Государственная поддержка
Настоящее изобретение было сделано при поддержке правительства по 1R01NS071835, присужденному Национальному институту неврологических расстройств и инсульта. Правительство имеет определенные права на данное изобретение.
Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам против тау и способам их применения.
Предпосылки создания изобретения
Агрегация связанного с микротрубочками белка тау связана с несколькими нейродегенеративными расстройствами, такими как болезнь Альцгеймера (AD) и лобно-височная деменция. При AD патологическая агрегация тау распространяется постепенно по всему мозгу, возможно, вдоль существующих нейронных сетей. AD является наиболее распространенной причиной слабоумия и растущей проблемой общественного здравоохранения. В настоящее время от данной болезни страдает примерно 5 миллионов человек в Соединенных Штатах с ожидаемым увеличением до 13 миллионов к 2050 году. Болезнь Альцгеймера приводит к потере памяти, когнитивной функции и, в конечном счете, потери независимости. Она приводит к тяжелым личностным и финансовым потерям для пациента и его семьи. Вследствие тяжести и увеличения распространенности AD и других нейродегенеративных заболеваний, связанных с агрегацией тау, в популяции, крайне важно разработать более эффективные способы лечения и диагностики.
Ссылка на цветные фигуры
Файл заявки содержит, по меньшей мере, одну фотографию, выполненную в цвете. Копии данной публикации патентной заявки с цветными фотографиями предоставляются Управлением после запроса и оплаты необходимой пошлины.
Краткое описание фигур
На фиг.1 показаны аминокислотные последовательности N-конца (A) и С-конца (B) человеческого белка тау (htau).
На фиг.2 приведены графики, демонстрирующие KD HJ8.1 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.3 приведены графики, демонстрирующие KD HJ8.2 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.4 приведены графики, демонстрирующие KD HJ8.3 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.5 приведены графики, демонстрирующие KD HJ8.4 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.6 приведены графики, демонстрирующие KD HJ8.5 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.7 приведены графики, демонстрирующие KD HJ8.7 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.8 приведены графики, демонстрирующие KD HJ8.8 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.9 приведены графики, демонстрирующие KD HJ9.1 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.10 приведены графики, демонстрирующие KD HJ9.2 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.11 приведены графики, демонстрирующие KD HJ9.3 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.12 приведены графики, демонстрирующие KD HJ9.4 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.13 приведены графики, демонстрирующие KD HJ9.5 против человеческого белка тау (А) и мышиного белка тау (B).
На фиг.14 приведены результаты иммуноблоттинга, демонстрирующие присутствие полноразмерного тау у мышей ISF дикого типа и P301S tg. (А) Лизаты гиппокампа мышей Tau KO (KO), дикого типа (WT) и Р301S tg (P301S tg) анализируют методом иммуноблоттинга с использованием антитела ВТ-2 против тау или антитела против актина. Тринадцать мкг белка загружают в лунку. Четыре полосы, соответствующие эндогенному мышиному тау, и одну полосу, соответствующую человеческому тау, обозначены белыми кружками и черными кружками, соответственно. Существует также полоса 39 кДа, соответствующая форме человеческого белка тау в лизате гиппокампа P301S tg. Такая форма может представлять собой продукт деградации тау. ISF тау мышей дикого типа (WT) и P301S tg (P301S tg) подвергают иммунопреципитации с использованием моноклональных антител против тау HJ9.3 (В) или HJ8.1 (С) и анализируют методом иммуноблоттинга. Полосы визуализируют с помощью биотинилированного антитела ВТ-2. Серые и черные стрелки обозначают эндогенный мышиный тау и человеческий тау, соответственно.
На фиг.15 (А) приведено схематическое изображение разных мутантных конструкций тау, используемых в данном исследовании, и на фиг.15 (B-D) приведены изображения, демонстрирующие, что белки Tau RD образуют фибриллярные агрегаты в трансфицированных клетках НЕК293. (A) В зависимости от схемы эксперимента, каждая форма мутантного тау несет на карбокси-конце маркер, представляющий собой голубой или желтый флуоресцентный белок (CFP или YFP), либо гемагглютинин (HA). (В) Анализ методом атомно-силовой микроскопии (AFM) SDS-нерастворимого вещества из клеток НЕК293, временно трансфицированных разными формами RD, показывает, что RD(ΔK)-HA и RD(LM)-HA продуцируют видимые фибриллярные частицы. Фибриллы не детектируются в устойчивых к агрегации RD(PP)-HA (n=2). Масштабная метка, 1 мкм. (С) Клетки НЕК293, временно трансфицированные различными формами RD-YFP и одним YFP, окрашивают X-34, амилоид-специфичным красителем. Включения, образованные RD(WT)-YFP, RD(ΔK)-YFP и RD(LM)-YFP, визуализируются с помощью конфокальной микроскопии и окрашиваются X-34. X-34-положительные клетки не обнаружены при экспрессии только YFP или RD(PP)-YFP. Стрелки указывают включения, окрашенные X-34. (n=3) (D) Нетрансфицированные клетки (NT) и разные формы RD-YFP/CFP трансфицируют в клетки HEK293 с последующей экстракцией тритоном/SDS и анализом методом вестерн-блоттинга с использованием антитела против участка RD. Обнаружены как мономеры, так и элементы с более высокой молекулярной массой. (S=растворимый белок, Р=осажденный нерастворимый белок). Процедуру повторяют три раза с получением одинаковых результатов.
На фиг.16 показаны агрегаты тау RD в клетках НЕК293, детектированные методом FRET. Чтобы количественно определить внутриклеточную агрегацию белка RD методом резонансного переноса энергии флуоресценции (FRET), разные мутанты RD (wt, ΔΚ, PP, LM), слитые с YFP и CFP, совместно трансфицируют в клетки HEK293. (А). Клетки НЕК293, совместно трансфицированные RD(LM)-CFP/YFP, обследуют и количественно определяют внутриклеточное образование агрегатов с использованием микроскопии с фотообесцвечиванием акцептора для FRET. Донорный сигнал до (Pre) и после (Post) фотообесцвечивания акцептора подтверждает, что включения RD(LM)-CFP/YFP продуцируют среднюю эффективность FRET 18,2%±0,058 SD (n=6). На верхних и нижних панелях изображены акцепторные и донорные каналы, соответственно, до и после фотообесцвечивания. В правом верхнем углу изображена тепловая карта рассчитанной эффективности FRET. Масштабная метка гистограммы показывает расчетную эффективность FRET на попиксельной основе. Эффективность FRET для агрегата тау RD составляет ~34% в данной ячейке. (В). Используя FPR, определяют относительный FRET для разных конструкций. В случае RD(PP)-CFP/YFP значимый уровень FRET не наблюдается. Но как RD(ΔK)-CFP/YFP, так и RD(LM)-CFP/YFP, продуцируют интенсивный сигнал FRET (n=3). (С). Клетки НЕК293, экспрессирующие RD(ΔK)-CFP/YFP, подвергают воздействию фибрилл RD(ΔK)-CFP/YFP в различных концентрациях (мономерные эквиваленты 0,01, 0,03, 0,1 и 0,3 мкΜ) в течение 9 ч. Внеклеточные фибриллы RD(wt)-HA в дозозависимой манере индуцируют агрегацию RD(ΔK)-CFP/YFP (n=3). (* обозначает р<0,05, ** обозначает р<0,001, планки погрешностей соответствуют SEM).
На фиг.17 приведены изображения и графики, демонстрирующие перенос агрегатов тау-RD между клетками и индукцию последующей агрегации. (А). Клетки НЕК293, трансфицированные RD(ΔK)-YFP, совместно культивируют в течение 48 ч с эквивалентным числом клеток, экспрессирующих RD(LM)-HA. Клетки фиксируют 4% параформальдегидом и окрашивают иммунофлюоресцентной меткой/X-34. В ряде клеток наблюдается совместная локализация RD(LM)-HA и RD(ΔK)-YFP во включениях. Эти включения также окрашиваются X-34, что свидетельствует о наличии бета-складчатой структуры (закрашенные стрелки). Кроме того, некоторые включения RD(LM)-HA окрашиваются X-34, но совместная локализация с включениями RD(ΔK)-YFP отсутствует (незакрашенные стрелки). (В). Две популяции клеток, одна из которых экспрессирует RD(ΔK)-CFP/YFP, и другая - RD(LM)-HA, совместно культивируют в течение 48 ч. В качестве контроля используют RD(PP)-HA или нетрансфицированные клетки, NT. FRET увеличивается в результате совместного культивирования с RD(LM)-HA, но не с RD(PP)-HA или клетками, трансфицированными суррогатом (n=3). (С). Чтобы анализировать гибель клеток, индуцированную агрегатами тау, как механизм высвобождения тау, клетки НЕК293 трансфицируют в течение 48 ч RD-HA (PP, ΔΚ или LM) или суррогатом. Трансфицированные суррогатом клетки обрабатывают различными концентрациями стауроспорина (1, 2, 4, 20 мкМ) в течение 30 минут при 37°С, чтобы индуцировать гибель клеток. Затем клетки подвергают воздействию пропидиййодида, 5 мкг/мл, и измеряют флуоресценцию с помощью планшет-ридера. Признаки гибели клеток в разных трансфицированных популяциях отсутствуют. (** обозначает р<0,001, планки погрешностей соответствуют SEM).
На фиг.18 приведены изображения и графики, демонстрирующие, что агрегаты RD способствуют передаче неправильного сворачивания от клетки к клетке. Клетки НЕК293 совместно трансфицируют разными конструкциями RD-CFP и RD-HA. Спустя 15 ч трансфицированные клетки культивируют совместно с клетками, экспрессирующими RD(ΔK)-YFP или RD(PP)-YFP, в течение 48 ч. (A) Для определения, происходит ли совместная агрегация в результате непосредственного контакта белков, проводят микроскопическое исследование FRET. Сигнал CFP измеряют до и после фотообесцвечивания YFP. Агрегаты RD(LM)-CFP и RD(LM)-YFP характеризуются средней эффективностью FRET 14,2%±0,053 SD (n=11), свидетельствующей о том, что RD(LM)-CFP и RD(LM)-YFP находятся в непосредственном контакте. Верхние и нижние панели изображают акцепторные и донорные каналы, соответственно, до (Pre) и после (Post) фотообесцвечивания. Типичная тепловая карта рассчитанной эффективности FRET приведена в правом верхнем углу. Гистограмма демонстрирует расчетную эффективность FRET на попиксельной основе. Эффективность FRET для агрегата тау RD составляет ~25% в данной ячейке. Отрицательные значения генерируются неспаренными CFP. (В) Сигнал FRET наблюдается, если клетки, экспрессирующие RD(ΔK)-CFP/RD-HA, культивируют совместно с клетками, экспрессирующими RD(ΔK)-YFP. Этот сигнал увеличивается, если агрегация RD(ΔK)-CFP индуцируется в результате совместной экспрессии склонных к агрегации форм тау, как ΔΚ, так и мутантов LM. RD-CFP или RD-YFP, содержащие мутацию PP, блокирующую образование β-складчатой структуры (n=3), не генерируют какого-либо значащего сигнала. (С) Чтобы подтвердить увеличение неправильной укладки, популяции клеток, экспрессирующих один CFP или RD(LM)-CFP, в течение 48 ч подвергают предварительному воздействию клеток, экспрессирующих RD-HA, содержащий одну из мутаций PP, ΔΚ или LM, чтобы инициировать неправильную укладку в разной степени. Затем указанные совместно культивируемые популяции делят на части и совместно культивируют в течение 48 ч с клетками, экспрессирующими RD(ΔΚ)-YFP, чтобы определить степень агрегации на основе переноса между клетками и FRET. Предварительное воздействие клеток RD(LM)-HA на клеточную популяцию RD(ΔΚ)-CFP индуцирует сигнал FRET, в 2,6 раза превышающий сигнал, индуцированный в результате предварительного воздействия на RD(PP)-HA. Введение клеток, экспрессирующих чистый CFP, во вторую популяцию клеток полностью блокирует влияние предварительного воздействия на склонных к агрегации мутантов RD-HA (n=3). (* указывает, что значение р<0,05, ** указывает, что значение р<0,001, планки погрешностей соответствуют SEM).
На фиг.19 приведены графики и результаты иммуноблоттинга, демонстрирующие распространение агрегатов тау через внеклеточную среду. (A) Клетки НЕК293, трансфицированные RD(LM)-HA, совместно культивируют в течение 48 ч с эквивалентным количеством клеток RD(ΔΚ)-CFP/YFP и затем проводят анализ FRET. Увеличение объема клеточной культуральной среды приводит к снижению эффективности трансклеточного перемещения агрегатов. (B) Перенос кондиционированной среды от клеток, экспрессирующих RD(LM)-HA, к клеткам, экспрессирующим RD(ΔΚ)-CFP/YFP, является достаточным для того, чтобы индуцировать агрегацию на 60%. (C) Добавление в среду антитела HJ9.3 приводит к уменьшению FRET, а также препятствует распространению агрегации. (D) Неспецифическое IgG не оказывает какого-либо влияния на распространение. (E) HJ9.3 не оказывает влияния на внутриклеточную агрегацию RD(ΔΚ)-CFP/YFP, совместно экспрессированного в той же клетке. (F) HJ9.3 блокирует эффект RD(LM)-HA, связанный с индукцией RD(ΔΚ)-YFP при совместном культивировании клеток, что определяют путем фракционирования в присутствии детергента и методом вестерн-блоттинга. (Т=общий белок, S=растворимый белок и Р=осадок нерастворимого белка). (G) Количественный анализ трех независимых исследований методом вестерн-блоттинга демонстрирует ~60% уменьшение фракции осадка после воздействия HJ9.3 по сравнению с общей фракцией. (H) Клетки, экспрессирующие RD(LM)-YFP и mCherry, совместно культивируют и анализируют методом проточной цитометрии. HJ9.3 уменьшает процент двойных положительных клеток от 2,07% до 1,31%. Клетки, смешанные непосредственно перед цитометрией, используют в качестве фонового контроля (* обозначает p<0,05, ** обозначает р<0,001, планки погрешностей соответствуют SEM).
На фиг.20 приведены изображения клеток НЕК293, трансфицированных RD(ΔΚ)-YFP (верхние панели) или суррогатом (нижние панели). HJ9.3 добавляют к культуральной среде и инкубируют в течение 48 ч. В конце эксперимента клетки фиксируют, пермеабилизируют и окрашивают вторичным антителом против мышиных антител (меченным Alexa 546). Локализацию комплексов HJ9.3/тау определяют методом конфокальной микроскопии. В верхних панелях показано, что многие комплексы детектируются при экспрессии RDΔ(K)-YFP, и в отсутствии указанной экспрессии комплексы не наблюдаются (нижние панели). Ортогональные анализы (правая панель) показывают, что комплексы в большинстве своем присутствуют на поверхности клетки, хотя иногда наблюдаются внутриклеточные комплексы.
На фиг.21 приведены изображения и график, демонстрирующие, что фибриллы тау опосредуют распространение от клетки к клетке. (А) Кондиционированную среду собирают из трансфицированных клеточных популяций, совместно культивированных с HJ9.3 или контрольным антителом IgG (1:1000) в течение 0 ч или 48 ч, и затем анализируют методом иммунопреципитации и вестерн-блоттинга. HJ9.3, в отличие от IgG, специфически улавливает частицы тау RD из клеточной среды. Агрегированные частицы высшего порядка присутствуют при экспрессии RD(ΔK)-YFP или RD(LM)-YFP, но не RD(PP)-YFP. (В) Количественный анализ результатов трех независимых исследований методом вестерн-блоттинга свидетельствует о ~10-кратном увеличении тау после 48 ч инкубации. (C) Клетки подвергают воздействию HJ9.3 в течение разных периодов времени. (D) Комплексы антитело/антиген, выделенные из среды, подвергшейся в течение 48 ч воздействию HJ9.3, помещают на чипы AFM и визуализируют. В среде клеток, экспрессирующих RD(ΔK)-HA и RD(LM)-HA, наблюдаются фибриллярные частицы, тогда как RD(PP)-HA продуцирует только аморфные агрегаты. Масштабная метка, 1 мкм.
На фиг.22 изображены схема и графики, демонстрирующие активность HJ8.5 и HJ9.4 в отношении рекомбинантного человеческого белка тау. (А) Схема, иллюстрирующая совместное культивирование клеток RD(LM)-CFP и RD(ΔK280)-YFP в присутствии и в отсутствии разных моноклональных полноразмерных антител против тау. (B) График, демонстрирующий, что HJ8.5, HJ9.3 и HJ9.4 способны блокировать распространение тау. (C) График, демонстрирующий, что HJ8.5, HJ9.3 и HJ9.4 можно использовать для детекции фибрилл RD-tau методом ELISA.
На фиг.23 приведена схема, иллюстрирующая экспериментальный план (A) интрацеребровентрикулярной инъекции и (B) имплантации осмотического насоса в боковой желудочек каждой мыши. (С) Изображение, подтверждающее размещение канюли путем окрашивания крезиловым фиолетовым.
На фиг.24 приведены изображения антител против тау после инфузии в течение 6 недель мышам P301S tg, полученных путем (A) окрашивания кумасси синим и (B) иммуноблоттинга против рекомбинантной самой длинной изоформы человеческого белка тау hTau40 с использованием антител, поступающих из насоса, до и после 6 недель инфузии.
На фиг.25 изображен график, показывающий отсутствие перекрестного взаимодействия вливаемых антител против тау в анализе общего тау методом HJ8.7-BT2B ELISA. Перед применением в методе ELISA антитела в указанных концентрациях предварительно инкубируют с рекомбинантным человеческим белком тау.
На фиг.26 приведены изображения коронарных срезов пириформной коры мышей P301S tg в возрасте 9 месяцев, получавших среду/PBS (верхние панели) или разные моноклональные антитела против тау (HJ8.5, HJ9.3, как указано в нижних панелях). Срезы окрашивают биотинилированным антителом AT8, которое распознает аномально фосфорилированную форму тау.
На фиг.27 приведены графики, демонстрирующие процент площади, покрытой нейрофибриллярными клубками, окрашенными AT8, в (А) гиппокампе CA2 и СА3, (В) миндалинах, (С) пириформной коре и (D) энторинальной коре.
На фиг.28 приведены графики, демонстрирующие детекцию фибрилл тау и мономера RD-tau методом ELISA с использованием антитела HJ9.3. Мономеры и фибриллы RD-wt tau в различных концентрациях помещают в планшет для ELISA. HJ9.3 используют в качестве первичного антитела. Для детекции используют антитела против мышиных антител, связанные с HRP.
На фиг.29 приведена схематическая иллюстрация трансклеточного распространения агрегации тау, происходящего посредством переноса фибрилл в клеточную среду. Белковый агрегат, присутствующий в донорной клетке, выходит из нее (А), поступает в клетку-реципиент (B) и непосредственно контактирует с белком, имеющим природную укладку (С), увеличивая неправильно уложенное состояние (D). Такое перемещение между клетками опосредуется фибриллами, которые высвобождаются непосредственно в среду. Данные фибриллы могут улавливаться в межклеточном пространстве антителом против тау (HJ9.3), что препятствует распространению между клетками (E).
Фиг.30. Характеристика антител против тау методами поверхностного плазмонного резонанса (SPR) и иммуноблоттинга. На фигуре изображены сенсограммы SPR, демонстрирующие связывание каждого антитела против тау с иммобилизованным рекомбинантным человеческим тау (самые длинные изоформы hTau40, 441 а.к.) и иммобилизованным мышиным тау (самые длинные изоформы mTau40, 432 а.к.). Каждое антитело используют в различных концентрациях (0,11, 0,23, 0,46, 0,90, 1,8, 3,7, 7,5 мкг/мл), и графики выделяют соответствующим цветом. (А) Сенсограммы SPR для связывания антитела HJ9.3 с иммобилизованным человеческим тау и иммобилизованным мышиным тау (B). (С) Сенсограммы SPR для связывания антитела HJ9.4 с иммобилизованным человеческим тау и иммобилизованным мышиным тау (D). Сенсограммы SPR для связывания антитела HJ8.5 с иммобилизованным (E) человеческим и (F) мышиным тау. (G) Растворимые фракции RAB мышей в возрасте 3 месяцев с нокаутом тау (KO), в возрасте 3 месяцев дикого типа (WT), в возрасте 3 месяцев Р301S (3mo) и в возрасте 9 месяцев Р301S (9mo) анализируют методом иммуноблоттинга с использованием указанных антител против тау.
Фиг.31. Сенсограммы SPR, иллюстрирующие взаимодействие антител против тау с иммобилизованными фибриллами человеческого белка тау. Сенсограммы SPR иллюстрируют взаимодействие антител против тау HJ9.3 (A), HJ9.4 (В) и HJ8.5 (С), использованных в различных концентрациях, и иммобилизованных фибрилл человеческого белка тау.
Фиг.32. Характеристика антител против тау в разных анализах. Иммуноокрашивание срезов головного мозга мышей в возрасте 3 месяцев с нокаутом тау (KO), в возрасте 3 месяцев дикого типа (WT), в возрасте 3 месяцев Р301S (3mo) и в возрасте 12 месяцев Р301S (12mo) из участка пириформной коры и лобной коры ткани, пораженной болезнью Альцгеймера (AD), проводят путем окрашивания биотинилированным антителом HJ8.5. Вставка в микрофотографию среза мыши P301S в возрасте 12 месяцев демонстрирует окрашивание тела клетки помимо диффузного окрашивания нейропиля. Черная стрелка указывает увеличение площади. Вставка в микрофотографию коры головного мозга человека с AD демонстрирует окрашивание нейрофибриллярных клубков (NFT) при большем увеличении. Черная стрелка указывает увеличение площади. Масштабная метка 250 мкм в панели с tau KO, изображения получены с таким же увеличением. Масштабная метка 50 мкм во вставках P301S 12mo и AD.
Фиг.33. Антитела против тау блокируют поглощение и затравочную активность агрегатов P301S tau, детектируемую методом FRET. Клетки НЕК293, экспрессирующие RD(ΔK280)-CFP/YFP, инкубируют с 2,5 мкг общего белка лизатов мозга 1×TBS в течение 24 ч. (A) Лизаты мозга мышей P301S в возрасте 12 месяцев вызывают гораздо большую затравочную активность (n=5), чем лизаты мышей с нокаутом (KO) (n=7), мышей дикого типа (WT) (n=6) или молодых мышей P301S в возрасте 3 месяцев (n=2) (****p<0,0001). (B) Клетки НЕК293 совместно трансфицируют с RD(ΔK280)-CFP и RD(ΔK280)-YFP. Через 18 ч к клеткам добавляют лизаты мозга Р301S, предварительно инкубированные в отсутствии или в присутствии антител против тау (HJ8.5, HJ9.3 и HJ9.4) или контрольного антитела (антитело HJ3.4 против Αβ). Все антитела против тау, инкубированные с лизатами головного мозга P301S, по существу блокируют затравочную активность. Статистическую значимость определяют с помощью однофакторного дисперсионного анализа с последующим апостериорным множественным сравнением Даннета с использованием программного обеспечения GraphPad Prism 5.0 (***p>0,001). (С) Титрование указанных антител в различных концентрациях (0,125 мкг/мл, 0,25 мкг/мл, 0,5 мкг/мл, 1 мкг/мл и 2 мкг/мл) проводят в присутствии фиксированного количества лизатов головного мозга Р301S. Через 24 часа проводят анализ методом FRET. Из всех использованных антител против тау HJ8.5 наиболее эффективно блокирует поглощение и затравочную активность лизатов головного мозга P301S. Статистическую значимость определяют с помощью двухфакторного дисперсионного анализа с последующим апостериорным множественным сравнением Бонферрони. (**p<0,0001, *p<0,01, результаты приведены в виде среднего значения±SEM).
Фиг.34. Поглощение клетками антител против тау, связанных с агрегатами P301S Tau, не обнаружено. Лизаты мозга P301S инкубируют с клетками НЕК293 в течение 3 ч. Чтобы детектировать тау, используют все 3 разных антитела против тау или контрольное антитело (антитело HJ3.4 против Αβ), с последующим окрашиванием антителами против мышиных IgG, меченных Alexa-fluor546. Кроме того, P301S лизаты мозга предварительно инкубируют в отсутствии и в присутствии 3-х разных антител против тау и антитела HJ3.4, после чего их добавляют к клеткам НЕК293, фиксируют и пермеабилизируют. Alexa-fluor546-меченые антитела против мышиных IgG используют для идентификации интернализованных антител. Для окрашивания ядра используют 4',6'-диамидино-2-фенилиндол (DAPI; показан синим цветом).
Фиг.35. Экспериментальная схема ICV инфузии антител и эффективность антитела в другом способе лечения. (А) Экспериментальный план инфузии антител или среды (PBS) путем интрацеребровентрикулярной инъекции в левый боковой желудочек головного мозга. (B) Показательное окрашивание крезиловым фиолетовым коронарных срезов участка мозга, чтобы подтвердить размещение имплантированного хирургическим способом зонда в левом боковом желудочке. В данном исследовании используют мышей с правильным размещением зонда в левом боковом желудочке.
Фиг.36. Антитела против тау сильно уменьшают окрашивание AT8 в мозге мыши P301S. Показательные коронарные срезы мышей P301S в возрасте 9 месяцев, обработанных PBS (A), антителом HJ3.4 (B), антителом HJ8.5 (C), антителом HJ9.3 (D) и антителом HJ9.4 (E), окрашенные биотинилированным антителом AT8, в участках, включающих пириформную кору и миндалину. Масштабная метка 250 мкм. Вставки в А-Е показывают окрашивание фосфорилированного тау биотинилированным антителом AT8 с более высоким увеличением, масштабная метка 50 мкм.
Фиг.37. Некоторые антитела против тау сильно уменьшают AT8-окрашивание мозга мышей P301S. Процент площади, покрытой аномально фосфорилированным тау, окрашенным биотинилированным AT8, в пириформной коре (А), энторинальной коре (B), миндалине (C) и участке гиппокампа СА1 (D) мышей Р301S в возрасте 9 месяцев, получавших антитела против тау HJ8.5 (N=13), HJ9.3 (N=15), HJ9.4 (N=13), антитело против Αβ, HJ3.4 (N=8) или PBS (N=16). Окрашивание AT8 в нескольких разных отделах головного мозга у мышей, получавших антитела против тау, ниже, чем у мышей, получавших PBS или антитело HJ3.4. HJ8.5 оказывает наибольшее воздействие. **p<0,01, *p<0,05, результаты приведены в виде среднего значения±SEM.
Фиг.38. Количественное определение окрашивания биотинилированным антителом AT8 у самцов и самок мышей Р301S. Процент площади, покрытой аномально фосфорилированным тау, окрашенным биотинилированным AT8, в пириформной коре (А и Е), энторинальной коре (B и F), миндалине (С и G) и участке гиппокампа CA1 (D и H) у самцов (А) и самок (B) мышей Р301S, получавших антитела против тау (HJ8.5, HJ9.3 и HJ9.4), контрольное антитело (HJ3.4) и PBS.
Фиг.39. Некоторые антитела против тау сильно уменьшают ThioS-окрашивание нейрофибриллярных клубков в головном мозге мышей Р301S. (A) Показательные изображения ThioS-окрашивания нейрофибриллярных клубков в пириформной коре мышей Р301S в возрасте 9 месяцев, получавших в течение 3 месяцев PBS и антитела HJ3.4, HJ8.5, HJ9.3 и HJ9.4. У мышей, получавших антитело HJ8.5, ThioS-окрашивание нейрофибриллярных клубков уменьшается по сравнению с мышами, получавшими PBS или антитело HJ3.4. Масштабная метка соответствует 100 мкм. (B) Полуколичественный анализ окрашивания ThioS с использованием оценок от 1 (отсутствие окрашивания) до 5 (максимальное окрашивание) проводят у всех мышей, получавших антитела против тау, и контрольных мышей. У мышей, получавших антитело HJ8.5, ThioS-окрашивание значительно уменьшается по сравнению с мышами, получавшими PBS или антитела HJ3.4. *p<0,05, **p<0,01.
Фиг.40. Корреляция между окрашиванием фосфорилированного тау и окрашиванием активированных клеток микроглии. (A) Окрашивание фосфорилированного тау биотинилированным AT8 у мышей Р301S в возрасте 9-месяцев, получавших HJ8.5 (N=6), HJ9.3 (N=6) и PBS (N=6 в каждой группе) в значительной степени коррелирует с PHF1-окрашиванием другого антитела против фосфорилированного тау. (B) Наблюдается значительная корреляция между CD68-окрашиванием активированных клеток микроглии и окрашиванием фосфорилированного тау биотинилированным AT8 во всех группах (n=6 в каждой группе) (C) Иммуноблоттинг типичных образцов фракции 70% FA (N=4) проводят с использованием поликлональных мышиных антител против тау (Abcam).
Фиг.41. CD68-окрашивание активированных клеток микроглии. Активацию микроглии анализируют у мышей Р301S. Типичные изображения CD68-окрашенных активированных клеток микроглии в пириформной коре 9-месячных мышей Р301S, получавших PBS (A), антитело HJ3.4 (B), антитело HJ8.5 (С), антитело HJ9.3 (D) и антитело HJ9.4 (Е).
Фиг.42. Уровень нерастворимого тау у мышей Р301S уменьшается в результате воздействия антител HJ8.5 и HJ9.3. Кору головного мозга всех обработанных мышей [получавших PBS (N=16) или антитела HJ3.4 (N=8), HJ8.5 (N=13), HJ9.3 (N=15), HJ9.4 (N=13)] последовательно экстрагируют RAB (A), RIPA (В) и 70% FA (С), после чего количественно определяют уровень тау с использованием метода ELISA. Среди разных групп отсутствуют статистические различия в уровнях растворимого тау во фракциях RAB и RIPA. Однако наблюдается значительное снижение уровня нерастворимого тау во фракциях 70% FA, полученных от мышей, получавших антитела против тау HJ8.5 и HJ9.3, по сравнению с группами, получавшими PBS или антитело HJ3.4. Уровень нерастворимого тау у мышей, получавших антитело HJ9.4, не отличается от уровня, наблюдающегося у контрольных групп (**p<0,01). Уровни человеческого белка тау (D), мышиного белка тау (E) и тау, фосфорилированного по Ser202 и Thr205 (F), определяют во фракциях 70% FA с использованием специфических антител против человеческого белка тау, против мышиного белка тау или против фосфорилированного тау методом ELISA (N=6 мышей на группу). Наблюдается снижение уровня человеческого белка тау у всех групп мышей, получавших антитела против тау, и изменения уровня мышиного белка тау отсутствуют. Авторы настоящего изобретения также обнаружили, путем детекции активности AT8, что во фракциях 70% FA уровень тау, фосфорилированного по Ser202 и Thr205, уменьшается у мышей, получавших антитела против тау по сравнению с контрольной группой, подобно уровню общего человеческого белка тау.
Фиг.43. У мышей Р301S, получавших антитело против тау, с помощью анализа FRET обнаруживают уменьшение затравочной активности тау в экстрактах коры головного мозга. (А) Затравочную активность тау измеряют с использованием растворимых фракций RAB, полученных от всех мышей, обработанных PBS (N=16), HJ3.4 (N=8), HJ8.5 (N=13), HJ9.3 (N=15) и HJ9.4 (N=13), на клетках HEK293 методом FRET. Клетки HEK293 совместно трансфицируют RD(ΔK280)-CFP и RD(ΔK280)-YFP. Через 18 ч к клеткам добавляют растворимые фракции RAB. У мышей, получавших антитела HJ8.5 и HJ9.3, затравочная активность гораздо ниже, чем у мышей, получавших PBS или HJ3.4. В растворимых фракциях RAB, полученных от мышей, обработанных антителом HJ9.4, не наблюдается снижение затравочной активности по сравнению с растворимыми фракциями RAB, полученными от мышей, обработанных PBS или антителом HJ3.4 (***p<0,001, результаты приведены в виде среднего значения±SEM). (B) Растворимые фракции RAB, полученные от мышей с нокаутом тау, обработанных PBS или антителом против тау, подвергают иммунопреципитации. Элюирование затравочной активности из комплексов антитело/гранулы измеряют методом FRET. У мышей, получавших HJ8.5 и HJ9.3, затравочная активность гораздо меньше, чем у мышей, получавших PBS (****р<0,0001, результаты приведены в виде среднего значения±SEM). (C) Результаты ELISA-анализа фракций 70% FA, полученных из участков коры головного мозга 9-месячных мышей Р301S всех экспериментальных групп, в значительной степени коррелируют с результатами анализа растворимых фракций RAB методом FRET. (D) Сравнивают уровни тау (X-ось) и затравочную активность (Y-ось) в растворимых фракциях RAB коры головного мозга всех мышей Р301S в возрасте 9 месяцев. Значимая корреляция между 2 указанными параметрами отсутствует. (E) Частицы тау, присутствующие в растворимых фракциях RAB, полученных от мышей с нокаутом в возрасте 3 месяцев (KO), мышей дикого типа в возрасте 3 месяцев (WT), мышей Р301S в возрасте 3 месяцев и мышей Р301S в возрасте 9 месяцев, получавших PBS, разделяют методом SDD-AGE с последующим анализом методом вестерн-блоттинга. Для детекции частиц тау используют поликлональные мышиные антитела против тау. Высокомолекулярные частицы присутствуют в растворимой фракции RAB и у мышей Р301S в возрасте 3 месяцев, и, в больших количествах, у мышей Р301S в возрасте 9 месяцев.
Фиг.44. Результаты тестирования условно-рефлекторной реакции на страх демонстрируют, что среди групп отсутствуют значимые различия в двигательной активности, сенсомоторных реакциях или компонентах слухового ориентира. Результаты rmANOVA не позволяют выявить значимые основные или интеракционные эффекты лечения, включающие влияние на общую способность к передвижению в тесте с использованием платформы с отверстиями (A), в тесте с прохождением мыши по рейке (B), или на любой другой показатель сенсомоторных реакций (не показано), или на способность удерживаться на вращающемся барабане (С). Данные, полученные в результате изменения базовых условий на 3-й день тестирования условно-рефлекторной реакции страха, демонстрируют наличие значимого эффекта лечения (*р=0,027), причем последующие сравнения показывают, что большая часть этого эффекта обусловлена значительными различиями между мышами, получавшими HJ9.4, и контрольной группой, получавшей PBS+HJ3.4 (р=0,0007). (D). Однако каких-либо существенных основных или интеракционных эффектов лечения не было обнаружено после анализа rmANOVA результатов тестирования слухового ориентира (мин 3-10), позволяя предположить, что уровни замирания существенно не различаются среди групп в течение указанного времени (E). Для определения, может ли уровень активности оказывать влияние на состояние замирания в процессе тестирования ситуативного страха на 2-й день, авторы рассчитали коэффициент корреляции Пирсона (r) между общей способностью к передвижению, измеренной с помощью теста на платформе с отверстиями, и % времени, проведенного в состоянии замирания во время тестирования ситуативного страха, и обнаружили, что достоверная корреляция отсутствует (р=0,39) (F).
Фиг.45. Дефицит условно-рефлекторной реакции на ситуативный страх у трансгенных мышей Р301S tau уменьшается в результате введения антител HJ8.5 и HJ9.4. (A) В 1 день тестирования условно-рефлекторной реакции на страх различие в уровне замирания среди групп не наблюдается ни во время 2-минутного исходного состояния, ни во время испытания звук/шок (T/S), о чем свидетельствует отсутствие значимых основных или интеракционных эффектов, связанных с введением антител, определяемое путем анализа полученных результатов методом rmANOVA. (B) И наоборот, значимый эффект введения антител (*р=0,019) и значимый эффект поминутного взаимодействия (**р=0,0001) определяют с помощью анализа rmANOVA результатов по уровням замирания, полученным на 2 день тестирования условно-рефлекторной реакции на страх. Только в группе, получавшей HJ9.4, наблюдается значительное привыкание от 1 до 8 минуты (#р=0,002). (C) Последующие запланированные сравнения демонстрируют, что замирание в группах, получавших антитела против тау HJ8.5 и HJ9.4, значительно увеличивается по сравнению с контрольной группой, получавшей PBS+HJ3.4, при усреднении значений, полученных в течение 8-минутного сеанса (**р=0,006 и *р=0,022, соответственно). Однако дальнейший анализ полученных данных свидетельствует о том, что наибольшее различие между группой, получавшей HJ9.4, и контрольной группой, получавшей PBS+HJ3.4, наблюдается в течение 2 минуты (†p=0,004), тогда как наибольшее различие между группой, получавшей HJ8.5, и контрольной группой наблюдается в течение 4-7 минут (††р<0,004), как изображено в "B".
На фиг.46 приведен график, демонстрирующий анализ тау методом сэндвич-ELISA, который можно использовать для различения образцов плазмы, положительных по затравочной активности, и образцов плазмы, отрицательных по затравочной активности. Затравочную активность определяют по способу, описанному Kfoury et al 2012 J Biol Chem 287(23). Количество агрегатов тау приводят в виде относительного связанного с изменением укладки возбуждения по сравнению с сигналом, полученным из плазмы здоровых молодых людей (т.е. фоновым сигналом анализа).
На фиг.47 приведены графики, демонстрирующие влияние антител настоящего изобретения против тау на распространение тау в клетке. В каждом графике первый столбец представляет среду без добавления антител, то есть, соответствует о фоновой эффективности распространения. (A) HJ8.1 и HJ8.2; (B) HJ8.3 и HJ8.4; (C) HJ8.5 и HJ8.7; (D) HJ8.8 и HJ9.1; (E) HJ9.2 и HJ9.3; (F) HJ9.4 и HJ9.5.
На фиг.48 приведен график, представляющий результаты клеточного анализа, демонстрирующие влияние отдельных антител против тау или эквимолярных смесей антител против тау на распространение тау.
На фиг.49 в разделе (А) приведен график, демонстрирующий, что антитело HJ9.3 не оказывает влияния на внутриклеточную агрегацию тау при одновременной экспрессии RD(ΔK)-CFP/YFP в той же клетке, и в разделе (В) приведен график, демонстрирующий, что неспецифический IgG не влияет на чресклеточное распространение агрегации тау.
На фиг.50 приведен график, демонстрирующий, что HJ9.3 ингибирует поглощение агрегатов тау по данным метода проточной цитометрии. Клетки подвергают воздействию рекомбинантных фибрилл RD, меченных флуоресцентным красителем химическим способом. После обработки трипсином и диспергирования клетки считают с использованием проточного цитометра. HJ9.3 дозозависимо уменьшает число клеток, несущих флуоресцентную метку, что указывает на ингибирование поглощения агрегатов.
Подробное описание изобретения
Общеизвестной минимальной связью между болезнью Альцгеймера и всеми видами таупатии является агрегированное состояние тау. При всех указанных болезненных состояниях мономерный тау, как известно, превращается в полимерные упорядоченные фибриллы. Нейрофибриллярные клубки (NFT), которые состоят из агрегатов фибриллярного тау, являются нейропатологическим признаком таупатии. Авторы настоящего изобретения обнаружили, что распространение тау-патологии в головном мозге может вызываться формой агрегата тау, которая высвобождается из "донорной" клетки и поступает во вторую "реципиентную" клетку, индуцируя дальнейшую неправильную укладку и агрегацию тау в реципиентной клетке путем непосредственного белок-белкового контакта. Специфическую форму агрегата тау, которая обеспечивает такое распространение агрегатов тау от клетки к клетке, называют "затравкой тау", и ее активность в настоящем описании называют "затравочной активностью", так как данная форма агрегатов тау может служить затравкой или центром образования агрегатов тау в клетке, в которую она проникает (т.е. в "реципиентной клетке").
Тау может существовать как в виде мономерной формы, так и в виде разных агрегированных форм. В настоящем описании термин "агрегат тау" относится к молекулярному комплексу, который содержит два или более мономеров тау. Не желая быть связанными с какой-либо теорией, авторы предположили, что агрегат тау может содержать практически неограниченное число мономеров, соединенных друг с другом. Например, агрегат тау может содержать два, три, четыре, пять, шесть, семь, восемь, девять, 10 или более мономеров тау. Альтернативно, агрегат тау может содержать 20, 30, 40, 50, 60, 70, 80, 90, 100 или более мономеров тау. Агрегат тау может также содержать 500, 1000, 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000 или более мономеров тау. Термины "фибриллярный агрегат тау" и "фибрилла тау" относятся к форме агрегатов тау и используются в данном описании как взаимозаменяемые. Фибриллярный агрегат тау представляет собой полимерное упорядоченное волокно, содержащее тау. Фибриллы тау, как правило, не растворяются, однако более короткие агрегаты, или олигомеры, могут быть растворимыми. Термин "агрегат тау" также относится к растворимым олигомерам и протофибриллам тау, которые могут служить промежуточными продуктами в процессе агрегации тау. Кроме того, определение "агрегат тау" охватывает термин "затравка тау", который относится к агрегату тау, который может выполнять роль центра образования или "затравки" внутриклеточных агрегатов тау после проникновения внутрь клетки, или при контакте с мономерным тау in vitro. Затравочную активность тау можно определить с помощью клеточного анализа агрегации тау, описанного в данном описании.
Кроме того, авторы разработали антитела, которые специфически связываются с тау, и способы их применения. В одном аспекте настоящее изобретение предоставляет антитела, которые специфически связываются с тау. В другом аспекте настоящее изобретение предоставляет способы эффективного замедления и/или уменьшения распространения агрегации тау от клетки к клетке. Антитела настоящего изобретения могут замедлять и/или уменьшать распространение агрегации тау путем инициации дезагрегации белковых фибрилл, блокировки превращения мономерного тау в агрегированный тау в клетке, стимуляции внутриклеточной деградации агрегатов тау, предотвращения проникновения агрегатов тау в соседние клетки, или путем комбинации указанных механизмов. В другом аспекте настоящее изобретение предоставляет способы детекции агрегатов тау в образце биологической жидкости, полученном от индивидуума. В другом аспекте настоящее изобретение предоставляет способы измерения количества агрегатов тау в образце биологической жидкости, полученной от индивидуума. В другом аспекте настоящее изобретение предоставляет способы отнесения индивидуума к определенной группе на основе количества агрегата тау, измеренного в образце биологической жидкости, полученной от индивидуума. Отнесение индивидуума к определенной группе на основе количества агрегата тау, измеренного в образце биологической жидкости, полученной от индивидуума, можно использовать для выявления индивидуумов, у которых может развиваться симптом и/или заболевание, связанные с агрегацией тау, на протяжении жизни индивидуума.
Настоящее изобретение охватывает открытие, заключающееся в том, что антитела против тау могут замедлять распространение фибриллярных агрегатов тау путем связывания внеклеточного тау, высвобождающегося из клеток, предотвращая тем самым проникновение агрегатов тау в соседние клетки и замедляя распространение агрегации тау. В одном аспекте настоящее изобретение предоставляет способы предотвращения проникновения агрегата тау в клетку. В другом аспекте настоящее изобретение предоставляет способы снижения внутриклеточной агрегации тау. В другом аспекте настоящее изобретение предоставляет способы уменьшения затравочной активности тау. Антитела настоящего изобретения можно использовать для предотвращения проникновения агрегатов тау в соседние клетки, в том числе антитела, которые связываются с эпитопом, присутствующим в тау.
I. Антитела, способные связываться с тау
У человека существуют шесть изоформ тау, которые образуются в результате альтернативного сплайсинга экзонов 2, 3 и 10. Изоформы содержат от 352 до 441 аминокислот. Каждый из экзонов 2 и 3 кодирует вставку из 29 аминокислот на N-конце (обозначенный N, и, следовательно, изоформы тау можно обозначать 2N (обе вставки), 1N (только экзон 2) или 0N (ни одной вставки). Все изоформы тау имеют три повтора микротрубочки-связывающего домена. Включение экзона 10 на С-конце приводит к включению четвертого микротрубочки-связывающего домена, кодируемого экзоном 10. Следовательно, изоформы тау могут состоять из четырех повторов микротрубочки-связывающего домена (экзон 10 входит в состав) или из трех повторов микротрубочки-связывающего домена (экзон 10 не входит в состав). Антитела настоящего изобретения против тау могут включать антитела, способные связываться с любой из изоформ тау. В иллюстративном варианте осуществления антитела настоящего изобретения против тау могут включать антитела, способные связываться с изоформой тау, которая содержит экзон 10.
Как указано выше, тау может присутствовать в растворимых и нерастворимых компартментах, в виде мономерных и агрегированных форм, упорядоченных и неупорядоченных структур, внутри клетки и вне клетки, и, кроме того, он может находиться в комплексе с другими белками или молекулами. Антитела настоящего изобретения против тау могут включать антитела, способные связываться с одной или несколькими из описанных выше форм тау. В некоторых вариантах осуществления антитело против тау связывается с мономером тау. В других вариантах осуществления антитело против тау связывается с агрегатом тау. В других вариантах осуществления антитело против тау связывается с фибриллами тау. В разных вариантах осуществления антитело против тау связывается с мономером тау и агрегатом тау. В альтернативных вариантах осуществления антитело против тау связывается с агрегатом тау и фибриллами тау. В разных вариантах осуществления антитело против тау связывается с фибриллами тау и мономером тау.
Используемые в настоящем изобретении антитела против тау также включают все антитела, способные специфически связывать агрегаты тау, присутствующие в биологическом образце. Используемые в настоящем изобретении антитела против тау включают все антитела, которые снижают распространение агрегации тау от клетки к клетке. Другими словами, подходящие для применения антитела замедляют проникновение тау в реципиентные клетки и/или уменьшают количество тау, проникающее в реципиентные клетки, по сравнению с количеством, которое могло бы проникнуть в реципиентную клетку в отсутствии антитела настоящего изобретения. Таким образом, подходящие для применения антитела уменьшают агрегацию тау в реципиентных клетках.
В одном аспекте используемые в настоящем изобретении антитела включают выделенные, охарактеризованные и очищенные функциональные антитела, которые были выделены (получены) для применения в функциональной терапевтической композиции, предназначенной для введения живому индивидууму. В другом аспекте используемые в настоящем изобретении антитела включают выделенные, охарактеризованные и очищенные функциональные антитела, которые были выделены (получены) для применения в анализе с целью детекции агрегатов тау в биологическом образце, полученном от живого индивидуума, и прогнозирования развития симптомов, связанных с агрегацией тау в течение всей жизни индивидуума. В другом аспекте используемые в настоящем изобретении антитела включают выделенные, охарактеризованные и очищенные функциональные антитела, которые были выделены (получены) для применения в анализе с целью детекции агрегатов тау в биологическом образце, полученном от живого индивидуума, и классификации индивидуума, как имеющего повышенный риск развития симптомов, связанных с агрегацией тау, в течение всей жизни индивидуума. В другом аспекте используемые в настоящем изобретении антитела включают выделенные, охарактеризованные и очищенные функциональные антитела, которые были выделены (получены) для применения, а также приведенные в таблице А и их варианты (например, гуманизированные формы, химерные формы и иммунологические фрагменты).
Антитела настоящего изобретения
Термин "антитело" включает термин "моноклональное антитело". "Моноклональное антитело" представляет собой антитело, полученное из одной копии или из одного клона, включающего, например, любой эукариотический, прокариотический или фаговый клон. Термин "моноклональное антитело" не ограничивается антителами, полученными с помощью гибридомной технологии. Моноклональные антитела можно получить с применением, например, хорошо известных в данной области гибридомных методов, а также рекомбинантных методов, методов фагового дисплея, синтетических методов или комбинаций указанных и других методов, хорошо известных в данной области техники. Кроме того, моноклональное антитело можно пометить детектируемой меткой, иммобилизовать на твердой фазе и/или конъюгировать с гетерологичным соединением (таким как фермент или токсин), применяя методы, известные в данной области техники.
Термин "антитело" также относится к функциональному моноклональному антителу или его иммунологически активному фрагменту; такому как фрагмент Fab, Fab' или F(аb')2. В некоторых контекстах данного описания фрагменты приведены специально для акцентирования; тем не менее, будет понятно, что независимо от того, указываются фрагменты или нет, термин "антитело" охватывает такие фрагменты, а также одноцепочечные формы. До тех пор, пока белок сохраняет способность специфически связываться со своей мишенью, он входит в определение термина "антитело". Термин "антитело" также включает, например, одноцепочечные формы, как правило, обозначаемые как фрагменты Fv антител, обладающих указанной специфичностью. Предпочтительно, но не обязательно, антитела, используемые в настоящем изобретении, получают рекомбинантным способом, включающим манипуляции, как правило, с мышиными или с другими нечеловеческими антителами, обладающими соответствующей специфичностью, с целью превращения их в гуманизированную форму. Антитела необязательно могут находиться в гликозилированном виде. Как известно, антитела можно соединять поперечными связями, такими как дисульфидные связи.
Основной формой используемого в настоящем изобретении антитела является тетрамер. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, причем каждая пара содержит одну "легкую" (размером примерно 25 кДа) и одну "тяжелую" цепь (примерно 50-70 кДа). Амино-концевой фрагмент каждой цепи содержит вариабельный участок, включающий примерно от 100 до 110 или более аминокислот, которые отвечают, главным образом, за распознавание антигена. Карбокси-концевой фрагмент каждой цепи содержит константный участок, ответственный, прежде всего, за эффекторную функцию.
Используемые в настоящем изобретении антитела против тау включают выделенные, охарактеризованные и очищенные функциональные антитела, которые были выделены (получены) с помощью способа их получения и, следовательно, доступны для применения в настоящем изобретении, в подходящей форме в количестве, достаточном для достижения терапевтических, медицинских или диагностических целей.
Легкие цепи подразделяют на типы гамма, мю, альфа и лямбда. Тяжелые цепи, которые подразделяют на типы гамма, мю, альфа, дельта или эпсилон, определяют изотип антитела, такой как IgO, IgM, IgA, IgD и IgE, соответственно. В легких и тяжелых цепях вариабельные и константные участки соединяются через "J"-участок, содержащий примерно 12 или более аминокислот, и тяжелая цепь также включает "D"-участок, содержащий примерно 10 или более аминокислот.
Вариабельные участки каждой пары легкой/тяжелой цепей образуют связывающий участок антитела. Следовательно, интактное антитело содержит два связывающих участка. Цепи имеют одинаковую общую структуру относительно консервативных каркасных участков (FR), соединенных тремя гипервариабельными участками, которые также называют участки, определяющие комплементарность (далее по тексту "CDR"). CDR из двух цепей выстраиваются в линию при посредстве каркасных участков, обеспечивая способность связываться со специфическим эпитопом. Как легкая, так и тяжелая цепь содержат, в направлении от N-конца к С-концу, домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4, соответственно. Отнесение аминокислот к конкретному домену осуществляют в соответствии с известными конвенциями (см. Kabat "Sequences of Proteins of Immunological Interest" National Institutes of Health, Bethesda, Md., 1987 and 1991; Chothia, et al, J. Mol. Bio. (1987) 196:901-917; Chothia, et al., Nature (1989) 342:878-883).
В одном аспекте моноклональные антитела против тау, обладающие соответствующей специфичностью, получают стандартными способами, включающими иммунизацию млекопитающих, получение гибридом из антитело-продуцирующих клеток указанных млекопитающих или иной способ иммортализации указанных клеток и культивирование гибридом или иммортализованных клеток с целью анализа их соответствующей специфичности. В соответствии с настоящим изобретением такие антитела можно получить путем иммунизации человека, кролика, крысы или мыши, например, пептидом, представляющим эпитоп, содержащий участок последовательности, кодирующей белок тау, или соответствующий его подучасток. Материалы для рекомбинантного манипулирования можно получить путем извлечения нуклеотидных последовательностей, кодирующих желательные антитела, из продуцирующей их гибридомы или другой клетки. При необходимости указанные нуклеотидные последовательности затем можно выделить, охарактеризовать, очистить, извлечь и подвергнуть манипуляциям с получением их гуманизированной формы для применения в настоящем изобретении.
Используемый в данном описании термин "гуманизированное антитело" включает антитело против тау, которое частично или полностью состоит из аминокислотных последовательностей, полученных из антител зародышевой линии человека путем изменения последовательности антитела, содержащего нечеловеческие участки, определяющие комплементарность ("CDR"). Простейшее из таких изменений может включать введение константного участка человеческого антитела вместо мышиного константного участка с получением химерной человеческой/мышиной молекулы, которая может обладать достаточно низкой иммуногенностью, позволяющей использовать молекулу для фармацевтического применения. Однако предпочтительно также гуманизировать вариабельный участок антитела и даже CDR с помощью методов, которые в настоящее время хорошо известны в данной области. Каркасные участки вариабельных доменов заменяют соответствующими человеческими каркасными участками, оставляя практически интактными нечеловеческие CDR, или даже CDR заменяют последовательностями, полученными из человеческого генома. CDR также могут быть случайно изменены так, что связывающая активность и аффинность в отношении тау поддерживаются или усиливаются в окружении полностью каркасных участков зародышевой линии человека или каркасных участков, которые являются по существу человеческими. По существу последовательности человеческих каркасных участков, по меньшей мере, на 90%, 95% или 99% идентичны известным человеческим каркасным последовательностям. Подходящие для применения полностью человеческие антитела также можно получить в генетически модифицированных мышах, чья иммунная система изменена так, чтобы соответствовать иммунной системе человека. Как указано выше, для применения в способах настоящего изобретения достаточно использовать иммунологически специфичные фрагменты антитела, в том числе фрагменты, представляющие собой одноцепочечные формы.
Кроме того, используемый в данном описании термин "гуманизированное антитело" относится к антителу против тау, содержащему человеческий каркасный участок, по меньшей мере, один CDR из нечеловеческого антитела, и константный участок, по существу идентичный константному участку иммуноглобулина человека, т.е. идентичный, по меньшей мере, примерно на 85-90%, предпочтительно, по меньшей мере, на 95%. Следовательно, все части гуманизированного антитела, за исключением, возможно, CDR, являются по существу идентичными соответствующим частям одной или нескольких последовательностей нативного иммуноглобулина человека.
При желании конструирование гуманизированных иммуноглобулинов можно проводить следующим образом. Если аминокислота попадает в нижеследующую категорию, аминокислоту каркасного участка используемого иммуноглобулина человека (акцепторный иммуноглобулин) заменяют аминокислотой каркасного участка из предоставляющего CDR нечеловеческого иммуноглобулина (донорного иммуноглобулина): (а) аминокислота в каркасном участке акцепторного иммуноглобулина человека является необычной для иммуноглобулина человека в данном положении, тогда как соответствующая аминокислота в донорном иммуноглобулине является типичной для иммуноглобулина человека в данном положении; (b) положение аминокислоты непосредственно примыкает к одному из CDR,; или (с) один из атомов боковой цепи аминокислоты каркасного участка находится на расстоянии 5-6 ангстрем (от центра до центра) от одного из атомов аминокислоты CDR в трехмерной модели иммуноглобулина (Queen, et al., op. cit., and Co, et al, Proc. Natl. Acad. Sci. USA (1991) 88:2869). Если одна из аминокислот каркасного участка акцепторного иммуноглобулина человека и соответствующая аминокислота донорного иммуноглобулина является необычной для иммуноглобулина человека в данном положении, такую аминокислоту заменяют аминокислотой, типичной для иммуноглобулина человека в данном положении.
Во всех случаях, антитело настоящего изобретения специфически связывает тау. В иллюстративных вариантах осуществления антитело настоящего изобретения специфически связывает человеческий тау. Фраза "специфически связывает" в данном описании означает, что антитело связывает белок с константой связывания или аффинностью взаимодействия (KD) в диапазоне от 0,1 пМ до 10 нМ, предпочтительно, в диапазоне от 0,1 пМ до 1 нМ. В данной области техники известны последовательности тау, полученные из разных видов, а также способы определения способности антитела связываться с тау. Например, их описание можно найти в разделе Примеры.
Антитела настоящего изобретения также можно использовать в виде слитых белков, известных как одноцепочечные вариабельные фрагменты (ScFv). Указанные scFv содержат вариабельные участки тяжелых и легких цепей, соединенные посредством линкера. В большинстве случаев, но не всегда, линкер может представлять собой пептид. Линкерный пептид предпочтительно содержит примерно от 10 до 25 аминокислот в длину. Предпочтительно линкерный пептид обогащен глицином, а также серином или треонином. ScFv можно использовать для проведения анализа методом фагового дисплея, проточной цитометрии или иммуногистохимии, или в качестве направляющих доменов. Способы получения и применения scFv известны в данной области.
В предпочтительном варианте осуществления scFv настоящего изобретения конъюгируют с человеческим константным доменом. В некоторых вариантах осуществления константный домен тяжелой цепи получают из домена IgG, такого как IgG1, IgG2, IgG3 или IgG4. В других вариантах осуществления константный домен тяжелой цепи можно получить из IgA, IgM или IgE.
Выделенное антитело настоящего изобретения, способное связываться с тау, предпочтительно распознает один из нескольких эпитопов. В одном варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп, указанный в таблице А. В другом варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп в аминокислотной последовательности SEQ ID NO:1 (DRKDQGGYTMHQD). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, три смежные аминокислоты SEQ ID NO:1, в том числе, по меньшей мере, 6 смежных аминокислот SEQ ID NO:1, по меньшей мере, 7 смежных аминокислот SEQ ID NO:1, по меньшей мере, 8 смежных аминокислот SEQ ID NO:1, по меньшей мере, 9 смежных аминокислот SEQ ID NO:1, по меньшей мере, 10 смежных аминокислот SEQ ID NO:1, по меньшей мере, 11 смежных аминокислот SEQ ID NO:1, по меньшей мере, 12 смежных аминокислот SEQ ID NO:1 и, по меньшей мере, 13 смежных аминокислот SEQ ID NO:1. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:1, представляет собой антитело HJ8.5. В другом иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:1, представляет собой антитело HJ8.1.1.
В другом варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп в аминокислотной последовательности SEQ ID NO:2 (KTDHGAE). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, три смежные аминокислоты SEQ ID NO:2, в том числе, по меньшей мере, 4 смежные аминокислоты SEQ ID NO:2, по меньшей мере, 5 смежных аминокислот SEQ ID NO:2, по меньшей мере, 6 смежных аминокислот SEQ ID NO:2 и, по меньшей мере, 7 смежных аминокислот SEQ ID NO:2. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:2, представляет собой антитело HJ8.1.2. В другом иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:2, представляет собой антитело HJ8.4.
В другом варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп внутри аминокислотной последовательности SEQ ID NO:3 (PRHLSNV). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, три смежные аминокислоты SEQ ID NO:3, в том числе, по меньшей мере, 4 смежные аминокислоты SEQ ID NO:3, по меньшей мере, 5 смежных аминокислот SEQ ID NO:3, по меньшей мере, 6 смежных аминокислот SEQ ID NO:3 и, по меньшей мере, 7 смежных аминокислот SEQ ID NO:3. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, который распознает эпитоп в последовательности SEQ ID NO:3, представляет собой антитело HJ8.2. В другом иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:3, представляет собой антитело HJ8.3. В следующем варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп в аминокислотной последовательности SEQ ID NO:4 (EPRQ). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, три смежные аминокислоты SEQ ID NO:4, в том числе, по меньшей мере, 4 смежные аминокислоты SEQ ID NO:4. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:4, представляет собой антитело HJ8.8.
В следующем варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп в аминокислотной последовательности SEQ ID NO:5 (AAGHV). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, три смежные аминокислоты SEQ ID NO:5, в том числе, по меньшей мере, 4 смежные аминокислоты SEQ ID NO:5 и, по меньшей мере, 5 смежных аминокислот SEQ ID NO:5. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп внутри последовательности SEQ ID NO:5, представляет собой антитело HJ8.7.
В другом варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп в аминокислотной последовательности SEQ ID NO:6 (TDHGAEIVYKSPVVSG). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, пять смежных аминокислот SEQ ID NO:6, в том числе, по меньшей мере, 6 смежных аминокислот SEQ ID NO:6, по меньшей мере, 7 смежных аминокислот SEQ ID NO:6, по меньшей мере, 8 смежных аминокислот SEQ ID NO:6, по меньшей мере, 9 смежных аминокислот SEQ ID NO:5, по меньшей мере, 9 смежных аминокислот SEQ ID NO:6, по меньшей мере, 10 смежных аминокислот SEQ ID NO:6, по меньшей мере, 11 смежных аминокислот SEQ ID NO:6, по меньшей мере, 12 смежных аминокислот SEQ ID NO:6, по меньшей мере, 13 смежных аминокислот SEQ ID NO:6, по меньшей мере, 14 смежных аминокислот SEQ ID NO:6, по меньшей мере, 15 смежных аминокислот SEQ ID NO:6 и, по меньшей мере, 16 смежных аминокислот SEQ ID NO:6. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:6, представляет собой антитело HJ9.1.
В другом варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп в аминокислотной последовательности SEQ ID NO:7 (EFEVMED). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, три смежные аминокислоты SEQ ID NO:7, в том числе, по меньшей мере, 4 смежные аминокислоты SEQ ID NO:6, по меньшей мере, 5 смежных аминокислот SEQ ID NO:7, по меньшей мере, 6 смежных аминокислот SEQ ID NO:7 и, по меньшей мере, 7 смежных аминокислот SEQ ID NO:7. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:7, представляет собой антитело HJ9.2. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:7, представляет собой антитело HJ9.4. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп в последовательности SEQ ID NO:7, представляет собой антитело HJ9.5.
В следующем варианте осуществления выделенное антитело настоящего изобретения, способное связываться с тау, распознает эпитоп в аминокислотной последовательности SEQ ID NO:8 (GGKVQIINKK). Предпочтительно, выделенное антитело распознает эпитоп, содержащий, по меньшей мере, три смежные аминокислоты SEQ ID NO:8, в том числе, по меньшей мере, 4 смежные аминокислоты SEQ ID NO:8, по меньшей мере, 5 смежных аминокислот SEQ ID NO:8, по меньшей мере, 6 смежных аминокислот SEQ ID NO:8, по меньшей мере, 7 смежных аминокислот SEQ ID NO:8, по меньшей мере, 8 смежных аминокислот SEQ ID NO:8, по меньшей мере, 9 смежных аминокислот SEQ ID NO:8 и, по меньшей мере, 10 смежных аминокислот SEQ ID NO:8. В иллюстративном варианте осуществления выделенное антитело настоящего изобретения, которое распознает эпитоп внутри последовательности SEQ ID NO:8, представляет собой антитело HJ9.3.
Предпочтительное антитело представляет собой гуманизированную форму мышиного антитела, полученного из гибридомы, обозначаемой HJ8.5. В настоящем описании термин "полученный из" означает, что "полученное" антитело содержит, по меньшей мере, один участок CDR из антитела, продуцируемого гибридомой HJ8.5. Иначе говоря, "полученное антитело" содержит, по меньшей мере, один участок CDR, аминокислотная последовательность которого выбрана из группы, состоящей из SEQ ID NO:16, 17, 18, 19, 20 и 21.
В одном варианте осуществления антитело настоящего изобретения может быть получено из гибридомы HJ8.5 и может кодироваться нуклеотидной последовательностью, на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичной последовательности, кодирующей вариабельный участок легкой цепи SEQ ID NO:12, или оно может кодироваться нуклеотидной последовательностью, на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичной последовательности, кодирующей вариабельный участок тяжелой цепи SEQ ID NO:13. В другом варианте осуществления антитело настоящего изобретения может быть получено из гибридомы HJ8.5 и может содержать аминокислотную последовательность, на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичную последовательности вариабельного участка легкой цепи SEQ ID NO:14, или оно может содержать аминокислотную последовательность, на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичную последовательности вариабельного участка тяжелой цепи SEQ ID NO:15. В каждом из приведенных выше вариантов осуществления антитело может быть гуманизированным.
В иллюстративном варианте осуществления антитело настоящего изобретения, способное связываться с тау, кодируется нуклеотидной последовательностью легкой цепи SEQ ID NO:12 и нуклеотидной последовательностью тяжелой цепи SEQ ID NO:13 [т.е. оно представляет собой моноклональное антитело, обозначаемое в данном описании HJ8.5]. В другом иллюстративном варианте осуществления антитело настоящего изобретения, способное связываться с тау, содержит аминокислотную последовательность легкой цепи SEQ ID NO:14 и аминокислотную последовательность тяжелой цепи SEQ ID NO:15 [т.е. оно представляет собой моноклональное антитело, обозначаемое в данном описании HJ8.5].
В одном варианте осуществления антитело настоящего изобретения может содержать CDR1 легкой цепи, например, как у антитела 1, указанного в таблице В. В другом варианте осуществления антитело настоящего изобретения может содержать CDR2 легкой цепи, например, как у антитела 4, указанного в таблице В. В следующем варианте осуществления антитело настоящего изобретения может содержать CDR3 легкой цепи, например, как у антитела 6, указанного в таблице В. В альтернативном варианте осуществления антитело настоящего изобретения может содержать комбинацию двух или трех CDR легкой цепи, например, как у антител 2, 3 и 5, указанных в таблице В.
Подобным образом, в одном варианте осуществления антитело настоящего изобретения может содержать CDR1 тяжелой цепи, например, как у антитела 7, указанного в таблице В. В другом варианте осуществления антитело настоящего изобретения может содержать CDR2 тяжелой цепи, например, как у антитела 10, указанного в таблице В. В следующем варианте осуществления антитело настоящего изобретения может содержать CDR3 тяжелой цепи, например, как у антитела 12, указанного в таблице В. В альтернативном варианте антитело настоящего изобретения может содержать комбинацию двух или трех CDR тяжелой цепи, например, как у антител 8, 9, 11, указанных в таблице В.
Альтернативно, антитело настоящего изобретения может содержать один или несколько CDR легкой цепи, и один или несколько CDR тяжелой цепи, например, как у антител 13-48, указанных в таблице В.
В разных вариантах осуществления антитело настоящего изобретения является гуманизированным. Например, в одном варианте осуществления гуманизированное антитело настоящего изобретения может содержать вариабельный участок легкой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:16, содержащей от нуля до двух аминокислотных замен, CDR2 с аминокислотной последовательностью SEQ ID NO:17, содержащей от нуля до двух аминокислотных замен, и CDR3 с аминокислотной последовательностью SEQ ID NO:18, содержащей от нуля до двух аминокислотных замен, или оно может содержать вариабельный участок тяжелой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:19, содержащей от нуля до двух аминокислотных замен, CDR2 с аминокислотной последовательностью SEQ ID NO:20, содержащей от нуля до двух аминокислотных замен, и CDR3 с аминокислотной последовательностью SEQ ID NO:21, содержащей от нуля до двух аминокислотных замен. В предпочтительном варианте осуществления гуманизированное антитело настоящего изобретения может содержать вариабельный участок легкой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:16, содержащей от нуля до двух аминокислотных замен, CDR2 с аминокислотной последовательностью SEQ ID NO:17, содержащей от нуля до двух аминокислотных замен, CDR3 с аминокислотной последовательностью SEQ ID NO:18, содержащей от нуля до двух аминокислотных замен, и вариабельный участок тяжелой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:19, содержащей от нуля до двух аминокислотных замен, CDR2 с аминокислотной последовательностью SEQ ID NO:20, содержащей от нуля до двух аминокислотных замен, и CDR3 с аминокислотной последовательностью SEQ ID NO:21, содержащей от нуля до двух аминокислотных замен. В иллюстративном варианте осуществления гуманизированное антитело настоящего изобретения может содержать вариабельный участок легкой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:16, CDR2 с аминокислотной последовательностью SEQ ID NO:17, CDR3 с аминокислотной последовательностью SEQ ID NO:18, и вариабельный участок тяжелой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:19, CDR2 с аминокислотной последовательностью SEQ ID NO:20 и CDR3 с аминокислотной последовательностью SEQ ID NO:21. Изобретение также охватывает соответствующие нуклеотидные последовательности SEQ ID NO:16, 17, 18, 19, 20 и 21, которые могут быть легко идентифицированы специалистом в данной области, и могут быть включены в вектор или другую большую молекулу ДНК, такую как хромосома, с целью экспрессии антитела настоящего изобретения.
II. Способ применения
В одном аспекте настоящее изобретение предоставляет антитела для применения в составе функциональной терапевтической композиции, предназначенной для введения живому индивидууму. В другом аспекте настоящее изобретение предоставляет антитела для применения в иммунологическом анализе, целью которого является детекция агрегатов тау в образце биологической жидкости, полученной из живого индивидуума. В другом аспекте настоящее изобретение предоставляет антитела для применения в иммунологическом анализе, целью которого является определение количества агрегатов тау в образце биологической жидкости, полученной из живого индивидуума. Количество агрегата тау в образце биологической жидкости, полученной от индивидуума, можно использовать для классификации индивидуума как имеющего высокое или низкое количество агрегата тау, и, кроме того, его можно использовать для прогнозирования риска развития симптомов и/или заболеваний, связанных с агрегацией тау, в течение всей жизни индивидуума.
Подходящие индивидуумы включают, но без ограничения, человека, домашний скот, домашнее животное, лабораторное животное и зоологическое животное. В одном варианте осуществления индивидуум представляет собой грызуна, такого как мышь, крыса, морская свинка и т.д. В другом варианте осуществления индивидуум может представлять собой домашний скот. Неограничивающие примеры подходящих сельскохозяйственных животных включают свиней, коров, лошадей, коз, овец, лам и альпак. В следующем варианте осуществления индивидуум может представлять собой домашнее животное. Неограничивающие примеры домашних животных включают таких животных, как собаки, кошки, кролики и птицы. В следующем варианте осуществления индивидуум может представлять собой зоологическое животное. В настоящем описании термин "зоологическое животное" относится к животному, которое можно найти в зоопарке. Такое животное может включать отличных от человека приматов, больших кошек, волков и медведей. В предпочтительных вариантах осуществления животное представляет собой лабораторное животное. Неограничивающие примеры лабораторных животных могут включать грызунов, собак, кошек и отличных от человека приматов. В некоторых вариантах осуществления животное представляет собой грызуна. Неограничивающие примеры грызунов могут включать мышей, крыс, морских свинок и т.д. В вариантах осуществления, где животное представляет собой мышь, мышь может относиться к штамму C57BL/6, Balb/c, 129sv или к любому другому лабораторному штамму. В иллюстративном варианте осуществления индивидуум представляет собой мышь C57BL/6J. В предпочтительном варианте осуществления индивидуумом является человек.
А. Способ лечения
В одном аспекте настоящее изобретение включает способ уменьшения распространения агрегации тау в головном мозге индивидуума. В другом аспекте настоящее изобретение включает способ уменьшения внутриклеточной агрегации тау, индуцированной затравкой тау. В каждом аспекте способ включает введение индивидууму фармакологически эффективного количества антитела против тау. Подходящие антитела описаны выше в разделе I. В предпочтительном варианте осуществления антитело выбрано из группы, включающей антитела, приведенные в таблице 1, и антитела, приведенные в таблице 2, в том числе гуманизированные антитела, химерные антитела или их иммунологические фрагменты.
Индивидуум может необязательно иметь симптом, связанный с агрегацией тау, до введения фармакологически эффективного количества антитела против тау. Иначе говоря, у индивидуума может присутствовать или отсутствовать симптом, связанный с агрегацией тау. Специалисту будет понятно, что патологическая агрегация тау, скорее всего, начинается до постановки диагноза или до появления симптомов, связанных с агрегацией тау. В некоторых вариантах осуществления у индивидуума присутствует симптом, связанный с агрегацией тау. В других вариантах осуществления у индивидуума отсутствует симптом, связанный с агрегацией тау. В других вариантах осуществления индивидуум имеет диагностируемую тау-патологию, но не имеет какого-либо другого симптома, связанного с агрегацией тау. Ограничение распространения агрегации тау в головном мозге индивидуума может уменьшить развитие и/или прогрессирование симптомов, связанных с патологической агрегацией тау.
Путем предотвращения распространения фибриллярных агрегатов тау можно лечить патологии, связанные с образованием и распространением агрегатов тау. В настоящем описании термины "лечить" или "лечение" включают профилактику, ослабление, обратное развитие или улучшение, по меньшей мере, одного симптома или признака заболевания, связанного с агрегацией тау. Одно из определений симптомов, связанных с агрегацией тау, включает любой симптом, вызванный образованием агрегатов тау, отчасти состоящих из фибрилл тау. Примеры расстройств, симптомы которых связаны с агрегацией тау, включают, но без ограничения, такие состояния, как прогрессирующий супрануклеарный паралич, деменция боксеров (хроническая травматическая энцефалопатия), лобно-височная деменция и паркинсонизм, связанный с хромосомой 17, болезнь Lytico-Bodig (паркинсонизм-деменция, комплекс острова Гуам), деменция с преобладанием клубков, ганглиоглиома и ганглиоцитома, менингоангиоматоз, подострый склерозирующий панэнцефалит, свинцовая энцефалопатия, туберозный склероз, болезнь Галлервордена-Шпатца, липофусциноз, болезнь Пика, кортикобазальная дегенерация, болезнь аргирофильных зерен (AGD), лобно-височная лобарная дегенерация, болезнь Альцгеймера и лобно-височная деменция. Методы диагностики таких заболеваний известны в данной области.
Примеры симптомов, связанных с агрегацией тау, могут включать нарушение когнитивной функции, изменение поведения, эмоциональное нарушение, судороги и нарушение структуры или функции нервной системы. Нарушение когнитивной функции включает, но без ограничения, проблемы с памятью, вниманием, концентрацией, языком, абстрактным мышлением, творчеством, способностью к целенаправленной деятельности, планированию и организации. Изменение поведения включает, но без ограничения, физическую или вербальную агрессию, импульсивность, снижение угнетения, апатию, снижение стимулирования, изменение личности, злоупотребление алкоголем, табаком или наркотиками, и другие, связанные с зависимостью, линии поведения. Эмоциональное нарушение включает, но без ограничения, депрессию, тревогу, манию, раздражительность и эмоциональную невоздержанность. Приступы включают, но без ограничения, генерализованные тонико-клонические судороги, комплексные парциальные судороги и неэпилептические психогенные судороги. Нарушение структуры или функции нервной системы включает, но без ограничения, гидроцефалию, паркинсонизм, нарушение сна, психоз, нарушение равновесия и координации. Такое нарушение включает моторные нарушения, такие как монопарез, гемипарез, тетрапарез, атаксия, баллизм и тремор. Оно также включает потерю или дисфункцию чувствительности, включающей обонятельные, осязательные, вкусовые, зрительные и слуховые ощущения. Кроме того, оно включает нарушения вегетативной нервной системы, такие как дисфункция кишечника и мочевого пузыря, половая дисфункция, нарушение регуляции кровяного давления и температуры. Наконец, оно включает гормональные нарушения, относимые к дисфункции гипоталамуса и гипофиза, такие как дефициты и нарушение регуляции гормона роста, тиреотропного гормона, лютеинизирующего гормона, фолликулостимулирующего гормона, гонадотропин-высвобождающего гормона, пролактина и многих других гормонов и модуляторов. Методы обнаружения и оценки симптомов, связанных с агрегацией тау, известны в данной области техники.
В некоторых вариантах осуществления симптом, связанный с агрегацией тау, относится к деменции. Деменция сама по себе не является конкретным заболеванием, и общим термином, который описывает широкий диапазон симптомов, связанных с ухудшением памяти или других навыков мышления, достаточно серьезных, чтобы уменьшить способность человека осуществлять повседневную деятельность. Деменция также является общим клиническим признаком многих заболеваний, связанных с агрегацией тау. Квалифицированному специалисту известны многочисленные методы определения степени тяжести деменции. Например, в данной области существует несколько когнитивных тестов и опросников для выявления деменции с разной степенью чувствительности и специфичности. Неограничивающие примеры включают краткую шкалу оценки психического статуса (MMSE), краткий тест на умственные способности (AMTS), модифицированную краткую шкалу оценки психического статуса (3MS), скрининговую шкалу для оценки когнитивных способностей (CASI), тест с построением маршрута, тест "рисования часов", опросник информанта о снижении когнитивной способности у пожилого человека, оценку когнитивных способностей врачом общего профиля, клиническую рейтинговую шкалу деменции (CDR), интервью информанта из восьми пунктов для дифференцирования старения и деменции (AD8).
В некоторых вариантах осуществления тяжесть симптомов деменции количественно определяют с использованием CDR. В соответствии с CDR оценка 0 указывает на отсутствие симптомов, оценка 0,5 указывает на очень слабые симптомы, оценка 1 указывает на легкие симптомы, оценка 2 указывает на умеренные симптомы, и оценка 3 указывает на тяжелые симптомы. Таким образом, любое увеличение оценки CDR указывает на ухудшение познавательной способности индивидуума и развитие деменции. Кроме того, изменение оценки CDR от 0 до значения, превышающего 0, указывает на развитие или появление деменции.
В некоторых вариантах осуществления симптом, связанный с агрегацией тау, относится к тау-патологии. Термин "тау-патология" относится к патологической агрегации белка тау. В некоторых вариантах осуществления тау-патология связана с наличием нейрофибриллярных клубков. В других вариантах осуществления тау-патология связана с гиперфосфорилированнием тау. В следующих вариантах осуществления тау-патология связана с повышенным уровнем агрегации тау в крови, плазме, сыворотке, CSF или ISF, который превышает уровень агрегации тау у лиц, не имеющих заболевания, примерно в 1,2-40 раз. Методы определения патологической агрегации тау известны в данной области и более подробно описаны в примерах.
В другом иллюстративном варианте осуществления способ уменьшения распространения агрегации тау в головном мозге индивидуума включает введение индивидууму фармакологически эффективного количества антитела против тау, где антитело выбрано из группы, включающей выделенное антитело, содержащее вариабельный участок легкой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:16, содержащей от нуля до двух аминокислотных замен, выделенное антитело, содержащее вариабельный участок легкой цепи, в состав которого входит CDR2 с аминокислотной последовательностью SEQ ID NO:17, содержащей от нуля до двух аминокислотных замен, выделенное антитело, содержащее вариабельный участок легкой цепи, в состав которого входит CDR3 с аминокислотной последовательностью SEQ ID NO:18, содержащей от нуля до двух аминокислотных замен, выделенное антитело, содержащее вариабельный участок тяжелой цепи, в состав которого входит CDR1 с аминокислотной последовательностью SEQ ID NO:19, содержащей от нуля до двух аминокислотных замен, выделенное антитело, содержащее вариабельный участок тяжелой цепи, в состав которого входит CDR2 с аминокислотной последовательностью SEQ ID NO:20, содержащей от нуля до двух аминокислотных замен, и выделенное антитело, содержащее вариабельный участок тяжелой цепи, в состав которого входит CDR3 с аминокислотной последовательностью SEQ ID NO:21, содержащей от нуля до двух аминокислотных замен.
В другом варианте осуществления способ уменьшения распространения агрегации тау в головном мозге индивидуума включает введение указанному индивидууму фармакологически эффективного количества антитела против тау, где антитело специфически связывается с тау и распознает эпитоп, содержащий последовательность SEQ ID NO:1 (DRKDQGGYTMHQD).
В другом варианте осуществления способ уменьшения распространения агрегации тау в головном мозге индивидуума включает введение индивидууму фармакологически эффективного количества антитела против тау, где антитело специфически связывается с тау и распознает эпитоп, содержащий последовательность SEQ ID NO:1 (DRKDQGGYTMHQD).
Антитела, которые можно вводить индивидууму в фармакологически эффективном количестве, предпочтительно имеют фармацевтическую степень чистоты и включают иммунологически активные фрагменты. Введение осуществляют с помощью стандартных эффективных методов, которые включают периферическое введение (т.е. не в центральную нервную систему) или локальное введение в центральную нервную систему. Периферическое введение включает, но без ограничения, внутривенное, внутрибрюшинное, подкожное, легочное, чрескожное, внутримышечное, интраназальное, трансбуккальное, сублингвальное введение или введение при помощи суппозиториев. Локальное введение, в том числе введение непосредственно в центральную нервную систему (ЦНС), включает, но без ограничения, введение с помощью люмбального, внутрижелудочкового или интрапаренхиматозного катетера или с использованием хирургически имплантированной композиции с контролируемым высвобождением.
Чтобы достичь эффективного введения, фармацевтические композиции специально разрабатывают для выбранного способа введения с использованием подходящих фармацевтически приемлемых эксципиентов, включающих совместимые диспергирующие агенты, буферы, поверхностно-активные вещества, консерванты, солюбилизирующие агенты, агенты, обеспечивающие изотоничность, стабилизирующие агенты и т.п. Специалистам в данной области хорошо известно, что описание способов получения композиций можно найти в сборнике Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton Pa, 16Ed ISBN:. 0-912734-04-3, последнее издание, включенное в данное описание посредством ссылки во всей полноте. Иногда желательно изменить характеристики растворимости антител, используемых в данном изобретении, с повышением их липофильности, например, путем заключения антител в липосомы или путем блокирования полярных групп.
Эффективная периферическая системная доставка путем внутривенной, внутрибрюшинной или подкожной инъекции является предпочтительным способом доставки лекарственного средства живому пациенту. Композиции для таких инъекций получают с использованием подходящих традиционных сред. Кроме того, введение также можно осуществлять через слизистые оболочки посредством назальных аэрозолей или суппозиториев. Композиции, подходящие для таких способов введения, хорошо известны и, как правило, включают поверхностно-активные вещества, которые облегчают перенос через мембрану. В качестве таких поверхностно-активных веществ часто используют производные стероидов или катионные липиды, включающие N-[1-(2,3-диолеоил)пропил]-N,N,N-триметилхлорид аммония (DOTMA) или такие соединения, как холестерин, фосфатидилхолин гемисукцинат глицерина и т.п.
Концентрация гуманизированного антитела в композициях, предназначенных для введения, представляет собой эффективное количество, варьирующее примерно от 0,1% по массе до 15 или 20% по массе, которое выбирают, прежде всего, с учетом объема, вязкости и других характеристик жидкости, в соответствии с конкретным выбранным способом введения, если это необходимо. Конкретная композиция для введения живому пациенту может содержать 1-5 мл стерильной буферированной воды, забуференным фосфатном солевым раствором, и примерно 1-5000 мг одного или комбинации гуманизированных антител настоящего изобретения. После получения композицию можно стерилизовать фильтрованием или сделать ее микробиологически приемлемой иным способом. Конкретная композиция для внутривенной инфузии может содержать 1-250 мл жидкости, такой как стерильный раствор Рингера, и антитело против тау в концентрации 1-100 мг на мл или более. Терапевтические средства настоящего изобретения можно хранить в замороженном или лиофилизированном виде и восстанавливать перед применением с использованием подходящего стерильного носителя. Лиофилизация и восстановление может привести к потере активности антител в различной степени (например, среди обычных иммуноглобулинов антитела IgM, как правило, теряют активность в большей степени, чем антитела IgG). Вводимые дозы, которые представляют собой эффективные дозы, можно корректировать. рН композиций, которые, как правило, имеют фармацевтическую степень чистоты, выбирают для достижения компромисса между стабильностью (химической и физической) антитела и комфортным состоянием пациента при введении. Как правило, рН в диапазоне от 4 до 8 хорошо переносится. Дозы варьируют от индивидуума к индивидууму в зависимости от размера, массы и других физико-биологических характеристик индивидуума, получающего композицию.
В настоящем описании термин "эффективное количество" означает количество вещества, такого как соединение, вызывающее измеримые и полезные эффекты у пациента при введении этого вещества, т.е. характеризующееся значительной эффективностью. Эффективное количество или дозу соединения, вводимого в соответствии с настоящим изобретением, определяют с учетом сопутствующих обстоятельств, включающих тип вводимого соединения, способ введения, состояние симптомов, подлежащих лечению, и подобные им параметры, касающиеся пациента и способа введения, в числе других параметров. В одном аспекте обычная доза описанного в данном описании антитела против тау составляет примерно от 0,01 мг/кг до 100 мг/кг. Дозы могут варьировать примерно от 0,05 мг/кг до 100 мг/кг, более предпочтительно, примерно от 0,1 мг/кг до 50 мг/кг или примерно от 0,5 мг/кг до 50 мг/кг. Частота введения дозы может оставлять один раз в день или один, два, три или более раз в неделю, или раз в месяц, в зависимости от того, какая частота требуется для эффективного лечения симптомов. Альтернативно, частота введения дозы может составлять, по меньшей мере, один раз в три месяца, если это требуется для эффективного лечения симптомов. Например, дозу можно вводить примерно каждые 5 недель, примерно каждые 6 недель, примерно каждые 7 недель, примерно каждые 8 недель, примерно каждые 9 недель, примерно каждые 10 недель, примерно каждые 11 недель или примерно каждые 12 недель.
Временная схема введения в процессе лечения самого заболевания и продолжительность лечения определяются сопутствующими обстоятельствами. Лечение можно начинать после диагностирования заболевания, связанного с агрегацией tau. Альтернативно, лечение можно начинать после клинического подтверждения симптома, связанного с агрегацией тау. Кроме того, лечение можно начинать после обнаружения тау-патологии. Лечение можно начинать сразу в больнице или клинике, или позже, после выписки из больницы, или после осмотра в поликлинике. Продолжительность лечения может варьировать от одной дозы вводимой однократно до пожизненного курса терапевтических процедур.
Хотя указанные выше способы являются наиболее удобными и наиболее подходящими и эффективными для введения таких белков, как гуманизированные антитела, после соответствующей адаптации можно использовать и другие эффективные способы введения, такие как внутрижелудочковое введение, трансдермальное введение и пероральное введение, при условии применения соответствующей композиции.
Кроме того, иногда желательно применять композиции с контролируемым высвобождением с использованием биоразлагаемых пленок и матриц, или осмотического мини-насоса, или систем доставки на основе декстрановых гранул, альгината или коллагена.
Конкретные уровни дозировки, которые можно определить и оптимизировать с помощью стандартных клинических методов, зависят от способа введения.
B. Способ детекции агрегатов тау в биологической жидкости
В одном аспекте настоящее изобретение предоставляет способы детекции агрегатов тау в образце биологической жидкости, полученной от индивидуума. В другом аспекте настоящее изобретение предоставляет способы измерения количества агрегатов тау в образце биологической жидкости, полученной от индивидуума. Способ, как правило, включает (i) получение образца биологической жидкости индивидуума, и (II) измерения количества агрегата тау в образце с использованием антитела, способного специфически связываться с tau. Подходящие антитела описаны выше в разделе I. Подходящие индивидуумы описаны выше.
В данном описании термин "биологическая жидкость" относится к жидкости, полученной от индивидуума. Можно использовать любую биологическую жидкость, содержащую агрегат тау. Неограничивающие примеры включают кровь, плазму, сыворотку, мочу, CSF и ISF. Жидкость можно использовать "как таковую", из жидкости можно выделить клеточные компоненты или из жидкости можно выделить белковую фракцию с использованием стандартных методов.
Как известно специалистам в данной области, способ получения образца биологической жидкости может и будет варьировать в зависимости от природы биологической жидкости и типа проводимого анализа. Для получения образца биологической жидкости можно использовать любой из ряда способов, широко известных в данной области техники. В общих чертах, способ предпочтительно поддерживает целостность образца таким образом, чтобы можно было точно детектировать агрегат тау и измерить его количество в соответствии с настоящим изобретением.
После получения образец обрабатывают in vitro, чтобы детектировать агрегат тау и измерить его количество с использованием антитела против тау. В некоторых вариантах осуществления перед проведением детекции и измерений концентрацию агрегата тау в образце увеличивают. В некоторых вариантах осуществления перед проведением детекции и измерений содержащийся в образце агрегат тау подвергают иммунопреципитации с использованием, по меньшей мере, одного выделенного антитела против тау. В других вариантах осуществления перед проведением детекции и измерений содержащийся в образце агрегат тау подвергают иммунопреципитации с использованием, по меньшей мере, двух выделенных антител против тау. В вариантах осуществления, в которых для иммунопреципитации агрегатов тау используют, по меньшей мере, два антитела, предпочтительно первое антитело связывается с первым эпитопом тау, и второе антитело связывается со вторым эпитопом тау, не перекрывающимся с первым эпитопом. Применение двух антител, которые связываются с двумя разными эпитопами тау, позволяет более эффективно улавливать все возможные конформеры агрегата тау. Неограничивающие примеры подходящих пар антител для иммунопреципитации приведены в таблице С. В предпочтительном варианте осуществления перед проведением детекции и измерения содержащийся в образце агрегат тау подвергают иммунопреципитации с использованием, по меньшей мере, двух выделенных антител против тау, где, по меньшей мере, первое антитело распознает эпитоп в последовательности SEQ ID NO:1, и, по меньшей мере, второе антитело распознает эпитоп в последовательности SEQ ID NO:8. Специалист в данной области посредством рутинного экспериментирования может определить, требуется ли проведение концентрирования или иммунопреципитации содержащегося в образце агрегата тау перед проведением детекции и измерения, и может осуществить указанные процедуры с помощью известных в данной области методов.
Способы детекции белка и измерения его количества с использованием антитела хорошо известны в данной области. Все подходящие способы детекции белка и измерения его количества с использованием антитела, известные специалистам в данной области, входят в объем настоящего изобретения. Неограничивающие примеры включают ELISA, сэндвич-иммуноанализ, радиоиммунный анализ, иммуноблоттинг или Вестерн-блоттинг, проточную цитометрию, иммуногистохимию и анализ с применением чипов.
Как правило, способ детекции агрегата тау и измерения его количества с помощью антител включает приведение в контакт части или всего образца, содержащего агрегат тау, с антителом против тау в условиях, обеспечивающих образование комплекса антитела с агрегатом тау. Обычно использование всего образца не требуется, что позволяет специалисту в данной области с течением времени многократно проводить детекцию агрегата тау и измерение его количества в образце. Способ можно проводить в растворе или антитело или агрегат тау можно иммобилизовать на твердой поверхности. Неограничивающие примеры подходящих поверхностей включают микротитрационные планшеты, пробирки, гранулы, смолы и другие полимеры. Присоединение к поверхности можно проводить с помощью широкого ряда способов, известных специалистам в данной области. Например, поверхность и антитело можно дериватизировать с использованием химических функциональных групп с последующим присоединением второго компонента. Например, поверхность можно дериватизировать химической функциональной группой, включающей, но без ограничения, аминогруппы, карбоксильные группы, оксогруппы или тиольные группы. В случае применения указанных функциональных групп, антитело можно присоединить к таким группам прямо или косвенно с использованием линкеров. Антитела против тау также можно присоединить к поверхности нековалентно. Например, биотинилированные антитела против тау могут связываться с поверхностью, к которой ковалентно присоединен стрептавидин. Альтернативно, антитело можно синтезировать на поверхности с использованием таких методов, как фотополимеризация и фотолитография.
Приведение в контакт образца с антителом в эффективных условиях и в течение периода времени, достаточного для образования комплекса, как правило, включает добавление композиции антитела против тау к образцу (или к иммунопреципитированному или концентрированному агрегату тау) и инкубацию смеси в течение периода времени, достаточного для связывания антитела против тау с любым присутствующим антигеном. По истечении этого времени комплекс промывают, детектируют и измеряют его количество с помощью любого хорошо известного в данной области способа. Способы детекции и измерения количества комплекса антитело-полипептид, как правило, основаны на обнаружении метки или маркера. Термин "метка" в соответствии с данным описанием относится к любому веществу, прикрепленному к антителу или другому субстратному материалу, где указанное вещество можно обнаружить с помощью способа детекции. Неограничивающие примеры подходящих меток включают люминесцентные молекулы, хемилюминесцентные молекулы, флуорохромы, средства, вызывающие гашение флуоресценции, окрашенные молекулы, радиоизотопы, сцинтилляторы, биотин, авидин, стрептавидин, белок A, белок G, антитела или их фрагменты, полигистидин, Ni2+, маркеры Flag, маркеры myc, тяжелые металлы и ферменты (включающие щелочную фосфатазу, пероксидазу и люциферазу). Способы детекции комплекса антитело-полипептид и измерения его количества, основанные на обнаружении метки или маркера, хорошо известны в данной области техники.
В предпочтительном варианте осуществления способ измерения количества агрегата тау в образце представляет собой иммуноанализ, включающий применение двух улавливающих антител и детекторного антитела, где каждое улавливающее антитело представляет собой выделенное антитело против тау, которое распознает эпитоп тау, отличный от эпитопа, распознаваемого другим улавливающим антителом, и детекторное антитело представляет собой выделенное антитело против тау, присоединенное к маркеру. Детекторное антитело может быть идентично одному из двух улавливающих антител или, альтернативно, детекторное антитело может распознавать эпитоп тау, не распознаваемый ни одним из улавливающих антител. Как правило, первое улавливающее антитело и второе улавливающее антитело можно использовать в соотношении, варьирующем примерно от 10:1 до 1:10, примерно от 5:1 до 1:5, примерно от 3:1 до 1:3 или примерно от 2:1 до 1:2. В некоторых вариантах осуществления первое улавливающее антитело и второе улавливающее антитело используют примерно в равных концентрациях. Неограничивающие примеры подходящих пар улавливающих антител включают антитела, описанные в таблице D и в таблице Е. Неограничивающие примеры подходящих детекторных антител включают антитела, перечисленные в таблице А, а также антитела, способные специфически связываться с тау и распознавать эпитоп в аминокислотной последовательности, выбранной из группы, включающей SEQ ID NO:1-11. В иллюстративном варианте осуществления первое улавливающее антитело представляет собой выделенное антитело, способное специфически связываться с тау и распознавать эпитоп в последовательности SEQ ID NO:7, второе улавливающее антитело представляет собой выделенное антитело, способное специфически связываться с тау и распознавать эпитоп в последовательности SEQ ID NO:8, и детекторное антитело представляет собой выделенное антитело, способное специфически связываться с тау и распознавать эпитоп в последовательности SEQ ID NO:8.
Первое и второе улавливающие антитела
Первое и второе улавливающие антитела: каждое антитело специфически связывается с тау и распознает эпитоп в аминокислотной последовательности, соответствующей указанному SEQ ID NO
В другом аспекте настоящее изобретение предоставляет способы отнесения индивидуума к определенной группе на основе размера агрегата тау, определенного в образце биологической жидкости, полученной от индивидуума. Способ, как правило, включает (i) получение образца биологической жидкости от индивидуума и измерение количества агрегата тау в образце с использованием антитела, способного специфически связываться с тау, (ii) сравнение количества агрегата тау в образце со стандартным значением, и (iii) отнесение индивидуума к группе, характеризующейся высоким или низким количеством агрегата тау, на основе количества агрегата тау, измеренного в образце. Способы получения образца биологической жидкости от индивидуума и измерения количества агрегата тау в образце с использованием антитела, способного специфически связываться с тау, подробно описаны выше и дополнительно описаны в разделе Примеры.
Можно использовать любое подходящее стандартное значение, известное в данной области. Например, в качестве подходящего стандартного значения можно использовать количество агрегата тау в образце биологической жидкости, полученной от индивидуума или группы индивидуумов одного вида, у которых отсутствуют клинически детектируемые симптомы агрегации тау. В другом примере в качестве подходящего стандартного значения можно использовать количество агрегата тау в биологическом образце жидкости, полученной от индивидуума или группы индивидуумов одного вида, у которых не диагностируется тау-патология. В другом примере в качестве подходящего стандартного значения можно использовать количество агрегата тау в биологическом образце жидкости, полученной от индивидуума или группы индивидуумов одного вида, у которых оценка по клинической рейтинговой шкале деменции равна нулю (CDR=0). В другом примере в качестве подходящего стандартного значения можно использовать фоновый сигнал анализа, определяемый с помощью известных в данной области техники способов. В другом примере в качестве подходящего стандартного значения можно использовать значение количества агрегата тау в стандартном образце, полученном от того же индивидуума. Стандартный образец представляет собой биологическую жидкость такого же типа, как и тестируемый образец, полученную от индивидуума, когда индивидуум не имел каких-либо клинически детектируемых симптомов агрегации тау. Специалисту в данной области известно, что не всегда возможно или желательно получить стандартный образец от индивидуума, когда индивидуум является в других отношениях здоровым. Например, при контролировании эффективности терапии стандартный образец может представлять собой образец, полученный от индивидуума до начала терапии. В таком примере индивидуум может иметь тау-патологию, но не иметь других симптомов агрегации тау (таких как деменция, ухудшение когнитивной способности и т.д.), или индивидуум может иметь тау-патологию и один или несколько других симптомов агрегации тау. В дополнительном примере в качестве подходящего стандартного значения можно использовать биологическую жидкость индивидуума или группы индивидуумов, у которых не обнаружены агрегаты тау.
Согласно изобретению, индивидуума можно отнести к определенной группе на основе количества агрегата тау, измеренного в образце. Отнесение индивидуума к определенной группе на основе количества агрегата тау, измеренного в образце биологической жидкости, полученном от индивидуума, можно использовать для выявления индивидуумов, у которых в течение жизни может развиться заболевание и/или симптом, связанный с агрегацией тау. В общих чертах, индивидуума можно отнести к определенной группе, характеризующейся высоким или низким количеством агрегата тау по сравнению со стандартным значением, где высокое количество агрегата тау представляет собой количество, превышающее стандартное значение, и низкое количество представляет собой количество, которое равно стандартному значению или ниже. В предпочтительных вариантах осуществления индивидуума относят к группе, характеризующейся высоким количеством агрегата тау, если количество агрегата тау в образце биологической жидкости превышает стандартное значение, по меньшей мере, в 2 раза. Например, количество агрегата тау в образце может превышать стандартное значение, по меньшей мере, в 2 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз, по меньшей мере, в 15 раз, по меньшей мере, в 20 раз, по меньшей мере, в 25 раз, по меньшей мере, в 30 раз, по меньшей мере, в 35 раз, по меньшей мере, в 40 раз, по меньшей мере, в 45 раз или, по меньшей мере, в 50 раз. Если количество агрегата тау в образце биологической жидкости, полученной от индивидуума, по меньшей мере, в 2 раза превышает стандартное значение, где стандартное значение измеряют в образце биологической жидкости того же типа, полученной от одного или нескольких индивидуумов, не имеющих заболевания или диагностированного симптома агрегации тау (или в эквивалентном ему образце), индивидуум характеризуется большой вероятностью развития заболевания и/или симптома, связанного с агрегацией тау, на протяжении жизни.
Определения
В настоящем описании термин "антитело" относится к молекуле, полученной из иммуноглобулина, которая способна специфически распознавать тау. Антитело настоящего изобретения может представлять собой полноразмерное антитело (IgM, IgG, IgA, IgE) или фрагмент антитела (Fab, F(аb')2, ScFv). Антитело может быть химерным или гуманизированным.
В настоящем описании "CDR" обозначает "участок, определяющий комплементарность". Термин "CDR" также может относиться к гипервариабельным участкам.
В настоящем описании термин "легкая цепь" относится к малой полипептидной субъединице антитела. Обычное антитело содержит две легкие цепи и две тяжелые цепи.
В настоящем описании термин "тяжелая цепь" относится к большой полипептидной субъединице антитела. Тяжелая цепь антитела содержит ряд доменов иммуноглобулина, в число которых входит, по меньшей мере, один вариабельный домен и, по меньшей мере, один константный домен.
Термин "гуманизация" в данном описании относится к способу получения моноклонального антитела с помощью технологии рекомбинантных ДНК, которая позволяет получить конструкцию, способную экспрессироваться в культуре клеток человека. Любые известные способы получения указанных конструкций можно использовать в целях настоящего изобретения.
В настоящем описании термины "одноцепочечные вариабельные фрагменты", или "ScFv", или "scFvs", относятся к гибридным белкам, содержащим вариабельные участки тяжелых и легких цепей иммуноглобулинов, соединенные посредством линкеров. В некоторых вариантах осуществления линкер представляет собой пептид, содержащий от 10 до 25 аминокислот.
Примеры
Следующие примеры приведены для демонстрации предпочтительных вариантов осуществления изобретения. Специалистам в данной области техники будет понятно, что способы, раскрытые в приведенных ниже примерах, представляют собой способы, предложенные авторами изобретения для успешного осуществления настоящего изобретения и, следовательно, могут рассматриваться как предпочтительные способы осуществления изобретения. Однако в свете настоящего описания специалистам в данной области техники будет понятно, что можно осуществить многочисленные изменения описанных конкретных вариантов осуществления и в результате все еще получить подобный или аналогичный результат без отклонения от сущности и объема настоящего изобретения.
Введение в примеры 1-8
Агрегация связанного с микротрубочками белка тау в нейронах и нейроглии связана более чем с 20 нейродегенеративными расстройствами, включающими болезнь Альцгеймера (AD), прогрессирующий надъядерный паралич и лобно-височную деменцию. Результаты последних исследований, проведенных на людях, позволяют предположить, что тау-патология не распространяется случайным образом через мозг, а связана с существующими сетями межнейронных связей. Фибриллярная тау-патология при AD развивается по всей протяженности известных анатомических связей, хотя механизмы дегенерации сетей неизвестны. Важно отметить, что последние патологические исследования позволяют предположить, что в головном мозге человека и мыши белковые агрегаты могут переходить из одной клетки в другую. Кроме того, связанные с болезнью человека фибриллярные формы рекомбинантных белков, таких как тау, SOD-1, α-синуклеин и полиглутамины, легко поглощаются из внеклеточного пространства, инициируя неправильную укладку белков внутри клетки. Указанные явления напоминают трансклеточное распространение прионов, которое предположительно происходит при посредстве экзосом и туннельных нанотрубочек. Вопрос, распространяется ли неправильная укладка белка, опосредованная агрегатами тау, от клетки к клетке через непосредственный контакт клетка-клетка или через межклеточное пространство, остается открытым. Кроме того, еще не определено, могут ли патологические виды тау опосредовать фактическое трансклеточное распространение агрегации, в результате которого агрегат высвобождается из "донорной" клетки, проникает во вторую "реципиентную" клетку и индуцирует дальнейшую неправильную укладку, посредством прямого контакта белок-белок или посредством более косвенных механизмов. Здесь проведены исследования для определения, способны ли фибриллы тау высвобождаться непосредственно во внеклеточное пространство и инициировать распространение агрегации по данному механизму.
Пример 1. Антитела против тау
С применением стандартных способов получают две серии антител против тау: серия HJ8 (мышиные моноклональные антитела против рекомбинантного человеческого белка тау) и серия HJ9 (мышиные моноклональные антитела против рекомбинантного мышиного белка тау) (таблица 1). Связывающие эпитопы картируют для разных антител (таблица A).
Антитела против человеческого и мышиного тау серии HJ8 и серии HJ9
Чтобы охарактеризовать аффинность связывания антител против мышиного тау и человеческого тау серий HJ8 и HJ9, используют технологию Biacore SPR. Сенсорный чип CM-5 Biacore (карбоксиметилированный декстран в качестве матрицы) активируют с использованием EDC (1-этил-3-(3-диметиламинопропил)карбодиимид) и NHS (N-гидроксисукцинимид) в соотношении 1:1. 2). Затем лиганд, мышиный тау или человеческий тау иммобилизуют (20 мкг/мл, в 10 мМ ацетате натрия, рН 3,5) на сенсорном чипе CM-5 Biacore при скорости потока 5 мкл/мин. Оставшиеся несвязанными участки на сенсорном чипе Biacore CM-5 дезактивируют путем пропускания 1М этаноламина рН 8,5.
После подготовки поверхности сенсорного чипа, анализируемые вещества (например, антитела) вводят в различных концентрациях (0,78 пМ-400 нМ) в отфильтрованном, дегазированном буфере 0,01М Hepes, который содержит 0,15М NaCl, 0,005% поверхностно-активного вещества Р20, рН 7,4, при скорости потока 10 мкл/мин. Все образцы анализируют в двух повторах. После каждого цикла/прогона с использованием одной концентрации антитела поверхность чипа регенерируют при использовании 10 мМ глицина, рН 1,7, удаляя связанное антитело/анализируемое вещество и оставляя присоединенные к поверхности мономер/фибриллу/лиганд.
Используя сенсограмму SPR (фиг.2-13), определяют скорость ассоциации, или скорость On (Ka), скорость диссоциации, или скорость Off (Kd), и константу аффинности или аффинность взаимодействия (KD, где KD=Kd/Ka) (таблицы 2 и 3).
Результаты анализа связывания антител серии HJ8 и серии HJ9 с мышиным тау
Результаты анализа связывания антител серии HJ8 и серии HJ9 с человеческим тау
Пример 2. Присутствие полноразмерного тау в ISF
Тау подвергают иммунопреципитации из образцов ISF мышей дикого типа и мышей, трансгенных по человеческому тау Р301S (мыши Р301S tg, подробно описанные в разделе Методы), с использованием антител против тау, распознающих как мышиный, так и человеческий тау. Используют два моноклональных антитела против тау, которые хорошо зарекомендовали себя в анализах методом иммунопреципитации, так как количество мономерного тау в ISF является относительно низким. После иммунопреципитации тау анализируют методом иммуноблоттинга. Эндогенные изоформы мышиного тау образуют полосы, соответствующие массе 48-62 кДа. Тау, содержащийся в лизатах мозга мышей дикого типа, образует четыре отдельные полосы в SDS-PAGE (фиг.14А). У мышей дикого типа в наибольшем количестве присутствует изоформа, соответствующая полосе 48 кДа. В мозге мышей Р301S tg помимо четырех эндогенных изоформ мышиного тау наблюдается сверхэкспрессия человеческого тау 1N4R, о чем свидетельствует интенсивная полоса, соответствующая 55 кД, также присутствует полоса, соответствующая 39 кДа, которая может быть образована продуктом деградации тау.
В отличие от общих лизатов головного мозга, ISF мышей дикого типа после иммунопреципитации дает одну полосу тау, детектируемого с помощью антитела HJ9.3, распознающего участок тау, связывающий микротрубочки (MTBR) (фиг.14В). Данная полоса соответствует наибольшей изоформе 2N4R, присутствующей в лизатах мозга мыши. Полоса, соответствующая человеческому тау, присутствующему в ISF мышей Р301S tg, совместно преципитируется с полосой вышеуказанного мышиного тау и соответствует несколько более низкой молекулярной массе (фиг.14В). Эти две полосы также преципитируются в присутствии другого мышиного моноклонального антитела против тау HJ8.1 (фиг.14С). Полученные результаты свидетельствуют о том, что основной тау-компонент, присутствующий в ISF, согласно анализу ELISA, скорее всего, представляет собой полноразмерный мономерный тау.
Пример 3. Белки тау RD образуют фибриллярные агрегаты в трансфицированных клетках НЕК293
Ген тау кодирует шесть изоформ белка, и совокупность мутаций вызывает доминантно наследуемое дегенеративное заболевание. В зависимости от сплайсинга белок тау имеет три или четыре повторяющихся участка, которые составляют склонное к агрегации ядро белка, называемое домен повторов (RD). Экспрессия тау RD вызывает патологию у трансгенных мышей, кроме того, существуют доказательства усечения полноразмерного тау с образованием фрагментов, составляющих фибриллы в организме пациента. Данную конструкцию используют вместо полноразмерного тау, поскольку она достоверно образует фибриллы в культивируемых клетках. В белок RD, содержащий четыре повтора, вводят различные мутации, которые заведомо увеличивают агрегацию тау: ΔΚ280 (обозначаемая ΔΚ), P301L и V337M. Мутации Р301L и V337M объединяют в одном белке (обозначаемом LM), с получением мутантной формы RD, характеризующейся значительно увеличенной склонностью к агрегации, подобно описанной выше. Такой "нефизиологичный" мутант облегчает проведение анализов событий передачи и трансклеточного распространения неправильной укладки, зависящих от эффективного образования внутриклеточных агрегатов, и дополняет аналогичные, но менее активные агрегационные фенотипы "физиологического" мутанта ΔΚ. В мутант ΔΚ также можно ввести две пролиновые замены, I227P и I308P (обозначаемые PP), которые ингибируют образование β-складчатой структуры и фибрилл, хотя и не блокируют образование аморфных агрегатов. Каждую форму мутантного тау гибридизуют по карбокси-концу с голубым или желтым флуоресцентным белком (CFP или YFP) или с маркером HA. Конструкции схематически изображены на фиг.15А.
Для определения характеристики внутриклеточных агрегатов тау RD, клетки HEK293 временно трансфицируют разными формами RD. Анализ SDS-нерастворимого вещества проводят методом атомно-силовой микроскопии (AFM). RD(ΔΚ)-HA и RD(LM)-HA продуцируют выраженные фибриллярные частицы (фиг.15В). Присутствующие в клетках агрегаты RD(ΔΚ)-HA и RD(LM)-HA также окрашиваются X-34, производным тиофлавина, которое взаимодействует с бета-складчатыми фибриллами и излучает в синей части спектра (фиг.15в). Кроме того, проводят детергентное фракционирование, чтобы проверить, имеют ли включения, видимые под световым микроскопом, биохимический аналог. В осадках, не растворимых в SDS (1% тритон Х-100 в 1×PBS, содержащий ингибиторы протеаз, используют для выделения растворимого осадка, после чего нерастворимую часть осадка экстрагируют SDS/RIPA), детектируют мономер и частицы с более высокой молекулярной массой, соответствующие олигомерам (фиг.15D).
Авторы настоящего изобретения ранее использовали метод резонансного переноса энергии флуоресценции (FRET) для количественного определения внутриклеточной агрегации белка хантингтина. Чтобы проверить, можно ли использовать данный метод для отслеживания агрегации тау RD, различные мутанты RD (wt, ΔΚ, PP, LM) гибридизуют с желтым флуоресцентным белком (YFP:акцептор для FRET) и голубым флуоресцентным белком (CFP:донор для FRET). Полученные гибриды совместно трансфицируют в клетки НЕК293 (обозначаемые как RD-CFP/RD-YFP) и количественно определяют образование внутриклеточных агрегатов, используя конфокальную микроскопию с фотообесцвечиванием акцептора для FRET и определение спектрального излучения FRET с использованием флуоресцентного планшет-ридера (FPR). С помощью конфокальной микроскопии визуализируют клетки, совместно экспрессирующие RD(LM)-CFP/RD(LM)YFP, и измеряют донорный сигнал до и после частичного и полного фотообесцвечивания акцептора. Увеличение донорного сигнала после фотообесцвечивания свидетельствует о средней эффективности FRET 18,2%±0,058 (n=6, значение±стандартное отклонение), подтверждающей наличие межмолекулярных взаимодействий между частицами RD, образующими пару для FRET (фиг.16а). Чтобы измерить агрегацию RD-CFP/YFP по спектральному излучению FRET с помощью FPR, используют стандартные методы. Чтобы максимизировать гашение донора в пределах детектирования сигнала, используют совместную трансфекцию RD-YFP и RD-CFP в соотношении 3:1. Значимый уровень FRET при использовании RD(PP)-CFP/YFP отсутствует. Однако как RD(ΔΚ)-CFP/YFP, так и RD(LM)-CFP/YFP продуцируют интенсивные сигналы FRET (фиг.16B), подтверждающие результаты микроскопии.
Ранее было обнаружено, что ряд клеток поглощают рекомбинантные фибриллы тау из внеклеточной среды. Такое поглощение инициирует внутриклеточную фибриллизацию полноразмерного белка тау с нативной укладкой, слитого с YFP. Данное наблюдение подтверждают методом FRET, позволяющим отслеживать агрегацию RD(ΔK)-CFP/YFP, индуцированную разными количествами рекомбинантных фибрилл RD. Клетки HEK293 совместно трансфицируют RD(ΔK)-CFP/YFP и культивируют в течение 15 ч. Затем к среде добавляют различные концентрации фибрилл RD-HA (0,01, 0,03, 0,1 и 0,3 мкΜ в пересчете на мономер) и инкубируют в течение 9 ч. Затем фибриллы удаляют путем замены среды, и клетки оставляют восстанавливаться в течение 4 ч, после чего их фиксируют и анализируют методом FRET. Наблюдают дозозависимое увеличение сигнала FRET, индуцированного рекомбинантными фибриллами, по сравнению с необработанными клетками RD(ΔK)-CFP/YFP (фиг.16C). Следовательно, можно сделать вывод, что существует корреляция между микроскопическими, молекулярными, биохимическими и биофизическими характеристиками агрегации тау RD и образованием фибрилл в клетках. В определенных пределах, особенно с использованием контролей по уровню экспрессии белка, анализ FRET, проводимый при использовании планшет-ридера, позволяет легко измерять параметры данного процесса.
Пример 4. Трансклеточная индукция агрегации RD
Авторы настоящего изобретения ранее определили, что включения тау из одной клетки переносятся в наивные клетки при совместном культивировании. Однако не было показано, что перенесенные агрегаты могут вызвать дальнейшую агрегацию в клетках-реципиентах, и что индукция агрегации основана на непосредственном белок-белковом взаимодействии. Сначала определяют, способны ли агрегаты RD(LM)-HA из одной популяции донорных клеток формировать включения RD(ΔK)-YFP в другой популяции реципиентных клеток при совместном культивировании. Одну группу клеток трансфицируют склонным к агрегации RD(LM)-HA, и другую группу трансфицируют RD(ΔK)-YFP. На следующий день популяции клеток повторно высевают и совместно культивируют в течение 48 ч. После фиксации клетки подвергают иммунному окрашиванию с использованием антитела против HA и контрастному окрашиванию X-34. Во многих клетках наблюдают включения с совместной локализацией RD(LM)-HA и RD(ΔK)-YFP (фиг.17а). Зачастую такие включения также окрашиваются X-34, что указывает на наличие бета-складчатой структуры. Данные исследования продолжают, применяя анализ FRET, позволяющий отслеживать агрегацию RD(ΔK)-CFP/YFP, индуцированную совместным культивированием с клетками, экспрессирующими RD(LM)-HA. В данном случае, совместно культивируют две популяции клеток. Донорная популяция экспрессирует RD(LM)-HA, и реципиентная популяция экспрессирует RD(ΔK)-CFP/YFP. В качестве отрицательных контролей используют устойчивую к образованию β-складчатой структуры форму тау RD(PP)-HA или клетки, трансфицированные суррогатом. Через 48 ч в клеточных монослоях измеряют сигнал FRET. Сильное увеличение сигнала FRET индуцируется в результате совместного культивирования с RD(LM)-HA по сравнению с RD(PP)-HA или клетками, трансфицированными суррогатом (фиг.17B). Небольшое увеличение сигнала FRET наблюдается после совместного культивирования клеток RD(LM)-HA с реципиентными клетками RD(WT)-CFP/YFP (данные не показаны). Полученные результаты свидетельствуют о перемещении склонной к агрегации разновидности тау от одной клетки к другой и совместной локализации во включениях, обогащенных бета-складчатыми структурами. Высвобождение агрегатов может происходить после гибели клеток, однако, каких-либо доказательств этому с помощью окрашивания пропидиййодидом разных трансфицированных популяций не получено (фиг.17C).
Пример 5. Распространение неправильной укладки в результате непосредственного контакта белков
Хотя полученные результаты с большой долей вероятности позволяют предположить, что совместная агрегация происходит в результате непосредственного контакта белков с образованием межмолекулярных связей между тау RD, полученным от донора, и соответствующим белком из реципиентной клетки, формально они этого не доказывают. Для решения данного вопроса используют FRET. Сначала RD(LM)-CFP совместно экспрессируют в популяции донорных клеток, в RD(LM)-YFP во второй популяции реципиентных клеток. Через 48 ч в клеточных монослоях измеряют сигнал FRET с использованием конфокальной микроскопии и FPR. С помощью конфокальной микроскопии сигнал CFP измеряют до и после фотообесцвечивания YFP. Регистрируют среднюю эффективность сигнала FRET ~14,2%, свидетельствующую о том, что включения содержат RD(LM)-CFP и RD(LM)-YFP, находящиеся в непосредственном контакте (фиг.18А). Затем относительные сигналы FRET сравнивают с помощью FPR, используя разные формы немеченого RD для индукции агрегации RD-CFP. Сначала RD(ΔK)-CFP и RD(LM)-HA совместно экспрессируют в донорной популяции клеток и RD(ΔK)-YFP во второй популяции реципиентных клеток. RD(LM)-HA служит усилителем агрегации и перемещения RD(ΔK)-CFP, стимулируя его последующий перенос в реципиентные клетки RD(ΔK)-YFP. Это приводит к небольшому, но воспроизводимому увеличению сигнала FRET в совместно культивируемых клетках. Данный сигнал исчезает, если CFP- или YFP-меченые конструкции RD содержат мутацию PP, которая блокирует образование β-складок (фиг.18), свидетельствуя о том, что оба члена пары должны иметь способность к образованию бета-складчатой структуры. Вместе с проведенными ранее экспериментами полученные результаты указывают на то, что распространение неправильной укладки происходит при непосредственном контакте, т.е. агрегат из одной клетки входит в контакт и запускает неправильную укладку естественно уложенного белка во второй клетке. Эти данные подразумевают, что усиление неправильной укладки также может происходить в серийных совместных культурах клеток. Было предсказано, что предварительное воздействие затравок агрегации на популяцию "донорных" клеток приведет к увеличению конечной агрегации в популяции реципиентных клеток. Это проверяют путем последовательного культивирования трех популяций клеток. Первая популяция экспрессирует различные формы нефлуоресцентного RD-HA с образованием "затравок" агрегации. Вторая популяция экспрессирует CFP или RD(ΔK)-CFP и либо не поддерживает (CFP), либо поддерживает (RD(ΔK)-CFP) агрегацию. Две указанные популяции совместно культивируют в течение 48 ч, чтобы обеспечить усиление неправильной укладки. Затем 50% совокупности первой и второй популяций совместно культивируют в течение 48 ч с третьей популяцией клеток, экспрессирующих RD(ΔK)-YFP. Третья реципиентная группа служит "репортером", указывая на степень внутриклеточной агрегации и распространения RD(ΔK)-CFP. Предварительное воздействие RD(LM)-HA на популяцию RD(АК)-CFP повышает конечный сигнал FRET в 2,6 раза по сравнению с клетками, которые не подвергались предварительному воздействию склонного к агрегации тау. Как и ожидалось, включение клеток, экспрессирующих чистый CFP, во вторую популяцию клеток полностью блокирует эффект предварительного воздействия "затравки" тау RD (фиг.18C). Вместе полученные данные свидетельствуют об усилении агрегации тау в серийно культивируемых клеточных популяциях.
Пример 6. Распространение от клетки к клетке опосредуется высвобождением агрегатов во внеклеточное пространство
Механизм, посредством которого белковые агрегаты перемещаются между клетками, неизвестен. Например, одни исследователи считают, что распространение белков прионов может происходить через туннелирующие нанотрубочки, а другие полагают, что такое распространение может происходить через экзосомы. Описанное ранее наблюдение, что антитела против белка тау уменьшают патологию in vivo, позволяет предположить, что агрегаты тау могут высвобождаться непосредственно во внеклеточное пространство. Хотя трансклеточное перемещение на основе межклеточного контакта не должно зависеть от объема внеклеточной среды, было высказано предположение, что трансклеточное перемещение тау может быть чувствительным к объему внеклеточной среды, как было описано для SOD1. Сначала тестируют эффект совместного культивирования в условиях разных объемов среды.
Было отмечено, что увеличение объема среды для культивирования клеток приводит к уменьшению эффективности трансклеточного перемещения агрегатов (фиг.19A). Кроме того, переноса кондиционированной среды от клеток, экспрессирующих RD(LM)-HA, было достаточно, чтобы вызвать агрегацию в клетках, экспрессирующих RD-CFP/YFP (фиг.19В). Указанные результаты свидетельствуют о том, что перемещение тау между клетками может происходить через межклеточное пространство, однако они не позволяют определить, заключается ли инкапсулированный белок в эндосому.
Разумно предположить, что доступ к инкапсулированному тау будет блокироваться липидной мембраной, тогда как свободный тау будет доступен для антитела. Поэтому проводят исследование для определения, будет ли мышиное моноклональное антитело (HJ9.3), способное вызывать иммунопреципитацию тау, блокировать трансклеточное распространение. Используют модификацию описанной выше клеточной модели распространения тау RD, в которой RD(LM)-HA и RD(ΔK)-CFP совместно экспрессируют в одной клеточной популяции, которую совместно культивируют в течение 48 ч с клетками, экспрессирующими RD(AK)-YFP, с последующим анализом методом FRET. HJ9.3, по сравнению с объединенным мышиным IgG, тестируют в течение 48-часового периода совместного культивирования. HJ9.3 вызывает дозозависимое уменьшение трансклеточного распространения, тогда как неспецифический IgG не влияет на такое распространение (фиг.19C и D). Важно отметить, что HJ9.3 не оказывает какого-либо эффекта на внутриклеточную агрегацию RD(ΔK)-CFP и RD(ΔK)-YFP, если два белка совместно экспрессируются в одной и той же клетке (фиг.19E), указывая на то, что антитело не участвует в непосредственном ингибировании внутриклеточной агрегации. Также анализируют роль свободного тау в трансклеточном распространении путем оценки индукции неправильной укладки тау с помощью биохимических методов. Индукцию агрегации подтверждают методами детергентного фракционирования и вестерн-блоттинга, которые позволяют обнаружить увеличение RD(ΔK)-YFP в нерастворимой фракции, индуцированное совместным культивированием с RD(LM)-HA. HJ9.3 блокирует эффект RD(LM)-HA, индуцируя нерастворимость RD-YFP в совместно культивируемых клетках (фиг.19F и G).
Эффективность добавления антитела позволяет предположить, что свободный тау непосредственно передается между клетками, но механизм ингибирования под действием антитела остается невыясненным. Было высказано предположение, что HJ9.3 блокирует поглощение фибрилл тау клетками. Чтобы проверить данное предположение, используют метод проточной цитометрии, позволяющий отслеживать воздействие антитела на трансклеточное перемещение агрегатов. Авторы настоящего изобретения ранее разработали цитометрическую схему, в которой одну популяцию клеток метят mCherry, и вторая популяция содержит гибриды тау-YFP. После совместного культивирования трансклеточное перемещение можно отслеживать по относительному проценту двойных положительных (YFP/mCherry) клеток. Популяцию клеток НЕК293 трансфицируют тау RD(LM)-YFP, и вторую популяцию трансдуцируют лентивирусом, экспрессирующим mCherry. После промывания и ресуспендирования клетки двух популяций совместно культивируют в течение 48 ч в присутствии или в отсутствии 10-кратных разведений HJ9.3 в среде. Клетки собирают и определяют относительное число двойных положительных клеток методом проточной цитометрии. Отрицательные контроли состоят из таких же клеточных популяций, смешанных до сортинга. Каждая экспериментальная точка получена с использованием биологических трипликатов. Популяция совместно культивированных клеток содержит значительно больше двойных положительных клеток RD(LM)-YFP/mCherry (2,07%), чем популяция предварительно перемешанных клеток (0,142%) (фон). HJ9.3 уменьшает процент двойных положительных клеток от 2,07% до 1,31% (фиг.19H). Это соответствует влиянию указанного антитела на трансклеточное распространение агрегации, измеренному методом FRET. Разница в способности данного антитела блокировать распространение по результатам измерения методами FRET и проточной цитометрии, скорее всего, обусловлена различиями между двумя методами, использованными для измерения указанного события.
Для дальнейшего мониторинга влияния антитела HJ9.3 на трансклеточное перемещение агрегатов используют прямой иммунофлюоресцентный метод, для определения, где откладываются комплексы HJ9.3/тау. Клетки RD(ΔK)-YFP или не трансфицированные клетки культивируют в присутствии HJ9.3 в течение 48 часов. Клетки фиксируют 4% PFA, пермеабилизируют 0,25% тритоном Х-100 и затем подвергают воздействию вторичного антитела, представляющего собой козлиное антитело против мышиных иммуноглобулинов, меченное Alexa 546. Внутри клеток присутствует очень небольшое количество комплексов HJ9.3/тау. Большинство комплексов обнаруживается вне клеток, в основном, связанными с клеточной мембраной. Такое окрашивание антителом отсутствует в нетрансфицированных клетках, свидетельствуя о том, что сигнал является специфичным для комплексов HJ9.3/тау (фиг.20). Таким образом, HJ9.3 блокирует поглощение агрегатов тау, улавливая агрегаты вне клетки.
Пример 7. Фибриллы тау опосредуют распространение от клетки к клетке
Активность HJ9.3 в анализе распространения создает возможность для определения отвечающих разновидностей тау. HJ9.3 используют для извлечения тау из клеточной среды. HJ9.3 или контрольный IgG добавляют в среду, используемую для культивирования клеток, экспрессирующих разные конструкции RD (wt, PP, ΔΚ, LM). Антитела добавляют в начале или в конце 48-часового периода культивирования. Среду собирают, и проводят аффинную очистку комплексов антитело/антиген с использованием агарозных гранул, содержащих белок G. Комплексы промывают и затем кипятят в SDS-содержащем буфере для загрузки, используемом в анализе методом Вестерн-блоттинга. HJ9.3 специфически улавливает частицы тау RD из клеточной среды, тогда как IgG не оказывает заметного эффекта (фиг.21A). ~10-Кратное увеличение количества белка тау, присутствующего в среде, наблюдают в случае присутствия HJ9.3 в течение всего периода культивирования, в отличие от случая, когда данное антитело добавляют в конце указанного периода (фиг.21B). В среде, используемой для культивирования клеток, трансфицированных RD(ΔK)-HA и RD(LM)-HA, также были обнаружены частицы с молекулярной массой высшего порядка, соответствующие агрегатам RD. В среде, использованной для культивирования клеток, трансфицированных тау RD(PP)-HA, содержится наименьшее количество белка, и частицы высшего порядка не обнаруживаются методом Вестерн-блоттинга. Динамика (0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 24 ч и 48 ч) описанного выше эксперимента демонстрирует зависимое от времени увеличение уровня тау в среде, подразумевая, что инкубация с HJ9.3 действительно приводит к повышению равновесного уровня белка тау, присутствующего в кондиционированной среде (фиг.21C). Вместе полученные результаты демонстрируют, что HJ9.3 блокирует распространение от клетки к клетке, препятствуя поглощению агрегатов клетками, и согласуются с данными по равновесным потокам агрегатов тау, направленным в клетки и из клеток.
Точная природа частиц тау, которые обеспечивают трансклеточное распространение, не известна. Поэтому HJ9.3 используют для улавливания этих частиц с целью визуализации методом AFM. Клетки НЕК293, трансфицированные разными мутантами тау, культивируют в присутствии HJ9.3. Через 48 ч комплексы антитело/антиген очищают с использованием агарозных гранул, содержащих белок G. Затем комплексы элюируют с гранул буфером с высоким содержанием соли и наносят на чипы AFM для визуализации. Видимые фибриллярные частицы детектируют в среде, использованной для культивирования клеток, экспрессирующих RD(ΔK)-HA и RD(LM)-HA, тогда как RD(PP)-HA продуцируют только аморфные агрегаты, (фиг.21D), и клетки, трансфицированные суррогатом, не продуцируют сигнал (данные не показаны). Полученные результаты согласуются с данными, свидетельствующими о том, что свободные фибриллы тау опосредуют трансклеточное распространение агрегации тау путем высвобождения в межклеточное пространство.
Пример 8. Влияние антител против тау на тау-патологию in vivo
Активность двух других антител против полноразмерного, рекомбинантного человеческого тау тестируют, применяя анализ распространения. Клетки RD(LM)-CFP и RD(ΔK)-YFP совместно культивируют в течение 48 часов в присутствии и в отсутствии различные моноклональных антител, направленных на различные эпитопы тау (HJ8.5, HJ9.3 и HJ9.4, фиг.22А). Антитело HJ3.4 против пептида Αβ используют в качестве отрицательного контроля. Все три антитела против тау блокируют трансклеточное распространение про-агрегационных мутантов RD-тау между клетками (фиг.22b). Отрицательный контроль HJ3.4 не блокирует трансклеточное распространение. С использованием HJ8.5, HJ9.3 и HJ9.4 также детектируют фибриллы RD-тау методом ELISA (фиг.22С).
Чтобы блокировать распространение агрегатов тау от клетки к клетке in vivo, используют способ пассивной вакцинации с использованием антител, предназначенных для различных эпитопов тау. Антитела против тау, HJ8.5 и HJ9.3 или среду вводят в боковой желудочек мышей Р301S tg в возрасте 6 месяцев, путем интрацеребровентрикулярной инъекции с использованием осмотических насосов Alzet (2006 модель, фиг.23а). Внутримозговую канюлю, прикрепленную к собранному насосу Alzet, хирургически имплантируют в левый боковой желудочек каждой мыши на расстоянии 0,4 мм от брегмы в переднезаднем направлении, 1,0 мм от средней линии в латеральном направлении и 2,5 мм в дорсовентральном направлении (фиг.23B).
После обработки размещение канюли подтверждают окрашиванием крезиловым фиолетовым (фиг.23C). Осмотический насос Alzet заменяют через 6 недель, и эксперимент заканчивают на 84 день. Для подтверждения того, что экспериментальная схема не вызывает деградации и/или дезактивации антител, антитела собирают из насоса Alzet после инфузии в мозг мыши в течение 6 недель и загружают на гель SDS-PAGE. Сначала гель окрашивают красителем кумасси голубым (фиг.24A), и затем анализируют вестерн-блоттингом с использованием антител, взятых из насоса до инфузии и через 6 недель после начала инфузии (фиг.24B). Все антитела являются стабильными и активными в течение 6 недель в насосе Alzet при физиологической температуре in vivo. Кроме того, было подтверждено, что блокировка рекомбинантного человеческого белка тау разными инфузионными антителами не препятствует проведению анализа ELISA с применением HJ8.7-BT2B для измерения общего тау (фиг.25).
Для определения, уменьшает ли обработка антителом окрашивание патологического тау, окрашивание тау анализируют в срезах тканей мышей Р301S tg в возрасте 9 месяцев, получавших среду/PBS или моноклональные антитела против тау. Коронарные срезы пириформной коры окрашиваются биотинилированным антителом AT8, которое распознает аномально фосфорилированную форму белка тау. Количественный анализ предварительных результатов иммуногистохимического анализа демонстрирует, что количество аномально фосфорилированного тау значительно уменьшается после инфузии HJ8.5 и HJ9.3 в мозг мыши (фиг.26 и 27). Биохимический анализ этих эффектов осуществляется в настоящее время. В случае успешного результата пассивная иммунизация против распространения и патологии тау может стать терапевтическим способом лечения болезни Альцгеймера, лобно-височной деменции или других таупатий.
Обсуждение примеров 3-7
Ранее было выдвинуто предположение, что прионо-подобные механизмы, включающие конформационное изменение матрицы и трансклеточное распространение агрегации, могут объяснить устойчивое прогрессирование таупатий и других нейродегенеративных заболеваний. Такой механизм включает высвобождение белкового агрегата из клетки-донора, проникновение его в реципиентную клетку и непосредственный контакт с белком, обладающим нативной укладкой, с увеличением форм, характеризующихся неправильной укладкой. Однако механистическое доказательство, поддерживающее данную модель таупатии, является неполным, и трансклеточное распространение неправильной укладки тау в этой модели не было показано ранее. Примеры 3-7 описывают трансклеточное распространение агрегации тау в культивируемых клетках посредством секретируемых агрегатов тау и предлагают возможный механизм. Сначала было описано спонтанное образование фибрилл тау RD в трансфицированных клетках, обнаруженное путем окрашивания экстрагированного вещества X-34 и его анализа методом AFM. Затем с помощью метода конфокальной микроскопии было обнаружено совпадение белков тау, присутствующих во внутриклеточных включениях двух разных клеток. Это было связано с повышением нерастворимости в детергенте тау RD(ΔK)-YFP при совместном культивировании с клетками, экспрессирующими склонную к агрегации форму белка, RD(LM)-HA. Данное увеличение агрегации также было продемонстрировано методом FRET с использованием RD(LM)-CFP/YFP, совместно экспрессированных в одних и тех же клетках. Для детекции используют фотообесцвечивание акцептора (микроскопия) и спектральные методы (FPR). Затем проводят FRET с использованием RD(LM)-CFP и RD(LM)-YFP, экспрессированных в раздельных клеточных популяциях, для подтверждения того, что распространение происходит в результате непосредственного контакта белков. Данный метод также используют для подтверждения увеличения неправильной укладки белка тау в клеточных популяциях при последовательном культивировании. Трансклеточное распространение агрегации тау опосредуется фибриллами, которые высвобождаются непосредственно в межклеточное пространство, и поскольку перенос чувствителен к объему внеклеточной среды, кондиционированная среда может увеличивать внутриклеточную агрегацию, а антитело против тау (HJ9.3) препятствует распространению от клетки к клетке и улавливает внеклеточные фибриллы тау. Таким образом, с помощью различных методов авторы подтверждают, что трансклеточное распространение агрегатов происходит посредством конформационного изменения матрицы, и предлагают простую модель для объяснения данного явления (фиг.29).
Трансклеточное распространение. Хотя спонтанное перемещение агрегированного тау между клетками было описано ранее, оставалось неизвестным, могут ли агрегаты белка тау распространять между клетками состояние, характеризующееся неправильной укладкой, путем непосредственного контакта белков, или они косвенно воздействуют на клетки. Исследования α-синуклеина в клеточной культуре также позволяют предположить распространение, однако неясно, какова природа частиц (например, агрегаты, димеры или мономеры), полученных из донорных клеток, и частиц, образующихся в реципиентных клетках. Кроме того, агрегаты SOD1 могут передаваться между клетками через среду, вызывая дальнейшую агрегацию, однако точная природа отвечающих конформеров белка остается невыясненной и непонятно, происходит ли непосредственный белок-белковый контакт. Введение очищенного Αβ42 и фибрилл тау в мозг трансгенных мышей вызывает агрегацию эндогенного тау с образованием поблизости фибрилл тау, однако трудно исключить индукцию агрегации под действием введенного белка. Исследование, проведенное в лаборатории авторов настоящего изобретения, и затем другие исследования подтверждают перемещение агрегатов тау извне внутрь клетки и индукцию агрегации под действием рекомбинантного белка. Однако ни одно предварительное исследование белка тау не продемонстрировало распространение в целом: перемещение агрегата от одной клетки к другой, непосредственный контакт с нативным белком, переход белка в реципиентной клетке в фибриллярное состояние и увеличение числа частиц с неправильной укладкой.
Данная работа демонстрирует указанные явления различными способами. Сначала было обнаружено, что совместное культивирование клеток, экспрессирующих склонную к агрегации форму тау RD(LM)-HA, с клетками, экспрессирующими RD(ΔK)-YFP, приводит к совместной локализации указанных молекул во включениях, положительных по β-складчатой структуре. Затем было отмечено, что совместное культивирование клеток, экспрессирующих RD(LM)-HA, с другой популяцией, экспрессирующей как RD(ΔK)-CFP, так и RD(ΔK)-YFP, приводит к увеличению сигнала FRET, позволяя предположить, что перемещение RD(LM)-HA в клетки, экспрессирующие пару молекул для FRET, индуцирует их агрегацию. Чтобы продемонстрировать непосредственный контакт и совместную агрегацию тау, агрегаты, перемещающиеся между клетками, RD(ΔK)-CFP и RD(ΔK)-YFP экспрессируют в отдельных популяциях. Это приводит к генерированию сигнала FRET в результате трансклеточного перемещения и совместной агрегации, который исчезает, если хотя бы одна из конструкций содержит двойную пролиновую мутацию, блокирующую образование β-складчатой структуры. Также наблюдается индукция агрегации полноразмерного тау-YFP в результате переноса агрегатов RD-CFP, но эффективность снижается (данные не показаны). Наконец, интенсивность сигнала FRET, индуцированного трансклеточным перемещением белковых агрегатов, значительно увеличивается в результате предварительного совместного культивирования клеток, экспрессирующих RD(LM)-HA, с клетками, экспрессирующими RD(ΔK)-CFP, свидетельствуя о том, что агрегированное состояние может увеличиваться в популяции клеток.
Модуляция распространения агрегатов тау под действием антитела. Антитела против пептида Αβ, который преимущественно является внеклеточным, позволяют предотвратить агрегацию Αβ в головном мозге и удалить существующие агрегаты. Хотя существуют потенциальные побочные эффекты, такие антитела являются перспективными лекарственными средствами. Однако успех вакцинации на мышиных моделях таупатии и синуклеинопатии является неожиданным с учетом того факта, что целевые белки преимущественно являются внутриклеточными. Было отмечено, что HJ9.3, мышиное моноклональное антитело против тау RD, ингибирует трансклеточное распространение агрегации тау. Однако это антитело не оказывает какого-либо влияния на внутриклеточную агрегацию тау. Периодическое воздействие клеточной среды на данное антитело сильно увеличивает равновесный уровень тау в среде. Это подтверждают анализы методом проточной цитометрии, которые демонстрируют, что HJ9.3 блокирует перемещение агрегатов из одной клетки в другую. И наконец, отсутствуют комплексы HJ9.3/tau, уловленные на поверхности клеток. Воздействие, оказываемое данным антителом, убедительно доказывает, что фибриллы тау высвобождаются во внеклеточное пространство и не распространяют неправильную укладку, главным образом, путем межклеточного переноса посредством экзосом или туннельных нанотрубочек, как в механизме, предложенном для прионов. Кроме того, если агрегаты, присутствующие вне клетки, не улавливаются HJ9.3, они, скорее всего, снова возвращаются в клетки. Может существовать несколько механизмов ингибирования под действием терапевтических антител, которые включают исчезновение белковых фибрилл, блокаду превращения в клетках и инициацию внутриклеточной деградации. Результаты, полученные авторами настоящего изобретения при использовании HJ9.3, наиболее согласуются с препятствованием поглощению клетками, как одним из механизмов блокирования таупатии, и позволяют предложить новые способы разработки и оптимизации терапевтических антител для лечения нейродегенеративных заболеваний.
Трансклеточное распространение посредством фибриллярного тау. Эффективная блокировка распространения агрегации тау под действием HJ9.3 позволяет использовать это антитело для улавливания отвечающих частиц. Иммуноаффинное выделение тау из кондиционированной среды демонстрирует наличие фибриллярного тау. Фибриллы тау отсутствуют в среде контрольных клеток, а также клеток, экспрессирующих RD(PP)-HA, устойчивый к образованию β-складчатой структуры, которые продуцируют аморфные агрегаты. Экспрессия же как RD(ΔK)-HA, так и RD(LM)-HA вызывает секрецию фибрилл во внеклеточное пространство. Было непонятно, как агрегация белков в одной клетке может повлиять на агрегацию в соседней клетке, и было выдвинуто предположение, что данный процесс могут опосредовать цитокины, экзосомы или непосредственные соединения между клетками. Эти возможности не могут быть полностью исключены. Однако полученные результаты наиболее согласуются с механизмом, в котором свободные фибриллярные частицы являются медиаторами распространения через внеклеточное пространство. Данное исследование предоставляет ответы на несколько важных вопросов, связанных с механизмами, посредством которых белковые агрегаты распространяются от одной клетки к другой в культуре и, следовательно, in vivo. В сочетании с описанными в данном описании способами контролирования трансклеточного распространения, данный процесс можно использовать в качестве мишени для фармакологических и биологических средств, обеспечивающих более эффективное лечение таупатий и других нейродегенеративных заболеваний.
Методы, использованные в примерах 1-8
Антитела
Самую длинную изоформу мышиного рекомбинантного тау mTau40 (432 а.к.) и самую длинную изоформу человеческого тау hTau40 (441 а.к.) получают в лаборатории Eva Mandelkow и используют в качестве стандартов в анализе тау методом ELISA. Моноклональное мышиное антитело Tau-5, которое распознает как человеческий, так и мышиный тау (эпитоп содержит остатки 218-225), получают из лаборатории L. Binder (LoPresti et al., 1995; Porzig et al., 2007). Моноклональные антитела HJ8.1 и HJ9.3 представляют собой мышиные моноклональные антитела, полученные путем иммунизации мышей с нокаутом тау против человеческого тау и мышиного тау, соответственно (Jackson Laboratory). Оба антитела распознают мышиный и человеческий тау в методах вестерн-блоттинга, иммунопреципитации и ELISA. HJ9.3 распознает участок тау, отвечающий за связывание микротрубочек (MTBR). Мышиное моноклональное антитело BT-2, которое также распознает человеческий и мышиный тау (эпитоп содержит остатки 194-198), получают от Pierce. Кроличьи поликлональные антитела против тау (ab64193, эпитоп расположен в участке домена повторов) получают от Abeam, Cambridge, MA. Мышиные моноклональные антитела против гемагглютинина HA (HA.11, клон 16B12) получают от Covance, Emeryville, СА. Кроличье поликлональное антитело GFP (sc-8334) получают от Santa Cruz Biotechnology.
Плазмиды
Последовательности, кодирующие домен четырех повторов (RD) связанного с микротрубочками белка тау, используют для экспрессии белка. Помимо формы дикого типа, получают различные мутанты тау: ΔΚ280 Δ(Κ); P301L/V337M (LM); ΔΚ280/I227P/I308P (PP). Указанные последовательности субклонируют либо в pcDNA3.1 (Invitrogen), с получением мутанта, содержащего на С-конце гемагглютинин (HA) в качестве маркера, либо в pEYFP-N1 или pECFP-N1 (Clontech), с получением гибридов, содержащих на C-конце флуоресцентные белки.
Животные
Мышей Р301S tg (линия PS19), характеризующихся сверхэкспрессией человеческой изоформы тау Р301S T34 (1N4R), получают с использованием B6C3 в качестве исходного штамма и заранее характеризуют. Мышей Р301S tg получают из Jackson Laboratory. Мышей с нокаутом тау получают из The Jackson Laboratory. Совпадающих по возрасту и генетическому фону детенышей нетрансгенных мышей используют в качестве мышей дикого типа. Во всех экспериментах данного исследования используют как самцов, так и самок.
Анализ с использованием иммунопреципитации и иммуноблоттинга
Анализ с использованием иммунопреципитации и иммуноблоттинга. Микродиализные образцы гиппокампа трансгенной по тау мыши Р301S и мышей дикого типа собирают со скоростью 1,0 л/мин в течение 15 ч. ISF подвергают иммунопреципитации с использованием Dynabeads (Invitrogen), покрытых антителом против тау HJ8.1 или HJ9.3 в соответствии с инструкциями производителя. Осажденные фракции загружают на восстанавливающий 4-12% мини-гель Bis-Tris (Invitrogen) и переносят на нитроцеллюлозную мембрану. Биотинилированное антитело ВТ-2 (Pierce) и поли-HRP-конъюгированный стрептавидин (Thermo Scientific) используют для устранения помех со стороны осажденных антител. Клетки HEK293 культивируют в модифицированной по Дульбекко среде Игла в (DMEM), содержащей 10% фетальной бычьей сыворотки, 100 мкг/мл пенициллина и 100 мкг/мл стрептомицина. Культуры поддерживают в увлажненной атмосфере, содержащей 5% СO2, при 37°С. Чтобы провести временную трансфекцию, клетки помещают в среду Optimem, трансфицируют, используя реагент Lipofectamine/PLUS и 600 нг соответствующих конструкций ДНК (Invitrogen, Carlsbad, CA, USA) в соответствии с рекомендациями производителя, собирают через 24 ч или 48 ч и используют для дальнейших анализов.
Анализы с использованием детергентного фракционирования и вестерн-блоттинга
Клетки HEK293 высевают в 12-луночный планшет с плотностью 400000 клеток/лунка. На следующий день клетки трансфицируют, используя 600 нг плазмиды. Через 48 ч клетки собирают путем обработки 0,05% трипсином в течение 3 минут при 37°C и кратковременного центрифугирования при 7000×g, после чего их лизируют в 100 мкл 1% раствора тритона в PBS, содержащего ингибиторы протеаз. Затем растворимые цитозольные белки собирают центрифугированием при 14000×g в течение 10 минут. Нерастворимые белки получают путем ресуспендирования осадка в буфере RIPA/SDS и центрифугирования при 20000×g в течение 15 минут, после чего нуклеиновые кислоты расщепляют нуклеазой бензоназой. Для проведения совместного культивирования используют равное число клеток, трансфицированных RD(LM)-HA и RD(ΔK)-YFP, которые культивируют вместе в течение 48 ч, затем клетки собирают и анализируют методом Вестерн-блоттинга. Эквивалентные количества белковых экстрактов клеток HEK293 из каждой фракции анализируют с использованием 4%-20% полиакриламидных гелей (BioRad); антитела против тау RD (распознающего эпитоп в участке RD) в разведении 1:2000 (ab64193, Abeam, Cambridge, MA) и/или антитела против GFP в разведении 1:1000 (SC-8334, Santa Cruz Biotechnology, Inc.). Проводят хемилюминесцентную реакцию вторичного антитела, конъюгированного с пероксидазой, используя для детекции рентгеновскую пленку. Количественное определение проводят с помощью программного обеспечения для анализа Image J.
Эксперименты с использованием совместного культивирования: измерение совместной агрегации RD-CFP/YFP методом FRET
Клетки HEK293 высевают в 12-луночный планшет с плотностью 300000 клеток/лунка. На следующий день клетки трансфицируют, используя 600 нг плазмиды, как описано выше. Совместно трансфицированные клетки получают комбинацию конструкций, включающих 150 нг RD-CFP и 450 нг RD-YFP. Через 15 ч клетки собирают путем обработки 0,05% трипсином в течение 3 минут при 37°C, затем часть клеток повторно высевают в 96-луночный планшет в четырехкратных повторах, или на μ-слайды ibidi (ibidi GmbH, Germany) для визуализации с помощью микроскопа. Затем клетки культивируют еще 48 ч, затем фиксируют 4% параформальдегидом и анализируют.
Эксперименты с использованием совместного культивирования: измерение индукции агрегации RD-YFP под действием RD-HA
Клетки НЕК293 трансфицируют RD(ΔK)-YFP или RD(LM)-HA в 12-луночных планшетах. Через 15 ч клетки снова высевают вместе на μ-слайды ibidi и культивируют в течение еще 48 ч. Затем их фиксируют, окрашивают антителом против HA и X-34 и анализируют с помощью микроскопии.
Эксперименты с использованием совместного культивирования: анализы распространения в совместной культуре
Две популяции клеток НЕК293 в 12-луночном планшете совместно трансфицируют, используя 300 нг RD(LM)-HA и 300 нг RD(ΔK)-CFP или RD(ΔK)-YFP. Через 15 ч равные части двух популяций совместно культивируют в течение 48 ч в формате 96-луночного планшета. Затем клетки фиксируют 4% параформальдегидом и анализируют методом FRET с использованием флуоресцентного планшет-ридера (FPR). Для проведения анализа методом FRET-микроскопии две популяции клеток НЕК293 в 12-луночном планшете трансфицируют, используя 600 нг RD(LM)-CFP или RD(LM)-YFP. Через 15 ч равные части двух популяций совместно культивируют в течение 48 ч на μ-слайдах ibidi. Затем клетки фиксируют 4% параформальдегидом и проводят FRET с фотообесцвечиванием акцептора.
Эксперименты с использованием совместного культивирования: увеличение агрегации тау при последовательном культивировании
Клетки НЕК293 трансфицируют в 12-луночном планшете с использованием 600 нг различных форм нефлуоресцентного RD-HA и культивируют в течение 24 ч. Вторую группу клеток трансфицируют CFP или RD(ΔK)-CFP. Затем равные части первой и второй популяций совместно культивируют в течение 48 ч. В этот момент 50% данной популяции высевают с популяцией клеток, трансфицированных RD(ΔK)-YFP, в 96-луночный планшет и культивируют в течение 48 ч. Затем клетки фиксируют 4% параформальдегидом и проводят анализ методом FRET с помощью FPR.
Перенос среды и эксперименты с использованием кондиционированной среды
Клетки НЕК293 трансфицируют в 12-луночном планшете либо 600 нг RD(LM)-HA, либо комбинацией 150 нг конструкции RD(ΔK)-CFP и 450 нг конструкции RD(ΔK)-YFP, спустя 15 ч клетки собирают путем обработки 0,05% раствором трипсина в течение 3 минут при 37°C. Равное количество клеток, экспрессирующих RD(ΔK)-YFP/CFP и RD(LM)-HA, совместно культивируют в течение 48 ч в различных количествах среды для культивирования клеток. Затем клетки фиксируют 4% параформальдегидом и проводят анализ FRET. Для экспериментов с использованием кондиционированной среды через 15 ч после трансфекции среду из клеток RD(LM)-HA, содержащих трансфицированные комплексы, заменяют свежей средой. Клетки, экспрессирующие RD(ΔK)-YFP/CFP, собирают путем обработки 0,05% раствором трипсина в течение 3 минут при 37°C и переносят в 96-луночный планшет. Через 24 ч кондиционированную среду, использованную для культивирования клеток, трансфицированных RD(LM)-HA, собирают и добавляют к клеткам, экспрессирующим RD(ΔK)-YFP/CFP. Спустя 48 ч клетки фиксируют 4% параформальдегидом и анализируют методом FRET.
Анализы с использованием резонансного переноса энергии флуоресценции (FRET): измерение FRET с использованием микроскопии с фотообесцвечиванием
Клетки НЕК293, трансфицированные для описанных выше экспериментов с совместной трансфекцией и совместным культивированием, используют для анализа FRET с помощью микроскопии с фотообесцвечиванием акцептора. Все изображения получают с использованием объектива C-Apochromat 40× 1,2 NA (Carl Zeiss Advanced Imaging Microscopy, 07740 Jena, Germany 100×(CFP). Цифровые изображения получают с использованием системы Zeiss LSM510 Meta NLO Multiphoton/конфокальный микроскоп с лазерным сканированием на Zeiss Axiovert 200M. Для визуализации используют следующие каналы: донорный CFP стимулируют с использованием аргонового лазера при 458 нм, и флуоресценцию регистрируют с использованием полосового фильтра 480-520 нм; акцепторный YFP стимулируют с использованием аргонового лазера при 514 нм, и флуоресценцию регистрируют с использованием длинноволнового пропускающего фильтра 560 нм. Для получения изображения, в котором интенсивность отражает оценку эффективности FRET, значение исходного изображения CFP вычитают из конечного изображения CFP, полученного после фотообесцвечивания на попиксельной основе, эту разность умножают на 100 и делят на конечную интенсивность изображения CFP: 100×(CFPfinal-CFPinitial)CFPfinal. Вносят соответствующие корректировки для частичного фотообесцвечивания акцептора. Преобразование изображений с арифметическим кодированием и черно-белых изображений в цветные осуществляют с помощью программного обеспечения NIH ImageJ 1,44.
Анализы FRET: флуоресцентный планшет-ридер
Спектральные измерения FRET (FRET/донор) проводят с использованием флуоресцентного планшет-ридера TecanM1000 в соответствии с описанными ранее способами. Если донор и акцептор не слиты с одним и тем же белком, спектральные измерения FRET зависят от тщательно подобранных относительных количеств донорных и акцепторных белков, экспрессированных в клетке. Из всех измеренных на ридере значений сначала вычитают фоновые значения, полученные для клеток, трансфицированных суррогатом. Сигнал YFP в каждой лунке (Smpl485ex/528em FRET) используют для оценки уровней экспрессии RD-YFP, допуская также, что в экспериментальных условиях не происходит независимого изменения RD-CFP/YFP. Это позволяет исключить возможность, что изменения кажущегося сигнала FRET возникают просто из-за различий в уровнях экспрессии RD. Относительный вклад активации акцептора (528 нм) под действием сигнала возбуждения донора (435 нм) в общий сигнал FRET корректируют путем определения доли "перекрестной активации" для акцептора, X, где X=сигнал RD-YFP, измеренный при 435ex/528em, и деления ее на сигнал, измеренный при 485ex/528em. "Перекрестная активация" является практически постоянной при различных уровнях экспрессии RD-YFP, встречающихся в экспериментах. "Измеренное" значение FRET в каждом образце регистрируют при 435ex/528em, "донорное" значение (CFP) регистрируют при 435ex/485em. Затем "фактическое" значение FRET/донор для каждой лунки рассчитывают по формуле:
FRETactual=(Smpl435ex/528em-X*(Smpl435ex/528em))/Smpl435ex/528em
Данный способ измерения агрегации белков методом FRET позволяет достоверно детектировать тонкие изменения в ответ на фармакологические средства, а также на генетические манипуляции с рецептором андрогенов и агрегацию белка хантингтина, которые подтверждаются визуальными и биохимическими анализами. Поскольку относительная величина измеренного спектрального сигнала FRET зависит от соотношения акцептор:донор, для RD-CFP и RD-YFP, совместно экспрессированных в одной клетке, используют постоянное соотношение 3:1. При измерении донорного сигнала такое соотношение обеспечивает близкую к максимальной эффективность FRET при приемлемом отношении сигнал:шум.
Атомно-силовая микроскопия (AFM)
RIPA-нерастворимые белки экстрагируют из трансфицированных клеток НЕК293 и инкубируют на чипах mica (Ted Pella, Inc) в течение 10 минут. Затем образцы промывают дважды 100 мкл ddH2O и оставляют сохнуть при комнатной температуре. На следующий день проводят атомно-силовую микроскопию с использованием атомно-силового микроскопа MFP-3D (Asylum Research).
Иммунофлуоресценция и конфокальная микроскопия
Клетки НЕК293, трансфицированные для описанных выше экспериментов с совместным культивированием, подготавливают для анализа методом иммунофлуоресценции и окрашивания X-34. После фиксации 4% параформальдегидом в течение 15 мин при комнатной температуре клетки дважды промывают PBS при комнатной температуре (КТ) в течение 5 мин и пермеабилизируют 0,25% раствором тритона X-100 в PBS при комнатной температуре в течение 10 минут. Клетки блокируют блокирующим раствором, содержащим 1% нормальной козьей сыворотки, 20 мг/мл BSA, 0,25% тритон X-100 в PBS, в течение 3 ч при комнатной температуре. Первичное мышиное моноклональное антитело против HA (Covance, Emeryville, CA) разводят 1:2000 блокирующим раствором, добавляют к клеткам и инкубируют в течение ночи при 4°С. Затем клетки промывают PBS, содержащим 0,1% тритон Х-100, 3 раза, каждый раз в течение 5 минут, и инкубируют с вторичным антителом против мышиных иммуноглобулинов, конъюгированным с Alexa546 (Invitrogen), разведенным в соотношении 1:400 блокирующим раствором. Затем клетки промывают PBS, содержащим 0,1% тритон Х-100 3 раза, каждый раз в течение 5 минут, и подвергают воздействию 1 мкΜ X-34 в растворе, содержащем 40% этанола, 60% PBS и 20 мМ NaOH, в течение 10 мин при комнатной температуре. Затем клетки промывают три раза, каждый раз в течение 2 мин, раствором, содержащим 40% EtOH и 60% PBS, и дважды промывают 1×PBS, каждый раз в течение 5 мин. Изображения получают с использованием конфокальной микроскопии (конфокальный микроскоп 405 Zeiss). Чтобы охарактеризовать механизм блокады распространения под действием антитела HJ9.3, клетки НЕК293 трансфицируют RD(ΔK)-YFP или суррогатом. После культивирования клеток, трансфицированных RD(ΔK)-YFP или суррогатом, в присутствии HJ9.3 в течение 48 часов, клетки фиксируют 4% PFA, пермеабилизируют 0,25% раствором тритона Х-100 и затем подвергают воздействию вторичного антитела против мышиных иммуноглобулинов, меченного Alexa 546. Изображения получают с использованием конфокальной микроскопии (конфокальный микроскоп Zeiss).
Анализ гибели клеток с использованием пропидиййодида (PI)
HEK293 клетки высевают в 96-луночный планшет с плотностью 75000 клеток/лунка. На следующий день клетки трансфицируют 100 нг разных форм нефлуоресцентных плазмид RD-НА в четырех повторах или подвергают воздействию трансфекционных комплексов в отсутствии ДНК. На следующий день среду, содержащую трансфекционные комплексы, удаляют и заменяют свежей средой. В качестве положительного контроля гибели клеток используют нетрансфицированные клетки, обработанные различными концентрациями стауроспорина (1, 2, 4, 20 мкΜ) в течение 30 минут при 37°С. Затем раствор стауроспорина удаляют и все клетки подвергают воздействию 5 мкг/мл пропидиййодида в течение 10 минут при 37°С. Затем раствор пропидиййодида заменяют на среду, не содержащую фенол, и считывают флуоресценцию на планшет-ридере при длине волны возбуждения 535 нм и длине волны испускания 617 нм.
Иммунопреципитация
Популяции трансфицированных клеток совместно культивируют либо сами по себе, либо в присутствии мышиного моноклонального антитела HJ9.3 (в разведении 1:1000, что эквивалентно концентрации антитела 2,5 нг/мкл), либо в присутствии общих мышиных антител IgG, в течение 3 ч, 6 ч, 9 ч, 12 ч, 24 ч или 48 ч. Кондиционированную среду собирают, к среде добавляют агарозные гранулы, содержащие белок G (100 мкл 50% суспензии гранул Pierce) и инкубируют, вращая, в течение ночи при 4°C. Через 18 часов к образцам добавляют 500 мкл буфера для связывания (Pierce) и центрифугируют при 2000×g в течение 3 минут. Супернатант отбрасывают и данную стадию промывки повторяют три раза. Белки, связанные с гранулами, элюируют с помощью буфера для элюирования с высоким содержанием соли (50 мкл) путем инкубации при комнатной температуре в течение 5 минут. Затем образцы центрифугируют при 2000×g в течение трех минут и собирают супернатант. Данную стадию элюирования повторяют один раз с получением общего объема элюата 100 мкл. Другой образец кондиционированной среды, изначально не подвергавшейся воздействию HJ9.3 или IgG, инкубируют с HJ9.3 (1:1000) или антителами IgG в течение ночи при 4°C с вращением, и затем подвергают описанному выше способу иммунопреципитации. Все образцы анализируют на 4-20% полиакриламидных гелях (BioRad), детектируя с помощью кроличьего поликлонального антитела против тау RD, разведенного 1:2000 5% раствором сухого молока в TBS/Tween (ab64193, Abeam, Cambridge, MA). Проводят хемилюминесцентную реакцию вторичного антитела, конъюгированного с пероксидазой, используя для детекции рентгеновскую пленку.
Проточная цитометрия
Клетки НЕК293 высевают в чашку размером 10 см и культивируют до ~80% слияния. Затем клетки трансфицируют, используя 24 мкг конструкции RD(LM)-YFP, или трансдуцируют лентивирусом mCherry. На следующий день клетки собирают путем обработки 0,05% раствором трипсина в течение 3 минут при 37°C, осаждают и ресуспендируют в свежей среде. Две популяции клеток культивируют совместно либо сами по себе, либо в присутствии мышиного моноклонального антитела HJ9.3 против тау RD в разведении 1:1000 или 1:10000 в течение 48 ч (разведение 1:1000 эквивалентно концентрации антитела 2,5 нг/мкл). По прошествии указанного времени клетки собирают и ресуспендируют в сбалансированной среде Хэнкса, содержащей 1% FBS и 1 мМ EDTA. Клетки, смешанные непосредственно перед цитометрией, используют в качестве отрицательного контроля. Клетки считают с использованием высокоскоростного клеточного сортера MoFlo (Beckman Coulter) и для каждого условия определяют процент двойных положительных клеток. Каждое условие реализуют в трех биологических повторах, используя для анализа каждого экспериментального условия 50000 клеток.
Интрацеребровентрикулярное (ICV) введение моноклональных антител против тау
В данном исследовании используют трансгенных мышей Р301S tau, которые экспрессируют изоформу Р301S человеческого T34 (1N4R). У данных мышей в возрасте 6 месяцев индуцируют тау-патологию. Антитела вводят в левый боковой желудочек путем церебровентрикулярной инъекции в возрасте 6 месяцев, повторяя такие вливания в течение 12 недель. После лечения мозг мышей обрабатывают для проведения иммуногистохимического и биохимического анализа с использованием ELISA и иммуноблоттинга.
Интрацеребровентрикулярные инъекции проводят с использованием осмотического насоса Alzet, модель 2006. Мозговую канюлю, прилагаемую к комплекту насоса Alzet, хирургически имплантируют в левый боковой желудочек каждой мыши на расстоянии 0,4 мм от брегмы в переднезаднем направлении, 1,0 мм от средней линии в латеральном направлении и 2,5 мм в дорсовентральном направлении. После обработки размещение канюли подтверждают окрашиванием крезиловым фиолетовым.
Введение к примерам 9-15
Тау представляет собой ассоциированный с микротрубочками белок, который образует внутриклеточные агрегаты при некоторых нейродегенеративных заболеваниях с общим названием таупатии. К ним относятся болезнь Альцгеймера (AD), прогрессирующий надъядерный паралич (PSP), кортикобазальная дегенерация (CBD) и лобно-височная деменция (FTD). Тау представляет собой хорошо растворимый и в нативном состоянии развернутый белок, который связывается с микротрубочками и активирует их сборку. При таупатиях тау накапливается в гиперфосфорилированных нейрофибриллярных клубках (NFT), которые визуализируются в дистрофических нейритах и телах клеток при соответствующем окрашивании. Количество патологии тау коррелирует с прогрессирующей дисфункцией нейронов и ухудшением синаптической функции, а также с функциональными расстройствами в организме человека и на моделях трансгенных мышей.
При таупатиях человека патология прогрессирует из одного участка мозга в другой по характерной для заболевания схеме, хотя механизм, лежащий в основе таких процессов, еще не выяснен. В соответствии с прионной гипотезой, агрегаты тау мигрируют из клеток, в которых они образовались, в соседние клетки, где они выполняют роль затравок дальнейшей агрегации тау с распространением патологии. Авторы настоящего изобретения ранее определили, что рекомбинантные фибриллы тау вызывают агрегацию полноразмерного внутриклеточного тау в культивируемых клетках, и что агрегированные формы тау передаются между клетками (Frost et al., 2009; Nat Rev Neurosci 11, 155-159). Кроме того, авторы настоящего изобретения обнаружили, что внутриклеточные фибриллы тау высвобождаются в свободном виде в среду, где они распространяют агрегацию путем непосредственного взаимодействия с нативным тау в реципиентных клетках. Антитело против тау (HJ9.3) блокирует этот процесс, предотвращая поглощение агрегатов тау реципиентными клетками (Kfoury et al., 2012; J Biol Chem 287, 19440-19451). В дополнение к подобным экспериментам с использованием рекомбинантного тау, было показано, что спаренные спиральные филаменты из мозга индивидуума с AD вызывают цитоплазматическую агрегацию тау. Введение экстракта мозга мышей, трансгенных по человеческому тау Р301S, в мозг мышей, экспрессирующих человеческий тау дикого типа, индуцирует сборку человеческого тау дикого типа в филаменты и распространение патологии. Похожие эффекты наблюдают после введения фибрилл рекомбинантного полноразмерного или усеченного тау, которые вызывают быструю индукцию NFT-подобных включений, распространяющихся из участков введения в связанные с ними участки мозга во время-зависимой манере. Наконец, селективная экспрессия тау в энторинальной коре вызывает позднюю патологию в концевых зонах аксонов клеток зубчатой извилины и гиппокампа, в соответствии с транс-синаптическим перемещением агрегатов. Таким образом, все большее число исследований подтверждает идею, заключающуюся в том, что агрегаты тау передаются между клетками и могут являться мишенями для терапевтических антител.
На мышиных моделях, которые имитируют признаки AD и болезни Паркинсона (PD), пассивная иммунизация с использованием антител против Αβ и синуклеина может уменьшить отложение Αβ и альфа-синуклеина в головном мозге, а также улучшить поведенческие нарушения. Активная иммунизация мышиных моделей таупатии с использованием фосфопептидов тау уменьшает тау-патологию и в некоторых исследованиях улучшает поведенческие нарушения. Однако в одном исследовании активная иммунизация мышей C57BL/6 дикого типа полноразмерным рекомбинантным тау индуцирует тау-патологию и неврологические нарушения. В двух исследованиях с пассивной вакцинацией наблюдают уменьшение тау-патологии и улучшение двигательной функции, если антитела вводят до появления патологии. Хотя в некоторых исследованиях с иммунизацией против тау наблюдают отдельные положительные эффекты, максимальная ожидаемая эффективность антител против тау, введенных после появления патологии, частицы тау, оптимальные для применения в качестве мишени, и механизм терапевтического эффекта остаются неизвестными.
Пример 9. Характеристика антител против тау
Авторы настоящего изобретения ранее обнаружили, что агрегаты тау, но не мономеры, поглощаются культивируемыми клетками, и что интернализованные агрегаты тау индуцируют внутриклеточную агрегацию тау в реципиентных клетках (Frost et al., 2009; Nat Rev Neurosci 11, 155-159; Kfoury et al., 2012; J Biol Chem 287, 19440-19451). Серию HJ8, включающую 8 мышиных моноклональных антител (полученных против полноразмерного человеческого белка тау), и серию HJ9, включающую 5 антител (полученных против полноразмерного мышиного тау), характеризуют с помощью адаптированной клеточной биосенсорной системы, описанной ранее в Kfoury et al. (2012; J Biol Chem 287, 19440-19451), которая позволяет измерить внутриклеточную агрегацию тау, индуцированную добавлением лизатов мозга, содержащих агрегаты тау. Эффекты антител при блокировании центров образования агрегатов являются непостоянными, несмотря на то, что все антитела эффективно связывают мономеры тау и окрашивают нейрофибриллярные клубки. Три антитела с разной активностью в отношении блокирования центров образования агрегатов выбирают для исследований, описанных в данном описании.
Перед тестированием in vivo определяют аффинность связывания и эпитопы антител, которые относятся к изотипу IgG2b. Человеческий и мышиный тау иммобилизуют на сенсорном чипе CM5 для проведения анализа методом поверхностного плазмонного резонанса (SPR) (фиг.30). Антитело HJ9.3, полученное против мышиного тау, распознает как человеческий (фиг.30А), так и мышиный (фиг.30B) тау с одинаковой константой связывания (KD=Kd/Ka=100 пМ) (фиг.30G). Константы ассоциации (Ka) и диссоциации (Kd) рассчитывают с помощью программного обеспечения BIAevaluation (Biacore AB), выбирая кинетические параметры Fit и одновременно Ka/Kd (общая подгонка) для модели взаимодействия 1:1 (Ленгмюра). Ka и Kd для комплексов HJ9.3 с человеческим (Ka=7,5×104 мсек-1, Kd=7,5×10-6 сек-1) и мышиным тау (Ka=8,6×104 мсек-1, Kd=9,1×10-6 сек-1) указывают на сильное связывание в обоих случаях. Эпитоп HJ9.3 находится в участке домена повторов (RD) между аминокислотами 306-320. HJ9.4, полученное против мышиного тау, обладает высокой аффинностью (KD=2,02 пМ) в отношении мышиного тау с высокой константой скорости ассоциации (Ka=2,28×105 мсек-1) и очень низкой константой диссоциации (Kd=5,1×10-7 сек-1) (фиг.30D и таблица 4). Однако то же антитело имеет значительно более низкую аффинность (KD=6,9 нМ) в отношении человеческого тау (фиг.30C и таблица 4) при почти такой же константе скорости ассоциации (Ka=1,5×105 мсек-1), как и у мышиного тау, но гораздо более высокой константе диссоциации (Kd=1,07×10-3 сек-1). Таким образом, взаимодействие HJ9.4 с человеческим тау менее стабильно, чем с мышиным тау. Эпитоп для этого антитела включает аминокислоты 7-13. HJ8.5 получают против человеческого тау. Данное антитело связывается с человеческим тау (фиг.30E), но не с мышиным тау (фиг.30F). KD (0,3 пМ) (фиг.30Е и таблица 4) и низкая скорость диссоциации (Kd=4,38×10-8 сек-1) свидетельствуют о том, что HJ8.5 связывается с человеческим тау с очень высокой аффинностью. Эпитоп HJ8.5 включает аминокислоты 25-30. SPR демонстрирует, что все 3 антитела против тау специфично распознают фибриллы человеческого тау (фиг.31). Поскольку фибриллы имеют несколько идентичных эпитопов, невозможно непосредственно рассчитать скорости ассоциации и диссоциации.
Константа скорости ассоциации (Ka), константа скорости диссоциации (Kd) и константа связывания (KD) для каждого антитела против человеческого и мышиного тау. Ka и Kd рассчитывают с помощью программного обеспечения BIAevaluation (Biacore AB), выбирая кинетические параметры Fit и одновременно Ka/Kd (общая подгонка) для модели взаимодействия 1:1 (Ленгмюра).
Mс-1=миллисекунда, M=молярный, с=секунда
Антитела анализируют методом иммуноблоттинга и иммуноокрашивания. Вестерн-блоттинг демонстрирует, что все 3 антитела связываются с человеческим тау (фиг.30H). HJ9.3 и HJ9.4 связываются с мышиным тау, а HJ8.5 не связывается с ним (фиг.30H). Согласно предыдущим результатам, полученным авторами настоящего изобретения (Yamada et al., 2011; J Neurosci 31, 13110-13117), у мышей Р301S в возрасте 9 месяцев меньше тау, растворимого в буфере для повторной сборки (RAB), чем у мышей Р301S в возрасте 3 месяцев. Кроме того, было обнаружено, что HJ8.5 окрашивают человеческий тау в мозге трансгенных мышей P301S в возрасте 3 месяцев и в возрасте 9-12 месяцев. Иммунореактивность тау присутствует во всех клеточных телах и процессах (фиг.32). У мышей Р301S в возрасте 9-12 месяцев, содержащих агрегаты тау, указанные агрегаты можно обнаружить с помощью HJ8.5 в клеточных телах (фиг.32A). Другие антитела дают аналогичные результаты (таблица 5). Все антитела связываются с нейрофибриллярными клубками и волокнами нейропиля в мозге, пораженном AD (фиг.32).
Относительная эффективность антител против тау в различных анализах
Пример 10. Антитела против тау блокируют поглощение и затравочную активность агрегатов тау Р301S
Чтобы оценить затравочную активность, присутствующую в лизатах головного мозга Р301S, используют адаптированную клеточную биосенсорную систему, ранее описанную авторами изобретения (Kfoury et al., 2012). Данная система основана на экспрессии домена повторов тау (а.к. 243-375), содержащего мутацию ΔΚ280 и слитого с голубым или желтым флуоресцентным белком (RD(ΔK)-CFP/YFP). Поглощение экзогенных агрегатов указанными клетками инициирует внутриклеточную агрегацию RD(ΔK)-CFP/YFP, которую детектируют методом резонансного переноса энергии флуоресценции (FRET), регистрируя результаты на флуоресцентном планшет-ридере. Очищенные лизаты мозга 12-месячных мышей Р301S после добавления к биосенсорной клеточной системе индуцируют интенсивную агрегацию репортера RD(ΔK)-CFP/YFP, указывая на наличие затравочной активности тау (фиг.33А). Затравочная активность в гомогенате мозга 12-месячных мышей Р301S соответствует примерно 50 нМ (в пересчете на мономер) рекомбинантных полноразмерных волокон (данные не показаны).
Лизаты мышей с нокаутом тау, мышей дикого типа или 3-месячных мышей Р301S, лишенных тау-патологии, практически не вызывают агрегацию или вызывают ее на низком уровне (фиг.33A). Антитела против тау (HJ8.5, HJ9.3 и HJ9.4) анализируют по способности блокировать поглощение и затравочную активность указанных лизатов. HJ3.4 (мышиное моноклональное антитело против Αβ) используют в качестве отрицательного контроля. Антитела против тау эффективно блокируют затравочную активность (фиг.33B). Для определения относительной эффективности антител, их титруют (0,125, 0,25, 0,5, 1, 2 мг/мл) против фиксированного количества лизата мозга Р301S (фиг.33C). Антитело HJ8.5 блокирует затравочную активность по сравнению с контрольной группой в такой низкой концентрации, как 0,25 мкг/мл. Антитела HJ8.5 и HJ9.3 в концентрации 0,5 мкг/мл в значительной степени блокируют поглощение и затравочную активность по сравнению с контролем. HJ9.4 наименее эффективно блокирует поглощение и затравочную активность в соответствии с его более высокой аффинностью в отношении мышиного тау. С помощью всех 3 антител против тау можно обнаружить интернализованные агрегаты тау после поглощения клетками НЕК293, где детекцию проводят путем анализа клеток после пермеабилизации и окрашивания методом post-hoc. Однако если указанные антитела предварительно инкубируют в присутствии и в отсутствии лизатов мозга Р301S, ни одно из этих антител не обнаруживается в клетках после окрашивания вторичным антителом против мышиных иммуноглобулинов (фиг.34). Хотя могут существовать и другие режимы ингибирования, полученные результаты согласуются с механизмом блокирования, основанным на поглощении клетками агрегатов тау.
Пример 11. Интрацеребровентрикулярная инфузия антител против тау
В мышиных колониях у мышей Р301S сначала развивают внутриклеточную тау-патологию, начиная с 5-месячного возраста. Для тестирования эффективности 3 антител путем хронического интрацеребровентрикулярного (ICV) введения, катетер хирургически имплантируют в левый боковой желудочек каждой мыши в возрасте 6 месяцев и осуществляют непрерывную инфузию антител против тау в течение 3 месяцев посредством подкожного осмотического мини-насоса Alzet (фиг.35A). В качестве отрицательных контролей используют антитело HJ3.4 против Αβ и забуференный фосфатом физиологический раствор (PBS). Через 6 недель каждый насос заменяют насосом, заполненным свежим раствором антитела или PBS. Во время вскрытия мозга размещение катетера в левом боковом желудочке каждой мыши подтверждают окрашиванием крезиловым фиолетовым (фиг.35B). В анализах используют только мышей с правильно размещенными катетерами. Для проверки стабильности антител in vivo через 6 недель (фиг.35A), остаточное содержимое насоса собирают после его удаления из животного, и антитела анализируют методом SDS-PAGE с окрашиванием Кумасси голубым. Легкие и тяжелые цепи являются неизменными, фрагментация отсутствует и, согласно результатам вестерн-блоттинга, антитела сохраняют тау-связывающую активность (данные не показаны). Для определения концентрации антител против тау в CSF и сыворотки во время инфузии, в течение 48 часов вводят биотинилированный HJ8.5 (HJ8.5B) (~7,2 мкг/сутки) (фиг.35А). Концентрация свободного HJ8.5B составляет 7,3 мкг/мл в CSF и 6,2 мкг/мл в сыворотке, свидетельствуя о значительном выведении антитела из ЦНС на периферию (таблица 6). HJ8.5B, связанный с человеческим тау, также детектируют как в CSF, так и в сыворотке, хотя его концентрация ниже, чем концентрация свободного антитела (таблица 6).
Уровни биотинилированного антитела HJ8.5 в свободном состоянии (не связанном с тау) и антитела HJ8.5, связанного с тау, в сыворотке и цереброспинальной жидкости (CSF) через 48 часов после в/б или ICV введения
Пример 12. Обработка антителом против тау уменьшает количество аномально фосфорилированного тау
Для определения степени тау-патологии у мышей Р301S после 3 месяцев лечения, проводят несколько окрашиваний на тау-патологию. Сначала анализируют срезы мозга путем иммунного окрашивания антителом АТ8 против фосфоридированного тау (фиг.36). AT8 связывает фосфорилированные остатки Ser202 и Thr205 как мышиного, так и человеческого тау (фиг.36). У мышей, обработанных PBS и HJ3.4, AT8 интенсивно окрашивает тела нейронных клеток и нейропиля в нескольких участках головного мозга, в частности, в пириформной коре, энторинальной коре, мозжечковой миндалине и гиппокампе (фиг.36A и 36B). Обработка HJ8.5 значительно уменьшает окрашивание AT8 (фиг.36С), особенно, в нейропиле. HJ9.3 и HJ9.4 также уменьшают окрашивание AT8, но с немного меньшей интенсивностью (фиг.36D и 36E). Количественный анализ окрашивания AT8 в пириформной коре (фиг.37 А), энторинальной коре (фиг.37В) и мозжечковой миндалине (фиг.37C) демонстрирует сильное, но не постоянное уменьшение фосфорилированного тау у всех мышей, обработанных антителом против тау. Антитело HJ8.5 заметно уменьшает окрашивание AT8 в пириформной коре, энторинальной коре и мозжечковой миндалине. HJ9.3 оказывает несколько сниженные эффекты по сравнению с HJ8.5, и HJ9.4 оказывает значительные эффекты в энторинальной коре и мозжечковой миндалине, но не в пириформной коре (фиг.37). В гиппокампе наблюдается гораздо более изменчивое окрашивание AT8 по сравнению с другими участками мозга, преимущественно в телах клеток, и, следовательно, обработка данным антителом не приводит к статистически значимому различию между экспериментальной и контрольной группами (фиг.37D). Поскольку было описано, что самцы мышей Р301S более склонны к тау-патологии, чем самки, также анализируют влияние обработки на оба пола (фиг.38). В дополнение к эффекту лечения отмечают, что у самцов мышей во всех участках мозга наблюдается значительно больше окрашивания AT8 (таблица 7). Тем не менее, влияние антител все же является весьма значительным и практически идентичным после корректировки по полу (таблица 8). В экспериментальных группах, сравниваемых с контрольными, также проводят отдельное сравнение между самцами и самками, причем эффект антитела HJ8.5 остается наиболее значительным (фиг.38A и 38B).
p-Значения для результатов анализа обработки/анализа влияния пола
Пример 13. Корреляция нескольких методов окрашивания
Для тестирования осаждения тау-амилоида, срезы головного мозга окрашивают тиофлавином S (ThioS) (фиг.39). Полуколичественный анализ окрашивания ThioS проводят со скрытием вида образца от оценивающего результат специалиста, который дает оценку от 1 (отсутствие окрашивания) до 5 (максимальное окрашивание) всем контрольным мышам и мышам, получавшим антитело против тау. Согласно результатам полуколичественного анализа, введение HJ8.5 значительно уменьшает окрашивание ThioS по сравнению с PBS и HJ3.4 (фиг.39А и 39B). Мышей, получавших PBS, HJ8.5 и HJ9.3 (n=6 в каждой группе), также подвергают окрашиванию моноклональным антителом PHF1, которое распознает фосфорилированные остатки тау Ser396 и Ser404. Окрашивания AT8 и PHF1 достоверно коррелируют (г=0,630, р=0,005) (фиг.40А), демонстрируя, что 2 антитела против фосфорилированного тау, направленные против разных эпитопов тау, дают сходные результаты.
Многие нейродегенеративные заболевания, в том числе таупатии, характеризуются активацией микроглии в участках мозга, окружающих участки агрегации белков и повреждения клеток. Активацию микроглии анализируют в экспериментальных группах с использованием антитела против CD68 (фиг.41). Введение HJ8.5 и HJ9.3 уменьшает активацию микроглии в пириформной коре, энторинальной коре и миндалине по сравнению с контрольной группой (фиг.41А-41D). HJ9.4 оказывает слабый эффект в пириформной коре по сравнению с HJ8.5 и HJ9.3 (фиг.41C-41E), что соответствует результатам окрашивания AT8 (фиг.37 A). Активация микроглии тесно коррелирует с окрашиванием AT8 во всех образцах (r=0,511, р=0,0038) (фиг.40В).
Пример 14. Антитела против тау уменьшают количество нерастворимого в детергенте тау и затравочную активность
Для определения уровня растворимого и нерастворимого тау в коре головного мозга, последовательно проводят биохимическую экстракцию RAB (водный буфер), радиоиммунопреципитационный анализ (RIPA) (детергент-содержащий буфер) и обработку 70% муравьиной кислотой (FA) для солюбилизации конечного осадка. Общий тау количественно определяют методом ELISA, используя антитело HJ8.7 против тау, которое распознает как человеческий, так и мышиный тау с одинаковыми значениями KD (0,34 пМ). Вероятность того, что используемые для обработки антитела будут препятствовать проведению ELISA, исключают путем добавления указанных антител к образцам положительного контроля до анализа, что показывает отсутствие помех (данные не показаны). Анализируют всех мышей, которых оценивали с использованием патологического анализа, приведенного на фиг.37. Уровни общего тау во фракциях, растворимых в RAB (фиг.42А) или RIPA (фиг.42В), были похожи во всех группах. Нерастворимые в детергенте/растворимые в 70% FA фракции анализируют путем нейтрализации образцов перед проведением ELISA и вестерн-блоттинга. В результате анализа каждого участвующего в исследовании животного было обнаружено, что HJ8.5 и HJ9.3 уменьшают количество нерастворимого в детергенте тау более чем на 50% по сравнению с контрольной группой (фиг.42C). Анализ типичных образцов (N=4 в каждой группе) методом вестерн-блоттинга демонстрирует уменьшение уровня нерастворимого тау у мышей, получавших HJ8.5 и HJ9.3 (фиг.40C). Уровень нерастворимого тау в группах, получавших HJ9.4, не отличается от уровня, наблюдающегося в группах, получавших PBS или HJ3.4. Человеческий и мышиный тау также специфически анализируют в детергент-нерастворимых/70% FA-растворимых фракциях, полученных от групп, содержащих по 6 мышей в каждой, с получением средних значений окрашивания AT8, отражающих средние значения результатов, приведенные на фиг.37. Во фракции, растворимой в 70% FA, присутствует значительно больше человеческого тау, чем мышиного тау, причем в данной фракции антитела значительно уменьшают уровень человеческого, но не мышиного тау (фиг.42D и 42E). В этих же образцах уровни иммунореактивного сигнала AT8 определяют методом ELISA. Сигнал AT8 ниже в образцах, обработанных антителом (фиг.42F), подобно эффекту, наблюдающемуся для общего тау в данной фракции.
Было выдвинуто предположение, что уменьшение агрегации тау в мозге может коррелировать со снижением затравочной активности. Таким образом, проводят клеточный биосенсорный анализ, позволяющий тестировать затравочную активность мозга Р301S в RAB-растворимых фракциях коры головного мозга, полученных от разных экспериментальных групп. Полученные ранее авторами настоящего изобретения результаты анализа тау в ISF мышей Р301S позволяют предположить присутствие внеклеточных агрегатов тау в равновесии как с биохимически растворимыми, так и нерастворимыми пулами тау (Yamada et al., 2011; J Neurosci 31, 13110-13117). Сначала анализируют внутриклеточную агрегацию RD(ΔK)-CFP/YFP после обработки клеток лизатами, полученными от мышей, получавших PBS или HJ3.4. Лизаты указанных групп сильно индуцируют сигнал FRET (фиг.43А). Заметно пониженную затравочную активность наблюдают в лизатах кортикальной ткани мышей, получавших HJ8.5 и HJ9.3 (фиг.43А). Это не является следствием присутствия остаточного антитела в лизатах головного мозга, поскольку иммунопреципитация лизатов головного мозга с последующим элюированием затравочной активности из комплексов антитело/гранула, дает такие же результаты (фиг.43В). Таким образом, HJ8.5 и HJ9.3 уменьшают затравочную активность в мозге трансгенных мыши Р301S тау. HJ9.4 не вызывает значительного снижения затравочной активности (фиг.43А). Затравочная активность тесно коррелирует с количеством нерастворимого в детергенте/растворимого в муравьиной кислоте тау, обнаруженном с помощью ELISA (по Пирсону r=0,529, р=0,0001) (фиг.43C), но не коррелирует с общим тау в RAB-фракциях (фиг.43D). Было выдвинуто предположение, что затравочная активность обусловлена агрегатами тау, присутствующими во фракции, растворимой в RAB. Для проверки данного предположения, проводят полуденатурирующий электрофорез в агарозном геле, содержащем детергент (SDD-AGE), с последующим вестерн-блоттингом. Помимо мономеров тау у 3-месячных мышей Р301S присутствуют частицы тау с более высокой молекулярной массой, число который увеличивается у мышей Р301S в возрасте 9 месяцев (фиг.43E). Компонент указанных частиц с более высокой молекулярной массой, по всей вероятности, вносит вклад в затравочную активность, детектируемую методом FRET, которая может находиться в равновесии с tau, присутствующим во фракции, нерастворимой в детергенте/растворимой в муравьиной кислоте.
Пример 15. Антитела против тау избавляют от дефицита ситуативного страха
Проводят исследования, в которых трансгенных мышей Р301S тау в возрасте 9 месяцев, получавших контрольное антитело и антитело против тау, сравнивают по разным параметрам поведения. Группы не различаются по двигательной активности, показателям исследовательской деятельности или сенсомоторной функции (фиг.44). Способность антитела против тау уменьшать когнитивный дефицит у мышей Р301S анализируют путем оценки поведения мышей на процедуре генерирования условно-рефлекторного страха. В 1 день все четыре экспериментальные группы мышей демонстрируют сходные фоновые уровни замирания в течение первых двух минут в тренировочной камере. Это подтверждают с помощью теста rmANOVA, который не позволяет выявить какие-либо значительные общие основные эффекты или взаимодействия, обусловленные лечением (фиг.45А). Кроме того, все четыре группы демонстрируют сходные уровни замирания во время действия пар условный раздражитель-безусловный раздражитель (CS-US) звук-шок (T/S) (фиг.45А). Общее отсутствие различий в уровне замирания среди групп при применении трех пар T/S подтверждает незначительное влияние лечения и незначительное влияние генотипа при минутных взаимодействиях.
В отличие от отсутствия различий между группами во время тестирования на 1 день, наблюдают надежные различия в уровнях замирания при тестировании ситуативного страха (форма ассоциативного обучения), проводимом на 2-й день, среди двух групп, включающих мышей, получавших антитело против тау, и контрольных мышей, получавших PBS+HJ3.4 (фиг.45B). Последующие плановые сравнения демонстрируют, что у мышей, получавших HJ8.5, наблюдается гораздо более высокий уровень замирания, усредненный для сеанса 8-минутного тестирования (фиг.45C), чем у мышей контрольной группы, получавших PBS+HJ3.4, [F(1,45)=8,30, р=0,006], аналогичный эффект, но в меньшей степени, наблюдается у мышей, получавших HJ9.4 [F(1,45)=5,60, р=0,022]. Таким образом, HJ8.5 оказывает более сильное влияние в целом на сохранение ассоциативного обучения.
Обсуждение результатов, полученных в примерах 9-15
В соответствии с одной моделью патогенеза таупатий агрегаты, образовавшиеся в клетке, покидают ее или высвобождаются в межклеточное пространство, индуцируя агрегацию в соседних клетках или в клетках, связанных с указанной клеткой. Было отмечено, что введение терапевтических антител, которые специфически блокируют затравочную активность тау в лизатах головного мозга, может приводить к интенсивным ответам in vivo, по меньшей мере, таким же интенсивным, если не более интенсивным, как и описанные ранее ответы на активную или пассивную вакцинацию тау. Эксперименты начинают с клеточного биосенсорного анализа, чувствительного к присутствию внеклеточных агрегатов тау. Обнаружено, что лизаты мозга трансгенных мышей Р301S содержат затравочную активность, которая может вызвать дальнейшую внутриклеточную агрегацию. После скрининга ряда антител против тау выбирают три антитела с разной способностью блокировать затравочную активность тау. Данные антитела вводят ICV в течение трех месяцев мышам Р301S с таупатией, начиная с момента инициации патологии (6 месяцев). Введение антител приводит к появлению заметной концентрации антител как в CSF, так и в сыворотке, в соответствии с предыдущими описаниями оттока антител из ЦНС к периферии. Обработка HJ8.5, наиболее активным антителом, in vitro, значительно уменьшает тау-патологию, сильно снижая ее в нейропиле. Этот эффект был обнаружен с применением нескольких независимых окрашиваний, биохимических анализов нерастворимого тау и анализа затравочной активности остаточного тау, присутствующего в лизатах мозга. Кроме того, указанная обработка улучшает один из поведенческих дефицитов, обнаруженных у данной модели. Все антитела блокируют поглощение агрегатов тау клетками, но ни одно из них не обнаружено в данных анализах внутри клеток в присутствии или в отсутствии внеклеточных агрегатов. Эффективность указанных антител позволяет предположить, что внеклеточный тау играет важную роль в патогенезе нейропатологии, который раньше считался независимым от клеток. Кроме того, данная работа является продолжением прежних исследований авторов изобретения, которые позволяют предположить, что поток агрегатов может происходить в условиях внутриклеточной патологии, увеличивая возможность применения терапий, которые могут способствовать выведению агрегатов путем направленности на внеклеточные частицы. И наконец, данная работа имеет важное значение для разработки терапевтических антител, позволяя предположить, что направленность на затравочную активность, в частности, может привести к разработке наиболее эффективных средств.
Механизм терапии антителами. Несколько разработанных ранее способов активной и пассивной периферической иммунотерапии против тау также уменьшают тау-патологию и улучшают поведенческие нарушения, но основной критерий выбора антител включал либо фосфо-эпитоп, реакционноспособность в отношении нейрофибриллярных клубков, либо не был установлен. В одном исследовании по тау-иммунизации, проводимом путем вакцинации мышей полноразмерным тау, индуцируют патологию у мышей дикого типа. Однако последующие способы активной иммунизации трансгенных по тау моделей фосфо-содержащими пептидами тау уменьшают тау-патологию и вызывают улучшение поведенческих нарушений. В исследовании с применением пассивной иммунизации трансгенным по тау мышам JNPL3 в возрасте 2-3 месяцев внутрибрюшинно вводят антитела PHF1 до появления таупатии. PHF-1 направлено против патологической формы аномально фосфорилированного тау. Лечение уменьшает тау-патологию и улучшает поведение. Однако, хотя оно уменьшает количество нерастворимого фосфорилированного тау, общее содержание нерастворимого тау не изменяется. В другом исследовании с применением пассивной иммунизации мышам JNPL3 и Р301S (в возрасте 2-3 месяцев, до появления таупатии) периферически вводят антитело PHF1 или МС1, направленное на связанный с агрегатами эпитоп. Оба способа лечения улучшают тау-патологию и задерживают появление двигательной дисфункции. В указанных предыдущих исследованиях механизм действия антител не был выяснен, и ни одно антитело не было протестировано явным образом. В действительности, некоторые исследователи предлагают внутриклеточный механизм. Кроме того, ни в одном исследовании не определяют величину уменьшения тау-патологии, описанной в приведенных в данном описании примерах, с указанием, что антитела вводят в ЦНС, тогда как в других исследованиях используют периферическую инфузию; и используют другие животные модели.
Настоящее исследование было в явной форме разработано для проверки предположения, что внеклеточные центры агрегации тау являются ключевым компонентом патогенеза. Исследование начинают с процесса селекции, выбирая антитела, способные блокировать образование центров агрегации тау in vitro, специальных тестирующих средств с диапазоном ожидаемой активности. Все антитела, тестированные in vivo, эффективно блокируют поглощение агрегатов и затравочную активность, что составляет основу их экспериментально определенной активности. Кроме того, корреляция аффинности антител, эпитопа, изотипа, гликозилирования и способности к связыванию фосфорилированных форм тау может иметь важное значение для анализа в будущих исследованиях. Это также первое исследование, описывающее эффекты инфузии антитела против тау непосредственно в ЦНС. Несмотря на то, что используемые антитела направлены против разных эпитопов тау и ни одно не направлено против фосфо-тау, 2 из 3 значительно снижают содержание аномального тау по данным иммуногистохимического и биохимического анализов, и два существенно улучшают память, один в большей степени, чем другой. Влияние на тау-патологию также хорошо коррелирует с уменьшением внутренней затравочной активности.
HJ8.5 и HJ9.3 значительно уменьшают патологические центры агрегации тау in vivo. Значительное снижение тау-патологии может происходить путем предотвращения индукции агрегации тау в соседних клетках. Хотя HJ9.4 не уменьшает патологию так эффективно, оно снижает тау-патологию в мозжечковой миндалине. Вариации среди антител по эффективности воздействия на разные участки головного мозга могут быть обусловлены образованием участок-специфичных агрегированных конформеров, значения аффинности связывания к которым у разных антител могут различаться.
Метаболическая судьба внеклеточных агрегатов тау после связывания с антителами против тау in vivo пока не ясна. Через 3 месяца после введения антитела наблюдается уменьшение активации микроглии, предположительно вследствие уменьшения тау-связанной патологии и нейродегенерации. Однако это может быть связано с более эффективным выведением внеклеточных агрегатов и связанным с ним уменьшением активации микроглии. Также было показано, что пассивная иммунизация антителами против Αβ в течение нескольких месяцев уменьшает микроглиоз. Механизм, посредством которого комплексы антитело/тау выводятся in vivo, и механизм, посредством которого они уменьшают тау-патологию, еще предстоит окончательно уточнить. Было высказано предположение, что иммунизация антителом против синуклеина способствует выведению агрегатов α-синуклеина путем стимуляции лизосомальной деградации. Проведенное в последнее время исследование с применением антител против α-синуклеина демонстрирует, что клиренс антител против α-синуклеина в основном осуществляется через микроглию, предположительно, посредством Fc-рецепторов. Нейроны экспрессируют рецепторы Fcγ и могут интернализовывать комплексы IgG с антигенами посредством высокоаффинного рецептора FcγRI. Интернализованные антитела против тау могут также связываться с тау в эндосомах и, в конечном итоге, индуцировать клиренс внутриклеточных агрегатов тау посредством эндосомально/аутофагально-лизосомальной системы. Хотя антитела против тау, использованные в описанном в данном описании исследовании, могут связывать внеклеточные скопления тау, доказательства значительной локализации в клетках отсутствуют. Однако это не исключает возможности того, что клетки поглощают комплексы антитело/тау in vivo, влияя на выведение агрегатов тау, а также на воспаление. Например, недавно было показано, что антитела в комплексе с вирусами могут связываться с цитозольным рецептором IgG TRIM21, направляющим комплексы антитело/вирус в протеасомы. Кроме того, было показано, что антитела, связанные с TRIM21, активируют иммунный сигнальный путь. Хотя данные по взаимодействию TRIM21 с комплексами антитело/неинфицирующий антиген до сих пор отсутствуют, интересно определить, имеет ли такой механизм отношение к антителам против тау. Важно отметить, что существуют свидетельства того, что на модели таупатии Р301S врожденная иммунная система активируется до заметного развития тау-патологии, и что ранняя иммуносупрессия уменьшает тау-патологию. Возможно, антитела улавливают агрегаты тау, индуцированные воспалением, уменьшая последующее индуцированное агрегатами воспаление и прогрессирование заболевания.
Внеклеточный тау и распространение тау-патологии. В представленной в данном описании работе косвенным образом определяют роль внеклеточного тау в патогенезе. Теперь совершенно ясно, что внеклеточные агрегаты тау могут вызывать образование фибрилл нативного тау внутри клеток, независимо от того, является ли их источник рекомбинантным белком или белком тау, выделенным из клеток млекопитающих. На основе предыдущей работы, проведенной авторами изобретения, в которой антитело HJ9.3, добавленное к клеточной среде, блокирует интернационализацию и иммунопреципитацию свободных фибрилл (Kfoury et al., 2012; J Biol Chem 287, 19440-19451), была выдвинута гипотеза о том, что свободные агрегаты тау (т.е. не заключенные в мембрану) могут выполнять роль посредников трансклеточного распространения.
На животных моделях, агрегаты тау, по-видимому, могут распространяется из одного участка в другой (например, из энторинальной коры в нижележащие нейроны зубчатой извилины и гиппокампа). Авторы настоящего изобретения обнаружили, что мономерный тау постоянно высвобождается in vivo в интерстициальную жидкость мозга даже при непатологических состояниях (Yamada et al., 2011; J Neurosci 31, 13110-13117). Авторы также обнаружили, что экзогенные агрегаты могут уменьшать уровень растворимого тау в ISF, свидетельствуя о том, что явления образования агрегатов и/или секвестрации могут происходить в этом пространстве (Yamada et al., 2011; J Neurosci 31, 13110-13117). Взятые вместе, многочисленные доказательства подтверждают концепцию, что внеклеточные агрегаты тау могут поглощаться соседними клетками, связанными клетками, или, возможно, теми же клетками, в которых они образовались, тем самым увеличивая уровень неправильной укладки белка. Данные доказательства позволяют сделать четкий прогноз: терапия, включающая улавливание внеклеточной затравочной активности должна приводить к улучшению при заболевании. Результаты, описанные в приведенных в данном описании примерах, соответствуют данной концепции.
Роль потока tau в патогенезе. Нельзя предсказать заранее, что на мышиной модели, такой как Р301S, у которой экспрессия мутанта тау находится под управлением промотора прионов практически во всех нейронах, может наблюдаться улучшение в результате лечения антителом, которое блокирует трансклеточное распространение агрегации. Теоретически патология может развиваться независимо во всех нейронах, которые экспрессируют указанный склонный к агрегации белок. Однако работа, проведенная ранее авторами изобретения на тканевой культуре, позволяет предположить, что поток агрегатов тау может играть некую роль, поскольку HJ9.3, добавленное к клеточной среде, повышает равновесный уровень агрегатов с течением времени. Хотя модель потока агрегатов требует дальнейшей проверки, представленные в данном описании результаты находятся в соответствии с этой концепцией, поскольку лечение антителом значительно уменьшает внутриклеточную тау-патологию. Было выдвинуто предположение, что антитела, которые блокируют поглощение тау, могут создавать "акцептор частиц" в межклеточном пространстве, способствуя выведению по другому механизму, возможно, с участием микроглии.
Терапевтические антитела и направленность на затравочную активность. Фармацевтическая промышленность уделяет все больше внимания разработке терапевтических антител, направленных на склонные к агрегации белки, которые накапливаются в клетках. Основным критерием является способность антител связываться с эпитопами, которые заведомо накапливаются в больном мозге. Данный подход необязательно может привести к получению антител с оптимальной активностью in vivo. Приведенные в данном описании примеры поддерживают новую модель развития терапевтического антитела, которая придает особое значение эффективности при блокировании затравочной активности, присутствующей в головном мозге. С помощью данного подхода были идентифицированы антитела с более высокой кажущейся эффективностью, чем описанная ранее. В дополнение к прионовой гипотезе также предполагают, что различные "штаммы" агрегатов тау могут преобладать у пациентов с различными типами таупатии, и они могут иметь уникальную чувствительность к разным антителам. В любом случае, применение чувствительных анализов эффективности антител in vitro, описанных в данном описании, может обеспечить более эффективное развитие и оптимизацию терапии на основе антител.
Сильный защитный эффект антител против тау, особенно антитела HJ8.5, позволяет предположить, что такой подход можно рассматривать как стратегию лечения таупатий у человека. Хотя возможно длительное ICV введение антител, в дальнейших исследованиях может иметь важное значение определение PK/PD ответа в случае периферического введения указанных антител при применении их как в профилактическом, так и в лечебном режиме. Кроме того, анализ затравочной активности тау можно использовать для мониторинга специфического взаимодействия с антителами.
Экспериментальные методы, использованные в примерах 9-15
Антитела. Мышиные моноклональные антитела HJ9.3 и HJ9.4 получают путем иммунизации мышей с нокаутом тау (The Jackson laboratory) против мышиного тау, и моноклональные антитела HJ8.5 и HJ8.7 получают путем иммунизации мышей с нокаутом тау против человеческого тау. Моноклональные антитела HJ9.3, HJ9.4 и HJ8.7 распознают как мышиный, так и человеческий тау. А моноклональное антитело HJ8.5 связывается только с человеческим тау (эпитоп включает остатки 25-30 [эталонная последовательность NCBI: NP_005901]). Антитело HJ9.3 распознает участок RD тау (эпитоп включает остатки 306-320). Антитело HJ9.4 распознает N-концевой участок тау (эпитоп включает остатки 7-13). В качестве контрольного антитела используют мышиное моноклональное антитело HJ3.4, которое распознает N-концевой участок последовательности человеческого Αβ (эпитоп включает остатки 1-16). Моноклональные антитела HJ8.5, 9,3 и 9,4 относятся к изотипу IgG2b. Кроличье поликлональное антитело против тау (ab64193, эпитоп расположен в области домена повторов) получают от Abeam. Мышиное моноклональное антитело BT-2, распознающее человеческий и мышиный тау (эпитоп включает остатки 194-198), получают от Pierce. Крысиное моноклональное антитело CD68 против мышиных иммуноглобулинов получают от ABD Serotec. Биотинилированное антитело AT8 получают от Thermo Scientific.
Поверхностный плазмонный резонанс. Эксперименты с применением поверхностного плазмонного резонанса проводят на приборе для измерения поверхностного плазмонного резонанса BIAcore 2000 (GE Healthcare, BIAcore). Сенсорный чип Biacore CM-5 активируют с использованием EDC (1-этил-3-(3-диметиламинопропил)карбодиимид) и NHS (N-гидроксисукцинимид) в соотношении 1:1 в течение 7 минут. Поверхность сенсорного чипа насыщают путем иммобилизации 5 мкг/мл рекомбинантного человеческого или мышиного белка тау или фибрилл человеческого тау в 10 мМ растворе ацетата натрия, рН 3,5, со скоростью потока 5 мкл/мин. Оставшиеся несвязанными участки блокируют 1М раствором этаноламина, рН 8,5. Для сравнения одну проточную кювету обрабатывают NHS и EDS, после чего блокируют 1М этаноламином. Затем все антитела против тау вводят в различных концентрациях (0,11, 0,23, 0,46, 0,9, 1,8, 3,7, 7,5 мкг/мл) в фильтрованном, дегазированном 0,01М буфере Hepes, содержащем 0,15М NaCl, 0,005% поверхностно-активного вещества P20, рН 7,4, при скорости потока 10 мкл/мин. Все образцы анализируют в двух повторах. После каждого цикла анализа с использованием одной концентрации антитела поверхность чипа полностью регенерируют с помощью 10 мМ глицина, рН 1,7, чтобы удалить связанное антитело против tau, не удаляя тау, иммобилизованный на чипе. Анализ данных выполняют с помощью программного обеспечения BIAevaluation (GE Healthcare, BIAcore).
Фибрилизация тау. 8 мкΜ раствор рекомбинантного человеческого полноразмерного тау предварительно инкубируют с 2 мМ дитиотреитолом в течение 45 мин при комнатной температуре, затем добавляют 10 мМ HEPES, 100 мМ NaCl и 8 мкΜ гепарин в общем объеме 200 мкл и затем инкубируют в течение 7 дней при 37°С с получением фибрилл. После образования фибрилл, оставшиеся в образце мономеры tau отделяют с использованием центрифужных фильтров 100 кДа Microcon в соответствии с инструкциями производителя (Millipore).
ВБ и ICV введение биотинилированного антитела HJ8.5. Мышиное моноклональное антитело HJ8.5 биотинилируют в соответствии с инструкциями производителя (набор Sulfo-NHS-LC-Biotin, Pierce). Биотинилированное HJ8.5 (HJ8.5B) вводят путем внутрибрюшинной инъекции (ВБ) в дозе 50 мг/кг мышам Р301S в возрасте 5-6 месяцев (n=3). Через 48 часов мышей умерщвляют. Сыворотку и CSF собирают и хранят при -80°C до использования. HJ8.5B также вводят путем интрацеребровентрикулярной (ICV) инъекции путем имплантации осмотических насосов в левый боковой желудочек 5-6 месячных мышей Р301S (n=3). Данное антитело вводят непрерывно путем инфузии в течение 48 ч. Через 48 часов мышей умерщвляют. Сыворотку и CSF собирают и хранят при -80°С до использования.
Методика интрацеребровентрикулярной (ICV) инъекции. ICV вливания осуществляют с использованием осмотических насосов ALZET 2006 модели (DURECT). На момент операции возраст мышей составляет 6 месяцев. Перед операцией L-образную канюлю прикрепляют к трубке (длиной 3 см), которую затем присоединяют к насосу Alzet, содержащему антитело или среду (забуференный фосфатом физиологический раствор - PBS, рН 7,4). Указанное собранное устройство предварительно инкубируют в PBS в течение 60 часов при 37°C, для активации насоса перед его размещением в боковом желудочке. Собранное устройство хирургически имплантируют с использованием стереотаксического аппарата (David Kopf Instruments) в левый боковой желудочек каждой мыши на расстоянии 0,4 мм от брегмы в переднезаднем направлении, 1,0 мм от средней линии в латеральном направлении и 2,5 мм в дорсовентральном направлении по отношению к поверхности мозга при анестезии изофлураном. Осмотические насосы ALZET помещают под кожу, делая подкожный карман с помощью изогнутых тупых ножниц. Каждую имплантированную канюлю укрепляют зубным цементом и небольшими анкерными винтиками из нержавеющей стали. После высушивания цемента кожу зашивают. Происходит постоянная инфузия антитела (2 мг/мл) или PBS из насоса в левый боковой желудочек мозга. Указанные осмотические насосы содержат максимум 200 мкл жидкости, которая откачивается со скоростью потока 3,6 мкл/день, обеспечивая вливание 7,2 мкг антитела в день. После 6 недель инфузии осмотические насосы у всех мышей заменяют. Раствор, оставшийся в насосе Alzet, собирают после его удаления из каждой мыши и хранят при -80°C. В возрасте 9 месяцев всех мышей умерщвляют. Всех мышей после хирургической имплантации держат в отдельных клетках.
Гистология. После 12 недель лечения мышей Р301S анестезируют внутрибрюшинно пентобарбиталом 200 мг/кг) и затем проводят перфузию 3 ед/мл гепарина в холодном PBS, забуференном фосфатом Дульбекко. Мозг удаляют и разрезают на два полушария. Левую сторону мозга фиксируют в течение 24 часов 4% раствором параформальдегида, затем переносят в 30% раствор сахарозы в PBS и хранят при 4°C до замораживания в порошкообразном сухом льде, затем хранят при -80°C. Из половинок мозга получают коронарные срезы 50 мкм с помощью замораживающего санного микротома, которые хранят в 24-луночных планшетах в криопротекторном растворе (0,2М забуференный фосфатом солевой раствор, 30% сахарозы, 30% этиленгликоля) при -20°С до использования. Гиппокамп и кору головного мозга выделяют из свежеперфузированного правого полушария каждого мозга и используют для биохимического анализа. Все выделенные ткани хранят при -80°C до проведения анализа. Размещение канюли в левом боковом желудочке подтверждают путем размещения участков головного мозга на расстоянии 300 мкм друг от друга и окрашивания крезиловым фиолетовым, как описано ранее (Holtzman et al., 1996; Ann Neurol 39, 114-122). Окрашенные ткани сканируют с помощью цифровой системы для анализа патологий NanoZoomer (Hamamatsu Photonics).
Клеточные культуры/анализ затравочной активности: лизаты мозга P301S и обработка антителами. Клетки HEK293 культивируют в модифицированной по Дульбекко среде Игла (DMEM), содержащей 10% фетальной бычьей сыворотки, 100 мкг/мл пенициллина и 100 мкг/мл стрептомицина. Культуры поддерживают во влажной атмосфере 5% СO2 при 37°С. Для проведения временной трансфекции клетки, HEK293 высевают в 12-луночный планшет с плотностью 250000 клеток/лунка в среде Optimem и трансфицируют с использованием реагента Lipofectamine 2000 и 600 нг соответствующих конструкций ДНК (Invitrogen) в соответствии с рекомендациями производителя. Совместно трансфицированные клетки получают комбинацию 150 нг конструкции RD(ΔK280)-CFP и 450 нг конструкции RD(ΔK280)-YFP. Через 15 ч клетки собирают путем обработки 0,05% раствором трипсина в течение 3 минут при 37°C, затем снова высевают в 96-луночный планшет в четырех повторах и инкубируют в течение 15 часов. Затем добавляют лизаты мозга Р301S [полученные в 1×TBS, содержащем ингибиторы протеазы (Roche) и фосфатазы (Roche)] в различных концентрациях (0,125 мкг/мл, 0,25 мкг/мл, 0,5 мкг/мл, 1 мкг/мл и 2 мкг/мл), которые предварительно инкубируют со всеми моноклональными антителами против тау (HJ8.5, 9,3 и 9,4) или антителом HJ3.4 (моноклональное антитело против Αβ), и инкубируют в течение 16 часов при 4°C при вращении. Для определения затравочную активность у мышей Р301S, получавших в течение 3 месяцев разные антитела, к клеткам также добавляют RAB-растворимые фракции всех мышей в различных концентрациях. Клетки культивируют еще 24 часа, затем фиксируют 4% параформальдегидом и анализируют методом FRET.
Иммунопреципитация. RAB-растворимые фракции мышей, получавших PBS или антитело, инкубируют в присутствии мышиных моноклональных антител против тау HJ9.3 и HJ8.5, конъюгированных с агарозными гранулами, содержащими белок G (набор Pierce Crosslink Immunoprecipitation, содержащий рекомендации), при 4°C и при вращении с переворачиванием в течение 24 часов. Кроме того, RAB-растворимые фракции мышей, получавших антитело, инкубируют в присутствии неконъюгированных агарозных гранул, содержащих белок G, при 4°C и при вращении с переворачиванием в течение 24 часов. Через 18 часов к образцам добавляют 500 мкл буфера для связывания/промывания (Pierce) и центрифугируют при 2000×g в течение 3 минут. Супернатант отбрасывают, и стадию промывания повторяют три раза. Затем белки, связанные с гранулами, элюируют, используя буфер для элюирования с низким значением рН (25 мкл) и инкубацию при комнатной температуре в течение 5 минут. Затем образцы центрифугируют при 2000×g в течение трех минут и собирают супернатант. Данную стадию элюирования повторяют один раз, с получением общего объема элюатов 50 мкл. Тау-иммунопреципитаты (IP), содержащие агрегаты тау, снова добавляют в эквивалентных количествах к клеткам, совместно трансфицированным RD(ΔK)-CFP/YFP, для проведения начальных экспериментов с применением лизатов головного мозга и последующего анализа затравочной активности.
Экстракция ткани мозга. Кору каждого мозга гомогенизируют в 30 мкл/мг буфера RAB [100 мМ MES, 1 мМ EDTA, 0,5 мМ MgSO4, 750 мМ NaCl, 20 мМ NaF, 1 мМ Na3VO4, содержащий ингибитор протеазы (Roche) и ингибитор фосфатазы (Roche)]. Коротко, образцы центрифугируют при 50000×g в течение 20 мин при 4°С при использовании ультрацентрифуги Optima MAX-TL (Beckman). Супернатанты собирают в виде RAB-растворимых фракций, осадки ресуспендируют в буфере RIPA [150 мМ NaCl, 50 мМ трис, 0,5% дезоксихолевая кислота, 1% тритон Х-100, 0,5% SDS-25 мМ EDTA, рН 8,0, содержащий ингибитор протеазы (Roche) и ингибитор фосфатазы (Roche)], 30 мкл/мг, и центрифугируют при 50000 г в течение 20 мин при 4°С. Супернатанты собирают в виде RIPA-растворимых фракций. Затем осадки ресуспендируют в 70% муравьиной кислоте, 10 мл/мг и центрифугируют при 50000×g в течение 20 мин при 4°С. Супернатанты собирают в виде фракций в 70% муравьиной кислоте. Все фракции хранят при -80°C до проведения анализа.
Электрофорез и иммунноблоттинг. Гель-электрофорез проводят в восстанавливающих условиях с использованием 4-12% гелей NuPAGE Bis-Tris (Invitrogen) с последующим переносом на мембрану PVDF с использованием устройства IBIot (Invitrogen). Фракции, полученные в 70% муравьиной кислоты, нейтрализуют перед загрузкой и проведением гель-электрофореза путем разбавления 1:3 смесью 10н NaOH и нейтрализующего буфера (1 моль/л трис-основания; 0,5 моль/л NaH4PO4) 1:1. В каждом цикле используют предварительно окрашенные белки-стандарты с известной молекулярной массой "SeeBlue" (Invitrogen). Мембраны блокируют 5% молоком в забуференном трисом физиологическом растворе (TBS), содержащем 0,1% твина 20. Затем мембраны промывают три раза в течение 5 минут. Кроличьи поликлональные антитела против тау (Abeam, 1:2000) используют в качестве первичных антител для детекции тау во фракциях, полученных в муравьиной кислоте. Обработанные мышиные антитела против тау, собранные до и после инфузии из осмотических насосов, также используют в качестве первичных антител. Затем мембраны инкубируют с козьим вторичным антителом против кроличьих или мышиных иммуноглобулинов (GE Healthcare, 1:2000). Окрашивание всех мембран проявляют с использованием первичного субстрата ECL (GE Healthcare). Полосы визуализируют с помощью хемилюминесцентного устройства для визуализации G:Box (Syngene).
Для определения иммунореактивности антител против тау в отношении тау из гомогенатов мозга, образцы RAB-растворимых фракций мышей Р301S в возрасте 9 месяцев, мышей Р301S в возрасте 3 месяца, мышей дикого типа в возрасте 3 месяцев и мышей с нокаутом тау в возрасте 3 месяцев подвергают анализу методом SDS-PAGE с последующим анализом методом вестерн-блоттинга. 1 мкг общего белка каждой RAB-растворимой фракции загружают на 4-12% гели NuPAGE Bis-Tris (Invitrogen) в восстанавливающих условиях с последующим переносом на нитроцеллюлозную мембрану с использованием устройства IBlot (Invitrogen). Мембраны блокируют 5% раствором молока в TBS, содержащем 0,05% твин 20 (TBST) и затем инкубируют с первичными антителами (HJ8.5, HJ9.3 и HJ9.4). HRP-конъюгированные ослиные антитела против мышиных IgG (1:2000, Santa Cruz) используют в качестве вторичных антител и окрашивание мембран проявляют с использованием Lumigen TMA6 (GE Healthcare).
Детекция свободного HJ8.5B и HJ8.5B, связанного с тау, методом ELISA. Концентрацию свободного HJ8.5B определяют в сыворотке крови и CSF мышей через 48 часов после ВБ или ICV введения. Девяносто шести-луночные планшеты для ELISA покрывают рекомбинантным человеческим белком тау, 50 нг/мл, при 4°C. Планшеты ELISA блокируют 4% BSA при 37°C в течение 1 часа. Затем планшеты промывают 5 раз и инкубируют с образцами сыворотки и CSF, разведенными буфером для образцов (0,25% BSA в PBS, 300 нМ трис, рН 7,4, с добавлением ингибиторов протеазы), при 4°С в течение ночи. На следующий день планшеты промывают 8 раз PBS, добавляют конъюгат стрептавидин-полипероксидаза хрена 40 (1:6000, Fitzgerald) и инкубируют в течение 1,5 ч в темноте при комнатной температуре. Затем планшеты промывают 8 раз PBS, развивают окраску с использованием Super Slow ELISA TMB (Sigma) и считывают при 650 нм. Для построения стандартной кривой используют различные концентрации HJ8.5B, которые анализируют в каждом планшете помимо образцов сыворотки и CSF.
Концентрацию HJ8.5B, связанного с тау, измеряют путем покрытия 96-луночных планшетов ELISA антителом HJ8.7 в концентрации 20 мкг/мл при 4°C. Планшеты ELISA блокируют 4% BSA при 37°C в течение 1 часа. Затем планшеты промывают 5 раз и инкубируют с образцами сыворотки и CSF, разведенными в буфере для образцов, при 4°С в течение ночи. На следующий день планшеты промывают 8 раз PBS и инкубируют с конъюгатом стрептавидин-поли-пероксидаза хрена 40 (1:6000, Fitzgerald) в течение 1,5 ч в темноте при комнатной температуре. Затем планшеты промывают 8 раз PBS, развивают окраску с использованием Super Slow ELISA TMB (Sigma) и считывают при 650 нм. Для построения стандартной кривой используют разные разведения очищенного HJ8.5B в комплексе с рекомбинантным тау, присутствующие в каждом планшете.
Анализ тау методом сэндвич-ELISA. Для определения уровня общего тау, 96-луночные планшеты ELISA (Costar) покрывают антителом HJ8.7 (20 мкг/мл) в карбонатном буфере, рН 9,6, и инкубируют при 4°С в течение ночи на шейкере. Планшеты ELISA промывают 5 раз PBS, содержащим средство для промывания планшетов BioTek EL×405, и блокируют 4% раствором BSA в PBS в течение 1 часа при 37°C. Затем планшеты промывают 5 раз, лунки инкубируют с RAB, RIPA или 70% FA, с помощью биохимических методов экстрагируют растворимые фракции тканей мозга, разводят их буфером для образцов (0,25% BSA в PBS, 300 нМ трис, рН 7,4, содержащий ингибиторы протеазы) и инкубируют при 4°C. Фракции, полученные в 70% FA, нейтрализуют путем разведения 1:20 1М раствором триса, рН 11, с последующим разведением буфером для образцов. На следующий день планшеты промывают 8 раз PBS, добавляют биотинилированное моноклональное мышиное антитело против тау BT-2 (0,3 мкг/мл, Pierce) в 0,5% растворе BSA в PBS и инкубируют в течение 1,5 ч при 37°C. Затем планшеты промывают 8 раз PBS с последующим добавлением конъюгата стрептавидин-поли-пероксидаза хрена 40 (1:4000) и инкубируют в течение 1,5 ч, в темноте, при комнатной температуре. Затем планшеты промывают 8 раз PBS, развивают окраску с использованием Super Slow ELISA TMB (Sigma) и считывают оптическую плотность при 650 нм на планшет-ридере BioTek Synergy 2. Каждый планшет содержит рекомбинантный человеческий тау, используемый в качестве стандарта. Каждый планшет также содержит лунки отрицательного контроля, в которых отсутствует первичное антитело. В качестве стандартов в анализах ELISA используют самые длинные изоформы рекомбинантного человеческого (hTau40, 441 а.к.) и мышиного тау (mTau40, 432 а.к.), полученные в лаборатории Eva-Maria Mandelkow.
Для определения уровня человеческого тау во фракциях 70% FA, 96-луночные планшеты ELISA покрывают моноклональным мышиным антителом Tau5 (20 мг/мл) и биотинилированным антителом HT7, специфичным к мышиному моноклональному антителу против человеческого белка тау (0,2 мкг/мл, Thermo Scientific), используемым для детекции. Для определения уровня мышиного тау во фракциях 70% FA, 96-луночные планшеты ELISA покрывают моноклональным специфичным антителом HJ9.2 против мышиного тау (20 мг/мл), и для детекции используют моноклональное антитело HJ8.7. Каждый планшет содержит рекомбинантные человеческие и мышиные белки тау, которые используют в качестве стандартов. Для определения уровней тау, фосфорилированного по положениям Ser202 и Thr205, половинные 96-луночные планшеты ELISA покрывают мышиным моноклональным антителом HJ8.7 (20 мг/мл), и биотинилированное антитело AT8 (0,2 мкг/мл, Thermo Scientific) используют в качестве детектирующего антитела.
Иммуногистохимия. Для детекции аномально фосфорилированного тау в головном мозге, анализируют три коронарных среза головного мозга толщиной 50 мкм на расстоянии 300 мкм друг от друга от каждой обработанной мыши. Срезы мозга блокируют 3% раствором молока в забуференном трисом физиологическом растворе (TBS) и 0,25% (объем/объем) тритоном-Х с последующей инкубацией с биотинилированным антителом AT8 (Thermo Scientific, 1:500) при 4°С в течение ночи, которое распознает тау, фосфорилированный по положению ser202 и thr205. Биотинилированное антитело PHF1 (1:200), которое распознает тау, аномально фосфорилированный по остаткам ser396 и ser404, также используют для определения корреляции между окрашиванием антителами AT8 и PHF1. Для корреляционных исследований мышей (n=6) выбирают случайным образом из групп, обработанных HJ8.5, HJ9.3 и PBS. Окрашенные ткани сканируют с помощью цифровой системы для анализа патологий NanoZoomer. Для определения корреляции между окрашиванием AT8 и окрашиванием активированной микроглии, срезы головного мозга мышей, выбранных из всех экспериментальных групп (n=6), блокируют 10% раствором нормальной козьей сыворотки в TBS, содержащим 0,25% (об/об) тритона X, и инкубируют с крысиным антителом против мышиного CD68 (ABD Serotec, 1:500) при 4°С в течение ночи. Затем срезы инкубируют с биотинилированным козьим антителом против крысиных IgG, адсорбирующих мышиные антитела (Vector, 1:2000). Все срезы сканируют с использованием слайдового сканера NanoZoomer (Hamamatsu Photonics). Все изображения экспортируют с помощью программы просмотра NDP и количественно анализируют с помощью программного обеспечения ImageJ (National Institutes of Health). Для окрашивания AT8 используют 3 среза головного мозга, полученных от каждой мыши, на расстоянии 300 мкм друг от друга, что соответствует примерно срезам, расположенным на расстоянии -1,4, -1,7 и -2,0 мм от брегмы в атласе мышиного мозга. Указанные срезы используют для определения процента площади, занимаемой окрашиванием аномально фосфорилированного биотинилированного антитела AT8. Во всех конвертированных изображениях единообразно устанавливают границы, для количественного определения окрашивания AT8, и среднее значение всех трех срезов используют для определения процента площади, окрашенной аномально фосфорилированным тау, для каждой мыши. Для окрашивания PHF-1 и CD68 используют два среза мозга от каждой мыши на расстоянии 300 мкм друг от друга, что соответствуют координатам -2,3 и -2,6 мм по отношению к брегме в атласе мозга мыши. Для определения окрашивания ThioS, срезы головного мозга случайно выбранных мышей из всех экспериментальных групп (n=6) окрашивают ThioS в 50% этаноле (0,25 мг/мл) в течение 3 мин с последующим промыванием 50% этанолом и дистиллированной водой. Затем срезы закрепляют, сушат и изображения анализируют с помощью микроскопии, используя Nanozoomer. Используют два среза мозга от каждой мыши, как описано, которые примыкают к срезам, использованным для окрашивания PHF-1 и CD68.
Полуденатурирующий электрофорез в агарозном геле (SDD-AGE). Для разделения частиц тау, присутствующих в разных RAB-растворимых фракциях мышей с нокаутом тау в возрасте 3 месяцев (KO), мышей дикого типа в возрасте 3 месяцев (WT), мышей Р301S в возрасте 3 месяцев и мышей Р301S в возрасте 9 месяцев, получавших PBS, используют ранее описанный метод полуденатурирующего электрофореза в агарозном геле, содержащем детергент (SDD-AGE), с незначительными изменениями. Образцы подвергают электрофорезу на горизонтальном 1,5% агарозном геле в буфере G (20 мМ трис, 200 мМ глицин), содержащем 0,2% SDS. Образцы инкубируют в буфере для образцов (60 мМ трис-HCl, рН 6,8, 0,2% SDS, 5% глицерин и 0,05% бромфеноловый синий) в течение 7 минут при комнатной температуре. После электрофореза белки переносят из гелей на пластины Immobilon-P PVDF (Millipore) при 4°C в буфере Лэммли (буфер G/0,1% SDS). Мембраны окрашивают с использованием специфических кроличьих поликлональных антител против тау (Abeam) в разведении 1:2000. Окрашивание пятен развивают с использованием системы GE ECL Plus.
Иммунофлуоресценция. Клетки HEK293 высевают при плотности 75000 клеток/лунка в 24-луночные планшеты, покрытые поли-D-лизином. Для определения, можно ли с помощью используемых антител против тау детектировать частицы тау, поглощенные клетками НЕК293, указанные клетки обрабатывают лизатами головного мозга Р301S в течение 2 часов, затем промывают 3 раза PBS, фиксируют 4% параформальдегидом в течение 15 мин при комнатной температуре и промывают еще 3 раза PBS при комнатной температуре. Клетки пермеабилизируют 0,1% тритоном Х-100 в течение 10 мин, промывают три раза PBS, затем блокируют 0,25% тритоном X-100 в PBS, содержащем 10% нормальной козьей сыворотки и 20 мг/мл BSA. Затем клетки инкубируют с вторичным антителом против мышиных иммуноглобулинов, конъюгированным с Alexa-fluor 546. Для определения, может ли антитело проникать в клетки, лизаты мозга Р301S предварительно инкубируют в присутствии и в отсутствии различных антител против тау, HJ8.5, HJ9.3 и HJ9.4, или антитела HJ3.4 против Αβ. После добавления лизатов клетки НЕК293 инкубируют в течение 2 ч, фиксируют и пермеабилизируют. Для идентификации антител используют вторичное антитело, конъюгированное с Alexa-fluor 546. Для окрашивания ядер используют 4',6'-диамидино-2-фенилиндол (DAPI; показан синим цветом). Все изображения получают с помощью конфокального микроскопа Zeiss LSM5 (Zeiss).
Статистический анализ результатов патологических и биохимических исследований. Все результаты представлены в виде среднего значения±SEM, результаты, полученные в разных условиях сравнивают с помощью однофакторного ANOVA с последующим использованием апостериорного теста Даннетта для сравнения контрольных групп с экспериментальными группами. Результаты считают статистически значимыми при p<0,05. Статистические анализы проводят с использованием GraphPad Prism 5.04 for Windows (GraphPad Software Inc.). При количественном определении окрашивания AT8 пол является важным фактором, поэтому результаты корректируют по полу с помощью программного обеспечения SAS, версия 9,2.
Статистический анализ, использующий обработку и пол в качестве факторов. Среднее значение, полученное в контрольной группе (PBS и HJ3.4), сравнивают со средним значением, полученным в каждой экспериментальной группе (среднее значение PBS+среднее значение HJ3.4)/2 сравнивают со средним значением в экспериментальной группе). Двухфакторный ANOVA используют для определения того, являются ли пол и обработка существенными факторами, где определение проводят с использованием PROC GLM в SAS версии 9.2, и значения р приведены в таблице 7. Контрастное условие используют в PROC GLM SAS версии 9.2 для доступа ко всем сравнениям. В двухфакторном ANOVA пол используют в качестве корректирующего фактора, и значения р до/после корректировки показаны на фиг.38D.
Поведенческие тесты. Мышей тестируют на двигательную активность и исследовательское поведение, а также на ряд сенсомоторных критериев и способность удерживаться на вращающемся барабане, для получения дополнительных контрольных данных, чтобы интерпретировать результаты тестирования условно-рефлекторного страха, которые используют для оценки когнитивной способности. Тестирование условно-рефлекторного страха проводят последним в серии испытаний, чтобы исключить влияние кратковременных ударов электрическим током по лапам на другие поведенческие показатели.
Исследование платформы с отверстиями, ряд сенсомоторных критериев и способность удерживаться на вращающемся барабане. Всех мышей тестируют на способность исследовать платформу с отверстиями, где общий объем перемещений (движений всего тела) и число тыканий в отверстия определяют в течение 30-минутного периода, с получением показателей двигательной активности и исследовательской способности. Протокол включает применение компьютеризированного устройства в виде платформы с отверстиями (41×41×38,5 см в высоту), содержащей 4 угловых и 4 боковых отверстия, причем боковые отверстия находятся на одинаковом расстоянии от угловых отверстий (Learning Holeboard; MotorMonitor, Kinder Scientific, LLC, Poway, CA). Светолучевые приборы используют для количественной оценки общего объема движений и исследования тыканий в отверстия во время сеанса тестирования. Данная процедура служит компонентом привыкания в общем тесте исследовательской способности/обонятельного предпочтения с использованием платформы с отверстиями. Мышей также тестируют по ряду из семи сенсомоторных критериев, которые используют для оценки способности удерживать равновесие (рейка, платформа), координации (шест, экраны с наклоном 60° и 90°), силы (перевернутый экран) и инициации движения из небольшой ограниченной области (начало ходьбы). Этот ряд критериев используется в предшествующих публикациях, и более подробное описание методики можно найти в (Wozniak et al. 2004; Neurobiol Dis 17, 403-414). Тест на удерживание на вращающемся барабане подобен ранее описанным методам и включает три вида испытаний: 1) неподвижный стержень (максимум 60 с); 2) барабан, вращающийся с постоянной скоростью (2,5 об/мин в течение максимум 60 с) и 3) барабан, вращающийся с ускорением (2,5-10,5 об/мин в течение 0-180 с). Протокол, использованный авторами настоящего изобретения, включает тестирование каждой мыши в одном испытании с применением неподвижного стержня, в двух испытаниях с применением барабана, вращающегося с постоянной скоростью, и в двух испытаниях с применением барабана, вращающегося с ускорением, которые входят в состав каждого из трех тестовых сеансов, проводимых с интервалом в 3 дня, чтобы ограничить усвоение двигательного навыка.
Условно-рефлекторный страх. Мышей анализируют путем тестирования на условно-рефлекторный страх, с помощью которого измеряют последний поведенческий критерий. Коротко, мышей подготавливают и тестируют в двух камерах для выработки условных рефлексов из плексигласа (26 см×18 см, высотой 18 см) (Med-Associates, St. Albans, VT), причем каждая камера содержит различные визуальные, обонятельные и тактильные сигналы. Каждую мышь помещают в камеру для выработки условных рефлексов для проведения 5-минутного испытания и количественно определяют замирание в течение 2-минутного фонового периода. Начиная с 3 мин, с 60-секундными промежутками мышей подвергают воздействию 3 пар звук-шок, где каждая пара включает 20-секундное воздействие тона 80 дБ (условный раздражитель; CS), состоящего из широкополосного белого шума, с последующим длительным ударом тока 1,0 мА по лапам (безусловный раздражитель; CS) на последней секунде воздействия звука. Широкополосный белый шум используют вместо звука с конкретной частотой, чтобы избежать возможных слуховых дефицитов, которые могут возникать с возрастом. На следующий день мышей помещают обратно в камеру для выработки условных рефлексов и количественно определяют замирание в течение 8-минутного периода, для оценки выработки условного рефлекса на ситуативный страх. Через двадцать четыре часа мышей помещают в другую камеру, содержащую различные сигналы и количественно определяют замирание в течение 2-минутного фонового периода "измененной ситуации" и в течение последующих 8 мин, в течение которых мышей подвергают воздействию звукового сигнала (звук; CS). Замирание количественно определяют с помощью программного обеспечения для анализа изображений FreezeFrame (Actimetrics, Эванстон), которое позволяет одновременно визуализировать поведение и корректировать "порог замирания", с помощью которого поведение классифицируют как замирание или отсутствие замирания с 0,75-секундными интервалами. Замирание определяют как отсутствие движения за исключением отсутствия движения, связанного с нормальным дыханием, и данные представляют в виде процента времени, проведенного в состоянии замирания. Чтобы оценить степень ситуативного условно-рефлекторного страха, авторы проводят анализ в каждой экспериментальной группе, который включает сравнение процента времени, проведенного в состоянии замирания, усредненного по 2-минутному фону, определенному в 1 день, с усредненным процентом времени, проведенного в состоянии замирания, в течение первых 2 мин тестирования ситуативного страха на 2 день, а также с уровнем замирания, усредненным по всему 8-минутному сеансу. Чувствительность к удару электрического тока оценивают после завершения тестирования условно-рефлекторного страха, в соответствии с методиками, описанными ранее в Khuchua et al. (2003; Neuroscience 119, 101-111).
Статистический анализ результатов поведенческого тестирования. Вариационный анализ (ANOVA) моделей, как правило, используют для анализа результатов поведенческих тестов (программное обеспечение Systat 12, Systat, Chicago, IL). Результаты тестирования условно-рефлекторного страха анализируют путем многократных измерений (rm) моделей ANOVA, включающих одну межсубъектную переменную (обработка) и одну внутрисубъектную переменную (многократные измерения) (в минутах). Корректировку Huynh-Feld альфа-уровней используют для всех внутрисубъектных эффектов, включающих более двух уровней защиты против нарушения допущений о сферичности/симметрии соединения, лежащих в основе моделей rmANOVA. Запланированные сравнения контрольной группы, получавшей PBS+HJ3.4, и каждой из трех экспериментальных групп, получавших другое антитело (то есть HJ8.5, HJ9.3, HJ9.4), проводят на моделях ANOVA для тестирования некоторых ключевых гипотез. В других случаях, проводят попарные сравнения релевантных, значимых общих эффектов ANOVA, которые подвергают коррекции Бонферрони, если это необходимо. Также рассчитывают коэффициент корреляции Пирсона (r) между общим числом движений, зарегистрированных в процессе тестирования на платформе с отверстиями, и процентом времени, проведенного в состоянии замирания во время тестирования ситуативного страха на 2-й день.
Пример 16. Анализ тау методом ELISA
Анализ ELISA разрабатывают для обнаружения присутствия патологических агрегатов тау в образцах плазмы пациентов. Антитела, использованные в данном анализе, включают мышиные моноклональные антитела против тау HJ9.3 и HJ9.2. HJ9.3 биотинилируют, используя набор для одностадийного биотинилирования антител (HJ9.3-Bio). В данном методе сэндвич-ELISA в качестве улавливающих антител используют HJ9.3 и HJ9.2 в эквивалентных концентрациях. 96-луночные планшеты с половинной площадью (Costar 3690) покрывают раствором антитела HJ9.2/HJ9.3 с концентрацией 20 мкг/мл в бикарбонатном буфере, рН 9,6 (50 мкл/лунка) и инкубируют при 4°C в течение ночи. После блокирования с использованием 4% BSA/PBS образцы плазмы (разведенные 1:4 буфером для образцов: 0,25% BSA/PBS, 300 нМ трис, рН 7,4-8,0, 1X ингибиторы протеаз) помещают в лунки в трех повторах (50 мкл/лунка). Затем планшеты инкубируют при 4°C в течение ночи. Перед проведением детекции в лунки добавляют раствор HJ9.3-Bio в 0,5% BSA/PBS с концентрацией 0,3 мкг/мл и инкубируют в течение 1,5 ч при 37°С. Конъюгат вторичное антитело-стрептавидин-полиHRP40, разведенный 1:4000 0,5% раствором BSA/PBS (50 мкл/лунка с инкубацией в течение 1,5 ч в темноте на шейкере при комнатной температуре), используют для конечной детекции посредством ферментативной реакции с применением сверхмедленного субстрата TMB. Метод ELISA разрабатывают так, чтобы оптимизировать детекцию редких частиц в плазме. Первоначальные варианты осуществления включают нанесение на поверхность планшета ELISA пар антител для оптимизации улавливания агрегатов. Однако для улавливания белков из образцов с большим объемом жидкости в равной степени можно использовать гранулы, покрытые антителом, что позволяет увеличить чувствительность анализа. Отрицательную плазму, полученную от здоровых молодых участников, используют для расчета фонового сигнала в анализе. Присутствие центров агрегации тау в экспериментальных образцах описывают в виде кратности индукции по сравнению с сигналом, полученным от отрицательной плазмы.
Набор образцов плазмы, полученных от предклинических пациентов и пациентов с болезнью Альцгеймера (AD), предварительно тестируют с помощью описанного анализа затравки образования агрегатов, используя метод сэндвич-ELISA. 12 контрольных пациентов (CDR 0), не содержащих затравочной активности (отрицательные), и 12 пациентов (CDR>0), содержащих затравочную активность (положительные), протестируют с использованием недавно разработанного анализа ELISA. Указанных пациентов заранее проверяют на присутствие затравочной активности в CSF и плазме с помощью биосенсорного клеточного анализа. В данном клеточном анализе используют фрагменты RD белка тау, содержащие мутацию ΔΚ280, слитые с голубым или желтым флуоресцентным белком. Это позволяет обнаружить агрегацию путем измерения резонансного переноса энергии флуоресценции методом FRET. Внеклеточные агрегаты проникают в клетку и инициируют внутриклеточную агрегацию FRET-репортерных белков тау.
Как показано на фиг.45, агрегаты тау не обнаружены в плазме пациентов с отрицательной затравочной активностью, по сравнению с очевидным присутствием центров агрегации тау в плазме пациентов с БА, характеризующихся положительной затравочной активностью. Данный бесклеточный анализ можно использовать в более клинических условиях в качестве неинвазивного диагностического инструмента для выявления многих таупатий, в том числе болезни Альцгеймера. Кроме того, с помощью данного анализа можно выявлять пациентов с начинающейся патологией, склонных к развитию слабоумия, облегчая проведение клинических испытаний путем обогащения выбранной популяцией. И, наконец, данный анализ можно использовать для мониторинга эффективности антител против тау или других лекарственных средств против деменции.
Пример 17
Анализ межклеточного распространения проводят для измерения распространения агрегатов тау из одной популяции в другую. Фрагмент tau, содержащий домен повторов (RD), используют как немеченую форму, содержащую либо две ассоциированные с болезнью мутации (LM: P301L/V337M), стимулирующие агрегацию CFP-меченой формы, либо одну ассоциированную с болезнью мутацию (ΔΚ: ΔΚ280). Одну группу клеток трансфицируют RD(LM) и RD(ΔK280)-CFP, и другую трансфицируют RD(ΔK280)-YFP. Сигналы FRET, полученные от клеток, выращенных в четырех повторах в 96-луночном формате, регистрируют на флуоресцентном планшет-ридере. Сигнал FRET генерируют агрегаты RD-CFP, перенесенные в клетки, содержащие RD-YFP, и наоборот. Несколько антител добавляют к среде при разных указанных разведениях. Исходная концентрация антитела составляет ~1 мг/мл. Например, разведение 10-3 указывает на то, что конечная концентрация составляет ~1 мкг/мл. Через 24 ч клетки фиксируют и регистрируют значения FRET. Результаты для отдельных антител приведены на фиг.47. Некоторые антитела являются очень активными в отношении предотвращения трансклеточного распространения агрегации (например, HJ8.2, HJ9.1). Другие обладают более умеренной активностью (например, HJ9.3), и некоторые из них являются практически неактивными (HJ8.7). В каждом графике первый столбец соответствует среде без добавления антител, то есть, он представляет фоновую эффективность распространения.
Чтобы тестировать синергическое действие антител, определяют их влияние на распространение в условиях отдельных антител, разведенных в соответствии с указанным диапазоном концентраций, или антител, смешанных в эквимолярном соотношении с последующим титрованием в том же диапазоне. Некоторые пары характеризуются высокой синергетической активностью (например, HJ9.3/9,4), тогда как другие препятствуют друг другу (HJ8.5/9,1) (фиг.48).
Влияние антитела на поглощение агрегатов тау можно измерить методом проточной цитометрии. Клетки подвергают воздействию рекомбинантных фибрилл RD, которые метят флуоресцентным красителем с помощью химического способа. После обработки трипсином и диспергирования, клетки подсчитывают с использованием проточного цитометра. HJ9.3 дозозависимо уменьшает количество флуоресцентномеченых клеток, что указывает на ингибирование поглощения агрегатов (фиг.50).
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПИЯ И ДИАГНОСТИКА НА ОСНОВЕ БЕЛКОВ ТАУ-ОПОСРЕДУЕМОЙ ПАТОЛОГИИ ПРИ БОЛЕЗНИ АЛЬЦГЕЙМЕРА | 2012 |
|
RU2645259C2 |
| ВЕЩЕСТВА И СПОСОБЫ ДЛЯ ПРИМЕНЕНИЯ ПРИ ПРЕДУПРЕЖДЕНИИ И/ИЛИ ЛЕЧЕНИИ БОЛЕЗНИ ГЕНТИНГТОНА | 2015 |
|
RU2742493C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К ГИПЕРФОСФОРИЛИРОВАННОМУ ТАУ-БЕЛКУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2727911C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К ГИПЕРФОСФОРИЛИРОВАННОМУ ТАУ-БЕЛКУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2760875C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ПРОТОФИБРИЛЛАМИ, И ИХ ПРИМЕНЕНИЕ В МЕТОДАХ ТЕРАПИИ И ДИАГНОСТИКИ БОЛЕЗНИ ПАРКИНСОНА, ДЕМЕНЦИИ, АССОЦИИРОВАННОЙ С ОБРАЗОВАНИЕМ ДИФФУЗНЫХ ТЕЛЕЦ ЛЕВИ, И ДРУГИХ АЛЬФА-СИНУКЛЕИНОПАТИЙ | 2011 |
|
RU2555526C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2825390C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2702108C2 |
| СКРИНИНГОВАЯ ПЛАТФОРМА CRISPR/CAS ДЛЯ ИДЕНТИФИКАЦИИ ГЕНЕТИЧЕСКИХ МОДИФИКАТОРОВ ЗАСЕВА ИЛИ АГРЕГАЦИИ ТАУ-БЕЛКА | 2020 |
|
RU2819350C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИ-ТАУ-АНТИТЕЛА | 2015 |
|
RU2743152C2 |
| ФОСФОСПЕЦИФИЧНЫЕ АНТИТЕЛА, РАСПОЗНАЮЩИЕ ТАУ | 2012 |
|
RU2639537C2 |








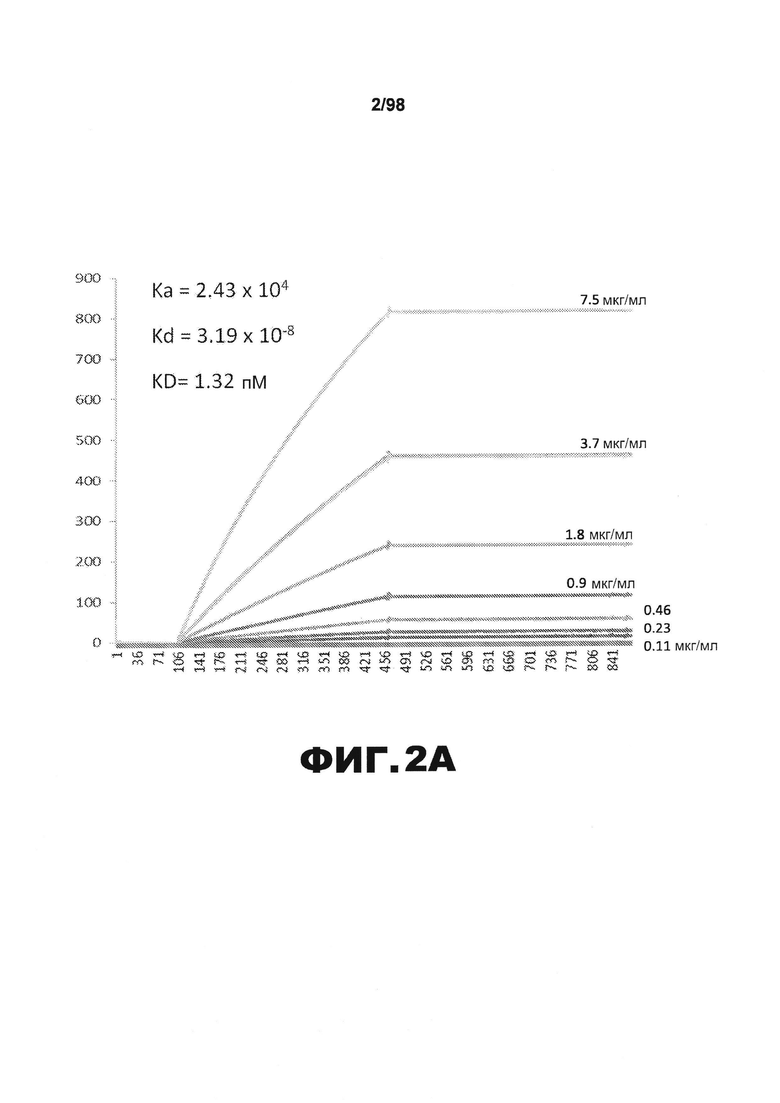
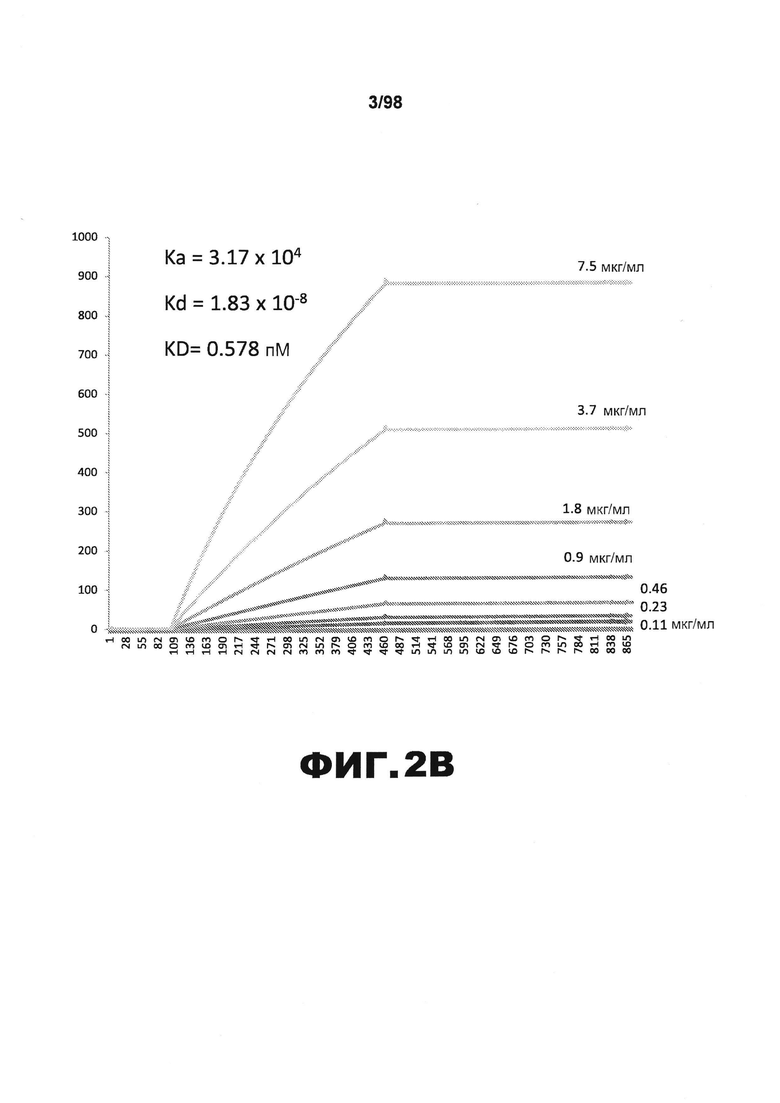
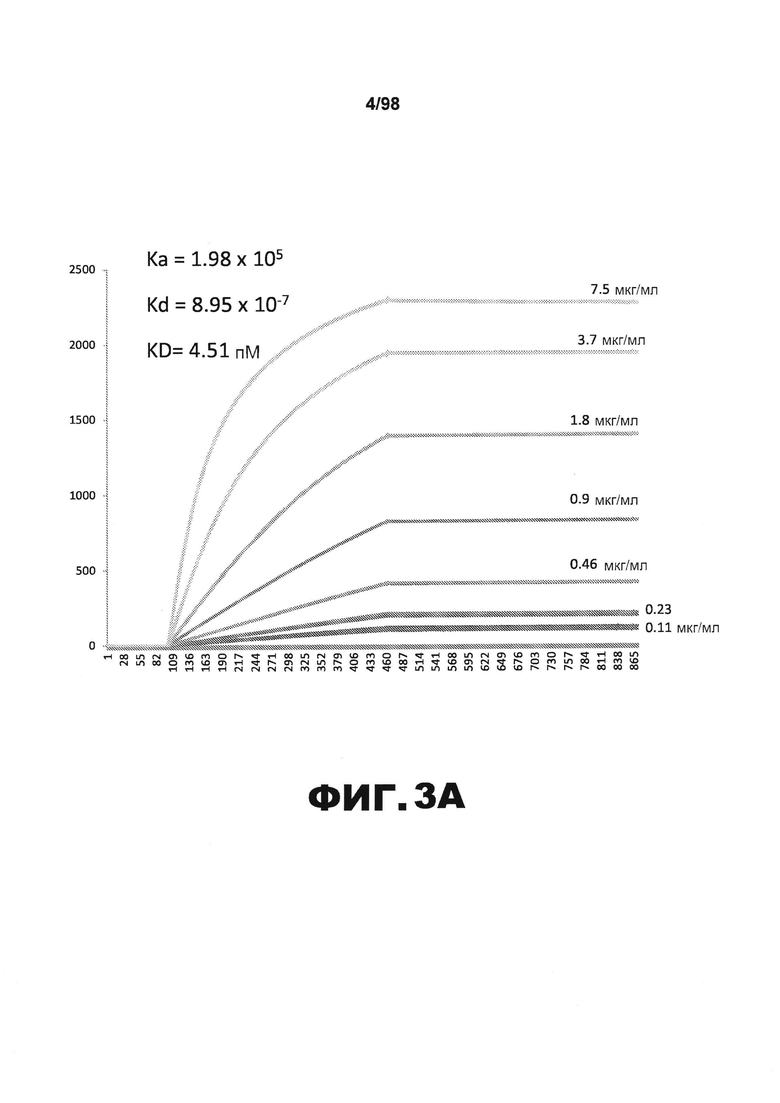
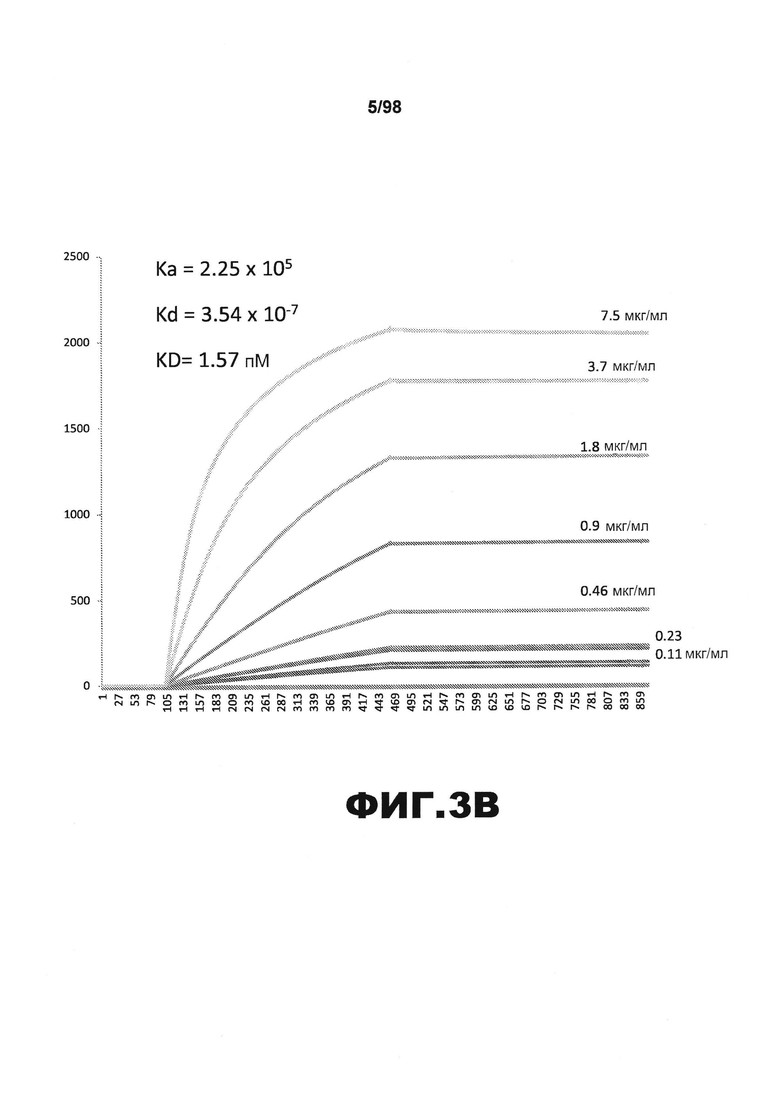
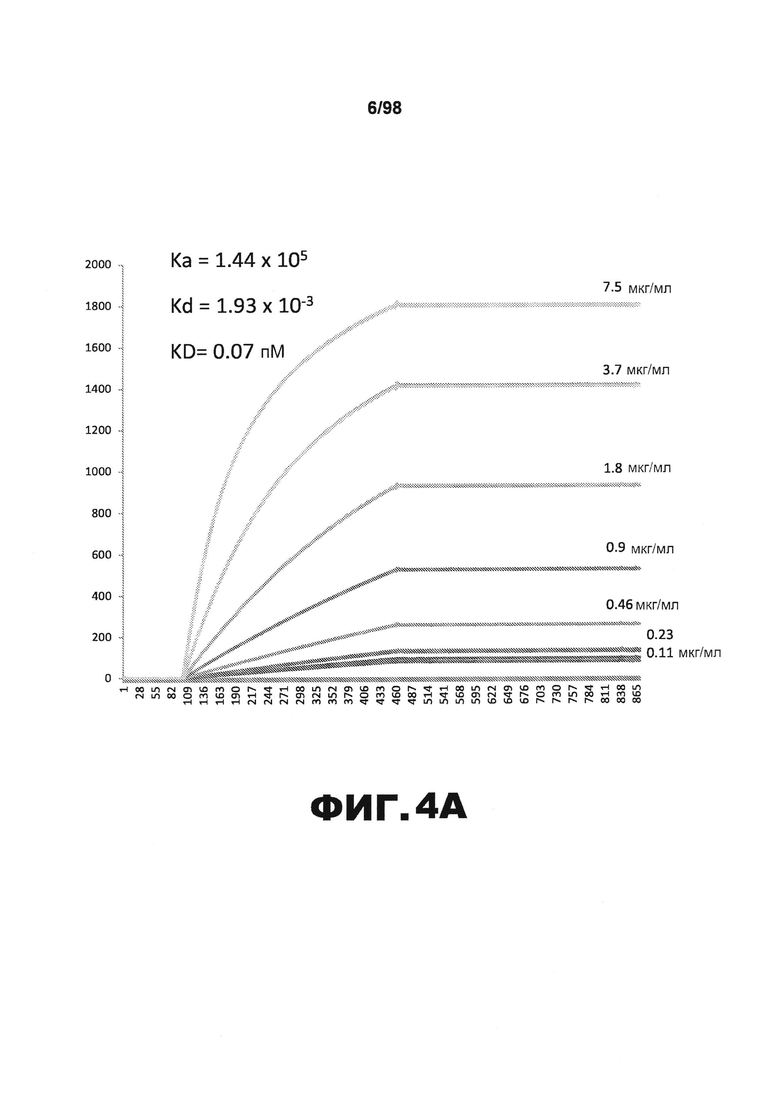
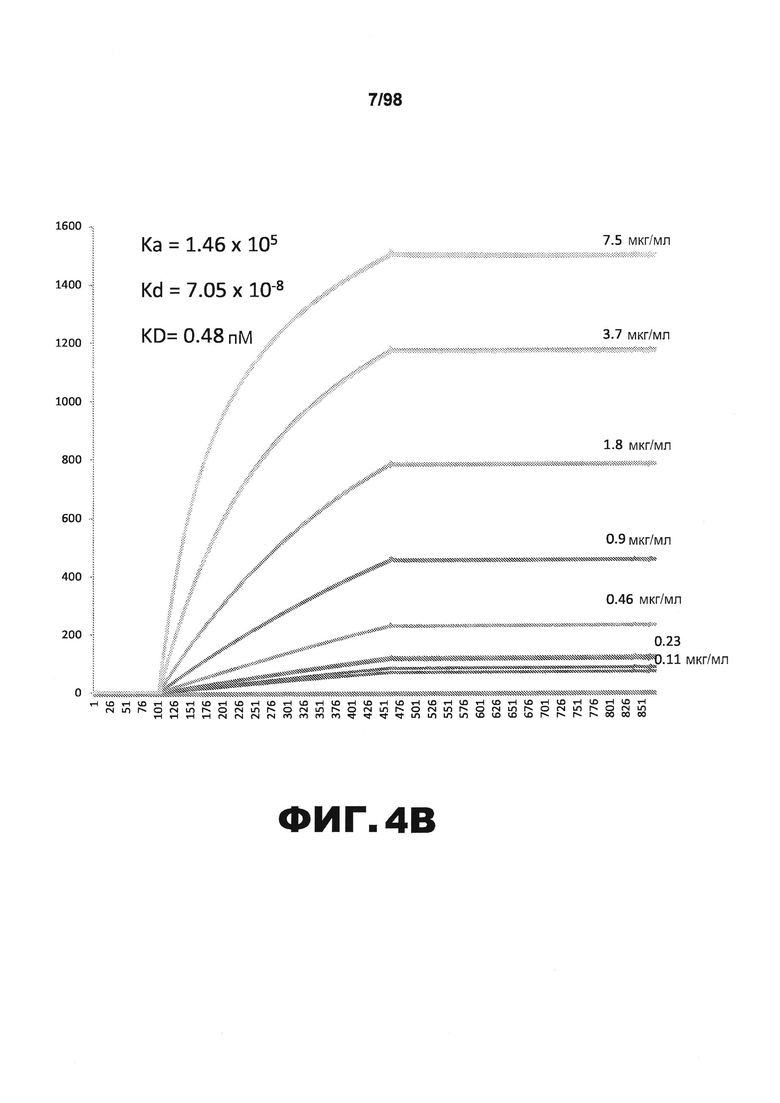
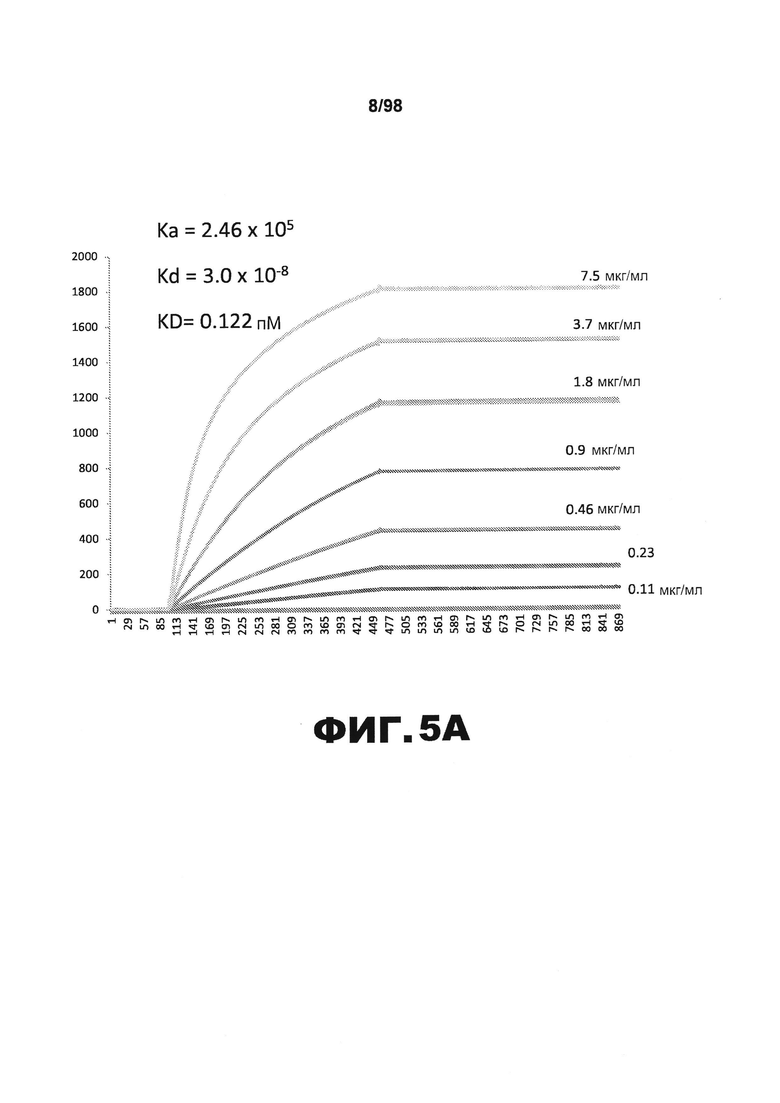
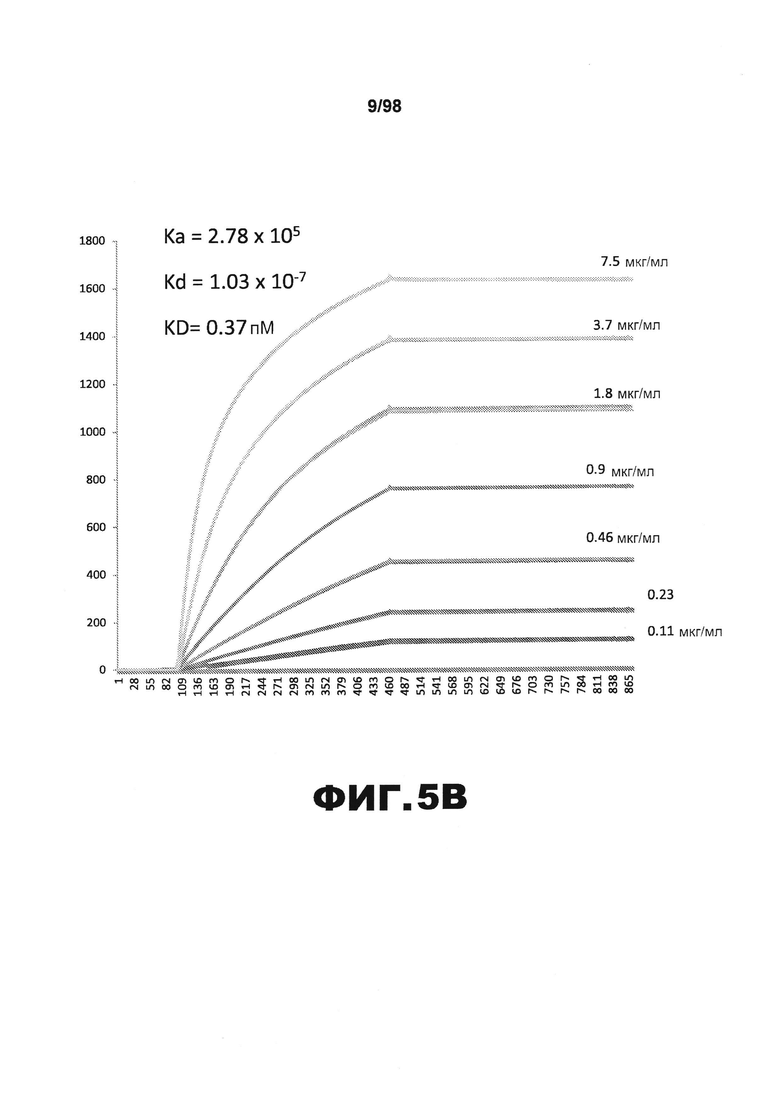
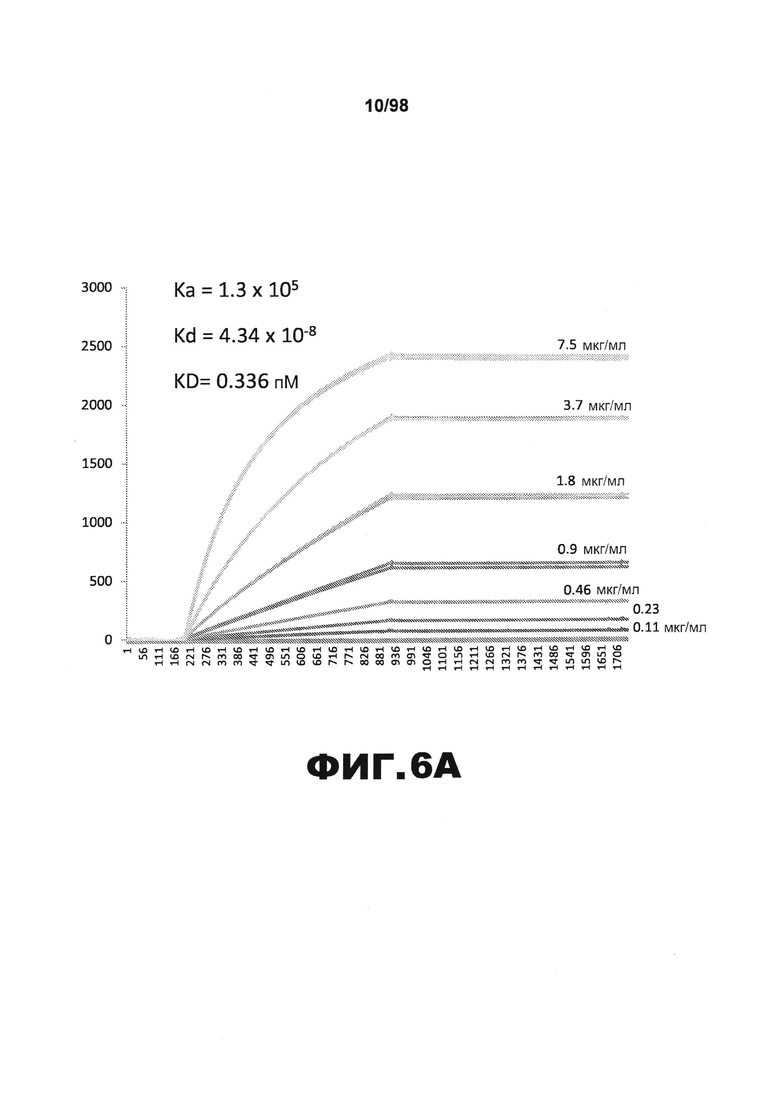
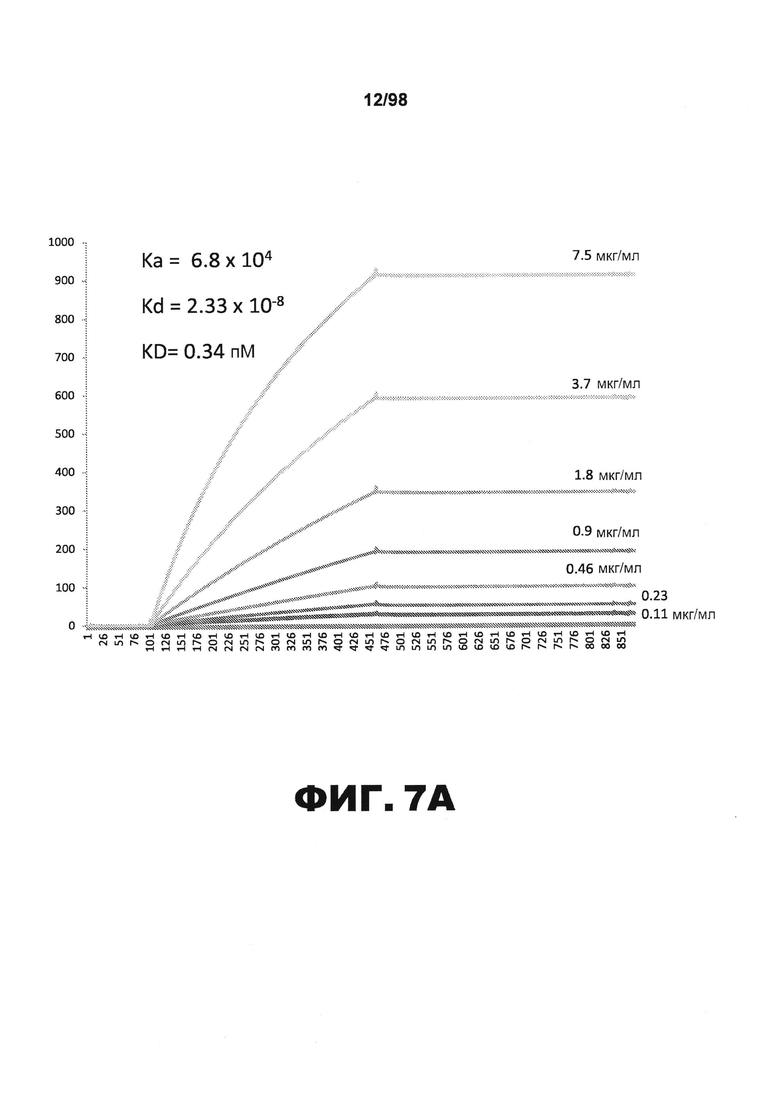
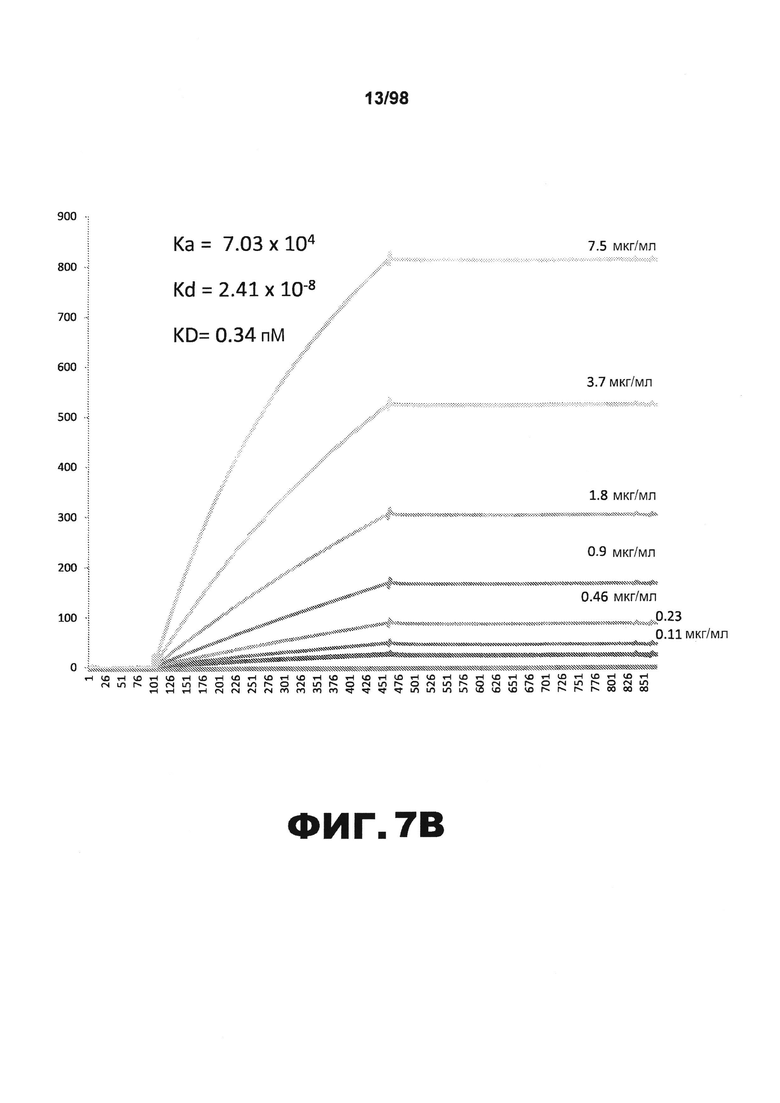
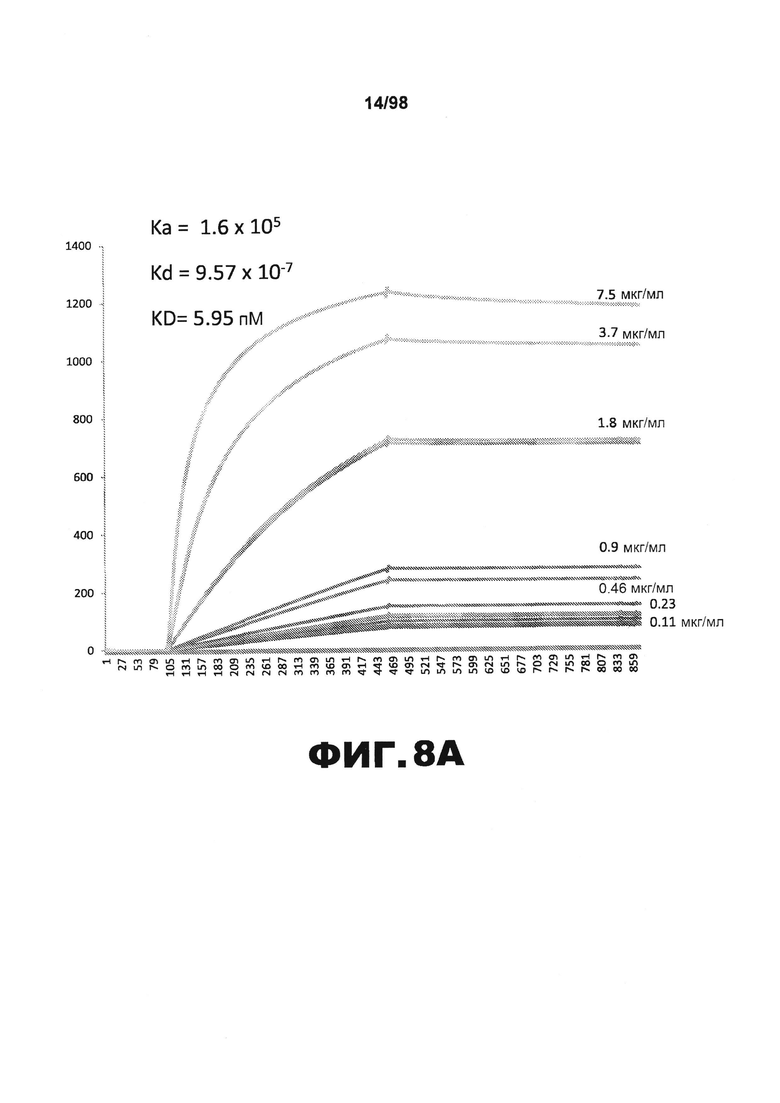
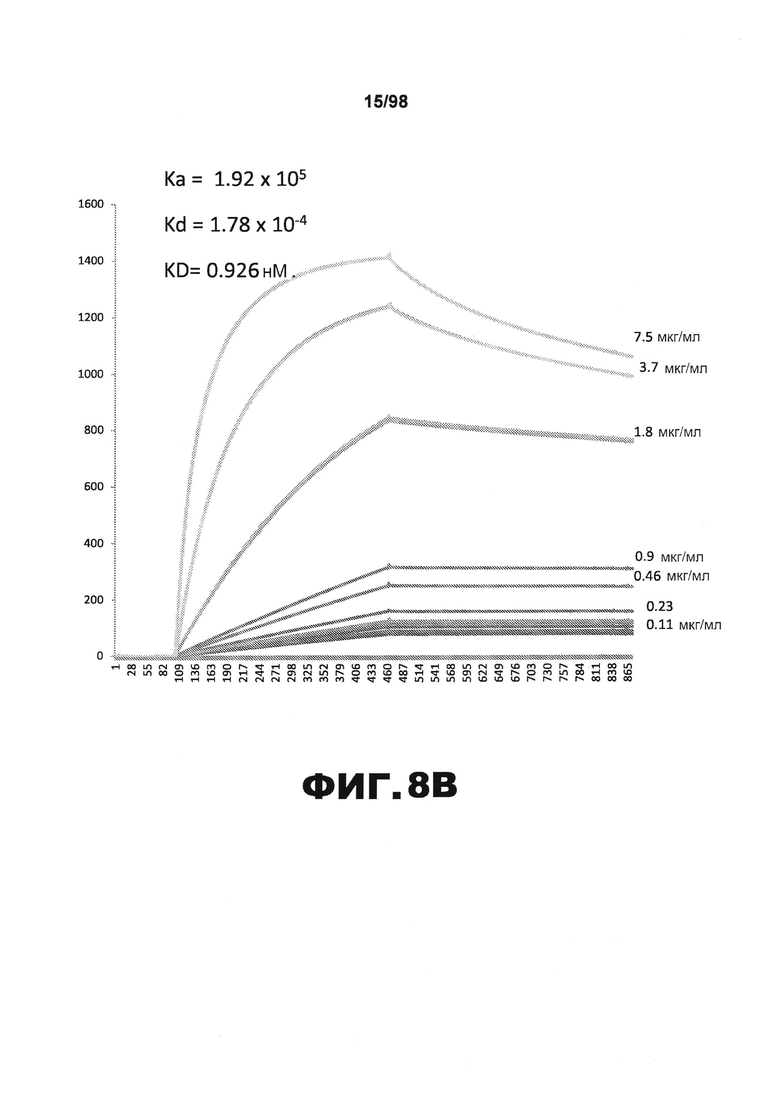
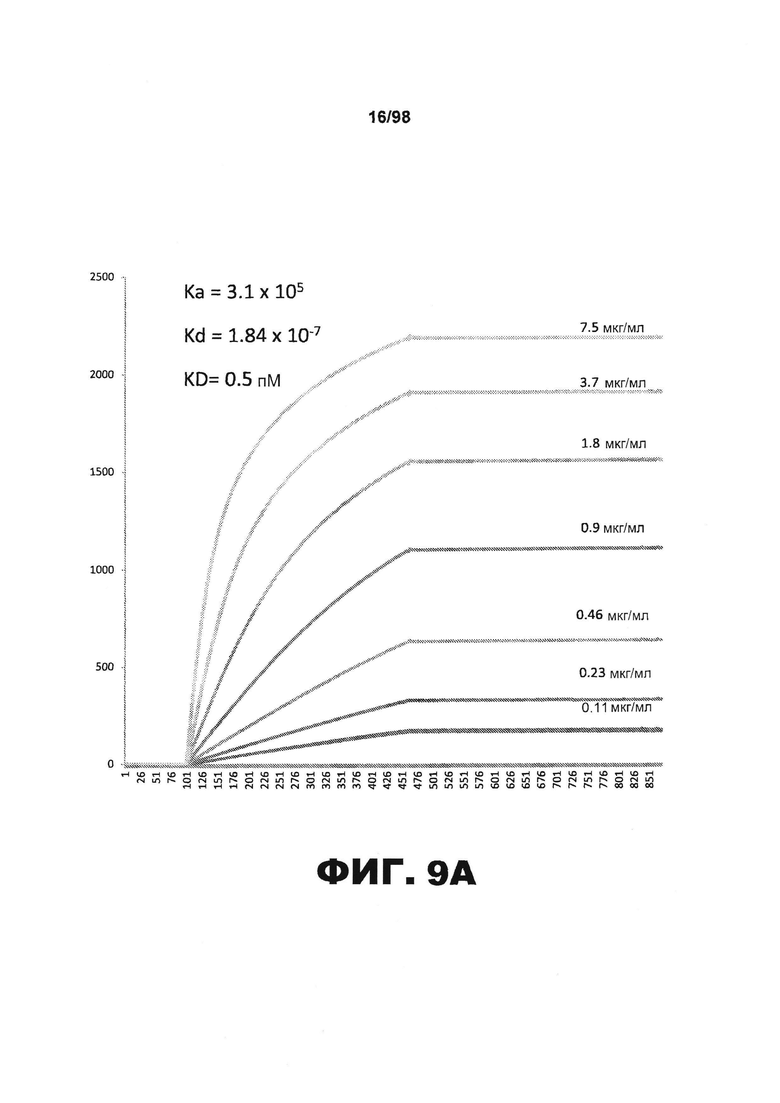
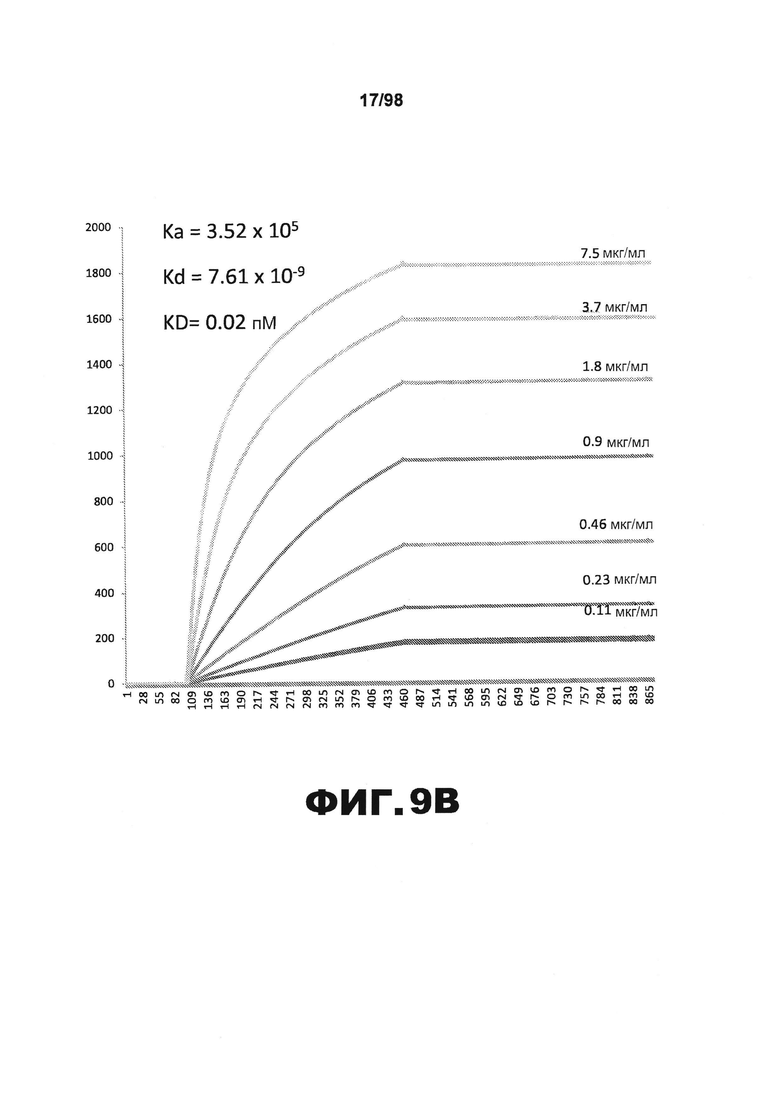
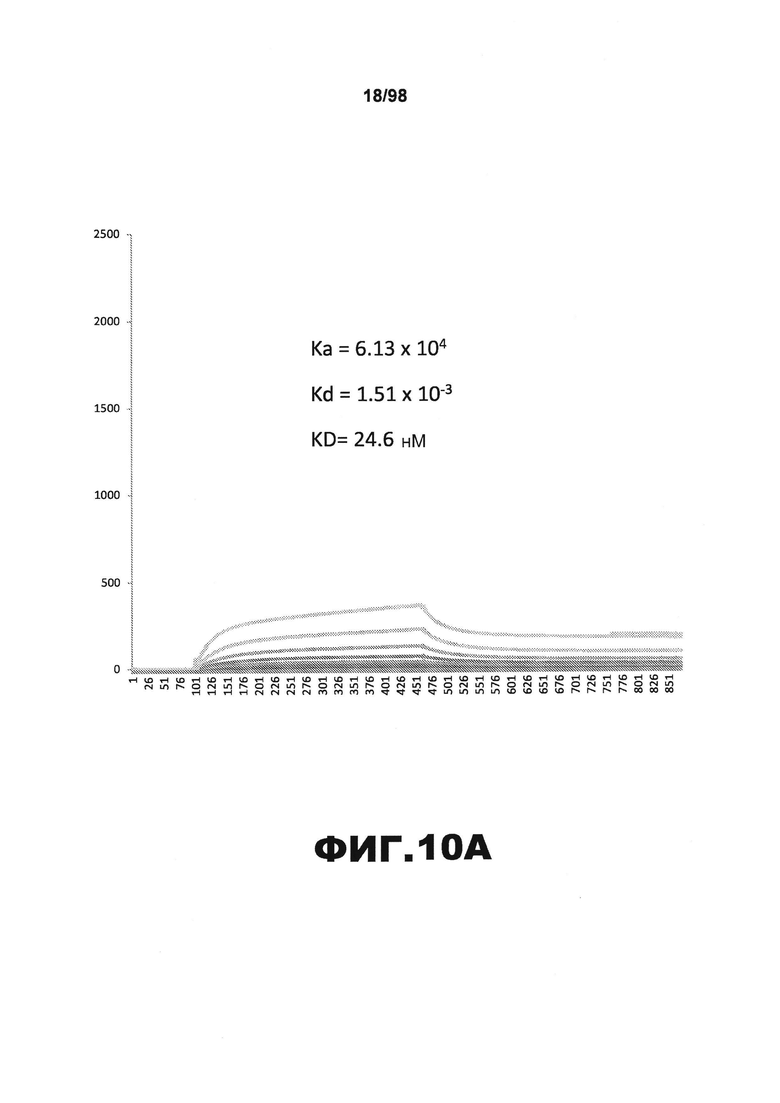
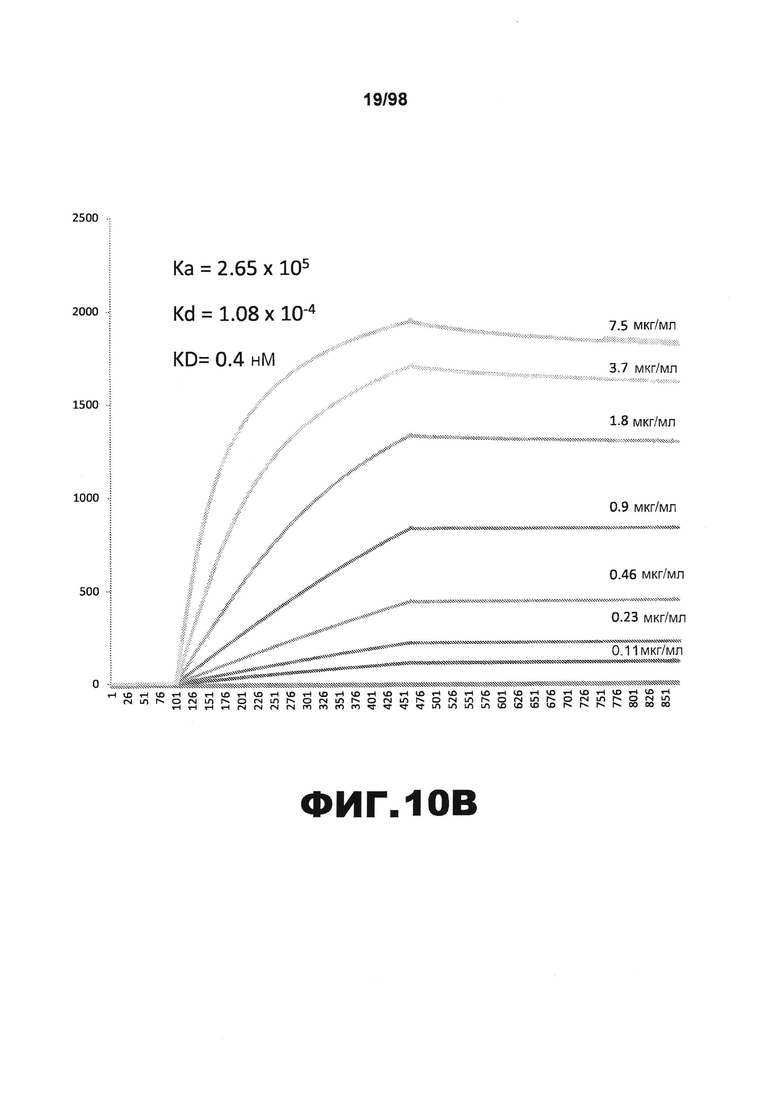
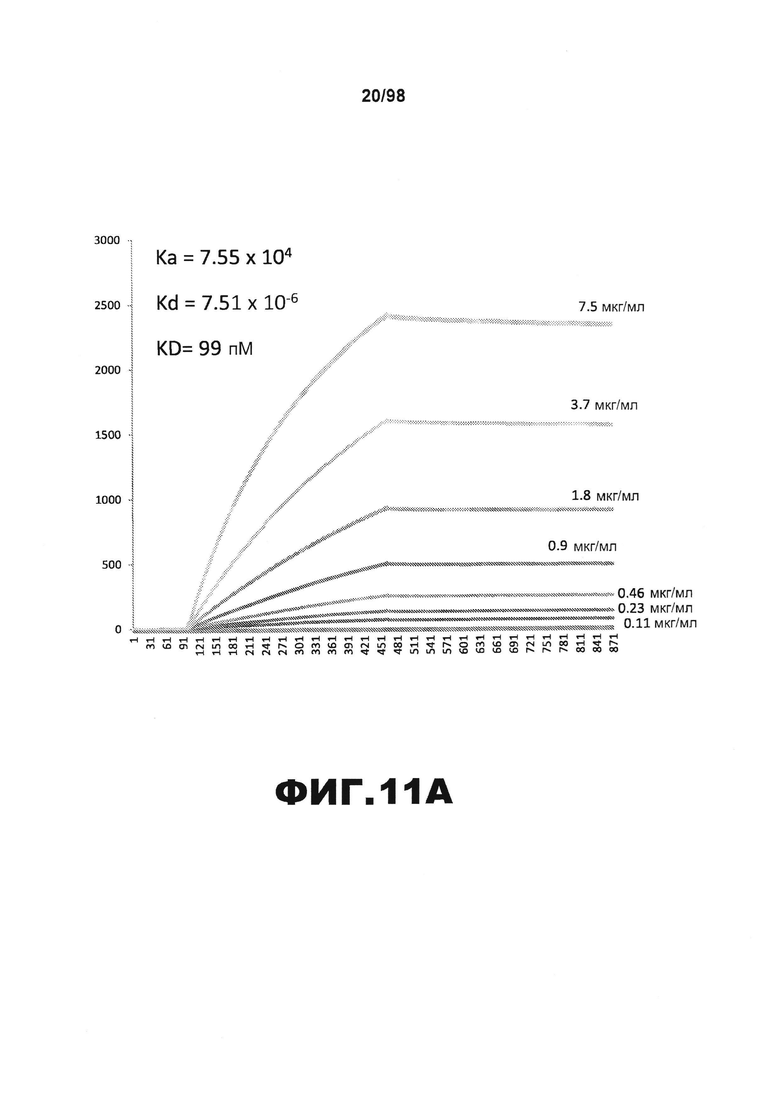
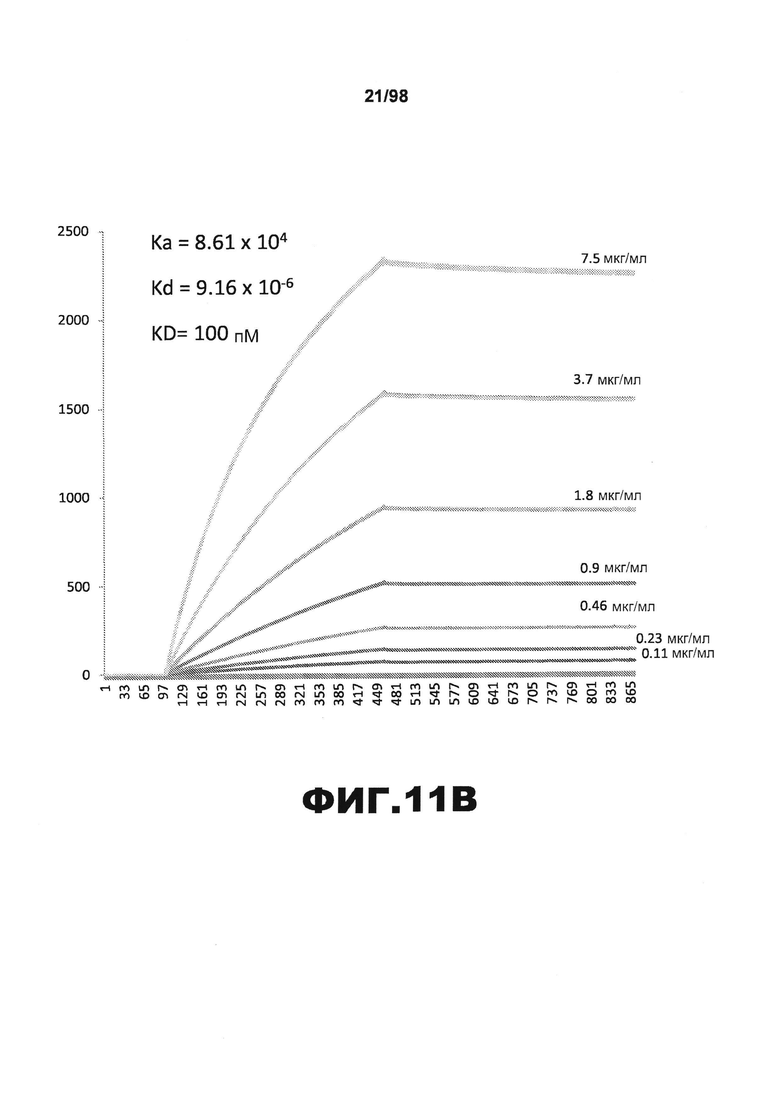
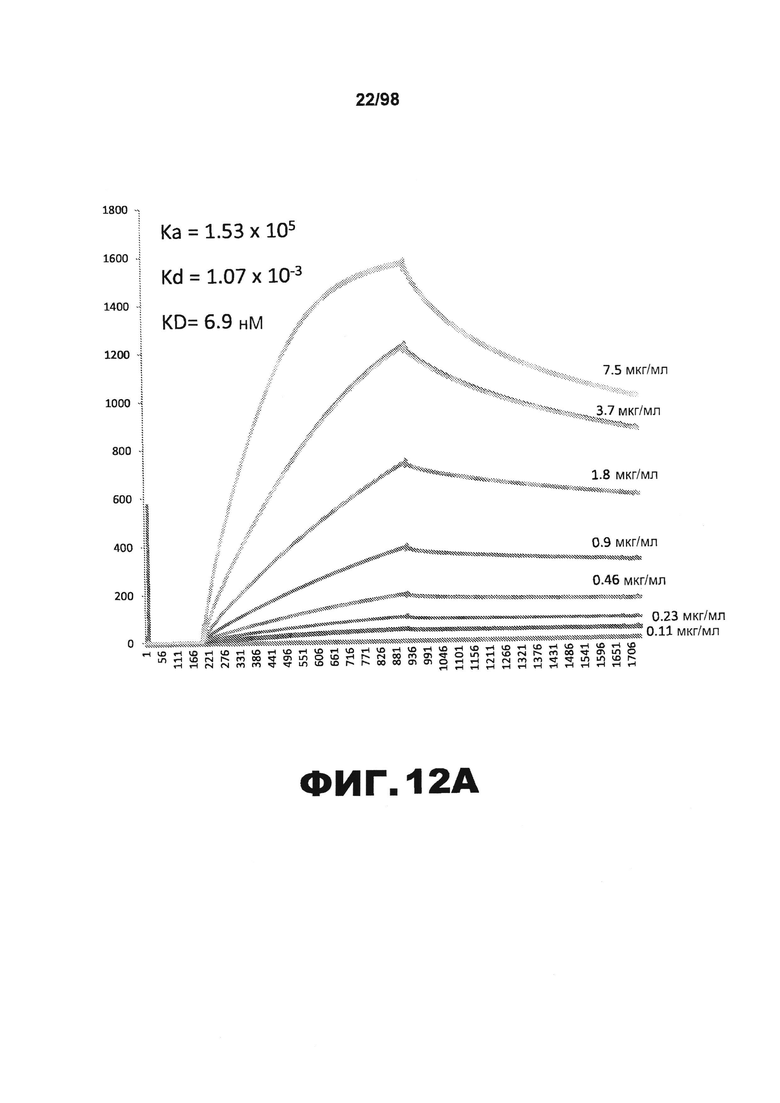
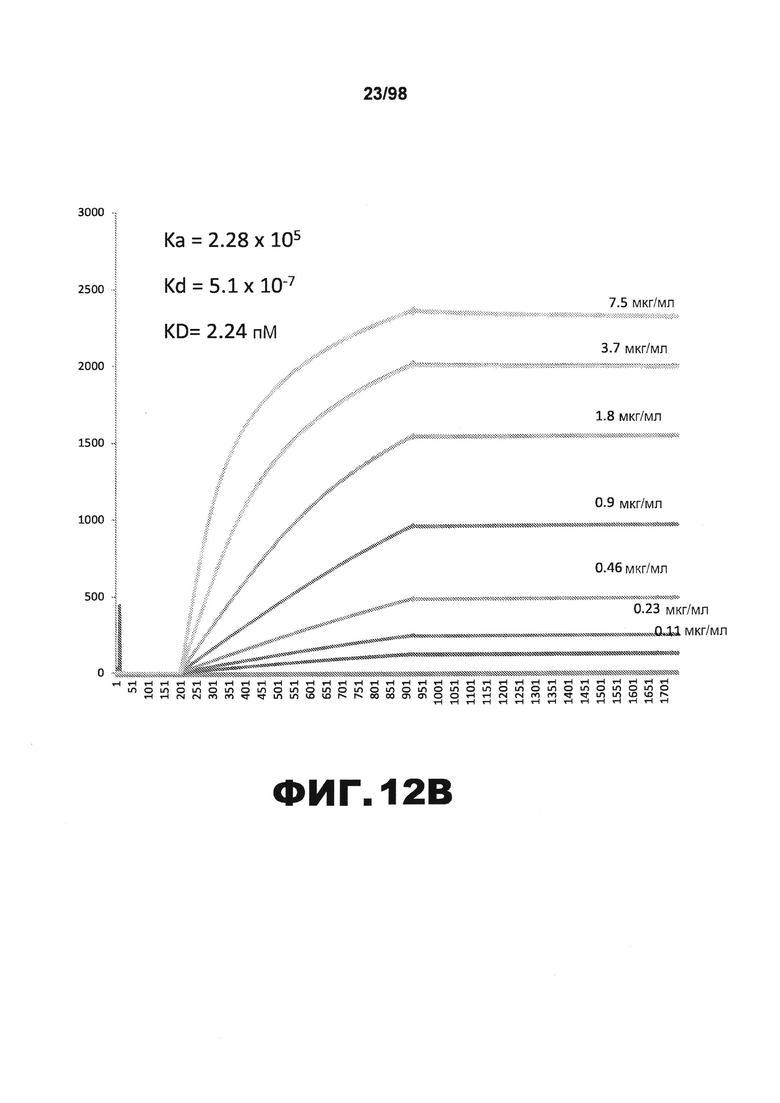
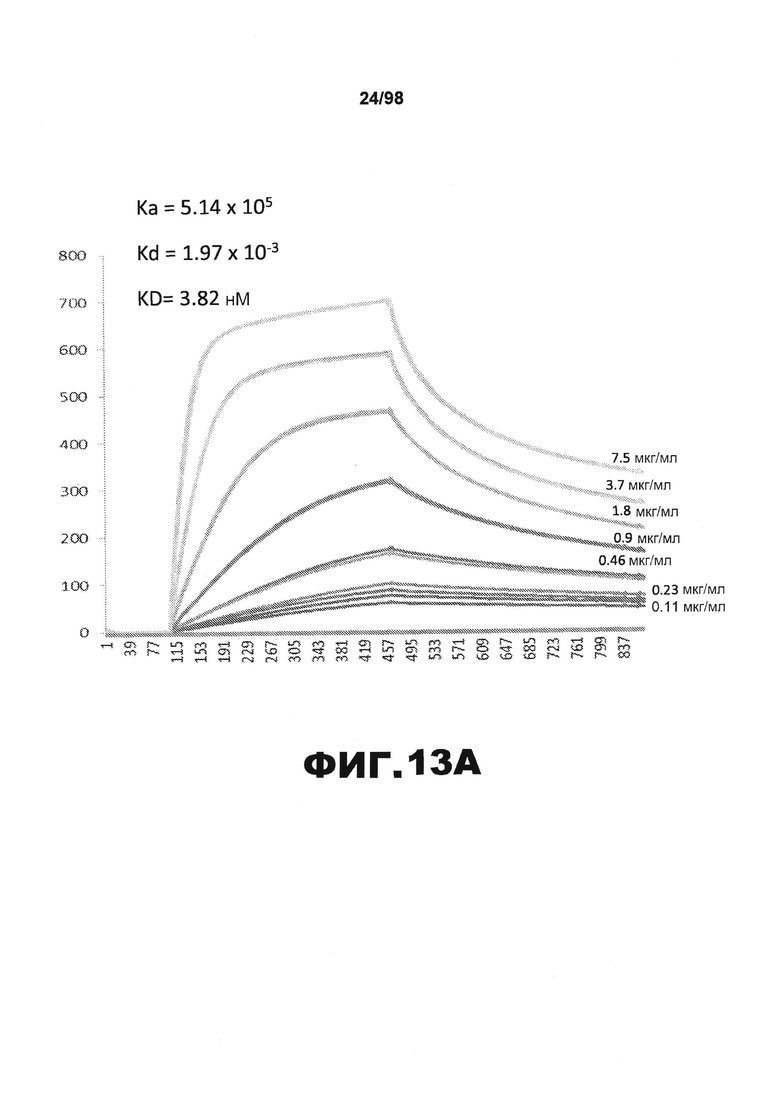
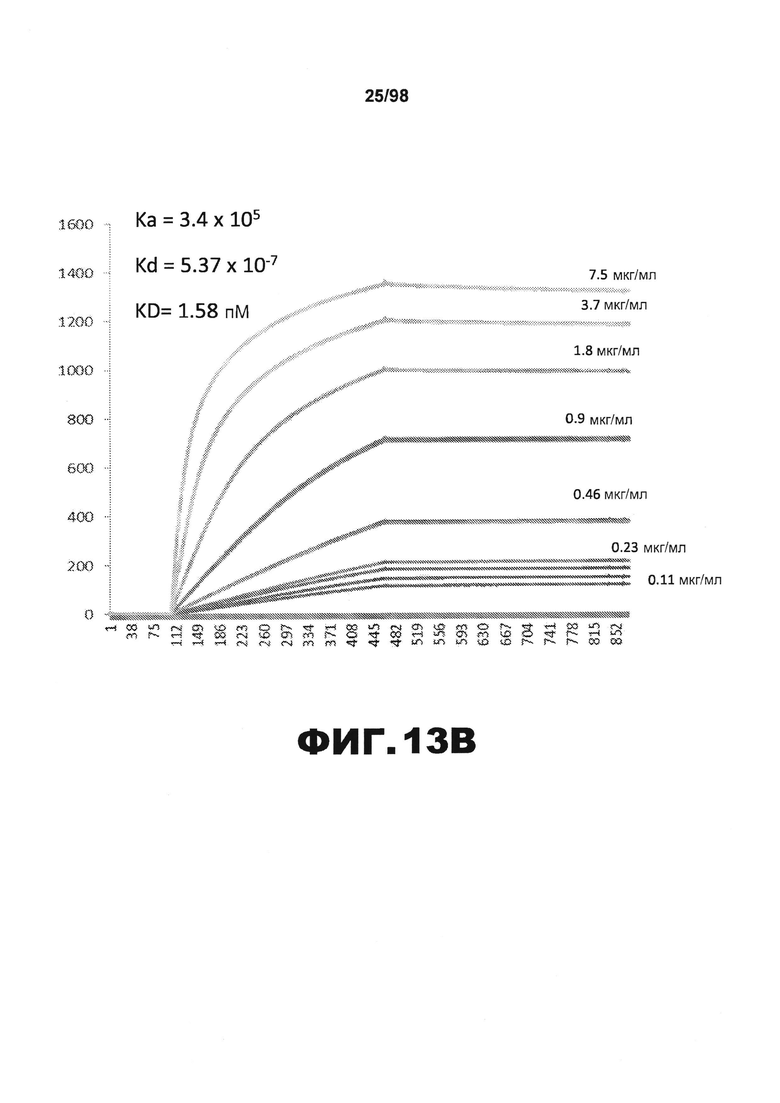
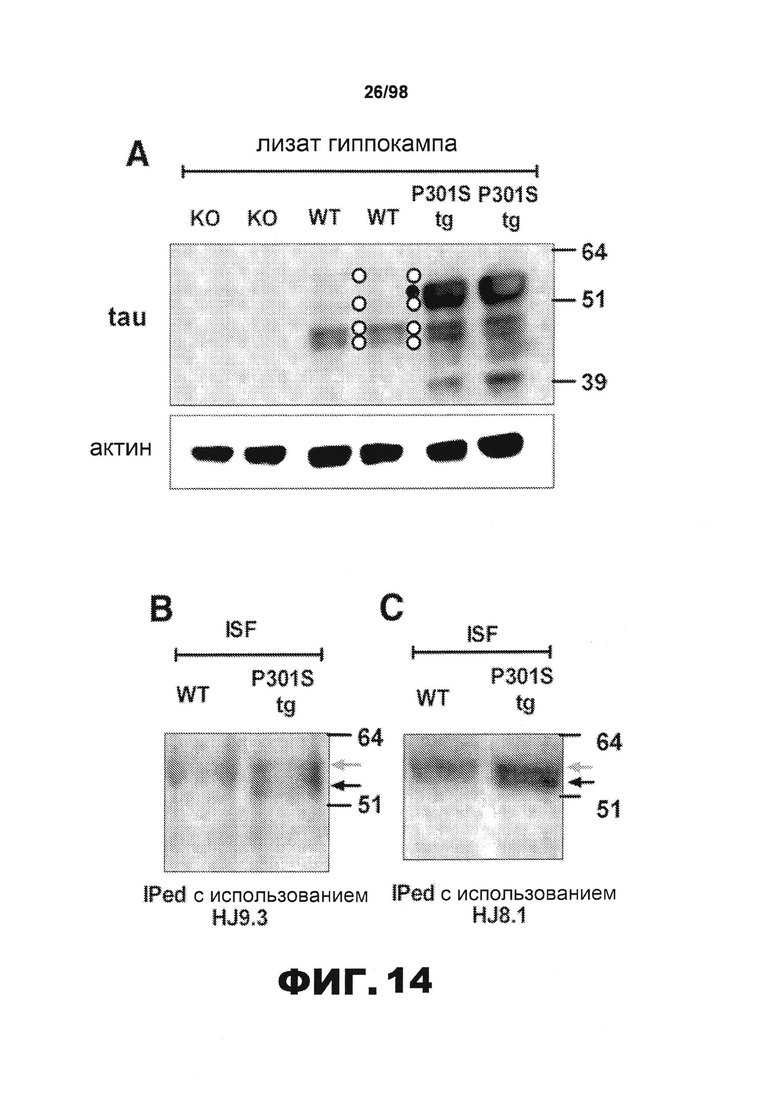
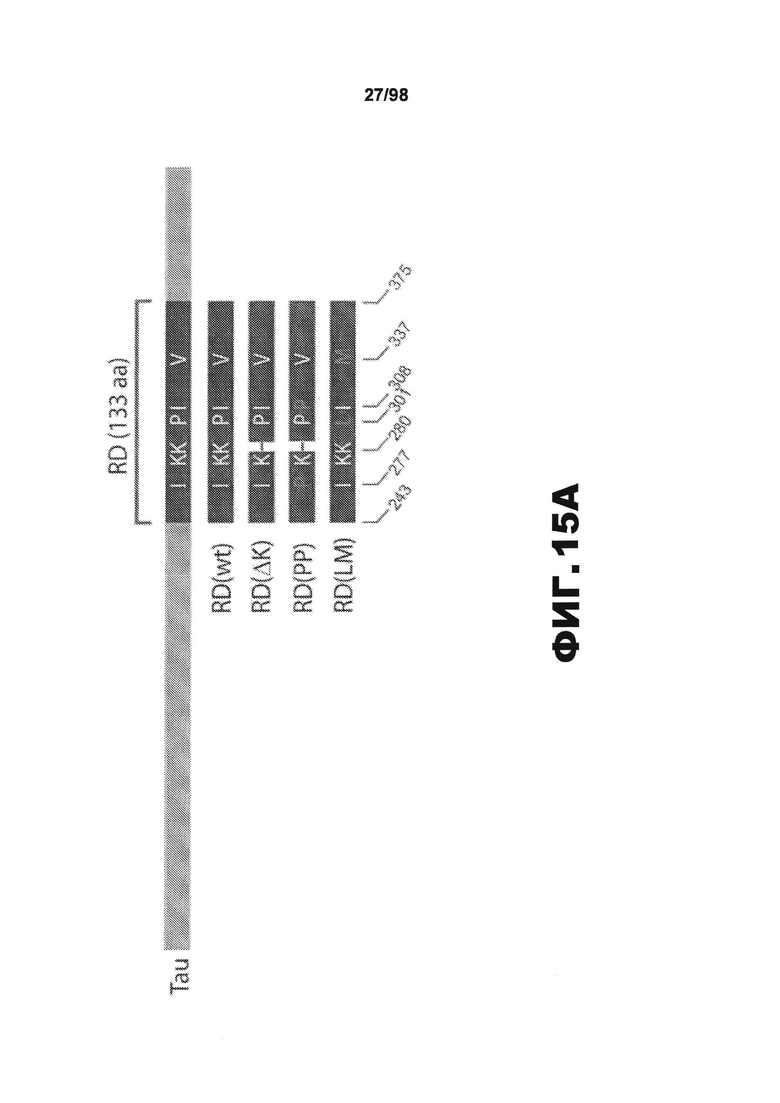

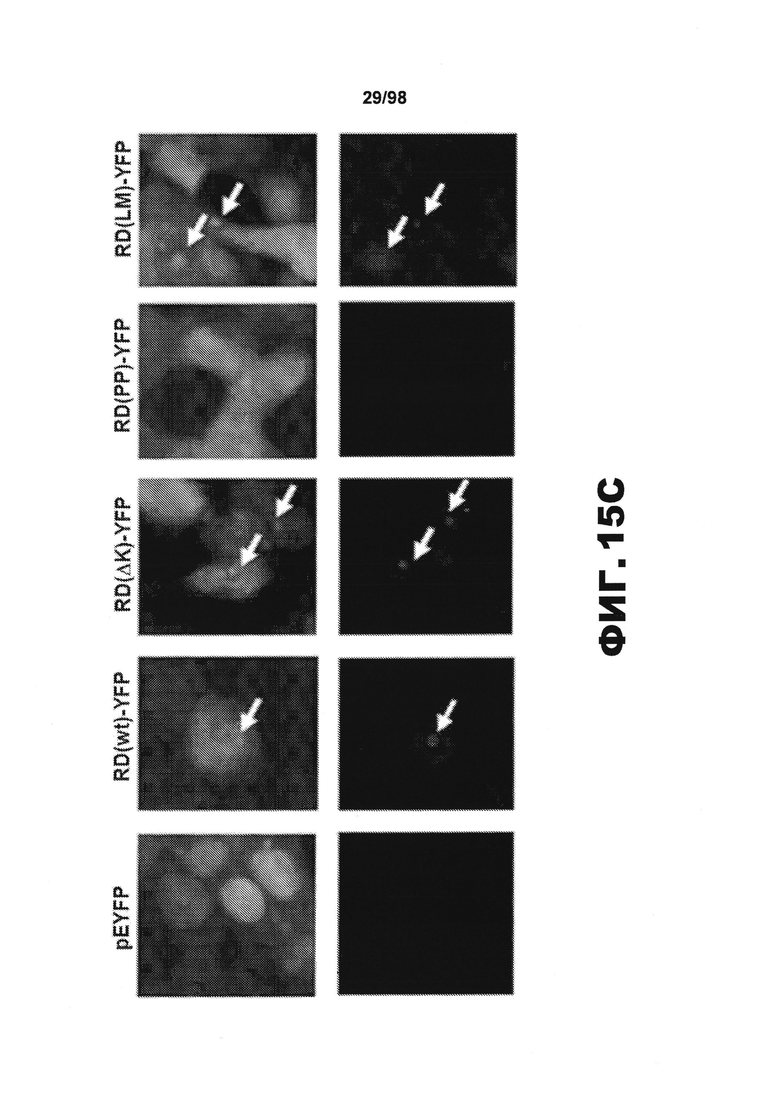
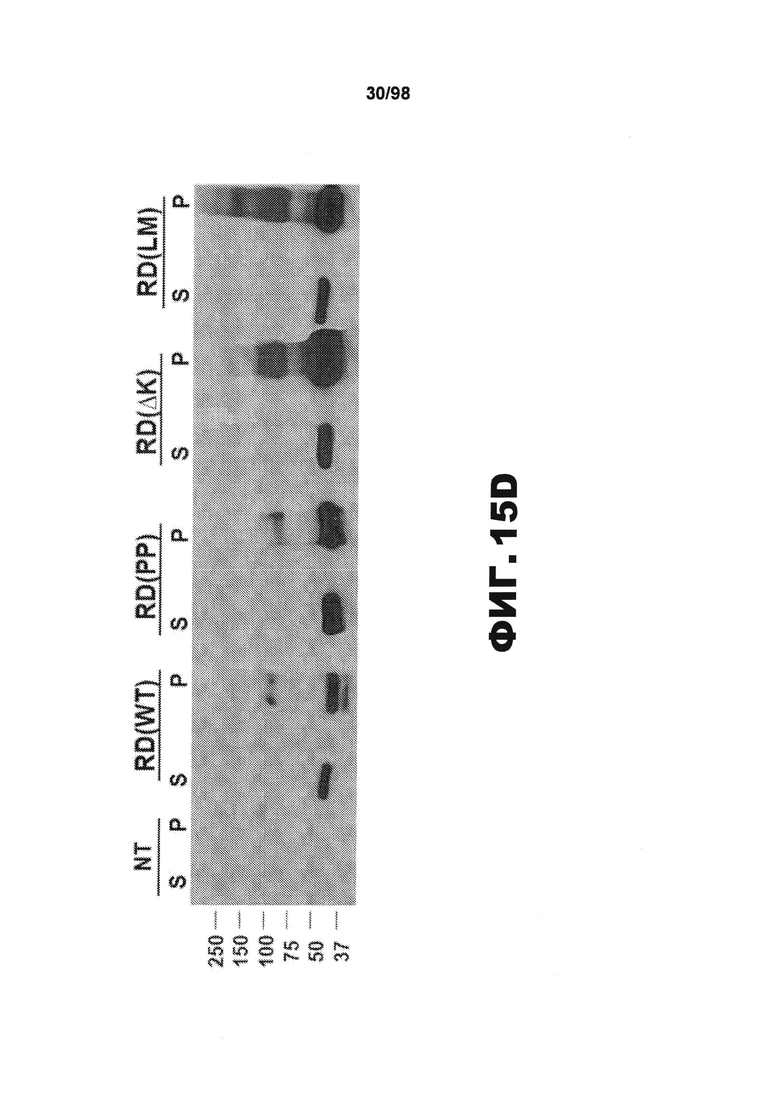
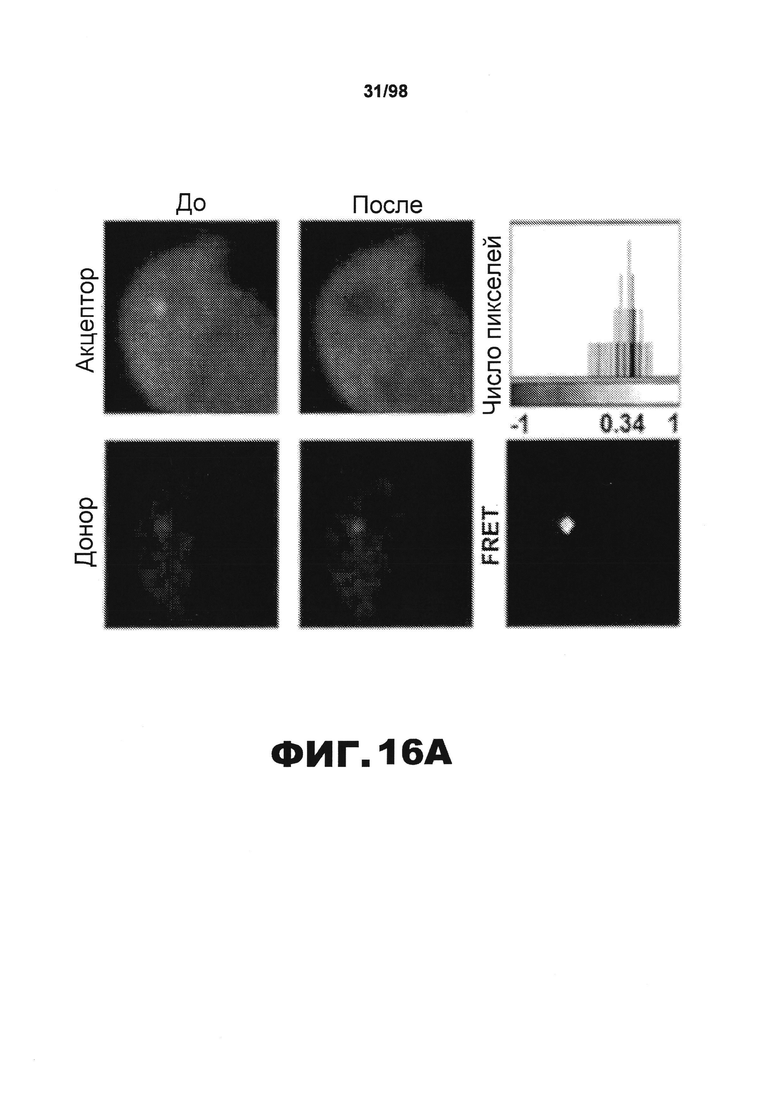
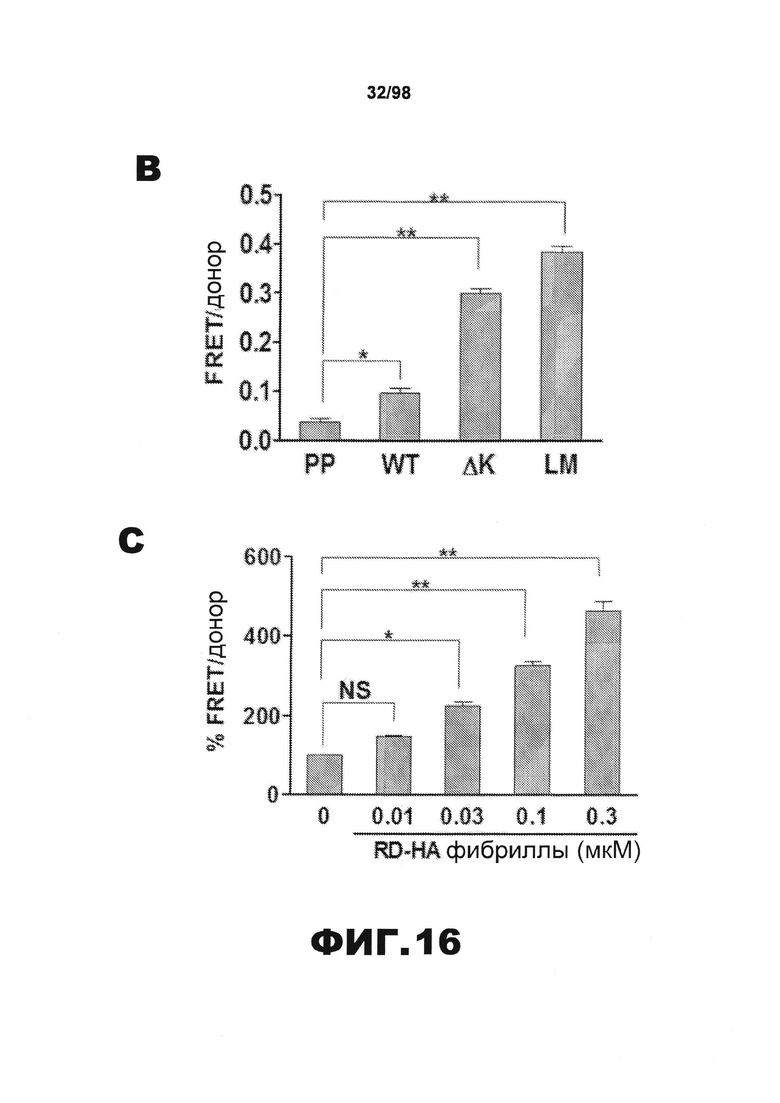
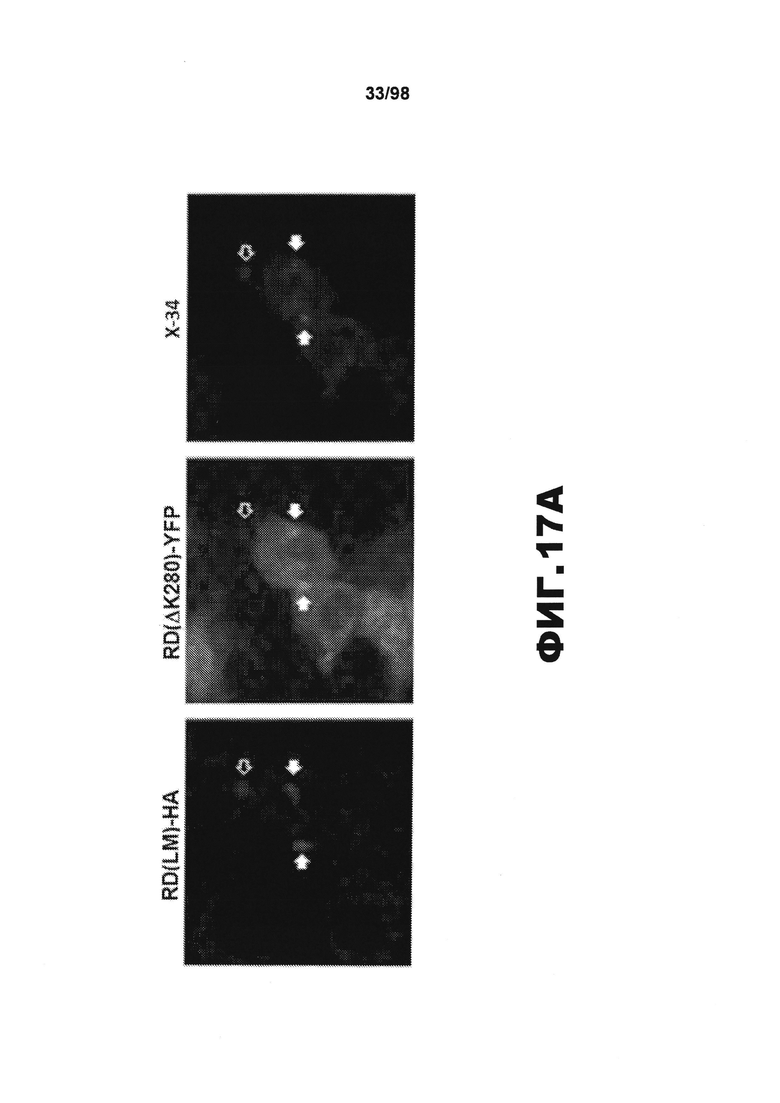
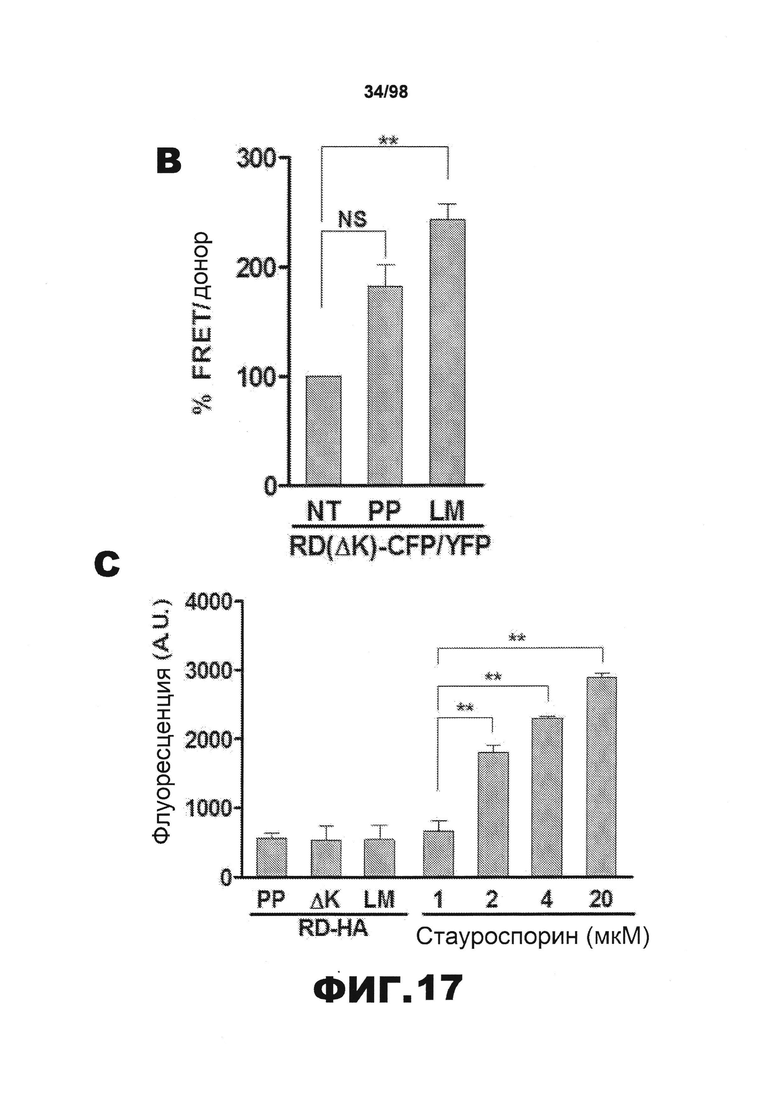
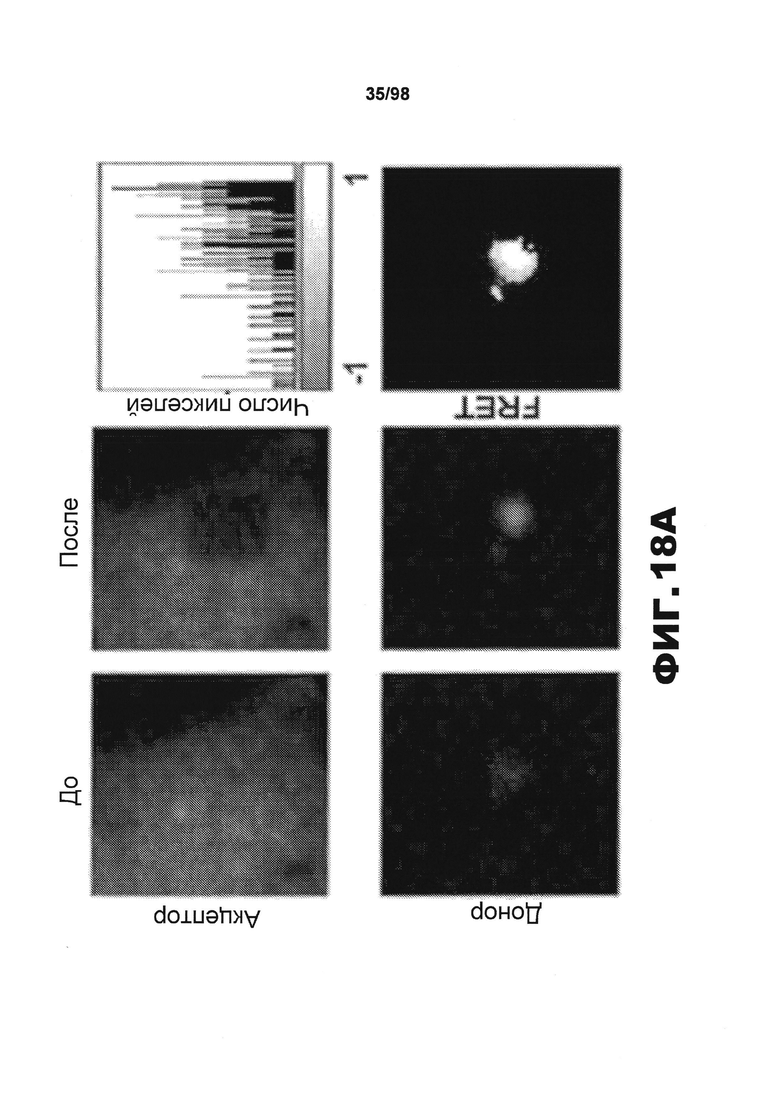
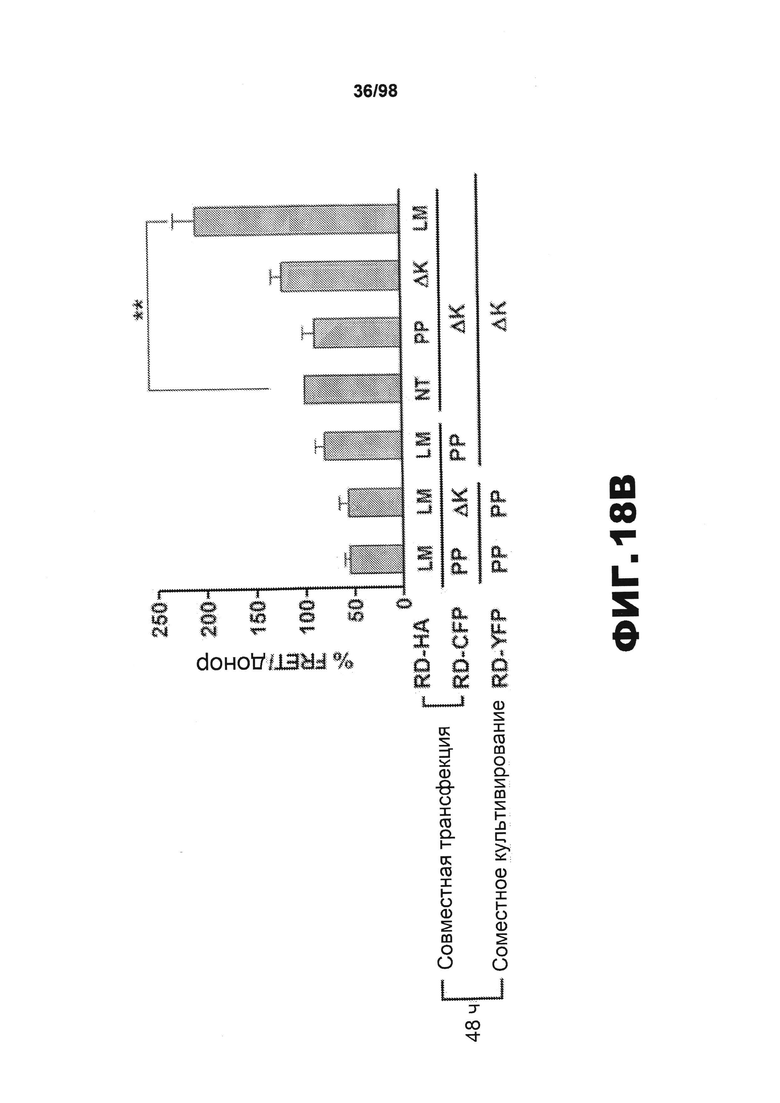
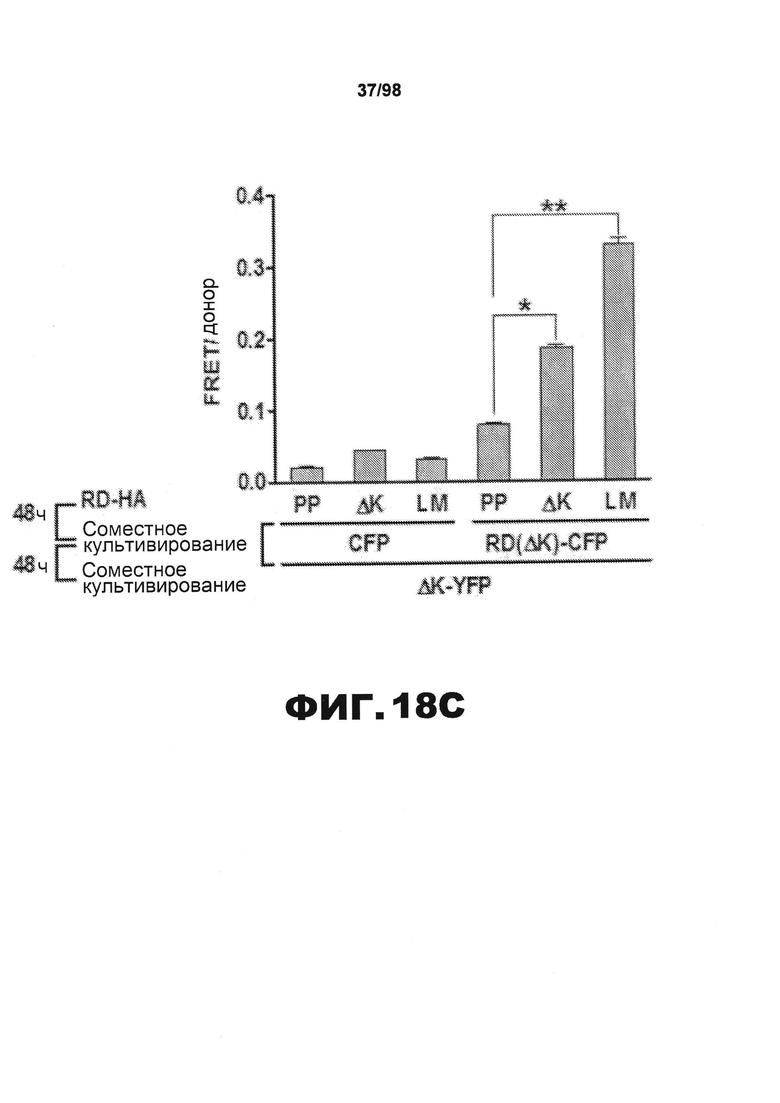
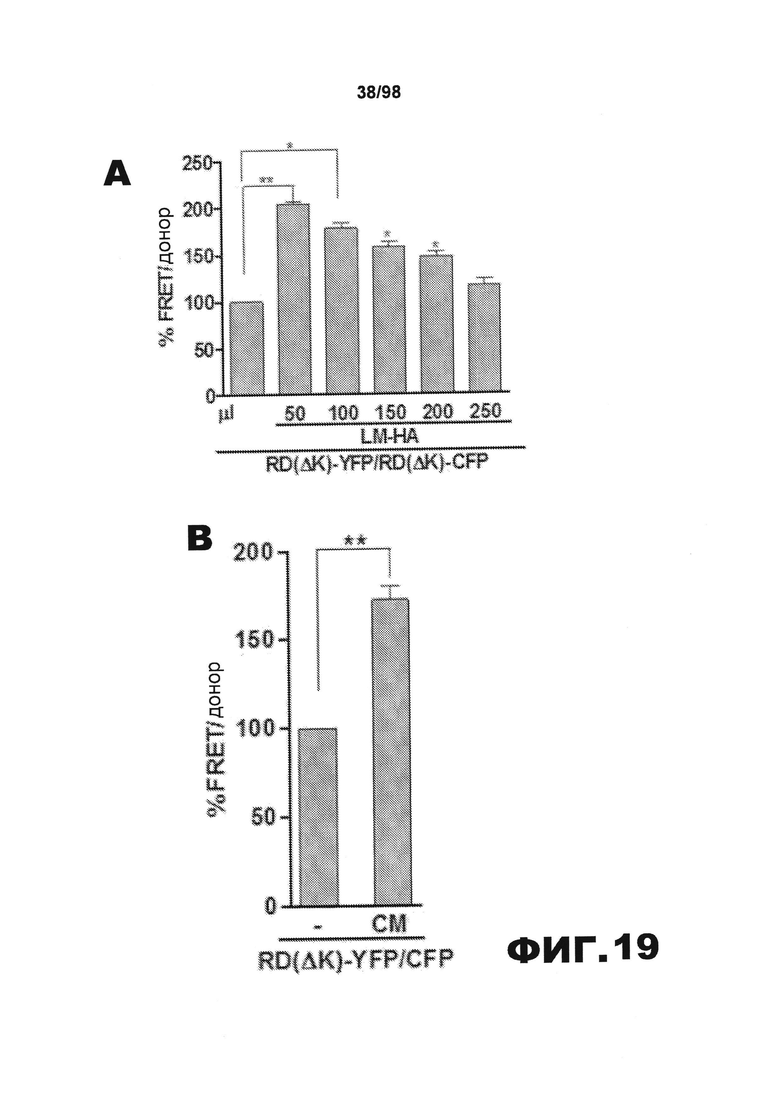
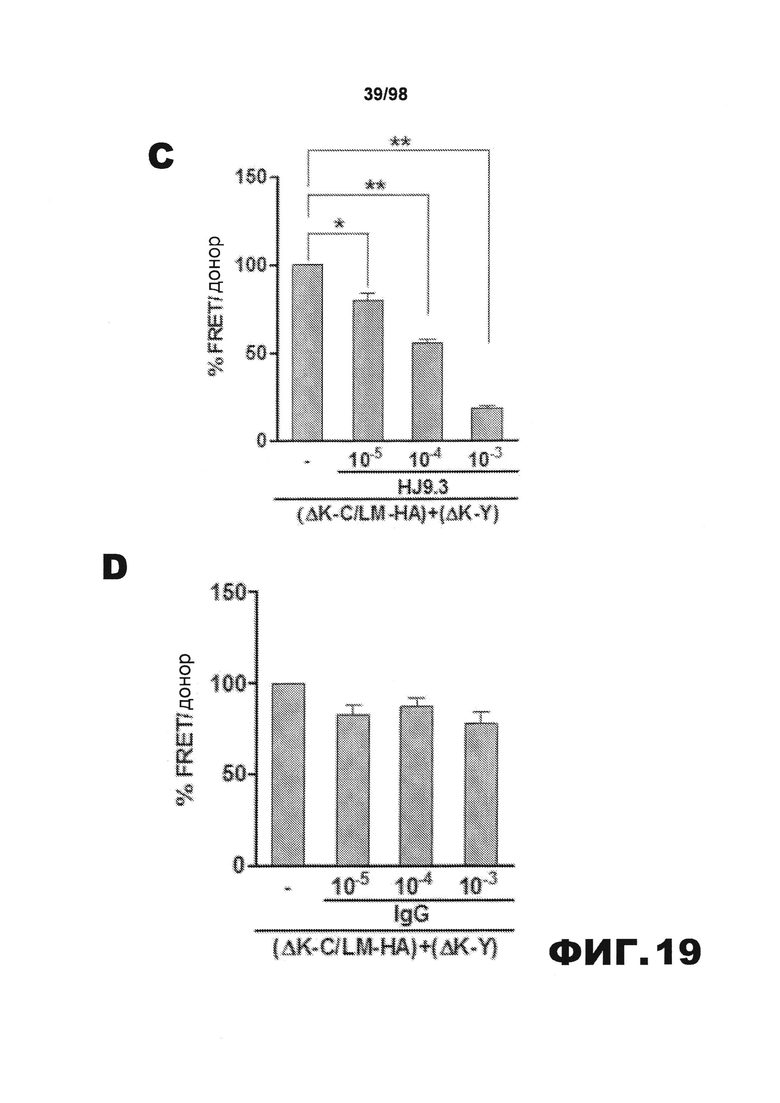
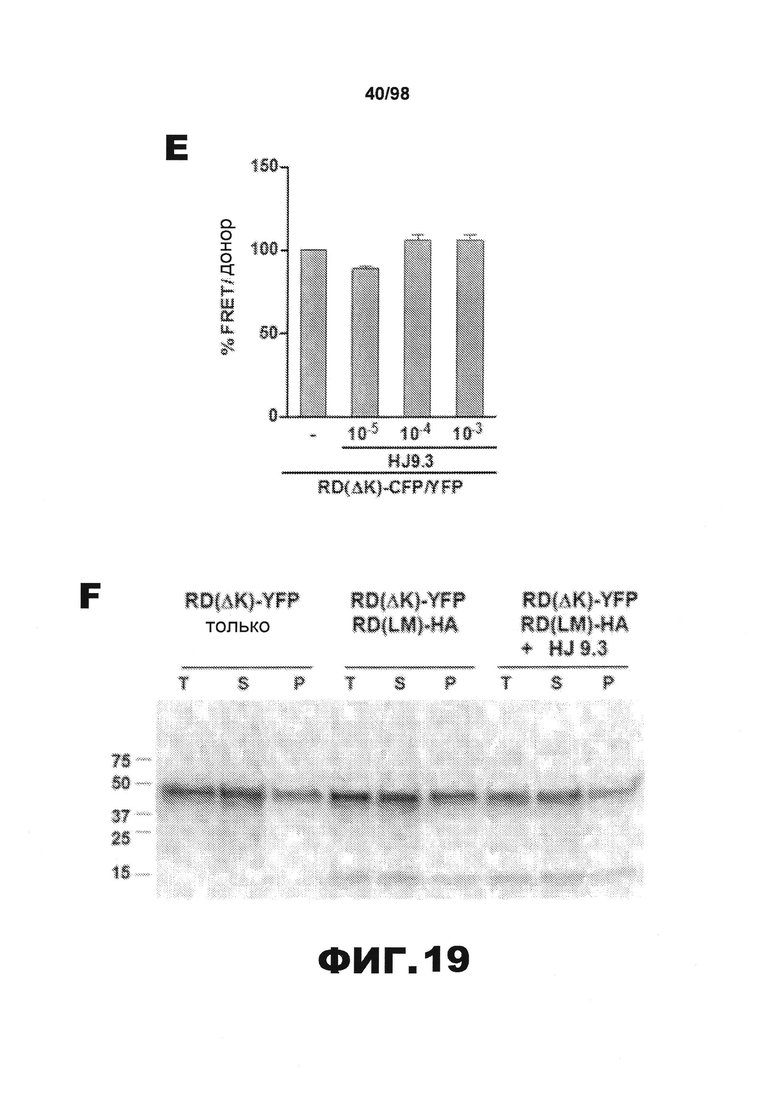
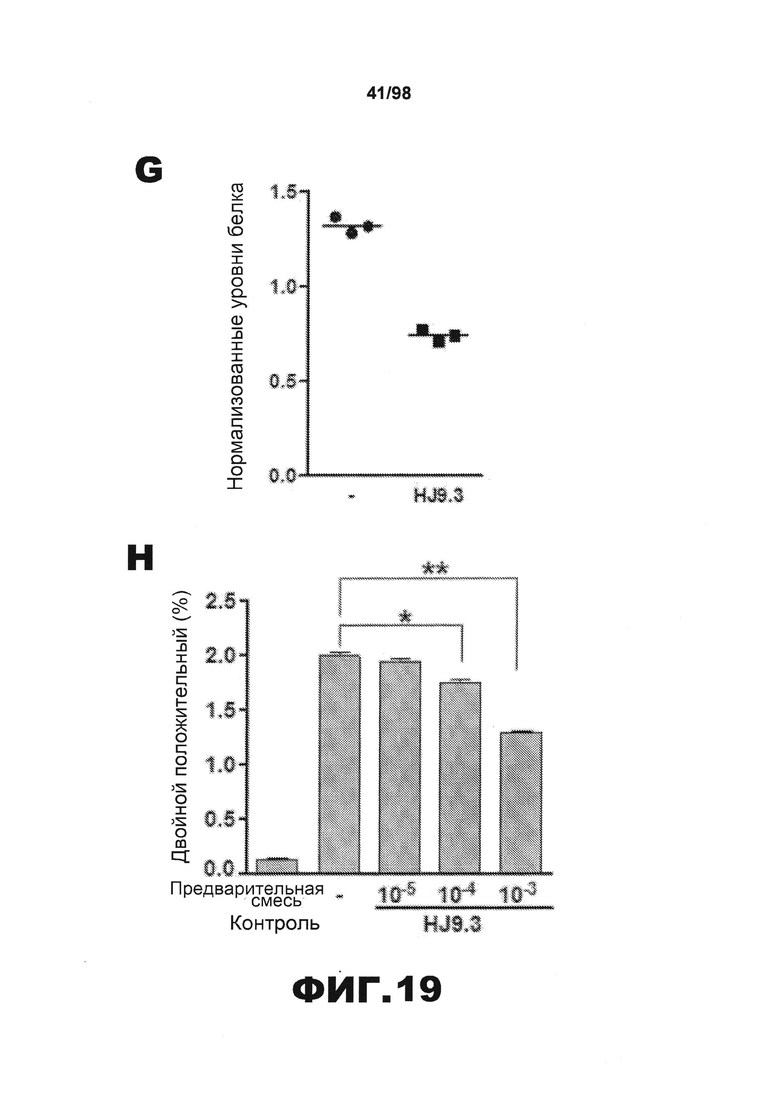
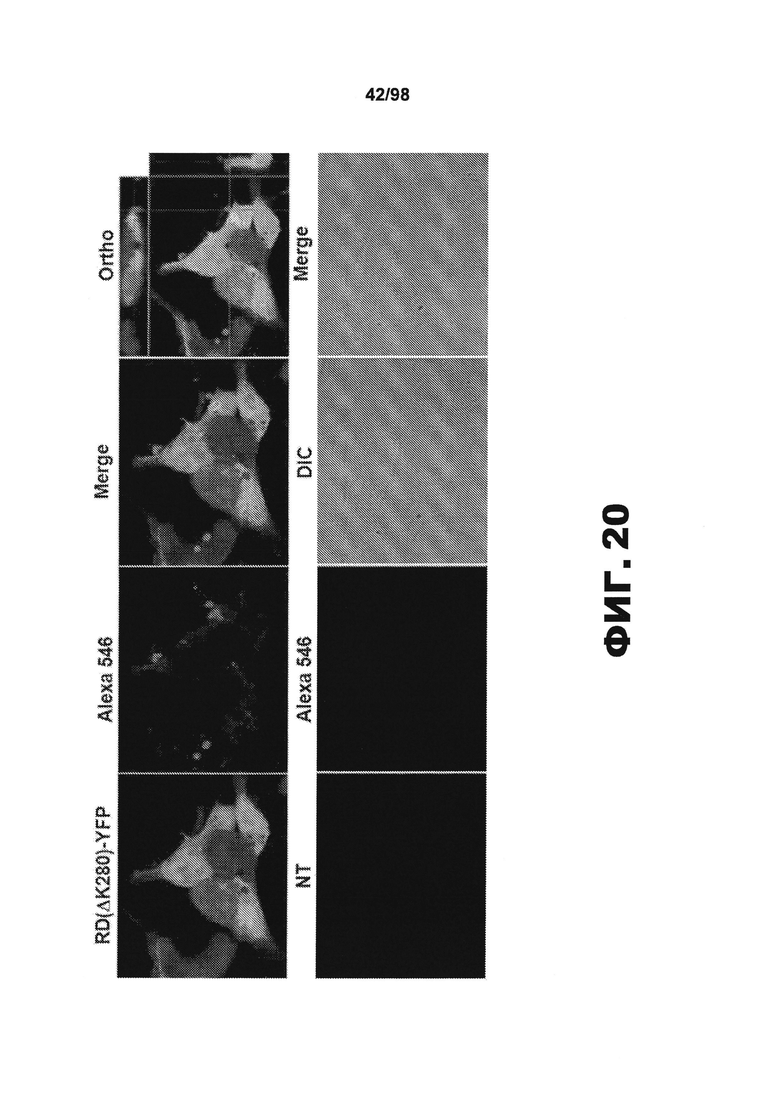
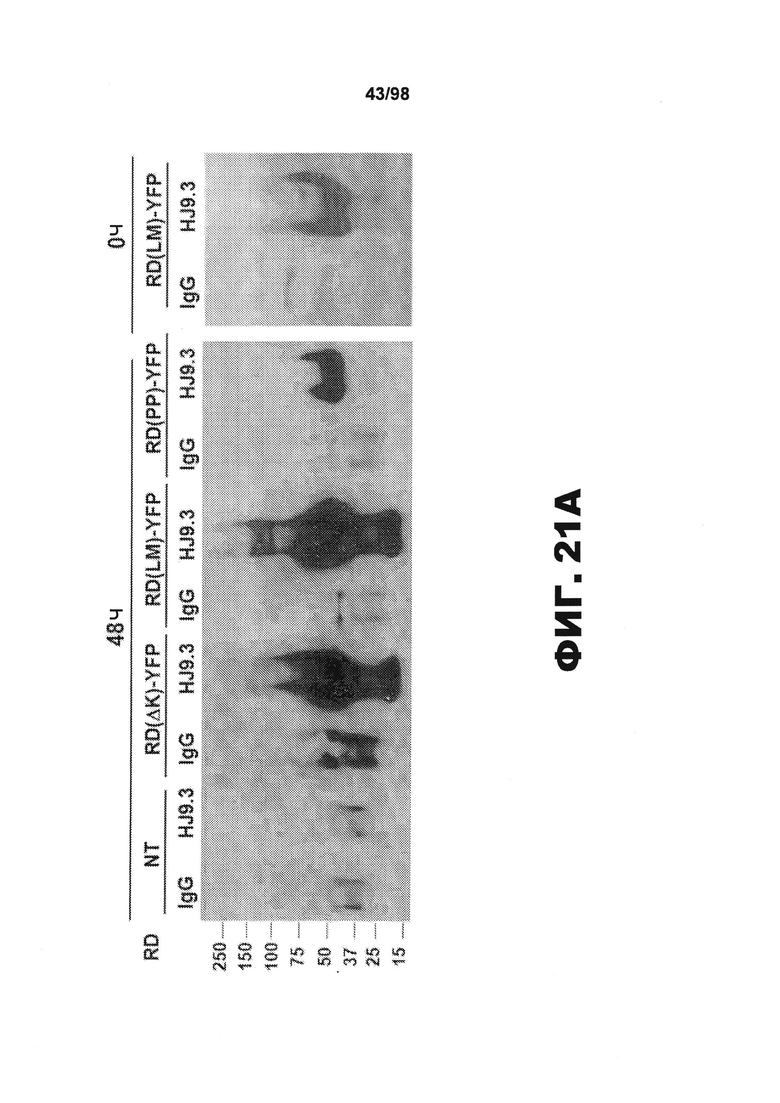
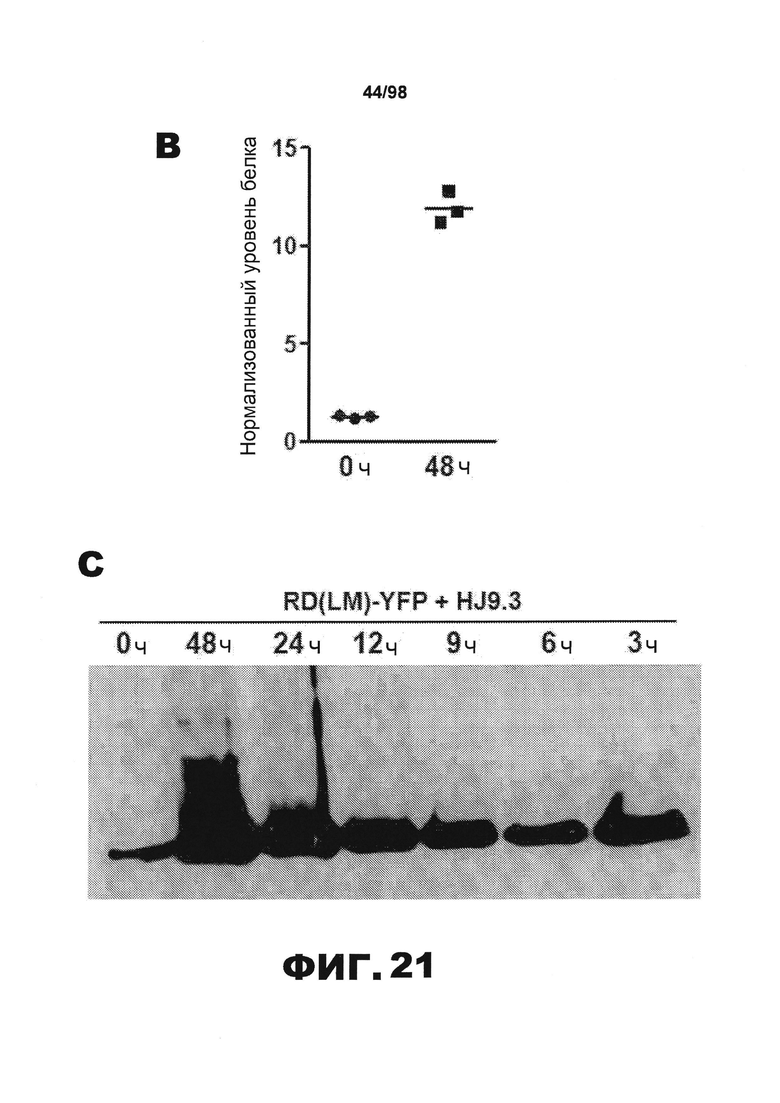

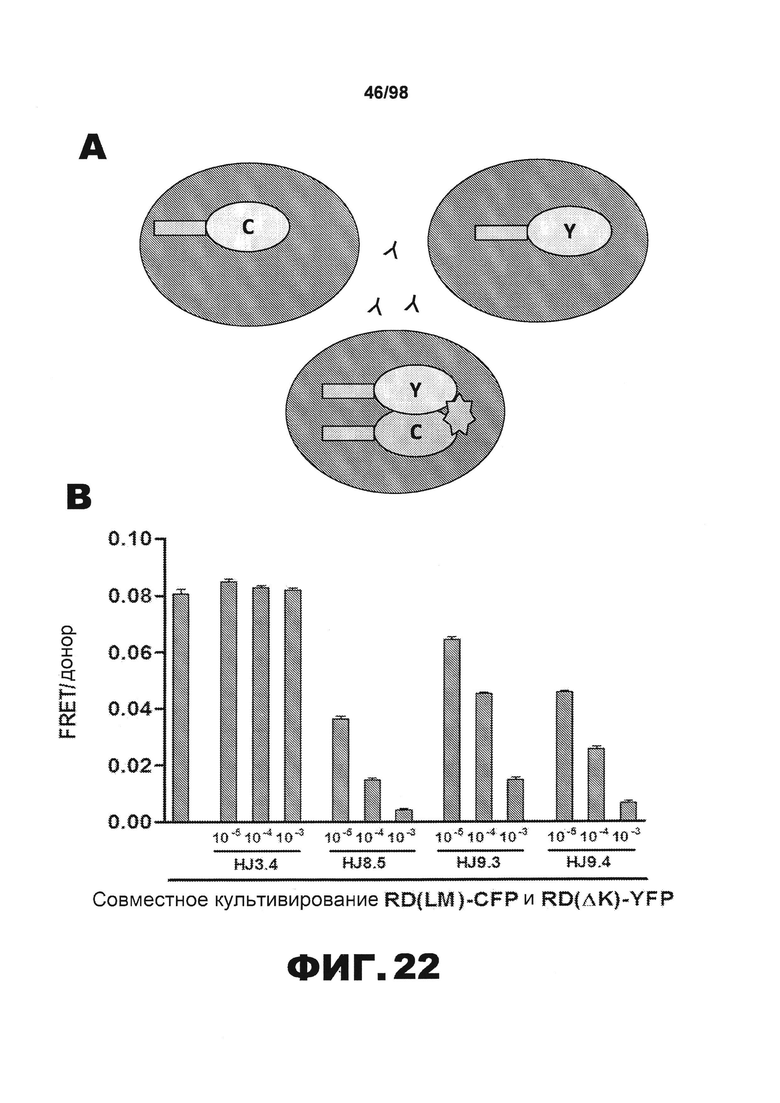
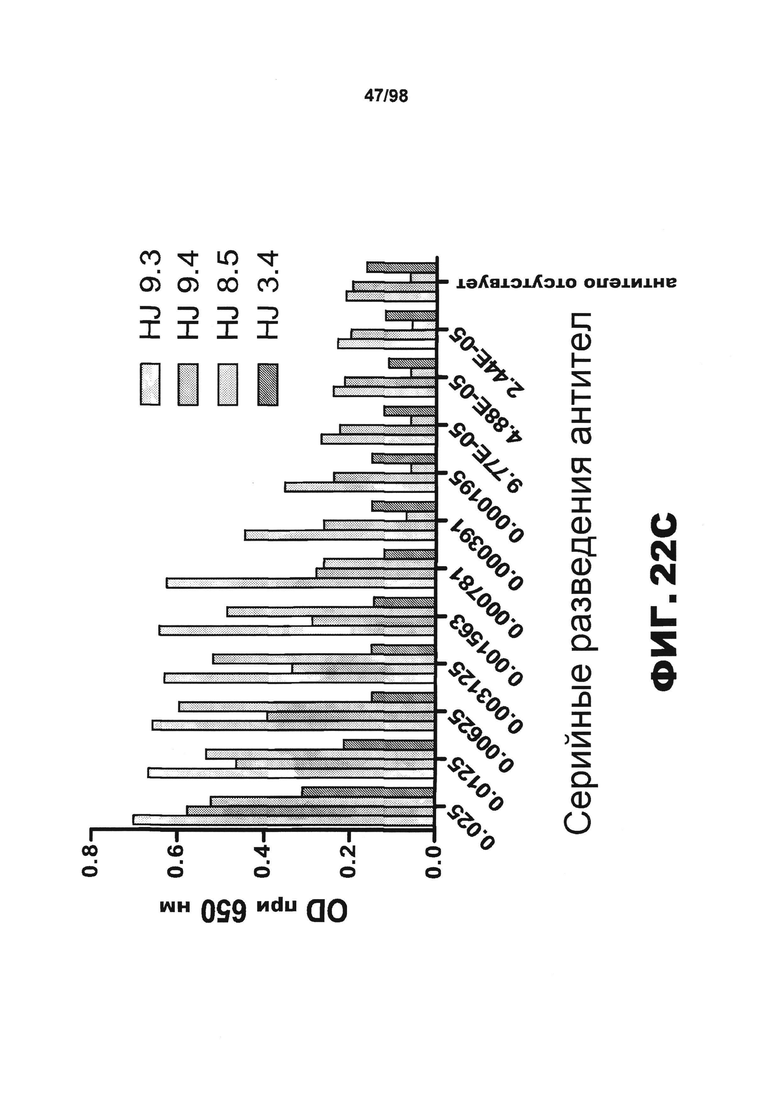

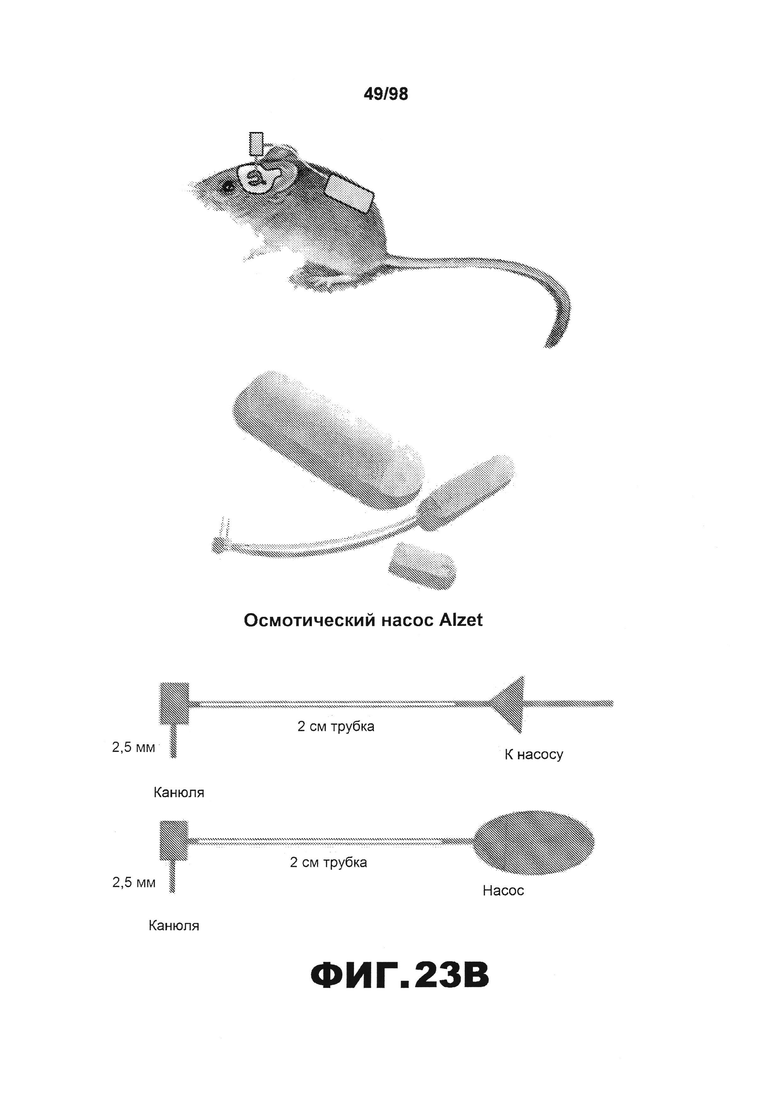
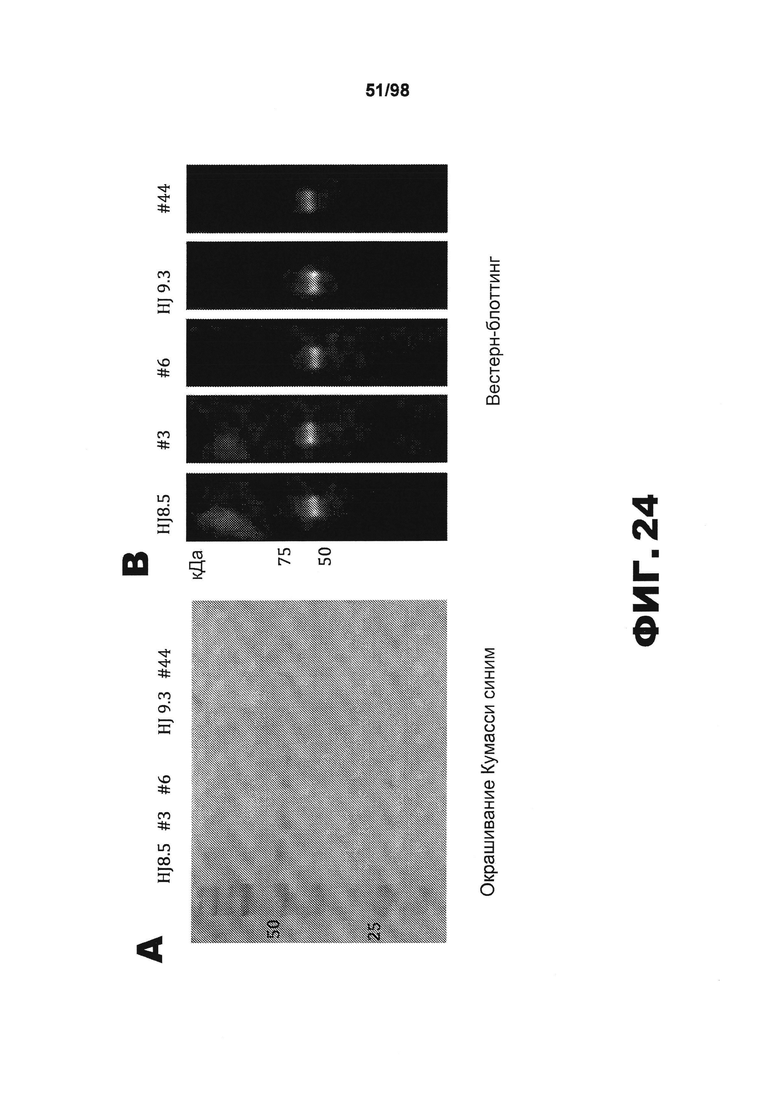
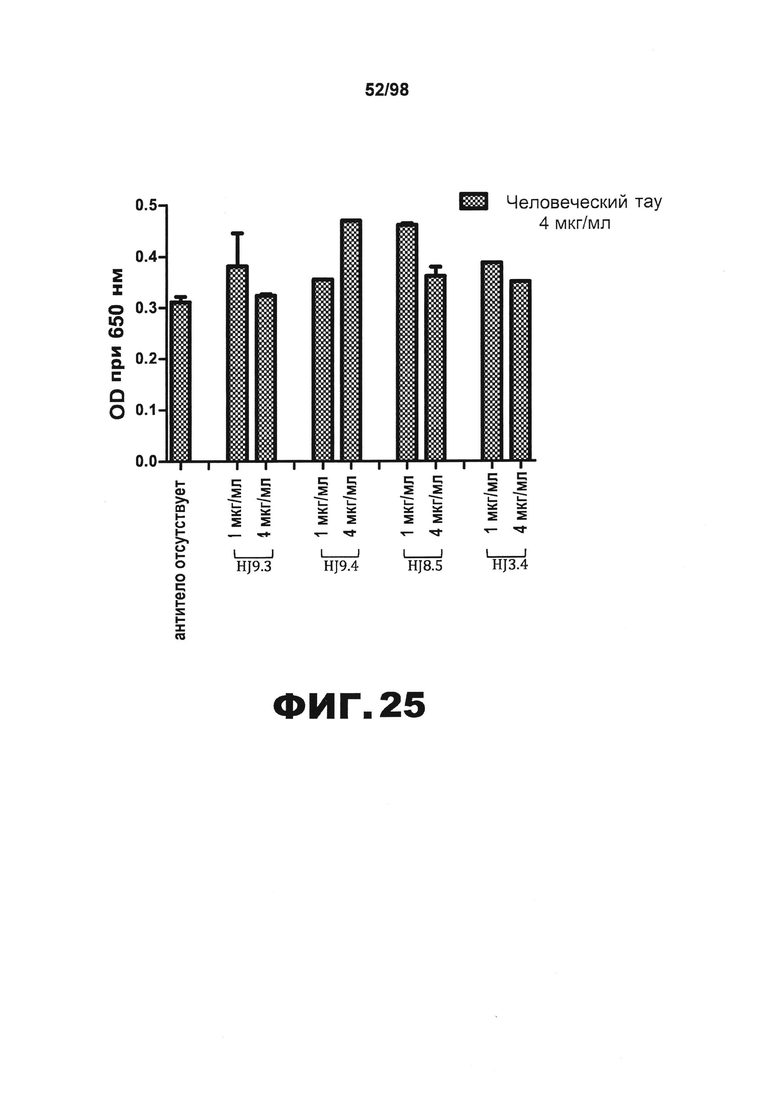
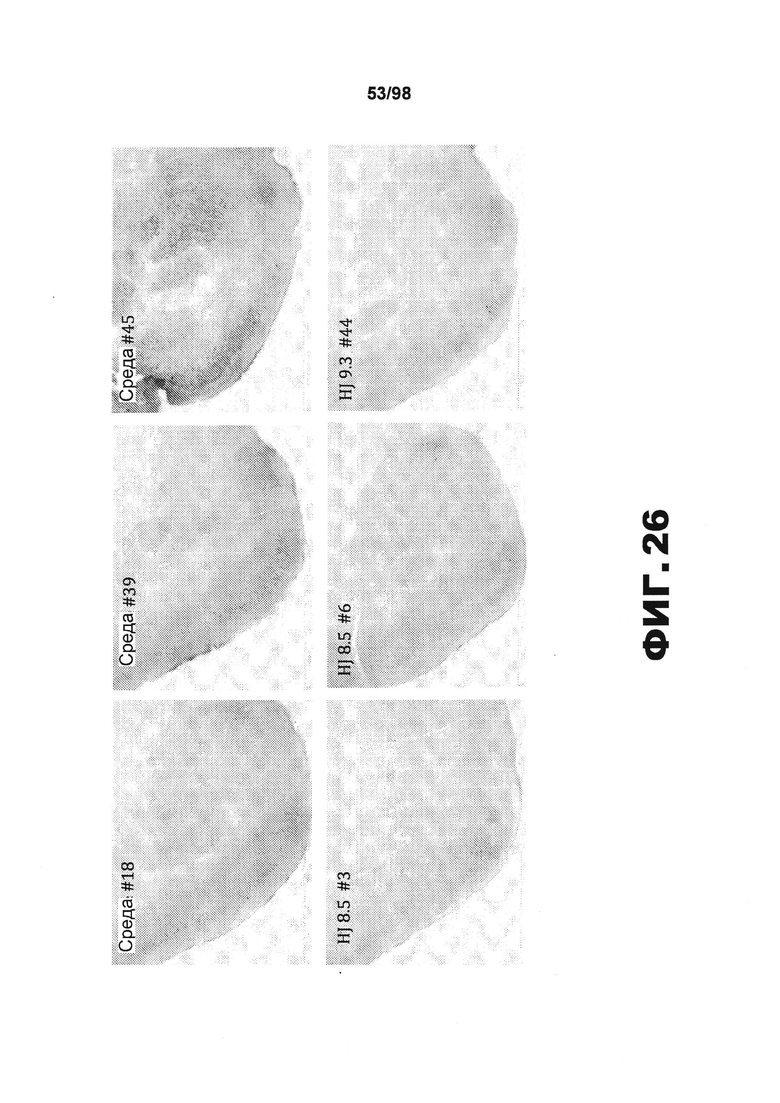
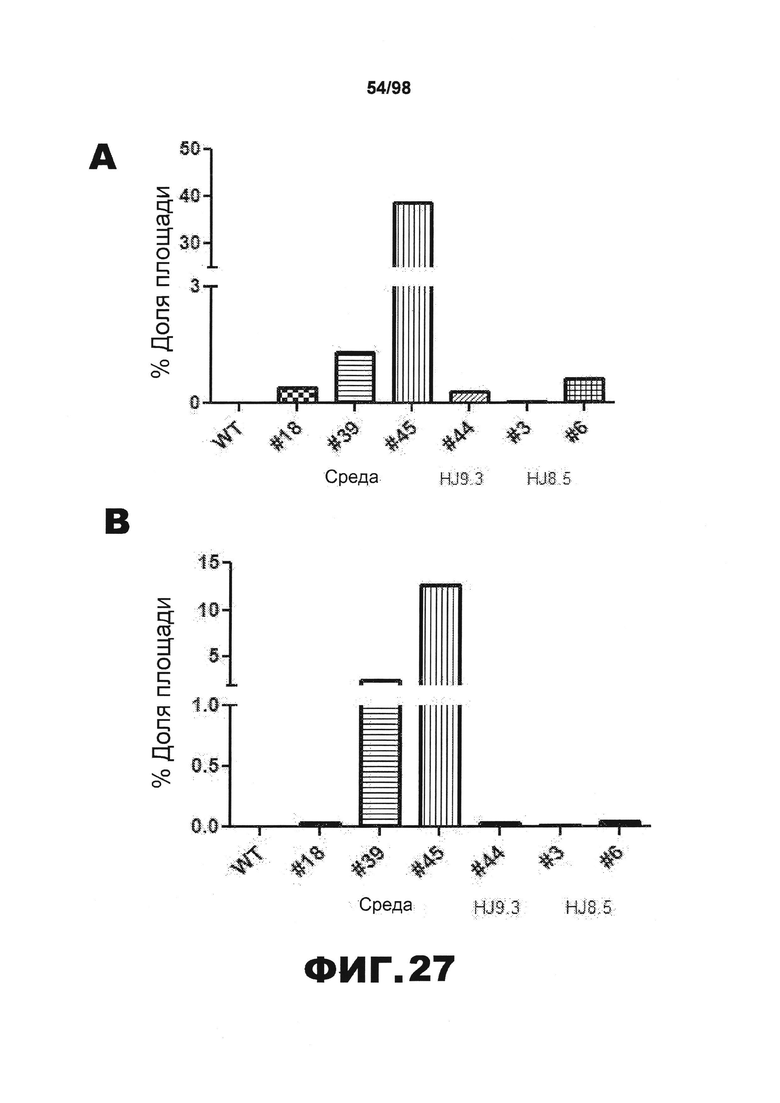
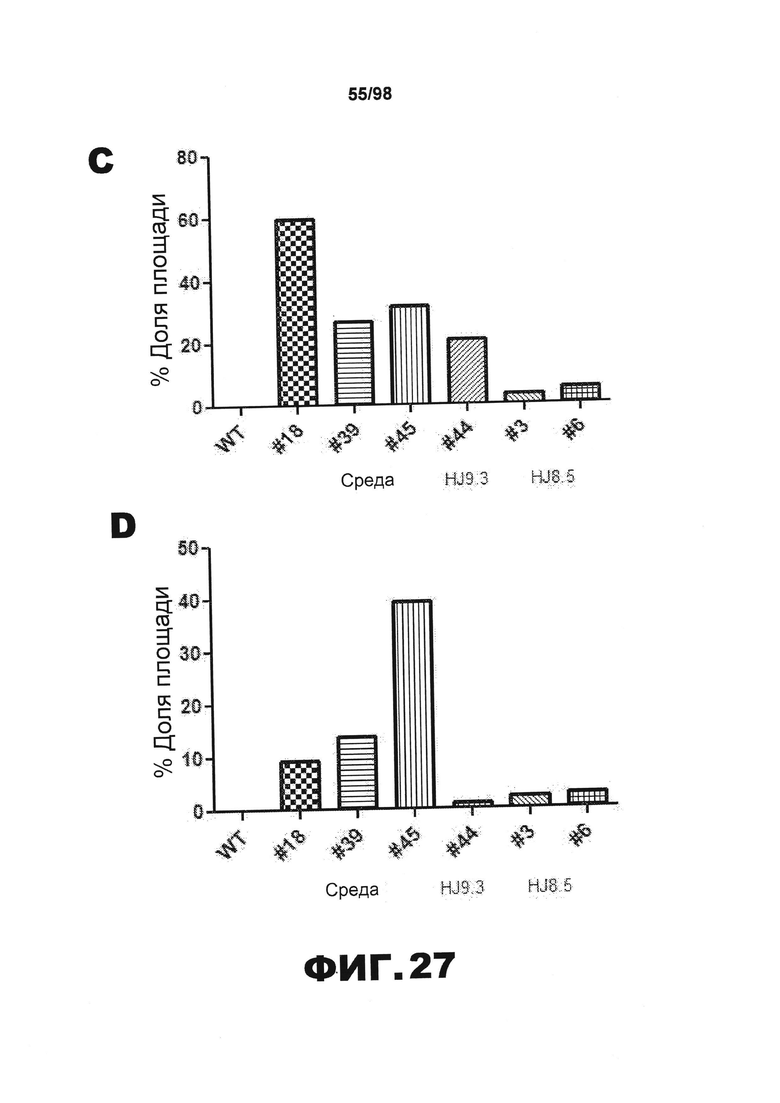
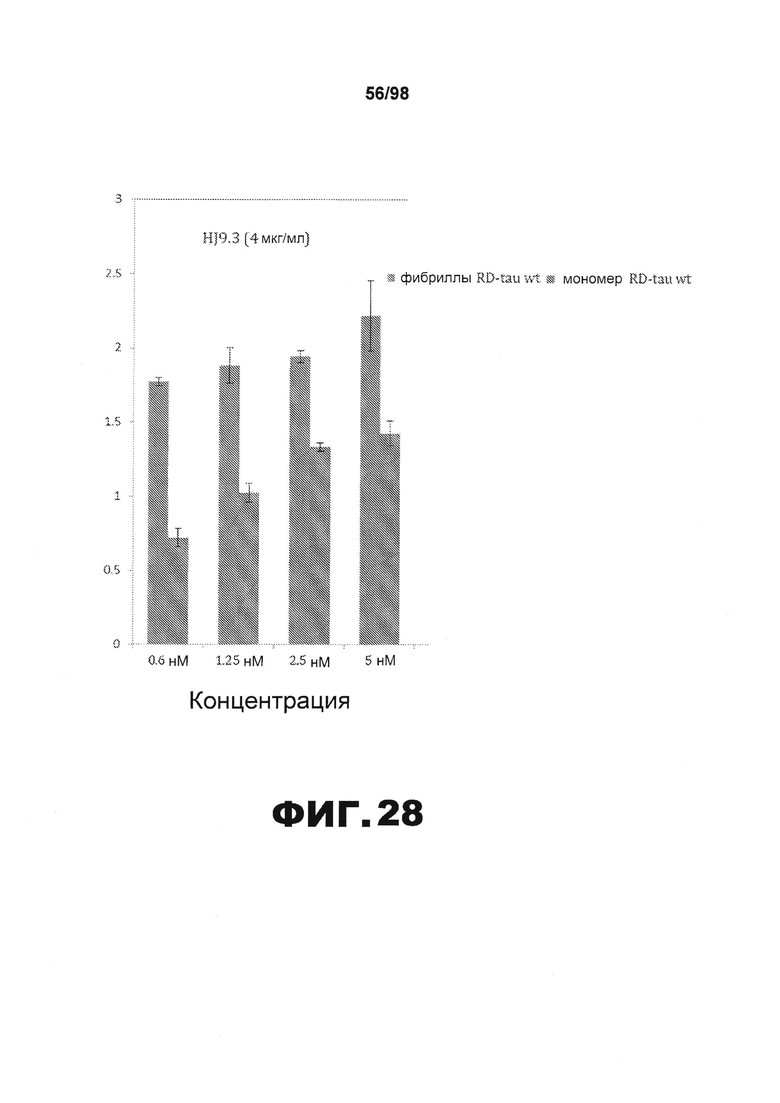
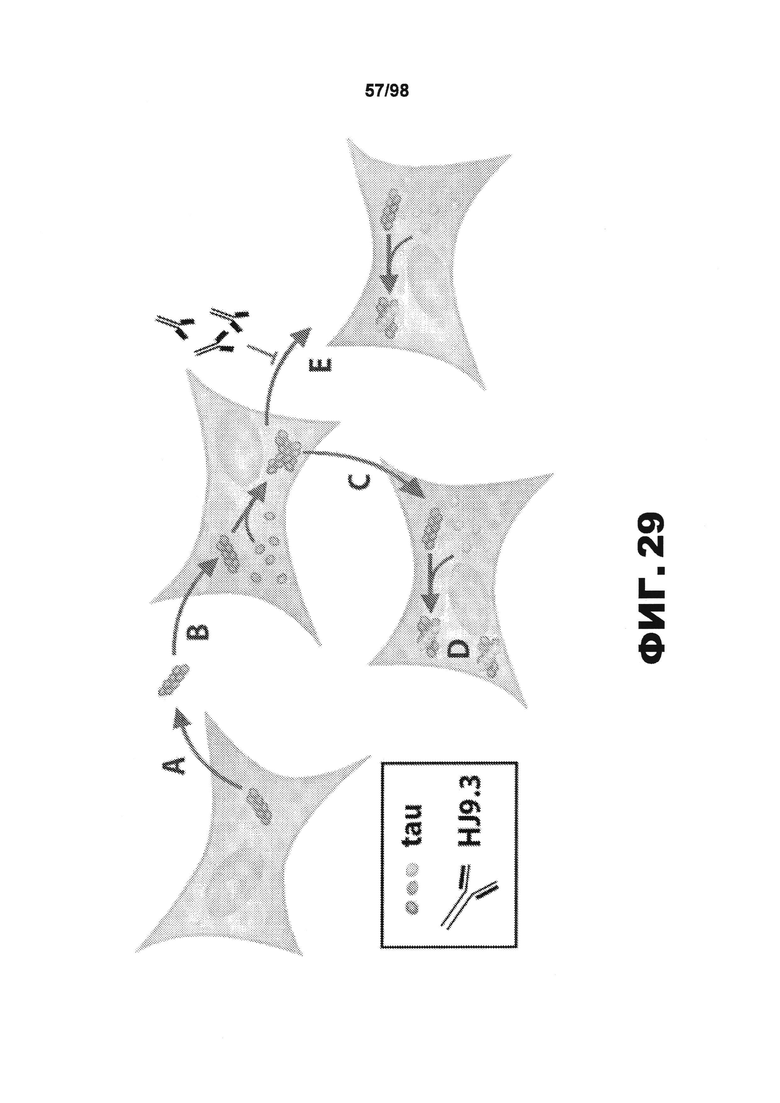
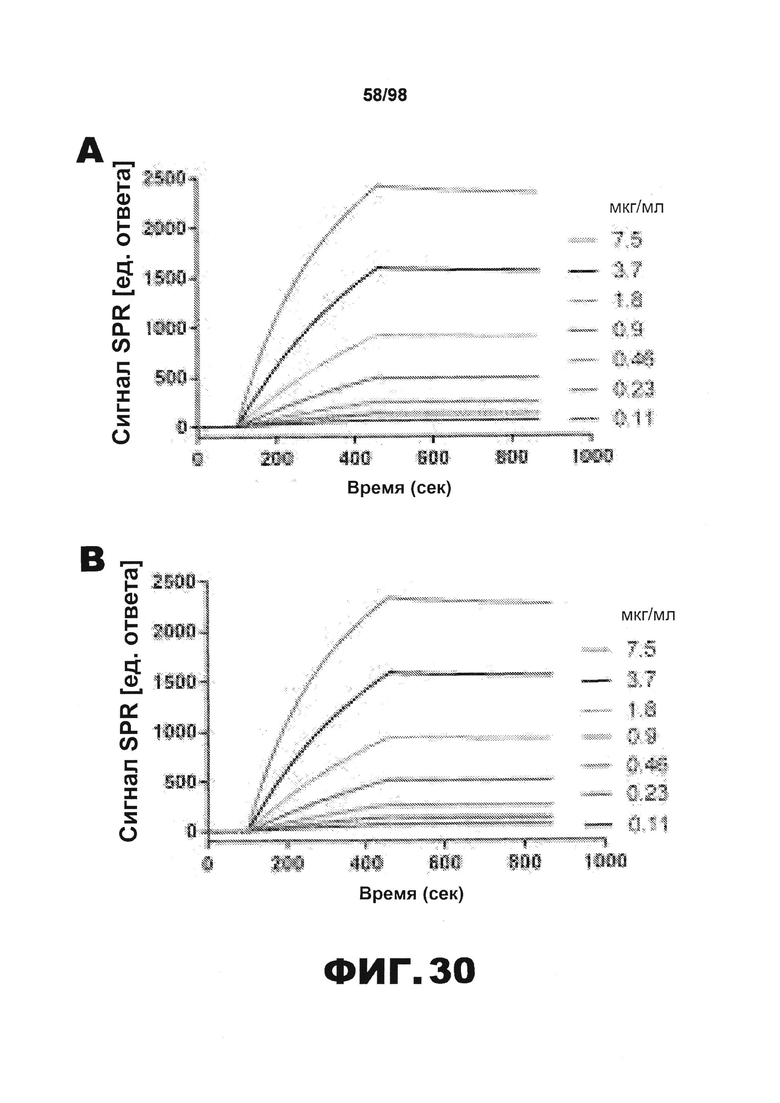
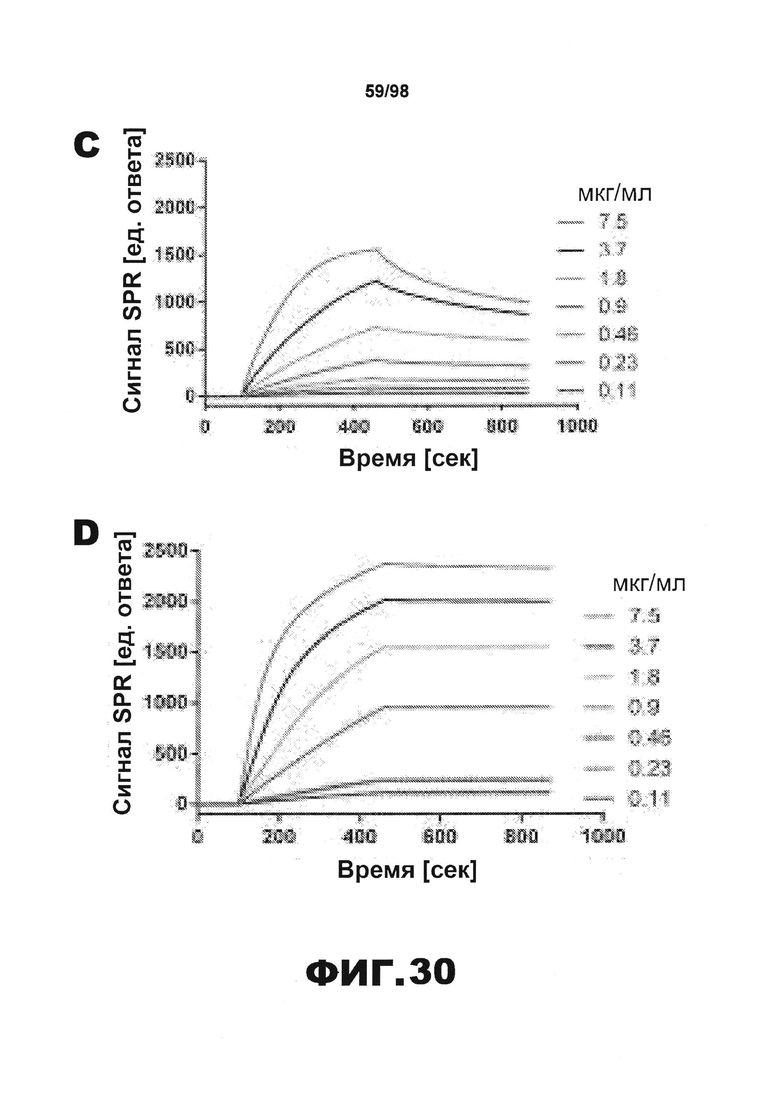
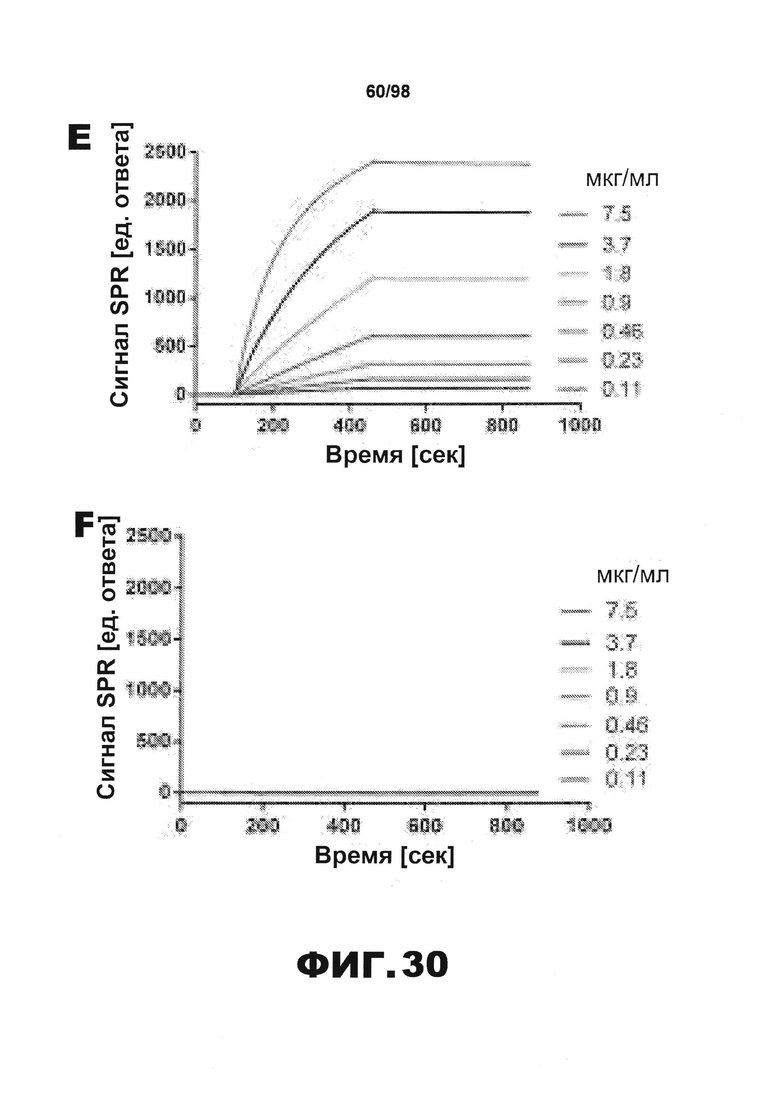
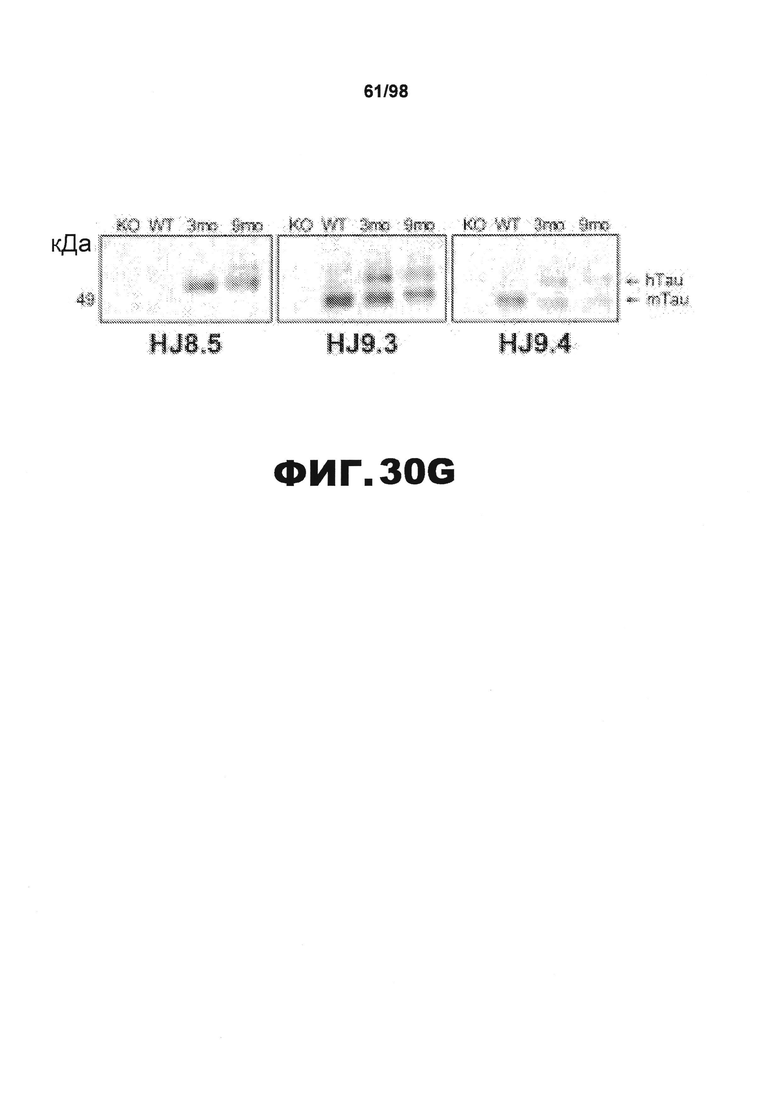
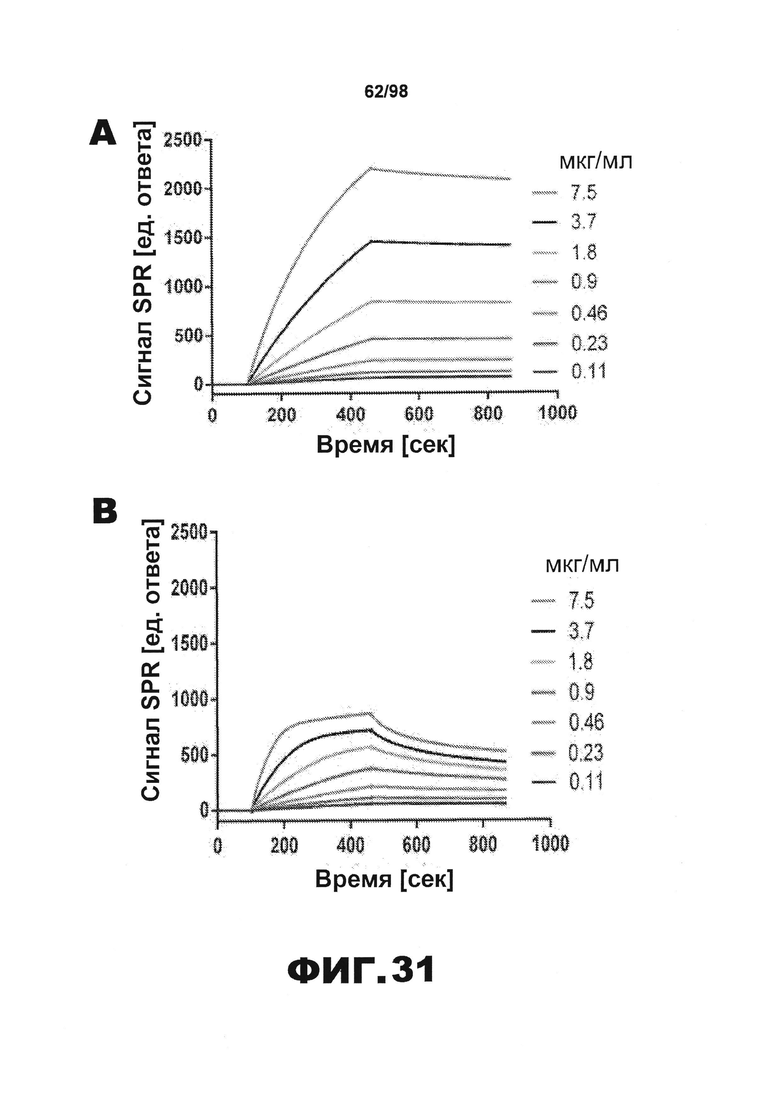
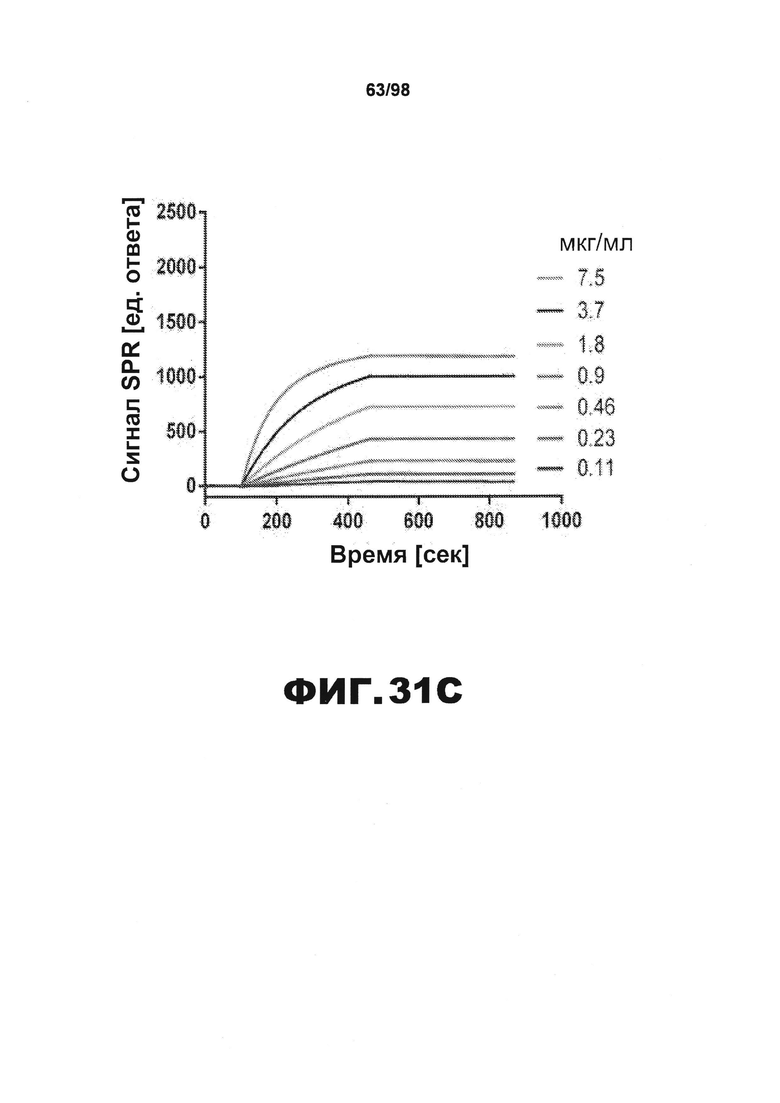

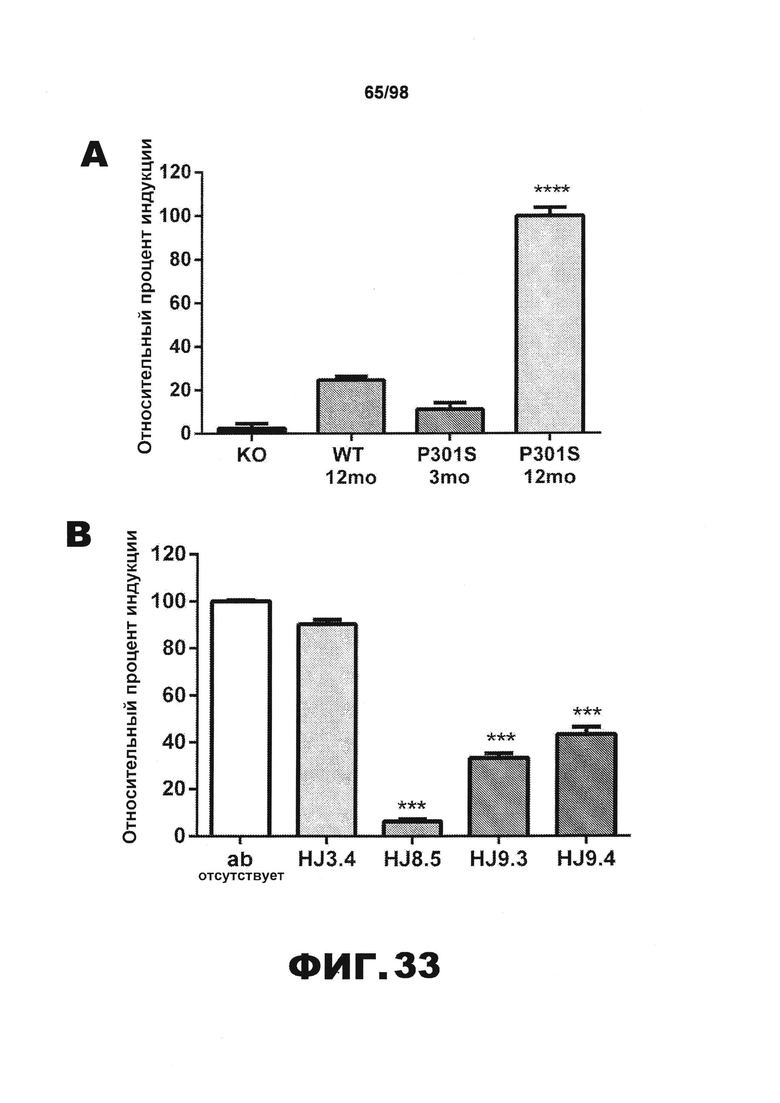
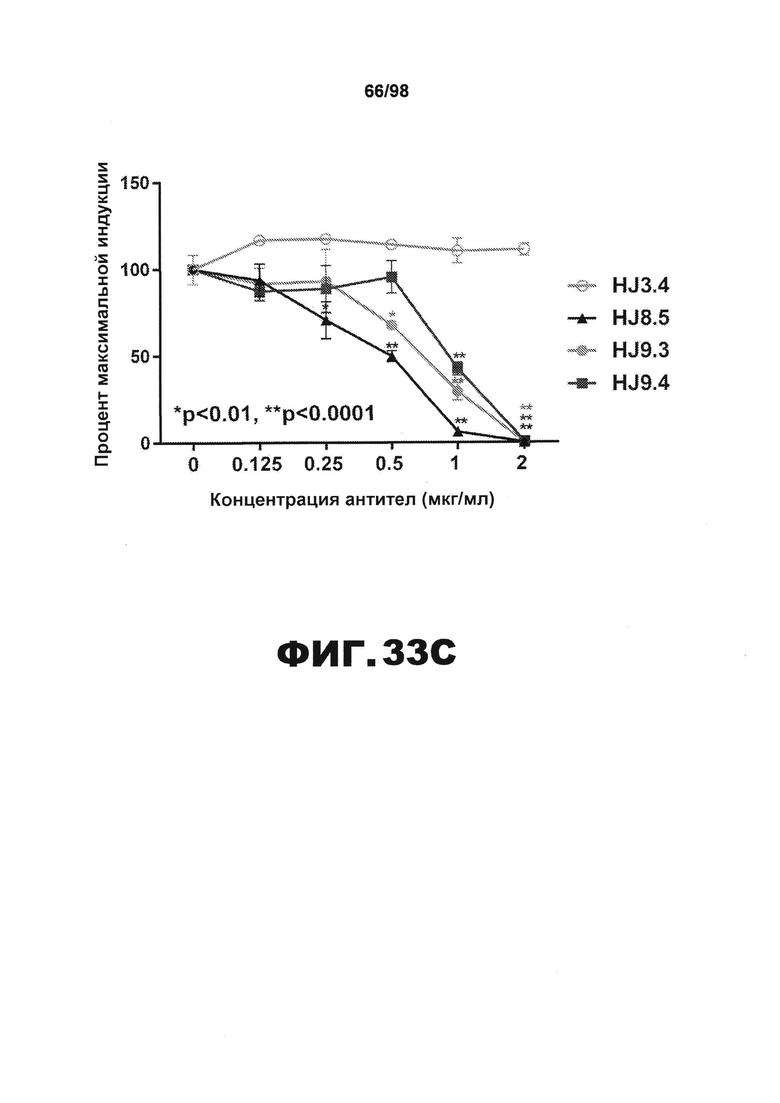

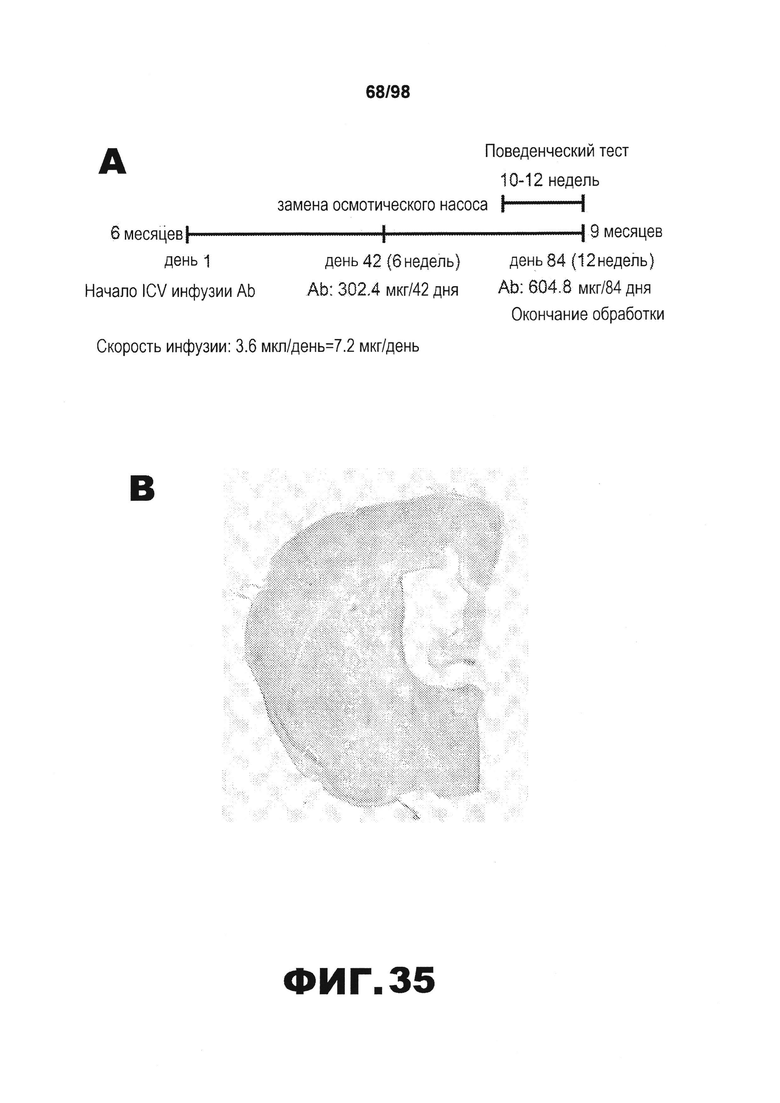
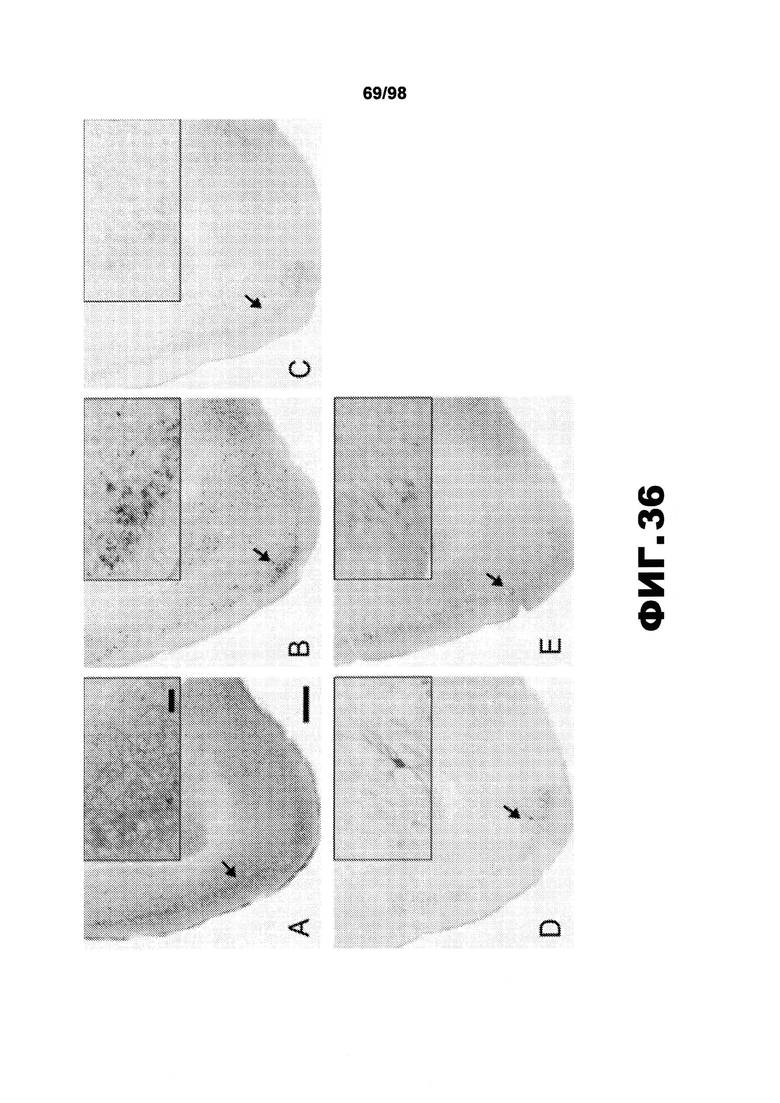
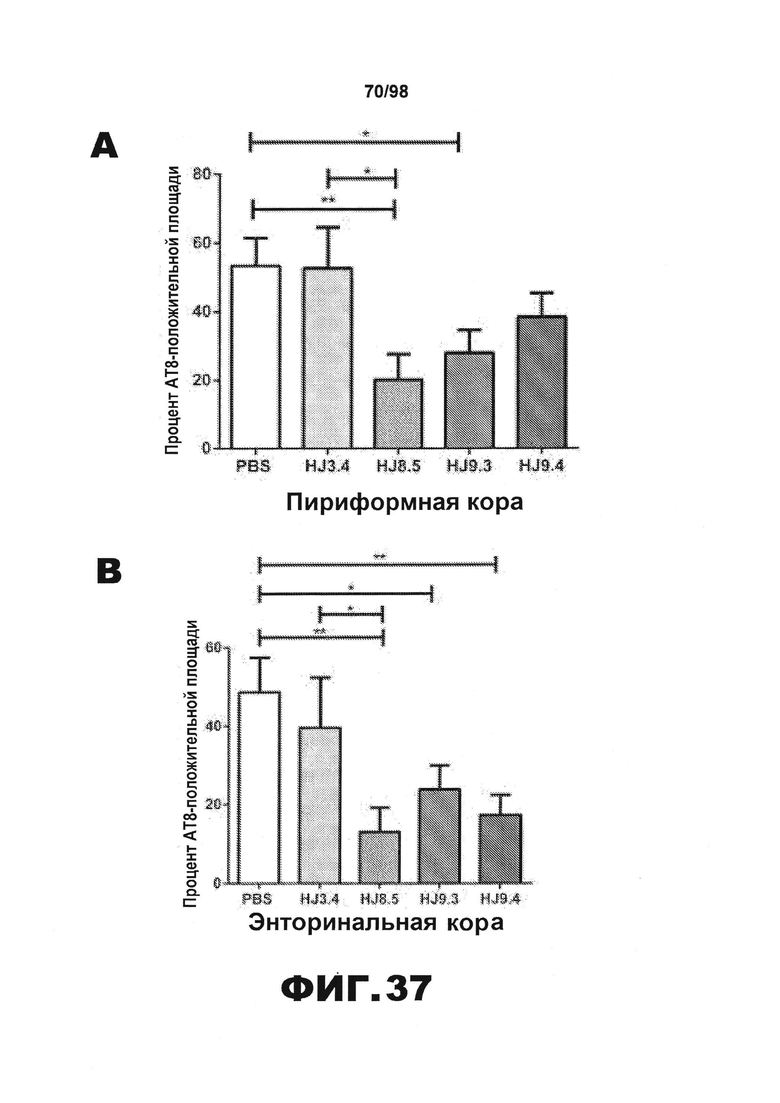

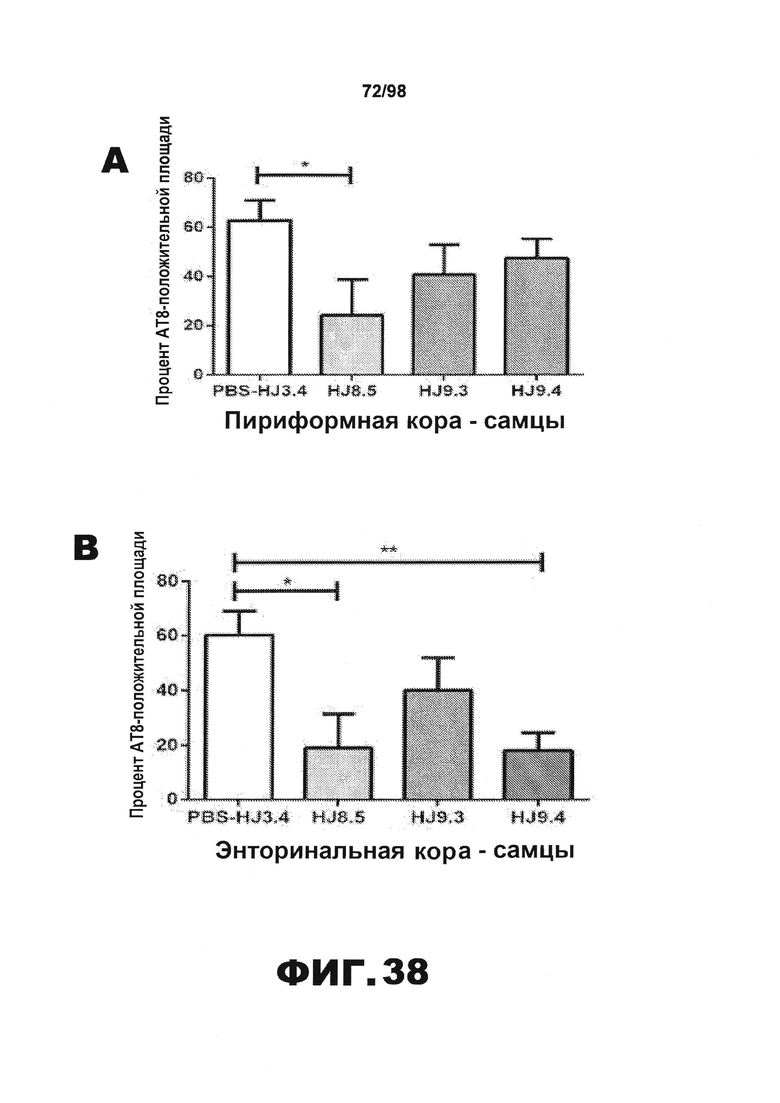
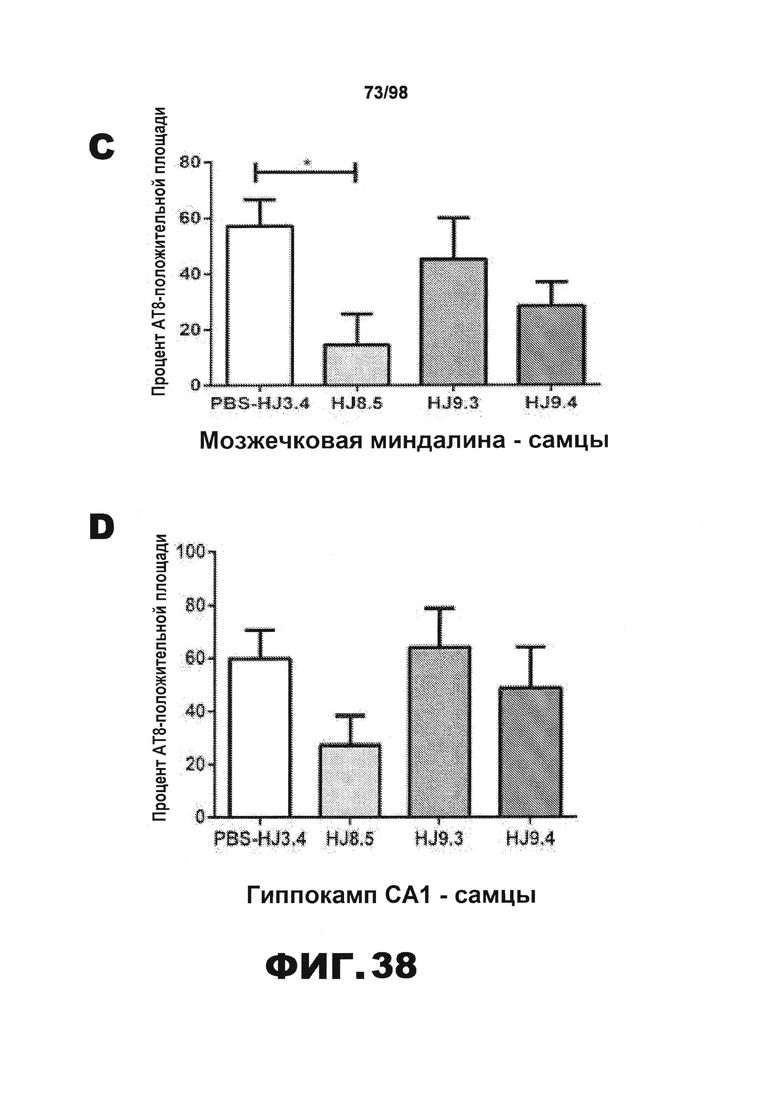
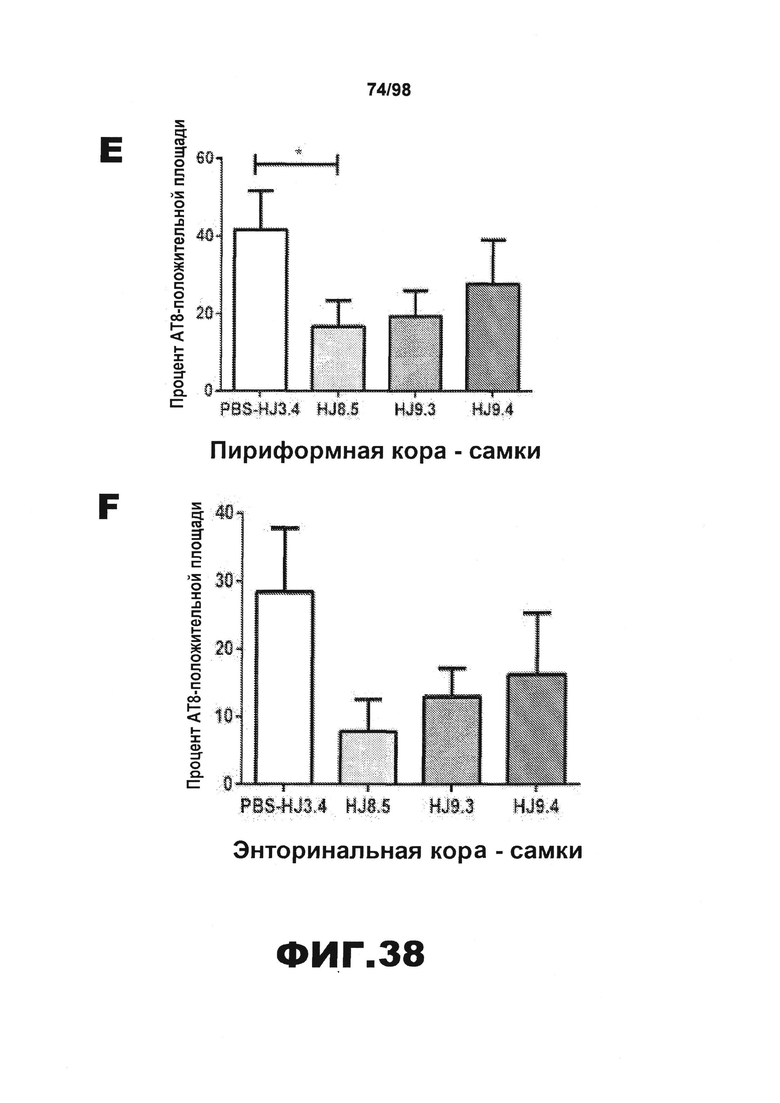
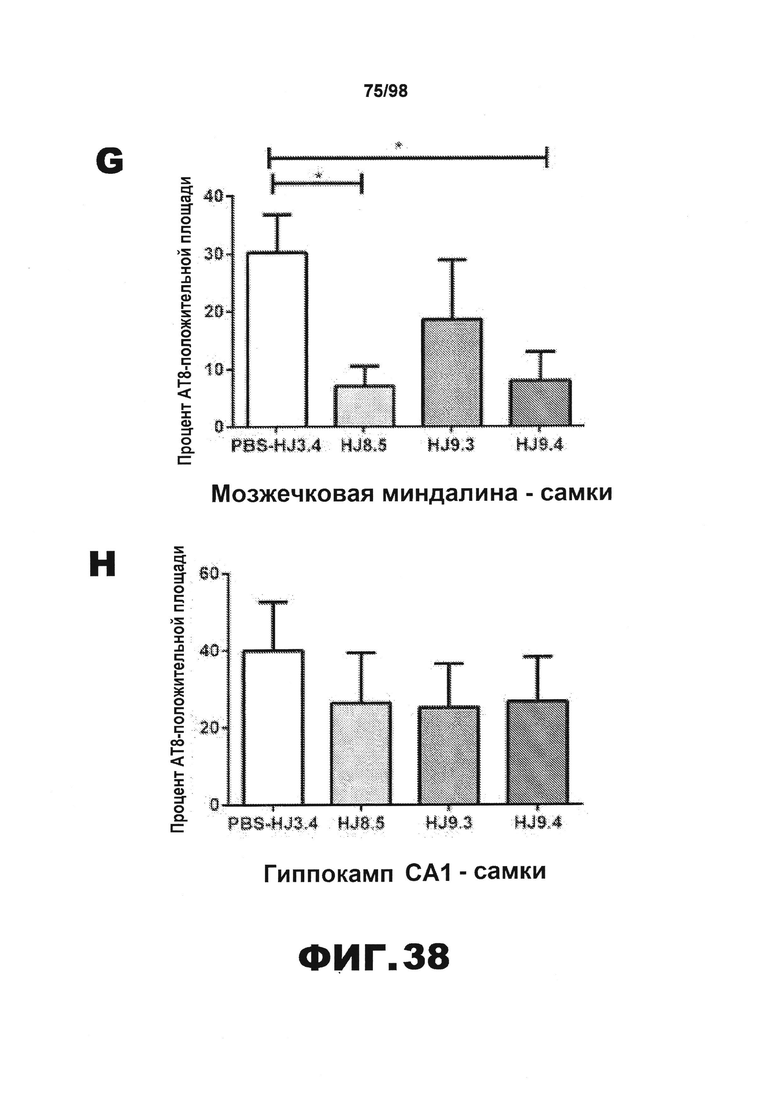

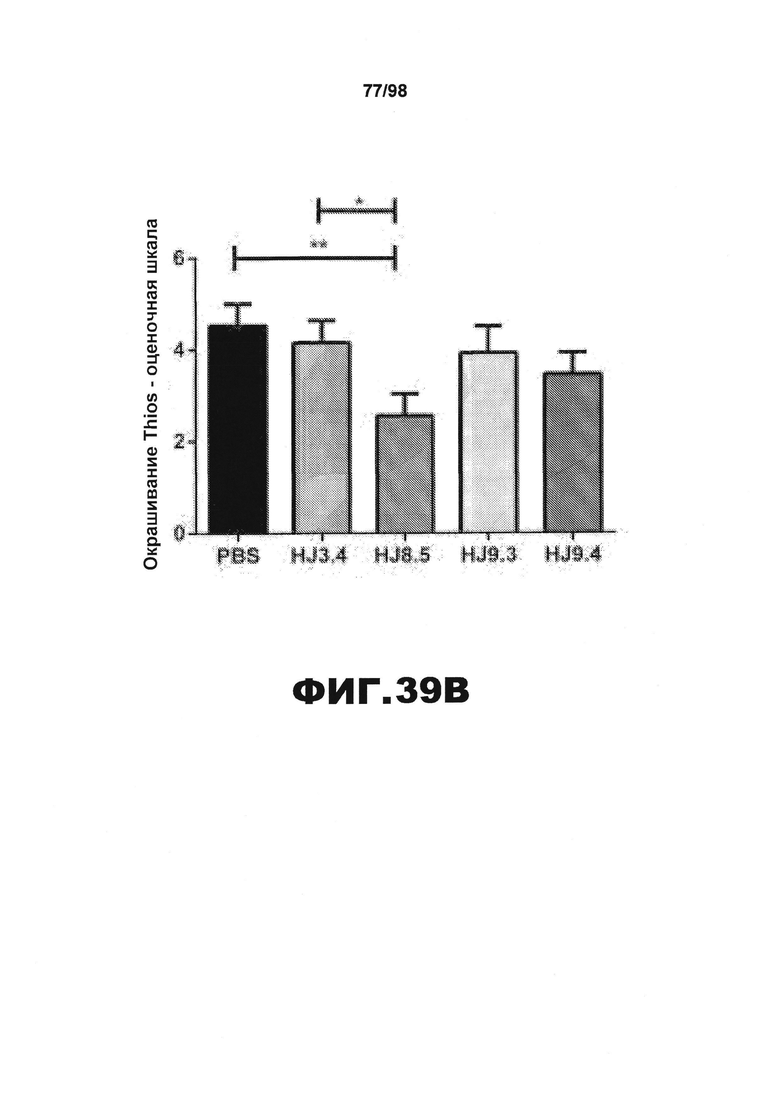
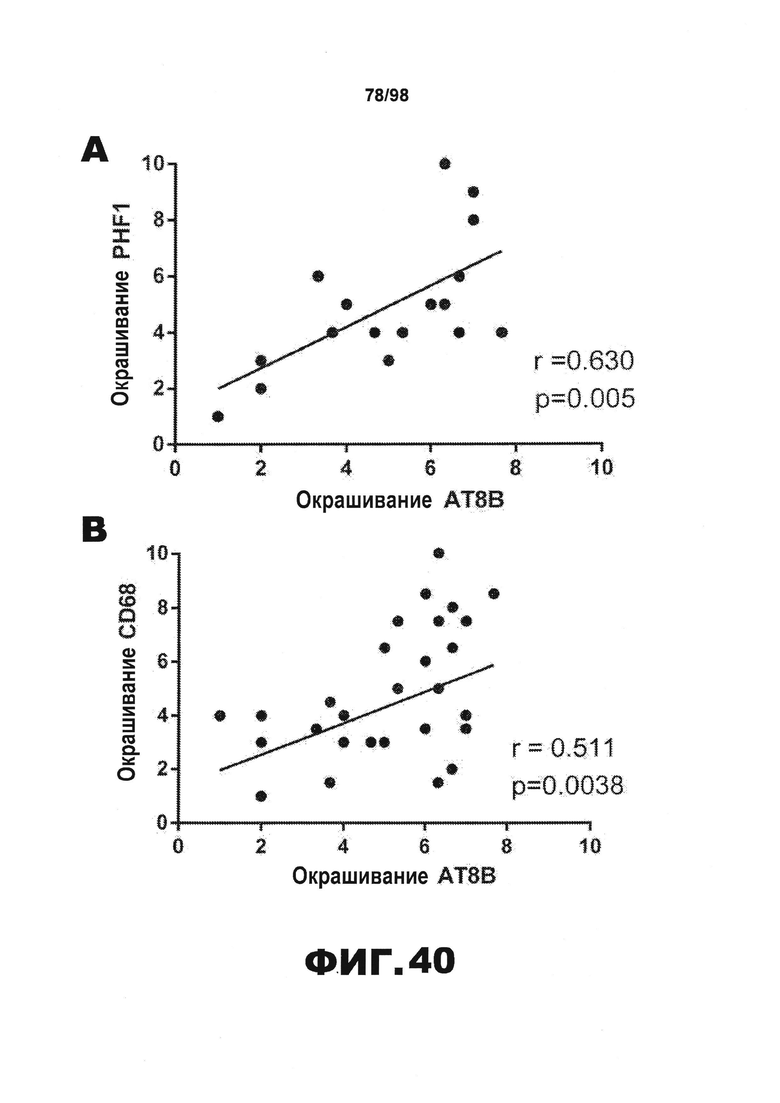

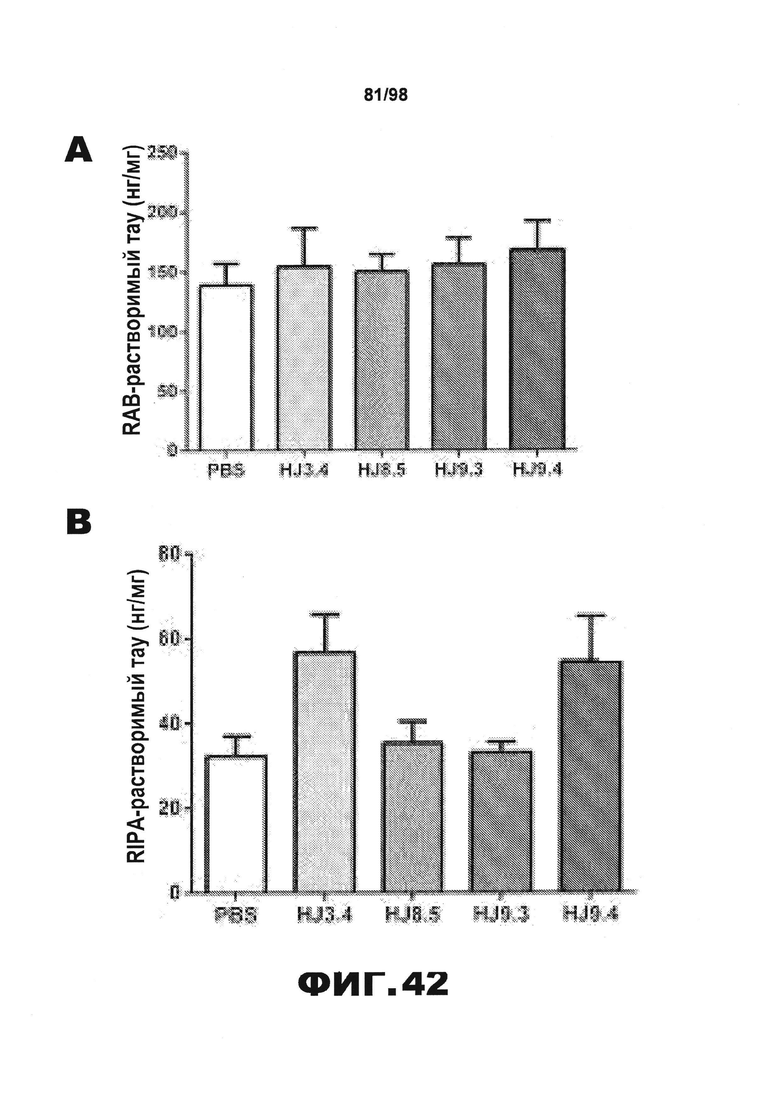
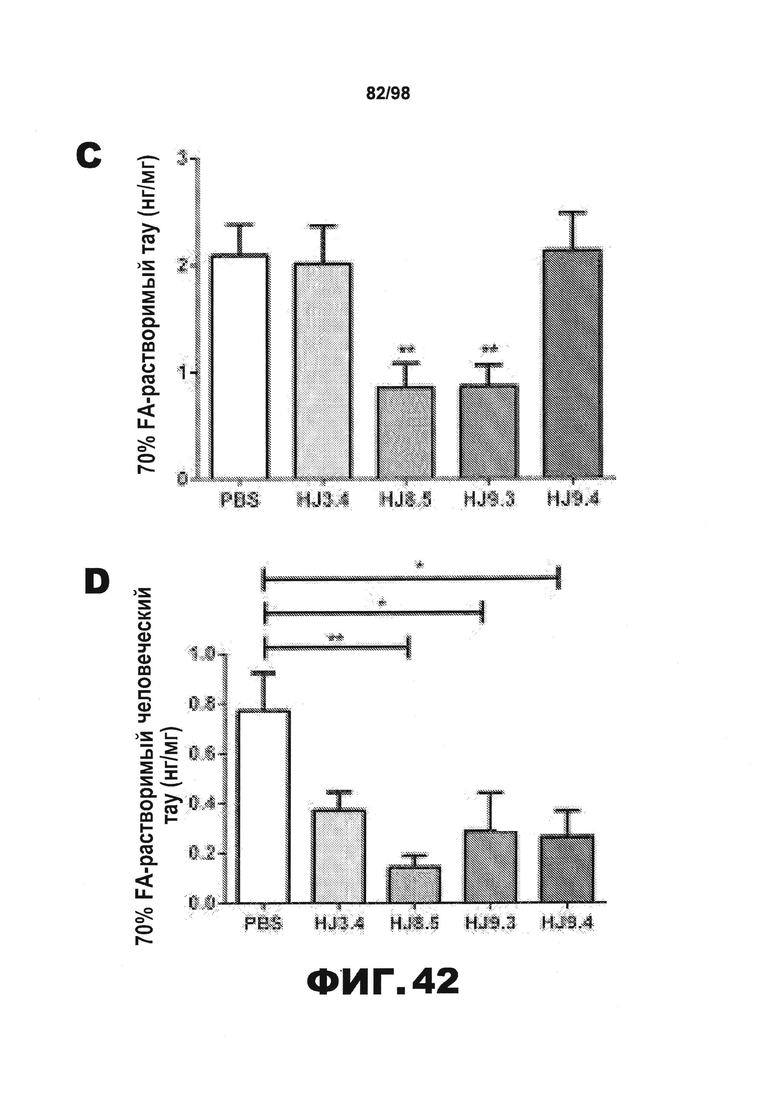
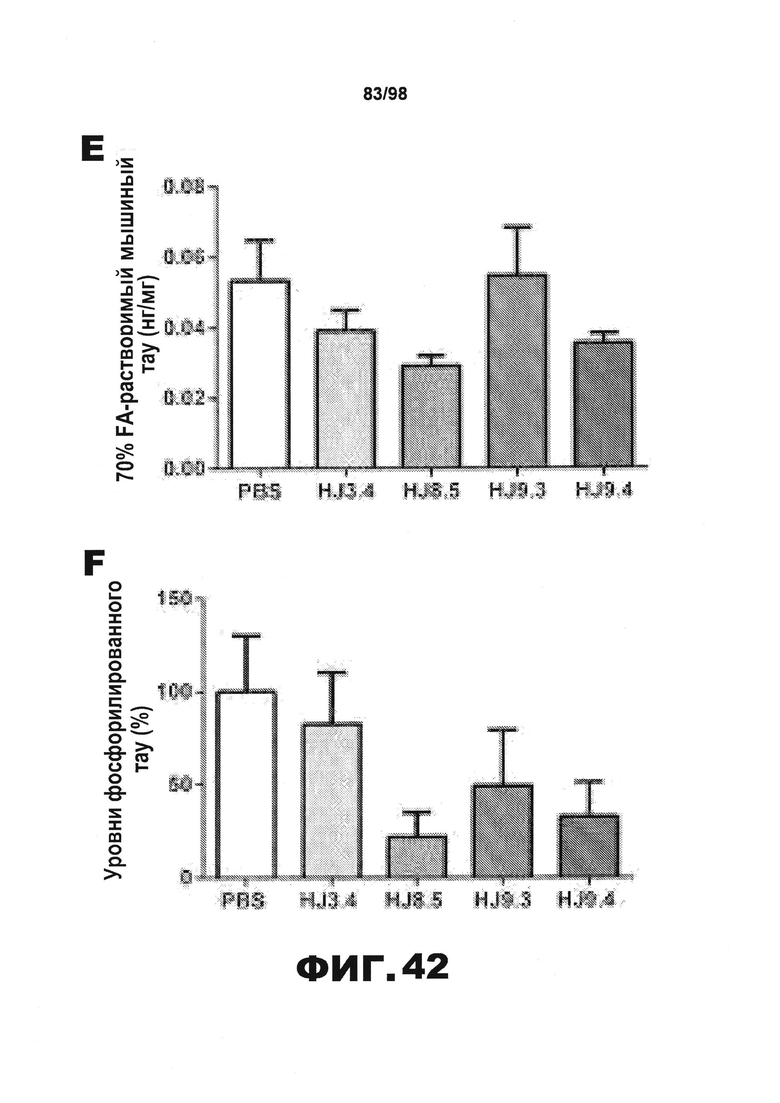
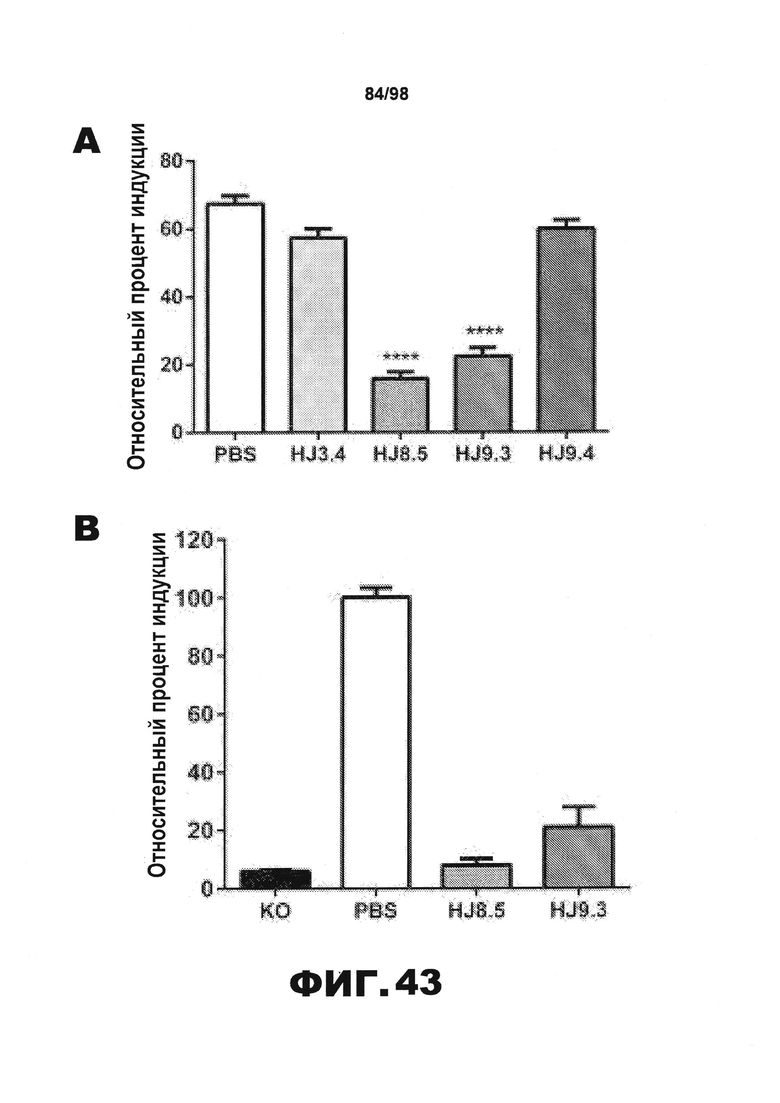
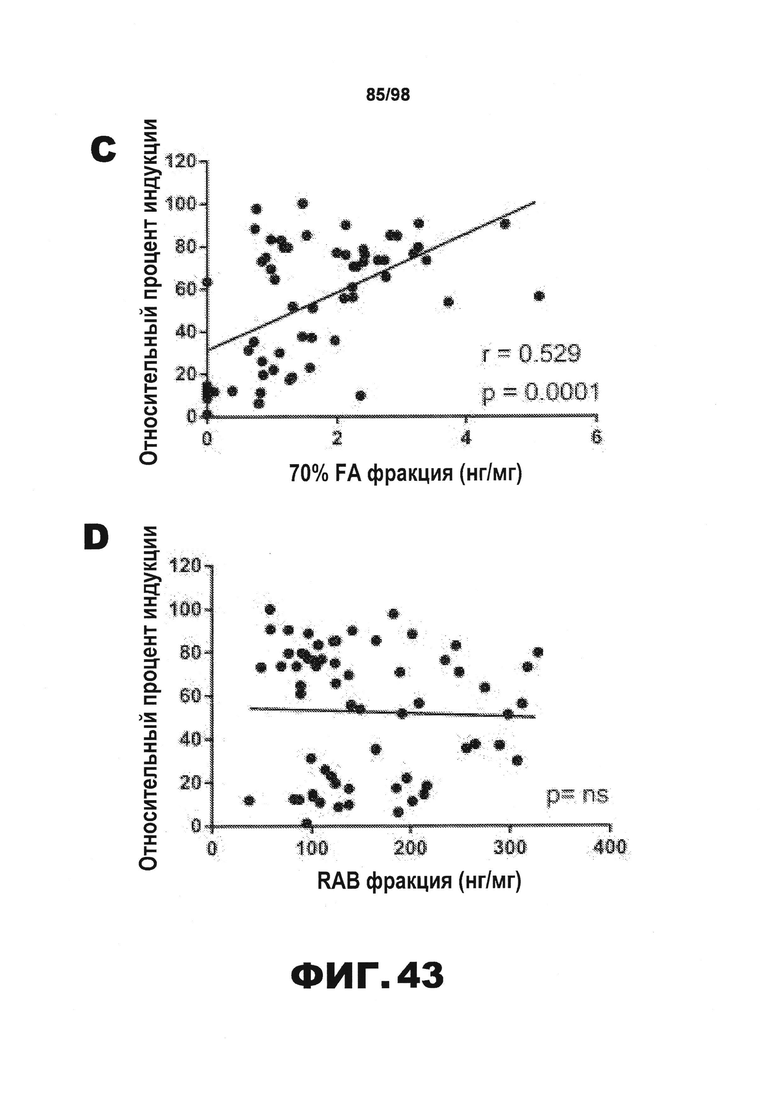
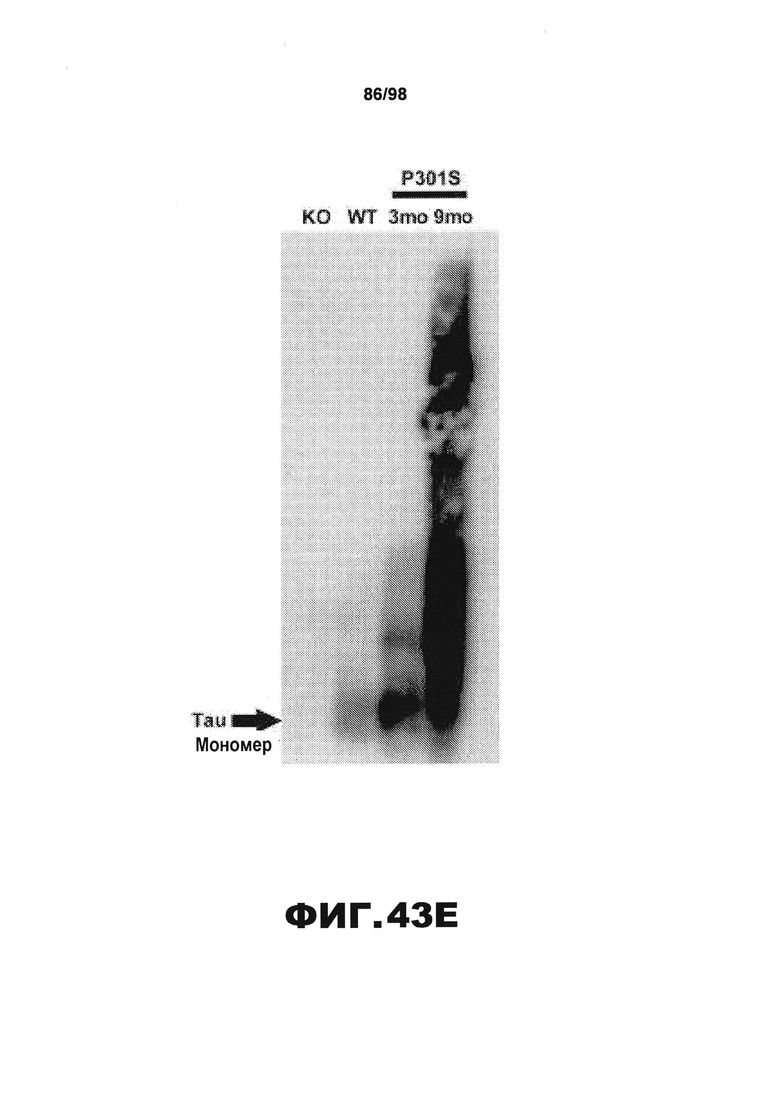
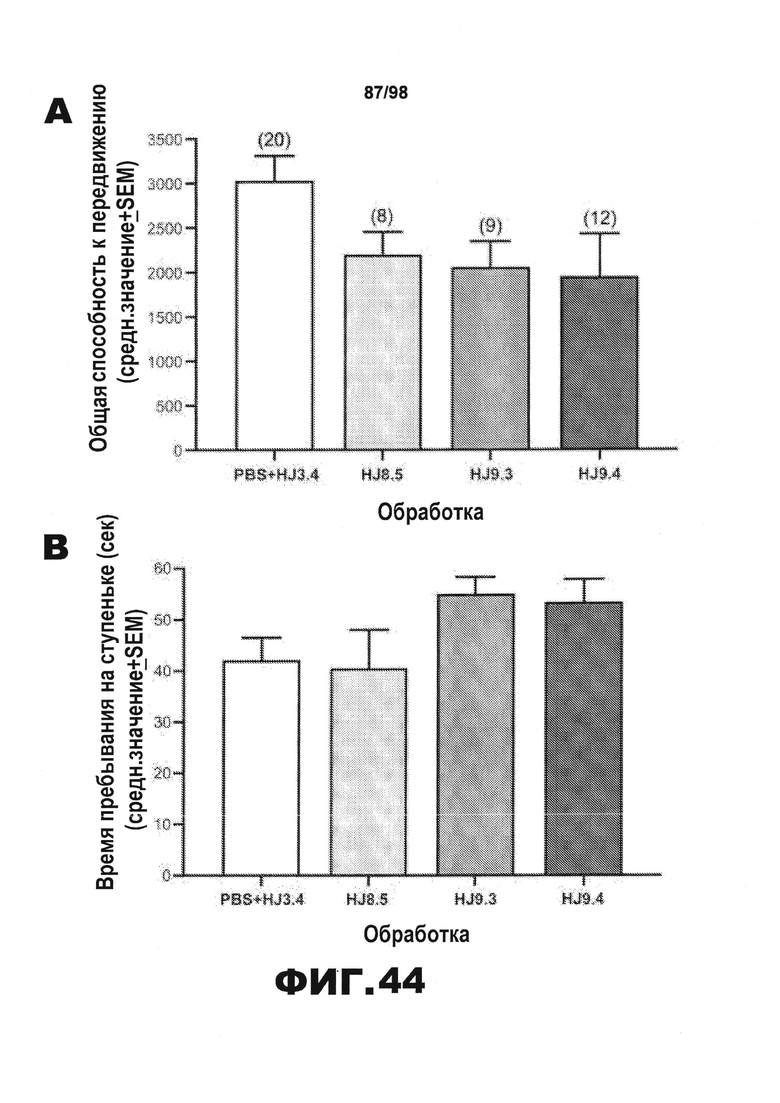
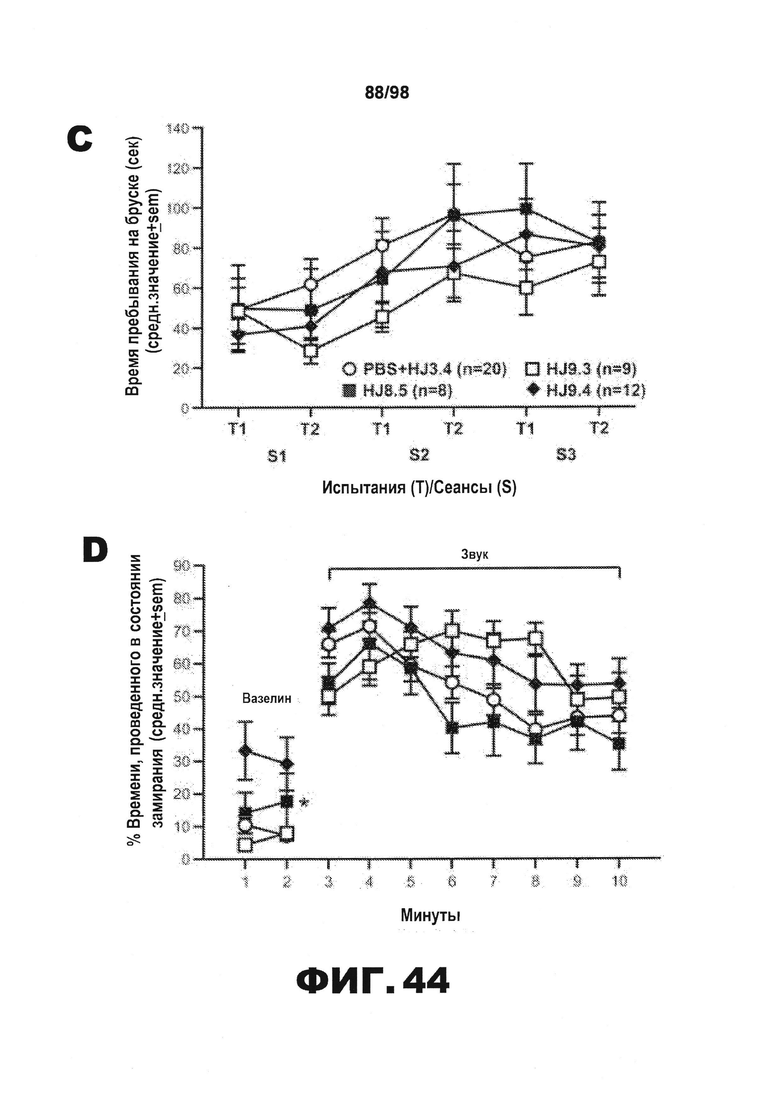
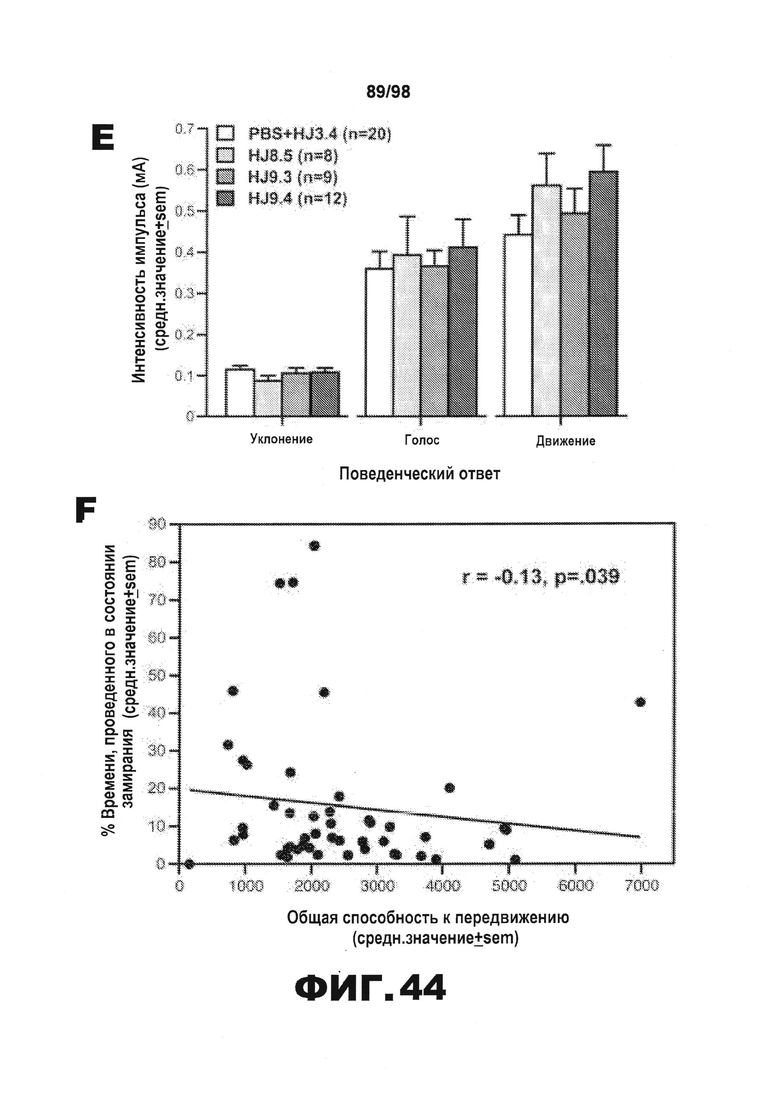
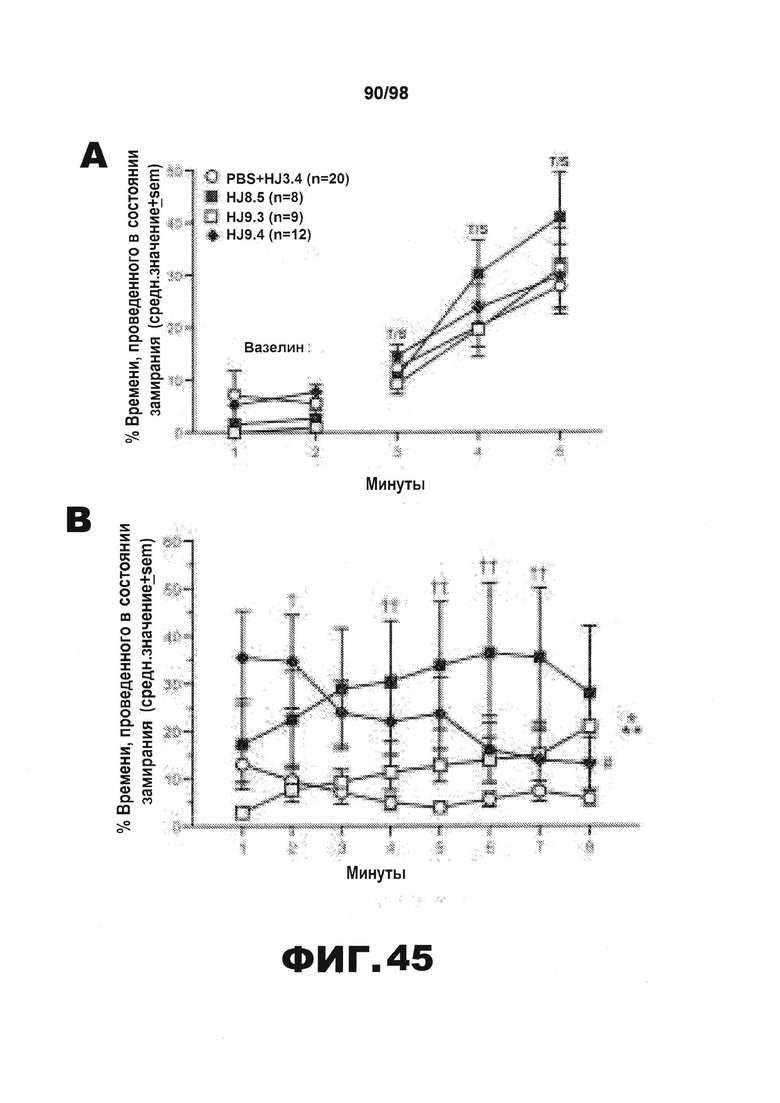
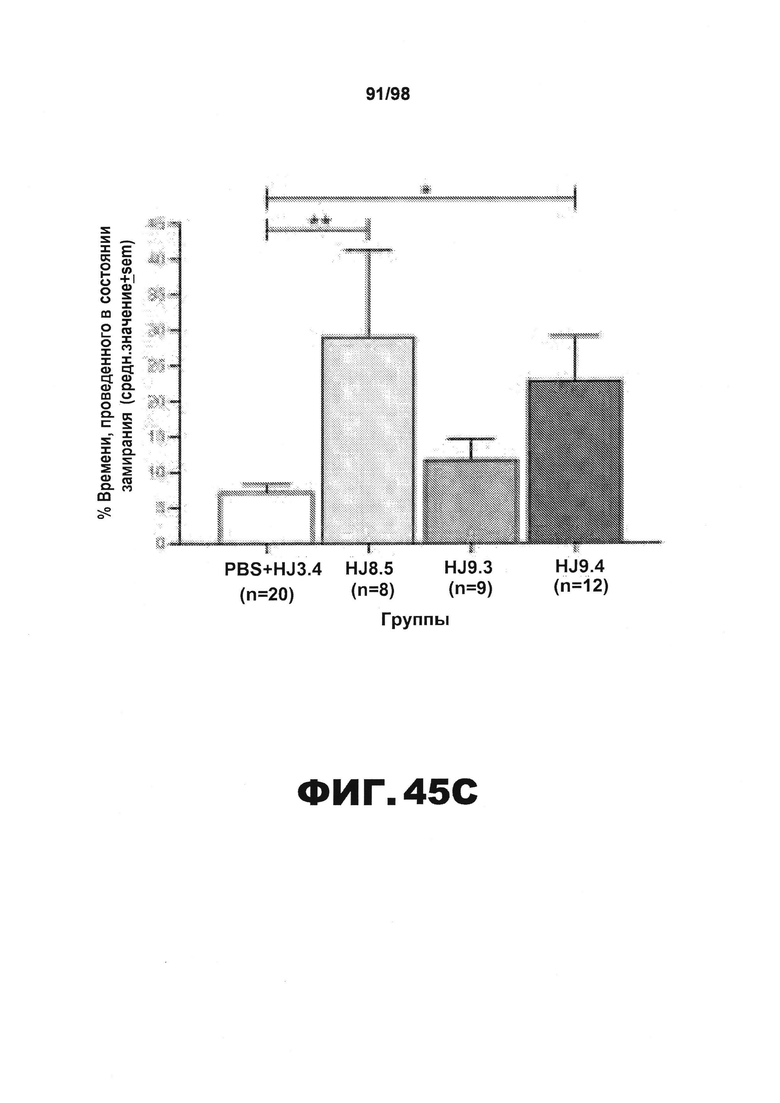
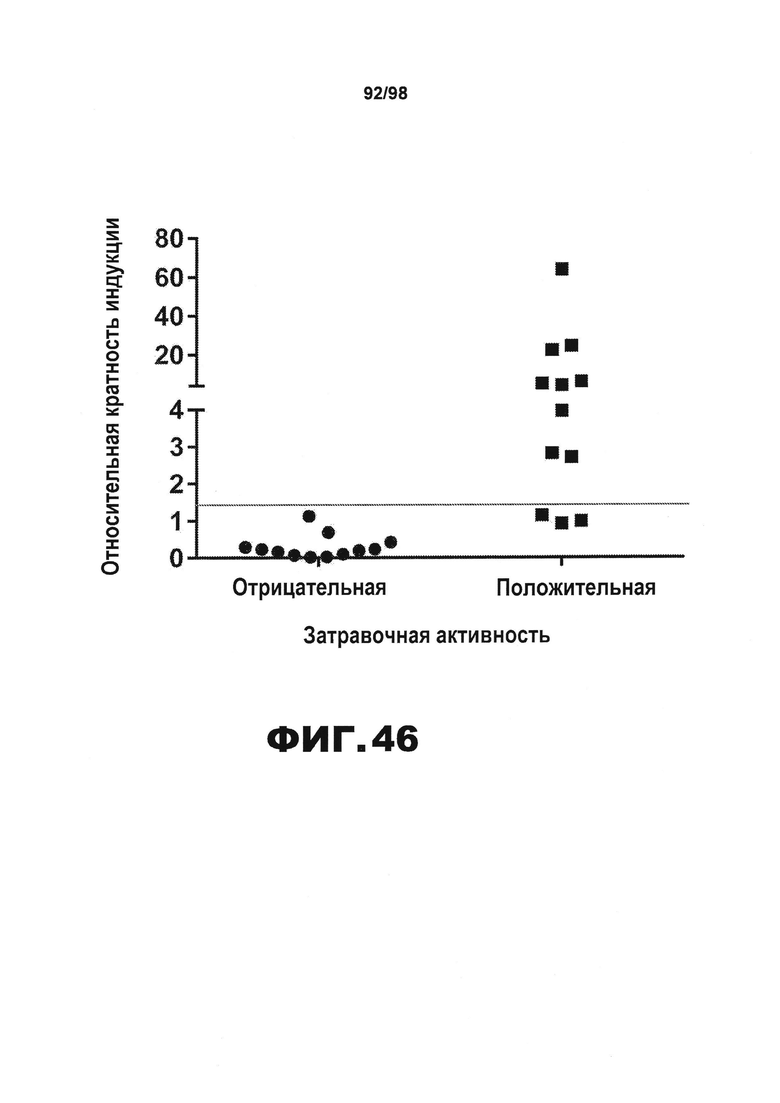
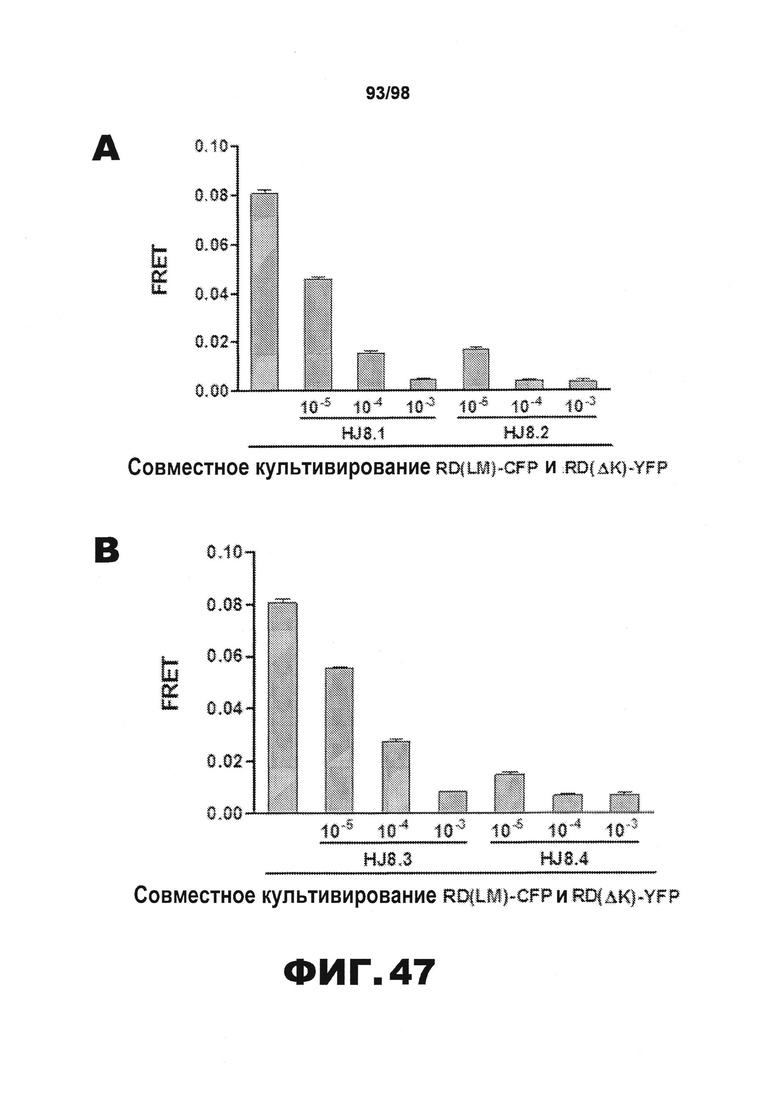

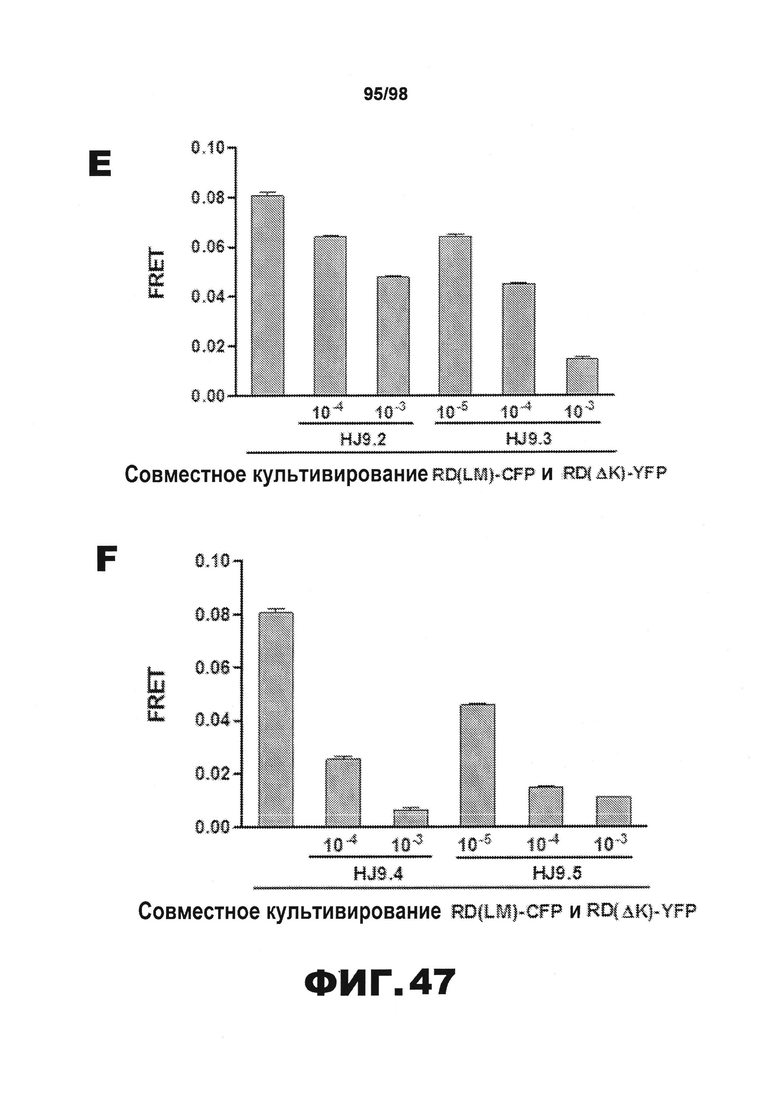
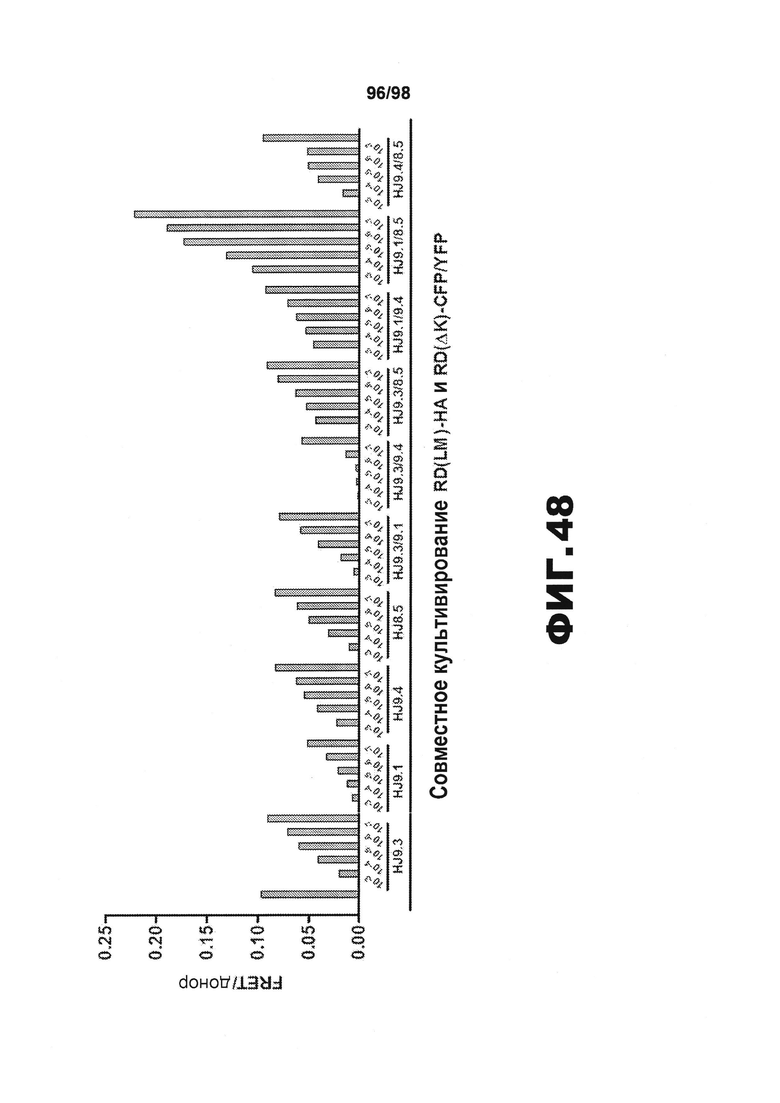
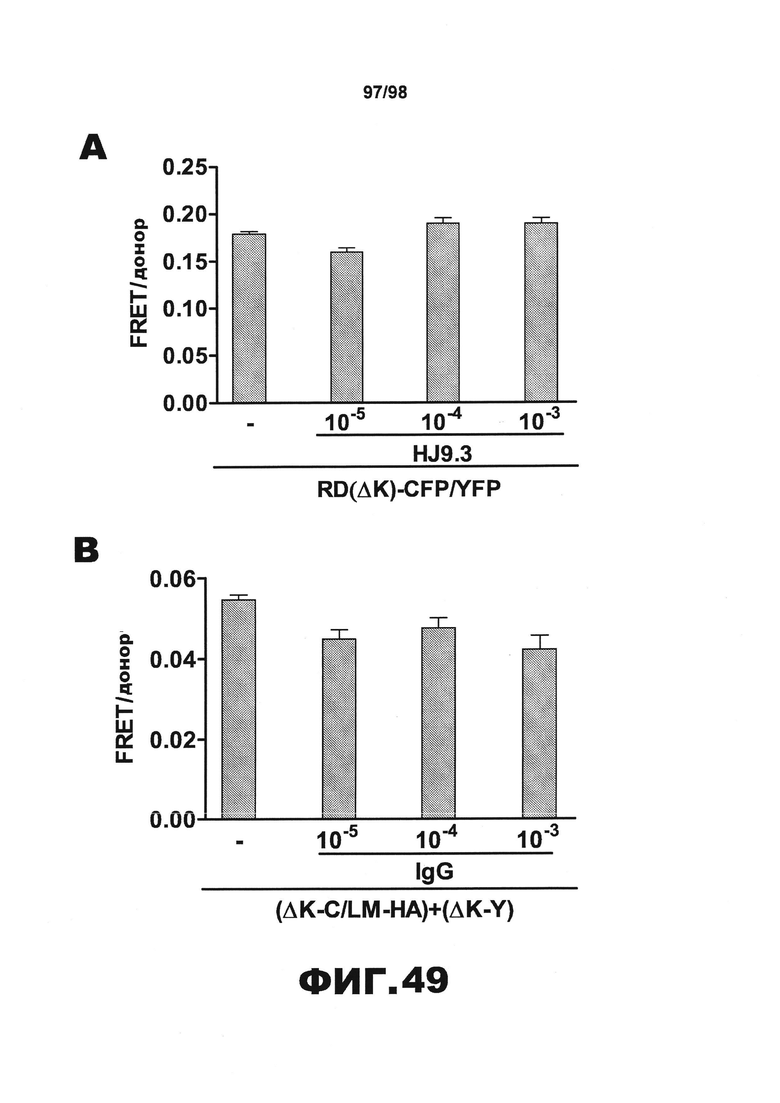
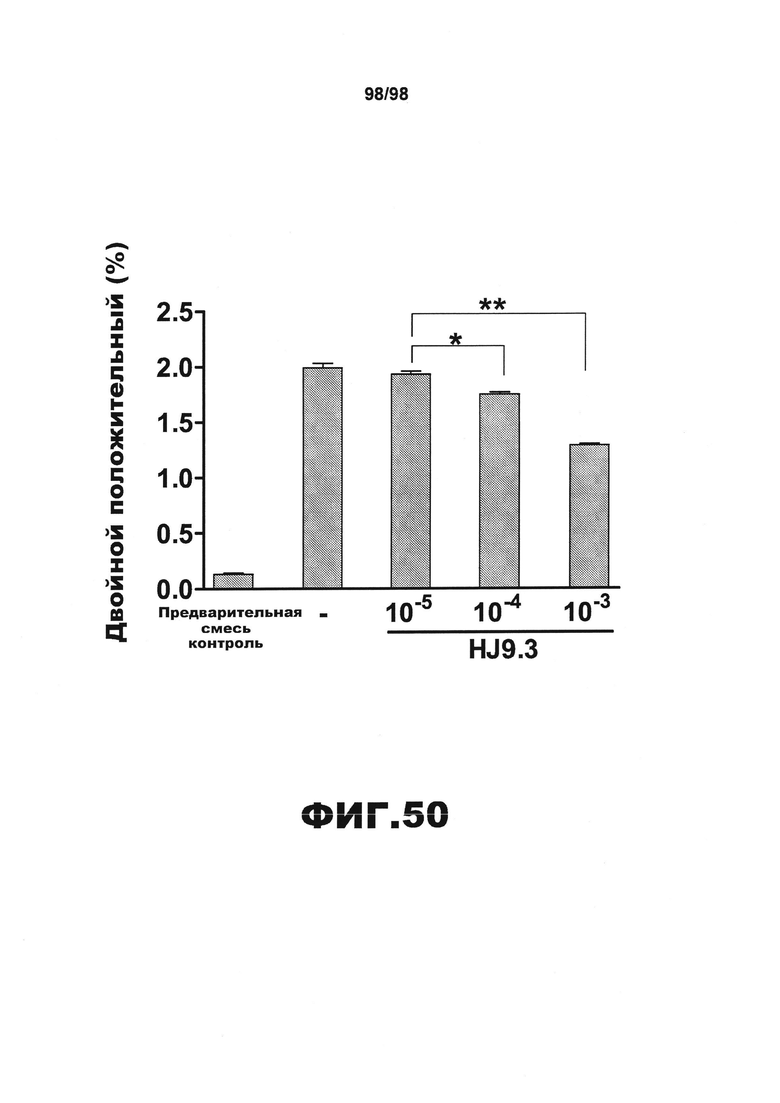
Изобретение относится к области биохимии, в частности к выделенному моноклональному антителу против ТАУ. Изобретение позволяет эффективно лечить заболевания, ассоциированные с ТАУ. 10 з.п. ф-лы, 50 ил., 13 табл., 17 пр.
1. Выделенное моноклональное антитело против тау, где антитело содержит
вариабельную область легкой цепи, содержащую аминокислотную последовательность CDR1 SEQ ID NO:16, аминокислотную последовательность CDR2 SEQ ID NO:17 и аминокислотную последовательность CDR3 SEQ ID NO:18, и
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность CDR1 SEQ ID NO:19, аминокислотную последовательность CDR2 SEQ ID NO:20 и аминокислотную последовательность CDR3 SEQ ID NO:21.
2. Выделенное моноклональное антитело против тау по п.1, где антитело специфически связывает тау с аффинностью взаимодействия (KD) от 0,1 пМ до 10 нМ.
3. Выделенное моноклональное антитело против тау по п.1, где антитело специфически связывает тау с аффинностью взаимодействия (KD) от 0,1 пМ до 1 нМ.
4. Выделенное моноклональное антитело против тау по п.1, где антитело специфически связывает тау человека и не связывает тау мыши.
5. Выделенное антитело по п. 1, где антитело способно специфически блокировать затравочную активность тау в клеточном анализе агрегации тау.
6. Выделенное моноклональное антитело против тау по п.1, где вариабельная область легкой цепи и вариабельная область тяжелой цепи содержат каркасные области.
7. Выделенное моноклональное антитело против тау по любому из пп.1 и 4-6, где антитело представляет собой химерное антитело.
8. Выделенное моноклональное антитело против тау по любому из пп.1 и 4-6, где антитело представляет собой гуманизированное антитело.
9. Выделенное моноклональное антитело против тау по п.7, где антитело является IgG изотипа.
10. Выделенное моноклональное антитело против тау по п.8, где антитело является IgG изотипа.
11. Выделенное моноклональное антитело против тау по п.10, где антитело является IgG4 изотипа.
| US 20100316564 A1, 16.12.2010 | |||
| НЕЙРОТРОФИЧЕСКИЙ ФАКТОР РОСТА ЧЕЛОВЕКА (ЭНОВИН), КОДИРУЮЩАЯ ЕГО ВЫДЕЛЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, ВЕКТОР (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ), АНТИТЕЛО, СПОСОБ ОБНАРУЖЕНИЯ ФАКТОРА РОСТА, НАБОР, СПОСОБ ИДЕНТИФИКАЦИИ АГОНИСТА ИЛИ АНТАГОНИСТА (ВАРИАНТЫ) | 1999 |
|
RU2238947C2 |
| WO 2010056754 A2, 20.05.2010 | |||
| WO 2000048621 A2, 24.08.2000. | |||

