Изобретение относится к иммунологии и биотехнологии, а именно к способам модуляции антиген-специфической иммунной реакции против антигенов вируса иммунодефицита человека (ВИЧ-1).
В основе развития нарушений иммунной защиты при проникновении ВИЧ в организм человека лежит поражение CD4+ Т-хелперов, дендритных клеток, что приводит к гибели иммуннокомпетентных клеток, дисрегуляции иммунной защиты и развития синдрома приобретенного иммунодефицита (СПИД).
В настоящее время большое внимание уделяется широконейтрализующим антителам как перспективным препаратам, способным нейтрализовать циркулирующие в крови вирусы. Однако реплицирующие формы вируса, находящиеся внутри клеток организма человека, не подвергаются воздействию гуморальных факторов защиты. В то же время важным звеном иммунитета, участвующим в элиминации вируса, находящегося внутри клетки, является Т-клеточный иммунитет (в основном цитотоксические Т-лимфоциты).
В настоящее время развиваются новые подходы профилактики и лечения больных, инфицированных ВИЧ, создаваемых на основе надежных и безопасных вакцин против ВИЧ, включающих в свой состав специально отобранные В- и Т-клеточные эпитопы [Кузнецова А.В., Данилова Т.И., Гладских О.П. и др. Дендритные клетки и их использование в иммунотерапии. // Молекулярная медицина. - 2003. - №3. - С.3-17; Bashan S.I., Belavin P.A., Seregin S.V., Danilyuk N.K., Babkina I.N., Karpenko L.I., NekrasovaN.A., Lebedev L.R., Ignatyev G.M., Agafonov A.P., Poryvaeva V.A., Abomeva I.V., Ilyichev A.A. Designing and engineering of DNA-vaccine construction encoding multiple CTL epitopes of major HIV-1 antigens // Vaccine. - 2004. - No 22. - P.1672-1682; Eroshkin A.M., Karginova E.A., Gileva I.P., Lomakin A.S., Lebedev L.R., Kamyinina T.P., Pereboev A.V., Ignat`ev G.M. Design of four-helix bundle protein as a candidate for HIV vaccine // Protein Eng. 1995. V.8. No2. P.167-173]. Такие вакцины содержат только те эпитопы, которые необходимы для формирования протективного специфического иммунитета. Данный подход был использован для создания искусственной полиэпитопной конструкции - белка TBI, созданного на базе ФГУН ГНЦ ВБ «Вектор» Роспотребнадзора [Лебедев Л.Р., Карпенко Л.И., Порываева В.А., Азаев М.Ш., Рябчикова Е.И., Гилева И.П., Ильичев A.A. Конструирование вирусоподобных частиц, экспонирующих эпитопы ВИЧ-1 // Молекулярная биология. - 2000. - Т. 34, №3. - С.480-485.; Eroshkin A.M., Karginova E.A., Gileva I.P., Lomakin A.S., Lebedev L.R., Kamyinina T.P., Pereboev A.V., Ignat`ev G.M. Design of four-helix bundle protein as a candidate for HIV vaccine // Protein Eng. 1995. V.8. No2. P.167-173; Karpenko L.I., Lebedev L.R., Ignatyev G.M., Agafonov A.P., Poryvaeva V.A., Pronyaeva T.R. et al. Construction of artificial virus-like particles exposing HIV epitopes, and the study of their immunogenic properties // Vaccine. - 2003. - No 21(5-6). - P.386-392].
Для достижения оптимальной активации Т-клеток против ВИЧ посредством ДК-иммунотерапии очень важно контролировать состояние созревания ДК, т.к. незрелые ДК слабо стимулируют Т-клетки и могут быть толерогены. Созревание ДК описывается как эффекторная функция, которая активирует антиген-специфические Т-клеточные иммунные реакции [Reis e Sousa С.Dendritic cells in a mature age. Nat. Rev. ImmunoL - 2006. - No.6. - P.476-483]. Это процесс включает экспрессию поверхностных молекул, таких как МНС.
В настоящее время активно исследуется возможность применения новых методов иммунокоррекции у больных ВИЧ, в частности использования дендритных клеток для модуляции специфического противовирусного иммунного ответа [Rinaldo C.R. Dendritic cell-based human immunodeficiency virus vaccine. // J.Intem Med. - 2009. - Vol.256. - No.1. - P.138-158]. Дендритные клетки (ДК) - гетерогенная по происхождению, морфологии и выполняемым функциям популяция иммунокомпетентных клеток, которые принимают участие практически во всех видах иммунных реакций. ДК являются основными антигенпрезентирующими клетками, что обусловливает их непосредственное и важнейшее участие в формировании эффективного специфического иммунного ответа. Кроме того, ДК обладают способностью продуцировать провоспалительные цитокины (ФНО-α, ИФН-γ, ИЛ-1, ИЛ-12 и др.), что приводит к активации функций других иммунокомпетентных клеток, участвующих в противовирусной защите.
По мере дифференцировки (созревания) изменяется функциональное состояние дендритных клеток. Так на стадии незрелых дендритных клеток они эффективно захватывают антигены, в основном посредством эндоцитоза и микропиноцитоза, но при этом их способность стимулировать Т-клетки ограничена. На стадии терминальной дифференцировки (зрелости) дендритные клетки способны презентировать антиген Т-лимфоцитам и активировать их, продуцируя соответствующие гуморальные факторы. То есть дендритные клетки, обладая уникальными механизмами захвата/представления антигена наивным Т-клеткам и активируя их, определяют направленность и эффективность иммунных реакций в организме.
Эффективность использования ДК объясняется, во-первых, тем, что на сегодня достаточно хорошо изучены молекулярные механизмы участия ДК в формировании иммунного ответа и, во-вторых, протоколы получения ДК и модуляции с их помощью иммунного ответа выглядят наиболее изученными и «прозрачными» с точки зрения контроля всех этапов этого метода клеточной терапии.
Эффективность модуляции ДК специфического иммунного ответа зависит от дополнительной стимуляции ДК in vitro. Оптимальное приготовление ДК для иммунотерапии дает возможность увеличения CD4+ и CD8+ Т-клеток непосредственно после инфузии. В данной модели созревание in vitro может активировать ДК для продукции ИЛ-12 при последующем взаимодействии с Тх1 клетками in vivo. ИЛ-12 продуцирующие ДК, которые ввели в организм, должны в последствии поляризовать CD4+ Т×1 клетки, которые преимущественно продуцируют ИФН-γ, который активирует CD8+цитотоксические Т-лимфоциты. Таким образом, посредством активированных in vitro ДК стимулируются CD8+ и преобладание Т×1 над Т×2 реакций in vivo, что позволит устранить дефицит Т-клеточной реактивности, наблюдаемой при ВИЧ-инфекции.
Было показано, что ВИЧ инфицирует как миелоидные, так и плазмацитоидные дендритные клетки, путем взаимодействия с CD4, DC-SIGN [Schimdt В., Fujimura S.H., Martin J.N., Levy J.A. Variations in plasmacytoid dendritic cell (PDC) and myeloid dendritic cell (MDC) levels in HIV-infected subjects on and off antiretroviral therapy. J. Clin. Immunol. 2006. - Vol.26. - P.55-64]. Репликация вируса в большей степени происходит в незрелых дендритных клетках по сравнению со зрелыми клетками. Миелоидные дендритные клетки у больных ВИЧ, не подвергавшиеся специфической терапии, имеют незрелый фенотип и индуцируют активность регуляторных Т-клеток, которые могут подавлять пролиферацию CD8+ Т-клеток и их функцию [Krathwohl M.D., Schacker T.W., Anderson J.L. Abnormal presence of semimature dendritic cells that induce regulatory Т cells in HIV-infected subjects. J. Infect Dis. - 2006. - Vol.193. - P.494-504].
Описан способ (прототип), при котором получали незрелые дендритные клетки из CD14+ мононуклеарных клеток периферической крови ВИЧ-позитивных (негативных) пациентов путем культивирования в полной среде с добавлением рекомбинантного человеческого гранулоцитарно-макрофагального колониестимулирующего фактора (рчГМ-КСФ) и интерлейкина-4 (рчИЛ-4) в течение 5-6 дней. Затем получали зрелые дендритные клетки путем добавления CD40L в культуральную среду и инкубации в течение 40 ч. В качестве ВИЧ-специфического антигена использовались эпитопы пептидов Nef и Gag. Антигены добавляли к незрелым или зрелым дендритным клеткам и инкубировали в течение 2 ч. Далее антиген-активированные клетки культивировали совместно с мононуклеарными клетками (МНК) в соотношении 1:10 в течение 16-18 ч («overnight»). Для оценки пролиферативной активности CD4- и CD8-позитивных клеток совместное культивирование продолжалось в течение 6 дней. Также оценивали внутриклеточную продукцию иммуннокомпетентными клетками IFN-γ, TNF-α методом проточной цитофлуорометрии. Наибольшее количество IFN-γ-положительных клеток было в группе МНК, сокультивированных со зрелыми ДК, к которым после созревания добавляли 15-мерный белок (эпитопы Nef и Gag), по сравнению с группой МНК, которые сокультивировали с ДК, к которым добавляли антигены на стадии незрелых клеток. В тоже время не было обнаружено стимуляции Т-клеточной реактивности в ответ на эпитопы белков Gag и Nef у трех ВИЧ-серонегативных пациентов. Эпитопы белка Nef стимулировали пролиферацию CD8-позитивных лимфоцитов в присутствии дендритных клеток, в культуре мононуклеарных клеток без сокультивирования с дендритными клетками белок Nef не стимулировал пролиферацию СD8-позитивных лимфоцитов. Белки Gag стимулировали пролиферацию CD4-и CD8-позитивных клеток. Наибольшее влияние белков на пролиферацию было при предварительной нагрузке ими дендритных клеток. Полученные результаты показали, что использование сочетания эпитопы белков Gag и Nef стимулирует продукцию множества иммунных медиаторов CD4- и СD8-позитивными клетками. При стимуляции МНК антиген-активированными дендритными клетками количество IFN-γ- и TNF-α-позитивных клеток достигало не более 2-3% от всей популяции лимфоцитов (Huang X.-L., Fan Z., Borowski L., Rinaldo C.R. Multiple T-cell responses to human immunodeficiency virus type 1 are enhanced by dendritic cell // Clinical and vaccine immunology. - 2009. - Vol.16. - N.10. - P.1504-1516).
Недостатком данного подхода является длительный срок культивирования дендритных клеток (5-6 дней). Создание полиэпитопного иммуногена, содержащего более 30 эпитопов ВИЧ, достаточно сложная в техническом плане процедура. Все это приводит к значительному удорожанию технологии. Кроме того, уровень стимуляции цитотоксических клеток, судя по количеству IFN-γ- и TNF-α-позитивных клеток, довольно низок и не превышает 2-3%.
Направленность развития протективного ответа зависит от многих факторов, в том числе от баланса про- и противовоспалительных цитокинов. Известно, что провоспалительный цитокин ИЛ-12 обладает широкой биологической активностью, стимулирует цитотоксические и НК-клетки, активирует дифференциацию CD4+ Т-лимфоцитов по 1 типу, индуцирует продукции NO макрофагами, тем самым потенцирует противовирусный иммунитет. Прововоспалительный цитокин ИЛ-18 является фактором, индуцирующим продукцию ИФН-γ Т- и НК-клетками, усиливая цитотоксический ответ. Показано, что при совместном применении ИЛ-12 и ИЛ-18 наблюдается наиболее выраженная стимуляция продукции ИНФ-γ и перфоринзависимой цитотоксичности НК-клеток, пролиферация Т-лимфоцитов, что позволяет максимально возможно стимулировать эффекторное звено иммунитета против ВИЧ [de Arquer G.R., Реnа R., Cabrere С., Coma G., Ruiz-Hernandez R., Guerola R., Clotet В., Ruiz L., Este J.A., Calle M.L., Bofill M. Skewed expression and up-regulation of the IL-12 and IL-18 receptors in resting and activated CD4 Т cells from HIV-1-infected patients // J. Leukoc. BioL - 2007. - Vol.82. - P.72-78].
Задачей настоящего изобретения является создание эффективного и экономичного способа генерации антиген-специфического иммунного ответа против антигенов вируса иммунодефицита человека.
Сущность предлагаемого изобретения заключается в следующем:
Мононуклеарные клетки неприлипающей фракции периферической крови условно здоровых доноров культивируют с антиген-активированными дендритными клетками, полученными из моноцитов прилипающей фракции мононуклеарных клеток. Антигенную активацию дендритных клеток проводят с помощью полиэпитопного вирусного белка TBI, а для созревания ДК используют рекомбинантный человеческий провоспалительный цитокин - фактор некроза опухоли (рчФНО-α). Для повышения модуляции потенциала цитотоксической активности мононуклеарных антиген-специфических клеток совместное культивирование неприлипающей фракции МНК и зрелых антиген-активированных ДК проводят в присутствии рчИЛ-12 и рчИЛ-18.
Принципиальным отличием разработанного способа от прототипа является использование рчИЛ-12 и рчИЛ-18 при сокультивировании ДК и МНК, что повышает эффективность необходимых для стимуляции и поддержания направленной дифференцировки наивных Т-клеток в Т-хелперы 1 типа, а также применение протоколов активации и созревания ДК, включающих использование антигенной вирусной конструкции - полиэпитопного белка TBI и рчФНО-α, что активирует мононуклеарные клетки против вирусных антигенов (увеличение содержания внутриклеточного перфорина в иммуннокомпетентных клетках), а также сокращает стадию получения зрелых антиген-активированных ДК до 3-х суток.
Предлагаемое изобретение повышает эффективность генерации цитотоксических клеток, а также позволяет сократить сроки сокультивирования ДК и МНК, что в итоге приводит к снижению стоимости процедуры.
Техническим результатом изобретения является генерация in vitro антиген-специфических клонов Т-клеток с активностью Т-хелперов 1 типа, необходимых для формирования эффективного противовирусного ответа.
Изобретение осуществляется следующим образом.
Выделенная из периферической крови условно здоровых доноров прилипающая фракция мононуклеарных клеток культивируется в концентрации 1 млн/мл в полной среде, содержащей 10% эмбриональной телячьей сыворотки, 2 мМ L-глютамина, 10 мМ HEPES, 5×10-5 мМ 2-меркаптоэтанола, 80 мкг/мл гентамицина, 100 мкг/мл ампициллина в атмосфере 5% СO2 при 37°С с добавлением рчГМ-КСФ (50 нг/мл) и рчИЛ-4 (100 нг/мл). Через 48 часов культивирования к полученным незрелым ДК добавляется антигенная вирусная конструкция (полиэпитопный белок TBI) в дозе 130 нг/мл, а еще через 24 часа для созревания незрелых дендритных клеток в течение последующих 24 часов вносится рчФ-НО-α в дозе 25 мкг/мл.
Полученные дендритные клетки культивируются с неприлипшей фракцией мононуклеарных клеток в соотношении 1:10 с добавлением рчИЛ-12 в дозе 10 нг/мл и рчИЛ-18 в дозе 100 нг/мл. Для оценки антиген-специфического ответа полученные группы культур клеток разделили на две подгруппы (спонтанная и TBI-стимулированная), к одной из них через 48 часов инкубации (5% СО2 и 37°С) добавляли белок TBI в количестве 130 нг/мл.
Для определения потенциальной цитотоксической активности Т-лимфоцитов через 48 часов оценивали количество перфоринсодержащих цитотоксических клеток и содержание ИЛ-4 и ИФН-γ в кондиционной среде культур клеток. Через 60 часов с момента добавления белка TBI в культуру мононуклеарных клеток оценивали внутриклеточную продукцию ИЛ-4 и ИНФ-γ иммуннокомпетентными клетками.
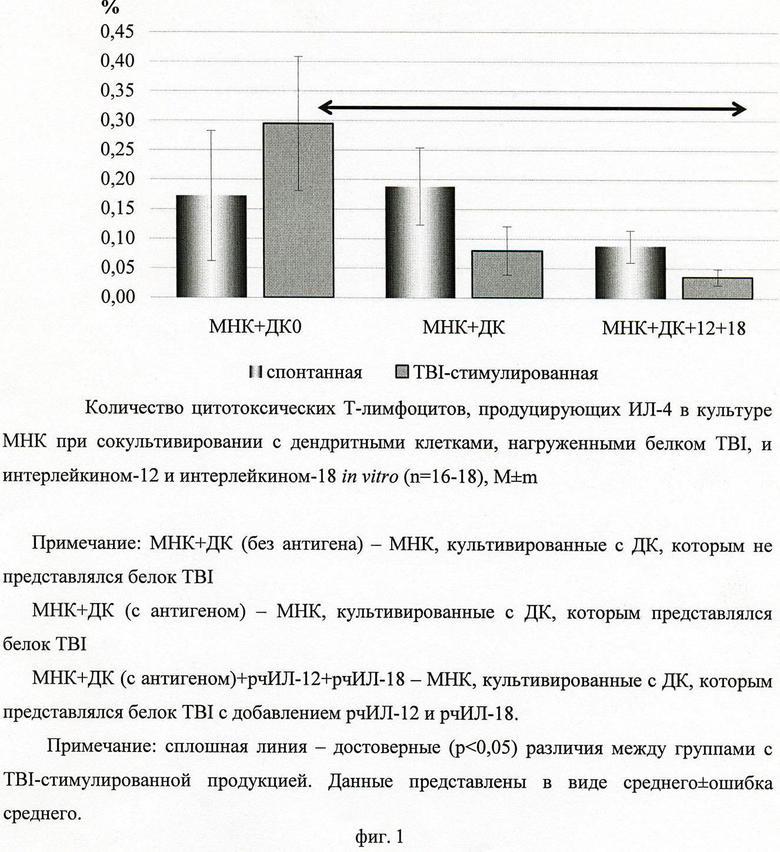
Исследованием количества цитотоксических Т-лимфоцитов было установлено, что при добавлении белка TBI в культуру МНК с дендритными клетками, нагруженными антигеном, происходит уменьшение количества цитотоксических Т-лимфоцитов, содержащих ИЛ-4 (фиг.1).
Также было показано, что добавление ИЛ-12 и ИЛ-18 к дендритным клеткам, нагруженным антигеном, приводит к дальнейшему снижению количества цитотоксических Т-лимфоцитов, содержащих ИЛ-4 (фиг.1).
Количество IFN-γ-содержащих цитотоксических Т-лимфоцитов достоверно не меняется как при инкубации с дендритными клетками, нагруженными белком TBI, так и в присутствии IL-12 и IL-18.
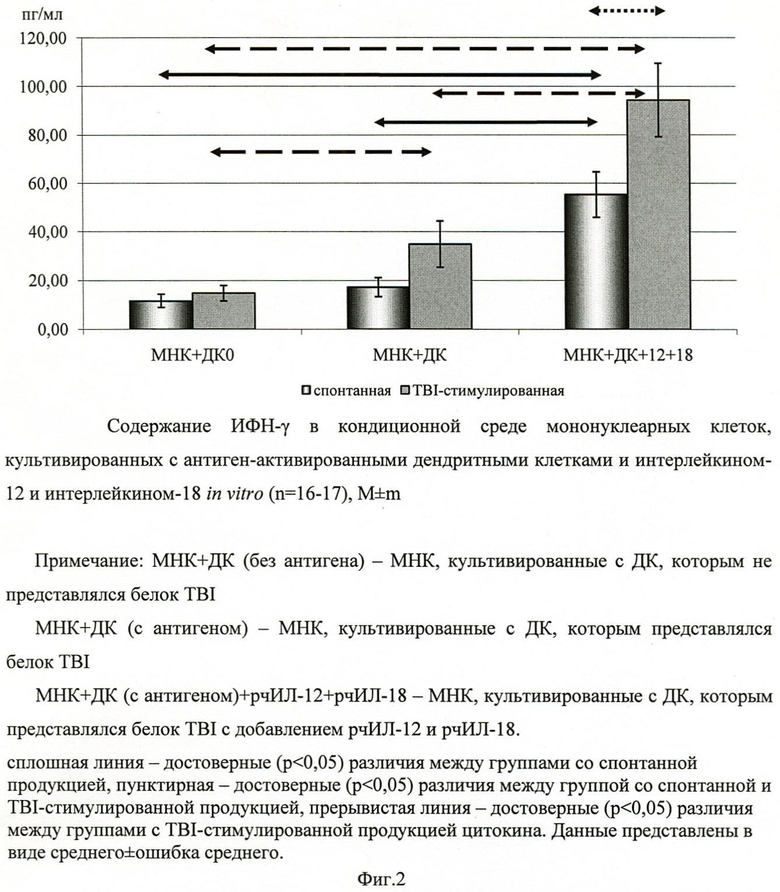
На следующем этапе оценивали уровень продукции ИЛ-4 и ИФН-γ мононуклеарными клетками при сокультивировании их с дендритными клетками, нагруженными белком TBI. Обнаружено, что сокультивирование МНК с дендритными клетками и цитокинами (ИЛ-12 и ИЛ-18) не влияет на продукцию ИЛ-4 В то же время дендритные клетки, нагруженные белком TBI, в сочетании с ИЛ-12 и ИЛ-18 достоверно увеличивают содержание ИФН-γ в культуре МНК, по сравнению с аналогичной культурой клеток, но без добавления ИЛ-12 и ИЛ-18 (фиг.2). Следует отметить, что внесение белка TBI в культуру МНК с дендритными клетками, нагруженными антигеном, вызывает увеличение содержания ИФН-γ в кондиционной среде. Этот эффект белка был наиболее выражен в культуре клеток, к которым добавляли ИЛ-12 и ИЛ-18.
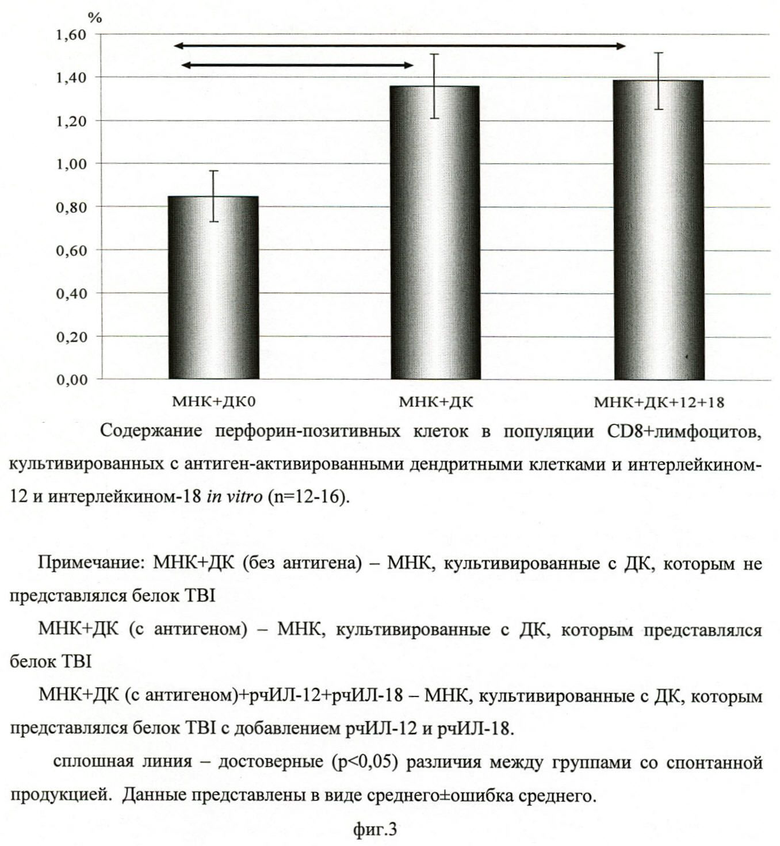
Для оценки влияния дендритных клеток, нагруженных белком TBI, на потенциальную цитотоксическую активность мононуклеарных клеток определяли содержание CD8-позитивных клеток, секретирующих внутриклеточный мономерный белок перфорин - фактор цитотоксичности, вызывающий образование пор в цитоплазматической мембране, приводящих к лизису инфицированных клеток. Установлено, что использование дендритных клеток, нагруженных белком TBI, достоверно увеличивает количество CD8-позитивных клеток, содержащих перфорин, в культуре МНК в отличие от дендритных клеток без нагрузки белком TBI, что свидетельствует о способности полиэпитопного белка (TBI) стимулировать цитотоксическую активность CD8-позитивных клеток (фиг.3).
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК, ОБЛАДАЮЩИХ АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2013 |
|
RU2596920C2 |
| СПОСОБ СТИМУЛЯЦИИ ЦИТОТОКСИЧЕСКОГО ИММУННОГО ОТВЕТА ПРОТИВ КЛЕТОК ОПУХОЛЕВОЙ ЛИНИИ АДЕНОКАРЦИНОМЫ МОЛОЧНОЙ ЖЕЛЕЗЫ, ЭКСПРЕССИРУЮЩИХ СПЕЦИФИЧЕСКИЕ АНТИГЕНЫ, С ПОМОЩЬЮ ДЕНДРИТНЫХ КЛЕТОК, ТРАНСФЕЦИРОВАННЫХ ПОЛИЭПИТОПНОЙ ДНК-КОНСТРУКЦИЕЙ | 2012 |
|
RU2520091C2 |
| СПОСОБ ГЕНЕРАЦИИ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК С АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК, ИНФИЦИРОВАННЫХ ВИРУСОМ ГЕПАТИТА В | 2008 |
|
RU2366707C1 |
| Способ получения in vitro популяций активированных антигенспецифических противоопухолевых цитотоксических Т-лимфоцитов, специфичных к эпитопам опухоль-ассоциированного антигена | 2016 |
|
RU2619186C1 |
| Полиэпитопная противоопухолевая вакцинная конструкция, содержащая эпитопы опухоль-ассоциированных антигенов, фармацевтическая композиция и ее применение для стимуляции специфического противоопухолевого иммунного ответа | 2016 |
|
RU2684235C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pCI-UB-POLYEPI, СОДЕРЖАЩАЯ ЭПИТОПЫ ОПУХОЛЬ-АССОЦИИРОВАННЫХ АНТИГЕНОВ ДЛЯ КОЛОРЕКТАЛЬНОГО РАКА, И СПОСОБ ЕЕ ПРИМЕНЕНИЯ ДЛЯ СТИМУЛЯЦИИ СПЕЦИФИЧЕСКОГО ПРОТИВООПУХОЛЕВОГО ИММУННОГО ОТВЕТА ПРОТИВ КЛЕТОК КОЛОРЕКТАЛЬНОГО РАКА | 2012 |
|
RU2507265C2 |
| СПОСОБ ГЕНЕРАЦИИ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК С ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ ПРИ РАКЕ МОЛОЧНОЙ ЖЕЛЕЗЫ | 2013 |
|
RU2521506C1 |
| Способ получения зрелых антиген-активированных дендритных клеток с трансфекцией РНК опухолевых клеток рака молочной железы | 2016 |
|
RU2631792C2 |
| СПОСОБ ГЕНЕРАЦИИ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК С АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК РАКА ЯИЧНИКА | 2012 |
|
RU2508298C2 |
| ПРОФИЛАКТИЧЕСКАЯ ИЛИ ТЕРАПЕВТИЧЕСКАЯ ПОЛИЭПИТОПНАЯ ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИННАЯ КОНСТРУКЦИЯ, ОБЕСПЕЧИВАЮЩАЯ ИНДУКЦИЮ КЛЕТОЧНОГО ИММУННОГО ОТВЕТА CD4+ ИЛИ CD8+ Т-ЛИМФОЦИТОВ | 2011 |
|
RU2539035C2 |
Изобретение относится к иммунологии и биотехнологии. Предложен способ модуляции специфического иммунного ответа против антигенов вируса иммунодефицита человека. Выделенные из периферической крови условно здоровых доноров дендритные клетки, презентирующие специфические антигены (полиэпитопный белок TBI), культивируются с неприлипшей фракцией мононуклеарных клеток в присутствии провоспалительных цитокинов рчИЛ-12 и рчИЛ-18. Способ позволяет получать in vitro антиген-специфические клоны Т-клеток с активностью Т-хелперов 1 типа, которые необходимы для формирования эффективного иммунного ответа на антигены ВИЧ. 3 ил.
Способ генерации специфического иммунного ответа против антигенов вируса иммунодефицита человека, заключающийся в совместном культивировании неприлипающей фракции мононуклеарных клеток (МНК), выделенных из периферической крови с дендритными клетками (ДК), активированными антигенами ВИЧ и полученными из моноцитов прилипающей фракции МНК, отличающийся тем, что для совместного культивирования берут зрелые ДК, полученные добавлением к антиген-активированным с помощью полиэпитопного вирусного белка ТВ1 незрелым ДК провоспалительного цитокина ФНО-α, а совместное культивирование неприлипающей фракции МНК и зрелых антиген-активированных ДК проводят в присутствии рекомбинантного человеческого интерлейкина - 12 и рекомбинантного человеческого интерлейкина - 18.
| HUANG X.L | |||
| et al., "Multiple T-cell responses to human immunodeficiency virus type 1 are enhanced by dendritic cells", Clin Vaccine Immunol | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pTBI-HBsAg, СОДЕРЖАЩАЯ ХИМЕРНЫЙ ГЕН TBI-HBsAg ПОД КОНТРОЛЕМ ПРОМОТОРА p7.5K ВИРУСА ОСПОВАКЦИНЫ, И ШТАММ РЕКОМБИНАНТНОГО ВИРУСА ОСПОВАКЦИНЫ, ВЫЗЫВАЮЩИЙ ИММУННЫЙ ОТВЕТ ПРОТИВ ВИЧ И ГЕПАТИТА B ЧЕЛОВЕКА В ОРГАНИЗМЕ ЖИВОТНЫХ | 2001 |
|
RU2194075C2 |
| СПОСОБ ПОДАВЛЕНИЯ ВИЧ-ИНФИЦИРОВАННЫХ КЛЕТОК МЛЕКОПИТАЮЩЕГО И БЕЛКОВЫЙ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1995 |
|
RU2165703C2 |

