Перекрестная ссылка на родственные заявки
Данная заявка испрашивает приоритет предварительной заявки на патент США №61/981,240, поданной 18 апреля 2014 года, раскрытие которой включено в данную заявку посредством отсылки.
Область техники, к которой относится изобретение
Настоящее изобретение, в общем, относится к гуманизированным антителам, которые распознают опухолевый антиген Томсена-Фриденрайха (TF) человека, и способам применения моноклональных антител.
Уровень техники
Во время канцерогенеза, изменения происходят в биосинтезе углеводных структур на поверхности клеток, и несколько различных углеводов, связанных или с белками, или с липидами, были признаны как такие, которые являются антигенами, ассоциированными с опухолью. Дисахарид Томсена-Фриденрайха (TF) (Galβ1-3GalNAcα) обычно обнаруживают О-связанным с остатками серина или треонина. TF-Ag (также известный как Т-антиген) ассоциировался с несколькими карциномами человека, включая те, которые найдены в поджелудочной железе, толстой кишке и молочной железе, и на этом основании был назван как пан-карциномный маркер. TF-Ag скрывается от иммунной системы в нормальных тканях взрослого организма путем расширения с большими гликановыми цепями. При раке, клеточный механизм гликозилирования может быть нарушен, что приводит к усечению этих цепей и воздействию TF антигена. Новые моноклональные антитела и их фрагменты антител, которые могут быть нацелены на TF-Ag, желательны и предусмотрены настоящим изобретением.
Сущность изобретения
Настоящее изобретение включает в различных вариантах осуществления композиции и способы лечения TF+ видов рака. В вариантах осуществления, настоящее изобретение включает частично гуманизированные моноклональные антитела (mAb) или их фрагменты, которые со специфичностью связываются с TF-Ag. mAb или его фрагмент содержит тяжелую цепь и легкую цепь, при этом тяжелая цепь содержит последовательность, выбранную из группы, состоящей из:
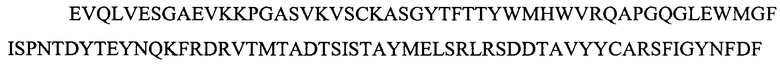
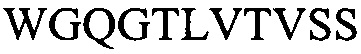 (SEQ ID NO: 7) (H1);
(SEQ ID NO: 7) (H1);
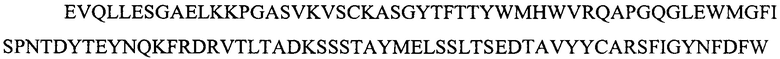
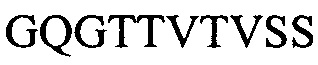 (SEQ ID NO: 8) (H2); и
(SEQ ID NO: 8) (H2); и
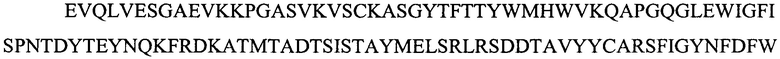
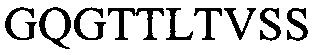 (SEQ ID NO: 9) (H3), и их комбинаций.
(SEQ ID NO: 9) (H3), и их комбинаций.
Легкая цепь содержит последовательность, выбранную из группы, состоящей из:
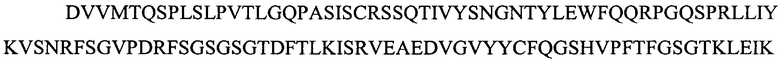
(SEQ ID NO: 10) (L1);
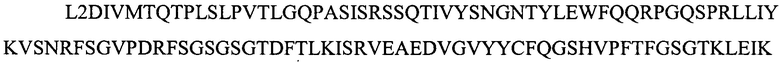
(SEQ ID NO: 11)(L2); и
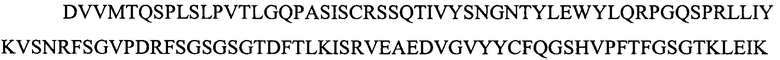
(SEQ ID NO: 12) (L3); и их комбинаций.
Дополнительные версии H2, H3, L2 и L3 были получены и испытаны. Они включают следующие:
Тяжелую вариабельную область Н2а:
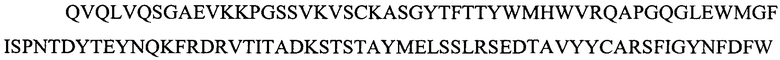
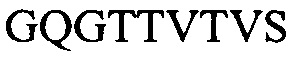 (SEQ ID NO: 13)
(SEQ ID NO: 13)
Тяжелую вариабельную область H3a:
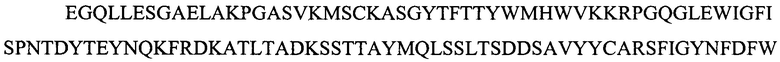
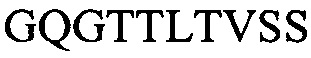 (SEQ ID NO: 14)
(SEQ ID NO: 14)
Легкую вариабельную область L2a:
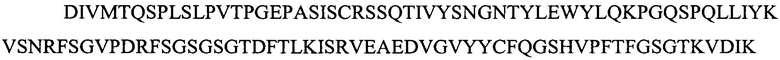
(SEQ ID NO: 15)
Легкую вариабельную область L3a:
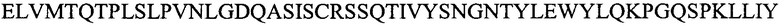
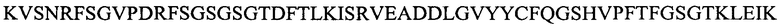
(SEQ ID NO: 16).
Комбинации тяжелых и вариабельных цепей, включенные в настоящее изобретение, представляют собой: H1-L1; H1-L2; H1-L2a; H1-L3; H1-L3a; H2-L1; Н2-L2; H2-L2a; H2-L3; H2-L3a; H3-L1; H3-L2; H3-L2a; H3-L3; H3-L3a; H2a-L1; H2a-L2; H2a-L2a; H2a-L3; H2a-L3a; H3a-L1; H3a-L2; H3a-L2a; H3a-L3; и H3a-L3a.
В вариантах осуществления, mAb содержит константную область IgG человека. В вариантах осуществления, mAb или его TF-Ag-связывающий фрагмент конъюгируется с агентом, выбранным из группы, состоящей из химиотерапевтических лекарственных средств, токсинов и радиоактивных изотопов.
В другом аспекте, настоящее изобретение включает способ профилактики и/или лечения рака у индивидуума, при этом рак содержит раковые клетки, экспрессирующие TF-Ag. Способ включает стадию, на которой вводят индивидууму одно или более mAb или фрагменты, как описано выше, при этом после введения ингибируется рост или выживание, или метастазы, или их комбинация, раковых клеток у индивидуума.
В другом аспекте, предусматриваются фармацевтические композиции, содержащие частично гуманизированное mAb или его фрагменты.
В другом аспекте, настоящее изобретение предусматривает in vitro клеточную культуру, при этом клетки в клеточной культуре экспрессируют частично гуманизированное mAb или его фрагменты.
В другом аспекте, настоящее изобретение предусматривает полинуклеотидные последовательности, кодирующие mAb и его TF-Ag-связывающие фрагменты, векторы экспрессии, содержащие такие полинуклеотиды, и in vitro клеточные культуры, содержащие такие векторы экспрессии. В вариантах осуществления, mAb или его TF-Ag-связывающий фрагмент кодируется более, чем одним вектором экспрессии.
В вариантах осуществления, предусматриваются способы получения mAb и их TF-Ag-связывающих фрагментов, и, как правило, включают стадии, на которых экспрессируют mAb или TF-Ag-связывающие фрагменты, или их комбинации в in vitro клеточной культуре, и отделяют mAb или TF-Ag-связывающие фрагменты, или их комбинации, от клеточной культуры.
Кроме того, предусматриваются наборы, содержащие mAb и их TF-Ag-связывающие фрагменты.
Краткое описание чертежей
На фигуре 1 представлены выравнивания аминокислотной последовательности тяжелой вариабельной области JAA-F11 и трех VH вариантных (H1, Н2 и H3) областей, которые включены в настоящее изобретение. Жирный шрифт означает идентичные аминокислоты между JAA-F11 и сконструированными, гуманизированными H цепями; выделенные жирным шрифтом и затененные аминокислоты означают различия между сконструированными гуманизированными цепями и мышиным JAA-F11. Алании в положении 72, показанный жирным шрифтом и курсивом, представляет собой мышиный остаток, который был сохранен для того, чтобы избежать стерических столкновений. Определяющие комплементарность области (CDR) показаны курсивом. Нумерация представлена в соответствии с системой Kabat. Последовательность mJAAF11 является SEQ ID NO: 17. Последовательность H1 является SEQ ID NO: 7; последовательность H2 является SEQ ID NO: 8; последовательность H3 является SEQ ID NO: 9.
Фигура 2 представляет выравнивания аминокислотных последовательностей легкой вариабельной области mJAA-F11 (JAA-F11VH) и трех VL вариантных (L1, L2 и L3) областей. Жирный шрифт означает мышиные аминокислоты или аминокислоты, идентичные мышиной последовательности, выделенные жирным шрифтом и затененные аминокислоты представляют собой различия между сконструированными гуманизированными вариантами JAA-F11 и мышиным JAA-F11. Лейцин в положении 51, показанный жирным шрифтом и курсивом, представляет собой мышиный остаток, который был сохранен для того, чтобы избежать стерических столкновений. Последовательность mJAAF11 является SEQ ID NO: 17. Последовательность L1 является SEQ ID NO: 10; последовательность L2 является SEQ ID NO: 11; последовательность L3 является SEQ ID NO: 12.
Фигура 3. Определение методом ELISA относительной аффинности с использованием ингибирования mJAA-F11 для того, чтобы оценить связывание каждого hJAA-F11 и химерного антитела с TF-Ag-BSA. Планки погрешностей представляют ±1 стандартную погрешность.
Фигура 4. Связывание hJAA-F11, химерного и мышиного JAA-F11 (50 мкг/мл) с 10 клеточными линиями рака молочной железы. Клеточные линии включают тройную отрицательную (НСС 70, ВТ 549, MDA-MB-231, MDA-MB-468, DU 4475), ER/PR-положительную (НСС 1419, AU 565, MDA-kb2) и HER2-положительную (САМА-1, НСС 1428). Для всех клеточных линий, испытанных выше, связывание с hJAA-F11, химерным или мышиным JAA-F11 (при 50 мкг/мл) было значительно выше (p≤0,05) (Т-критерий Стьюдента), чем у TF-Ag отрицательного контроля миеломы. Каждая планка погрешности представляет ±1 стандартное отклонение.
Фигура 5. Химерное антитело и гуманизированные антитела JAA-F11, кроме H3L3, вызывали небольшое (~6-11%), но статистически значимое ингибирование роста опухолевых клеток. Не было обнаружено никакого усиления пролиферации клеток. Не было обнаружено никакого значимого результата для какого-либо из антител в концентрации 2 или 1 мкг/мл.
Фигура 6А. ADCC активность мышиных, химерных и 4 конструктов hJAA-F11 против TF-Ag положительной клеточной линии рака молочной железы человека (ВТ 549). Результаты представлены в виде процента лизиса клеток в клетках, обработанных антителами, по сравнению с 100% лизисом клеток. H2L2 и H3L3 показали статистически большую ADCC активность, чем химерные или мышиные антитела (p<0,05) во всех случаях в клеточной линии человека. Это представлено как среднее значение всех экспериментов на РВМС от донора №1 и донора №2 (более, чем 3 независимых эксперимента). Планки погрешности представляют ±1 стандартную погрешность.
Фигура 6B. ADCC активность мышиных, химерных и 4 конструктов hJAA-F11 против TF-Ag положительной клеточной линии рака молочной железы человека (ВТ 549) на РВМС донора №1. Результаты представлены в виде процента лизиса клеток в клетках, обработанных антителами, по сравнению с 100% лизисом клеток. H2L2 и H3L3 показали статистически большую ADCC активность, чем химерные или мышиные антитела (p<0,05) во всех случаях в клеточной линии человека. Это представлено как среднее значение, по меньшей мере, 3 независимых экспериментов. Планки погрешности представляют ±1 стандартную погрешность.
Фигура 6C. ADCC активность мышиных, химерных и 4 конструктов hJAA-F11 против TF-Ag положительной клеточной линии рака молочной железы человека (ВТ 549) на РВМС от донора №2. Результаты представлены в виде процента лизиса клеток в клетках, обработанных антителами, по сравнению с 100% лизисом клеток. H2L2 и H3L3 показали статистически большую ADCC активность, чем химерные или мышиные антитела (p<0,05) во всех случаях в клеточной линии человека. Это представлено как среднее значение, по меньшей мере, 3 независимых экспериментов. Планки погрешности представляют ±1 стандартную погрешность.
Фигура 6D. ADCC активность мышиных, химерных и 3 конструктов hJAA-F11 против 4Т1 TF-Ag положительной мышиной клеточной линии рака молочной железы. Результаты представлены в виде процента лизиса клеток в клетках, обработанных антителами, по сравнению с 100% лизисом клеток. H3L3 и химерный показали статистически большую ADCC активность, чем мышиное антитело (p<0,05). Планки погрешности представляют ±1 стандартную погрешность. ADCC проводили только один раз с РВМС от донора №3.
Фигура 7. Гуманизированное, химерное и мышиное JAA-F11 не вызывают комплемент-зависимую цитотоксичность (CDC). Испытывали активность CDC мышиных, химерных и 2 конструктов hJAA-F11 против клеточной линии рака молочной железы НСС 1428 человека. Результаты представлены в виде процента лизиса клеток в клетках, обработанных антителами, по сравнению с 100% лизисом клеток. Положительный контроль: LDH положительные клетки, включенные в набор реагентов, показали лизис. Планки погрешности представляют ±1 стандартную погрешность.
Фигура 8. Интернализация мышиных, химерных и 4 антител hJAA-F11 с использованием иммуноферментного анализа для того, чтобы измерить поверхностное связывание. Интернализацию анализировали путем инкубирования мышиных 4Т1 клеток рака молочной железы с антителами при 37°C или 4°C. Как ожидалось, картина была противоположной картине ADCC, показанной для H3L3 на фигуре 20, но была неожиданной для H2L2, которое индуцировало ADCC и интернализовало. Мышиное, H2L2, химерное Ab, H2L3 и H1L1 значительно интернализовали (p<0,05). Планки погрешности представляют ±1 стандартную погрешность.
Фигура 9. МикроРЕТ визуализация, показывающая иммунолокализацию антитела hJAA-F11 у мыши, которая имеет 4Т1 опухоль.
Фигура 10. МикроРЕТ визуализация, показывающая локализацию свободного 124 йода в щитовидной железе у мыши, которая имеет 4Т1 опухоль.
Подробное описание изобретения
Настоящее изобретение включает различные гуманизированные моноклональные антитела (mAb) и их фрагменты, которые специфически распознают TF-Ag, способы получения mAb и способы применения mAb для профилактических и/или лечебных целей, и для диагностической визуализации. Аминокислотные последовательности mAb, предусмотренные настоящим изобретением, были разработаны с использованием нового подхода для того, чтобы изменить аминокислотные последовательности в mAb, продуцированные гибридомой, депонированной под номером CRL-2381 каталога АТСС, таким образом, что каркасные области mAb включают как мышиные, так и человеческие последовательности иммуноглобулина (Ig) в управляемом способе, что сохраняет специфичность и уменьшает иммуногенность. Гибридома продуцирует mAb, известное как JAA-F11. Изменения, внесенные в последовательность JAA-F11 в контексте мышиных каркасных областей в результате приводят к ансамблю из трех различных тяжелых и трех различных легких вариабельных цепей, которые могут быть скомбинированы для того, чтобы продуцировать двадцать пять различных mAb, которые являются приемлемыми для борьбы с любым из различных видов рака, которые содержат раковые клетки, которые экспрессируют TF-Ag. Кроме того, mAb имеют желаемые характеристики, которые делают их особенно приемлемыми для предпочтительного вызывания различных противораковых механизмов у индивидуума. Например, в зависимости от выбора тяжелой и легкой цепи, настоящее изобретение включает обеспечение стимулирования усиленной антитело-зависимой цитотоксичности, или комплемент-зависимой цитотоксичности, или интернализации mAb.
Варианты осуществления настоящего изобретения включают CDR последовательности из mAb JAA-F11. Они представляют собой: VH цепь: CDR1: 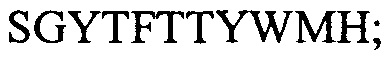 (SEQ ID NO: 1); CDR2:
(SEQ ID NO: 1); CDR2:  (SEQ ID NO: 2); CDR3:
(SEQ ID NO: 2); CDR3: 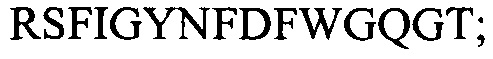 (SEQ ID NO: 3); и VL цепь: CDR1:
(SEQ ID NO: 3); и VL цепь: CDR1:  (SEQ ID NO: 4); CDR2:
(SEQ ID NO: 4); CDR2:  (SEQ ID NO: 5); и CDR3:
(SEQ ID NO: 5); и CDR3: 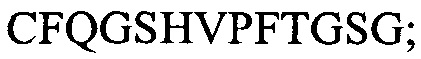 (SEQ ID NO: 6).
(SEQ ID NO: 6).
CDR последовательности располагаются в контексте модифицированных каркасных последовательностей. Как следствие, mAb и их TF-Ag-связывающие фрагменты содержат тяжелую цепь, выбранную из:
H1 -

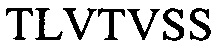 (SEQ ID NO: 7);
(SEQ ID NO: 7);
H2 -

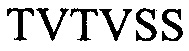 (SEQ ID NO: 8);
(SEQ ID NO: 8);
H3 -
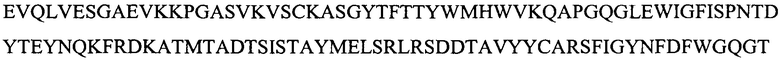
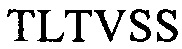 (SEQ ID NO: 9) и и их комбинаций;
(SEQ ID NO: 9) и и их комбинаций;
и легкую цепь, выбранную из группы, состоящей из:
L1 -
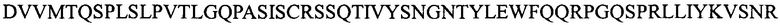
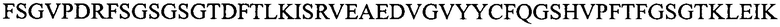 (SEQ ID NO: 10);
(SEQ ID NO: 10);
L2 -
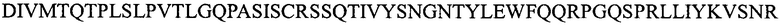
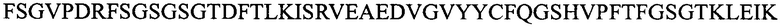 (SEQ ID NO: 11);
(SEQ ID NO: 11);
L3 -
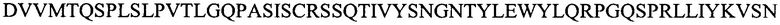
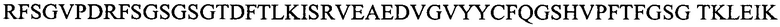 (SEQ ID NO: 12) и и их комбинаций.
(SEQ ID NO: 12) и и их комбинаций.
В дополнение к указанным выше последовательностям, были получены и испытаны в определенных комбинациях следующие тяжелые и легкие цепи:
Тяжелая вариабельная область Н2а:
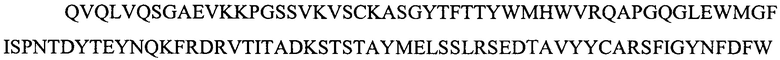
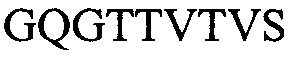 (SEQ ID NO: 13)
(SEQ ID NO: 13)
Тяжелая вариабельная область H3a:

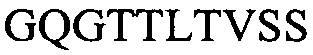 (SEQ ID NO: 14)
(SEQ ID NO: 14)
Легкая вариабельная область L2a:

(SEQ ID NO: 15)
Легкая вариабельная область L3a:
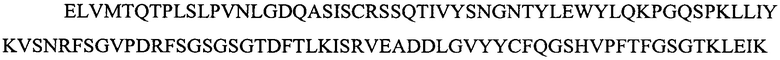
(SEQ ID NO: 16).
Настоящее изобретение охватывает mAb и их TF-Ag-связывающие фрагменты, содержащие аминокислотные последовательности, которые содержат или состоят из указанных выше последовательностей. В вариантах осуществления, настоящее изобретение включает mAb и их TF-Ag-связывающие фрагменты, при этом каркасные и CDR последовательности состоят из SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ NO: 13, SEQ ID NO: 14, SEQ ID NO: 15 и SEQ ID NO: 16.
Таким образом, следующие 25 комбинаций тяжелой и легкой цепи охвачены настоящим изобретением: H1-L1; H1-L2; H1-L2a; H1-L3; H1-L3a; H2-L1; H2-L2; Н2-L2a; H2-L3; H2-L3a; H3-L1; H3-L2; H3-L2a; H3-L3; H3-L3a; H2a-L1; H2a-L2; H2a-L2a; H2a-L3; H2a-L3a; H3a-L1; H3a-L2; H3a-L2a; H3a-L3; и H3a-L3a. В вариантах осуществления, настоящее изобретение включает mAb и их TF-Ag-связывающие фрагменты, содержащие комбинацию из H1L1, H2L2, H3L3, H2L3, Н2а и L2a, и H3a и L3a. В настоящем изобретении, в дополнение к результатам для комбинаций Н и L цепей, как описано ниже, H2a-L2a и H3aL3a были получены и испытаны, и результаты для данных двух комбинаций были аналогичны тем, которые сообщены для H2-L2 и H3-L3, соответственно. Комбинации H2a-L2a и H3a-L3a были получены в результате процедур субклонирования, что позволяет выбор дополнительных субклонов с дополнительными улучшенными характеристиками связывания для TF-Ag, как определяется самой высокой реакционной способностью в иммуноферментном анализе (EIA). Из данных, наилучшими для интернализации являются H2L2 и H2aL2a, в то время как наилучшими для ADCC являются H2L2, H2aL2a, H3L3 и H3aL3a.
Репрезентативные VH и VL последовательности дополнительно описаны с помощью фигур 1 и 2. В частности, на фигуре 1 представлены выравнивания аминокислотной последовательности JAA-F11 тяжелой вариабельной и трех VH вариантных (H1, H2 и H3) областей, которые включены в настоящее изобретение. Жирный шрифт означает идентичные аминокислоты между JAA-F11 и сконструированными, гуманизированными Н цепями; выделенные жирным шрифтом и затененные аминокислоты означают различия между сконструированными гуманизированными цепями и мышиным JAA-F11. Алании в положении 72, показанный жирным шрифтом и курсивом, представляет собой мышиный остаток, который был сохранен для того, чтобы избежать стерических столкновений. CDR показаны курсивом. Нумерация представлена в соответствии с системой Kabat. Таким образом, следует признать, что, в то время как mAb, описанные в данной заявке, упоминаются как «гуманизированные», они содержат определенные мышиные остатки, и таким образом могут считаться частично гуманизированными.
Фигура 2 представляет выравнивания аминокислотных последовательностей mJAA-F11 легкой вариабельной (JAA-F11VH) и трех VL вариантных (L1, L2 и L3) областей. Жирный шрифт означает мышиные аминокислоты или аминокислоты идентичные мышиной последовательности, выделенные жирным шрифтом и затененные аминокислоты представляют собой различия между сконструированными гуманизированными JAA-F11 вариантами и мышиным JAA-F11. Лейцин в положении 51, показанный жирным шрифтом и курсивом, представляет собой мышиный остаток, который был сохранен для того, чтобы избежать стерических столкновений. Как будет видно из фигур 1 и 2, которые представлены в соответствии с настоящим изобретением, гуманизированные Н и L цепи содержат отличия от JAA-F11 мышиного антитела и различия между собой, но определенные мышиные аминокислоты были сохранены для поддержания специфичности гуманизированных mAb.
Фрагменты гуманизированных mAb, описанных в данной заявке, также включены в настоящее изобретение. Примеры приемлемых фрагментов антитела включают Fab, Fab', F(ab')2, ScFv и Fv фрагменты. Различные методики были разработаны для получения фрагментов антитела и являются включенными в объем настоящего изобретения. В вариантах осуществления, mAb или их фрагменты продуцируются клетками-хозяивами посредством рекомбинантных векторов экспрессии. Настоящее изобретение включает все полинуклеотидные последовательности, кодирующие аминокислотные последовательности, описанные в данной заявке, векторы экспрессии, содержащие такие полинуклеотидные последовательности, и in vitro клеточные культуры, содержащие такие векторы экспрессии. В вариантах осуществления, клеточные культуры представляют собой эукариотные клетки. В вариантах осуществления, клеточные культуры представляют собой клетки млекопитающих. В вариантах осуществления, клетки представляют собой СНО клетки. Наборы, содержащие mAb и/или их TF-Ag-связывающие фрагменты, и/или клеточные культуры, экспрессирующие mAb и/или их TF-Ag-связывающие фрагменты, предусматриваются настоящим изобретением. В общем, наборы содержат один или более герметично закрытых контейнеров, которые содержат mAb и/или их TF-Ag-связывающие фрагменты, или клетки, экспрессирующие их. Инструкции по использованию mAb и/или TF-Ag-связывающих фрагментов для лечебных целей и/или целей визуализации могут быть включены в наборы.
В вариантах осуществления, настоящее изобретение включает способ получения mAb или их TF-Ag-связывающих фрагментов, который включает стадии, на которых культивируют клетки, содержащие вектор экспрессии или другую полинуклеотидную последовательность, кодирующую mAb или их TF-Ag-связывающие фрагменты, позволяют экспрессирование mAb или их TF-Ag-связывающих фрагментов, и отделяют mAb или их TF-Ag-связывающие фрагменты от клеточной культуры. Нуклеотидные последовательности, кодирующие mAb или их TF-Ag-связывающие фрагменты, могут быть экспрессированными с использованием любого приемлемого вектора экспрессии, многие из которых известны в данной области с уровня техники и/или являются коммерчески доступными. В одном варианте осуществления, тяжелая и легкая цепи экспрессируются на одном векторе экспрессии, таком как плазмида. В другом варианте осуществления, тяжелая и легкая цепи экспрессируются на разных плазмидах в одной и той же клетке, после чего экспрессированные тяжелая и легкая цепи образуют обычную архитектуру mAb. mAb или их TF-Ag-связывающие фрагменты могут быть выделены и/или очищены с использованием общепринятых методик, принимая во внимание преимущество настоящего изобретения.
В другом аспекте, настоящее изобретение предусматривает способ ингибирования роста раковых клеток у индивидуума, и/или ингибирования метастаз раковых клеток у индивидуума, где раковые клетки экспрессируют TF-Ag молекулы. Способ включает стадию, на которой вводят индивидууму терапевтическое количество гуманизированных mAb и/или их фрагментов, при этом введение ингибирует рост и/или ингибирует метастазы TF-Ag экспрессирующих раковых клеток. В вариантах осуществления, исполняемый на практике способ в соответствии с настоящим изобретением уменьшает объем опухоли, и/или уменьшает образование метастатических очагов, или вторичных опухолей. В вариантах осуществления, способ предусматривается для индивидуума, который в этом нуждается. В вариантах осуществления, у индивидуума, который в этом нуждается, диагностируется, подозревается наличие, или находится в группе риска развития или наличия рецидива рака. В вариантах осуществления, используют терапевтически эффективное количество mAb или его TF-Ag-связывающего фрагмента. Термин «терапевтически эффективное», как используется в данной заявке, означает, что количество введенного mAb или его TF-Ag-связывающего фрагмента является достаточным количеством для ингибирования роста, выживания и/или метастаз TF+ раковых клеток.
В различных вариантах осуществления, гуманизированные mAb и/или их фрагменты могут быть конъюгированы с химиотерапевтическим агентом для возможности локализации химиотерапевтического агента в раковых клетках посредством связывания с клетками, экспрессирующими TF-Ag. Химиотерапевтические агенты, полезные для получения таких конъюгатов антител, включают ферментативно активные токсины и их фрагменты. Приемлемые ферментативно активные токсины включают дифтерийную А цепь, несвязывающие активные фрагменты дифтерийного токсина, А-цепь экзотоксина (от Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модекцина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Химиотерапевтические агенты могут быть ковалентно связаны с mAb или его TF-Ag-связывающим фрагментом с использованием любого приемлемого подхода химического конъюгирования. В вариантах осуществления, химиотерапевтический агент может содержать сегмент слитого белка с mAb или TF-Ag-связывающим фрагментом.
В другом аспекте, настоящее изобретение предусматривает способ определения у индивидуума метастатических очагов, опухолей или и их комбинаций, при этом метастатические очаги или опухоли содержат клетки, экспрессирующие TF-Ag. Способ включает стадии, на которых вводят индивидууму гуманизированные mAb и/или их фрагменты, при этом гуманизированные mAb и/или их фрагменты были конъюгированы с обнаруживаемой меткой, и обнаруживают обнаруживаемую метку для того, чтобы определить метастатические очаги, опухоли или и их комбинации. Таким образом, гуманизированные mAb и/или их фрагменты могут быть конъюгированы с обнаруживаемой меткой, такой как радиоактивный агент. Множество радиоактивных изотопов являются доступными для конъюгирования с JAA-F11 mAb таким образом, что клетки, с которыми JAA-F11 mAb связываются, могут быть визуализированы или выборочно разрушены. Для выборочного разрушения клеток, экспрессирующих TF-Ag, JAA-F11 mAb могут быть конъюгированы с высоко радиоактивным атомом, таким как In111, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивными изотопами Lu.
Когда гуманизированные mAb и/или их фрагменты используются для идентификации клеток, экспрессирующих TF-Ag в метастатических очагах или в опухолях, то они могут содержать радиоактивный атом для сцинтиграфических исследований, например, Tc99m (метастабильный технеций-99), I123 или спиновую метку для томографии ядерного магнитного резонанса (ЯМР) (также известной как магнитно-резонансная томография или «МРТ»), такую как I123, I131, I124, F19, С13, N15, O17 или гадолиний (III) или марганец (II).
Меченные гуманизированные mAb и/или их фрагменты могут быть инъекционно введены пациентам, у которых диагностировано или подозревается наличие метастатического заболевания для того, чтобы определить метастатические очаги и/или опухоли. Информация от такой визуализации может быть использована для диагностирования или определения стадии состояния болезни пациента. Используемая метка может быть выбрана в соответствии с системой визуализации, которую используют. Например, индий 111, технеций 99 или йод 131 могут быть использованы для плоских сканирований или однофотонной эмиссионной компьютерной томографии (SPECT). Позитронные излучающие метки, такие как фтор 19, йод 123 и йод 124 могут быть использованы в позитронной эмиссионной томографии. Парамагнитные ионы, такие как гадолиний (III) или марганец (II) могут быть использованы в магнитно-резонансной томографии (МРТ). Локализация метки в пределах определенной ткани индивидуума позволяет определить локализацию метастатических очагов или опухолей, которые содержат клетки, экспрессирующие TF-Ag. Концентрация метки в определенном местоположении большая, чем фоновая, позволяет определить наличие метастазированных клеток. В предпочтительном варианте осуществления, после введения меченных гуманизированных mAb и/или их фрагментов, дается приемлемый период времени для прохождения таким образом, что несвязанные гуманизированные mAb и/или их фрагменты выводятся из индивидуума, таким образом, что фоновая метка значительно снижается.
Терапевтические составы, содержащие конъюгированные или неконъюгированные гуманизированные mAb и/или их фрагменты, могут быть получены за счет смешивания фармацевтически приемлемых носителей, эксципиентов или стабилизаторов (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в виде лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в применяемых дозировках и концентрациях, и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее, чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; образующие соль противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (ПЭГ). Гуманизированные mAb и/или их фрагменты могут быть объединены с другими химиотерапевтическими агентами в фармацевтических композициях.
Гуманизированные mAb и/или их TF-связывающие фрагменты могут быть введены любыми приемлемыми способами, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное и назальное введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное, внутрилимфатическое или подкожное введение. Кроме того, гуманизированные mAb и/или их фрагменты соответствующим образом могут быть введены путем импульсной инфузии, например, с помощью снижения доз антитела. Предпочтительно, дозирование осуществляется с помощью инъекций, наиболее предпочтительно внутривенных или подкожных инъекций, отчасти в зависимости от того, является ли введение кратким или продолжительным. В некоторых вариантах осуществления, гуманизированные mAb и/или их фрагменты вводят индивидууму, у которого диагностирован или подозревается наличие рака молочной железы, толстой кишки, предстательной железы, яичников, мочевого пузыря или других TF Ag+ видов рака, для того чтобы ингибировать метастазы или ингибировать рост раковых клеток.
Другие соединения, такие как химиотерапевтические агенты, иммунодепрессивные агенты и/или цитокины также могут быть введены. Комбинированное введение может включать совместное введение, с использованием отдельных составов или одного фармацевтического состава, и также может включать последовательное введение в любом порядке, при этом предпочтительно существует период времени, в течение которого оба (или все) активные агенты одновременно проявляют свои биологические активности.
Гуманизированные mAb и/или их фрагменты могут быть введены человеку или другому животному в соответствии с упомянутыми выше способами лечения в количестве, достаточном для ингибирования метастаз и/или роста клеток, экспрессирующих TF-Ag. Квалифицированным специалистом в данной области техники будет установлено, что форма и характер фармацевтически приемлемого носителя или разбавителя определяются количеством активного ингредиента, с которым он должен быть скомбинирован, путем введения и другими хорошо известными переменными, такими как размер индивидуума и стадия заболевания.
Пример 1
Следующий пример описывает получение и использование гуманизированных mAb в соответствии с настоящим изобретением, а также физические и функциональные свойства mAb.
Создание и конструирование гуманизированного JAA-F11
После того, как последовательности вариабельных областей мышиного JAA-F11 были верифицированы, CDR были определены таким образом, чтобы сохранять специфичность и аффинность для TF-Ag. Инклюзивные CDR были определены в соответствии с как Kabat et al [139], так и Chothia et al [141], на основе последовательности и структурной вариабельности, соответственно. Две аминокислоты, как считается, контактируют друг с другом в структуре белка, если обе содержат пару атомов, разделенных 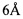 или меньше [138]. Рентгеновская кристаллография и компьютерные исследования углеводного протягивания на мышиных JAA-F11 [142] идентифицировали аминокислотные остатки в пределах
или меньше [138]. Рентгеновская кристаллография и компьютерные исследования углеводного протягивания на мышиных JAA-F11 [142] идентифицировали аминокислотные остатки в пределах 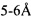 от сайта связывания. Данные аминокислоты, а также два цистеиновых остатка в положении L23 и L88 легкой цепи, которые являются важными для конформационной структуры, были включены в окончательное инклюзивное определение CDR, использованных в гуманизации JAA-F11.
от сайта связывания. Данные аминокислоты, а также два цистеиновых остатка в положении L23 и L88 легкой цепи, которые являются важными для конформационной структуры, были включены в окончательное инклюзивное определение CDR, использованных в гуманизации JAA-F11.
Выбор человеческих каркасов
Три различных подхода применяли для выбора каркасов человеческого акцепторного антитела для тяжелой и легкой JAAF11 цепей вариабельной области. Три варианта тяжелой цепи (H1, H2, H3) и 3 варианта легкой цепи (L1, L2, L3) были сконструированы. Любая из 3 тяжелых цепей (VH) может создавать пару с любой из 3 легких цепей (VL), чтобы создать в общей сложности 9 возможных hJAA-F11 VH/VL комбинационных вариантов.
Для того, чтобы создать эти варианты, поиск BLAST осуществляли, отдельно сравнивая VH и VL последовательность mJAA-F11 (мышиное антитело) по отношению к человеческим последовательностям иммуноглобулина. Первые 10 наиболее гомологичных человеческих VH и VL последовательностей выравнивали в соответствии с вариабельными областями mJAA-F11, используя программу выравнивания последовательности Seaview.
Для первых вариантов, H1 и L1, аминокислотная последовательность каркасных областей была выбрана на основе аминокислоты, наиболее часто проявляющейся в каждом положении в первых 10 наиболее гомологичных человеческих IgG вариабельных последовательностях. Для вариантов Н2 и L2, аминокислотные последовательности каркасных областей выбирали следующим образом: если аминокислота в любой из первых 10 человеческих последовательностей подходила под пару соответствующей аминокислоте в мышином иммуноглобулине, то была выбрана такая аминокислота, в остальных положениях была выбрана аминокислота, наиболее часто встречающаяся в человеческих последовательностях. Для третьего варианта, H3 и L3, аминокислотная последовательность каркасной области содержала 3 мышиные аминокислоты на какой-либо стороне каждой CDR, в то время как остальная часть последовательности состоит из аминокислот, которые наиболее часто встречается в первых 10 человеческих последовательностях.
Кроме того, было сконструировано химерное JAA-F11, которое состояло из всей мышиной JAA-F11 вариабельной области, присоединенной к человеческим константным областям. Химерное получали для верификации того, что правильные мышиные вариабельные области были клонированы и секвенированы, а также для того, чтобы служить в качестве положительного контроля при оценке гуманизированных JAA-F11 антител. Химерное, как ожидается, сохраняет те же характеристики связывания, что и мышиное JAA-F11, при этом имея человеческую константную область.
Оценка моделей hJAA-F11
Первоначально предложенные hJAA-F11 конструкты оценивали касательно конформационных эффектов на сайте связывания. Три тяжелые цепи вариантов (H1, H2, H3) имели замену на аланин аргинина в положении 72, что возможно могло привести к серьезному стерическому взаимодействию с окружающими аминокислотами. Лейцин в положении 51 для легкой цепи вариантов (L1, L2, L3) был заменен на аргинин, что в результате могло привести к стерическим столкновениям с окружающими аминокислотными боковыми цепями. Остаток аргинина был удален и заменен на исходный мышиный JAA-F11 каркасный остаток, аланин 72 (VH) и лейцин 51 (VL), как показано на фигурах 1 и 2. Сравнения выравнивания между мышиными JAA-F11 тяжелыми и легкими вариабельными аминокислотными последовательностями и hJAA-F11 конструктами являются такими, как показано на фигурах 1 и 2. Квалифицированные специалисты в данной области техники также признают различия в аминокислотных последовательностях, представленных в Н2а, H3a, L2a и L3a, по сравнению с другими Н и L последовательностями.
Прогнозирование иммуногенности hJAA-F11 вариантов
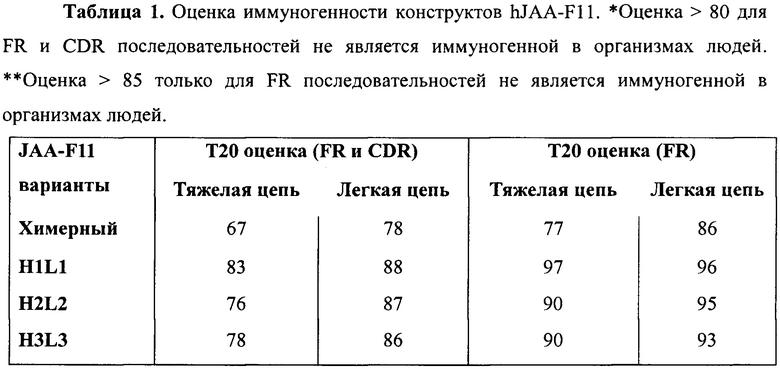
Прогнозируемая иммуногенность с использованием Т20 оценки наших гуманизированных вариантов (CDR-привитых), как ожидается, является очень низкой (Таблица 1). Т20 оценка используется для измерения «гуманизированности» последовательностей вариабельной области моноклонального антитела. Данная система оценки была разработана Gao et al [Monoclonal antibody humanness score and its applications. 2013. BMC Biotechnology, 13:55], используя базу данных более 38000 последовательностей человеческого антитела. В данном способе функционирует белок BLAST из этой базы данных и испытуемое гуманизированное Ab сравнивают с этими человеческими последовательностями. Гуманизированное антитело сравнивают с первыми 20 человеческими Ab BLAST парами и оценивают касательно сходства с этими последовательностями. Наиболее высокая возможная оценка составляет 100 (наиболее подобные человеку).
Валидация данного способа показана, когда Gao et al испытывали данный способ оценки иммуногенности на более, чем 90 антителах, которые были использованы клинически в организмах людей, и обнаружили, что антитела с Т20 оценками FR и CDR последовательностей выше 80 не были иммуногенными, в то время как Т20 оценки только для FR последовательностей, которые были выше 85, не были иммуногенными. Используя значения Т20 нынешних вариантов hJAA-F11, очень низкая иммуногенность ожидается у пациентов. Это показывает, что CDR-привитые антитела являются более человеческими и, как ожидается, являются менее иммуногенными, чем химерные, что H1L1 вариант является наиболее человеческим CDR-привитым вариантом, и что все hJAA-F11 варианты, как ожидается, имеют низкую иммуногенность.
Экспрессия и продуцирование гуманизированных и химерных JAA-F11 вариантов
hJAA-F11 и химерные VH и VL гены клонировали в двух различных векторах экспрессии млекопитающих, содержащих константную область человеческой IgG1 тяжелой цепи (6307 pAH) и константную область человеческой каппа-легкой цепи (6714 pAN), соответственно, для того, чтобы продуцировать плазмиды, содержащие полный IgG1 и каппа-гены. Правильная последовательность и направленность каждой из клонированных VH и VL областей в 6307 pAH и 6714 pAN векторах экспрессии были верифицированы путем секвенирования.
После стабильной совместной трансфекции двух векторов в СНО-K1 клетках, стабильные клоны, экспрессирующие hJAA-F11 или химерные, были выбраны на основе устойчивости к неомицину с использованием антибиотика G418 и экспрессии химерного или hJAA-F11. Скрининг hJAA-F11 кандидатов проводили с использованием ELISA для антитела к антигену TF-Ag с использованием установленных методик. Клоны из совместной трансфекции каждого из гуманизированных JAA-F11 H/L комбинационных вариантов и химерных JAA-F11, которые показали самую высокую реакционную способность с TF-Ag, были выбраны для дальнейших характеристик как описано в данной заявке. Гуманизированные JAA-F11 конструкты и химерные JAA-F11 индивидуально очищали от культуральных супернатантов с помощью белковой А колоночной хроматографии. Продуцированным было химерное антитело, а также H1L1, H2L2, H3L3 и H2L3, H2aL3a и H3aL3a. С учетом преимуществ настоящего изобретения квалифицированный специалист может легко получить остальные комбинации Н и L цепей антитела, которые охватываются в настоящем изобретении.
Анализ химической специфичности по гликановому массиву
После первоначального скрининга, химическую специфичность hJAA-F11 и химерных JAA-F11 вариантов определяли с использованием гликанового массива от Consortium for Functional Glycomics. Эти данные сравнивали с данными, полученными для мышиных JAA-F11. Анализ гликанового массива является непрямым иммунофлуоресцентным способом для определения гликан-связывающей реакционной способности лектинов и антител. Ранее гликановый массив использовали для демонстрации химической специфичности мышиных JAA-F11. Этот способ использовали для анализа реакционной способности кандидатов антител с 610 различными гликанами. hJAA-F11 H1L1, H2L2, H3L3 и химерное антитело показали такую же тонкую специфичность связывания как мышиное антитело, включая ограничение связывания с Galβ1-3GalNAc-α (TF-Ag) связанными структурами, и отсутствие связывания с α2-3 сиалилированными структурами. Гликановый массив показывает, что из 610 гликанов, гуманизированные антитела, химерное и мышиное JAA-F11 связывают только TF-Ag и четыре других TF-Ag содержащих сахаридных структуры. Четыре дополнительных сахарида, чьи мышиные JAA-F11, химерные и bJAA-F11 конструкты связываются с 440 или 610 испытуемыми сахаридами, не должны быть биологически проблематичными для нацеливания на опухоль JAA-F11. Незамещенные Neu5Acα2-6(Galβ1-3)GalNAcβ, GlcNAcβ1-6(Galβ1-3)GalNAcβ и Galβ1-4GlcNAcβ1-6(Galβ1-3)GalNAcβ были найдены только в выделениях из организма при раке или других болезненных состояниях. Neu5Acβ2-6 Galβ1-3GalNAc не является естественной структурой и и не встречается у людей.
Специфичность гуманизированных JAA-F11 конструктов, как представляется, является такой же самой или даже улучшенной по сравнению с химерными и мышиными антителами. Например, H2L3 показывает статистически (p<0,05 согласно ANOVA) меньшее связывание с любым дополнением к TF-Ag, и H1L1 и H2L2 не позволяют дисахариду быть добавленным к С-6 гидроксилу GalNAc и это представляет собой статистическое различие (p<0,05) от других антител.
Отсутствие связывания с более, чем 600 отрицательными сахаридами на массиве указывает, вероятно, на способность нацеливания всех этих антител. Важные примеры тесно связанных сахаридов, которые не связываются, представляют собой Galβ1-3GalNAc-бета-связанные структуры, указывающие на то, что антитело будет связываться с опухолевыми тканями, а не с GM1 ганглиозидом центральной нервной системы, асиало-GM1 NK клеток, GD1 гликолипидов или с асиало-GM1 периферической нервной ткани (данные являются бета-связанными). Хорошо известное удлинение TF-Ag на нормальных тканях осуществляется с помощью β1-3N-ацетилглюкозаминилтрансферазы, добавляя GlcNAcβ1-3 на Gal, образуя GlcNAcβ1-3Galβ1-3GalNAcα-Sp8, или с помощью 2-3 сиалилтрансферазы, образуя Neu5Acα2-3Galβ1-3GalNAc (сиалил-TF), или с помощью второй сиалилтрансферазы, образуя Neu5Ac2-3Galβ1-3(Neu5Acα2-6)GalNAc (дисиалил-TF), все из которых не связывают мышиные, химерные или гуманизированные JAA-F11. Таким образом, связывание нормальной ткани не ожидается для JAA-F11 или the hJAA-F11 конструктов.
Анализ относительной аффинности
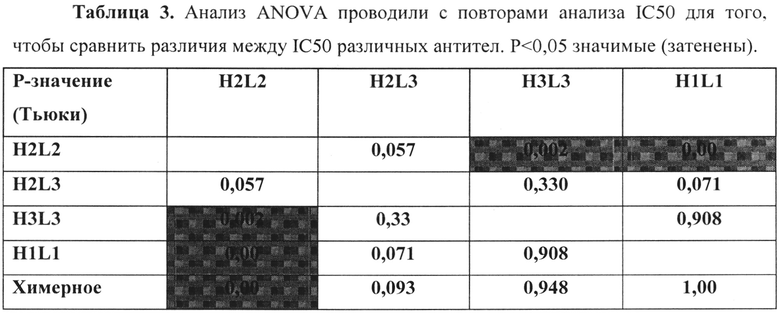
Относительные аффинности связывания четырех hJAA-F11 и химерных антител с TF-Ag определяли путем сравнения способности мышиного антитела конкурировать с гуманизированными антителами в иммуноферментном анализе. В этом анализе, 3 мкг каждого гуманизированного Ab смешивают с серийными разбавлениями мышиного антитела к антигену TF-Ag (mJAA-F11). Связывание человеческого антигена TF-Ag с планшетом, покрытым TF-Ag, измеряют с использованием видов специфического античеловеческого IgG. Количество мышиного mJAA-F11, необходимое для ингибирования 1 мкг hJAA-F11 до 50%, экстраполируется и принимается в качестве меры относительной аффинности. Чем больше количество мышиного антитела, требуемое для ингибирования, тем выше относительная аффинность антитела.
Результаты представлены на фигуре 3 и в таблицах 2 и 3. Таблица 2 показывает половину максимальной ингибирующей концентрации мышиного JAA-F11, требуемой (IC50) для различных антител, чтобы конкурировать с 1 мкг данного гуманизированного антитела. H2L2 показан с самой высокой относительной аффинностью к TF-Ag среди антител, с IC50 2,31 мкг мышиного антитела. Таблица 7 показывает p-значения, рассчитанные для каждого антитела по отношению друг к другу по ретроспективному анализу ANOVA по способу Тьюки. Можно видеть, что эта улучшенная разница аффинности H2L2 антитела является статистически значимой (p<0,05), при сравнении с каждым из hJAA-F11 и химерным антителом, за исключением для H2L3, где она приближалась к значимости (p=0,057) (Таблица 3). H2L3 значительно не отличается от H1L1, H2L2, H3L3 и химерного антитела. Относительные аффинности H1L1, H3L3 и химерного антитела значительно не отличаются друг от друга.

Анализ биологической реакционной способности, специфичности и активности hJAA-F11
3hJAA-F11 и химерное JAA-F11 антитела связываются с клеточными линиями опухоли человека.
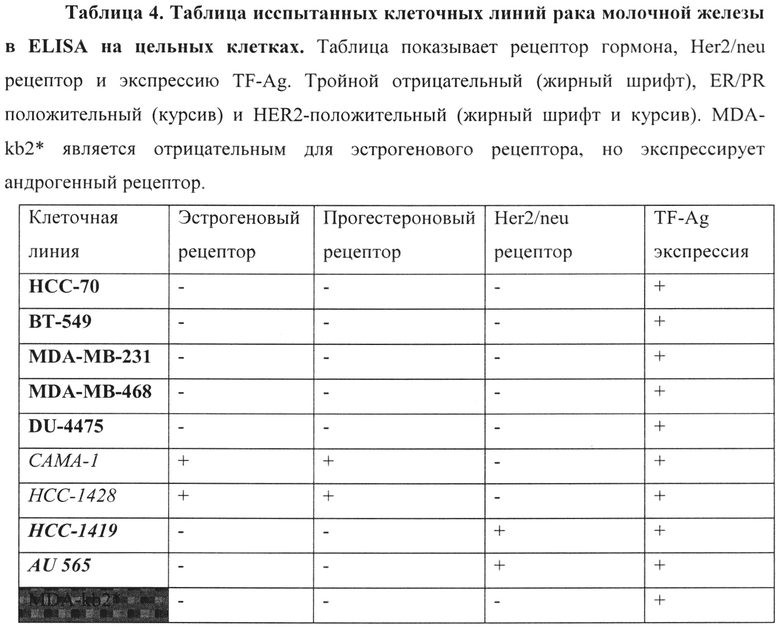
Связывание hJAA-F11 и химерного JAA-F11 антител с различными клеточными линиями опухоли молочной железы человека оценивали с использованием ELISA на цельных клетках. Клеточная линия TF-Ag положительной мышиной опухоли молочной железы 4Т1 служила в качестве положительного контроля, в то время как миеломная клеточная линия P3-X63-Ag8 служила в качестве TF-Ag отрицательного контроля. Анализы проводили в течение трех разных дней и анализировали от 3 до 4 клеточных линий за один раз на четырех одинаковых экземплярах с контрольными клеточными линиями. Клеточная линия считалась положительной, если относительное связывание возникшее с hJAA-F11 или химерным JAA-F11 (при 50 мкг/мл) было значительно выше (p≤0,05), чем у TF-Ag отрицательного контроля миеломы. Результаты показаны на фигуре 4.
Испытанные раковые клеточные линии состоят из различных подгрупп рака молочной железы, гормональных положительных или отрицательных, Her2-положительных или отрицательных, и тройных отрицательных рецепторов. Испытанные положительные клеточные линии эстрогенового рецептора (ER) и прогестеронового рецептора (PR) представляли собой САМА-1 и НСС-1428. Испытывали две положительные клеточные линии Her2/neu рецептора, НСС-1419 и AU-565. Испытанные клеточные линии тройного отрицательного рака молочной железы (TNBC) представляли собой НСС-70, MDA-MB-231, MDA-MB-468, DU-4475 и ВТ-549. Одна клеточная линия, MDA-kb2, экспрессирует андрогенный рецептор, но является отрицательным эстрогенновым рецептором. Эти клеточные линии рака молочной железы были рассмотрены ранее, используя мышиные JAA-F11, и все были положительными для TF-Ag экспрессии. В настоящем изобретении, результаты показывают, что hJAA-F11 и химерные JAA-F11 антитела связываются со всеми 10 рассмотренными клеточными линиями рака молочной железы, что подтверждает экспрессию TF-Ag на данных клеточных линиях. Среди различных подгрупп рака молочной железы, в настоящее время не существует целевой терапии для лечения пациентов с TNBC. Полученные данные свидетельствуют о терапевтической роли TF-Ag в нацеливании на агрессивный TNBC и использовании hJAA-F11 антител для лечения и увеличения выживаемости пациентов с раком молочной железы независимо от статуса рецепторов. В таблице 4 приведены виды рака молочной железы и экспрессия TF-Ag каждой испытанной клеточной линии.
Гуманизированное JAA-F11 ингибирует пролиферацию раковых клеток in vitro
Поскольку некоторые антитела к антигену TF-Ag, как было показано, вызывают пролиферацию опухолевых клеток, важным было определить влияние различных конструктов на пролиферацию клеток. МТТ анализ, который измеряет метаболическую активность, был выбран в качестве замены для прямого пролиферативного анализа, меченного тритием тимидина. Влияние гуманизированных, химерных и мышиных JAA-F11 антител на рост раковых клеток по сравнению с контролем без антитела определяли при 4 мкг/мл антител, используя известные способы. Данные представлены на фигуре 5. Химерное антитело и гуманизированные JAA-F11 антитела, за исключением H3L3, вызвали небольшое (~6-11%), но статистически значимое ингибирование роста опухолевых клеток. Не было обнаружено никакого усиления пролиферации клеток.
Гуманизированное JAA-F11 вызывает ADCC
Опосредованная антитело-зависимыми клетками цитотоксичность (ADCC) была проведена на клеточных линиях опухоли молочной железы человека с использованием CytoTox 96 нерадиоактивного анализа на цитотоксичность (Promega, Madison, WI), основанного на анализе высвобождения лактатдегидрогеназы (LDH) в соответствии со стандартными методиками со следующими приспособлениями. Свежие мононуклеарные клетки периферической крови человека были выделены из EDTA некоагулированной цельной крови для использования с гуманизированным антителом и 4Т1 мышиными клетками рака молочной железы при соотношении эффектора к мишени 100:1. Высвобождение LDH использовали для количественной оценки цитотоксичности.
Фигура 6А показывает количество ADCC, уменьшенное с помощью мышиных, химерных, H1L1 и H3L3 антител по сравнению с клеточными линиями рака молочной железы человека, ВТ 549. Эксперименты повторяли больше, чем три раза. Во всех экспериментах, H2L2 и H3L3 показали значительно большую (p<0,5) ADCC, чем или химерное, или мышиное антитело в клеточных линиях человека. Более, чем 20% TF-Ag, имеющих ВТ549 и НСС70 клетки-мишени, были лизированы с помощью H2L2 и H3L3 антитела в концентрации 200 мкг/мл при соотношении 100:1 эффектора к мишени (Е:Т). Кроме того, во всех экспериментах на мышиной 4Т1 клеточной линии, хотя все испытанные антитела вызывали менее, чем 10% ADCC, H3L3 антитело показало значительно большую (p<0,05) ADCC, чем или химерное, или мышиное антитело. Это указывает на то, что в настоящее время H2L2 и H3L3 антитело представляет собой наилучший выбор для иммунотерапии. В отличие от этого, мышиное JAA-F11 не показало статистически значимые возможности ADCC в каком-либо из испытаний. ADCC результаты от трех отдельных доноров РВМС являются такими, как показано на фигурах 6B, 6С и 6D.
Гуманизированное JAA-F11 не вызывает CDC
Способность hJAA-F11 и химерных антител опосредовать комплемент-зависимую цитотоксичность (CDC) определяли, используя анализ высвобождения лактатдегидрогеназы (LDH), используя клетки рака молочной железы человека НСС-1428 в качестве клеток-мишеней. LDH положительный контроль, обеспеченный набором реагентов, был использован в качестве положительного контроля, и показал лизис. Мышиные JAA-F11, химерные, H1L1, H3L3, H2L3 антитела не вызывали комплемент-зависимой цитотоксичности, поскольку не происходил лизис как показано на фигуре 7.
Гуманизированное JAA-F11 интернализирует в раковых клетках
Интернализацию гуманизированного и химерного JAA-F11 в 4Т1 клетках опухоли молочной железы определяли двумя способами, иммуноферментным анализом со связыванием с поверхностью, измеренным и сравненным после инкубирования клеток при или 4°C, или 37°C, и способом иммунофлуоресцентной микроскопии с LAMP-1 (лизосомальный ассоциированный мембранный белок, лизосомальный маркер) и DAPI окрашиванием для ядерного окрашивания. В иммуноферментном анализе, как показано на фигуре 8, мышиное, H2L2, химерное, H2L3 и H1L1 антитела значительно интернализовали с p-значениями 0,001, 0,002, 0,001, 0,002, и 0,014, соответственно. Однако, H3L3 антитело не показало значимой интернализации (p=0,16), и, как ожидается, H3L3 показало значительно большую (p<0,5) ADCC, чем химерное или мышиное JAA-F11.
В иммунофлуоресцентных экспериментах, в согласии с иммуноферментным анализом, мышиное, химерное, H2L2, H2L3 антитела показали интернализацию, и совместная локализация с LAMP-1 и H3L3 показала мембранное окрашивание, которое не интернализовало. Эти данные подтвердили, что получены с использованием поверхностного иммуноферментного анализа. Можно было бы ожидать, что антитела, которые показали более высокую ADCC активность, покажут более низкую интернализацию, поскольку присутствие антитела на поверхности клетки является необходимым для функции ADCC. Анализы интернализации были проведены на hJAA-F11, химерном и mJAA-F11. H3L3 показало согласие с этим ожиданием, имея небольшие количества интернализации, но относительно высокую ADCC. H1L1, H2L3, химерное и mJAA-F11, все показали статистически значимую интернализацию и не выполняли ADCC хорошо. Неожиданный результат наблюдался для H2L2 антитела, которое вызывало высокую ADCC, подобную H3L3, но показало самый высокий процент активности интернализации по сравнению с химерными и другими гуманизированными антителами.
Эксперимент иммунофлуоресцентной интернализации первоначально проводили с использованием 5 мкг/мл антитела и повторяли с использованием концентраций антитела 0,1 мкг/мл, и антител, которые интернализировали таким образом даже при данной низкой концентрации. Приведенные выше данные показывают, что из-за высокого процента интернализации, и высокой скорости этой интернализации, химерные, H2L2 и H2L3 конструкты имеют потенциал для того, чтобы быть использованными в качестве конъюгатов антитело-лекарственное средство. Данные гликановой специфичности, относительной аффинности, интернализации, ADCC и CDC приведены в таблице 5 ниже.

МикроРЕТ визуализация
МикроРЕТ визуализацию проводили поочередно на одной мыши, которой каждый раз инъекционно вводили 124 йод-hJAA-F11 H2L2 антитело, и свободный 124 йод. Визуализацию проводили через 24, 48, 72, 96, 168 и 192 часов. Фигура 9 показывает корональный вид мыши, которой инъекционно вводили 124 йод-hJAA-F11 H2L2 антитело в различные моменты времени, в то время как фигура 10 представляет контрольную мышь, которой инъекционно вводили свободный 124 йод. Поглощение радиоактивно меченного антитела опухолью наблюдали через 24, 48, 72 и 96 часов после инъекции. Поглощение антитела также наблюдалось в селезенке через 48 часов (фигура 9). Отрицательный контроль мыши, имеющей 4Т1 TF-Ag положительную опухоль, которая получала только свободный 124 йод, показал, что не было никакой локализации в любых органах или опухоли за исключением щитовидной железы на протяжении всего исследования (Фигура 10).
Полностью мышиное JAA-F11 имеет потенциал для пассивной гуморальной иммунотерапии и лечения конъюгатом лекарственного средства у пациентов с раком молочной железы. Однако, поскольку было показано, что применение мышиных антител у людей ограничивается развитием ответов на человеческие анти-мышиные антитела (НАМА) и коротким периодом полувыведения у пациентов, гуманизация является желательной для того, чтобы уменьшить иммуногенность мышиного JAA-F11 антитела, а также позволяет ему оставаться в обращении в течение более длительного времени.
Будет признано из приведенного выше описания результатов, что настоящее изобретение предусматривает, среди других аспектов, описание нового подхода для гуманизации мышиного JAA-F11 mAb, а также сравнения и различия между мышиными, химерными и гуманизированными JAA-F11 антителами. mJAA-F11 было гуманизировано частично путем CDR-привития. Как будет признано квалифицированным специалистом в данной области техники, традиционно, гуманизация путем CDR-привития использует один каркас человеческого акцепторного антитела [Jones, et al. Replacing the complementarity-determining regions in a human antibody with those from a mouse. 1986. Nature 321: 522-525]. Таким образом, человеческие каркасные последовательности выбирают из существующих генов зародышевой линии человека. В противоположность этому, в настоящем изобретении, три новых и разных подхода применяли для выбора каркаса человеческого акцепторного антитела для тяжелой и легкой JAAF11 цепей вариабельной области, образуя три варианта тяжелой цепи (H1, H2, H3) и 3 варианта легкой цепи (L1, L2, L3). Как уже обсуждалось выше, любая из 3 тяжелых цепей (VH) может создавать пару с любой из 3 легких цепей (VL), чтобы создать в общей сложности 9 возможных hJAA-F11 VH/VL комбинационных вариантов. Химерное JAA-F11, которое имеет цельную мышиную вариабельную область и человеческий IgG1 и каппа константные области, также образовывалось и использовалось в качестве контроля.
Одна проблема представляла собой подход, использованный при определении CDR и FR областей, который уменьшает иммуногенность гуманизированных антител. Способ Т20 оценки, разработанный Gao et al [Monoclonal antibody humanness score and its applications. 2013. BMC Biotechnology, 13:55], и валидированный анализом 90 антител, которые применяли у пациентов, использовали для прогнозирования иммуногенности гуманизированных JAA-F11 вариантов. Т20 оценки гуманизированных JAA-F11 вариантов являются все более высокими (улучшенными) (>85), чем химерного JAA-F11 (<85), что указывает на гуманизацию тяжелых и легких вариабельных цепей всеми тремя из способов в соответствии с настоящим изобретением, созданных hJAA-F11 вариантов, которые будут менее иммуногенными, чем химерный вариант.
Мы гуманизировали мышиные JAA-F11, и пять антител были получены с использованием СНО-K1 клеток, химерного и 4 hJAA-F11 конструктов, H1L1, H2L2, H3L3 и H2L3. Таким образом, принимая во внимание преимущество настоящего изобретения, квалифицированный специалист сможет получить любое применение других четырех возможных VH и VL комбинаций.
Гуманизация антитела путем CDR-привития может привести к снижению аффинности или потере связывания антигена, которая может происходить из-за конформации CDR, и сайта связывания антигена, который отрицательно сказывается на изменениях в некоторых каркасных аминокислотных остатках в гуманизированном антителе. CDR состоят из остатков, которые взаимодействуют с каркасными областями и другими CDR. В дополнение к взаимодействию CDR аминокислот с последовательно расположенными рядом аминокислотами, некоторые каркасные остатки, которые могут влиять на связывание с антигеном непосредственно или опосредовано, включают остатки зоны Верньера и остатки на разделе VL/VH. Остатки зоны Верньера представляют собой остатки в каркасе β-листа, лежащие в основе CDR, которые обеспечивают основу для конформации структур петель. Остатки Верньера на легкой цепи находятся в положении 2, 4, 35, 36, 46, 48, 49, 64, 67, 69 и 71, в то время как на тяжелой цепи они определены в положении 2, 27, 28, 29, 30, 47, 48, 49, 67, 69, 71, 73, 78, 93, 94 и 103. Остатки на разделе VL/VH были определены Chothia и его сотрудниками [Chothia С, et al. The packing of variable domains. 1989. J. Mol. Biol. 186:651-63], и находятся в положении 34, 36, 38, 44, 46, 87, 89, 91, 96 и 98 легкой цепи, и в положении 35, 37, 39, 45, 47, 91, 93, 95, 100-100K и 103 для тяжелой цепи.
Мы показали, что подходы, используемые в настоящем изобретении для выбора человеческих FR и CDR отрицательно не изменяли аффинность гуманизированного JAA-F11 для TF-Ag, по сравнению с химерными и мышиными JAA-F11. При определении CDR, остатки в пределах от 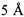 до
до 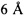 сайта связывания были включены для того, чтобы гарантировать, что аффинность и специфичность к TF-Ag сохраняется. Данные остатки находятся в положении Н31, Н32, H33, Н35, Н50, Н52, Н53, Н54, Н95, Н96, Н97, Н98 и Н100 на тяжелой цепи, и в положениях L27d, L28, L30, L32, L34, L50, L89, L91, L92 и L96 на легкой цепи. Однако, из исследований относительной аффинности, в то время как все гуманизированные антитела сохраняют аффинность для TF-Ag, мы обнаружили различия в аффинности между антителами. Исследования относительной аффинности показали, что H2L2 антитело обладает самой высокой аффинностью антитела, с последующим H2L3 антителом. H2L2 антитело обладает самой высокой относительной аффинностью к TF-Ag среди антител, требуя более 2 мг мышиного антитела для ингибирования 1 мг H2L2 антитела. H2L3 антитело также обладало более высокой аффинностью, чем H1L1 и химерное антитело, хотя и не значительно, в то время как относительные аффинности H1L1, H3L3 и химерного антитела к TF-Ag значительно не отличаются друг от друга.
сайта связывания были включены для того, чтобы гарантировать, что аффинность и специфичность к TF-Ag сохраняется. Данные остатки находятся в положении Н31, Н32, H33, Н35, Н50, Н52, Н53, Н54, Н95, Н96, Н97, Н98 и Н100 на тяжелой цепи, и в положениях L27d, L28, L30, L32, L34, L50, L89, L91, L92 и L96 на легкой цепи. Однако, из исследований относительной аффинности, в то время как все гуманизированные антитела сохраняют аффинность для TF-Ag, мы обнаружили различия в аффинности между антителами. Исследования относительной аффинности показали, что H2L2 антитело обладает самой высокой аффинностью антитела, с последующим H2L3 антителом. H2L2 антитело обладает самой высокой относительной аффинностью к TF-Ag среди антител, требуя более 2 мг мышиного антитела для ингибирования 1 мг H2L2 антитела. H2L3 антитело также обладало более высокой аффинностью, чем H1L1 и химерное антитело, хотя и не значительно, в то время как относительные аффинности H1L1, H3L3 и химерного антитела к TF-Ag значительно не отличаются друг от друга.
Данные улучшения в аффинности могут относится к изменениям в некоторых остатках между различными гуманизированными вариантами. Различия между вариантами перечислены ниже в таблице 6A и 6B.
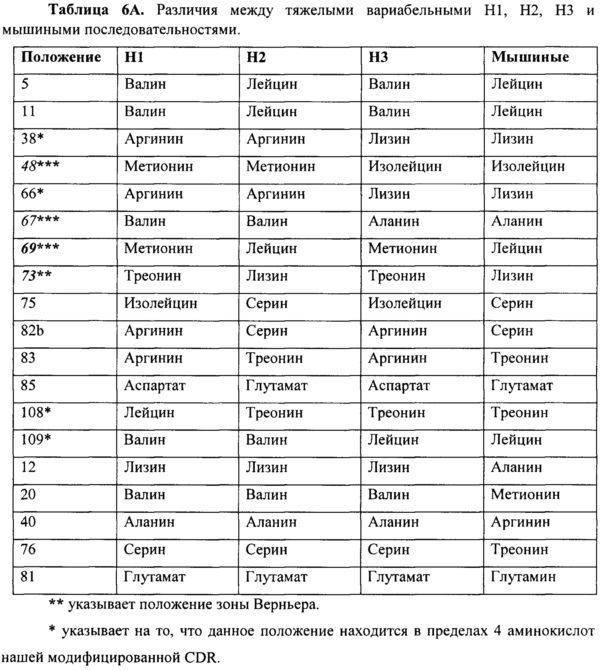
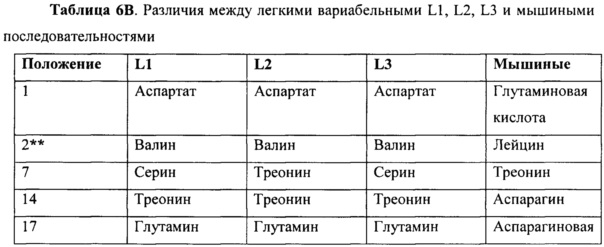
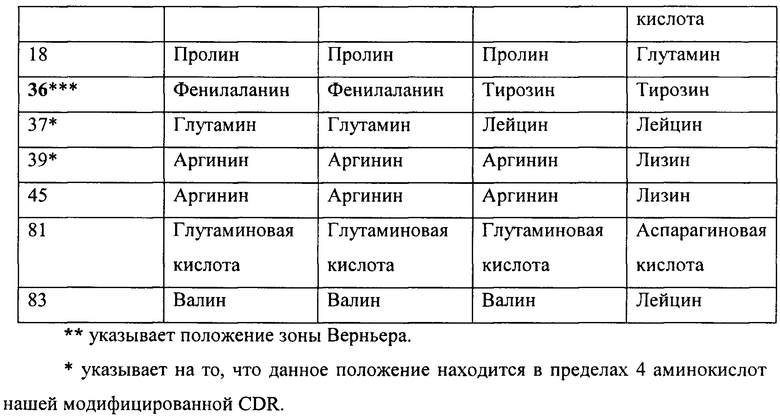
Как было упомянуто выше, каркасные остатки, влияющие на связывание антигена, включают остатки зоны Верньера и остатки на разделе VL/VH. При сравнении гуманизированных конструктов, mAb в соответствии с настоящим изобретением сохраняют большую часть остатков зоны Верньера и остатков на разделе VL/VH, которые могут иметь важное значение для конформации сайта связывания, однако некоторые изменения сделаны для создания менее иммуногенных гуманизированных вариантов. Аминокислоты, которые были изменены, которые находились в зоне Верньера или на разделе VH/VL, и изменения, сделанные с аминокислотами в пределах 4 аминокислот (последовательно) для CDR, являются такими, как показано звездочками. Существует четыре положения зоны Верньера на последовательности тяжелой цепи, которые отличаются между тремя конструктами, положения 48, 67, 69 и 73. H2 сохраняет мышиные остатки, но отличается между H1 и H3 конструктами в восьми положениях 5, 11, 69, 73, 75, 82b, 83 и 85. Из этих восьми остатков, два представляют собой остатки зоны Верньера, в положениях 69 и 73. В тоже время, H2 и H1 имеют замещения в двух положениях зоны Верньера, то есть, в остатке 48 и 67, которые сохраняются как мышиные остатки на H3. Поскольку не существует никакого различия в аффинности между H1L1 и H3L3, это предполагает, что положения зоны Верньера 48 и 67 не могут, вероятно, способствовать различию аффинности, поскольку они сохраняются в H3, но изменяются в H1. H2, однако, сохраняет мышиные остатки зоны Верньера в положениях 69 и 73, в отличие от H1 и H3, предполагая, что изменения в этих двух положениях зоны Верньера могут способствовать различиям в аффинности. Эти два остатка зоны Верньера, которые лежат в основе CDR2, могут иметь повышенное H2 связывание по сравнению с H1 и H3. Из аминокислотного различия между гуманизированными конструктами, положения 69 и 108 являются наиболее близкими к сайту связывания. Аминокислота в положении 69 в H2 (например, в мышином антителе) представляет собой лейцин, в то время как оба других гуманизированных конструкта имеют метионин. Из различий между H2 и мышиным антителом, изменения в аминокислотах 38, 48, 66, 67 и 109 находятся ближе всего к CDR и также могут влиять на аффинность.
Поскольку H2L2 имеет более высокую аффинность, чем H2L3, сравнение L2 и L3 может помочь дополнительно разграничить причину этого различия. Не было никаких изменений для остатков VL/VH и большинство из остатков зоны Верньера на легкой цепи были сохранены среди 3 гуманизированных конструктов, за исключением положений Верньера 2 и 36. В отличие от L3, но подобно мышиному антителу, более высокую аффинность L2 имеет фенилаланин, а не тирозин в положении Верньера 36. Аминокислоты в положении 36 и 37 могут влиять на конформацию благоприятным образом, так как они находятся в пределах 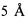 сайта связывания, и являются частью аминокислот в положениях 24-35, которые, как было найдено, являются важными в сохранении требования альфа связывания при уменьшении конечной GalNAc.
сайта связывания, и являются частью аминокислот в положениях 24-35, которые, как было найдено, являются важными в сохранении требования альфа связывания при уменьшении конечной GalNAc.
Многие нормальные ткани имеют неидентичные, но очень схожие структуры с TF-Ag, такие как Galβ1-3GalNAc-бета связанные структуры, которые обычно встречаются в центральной нервной системе, в клетках NK, и периферической нервной ткани, таким образом, сохранение хорошей специфичности является ключевым признаком в данной гуманизации. hJAA-F11 H1L1, H2L2, H3L3 и химерное антитело сохраняли такую же или улучшенную хорошую специфичность связывания с антигеном по сравнению с мышиным антителом, включая ограничение связывания с Galβ1-3GalNAc-α (TF-Ag) связанными структурами, и отсутствие связывания с α2-3 сиалилированными структурами на основе гликанового массива. Специфичность гуманизированных конструктов JAA-F11, как представляется, является такой же или даже улучшенной при испытании по отношению к более, чем 600 различным сахаридам. Мышиное JAA-F11 и все гуманизированные конструкты реагируют только с 5 сахаридами, TF-Ag и 3 трисахаридами с одним дополнительным сахаром 1-6, связанным с GlcNAc (NeuAc в 2 и GlcNAc в 3-ем) или с одним с 2 дополнительными сахарами GalGlcNAc, связанными 1-6 с GalNAc. Например, H2L3 показал меньшее связывание на всех 4 других сахаридах, которые связываются с JAA-F11, в то время как H1L1 и H2L2 не имеют никакого связывания с тетрасахаридом, который связывается с JAA-F11, по сравнению с химерными и мышиными антителами. Хотя эти 4 других сахарида, которые связываются с JAA-F11, как известно, не экспрессируются в нормальных тканях, повышенная специфичность для целевой структуры всегда является желаемой. Это вероятно демонстрирует способность нацеливания для всех этих антител в лечении человека. Поскольку H2L3 увеличил специфичность связывания, как показано ниже за счет меньшего связывания с 3 из 4 сахаридов по сравнению с другими конструктами, различие между H2 и H3 может вызывать это различие. Различия между вариантами, которые уже обсуждались в разделе аффинность, могут быть причиной для этой улучшенной специфичности. H2L3 отличается от H2L2 своим сниженным связыванием с NeuAcalpha2-6 (TF-Ag) и GlcNAcbeta2-6 (TF-Ag).
Не желая ограничиваться какой-либо теорией, полагают, что причина того, что H1L1 и H2L2 не связываются вообще с тетрасахаридом, может состоять в общем различии между данными антителами, при сравнении с H3L3 и мышиными антителами. H1 и H2 оба имеют метионин в положении 48, который одновременно находится в положении зоны Верньера и находится последовательно в пределах 4 аминокислот CDR. Данный метионин находится вместо изолейцина мышиного и H3 антител.
В настоящем изобретении, результаты EIA на цельных клетках указывают на способность hJAA-F11 и химерных JAA-F11 антител связывать тройные отрицательные клеточные линии рака молочной железы. Полученные данные свидетельствуют о возможной лечебной роли TF-Ag в нацеливании на все виды рака молочной железы, но особенно на агрессивные тройные отрицательные виды рака молочной железы (TNBC), поскольку не существует целевого лечения для этих видов рака в настоящее время.
Некоторые лектины и антитела, которые связываются с TF-Ag, вызывают повышенную пролиферацию TF-Ag экспрессирующих клеток опухоли. Полагают, что это, как правило, связано с тем, что или лектин или антитело связывается как с альфа, так и с бета аномерами на уменьшенном конце (вызывание пролиферации) или только с альфа аномером (ингибирование). В то время как мышиное JAA-F11 связывается только с альфа аномером и не вызывает пролиферацию клеток опухоли, это является важным для определения, имели ли гуманизированные конструкты имели такой эффект на клетки опухоли. Гуманизированные JAA-F11 и химерные антитела не вызывают пролиферацию раковых клеток in vitro, и наоборот, наблюдали эффект, подобный эффекту мышиного JAA-F11 на небольшое (~6-11%), но значимое ингибирование роста мышиных 4Т1 и человеческих раковых клеток.
Поскольку ADCC представляет собой один из основных механизмов, с помощью которого антитела уничтожают клетки опухоли, и является важной для антител, которые применяют для лечения рака, исследовали способность гуманизированных JAA-F11 вариантов вызывать ADCC. ADCC активность была замечена с некоторыми, но не всеми гуманизированными конструктами, и была замечена с РВМС трех индивидуумов. Оба H2L2 и H3L3 антитела вызывали значительно большую ADCC, чем или химерное или мышиное антитело как мышиной 4Т1 клеточной линии рака молочной железы, так и клеточной линии рака молочной железы человека, показывая, что на данный момент H2L2 и H3L3 антитело может быть предпочтительным выбором для прямой пассивной иммунотерапии. Мышиное JAA-F11 не показало каких-либо статистически значимых ADCC способностей в каких-либо испытаниях. Другая эффекторная функция, которая использует антитела для уничтожения клеток опухоли, представляет собой CDC. Никакие из гуманизированных, химерных и мышиных антител JAA-F11 не вызывали CDC, но данная потеря CDC активности не исключает ее использование в качестве иммунотерапевтического средства, более того герцептин, как найдено, вызывает лишь номинальное количество CDC, и это не единственный из способов его действия. Данные с ритуксаном фактически показывают, что комплементарное связывание ингибирует связывание NK клеток и снижает эффективность.
Антитела потенциально могут быть использованы для переноса лекарственных средств или токсинов в форме конъюгатов антитело-лекарственное средство в клетку. Таким образом, мы оценивали способность гуманизированных JAA-F11 вариантов к интернализации после связывания с TF-Ag на раковых клетках. Мышиное JAA-F11, как было показано ранее, интернализирует в пределах 1 часа. Используя иммуноферментный анализ, мы показали, что мышиные, H2L2, химерные, H2L3 и H1L1 антитела значительно интернализируют в мышиных 4Т1 клетках рака молочной железы, тогда как H3L3 антитело не показало значительной интернализации. Способ флуоресцентной микроскопии живых клеток подтвердил результаты, полученные в иммуноферментном анализе. Мы ожидали, что антитела, которые показали более высокую ADCC активность, покажут более низкую интернализацию, и результаты от обоих анализов интернализации показывают согласие с данным ожиданием, в котором H3L3 антитело продемонстрировало большую ADCC, чем или мышиные, или химерные антитела, но не сильно интернализирует. Однако, H2L2, даже если оно хорошо зарекомендовало себя в анализе ADCC, интернализирует хорошо. Механизм того, как оба из них могут происходить с тем же антителом еще не ясен, но одна возможность заключается в том, что различные скорости продуцирования в СНО клетках могут в результате приводить к различным скоростям фукозилирования, таким образом, если H2L2 является менее фукозилированным, оно может работать лучше в ADCC в течение того времени, когда оно находится на поверхности клетки, даже если оно интернализируется. Данные результаты показывают, что химерный, H2L2 и H2L3 конструкты имеют потенциал для использования в качестве конъюгатов антитело-лекарственное средство.
Мышиное антитело 124I-JAA-F11, как было показано, локализуется в TF-Ag опухоли молочной железы у мышей. Меченное антитело JAA-F11 остается связанным с опухолью 4Т1 в течение, по меньшей мере, 20 дней и 24 дней для тройной отрицательной опухоли молочной железы человека, подразумевая, что JAA-F11 может быть использовано для обнаружения метастаз и лечения опухоли, имеющей TF-Ag. Для того, чтобы исследовать, где меченное йодом-124 гуманизированное антитело будет локализоваться в человеческой опухоли молочной железы у мышей, использовали гуманизированное антитело с самой высокой аффинностью, H2L2 вариант, (который интернализирует и выполняет ADCC). Визуализация показала преимущественное поглощение опухоли, с поглощением в щитовидной железе и в селезенке, которое было заблокировано в более позднем эксперименте с холодным кроличьим иммуноглобулином. МикроРЕТ визуализация показала, что радиоактивно меченное гуманизированное антитело было поглощено опухолью в пределах 24 часов и могло быть видным до 96 часов. Такая радиоактивная локализация у пациентов может быть использована для обнаружения метастаз, и также используется для определения существует ли что-либо от нацеленного связывания у конкретного пациента перед или прямой пассивной иммунотерапией или лечением конъюгатом антитело-лекарственное средство.
Антиген Томсена-Фриденрайха (TF-Ag) присутствует в более, чем 80% нескольких человеческих карцином, включая виды рака молочной железы. Он имеет функциональную роль в адгезии опухоли и метастазах, поэтому способность антитела к нацеливанию на TF-Ag предполагает возможное применение в качестве иммунотерапевтического средства, в качестве конъюгата антитело-лекарственное средство для уничтожения раковых клеток и для ингибирования метастаз. Антитело JAA-F11 с его уникальной высокой специфичностью для TF-Ag имеет большой потенциал в качестве пассивного ответа антигена TF-Ag для лечения рака молочной железы и других видов рака. В нашем подходе гуманизации, мы сохранили эту уникальную специфичность и вероятно улучшили ее с помощью гуманизированных вариантов JAA-F11.
Пример 2
Данный пример предусматривает описание материалов и способов, используемых для получения результатов, описанных в данной заявке.
JAA-F11 CDR ранее были предсказаны. Мы проводили клонирование и секвенирование мышиного антитела JAA-F11, чтобы подтвердить аминокислотные последовательности вариабельных областей как тяжелой, так и легкой цепи.
Для того, чтобы иметь терапевтический полезный эффект в организме человека, антитело должно сохранять свою специфичность для его антигена-мишени, и в то же время не образовывать анти-мышиные иммунные реакции. Мы выбрали подход CDR-привития, чтобы сохранить CDR мышиного JAA-F11 таким образом, чтобы сохранить специфичность и аффинность для TF-Agα. CDR выбирали, используя способы Chothia и Kabat, а также рентгеновскую кристаллическую структуру и компьютерное углеводное протягивание.
Для того, чтобы выбрать каркас человеческого акцепторного антитела для каждой вариабельной области из тяжелой и легкой цепи JAAF-11, Поиск белка BLAST® (BLASTP) проводили на blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins по отношению ко всей нерезервированной человеческой (Homo sapiens) базе данных Genbank, для того, чтобы определить гомологичные человеческие антитела к мышиному JAA-F11. Все белковые последовательности не Homo sapiens, гуманизированные антитела и последовательности фагового дисплея были исключены из результатов BLASTP. Полученная первая десятка наиболее гомологичных последовательностей для каждой последовательности тяжелой и легкой вариабельной цепи JAA-F11 была выбрана в качестве возможных каркасов человеческих акцепторных антител для гуманизированного JAA-F11 (hJAA-F11). Программа выравнивания последовательности SeaView была использована для выравнивания десяти возможных человеческих акцепторных вариабельных тяжелых или легких каркасных последовательностей с мышиными JAA-F11 тяжелыми или легкими последовательностями вариабельной области.
Три вариабельные области тяжелой цепи (H1, H2 и H3) и три области легкой цепи (L1, L2 и L3) были сконструированы на основе 10 наиболее подобных последовательностей человеческого иммуноглобулина, выбранных при помощи белкового BLAST. Конечные CDR из мышиного JAA-F11 потом были привиты на человеческие каркасные области с помощью одного из трех различных способов, создавая 3 различные тяжелые цепи и 3 различные легкие цепи. В первом способе, (а) последовательности тяжелой H1 и легкой L1 FR были сконструированы таким образом, что содержат наиболее часто встречающуюся человеческую аминокислоту в каждом сайте среди первых десяти выбранных человеческих FR последовательностей. Во втором способе, (b) тяжелая H2 и легкая L2 были сконструированы таким образом, что содержат наиболее часто встречающуюся человеческую аминокислоту среди первых десяти выбранных человеческих FR последовательностей в каждом сайте, за исключением случаев, когда аминокислота присутствовала в каком-либо из первых десяти человеческих FR, что соответствует тому, что обнаружено в мышином JAA-F11, затем аминокислоту сохраняли в качестве мышиного JAA-F11. В третьем способе, (с) в тяжелой H3 или легкой L3, три FR остатка перед и после CDR1, CDR2 и CD3 мышиного JAA-F11 были сохранены, и остальные остатки представляли собой наиболее часто встречающиеся аминокислоты в человеческих последовательностях как в вариантах H1 или L1. Любая одна из данных сконструированных тяжелых вариабельных цепей может создать пару с любой подобным образом сконструированной легкой вариабельной цепью, например, H1/L1, H1/L2, H1L3, H2L1, H2L3, и т.д.
Кроме того, сконструированым было химерное JAA-F11, в котором вся вариабельная область мышиного JAA-F11 была присоединена к константной области IgG1 человека. Это должно было обеспечить базовый уровень, который должен иметь исходную специфичность и аффинность мышиного антитела, в то время как, имея человеческую константную область, будет использоваться в качестве положительного контроля для оценки гуманизированных вариантов.
Оценка моделей hJAA-F11
Различные hJAA-F11 конструкты оценивали касательно конформационных эффектов. В кратком изложении, то предложенные последовательности тяжелой и легкой вариабельной цепи различных конструктов hJAA-F11 выравнивали с последовательностью JAA-F11. Любой аминокислотный остаток в каркасе для каждого гуманизированного варианта, который потенциально может вызывать серьезное стерическое взаимодействие с окружающими аминокислотами, был удален и замещен на исходный каркасный остаток мышиного JAA-F11.
Предсказание иммуногенности hJAA-F11 вариантов с использованием Т20 оценки.
Несмотря на то, что гуманизированные или полностью человеческие антитела считаются неиммуногенными и безопасными для использования у человека, иммуногенность полностью человеческого и гуманизированного антител была зарегистрирована у пациентов. Для того, чтобы определить, были ли выбранные последовательности с низкой иммуногенностью, анализ иммуногенности проводили с помощью способа Т20 оценки. Анализатор Т20 оценки, разработанный Gao et al, вычисляет «гуманизированность» последовательностей вариабельной области моноклонального антитела, используя базу данных из более, чем 38000 последовательностей человеческого антитела.
В этом способе проводят поиск на основе белкового BLAST, и испытуемое гуманизированное Ab сначала сравнивают со всеми данными человеческими последовательностями. Гуманизированное антитело затем сравнивают с 20 человеческими Ab, демонстрирующими наилучшее совпадение в BLAST, и оценивают в отношении подобия этих последовательностей. Т20 оценку для гуманизированного антитела получают из среднего значения процента идентичности 20 лучших подобранных человеческих последовательностей. Наиболее высокая возможная оценка составляет 100 (наиболее подобные человеку). В качестве доказательства концепции взаимосвязи этого способа с иммуногенностью у пациента, Gao et al сравнивали результаты иммуногенности in vivo у пациента более, чем 90 антител, которые либо одобрены для клинического применения, или на различных стадиях клинической разработки (последовательности мышиного, химерного (из мышиного), гуманизированного (n=22) и полностью человеческого антитела) с помощью Т20 оценки, и нашли, что Ab с Т20 оценками FR и CDR последовательностей более 80 не были иммуногенными, и используя Т20 оценки только для FR последовательностей, данные более 85 не были иммуногенными. Т20 оценки для гуманизированных JAA-F11 вариантов были рассчитаны с использованием базы данных Т20 Cutoff Human Databases на abanalyzer.lakepharma.com/.
Оптимизация кодонов и синтез генов. После конструирования гуманизированных JAA-F11 VH и VL вариантов аминокислотных последовательностей, соответствующие нуклеотидные последовательности были выбраны вручную для использования кодонов, оптимальных для продуцирования клеточного белка (СНО) Cricetulus griseus, и синтезированы и вставлены в pUC57 плазмиду, обычно используемый клонирующий вектор.
Субклонирование и секвенирование hJAA-F11 и химерного JAA-F11 вариабельного тяжелого (VH) и легкого (VL) генов в векторах экспрессии млекопитающих. hJAA-F11 VH гены были субклонированы и вставлены в вектор экспрессии (рАН6307), содержащий лидерную/константную область тяжелой цепи человеческого IgG1 при контроле промотора человеческого цитомегаловируса (CMV) и кассеты ампицилина (Amp) и гистидинолдегидрогеназы (hisD). VL гены были вставлены в вектор экспрессии (pAN 6714), содержащий лидерную/константную область человеческой каппа-легкой цепи при контроле промотора человеческого CMV и кассеты неомицинфосфотрансферазы (neoR). Аналогично, были экспрессированы VH и VL химерного JAA-F11 и использованы в качестве положительного контроля в последующих анализах.
Экспрессия и продуцирование вариантов гуманизированного и химерного JAA-F11.
Совместная трансфекция в СНО-K1 клетках.
Адгезивные клетки яичника китайского хомяка (СНО-K1; АТСС № CCL-61, Manassas, VA) культивировали в течение 18 часов перед трансфекцией в среде F12 Хама (Corning Cellgro, Manassas, VA), дополненной 10% фетальной телячей сыворотки (FCS; Hyclone) при 37°C и 5% диоксида углерода (CO2) в увлажненном воздухе и собирали при 50-80% конфлюентности. 6307 рАН (VH) и 6714 pAN (VL) векторы экспрессии млекопитающих совместно трансфицировали в СНО-K1 клетках с помощью электропорации (Gene Pulser System (Bio-Rad, Hercules, CA). В кратком изложении, каждую плазмиду (VH и VL) линеаризовали, используя фермент рестрикции PvuI (Promega, Madison, WI), и 5 мкг каждой плазмиды добавляли к 5×106 СНО-K1 клеткам в холодной среде F12 Хэма (Corning Cellgro, Manassas, VA) в общем объеме 500 мкл в 0,4 см кювете для электропорации (Bio-Rad, Hercules, СА). Кювету, содержащую смесь, электропорировали на 960 мкФ и 250 мВ. Смесь трансфекции затем разбавляли предварительно нагретой неселективной средой (F12 Хэма, дополненной 10% FCS) до концентрации 1×105 клеток/мл. Двести микролитров затем высевали в 96-луночный планшет(ы) для тканевых культур (BD Bioscience, San Jose, California) с плотностью 1×104 клеток на лунку. Трансфицированные клетки инкубировали при 37°C и 5% CO2 в инкубаторе для тканевых культур. Через 72 часа, неселективную среду удаляли из 96-луночного планшета для тканевой культуры и заменяли на 200 мкл селективной среды F12 Хэма (плюс 10% FCS), содержащей 700 мкг/мл G418 (Gibco, Life Techologies, Grand Island, NY), 5 мМ гистидинола (Sigma-Aldrich, St Louis, МО) и 10 мМ буфера HEPES (Corning Cellgro, Manassas, VA), и колонии оставляли расти на срок до 3 недель. СНО-K1 клетки, трансфицированные плазмидами 6307 рАН и 6714 pAN будут продолжать расти, в то время как нетрансфицированные СНО-K1 клетки расти не будут. Каждые 2-4 дня в течение следующих от 14 до 21 дней, меняли селективную среду для того, удалить дебрис мертвых клеток, и колониям устойчивых клеток позволяли расти в селективной среде.
Анализ культуральных супернатантов с использованием иммуноферментного анализа (ELISA)
В период 14-21 день после отбора, СНО-К1 культуральные супернатанты из 96-луночного планшета(ов) собирали и анализировали касательно антител с использованием иммуноферментного анализа (ELISA). В кратком изложении, 100 мкл культуральных супернатантов добавляли в Immulon 1 В-среду связывания в 96-луночный планшет(ы) для микротитрования (Thermo Scientific, Milford, MA), который был покрыт 1,25 мкг/мл конъюгата TF-Ag-BSA в буфере для покрытия (0,1 М Na2CO3 при pH 9,6) и промывали пять раз TBS-Brij (pH 7,2). Разбавление в соотношении 1:500 очищенного мышиного JAA-F11 (1 мг/мл) в стерильном PBS также добавляли в 2-3 лунки в каждом планшете, чтобы служили в качестве положительных контролей. После 2-часового инкубирования при 37°C, планшеты промывали пять раз, и 100 мкл конъюгированного вторичного антитела анти-человеческого IgG-щелочной фосфатазы (Sigma-Aldrich, St Louis, МО) в 1% буфере BSA-PBST (1:10000) добавляли в каждую лунку для трансфекции клеток, в то время как разбавление 1:1000 конъюгированного вторичного антитела анти-мышиного IgG-щелочной фосфатазы (Sigma-Aldrich, St Louis, МО) в 1% буфере BSA-PBST использовали для контрольной лунки мышиного JAA-F11. После 1-часового инкубирования при комнатной температуре, планшеты промывали пять раз, а затем 100 мкл субстрата фосфатазы (п-нитрофенилфосфата (pNPP)) (Sigma-Aldrich, St Louis, МО) добавляли в каждую лунку. Планшеты считывали при 405 нм, используя планшетный считыватель после часового инкубирования при комнатной температуре для скрининга на наличие IgG антител к TF-Ag. Как холостую пробу, среду F12 Хэма (плюс 10% FCS) использовали во время инкубирования первичных антител, с последующим инкубированием вторичного антитела и инкубированием субстрата.
Образование стабильных клонов и продуцирование
После первоначального скрининга для антитела к антигену TF-Ag через 14-21 дней после начала отбора, выбрали 10-12 клонов с наиболее высокими показаниями абсорбции при считывании и перенесли в 24-луночные планшеты для тканевых культур для распространения. Культуральные супернатанты из этих клонов затем анализировали с помощью ELISA. 4 клона, которые давали наиболее высокие показания абсорбции при считывании переносили в колбы Т25 для тканевой культуры, распространяли и снова подвергали скринингу для антитела к антигену TF-Ag с помощью ELISA, чтобы убедиться, что клоны все еще продолжали продуцировать антитела, реакционноспособные с TF-Ag. Эти 4 клона затем переносили в колбы Т75 для тканевой культуры, распространяли, снова подвергали скринингу для антитела к антигену TF-Ag, и клетки хранили в замороженном виде в жидком азоте. Клон с наиболее высоким результатом антигена TF-Ag затем дополнительно распространяли и пассировали 2-3 раза, чтобы убедиться, что клеточная линия является стабильной.
Стабильные отдельные клеточные клоны для каждой комбинации тяжелой и легкой цепи hJAA-F11 из наилучшего клона, полученного выше, получали путем субклонирования с помощью лимитирующего разбавления. В кратком изложении, клетки выращивали до конфлюентности в колбе Т75 для тканевой культуры, собирали и подсчитывали, и высевали при разбавлении 0,3 клеток/лунку в каждую лунку 96-луночных планшетов в селективной среде. Планшеты тщательно исследовали через 7-10 дней под микроскопом, и лунки, которые показали один фокус клеток, были отмечены, и их контролировали пока не наблюдалась достаточная плотность роста. Супернатанты из этих отмеченных лунок испытывали на антитело к антигену TF-Ag с использованием ELISA. 4 клона, которые давали наиболее высокие показания абсорбции при считывании распространяли, как описано выше, для получения родительских клонов. Субклон, который дал наиболее высокую абсорбцию, размножали и использовали для продуцирования супернатанта в Т200 NuncTM клеточной культуре Tripleflasks (Thermo Fisher Scientific, Inc.).
Для продуцирования в колбах с культурой Tripleflasks для каждого гуманизированного варианта JAA-F11, клетки культивировали в течение 3-х недель перед тем, как собирали 1 литр культурального супернатанта. Клон, продуцирующий химерное JAA-F11, был обработан таким же образом, как гуманизированные JAA-F11 варианты.
Очистка гуманизированных и химерных JAA-F11 вариантов
Супернатанты гуманизированных и химерных JAA-F11 вариантов очищали с использованием Protein A-Sepharose® 4B, колонки с аффинностью при быстром потоке (Sigma-Aldrich, St Louis, МО). Для того, чтобы получить колонку Protein А, 1:1 суспензию смолы в буфере А (0,02 М NaH2PO4, 0,15 М NaCl, pH до 8,0) выливали в колонку. После того, как колонка осела, ее промывали 20 объемами колонки (CV) буфера А. Один литр СНО-K1 супернатанта клеточной культуры центрифугировали при 3500 оборотах в минуту в течение 30 минут, чтобы очистить от мертвых клеток или дебриса, а затем фильтровали. Отфильтрованный супернатант загружали в колонку Protein А и позволяли стечь под действием силы тяжести при скорости потока 1 мл/мин. Затем колонку промывали 10 CV буфера А. Антитело элюировали из колонки Protein А, используя 3 CV буфера В (0,2 М Na2HPO4, 0,1 М лимонной кислоты, pH 3,9). Элюат осторожно нейтрализовали 0,1 М NaOH, чтобы минимизировать влияние низкого pH на антитело. Колонку Protein А повторно уравновешивали 20-30 CV буфера А и хранили при 2-8°C. Каждую колонку использовали до пяти раз для одного и того же антитела. Очищенное антитело подвергали диализу в течение ночи при 4°C (Slide-A-Lyzer диализная кассета; Thermo Scientific) по отношению к или 1X фосфатному буферному солевому раствору (PBS) или RPMI среде с или без фенола красного как это требуется для последующих анализов. После диализа антитело фильтровали с использованием 0,22 мкм фильтра (Corning) и определяли концентрацию белка с помощью Bio-Rad белкового анализа, как детально описано ниже. Антитело хранили при 4°C и проверяли на связывание с TF-Ag, используя ELISA.
Bio-Rad белковый анализ
Концентрацию антитела определяли с использованием Bio-Rad белкового анализа, который основан на методе Брэдфорда связывания красителя. В кратком изложении, красящий реагент разбавляли дистиллированной водой при соотношении 1:4 и фильтровали с использованием бумажного фильтра Ватман №1. Серийные разбавления известного стандарта белка (1,44 мг/мл гамма-глобулина, Bio-Rad, Hercules, СА) и образец гуманизированного антитела получали с использованием 1X PBS буфера. 10 мкл каждого раствора стандарта и образца помещали в трех повторах в лунки планшета для микротитрования. Двести микролитров разбавленного красящего реагента затем добавляли во все лунки с помощью многоканальной пипетки и тщательно перемешивали пипетированием вверх и вниз, не создавая пузырьков. Планшет инкубировали при комнатной температуре в течение, по меньшей мере, от 5 минут до 1 часа. Затем считывали абсорбцию при 595 нм на планшетном считывателе (Bio-Tek Instruments).
Анализ химической специфичности и аффинности гуманизированных и химерных JAA-F11 антител
Определение химической специфичности
Химическую специфичность различных hJAA-F11 конструктов и химерного JAA-F11 определяли с использованием напечатанного гликанового массива. Их сравнивали с данными, полученными ранее для мышиного JAA-F11. Гликановый массив является непрямым иммунофлуоресцентным способом, описанным на вебсайте Consortium for Functional Glycomics. Каждый конструкт анализировали на реакционную способность с 610 различными гликанами. В кратком изложении, напечатанный массив инкубировали последовательно с антителом, промывали и инкубировали с вторичным антителом, меченным FITC. После промывания изображение считывали на Perkin Elmer Microscanarray XL4000 и tiff-файл изображения сохраняли, и анализ изображения проводили с использованием программного обеспечения для анализа изображений Imagene V.6. Относительное связывание каждого связывания гликана выражали и нормализировали до связывания исходного гликана TF-Ag. Сравнительный и статистический анализ относительных способностей связывания проводили с использованием ANOVA.
Определение относительной аффинности
Относительную аффинность связывания hJAA-F11 с TF-Ag анализировали с использованием конкурентного ингибирования ELISA с помощью мышиного JAA-F11 антитела и химерного JAA-F11 антитела. В кратком изложении, 3 мкг/мл hJAA-F11 или химерного JAA-F11 инкубировали в присутствии различных концентраций мышиного JAA-F11 (10, 8, 6, 4, 2 мкг/мл) в 96-луночных планшетах, покрытых 1,25 мкг/мл TF-Ag (Thermo Scientific, Milford, MA). Связанное антитело обнаруживали путем инкубирования с анти-человеческим IgG вторичным антителом и субстратом и показатели абсорбции при считывании получали как описано в данных материалах и способах. Определяли и сравнивали количество мышиного антитела, требуемое для того, чтобы ингибировать связывание 3 мкг/мл гуманизированного или химерного антител на 50%. Чем большее количество мышиного антитела требуется для ингибирования, тем более высокая относительная аффинность антитела. Относительную аффинность каждого из гуманизированных антител и химерного антитела сравнивали с использованием ANOVA.
Анализ биологической эффективности hJAA-F11
Оценка связывания с клеточными линиями опухоли молочной железы человека
Связывание hJAA-F11 с различными клеточными линиями опухоли молочной железы человека анализировали с использованием ELISA на цельных клетках. Клеточная линия мышиной опухоли молочной железы 4Т1, имеющая TF-Ag, служила в качестве положительного контроля, в то время как клеточная линия миеломы Р3-Х63-Ag8 (АТСС Number: CRL-1580), которая была партнером по слиянию для продуцирования гибридомы JAA F11, служила в качестве TF-антигенного отрицательного контроля. Положительное связывание определяли путем сравнения реакционной способности каждого антитела с каждой клеточной линией с реакционной способностью этого антитела с миеломной клеточной линией с использованием Т-критерия Стьюдента. Для стандартизации данных, связывание с каждой клеточной линией было выражено как деление показания абсорбции при считывании испытуемой клеточной линии на показание абсорбции при считывании миеломных клеток.
Подготовка клеток
Клеточные линии 4Т1 опухоли молочной железы и миеломы собирали с использованием неферментативного Cellstripper (Mediatech, Inc. VA, USA). Для предотвращения слипания клеток, среду, буфер и реагенты предварительно нагревали перед использованием. Для адгезивных клеток, среду удаляли из емкостей для культивирования, и клетки промывали IX фосфатно-солевым буфером Дульбекко (DPBS) без кальция и магния (Mediatech Inc., Cellgro). Пять миллилитров раствора для диссоциации клеток добавляли в каждую колбу, и затем инкубировали при 37°C в течение 10 минут. Колбы встряхивали, чтобы выбить клетки. Для неадгезивных клеток миеломы колбы хорошо встряхивали, и культуральный супернатант центрифугировали при 1000 х G в течение 10 мин. Клеточный осадок снова суспендировали в растворе для диссоциации клеток и инкубировали при 37°C в течение 10 мин. После инкубирования 20 мл 1X DPBS добавляли к клеткам и пипетировали несколько раз, чтобы удалить комки. Клеточные суспензии затем центрифугировали при 1000 х G в течение 10 минут. Супернатанты декантировали и осадок снова суспендировали в 5 мл DPBS. Клетки подсчитывали с использованием трипанового синего красителя и гемоцитометра. Клетки разбавляли, чтобы получить 1×106 жизнеспособных клеток/мл. Двести микролитров суспензии клеток (2×105 клеток) помещали в 5 мл полистирольные пробирки квадруплетов. Двести микролитров 4% раствора формальдегида добавляли в каждую пробирку и инкубировали в течение 20 мин при комнатной температуре перед центрифугированием при 1500 х G в течение 10 минут. Супернатант осторожно декантировали в один прием, клетки промывали в DPBS с последующим центрифугированием и декантацией. Двести микролитров PBS-Tween 1% BSA (масс./об.) добавляли в каждую пробирку и хранили при 4°C в течение ночи или до двух недель.
Иммуноферментный анализ на клетках
Двести микролитров 50 мкг/мл мышиного JAA-F11, hJAA-F11 или химерного JAA-F11 антитела добавляли в пробирки, содержащие различные клеточные линии, испытанные в квадруплете, и инкубировали при 37°C в течение двух часов. Один набор пробирок, обработанных 200 мкл 1X PBS-0,1% Tween 20-1% BSA, служил в качестве отрицательного контроля для каждой испытанной клеточной линии. Пробирки трижды промывали 3 мл промывочного буфера (1X PBS Tween, без азида), а затем центрифугировали в течение 10 минут при 1500 х G. Супернатант осторожно декантировали между каждым промыванием. Двести микролитров вторичного антитела анти-мышиного IgG (γ-цепь-специфический) с пероксидазой хрена (1:1000, Sigma-Aldrich, St. Louis, МО) или вторичного антитела анти-человеческого IgG (γ-цепь-специфический) с пероксидазой хрена (1:2000, Sigma Aldrich, St. Louis, МО) в PBS-Tween-1% BSA добавляли в соответствующие пробирки, а затем инкубировали в течение 1 часа при комнатной температуре (к.т.). После инкубирования пробирки декантировали и промывали три раза, и центрифугировали при 1500 х G между промываниями. Двести микролитров раствора субстрата О-фенилендиамина дигидрохлорида (OPD; Sigma, St. Louis, МО) затем добавляли в каждую пробирку и инкубировали в течение 1 часа при комнатной температуре. После инкубирования, реакцию останавливали добавлением 100 мкл стоп-раствора (1 N H2SO4) и центрифугировали в течение 10 минут при 1500 х G. Далее, 200 мкл супернатанта удаляли из каждой пробирки и переносили в соответствующие лунки в планшете для микротитрования. Абсорбцию считывали при 490 нм с использованием микропланшетного считывателя (Microplate Autoreader, Model EL311, Bio-Tek Instruments, Inc.) и непрореагировавший субстрат OPD был использован в качестве холостой пробы. Для различных клеточных линий, каждую соответствующую среднюю холостую пробу (пробирки только с PBS-Tween-1% BSA), вычитали из их соответствующего среднего OD для того, чтобы получить конечные оптические показания при считывании. Каждый эксперимент повторяли 3 раза.
Влияние hJAA-F11 на пролиферацию раковых клеток in vitro
Для изучения влияния гуманизированных и химерных JAA-F11 антител на рост раковых клеток in vitro проводили анализы пролиферации 3-[4,5-диметилтиазол-2-ил]-2,5-дифенилтетразолий бромида (тиазолил синий, МТТ). 4Т1 клетки мышиной опухоли молочной железы и клетки опухоли молочной железы человека высевали в количестве 1×104 клетки/лунку в 10 повторах в 96-луночные планшеты в присутствии различных количеств JAA-F11, гуманизированных и химерных JAA-F11 (4, 2 и 1 мкг/мл). Когда высевали при этой плотности клеток, клетки находятся в линейной части их кривых роста в течение 72 часов. Клетки, которые выращивали в культуральной среде без антител, служили в качестве контролей нормального роста. Сама культуральная среда была использована как холостая проба. После 68-часового роста клеток при 37°C, 10 мкл соли тетразолия МТТ (5 мг/мл) добавляли в каждую лунку, и планшеты были возвращены в инкубатор еще на 4 часа. В конце инкубирования, образовавшийся в результате продукт формазана в каждой лунке солюбилизировали добавлением 120 мкл диметилсульфоксида (ДМСО, Fisher Scientific), и затем измеряли абсорбцию каждой лунки при 570 нм (MicroplateAutoreader, модель EL311, Bio-Tek Instruments, Inc.).
Анализ антитело-зависимой клеточной цитотоксичности
Для изучения эффекторных функций hJAA-F11 антител, проводили анализы антитело-зависимой клеточной цитотоксичности (ADCC) и комплемент-зависимой цитотоксичности (CDC).
ADCC определяли с помощью анализа высвобождения лактатдегидрогеназы (LDH) (CytoTox 96 анализ нерадиоактивной цитотоксичности; Promega, Madison, WI) с использованием человеческих мононуклеарных клеток периферической крови (РВМС) в качестве эффекторных клеток и клеточных линий опухоли молочной железы человека в качестве клеток-мишеней при соотношении эффектора к мишени (Е:Т) 100:1.
РВМС получали из цельной крови путем центрифугирования в градиенте плотности с использованием Ficoll-Paque. Цельную кровь, взятую в пурпурный верхний EDTA вакуумный контейнер смешивали с равным объемом предварительно нагретого стерильного DPBS. Двадцать миллилитров разбавленной крови затем осторожно наслаивали на 15 мл Ficoll-Paque Plus (GE Healthcare) в 50 мл конических пробирках. Образцы центрифугировали при 1300 оборотах в минуту в течение 30-40 мин при комнатной температуре, без остановки. Затем слой РВМС собирали и стерильный PBS добавляли в РВМС для общего объема 40 мл. Затем смесь центрифугировали при 1000 оборотах в минуту в течение 10 мин при 18°C-22°C, чтобы удалить любые загрязняющие Ficoll и тромбоциты/белки плазмы. Супернатант отбрасывали, клетки снова суспендировали в свежем стерильном PBS, и стадию центрифугирования повторяли. Клетки снова суспендировали в культуральной среде RPMI 1640 (10% FCS) и подсчитывали с использованием гемоцитометра и трипанового синего.
Клетки-мишени (1×104; 30 мкл) и РВМС (1×106; 30 мкл) добавляли в 96-луночные с U-образным дном планшеты и инкубировали с hJAA-F11 или химерными антителами (200 мкг/мл; 30 мкл) в течение 17 часов при 37°C в инкубаторе тканевых культур. За сорок пять минут до завершения 17-часового инкубирования, 10 мкл лизисного раствора (х10) добавляли в лунки, содержащие клетки-мишени с максимальным контролем высвобождения LDH (клетки-мишени и среда) и контроль коррекции объема (только среда). В конце инкубирования, планшет центрифугировали в течение 4 минут при 1000 оборотах в минуту. Пятьдесят микролитров аликвоты переносили из всех лунок в свежий 96-луночный планшет с плоским дном. Затем 50 мкл смеси повторно растворенного субстрата добавляли в каждую из этих лунок. Планшет покрывали фольгой и инкубировали в течение 30 минут при комнатной температуре. Реакцию останавливали добавлением 50 мкл стоп-раствора в каждую лунку, и абсорбцию регистрировали при 490 нм с использованием микропланшетного считывателя (MicroplateAutoreader, Model EL311, Bio-Tek Instruments, Inc.). Цитотоксичность рассчитывается по следующей формуле:
Цитотоксичность (%)=100×[(Е-SE)]/[(М-SE)],
где для каждого условия измеряют абсорбцию субстрата. Условия являются следующими: E представляет собой экспериментальную лунку, SE представляет собой спонтанное высвобождение без контрольного антитела (клетки-мишени инкубировали с эффекторными клетками и PBS), М представляет собой максимальное высвобождение, которое определяли с помощью клеток-мишеней, лизированных 10Х лизисным раствором.
Анализ комплемент-зависимой цитотоксичности (CDC)
CDC определяли с помощью анализа высвобождения лактатдегидрогеназы (LDH) (CytoTox 96 анализ нерадиоактивной цитотоксичности; Promega, Madison, WI) с использованием НСС 1428 клеток опухоли молочной железы (АТСС® CRL-2327, Manassas, VA) в качестве популяции клеток-мишеней. LDH положительный образец, который содержит LDH бычьго сердца, предусмотренный с комплектом реагентов, был использован в качестве положительного контроля. После сравнения комплемента кролика, крольченка и морской свинки для фоновых уровней цитотоксичности, лиофилизированная сыворотка морской свинки (CL3112, Cedarlane Laboratories, Burlington, NC) была использована в качестве источника комплемента и восстановлена в соответствии с инструкциями изготовителя. Оптимальная концентрация клеток, использованная в анализе, составляла 1×104 клеток/50 мкл на лунку. Используемые антитела диализовали по отношению к RPMI средам, свободным от фенола красного. Для экспериментальных лунок, 50 мкл НСС 1428 клеточной суспензии, 20 мкл разбавления комплемента (конечное разбавление 1:20) и 30 мкл антитела (при конечной концентрации 100 мкг/мл) смешивали в каждой лунке в наборах из четырех экземпляров в 96-луночном планшете для культуры с круглым дном. Включенными были контрольные лунки, содержащие клетки-мишени и комплемент без антитела для контроля за любой опосредованной цитотоксичностью с помощью сыворотки, используемой в качестве источника комплемента и спонтанного высвобождения LDH. Планшет центрифугировали при 1000 оборотах в минуту в течение 4 минут, а затем инкубировали в течение 2 часов при 37°C с 5% диоксида углерода. За сорок пять минут до завершения 2-часового инкубирования, 10 мкл лизисного раствора (х10) добавляли в лунки, содержащие клетки-мишени с максимальным высвобождением контроля LDH и контроль коррекции объема.
В конце инкубирования, планшет центрифугировали в течение 4 минут при 1000 оборотах в минуту. Пятьдесят микролитров аликвоты затем переносили из всех лунок в свежий 96-луночный планшет с плоским дном. Затем 50 мкл восстановленной субстратной смеси добавляли в каждую лунку. Планшет накрывали фольгой и инкубировали в течение 30 минут при комнатной температуре. Реакцию останавливали добавлением 50 мкл стоп-раствора в каждую лунку, и затем регистрировали абсорбцию при 490 нм с использованием микропланшетного считывателя (MicroplateAutoreader, Model EL311, Bio-Tek Instruments, Inc.). Цитотоксичность рассчитывается по следующей формуле:
Цитотоксичность (%)=100×[(Е-SE)]/[(М-SE)],
где для каждого условия измеряют абсорбцию субстрата. Условия являются следующими: Е представляет собой абсорбцию экспериментальной лунки, SE представляет собой спонтанное высвобождение с комплементом, но без антитела (клетки-мишени инкубировали с комплементом и средой), М является таким как у клеток-мишеней, лизированных 10Х лизисным раствором.
Анализы клеточной интернализации
Интернализацию гуманизированного и химерного JAA-F11 в 4Т1 клетках опухоли молочной железы анализировали двумя способами, иммуноферментным анализом с поверхностным связыванием, измеренным при 2 температурах инкубирования, 4°C и 37°C, и способом иммунофлуоресцентной микроскопии.
Способ иммуноферментного анализа интернализации
Клеточную линию 4Т1 мышиного рака молочной железы испытывали касательно интернализации JAA-F11, hJAA-F11 или химерного JAA-F11. Пятьсот тысяч (5×105) клеток высевали во всех шести лунках 6-луночного планшета и культивировали до конфлюентности. Для каждого испытуемого антитела подготовили два планшета. Среду удаляли из каждого планшета и 1 мл антитела (200 мкг/мл) добавляли в 3 лунки каждого планшета, и 1 мл PBS разбавления добавляли к оставшимся 3 лункам. Один набор планшетов инкубировали при 37°C, чтобы позволить интернализацию, а другой планшет инкубировали при 4°C. После 1-часового инкубирования среду удаляли и лунки промывали 4 раза 1 мл среды RPMI, свободной от фенолового красного. Затем добавляли 1 мл 2% параформальдегида добавляли в каждую лунку, и планшеты инкубировали при комнатной температуре в течение 20 минут, чтобы зафиксировать клетки. Затем планшеты промывали 4 раза средой RPMI, свободной от фенолового красного. Затем один мл вторичного антитела антимышиного или анти-человеческого IgG (γ-цепь-специфический)-щелочной фосфатазы (1:5000, Sigma, St. Louis, МО) в 1% BSA/PBS добавляли и планшеты инкубировали при 37°C в течение 1 часа. Затем планшеты 4 раза промывали средой RPMI, свободной от фенолового красного, и добавляли 1 мл п-нитрофенилфосфатного субстрата (pNPP) и инкубировали в течение 1 часа в темноте. После 1 ч инкубирования, 200 мкл из каждой лунки переносили в соответствующие лунки 96-луночного планшета и абсорбцию регистрировали при 405 нм с использованием микропланшетного считывателя (MicroplateAutoreader, Model EL311, Bio-Tek Instruments, Inc.). Непрореагировавший субстрат использовали в качестве холостой пробы. Эти тройные повторы лунки были усреднены, среднюю оптическую плотность сред только холостых проб вычитали из лунок, содержащих антитело. Процент интернализации рассчитывали по формуле:
% интернализации=100*[1-(37°C образец-37°C холостая проба)/(4°C образец-4°C холостая проба)]
Способ иммунофлуоресцентной микроскопии
В этом способе, 4Т1 клетки рака молочной железы высевали на покровные стекла в лунки двух 6-луночных планшетах для культур при плотности 3×105 клеток в 10% среде FCS RPMI 1640 и инкубировали при 37°C и 5% CO2 в течение 24 часов. Среду удаляли из планшетов и 1,5 мл испытуемых антител (5 мкг/мл), разбавленных безсывороточной средой RPMI 1640 добавляли в соответствующие лунки. Оба планшета были размещены при 4°C в течение 20 минут, чтобы позволить поверхностное связывание антител. После 20-минутного инкубирования, покровные стекла переносили из одного планшета в новый 6-луночный планшет. Разбавления антитела удаляли из второго планшета и добавляли предварительно нагретую безсывороточную среду RPMI 1640 и планшет инкубировали при 37°C в течение 1 часа в инкубаторе для тканевых культур. Покровные стекла из первого планшета дважды промывали охлажденным до ледяного 5% BSA/PBS, один раз ополаскивали 1X PBS, а затем фиксировали 4% раствором параформальдегида (Affymetrix) в течение 15 мин при комнатной температуре. После 1-часового инкубирования при 37°C, покровные стекла из второго планшета переносили в новый 6-луночный планшет, промывали, ополаскивали и фиксировали.
После трех промываний в 1X PBS, клетки в обоих планшетах пермеабилизировали 0,1% Triton Х-100, 0,1% дезоксихолатом натрия в PBS в течение 10 мин при комнатной температуре. Затем клетки ополаскивали три раза 1X PBS, а затем инкубировали в 5% BSA/PBS в течение 30 минут при комнатной температуре. Покровные стекла затем инкубировали с кроличьим анти-лизосомальным мембранным белком 1 (LAMP1; Abcam, 24170) антитела при разбавлении 1 мкг/мл в 5% BSA/PBS в течение 1 часа при комнатной температуре. Покровные стекла затем ополаскивали три раза 1X PBS и инкубировали с анти-мышиным IgG-Alexa 647 и анти-кроличьим IgG Alexa 488 (мышиные JAA-F11 покровные стекла) и анти-человеческим IgG-Alexa 647 и анти-кроличьим IgG Alexa 488 (химерное и hJAA-F11) вторичными антителами (Molecular Probes, Invitrogen), при разбавлении 1:500 в 5% BSA/PBS. Анти-кроличье IgG Alexa 488 вторичное антитело использовали для обнаружения анти-LAMP 1 антитела. Покровные стекла инкубировали в течение 1 часа при комнатной температуре в темноте. После инкубирования, клетки ополаскивали три раза 1X PBS в темноте. Покровные стекла помещали с клетками лицевой частью вниз на Slowfade Gold реагент (Molecular Probes, Life Technologies) с DAPI средой на микроскопических слайдах и герметично скрепляли лаком для ногтей. Клетки анализировали с помощью AxioImager флуоресцентного микроскопа (Zeiss). Получали изображения и анализировали, используя программное обеспечение AxioVision Release 4.8.2.
In vivo испытания для того, чтобы испытать эффективность гуманизированного JAA-F11 для обнаружения опухолей молочной железы, имеющих TF-Ag.
Используя H2L2 гуманизированное JAA-F11 антитело, полученное из указанных выше биологических исследований in vitro, проводили исследование иммунолокализации у мышей для того, чтобы испытать, будет ли меченное йодом-124 гуманизированное антитело локализоваться в TF-Ag опухолях молочной железы у мышей.
Введение метки [124]йод в антитело hJAA-F11 (H2L2)
Введение метки [124]йод в гуманизированное антитело JAA-F11 осуществляли, используя метод Болтона-Хантера. Перед йодированием, гуманизированное антитело сначала модифицировали, используя растворимый в воде реагент Болтона-Хантера (Sulfo-SHPP) (Thermo Scientific, Rockford IL, USA). В кратком изложении, 1,9 мг hJAA-F11 антитела растворяли в модифицированном буфере (200 мМ боратного буфера, pH 9,0). Пять мг растворимого в воде реагента Болтона-Хантера растворяли в 1 мл модифицированного буфера непосредственно перед использованием. 100 мкл раствора растворимого в воде реагента Болтона-Хантера затем добавляли в образец антитела и инкубировали на льду в течение 3 часов при периодическом перемешивании. Непрореагировавший растворимый в воде реагент Болтона-Хантера удаляли диализом против фосфатного буферного солевого раствора (PBS: 0,1 М фосфата натрия, 150 мМ хлорида натрия). На этой промежуточной стадии гуманизированное антитело содержит связывающий агент, прикрепленный к некоторым лизинам антитела, и он стабилен в течение 2 недель.
Модифицированное антитело hJAA-F11 затем метили с использованием способа непрямого введения метки Chizzonite [158]. В кратком изложении, предварительно покрытую пробирку для йодирования Pierce (Thermo Fisher, Rockford, IL, USA) предварительно смачивали 1 мл HighTris буфера для йодирования (0,125 М Tris-HCl, pH 6,8, 0,15 М NaCl). Буфер декантировали и затем 100 мкл HighTris буфера для йодирования добавляли непосредственно на дно пробирки с последующим добавлением 280 мкл (4,59 мКи) 124I натрия йодида в 0,02 М растворе NaOH (IBA Molecular, Richmond, VA). Первоначальный подсчет брали, используя радиоизотопный калибратор CRC 12 (Capintec). Измеряли pH смеси, чтобы убедиться, что она была при pH 7, нейтральной. После 6 минутной активации при комнатной температуре, активированный йодид добавляли к раствору предварительно модифицированного huJAA-F11. После 9 минутного периода инкубирования, 50 мкл буфера для остановки реакции (10 мг тирозина/мл в Tris буфере для йодирования; 25 мМ Tris, pH 7,5, 0,4 М NaCl) и смесь инкубировали в течение 5 минут. Цель буфера для остановки реакции состоит в удалении свободного йода, который вступает в реакцию с тирозином в буфере.
1 мл Tris/NaCl/EDTA буфера (25 мМ Tris-HCl, pH 7,5, 0,4 М NaCl, 5 мМ EDTA, 0,05% азида натрия) затем добавляли к реакционной смеси. Образец затем добавляли в 10 мл обессоливающую колонку, которая была предварительно уравновешена Tris/NaCl/EDTA буфером. Пробирку с образцом промывали 0,5 мл Tris/NaCl/EDTA буфера и промывную воду добавляли в колонку. Образец элюировали на пятнадцать фракций по 500 мкл каждая, используя Tris/NaCl/EDTA буфер, и испытывали на радиоактивность. Эффективность введения радиоактивной метки определяли на фракции с наиболее высокой активностью с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). Радиоактивно меченое антитело инъекционно вводили животным в пределах 2 часов введения метки. Для того, чтобы убедиться, что меченое гуманизированное JAA-F11 сохраняет свою TF-Ag реакционную способность, кроме того, проводили радиоиммунный анализ.
Животные и модели опухолей
Животных в этом исследовании, размещали и использовали в соответствии с нормативными актами Institutional Animal Care and Use Committees (IACUC). Все протоколы были одобрены IACUC университета в Буффало. 4Т1 клетки рака молочной железы мыши имплантируют 7-8 недельным самкам мышей линии Balb/C путем инъекционного введения 5×104 клеток в 0,1 мл D-PBS подкожно под один из правых сосков. Мыши были разделены на 2 группы и им инъекционно вводили [124I]-hJAA-F11 (n=13) и свободный 124124 (n=7) через 10-14 дней после имплантации опухоли в хвостовую вену и затем подвергали исследованиям биораспределения и микро-РЕТ визуализации. Одну мышь из каждой из 2 групп обследовали и наблюдали на протяжении всего исследования. Все мыши получали воду с 0,2 г/л йодида калия после инъекции с меченым антителом, и наблюдали на протяжении всего исследования в щитовидно блокирующем режиме.
Исследования биораспределения
Мышей умерщвляли с помощью инъекции 0,1 мл пентобарбитала натрия (Fatal Plus) внутрибрюшинно через 72, 96, 168 и 192 часов после инъекции радиомеченого антитела. В каждый момент времени, три мыши, которые получали меченые антитела, и две мыши, которые получали свободный йод, были умерщвлены. Кровь, мышцы, селезенку, легкие, почки, сердце, печень, тонкий и толстый кишечник, желудок, головной мозг, кожу, опухолевые ткани, кости, хвост, пищевод, щитовидную железу и яичники собирали и помещали в предварительно взвешенные 5 мл полипропиленовые пробирки. Эти пробирки были повторно взвешены, чтобы получить фактический вес каждой ткани/органа. Все пробирки закрывали и измеряли радиоактивность с помощью гамма-счетчика. Поглощение радиоактивности для каждой ткани рассчитывали, как процент от инъекционно введенной дозы на грамм ткани (% ID/г) в соответствии со следующей формулой, как и в Приложении 2.
Микро-РЕТ визуализация
Локализацию меченого антитела в одной мыши каждой из двух групп определяли с помощью микроРЕТ визуализации с использованием микроРЕТ камеры, Focus 120® (Siemens Concorde Microsystems) через 24, 48, 72, 96, 168 и 192 часа после инъекции с использованием известных методик. В кратком изложении, перед сканированием мышей анестезировали O2/изофлураном (1%-3% изофлурана) и затем визуализировали в положении лежа в гентри микроРЕТ сканера. Окно сканирования излучения было установлено от 350 до 750 кэВ. Сканирование проводили в течение 30 минут для каждой мыши.
Радиоиммунный анализ
Для определения иммунореактивности радиоактивно меченого антитела, проводили радиоиммунный анализ (РИА). Планшет для микротитрования покрывали 100 мкл 1,25 мкг/мл конъюгата TF-Ag-BSA в покрывающем буфере. Буфер удаляли из лунок и промывали 1% BSA/PBS-Tween. Сто микролитров серийных разбавлений радиоактивно меченного hJAA-F11 в 1% BSA/PBS-Tween добавляли в лунки и позволяли связываться при комнатной температуре в течение 1 часа. После инкубирования, несвязанное антитело удаляли путем промывания лунок 1% BSA/PBS-Tween три раза вручную, с помощью многоканальный пипетки. Связанное hJAA-F11 удаляли из планшета путем инкубирования с 200 мкл буфера 1 М уксусной кислоты/0,15 М NaCl (pH 2,4) в течение 30 минут при комнатной температуре. После инкубирования, 100 мкл раствора из каждой лунки добавляли в отдельные полипропиленовые тестовые пробирки и радиоактивность измеряли с использованием гамма-счетчика.
В то время как настоящее изобретение было описано с помощью конкретных вариантов осуществления, общепринятые модификации будут очевидны специалистам в этой области техники, и такие модификации, как подразумевается, находятся в пределах объема настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕКОНКУРЕНТНЫЕ В ОТНОШЕНИИ НЕЙРЕГУЛИНА АЛЛОСТЕРИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО HER3 И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2704228C2 |
| АНТИТЕЛА ПРОТИВ IL2 | 2006 |
|
RU2425054C2 |
| Конъюгаты "антитело-лекарственное средство" | 2012 |
|
RU2624141C2 |
| Антитело, связывающееся с карбоангидразой, и его применение | 2017 |
|
RU2727682C1 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АЛЬФА-ЭНОЛАЗЕ, И СПОСОБЫ ПРИМЕНЕНИЯ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2014 |
|
RU2761662C2 |
| АНТИТЕЛО ПРОТИВ КСФ-1R | 2012 |
|
RU2621859C2 |
| АНТИТЕЛА К HER3 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2560583C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| Гуманизированные антитела к LIV-1 и их применение для лечения рака | 2011 |
|
RU2608646C2 |









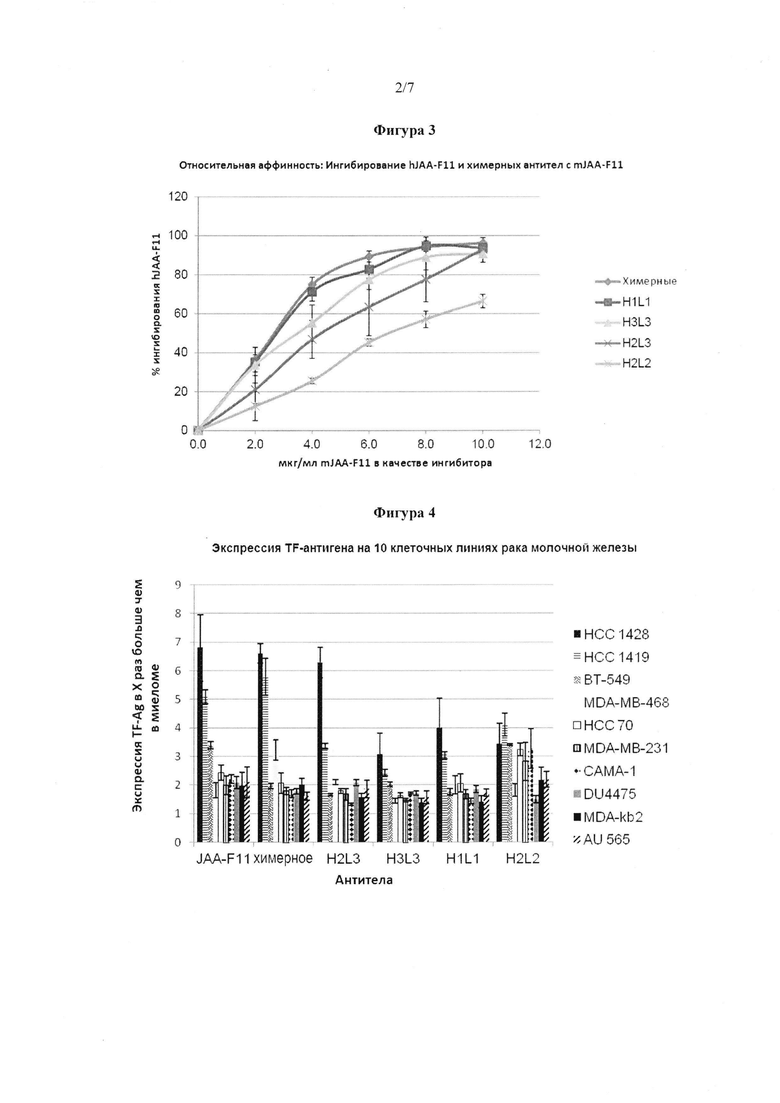
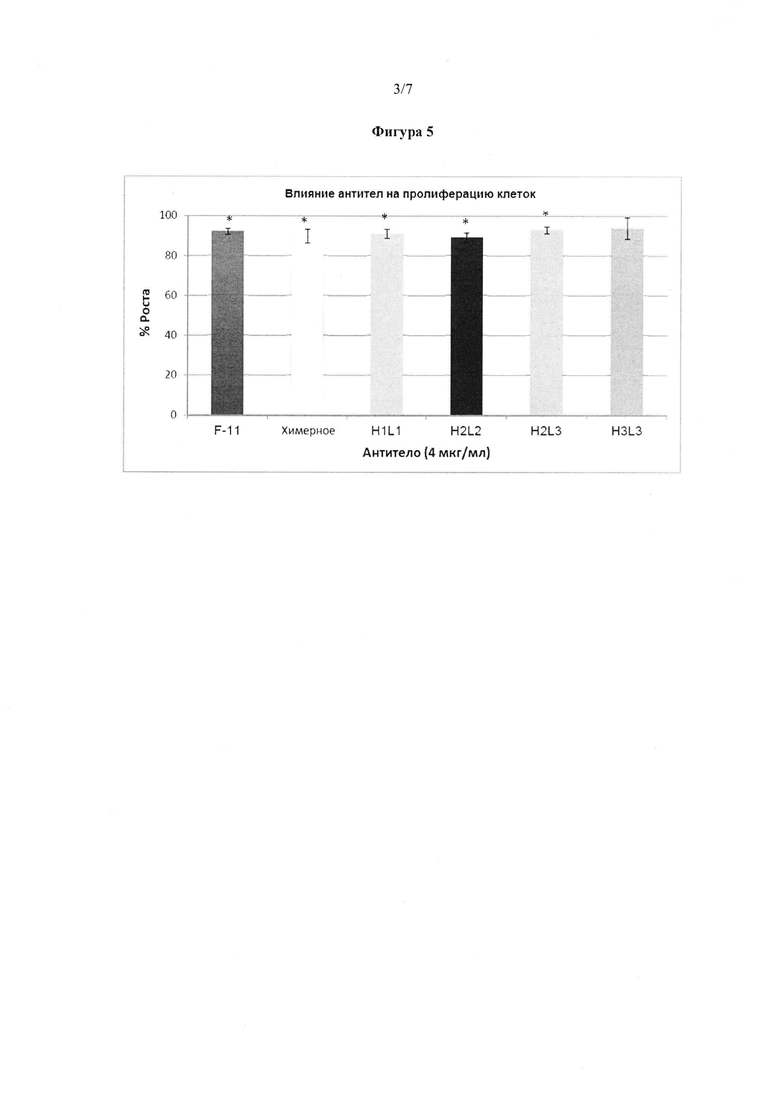
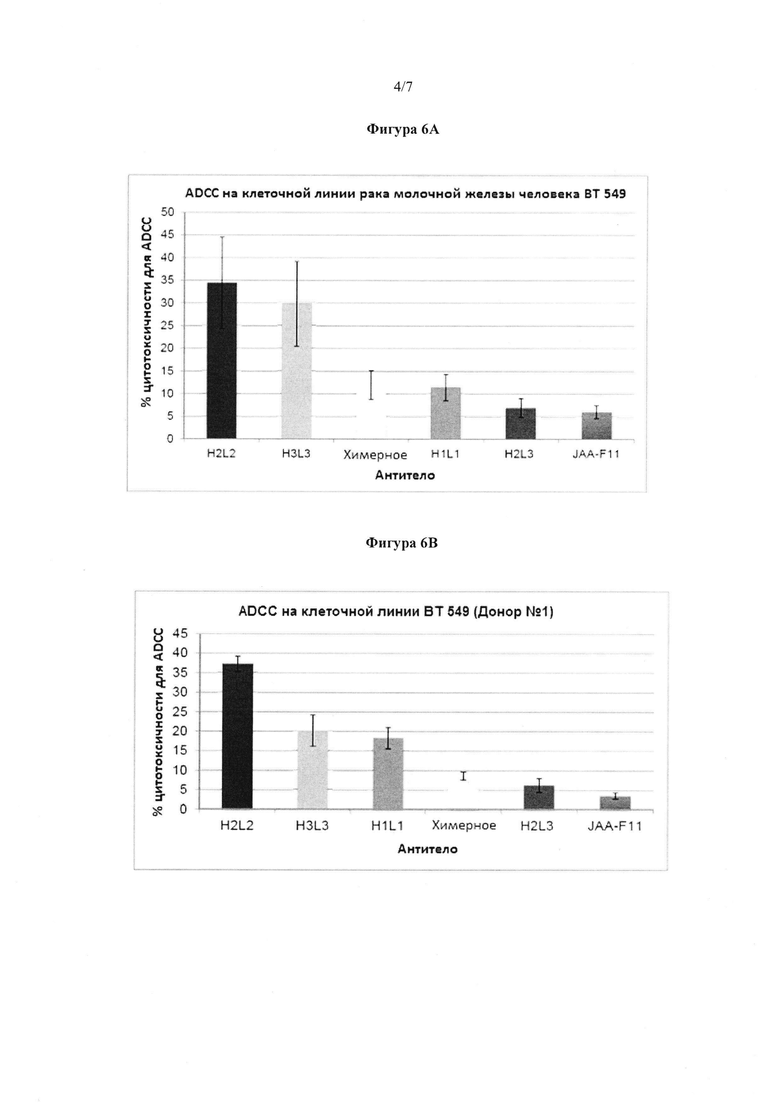
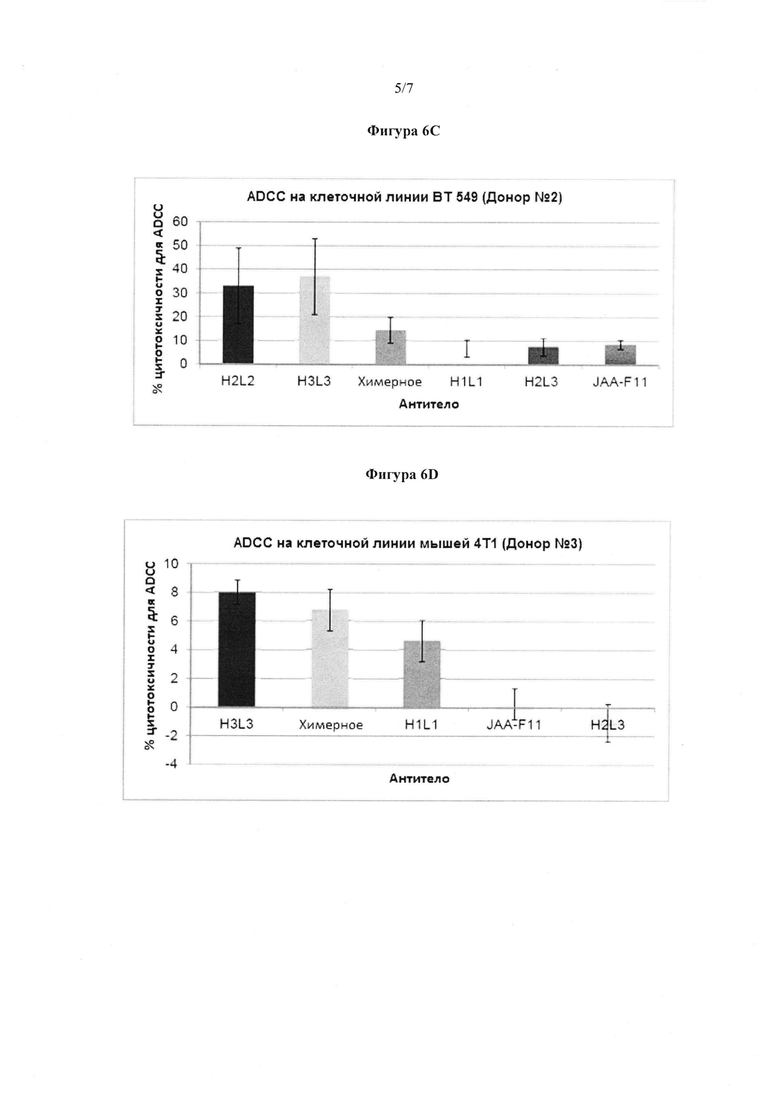
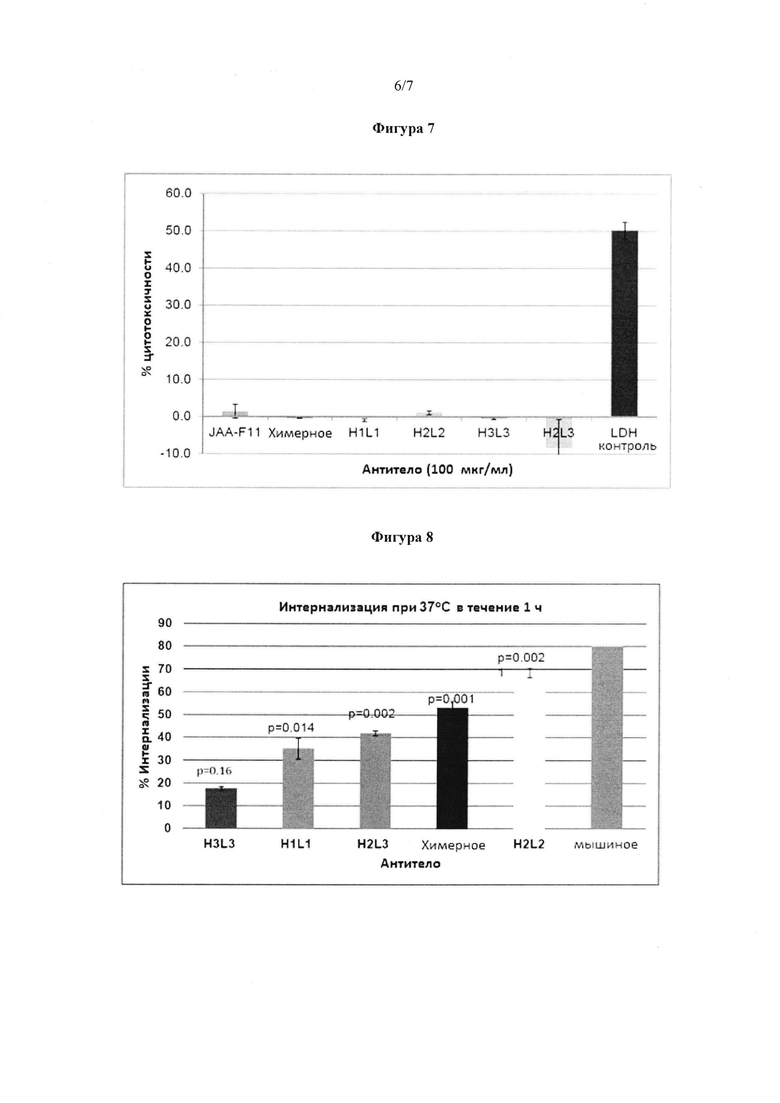
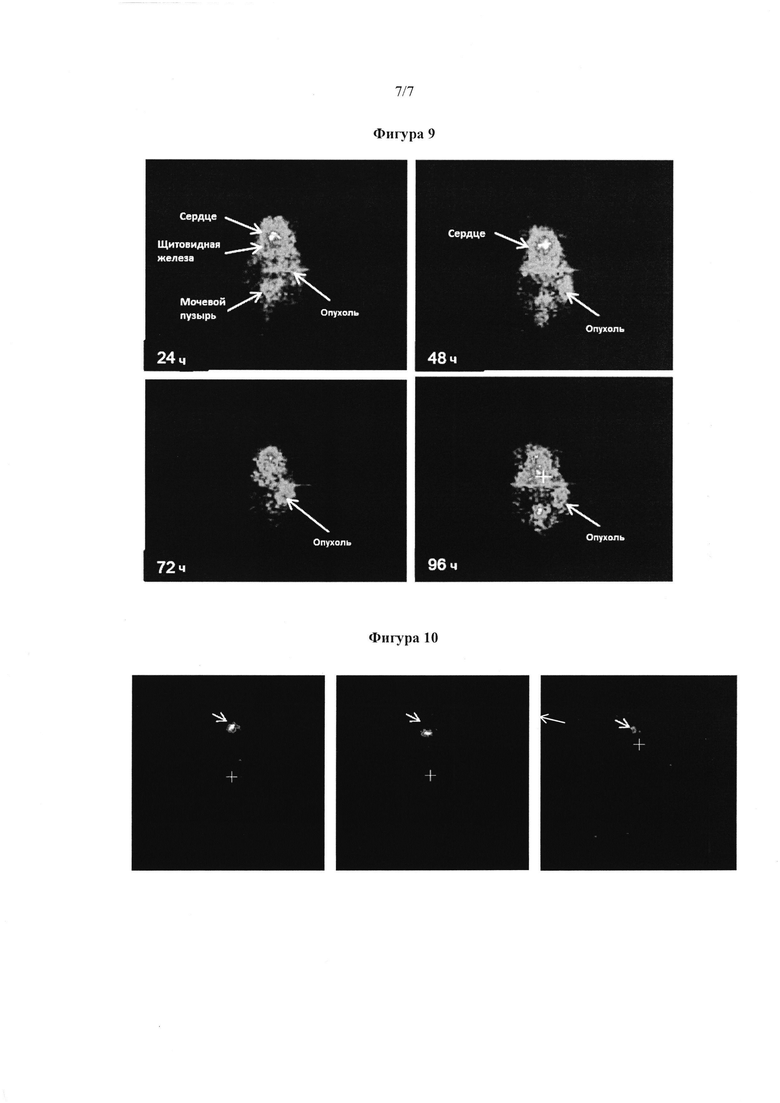
Изобретение относится к области биохимии. Описана группа изобретений, включающая гуманизированное моноклональное антитело или его антиген-связывающий фрагмент, которое связывается со специфичностью с антигеном Томсена-Фриденрайха (TF-Ag), конъюгат, связывающийся с раковыми клетками, которые экспрессируют TF-Ag, способ профилактики или лечения рака у индивидуума, при этом рак содержит раковые клетки, экспрессирующие TF-Ag, фармацевтическая композиция для лечения TF+ видов рака, содержащая вышеуказанное антитело или его антиген-связывающий фрагмент в терапевтически эффективном количестве, клетку-хозяин млекопитающего, предназначенную для экспрессии вышеуказанного антитела или его антиген-связывающего фрагмента. В одном из вариантов изобретения вышеуказанное антитело содержит константную область IgG человека. Представленное изобретение расширяет арсенал средств для связывания с антигеном Томсена-Фриденрайха. 5 н. и 2 з.п. ф-лы, 7 табл., 13 ил., 2 пр.
1. Гуманизированное моноклональное антитело (mAb), которое связывается со специфичностью с антигеном Томсена-Фриденрайха (TF-Ag), или его антиген-связывающий фрагмент, при этом моноклональное антитело или его антиген-связывающий фрагмент содержат тяжелую цепь и легкую цепь, причем тяжелая цепь содержит последовательность, выбранную из группы, состоящей из:
a) последовательности, состоящей из:
EVQLVESGAEVKKPGASVKVSCKASGYTFTTYWMHWVRQAPGQGLEWMGFISPNTDYTEYNQKFRDRVTMTADTSISTAYMELSRLRSDDTAVYYCARSFIGYNFDFWGQGTLVTVSS (SEQ ID NO:7) (H1);
b) последовательности, состоящей из:
EVQLLESGAELKKPGASVKVSCKASGYTFTTYWMHWVRQAPGQGLEWMGFISPNTDYTEYNQKFRDRVTLTADKSSSTAYMELSSLTSEDТАVYYCARSFIGYNFDFWGQGTTVTVSS (SEQ ID NO:8) (H2);
c) последовательности, состоящей из:
EVQLVESGAEVKKPGASVKVSCKASGYTFTTYWMHWVKQAPGQGLEWIGFISPNTDYTEYNQKFRDKATMTADTSISTAYMELSRLRSDDTAVYYCARSFIGYNFDFWGQGTTLTVSS (SEQ ID NO:9) (H3),
d) последовательности, состоящей из:
QVQLVQSGAEVKKPGSSVKVSCKASGYTFTTYWMHWVRQAPGQGLEWMGFISPNTDYTEYNQKFRDRVTITADKSTSTAYMELSSLRSEDТАVYYCARSFIGYNFDFWGQGTTVTVS (SEQ ID NO:13) (H2a);
e) последовательности, состоящей из:
EGQLLESGAELAKPGASVKMSCKASGYTFTTYWMHWVKKRPGQGLEWIGFISPNTDYTEYNQKFRDKATLTADKSSTTAYMQLSSLTSDDSAVYYCARSFIGYNFDFWGQGTTLTVSS (SEQ ID NO:14) (H3a); и
их комбинаций;
и при этом легкая цепь содержит последовательность, выбранную из группы, состоящей из:
d) последовательности, состоящей из:
DVVMTQSPLSLPVTLGQPASISCRSSQTIVYSNGNTYLEWFQQRPGQSPRLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPFTFGSGTKLEIK (SEQ ID NO:10) (L1);
e) последовательности, состоящей из:
DIVMTQTPLSLPVTLGQPASISCRSSQTIVYSNGNTYLEWFQQRPGQSPRLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPFTFGSGTKLEIK (SEQ ID NO:11) (L2);
f) последовательности, состоящей из:
DVVMTQSPLSLPVTLGQPASISCRSSQTIVYSNGNTYLEWYLQRPGQSPRLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPFTFGSGTKLEIK (SEQ ID NO:12) (L3);
g) последовательности, состоящей из:
DIVMTQSPLSLPVTPGEPASISCRSSQTIVYSNGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPFTFGSGTKVDIK (SEQ ID NO:15) (L2a);
h) последовательности, состоящей из:
ELVMTQTPLSLPVNLGDQASISCRSSQTIVYSNGNTYLEWYLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEADDLGVYYCFQGSHVPFTFGSGTKLEIK (SEQ ID NO:16) (L3a);
и их комбинаций.
2. mAb по п. 1, при этом mAb содержит константную область IgG человека.
3. Конъюгат, связывающийся с раковыми клетками, экспрессирующими TF-Ag, в котором mAb или его антиген-связывающий фрагмент по п. 1 конъюгирован с агентом, выбранным из группы, состоящей из химиотерапевтических лекарственных средств, токсинов и радиоактивных изотопов.
4. Способ профилактики или лечения рака у индивидуума, при этом рак содержит раковые клетки, экспрессирующие TF-Ag, причем способ включает стадию, на которой вводят индивидууму терапевтически эффективное количество одного или более mAb или их антиген-связывающих фрагментов по п. 1, или конъюгата по п. 3, при этом рост, или выживание, или метастазирование, или их комбинация, раковых клеток у индивидуума ингибируется после введения.
5. Способ по п. 4, в котором mAb содержит константную область IgG человека.
6. Фармацевтическая композиция для лечения TF+ видов рака, содержащая гуманизированное mAb или его антиген-связывающий фрагмент по п. 1 в терапевтически эффективном количестве и, необязательно, фармацевтически приемлемые носители, эксципиенты или стабилизаторы.
7. Клетка-хозяин млекопитающего, предназначенная для экспрессии гуманизированного mAb или его антиген-связывающего фрагмента по п. 1, причем указанная клетка-хозяин трансформирована одним или более рекомбинантных векторов экспрессии млекопитающих, который(е) кодирует(ют) тяжелую цепь и легкую цепь, охарактеризованные в независимом п. 1.
| US 20060018913 A1, 26.01.2006 | |||
| US 20050260680 A1, 24.11.2005 | |||
| US 20070253950 A1, 01.11.2007 | |||
| US 20110117086 A1, 19.05.2011 | |||
| АНТИ-CD 16 СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2006 |
|
RU2491294C2 |

