Изобретение относится к области медицины и биотехнологии и может быть использовано в фармацевтической промышленности. Раскрыт способ контролируемого дегликозилирования и очистки рекомбинантного белка глюкоцереброзидазы, который применяется при длительной фермент-замещающей терапии у пациентов с диагнозом болезнь Гоше.
Болезнь Гоше - наследственное аутосомно-рецессивное метаболическое заболевание, возникающее вследствие дефицита лизосомального фермента бета-глюкоцереброзидазы (GBA). Это самое частое лизосомальное заболевание. Наибольшая распространенность отмечается среди евреев Ашкенази (1 на 855 человек), тогда как среди других популяций она составляет 1 на 100 тыс. В России распространенность составляет 1 на 50 тыс., то есть, приблизительно, около трех тысяч пациентов (1).
Функциональная недостаточность GBA приводит к накоплению субстрата этого фермента, глюкоцереброзида, в организме, преимущественно в печени, селезенке и костном мозге. Наиболее частыми осложнениями болезни Гоше являются костно-суставные поражения, которые являются наиболее частой причиной инвалидности и смертности при болезни Гоше. Существует три типа болезни Гоше. Наиболее распространенный - тип 1, характеризуется широким спектром проявлений, обусловленных цитопенией. Тип 2 и тип 3 - редкие варианты, проявляются в детском возрасте и сопровождаются неврологическими расстройствами. Наиболее эффективна при данном заболевании заместительная терапия глюкоцереброзидазой, которая, в сравнении с другими препаратами, приносит наиболее существенные результаты.
Первым препаратом для ферментативной заместительной терапии была альглюцераза (GBA, выделенная из плацентарного материала человека). Препарат выпускался под названием Глуцераза. Следующим шагом было создание рекомбинантного белка, мутантной формы бета-глюкоцереброзидазы человека с одной аминокислотной заменой Arg495His (имиглюцераза), экспрессированного в трансформированной линии СНО (клетки яичников китайского хомячка). Лекарственное средство на основе этого белка известно под названием Церезим.
Основной проблемой при получении имиглюцеразы является ее сложный профиль гликозилирования. В природной глюкоцереброзидазе имеется 4 сайта гликозилирования, при этом, боковые цепи не содержат концевых манноз. В связи с этим, природная глюкоцереброзидаза практически не способна к проникновению в макрофаги по механизму взаимодействия с их маннозными рецепторами. Рекомбинантная имиглюцераза имеет от одной до девяти структур Man3→Man9 и обладает большим сродством к маннозным рецепторам на поверхности макрофагов. Однако, указанные остатки маннозы «скрыты» другими сахарами и поэтому, чтобы получить активный белок, СНО-экспрессированную имиглюцеразу подвергают в ходе выделения и очистки дальнейшему процессингу с использованием ферментов, изменяющих ее паттерн гликозилирования.
Для решения этой задачи в статье Furbish с сотр. (BiochimicaetBiophysicaActa, 1981, 673, 425-434) применялась последовательная обработка белка нейраминидазой, галактозидазой и β-N-ацетилглюкозаминидазой, для удаления, соответственно, остатков сиаловой кислоты, галактозы и N-ацетилглюкозамина. Эти несколько этапов ферментативной обработки позволяют высвободить значительное число маннозных остатков таким образом, чтобы боковые цепи гликанов преимущественно оканчивались остатками маннозы. Результирующая углеводная структура имиглюцеразы отличается от таковой глюкоцереброзидазы, что обеспечивает существенно большее время полужизни in vivo и более высокое сродство к маннозным рецепторам на поверхности человеческих клеток.
Однако такая последовательная обработка ферментами, хотя и позволяет получить активный белок, не очень эффективна, поскольку существует риск неполного гидролиза соответствующих моносахаридных звеньев одним из ферментов, что блокирует функцию последующих ферментов. Таким образом, данный подход требует продолжительного времени, больших количеств ферментов и в итоге оказывается весьма дорогостоящим, что, в конечном счете, значительно увеличивает стоимость полученного белка. Поэтому задача разработки новых, более экономичных и эффективных, способов выделения и очистки биологически активной глюкоцереброзидазы сохраняется.
Известны различные способы очистки глюкоцереброзидазы с использованием комбинаций хроматографических методов, а также стадий осаждения и экстракции. Однако часть из них относится к очистке белка, выделенного из плаценты (например, J. Biol. Chem. 1973, 248, 5256-5261; P. N. A. S.1977, 74, 3560-3563; ANALYTICAL BIGCHEMISTRY 1986, 154,655-663;CAN. J. BIOCHEM.1982, 60,1025-1031 и т.п.), то есть такие способы не учитывают особенностей белка, полученного рекомбинантным способом, и, как правило, имеют довольно низкие выходы (около 30%). Более современные способы очистки разработаны для глюкоцереброзидазы, в которой в силу различных причин не требуется проводить контролируемое дегликозилирование, например, для белков, содержащих мутации (как в способе, описанном в WO 2011119115) или полученных в специально созданной линии клеток (например, Plant Biotechnology Journal 2007,5, 579-590).
Наиболее близким к заявленному можно считать способ выделения и очистки глюкоцереброзидазы, использованный в патенте US 5549892. Способ включает очистку полученного в клетках СНО белка и последовательную обработку его ферментами, в соответствии с методикой, описанной в статье Furbish (см. выше). Однако детали способа очистки в патенте не раскрыты.
Таким образом, хотя известны различные способы очистки глюкоцереброзидазы и способ ферментативного отщепления остатков сахаров, из уровня техники не известен удобный и эффективный метод, объединяющий эти этапы и позволяющий получить продукт, пригодный далее для создания лекарственного препарата.
Задача заявленного технического решения - получение имиглюцеразы фармакопейного качества путем оптимизации этапов ферментативной обработки белка и многостадийной схемы хроматографической очистки.
Поставленная задача решается за счет разработки способа очистки имиглюцеразы, который совмещает этап гидрофобной хромотографии с дегликозилированием целевого белка за счет ферментативной обработки нейраминидазой, β-1,4-галактозидазой и β-N-ацетилглюкозаминидазой, проходящей в одну стадию.
В одном из вариантов осуществления настоящее изобретение относится к способу дегликозилирования имиглюцеразы, полученной в клетках млекопитающих, отличающемуся тем, что обработка нейраминидазой, β-1,4-галактозидазой и β-N-ацетилглюкозаминидазой проводится в одну стадию.
В предпочтительном варианте соотношение ферментов нейраминидазы: β-1,4-галактозидазы: β-N-ацетилглюкозаминидазыв смеси составляет: 6Е : 1Е : 14Е в расчете на 1 г белка.
В предпочтительном варианте дегликозилирование проводится в течение 22-36 часов при комнатной температуре.
В предпочтительном варианте дегликозилирование проводится в ходе хроматографической очистки целевого белка.
В одном из вариантов осуществления настоящее изобретение относится к способу очистки имиглюцеразы, полученной в клетках млекопитающих, при котором в ходе хроматографической очистки проводится дегликозилирование имиглюцеразы.
В предпочтительном варианте способ очистки имиглюцеразы включает следующие стадии:
а) гидрофобную хроматографию на Butyl-сефарозе,
б) катионный обмен на SP-сефарозе,
в) металло-хелатную хроматографию на IMAC-сефарозе,
г) гидрофобную хроматографию на Phenyl-сефарозе, совмещенную с дегликозилированием целевого белка по любому из пп. 1-4,
д) фильтрацию через Q-сефарозу,
е) катионный обмен на СМ-сефарозе.
Используемый в настоящем документе термин «имиглюцераза» обозначает рекомбинантную форму бета-глюкоцереброзидазы.
Используемый в настоящем документе термин «белок фармакопейного качества» обозначает следующие критерии его чистоты: не менее 99% основного вещества (по обрашенно-фазовой ВЭЖХ), не более 4% родственных примесей (по эксклюзионной ВЭЖХ), удельная активность не менее 35 МЕ/мг, содержание остаточных белков клетки-продуцента не более 50 нг/мг имиглюцеразы.
Предлагаемый способ выделения и очистки имиглюцеразы, синтезируемой в клетках СНО, состоит из нескольких последовательных стадий.
После культивирования рекомбинантного клона-продуцента культуральную жидкость (КЖ), содержащую целевой белок, наносят на колонку с Butyl-сефарозой (гидрофобная хроматография). На данной стадии происходит первичный захват белка, а применение нескольких промывок, содержащих этиленгликоль, позволяет удалить основные родственные примеси имиглюцеразы. Из уровня техники (ANALYTICAL BIGCHEMISTRY 1986, 154, 655-663) известно применение аффинного сорбента для захвата белка из культуральной жидкости, однако невозможность отделения родственных примесей при его использовании, а также высокая стоимость изготовления такого сорбента, подтверждают очевидные преимущества применения гидрофобной хроматографии на первой стадии выделения.
На второй стадии очистки проводится катионообменная хроматография на SP-сефарозе, позволяющая удалить основную часть остаточных примесных белков клеток-продуцентов за счет солевой промывки и щадящей элюции целевого белка с сорбента путем увеличения pH. Хроматографическая чистота целевого белка после первых двух стадий очистки достигает 96-98% как на обращенно-фазовой, так и на эксклюзионной ВЭЖХ.
В целях дальнейшего увеличения чистоты целевого белка до 98-99%, для его использования в качестве фармацевтической субстанции, на третьем этапе очистки используется цинк-хелатная хроматография на IMAC-сефарозе со специфической элюцией имиглюцеразы комбинированным буферным раствором с высоким содержанием хлорида натрия и этиленгликоля. В результате получается высокоочищенный белок, пригодный для дальнейшей обработки дегликозилирующими ферментами с целью увеличения его функциональной активности.
Фракция целевого белка с третьей стадии очистки наносится на сорбент Phenyl-сефарозу. На данном этапе (гидрофобная хроматография) проводится контролируемое дегликозилирование целевого белка при помощи смеси трех ферментов (нейраминидаза, β-1,4-галактозидаза, β-N-ацетилглюкозаминидаза) для укорачивания полисахаридных цепей белка до концевых маннозных остатков. Подобраны минимальные концентрации гликозидаз в реакционной смеси (снижение более чем в 10 раз по сравнению с известными из уровня техники), обеспечивающие полноту прохождения реакции и приемлемую для коммерческого производства скорость процесса. Более того, подтверждена возможность многократного использования оптимизированной ферментной смеси без потери эффективности дегликозилирования. В связи с высокой стоимостью коммерчески доступных гликозидаз, проведенная оптимизация позволяет значительно снизить затраты на получение целевой субстанции.
При использовании в производстве коммерческих ферментов, произведенных в бактериальных клеточных линиях и не прошедших достаточной очистки, может быть введен дополнительный этап удаления эндотоксинов. В таком случае целевой белок после дегликозилирования промывается буферным раствором, содержащим изопропанол, перед элюцией с сорбента.
Полученный частично дегликозилированный белок далее подвергают фильтрации через анионообменный сорбент Q-сефарозу для удаления остаточных эндотоксинов и ДНК клеток-продуцентов.
Последнюю стадию хроматографической очистки проводят на СМ-сефарозе, осуществляя финальную доочистку от примесных белков клеток-продуцентов и перевод белка в буфер готовой формы. В результате получают субстанцию с хроматографической чистотой не менее 99% по обращенно-фазовой ВЭЖХи специфической активностью около 40 МЕ/мг, далее используемую для получения готовой формы лекарственного средства. Общий выход продукта после всех стадий выделения составляет приблизительно 40%.
Ключевые преимущества данного способа выделения и очистки имиглюцеразы состоят в усовершенствовании и оптимизации наиболее сложного и дорогостоящего этапа дегликозилирования белка за счет подтвержденной возможности использования смеси трех гликозидаз, что позволяет непосредственно в ходе хроматографической очистки модифицировать профиль гликозилирования белка для активации его функциональных свойств. Разработанная методика дает возможность получения активного целевого белка высочайшей степени чистоты пригодного для фармацевтического применения. Кроме того, возможность использовать оптимизированную ферментную смесь повторно позволяет существенно снизить затраты на производство конечного продукта.
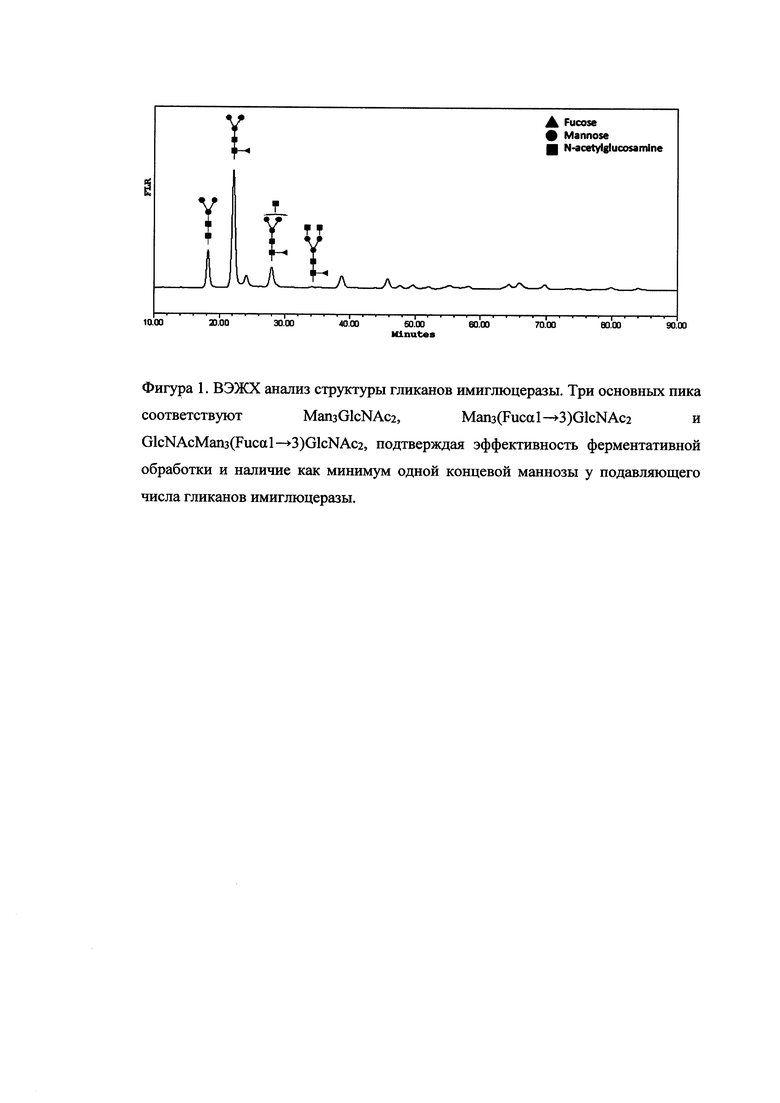
После завершения хроматографической очистки проводили контроль углеводной структуры субстанции. Аспарагин-связанные олигосахариды извлекались путем инкубации имиглюцеразы с N-гликаназой с последующим флуоресцентным мечением и анализом полученной смеси на гидрофильной ВЭЖХ-колонке высокого разрешения (фиг. 1). Методом масс-спектрометрического анализа показано, что три основных пика олигосахаридов соответствуют Man3GlcNAc2, Man3(Fucα1→3)GlcNAc2 и GlcNAcMan3(Fucα1→3)GlcNAc2, то есть подавляющее число углеводных цепей полученного гликопротеина содержат один или два концевых маннозных остатка необходимых для его биологической активности.
Ферментативную активность имиглюцеразы измеряли при помощи хромогенного метода с использованием 4-нитрофенил-β-D-глюкопиранозида в качестве субстрата. Биологическую активность субстанции подтверждали по способности имиглюцеразы к интернализации в иммунные клетки крови мыши с сохранением ферментативной активности, детектируемой более чувствительным методом с флуорогенным субстратом (4-метилумбеллиферил-β-D-глюкопиранозид). Показано, что целевой белок обладает требуемой специфической активностью (около 40 МЕ/мг) и эффективно проникает в клетку, подтверждая тем самым свою полную функциональность.
Полученный белок характеризуется следующими параметрами: не менее 99% основного вещества (по обрашенно-фазовой ВЭЖХ), не более 4% родственных примесей (по эксклюзионной ВЭЖХ), удельная активность не менее 35 МЕ/мг, содержание остаточных белков клетки-продуцента не более 50 нг/мг имиглюцеразы. Изобретение иллюстрируется, но не ограничивается приведенными ниже примерами.
ПРИМЕРЫ
Пример 1. Создание продуцента имиглюцеразы.
Ген, кодирующий имиглюцеразу, был сконструирован синтетическим способом, используя кодоны, оптимизированные для экспрессии в клетках грызунов (китайского хомячка). Аминокислотная последовательность белка была взята из базы данных UniProt (UniProtKB - Р04062 (GLCM_HUMAN). Для обеспечения секреции целевого белка в культуральную среду, ген имиглюцеразы содержал сигнальный пептид, обеспечивающий направление белковой молекулы по секреторному пути. Сигнальный пептид был взят из последовательности человеческого фактора свертываемости крови FVII. После прохождения через эндоплазматический ретикулум сигнальный пептид отщепляется от полипептида имиглюцеразы.
Ген имиглюцеразы был клонирован в плазмидные векторы для экспрессии в клетках млекопитающих под контролем сильного промотора и экспрессировался в клетках СНО (линия клеток яичников китайского хомячка). Два сконструированных вектора содержали идентичный ген, но отличались устойчивостью к селективному антибиотику. Вектор pMS590 нес ген устойчивости к пуромицину, а вектор pMS591 нес ген устойчивости к гигромицину, что обеспечивало устойчивость трансфецированной клеточной линии к селективным антибиотикам.
Экспрессия осуществлялась путем введения экспрессионных плазмид pMS590 pMS591, созданных на основе модифицированной плазмиды pUC18, в клеточную линию-хозяина посредством трансфекции клеток. Клеточная линия-хозяин представляла собой субклон линии клеток яичников китайского хомячка СНО-К1, приспособленный к выращиванию в виде суспензии в питательной среде, не содержащей сыворотки крови животных. В качестве способа трансфекции применяли электропорацию, в результате чего, введенная последовательность стабильно интегрировалась в геном хозяина.
Для идентификации и выделения клеток, продуцирующих имиглюцеразу, после трансфекции и добавления селективного антибиотика в культуральную среду, суспензия клеток разбавлялась и рассевалась таким образом, чтобы изолировать популяции, выросшие из единичных клеток, то есть клеточные клоны. После выделения клоны выращивались до состояния конфлюэнтности, и содержание целевого белка в супернатанте оценивалось методом иммуноферментного анализа, а также Вестерн-блоттинга. Клеточные клоны, секретирующие β-глюкоцереброзидазу на высоком уровне, размножали и хранили с помощью криоконсервации.
Пример 2. Культивирование клеток-продуцентов.
Для производства имиглюцеразы культивирование клеточного клона, стабильно продуцирующего целевой белок, проводили в течение не менее 20 суток в непрерывном (перфузионном) режиме в одноразовом биореакторе волнового типа со встроенной перфузионной мембраной. Для культивирования использовали базовую питательную среду SFM4CHO (HyClone), не содержащую сыворотки крови крупного рогатого скота и иных компонентов животного происхождения, дополненную нутриентной добавкой CHOFeedBioreactorSupplement (SAFC) в количестве 5 мл/л. Температура культивирования составляла 37°С в фазе экспоненциального роста и 32°С в продукционной фазе. Скорость протока питательной среды достигала одного объема биореактора в сутки. Культуральная жидкость, отделенная от клеток, раз в сутки передавалась на хроматографическую очистку. Процесс культивирования завершали после снижения жизнеспособности клеток ниже 80%.
Пример 3. Хроматография на сорбенте Butyl Sepharose.
К культуральной жидкости, содержащей рекомбинантную имиглюцеразу, добавляли хлорид натрия до концентрации 0,5 М. Сорбент уравновешивали буферным раствором состава 50 мM Tris-HCl, 0,5 М NaCl, 10% этиленгликоль (pH 7,2), затем наносили культуральную жидкость. По окончании нанесения сорбент последовательно промывали несколькими объемами буферных растворов следующего состава:
- 50 мМ Tris-HCl, 0,5 М хлорид натрия, 10% этиленгликоль (pH 7,2);
- 10 мМ Tris-HCl, 0,1 М хлорид натрия, 10% этиленгликоль (pH 7,2);
- 10 мМ ацетат натрия, 0,1 М хлорид натрия, 10% этиленгликоль (pH 5,0).
Целевой белок элюировали буферным раствором, содержащим 10 мМ ацетат натрия, 0,15 М хлорид натрия, 70% этиленгликоль (pH 5,0). Элюат собирали и передавали на следующую стадию очистки.
Пример 4. Хроматография на сорбенте SP Sepharose.
Элюат с Butyl-сефарозы разбавляли в два раза раствором 20 мМ ацетата натрия (pH 5,0) и наносили на сорбент, предварительно уравновешенный буферным раствором состава 20 мМ ацетат натрия, 25 мМ хлорид натрия, 10% этиленгликоль (pH 5,0).
По окончании нанесения сорбент последовательно промывали буферными растворами следующего состава:
- 20 мМ ацетат натрия, 25 мМ хлорид натрия, 10% этиленгликоль (pH 5,0);
- 20 мМ ацетат натрия, 100 мМ хлорид натрия, 10% этиленгликоль(pH 5,0).
Целевой белок элюировали буферным раствором, содержащим 20 мМ фосфат натрия, 100 мМ хлорид натрия, 20% этиленгликоль (pH 7,0). Элюат собирали и передавали на следующую стадию очистки.
Пример 5. Хроматография на сорбенте IMAC Sepharose.
Перед нанесением белка сорбент насыщали ионами Zn2+ путем последовательных промывок водой, раствором 0,2 М хлорида цинка, водой, а затем уравновешивали буферным раствором состава 20 мМ фосфат натрия, 0,1М хлорид натрия, 20% этиленгликоль (pH 7,0). Элюат с SP-сефарозы наносили на сорбент и по окончании нанесения проводили последовательные промывки буферными растворами следующего состава:
- 20 мМ фосфат натрия, 0,1 М хлорид натрия, 20% этиленгликоль (pH 7,0)
- 20 мМ фосфат натрия (pH 7,0);
- 20 мМ фосфат натрия, 1 М хлорид натрия (pH 7,0);
- 20 мМ фосфат натрия, 1 М хлорид натрия, 20% этиленгликоль (pH 7,0).
Целевой белок элюировали буферным раствором, содержащим 50 мМ фосфат натрия, 2,8 М хлорид натрия, 30% этиленгликоль (pH 7,0). Элюат собирали и передавали на следующую стадию очистки.
Пример 6. Хроматография на сорбенте Phenyl Sepharose и ферментативная обработка.
Элюат с IMAC-сефарозы разбавляли в 5 раз раствором 50 мМ фосфата натрия (pH 7,5) и наносили на сорбент, предварительно уравновешенный буферным раствором состава 50 мМ фосфат натрия, 0,5 М хлорид натрия, 5% этиленгликоль (pH 7,5). По окончании нанесения сорбент последовательно промывали буферными растворами следующего состава:
- 50 мМ фосфат натрия, 0,5 М хлорид натрия, 5% этиленгликоль (pH 7,5);
- 50 мМ цитрат натрия, 10% этиленгликоль (pH 5,7).
Последний буферный раствор также применялся для приготовления реакционной смеси ферментов для контролируемого дегликозилирования иммобилизованной на сорбенте имиглюцеразы. Объем реакционной смеси составлял 110% от колоночного объема; ферментный состав смеси рассчитывался из следующих соотношений: 6Е нейраминидазы (Worthington), 1E β-1,4-галактозидазы (Calbiochem) и 14Е β-N-ацетилглюкозаминидазы (Sigma) на 1 г имиглюцеразы.
Реакционную смесь для дегликозилирования пропускали через сорбент в режиме рециркуляции с линейной скоростью 90 см/ч в течение ~22-36 часов. После гидролиза олигосахаридных цепей реакционную смесь собирали и хранили при температуре 4°С (показана стабильность в течение минимум двух недель) для повторного использования. Сорбент промывали буферным раствором состава 50 мМ цитрат натрия, 10% изопропанол (pH 5,7). Целевой белок с оптимизированным профилем гликозилирования элюировали буферным раствором, содержащим 40 мМ фосфат натрия, 55% пропиленгликоль (pH 7,5). Элюат собирали и передавали на следующую стадию очистки.
Пример 7. Хроматография на сорбенте Q Sepharose.
Элюат с Phenyl-сефарозы разбавляли в 2 раз раствором 100 мМ фосфата натрия (pH 7,5) и наносили на сорбент, предварительно уравновешенный буферным раствором состава 50 мМ фосфат натрия, 10% этиленгликоль (pH 7,5). Данную стадию проводили в режиме фильтрации; после нанесения фракции целевого белка сорбент промывали буфером для уравновешивания. Фильтрат собирали и передавали на следующую стадию очистки.
Пример 8. Хроматография на сорбенте CM Sepharose.
К фильтрату с Q-сефарозы добавляли раствор 1 М лимонной кислоты до значения pH 5,0 и наносили на сорбент, предварительно уравновешенный раствором 25 мМ цитрата натрия (pH 5,0). По окончании нанесения сорбент последовательно промывали буферными растворами следующего состава:
- 25 мМ цитрат натрия (pH 5,0);
- 25 мМ цитрат натрия, 10% изопропанол (pH 5,0).
Затем, для удаления фракции белка с высоким содержанием остаточных белков СНО, проводили промывку линейным градиентом буфера состава 25 мМ цитрат натрия, 10% изопропанол, 200 мМ хлорид натрия (pH 5,0) с остановкой градиента при достижении проводимости 7-8 mS/cm. После этого сорбент последовательно промывали буферными растворами следующего состава:
- 25 мМ цитрат натрия, 10% изопропанол (pH 5,0)
- 25 мМ цитрат натрия, 0,01% полисорбат-80 (pH 5,0).
Целевой белок элюировали буферным раствором, содержащим 55 мМ цитрат натрия, 0.01% полисорбат-80 (pH 6,3). Для приготовления готовой субстанции в полученный раствор имиглюцеразы вносили в сухом виде недостающие компоненты финальной формуляции.
Пример 9. Определение углеводной структуры имиглюцеразы.
Аликвоту 0,5 мг субстанции имиглюцеразы переводили в 0,2 мл гликозидазного буфера (50 мМ Tris-HCl, pH 8,0) при помощи 250-кратного разбавления на микроконцентраторе Amicon-10К (Millipore). Затем к образцу добавляли около 20 единиц активности N-glycanase (Prozyme) и термостатировали при +37°С в течение 8 ч. Олигосахариды отделяли от белка ультрафильтрацией на Amicon-10К с дополнительными промывками деионизованной водой. Собранный проскок (99% материала) упаривали досуха на вакуумном концентраторе CentriVap (Labconco).
Реагент для флуоресцентного мечения олигосахаридов получали последовательным растворением 1 мкг 2-аминобензойной кислоты и 1,25 мкг цианоборогидрида натрия в 21 мкл смеси диметилсульфоксида с уксусной кислотой. Образцы смеси олигосахаридов растворяли в 3 мкл деионизованной воды и затем добавляли 10 мкл реагента для мечения. Реакционную смесь инкубировали в течение 3 ч при +45°С. Избыточный краситель удаляли методом твердофазной экстракции на колонках LudgerClean™ T1 Cartridge Kit (Ludger) согласно рекомендуемому протоколу. Пробы упаривали до объема около 50 мкл и использовали для анализа.
Хроматографический анализ (фиг. 1) осуществляли на приборе Alliance HPLC (Waters) с применением гидрофильной ВЭЖХ-колонки высокого разрешения TSKgel Amide-80 (Tosoh Bioscience) с предколонкой TSKguardgel Amide-80. Разделение проводили при температуре колонки 45°С в градиенте подвижной фазы (раствор формиата аммония с ацетонитрилом). Сигнал детектировали по флуоресценции при 425 нм при возбуждении светом 360 нм.
Пример 10. Определение биологической активности полученного белка и способности его интернализации в клетку.
Ферментативную активность имиглюцеразы измеряли хромогенным методом с 4-нитрофенил-β-D-глюкопиранозидом в качестве субстрата. В исследуемый образец в нескольких разведениях вносили четырехкратный объем 10 мМ раствора субстрата и инкубировали 15 мин при 37°С, после чего реакцию останавливали прибавлением восьми объемов 0,1 М раствора глицина. Измеряли оптическую плотность растворов на спектрофотометре в максимуме поглощения при длине волны 400 нм. Активность определяли, исходя из того, что одна единица активности соответствует гидролизу одного мкмоля субстрата/мин при температуре 37°С.
Биологическую функциональность имиглюцеразы определяли по способности исследуемых образцов белка интернализироваться в перитонеальные макрофаги (моноциты) мыши в присутствии или в отсутствие дрожжевого маннана с последующей детекцией ферментативной активности интернализированной имиглюцеразы в клеточном лизате.
Пул мышиных клеток, содержащих перитонеальные макрофаги, осаждали центрифугированием с последующим ресуспендированием клеточного осадка в питательной среде DMEM/F12 с 10% бычьей сывороткой, 2 мМ глутамином и пенициллин-стрептомицином. Полученные клетки высевали в 6 луночный планшет в количестве 7 млн/лунку. По окончании полуторачасовой инкубации прикрепившиеся клетки перитонеальных макрофагов двукратно промывали средой с последующим добавлением 2 мл свежей среды в каждую лунку. К клеткам добавляли аликвоты исследуемых образцов имиглюцеразы до концентрации 100 мкг/мл. В качестве контроля использовали клетки, к которым предварительно добавляли маннан (до концентрации 800 мкг/мл), блокирующий маннозные рецепторы. После 3-х часовой инкубации при 37°С макрофаги промывали фосфатно-солевым буфером; смывы после 4 и 5 промывок собирались для анализа. По окончании последней промывки клетки каждой лунки смывали 500 мкл лизирующего буфера. Полученные образцы клеточных лизатов и промывочных растворов были использованы для детектирования активной формы белка.
Определение ферментной активности имиглюцеразы в клеточном лизате проводили флюорогенным методом: под действием имиглюцеразы происходит расщепление синтетического субстрата (4-метилумбеллиферил-β-D-глюкопиранозид) с образованием флюоресцентного вещества 4-метилумбеллиферона.
К 20 мкл образцов имиглюцеразы, разведенных в фосфатном буфере, добавляли 80 мкл 4-метилумбеллиферил-β-D-глюкопиранозида (Sigma) и инкубировали 30 мин при 37°С. Реакцию останавливали добавлением 800 мкл 0,1 М раствора глицина, pH 10,5. Флюоресценцию испытуемых растворов и раствора сравнения, не содержащего белка, измеряли спектрофотометрически при длине волны возбуждения 365 нм и длине волны эмиссии 440 нм. Флюоресценция продукта реакции количественно оценивается с помощью калибровочной кривой, построенной для заданных концентраций 4-метилумбеллиферона (Sigma).
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ α-ГАЛАКТОЗИДАЗНОЙ А НЕДОСТАТОЧНОСТИ | 2000 |
|
RU2248213C2 |
| ЛЕЧЕНИЕ В СЛУЧАЕ ДЕФИЦИТА α-ГАЛАКТОЗИДАЗЫ А | 1997 |
|
RU2179034C2 |
| АНТИТЕЛА ДЛЯ СВЯЗЫВАНИЯ ИМИГЛЮЦЕРАЗЫ И ИХ ПРИМЕНЕНИЕ В АФФИННОЙ ХРОМАТОГРАФИИ | 2017 |
|
RU2676956C1 |
| СПОСОБ ВЫДЕЛЕНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОЙ ЧЕЛОВЕЧЕСКОЙ ПРОУРОКИНАЗЫ М5 | 2013 |
|
RU2553533C2 |
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2004 |
|
RU2385928C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕННО-ИНЖЕНЕРНОГО ИНСУЛИНА ЧЕЛОВЕКА | 2006 |
|
RU2322504C1 |
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2009 |
|
RU2535342C2 |
| Синтетическая ДНК, кодирующая интерлейкин-7 человека, содержащий ее экспрессионный вектор (варианты), штамм-продуцент интерлейкина-7 человека и способ получения интерлейкина-7 человека | 2015 |
|
RU2615447C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ В-1,4-ГАЛАКТОЗИДАЗЫ BGAA ИЗ STREPTOCOCCUS PNEUMONIAE | 2019 |
|
RU2698460C1 |
| СПОСОБ КРУПНОМАСШТАБНОГО ПОЛУЧЕНИЯ, ВЫДЕЛЕНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО ГРАНУЛОЦИТАРНОГО КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА ЧЕЛОВЕКА | 2010 |
|
RU2487885C2 |
Группа изобретений относится к медицине и биотехнологии и может быть использована в фармацевтической промышленности. Предложены способ дегликозилирования имиглюцеразы и очистки имиглюцеразы, включающий этап дегликозилирования имиглюцеразы. Способ дегликозилирования включает выделение полученной в клетках СНО имиглюцеразы и ее последующую обработку нейраминидазой, β-1,4-галактозидазой и β-N-ацетилглюкозаминидазой в одну стадию. Соотношение ферментов нейраминидазы: β-1,4-галактозидазы: β-N-ацетилглюкозаминидазы в смеси составляет: 6 Е:1 Е:14 Е в расчете на 1 г белка. Способ очистки имиглюцеразы включает гидрофобную хроматографию на Butyl-сефарозе, катионный обмен на SP-сефарозе, металло-хелатную хроматографию на IMAC-сефарозе, гидрофобную хроматографию на Phenyl-сефарозе, совмещенную с дегликозилированием целевого белка, фильтрацию через Q-сефарозу и катионный обмен на СМ-сефарозе. Изобретения позволяют получать продукт фармакопейного качества с содержанием не менее 99% имиглюцеразы (по обрашенно-фазовой ВЭЖХ), не более 4% родственных примесей (по эксклюзионной ВЭЖХ), с удельной активностью не менее 35 МЕ/мг и содержанием остаточных белков клетки-продуцента не более 50 нг/мг. 2 н. и 2 з.п. ф-лы, 1 ил., 10 пр.
1. Способ дегликозилирования имиглюцеразы, полученной в клетках млекопитающих, включающий выделение полученной в клетках СНО имиглюцеразы и ее последующую обработку нейраминидазой, β-1,4-галактозидазой и β-N-ацетилглюкозаминидазой, отличающийся тем, что обработка нейраминидазой, β-1,4-галактозидазой и β-N-ацетилглюкозаминидазой проводится в одну стадию, и соотношение ферментов нейраминидазы: β-1,4-галактозидазы: β-N-ацетилглюкозаминидазы в смеси составляет: 6 Е:1 Е:14 Е в расчете на 1 г белка.
2. Способ дегликозилирования имиглюцеразы по п. 1, отличающийся тем, что дегликозилирование проводится в течение 22-36 часов при комнатной температуре.
3. Способ дегликозилирования имиглюцеразы по любому из пп. 1, 2, отличающийся тем, что дегликозилирование проводится в ходе хроматографической очистки.
4. Способ очистки имиглюцеразы из культуральной жидкости, полученной при культивировании клеток млекопитающих, включающий следующие стадии:
а) гидрофобную хроматографию на Butyl-сефарозе,
б) катионный обмен на SP-сефарозе,
в) металло-хелатную хроматографию на IMAC-сефарозе,
г) гидрофобную хроматографию на Phenyl-сефарозе, совмещенную с дегликозилированием целевого белка по любому из пп. 1-3,
д) фильтрацию через Q-сефарозу,
е) катионный обмен на СМ-сефарозе.
| US 5549892 A1, 27.08.1996 | |||
| DOEBBER T | |||
| et al | |||
| "Enhanced macrophage uptake of synthetically glycosylated human placental beta-glucocerebrosidase" | |||
| J | |||
| Biol | |||
| Chem | |||
| Устройство для видения на расстоянии | 1915 |
|
SU1982A1 |
| FURBISH F | |||
| S | |||
| et al | |||
| "Uptake and Distribution of Placental Glucocerebrosidase in Rat Hepatic Cells and Effects of Sequential Deglycosylation" Biochimica et Biophysica Acta, 1981, 673, p.425-434 | |||
| FURBISH F | |||
| S | |||
| et al | |||
| "Enzyme replacement therapy in Gaucher's disease: Large-Scale purification of glucocerebrosidase suitable for human administration" Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| Шеститрубный элемент пароперегревателя в жаровых трубках | 1918 |
|
SU1977A1 |
| FRIEDMAN B | |||
| et al | |||
| "A comparison of the pharmacological properties of carbohydrate remodeled recombinant and placentalderived beta-glucocerebrosidase: implications for clinical efficacy in treatment of Gaucher disease" | |||
| Blood, 1999, 93, 2807-2816 | |||
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2009 |
|
RU2535342C2 |

