Изобретение относится к области биотехнологии, а именно к способам производства терапевтических препаратов, выделения и очистки рекомбинантных белков, в частности к получению высокоочищенного препарата рекомбинантной β-1,4-галактозидазы (BgaA). Основными сферами применения полученной предлагаемым способом бактериальной β-1,4-галактозидазы BgaA является включение ее в технологический процесс получения ряда медицинских лекарственных препаратов, а также биомедицинские исследования в области гликобиологии.
Изобретение решает задачу создания способа получения в промышленных объемах высокоочищенной ферментативно активной рекомбинантной β-1,4-галактозидазы BgaA, пригодной для использования на стадии ремоделирования для получения терапевтически функциональных препаратов гликопротеинов с заданными свойствами, для разработки инструментов для высокочувствительного анализа гликановых цепей гликопротеинов тканей растений, животных и микроорганизмов. Изобретение относится также к способу получения препарата рекомбинантной β-1,4-галактозидазы BgaA, имеющего более высокую степень чистоты по сравнению с коммерческими препаратами, гомогенного по целевому продукту, с более низким содержанием эндотоксинов, белков и ДНК продуцента в сравнении с коммерческими препаратами.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
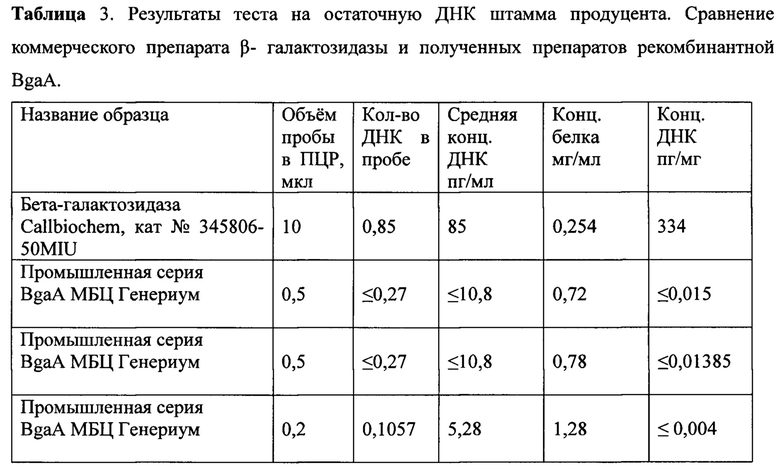
Фиг. 1. Схема строения N-гликозилированного человеческого гликопротеина. Названия сахарных остатков расположены под соответствующими им символами. Линии - связи между сахарными остатками, числа наверху - специфичность связи. Стрелки - связи, которые потенциально расщепляются гликозидазами, обозначенными вверху схемы;
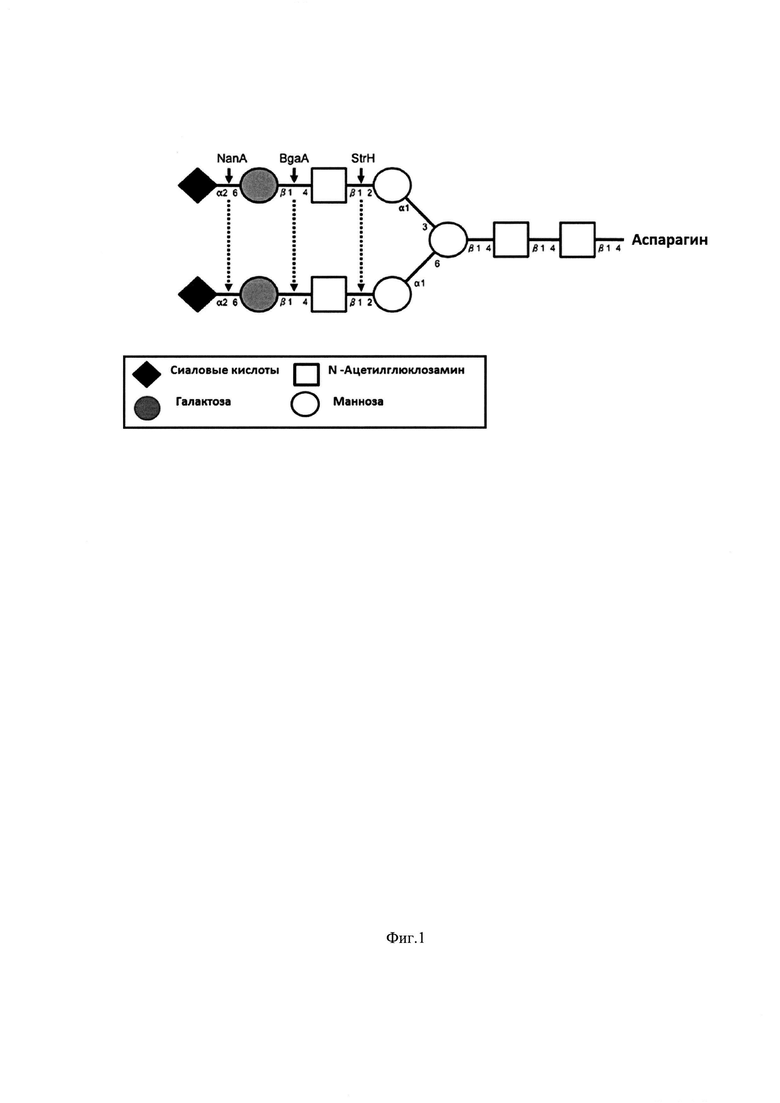
Фиг. 2. Плазмидная карта рМТ1620 (экспрессионный вектор на основе pET15b для получения стрептококковой β-галактозидазы BgaA)
Обозначения:
Т7 promoter - промотор Т7 РНК-полимеразы; N-terminal His-tagged BGAA_STRPN - ген белка His-тэгированного (N-конец) BGAA_STRPN, составной частью которого являются последовательности 20 аминокислотных остатков, содержащих гексагистидиновый тэг, и 2053 аминокислотных остатков (без сигнального пептида, G5 домена и якорного пептида) β-галактозидазы (bgaA) Streptococcus pneumoniae; Т7 terminator - терминатор Т7 РНК-полимеразы; AmpR promoter - промотер гена устойчивости к ампициллину; AmpR - ген устойчивости к ампициллину; ori - ориджин репликации; lacI -ген лактозного репрессора; lacI promoter - промотор гена лактозного репрессора; NdeI - сайт узнавания эндонуклеазы рестрикции NdeI; BamHI - сайт узнавания эндонуклеазы рестрикции BamHI
Фиг. 3. Динамика накопления биомассы при культивировании штамма-продуцента E. coli 12746 в объеме 15 л по предлагаемой схеме. По оси абсцисс - часы культивирования, по оси ординат - значения параметров культивирования.
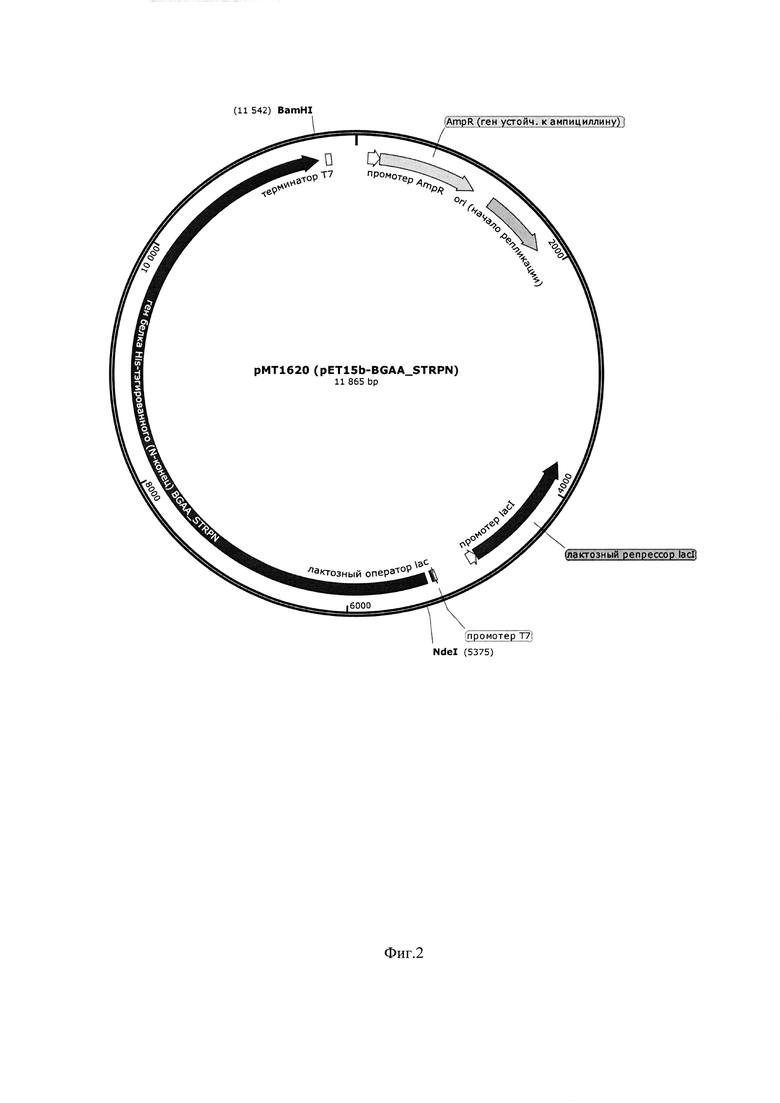
Фиг. 4. Параметры культивирования штамма-продуцента E. Coli ВКПМ 12746 в объеме 15 л по предлагаемой схеме. По оси абсцисс - часы культивирования, по оси ординат - значения параметров культивирования.
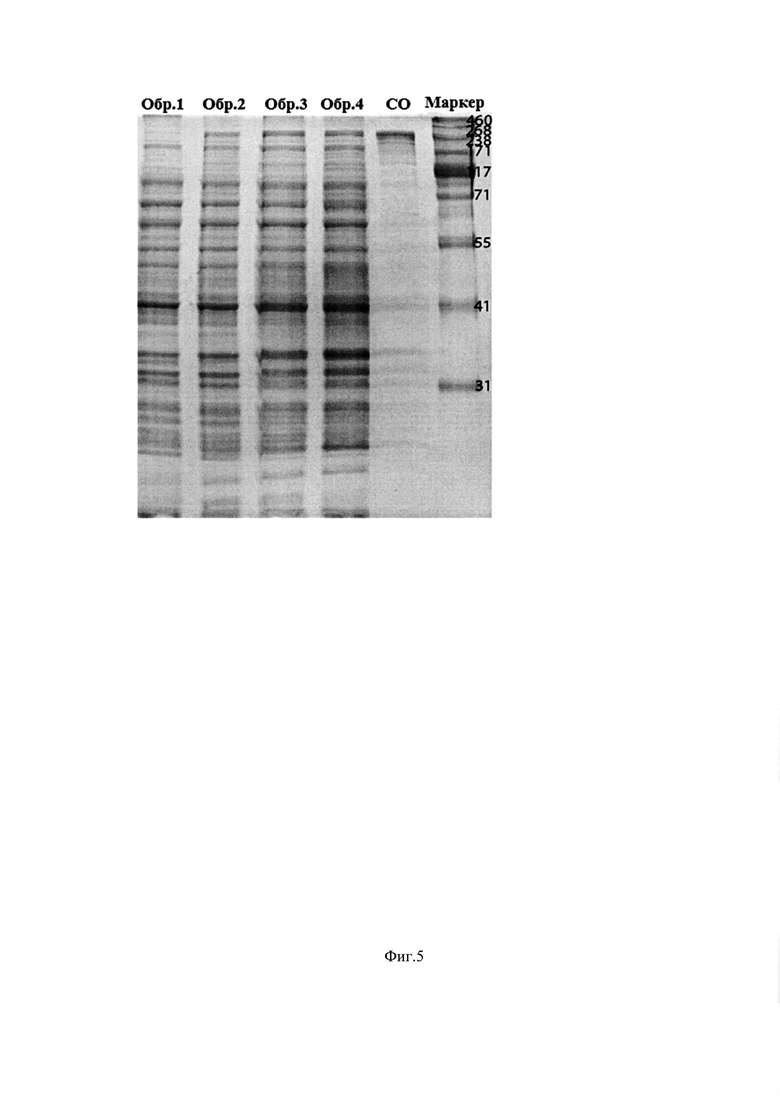
Фиг. 5. Анализ накопления белка β-галактозидазы BgaA методом электрофореза в восстанавливающих условиях в 8% ПААГ при культивировании штамма-продуцента E. Coli ВКПМ В-12746 в объеме 15 л по предлагаемой схеме. По оси абсцисс - часы культивирования, по оси ординат - накопление белка. Маркер - маркеры молекулярного веса, Обр. 1 - 2,5 час роста, Обр. 2 - 3 час роста, Обр. 3-4 час роста, Обр. 4 - 6 часов роста, СО - стандартный образец β-галактозидазы BgaA.
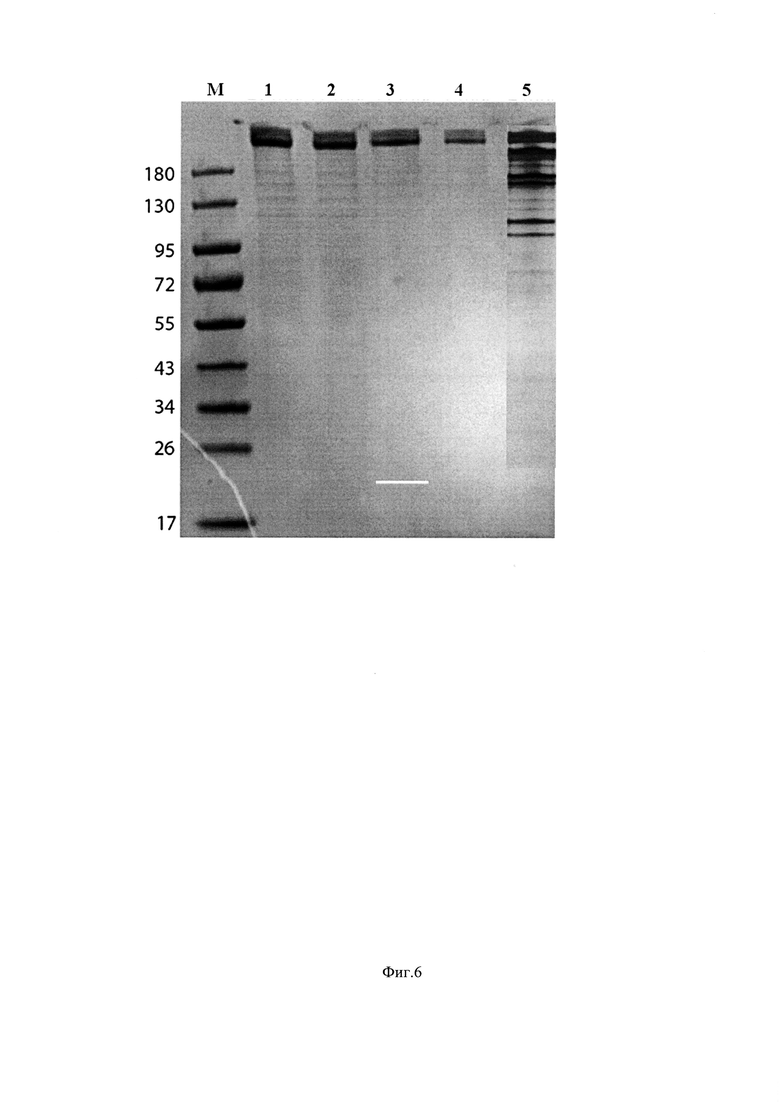
Фиг. 6. Результат выделения BgaA. Анализ электрофорезом в 4 - 20% градиентном ПААГ. Сравнение коммерческого препарата β-галактозидазы и полученного препарата BgaA.
Дорожка 1 - маркеры молекулярной массы Thermo Scientific;
Дорожки 2-4 - очищенный препарат BgaA с различным количеством внесенного в лунку препарата.
Дорожка 5 - коммерческая β-галактозидаза;
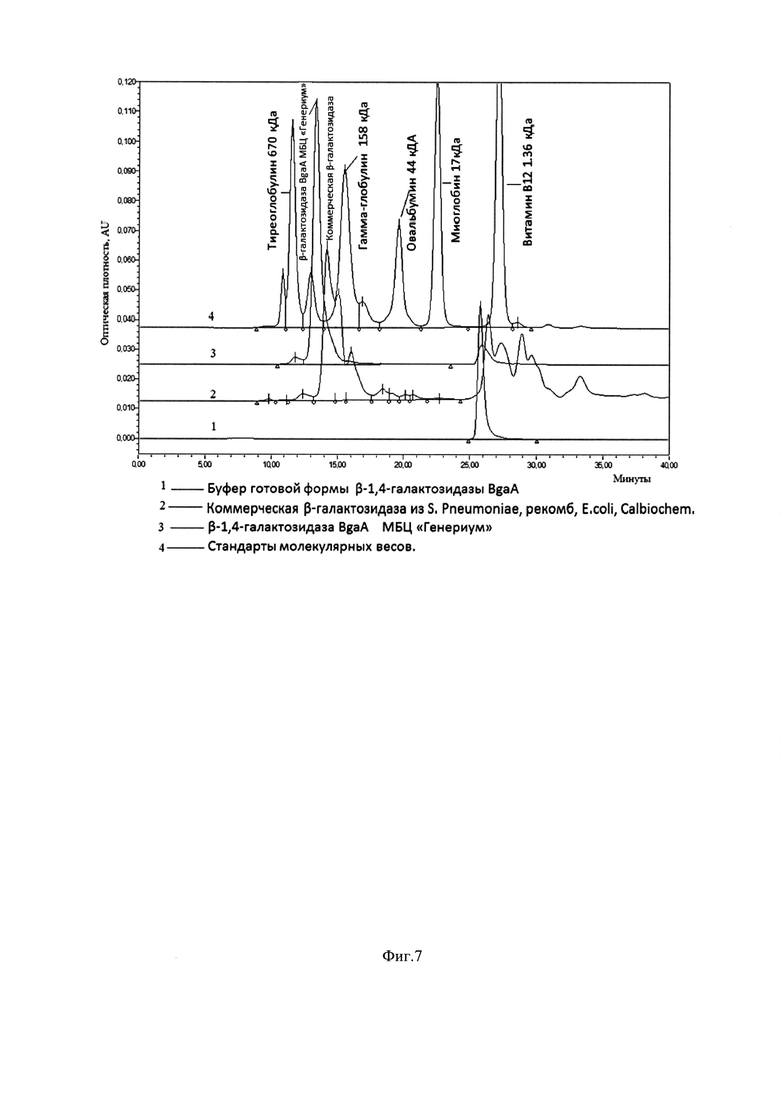
Фиг. 7. Сравнение хроматографических профилей β-галактозидазы из Streptococcus pneumoniae, рекомб, E.coli, Calbiochem, и β-галактозидазы BgaA МБЦ «Генериум» при помощи гель-фильтрации. Линия 1 - Плацебо; Линия 2 - Коммерческая β-галактозидаза из S. Pneumoniae, рекомб, E.coli, Calbiochem, 3 - β-галактозидаза BgaA МБЦ «Генериум»; Линия 4 - Стандарты молекулярных весов. Колонка: Superdex200 Increase 10/300 GL, GE Healthcare; Скорость потока: 0,75 мл/мин; Объем инжекции: 100 мкл; Время и параметры регистрации хроматограммы: 40 мин, tкол=30°С; tобр=5°С; λ=280/214 нм; ПФ: 0,02 М NaH2PO4 0,3 М NaCl рН=7.0.
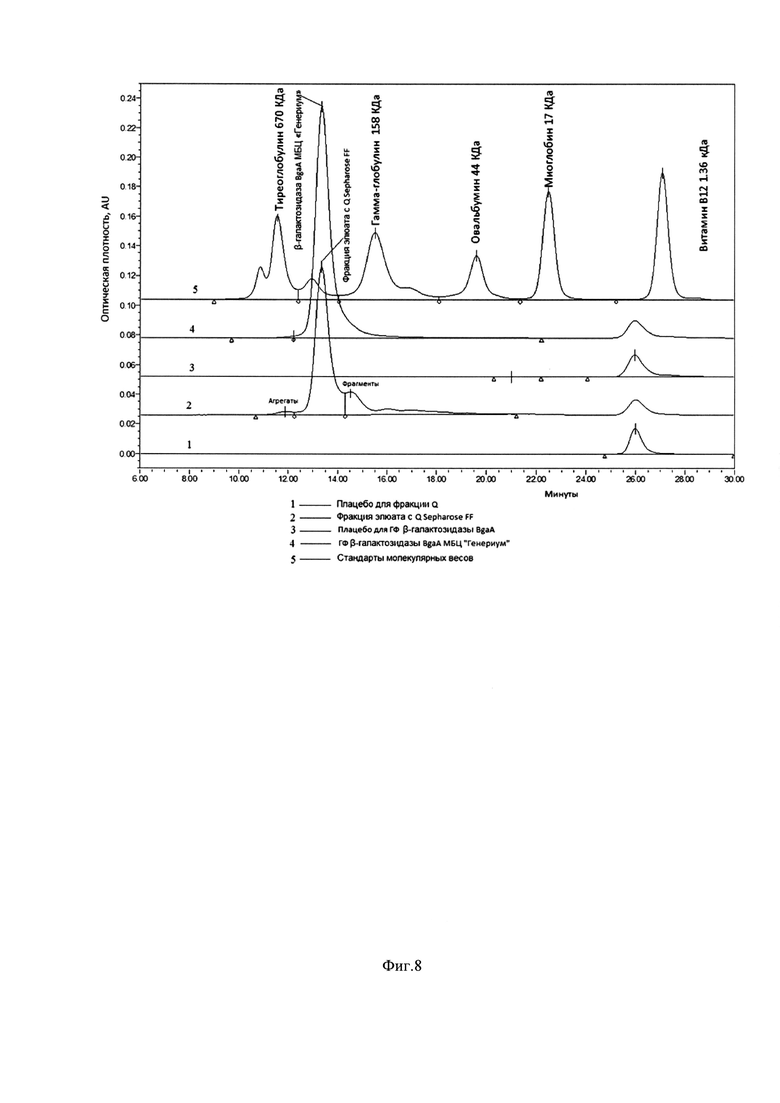
Фиг. 8. Результат анализа ГФ ВЭЖХ. Анализ фракций с промежуточных серий и готовая форма промышленной серии BgaA.
Линия 1 - Плацебо для фракции Q Sepharose FF; Линия 2 - Фракция элюата с Q Sepharose FF; Линия 3 - плацебо для ГФ BgaA; Линия 4 - ГФ BgaA; Линия 5 - Стандарты молекулярных весов. Колонка: Superdex200 Increase 10/300 GL, GE Healthcare; Скорость потока: 0,75 мл/мин; Объем инжекции: 100 мкл; Время и параметры регистрации хроматограммы: 40 мин, tкол=30°С; tобр=5°С; λ=280/214 нм; ПФ: 0,02 М NaH2PO4 0,3 М NaCl рН=7.0.
Отсутствие отечественных коммерчески доступных препаратов полноразмерной рекомбинантной β-1,4-галактозидазы BgaA из Streptococcus pneumoniae, экспрессированных в E.coli, и чрезвычайно высокая стоимость препаратов у зарубежных производителей, таких как QA-Bio, NEB, Calbiochem, делает коммерчески невыгодным использование этих препаратов при производстве лекарственных препаратов. Препараты данных фирм производятся не в соответствии со стандартами «Надлежащей производственной практики»/«Good Manufacturing Practice» (GMP) и предназначены только для аналитических исследований.
Подробные характеристики β-1,4-галактозидазы BgaA, принадлежащей к большому семейству галактозидаз (лактаз, β-галактозид-галактогидралаз), которые действуют на N/O-гликозильные соединения и отщепляют концевой нередуцированный остаток β-D-галактозы в β-галактозидах, включая лактозу, с образованием свободных моносахаридов, либо переносят остаток β-D-галактозы на молекулу лактозы или других β-D-галактозидов с образованием галактоолигосахаридов, даны в публикациях А.А. Костеневич, Л.И. Сапунова, Бактериальные β-галактозидазы, биохимическое и генетическое разнообразие // Труды БГУ. -2013 - Т. 8. 4.1 - с. 52-63, А.Е. Браунштейн, Номенклатура ферментов / Рекомендации Международного биохимического союза по номенклатуре и классификации ферментов, а также по единицам ферментов и символам кинетики ферментативных реакций, // - 1979 - ВИНИТИ. - с. 320, Panesar P.S.et al, Microbial production, immobilization and applications of β-D-galactosidase, // J. Chem. Technol. Biotechnol. - 2006. - V. 81 - p. 530-543, Ah-Reum Park, Deok-Kun Oh, Galacto-oligosaccharide production using microbial β-galactosidase: current state and perspectives, // - 2010 - Appl Microbiol Biotechnol - V. 85 - p. 1279-1286).
Эти ферменты широко распространены в природе и представляют большой интерес для биотехнологической и фармацевтической промышленности, их гидролитическая активность в течение десятилетий применяется в пищевой промышленности для уменьшения содержания лактозы в молочных продуктах, чтобы обойти лактозную непереносимость, преобладающую в более чем половине населения мира (Alejandro Vian, Alfonso V. Carrascosa, Jose L. Garcia, Estrella Cortes, Structure of the b-Galactosidase Gene from Thermus sp.Strain T2: Expression in Escherichia coli and Purification in a Single Step of an Active Fusion Protein, // Applied and Environmental Microbiolology - 1998, V. 64 - N.6 - p. 2187-2191).
Некоторые из β-галактозидаз были очищены, секвенированы и широко охарактеризованы. Препараты β-галактозидаз, имеющиеся в продаже, получают главным образом из дрожжей Kluyveromyces fragilis, Kluyveromyces lactis и Candida pseudotropicalis, грибов Aspergillus niger и Aspergillus oryzae и Bacillus stearothermophilus (Parmjit S. Panesar, Reeba Panesar, Ram S. Singh, John F Kennedy and Harish Kumar, Review.Microbial production, immobilization and applications of β-D-galactosidase // Journal of Chemical Technology and Biotechnology- 2006 - V.81 - p. 530-543).
Известны работы по выделению и клонированию генов, ответственных за синтез β-галактозидазы у различных бактерий. Наиболее изучен ген β-галактозидазы Е. coli - lacZ, который кодирует ферментный белок-тетрамер (540 кДа), состоящий из идентичных субъединиц (A. Kalnins et al, lacZ Sequence of the gene of Escherichia coli // EMBO J. - 1983. - V. 2, N. 4. - p. 593-597).
Несмотря на многолетнюю историю применения β-галактозидаз в различных отраслях промышленности и научных исследованиях, не все β-галактозидазы могут служить в качестве инструментов для анализа и ремоделирования гликанов, а также для получения олигосахаридов из природных материалов или субстратов для гликозилтрансфераз. Например, фермент из Escherichia coli используется во многих областях биохимии, но не для гликоконъюгатов (Reinhard Zeleny, Friedrich Altmann and Werner Praznik, A Capillary Electrophoretic Study on the Specificity of β-Galactosidases from Aspergillus oryzae, Escherichia coli, Streptococcus pneumoniae, and Canavalia ensiformis (Jack Bean) // Analitical Biochemistry - 1997 - V. 246. - p. 96-101).
В работе Kyoko Kojima, Masao Iwamori, Sei-Ichi Takasaki, Shirou Nozawa, Rihachi Iizuka, Yoshitaka Nagalt, Diplococcal β- Galactosidase with a Specificity Reacting to β 1-4 Linkage but Not to β-1-3 Linkage as a Useful Exoglycosidase for the Structural Elucidation of Glycolipids, // Analytical Biochemistrty - 1987 - V. 165 - p. 465-469, было показано, что фермент β-1,4-галактозидаза BgaA из Streptococcus pneumoniae (ранее Diplococcus pneumoniae) имеет довольно строгую специфичность для связей β-1-4 и, следовательно, может служить для удаления или выяснения положения галактозного остатка, кроме того, в публикации Dominique Н. Limoli, Julie A. Sladek, Lindsey A. Fuller, Anirudh К. Singh and Samantha J. King, BgaA acts as an adhesin to mediate attachment of some pneumococcal strains to human epithelial cells,// Microbiology - 2011 - V.157 - p. 2369-2381 было сообщено, что в отличие от других β-галактозидаз, основная роль BgaA заключается не в метаболизме лактозы, а в дегликозилировании гликоконъюгатов человека. Необычно высокая специфичность фермента BgaA для β-1,4-гликозидных связей и более чем 10-кратная высокая специфичность для N-ацетилгалактозамина (Ga1β1-4GlcNAc), чем для лактозы (Zeleny, R, F. Altmann, and W. Praznik, A capillary electrophoretic study on the specificity of β-galactosidases from Aspergillus oryzae, Escherichia coli, Streptococcus pneumoniae, and Canavalia ensiformis (jack bean) // Anal. Biochem - 1997 - V.246 - p - 96-101), позволяют считать фермент BgaA идеальным кандидатом для атаки полисахаридов, конъюгированных с поверхностными компонентами эукариотических клеток (Yang, Y, and R. Orlando, Simplifying the exoglycoside digestion / MALDI-MS procedure for sequencing N-linked carbohydrate side chains, // Anal. Chem - 1996 - V. 68 - p. 570-572, Vanessa S. Terra, Karen A. Homer, Susmitha G. Rao, Peter W. Andrew and Hasan Yesilkaya, Characterization of Novel - Galactosidase Activity That Contributes to Glycoprotein Degradation and Virulence in Streptococcus pneumoniae. // Infection and Immunity - 2010 - V. 78 - N. 1 - p. 348-357), что делает фермент BgaA чрезвычайно привлекательным для использования его в области гликобиологии. В работе Amanda М. Burnaugh, Laura J. Frantz, and Samantha J. King, Growth of Streptococcus pneumoniae on Human Glycoconjugates Is Dependent upon the Sequential Activity of Bacterial Exoglycosidases // Journal of Bactereology - 2008 - V. 230 - N. 1 - p. 221-230 было продемонстрировано, что BgaA может совместно с нейраминидазой NanA и β-N-ацетилглюкозаминидазой StH последовательно удалять сахара на N-связанных гликанах гликоконъюгатов человека (Фиг. 1).
Впервые фермент BgaA был очищен из фильтрата культуры Streptococcus 6646 и охарактеризован в работах Takao Kiyohara, Tadao Terao, Kohei Shioiri-Nakano and Toshiaki Osawa, Purification and Characterization of β-N-Acetylhexosaminidases and β-Galactosidase from Streptococcus 6646 K1, // - J. Biochem - 1976 - V. 80 - p. 9-17, и Lowrie R. Glasgows, S. James, C. Paulson and Robert L. Hill, Systematic Purification of Five Glycosidases from Streptococcus (Diplococcus) pneumoniae", // The Journal of Biological Chemistry-1977 - V. 252 - N. 23 - p. 8615-3623. В исследовании Dorothea Zahner and Regine Hakenbeck, The Streptococcus pneumoniae Beta-Galactosidase Is a Surface Protein // Journal of Bacteriology - 2000, - V. 182 - N. 20 - p. 5919-5921, был идентифицирован ген, кодирующий β-галактозидазу BgaA из S. Pneumoniae. В отличие от типичных галактозидаз, которые обычно представляют собой цитоплазматические белки, ген BgaA S. Pneumoniae кодирует связанную с поверхностью β-1,4-галактозидазу, состоящую из 2 235 аминокислотных остатков с молекулярным весом более 200 кДа. Полный сиквенс генома S. Pneumoniae представлен в работе Herve Tettelin et al, Complete Genome Sequence of a Virulent Isolate of Streptococcus pneumonia // - Science - 2001 - V. 293 - p. 498-506. В исследовании Anirudh K. Singh et al, Unravelling the Multiple Functions of the, Architecturally Intricate Streptococcus pneumoniae β-galactosidase BgaA, // PLOS Pathogens/ www.plospathogens.org - 2014 - V. 10 - I.9 - e1004364, благодаря детальному структурному и функциональному анализу, раскрыта сложная архитектура и разнообразие функций BgaA.
В настоящее время исследованы полные геномные последовательности семи штаммов S. Pneumoniae: S. Pneumoniae АТСС 700669, S. Pneumoniae G54, S. Pneumoniae CGSP14, S. Pneumoniae Hungary 19A-6, S. PneumoniaeD39, S. Pneumoniae R6 и S. Pneumoniae TIGR4, доступны в публичных базах данных (Lanie, J.A, W.L. Ng, K.M. Kazmierczak, Т.M. Andrzejewski, Т.М. Davidsen, K.J. Wayne, Н. Tettelin, J.I. Glass, and M.E. Winkler. Genome sequence of Avery's virulent serotype 2 strain D 39 of Streptococcus pneumoniae and comparison with that of unencapsulated laboratory strain R6. // - J. Bacteriol. - 2007. - V. 189. - p. 38-51).
В работе Jae Кар Jeong, Ohsuk Kwon, Yun Mi Lee, 1 Doo-Byoung Oh, Jung Mi Lee, Seonghun Kim, Eun-Hye Kim, Tu Nhat Le, Dong-Kwon Rhee, and Hyun Ah Kang, Characterization of the Streptococcus pneumoniae BgaC Protein as a Novel Surface - Galactosidase with Specific Hydrolysis Activity for the Gal1-3GlcNAc Moiety of Oligosaccharides. // -Journal of Bacteriology. - 2009. - V. 191. - N. 9.- p. 3011-3023 сообщается еще об одной β-галактозидазе BgaC Streptococcus pneumoniae, которая представляет собой так же поверхностно-ассоциированную β-галактозидазу с высокой специфической гидролизной активностью для Gal1-3GlcNAc-фрагмента олигосахаридов, для β-1,3-гликозидной связи без каких-либо модификаций, состоящую из 595 аминокислотных остатков с молекулярным весом около 70 кДа. В патенте Kang et al. Recombinant beta-Galactosidase derived from Streptococcus Pneumoniae, // US 0287463 A1., - 2011, раскрыто получение рекомбинантной β-галактозидазы BgaC с молекулярным весом 69 кДа, экспрессируемой в растворимой фракции клеток Е. Coli с дальнейшим разрушением клеток ультразвуком и последующим центрифугированием и получением целевого белка очищением супернатанта аффинной хроматографией на колонке с Ni-NTA агарозой. Данная β-1-3 галактозидаза обладает чрезвычайно узкой субстратной специфичностью и не применяется для ремоделирования гликоконьюгатов. Получение β-1-4 галактозидазы из Bacteroides fragilis раскрыто в патенте Sharon T.Wong - Madden, Ellen P.Guthrie et al, Isolation and compositionof novel glycosidases, // US 7094563 B2, - 2006, фермент выделяли из грубого лизата клеток Bacteroides fragilis хроматографической очисткой на DEAE Sepharose FF в режиме проскока с последующей хроматографией на Phenyl Sepharose FF, диализом активных фракций, последующей хроматографией на Heparin-Sepharose и диализом активных фракций, хроматографией на Mono Q с финальной хроматографией на Heparin TSK.
Хотя в настоящее время доступны некоторые коммерческие препараты β-1,4 галактозидаз, как произведенные из природных источников, так и рекомбинантные, экспрессированные в Е. Coli, существует потребность в новых в гораздо более чистых препаратах β-1,4 галактозидаз со строго определенной субстратной специфичностью и воспроизводимой энзиматической активностью.
В публикации S. Campuzano, В. Serra, D. Llull, J.L. Garcia, E. Garcia, P. Garcia, Cloning, Expression, and Characterization of a Peculiar Choline-Binding-Galactosidase from Streptococcus mitis, // Applied and Environmental Microbiology, - 2009. - V.75, - N.18. - p.5972-5980, описан наиболее близкий к заявленному изобретению способ получения ортологичной β-1,4 галактозидазы из Streptococcus mitis. Геномный фрагмент ДНК Streptococcus mitis, несущий ген SMT1224, кодирующий β-галактозидазу, идентифицировали, клонировали и экспрессировали в Escherichia coli. Этот ген кодирует белок длиной 2411 аминокислот с предсказанной молекулярной массой 268 кДа. Рекомбинантный белок содержит N-концевой сигнальный пептид и С-концевой холинсвязывающий домен. Холинсвязывающая способность белка облегчает его очистку с использованием аффинной хроматографии на DEAE-целлюлозе, хотя полная очистка целевого белка была достигнута путем конструирования в виде фьюжен белка с полигистидиновой последовательностью.
Вариант рекомбинантного белка с холинсвязывающим доменом очищали с помощью аффинной хроматографии на DEAE-целлюлозе, активные фракции, полученные после DEAE-аффинной хроматографии, объединяли, диализовали и концентрировали 10 раз в диализном мешке с полиэтиленгликолем 6000. Концентрированный раствор фермента доочищали гель - фильтрационной хроматографией на колонке Sephacryl S-300. Данным способом получали 0.36 мг целевого белка с 1 л культуральной жидкости.
Вариант рекомбинантного белка с полигистидиновой последовательностью экспрессированный в E.coli МС4100 (pHGM02), очищали за одну стадию аффинной хроматографией на IMAC картриджах (Bio-Rad) с выходом 0,004 г на 1 л культуральной жидкости.
Данные схемы очистки являются неприемлемыми для масштабного производства препарата, так как авторы получали препарат с низким выходом и только для аналитических целей. Кроме того, авторы не сообщают об уровне содержания в препарате бактериальных эндотоксинов. Удаление бактериальных эндотоксинов является принципиальным и необходимым требованием, как с точки зрения российских, так и международных стандартов (ГФ XIII, Document Q AS/ 11.452 FINAL July 2012). Однако часто остается технически трудновыполнимой задачей, поэтому содержание бактериальных эндотоксинов в промежуточных продуктах, применяемых при производстве ЛС, является критическим параметром. При оценке качества таких промежуточных продуктов, как гликозидазы, помимо соответствия субстратной специфичности, активности, особо должны оцениваться такие потенциально опасные контаминанты, как бактериальные эндотоксины, клеточная ДНК, в том числе потенциально онкогенная, также количество белков хозяина штамма-продуцента и гомогенность препарата. И, как правило, при производстве препаратов из природных источников существуют трудности с получением чистых препаратов. Обычно они загрязнены примесями других гликозидаз, протеаз.
Для решения задачи получения высокоочищенной BgaA в промышленных объемах авторы изобретения предлагают новый способ получения рекомбинантной β-1,4 галактозидазы BgaA из Streptococcus pneumoniae, включающий синтез оптимизированной для трансляции в E.coli последовательности ДНК, кодирующий белок BgaA, нуклеотидная последовательность которого приведена в составе SEQ ID NO1, конструирование экспрессионного плазмидного вектора, кодирующего химерный белок с N-концевой химеризацией, составленного пептидными последовательностями шести гистидинов, сайт узнавания тромбина а также 2053 аминокислотных остатков (без сигнального пептида, G5 домена и якорного пептида) β-1,4-галактозидазы (BgaA) Streptococcus pneumoniae с молекулярным весом 231 кДа (аминокислотная последовательность представлена SEQ ID NO 2), получение рекомбинантного белка продукцией в E.coli, и далее хроматографичекую очистку белка из растворимой фракции клеточного лизата, где первые две стадии проводят на одном и том же Ni+ металлохелатном сорбенте при разных разных условиях элюции, псевдоаффинной хроматографии в режиме проскока на Blue Sepharose анионобменной хроматографии на Toyopearl Giga Cap Q- 650М, гидрофобной хроматографии в режиме проскока на Butyl Capto, концентрирующей анионобменной хроматографию на Q Sepharose FF с последующей гель-фильтрацией на Superdex 200 с выходом высокоочищенного активного препарата BgaA, пригодного для использования на производстве ЛС и гликобиологических исследований, не менее 0,01 г с литра бактериальной культуры.
Основным техническим результатом изобретения является получение в промышленных масштабах высокоочищенного активного препарата ферментативно активной рекомбинантной β-1,4-галактозидазы BgaA, с содержащей минимальные остаточные концентрации белков и ДНК клетки хозяина, что делает ее пригодной для использования на стадии ремоделирования в производстве терапевтически функциональных препаратов гликопротеинов с заданными свойствами, а также для разработки инструментов высокочувствительного анализа гликановых цепей гликопротеинов тканей растений, животных и микроорганизмов.
Для осуществления изобретения был создан штамм генно-инженерно-модифицированного микроорганизма E.coli ВКПМ В- 12746: реципиентного штамма-хозяина BL21(DE3) плазмидами pLysS, несущей ген Т7 лизоцима, являющегося ингибитором Т7 РНК полимеразы, рМТ1620 (pET15b-BGAA_STRPN), несущей ген белка BGAA_STRPN, составной частью которого являются последовательности 20 аминокислотных остатков, содержащих гексагистидиновый тэг, и 2053 аминокислотных остатков (без сигнального пептида, G5 домена и якорного пептида) β-галактозидазы BgaA Streptococcus pneumoniae с расчетной молекулярной массой 231 кДа. (Фиг. 2).
Рефолдинг продукта не требуется, так как он синтезируется в бактериальной цитоплазме преимущественно в растворенном состоянии при культивировании E.coli ВКПМ В-12746 с уровнем биосинтеза вариантов целевого белка не менее 10 мг/л культуральной жидкости.
Высокоэффективное извлечение целевого белка BgaA из клеток достигается в результате процедуры лизиса бактериальных клеток с использованием гомогенизатора высокого давления.
Получение активного, хроматографически гомогенного с низким содержанием бактериальных эндотоксинов, ДНК и белков хозяина препарата BgaA достигается благодаря схеме хроматографической очистки, которая, в совокупности с применением некоторых стандартных приемов очистки белков (анионобменная хроматография на высокомолекулярном анионообменном сорбенте, гидрофобная хроматография в режиме проскока, концентрирующая анионобменная хроматография и полирующая гель - фильтрация), включает в себя две стадии хроматографии на одном и том же Ni+ металлхелатном сорбенте в разных условиях элюции и далее псевдоаффинную хроматографию в режиме проскока с использованием матрицы сорбента, к которой ковалентно пришит Cibacron Blue 3G, анионный антрахиноновый краситель. Обычно подобный сорбент используют для очистки альбумина и интерферона как в лабораторном, так и в технологическом масштабе. Также используется для выделения групп ферментов и удаления альбумина, так как синий краситель связывает многие белки, такие как альбумин, интерферон, липопротеины и факторы свертывания крови. Он также связывает несколько ферментов, включая киназы, дегидрогеназы и большинство ферментов, требующих аденилсодержащих кофакторов, например, NAD+. В заявленном изобретении использовали данный тип сорбента для удаления некоторых клеточных белков E.coli, в связи с неожиданным открытием авторами того факта, что при очистке β-галактозидаза BgaA, не сорбируясь, проходит через сорбент, в то время как клеточные белки задерживаются. Обычно данный сорбент не используется в подобных целях. Таким образом, в отличие от известных ранее методов, технический результат достигается введением в схему очистки от клеточных белков стадии псевдоаффинной хроматографии на сорбентах с ковалентно связанным Cibacron Blue 3G.
Представленный в изобретении пример иллюстрирует использование двух Ni2+ металлохелатных хроматографий на IMAC Sepharose 6 FF с разными условиями элюирования целевого белка, что позволило максимально отделить целевой белок от мажорных контаминирующих примесей клеточных белков, имеющих сродство к IMAC Sepharose. Данный технический результат имеет одно из решающих значений для качества дальнейшей очистки за счет снижения нагрузки контаминирующими белками на все последующие сорбенты. На рынке доступны и другие сорбенты, которые также могут быть использованы для металлохелатной хроматографии в предложенном способе, например, можно использовать IMAC Sepharose High Performance. Сорбент Chelating Sepharose Fast Flow является менее предпочтительным, так как обладает более низкой емкостью и меньшим аффинитетом белка интереса. Для обеих стадий хроматографии приемлемым является единый уравновешивающий буфер, рН которого варьирует в диапазоне 7,5 - 7,8. Элюцию на первой стадии проводят содержащим NaCl натрий -ацетатным буфером, имеющим рН в диапазоне 4,2-4,3 единицы, на второй стадии целевой белок элюируют 150 мМ - 200 мМ имидазола. Предпочтительный рН буфера отмывки, применяемого на второй стадии металлхелатной хроматографии, находится в диапазоне 6,5-6,6.
Введение в схему очистки стадии псевдоаффинной хроматографии на сорбенте с ковалентно связанным Cibacron Blue 3G в режиме проскока является еще одной ключевой стадией, позволяющей значительно очистить целевой белок от контаминирующих примесей, коэлюирующихся с целевым белком на всех предыдущих стадиях. На рынке помимо использованной авторами изобретения Blue Sepharose 6 Fastp Flow доступен и ряд сорбентов, подходящих для использования в заявленном изобретении: Capto Blue (high sub), Capto Blue и другие с аналогичными свойствами. Предпочтительный состав уравновешивающего буфера на данной стадии 20 mM Tris-HCl, 80 мМ NaCl, 5% изопропанол, рН 8.2-8.3.
После проведения двух металлхелатных хроматографий и псевдоаффинной хроматографии в режиме проскока доочистка белка может быть проведена с использованием различных видов хроматографии, рутинно применяемых в очистке белков: гидрофобной, анионобменной, концентрирующей хроматографии и полирующей гель-фильтрации. Выбор сорбентов и условий может быть проведен специалистом на основании общих знаний уровня техники и рекомендаций производителей сорбентов. В заявленном изобретении авторы привели примеры применения конкретных марок сорбентов в конкретные условия применения, однако со сменой сорбента условия могут быть изменены в соответствии с рекомендациями производителя и общими правилами. Наиболее предпочтительными сорбентами для последующей анионобменной хроматографии являются сорбенты TOYOPEARL GigaCap Q-650M (Toson) и MacroCap Q (GE), оптимизированные для сорбции и очистки белков с большим молекулярным весом. Предпочтительный рН уравновешивающего буфера для этой стадии находится в диапазоне 7,5-7,7. Солевой состав буфера элюции представляет собой градиент от 5 мМ до 260 мМ NaCl в стартовом буфере. Предлагаемые условия посадки и съема целевого белка позволяют максимально очистить целевой белок от контаминирующих примесей клеточных белков и эндотоксинов. Применение гидрофобной хроматографии в режиме проскока позволяет удалить контаминирующие примеси гидрофобных белков и липополисахаридов. В качестве сорбента для предколонки можно использовать целый ряд присутствующих на рынке сорбентов: Butyl Capto, Butyl Sepharose High Performance Capto Butyl ImpRes, SuperButyl-550C и др. В качестве концентрирующего сорбента помимо Q Sepharose FF, фигурирующей в примере осуществления данного изобретения, могут быть использованы Capto Q, Capto Q ImpRes, Capto Q XP. Данные сорбенты обладают большей емкостью, что следует учитывать при выборе концентрации соли в элюирующем буфере. Предпочтительный рН уравновешивающего буфера находится в диапазоне 7,5-7,7. Состав предпочтительного буфера элюции для данной стадии- 350 mM - 360 мМ NaCl в 20 mM Tris - HCl, рН 7,5-7,7. Заключительная стадия гель-фильтрационной хроматографии является полирующей, позволяющей получить гомогенный препарат в буфере готовой формы. Предпочтительным является использование преднабитой колонки HiLoad Superdex 200 26/600 (GE), возможно использование также Superdex 200, HiPrep 26/60 Sephacryl S-400 HR, сорбент Sephacryl S-400 HR. Предпочтительный буфер для готовой формы - 20 mM Tris HCl, 0.05 М NaCl, 1 mM EDTA рН 7.5-7,6.
Материалы и методы
Для экспрессии BgaA получают генно-инженерные конструкции (ГИК) рМТ1620. Для получения ГИК, кодирующих BgaA, была разработана схема клонирования на базе вектора pET-15b (Novagen), предполагающая получение полной версии, имеющей 6×His-tag на N-конце. Для получения соответствующей ГИК в компании TOP Gene Technologies был заказан синтетический ген, кодирующий BgaA (геномная последовательность бактерии Streptococcus pneumoniae, кодирующая 2053 аминокислотных остатков (без сигнального пептида, одного из двух G5 доменов и якорного пептида) β-1,4-галактозидазы (BgaA) Streptococcus pneumoniae, слитая на 5'-конце с частью векторной последовательности материнской плазмиды pET15b(+), кодирующей 20 аминокислотных остатков, содержащих гексагистидиновый тэг), фланкированный сайтами рестрикции NdeI, BamHI, содержащий внутренние сайты рестрикции NheI для получения «каталитического» домена и стоп-кодоны на 3'-конце последовательности (TGATAA). В TOP Gene Technologies была проведена кодон-оптимизация последовательности для экспрессии белка в E.coli, принимая во внимание требование об отсутствии в последовательности «внутренних» сайтов эндонуклеаз рестрикции NdeI, BamHI, XhoI и NheI. Кодон-оптимизация полученной последовательности BgaA проверена на сайте http://gcua.schoedl.de/sequential_v2.html. Соответствие транслируемой аминокислотной последовательности, заказанной подтверждено с помощью выравнивания в программе MEGA 6.0. Отсутствие нежелательных сайтов рестрикции подтверждено с помощью программного обеспечения CloneManager 9.0. Для получения ГИК рМТ1620 (pET15b-BGAA_STRPN) проведена наработка и рестрикционное картирование синтетического гена pAPG110-BgaA_STRPN была проведена наработка и рестрикционное картирование синтетического гена pAPG110-BgaA_STRPN. Согласно разработанной схеме было проведено клонирование фрагмента, кодирующего стрептококковую BgaA из плазмиды pAPG110-BgaA_STRPN, в векторную плазмиду pET15b по сайтам рестрикции NdeI, BamHI. Полученные в ходе рестрикции фрагменты очищали и использовали для лигирования. Лигазной смесью трансформировали химически компетентные клетки E.coli XL10-GOLD. Клетки высевали для получения трансформированных клонов.
Затем выделяли плазмидную ДНК и проводили рестрикционное картирование с использованием эндонуклеазы рестрикции BglII. Образец плазмидной ДНК передавали на секвенирование в "Евроген". Полученная последовательность была выровнена с «теоретической» последовательностью рМТ1620.
Для проведения аналитической экспрессии плазмидную ДНК ГИК рМТ1620 трансформировали в штаммы BL21(DE3) pLysS. На основании аналитической экспрессии отбирали клоны-суперпродуценты BgaA. Затем получали штамм-суперпродуцент BgaA. Штамм был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под инвентарным номером - ВКПМ В-12746.
Для получения целевого белка BgaA проводили препаративную экспрессию путем культивирования E.Coli ВКПМ В-12746 в полноценной питательной среде с относительным содержанием растворенного кислорода не менее 10% при температуре 37оС, значении рН - 6,5-7,5. Длительность процесса культивирования составляет 5-7 часов.
Индукцию экспрессии целевого продукта на этапе биосинтеза обеспечивали внесением раствора Изопропил-β-D-1-тиогалактопиранозида до конечной концентрации 1 мМ в питательной среде на 2,5-3,0 ч роста по достижении оптической плотности культуры 2,5-5,0 о.е
В течение первого этапа клетки активно растут благодаря обогащенной культуральной среде. Присутствие в среде глюкозы подавляет экспрессию рекомбинантного белка, а, следовательно, снижает нагрузку на клетку штамма-продуцента, что приводит к увеличению количества накапливаемой биомассы.
Непосредственно перед внесением индуктора в среду добавляли глицерин, который служит источником углерода и энергии, не подавляя экспрессию рекомбинантного белка. Это приводит к увеличению количества накапливаемого рекомбинантного белка β-галактозидазы.
С 3-3,5 часа культивирования по достижении оптической плотности начинается этап биосинтеза гибридного белка. Экспрессия белка BgaA находится под контролем промотора, который дерепрессируется внесением раствора ИПТГ до конечной концентрации 0,0238 мас. %.
Культивирование проводили в течение 5-7 ч.
Уровень биосинтеза вариантов целевого белка BgaA составляет не менее 5 мг/л культуральной жидкости. Бактериальную биомассу собирали центрифугированием и хранили при - 70°С. Клеточную биомассу лизировали с помощью гомогенизатора высокого давления Panda Plus 2000(GEA Niro Soavi), лизат осветляли центрифугированием и проводили выделение белка BgaA из лизата путем очистки целевого белка от примесных белков двумя металлохелатными хроматографиями с разными условиями элюирования на колонках с Ni2+ IMAC Sepharose FF. Доочищали фермент последовательными хроматографиями на Blue Sepharose FF в режиме проскока, анионообменной Toyopearl Giga Cap Q- 650M, Butyl Capto в режиме проскока, хроматографией на Q Sepharose FF, концентрированием и гель-фильтрацией. На каждом этапе контроль степени очистки проводили электрофорезом в ПААГ.
Выход белка составляет не менее 0,01 мг на литр бактериальной культуры.
Измерения параметров различных партий полученного целевого белка BgaA показали, что в результате очистки он имеет следующие характеристики:
- содержание белка (УФ-спектрофотометрия) - не менее 0,5 мг/мл;
- содержание бактериальных эндотоксинов (ГФ XIII, гель-тромб тест с ЛАЛ-реактивом ОФС.1.2.4.0006.15) не более 2 ЭЕ/мг;
- Хроматографическая чистота (ГФ ВЭЖХ) не менее 94%;
- Ферментативная активность (Колориметрический метод, синтетический субстрат ONPG (ortho-nitrophenyl-β-D-galactopyranoside), «β-Gal Assay Kit» (Invitrogen)) не менее 13000 U/мг;
- Остаточная ДНК штамма-продуцента (Количественный ПЦР в реальном времени) не более 0,015 пг/мг;
- Остаточные белки штамма-продуцента не более 4 нг/мг;
Таким образом, показано, что с использованием предложенного в заявленном изобретении способа очистки получается препарат BgaA с хроматографической чистотой, определенной с помощью гель-фильтрационной ВЭЖХ - более 94%-ной гомогенности, удельная активность составляет от 13000 единиц/мг белка и выше, содержание бактериальных эндотоксинов составляет не более 2 ЕЭ /мг белка, содержание остаточной ДНК штамма-продуцента составляет не более 0,015 пг/мг белка, содержание остаточных белков штамма - продуцента составляет не более 4 нг/мг белка.
Получение BgaA на основе оптимизированной синтетической последовательности ДНК с короткой последовательностью из шести гистидинов на N-конце рекомбинантного белка происходит без удаления гексагистидинового тэга, что позволяет упростить очистку и удалить из схемы очистки применение дорогостоящих препаратов для удаления тэга. Аминокислотная последовательность BgaA представлена SEQ ID NO 2. Эксперименты показали, что, несмотря на присутствие гексагистидинового тэга, обладая большим молекулярным весом (231 кДа), целевой белок BgaA плохо проникает в поры сорбента, сорбируясь на его поверхности. В то же время глубокие слои сорбента занимают примеси, обладающие разным, в том числе и более высоким сродством к сорбенту, чем целевой белок. Для удаления этих примесей, без которого невозможна эффективная очистка белка, в заявленном изобретении предложен неожиданный и оригинальный подход, заключающийся в проведении двух стадий металлохелатной хроматографии с различными условиями элюирования, позволяющие удалить примеси, мешающие дальнейшей очистке. Таким образом, авторы изобретения считают введение этих двух стадий очистки наиболее существенными для получения технического результата.
На первой стадии хроматографии на Ni+металлхелатном сорбенте авторы изобретения применяют элюцию целевого белка буфером с низким рН и с высокой ионной силой, на второй стадии хроматографии на этом же сорбенте авторы применяют отмывки буфером со слабокислым водородным показателем среды, предпочтительно рН 6,5, и высокой ионной силой, и далее элюцию буфером, содержащим имидазол в невысокой концентрации. Применение псевдоаффинной хроматографии на Blue Sepharose в режиме проскока позволяет очистить целевой белок от примесей клеточных белков, коэлюирующихся с целевым белком на всех стадиях. Анионообменную хроматографию предпочтительно проводят на высокомолекулярном анионообменном сорбенте с высоким разрешением, оптимизированным для захвата и очистки белков с большим молекулярным весом. Далее анионообменная хроматография позволяет практически без потерь сконцентрировать целевой белок без процесса мембранных концентрирования и диафильтраци. После проведения итоговой полирующей гель-фильтрации получают высокоочищенный активный препарат BgaA, пригодный для использования на производстве ЛС и гликобиологических исследований, с выходом не менее 0,01 г с литра бактериальной культуры.
Заявленный способ позволяет получить препарат рекомбинантной BgaA, имеющий более высокую степень чистоты по сравнению с коммерческими аналогами, результаты сравнения представлены в Таблице 1.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного исполнения:
Пример 1. Микробиологический синтез белка BgaA
Заявляемый способ реализован для штамма-продуцента E.Coli ВКПМ В-12746 при культивировании в ферментере (Biotron LiFlus SP/SL-20L, Юж. Корея) в объеме питательной среды 15 л. рН поддерживают на уровне 6,9-7,1. Культивирование ведут при температуре 37°С. Относительное содержание растворенного кислорода поддерживают на уровне 5-100% путем перемешивания со скоростью 800 об/мин и подачи воздуха с объемным расходом 15 л/мин. Перед началом процесса в охлажденную питательную среду асептически вносят 50% раствор глюкозы до конечной концентрации 0,5 мас. %. Инокулят в объеме 0,75 л, выращенный до оптической плотности, измеренной при длине волны 600 нм (ОД600), равной 1,8 о.е. (оптических единиц), вносят в ферментер со стерильной питательной средой, содержащей гидролизат казеина, дрожжевой экстракт, дигидрофосфат калия, сульфат аммония, гидрофосфат натрия.
В ходе процесса культивирования производят мониторинг основных параметров роста культуры: ОД600, рН, морфология клеток, относительное содержание растворенного кислорода. На 2,5 час культивирования, когда оптическая плотность культуры достигает 3,0 о.е, вносят последовательно стерильный 50% раствор глицерина - до конечной концентрации 0,5 мас. % и индуктор - 23,8% стерильный растворизопропил-β-D-1-тиогалактопиранозида - до конечной концентрации 0,0238 мас. % (1 мМ). Культивирование проводят на протяжении 6 часов. Конечная оптическая плотность -20,3 о.е, выход биомассы - 28 г/л. Продуктивность по целевому продукту штамма E.Coli ВКПМ В-12746 составляет 10,0 мг/л культуральной жидкости.
На Фиг. 4. представлена динамика накопления биомассы при культивировании штамма-продуцента E.coli ВКПМ В-12746.
Пример 2. Выделение и очистка белка BgaA из бактериальной биомассы. Получение промышленной серии препарата BgaA.
2.1 Дезинтеграция биомассы.
Буфер А - 20 mM Tris-HCl, 0.5 М NaCl, рН 7.8;
Замороженную биомассу клеток 288 г, суспендируют в 1400 мл буфера А с PMSF до 0,003% в течение 45 мин на льду и разрушают в проточном гомогенизаторе высокого давления Panda Plus 2000(GEANiro Soavi). Гомогенизатор промывают охлажденной до 0°С водой очищенной, настраивают по охлажденному до 0°С буферу А, настройка II ступени - 130 бар, настройка I ступени - 750 бар, затем лизируют суспензию клеток в этом режиме, собирая лизат в емкость, помещенную в ледяную баню, промывают гомогенизатор буфером А. Лизис повторяют еще 2 раза в том же режиме. Лизат объемом 2200 мл осветляют центрифугированием 33250 × g (JLA-16.250) при 4°С в течение 90 мин.
Хроматографии проводят при 4°С, хроматографическая система  Purifer 100 (GE).
Purifer 100 (GE).
2.2. Хроматография на Ni2+ IMAC Sepharose FF - стадия 1.
Супернатант 2200 мл доводят рН до 7,8 2М NaOH, фильтруют через глубинный фильтр Sartopor 2 300, разбавляют буфером А до 3000 л и наносят на колонку с Ni2+ IMAC Sepharose, объемом 300 мл, уравновешенную буфером А. После нанесения колонку промывают буфером А, целевой белок элюируют натрий - ацетатным буфером с 1 М NaCl, рН 4.3.
2.3. Хроматография на Ni2+ IMAC Sepharose FF - стадия 2.
Элюат с Ni2+ IMAC Sepharose FF - 1 разбавляют охлажденным до 4°С 20 mM Трис-HCl буфером, рН 7,8, доводят рН до 7,8 2М NaOH и наносят на колонку с Ni2+ IMAC Sepharose FF объемом 150,0 ml, уравновешенную буфером А. После нанесения колонку промывают 20 mM натрий фосатным буфером с 1 М NaCl, рН 6.5. Целевой белок элюируют 150 мМ имидазола.
2.4. Псевдоаффинная хроматография
Элюат с Ni2+ IMAC Sepharose разбавляли стартовым буфером (20 mM Tris-HCl, 80 мМ NaCl, 5% изопропанол, рН 8.3) до проводимости 2,3 mSm/sm, доводят рН до 8,3, и наносят на колонку с Blue Sepharose 6 FF 50 мл, уравновешенную стартовым буфером. Целевой белок собирают в режиме проскока.
2.5. Анионобменная хроматография
Проскок с Blue Sepharose 6 FF разбавляют охлажденной до 4°С водой очищенной до проводимости менее 1,0 mSm/sm, доводят рН до 7,5 и наносят на колонку с Toyopearl Giga Cap Q- 650M, объемом 50 мл, уравновешенную буфером 20 mM Tris-HCl, рН 7.5. Целевой белок элюируют градиентом от 5 до 0,25 М NaCl в стартовом буфере, собирая по фракциям.
2.6. Концентрирование с предколонкой, содержащей гидрофобный сорбент. Фракции элюата, содержащие целевой белок, с Toyopearl Giga Cap Q- 650М разбавляют водой очищенной до проводимости 2,3 mSm/sm, доводят рН до 7,5 и наносят на колонки с Q Sepharose FF с предколонкой Butyl Capto, уравновешенные буфером 20 mM Tris - HCl, рН 7.5. Отмывают стартовым буфером до выхода элюата не сорбировавшихся компонентов, колонку с Butyl Capto отсоединяют, отмывают стартовым буфером и элюируют 350 mM NaCl в стартовом буфере
2.7. Полирующая гель - фильтрация.
Элюат с Q Sepharose FF наносят на колонку с Superdex 200 26/600, уравновешенную буфером В - 20 mM Tris HCl, 0.05 М NaCl, 1 mM EDTA рН 7.5. Элюируют с колонки в том же буфере.
2.8. Разведение, стерилизующая фильтрация и фасовка.
Элюат с Superdex 200 разбавляют буфером В до получения готовой формы и стерильно фильтруют через FILTER-MAX, "rapid" - 150 ml, 0.22 μm, аликвотируют и хранят при минус 20°С.
Получено:
122,46 мг, 157,0 мл, ε280=1,17, С=0.78 мг/мл,
Содержание бактериальных эндотоксинов: менее 0,6ЕЭ/мл.
Активность на синтетическом субстрате: 17061 U/мг.
Чистота образца по ГФ ВЭЖХ составляет: 99,5%.
Остаточная ДНК штамма продуцента (E.colli): ≤0,01385пг/мг.
Остаточные белки штамма продуцента (E.colli): ≤4,0 нг/мг.
Пример 3. Анализ активности рекомбинантной BgaA
Определение активности рекомбинантной β-галактозидазы BgaA проводят постановкой теста на синтетическом субстрате ONPG (ortho-nitrophenyl-β-D-galactopyranoside), используя набор «β-Gal Assay Kit» (Invitrogen). В результате гидролиза субстрата ONPG ферментом β-галактозидаза образуется продукт ortho-nitrophenol (ONP), содержание которого измеряется колориметрически при длине волны 420 нм.
Для проведения теста готовят однократные растворы PBS и Cleavage Buffer, с помощью разбавления стоковых растворов водой очищенной (10 мл раствора «10xPBS» добавляют к 90 мл воды очищенной и 10 мл раствора «10xCleavage Buffer» добавиляют к 90 мл воды очищенной). Перед использованием вносят 270 мкл β-меркаптоэтанола в 100 мл однократного раствора Cleavage Buffer. Образцы испытуемых растворов используются в реакции с предварительным разбавлением в однократном растворе PBS в 10, 20, 40, 80 раз. Полученные разведения образцов в объеме 2 мкл вносят в лунки 96-луночного планшета, содержащие 50 мкл однократного раствора «Cleavage Buffer» с внесенным β-меркаптоэтанолом и 17 мкл субстрата ONPG. Планшет закрывают пленкой и инкубируют при температуре 37°С в течение 30 минут. Вносят в лунки по 125 мкл раствора «Stop Buffer» и измеряют оптическую плотность образцов при длине волны 420 нм. Финальный объем реакционной смеси составляет 192 мкл.
Количество гидролизованного субстрата ONPG (нмоль) вычисляли по формуле:
(OD420)×(1.92×105нл)/(4500нл/нмоль-cm)×(1cm), где
OD420 - оптическая плотность раствора образцов при длине волны 420 нм
1.92×105 нл - объем реакционной смеси, нл
4500 - коэффициент экстинкции субстрата
Для определения специфической активности β-Galactosidase, выраженной в единицах нмоль/мин/мг, количество нмоль гидролизованного ONPG, вычисленное по вышеприведенной формуле, делят на время инкубации (30 минут) и на количество белка β-галактозидазы, содержащегося в реакционной смеси.
Пример 4. Электрофоретическое разделение белков
Электрофорез проводят в 4-20% градиентном ПААГ в восстанавливающих условиях, окрашивают солями серебра (Thermo Scientific).
Пробоподготовку образцов проводят разведением образцов до концентрации ~50 мкг/мл добавлением буфера для нанесения проб (S.b) с добавлением 2 М раствора дитиотреитола (ДТТ). Прогревают в течение 5 минут при (100±2) 0С. Центрифугируют 1 мин, 13000 об/мин и наносят пробы (растворы, полученные при разведении образцов) по 10 мкл в лунку геля, маркеры молекулярной массы - 5 мкл в лунку геля.
Электрофорез проводят при напряжении - 400 В, силе тока - 30 мА, время - до выхода красителя. Затем окрашивают солями серебра по инструкции к набору Thermo Scientific. Результаты представлены на Фиг. 6.
Пример 5. Тестирование очищенных препаратов рекомбинантной BgaA в сравнении с коммерческим препаратом β-галактозидазы на содержание бактериальных эндотоксинов.
Анализ на содержание бактериальных эндотоксинов проводят методом гель-тромб теста в соответствии с ГФ XIII, гель-тромб тест с ЛАЛ-реактивом ОФС.1.2.4.0006.15. Образцы разводят водой для лал-теста («Пиротест», Россия). Используют для работы: LAL-реактив Endosafe (Charles River, USA) с чувствительностью 0,03 ЕЭ/мл; Контрольный стандартный образец эндотоксина (Charles River, USA) с содержанием 20 ЕЭ/мл. В пробирки для анализа помещают по 100 мкл образцов и по 100 мкл LAL-реактива и инкубируют в течение 60 мин при температуре 37 0С. Положительный результат: образование плотного сгустка геля. При переворачивании пробирки на 180 градусов гель не разрушается. Отрицательный результат: гель не образуется или разрушается при переворачивании пробирки.
Содержание эндотоксинов в конечной точке выражается в ЕЭ/мл и рассчитывается по формуле: «С»=(разведение в конечной точке)×(чувствительность LAL-реактива). Полученные результаты представлены в Таблице 2.
Примечания к таблице:
«+» - положительный результат; «-» - отрицательный результат.
Положительный результат: образование плотного сгустка геля. При переворачивании пробирки на 180 градусов гель не разрушается. Отрицательный результат: гель не образуется или разрушается при переворачивании пробирки.
Содержание эндотоксинов в конечной точке выражается в ЕЭ/мл и рассчитывается по формуле: «С»=(разведение в конечной точке)×(чувствительность LAL-реактива).
Пример 6. Тестирование хроматографической чистоты очищенных препаратов рекомбинантной BgaA в сравнении с коммерческим препаратом β-галактозидазы.
Тестирование производят методом ВЭЖХ гель-фильтрации на колонке с Superdex200 Increase 10/300 GL (GE) на хроматографической системе ВЭЖХ Alliance 2695, UV/Visible Detector 2487 (Waters). В качестве подвижной фазы используют буфер 0,02 М NaH2PO4; 0,3 M NaCl; рН=7.0. Препарат BgaA МБЦ «Генериум» и коммерческий препарат β1,4-Galactosidase, Streptococcus pneumoniae, Recombinant, E. Coli, Calbiochem Cat. №345806-50MIU Lot 2625690, 3 U/ml, 11.8 U/mg, 0,254 mg/ml, M.w. 350000 до внесения в систему разводят в 2 раза подвижной фазой. Сравнение хроматографических профилей испытуемых образцов показывает (Фиг. 7), что BgaA МБЦ «Генериум» существенно более однородна по составу, тогда как коммерческая β-галактозидаза более гетерогенна, что вызывает трудности в определении основного пика на хроматограмме коммерческого образца. Тестирование промышленных серий препарата рекомбинантной BgaA проводят таким же образом. На Фиг. 8 представлен типичный хроматографический профиль тестируемой промышленной серии BgaA.
Пример 7. Тестирование препаратов рекомбинантной BgaA в сравнении с коммерческим препаратом β-галактозидазой на остаточную ДНК штамма продуцента.
Для определения остаточной ДНК штамма продуцента используют метод количественного ПЦР в реальном времени. Для дизайна количественной полимеразной цепной реакции (The quantitative polymerase chain reaction - QPCR) была выбрана система Taqman, в которой при размножении специфической последовательности ДНКа, зонд деградируется ДНКа-полимеразой. Для детекции ДНК штамма продуцента E.coli был использован набор праймеров и зонда 16SER2.
Подготовка образцов: Образцы были очищены с использованием набора QIAamp DNA Blood Mini Kit (Qiagen). Объем образца для выделения составлял 0,5 мл. Конечный объем при элюции - 0,1 мл. В реакцию qPCR (пробу) добавляли 0,005 мл образца. Количество повторов для каждого образца (пробы) - 3.
Результаты анализа:
Концентрация ДНК в образцах вычисляются по формуле:
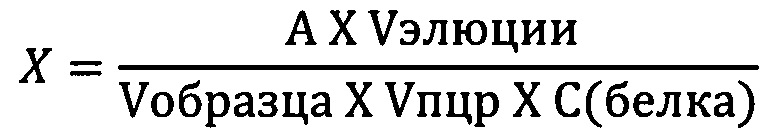
X - концентрация ДНК относительно белка в испытуемом образце (пг/мг белка);
С(белка) - концентрация белка в испытуемом образце (мг/мл); А - количество ДНК в образце, пг; Vобразца - объем образца для выделения взятый для выделения, мл (0,5 мл); Vпцр - объем элюата внесенного в ПЦР пробирку, мл (0,005 мл); Vэлюции - объем элюции, мл (0,1 мл).
Анализ каждого образца проведен в трех проворностях. Полученные результаты (А) усреднены.
*Если полученные значения ниже 0,27 пг в пробе, то количество ДНК ниже ПКО (Предел Количественного Определения) и принимается значение менее 0,27 пг (≤0,27 пг). Результаты анализа представлены в Таблице 3.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ БЕТА-N-АЦЕТИЛГЛЮКОЗАМИНИДАЗЫ STRH ИЗ STREPTOCOCCUS PNEUMONIAE | 2018 |
|
RU2693660C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ НЕЙРАМИНИДАЗЫ NANH ИЗ CLOSTRIDIUM PERERINGENS | 2018 |
|
RU2698774C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ЭНДОНУКЛЕАЗЫ SERRATIA MARCESCENS | 2021 |
|
RU2795623C2 |
| СПОСОБ ОЧИСТКИ РЕКОМБИНАНТНОГО ТКАНЕВОГО АКТИВАТОРА ПЛАЗМИНОГЕНА ДЛЯ МЕДИЦИНСКОГО ПРИМЕНЕНИЯ | 2017 |
|
RU2683950C1 |
| СПОСОБ ВЫДЕЛЕНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОЙ ЧЕЛОВЕЧЕСКОЙ ПРОУРОКИНАЗЫ М5 | 2013 |
|
RU2553533C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВЫСОКООЧИЩЕННОГО РЕКОМБИНАНТНОГО ИНГИБИТОРА С1-ЭСТЕРАЗЫ ЧЕЛОВЕКА ДЛЯ МЕДИЦИНСКОГО ПРИМЕНЕНИЯ | 2020 |
|
RU2769201C2 |
| Способ выделения и очистки рекомбинантного белка, аналога фрагмента каппа-казеина человека, обладающего цитотоксической активностью по отношению к раковым клеткам человека | 2018 |
|
RU2693251C1 |
| СПОСОБ ПРОМЫШЛЕННОЙ ОЧИСТКИ РОМИПЛОСТИМА | 2021 |
|
RU2804622C2 |
| Способ получения препарата рекомбинантной нуклеазы CAS, по существу, свободного от бактериальных эндотоксинов, полученный данным способом препарат и содержащий его набор для использования в системе CRISPR/Cas | 2019 |
|
RU2707542C1 |
| СЛИТЫЙ БЕЛОК ТИОРЕДОКСИНА И ДОМЕНА 4 ИНФЕСТИНА, СПОСОБ ЕГО ПОЛУЧЕНИЯ, ЭКСПРЕССИОННАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СЛИТЫЙ БЕЛОК, И БАКТЕРИЯ РОДА Escherichia coli, ТРАНСФОРМИРОВАННАЯ ТАКОЙ ПЛАЗМИДНОЙ ДНК | 2012 |
|
RU2528251C2 |









Изобретение относится к области биотехнологии и решает задачу получения в промышленных объемах высокоочищенной ферментативно активной рекомбинантной β-1,4-галактозидазы BgaA, пригодной для использования на стадии ремоделирования для получения терапевтически функциональных препаратов гликопротеинов с заданными свойствами, для разработки инструментов для высокочувствительного анализа гликановых цепей гликопротеинов тканей растений, животных и микроорганизмов. Способ предусматривает применение металлохелатной, псевдоаффинной, гидрофобной, анионобменной, концентрирующей хроматографии с последующей полирующей гель-фильтрацией, при этом белок из растворимой фракции клеточного лизата очищают с помощью двух стадий хроматографии на одном и том же Ni2+ металлохелатном сорбенте в различных условиях элюции и псевдоаффинной хроматографией в режиме проскока на Blue Sepharose. Применение вышеуказанных трех стадий помогает максимально очистить препарат от мажорных контаминирующих примесей. 9 з.п. ф-лы, 8 ил., 3 табл., 6 пр.
1. Способ очистки в промышленных объемах рекомбинантной β-1,4-галактозидазы BgaA в E. coli с применением металлохелатной, псевдоаффинной, гидрофобной, анионобменной, концентрирующей хроматографии и полирующей гель-фильтрации, отличающийся тем, что белок из растворимой фракции клеточного лизата очищают с помощью двух стадий хроматографии на одном и том же Ni2+ металлохелатном сорбенте, где элюцию на первой стадии проводят содержащим NaCl натрий-ацетатным буфером, имеющим рН в диапазоне от 4,2 до 4,3, на второй стадии целевой белок элюируют буфером, содержащим имидазол, и элюат со второй стадии используют для проведения псевдоаффинной хроматографии в режиме проскока на сорбенте с ковалентно связанным Cibacron Blue 3G.
2. Способ по п. 1, где предпочтительный рН буфера отмывки на второй стадии металлхелатной хроматографии находится в диапазоне от 6,5 до 6,6.
3. Способ по п. 1, где обе стадии металлхелатной хроматографии проводят на IMAC Sepharose 6 FF.
4. Способ по п. 1, где обе стадии металлхелатной хроматографии проводят на IMAC Sepharose High Performance.
5. Способ по п. 1, где на первой стадии металлохелатной хроматографии предпочтительный уровень рН буфера элюции составляет 4,3.
6. Способ по любому из пп. 1-5, где промывка на второй стадии металлхелатной хроматографии проводится буфером с рН 6,5 и высокой ионной силой.
7. Способ по п. 1, где на второй стадии металлохелатной хроматографии буфер элюции содержит от 150 мМ до 200 мМ имидазола.
8. Способ по п. 1, где псевдоаффинная хроматография проводится на Blue Sepharose.
9. Способ по п. 1, где на псевдоаффинной хроматографии при уравновешивании используют 5% изопропанол и 80 мМ NaCl, рН 8,3.
10. Способ по п. 1, где аминокислотная последовательность рекомбинантной β-1,4-галактозидазы BgaA с гексагистидиновым тагом идентична последовательности SEQ ID NO 2.
| US 8236520 B2, 07.08.2012 | |||
| US 6555348 B2, 2003.04.29 | |||
| БЕТА-ГАЛАКТОЗИДАЗА С ТРАНСГАЛАКТОЗИЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2007 |
|
RU2435856C2 |
| Campuzano S., Cloning, expression, and characterization of a peculiar choline-binding beta-galactosidase from Streptococcus mitis, Appl Environ Microbiol | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Zahner and Hakenberck, The Streptococcus pneumoniae Beta-Galactosidase Is a Surface Protein, J | |||
| Bacteriol, J Bacteriol., 2000 Oct; 182(20): 5919-5921, найдено онлайн, найдено в Интернет: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC94720/. | |||

