Настоящее изобретение относится к способам и композициям для лечения α -галактозидазной А недостаточности.
Предшествующий уровень техники
Болезнь Фабри представляет собой Х-сцепленную наследственную болезнь, связанную с накоплением лизосом, характеризующуюся тяжелым поражением почек, ангиокератомами и сердечно-сосудистыми нарушениями, включая увеличение желудочка и недостаточность митрального клапана. Болезнь Фабри также воздействует на периферическую нервную систему, вызывая приступы мучительных жгучих болей в конечностях. Причиной развития болезни Фабри является дефицит фермента α -галактозидазы A (α -Gal A). α -Gal А представляет собой лизосомную гликогидролазу, которая отщепляет концевые α -галактозильные группы различных гликоконъюгатов. Болезнь Фабри приводит к блокированию катаболизма нейтрального гликосфинголипида, керамид-тригексозида (СТН) и к аккумуляции субстрата фермента в клетках и в кровотоке.
Вследствие того что данное заболевание имеет X-сцепленный тип наследования, то большинство пациентов с болезнью Фабри составляют мужчины. Хотя у гетерозиготных женщин наблюдаются серьезные поражения, однако, в большинстве случаев, эти гетерозиготные женщины не обнаруживают каких-либо симптомов поражения или обнаруживают относительно слабые симптомы (такие как характерное помутнение роговицы). Атипический вариант болезни Фабри, проявляющийся в низкой остаточной активности α -Gal А и либо обнаруживающий очень слабые симптомы, либо не обнаруживающий никаких других явных симптомов, характерных для болезни Фабри, коррелирует с гипертрофией левого желудочка и заболеванием сердца. Nakano et al. New Engl. J.Med. 333:288-293 (1995). Причиной таких патологий сердца может быть снижение активности α -Gal A.
Были выделены и секвенированы кДНК и ген, кодирующие α -Gal А человека. α -Gal А человека экспрессируется в виде полипептида из 429 аминокислот, где N-концевые 31 аминокислота представляют собой сигнальный пептид. Этот человеческий фермент был экспрессирован в клетках яичника китайского хомячка (СНО) (Desnick et al., патент США №5356804; loannou et al., J.Cell.Biol. 119: 1137 (1992)) и в клетках насекомых (Calhoun et al., WO 90/11353).
Однако современные препараты α -Gal А имеют ограниченную эффективность. Методы получения α -Gal А с относительно высокой чистотой зависят от использования аффинной хроматографии с применением комбинации аффинной хроматографии на лектине (конкавалине A (Con А)-Сефарозе®) и аффинной хроматографии, основанной на связывании α -Gal А с аналогом субстрата N-6-аминогексаноил-α -В-галактозиламином, связанным с сефарозной матрицей (Sepharose®). См., например, Bishop et al., J. Biol. Chem. 256:1307-1316 (1981). Использование смол, обладающих аффинностью к белку лектину, и смол, являющихся аналогом субстрата, обычно ассоциировано с непрерывным выщелачиванием аффинного агента из твердого носителя (Marikar et al., Anal. Biochem. 201:306-310 (1992)), что приводит к загрязнению очищенного продукта аффинным агентом либо в свободной форме в растворе, либо в форме, связанной с проэлюированным белком. Такие загрязнения делают этот продукт неприемлемым для использования в фармацевтических препаратах. Связанные аналоги субстратов и лектины могут также оказывать значительные негативные эффекты на ферментативные, функциональные и структурные свойства белков. Более того, α -Gal А, получаемая известными методами, быстро выводится печенью.
Таким образом, разработка схемы очистки с использованием стандартных хроматографических смол, являющихся коммерчески доступными и обладающих качеством, подходящим для их широкомасштабного коммерческого использования, которые позволят получить препарат α -Gal А, не содержащий аффинного агента, остается актуальной. Кроме того, также остается актуальной необходимость в получении препаратов α -Gal А с более продолжительным временем полужизни в кровотоке и с повышенным уровнем их поглощения в конкретных тканях, не относящихся к печени.
Краткое описание изобретения
Настоящее изобретение относится к высокоочищенным препаратам α -Gal А и к различным способам очистки гликоформ α -Gal
А. Настоящее изобретение также относится к препаратам α -Gal A с измененным зарядом и к способам получения таких препаратов. Изменение заряда достигается увеличением содержания сиаловой кислоты в α -Gal А и/или увеличением уровня фосфорилирования α -Gal А. Кроме того, настоящее изобретение относится к препаратам α -Gal А, имеющим более продолжительное время полужизни в кровотоке млекопитающего-хозяина и к способам их получения. И наконец, настоящее изобретение относится к способам и к дозам введения препарата α -Gal А индивидууму. Препараты α -Gal А настоящего изобретения могут быть использованы для лечения индивидуумов с болезнью Фабри или с атипическим вариантом болезни Фабри, например, конкретных групп пациентов, страдающих болезнью Фабри с преобладающими сердечно-сосудистыми патологиями, такими как увеличение желудочков, например гипертрофия левого желудочка (ГЛЖ) и/или недостаточность митрального клапана, или групп пациентов, страдающих болезнью Фабри с преобладающими почечными нарушениями.
Краткое описание чертежей
На фиг.1 представлен 210 п.н. - зонд, который был использован для выделения кДНК α -Gal А из библиотеки кДНК фибробластов человека (SEQ ID NО:1). Эта последовательность происходит от экзона 7 гена α -Gal А. Зонд был выделен из геномной ДНК человека с помощью полимеразной цепной реакции (ПЦР). Подчеркнутые на фиг.1 области соответствуют последовательностям праймеров для амплификации.
На фиг.2 представлена последовательность ДНК-фрагмента, которую завершает 5'-конец кДНК-клона α -Gal A (SEQ ID NО:2). Этот фрагмент был амплифицирован из геномной ДНК человека с помощью ПЦР. Подчеркнутые области соответствуют последовательностям праймеров. Также показаны положения NcoI- и SacII-сайтов рестриктирующих эндонуклеаз, которые были использованы для субклонирования, как описано в Примере 1.
На фиг.3 представлена последовательность кДНК α -Gal A, включающая последовательность, которая кодирует сигнальный пептид (SEQ ID NO:3).
На фиг.4 схематически представлена карта pXAG-16, α -Gal А-экспрессирующей конструкции, которая включает промотор CMV (цитомегаловируса), экзон 1 и первый интрон, последовательность, кодирующую сигнальный пептид чГР и первый интрон, кДНК, кодирующую α -Gal А (в которой отсутствует последовательность сигнального пептида α -Gal А) и 3'UTS чГР. pcDNeo указывает на положение гена nео, происходящего от плазмиды pcDNeo.
На фиг.5 схематически представлена карта pXAG-28 конструкции, экспрессирующей α -Gal А, которая включает промотор коллагена Iα 2 и первый экзон 1, интрон β -актина, последовательность, кодирующую сигнальный пептид чГР и первый интрон, кДНК, кодирующую α -Gal А (в которой отсутствует последовательность сигнального пептида α -Gal А) и 3'UTS чГР. pcDNeo указывает на положение гена nео, происходящего от плазмиды pcDNeo.
На фиг.6 представлена аминокислотная последовательность α -Gal А человека (SEQ ID NO:4).
На фиг.7 представлена кДНК-последовательность, кодирующая α -Gal А человека (без сигнального пептида) (SEQ ID NO:5).
На фиг.8 представлена ахроматограмма стадии очистки α -Gal А с использованием бутилсефарозной смолы (Butyl Sepharose®). Показано поглощение при 280 нм (сплошная линия) и активность α -Gal А (пунктирная линия) выбранной фракции.
На фиг.9 схематически представлена карта pGA213C.
На фиг.10, в виде диаграммы, представлена конструкция для целевой доставки, pGA213C, и гомологичная рекомбинация с эндогенным локусом α -галактозидазы А. pGA213C изображена в виде последовательностей для доставки, расположенных выше соответствующих последовательностей в локусе α -галактозидазы А на хромосоме X. Положения по отношению к инициирующему кодону метионина ATG показаны номерами, обозначенным выше линейных карт. Активационный компонент, содержащий последовательности мышиного гена dhfr, бактериального гена nео и промотора CMV/интрона альдолазы, показан выше положения (-221), в которые они были встроены путем клонирования ДНК. Последовательности, кодирующие α -галактозидазу А, показаны затемненными рамками. Некодирующие геномные последовательности α -галактозидазы показаны слегка заштрихованными рамками. Крупные стрелки указывают на направление транскрипции комет экспрессии dhfr и nео. Сплайсинг мРНК GA-GAL после успешной доставки и активации гена показан сегментированной линией, расположенной ниже карты локуса активированной α -галактозидазы А (GA-GAL).
Подробное описание изобретения
Введение
Описанное здесь изобретение относится к некоторым новым препаратам α -Gal А и к способам их получения, а также к способам лечения пациентов с болезнью Фабри или с атипическими вариантами болезни Фабри с использованием данных препаратов. Некоторые рассматриваемые репрезентативные варианты осуществления изобретения систематизированы и подробно описаны ниже.
В настоящем изобретении рассматривается использование α -Gal А, продуцируемой в любой клетке (в клетке, продуцирующей α -Gal A), для лечения болезни Фабри. В предпочтительном варианте осуществления изобретения используется человеческая α -Gal А, продуцируемая стандартными методами генной инженерии (на основе введения клонированного гена или кДНК α -Gal А в клетку-хозяина) или путем активации гена.
Настоящее изобретение относится к препаратам и к способам получения препаратов, которые содержат α -Gal А, имеющую более высокую степень чистоты по сравнению с α -Gal А, полученной ранее. С использованием методов очистки настоящего изобретения композиции препаратов α -Gal А человека предпочтительно очищают, по крайней мере, до 98%-ной гомогенности, более предпочтительно, по крайней мере, до 99%-ной гомогенности, а наиболее предпочтительно, по крайней мере, до 99,5%-ной гомогенности, как было определено с помощью электрофореза в ПААГ с ДСН или с помощью обращенно-фазовой ВЭЖХ. Удельная активность препаратов α -Gal А настоящего изобретения предпочтительно составляет, по крайней мере, 2,0× 106 единиц/мг белка, более предпочтительно, по крайней мере, 3,0× 106 единиц/мг белка и наиболее предпочтительно, по крайней мере, 3,5× 106 единиц/мг белка.
В одном из вариантов осуществления настоящего изобретения препарат α -Gal А очищают путем отделения различных гликоформ α -Gal А от других компонентов на смоле для гидрофобного взаимодействия, но не проводят стадию хроматографии на лектине. В предпочтительном варианте осуществления настоящего изобретения, функциональная группа смолы для гидрофобного взаимодействия включает бутильную группу.
В альтернативном варианте осуществления настоящего изобретения, препарат α -Gal А очищают сначала путем связывания различных гликоформ α -Gal А с катионнообменной смолой на колонке при кислотном рН в уравновешивающем буфере. Затем эту колонку промывают уравновешивающим буфером для элюирования несвязанного материала и указанные различные гликоформы α -Gal A элюируют с использованием в качестве элюирующего раствора солевого раствора при 10-100 мМ, буферного раствора с рН 4-5 или их комбинации. В предпочтительном варианте осуществления настоящего изобретения буфер для уравновешивания имеет рН около 4,4.
В другом альтернативном варианте осуществления настоящего изобретения, препарат α -Gal А очищают путем отделения различных гликоформ α -Gal А в образце от других компонентов данного образца с использованием процедуры очистки, включающей стадию проведения, по крайней мере, одной из хроматографий, таких как хроматофокусирующая хроматография, аффинная хроматография на хелате металла или иммунноаффинная хроматография.
Кроме того, настоящее изобретение относится к препаратам α -Gal А и к способам получения препаратов α -Gal А, которые содержат α -Gal А с измененным зарядом. Данные препараты могут включать различные гликоформы α -Gal А. Изменение заряда достигаются путем увеличения содержания сиаловой кислоты в препаратах α -Gal А и/или путем повышения уровня фосфорилирования препаратов α -Gal A.
Содержание сиаловой кислоты в препаратах α -Gal А увеличивают (i) путем выделения высоко заряженных и/или более высокомолекулярных гликоформ α -Gal А в процессе очистки или после очистки; (ii) путем присоединения остатков сиаловой кислоты с использованием клеток, генетически модифицированных (либо стандартными методами генной инженерии, либо активацией гена) для экспрессии гена или кДНК сиалилтрансферазы, или (iii) путем ферментации или культивирования клеток, экспрессирующих данный фермент в среде с низким содержанием аммония.
Уровень фосфорилирования препаратов α -Gal А увеличивают (i) путем присоединения фосфатных остатков с использованием клеток, генетически модифицированных (либо стандартными методами генной инженерии, либо путем активации гена) для экспрессии гена или кДНК фосфорилтрансферазы, или (ii) путем добавления ингибиторов фосфатазы к культивированным клеткам.
С использованием способов настоящего изобретения получают препараты гликозилированной α -Gal А человека, где заряженными являются от 35 до 85% олигосахаридов. В предпочтительном варианте осуществления настоящего изобретения заряженными являются, по крайней мере, 35% олигосахаридов. В более предпочтительном варианте осуществления настоящего изобретения заряженными являются, по крайней мере, 50% олигосахаридов.
Альтернативные предпочтительные препараты человеческой гликозилированной α -Gal А имеют множество гликоформ α -Gal А, содержащих предпочтительно, по крайней мере, 20%, более предпочтительно, по крайней мере, 50%, а наиболее предпочтительно, по крайней мере, 70% сложных гликанов с 2-4 остатками сиаловой кислоты. В альтернативном предпочтительном варианте осуществления настоящего изобретения препараты человеческой гликозилированной α -Gal А с множеством гликоформ имеют олигосахаридный заряд, измеряемый числом Z, которое превышает 100, предпочтительно превышает 150, а более предпочтительно превышает 170. В другом альтернативном предпочтительном варианте осуществления настоящего изобретения препараты человеческой гликозилированной α -Gal А с множеством гликоформ имеют, в среднем, по крайней мере, 16-50%, предпочтительно 25-50%, а более предпочтительно, по крайней мере, 30% фосфорилированных гликоформ. В другом альтернативном варианте осуществления настоящего изобретения в указанных препаратах с множеством гликоформ, 50-75%, а предпочтительно 60% гликанов от общего числа гликанов являются сиалилированными.
В одном из вариантов осуществления настоящего изобретения препарат гликозилированной α -Gal А, имеющий повышенный олигосахаридный заряд, получают сначала путем введения полинуклеотида, кодирующего трансферазу GlcNAc III (GnT-III), в клетку, продуцирующую α -Gal А, или путем введения, посредством гомологичной рекомбинации, регуляторной последовательности, контролирующей экспрессию эндогенного гена GnT-III. Затем клетку, продуцирующую α -Gal А, культивируют в условиях культивирования, способствующих экспрессии α -Gal А и GnT-III. Конечная стадия предусматривает выделение препарата α -Gal А с увеличенным олигосахаридным зарядом.
В альтернативном варианте осуществления настоящего изобретения препарат гликозилированной α -Gal А, имеющий повышенный олигосахаридный заряд, продуцируют сначала путем введения полинуклеотида, кодирующего сиалилтрансферазу, в клетку, продуцирующую α -Gal А, или путем введения, посредством гомологичной рекомбинации, регуляторной последовательности, контролирующей экспрессию эндогенного гена сиалилтрансферазы. Затем клетку, продуцирующую α -Gal А, культивируют в условиях культивирования, способствующих экспрессии α -Gal А и сиалилтрансферазы. Конечная стадия предусматривает выделение препарата α -Gal А с повышенным олигосахаридным зарядом. Предпочтительными сиалилтрансферазами являются α 2,3-сиалилтрансфераза и α 2,6-сиалилтрансфераза. В предпочтительном варианте осуществления настоящего изобретения данный способ включает дополнительную стадию отбора гликоформ α -Gal А с увеличенным размером или увеличенным зарядом путем фракционирования или очистки данного препарата.
В другом варианте осуществления изобретения препарат гликозилированной α -Gal А с повышенным уровнем сиалилирования получают путем контактирования клетки, продуцирующей α -Gal А, с культуральной средой, содержащей концентрацию аммония ниже 10 мМ, а более предпочтительно ниже 2 мМ. В предпочтительном варианте осуществления изобретения среду с низким содержанием аммония получают путем добавления глутаминсинтетазы к указанной культуральной среде. В альтернативном предпочтительном варианте осуществления настоящего изобретения среду с низким содержанием аммония получают путем непрерывной или периодической перфузии α -Gal А-продуцирующей клетки свежей культуральной средой для поддержания концентрации аммония ниже 10 мМ, а более предпочтительно ниже 2 мМ.
В еще одном варианте осуществления настоящего изобретения препарат гликозилированной α -Gal А с повышенным уровнем фосфорилирования получают сначала путем введения в α -Gal А-продуцирующую клетку полинуклеотида, кодирующего фосфорилтрансферазу, или путем введения регуляторной последовательности посредством гомологичной рекомбинации, регулирующей экспрессию эндогенного гена фосфорилтрансферазы. Затем клетку, продуцирующую α -Gal А, культивируют в условиях культивирования, способствующих экспрессии α -Gal А и фосфорилтрансферазы. Затем выделяют препарат α -Gal А, который имеет повышенный уровень фосфорилирования по сравнению с препаратами α -Gal А, продуцированной в данной клетке без полинуклеотида. В предпочтительном варианте осуществления настоящего изобретения препараты α -Gal А, продуцируемые способами настоящего изобретения, имеют множество гликоформ, из которых 16-50%, предпочтительно 25-50%, а более предпочтительно, по крайней мере, 30% являются фосфорилированными. В предпочтительном варианте осуществления настоящего изобретения данный способ включает дополнительную стадию отбора гликоформ α -Gal А с увеличенным размером или увеличенным зарядом путем фракционирования или очистки данного препарата.
В еще одном варианте осуществления изобретения препарат гликозилированной α -Gal А с повышенным уровнем фосфорилирования получают путем добавления к культивированным клеткам ингибитора фосфатазы, например бромтетрамизола. Низкие уровни щелочной фосфатазы бычьей плазмы могут присутствовать в фетальной сыворотке теленка, используемой в качестве ростовой добавки для культивированных клеток. Это приводит к вероятности того, что экспонированные эпитопы Man-6-P на секретированной α -Gal А могут быть субстратом для щелочной фосфатазы сыворотки. Было также показано, что бромтетрамизол является сильным ингибитором щелочной фосфатазы, Ki=2,8 мМ (Metaye et al., Biochem. Pharmacol. 15:4263-4268 (1988)), и полное ингибирование достигается при концентрации 0,1 мМ (Borgers & Thone, Histochemistry 44:277-280 (1975)). Следовательно, в одном из вариантов осуществления изобретения ингибитор фосфатазы, например бромтетрамизол, может быть добавлен к культивированным клеткам для максимизации содержания форм α -Gal А с высоким поглощением, присутствующих в культуральной среде, благодаря предупреждению гидролиза сложноэфирных групп Man-6-P.
Настоящее изобретение, кроме того, относится к препаратам α -Gal А и к способам получения таких препаратов, которые имеют увеличенное время полужизни в кровотоке млекопитающего-хозяина. Время полужизни в кровотоке и клеточное поглощение увеличивают путем (i) повышения содержания сиаловой кислоты в α -Gal А (достигаемого, как описано выше); (ii) повышения уровня фосфорилирования α -Gal А (достигаемого, как описано выше); (iii) ПЭГилирования α -Gal А или (iv) последующего удаления сиаловой кислоты и концевых остатков галактозы, или удаления концевых остатков галактозы на олигосахаридных цепях α -Gal А.
Повышение уровня сиалилирования препаратов α -Gal А приводит к увеличению времени полужизни экзогенной α -Gal А в кровотоке. Кроме того, по сравнению с поглощением в гепатоцитах, повышенный уровень сиалилирования α -Gal А приводит к повышению уровня его поглощения в не-гепатоцитах, таких как эндотелиальные клетки печени, синусоидальные клетки печени, клетки легких, клетки почек, нервные клетки, эндотелиальные клетки или клетки сердца. Препарат человеческой гликозилированной α -Gal A с повышенным содержанием сиаловой кислоты предпочтительно включает множество гликоформ, где, по крайней мере, 20% сложных гликанов имеют 2-4 остатка сиаловой кислоты. Альтернативный предпочтительный препарат человеческой гликозилированной α -Gal А имеет множество гликоформ, где 50-75%, а предпочтительно, по крайней мере, 60% от всех гликанов являются сиалилированными.
Фосфорилирование препаратов α -Gal А также увеличивает уровень вхождения α -Gal А в клетки. Фосфорилирование происходит в клетках, экспрессирующих данную α -Gal А. Один предпочтительный препарат человеческой гликозилированной α -Gal А настоящего изобретения предпочтительно включает множество гликоформ, где, в среднем, по крайней мере, 16-50%, предпочтительно 25-50%, а более предпочтительно, по крайней мере, 30% данных гликоформ являются фосфорилированными.
В альтернативном варианте осуществления изобретения время полужизни в кровотоке препарата человеческой α -Gal А увеличивают путем образования комплекса α -Gal А с полиэтиленгликолем. В предпочтительном варианте осуществления изобретения данный препарат α -Gal А подвергают реакции образования комплекса с использованием трезилмонометокси-ПЭГ (TMPEG) с получением ПЭГилированной α -Gal А. Затем комплекс ПЭГ-α -Gal А очищают и выделяют препарат ПЭГ-α -Gal А. ПЭГилирование α -Gal A увеличивает время полужизни в кровотоке и повышает in vivo-эффективность данного белка.
Сиалилирование влияет на время полужизни в кровотоке и биологическое распределение белков. Белки с минимальным количеством сиаловой кислоты или без сиаловой кислоты легко интернализуются посредством рецептора асиалогликопротеина (рецептора Эшвелла) на гепатоцитах благодаря экспонированным галактозным остаткам на белке. Время полужизни в кровотоке α -Gal А с галактозным концом может быть увеличено путем проведения последовательных стадий (1) удаления сиаловой кислоты посредством контактирования α -Gal А с нейраминидазой (сиалидазой), в результате чего концевые галактозные группы становятся экпонируемыми, и (2) удаления концевых галактозидных остатков посредством контактирования десиалилированной α -Gal А с β -галактозидазой. Полученный препарат α -Gal А имеет меньшее число концевых остатков сиаловой кислоты и/или концевых галактозидных остатков на олигосахаридных цепях по сравнению с препаратами α -Gal А, которые не были последовательно подвергнуты контактированию с нейраминидазой и β -галактозидазой. Альтернативно, время полужизни в кровотоке β -Gal А с галактозным концом может быть увеличено лишь путем удаления концевых галактозидных остатков посредством контактирования десиалилированной α -Gal А с β -галактозидазой. Полученный α -Gal А-препарат имеет меньшее число концевых галактозидных остатков на олигосахаридных цепях по сравнению с α -Gal А-препаратами, которые не были подвергнуты контактированию с β -галактозидазой. В предпочтительном варианте осуществления изобретения, после контактирования нейраминидазы и β -галактозидазы, полученные α -Gal А - препараты затем подвергают контактированию с β -гексозаминидазой, что приводит к отщеплению олигосахарида с получением триманнозной сердцевины.
Кроме того, уровни сиалилирования могут зависеть от типа используемой клетки. Поэтому, в другом предпочтительном варианте осуществления изобретения, уровень сиалилирования α -Gal A может быть увеличен благодаря скринингу на клетки млекопитающих, например клетки человека, которые имеют относительно высокий уровень сиалилтрансферазной активности, и благодаря использованию таких клеток в качестве α -Gal А-продуцирующих клеток.
Кроме того, настоящее изобретение относится к композициям α -Gal А-препаратов, которые, в основном, не содержат белков α -Gal А, таких как альбумин; белков, не относящихся к α -Gal А и продуцируемых клетками-хозяевами, или белков, выделенных из тканей или физиологических жидкостей животного. В одном из предпочтительных вариантов осуществления изобретения данная композиция, кроме того, содержит наполнитель. Предпочтительными наполнителями являются маннит, сорбит, глицерин, аминокислоты, липиды, EDTA, EGTA, хлорид натрия, полиэтиленгликоль, поливинилпироллидон, декстран или комбинации любых из указанных наполнителей. В другом предпочтительном варианте осуществления изобретения композиция, кроме того, содержит неионный детергент. Предпочтительными неионогенными детергентами являются полисорбат 20, полисорбат 80, тритон Х-100, тритон Х-114, нонидет Р-40, октил-а-глюкозид, октил-b-глюкозид, Brij 35, плюроник и твин 20. В предпочтительном варианте осуществления изобретения неионогенный детергент включает полисорбат 20 или полисорбат 80. Предпочтительная композиция, кроме того, содержит забуференный фосфатом физиологический раствор, предпочтительно, при рН 6.
Кроме того, настоящее изобретение относится к способам введения препарата α -Gal А индивидууму. В предпочтительном варианте осуществления изобретения препаратом α -Gal А является препарат α -Gal А с измененным зарядом, например с повышенным зарядом олигосахарида и/или с более продолжительным временем полужизни, как описано в настоящей заявке. Доза введения предпочтительно составляет 0,05-5,0 мг, а более предпочтительно 0,1-0,3 мг препарата α -Gal А на килограмм массы тела, и вводится один раз в неделю или один раз в две недели. В предпочтительном варианте осуществления изобретения доза введения составляет приблизительно 0,2 мг на килограмм массы тела и вводится раз в две недели. В указанных способах эта доза может быть введена внутримышечно, перорально, ректально, подкожно, внутриартериально, внутрибрюшинно, интрацеребрально, интраназально, внутрикожно, внутриоболочечно, через слизистую, чрескожно или путем ингаляции. В одном из вариантов осуществления изобретения способ доставки препарата α -Gal А индивидууму предусматривает подкожное введение дозы, составляющей от 0,01 до 10,0 мг, а предпочтительно 0,1-5,0 мг препарата α -Gal А на килограмм массы тела, один раз в неделю или один раз в две недели. Данный препарат α -Gal А может быть также введен внутривенно, например путем внутривенной инъекции болюса, внутривенной инъекции посредством медленной интубации или путем непрерывной внутривенной инфузии. В любом из вышеуказанных способов доставка препарата α -Gal А может быть осуществлена с использованием такой системы доставки, как насос, инкапсулированная клетка, липосомы, инъекции с использованием игл, безыгольное впрыскивание, ингалятор, аэрозольный распылитель, электропорация и пластырь для чрескожной доставки. Указанными способами может быть введен любой из вышеописанных препаратов α -Gal А.
Индивидуум с подозрением на болезнь Фабри или с уже установленным диагнозом болезни Фабри может быть подвергнут лечению путем введения вышеописанного препарата α -Gal А с использованием вышеописанных способов введения и доз. В настоящем изобретении рассматривается лечение индивидуумов, в основном, с болезнью Фабри, а также с атипическими вариантами болезни Фабри, например конкретных групп пациентов, страдающих болезнью Фабри с преобладающими сердечно-сосудистыми нарушениями, такими как увеличение желудочков, например гипертрофия левого желудочка (ГЛЖ), и/или недостаточность митрального клапана, или групп пациентов, страдающих болезнью Фабри с преобладающими почечными нарушениями.
α -Gal A
α -Gal А представляет собой гомодимерный гликопротеин, который отщепляет концевые α -галактозильные группы от гликолипидов и гликопротеинов.
Термины зрелая "α -Gal A", "GA-GAL" и "SEQ ID NO:5" (см.фиг.7) относятся к α -Gal А без сигнального пептида (для α -Gal А с сигнальным пептидом см. фиг.3 и SEQ ID NO:3). Определенный здесь термин "препарат α -Gal А" и термин "препарат гликозилированной α -Gal А" являются взаимозаменяемыми и включают различные гликоформы гликолизированной α -Gal А.
Термин "сигнальный пептид" означает пептидную последовательность, которая направляет новосинтезированный полипептид, к которому присоединен данный сигнальный пептид, в эндоплазматический ретикулум (ЭР) для последующего посттрансляционного процессинга и распределения.
Термин "гетерологичный сигнальный пептид", используемый здесь в связи с α -Gal А, означает сигнальный пептид, который не является сигнальным пептидом α -Gal А человека, а обычно представляет собой сигнальный пептид белка некоторых млекопитающих, не являющегося α -Gal А.
Специалистам известно, что ДНК-последовательность (либо кДНК [SEQ ID NO:5], либо геномная ДНК) человеческой α -Gal A или последовательности, которые отличаются от ДНК α -Gal А либо заменами молчащего кодона, либо заменами кодона, приводящими к заменам консервативных аминокислот, могут быть использованы для генетической модификации культивированных клеток человека так, чтобы они сверхпродуцировали и секретировали указанный фермент. Некоторые мутации в ДНК-последовательности α -Gal A могут кодировать полипептиды, которые сохраняют ту же ферментативную активность или обладают улучшенной ферментативной активностью α -Gal А. Так, например, ожидается, что замены консервативных аминокислот будут иметь незначительное влияние или вообще не будут оказывать влияния на биологическую активность, а особенно, если они представляют менее чем 10% от общего числа остатков в данном белке. Консервативными заменами обычно являются замены внутри следующих групп: глицин, аланин; валин, изолейцин, лейцин; аспарагиновая кислота, глутаминовая кислота; аспарагин, глутамин; серин, треонин; лизин, аргинин; фенилаланин, тирозин. См., например, патент США 5356804, который вводится в настоящее описание посредством ссылки.
Болезнь Фабри
Болезнь Фабри представляет собой наследственное заболевание, вызываемое дефицитом активности фермента α -Gal А. Термин "дефицит α -Gal А" означает любой дефицит количества или активности этого фермента у пациента, приводящий к аномальной аккумуляции нейтральных гликолипидов (например, глоботриаозилцерамида) в гистиоцитах в стенках кровеносных сосудов с образованием ангиокератом в области бедер, ягодиц и гениталий, к гипогидрозу, парестезии в конечностях, извитости роговицы и к спицеобразной задней субкапсулярной катаракте. Отложение этого материала может приводить к болям, тяжелому почечному и сердечно-сосудистому заболеванию и к инсульту. Аккумуляция гликолипида может приводить к тяжелым симптомам, обычно наблюдаемым у мужчин, страдающих болезнью Фабри. Альтернативно, такая аккумуляция может индуцировать относительно умеренные симптомы, которые могут иногда наблюдаться у женщин - гетерозиготных носителей дефектного гена. Страдающие этим заболеванием индивидуумы имеют прогнозируемую короткую продолжительность жизни; смерть обычно наступает в результате осложнений на почки, сердце или сердечно-сосудистую систему в возрасте приблизительно 40 лет. Какого-либо конкретного способа лечения этой болезни не существует. Болезнь Фабри классифицируется как расстройство, связанное с накоплением лизосом, которым страдают более чем 15000 человек во всем мире.
Болезнь Фабри, описанная выше, представляет собой сложный клинический синдром, характеризующийся вовлечением в патологический процесс множества органов и систем. Пациентов, у которых наблюдается комбинация признаков дистрофии роговицы, поражений кожи (ангиокератомы), болезненной невропатии, сердечно-сосудистого заболевания, кардиомиопатии и почечной дисфункции, относят к категории, рассматриваемой как "классический" фенотип. Однако имеются пациенты, у которых проявляются некоторые, но не все аспекты указанного классического фенотипа. Эти пациенты классифицируются как пациенты с "атипическими вариантами болезни Фабри". Существует несколько фенотипов атипического варианта, ассоциированных с дефицитом α -галактозидазы А. Так, например, некоторые пациенты с дефицитом α -галактозидазы А страдают вариантом болезни Фабри, сопровождающимся лишь поражением сердца, например гипертрофией левого желудочка (ГЛЖ).
Имеется также другой фенотипический вариант, при котором пациенты страдают лишь поражением почек. Хотя оба этих фенотипических варианта были определены в мужских гемизиготах, однако, вариантные формы болезни Фабри были также описаны в женских гетерозиготах.
У пациентов, страдающих атипическим вариантом с поражением сердца, симптомы этой болезни проявляются в более позднем возрасте. Средний возраст диагностики этой болезни у пациентов с фенотипическим вариантом, проявляющимся поражением сердца, составляет приблизительно 52 года, тогда как для классического фенотипа, этот возраст составляет приблизительно 29 лет (Desnick et al., In The Metabolic and Molecular Bases of Inherited Disease, 6th edition (1996), Scriver et al. (eds), McGraw-Hill (New York), pp. 2741-2784; Meikle et al., J.Am.Med. Assoc. 281:249-254 (1999)). Среди пациентов с этим синдромом часто встречаются пациенты со слабыми симптомами нарушения сердечной функции, такими как одышка при физической нагрузке. Обычно стандартный эхокардиографический анализ показывает, что у пациентов с сердечным фенотипическим вариантом наблюдается гипертрофия левого желудочка (ГЛЖ) или асимметрическая гипертрофия перегородки. Однако могут также встречаться пациенты с инфарктом миокарда или кардиомиопатией (Scheidt et al., New Engl. J.Med. 324:395-399 (1991); Nakao et al., New Engl. J.Med. 333:288-293 (1995)). Этих пациентов в большинстве случаев подвергают миокардиальной биопсии, которая показывает, что патология с такой разновидностью синдрома, в основном, аналогична классической болезни Фабри, т.е. миокардиальной инфильтрации в результате отложения гликолипида. Ферментные анализы α -галактозидазы А у этих пациентов выявили наличие широкого диапазона уровней фермента. Так, например, сообщалось, что пациенты с "сердечным" вариантом имеют ферментативную активность, составляющую до 30% от нормальных уровней активности фермента α -галактозидазы А, и, таким образом, до настоящего времени эти пациенты рассматривались как кандидаты на заместительную терапию α -Gal A.
Авторами настоящего изобретения было неожиданно обнаружено, что, хотя пациенты с атипическим сердечным вариантом или атипическим почечным вариантом могут иметь уровни активности фермента α -галактозидазы А, которые являются относительно высокими по сравнению с уровнями у пациентов с классическим фенотипом болезни Фабри, однако, у этих пациентов может также наблюдаться благоприятный исход в результате проведения терапии с использованием фермента α -галактозидазы А. Так, например, пациенты могут иметь мутацию, продуцирующую кинетически нестабильный фермент α -Gal А в клетке, и у этих пациентов уровни фермента α -Gal А могут быть значительно увеличены путем введения препаратов α -Gal А настоящего изобретения. Кроме того, сообщалось, что у некоторых пациентов, страдающих атипичным фенотипическим сердечным вариантом, имеется точковая мутация в аминокислоте 215 α -галактозидазы А. Эта аминокислота в немутированном белке представляет собой аспарагин, который является гликозилированным (Eng et al., Am. J. Hum. Genet. 53:1186-1197 (1993)). Таким образом, заместительная терапия ферментом α -Gal А с использованием препаратов на основе соответствующим образом гликозилированной α -галактозидазы А настоящего изобретения может оказаться эффективной для этих пациентов. Более того, сообщалось, что у пациентов с атипическим почечным вариантом, у которых наблюдалось лишь клиническое проявление болезни Фабри, имеется слабая протеинурия. Однако биопсия почек выявила типичные для болезни Фабри гликолипидные включения, а анализ на фермент α -Gal А выявил более низкие уровни α -Gal А по сравнению с нормальными уровнями α -Gal А. Однако, поскольку отложения церамидтригексозида в почках могут быть обнаружены в клетках отводящих почечных канальцев при осаждении мочи у этих пациентов, то введение препаратов α -Gal А настоящего изобретения может значительно снижать эти уровни. Лизосомные ферменты, такие как α -Gal А, доставляются в лизосомный компартмент клетки посредством взаимодействия с рецептором маннозо-6-фосфата (М6Р), который связывается с остатками М6Р, присутствующими в олигосахаридных группах ферментов, нацеленных на лизосомный компартмент. Kornfeld & Mellman, Ann. Rev. Cell. Biol. 5:483-525 (1989). Первичное взаимодействие происходит в аппарате Гольджи, где ферменты, связанные с рецепторами М6Р Гольджи, отделяются для транспорта в лизосомы. Очевидно, что второй тип взаимодействия происходит между внеклеточной α -Gal А и рецепторами М6Р на клеточной поверхности. Ферменты, которые "ускользают" из системы обычного "маршрута", секретируются клетками по конститутивному секреторному пути и часто снова захватываются рецепторами М6Р клеточной поверхности, которые возвращают α -галактозидазу А в лизосому по эндоцитному пути. Внеклеточные вещества, интернализованные клетками, транспортируются через цитоплазму в эндоцитные везикулы, которые сливаются с первичными лизосомами и переносят свое содержимое в лизосомы. В этом процессе рецепторы М6Р клеточной поверхности также включаются в эндоцитные везикулы и транспортируются в лизосомы. В частности, препараты α -Gal А настоящего изобретения, в которых присутствуют высокие уровни сиалилирования и/или фосфорилирования, являются предпочтительными для лечения пациентов с атипическими вариантами болезни Фабри. Такие препараты, например, минимизируют фракцию инъецированной α -Gal А, которая удаляется гепатоцитами, и позволяют поглощать высокие уровни α -Gal А клетками, не являющимися клетками печени, такими как клетки почек, клетки сосудов, клетки канальцев, гломерулярные клетки, миоциты сердца и клетки сердечно-сосудистой системы.
Внеклеточная α -Gal А, несущая остатки М6Р, может связываться с рецепторами М6Р клеточной поверхности и транспортироваться в лизосомный компартмент. После транспортировки в лизосомный компартмент α -Gal А может выполнять соответствующую функцию. Это является тем аспектом транспортировки лизосомного фермента, который делает заместительную терапию с использованием фермента α -галактозидазы А подходящей для эффективного терапевтического лечения пациентов с болезнью Фабри. Таким образом, даже если клетка является генетически дефицитной по продуцированию α -Gal А, эта клетка может поглощать внеклеточную α -Gal А, если указанная α -Gal А является соответствующим образом гликозилированной, а дефицитная клетка несет рецепторы М6Р. У пациентов с болезнью Фабри сосудистые эндотелиальные клетки почек и сердца обнаруживают серьезные гистопатологические аномалии и вносят определенный вклад в клиническую патологию данного заболевания. Такие клетки, которые несут рецепторы М6Р, являются конкретной терапевтической мишенью для α -Gal А. Целью настоящего изобретения является получение препарата α -Gal А, в котором М6Р присутствует в N-связанных олигосахаридах.
Степень, с которой N-связанные олигосахариды α -Gal А модифицируются путем сиалилирования, оказывает значительное влияние на фармакокинетику и биологическое распределение α -Gal А. В отсутствие соответствующего сиалилирования α -Gal А быстро выводится из кровотока благодаря связыванию печеночных асиалогликопротеиновых рецепторов (рецепторов Эшвелла) с последующей интернализацией и деградацией гепатоцитами. Ashwell & Harford, Ann.Rev.Biochem. 51:531-554 (1982). Это приводит к снижению количества α -Gal А, имеющегося в кровотоке для связывания с рецепторами М6Р на клетках, которые вносят свой вклад в клиническую патологию болезни Фабри, таких как сосудистые эндотелиальные клетки почек и сердца. α -Gal А, секретируемая генетически модифицированными клетками человека, обладает гликозилирующими свойствами, которые являются подходящими для лечения болезни Фабри, либо путем стандартного фармацевтического введения очищенного секретированного белка, либо посредством генной терапии, и не требует каких-либо дополнительных ферментных модификаций, которые, как сообщалось, необходимы для лизосомного фермента, глюкоцереброзидазы, где поглощение очищенного глюкоцереброзидазного фермента клинически релевантными клетками требует проведения сложных ферментативных модификаций данного фермента после очистки от плаценты человека. Beutler, New Engl. J.Med. 325:1354-1360 (1991).
Клетки, подходящие для продуцирования α -Gal A
Индивидуум с подозрением на дефицит α -Gal А, такой как болезнь Фабри, может быть подвергнут лечению с использованием очищенной человеческой α -Gal А, полученной из культивированных, генетически модифицированных клеток, предпочтительно клеток человека.
Если для лечения болезни Фабри необходима генетическая модификация клеток, то эти клетки могут быть модифицированы стандартными методами генной инженерии или путем активации гена.
В соответствии со стандартными методами молекула ДНК, содержащая кДНК или геномную ДНК-последовательность α -Gal А, может находиться в экспрессионной конструкции и трансфецироваться в первичные, вторичные или иммортализованные клетки стандартными методами, включая, но не ограничиваясь ими, трансфекцию, опосредованную липосомами, полибреном или DEAE-декстраном, электропорацию, осаждение фосфатом кальция, микроинъекцию или бомбардировку микрочастицами с регулируемой скоростью ("биобаллистика") (см. например, совместно рассматриваемую заявку, USSN 08/334797, которая вводится в настоящее описание посредством ссылки). Альтернативно, может быть использована система, которая доставляет генетическую информацию с помощью вирусного вектора. Известными вирусами, которые могут быть использованы для переноса генов, являются аденовирусы, аденоассоциированный вирус, вирус герпеса, вирус свинки, полиовирус, ретровирусы, вирус Синдбис и вирус коровьей оспы, такой как поксвирус канареек.
Альтернативно, указанные клетки могут быть модифицированы методом активации гена ("GA"), таким как метод, описанный в патентах Соединенных Штатов №5733761 и 5750376, каждый из которых вводится в настоящее описание посредством ссылки. α -Gal А, полученный путем активации гена, обозначен в настоящем описании GA-GAL.
В соответствии с этим термин "генетически модифицированный", используемый в данном описании применительно к клеткам, относится к клеткам, которые экспрессируют конкретный генный продукт после введения ДНК-молекул, кодирующих этот генный продукт и/или регуляторные элементы, контролирующие экспрессию последовательности, кодирующей данный генный продукт. Указанная ДНК-молекула может быть введена посредством доставки гена или посредством гомологичной рекомбинации, т.е. введения этой ДНК-молекулы в конкретный геномный сайт. Гомологичная рекомбинация может быть использована для замены самого дефектного гена (дефектный ген α -Gal А или его часть могут быть заменены в клетках пациента с болезнью Фабри на целый ген или его часть).
Используемый здесь термин "первичная клетка" означает клетку, присутствующую в суспензии клеток, выделенных из тканевого источника позвоночных (перед их высеванием, т.е. прикреплением к субстрату тканевой культуры, такому как чашка или колба); клетку, присутствующую в эксплантате, происходящем от ткани; клетки обоих вышеупомянутых типов, высеваемые в первый раз, и клеточные суспензии, полученные от указанных засеянных клеток.
Термин "вторичные клетки" означает клетки во всех последующих стадиях культивирования. То есть в первый раз засеянная первичная клетка, удаляемая из субстрата в культуре и засеиваемая снова (пассированная), называется вторичной клеткой, как и все клетки в последующих пассажах.
"Клеточный штамм" состоит из вторичных клеток, которые были пассированы один или несколько раз; имеют определенное число удвоения средней популяции в культуре; обладают способностью к контактно ингибируемому росту на "якорной" подложке (за исключением клеток, размноженных в суспензионной культуре) и не являются иммортализованными.
Термин "иммортализованная клетка" означает клетку, происходящую от стабильной клеточной линии, которая обладает очевидной неограниченной продолжительностью жизни в культуре.
Примерами первичных или вторичных клеток являются фибробласты, эпителиальные клетки, включая клетки млекопитающих и эпителиальные кишечные клетки, эндотелиальные клетки, форменные элементы крови, включая лимфоциты и клетки костного мозга, глиальные клетки, гепатоциты, кератиноциты, мышечные клетки, нервные клетки или предшественники клеток этого типа. Примерами иммортализованных клеточных линий человека, которые могут быть использованы в способах настоящего изобретения, являются, но не ограничиваются ими, клетки меланомы Боуэса (номер доступа АТСС CRL 9607), клетки Дауди (номер доступа АТСС CCL 213), клетки HeLa и производные клеток HeLa (номер доступа АТСС CCL 2, CCL 2.1 и CCL 2.2), клетки HL-60 (номер доступа АТСС CCL 240), клетки НТ-1080 (номер доступа АТСС CCL 121), клетки Jurkat (номер доступа АТСС TIB 152), клетки карциномы KB (номер доступа АТСС CCL 17), клетки лейкоза К-562 (номер доступа АТСС CCL 243), клетки рака молочной железы MCF-7 (номер доступа АТСС ВТН 22), клетки MOLT-4 (номер доступа АТСС 1582), клетки Намальвы (номер доступа АТСС CRL 1432), Raji-клетки (номер доступа АТСС CCL 86), клетки RPMI 8226 (номер доступа АТСС CCL 155), клетки U-937 (номер доступа АТСС CRL 1593), клетки 2R4 сублинии WI-38VA13 (номер доступа АТСС CLL 75,1), клетки CCRF-CEM (номер доступа АТСС CCL 119) и клетки карциномы яичника 2780AD (Van der Blick et al., Cancer Res. 48:5927-5932, 1988), а также клетки гетерогибридомы, продуцируемые путем слияния клеток человека и клеток другого вида.
После генетической модификации клеток человека для продуцирования клетки, секретирующей α -Gal А, может быть генерирован клональный клеточный штамм, состоящий, в основном, из множества генетически идентичных культивированных первичных клеток человека, либо, если эти клетки являются иммортализованными, то может быть генерирована клональная клеточная линия, состоящая, в основном, из множества генетически идентичных иммортализованных человеческих клеток. В одном варианте осуществления настоящего изобретения клетками клонального клеточного штамма или клональной клеточной линии являются фибробласты. В предпочтительном варианте осуществления изобретения такими клетками являются вторичные фибробласты человека, например клетки BRS-11.
После генетической модификации указанные клетки культивируют в условиях, способствующих секреции α -Gal А. Данный белок выделяют из культивированных клеток путем сбора среды, в которой были выращены указанные клетки, и/или путем лизиса клеток с высвобождением их содержимого, с последующей их обработкой методами очистки белков.
Очистка α -Gal А из кондиционированной среды стабильно трансфецированных клеток
В соответствии со способами настоящего изобретения белок α -Gal А выделяют из культивированных клеток ("клеток, продуцирующих α -Gal А") путем сбора среды, в которой были выращены эти клетки, или лизиса клеток с высвобождением их содержимого, а затем белки очищают методами очистки белков без использования аффинной хроматографии на лектине. Предпочтительный способ очистки описан ниже в примере 2.
Для очистки α -Gal А могут быть также использованы альтернативные смолы гидрофобного взаимодействия, такие как Source Iso (Pharmacia), метиловая подложка Macro-Prep® Methyl Support (Bio-Rad), TSK-бутил (Tosohaas) или фенилсефароза® (Pharmacia). Колонка может быть уравновешена в относительно высокой концентрации соли, например в 1 М сульфате аммония или 2 М хлориде натрия, в буфере при рН 5,6. Очищаемый образец получают путем доведения рН и концентрации соли до рН и концентрации уравновешивающего буфера. Образец наносят на колонку и эту колонку промывают уравновешивающим буфером для удаления несвязанного материала. α -Gal А элюируют из колонки буфером с более низкой ионной силой, водой или органическим растворителем в воде, например 20% этанолом или 50% пропиленгликолем. Альтернативно, α -Gal А можно заставить проходить через колонку с использованием более низкой концентрации соли в уравновешивающем буфере и в пробе или с использованием другого рН. Другие белки могут быть связаны с колонкой, что приводит к очистке α -Gal А-содержащего образца, который не был связан с колонкой. Первая стадия очистки, предпочтительно, предусматривает использование колонки с гидроксиапатитом.
В альтернативной стадии очистки α -Gal А могут быть использованы катионообменные смолы, например сефарозы: SP Sepharose® 6 Fast Flow (Pharmacia), Source 30S (Pharmacia), CM Sepharose® Fast Flow (Pharmacia), Macro-Prep® Methyl Support (Bio-Rad) или Macro-Prep® High S Support (Bio-Rad). Термин "первая стадия хроматографии" означает первое нанесение образца на хроматографическую колонку (все стадии, связанные с получением образца исключаются). α -Gal А может быть связана с колонкой при рН 4,4. Для уравновешивания колонки может быть использован буфер, такой как 10 мМ ацетат натрия, рН 4,4, 10 мМ цитрат натрия, рН 4,4, или другой буфер с аналогичной забуферивающей способностью, приблизительно при рН 4,4. Образец, предназначенный для очистки доводят до рН и ионной силы уравновешивающего буфера. Этот образец наносят на колонку и после загрузки эту колонку промывают для удаления несвязанного материала. Для элюирования α -Gal А с колонки может быть использована соль, такая как хлорид натрия или хлорид калия. Альтернативно, α -Gal А может быть проэлюирован с колонки буфером с более высоким рН или с буфером с более высокой концентрацией соли и с более высоким рН. α -Gal А можно также заставить проходить через колонку во время загрузки путем увеличения концентрации соли в уравновешивающем буфере и в загрузке пробы при проведении хроматографии на колонке при более высоком рН или путем комбинации повышения концентрации соли и увеличения рН.
В другой стадии очистки α -Gal А может быть использована сефароза® Q (Q Sepharose® 6 Fast Flow). Указанная Сефароза® Q является относительно сильной анионообменной смолой. Для очистки α -Gal А может быть использована и более слабая анионообменная смола, такая как DEAE-сефароза® (DEAE Sepharose® 6 Fast Flow; Pharmacia) или Macro-Prep® DEAB (Bio-Rad). Колонку уравновешивают в буфере, например в 10 мМ фосфате натрия, рН 6. рН образца доводят до рН 6, а низкая ионная сила достигается разведением или диафильтрацией образца. Этот образец наносят на колонку в условиях, благоприятствующих связыванию α -Gal А. Колонку промывают уравновешивающим буфером для удаления несвязанного материала. α -Gal А элюируют с использованием соли, например хлорида натрия или хлорида калия, или с использованием буфера с более низким рН, или с использованием комбинации более высокой концентрации соли и более низким рН. α -Gal А можно также заставить проходить через колонку во время загрузки путем увеличения концентрации соли в уравновешивающем буфере и в загрузке пробы при проведении хроматографии на колонке при более высоком рН или путем комбинации повышения концентрации соли и увеличения рН.
В другой стадии очистки α -Gal А может быть использована вытеснительная хроматография на супердексе® (Superdex® 200; Pharmacia). Для очистки α -Gal А могут быть использованы и другие смолы для вытеснительной хроматографии, такие как сефакрил® (Sephacryl® S-200 HR) или Bio-Gel® А-1,5 м. Предпочтительным буфером для вытеснительной хроматографии является 25 мМ фосфат натрия, рН 6,0, содержащий 0,15 М хлорида натрия. Могут быть использованы и другие, совместимые с данной композицией буферы, например 10 мМ цитрат натрия или калия. Данный буфер может иметь рН от 5 до 7 и должен содержать соль, например хлорид натрия или смесь хлорида натрия и хлорида калия.
В другой стадии для очистки α -Gal А может быть использована хроматофокусирующая смола, такая как Polybuffer Exchanger РВЕ 94 (Pharmacia). Колонку уравновешивают при относительно высоких рН (например, рН 7 или выше), рН образца, подвергаемого очистке, доводят до того же самого рН и данный образец наносят на колонку. Белки элюируют с уменьшающимся градиентом рН до, например, рН 4 с использованием буферной системы, например Polybuffer 74 (Pharmacia), которая была доведена до рН 4.
Альтернативно, для очистки α -Gal А может быть использована иммуноаффинная хроматография. Соответствующее поликлональное или моноклональное антитело против α -Gal А (генерированное путем иммунизации α -Gal А или пептидом, полученным из последовательности α -Gal А стандартными методами) может быть иммобилизовано на активированной связывающей смоле, например NHS-активированной сефарозе® (Sepharose® 4 Fast Flow, Pharmacia) или на CNBr-активированной сефарозе® (Sepharose® 4 Fast Flow; Pharmacia). Предназначенный для очистки образец может быть нанесен на колонку с иммобилизованным антителом примерно при рН 6 или рН 7. Эту колонку промывают для удаления несвязанного материала. α -Gal А элюируют с колонки типичными реагентами, используемыми для элюирования на аффинной колонке, такими как денатурирующий агент с низким рН, например рН 3, например гуанидин-НСl или тиоцианат, либо органический растворитель, например 50% пропиленгликоль в буфере с рН 6. В процедуре очистки α -Gal А может быть также использована аффинная смола, образующая хелатный комплекс с металлом, например хелатообразующая сефароза (Sepharose® Fast Flow; Pharmacia). Колонку предварительно загружают ионами металла, например Сu2+, Zn2+, Са2+, Мg2+ или Сd2+. Очищаемый образец наносят на колонку при соответствующем рН, например рН 6-7,5, и эту колонку промывают для удаления несвязанных белков. Связанные белки элюируют путем конкурентного элюирования имидазолом или гистидином, либо путем снижения рН с использованием цитрата натрия или ацетата натрия до рН менее 6, либо путем введения хелатообразующих агентов, таких как EDTA или EGTA.
В соответствии с вышеуказанными протоколами настоящее изобретение относится к препаратам с более высокой чистотой, чем полученные ранее препараты α -Gal А, то есть к препаратам, очищенным, по крайней мере, до 98%-ной гомогенности, более предпочтительно, по крайней мере, до 99%-ной гомогенности, а наиболее предпочтительно, по крайней мере, до 99,5%-ной гомогенности, измеренной с помощью электрофореза в ПААГ с ДСН или обращенно-фазовой ВЭЖХ. Препараты α -Gal А настоящего изобретения могут содержать большое количество гликоформ α -Gal А. В соответствии с этим термин "гомогенность", используемый здесь по отношению к препаратам α -Gal А, относится к препаратам, которые, в основном, не содержат белков (<2% от общего числа белков), не являющихся α -Gal А. Примерами таких не-α -Gal А белков являются альбумин, белки, не относящиеся к α -Gal А, продуцируемые клеткой-хозяином, и белки, не относящиеся к α -Gal А, выделенные из ткани или физиологической жидкости животного. Удельная активность препаратов α -Gal А настоящего изобретения предпочтительно составляет, по крайней мере, 2,0× 106 единиц/мг белка, более предпочтительно, по крайней мере, 3,0× 106 единиц/мг белка, а наиболее предпочтительно, по крайней мере, 3,5× 106 единиц/мг белка.
Увеличение времени полужизни в кровотоке препаратов α -Gal A путем гликанового ремоделирования в целях увеличения заряда олигосахаридов
Настоящее изобретение относится к программе модификации гликопротеина для усиления поглощения терапевтического фермента конкретными тканями, не относящимися к печени, и макрофагами. С использованием методов настоящего изобретения были получены препараты гликозилированной человеческой α -Gal А, где от 35 до 85% олигосахаридов, а предпочтительно, по крайней мере, 50% олигосахаридов, являются заряженными.
N-Гликозилирование белка осуществляют путем модификации соответствующих аспрагиновых остатков белков олигосахаридными структурами, что влияет на их свойства и биоактивность. Kukuruzinska & Lennon, Crit.Rev.Oral.Biol.Med. 9: 415-48 (1998). Настоящее изобретение относится к выделенному препарату α -Gal А, в котором высокий процент олигосахаридов является отрицательно заряженным, главным образом, путем присоединения от одного до четырех остатков сиаловой кислоты к сложным гликанам, либо одной или двух фосфатных групп к гликанам с высоким содержанием маннозы, либо одного фосфата и одной сиаловой кислоты к гибридным гликанам. Могут также присутствовать сульфатированные сложные гликаны в меньших количествах. Высокое содержание заряженных структур имеет две основные функции. Во-первых, кэпирование предпоследних галактозных остатков 2,3- или 2,6-связанной сиаловой кислотой предупреждает их преждевременное удаление из кровотока под действием асиалогликопротеинового рецептора, присутствующего на гепатоцитах. Этот рецептор узнает гликопротеины с концевыми галактозными остатками. Увеличение времени полужизни α -Gal А в кровотоке дает возможность таким важным органам-мишеням, как сердце и почки, осуществлять эндоцитоз больших количеств фермента из плазмы после инфузии фермента. Во-вторых, присутствие ман-6-фосфата на гликанах с высоким содержанием маннозы или на гибридных гликанах позволяет осуществлять опосредованное рецептором поглощение с помощью катиононезависимого ман-6-фосфатного рецептора (CI-MPR). Это опосредованное рецептором поглощение происходит на поверхности многих клеток, включая эндотелиальные клетки сосудов, которые являются главным местом накопления СТН у пациентов с болезнью Фабри. Молекулы фермента с двумя ман-6-фосфатными остатками обладают гораздо большей аффинностью по отношению к CI-MPR, чем ферменты с одним ман-6-фосфатом. Характерные гликановые структуры представлены в таблице 1.

В биосинтезе N-гликопротеина участвует множество ферментов, гликозилтрансфераз и гликозидаз. Большинство из этих ферментов функционирует в эндоплазматическом ретикулуме (ЭР) и аппарате Гольджи упорядоченным и хорошо организованным образом. Сложность N-гликозилирования возрастает вследствие того факта, что другие аспарагиновые остатки в тех же самых полипептидах могут быть модифицированы другими олигосахаридными структурами, и различные белки отличаются друг от друга своими углеводными группами. Прогресс, достигнутый за последнее время в молекулярной генетике, ускорил идентификацию, выделение и характеризацию генов N-гликозилирования. В результате этого была получена информация относительно взаимосвязи между гликозилированием и другими клеточными функциями.
Процессинг N-связанного гликопротеина в клетке начинается в том случае, когда олигосахаридная цепь с Glc3Man9GlcNAc2 присоединяется к акцептору аспарагина на растущей пептидной цепи в просвете ЭР в виде одного звена. Олигосахаридную цепь из четырнадцати сахаров, состоящую из Glc3Man9GlcNAc2, строят на долихоле, алифатическом спирте с очень длинной цепью:

Этот олигосахарид переносят в виде одного звена к остатку аспарагина-акцептора на растущей пептидной цепи в просвете ЭР. Гликан, имеющий большой размер по отношению к пептиду, может управлять укладкой белка. Эти три глюкозных остатка служат в качестве сигнала, завершающего олигосахарид, в результате чего данный олигосахарид становится готовым для переноса олигосахарилтрансферазой. Этот фермент будет также переносить неглюкозилированные олигосахариды, но только фракцию с размером, равным завершенной цепи, поскольку они являются субоптимальными субстратами. Было показано, что одна из форм синдрома углеводного дефицита гликопротеина у человека обусловлена дефицитом долихол-P-Glc:Man9GlcNАс2-РР-долихол-гликозилтрансферазы, первого фермента в пути присоединения глюкозы, что приводит к гипогликозилированию белков сыворотки. Korner et al., Proc.Natl. Acad. Sci., USA, 95:13200-13205 (1998). После удаления трех глюкозных остатков и достижения правильной конформации вновь синтезированный гликопротеин экспортируется в аппарат Гольджи. В зависимости от доступности гликана маннозидазам аппарата Гольджи после укладки белка гликановая цепь может оставаться в виде цепи с высоким содержанием маннозы с 5-9 маннозными остатками. Альтернативно, гликановая цепь может быть затем процессирована в триманнозиловую сердцевину и стать акцептором для других гликозилтрансфераз, которые образуют сложные цепи путем присоединения других остатков GlcNAc, а затем Gal, NeuAc и Fuc. Третьей возможностью, в том случае, если белок имеет два лизиновых остатка на расстоянии точно в 34 ангстрема и в правильной пространственной взаимосвязи с цепью с высоким содержанием маннозы, является присоединение GlcNAcα -1-РO4 к углероду 6 одного, а иногда двух маннозных остатков. Cuozzo et аl., J.Biol.Chem. 273:21069-21076 (1998). После удаления α -связанного GlcNAc специфическим ферментом, генерируется концевой эпитоп М6Р, который узнается рецептором М6Р в транс-сети Гольджи, которая затем направляет эти ферменты в лизосомы клеток, происходящих из мезенхимы.
Для доставки α -Gal А по возможности в большее число тканей могут быть использованы многие другие углеводные структуры (гликоформы). Matsuura et al., Glycobiology 8:329-339 (1998), сообщают, что гликановые структуры на человеческой α -Gal А, продуцированные в клетках СНО, имеют 41% гликанов с высоким содержанием маннозы, а уровень фосфорилирования составляет 24%. Однако уровень сиалилированных сложных гликанов составлял лишь 11%. Таким образом, 2/3 указанных сложных цепей не были сиалилированы, что приводило к быстрому удалению α -Gal А печенью. α -Gal А, продуцированная в человеческих клетках настоящего изобретения, имеет более высокий процент заряженных олигосахаридов, чем α -Gal А, продуцированная ранее в клетках СНО. Так, например, α -Gal А, продуцированная в описанных здесь клетках НТ-1080, является особенно подходящей, поскольку α -Gal А, продуцированная в клетках НТ-1080, содержит приблизительно 15% нейтральных структур (с высоким содержанием маннозы и гибридов), приблизительно 16% фосфорилированных гликанов и приблизительно 67% сложных гликанов в 2-4 остатками сиаловой кислоты. Таким образом, в основном, все эти сложные цепи являются сиалилированными по сравнению с α -Gal А, продуцируемой в клетках СНО. α -Gal А в клетках НТ-1080 имеет три сайта гликозилирования по N-связям. Два сайта процессируются в сложные гликаны в аппарате Гольджи, а третий сайт занят гликаном с высоким содержанием маннозы, 50% которого модифицировано лизосомным фермент-специфическим фосфорилированием с образованием как монофосфорилированных, так и дифосфорилированных молекул.
Существует четыре способа углеводного ремоделирования на белке, содержащем N-связанные гликановые цепи. В первом способе соотношение заряженной α -Gal А может быть увеличено путем селективного выделения гликоформ в процессе очистки. Настоящее изобретение относится к увеличению содержания в высокой степени заряженных и высокомолекулярных гликоформ α -Gal А путем фракционирования молекул α -Gal А на хроматографической колонке со смолой во время и/или после очистки. Молекулы гликоформ α -Gal А с большим зарядом содержат большее количество сиаловой кислоты и/или большее количество фосфата, а более высокомолекулярные гликоформы также должны содержать полностью гликозилированные, наиболее высокоразветвленные и высокозаряженные молекулы. Отбор заряженных молекул или удаление негликозилированных, слабо гликозилированных или слабо сиалилированных и/или фосфорилированных молекул α -Gal А должно приводить к получению популяции гликоформ α -Gal А с 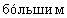 содержанием сиаловой кислоты и/или с
содержанием сиаловой кислоты и/или с  содержанием фосфата, а следовательно, к получению препарата α -Gal А с более высоким временем полужизни и с более высокой потенциальной терапевтической эффективностью.
содержанием фосфата, а следовательно, к получению препарата α -Gal А с более высоким временем полужизни и с более высокой потенциальной терапевтической эффективностью.
Этот процесс фракционирования может происходить, но не ограничиваться ими, на подходящих смолах для хроматографических колонок, используемых для очистки или для выделения α -Gal А. Так, например, фракционирование может происходить, но не ограничиваться ими, на катионнообменных смолах (таких как SP-Sepharose®), анионнообменных смолах (Q-Sepharose®), аффинных смолах (Heparin Sepharose®, колонки с лектином), смолах на колонках для вытеснительной хроматографии (Superdex® 200) и смолах на колонках для гидрофобного взаимодействия (Butyl Sepharose®) и на других смолах для хроматографических колонок, известных специалистам.
Поскольку α -Gal А продуцируется в клетках в виде гетерогенной смеси гликоформ, которые отличаются по своей молекулярной массе и заряду, то α -Gal А имеет тенденцию к элюированию с хроматографических смол в относительно широких пиках. При таком элюировании указанные гликоформы имеют конкретное распределение в зависимости от природы используемой смолы. Так, например, при вытеснительной хроматографии самые крупные гликоформы элюируются раньше на профиле элюции, чем более мелкие гликоформы.
При ионообменной хроматографии наиболее отрицательно заряженные гликоформы будут связываться с положительно заряженной смолой (такой как Q-Sepharose®) с более высокой аффинностью, чем менее отрицательно заряженные гликоформы, а следовательно, они будут позднее элюироваться на профиле элюции. В противоположность этому указанные в высокой степени отрицательно заряженные гликоформы могут менее жестко связываться с отрицательно заряженной смолой, такой как SP Sepharose®, чем менее отрицательно заряженные молекулы, либо они даже могут вообще не связываться со смолой.
Фракционирование молекул гликоформ на хроматографических смолах может зависеть от рН, ионной силы, выбора буферной соли, вязкости и/или других параметров, таких как тип выбранной смолы. Использование различных типов градиентного элюирования (линейных градиентов, нелинейных градиентов, например экспоненциальных градиентов) или использование серии коротких стадий элюции для селективного элюирования молекул α -Gal А с хроматографической колонки может быть оптимизировано для фракционирования α -Gal А. Все эти факторы, взятые отдельно или в комбинации, могут быть оптимизированы для достижения эффективного фракционирования гликоформ. Фракционирование может быть осуществлено после завершения процесса очистки на конкретной хроматографической смоле, селективно оптимизированной для фракционирования и отбора нужной популяции гликоформ.
Отбор популяции гликоформ из фракционированных молекул α -Gal А может быть осуществлен после анализа элюированных гликоформ α -Gal А. Пик элюции может быть проанализирован различными способами, но не ограничиваясь ими, такими как электрофорез в ДСН-ПААГ, изоэлектрическое фокусирование, капиллярный электрофорез, аналитическая ионообменная ВЭЖХ и/или аналитическая вытеснительная ВЭЖХ. Могут быть отобраны конкретные фракции, которые имеют желаемый размер или профиль заряда. Отбор может быть осуществлен в каждой хроматографической стадии данного процесса, что позволяет постепенно достичь получения нужной популяции гликоформ, или, в случае высокой эффективности фракционирования в данной стадии (стадиях), этот отбор может быть ограничен этой конкретной стадией или стадиями. Фракционирование может быть также осуществлено после завершения процесса очистки на конкретной хроматографической смоле, селективно оптимизированной для фракционирования и отбора нужной популяции гликоформ.
Фракционирование и отбор высокозаряженных и/или высокомолекулярных гликоформ α -Gal А могут быть осуществлены на любом препарате α -Gal А, таком как препарат, полученный из генетически модифицированных клеток, таких как клетки, модифицированные стандартными методами генной инженерии или путем активации гена (АГ). Они могут быть осуществлены на клеточных линиях, культивированных в оптимизированных системах с достижением высокой степени сиалилирования и фосфорилирования, как описано выше, либо на ПЭГилированных α -Gal А, как описано ниже.
Так, например, в описанном здесь процессе очистки α -Gal А фракционирование гликоформ α -Gal А может быть осуществлено на различных стадиях этого процесса. На гидрофобной смоле, бутилсефарозе, Butyl Sepharose® Fast Flow, первыми элюируются наиболее заряженные гликоформы α -Gal А, а за ними - молекулы с меньшим зарядом. На гепарин-сефарозе, Heparin Sepharose®, молекулы с наиболее высоким зарядом также элюируются первыми в пике элюции, а менее заряженные молекулы элюируются позже. Это отличается от элюирования на Q-серфарозе®, где наименее заряженные молекулы элюируются первыми, а затем элюируются наиболее высоко заряженные гликоформы. При вытеснительной хроматографии на супердексе (Superdex® 200), наиболее высокомолекулярные гликоформы элюируются первыми, а за ними элюируются молекулы гликозилированной α -Gal А с меньшей молекулярной массой. Для эффективного фракционирования конкретных популяций гликоформ α -Gal А может быть объединено множество хроматографических стадий, и все эти гликоформы фракционируются разными физическими методами. Так, например, для получения гликоформ α -Gal А, содержащих наименьший рI (то есть, содержащих наибольший отрицательный заряд), ограничивание сбором рано элюирующихся бутильных фракций должно приводить к увеличению α -Gal А с большим зарядом. Осуществляя хроматографию этого отобранного пула на гепариновой колонке и снова ограничиваясь сбором пула рано элюирующихся молекул α -Gal А с более высоким отрицательным зарядом, можно достичь еще большего увеличения фракции гликоформ α -Gal А с низким рI в данном пуле. Последующая более тонкая корректировка популяции гликоформ может быть проведена в различных стадиях процесса очистки путем мониторинга распределения размера и заряда пулов элюции с помощью электрофореза в ДСН-ПААГ и изоэлектрического фокусирования. Пример фракционирования по размерам и по заряду показан ниже в Примере 2.4.
Вторым способом ремоделирования углеводов является модификация некоторых гликоформ очищенной α -Gal А путем присоединения дополнительного концевого сахарного остатка с использованием очищенной гликозилтрансферазы и соответствующего донора нуклеотидного сахара. Такая обработка воздействует лишь на те гликоформы, которые имеют соответствующий свободный концевой сахарный остаток, действующий как акцептор для используемой гликозилтрансферазы. Так, например, α 2,6-сиалилтрансфераза добавляет сиаловую кислоту к 2,6-связи на акцептор концевого Galβ 1, 4GlcNac-R с использованием СМР-сиаловой кислоты в качестве донора нуклеотидного сахара. Коммерчески доступными ферментами и молекулами, от которых они происходят, являются фукозо-α 1,3-трансферазы III, V и VI (человека); галактозо-α 1,3-трансфераза (свиньи); галактозо-β 1,4-трансфераза (коровы); маннозо-α 1,2-трансфераза (дрожжей); сиаловая кислота-α 2,3-трансфераза (крысы) и сиаловая кислота-α 2,6-трансфераза (крысы). После завершения реакции гликозилтрансфераза может быть удалена из реакционной смеси с использованием гликозилтрансферазо-специфической аффинной колонки, включающей соответствующий нуклеотид, связанный с гелем через 6-углеродный спейсер посредством пирофосфатной (GDP, UDP) или фосфатной (СМР) связи, или с использованием других хроматографических известных методов. Для ферментной заместительной терапии человека, из вышеперечисленных гликозилтрансфераз, особенно подходящими для модификации ферментов, таких как α -Gal А, являются сиалилтрансферазы. Использование любой сиалилтрансферазы с СМР-5-флуоресцеин-нейраминовой кислотой в качестве донора нуклеотидного сахара позволяет получить флуоресцентно меченный гликопротеин, поглощение и тканевая локализация которого может быть легко прослежена.
Третий способ углеводного ремоделирования, предусматривающий осуществление глико-инженерии, например введение генов, которые воздействуют на клеточные механизмы продуцирования α -Gal А с модификацией посттрансляционного процессинга в аппарате Гольджи, является предпочтительным способом.
Четвертый способ углеводного ремоделирования предусматривает обработку α -Gal А соответствующими гликозидазами для снижения числа других присутствующих гликоформ. Так, например, последующая обработка сложных гликановых цепей нейраминидазой, β -галактозидазой и β -гексозаминидазой приводит к отщеплению олигосахарида до триманнозной сердцевины.
Структура N-связанного гликана зависит от доступности гликановой цепи для маннозидаз, осуществляющих процессинг в аппарате Гольджи после укладки белка, и от присутствия в аппарате Гольджи семейства гликозилтрансфераз и соответствующих доноров нуклеотидного сахара. Многие гликозил-трансферазы катализируют конкурентные реакции, которые могут приводить к удлинению гликановой цепи некоторыми другими и совместимыми путями в зависимости от того, какой фермент реагирует первым. Это приводит к микрогетерогенности и образованию семейства сложных гликоформ. Некоторые структуры являются уникальными для одной ткани, такие как структуры, образованные в результате модификации некоторых гормонов гипофиза путем присоединения GalNAc-4-SO4, или ограничиваются несколькими органами.
Примером последней из вышеупомянутых структур является так называемая разделяющая пополам (бисекторная) структура GlcNAc (GlcNAc, β 1,4-связанная с коровым β -маннозным остатком) на сложных гликанах глутамилтранспептидазы в почках, но не в печени. Разделенная пополам биантеннальная структура на γ ~ глутамилтранспептидазе показана ниже:

У млекопитающих ответственный фермент, GlcNAc-трансфераза III (GnT-III), обнаружен в некоторых клетках головного мозга и почек и в некоторых клетках печени у пациентов с гепатокарциномой. GnT-III катализирует присоединение N-ацетилглюкозамина в β -1-4-связи к β -связанной маннозе триманнозиловой сердцевины N-связанных сахарных цепей с продуцированном бисекторного остатка GlcNAc. Были клонированы гены GnT-III мыши, крысы и человека. lhara et al., J. Biochem. (Tokyo) 113:692-698 (1993).
Присутствие дополнительной активности GlcNAc T-III в клетках человека может продуцировать увеличение уровня монофосфорилированных гибридных гликанов за счет би-, три- и тетраантеннальных сложных гликанов. Это не должно негативно влиять на время полужизни в плазме, но может приводить к увеличению уровня доставки в сосудистые эндотелиальные клетки. Характерная структура представлена ниже:

Некоторые из α -Gal А поглощаются почками и приводят к значительному их снижению накопления гликолипидов. Поскольку почки могут образовывать N-гликаны с бисекторными остатками GlcNAc, почечные эпителиальные клетки могут распознавать гликопротеины с этим эпитопом с особенно высокой специфичностью.
Повышенная активность GnT-III может вызывать дисбаланс в разветвлении на триманнозильной сердцевине путем ингибирования последующего разветвления посредством GnT-II, IV, V и Gal-β -1,4-трансферазы на уровне субстрата. Недавно путем сверхэкспрессии рекомбинантного GnT-III была сконструирована клеточная линия яичника китайского хомячка (СНО), способная продуцировать бисекторные олигосахариды на гликопротеинах. Sburtali et al., Biotechnol. Progr. 14:189-192 (1998). Интерферон β (IFN-β ) был выбран как модель и потенциальный терапевтический секретированный гетерологичный белок, на котором может быть оценено влияние GnT-III-экспрессии на гликозилирование продукта. IFN-β с бисекторными олигосахаридами был продуцирован клетками СНО, сконструированными с использованием GnT-III, но не клетками, представляющими собой немодифицированную родительскую клеточную линию.
Для продуцирования гликопротеиновых терапевтических средств необходима характеризация гликозилирования в отношении консистенции от партии к партии. "Гипотетический заряд Z N-гликана" был использован в качестве параметра для характеризации гликозилирования белка простым эффективным способом. Определение Z было осуществлено во многих повторных экспериментах и было подтверждено, что он имеет высокую степень точности и надежности. Hermentin et al. Glycobiology 6:217-230 (1996). Гипотетический заряд N-гликана данного гликопротеина выводят из профиля картирования N-гликана, полученного с помощью высокоэффективной анионообменной хроматографии (ВЭАОХ)/импульсной амперометрической детекции (ИАД). В ВЭАОХ N-гликаны четко разделяются в соответствии с их зарядом, например в соответствии с их числом остатков сиаловой кислоты, что позволяет получить четкие области для нейтральных структур, а также для моно-, ди-, три- и тетрасиалилированных N-гликанов. Z определяют как сумму продуктов соответствующих площадей (А) в асиало-, моносиало-, дисиало-, трисиало-, тетрасиало- и пентасиало-области, каждую из которых умножают на соответствующий заряд:
Z=A(aсиaлo)·+A(MS)·1+A(Dis)·2+A(TriS)·3+A(TetraS)·4[+A(PentaS)·5]
Z=Σ А(i)·(i)
где i равно нулю в асиало-области, 1 в моносиало (MS) - области, 2 в дисиало (DiS) - области, 3 в трисиало (TriS) - области, 4 в тетрасиало (TetraS) - области и 5 в пентасиало (PentaS) - области.
Таким образом, гликопротеин, имеющий, в основном, структуры С4-4*, будет иметь Z ≅ 400, гликопротеин, несущий множество структур С2-2*, будет иметь Z ≅ 200, а гликопротеин, обладающий лишь структурой с высоким содержанием маннозы или усеченной структурой, будет иметь Z ≅ 0.
Препараты человеческой гликозилированной α -Gal А настоящего изобретения имеют олигосахаридный заряд, выраженный числом Z, превышающим 100, предпочтительно превышающим 150, а более предпочтительно превышающим 170.
Изменение времени полужизни α -Gal А в сыворотке путем фосфорилирования
Фосфорилирование α -Gal А может быть модифицировано в целях изменения времени полужизни в кровотоке α -Gal А и уровня α -Gal А, входящих в клетки. Такое фосфорилирование, предпочтительно, достигается в клетке, экспрессирующей α -Gal А. В данной заявке конкретно рассматривается получение гликозилированного препарата α -Gal А с повышенным уровнем гликозилирования путем первоначального введения в α -Gal А-продуцирующую клетку ДНК-последовательности, кодирующей фосфорилилтрансферазу, или путем введения регуляторной последовательности посредством гомологичной рекомбинации, которая регулирует экспрессию эндогенного гена фосфорилтрансферазы. Затем клетку, продуцирующую α -Gal А, культивируют в соответствующих условиях культивирования, которые способствуют экспрессии α -Gal А и фосфорилтрансферазы. После этого осуществляют выделение препарата α -Gal А, который по сравнению с α -Gal А, продуцированной в клетке без полинуклеотида, имеет повышенный уровень фосфолилирования. Такие фосфорилтрансферазы хорошо известны специалистам. См., например, патенты США 5804413 и 5789247, каждый из который вводится в настоящее описание посредством ссылки.
Для генерирования маркера распознавания ман-6-фосфата на лизосомном проферменте необходимы согласованные действия двух мембраноассоциированных ферментов Гольджи. Первый фермент, UDP-N-ацетилглюкозамин: гликопротеин-N-ацетилглюкозамин-1-фосфотрансфераза (GlcNAc-фосфотрансфераза) требует присутствия белокраспознающей детерминанты на лизосомном ферменте, которая состоит из двух лизиновых остатков, находящихся на расстоянии точно 34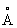 и в соответствующем правильном пространственном соотношении с цепью, имеющей высокое содержание маннозы. Второй фермент, N-ацетилглюкозамин-1-фосфодиэфир-а-N- ацетилглюкозаминидаза(фосфодиэфир-а-GlсNАс-аза) гидролизует α -GlcNAc-фосфатную связь, что приводит к экспонированию сайта распознавания ман-6-фосфата.
и в соответствующем правильном пространственном соотношении с цепью, имеющей высокое содержание маннозы. Второй фермент, N-ацетилглюкозамин-1-фосфодиэфир-а-N- ацетилглюкозаминидаза(фосфодиэфир-а-GlсNАс-аза) гидролизует α -GlcNAc-фосфатную связь, что приводит к экспонированию сайта распознавания ман-6-фосфата.
В соответствии со способами настоящего изобретения препараты α -Gal А, полученные способами настоящего изобретения, имеют множество гликоформ, из которых 16-50%, предпочтительно 25-50%, а более предпочтительно, по крайней мере, 30% гликоформ являются фосфорилированными.
Изменение времени полужизни α -Gal А в сыворотке путем увеличения уровня сиалилирования
Увеличение уровня сиалилирования недостаточно сиалилированных гликанов с концевыми галактозными остатками может быть осуществлено путем трансфекции клеток млекопитающих, а предпочтительно, клеток человека, геном сиалилтрансферазы.
Настоящее изобретение относится к получению гликозилированной α -Gal А, имеющей более высокозаряженные олигосахариды, продуцированные благодаря предварительному введению в α -Gal А-продуцирующую клетку полинуклеотида, кодирующего сиалилтрансферазу, или благодаря введению регуляторной последовательности посредством гомологичной рекомбинации, которая регулирует экспрессию эндогенного гена сиалилтрансферазы. Затем клетку, продуцирующую α -Gal А, культивируют в соответствующих культуральных условиях, которые приводят к экспрессии α -Gal А и сиалилтрансферазы. Последующая стадия предусматривает выделение препарата α -Gal А с повышенным зарядом олигосахаридов. Предпочтительными сиалилтрансферазами являются α 2,3-сиалилтрансфераза и α 2,6-сиалилтрансфераза. Указанные сиалилтрансферазы хорошо известны специалистам. См., например, патент США 5858751, который вводится в настоящее описание посредством ссылки.
В предпочтительном варианте осуществления изобретения указанный способ увеличения уровня сиалилирования включает дополнительную стадию отбора гликоформ α -Gal А с увеличенным размером или с увеличенным зарядом путем фракционирования или очистки данного препарата (как обсуждается ниже).
Альтернативно, настоящее изобретение относится к увеличению степени сиалилирования путем выдерживания клеток в среде с низким содержанием аммония. В частности, препарат гликозилированной α -Gal А с повышенным уровнем сиалилирования получают посредством контактирования клетки, продуцирующей α -Gal А, с культуральной средой, имеющей концентрацию аммония ниже 10 мМ, а более предпочтительно ниже 2 мМ. Увеличение уровня сиалилирования может быть осуществлено путем перфузии продуктивных клеток, при которой токсичные метаболиты, такие как аммоний, периодически удаляются из культуральной среды. В предпочтительном варианте осуществления изобретения среду с низким содержанием аммония получают путем добавления в продуктивные клетки гена или кДНК глутаминсинтетазы. Альтернативно, среду с низким содержанием аммония получают путем перфузии α -Gal А-продуцирующих клеток свежей культуральной средой с поддержанием концентрации аммония ниже 10 мМ, более предпочтительно ниже 2 мМ. Продуктивные клетки могут быть подвергнуты непрерывной перфузии свежей культуральной средой с концентрацией аммония ниже 10 мМ, а более предпочтительно ниже 2 мМ. Альтернативно, продуктивные клетки могут быть периодически перфузированы свежей культуральной средой. Используемая здесь периодическая перфузия означает перфузию, осуществляемую регулярно, через определенные интервалы времени, или после измерения концентрации аммония с достижением целевой концентрации (то есть, 10 мМ, более предпочтительно ниже 2 мМ). Периодические перфузии должны быть осуществлены через интервалы времени с достаточной частотой так, чтобы концентрация аммония никогда не превышала целевую концентрацию. Продуктивные клетки подвергают перфузии в течение периода времени, необходимого для получения препарата α -Gal А, в котором 50-70%, а предпочтительно 60% гликанов от всего количество гликанов, являются сиалилированными.
Увеличение времени полужизни α -Gal А в сыворотке путем ПЭГилирования α -Gal A
Кроме того, в соответствии с настоящим изобретением, время полужизни препарата человеческой гликозилированной α -Gal A в кровотоке, увеличивают посредством образования комплекса α -Gal А с полиэтиленгликолем. Полиэтиленгликоль (ПЭГ) представляет собой водорастворимый полимер, который, при его ковалентном связывании с белками, изменяет их свойства так, что это приводит к расширению возможностей их использования. Модификация полиэтиленгликолем ("ПЭГилирование") представляет собой хорошо разработанную методику, которая позволяет решить или минимизировать многие проблемы, связанные с получением белковых и пептидных фармацевтических средств.
Улучшенное фармакологическое качество ПЭГ-белков, по сравнению с их немодифицированными аналогами, позволяет получать конъюгаты такого типа в качестве терапевтических агентов. Состояния, связанные с дефицитом ферментов, для лечения которых терапия нативным ферментом является недостаточной (вследствие быстрого их выведения и/или иммунологических реакций), могут быть в настоящее время подвергнуты лечению эквивалентными ПЭГ-ферментами. Так, например, ПЭГ-аденозиндезаминаза уже была утверждена FDA (Управлением по контролю за качеством пищевых продуктов и медикаментов). Delgado et al., Crit.Rev.Ther.Drug Carrier Syst. 9: 249-304 (1992).
Ковалентное связывание ПЭГ с α -галактозидазой зеленых кофейных бобов вызывает изменение каталитических свойств данного фермента путем маскировки сайтов специфических детерминант на данной молекуле. Это приводит к увеличению величин Km и снижению величины Vmax по сравнению с аналогами п-нитрофениловых субстратов. Wieder & Davis, J.AppI.Biochem. 5:337-47 (1983). α -Галактозидаза еще обладала способностью отщеплять концевые галактозные остатки от вещества группы крови В в слюне человека. ПЭГ-α -галактозидаза теряла способность к специфическому связыванию с антителом и лектином. Антитела, генерированные нативной α -галактозидазой, могут блокировать ферментативную активность, и способность к такому ингибированию постепенно терялась при тестировании по сравнению с препаратами ферментов, содержащих постепенно возрастающие количества ПЭГ. В противоположность этому, антисыворотка, полученная от животных, иммунизованных ПЭГ-α -галактозидазой, не ингибировала ферментативную активность в любом из α -галактозидазных или ПЭГ-α -галактозидазных препаратов. Эти результаты показали, что ПЭГ имеет тенденцию покрывать лектин-специфические углеводные группы и антигенные детерминанты и что эти сайты, вероятно, остаются "скрытыми" во время in vivo-процессинга ПЭГ-ферментов.
Ковалентное связывание ПЭГ с белками требует активации гидроксильной концевой группы полимера подходящей уходящей группой, которая может быть удалена нуклеофильной атакой ε -аминоконца лизина и α -аминогруппы N-конца. Для активации ПЭГ было использовано несколько химических групп. Для каждого конкретного применения разные методы имеют свои определенные преимущества. Разные методы ПЭГилирования обнаруживали неожиданное и резкое воздействие на такие факторы, как сохранение биологической активности, стабильность и иммуногенность полученных ПЭГилированных белков и пептидов. Francis et al., Int.J.Hematol. 68(1): 1-18 (1998). Так, например, в методах ПЭГилирования без использования линкера к молекуле-мишени присоединяется лишь ПЭГ. Более конкретно, применение методов биологически оптимизированного ПЭГилирования с использованием трезилмонометокси-ПЭГ (ТМПЭГ) к ряду белков-мишеней показало, что при этом, как описано Francis et al., Int.J.Hematol. 68(1): 1-18 (1998), абсолютно сохраняется биологическая активность данной мишени. Этот факт и преимущество добавления к данному белку лишь только одного ПЭГ (которое, как было показано, безопасно для использования в терапевтических препаратах для лечения человека) делает этот способ идеальным для модификации α -Gal A.
Для присоединения ПЭГ к белкам имеются четыре возможных сайта: (1) аминогруппы (N-конец и лизин); (2) карбоксильные группы (аспарагиновая кислота и глутаминовая кислота); (3) сульфгидрильные группы (цистеин) и (4) углеводные группы (альдегиды, продуцируемые после обработки периодатом). Присоединение к карбоксильным группам белков и к альдегидным группам на углеводах требует использования реагента ПЭГ с нуклеофильной аминогруппой. Этот химический процесс меняет рI фермента α -Gal А после связывания отрицательно заряженных карбоксильных групп с ПЭГ. Любые изменения рI могут влиять на биологическую активность α -Gal А. Более того, связывание ПЭГ с углеводными цепями может влиять на поглощение α -Gal А посредством рецептора М6Р, который играет существенную роль в биологической активности. Химический процесс с использованием сульфгидрила также влияет на физическую структуру данной молекулы, а поэтому его использовать не рекомендуется.
Общеизвестные методы ПЭГилирования позволяют образовывать амидную связь между аминогруппами белка и метоксигруппой на монометокси-ПЭГ. NHS-ПЭГ является коммерчески доступным и позволяет образовывать амидную связь между белком и ПЭГ. Однако образование амидной связи изменяет рI, что обусловлено потерей положительного заряда группы –NH2.
Способ присоединения ПЭГ к α -Gal А без влияния на его рI предусматривает использование трезил-ПЭГ. Трезил-ПЭГ присоединяется через аминогруппы и образует стабильный вторичный амин. Вторичные амины имеют то преимущество, что они сохраняют положительный заряд аминогруппы. Реагент трезил-ПЭГ является коммерчески доступным и стабильным в качестве лиофилизованного и высушенного порошка. Трезил-ПЭГ был тщательно охарактеризован и были хорошо определены его реакция и побочные продукты. В соответствии с этим, в предпочтительном варианте осуществления настоящего изобретения, препарат α -Gal А подвергают комплексообразованию с использованием трезилмонометокси-ПЭГ (ТМПЭГ), в результате чего получают ПЭГилированный-α -Gal А. Затем ПЭГилированный-α -Gal А очищают с получением выделенного ПЭГилированного α -Gal A.
Схематическая реакция

α -Gal А содержит 18 аминогрупп, 17 ε -аминогрупп (лизин) и одну α -аминогруппу (N-конец). Эта реакция может быть отрегулирована для продуцирования α -Gal А с минимальными заменами, а затем молекулы с одним ПЭГ на молекулу или с наименьшим средним числом групп ПЭГ на молекулу могут быть очищены от незамещенных форм и форм с множеством замен. Множество замен на α -Gal А может незначительно влиять на биологическую активность, а поэтому конечный продукт может содержать гетерогенную смесь из 1-18 присоединенных молекул ПЭГ. Уровень замены зависит от уровня остаточной ферментативной активности. Следует отметить, что снижение ферментативной активности может быть компенсировано повышенным терапевтическим эффектом, достигнутым в результате увеличения времени полужизни в кровотоке и снижения способности к иммунному распознаванию α -Gal А. Таким образом, при разработке продукта ПЭГ-α -Gal А отношение ПЭГ к α -Gal A должно зависеть от биологической активности, а не только от ферментативной активности.
Реакция ПЭГилирования требует контроля за рН, составом буфера и концентрацией белка. Нужные реакционные условия могут быть достигнуты путем проведения стадии ультрафильтрации/диафильтрации, которая в настоящее время используется в промышленном производстве. Непосредственно перед реакцией трезил-ПЭГ быстро солюбилизируется в воде при непрерывном перемешивании. Затем этот раствор добавляют к полученной α -Gal A и оставляют для реакции в течение контролируемого периода времени и при регулируемой температуре (например, на 2 часа при 250° С). ПЭГилирование может быть осуществлено перед конечной процедурой очистки, которая позволяет избежать проведения стадий присоединения до процедуры очистки. После завершения присоединения ПЭГ-α -Gal А обрабатывают путем проведения остальных стадий очистки. Осуществление реакции перед хроматографией на колонке Q (анионообменной хроматографии) позволяет провести две стадии очистки для удаления побочных продуктов реакции. Поскольку ПЭГ не содержит какого-либо отрицательного заряда, он не будет удерживаться Q-сефарозой® и будет элюироваться в пустом объеме.
Степень ПЭГилирования может быть определена известными методами. Так, например, флуорескамин флуоресцирует при связывании с α -амино- и ε -аминогруппами белков. Процент потери флуоресценции после ПЭГилирования соответствует процентному отношению связанного ПЭГ к α -Gal А. Для определения концентрации белка может быть использован ВСА-анализ Пирса (Pierce) на общий белок. Для оценки эффекта ферментативной активности ПЭГ-α -Gal А используют анализ на активность метилумбеллиферил-α -D-галактопиранозида (4-MUF-α -Gal). α -Gal А содержит М6Р, который необходим для поглощения лизосомами. Влияние ПЭГ на узнавание М6Р-рецептором может быть оценено с использованием клеточного анализа для мониторинга поглощения ПЭГ-α -Gal А лизосомами клеток.
Способы введения препарата α -Gal A
Композиции настоящего изобретения (то есть содержащие различные гликоформы α -Gal А) могут быть введены любым способом, совместимым с данным препаратом α -Gal А. Очищенный препарат α -Gal А может быть введен индивидуумам, у которых вырабатывается недостаточное количество белка α -Gal А или дефектный белок α -Gal А, либо индивидуумам, на которых α -Gal А-терапия оказывает благоприятное воздействие. Терапевтические препараты настоящего изобретения могут быть введены индивидууму любым подходящим способом, непосредственно (например, местно, путем инъецирования, имплантации или наружного нанесения на участок ткани) или системно (например, перорально или парентерально).
Способ введения может быть пероральным или парентеральным, включая внутривенное, подкожное, внутриартериальное, внутрибрюшинное, внутриглазное, внутримышечное, трансбуккальное, ректальное, вагинальное, интраорбитальное, интрацеребральное, чрескожное, внутричерепное, интраспинальное, интравентрикулярное, внутриоболочечное, интрацистернальное, интракапсулярное, внутрилегочное, интраназальное, чрезслизистое и трансдермальное введение или введение путем ингаляции. Способы, аппарат и лекарственные препараты для внутрилегочной доставки описаны, например, в патентах США 5785049, 5780019 и 5775320, каждый из которых вводится в настоящее описание посредством ссылки. Предпочтительным способом чрескожной доставки является ионофоретическая доставка лекарственного средства с использованием пластырей; пример такой доставки описан в патенте США 5843015, который вводится в настоящее описание посредством ссылки.
Особенно эффективным является введение путем подкожного инъецирования. Препарат α -Gal А настоящего изобретения изготавливают так, чтобы вся требуемая доза могла быть введена в виде одной инъекции в количестве одного или двух миллилитров.
Для получения объема инъекции в один или два миллилитра препарат α -Gal А настоящего изобретения может быть изготовлен в концентрации, при которой предпочтительная доза доставляется в объеме от одного до двух миллилитров, либо этот препарат α -Gal А может быть изготовлен в лиофилизованной форме, которую разводят в воде или в соответствующем физиологически совместимом буфере непосредственно перед введением. Подкожные инъекции препаратов α -Gal А имеют то преимущество, что они удобны для пациента, а в частности позволяют ему вводить инъекции самостоятельно, что также приводит к более продолжительному времени полужизни в плазме по сравнению, например, с внутривенным введением. Увеличение продолжительности полужизни в плазме приводит к поддержанию эффективных уровней α -Gal А в плазме в течение более продолжительных периодов времени, благоприятный эффект которых заключается в увеличении воздействия инъецированного α -Gal А на клинически пораженные ткани и тем самым в увеличении поглощения α -Gal А указанными тканями. Это оказывает более благоприятное воздействие на пациента и/или позволяет снизить частоту введения. Более того, вместе с препаратами α -Gal А настоящего изобретения, обсуждаемыми в настоящем описании, может быть использован ряд устройств, предназначенных для удобства пациента, таких как устройство для инъекций в виде перьевой ручки для инъекции с заправкой и устройство для безыгольного введения.
Введение может быть осуществлено путем периодических инъекций болюса данного препарата либо оно может быть осуществлено путем внутривенного или внутрибрюшинного введения из резервуара, который может быть внешним (например, баллон для i.v.) или внутренним (например, биологически разлагаемый имплантат, биологический искусственный орган или популяция имплантированных α -Gal А-продуцирующих клеток). См., например, патенты США 4407957 и 5798113, каждый из которых вводится в настоящее описание посредством ссылки. Способы и аппарат для внутрилегочного введения описаны, например, в патентах США 5654007, 5780014 и 5814607, каждый из которых вводится в настоящее описание посредством ссылки. Другие эффективные системы для парентеральной доставки предусматривают использование частиц из сополимера этилена-винилацетата, осмотических насосов, имплантируемых систем для инфузий, насосов для доставки, инкапсулированных клеток, липосом, инъекций с применением игл, безыгольного впрыскивания, ингалятора, аэрозольного распылителя, электропорации и пластыря для чрескожной доставки. Устройство для безыгольного впрыскивания описано в патентах США 5879327; 5520639, 5846233 и 5704911, содержание которых вводится в настоящее описание посредством ссылки. Указанными способами может быть введен любой из вышеописанных препаратов α -Gal A.
Способ введения и количество доставляемого белка может определяться факторами, хорошо известными специалистам. Кроме того, специалистам известно, что способ введения и доза терапевтического белка могут варьироваться для данного пациента до тех пор, пока не будет получен терапевтический уровень доз.
Фармацевтическая композиция белка α -Gal А
Настоящее изобретение, кроме того, относится к новым композициям препарата α -Gal А, которые, в основном, не содержат белков, не являющихся α -Gal А, таких как альбумин, белки не-α -Gal А, продуцируемые клеткой-хозяином, или белки, выделенные из ткани или физиологической жидкости животного.
Данный препарат предпочтительно включает часть водной или физиологически совместимой жидкой суспензии или раствора. Носитель или наполнитель являются физиологически совместимыми так, чтобы при их добавлении для доставки нужного препарата пациенту они не оказывали какого-либо негативного влияния на электролитный и/или объемный баланс пациента. Используемые растворы для парентерального введения могут быть получены любыми методами, хорошо известными в фармацевтической практике. См., например, Remingtons Pharmaceutical Sciences (Gennaro A. ed.), Mack Pub., 1990. Могут быть также использованы композиции, не предназначенные для парентерального введения, такие как суппозитории и композиции для перорального введения.
Предпочтительно, чтобы данная композиция содержала наполнитель. Фармацевтически приемлемыми наполнителями для α -Gal А, которые могут быть включены в указанную композицию, являются буферы, такие как цитратный буфер, фосфатный буфер, ацетатный буфер и бикарбонатный буфер, аминокислоты, мочевина, спирты, аскорбиновая кислота, фосфолипиды; белки, такие как сывороточный альбумин, коллаген и желатин; соли, такие как EDTA или EGTA и хлорид натрия; липосомы; поливинилпирролидон; сахара, такие как декстран, маннит, сорбит и глицерин; пропиленгликоль и полиэтиленгликоль (например, ПЭГ-4000, ПЭГ-6000); глицерин; глицин или другие аминокислоты; липиды. Буферными системами, используемыми для препаратов α -Gal А, являются цитратные, ацетатные, бикарбонатные и фосфатные буферы (все эти буферы поставляются фирмой Sigma). Предпочтительным буфером для осуществления настоящего изобретения является фосфатный буфер. Предпочтительным рН для препаратов α -Gal А является рН 4,5-7,4.
Указанная композиция также предпочтительно содержит неионогенный детергент. Предпочтительными неионогенными детергентами являются полисорбат 20, полисорбат 80, тритон Х-100, тритон Х-114, нонидет Р-40, октил-α -глюкозид, октил-β -глюкозид, Brij 35, плюроник и твин 20 (все они поставляются фиромой Sigma).
Особенно предпочтительная композиция содержит неионогенный детергент полисорбат 20 или полисорбат 80 и забуференный фосфатом физиологический раствор, предпочтительнее всего при рН 6.
Для лиофилизации препаратов α -Gal А концентрация белка может составлять 0,1-10 мг/мл. К смеси для лиофилизации могут быть добавлены агенты, придающие объем, такие как глицин, маннит, альбумин и декстран. Кроме того, к смеси для лиофилизации могут быть добавлены возможные криозащитные вещества, такие как дисахариды, аминокислоты и ПЭГ. Могут быть также добавлены любые буферы, наполнители и детергенты, перечисленные выше.
В предпочтительной композиции концентрация α -Gal А для инъекции составляет 1 мг/мл.
Для поддержания данного агента в нужном участке композиции для введения могут включать глицерин и другие композиции с высокой вязкостью. Для регуляции высвобождения агента in vivo могут быть использованы наполнители, такие как биологически совместимые полимеры, а предпочтительно биологически ресорбируемые, биологически совместимые полимеры (включая, например, полимеры гиалуроновой кислоты, коллагена, полибутирата, лактида и гликолида и сополимеры лактида/гликолида). Композиции для парентерального введения могут включать гликохолат для транс-буккального введения, метоксисалицилат для ректального введения или лимонную кислоту для вагинального введения. Суппозитории для ректального введения могут быть получены путем смешивания препаратов α -Gal А настоящего изобретения с нераздражающим наполнителем, таким как масло какао или с другими композициями, которые являются твердыми при комнатной температуре и жидкими при температуре тела.
Композиции для введения путем ингаляции могут содержать лактозу или другие наполнители, либо они могут представлять собой водные растворы, которые могут содержать полиоксиэтилен-9-лауриловый эфир, гликохолат или дезоксикохолат. Предпочтительным аэрозолем для ингаляции является аэрозоль, характеризующийся тем, что он имеет частицы с небольшой плотностью по массе и крупного размера. Частицы с плотностью по массе менее чем 0,4 грамма на кубический сантиметр и средним диаметром, превышающим 5 мкм, способствуют эффективной доставке терапевтических средств в кровоток путем ингаляции. Такие частицы вдыхаются вглубь легких и удаляются из легких в соответствии с механизмами природного клиренса до тех пор, пока вдыхаемые частицы не доставят свое ценное терапевтическое содержимое (Edwards et al., Science 276: 1868-1872 (1997)). Препараты α -Gal А настоящего изобретения могут быть введены в форме аэрозолей, например, методами получения и изготовления композиций, как описано в патентах США 5654007, 5780014 и 5814607, каждый из которых вводится в настоящее описание посредством ссылки. Композиция для интраназального введения может включать масляные растворы для введения в виде капель в нос или в виде геля для интраназального введения.
Композиции для местного нанесения на поверхность кожи могут быть получены путем диспергирования препарата α -Gal А в дерматологически приемлемом носителе, таком как лосьон, крем, мазь или мыло. Особенно полезными являются носители, способные образовывать пленку или слой на коже для локализации в месте нанесения и предупреждения удаления лекарственного средства. Для локального введения во внутренние поверхности тканей препарат α -Gal А может быть диспергирован в жидком тканевом клее или в другом веществе, о котором известно, что он способствует усилению адсорбции на поверхности ткани. Так, например, для доставки лекарственного средства через слизистую оболочку были описаны некоторые адгезивы для слизистых и таблетки для растворения в щечном кармане, такие как препараты, описанные в патентах США 4740365, 4764378 и 5780045, каждый из которых вводится в настоящее описание посредством ссылки. Могут быть также включены растворы гидроксипропилцеллюлазы или фибриногена/тромбина. Альтернативно, могут быть использованы растворы для нанесения на ткань, такие как пектинсодержащие композиции.
Препараты настоящего изобретения могут быть изготовлены в контейнерах, подходящих для поддержания стерильности, защиты активности активных ингредиентов в процессе соответствующего распределения и хранения, и обеспечения удобства и доступности данного препарата для введения пациенту. Инъецируемая композиция препарата α -Gal А может быть помещена в закрывающийся сосуд, подходящий для извлечения содержимого с использованием иглы и шприца. Данный сосуд может быть предназначен либо для одноразового, либо для многократного использования. Указанный препарат может быть также изготовлен в виде предварительно заполненного шприца. В некоторых случаях содержимое может присутствовать в виде жидкой композиции, а иногда оно может находиться в сухом или лиофилизованном состоянии, при котором, в некоторых случаях, может потребоваться разведение стандартным или готовым разбавителем до жидкого состояния. Если данный препарат поставляется в виде жидкости для внутривенного введения, то он может быть изготовлен в виде стерильного пакета или контейнера, подходящего для подсоединения к трубке или к катетеру для внутривенного введения. В предпочтительных вариантах осуществления настоящего изобретения препараты настоящего изобретения поставляются либо в виде жидких, либо в виде порошкообразных композиций в устройствах, которые обеспечивают удобное введение заранее установленной дозы данного препарата; примерами таких устройств являются безыгольный впрыскиватель для подкожной или внутримышечной инъекции и аэрозольное устройство с дозируемой подачей лекарственного средства. В других случаях данный препарат может быть изготовлен в форме, подходящей для пролонгированного высвобождения, такой как пластырь или перевязочный материал, применяемый для нанесения на кожу при чрескожном введении, или в форме разлагаемых приспособлений для введения через слизистую. В случаях, когда данный препарат вводят перорально в виде таблетки или драже, указанный препарат может быть изготовлен в виде сосуда с удаляемой верхней крышкой. Эти контейнеры могут быть снабжены информацией, такой как тип препарата, имя производителя или распространителя, показания, предполагаемые дозы, инструкции по правильному хранению или инструкции по введению.
Дозы для введения препарата α -Gal A
Настоящее изобретение также относится к способам введения препарата α -Gal А пациенту, страдающему болезнью Фабри, атипическим вариантом болезни Фабри или любым состоянием, характеризующимся присутствием пониженных уровней или мутантных форм α -Gal А. Дозы для введения составляют предпочтительно 0,05-5,0 мг, а более предпочтительно 0,1-0,3 мг препарата α -Gal A на килограмм массы тела и вводятся раз в неделю или раз в две недели. В предпочтительном варианте осуществления изобретения дозу примерно 0,2 мг/кг вводят раз в две недели. Повторные дозы белка необходимо регулярно вводить в течение всей жизни пациента. Для поддержания более продолжительного системного воздействия лекарственного средства могут быть использованы подкожные инъекции. Подкожная доза может составлять в пределах от 0,01 до 10,0 мг, а предпочтительно 0,1-5,0 мг препарата α -Gal А на один килограмм массы тела, один раз в неделю или раз в две недели. Дозы препаратов α -Gal А для внутримышечного введения могут быть такими же или отличаться от доз для подкожного введения, а в предпочтительном варианте осуществления изобретения дозы для внутримышечного введения могут быть меньше и вводиться с меньшей частотой. Препарат α -Gal А может быть также введен внутривенно, например путем внутривенной инъекции болюса, внутривенной инъекции посредством медленной интубации или путем непрерывной внутривенной инфузии. Непрерывная внутривенная (i.v.) инфузия (например, в течение 2-6 часов) позволяет поддерживать конкретные уровни в кровотоке.
Альтернативный предпочтительный способ введения препарата α -Gal А пациенту предусматривает введение предпочтительной дозы препарата α -Gal А раз в неделю или раз в две недели в течение нескольких лет, например вплоть до трех лет, во время которых пациент находится под непрерывным наблюдением для оценки статуса его или ее болезни. Клиническое улучшение состояния, определяемое, например, улучшением почечной или сердечной функции или общего состояния пациента (например, болевых ощущений), и улучшение лабораторных показателей, определяемое, например, снижением уровней СТН в моче, в плазме или в ткани, может свидетельствовать о состоянии здоровья пациента. В случае клинического улучшения, наблюдаемого после указанного курса лечения и периода мониторинга, частота введения α -Gal А может быть снижена. Так, например, пациент, получающий еженедельные инъекции препарата α -Gal А, может быть переведен на инъекции раз в две недели. Альтернативно, пациент, получающий инъекции препарата α -Gal А раз в две недели, может быть переведен на ежемесячные инъекции. После такой замены частоты введения доз пациент должен проходить обследование в течение последующих нескольких лет, например в течение трех лет, для оценки клинических и лабораторных параметров, связанных с болезнью Фабри. В предпочтительном варианте осуществления изобретения вводимую дозу не меняют, если была изменена частота введения доз. Это дает гарантию того, что некоторые фармакокинетические параметры (например, максимальная концентрация в плазме [Cmax], время до достижения максимальной концентрации в плазме [tmax], время полужизни в плазме [t1/2] и эффект, измеряемый по площади под кривой [AUC]) будут оставаться относительно постоянными после каждой вводимой дозы. Поддержание этих фармакокинетических параметров будет приводить к относительно постоянным уровням опосредованного рецептором поглощения α -Gal А в тканях по мере изменения частоты введения доз.
Пациента с атипическим вариантом болезни Фабри, у которого, например, обнаруживаются преимущественно сердечно-сосудистые патологии или поражения почек, подвергают лечению с применением аналогичных схем введения доз, например, от 0,05 до 5 мг/кг раз в неделю или раз в две недели. При необходимости эту дозу корректируют. Так, например, у пациента с фенотипическим вариантом сердечной патологии, которого подвергают лечению заместительной терапией ферментом α -галактозидазой А, будет наблюдаться изменение химического состава сердца и улучшение сердечной функции после проведения терапии. Это изменение может быть определено с помощью стандартной эхокардиографии, которая позволяет определить увеличенную толщину стенок левого желудочка у пациентов с болезнью Фабри (Goldman et al., J.Am. Coil. Cardiol. 7:1157-1161 (1986)). Серийные эхокардиографические измерения толщины стенок левого желудочка могут быть проведены в процессе терапии, а снижение размера стенок желудочка является показателем терапевтического ответа. У пациентов, подвергаемых заместительной терапии ферментом α -Gal А, может быть также осуществлен ядерно-магнитный резонанс (ЯМР) сердца. ЯМР позволяет проводить оценку относительного состава данной ткани. Так, например, ЯМР сердца у пациентов с болезнью Фабри позволяет выявить отложения липидов в миокарде по сравнению с контрольными пациентами (Matsui et al. Am. Heart J. 117:472-474 (1989)). Серийные оценки ЯМР сердца у пациентов, подвергаемых заместительной терапии ферментами, могут выявить изменения в отложении липидов в сердце пациента. У пациентов с почечным фенотипическим вариантом могут также проявляться благоприятные эффекты от лечения заместительной терапией ферментом α -галактозидазой А. Эффект терапии может быть определен в соответствии со стандартными тестами на функцию почек, такую как 24-часовой уровень белка в моче, клиренс креатинина и скорость гломерулярной фильтрации. Нижеследующие примеры представлены для более полной иллюстрации предпочтительных вариантов осуществления настоящего изобретения. Эти примеры не должны рассматриваться как ограничение объема настоящего изобретения, определенного в прилагаемой формуле изобретения.
Пример 1
Получение и использование конструкций, предназначенных для доставки и экспрессии α -Gal A
Были сконструированы две экспрессионные плазмиды, pXAG-16 и pXAG-28. Эти плазмиды содержат кДНК α -Gal А человека, кодирующую 398 аминокислот фермента α -Gal А (без сигнального пептида α -Gal A); геномную ДНК-последовательность сигнального пептида гормона роста человека (чГР), которая прерывается первым интроном гена чГР, и З'-нетранслируемую последовательность (UTS) гена чГР, содержащую сигнал полиаденилирования. Плазмида pXAG-16 имеет предранний промотор цитомегаловируса человека (IE CMV) и первый интрон (фланкированный некодирующими последовательностями экзона), a pXAG-28 находится под контролем промотора коллагена Iα 2 и экзона 1 и также содержит 5'UTS гена β -актина, содержащую первый интрон гена β -актина.
1.1. Клонирование полной кДНК α -Gal А и конструирование экспрессионной плазмиды pXAG-16 α -Gal A
кДНК α -Gal А человека клонировали из кДНК-библиотеки фибробластов человека, которая была сконструирована методом, описанным ниже. мРНК Poly-A+ выделяли из полной РНК и синтез кДНК осуществляли с использованием реагентов для системы лямбда ZapII® в соответствии с инструкциями производителей (Stratagene Inc., LaJolla, CA). Для этого "первую цепь" кДНК генерируют посредством обратной транскрипции в присутствии олиго-dТ-праймера, содержащего внутренний сайт рестриктирующей эндонуклеазы Xhol. После обработки РНКазой Н кДНК подвергали никтрансляции под действием ДНК-полимеразы I с генерированием двухцепочечной кДНК. Эту кДНК затупляли по концам посредством ДНК-полимеразы Т4 и лигировали с EcoRI-адапторами. Продукты этого лигирования обрабатывали ДНК-киназой Т4 и гидролизовали Xhol. кДНК фракционировали с помощью хроматографии на Сефакриле®-400. Фракции большого и среднего размера объединяли и кДНК лигировали с EcoRI- и Xhol-расщепленными плечами лямбда ZapII. Продукты этого лигирования упаковывали и титровали. Первичная библиотека имела титр 1,2× 107 б.о.е./мл, а средний размер вставки составлял 925 п.н.
Для выделения кДНК использовали 210 п.н.- зонд, полученный от экзона 7 гена α -Gal А человека (фиг.1, SEQ ID NO:1). Сам зонд был выделен из геномной ДНК путем полимеразной цепной реакции (ПЦР) с использованием нижеследующих олигонуклеотидов:
5'-CTGGGCTGTAGCTATGATAAAC-3' (олиго 1; SEQ ID NO:6) и
5'-TCTAGCTGAAGCAAAACAGTG-3' (олиго 2; SEQ ID NO:7).
Затем этот ПЦР-продукт был использован для скрининга библиотеки кДНК фибробластов, и положительные клоны были выделены, а затем охарактеризованы. Один положительный клон, фаг ЗА, вырезали согласно протоколу вырезания с использованием системы лямбда ZapII (Stratagene, Inc., La Jolla, CA) в соответствии с инструкциями производителей. Эта процедура позволяла получить плазмиду pBSAG3A, которая содержит кДНК-последовательность α -Gal А в плазмидном скелете Bluescript II SK™ . Секвенирование ДНК выявило, что эта плазмида не содержит полного 5'-конца кДНК-последовательности. Следовательно, 5'-конец был снова сконструирован с использованием ПЦР-фрагмента, амплифицированного из геномной ДНК человека. Для осуществления этой процедуры 268 п.н. - фрагмент геномной ДНК (фиг.2, SEQ ID NO:2) амплифицировали с использованием нижеследующих олигонуклеотидов:
5'-ATTGGTCCGCCCCTGAGGT-3' (олиго 3; SEQ ID NO:8) и 5'-TGATGCAGGAATCTGGCTCT-3' (олиго 4; SEQ ID NO:9).
Этот фрагмент субклонировали в клонирующую плазмиду "ТА" (Invitrogen, Corp. San Diego, CA) с образованием плазмиды pTAAGEI. Плазмиду рВSАG3А, содержащую 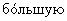 часть кДНК-последовательности α -Gal А, и плазмиду pTAAGEI, содержащую 5'-конец кДНК α -Gal A, расщепляли SacII и Ncol. Положения соответствующих SacII-Ncol-сайтов в амплифицированном ДНК-фрагменте показаны на фиг.2. 0,2 т.п.н. SacII-NcoI-фрагмент из pTAAGEI выделяли и лигировали с эквивалентно расщепленной pBSAG3A. Эта плазмида pAGAL содержит полную кДНК-последовательность α -Gal А, включая последовательность, кодирующую сигнальный пептид α -Gal А. кДНК была полностью секвенирована (как показано на Фиг.3, включая сигнальный пептид α -Gal A; SEQ ID NO:3) и было обнаружено, что она идентична опубликованной последовательности кДНК α -Gal A человека (последовательность Genbank HUMGALA).
часть кДНК-последовательности α -Gal А, и плазмиду pTAAGEI, содержащую 5'-конец кДНК α -Gal A, расщепляли SacII и Ncol. Положения соответствующих SacII-Ncol-сайтов в амплифицированном ДНК-фрагменте показаны на фиг.2. 0,2 т.п.н. SacII-NcoI-фрагмент из pTAAGEI выделяли и лигировали с эквивалентно расщепленной pBSAG3A. Эта плазмида pAGAL содержит полную кДНК-последовательность α -Gal А, включая последовательность, кодирующую сигнальный пептид α -Gal А. кДНК была полностью секвенирована (как показано на Фиг.3, включая сигнальный пептид α -Gal A; SEQ ID NO:3) и было обнаружено, что она идентична опубликованной последовательности кДНК α -Gal A человека (последовательность Genbank HUMGALA).
Плазмида pXAG-16 была сконструирована с получением нескольких промежуточных конструкций следующим образом. Сначала pAGAL расщепляли SacII и Xhol и затупляли по концам. Затем концы полноразмерной кДНК α -Gal А лигировали с ХbаI-линкерами и субклонировали в Xbal-расщепленную pEF-BOS (Mizushima et al., Nucl. Acids Res. 18:5322, 1990) с образованием pXAG-1. Эта конструкция содержит 3’-UTS гранулоцитарного колониестимулирующего фактора человека (Г-КСФ) и промотор человеческого фактора элонгации 1a (EF-1a), фланкирующий кДНК, кодирующую α -Gal А плюс сигнальный пептид α -Gal А так, что 5' -конец кДНК α -Gal А присоединен к промотору EF-la. Для создания конструкции, содержащей IE-промотор CMV и первый интрон, кДНК α -Gal A и 3'-UTS Г-КСФ удаляли из pXAG-1 в виде 2 т.п.н.-ХbаI-ВаmНI-XbaI-BamHI-фрагмента. Этот фрагмент затупляли по концам, лигировали с BamHI-линкерами и встраивали в BamHI-расщепленную плазмиду pCMVflpNeo (которая была сконструирована, как описано ниже). Ориентация была такой, что 5'-конец кДНК α -Gal А был присоединен к области IE-промотора CMV.
pCMVflpNeo конструировали следующим образом. Фрагмент IE-промотора гена CMV амплифицировали с помощью ПЦР с использованием геномной ДНК CMV в качестве матрицы и олигонуклеотидов:
5'-TTTTGGATCCCTCGAGGACATTGATTATTGACTAG-3' (SEQ ID NO:10) и 5'-TTTTGGATCCCGTGTCAAGGACGGTGAC-3' (SEQ ID NO:11).
Полученный продукт (1,6 т.п.н.-фрагмент) расщепляли ферментом BamHI с получением фрагмента, содержащего промотор CMV и имеющего липкие BamHI-расщепленные концы. Единицу, экспрессирующую nео, выделяли из плазмиды pMC1neopA (Stratagene, La Jolla, CA) в виде 1,1 т.п.н.-XhoI-BamHI-фрагмента. Фрагменты, содержащие промотор CMV и пео, встраивали в BamHI-, XhoI-расщепленную плазмиду (pUC12). Следует отметить, что pCMVflpNeo содержит область IE-промотора CMV, начинающуюся у нуклеотида 546 и кончающуюся у нуклеотида 2105 (последовательность Genbank HS5MIEP), а ген резистентности к неомицину, находящийся под контролем промотора тимидинкиназы вируса простого герпеса (HSV) (гена ТКnео), расположен непосредственно перед фрагментом IE-промотора CMV с 5'-стороны. Направление транскрипции гена nео то же самое, что и у фрагмента промотора CMV. Эта промежуточная конструкция была обозначена pXAG-4.
Для присоединения 3'UTS чГР из pXAG-4 удаляли 3'UTS Г-КСФ в виде XbaI-SmaI-фрагмента и концы pXAG-4 затупляли. 3'UTS чГР удаляли из pXGH5 (Selden et al., Mol.Cell.Biol. 6:3173-3179, 1986) в виде 0,6 т.п.н.-SmaI-EcoRI-фрагмента. После затупления концов этого фрагмента его лигировали в pXAG-4 непосредственно после затупленного по концам Xbal-сайта pXAG-4. Эту промежуточную конструкцию обозначали pXAG-7. Из этой плазмиды удаляли ТКnео-фрагмент в виде HindIII-ClaI-фрагмента и концы этой плазмиды затупляли путем "достраивания" фрагментом Кленова ДНК-полимеразы I. Ген резистентности к неомицину, находящийся под контролем раннего промотора SV40, лигировали в затупленный по концам ClaI-BsmBI-фрагмент в результате расщепления pcDNeo (Chen et al., Mol. Cell. Biol. 7:2745-2752, 1987), при этом транскрипционная единица nео находилась в той же ориентации, что и транскрипционная единица α -Gal А. Эту промежуточную конструкцию обозначали pXAG-13.
Для завершения конструирования pXAG-16, которая содержит последовательность, кодирующую сигнальный пептид чГЗ, состоящий из 26 аминокислот, и первый интрон гена чГР, удаляли 2,0 т.п.н.-EcoRI-BamHI-фрагмент pXAG-13. Этот фрагмент включал кДНК α -Gal А и 3'UTS чГР. Этот крупный фрагмент заменяли 3 фрагментами. Первый фрагмент состоит из 0,3 т.п.н.-ПЦР-продукта pXGH5, который содержит последовательность, кодирующую сигнальный пептид чГР, и включает последовательность первого интрона чГР, от синтезированного BamHI-сайта, локализованного непосредственно выше от консенсусной последовательности Kozak, до конца последовательности, кодирующей сигнальный пептид чГР. Для амплификации этого фрагмента (фрагмента 1) использовали следующие олигонуклеотиды:
5'-TTTTGGATCCACCATGGCTA-3' (олиго HGH101; SEQ ID NO:12) и
5'-TTTTGCCGGCACTGCCCTCTTGAA-3' (олиго HGH102; SEQ ID NO:13).
Второй фрагмент состоял из 0,27 т.п.н.-ПЦР-продукта, содержащего последовательности, соответствующие началу кДНК, кодирующей фермент α -Gal А, состоящий из 398 аминокислот (то есть в нем отсутствовал сигнальный пептид α -Gal А) до Nhel-сайта. Для амплификации этого фрагмента (фрагмента 2) использовали следующие олигонуклеотиды: 5'-TTTTCAGCTGGACAATGGATTGGC-3' (олиго AG10; SEQ ID NO:14) и 5'-TTTTGCTAGCTGGCGAATCC-3' (олиго AG11; SEQ ID NO:15). Третий фрагмент состоял из NheI-EcoRI-фрагмента pXAG-7, содержащего остальную последовательность α -Gal А, а также 3'UTS чГР (фрагмент 3).
Фрагмент 1 (расщепленный BamHI и Nhel), фрагмент 2 (расщепленный PvuII и Nhel) и фрагмент 3 смешивали с 6,5 т.п.н.-BamHI-EcoRI-фрагментом pXAG-7, содержащим ген nео и IE-промотор CMV, и лигировали вместе, в результате чего получали плазмиду pXAG-16 (фиг.4).
1.2. Конструирование α -Gal А-экспрессирующей плазмиды pXAG-28
Промотор коллагена Iα 2 человека, используемый для конструирования α -Gal А-экспрессирующей конструкции pXAG-28, выделяли следующим образом. 408 п.н.-ПЦР-фрагмент человеческой геномной ДНК, содержащей часть промотора человеческого коллагена Iα 2, выделяли с использованием следующих олигонуклеотидов: 5’-TTTTGGATCCGTGTCCCATAGTGTTTCCAA-3' (олиго 72; SEQ ID NO: 16) и 5’-TTTTGGATCCGCAGTCGTGGCCAGTACC' (олиго 73; SEQ ID NO:17).
Этот фрагмент использовали для скрининга библиотеки человеческих лейкоцитов в EMBL3 (Clontech Inc., Palo Alto, CA). Один позитивный клон (фаг 7Н), содержащий 3,8 т.п.н.-EcoRI-фрагмент выделяли и клонировали в pBSIISK+ (Stratagene, La Jolla, CA) в EcoRI-сайт (с образованием pDS/7H.2). AvrII-сайт вводили в pBSIISK+ путем расщепления ферментом Spel, который расщепляет последовательность внутри pBSIISK+полилинкера, с "достраиванием" концов фрагментом Кленова ДНК-полимеразы I и с встраиванием олигонуклеотида 5'-CTAGTCCTAGGA-3' (SEQ ID NO:18). Этот вариант pBSIISK+ расщепляли ферментами BamHI и AvrII и лигировали с 121 п.н.-BamHI-AvrII-фрагментом исходного 408 п.н.-ПЦР-фрагмента промотора коллагена Iα 2, описанного выше, в результате чего получали pBS/121COL.6.
Указанную плазмиду pBS/121COL.6 расщепляли ферментом XbaI, который расщепляет последовательность внутри pBSIISK+полилинкера, с "достраиванием" концов фрагментом Кленова ДНК-полимеразы I и расщепляли AvrII. 3,8 т.п.н.-BamHI-AvrII-фрагмент pBS/7H.2 выделяли и BamHI-сайт затупляли по концам путем обработки ферментом Кленова. Затем этот фрагмент расщепляли AvrII и лигировали с AvrII-расщепленным вектором, в результате чего получали плазмиду pBS/121COL.7Н.18, содержащую промотор коллагена.
Затем промотор коллагена сливали с 3'UTS человеческого гена β -актина, который содержит первый интрон человеческого гена β -актина. Для выделения этой последовательности 2 т.п.н. - ПЦР-фрагмент выделяли из геномной ДНК человека с использованием следующих олигонуклеотидов:
5’-TTTTGAGCACAGAGCCTCGCCT-3' (олиго ВА1; SEQ ID NO:19) и
5'-TTTTGGATCCGGTGAGCTGCGAGAATAGCC-3' (олиго ВА2; SEQ ID NO:20).
Этот фрагмент расщепляли BamHI и BsiHKAl с высвобождением 0,8 т.п.н.-фрагмента, содержащего 5'UTS β -актина и интрон. Затем из плазмиды pBS/121COL7H.18, содержащей промотор коллагена, выделяли 3,6 т.п.н.-Sall-SrfI-фрагмент следующим образом. pBS/121COL7H.18 частично расщепляли BamHI (BamHI-сайт расположен у 5'-конца фрагмента промотора коллагена Iα 2), затупляли по концам обработкой фрагментом Кленова и лигировали с Sall-линкером (5'-GGTCGACC-3'), в результате чего Sall-сайт был локализован выше от промотора коллагена Iα 2. Затем эту плазмиду расщепляли ферментами Sall и Srfl (Srfl-сайт расположен на 110 п.н. выше от САР-сайта промотора коллагена Iα 2) и выделяли 3,6 т.п.н.-фрагмент. 0,8-и 3,6 т.п.н.-фрагменты объединяли с Sall- и BamHI-расщепленным pBSIISK (Stratagene, La Jolla, CA) и фрагмент, состоящий из нижеследующих четырех олигонуклеотидов, отжигали вместе (с образованием фрагмента с тупым концом и BSiHKAI-концом):

Эти четыре олигонуклеотида, после их отжига, соответствовали области, начинающейся у Srfl-сайта промотора коллагена и охватывающей BSiHKAI-сайт промотора β -актина. Полученную плазмиду обозначали "pCOL/β -актин".
Для завершения конструирования pXAG-28 выделяли Sall-BamHI-фрагмент pCOL/β -актина, содержащего промотор коллагена Iα 2 и 3'UTS β -актина. Этот фрагмент лигировали с двумя фрагментами из pXAG-16 (см., пример 1,1 и фиг.4): (1) 6,0 т.п.н.-BamHI-фрагментом (содержащим ген nео, скелет плазмиды, кДНК, кодирующую фермент α -Gal А, состоящий из 398 аминокислот, и 3'UTS чГР); (2) 0,3 т.п.н.-BamHI-XhoI-фрагментом (содержащим последовательность поли-А SV40 из pcDneo). pXAG-28 содержит промотор коллагена Iα 2, слитый с 5'UTS β -актином человека, сигнальным пептидом чГР (который прерывается первым интроном чГР), кДНК, кодирующей фермент α -Gal А и 3'UTS чГР. Карта законченной экспрессионной конструкции pXAG-28 представлена на фиг.5.
1.3. Трансфекция и отбор фибробластов, в которые путем электропорации были введены α -Gal А-экспрессирующие плазмиды
Для экспрессии α -Gal А в фибробластах вторичные фибробласты культивировали и трансфецировали в соответствии с опубликованными процедурами (Selden et al., WO 93/09222).
Плазмиды pXAG-13, pXAG-16 и pXAG-28 трансфецировали путем электропорации в фибробласты крайней плоти человека в целях генерирования штаммов стабильно трансфецированных клональных клеток и проводили мониторинг полученных уровней экспрессии α -Gal А, как описано в примере 1.4. Уровень секреции α -Gal A нормальными фибробластами крайней плоти человека составляет в пределах 2-10 единиц/106 клеток/24 часа. Для сравнения средние наблюдаемые уровни экспрессии трасфецированных фибробластом показаны в таблице 2.

Эти данные указывают на то, что все три экспрессионные конструкции способны по сравнению с нетрансфецированными фибробластами во много раз усиливать экспрессию α -Gal А. Уровни экспрессии фибробластами, стабильно трансфецированными конструкцией pXAG-13, кодирующей фермент α -Gal А, присоединенный к сигнальному пептиду α -Gal А, были значительно ниже, чем уровни экспрессии фибробластами, трансфецированными плазмидой pXAG-16, которая отличается только тем, что в ней сигнальным пептидом является сигнальный пептид чГР, кодирующая последовательность которого прерывается первым интроном гена чГР.
Каждый раз, трансфецированные клетки пассировали, определяли активность секретированной α -Gal А, указанные клетки подсчитывали и вычисляли клеточную плотность. Исходя из числа собранных клеток и времени, требующегося для секреции α -Gal А, определяли уровень специфической экспрессии α -Gal А, и полученные данные были представлены в таблицах 3 и 4 в виде секретированных единиц (α -Gal А) на 106 клеток за период времени 24 часа. Клеточные штаммы, необходимые для генной терапии или для использования в получении материала для очистки α -Gal А, должны демонстрировать стабильный рост и экспрессию на протяжении нескольких пассажей. Данные, полученные для клеточных штаммов, показанных в таблицах 3 и 4, которые были стабильно трансфецированы α -Gal А-экспрессирующей конструкцией pXAG-16, иллюстрируют тот факт, что экспрессия α -Gal А стабильно поддерживается в процессе всего серийного пассирования.
Рост и экспрессия клеток BRS-11, содержащих конструкцию pXAG-16, экспрессирующую α -Gal A
Рост и экспрессия клеток HF503-242, содержащих конструкцию pXAG-16, экспрессирующую α -Gal А
1.4. Количественная оценка экспрессии α -Gal A
Активность α -Gal А измеряли с использованием водорастворимого субстрата 4-метилумбеллиферил-α -D-галактопиранозида (4-MUF-gal; Reserch Products, Inc) в соответствии с измененным протоколом, описанным loannou et al., J.Cell.Biol. 119: 1137-1150 (1992). Этот субстрат растворяли в буфере для субстрата (0,1 М цитратно-фосфатного, рН 4,6) до концентрации 1,69 мг/мл (5 мМ). Обычно 10 мл супернатанта культуры добавляли к 75 мл субстратного раствора. Пробирки накрывали и оставляли для инкубирования на 60 минут в водяной бане при 37° С. По окончании периода инкубирования добавляли 2 мл глицин-карбонатного буфера (130 мМ глицин, 83 мМ карбонат натрия, при рН 10,6) для прекращения реакции. Относительную флуоресценцию каждого образца измеряли с использованием флуориметра модели ТК0100 (Hoefer Scientific Instruments), который имел фиксированную длину волны возбуждения 365 нм, и осуществляли детекцию при фиксированной длине волны излучения 460 нм. Данные, полученные при считывании образцов, сравнивали со стандартами, полученными от 1 мМ маточного раствора метилумбеллиферона (Sigma Chemical Co.), и подсчитывали количество расщепленного субстрата. Активность α -Gal А выражали в единицах; при этом одна единица активности α -Gal А эквивалентна одному наномолю субстрата, расщепленного за 1 час при 37° С. Данные экспрессии клетками, в основном, выражали как единицы секретированной α -Gal А-активности/106 клеток/24 часа. Этот анализ был также использован для измерения количества α -Gal А-активности в клеточных лизатах и в образцах из различных стадий очистки α -Gal А, как обсуждается ниже.
1.5. Получение α -Gal А, активированной геном (GA-GAL)
Продуцирование активированной геном α -Gal A (GA-GAL) осуществляли путем встраивания регуляторной и структурной ДНК-последовательностей выше α -Gal А-кодирующей последовательности человека с использованием GA-техники, в основном, описанной в патенте США 5733761, который вводится в настоящее описание посредством ссылки. Точное встраивание ген-активирующей последовательности происходит в результате гомологичной рекомбинации между ДНК, присутствующей на фрагменте трансфецированной ДНК, и последовательностями геномной ДНК, расположенными выше локуса α -Gal А в клетке человека. Сама ген-активирующая последовательность содержит кодирующую последовательность α -Gal A вплоть до, но не включительно, сайта отщепления сигнального пептида. Клетки, содержащие активированный локус α -Gal А, выделяли и подвергали отбору с использованием лекарственного средства в целях выделения клеток с повышенным продуцированием GA-GAL.
Обеспечивающий доставку ДНК-фрагмент, содержащий соответствующую ген-активирующую последовательность, вводили в клеточные линии человека путем электропорации. Одной такой клеточной линией является сертифицированная клеточная линия НТ-1080, депонированная в АТСС (Rockville, Maryland). Ген-активирующая плазмида (конструкция для доставки) pGA213C, содержащая такой ДНК-фрагмент, показана на фиг.9. Эта плазмида содержит последовательности, предназначенные для активации части эндогенного локуса α -Gal А в клеточной линии хозяина, а также содержит последовательности, кодирующие сигнальный пептид, но не α -Gal А человека. Конструкция для доставки также содержит экспрессионные кассеты для бактериального гена пео и мышиного гена dhfr. Это позволяет проводить отбор стабильно интегрированных фрагментов для доставки (посредством гена пео) и последующий отбор гена dhfr путем постадийного отбора с использованием метотрексата (МТХ).
Кроме того, pGA213C содержит последовательности, сконструированные для доставки хромосомных последовательностей выше эндогенного локуса α -Gal А путем гомологичной рекомбинации. Гомологичная рекомбинация между эндогенным локусом α -Gal А и 9,6 т.п.н.-ДНК-фрагментом pGA213C показана на фиг.10.
pGA213C была сконструирована для делеции 962 п.н. геномных последовательностей, простирающихся от положений от –1183 до -222 по отношению к метиониновому кодону инициации α -Gal А, после гомологичной рекомбинации фрагмента pGA213C с X-хромосомным локусом α -Gal А. Активация транскрипции локуса α -Gal А происходит путем точной доставки экзогенных регуляторных последовательностей выше кодирующей области α -Gal А. Полученный локус GA-GAL вызывает транскрипцию с инициацией от промотора CMV и с протеканием реакции через экзон 1 CMV, интрон альдолазы и семь экзонов и шесть интронов кодирующей последовательности α -Gal А. Сплайсинг крупного предшественника мРНК соединяет экзогенный экзон CMV (встроенный путем доставки) с целым эндогенным первым экзоном транскрипта α -Gal А. Трансляция мРНК GA-GAL приводит к образованию пре-GA-GAL с сигнальным пептидом из 31 аминокислоты. После секреции из клетки-хозяина сигнальный пептид удаляется. Клеточные линии с точной доставкой сначала индентифицируют путем скрининга посредством полимеразной цепной реакции на присутствие мРНК GA-GAL. Также было обнаружено, что клоны, продуцирующие мРНК GA-GAL, секретируют ферментативно активный α -Gal А в культуральную среду. Последующее подтверждение событий доставки осуществляют путем расщепления рестриктазой и анализа геномной ДНК посредством Саузерн-блот-гибридизации.
Клетки были экспонированы путем постадийного отбора с использованием метотрексата ("МТХ"). После отбора в 0,05 мкМ МТХ клеточные клоны выделяли и подвергали отбору с использованием 0,1 мкМ МТХ. В результате этого процесса был выделен пул клеток, резистентных к 0,1 мкМ МТХ (клеточной линии RAG001), которые были размножены в культуре и охарактеризованы.
Пример 2
Очистка α -Gal A
Ниже описан предпочтительный метод продуцирования, очистки и тестирования α -Gal А. В процессе очистки α -Gal А поддерживался в растворимой, активной, нативной форме в течение всего процесса очистки. Белки не подвергались воздействию экстремальных величин рН, органических растворителей или детергентов, не подвергались протеолитическому расщеплению в процессе очистки и не образовывали агрегаты. Процесс очистки не оказывал влияния на распределение гликоформ α -Gal A.
2.1. Очистка α -Gal A
В примере 2.1 показано, что α -Gal А может быть выделен в почти гомогенной форме из кондиционированной среды культивированных штаммов клеток человека, которые были стабильно трансфецированы с продуцированном фермента. α -Gal А выделяли из α -Gal А-содержащей среды посредством проведения серии из пяти хроматографических стадий. В этих пяти стадиях, для выделения α -Gal А из материала с примесями, использовали разные методы выделения, которые имеет преимущества в отношении разных физических свойств фермента. Эти стадии включали хроматографию на основе гидрофобного взаимодействия с использованием бутилсефарозы®, ионообменную хроматографию на гидроксиапатите, анионообменную хроматографию на Q Сефарозе® и вытеснительную хроматографию на Супердексе® 200. Помимо этого, конечная стадия в данном процессе очистки, т.е. вытеснительная хроматография, также служит в качестве эффективного средства для обмена очищенного белка в буфере, совместимом для данного препарата.
А. Использование хроматографии на бутилсефарозе а качестве первой стадии очистки α -Gal A
Холодную кондиционированную среду (1,34 литра) осветляли путем центрифугирования и фильтровали через фильтр, содержащий 0,45 мкм ацетата целлюлозы, с использованием предварительных стекловолоконных фильтров. При перемешивании рН холодной отфильтрованной среды доводили до 5,6 путем добавления по каплям 1 н. НСl и добавляли сульфат аммония до конечной концентрации 0,66 М путем добавления по каплям маточного раствора (комнатной температуры) 3,9 М сверхчистого сульфата аммония. Эту среду перемешивали еще в течение 5 минут при 4° С, фильтровали, как описано выше, и наносили на колонку с бутилсефарозой (Butyl Sepharose® 4 Fast Flow (объем колонки 81 мл, 2,5× 16,5 см; Pharmacia, Uppsala, Sweden), которую уравновешивали в 10 мМ MES-Трис, рН 5,6, содержащем 0,66 М сульфата аммония (буфер А). Хроматографию проводили при 4° С на системе Gradi-Frac™ (Pharmacia, Uppsala, Sweden), снабженной встроенным УФ-монитором (280 нМ) и монитором для определения проводимости в целях оценки общего белка и концентрации соли, соответственно. После нанесения пробы при скорости потока 10 мл/мин колонку промывали 10 колоночными объемами буфера A. α -Gal А элюировали с колонки на бутилсефарозе (Butyl Sepharose®) 14 колоночными объемами линейного градиента буфера А (содержащего сульфат аммония) - 10 мМ MES-Трис, рН 5,6 (не содержащего сульфата аммония). Фракции оценивали на α -Gal А-активность посредством анализа с использованием 4-MUF-gal, и те фракции, которые содержали заметную ферментативную активность, собирали. Как видно на фиг.8 и из систематизированных данных очистки (таблица 5), эта стадия позволяет удалить приблизительно 99% примесного белка (образец на пред-колонке=8,14 г общего белка; образец на пост-колонке=0,0638 г общего белка).
Очистка α -Gal А из кондиционированной среды со стабильно трансфецированными фибробластами человека
В. Использование хроматографии на гепарин-сефарозе® в качестве стадии очистки α -Gal A
Фракции пика, собранные с колонки на бутилсефарозе® диализовали при 4° С против (4 литра) 10 мМ МЕS-трис, рН 5,6 (с одной заменой). Проводимость диализата доводили до 1,0 мМ/Ом при 4° С добавлением H2O или NaCl, если это необходимо. Пробу наносили на колонку с гепарин-сефарозой® (Sepharose® 4 Fast Flow, Pharmacia, Uppsala, Sweden; объем колонки 29 мл, 2,5× 6 см), которую уравновешивали в 10 мМ МЕS-трис, рН 5,6, содержащего 9 мМ NaCl (буфер В). Это осуществляли при 4° С и скорости потока 10 мл/мин. На встроенном УФ-мониторе (280 нм) и мониторе для измерения проводимости определяли количество общего белка и концентрацию соли. После нанесения образца колонку промывали 10 колоночными объемами буфера В, а затем 3 колоночными объемами линейного градиента 8% буфера С/92% буфера В (где буфер С представляет собой 10 мМ МЕS-трис, рН 5,6, содержащий 250 мМ NaCl) и 10 колоночными объемами 8% буфера С. После этого α -Gal А элюировали 1,5 колоночными объемами линейного градиента до 29% буфера С, а затем 10 колоночными объемами линейного градиента до 35% буфера С. Фракции анализировали на α -Gal А-активность и те фракции, которые содержали заметную активность, собирали.
С. Использование хроматографии на гидроксиапатите в качестве стадии очистки α -Gal A
Гепариновый пул фильтровали и наносили непосредственно на колонку с керамическим гидроксиапатитом НС (40 мкм; American International Chemical, Natick, MA; объем колонки 12 мл, 1,5× 6,8 см), которая была уравновешена в 1 мМ фосфате натрия, рН 6,0 (буфер D). Хроматографию осуществляли при комнатной температуре на гибридной системе Gradi-Frac™ /FPLC® (Pharmacia, Uppsala, Sweden), снабженной встроенным УФ-монитором (280 нм) и монитором для измерения проводимости. После нанесения проб (5 мл/мин) колонку промывали 10 колоночными объемами буфера D. α -Gal А элюировали 7 колоночными объемами линейного градиента до 42% буфера Е/58% буфера D (где буфером Е является 250 мМ фосфат натрия, рН 6,0), а затем 10 колоночными объемами градиента до 52% буфера Е. Фракции анализировали на активность α -Gal А и те фракции, которые обладали значительной активностью, собирали.
D. Использование анионообменной хроматографии на Q-сефарозе® в качестве стадий очистки α -Gal A
Гидроксиапатитный пул приблизительно 1,5-кратно разводили H2O до конечной проводимости 3,4-3,6 мМ/Ом при комнатной температуре. После фильтрации пробу наносили на колонку с Q-сефарозой® HP (Pharmacia, Uppsala, Sweden; объем колонки 5,1 мл; 1,5× 2,9 см), уравновешенную 10% буфером G/90% буфером F, где буфер F представляет собой 25 М фосфат натрия, рН 6,0, а буфер G представляет собой 25 мМ фосфат натрия, рН 6,0, 250 мМ NaCl. Хроматографию осуществляли при комнатной температуре на гибридной системе Gradi-Frac™ /FPLC® (Pharmacia, Uppsala, Sweden) и полные концентрации белка и соли прослеживали с помощью встроенных мониторов. Пробу наносили при скорости потока 5 мл/мин, а затем осуществляли следующие стадии: (1) промывки 5 колоночными объемами 10% буфера G, (2) промывки 7 колоночными объемами 12% буфера G, (3) промывки 3 колоночными объемами линейного градиента до 50% буфера G, (4) промывки 10 колоночными объемами линейного градиента до 53% буфера G, (5) промывки 3 колоночными объемами градиента до 100% буфера G и (6) промывки 10 колоночными объемами 100% буфера G. α -Gal А элюировался, главным образом, в стадиях 3 и 4. Фракции, обладающие значительной активностью, собирали ("О-пул").
Е. Использование гель-фильтрации на супердексе®-200 в качестве стадии очистки α -Gal A
Q-пул концентрировали приблизительно 5-кратно с использованием центрифужных концентраторов Centriprep®-10 (Amicon, Beverly, MA) и наносили на колонку с супердекс® 200 (Pharmacia, Uppsala, Sweden; объем колонки 189 мл; 1,6× 94 см). Колонку уравновешивали и элюировали 25 мМ фосфатом натрия, рН 6,0, содержащим 150 мМ NaCl. Хроматографию осуществляли при комнатной температуре на системе FPLC® (Pharmacia, Uppsala, Sweden) с использованием встроенного УФ-монитора (280 нм) с последующим элюированием белка. Объем пробы, наносимый на колонку, составлял ≤ 2 мл, скорость потока составляла 0,5 мл/мин, а размер фракции составлял 2 мл. Выполняли множество прогонов через колонку; фракции анализировали на α -Gal А-активность и фракции, обладающие значительной активностью, собирали.
Фракции, собранные с колонки с супердексом® 200, концентрировали с использованием концентратора Centriprep 10, разделяли на аликвоты, подвергали мгновенному замораживанию и хранили при -80° С в течение коротких промежутков времени. Результаты очистки α -Gal А, полученные в данном примере, систематизированы в таблице 5. Конечный выход α -Gal А составлял 59% от исходной активности материала, а удельная активность очищенного продукта составляла 2,92× 106 единиц/мг белка. Полученный продукт обнаруживал высокий уровень чистоты после электрофореза, проводимого в условиях восстановления на 4-15% ДСН-полиакриламидном геле, который был затем окрашен серебром.
Выводы
В результате очистки получали в высокой степени очищенную α -Gal А. Наибольшая очистка происходила в первые две стадии данного процесса, тогда как последние три стадии служили для более тонкой очистки материала в целях удаления оставшихся следовых количеств примесей. Последняя стадия вытеснительной хроматографии на супердексе® 200 также служила для обмена α -Gal А в совместимом для композиции буфере.
2.2. Размер α -Gal А, продуцированной стабильно трансфецированными клетками человека в культуре
Были проведены исследования структурных и функциональных свойств очищенной α -Gal А человека. Полученный продукт обнаруживал высокий уровень чистоты после электрофореза, проводимого в условиях восстановления на 4-15% ДСН-полиакриламидном геле, который затем окрашивали серебром.
Молекулярную массу α -Gal А оценивали с помощью масс-спектрометрии MALDI-TOF. Полученные результаты показали, что молекулярная масса димера составляет 102353 Да, а молекулярная масса мономера составляет 51002 Да. Ожидаемая молекулярная масса мономера, исходя из аминокислотного состава, составляет 45400 Да. Следовательно, углеводное содержание данного фермента составляет вплоть до 5600 Да по его молекулярной массе.
2.3. Углеводная модификация α -Gal А, продуцируемого стабильно трансфецированными клетками человека
Был также оценен характер гликозилирования α -Gal А, продуцируемого в соответствии с настоящим изобретением. Правильное гликозилирование является важным фактором для оптимальной in vivo-активности α -Gal A; α -Gal А, экспрессируемая в не-гликозилирующих системах, является неактивной или нестабильной. Hantzopolous et al., Gene 57:159 (1987). Гликозилирование также играет важную роль для интернализации α -Gal А в нужные клетки-мишени и влияет на время полужизни фермента в кровотоке in vivo. На каждой субъединице α -Gal А имеются четыре сайта, подходящие для присоединения связанных с аспарагином углеводных цепей, из которых три являются занятыми. Desnick et al., In The Metabolic and Molecular Bases of Inherited Disease (McGraw Hill, New York, 1995), pp.2741-2780.
Образец α -Gal А, продуцируемый стабильно трансфецированными клетками, был обработан нейраминидазой, выделенной из A.urafaciens (Boehringer-Mannheim, Indianapolis, IN), для удаления сиаловой кислоты. Эту реакцию осуществляли путем обработки 5 мг α -Gal А в течение ночи 10 мЕ нейраминидазой при комнатной температуре в полном объеме 10 мл забуференного ацетатом физиологического раствора (ABS, 20 мМ ацетат натрия, рН 5,2, 150 мМ NaCl).
Очищенную α -Gal А, продуцируемую стабильно трансфецированными клетками, также дефосфорилировали с использованием щелочной фосфатазы (щелочная фосфатаза кишечника теленка, Boehringer-Mannheim, Indianapolis, IN) путем обработки 5 мг α -Gal А в течение ночи при комнатной температуре 15 единицами щелочной фосфатазы в ABS (рН поднимали до 7,5 с использованием 1 М Трис).
Образцы анализировали с помощью электрофореза в ПААГ с ДСН и/или с помощью изоэлектрического фокусирования, а затем проводили Вестерн-блоттинг с использованием специфического антитела против α -Gal А. Используемое антитело представляло собой кроличье поликлональное антитело против пептида, которое было продуцировано с использованием пептида, представляющего собой аминокислоты 68-81 α -Gal А в качестве иммуногена. После переноса белка на PVDF (Millipore, Bedford, MA) мембрану зондировали при разведении 1:2000 антисывороткой в 2,5% блотто (обезжиренное сухое молоко в 20 мМ Трис-HCl, рН 7,5, 0,05% Твин-20). Затем проводили детекцию с использованием козьего антитела против IgG кролика, конъюгированного с пероксидазой хрена (Organo Technique/Cappella, Durham, NC; разведение 1:5000), и реагентов набора для электрохемилюминисценции (ЭХЛ) (Amersham, Arlington Heights, IN).
Обработка α -Gal А нейраминидазой с последующим анализом с помощью электрофореза в ПААГ с ДСН приводит к сдвигу молекулярной массы (приблизительно 1500-2000 Да или 4-6 сиаловых кислот/мономер), что дает основание предположить о наличии интенсивной модификации α -Gal А сиаловой кислотой. Для справки, плазменная форма α -Gal А имеет 5-6 остатков сиаловой кислоты на мономер, а плацентарная форма имеет 0,5-1,0 остатков сиаловой кислоты на мономер. Bishop et al., J. Biol. Chem. 256:1307 (1981).
Другим методом, используемым для оценки модификаций α -Gal А сиаловой кислотой и М6Р, является изоэлектрическое фокусирование (ИЭФ), где данные образцы разделяют на основе их изоэлектрической точки (рI) или суммарного заряда. Таким образом, предполагается, что удаление заряженных остатков, таких как сиаловая кислота или фосфат, из α -Gal А будет приводить к изменению подвижности белка в системе ИЭФ.
Для осуществления ИЭФ-эксперимента образцы α -Gal А, продуцируемые в соответствии с настоящим изобретением, обрабатывали нейраминидазой и/или щелочной фосфатазой, смешанными 1:1 с 2Х буфером для образца Novex (с 8 М мочевиной, рН 3,0-7,0), и загружали на ИЭФ-гель (5,5% полиакриламидный) с 6 М мочевиной, изготовленный с использованием Pharmalyte® (Pharmacia, Uppsala, Sweden; pH 3,0-6,5; Pharmalyte® 4-6,5 и 2,5-5,5, 0,25 мл каждого на гель). Были также включены стандарты изоэлектрической точки (Bio-Rad). После электрофореза гель переносили на PVDF и проводили Вестерн-блот-анализ, как описано выше.
Нейраминидазная обработка фермента приводила к увеличению рI всех трех изоформ, что указывало на то, что все они были до некоторой степени модифицированы сиаловой кислотой. Эти данные дают основание предполагать, что α -Gal А-препараты, полученные, как описано в данной заявке, должны иметь желаемое время полужизни в плазме, что означало бы, что данный материал является в достаточной степени подходящим для фармакологического использования. Кроме того, обработка щелочной фосфатазой фермента α -Gal А, уже обработанного нейраминидазой, приводит к дополнительному увеличению рI части данного белка приблизительно до 5,0-5,1, что указывает на то, что данный фермент несет один или несколько остатков М6Р. Такая модификация необходима для эффективной интернализации α -Gal А клетками-мишенями.
N-Связанные углеводные цепи α -Gal А были проанализированы с помощью ионообменной ВЭЖХ (Glyco-Sep С) и мечения невосстанавливающего конца флуоресцентным соединением 2-аминобензамидом (АВ). Результаты анализа АВ-гликанов от трех отдельных α -Gal А-препаратов систематизированы в таблице 6. Все три препарата имели число Z, превышающее 170. Кроме того, более чем 67% гликанов были сиалилированы, более чем 16% гликанов были фосфорилированы, а менее чем 16% были нейтральными. Эти данные указывают на значительно лучшие результаты по сравнению с известными результатами. Так, например, Desnick et аl. (патент США 5356804) сообщали, что свыше 60% гликанов были нейтральными и только 11% были сиалилированы.
Результаты анализа АВ-гликанов от GA-GAL
Дополнительная подробная характеристика очищенных препаратов GA-GAL представлена в таблице 7.
Объем очищенного препарата GA-GAL
2.4. Увеличение содержания заряженной α -Gal А путем фракционирования молекул α -Gal A
Как обсуждалось выше, фракционирование гликоформ α -Gal A может быть осуществлено в различных стадиях в процессе очистки, как описано в настоящей заявке. В настоящем примере гликоформы α -Gal А были фракционированы по размеру и по заряду. Фракционировать α -Gal А можно также с использованием комбинации этих и других хроматографических методов, описанных выше.
Для фракционирования по размеру гликоформ α -Gal А осуществляли вытеснительную хроматографию на колонке с супердексом® 200 (Pharmacia, 1,6 см × 94,1 см), уравновешенной в забуференном фосфатом физиологическом растворе при рН 6. α -Gal A (2,6 мг в 1 мл) загружали на колонку и эту колонку элюировали при 0,35 мл/мин. Фракции собирали по всему профилю элюции и фракции, содержащие широкий пик элюирования α -Gal А, анализировали с помощью электрофореза в ПААГ с ДСН, а затем визуализировали окрашиванием серебром. Фракции ведущего края пика содержали α -Gal А с наиболее высокой молекулярной массой, и по мере продвижения фракций по пику кажущаяся молекулярная масса α -Gal A постепенно снижалась. Затем фракции α -Gal А собирали и объединяли с полученим препарата α -Gal А с нужной молекулярной массой.
Для фракционирования гликоформ α -Gal А по заряду α -Gal A фракционировали с помощью хроматографии на Q-Сефарозе®. Колонку с Q-Сефарозой® (1,6 см × 94,1 см) уравновешивали в 20 мМ фосфате натрия при рН 6, содержащем 30 мМ NaCl, а скорость потока поддерживали при 5 мл/мин. α -Gal A (130 мг в 166 мл) загружали на колонку, промывали уравновешивающим буфером, а затем элюировали 20 мМ фосфатом натрия, рН 6,0, содержащим 130 мМ NaCl. Для более интесивного фракционирования может быть использовано градиентное элюирование (например, 10 колоночными объемами) уравновешивающим буфером - элюирующим буфером. Фракции собирали по всему профилю элюирования и фракции, содержащие пик элюирования α -Gal А, анализировали с помощью электрофореза в ПААГ с ДСН, а затем визуализировали окрашиванием серебром. Молекулы с наименьшей молекулярной массой наблюдались на геле, элюированном в промывке и в ведущем крае пика, а гликоформы с наиболее высокой молекулярной массой элюировались ближе к концу пика. Молекулы с более низкой молекулярной массой соответствовали менее отрицательно заряженным гликоформам α -Gal А, которые менее жестко связывались с колонкой с положительно заряженной Q-Сефарозой® (состоящей из смолы, замещенной четвертичным амином). Молекулы α -Gal А с наиболее высоким отрицательным зарядом элюировались позднее на профиле элюции и имели более высокую молекулярную массу, как было проанализировано с помощью электрофореза в ДСН-ПААГ. Фракционирование по заряду было подтверждено с помощью изоэлектрического фокусирования проэлюированных фракций или отобранных пулов.
Таким образом, как фракционирование по размеру, так и фракционирование по заряду позволяло проводить отбор в высокой степени заряженных гликоформ α -Gal A.
2.5. Интернализация α -Gal А, опосредованная маннозой или маннозо-6-фосфатом (М6Р)
Что касается α -Gal А, продуцированной стабильно трансфецированными клетками и используемой в качестве эффективного терапевтического агента для лечения состояний с дефицитом α -Gal А, то указанный фермент должен быть интернализирован пораженными клетками. α -Gal А является минимально активным при физиологических уровнях рН, например, в крови и в тканевых жидкостях. α -Gal А оптимально метаболизирует аккумулированные липидные субстраты лишь при интернализации в кислотном окружении лизосомы. Эта интернализация опосредуется связыванием α -Gal A с рецепторами М6Р, которые экспрессируются на клеточной поверхности и доставляют данный фермент в лизосомы по эндоцитному пути. Рецептор М6Р экспрессируется почти везде и в той или иной степени М6Р экспрессируется большинством соматических клеток. Маннозный рецептор, который является специфическим для экспонированных маннозных остатков на гликопротеинах, является менее распространенным. Маннозные рецепоры, в основном, обнаруживаются только на макрофагах или макрофагподобных клетках и являются дополнительным средством для вхождения α -Gal А в эти клетки.
Для иллюстрации МбР-опосредованной интернализации α -Gal А фибробласты кожи, взятые от пациента с болезнью Фабри (NIGMS Human Genetic Mutant Cell Repository), культивировали в течение ночи в присутствии возрастающих концентраций очищенного α -Gal A настоящего изобретения. Некоторые из образцов содержали 5 мМ растворимого М6Р, который конкурентно ингибирует связывание с рецептором М6Р и опосредованную им интернализацию. Другие образцы содержали 30 мг/мл маннана, который ингибирует связывание с маннозным рецептором и опосредованную им интернализацию. После инкубирования клетки промывали и собирали путем соскабливания в буфер для лизиса (10 мМ трис, рН 7,2, 100 мМ NaCl, 5 мМ EDTA, 2 мМ Pefabloc™ (Boehringer-Mannhiem, Indianapolis, IN) и 1% NP-40). Затем лизированные образцы анализировали на концентрацию белка и α -Gal А-активность. Результаты выражали как единицы α -Gal А-активности/мг клеточного белка. α -Gal A интернализировался в клетки Фабри дозо-зависимым образом. Указанная интернализация ингибировалась М6Р, но не ингибировалась маннаном. Поэтому интернализация α -Gal А в фибробласты Фабри опосредуется рецептором М6Р, но не маннозным рецептором.
α -Gal А также интернализируется in vitro эндотелиальными клетками, которые являются важными клетками-мишенями при лечении болезни Фабри. Эндотелиальные клетки пупочной вены человека (HUVEC) культивировали в течение ночи вместе с 7500 единицами α -Gal А, причем некоторые лунки содержали М6Р. После инкубирования клетки собирали и анализировали на α -Gal А, как описано выше. Клетки, инкубированные с α -Gal А, содержали уровни фермента, которые почти в 10 раз превышали уровни контрольных клеток (не инкубированных с α -Gal А). М6Р ингибировал внутриклеточную аккумуляцию α -Gal А, что дает основание предположить, что интернализация α -Gal А клетками HUVEC опосредуется рецептором М6Р. Таким образом, человеческая α -Gal А настоящего изобретения интернализируется клинически важными клетками.
Существует несколько культивированных клеточных линий человека, которые, как известно, экспрессируют маннозный рецептор. Однако для того, чтобы определить, интернализируется ли очищенная α -Gal А посредством маннозного рецептора, может быть использована мышиная макрофагподобная клеточная линия (J774.E), которая несет маннозные рецепторы, но имеет очень мало, а то и вообще не имеет рецепторов М6Р. Diment et al., J. Leukocyte Biol. 42:485-490 (1987). Клетки J774.E культивировали в течение ночи в присутствии 10000 единиц/мл α -Gal А. Выбранные образцы также содержали 2мМ М6Р, а другие содержали 100 мг/мл маннана. Эти клетки промывали и собирали, как описано выше, и для каждого образца определяли содержание общего белка и α -Gal А-активность. М6Р не ингибировал поглощение α -Gal A этими клетками, тогда как маннан способствовал снижению уровней аккумулированного α -Gal А на 75%. Таким образом, α -Gal A настоящего изобретения может быть интернализирован маннозным рецептором в клетки тех типов, которые экспрессируют этот конкретный рецептор клеточной поверхности.
Пример 3
Фармацевтическая композиция
Получение буферных растворов и композиций
Массу очищенного α -Gal А разводили до конечной концентрации разбавителем для α -Gal А. Исходя из объема очищенной массы, используемой для изготовления композиции, определяли требуемые концентрацию α -Gal А (мг/мл) и нужную концентрацию α -Gal А для конечной композиции, а также объем разбавителя для α -Gal А. Разбавитель для α -Gal А получали не более чем за 24 часа до использования путем смешивания соответствующих количеств WFI, хлорида натрия и моноосновного фосфата натрия, а рН доводили до 6,0 раствором гидроксида натрия. Состав разбавителя для α -Gal А приводится в таблице 8.
Состав разбавителя для α -Gal А (на литр)
Один литр или меньшие объемы разбавителя для α -Gal A фильтровали путем вакуумной фильтрации с использованием стерильных найлоновых 0,2 мм-фильтров (Nalge Nunc International, Rochester, NY). 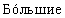 объемы фильтровали при положительном давлении с использованием перистальтического насоса и капсульных 0,2 мм-фильтров Supor® (Pall, Port Washington, NY). Все фильтры были подвергнуты тесту на целостность при температуре начала кипения после фильтрации. Стадии смешивания и фильтрации осуществляли в вытяжном шкафу с ламинарным потоком, сертифицированном как класс 100. Разбавитель α -Gal А добавляли к массе очищенного α -Gal А в сосуде для смешивания, в результате чего получали 1 мг/мл конечного раствора. Затем добавляли соответствующий объем полисорбата 20 (твин 20, Spectrum) до достижения концентрации 0,02%.
объемы фильтровали при положительном давлении с использованием перистальтического насоса и капсульных 0,2 мм-фильтров Supor® (Pall, Port Washington, NY). Все фильтры были подвергнуты тесту на целостность при температуре начала кипения после фильтрации. Стадии смешивания и фильтрации осуществляли в вытяжном шкафу с ламинарным потоком, сертифицированном как класс 100. Разбавитель α -Gal А добавляли к массе очищенного α -Gal А в сосуде для смешивания, в результате чего получали 1 мг/мл конечного раствора. Затем добавляли соответствующий объем полисорбата 20 (твин 20, Spectrum) до достижения концентрации 0,02%.
Пример 4
Десиалилированная дегалактозилированная α -Gal A
Для исследования влияния гликозилирования на биологическое распределение α -Gal А очищенный препарат α -Gal А последовательно дегликозилировали и каждую форму инъецировали мышам. Через четыре часа после инъекции органы мышей собирали и проводили иммуногистохимический анализ этих тканей для визуализации возможных изменений в биологическом распределении белка.
Сначала α -Gal А обрабатывали нейраминидазой (сиалидазой) для удаления остатков сиаловой кислоты, в результате чего галактозные части оставались экспонируемыми. Затем часть этого обработанного сиалидазой фермента подвергали реакции с β -галактозидазой для удаления галактозных остатков; при этом оставались экспонируемыми N-ацетилглюкозаминидазные остатки (GlcNAC). Эти GlcNAC затем удаляли с помощью N-ацетилглюкозаминидазы, в результате чего на белке оставались маннозные группы сердцевины. Необработанную α -Gal А (контроль) или одну из обработанных форм белка инъецировали мышам в хвостовую вену. Через четыре часа после инъекций печень, селезенку, сердце, почки и легкие мышей собирали, обрабатывали консервантом и подвергали иммуноокрашиванию для детекции α -Gal А.
Сравнение с контрольными животными, получавшими необработанный белок, показало, что мыши, которым был инъецирован обработанный сиалидазой фермент (с экспонированными галактозными остатками), имели преимущественную локализацию α -Gal А в печени и соответственно меньшее количество этого фермента в других исследованных органах. Кроме того, картина окрашивания в печени у этих животных очень сильно отличалась. У контрольных животных α -Gal А локализировался, главным образом, в клетках Купффера и в эндотелиальных клетках с умеренным окрашиванием в гепатоцитах. У животных, получавших обработанный сиалидазой α -Gal А, этот фермент локализировался только в гепатоцитах, что соответствовало известному биологическому распределению рецептора асиалогликопротеина. Это влияние дегликозилирования на биологическое распределение было обратным при удалении галактозных остатков под действием β -галактозидазы. Картина окрашивания, наблюдаемая в печени мышей, получавших данный белок без галактозных остатков, была аналогична картине окрашивания для контрольных животных; при этом наибольшее окрашивание наблюдалось в клетках Купффера и в эндотелиальных клетках с минимальным окрашиванием гепатоцитов. Последующая обработка α -Gal А N-ацетилглюкозаминидазой не изменяла характера окрашивания, который наблюдался для белка, обработанного β -галактозидазой, то есть удаление N-ацетилглюкозаминовых остатков, очевидно, оказывает незначительное влияние на биологическое распределение α -Gal A.
Пример 5
Коррекция фибробластов Фабри путем экспрессии α -Gal A фибробластами человека
Для генной терапии имплантация аутологичных клеток, продуцирующих α -Gal А, должна приводить к продуцированию фермента в форме, соответствующим образом модифицированной для "коррекции" дефицита α -Gal А в клетках-мишенях. Для оценки влияния продуцирования α -Gal А трансфецированными фибробластами человека на клетки Фабри, фибробласты, взятые у пациентов с болезнью Фабри (NIGMS Human Genetics Mutant Cell Repository), были совместно культивированы с клеточным штаммом, продуцирующим α -Gal A (BRS-11) в Transwells® (Costar, Cambridge, MA). Клетки Фабри культивировали в 12-луночных планшетах для культивирования ткани, некоторые из которых содержали вставки (Transwells®, размер пор 0,4 мм), имеющие поверхность, на которой могут быть выращены клетки. Матрица вставки для роста клеток является пористой и позволяет макромолекулам проходить от верхней до нижней части среды. Одна серия вставок содержала нормальные фибробласты крайней плоти человека (HF), которые секретируют минимальные уровни α -Gal А, а другая серия содержала стабильно трансфецированный штамм фибробластов человека, BRS-11, которые секретируют большое количество α -Gal А. В лунках, культивированных вместе с клетками, продуцирующими α -Gal А, α -Gal А может проникать в среду, заливающую клетки Фабри, и потенциально интернализироваться клетками Фабри.
Данные, представленные в таблице 9, показывают, что клетки Фабри интернализируют секретированную α -Gal А. Мониторинг внутриклеточных уровней α -Gal А проводили в течение 3 дней. Эти клетки, культивированные отдельно (без вставки) или в присутствии нетрансфецированных фибробластов крайней плоти (вставка HF) имели очень низкие внутриклеточные уровни α -Gal А-активности. Однако клетки Фабри, культивированные с клетками, продуцирующими α -Gal А (вставка BSR-11), обнаруживали уровни фермента, аналогичные уровням фермента в нормальных клетках в конце дня 2 (нормальные фибробласты имели 25-80 единиц α -Gal А/мг белка). Такая коррекция, обусловленная α -Gal А, поглощаемого посредством рецептора М6Р, была продемонстрирована благодаря ингибированию М6Р (вставка BSR-11 + М6Р).
"Коррекция" фибробластов Фабри нормальными фибробластами человека, экспрессирующими α -Gal А-активность (единиц/мг общего белка)
Вышеприведенное описание изобретения представлено лишь в иллюстративных целях и настоящее изобретение не ограничивается конкретно описанной формой, а ограничивается лишь прилагаемой формулой изобретения. В настоящем описании и в прилагаемой формуле изобретения отдельные формы включают множество известных форм, если это не указано конкретно. Все патенты и публикации, цитированные в настоящем описании, вводятся в описание посредством ссылки.


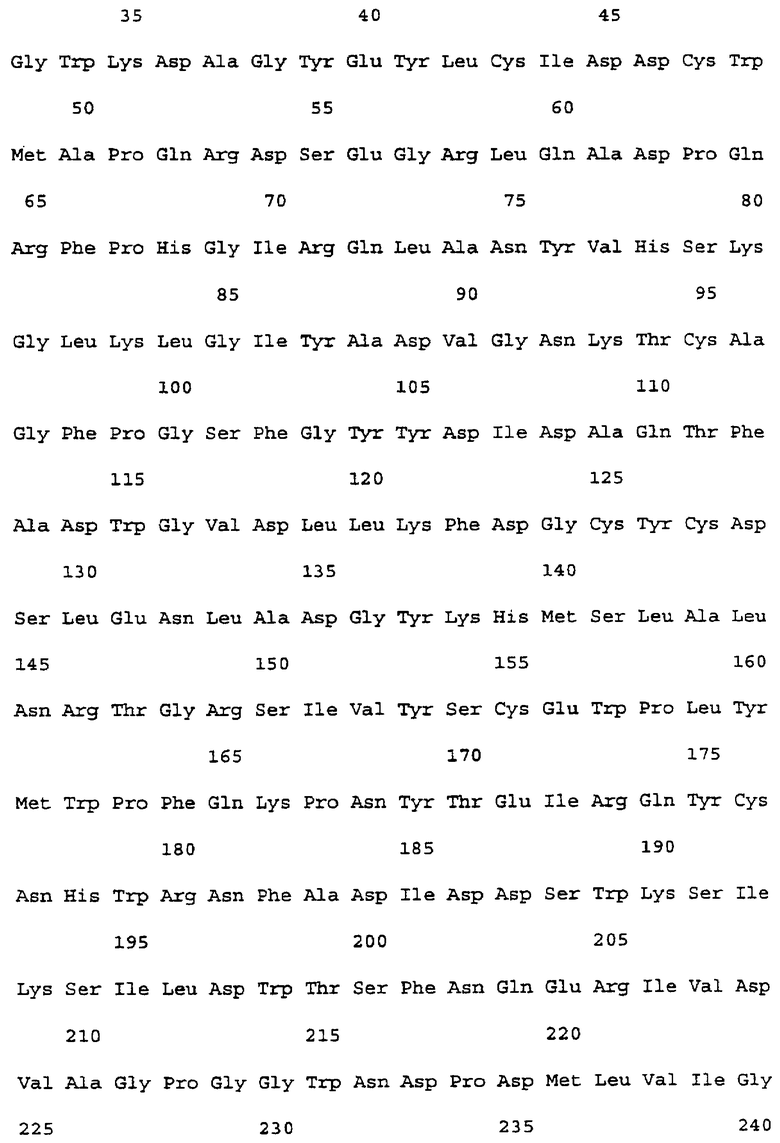






| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ В СЛУЧАЕ ДЕФИЦИТА α-ГАЛАКТОЗИДАЗЫ А | 1997 |
|
RU2179034C2 |
| СТАБИЛИЗИРОВАННАЯ АЛЬФА-ГАЛАКТОЗИДАЗА И ЕЕ ПРИМЕНЕНИЕ | 2011 |
|
RU2604795C2 |
| ВЕКТОР ДЛЯ ЭКСПРЕССИИ ПОЛИПЕПТИДОВ С СИАЛИДАЗНОЙ АКТИВНОСТЬЮ, СПОСОБ ОБЕСПЕЧЕНИЯ СИАЛИДАЗНОЙ АКТИВНОСТИ В КУЛЬТУРЕ КЛЕТОК И СПОСОБ РЕГУЛИРОВАНИЯ СВОЙСТВ Fc-СОДЕРЖАЩИХ МОЛЕКУЛ, ЭКСПРЕССИРУЕМЫХ В ЛИНИИ КЛЕТОК | 2007 |
|
RU2466189C2 |
| КОНЪЮГАТ ПОЛИПЕПТИДА ФАКТОРА VII, СПОСОБ ЕГО ПОЛУЧЕНИЯ, ЕГО ПРИМЕНЕНИЕ И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2003 |
|
RU2362807C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2017 |
|
RU2788133C2 |
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2004 |
|
RU2385928C2 |
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2009 |
|
RU2535342C2 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2011 |
|
RU2604811C2 |
| ПРОДУКЦИЯ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ФУКОЗИЛИРОВАНИЕМ | 2008 |
|
RU2479629C2 |
| СХЕМА ЛЕЧЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ С ИСПОЛЬЗОВАНИЕМ СТАБИЛИЗИРОВАННОЙ α-ГАЛАКТОЗИДАЗЫ | 2018 |
|
RU2793184C2 |
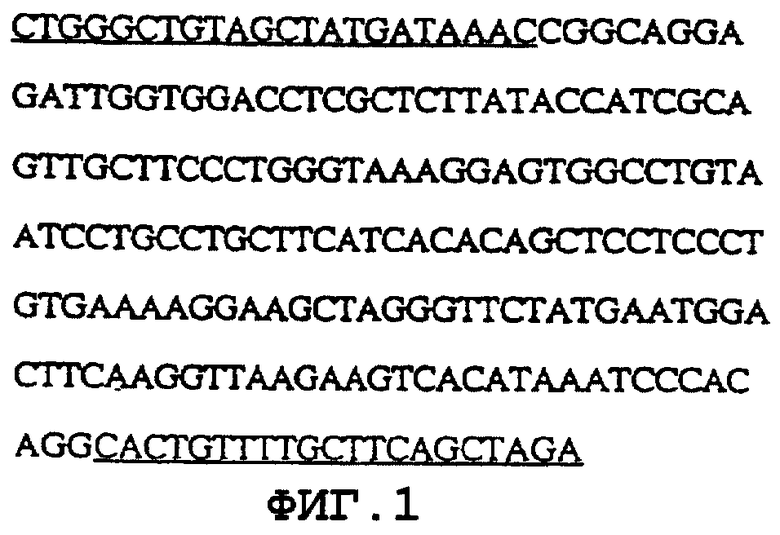


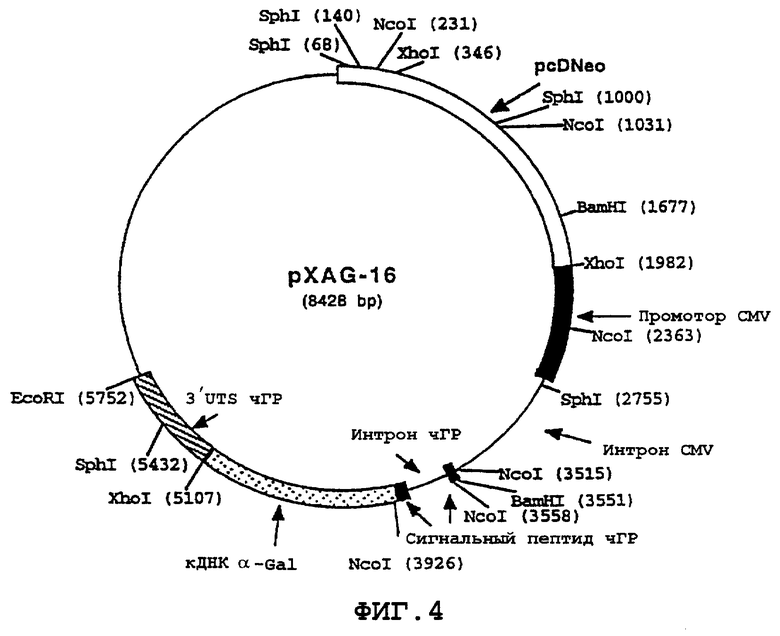
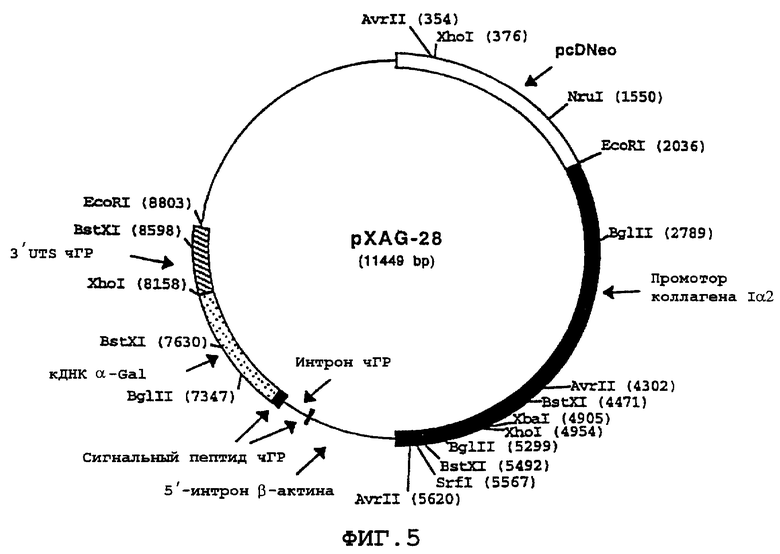
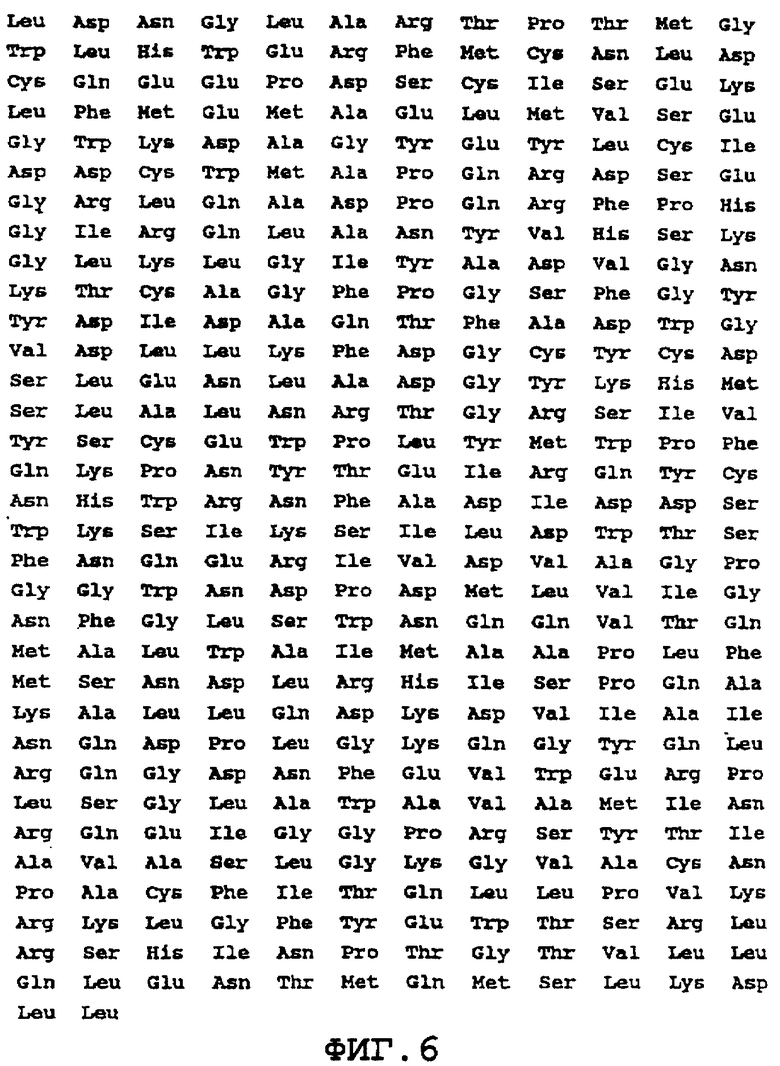

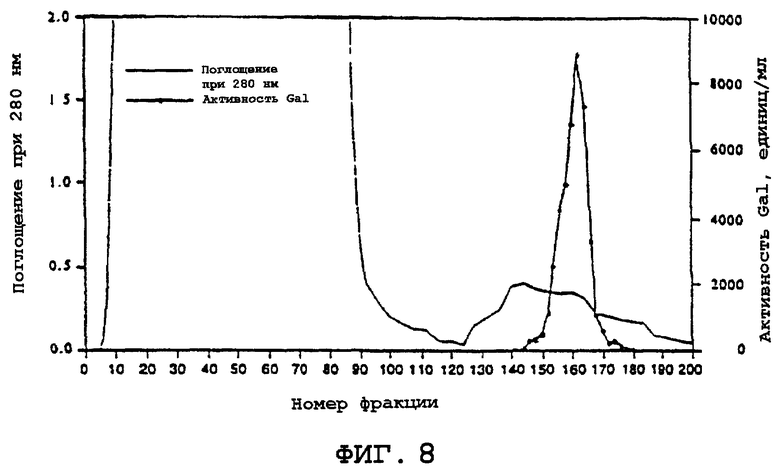
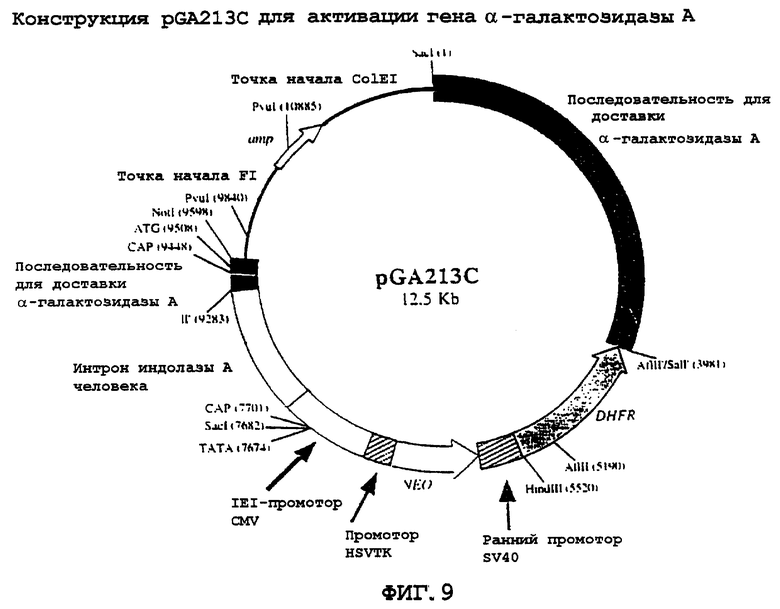
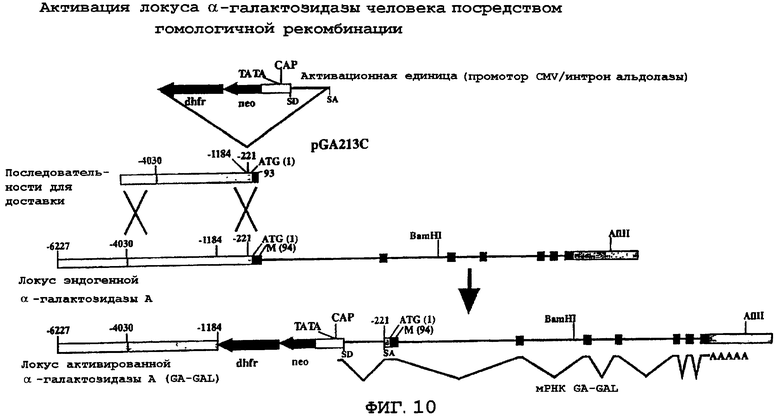
Настоящее изобретение относится к области фармакологии, в частоности к высокоочищенной α-Gal А и к различным способам ее очистки. Сущность состоит в создании препаратов α-Gal А с измененным зарядом и к способам получения таких препаратов, а также к препаратам α-Gal A, имеющим увеличенное время полужизни в кровотоке млекопитающего-хозяина и к способам их получения; к способам и дозам введения препарата α-Gal А индивидууму. Технический результат - расширение арсенала средств борьбы с α-галактозидазной А недостаточностью. 2 н. и 13 з.п. ф-лы, 9 табл., 10 фиг.
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Экономайзер | 0 |
|
SU94A1 |
| Э.Маршелл “Биофизическая химия”, т.2 | |||
| Приспособление для изготовления в грунте бетонных свай с употреблением обсадных труб | 1915 |
|
SU1981A1 |

