Область техники, к которой относится изобретение
Настоящее изобретение относится, главным образом, к созданию и характеристике нейтрализующих анти-IFN-α моноклональных антител с широкой реактивностью к различным подтипам IFN-α. Настоящее изобретение относится также к использованию таких анти-IFN-α антител в диагностике и лечении нарушений, связанных с повышенной экспрессией IFN-α, в частности аутоиммунных нарушений, таких как инсулинозависимый сахарный диабет (IDDM) и системная красная волчанка (SLE).
Описание близкого уровня техники
Интерферон-α (IFN-α)
Хотя интерфероны поначалу были открыты по их противовирусным активностям, при последующем изучении обнаружились многочисленные регуляторные активности, ассоциированные с этими сильнодействующими цитокинами. Интерфероны типа I образуют древнее семейство цитокинов, которое включает IFN-α, IFN-β, IFN-δ, IFN-ω и IFN-τ (Roberts et al., J. Interferon Cytokine Res. 18:805-816 [1998]). Они кодируются безынтронными генами и широко распространены среди позвоночных. Несмотря на то что IFN-β кодируется у приматов и грызунов единственным геном, у человека и мышей обнаруживается более 10-15 разных подтипов IFN-α. Другие интерфероны типа I менее распространены, например IFN-δ присутствует у свиней, IFN-τ - у крупного рогатого скота и овец, а IFN-ω - у крупного рогатого скота и людей. Таким образом, интерфероны типа I человека включают множество членов семейства IFN-α и по одному члену из семейств IFN-β и IFN-ω. Все IFN типа I связываются с единственным рецептором, который включает, по меньшей мере, два белка, пронизывающих мембрану. С другой стороны, интерфероны типа II представлены одним членом, IFN-γ, который связывается с отдельным рецептором.
Хотя все IFN типа I, в том числе и IFN-α, проявляют противовирусное и антипролиферативное действие и, тем самым, помогают бороться с вирусными инфекциями и злокачественными опухолями (Lefevre et al., Biochimie 80:779-788 [1998]; Horton et al., Cancer Res. 59:4064-4068 [1999]; Alexenko et al., J. Interferon Cytokine Res. 17:769-779 [1997]; Gresser, J. Leukoc. Biol. 61:567-574 [1997]), существует несколько аутоиммунных заболеваний, которые ассоциируются с повышенной экспрессией IFN-α, это прежде всего инсулинозависимый сахарный диабет (IDDM) и системная красная волчанка (SLE).
Диабет тип I, известный также как аутоиммунный диабет или инсулинозависимый сахарный диабет (IDDM), представляет собой аутоиммунное заболевание, характеризующееся избирательным разрушением панкреатических β-клеток аутореактивными Т-лимфоцитами (Bach, Endocr. Rev. 15:516-542 [1994]; Castano and Eisenbarth, Annu. Rev. Immunol. 8:647-679 [1990]; Shehadeh and Lafferty, Diabetes Rev. 1:141-151 [1993]). Патология IDDM является очень сложной, включающей взаимодействие между эпигенетическим событием (возможно, вирусная инфекция), панкреатическими β-клетками и иммунной системой у генетически чувствительного хозяина. Ряд цитокинов, в том числе IFN-α и IFN-γ, вовлечены у человека в патогенез IDDM и в животных моделях данного заболевания (Campbell et al., J. Clin. Invest 87:739-742 [1991]; Huang et al., Diabetes 44:658-664 [1995]; Rhodes and Taylor, Diabetologia 27:601-603 [1984]). Например, сообщается об экспрессии панкреатической мРНК Ifn-α и о наличии иммунореактивных IFN-α в β-клетках пациентов с IDDM (Foulis et al., Lancet 2:1423-1427 [1987]; Huang et al., [1995] выше; Somoza et al., J. Immunol. 153:1360-1377 [1994]). Экспрессия IFN-α ассоциируется в островках Лангерганса человека со сверхэкспрессией антигенов главного комплекса гистосовместимости (МНС) класса IA (Foulis et al., [1978] выше; Somoza et al., [1994] выше). В двух моделях аутоиммунного заболевания грызунов, подверженных диабету DP-BB-крыс и обработанных стрептозотоцином мышей, экспрессия мРНК Ifn-α в островках Лангерганса предшествует инсулиновой недостаточности и диабету (Huang et al., Immunity 1:469-478 [1994]). Кроме того, у трансгенных мышей, содержащих гибридную конструкцию человеческий инсулиновый промотор-Inf-α, развивается гипоинсулинемический диабет, сопровождаемый инсулиновой недостаточностью (Stewart et al., Science 260:1942-1946 [1993]).
Оказалось, что локальная экспрессия IFN-α клетками панкреатических островков в ответ на возможные диабетогенные стимулы, такие как вирусы, может запускать процесс развития инсулиновой недостаточности. Сообразно его роли в качестве инициирующего агента показано, что IFN-α индуцирует в эндотелиальных клетках островков Лангерганса человека фактор межклеточной адгезии-I (ICAM-1) и HLA класса IA, что может способствовать лейкоцитарной инфильтрации в период инсулиновой недостаточности (Chakrabarti et al., J. Immunol. 157:522-528 [1996]). Кроме того, IFN-α способствует Т-клеточной стимуляции в результате индукции ко-стимулирующих молекул ICAM-1 и В7.2 в антиген-презентирующих клетках островков Лангерганса (Chakrabarti et al., Diabetes 45:1336-1343 [1996]). Вместе эти исследования свидетельствуют о том, что ранняя экспрессия β--клетками IFN-α может являться ключевым событием в инициации аутоиммунного диабета. Несмотря на имеющийся ряд сообщений об участии IFN-γ в развитии IDDM в моделях грызунов корреляция между экспрессией данного цитокина и IDDM человека мала. Таким образом, клетки, экспрессирующие IFN-γ, можно обнаружить в островках Лангерганса в подгруппе пациентов-людей, подобранных по существенной лимфоцитарной инфильтрации в указанных островках. В группе пациентов, которых не подбирали по этому критерию, ассоциация между экспрессией IFN-γ и IDDM человека оказалась неочевидной.
Основываясь на повышенном уровне экспрессии IFN-α у пациентов с системной красной волчанкой (SLE), считается, что IFN-α также участвует в патогенезе SLE (Ytterberg and Schnitzer, Arthritis Rheum 25:401-406 [1982]; Shi et al., Br. J. Dermatol. 117:155-159 [1987]). Интересно отметить, что в настоящее время IFN-α используют для лечения рака, а также вирусной инфекции, такой как хронический гепатит, вызванный инфицированием вируса гепатита В и вирусом гепатита С. В соответствии с наблюдениями о повышенных уровнях IFN-α, запускающих аутоиммунность, сообщается о значительном повышении частоты возникновения аутоиммунных нарушений, таких как IDDM, SLE, тиреоидит, у пациентов, подвергающихся IFN-α-терапии. Например, было показано, что продолжительное использование IFN-α в качестве противовирусной терапии индуцирует IDDM (Waguri et al., Diabetes Res. Clin. Pract. 23:33-36 [1994]; Fabris et al., J. Hepatol. 28:514-517 [1998] или SLE (Garcia-Porrua et al., Clin. Exp. Rheumatol. 16:107-108 [1998]). Лечение инфекции, вызванной вирусом коксаки В (CBV), с помощью IFN-α-терапии также ассоциируется с индукцией IDDM (Chehadeh et al., J. Infect. Dis. 181:1929-1939 [2000]). Также сообщается о множестве случаев, свидетельствующих о развитии IDDM или SLE у пациентов с раком, которых лечили IFN-α (Ronnblom et al., J. Intern. Med. 227:207-210 [1990]).
Терапия антителами
Использование моноклональных антител в качестве терапевтических средств было признано для некоторых моноклональных антител (mAb), либо одобренных для использования на человеке, либо находящихся на последней стадии клинических испытаний. Первые mAb для лечения отторжения аллотрансплантата, одобренные в 1986 г. Управлением по контролю за продуктами и лекарствами (FDA), были антителами против CD3 (OKT3). С того времени достижения в области mAb значительно выросли, особенно с 1994 г., что привело к одобрению еще семи видов mAb для лечения человека. Они включают ReoPro® для терапии осложнений коронарной ангиопластики в 1994 г., Zenapax® (анти-CD25) для предотвращения отторжения аллотрансплантата в 1997 г., Rituxan® (анти-CD20) для лечения В-клеточной лимфомы не-Ходжкина в 1997 г., Infliximab® (анти-IFN-α) первоначально для лечения болезни Крона в 1998 г., а впоследствии для лечения ревматоидного артрита в 1998 г., Simulect® (анти-CD25) для предотвращения отторжения аллотрансплантата в 1998 г., Synagis® (антитела против F-белка респираторно-синцитиального вируса) для лечения респираторных инфекций в 1998 г., и Herceptin® (анти-HER2/neu) для лечения сверхэкспрессии HER2 метастазирующих злокачественных опухолей молочной железы в 1998 г. (Glennie and Johnson, Immunol. Today 21:403-410 [2000]).
Антитела против IFN-α
Болезненные состояния, которые поддаются воздействию mAb, включают все состояния, в которых наблюдается патологический уровень антигена-мишени. Например, антитело, которое нейтрализует IFN-α, присутствующий в сыворотке крови пациентов, больных SLE, и экспрессирующийся панкреатическими островками у больных IDDM, является потенциальным кандидатом для терапевтического воздействия на эти заболевания. Его можно было бы также использовать для терапевтического воздействия на другие аутоиммунные заболевания с подразумеваемым повышением причинно-обусловленной роли экспрессии IFN-α. Оказалось, что при заболевании человека IDDM (Foulis et al., Lancet 2:1423-1427 [1987]; Huang et al., Diabetes 44:658-664 [1995]; Somoza et al., J. Immunol. 153:1360-1377 [1994] и SLE (Hooks et al., Arthritis & Rheumatism 25:396-400 [1982]; Kim et al., Clin. Exp. Immunol. 70:562-569 [1987]; Lacki et al., J. Med. 28:99-107 [1997]; Robak et al., Archivum Immunologiae et Therapiae Experimentalis 46:375-380 [1998]; Shiozawa et al., Arthritis & Rheumatism 35:417-422 [1992]; von Wussow et al., Rheumatology International 8:225-230 [1988]), существует корреляция между заболеванием и IFN-α, но не IFN-β или IFN-γ. Таким образом, при антиинтерфероновом воздействии mAb при IDDM или SLE потребуется специфическая нейтрализация большинства, если не всех, подтипов IFN-α, без какой-либо существенной нейтрализации IFN-β или IFN-γ. Остаточная активность этих последних двух интактных интерферонов может также иметь преимущество, обеспечивая сохранение значительной противовирусной активности.
В то же время обнаружено, что несколько mAb, которые демонстрируют реактивность в отношении описанных подтипов рекомбинантного человеческого IFN-α, нейтрализуют лишь ограниченную проанализированную подгруппу подтипов рекомбинантного IFN-α или оказались неспособны нейтрализовать смесь подтипов IFN-α, полученную с помощью стимулированных лейкоцитов периферической крови (Tsukui et al., Microbiol. Immunol. 30:1129-1139 [1986]; Berg, J. Interferon Res. 4:481-491 [1984]; Meager and Berg, J. Interferon Res. 6:729-736 [1986]; Патент США №4902618; и EP публикация №0139676 В1).
Поэтому существует большая потребность в анти-IFN-α антителах, которые не только связываются с большинством подтипов IFN-α, предпочтительно всеми, но также нейтрализуют такие подтипы, которые не влияют на указанную биологическую функцию других интерферонов.
Краткое изложение существа изобретения
Настоящее изобретение основано на создании моноклональных антител, которые, что было экспериментально установлено, нейтрализуют все семь анализируемых разных подтипов рекомбинантного человеческого IFN-α и два независимых пула природных подтипов человеческого IFN-α.
Первой целью настоящего изобретения является создание анти-IFN-α моноклонального антитела человека, которое связывается и нейтрализует биологическую активность, по меньшей мере, IFN-α человека подтипов IFN-α1, IFN-α2, IFN-α4, IFN-α5, IFN-α8, IFN-α10, и IFN-α21. Еще одной целью настоящего изобретения является создание анти-IFN-α моноклонального антитела человека, которое связывается со всеми подтипами IFN-α человека и нейтрализует их биологическую активность. Антитело настоящего изобретения может в значительной мере уменьшить или подавить биологическую активность рассматриваемого IFN-α человека. В одном из вариантов осуществления настоящего изобретения антитело настоящего изобретения способно нейтрализовать, по меньшей мере, 60% или, по меньшей мере, 70%, предпочтительно, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 80%, еще более предпочтительно, по меньшей мере, 85% и еще более предпочтительно 90%, еще более предпочтительно, по меньшей мере, 95% и наиболее предпочтительно, по меньшей мере, 99% биологической активности IFN-α человеческого индивида. В другом варианте осуществления настоящего изобретения моноклональное антитело, нейтрализующее биологическую активность IFN-α человека, не нейтрализует соответствующую биологическую активность IFN-∃ человека.
Биологическая активность IFN-α человеческого индивида может представлять собой IFNAR2-связывающую активность. В конкретном варианте осуществления настоящего изобретения представлено моноклональное антитело против IFN-∀ человека, способное связывать и блокировать, по меньшей мере, 60% или, по меньшей мере, 70%, предпочтительно, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 80%, еще более предпочтительно, по меньшей мере, 85%, еще более предпочтительно, по меньшей мере, 90%, еще более предпочтительно, по меньшей мере, 95%, наиболее предпочтительно, по меньшей мере, 99% IFNAR2-связывающей активности всех или практически всех подтипов IFN-α человека. В другом варианте осуществления настоящего изобретения представлено моноклональное антитело против IFN-∀ человека, которое способно связывать и блокировать, по меньшей мере, 60% или, по меньшей мере, 70%, предпочтительно, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 80%, еще более предпочтительно, по меньшей мере, 85%, еще более предпочтительно, по меньшей мере, 90%, еще более предпочтительно, по меньшей мере, 95%, наиболее предпочтительно, по меньшей мере, 99% IFNAR2-связывающей активности каждого подтипа IFN-∀ человека - 1, 2, 4, 5, 8, 10 и 21. В другом варианте осуществления настоящего изобретения моноклональное антитело против IFN-∀ человека не обладает перекрестной реактивностью с IFN-∃ человека.
Биологическая активность IFN-α человеческого индивида может представлять собой противовирусную активность. В одном из вариантов осуществления настоящего изобретения моноклональное антитело против IFN-∀ человека способно связываться со всеми или практически со всеми подтипами IFN-α человека и нейтрализовать их противовирусную активность. В другом варианте осуществления настоящего изобретения моноклональное антитело против IFN-∀ человека способно связываться и нейтрализовать противовирусную активность каждого подтипа IFN-∀ человека - 1, 2, 4, 5, 8, 10 и 21. В конкретном варианте осуществления настоящего изобретения создано моноклональное антитело против IFN-∀ человека, которое способно связываться и нейтрализовать, по меньшей мере, 60% или, по меньшей мере, 70%, предпочтительно, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 80%, еще более предпочтительно, по меньшей мере, 85%, еще более предпочтительно, по меньшей мере, 90%, еще более предпочтительно, по меньшей мере, 95%, наиболее предпочтительно, по меньшей мере, 99% противовирусной активности всех или практически всех подтипов IFN-α человека. Еще в одном варианте осуществления настоящего изобретения создано моноклональное антитело против IFN-∀ человека, которое связывает и нейтрализует, по меньшей мере, 60% или, по меньшей мере, 70%, предпочтительно, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 80%, еще более предпочтительно, по меньшей мере, 85%, еще более предпочтительно, по меньшей мере, 90%, еще более предпочтительно, по меньшей мере, 95%, наиболее предпочтительно, по меньшей мере, 99% противовирусной активности каждого подтипа IFN-∀ человека - 1, 2, 4, 5, 8, 10 и 21. Еще в одном варианте осуществления настоящего изобретения моноклональное антитело, нейтрализующее противовирусную активность IFN-α человека, не нейтрализуют противовирусную активность IFN-∃ человека.
Антитело может быть мышиным, гуманизированным или антителом человека. Антитело может представлять собой мышиное моноклональное антитело 9F3 против IFN-α человека или его гуманизированный вариант, такой, например, как вариант 13 (V13), или его химерную форму. В рамки настоящего изобретение включено также антитело, которое существенно связывает тот же эпитоп IFN-α, что и эпитоп мышиного моноклонального антитела 9F3 против IFN-α человека, или эпитоп его гуманизированной или химерной формы. Например, стандартным антителом для этой цели является анти-IFN-α антитело, полученное с помощью мышиной гибридомной клеточной линии 9F3.18.5, депонированной в АТСС 18 января 2001 г. с инвентарным №РТА-2917. В другом варианте осуществления настоящего изобретения представлено мышиное или химерное, мышь/человек, моноклональное антитело против IFN-α человека, включающее аминокислотную последовательность вариабельного домена (вариабельной области) легкой цепи, представленной на Фиг.5А (SEQ ID NO:1), и/или аминокислотную последовательность вариабельного домена (вариабельной области) тяжелой цепи, представленной на Фиг.5В (SEQ ID NO:2). Еще в одном варианте осуществления настоящего изобретения представлено гуманизированное моноклональное антитело против IFN-α человека, включающее аминокислотную последовательность гуманизированной легкой цепи вариабельного домена, представленную на Фиг.5А (SEQ ID NO:3), и/или аминокислотную последовательность вариабельного домена гуманизированной тяжелой цепи, представленной на Фиг.5В (SEQ ID NO:5).
Кроме того, представлено моноклональное антитело против IFN-α человека, которое существенно связывает тот же эпитоп в подтипах 1, 2, 4, 5, 8, 10 и 21 IFN-α человека, который связывается мышиным моноклональным антителом 9F3 против IFN-α человека, либо гуманизированным или химерным его вариантом. Также представлено моноклональное антитело против IFN-α человека, которое конкурирует с мышиным моноклональным антителом 9F3 против IFN-α человека за связывание с каждым из подтипов 1, 2, 4, 5, 8, 10 и 21 IFN-α человека.
Кроме того, представлена выделенная молекула нуклеиновой кислоты, кодирующая любое из описанных здесь антител, вектор, включающий выделенную молекулу нуклеиновой кислоты, клетку-хозяина, трансформированную данной молекулой нуклеиновой кислоты, и способ получения антитела, включающий культивирование клетки-хозяина в условиях, в которых экспрессируется указанная молекула нуклеиновой кислоты с получением антитела, и необязательно выделение антитела из клетки-хозяина. Антитело может происходить из IgG-класса и представлять изотип, такой как IgG1, IgG2; IgG3 или IgG4. Рамки настоящего изобретения включают также фрагменты антител, такие как фрагменты Fv, scFv, Fab, F(ab′)2, и Fab′.
В соответствии с другой целью в настоящем изобретении представлена легкая цепь моноклонального антитела против IFN-α человека или ее фрагмент, включающая следующие гипервариабельные области (CDR) (как указано у Kabat, et al., Sequences of Proteins of Immune-logical Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD [1991]. vols 1-3): (a) LI формулы RASQSVSTSSYSYMH (SEQ ID NO:7); (b) L2 формулы YASNLES (SEQ ID NO:8); и (с) L3 формулы QHSWGIPRTF (SEQ ID NO:9). Рамки настоящего изобретения включают также вариабельный домен фрагмента легкой цепи моноклонального антитела против IFN-α человека. Кроме того, рамки настоящего изобретения включают полипептид легкой цепи моноклонального антитела против IFN-α человека, включающий аминокислотную последовательность вариабельного домена химерной легкой цепи мыши/человека, либо аминокислотную последовательность полипептида всей химерной легкой цепи, кодируемой вектором XAIFN-ChLpDR1, депонированным в АТСС 9 января 2001 г. с инвентарным №РТА-2880. Рамки настоящего изобретения включают также полипептид легкой цепи моноклонального антитела против IFN-α человека, включающий аминокислотную последовательность вариабельного домена гуманизированной легкой цепи, или аминокислотную последовательность полипептида всей гуманизированной легкой цепи, кодируемой вектором VLV30-IgG, депонированным в АТСС 9 января 2001 г. с инвентарным №РТА-2882.
Еще одной целью настоящего изобретения является создание тяжелой цепи моноклонального антитела против IFN-α человека, или ее фрагмента, включающей следующие CDR: (а) Н1 формулы GYTFTEYIIH (SEQ ID NO:10); (b) H2 формулы SINPDYDITNYNQRFKG (SEQ ID NO:11) и (с) Н3 формулы WISDFFDY (SEQ ID NO:12). Рамки настоящего изобретения включают также вариабельный домен такого фрагмента тяжелой цепи моноклонального антитела против IFN-α человека. Рамки настоящего изобретения дополнительно включают полипептид тяжелой цепи моноклонального антитела против IFN-α человека, включающий аминокислотную последовательность вариабельного домена химерной тяжелой цепи мыши/человека, или аминокислотную последовательность полипептида всей химерной тяжелой цепи, кодируемой вектором XAIFN-ChHpDR2, депонированным в АТСС 9 января 2001 г. с инвентарным №РТА-2883. Дополнительно включен полипептид тяжелой цепи моноклонального антитела против IFN-α человека, включающий аминокислотную последовательность вариабельного домена гуманизированной тяжелой цепи, или аминокислотную последовательность полипептида всей гуманизированной тяжелой цепи, кодируемой вектором VHV30-IgG2, депонированным в АТСС 9 января 2001 г. с инвентарным №РТА-2881.
В соответствии с дополнительной целью в настоящем изобретении представлено моноклональное антитело против IFN-α человека, включающее (А) по меньшей мере, одну легкую цепь или ее фрагмент, включающие следующие CDR: (а) L1 формулы RASQSVSTSSYSYMH (SEQ ID NO:7); (b) L2 формулы YASNLES (SEQ ID NO:8); и (с) L3 формулы QHSWGIPRTF (SEQ ID NO:9); и (В) по меньшей мере, одну тяжелую цепь, или ее фрагмент, включающую следующие CDR: (а) Н1 формулы GYTFTEYIIH (SEQ ID NO:10); (b) H2 формулы SINPDYDITNYNQRFKG (SEQ ID NO:11); и (с) Н3 формулы WISDFFDY (SEQ ID NO:12). Антитело может обладать гомотетрамерной структурой, состоящей из двух пар цепей, тяжелой и легкой, антитела, соединенных дисульфидной связью. Рамки настоящего изобретения включают, в частности, линейное антитело, мышиное антитело, химерное антитело, гуманизированное антитело или антитело человека. Дополнительно представлено химерное антитело, включающее (1) аминокислотную последовательность вариабельного домена химерной легкой цепи мыши/человека или аминокислотную последовательность полипептида всей химерной легкой цепи, кодируемой вектором XAIFN-ChLpDR1, депонированным в ATCC 9 января 2001 г. с инвентарным №РТА-2880; и (2) аминокислотную последовательность вариабельного домена химерной тяжелой цепи мыши/человека или аминокислотную последовательность полипептида всей химерной тяжелой цепи, кодируемой вектором XAIFN-ChHpDR2, депонированным в ATCC 9 января 2001 г. с инвентарным №РТА-2883. Дополнительно представлено гуманизированное антитело, включающее (1) аминокислотную последовательность вариабельного домена гуманизированной легкой цепи или аминокислотную последовательность полипептида всей гуманизированной легкой цепи, кодируемой вектором VLV30-IgG, депонированным в ATCC 9 января 2001 г. с инвентарным №РТА-2882; и (2) аминокислотную последовательность вариабельного домена гуманизированной тяжелой цепи или аминокислотную последовательность полипептида всей гуманизированной тяжелой цепи, кодируемой вектором VHV30-IgG2, депонированным в АТСС 9 января 2001 г. с инвентарным №РТА-2881.
Еще одной целью настоящего изобретения является представление фармацевтической композиции, включающей эффективное количество антитела настоящего изобретения в смеси с фармацевтически приемлемым носителем.
Другой целью настоящего изобретения является представление способа диагностики состояния, ассоциированного с экспрессией IFN-α в клетке, включающего контактирование клетки с анти-IFN-α антителом и детектирование присутствия IFN-α.
Еще одной целью настоящего изобретения является представление способа лечения заболевания или состояния, ассоциированного с экспрессией у пациента IFN-α, включающего введение пациенту эффективного количества анти-IFN-α антитела. Пациент является пациентом-млекопитающим, предпочтительно пациентом-человеком. Заболевание является аутоиммунным заболеванием, таким как инсулинозависимый сахарный диабет (IDDM); системная красная волчанка (SLE); или аутоиммунный тиреоидит.
Краткое описание чертежей
На Фиг.1 представлена принципиальная схема способа, используемого при создании моноклональных антител против IFN-α человека.
На Фиг.2 показано, каким образом мышиное mAb против IFN-α человека (9F3) способно нейтрализовать спектр подтипов рекомбинантного IFN-α, но не рекомбинантного IFN-β. Указанные IFN оценивают по ингибированию в клетках А549 роста вируса энцефаломиокардита (EMC) в присутствии увеличивающихся концентраций mAb 9F3. Данные, представленные в виде процентов ингибирования активности вирусного роста, получены для указанного IFN в отсутствие mAb 9F3.
На Фиг.3А-3В показана нейтрализация лейкоцитарного интерферона (Sigma) (Фиг.3А) и лимфобластоидного интерферона (NIH-стандарт Ga23-901-532) (Фиг.3В). На Фиг.3А, 20000 МЕ/мл (заштрихованные столбики) или 5000 МЕ/мл (незаштрихованные столбики) лейкоцитарного интерферона (Sigma Product No. I-2396) инкубируют с пустым контролем (только буфер) (обозначен в виде "-"), 10 г/мл IgG контрольной мыши (обозначен в виде "mIgG"), или 10 г/мл mAb 9F3 (обозначен в виде "9F3"). Использовали различные разведения и показана величина остаточной активности. Полученные результаты представляют собой средние значения повторных определений. На Фиг.3В лимфобластоидный интерферон (NIH-стандарт Ga23-901-532) анализируют при 10 (заштрихованные колонки) или 3 (незаштрихованные колонки) МЕ/мл в присутствии или в отсутствие указанных концентраций mAb 9F3. Наибольший цитопатический эффект является свидетельством уменьшения активности интерферона. Полученные результаты представлены в виде средних значений повторных определений.
На Фиг.4 изображены результаты анализа изменений электрофоретической подвижности (EMSA), показывающие индукцию ISGF3/ISRE-комплекса с помощью IFN-α и способность 9F3 mAb предотвращать образование данного комплекса. EMSA осуществляют в присутствии или в отсутствие либо IFN-α2 человека (обозначен в виде "α2"), либо IFN-∃ (обозначен в виде "∃") в концентрации 25 нг/мл для mAb 9F3 (обозначен в виде "9F3") или мышиного контрольного антитела IgG (обозначен в виде "IgG") в концентрации 10 г/мл.
На Фиг.5A показано сопоставление аминокислотных последовательностей вариабельного домена легкой цепи мышиной 9F3 (мышиная, SEQ ID NO:1), варианта 13 гуманизированной 9F3 (V13, SEQ ID NO:3) и консенсусной вариабельной области легкой κ-цепи подгруппы I человека (huκI, SEQ ID NO:4). CDR (L1, SEQ ID NO:7; L2, SEQ ID NO:8; и L3, SEQ ID NO:9) выделены подчеркиванием. Нумерация аминокислотных остатков по Kabat et al., (1991) выше. Различия между мышиными последовательностями 9F3 и V13, а также различия между последовательностями 9F3 и huκI указаны звездочками.
На Фиг.5В показано сопоставление аминокислотных последовательностей вариабельной области тяжелой цепи мышиной 9F3 (мышиная, SEQ ID NO:2), варианта 13 гуманизированной 9F3 (V13, SEQ ID NO:5) и консенсусной вариабельной области подгруппы III тяжелой цепи человека (huIII, SEQ ID NO:6). CDR (H1, SEQ ID NO:10; H2, SEQ ID NO:11; и H3, SEQ ID NO:12) выделены подчеркиванием. Нумерация остатков по Kabat et al., (1991) выше. Различия между мышиными последовательностями 9F3 и V13, а также различия между последовательностями 9F3 и huIII указаны звездочками.
На Фиг.6 представлена картина нейтрализационной активности исходных mAb 9F3 (левая панель) и химерного белка СН8-2 (правая панель) по отношению к ингибированию проявляемого вирусного роста рекомбинантными подтипами IFN-α в клетках А549, зараженных вирусом энцефаломиокардита (EMC).
На Фиг.7 изображена модель гуманизированного варианта 13 9F3. Каркас областей VL и VH представлен в виде ленты. CDR показаны белым цветом и промаркированы (L1, L2, L3, H1, H2, H3). Структура боковых цепей, изменяемая от человека к мыши, представлена светлым изображением и промаркирована оцифрованным остатком.
Подробное описание предпочтительных вариантов осуществления настоящего изобретения
А. Определения
Если не оговорено иначе, используемые здесь технические и научные термины имеют то же значение, что и очевидное в большинстве случаев специалисту в данной области техники, к которой относится и настоящее изобретение. См., например, Singleton et al., Dictionary of Microbiology and Molecular Biology 2nd ed., J. Wiley & Sons (New York, NY 1994); Sambrook et al., Molecular Cloning, A Laboratory Manual Cold Springs Harbor Press (Cold Springs Harbor, NY 1989). Для целей настоящего изобретения ниже приводятся определения следующих терминов.
Используемый здесь термин "интерферон типа I" определяют в качестве термина, который включает все подтипы нативной последовательности интерферона типа I любого вида млекопитающих, в том числе интерферон-α, интерферон-β, интерферон-δ, интерферон-ω и интерферон-τ. Подобным образом термин "интерферон человека типа I" определяют в качестве термина, который включает все подтипы нативной последовательности интерферонов типа I человека, в том числе классы интерферона-α, интерферона-β и интерферона-ω, которые связываются с обычными клеточными рецепторами.
Если не предусмотрено специально, используемые здесь термины "интерферон-α", "IFN-α", и "интерферон-α человека", "IFN-α человека" и "IFN-∀" относятся ко всем видам нативной последовательности интерферона-альфа человека, в том числе ко всем подтипам нативной последовательности интерферона-α человека. Природный (нативная последовательность) интерферон-α человека включает 23 или более близкородственных белков, кодируемых отдельными генами с высокой степенью структурной гомологии (Weissmann and Weber, Prog. Nucl. Acid. Res. Mol. Biol., 33:251 [1986]; J. Interferon Res., 13:443-444 [1993]; Roberts et al., J. Interferon Cytokine Res. 18:805-816 [1998]). Локус IFN-α человека включает два подсемейства. Первое подсемейство состоит, по меньшей мере, из 14 функциональных, неаллельных генов, в том числе генов, кодирующих IFN-αА (IFN-α2), IFN-αВ (IFN-α8), IFN-αС (IFN-α10), IFN-αD (IFN-α1), IFN-αЕ (IFN-α22), IFN-αF (IFN-α21), IFN-αG (IFN-α5) и IFN-αН (IFN-α14), и псевдогенов, обладающих, по меньшей мере, 80% гомологией. Второе подсемейство, αII или Т, содержит, по меньшей мере, 5 псевдогенов и один функциональный ген (обозначаемый здесь в виде "IFN-αII1" или "IFN-Т"), который проявляет 70% гомологию с генами IFN-α (Weissmann and Weber [1986] выше).
Используемые здесь термины "первый рецептор интерферона-∀, (hIFN-∀) человека", "IFN-∀R", hIFNАR1", "IFNАR1", и "Uze-цепь" определены в виде белкового рецептора из 557 аминокислот, клонированного Uze et al., Cell, 60:225-234 (1990), включающего внеклеточный домен из 409 остатков, трансмембранный домен из 21 остатка и внутриклеточный домен из 100 остатков, как показано на Фиг.5 стр. 229 публикации Uze et al. Включены также вышеизложенные термины и фрагменты IFNAR1, которые содержат внеклеточный домен (ECD) (или фрагменты ECD) IFNAR1.
Используемые здесь термины "второй рецептор интерферона-∀ (hIFN-∀) человека", "IFN-∀∃R", hIFNAR2", "IFNAR2", и "Novick-цепь" определены в виде рецепторного белка из 515 аминокислот, клонированного Domanski et al., J. Biol. Chem., 37:21606-21611 (1995), включающего внеклеточный домен из 217 остатков, трансмембранный домен из 21 остатка и внутриклеточный домен из 250 остатков, как показано на Фиг.1 стр. 21608 публикации Domanski et al. Включены также вышеизложенные термины и фрагменты IFNAR2, которые содержат внеклеточный домен (ECD) (или фрагменты ECD) IFNAR2, и растворимые формы IFNAR2, такие как ECD IFNAR2, слитый с иммуноглобулиновой последовательностью, например, IFNAR2 ECD-Fc IgG, которая описана ниже.
Термин "нативная последовательность", в связи с интерфероном типа I, IFN-α или любым другим полипептидом, относится к полипептиду, который обладает той же аминокислотной последовательностью, что и соответствующий полипептид, полученный в природе, независимо от способа его получения. Такую нативную последовательность полипептида можно выделить из природного источника или можно изготовить рекомбинантным и/или синтетическим способами или любым их сочетанием. Термин "нативная последовательность" включает, в частности, естественно встречаемые усеченную или секретируемую формы (например, последовательность внеклеточного домена), естественно встречаемые вариантные формы (например, альтернативно сплайсированные формы) и естественно встречаемые аллельные варианты полноразмерных полипептидов.
"Полимеразная цепная реакция" или "ПЦР" относится к технологическому процессу или методу, в котором амплифицируют небольшие количества отдельного участка нуклеиновой кислоты, РНК и/или ДНК, как описано в Патенте США №4683195, выданном 28 июля 1987 г. Как правило, необходима информация о последовательности на концах области, представляющей интерес, или за ее пределами, так, чтобы сконструировать олигонуклеотидные праймеры; эти праймеры должны быть идентичны или подобны последовательности противоположных цепей матрицы, которая амплифицируется. 5'-Концевые нуклеотиды двух праймеров могут совмещаться с концами амплифицируемого материала. ПЦР можно использовать для амплификации специфичных РНК-последовательностей, специфичных ДНК-последовательностей из тотальной геномной ДНК и кДНК, транскрибируемой с тотальной клеточной РНК, бактериофаговых или плазмидных последовательностей, и т.д. и т.п. См. в основном Mullis et al., Cold Spring Harbor Symp. Quant. Biol. 51:263 (1987); Erlich, ed., PCR Technology (Stockton Press, NY, 1989). Используемая здесь ПЦР рассматривается в качестве одного, но не единственного примера способа полимеразной реакции нуклеиновой кислоты для амплификации тестируемого образца нуклеиновой кислоты, включающего использование известной нуклеиновой кислоты в качестве праймера и полимеразы нуклеиновой кислоты для амплификации или образования специфического участка нуклеиновой кислоты.
"Антитела" (Ab) и "иммуноглобулины" (Ig) являются гликопротеинами, обладающими одними и теми же структурными характеристиками. В то время как антитела проявляют специфичность связывания к специфичному антигену, иммуноглобулины включают в себя и антитела, и другие антитело-подобные молекулы, у которых отсутствует антигенная специфичность. Полипептиды последнего типа, например, продуцируются в малых количествах лимфатической системой и в повышенных количествах при миеломе.
"Нативные антитела и иммуноглобулины" представляют собой, как правило, гетеротетрамерные гликопротеины около 150000 дальтон, состоят из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь присоединена к тяжелой цепи одной ковалентной дисульфидной связью, причем число дисульфидных связей между тяжелыми цепями различных иммуноглобулиновых изотипов варьирует. Каждая тяжелая и легкая цепь обладает также расположенными с равными интервалами межцепочечными дисульфидными мостиками. Каждая тяжелая цепь содержит на одном конце вариабельную область (домен) (VH), после которой следует ряд константных доменов. Каждая легкая цепь обладает вариабельной областью на одном конце (VL) и константной областью на своем другом конце; константную область легкой цепи сопоставляют с первой константной областью тяжелой цепи, а вариабельную область легкой цепи сопоставляют с вариабельной областью тяжелой цепи. Предполагают, что отдельные аминокислотные остатки образуют стык (область контакта) между вариабельными областями легкой и тяжелой цепи (Chothia et al., J. Mol. Biol. 186:651 [1985]; Novotny and Haber, Proc. Natl. Acad. Sci. U.S.A. 82:4592 [1985]; Chothia et al., Nature 342: 877-883 [1989]).
Термин "вариабельный" относится к тому обстоятельству, что определенные части вариабельных доменов существенно отличаются у антител и используются для связывания и специфичности каждого отдельного антитела в отношении своего конкретного антигена. Однако вариабельность не распределена равномерно на всем протяжении вариабельных доменов антител. Она концентрируется в трех сегментах, именуемых определяющими комплементарность областями (CDR) или гипервариабельными областями в вариабельных доменах легкой и тяжелой цепи. Более высококонсервативные участки вариабельных доменов называют каркасными (FR). Вариабельные домены каждой нативной тяжелой и легкой цепи включают четыре FR-области, в большой степени принимающих β-складчатую конфигурацию, связанных с помощью трех CDR, которые образуют соединительные петли, и в некоторых случаях образующих отчасти структуру β-складки. В каждой цепи CDR удерживаются вместе в непосредственной близости от FR-областей и CDR другой цепи, способствуя образованию антиген-связывающего сайта антител (см. Kabat et al. (1991) выше). Константные домены прямо не участвуют в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антитело-зависимой клеточной токсичности.
Расщепление антител папаином приводит к образованию двух идентичных антиген-связывающих фрагментов, названных "Fab"-фрагментами, каждый с единственным антиген-связывающим сайтом, и остаточным "Fc"-фрагментом, наименование которого отражает его способность легко кристаллизоваться. Обработка пепсином дает F(ab')2-фрагмент, который обладает двумя антиген-связывающими сайтами и все еще способен перекрестно взаимодействовать с антигеном.
"Fv" является наименьшим фрагментом антитела, который содержит антиген-распознающий и -связывающий сайт. В двухцепочечных Fv-видах данная область состоит из димера одной тяжелой и одной легкой цепи вариабельного домена, находящихся в тесной нековалентной ассоциации. В одноцепочечных Fv-видах тяжелая и легкая цепь вариабельного домена могут соединиться ковалентно с помощью гибкого пептидного линкера так, чтобы легкая и тяжелая цепи смогли ассоциировать в "димерную" структуру, аналогичную структуре двухцепочечных Fv-видов. Отметим, что в данной конфигурации три CDR каждого вариабельного домена взаимодействуют с определенным антиген-связывающим сайтом на поверхности VH-VL-димера. Совместно эти шесть CDR придают данному антителу антиген-связывающую специфичность. Однако даже одиночная вариабельная область (или половина Fv, включающая лишь три CDR, специфичных для антигена) обладает способностью распознавать и связывать антиген несмотря на пониженное сродство в сравнении с полным связывающим сайтом.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов из-за добавления нескольких остатков по карбоксиконцу тяжелой цепи CH1-домена, в том числе одного или более цистеинов шарнирной области антитела. Fab'-SH обозначает здесь Fab', в котором цистеиновый остаток(и) константных доменов несут свободную тиольную группу. F(ab')2-фрагменты антитела исходно получают в виде пары Fab'-фрагментов, которые содержат между собой шарнирные цистеины. Известны также и другие химические соединения фрагментов антител.
Термин "легкие цепи" антител (иммуноглобулинов) для любых видов позвоночных можно отнести на основании аминокислотных последовательностей их константных доменов к одному из двух четко отличающихся типов, названных κ и λ.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей иммуноглобулины можно отнести к разным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно еще разделить на подклассы (изотипы), напр., IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют разным классам иммуноглобулинов, называют соответственно α, β, ε, γ и μ. Структура субъединиц и пространственная конфигурация разных классов иммуноглобулинов хорошо известны.
Термин "антитело" включает все классы и подклассы интактных иммуноглобулинов. Термин "антитело" включает также фрагменты антител. Термин "антитело" в частности включает моноклональные антитела, в том числе клоны фрагментов антител.
"Фрагменты антител" включают часть интактного антитела, которая содержит антиген-связывающую или вариабельную область интактного антитела. Примеры фрагментов антител включают фрагменты Fab, Fab', F(ab')2 и Fv; тельца включения; одноцепочечные молекулы антител, в том числе одноцепочечные молекулы Fv (scFv); и мультиспецифичные антитела, образуемые из фрагментов антител.
Используемый здесь термин "моноклональное антитело" относится к антителу (или к фрагменту антитела), полученному из популяции практически гомогенных антител, т.е. составляющих популяцию идентичных индивидуальных антител, если не учитывать возможные естественно встречаемые мутации, которые могут быть представлены в небольших количествах. Моноклональные антитела являются высокоспецифичными, направленными против единственного антигенного сайта. Кроме того, в противоположность препаратам традиционных (поликлональных) антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против единственной детерминанты в антигене. Помимо специфичности моноклональные антитела обладают тем преимуществом, что они синтезируются с помощью гибридомной культуры и не загрязнены примесью других иммуноглобулинов. Определение "моноклональный" указывает на признак данного антитела, которое получают из практически гомогенной популяции антител, и не истолковывается в качестве требования получения антитела с помощью какого-либо конкретного способа. Например, моноклональные антитела, используемые в соответствии с настоящим изобретением, можно создать с помощью гибридомного способа, впервые описанного Kohler et al., Nature, 256:495 (1975), или с помощью методов рекомбинантной ДНК (см., например, Патент США №4816567). Термин "моноклональные антитела" включает также клоны антиген-распознающего и антиген-связывающего сайта, содержащего фрагменты антител (Fv-клоны), выделяемые из фаговых библиотек антител с использованием методик, описанных, например, у Clackson et al., Nature, 352:624-628 (1991) и Marks et al., J. Mol. Biol. 222:581-597 (1991).
При этом моноклональные антитела включают, в частности, "химерные" антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных от конкретных видов или относящихся к определенному классу или подклассу антител, а остальная цепь(и) идентична или гомологична соответствующим последовательностям в антителах, полученных от других видов или относящихся к другому классу или подклассу антител, а также к фрагментам таких антител, при условии, что они проявляют требуемую биологическую активность (Патент США №4816567 Cabilly et al.; Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 [1984]).
"Гуманизированные" формы антител, не относящихся к человеку (например, мышиные), представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антиген-связывающие субпоследовательности антител), которые содержат наименьшую последовательность, полученную из иммуноглобулина, не относящегося к человеку. Большей частью гуманизированные антитела представляют собой иммуноглобулины человека (реципиентные антитела), в которых остатки, частично или все гипервариабельной области (CDR) реципиента, заменяют остатками CDR видов, не относящихся к человеку (донорные антитела), таких как мышь, крыса или кролик, обладающие требуемой специфичностью, сродством и производительностью. В некоторых случаях остатки каркасной области Fv (FR) иммуноглобулина человека заменяют на соответствующие остатки, не принадлежащие человеку. Более того, гуманизированные антитела могут включать остатки, которые не обнаруживаются ни в реципиентных антителах, ни в чужеродной CDR или в каркасных последовательностях. Эти модификации осуществляются для дополнительного усовершенствования и оптимизации функционирования антител. Вообще гуманизированное антитело должно включать в сущности все или, по меньшей мере, один или, как правило, два вариабельных домена, в которых все или практически все CDR-области соответствуют таковым в иммуноглобулине, не относящемся к человеку, или все, или практически все FR-области соответствуют таковым в последовательности иммуноглобулина человека. Оптимальное гуманизированное антитело должно включать также, по меньшей мере, часть константной области (Fc) иммуноглобулина, как правило, иммуноглобулина человека. Более подробно см. Jones et al., Nature, 321:522-525 (1986); Reichmann et al., Nature, 332:323-329 (1988); Presta, Curr. Op. Struct. Biol., 2:593-596 (1992); и Clark, Immunol. Today 21:397-402 (2000). Гуманизированное антитело включает приматизированное (Primatized™) антитело, в котором антиген-связывающая область данного антитела получена из антитела, полученного путем иммунизации обезьян-макак представляющим интерес антигеном.
"Одноцепочечные Fv" или "scFv" фрагменты антител включают VH- и VL-домены антител, которые представлены одиночной полипептидной цепью. Как правило, scFv-полипептид дополнительно включает полипептидный линкер между VH- и VL-доменами, который позволяет scFv образовать требуемую структуру для связывания антигена. Обзор по scFV см. Pluckthun, в The Pharmacology of Monoclonal Antibodies, vol.113, Rosenburg and Moore eds., Springer-Verlag, New York, pp.269-315 (1994), Dall'Acqua and Carter, Curr. Opin. Struct. Biol. 8:443-450 (1998), и Hudson, Curr. Opin. Immunol. 11:548-557 (1999).
Термин "диательца" относится к небольшим фрагментам антител с двумя антиген-связывающими сайтами, которые включают вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в той же полипептидной цепи (VH-VL). С использованием линкера, который настолько короткий, что обеспечивает спаривание двух доменов в той же цепи, указанные домены вынуждены спариваться с комплементарными доменами другой цепи и создавать два антиген-связывающих сайта. Более полно "диательца" описаны, например, в ЕР 404097; WO 93/11161; и у Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993).
"Выделенное" антитело является антителом, которое было идентифицировано и отделено и/или извлечено из составляющего его естественного окружения. Загрязняющими компонентами его естественного окружения являются вещества, которые препятствуют диагностическому или терапевтическому использованию данного антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления настоящего изобретения антитело следует выделять очисткой (1) до более чем 95% массы антитела, что определяют методом Лоури, и наиболее предпочтительно до более чем 99% массы, (2) до степени, достаточной для получения, по меньшей мере, N-концевой или внутренней аминокислотной последовательности из 15 остатков путем использования высокоскоростного секвенатора, или (3) до гомогенности с помощью SDS-PAGE в восстанавливающих или в невосстанавливающих условиях с использованием окрашивания Кумасси голубым или предпочтительно серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, поскольку, по меньшей мере, ни один компонент из естественного окружения антитела не должен присутствовать. Но обычно выделенное антитело получают с помощью, по меньшей мере, одной стадии операции очистки.
Под "нейтрализующим антителом" подразумевают молекулу антитела, которая способна подавить или существенно снизить эффекторную функцию антигена-мишени, с которым оно связывается. В соответствии с этим "нейтрализующее" анти-IFN-α антитело способно подавить или существенно снизить эффекторную функцию IFN-α, например, связыванием рецептора и/или вызыванием клеточного ответа.
Для целей настоящего изобретения, например, способность анти-IFN-α антитела нейтрализовать рецепторную стимулирующую активность IFN-α в Анализе Стимуляции Рецептора Киназы (KIRA), как описано в WO 95/14930, опубликованной 1 июня 1995 г., можно контролировать путем измерения способности антитела-кандидата уменьшать фосфорилирование тирозина (возникающее при связывании лиганда) рецепторного комплекса IFNAR1/R2.
Для целей настоящего изобретения способность анти-IFN-α антител нейтрализовать проявление клеточного ответа с помощью IFN-α предпочтительно определяется путем контролирования нейтрализации противовирусной активности IFN-α, как описано у Kawade, J. Interferon Res. 1:61-70 (1980), или у Kawade and Watanabe, J. Interferon Res. 4:571-584 (1984), или у Yousefi, et al., Am. J. Clin. Pathol. 83:735-740 (1985), или путем тестирования способности анти-IFN-α антитела нейтрализовать способность IFN-α активировать связывание сигнальной молекулы, интерферон-стимулирующий фактор 3 (ISGF3), с олигонуклеотидом, полученным в интерферон-стимулированном элементе ответа (ISRE), в анализе изменения электрофоретической подвижности, как описано у Kurabayashi et al., Mol. Cell Biol., 15:6386 (1995).
"Существенное" уменьшение подразумевает, по меньшей мере, около 60% или, по меньшей мере, около 70%, предпочтительно, по меньшей мере, около 75%, более предпочтительно, по меньшей мере, около 80%, еще более предпочтительно, по меньшей мере, около 85%, еще более предпочтительно, по меньшей мере, около 90%, еще более предпочтительно, по меньшей мере, около 95%, наиболее предпочтительно, по меньшей мере, около 99% уменьшения эффекторной функции антигена-мишени (например IFN-α), например, связыванием рецептора (например IFNAR2) и/или вызыванием клеточного ответа. Предпочтительно "нейтрализующие" антитела, как определено здесь, способны нейтрализовать, по меньшей мере, около 60% или, по меньшей мере, около 70%, предпочтительно, по меньшей мере, около 75%, более предпочтительно, по меньшей мере, около 80%, еще более предпочтительно, по меньшей мере, около 85%, еще более предпочтительно, по меньшей мере, около 90%, еще более предпочтительно, по меньшей мере, около 95%, наиболее предпочтительно, по меньшей мере, около 99% противовирусной активности IFN-α, как определено с помощью противовирусного анализа у Kawade (1980), выше, или у Yousefi (1985), выше. В другом предпочтительном варианте осуществления настоящего изобретения "нейтрализующие" антитела способны снижать фосфорилирование тирозина из-за связывания IFN-α рецепторным комплексом IFNAR1/IFNAR2 на, по меньшей мере, около 60% или, по меньшей мере, около 70%, предпочтительно, по меньшей мере, около 75%, более предпочтительно, по меньшей мере, около 80%; еще более предпочтительно, по меньшей мере, около 85%, еще более предпочтительно, по меньшей мере, около 90%, еще более предпочтительно, по меньшей мере, около 95%, наиболее предпочтительно, по меньшей мере, около 99%, как определено в вышеприведенном анализе KIRA. В особенно предпочтительном варианте осуществления настоящего изобретения нейтрализующие анти-IFN-α антитела способны нейтрализовать все или практически все подтипы IFN-α и неспособны нейтрализовать IFN-∃. В данном контексте термин "практически все" подразумевает, что нейтрализующее анти-IFN-α антитело должно нейтрализовать, по меньшей мере, IFN-α1, IFN-α2, IFN-α4, IFN-α5, IFN-α8, IFN-α10 и IFN-α21.
Для целей настоящего изобретения способность анти-IFN-α антитела блокировать связывание IFN-α с рецептором определяется как свойство или способность определенной концентрации антитела уменьшить или подавить связывание IFN-α с IFNAR2 в конкурентном анализе связывания по сравнению с действием эквивалентной концентрации нерелевантного контрольного антитела в анализе связывания IFN-α с IFNAR2. Предпочтительно блокирование анти-IFN-α антитела снижает связывание IFN-α с IFNAR2, по меньшей мере, на около 50% или, по меньшей мере, около 55%, или, по меньшей мере, около 60%, или, по меньшей мере, около 65%, или, по меньшей мере, около 70%, или, по меньшей мере, около 75%, или, по меньшей мере, около 80%, или, по меньшей мере, около 85%, или, по меньшей мере, около 90%, или, по меньшей мере, около 95%, или, по меньшей мере, около 99% по сравнению с нерелевантным контрольным антителом.
Для цели настоящего изобретения способность анти-IFN-α антитела блокировать связывание IFN-α с IFNAR2 можно определить в стандартном конкурентном анализе, таком, который описан в Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). Например, метод IFN-α-связывания в ELISA, описанный ниже в Примере 2, можно модифицировать для использования конкурентного связывания анти-IFN-α антитела и растворимого IFNAR2. Такой анализ можно осуществить путем наслоения IFN-α в планшеты для микротитрования, инкубирования этих наслоений в планшетах с последовательными разведениями немеченого анти-IFN-α антитела или немеченого контрольного антитела, смешанного с выбранной концентрацией меченого IFNAR2 слитого белка ECD-Fc IgG человека, детектируя и измеряя сигнал в каждой инкубационной смеси, а затем сравнивая измерения сигнала при разных разведениях антитела.
В особенно предпочтительном варианте осуществления настоящего изобретения блокирующие анти-IFN-α антитела должны обладать способностью блокировать IFNAR2-связывание всех или практически всех подтипов IFN-α и не реагировать перекрестно с IFN-β. В этом смысле термин "практически все" означает, что блокирующее анти-IFN-α антитело должно блокировать IFNAR2-связывание, по меньшей мере, IFN-α1, IFN-α2, IFN-α4, IFN-α5, IFN-α8, IFN-α10 и IFN-α21. В особенно предпочтительном варианте осуществления настоящего изобретения блокирующие анти-IFN-α антитела настоящего изобретения будут блокировать IFNAR2-связывание всех известных подтипов IFN-α.
Используемый термин "эпитоп" относится к связывающим сайтам (моноклональных или поликлональных) антител в белковых антигенах.
Антитела, которые связываются с конкретным эпитопом, можно идентифицировать путем "эпитопного картирования". Существует много способов, известных в данной области техники, для картирования и характеристики расположения эпитопов в белках, включая растворение кристаллической структуры комплекса антитело-антиген, конкурентные методы, методы экспрессии генного фрагмента, и методы, основанные на пептидном синтезе, как описано, например, в Главе 11 у Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999. Конкурентные методы обсуждаются выше и ниже. В соответствии с анализом экспрессии генного фрагмента открытая рамка считывания, кодирующая белок, фрагментируется либо случайным образом, либо с помощью специфичных генетических конструкций, и определяется реактивность экспрессируемых фрагментов белка с тестируемым антителом. Генные фрагменты можно, к примеру, получить с помощью ПЦР, а затем транскрибировать и транслировать в белок in vitro в присутствии радиоактивных аминокислот. Связывание антитела с радиоактивно меченными белковыми фрагментами определяют затем с помощью иммунопреципитации и гель-электрофореза. Некоторые эпитопы можно также идентифицировать путем использования больших библиотек случайных пептидных последовательностей, обнаруживаемых на поверхности фаговых частиц (фаговые библиотеки). Альтернативно определенную библиотеку перекрывающихся пептидных фрагментов можно тестировать на связывание с испытуемым антителом в простых анализах связывания. Последний подход пригоден для определения линейных эпитопов примерно из 5-15 аминокислот.
Антитело связывает "практически тот же эпитоп" в качестве эталонного антитела, если два антитела распознают идентичные или стерически перекрывающиеся эпитопы. Наиболее широко используемые и быстрые способы для определения того, являются ли два эпитопа идентично связанными или стерически перекрывающимися эпитопами, представляют собой методы конкурентного анализа, которые могут использоваться для обнаружения любого показателя в любой форме с использованием либо меченого антигена, либо меченого антитела. Как правило, антиген иммобилизуют в 96-луночный планшет, а способность немеченых антител блокировать связывание меченых антител измеряют с использованием радиоактивных или ферментных меток.
Используемый здесь термин аминокислота или аминокислотный остаток относится к природным L-аминокислотам или к D-аминокислотам, которые описаны далее ниже по вариантам. Для аминокислот здесь используются обычно применяемые одно- и трехбуквенные сокращения (Bruce Alberts et al., Molecular Biology of the Cell, Garland Publishing, Inc., New York (3d ed. 1994)).
"Процент (%) идентичности аминокислотной последовательности" в отношении вышеуказанных полипептидных последовательностей определяют в виде процента аминокислотных остатков в кандидатной последовательности, которые идентичны с аминокислотными остатками с последовательностью, после сопоставления последовательностей, и при необходимости введены пробелы для достижения максимального процента идентичности последовательностей, и без учета консервативных замен как части элемента идентичности последовательности. Сопоставление для определения процента идентичности последовательности аминокислот может быть достигнуто разными способами, которые находятся в пределах компетенции специалистов в данной области, например с использованием компьютерного программного обеспечения, находящегося в открытом доступе, такого как BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области техники могут установить соответствующие параметры для осуществления сопоставления, в том числе любые алгоритмы, необходимые для достижения максимального сопоставления по всем полноразмерным последовательностям, которые сравниваются. Однако для используемых здесь целей % идентичности аминокислотной последовательности получают, как описано ниже, путем использования компьютерной программы ALIGN-2 для сравнения последовательностей. Авторство компьютерной программы ALIGN-2 для сравнения последовательностей принадлежит Genentech, Inc., и ее исходный код был оформлен в виде инструкции для пользователя в U.S. Copyright Office, Washington D.C., 20559, где ее зарегистрировали под защищенным авторским правом США инвентарным №TXU510087. Программа ALIGN-2 находится в открытом доступе от Genentech, Inc., South San Francisco, California, а исходный код для программы ALIGN-2 и инструкции для ее использования раскрыты в изданной Международной Заявке №WO2000/39297, опубликованной 6 июля 2000 г. Программу ALIGN-2 следует компилировать для использования в операционной системе UNIX, предпочтительно цифровой UNIX V4.0D. Все параметры сравниваемых последовательностей устанавливаются с помощью программы ALIGN-2 и не меняются.
Для целей настоящего изобретения % идентичности аминокислотной последовательности данной аминокислотной последовательности А к, с или по сравнению с данной аминокислотной последовательностью В (которая может быть составлена в обратном направлении в виде данной аминокислотной последовательности А, которая обладает или включает определенный % идентичности аминокислотной последовательности к, с или по сравнению с данной аминокислотной последовательностью В) вычисляют следующим образом:
100 умножить на дробь X/Y
где Х равно числу аминокислотных остатков, подсчитанных в виде идентичных совпадений с помощью программы сопоставления последовательностей ALIGN-2, в которой сопоставляют А и В, и где Y равен общей численности аминокислотных остатков в последовательности В. Следует иметь в виду, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то % идентичности аминокислотной последовательности А с В не равен % идентичности аминокислотной последовательности В с А. Если не оговорено особо, все значения % идентичности аминокислотной последовательности, используемые здесь, получают, как описано выше, с использованием компьютерной программы ALIGN-2 для сравнения последовательностей. Однако % идентичности аминокислотной последовательности можно также определить с использованием программы NCBI-BLAST2 для сравнения последовательностей (Altschul et al., Nucleic Acids Res. 25:3389-3402 (1997)). Программу NCBI-BLAST2 для сравнения последовательностей можно загрузить с http://www.ncbi.nlm.nih.gov. NCBI-BLAST2 использует несколько поисковых параметров, в которой все эти поисковые параметры устанавливаются по умолчанию, в том числе, например, обнаружение = да, цепь = все, ожидаемые вхождения = 10, минимальная низкая сложность длины = 15/5, e-величина неоднократного прохода = 0,01, константа для неоднократного прохода = 25, выход для конечного сопоставления пробелов = 25 и матрица оценок = BLOSUM62.
В ситуациях, в которых для сравнения аминокислотных последовательностей используют NCBI-BLAST2, % идентичности аминокислотной последовательности данной аминокислотной последовательности А к, с или по сравнению с данной аминокислотной последовательности В (что можно иначе сформулировать как данная аминокислотная последовательность А, которая обладает или включает определенный % идентичности аминокислотной последовательности к, с или по сравнению с данной аминокислотной последовательностью В) вычисляют следующим образом:
100 умножить на дробь X/Y
где Х равно числу аминокислотных остатков, подсчитанных в виде идентичных совпадений с помощью программы сопоставления последовательностей NCBI-BLAST2, в которой сопоставляют А и В, и где Y равно общей численности аминокислотных остатков в В. Следует иметь в виду, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то % идентичности аминокислотной последовательности А с В не равен % идентичности аминокислотной последовательности В с А.
Нуклеиновая кислота "функционально соединена", если она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК последовательности-предшественника или секреторной лидерной функционально соединяют с ДНК полипептида, когда он экспрессируется в виде пребелка, который участвует в секреции полипептида; промотор или энхансер функционально соединен с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или рибосом-связывающий сайт функционально соединен с кодирующей последовательностью, если он расположен таким образом, чтобы способствовать трансляции. Вообще "функционально соединен" означает, что присоединяемые ДНК-последовательности представляют собой контиги, а в случае секреторного лидера представляют собой контиги и находятся в фазе считывания. Однако энхансеры не могут представлять собой контиги. Присоединение осуществляют путем лигирования по подходящим сайтам рестрикции. Если таких сайтов не существует, то в соответствии с общепринятой практикой используют синтетические олигонуклеотидные адаптеры или линкеры.
Термин "болезненное состояние" относится к физиологическому состоянию клетки или целому организму млекопитающего, в котором происходит сбой, прекращаются или нарушаются функции клеток или целого организма, систем или органов.
Термин "эффективное количество" относится к количеству лекарственного средства, эффективному для лечения (включая профилактику) заболевания, нарушения или нежелательных физиологических состояний у млекопитающего. В настоящем изобретении "эффективное количество" анти-IFN-α антитела может уменьшать, замедлять или приостанавливать аутоиммунное нарушение, такое как IDDM или SLE; снижать, предотвращать или подавлять (т.е. до некоторой степени замедлять или предпочтительно останавливать) развитие аутоиммунного нарушения, такого как IDDM или SLE; и/или до определенного уровня ослаблять один или более симптомов, ассоциированных с аутоиммунными нарушениями, такими как IDDM или SLE.
В способах настоящего изобретения термин "контроль" и его грамматические варианты используют для обозначения предупреждения, частичного или полного ингибирования, уменьшения, приостановки или замедления нежелательного явления, например физиологического состояния, такого как образования аутореактивных Т-клеток и развитие аутоиммунности.
"Лечение" относится к терапевтическому лечению, а также к профилактике или предупредительным мерам. Нуждающиеся в лечении включают индивидов уже с определенным нарушением, а также индивидов со склонностью к нарушению или тех, кому требуется профилактика нарушения. Для целей настоящего изобретения благоприятные или требуемые клинические результаты включают, но не ограничиваются, облегчение симптомов, ослабление заболевания, стабилизацию (т.е. неухудшение) состояния заболевания, приостановку или замедление развития заболевания, улучшение или временное облегчение болезненного состояния и ремиссию (частичную или общую), явную или скрытую. "Лечение" может также подразумевать повышенную выживаемость в сравнении с ожидаемой выживаемостью в отсутствие лечения. Нуждающиеся в лечении включают также индивидов уже с болезненным состоянием или нарушением, а также со склонностью к состоянию или нарушению, или кому требуется профилактика состояния или нарушения.
"Фармацевтически приемлемые" носители, наполнители или стабилизаторы - это те, которые нетоксичны для клетки или млекопитающего, которые подвергаются их воздействию в используемых дозах и концентрациях. Зачастую физиологически приемлемый носитель представляет забуференный водный раствор. Примеры физиологически приемлемых носителей включают буферы, такие как фосфатный, цитратный, и других органических кислот; антиоксиданты, в том числе аскорбиновую кислоту; низкомолекулярные (менее чем около 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон, аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу, или декстрины; хелатообразующие агенты, такие как ЭДТА; сахарные спирты, такие как манит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как Tween™, полиэтиленгликоль (PEG) и Pluronics™.
"Млекопитающее" для целей лечения относится к любому животному, классифицируемому в качестве млекопитающего, включая человека, домашних или сельскохозяйственных животных, а также предназначенных для зоопарков и спорта, или домашних животных, таких как собаки, лошади, кошки, коровы и т.п. Предпочтительным млекопитающим является человек.
В. Способы осуществления настоящего изобретения
1. Получение антител
(i) Поликлональные антитела
Способы получения поликлональных антител известны в области техники. Поликлональные антитела можно индуцировать у млекопитающего, например, одной или более инъекциями иммунизирующего агента и, если необходимо, адъюванта. Как правило, иммунизирующий агент и/или адъювант следует инъецировать млекопитающему путем неоднократных подкожных или внутрибрюшинных инъекций. Это может оказаться пригодным для получения конъюгата данного иммунизирующего агента с белком, известным в качестве иммуногенного для иммунизируемого млекопитающего, таким как сывороточный альбумин или ингибитор трипсина в виде соевого масла. Примеры адъювантов, которые можно использовать, включают полный адъювант Фрейнда и MPL-TDM.
В другом предпочтительном варианте осуществления настоящего изобретения животных иммунизируют смесью разных, предпочтительно всех, подтипов IFN-α с целью получения анти-IFN-α антител с широкой реактивностью против подтипов IFN-α. В другом предпочтительном варианте осуществления настоящего изобретения животных иммунизируют смесью подтипов IFN-α человека, которые представлены у человека лимфобластоидными интерферонами, секретируемые клетками лимфомы Беркитта (Namalva-клетки), индуцируемые вирусом Сендай, как описано ниже в Примере 1. Соответствующий препарат таких лимфобластоидных интерферонов человека можно купить (Product No. I-9887) у Sigma Chemical Company, St. Louis, MO.
(ii) Моноклональные антитела
Моноклональные антитела можно создать с использованием гибридомного способа, впервые описанного Kohler et al., Nature 256:495 (1975), или можно получить методами рекомбинантной ДНК (Патент США №4816567).
В гибридомном способе мышь или иное подходящее животное-хозяин, такое как хомячок или обезьяна макака, иммунизируют, как описано выше, для получения лимфоцитов, которые продуцируют или способны продуцировать антитела, которые должны специфически связываться с белком, используемым для иммунизации. Альтернативно лимфоциты могут быть иммунизированы in vitro. Затем лимфоциты сливают с миеломными клетками с использованием подходящего агента для слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103, [Academic Press, 1996]).
Полученные таким образом гибридомные клетки высевают и выращивают в соответствующей культуральной среде, которая предпочтительно содержит одно или более веществ, ингибирующих рост или выживаемость неслитых родительских миеломных клеток. Например, если у родительских миеломных клеток отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), то культуральная среда для данных гибридом должна, как правило, включать гипоксантин, аминоптерин и тимидин (HAT-среда), вещества, которые предотвращают рост клеток, дефицитных по HGPRT.
Предпочтительными миеломными клетками являются те клетки, которые эффективно сливают, обеспечивая стабильное на высоком уровне образование антител с помощью селективных антителопродуцирующих клеток, чувствительных к такой среде, как HAT-среда. Из них предпочтительными миеломными клеточными линиями являются мышиные миеломные линии, такие, которые создаются из мышиных опухолей MOP-21 и МС-11, доступных от Salk Institute Cell Distribution Center, San Diego, California USA, и клеток SP-2 или X63-Ag8-653, доступных от American Type Culture Collection, Rockville, Maryland USA. Человеческая миеломная и гетеромиеломная, мышь-человек, клеточные линии также описаны в качестве продуцентов моноклональных антител человека (Kozbor, J. Immunol. 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63, Marcel Dekker, Inc. New York, [1987]).
Культуральную среду, в которой выращивают гибридомные клетки, оценивают по производительности моноклональных антител против антигена. Предпочтительно специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, определяют с помощью иммунопреципитации или с помощью in vitro связывающего анализа, такого как радиоиммуноанализ (RIA) или иммуноферментный анализ (ELISA).
Сродство связывания моноклонального антитела можно, например, определить с помощью анализа по Скэтгарду, описанному Munson et al., Anal. Biochem 107:220 (1980).
Затем гибридомные клетки, которые продуцируют антитела требуемой специфичности, сродства и/или активности, идентифицируют, после чего эти клетки можно субклонировать методами лимитирующего разведения и выращивать стандартными способами (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103, Academic Press, 1996). Для этой цели подходящая культуральная среда включает, например, среду DMEM или RPMI-1640. Кроме того, такие гибридомные клетки могут расти in vivo в виде асцитных опухолей у животных.
Моноклональные антитела, секретируемые субклонами, соответствующим образом отделяют от культуральной среды, асцитной жидкости или сыворотки традиционными методами очистки иммуноглобулинов, такими, например, как хроматография на белок А-Сефарозе, на гидроксилапатите, гелевый электрофорез, диализ или хроматография по сродству.
ДНК, кодирующую моноклональные антитела, легко выделить и секвенировать с использованием традиционных методов (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи моноклональных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно поместить в экспрессионные векторы, которые затем переносят в клетки-хозяева, такие как клетки E. coli, COS-клетки обезьяны, клетки яичника китайского хомячка (CHO) или миеломные клетки, которые в иных обстоятельствах не продуцируют иммуноглобулиновый белок, для осуществления синтеза моноклональных антител в рекомбинантных клетках-хозяевах. ДНК можно также модифицировать, например, заменяя последовательности, кодирующие константные домены тяжелой и легкой цепи человека, на гомологичные мышиные последовательности (Morrison, et al., Proc. Nat. Acad. Sci 81:6851 [1984], или путем ковалентного присоединения к последовательности, кодирующей иммуноглобулин, всей или части кодирующей последовательности неиммунноглобулинового полипептида. Этим способом получают "химерные" или "гибридные" антитела, которые обладают в настоящем изобретении специфичностью связывания анти-IFN-α моноклонального антитела.
Обычно такие неиммуноглобулиновые полипептиды заменяют на константные домены антител настоящего изобретения или же их заменяют на вариабельные домены одного из антител настоящего изобретения с антиген-связывающим сайтом для создания химерного бивалентного антитела, включающего один антиген-связывающий сайт, обладающий специфичностью к IFN-α, и другой антиген-связывающий сайт, обладающий специфичностью к другому антигену.
Химерные или гибридные антитела можно также получить in vitro с использованием известных способов в химии белкового синтеза, в том числе c использованием перекрестносшивающих агентов. Например, иммунотоксины можно сконструировать с использованием реакции дисульфидного обмена или путем образования тиоэфирной связи. Примеры соответствующих реагентов для этой цели включают иминотиоэфир и метил-4-меркаптобутиримидат.
Образование рекомбинантных антител более подробно будет описано ниже.
(iii) Гуманизированные антитела
Как правило, гуманизированное антитело обладает одним или более аминокислотными остатками, введенными в него из не относящегося к человеку источника. Эти не относящиеся к человеку аминокислотные остатки часто именуются в качестве "импортируемых" остатков, которые обычно извлекают из "импортируемого" вариабельного домена. Гуманизация может, в сущности, производиться по способу Winter и соавторов путем замещения CDR грызунов или CDR-последовательностей, на соответствующие последовательности антитела человека (Jones et al., Nature 321:522-525 [1986]; Riechmann et al., Nature 332:323-327 [1988]; Verhoeyen et al., Science 239:1534-1536 [1988]; обзор у Clark, Immunol. Today 21:397-402 [2000]).
Следовательно, такие "гуманизированные" антитела являются химерными антителами (Cabilly, выше), которые существенно меньше, чем интактный вариабельный домен человека, замещенный соответствующей последовательностью из видов, не принадлежащих человеческому роду. На практике гуманизированные антитела обычно являются антителами человека, в которых некоторые CDR-остатки, а возможно и некоторые FR-остатки замещены остатками из аналогичных участков антител грызунов.
Важно, чтобы антитела, подвергнутые гуманизации, сохраняли высокое сродство к антигену, а также другие подходящие биологические свойства. Для достижения данной цели в соответствии с предпочтительным способом гуманизированные антитела получают в ходе анализа родительских последовательностей и различных концептуальных гуманизированных продуктов с использованием пространственных моделей родительских и гуманизированных последовательностей. Пространственные иммуноглобулиновые модели в большинстве случаев доступны и хорошо знакомы специалистам в данной области. Доступны и компьютерные программы, которые иллюстрируют и демонстрируют вероятностные трехмерные конформационные структуры выбранных кандидатных иммуноглобулиновых последовательностей. Обследование этих изображений позволяет проанализировать возможную роль остатков для функционирования кандидатной иммуноглобулиновой последовательности, т.е. проанализировать остатки, которые влияют на способность кандидатного иммуноглобулина связывать его антиген. Таким способом можно выбрать FR-остатки и скомбинировать их с консенсусной и "импортируемой" последовательностью так, чтобы получить антитело с требуемой характеристикой, такой как повышенное сродство к антигену(ам)-мишени. Вообще CDR-остатки непосредственно и больше всего оказывают влияние на связывание антигена. Более подробно см. патент США №5821337.
(iv) Антитела человека
Попытки использовать такую же технологию для создания человеческих mAb натолкнулись на отсутствие подходящей человеческой миеломной клеточной линии. Наилучшие результаты были получены с использованием гетеромиелом (гибридные миеломы мышь х человек) в качестве партнеров слияния (Kozbor, J. Immunol. 133:3001 (1984); Brodeur, et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63, Marcel Dekker, Inc. New York, [1987]). Альтернативно человеческие антителосекретирующие клетки можно иммортализовать инфицированием вирусом Эпштейна-Барра (EBV). Однако EBV-инфицированные клетки трудно клонировать, и в большинстве случаев они дают относительно низкий выход иммуноглобулина (James and Bell, J. Immunol. Methods 100:5-40 [1987]). В будущем иммортализацию В-клеток человека можно, вероятно, осуществить путем введения определенного сочетания трансформирующих генов. Такая возможность выдвигается на первый план благодаря недавней демонстрации того, что экспрессия каталитической субъединицы теломеразы вместе с большим Т-онкопротеином SV40 и онкогенным аллелем H-ras приводит к канцерогенному превращению нормальных эпителиальных клеток и фибробластов человека (Hahn et al., Nature 400:464-468 [1999]).
В настоящее время можно создавать трансгенных животных (например мышей), способных после иммунизации продуцировать набор человеческих антител при отсутствии эндогенного продуцирования иммуноглобулинов (Jakobovits et al., Nature 362:255-258 [1993]; Lonberg and Huszar, Int. Rev. Immunol. 13:65-93 [1995]; Fishwild et al., Nat. Biotechnol. 14:845-851 [1996]; Mendez et al., Nat. Genet. 15:146-156 [1997]; Green, J. Immunol. Methods 231:11-23 [1999]; Tomizuka et al., Proc. Natl. Acad. Sci USA 97:722-727 [2000]; обзор у Little et al., Immunol. Today 21:364-370 [2000]). Например, описано, что гомозиготное делетирование из гена соединительной области (JH) тяжелой цепи антитела в химерной и эмбриональной линии мутантных гомозиготных мышей приводит к полному ингибированию продуцирования эндогенного антитела (Jakobovits et al., Proc. Natl. Acad. Sci. USA 90:2551-2555 [1993]). Перенос иммуноглобулинового генного набора из человеческих клеток зародышевой линии в клетки зародышевой линии мутантных мышей ведет к образованию в ней человеческих антител после стимуляции антигеном (Jakobovits et al., Nature 362:255-258 [1993]).
Mendez et al. (Nature Genetics 15:146-156 [1997]) создали линию трансгенных мышей, названную "XenoMouse® II", которые при стимуляции антигеном образуют полностью человеческие антитела с высоким сродством. Это осуществляется путем интеграции локусов с мега парами оснований зародышевой линии тяжелой цепи и легкой цепи человека мышам с делецией в эндогенный JH-сегмент, как описано выше. Клетки линии XenoMouse® II несут 1020 т.п.н. локус тяжелой цепи человека, содержащий приблизительно 66 генов VH, полные области DH и JH и три разные константные области (μ, δ и γ), а также несут 800 т.п.н. человеческий κ-локус, содержащий 32 Vκ-гена, Jκ-сегменты и Сκ-гены. Антитела, продуцируемые этими мышами, во всех отношениях очень схожи с антителами человека, включая генную реаранжировку, упорядоченную структуру и набор. Предпочтительно экспрессируемые человеческие антитела превышают экспрессию эндогенных антител вследствие делеции в эндогенном JH-сегменте, который препятствует генной перестройке в мышином локусе.
Tomizuka et al. (Proc. Natl. Acad. Sci USA 97:722-727 [2000] недавно описали образование двойных транс-хромосомных (Tc) мышей после встраивания двух отдельных фрагментов хромосомы человека (hCF), один из которых содержит полноразмерный локус Ig-тяжелой цепи (IgH, ˜1,5 Mb), а другой - полноразмерный κ-локус легкой цепи (Igκ, ˜2 Mb), в мышиную линию, эндогенные локусы которой IgH и Igκ были инактивированы. У этих мышей закрепляется антиген-специфичный ответ человеческих антител при отсутствии мышиных антител. Tc-технология делает возможной гуманизацию комплексных локусов или генных кластеров с размером, превышающим мега пар оснований (таких, которые кодируют Т-клеточные рецепторы, главный комплекс гистосовместимости, кластер Р450 и т.п.) мышей или иных животных. Другое преимущество данного способа заключается в исключении необходимости клонирования больших локусов. Это является большим преимуществом, так как клонирование ДНК-фрагментов с размером, превышающим мега пар оснований, включающих целые Ig-локусы, остается трудным даже при использовании дрожжевых искусственных хромосом (Peterson et al., Trends Genet. 13:61-66 [1997]; Jacobovits, Curr. Biol. 4:761-763 [1994]). Кроме того, известно, что константная область человеческого IgH-локуса содержит последовательности, которые трудны для клонирования (Kang and Cox, Genomics 35:189-195 [1996]).
Альтернативно для получения in vitro человеческих антител и антительных фрагментов из генных наборов вариабельного (V) домена иммуноглобулина от иммунизированных доноров можно использовать технику фагового дисплея (McCafferty et al., Nature 348:552-553 [1990]; reviewed in Kiprianov and Little, Mol. Biotechnol. 12:173-201 [1999]; Hoogenboom and Chames, Immunol. Today 21:371-378 [2000]). В соответствии с данным методом гены V-домена антитела клонируют в рамку считывания либо основного, либо минорного белкового гена оболочки нитевидного бактериофага, такого как М13 или fd, и обнаруживают в виде функциональных антительных фрагментов на поверхности фаговой частицы. Поскольку нитевидная частица содержит одноцепочечную ДНК-копию фагового генома, селекция по функциональным свойствам антитела также ведет к отбору гена, кодирующего антитело, проявляющего те же свойства. Таким образом, фаг имитирует некоторые свойства В-клетки. Фаговый дисплей можно осуществить в разнообразном виде (reviewed in Johnson and Chiswell, Current Opinion in Structural Biology 3:564-571 [1993]; Winter et al., Annu. Rev. Immunol. 12:433-455 [1994]; Dall'Acqua and Carter, Curr. Opin. Struct. Biol. 8:443-450 [1998]; Hoogenboom and Chames, Immunol. Today 21:371-378 [2000]). Для фагового дисплея могут быть использованы несколько источников V-генных сегментов. Clackson et al., (Nature 352:624-628 [1991]) выделили из небольшой случайной комбинаторной библиотеки V-генов, полученной из селезенок иммунизированных мышей, ряд разнообразных антител против оксазолона. Можно сконструировать набор V-генов от иммунизированных человеческих доноров и можно по существу выделить антитела к разнообразным антигенам (в том числе и к собственному антигену) по методам, описанным Marks et al., J. Mol. Biol. 222:581-597 (1991), или Griffiths et al., EMBO J. 12:725-734 (1993). При естественном иммунном ответе гены антител накапливают мутации с высокой скоростью (соматическое гипермутирование). Некоторые из включенных изменений будут придавать высокое сродство, и В-клетки, проявляющие высокое сродство поверхности иммуноглобулина, предпочтительно реплицируются и дифференцируются во время последующей стимуляции антигеном. Этот естественный процесс можно имитировать при использовании метода, известного как "перетасовка цепей" (Marks et al., Bio/Technol. 10:779-783 [1992]). В данном способе сродство "первичных" человеческих антител, получаемых с помощью фагового дисплея, можно повысить путем последовательного замещения тяжелой и легкой цепи генов V-области наборами естественно встречаемых вариантов (наборов) генов V-области, получаемых от иммунизированных доноров. Данная методика позволят продуцировать антитела и фрагменты антител со сродством в нМ диапазоне. Методология создания очень больших фаговых наборов антител (известных также как "the mother-of-all libraries") была описана Waterhouse et al., Nucl. Acids Res. 21:2265-2266 (1993), а выделение человеческого антитела с высоким сродством непосредственно из такой большой фаговой библиотеки описано Griffiths et al., EMBO J. 13:3245-3260 (1994). Перетасовка генов может быть также использована для получения человеческих антител из антител грызунов, поскольку человеческие антитела обладают аналогичным сродством и специфичностью с исходным антителом грызуна. В соответствии с данным способом, который также называется "эпитопный импринтинг", тяжелую или легкую цепь гена V-домена антител грызунов, полученную методом фагового дисплея, замещают набором человеческих генов V-домена, создавая химеры грызун-человек. Отбор по антигену влияет на выделение человеческого вариабельного домена, способного сохранять функциональный антиген-связывающий сайт, т.е. эпитоп, определяет (импринтирует) выбор партнера. Если данный процесс повторяют с целью заменить остающийся V-домен грызуна, то получают человеческое антитело (см. патентную РСТ-заявку WO 93/06213, опубликованную 1 апреля 1993 г.). В отличие от традиционной гуманизации антител грызунов с помощью CDR-пересадки данный метод позволяет получить полноразмерные человеческие антитела, которые не имеют остатков каркасной области или CDR, происходящих от грызунов.
(v) Биспецифичные антитела
Биспецифичными антителами являются моноклональные антитела, предпочтительно человеческие или гуманизированные, которые обладают специфичностью связывания, по меньшей мере, двух разных антигенов. В данном случае одна из специфичностей связывания относится к IFN-α для создания нейтрализующего антитела, а вторая относится к любому другому антигену.
Способ создания биспецифичных антител известен в данной области техники. Обычно рекомбинантное получение биспецифичных антител основывается на коэкспрессии двух пар тяжелой и двух пар легкой цепи иммуноглобулина, причем две тяжелые цепи обладают разной специфичностью (Millstein and Cuello, Nature 305:537-539 [1983]). Поскольку выбор тяжелой и легкой цепи иммуноглобулина случаен, то эти гибридомы (квадромы) образуют потенциальную смесь из 10 различных антительных молекул, из которых лишь одна обладает правильной биспецифичной структурой. Очистка правильной молекулы, которую обычно осуществляют с помощью стадий хроматографии по сродству, является довольно обременительной, а выход продукта низкий. Аналогичные методы раскрыты в РСТ-заявке, опубликованной под №WO 93/08829 (опубликованной 13 мая 1993 г.) и у Traunecker et al., EMBO J. 10:3655-3659 (1991).
В соответствии с другим и более предпочтительным подходом вариабельные домены антител с требуемой специфичностью связывания (участок соединения антитела с антигеном) сливают с последовательностями константных доменов иммуноглобулина. Слияние предпочтительно осуществляют с тяжелой цепью константного домена иммуноглобулина, включающей, по меньшей мере, часть шарнирной, СН2 и СН3 областей. Целесообразно, чтобы константная область первой тяжелой цепи (СН1), содержащая участок, необходимый для связывания легкой цепи, была представлена хотя бы в одном из слияний. ДНК, кодирующие слияние тяжелых цепей иммуноглобулина и при необходимости легкой цепи иммуноглобулина, встраивают в отдельные экспрессионные векторы и котрансфицируют в соответствующий организм-хозяин. Это обеспечивает большую гибкость для взаимной подгонки этих интегрируемых трех полипептидных фрагментов в изобретении при неравном соотношении трех полипептидных цепей, используемых в конструкции для создания оптимального выхода продукта. Тем не менее представляется возможным встроить кодирующие последовательности двух или всех трех полипептидных цепей в один экспрессионный вектор, в случае когда экспрессия, по меньшей мере, двух полипептидных цепей дает продукцию с большим выходом в равном соотношении, или в случае, когда данные соотношения не являются существенно значимыми. В предпочтительном варианте осуществления настоящего подхода биспецифичные антитела скомбинированы из гибридной тяжелой цепи иммуноглобулина с первой связывающей специфичностью на одном плече и гибридной пары тяжелая цепь - легкая цепь иммуноглобулина (создающей вторую связывающую специфичность) на другом плече. Установлено, что данная асимметричная структура облегчает отделение требуемого биспецифичного соединения от нежелательных комбинаций иммуноглобулиновой цепи, так как присутствие легкой иммуноглобулиновой цепи в одной только половине биспецифичной молекулы обеспечивает более легкий способ разделения.
Более подробно об образовании биспецифичных антител см., например, Suresh et al., Methods in Enzymology 121, 210 (1986).
(vi) Гетероконъюгатные антитела
Гетероконъюгатные антитела также находятся в рамках настоящего изобретения. Гетероконъюгатные антитела состоят из двух ковалентно соединенных антител. Такие антитела были предложены, например, для нацеливания клеток иммунной системы на нежелательные клетки (Патент США №4676980), и для лечения ВИЧ-инфекции (РСТ-заявки, опубликованные под №№WO 91/00360 и WO 92/200373; EP 03089). Гетероконъюгатные антитела можно создать с использованием любого удобного способа сшивки. Подходящие сшитые агенты, а также ряд способов сшивки хорошо известны в данной области и раскрыты в Патенте США №4676980.
(vii) Фрагменты антител
В некоторых вариантах осуществления настоящего изобретения нейтрализующее анти-IFN-α антитело (в том числе мышиные, человеческие и гуманизированные антитела и антительные варианты) представляет собой антительный фрагмент. Для получения антительных фрагментов разработаны разнообразные методы. Обычно эти фрагменты получают путем протеолитического расщепления интактных антител (см., например, Morimoto et al., J. Biochem. Biophys. Methods 24:107-117 [1992] and Brennan et al., Science 229:81 [1985]). Но в настоящее время данные фрагменты можно получить прямо с помощью рекомбинантных клеток-хозяев (reviewed Hudson, Curr. Opin, Immunol. 11:548-557 [1999]; Little et al., Immunol. Today 21:364-370 [2000]). Например, Fab'-SH-фрагменты можно непосредственно извлечь из E. coli и химически соединить с образованием F(ab')2-фрагментов (Carter et al., Bio/Technology 10:163-167 [1992]). В другом варианте осуществления настоящего изобретения F(ab')2 создают с использованием лейциновой застежки GCN4, которая ускоряет сборку F(ab')2-молекулы. В соответствии с другим подходом фрагменты Fv, Fab или F(ab')2 можно выделить непосредственно из рекомбинантной культуры клетки-хозяина. Другие методы получения фрагментов антител очевидны специалистам-практикам.
(viii) Варианты аминокислотных последовательностей антител
Рассматривается описанная здесь модификация(и) аминокислотных последовательностей анти-IFN-α антител. Например, может возникнуть необходимость улучшить для антитела сродство связывания и/или другую характеристику. Варианты аминокислотных последовательностей анти-IFN-α антител получают путем введения надлежащих нуклеотидных изменений в нуклеиновую кислоту, кодирующую цепи анти-IFN-α антитела, или с помощью пептидного синтеза. Такие модификации включают, например, делеции, и/или вставки, и/или замены остатков в аминокислотных последовательностях анти-IFN-α антитела. Для создания конечной конструкции осуществляют любое сочетание делеции, вставки и замены при условии, что конечная конструкция обладает требуемыми характеристиками. Аминокислотные замены могут также изменять посттрансляционные процессы анти-IFN-α антитела, такие как изменение числа или положения сайтов гликозилирования.
Полезный способ идентификации некоторых остатков или областей нейтрализующего анти-IFN-α антитела, который предпочтительно предназначается для мутагенеза, называется "аланин-сканирующий мутагенез", который описан у Cunningham and Wells, Science 244:1081-1085 (1989). При этом идентифицируют остаток или группу целевых остатков (например, заряженные остатки, такие как arg, asp, his, lys и glu) и замещают нейтральной или отрицательно заряженной аминокислотой (наиболее предпочтителен аланин или полиаланин), которая влияет на взаимодействие аминокислот с антигеном. Затем уточняют местоположения аминокислот, демонстрирующих функциональную чувствительность к заменам, путем введения в участки замещения дополнительных или иных вариантов. Поэтому несмотря на то что участок для введения разновидности аминокислотной последовательности установлен заранее, природа мутации как таковой не нуждается в предопределении. Например, для анализа функционирования мутации по данному сайту ala-сканирующий или случайный мутагенез осуществляют по целевому кодону или участку и экспрессируемые варианты анти-IFN-α антитела скринируют на нужную активность.
Вставки аминокислотных последовательностей включают слияния амино- и/или карбоксиконцов, имеющие в длину от одного остатка до полипептида, содержащего сто и более остатков, а также вставки внутри последовательности одиночных или множественных аминокислотных остатков. Примеры терминальных вставок включают нейтрализующее анти-IFN-α антитело с N-концевым метионильным остатком или данное антитело, слитое с эпитопным маркером. Другие варианты вставок молекулы анти-IFN-α антитела включают слияние с N- или С-концом антитела фермента или полипептида, что увеличивает время полужизни данного антитела в сыворотке.
Другой тип варианта представляет собой вариант аминокислотного замещения. Эти варианты обладают, по меньшей мере, одним удаляемым аминокислотным остатком в молекуле нейтрализующего анти-IFN-α антитела и другим остатком, встроенным на его место. Участки, представляющие наибольший интерес для мутагенеза путем замещения, включают гипервариабельные области, хотя рассматриваются также и FR-изменения. Консервативные замены показаны в Таблице 1, озаглавленной "предпочтительные замены". Если такие замены приводят к изменению биологической активности, тогда можно ввести более существенные изменения, обозначенные в Таблице 1 "типичные замены", или в качестве дополнительных, описанных ниже, что касается классов аминокислот, и полученные продукты проскринировать.
Существенные модификации биологических характеристик антитела осуществляют путем избирательных замен, которые существенно отличаются своим влиянием на поддержание (а) структуры полипептидного скелета области замен, например, складчатая или спиральная конформация, (b) заряда или гидрофобности молекулы в данном целевом сайте, или (с) структуры боковой цепи. Естественно встречаемые остатки распределяют в группы на основании обычных характеристик боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr;
(3) кислотные: asp, glu;
(4) основные: asn, gln, his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены повлекут за собой замену члена одного из этих классов на представителя другого класса.
Любой цистеиновый остаток, не участвующий в поддержании надлежащей конформации нейтрализующего анти-IFN-α антитела, может быть также заменен, обычно на серин, для улучшения окислительной стабильности его молекулы и предотвращения аберрантного сшивания. С другой стороны, к антителу можно добавить цистеиновую связь(и) для повышения его стабильности (особенно в том случае, когда антитело представляет собой антительный фрагмент, такой как Fv-фрагмент).
Особенно предпочтительный тип варианта замены включает замену одного или более остатков гипервариабельной области родительского антитела (например, гуманизированного или человеческого антитела). Как правило, получаемый вариант(ы), выбранный для дальнейшего усовершенствования, должен обладать улучшенными биологическими характеристиками относительно родительского антитела, от которого он происходит. Удобный способ для образования таких вариантов замены представляет собой созревание по сродству с использованием фагового дисплея. Вкратце, несколько участков гипервариабельной области (например, 6-7 участков) мутируют с образованием всех возможных аминокислотных замен в каждом участке. Получаемые таким образом варианты антител "выставляются" в моновалентной форме на нитевидных фаговых частицах в виде слияний с продуктом гена III каждой частицы, упакованной в М13. Затем варианты фагового дисплея скринируют на их биологическую активность (например, антагонистическую активность) так, как здесь раскрыто. Для того чтобы идентифицировать кандидатные сайты гипервариабельной области для модификации, осуществляют аланин-сканирующий мутагенез для идентификации остатков гипервариабельной области, существенно содействуя связыванию антигена. Альтернативно, или в дополнение, это может благоприятствовать анализу кристаллической структуры комплекса антиген-антитело, чтобы идентифицировать точки контактирования между антителом и IFN-α. Такие контактирующие остатки и прилегающие остатки являются кандидатами для замены в соответствии с детально разработанными здесь методами. После того как такие варианты созданы, полученную панель вариантов подвергают скринированию, как здесь описано, и антитела с наилучшими характеристиками в одном или более соответствующих анализах могут быть выбраны для последующего совершенствования.
(ix) Варианты гликозилирования
Антитела гликозилируют по консервативным положениям в их константных областях (Jefferis and Lund, Chem. Immunol. 65:111-128 [1997]; Wright and Morrison, Trends Biotechnol. 15:26-32 [1997]). Олигосахаридные боковые цепи иммуноглобулинов влияют на функцию белка (Boyd et al., Mol. Immunol. 32:1311-1318 [1996]; Wittwe and Howard, Biochem. 29:4175-4180 [1990]), а внутримолекулярное взаимодействие между частями гликопротеина может влиять на конформацию и представленную трехмерную поверхность этого гликопротеина (Jefferis and Lund, выше; Wyss and Wagner, Current Opin. Biotech. 7:409-416 [1996]). Олигосахариды могут также служить для нацеливания данного гликопротеина на определенные молекулы, основанного на специфическом распознавании структур. Например, сообщается о том, что в агалактозилированном IgG олигосахаридная составляющая "выпрыгивает" из СН2-промежутка и концевой остаток N-ацетилглюкозамина становится доступным для связывания связывающего белка маннозы (Malhorta et al., Nature Med. 1:237-243 [1995]). Удаление с помощью гликопептидазы олигосахаридов из CAMPATH-1H (рекомбинантное гуманизированное мышиное моноклональное антитело IgG1, которое распознает CDw52-антиген человеческих лимфоцитов), производимого в клетках яичника китайского хомячка (СНО), приводит к полному снижению лизиса, опосредованного комплементом (CMCL) (Boyd et al., Mol. Immunol. 32:1311-1318 [1996], в то время как избирательное удаление остатков сиаловой кислоты с использованием нейраминидазы не приводит к утрате DMCL. Сообщается также о том, что гликозилирование антител влияет на антителозависимую клеточную цитотоксичность (ADCC). В частности, сообщается о том, что регулируемая тетрациклином экспрессия СНО-клетками β(1,4)-N-ацетилглюкозаминтрансферазы III (GnTIII), катализируемое гликозилтрансферазой образование бисекторной GlcNAc, повышает активность ADCC (Umana et al., Mature Biotech. 17:176-180 [1999]).
Гликозилирование антител является обычно N- или О-сцепленным. N-сцепленное гликозилирование относится к присоединению углеводной составляющей к боковой цепи аспарагинового остатка. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где Х представляет собой любую аминокислоту за исключением пролина, являются распознающими последовательностями для ферментного присоединения углеводной составляющей к боковой цепи аспарагина. Поэтому присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. О-сцепленное гликозилирование относится к присоединению одного из сахаров N-галактозамина, галактозы или ксилозы к оксииминокислоте, чаще всего сериновой или треониновой, хотя можно использовать и 5-гидроксипролин или 5-гидроксилизин.
Варианты гликозилирования антител представляют собой варианты, в которых паттерн гликозилирования антитела изменяется. Под изменением подразумевают удаление одной или более углеводных составляющих, обнаруживаемых в антителе, добавление одной или более углеводных составляющих к данному антителу, изменение состава гликозилирования (паттерна гликозилирования), степени гликозилирования и т.п.
Добавление числа сайтов гликозилирования к антителу удобно осуществлять путем изменения аминокислотной последовательности таким образом, чтобы он содержал одну или более вышеописанных трипептидных последовательностей (для N-сцепленных сайтов гликозилирования). Изменение можно осуществить путем добавления или замещения одного или более остатков серина или треонина в последовательности исходного антитела (что касается О-сцепленных сайтов гликозилирования). Подобным образом удаление сайтов гликозилирования можно осуществить путем аминокислотного изменения в нативных сайтах гликозилирования антитела.
Обычно аминокислотную последовательность изменяют в результате изменения основной последовательности нуклеиновой кислоты. Молекулы нуклеиновой кислоты, кодирующие варианты аминокислотной последовательности анти-IFNα антитела, получают разнообразными способами, известными в данной области техники. Данные способы включают, но не ограничиваются, выделение из естественного источника (в случае естественно встречаемых вариантов аминокислотных последовательностей) или получение с помощью одигонуклеотид-опосредованного (или сайт-направленного) мутагенеза, ПЦР-мутагенеза, а также кассетного мутагенеза ранее полученного варианта или безвариантной версии анти-IFN-α антитела.
Гликозилирование (включая паттерн гликозилирования) антител можно также изменить без изменения аминокислотной последовательности или основной нуклеотидной последовательности. Гликозилирование в значительной степени зависит от клетки-хозяина, используемой для экспрессии антитела. Поскольку тип клетки, используемой для экспрессии рекомбинантных гликопротеинов, например антител, в качестве возможных терапевтических средств, редко представляет собой нативную клетку, можно ожидать значительную изменчивость в паттерне гликозилирования антител (см., например,. Hse et al., J. Biol. Chem. 272:9062-9070 [1997]). Кроме выбора клеток-хозяев факторы, которые влияют на гликозилирование в ходе получения рекомбинантных антител, включают характер роста, состав среды, плотность клеток в культуре, кислородонасыщение, рН, схемы очистки и т.п. Для изменения паттерна гликозилирования, осуществляемого в конкретном организме-хозяине, были предложены разнообразные способы, в том числе введение или сверхэкспрессирование некоторых ферментов, участвующих в образовании олигосахарида (Патенты США №№5047335; 5510261 и 5278299). Гликозилирование или некоторые виды гликозилирования, можно ферментативно исключить для данного гликопротеина, например, используя эндогликозидазу Н (Endo H). Кроме того, рекомбинантную клетку-хозяина можно сконструировать генно-инженерным способом, например, сделать дефектной для процессирования некоторых видов полисахаридов. Эти и аналогичные методы хорошо известны в данной области техники.
Гликозилированную структуру антител можно легко проанализировать с помощью удобных методов углеводного анализа, включая лектин-хроматографию, ЯМР, масс-спектрометрию, HPLC, GPC, анализ состава моносахаридов, последовательное ферментативное расщепление и HPAEC-PAD, в котором используют анионообменную хроматографию с высоким рН для разделения олигосахаридов по заряду. Известны также способы высвобождения олигосахаридов для аналитических целей, которые включают, без ограничения, ферментативную обработку (обычно осуществляемую с использованием пептид-N-гликозидазы F/эндо-β-галактозидазы), исключающую применение жесткой щелочной среды для высвобождения, в основном O-сцепленных структур, и способы, использующие безводный гидразин для высвобождения N- и O-сцепленных олигосахаридов.
(x) Другие модификации антител
Раскрытые здесь нейтрализующие анти-IFN-α антитела можно создать в виде иммунолипосом. Липосомы, содержащие антитело, получают с помощью известных в данной области техники способов, которые описаны, например, у Epstein et al., Proc. Natl. Acad. Sci. USA 82:3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA 77:4030 (1980); и в Патентах США №№4485045 и 4544545. Липосомы с увеличенным временем циркуляции раскрыты в Патенте США №5013556.
Особенно полезные липосомы можно создать с помощью способа обратнофазового выпаривания липидной композиции, включающей фосфатидилхолин, холестерин и PEG-производный фосфатидилэтаноламин (PEG-PE). Липосомы продавливают через фильтры с определенным размером пор с получением липосом требуемого диаметра. Fab'-фрагменты антитела настоящего изобретения можно конъюгировать с липосомами, как описано у Martin et al., J. Biol. Chem. 257:286-288 (1982), с помощью реакции взаимного дисульфидного обмена. Химиотерапевтический агент (такой как доксорубицин) не обязательно содержится в липосоме. См. Gabizon et al., J. National Cancer Inst. 81(19):1484 (1989).
Антитело настоящего изобретения можно также использовать в ADEPT путем конъюгации антитела c ферментом, активирующим пролекарства, который превращает пролекарство (например, пептидильный химиотерапевтический агент, см. WO 81/01145) в действующее лекарство. См., например, WO 88/07378 и Патент США №4975278.
Ферментный компонент иммуноконъюгата, пригодного для ADEPT, включает любой фермент, способный подействовать на пролекарство таким образом, чтобы превратить его в более активную форму, проявляющую требуемые биологические характеристики.
Ферменты, которые пригодны для данного способа настоящего изобретения, включают, но не ограничиваются, щелочную фосфатазу, пригодную для преобразования фосфатсодержащих пролекарств в не связанные с белками плазмы крови лекарственные вещества; арилсульфатазу, пригодную для преобразования сульфатсодержащих пролекарств в не связанные с белками плазмы крови лекарственные вещества; цитозиндезаминазу, пригодную для преобразования нетоксичного 5-фторцитозина в противоопухолевое лекарственное вещество 5-фторурацил; протеазы, такие как пилообразная протеаза, термолизин, субтилизин, карбоксипептидазы и катепсины (такие как катепсины В и L), которые пригодны для преобразования пептидсодержащих пролекарств в не связанные с белками плазмы крови лекарственные вещества; D-аланилкарбоксипептидазы, пригодные для преобразования пролекарств, которые содержат D-аминокислотные заместители; расщепляющие углеводы ферменты, такие как β-галактозидаза и нейраминидаза, пригодные для преобразования гликозилированных пролекарств в не связанные с белками плазмы крови лекарственные вещества; β-лактамазу, пригодную для преобразования лекарственных веществ, производных ∃-лактамов, в не связанные с белками плазмы крови лекарственные вещества; и пенициллиновые амидазы, такие как пенициллин-V-амидаза или пенициллин-G-амидаза, пригодные для преобразования лекарственных веществ, производных по их аминонитрогруппам с феноксиацетильными или фенилацетильными группами, соответственно в не связанные с белками плазмы крови лекарственные вещества. Альтернативно антитела с ферментативной активностью, известные также как "абзимы", можно использовать для преобразования пролекарств настоящего изобретения в не связанные с белками плазмы крови лекарственные вещества (см., например, Massey, Nature 328:457-458 (1987)). Конъюгаты антитело-абзим можно получить, как описано здесь, для доставки абзима в нужную популяцию клеток.
Данные ферменты могут ковалентно связываться с нейтрализующими анти-IFN-α антителами с помощью методов, хорошо известных в данной области техники, таких, которые используют вышерассмотренные реагенты для гетеробифункционального перекрестного сшивания. Альтернативно слитые белки, включающие, по меньшей мере, антиген-связывающую область антитела настоящего изобретения, присоединенные, по меньшей мере, к функционально активному участку фермента настоящего изобретения, можно сконструировать с использованием методов рекомбинантной ДНК, хорошо известных в данной области техники (см., например, Neuberger et al., Nature 312:604-608 [1984]).
В некоторых вариантах осуществления настоящего изобретения желательно использовать фрагмент антитела, а не интактное антитело. В этом случае требуется модифицировать фрагмент антитела с тем, чтобы повысить его время полужизни в сыворотке. Это можно осуществить, например, путем встраивания в антительный фрагмент улучшенного связывающегося с рецептором эпитопа (например, путем изменения соответствующей области в антительном фрагменте или путем встраивания эпитопа в пептидный маркер, который затем сливают с антительным фрагментом либо на конце, либо в средине, например, с помощью ДНК или пептидного синтеза). См. WO 96/32478, опубликованную 17 октября 1996 г.
Улучшенный связывающийся рецептором эпитоп, как правило, представляет собой область, в которой любой один или более аминокислотных остатков из одной или двух петель Fc-области переносят в аналогичное положение в антительном фрагменте. Еще предпочтительней, если переносят три или более остатков из одной или двух петель Fc-области. Более предпочтительно, если указанный эпитоп берут из СН2-домена Fc-области (например, IgG) и переносят в СН1, СН3 или VH-область, или в более чем одну такую область антитела. Или же данный эпитоп берут из СН2-домена Fc-области и переносят в CL-область или в VL-область или в обе области антительного фрагмента.
Ковалентные модификации нейтрализующих анти-IFN-α антител также включены в рамки настоящего изобретения. Их можно осуществить с помощью химического синтеза или, насколько применимо, с помощью ферментативного или химического расщепления антитела. Другие виды ковалентных модификаций антитела вводят в молекулу путем взаимодействия целевых аминокислотных остатков антитела с органическим преобразующим агентом, который способен взаимодействовать с селективными боковыми цепями или с N- либо С-концевыми остатками. Типичные ковалентные модификации полипептидов, которые описаны в Патенте США №5534615, специфически включены здесь путем ссылки. Предпочтительный тип ковалентной модификации антитела включает связывание антитела с одним из многих небелковых полимеров, например с полиэтиленгликолем, пропиленгликолем или полиоксиалкенами, способом, изложенным в Патентах США №№4640835; 4496689; 4301144; 4670417; 4791192 или 4179337.
2. Скрининг антител с нужными характеристиками
Методы создания антител описаны выше. Анти-IFN-α антитела с требуемыми широкого диапазона нейтрализующими характеристиками можно в последующем идентифицировать с помощью способов, известных в данной области техники.
(i) Анализ связывания
Так, нейтрализующие анти-IFN-α антитела настоящего изобретения можно, например, идентифицировать в анализе связывания IFN-α, путем инкубации антитела-кандидата с одним или более индивидуальными подтипами IFN-α, или с набором или со смесью разных подтипов IFN-α, и проконтролировать связывание и нейтрализацию биологической активности IFN-α. Анализ связывания можно осуществить с очищенным полипептидом(ами) IFN-α. В одном из вариантов осуществления настоящего изобретения анализ связывания представляет собой конкурентный анализ связывания, в котором оценивают способность антитела-кандидата конкурировать с известным анти-IFN-α антителом за связывание IFN-α. Анализ можно осуществить в разных форматах, включая ELISA-формат, который проиллюстрирован в Примерах ниже. Связывание IFN-α антителом-кандидатом можно также контролировать в BIAcoreTM Biosensor-анализе, который описан ниже.
Чтобы охарактеризовать конкурентную способность анти-IFN-α моноклонального антитела-кандидата с мышиным моноклональным антителом 9F3 против IFN-α человека за связывание с конкретным видом IFN-α, можно использовать любой подходящий конкурентный анализ связывания, известный в данной области техники. Общепринятый конкурентный анализ описан в Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Ed.Harlow and David Lane (1988). В другом варианте осуществления настоящего изобретения ELISA-анализ связывания IFN-α, описанный ниже в Примере 2, можно было бы модифицировать с тем, чтобы использовать конкуренции между антителом-кандидатом и антителом 9F3 за связывание IFN-α. Такой анализ можно было бы осуществить путем наслаивания IFN-α на плашки для микротитрования, инкубации наслоенных плашек в последовательных разведениях немеченого анти-IFN-α антитела и немеченого контрольного антитела, смешиваемых с выбранной концентрацией меченого 9F3-антитела, детектирования и измерения сигнала метки антитела 9F3 и последующего сравнения полученных измерений сигнала, проявляемых в разных разведениях антитела.
(ii) Противовирусный анализ
Способность антитела-кандидата нейтрализовать биологическую активность IFN-α можно выяснить, например, с помощью мониторинга нейтрализации противовирусной активности IFN-α, как описано у Kawade, J.Interferon Res. 1:61-70 (1980), или у Kawade and Watanabe, J.Interferon Res. 4:571:584 (1984). Вкратце, фиксированную концентрацию IFN-α, предварительно смешанного с разными разведениями антитела-кандидата, добавляют к FL-клеткам, полученным из амниона человека, и определяют способность антитела-кандидата нейтрализовать противовирусную активность IFN-α, используя соответствующий вирус, к примеру, вирус Синдбис. Полученные титры выражают в международных единицах (IU), как определено для международного эталона человеческого IFN-α (NIH Ga23-901-527).
Анти-IFN-α антитело-кандидат считают способным ингибировать противовирусную активность выбранного IFN-α-подтипа, если определенная концентрация антитела подавляет противовирусную активность в большей мере по сравнению с базовым уровнем ингибирования противовирусной активности, измеренной в присутствии эквивалентной концентрации контрольного антитела. Выбранная определенная концентрация анти-IFN-α антитела-кандидата необязательно должна подавлять, по меньшей мере, или около 60%, или, по меньшей мере, или около 70%, предпочтительно, по меньшей мере, или около 75%, или более предпочтительно, по меньшей мере, или около 80%, или даже более предпочтительно, по меньшей мере, или около 85%, или еще более предпочтительно, по меньшей мере, или около 90%, или еще более предпочтительно, по меньшей мере, или около 95%, или наиболее предпочтительно, по меньшей мере, или около 99% противовирусной активности выбранного подтипа IFN-α в анализе на противовирусную активность по сравнению с базовой активностью, измеренной в присутствии эквивалентной концентрации контрольного антитела. Анти-IFN-α антитело-кандидат считают неспособным подавлять противовирусную активность выбранного подтипа IFN-α, если имеющаяся концентрация данного антитела не проявляет большей противовирусной ингибирующей активности по сравнению с базовым уровнем ингибирования противовирусной активности, измеренной в присутствии эквивалентной концентрации контрольного антитела.
В предпочтительном варианте осуществления настоящего изобретения каждый вид интерферона, используемый в анализе на инфекционность вируса, титруют до концентрации, которая обеспечивает один и тот же уровень ингибирования вирусного роста, который индуцируется ранее выбранным числом единиц стандарта IFN-α. Данная концентрация пригодна для того, чтобы стандартизовать единицы, представляющие вид данного интерферона. Чтобы оценить способность анти-IFN-α антитела подавлять противовирусную активность различных подтипов IFN-α, эффективную концентрацию (ЕС50) анти-IFN-α антитела для 50% ингибирования противовирусной активности отдельного подтипа IFN-α (в концентрации, титруемой для получения стандартизованных единиц активности) определяют для каждого тестируемого подтипа IFN-α.
В одном из вариантов осуществления настоящего изобретения анализ нейтрализации противовирусной активности осуществляют, как описано ниже в Примере 1. Вкратце, клетки А549 выращивают в 96-луночных планшетах для титрования до плотности 5×105 А549-клеток/лунку. Последовательные разведения анти-IFN-α антитела-кандидата инкубируют с 0,2 единицы/1 выбранного подтипа IFN-α (стандартизованные до 0,2 единицы/1 стандартного эталона NIH рекомбинантного человеческого IFN-α2) в общем объеме 100:1 при 37°С в течение одного часа. Затем в индивидуальную лунку планшета для микротитрования к 5×105 клеток А549 добавляют по 100:1 объема инкубационной смеси антитело/интерферон культуральной среды, получая конечную концентрацию IFN-α в каждой лунке, равную 100 единицам/мл. Полученные смеси культуры клеток инкубируют в течение 24 часов при 37°С. После чего клетки заражают 2×105 б.о.е./мл вируса энцефаломиокардита (ЕМС) и инкубируют еще в течение 24 часов при 37°С. По окончании инкубации определяют жизнеспособность клеток путем визуальной проверки под микроскопом или окрашиванием кристалвиолетом. Для каждого тестируемого подтипа IFN-α в данном анализе определяют эффективную концентрацию (ЕС50) анти-IFN-α антитела-кандидата для 50% ингибирования противовирусной активности отдельного подтипа IFN-α.
В соответствии с одним из аспектов настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 до или около 20 г/мл, либо до или около 10 г/мл, либо до или около 5 г/мл, либо до или около 4 г/мл, либо до или около 3 г/мл, либо до или около 2 г/мл, либо до или около 1 г/мл в отношении каждого из представленных подтипов IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,1 г/мл до или около 20 г/мл, от или около 0,1 г/мл до или около 10 г/мл, от или около 0,1 г/мл до или около 5 г/мл, от или около 0,1 г/мл до или около 4 г/мл, от или около 0,1 г/мл до или около 3 г/мл, от или около 0,1 г/мл до или около 2 г/мл, от или около 0,1 г/мл до или около 1 г/мл в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,2 г/мл до или около 20 г/мл, от или около 0,2 г/мл до или около 10 г/мл, от или около 0,2 г/мл до или около 5 г/мл, от или около 0,2 г/мл до или около 4 г/мл, от или около 0,2 г/мл до или около 3 г/мл, от или около 0,2 г/мл до или около 2 г/мл, от или около 0,2 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,3 г/мл до или около 20 г/мл, от или около 0,3 г/мл до или около 10 г/мл, от или около 0,3 г/мл до или около 5 г/мл, от или около 0,3 г/мл до или около 4 г/мл, от или около 0,3 г/мл до или около 3 г/мл, от или около 0,3 г/мл до или около 2 г/мл, от или около 0,3 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,4 г/мл до или около 20 г/мл, от или около 0,4 г/мл до или около 10 г/мл, от или около 0,4 г/мл до или около 5 г/мл, от или около 0,4 г/мл до или около 4 г/мл, от или около 0,4 г/мл до или около 3 г/мл, от или около 0,4 г/мл до или около 2 г/мл, от или около 0,4 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,5 г/мл до или около 20 г/мл, от или около 0,5 г/мл до или около 10 г/мл, от или около 0,5 г/мл до или около 5 г/мл, от или около 0,5 г/мл до или около 4 г/мл, от или около 0,5 г/мл до или около 3 г/мл, от или около 0,5 г/мл до или около 2 г/мл, от или около 0,5 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,6 г/мл до или около 20 г/мл, от или около 0,6 г/мл до или около 10 г/мл, от или около 0,6 г/мл до или около 5 г/мл, от или около 0,6 г/мл до или около 4 г/мл, от или около 0,6 г/мл до или около 3 г/мл, от или около 0,6 г/мл до или около 2 г/мл, от или около 0,6 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,7 г/мл до или около 20 г/мл, от или около 0,7 г/мл до или около 10 г/мл, от или около 0,7 г/мл до или около 5 г/мл, от или около 0,7 г/мл до или около 4 г/мл, от или около 0,7 г/мл до или около 3 г/мл, от или около 0,7 г/мл до или около 2 г/мл, от или около 0,7 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 0,8 г/мл до или около 20 г/мл, от или около 0,8 г/мл до или около 10 г/мл, от или около 0,8 г/мл до или около 5 г/мл, от или около 0,8 г/мл до или около 4 г/мл, от или около 0,8 г/мл до или около 3 г/мл, от или около 0,8 г/мл до или около 2 г/мл, от или около 0,8 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность представленного подтипа IFN-α, должно проявлять ЕС50 от или около 0,9 г/мл до или около 20 г/мл, от или около 0,9 г/мл до или около 10 г/мл, от или около 0,9 г/мл до или около 5 г/мл, от или около 0,9 г/мл до или около 4 г/мл, от или около 0,9 г/мл до или около 3 г/мл, от или около 0,9 г/мл до или около 2 г/мл, от или около 0,9 г/мл до или около 1 г/мл, в отношении каждого представленного подтипа IFN-α в вышеописанном анализе ингибирования вирусных клеток А549 ЕМС.
В соответствии с другим аспектом настоящего изобретения анти-IFN-α антитело, которое проявляет нейтрализующую противовирусную активность против представленных подтипов IFN-α, должно проявлять ЕС50 от или около 1 г/мл до или около 20 г/мл, от или около 1 г/мл до или около 10 г/мл, от или около 1 г/мл до или около 5 г/мл, от или около 1 г/мл до или около 4 г/мл, от или около 1 г/мл до или около 3 г/мл, от или около 1 г/мл до или около 2 г/мл, в отношении каждого представленного подтипа IFN-α в описанном выше опыте по ингибированию вируса ЕМС в клетках А549.
(iii) Анализ с перекрестным блокированием
Скринирование антител, которые связываются с эпитопом IFN-α, связываемым представляющим интерес антителом, можно осуществить с помощью стандартного анализа перекрестного блокирования, который описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). Альтернативно, или дополнительно, эпитопное картирование можно осуществить с помощью способов, известных в данной области техники. Например, IFN-α-эпитоп, связываемый моноклональным антителом настоящего изобретения, можно определить с помощью анализа конкурентного связывания, как описано у Fendly et al., Cancer Research 50:1550-1558 (1990). В другом примере исследования по перекрестному блокированию можно осуществить вместе с прямой флуоресценцией в планшетах для микротитрования. В данном способе представляющее интерес моноклональное антитело конъюгируют с флуоресцеинизотиоцианатом (FITC), используя признанные методы (Wofsy et al., Selected Methods in Cellular Immunology, p.287, Mishel and Schiigi (eds.) San Francisco; W.J.Freeman Co. (1980)). Выбранный IFN-α наслаивают в лунки планшетов для микротитрования, эти наслоенные лунки инкубируют со смесью, состоящей из (1) FITC-меченного моноклонального антитела, представляющего интерес, и (2) немеченого тестируемого моноклонального антитела, и в каждой лунке количественно оценивают флуоресценцию для определения уровня перекрестного блокирования, проявляемого антителами. Считается, что моноклональные антитела используют совместно эпитоп, если каждый блокирует связывание другого на 50% или больше в сравнении c нерелевантным контрольным моноклональным антителом.
Данные результаты, полученные в биологическом анализе на клетках, можно затем проверить на животном, например, мышиных моделях, и в клинических испытаниях на человеке. При необходимости мышиные моноклональные антитела, обладающие требуемыми характеристиками, можно преобразовать в химерные антитела или в гуманизированные антитела с помощью методик, хорошо известных в данной области техники, в том числе методом "генного конверсионного мутагенеза", как описано в Патенте США №5821337, полнота раскрытия которого включена, таким образом, путем ссылки. Ниже, здесь в Примерах, описана также гуманизация конкретного анти-IFN-α антитела.
(iv) Способ фагового дисплея
Анти-IFN-α антитела настоящего изобретения можно идентифицировать путем использования комбинаторных библиотек для скринирования клонов синтетических антител с нужной активностью или активностями. В принципе, клоны синтетических антител выбирают путем скринирования фаговых библиотек, содержащих фаг, который обнаруживает различные фрагменты (например, Fab, F(ab')2, и т.п.) вариабельной области антитела (Fv), слитого с белком оболочки фага. Такие фаговые библиотеки создают с помощью хроматографии по сродству против требуемого антигена. Клоны, экспрессирующие Fv-фрагменты, способны связываться с требуемым антигеном и адсорбироваться на этом антигене и, таким образом, отделяться от несвязывающих клонов данной библиотеки. Затем связывающиеся клоны элюируют с антигена и в дальнейшем их можно обогатить дополнительными циклами антигенной адсорбции/элюции. Любые анти-IFN-α антитела настоящего изобретения можно получить путем создания соответствующей антигенной скринирующей методики для отбора представляющего интерес фагового клона с последующим конструированием полноразмерного клона анти-IFN-α антитела с использованием Fv-последовательностей из представляющего интерес фагового клона и последовательностей соответствующей константной области (Fc), описанной Kabat et al. (1991), выше.
Конструирование библиотек фагового дисплея
Антиген-связывающий участок антитела образуется из двух вариабельных (V) областей со 110 аминокислотами каждая, а также из легкой (VL) и тяжелой (VH) цепей, обе из которых представлены тремя гипервариабельными петлями или областями (CDR). Вариабельные области могут функционально обнаруживаться на фаге либо в виде одноцепочечных фрагментов Fv (scFv), в которых VH и VL ковалентно соединены с помощью короткого гибкого пептида, или в виде фрагментов D(ab')2, в которых каждый из них слит с константной областью и нековалентно взаимодействуют, как описано у Winter et al., Ann. Rev. Immunol., 12:433-455 (1994). Как использовано здесь, фаговые клоны, кодирующие scFv, и фаговые клоны, кодирующие Fab или F(ab')2, в совокупности именуются как "фаговые клоны Fv" или "Fv-клоны".
"Наивный" набор животного (набор до антигенной стимуляции) представлен антителами, которые могут связывать, с умеренным сродством (Kd -1 около 106-107 М-1), практически любую не свою молекулу. Разнообразие последовательностей антиген-связывающих сайтов не кодируется непосредственно в зародышевой линии, а собирается комбинаторным способом из сегментов V-гена. В человеческих тяжелых цепях первые две гипервариабельные петли (Н1 и Н2) извлекаются из менее чем 50 сегментов VH-гена, которые объединяются с D-сегментами и JH-сегментами, чтобы создать третью гипервариабельную петлю (Н3). В человеческих легких цепях первые две гипервариабельные петли (L1 и L2), а часто и третья (L3) извлекаются из приблизительно менее чем 30 V8- и приблизительно менее чем 30 V6-сегментов для завершения третьей гипервариабельной петли (L3).
Каждая комбинаторная перестройка сегментов V-гена в стволовых клетках дает начало В-клетке, которая экспрессирует одиночное сочетание VH-VL. Иммунизация, запускаемая любой В-клеткой, создает объединение, связывающее иммуноген для пролиферации (клональная экспансия) и секреции соответствующего антитела. Затем "наивные" антитела за счет мутагенеза и селекции созревают до высокого сродства (Kd -1 в 109-1010 М-1), известного в качестве процесса созревания по сродству. После прохождения этой стадии данные клетки обычно удаляют для получения гибридом и образования моноклональных антител с высоким сродством.
На трех стадиях данного процесса наборы VH- и VL-генов можно раздельно клонировать с помощью полимеразной цепной реакции (ПЦР) и рекомбинировать случайным образом в фаговые библиотеки, которые затем могут служить для поиска антиген-связывающих клонов, как описано у Winter et al., Ann. Rev. Immunol., 12:433-455 (1994). Библиотеки из иммунизированных источников обеспечивают антитела с высоким сродством к иммуногену без необходимости конструировать гибридомы. Альтернативно "наивный" набор можно клонировать для создания единственного источника человеческих антител для широкого ряда не своих и собственных антигенов без какой-либо иммунизации, как описано у Griffiths et al., EMBO J., 12:725-734 (1993). Наконец, "наивные" библиотеки можно также создавать синтетически путем клонирования нереаранжированных V- или VIII-генных сегментов стволовых клеток и используя ПЦР-праймеры, содержащие случайную последовательность для кодирования высоковариабельных CDR3-областей и для осуществления перестройки in vitro, как описано у Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992).
Фаговый дисплей имитирует В-клетку. Нитевидный фаг используют для обнаружения фрагментов антител путем слияния с минорным pIII-белком оболочки. Антительные фрагменты могут выявляться в виде одноцепочечных Fv-фрагментов, в которых VH- и VL-домены соединяются в одну полипептидную цепь с помощью гибкого полипептидного спейсера, как, например, описано Marks et al., J. Mol. Biol., 222:581-597 (1991), или в виде Fab- (включая F(ab')2) фрагментов, у которых одна цепь сливается с pIII, а другая секретируется в периплазму бактериальной клетки-хозяина, где собирается структура Fab-белка оболочки, которая выставляется на поверхности фага в результате вытеснения некоторых оболочечных белков дикого типа, как описано, например, у Hoogenboom et al., Nucl. Acids Res., 19:4133-4137 (1991). При слиянии фрагментов антитела с N-концевым доменом pIII данный фаг становится инфекционным. Однако если N-концевой домен pIII вырезать и слияние осуществить со вторым доменом, фаг не становится инфекционным, и pIII дикого типа следует создать при помощи фага-хелпера.
Слияние pIII и других белков фага может кодироваться полностью в одном и том же фаговом репликоне или в разных репликонах. При использовании двух репликонов слияние pIII кодируется фагемидой, плазмидой, содержащей ориджин репликации фага. Фагемиды можно упаковать в фаговые частицы путем "спасения" с помощью фага-хелпера, такого как M13K07, который создает все фаговые белки, в том числе pIII, но вследствие дефектного ориджина сам плохо упаковывается из-за конкуренции с фагемидами, как описано у Vieira and Messing, Meth. Enzymol., 153:3-11 (1987). В предпочтительном способе система фагового дисплея создается так, чтобы рекомбинантный фаг мог расти в клетке-хозяине в условиях, позволяющих лишь небольшому количеству фаговых частиц выставлять на поверхности фаговой частицы более одной копии слитой конструкции Fv-оболочечный белок, как описано у Bass et al., Proteins, 8:309-314 (1990) и в WO 92/09690 (PCT/US91/09133, опубликованной 11 июня 1992 г.).
Вообще, нуклеиновые кислоты, кодирующие генные фрагменты антител, получают из иммунных клеток, собранных от людей или животных. Если требуется смещение библиотеки в пользу клонов анти-IFN-α, индивида иммунизируют IFN-α с получением гуморального ответа, и для конструирования библиотеки извлекают клетки селезенки и/или циркулирующие В-клетки других лимфоцитов периферической крови (PBL). В предпочтительном варианте осуществления настоящего изобретения библиотеку генного фрагмента человеческого антитела, смещаемую в пользу анти-IFN-α клонов человека, получают в результате образования гуморального ответа анти IFN-α антитела человека у трансгенных мышей, несущих последовательность функционального гена иммуноглобулина человека (и не имеющих системы образования функционального эндогенного антитела), так что IFN-α-иммунизация дает начало В-клеткам, продуцирующим последовательности анти-IFN-α антител человека.
В другом предпочтительном варианте осуществления настоящего изобретения животных иммунизируют смесью разных, предпочтительно всех, подтипов IFN-α с целью получить гуморальный ответ, который включает продуцирующие В-клетками анти-IFN-α антитела c широкой реактивностью против подтипов IFN-α. В другом предпочтительном варианте осуществления настоящего изобретения животных иммунизируют смесью подтипов IFN-α человека, которые представлены лимфобластоидными интерферонами человека, секретируемыми клетками лимфомы Бэркитта (Namalva-клетки), индуцированных вирусом Сендай, как описано ниже в Примере 1. Соответствующий препарат таких лимфобластоидных интерферонов человека можно купить (Product No. I-9887) у Sigma Chemical Company, St. Louis, MO.
Дополнительно обогащенные популяции анти-IFN-α клеток можно получить с использованием соответствующего метода скрининга для выделения В-клеток, экспрессирующих специфичное мембраносвязанное анти-IFN-α антитело, например, путем разделения клеток с помощью хроматографии по сродству с IFN-α или адсорбцией клеток меченного флуорохромом IFN-α с последующей активируемой флуоресценцией сортировкой клеток (FACS).
Альтернативно использование клеток селезенки и/или В-клеток или иных PBL от иммунизированного донора обеспечивает лучшее представление возможного набора антител, а также позволяет конструировать библиотеку антител с использованием любых видов животных (человека или отличных от человека), у которых IFN-α не является антигенным. Для библиотек со встроенной in vitro генной конструкцией антитела стволовые клетки собирают от субъекта с получением нуклеиновых кислот, кодирующих нереаранжированные генные сегменты антитела. Представляющие интерес иммунные клетки можно получить от разнообразных видов животных, таких как человек, мышь, крыса, зайцеобразные, волк, собака, кошка, свинья, корова, лошадь, а также виды птиц, и т.п.
Нуклеиновую кислоту, кодирующую вариабельные генные сегменты антитела (включая сегменты VH и VL), извлекают из представляющих интерес клеток и амплифицируют. В случае реаранжированных библиотек генов VH и VL нужную ДНК можно получить путем выделения геномной ДНК или мРНК из лимфоцитов с последующей полимеразной цепной реакцией (ПЦР) с праймерами, соответствующими 5'- и 3'-концам реаранжированных генов VH и VL, как описано у Orlandi et al., Proc. Natl. Acad. Sci. (USA), 86:3833-3837 (1989), создавая, таким образом, разнообразные V-генные наборы для экспрессии. V-гены могут быть амплифицированы из кДНК и геномной ДНК с помощью обратных праймеров по 5'-концу экзона, кодирующего зрелую V-область, и с помощью прямых праймеров, на основе J-сегмента, как описано у Orlandi et al. (1989) и у Ward et al., Nature, 341:544-546 (1989). Однако для амплификации из кДНК обратные праймеры можно также создать на основе лидерного экзона, как описано у Jones et al., Biotechnol., 9:88-89 (1991), а прямые праймеры - на основе константной области, как описано у Sastry et al., Proc. Natl. Acad. Sci. (USA), 86:5728-5732 (1989). Для максимального повышения комплементарности в данные праймеры можно ввести вырожденность, как описано у Orlandi et al. (1989) или у Sastry et al. (1989). Предпочтительно разнообразие библиотеки увеличивают до предела путем использования ПЦР-праймеров, нацеленных на каждое семейство V-гена с целью амплифицировать все доступные VH- и VL-структуры, представленные в образце нуклеиновой кислоты иммунной клетки, например, как описано в способе Marks et al., J. Mol. Biol., 222:581-597 (1991) или как описано в способе Orum et al., Nucleic Acids Res., 21:4491-4498 (1993). Для клонирования амплифицированной ДНК в экспрессионные векторы в данный ПЦР-праймер можно ввести в качестве метки по одному концу редкий сайт рестрикции, как описано у Orlandi et al. (1989), или путем дополнительной ПЦР-амплификации c маркированным праймером, как описано у Clackson et al., Nature, 352:624-628 (1991).
Наборы синтетически реаранжированных V-генов можно создать in vitro из сегментов V-гена. Большинство человеческих сегментов VH-гена клонируют и секвенируют (reported in Tomlinson et al., J. Mol. Biol., 227:776-798 (1992)), а также картируют (reported in Matsuda et al., Nature Genet., 3:88-94 (1993); эти клонированные сегменты (в том числе все главные конформации Н1- и Н2-петли) можно использовать для получения разнообразных наборов VH-гена с помощью ПЦР-праймеров, кодирующих H3-петли иной последовательности и длины, как описано у Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992). VH-наборы можно также создать с помощью всего разнообразия последовательностей, собранных в большой H3-петле одной длины, как описано у Barbas et al., Proc. Natl. Acad. Sci. USA, 89:4457-4461 (1992). Человеческие V6- и V8-сегменты клонируют и секвенируют (reported in Williams and Winter, Eur. J. Immunol., 23:1456-1461 (1993), после чего их можно использовать для создания синтетических наборов легкой цепи. Синтетические наборы V-гена, созданные на основе ряда укладок VH и VL, а также на основе ряда длин L3 и H3, будут кодировать антитела со значительным структурным разнообразием. После амплификации V-гена, кодирующего ДНК, сегменты V-гена клеток зародышевого пути можно реаранжировать in vitro в соответствии со способами Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992).
Наборы антительных фрагментов можно сконструировать путем объединения генных наборов VH и VL несколькими способами. Каждый набор можно создать в разных векторах и данные векторы рекомбинировать in vitro, как описано, например, у Hogrefe et al., Gene, 128:119-126 (1993), или in vivo с помощью комбинаторного инфицирования, например, системой loxP, описанной у Waterhouse et al., Nucl. Acids Res., 21:2265-2266 (1993). Метод рекомбинации in vivo использует двухцепочечный характер Fab-фрагментов, чтобы преодолеть ограничение в размере библиотеки, продиктованное эффективностью E. coli-трансформации. Исходные наборы VH и VL клонируют раздельно, один в фагемиду, а другой в фаговый вектор. Затем эти две библиотеки объединяют с помощью фагового инфицирования фагемид-содержащими бактериями так, что каждая клетка содержит разную комбинацию, а размер библиотеки ограничен лишь числом представленных клеток (около 1012 клонов). Оба вектора содержат сигналы рекомбинации in vivo так, что гены VH и VL рекомбинируют в единственный репликон и вместе упаковываются в фаговые вирионы. Эти гигантские библиотеки создают большое многообразие антител с хорошим сродством (Kd -1 около 10-8 М).
Альтернативно наборы можно клонировать последовательно в один и тот же вектор, например, как описано у Barbas et al., Proc. Natl. Acad. Sci. USA, 88:7978-7982 (1991), или собрать вместе с помощью ПЦР, а затем клонировать, например, как описано у Clackson et al., Nature, 352:624-628 (1991). ПЦР-сборку можно также использовать для присоединения VH-ДНК и VL-ДНК к ДНК, кодирующей гибкий пептидный спейсер для получения одноцепочечных наборов Fv (scFv). Еще один метод, "клеточная ПЦР-сборка", используют для объединения генов VH и VL в лимфоцитах с помощью ПЦР, а затем клонируют наборы сцепленных генов, как описано у Embleton et al., Nucl. Acids Res., 20:3831-3837 (1992).
Антитела, полученные с помощью первичных библиотек (естественных или синтетических), могут обладать умеренным сродством (Kd -1 около 106-107 М-1), но созревание по сродству можно также имитировать in vitro путем создания и повторной селекции вторичных библиотек, как описано у Winter et al. (1994), выше. Например, мутацию можно встроить in vitro случайным образом с использованием "ошибающейся" полимеразы (reported in Leung et al., Technique, 1:11-15 (1989) в способе Hawkins et al., J. Mol. Biol., 226:889-896 (1992) или в способе Gram et al., Proc. Natl. Acad. Sci USA, 89:3576-3580 (1992). Кроме того, созревание по сродству можно осуществить путем случайного мутирования одной или более CDR в выбранных индивидуальных Fv-клонах, например, с использованием ПЦР и праймеров, несущих случайную последовательность, представляющей интерес, пронизывающей CDR, и скринирования клонов по наибольшему сродству. WO 9607754 (опубликована 14 марта 1996 г.) описывает способ индукции мутагенеза в гипервариабельной области легкой цепи иммуноглобулина для создания библиотеки генов легкой цепи. Другой эффективный метод заключается в рекомбинации доменов VH или VL, выбранных с помощью фагового дисплея и наборов естественно встречаемых вариантов V-домена, полученных от неиммунизированных доноров, и скринировании на наибольшее сродство в нескольких циклах повторной перетасовки цепи, как описано у Marks et al., Biotechnol., 10:779-783 (1992). Данный метод позволяет получать антитела и фрагменты антител со сродством в диапазоне 10-9 М.
Приготовление библиотек фагового дисплея для анти-IFN-α клонов
a. Синтез IFN-α
Используемую здесь последовательность нуклеиновой кислоты, кодирующую подтипы IFN-α, можно создать с использованием опубликованных аминокислотных и нуклеотидных последовательностей интерферонов, см., например, J. Interferon Res., 13:443-444 (1993) компиляцию ссылок, содержащих геномные и кДНК-последовательности различных интерферонов типа I, а также цитируемые в ней ссылки. Аминокислотные последовательности для IFN-αА (IFN-α2), IFN-αВ (IFN-α8), IFN-αС (IFN-α10), IFN-αD (IFN-α1), IFN-αЕ (IFN-α22), IFN-αF (IFN-α21), IFN-αG (IFN-α5) и IFN-αH (IFN-α14) или кДНК-последовательности показаны на Фиг.3 и 4 на стр. 23-24 у Goeddel et al., Nature, 290:20-26 (1981). кДНК, кодирующая аминокислотную последовательность IFN-α7 (IFN-αJ), показана Cohen et al., Dev. Biol. Standard, 60: 111-122 (1985). ДНК, кодирующие представляющие интерес интерфероны, можно получить разнообразными способами, известными в данной области техники. Эти способы включают, но не ограничиваются, химический синтез с помощью любого способа, описанного у Engels et al., Agnew. Chem. Int. Ed. Engl., 716-734 (1989), такого как триэфирный, фосфитный, фосфороамидитный и Н-фосфонатный способ. В одном из вариантов осуществления настоящего изобретения кодоны, предпочтительные благодаря экспрессирующей клетке-хозяину, используют для конструирования ДНК, кодирующей интерферон. Альтернативно кодирующую интерферон ДНК можно выделить из геномной или из кДНК-библиотеки.
После конструирования молекулы ДНК, кодирующей представляющий интерес интерферон, полученную молекулу ДНК функционально присоединяют к последовательности, контролирующей экспрессию в экспрессирующем векторе, таком как плазмида, где контролирующая последовательность распознается клеткой-хозяином, трансформированной вектором. Вообще плазмидные векторы содержат реплицирующие и контролирующие последовательности, которые получены из видов, сопоставимых с клеткой-хозяином. Вектор обычно несет сайт репликации, а также последовательности, которые кодируют белки, обеспечивающие отбор по фенотипам в трансформированных клетках.
Для экспрессии в прокариотических хозяевах подходящие векторы включают pBR322 (ATCC No.37017), phGH107 (ATCC No.40011), pBO475, pSO132, pRIT5, любой вектор серии pRIT20 или pRIT30 (Nilsson and Abrahmsen, Meth. Enzymol., 185:144-161 (1990)), pRIT2T, pKK233-2, pDR540 и pPL-лямбда. Прокариотические клетки-хозяева, содержащие экспрессионные векторы, пригодные для использования в настоящем изобретении, включают E.coli K12 штамм 294 (ATCC NO.31446), E.coli штамм JM101 (Messing et al., Nucl. Acid Res., 9:309 (1981)), E.coli штамм В, E.coli штамм Π1776 (ATCC No.31537), E.coli c600 (Appleyard, Genetics, 39:440 (1954)), E.coli W3110 (F-, гамма, прототрофный, ATCC No.27325), E.coli штамм 27С7 (W3110, tonA, phoA E15, (argF-lac)169, ptr3, degP41, ompT, kanr) (Патент США №5288931, ATCC No.55244), Bacillus subtilis, Salmonella typhimurium, Serratia marcesans и виды Pseudomonas.
Помимо прокариот в качестве клеток-хозяев могут использоваться эукариотические организмы, такие как дрожжи, или клетки, полученные из многоклеточных организмов. Для экспрессии в дрожжевых клетках-хозяевах, таких как обычные пекарские дрожжи или Saccharomyces cerevisiae, подходящие векторы включают эписомно реплицирующиеся векторы на основе 2-микронной плазмиды, интегрирующие векторы и векторы искусственной дрожжевой хромосомы (YAC). Для экспрессии в клетках-хозяевах насекомых, таких как клетки Sf9, подходящие векторы включают бакуловирусные векторы. Для экспрессии в клетках-хозяевах растений, особенно двудольных растений-хозяев, таких как табак, соответствующие экспрессионные векторы включают векторы, созданные из Agrobacterium tumefaciens Ti-плазмиды.
Однако наиболее интересными являются клетки-хозяева позвоночных. Примеры пригодных клеток-хозяев млекопитающих включают CV1-линию клеток почки обезьяны, трансформированную SV40 (COS-7, ATCC CRL 1651); линию эмбриональных клеток почки человека (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol., 36:59 (1977); почечные клетки детеныша хомячка (BHK, ATCC CCL 10); клетки яичника/-DHER китайского хомячка (CHO, Urlaub and Chasin, Proc. Natl. Acad. Sci. USA, 77:4216 (1980)); мышиные клетки Сертоли (TM4, Mather, Biol. Reprod., 23:243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки Африканской зеленой мартышки (VERO-76, ATCCC CRL-1587); раковые клетки шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени buffalo rat (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); TRI-клетки (Mather et al., Annals N.Y. Acad. Sci., 383:44-68 (1982)); клетки MRC 5; FS4-клетки; и линия клеток гепатомы человека (Hep G2). Для экспрессии в клетках-хозяевах млекопитающих пригодные векторы включают векторы, получаемые из SV40, векторы, получаемые из цитомегаловируса, такие как pRK-векторы, в том числе векторы pRK5 и pRK7 (Suva et al., Science, 237:893-896 (1987), EP 307247 (3/15/89), EP 278776 (8/17/88), векторы, получаемые из вируса коровьей оспы или из других поксвирусов, а также ретровирусные векторы, такие как векторы, получаемые из вируса лейкоза мышей Молони (MoMLV).
Необязательно, ДНК, кодирующую интересующий интерферон, функционально присоединяют к секреторной лидерной последовательности для секреции экспрессируемого продукта клеткой-хозяином в культуральную среду. Примеры секреторных лидерных последовательностей включают stII, экотин, lamB, герпес GD, lpp, щелочной фосфатазы, инвертазы и альфа-фактора. Пригодной для использования в настоящем изобретении является также 36 аминокислотная лидерная последовательность белка А (Abrahmsen et al., EMBO J., 4:3901 (1985)).
Клетки-хозяева трансфицируют и предпочтительно трансформируют с помощью вышеописанной экспрессии или клонирующими векторами настоящего изобретения и культивируют в стандартной питательной среде, модифицированной для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих требуемые последовательности.
Трансфекция относится к помещению экспрессионного вектора в клетку-хозяина независимо от того, какая из кодирующих последовательностей экспрессируется в действительности. Специалистам в данной области известно множество способов трансфекции, например осаждение CaPO4 и электропорация. Об успешной трансфекции обычно узнают по какому-либо функционированию данного вектора в клетке-хозяине.
Трансформация подразумевает введение ДНК в организм таким образом, чтобы эта ДНК реплицировалась, либо в качестве внехромосомного элемента, либо в виде хромосомного компонента. В зависимости от используемой клетки-хозяина трансформацию осуществляют с использованием стандартных методов, пригодных для таких клеток. Обработку кальцием с использованием хлорида кальция, как описано в разделе 1.82 у Sambrook et al., Molecular Cloning (2nd ed.), Cold Spring Harbor Laboratory, NY (1989), как правило, применяют для прокариот или других клеток, которые содержат существенные препятствия в виде клеточных стенок. Инфицирование Agrobacterium tumefaciens используют для трансформации некоторых растительных клеток, как описано у Shaw et al., Gene, 23:315 (1983) и WO 89/05859, опубликованной 29 июня 1989 г. Для клеток млекопитающих, не имеющих таких клеточных стенок, способ осаждения фосфатом кальция, описанный в разделах 16.30-16.37 у Sambrook et al., выше, является предпочтительным. Общие аспекты системной трансформации клетки-хозяина млекопитающего описаны Axel в патенте США №4399216, выданном 16 августа 1983 г. Трансформацию в дрожжах обычно осуществляют в соответствии со способом Van Solingen et al., J. Bact., 130:946 (1977), а также Hsiao et al., Proc. Natl. Acad. Sci. (USA), 76:3829 (1979). Но можно также использовать и другие способы введения ДНК в клетки, такие как инъецирование ядра, электропорация, или с помощью слияния протопластов.
Прокариотические клетки-хозяева, используемые для получения интересующего интерферона, можно культивировать, как описано в основном у Sambrook et al., выше.
Клетки-хозяева млекопитающих, используемые для получения интересующего интерферона, можно культивировать в разнообразных средах. Имеющиеся в продаже среды, такие как среда Хэма F10 (Sigma), минимальная основная среда ((MEM), Sigma), RPMI-1640 (Sigma), среда Игла в модификации Дюльбекко ((DMEM), Sigma) пригодны для культивирования клеток-хозяев. Кроме того, любая среда, описанная у Ham and Wallace, Meth. Enz., 58:44 (1979), Barnes and Sato, Anal. Biochem., 102:255 (1980), в Патентах США №№4767704; 4657866; 4927762; или 4560655; WO 90/03430; WO 87/00195; Патент США (в деле) №30985; или Патент США №5122469, раскрытие которых, всех, включено здесь путем ссылки, может использоваться с качестве культуральной среды для клеток-хозяев. Любая из этих сред может содержать добавки в виде гормонов и/или других ростовых факторов (таких как фактор роста инсулина, трансферрина или эпидермиса), соли (такие как хлорид натрия, соли кальция, магния и фосфата), буферы (такие как HEPES), нуклеозиды (такие как аденозин или тимидин), антибиотики (такие как лекарственное средство Gentamycin™), микроэлементы (указанные в виде неорганических соединений, представленные обычно в конечных концентрациях, измеряемых в микромолях), и глюкоза или эквивалентный источник энергии. Любые другие необходимые добавки могут быть также включены в надлежащих концентрациях, которые должны быть известны специалистам в данной области техники. Условия культивирования, такие как температура, рН и тому подобное, представляют собой условия, которые раньше использовали для клеток-хозяев, селектируемых для экспрессии, и должны быть очевидны специалистам в данной области.
Указанные в настоящем описании клетки-хозяева включают клетки для in vitro-культуры, а также клетки, которые находятся в животном-хозяине.
При внутриклеточной экспрессионной системе или системе, секретирующей в периплазматическое пространство, рекомбинантно экспрессируемый белок интерферон можно извлечь из культуры клеток в результате разрыва клеточной мембраны/клеточной стенки хозяина (например, с помощью осмотического шока или солюбилизации мембраны клетки-хозяина в детергенте). Альтернативно рекомбинантный белок можно извлекать из культуральной среды при использовании внеклеточной секретирующей системы. На первой стадии культуральную среду или лизат центрифугируют для удаления любых частиц клеточного дебриса. Затем разделяют мембраны и фракции растворимого белка. Обычно интерферон выделяют очисткой из фракции растворимого белка. Если IFN-α экспрессируется в мембранно-связанном виде, то мембранно-связанный пептид можно извлечь из мембранной фракции солюбилизацией с помощью детергентов. Затем неочищенный пептидный экстракт можно еще раз подвергнуть выделению очисткой с помощью соответствующих методик, таких как фракционирование на иммунноаффинных или ионообменной колонках; этанольное осаждение; обращенно-фазовая HPLC; хроматография на силикагеле или на анионообменной смоле, такой как DEAE; хроматофокусировка; SDS-PAGE; осаждение сульфатом аммония; гель-фильтрация с использованием, например, Сефадекс G-75; гидрофобноаффиные смолы и сродство к лиганду с использованием интерферонового рецептора, иммобилизованного на матрице.
Многие используемые здесь IFN-α человека можно получить из коммерческих источников, например, от Sigma (St. Louis, MO), Calbiochem-Novabiochem Corporation (San Diego, CA) или ACCURATE Chemical & Scientific Corporation (Westbury, NY).
Для конструирования плазмид, которые управляют транслокацией разных видов hIFN-α в периплазматическом пространстве E.coli, используют общепринятые методы клонирования, описанные у Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989). ПЦР-реакцию осуществляют на кДНК-клонах различных подвидов hIFN-α, раскрытых у Goeddel et al., Nature 290:20-26 (1981), с помощью добавляемых в соответствующие праймеры сайтов рестрикции Nsil и StyI. Затем полученные ПЦР-продукты субклонируют в соответствующие сайты экспрессионного вектора pB0720, описанного у Cunningham et al., Science 243:1330-1336 (1989). Полученные плазмиды помещают для продуцирования подтипов hIFN-α контролем промотора phoA E.coli и теплоустойчивого сигнального пептида энтеротоксина II, как описано у Chang et al., Gene 55:189-196 (1987). Правильность ДНК-последовательности каждого гена подтверждают с использованием набора United States Biochemical Sequenase Kit version 2.0. Каждую плазмиду трансформируют в E.coli-штамме 27С7 (ATCC #55244) и выращивают в 10-литровых ферментерах, как описано у Carter et al., Bio/Technology 10:163-167 (1992). Человеческие hIFN выделяют очисткой из E. coli-клеточной пасты, содержащей каждый IFN-α, с помощью хроматографии по сродству. Бактериальные клетки лизируют и лизат центрифугируют при 10000×g для удаления дебриса. Полученный супернатант наносят на иммуноаффинную колонку, содержащую мышиные анти-hIFN-αB антитела (LI-1), которые получают, как описано у Staehelin et al., Proc. Natl. Acad. Sci. 78:1848-1852 (1981). LI-1 иммобилизуют на стекло с контролируемыми порами с помощью модифицированного способа Roy et al., Journal of Chromatography, 303:225-228 (1984). Связанный интерферон элюируют из колонки 0,1 М цитратом, рН 3,0, содержащим 20% (мас./об.) глицерина. Выделенный очисткой IFN анализируют с помощью SDS-PAGE и иммуноблоттинга, после чего оценивают биоактивность в hIFN-индуцированном противовирусном анализе, как здесь описано.
Человеческую IFN-α2/1-гибридную молекулу (IFN-α21-62/α64-166 получают, как описано у Rehberg et al., J. Biol. Chem., 257:11497-11502 (1992) или у Horisberger and Marco, Pharmac. Ther., 66:507-534 (1995).
b. Иммобилизация IFN-α
Выделенный очисткой IFN-α можно присоединить к подходящей матрице, такой как гранулы агарозы, акриламидные гранулы, стеклянные бусинки, целлюлоза, различные акриловые сополимеры, гидроксилметакрилатные гели, полиакриловые или полиметакриловые сополимеры, найлон, нейтральные и ионные носители и т.п., для использования в разделительной хроматографии по сродству клонов фагового дисплея. Присоединение белка IFN-α к определенному матриксу можно осуществить с помощью способов, описанных в Methods in Enzymology, vol.44 (1976). Обычно используемый метод для присоединения белковых лигандов к полисахаридным матрицам, например, к агарозе, декстрану или целлюлозе, включает активацию носителя с помощью цианистых галогенидов и последующего связывания первичных алифатических или ароматических аминов пептидного лиганда с активированной матрицей.
Альтернативно IFN-α можно использовать для покрытия лунок адсорбционных планшетов экспрессируемыми клетками-хозяевами, прикрепляемыми к адсорбционным планшетам или используемыми для сортировки клеток, или конъюгированными с биотином для улавливания гранулами, покрытыми стрептавидином, или используемыми в любом ином хорошо известном в данной области техники способе для укладки библиотек фагового дисплея.
с. Методы пэннинга
Образцы фагового дисплея приводят в соприкосновение с иммобилизованным IFN-α в условиях, пригодных для связывания, по меньшей мере, части фаговых частиц с адсорбентом. Условия, включающие рН, ионную силу, температуру и т.п., обычно подбирают так, чтобы имитировать физиологические условия. Связанный с твердой фазой фаг промывают и затем элюируют кислотой, например, как описано у Barbas et al., Proc. Natl. Acad. Sci. USA 88:7978-7982 (1991), или щелочью, например, как описано у Marks et al, J. Mol. Biol. 222:581-597 (1991), или IFN-α-антигеном, например, по методу, аналогичному антиген-конкурентному способу Clackson et al., Nature, 352:624-628 (1991). Фаг можно обогатить 20-1000-кратно за один цикл селекции. Кроме того, обогащенный фаг можно вырастить в бактериальной культуре и подвергнуть последующим циклам селекции.
В предпочтительном варианте осуществления настоящего изобретения фаг, последовательно инкубированный с разными подтипами IFN-α, иммобилизуют для его идентификации и последующей характеристики фаговых клонов, которые проявляют существенное связывание с большинством, предпочтительно со всеми, подтипами IFN-α. В данном способе фаг вначале инкубируют с одним специфичным подтипом IFN-α. Связанный с данным подтипом фаг элюируют и подвергают селекции с помощью другого подтипа IFN-α. Затем данный процесс связывания и элюирования повторяют со всеми подтипами IFN-α. В самом конце данная операция дает популяцию фага, воспроизводящего антитела с широкой реактивностью против всех подтипов IFN-α. Затем эти фаги можно тестировать против других видов IFN, т.е. иных, чем IFN-α, с целью отобрать те клоны, которые не демонстрируют существенного связывания с другими видами IFN. Наконец, отобранные фаговые клоны можно проверить на их способность нейтрализовать биологическую активность, например, противовирусную активность, различных подтипов IFN-α, после чего клоны, представляющие антитела с широкой нейтрализующей активностью против большинства, предпочтительно против всех подтипов IFN-α, отбирают окончательно.
Эффективность отбора зависит от многих факторов, в том числе от кинетик диссоциации во время промывки и от того, может ли множество антительных фрагментов одного фага одновременно связаться с антигеном. Антитела с быстрой кинетикой диссоциации (и слабым сродством связывания) можно удержать путем применения коротких промывок, мультивалентного фагового дисплея и высокой плотности покрывающего антигена на твердой фазе. Высокая плотность не только стабилизирует фаг вследствие мультивалентных взаимодействий, но и благоприятствует повторному связыванию диссоциированного фага. Отбор антител с медленной кинетикой диссоциации (и хорошим сродством связывания) может способствовать применению длительных промывок и моновалентного фагового дисплея, как описано у Bass et al., Proteins, 8:309-314 (1990) и в WO 92/09690, и низкой плотности покрытия антигеном, как описано у Marks et al., Biotechnol., 10:779-783 (1992).
Представляется возможным выбирать между фаговыми антителами IFN-α с разным сродством, даже с незначительным сродством. Однако случайная мутация в выбранном антителе (например, осуществляемая в некоторых вышеописанных методах созревания по сродству), вероятно, дает начало многим мутантам, в большинстве связывающимися с антигеном, и лишь немногие из которых - с более высоким сродством. При ограничении IFN-α фаг с высоким сродством может оказаться вне конкуренции. Чтобы сохранить все мутанты с наибольшим сродством, фаг можно инкубировать с избытком биотинилированного IFN-α, однако с биотинилированным IFN-α - в концентрации с пониженной молярностью по сравнению с целевой молярной константой сродства для IFN-α. Связывающий с высоким сродством фаг можно затем уловить с помощью парамагнитных шариков, покрытых стрептавидином. Такой "равновесный захват" позволяет антителам селектироваться в соответствии с их сродством связывания и с чувствительностью, которая позволяет выделять мутантные клоны как минимум с двукратно превышающим сродством из большого числа фагов с более низким сродством. Условия, используемые для промывки фага, связанного с твердой фазой, можно также регулировать для разделения на основе кинетик диссоциации.
В одном из вариантов осуществления настоящего изобретения фаг последовательно инкубируют с разными подтипами IFN-α, иммобилизованными на твердом носителе, таком как вышеописанная матрица из полимерных гранул для хроматографии. В данном способе фаг вначале инкубируют с одним специфичным подтипом IFN-α. Связанный с этим подтипом фаг элюируют из твердой фазы подходящим элюентом, например любой солью или кислым буфером, способными высвободить связанный фаг в раствор. Затем элюированные фаговые клоны подвергают селекции с другим подтипом IFN-α. Чтобы обогатить популяцию клонов, которые конкурируют с растворенным IFNAR2 за связывание с IFN-α, фаговые клоны, извлеченные в нескольких циклах хроматографического разделения подтипов IFN-α, инкубируют с комплексом иммобилизованных IFNAR2, предварительно адсорбированных с IFN-α, а неадсорбированные фаговые клоны извлекают из инкубационной реакционной смеси.
Процедуры отбора можно разработать для использования в любом подходящем методе ступенчатой хроматографии. В одном из вариантов осуществления настоящего изобретения фаговые клоны адсорбируют на матрице с IFN-α в суспензии полимерных гранул, данные адсорбированные гранулы извлекают центрифугированием, извлеченные гранулы ресуспендируют и инкубируют в соответствующем буфере для элюции, как например, в любом солевом или кислом буфере, способном высвободить связанный фаг в раствор, после чего полученную элюционную смесь центрифугируют, а элюированные фаговые клоны извлекают из полученного супернатанта и затем операцию адсорбция/элюция повторяют для каждого очередного подтипа IFN-α. Чтобы обогатить популяцию клонов, которые конкурируют с растворимым IFNAR2 за связывание с IFN-α, фаговые клоны извлекают из хроматографически отделенного подтипа IFN-α и инкубируют с суспензией IFNAR2 в форме производных полимерных гранул, предварительно адсорбированных с IFN-α, эту инкубационную смесь центрифугируют и неадсорбированные фаговые клоны извлекают из супернатанта.
В другом варианте осуществления настоящего изобретения процедура отбора предназначается для обогащения популяции фага, способной ингибировать связывание IFN-α с IFNAR2 в течение каждого хроматографического разделения по сродству. В данном способе фаг последовательно инкубируют с каждым специфичным подтипом IFN-α, иммобилизованным на твердом носителе, а затем элюируют с твердой фазы с помощью элюента, включающего избыток растворимого IFNAR2, такого, например, как IFNAR2 ECD-IgG Fc, в условиях, где растворимый IFNAR2 способен вытеснять любой фаговый клон, который конкурирует с IFNAR2 за связывание с иммобилизованным IFN-α. Затем этот процесс по связыванию и элюции повторяют с каждым специфичным подтипом IFN-α.
В другом варианте осуществления настоящего изобретения фаговые клоны адсорбируют с суспензией IFN-α в форме производных полимерных гранул, адсорбированные гранулы извлекают центрифугированием, извлеченные гранулы ресуспендируют и инкубируют в подходящем элюирующем буфере, включающем избыток растворимого IFNAR2 (такого как IFNAR2 ECD-IgG Fc) в условиях, где растворимый IFNAR2 способен вытеснить любой фаговый клон, который конкурирует с IFNAR2 за связывание с иммобилизованным IFN-α, и высвобождает связанный фаг в раствор, полученную элюционную смесь центрифугируют, элюированные фаговые клоны извлекают из супернатанта, и затем операцию адсорбция/элюция повторяют для каждого очередного подтипа IFN-α.
В конце процедура дает популяцию фага, обнаруживающего антитела с активностью, ингибирующей связывание IFNAR2 в широком диапазоне подтипов IFN-α. Эти фаги можно затем тестировать против других видов IFN (иных, чем виды IFN-α), такого как IFN-∃, с тем, чтобы отобрать те клоны, которые не проявляют существенного связывания с другими видами IFN. Наконец, отобранные фаговые клоны можно проверить на их способность нейтрализовать биологическую активность, например, противовирусную активность, разных подтипов IFN-α, и клоны, представляющие антитела с широкой нейтрализующей активностью против большинства, предпочтительно всех подтипов IFN-α, отбирают окончательно.
Селекционная активность анти-IFN-α клонов
В одном из вариантов осуществления настоящего изобретения созданы анти-IFN-α антитела, которые связывают, а также нейтрализуют активность большинства, предпочтительно всех подтипов IFNα и существенно не связывают или существенно не нейтрализуют активность любых других видов интерферона. Например, можно протестировать способность различных фаговых клонов нейтрализовать противовирусные активности различных подтипов IFN-α практически тем же способом, который описан раньше для антител.
4. Получение растворимого IFNAR2-IgG
кДНК, кодирующую человеческие иммуноглобулиновые слитые белки (иммуноадгезины), на основе внеклеточного домена (ECD) hIFNAR2 (клон pRK5 hIFNAR2-IgG), можно создать с использованием способов, аналогичных способам, которые описаны Haak-Frendscho et al., Immunology 79:594-599 (1993) для конструирования мышиного IFN-α рецепторного иммуноадгезина. Вкратце, плазмиду pRKCD42Fc1 конструируют так, как описано в Примере 4 WO 89/02922 (PCT/US88/03414, опубликованная 6 апреля 1989 г.). кДНК, кодирующую последовательность первых 216 остатков зрелого hIFNAR2 ECD, получают из опубликованной последовательности (Novick et al., Cell, 77:391-400 [1994]). Последовательность, кодирующую CD4 в pRKCD4Fc1, замещают кДНК, кодирующей hIFNAR2 ECD, с образованием клона pRK5hIFNAR2-IgG. hIFNAR2-IgG экспрессируется в клетках 293 почки эмбриона человека путем временной трансфекции с использованием метода осаждения фосфатом кальция. Полученный иммуноадгезин выделяют очисткой из супернатантов бессывороточных клеточных культур одной стадией хроматографией по сродству на колонке с белком А-сефарозой, как описано у Haak-Frendscho et al. (1993), выше. Связанный hIFNAR2-IgG элюируют 0,1 М цитратным буфером, рН 3,0, содержащим 20% (мас./об.) глицерина. hIFNAR2-IgG выделяют очисткой, достигающей 95% чистоты, если судить по SDS-PAGE.
5. Использование анти-IFN-α антител в диагностике
Анти-IFN-α антитела настоящего изобретения являются уникальными аналитическими реагентами в диагностических анализах экспрессии IFN-α. Как указано ранее, экспрессия IFN-α повышается при некоторых аутоиммунных заболеваниях, таких как IDDM, SLE, и аутоиммунном тиреоидите. Повышенную экспрессию различных подтипов IFN-α в таких заболеваниях можно выявить и количественно оценить с использованием анти-IFN-α антител настоящего изобретения с широкой реактивностью против большинства подтипов IFN-α. Анти-IFN-α антитела пригодны также для выделения очисткой по сродству различных подтипов IFN-α из рекомбинантных клеточных культур или из природных источников.
Анти-IFN-α антитела можно использовать для детекции IFN-α в любом из многочисленных хорошо известных диагностических способов анализа. Например, биологический образец можно проанализировать на наличие IFN-α после получения такого образца из требуемого источника, смешивания данного образца с анти-IFN-α антителом, что позволяет антителу образовать комплекс антитело/IFN-α с любым подтипом IFN-α, присутствующим в смеси, и детектировать любой комплекс антитело/IFN-α, представленный в смеси. Исследуемый биологический образец можно подготовить для анализа с помощью способов, известных в данной области техники и подходящих для конкретного образца. Способы смешивания биологического образца с антителами и способы детектирования комплекса антитело/IFN-α выбирают в соответствии с видом применяемого анализа. Такие анализы включают конкурентный и сэндвич-анализ, а также анализ стерического ингибирования. В конкурентном способе и сэндвич-способе используют стадию фазоразделения в качестве составляющей данного способа, тогда как анализ стерического ингибирования осуществляют в одной реакционной смеси.
В аналитических способах для всех IFN-α используют один или более из следующих реагентов: меченый аналог IFN-α, иммобилизованный аналог IFN-α, меченое анти-IFN-α антитело, иммобилизованное анти-IFN-α антитело и стерические конъюгаты. Меченые реагенты известны также в качестве "трейсеров" ("радиометок").
Используемая метка представляет собой любую детектируемую функциональность, которая не влияет на связывание IFN-α с анти-IFN-α антителом. Известно множество меток, которые используют в иммунологическом анализе, примеры включают составляющие, которые можно детектировать непосредственно, такие как флуорохром, хемилюминесцентная составляющая и радиоактивные метки, а также такие составляющие, как ферменты, которые должны взаимодействовать или дериватизироваться, чтобы детектироваться. Примеры таких меток включают радиоизотопы Р32, С14, I125, Н3 и I131, флуорофоры, такие как редкоземельные хелаты или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люцифераза светлячка и бактериальная люцифераза (Пат. США №4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидаза хрена (HRP), щелочная фосфатаза, β-галактозидаза, глюкоамилаза, лизоцим, сахаридоксидазы, например, глюкооксидаза, галактооксидаза и глюкозо-6-фосфатдегидрогеназа, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, конденсированные с ферментом, который использует перекись водорода для окисления красителя-предшественника, такого как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спин-метки, метки бактериофага, стабильные свободные радикалы и т.п.
Для ковалентного связывания этих меток с белками в распоряжении имеются традиционные способы. Например, конденсирующие агенты, такие как диальдегиды, карбодиимиды, дималеимиды, бис-имидаты, бис-диазотированный бензидин и им подобные, можно использовать для маркирования антител с помощью вышеуказанных флуоресцентных, хемилюминесцентных и ферментативных меток. См., например, патент США №№3940475 (флуориметрия) и 3645090 (ферменты); Hunter et al., Nature, 144:945 (1962); David et al., Biochemistry, 13:1014-1021 91974); Pain et al., J. Immunol. Methods, 40:219-230 (1981); и Nygren, J. Histochem. and Cytochem., 30:407-412 (1982). Предпочтительными метками здесь являются ферменты, такие как пероксидаза хрена и щелочная фосфатаза.
Конъюгирование такой метки, в том числе и ферментов, с антителом является стандартной рабочей операцией для специалистов в области иммунологического анализа. См., например, O'Sullivan et al., "Methods for the Preparation of Enzyme-antibody Conjugates for Use in Enzyme Immunoassay", in Methods Enzymology, ed. J.J.Langone and H.Van Vunakis, Vol.73 (Academic Press, New York, New York, 1981), pp.147-166.
Для некоторых способов анализа необходимы реагенты для иммобилизации. Иммобилизация влечет за собой отделение анти-IFN-α антитела от любого IFN-α, который остается свободным в растворе. Это обычно происходит либо в результате нерастворения анти-IFN-α антитела или аналога IFN-α до процедуры анализа, а также в результате адсорбции на водонерастворимую матрицу или поверхность (Bennich et al., Пат. США №3720760) вследствие ковалентного присоединения (например, с использованием сшивания глутаральдегидом) либо из-за наступившего позднее нерастворения анти-IFN-α антитела или аналога IFN-α, например вследствие иммунопреципитации.
Другие способы анализа, известные в качестве конкурентного или сэндвич-анализа, весьма укоренились и широко используются в коммерческой диагностике.
Конкурентные анализы зависят от способности индикаторного аналога IFN-α конкурировать с тестируемым образцом IFN-α за ограниченное количество антиген-связывающих сайтов анти-IFN-α антитела. Анти-IFN-α антитело обычно инсолюбилизируют до или после конкуренции и радиометку и IFN-α, связанные с анти-IFN-α антителом, отделяют от несвязанных радиометки и IFN-α. Разделение осуществляют путем декантирования (причем связывающий партнер заранее инсолюбилизируют) или путем центрифугирования (когда партнер связывания осаждают после конкурентной реакции). Количество тестируемого образца IFN-α обратно пропорционально количеству связанной радиометки, что измеряют по количеству маркерного вещества. Получают дозозависимые кривые с известными количествами IFN-α и сравнивают с результатами тестирования количественного определения присутствующего в тест-образце содержания IFN-α. При использовании ферментов в качестве детектируемых маркеров эти анализы именуются ELISA-системами.
Другие виды конкурентного анализа, называемые "гомогенным" анализом, не нуждаются в фазоразделении. При этом получают конъюгат фермента с IFN-α и используют его так, что при связывании анти-IFN-α антитела с IFN-α присутствие анти-IFN-α антитела модифицирует ферментативную активность. В данном случае IFN-α или его иммунологически активные фрагменты конъюгируют с помощью бифункционального органического мостика с ферментом, таким как пероксидаза. Конъюгаты отбирают для использования с анти-IFN-α антителом так, что связывание анти-IFNα антитела ингибирует или потенцирует ферментативную активность метки. Этот способ per se широко практикуется под названием EMIT.
Стерические конъюгаты используют для способов стерического препятствования в гомогенном анализе. Эти конъюгаты синтезируют путем ковалентного связывания низкомолекулярных гаптенов с небольшим фрагментом IFN-α так, чтобы антитело с гаптеном оказались фактически неспособными связать конъюгат одновременно с анти-IFN-α антителом. Во время данной аналитической процедуры IFN-α, присутствующий в данном тестируемом образце, должен связать анти-IFNα антитело, позволяя тем самым анти-гаптену связать конъюгат, приводя к изменению свойств конъюгированного гаптена, например, к изменению флуоресценции, если данный гаптен является флуорофором.
Сэндвич-анализ особенно пригоден для определения IFN-α или анти-IFN-α антител. В последовательных сэндвич-анализах иммобилизованное анти-IFN-α антитело используют для адсорбции тестируемого образца IFN-α, данный тестируемый образец удаляют путем промывки, связанный IFN-α используют для адсорбции второго меченого анти-IFN-α антитела и затем связанный материал отделяют от оставшейся радиометки. Количество связанной радиометки прямо пропорционально тестируемому образцу IFN-α. В "одновременных" сэндвич-анализах тестируемый образец не отделяют перед добавлением меченого анти-IFN-α. Последовательный сэндвич-анализ с использованием анти-IFN-α моноклонального антитела в качестве первого антитела и поликлонального анти-IFN-α антитела в качестве другого пригоден для тестирования образцов на IFN-α.
Вышеприведенное представляет собой простой типовой диагностический анализ на IFN-α. Другие современные способы или создаваемые в дальнейшем, которые используют анти-IFN-α антитело для определения IFN-α, включены в рамки настоящего изобретения, в том числе и вышеописанные биоаналитические методы.
6. Терапевтические композиции и введение анти-IFN-α антител
Терапевтические композиции анти-IFN-α антител настоящего изобретения получают для хранения путем смешивания антитела, обладающего требуемой степенью очистки, с необязательно физиологически приемлемыми носителями, наполнителями или стабилизаторами (Remington: The Science and Practice of Pharmacy, 19th Edition, Alfonso, R., ed, Mack Publishing Co. (Easton, PA: 1995)), в виде лиофилизованного брикета или водных растворов. Приемлемые носители, наполнители или стабилизаторы не являются токсичными для реципиентов в используемых дозах и концентрациях и включают буферы, такие как фосфатный, цитратный, и другие органические кислоты; антиоксиданты, в том числе аскорбиновую кислоту; низкомолекулярные (менее чем около 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и иные углеводы, включая глюкозу, маннозу или декстрины; хелатообразующие (комплексообразующие) агенты, такие как ЭДТА; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как Твин, Плуроник или полиэтиленгликоль (PEG).
Анти-IFN-α антитело, используемое для введения in vivo, должно быть стерильным. Это легко осуществляется путем фильтрации через стерильные фильтрующие мембраны до или после лиофилизации и восстановления влагосодержания. Анти-IFN-α антитело обычно хранят в лиофилизованной форме или в растворе.
Терапевтические композиции анти-IFN-α антитела, как правило, помещают в контейнер, обладающий стерильным входным отверстием, например, мягкий резервуар для внутривенного раствора или флакон, обладающий пробкой, прокалываемой подкожной инъекционной иглой.
Путь введения анти-IFN-α антитела согласуется с известными способами, например инъекция или инфузия с помощью внутривенного, внутрибрюшинного, внутричерепного, подкожного, внутримышечного, внутриглазного, внутриартериального, внутрицереброспинального или внутриочагового пути, или, как указано выше, с помощью систем замедленного высвобождения. Предпочтительно данное антитело дают системно.
Соответствующие примеры препаратов с замедленным высвобождением включают полимерные полупроницаемые матрицы в виде профилированных изделий, например, пленок или микрокапсул. Матрицы с замедленным высвобождением включают полиэфиры, гидрогели, полилактиды (США №3773919, ЕР 58481), сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамат (Sidman et al., Biopolymers, 22:547-556 (1983)), поли(2-гидроксиэтилметакрилат) (Langer et al., J. Biomed. Mater. Res., 15:167-277 (1981) и Langer, Chem. Tech., 12:98-105 (1982)), этиленвинилацетат (Langer et al., выше) или поли-D-(-)-3-гидроксимасляная кислота (ЕР 133988). Композиции анти-IFNAR2 антител с замедленным высвобождением включают также захваченное в липосому антитело. Содержащие антитело липосомы получают способом, известным per se: DE 3218121; Epstein et al., Proc. Natl. Acad. Sci. USA, 82:3688-3692 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA, 77:4030-4034 (1980); EP 52322; EP 36676; EP 88046; EP 143949; EP 142641; Японская патентная заявка 83-118008; США 4485045 и 4544545; а также ЕР 102324. Обычно эти липосомы представляют собой небольшие (около 200-800 ангстрем) однослойные липосомы, в которых липидное содержание составляет более чем около 30 мол.% холестерина, выбранную пропорцию которого доводят для оптимальной антительной терапии.
Анти-IFN-α антитело можно также вводить путем ингаляции. Имеющиеся в продаже ингаляторы для жидких композиций, включая струйные ингаляторы и ультразвуковые ингаляторы, пригодны для введения. Жидкие композиции можно распылять сразу, а лиофилизированный порошок можно распылять после восстановления влагосодержания. Альтернативно анти-IFN-α антитело можно распылять с использованием фторированного углеводородного газа и дозируемого ингалятора, или распылять в виде лиофилизированного и измельченного порошка.
"Эффективное количество" терапевтически используемого анти-IFN-α антитела зависит, например, от терапевтических целей, пути введения, используемого типа анти-IFN-α антитела и состояния пациента. В соответствии с этим терапевту необходимо учесть титр дозы и подобрать путь введения, необходимого для получения оптимального терапевтического эффекта. Обычно лечащий врач вводит анти-IFN-α антитело в дозе, достаточной для получения требуемого эффекта. За ходом лечения нетрудно следить с помощью традиционных анализов.
Пациенты, которых лечат анти-IFN-α антителом настоящего изобретения, включают доклинических пациентов или пациентов, у которых недавно проявились иммуноопосредованные нарушения и, в частности, аутоиммунные нарушения. Пациенты являются кандидатами для лечения согласно настоящему изобретению до того момента, пока больная ткань остается незащищенной от иммуноопосредованного разрушения. Например, пациенту, страдающему инсулинозависимым сахарным диабетом (IDDM), лечение анти-IFN-α антителом настоящего изобретения может приносить пользу до тех пор, пока панкреатические островковые клетки не утратят своей жизнеспособности. При развитии иммуноопосредованного или аутоиммунного нарушения желательно вводить анти-IFN-α антитело настоящего изобретения как можно раньше и при необходимости продолжать лечение как можно дольше, чтобы защитить здоровую ткань от разрушения со стороны иммунной системы пациента. Например, пациента с IDDM лечат до тех пор, пока инсулиновый мониторинг не станет демонстрировать адекватный ответ островков, а другие показатели некроза островков не уменьшатся (например, снижение титра анти-островковых антител), после чего этому пациенту можно отменить лечение анти-IFN-α антителом на испытательный срок, в течение которого контролируют инсулиновый ответ и уровень анти-островковых антител до рецидива болезни.
Для лечения и профилактики иммуноопосредованного или аутоиммунного нарушения анти-IFN-α антителом составляют композицию для данного антитела, дозируют и вводят ее способом, согласующимся с хорошей медицинской практикой. Факторы, рассматриваемые в этой связи, включают конкретное нарушение, которое лечат, конкретное млекопитающее, которое лечат, клиническое состояние отдельного пациента, причину нарушения, участок доставки антитела, конкретный тип антитела, способ введения, составление графика введения и другие факторы, известные лечащему врачу. "Терапевтически эффективное количество" антитела, которое нужно ввести, должно определяться такими соображениями и равно минимальному количеству, необходимому для профилактики, улучшения или лечения данного нарушения, включая лечение хронических аутоиммунных состояний и поддержание иммуносупрессии у реципиентов с трансплантатами. Такое количество предпочтительно ниже количества, которое токсично для хозяина или придает хозяину гораздо большую чувствительность к инфицированию.
В качестве общего высказывания, начальное фармацевтически эффективное количество парентерально вводимого антитела колеблется в диапазоне от около 0,1 до 50 мг/кг массы тела пациента в день, с обычным исходным диапазоном для используемого антитела, составляющим 0,3-20 мг/кг/день, более предпочтительно 0,3-15 мг/кг/день. Требуемую дозу доставляют с помощью однократного болюсного введения, с помощью множественных болюсных введений или путем непрерывной инфузии антитела в зависимости от характера фармакокинетического ослабления, которого желает достигнуть практикующий врач.
Однако, как указано выше, эти предполагаемые количества антитела зависят во многом от лечения по усмотрению врача. Решающим фактором в выборе надлежащей дозы и графика ее ведения является получаемый результат, как указано выше.
Антитело можно, но необязательно, готовить и сочетании с одним или более агентами, используемыми в настоящее время для профилактики или лечения иммуноопосредуемого или аутоиммунного нарушения, о котором идет речь. Например, при ревматоидном артрите антитело можно назначать в сочетании с глюкокортикостероидом. Эффективное количество таких других агентов зависит от количества анти-IFN-α антитела, присутствующего в композиции, вида нарушения или лечения и иных указанных выше факторов. Их, как правило, используют в таких же дозах и вводят теми же путями, что и используемые выше, или около от 1 до 99% от доз, используемых до этого.
Дополнительные подробности настоящего изобретения можно найти в нижеследующих примерах, которые также приведены в рамках настоящего изобретения. Все ссылки, указанные в настоящем описании, а также процитированные ссылки, представленные таким образом, включены путем ссылки в их полноте.
ПРИМЕРЫ
Нижеследующие примеры предлагаются в качестве иллюстрации, а не в качестве ограничения. Данные примеры составлены таким образом, чтобы обеспечить специалистов в данной области техники полнотой раскрытия и описанием создания и использования данных соединений, композиций, а также способов настоящего изобретения, и они не подразумевают ограничить все то, что заявители относят к своему изобретению. Предприняты усилия обеспечить точность в отношении используемых чисел (например, количественные, температурные и т.п.), хотя нельзя и исключить некоторых экспериментальных ошибок и отклонений. Если не указано иначе, доли представляют собой доли массы, температура представлена в градусах С, а давление равно или близко атмосферному. Раскрытие всех цитирований в настоящем описании включено здесь путем ссылки.
Пример 1. Получение и характеристика анти-IFN-α моноклонального антитела мыши с широкой реактивностью
Материал и методы
Мышиное моноклональное антитело с широкой реактивностью против IFN-α-подтипов
Общее антитело, нейтрализующее IFN-α, было создано путем последовательной иммунизации мышей смесью подтипов IFN-α человека, образования большого числа кандидатных mAb и последующего их скринирования на связывание и активность. В частности, мышей Balb/c иммунизируют в подушечку задней лапы 9 раз (с интервалом в две недели) 2,5 г лимфобластоидного hIFN-∀ (Product No.I-9887 от Sigma, St. Louis, MO), ресуспендированного в MPL-TDM (Ribi Immunochemical Research, Inc., Hamilton, MT). Через три дня после последней иммунизации клетки подколенного лимфатического узла сливают с мышиными миеломными клетками P3X63Ag8.U.1 (ATCC CRL1597) с использованием 35% полиэтиленгликоля. Гибридомы селектируют в HAT-среде. Через десять дней после слияния культуральные супернатанты гибридом впервые скринируют на mAb-связывание с разными видами hIFN-α в ELISA. Отобранные супернатанты гибридомных культур тестируют на их способность подавлять противовирусный цитопатический эффект IFN в клеточной линии А549 клеток рака легкого человека, как описано ниже. Как указано на Фиг.1, три mAb, полученные в 1794 луночных слияниях, оказались способны нейтрализовать разнообразный набор подтипов IFN-α. Эти три mAb были субклонированы и проанализированы повторно.
Нейтрализация противовирусной активности IFN-α
Способность кандидатного антитела нейтрализовать противовирусную активность IFN-α анализируют, как описано у Yousefi, S., et al., Am. J. Clin. Pathol., 83:735-740 (1985). Вкратце, данный анализ осуществляют с использованием А549-клеток рака легкого человека, зараженных вирусом энцефаломиокардита (EMC). Последовательные разведения mAb инкубируют с различными представителями интерферонов типа I в течение одного часа при 37°С в общем объеме 100:1. Затем эти смеси инкубируют с 5×105 клеток А549 в соотношении 100:1 в клеточной культуральной среде в течение 24 часов. После этого клетки заражают с помощью 2×105 б.о.е. вируса ЕМС в последующие 24 часа. В конце инкубации путем визуального микроскопирования или окрашиванием кристалвиолетом определяют клеточную жизнеспособность. Титр (ЕС50) нейтрализующего антитела определяют в виде концентрации антитела, которая нейтрализует 50% противовирусного цитопатического эффекта, обусловленного IFN типа I, 100 единиц/мл. Представителей IFN типа I, используемых в данном исследовании, определяют с использованием NIH-эталонного рекомбинантного человеческого IFN-∀2 в качестве стандарта. Были протестированы специфические активности следующих разных IFN типа I: IFN-∀2/-∀1 (IFN-∀2 остатки 1-62/-∀1 остатки 64-166) (2×107 МЕ/мг), IFN-∀1 (3×107 МЕ/мг), IFN-∀2 (2×107 МЕ/мг), IFN-∀5 (8×107 МЕ/мг), IFN-∀8 (19х107) и IFN-∀10 (1,5×105 МЕ/мг). Тестируемый IFN лейкоцитов был представлен Sigma Product No. I-2396. Тестируемый лимфобластоидный IFN был представлен NIH-эталонным стандартом Ga23-901-532. Данные, приведенные на Фиг.3В, были получены с использованием вышеописанной формы анализа в экспериментах, осуществленных в Access Biomedical (San Diego, CA) по просьбе заявителя.
Анализ изменения электрофоретической подвижности
Большинство непосредственных действий IFN связано с активацией латентных цитоплазматических сигнальных трансдукторов и активаторов транскрипции (STAT) белков, которые образуют мультибелковый комплекс, интерферон-стимулированный генный фактор-3 (ISGF3), индуцирующий транскрипцию с целевого промотора интерферон-стимулированного элемента ответа (ISRE). ISGF3 состоит из трех белковых субъединиц: STAT1, STAT2 и p48/ISGF3γ. Белок р48 относится к семейству факторов, регулирующих интерферон (IFR), и представляет собой белок, связывающий ДНК, который непосредственно взаимодействует с ISRE. Таким образом, мониторинг ISRE, специфичного клеточного комплекса, связывающего ДНК в ответ на активирование IFN, дает простой, быстрый и удобный способ оценки влияния IFN на клетки-мишени. Одной из удобных форм осуществления такого исследования является анализ изменения электрофоретической подвижности (EMSA), в котором индукция ISRE-связывающей активности обработкой IFN приводит к изменению электрофоретической подвижности радиоактивно меченного двухцепочечного олигонкулеотидного зонда, соответствующего консенсусной последовательности ISRE.
Данный анализ осуществляют практически так, как описано Kurabayashi et al., Mol. Cell Biol. 15:6386 (1995). Вкратце, 5 нг специфичного подтипа IFN-α и разные концентрации (5-100 мкг/мл) анти-IFN-α mAb инкубируют с 5х105 клеток HeLa в 200 мкл DMEM в течение 30 минут при 37°С. Клетки преинкубируют с антителом в течение 15 минут при 4°С перед добавлением hIFN-∀. Клетки промывают в PBS и ресуспендируют в 125 мкл буфера А (10 мМ HEPES, рН 7,9, 10 мМ KCl, 0,1 мМ ЭДТА, 1 мМ DTT, 1 мМ фенилметилсульфонилфторида, 10 мкг/мл лейпептина, 10 мг/мл апротинина). После 15 минут инкубации на льду клетки лизируют добавлением 0,025% NP40. Осадок ядер получают путем центрифугирования и ресуспендируют в 50 мкл буфера В (20 мМ HEPES, рН 7,9, 400 мМ NaCl, 0,1 мМ ЭДТА, 1 мМ DTT, 1 мМ фенилметилсульфонилфторида, 10 мкг/мл лейпептина, 10 мкг/мл апротинина) и выдерживают на льду в течение 30 мин. Полученную ядерную фракцию осветляют центрифугированием, а полученный супернатант хранят при -70°С перед использованием. Двухцепочечные зонды получают из одноцепочечных олигонуклеотидов (ISG15 верхний: 5'-GATCGGGAAAGGGAAACCGAAACTGAAGCC-3' [SEQ ID NO.13], ISG15 нижний: 5'-GATCGGCTTCAGTTTCGGTTTCCCTTTC CC-3' [SEQ ID NO.14]), используя ДНК-полимеразу I Кленова и реакцию включения Р32-dATP (3000 Ки/мМ, Amersham). Меченые олигонуклеотиды выделяют очисткой из невключенных радиоактивно меченных нуклеотидов с использованием колонок BIO-Spin 30 (Bio-Rad). Реакции связывания, содержащие 5 мкл ядерного экстракта, 25000 cpm меченого зонда и 2 мкг неспецифичного конкурента поли(dI-dC)-поли (dI-dC) в 15 мкл буфера для связывания (10 мМ Трис-HCl, рН 7,5, 50 мМ NaCl, 1 мМ ЭДТА, 1 мМ DTT, 1 мМ фенилметилсульфонилфторида и 15% глицерина), инкубируют при комнатной температуре в течение 30 минут. Комплексы ДНК-белок растворяют в 6% неденатурирующих полиакриламидных гелях и анализируют с помощью ауторадиографа. Специфичность анализа определяют добавлением в отдельные реакционные смеси 350 нг немеченого зонда ISG15. Образование специфичного комплекса ISGF3 подтверждают путем анализа резкого изменения подвижности анти-STAT1-антитела.
Клонирование гена, кодирующего моноклональное антитело 9F3 против IFN-α
Получают мышиное моноклональное антитело 9F3 против IFN-α человека, клонируют его и секвенируют. Плазмида pEMX1, используемая для экспрессии и мутагенеза F(ab)s в Е.coll, была описана раньше (Werther et а1., J.Immunol. 157:4986-4995 [1996]). Вкратце, плазмида содержит фрагмент ДНК, кодирующий консенсус легкой κ-цепи подгруппы 1 человека (VLκ1-CL) и консенсус тяжелой цепи подгруппы III человека (VHIII-CH1), а также промотор щелочной фосфатазы. Использование консенсусных последовательностей для VL и VH было описано раньше (Carter et а1., Proc. Natl. Acad. Sci. USA 89:4285-4289 [1992]).
Результаты
Авторы изобретения ранее показали, что существует широкий спектр подтипов IFN-α, экспрессируемых островками Лангерганса у пациентов с IDDM (Huang et al., Diabetes 44:658-664 [1995]). Авторы также продемонстрировали, что не существует явной взаимосвязи между IDDM и экспрессией IFN-β или IFN-γ (Huang et al., [1995] выше). Несмотря на то что не было показано, что специфичные подтипы IFN-α экспрессируются как часть SLE-патологии, а также IDDM, взаимосвязь существует с IFN-α, но не с IFN-β или IFN-γ (Hooks, et al., Arthritis & Rheumatism 25:396-400 [1982]; Kirn, et al., Clin. Exp. Immunol. 70:562-569 [1987]; Lacki, et al., J. Med. 28:99-107 [1997]; Robak, et al., Archivum Immunologiae et Theraplae Experimental is 46:375-380 [1998]; Shiozawa, et al., Arthritis & Rheumatism 35:417-422 [1992]; von Wussow, et al., Rheumatology International 8:225-230 [1988]). Эти наблюдения привели авторов к предположению о том, что кандидатное антитело для терапевтического вмешательства при IDDM или SLE необходимо для нейтрализации большинства подтипов IFN-α, тогда как остальные интактные активности других интерферонов (β, γ и ω) и интерлейкинов могут понадобиться для защиты хозяина.
Одно из них (9F3) способно нейтрализовать широкий спектр подтипов рекомбинантного интерферона α и было дополнительно охарактеризовано. Как показано на Фиг.2, 9F3 способно нейтрализовать противовирусную активность семи рекомбинантных интерферонов IFN-α - 2, 4, 5, 8 и 10 (Фиг.2), а также IFN-α 1 и 21 (Таблица 2 и Фиг.6). Эти подтипы IFN-α охватывают весь спектр последовательностей, в виде прогнозируемой дендрограммы последовательностей для интерферона типа I. Более того, mAb 9F3, которое нейтрализует подтипы IFN-α, оказалось неспособно нейтрализовать IFN-β (Фиг.2, Таблица 2) или IFN-γ. Небольшое увеличение активности для IFN-β, показанное на Фиг.2, не воспроизводится в других анализах и возникает в результате погрешности анализа.
Были созданы и другие mAb, которые являются нейтрализующими в отношении IFN-α (Tsukui et al., Microbiol. Immunol. 30:1129-1139 [1986]; Berg, J. Interferon Res. 4:481-491 [1984]; Meager and Berg, J. Interferon Res. 6:729-736 [1986]; Патент США №4902618; и ЕР-публикация №0139676 В1). Однако эти антитела нейтрализуют лишь ограниченное число рекомбинантных IFN-α-подтипов и неспособны нейтрализовать широкий спектр подтипов IFN-α, таких, которые продуцируются активированными лейкоцитами. Напротив, mAb 9F3 способно нейтрализовать, по меньшей мере, 95% противовирусной активности в коллекции различных подтипов IFN-α, продуцируемых активированными лейкоцитами (Фиг.3А). Подобным образом mAb 9F3 также способно блокировать противовирусную активность независимого препарата лимфобластоидного IFN (эталонный стандарт NIH), что установлено в независимом эксперименте (Фиг.3В).
Способность mAb 9F3 нейтрализовать IFN-α тестировали также с использованием альтернативного биоаналитического подхода. Данный анализ основан на способности IFN-α активировать связывание сигнальной молекулы интерферон-стимулируемого генного фактора 3 (ISGF3) с олигонуклеотидом, полученным из интерферон-стимулируемого ответного элемента (ISRE) в анализе по связыванию ДНК, известном в качестве анализа изменения электрофоретической подвижности (Horvath et al., Genes Dev. 9:984-994 [1995]). Трансдукция сигналов интерферона типа I в ядро зависит от активации белкового комплекса, ISGF3, с двумя сигнальными трансдукторами и активаторами транскрипции (STAT) белков, STAT1 и STAT2, а также белкового фактора, регулирующего интерферон (IRF), p48/ISGF3γ (Wathelet et al., Mol. Cell 1:507-518 [1998]). Последний представляет собой ДНК-последовательность, распознающую субъединицу ISGF3, и непосредственно взаимодействует с ISRE (McKendry et al., Proc. Natl. Acad. Sci. USA 88:11455-11459 [1991]; John et al., Mol. Cell. Biol. 11:4189-4195 [1991]). Обработка COS-клеток IFN-α или IFN-β приводит к появлению комплекса, соответствующего связыванию ISGF3 с полученным ISRE-зондом. Появление комплекса, индуцируемого IFN-α, но не IFN-β, блокируется mAb 9F3 (Фиг.4). Кроме того, mAb 9F3 способно нейтрализовать активность шести рекомбинантных IFN-α-подтипов, которые протестированы в данном анализе (Таблица 2).
Ингибирование mAb 9F3 образования ISGF3, индуцированного IFN тип I
Обладая установленным фактом, что 9F3 способно нейтрализовать и широкое разнообразие рекомбинантных IFN-α-подтипов и смесь подтипов IFN-α-подтипов, продуцируемых активированными лейкоцитами, авторы клонировали и секвенировали кДНК, кодирующие тяжелую и легкую цепи mAb 9F3. Тяжелая и легкая цепи были выделены очисткой, а полученные N-концевые аминокислотные последовательности используют для конструирования вырожденных 5'-праймеров, соответствующих N-концу, тогда как вырожденные 3'-праймеры были сконструированы для соответствия константной области мышиной легкой κ-цепи и тяжелой цепи IgG2. Соответствующие кДНК клонируют с использованием удобного ПЦР-метода и определяют нуклеотидные последовательности вставок. На Фиг.5 представлено сопоставление последовательности доменов VL (5A) и VH (5B) мышиного моноклонального антитела 9F3, гуманизированного варианта (V13) и консенсусной последовательности тяжелой цепи подгруппы III человека и легкой κ-цепи подгруппы III. Для того чтобы гарантировать, что клонированные кДНК кодируют правильное Mab, отражающее специфичность и параметры mAb 9F3, были созданы рекомбинантные химерные белки, которые используют мышиные кДНК-последовательности и СН1-домен человека, представленные на Фиг.5. Полученная химера (СН8-2) оказалась способна полностью нейтрализовать различные рекомбинантные IFN-α-подтипы (Фиг.6). Аминокислотные последовательности тяжелой и легкой цепей были затем использованы для создания гуманизированного антитела.
Пример 2. Гуманизация моноклонального антитела 9F3, нейтрализующего пан-IFN-α
Материал и методы
Конструирование гуманизированных F(ab)
Чтобы сконструировать первый F(ab)-вариант гуманизированного 9F3, осуществляют сайт-направленный мутагенез (Kunkel, Proc. Natl. Acad. Sci. USA 82:488-492 [1985]) на дезоксиуридин-содержащей матрице pEMX1. Шесть CDR были заменены мышиной последовательностью 9F3 (Фиг.5); остатки, включенные в каждую CDR, происходят из CDR, сформированной на основе данной последовательности (Kabat et al., (1991) выше). Следовательно, F-1 состоит полностью из человеческой каркасной области (подгруппа 1 VL κ и подгруппа III VH) с шестью полными мышиными последовательностями CDR. Плазмиды для всех других F(ab)-вариантов конструируют из плазмидной матрицы F-1. Плазмиды трансформируют в E.coli штамма XL-1 Blue (Stratagene, San Diego, CA) для получения двух- и одноцепочечной ДНК с использованием имеющихся в продаже наборов (Qiagen, Valencia, CA). Для каждого варианта ДНК, кодирующую легкую и тяжелую цепи, полностью секвенируют с использованием способа терминации цепи дидезоксинуклеотидом (Sequenase, U.S. Biochemical Corp., Clevalend, OH). Плазмиды трансформируют в E. coli штамм 16С9, полученный из ММ294, высевают в чашки на среду Луриа, содержащую 50 мкг/мл карбенициллина, и отдельную колонию отбирают для экспрессии белка. Отдельную колонию выращивают в 5 мл среды Луриа со 100 мкг/мл карбенициллина в течение 5-8 ч при 37°С. 5 мл культуры вносят в 500 мл среды АР5, содержащей 50 мкг/мл карбенициллина, и оставляют расти в течение 20 ч в 4-литровой встряхиваемой колбе при 30°С. Среда АР5 состоит из: 1,5 г глюкозы, 11,0 г Hycase SF, 0,6 г дрожжевого экстракта (сертифицированный), 0,19 г MgSO4 (безводный), 1,07 г NH4Cl, 3,73 г KCl, 1,2 г NaCl, 120 мл 1 М триэтаноламина, рН 7,4, воды до 1 л, после чего стерильно фильтруют через 0,1 мкм Sealkeen-фильтр. Клетки собирают центрифугированием в 1 л центрифужной бутыли при 3000×g и супернатант удаляют. После замораживания в течение 1 ч осадок ресуспендируют в 25 мл холодного раствора, содержащего 10 мМ Трис-1 мМ ЭДТА-20% сахарозы, рН 7,5, к которому для подавления протеолиза добавляют 250 мкл 0,1 М бензамидина (Sigma, St. Louis, MO). После осторожного перемешивания на льду в течение 3 ч образец центрифугируют при 40000хg в течение 15 мин. Затем полученный супернатант наносят на колонку (чистый объем 0,5 мл) с белком G-Сефарозой CL-4B (Pharmacia, Uppsala, Sweden), уравновешенной 10 мМ Трис-1 мМ ЭДТА, рН 7,5. Колонку промывают 10 мл 10 мМ Трис-1 мМ ЭДТА, рН 7,5, и элюируют 3 мл 0,3 М глицина, рН 3,0, в 1,25 мл 1 М Триса, рН 8,0. Затем F(ab)-буфер меняют на PBS, применяя Centricon-30 (Amicon, Beverly, MA) и концентрируют до конечного объема 0,5 мл. Гели SDS-PAGE, содержащие все F(ab), разгоняют, чтобы выяснить степень очистки, а молекулярную массу каждого варианта проверяют с помощью электрораспылительной масс-спектрометрии. Концентрации F(ab) определяют с использованием количественного аминокислотного анализа.
Конструирование химерного и гуманизированного IgG
Для образования вариантов IgG2 химерного и гуманизированного 9F3 человека соответствующие мышиные или гуманизированные домены VL и VH (F-13, Таблица 3) субклонируют в отдельные ранее описанные pRK-векторы (Eaton et al., Biochemistry 25:8343-8347 [1986]), которые содержат ДНК, кодирующую CH1-Fc IgG2 человека или CL-домен легкой цепи человека. ДНК, кодирующую полную легкую и полную тяжелую цепь каждого варианта проверяют с помощью дидезоксинуклеотидного секвенирования. Химерный IgG состоит из полного VH-домена мышиного 9F3, слитого с СН1-доменом человека по аминокислоте SerH113, и полного VL-домена мышиного 9F3, слитого с CL-доменом человека по аминокислоте LysL 107.
Тяжелую и легкую цепь плазмид котрансфицируют в трансформированную аденовирусом линию эмбриональных клеток почки человека, 293 (Graham et al., J. Gen. Virol. 36:59-74 [1977]), с использованием высокоэффективного метода (Gorman et al., DNA Prot. Eng. Tech. 2:3-10 [1990]). Ежедневно меняют среды на бессывороточные и в течение всех пяти дней собирают клетки. Антитела выделяют очисткой из объединенных супернатантов с использованием белок А-Сефарозы CL-4B (Pharmacia). Элюированному антителу меняют буфер на PBS с использованием Centricon-30 (Amicon), концентрируемого до 0,5 мл, стерильно фильтруют с использованием Millex-GV (Millipore, Bedford, MA) и хранят при 4°С. Концентрации IgG2 определяют с использованием количественного аминокислотного анализа.
Анализ связывания IFN-α
Для осуществления ELISA 96-луночные планшеты для микротитрования (Nunc) покрывают внесением в каждую лунку 50 мкл 0,1 мкг/мл IFN-α в PBS и инкубируют при 4°С в течение ночи. Затем планшеты трижды промывают буфером для промывки (PBS c 0,05 % Твин-20). После этого лунки планшета для микротитрования блокируют с помощью 200 мкл SuperBlock (Pierce) и инкубируют при комнатной температуре в течение 1 часа. Планшеты вновь промывают три раза промывочным буфером. После стадии промывки в указанные лунки вносят 100 мкл серийно разведенного гуманизированного mAb с исходной концентрацией 10 мкг/мл. Планшеты инкубируют при комнатной температуре в течение 1 часа на аппарате для встряхивания и затем три раза промывают промывочным буфером. Далее в каждую лунку вносят конъюгированные с пероксидазой хрена (HRP) козьи антитела, специфичные к Fab человека (Cappel), разведенные 1:1000 в аналитическом буфере (0,5 % бычьего сывороточного альбумина, 0,05% Твин-20 в PBS). Планшеты инкубируют при комнатной температуре на аппарате для встряхивания и затем промывают три раза промывочным буфером с последующим добавлением в каждую лунку 100 мкл субстрата (TMB, 3,3',5,5'-тетраметилбензидин; Kirkegaard & Perry) и инкубируют при комнатной температуре в течение 10 минут. Реакцию останавливают добавлением в каждую лунку 100 мкл стоп-раствора (от Kirkegaard & Perry) и с помощью автоматизированного считывающего устройства для микротитровальных планшетов считывают поглощение при 450 нм.
BIAcore™-биосенсорный анализ
Связывание IFN-α гуманизированными F(ab), химерными и гуманизированными IgG2-антителами измеряют с использованием BIAcore™-биосенсора (Karlsson et al., Methods: A companion to Methods in Enzymology 6:97-108 [1994]). IFN-α иммобилизуют на сенсорном чипе в концентрации 60 мкг/мл в 50 мМ MES-буфере, рН 6,3. Антитела экспонируют на чипе в концентрации 75 мкг/мл (500 нм) в фосфатно-солевом буферном растворе/1% Твин-20. Измеряют антительный on-rate (kon).
Методы компьютерной графики мышиных и гуманизированных F(ab)
Последовательности VL и VH доменов (Фиг.5А и В) используют для построения компьютерной графикой модели доменов VL-VH мышиных 9F3 (Фиг.7). Данную модель используют для определения остатков каркасных областей, которые должны быть встроены в гуманизированное антитело. Создают также модель гуманизированного F(ab), чтобы проверить правильность отбора остатков каркасной области мыши. Построение моделей осуществляют так, как описано раньше (Carter et al., [1992] выше; Werther et al., [1996] выше).
Результаты
Консенсусную последовательность тяжелой цепи подгруппы III и легкой цепи подгруппы I человека используют в качестве каркасной области для гуманизации, как показано на Фиг.5 (Kabat et al., (1991), выше). Данную каркасную область успешно используют для гуманизации других мышиных антител (Carter et al., Proc. Natl. Acad. Sci. USA 89:4285-4289 [1992]; Presta et al., J. Immunol. 151:2623-2632 [1993]; Eigenbrot et al., Proteins 18:49-62 [1994]; Werther et al., J. Immunol. 157:4986-4995 [1996]). Все гуманизированные варианты первоначально были созданы и скринированы для связывания F(ab), экспрессируемых в E.coli. Обычный выход для 500 мл колб составляет 0,1-0,4 мг F(ab).
Остатки гипервариабельной области (CDR) идентифицируют либо на основе гипервариабельности последовательности (Kabat et al., (1991) выше), либо по кристаллической структуре F(ab)-антигенных комплексов (Chothia et al., Nature 342:877-883 [1989]). Хотя идентификация CDR на основе последовательности больше, чем идентификация CDR на основе структуры, обе они согласуются между собой за исключением CDR-H1. В соответствии с идентификацией на основе последовательности CDR-H1 включает остатки H31-H35, тогда как система на основе структуры идентифицирует в качестве CDR-H1 остатки H26-H32 (остатки легкой цепи количественно присоединяются к L; остатки тяжелой цепи количественно присоединяются к H). В настоящем исследовании CDR-H1 идентифицируют в виде сочетания из двух, т.е. включающие остатки H26-H35. Другие CDR определяют с использованием идентификации на основе последовательности (Kabat et al., (1991) выше).
В первоначальном варианте F-1, CDR-остатки переносят из мышиного антитела в каркасную область человека. Кроме того, создают и тестируют на связывание F(ab), которые состоят из химерной тяжелой цепи и F-1-легкой цепи (Ch-1), а также F-1-тяжелой цепи и химерной легкой цепи (Ch-2). F-1 плохо связывает IFN-α (Таблица 3). Сравнение сродства связывания Ch-1 и Ch-2 (Таблица 3) дает основание полагать, что остатки каркасной области в VH-домене F-1 нужно заменить для улучшения связывания.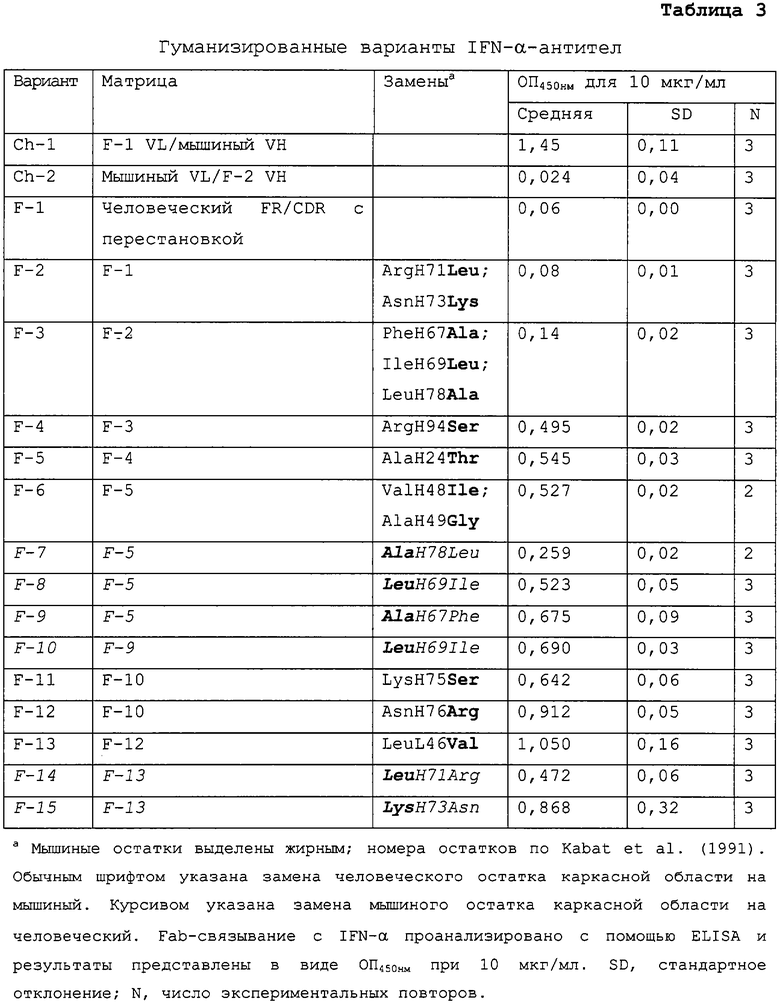
В предшествующих экспериментах по гуманизации (Xiang et al., J. Mol. Biol. 253:385-390 [1995]; Werther et al., [1996] выше), а также в исследованиях F(ab)-антигенных кристаллических структур (Chothia et al., [1989] выше; Tramontano et al., J. Mol. Biol. 215:175-182 [1990]) показано, что остатки Н71 и Н73 могут обладать абсолютным влиянием на связывание, возможно, из-за влияния конформаций CDR-H1 и CDR-H2. Замена человеческих остатков в положениях Н71 и Н73 их мышиными копиями лишь незначительно увеличивает связывание (вариант F-2, Таблица 3). Дальнейшие одновременные замены в положениях Н67, Н69 и Н78 (вариант F-3), а впоследствии и замены ArgH94Ser (вариант F-4) и AlaH24Thr (вариант F-5) существенно увеличивают связывание (Таблица 3). После того как положения Н67, Н69 и Н78 были одновременно заменены, каждое положение вновь заменяют на человеческий консенсусный остаток каркасной области; варианты F-7, F-8, F-9 и F-10 свидетельствуют, что человеческий остаток предпочтительно находится в положении Н67, а положение Н69 не демонстрирует какого-либо предпочтения для человеческого или мышиного остатка, но мышиный остаток предпочтителен в положении Н78.
Во время прежних гуманизаций авторами было обнаружено, что остатки в каркасной петле, FR-3 (Kabat et al., (1991) выше), примыкающие к CDR-H1 и CDR-H2, могут влиять на связывание (Eigenbrot et al., (1994) выше). В соответствии с этим два остатка в этой петле были заменены на соответствующие участки мыши: LysH75 на мышиный Ser (вариант F-11), а AsnH76 на мышиный Arg (вариант F-12). Лишь замена AsnH76Arg улучшает связывание (Таблица 3).
Исследование моделей мышиных и гуманизированных F(ab) дает основание полагать, что остаток L46, погруженный в область контакта VL-VH и взаимодействующий с CDR-H3, может также играть роль в определении конформации CDR-H3 и/или влиять на взаимодействие доменов VL и VH. Подобным образом, положение L49, которое примыкает к CDR-L2, отличается между человеческим консенсусом (Tyr) и последовательности 9F3 (Ser). Поэтому остатки LeuL46Val и TyrL49Ser были заменены одновременно, что дает вариант (F-13) с дополнительным улучшением связывания (Таблица 3). На основании самой лучшей способности связывания среди всех полученных вариантов F-13 был выбран в качестве окончательного гуманизированного варианта.
Гуманизированное рекомбинантное анти-IFN-α моноклональное антитело (V13IgG2) создают путем слияния доменов VH и VL, полученных из F-13, соответственно для CH1-Fc человеческого IgG2 и человеческих CL-доменов. Затем уровни KON и величины KD V13IgG2 сравнивают с химерным IgG2 или мышиным 9F3. BIACore™-измерение связывания V13IgG2 и химерного IgG2 с иммобилизованным IFN-α свидетельствует, что их KON-уровни сходны (Таблица 4). Измерение сродства с использованием Kinexa™-технологии показывает, что сродство V13IgG2 с IFN-α снижено в 2 раза по сравнению с родительским мышиным антителом 9F3 (Таблица 4).

Депонирование материала
Нижеследующие материалы были депонированы в Американской коллекции типовых культур, 10801 University Bvld., Manassas, VA 20110-2209, США (ATCC):
(ID. Ref.: 9F3.18.5)
(ID. Ref.: XAIFN-ChHpDR2)
(ID. Ref.: XAIFN-ChLpDR1)
(ID. Ref.: VHV30-IgG2)
(ID. Ref.: VLV30-IgG)
Данное депонирование осуществляли в соответствии с положениями Будапештского Договора о международном признании депонирования микроорганизмов для целей патентной процедуры и Правил на его основе (Будапештский Договор). Это гарантирует сохранение жизнеспособности культуры в течение 30 лет с даты депонирования. Депонированный материал будет доступен в ATCC в рамках Будапештского Договора и в соответствии с соглашением между Genentech, Inc. и ATCC, которое обеспечивает постоянный и неограниченный общественный доступ к потомству культуры-депонента с момента выдачи соответствующего патента США, или после опубликования патентной заявки США или иностранной заявки в зависимости от того, какое событие наступит раньше, и обеспечивает доступность потомства, как это определено для Уполномоченного по патентам и товарным знакам, имеющим на это право, в соответствии со статьей 35 §122 свода законов США и в соответствии с правилами для Уполномоченного (включая ст. 37 § 1.14 C.F.R. (Свод федеральных правил) c особой ссылкой на 886 OG (наименование официального периодического издания, выпускаемого Бюро Патентов США) 638).
Назначенное лицо по настоящей заявке согласно с тем, что в случае гибели или утраты или уничтожения материалов депонированной культуры при культивировании в подходящих условиях, материалы следует сразу же заменить на другие такие же по получении уведомления. Доступность депонированного материала нельзя рассматривать в качестве лицензии на использование настоящего изобретения в нарушение прав в соответствии с доверенностью от любого правительства, согласно его патентным законам.
Вышеизложенное в письменной форме описание изобретения считается достаточным для возможности осуществления настоящего изобретения специалистом в данной области. Настоящее изобретение не ограничено депонированным материалом, который проиллюстрирован в качестве предпочтительного варианта осуществления настоящего изобретения лишь в одном аспекте, при этом любой функционально эквивалентный материал также подпадает под рамки настоящего изобретения. При этом депонированный материал не является признанием того, что представленное здесь письменное описание является недостаточным для осуществления любого аспекта настоящего изобретения, в том числе лучшего способа осуществления изобретения, а также не является основанием ограничения широты притязаний конкретными представленными примерами. В действительности, различные модификации настоящего изобретения помимо представленных и описанных здесь должны быть очевидны специалистам в данной области техники на основании вышеизложенного описания, которое находится в рамках заявленной формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ АЛЬФА-ИНТЕРФЕРОНА ЧЕЛОВЕКА | 2009 |
|
RU2532832C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ИНТЕРФЕРОНА-АЛЬФА И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2431638C2 |
| АНТИТЕЛА РЕЦЕПТОРА 1 ИНТЕРФЕРОНА АЛЬФА И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2600884C2 |
| АНТИТЕЛА РЕЦЕПТОРА 1 ИНТЕРФЕРОНА АЛЬФА И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2412202C2 |
| Гуманизированное нейтрализующее антитело к интерферону-бета человека | 2019 |
|
RU2737466C1 |
| Вариабельные домены лёгкой и тяжёлой цепи мышиного моноклонального антитела против интерферона альфа (IFN-α) человека, антигенсвязывающий фрагмент (Fab) против IFN-α человека, содержащий указанные домены | 2018 |
|
RU2694672C1 |
| СПОСОБ ЛЕЧЕНИЯ РАССЕЯННОГО СКЛЕРОЗА | 2005 |
|
RU2384345C2 |
| АНТИ-PD-1 АНТИТЕЛА | 2017 |
|
RU2752832C2 |
| АНТИТЕЛА К PD-L1 И ИХ ПРИМЕНЕНИЕ ДЛЯ УСИЛЕНИЯ ФУНКЦИИ Т-КЛЕТОК | 2009 |
|
RU2636023C2 |
| АНТИ-IFNAR1 АНТИТЕЛА С ПОНИЖЕННЫМ СРОДСТВОМ С ЛИГАНДОМ Fc | 2009 |
|
RU2556125C2 |
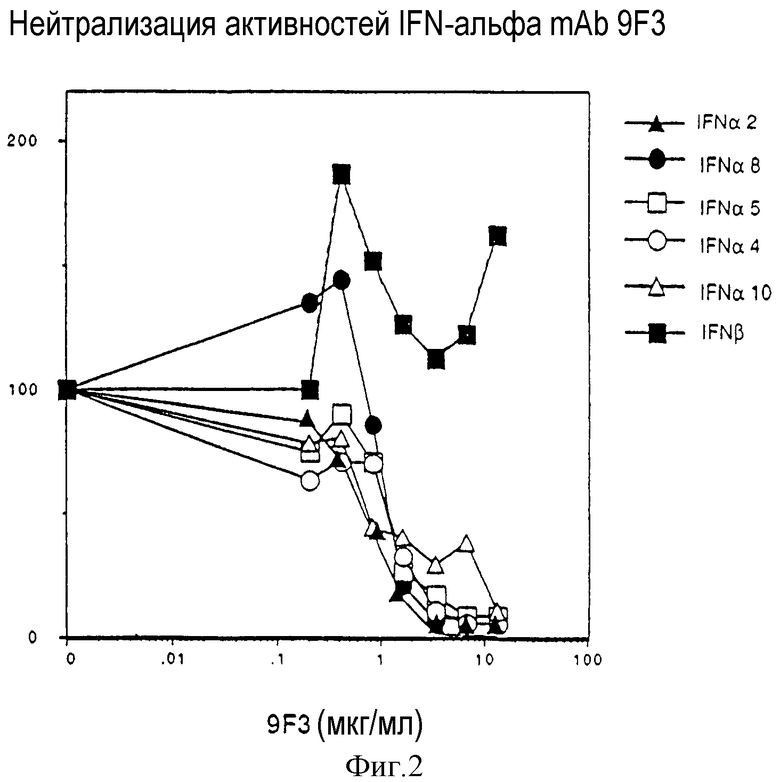
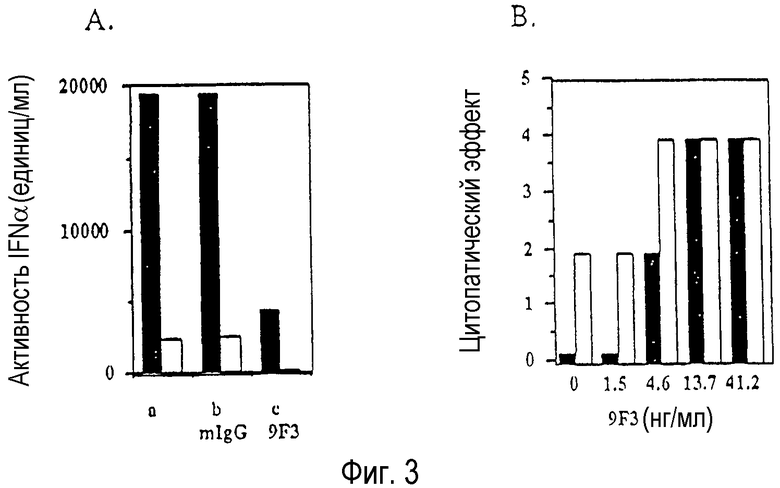
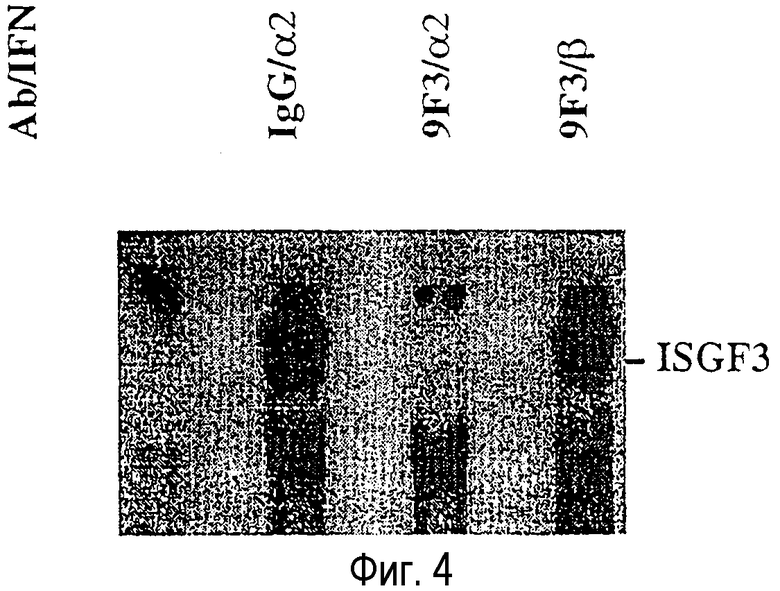


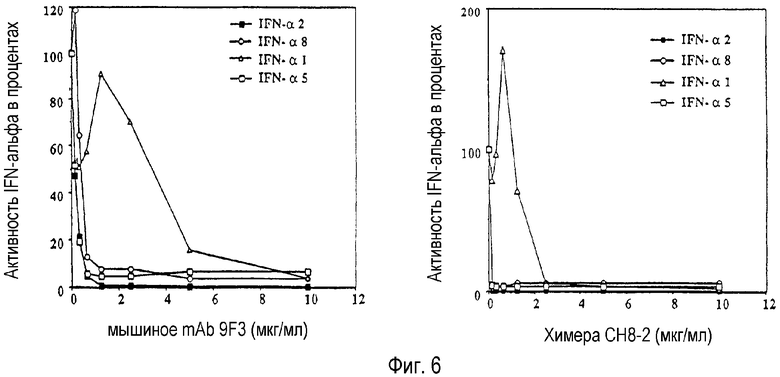

Изобретение относится к биотехнологии и иммунологии. Описано моноклональное анти-IFNα антитело, которое связывается с подтипами IFNα: IFNα1, IFNα2, IFNα4, IFNα5, IFNα8, IFNα10, IFNα21 и содержит три CDR участка тяжелой цепи. Аминокислотная последовательность в описании. Раскрыта тяжелая цепь анти-IFNα антитела или ее фрагмент, которые также содержат указанные CDR участки. Описано анти-IFNα антитело, которое содержит, по-меньшей мере, одну легкую цепь и одну тяжелую. Раскрыты варианты нуклеиновых кислот, кодирующих указанные антитела, и варианты векторов для экспрессии нуклеиновых кислот, а также варианты трансформированных клеток-хозяина. Среди векторов экспрессии описаны также векторы, депонированные под №2881 и под №2882, несущие соответственно тяжелую и легкую цепь антитела. Описан способ получения антитела из указанных клеток. Раскрыта гибридомная линия клеток мыши, депонированная в АТСС под номером №РТА-2917, и антитело, продуцируемое указанной линией. Описаны также варианты фармацевтической композиции на основе антитела и способ диагностики аутоиммунного заболевания. Раскрыто также применение антител для лечения заболевания или состояния, ассоциированного у пациента с повышенными уровнями IFNα. Использование изобретения позволяет одновременно подавлять биологическую активность, по крайней мере, семи подтипов IFNα человека, а именно IFNα1, IFNα2, IFNα4, IFNα5, IFNα8, IFNα10, IFNα21, что может найти применение в диагностике и терапии различных заболеваний человека, опосредуемых IFNα, таких как инсулинозависимый сахарный диабет или системная красная волчанка. 24 н. и 29 з.п. ф-лы. 5 табл., 7 ил.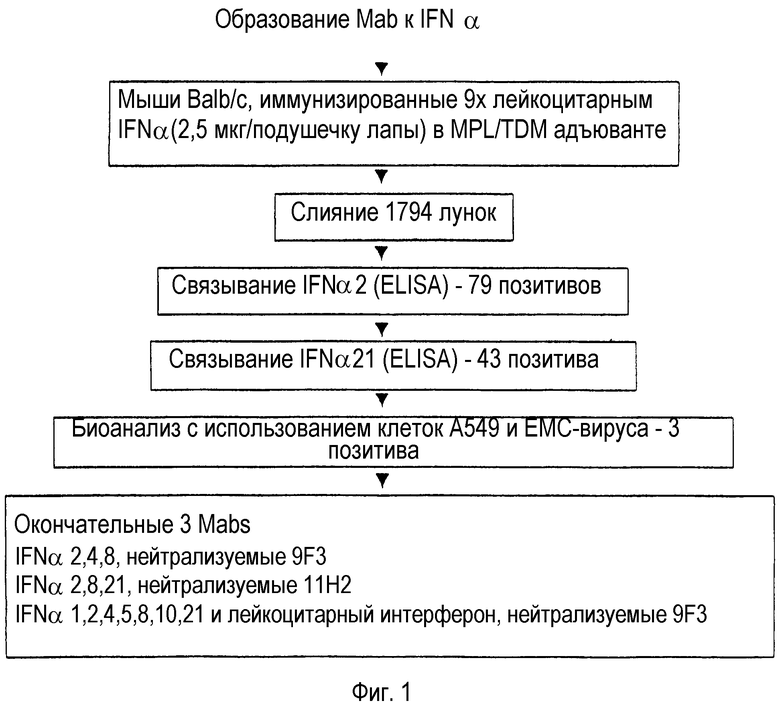
(a) Н1 формулы GYTFTEYIIH (SEQ ID NO:10);
(b) H2 формулы SINPDYDITNYNQRFKG (SEQ ID NO:11);
(c) НЗ формулы WISDFFDY (SEQ ID NO:12).
(a) L1 формулы RASQSVSTSSYSYMH (SEQ ID NO:7);
(b) L2 формулы YASNLES (SEQ ID NO:8); и
(c) L3 формулы QHSWGIPRTF (SEQ ID NO:9); и (В) по меньшей мере, одну тяжелую цепь или ее фрагмент, включающую следующие определяющие комплементарность области:
(a) Н1 формулы GYTFTEYIIH (SEQ ID NO:10);
(b) H2 формулы SINPDYDITNYNQRFKG (SEQ ID NO:11); и
(c) НЗ формулы WISDFFDY (SEQ ID NO:12).
(a) L1 формулы RASQSVSTSSYSYMH (SEQ ID NO:7);
(b) L2 формулы YASNLES (SEQ ID NO:8); и
(c) L3 формулы QHSWGIPRTF (SEQ ID NO:9).
(a) Н1 формулы GYTFTEYIIH (SEQ ID NO:10);
(b) H2 формулы SINPDYDITNYNQRFKG (SEQ ID NO:11); и
(c) НЗ формулы WISDFFDY (SEQ ID NO:12).
(a) L1 формулы RASQSVSTSSYSYMH (SEQ ID NO:7);
(b) L2 формулы YASNLES (SEQ ID NO:8);
(c) L3 формулы QHSWGIPRTF (SEQ ID NO:9);
(d) HI формулы GYTFTEYIIH (SEQ ID NO:10);
(e) H2 формулы SINPDYDITNYNQRFKG (SEQ ID NO:11); и
(f) НЗ формулы WISDFFDY (SEQ ID NO:12), депонированная в Американской Коллекции Типовых Культур с инвентарным №РТА-2917.
(a) L1 формулы RASQSVSTSSYSYMH (SEQ ID NO:7);
(b) L2 формулы YASNLES (SEQ ID NO:8); и
(c) L3 формулы QHSWGIPRTF (SEQ ID NO:9),
депонированный в АТСС с инвентарным №РТА-2882.
(a) Н1 формулы GYTFTEYIIH (SEQ ID NO:10);
(b) H2 формулы SINPDYDITNYNQRFKG (SEQ ID NO:11); и
(c) Н3 формулы WISDFFDY (SEQ ID NO:12),
депонированный в АТСС с инвентарным №РТА-2881.
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| RU 98117079 A, 27.06.2000. | |||

