ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0001] Настоящее изобретение относится к новому классу моноклональных антител, которые специфично связываются с фосфорилированным сериновым остатком 396 в патологически гиперфосфорилированном (PHF) тау-белке (pS396), а также к способам применения этих молекул и их тау-связывающих фрагментов в лечении болезни Альцгеймера и таупатий.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит один или несколько перечней последовательностей в соответствии с параграфами 1.821 и последующими раздела 37 C.F.R., которые раскрыты на машиночитаемых носителях (название файла: 0995.txt, создан 23 июня 2016 г. и имеет размер 40 кБ), и этот файл включен в данный документ посредством ссылки во всей своей полноте.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0003] Возрастные нейродегенеративные заболевания, такие как болезнь Альцгеймера (AD) и деменция, в настоящее время являются одной из крупнейших общественных проблем. По оценкам Всемирной организации здравоохранения стоимость лечения лиц старшего возраста будет продолжать повышаться, и количество диагностированных случаев деменции утроится к 2050 году (World Health Organization and Alzheimer's Disease International - Status Report (2012) DEMENTIA: A public health priority, WHO). Первыми средствами лечения AD были модуляторы высвобождения нейромедиаторов, такие как ингибиторы ацетилхолинэстеразы, и модуляторы NMDA-рецепторов. Эти терапевтические средства стали доступными на рубеже тысячелетий и по-прежнему образуют краеугольный камень облегчения симптомов дефицитов памяти, связанных с деменцией и AD. Тем не менее, эти лекарственные средства не воздействуют целенаправленно на первопричины AD - накопление агрегатов β-амилоидного (Аβ) пептида и тау-белка и ассоциированную с ним утрату нейронных синапсов и в конечном счете нейронов.
[0004] Долгосрочные исследования в широком общественном масштабе с участием лиц старшего возраста (Weiner, M.W. et al. (2014) ADNI online: http://www.adni-info.org/; Breteler, M.M. et al. (1992) Neuroepidemiology 11 Suppl 1, 23-28; Launer, L.J. (1992) Neuroepidemiology 11 Suppl 1, 2-13) наряду с крупными полногеномными исследованиями ассоциаций (Lambert, J.C. et al. (2013) Nat. Genet. 45, 1452-1458) показали, что AD представляет собой неоднородную комбинацию форм деменции, где до 10 процентов пациентов с запущенной формой AD не имеют патологии амилоидного пептида (Crary, J.F. et al. (2014) Acta Neuropathol. 128, 755-766). Дополнительно, фундаментальные патологоанатомические исследования, проведенные Braak & Braak (Braak, Н. and Braak, E. (1996) Acta Neurol. Scand. Suppl 165, 3-12), продемонстрировали четкую корреляцию между степенью патологии нейрофибриллярных клубков и когнитивным статусом перед вскрытием. Эти наблюдения подкреплялись несколькими исследователями (Nelson, Р.Т. et al. (2012) J. Neuropathol. Exp. Neurol. 71, 362-381) и в недавних долгосрочных исследованиях биомаркеров, которые указывают на то, что уровни тау-белка и фосфорилированного тау-белка в цереброспинальной жидкости (CSF) повышаются на протяжении ранних и поздних стадий заболевания (Jack, C.R., Jr. et al. (2013) Lancet Neurol. 12, 207-216).
[0005] Как указано выше, тау-белок, ассоциированный с микротрубочками, и его гиперфосфорилированный вариант образуют основную составляющую внутриклеточных нейрофибриллярных клубков, которые являются одной из главных характерных особенностей AD. Дополнительно, определенные варианты гена тау-белка ассоциированы с семейными формами лобно-височной деменции (FTD). Появление патологии тау-белка при AD происходит в соответствии с четко выраженным пространственным паттерном, начиная с энторинальной коры с последующим переходом в гиппокампальные и корковые зоны (Braak, Н. and Braak, Е. (1996) Acta Neurol. Scand. Suppl 165, 3-12). Конкретная стадия патологии тау-белка также хорошо коррелирует с когнитивными способностями (Nelson, Р.Т. et al. (2012) J. Neuropathol. Exp. Neurol. 71, 362-381; Braak, E. et al. (1999) Eur. Arch. Psychiatry Clin. Neurosci. 249 Suppl 3, 14-22). Взятые вместе, эти данные составляют основу гипотезы, опирающейся на то, что тау-белок играет определенную роль в AD. Она подразумевает, что внутриклеточное накопление тау-белка приводит к разрушению микротрубочек и коллапсу дендритных шипиков. В результате этого нарушается коммуникация между нейронами и наступает гибель клеток. Недавно также было показано, что тау-белок сам по себе может образовывать эндопатогенные молекулы, которые могут способствовать распространению нейродегенерации от одной клетки к следующей (Clavaguera, F. et al. (2009) Nat. Cell Biol. 11, 909-913).
I. Тау-белок в качестве эндопатогена
[0006] Clavaguera и соавторы продемонстрировали, что тау-белок сам по себе может действовать в качестве эндопатогена (Clavaguera, F. et al. (2009) Nat. Cell Biol. 11, 909-913). Экстракты головного мозга, получаемые при низкой скорости центрифугирования, выделяли из трансгенных по тау-белку мышей P301S (Allen, В. et al. (2002) J. Neurosci. 22, 9340-9351), разбавляли и инъецировали в гиппокамп и корковые зоны молодых мышей ALZ17. Мыши ALZ17 представляют собой трансгенную по тау-белку линию мышей, у которых развивается только поздняя патология (Probst, A. et al. (2000) Acta Neuropathol. 99, 469-481). У подвергнутых инъекции мышей ALZ17 быстро развивалась патология плотных филаментов, и введение иммуноистощенных экстрактов головного мозга от мышей P301S или экстрактов от мышей дикого типа не индуцировало патологию тау-белка. Фракционирование экстрактов головного мозга на растворимый (S1) и нерастворимый в саркозиле (Р3) тау-белок (Sahara, N. et al. (2013) J. Alzheimer's. Dis. 33, 249-263) и их инъекция мышам ALZ17 продемонстрировали, что фракция Р3 в наибольшей степени способна индуцировать патологию. Она содержит большую часть внутриклеточного гиперфосфорилированного филаментозного тау-белка. Патологию в большинстве случаев также можно было индуцировать при инъекции экстрактов P301S в головной мозг мышей дикого типа, но NFT не образовывались. В последующих исследованиях Clavaguera и соавт. показали, что человеческий тау-белок, экстрагированный посмертно из ткани головного мозга с другими таупатиями (болезнью аргирофильных зерен (AGD), прогрессирующим надъядерным параличом (PSP) и кортикобазальной дегенерацией (CBD)), также может индуцировать патологию тау-белка в модели ALZ17 (Clavaguera, F. et al. (2013) Proc. Natl. Acad. Sci. U.S.A. 110, 9535-9540). С момента представления этих данных сообщалось о некоторых других моделях затравочного действия и разнесения тау-белка (Ahmed, Z. et al. (2014) Acta Neuropathol. 127, 667-683; Walker, L.C. et al. (2013) JAMA Neurol. 70, 304-310). Основной вывод этих исследований указывает на механизм, посредством которого патогенный тау-белок во внутриклеточных включениях секретируется из клетки в периплазматическое пространство. Патологический материал тау-белка затем транспортируется по оболочке синаптических пузырьков как в антероградном, так и в ретроградном направлении и впоследствии поглощается соседними клетками путем объемного эндоцитоза. Этот механизм объясняет, почему разнесение патологии, наблюдаемое при заболевании у человека, следует четко выраженному анатомическому паттерну. Весьма любопытно, что периферическое введение патологического тау-белка может ускорять формирование патологии тау-белка у мышей ALZ17 (Clavaguera, F. et al. (2014) Acta Neuropathol. 127, 299-301). Этот механизм разнесения может объяснять распространение заболевания при других протеинопатиях (Goedert, М. etal. (2010) Trends Neurosci. 33, 317-325; Sigurdsson, E.M. et al. (2002) Trends Mol. Med. 8, 411-413).
II. Молекулы тау-белка
[0007] Обнаружение того, что тау-белок может действовать в качестве эндопатогена, побудило к поиску «патогенных молекул», на которые можно было целенаправленно воздействовать в рамках потенциальных методов интервенционной терапии.
[0008] Ген тау-белка, ассоциированного с микротрубочками (МАРТ), расположен в хромосоме 17 человеческого генома и экспрессирует шесть изоформ тау-белка в головном мозге взрослого человека. Эти изоформы образуются в результате альтернативного сплайсинга экзонов 2, 3 и 10 из 16 экзонов в гене МАРТ. Экзоны 2 и 3 экспрессируют повтор из 29 аминокислот, а экзон 10 экспрессирует дополнительный домен, связывающийся с микротрубочками. В результате этого изоформы тау-белка будут содержать 0, 1 или 2 N-концевых повтора и 3 или 4 С-концевых домена, связывающихся с микротрубочками (тау-белок 3R или 4R). Обычно экспрессируются шесть изоформ тау-белка. Наиболее длинная (2N4R) и наиболее короткая (0N3R) изоформы состоят из 441 и 352 аминокислот соответственно (Kolarova, М. et al. (2012) Int. J. Alzheimers. Dis. 2012, 731526). N-концевой выступающий домен тау-белка (2N4R) состоит из богатого глицином "хвоста" из 44 аминокислот и остатков 45-102, охватывающих две высококислые области (N1, N2-домены). В остатках 151-243 обнаружены две области, богатые пролином (Р1, Р2-домены). Остальную часть белка составляют четыре домена, связывающихся с микротрубочками (R1-R4), за которыми расположена короткая С-концевая область.
[0009] Тау-белок является весьма растворимым и высоколабильным в отношении фосфорилирования белком. Примерно 20 процентов, или 85, аминокислотных остатков в наиболее длинной изоформе тау-белка являются потенциальными (Ser, Thr или Tyr) сайтами фосфорилирования. Порядка половины из них наблюдались экспериментально (Hanger, D.P. et al. (2009) Trends Mol. Med. 15, 112-119; Hasegawa, M. et al. (1992) J. Biol. Chem. 267, 17047-17054) и группируются вокруг концевых остатков доменов, связывающихся с микротрубочками. Тау-белок подвергается динамичному фосфорилированию и дефосфорилированию в ходе клеточного цикла. Он должен диссоциировать из микротрубочек, чтобы обеспечить прохождение мейоза. Его основной ролью в постмитотических клетках (дифференцированных нейронах) является действие в качестве стабилизатора микротрубочек, что обеспечивает оптимальный аксональный транспорт. Он может ассоциировать с микротрубочками только в своей главным образом дефосфорилированной форме, поэтому фосфорилирование выступает в качестве непосредственного переключателя ассоциации с микротрубочками/диссоциации из микротрубочек в нейроне. В нормальных условиях цитозольный тау-белок в среднем содержит два фосфорилированных сайта. В материале парных спиральных филаментов по меньшей мере 7-8 сайтов являются фосфорилированными (Hanger, D.P. et al. (2009) Trends Mol. Med. 15, 112-119; Hasegawa, M. et al. (1992) J. Biol. Chem. 267, 17047-17054). Парные спиральные филаменты гиперфосфорилированного тау-белка являются ключевой характерной особенностью болезни Альцгеймера (Kosik et. al. (1986) PNAS, 86, 4044-4048), в иммуноцитохимическом анализе материала головного мозга человека с AD наблюдается четко выраженный сдвиг подвижности гиперфосфорилированного тау-белка.
[0010] Исследование тау-белка с помощью традиционных структурных методик, таких как рентгеновская кристаллография или спектроскопия ЯМР, было затруднительным, что отражает его метастабильную природу. Такие исследования проводились главным образом в отношении фрагментов доменов нефосфорилированного белка. Единственное до настоящего времени структурное исследование полноразмерного тау-белка (2N4R) с помощью спектроскопии ЯМР выявило, что белок содержит только редкие фрагменты стабильной вторичной структуры (Mukrasch, M.D. et al. (2009) PLoS. Biol. 7, e34). Этот анализ указывает на то, что вторичная структура пептидного остова обладает высокой склонностью принимать структуру β-листа. Первые 200 остатков остова являются значительно более упорядоченными, чем С-конец, охватывающий домены, связывающиеся с микротрубочками. Наличие большого количества специфических взаимодействий дальнего порядка в белке в растворе указывает на то, что он существует в сильно разупорядоченном состоянии расплавленной глобулы (Ohgushi, М. and Wada, А. (1983) FEBS Lett. 164, 21-24).
[0011] В материале клубков были идентифицированы продукты расщепления тау-белка протеазами, образующиеся, в частности, под действием каспазы и кальпаина (Asp13, Glu391 и Asp421) (Gamblin, Т.С. et al. (2003) Proc. Natl. Acad. Sci. U.S.A. 100, 10032-10037). В частности, усечение по Asp421 подробно изучалось с применением антитела С3 к тау-белку, которое связывается со свободным концом Asp421. Было предположено, что это усечение является ранним событием в патогенезе AD, ассоциированным с индукцией апоптоза (deCalignon A. et al. (2010) Nature 464, 1201-1204). N-концевое расщепление по Asp 13 и С-концевое расщепление по Glu391 считаются поздними событиями в патогенезе (deCalignon A. et al. (2010) Nature 464, 1201-1204; Delobel, P. et al. (2008) Am. J. Pathol. 172, 123-131). Недавно в CSF от пациентов с AD и PSP был идентифицирован дополнительный N-концевой фрагмент (остатки 1-224), и была выдвинута гипотеза, что он является ранним маркером заболевания и особенно патогенным (US 14/092539; Bright, J. et al. (2014) Neurobiol. Ageing, 1-17). Об аналогичном фрагменте, образующемся в результате расщепления кальпаином, сообщали другие группы (Ferreira, A. and Bigio, Е.Н. (2011) Mol. Med. 17, 676-685; Reinecke, J.B. et al. (2011) PLoS. One. 6, e23865).
[0012] Было выдвинуто предположение, что, помимо гиперфосфорилирования и фрагментации тау-белка, посттрансляционное ацетилирование (Cohen, T.J. et al. (2011) Nat. Commun. 2, 252; Min, S.W. et al. (2010) Neuron 67, 953-966) и O-GlcNAc-илирование (Zhu, Y. et al. (2014) J. Biol. Chem.) являются процессами, определяющими патологию, при формировании патологии клубков, ассоциированной с AD.
III. Виды иммунотерапии с применением тау-белка
[0013] Виды иммунотерапии традиционно разделяются на пассивные и активные подходы к вакцинации. При активном подходе к вакцинации патогенное средство вводят пациенту путем инъекции, и система врожденного иммунитета вызывает иммунный ответ. Это запускает созревание В-клеток, вырабатывающих высокоаффинные антитела к введенному антигену. При пассивном подходе к вакцинации запуска системы врожденного иммунного ответа избегают путем инфузии антитела, специфичного к антигену. Собственная система очищения организма затем удаляет связанный с антителом лиганд.
[0014] AC Immune предоставляет моноклональное мышиное антитело к фосфосерину-409 тау-белка. Определяли профиль антител к антигенам ткани головного мозга человека с AD и контрольной ткани головного мозга, и их отбирали на основании их способности к распознаванию патологии клубков. Оба гуманизированных варианта двух антител hACI-36-2B6-Ab1 и hACI-36-3A8-Ab1 связываются с эпитопом тау-белка в пределах аминокислот 401-418 (WO 2013/151762).
[0015] Группа Roger Nitsch выделила аутоантитела к тау-белку от пожилых здоровых индивидуумов без признаков дегенеративной таупатии. Был выделен ряд антител с применением полноразмерного рекомбинантного человеческого тау-белка (2N4R) для обнаружения антител, специфичных к тау-белку. Их потом подвергали скринингу в отношении их способности к проведению различий между изолятами тау-белка от больных и здоровых индивидуумов. Три лидерных антитела 4Е4, 4А3 и 24В2 были описаны в патентной литературе (WO 2012049570; US 2012087861). Картирование их эпитопов указывает на то, что все они распознают аминокислоты в пределах области связывания с микротрубочками и в С-концевом направлении от него в положениях от V339 до K369. Эти антитела не проявляют какую-либо фосфоспецифичность.
[0016] C2N Diagnostics фокусируется главным образом на разработке средств диагностики для раннего выявления нейродегенеративного заболевания. Получали антитела к полноразмерному человеческому и мышиному тау-белку. Идентифицировали восемь и пять антител, соответственно распознающих человеческий и мышиный тау-белок (Yanamandra, K. et al. (2013) Neuron 80, 402-414). Три антитела с разными показателями кинетики связывания отбирали для оценивания in vivo. А именно это были HJ9.3, HJ9.4 и HJ8.5, распознающие соответственно остатки 306-320, 7-13 и 25-30 тау-белка, при этом последнее является специфичным к человеческому тау-белку. Антитела также отбирали на основании их способности к предупреждению переноса патологии в оригинальном механизированном репортерном анализе трансцеллюлярного распространения патологии тау-белка (Sanders, D.W. et al. (2014) Neuron 82, 1271-1288; Kfoury, N. etal. (2012) J. Biol. Chem. 287, 19440-19451). Их оценивание в исследованиях с длительной i.c.v. инъекцией у трансгенных мышей P301S продемонстрировало их способность к уменьшению уровней гиперфосфорилированного тау-белка, определенную по окрашиванию АТ8 в иммуногистохимическом анализе обработанных мышей.
[0017] Антитела Peter Davies изначально разрабатывали как средства диагностики, которые могут обеспечивать установление различий между патологическим и нормальным тау-белком в материале головного мозга с AD и контрольном материале головного мозга (Greenberg, S.G. and Davies, P. (1990) Proc. Natl. Acad. Sci. U.S.A. 87, 5827-5831). Оценивание терапевтической полезности антител PHF1 и MC1 было продемонстрировано у мышей P301S и JPNL3 (P301L) (Boutajangout, A. et al. (2011) J. Neurochem. 118, 658-667; Chai, X. et al. (2011) J. Biol. Chem. 286, 34457-34467; D'Abramo, C. et al. (2013) PLoS. One. 8, e62402). PHF1 распознает линейный эпитоп фосфорилированного тау-белка (pS396, pS404), тогда как MC1 представляет собой конформационно-зависимое антитело, которое распознает структурный эпитоп тау-белка, для которого требуются две отдельные части линейной последовательности, эпитоп в пределах остатков 46-202 и С-концевой эпитоп в остатках 312-342 (Jicha, G.A. et al. (1997) J. Neurosci. Res. 48, 128-132). Инъекция этих двух антител в исследованиях с длительной 12-13-недельной иммунизацией приводила к существенному уменьшению патологии спинного мозга и ствола головного мозга среди других областей головного мозга, что преобразовывалось в ослабление двигательного дефицита, наблюдаемого у этих мышей. (D'Abramo, С. et al. (2013) PLoS. One. 8, e62402).
[0018] iPerian/Bristol Meyers Squibb разработали антитела к тау-белку, направленные на предполагаемые патологические молекулы тау-белка, состоящие из N-концевого фрагмента тау-белка (eTau: остатки 1-224), которые способствовали гиперактивности в культурах нейронов из индуцированных плюрипотентных стволовых клеток. Был разработан портфель антител, но получение характеристик было сфокусировано на антителах IPN001 и IPN002, которые распознают N-концевой эпитоп в пределах остатков 9-18. Соответственно, эти антитела выявляют повышенные уровни тау-белка в CSF от пациентов с установленной стадией AD и PSP, которые могут являться ранним признаком заболевания. Инъекции антител мышам JPNL3 (P301L) in vivo приводили к частичному устранению прогрессирующих двигательных дефицитов (US 14/092539).
[0019] Einar Sigurdsson разработал первую программу, демонстрирующую эффективность иммунотерапии на основе применения тау-белка. Для иммунизации мышей JPNL3 (P301L) применяли активную вакцину, состоящую из пептида тау-белка 379-408 [pS396, pS404] вместе с адъювантом Adju-Phos. В этом исследовании наблюдали выраженное уменьшение патологии тау-белка у мышей, обработанных вакциной, по сравнению с контрольными животными. Также выявляли ослабление двигательного фенотипа, связанного с таупатией. Его эффективность подтверждали в другой мышиной модели (htau/PS1), функционирующей без воздействия мутантного тау-белка (Boutajangout, A. et al. (2011) AAIC 2011 (7, issue 4, Supplement edn) p. s480-s431; Congdon, E.E. et al. (2013) J. Biol. Chem. 288, 35452-35465; Gu, J. et al. (2013) J. Biol. Chem. 288, 33081-33095).
[0020] В Prothena оценили три антитела к тау-белку у трансгенных по тау-белку мышей K369I (K3) и в мышиной модели P301L. Антитела с различными свойствами отбирали для оценивания in vivo. Два антитела к pS404 с разными изотипами (IgG1/k и IgG2a/k) или общее (панспецифическое) антитело к тау-белку (IgG1/k) инъецировали в рамках парадигмы длительного исследования. Мышей K369I обрабатывали посредством еженедельных инъекций в течение 21 недели, начиная с возраста 3 недель, а мышей P301L обрабатывали в течение 7 месяцев посредством еженедельных инъекций, начиная с возраста 4 месяцев. У мышей K3, обработанных антителом IgG2a/k к pS404, наблюдали уменьшение количества положительных по тау-белку нейрофибриллярных включений. Оба антитела к pS404 были способны уменьшать уровни pS422-положительного тау-белка, тогда как у мышей, обработанных панспецифическим антителом к тау-белку, уменьшение не наблюдалось. Эти исследования позволяют предположить, что: 1) очищение от тау-белка может зависеть от изотипа, и 2) может быть важным целенаправленное воздействие на молекулы тау-белка, имеющие отношение к заболеванию, поскольку общее антитело к тау-белку было не способно уменьшать уровни гиперфосфорилированного тау-белка (PCT/US 2014/025044).
[0021] Авторы настоящего изобретения неожиданно обнаружили антитела, специфичные к фосфорилированному сериновому остатку 396 тау-белка (pS396); это входит в противопоставление с антителами из предшествующего уровня техники, которые распознают главным образом тау-белки, фосфорилированные как по остатку 396, так и по остатку 404, фосфорилированные только по остатку 404 или по другим остаткам тау-белка.
[0022] Авторы настоящего изобретения разработали антитела, которые дополнительно характеризуются значительной специфичностью и селективностью в отношении патологического человеческого тау-белка. Существует необходимость в антителах, характеризующихся высокой степенью селективности и специфичности в отношении патогенного тау-белка. Антитела по настоящему изобретению демонстрируют намного более высокую степень специфичности и селективности в отношении патологического человеческого тау-белка по сравнению с непатологическим тау-белком, чем антитела из WO 2013/050567 (см. фигуру 1 из WO 2013/050567). Образование антител из WO 2012/045882, которые, как сообщалось, характеризуются специфичным связыванием, вызывалось в ответ на аминокислотные последовательности с 6-9 остатками из аминокислот 393-401, 396-401, 394-400 и 393-400 тау-белка. Это входит в противопоставление с антителами по настоящему изобретению, образование которых происходит в ответ на патогенный гиперфосфорилированный тау-белок, содержащий более длинную аминокислотную последовательность, описанную в данном документе.
[0023] Как показано в примерах, сравнение с пятью опубликованными антителами к тау-белку hACI-2B6 (описанным в WO 2013151762); IPN002 (описанным в WO 2014028777); HJ8.5 (описанным в WO 2014008404); моноклональным антителом 2.10.3 к тау-белку pS422 (описанным в US 8609097); PHF13 (коммерчески доступным антителом (например, Sigma-Aldrich), рекомендованным для выявления тау-белка, фосфорилированного по Ser 396, мышиного, крысиного и человеческого происхождения и обсуждаемым Sankaranarayanan (PLOSONE, DOI:10.1371/journal.pone.0125614 May 1, 2015 и Otvos (Biochemistry 1997, 36, 8114-8124); а также антителом 4E4 (описанным как связывающееся с V339, Е342, D387, Е391 и K395 в US 8940272) показало, что антитела и их эпитопсвязывающие фрагменты по настоящему изобретению проявляют более высокую степень специфичности и селективности в отношении патологического человеческого тау-белка, чем любые из сравниваемых антител.
[0024] Дополнительно, антитела и их эпитопсвязывающие фрагменты по настоящему изобретению демонстрируют множество преимущественных признаков, таких как способность к проведению различий между патологическим и непатологическим человеческим тау-белком и, в частности, к связыванию с тау-белком, ассоциированным с альцгеймеровской (AD) патологией. В электрофизиологических исследованиях антитела и их эпитопсвязывающие фрагменты по настоящему изобретению были дополнительно способны устранять уменьшенные парное облегчение и самопроизвольный миниатюрный возбуждающий синаптический ток (mEPSC).
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0025] Настоящее изобретение относится к моноклональным антителам и их эпитопсвязывающим фрагментам, способным специфично связываться с фосфорилированным остатком серином-396 человеческого тау-белка (изоформы 2N4R) (SEQ ID NO: 33), и к тем антителам, которые были получены с помощью нового способа, обеспечивающего такое специфичное выделение и извлечение. Антитела дополнительно характеризуются своей способностью различать фосфорилированные остатки 396 и 404, так что они практически не связываются с фосфорилированным остатком 404.
[0026] Без ограничения какой-либо конкретной теорией данные авторов настоящего изобретения демонстрируют, что различительная способность и селективность антител по настоящему изобретению в отношении человеческого тау-белка, фосфорилированного по остатку 396, в присутствии тау-белка, фосфорилированного по остатку 404, но не по 396, является значимой с точки зрения патологической и терапевтической перспективы. Антитела по настоящему изобретению являются селективными в отношении патологического тау-белка в присутствии непатологического, но при этом фосфорилированного, тау-белка. Антитела по настоящему изобретению способны вызывать истощение клубков тау-белка из патологического тау-белка в присутствии нормального тау-белка. Без ограничения какой-либо конкретной теорией полагают, что истощение клубков тау-белка, содержащих тау-белок, который был фосфорилирован в положении 396 тау-белка, предупреждает затравочное действие патологического тау-белка с образованием клубков тау-белка. Соответственно, один аспект настоящего изобретения относится к антителу, способному селективно связываться с фосфорилированным в положении 396 тау-белком даже в тех случаях, когда такие молекулы находятся в присутствии тау-белка, который был фосфорилирован в положении 404 тау-белка. Связанный аспект настоящего изобретения относится к антителу, способному селективно связываться с фосфорилированным в положении 396 тау-белком даже в тех случаях, когда такие молекулы находятся в присутствии непатогенного тау-белка. Как определено дополнительно, настоящее изобретение относится к антителу, селективному в отношении патологического тау-белка, при этом указанный патологический тау-белок является гиперфосфорилированным тау-белком, появляющимся в виде полосы, соответствующей 64 кДа (в вестерн-блот-анализе), у трансгенных мышей, сверхэкспрессирующих изоформу 2N4R человеческого тау-белка.
[0027] Один аспект настоящего изобретения направлен на антитело к тау-белку, удовлетворяющее следующим критериям тестирования: i) антитело не связывается с нефосфорилированным тау-белком; ii) антитело не связывается с тау-белком, фосфорилированным в 404 и не фосфорилированным в 396; iii) антитело связывается с тау-белком, фосфорилированным в 396; и iv) антитело связывается с тау-белком, фосфорилированным как в 396, так и в 404. Авторы настоящего изобретения обнаружили, что связывание в соответствии с критериями тестирования iii) и iv) имеет один и тот же порядок величины, и предположили, что фосфорилирование в положении 404 не препятствует процессу связывания, равно как и не усиливает его. Авторы настоящего изобретения дополнительно обнаружили, что, в противоположность критерию тестирования ii), связывание с тау-белком, который не является фосфорилированным в 396, но является фосфорилированным в 404, не вызывает истощение клубков или очищение от патологического тау-белка в тестовых моделях.
[0028] Один аспект настоящего изобретения направлен на антитело к тау-белку, которое при применении с иммуноистощенными экстрактами от трансгенных мышей rTg4510 специфично уменьшает полосы гиперфосфорилированного тау-белка размером 64 и 70 кДа по меньшей мере на 90%, уменьшая при этом полосу тау-белка размером 55 кДа не более чем на 10%. Дополнительный аспект настоящего изобретения направлен на антитело к тау-белку, которое специфично уменьшает полосы гиперфосфорилированного тау-белка размером 64 и 70 кДа по меньшей мере на 90%, уменьшая при этом полосу тау-белка размером 55 кДа не более чем на 10%; или которое при применении согласно описанному в данном документе с посмертными экстрактами головного мозга людей с AD обладает способностью к специфичному уменьшению полос фосфорилированного в S396 гиперфосфорилированного тау-белка по меньшей мере на 90% без уменьшения при этом полос негиперфосфорилированного тау-белка более чем на 10%.
[0029] Другой аспект настоящего изобретения направлен на способ лечения пациента с таупатией, такой как болезнь Альцгеймера, включающий истощение клубков или ослабление прогрессирования образования указанных клубков, при этом указанные клубки содержат гиперфосфорилированный тау-белок, при этом указанный способ включает приведение гиперфосфорилированного тау-белка в контакт с антителом по настоящему изобретению таким образом, что клубки истощаются, содержание в них гиперфосфорилированного тау-белка уменьшается или прогрессирование образования клубков ослабляется.
[0030] Как определено в качестве альтернативы, настоящее изобретение относится к способу лечения пациента с таупатией, такой как болезнь Альцгеймера, при этом указанный способ включает приведение клубков в контакт с антителом, селективным в отношении тау-белка, имеющего фосфорилированный остаток 396, таким образом, что в клубках происходит истощение по гиперфосфорилированному тау-белку.
[0031] Более конкретно, настоящее изобретение относится к любому из четырех моноклональных антител, выбранных из группы, включающей:
антитело С5.2,
где антитело С5.2 содержит:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 17;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 18;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 19;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 20;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 21; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 22;
антитело С8.3,
где антитело С8.3 содержит:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 25;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 26;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 27;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 28;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 29; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 30;
антитело С10-2,
где антитело С10-2 содержит:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 9;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 10;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 11;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 13; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 14;
и
антитело D1.2,
где антитело D1.2 содержит:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 1;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 2;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 3;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 4;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 5; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 6.
[0032] Аминокислотные последовательности полных легкой и тяжелой цепей приводимого в качестве примера антитела С5.2, в том числе их константных доменов, показаны соответственно под SEQ ID NO: 23 и SEQ ID NO: 24 (как используется в примерах).
[0033] Аминокислотные последовательности полных легкой и тяжелой цепей приводимого в качестве примера антитела С8.3, в том числе их константных доменов, показаны соответственно под SEQ ID NO: 31 и SEQ ID NO: 32 (как используется в примерах).
[0034] Аминокислотные последовательности полных легкой и тяжелой цепей представляющего интерес антитела С10-2, в том числе их константных доменов, показаны соответственно под SEQ ID NO: 15 и SEQ ID NO: 16 (как используется в примерах). Аминокислотная последовательность тяжелой цепи гуманизированного антитела С10-2 показана под SEQ ID NO: 35. Аминокислотная последовательность легкой цепи гуманизированного антитела С10-2 показана под SEQ ID NO: 36. Один аспект настоящего изобретения относится к антителу по настоящему изобретению, содержащему SEQ ID NO: 35 или SEQ ID NO: 36 или их обе.
[0035] Аминокислотные последовательности полных легкой и тяжелой цепей приводимого в качестве примера антитела D1.2, в том числе их константных доменов, показаны соответственно под SEQ ID NO: 7 и SEQ ID NO: 8 (как используется в примерах).
[0036] В альтернативном варианте осуществления антитело D1.2 содержит легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 34, где аминокислота в положении 3 представляет собой валин (тогда как в приводимой в качестве примера легкой цепи под SEQ ID NO: 7 эта аминокислота представляет собой метионин). Эта легкая цепь может находиться в паре с тяжелой цепью, описанной выше, т.е. имеющей CDR под SEQ ID NO: 4, 5 и 6. Например, антитело может содержать легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 34, вместе с тяжелой цепью, имеющей аминокислотную последовательность SEQ ID NO: 8 (антитело «D 1.2* »).
[0037] Один аспект настоящего изобретения направлен на антитело, содержащее:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 9;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 10; и/или
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 11.
[0038] Дополнительный аспект настоящего изобретения направлен на антитело, содержащее или дополнительно содержащее:
(a) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12;
(b) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 13; и/или
(c) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 14.
[0039] Антитела и их эпитопсвязывающие фрагменты по настоящему изобретению можно применять в лечении таупатий, таких как болезнь Альцгеймера (AD), болезнь аргирофильных зерен (AGD), прогрессирующий надъядерный паралич (PSP), кортикобазальная дегенерация (CBD), TBI (травматическое повреждение головного мозга, легкое, острое или хроническое) и хроническая травматическая энцефалопатия (СТЕ).
[0040] Антитела и их эпитопсвязывающие фрагменты по настоящему изобретению дополнительно предназначены для применения в лечении психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фигура 1. Дот-блот-анализ связывания с патологическим материалом
[0041] На фигуре 1 (панели А-В) представлены результаты дот-блот-анализа, отображающие 500 нг фракций S1 и Р3 (образование фракций S1 и Р3 раскрыто в примере 3), полученных из головного мозга пациентов с AD (AD) и пожилых здоровых индивидуумов (con) или из 32-недельных однопометных rTg4510 и нетрансгенных животных (wt), которые зондировали 1 мкг/мл D1.2 или С10-2 для оценки выявления патологического тау-белка (пример 3). На точечной диаграмме показано, что D1.2 (панель А) или С10-2 (панель В) специфично реагирует с пораженным заболеванием материалом от пациентов с AD или человеческим тау-белком (P301L), экспрессируемым у трансгенных мышей (Tg4510).
Фигура 2. Вестерн-блот-анализ антител D1.2 и С10-2
[0042] На фигуре 2 (панели А-В) представлены результаты вестерн-блот-анализа, отображающие 2 мкг фракций S1 и Р3, полученных из головного мозга 32-недельных однопометных rTg4510 и нетрансгенных животных (wt), или 20 мкг фракций S1 и Р3, полученных из головного мозга пациентов с AD (AD) и пожилых здоровых индивидуумов (con), которые зондировали 1 мкг/мл D1.2 (панель А) или С10-2 (панель В). Фракции S1 и Р3, которые получали из 0,01 мг ткани, загружали в соотношении 1:50 (по массе ткани). В вестерн-блот-анализе нормальный человеческий тау-белок 4R0N с мутацией P301L отображается при 55 кДа, а гиперфосфорилированная молекула человеческого тау-белка 4R0N с мутацией P301L отображается при 64 и 70 кДа. Во фракциях Р3 из материала с AD гиперфосфорилированный тау-белок отображается при 54, 64, 69 и 74 кДа (пример 3). На фигуре проиллюстрировано, что антитела специфично связываются с гиперфосфорилированным тау-белком со сдвигом подвижности.
Фигура 3. Связывание с патологическим материалом Р3 на MSD
[0043] На фигуре 3 (панели A-D) представлены результаты связывания D1.2 (панель А), С5-2 (панель В), С10-2 (панель С) и С8-3 (панель D) в ходе ELISA на платформе Meso Scale Discovery (MSD) с тау-белком, выделенным из головного мозга людей с AD и не пораженного заболеванием контрольного головного мозга (пример 4). Аналогично продемонстрированному на фигуре 1, иммобилизацию тау-белка, выделенного из пораженного заболеванием (AD) и здорового контрольного головного мозга, на планшетах для ELISA можно применять для демонстрирования того, что антитела в настоящем изобретении специфично связываются с патологическими молекулами тау-белка. Повышение концентраций антитела приводит к насыщающему связыванию. Количество связанного антитела выявляют с помощью вторичного антитела к мышиным иммуноглобулинам.
Фигура 4. Аффинность к пептидам и селективность в отношении pS396 (связывание с пептидами)
[0044] На фигуре 4 (панели A-D) представлены результаты анализа специфичного связывания С10-2 (панель А) и D1.2 (панель В) с пептидами тау-белка (386-409) со всеми комбинациями фосфорилирования в положениях S396 и S404 (пример 5). Специфичную аффинность в отношении патологического материала человека трудно оценить, и по этой причине авторы настоящего изобретения использовали специфичное связывание с пептидами для определения точной аффинности к эпитопу с применением специфических фосфорилированных и нефосфорилированных пептидов. Специфические кривые зависимости доза-ответ показаны для связывания антител С10-2 (панель С) и D1.2 (панель D) с пептидом TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37) (pS396/pS404), фосфорилированным по остаткам Ser396 и Ser404. Конкурентное связывание проводили с нефосфорилированным пептидом (NP) и монофосфорилированными пептидами (pS396 и pS404). Дополнительно был включен контрольный пептид, соответствующий фосфорилированному серину-262. Конкурентное связывание демонстрирует, что во всех случаях связывание достигается посредством фосфорилированного серинового остатка 396. Дополнительно, данные демонстрируют, что фосфорилирование по остатку 404 не препятствует связыванию антител с фосфосерином-396.
Фигура 5. Получение характеристик специфических для патологии антител гистологическими методами
[0045] На фигуре 5, панели А показано, что антитела С10-2 (левый столбец) и D1.2 (правый столбец) связываются с молекулами pTau в клеточных телах и нейропилях Tg4510 (верхний ряд). В срезах головного мозга животных, отличных от Tg, иммунореактивность не выявлена (нижний ряд). На фигуре 5, панели В показано, что антитела С10-2 (левый столбец) и D1.2 (правый столбец) связываются с молекулами pTau в клеточных телах и нитях нейропиля в донорном материале с AD (AD) (верхний ряд). В контрольном донорном головном мозге отсутствует иммунореактивность (нижний ряд) (пример 6).
Фигура 6. Связывание с патологическим и непатологическим Р3 для С10-2 и эталонных антител
[0046] На фигуре 6 (панели А-Е) представлены результаты, демонстрирующие превосходство антитела С10-2 (панель С) по настоящему изобретению в распознавании патологического материала по сравнению с антителами 2-10-3 (панель A), HACI-2B6 (панель В), IPN 002 (панель D) и HJ8.5 (панель Е) из предшествующего уровня техники. На фигуре показано специфичное связывание С10-2 с тау-белком из здорового (в качестве контроля) и пораженного заболеванием (AD) головного мозга людей наряду со связыванием с тау-белком из мышей Tg4510 в возрасте 10 месяцев, экспрессирующих человеческий тау-белок с мутацией P301L. К материалу тау-белка Р3, иммобилизованному на планшетах для ELISA, добавляют антитело в повышающихся концентрациях. Степень селективности в отношении патологического тау-белка определяют при полном насыщении активными молекулами. Кратность селективности для каждого из антител из предшествующего уровня техники показана на фигуре (пример 7).
Фигура 7. Предупреждение затравочного действия в клетках HEK293 и in vitro
[0047] На фигуре 7 (панели А-С) представлена количественная оценка агрегации тау-белка с помощью анализа Cisbio. Клетки HEK293, трансфицированные pcDNA для введения затравки, не продемонстрировали сигнал, что подтверждало отсутствие выявления входящего затравочного материала. Затравочный материал Wt (дикого типа) (WW) не продемонстрировал затравочного действия, однако, в отличие от этого, гомогенаты rTg4510 (СС) характеризовались эффективным затравочным действием по сравнению с лишенными затравки. На этот затравочный эффект не влияла обработка с помощью HEL, но он частично устранялся путем обработки антителами к тау-белку (С10-2 > D1.2 > hACI36-2B6-Ab1). На графических изображениях (панели А-С) представлены три независимых набора образцов, и они выполнены в виде диаграммы относительной агрегации тау-белка (кратности сигнала относительно фонового уровня, нормализованной к общему белку) (пример 8).
Фигура 8. Устранение электрофизиологического дефицита
[0048] На фигуре 8 показано устранение антителами дефицитов парного облегчения (панели В и D) и базальной синаптической передачи (панели А и С) в вызванных полевых потенциалах в СА1 (С 10-2, панель A; D1.2, панель В), где проиллюстрированы вызванные полевые потенциалы при обработке СА1 средней длительности с помощью С10-2 у мышей Tg4510 с и мышами tTA в качестве контроля. Животных обрабатывали дважды в неделю дозой 15 мг/кг антитела в течение двух недель (см. пример 9). На панели А (для С10-2) и панели С (для D1.2) градиент полевых потенциалов (fEPSP) представлен на диаграмме в зависимости от интенсивности стимуляции. На панелях А и С проиллюстрировано, что электрофизиологическая оценка синаптической передачи и пластичности в CA1-зоне гиппокампа у мышей rTg4510 (нижние 2 кривые) и контрольных мышей tTA (верхние 2 кривые) в возрасте 4,5-5,5 месяцев in vivo показала i) значительное ухудшение базальной синаптической передачи у мышей rTg4510 по сравнению с tTA и ii) значительное уменьшение парного облегчения у мышей rTg4510 по сравнению с tTA.
[0049] Парное облегчение - краткосрочную синаптическую пластичность, в основе которой, как полагают, лежат пресинаптические механизмы - дополнительно измеряли у мышей rTg4510 и tTA (панель В для С10-2 и панель D для D1.2). Вкратце, в отношении коллатерали Шеффера прилагали пару стимулов с интервалом между стимулами (ISI), варьирующимся от 25 до 1000 мс, и градиент второго fEPSP сравнивали с градиентом первого fEPSP. Облегчение наблюдали при всех ISI с максимальным облегчением при ISI, составляющем 50 и 75 мс. Интересно, что у мышей rTg4510 наблюдали значительно более низкое PPF (вторые 2 столбца) по сравнению с мышами tTA (первые 2 столбца).
Фигура 9. Краткий обзор скрининга, в общих чертах представленного на фигурах 1-8
[0050] Индуцировали выработку антител к дифосфорилированному пептиду TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающему остатки 386-410 тау-белка 2N4R. Гибридомы подвергали скринингу с помощью дот-блот-анализа и MSD-ELISA с иммобилизованным патологическим и непатологическим человеческим тау-белком (пример 4) для выделения клонов, которые были высокоспецифичными в отношении любого из фосфорилированных эпитопов S396 и/или S404 и в то же время специфично распознавали гиперфосфорилированный тау-белок из головного мозга людей с болезнью Альцгеймера. Способность к проведению различий между патологическим и непатологическим человеческим тау-белком в ходе дот-блот-анализа и вестерн-блот-анализа применяли для отбора гибридом. Отбирали 16 клонов, из которых четыре клона (D1.2, С10-2, С5.2 и С8.3) проявляли чрезвычайно высокие способности к связыванию с патологическим материалом человека. В случае применения специфического протокола иммунизации и скрининга вырабатываются высокоспецифичные антитела к фосфосерину-396 (pS396).
Фигура 10. Остаток pSer396 связывается в центре антигенсвязывающего участка mAb С5.2
[0051] Представлена кристаллическая структура mAb С5.2 в комплексе с фосфорилированным пептидом 386-410 при разрешении 1,9  . В этой структуре разрешена электронная плотность для остатков 392-398. Остаток {p} Ser396 связывается в центре антигенсвязывающего участка mAb С5.2. В этом структурном исследовании mAb к тау-белку эпитоп связывается между тяжелой цепью (справа внизу) и легкими (слева внизу) цепями.
. В этой структуре разрешена электронная плотность для остатков 392-398. Остаток {p} Ser396 связывается в центре антигенсвязывающего участка mAb С5.2. В этом структурном исследовании mAb к тау-белку эпитоп связывается между тяжелой цепью (справа внизу) и легкими (слева внизу) цепями.
Фигура 11. Взаимодействие антитела С5.2 с пептидом тау-белка (292-298) с фосфосерином
[0052] На фигуре 11 представлено взаимодействие между антителом С5.2 и пептидом тау-белка (292-298) с фосфосерином. Показана структура Ile(392)-Val(393)-Tyr(394)-Lys(395)-P-Ser(396)-Pro(397)-Val(398). В основном взаимодействии участвует гидрофобный карман, образуемый L3:H3, L3:F8*, Н1:Н13, H2:Y1, H2:Y3 и Y(394) пептида тау-белка. Существует разветвленная сеть водородных связей, образующаяся между сольватированным {p}S(396) и L3:T4, H1:R10, Н1:Т11, H3:R1, H3:T3. В используемой номенклатуре первая буква (например, «L» в L3:H3) означает, относится ли участвующий остаток CDR к CDR легкой цепи или CDR тяжелой цепи, первая цифра означает, какая CDR такой цепи участвует (например, «L3 » означает CDR3 легкой цепи), остальные обозначения (например, «Н3» в L3:H3) означают название и положение участвующей аминокислоты (например, «Н3» означает гистидин в третьем положении остатка CDR); таким образом, «L3:H3» означает гистидиновый остаток в третьем положении CDR3 легкой цепи. Существуют образование сильных водородных связей и взаимодействия зарядов/полярные взаимодействия между Y(394) боковой цепи и остовом, при этом фосфонат {p}S396 образует изгиб в пептидном остове. (*) L3:F8 представляет собой С-концевой остаток каркасной области, фланкирующий CDR L3.
[0053] Последовательности CDR С5.2 представляют собой:
CDR L1: QASQDTSINLN (SEQ ID NO: 17)
CDR L2: GASNLED (SEQ ID NO: 18)
CDR L3: LQHTYLP (SEQ ID NO: 19)
CDR H1: KASGYTFTDRTIH (SEQ ID NO: 20)
CDR H2: YIYPGDDSTKYNDNFKG (SEQ ID NO: 21)
CDR H3: RGTMDY (SEQ ID NO: 22)
Фигура 12. Истощение по тау-белку для анализа затравочного действия (HEK293)
[0054] На фигуре 12 (панели А-В) показано иммуноистощение гомогенатов головного мозга rTg4510 с помощью мышиного С10-2 (mC10-2) и гуманизированного С10-2 (hC10-2). Выявление в истощенных гомогенатах в ходе вестерн-блоттинга проводили с помощью Е1 (общий тау-белок; панель А; нижняя часть) и С10-2 (тау-белок pS396; панель А; верхняя часть), и как mC10-2, так и hC10-2 осуществляли эффективное истощение по гиперфосфорилированному тау-белку (верхние полосы на картине блоттинга для Е1 и все полосы на картине блоттинга для С10-2). Истощенные гомогенаты также анализировали в отношении истощения по агрегированному тау-белку с помощью анализа Cisbio. На панели В показано изменение количества агрегированного тау-белка в образцах. В исследованиях истощения с использованием mC10-2 и hC10-2 агрегаты тау-белка были удалены соответственно на 99 и 99,5% (панель В).
Фигура 13. Анализ затравочного действия (HEK293) с истощенным материалом
[0055] На панелях А-С показаны истощенные гомогенаты, применяемые для осуществления затравочного действия P301L-hTau в клетках HEK293. Гомогенаты от контрольных животных (WW) не демонстрировали затравочного действия, тогда как гомогенаты rTg4510 (СС) характеризовались эффективным затравочным действием, согласно измерениям в ходе анализа агрегации Cisbio в общих клеточных лизатах или в ходе фракционирования клеток HEK293 в 1% Triton-X (по количественной оценке нерастворимого гиперфосфорилированного D1.2 и тау-белка (панель А, верхняя и нижняя части)). Истощение с помощью антител HEL и hHEL не влияло на затравочное действие, тогда как истощение с помощью mC10-2 и hC10-2 предупреждало агрегацию тау-белка на 88% и 96% (панель С), а нерастворимого тау-белка на 97% и 100% (панель В) соответственно.
Фигура 14. Иммуноистощенный материал rTg4510 (применяемый для исследований затравочного действия in vivo)
[0056] На панелях А-С продемонстрирован вестерн-блот-анализ (панель А; верхняя, нижняя части) иммуноистощенных экстрактов головного мозга rTg4510. С10-2 и D1.2 специфично уменьшают полосу гиперфосфорилированного человеческого белка размером 64 кДа на 90% и не оказывают эффект на тау-белок размером 55 кДа. Tau5, коммерческое антитело к общему тау-белку, в отличие от этого, уменьшает полосу нормального тау-белка размером 55 кДа на 74% и также не оказывает эффект на человеческий тау-белок размером 64 кДа (панели В-С).
Фигура 15. Иммуноистощенный материал с AD (применяемый для исследований затравочного действия in vivo)
[0057] На фигуре 15 (панели А-С) изображен вестерн-блот-анализ (панель А) иммуноистощенных экстрактов головного мозга с болезнью Альцгеймера. Иммуноистощение с помощью С10-2 и D1.2 не приводит к уменьшению уровней общего тау-белка более чем на 10%, но обуславливает специфичное снижение уровня гиперфосфорилированного тау-белка (90% уменьшение) (панели В-С).
Фигура 16 (панель А). Патология тау-белка в гиппокампах у мышей rTg4510 с затравкой иммуноистощенным материалом rTg4510
[0058] На фигуре 16 (панель А) проиллюстрирована количественная оценка патологии тау-белка в головном мозге rTg4510 с затравкой гомогенатами головного мозга rTg4510 или с AD. Перед введением затравки уровень гиперфосфорилированного тау-белка, но не нормального тау-белка, был уменьшен в гомогенатах на 90-95% путем применения С10-2 или D1.2. При удалении гиперфосфорилированного тау-белка из гомогенатов гомогенаты больше не индуцируют затравочное действие с формированием патологии тау-белка.
Фигура 16 (панель В). Патология клубков в гиппокампах мышей rTg4510 с затравкой иммуноистощенным материалом с AD
[0059] На фигуре 16 (панель В) проиллюстрирована количественная оценка патологии тау-белка в головном мозге rTg4510 с затравкой гомогенатами головного мозга rTg4510 (А) или с AD (В). Перед введением затравки уровень гиперфосфорилированного тау-белка, но не нормального тау-белка, был уменьшен в гомогенатах на 90-95% путем применения С10-2 или D1.2. При удалении гиперфосфорилированного тау-белка из гомогенатов гомогенаты больше не индуцируют затравочное действие с формированием патологии тау-белка.
Фигура 17. Патология клубков в гиппокампах мышей rTg4510 с затравкой, обработанных D1.2
[0060] На фигуре 17 изображена количественная оценка нейронов, содержащих клубки, в гиппокампах мышей rTg4510 с затравкой. Данная патология увеличивается с течением времени (IgG - 1 месяц; IgG - 2 месяца; IgG - 3 месяца). Однако после обработки мышей с помощью D1.2 патология значительно уменьшалась через 1, 2 и 3 месяца после введения затравки. (D1.2-1 месяц; D1.2-2 месяца; D1.2-3 месяца).
Фигура 18. Истощение по тау-белку для анализа затравочного действия (HEK293)
[0061] На панели А показано иммуноистощение гомогенатов головного мозга rTg4510 с помощью мышиного С10-2 (mC10-2) и гуманизированного С10-2 (hC10-2). Выявление в истощенных гомогенатах в ходе вестерн-блоттинга проводили с помощью Е1 (общий тау-белок) и С10-2 (тау-белок pS396), и как mC10-2, так и hC10-2 осуществляли эффективное истощение по гиперфосфорилированному тау-белку (верхние полосы на картине блоттинга для Е1 и все полосы на картине блоттинга для С10-2). Истощенные гомогенаты анализировали в отношении истощения по агрегированному тау-белку с помощью анализа Cisbio. На панели В показано истощение с использованием mC10-2 и hC10-2, в ходе которого агрегаты тау-белка были удалены соответственно на 99 и 99,5%.
Фигура 19. Анализ затравочного действия (HEK293) с истощенным материалом
[0062] На панели А показано фракционирование тау-белка (вестерн-блоттинг нерастворимой фракции. На панели В показана количественная оценка вестерн-блоттинга. На панели С показан агрегированный тау-белок в клеточных лизатах. Истощенные гомогенаты применяли для осуществления затравочного действия P301L-hTau в клетках HEK293. Гомогенаты от контрольных животных (WW) не демонстрировали затравочного действия, тогда как гомогенаты rTg4510 (СС) характеризовались эффективным затравочным действием, согласно измерениям в ходе анализа агрегации Cisbio в общих клеточных лизатах или в ходе фракционирования клеток HEK293 в 1% Triton-X (по количественной оценке нерастворимого гиперфосфорилированного D1.2+ тау-белка). Истощение с помощью антител HEL и hHEL не влияло на затравочное действие, тогда как истощение с помощью mC10-2 и hC10-2 (панель С) предупреждало агрегацию тау-белка на 88% и 96%, а нерастворимого тау-белка на 97% и 100%» соответственно (панель В).
Фигура 20. Иммуноселективность С10-2 и D1.2 в отношении гиперфосфорилированного тау-белка по сравнению с нормальным тау-белком
[0063] Иммуноистощенный материал rTg4510 применяли для исследований затравочного действия in vivo. На панели А показан вестерн-блот-анализ иммуноистощенных экстрактов головного мозга rTg4510. На панели В показано, что С10-2 и D1.2 специфично уменьшают полосу гиперфосфорилированного белка размером 64 кДа, фосфорилированного по серину-396, по сравнению с полосой тау-белка размером 55 кДа, который не содержит значительное количество р396. В отличие от этого, Tau5, коммерческое антитело к общему тау-белку, уменьшает полосу нормального тау-белка размером 55 кДа и является неэффективным в связывании с тау-белком размером 64 кДа.
Фигура 21. Иммуноселективность С10-2 и D1.2 в отношении гиперфосфорилированного тау-белка по сравнению с нормальным тау-белком
[0064] Иммуноистощенный материал с AD применяли для исследований затравочного действия in vivo. На панели А показан вестерн-блот-анализ иммуноистощенных экстрактов головного мозга с болезнью Альцгеймера. Иммуноистощение с помощью mC10-2 и D1.2 не приводит к уменьшению уровней общего тау-белка более чем на 10% (панель В), но обуславливает специфичное снижение уровня гиперфосфорилированного тау-белка (90% уменьшение) (панель С).
Фигура 22. Патология тау-белка в гиппокампах мышей rTg4510
[0065] На панели А показана патология тау-белка в гиппокампах у мышей rTg4510 с затравкой иммуноистощенным материалом rTg4510. На панели В показана патология клубков в гиппокампах у мышей rTg4510 с затравкой иммуноистощенным материалом с AD. Показана количественная оценка патологии тау-белка в головном мозге rTg4510 с затравкой гомогенатами головного мозга rTg4510 (А) или с AD (В). Перед введением затравки уровень гиперфосфорилированного тау-белка, но не нормального тау-белка, был уменьшен в гомогенатах на 90-95% путем применения антител С10-2 или D1.2. При удалении гиперфосфорилированного тау-белка из гомогенатов гомогенаты больше не индуцируют затравочное действие с формированием патологии тау-белка.
Фигура 23. Патология клубков в гиппокампах мышей rTg4510 с затравкой, обработанных D1.2
[0066] Показана количественная оценка нейронов, содержащих клубки, в гиппокампах мышей rTg4510 с затравкой. На фигуре показано, что патология увеличивается с течением времени, и при обработке мышей с помощью D1.2 патология значительно уменьшается через 2 и 3 месяца после введения затравки.
Фигура 24. Вестерн-блот-анализ иммуноистощенных экстрактов от людей с AD
[0067] На фигуре проиллюстрировано, что гуманизированный вариант С10-2 (hC10-2), а также mC10-2 отличаются от антитела 2.10.3 (P-S422) тем, что, хотя уровень остающегося общего тау-белка несущественно отличается (левая панель) от такового в случае с 2.10.3, С10-2 (hC10-2), а также mC10-2 удаляют большее количество гиперфосфорилированного тау-белка, присутствующего в экстрактах головного мозга с болезнью Альцгеймера, с помощью способов иммуноистощения. Это подтверждается на фигуре 25 путем количественной оценки.
Фигура 25. Количественная оценка агрегированного тау-белка после иммуноистощения
[0068] Антитела hC10-2 и mC10-2 отличаются от антитела 2.10.3 своей способностью к удалению большего количества агрегированного тау-белка, присутствующего в экстрактах головного мозга с болезнью Альцгеймера, с помощью способов иммуноистощения.
Фигура 26. Общий тау-белок, остающийся после иммуноистощения
Показана количественная оценка сигналов вестерн-блоттинга после иммуноистощения экстрактов материала с болезнью Альцгеймера с помощью различных количеств гуманизированного С10-2  и антитела 2.10.3
и антитела 2.10.3  . На фигуре 26 показана количественная оценка сигнала общего тау-белка с применением Tau5 (все изоформы тау-белка были включены в анализ). Оба антитела удаляли небольшую долю тау-белка из препарата головного мозга субъекта с болезнью Альцгеймера. 2.10.3, сконструированное таким образом, что оно обладает специфичностью к тау-белку Р-S422, удаляет до 24% от общего количества тау-белка, а С10-2 удаляет до 15% общего тау-белка (см. фигуру 26).
. На фигуре 26 показана количественная оценка сигнала общего тау-белка с применением Tau5 (все изоформы тау-белка были включены в анализ). Оба антитела удаляли небольшую долю тау-белка из препарата головного мозга субъекта с болезнью Альцгеймера. 2.10.3, сконструированное таким образом, что оно обладает специфичностью к тау-белку Р-S422, удаляет до 24% от общего количества тау-белка, а С10-2 удаляет до 15% общего тау-белка (см. фигуру 26).
Фигура 27. Общий тау-белок, остающийся после иммуноистощения по гиперфосфорилированному тау-белку
[0069] На фигуре 27 проиллюстрирована количественная оценка гиперфосфорилированного тау-белка, фосфорилированного по серину-422 (все полосы и высокомолекулярный шмер были включены в анализ). Как 2.10.3  , так и С10-2
, так и С10-2  удаляют более 90% тау-белка, фосфорилированного по серину-422. Однако количество антитела, необходимого для удаления 50% тау-белка, различается: в случае с антителом 2.10.3 необходимо 0,42 мкг антитела, тогда как в случае с С10-2 для того же эффекта необходимо 0,27 мкг.
удаляют более 90% тау-белка, фосфорилированного по серину-422. Однако количество антитела, необходимого для удаления 50% тау-белка, различается: в случае с антителом 2.10.3 необходимо 0,42 мкг антитела, тогда как в случае с С10-2 для того же эффекта необходимо 0,27 мкг.
Фигура 28. Общий тау-белок, остающийся после иммуноистощения по гиперфосфорилированному тау-белку
[0070] Показана количественная оценка гиперфосфорилированного тау-белка, фосфорилированного по серину-396 (все полосы и высокомолекулярный шмер были включены в анализ). С10-2  эффективно удаляет тау-белок, фосфорилированный по серину-396 (максимальный эффект: 88%, и половина этого эффекта достигается при применении 0,30 мкг антитела). 2.10.3
эффективно удаляет тау-белок, фосфорилированный по серину-396 (максимальный эффект: 88%, и половина этого эффекта достигается при применении 0,30 мкг антитела). 2.10.3  удаляет меньшую долю тау-белка, фосфорилированного по серину-396 (максимальный эффект: 60%, и половина этого эффекта достигается при применении 0,63 мкг антитела). Это указывает на то, что весь тау-белок, фосфорилированный по серину-422, также является фосфорилированным по серину-396, но существует часть гиперфосфорилированного тау-белка, фосфорилированного по серину-396, в которой отсутствует фосфорилированный серии в положении 422.
удаляет меньшую долю тау-белка, фосфорилированного по серину-396 (максимальный эффект: 60%, и половина этого эффекта достигается при применении 0,63 мкг антитела). Это указывает на то, что весь тау-белок, фосфорилированный по серину-422, также является фосфорилированным по серину-396, но существует часть гиперфосфорилированного тау-белка, фосфорилированного по серину-396, в которой отсутствует фосфорилированный серии в положении 422.
Фигура 29. Общий тау-белок, остающийся после иммуноистощения по гиперфосфорилированному тау-белку
[0071] Показана количественная оценка гиперфосфорилированного тау-белка, фосфорилированного по серину-199/202 (все полосы и высокомолекулярный шмер были включены в анализ). Значительная часть тау-белка, удаляемая С10-2  , также является фосфорилированной по серину-199/202, поскольку 69% тау-белка, имеющего такое фосфорилирование, подвергается влиянию иммуноистощения (50% эффект при применении 0,34 мкг антитела). Иммуноистощение с помощью 2.10.3
, также является фосфорилированной по серину-199/202, поскольку 69% тау-белка, имеющего такое фосфорилирование, подвергается влиянию иммуноистощения (50% эффект при применении 0,34 мкг антитела). Иммуноистощение с помощью 2.10.3  не давало сигмоидальную кривую зависимости доза-ответ для тау-белка P-S199/202, хотя при повышении количества антитела наблюдается снижение интенсивности сигнала (максимальное уменьшение 52% при применении максимального количества антитела (5 мкг). Эти данные указывают на то, что антитело С10-2, нацеливающееся на фосфорилированный серин-396, связывается с более крупным пулом гиперфосфорилированных тау-белков, чем антитело 2.10.3, нацеливающееся на фосфорилированный серии в положении 422.
не давало сигмоидальную кривую зависимости доза-ответ для тау-белка P-S199/202, хотя при повышении количества антитела наблюдается снижение интенсивности сигнала (максимальное уменьшение 52% при применении максимального количества антитела (5 мкг). Эти данные указывают на то, что антитело С10-2, нацеливающееся на фосфорилированный серин-396, связывается с более крупным пулом гиперфосфорилированных тау-белков, чем антитело 2.10.3, нацеливающееся на фосфорилированный серии в положении 422.
Фигура 30. Ингибирование mD1.2 и mC10-2 захвата антигенного тау-белка в планшетах, покрытых mC10-2
[0072] Это происходило в рамках жидкофазного ELISA, где смесь препарата Р3 rTg4510 и различных количеств антител С10-2 или D1.2 добавляли на планшеты, покрытые С10-2. Чем больше антител связывалось с тау-белком Р3 в растворе, тем меньше доступных эпитопов тау-белка могло связываться с планшетами. Величину связывания тау-белка с планшетами определяют с помощью антитела к человеческому тау-белку, меченного SULFO-TAG. С10-2  и D1.2 (□) характеризуются разным связыванием с тау-белком в растворе, при этом связывание с С10-2 может полностью превосходить связывание с планшетами (IC50-20 нМ). В то же время D1.2 демонстрирует очень низкий уровень связывания с тау-белком в растворе.
и D1.2 (□) характеризуются разным связыванием с тау-белком в растворе, при этом связывание с С10-2 может полностью превосходить связывание с планшетами (IC50-20 нМ). В то же время D1.2 демонстрирует очень низкий уровень связывания с тау-белком в растворе.
Фигура 31. Ингибирование PHF13 и mC10-2 захвата антигенного тау-белка в планшетах, покрытых mC10-2
[0073] В рамках жидкофазного ELISA смесь препарата Р3 из материала с AD и различных количеств антител С10-2 и PHF13 добавляли на планшеты, покрытые С10-2. Чем больше антител связывалось с тау-белком Р3 в растворе, тем меньше доступных эпитопов тау-белка могло связываться с планшетами. Величину связывания тау-белка с планшетами определяют с помощью антитела к человеческому тау-белку, меченного SULFO-TAG. С10-2 и PHF13 характеризуются разным связыванием с тау-белком в растворе, при этом связывание с С10-2 может полностью превосходить связывание с планшетами (IC50=3 нМ), тогда как в случае с PHF13 это не происходит.
Фигура 32. Как mC10-2, так и PFH-13 связываются с pTau 386-408 (pS396/pS404) дозозависимым образом
[0074] На фигуре 32 показано, что mC10-2 и PHF-13 в равной степени хорошо связываются в планшетах MSD, покрытых 100 нг/мл pTau 386-408 (pS396/pS404. Антитела в повышающихся концентрациях (указанных по оси х) инкубировали в лунках в течение 2 ч. с последующим промыванием и выявлением связанных антител с помощью антител к человеческому IgG, меченных SULFO-TAG. Это указывает на то, что полученное PHF-13, применяемое в последующих примерах, является активным.
Фигура 33. Сравнение связывания mD1.2 и mC10-2 с AD-P3
[0075] На фигуре 33 показано, что mD1.2 и mC10-2 в равной степени хорошо связываются в планшетах MSD, покрытых 1 мкг/мл AD-P3. Антитела в повышающихся концентрациях (указанных по оси х) инкубировали в присутствии и в отсутствие 10 мкМ пептида pTau 386-408 (pS396/pS404) в течение 1 часа при комнатной температуре с последующим инкубированием в лунках в течение 2 часов, протекавших перед выявлением связанных антител с помощью антител к человеческому IgG, меченных SULFO-TAG. Значения IC50 составляли 320 нМ и 11 нМ для захвата AD-P3 и AD-S1(p). В отличие от этого, mD1.2 продемонстрировало значительно более слабое ингибирование захвата антигенного тау-белка при значениях IC50, составляющих 589 и 503 нМ, что позволяет предположить значительно более низкую аффинность связывания с растворимыми антигенами.
[0076] Анализ выполняли в две стадии. А: 1 мкг/мл AD-P3 и 20 нг/мл AD-S1(p) соответственно инкубировали с mD1.2 и mC10-2 в повышающихся концентрациях и инкубировали в течение 1 часа при комнатной температуре для обеспечения повышения связывания антитела и антигена (занятости). В: образцы инкубировали на планшетах MSD, покрытых AD-P3 (1 мкг/мл), в течение 2 часов с последующим промыванием и выявлением захваченных антигенных тау-белков с помощью антител к общему тау-белку, меченных SULFO-TAG.
Фигура 34. mC10-2, но не PHF-13, эффективно связывается с антигенами AD-P3, представленными на твердой фазе
Высокоспецифичное связывание mC10-2, но не PHF-13: на фигуре 34 показано, что mC10-2 эффективно связывается с планшетами MSD, покрытыми антигенами AD-P3 (1 мкг/мл). Для сравнения, низкая активность связывания PHF-13 указывает на более низкую аффинность к физиологическим антигенам pTau. Дополнительно, PHF-13 демонстрировало значительно более высокую степень неспецифичного связывания по сравнению с mC10-2 (см. таблицу 6). Антитела в повышающихся концентрациях (указанных по оси х) инкубировали в течение 2 часов, протекавших перед выявлением связанных антител с помощью антител к человеческому IgG, меченных SULFO-TAG. В сигнал связывания вносили поправку на активность неспецифического связывания (определенную как сигналы, измеренные в присутствии 10 мкМ пептида pTau 386-408 (pS396/pS404). Значение IC50 составляло 3 нМ для захвата mC10-2 AD-P3. В отличие от этого, PHF-13 практически не продемонстрировало ингибирование.
[0077] Анализ выполняли в две стадии. А: 1 мкг/мл AD-P3 инкубировали с mC10-2 и PHF-13 в повышающихся концентрациях и инкубировали в течение 1 часа при комнатной температуре для обеспечения повышения связывания антитела и антигена (занятости). В: образцы инкубировали на планшетах MSD, покрытых AD-P3 (1 мкг/мл), в течение 2 часов с последующим промыванием и выявлением захваченных антигенных тау-белков с помощью антител к общему тау-белку, меченных SULFO-TAG.
ПОСЛЕДОВАТЕЛЬНОСТИ, ВКЛЮЧЕННЫЕ ПОСРЕДСТВОМ ССЫЛКИ
SEQ ID NO: 1 CDR1 легкой цепи D1.2
SEQ ID NO: 2 CDR2 легкой цепи D1.2
SEQ ID NO: 3 CDR3 легкой цепи D1.2
SEQ ID NO: 4 CDR1 тяжелой цепи D1.2
SEQ ID NO: 5 CDR2 тяжелой цепи D1.2
SEQ ID NO: 6 CDR3 тяжелой цепи D1.2
SEQ ID NO: 7 легкая цепь D1.2
SEQ ID NO: 8 тяжелая цепь D1.2
SEQ ID NO: 9 CDR1 легкой цепи C10-2
SEQ ID NO: 10 CDR2 легкой цепи С10-2
SEQ ID NO: 11 CDR3 легкой цепи С10-2
SEQ ID NO: 12 CDR1 тяжелой цепи С10-2
SEQ ID NO: 13 CDR2 тяжелой цепи С10-2
SEQ ID NO: 14 CDR3 тяжелой цепи С10-2
SEQ ID NO: 15 легкая цепь С10-2
SEQ ID NO: 16 тяжелая цепь С10-2
SEQ ID NO: 17 CDR1 легкой цепи C5.2
SEQ ID NO: 18 CDR2 легкой цепи C5.2
SEQ ID NO: 19 CDR3 легкой цепи C5.2
SEQ ID NO: 20 CDR1 тяжелой цепи C5.2
SEQ ID NO: 21 CDR2 тяжелой цепи C5.2
SEQ ID NO: 22 CDR3 тяжелой цепи C5.2
SEQ ID NO: 23 легкая цепь C5.2
SEQ ID NO: 24 тяжелая цепь C5.2
SEQ ID NO: 25 CDR1 легкой цепи C8.3
SEQ ID NO: 26 CDR2 легкой цепи C8.3
SEQ ID NO: 27 CDR3 легкой цепи C8.3
SEQ ID NO: 28 CDR1 тяжелой цепи C8.3
SEQ ID NO: 29 CDR2 тяжелой цепи C8.3
SEQ ID NO: 30 CDR3 тяжелой цепи C8.3
SEQ ID NO: 31 легкая цепь C8.3
SEQ ID NO: 32 тяжелая цепь C8.3
SEQ ID NO: 33 человеческий тау-белок
SEQ ID NO: 34 легкая цепь D1.2*
SEQ ID NO: 35 тяжелая цепь гуманизированного С10-2
SEQ ID NO: 36 легкая цепь гуманизированного С10-2
SEQ ID NO: 37 остатки 386-408 тау-белка (pS396, pS404)
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0078] Используемый в данном документе термин «тау» является синонимом «тау-белка» и относится к любой из изоформ тау-белка (идентифицированных, например, в UniProt как Р10636, 1-9). Нумерация аминокислот тау-белка, используемая в данном документе, приведена применительно к изоформе 2 (SEQ ID NO: 33), показанной ниже, при этом метионин (М) является аминокислотным остатком 1. SEQ ID NO: 33:

[0079] Настоящее изобретение относится к антителам и их эпитопсвязывающим фрагментам, которые способны специфично связываться с тау-белком, и в частности, с человеческим тау-белком, и в одном варианте осуществления проявляют способность к специфичному связыванию с фосфорилированным остатком S396 (pS396) человеческого тау-белка. Антитела и их эпитопсвязывающие фрагменты по настоящему изобретению дополнительно характеризуются тем, что они не способны или практически не способны специфично связываться с фосфорилированным остатком 404 (pS404) в человеческом тау-белке, например, в условиях ограниченного количества антител или ненасыщающих условиях. Дополнительно, фосфорилирование в pS404 не препятствует специфичному связыванию с pS396. Используемые в данном документе обозначения «pS» и «{p}S» означают фосфосериновый аминокислотный остаток. Как используется в данном документе, антитело «практически» не способно связываться с эпитопом, если относительно другого эпитопа такое связывание составляет менее 20%, менее 10%, менее 5%, менее 2% и более предпочтительно менее 1% от связывания, наблюдаемого для такого другого эпитопа.
[0080] Термин «антитело» (Ab) в контексте настоящего изобретения относится к молекуле иммуноглобулина или, согласно некоторым вариантам осуществления настоящего изобретения, фрагменту молекулы иммуноглобулина, которые обладают способностью к специфичному связыванию с эпитопом молекулы («антигена»). Встречающиеся в природе антитела в типичном случае являются тетрамерами, которые обычно состоят по меньшей мере из двух тяжелых (Н) цепей и по меньшей мере двух легких (L) цепей. Каждая тяжелая цепь состоит из вариабельного домена тяжелой цепи (сокращенно называемого в данном документе VH) и константного домена тяжелой цепи, обычно состоящего из трех доменов (CH1, СН2 и СН3). Тяжелые цепи могут относиться к любому изотипу, включая IgG (подтипы IgG1, IgG2, IgG3 и IgG4). Каждая легкая цепь состоит из вариабельного домена легкой цепи (сокращенно называемого в данном документе VL) и константного домена легкой цепи (CL). Легкие цепи включают каппа-цепи и лямбда-цепи. Вариабельный домен тяжелой и легкой цепей в типичном случае отвечает за распознавание антигена, а константный домен тяжелой и легкой цепей может опосредовать связывание иммуноглобулина с тканями или факторами реципиента, в том числе с различными клетками иммунной системы (например, эффекторными клетками) и первым компонентом (C1q) классического пути активации системы комплемента. VH- и VL-домены могут дополнительно подразделяться на гипервариабельные домены, называемые «областями, определяющими комплементарность», которые перемежаются с доменами с более консервативной последовательностью, называемыми «каркасными областями» (FR). Каждый VH и VL состоит из трех CDR-доменов и четырех FR-доменов, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4. Вариабельные домены тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Особенно значимыми являются антитела и их эпитопсвязывающие фрагменты, которые были «выделены» таким образом, чтобы они существовали в физической среде, отличающейся от той, в которой они могут встречаться в природе, или которые были модифицированы таким образом, чтобы они отличались по аминокислотной последовательности от встречающегося в природе антитела.
[0081] Термин «эпитоп» означает антигенную детерминанту, способную специфично связываться с антителом. Эпитопы обычно состоят из поверхностных групп молекул, таких как аминокислоты или сахарные боковые цепи, и обычно имеют специфические характеристики трехмерной структуры, а также специфические характеристики заряда. Конформационные и линейные эпитопы отличаются тем, что связывание с первыми, но не с последними, всегда утрачивается в присутствии денатурирующих растворителей. Эпитоп может содержать аминокислотные остатки, непосредственно участвующие в связывании, и другие аминокислотные остатки, которые непосредственно не участвуют в связывании, такие как аминокислотные остатки, эффективно блокируемые пептидом, специфично связывающимся с эпитопом (другими словами, аминокислотный остаток находится в пределах области узнавания для пептида, специфично связывающегося с эпитопом).
[0082] Используемый в данном документе термин «эпитопсвязывающий фрагмент антитела» означает фрагмент, часть, область или домен антитела (независимо от того, как они получены (например, путем расщепления, рекомбинантным путем, синтетическим путем и т.п.)), которые способны специфично связываться с эпитопом. Эпитопсвязывающий фрагмент может содержать 1, 2, 3, 4, 5 или все 6 CDR-доменов такого антитела и, несмотря на то, что способен специфично связываться с таким эпитопом, может проявлять специфичность, аффинность или селективность в отношении того эпитопа, который отличается от эпитопа для такого антитела. Предпочтительно, однако, чтобы эпитопсвязывающий фрагмент содержал все 6 CDR-доменов такого антитела. Эпитопсвязывающий фрагмент антитела может представлять собой или содержать часть одной полипептидной цепи (например, в scFv) или может представлять собой или содержать часть двух илиболее полипептидных цепей, каждая из которых имеет амино-конец и карбоксильный конец (например, в диателе, Fab-фрагменте, Fab2-фрагменте и т.п.). Фрагменты антител, которые проявляют способность к связыванию с эпитопом, можно получить, например, путем расщепления интактных антител протеазами. Более предпочтительно, чтобы, несмотря на то, что два домена Fv-фрагмента, VL и VH, в естественных условиях кодируются отдельными генами, полинуклеотиды, которые кодируют такие последовательности генов (например, их кодирующую кДНК), можно было с помощью рекомбинантных способов соединить гибким линкером, который обеспечивает возможность их получения в виде одной белковой цепи, в которой VL- и VH-области ассоциируют друг с другом с образованием одновалентных эпитопсвязывающих молекул (известных как одноцепочечные Fv (scFv); см., например, Bird et al., (1988) Science 242: 423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. (U.S.A.) 85: 5879-5883). В качестве альтернативы, путем использования гибкого линкера, который является слишком коротким (например, имеет длину менее чем приблизительно 9 остатков) для того, чтобы обеспечить возможность ассоциации VL- и VH-доменов одной полипептидной цепи друг с другом, можно образовать биспецифическое антитело, диатело или аналогичную молекулу (в которой две такие полипептидные цепи ассоциируют друг с другом с образованием двухвалентной эпитопсвязывающей молекулы) (см., например, PNAS USA 90(14), 6444-8 (1993) в отношении описания диател). Примеры эпитопсвязывающих фрагментов, охватываемых настоящим изобретением, включают (i) Fab'- или Fab-фрагмент - одновалентный фрагмент, состоящий из VL-, VH-, CL- и СН1-доменов, или одновалентное антитело, описанное в WO 2007059782; (ii) Р(ab')2-фрагменты -двухвалентные фрагменты, содержащие два Fab-фрагмента, соединенных дисульфидным мостиком в шарнирном домене; (iii) Fd-фрагмент, по сути состоящий из VH- и СН1-доменов; (iv) Fv-фрагмент, по сути состоящий из VL- и VH-доменов, (v) dAb-фрагмент (Ward et al., Nature 341, 544-546 (1989)), который по сути состоит из VH-домена и также называется доменным антителом (Holt et al; Trends Biotechnol. 2003 Nov; 2i(II): 484-90); (vi) антитела верблюдовых или нанотела (Revets et al; Expert Opin Biol Ther. 2005 Jan; 5_(I): I II-24) и (vii) выделенную область, определяющую комплементарность (CDR). Дополнительно, несмотря на то, что два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, их с помощью рекомбинантных способов можно соединить синтетическим линкером, который обеспечивает возможность их получения в виде одной белковой цепи, в которой VL- и VH-домены находятся в паре, с образованием одновалентных молекул (известных как одноцепочечные антитела или одноцепочечные Fv (scFv), см., например, Bird et al., Science 242, 423-426 (1988), и Huston et al., PNAS USA 85, 5879-5883 (1988)). Эти и другие фрагменты антител, применимые в контексте настоящего изобретения, дополнительно обсуждаются в данном документе. Также следует понимать, что термин «антитело», если не указано иное, также включает антителоподобные полипептиды, такие как химерные антитела и гуманизированные антитела, и фрагменты антител, сохраняющие способность к специфичному связыванию с антигеном (эпитопсвязывающие фрагменты), получаемые с помощью любой известной методики, такой как ферментативное расщепление, синтез пептидов и рекомбинантные методики. Полученное антитело может иметь любой изотип. Как используется в данном документе, «изотип» относится к классу иммуноглобулина (например, IgG1, IgG2, IgG3 или IgG4), кодируемому генами константных доменов тяжелых цепей. Такие фрагменты антител получают с помощью традиционных методик, известных специалистам в данной области; подходящие фрагменты, способные связываться с желаемым эпитопом, можно без труда подвергнуть скринингу в отношении полезности таким же образом, как и интактное антитело.
[0083] Термин «биспецифическое антитело» относится к антителу, содержащему два независимых эпитопсвязывающих фрагмента, каждый из которых нацеливается на независимые мишени. Эти мишени могут представлять собой эпитопы, присутствующие в различных белках, или различные эпитопы, присутствующие в одной и той же мишени. Молекулы биспецифических антител можно получать с помощью компенсаторных изменений аминокислот в константных доменах НС исходных молекул моноспецифических двухвалентных антител. Полученное в результате гетеродимерное антитело содержит один Fab, вклад в образование которого вносят два разных исходных моноспецифических антитела. Изменения аминокислот в Fc-домене приводят к повышению стабильности гетеродимерного антитела с биспецифичностью, стабильной с течением времени. (Ridgway et al., Protein Engineering 9, 617-621 (1996), Gunasekaran et al., JBC 285, 19637-1 (2010), Moore et al., MAbs 3:6 546-557 (2011), Strop et al., JMB 420, 204-219 (2012), Metz et al., Protein Engineering 25:10 571-580 (2012), Labrijn et al., PNAS 110:113, 5145-5150 (2013), Spreter Von Kreudenstein et al., MAbs 5:5 646-654 (2013)). Биспецифические антитела также могут включать молекулы, получаемые с помощью слияний scFv. Два моноспецифических scFv затем независимо соединяют с Fc-доменами, способными образовывать стабильные гетеродимеры, с получением одной биспецифической молекулы (Mabry et al., PEDS 23:3 115-127 (2010)). Биспецифические молекулы обладают способностями к двойному связыванию.
[0084] Термин «антитело D1.2» подразумевается как означающий антитело или его эпитопсвязывающий фрагмент, содержащие вариабельный домен легкой цепи антитела, имеющий:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: l;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 2; и
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 3;
и вариабельный домен тяжелой цепи антитела, имеющий:
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 4;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 5; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 6, или состоящие из них.
[0085] В одном варианте осуществления антитело D1.2 или его эпитопсвязывающий фрагмент могут содержать тяжелую цепь под SEQ ID NO: 8 и/или легкую цепь под SEQ ID NO: 7 или состоять из них.
[0086] В связанном варианте осуществления антитело D1.2* или его эпитопсвязывающий фрагмент могут содержать тяжелую цепь SEQ ID NO: 8 и/или легкую цепь SEQ ID NO: 34 или состоять из них.
[0087] Термин «антитело С10-2» подразумевается как означающий антитело или его эпитопсвязывающий фрагмент, содержащие вариабельный домен легкой цепи антитела, имеющий:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 9;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 10; и
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 11;
и вариабельный домен тяжелой цепи антитела, имеющий:
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 13; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 14, или состоящие из них.
[0088] В одном варианте осуществления антитело С10-2 или его эпитопсвязывающий фрагмент могут содержать тяжелую цепь под SEQ ID NO: 16 и/или легкую цепь под SEQ ID NO: 15 или состоять из них.
[0089] В дополнительном варианте осуществления гуманизированное антитело С10-2 или его эпитопсвязывающий фрагмент могут содержать тяжелую цепь SEQ ID NO: 35, легкую цепь SEQ ID NO: 36 или их обе или состоять из них. Один вариант осуществления настоящего изобретения направлен на антитело или его эпитопсвязывающий фрагмент, содержащий тяжелую цепь SEQ ID NO: 35, легкую цепь SEQ ID NO: 36 или состоящий из них.
[0090] Термин «антитело С5.2» подразумевается как означающий антитело или его эпитопсвязывающий фрагмент, содержащие вариабельный домен легкой цепи антитела, имеющий:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 17;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 18; и
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 19;
и вариабельный домен тяжелой цепи антитела, имеющий:
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 20;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 21; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 22, или состоящие из них.
[0091] В одном варианте осуществления антитело С5.2 или его эпитопсвязывающий фрагмент могут содержать тяжелую цепь под SEQ ID NO: 24 и/или легкую цепь под SEQ ID NO: 23 или состоять из них.
[0092] Термин «антитело С8.3» подразумевается как означающий антитело или его эпитопсвязывающий фрагмент его эпитопсвязывающий фрагмент, содержащие вариабельный домен легкой цепи антитела, имеющий:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 25;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 26; и
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 27;
и вариабельный домен тяжелой цепи антитела, имеющий:
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 28;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 29; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 30, или состоящие из них.
[0093] В одном варианте осуществления антитело С8.3 или его эпитопсвязывающий фрагмент могут содержать тяжелую цепь под SEQ ID NO: 32 и/или легкую цепь под SEQ ID NO: 31 или состоять из них.
[0094] «Антитело к тау-белку» представляет собой антитело или его эпитопсвязывающий фрагмент, специфично связывающиеся с тау-белком или фрагментом тау-белка.
[0095] Термины «моноклональное антитело» или «композиция на основе моноклональных антител», используемые в данном документе, относятся к препарату из молекул антител единого молекулярного состава. Традиционная композиция на основе моноклональных антител демонстрирует единую специфичность связывания и аффинность к конкретному эпитопу. В определенных вариантах осуществления моноклональное антитело может состоять из более чем одного Fab-домена с повышением таким образом специфичности более чем к одной мишени. Термины «моноклональное антитело» или «композиция на основе моноклональных антител» не подразумеваются как ограниченные каким-либо конкретным способом получения (например, рекомбинантным, трансгенным, гибридомным и т.п.).
[0096] Антитела по настоящему изобретению и их эпитопсвязывающие фрагменты предпочтительно являются «гуманизированными», в частности, если они используются для терапевтических целей. Термин «гуманизированный» относится к молекуле, обычно получаемой с помощью рекомбинантных методик, которая имеет эпитопсвязывающий участок, полученный из иммуноглобулина от вида, отличного от человека, и остальную часть структуры иммуноглобулина, в основе которой лежит структура и/или последовательность человеческого иммуноглобулина. Эпитопсвязывающий участок может содержать либо полные вариабельные домены антитела, отличного от человеческого, слитые с человеческими константными доменами, либо только области, определяющие комплементарность (CDR), таких вариабельных доменов, трансплантированные на соответствующие человеческие каркасные области человеческих вариабельных доменов. Остатки каркасных областей таких гуманизированных молекул могут быть дикого типа (например, полностью человеческими), или они могут быть модифицированы таким образом, чтобы содержать одну или несколько аминокислотных замен, не обнаруживаемых в человеческом антителе, последовательность которого служила в качестве основы для гуманизации. Гуманизация уменьшает или устраняет вероятность того, что константный домен молекулы будет действовать в качестве иммуногена у индивидуумов-людей, однако возможность выработки иммунного ответа на чужеродный вариабельный домен сохраняется (LoBuglio, A.F. et al. (1989) "Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response" Proc. Natl. Acad. Sci. (U.S.A.) 86: 4220-4224). Другой подход фокусируется не только на получении константных доменов человеческого происхождения, но также и на модификации вариабельных доменов таким образом, чтобы реконструировать их как можно более близко к человеческой форме. Известно, что вариабельные домены как тяжелой, так и легкой цепей содержат три области, определяющие комплементарность (CDR), которые различаются по ответу на рассматриваемые антигены и определяют способность к связыванию, фланкированные четырьмя каркасными областями (FR), которые являются относительно консервативными у данного вида и которые предположительно обеспечивают остов для CDR. В тех случаях, когда антитела, отличные от человеческих, получают в отношении конкретного антигена, вариабельные домены можно «реконструировать» или «гуманизировать» путем трансплантации CDR, полученных из антитела, отличного от человеческого, на FR, присутствующие в человеческом антителе, подлежащем модификации. О применении данного подхода к различным антителам сообщалось в Sato, K. et al. (1993) Cancer Res 53: 851-856. Riechmann, L. et al. (1988) "Reshaping Human Antibodies for Therapy", Nature 332: 323-327; Verhoeyen, M. etal. (1988)" Reshaping Human Antibodies: Grafting An Antilysozyme Activity", Science 239: 1534-1536; Kettleborough, C. A. et al. (1991) "Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation", Protein Engineering 4: 773-3783; Maeda, H. et al. (1991) "Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity", Human Antibodies Hybridoma 2: 124-134; Gorman, S. D. et al. (1991) "Reshaping A Therapeutic CD4 Antibody", Proc. Natl. Acad. Sci. (U.S.A.) 88: 4181-4185; Tempest, P.R. et al. (1991) "Reshaping A Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo", Bio/Technology 9: 266-271; Co, M. S. et al. (1991) " Humanized Antibodies For Antiviral Therapy", Proc. Natl. Acad. Sci. (U.S.A.) 88: 2869-2873; Carter, P. et al. (1992) "Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy", Proc. Natl. Acad. Sci. (U.S.A.) 89: 4285-4289; и Co, M.S. et al. (1992) "Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen", J. Immunol. 148: 1149-1154. В некоторых вариантах осуществления в гуманизированных антителах сохраняются все последовательности CDR (например, в гуманизированном мышином антителе, которое содержит все шесть CDR мышиных антител). В других вариантах осуществления гуманизированные антитела имеют одну или несколько CDR (одну, две, три, четыре, пять, шесть), измененных по сравнению с исходным антителом, которые также называют одной или несколькими CDR, «полученными из» одной или нескольких CDR исходного антитела. Возможность гуманизации антигена хорошо известна (см., например, патенты США №№5225539; 5530101; 5585089; 5859205; 6407213; 6881557).
[0097] Термин «антитело «XX» подразумевается как означающий антитело или его эпитопсвязывающий фрагмент (например, антитело "С10-2"), содержащие легкую цепь, вариабельный домен легкой цепи или CDR1-3 вариабельного домена легкой цепи, определенные по их соответствующему SEQ ID NO, и тяжелую цепь, вариабельный домен тяжелой цепи или CDR1-3 вариабельного домена тяжелой цепи, определенные по их соответствующему SEQ ID NO, или состоящие из них. В определенных вариантах осуществления антитело или его эпитопсвязывающий фрагмент определены полным вариабельным доменом тяжелой цепи, содержащимся в них, определенным по его SEQ ID NO, и вариабельным доменом легкой цепи, определенным по его SEQ ID NO.
[0098] Если в данном документе не указано иное, нумерация аминокислотных остатков в Fc-области или константном домене антитела соответствует системе нумерации EU, также называемой EU-индексом, которая описана в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
[0099] Как используется в данном документе, говорят, что антитело или его эпитопсвязывающий фрагмент «специфично» связываются с областью другой молекулы (т.е. эпитопом), если они реагируют или ассоциируют с этим эпитопом чаще, быстрее, с большей длительностью и/или с большей аффинностью или авидностью по сравнению с альтернативными эпитопами. Также при прочтении этого определения следует понимать, что, например, антитело или его эпитопсвязывающий фрагмент, которые специфично связываются с первой мишенью, могут специфично или преимущественно связываться со второй мишенью или могут не делать этого. Используемый в данном документе термин «связывание» применительно к связыванию антитела с предварительно определенным антигеном в типичном случае относится к связыванию с аффинностью, соответствующей KD, составляющей приблизительно 10-7 М или меньше, как, например, приблизительно 10-8 М или меньше, как, например, приблизительно 10-9 М или меньше, при определении с помощью, например, технологии поверхностного плазмонного резонанса (SPR) на приборе BIAcore® 3000 с применением антигена в качестве лиганда и антитела в качестве анализируемого вещества, и к связыванию с предварительно определенным антигеном с аффинностью, соответствующей KD, по меньшей мере в десять раз более низкой, как, например, по меньшей мере в 100 раз более низкой, например, по меньшей мере в 1000 раз более низкой, как, например, по меньшей мере в 10000 раз более низкой, например, по меньшей мере в 100000 раз более низкой, чем для его аффинности связывания с неспецифичным антигеном (например, BSA, казеином), отличным от предварительно определенного антигена, или близкородственным антигеном. Величина, на которую аффинность является более низкой, зависит от KD антитела, так что если KD антитела является очень низкой (то есть антитело является высокоспецифичным), то величина, на которую аффинность к антигену является более низкой, чем аффинность к неспецифичному антигену, может быть по меньшей мере 10000-кратной.
[00100] Термин «kd» (с-1 или 1/с), используемый в данном документе, относится к константе скорости диссоциации для конкретного взаимодействия антитела и антигена. Указанное значение также называют значением koff.
[00101] Термин «ka» (М-1 х с-1 или 1/М⋅с), используемый в данном документе, относится к константе скорости диссоциации для конкретного взаимодействия антитела и антигена.
[00102] Термин «KD» (М), используемый в данном документе, относится к равновесной константе диссоциации для конкретного взаимодействия антитела и антигена, получаемой путем деления kd на ka.
[00103] Термин «KA» (М-1 или 1/М), используемый в данном документе, относится к равновесной константе ассоциации для конкретного взаимодействия антитела и антигена, получаемой путем деления ka на kd.
[00104] В одном варианте осуществления настоящее изобретение относится к антителу к тау-белку или его эпитопсвязывающему фрагменту, которые проявляют одно или несколько следующих свойств:
(i) практически отсутствующую способность к связыванию с нефосфорилированным тау-белком;
(ii) практически отсутствующую способность к связыванию с тау-белком, фосфорилированным в S404 и не фосфорилированным в S396;
(iii) способность к связыванию с тау-белком, фосфорилированным в S396;
(iv) способность к связыванию с тау-белком, фосфорилированным как в S396, так и в S404;
(v) способность к селективному проведению различий между фосфорилированными остатками S396 и S404 тау-белка, так что они практически не способны связываться с фосфорилированным остатком 404 (pS404);
(vi) способность к связыванию с гиперфосфорилированным тау-белком из головного мозга людей с болезнью Альцгеймера;
(vii) способность к проведению различий между патологическим и непатологическим человеческим тау-белком и/или
(viii) способность к специфичному уменьшению полос гиперфосфорилированного тау-белка размером 64 кДа и 70 кДа по меньшей мере на 90% без уменьшения при этом полосы тау-белка размером 55 кДа более чем на 10% при применении согласно описанному в данном документе с иммуноистощенными экстрактами от трансгенных мышей rTg4510 или способность к специфичному уменьшению полос фосфорилированного в S396 гиперфосфорилированного тау-белка по меньшей мере на 90% без уменьшения при этом полос негиперфосфорилированного тау-белка более чем на 10% при применении согласно описанному в данном документе с посмертными экстрактами головного мозга людей с AD.
[00105] Дополнительный вариант осуществления настоящего изобретения относится к антителу, получаемому с помощью способа получения высокоспецифичных высокоаффинных антител, которые являются иммуноспецифичными в отношении патогенного гиперфосфорилированного тау-белка, содержащего фосфорилированный остаток S396, где указанный способ включает стадии:
(A) проведения инъекции иммуногена млекопитающему, при этом указанный иммуноген содержит дифосфорилированный пептид, содержащий 18-40, как, например, 18-30, как, например, 20-30, последовательных аминокислотных остатков, содержащих TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающий остатки 386-410 тау-белка 2N4R, с иммунизацией таким образом указанного млекопитающего;
(B) повторения указанной иммунизации указанного млекопитающего два или более раз;
(C) проведения скрининга образца сыворотки крови от указанного многократно иммунизированного млекопитающего в отношении наличия высокоспецифичных высокоаффинных антител, способных связываться с патогенным гиперфосфорилированным тау-белком, содержащим фосфорилированный остаток S396, но в значительно меньшей степени способных связываться с непатогенным тау-белком; и
(D) извлечения указанных высокоспецифичных высокоаффинных антител.
[00106] Как используется в данном документе, «практически отсутствующая способность» к связыванию с молекулой тау-белка означает более чем 20% разницу, более чем 40% разницу, более чем 60% разницу, более чем 80% разницу, более чем 100% разницу, более чем 150% разницу, более чем 2-кратную разницу, более чем 4-кратную разницу, более чем 5-кратную разницу или более чем 10-кратную разницу в функциональных характеристиках по сравнению с выявляемым связыванием, опосредованным эталонным антителом.
[00107] Термины «селективный» и «иммуноселективный», когда речь идет о способностях к связыванию, которыми обладает антитело к тау-белку в отношении двух эпитопов, подразумеваются как означающие то, что для наблюдаемого связывания в насыщающих условиях проявляется по меньшей мере 80% разница, по меньшей мере 95% разница и наиболее предпочтительно 100% разница (т.е. отсутствие выявляемого связывания с одним эпитопом). Термины «селективный» и «иммуноселективный», когда речь идет об антителе к тау-белку, дополнительно подразумеваются как означающие то, что антитело связывается с гиперфосфорилированным тау-белком из головного мозга людей с болезнью Альцгеймера и способно различать патологический и непатологический человеческий тау-белок.
[00108] Термины «экстрагируемая в TBS» (S1), «экстрагируемая в высокосолевом растворе/саркозиле» (S3) и «нерастворимая в саркозиле» (Р3) фракции означают фракции, получаемые с помощью биохимического фракционирования тау-белка, описанного в данном документе.
[00109] В некоторых антителах только часть CDR, а именно подмножество остатков CDR, требуемых для связывания, называемое SDR, необходима для сохранения связывания в гуманизированном антителе. Остатки CDR, не контактирующие с соответствующим эпитопом и не содержащиеся в SDR, можно идентифицировать на основании предыдущих исследований (например, остатки Н60-Н65 в CDR Н2 часто не требуются), из областей CDR по Kabat, лежащих за пределами гипервариабельных петель по Chothia (см. Kabat et al. (1992) Sequences of Proteins of Immunological Interest, National Institutes of Health Publication No. 91-3242; Chothia, C. et al. (1987) "Canonical Structures For The Hypervariable Regions Of Immunoglobulins", J. Mol. Biol. 196: 901-917), с помощью молекулярного моделирования и/или эмпирическим путем или согласно описанному в Gonzales, N.R. et al. (2004) "SDR Grafting Of A Murine Antibody Using Multiple Human Germline Templates To Minimize Its Immunogenicity", Mol. Immunol. 41: 863-872. В таких гуманизированных антителах в положениях, в которых отсутствуют один или несколько остатков донорного CDR или в которых весь донорный CDR пропущен, аминокислота, занимающая положение, может представлять собой аминокислоту, занимающую соответствующее положение (согласно нумерации по Kabat) в акцепторной последовательности антитела. Количество таких замен акцепторных аминокислот на донорные в CDR, которые нужно включить, отражает баланс альтернативных мнений. Такие замены являются потенциально преимущественными для снижения количества мышиных аминокислот в гуманизированном антителе и снижения вследствие этого потенциальной иммуногенности. Тем не менее, замены также могут обуславливать изменения аффинности, и значительного уменьшения значений аффинности предпочтительно избегают. Положения для замены в пределах CDR и аминокислоты, необходимые для замены, также можно выбрать эмпирическим путем.
[00110] Тот факт, что изменение одной аминокислоты в остатке CDR может приводить к утрате функционального связывания (Rudikoff, S. etc.. (1982) "Single Amino Acid Substitution Altering Antigen-Binding Specificity," Proc. Natl. Acad. Sci. (USA) 79(6): 1979-1983), обеспечивает возможность систематической идентификации альтернативных функциональных последовательностей CDR. В одном предпочтительном способе получения таких вариантов CDR полинуклеотид, кодирующий CDR, подвергают мутагенезу (например, посредством случайного мутагенеза или с помощью сайт-направленного способа (например, амплификации, опосредованной полимеразной цепной реакцией, с праймерами, кодирующими мутантный локус)) с получением CDR, имеющего замененный аминокислотный остаток. Путем сравнения по соответствующему остатку идентичности исходной (функциональной) последовательности CDR с идентичностью варианта последовательности CDR с заменой (нефункционального) для этой замены можно определить вес замены согласно BLOSUM62.iij. В системе BLOSUM представлена матрица аминокислотных замен, создаваемая путем анализа базы данных последовательностей в отношении достоверных выравниваний (Eddy, S.R. (2004) "Where Did The BLOSUM62 Alignment Score Matrix Come From?", Nature Biotech. 22(8): 1035-1036; Henikoff, J.G. (1992) "Amino acid substitution matrices from protein blocks", Proc. Natl. Acad. Sci. (USA) 89: 10915-10919; Karlin, S. et al. (1990) "Methods For Assessing The Statistical Significance Of Molecular Sequence Features By Using General Scoring Schemes, " Proc. Natl. Acad. Sci. (USA) 87: 2264-2268; Altschul, S.F. (1991) "Amino Acid Substitution Matrices From An Information Theoretic Perspective", J. Mol. Biol. 219, 555-565. В настоящее время наиболее совершенной базой данных BLOSUM является база данных BLOSUM62 (BLOSUM62.iij). В таблице 1 представлены значения веса замен согласно BLOSUM62.iij (чем больше вес, тем более консервативной является замена и, таким образом, тем более вероятно, что замена не будет влиять на функцию). Если эпитопсвязывающий фрагмент, содержащий полученную в результате CDR, не может связываться с тау-белком, например, то полагают, что вес замены согласно BLOSUM62.iij свидетельствует о ее недостаточной консервативности, и выбирают и производят новую замену-кандидата, имеющую больший вес замены. Таким образом, например, если исходный остаток представлял собой глутамат (Е), а нефункциональный заменяющий остаток представлял собой гистидин (Н), то вес замены согласно BLOSUM62.iij равен 0, и предпочтительными являются более консервативные изменения (как, например, на аспартат, аспарагин, глутамин или лизин).
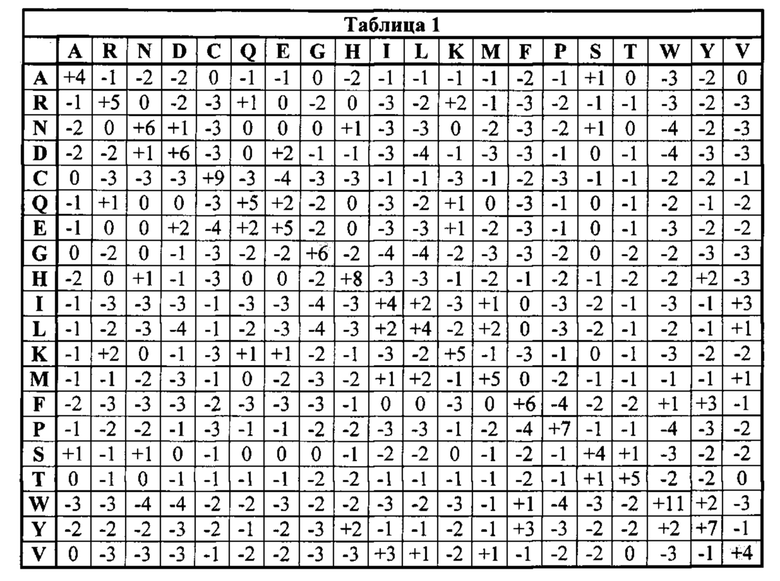
[00111] Настоящее изобретение, таким образом, предполагает применение случайного мутагенеза для идентификации улучшенных CDR. В контексте настоящего изобретения консервативные замены могут быть определены как замены в пределах классов аминокислот, отраженных в одной или нескольких из таблиц 2, 3 или 4.
Классы аминокислотных остатков для консервативных замен

Альтернативные классы аминокислотных остатков для консервативных замен
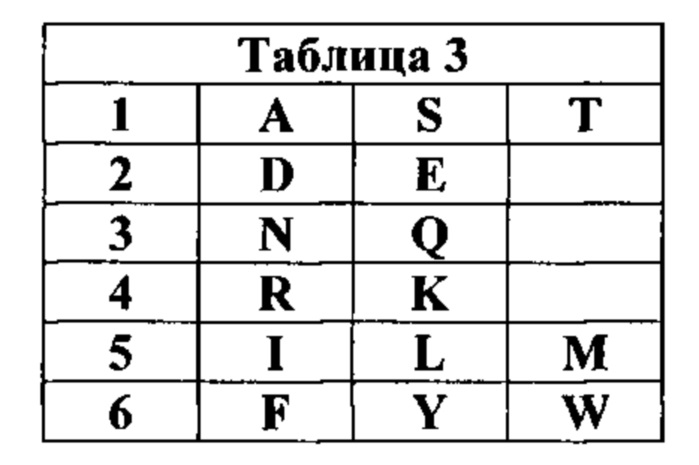
Альтернативные физические и функциональные классификации аминокислотных остатков

[00112] Группы более консервативных замен включают валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин и аспарагин-глутамин.
[00113] Дополнительные группы аминокислот также могут быть составлены с использованием принципов, описанных, например, в Creighton (1984) Proteins: Structure and Molecular Properties (2d Ed. 1993), W. H. Freeman and Company.
[00114] В качестве альтернативы можно применять технологию фагового дисплея для повышения (или снижения) аффинности CDR. В этой технологии, называемой созреванием аффинности, используется мутагенез или «прогулка по CDR» и повторный отбор с применением антигена-мишени или его антигенного эпитопсвязывающего фрагмента для идентификации антител, имеющих CDR, которые связываются с более высокой (или более низкой) аффинностью с антигеном по сравнению с первоначальным или исходным антителом (см., например, Glaser et al. (1992) J. Immunology 149:3903). Мутагенез целых кодонов, а не отдельных нуклеотидов, приводит к получению полу случайного набора аминокислотных мутаций. Можно конструировать библиотеки, состоящие из пула вариантов клонов, каждый из которых отличается одним изменением аминокислоты в одной CDR, который также содержит варианты, представляющие каждую возможную аминокислотную замену для каждого остатка CDR. Мутантные формы с повышенной (или пониженной) аффинностью связывания с антигеном можно подвергнуть скринингу путем приведения иммобилизованных мутантных форм в контакт с меченым антигеном. Можно применять любой способ скрининга, известный из уровня техники, для идентификации мутантных антител с повышенной или пониженной аффинностью к антигену (например, ELISA) (см. Wu et al. 1998, Proc. Natl. Acad. Sci. (U.S.A.) 95:6037; Yelton etal, 1995, J. Immunology 155:1994). Возможно также применять прогулку по CDR, в ходе которой легкая цепь подвергается случайному мутагенезу (см. Schier et al., 1996, J. Mol. Bio. 263:551).
[00115] Способы выполнения такого созревания аффинности описаны, например, в Krause, J.C. et al. (2011) "An Insertion Mutation That Distorts Antibody Binding Site Architecture Enhances Function Of A Human Antibody," MBio. 2(1) pii: e00345-10. doi: 10.1128/mBio.00345-10; Kuan, C.T. et al. (2010) "Affinity-Matured Anti-Glycoprotein NMB Recombinant Immunotoxins Targeting Malignant Gliomas And Melanomas", Int. J. Cancer 10.1002/ijc.25645; Hackel, B.J. et al. (2010) "Stability And CDR Composition Biases Enrich Binder Functionality Landscapes", J. Mol. Biol. 401(1): 84-96; Montgomery, D.L. et al. (2009) "Affinity Maturation And Characterization Of A Human Monoclonal Antibody Against HIV-1 gp41", MAbs 1(5):462-474; Gustchina, E. et al. (2009) "Affinity Maturation By Targeted Diversification Of The CDR-H2 Loop Of A Monoclonal Fab Derived From A Synthetic  Human Antibody Library And Directed Against The Internal Trimeric Coiled-Coil Of Gp41 Yields A Set Of Fabs With Improved HIV-1 Neutralization Potency And Breadth", Virology 393(1):112-119; Finlay, W.J. et al. (2009) "Affinity Maturation Of A Humanized Rat Antibody For Anti-RAGE Therapy: Comprehensive Mutagenesis Reveals A High Level Of Mutational Plasticity Both Inside And Outside The Complementarity-Determining Regions", J. Mol. Biol. 388(3):541-558; Bostrom, J. et al. (2009) "Improving Antibody Binding Affinity And Specificity For Therapeutic Development", Methods Mol. Biol. 525:353-376; Steidl, S. et al. (2008) "In Vitro Affinity Maturation Of Human GM-CSF Antibodies By Targeted CDR-Diversification", Mol. Immunol. 46(1):135-144; и Barderas, R. et al. (2008) "Affinity Maturation Of Antibodies Assisted By In Silico Modeling," Proc. Natl. Acad. Sci. (USA) 105(26):9029-9034.
Human Antibody Library And Directed Against The Internal Trimeric Coiled-Coil Of Gp41 Yields A Set Of Fabs With Improved HIV-1 Neutralization Potency And Breadth", Virology 393(1):112-119; Finlay, W.J. et al. (2009) "Affinity Maturation Of A Humanized Rat Antibody For Anti-RAGE Therapy: Comprehensive Mutagenesis Reveals A High Level Of Mutational Plasticity Both Inside And Outside The Complementarity-Determining Regions", J. Mol. Biol. 388(3):541-558; Bostrom, J. et al. (2009) "Improving Antibody Binding Affinity And Specificity For Therapeutic Development", Methods Mol. Biol. 525:353-376; Steidl, S. et al. (2008) "In Vitro Affinity Maturation Of Human GM-CSF Antibodies By Targeted CDR-Diversification", Mol. Immunol. 46(1):135-144; и Barderas, R. et al. (2008) "Affinity Maturation Of Antibodies Assisted By In Silico Modeling," Proc. Natl. Acad. Sci. (USA) 105(26):9029-9034.
[00116] Таким образом, последовательность вариантов CDR охватываемых антител или их эпитопсвязывающих фрагментов может отличаться от последовательности CDR исходного антитела D1.2, С10-2, С5.2 или С8.3 благодаря заменам; например, замененным 4 аминокислотным остаткам, 3 аминокислотным остаткам, 2 аминокислотным остаткам или 1 аминокислотному остатку. Согласно варианту осуществления настоящего изобретения дополнительно предусматривается, что аминокислоты в CDR-областях можно заменять посредством консервативных замен, определенных в 3 таблицах выше. Например, кислый остаток Asp можно заменить на Glu без значительного влияния на характеристики связывания антитела.
[00117] Термин «нормальный тау-белок» относится к нормальному тау-белку головного мозга, содержащему 2-3 моля фосфата на моль белка.
[00118] Термин «гиперфосфорилированный тау-белок» относится к полифосфорилированным молекулам тау-белка, соответствующим индуцированному полианионными молекулами сдвигу подвижности в вестерн-блоттинге, или к молекулам тау-белка, имеющим более пяти, шести или семи фосфорилированных сериновых, треониновых или тирозиновых сайтов.
[00119] Термин «тау-белок, имеющий фосфорилированный остаток 396» относится к гиперфосфорилированному тау-белку, в котором остаток 396 является фосфорилированным, а остаток 404 является или не является фосфорилированным.
[00120] Термин «трансгенное животное, отличное от человека» относится к животному, отличному от человека, имеющему геном, содержащий один или несколько человеческих трансгенов или трансхромосом, кодирующих тяжелую и/или легкую цепь (интегрированных или не интегрированных в природную геномную ДНК животного), которое способно экспрессировать полностью гуманизированные антитела. Например, трансгенная мышь может иметь трансген, кодирующий гуманизированную легкую цепь, и либо трансген, кодирующий гуманизированную тяжелую цепь, либо трансхромосому, кодирующую гуманизированную тяжелую цепь, так что у мыши вырабатывается гуманизированное антитело к тау-белку при иммунизации антигенным тау-белком и/или клетками, экспрессирующими тау-белок. Трансген, кодирующий гуманизированную тяжелую цепь, может быть интегрирован в хромосомную ДНК мыши, как в случае с трансгенными мышами, например, мышами HuMAb, такими как мыши НСо7 или НСо12, или трансген, кодирующий гуманизированную тяжелую цепь, может сохраняться внехромосомно, как в случае с трансхромосомными мышами KM, описанными в WO 02/43478. Такие трансгенные и трансхромосомные мыши (в совокупности называемые в данном документе «трансгенными мышами») способны вырабатывать несколько изотипов гуманизированных моноклональных антител к данному антигену (таких как IgG, IgA, IgM, IgD и/или IgE), подвергаясь V-D-J-рекомбинации и переключению изотипа.
[00121] Трансгенное животное, отличное от человека, также можно использовать для получения антител к специфическому антигену путем введения генов, кодирующих такое специфичное антитело, например, путем формирования функциональной связи генов с геном, экспрессируемым в молоке животного.
[00122] Термин «лечение» или «осуществление лечения», используемый в данном документе, означает облегчение, замедление, ослабление или устранение прогрессирования или тяжести заболевания или нарушения или облегчение, замедление, ослабление или устранение одного или нескольких симптомов или побочных эффектов такого заболевания или нарушения. Для целей настоящего изобретения «лечение» или «осуществление лечения» дополнительно означает подход к получению благоприятных или желаемых клинических результатов, где «благоприятные или желаемые клинические результаты» включают без ограничения частичные либо полные выявляемые или невыявляемые уменьшение интенсивности проявления симптома, снижение степени тяжести нарушения или заболевания, стабилизацию (т.е. отсутствие усугубления) состояния заболевания или нарушения, задержку или замедление прогрессирования состояния заболевания или нарушения, облегчение или смягчение состояния заболевания или нарушения и ремиссию заболевания или нарушения.
[00123] «Эффективное количество», применяемое в отношении антитела или его эпитопсвязывающего фрагмента по настоящему изобретению, относится к количеству, которое при введении в необходимых дозах и в течение необходимых периодов времени является достаточным для достижения намеченного биологического эффекта или желаемого терапевтического результата, в том числе без ограничения клинических результатов. Фраза «терапевтически эффективное количество», применяемая в отношении антитела или его эпитопсвязывающего фрагмента по настоящему изобретению, подразумевается как означающая количество антитела или его эпитопсвязывающего фрагмента, которое является достаточным для облегчения, смягчения, стабилизации, устранения, замедления, ослабления или задержки прогрессирования состояния нарушения или заболевания или симптома нарушения или заболевания. В варианте осуществления способ по настоящему изобретению предусматривает введение антитела или его эпитопсвязывающего фрагмента в комбинациях с другими соединениями. В таких случаях «эффективное количество» представляет собой количество комбинации, достаточное для того, чтобы вызывать намеченный биологический эффект.
[00124] Терапевтически эффективное количество антитела к тау-белку или его эпитопсвязывающего фрагмента по настоящему изобретению может варьироваться в зависимости от таких факторов, как состояние заболевания, возраст, пол и масса индивидуума, а также от способности антитела к тау-белку или его эпитопсвязывающего фрагмента вызывать желаемый ответ у индивидуума. Терапевтически эффективным также является такое количество, при котором любые токсические или вредные эффекты антитела или части антитела перевешиваются терапевтически благоприятными эффектами.
[00125] Как указано выше, настоящее изобретение, в частности, относится к моноклональным антителам или их эпитопсвязывающим фрагментам и к абсолютно новому способу получения таких молекул (и, следовательно, таких их эпитопсвязывающих фрагментов). Этот способ в общих чертах представлен на фигуре 9. Эта возможность выделять моноклональные антитела с помощью нового способа представлена в данном документе на примере его применения для выделения моноклональных антител, способных специфично связываться с фосфорилированным остатком серином-396 ({p}S396) человеческого тау-белка (SEQ ID NO: 33). Эти антитела дополнительно характеризуются своей способностью к проведению различий между фосфорилированными остатками серином-396 и серином-404 (pS404), так что они не связываются с тау-белком с фосфорилированным серином 404, за исключением случаев, когда тау-белок также фосфорилирован по остатку 396.
[00126] Антитела по настоящему изобретению или их эпитопсвязывающий фрагмент были получены и выделены путем применения нового способа (фигура 9), который способствует отбору {p}S396-специфичных антител (фигура 9). Дополнительно, посредством применения этой весьма строгой процедуры отбора клонов антител были получены антитела, которые являются не только высокоспецифичными в отношении S396, но также и высокоселективными в отношении фосфорилированного эпитопа {p}S396. Эти антитела уникальным образом распознают тау-белок из головного мозга с болезнью Альцгеймера. Авторы настоящего изобретения также демонстрируют, что процедура скрининга, в общих чертах представленная на фигуре 9, обеспечивает идентификацию антител, обладающих функциональной и терапевтической полезностью.
[00127] Индуцировали выработку антител к дифосфорилированному пептиду TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающему остатки 386-408 тау-белка 2N4R (пример 1). Мышей иммунизировали фосфорилированным пептидом. После получения достаточных титров антител мышей умерщвляли, и получали гибридомы (пример 2). Гибридомы подвергали скринингу с помощью дот-блот-анализа (пример 3) и MSD-ELISA с иммобилизованным патологическим и непатологическим человеческим тау-белком (пример 4). Способность к проведению различий между патологическим и непатологическим человеческим тау-белком в ходе дот-блот-анализа и вестерн-блот-анализа применяли для отбора гибридом. Отбирали шестнадцать клонов, из которых извлекали четыре гибридомных клона, которые вырабатывали антитела, характеризующиеся чрезвычайно высокими способностями к связыванию с патологическим материалом человеческого тау-белка.
[00128] Специфичное связывание с патологическим и непатологическим тау-белком также определяли путем выделения тау-белка из пораженного заболеванием и не пораженного заболеванием головного мозга людей с AD и иммобилизации этого материала на планшетах для MSD-ELISA (пример 4).
[00129] Дополнительный аспект настоящего изобретения относится к моноклональному антителу или его эпитопсвязывающему фрагменту, образование которых происходит в ответ на дифосфорилированный пептид, содержащий по меньшей мере 18, как, например, по меньшей мере 20, последовательных аминокислотных остатков в пределах TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающего остатки 386-410 тау-белка 2N4R. В этом аспекте настоящего изобретения образование моноклонального антитела или его эпитопсвязывающего фрагмента в типичном случае вызывается в ответ на дифосфорилированный пептид, содержащий 18-40, как, например, 18-30, как, например, 20-30, последовательных аминокислотных остатков, содержащих TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающий остатки 386-410 тау-белка 2N4R.
[00130] Дополнительный аспект настоящего изобретения направлен на моноклональное антитело или его эпитопсвязывающий фрагмент по настоящему изобретению, обладающие специфичностью в отношении фосфорилированного тау-белка (pTau) от пациентов, пораженных AD, по сравнению со здоровыми контрольными субъектами в соответствующей возрастной группе, так что указанные моноклональное антитело или его эпитопсвязывающий фрагмент характеризуются разницей в специфичности в отношении фосфорилированного тау-белка (pTau) от пациентов, пораженных AD, по сравнению с тау-белком от контрольных субъектов в соответствующей возрастной группе здоровых людей, представляющей собой более чем 50-кратное, как, например, более чем 100-кратное повышение специфичности в отношении материала, пораженного AD, по сравнению с материалом от здоровых контрольных субъектов в основанном на ELISA анализе для выявления фосфорилированного тау-белка (pTau) в гомогенатах головного мозга от пораженных AD и от здоровых контрольных субъектов с применением специфической схемы 1 ELISA для фосфорилированных и мультимерных молекул, описанной в данном документе.
[00131] Связанный аспект настоящего изобретения направлен на моноклональное антитело или его эпитопсвязывающий фрагмент по настоящему изобретению, обладающие специфичностью в отношении тау-белка из материала, пораженного AD, так что указанные моноклональное антитело или его эпитопсвязывающий фрагмент характеризуются разницей в специфичности в отношении материала, пораженного AD, по сравнению с материалом от контрольных субъектов в соответствующей возрастной группе здоровых людей, представляющей собой более чем 50-кратное, как, например, более чем 100-кратное повышение специфичности в отношении материала, пораженного AD, по сравнению с материалом от здоровых контрольных субъектов в основанном на ELISA анализе для выявления фосфорилированного тау-белка (pTau) в гомогенатах головного мозга от пораженных AD и от здоровых контрольных субъектов с применением специфической схемы 1 ELISA для фосфорилированных и мультимерных молекул.
[00132] Способ выполнения схемы 1 ELISA включает стадии А) захвата патологических антигенных человеческих тау-белков из головного мозга с AD с помощью планшетов, покрытых С10-2; В) инкубирования антигенных тау-белков с pS396-специфичными антителами в повышающихся концентрациях и С) выявления захвата антигенных тау-белков и его антителоопосредованного ингибирования с помощью антител к человеческому (общему) тау-белку от MSD, меченных SULFO-TAG.
[00133] Настоящее изобретение дополнительно относится к антителу, получаемому с помощью способа получения высокоспецифичных высокоаффинных антител, которые являются иммуноспецифичными в отношении патогенного гиперфосфорилированного тау-белка, содержащего фосфорилированный остаток S396, где указанный способ включает стадии:
(A) проведения инъекции иммуногена млекопитающему, при этом указанный иммуноген содержит дифосфорилированный пептид, содержащий 18-40, как, например, 18-30, как, например, 20-30, последовательных аминокислотных остатков, содержащих TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающий остатки 386-410 тау-белка 2N4R, с иммунизацией таким образом указанного млекопитающего;
(B) повторения указанной иммунизации указанного млекопитающего два или более раз;
(C) проведения скрининга образца сыворотки крови от указанного многократно иммунизированного млекопитающего в отношении наличия высокоспецифичных высокоаффинных антител, способных связываться с патогенным гиперфосфорилированным тау-белком, содержащим фосфорилированный остаток S396, но в значительно меньшей степени способных связываться с непатогенным тау-белком;
и
(D) извлечения указанных высокоспецифичных высокоаффинных антител.
[00134] Более конкретно, стадия А включает: покрытие планшетов MSD (в типичном случае в течение ночи при 4 С) антителом С10-2, в типичном случае при 0,5 мкг/мл (захватывающим антителом) в покрывающем буфере, блокирование (в типичном случае в течение 1 часа при комнатной температуре) и промывание, в типичном случае 3 раза. Стадия В включает: смешивание образцов лизата Р3 (в типичном случае при 1:1000=2-4 мкг/мл общего белка) и/или S1(p) (в типичном случае при 1:300=20-40 нг/мл общего белка) из материала от субъектов с AD (объединенных от 3 пациентов) с антителом, специфичным к пептидному эпитопу pS396, в ступенчато изменяющихся концентрациях и инкубирование (в типичном случае в течение 1 часа при комнатной температуре). Реакционные смеси затем инкубируют в течение 2 часов на планшетах, подготовленных на стадии А. Стадия С включает выявление тау-белка, захваченного С10-2, с помощью антитела к человеческому тау-белку, меченного SULFO-TAG. Антитело к тау-белку (в типичном случае 1:50) от MSD, соответствующее инструкциям производителя. Планшеты анализируют на SECTOR® S600 от MSD. Р3 из материала с AD и S1(p) из материала с AD тестируют согласно аналогичной схеме.
[00135] Дополнительный вариант осуществления направлен на антитело или его антигенсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33), которые были получены или произведены в линии клеток, такой как линия человеческих клеток, линия клеток млекопитающего, отличного от человека, линия клеток насекомого, дрожжей или бактерии.
[00136] Антитело или его антигенсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33), получают в линии клеток СНО, линии клеток HEK, линии клеток BHK-21, линии мышиных клеток (такой как линия клеток миеломы), линии клеток фибросаркомы, линии клеток PER.C6, линии клеток HKB-11, линии клеток CAP и линии человеческих клеток HuH-7.
[00137] Характеристики специфичной аффинности и свойств связывания D1.2 и С10-2 получали с применением пептидов 386-410 тау-белка (2R4N), фосфорилированных либо не фосфорилированных в положении 396 или 404 (SEQ ID NO: 29-32). Путем применения специфического протокола иммунизации и скрининга (фигура 9), в общих чертах представленного в настоящей заявке, получают высокоспецифичные антитела к фосфосерину-396 (pS396), продемонстрированные на фигуре 4.
[00138] Чтобы продемонстрировать, что антитела являются специфичными в отношении патологического тау-белка, также получали характеристики антител D1.2 и С10-2 с помощью иммуногистохимического анализа (пример 6). Антитела проявляют высокоспецифичное связывание с нейрофибриллярными клубками в головном мозге и в срезах с болезнью Альцгеймера от трансгенных по тау-белку мышей Tg4510, экспрессирующих человеческий (P301L) мутантный тау-белок (фигура 5). Связывание с тканью из контрольного головного мозга людей и из головного мозга нетрансгенных мышей не наблюдается, что демонстрирует, что антитела специфично связываются с человеческим тау-белком, и в частности, с тау-белком, ассоциированным с альцгеймеровской патологией.
[00139] Уникальная способность этих антител к распознаванию тау-белка, ассоциированного с болезненной патологией, продемонстрирована в данном документе в примере 7. Авторы настоящего изобретения сравнивают связывание патологического и непатологического тау-белка в анализе, описанном в примере 3. Сравнение проводят в отношении пяти опубликованных антител к тау-белку: hACI-2B6, IPN002, HJ8.5,2.10.3 и 4Е4. На фигуре 6 проиллюстрировано связывание каждого из эталонных антител с тау-белком из здорового и пораженного заболеванием головного мозга и связывание с человеческим тау-белком с мутацией P301L, выделенным из мышей Tg4510 в возрасте 10 месяцев, трансгенных по тау-белку. Это демонстрирует, что выделенные антитела демонстрируют исключительно высокую степень специфичности и селективности в отношении человеческого патологического тау-белка. Эта селективность превосходит селективность любого из сравниваемых антител, показанных в таблице 5.


[00140] При насыщающем связывании антитела D1.2 и С10-2 проявляют более чем 100-кратную селективность в отношении тау-белка Р3, выделенного из головного мозга людей с AD.
[00141] Чтобы продемонстрировать, что отобранные антитела обладают функциональной и терапевтической полезностью, антитела тестировали в анализах агрегации тау-белка in vitro и в клетке (пример 8). Эти анализы представляют собой функциональные анализы, в которых демонстрируется, что антитела способны препятствовать процессу патологической агрегации тау-белка. Клетки HEK293 транзиентно трансфицируют человеческим тау-белком P301L-FLAG (4R0N). Затем клетки подвергают воздействию экстрактов тау-белка из головного мозга людей с AD или из головного мозга трансгенных Tg4510. Это воздействие патологического тау-белка способствует поглощению тау-белка клетками и его внутриклеточной агрегации. Как иммуноистощение препаратов тау-белка с помощью антител D1.2 и С10-2, так и непосредственная обработка клеток этими антителами способны обеспечивать существенное уменьшение образования агрегатов тау-белка (фигура 7).
[00142] Терапевтическую полезность антител D1.2 и С10-2 также оценивали у мышей, экспрессирующих человеческий тау-белок/PS1 (пример 9). Эта мышиная модель является более подходящей животной моделью заболевания AD, в которой патология AD образуется лишь в позднем возрасте (в возрасте 12-18 месяцев). Тем не менее, у мышей проявляется гиперфосфорилирование тау-белка до появления патологии плотных клубков. Мышам инъецировали дозу 15 мг/кг два раза в неделю в течение длительного периода 13 недель. У мышей, обработанных антителом, проявляется существенное уменьшение уровня фосфорилированного тау-белка, продемонстрированное на фигуре 9, что указывает на то, что при длительной обработке антителами D1.2 и С10-2 будет уменьшаться патология клубков и, таким образом, последующая нейро дегенерация in vivo.
[00143] Антитела по настоящему изобретению специфично удаляют гиперфосфорилированный тау-белок из экстрактов головного мозга мышей rTg4510 с помощью способов иммуноистощения. Кроме того, антитела по настоящему изобретению не удаляют нормальный тау-белок из гомогенатов, тогда как коммерчески доступное антитело Tau5 делает это. В отличие от коммерческих антител, связывающихся с тау-белками, в которых имеет место фосфорилирование по остатку 404 или как по остатку 404, так и по остатку 396, антитела по настоящему изобретению специфично удаляют на 95% гиперфосфорилированный тау-белок, который фосфорилирован по серину-396. Эксперименты (пример 12) демонстрируют, что несмотря на то, что антитело по настоящему изобретению удаляет лишь очень небольшую долю общего тау-белка в гомогенате головного мозга (8%), антитела, тем не менее, специфично удаляют гиперфосфорилированный тау-белок (на 90%). Соответственно, один аспект настоящего изобретения направлен на моноклональное антитело или его эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с патогенным гиперфосфорилированным тау-белком. Дополнительно, в экспериментах, в которых гиперфосфорилированный тау-белок удаляли с помощью антитела по настоящему изобретению, активность затравочного действия устраняется. При удалении гиперфосфорилированного тау-белка из гомогенатов гомогенаты больше не индуцируют затравочное действие с формированием патологии тау-белка. Было выдвинуто предположение, что при уменьшении затравочного действия уменьшается развитие образования клубков и прогрессирование таупатий, в том числе болезни Альцгеймера. Соответственно, дополнительный аспект настоящего изобретения направлен на антитело по настоящему изобретению для применения в уменьшении прогрессирования AD или интенсивности симптомов AD.
[00144] Более конкретно, как подробно описано выше, настоящее изобретение относится к любому из четырех моноклональных антител, выбранных из группы, включающей:
Антитело D1.2
(a) CDR1 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 1;
(b) CDR2 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 2;
(c) CDR3 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 3;
(d) CDR1 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 4;
(e) CDR2 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 5; и
(f) CDR3 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 6.
[00145] Антитело или его эпитопсвязывающий фрагмент могут содержать вариабельный домен тяжелой цепи под SEQ ID NO: 8 и/или вариабельный домен легкой цепи под SEQ ID NO: 7 или состоять из них.
[00146] В связанном варианте осуществления антитело D1.2 или его эпитопсвязывающий фрагмент могут содержать тяжелую цепь SEQ ID NO: 8 и/или легкую цепь SEQ ID NO: 34 или состоять из них.
Антитело С10-2
(a) CDR1 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 9;
(b) CDR2 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 10;
(c) CDR3 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 11;
(d) CDR1 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 12;
(e) CDR2 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 13; и
(f) CDR3 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 14.
[00147] Антитело или его эпитопсвязывающий фрагмент могут содержать вариабельный домен тяжелой цепи под SEQ ID NO: 15 и/или вариабельный домен легкой цепи под SEQ ID NO: 16 или состоять из них.
[00148] Аминокислотная последовательность тяжелой цепи гуманизированного антитела С10-2 показана под SEQ ID NO: 35. Аминокислотная последовательность легкой цепи гуманизированного антитела С10-2 показана под SEQ ID NO: 36.
[00149] В совокупности примеры показывают, что антитела по настоящему изобретению, в том числе С10-2, эффективно связываются с планшетами MSD, покрытыми антигенами AD-P3. Для сравнения, коммерческие антитела, такие как PHF-13, обладают низкой активностью связывания. Дополнительно, PHF-13 демонстрировало значительно более высокую степень неспецифичного связывания по сравнению с антителами по настоящему изобретению (см. таблицы 6A-6D). В таблице 6 показано, что жидкофазное ингибирование mC10-2 захвата антигена pTau в планшете, покрытом С10-2, является эффективным IC50=10-20 нМ), тогда как в случае с mD1.2 оно является неэффективным (IC50=500-1000 нМ). Жидкофазное ингибирование mC10-2 захвата антигена pTau в планшете, покрытом mC10-2, является эффективным при IC50=10-20 нМ, тогда как в случае с PHF-13 оно является неэффективным (IC50=500-1000 нМ).
[00150] Один аспект настоящего изобретения направлен на антитело, содержащее:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 9;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 10;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 11.
[00151] Дополнительный аспект настоящего изобретения направлен на антитело, содержащее:
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 13; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 14.
[00152] Дополнительный аспект настоящего изобретения направлен на антитело, содержащее:
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 13; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 14, а также одну, две или три из
(a) CDR1 легкой цепи, имеющей аминокислотную последовательность SEQ ID NO:9;
(b) CDR2 легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 10; и
(c) CDR3 легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 11.
Антитело С5.2
(a) CDR1 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 17;
(b) CDR2 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 18;
(c) CDR3 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 19;
(d) CDR1 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 20;
(e) CDR2 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 21; и
(f) CDR3 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 22.
[00153] Антитело или его эпитопсвязывающий фрагмент могут содержать вариабельный домен тяжелой цепи под SEQ ID NO: 23 и/или вариабельный домен легкой цепи под SEQ ID NO: 24 или состоять из них.
[00154] Как можно видеть из кристаллической структуры на фигуре 10, эпитоп связывается между тяжелой цепью и легкими цепями С5.2. Соответственно, в связанном варианте осуществления антитело по настоящему изобретению или его эпитопсвязывающий фрагмент содержат:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 17; или
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 18; или
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 19; и
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 20; или
(d) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 21; или
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 22.
Антитело С8.3
(a) CDR1 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 25;
(b) CDR2 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 26;
(c) CDR3 легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 27;
(d) CDR1 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 28;
(e) CDR2 тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO; 29; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 30.
[00155] Антитело или его эпитопсвязывающий фрагмент могут содержать вариабельный домен тяжелой цепи под SEQ ID NO: 31 и/или вариабельный домен легкой цепи под SEQ ID NO: 32 или состоять из них.
[00156] Антитело или его эпитопсвязывающий фрагмент предпочтительно представляют собой человеческое или гуманизированное антитело.
[00157] Антитела и их эпитопсвязывающие фрагменты, упомянутые выше, согласно одному варианту осуществления могут дополнительно содержать вариант такой CDR1, CDR2 или CDR3 легкой и/или тяжелой цепи (не более чем с 4 отличиями в аминокислотах, или не более чем с 3 отличиями в аминокислотах, или не более чем с 2 отличиями в аминокислотах, или не более чем с 1 отличием в аминокислотах.
[00158] Как можно видеть из фигуры 11, CDR1 НС, CDR2 НС, CDR3 НС и CDR3 LC по меньшей мере в одном варианте осуществления важны для связывания с областью 392-398 тау-белка. В одном варианте осуществления настоящего изобретения антитело по настоящему изобретению или его эпитопсвязывающий фрагмент содержат:
a) CDR1 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 12, SEQ ID NO: 20 и SEQ ID NO: 28;
b) CDR2 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, SEQ ID NO: 13, SEQ ID NO: 21 и SEQ ID NO: 29; и
c) CDR3 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 14, SEQ ID NO: 22 и SEQ ID NO: 30; и
d) CDR3 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 11, SEQ ID NO: 19 и SEQ ID NO: 27.
[00159] Антитело по настоящему изобретению или его эпитопсвязывающий фрагмент могут содержать:
a) CDR1 тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 20;
b) CDR2 тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 21;
c) CDR3 тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 22; и
d) CDR3 легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 19.
[00160] В одном аспекте настоящего изобретения настоящее изобретение направлено на антитело или его эпитопсвязывающие фрагменты, образующие гидрофобный карман, образуемый L3:H3, L3:F8*, H1:H13, H2:Y1, H2:Y3 с Y394 пептида тау-белка. В варианте осуществления настоящее изобретение направлено на антитело, которое конкурирует с антителом, дополнительно описанным в данном документе, за образование сети водородных связей между сольватированным {p}S396 и L3:T4, H1:R10, H1:T11, H3:R1, Н3:Т3; (*) L3:F8 представляет собой С-концевой остаток каркасной области, фланкирующий CDR L3 (см. фигуру 11).
[00161] Как можно видеть из кристаллической структуры, определенной с помощью рентгенографического анализа, антитело по настоящему изобретению связывается на двух уровнях селективности. Первый уровень селективности представляет собой селективность в отношении гиперфосфорилированного патологического тау-белка, а второй уровень селективности представляет собой селективность в отношении фосфорилированного серинового остатка, где фосфат указанного фосфорилированного серина связан водородной связью с боковой цепью тирозинового остатка, удаленного на два остатка от указанного фосфорилированного серина. Соответственно, представляющий интерес аспект настоящего изобретения направлен на антитело или его эпитопсвязывающий фрагмент, селективные в отношении аминокислотного мотива гиперфосфорилированного тау-белка, содержащего фосфорилированный серин, удаленный на два остатка от тирозинового остатка. В типичном случае аминокислотный мотив имеет следующую последовательность:
Y - Х - S(фосфорилированный) - Р -,
где Y представляет собой тирозин, Х представляет собой встречающуюся в природе аминокислоту, Р представляет собой пролин, а S(фосфорилированный) представляет собой серин с фосфорилированной гидроксильной группой боковой цепи.
[00162] Подобным образом, представляющий интерес аспект настоящего изобретения направлен на антитело или его эпитопсвязывающий фрагмент, которые связываются с фосфорилированным тау-белком, предпочтительно с гиперфосфорилированным тау-белком, где указанные антитело или его эпитопсвязывающий фрагмент являются селективными в отношении мотива IA из аминокислотных остатков, где R представляет собой боковую цепь встречающейся в природе аминокислоты.
[00163] Без ограничения какой-либо конкретной теорией полагают, что антитело по настоящему изобретению является селективным в отношении аминокислотного мотива IA, где указанный мотив имеет конформацию, которую принимает патологический тау-белок. Соответственно, аминокислотный мотив IA в типичном случае представляет собой последовательность, селективно распознаваемую антителом по настоящему изобретению. Соответственно, представляющий интерес аспект настоящего изобретения направлен на антитело или его эпитопсвязывающий фрагмент, которые связываются с фосфорилированным тау-белком, предпочтительно с гиперфосфорилированным тау-белком, где указанные антитело или его эпитопсвязывающий фрагмент являются селективными в отношении мотива IB из аминокислотных остатков, где R представляет собой боковую цепь встречающейся в природе аминокислоты.
[00164] В типичном варианте осуществления данного аспекта настоящего изобретения настоящее изобретение направлено на антитело или его эпитопсвязывающий фрагмент, которые связываются с фосфорилированным тау-белком, предпочтительно с гиперфосфорилированным тау-белком, где указанные антитело или его эпитопсвязывающий фрагмент являются селективными в отношении мотива IB из аминокислотных остатков, такого как, без ограничения, IC или ID, где R представляет собой боковую цепь встречающейся в природе аминокислоты.
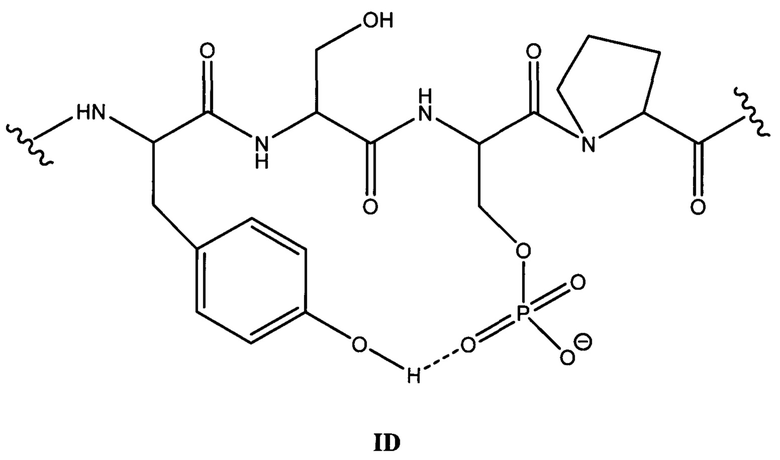
[00165] Настоящее изобретение также предусматривает способ уменьшения образования клубков тау-белка у пациента, включающий введение пациенту, нуждающемуся в таком лечении, терапевтически эффективного количества антитела по настоящему изобретению или его эпитопсвязывающих фрагментов.
[00166] Один аспект настоящего изобретения направлен на способ лечения таупатии с помощью антитела по настоящему изобретению или его эпитопсвязывающих фрагментов. В типичном случае таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, психиатрических симптомов у пациентов с деменцией с тельцами Леви, прогрессирующего надъядерного паралича (PSP), лобно-височной деменции (FTD или ее вариантов), TBI (травматического повреждения головного мозга, острого или хронического), кортикобазальной дегенерации (CBD), болезни Пика, первичной возрастной таупатии (PART), сенильной деменции с преобладанием нейрофибриллярных клубков, деменции боксеров, хронической травматической энцефалопатии, инсульта, восстановления после инсульта, нейродегенерации, связанной с болезнью Паркинсона, паркинсонизма, сцепленного с хромосомой, болезни Литико-Бодига (гуамского комплекса паркинсонизм-деменция), ганглиоглиомы и ганглиоцитомы, менингоангиоматоза, постэнцефалитического паркинсонизма, подострого склерозирующего панэнцефалита, болезни Хантингтона, свинцовой энцефалопатии, туберозного склероза, болезни Галлервордена-Шпатца и липофусциноза. В более типичном случае таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, психиатрических симптомов у пациентов с деменцией с тельцами Леви, прогрессирующего надъядерного паралича (PSP), лобно-височной деменции (FTD или ее вариантов), TBI (травматического повреждения головного мозга, острого или хронического), кортикобазальной дегенерации (CBD) и болезни Пика. В частности, таупатии могут быть выбраны из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, обусловленного AD, или психоза у пациентов с AD и психиатрических симптомов у пациентов с деменцией с тельцами Леви.
[00167] Соответственно, дополнительный аспект настоящего изобретения направлен на антитело по настоящему изобретению или его эпитопсвязывающие фрагменты для применения в лечении таупатии. В типичном случае таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, психиатрических симптомов у пациентов с деменцией с тельцами Леви, прогрессирующего надъядерного паралича (PSP), лобно-височной деменции (FTD или ее вариантов), TBI (травматического повреждения головного мозга, острого или хронического), кортикобазальной дегенерации (CBD), болезни Пика, первичной возрастной таупатии (PART), сенильной деменции с преобладанием нейрофибриллярных клубков, деменции боксеров, хронической травматической энцефалопатии, инсульта, восстановления после инсульта, нейродегенерации, связанной с болезнью Паркинсона, паркинсонизма, сцепленного с хромосомой, болезни Литико-Бодига (гуамского комплекса паркинсонизм-деменция), ганглиоглиомы и ганглиоцитомы, менингоангиоматоза, постэнцефалитического паркинсонизма, подострого склерозирующего панэнцефалита, болезни Хантингтона, свинцовой энцефалопатии, туберозного склероза, болезни Галлервордена-Шпатца и липофусциноза. В более типичном случае таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, психиатрических симптомов у пациентов с деменцией с тельцами Леви, прогрессирующего надъядерного паралича (PSP), лобно-височной деменции (FTD или ее вариантов), TBI (травматического повреждения головного мозга, острого или хронического), кортикобазальной дегенерации (CBD) и болезни Пика. В частности, таупатии могут быть выбраны из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, обусловленного AD, или психоза у пациентов с AD и психиатрических симптомов у пациентов с деменцией с тельцами Леви.
[00168] Дополнительный аспект настоящего изобретения направлен на антитело по настоящему изобретению или его эпитопсвязывающие фрагменты в композиции вместе с фармацевтически приемлемым носителем, разбавителем, вспомогательным средством и/или стабилизатором. Антитела по настоящему изобретению или их эпитопсвязывающие фрагменты можно применять в терапии для лечения таупатии. В типичном случае таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, психиатрических симптомов у пациентов с деменцией с тельцами Леви, прогрессирующего надъядерного паралича (PSP), лобно-височной деменции (FTD или ее вариантов), TBI (травматического повреждения головного мозга, острого или хронического), кортикобазальной дегенерации (CBD), болезни Пика, первичной возрастной таупатии (PART), сенильной деменции с преобладанием нейрофибриллярных клубков, деменции боксеров, хронической травматической энцефалопатии, инсульта, восстановления после инсульта, нейродегенерации, связанной с болезнью Паркинсона, паркинсонизма, сцепленного с хромосомой, болезни Литико-Бодига (гуамского комплекса паркинсонизм-деменция), ганглиоглиомы и ганглиоцитомы, менингоангиоматоза, постэнцефалитического паркинсонизма, подострого склерозирующего панэнцефалита, болезни Хантингтона, свинцовой энцефалопатии, туберозного склероза, болезни Галлервордена-Шпатца и липофусциноза. В более типичном случае таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, психиатрических симптомов у пациентов с деменцией с тельцами Леви, прогрессирующего надъядерного паралича (PSP), лобно-височной деменции (FTD или ее вариантов), TBI (травматического повреждения головного мозга, острого или хронического), кортикобазальной дегенерации (CBD) и болезни Пика. В частности, таупатии могут быть выбраны из болезни Альцгеймера, болезни аргирофильных зерен (AGD), психоза, обусловленного AD, или психоза у пациентов с AD и психиатрических симптомов у пациентов с деменцией с тельцами Леви.
[00169] Лечение, предусматриваемое настоящим изобретением, может быть длительным, и пациент может получать лечение по меньшей мере в течение 2 недель, как, например, по меньшей мере в течение 1 месяца, 6 месяцев, 1 года или дольше.
[00170] Антитела по настоящему изобретению могут, например, представлять собой моноклональные антитела, получаемые с помощью гибридомного способа, впервые описанного в Kohler et al., Nature 256, 495 (1975), или могут представлять собой моноклональные антитела, получаемые с помощью рекомбинантных ДНК или других способов, или более предпочтительно могут быть получены с помощью нового способа, раскрытого в данном документе (фигура 9). Моноклональные антитела также можно выделять из фаговых библиотек антител с помощью методик, описанных, например, в Clackson et al., Nature 352, 624-628 (1991), и Marks et al., J. Mol. Biol. 222, 581-597 (1991). Моноклональные антитела можно получать из любого подходящего источника. Таким образом, например, моноклональные антитела можно получать из гибридом, полученных из мышиных В-лимфоцитов селезенки, полученных от мышей, иммунизированных антигеном, представляющим интерес, например, в виде клеток, экспрессирующих антиген на поверхности, или нуклеиновой кислоты, кодирующей антиген, представляющий интерес. Моноклональные антитела также можно получать из гибридом, происходящих из клеток, экспрессирующих антитела, иммунизированных людей или млекопитающих, отличных от человека, таких как крысы, кролики, собаки, овцы, козы, приматы и т.п.
[00171] В одном варианте осуществления антитело по настоящему изобретению представляет собой гуманизированное антитело. Гуманизированные моноклональные антитела, направленные против тау-белка, можно получать с использованием трансгенных или трансхромосомных мышей, несущих части человеческой иммунной системы, а не мышиной системы. Такие трансгенные и трансхромосомные мыши включают мышей, называемых в данном документе соответственно мышами HuMAb (от слов «гуманизированное моноклональное антитело») и мышами KM, и в совокупности называются в данном документе «трансгенными мышами».
[00172] Мышь HuMAb содержит минилокус гена человеческого иммуноглобулина, который кодирует переаранжированные последовательности вариабельных и константных доменов тяжелых цепей (μ, и γ) и вариабельных и константных доменов легких цепей (κ) человеческого иммуноглобулина, вместе с целевыми мутациями, которые инактивируют эндогенные локусы, кодирующие μ- и κ-цепи (Lonberg, N. et al., Nature 368, 856-859 (1994)). Соответственно, у мышей проявляется уменьшенная экспрессия мышиного IgM или IgK, и в ответ на иммунизацию введенные трансгены, кодирующие человеческую тяжелую и легкую цепь, подвергаются переключению класса и соматической мутации с образованием высокоаффинных человеческих моноклональных антител IgG κ-изотипа (Lonberg, N. et al. (1994), выше, обзор которых приведен в Lonberg, N., Handbook of Experimental Pharmacology 113, 49-101 (1994), Lonberg, N. and Huszar, D., Intern. Rev. Immunol. Vol. 13 65-93 (1995) и Harding, F. and Lonberg, N., Ann. N. Y. Acad. Sci 764 536-546 (1995)). Получение мышей HuMAb подробно описано в Taylor, L. et al., Nucleic Acids Research 20, 6287-6295 (1992), Chen, J. et al.. International Immunology 5, 647-656 (1993), Tuaillon et al., J. Immunol. 152, 2912-2920 (1994), Taylor, L. et al., International Immunology 6, 579-591 (1994), Fishwild, D. et al., Nature Biotechnology 14, 845-851 (1996). См. также US 5545806, US 5569825, US 5625126, US 5633425, US 5789650, US 5877397, US 5661016, US 5814318, US 5874299, US 5770429, US 5545807, WO 98/24884, WO 94/25585, WO 93/1227, WO 92/22645, WO 92/03918 и WO 01/09187.
[00173] Мыши НСо7, НСо12, НСо17 и НСо20 имеют JKD-разрушение в своих эндогенных генах легких цепей (каппа) (описанное в Chen et al., EMBO J. 12, 811-820 (1993)), CMD-разрушение в своих эндогенных генах тяжелых цепей (описанное в примере 1 WO 01/14424) и трансген KCo5, кодирующий человеческую легкую каппа-цепь (описанный в Fishwild et al.. Nature Biotechnology 14, 845-851 (1996)). Дополнительно, мыши НСо7 имеют трансген НСо7, кодирующий человеческую тяжелую цепь (описанный в US 5770429), мыши НСо12 имеют трансген НСо12, кодирующий человеческую тяжелую цепь (описанный в примере 2 WO 01/14424), мыши НСо17 имеют трансген НСо17, кодирующий человеческую тяжелую цепь (описанный в примере 2 WO 01/09187), а мыши НСо20 имеют трансген НСо20, кодирующий человеческую тяжелую цепь. Полученные в результате мыши экспрессируют трансгены, кодирующие тяжелые цепи и легкие каппа-цепи человеческих иммуноглобулинов, в генетическом окружении, гомозиготном по разрушению эндогенных локусов, кодирующих мышиные тяжелые цепи и легкие каппа-цепи.
[00174] У мышей линии KM эндогенный ген мышиной легкой канна-цепи был подвергнут гомозиготному разрушению согласно описанному в Chen et al., EMBO J. 12, 811-820 (1993), и эндогенный ген мышиной тяжелой цепи был подвергнут гомозиготному разрушению согласно описанному в примере 1 WO 01/09187. Данная линия мышей несет трансген KCo5, кодирующий человеческую легкую каппа-цепь, который описан в Fishwild et al.. Nature Biotechnology 14, 845-851 (1996). Данная линия мышей также несет трансхромосому, кодирующую человеческую тяжелую цепь, состоящую из hCF-фрагмента хромосомы 14 (SC20), описанного в WO 02/43478. Мышей HCo12-Balb/c, HCo17-Balb/c и НСо20-Balb/с можно получать путем скрещивания НСо12, НСо17 и НСо20 с KCo5[J/K](Balb) согласно описанному в WO 09/097006.
[00175] Мыши rTg4510 представляют собой известную модель таупатии, обеспечивающую временной и пространственный контроль над экспрессией трансгена, кодирующего мутантный тау-белок. У мышей линии KM эндогенный ген мышиной легкой каппа-цепи был подвергнут гомозиготному разрушению согласно описанному в Chen et al., EMBO J. 12, 811-820 (1993), и эндогенный ген мышиной тяжелой цепи был подвергнут гомозиготному разрушению согласно описанному в примере 1 WO 01/09187. Данная линия мышей несет трансген KCo5, кодирующий человеческую легкую каппа-цепь, который описан в Fishwild et al., Nature Biotechnology 14, 845-851 (1996). Данная линия мышей также несет трансхромосому, кодирующую человеческую тяжелую цепь, состоящую из отвечающего за связывание с эпитопом hCF-фрагмента хромосомы 14 (SC20), описанного в WO 02/43478.
[00176] Спленоциты из этих трансгенных мышей можно использовать для образования гибридом, секретирующих гуманизированные моноклональные антитела, согласно хорошо известным методикам. Гуманизированные моноклональные или поликлональные антитела по настоящему изобретению, или антитела по настоящему изобретению, происходящие из других видов, также можно получать трансгенным путем посредством получения другого млекопитающего, отличного от человека, или растения, которое является трансгенным по последовательностям тяжелых и легких цепей иммуноглобулинов, представляющим интерес, и получения из него антитела в извлекаемой форме. Что касается получения антител у млекопитающих трансгенным путем, их можно получать в молоке коз, коров или других млекопитающих и извлекать из него. См., например, US 5827690; US 5756687; US 5750172 и US 5741957.
[00177] Антитело по настоящему изобретению может относиться к любому изотипу. При выборе изотипа в типичном случае будут исходить из желаемых эффекторных функций, таких как индукция ADCC. Приводимыми в качестве примера изотипами являются IgG1, IgG2, IgG3 и IgG4. Можно использовать константные домены любой из человеческих легких цепей каппа- или лямбда-типа. При желании класс антитела к тау-белку по настоящему изобретению можно переключить с помощью известных способов. Например, антитело по настоящему изобретению, которое изначально представляло собой IgM, можно подвергнуть переключению класса на антитело IgG по настоящему изобретению. Дополнительно, методики переключения класса можно применять для превращения одного подкласса IgG в другой, например, из IgGI в IgG2. Таким образом, эффекторную функцию антител по настоящему изобретению можно изменить путем переключения изотипа, например, на антитело IgG1, IgG2, IgG3, IgG4, IgD, IgA, IgE или IgM для различных терапевтических путей применения. В одном варианте осуществления антитело по настоящему изобретению представляет собой антитело IgG1, например, IgG1 κ-изотипа. Говорят, что антитело относится к конкретному изотипу, если его аминокислотная последовательность в наибольшей степени гомологична этому изотипу по сравнению с другими изотипами.
[00178] В одном варианте осуществления антитело по настоящему изобретению представляет собой полноразмерное антитело, предпочтительно антитело IgG, в частности, антитело IgG1 к-изотипа. В другом варианте осуществления антитело по настоящему изобретению представляет собой эпитопсвязывающий фрагмент антитела или одноцепочечное антитело.
[00179] Антитела и их эпитопсвязывающие фрагменты можно, например, получать путем разделения на эпитопсвязывающие фрагменты с помощью традиционных методик, а эпитопсвязывающие фрагменты подвергать скринингу в отношении полезности таким же образом, как описано в данном документе для полных антител. Например, эпитопсвязывающие F(ab')2-фрагменты можно получать путем обработки антитела пепсином. Полученный в результате эпитопсвязывающий F(ab')2-фрагмент можно обработать для восстановления дисульфидных мостиков с получением эпитопсвязывающих Fab'-фрагментов. Эпитопсвязывающие Fab-фрагменты можно получать путем обработки антитела IgG папаином; эпитопсвязывающие Fab'-фрагменты можно получать путем расщепления антитела IgG пепсином. Эпитопсвязывающий F(ab')-фрагмент также можно получить посредством связывания Fab', описанных ниже, с помощью тиоэфирной связи или дисульфидной связи. Эпитопсвязывающий Fab'-фрагмент представляет собой Эпитопсвязывающий фрагмент антитела, полученный путем разрезания дисульфидной связи в шарнирном домене F(ab')2. Эпитопсвязывающий Fab'-фрагмент можно получить путем обработки эпитопсвязывающего F(ab')2-фрагмента восстановителем, таким как дитиотреитол. Эпитопсвязывающий фрагмент антитела также можно получить посредством экспрессии нуклеиновых кислот, кодирующих такие эпитопсвязывающие фрагменты, в рекомбинантных клетках (см., например, Evans et al., J. Immunol. Meth. 184, 123-38 (1995)). Например, химерный ген, кодирующий часть эпитопсвязывающего F(ab')2-фрагмента, может содержать последовательности ДНК, кодирующие СН1-домен и шарнирный домен Н-цепи, за которыми расположен стоп-кодон для остановки трансляции, для получения такой усеченной молекулы эпитопсвязывающего фрагмента антитела.
[00180] В одном варианте осуществления антитело к тау-белку представляет собой одновалентное антитело, предпочтительно одновалентное антитело, описанное в WO 2007059782 (включенной в данный документ посредством ссылки во всей своей полноте), имеющее делецию в шарнирной области. Соответственно, в одном варианте осуществления антитело представляет собой одновалентное антитело, где указанное антитело к тау-белку сконструировано с помощью способа, включающего: i) обеспечение конструкции нуклеиновой кислоты, кодирующей легкую цепь указанного одновалентного антитела, при этом указанная конструкция содержит нуклеотидную последовательность, кодирующую VL-область выбранного антиген-специфичного антитела к тау-белку, и нуклеотидную последовательность, кодирующую константную CL-область Ig, где указанная нуклеотидная последовательность, кодирующая VL-область выбранного антиген-специфичного антитела, и указанная нуклеотидная последовательность, кодирующая CL-область Ig, функционально связаны друг с другом, и где в случае с подтипом IgG1 нуклеотидная последовательность, кодирующая CL-область, была модифицирована таким образом, чтобы CL-область не содержала каких-либо аминокислот, способных образовывать дисульфидные связи или ковалентные связи с другими пептидами, содержащими идентичную аминокислотную последовательность CL-области, в присутствии поликлонального человеческого IgG или при введении животному или человеку; ii) обеспечение конструкции нуклеиновой кислоты, кодирующей тяжелую цепь указанного одновалентного антитела, при этом указанная конструкция содержит нуклеотидную последовательность, кодирующую VH-область выбранного антиген-специфичного антитела, и нуклеотидную последовательность, кодирующую константную СН-область человеческого Ig, где нуклеотидная последовательность, кодирующая СН-область, была модифицирована таким образом, чтобы область, соответствующая шарнирной области и, как того требует подтип Ig, другим областям СН-области, таким как СН3-область, не содержала каких-либо аминокислотных остатков, принимающих участие в образовании дисульфидных связей или ковалентных или стабильных нековалентных связей между тяжелыми цепями с другими пептидами, содержащими идентичную аминокислотную последовательность СН-области человеческого Ig, в присутствии поликлонального человеческого IgG или при введении животному, человеку, где указанная нуклеотидная последовательность, кодирующая VH-область выбранного антиген-специфичного антитела, и указанная нуклеотидная последовательность, кодирующая СН-область указанного Ig, функционально связаны друг с другом; iii) обеспечение клеточной системы экспрессии для получения указанного одновалентного антитела; iv) получение указанного одновалентного антитела путем совместной экспрессии конструкций нуклеиновых кислот из (i) и (ii) в клетках клеточной системы экспрессии из (iii).
[00181] Подобным образом, в одном варианте осуществления антитело к тау-белку по настоящему изобретению представляет собой одновалентное антитело, которое содержит:
(i) вариабельный домен антитела по настоящему изобретению, описанный в данном документе, или эпитопсвязывающую часть указанного домена, и
(ii) СН-домен иммуноглобулина или его домен, содержащий СН2- и СН3-домены, где СН-домен или его домен был модифицирован таким образом, чтобы домен, соответствующий шарнирному домену и, если иммуноглобулин не относится к подтипу IgG4, другим доменам СН-домена, таким как СН3-домен, не содержал каких-либо аминокислотных остатков, способных образовывать дисульфидные связи с идентичным СН-доменом или другие ковалентные или стабильные нековалентные связи между тяжелыми цепями с идентичным СН-доменом в присутствии поликлонального человеческого IgG.
[00182] В дополнительном варианте осуществления тяжелая цепь одновалентного антитела по настоящему изобретению была модифицирована таким образом, чтобы вся шарнирная область была подвергнута делеции.
[00183] В другом дополнительном варианте осуществления последовательность одновалентного антитела была модифицирована таким образом, чтобы она не содержала каких-либо акцепторных сайтов N-связанного гликозилирования.
[00184] Настоящее изобретение также включает «биспецифические антитела», где область связывания антитела с тау-белком (например, область связывания с тау-белком моноклонального антитела к тау-белку) является частью двухвалентного или поливалентного биспецифического остова, который нацеливается более чем на один эпитоп (например, второй эпитоп может включать эпитоп рецептора, участвующего в активном транспорте, так что биспецифическое антитело будет проявлять улучшенный трансцитоз через биологический барьер, такой как гематоэнцефалический барьер). Таким образом, в другом дополнительном варианте осуществления одновалентный Fab антитела к тау-белку может быть соединен с дополнительным Fab или scFv, который нацеливается на другой белок, с получением биспецифического антитела. Биспецифическое антитело может иметь двойную функцию, например, терапевтическую функцию, придаваемую доменом антитела, связывающимся с тау-белком, и транспортную функцию, благодаря которой оно может связываться с молекулой рецептора для усиления переноса через биологический барьер, такой как гематоэнцефалический барьер.
[00185] Антитела и их эпитопсвязывающие фрагменты по настоящему изобретению также включают одноцепочечные антитела. Одноцепочечные антитела представляют собой пептиды, в которых Fv-домены тяжелой и легкой цепи соединены друг с другом. В одном варианте осуществления настоящее изобретение предусматривает одноцепочечный Fv (scFv), где тяжелая и легкая цепи в Fv антитела к тау-белку по настоящему изобретению соединены с помощью гибкого пептидного линкера (в типичном случае длиной приблизительно 10, 12, 15 или более аминокислотных остатков) в одну пептидную цепь. Способы получения таких антител описаны, например, в US 4946778, Pluckthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315 (1994), Bird et al., Science 242, 423-426 (1988), Huston et al., PNAS USA 85, 5879-5883 (1988) и McCafferty et al. Nature 348, 552-554 (1990). Одноцепочечное антитело может быть одновалентным, если используется только один VH и VL, двухвалентным, если используются два VH и VL, или поливалентным, если используется более двух VH и VL.
[00186] Антитела и их эпитопсвязывающие фрагменты, описанные в данном документе, можно модифицировать путем включения, можно модифицировать путем включения любого пригодного количества модифицированных аминокислот и/или связей с такими конъюгированными заместителями. Пригодность в данном контексте обычно определяется способностью по меньшей мере в значительной степени сохранять селективность в отношениитау-белка и/или специфичность в отношении тау-белка, присущую недериватизированному исходному антителу к тау-белку. Включение одной или нескольких модифицированных аминокислот может быть преимущественным, например, для увеличения периода полувыведения полипептида из сыворотки крови, уменьшения антигенности полипептида или увеличения стабильности полипептида при хранении. Аминокислоту (аминокислоты) модифицируют, например, котрансляционно или посттрансляционно в ходе получения рекомбинантным путем (например, посредством N-связанного гликозилирования в мотивах N-X-S/T в ходе экспрессии в клетках млекопитающих) или модифицируют с помощью синтетических способов. Неограничивающие примеры модифицированной аминокислоты включают гликозилированную аминокислоту, сульфатированную аминокислоту, пренилированную (например, фарнезилированную, геранилгеранилированную) аминокислоту, ацетилированную аминокислоту, ацилированную аминокислоту, пегилированную аминокислоту, биотинилированную аминокислоту, карбоксилированную аминокислоту, фосфорилированную аминокислоту и т.п. В литературе обильно представлены надлежащие источники для обеспечения специалиста руководством по модификации аминокислот. Приводимые в качестве примера протоколы находятся в Walker (1998) Protein Protocols On CD-Rom, Humana Press, Totowa, NJ. Модифицированная аминокислота может, например, быть выбрана из гликозилированной аминокислоты, пегилированной аминокислоты, фарнезилированной аминокислоты, ацетилированной аминокислоты, биотинилированной аминокислоты, аминокислоты, конъюгированной с липидным компонентом, или аминокислоты, конъюгированной с органическим дериватизирующим средством.
[00187] Антитела и их эпитопсвязывающие фрагменты по настоящему изобретению также можно модифицировать химическим путем посредством ковалентного конъюгирования с полимером, например, для увеличения их периода полувыведения из кровотока. Приводимые в качестве примера полимеры и способы присоединения их к пептидам проиллюстрированы, например, в US 4766106; US 4179337; US 4495285 и US 4609546. Дополнительные иллюстративные полимеры включают полиоксиэтилированные полиолы и полиэтиленгликоль (PEG) (например, PEG с молекулярной массой от приблизительно 1000 до приблизительно 40000, как, например, от приблизительно 2000 до приблизительно 20000, например, приблизительно 3000-12000 г/моль).
[00188] Антитела и их эпитопсвязывающие фрагменты по настоящему изобретению можно дополнительно применять в способе диагностики или в качестве диагностического визуализирующего лиганда.
[00189] В одном варианте осуществления предусмотрены антитела и их эпитопсвязывающие фрагменты по настоящему изобретению, содержащие одну или несколько аминокислот, меченных радиоактивными изотопами. Антитело к тау-белку, меченное радиоактивным изотопом, можно применять как в диагностических, так и в терапевтических целях (конъюгирование с молекулами, меченными радиоактивными изотопами, является другой возможной характерной особенностью). Неограничивающие примеры таких меток включают без ограничения висмут (213Bi), углерод (11С, 13С, 14С), хром (51Cr), кобальт (57Со, 60Со), медь (64Cu), диспрозий (165Dy), эрбий (169Er), фтор (18F), гадолиний (153Gd, 159Gd), галлий (68Ga, 67Ga), германий (68Ge), золото (198Au), гольмий (166Ho), водород (3Н), индий (111In, 112In, 113In, 115In), йод (121I,123I, 125I, 131I), иридий (192Ir), железо (59Fe), криптон (81mKr), лантан (140La), лютеций (177Lu), марганец (54Mn), молибден (99Мо), азот (13N, 15N), кислород (15О), палладий (103Pd), фосфор (32Р), калий (42K), празеодим (142Pr), прометий (149Pm), рений (186Re, 188Re), родий (105Rh), рубидий (81Rb, 82Rb), рутений (82Ru, 97Ru), самарий (153Sm), скандий (47Sc), селен (75Se), натрий (24Na), стронций (85Sr, 89Sr, 92Sr), серу (35S), технеций (99Tc), таллий (201Tl), олово (113Sn, 117Sn), ксенон (133Хе), иттербий (169Yb, 175Yb, 177Yb), иттрий (90Y), цинк (65Zn) и цирконий (89Zr). Цирконий (89Zr) представляет особенный интерес. Способы получения аминокислот, меченных радиоактивными изотопами, и соответствующих производных пептидов известны из уровня техники (см., например, Junghans et al., в Cancer Chemotherapy and Biotherapy 655-686 (2nd edition, Chafher and Longo, eds., Lippincott Raven (1996)) и US 4681581; US 4735210; US 5101827; US 5102990 (US RE35500), US 5648471 и US 5697902. Например, радиоактивный изотоп можно конъюгировать с помощью способа с использованием хлорамина Т (Lindegren, S. et al. (1998) "Chloramine-T In High-Specific-Activity Radioiodination Of Antibodies Using N-Succinimidyl-3-(Trimethylstannyl)Benzoate As An Intermediate", Nucl. Med. Biol. 25(7):659-665; Kurth, M. et al. (1993) "Site-Specific Conjugation Of A Radioiodinated Phenethylamine Derivative To A Monoclonal Antibody Results In Increased Radioactivity Localization In Tumor", J. Med. Chem. 36(9):1255-1261; Rea, D.W. et al. (1990) "Site-specifically radioiodinated antibody for targeting tumors", Cancer Res. 50(3 Suppl):857s-861s).
[00190] Настоящее изобретение также предусматривает антитела к тау-белку и их эпитопсвязывающие фрагменты, меченные с помощью выявляемых флуоресцентной метки (такой как хелат редкоземельного элемента (например, хелат европия)), метки флуоресцеинового типа (например, флуоресцеина, изотиоцианата флуоресцеина, 5-карбоксифлуоресцеина, 6-карбоксифлуоресцеина, дихлортриазиниламинофлуоресцеина), метки родаминового типа (например, ALEXA FLUOR® 568 (Invitrogen), TAMRA® или дансилхлорида), VIVOTAG 680 XL FLUOROCHROME™ (Perkin Elmer), фикоэритрина; умбеллиферона, лиссамина; цианина; фикоэритрина, техасского красного, BODIPY FL-SE® (Invitrogen) или его аналога, все из которых подходят для оптического выявления. Можно использовать хемилюминесцентные метки (например, люминол, люциферазу, люциферин и экворин). Такую диагностику и выявление также можно выполнять путем присоединения диагностической молекулы по настоящему изобретению к выявляемым веществам, включающим без ограничения различные ферменты, при этом ферменты включают без ограничения пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу, или к комплексам простетических групп, таким как, без ограничения, стрептавидин/биотин и авидин/биотин.
[00191] Можно использовать хемилюминесцентные метки (например, люминол, люциферазу, люциферин и экворин). Такую диагностику и выявление также можно выполнять путем присоединения диагностической молекулы по настоящему изобретению к выявляемым веществам, включающим без ограничения различные ферменты, при этом ферменты включают без ограничения пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу, или к комплексам простетических групп, таким как, без ограничения, стрептавидин/биотин и авидин/биотин. Также можно использовать парамагнитные метки, и их предпочтительно выявляют с помощью позитронно-эмиссионной томографии (PET) или однофотонной эмиссионной компьютерной томографии (SPECT). Такие парамагнитные метки включают без ограничения соединения, содержащие парамагнитные ионы алюминия (А1), бария (Ва), кальция (Са), церия (Се), диспрозия (Dy), эрбия (Er), европия (Eu), гадолиния (Gd), гольмия (Но), иридия (Ir), лития (Li), магния (Mg), марганца (Mn), молибдена (М), неодима (Nd), осмия (Os), кислорода (О), палладия (Pd), платины (Pt), родия (Rh), рутения (Ru), самария (Sm), натрия (Na), стронция (Sr), тербия (Tb), тулия (Tm), олова (Sn), титана (Ti), вольфрама (W) и циркония (Zi) и, в частности, Со+2, Cr+2, Cr+3, Cu+2, Fe+2, Fe+3, Ga+3, Mn+3, Ni+2, Ti+3, V+3 и V+4, позитрон-излучающие изотопы металлов при использовании различных видов позитронно-эмиссионной томографии и нерадиоактивные парамагнитные ионы металлов.
[00192] Таким образом, в одном варианте осуществления антитело к тау-белку или его фрагмент, связывающийся с тау-белком, по настоящему изобретению могут быть меченными флуоресцентной меткой, хемилюминесцентной меткой, парамагнитной меткой, радиоизотопной меткой или ферментной меткой. Меченое антитело или его фрагмент можно применять для выявления наличия или измерения количества указанного тау-белка в головном мозге субъекта. Этот способ может включать выявление или измерение количества антитела к тау-белку или фрагмента, связывающегося с тау-белком, связанного с указанным тау-белком, посредством визуализации in vivo и может включать визуализацию ex vivo указанного антитела к тау-белку или фрагмента, связывающегося с тау-белком, связанного с таким тау-белком.
[00193] В дополнительном аспекте настоящее изобретение относится к вектору экспрессии, кодирующему одну или несколько полипептидных цепей антитела по настоящему изобретению или его фрагмента, связывающегося с тау-белком. Такие векторы экспрессии можно применять для получения антител или их эпитопсвязывающих фрагментов по настоящему изобретению рекомбинантным путем.
[00194] Вектор экспрессии в контексте настоящего изобретения может представлять собой любой подходящий ДНК- или РНК-вектор, включая хромосомные, нехромосомные и синтетические векторы на основе нуклеиновых кислот (последовательность нуклеиновой кислоты, содержащую подходящий набор элементов, контролирующих экспрессию). Примеры таких векторов включают производные SV40, бактериальные плазмиды, фаговую ДНК, бакуловирус, дрожжевые плазмиды, векторы, полученные в результате объединения плазмид и фаговой ДНК, и векторы на основе вирусной нуклеиновой кислоты (РНК или ДНК). В одном варианте осуществления нуклеиновая кислота, кодирующая антитело к тау-белку, содержится в векторе на основе «голой» ДНК или РНК, включающем в себя, например, линейный экспрессионный элемент (описанный, например, в Sykes and Johnston, Nat Biotech 12, 355-59 (1997)), вектор на основе плотно упакованной нуклеиновой кислоты (описанный, например, в US 6077835 и/или WO 00/70087), плазмидный вектор, такой как pBR322, pUC 19/18 или pUC 118/119, вектор минимального размера MIDGE на основе нуклеиновой кислоты (описанный, например, в Schakowski et al., Mol Ther 3, 793-800 (2001)), или представлена в виде осаждаемой векторной конструкции нуклеиновой кислоты, такой как конструкция, осаждаемая CaPO4 (описанная, например, в WO 00/46147, Benvenisty and Reshef, PNAS USA 83, 9551-55 (1986), Wigler et al., Cell 14, 725 (1978), и Coraro and Pearson, Somatic Cell Genetics 2,603 (1981)). Такие векторы на основе нуклеиновых кислот и их применение хорошо известны из уровня техники (см., например, US 5589466 и US 5973972).
[00195] В одном варианте осуществления вектор подходит для экспрессии антител к тау-белку или их эпитопсвязывающих фрагментов по настоящему изобретению в бактериальной клетке. Примеры таких векторов включают векторы экспрессии, такие как BlueScript (Stratagene), векторы pIN (Van Heeke & Schuster, J. Biol. Chem. 264, 5503-5509 (1989), векторы pET (Novagen, Мэдисон, Висконсин) и т.п.
[00196] Вектор экспрессии может также или в качестве альтернативы представлять собой вектор, подходящий для экспрессии в дрожжевой системе. Можно использовать любой вектор, подходящий для экспрессии в дрожжевой системе. Подходящие векторы включают в себя, например, векторы, содержащие конститутивные или индуцируемые промоторы, такие как промоторы генов фактора альфа, алкогольоксидазы и PGH (обзор которых приведен в: F. Ausubel et al., ed. Current Protocols in Molecular Biology, Greene Publishing and Wiley InterScience New York (1987), Grant et al.. Methods in Enzymol 153, 516-544 (1987), Mattanovich, D. et al. Methods Mol. Biol. 824, 329-358 (2012), Celik, E. et al. Biotechnol. Adv. 30(5), 1108-1118 (2012), Li, P. et al. Appl. Biochem. Biotechnol. 142(2), 105-124 (2007),  , E. et al. Appl. Microbiol. Biotechnol. 77(3), 513-523 (2007), van der Vaart, J.M. Methods Mol. Biol. 178, 359-366 (2002), и Holliger, P. Methods Mol. Biol. 178, 349-357 (2002)).
, E. et al. Appl. Microbiol. Biotechnol. 77(3), 513-523 (2007), van der Vaart, J.M. Methods Mol. Biol. 178, 359-366 (2002), и Holliger, P. Methods Mol. Biol. 178, 349-357 (2002)).
[00197] В векторе экспрессии по настоящему изобретению нуклеиновые кислоты, кодирующие антитело к тау-белку, могут содержать любой подходящий промотор, энхансер и другие элементы, способствующие экспрессии, или быть связанными с ними. Примеры таких элементов включают сильные экспрессионные промоторы (например, IE-промотор/энхансер CMV человека, а также промоторы RSV, SV40, SL3-3, MMTV и LTR HIV), эффективные поли-(А)-последовательности терминации, точку начала репликации для продукта плазмиды в E.coli, ген устойчивости к антибиотику в качестве селектируемого маркера и/или подходящий сайт клонирования (например, полилинкер). Нуклеиновые кислоты также могут содержать индуцируемый промотор, а не конститутивный промотор, такой как IE CMV (специалисту в данной области будет понятно, что такие термины фактически описывают степень экспрессии гена в определенных условиях).
[00198] В еще одном дополнительном аспекте настоящее изобретение относится к рекомбинантной эукариотической или прокариотической клетке-хозяину, такой как трансфектома, которая вырабатывает антитело или его эпитопсвязывающий фрагмент по настоящему изобретению, определенные в данном документе, или биспецифическую молекулу по настоящему изобретению, определенную в данном документе. Примеры клеток-хозяев включают клетки дрожжей, бактерий и млекопитающих, такие как клетки СНО или HEK. Например, в одном варианте осуществления настоящее изобретение предусматривает клетку, содержащую нуклеиновую кислоту, стабильно интегрированную в клеточный геном, которая содержит последовательность, кодирующую экспрессируемое антитело к тау-белку по настоящему изобретению или его эпитопсвязывающий фрагмент. В другом варианте осуществления настоящее изобретение предусматривает клетку, содержащую неинтегрированную нуклеиновую кислоту, такую как плазмида, космида, фагмида или линейный экспрессионный элемент, которая содержит последовательность, кодирующую экспрессируемое антитело к тау-белку или его эпитопсвязывающий фрагмент по настоящему изобретению.
[00199] В дополнительном аспекте настоящее изобретение относится к способу получения антитела к тау-белку по настоящему изобретению, при этом указанный способ включает стадии а) культивирования гибридомы или клетки-хозяина по настоящему изобретению, описанных в данном документе выше, и b) очистки антитела по настоящему изобретению от культуральной среды.
[00200] В одном варианте осуществления настоящее изобретение относится к препарату в том смысле, в котором этот термин используется в данном документе, который содержит антитело к тау-белку, определенное в данном документе, и который практически не содержит антител, образующихся в естественных условиях, которые либо не способны связываться с тау-белком, либо существенным образом не изменяют функциональные характеристики антитела к тау-белку в препарате. Таким образом, такой препарат не охватывает сыворотку крови, образующуюся в естественных условиях, или очищенный производный продукт такой сыворотки крови, который содержит смесь антитела к тау-белку и другого антитела, которое не изменяет функциональные характеристики антитела к тау-белку в препарате, где такие функциональные характеристики представляют собой:
(i) практически отсутствующую способность к связыванию с нефосфорилированным тау-белком;
(ii) практически отсутствующую способность к связыванию с тау-белком, фосфорилированным в S404 и не фосфорилированным в S396;
(iii) способность к связыванию с тау-белком, фосфорилированным в S396;
(iv) способность к связыванию с тау-белком, фосфорилированным как в S396, так и в S404;
(v) способность к селективному проведению различий между фосфорилированными остатками S396 и S404 тау-белка, так что они практически не способны связываться с фосфорилированным остатком 404;
(vi) способность к связыванию с гиперфосфорилированным тау-белком из головного мозга людей с болезнью Альцгеймера;
(vii) способность к проведению различий между патологическим и непатологическим человеческим тау-белком и/или
(viii) способность к специфичному уменьшению полос гиперфосфорилированного тау-белка размером 64 кДа и 70 кДа по меньшей мере на 90% без уменьшения при этом полосы тау-белка размером 55 кДа более чем на 10% при применении согласно описанному в данном документе с иммуноистощенными экстрактами от трансгенных мышей rTg4510 или способность к специфичному уменьшению полос фосфорилированного в S396 гиперфосфорилированного тау-белка по меньшей мере на 90% без уменьшения при этом полос негиперфосфорилированного тау-белка более чем на 10% при применении согласно описанному в данном документе с посмертными экстрактами из головного мозга людей с AD.
[00201] Настоящее изобретение, в частности, относится к препаратам такого антитела к тау-белку, имеющего структурное изменение в своей аминокислотной последовательности (в любых своих CDR, вариабельных доменах, остатках каркасной области и/или константных доменах) по сравнению со структурой встречающегося в природе антитела к тау-белку, где указанное структурное изменение обуславливает проявление антителом к тау-белку значительно измененных функциональных характеристик (т.е. более чем с 20% разницей, более чем с 40% разницей, более чем с 60% разницей, более чем с 80% разницей, более чем со 100% разницей, более чем со 150% разницей, более чем с 2-кратной разницей, более чем с 4-кратной разницей, более чем с 5-кратной разницей или более чем с 10-кратной разницей в функциональных характеристиках) по сравнению с функциональными характеристиками, проявляемыми указанным встречающимся в природе антителом к тау-белку; где указанные функциональные характеристики представляют собой:
(i) практически отсутствующую способность к связыванию с нефосфорилированным тау-белком;
(ii) практически отсутствующую способность к связыванию с тау-белком, фосфорилированным в S404 и не фосфорилированным в S396;
(iii) способность к связыванию с тау-белком, фосфорилированным в S396;
(iv) способность к связыванию с тау-белком, фосфорилированным как в S396, так и в S404;
(v) способность к селективному проведению различий между фосфорилированными остатками S396 и S404 тау-белка, так что они практически не способны связываться с фосфорилированным остатком 404;
(vi) способность к связыванию с гиперфосфорилированным тау-белком из головного мозга людей с болезнью Альцгеймера;
(vii) способность к проведению различий между патологическим и непатологическим человеческим тау-белком и/или
(viii) способность к специфичному уменьшению полос гиперфосфорилированного тау-белка размером 64 кДа и 70 кДа по меньшей мере на 90% без уменьшения при этом полосы тау-белка размером 55 кДа более чем на 10% при применении согласно описанному в данном документе с иммуноистощенными экстрактами от трансгенных мышей rTg4510 или способность к специфичному уменьшению полос фосфорилированного в S396 гиперфосфорилированного тау-белка по меньшей мере на 90% без уменьшения при этом полос негиперфосфорилированного тау-белка более чем на 10% при применении согласно описанному в данном документе с посмертными экстрактами головного мозга людей с AD.
[00202] Термин «практически не содержит антител, образующихся в естественных условиях» относится к полному отсутствию таких антител, образующихся в естественных условиях, в таких препаратах или к включению таких антител, образующихся в естественных условиях, в такие препараты в концентрации, в которой они существенным образом не влияют на свойства связывания с тау-белком данных препаратов. Говорят, что антитело является «выделенным», если оно не имеет эквивалента, образующегося в естественных условиях, или было отделено или очищено от компонентов, которые в естественных условиях сопутствуют ему.
[00203] Термин «антитела, образующиеся в естественных условиях», используемый в отношении таких препаратов, относится к антителам (в том числе аутоантителам, образующимся в естественных условиях), образование которых происходит в организме живых людей или других животных в качестве естественного следствия функционирования их иммунных систем.
[00204] Таким образом, препараты по настоящему изобретению не исключают, а в действительности явным образом охватывают препараты, которые содержат антитело к тау-белку и преднамеренно добавленное дополнительное антитело, способное связываться с эпитопом, не содержащимся в тау-белке. Такие препараты, в частности, включают их варианты осуществления, в которых препарат проявляет повышенную эффективность в лечении болезни Альцгеймера (AD), болезни аргирофильных зерен (AGD), прогрессирующего надъядерного паралича (PSP) и кортикобазальной дегенерации (CBD). Дополнительно, настоящее изобретение направлено на препараты, которые содержат антитуло, антитела к тау-белку или их эпитопсвязывающие фрагменты, предназначенные для применения в лечении психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, и психиатрических симптомов у пациентов с деменцией с тельцами Леви. Дополнительно, препараты по настоящему изобретению содержат антитело, антитела к тау-белку или их эпитопсвязывающие фрагменты, которые можно применять в лечении инсульта, восстановления после инсульта, нейродегенерации, связанной с болезнью Паркинсона.
[00205] В еще одном дополнительном аспекте настоящее изобретение относится к фармацевтической композиции, содержащей:
(i) антитело к тау-белку или его эпитопсвязывающий фрагмент, оба из которых определены в данном документе, или препарат в том смысле, в котором этот термин определен в данном документе, который содержит такое антитело к тау-белку или его эпитопсвязывающий фрагмент; и
(ii) фармацевтически приемлемый носитель.
[00206] Фармацевтические композиции можно составлять с фармацевтически приемлемыми носителями или разбавителями, а также с любыми другими известными вспомогательными средствами и наполнителями согласно традиционным методикам, таким как раскрытые в Remington: The Science and Practice of Pharmacy, 22nd Edition, Gennaro, Ed., Mack Publishing Co., Easton, PA, 2013.
[00207] Фармацевтически приемлемые носители или разбавители, а также любые другие известные вспомогательные средства и наполнители должны быть пригодными для выбранного соединения по настоящему изобретению и выбранного способа введения. Пригодность носителей и других компонентов фармацевтических композиций определяют на основании отсутствия значительного отрицательного влияния на желаемые биологические свойства выбранного соединения или фармацевтической композиции по настоящему изобретению (например, менее чем существенного влияния (10% или меньшего относительного ингибирования, 5% или меньшего относительного ингибирования и т.п.)) на связывание с эпитопом.
[00208] Фармацевтическая композиция по настоящему изобретению также может содержать разбавители, заполнители, соли, буферы, детергенты (например, неионогенный детергент, такой как Tween-20 или Tween-80), стабилизаторы (например, сахара или белковые свободные аминокислоты), консерванты, фиксаторы для тканей, солюбилизаторы и/или другие материалы, подходящие для включения в фармацевтическую композицию. Разбавитель выбирают таким образом, чтобы он не влиял на биологическую активность комбинации. Примерами таких разбавителей являются дистиллированная вода, физиологический раствор с фосфатным буфером, раствор Рингера, раствор декстрозы и раствор Хэнкса. В дополнение, фармацевтическая композиция или состав также могут содержать другие носители или нетоксичные нетерапевтические неиммуногенные стабилизаторы и т.п. Композиции также могут содержать крупные, медленно метаболизируемые макромолекулы, такие как белки, полисахариды, такие как хитозан, полимолочные кислоты, полигликолевые кислоты и их сополимеры (например, функционализированную латексом сефарозу, агарозу, целлюлозу и т.п.), полимеры аминокислот, сополимеры аминокислот и липидные агрегаты (например, капли масла или липосомы).
[00209] Фактические уровни доз активных ингредиентов в фармацевтических композициях по настоящему изобретению можно изменять таким образом, чтобы получить количество активного ингредиента, эффективное для достижения желаемого терапевтического ответа для конкретных пациента, композиции и способа введения. Выбранный уровень дозы будет зависеть от ряда фармакокинетических факторов, включающих активность конкретных используемых композиций по настоящему изобретению или их амидных форм, путь введения, время введения, скорость экскреции конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, применяемые в комбинации с конкретными используемыми композициями, возраст, пол, массу, состояние, общее состояние здоровья и анамнез пациента, подвергаемого лечению, и подобные факторы, хорошо известные в области медицины.
[00210] Фармацевтическую композицию можно вводить любым подходящим путем и способом, в том числе парентеральным, местным, пероральным или интраназальным путями, для профилактического и/или терапевтического лечения. В одном варианте осуществления фармацевтическую композицию по настоящему изобретению вводят парентерально. Фразы «парентеральное введение» и «вводимый парентерально», используемые в данном документе, означают способы введения, отличные от энтерального и местного введения, обычно посредством инъекции, и включают эпидермальную, внутривенную, внутримышечную, внутриартериальную, интратекальную, внутрикапсульную, внутриглазничную, внутрисердечную, внутрикожную, внутрибрюшинную, внутрисухожильную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, подкапсульную, субарахноидальную, интраспинальную, внутричерепную, интраторакальную, эпидуральную и внутригрудинную инъекцию и инфузию.
[00211] Дополнительные подходящие пути введения соединения по настоящему изобретению in vivo и in vitro хорошо известны из уровня техники и могут быть выбраны специалистами обычной квалификации в данной области.
[00212] В одном варианте осуществления эту фармацевтическую композицию вводят посредством внутривенной или подкожной инъекции или инфузии.
[00213] Фармацевтически приемлемые носители включают любые возможные подходящие растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические средства, антиоксиданты и средства, замедляющие всасывание, и т.п., которые являются физиологически совместимыми с соединением по настоящему изобретению.
[00214] Примеры подходящих водных и неводных носителей, которые можно использовать в фармацевтических композициях по настоящему изобретению, включают воду, солевой раствор, фосфатно-солевой буферный раствор, этанол, декстрозу, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и их подходящие смеси, растительные масла, такие как оливковое масло, кукурузное масло, арахисовое масло, хлопковое масло и кунжутное масло, коллоидные растворы карбоксиметилцеллюлозы, трагакантовую камедь и инъекционные органические сложные эфиры, такие как этилолеат, и/или различные буферы. Другие носители хорошо известны в области фармацевтики.
[00215] Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для индивидуального приготовления стерильных инъекционных растворов или дисперсий. Применение таких сред и средств для фармацевтически активных веществ известно из уровня техники. За исключением случаев, когда какие-либо традиционные среда или средство являются несовместимыми с активным соединением, предполагается их применение в фармацевтических композициях по настоящему изобретению.
[00216] Надлежащую текучесть можно поддерживать, например, путем применения материалов для покрытия, таких как лецитин, путем поддержания требуемого размера частиц в случае дисперсий и путем применения поверхностно-активных веществ.
[00217] Фармацевтические композиции по настоящему изобретению также могут содержать фармацевтически приемлемые антиоксиданты, например, (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфит натрия и т.п.; (2) жирорастворимые антиоксиданты, такие как аскорбилпальмитат, бутилированный гидроксианизол (ВНА), бутилированный гидрокситолуол (ВНТ), лецитин, пропилгаллат, альфа-токоферол и т.п.; и (3) металлохелаторы, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбит, винная кислота, фосфорная кислота и т.п.
[00218] Фармацевтические композиции по настоящему изобретению также могут содержать изотонические средства, такие как сахара, многоатомные спирты, такие как маннит, сорбит, глицерин, или хлорид натрия, в композициях.
[00219] Фармацевтические композиции по настоящему изобретению также могут содержать одно или несколько вспомогательных средств, подходящих для выбранного пути введения, таких как консерванты, смачивающие средства, эмульгаторы, диспергирующие средства, консерванты или буферы, которые могут повышать сохраняемость или эффективность фармацевтической композиции. Соединения по настоящему изобретению можно получать с носителями, которые будут защищать соединение от быстрого высвобождения, как, например, в составе с контролируемым высвобождением, включающем имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Такие носители могут включать желатин, глицерилмоностеарат, глицерилдистеарат, биоразлагаемые биосовместимые полимеры, такие как сополимер этилена и винилацетата, полиангидриды, полигликолевую кислоту, коллаген, сложные полиортоэфиры и полимолочную кислоту отдельно или вместе с воском или другие материалы, хорошо известные из уровня техники. Способы получения таких составов общеизвестны для специалистов в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
[00220] В одном варианте осуществления соединения по настоящему изобретению можно составлять для обеспечения надлежащего распределения in vivo. Фармацевтически приемлемые носители для парентерального введения включают стерильные водные растворы или дисперсии и стерильные порошки для индивидуального приготовления стерильных инъекционных растворов или дисперсий. Применение таких сред и средств для фармацевтически активных веществ известно из уровня техники. За исключением случаев, когда какие-либо традиционные среда или средство являются несовместимыми с активным соединением, предполагается их применение в фармацевтических композициях по настоящему изобретению. Дополнительные активные соединения также можно включать в состав композиций.
[00221] Фармацевтические композиции для инъекции в типичном случае должны быть стерильными и стабильными в условиях производства и хранения. Композицию можно составлять в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Носитель может представлять собой водный или неводный растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и их подходящие смеси, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Надлежащую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Во многих случаях будет предпочтительным включение изотонических средств, например, сахаров, многоатомных спиртов, таких как глицерин, маннит, сорбит, или хлорида натрия, в композицию. Длительного всасывания инъекционных композиций можно достичь путем включения в композицию средства, замедляющего всасывание антитела, например, моностеаратных солей и желатина. Стерильные инъекционные растворы можно получать путем помещения активного соединения в требуемом количестве в соответствующий растворитель с одним ингредиентом или их комбинацией, например, перечисленными выше, в соответствии с требованиями с последующей стерилизующей микрофильтрацией. Обычно дисперсии получают путем помещения активного соединения в стерильную основу, которая содержит основную дисперсионную среду и другие требуемые ингредиенты, например, из перечисленных выше. В случае стерильных порошков для получения стерильных инъекционных растворов примерами способов получения являются вакуумная сушка и сублимационная сушка (лиофилизация), в результате которой получают порошкообразный активный ингредиент вместе с любым дополнительным желаемым ингредиентом из их раствора, предварительно подвергнутого стерилизующей фильтрации.
[00222] Стерильные инъекционные растворы можно получать путем помещения активного соединения в требуемом количестве в соответствующий растворитель с одним ингредиентом или их комбинацией, перечисленными выше, в соответствии с требованиями с последующей стерилизующей микрофильтрацией. Обычно дисперсии получают путем помещения активного соединения в стерильную основу, которая содержит основную дисперсионную среду и другие требуемые ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных инъекционных растворов примерами способов получения являются вакуумная сушка и сублимационная сушка (лиофилизация), в результате которой получают порошкообразный активный ингредиент вместе с любым дополнительным желаемым ингредиентом из их раствора, предварительно подвергнутого стерилизующей фильтрации.
[00223] Режимы дозирования в вышеуказанных способах лечения и путях применения, описанных в данном документе, корректируют для получения оптимального желаемого ответа (например, терапевтического ответа). Например, можно вводить однократную болюсную дозу, можно вводить несколько разделенных доз в течение некоторого времени, или дозу можно пропорционально уменьшать или повышать, как определяется потребностями терапевтической ситуации. Композиции для парентерального введения можно составлять в виде единичной лекарственной формы для простоты введения и равномерности дозирования. Единичная лекарственная форма, используемая в данном документе, относится к физически дискретным единицам, подходящим в качестве единичных доз для субъектов, подлежащих лечению; при этом каждая единица содержит предварительно определенное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта, совместно с требуемым фармацевтическим носителем. Технические требования к единичным лекарственным формам по настоящему изобретению продиктованы (а) уникальными характеристиками активного соединения и конкретного терапевтического эффекта, которого необходимо достичь, и (b) ограничениями, присущими области составления такого активного соединения для лечения чувствительности у индивидуумов, и непосредственно зависят от них.
[00224] Эффективные дозы и режимы дозирования антител или их эпитопсвязывающих фрагментов по настоящему изобретению зависят от заболевания или состояния, подлежащего лечению, и могут быть определены специалистами в данной области. В любой данный день, в который дают дозу, доза может находиться в диапазоне от приблизительно 0,0001 до приблизительно 100 мг/кг и в более обычном случае от приблизительно 0,01 до приблизительно 5 мг/кг массы тела реципиента. Например, дозы могут составлять 1 мг/кг массы тела или 10 мг/кг массы тела или находиться в диапазоне 1-10 мг/кг массы тела. Приводимые в качестве примера дозы, таким образом, включают в себя: от приблизительно 0,1 до приблизительно 10 мг/кг массы тела, от приблизительно 0,1 до приблизительно 5 мг/кг массы тела, от приблизительно 0,1 до приблизительно 2 мг/кг массы тела, от приблизительно 0,1 до приблизительно 1 мг/кг массы тела, например, приблизительно 0,15 мг/кг массы тела, приблизительно 0,2 мг/кг массы тела, приблизительно 0,5 мг/кг массы тела, приблизительно 1 мг/кг массы тела, приблизительно 1,5 мг/кг массы тела, приблизительно 2 мг/кг массы тела, приблизительно 5 мг/кг массы тела или приблизительно 10 мг/кг массы тела.
[00225] Врач, являющийся специалистом обычной квалификации в данной области, может без труда определить и назначить эффективное количество требуемой фармацевтической композиции. Например, врач может начинать с доз антитела или его эпитопсвязывающего фрагмента по настоящему изобретению, используемых в фармацевтической композиции, на более низких уровнях, чем требуется для достижения желаемого терапевтического эффекта, и постепенно повышать дозу до достижения желаемого эффекта. Как правило, подходящая суточная доза композиции по настоящему изобретению будет представлять собой такое количество соединения, которое является наиболее низкой дозой, эффективной для получения терапевтического эффекта. Такая эффективная доза обычно будет зависеть от факторов, описанных выше. Введение может, например, быть внутривенным, внутримышечным, внутрибрюшинным или подкожным. При желании эффективную суточную дозу фармацевтической композиции можно вводить в виде двух, трех, четырех, пяти, шести или более частей дозы, вводимых по отдельности с соответствующими интервалами в течение всего дня, необязательно в виде единичных лекарственных форм. Хотя соединение по настоящему изобретению можно вводить отдельно, предпочтительно вводить соединение в виде фармацевтической композиции, описанной выше.
[00226] Меченые антитела или их эпитопсвязывающие фрагменты по настоящему изобретению можно применять в диагностических целях для выявления, диагностики или мониторинга заболеваний или нарушений. Настоящее изобретение предусматривает выявление или диагностику нейродегенеративного или когнитивного заболевания или нарушения, в том числе без ограничения болезни Альцгеймера, болезни аргирофильных зерен (AGD), прогрессирующего надъядерного паралича (PSP) и кортикобазальной дегенерации (CBD), включающие: (а) проведение анализа наличия пироглутамилированных Аβ-фрагментов в клетках или образцах тканей субъекта с помощью одного или нескольких антител, специфично связывающихся с тау-белком; и (b) проведение сравнения уровня антигена с контрольным уровнем, например, с уровнями в нормальных образцах тканей, при котором повышение анализируемого уровня антигена по сравнению с контрольным уровнем антигена свидетельствует о заболевании или нарушении или свидетельствует о тяжести заболевания или нарушения.
[00227] Антитела или их эпитопсвязывающие фрагменты по настоящему изобретению можно применять для анализа тау-белка или фрагментов тау-белка в биологическом образце с помощью иммуногистохимических способов, хорошо известных из уровня техники. Другие способы с использованием антител, применимые для выявления белка, включают иммунологические анализы, такие как иммуноферментный анализ (ELISA) и радиоиммунологический анализ (RIA), а также анализы с использованием платформы Meso Scale Discovery (MSD). В таких наборах и способах можно применять подходящие метки для антител, и метки, известные из уровня техники, включают ферментные метки, такие как щелочная фосфатаза и глюкозооксидаза; радиоизотопные метки, такие как йод (125I, 131I), углерод (14С), сера (35S), тритий (3Н), индий (121In) и технеций (99mTc); а также люминесцентные метки, такие как люминол и люцифераза; и флуоресцентные метки, такие как флуоресцеин и родамин.
[00228] Наличие меченых антител к тау-белку или их фрагментов, связывающихся с тау-белком, можно выявлять in vivo в диагностических целях. В одном варианте осуществления диагностика включает: а) введение субъекту эффективного количества такой меченой молекулы; b) выжидание в течение определенного временного интервала после введения для обеспечения концентрирования меченой молекулы в местах отложения Aβ (при наличии таковых) и для обеспечения очищения от несвязанной меченой молекулы до фонового уровня; с) определение фонового уровня и d) выявление меченой молекулы у субъекта таким образом, что выявление меченой молекулы на уровне, превышающем фоновый, свидетельствует о том, что субъект имеет заболевание или нарушение, или свидетельствует о тяжести заболевания или нарушения. В соответствии с таким вариантом осуществления молекулу метят визуализирующим компонентом, подходящим для выявления, с помощью конкретной системы визуализации, известной специалистам в данной области. Фоновые уровни можно определить с помощью различных способов, известных из уровня техники, в том числе путем сравнения количества выявленного меченого антитела со стандартным значением, предварительно определенным для конкретной системы визуализации. Способы и системы, которые можно применять в способах диагностики по настоящему изобретению, включают без ограничения компьютерную томографию (СТ), сканирование всего тела, такое как позитронно-эмиссионная томография (PET), магнитно-резонансную визуализацию (MRI) и ультразвуковое исследование.
[00229] В дополнительном аспекте настоящее изобретение предусматривает моноклональное антитело или его эпитопсвязывающий фрагмент, определенные в данном документе, для применения в терапии.
[00230] В дополнительном аспекте настоящее изобретение предусматривает моноклональное антитело или его эпитопсвязывающий фрагмент, определенные в данном документе, для применения в лечении, диагностике или визуализации таупатий.
[00231] В дополнительном аспекте настоящее изобретение предусматривает моноклональное антитело или его эпитопсвязывающий фрагмент, определенные в данном документе, для применения в лечении болезни Альцгеймера, болезни аргирофильных зерен (AGD), прогрессирующего надъядерного паралича (PSP) и кортикобазальной дегенерации (CBD).
[00232] В дополнительном аспекте настоящее изобретение предусматривает моноклональное антитело или его эпитопсвязывающий фрагмент, определенные в данном документе, для применения в производстве лекарственного препарата для лечения, диагностики или визуализации таупатий.
[00233] Лекарственный препарат предпочтительно предназначен для лечения болезни Альцгеймера (AD), болезни аргирофильных зерен (AGD), прогрессирующего надъядерного паралича (PSP) и кортикобазальной дегенерации (CBD), наиболее предпочтительно болезни Альцгеймера (AD). Лекарственный препарат также предпочтительно предназначен для лечения психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, и психиатрических симптомов у пациентов с деменцией с тельцами Леви.
[00234] В дополнительном аспекте настоящее изобретение предусматривает способ лечения, диагностики или визуализации болезни Альцгеймера или других таупатий у субъекта, при этом указанный способ включает введение лекарственного препарата на основе моноклонального антитела или его эпитопсвязывающего фрагмента, определенных в данном документе, указанному субъекту в эффективном количестве.
[00235] В предпочтительном варианте осуществления лечение является длительным и предпочтительно продолжается по меньшей мере в течение 2 недель, как, например, по меньшей мере в течение 1 месяца, 6 месяцев, 1 года или дольше.
[00236] В дополнительном аспекте настоящее изобретение предусматривает набор, содержащий антитело или его фрагмент, определенные в данном документе, для применения в терапии.
Варианты осуществления
1. Моноклональное антитело или его эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка, таким как фосфорилированный остаток 396 SEQ ID NO: 33.
2. Антитело в соответствии с вариантом осуществления 1, состоящее из интактного антитела.
3. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантами осуществления 1 или 2, содержащие эпитопсвязывающий фрагмент, выбранный из группы, состоящей из Fv-фрагмента (например, одноцепочечного Fv и Fv, стабилизированного дисульфидными связями); Fab-подобного фрагмента (например, Fab-фрагмента, Fab'-фрагмента и F(ab)2-фрагмента); (Fv)2-CH3-домена миниантитела и доменного антитела (например, отдельного вариабельного VH-домена или вариабельного VL-домена), или состоящие из него.
4. Антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих вариантов осуществления, где антитело выбрано из группы, состоящей из антител подтипа IgG1, IgG2, IgG3 или IgG4.
5. Моноклональное антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих вариантов осуществления, которые являются человеческими или гуманизированными.
6. Моноклональное антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих вариантов осуществления, где антитело или эпитопсвязывающий фрагмент проявляют одно или несколько из следующих свойств:
(a) селективность и специфичность в отношении человеческого патологического тау-белка;
(b) аффинность связывания (KD) с pTau 386-408 (pS396/pS404) (SEQ ID NO: 33), составляющую 0,5-10 нМ, как, например, 1-5 нМ или 1-2 нМ.
7. Моноклональное антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих вариантов осуществления, где указанное антитело практически не связывается с фосфорилированным остатком 404 тау-белка (SEQ ID NO: 33).
8. Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащие:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 2 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 4 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 5 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 6 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
9. Моноклональное антитело в соответствии с вариантом осуществления 8, содержащее вариабельный домен тяжелой цепи под SEQ ID NO: 8 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах, и/или вариабельный домен легкой цепи под SEQ ID NO: 7, имеющий не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
10. Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащие:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 9 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 10 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(с) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 11 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 13 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 14 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
11. Моноклональное антитело в соответствии с вариантом осуществления 10, содержащее вариабельный домен тяжелой цепи под SEQ ID NO: 16 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах, и или вариабельный домен легкой цепи под SEQ ID NO: 15 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
12. Моноклональное антитело, где эпитопсвязывающий фрагмент содержит:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 17 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 18 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(с) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 19 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 20 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 21 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 22 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
13. Моноклональное антитело в соответствии с вариантом осуществления 12, содержащее вариабельный домен тяжелой цепи под SEQ ID NO: 24 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах, и/или вариабельный домен легкой цепи под SEQ ID NO: 23 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
14. Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащие:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 25 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 26 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(с) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 27 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 28 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 29 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 30 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
15. Моноклональное антитело в соответствии с вариантом осуществления 14, содержащее вариабельный домен тяжелой цепи под SEQ ID NO: 32 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах, и/или вариабельный домен легкой цепи под SEQ ID NO: 31 или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
16. Антитело или его эпитопсвязывающий фрагмент в соответствии с любым из вариантов осуществления 1-7, где указанные антитело или его фрагмент конкурируют с антителом или его эпитопсвязывающим фрагментом, определенными в вариантах осуществления 8-15, за связывание с человеческим тау-белком.
17. Антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих вариантов осуществления, содержащие Fc-домен.
18. Антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих вариантов осуществления, дополнительно содержащие компонент для увеличения периода полувыведения in vivo.
19. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантом осуществления 18, где компонент для увеличения периода полувыведения in vivo выбран из группы, состоящей из полиэтиленгликоля (PEG), человеческого сывороточного альбумина, гликозилирующих групп, жирных кислот и декстрана.
20. Антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих вариантов осуществления, где антитело дополнительно содержит выявляемый компонент.
21. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантом осуществления 20, где выявляемый компонент выбран из группы, состоящей из флуоресцентной метки, хемилюминесцентной метки, парамагнитной метки, радиоизотопной метки или ферментной метки.
22. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантами осуществления 20 или 21, где выявляемый компонент содержит радиоактивный изотоп или состоит из него.
23. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантом осуществления 22, где радиоактивный изотоп выбран из группы, состоящей из 99mTc, 111In, 67Ga, 68Ga, 72As, 89Zr, 123I и 201Tl.
24. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантом осуществления 21, где выявляемый компонент содержит парамагнитный изотоп или состоит из него.
25. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантом осуществления 24, где парамагнитный изотоп выбран из группы, состоящей из 157Gd, 55Mn, 162Dy, 52Cr и 56Fe.
26. Антитело или его эпитопсвязывающий фрагмент в соответствии с любым из вариантов осуществления 20-25, где выявляемый компонент выявляется с помощью методики визуализации, такой как SPECT, PET, MPI, оптическая или ультразвуковая визуализация.
27. Антитело или его эпитопсвязывающий фрагмент в соответствии с любым из вариантов осуществления 20-26, где выявляемый компонент опосредованно соединен с антителом или его эпитопсвязывающим фрагментом с помощью линкерного компонента.
28. Антитело или его эпитопсвязывающий фрагмент в соответствии с вариантом осуществления 27, где линкерный компонент выбран из группы, состоящей из производных 1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусной кислоты (DOTA), дефероксамина (DFO), производных диэтилентриаминпентауксусной кислоты (DTPА), производных S-2-(4-изотиоцианатобензил)-1,4,7-триазациклононан-1,4,7-триуксусной кислоты (NOTA) и производных 1,4,8,11-тетраазациклододекан-1,4,8,11-тетрауксусной кислоты (ТЕТА).
29. Моноклональное антитело или его эпитопсвязывающий фрагмент, где тяжелая цепь выбрана из группы, состоящей из SEQ ID NO: 8, SEQ ID NO: 16, SEQ ID NO: 24, SEQ ID NO: 32 и SEQ ID NO: 35, а легкая цепь выбрана из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 15, SEQ ID NO: 23 и SEQ ID NO: 36.
30. Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащие:
(a) CDR1 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 12, SEQ ID NO: 20 и SEQ ID NO: 28;
(b) CDR2 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, SEQ ID NO: 13, SEQ ID NO: 21 и SEQ ID NO: 29; и
(c) CDR3 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 14, SEQ ID NO: 22 и SEQ ID NO: 30; и
(d) CDR3 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 11, SEQ ID NO: 19 и SEQ ID NO: 27.
31. Антитело по настоящему изобретению или его эпитопсвязывающий фрагмент в соответствии с любым из вариантов осуществления 1-7, содержащие:
a) CDR1 тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 20;
(b) CDR2 тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 21;
(c) CDR3 тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 22; и
(d) CDR3 легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 19.
32. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его эпитопсвязывающий фрагмент, определенные в любом из вариантов осуществления 1-31.
33. Молекула нуклеиновой кислоты в соответствии с вариантом осуществления 32, где молекула представляет собой молекулу кДНК.
34. Вектор, содержащий молекулу нуклеиновой кислоты, определенную в вариантах осуществления 32 или 33.
35. Рекомбинантная клетка-хозяин, содержащая молекулу нуклеиновой кислоты, определенную в любом из вариантов осуществления 32-34.
36. Способ получения антитела или эпитопсвязывающего фрагмента, определенных в любом из вариантов осуществления 1-31, при этом способ включает культивирование клетки-хозяина, определенной в варианте осуществления 35, в условиях, обеспечивающих возможность экспрессии кодируемого антитела или его эпитопсвязывающего фрагмента.
37. Препарат, содержащий антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих пунктов, где указанный препарат практически не содержит антител, образующихся в естественных условиях, которые либо не способны связываться с тау-белком, либо существенным образом не изменяют функциональные характеристики антитела к тау-белку в препарате, где указанные функциональные характеристики выбраны из группы, состоящей из:
(i) практически отсутствующей способности к связыванию с нефосфорилированным тау-белком;
(ii) практически отсутствующей способности к связыванию с тау-белком, фосфорилированным в S404 и не фосфорилированным в S396;
(iii) способности к связыванию с тау-белком, фосфорилированным в S396;
(iv) способности к связыванию с тау-белком, фосфорилированным как в S396, так и в S404;
(v) способности к селективному проведению различий между фосфорилированными остатками S396 и S404 тау-белка, так что они практически не способны связываться с фосфорилированным остатком 404;
(vi) способности к связыванию с гиперфосфорилированным тау-белком из головного мозга людей с болезнью Альцгеймера;
(vii) способности к проведению различий между патологическим и непатологическим человеческим тау-белком и/или
(viii) способности к специфичному уменьшению полос гиперфосфорилированного тау-белка размером 64 кДа и 70 кДа по меньшей мере на 90% без уменьшения при этом полосы тау-белка размером 55 кДа более чем на 10% % при применении согласно описанному в данном документе с иммуноистощенными экстрактами от трансгенных мышей rTg4510 или способности к специфичному уменьшению полос фосфорилированного в S396 гиперфосфорилированного тау-белка по меньшей мере на 90% без уменьшения при этом полос негиперфосфорилированного тау-белка более чем на 10% при применении согласно описанному в данном документе с посмертными экстрактами из головного мозга людей с AD.
38. Препарат, содержащий антитело или его эпитопсвязывающий фрагмент в соответствии с любым из предыдущих пунктов, где указанное антитело или его указанный эпитопсвязывающий фрагмент имеют структурное изменение в своей аминокислотной последовательности по сравнению со структурой встречающегося в природе антитела к тау-белку, где указанное структурное изменение обуславливает проявление указанным антителом или указанным фрагментом измененных функциональных характеристик по сравнению с функциональными характеристиками, проявляемыми указанным встречающимся в природе антителом к тау-белку, где указанные функциональные характеристики выбраны из группы, состоящей из:
(i) практически отсутствующей способности к связыванию с нефосфорилированным тау-белком;
(ii) практически отсутствующей способности к связыванию с тау-белком, фосфорилированным в S404 и не фосфорилированным в S396;
(iii) способности к связыванию с тау-белком, фосфорилированным в S396;
(iv) способности к связыванию с тау-белком, фосфорилированным как в S396, так и в S404;
(v) способности к селективному проведению различий между фосфорилированными остатками S396 и S404 тау-белка, так что они практически не способны связываться с фосфорилированным остатком 404;
(vi) способности к связыванию с гиперфосфорилированным тау-белком из головного мозга людей с болезнью Альцгеймера;
(vii) способности к проведению различий между патологическим и непатологическим человеческим тау-белком и/или
(viii) способности к специфичному уменьшению полос гиперфосфорилированного тау-белка размером 64 кДа и 70 кДа по меньшей мере на 90% без уменьшения при этом полосы тау-белка размером 55 кДа более чем на 10% при применении согласно описанному в данном документе с иммуноистощенными экстрактами от трансгенных мышей rTg4510 или способности к специфичному уменьшению полос фосфорилированного в S396 гиперфосфорилированного тау-белка по меньшей мере на 90% без уменьшения при этом полос негиперфосфорилированного тау-белка более чем на 10% при применении согласно описанному в данном документе с посмертными экстрактами из головного мозга людей с AD.
39. Фармацевтическая композиция, содержащая моноклональное антитело или его эпитопсвязывающий фрагмент, определенные в любом из вариантов осуществления 1-31, или препарат, определенный в любом из вариантов осуществления 37-38; и фармацевтически приемлемый носитель.
40. Моноклональное антитело или его фрагмент согласно любому из вариантов осуществления 1-31, препарат согласно любому из вариантов осуществления 37-38 или фармацевтическая композиция согласно варианту осуществления 39 для применения в медицине.
41. Моноклональное антитело или его фрагмент согласно любому из вариантов осуществления 1-31, препарат согласно любому из вариантов осуществления 37-38 или фармацевтическая композиция согласно варианту осуществления 39 для применения в лечении таупатии.
42. Моноклональное антитело или его фрагмент, препарат или фармацевтическая композиция в соответствии с вариантом осуществления 41, где таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), прогрессирующего надъядерного паралича (PSP), кортикобазальной дегенерации (CBD), психоза, в частности, психоза, обусловленного AD, или психоза у пациентов с AD, и психиатрических симптомов у пациентов с деменцией с тельцами Леви.
43. Применение моноклонального антитела или его фрагмента согласно любому из вариантов осуществления 1-31, препарата согласно любому из вариантов осуществления 37-38 или фармацевтической композиции согласно варианту осуществления 39 в производстве лекарственного препарата для лечения таупатии.
44. Применение моноклонального антитела или его фрагмента, препарата или фармацевтической композиции в соответствии с вариантом осуществления 43, где таупатия выбрана из группы, состоящей из болезни Альцгеймера, болезни аргирофильных зерен (AGD), прогрессирующего надъядерного паралича (PSP), кортикобазальной дегенерации (CBD, психоза, обусловленного AD, или психоза у пациентов с AD, и психиатрических симптомов у пациентов с деменцией с тельцами Леви.
45. Способ лечения болезни Альцгеймера или других таупатий у субъекта, при этом указанный способ включает введение моноклонального антитела или его фрагмента согласно любому из вариантов осуществления 1-31, препарата согласно любому из вариантов осуществления 37-38 или фармацевтической композиции согласно варианту осуществления 39 указанному субъекту в эффективном количестве.
46. Способ в соответствии с вариантом осуществления 45, где лечение является длительным.
47. Способ в соответствии с вариантом осуществления 46, где длительное лечение продолжают по меньшей мере в течение 2 недель, как, например, по меньшей мере в течение 1 месяца, 6 месяцев, 1 года или дольше.
48. Способ в соответствии с любым из вариантов осуществления 45-47, где субъектом является человек.
49. Набор, содержащий моноклональное антитело или его фрагмент согласно любому из вариантов осуществления 1-31, препарат согласно любому из вариантов осуществления 37-38 или фармацевтическую композицию согласно варианту осуществления 39, для применения в медицине.
50. Моноклональное антитело или его фрагмент согласно любому из вариантов осуществления 1-31, препарат согласно любому из вариантов осуществления 37-38 или фармацевтическая композиция согласно варианту осуществления 39 для применения в выявлении наличия или измерении количества указанного тау-белка в головном мозге субъекта.
51. Моноклональное антитело или его фрагмент, препарат или фармацевтическая композиция согласно варианту осуществления 50, где указанное выявление или измерение включает визуализацию in vivo указанного антитела к тау-белку, связанного с указанным тау-белком.
52. Моноклональное антитело или его фрагмент, препарат или фармацевтическая композиция согласно варианту осуществления 50, где указанное выявление или измерение включает визуализацию ex vivo указанного антитела к тау-белку или его указанного фрагмента, связанных с указанным тау-белком.
53. Моноклональное антитело или его эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33) в присутствии человеческого тау-белка, фосфорилированного по остатку 404, но не фосфорилированного по остатку 396.
54. Моноклональное антитело или его эпитопсвязывающий фрагмент, характеризующиеся иммуноспецифичным связыванием с человеческим тау-белком, содержащим фосфорилированный остаток 396, в соответствии со следующими критериями тестирования: i) антитело практически не связывается с нефосфорилированным тау-белком; ii) антитело практически не связывается с тау-белком, фосфорилированным в 404, если он при этом не фосфорилирован в 396; iii) антитело связывается с тау-белком, фосфорилированным в 396; и iv) антитело связывается с тау-белком, если как 396, так и 404 являются фосфорилированными.
55. Моноклональное антитело, выработка которого индуцируется в ответ на дифосфорилированный пептид TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающий остатки 386-410 тау-белка 2N4R, или его эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33).
56. Моноклональное антитело в соответствии с вариантом осуществления 55, где гибридомы подвергаются скринингу с использованием человеческого патологического и непатологического тау-белка для выделения клонов, которые как i) являются иммуноспецифичными в отношении любого из фосфорилированных эпитопов S396, так и ii) специфично распознают гиперфосфорилированный тау-белок из головного мозга людей с болезнью Альцгеймера, где указанные антитела способны различать патологический и непатологический человеческий тау-белок.
57. Способ удаления гиперфосфорилированного тау-белка из клубка, при этом указанный клубок содержит гиперфосфорилированный тау-белок, при этом указанный способ включает приведение гиперфосфорилированного тау-белка в контакт с антителом, при этом указанное антитело является селективным в отношении тау-белка, имеющего фосфорилированный остаток 396, таким образом, чтобы в результате получить 90% истощение клубка по гиперфосфорилированному тау-белку.
58. Способ задержки прогрессирования болезни Альцгеймера у пациента, при этом указанный способ включает уменьшение или ослабление накопления патологического тау-белка у указанного пациента, при этом указанный способ включает введение антитела, которое удаляет тау-белок с фосфорилированным остатком 396.
59. Способ задержки прогрессирования болезни Альцгеймера у пациента, при этом указанный способ включает удаление тау-белков, из которых посредством затравочного действия образуются патологические тау-белки, при котором удаляют тау-белки, имеющие фосфорилированный остаток 396.
60. Способ лечения пациента с болезнью Альцгеймера, включающий удаление гиперфосфорилированного тау-белка из клубка, при этом указанный клубок содержит гиперфосфорилированный тау-белок и нормальный тау-белок, путем приведения гиперфосфорилированного тау-белка в контакт с антителом, селективным в отношении тау-белка, имеющего фосфорилированный остаток 396.
61. Способ в соответствии с любым из вариантов осуществления 57-59, включающий применение антитела, определенного в любом из вариантов осуществления 1-31, 40-42 или 50-56.
62. Выделенное моноклональное антитело или его выделенный эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33).
63. Рекомбинантное человеческое или рекомбинантное гуманизированное моноклональное антитело или его выделенный эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33).
64. Рекомбинантное моноклональное антитело или его эпитопсвязывающий фрагмент, выработка которых индуцируется в ответ на дифосфорилированный пептид TDHGAEIVYK{p}SPVVSGDT{p}SPRHL (SEQ ID NO: 37), охватывающий остатки 386-410 тау-белка 2N4R, где указанное рекомбинантное моноклональное антитело или его эпитопсвязывающий фрагмент способны иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33).
65. Фармацевтическая композиция, содержащая выделенное моноклональное антитело или его выделенный эпитопсвязывающий фрагмент, где указанное выделенное моноклональное антитело или его выделенный эпитопсвязывающий фрагмент определены в любом из вышеприведенных вариантов осуществления.
66. Химерное моноклональное антитело или его выделенный эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33).
67. Антитело или его антигенсвязывающий фрагмент, определенные в любом из вариантов осуществления 1-31 и 51-56, которые были получены или произведены в линии клеток, такой как линия человеческих клеток, линия клеток млекопитающего, отличного от человека, линия клеток насекомого, дрожжей или бактерии.
68. Антитело или его антигенсвязывающий фрагмент в соответствии с вариантом осуществления 67, полученные в линии клеток СНО, линии клеток HEK, линии клеток BHK-21, линии мышиных клеток (такой как линия клеток миеломы), линии клеток фибросаркомы, линии клеток PER.C6, линии клеток HKB-11, линии клеток САР и линии человеческих клеток HuH-7.
ПРИМЕРЫ
Пример 1. Иммунизация мышей фосфорилированными пептидами 396/404
[00237] Мышей (C56/BL6 и FVB иммунизировали 10 мкг фосфорилированного тау-белка 386-408 (pS396/pS404) (SEQ ID NO: 37), конъюгированного с Р30, составленного в адъюванте TiterMax.
[00238] Мыши были самками и самцами из линий C56/BL6 и FVB. Мышей в возрасте 2-3 месяцев иммунизировали фосфорилированным тау-белком 386-408, конъюгированным с пептидным эпитопом Р30.
[00239] Иммуногенный фосфорилированный пептид тау-белка 386-408 (pS396/pS404), конъюгированный с Р30, составляли в TiterMax (400 мкг/мл пептида в смеси 1:1 об.:об.) согласно протоколу поставщика TiterMax, и мышам подкожно инъецировали 20 мкг антигенного пептида (100 мкл). Контрольным мышам инъецировали только адъювант. Всех мышей, иммунизированных пептидом, подвергали повторной иммунизации 0,5 мкг пептида/TiterMax (10 мкг/мл пептида, составленного согласно описанному выше и инъецированного) с месячными интервалами. В заключение мышей подвергали повторной иммунизации фосфорилированным тау-белком 386-408 (pS396/pS404), конъюгированным с Р30, без TiterMax за 3 дня до слияния спленоцитов с клетками SP-2. Гибридомы отбирали для циклов повторного клонирования после того, как они проявляли положительное связывание с планшетами для ELISA, которые были покрыты 1 мкг/мл фосфорилированного тау-белка 386-408 (pS396/pS404), и проявляли активность предпочтительного связывания с антигенами S1 и Р3 из лизата головного мозга субъектов с AD и TG4510 (как описано ниже в примере 3). Такое связывание сравнивали с активностью связывания таких антител с лизатом головного мозга от контрольных субъектов с помощью дот-блот-анализов и планшетов для ELISA или MSD, покрытых лизатом головного мозга.
Пример 2. Образование гибридом
[00240] Мышей подвергали повторной иммунизации фосфорилированным тау-белком 386-408 (pS396/pS404), конъюгированным с Р30, без TiterMax за 3 дня до слияния спленоцитов с клетками SP-2. Гибридомы отбирали для циклов повторного клонирования после положительного связывания в планшетах для ELISA, покрытых 1 мкг/мл фосфорилированного тау-белка 386-408 (pS396/pS404), и проявления активности предпочтительного связывания с антигенами S1 и Р3 из лизата головного мозга субъектов с AD и TG4510 по сравнению с лизатом головного мозга от контрольных субъектов с помощью дот-блот-анализов и планшетов для ELISA или MSD, покрытых лизатом головного мозга.
Пример 3. Вестерн-блот-анализ и дот-блот-анализ специфичных антител
Биохимическое фракционирование тау-белка
[00241] Ткани головного мозга людей или мышей rTg4510, сверхэкспрессирующие человеческий тау-белок с мутацией P301L, гомогенизировали в 10 объемах Tris-солевого буферного раствора, содержащего ингибиторы протеаз и фосфатаз, как указано далее: 50 мМ Tris/HCl (рН 7,4); 274 мМ NaCl; 5 мМ KCl; 1% смесь ингибиторов протеаз (Roche); 1% коктейль ингибиторов фосфатаз I и II (Sigma) и 1 мМ фенилметилсульфонилфторид (PMSF; Sigma). Гомогенаты центрифугировали при 27000 × g в течение 20 мин. при 4°С с получением фракций надосадочной жидкости (S1) и осадка. Образцы осадка повторно гомогенизировали в 5 объемах буфера с высокой концентрацией солей/сахарозы (0,8 М NaCl, 10% сахароза, 10 мМ Tris/HCl, [рН 7,4], 1 мМ EGTA, 1 мМ PMSF) и центрифугировали согласно описанному выше. Образцы надосадочной жидкости собирали и инкубировали с саркозилом (конечная концентрация 1%; Sigma) в течение часа при 37°С с последующим центрифугированием при 150000 × g в течение часа при 4°С с получением образцов осадка, нерастворимых в саркозиле, называемых фракцией Р3. Осадок Р3 ресуспендировали в буфере ТЕ (10 мМ Tris/HCl [рН 8,0], 1 мМ EDTA) до объема, равного половине исходного объема, используемого для гомогенатов головного мозга.
Вестерн-блот- и дот-блот-анализы
[00242] Фракционированные экстракты тканей S1 и Р3 растворяли в SDS-буфере для образца, содержащем 0,1 М DTT. Подвергнутые тепловой обработке образцы (95°С в течение 10 мин.) разделяли с помощью гель-электрофореза в 4-12% Bis-Tris-гелях для SDS-PAGE (Invitrogen) и переносили на мембраны из PVDF (BioRad Laboratories, Геркулес, Калифорния). Образцы для дот-блоттинга наносили пятнами непосредственно на нитроцеллюлозные мембраны (Amersham, Питтсбург, Пенсильвания) в известных концентрациях для всех образцов. Мембраны как для вестерн-блоттинга, так и для дот-блоттинга блокировали в 5% обезжиренном сухом молоке в TBS-Tween (0,5%) с рН 7,4 с последующим инкубированием в 1 мкг/мл D1.2 или С10-2 в течение ночи при 4°С.
Мембраны промывали и инкубировали с антителом к мышиному IgG, конъюгированным с пероксидазой (1:5000; Jackson ImmunoResearch, Вест Гроув, Пенсильвания). Связанные антитела выявляли с помощью системы усиленной хемилюминесценции (набор ECL PLUS; PerkinElmer). Количественную оценку и визуальный анализ иммунореактивности в рамках вестерн-блоттинга и дот-блоттинга выполняли с помощью подключенной к компьютеру биовизуализационной аналитической системы LAS-4000 (Fujifilm, Токио, Япония) и программного обеспечения Multi Gauge v3.1 (Fujifilm). Белковую нагрузку корректировали по объему исходных фракций, и ее можно преобразовать в исходную сырую массу ткани. Результаты показаны на фигуре 1 и фигуре 2.
Пример 4. Скрининг и отбор антител к 396/404 с применением иммобилизованного человеческого патологического материала
[00243] Образцы надосадочной жидкости культур гибридом подвергали скринингу в отношении связывания антител в планшетах Nunc, покрытых 1 мкг/мл фосфорилированного пептида тау-белка 386-408 (pS396/pS404), с применением 0,1 М карбонатного буфера с рН 9.
[00244] Положительные образцы надосадочной жидкости затем разбавляли 1:50-1:800 в PBS, 0,1% BSA и 0,1% NP40 для связывания в планшетах для ELISA или MSD, покрытых соответственно антигенами лизата головного мозга (осадком Р3, см. пример 3) от пораженных AD и здоровых контрольных субъектов (НС). Антигены лизата головного мозга разбавляли в 1500 раз в 0,1 М карбонатном буфере с рН 9 перед инкубированием/покрытием планшетов для ELISA или MSD. Лунки затем блокировали в течение 2 ч. при комнатной температуре (PBS, 3 мг/мл BSA, 0,1% NP-40), и активность связывания антител выявляли с помощью антитела к мышиному IgG, конъюгированного с HRP (DAKO) и SULFO-TAG (MSD, № продукта), согласно протоколу поставщика. Выбранные антитела (D1-2, С5-2, С8-3 и С10-2), разбавленные в PBS с 0,1% BSA, характеризовались дозозависимым эффектом и демонстрировали активность связывания с планшетами, покрытыми антигеном AD-P3, от субнаномолярной до наномолярной, и дополнительно характеризовались активностью связывания с выбранными специфическими и контрольными пептидами. Результаты показаны на фигуре 3.
Пример 5. Специфичность в отношении пептидов и аффинность связывания с ними
[00245] Антитела, характеризующиеся положительным связыванием с патологическим тау-белком, дополнительно характеризовали в отношении кажущейся аффинности (IC50) и селективности/специфичности в отношении ряда фосфорилированных пептидных эпитопов (р). Планшеты для MSD покрывали 100 нг/мл фосфорилированного тау-белка 386-408 (pS396/pS404), описанного выше. Антитела к фосфорилированному тау-белку анализировали в анализах зависимости доза-ответ для определения концентраций антител, обеспечивающих надлежащий уровень аналитического сигнала (в типичном случае 5000-20000 RU в MSD, что соответствует 0,5-2% от максимального аппаратного сигнала, или сигналы OD 1,0-1,5 при 450 нм в ELISA. Выбранные антитела инкубировали с фосфорилированным тау-белком 386-408 (pS396/404) в ступенчато изменяющихся концентрациях (0-1000 нМ) в течение 2 ч. при комнатной температуре. Реакционные смеси затем наносили на покрытые пептидом планшеты для MSD, покрытые 100 нг/мл фосфорилированного пептида тау-белка 386-408 (pS396/pS404), описанного выше, и измеряли активность связывания. Значения IC50 из анализов ингибирования соответствуют значениям кажущейся аффинности (KD), составляющим 10-100 нМ.
[00246] Специфичность и фосфоселективность. Моноклональное антитело в надлежащей концентрации инкубировали со 100 нМ дифосфорилированного (pS396/pS404), нефосфорилированного или монофосфорилированного (pS396 или pS404) фосфорилированного тау-белка 386-408 и анализировали в отношении активности связывания (в анализах ингибирования). Контрольные фосфорилированные пептиды тау-белка (фосфорилированный тау-белок 260-270 (pS262) или фосфорилированный тау-белок 240-270 (pS262)) и рекомбинантный нефосфорилированный тау-белок анализировали для сравнения. Все антитела, положительно реагирующие с антигеном AD-P3, демонстрировали сильное предпочтение к фосфорилированному пептиду тау-белка 386-408 (pS396/pS404) и монофосфорилированному пептиду фосфорилированного тау-белка 386-408 (pS396) и отсутствие активности связывания с монофосфорилированным пептидом фосфорилированного тау-белка 386-408 (pS404) и нефосфорилированным пептидом тау-белка 386-408. Фосфорилированные пептиды тау-белка 240-270 и фосфорилированный тау-белок были контрольными. Результаты показаны на фигуре 4 и фигуре 32.
Пример 6. Получение характеристик антител гистологическими методами с помощью иммуногистохимического исследования
[00247] Ткани головного мозга мышей собирали от мышей rTg4510 в возрасте 8 месяцев (сверхэкспрессирующих человеческий тау-белок P301L под контролем промотора CamKII) и нетрансгенных однопометников (отличных от Tg), фиксировали в 4% параформальдегиде и заливали в парафин. Залитые в парафин образцы лобной доли коры головного мозга человека приобретали у Tissue Solutions (Глазго, Великобритания). Ткань от доноров с диагностированной конечной стадией болезни Альцгеймера сравнивали с тканью от не пораженных деменцией контрольных доноров в соответствующей возрастной группе. Срезы толщиной 4 мкм депарафинировали и подвергали демаскированию антигена путем микроволновой обработки срезов в 10 мМ цитратном буфере, рН 6, в течение 10 минут. Эндогенные пероксидазы блокировали с помощью 1% гидропероксидазы, а затем 5% нормальной свиной сыворотки крови в PBS/1% BSA/0,3% Triton Х-100 (PBS-BT). Срезы инкубировали в течение ночи при 4°С с антителами D1.2 и С10-2, разбавленными в PBS-BT в диапазоне концентраций. Срезы промывали в PBS, 0,25% BSA, 0,1% Triton Х-100 перед инкубированием с биотинилированным вторичным свиным антителом к мышиным иммуноглобулинам (Е0464; DAKO, Глоструп, Дания) при 1:500 в течение 1 часа. После дополнительного промывания применяли набор для образования комплекса стрептавидин-биотин (Vector Laboratories, Бурлингейм, Калифорния), и иммунореактивность визуализировали с помощью диаминобензидина. Срезы подвергали контрастному окрашиванию гематоксилином. Результаты показаны на фигуре 5.
Пример 7. Селективность антител в отношении патологического тау-белка
[00248] Планшеты для MSD покрывали солюбилизированными антигенами Р3 из головного мозга субъекта с AD (разбавленными 1:1500) или головного мозга TG4510 (разбавленными 1:3000). Результаты показаны на фигуре 6.
[00249] Выявление выполняют согласно описанному в примере 4 выше.
Пример 8. Анализ затравочного действия в клетках HEK
[00250] Клетки HEK293 подвергали транзиентной трансфекции человеческим тау-белком P301L-FLAG в 6-луночных планшетах через 24 ч. после посева с последующим спустя 24 ч. инкубированием с гомогенатом головного мозга в течение 24 ч. с последующим разделением и пересевом клеток и сбором через дополнительные 24 ч. Клетки лизировали и подвергали ультразвуковой обработке в PBS, дополненном буфером с 1% Triton-X, ингибитором фосфатаз PhosSTOP и ингибитором протеаз cOmplete (Roche), и ультрацентрифугировали при 100000 × g в течение 30 мин. Осадок ресуспендировали в SDS, подвергали ультразвуковой обработке и ультрацентрифугировали в течение 30 мин. при 100000 × g. Образцы надосадочной жидкости анализировали с помощью вестерн-блоттинга. Клетки, экспрессирующие человеческий тау-белок P301L, продемонстрировали наличие нерастворимого (фракция SDS, выявление с помощью E1/FLAG) гиперфосфорилированного (выявление с помощью D1.2}pS396) тау-белка при затравке общими гомогенатами головного мозга от мышей rTg4510, трансгенных по тау-белку. Клетки, обработанные гомогенатом контрольного головного мозга мышей, продемонстрировали отсутствие агрегированного гиперфосфорилированного человеческого тау-белка. Дополнительно, общие клеточные лизаты клеток HEK293 анализировали с помощью анализа агрегации тау-белков от Cisbio. В основе этого анализа лежит флуоресценция с временным разрешением с применением одного и того же антитела в качестве как донорного (конъюгированного с Tb3+), так и акцепторного (конъюгированного с d2) антитела в FRET. Образец объемом 10 мкл смешивали со смесью антител объемом 10 мкл и инкубировали в течение 20 ч. Планшет прочитывали на планшет-ридере PHERAstar для оценки флуоресценции с временным разрешением (измеренного/интегрированного после включения возбуждающего света сигнала FRET). В анализе измеряют уровень агрегированного тау-белка как в человеческом секционном материале, так и у мышей rTg4510 и в клетках HEK с затравкой с высокой специфичностью и чувствительностью. Результаты показаны на фигуре 7 и демонстрируют, что на затравочный эффект не влияла обработка с помощью HEL, но он частично устранялся путем обработки антителами к тау-белку (С10-2 > D1.2 > hACI36-2B6-Ab1).
Пример 9. Устранение функциональной (электрофизиологической (эл.-физ.)) реакции in vivo для D1.2 и С10-2
[00251] Электрофизиологическая оценка синаптической передачи и пластичности в CA1-зоне гиппокампа у мышей rTg4510 и контрольных мышей tTA в возрасте 4,5-5,5 месяцев in vivo показала i) значительное ухудшение базальной синаптической передачи у мышей rTg4510 по сравнению с tTA и ii) значительное уменьшение парного облегчения у мышей rTg4510 по сравнению с tTA.
[00252] Все эксперименты проводили в соответствии с Директивой Совета Европейского Союза (86/609/ЕЕС) об уходе за лабораторными животными и их использовании и законодательством Дании, регулирующим проведение экспериментов на животных.
[00253] В настоящем исследовании использовали самцов мышей rTg4510 и tTA (Taconic Europe A/S) в возрасте 5-5,5 месяцев на момент регистрации данных. Мышей содержали в группах в условиях контролируемой температуры (22±1,5°С) и влажности (55-65%) и выдерживали при цикле чередования света и темноты 12:12 часов (свет включали в 06:00 ч.). Пища и вода были доступны без ограничений.
[00254] Животные получали анестезию путем внутрибрюшинной (i.p.) инъекции уретана (1,2 г/кг). Мышей затем фиксировали в стереотаксической рамке, их температуру доводили до 37,5°С с помощью электрической грелки, и обнажали череп. На лобную кость помещали платиновую проволоку, выступающую в качестве эталонного электрода, и просверливали дополнительное отверстие для введения регистрирующего и стимулирующего электродов в гиппокамп по следующим координатам согласно атласу Paxinos и Franklin (Paxinos and Franklin, 2001): регистрирующий - на 1,5-1,7 мм кзади от брегмы, на 1,0-1,2 мм латеральнее от срединной линии, на 1,4-1,7 мм ниже поверхности головного мозга; стимулирующий - на 1,8-2,0 мм кзади от брегмы, на 1,5-1,7 мм латеральнее от срединной линии, на 1,3-1,7 мм ниже поверхности головного мозга. Животных оставляли в стереотаксической рамке в течение всего периода регистрации данных, и их уровень анестезии регулярно проверяли.
[00255] Полевые потенциалы (fEPSP) вызывали в СА1 путем электростимуляции коллатерали Шеффера каждые 30 с, и глубину расположения регистрирующего электрода корректировали до тех пор, пока в ответ на монополярный прямоугольный импульс не был зарегистрирован отрицательный fEPSP. Измеренный градиент вызванного fEPSP в типичном случае составлял от 30 до 70% максимальной амплитуды fEPSP.
[00256] После индукции оптимального fEPSP оценивали базальную синаптическую передачу по зависимости между интенсивностью стимуляции и градиентом вызванного fEPSP (зависимости «вход-выход»). Различные значения интенсивности стимуляции составляли 0, 25, 50, 75, 100, 150, 200, 300, 400 и 500 мкА, и их последовательно прилагали в порядке возрастания с 2-3 повторами для каждого значения интенсивности. Было обнаружено, что базальная синаптическая передача значительно ухудшалась у мышей rTg4510 по сравнению с tTA.
[00257] Парное облегчение - краткосрочную синаптическую пластичность, в основе которой, как полагают, лежат пресинаптические механизмы - дополнительно измеряли у мышей rTg4510 и tTA. Вкратце, в отношении коллатерали Шеффера прилагали пару стимулов с интервалом между стимулами (ISI), варьирующимся от 25 до 1000 мс, и градиент второго fEPSP сравнивали с градиентом первого fEPSP. Облегчение наблюдали при всех ISI с максимальным облегчением при ISI, составляющем 50 и 75 мс. Интересно, что у мышей rTg4510 наблюдали значительно более низкое PPF по сравнению с мышами tTA.
[00258] Идентифицированные ухудшения базальной синаптической передачи и парного облегчения у мышей rTg4510 дополнительно использовали в качестве регистрируемых показаний для тестирования эффективности антител.
[00259] Регистрацию данных выполняли во всех экспериментах через 2-4 дня после введения 4 доз антитела дважды в неделю в течение 2 недель i.p. У каждого животного в гиппокампе при наличии возможности регистрировали как базальную синаптическую передачу, так и парное облегчение, которые дополнительно использовали в отдельных экспериментах. Результаты показаны на фигуре 8 и демонстрируют устранение антителами дефицитов парного облегчения и базальной синаптической передачи в вызванных полевых потенциалах в СА1.
Пример 10. Иммуноистощение экстрактов из головного мозга rTg4510 по тау-белку
[00260] 60 мкг мышиного и гуманизированного антитела С10-2 иммобилизовали на магнитных гранулах Dynabeads в 300 мкл суспензии (набор для иммунопреципитации Dynabeads с белком G Novex, № по кат. 10007D). После тщательного промывания гранулы смешивали с 60 мкл экстракта из головного мозга rTg4510 и инкубировали при комнатной температуре в течение 10 минут. Магнитные гранулы отделяли от экстракта, и экстракты анализировали с помощью вестерн-блоттинга. При истощении с помощью mC10-2 и hC10-2 было удалено соответственно 99% и 99,5% агрегатов тау-белка. Результаты показаны на фигуре 12.
Пример 11. Анализ затравочного действия в клетках HEK с применением иммуноистощенных экстрактов
[00261] Клетки HEK293 транзиентно трансфицировали человеческим тау-белком P301L-FLAG в 6-луночных планшетах. Спустя 24 ч. клетки инкубировали с гомогенатом головного мозга, который был подвергнут иммуноистощению с помощью гуманизированного или мышиного С10-2. Через 24 ч. клетки пересевали и собирали через дополнительные 24 ч. Клетки лизировали и подвергали ультразвуковой обработке в TBS, дополненном 1% Triton-X, ингибиторами фосфатаз и протеаз (Roche), и ультрацентрифугировали при 100000 × g в течение 30 мин. Осадок ресуспендировали в 1% SDS, подвергали ультразвуковой обработке и ультрацентрифугировали в течение 30 мин. при 100000 × g. Образцы надосадочной жидкости анализировали с помощью вестерн-блоттинга. Клетки, экспрессирующие человеческий тау-белок P301L, продемонстрировали наличие нерастворимого (фракция SDS, выявление с помощью E1/FLAG) гиперфосфорилированного тау-белка (D1.2/тау-белок pS396, прогон при более высокой молекулярной массе) при затравке общими гомогенатами головного мозга от мышей rTg4510, трансгенных по тау-белку. Клетки, обработанные гомогенатом контрольного головного мозга мышей tTA, продемонстрировали отсутствие агрегированного гиперфосфорилированного человеческого тау-белка. Дополнительно, общие клеточные лизаты клеток HEK293 анализировали с помощью анализа агрегации тау-белков от Cisbio. Истощение с помощью антител HEL и hHEL не влияло на затравочное действие, тогда как истощение с помощью mC10-2 и hC10-2 предупреждало агрегацию тау-белка на 88% и 96%, а нерастворимого тау-белка на 97% и 100% соответственно. Результаты показаны на фигуре 13.
Пример 11. Иммуноистощение экстрактов из головного мозга rTg4510 по тау-белку
[00262] 100 мкг мышиных антител С10-2, D1.2 и Tau5 (Invitrogen) иммобилизовали на магнитных гранулах Dynabeads в 500 мкл суспензии (набор для иммунопреципитации Dynabeads с белком G Novex, № по кат.10007D). После тщательного промывания гранулы смешивали со 100 мкл экстракта из головных мозгов rTg4510 и инкубировали при комнатной температуре в течение 10 минут. Магнитные гранулы отделяли от экстракта, и экстракты анализировали с помощью вестерн-блоттинга. С10-2 и D1.2 не удаляют нормальный тау-белок из гомогенатов, как это делает коммерчески доступное антитело Tau5. В отличие от этого, два антитела по настоящему изобретению специфично удаляют гиперфосфорилированный тау-белок (64 кДа), то есть тау-белок, фосфорилированный по серину-396, на 95%. Результаты показаны на фигуре 14.
Пример 12. Иммуноистощение экстрактов из головного мозга субъектов с болезнью Альцгеймера по тау-белку
[00263] 100 мкг мышиных антител С10-2 и D1 иммобилизовали на магнитных гранулах Dynabeads в 500 мкл суспензии (набор для иммунопреципитации Dynabeads с белком G Novex, № по кат. 10007D). После тщательного промывания гранулы смешивали со 100 мкл экстракта из головного мозга субъектов с болезнью Альцгеймера и инкубировали при комнатной температуре в течение 10 минут. Магнитные гранулы отделяли от экстракта, и экстракты анализировали с помощью вестерн-блоттинга. D1.2 и С10-2 удаляют лишь очень небольшую долю общего тау-белка в гомогенате головного мозга (8%). Антитела, тем не менее, специфично удаляют гиперфосфорилированный тау-белок (90%), специфичный для пациентов с AD. Результаты показаны на фигуре 15.
Пример 13. Затравочное действие у мышей rTg4510 с применением иммуноистощенных экстрактов
[00264] Использовали трансгенных мышей, экспрессирующих человеческий мутантный тау-белок (P301L 0N4R) под контролем элемента ответа Tet-Off в CamK2-положительных нейронах (rTg4510). В этой модели патология тау-белка обычно начинает развиваться в возрасте 3 месяцев, но при скармливании матерям доксициклина во время беременности и в течение первых 3 недель жизни детеныша патология развивается на более поздней стадии (начиная с возраста 6 месяцев). Используемые в данных исследованиях мыши, предварительно обработанные доксициклином, имели возраст 2,5 месяца в момент времени, соответствующий инъекции. Мыши получали анестезию, будучи зафиксированными в стереотаксической рамке, путем вдыхания изофлурана. Череп обнажали, и его положение выравнивали, пока брегма и лямбда не расположились на одном уровне. В черепе просверливали отверстие на 2 мм латеральнее (справа) и на 2,4 мм кзади от брегмы. Шприц со скошенным наконечником на 10 мкл (SGE) применяли для инъекции затравочного материала на 1,4 мм вентральнее поверхности головного мозга по вышеуказанным координатам. 2 мкл иммуноистощенных экстрактов, описанных в примерах 11 и 12, медленно инфундировали в необходимое место (1 мкл/минута), и шприц оставляли на 5 минут перед его извлечением. Рану закрывали швами, и мышей согревали, пока они пробуждались. Мышей содержали в течение 3 месяцев, а затем умерщвляли и подвергали перфузионной фиксации с помощью 4% PFA.
[00265] Иммуногистохимическое исследование. Фиксированные головной мозг разрезали на коронарные срезы толщиной 35 мкм в NSA, и каждый 6-й срез окрашивали для выявления клубков тау-белка (серебряный краситель по Gallyas) и для выявления гиперфосфорилированного тау-белка (АТ8). Положительно окрашенные нейроны (сомы) подсчитывали на ипси- и контралатеральной сторонах гиппокампа во всех головных мозгах. Были включены все подобласти гиппокампа. Подсчет проводили в восьми срезах на головной мозг. Результаты отражают суммарное количество положительных нейронов из 8 срезов. У 2 мышей, не подвергнутых инъекции, определяли фоновый сигнал. При удалении гиперфосфорилированного тау-белка из гомогенатов гомогенаты больше не индуцируют затравочное действие с формированием патологии тау-белка. Результаты показаны на фигуре 16. Показана количественная оценка патологии тау-белка в головном мозге rTg4510 с затравкой гомогенатами головного мозга rTg4510 (А) или с AD (В). Перед введением затравки уровень гиперфосфорилированного тау-белка, но не нормального тау-белка, был уменьшен в гомогенатах на 90-95% путем применения С10-2 или D1.2. При удалении гиперфосфорилированного тау-белка из гомогенатов гомогенаты больше не индуцируют затравочное действие с формированием патологии тау-белка.
[00266] Гомогенаты из головного мозга rTg4510 или субъектов с болезнью Альцгеймера могут индуцировать затравочное действие с формированием патологии тау-белка у мышей rTg4510 на стадии, на которой эндогенная патология тау-белка не развивалась. При удалении гиперфосфорилированного тау-белка из гомогенатов путем применения D1.2 или С10-2, как описано в примерах 11 и 12, активность затравочного действия устраняется.
Пример 14. Обработка антителами у мышей rTg4510 с затравкой
[00267] Мышей rTg4510, обработанных доксициклином (как описано в примере 13), подвергали длительной обработке мышиным антителом D1.2 или контрольным антителом при 15 мг/кг/неделю, начиная с возраста 2 месяцев. В возрасте 2,5 месяцев экстракт из головного мозга rTg4510 инфундировали в гиппокамп (как описано в примере 13). Мышей умерщвляли через 1, 2 и 3 месяца после инфузии в головной мозг, и иммуногистохимическое исследование и последующий анализ выполняли согласно описанному в примере 13. При обработке с помощью D1.2 патология тау-белка значительно уменьшалась через 2 и 3 месяца после начала затравочного действия. Результаты показаны на фигуре 17.
[00268] Показана количественная оценка нейронов, содержащих клубки, в гиппокампах мышей rTg4510 с затравкой. Патология увеличивается с течением времени, и при обработке мышей с помощью D1.2 патология значительно уменьшается через 2 и 3 месяца после введения затравки. Показана количественная оценка нейронов, содержащих клубки, в гиппокампах мышей rTg4510 с затравкой. Патология увеличивается с течением времени, и при обработке мышей с помощью D1.2 патология значительно уменьшается через 2 и 3 месяца после введения затравки.
[00269] Гомогенаты из головного мозга rTg4510 или субъектов с болезнью Альцгеймера могут индуцировать затравочное действие с формированием патологии тау-белка у мышей rTg4510 на стадии, на которой эндогенная патология тау-белка не развивалась. Путем системной обработки мышей с помощью D1.2 можно значительно уменьшить развитие патологии тау-белка.
Пример 15. Иммуноистощение экстрактов из головного мозга субъектов с болезнью Альцгеймера по тау-белку с помощью гуманизированных антител к тау-белку
[00270] 100 мкг мышиного и гуманизированного С10-2, а также антител 2.10.3 и HJ8.5 из предшествующего уровня техники (источников) иммобилизовали на магнитных гранулах Dynabeads в 500 мкл суспензии (набор для иммунопреципитации Dynabeads с белком G Novex, № по кат. 10007D). После тщательного промывания гранулы смешивали со 100 мкл экстракта из головного мозга субъектов с болезнью Альцгеймера и инкубировали при комнатной температуре в течение 10 минут. Магнитные гранулы отделяли от экстракта, и экстракты анализировали с помощью вестерн-блоттинга и анализа Cisbio. Мышиное и гуманизированное С10-2 эффективно удаляли 93% и 97% фосфорилированного тау-белка pS396, но только 22% и 17% общего тау-белка по сравнению с контрольным антителом hHEL. Антитело 2.10.3 удаляло 91% тау-белка, фосфорилированного по серину-396, и 10% общего тау-белка. По всей видимости, 2.10.3 является менее эффективным в удалении одной из полос гиперфосфорилированного белка по сравнению с антителами С10-2 (средняя полоса, соответствующая 64 кДа). Антитело HJ8.5 имеет совершенно иной профиль по сравнению с антителами как С10-2, так и 2.10.3, удаляя большую часть тау-белка, 89% общего тау-белка и 88% тау-белка pS396. Результаты показаны на фигурах 23-24.
Пример 16. Иммуноистощение экстрактов из головного мозга субъектов с болезнью Альцгеймера по агрегированному тау-белку с помощью гуманизированных антител к тау-белку
[00271] Иммуноистощенные экстракты от субъектов с AD, описанные в примере 13, также анализировали путем применения анализа агрегации тау-белков, описанного в примере 10. Антитела С10-2 и HJ8.5 более эффективно удаляют агрегированный тау-белок из материала от субъектов с AD, чем антитело 2.10.3. В порядке убывания эффективности: HJ8.5 (99%), hC10-2 (98%), mC10-2 (96%) и 2.10.3 (90%), во всех случаях по сравнению с антителом hHEL. Результаты показаны на фигуре 25.
Пример 17. Иммуноистощение по тау-белку
[00272] 25 мкг антитела (гуманизированного С10-2 или 2.10.3) иммобилизовали на магнитных гранулах Dynabeads в 125 мкл суспензии (набор для иммунопреципитации Dynabeads с белком G Novex, № по кат. 10007D). После тщательного промывания покрытые гранулы смешивали с различными количествами непокрытых промытых гранул. Начинали со 100% гранул, покрытых Ab, что соответствовало 5 мкг антитела, со снижением до 100% непокрытых гранул. Общее количество гранул было одинаковым во всех образцах. Гранулы смешивали с 20 мкл экстракта от субъектов с AD и инкубировали при комнатной температуре в течение 10 минут. Магнитные гранулы отделяли от экстракта, и экстракты разделяли на аликвоты, подвергали мгновенной заморозке и выдерживали при -80 С до применения.
Анализ истощения с помощью вестерн-блоттинга
[00273] Образцы кипятили в 1х загрузочном LDS-буфере и 100 мМ DTT. Объем, соответствующий 3 мкл экстрактов, загружали в 4-12% Bis-Tris-гель NuPAGE (Life Tech, Novex). После электрофореза белки блотировали на мембрану Immobilon-FL из PVDF (0,45 мкм, IPFL10100, Millipore). Мембрану блокировали с помощью блокирующего буфера SEA (№ продукта 37527, Thermo). Уровни тау-белка и pTau оценивали в образцах с помощью Tau5 (ab80579, Abcam, 1:2000), мышиного С10-2 (1 мкг/мл), P-S199/202 (44768G, Invitrogen, 1:1000), P-S422 (ab79415, Abcam, 1:750), человеческого IPN (1 мкг/мл). GAPDH и актин применяли в качестве контролей нагрузки (ab9484, Abcam, 1:2000, А5441, Sigma, 1:20000). Применяли вторичные антитела IgG, конъюгированные с флуорофором (козье антитело к человеческим иммуноглобулинам, конъюгированное с IRDye 800CW, козье антитело к кроличьим иммуноглобулинам, конъюгированное с IRDye 800CW, козье антитело к мышиным иммуноглобулинам, конъюгированное с IRDye 680, LI-COR Biosciences), и сигнал оценивали количественно с помощью Odyssey CLx и программного обеспечения Image Studio (LI-COR Biosciences).
Проводили количественную оценку отдельных полос, а также сигнала в целых дорожках, и на ее основании строили сигмоидальные кривые зависимости доза-ответ и при наличии возможности оценивали максимальный эффект и значения ЕС50.
Результаты
[00274] Оба антитела удаляли небольшую долю тау-белка из препарата головного мозга субъекта с болезнью Альцгеймера. 2.10.3, сконструированное таким образом, что оно обладает специфичностью к тау-белку P-S422, удаляет до 24% от общего количества тау-белка, а С10-2 удаляет до 15% общего тау-белка (см. фигуру 26).
[00275] Как 2.10.3, так и С10-2 удаляют более 90% тау-белка, фосфорилированного по серину-422, хотя количество антитела, необходимое для удаления 50% тау-белка Р-S422, различается: в случае с 2.10.3 необходимо 0,42 мкг антитела, а в случае с С10-2 для достижения того же самого эффекта необходимо 0,27 мкг (см. фигуру 27).
[00276] С10-2 эффективно удаляет тау-белок, фосфорилированный по серину-396 (максимальный эффект: 88%, и половина этого эффекта достигается при применении 0,30 мкг антитела). 2.10.3 удаляет меньшую долю тау-белка, фосфорилированного по серину-396 (максимальный эффект: 60%, и половина этого эффекта достигается при применении 0,63 мкг антитела) (см. фигуру 28). Это указывает на то, что весь тау-белок, фосфорилированный по серину-422, также является фосфорилированным по серину-396, но существует часть гиперфосфорилированного тау-белка, фосфорилированного по серину-396, в которой отсутствует фосфорилированный серин в положении 422.
[00277] Значительная часть тау-белка, удаляемая С10-2, также является фосфорилированной по серину-199/202, поскольку 69% тау-белка, имеющего такое фосфорилирование, подвергается влиянию иммуноистощения (50% эффект при применении 0,34 мкг антитела) (см. фигуру 29). Иммуноистощение с помощью 2.10.3 не дает сигмоидальную кривую зависимости доза-ответ для тау-белка P-S199/202, хотя при повышении количества антитела наблюдается снижение интенсивности сигнала (максимальное уменьшение 52% при применении максимального количества антитела (5 мкг) (см. фигуру 29).
[00278] Эти данные указывают на то, что антитело С10-2, нацеливающееся на фосфорилированный серин-396, связывается с более крупным пулом гиперфосфорилированных тау-белков, чем антитело 2.10.3, нацеливающееся на фосфорилированный серин в положении 422.
Пример 18. Антителоопосредованное ингибирование специфичного захвата mС10-2 патологических антигенных тау-белков в лизатах головного мозга от субъектов с AD
Материалы и способы
[00279] Материал
Покрывающий буфер: карбонатный буфер, рН 8,5, 150 мМ NaCl. Блокирующий буфер: 3% BSA (фракция V), 0,1% NP40 в PBS, рН 7,4. Промывочный буфер: 0,1% BSA (фракция V), 0,1% NP40 в PBS, рН 7,4. Гуманизированное общее козье антитело к тау-белку, меченное SULFO-TAG (D221LA-1 MSD, 50 мкг/мл).
[00280] Способ направлен на измерение захвата патологических человеческих антигенных тау-белков из головного мозга субъектов с AD с применением планшетов, покрытых С10-2 (стадия А), после инкубирования антигенных тау-белков с pS396-специфичными антителами в повышающихся концентрациях (стадия В). Захват антигенного тау-белка и антителоопосредованное ингибирование выявляли с помощью антител к человеческому (общему) тау-белку от MSD, меченных SULFO-TAG.
[00281] А: планшеты для MSD покрывали (в течение ночи при 4°С) 0,5 мкг/мл mC10-2 (захватывающего антитела) в покрывающем буфере и затем блокировали в течение 1 часа при комнатной температуре) и промывали 3 раза (фигура 30).
[00282] В: образцы лизата Р3 (1:1000=2-4 мкг/мл общего белка) и/или S1(p) (1:300=20-40 нг/мл общего белка) из материала от субъектов с AD (объединенного от 3 пациентов) смешивали с антителом, специфичным к пептидному эпитопу pS396, в ступенчато изменяющихся концентрациях и инкубировали в течение 1 часа при комнатной температуре. Реакционные смеси затем инкубировали в течение 2 часов в планшетах, подготовленных на стадии А (фигура 31).
[00283] С: тау-белок, захваченный С10-2, выявляли с помощью антитела к человеческому тау-белку, меченного SULFO-TAG. Антитело к тау-белку (1:50) от MSD, соответствующее инструкции производителя. Планшеты анализируют на SECTOR® S 600 от MSD. Р3 из материала от субъектов с AD и S1(p) из материала от субъектов с AD тестируют согласно аналогичной схеме (фигура 33/34).


--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> H. Lundbeck A/S
<120> Антитела к тау-белку (396)
<130> 0995-WO-PCT
<160> 37
<170> PatentIn версия 3.5
<210> 1
<211> 16
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 легкой цепи D1.2
<400> 1
Arg Ser Ser Gln Ser Leu Val His Ser Asn Gly Asn Thr Tyr Leu His
1 5 10 15
<210> 2
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 легкой цепи D1.2
<400> 2
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 3
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 легкой цепи D1.2
<400> 3
Ser Gln Ser Thr His Val Pro
1 5
<210> 4
<211> 13
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 тяжелой цепи D1.2
<400> 4
Lys Ala Ser Gly Asn Thr Phe Thr Asp Tyr Glu Ile His
1 5 10
<210> 5
<211> 17
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 тяжелой цепи D1.2
<400> 5
Ala Ile Asp Pro Glu Thr Gly Asn Thr Ala Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 6
<211> 6
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 тяжелой цепи D1.2
<400> 6
Ser Arg Gly Phe Asp Tyr
1 5
<210> 7
<211> 219
<212> БЕЛОК
<213> Искусственная
<220>
<223> Легкая цепь D1.2
<400> 7
Asp Val Met Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp His Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Phe Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 8
<211> 451
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь D1.2
<400> 8
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Asn Thr Phe Thr Asp Tyr
20 25 30
Glu Ile His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asn Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Arg Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser Arg Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
100 105 110
Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro
115 120 125
Gly Cys Gly Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val
130 135 140
Lys Gly Tyr Phe Pro Glu Ser Val Thr Val Thr Trp Asn Ser Gly Ser
145 150 155 160
Leu Ser Ser Ser Val His Thr Phe Pro Ala Leu Leu Gln Ser Gly Leu
165 170 175
Tyr Thr Met Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser
180 185 190
Gln Thr Val Thr Cys Ser Val Ala His Pro Ala Ser Ser Thr Thr Val
195 200 205
Asp Lys Lys Leu Glu Pro Ser Gly Pro Ile Ser Thr Ile Asn Pro Cys
210 215 220
Pro Pro Cys Lys Glu Cys His Lys Cys Pro Ala Pro Asn Leu Glu Gly
225 230 235 240
Gly Pro Ser Val Phe Ile Phe Pro Pro Asn Ile Lys Asp Val Leu Met
245 250 255
Ile Ser Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Val Ser Glu
260 265 270
Asp Asp Pro Asp Val Arg Ile Ser Trp Phe Val Asn Asn Val Glu Val
275 280 285
His Thr Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Ile
290 295 300
Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly
305 310 315 320
Lys Glu Phe Lys Cys Lys Val Asn Asn Lys Asp Leu Pro Ser Pro Ile
325 330 335
Glu Arg Thr Ile Ser Lys Ile Lys Gly Leu Val Arg Ala Pro Gln Val
340 345 350
Tyr Ile Leu Pro Pro Pro Ala Glu Gln Leu Ser Arg Lys Asp Val Ser
355 360 365
Leu Thr Cys Leu Val Val Gly Phe Asn Pro Gly Asp Ile Ser Val Glu
370 375 380
Trp Thr Ser Asn Gly His Thr Glu Glu Asn Tyr Lys Asp Thr Ala Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Tyr Phe Ile Tyr Ser Lys Leu Asp Ile
405 410 415
Lys Thr Ser Lys Trp Glu Lys Thr Asp Ser Phe Ser Cys Asn Val Arg
420 425 430
His Glu Gly Leu Lys Asn Tyr Tyr Leu Lys Lys Thr Ile Ser Arg Ser
435 440 445
Pro Gly Lys
450
<210> 9
<211> 11
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 легкой цепи C10.2
<400> 9
Gln Ala Ser Gln Gly Thr Ser Ile Asn Leu Asn
1 5 10
<210> 10
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 легкой цепи C10.2
<400> 10
Gly Ala Ser Asn Leu Glu Asp
1 5
<210> 11
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 легкой цепи C10.2
<400> 11
Leu Gln His Thr Tyr Leu Pro
1 5
<210> 12
<211> 13
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 тяжелой цепи C10.2
<400> 12
Lys Ala Ser Gly Tyr Thr Phe Thr Asp Arg Thr Ile His
1 5 10
<210> 13
<211> 17
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 тяжелой цепи C10.2
<400> 13
Tyr Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Asn Glu Asn Phe Lys
1 5 10 15
Gly
<210> 14
<211> 6
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 тяжелой цепи C10.2
<400> 14
Arg Gly Ala Met Asp Tyr
1 5
<210> 15
<211> 214
<212> БЕЛОК
<213> Искусственная
<220>
<223> Легкая цепь C10.2
<400> 15
Asp Val Gln Met Ile Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Ile Val Thr Met Thr Cys Gln Ala Ser Gln Gly Thr Ser Ile Asn
20 25 30
Leu Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Glu Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Asp
65 70 75 80
Glu Asp Met Ala Thr Tyr Phe Cys Leu Gln His Thr Tyr Leu Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Ala Asp Ala Ala
100 105 110
Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly
115 120 125
Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu
145 150 155 160
Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser
165 170 175
Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr
180 185 190
Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser
195 200 205
Phe Asn Arg Asn Glu Cys
210
<210> 16
<211> 439
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь C10.2
<400> 16
Gln Val Gln Leu Gln Gln Ser Asp Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Arg
20 25 30
Thr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Asn Glu Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
100 105 110
Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro
115 120 125
Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val
130 135 140
Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser
145 150 155 160
Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu
165 170 175
Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser
180 185 190
Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val
195 200 205
Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys
210 215 220
Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
225 230 235 240
Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val
245 250 255
Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp
260 265 270
Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe
275 280 285
Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp
290 295 300
Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe
305 310 315 320
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys
325 330 335
Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys
340 345 350
Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp
355 360 365
Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys
370 375 380
Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser
385 390 395 400
Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr
405 410 415
Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser
420 425 430
Leu Ser His Ser Pro Gly Lys
435
<210> 17
<211> 11
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 легкой цепи C5.2
<400> 17
Gln Ala Ser Gln Asp Thr Ser Ile Asn Leu Asn
1 5 10
<210> 18
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 легкой цепи C5.2
<400> 18
Gly Ala Ser Asn Leu Glu Asp
1 5
<210> 19
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 легкой цепи C5.2
<400> 19
Leu Gln His Thr Tyr Leu Pro
1 5
<210> 20
<211> 13
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 тяжелой цепи C5.2
<400> 20
Lys Ala Ser Gly Tyr Thr Phe Thr Asp Arg Thr Ile His
1 5 10
<210> 21
<211> 17
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 тяжелой цепи C5.2
<400> 21
Tyr Ile Tyr Pro Gly Asp Asp Ser Thr Lys Tyr Asn Asp Asn Phe Lys
1 5 10 15
Gly
<210> 22
<211> 6
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 тяжелой цепи C5.2
<400> 22
Arg Gly Thr Met Asp Tyr
1 5
<210> 23
<211> 214
<212> БЕЛОК
<213> Искусственная
<220>
<223> Легкая цепь C5.2
<400> 23
Asp Val Gln Met Ile Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Ile Val Thr Met Thr Cys Gln Ala Ser Gln Asp Thr Ser Ile Asn
20 25 30
Leu Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Glu Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Asp
65 70 75 80
Glu Asp Met Ala Thr Tyr Phe Cys Leu Gln His Thr Tyr Leu Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Ala Asp Ala Ala
100 105 110
Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly
115 120 125
Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu
145 150 155 160
Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser
165 170 175
Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr
180 185 190
Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser
195 200 205
Phe Asn Arg Asn Glu Cys
210
<210> 24
<211> 439
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь C5.2
<400> 24
Gln Val Gln Leu Gln Gln Ser Asp Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Arg
20 25 30
Thr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Pro Gly Asp Asp Ser Thr Lys Tyr Asn Asp Met Phe
50 55 60
Lys Ala Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Asp Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Thr Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
100 105 110
Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro
115 120 125
Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val
130 135 140
Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser
145 150 155 160
Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu
165 170 175
Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser
180 185 190
Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val
195 200 205
Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys
210 215 220
Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
225 230 235 240
Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val
245 250 255
Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp
260 265 270
Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe
275 280 285
Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp
290 295 300
Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe
305 310 315 320
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys
325 330 335
Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys
340 345 350
Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp
355 360 365
Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys
370 375 380
Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser
385 390 395 400
Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr
405 410 415
Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser
420 425 430
Leu Ser His Ser Pro Gly Lys
435
<210> 25
<211> 11
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 легкой цепи C8.3
<400> 25
Gln Ala Ser Gln Gly Thr Ser Ile Asn Leu Asn
1 5 10
<210> 26
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 легкой цепи C8.3
<400> 26
Gly Ser Ser Asn Leu Glu Asp
1 5
<210> 27
<211> 7
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 легкой цепи C8.3
<400> 27
Leu Gln His Ser Tyr Leu Pro
1 5
<210> 28
<211> 13
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 1 тяжелой цепи C8.3
<400> 28
Lys Ala Ser Gly Tyr Thr Phe Thr Asp Arg Thr Ile His
1 5 10
<210> 29
<211> 17
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 2 тяжелой цепи C8.3
<400> 29
Tyr Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Asn Glu Asn Phe Lys
1 5 10 15
Gly
<210> 30
<211> 6
<212> БЕЛОК
<213> Искусственная
<220>
<223> CDR 3 тяжелой цепи C8.3
<400> 30
Arg Gly Ala Met Asp Tyr
1 5
<210> 31
<211> 214
<212> БЕЛОК
<213> Искусственная
<220>
<223> Легкая цепь C8.3
<400> 31
Asp Val Gln Met Ile Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Ile Val Thr Met Thr Cys Gln Ala Ser Gln Gly Thr Ser Ile Asn
20 25 30
Leu Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ser Ser Asn Leu Glu Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Asp
65 70 75 80
Glu Asp Met Ala Thr Tyr Phe Cys Leu Gln His Ser Tyr Leu Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Ala Asp Ala Ala
100 105 110
Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly
115 120 125
Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu
145 150 155 160
Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser
165 170 175
Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr
180 185 190
Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser
195 200 205
Phe Asn Arg Asn Glu Cys
210
<210> 32
<211> 439
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь C8.3
<400> 32
Gln Val Gln Leu Gln Gln Ser Asp Ala Glu Leu Val Asn Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Arg
20 25 30
Thr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Asn Glu Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
100 105 110
Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro
115 120 125
Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val
130 135 140
Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser
145 150 155 160
Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu
165 170 175
Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser
180 185 190
Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val
195 200 205
Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys
210 215 220
Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
225 230 235 240
Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val
245 250 255
Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp
260 265 270
Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe
275 280 285
Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp
290 295 300
Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe
305 310 315 320
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys
325 330 335
Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys
340 345 350
Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp
355 360 365
Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys
370 375 380
Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser
385 390 395 400
Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr
405 410 415
Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser
420 425 430
Leu Ser His Ser Pro Gly Lys
435
<210> 33
<211> 441
<212> БЕЛОК
<213> Искусственная
<220>
<223> Человеческий тау-белок, № 33
<400> 33
Met Ala Glu Pro Arg Gln Glu Phe Glu Val Met Glu Asp His Ala Gly
1 5 10 15
Thr Tyr Gly Leu Gly Asp Arg Lys Asp Gln Gly Gly Tyr Thr Met His
20 25 30
Gln Asp Gln Glu Gly Asp Thr Asp Ala Gly Leu Lys Glu Ser Pro Leu
35 40 45
Gln Thr Pro Thr Glu Asp Gly Ser Glu Glu Pro Gly Ser Glu Thr Ser
50 55 60
Asp Ala Lys Ser Thr Pro Thr Ala Glu Asp Val Thr Ala Pro Leu Val
65 70 75 80
Asp Glu Gly Ala Pro Gly Lys Gln Ala Ala Ala Gln Pro His Thr Glu
85 90 95
Ile Pro Glu Gly Thr Thr Ala Glu Glu Ala Gly Ile Gly Asp Thr Pro
100 105 110
Ser Leu Glu Asp Glu Ala Ala Gly His Val Thr Gln Ala Arg Met Val
115 120 125
Ser Lys Ser Lys Asp Gly Thr Gly Ser Asp Asp Lys Lys Ala Lys Gly
130 135 140
Ala Asp Gly Lys Thr Lys Ile Ala Thr Pro Arg Gly Ala Ala Pro Pro
145 150 155 160
Gly Gln Lys Gly Gln Ala Asn Ala Thr Arg Ile Pro Ala Lys Thr Pro
165 170 175
Pro Ala Pro Lys Thr Pro Pro Ser Ser Gly Glu Pro Pro Lys Ser Gly
180 185 190
Asp Arg Ser Gly Tyr Ser Ser Pro Gly Ser Pro Gly Thr Pro Gly Ser
195 200 205
Arg Ser Arg Thr Pro Ser Leu Pro Thr Pro Pro Thr Arg Glu Pro Lys
210 215 220
Lys Val Ala Val Val Arg Thr Pro Pro Lys Ser Pro Ser Ser Ala Lys
225 230 235 240
Ser Arg Leu Gln Thr Ala Pro Val Pro Met Pro Asp Leu Lys Asn Val
245 250 255
Lys Ser Lys Ile Gly Ser Thr Glu Asn Leu Lys His Gln Pro Gly Gly
260 265 270
Gly Lys Val Gln Ile Ile Asn Lys Lys Leu Asp Leu Ser Asn Val Gln
275 280 285
Ser Lys Cys Gly Ser Lys Asp Asn Ile Lys His Val Pro Gly Gly Gly
290 295 300
Ser Val Gln Ile Val Tyr Lys Pro Val Asp Leu Ser Lys Val Thr Ser
305 310 315 320
Lys Cys Gly Ser Leu Gly Asn Ile His His Lys Pro Gly Gly Gly Gln
325 330 335
Val Glu Val Lys Ser Glu Lys Leu Asp Phe Lys Asp Arg Val Gln Ser
340 345 350
Lys Ile Gly Ser Leu Asp Asn Ile Thr His Val Pro Gly Gly Gly Asn
355 360 365
Lys Lys Ile Glu Thr His Lys Leu Thr Phe Arg Glu Asn Ala Lys Ala
370 375 380
Lys Thr Asp His Gly Ala Glu Ile Val Tyr Lys Ser Pro Val Val Ser
385 390 395 400
Gly Asp Thr Ser Pro Arg His Leu Ser Asn Val Ser Ser Thr Gly Ser
405 410 415
Ile Asp Met Val Asp Ser Pro Gln Leu Ala Thr Leu Ala Asp Glu Val
420 425 430
Ser Ala Ser Leu Ala Lys Gln Gly Leu
435 440
<210> 34
<211> 219
<212> БЕЛОК
<213> Искусственная
<220>
<223> Легкая цепь D1.2*
<400> 34
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp His Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Phe Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 35
<211> 444
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь hC10.2
<400> 35
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Arg
20 25 30
Thr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Ser Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 36
<211> 214
<212> БЕЛОК
<213> Искусственная
<220>
<223> LC hC10.2
<400> 36
Asp Val Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Met Thr Cys Gln Ala Ser Gln Asp Thr Ser Ile Asn
20 25 30
Leu Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Met Ala Thr Tyr Tyr Cys Leu Gln His Thr Tyr Leu Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 37
<211> 23
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тау-белок 386-408 с фосфорилированными S396 и S404
<400> 37
Thr Asp His Gly Ala Glu Ile Val Tyr Lys Ser Pro Val Val Ser Gly
1 5 10 15
Asp Thr Ser Pro Arg His Leu
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К ГИПЕРФОСФОРИЛИРОВАННОМУ ТАУ-БЕЛКУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2760875C1 |
| Моноклональные антитела к альфа-синуклеину для предотвращения агрегации тау-белка | 2017 |
|
RU2760334C2 |
| АНТИТЕЛА ПРОТИВ ТАУ-БЕЛКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2732122C2 |
| Иммунногенный слитый белок | 2016 |
|
RU2757426C2 |
| БИСПЕЦИФИЧНЫЕ МОЛЕКУЛЫ, ИММУНОРЕАКТИВНЫЕ С ИММУННЫМИ ЭФФЕКТОРНЫМИ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ АКТИВИРУЮЩИЙ РЕЦЕПТОР | 2014 |
|
RU2721707C2 |
| СПОСОБЫ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2789485C2 |
| АНТИТЕЛА И КОМПОЗИЦИИ ПРОТИВ FLT3 | 2021 |
|
RU2836004C1 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
| АНТИТЕЛА ПРОТИВ PD-1 И КОМПОЗИЦИИ | 2016 |
|
RU2750675C1 |
| КОМБИНИРОВАННЫЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА, НАЦЕЛЕННЫЕ НА PD-1, TIM-3 И LAG-3 | 2018 |
|
RU2795232C2 |

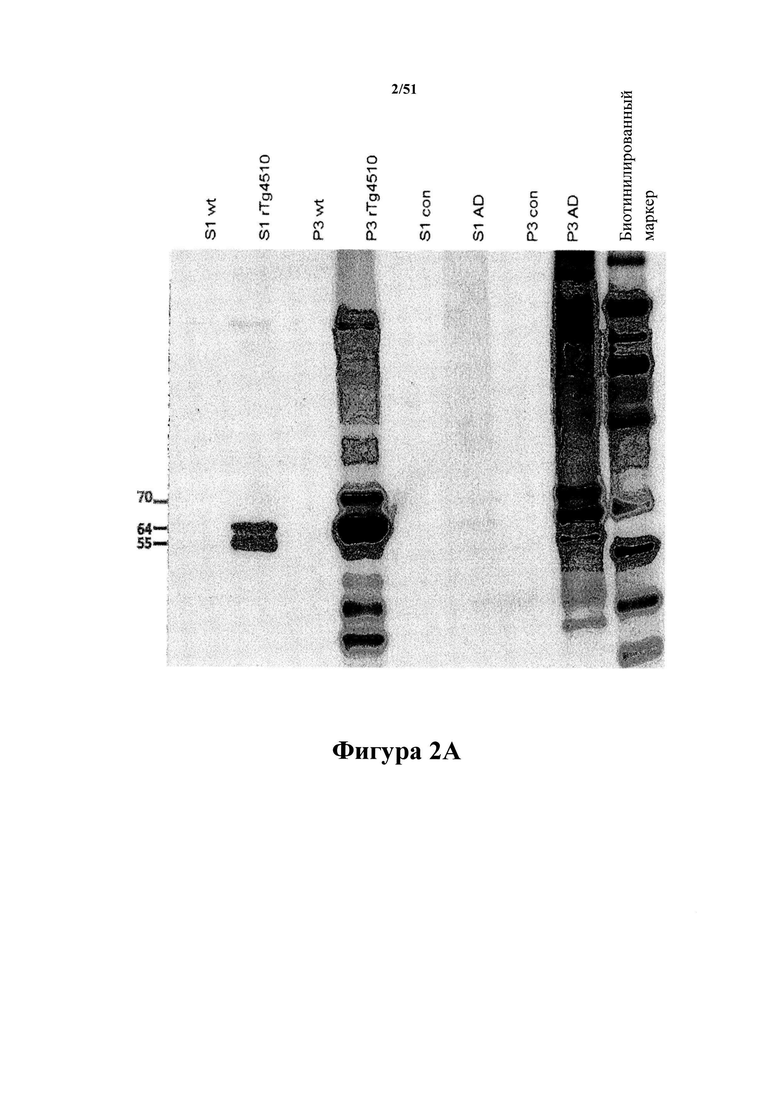
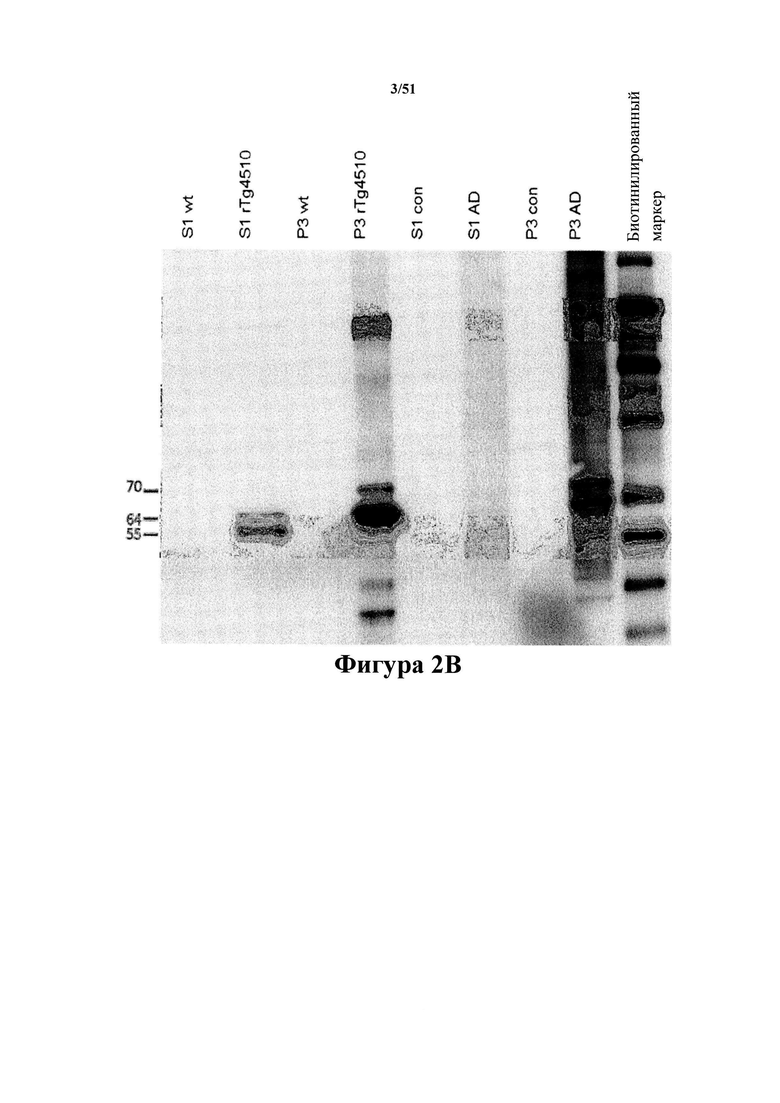
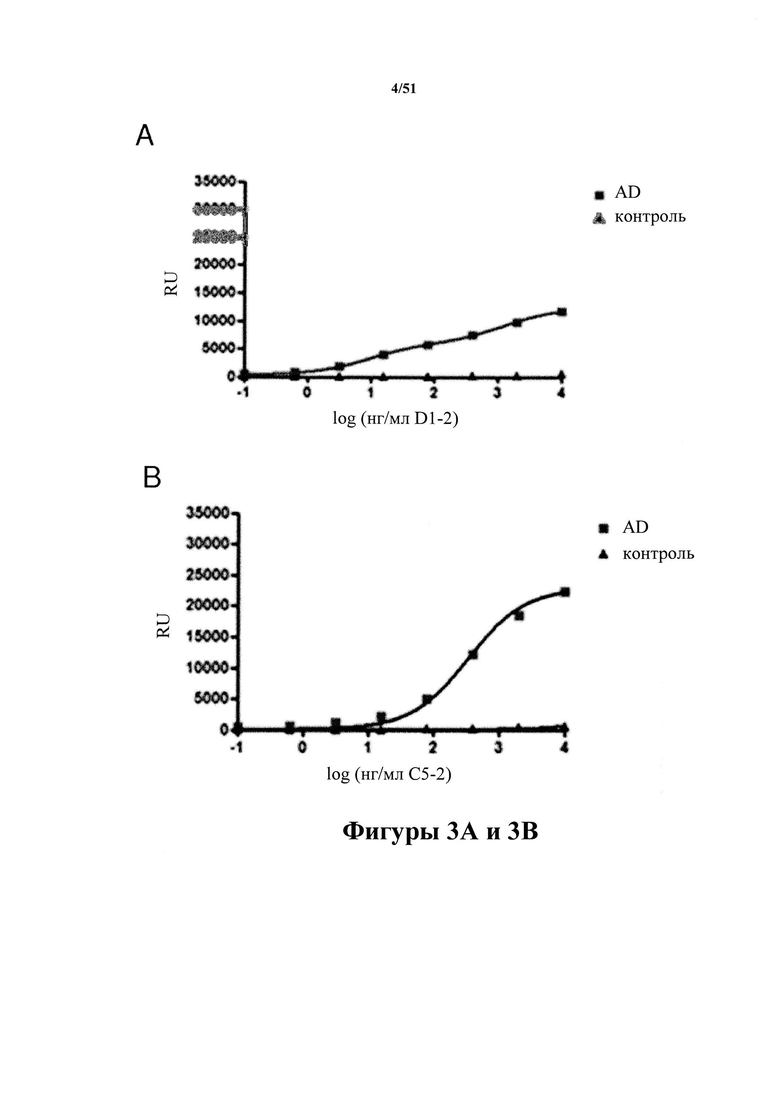
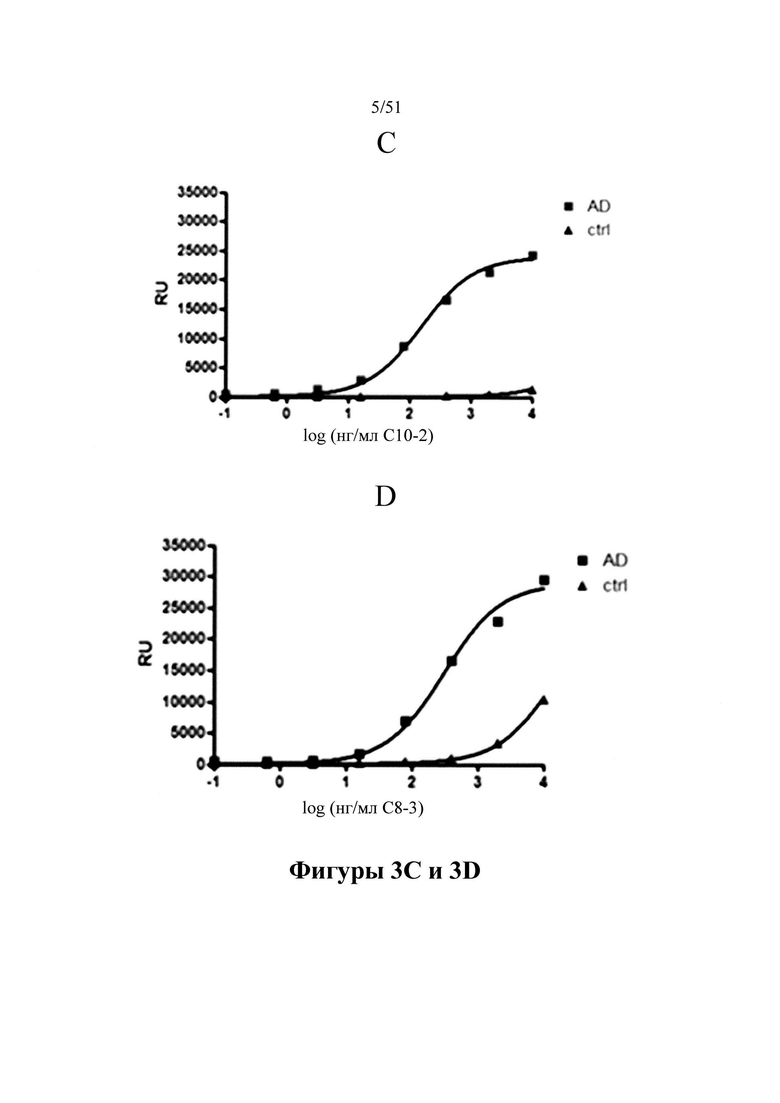
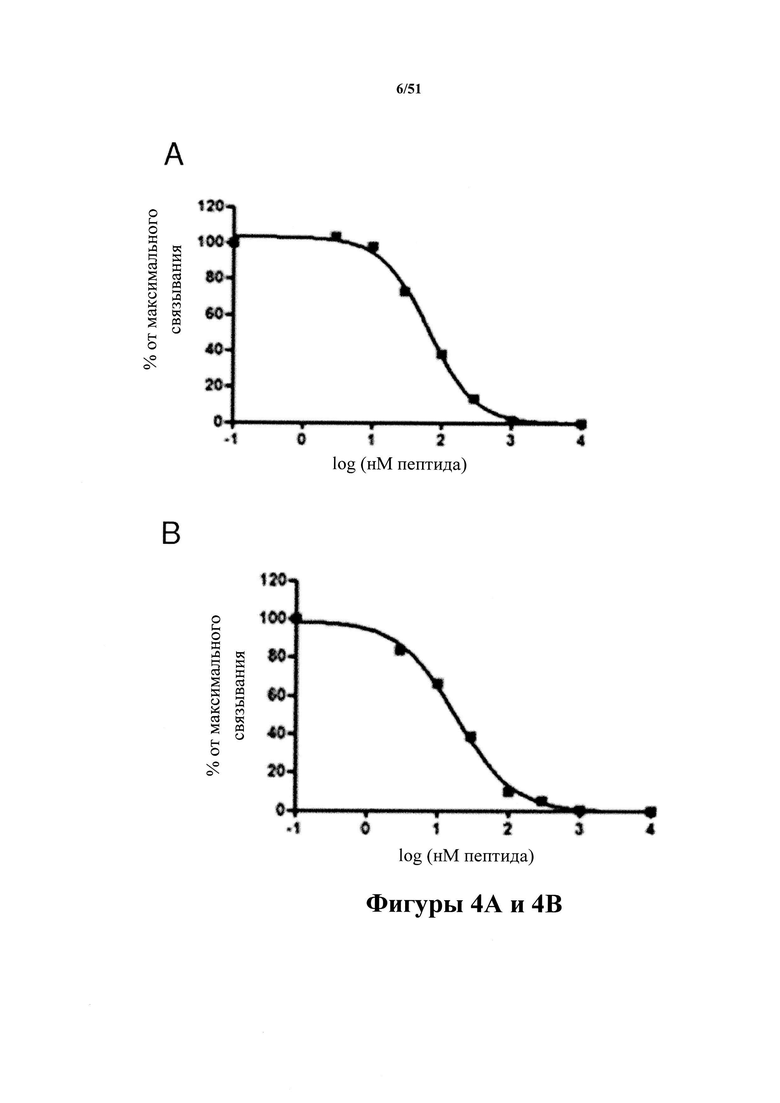

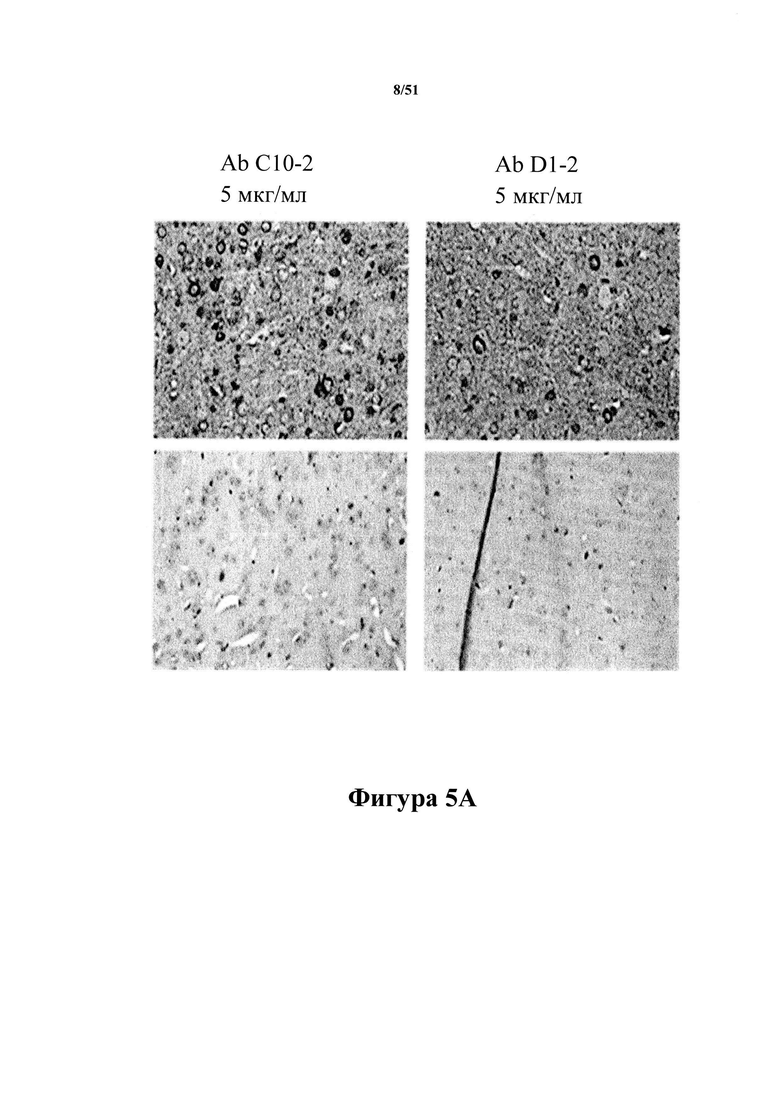
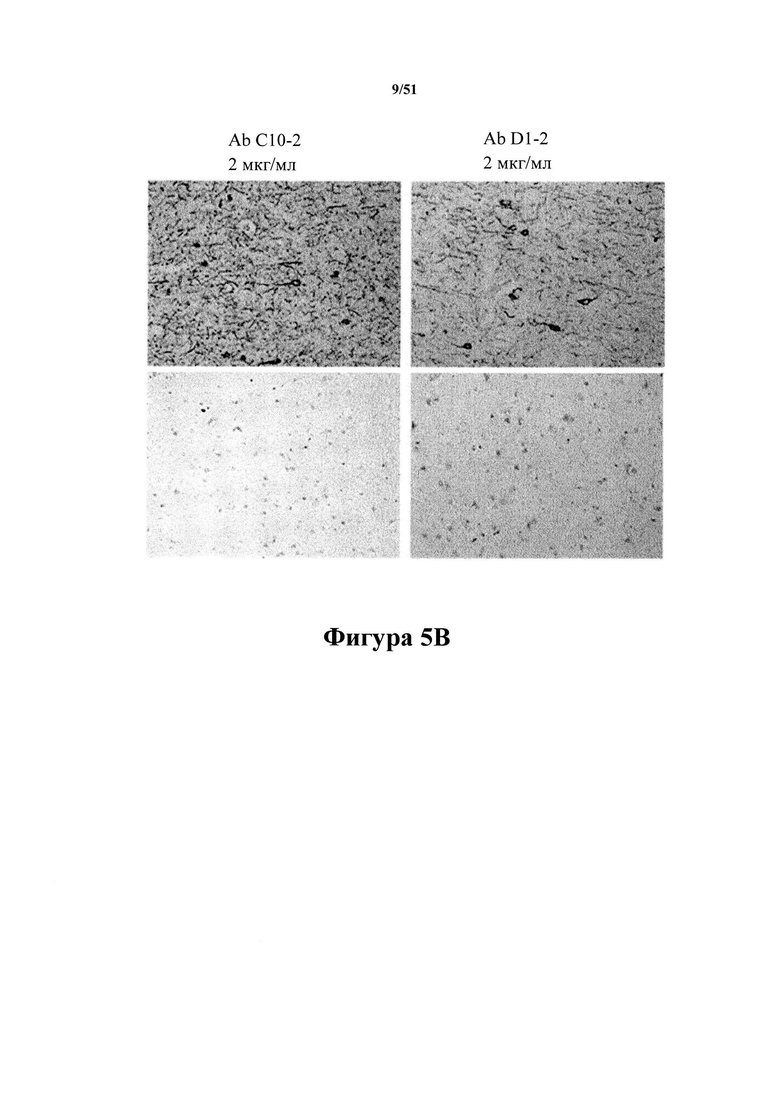

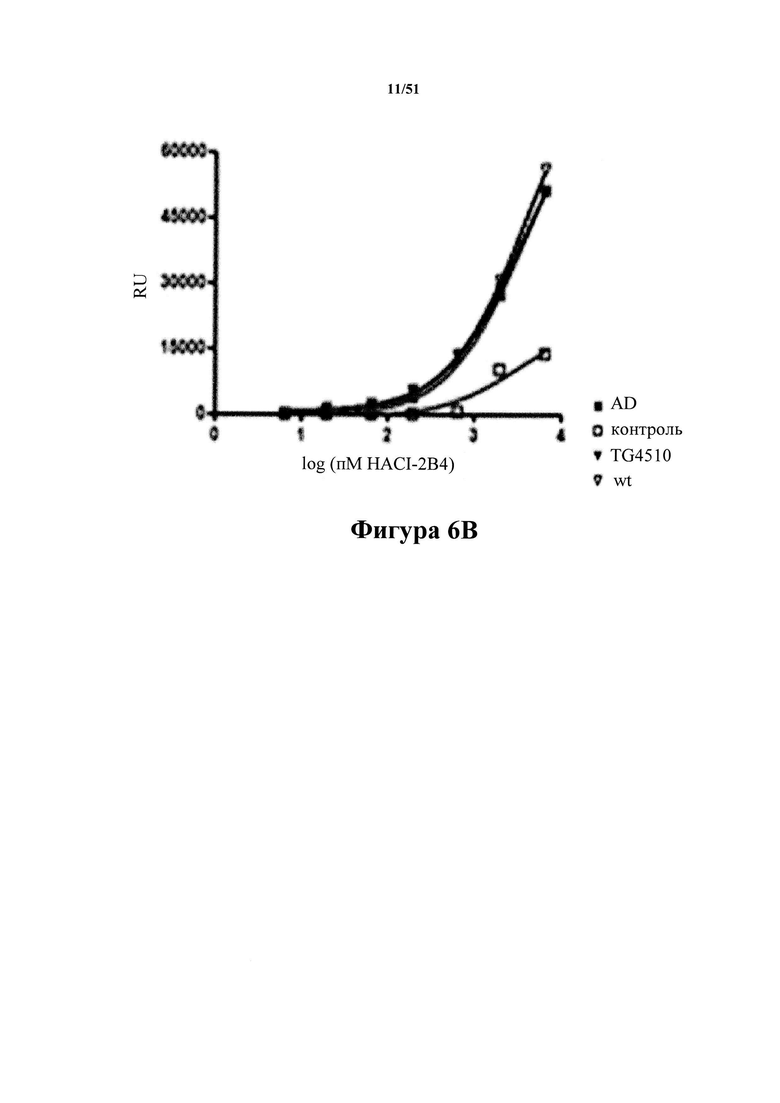
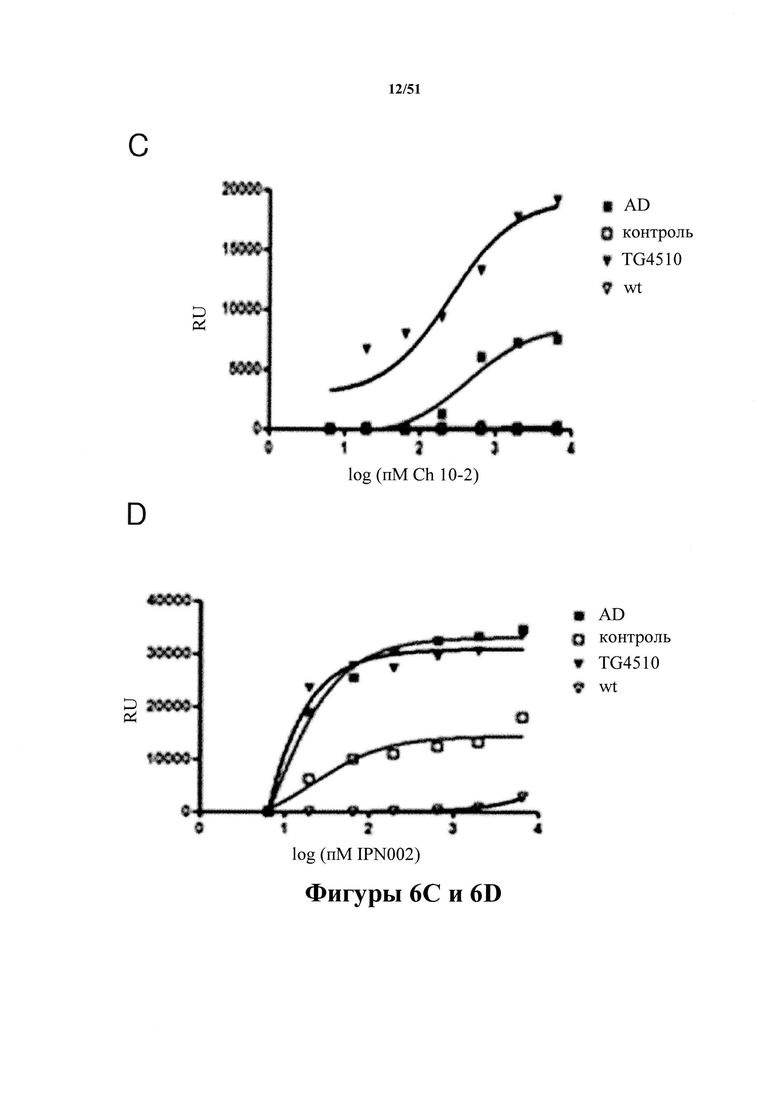
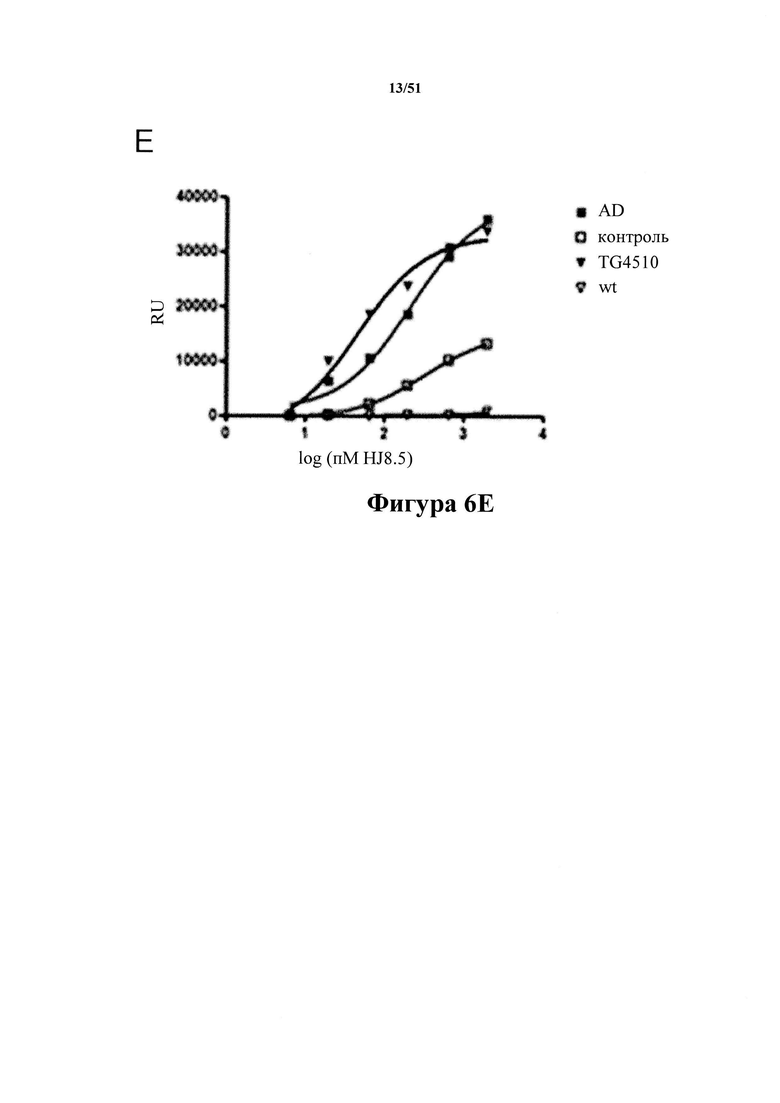


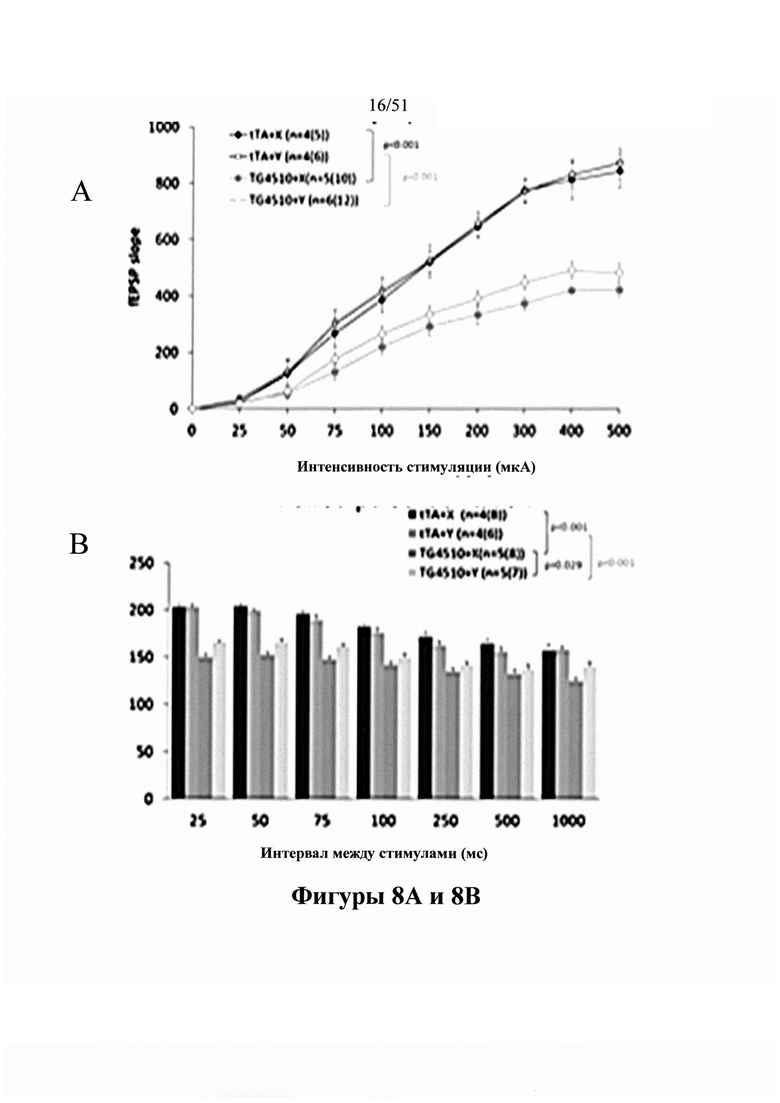

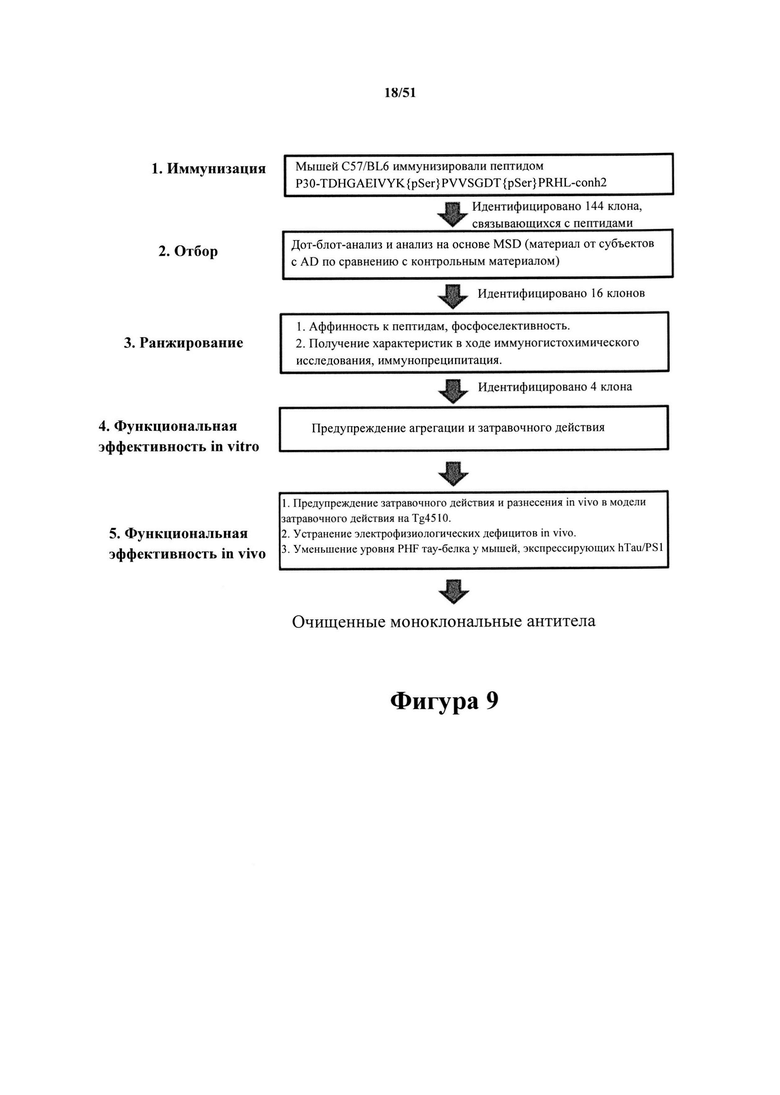
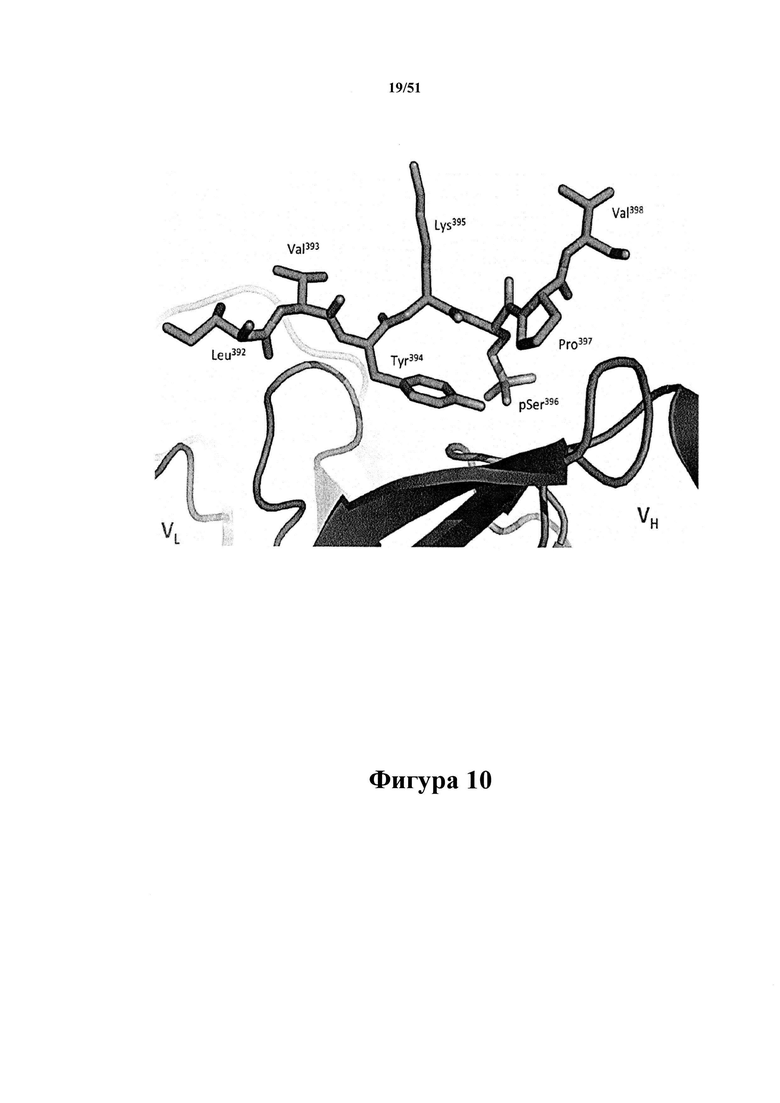
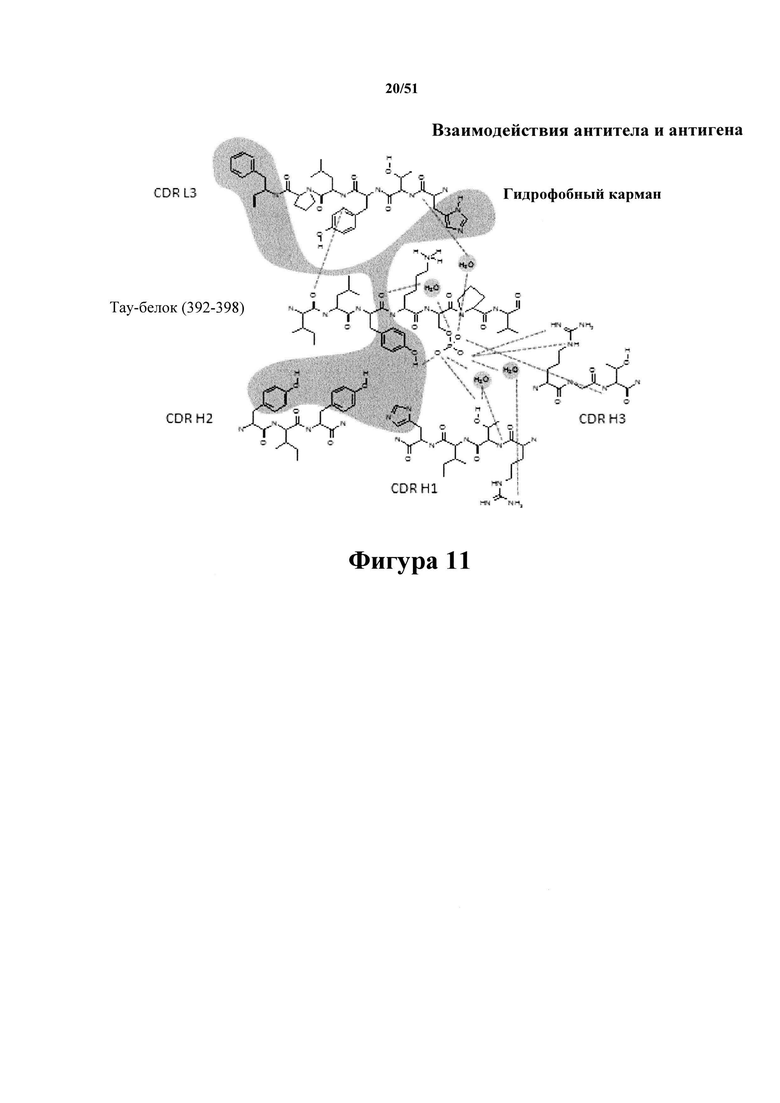
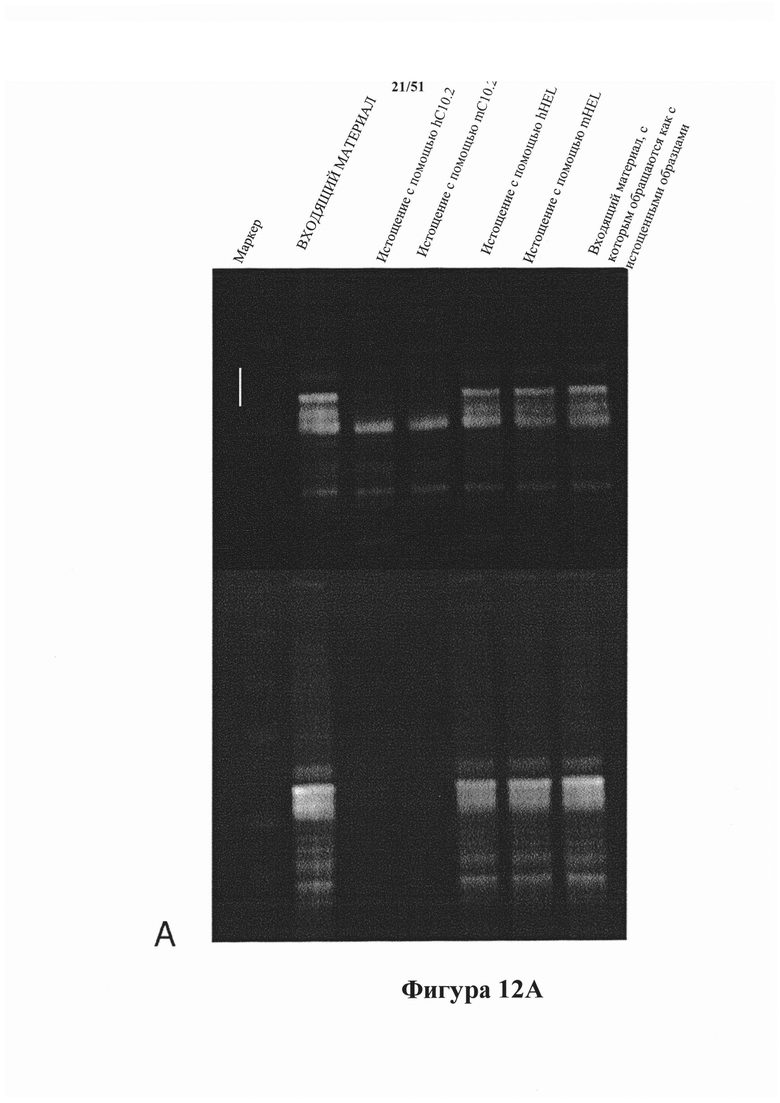
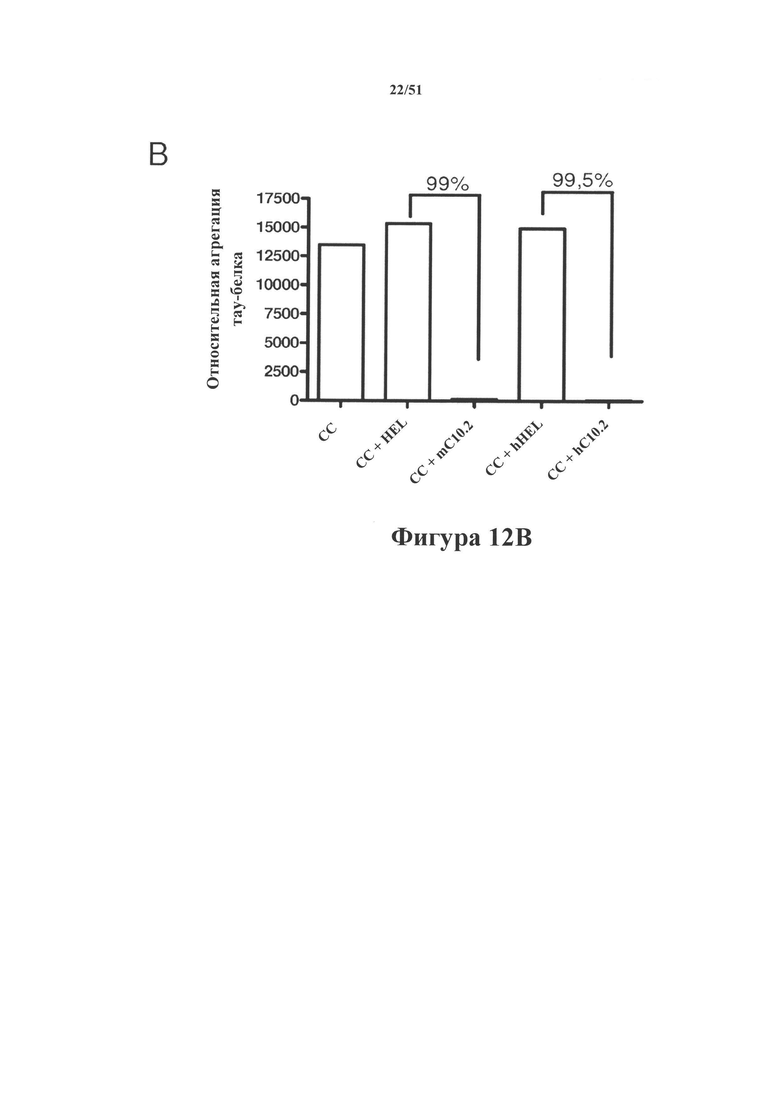
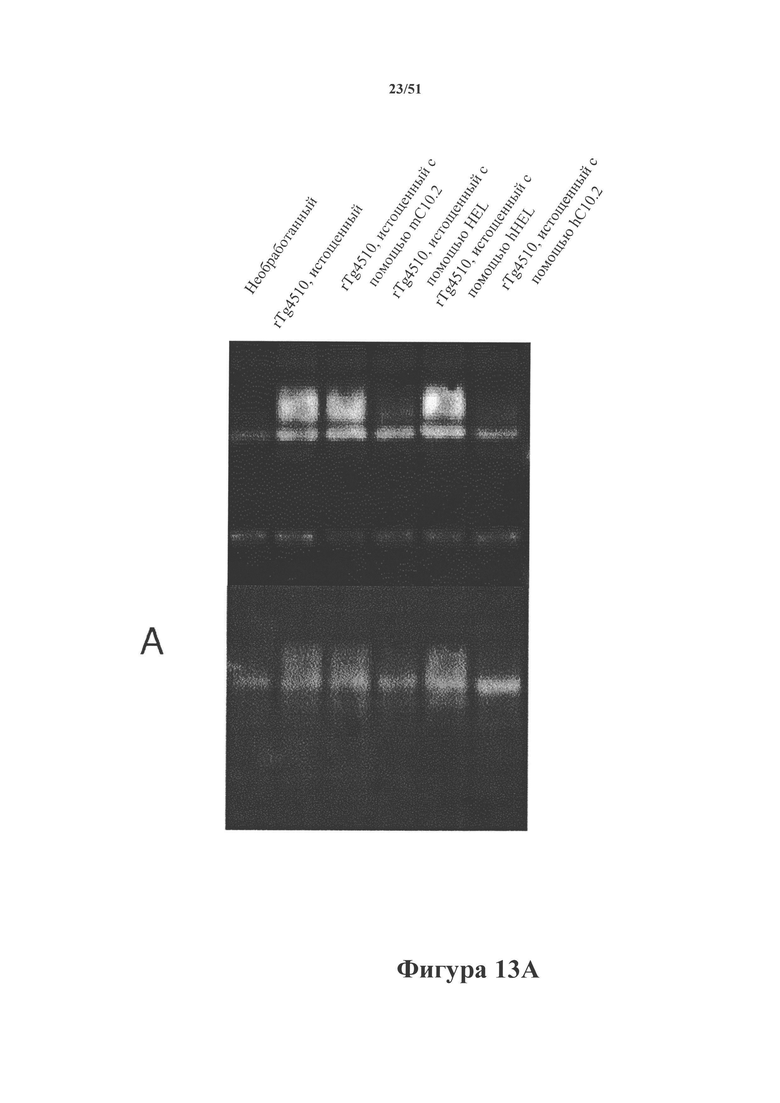



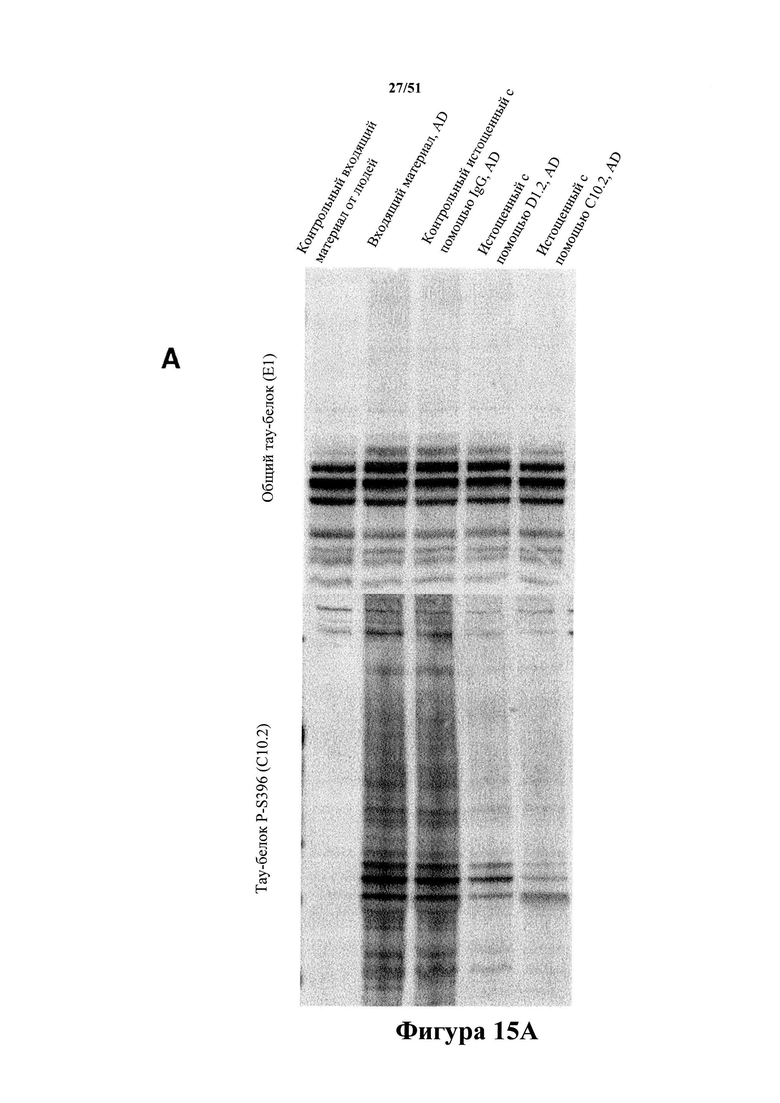

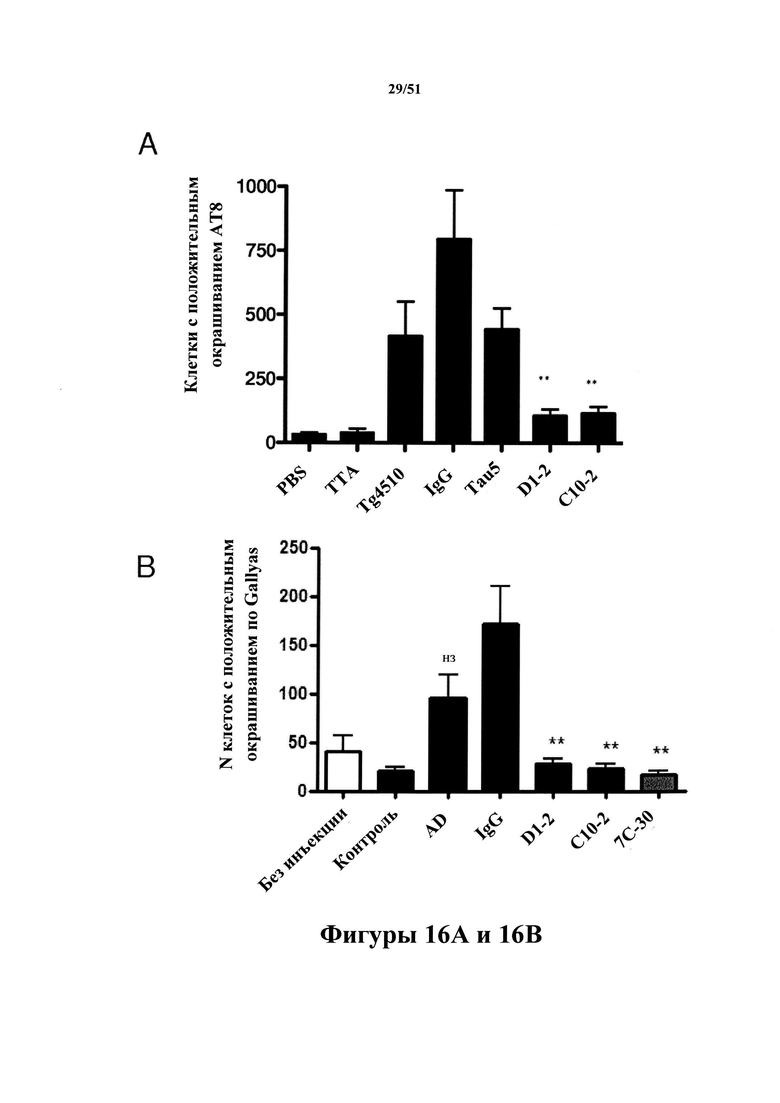
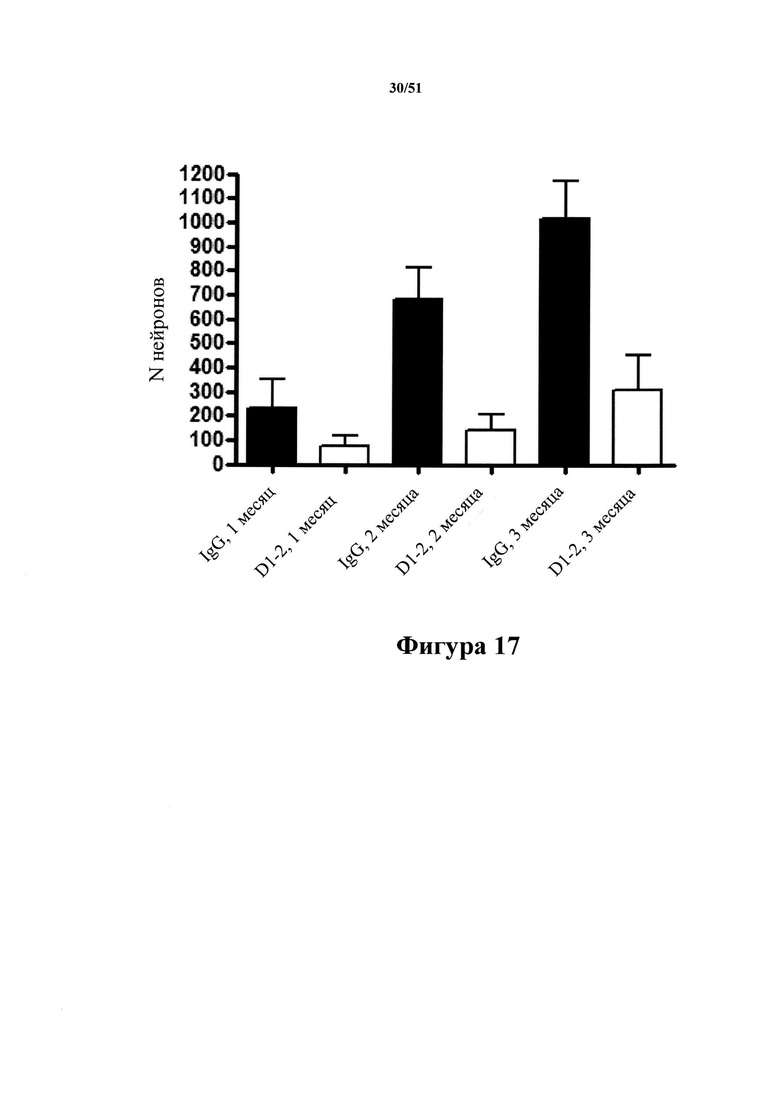
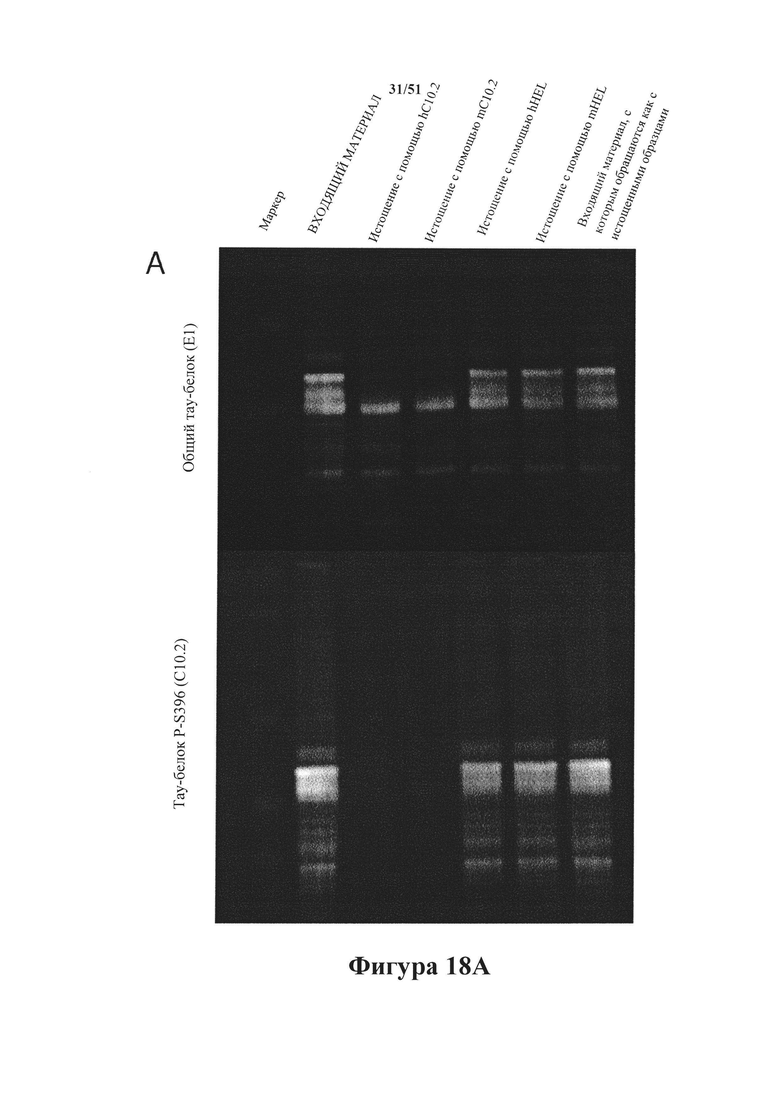

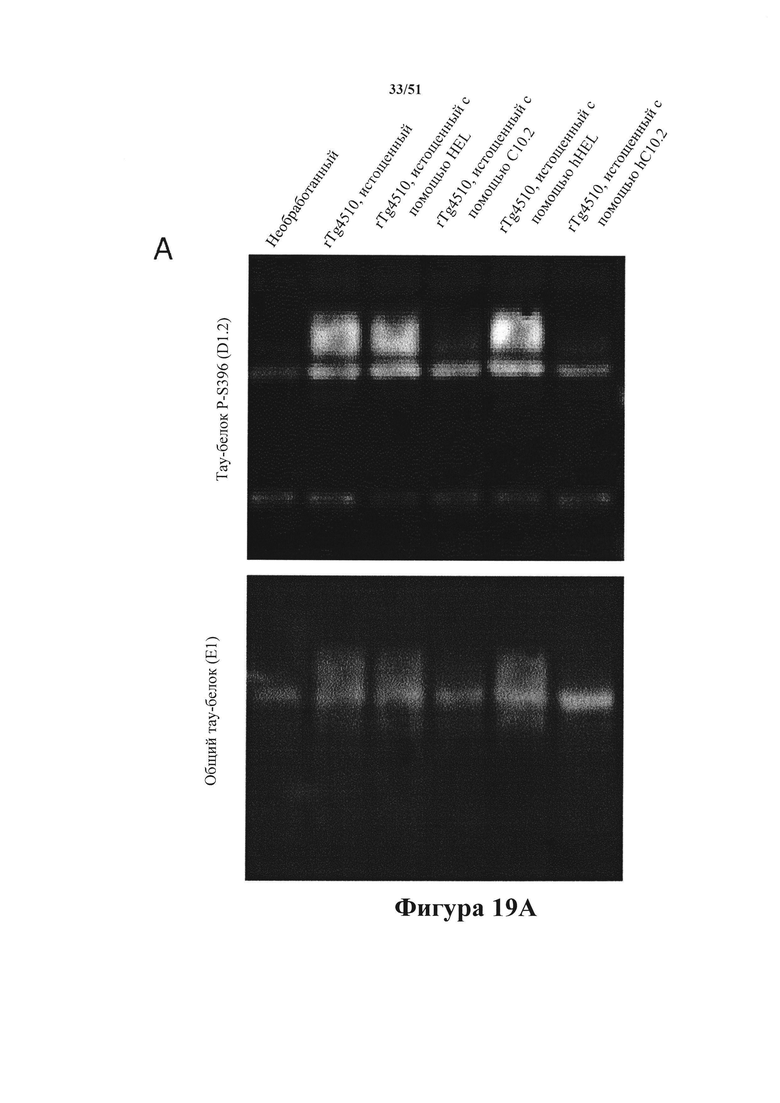
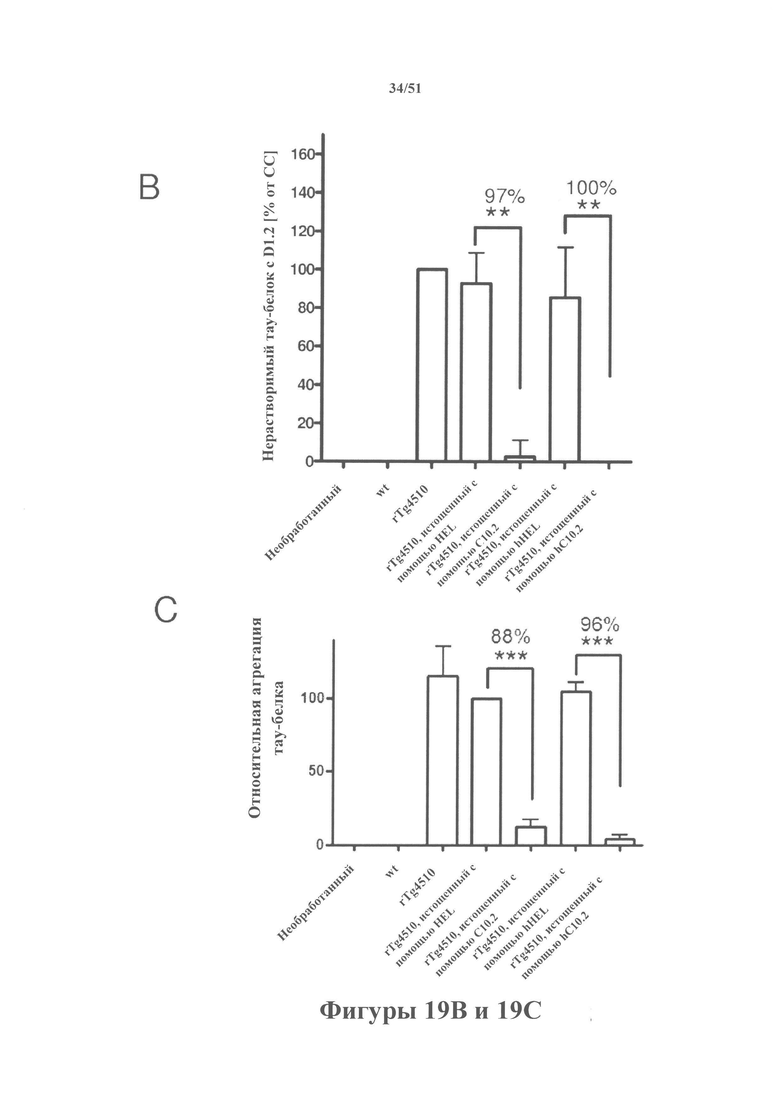


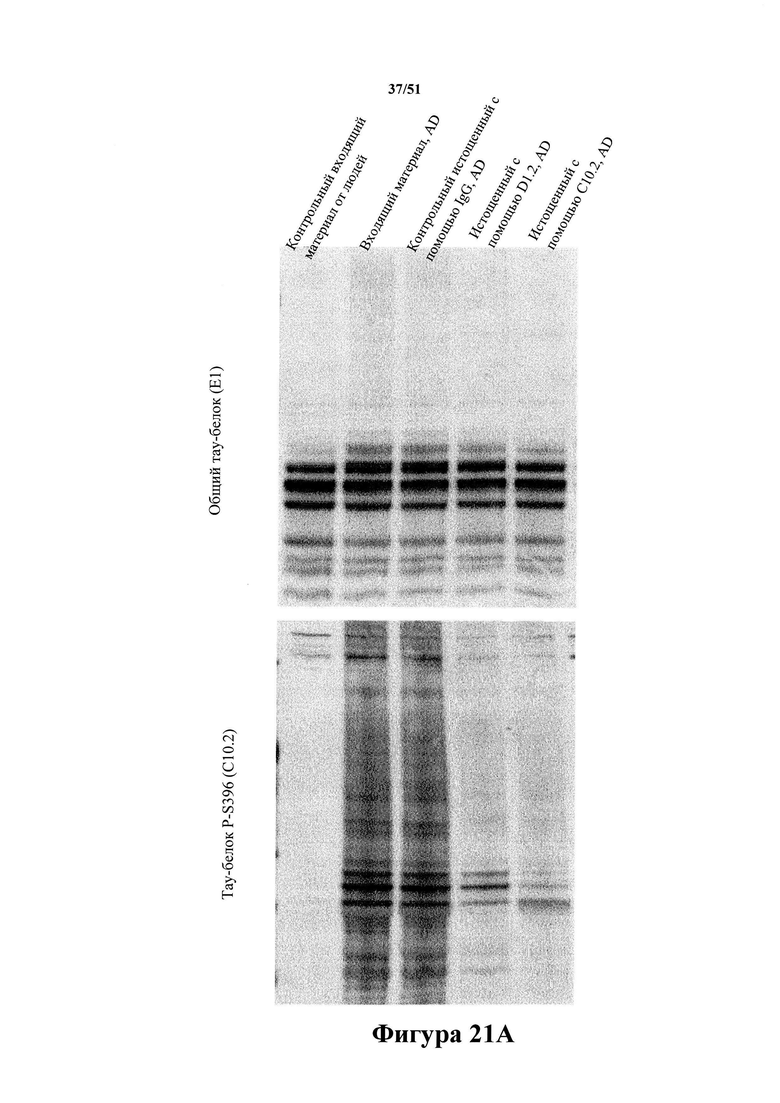
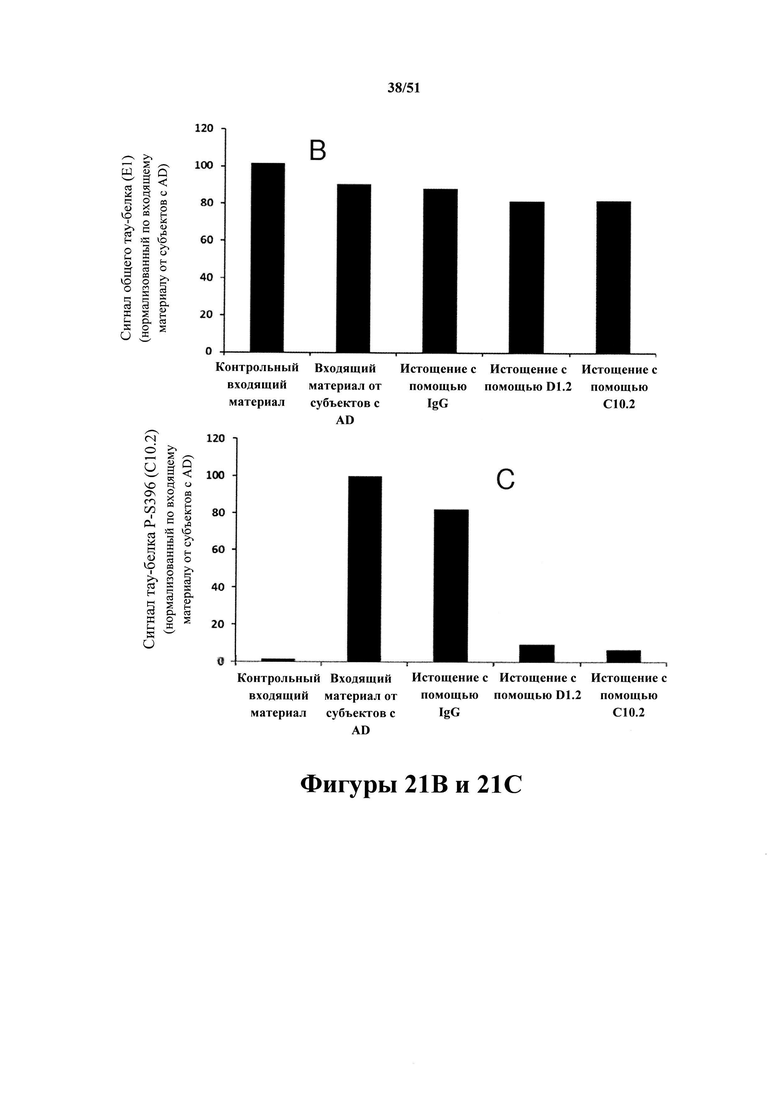

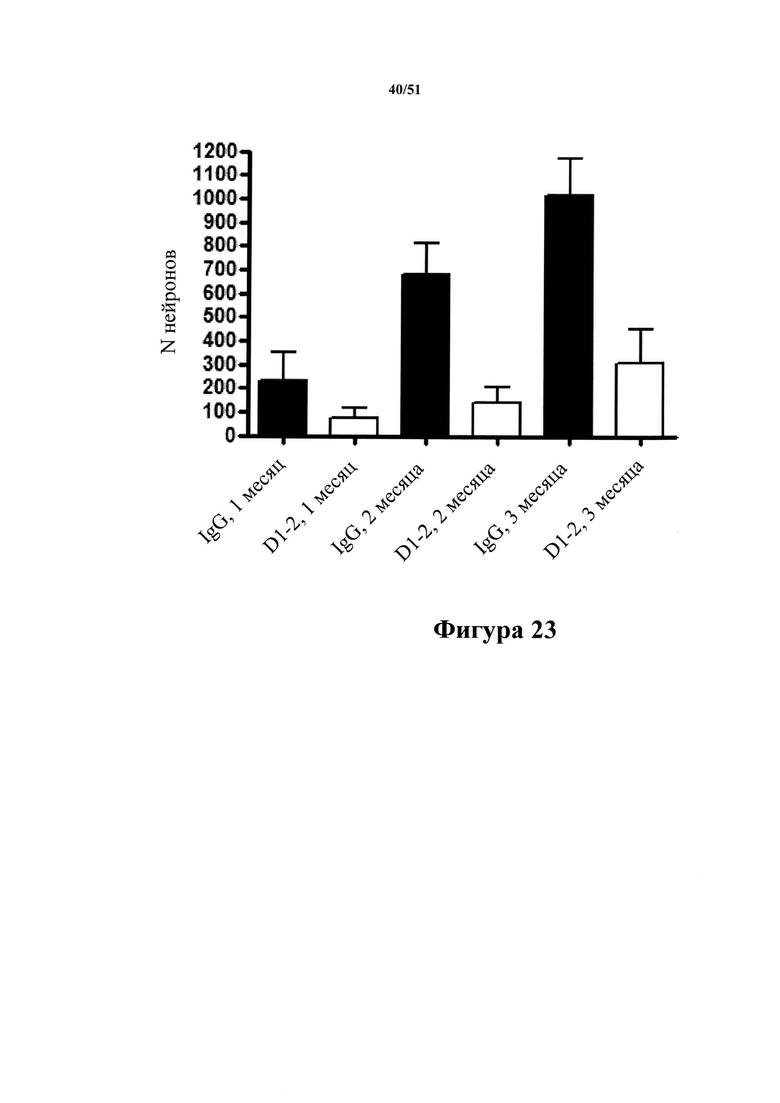
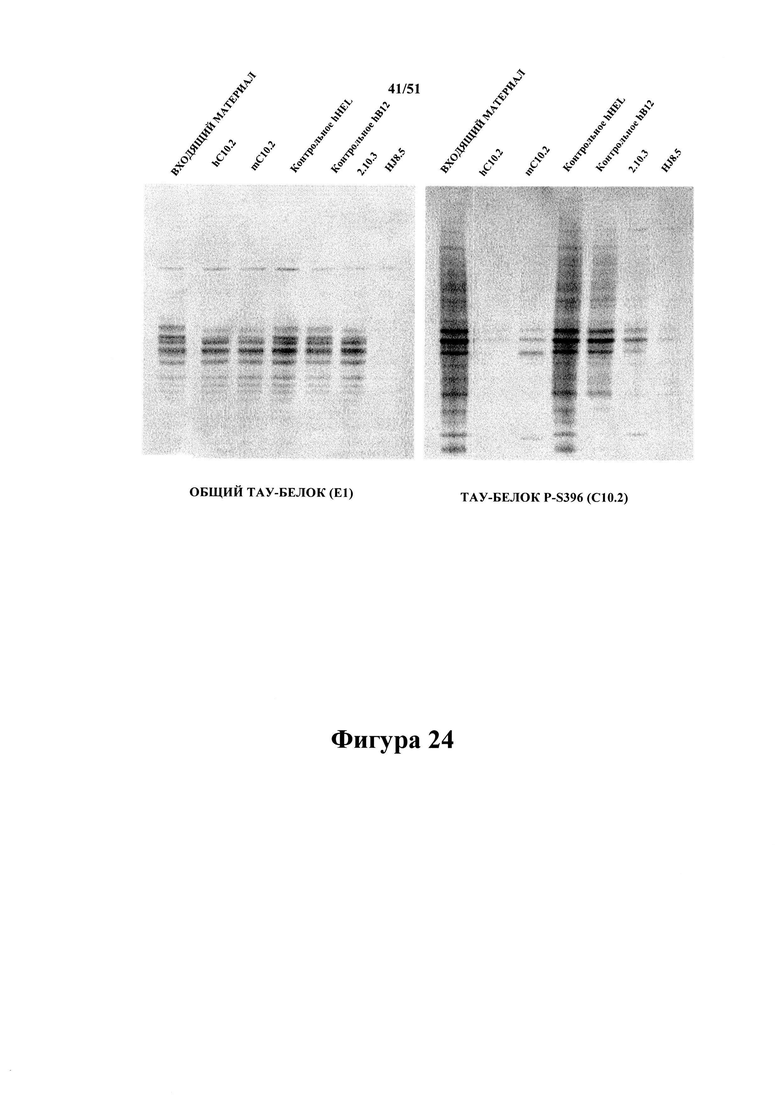
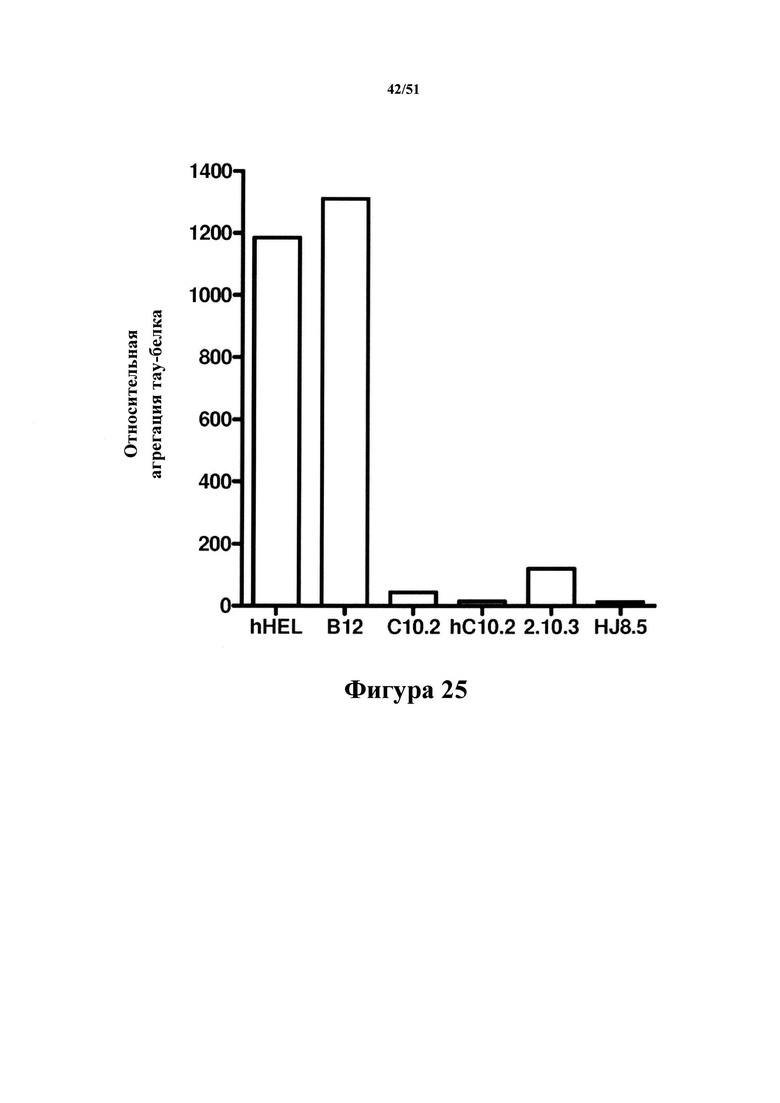
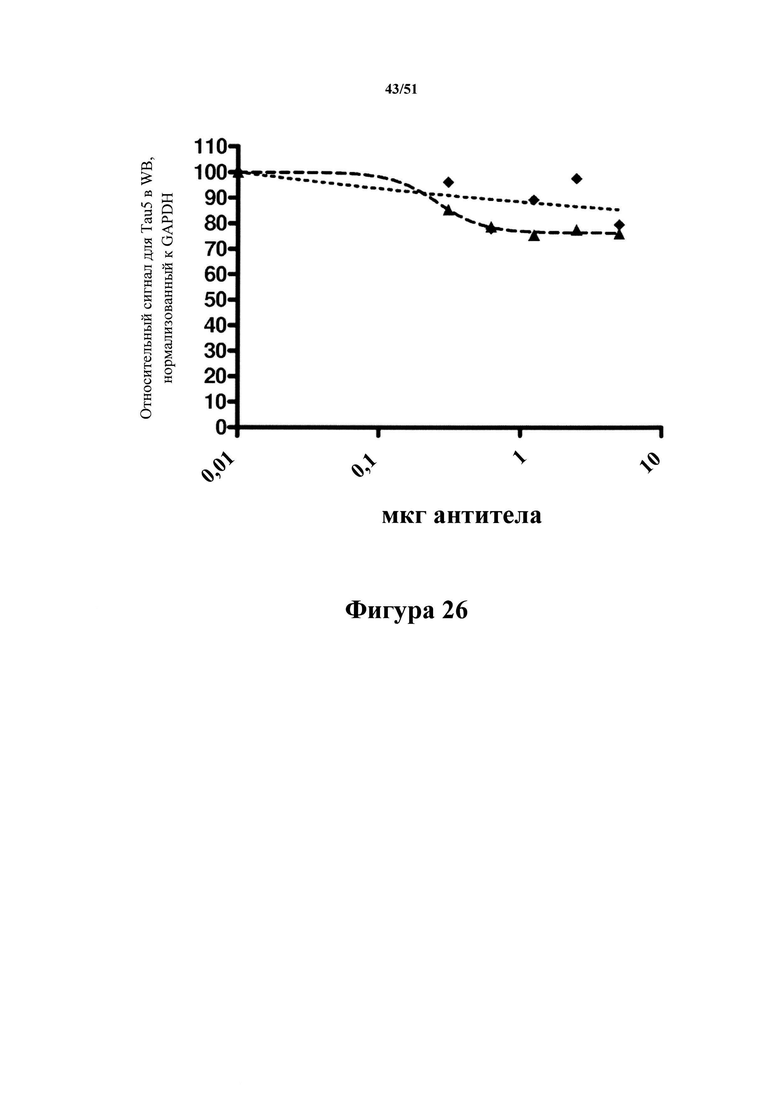



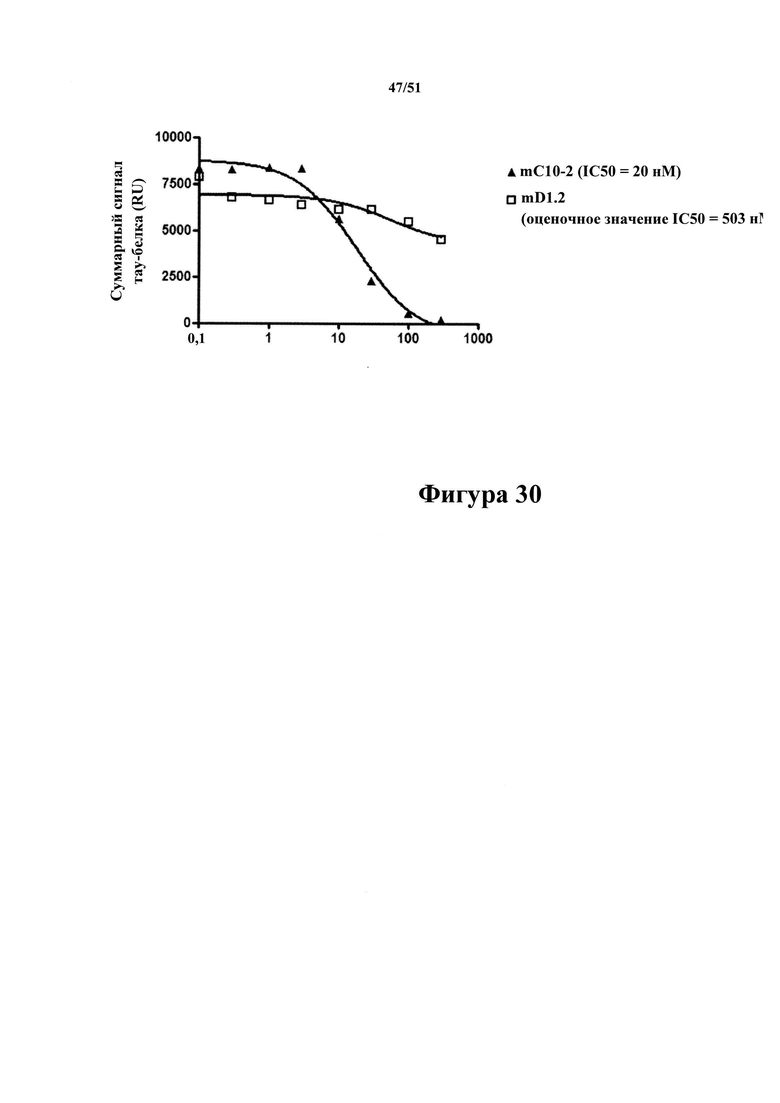
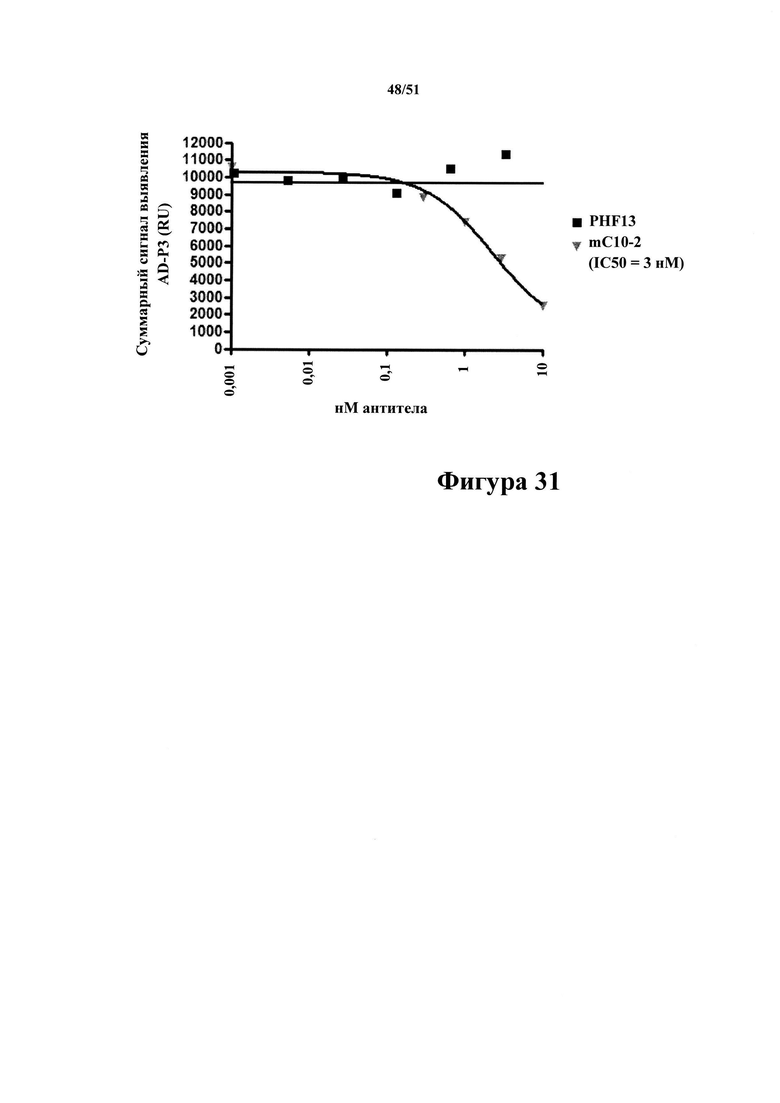
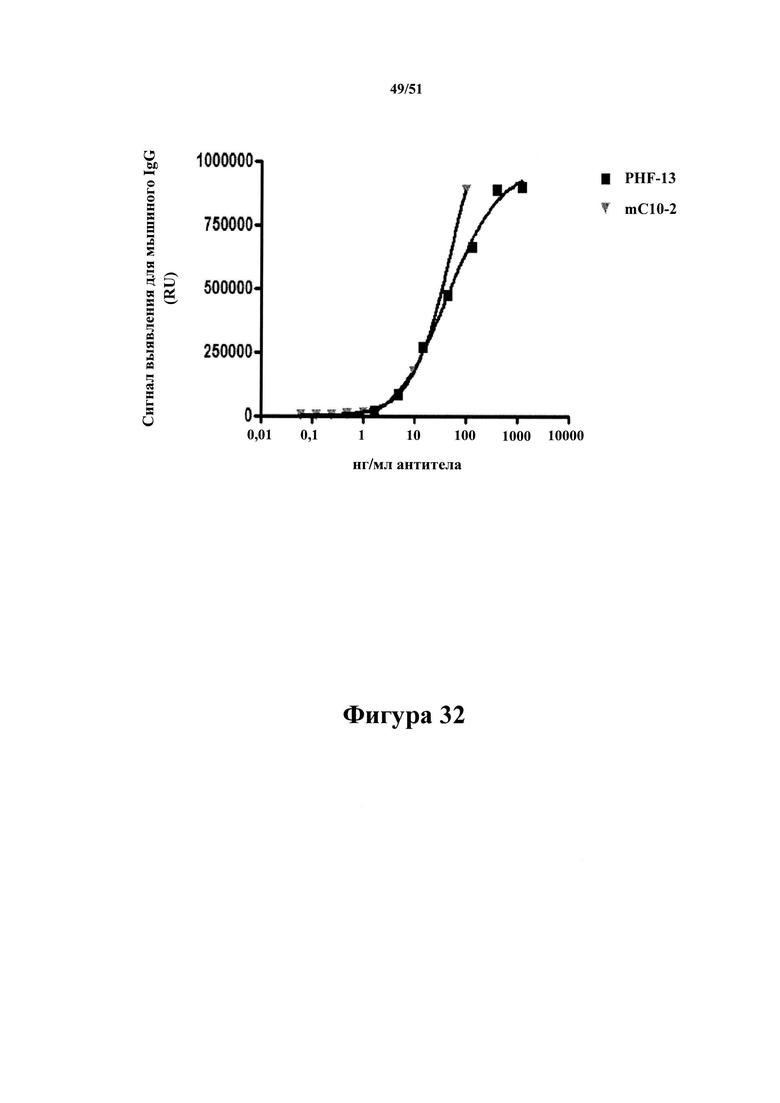
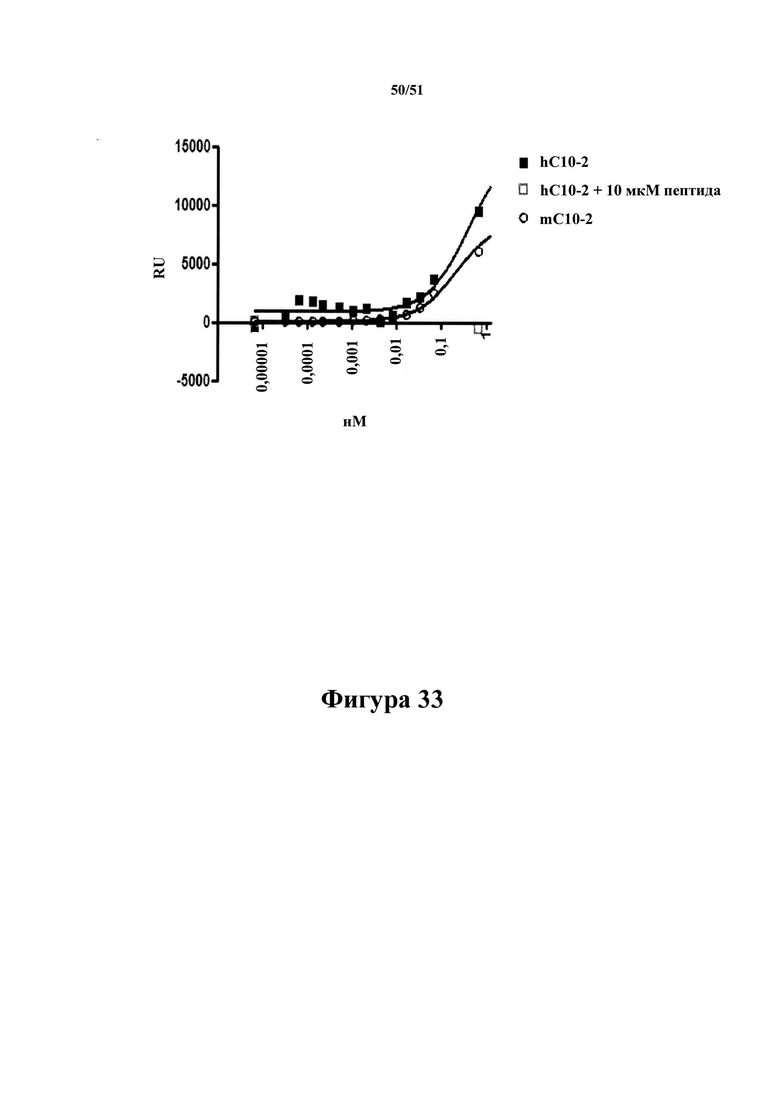
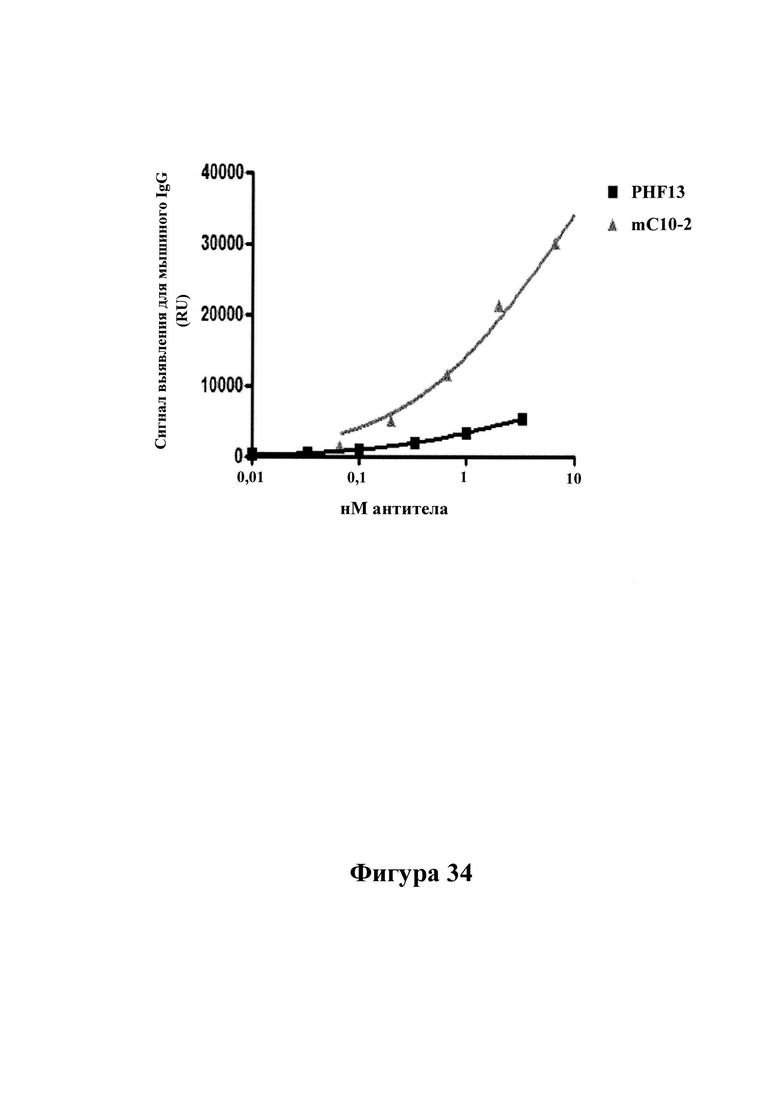
Изобретение относится к области биохимии, в частности к моноклональному антителу или его эпитопсвязывающему фрагменту, способному иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка, а также к композиции, его содержащей. Изобретение позволяет эффективно осуществлять лечение таупатии. 2 н. и 1 з.п. ф-лы, 34 ил., 6 табл., 18 пр.
1. Моноклональное антитело или его эпитопсвязывающий фрагмент, способные иммуноспецифично связываться с фосфорилированным остатком 396 человеческого тау-белка (SEQ ID NO: 33), выбранное из группы, включающей:
i). Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 1;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 2;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 3;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 4;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 5; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 6, или
ii). Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 9;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 10;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 11;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 13; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 14, или
iii). Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащие:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 17;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 18;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 19;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 20;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 21; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 22, или
iv). Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее:
(a) CDR1 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 25;
(b) CDR2 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 26;
(c) CDR3 легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 27;
(d) CDR1 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 28;
(e) CDR2 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 29; и
(f) CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 30.
2. Моноклональное антитело или его эпитопсвязывающий фрагмент по п. 1, выбранное из группы, включающей
i) Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее тяжелую цепь под SEQ ID NO: 8 и легкую цепь под SEQ ID NO: 7;
ii) Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее тяжелую цепь под SEQ ID NO: 8 и легкую цепь под SEQ ID NO: 34;
iii) Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее тяжелую цепь под SEQ ID NO: 16 и/или легкую цепь под SEQ ID NO: 15;
iv) Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее тяжелую цепь под SEQ ID NO: 24 и/или легкую цепь под SEQ ID NO: 23;
v) Моноклональное антитело или его эпитопсвязывающий фрагмент, содержащее тяжелую цепь под SEQ ID NO: 32 и/или легкую цепь под SEQ ID NO: 31.
3. Фармацевтическая композиция для лечения таупатии, содержащая терапевтически эффективное количество моноклонального антитела или его эпитопсвязывающий фрагмент по любому из пп. 1, 2 и фармацевтически приемлемый носитель.
| WO2013050567 A1, 11.04.2013 | |||
| WO2013041962 A1, 28.03.2013 | |||
| FORNO L | |||
| et al., Ubiquitin-positive neuronal and tau 2-positive glial inclusions in frontotemporal dementia of motor neuron type, Acta Neuropathol, 21.02.2002, Vol | |||
| Клапанный регулятор для паровозов | 1919 |
|
SU103A1 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ TAU PS422 ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ГОЛОВНОГО МОЗГА | 2010 |
|
RU2536247C2 |

