебаниями (υC-O-C) гликозидных связей углеводов, лактонов, С-О-С-связями циклических и алифатических простых эфиров, при этом присутствие С-O связей полисахаридов также подтверждается колебаниями в области 830-1100 см-1. В области волновых чисел от 800 до 600 см-1 наблюдаются слабые полосы поглощения, возможно обусловленные внеплоскостными деформационными колебаниями (бон) в ароматических кольцах, имеющих два и более незамещенных атомов водорода, и в том числе присутствием конденсированных многоядерных аренов (755-760 см-1).
Спектральные коэффициенты являются характерным диагностическим показателем структуры ГК. Высокая интенсивность полос поглощения говорит о большей обогащенности функциональными группами, т.е. чем выше интенсивность поглощения, тем больше содержание этой группы или фрагмента. Относительная количественная оценка содержания функциональных групп в молекулах ГК по данным ИК-спектроскопии дана на основании отношений оптических плотностей полос поглощения (ОППП) кислородсодержащих функциональных групп (υОН3400 см-1, υС=О 1720 см-1, υC-O, С-О-С 1225 см-1, υC-O 1035 см-1,) к оптическим плотностям полос поглощения, соответствующим ароматическим (υС=С1610 см-1) и алифатическим (υAlif2920 см-1) фрагментам структуры, и представлена в таблице 3.


Одними из основных кислородсодержащих функциональных групп в молекуле ГК верхового сосново-сфагново-пушицевого торфа являются: гидроксильные (υOH3400-3300 см-1, υС-О 1150-1000 см-1) группы, карбоксильные группы и их функциональные производные (υС=О 1725-1700 см-1, υС-О 1260-1225 см-1), а также простые эфирные группы (υC-O-C 1050-1035 см-1). Рассматривая отношения ОППП алифатических фрагментов структуры к ароматическим (AAlif2920/AC=C 1610) можно отметить, что наблюдается преобладание ароматических структур над алкильными и над кислородсодержащими функциональными группами. Также наблюдается обратная зависимость в отношениях ОППП кислородсодержащих функциональных групп к алифатическим фрагментам структуры, где в основном преобладают группы карбоновых кислот и сложных эфиров.
4. Молекулярную массу определяли методом гель-проникающей хроматографии на приборе Ultimate 3000 (Thermo Scientific, США) с использованием колонки Ultrahydrogel 250, 300×7,8 мм, подвижная фаза - 0,1 М NaNO3, 0,01% NaN3 в воде, скорость потока 1 мл/мин. Детектирование спектрофотометрическое при длине волны 200 нм. Расчет молекулярной массы проводили по калибровочному графику log Mw=f(tR) построенному по стандартам PSS (polystyrene sulfonate) 891 - 976000 Da (Polymer Standarts Service Service GmbH, Германия). Среднечисленная молекулярная масса ГК составила 6110,2 Да; среднемассовая молекулярная масса составила 22783,9; полидисперсность - 3,7; медиана - 11798,9 Да.
Биологическую активность ГК оценивали на линейных мышах C57BL/6 [14]. Животные в возрасте 8-10 недель были получены из отдела экспериментальных биологических моделей НИИФиРМ им. Е.Д. Гольдберга Томский НИМЦ РАН (ветеринарное свидетельство 270 №0008633 15 июля 2015 г.). Все процедуры (содержание, введение исследуемых веществ, умерщвление) были проведены в соответствии с Директивой 2010/63/EU Европейского Парламента и Совета ЕС по охране животных используемых в научных целях, ГОСТ 33216-2014 «Руководство по содержанию и уходу за лабораторными животными». В качестве контроля в экспериментах in vivo использовали ликопид и пирогенал, in vitro - мурамилдипептид.
Макрофаги получали из суспензии перитонеальных клеток, для чего животных забивали дислокацией шейного отдела позвоночника, брюшную полость промывали ледяным изотоническим раствором хлорида натрия (ФР), клетки осаждали, ресуспендировали в культуральной среде и оценивали их жизнеспособность в тесте с 0,1% трипановым синим. В экспериментах использовали суспензии, содержащие не менее 95% жизнеспособных клеток. Далее клетки (1,5-2,0×106/мл) помещали в пластиковые чашки Петри, культивировали 2 ч при 37°C (в атмосфере 5% СО2 и абсолютной влажности) в среде RPMI 1640 («Sigma»), 10% ЭТС («Hyclone»), 20 мМ HEPES («Sigma»), 0,05 мМ 2-меркаптоэтанол («Sigma»), 50 мкг/мл гентамицин («Sigma»), 2 мМ L-глютамин («Sigma»)). Полученные после прилипания макрофаги переносили в плоскодонные 96-луночные планшеты (2,5-3,0×106 клеток/мл) и культивировали в указанных выше условиях в присутствии ГК (100 мкг/мл); 1 мкг/мл ЛПС (серотип O111:В4, «Sigma»); 10 мкг/мл мурамилдипептида (МДП) (N-ацетилмурамил-L-аланил-D-изоглютамин, «Calbiochem»). Продукцию монокинов стимулировали добавлением ЛПС (1 мкг/мл), лимфокинов - конканавалином А (4 мкг/мл) или митогеном лаконоса (5 мкг/мл). Через 24 ч от начала культивирования собирали из лунок надосадок и замеряли в нем концентрацию цитокинов твердофазным иммуноферментным методом при помощи тест-систем согласно прилагаемым протоколам: ИЛ-2, ИЛ-4, ИЛ-10, ИЛ-12 и ИФН-γ («eBioscience»).
Количество цитокинов, вырабатываемых мононуклеарами периферической крови человека (Мн), определяли в супернатантах клеток, получаемых следующим образом. Жидкость для сепарации клеток «Histopaque-1077» («Sigma-Aldrich») с плотностью 1,077 помещали в пробирки, затем осторожно наслаивали цельную кровь здоровых доноров с добавлением гепарина (10 ЕД/мл). После 15-минутного центрифугирования при 400 g собирали клетки, сформировавшие кольцо на градиенте плотности, трижды отмывали их холодным ФР, ресуспендировали в культуральной среде, оценивали жизнеспособность. Далее Мн (2,5-3,0×106 клеток/мл) помещали в 96-луночный планшет и вносили ГК (100 мкг/мл), продукцию цитокинов стимулировали добавлением митогенов, как указано выше. Через 24 ч инкубации собирали бесклеточный супернатант, в котором иммуноферментным методом при помощи тест-систем определяли количество цитокинов согласно прилагаемым протоколам: ИЛ-10 и ИФН-γ («R@D Systems»), ФНО-α - («Вектор-Бэст»).
ГК вводили мышам ежедневно внутрибрюшинно в течение 10 дней. В предварительных экспериментах in vivo было выявлено, что для проявления иммуномодулирующего эффекта оптимальной суточной дозой ГК является концентрация 1 мг/кг массы тела, пирогенала (Филиал «Медгамал» ФНИЦЭМ им. Н.Ф. Гамалеи, Россия) - 5 мкг/кг и ликопида (ЗАО «Пептек», Россия) - 2 мг/кг. Для индукции Th1-зависимого типа иммунного ответа через 5 дней от начала введения ПС животных иммунизировали внутрибрюшинной инъекцией эритроцитов барана (5×107).
Для оценки действия ГК на клеточное звено иммунитета использовали реакцию гиперчувствительности замедленного типа. Для этого на 5-е сутки после иммунизации животным проводили вторую (разрешающую) инъекцию эритроцитов барана в подушечку задней лапы - «опытная лапа» (108 эритроцитов барана в 0,05 мл изотонического раствора хлорида натрия). В контрлатеральную лапу вводили 0,05 мл стерильного изотонического раствора хлорида натрия («контрольная лапа»). Через 24 часа животных забивали, обе лапы отрезали по выступу кости ниже сочленения мало- и большеберцовой кости и выше пяточного сустава, местную воспалительную реакцию оценивали по разнице массы опытной и контрольной лап.
Влияние ГК на гуморальное звено иммунитета оценивали по количеству антителообразующих клеток (АОК) в селезенке, для чего животных, получавших курс ПС, забивали на 5-е сутки после иммунизации эритроцитами барана (на пике IgM-AOK и IgG-АОК, соответственно).
Полученные в ходе исследования данные будут обрабатываться с помощью пакета статистических программ Statistica 8,0. Для каждой выборки будет вычисляться среднее арифметическое (X), ошибка среднего арифметического (m), среднее арифметическое отклонение (σ). Проверка на нормальность распределения будет проводиться с помощью критерия Шапиро-Уилка. Сравнение выборочных средних будет осуществляться по критерию Даннета для сравнения нескольких экспериментальных выборок с одной контрольной в случае нормального распределения или по критерию Крускалла-Уоллиса для к-несвязанных выборок (к>2) и критерия Данна в случае распределения, отличающегося от нормального.
Пример 1.
ГК, выделенные из верхового сосново-сфагново-пушицевого вида торфа с глубины 10-50 см торфяного месторождения «Васюганское» (участок Высокий рям) Бакчарского района Томской области, при культивировании с интактными МФ значительно, в 28 раз, увеличивали концентрацию ИЛ-12 - ключевого цитокина, ответственного за эффективность Т-клеточного иммунного ответа (Таблица 4) - с 0,166±0,018 пг/мл в контроле до 4,480±0,702 пг/мл. При культивировании макрофагов без добавления каких-либо стимуляторов (среда) сколько-нибудь значимых количеств данного цитокина в супернатанте обнаружено не было, а ЛПС даже снижал показатель в 2,1 раза. Активирующее действие мурамилдипептид (МДП) также было значительным относительно контроля и существенно не отличалось от ГК.
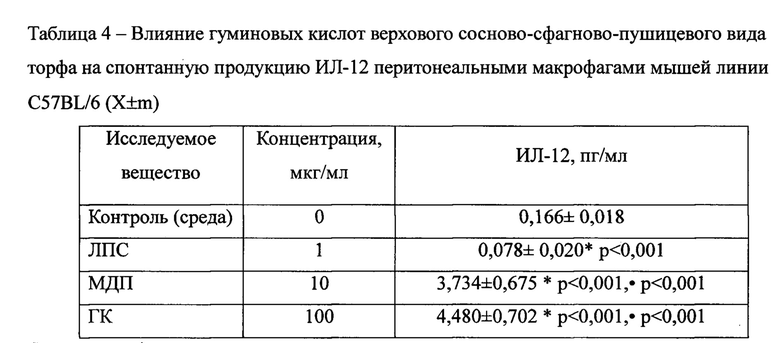
Примечания:* - достоверные различия с контролем; • - достоверные различия с митогеном; n=6.
ГК значительно, в 2,5 раза, усиливали секрецию другого провоспалительного цитокина ИЛ-2 митогенстимулированными спленоцитами мышей с 105,453±16,797 пг/мл при стимуляции Кон А до 268,310±3,170 пг/мл при добавлении ГК (Таблица 5). Кроме того, под влиянием ГК концентрация ИФН-γ увеличивалась в 5,9 раза с 2,901±0,543 в контроле до 17,101±2,521 пг/мл в опыте.
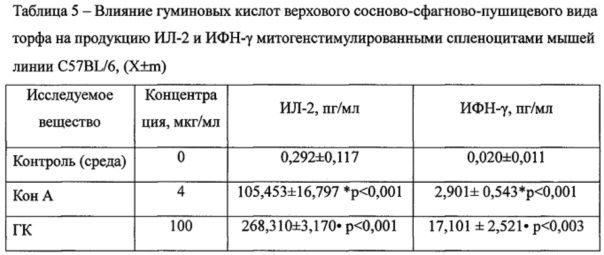
Примечания:* - достоверные различия с контролем; • - достоверные различия с митогеном. n=6.
В отсутствии каких-либо стимуляторов макрофаги продуцировали 20,048±5,036 пг/мл ИЛ-10. При добавлении стандартного активатора макрофагов ЛПС концентрация цитокина в супернатанте увеличилась в 6,9 раза до 138,770±7,635 пг/мл и в 4,6 (до 91,805±12,882 пг/мл) и 5 (до 100,567±5,485 пг/мл) раз в присутствии МДП и ГК, соответственно. Однако в условиях экспериментального воспаления (ЛПС стимуляция) ГК резко, в 1,5 раза, снижали митогенстимулированную продукцию ИЛ-10, в отличие от МДП, сохранявшего свои активирующие свойства (Таблица 6).
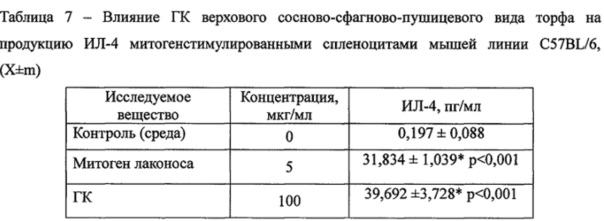
ГК не влияли на продукцию ИЛ-4 спленоцитами мышей, стимулированную митогеном лаконоса (Таблица 7).
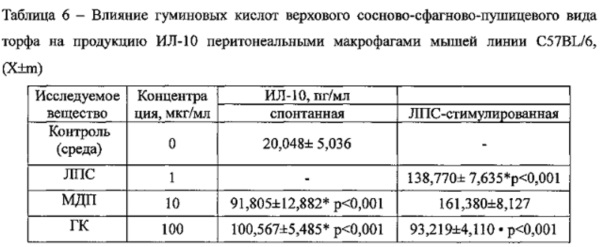
Примечания:* - достоверные различия с контролем; • - достоверные различия с митогеном. n=6.
Примечания: * - достоверные различия с контролем. n=6.
На модели моноцитов периферической крови человека показано (Таблица 8), что при добавлении водорастворимых гуминовых кислот верхового сосново-сфагново-пушицевого вида торфа концентрация ФНО-α резко увеличивалась с 1,486±0,92 пг/мл в контроле до 92,176±3,354 пг/мл как при культивировании с ГК, так и с МДП до 151,070±11,263 пг/мл. Активирующее действие ГК сохранялось и при оценке их влияния на продукцию ИФН-γ лимфоцитами: концентрация цитокина увеличивалась в 66,9 раза с 0,042±0,007 до 2,809±2,144 пг/мл.
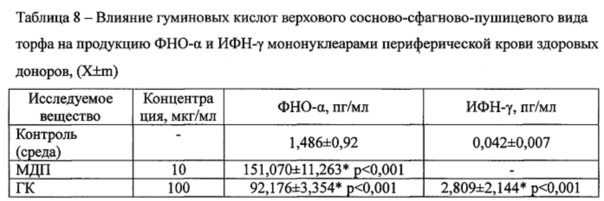
Примечание: * - различия с контролем достоверны. n=6.
Оценка влияния исследуемых веществ на секрецию ИЛ-10 показала, что ГК не влияют на спонтанную продукцию цитокина в отличие от МДП, который ее значительно, в 7,1 раза, усиливал. Более того, инкубация мононуклеаров с ГК в условиях ЛПС-моделированного воспаления приводила к существенному снижению в 3,8 раза концентрации цитокина (Таблица 9). ГК не усиливали митогенстимулированную продукцию ИЛ-4 (Таблица 10).
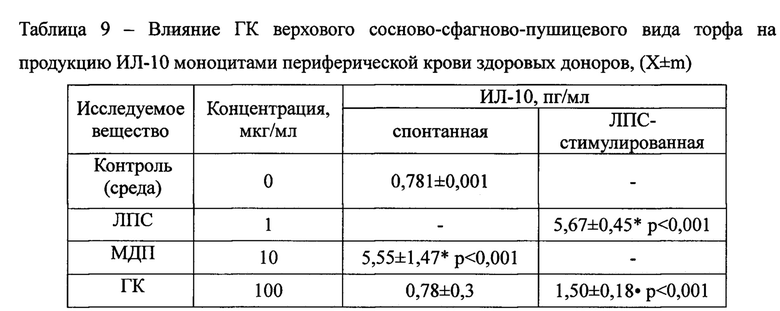
Примечания: * - достоверные различия с контролем; • - достоверные различия с митогеном. n=6.
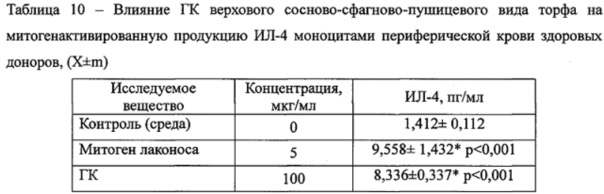
Примечания:* - достоверные различия с контролем. n=6.
Таким образом, экспериментально установлено, что ГК, выделенные из верхового сосново-сфагново-пушицевого вида торфа, снижают продукцию ИЛ-10 на фоне стимуляции выработки антигенпрезентирующими клетками ключевых провоспалительных цитокинов ИЛ-2, ИЛ-12, ФНО-α и ИФН-γ лимфоцитами. Следует отметить, что по своему цитокин-стимулирующему действию ГК значительно превосходят препарат сравнения МДП.
Пример. 2.
Курсовое введение ГК верхового сосново-сфагново-пушицевого вида торфа животным на фоне развития у них Th1-зависимого иммунного ответа, индуцированного введением эритроцитов барана, привело к подавлению реакции ГЗТ - маркерной реакции клеточного Тh1 иммунного ответа (Таблица 11). Курсовое введение препарата сравнения - ликопида - усиливало показатель в 1,3 раза. Величина отека при применении ГК и пирогенала снижалась в 1,6 по сравнению с контролем и в 2-2,2 раза относительно ликопида.
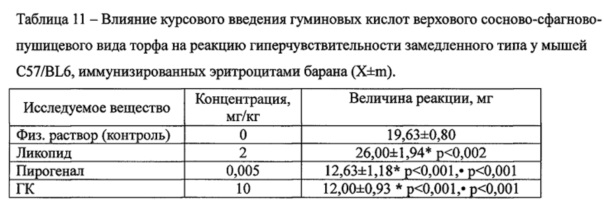
Примечание: * - различия с контролем достоверны. n=8.
Пример 3.
Влияние исследуемых гуминовых кислот на гуморальное звено Th1-зависимого иммунного ответа оценивали по количеству АОК. Показано, что курсовое введение ГК, выделенных при экстракции пирофосфатом натрия из верхового сосново-сфагново-пушицевого торфа, животным на фоне развития у них Th1-зависимого иммунного ответа, приводило к достоверному повышению числа АОК (Таблица 12). Показатель увеличивался как у мышей, получавших ликопид, так и ГК в 1,2 и 1,7 раза, соответственно. Пирогенал, напротив, значительно, в 3,5 раза, снижал интенсивность антителообразования. При этом титр гемаагглютининов неизменно увеличивался при курсовом введении всех исследуемых веществ в 1,8 (ликопид), 1,7 (пирогенал) и в 2,8 (ГК) раза. Более того, ГК в среднем в 1,5 раза усиливали РГА по сравнению с ликопидом и пирогеналом.
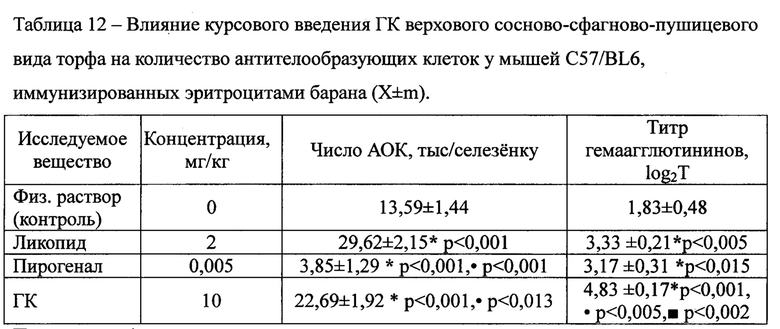
Примечание: * - различия показателя с контролем достоверны; • - различия показателя со ликопидом достоверны; ■ - различия показателя с пирогеналом достоверны. n=6.
Таким образом, экспериментально установлено, что гуминовые кислоты, выделенные из верхового сосново-сфагново-пушицевого вида торфа, снижают продукцию ИЛ-10 на фоне стимуляции выработки ключевых провоспалительных цитокинов антигенпрезентирующими клетками - ИЛ-2, ИЛ-12, ФНО-α и лимфоцитами - ИФН-γ. Курсовое введение гуминовых кислот снижает интенсивность реакции клеточного иммунитета и усиливает показатели гуморального иммунитета. Гуминовые кислоты, выделенные из верхового сосново-сфагново-пушицевого вида торфа, являются активаторами воспалительных свойств макрофагов по Th1-зависимому типу иммунного ответа и могут расширить арсенал малотоксичных средств растительного происхождения, способных стимулировать иммунный ответ при инфекционно-воспалительных процессах, хронических и онкологических заболеваниях.
Список литературы
1. Бакина, Л.Г. Особенности извлечения гумусовых кислот из почв растворами пирофосфата натрия различной щелочности / Л.Г. Бакина, Н.Е. Орлова // Почвоведение. - 2012. - №4. - С. 445-452.
2. Бузлама, А.В. Анализ фармакологических свойств, механизмов действия и перспектив применения гуминовых веществ в медицине / А.В. Бузлама, Ю.Н. Чернов // Экспер. и клин. фармакол. - 2010. - Т. 73, - №9. - С. 43-48.
3. Ваксман, С.А. Гумус, происхождение, химический состав и значение в природе / С.А. Ваксман // М.: Сельхозгиз. - 1937. - 437 с.
4. Василевич, Р.С. Строение высокомолекулярных органических веществ тундровых бугристых торфяников / Р.С. Василевич, В.А. Безносиков, Е.Д. Лодыгин // Материалы VI Всероссийской научной конференции с международным участием «Гуминовые вещества в биосфере». - Сыктывкар. - 2014. - c. 49-52.
5. Влияние гуминовых кислот торфа различного генеза на продукцию оксида азота in vitro (скрининговое исследование) / Е.С. Трофимова [и др.] // Бюлл. эксперим. биол. и медицины. - 2016. - №5. - С. 626-636.
6. Гостищева, М.В. Характеристика органического вещества торфяных почв эвтрофного болота Таган Томской области / М.В. Гостищева, Л.И. Инишева, А.И. Щеголихина // Вестник ТГПУ. - 2010. - №3. - С. 114-119.
7. Гуминовые кислоты торфа и препараты на их основе / И.И. Лиштван [и др.] // Природопользование. - 2004. - Вып. 10. - С. 114-119.
8. Исматова, Р.Р. Низкая токсичность и противовоспалительная активность гуматов, выделенных из торфа и сапропеля Томской области / Р.Р. Исматова, А.У. Зиганшин, С.Е. Дмитрук, И.В. Федько // Казанский медицинский журнал. - 2006. - Т. 87. - №6. - С. 454-455.
9. Исследование гепатозащитных свойств нативных гуминовых кислот низинного торфа Томской области / М.В. Белоусов [и др.] // Хим.-фарм. журн. - 2014. Т. 48, №4. С. 28-31.
10. Комиссаров, И.Д. Влияние способа извлечения гуминовых кислот из сырья на химический состав полученных препаратов / И.Д. Комиссаров, И.Н. Стрельцова // Гуминовые препараты: Научный трактат. Т. XIV. Тюмень. - 1974. - С. 48-62.
11. Кухаренко, Т.А. Об определении понятия и классификации гуминовых кислот / Т.А. Кухаренко // Химия твердого топлива. - 1979. - №5. - С. 3-11.
12. Левашенко, Д.В. Климатический оптимум голоцена в дельте Печоры / Д.В. Левашенко, Е.С. Малясова // Известия РАН. Сер. географическая. - 2007. - №4. - С. 125-132.
13. Орлов, Д.С. Гумусовые кислоты почв и общая теория гумификации. - М.: МГУ, 1990. - 325 с.
14. Руководство по проведению доклинических исследований лекарственных средств. 4.1. - М.: Гриф и К. 2013. - С. 64-79.
15. Сартаков, М.П. Графостатистический анализ и спектроскопия 13С-ЯМР молекул гуминовых кислот торфов Среднего Приобья / М.П. Сартаков, В.Д. Тихова // Вестник КрасГАУ. - 2009. - №6. - С. 76-80.
16. Хричтева, Л.А. Роль гуминовой кислоты в питании высших растений и гуминовые удобрения / Л.А. Хричтева // Труды почвенного института им. В.В. Докучаева, М.: 1951. - Т. 38. - 314 с.
17. In vitro evaluation of the antiviral properties of Shilajit and investigation of its mechanisms of action / V. Cagno [et al.] // Journal of Ethnopharmacology. - 2015. -, V. 166. P. - 129-134.
18. Inglot, A.D. A. A method to assess the immunomodulating effects of the Tolpa Torf Preparation (TTP) by measuring the hyporeactivity to interferon induction and tumor necrosis factor response / A.D. Inglot,  A.Sypula // Archivum Immunologiae et Therapiae Experimentalis. - 1993. - V. 41. - Is. 1. - P. - 87-93.
A.Sypula // Archivum Immunologiae et Therapiae Experimentalis. - 1993. - V. 41. - Is. 1. - P. - 87-93.
19. Li, H.H. Humic Acid Increases Amyloid β-Induced Cytotoxicity by Induction of ER Stress in Human SK-N-MC Neuronal Cells / H.H. Li [et al.] // International Journal of Molecular Sciences. - 2015. -V. 16. - Is. 5. - P. - 10426-10442.
20. Mosmann, T.R. TH1 and TH2 cells: different patterns of lymfokine secrecion lead to different functional properties / T.R. Mosmann, R.L. Coffman // Ann. Rev. Immunol. - 1989. - V. 7. - P. 145-173.
21. Prion protein interaction with soil humic substances: environmental implications / G. Giachin [et al.] // PloS one. - 2014. - V. 9. - Is. 6. - P. - e100016. (DOI: 10.1371/journal.pone.0100016).
22.  D.M. Angiomodulatory properties of some antibiotics and Tolpa Peat Preparation / D.M. [et al.] // Central-European Journal of Immunology. - 2016. -V. 41. - Is. 1. - P. - 19-24.
D.M. Angiomodulatory properties of some antibiotics and Tolpa Peat Preparation / D.M. [et al.] // Central-European Journal of Immunology. - 2016. -V. 41. - Is. 1. - P. - 19-24.
23. Rivera, A. Innate cell communication kick-starts pathogen-specific immunity / A. Rivera, [et al.] // Nature Immunology. - 2016. - V. 17. - Is.4. - P. - 356-363.
24. Van Rensburg, C. J.In vitro and in vivo assessment of humic acid as an aflatoxin binder in broiler chickens / C. J. van Rensburg [et al.] // Poultry Science - 2006. - V. 85. - Is.9. - P. - 1576-1583.
25. Van Rensburg, C.E. Potassium humate inhibits complement activation and the production of inflammatory cytokines in vitro / C.E. Van Rensburg, P.J. Naude // Inflammation - 2009. - V. 32. - Is. 4. - P. - 270-276.
26. Van Rensburg, С.E. Topical application of oxifulvic acid suppresses the cutaneous immune response in mice / С. E. J. Van Rensburg, S. С. K. Malfeld, J. Dekker // Chemotherapy. - 2002. - V. 48. - P. - 138-143.
27. Vetvicka, V. Prophylactic effects of humic acid-glucan combination against experimental liver injury / V. Vetvicka, J.M. Garcia-Mina, J.C. Yvin // Journal of Intercultural Ethnopharmacology. - 2015. - V. 4. - Is. 3. - P. - 249-255.
28. Vetvicka, V. The relative abundance of oxygen alkyl-related groups in aliphatic domains is involved in the main pharmacological-pleiotropic effects of humic acids / V. Vetvicka [et al.] // Journal of Medicinal Food. - 2013. - V. 16. - Is. 7. - P. - 625-632.
29. Worall, F. Can climate change explain increases in DOC flux from upland peat catchments? / F. Worall, T. Burt, J. Adamson // Science of the Total Enviroment. - 2004. - N. 326. - P. - 95-112.
30. Welte, E. Zur Konzentrationsmessung von Huminsauren / E. Welte // Z. Pflanzenernahr., Dung., Bodenkunde. - 1956. - V. 74. - №3.
| название | год | авторы | номер документа |
|---|---|---|---|
| Средство гуминовой природы, обладающее иммуномодулирующим действием | 2022 |
|
RU2783772C1 |
| Средство, гуминовой природы, обладающее иммуномодулирующей активностью | 2019 |
|
RU2716504C1 |
| Средство гуминовой природы, обладающее противоаллергической активностью | 2020 |
|
RU2750098C1 |
| СРЕДСТВО ГУМИНОВОЙ ПРИРОДЫ, УСИЛИВАЮЩЕЕ ФАГОЦИТИРУЮЩУЮ АКТИВНОСТЬ КЛЕТОК | 2023 |
|
RU2834961C1 |
| ИММУНОМОДУЛИРУЮЩЕЕ ГУМИНОВОЕ СРЕДСТВО | 2020 |
|
RU2756353C1 |
| Средство, обладающее иммуномодулирующей активностью | 2018 |
|
RU2697526C1 |
| СРЕДСТВО, ПОВЫШАЮЩЕЕ ПРОДУКЦИЮ ОКСИДА АЗОТА МАКРОФАГАМИ in vitro, НА ОСНОВЕ ГУМИНОВЫХ КИСЛОТ ИЗ ТОРФА БОЛОТ ТОМСКОЙ ОБЛАСТИ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2015 |
|
RU2610446C2 |
| Средство, обладающее иммуномодулирующей активностью | 2017 |
|
RU2657819C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ИММУНОМОДУЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2011 |
|
RU2470656C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ИММУНОМОДУЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2010 |
|
RU2423141C1 |
Изобретение относится к медицине, конкретно к фармакологии, и касается водорастворимых гуминовых кислот, выделенных из верхового сосново-сфагново-пушицевого вида торфа для коррекции нарушений в иммунной системе при патологических состояниях, связанных с недостаточностью Th1-зависимого иммунного ответа. Изобретение обеспечивает расширение арсенала таргетных нетоксичных иммуномодулирующих средств растительного происхождения за счет селективного повышения продукции провоспалительных цитокинов интерлейкина-2, интерлейкина-12, фактора некроза опухоли-альфа макрофагами и интерферона-гамма лимфоцитами, сопровождающееся отсутствием влияния на продукцию интерлейкина-4 и интерлейкина-10. 2 ил., 12 табл., 2 пр.
Применение водорастворимых гуминовых кислот, выделенных из верхового сосново-сфагново-пушицевого вида торфа экстракцией пирофосфатом натрия, в качестве средства, обладающего иммуномодулирующей активностью и стимулирующего развитие реакций Th1-зависимого типа иммунного ответа.
| СПОСОБ ЛЕЧЕНИЯ ЯЗВЕННОЙ БОЛЕЗНИ ЖЕЛУДКА И ДВЕНАДЦАТИПЕРСТНОЙ КИШКИ | 2007 |
|
RU2357741C1 |
| И.И | |||
| ЛИШТВАН и др | |||
| Гуминовые кислоты торфа и препараты на их основе // Природопользование | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| - Вып | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| - С | |||
| Способ получения борнеола из пихтового или т.п. масел | 1921 |
|
SU114A1 |
| JUNEK R., et al | |||
| Индукционный выпрямитель переменного тока в постоянный | 1924 |
|
SU937A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| - Vol | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| - P | |||
| РУЧНОЙ СТАНОК ДЛЯ ФОРМОВКИ ПУСТОТЕЛЫХ КАМНЕЙ РАЗЛИЧНОЙ ФОРМЫ | 1922 |
|
SU470A1 |

