Область изобретения
Настоящее изобретение относится к трансформированным клеткам-хозяевам для продуцирования белков с высоким содержанием маннозы и способу и системе продуцирования таких белков, в частности, в растительной культуре.
Предпосылки создания изобретения
Болезнь Гоше представляет собой наиболее распространенную лизосомальную болезнь накопления. Она вызвана рецессивным генетическим нарушением (хромосома 1 q21-q31), приводящим к дефициту глюкоцереброзидазы, также известной как глюкозилкерамидаза, которая представляет собой мембранно-связанный лизосомальный фермент, который катализирует гидролиз глюкозфинголипидного глюкоцереброзида (глюкозилкерамид, GlcCer) до глюкозы и керамида. Болезнь Гоше вызывается точечными мутациями в гене (GBA) hGCD (глюкоцереброзидазы человека), что приводит к накоплению GlcCer в лизосомах макрофагов. Характерные клетки накопления, называемые клетками Гоше, находятся в печени, селезенке и костном мозге. Связанные с этим клинические симптомы включают серьезную гепатоспленомегалию, анемию, тромбоцитопению и повреждение скелета.
Ген, кодирующий GCD человека, был впервые секвенирован в 1985 г. (6). Белок, состоящий из 497 аминокислот, получен из 536-mer пробелка. Зрелый GCD содержит пять N-гликозилированных согласованных последовательностей аминокислот (Asn-X-Ser/Thr). Четыре из этих сайтов гликозилированы обычным образом. Гликозилирование пятого сайта является необходимым для продуцирования активного белка. Были идентифицированы как высокоманнозные, так и комплексные олигосахаридные цепи (7). HGCD плаценты содержит 7% углевода, 20% которого относится к высокоманнозному типу (8). Биохимические исследования и исследования сайт-направленного мутагенеза обеспечили первоначальную карту областей и остатков, важных для складчатости, активаторного взаимодействия и расположения активного сайта.
Обработка плацентарной hGCD нейраминидазой (приводящая к азиало-ферменту) приводит к повышенным скоростям выведения и поглощения в клетках печени крысы с сопутствующим увеличением гепатической ферментативной активности (Furbish et al., 1981, Biochim.Biophys. Acta 673:425-434). Такая гликан-модифицированная плацентарная hGC обычно используется в качестве терапевтического агента при лечении болезни Гоше. Биохимические исследования и исследования сайт-направленного мутагенеза обеспечили первоначальную карту областей и остатков, важных для складчатости, активаторного взаимодействия и расположения активного сайта [Grace et al., J.Biol.Chem., 269:228302291 (1994)].
Существует три различных типа болезни Гоше, каждый из которых определяется уровнем активности hGC. Основными клетками, поражаемыми заболеванием, являются макрофаги, которые являются сильно увеличенными благодаря накоплению GlcCer, и, соответственно, упоминаются как «клетки Гоше».
Идентификация дефекта в GCD в качестве первичной причины болезни Гоше привела к развитию ферментативной заместительной терапии в качестве терапевтической стратегии для данного заболевания.
De Duve первым предположит, что замещение недостающего лизосомального фермента экзогенным биологически активным ферментом могло бы оказаться подходом, приносящим хорошие результаты при лечении лизосомальных болезней накопления. [Fed Proc. 23:1045 (1964)].
С этого времени в различных исследованиях предполагалось, что ферментативная заместительная терапия может оказаться выигрышной для лечения различных лизосомальных болезней накопления. Максимальный успех был достигнут в случае индивидуумов, имеющих болезнь Гоше типа I, которых лечили экзогенным ферментом (β-глюкоцереболзидазой), полученным из плаценты (Цередаза (CeredaseTM)), или, позднее, полученным рекомбинантным образом (Церезим (CerezymeTM)).
Немодифицированная глюкоцереброзидаза, полученная из природных источников, представляет собой гликопротеин с четырьмя углеводными цепями. Данный белок не направлен на фагоцитные клетки тела и, следовательно, имеет ограниченное терапевтическое значение. При развитии современной терапии болезни Гоше концевые сахара на углеводных цепях глюкоцереброзидазы последовательно удаляли с помощью обработки тремя различными гликозидазами. Такая обработка гликозидазами приводит к гликопротеину, концевые сахара которого состоят из остатков маннозы. Поскольку фагоциты имеют рецепторы маннозы, которые распознают гликопротеины и гликопептиды с олигосахаридными цепями, на конце которых расположены остатки маннозы, углеводное модифицирование глюкоцереброзидазы улучшило нацеливание фермента на данные клетки [Furbish et al., Biochim.Biophys. Acta 673: 425, (1981)].
Как указано в данном описании, гликозилирование играет решающую роль в активности hGCD, следовательно, дегликозилирование hGCD, экспрессируемой клеточными линиями, с использованием либо туникамицина (клетки Sf9), либо точечных мутаций, разрушающее все сайты гликозилирования (как клетки Sf9, так и COS-1), приводит к полной потере ферментативной активности. Кроме того, было установлено, что hGCD, экспрессируемый в E.coli, не является активным. Дополнительное исследование указало на значимость различных сайтов гликозилирования в активности белка. В дополнение к роли гликозилирования в фактической активности белка коммерчески получаемый фермент содержит модификации последовательности гликана, которые облегчают специфическую доставку лекарственного средства. Гликозилированные белки модифицируют после экстракции так, чтобы они включали только содержащие маннозу последовательности гликана.
Фермент GCD человека содержит 4 сайта гликозилирования и 22 лизина. Получаемый рекомбинантным образом фермент (ЦерезимТМ) отличается от плацентарного фермента (Цередаза ТМ) в положении 495, где аргинин был замещен на гистидин. Кроме того, олигосахаридный состав различается между рекомбинантной и плацентарной GCD, поскольку первый содержит больше фукозных и N-ацетилглюкозаминных остатков, тогда как последний сохраняет только одну цепь с высоким содержанием маннозы. Как отмечено выше, оба типа GCD обрабатывают тремя различными гликозидазами (нейраминидазой, галактозидазой и P-N ацетилглюкозаминидазой), чтобы оставить незащищенными концевые маннозы, которые дают возможность целеуказания в отношении фагоцитных клеток. Фармацевтический препарат, содержащий полученный рекомбинантным образом фермент, описан в патенте США 5549892. Следует отметить, что все отмеченные в данном описании публикации включены посредством ссылки во всей своей полноте.
Один из недостатков, связанных с существующей в настоящее время лизосомальной заместительной терапией, заключается в том, что биоактивность фермента in vivo является нежелательно низкой, например, вследствие низкого поглощения, пониженного целеуказания на лизосомы специфических клеток, где аккумулируется субстрат, и короткого функционального времени полураспада в лизосомах in vivo.
Другим основным недостатком существующих рекомбинантных ферментов GCD является их стоимость, которая может лечь тяжелым экономическим бременем на системы здравоохранения. Высокая стоимость данных рекомбинантных ферментов связана со сложным протоколом их очистки и относительно большими количествами лекарственных средств, требуемых для существующего в настоящее время лечения. Следовательно, существует неотложная необходимость в снижении стоимости GCD так, чтобы данное сохраняющее жизнь людей лечение могло быть более доступно по средствам для всех, кто в нем нуждается.
Белки для фармацевтического применения традиционно продуцируются в системах экспрессии млекопитающих и бактерий. В последнее десятилетие была разработана новая система экспрессии в растениях. В данной методологии используется Agrobacterium, бактерия, способная встраивать молекулы отдельных цепей ДНК (Т-ДНК) в растительный геном. Вследствие относительной простоты введения генов для массового продуцирования белков и пептидов данная методология становится все более популярной в качестве альтернативной системы экспрессии белка (1).
Хотя посттрансляционные модификации не существуют в бактериальных системах экспрессии, системы экспрессии, полученные из растений, действительно облегчают данные модификации, известные как решающие для экспрессии и активности белка. Одним из основных различий между системой экспрессии млекопитающих и растений является изменение боковых цепей сахаров белка, вызванное разницей в биосинтетических путях. Показано, что гликозилирование оказывает чрезвычайное влияние на активность, складчатость, стабильность, растворимость, восприимчивость к протеазам, скорость выведения из крови и антигенный потенциал белков. Следовательно, при любом продуцировании белка в растениях следует принимать во внимание потенциальные разветвления растительного гликозилирования.
Гликозилирование белка подразделяют на две категории: N-связанные и О-связанные модификации (2). Эти два типа отличаются аминокислотой, к которой присоединен гликановый фрагмент: N-связанный присоединен к остаткам Asn, тогда как О-связанный присоединен к остаткам Ser или Thr. Кроме того, последовательность гликанов каждого типа содержит уникальные отличительные особенности. Среди этих двух типов N-связанное гликозилирование является более частым, и его влияние на функцию белка интенсивно исследовалось. О-связанные гликаны, с другой стороны, являются относительно редкими, и относительно их влияния на белки доступно меньшее количество информации.
Краткое изложение сущности изобретения
В предшествующем уровне развития данной области не указаны или не предложены устройство, система или способ селективного продуцирования гликозилированных белков в растительной культуре. Также в предшествующем уровне данной области не указываются или не предлагаются такие устройство, система или способ продуцирования высокоманнозных белков в растительной культуре. Кроме того, в предшествующем уровне данной области не указывается или не предлагается устройство, система или способ продуцирования белков в растительной культуре посредством эндоплазматической сети (ER). Также в предшествующем уровне данной области не указывается или не предлагается устройство, система или способ продуцирования белков в растительной культуре посредством эндоплазматической сети (ER), при этом в обход аппарата Гольджи. Также в предшествующем уровне данной области не указывается или не предлагается устройство, система или способ продуцирования белков в растительной культуре с использованием ER-сигнала для обхода аппарата Гольджи.
В настоящем изобретении преодолены данные недостатки предшествующего уровня техники посредством разработки устройства, системы или способа продуцирования гликозилированных белков в растительной культуре, в частности белков, имеющих высокоманнозное гликозилирование, в то время как необязательно и предпочтительно для таких белков проводят целеуказание (и/или в других обстоятельствах манипулируют их переработкой) с помощью ER-сигнала. Не желая ограничиваться какой-либо одной гипотезой, авторы настоящего изобретения полагают, что такое целеуказание вызывает обход белками аппарата Гольджи и, следовательно, сохраняет желаемое гликозилирование, в частности высокоманнозное гликозилирование. Следует отметить, что термин «растительная культура», как он использован в данном описании, включает любой тип трансгенных и/или иначе генетически сконструированных клеток растений, которые выращивают в культуре. Генная инженерия может быть постоянной или временной. Предпочтительно отличительной особенностью культуры является то, что клетки не группируются, образуя полное растение, так что, по крайней мере, не присутствует одна биологическая структура растения. Необязательно и предпочтительно культура может отличаться множеством различных типов растительных клеток, но предпочтительно в культуре используют конкретный тип растительной клетки. Следует отметить, что необязательно растительные культуры, отличительной особенностью которых является определенный тип растительной клетки, могут первоначально происходить из множества различных типов таких растительных клеток.
Растительные клетки можно выращивать в соответствии с любым типом подходящего способа культивирования, включая, но не ограничиваясь указанным, культивирование на твердой поверхности (такой, как пластиковые сосуды для культивирования или, например, чашки Петри) или в суспензии.
Кроме того, отличительной особенностью изобретения являются векторы и способы экспрессии и продуцирования ферментативно активных высокоманнозных лизосомальных ферментов с использованием корней трансгенных растений, в частности клеток моркови. Более конкретно, отличительной особенностью изобретения являются клетки-хозяева, в частности трансгенные суспендированные клетки-моркови, векторы и способы экспрессии и продуцирования с высоким выходом биологически активной высокоманнозной глюкоцереброзидазы (GCD). Кроме того, изобретение относится к композициям и способам лечения лизосомальных болезней накопления.
Настоящее изобретение также относится к устройству, системе и способу обеспечения дефицитных клеток достаточными количествами биологически активных лизосомальных ферментов и, в частности, GCD человека. Настоящее изобретение также относится к клеткам-хозяевам, включающим композиции новых векторов, которые дают возможность эффективного продуцирования генов, кодирующих лизосомальные ферменты, такие как GCD.
Таким образом, настоящее изобретение решает продолжительную необходимость в экономически жизнеспособной технологии получения белков, соответствующих определенным требованиям в отношении гликозилирования, таким как высокоманнозное гликозилирование лизосомальных ферментов, таких, например, как GCD. Настоящее изобретение способно решить эту продолжительную необходимость посредством использования культуры клеток растений.
Для дальнейшего объяснения настоящего изобретения будет приведено краткое объяснение биосинтетического пути для белков с высоким содержание маннозы. Основной путь биосинтеза высокоманнозных и комплексных N-связанных гликанов является высококонсервативным для всех эукариотов. Биосинтез начинается в эндоплазматической сети (ER) с переносом предшественника гликана от длинного липидного носителя к специфическому остатку Asn на белке с помощью олигосахарилтрансферазы. Предшественник впоследствии модифицируется в ER гликозидазами I и II и предположительной маннозидазой, давая высокоманнозные структуры, аналогично процессу, происходящему у млекопитающих.
Последующие модификации последовательности гликана в комплексные и гибридные структуры происходят в аппарате Гольджи. Такие модификации включают удаление одного из четырех маннозных остатков с помощью α-маннозидазы I, присоединение остатка N-ацетилглюкозамина, удаление двух дополнительных остатков маннозы с помощью α-маннозидазы II, присоединение N-ацетилглюкозамина, и, необязательно, на данной стадии могут присоединяться остатки ксилозы и фукозы, давая растительно-специфические N-связанные гликаны. После переноса ксилозы и фукозы в ядро N-гликаны комплексного типа могут в дальнейшем перерабатываться путем присоединения концевой фукозы и галактозы. Последующие модификации также могут происходить во время транспорта гликопротеина.
В данной области в настоящее время используется несколько подходов для контроля и приспособления для определенных целей гликозилирования белков в растениях, все из которых имеют заметные недостатки, в частности в сравнении с настоящим изобретением. Модификации Гросса, такие как полное ингибирование гликозилирования или удаление сайтов гликозилирования из пептидной цепи, представляют собой одну из стратегий. Однако данный подход может привести к структурным дефектам. Дополнительный подход включает сильный удар и введение специфических ферментов переработки углеводов. Опять же данный подход труден и также может оказать вредное влияние на сами клетки растений.
Настоящее изобретение преодолевает такие недостатки этих подходов предшествующего уровня техники посредством использования ER-сигнала и/или путем блокирования секреции из эндоплазматической сети в аппарат Гольджи. Не желая ограничиваться одной гипотезой, поскольку предпочтительной является высокоманнозная структура лизосомальных ферментов, если секреция может быть блокирована и белок может сохраняться в эндоплазматической сети, природные высокоманнозные структуры получают без необходимости в последующей трансформации.
Как указано выше, белки, транспортируемые через эндомембранную систему, сначала проходят в эндоплазматическую сеть. Необходимый транспортный сигнал для данной стадии представлен сигнальной последовательность на N-конце молекулы так называемым сигнальным пептидом. Как только данный сигнальный пептид выполнил свою функцию, которая заключается во введении присоединенного к нему белка-предшественника в эндоплазматическую сеть, он протеолитически отщепляется от белка-предшественника. На основании его специфической функции последовательность данного типа сигнального пептида оставалась в высокой степени консервативной во время эволюции во всех живых клетках вне зависимости от того, являются ли они бактериями, дрожжами, грибами, животными или растениями.
Многие растительные белки, которые вводятся в эндоплазматическую сеть с помощью сигнального пептида, не находятся в ER, а транспортируются из эндоплазматической сети в аппарат Гольджи и продолжают движение из аппарата Гольджи в вакуоли. Одним из классов таких сортирующих сигналов для данного перемещения являются сигналы, которые находятся на С-концевой части белка-предшественника [Neuhas and Rogers, (1998) Plant Mol. Biol. 38: 127-144]. Предполагается, что белки, содержащие как N-концевой сигнальный пептид для введения в эндоплазматическую сеть, так и С-вакуолярный нацеливающий сигнал, содержат комплексные гликаны, которые присоединяются к ним в аппарате Гольджи [Lerouge et al., (1998) Plant Mol.Biol., 38:31-48]. Природа таких С-концевых сортирующих сигналов может варьироваться очень широко. В патенте США 6054637 описаны пептидные фрагменты, полученные из области основной хитиназы табака, которая представляет собой вакуолярный белок, которые действуют как вакуолярные нацеливающие пептиды. Примером вакуолярного белка, содержащего С-концевой нацеливающий сигнал и комплексные гликаны, является фазеолинхранящий белок семян бобов [Frigerio et al., (1998) Plant Cell, 10:1031-1042; Frigerio et al., (2001) Plant Cell 13:1109-1126].
Принципиально то, что во всех эукариотических клетках вакуолярные белки проходят через ER и аппарат Гольджи перед изолированием в вакуоле в качестве их конечного места назначения. Неожиданно трансформированные клетки корней растений по настоящему изобретению продуцировали GCD с необычно высоким содержанием маннозы. Было установлено, что преимущественно такой высокоманнозный продукт является биологически активным и, следовательно, нет необходимости в дополнительных стадиях для его активации. Не желая ограничиваться единственной гипотезой, будет очевидно, что применение ER-сигнала вместе с рекомбинантным белком, продуцируемым в культуре растительных клеток, способно преодолеть транспортирование через аппарат Гольджи и, следовательно, сохранять желаемое высокоманнозное гликозилирование. Необязательно любой тип механизма, который способен продуцировать высокоманнозное гликозилирование, включая любой тип механизма обхода аппарата Гольджи, можно использовать в соответствии с настоящим изобретением.
В первом аспекте настоящее изобретение относится к клетке-хозяину, продуцирующей высокоманнозный рекомбинантный белок, представляющий интерес. Данная клетка может быть трансформирована или трансфицирована молекулой рекомбинантной нуклеиновой кислоты, кодирующей представляющий интерес белок, или вектором экспрессии, включающим молекулу нуклеиновой кислоты. Такая молекула нуклеиновой кислоты включает первую последовательность нуклеиновой кислоты, кодирующую целевой белок, функционально связанную со второй последовательность нуклеиновой кислоты, кодирующей вакуолярный нацеливающий сигнальный пептид. Первая последовательность нуклеиновой кислоты может быть необязательно дополнительно функционально связана с третьей последовательностью нуклеиновой кислоты, кодирующей ER(эндоплазматическая цепь)-нацеливающий сигнальный пептид. Клетка-хозяин по изобретению отличается тем, что представляющий интерес белок продуцируется клеткой в высокоманнозилированной форме.
Клетка-хозяин по изобретению может представлять собой эукариотическую или прокариотическую клетку.
В одном варианте осуществления клетка-хозяин по изобретению представляет собой прокариотическую клетку, предпочтительно бактериальную клетку, наиболее предпочтительно клетку Agrobacterium tumefaciens. Данные клетки используются для инфицирования предпочтительных растительных клеток-хозяев, описанных далее.
В другом предпочтительном варианте осуществления клетка-хозяин по изобретению может представлять собой эукариотическую клетку, предпочтительно растительную клетку и наиболее предпочтительно клетку корня растений, выбранную из группы, включающей трансформированную Agrobacterium rihzogenes клетку корня, клетку сельдерея, клетку имбиря, клетку хрена и клетку моркови.
В предпочтительном варианте осуществления клетка корня растения представляет собой клетку моркови. Следует отметить, что трансформированные клетки моркови по изобретению выращивают в суспензии. Как отмечено выше и описано в примерах, данные клетки были трансформированы с использованием клеток Agrobacterium tumefaciens.
В другом варианте осуществления молекула рекомбинантной нуклеиновой кислоты, включенная в клетку-хозяина по изобретению, включает первую последовательность нуклеиновой кислоты, кодирующую лизосомальный фермент, которая функционально связана со второй последовательностью нуклеиновой кислоты, кодирующей вакуолярный нацеливающий сигнальный пептид, полученный из основного гена хитиназы А табака. Данный вакуолярный сигнальный пептид имеет последовательность аминокислот, представленную в SEQ ID NO:2. Первая последовательность нуклеиновой кислоты необязательно может быть дополнительно связана с третьей последовательностью нуклеиновой кислоты, кодирующей нацеливающий сигнальный пептид ER (эндоплазматической сети), обозначенный как SEQ ID NO:1. В одном варианте осуществления рекомбинантная молекула нуклеиновой кислоты, включенная в клетку-хозяина по изобретению, дополнительно включает промотор, который является функциональным в клетках растений. Данный промотор должен быть функционально связанным с рекомбинантной молекулой по изобретению.
В другом варианте осуществления данная рекомбинантная молекула нуклеиновой кислоты необязательно дополнительно может включать функционально связанный терминатор, который предпочтительно является функциональным в клетках растений. Рекомбинантная молекула нуклеиновой кислоты по изобретению необязательно может дополнительно включать дополнительные контрольные, промотирующие и регулирующие элементы и/или селектируемые маркеры. Следует отметить, что данные регуляторные элементы являются функционально связанными с рекомбинантной молекулой.
В предпочтительном варианте осуществления представляющий интерес высокоманнозный белок, продуцируемый клеткой-хозяином по изобретению, может представлять собой высокоманнозный гликопротеин, имеющий незащищенные концевые остатки маннозы.
Такой высокоманнозный белок может в соответствии с другим предпочтительным вариантом осуществления представлять собой лизосомальный фермент, выбранный из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. В предпочтительном варианте осуществления лизосомальный фермент может представлять собой глюкоцереброзидазу человека (GCD). Далее рекомбинантный GCD, rGCD, rhGCD все упоминаются как различные формы рекомбинантной GCD человека, если не указано другое.
Как описано ранее, болезнь Гоше, наиболее распространенная лизосомальная болезнь накопления, вызвана точечными мутациями гена (GBA) hGCD (глюкоцереброзидаза человека, что приводит к накоплению GlcCer в лизосомах макрофагов. Идентификация дефицита GCD в качестве первичной причины болезни Гоше привела к развитию ферментозаместительной терапии как терапевтической стратегии для данного заболевания. Однако гликозилирование играет решающую роль в активности hGCD и поглощении целевыми клетками.
Следовательно, в соответствии с другими предпочтительными вариантами настоящего изобретения предпочтительно разработан подходящим образом гликозилированный hGCD посредством контролирования экспрессии hGCD в клетках растительной культуры и более предпочтительно путем обеспечения сигнала ER и/или в других обстоятельствах посредством, необязательно и более предпочтительно, блокирования транспортировки в аппарат Гольджи.
Необязательно и предпочтительно hGCD имеет по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы для лечения или профилактики болезни Гоше.
В дальнейшем в конкретном варианте осуществления данную предпочтительную клетку-хозяина трансформируют или трансфектируют молекулой рекомбинантной нуклеиновой кислоты, которая дополнительно включает 35S промотор вируса мозаики цветной капусты, терминатор октопинсинтазы Agrobacterium tumefaciens и омега-трансляционный элемент-энхансер регуляторный элемент представляет собой TMV (вируса мозаики табака). В соответствии с предпочтительным вариантом осуществления данная рекомбинантная молекула нуклеиновой кислоты включает последовательность нуклеиновой кислоты по существу такую, как представлено в SEQ ID NO:13, и кодирует высокоманнозную GCD, имеющую последовательность аминокислот по существу такую, как представлено в SEQ ID NO: 14.
Следует понимать, что настоящее изобретение дополнительно обеспечивает вектор экспрессии, включающий молекулу нуклеиновой кислоты, кодирующую биологически активный лизосомальный фермент.
В одном предпочтительном варианте осуществления вектор экспрессии по изобретению включает молекулу нуклеиновой кислоты, кодирующую биологически активную высокоманнозную глюкоцереброзидазу человека (GCD). Предпочтительно предпочтительный вектор экспрессии включает нуклеиновую рекомбинантную молекулу нуклеиновой кислоты, которая имеет последовательность нуклеиновой кислоты по существу такую, как представлено в SEQ ID NO:13.
Во втором аспекте настоящее изобретение относится к рекомбинантному высокоманнозному белку, продуцируемому клеткой-хозяином по изобретению.
В предпочтительном варианте осуществления данный высокоманнозный белок может представлять собой биологически активный высокоманнозный лизосомальный фермент, выбранный из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. Наиболее предпочтительно данный лизосомальный фермент может представлять собой глюкоцереброзидазу человека (GCD).
В дальнейшем изобретение относится к рекомбинантному биологически активному высокоманнозному лизосомальному ферменту, имеющему по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы.
В соответствии с предпочтительным вариантом осуществления рекомбинантный лизосомальный фермент по изобретению может связываться с рецептором маннозы на клетке-мишени в сайте-мишени. Предпочтительно данный сайт может находиться в организме пациента, страдающего от лизосомальной болезни накопления.
Следует отметить, что рекомбинантный лизосомальный фермент обладает повышенной аффинностью по отношению к клетке-мишени по сравнению с соответствующей аффинностью природного лизосомального фермента по отношению к клетке-мишени. В конкретном варианте осуществления клетка-мишень с сайтом-мишенью может представлять собой клетку Купфера печени пациента.
В предпочтительном варианте осуществления рекомбинантный лизосомальный фермент может быть выбран из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы.
Наиболее предпочтительно рекомбинантный лизосомальный фермент представляет собой глюкоцереброзидазу (GCD).
В третьем аспекте изобретение относится к способу продуцирования высокоманнозного белка. Соответственно, способ изобретения включает стадии: (а) получение культуры рекомбинантных клеток-хозяев, трансформированных или трансфицированных рекомбинатными молекулами нуклеиновых кислот, кодирующими целевой рекомбинантный белок, или вектором экспрессии, включающим рекомбинантные молекулы нуклеиновых кислот; (b) культивирование данных культур клеток-хозяев, полученных на стадии (а) в условиях, допускающих экспрессию белка, где клетки-хозяева продуцируют данный белок в высокоманнозилированной форме; (с) выделение белка из клеток и сбор клеток из культуры, обеспеченных на стадии (а); и (d) очистку белка со стадии (с) с помощью подходящего способа очистки белков.
В соответствии с предпочтительным вариантом осуществления клетка-хозяин, используемая в данном способе, представляет собой клетку-хозяина по изобретению.
В другом предпочтительном варианте осуществления высокоманнозный белок, продуцируемый способом по изобретению, может представлять собой биологически активный высокоманнозный лизосомальный фермент, имеющий по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы.
Данный рекомбинантный фермент может связываться с рецептором маннозы на клетке-мишени в сайте-мишени. Более конкретно, рекомбинантный фермент, продуцируемый способом по изобретению, обладает повышенной аффинностью по отношению к клетке-мишени по сравнению с соответствующей аффинностью природного лизосомального фермента по отношению к клетке-мишени. Соответственно клетка-мишень в сайте-мишени может представлять собой клетку Купфера печени пациента.
В конкретном варианте осуществления лизосомальный фермент может быть выбран из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. Наиболее предпочтительно данный лизосомальный фермент может представлять собой глюкоцереброзидазу (GCD).
В другом предпочтительном варианте осуществления клетка-хозяин, используемая в способе по изобретению, может представлять собой клетку корней растений, выбранную из трансформированной Agrobacterium rihzogenes клетки корня, клетки сельдерея, клетки имбиря, клетки хрена и клетки моркови. Наиболее предпочтительно клетка корня растений представляет собой клетку моркови. Следует особо отметить, что в способе по изобретению трансформированные клетки-хозяева моркови выращивают в суспензии.
В следующем аспекте настоящее изобретение относится к способу лечения пациента, имеющего лизособную болезнь накопления с использованием экзогенного рекомбинантного лизосомального фермента, включающему: (а) получение биологически активной формы рекомбинантного лизосомального фермента, полученного очисткой из трансформированных клеток корней растений и способного эффективно направляться к клеткам с аномальным дефицитом лизосомального фермента. Данный рекомбинантный лизосомальный фермент имеет незащищенные концевые остатки маннозы на присоединенных олигосахаридах; и (b) введение пациенту терапевтически эффективного количества рекомбинантного биологически активного лизосомального фермента. В предпочтительном варианте осуществления рекомбинантный высокоманнозный фермент, используемый в способе по изобретению, может продуцироваться клетками-хозяевами по изобретению. Предпочтительно данная клетка-хозяин представляет собой клетку моркови.
В другом предпочтительном варианте осуществления лизосомальный фермент, используемый в способе по изобретению, может представлять собой высокоманнозный фермент, включающий по крайней мере одну олигосахаридную цепь, имеющую незащищенный остаток маннозы. Данный рекомбинантный фермент может связываться с рецептором маннозы на клетке-мишени в сайте-мишени в организме пациента. Более предпочтительно данный рекомбинантный лизосомальный фермент обладает повышенной аффинностью к данным клеткам-мишеням по сравнению с соответствующей аффинностью по отношению к клетке-мишени природного лизосомального фермента.
Более конкретно, лизосомальный фермент, используемый в способе по изобретению, может быть выбран из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. Предпочтительно данный лизосомальный фермент представляет собой глюкоцереброзидазу (GCD).
В соответствии с предпочтительным вариантом осуществления способ по изобретению, следовательно, предназначен для лечения лизосомальной болезни накопления, в частности болезни Гоше.
В таком случае клетка-мишень у сайта-мишени может представлять собой клетку Купфера печени пациента.
Изобретение, кроме того, относится к фармацевтической композиции для лечения лизосомальной болезни накопления, включающей в качестве активного ингредиента рекомбинантный биологически активный высокоманнозный лизосомальный фермент, как определено в данном изобретении. Композиция по изобретения необязательно может включать фармацевтически приемлемый разбавитель, носитель или эксципиент.
В конкретном варианте осуществления композиция по изобретению предназначена для лечения болезни Гоше. Такая композиция предпочтительно может включать в качестве эффективного ингредиента биологически активную высокоманнозую глюкоцереброзидазу человека (GCD), как определено в изобретении.
Изобретение дополнительно относится к применению рекомбинантного биологически активного высокоманнозного лизосомального фермента по изобретению для получения лекарственного средства для лечения или профилактики лизосомальной болезни накопления. Более конкретно, данное заболевание может представлять собой болезнь Гоше.
Соответственно данный биологически активный лизосомальный фермент представляет собой биологически активную высокоманнозую глюкоцереброзидазу человека (GCD), как определено в изобретении.
Согласно настоящему изобретению разработана клетка-хозяин, продуцирующая высокоманнозный рекомбинантный белок, включающая полинуклеотид, кодирующий рекомбинантный белок и сигнал, вызывающий продуцирование рекомбинантного белка в виде высокоманнозного белка. Предпочтительно полинуклеотид включает первую последовательность нуклеиновой кислоты, кодирующую целевой белок, функционально связанную со второй последовательностью нуклеиновой кислоты, кодирующей сигнальный пептид. Необязательно сигнальный пептид включает нацеливающий сигнальный пептид ER (эндоплазматической сети). Предпочтительно полинуклеотид дополнительно включает третью последовательность нуклеиновой кислоты для кодирования вакуолярного нацеливающего сигнального пептида.
Предпочтительно сигнал вызывает нацеливание рекомбинантного белка в ER. Более предпочтительно сигнал включает сигнальный пептид, вызывающий нацеливание рекомбинантного белка в ER. Наиболее предпочтительно полинуклеотид включает сегмент нуклеиновой кислоты для кодирования сигнального пептида.
Необязательно и предпочтительно сигнал вызывает обход рекомбинантным пептидом аппарата Гольджи. Предпочтительно сигнал включает сигнальный пептид, который вызывает то, что рекомбинантный белок не направляется в аппарат Гольджи. Более предпочтительно полинуклеотид включает сегмент нуклеиновой кислоты для кодирования сигнального пептида.
Необязательно и предпочтительно клетка-хозяин является любой эукариотической и прокариотической клеткой. Необязательно прокариотическая клетка представляет собой бактериальную клетку, предпочтительно клетку Agrobacterium tumefaciens. Предпочтительно эукариотическая клетка представляет собой растительную клетку. Более предпочтительно растительная клетка представляет собой клетку корня растений, выбранную из группы, состоящей из трансформированной Agrobacterium rihzogenes клетки корня, клетки сельдерея, клетки имбиря, клетки хрена и клетки моркови. Наиболее предпочтительно клетка корня растения представляет собой клетку моркови.
Предпочтительно рекомбинантный полинуклеотид включает первую последовательность нуклеиновых кислот, кодирующую целевой белок, которая функционально связана со второй последовательностью нуклеиновой кислоты, кодирующей вакуолярный сигнальный пептид, полученный из основного гена хитиназы А табака, при этом вакуолярный сигнальный пептид имеет последовательность аминокислот, обозначенную как SEQ ID NO: 2, где первая последовательность нуклеиновой кислоты необязательно дополнительно функционально связана с третьей последовательность нуклеиновой кислоты, кодирующий нацеливающий сигнальный пептид ER (эндоплазматической сети), обозначенный как SEQ ID NO: 1.
Более предпочтительно рекомбинантный полинуклеотид дополнительно включает промотор, который является функциональным в растительных клетках, где промотор функционально связан с рекомбинантной молекулой.
Наиболее предпочтительно рекомбинантный полинуклеотид дополнительно включает терминатор, который является функциональным в растительных клетках, где терминатор функционально связан с рекомбинантной молекулой.
Также наиболее предпочтительно рекомбинантный полинуклеотид дополнительно включает дополнительные контрольные, промотирующие и регулирующие элементы и/или селектируемые маркеры, где регулирующие элементы функционально связаны с рекомбинантной молекулой.
Предпочтительно высокоманнозный белок представляет собой высокоманнозный гликопротеин, гликозилированный по крайней мере одним незащищенным остатком маннозы. Более предпочтительно высокоманнозный белок представляет собой биологически активный высокоманнозный лизосомальный фермент, выбранный из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы.
Наиболее предпочтительно данный лизосомальный фермент представляет собой глюкоцереброзидазу человека (GCD).
Предпочтительно GCD включает последовательность аминокислот по существу такую, как последовательность, обозначенная как SEQ ID NO: 8, кодируемую последовательностью нуклеиновой кислоты, обозначенной как SEQ ID NO: 7.
Более предпочтительно клетку трансформируют или трансфицируют рекомбинантный полинуклеотидом или вектором экспрессии, включающим молекулу, при этом рекомбинантный полинуклеотид дополнительно включает 35S промотор вируса мозаики цветной капусты, терминатор октопинсинтазы Agrobacterium tumefaciens и регуляторный элемент представляет собой омега-трансляционный элемент-энхансер TMV (вируса мозаики табака), и имеет последовательность нуклеиновой кислоты по существу такую, как последовательность, обозначенная SEQ ID NO:13, кодирующая GCD, имеющую последовательность аминокислот по существу такую, как представлено в SEQ ID NO: 14.
Согласно предпочтительным вариантам осуществления разработан рекомбинантный высокоманнозный белок, продуцируемый описанной выше клеткой-хозяином.
Предпочтительно высокоманнозный белок представляет собой биологически активный высокоманнозный лизосомальный фермент, выбранный из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы.
Более предпочтительно лизосомальный фермент представляет собой глюкоцереброзидазу человека (GCD).
В соответствии с другими предпочтительными вариантами осуществления настоящего изобретения разработан рекомбинантный биологически активный высокоманнозный фермент, имеющий по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы.
В соответствии со следующими предпочтительными вариантами осуществления разработан рекомбинантный белок, включающий первую часть, обладающую активностью сигнального пептида, и вторую часть, обладающую активностью лизосомального фермента, при этом первая часть вызывает переработку второй части в растительной клетке так, чтобы она включала по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы.
Предпочтительно лизосомальный фермент включает белок для лечения или профилактики болезни Гоше.
Более предпочтительно белок включает hGCD.
Предпочтительно первая часть включает нацеливающий сигнальный пептид ER растительной клетки. Более предпочтительно рекомбинантный фермент может связываться с рецептором маннозы на клетки-мишени в сайте-мишени в организме пациента, который страдает от лизосомальной болезни накопления. Наиболее предпочтительно рекомбинантный лизосомальный фермент обладает повышенной аффинностью по отношению к клетке-мишени по сравнению с соответствующей аффинностью природного лизосомального фермента по отношению к клетке-мишени.
Также наиболее предпочтительно рекомбинантный лизосомальный фермент выбирают из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы.
Предпочтительно рекомбинантный лизосомальный фермент представляет собой глюкоцереброзидазу (GCD).
Также предпочтительно клетка-мишень при сайте-мишени представляет собой клетку Купфера печени пациента.
В соответствии с другими предпочтительными вариантами разработан рекомбинантный высокоманнозный белок, продуцируемый культурой растительных клеток. Предпочтительно отличительной особенностью белка является наличие растительного сигнального пептида для нацеливания белка в эндоплазматическую сеть.
Более предпочтительно растительный сигнальный пептид включает пептид для нацеливания белка в эндоплазматическую сеть в культуре клеток корня растений. Наиболее предпочтительно культура клеток корней растения включает клетки моркови.
Еще в соответствии с другими предпочтительными вариантами осуществления разработан рекомбинантный высокоманнозый белок hGCD, продуцируемый в культуре растительных клеток.
В соответствии со следующими предпочтительными вариантами осуществления разработано применение культуры растительных клеток для продуцирования высокоманнозного белка.
В соответствии с другими предпочтительными вариантами осуществления разработан способ продуцирования высокоманнозного белка, включающий: получение культуры рекомбинантных клеток-хозяев, трансформированных или трансфицированных рекомбинантным полинуклеотидом, кодирующим рекомбинантный белок; культивирование культуры клеток-хозяев в условиях, дающих возможность экспрессии белка, где клетки-хозяева продуцируют белок в высокоманнозной форме.
Предпочтительно культуру клеток-хозяев культивируют в суспензии. Наиболее предпочтительно способ дополнительно включает очистку белка.
В соответствии с другими предпочтительными вариантами осуществления способ осуществляют с использованием клеток-хозяев, как описано ранее. Предпочтительно высокоманнозный белок представляет собой биологически активный высокоманнозный лизосомальный фермент, имеющий по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы. Более предпочтительно рекомбинантный фермент связывается с рецептором маннозы на клетки-мишени в сайте-мишени. Наиболее предпочтительно рекомбинантный фермент обладает повышенной аффинностью по отношению к клетке-мишени по сравнению с соответствующей аффинностью природного лизосомального фермента по отношению к клетке-мишени.
Предпочтительно лизосомальный фермент выбирают из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы.
Более предпочтительно лизосомальный фермент представляет собой глюкоцереброзидазу (GCD). Наиболее предпочтительно клетка-мишень при сайте-мишени представляет собой клетку Купфера печени пациента.
Предпочтительно клетка-хозяин представляет собой клетку корней растений, выбранную из группы, состоящей из трансформированной Agrobacterium rihzogenes клетки корня, клетки сельдерея, клетки имбиря, клетки хрена и клетки моркови.
Более предпочтительно клетка корня растений представляет собой клетку моркови.
Наиболее предпочтительно трансформированные клетки-хозяева моркови выращивают в суспензии.
В соответствии со следующими предпочтительными вариантами осуществления разработан способ лечения пациента, имеющего лизосомальную болезнь накопления, с использованием экзогенного рекомбинантного лизосомального фермента, включающий: получение рекомбинантной биологически активной формы лизосомального фермента, полученного очисткой из трансформированных клеток корня растений и способного эффективно поступать в клетки с аномальным дефицитом лизосомального фермента, где рекомбинантный биологически активный фермент имеет незащищенный концевой остаток маннозы на присоединенных олигосахаридах; и введение пациенту терапевтически эффективного количества рекомбинантного биологически активного лизосомального фермента. Данный способ необязательно может быть осуществлен с использованием любой клетки-хозяина и/или белка, как описано выше.
Предпочтительно рекомбинантный фермент может связываться с рецептором маннозы на клетке-мишени в сайте-мишени в организме пациента. Более предпочтительно рекомбинантный лизосомальный фермент обладает повышенной аффинностью по отношению к клетке-мишени по сравнению с соответствующей аффинностью природного лизосомального фермента по отношению к клетке-мишени. Наиболее предпочтительно лизосомальный фермент выбирают из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. Также наиболее предпочтительно лизосомальный фермент представляет собой глюкоцереброзидазу (GCD).
Также наиболее предпочтительно лизосомальная болезнь накопления представляет собой болезнь Гоше. Также наиболее предпочтительно клетка-мишень при сайте-мишени представляет собой клетку Купфера печени пациента.
В соответствии с другими предпочтительными вариантами осуществления разработана фармацевтическая композиция для лечения лизосомальной болезни накопления, включающая в качестве активного ингредиента рекомбинантный биологически активный высокоманнозный лизосомальный фермент, как описано выше, данная композиция необязательно дополнительно включает фармацевтически приемлемый разбавитель, носитель или эксципиент. Предпочтительно лизосомальная болезнь накопления представляет собой болезнь Гоше. Более предпочтительно рекомбинантный лизосомальный фермент представляет собой биологически активную высокоманнозную глюкоцереброзидазу человека (GCD).
В соответствии со следующими предпочтительными вариантами осуществления предложено применение рекомбинантного биологически активного высокоманнозного лизосомального фермента, как описано выше, для получения лекарственного средства для лечения или профилактики лизосомальной болезни накопления. Предпочтительно болезнь представляет собой болезнь Гоше. Более предпочтительно биологически активный лизосомальный фермент представляет собой биологически активную высокоманнозную глюкоцереброзидазу человека (GCD).
Изобретение будет дополнительно описано с использованием следующих фигур, которые являются только иллюстративными и не ограничивают объем изобретения, который также определен прилагаемой формулой изобретения.
Краткое описание чертежей
Изобретение описано только в качестве примера со ссылкой на прилагаемые чертежи.
На фиг.1А показана полученная кассета экспрессии, включающая 35S промотор вируса мозаики цветной капусты, омега-трансляционный элемент-энхансер TMV (вируса мозаики табака), ER нацеливающий сигнал, последовательность GCD человека (также обозначенную SEQ ID NO: 7), вакуолярный сигнал и терминаторную последовательность октопинсинтазы из Agrobacterium tumefaciens.
На фиг.1В показана схематическая карта скелета плазмиды pGreenII.
На фиг.2 показан вестерн-блот анализ экстракта трансформированных hGCD клеток с использованием специфических против hGCD антител. В качестве положительного контроля использовали стандарт Церезим (полоса 1), нетрансформированный каллюс использовали в качестве отрицательного контроля (полоса 2), различные выбранные экстракты каллюса показаны на полосах 3-8.
На фиг.3А-3С показана первая стадия очистки rhGCD на сильной катионообменной смоле (Macro-Prep high-S носитель, Bio-Rad), набитой в колонку ХК (2,6×20 см). Колонка была объединена с АКТА основной системой (Amersham Pharmacia Biotech), что позволяет проводить мониторинг проводимости, рН и поглощения при 280 нм. Элюирование rh-GCD проводили уравновешивающим буфером, содержащим 600 мМ NaCl. На фиг.3А представлен стандартный эксперимент при проведении данной стадии очистки. Фракции, собранные во время прохождения эксперимента, контролируют с помощью анализа ферментативной активности, как показано на фиг.3В, и объединяют пробирки, содержимое которых проявляет ферментативную активность (в пиках элюирования). На фиг.3С показано окрашивание кумассином синим элюированных фракций, проанализированных на активность.
На фиг. 3D-3F показаны соответствующие графики, как на фиг.3А-3С, но для второй колонки.
На фиг.4А-С показана конечная стадия очистки рекомбинантного hGCD на смоле с гидрофобным взаимодействием (гель TSK, Toypearl Phenyl-650C, Tosoh Corp.), набитой в ХК колонку (2,6×20 см). Колонка была объединена с АКТА основной системой (Amersham Pharmacia Biotech), что позволяет проводить мониторинг, рН и поглощения при 280 нм. Объединенную фракцию GCD из предшествующей колонки загружали при скорости 6 мл/мин с последующей промывкой уравновешивающим буфером до тех пор, пока поглощение в УФ не достигало базисной линии. Чистую GCD элюировали 10мМ цитратным буфером, содержащим 50% этанола.
На фиг.4А показан стандартный эксперимент данной стадии очистки.
На фиг.4В показаны фракции, собранные во время эксперимента, которые контролировали с помощью анализа ферментативной активности.
На фиг.4С показано окрашивание кумассином синим элюированных фракций, проанализированных на активность.
На фиг.5 показана активность рекомбинантного hGCD после поглощения перитонеальными макрофагами (фиг.5А-5С), тогда как на фиг.5D показан вестерн-блот анализ рекомбинантной GCD согласно настоящему изобретению.
На фиг.6 показаны сравнительные гликозилированные структуры rGCD в соответствии с настоящим изобретением.
На фиг.8 показаны дополнительные N-гликан-гликозилированные структуры rGCD в соответствии с настоящим изобретением.
Подробное описание изобретения
Белки для фармацевтического применения традиционно продуцируют в системах экспрессии млекопитающих и бактерий. В последние несколько лет была выявлена обещающая новая система экспрессии в растениях. Вследствие относительной простоты введения новых генов и потенциала для массового производства белков и пептидов «молекулярная фармацевтика» становится все более популярной в качестве системы экспрессии белков.
Одним из основных отличий между относящейся к млекопитающим и растительной системами экспрессии белка является изменение в последовательностях гликозилирования белка, вызванное различием биосинтетических путей. Показано, что гликозилирование оказывает огромное влияние на активность, складчатость, стабильность, растворимость, восприимчивость к протеазам, скорость выведения из крови и антигенный потенциал белков. Следовательно, при любом продуцировании белка в растениях следует принимать во внимание потенциальные разветвления растительного гликозилирования.
Углеводный фрагмент представляет собой одну из наиболее обычных посттрансляционных модификаций белков. Гликозилирование белков подразделяют на две категории: N-связанное и О-связанное. Эти два типа отличаются аминокислотой, к которой присоединен гликановый фрагмент в белке: N-связанный присоединен к остаткам Asn, тогда как О-связанный присоединен к остаткам Ser или Thr. Кроме того, последовательность гликанов каждого типа содержит уникальные отличительные особенности. Среди этих двух типов N-связанное гликозилирование является более частым, и его влияние на белки было широко исследовано. О-связанные гликаны, с другой стороны, являются относительно редкими, и относительно их влияния на белки доступно меньшее количество информации. Большинство доступных данных по гликозилированию белка в растениях сфокусировано на N-связанных, а не на О-связанных гликанах.
В настоящем изобретении описана растительная система экспрессии, основанная на клетках трансгенных растений, которые предпочтительно представляют собой клетки корней, необязательно и предпочтительно выращенные в суспензии. Такая система экспрессии специально разработана для эффективного продуцирования высокоманнозного белка, представляющего интерес. Термин «высокоманнозный» включает гликозилирование, имеющее по крайней мере один незащищенный остаток маннозы.
Таким образом, в первом аспекте настоящее изобретение относится к клетке-хозяину, продуцирующей высокоманнозный рекомбинантный белок, представляющий интерес. Предпочтительно рекомбинантный белок характеризуется сигнальным пептидом ER (эндоплазматической сети), более предпочтительно ER-направляющим сигнальным пептидом. Альтернативно или дополнительно, рекомбинантный белок характеризуется сигналом, который вызывает обход белком аппарата Гольджи. Сигнал предпочтительно дает возможность получения рекомбинантного белка, который отличается высокоманнозным гликозилированием, более предпочтительно путем сохранения такого гликозилирования и наиболее предпочтительно путем направления в ER и/или путем обхода аппарата Гольджи. Как описано здесь более подробно, такой сигнал предпочтительно осуществляет выполнение как сигнальный пептид, который более предпочтительно образует часть последовательности белка, необязательно и более предпочтительно путем конструирования белка, отличительной особенностью которого является сигнальный пептид как часть белка. Следует отметить, что сигнал необязательно может представлять собой нацеливающий сигнал, сигнал сохранения, сигнал уклонения (обхода) или любую их комбинацию или любой другой тип сигнала, способного обеспечивать желаемую структуру с высокоманнозным гликозилированием.
Не желая ограничиваться единственной гипотезой, следует понимать, что применение ER-направляющего сигнала вместе с рекомбинантным белком, продуцируемым в культуре растительных клеток, способно преодолеть транспортировку в аппарат Гольджи и, следовательно, сохранить желаемое высокоманнозное гликозилирование. Необязательно любой тип механизма, который способен продуцировать высокоманнозное гликозилирование, включая любой тип механизма обхода аппарата Гольджи, можно использовать в соответствии с настоящим изобретением. ER-нацеливающие сигнальные пептиды хорошо известны в данной области, и они представляют собой N-концевые сигнальные пептиды. Необязательно любой подходящий ER-нацеливающий сигнальный пептид можно использовать в соответствии с настоящим изобретением.
Клетка-хозяин в соответствии с настоящим изобретением необязательно может быть трансформирована или трансфицирована (постоянно и/или временно) молекулой рекомбинантной нуклеиновой кислоты, кодирующей представляющий интерес белок, или вектором экспрессии, включающим молекулу нуклеиновой кислоты. Такая молекула нуклеиновой кислоты включает первую последовательность нуклеиновой кислоты, кодирующую целевой белок, необязательно и предпочтительно функционально связанную со второй последовательностью нуклеиновой кислоты, кодирующей вакуолярный направляющий сигнальный пептид. Следует отметить, что, как он использован в данном описании, термин «функционально» связанный не относится в обязательном порядке к физической связи. Первая последовательность нуклеиновой кислоты может быть необязательно и предпочтительно дополнительно быть функционально связанной с третьей последовательностью нуклеиновой кислоты, кодирующей ER(эндоплазматическая сеть)-нацеливающий сигнальный пептид. Клетка-хозяин по изобретению отличается тем, что представляющий интерес белок продуцируется клеткой в форме, которая включает по крайней мере одни незащищенный остаток маннозы, но предпочтительно в высокоманнозилированной форме.
Термины «клетки», «клетки-хозяева» или «рекомбинантные клетки-хозяева» используются в данном описании взаимозаменяемо. Понятно, что такие термины относятся не только к конкретным рассматриваемым клеткам, но и к потомству или потенциальному потомству таких клеток. Поскольку определенные модификации могут происходить в последующих поколениях либо за счет мутации, или под влиянием окружающей среды, такое потомство может фактически не быть идентичным исходной клетке, но все еще включено в объем термина, как он используется в данном описании. Термин «клетка-хозяин», как он используется в данном описании, относится к клеткам, которые могут быть рекомбинантно трансформированы изолированной ДНК или вектором экспрессии, сконструированным с использованием рекомбинантных ДНК технологий. Как использовано в данном описании, термин «трансфекция» означает введение нуклеиновой кислоты, например изолированной ДНК или вектора экспрессии, в клетки-реципиенты посредством опосредованного нуклеиновой кислотой переноса гена. Термин «трансформация», как он использован в данном описании, относится к процессу, в котором генотип клетки изменяется в результате клеточного поглощения экзогенной ДНК или РНК, и, например, трансформированная клетка экспрессирует рекомбинантную форму желаемого белка.
Следует понимать, что лекарственно-устойчивый или другой селектируемый маркер предназначен частично для облегчения селекции трансформантов. Кроме того, присутствие селектируемого маркера, например маркера устойчивости в отношении лекарственных препаратов, может использоваться для предотвращения размножения загрязняющих микроорганизмов в культуральной среде. Такая чистая культура трансформированных клеток-хозяев была бы получена путем культивирования клеток в условиях, которые необходимы для выживания индуцированного фенотипа.
Как указано выше, клетки-хозяева по изобретению могут быть трансфицированы или трансформированы молекулой нуклеиновой кислоты. Как использовано в данном описании, термин «нуклеиновая кислота» относится к полинуклеотидам, таким как дезоксирибонуклеиновая кислота (ДНК) и, где это является подходящим, рибонуклеиновая кислота (РНК). Следует также понимать, что термины включают в качестве эквивалентов аналоги либо РНК, либо ДНК, полученные из аналогов нуклеотидов и, как применимо к описываемому варианту осуществления, полинуклеотиды с единичной цепью (такой, как смысловая или антисмысловая) и с двойной цепью.
Еще в другом варианте осуществления клетка-хозяин по изобретению может быть трансфицирована или трансформирована вектором экспрессии, включающим рекомбинантную молекулу нуклеиновой кислоты. Термин «векторы экспрессии», как он использован в данном описании, охватывает векторы, такие как плазмиды, вирусы, бактериофаги, интегрируемые фрагменты ДНК и другие переносчики, которые способны интегрировать фрагменты ДНК в геном хозяина. Векторы экспрессии обычно представляют собой самореплицирующие конструкции ДНК или РНК, содержащие желаемый ген или его фрагменты, и функционально связанные генные контрольные элементы, которые распознаются в подходящей клетке-хозяине и выполняют экспрессию желаемых генов. Данные контрольные элементы способы выполнять экспрессию в подходящем хозяине. Обычно генные контрольные элементы могут включать систему прокариотического промотора и контрольную систему экспрессии эукариотического промотора. Такие системы обычно включают промотор транскрипции, необязательный оператор для контроля начала транскрипции, энхансеры транскрипции для повышения уровня экспрессии РНК, последовательность, которая кодирует подходящий сайт связывания рибосома, экзон-интронные сочленения РНК, последовательности, которые заканчивают транскрипцию и трансляцию и так далее. Векторы экспрессии обычно содержат источник репликации, который дает возможность вектору реплицировать независимо от клетки-хозяина.
Плазмиды представляют собой наиболее распространенную форму вектора, но другие формы векторов, которые выполняют эквивалентную функцию и которые являются или станут известными в данной области, подходят для применения в данном изобретении. См., например, Pouwels et al., Cloning Vectors: a Laboratory Manual (1985 и приложения), Elsevier N.Y.; и Rodriquez et al (eds.), Vectors: a Survey of Molecular Cloning Vectors and Their Uses, Buttersworth, Boston, Mass (1988), которые включены в настоящее описание путем ссылки.
Обычно такие векторы содержат, кроме того, специфические гены, которые способны обеспечить фенотипическую селекцию в трансформированных клетках. Также предполагается применение прокариотическиих и эукариотических вирусных векторов экспрессии для экспрессии генов, кодирующих полипептиды по настоящему изобретению.
Необязательно вектор может представлять собой общий вектор растения (как описано в приведенных ниже примерах). Альтернативно, вектор необязательно может быть специфическим для клеток корней.
В одном предпочтительном варианте осуществления клетка-хозяин по изобретению может представлять собой эукариотическую или прокариотическую клетку.
В конкретном варианте осуществления клетка-хозяин по изобретению представляет собой прокариотическую клетку, предпочтительно бактериальную клетку, наиболее предпочтительно клетку Agrobacterium tumefaciens. Данные клетки используются для инфицирования предпочтительных растительных клеток-хозяев, описанных далее.
В другом предпочтительном варианте осуществления клетка-хозяин по изобретению может представлять собой эукариотическую клетку, предпочтительно растительную клетку и наиболее предпочтительно клетку корня растений, выбранную из группы, включающей трансформированную Agrobacterium rihzogenes клетку корня, клетку сельдерея, клетку имбиря, клетку хрена и клетку моркови.
В предпочтительном варианте осуществления клетка корня растения представляет собой клетку моркови. Следует отметить, что трансформированные клетки моркови по изобретению выращивают в суспензии. Как отмечено выше и описано в примерах, данные клетки были трансформированы с использованием клеток Agrobacterium tumefaciens по изобретению.
Векторы экспрессии или молекулы рекомбинантных нуклеиновых кислот, используемые для транфекции или трансформации клеток-хозяев по изобретению, могут быть дополнительно модифицированы в соответствии со способами, известными специалистам в данной области, для добавления, удаления или модификации каким-либо другим образом последовательностей сигнальных пептидов для изменения расщепления сигнального пептида или для увеличения или изменения целеуказания экспрессируемого лизосомального фермента через эндомембранную систему растения. Например, но не в качестве ограничения, конструкция экспрессии может быть специально разработана для целеуказания лизосомального фермента для секреции, или вакуолярной локализации, или сохранения в эндоплазматической сети (ER).
В одном варианте осуществления вектор экспрессии или молекула рекомбинантной нуклеиновой кислоты могут быть сконструированы для введения последовательности нуклеотидов, которая кодирует сигнал, направляющий лизосомальный фермент в вакуоль растения. Например, но не в качестве ограничения, молекула рекомбинантной нуклеиновой кислоты, заключающаяся в клетке-хозяине по изобретению, включает первую последовательность нуклеиновой кислоты, кодирующую лизосомальный фермент, которая функционально связана со второй последовательностью нуклеиновой кислоты, кодирующей вакуолярный нацеливающий сигнальный пептид, полученной из основного гена хитиназы А табака. Данный вакуолярный сигнальный пептид имеет последовательность аминокислот, обозначенную как SEQ ID NO: 2. Первая последовательность нуклеиновой кислоты необязательно может быть дополнительно связана с помощью функциональной связи с третьей последовательностью нуклеиновой кислоты, кодирующей ER (эндоплазматическая сеть)-нацеливающий сигнальный пептид, обозначенный SEQ ID NO: 1. В одном варианте осуществления молекула рекомбинантной нуклеиновой кислоты, заключающаяся в клетке-хозяине по изобретению, дополнительно включает промотор, который является функциональным в растительных клетках. Такой промотор должен быть функционально связан с рекомбинантной молекулой по изобретению.
Термин «функционально связан» используется в данном описании для указания того, что первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты помещена в функциональную взаимосвязь со второй последовательностью нуклеиновой кислоты. Например, промотор является функционально связанным с кодирующей последовательностью, если промотор оказывает влияние на транскрипцию или экспрессию кодирующей последовательности. Необязательно и предпочтительно функционально связанные последовательности ДНК являются прилегающими (например, физически связаны) и, когда необходимо объединить две протеин-кодирующие области, находятся в одной и той же рамке считывания. Таким образом, последовательность ДНК и регуляторная(ые) последовательность(и) связаны таким образом, чтобы допускать экспрессию гена, когда подходящие молекулы (например, транскрипционные активирующие белки) связаны с регуляторной(ыми) последовательностью(ями).
В другом варианте осуществления такая молекула рекомбинантной нуклеиновой кислоты необязательно может дополнительно включать функционально связанный терминатор, который предпочтительно является функциональным в растительных клетках. Рекомбинантная молекула нуклеиновой кислоты по изобретению необязательно может дополнительно включать дополнительные контрольные, промотирующие и регуляторные элементы и/или селектируемые маркеры. Следует отметить, что данные регуляторные элементы являются функционально связанными с рекомбинантной молекулой.
Регуляторные элементы, которые можно использовать в конструкциях экспрессии, включают промоторы, которые могут быть или гетерологичными, или гомологичными по отношению к клеткам растений. Промотор может представлять собой растительный промотор или не растительный промотор, который способен вызывать высокие уровни транскрипции связанной последовательности в растительных клетках и растениях. Неограничивающие примеры растительных промоторов, которые можно эффективно использовать при практической реализации изобретения, включают вирус мозаики цветной капусты (CaMV) 35S, rbcS, промотор для хлорофилл a/b связывающего белка, AdhI, NOS и HMG2, или их модификации или производные. Промотор может быть или конструктивным, или индуцируемым. Например, но не в качестве ограничения, индуцируемый промотор может представлять собой промотор, который стимулирует или повышает экспрессию нуклеотидной последовательности лизосомального фермента после механической активации гена (MGA) растения, ткани растения или клетки растения.
Векторы экспрессии, используемые для трансфицирования или трансформирования клеток-хозяев по изобретению, могут быть модифицированы дополнительно с использованием способов, известных специалистам в данной области, для усиления или оптимизации экспрессии гетерологичного гена в растениях и растительных клетках. Такие модификации включают, но не ограничены указанным, мутирование регуляторных элементов ДНК для увеличения эффективности промотора или для изменения целевого белка.
В предпочтительном варианте осуществления целевой высокоманнозный белок, продуцируемый клетками-хозяевами по изобретению, может представлять собой высокоманнозный гликопротеин, имеющий по крайней мере один незащищенный остаток маннозы (по крайней мере один концевой остаток маннозы).
Такой высокоманнозный белок может представлять собой в соответствии с другим предпочтительным вариантом осуществления лизосомальный фермент, выбранный из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы.
Термин «лизосомальный фермент», как он использован в данном описании по отношению к любому такому ферменту и продукту, продуцируемому в описанной в изобретении растительной систем экспрессии, относится к рекомбинантному пептиду, экспрессируемому в трансгенной растительной клетке из нуклеотидной последовательности, кодирующей лизосомальный фермент человека или животного, модифицированный лизосомальный фермент человека или животного, или фрагмент, производное, или модификацию такого фермента. Полезные модифицированные лизосомальные ферменты человека или животного включают, но не ограничиваются указанным, лизосомальные ферменты человека или животного, содержащие добавление, удаление (делецию) и/или замещение одной или нескольких природных или искусственно введенных аминокислот.
Растворимые лизосомальные ферменты имеют такие первоначальные стадии биосинтеза, что и секреторные белки, т.е. синтез на рибосоме, связывание N-концевого сигнального пептида с поверхностью неровности эндоплазматической сети (ER), транспорт в просвет ER, где отщепляется сигнальный пептид, и присоединение олигосахаридов к специфическим аспарагиновым остаткам (N-связывание) с последующей дальнейшей модификацией образующегося белка в аппарате Гольджи [von Figura and Hasilik, Annu. Rev. Biochem., 55:167-193 (1986)]. N-связанные олигосахариды могут быть комплексными, различными и гетерогенными и могут содержать высокоманнозные остатки. Белки подвергаются дальнейшей переработке в пост-ER, предшествующем аппарату Гольджи отделе и в цис-аппарате Гольджи, образуя или N-связанный манноза-6-фосфат (М-6-Р) олигосахарид-зависимый или N-связанный (М-6-Р) олигосахарид-независимый сигнал распознавания для расположенных в лизосомах ферментов [Kornfeld & Mellman, Ann. Rev.Cell.Biol., 5:483-525 (1989); Kaplan et al., natl.Acad.Sci. USA, 74:2026 (1977)]. Присутствие сигнала распознавания М-6-Р приводит к связыванию фермента с рецепторами М-6-Р (MPR). Такие связанные ферменты, оставшиеся в клетке, в конце концов, упаковываются в лизосомах и таким образом отделяются от белков, нацеленных для секреции или к плазматической мембране.
В предпочтительном варианте осуществления лизосомальный фермент может представлять собой глюкоцереброзидазу человека (GCD).
В последующем в конкретном варианте осуществления данную предпочтительную клетку-хозяина трансформируют или трансфицируют рекомбинантной молекулой нуклеиновой кислоты, которая дополнительно включает 35S промотор вируса мозаики цветной капусты, предпочтительно имеющий последовательность нуклеиновой кислоты, обозначенную как SEQ ID NO: 9, терминатор октопинсинтазы Agrobacterium tumefaciens, предпочтительно имеющий последовательность нуклеиновой кислоты, обозначенную как SEQ ID NO: 12, и омега-трансляционный элемент-энхансер TMV (вируса мозаики табака). В соответствии с предпочтительным вариантом осуществления данная рекомбинантная молекула нуклеиновой кислоты включает последовательность нуклеиновой кислоты по существу такую, которая обозначена SEQ ID NO:13, и кодирует высокоманнозную GCD, имеющую последовательность аминокислот по существу такую, как представлено в SEQ ID NO: 14.
Следует понимать, что настоящее изобретение дополнительно относится к вектору экспрессии, включающему молекулу нуклеиновой кислоты, кодирующую биологически активный высокоманнозный лизосомальный фермент.
В одном предпочтительном варианте осуществления данного аспекта вектор экспрессии по изобретению включает молекулу нуклеиновой кислоты, кодирующую биологически активную высокоманнозную глюкоцереброзидазу человека (GCD). Предпочтительно данный предпочтительный вектор экспрессии включает рекомбинантную молекулу нуклеиновой кислоты, которая имеет последовательность нуклеиновой кислоты по существу такую, которая обозначена SEQ ID NO: 13. В соответствии с конкретным вариантом осуществления в предпочтительном векторе экспрессии используется плазмида pGREEN II, как описано далее в примере 1.
Далее следует отметить, что изобретение относится к кассете экспрессии, включающей в себя описанный выше вектор экспрессии.
Во втором аспекте настоящее изобретение относится к рекомбинантному высокоманнозному белку, продуцируемому клеткой-хозяином по изобретению.
В предпочтительном варианте осуществления такой высокоманнозный белок может представлять собой биологически активный высокоманнозный лизосомальный фермент, выбранный из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. Наиболее предпочтительно данный лизосомальный фермент может представлять собой глюкоцереброзидазу человека (GCD).
Термин «биологически активный» используется в данном описании в отношении любого рекомбинантного лизосомального фермента, продуцируемого в растительной системе экспрессии, для обозначения того, что данный рекомбинантный лизосомальный фермент способен гидролизовать либо природный субстрат, или аналог, или синтетический субстрат соответствующего лизосомального фермента человека или животного на обнаруживаемом уровне.
Далее изобретение относится к рекомбинантному биологически активному высокоманнозному лизосомальному ферменту, имеющему по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы.
В соответствии с предпочтительным вариантом осуществления рекомбинантный лизосомальный фермент по изобретению может связываться с рецептором маннозы на клетке-мишени в сайте-мишени. Предпочтительно данный сайт может находиться в организме пациента, страдающего от лизосомальной болезни накопления.
Необязательно и предпочтительно данный рекомбинантный лизосомальный фермент обладает повышенной аффинностью по отношению к клетке-мишени по сравнению с соответствующей аффинностью природного лизосомального фермента по отношению к клетке-мишени. В конкретном варианте осуществления клетка-мишень с сайтом-мишенью может представлять собой клетку Купфера печени пациента.
В предпочтительном варианте осуществления рекомбинантный лизосомальный фермент может быть выбран из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы.
Наиболее предпочтительно данный лизосомальный фермент представляет собой глюкоцереброзидазу (GCD).
В третьем аспекте изобретение относится к способу продуцирования высокоманнозного белка. Соответственно, способ по изобретению включает следующие стадии: (a) получение культуры рекомбинантных клеток-хозяев, трансформированных или трансфицированных рекомбинантными молекулами нуклеиновых кислот, кодирующими представляющий интерес рекомбинантный белок, или вектором экспрессии, включающим рекомбинантные молекулы нуклеиновых кислот; (b) культивирование указанной культуры клетки-хозяина, полученной на стадии (а) в суспензии в условиях, допускающих экспрессию высокоманнозного белка, где указанные клетки-хозяева продуцируют белок в высокоманнозилированной форме; (с) сбор клеток культуры, обеспеченной на стадии (а), и выделение белка из клеток; и (d) очистка белка со стадии (с) с использованием подходящего способа очистки белка.
Необязательно и предпочтительно рекомбинантный белок может продуцироваться растительными клетками по изобретению при культивировании в устройстве, описанном в патенте США 6391638, выданном 21 мая 2002 г. и включенном в данное описание путем ссылки, как если бы оно было полностью изложено в данном описании. Что касается условий культивирования клеток растений в суспензии с использованием данного устройства, то они описаны в патентной заявке США, озаглавленной «Устройство, система и способ культивирования клеток/тканей» (“Cell/tissue culturing device, system and method”), автором которой является один из авторов настоящего изобретения и признанной одновременно вместе с настоящей заявкой, которая включена в настоящее описание путем ссылки, как если бы она была полностью изложена здесь, и которая была подана в тот же день, что и настоящая заявка.
Конкретный и неограничивающий пример выделения и очистки представляющего интерес высокоманнозного белка, получаемого с использованием способа по изобретению, можно найти в приведенных далее примерах. В примере показано, что рекомбинантная h-GCD, продуцируемая по изобретению, неожиданно оказалась связанной с внутренней мембраной трансформированных клеток моркови по изобретению и не секретировалась в среду. Растворимая rh-GCD может быть отделена от дебриса клеток и других нерастворимых компонентов в соответствии со способами, известными в данной области, такими как фильтрование или осаждение. Например, после цикла замораживание-оттаивание клетки разрушают и высвобождают внутриклеточные растворимые белки, тогда как rh-GCD остается связанной с нерастворимым дебрисом мембраны. Данную смесь растворимых компонентов и нерастворимого дебриса мембраны затем центрифугируют и растворимую фракцию удаляют, упрощая таким образом очистку. Связанная с мембраной h-GCD затем может быть растворена путем механического разрушения в присутствии мягкого детергента, ингибиторов протеазы и нейтрализующего окислителя. Растворимый фермент далее может быть очищен с использованием хроматографических способов, например, на катионообменных колонках и хроматографических колонках гидрофобного взаимодействия. Во время продуцирования rh-GCD в биореакторе и процесса очистки идентичность, выход, чистота и ферментативная активность могут быть определены с помощью одного или нескольких биохимических анализов, включая, но не ограничиваясь указанным, определение гидролиза субстрата фермента или аналога субстрата, анализ с использованием электрофореза на SDS-полиакриламидном геле и иммунологические анализы, такие как ELISA и вестерн-блот анализ.
В соответствии с предпочтительным вариантом осуществления клетка-хозяин, использованная в данном способе, включает клетку-хозяина по изобретению.
В другом предпочтительным варианте осуществления высокоманнозный белок, продуцируемый способом по изобретению, может представлять собой биологически активный высокоманнозный фермент, имеющий по крайней мере одну олигосахаридную цепь, включающую незащищенный остаток маннозы.
Данный рекомбинантный фермент может связываться с рецептором маннозы на клетке-мишени в сайте-мишени. Более конкретно, рекомбинантный фермент, продуцируемый способом по изобретению, обладает повышенной аффинностью по отношению к клетке-мишени по сравнению с соответствующей аффинностью природного лизосомального фермента по отношению к клетке-мишени. Соответственно клетка-мишень при сайте-мишени может представлять собой клетку Купфера печени пациента.
В конкретном варианте осуществления данный лизосомальный фермент может быть выбран из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. Наиболее предпочтительно данный лизосомальный фермент может представлять собой глюкоцереброзидазу (GCD).
В другом предпочтительном варианте осуществления клетка-хозяин, используемая в способе по изобретению, может представлять собой клетку корня растения, выбранную из группы, состоящей из трансформированной Agrobacterium rihzogenes клетку корня, клетку сельдерея, клетку имбиря, клетку хрена и клетку моркови. Наиболее предпочтительно клетка корня растения представляет собой клетку моркови. Следует особо отметить, что трансформированные клетки-хозяева моркови выращивают в суспензии.
В следующем аспекте настоящее изобретение относится к способу лечения пациента, предпочтительно пациента, относящегося к группе млекопитающих, имеющего лизосомальную болезнь накопления, с использованием экзогенного рекомбинантного лизосомального фермента.
Следует понимать, что раскрытое и описанное данной изобретение не ограничено конкретными примерами, стадиями способов и раскрытыми здесь материалами, поскольку такие стадии способа и материалы могут в определенной степени изменяться. Также следует понимать, что использованная в данном описании терминология используется только в целях описания конкретных вариантов осуществления и не предназначена для ограничения, поскольку объем настоящего изобретения будет ограничен только прилагаемой формулой изобретения и ее эквивалентами.
По всему тексту в описании и в последующей формуле изобретения, если контекст не предполагает другого, следует понимать, что слово «включать» и его вариации, такие как «включает» и «включающий», подразумевают включение указанного целого или стадии или группы целых или стадий, но не включение любого другого целого или стадии или группы целых или стадий.
Следует отметить, что, как это использовано в данном описании и прилагаемой формуле изобретения, использованная неопределенная и определенная форма единственного числа включают ссылку на множественную форму, если контекст четко не указывает другого.
Следующие примеры являются иллюстрацией способов, использованных авторами данного изобретения для осуществления аспектов настоящего изобретения. Следует понимать, что, хотя данные способы являются иллюстрацией предпочтительных вариантов осуществления для практической реализации изобретения, в свете настоящего описания специалистам в данной области будет очевидно, что можно провести многочисленные модификации, не отступая от сути и предполагаемого объема изобретения.
Примеры
Экспериментальные методики:
Векторы плазмид
* CE-T-Was сконструирована из плазмиды СЕ, полученной от профессора Галили (Prof. Galili) [патент Соединенный Штатов Америки 5367110, 22 ноября (1994)].
Плазмиду СЕ расщепляли с помощью SalI.
Сцепленный с SalI конец притупляли с использованием большого фрагмента ДНК полимеразы I. Затем плазмиду расщепляли с помощью PstI и лигировали к фрагменту ДНК, кодирующему ER-направляющий сигнал, от гена основной эндохитиназы [Arabidopsis thaliana] ATGAAGACTAATCTTTTTCTCTTTCTCATCTTTTCA
CTTCTCCTATCATTATCCTCGGCCGAATTC и вакуолярный нацеливающий сигнал от хитиназы А табака: GATCTTTTAGTCGATACTATG расщепляли SmaI и PstI.
pGREENII получали от Dr.P.Mullineaux [Roger P.Hellens et al., (2000) Plant.Mol.Bio., 42:819-832] Экспрессию от вектора pGREEN II контролировали с использованием 35S промотора вируса мозаики цветной капусты, омега-трансляционного элемента-энхансера TMV (вируса мозаики табака) и терминаторной последовательности октопинсинтазы Agrobacterium tumefaciens.
кДНК
hGCD - получен от АТСС (каталожный № 65696)б GC-2.2 [GCS-2 т.п.о., лямбда-EZZ-гамма3 Homo sapiens], содержащий глюкозидаза-бета-кислоту [глюкоцереброзидаза]. Длина вставок (т.п.о.): 2,20; ткань: фибробласт клетки WI-38.
Конструкция плазмиды экспрессии
кДНК, кодирующую hGCD, амплифицировали с использованием прямого 5' CAGAATTCGCCCGCCCCTGCA 3' и обратного 5' CTCAGATCTTGGCGATGCCACA 3' праймеров. Очищенный с помощью ПЦР продукт ДНК расщепляли эндонуклеазами EcoRI и BglII (см. распознающие последовательности, подчеркнутые в праймерах) и лигировали в промежуточный вектор, имеющий кассету экспрессии CE-T, расщепленную теми же ферментами. Кассету экспрессии вырезали и элюировали из промежуточного вектора и лигировали в бинарный вектор pGREENII с использованием ферментов рестрикции SmaI и XbaI, образуя конечный вектор экспрессии. Устойчивость к канамицину придается с помощью гена NPTII, управляемого с помощью nos промотора, полученного вместе с вектором pGREEN (фиг.1В). Полученная кассета экспрессии представлена на Фиг.1А.
Полученную плазмиду секвенировали для того, чтобы гарантировать корректное слияние сигналов в рамке считывания с использованием следующих секвенирующих праймеров: 5' 35S промотор: 5' CTCAGAAGACCAGAGGGC 3' и 3' терминатор: 5' CAAAGCGGCCATCGTGC 3'.
Создание каллюса моркови и суспензионных культур клеток
Создание каллюса моркови и суспензионных культур клеток авторы осуществляли аналогично описанному ранее Torres K.C. (Tissue culture techniques for horticular crops, p.p. 111, 169).
Трансформация клеток моркови и выделение трансформированных клеток.
Трансформацию клеток моркови предварительно проводили с использованием трансформации Argrobacterium, путем приспосабливания ранее описанного способа [Wurtele E.S. and Bulka K. Plant. Sci., 61:253-262 (1989)]. Клетки, выращенные в жидкой среде, использовали на протяжении всего способа вместо каллюса. Время инкубации и роста адаптировали для трансформации клеток в жидкой культуре. Argrobacterium трансформировали с использованием вектора pGREEN II путем электропорации [den Dulk-ra, A. And Hooykaas P.J. (1995), Methods Mol.Biol., 55: 63-72], а затем отбирали с использованием 30 мг/мл антибиотика паромомицина. Клетки моркови трансформировали с использованием Argrobacterium и отбирали с использованием 60 мг/мл антибиотика паромомицина в жидкой среде.
Скрининг трансформированных клеток моркови на выделение каллюса, экспрессирующего высокие уровни GCD
Через 14 дней после трансформации клетки культуры помещали на твердую среду при разведении 3% упакованного объема клеток для трансформации каллюса от индивидуальных кластеров клеток. Когда индивидуальный каллюс достигал 1-2 см в диаметре, клетки гомогенизировали в буфере для SDS образцов и полученные экстракты белка делили на SDS-PAGE [Laemmli U., (1970) Nature 227: 680-885] и переносили на нитроцеллюлозную мембрану (нитроцеллюлоза hybond C, 0,45 микрон. Каталожный № RPN203C от Amersham Life Science). Вестерн-блот анализ для обнаружения GCD проводили с использованием поликлональных антител против hGCD (описано далее). Каллюсы, экспрессирующие значительные уровни GCD, увеличивали в размерах и переносили для роста в жидкую среду для масштабирования, очистки белка и анализа.
Получение поликлональных антител
75 микрограммов рекомбинантного GCD (ЦерезимТМ) суспендировали в полном адъюванте Фрейнда и вводили в виде инъекции каждому из двух кроликов. Каждому кролику проводили повторную инъекцию через две недели. У кроликов отбирали кровь примерно через 10 дней после второй инъекции и еще раз с недельными интервалами до тех пор, пока титр антител не начинал падать. После удаления сгустка крови сыворотку крови делили на аликвоты и хранили при -20°С.
Масштабированное выращивание высококачественной культуры в биореакторах.
Примерно 1 см (в диаметре) каллюс клеток генетически модифицированной моркови, содержащий ген rh-GCD, помещали в агаровые чашки диаметром 9 см Murashige and Skoog, содержащие 4,4 г/л среды MSD (Duchefa), 9,9 мг/л тиамина HCl (Duchefa), 0,5 мг фолиевой кислоты (Sigma), 0,5 мг/л биотина (Duchefa), 0,8 г/л гидролизата казеина (Ducifa), сахар 30 г/л и гормоны 2-4 D (Sigma). Каллюс выращивали в течение 14 дней при 25°С.
Суспензионную культуру клеток получали путем субкультивирования трансформированного каллюса в жидкой среде MSD (Murashige & Skoog (1962), содержащей 0,2 мг/л 2,4-дихлоруксусной кислоты), как хорошо известно в данной области. Суспензию клеток культивировали в колбе Эрленмейера объемом 250 мл (рабочий объем начинается с 25 мл, через 7 дней увеличивается до 50 мл) при 25°С при скорости встряхивания 60 об/мин. Впоследствии объем клеточной культуры увеличивали до использования 1 л колбы Эрленмейера путем добавления рабочего объема до 300 мл при тех же условиях. Инокуляцию небольшого биореактора (10 л) [смотри WO 98/13469], содержащего 4 л среды MSD, проводили, добавляя 400 мл суспензии клеток, полученной из двух колб Эрленмейера, объемом 1 л, которые культивировали в течение семи дней. После недели культивирования при 25°С с использованием 1 л/мин потока воздуха среду MSD добавляли до 10 л и культивирование продолжали при тех же условиях. После дополнительного культивирования в течение 5 дней большую часть клеток процеживали и собирали, пропуская клеточную среду через сеть с ячейками 80 мкм. Избыточную среду отдавливали прессованием и упакованный осадок клеток хранили при -70°С.
Дополнительные подробности устройства биореактора можно найти в патенте США 6391638, выданном 21 мая 2002 г., и процитированном ранее путем ссылки.
Очистка белка
Для отделения среды от нерастворимой GCD замороженный осадок клеток, содержащий примерно 100 г сырого веса клеток, оттаивали с последующим центрифугированием при 17000xg в течение 20 минут при 4°С. Нерастворимые материалы и неразрушенные клетки промывали с помощью повторного суспендирования в 100 мл промывочного буфера (20 мМ фосфата натрия, рН 7,2, 20 мМ EDTA) и затем осаждали центрифугированием при 17000xg в течение 20 минут при 4°С. rh-GCD (рекомбинантная GCD человека) экстрагировали и солюбилизировали путем гомогенизации осадка в 200 мл экстрагирующего буфера (20 мМ фосфата натрия, рН 7,2, 20 мМ EDTA, 1 мМ PMSF, 20 мМ аскорбиновой кислоты, 3,8 г поливинилпирролидона (PVPP), 1 мМ DTT, 1% Triton-x-100). Гомогенат затем встряхивали в течение 30 минут при комнатной температуре и осветляли центрифугированием при 17000xg в течение 20 минут при 4°С. Осадок отбрасывали и рН супернатанта доводили до рН 5,5 добавлением концентрированной лимонной кислоты. Мутность, возникающую после доведения рН, осветляли центрифугированием в тех же условиях, как описано выше.
Дальнейшую очистку проводили с помощью хроматографических колонок следующим образом: на первой стадии 200 мл осветленного экстракта загружали на 20 мл сильной катионообменной смолы (Macro-Prep high-S носитель, Bio-Rad), уравновешенной в 25 мМ буфере цитрата натрия, рН 5,5 и набитой в колонку ХК (2,6×20 см). Колонка была объединена с АКТА основной системой (Amersham Pharmacia Biotech), что позволяет проводить мониторинг проводимости, рН и поглощения при 280 нм. Образец загружали при скорости 20 мл/мин, после чего колонку промывали уравновешивающим буфером (25 мМ буфер цитрата натрия, рН 5,5) при скорости потока 12 мл/мин до тех пор, пока поглощение в УФ не достигало базисной линии. Предварительное элюирование rh-GCD проводили с использованием уравновешивающего буфера, содержащего 200мМ NaCl, и элюирование проводили уравновешивающим буфером, содержащим 600 мМ NaCl. Фракции, собранные во время прохождения эксперимента, контролировали с помощью анализа ферментативной активности и объединяли пробирки, содержимое которых проявляло ферментативную активность (в пиках элюирования). Собранные образцы разбавляли в воде (1:5), содержащей 5% этанола, и рН доводили до 6,0 с использованием NaOH. Элюированные фракции, содержащие rGCD, наносили на вторую колонку ХК (1,6×20 см), набитую 10 мл той же резины, что и предшествующая колонка. Смолу на данной колонке уравновешивали 20 мМ цитратного буфера, рН 6,0, содержащего 5% этанола. После загрузки образца колонку промывали уравновешивающим буфером и GCD элюировали из колонки с помощью элюирующего буфера (20 мМ цитратного буфера, рН 6,0, 5% этанола и 1 М NaCl). Фракции с пиком поглощения на стадии элюирования собирали и наносили на третью колонку. Конечную стадию очистки проводили на колонке ХК (1,6×20 см), набитой 8 мл смолы с гидрофобным взаимодействием (гель TSK, Toypearl Phenyl-650C, Tosoh Corp.). Смолу уравновешивали 10 мМ цитратного буфера, рН 6,0, содержащего 5% этанола. Объединенную фракцию GCD, элюированную из предшествующей колонки, загружали при скорости 6 мл/мин с последующей промывкой уравновешивающим буфером до тех пор, пока поглощение в УФ не достигало базисной линии. Чистую GCD элюировали 10 мМ цитратным буфером, содержащим 50% этанола, объединяли и хранили при -20°С.
Определение концентрации белка
Концентрации белка в экстрактах и фракциях клеток анализировали способом Lowry/Bradford (анализ белка Bio Rad) [Bradford M., Anal.Biochem. (1976) 72:248] с использованием стандарта бычьего сывороточного альбумина (фракция V Sigma). Альтернативно, концентрацию в гомогенных образцах белка определяли по поглощению при 280 нм, 1 мг/мл=1,4 ОП280. Чистоту определяли по соотношению 280/260 нм.
Анализ ферментативной активности GCD
Ферментативную активность GCD определяли с использованием п-нитрофенил-β-D-глюкопиранозида (Sigma) в качестве субстрата. Буфер для анализа содержал 60 мМ фосфат-цитратного буфера, рН 6, 4 мМ β-меркаптоэтанола, 1,3 мМ EDTA, 0,15% Triton X-100, 0,125% таурохолята натрия. Анализ предворительно формировали в 96-луночных планшетах ELISA, 0-50 микролитра образца инкубировали с 250 микролитрами буфера для анализа и субстрат добавляли до конечной концентрации 4 мМ. Реакционную смесь инкубировали при 37°С в течение 60 минут. Образование продукта (п-нитрофенил, pNP) определяли по поглощению при 405 нм. Поглощение при 405 нм контролировали в момент времени t=0 и в конечной точке. Через 60 минут 6 микролитров 5 н. NaOH добавляли в каждую лунку и опять контролировали поглощение при 405 нм. Параллельно анализируемую кривую ссылочного стандарта использовали для количественной оценки концентраций GCD в тестируемых образцах [Friedman et al., (1999), Blood, 93(9): 2807-16].
Биохимические анализы
Протеолиз в геле и масс-спектрометрический анализ
Окрашенные полосы белка в геле вырезали с помощью чистого лезвия бритвы и белки в геле восстанавливали с использованием 10 мМ DTT и модифицировали 200 мМ иодацетамида в 10 мМ бикарбонате аммония. Куски геля обрабатывали 50% ацетонитрилом в 10 мМ бикарбонате аммония для удаления красителя из белков с последующим высушиванием кусков геля. В высушенных кусках геля восстанавливали влагосодержание с использованием 10% ацетонитрила в 10 мМ бикарбонате аммония, содержащем примерно 0.1 мкг трипсина на образец. Куски геля инкубировали в течение ночи при 37°С и полученные пептиды выделяли 60%-ным ацетонитрилом с добавлением 0,1% трифторацетата.
Расщепленные трипсином пептиды разделяли с помощью хроматографии на обращенной фазе на 0,1Х 300 мм кварцевых капиллярных колонках (J&W, внутренний диаметр 100 микрометров), наполненных самостоятельно пористым R2 (Perspective). Пептиды элюировали c использованием линейного градиента в течение 80 минут, начиная от 5 до 95% ацетонитрила с добавлением 0,1% уксусной кислоты в воде при скорости потока примерно 1 мкл/мин. Жидкость из колонки подвергали электрораспылению в масс-спектрометре с ионной ловушкой (LCQ, Finnegan, San Jose, CA). Масс-спектрометрический анализ проводили в режиме положительных ионов с использованием повторяющегося полного МС-сканирования с последующей индуцированной столкновением диссоциацией (CID) наиболее доминирующего иона, выбранного из первого МС-скана. Масс-спектрометрические данные сравнивали с моделированным протеолизом и CID белков в NR-NCBI базе данных с использованием программного обеспечения Sequest [J.Eng and J.Yates, University of Washington and Finnegan, San Jose].
Аминные концы белка секвенировали на пептидном секвенаторе Peptide Sequencer 494A (Perkin Elmer) в соответствии с инструкциями производителя.
Поглощение GCD перитонеальными макрофагами
Известно, что направление GCD к макрофагам и поглощение макрофагами опосредуется манноза/N-ацетилглюкозиминным рецептором и может быть определено с использованием триогликолят-извлеченных перитонеальных макрофагов, полученных у мышей, как описано Stahl P. и Gordon S. [J.cell Biol. (1982) 93(1): 49-56]. Кратко, мышам (самки, штамм С57-В6) внутрибрюшинно вводили 2,5 мл 2,4% Bacto-тиогликолятной среды вес./об. в декстрозе (Difco Cat. No. 0363-17-2). Через 4-5 дней обработанных мышей умерщвляли путем смещения шейных позвонков и полость брюшины ополаскивали забуференным фосфатом физиологическим раствором. Клетки осаждали центрифугированием (1000xg, 10 минут) и повторно суспендировали в DMEM (Beit Haemek, Israel), содержащей 10% фетальной телячьей сыворотки. Затем клетки помещали из расчета 1-2×105 клеток/лунку в 96-луночных планшетах для культур тканей и инкубировали при 37°С. Через 90 минут не прилипшие клетки промывали три раза с использованием PBS и прилипшие макрофаги инкубировали в течение 90 минут при 37°С в культуральной среде, содержащей определенные количества rhGCD, колеблющиеся от 0 до 40 микрограммов в конечном объеме 200 микролитров, в отсутствие и в присутствие дрожжевой маннаны (2-10, 5 мг/мл). После инкубации среду, содержащую избыток rGCD, удаляли и клетки промывали три раза PBS, а затем лизировали с использованием лизисного буфера (10 мМ Tris, pH 7,3, 1 мМ MgCl2, 0,5% NP-40 и ингибиторы протеазы). Активность rGCD, поглощенной клетками, определяли, подвергая клеточные лизаты гликозидазному анализу in vitro, как описано выше.
Пример 1
Конструирование плазмиды экспрессии
В данном примере более подробно описано конструирование иллюстративной плазмиды экспрессии, использованной в дальнейших примерах.
кДНК, кодирующую hGCD (ATTC. Номер клона 65696), амплифицировали с использованием прямого 5' CAGAATTCGCCCGCCCCTGCA 3' (также обозначенного как SEQ ID NO: 1) и обратного 5' CTCAGATCTTGGCGATGCCACA 3' (также обозначенного как SEQ ID NO: 2) праймеров.
Очищенный ПЦР продукт ДНК расщепляли эндонуклеазами EcoRI и BglII (см. распознающие последовательности, подчеркнутые в праймерах) и лигировали в промежуточный вектор, имеющий кассету экспрессии CE-T, расщепленную теми же ферментами. СЕ-Т включает ER нацеливающий сигнал MKTNLFLFLIFSLLLSLSSAEA (также обозначенный как SEQ ID NO: 3) из основного гена эндохитиназы [Arabidopsis thaliana] и вакуолярный нацеливающий сигнал хитиназы А табака: DLLVDTM* (также обозначенный как SEQ ID NO: 4).
Кассету экспрессии вырезали и элюировали из промежуточного вектора и лигировали в бинарный вектор pGREENII с использованием ферментов рестрикции SmaI и XbaI, образуя конечный вектор экспрессии. Устойчивость к канамицину придается с помощью гена NPTII, управляемого с помощью nos промотора вместе с вектором pGREEN (фиг.1В). Полученная кассета экспрессии представлена на фиг.1А.
Полученную плазмиду секвенировали для того, чтобы гарантировать корректное слияние сигналов в рамке считывания с использованием следующих секвенирующих праймеров:
Праймер из 5' 35S промотора: 5' CTCAGAAGACCAGAGGGC 3' (также обозначенный как SEQ ID NO: 5) и 3' терминатор: 5' CAAAGCGGCCATCGTGC 3', также обозначенный как SEQ ID NO: 6). Верифицированная клонированная кодирующая последовательность hGCD обозначена как SEQ ID NO: 7.
Пример 2
Трансформация клеток моркови и скрининг для выявления трансформированных клеток, экспрессирующих rhGCD
В данном примере описывается иллюстративный способ трансформирования клеток моркови согласно настоящему изобретению, как он использован в примере ниже.
Трансформацию клеток моркови предварительно проводили с использованием трансформации Argrobacterium, как описано ранее [Wurtele and Bulka (1989), там же]. Генетически модифицированные клетки моркови высевали на агаровой среде Murashige и Skoog, содержащей антибиотики для отбора трансформантов. Как показано на фиг.2, экстракты, полученные от растущего каллюса, тестировали на экспрессию GCD с помощью вестерн-блот анализа с использованием поликлональных антител против hGCD и сравнивали с церезимом в качестве стандарта (положительный контроль) и экстрактами нетрансформированных клеток (отрицательный контроль). Среди различных протестированных каллюсов один каллюс (номер 22) был отобран для масштабированного роста и очистки белка.
Вестерн-блот анализ проводили следующим образом.
Для данного анализа белки из полученного образца делили электрофорезом на SDS полиакриламидном геле и переносили на нитроцеллюлозу. Для этой цели SDS полиакриламидные гели получали следующим образом. SDS гели состояли из концентрирующего геля и разделяющего геля (в соответствии с Laemmli, UK, 1970, Cleavage of structural proteins during assembly of the head of bacteriphage T4, Nature 227, 680-685). Состав разделяющих гелей был следующим: 12% акриламида (Bio-Rad), 4 микролитра TEMED (N,N,N',N'-тетраметилэтилендиамин, каталожный номер Sigma T9281) на 10 мл раствора геля, 0,1% SDS, 375 мМ Tris-HCl, pH 8,8 и персульфат аммония (APS), 0,1% TEMED и персульфат аммония использовали в данном контексте в качестве источников свободных радикалов для полимеризации. Примерно через 20 минут после инициирования полимеризации концентрирующий гель (3% акриламида, 0,1% SDS, 126 мМ Tris-HCl, pH 6,8, 0,1% APS и 5 микролитров TEMED на 5 мл раствора концентрирующего геля) выливали на разделяющий гель и вводили гребенку с 12 или 18 промежутками для создания лунок для образцов.
Анодную и катодную камеры наполняли идентичными буферными растворами: Tris-глициновый буфер, содержащий SDS (Biorad, каталожный номер 161-0772), рН 8,3. Содержащий антиген материал обрабатывали 0,5 объема буфера для загрузки образца (30 мл глицерина (Sigma, каталожный номер G9012), 9% SDS, 14% меркаптоэтанола (Sigma, каталожный номер М6250), 187,5 мМ Tris-HCl, рН 6,8, 500 микролитров бромфенола синего, все объемы на 100 мл буфера для образцов) и затем смесь нагревали при 100°С в течение 5 минут и загружали на концентрирующий гель.
Электрофорез проводили при комнатной температуре в течение подходящего промежутка времени, например 45-60 минут, с использованием постоянной силы тока в 50-70 вольт, после чего в течение 54-60 минут при 180-200 вольт для гелей размером 13 на 9 см. Затем антигены переносили на нитроцеллюлозу (Schleicher and Schuell, Dassel).
Перенос белка осуществляли по существу, как описано в данном описании. Гель размещали вместе с прилегающей нитроцеллюлозой между листами фильтровальной бумаги Whatmann 3 MM, проводящим вспененным материалом толщиной 0,5 см и проволочными электродами, которые проводили ток с помощью платиновых электродов. Фильтровальную бумагу, вспененный материал и нитроцеллюлозу тщательно вымачивали в буфере для переноса (буфер TG от Biorad, каталожный номер 161-0771, разведенный в 10 раз метанолом и водным буфером (20%-ный метанол)). Перенос был осуществлен при 100 вольтах в течение 90 минут при 4°С.
После переноса свободные сайты связывания на нитроцеллюлозе насыщали при 4°С в течение ночи с использованием блокирующего буфера, содержащего 1% сухого молока (Dairy America) и 0,1% Tween 20 (Sigma Cat P1379), разведенного фосфатным буфером (Riedel deHaen, каталожный номер 30435). Полосы блота инкубировали с антителом (разведение 1:6500 в фосфатном буфере, содержащем 1% сухого молока и 0,1% Tween 20 как указано выше, рН 7,5) при 37°С в течение 1 часа.
После инкубации с антителом блот промывали три раза в каждом случае по 10 минут с использованием PBS (забуференный фосфатом буфер фосфата натрия (Riedel deHaen, каталожный номер 30435). Затем полосы блота инкубировали при комнатной температуре в течение 1 часа с подходящим вторичным антителом (козье антикроличье антитело (целая молекула) HRP (Sigma, каталожный номер А-4914)), разведение 1:3000 в буфере, содержащем % сухого молока (Dairy America) и 0,1% Tween 20 (Sigma Cat P1379), разведенном фосфатным буфером (Riedel deHaen, каталожный номер 30435). После промывания несколько раз с использованием PBS полосы блота окрашивали проявляющими реагентами ELC (Amersham RPN 2209).
После погружения блотов в реагенты ELC блоты подвергали воздействию рентгеновской пленки FUJI SUPER RX 18х24 и проявляли с использованием проявителя FUJI-ANATOMIX и фиксажа (FUJI-X fix каталожный номер FIXRTU 1 из 2). Полосы, характеризующие белки, которые были связаны антителом, становились видимыми после такой обработки.
Масштабированное выращивание высококачественной культуры в биореакторах.
Суспензионные культуры каллюса 22 получали путем субкультивирования трансформированного каллюса в жидкой среде. Клетки культивировали при встряхивании в колбах Эрленмейера до тех пор, пока общий объем не оказывался достаточным для инокулирования биореактора (как описано в экспериментальных методиках). Генетически модифицированные клетки моркови можно культивировать месяцами и сбор клеток можно проводить с цикличностью в 5-7 дней (данные не показаны). На семнадцатый день культивации, когда количество продуцирования rh-GCD в клетках моркови достигает максимума, клетки собирают, пропуская культуру через сетку с диаметром ячеек 100 меш. Следует отметить, что клетки можно собирать с помощью способов, известных в данной области, таких как фильтрование или центрифугирование. Запакованный осадок клеток, который обеспечивает материал для очистки h-GCD до гомогенности, можно хранить при температуре замораживания.
Пример 3
Очистка рекомбинантного активного белка HGCD из трансформированных клеток моркови
Установлено, что рекомбинантная h-GCD, экспрессируемая в трансформированных клетках моркови, связана с внутренними мембранами клеток и не выделяется в среду. Механически разрушенные клетки оставляют rGCD связанную с нерастворимым дебрисом мембраны (данные не показаны). Затем rGCD растворяли с использованием мягких детергентов и отделяли от клеточного дебриса и других нерастворимых компонентов. Растворимый фермент далее очищали с использованием хроматографических методов, включая катионообменные хроматографические колонки и колонки гидрофобного взаимодействия, как описано в экспериментальных методиках.
Для отделения среды от нерастворимой GCD замороженный осадок клеток, содержащий примерно 100 г сырого веса клеток, оттаивали с последующим центрифугированием при 17000xg в течение 20 минут при 4°С. Нерастворимые материалы и неразрушенные клетки промывали с помощью повторного суспендирования в 100 мл промывочного буфера (20 мМ фосфата натрия, рН 7,2, 20 мМ EDTA) и осаждения центрифугированием при 17000xg в течение 20 минут при 4°С. rGCD экстрагировали и солюбилизировали путем гомогенизации осадка в 200 мл экстрагирующего буфера (20 мМ фосфата натрия, рН 7,2, 20 мМ EDTA, 1 мМ PMSF, 20 мМ аскорбиновой кислоты, 3,8 г поливинилпирролидона (PVPP), 1 мМ DTT, 1% Triton-x-100 (Sigma)). Гомогенат встряхивали в течение 30 минут при комнатной температуре и осветляли центрифугированием при 17000xg в течение 20 минут при 4°С. Осадок отбрасывали и рН супернатанта доводили до рН 5,5 добавлением концентрированной лимонной кислоты. Мутность, возникающую после доведения рН, осветляли центрифугированием в тех же условиях, как описано выше.
Дальнейшую очистку проводили с помощью хроматографических колонок следующим образом: на первой стадии 200 мл осветленного экстракта загружали на 20 мл сильной катионообменной смолы (Macro-Prep high-S носитель, Bio-Rad), уравновешенной в 25 мМ буфере цитрата натрия, рН 5,5, и набитой в колонку ХК (2,6 Х 20 см). Колонка была объединена с АКТА основной системой (Amersham Pharmacia Biotech), что позволяет проводить мониторинг проводимости, рН и поглощения при 280 нм. Образец загружали при скорости 20 мл/мин, после чего колонку промывали уравновешивающим буфером (25 мМ буфер цитрата натрия, рН 5,5) при скорости потока 12 мл/мин до тех пор, пока поглощение в УФ не достигало базисной линии. Предварительное элюирование rh-GCD проводили с использованием уравновешивающего буфера, содержащего 200 мМ NaCl, и элюирование проводили уравновешивающим буфером, содержащим 600 мМ NaCl. Фракции, собранные во время прохождения эксперимента, контролировали с помощью анализа ферментативной активности и объединяли пробирки, содержимое которых проявляло ферментативную активность (в пиках элюирования). Собранные образцы разбавляли в воде (1:5), содержащей 5% этанола, и рН доводили до 6,0 с использованием NaOH.
На фиг.3А показан стандартный эксперимент данной стадии очистки. Фракции, собранные по время эксперимента, контролировали с помощью анализа ферментативной активности, как показано на фиг.3В и фиг.3С, демонстрирующих окрашивание кумассином синим элюированных фракций, проанализированных на активность.
Элюированные фракции, содержащие rGCD, наносили на вторую колонку ХК (1,6×20 см), набитую 10 мл той же резины, что и предшествующая колонка, для проведения второй стадии очистки. Смолу на данной колонке уравновешивали 20 мМ цитратного буфера, рН 6,0, содержащего 5% этанола. После загрузки образца колонку промывали уравновешивающим буфером и rGCD элюировали из колонки с помощью элюирующего буфера (20 мМ цитратного буфера рН 6,0, 5% этанола и 1 М NaCl). На фиг.3D показан стандартный эксперимент данной стадии очистки. Фракции, собранные по время эксперимента, контролировали с помощью анализа ферментативной активности, как показано на фиг.3Е и фиг.3F, демонстрирующих окрашивание кумассином синим элюированных фракций, проанализированных на активность.
Фракции, имеющие пик поглощения на стадии элюирования, собирали и наносили на третью колонку для проведения третьей стадии очистки. Третью стадию очистки проводили на колонке ХК (1,6×20 см), набитой 8 мл смолы с гидрофобным взаимодействием (гель TSK, Toypearl Phenyl-650C, Tosoh Corp.). Смолу уравновешивали 10 мМ цитратного буфера, рН 6,0, содержащего 5% этанола. Объединенную фракцию GCD из предшествующей колонки загружали при скорости 6 мл/мин с последующей промывкой уравновешивающим буфером до тех пор, пока поглощение в УФ не достигало базисной линии. Чистую GCD элюировали 10 мМ цитратным буфером, содержащим 50% этанола, объединяли и хранили при -20°С.
На фиг.4А показан стандартный эксперимент данной стадии очистки. Фракции, собранные во время эксперимента, контролировали с помощью анализа ферментативной активности (фиг.4В), на фиг.4С показано окрашивание кумассином синим элюированных фракций, проанализированных на активность.
Для обрабатываемой порции клеток, предназначенной для очистки, белок rGCD очищали до уровня, превышающего 95%; если проводили только первую и третью стадии, чистота достигала уровня выше 80% (результаты не представлены).
Биохимический анализ
Для подтверждения идентичности очищенного rGCD проводили Mass-Spec Mass-Spec (MSMS) анализ. Полученные результаты показали 49% границы последовательности белка, которая соответствует прогнозируемой последовательности аминокислот, на основании кассеты экспрессии ДНК, включая лидерный пептид и нацеливающие последовательности.
Поглощение и активность рекомбинантной hGCD в перитонеальных макрофагах.
Для того чтобы определить, была ли продуцированная в моркови rhGCD гликозилирована правильным образом и может ли она поглощаться клетками-мишенями и, следовательно, использоваться для лечения болезни Гоше, далее анализировали способность rhGCD связываться и поглощаться макрофагами. Нацеливание rhGCD на маркофаги опосредуется рецептором маннозы/N-ацетилглюкозамиина (Man/GlcNAc) и может быть определено с использованием тиогликолят-выявляемыми перитонеальными макрофагами. Как показано на фиг.5, rGCD поглощается клетками на высоком уровне. На фиг.5А показано поглощение клетками rGCD согласно настоящему изобретению относительно концентрации маннана.
На фиг.5А показано поглощение на сравнимых уровнях церезимаТМ (данный препарат был получен с 80%-ной чистотой с использованием только первой и третьей описанных выше стадий очистки).
На фиг.5В и 5С показано, что поглощение rGCD находится на более высоком уровне, чем для церезимаТМ, поскольку данный препарат был получен с чистотой более 95% с использованием всех трех описанных выше стадий очистки.
Что касается фиг.5С, ясно, что процент специфической активности от общей активности, ингибируемой 4 мг/мл маннана, выше для GCD по настоящему изобретению (rGCD или рекомбинантная GCD человека), чем для доступного в настоящее время на рынке продукта, следующим образом: GCD (СВ-mix1, которая представляет собой rGCD по настоящему изобретению) - 75%, церезим - 65%.
Кроме того, как показано на фигурах, добавление маннана четко ингибирует связывание rGCD клетками. При концентрации в 2 мг/мл маннана связывание rGCD ингибировалось на 50%.
Данные результаты показывают, что даже без модификации структур гликана, rhGCD, экспрессированный и очищенный из трансформированных клеток моркови специфически поглощается клетками-мишенями макрофагов посредством Man/GlcNAc рецепторов. Более того, данный рекомбинантный rhGCD является ферментативно активным.
На фиг.5D показано, что rhGCD также распознается антителами против GCD в вестерн-блот анализе; rGCD относится к белку согласно настоящему изобретению, тогда как стандарт GCD (показан при 5, 10 и 25 нг на полосу) представляет собой полученную из коммерческого источника GCD (Церезим ТМ).
Пример 4
Токсикологическое исследование
Материал, полученный в соответствии с вышеуказанным способом очистки, тестировали в соответствии со стандартными протоколами токсикологического тестирования (Guidance for Industry on Single Dose Acute Toxicity Testing for Pharmaceuticals (Руководство для промышленности по тестированию острой токсичности однократной дозы фармацевтических средств), Center for Drug Evaluation and Research (CDER), PT1 (61 Fr 43934, August 26, 1996) и ICH M3(M) Non-clinical Safety Studies for the Conduct of Human Clinical trials for Pharmaceuticals (Исследование неклинической безопасности при проведении клинических испытаний фармацевтических препаратов для людей) CPMP/ICH/286/95 модификация, 16 ноября 2000 г.).
Мышей инъецировали следующим образом. После первоначальной дозы 1,8 мг/кг (клиническая доза) вводили дозы 9 и 18 мг/кг. Группы тестирования включали по шесть мышей (ICR CD01; 3 самца и 3 самки), получавших rGCD согласно настоящему изобретению (в жидком носителе, содержащем 25 мМ цитратного буфера, 150 мМ NaCl, 0,01 Tween 80, 5% этанола), и другие шесть мышей обрабатывали только носителем в качестве контрольной группы. Затем мышей наблюдали в течение 14 дней и проводили эвтаназию. Ни одна из мышей не умерла перед проведением эвтаназии по расписанию. Ни одна из мышей не продемонстрировала каких-либо заметных эффектов от обработки. Ни у одной из мышей не обнаружили бросающейся в глаза патологии и/или изменений веса тела.
Пример 5
Анализ гликозилирования
Был проведен анализ структур гликанов, присутствующих на rGCD, продуцированном, как описано в предшествующих примерах. Как подробно описано далее, результаты показывают, что большинство гликанов содержит терминальные остатки маннозы, а также имеют высокоманнозные структуры. Установлено, что преимущественно высокоманнозный продукт является биологически активным и, следовательно, нет необходимости в последующих стадиях для его активации.
Следующие способы были использованы для определения структуры гликозилирования рекомбинантного hGCD, продуцированного в соответствии с приведенными выше примерами. Кратко, моносахаридные связи как в N-, так и в О-связанных гликанах определяли с помощью гидролиза и ГХ-масс-спектрометрической стратегии. В данном способе оценивается тип связи углеводов с пептидом и общий моносахаридный состав гликопротеина. На основании предшествующих знаний и также соотношений между различными моносахаридами с помощью данного способа можно предложить тип гликанов на гликопротеине. Данная информация важна для оценки возможных структур гликанов, присутствующих в белке.
С использованием другого способа проводили олигосахаридный анализ популяции N-гликана. Проводили масс-спектральный FAB и MALDI-TOF анализы после расщепления аликвот образцов трипсином и пептид-N-гликозидазой F (PNGaseF) и исчерпывающего метилирования гликанов. Данный способ используется для разъединения и выделения N-связанных углеводов от ферментативно расщепленного гликопротеина. Определяли массы популяции гликанов в выделенной смеси гликанов и их массы сравнивали с массами известных структур по базам данных с точки зрения анализа композиции моносахаридов. Предлагаемые структуры основаны также на схеме гликозилирования организма-источника.
Другой способ включал анализ популяции О-гликанов после восстановительного элиминирования обработанных трипсином и (PNGaseF) гликопептидов, обессоливания и исчерпывающего метилирования. О-гликаны не высвобождаются PNGaseF, следовательно, гликаны, остающиеся связанными с пептидами, наиболее вероятно представляют собой О-связанные гликаны. Данные гликаны затем высвобождают путем восстановительного элиминирования и анализируют их массы.
Анализ композиции моносахаридов (суммировано ниже) выявил характеристическое распределение гексоз, гексозаминов и пентоз, характерных для растительного гликозилирования. Соотношение между GlcNac и маннозой позволяет предположить, что характерные N-связанные структуры являются доминирующими в популяции гликанов.
Масс-спектрометрический анализ N-гликанов из hGCD, продуцированной, как описано выше, указывает, что доминирующая популяция N-гликанов имеет состав моносахаридов Pent.deoxyHex.Hex3.HexNAc2.
Материалы и методы
Анализ проводили с использованием комбинации газовой хроматографии - масс-спектрометрии (ГХ-МС), масс-спектрометрии с бомбардировкой быстрыми атомами (FAB-MS) и масс-спектрометрии с использованием замедленной экстракции - матрицевспомогательной лазерной десорбционной ионизации - времени пролета (DE-MALDI-TOF MS).
Для анализа олигосахаридов, популяцию N-гликанов анализировали с помощью масс-спектрометрии FAB-MS и MALDI-TOF после расщепления аликвот образцов трипсином и пептид-N-гликозидазой F (PNGaseF) и исчерпывающего метилирования гликанов. Популяцию О-гликанов анализировали после восстановительного элиминирования обработанных трипсином и (PNGaseF) гликопептидов, обессоливания и исчерпывающего метилирования.
Моносахаридные связи как для N-, так и для О-гликанов определяли с использованием гидролиза и ГХ-МС стратегии для полученных производных.
Экспериментальное описание
Образец
Полученным ампулам с образцами присваивали уникальный номер образца следующим образом (таблица 1)
62996
62997
62998
Образцы хранили при температуре между -10 и -30°С до момента, когда они потребуются.
Химия белка
Диализ цельных образцов
Одну ампулу (содержащую 1 мл белка в установленной концентрации 0,8 мг/мл) вводили в кассету для диализа Slide-A-Lyzer (отсекание по молекулярной массе 10 кДа) и проводили диализ при 4°С в течение 24 часов против воды, при этом воду заменяли 3 раза. После диализа образец удаляли из кассеты и лиофилизовали.
Расщепление трипсином цельных образцов
Диализованные, лиофилизованные образцы повторно суспендировали в 50 мМ аммоний-бикарбонатном буфере, доведенном до рН 8,4 10% водным аммиаком, и расщепляли обработанным ТРСК трипсином в течение 4 часов при 37°С в соответствии с SOPs B001 и В003. Реакцию заканчивали путем помещения образца на 2 минуты в нагревательный блок при температуре 95°С с последующей лиофилизацией.
Углеводная химия
Расщепление пептид-N-Гликозидазой А
Расщепленную трипсином смесь пептид/гликопетид из образца гликопротеина обрабатывали ферментом пептид-N-Гликозидазой А (PNGaseF) в аммонийацетатном буфере, рН 5,5 при 37°С в течение 15 часов. Реакцию останавливали путем лиофильной сушки. Полученные продукты очищали с использованием катриджа С18 Sep-Pak.
Восстановительное элиминирование
Фракцию Sep-Pak, содержащую потенциальные О-связанные гликопептиды, растворяли в растворе, содержащем 10 мг/мл боргидрида натрия в 0,05 М гидроксиде натрия и инкубировали при 45°С в течение 16 часов. Реакцию заканчивали прибавлением ледяной уксусной кислоты.
Высаливание восстановительно элиминированного материала.
Высаливание с использованием шариков смолы Dowex проводили в соответствии с SOP B022. Образец загружали на колонку и элюировали с использованием 4 мл 5%-ной водной уксусной кислоты. Собранную фракцию лиофилизовали.
Исчерпывающее метилирование высвобожденных углеводов
Для N-связанных углеводов, элюированных 5%-ной уксусной кислотой из фракции Sep-Pak и потенциальных О-связанных гликанов, высвобожденных с помощью восстановительного элиминирования, проводили исчерпывающее метилирование с использованием методики (SOP B018) с применением гидроксида натрия (NaOH)/иодистого метила (MeI). Часть перметилированной смеси N-гликанов анализировали с использованием масс-спектрометических анализов FAB-MS и MALDI-TOF MS, а остаток подвергали анализу на связывание.
Анализ связывания N-связаннного углевода
Получение производных
Образец смеси перметилированных гликанов, полученной после расщепления трипсином и PNGase или восстановительного элиминирования, гидролизовали (2 М ТФУК, 2 часа при 120°С) и восстанавливали (бордейтерид натрия (NaBD4) в 2 М NH4OH, 2 часа при комнатной температуре, SOP B025). Борат, образующийся при разрушении бордейтерида, удаляли, 3 раза добавляя смесь метанола с ледяной уксусной кислотой (90:10) с последующей лиофилизацией. Затем образцы ацетилировали с использованием уксусного ангидрида (1 час при 100°). Ацетилированные образцы очищали экстракцией в хлороформ. Частично метилированные альдитолацетаты затем исследовали с помощью газовой хроматографии/масс-спектрометрии (ГЖ/МС). Стандартные смеси частично метилированных альдитолацетатов и пустой образец также анализировали при тех же условиях.
Газожидкостная хроматография/ масс-спектрометрия (ГХ/МС)
Аликвоту (1 мкл) образца углеводов после получения производных, растворенную в гексане, анализировали методом ГХ/МС с использованием Perkin Elmer Turbomass Gold масс-спектрометра с газовым хроматографом Autosystem XL и системой данных Dell при следующих условиях:
Газовая хроматография
Колонка: DB5
Впрыскивание: прямо в колонку
Температура инжектора: 40°С
Программа: 1 минута при 40°С, затем 70°С/минуту до 100°С, выдерживание при 100°С в течение 1 минуты, затем 8°С/минуту до 290°С, наконец выдерживание при 290°С в течение 5 минут
Газ-носитель: гелий
Масс-спектрометрия
Напряжение ионизации: 70 эВ
Способ получения данных: сканирование
Диапазон масс: 35-450 дальтон
Разрешение масс-спектра: единица
Анализ сахаров незатронутой глюкоцереброзидазы
Получение производных
Аликвоту, эквивалентную 500 мкг глюкоцереброзидазы, лиофилизовали с использованием 10 мкг арабитола в качестве внутреннего стандарта. Затем ее метанолизовали в течение ночи при 80°С и сушили в атмосфере азота. Высвободившиеся моносахариды затем повторно N-ацетилировали с использованием раствора метанола, пиридина и уксусного ангидрида, сушили опять в атмосфере азота и преобразовывали в триметилсилильные (ТМС) производные в соответствии с SOP B023. ТМС-производные восстанавливали в объеме в атмосфере азота, растворенного в 2 мл гексана, и обрабатывали ультразвуком в течение 3 минут. Образцы затем оставляли уравновешиваться при 4°С в течение ночи. Параллельно получали пустой образец, содержащий 10 мкг арабитола, и стандартную смесь моносахаридов, содержащую по 10 мкг фукозы, ксилозы, маннозы, галактозы, глюкозы, N-ацетилгалактозамина, N-ацетилглюкозамина, N-ацетилнеураминовой кислоты и арабитол. ТМС-производные затем исследовали методом газовой хроматографии/масс-спектрометрии (ГЖ/МС).
Газожидкостная хроматография/ масс-спектрометрия (ГХ/МС)
Аликвоту (1 мкл) образца углеводов после получения производных, растворенную в гексане, анализировали методом ГХ/МС с использованием Perkin Elmer Turbomass Gold масс-спектрометра с газовым хроматографом Autosystem XL и системой данных Dell при следующих условиях:
Газовая хроматография
Колонка: DB5
Впрыскивание: прямо в колонку
Температура инжектора: 40°С
Программа: 1 минута при 90°С, затем 25°С/минуту до 140°С, 5°С/минуту до 220°С, наконец 10°С/минуту до 300°С и выдерживание при 300°С в течение 5 минут
Газ-носитель: гелий
Масс-спектрометрия
Напряжение ионизации: 70 эВ
Способ получения данных: сканирование
Диапазон масс: 50-620 дальтон
Разрешение масс-спектра: единица
Масс-спектрометрия замедленной экстракции - матрицевспомогательной лазерной десорбционной ионизации (DE-MALDI-MC) и масс-спектрометрия при бомбардировке быстрыми атомами (FAB-МС)
MALDI-TOF масс-спектрометрию осуществляли с использованием лазерно-десорбционного масс-спектрометра Voyager STR Biospectrometry Research, связанного с замедленной экстракцией (DE).
Высушенные перметилированные гликаны растворяли в смеси метанол:вода (80:20) и анализировали с использованием матрицы 2,5-дигидроксибензойной кислоты. В качестве внешних калибрантов использовали брадикинин, ангиотензин и АСТН.
Масс-спектрометрический анализ положительных ионов при бомбардировке быстрыми атомами проводили на масс-спектрометре M-Scan's VG AutoSpecE, функционирующем при Vacc=8 кВ для 4500 массового диапазона при полной чувствительности с разрешением приблизительно 2500. Для генерации спектра использовали цезиевую ионную пушку, работающую при 30 кВ. Спектры регистрировали на системе данных VAX 3100 M76 с использованием программного обеспечения Opus.
Высушенные перметилированные гликаны растворяли в метаноле и загружали на мишень, предварительно намазанную 2-4 мкл тиоглицерина в качестве матрицы перед введением в источник.
Результаты и обсуждение
TMS анализ сахаров глюкоцереброзидазы
Скрининг N-связанных олигосахаридов
Цельный гликопротеин подвергали диализу с последующим расщеплением трипсином и лиофилизованные продукты расщепляли в использованием PNGase A, а затем очищали с использованием С18 Sep-Pak. Фракцию 5% водной уксусной кислоты (содержащую N-связанные олигосахариды) подвергали исчерпывающему метилированию и получали FAB масс-спектры с использованием части дериватизованного олигосахарида в диапазоне низких масс для получения фрагментированных ионов и DE-MALDI-TOF масс-спектры получали с использованием части дериватизованных олигосахаридов в диапазоне высоких масс для получения молекулярных ионов.
Анализ N-гликанов глюкоцереброзидазы
В таблице 1 перечислены доминирующие фрагменты ионов, присутствующих в FAB спектре, и молекулярные ионы, присутствующие в MALDI спектре. Область молекулярного иона (показана в приложении III) содержит доминирующий сигнал при m/z 1505,8 (соответствует [M+Na]+ квазимолекулярному иону для структуры, имеющей состав Pent.deoxyHex.Hex3.HexNAc2). Также обнаруживали диапазон квазимолекулярных ионов меньшей интенсивности, соответствующий комплексу и высокоманнозным структурам. Обнаруженные высокоманнозные структуры колеблются в размере от Hex5HexNAc2 при m/z 1579,8 до Hex8HexNAc2 при m/z 2193,0. Сигнальные комплексы образуются от менее сильно обработанных N-гликанов, как, например, m/z 1331,7 (соответствует [M+Na]+ квазимолекулярному иону для структуры, имеющей состав Pent.Hex3.HexNAc2) или от больших N-гликанов, например, имеющих m/z 1751,0 (соответствует [M+Na]+ квазимолекулярному иону для структуры, имеющей состав Pent.deoxyHex Hex3.HexNAc3), m/z 2375,4 (соответствует [M+Na]+ квазимолекулярному иону для структуры, имеющей состав Pent.deoxyHex2Hex4.HexNAc4) и m/z 2753,6 (соответствует [M+Na]+ квазимолекулярному иону для структуры, имеющей состав Pent.deoxyHex3 Hex5.HexNAc4).
FAB масс-спектр обеспечивает информацию, касающуюся антеннообразной структуры, путем визуализации фрагментарных ионов в области низких масс спектра (данные не показаны). Сигналы обнаруживали, идентифицируя гексозу (при m/z 219) и HexNAc (при m/z 260) в качестве невосстанавливающихся концевых моносахаридов в N-гликанах.
Все массы в первой колонке являются моноизотопными, если не указано другое. Массовые числа могут не относиться непосредственно к необработанным данным, поскольку программное обеспечение часто относит массовые числа к пикам 13С изотопа, в особенности для масс, превышающих 1700 Да.
Анализ связывания N-гликанов гликоцереброзидазы
Анализ связывания проводили на N-связанных углеводах, высвобождающихся после расщепления PNGase А, очисткой на Sep-Pak и исчерпывающего метилирования.
Комплексная хроматограмма была получена вместе с несколькими пиками примесей, возникающими при дериватизации реагентов. Сравнение времени удерживания и спектра со стандартными смесями позволило провести условное отнесение пиков, содержащих сахар, перечисленных в таблице 3.
Времена удерживания различным образом сшитых моносахаридов, определяемых в виде из частично метилированных альдитолацетатов в ГХ-масс-спектрометрическом анализе глюкоцереброзидазы (ссылочный номер 62996) после расщепления трипсином и пептид N-гликозидазой А
Ксилоза
Фукоза
Манноза
Галактоза
Манноза
Глюкоза
Манноза
Манноза
Манноза
GlcNAc
GlcNAc
4.3. Скрининг О-связанных олигосахаридов
Проводили восстановительное элиминирование на фракции 60%-ного 2-пропанола (потенциальная фракция О-сшитого гликопротеина), полученной при очистке на Sep-Pak глюкоцереброзидазы после расщепления трипсином и PNGase А. Образец высаливали после окончания реакции и после удаления бората проводили исчерпывающее метилирование (перметилирование). Получали FAB-масс-спектры, используя порцию производного олигосахарида, в диапазоне низких масс с целью получения ионов фрагментации, и DE-MALDI-TOF масс-спектр получали, используя порцию производного олигосахарида, в диапазоне высоких масс с целью получения молекулярных ионов. Сигналы, согласующиеся с наличием О-сшитых гликанов, не наблюдались (данные не представлены).
Анализ связывания О-гликанов глюкоцереброзидазы
Анализ связывания проводили для продуктов восстановительного элиминирования после исчерпывающего метилирования. Сигналов, соответствующих присутствию типичных О-связанных гликанов, не наблюдалось (данные не показаны).
На фиг.6 показаны некоторые иллюстративные структуры гликанов для сравнения между GCD, полученной из клеток СНО (яичник китайского хомячка), которые являются клетками млекопитающих (ЦерезимТМ), и GCD по настоящему изобретению из клеток моркови. Как показано, для ЦерезимаТМ необходима модификация данных структур для получения незащищенных остатков маннозы. Например, такие незащищенные остатки маннозы непосредственно получают в случае GCD, полученной из клеток растений в соответствии с настоящим изобретением, где не требуются дальнейшие манипуляции, например, с использованием гликозилаз.
На фиг.7 представлена основная структура гликана, установленная для rGCD. На фиг.7 показаны предполагаемые структуры: а) доминантной популяции олигосахаридов, выявленной на hGCD, экспрессируемой в суспензии клеток моркови (1505,7 m/z); b) типичное N-связанное ядро; с) фукозилированное растительное N-связанное ядро. N-связанные гликаны сшиты с белком посредством аспарагина и через восстанавливающий конец остатка GlcNac (GN) с правой стороны диаграммы. N-растительные схемы гликозилирования, фукозные остатки могут быть частью структуры ядра, связанной с первым GlcNac с использованием альфа(1-3) гликозидной связи, тогда как в структурах, относящихся к млекопитающим, обычно используется альфа(1-6) гликозидная связь.
На фиг.8А-8D показаны все возможные структуры для N-гликанов, обнаруживаемые на rGCD белке согласно настоящему изобретению. Доминантная структура гликана, которая была идентифицирована, представляет собой ядерную гликановую структуру, выявленную в большинстве растительных гликопротеинов из риса, маиса и других съедобных растений. Данная структура содержит ядерный остаток ксилозы, а также ядерную альфа-(1,3)-фукозу. Исследование, проведенное Bardor и др. (33), показывает, что 50% неаллергических доноров крови имеют специфические антитела к ядерной ксилозе в сыворотке крови и 25% имеет специфические антитела к ядерной альфа-(1,3)-фукозе. Однако все еще требуется исследование, могут ли такие антитела вводить ограничения на применение полученных из растений биофармацевтических гликопротеинов.
Минорная популяция гликанов hGCD, продуцируемой, как описано выше, представляли собой высокоманнозные структуры от Hex4HexNAc2 до Hex8HexNAc2. Среди комплексных структур проявлялись структуры, такие как Pent.deoxyHex2 Hex4.HexNAc3 и Pent.deoxyHex3 Hex5.HexNAc3. Структура Pent.deoxyHex3 HexNAc2 обнаруживалась в меньших долях.
Основными концевыми моносахаридами являются гексоза (манноза или галактоза) и N-ацетилгексозамин, что соответствует присутствию высокоманнозных структур и частично переработанных комплексных структур.
Что касается скрининга О-связанных олигосахаридов, сигналов, которые соответствовали бы типичным О-связанным гликанам, не наблюдалось. Известно, что GCD не содержит О-связанных олигосахаридов, так что данные результаты соответствуют известному гликозилированию GCD в других клеточных системах, включая природную GCD и рекомбинантную GCD, продуцированную в культуральных системах, относящихся к млекопитающим. Однако в композиции моносахаридов обнаруживались сигналы, соответствующие арабинозе.
Важным моментом, касающимся настоящего изобретения, является то, что анализ композиции N-гликанов белка hGCD показал, что большинство N-гликанов имеет на конце маннозные остатки. Это согласуется с требованиями для концевых маннозных групп в N-гликанах, способствующих поглощению терапевтической hGCD рецептором маннозы макрофагов. Однако ни природная GCD, ни рекомбинантная GCD, продуцируемая клетками млекопитающих, не являются высокоманнозными. Следовательно, настоящее изобретение преодолевает заметный недостаток коммерчески производимых hGCD белков, который заключается в том, что данные белки модифицируют для того, чтобы они содержали и концевые маннозные сахара, в отличие от белка, продуцируемого, как описано выше.
Пример 6
Лечение с использованием настоящего изобретения
Рекомбинантный белок, продуцируемый в соответствии с настоящим изобретением, предпочтительно включает подходящим образом гликозилированный белок, продуцируемый культурой растительных клеток, который предпочтительно представляет собой, например, лизосомальный фермент и/или высокоманнозный гликозилированный белок.
В соответствии с предпочтительными вариантами осуществления белок, продуцируемый в соответствии с настоящим изобретением, является подходящим для лечения лизосомно-связанных заболеваний, таких, например, как лизосомальные болезни накопления.
Способ лечения необязательно и предпочтительно включает: (а) получение рекомбинантной биологически активной формы лизосомального фермента, полученного очисткой из трансформированных клеток корней растений и способного эффективно поступать в клетки с аномальным дефицитом лизосомального фермента. Данный рекомбинантный биологически активный фермент имеет незащищенные концевые остатки маннозы на прикрепленных олигосахаридах; и (b) введение пациенту терапевтически эффективного количества рекомбинантного биологически активного лизосомального фермента или включающей его композиции. В предпочтительном варианте осуществления рекомбинантный высокоманнозный лизосомальный фермент, используемый в способе по изобретению, может продуцироваться клетками-хозяевами по изобретению. Предпочтительно данная клетка-хозяин представляет собой клетку моркови.
Термин «субъект-млекопитающее» или «пациент-млекопитающее» означает любое млекопитающее, для которого желательна генная терапия, включая субъектов, представляющих собой людей, коров, лошадей, субъектов семейства псовых и семейства кошачьих, наиболее предпочтительно людей.
Следует понимать, что термин «лечение» также включает улучшение или облегчение патологического состояния и/или одного или нескольких его симптомов, излечение такого состояния или предотвращение возникновения такого состояния.
В другом предпочтительном варианте осуществления лизосомальный фермент, используемый с помощью способа по изобретению, может представлять собой высокоманнозный фермент, включающий оп крайней мере одну олигосахаридную цепь, имеющую незащищенный остаток маннозы. Такой рекомбинантный фермент может связываться с рецептором маннозы на клетки-мишени в сайте-мишени в организме пациента. Более предпочтительно данный рекомбинантный лизосомальный фермент обладает повышенной аффинностью к данным клеткам-мишеням по сравнению с соответствующей аффинностью к клеткам-мишеням природного лизосомального фермента. Следовательно, каждая доза зависит от эффективного определения клеток с аномальным дефицитом GCD, и каждая доза такой формы GCD по существу меньше, чем доза природного GCD, который был бы в других обстоятельствах введен аналогичным образом для достижения терапевтического эффекта.
В соответствии с предпочтительными вариантами осуществления настоящего изобретения белок является подходящим для лечения лизосомальных болезней накопления, так что настоящее изобретение также включает способ лечения таких заболеваний. Лизосомальные болезни накопления представляют собой группу из более чем 40 заболеваний, которые возникают в результате дефекта генов, кодирующих ферменты, которые разрушают гликолипидные или полисахаридные побочные продукты в лизосомах клеток. Ферментативные продукты, например сахара и липиды, затем превращаются в новые продукты. Каждое из этих нарушений является результатом наследственной аутосомной или Х-связанной рецессивной черты, которая оказывает влияние на уровни ферментов в лизосоме. Обычно затронутые ферменты в клетках и тканях пораженных индивидуумов не проявляют биологической или функциональной активности. При таких заболеваниях дефицит ферментативной функции создает прогрессирующее системное отложение жира или углеводного субстрата в лизосомах клетках тела, в конечном счете вызывая потерю функции органа и смерть. Генетическая этиология, клинические проявления, молекулярная биология и возможность лизосомных болезней накопления подробно описаны Scriver и др. [Scriver et al., eds., The Metablic and Molecular Basis of Inherited Disease, 7th Ed., Vol.II, McGraw Hill, (1995)].
Примеры лизосомальных болезней накопления (и связанных с ними дефицитных ферментов) включают, но не ограничиваются указанным болезнь Фабри (α-галактозидаза), болезнь Фарбера (керамидаза), болезнь Гоше (глюкоцереброзидаза), Gml ганглиозидоз (β-галактозидаза), болезнь Тея-Сакса (β-гексозаминидаза), болезнь Ниманна-Пика (сфингомиелиназа), болезнь Шиндлера (α-N-ацетилгалактозаминидаза), синдром Гунтера (идуронат-2-сульфатаза), синдром Слая (β-глюкуронидаза), синдромы Гурлера и Гурлера/Шейе (идуронидаза) и синдром I-клетка/Сан-Филипо (транспортер манноза-6-фосфатазы).
Болезнь Гоше является наиболее распространенной лизосомальной болезнью накопления у людей, при этом с наибольшей частотой она встречается у популяции евреев ашкенази. Примерно от 5000 до 10000 людей в Соединенных штатах Америки поражены данным заболеванием [Grabowski, Adv. Hum.Genet. 21:377-441 (1993)]. Болезнь Гоше является результатом дефицита глюкоцереброзидазы (hGCD; глюкозилкерамидаза). Этот дефицит приводит к накоплению субстрата фермента, глюкоцереброзида, в ретикулоэндотелиальных клетках костного мозга, селезенки и печени, приводя к значительным скелетным осложнениям, таким как расширение костного мозга и повреждение костей, а также к спленомегалии, гепатомегалии, тромбоцитопении, анемии и осложнениям легких [Grabowski, (1993) ibid.; Lee, Prog. Clin.Biol.Res. 95:177-217 (1982)].
Более конкретно, лизосомальный фермент, используемый в способе по изобретению, может быть выбран из группы, состоящей из глюкоцереброзидазы (GCD), кислотной сфингомиелиназы, гексозаминидазы, α-N-ацетилгалактозаминидизы, кислотной липазы, α-галактозидазы, глюкоцереброзидазы, α-L-идуронидазы, идуронатсульфатазы, α-маннозидазы или зиалидазы. Предпочтительно, когда заболевание, подвергаемое лечению, представляет собой болезнь Гоше, лизосомальный фермент, используемый в способе по изобретению, представляет собой глюкоцереброзидазу (GCD).
Белок по настоящему изобретению можно использовать для получения фармацевтической композиции. Таким образом, в соответствии с другим аспектом настоящего изобретения, разработана фармацевтическая композиция, которая включает в качестве активного ингредиента белок и фармацевтически приемлемый носитель. Как использовано в данном описании, термин «фармацевтическая композиция» относится к препарату одного или нескольких описанных здесь активных ингредиентов, таких как рекомбинантный белок, вместе с другими химическими компонентами, такими как традиционные лекарственные средства, физиологически подходящие носители и эксципиенты. Целью фармацевтической композиции является облегчение введения белка или клетки в организм. Фармацевтически композиции по настоящему изобретению могут быть изготовлены способами, хорошо известными в данной области, например с помощью обычных способов смешивания, растворения, гранулирования, изготовления драже, растирания в порошок, эмульгирования, инкапсулирования, захвата или лиофилизации.
В предпочтительном варианте осуществления термин «фармацевтически приемлемый» означает одобренный органами государственного регулирования Федерального правительства или правительства государства или перечисленный в Фармакопее США или других общепринятых фармакопеях для применения для животных и, более конкретно, для человека. Далее фразы «физиологически подходящий носитель» и «фармацевтически приемлемый носитель» используются взаимозаменяемо и относятся к одобренному носителю или разбавителю, который не вызывает заметного раздражения организма и не аннулирует биологическую активность и свойства вводимого конъюгата.
Термин «носитель» относится к разбавителю, адьюванту (вспомогательному агенту, усиливающему действие лекарственного средства), эксципиенту или переносчику, вместе с которым вводится лекарственное средство. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла, включая масла нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобные. Вода представляет собой предпочтительный носитель, когда фармацевтическую композицию вводят внутривенно. Физиологические растворы и водные растворы декстрозы и глицерина также можно использовать в качестве жидких носителей, в особенности для инъецируемых растворов. Подходящие фармацевтические эксципиенты включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, глицеринмоностеарат, тальк, хлорид натрия, высушенное снятое молоко, глицерин, пропилен, гликоль, воду, этанол и тому подобные. Композиция, при желании, также может содержать незначительные количества смачивающих или эмульгирующих агентов или рН-буферных агентов. Данные композиции могут иметь вид растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, препаратов замедленного высвобождения и тому подобного. Композиция может быть изготовлена в виде суппозитория с традиционными связующими веществами и носителями, какими как триглицериды. Пероральные препараты могут включать стандартные носители, такие как имеющие фармацевтическое качество маннит, лактоза, крахмал, стеарат магния, сахаринат натрия, целлюлоза, карбонат магния и т.д. Примеры подходящих фармацевтических носителей описаны в справочнике E.W.Martin “Remington's Pharmaceutical Science”. Такие композиции будут содержать терапевтически эффективное количество белка, предпочтительно в очищенном виде, вместе с подходящим количеством носителя таким образом, чтобы обеспечить форму для надлежащего введения пациенту. Препарат должен быть подходящим для способа введения.
В данном описании термин «эксципиент» относится к инертному веществу, добавляемому к фармацевтической композиции для дополнительного облегчения процессов и введения активных ингредиентов. Примеры эксципиентов, без ограничения, включают карбонат кальция, фосфат кальция, различные сахара и типы крахмалов, производные целлюлозы, желатин, растительные масла и полиэтиленгликоли.
Дополнительные способы получения рецептур лекарственных препаратов и введения активных ингредиентов можно найти в книге Remington's Pharmaceutical Science”, Mark Publishing Co., Easton PA, последнее издание, которое включено в данное описание во всей полноте путем ссылки.
Описанные здесь фармацевтические композиции также могут включать подходящие твердые или гелеобразные носители или эксципиенты. Примеры таких носителей или эксципиентов включают, но не ограничиваются указанным, карбонат кальция, фосфат кальция, различные сахара, крахмалы, производные целлюлозы, желатин и полимеры, такие как полиэтиленгликоли.
Подходящие пути введения могут, например, включать пероральный, ректальный, чресслизистый, чрескожный, кишечный или парентеральный пути доставки, включая внутримышечные, подкожные и интрамедуллярные инъекции, а также подоболочечные, прямые внутрижелудочковые, внутривенные, внутрибрюшинные, внутриназальные или внутриглазные инъекции.
Таким образом, фармацевтические композиции для применения в соответствии с настоящим изобретением могут быть получены обычным образом с использованием одного или нескольких фармацевтически приемлемых носителей, включая эксципиенты и вспомогательные добавки, которые облегчают переработку активных ингредиентов в препараты, которые могут фармацевтически использоваться. Надлежащий препарат зависит от выбранного пути введения.
Для инъекции активные ингредиенты по изобретению могут быть изготовлены в виде лекарственных препаратов в водных растворах, предпочтительно в физиологически совместимых буферных растворах, таких как раствор Хэнка, раствор Рингера или физиологический буферный раствор. Для введения через слизистую оболочку в препаратах используются агенты, способствующие проницаемости. Такие агенты, способствующие проницаемости, обычно известны в данной области.
Для перорального введения активные ингредиенты необязательно могут быть изготовлены в виде лекарственных препаратов для введения цельных клеток, продуцирующих белок по настоящему изобретению, такой, например, как GCD. Активный ингредиент также может быть введен в состав лекарственных препаратов путем объединения активных ингредиентов и/или клеток с фармацевтически приемлемыми носителями, хорошо известными в данной области. Такие носители дают возможность введения активных ингредиентов по изобретению в состав таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, гидросмесей, суспензий, и тому подобного для перорального всасывания у пациента. Фармакологические препараты для перорального использования могут быть получены с использованием твердого эксципиента, необязательно путем измельчения полученной смеси и переработки смеси в гранулы, после добавления, при желании, подходящих вспомогательных добавок, для получения ядра таблеток или драже. Подходящими эксципиентами, в частности, являются наполнители, такие как сахара, включая лактозу, сахарозу, маннит или сорбит; препараты целлюлозы такие как, например, кукурузный крахмал, пшеничный крахмал, рисовый крахмал, картофельный крахмал, желатин, трагакантовая камедь, метилцеллюлоза, гидроксипропилметилцеллюлоза, карбоксиметилцеллюлоза натрия; и/или физиологически приемлемые полимеры, такие как поливинилпирролидон (ПВП). При желании могут быть добавлены дезинтегрирующие агенты, такие как сшитый поливинилпирролидон, агар или альгиновая кислота или ее соли, такие как альгинат натрия.
Ядро драже обеспечивают подходящими покрытиями. С этой целью можно использовать концентрированные растворы сахара, которые необязательно могут содержать аравийскую камедь, тальк, поливинилпирролидон, карбополовый гель, полиэтиленгликоль, диоксид титана, растворы глазури и подходящие органические растворители или смеси растворителей. В покрытия таблеток или драже могут быть добавлены красители или пигменты для идентификации или охарактеризовывания различных комбинаций доз активных ингредиентов.
Фармацевтические композиции, которые можно использовать перорально, включают капсулы с плотной посадкой, изготовленные из желатина, а также мягкие герметично закрытые капсулы, изготовленные из желатина и пластификатора, такого как глицерин или сорбит. Капсулы с плотной посадкой могут содержать активные ингредиенты в смеси с наполнителем, таким как лактоза, связующими веществами, такими как крахмалы, лубрикантами, такими как тальк или стеарат магния и, необязательно, стабилизаторами. В мягких капсулах активные ингредиенты могут быть растворены или суспендированы в подходящих жидкостях, таких как жирные масла, жидкий парафин или жидкие полиэтиленгликоли. Кроме того, могут быть добавлены стабилизаторы. Все препараты для перорального введения должны быть в дозировках, подходящих для выбранного пути введения.
Для буккального введения композиция может иметь вид таблеток или пастилок, полученных обычным образом.
При введении путем ингаляции активные ингредиенты для применения в соответствии с настоящим изобретением обычно доставляют с помощью аэрозольного спрея из упаковки под давлением или небулайзера с использованием подходящего пропеллента, например дихлордифторметана, трихлорфторметана, дихлортетрафторметана или диоксида углерода. В случае аэрозоля под давлением единица дозировки может определяться за счет обеспечения клапана для доставки отмеренного количества. Могут быть получены капсулы и катриджи, например, из желатина, содержащие порошкообразную смесь активного ингредиента и подходящей порошкообразной основы, такой как лактоза или крахмал, для применения в ингаляторе или аппарате для вдувания.
Описанные здесь активные ингредиенты могут быть использованы для получения препаратов для парентерального введения, например, посредством болюсной инъекции или непрерывного вливания. Препараты для инъекции могут быть представлены в виде единичной препаративной лекарственной формы, например в ампулах или в многодозовых контейнерах, необязательно с добавлением консерванта. Композиции могут представлять собой суспензии, растворы или эмульсии в маслянистых или водных носителях и могут содержать агенты для получения лекарственных препаратов, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты.
Фармацевтические композиции для парентерального введения включают водные растворы активного препарата в водорастворимой форме. Кроме того, могут быть получены суспензии активных ингредиентов в виде подходящих масляных суспензий для инъекций. Подходящие липофильные растворители или переносчики включают жирные масла, такие как кунжутное масло, или синтетические эфиры жирных кислот, такие как этилолеат, триглицериды или липосомы. Водные инъекционные суспензии могут содержать вещества, которые увеличивают вязкость суспензии, такие как карбоксиметилцеллюлоза натрия, сорбит или декстран. Необязательно суспензия также может содержать подходящие стабилизаторы или агенты, которые увеличивают растворимость активных ингредиентов, чтобы иметь возможность получения растворов с высокой концентрацией.
В предпочтительном варианте осуществления композицию получают в соответствии с обычными способами в виде фармацевтической композиции, адаптированной для внутривенного введения людям. Обычно фармацевтические композиции для внутривенного введения представляют собой растворы в стерильном изотоническом водном буфере. Обычно ингредиенты обеспечивают или по отдельности или смешивают их вместе в единичной препаративной лекарственной форме, например в виде сухого лиофилизованного порошка или не содержащего воды концентрата в герметически закупоренном контейнере, таком как ампула или саше, с указанием количества активного агента. Когда композиция предназначена для введения путем вливания, ее можно отпускать вместе с бутылкой для инфузии, содержащей воду или физиологический раствор фармацевтического качества. Когда композиции вводят путем инъекции, может обеспечиваться ампула со стерильной водой для инъекции или с физиологическим раствором так, чтобы ингредиенты могли быть смешаны перед введением.
Фармацевтические композиции по изобретению могут быть получены в нейтральном виде или в виде соли. Фармацевтически приемлемые соли включают те, которые образованы анионами, такими как полученные из хлористоводородной, фосфорной, уксусной, щавелевой, винной кислот и т.д., и те, которые образованы катионами, такими как полученные из гидроксидов натрия, калия, аммония, кальция, железа, изопропиламина, триэтиламина, 2-этиламминоэтанола, гистидина, прокаина, и т.д.
Активные ингредиенты по настоящему изобретению также могут быть введены в состав ректальных композиций, таких как суппозитории или удерживающие клизмы с использованием, например, обычных основ для суппозиториев, таких как масло какао или другие глицериды.
Описанные здесь фармацевтические композиции также могут включать подходящие твердые или гелеобразные носители или эксципиенты. Примеры таких носителей или эксципиентов включают, но не ограничиваются указанным, карбонат кальция, фосфат кальция, различные сахара, крахмалы, производные целлюлозы, желатин и полимеры, такие как полиэтиленгликоли.
Введение необязательно проводят наружным (местным) путем, что сопровождается применением носителя для наружного применения. Носитель для наружного применения представляет собой такой носитель, который, в общем, является подходящим для местного введения активного ингредиента и включает любые подобные материалы, известные в данной области. Носитель для местного применения выбирают таким образом, чтобы обеспечить желаемую форму композиции, например, как жидкий или не жидкий носитель, лосьон, крем, паста, гель, порошок, мазь, растворитель, жидкий разбавитель, капли и тому подобное, и он может включать материалы либо природного, либо синтетического происхождения. Ясно, что существенным являются, чтобы выбранный носитель не оказывал неблагоприятного действия на активный ингредиент или другие компоненты местного препарата и был стабильным в отношении всех компонентов местного препарата. Примеры подходящих носителей для местного применения включают воду, спирты и другие нетоксичные органические растворители, глицерин, минеральное масло, силикон, вазелин, ланолин, жирные кислоты, растительные масла, парабены, воски и тому подобное. Предпочтительные препараты в данном описании представляют собой бесцветные, не имеющие запаха мази, жидкости, лосьоны, кремы и гели.
Мази представляют собой полутвердые препараты, которые обычно основаны на вазелине или других производных вазелина. Конкретная используемая основа мази, как это будет очевидно специалистам в данной области, будет представлять собой ту, которая будет обеспечивать оптимальную доставку активных ингредиентов и, предпочтительно будет обеспечивать другие желаемые характеристики, например смягчающие свойства или тому подобные. Как и в случае других носителей или переносчиков, основа мази должна быть инертной, стабильной, не раздражающей и не чувствительной. Как объясняется в справочнике Remington: The Science and Practice of Pharmacy, 19th Ed. (Easton, Pa.: Mark Publishing Co., 1995) на страницах 1399-1404, основы мазей могут быть сгруппированы в четыре класса: маслянистые основы; эмульгируемые основы; эмульсионные основы и водорастворимые основы. Маслянистые основы мазей включают, например, растительные масла, жиры, полученные из животных, и полутвердые углеводороды, полученные из нефти. Эмульгируемые основы мазей, также известные как абсорбирующие основы мазей, содержат незначительное количество воды или не содержат воды и включают, например, гидростеаринсульфат, безводный ланолин и гидрофильный вазелин. Эмульсионные основы мазей представляют собой или эмульсии вода-в-масле (В/М) или эмульсии масло-в-воде (М/В) и включают, например, цетиловый спирт, глицерилмоностеарат, ланолин и стеариновую кислоту. Предпочтительные водорастворимые основы мазей получают из полиэтиленгликолей с различной молекулярной массой; опять же для получения дополнительной информации можно сослаться на Remington: The Science and Practice of Pharmacy.
Лосьоны представляют собой препараты для нанесения на поверхность кожи без трения, и обычно они представляют собой жидкие или полужидкие препараты, в которых твердые частицы, включая активный агент, присутствуют в воде или в спиртовой основе. Лосьоны обычно представляют собой суспензии твердых веществ и могут включать жидкую маслянистую эмульсию типа масло-в-воде. Лосьоны являются предпочтительными препаратами в данном изобретении для лечения больших площадей тела вследствие легкости нанесения большого количества жидкой композиции. Обычно необходимо, чтобы нерастворимое вещество в растворе было тонкодисперсным. Лосьоны обычно будут содержать суспендирующие агенты для получения лучших дисперсий, а также активные ингредиенты, полезные для локализации и поддерживания контакта активного агента с кожей, например метилцеллюлозу, карбоксиметилцеллюлозу и тому подобное.
Кремы, содержащие выбранные активные ингредиенты, представляют собой, как известно в данной области, вязкие жидкие и полутвердые эмульсии или масло-в-воде, или вода-в-масле. Основы крема являются смываемыми водой и содержат масляную фазу, эмульгатор и водную фазу. Масляная фаза, также иногда называемая «внутренней фазой», обычно включает вазелин и жирный спирт, такой как цетиловый или стеариловый спирт; водная фаза обычно, хотя и не обязательно, превышает масляную фазу по объему и обычно содержит увлажнитель. Эмульгатор в препарате крема, как объяснено в справочнике Remington, процитированном выше, обычно представляет собой неионное, анионное, катионное или амфотерное поверхностно-активное вещество.
Препараты в виде геля являются предпочтительными для нанесения на кожу головы. Как будет очевидно работающим в области препаратов активных ингредиентов для наружного применения, гели представляют собой полутвердые системы типа суспензий. Однофазные гели содержат органические макромолекулы, по существу равномерно распределенные в жидком носителе, который обычно является водным, но также предпочтительно содержит спирт и, необязательно, масло.
В препараты по изобретению для наружного применения могут быть включены различные вспомогательные добавки, известные специалистам в данной области. Например, растворители можно использовать для солюбилизации некоторых активных веществ-ингредиентов. Другие необязательные добавки включают усилители проницаемости кожи, глушители, антиоксиданты, желирующие агенты, загустители, стабилизаторы и том подобные.
Композиции для наружного применения по настоящему изобретению также могут доставляться к коже с помощью обычных пластырей дермального типа или изделий, в которых композиции активных ингредиентов содержатся внутри ламинированной структуры, которая служит устройством доставки лекарственного средства, прикрепляемым к коже. В такой структуре композиция активных ингредиентов содержится в слое или «резервуаре», расположенном под верхним защитным слоем. Ламинированная структура может содержать один резервуар или же она может содержать множество резервуаров. В одном варианте осуществления резервуар включает полимерную матрицу из фармацевтически приемлемого контактного прилипающего материала (адгезива), который служит для прикрепления системы к коже во время доставки активных ингредиентов. Примеры подходящих для кожи контактных прилипающих материалов включают, но не ограничиваются указанным, полиэтилены, полисилоксаны, полиизобутилены, полиакрилаты, полиуретаны и тому подобное. Конкретные полимерные выбираемые прилипающие материалы будут зависеть от конкретных активных ингредиентов, носителя и т.д., т.е. прилипающий материал должен быть совместимым со всеми компонентами композиции, содержащей активные ингредиенты. Альтернативно, резервуар, содержащий активные компоненты, и контактирующий с кожей прилипающий материал могут присутствовать в виде отдельных и индивидуальных слоев, при этом прилипающий материал расположен ниже резервуара, который в таком случае может или представлять собой полимерную матрицу, как описано выше, или может представлять собой жидкий или гидрогелевый резервуар, или может иметь какую-либо другую форму.
Защитный слой в таких ламинатах, который служит верхней поверхностью устройства, функционирует в качестве первичного структурного элемента ламинированной структуры и обеспечивает устройством большей частью его гибкости. Материал, выбираемый для защитного слоя, следует выбирать таким образом, чтобы он был по существу непроницаемым для активных ингредиентов и для любых других компонентов композиции, содержащей активные ингредиенты, предотвращая таким образом потерю любых компонентов через верхнюю поверхность устройства. Защитный слой может быть или герметичным или не герметичным, в зависимости от того, является ли желательным, чтобы кожа увлажнялась во время доставки активных ингредиентов. Защитный слой обычно изготавливают из листа или плетки предпочтительно гибкого эластомерного материала. Примеры полимеров, которые являются подходящими для защитного слоя, включают полиэтилен, полипропилен и сложные полиэфиры.
Во время хранения и перед использованием ламинированная структура включает снимаемую прокладку. Непосредственно перед употреблением данный слой удаляют из устройства, оставляя открытой основную структуру, или резервуар с активными ингредиентами, или отдельный контактный адгезивный слой, таким образом, чтобы можно было прикрепить систему к коже. Такая снимаемая прокладка должна быть изготовлена из материала, не проницаемого для активных ингредиентов/носителя.
Такие устройства могут быть изготовлены с использованием обычных технологий, известных в данной области, например, путем выливания жидкой смеси адгезива, активных ингредиентов и носителя на защитный слой с последующим ламинированием с использованием снимаемой прокладки. Аналогично, адгезивную смесь можно выливать на снимаемую прокладку с последующим ламинированием с использованием защитного слоя. Альтернативно, резервуар для активных ингредиентов может быть получен в отсутствие активных ингредиентов или эксципиента, а затем нагружен ими путем «пропитки» смесью активных ингредиентов и носителя.
Что касается лекарственных форм для наружного (местного) применения по изобретению, композиции активных ингредиентов, содержащиеся в резервуарах для активных ингредиентов данных ламинированных систем, могут содержать ряд компонентов. В некоторых случаях активные ингредиенты могут доставляться «неразбавленными», т.е. в отсутствие дополнительной жидкости. Однако в большинстве случаев активные ингредиенты будут растворены, диспергированы или суспендированы в подходящем фармацевтически приемлемом носителе, обычно в растворителе или геле. Другие компоненты, которые могут присутствовать, включают консерванты, стабилизаторы, поверхностно-активные вещества и тому подобное.
Следует отметить, что белок по изобретению, такой как высокоманнозный лизосомальный фермент, предпочтительно вводят нуждающемуся в этом пациенту в эффективном количестве. Как использовано в данном описании, термин «эффективное количество» означает количество, необходимое для достижения выбранного результата. Например, эффективное количество композиции по изобретению может быть выбрано как используемое при лечении лизосомальной болезни накопления.
Фармацевтические композиции, подходящие для применения в контексте настоящего изобретения, включают композиции, в которых активные ингредиенты содержатся в количествах, эффективных для достижения предполагаемой цели. Более конкретно, выражение «терапевтически эффективное количество» означает количество активного ингредиента, эффективного для профилактики, облегчения или улучшения симптомов заболевания или продления жизни пациента, подвергаемого лечению.
Определение терапевтически эффективного количества находится в пределах способностей специалиста в данной области, особенно в свете представленного здесь подробного описания.
Для любого активного ингредиента, используемого в способах изобретения, терапевтически эффективное количество или доза могут вначале быть оценены с помощью анализов активности на животных. Например, доза может быть разработана на животных моделях для достижения диапазона циркулирующей концентрации, который включает значение IC50, определенное с помощью анализов активности.
Токсичность и терапевтическая эффективность описанных здесь активных ингредиентов может быть определена с помощью стандартных фармацевтических способов на экспериментальных животных, например, путем определения IC50 и LD50 (летальная доза, вызывающая смерть 50% тестируемых животных) для рассматриваемого активного ингредиента. Данные, полученные в данных анализах активности и при исследованиях на животных, можно использовать при разработке диапазона дозировки для применения человеком. Например, терапевтически эффективные дозы, подходящие для лечения генетических заболеваний, могут быть определены из экспериментов на животных моделях для данных заболеваний.
Дозировка может изменяться в зависимости от используемой препаративной лекарственной формы и используемого пути введения. Точная рецептура, путь введения и дозировка могут быть выбраны отдельным лечащим врачом, исходя из состояния пациента. (См., например, Fingl et al., 1975, в “The Pharmacological Basis of Therapeutics”, часть 1, стр.1.)
Количества и диапазон дозировки могут регулироваться индивидуально для обеспечения уровней активного фрагмента в плазме крови, которые являются достаточными для поддержания модулирующего действия, выраженные в минимальной эффективной концентрации (МЕС). МЕС будет изменяться для каждого препарата, но необязательно может быть оценена по данным для животного в целом.
Интервалы дозировки также могут быть определены с использованием величины МЕС. Препараты необязательно можно вводить с использованием режима, который поддерживает уровни в плазме крови, превышающие МЕС на 10-90% во времени, предпочтительно между 30-90% и наиболее предпочтительно между 50-90%.
В зависимости от серьезности и способности к реагированию состояния, подвергаемого лечению, доза также может представлять собой единственное введение описанной выше композиции медленного высвобождения при курсе лечения, продолжающемся от нескольких дней до нескольких недель, или до тех пор, пока излечение или уменьшение болезненного состояния не будут достигнуты.
Композиции по настоящему изобретению могут, при желании, быть представлены в упаковке или в дозирующем устройстве, таком как набор, утвержденный Управлением по контролю за продуктами и лекарствами США, который может содержать одну или несколько единичных препаративных лекарственных форм, содержащих активный ингредиент. Упаковка, например, может включать металлическую или пластиковую фольгу, представляя собой, например, блистерную упаковку. Упаковка или раздаточное устройство могут сопровождаться инструкцией по введению. Упаковка или раздаточное устройство также могут сопровождаться уведомлением, связанным с контейнером, в форме, установленной правительственным агентством, регулирующим производство, применение или продажу фармацевтических препаратов, указанное предупреждение отражает одобрение агентством формы композиции или введения людям или животным. Такое уведомление, например, может быть с пометкой одобрения Управлением по контролю за продуктами и лекарствами США для лекарств, отпускаемых по рецепту, или в виде вкладыша одобренного продукта. Композиции, включающие активный ингредиент по изобретению, содержащийся в совместимом фармацевтическом носителе, также могут быть получены, помещены в подходящий контейнер и помечены как предназначенные для лечения указанного состояния.
Как использовано в данном описании, термин «модулировать» включает по существу ингибирование, замедление или обращение прогрессирования заболевания, по существу улучшение клинических симптомов заболевания или состояния или по существу предотвращение появления клинических симптомов заболевания или состояния. Термин «модулятор», следовательно, относится к агенту, который может модулировать заболевание или состояние.
Список последовательностей
SEQ ID NO: 1
Последовательность аминокислот сигнального пептида ER
MKTNLFLFLIFSLLLSLSSAEF
SEQ ID NO: 2
Последовательность аминокислот вакуолярного нацеливающего сигнала хитиназы А табака:
DLLVDTM
SEQ ID NO: 3
Последовательность нуклеиновой кислоты прямого праймера:
cagaattcgcccgcccctgca
SEQ ID NO: 4
Последовательность нуклеиновой кислоты обратного праймера:
ctcagatcttggcgatgccaca
SEQ ID NO: 5
Последовательность нуклеиновой кислоты прямого праймера промотора 35S:
ctcagaagaccagagggct
SEQ ID NO: 6
Последовательность нуклеиновой кислоты обратного праймера терминатора:
caaagcggccatcgtgc
SEQ ID NO: 7
Последовательность нуклеиновой кислоты кДНК GCD человека, используемая в конструктах по изобретению

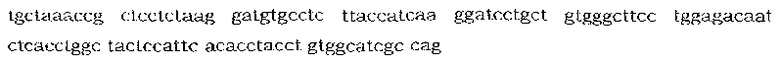
SEQ ID NO: 8
Последовательность аминокислот глюкоцереброзидазы

SEQ ID NO: 9
Последовательность нуклеиновой кислоты промотора 35S

SEQ ID NO: 10
Последовательность нуклеиновой кислоты, кодирующая сигнальный ER-пептид
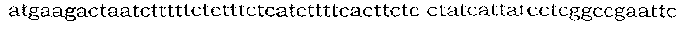
SEQ ID NO: 11
Последовательность нуклеиновой кислоты, кодирующая вакуолярную нацеливающую последовательность
gatcttttagtcgatactatg
SEQ ID NO: 12
Последовательность нуклеиновой кислоты терминатора
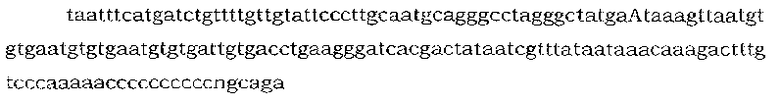
SEQ ID NO: 13
Последовательность нуклеиновой кислоты кассеты экспрессии по изобретению



SEQ ID NO: 14
Последовательность аминокислот рекомбинантного белка по изобретению
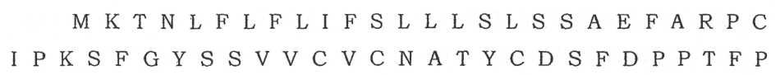

Другие варианты осуществления
Следует понимать, что, хотя изобретение было описано в сочетании с его подробным описание, вышеуказанное описание предназначено для иллюстрации, а не для ограничения объема изобретения, который определен объемом прилагаемой формулы изобретения. Другие аспекты, преимущества и модификации входят в объем следующей формулы изобретения.
Список литературы
1. Ma, J.K.C, Drake, P.M.W., and Christou, P. (2003) Nature reviews 4, 794-805.
2. Lerouge, P., Cabanes-Macheteau, M., Rayon, C., Fischette-Laine, A.C., Gomord, V., Faye, L. (1998) Plant Mol Biol 38, 31-48.
3. Lee, R.E. (1982) Prog Clin Biol Res 95, 177-217.
4. Grabowski, G. (1993) Adv Hum Genet. 21, 377-441.
5. Grabowski, G.A., and Hopkin, R.J. (2003) Annual Review of Genomics and Human Genetics 4, 403-436.
6. Sorge, J.W., C., Westwood, B., Beutler, E. (1985) Proc Natl Acad Sci USA. 82, 7289-7293.
7. Berg-Fussman, A., Grace, M., Ioannou, Y., and Grabowski, G. (1993) J. Biol. Chem. 268, 14861-14866.
8. Grace, M., Grabowski, GA. (1990) Biochem Biophys Res Commun 168, 771-777.
9. Grace, M., Newman, K., Scheinker, V., Berg-Fussman, A., and Grabowski, G. (1994) J. Biol. Chem. 269, 2283-2291.
10. Barton, N.W., Brady, R.O., Dambrosia, J.M., Di Bisceglie, A.M., Doppelt, S.H., Hill, S.C., Mankin, H.J., Murray, G.J., Parker, R.I., Argoff, C.E., et al. (1991) N Engl J Med. 324, 1464-1470.
11. Grabowski, G.A., Barton, N.W., Pastores, G., Dambrosia, J.M., Banerjee, T.K., McKee, M.A., Parker, C., Schifrmann, R., Hill, S.C., and Brady, R.O. (1995) Ann Intern Med 122, 33-39.
12. Pastores, G.M., Sibille, A.R., Grabowski, G.A. (1993) Blood 82, 408-416.
13. Weinreb, N.J., Charrow, J., Andersson, H.C., Kaplan, P., Kolodny, E.H., Mistry, P., Pastores, G., Rosenbloom, BE., Scott, C.R., Wappner, R.S., Zimran, A. (2002) Am J Med 113, 112-119.
14. Bijsterbosch, M.K., Donker, W., van de Bilt, H., van Weely, S., van Berkel, T.J., Aerts, J.M. (1996) Eur J Biochem 237 , 344-349.
15. Friedman, B., Vaddi, K., Preston, C., Mahon, E., Cataldo, J.R., and McPherson, J.M. (1999) Blood 93 , 2807-2816.
16. Furbish, F.S., Steer, C.J., Krett, N.L., Barranger, J.A. (1981) Biochim Biophys Acta 673, 425-434.
17. Doebber, T., Wu, M., Bugianesi, R., Ponpipom, M., Furbish, F., Barranger, I., Brady, R., and Shen, T. (1982) J. Biol. Chem. 257 , 2193-2199.
18. Dwek, R.A., Butters, T.D., Platt, F.M., Zitzmann, N. (2002) Nature reviews 1, 65-75.
19. Neuhaus, J.M., Rogers, J.C. (1998) Plant Mol Biol 38, 127-144.
20. Vitale, A., and Galili, G. (2001) Plant Physiol. 125, 115-118.
21. Hellens, R., Edwards, E.A., Leyland, N.R., Bean, S., Mullineaux, P.M. (2000) Plant Mol Biol 42, 819-832.
22. Wurtele, E.S., Bulka, K. (1989) Plant Sci 61, 253-262.
23. den Dulk-Ras, A., Hooykaas, P.J. (1995) Methods Mol Biol. 55, 63-72.
24. Laemmli, U.K. (1970) Nature reviews 227, 680-685.
25. Bradford, M.M. (1976) Anal Biochem 72, 248-254.
26. Stahl, P.G.S. (1982) J Cell Biol 93, 49-56.
27. Takasaki, S., Murray, G., Furbish, F., Brady, R., Barranger, J, and Kobata, A. (1984) J. Biol. Chem. 259, 10112-10117.
28. Lerouge, P., Cabanes-Macheteau, M., Rayon, C., Fitchette-Laine, A. C., Gomord, V., and Faye, L. (1998) Plant Mol. Biol. 38, 31-48.
29. Frigerio, L., Pastres, A., Prada, A., and Vitaie, A. (2001) Plant Cell 13, 1109-1126.
30. Frigerio, L., de Virgilio, M., Prada, A., Faoro, F., and Vitaie, A. (1998) Plant Cell 10, 1031-1042.
31. Hadlington JL, D. J. (2000) Curr Opin Plant Biol. 3, 461-68.
32. Okamoto, T., Shimada, T., Hara-Nishimura, I., Nishimura, M., and Minamikawa, T. (2003) Plant Physiol. 132, 1892-1900.
33. Bardor, M., Faveeuw, C. Fitchette, A.-C., Gilbert, D., Galas, L., Trottein, F., Faye, L., and Lerouge, P. (2003) Glycobiology 13, 427-434.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2004 |
|
RU2385928C2 |
| ПРОДУКЦИЯ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ФУКОЗИЛИРОВАНИЕМ | 2008 |
|
RU2479629C2 |
| ИЗГОТОВЛЕНИЕ АКТИВНЫХ ВЫСОКОФОСФОРИЛИРОВАННЫХ ЛИЗОСОМАЛЬНЫХ ФЕРМЕНТОВ СУЛЬФАТАЗ ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2510820C2 |
| ФЕРМЕНТ ЛИЗОСОМАЛЬНОЙ БОЛЕЗНИ НАКОПЛЕНИЯ | 2017 |
|
RU2741116C2 |
| Лекарственное средство на основе бифункционального антитела для лечения мукополисахаридоза II типа | 2016 |
|
RU2673038C2 |
| ФЕРМЕНТ ЛИЗОСОМАЛЬНОЙ БОЛЕЗНИ НАКОПЛЕНИЯ | 2011 |
|
RU2575074C2 |
| Способ контроля гликозилирования рекомбинантного гликопротеина | 2015 |
|
RU2670889C9 |
| СПОСОБ ОЧИСТКИ ИМИГЛЮЦЕРАЗЫ | 2017 |
|
RU2668158C1 |
| ПРОИЗВОДСТВО АКТИВНОЙ ВЫСОКОФОСФОРИЛИРОВАННОЙ N-АЦЕТИЛГАЛАКТОЗАМИН-6-СУЛЬФАТАЗЫ ЧЕЛОВЕКА И ЕЕ ПРИМЕНЕНИЕ | 2011 |
|
RU2607376C2 |
| СПОСОБЫ И МАТЕРИАЛЫ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ПОМПЕ | 2013 |
|
RU2644258C2 |










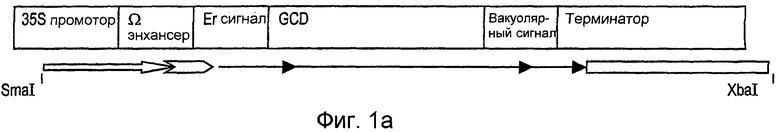
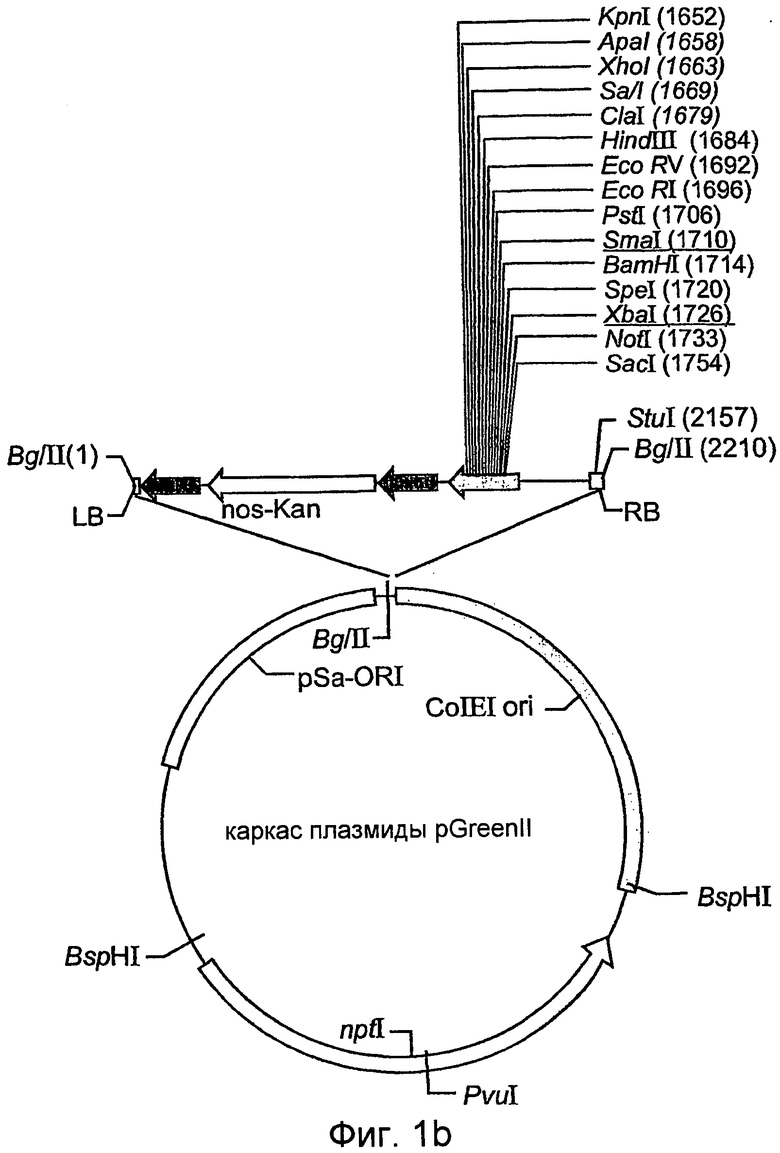

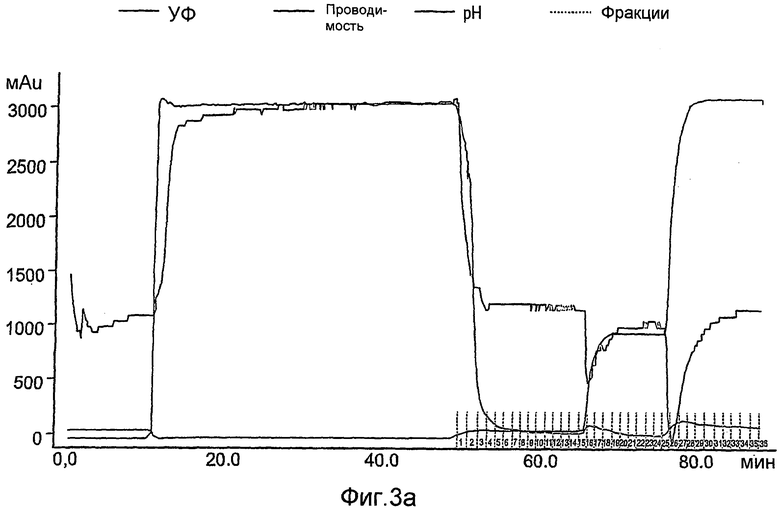
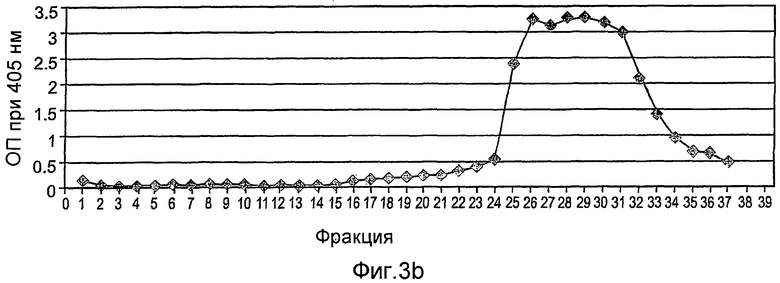
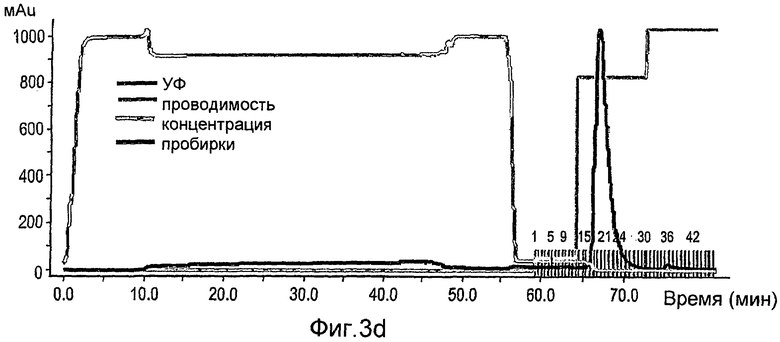
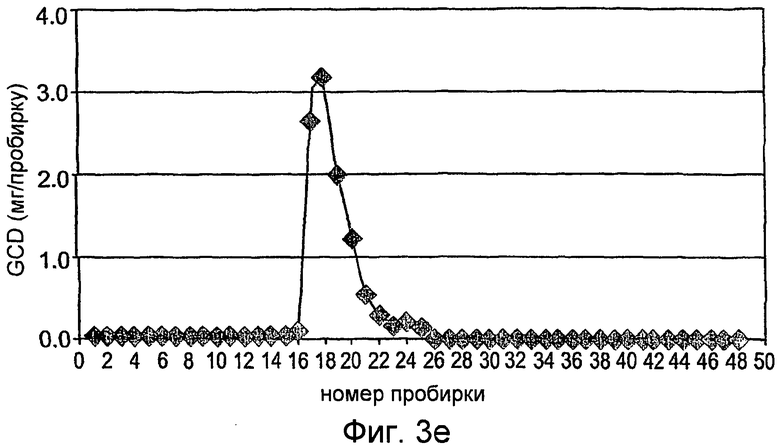

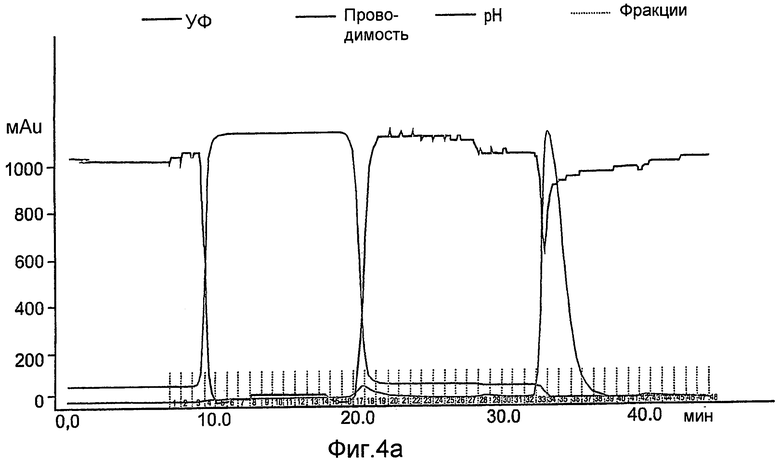
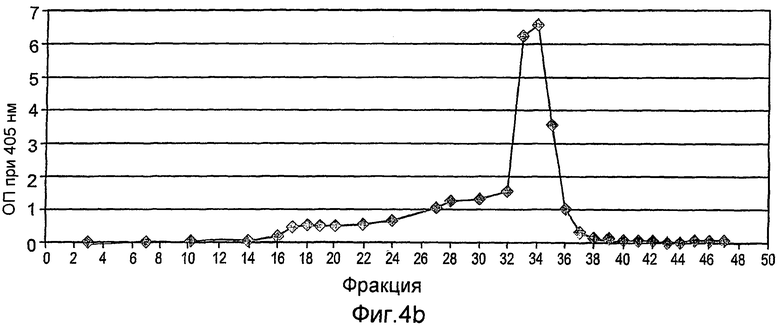
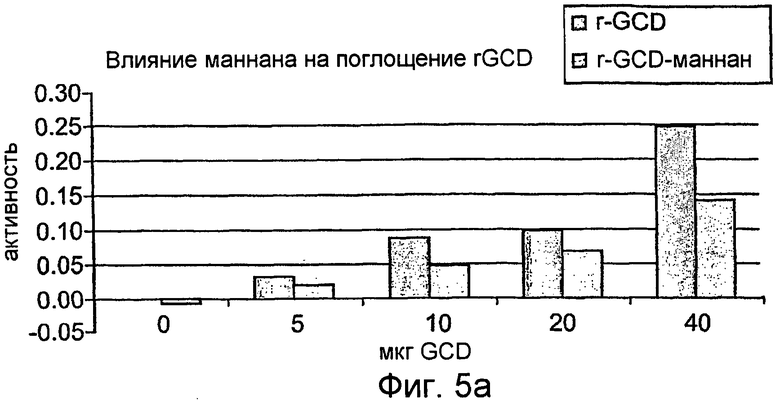
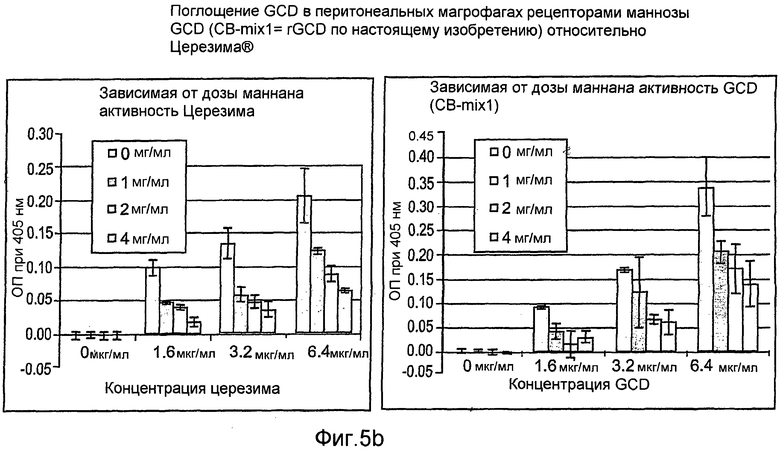
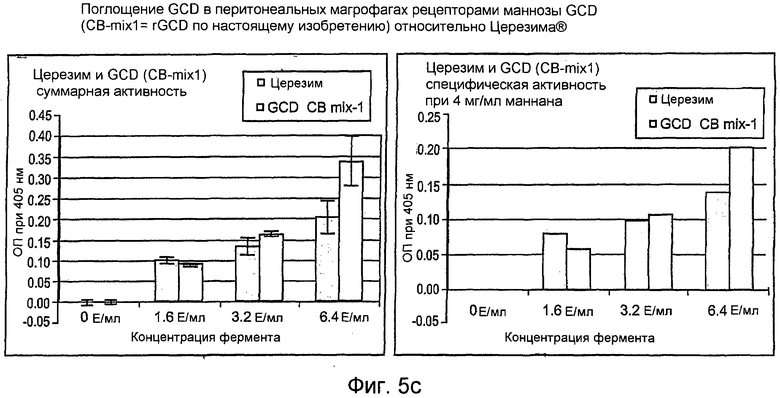
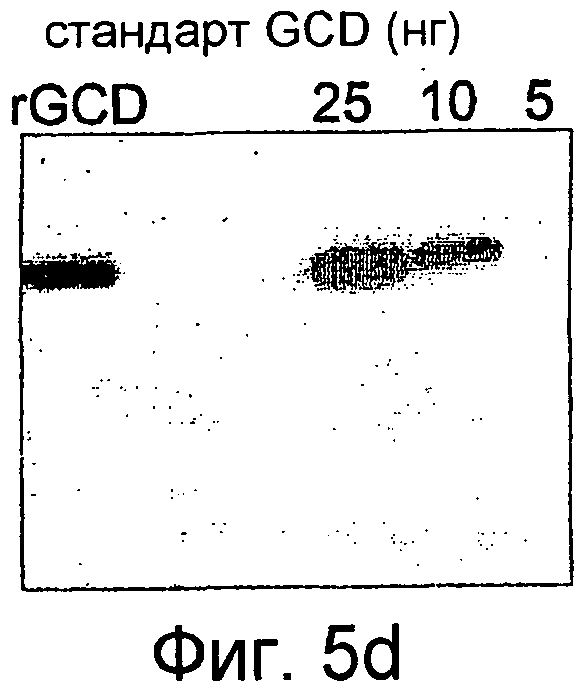
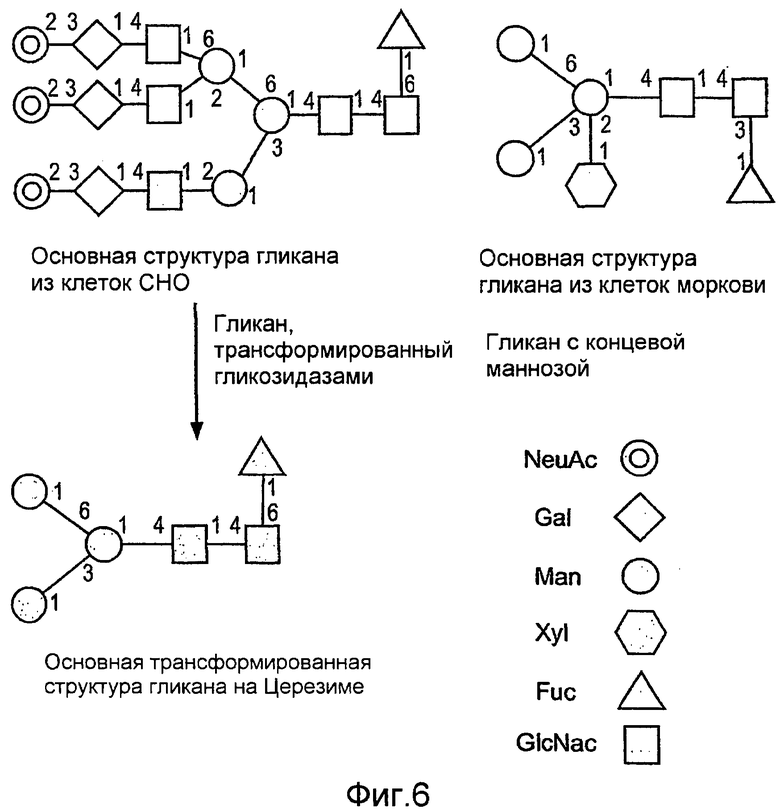
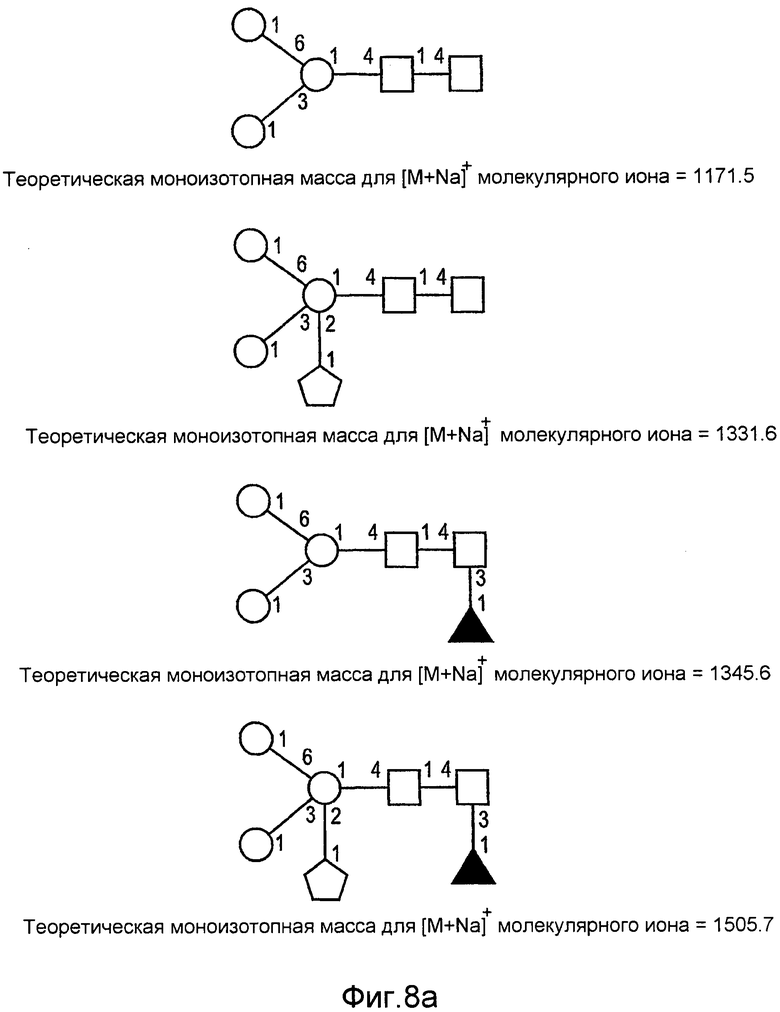
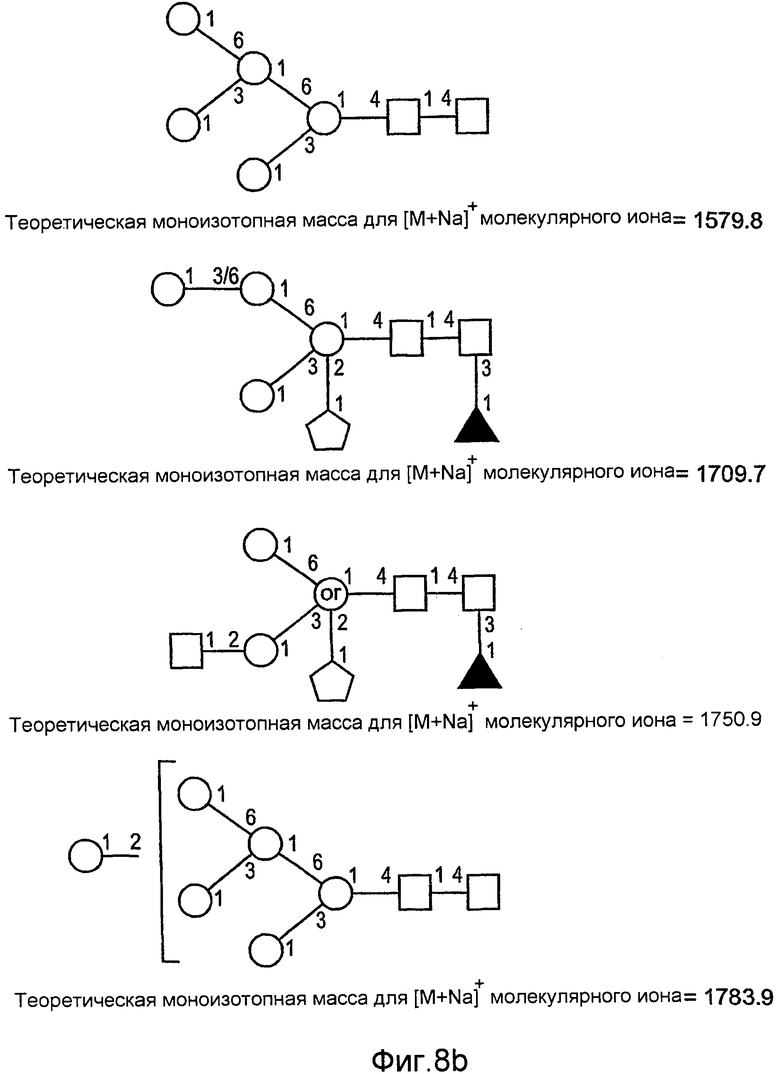
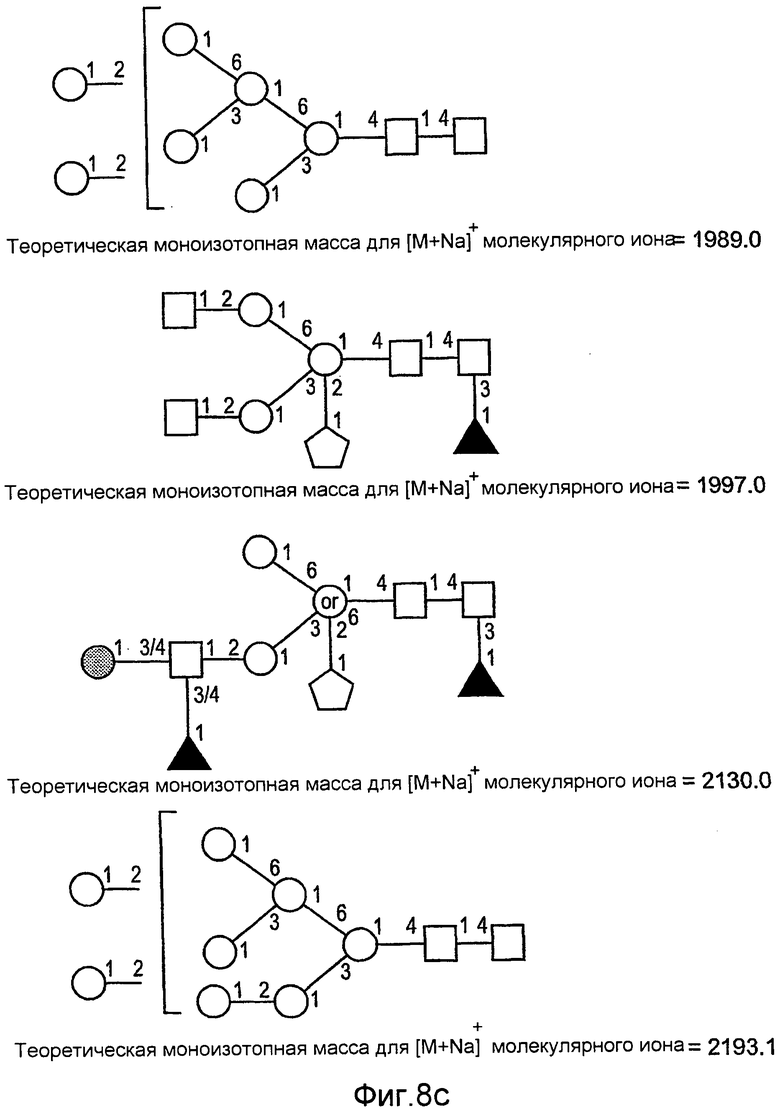
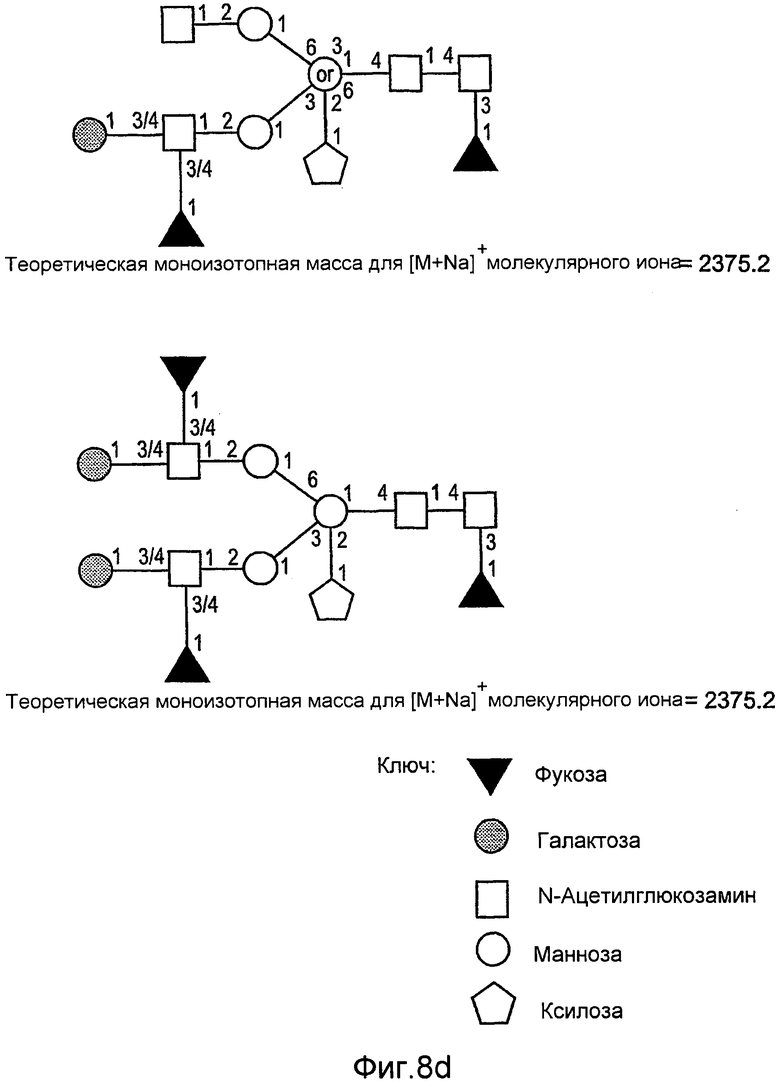
Изобретение относится к биохимии и представляет собой фармацевтическую композицию для лечения болезни Гоше, содержащую в качестве активного ингредиента лиофилизированные растительные клетки, экспрессирующие рекомбинантную человеческую глюкоцереброзидазу, и фармацевтически приемлемый носитель, где указанная фармацевтическая композиция составлена для перорального введения и где указанная человеческая глюкоцереброзидаза имеет высокоманнозное гликозилирование. Изобретение позволяет получить эффективную фармацевтическую композицию для лечения болезни Гоше с высокой биоактивностью фермента вследствие его высокоманнозного гликозилирования. 2 н. и 14 з.п. ф-лы, 21 ил., 3 табл., 6 пр.
1. Фармацевтическая композиция для лечения болезни Гоше, содержащая в качестве активного ингредиента лиофилизированные растительные клетки, экспрессирующие рекомбинантную человеческую глюкоцереброзидазу, и фармацевтически приемлемый носитель, где указанная фармацевтическая композиция составлена для перорального введения и где указанная человеческая глюкоцереброзидаза имеет высокоманнозное гликозилирование.
2. Фармацевтическая композиция по п.1, где указанный рекомбинантный человеческий лизосомальный протеин содержит по меньшей мере одну фукозу, имеющую альфа (1-3)гликозидную связь и по меньшей мере одну ксилозу.
3. Фармацевтическая композиция по п.1, где основная структура гликана указанного лизосомального человеческого протеина указанных растительных клеток содержит незащищенную маннозу.
4. Фармацевтическая композиция по п.1, где указанная растительная клетка является клеткой корня растений, выбранная из группы, состоящей из трансформированной Agrobacterium rihzogenes клетки корня, клетки сельдерея, клетки имбиря, клетки хрена и клетки моркови.
5. Фармацевтическая композиция по п.1, где указанная растительная клетка является клеткой моркови.
6. Фармацевтическая композиция по п.1, где указанные растительные клетки выращивают в культуре клеток.
7. Фармацевтическая композиция по п.5, где указанные растительные клетки выращивают как часть целого растения.
8. Фармацевтическая композиция по п.6, где указанную культуру клеток культивируют в суспензии.
9. Применение лиофилизированных растительных клеток, экспрессирующих рекомбинантную человеческую глюкоцереброзидазу, для производства лекарственного средства для лечения болезни Гоше путем перорального введения, где указанная человеческая глюкоцереброзидаза имеет высокоманнозное гликозилирование.
10. Применение по п.9, где указанный рекомбинантный человеческий лизосомальный фермент содержит по меньшей мере одну фукозу, имеющую альфа (1-3)гликозидную связь и по меньшей мере одну ксилозу.
11. Применение по п.9, где основная структура гликана указанного лизосомального человеческого протеина указанных растительных клеток содержит незащищенную маннозу.
12. Применение по п.9, где указанная растительная клетка является клеткой корня растений, выбранная из группы, состоящей из трансформированной Agrobacterium rihzogenes клетки корня, клетки сельдерея, клетки имбиря, клетки хрена и клетки моркови.
13. Применение по п.9, где указанная растительная клетка является клеткой моркови.
14. Применение по п.13, где указанные растительные клетки выращивают в культуре клеток.
15. Применение по п.13, где указанные растительные клетки выращивают как часть целого растения.
16. Применение по п.14, где указанную культуру клеток культивируют в суспензии.
| СПОСОБ ПОЛУЧЕНИЯ МОНО- ИЛИ БИС-β,β-ДИГАЛОИДВИНИЛОВЫХ ЭФИРОВ НЕПРЕДЕЛЬНЫХ СПИРТОВ И ГЛИКОЛЕЙ | 1967 |
|
SU215927A1 |
| US2002127219 A1, 12.09.2002 | |||
| RU 99123374 A, 20.12.2001 | |||

