Перекрестная ссылка на родственные заявки
Данная заявка заявляет приоритет перед заявками US 61/720,218, поданной 30 октября 2012, US 61/870,418, поданной 27 августа 2013 г., US 61/870,435, поданной 27 августа 2013 г., которые включены в данный документ во всей полноте посредством ссылки.
Область техники
Область изобретения в целом относится к соединениям, композициям и способам лечения рака.
Предшествующий уровень техники
Рак является ведущей причиной смертности в Канаде. По оценке Канадского общества рака в 2011 г. ожидается 170000 новых случаев рака, и приблизительно 75000 смертей в результате рака.
Перспективный подход к лечению рака связан с концепцией синтетической летальности. Два гена (или продукты двух генов) являются синтетически летальными, если мутация какого-либо одного из них совместима с жизнью, а мутация обоих приводит к смерти. Другими словами, «синтетическая летальность» описывает ситуацию, при которой мутация и лекарственное средство (например) вместе вызывают гибель раковых клеток, при этом только мутация или только лекарственное средство не приводят к гибели клеток. Нацеливание на ген (или продукт гена), являющийся синтетически летальным в отношении мутации связанной с раком, должно приводить к гибели только раковых клеток и не затрагивать нормальные клетки. Таким образом, синтетическая летальность обеспечивает фундамент для развития специфичных антираковых агентов.
Подход синтетической летальности к лечению рака является развивающимся, а не рутинным подходом, в основном из-за отсутствия идентификации генов синтетической летальности (и продуктов генов).
N-миристоилирование белков представляет собой модификацию, при которой миристат (14-углеродная насыщенная жирная кислота) ковалентно прикреплен к NH2 терминальному глицину различных клеточных, вирусных и онко-белков (например, онкогенные Src-связанные тирозинкиназы, гетеротримерные G альфа субъединицы, и т.д.).
Клеточные миристоилированные белки выполняют различные биологические функции в сигнальной трансдукции и онкогенезе. Модификация белков посредством миристоилирования необходима для внутриклеточного нацеливания, конформации белка и биологической активности многих важных белков в эукариотических клетках, включая таковые необходимые для сигнальной трансдукции и регуляторных функций, важных для клеточного роста. Тирозинкиназы семейства Src (протоонкогены) являются наиболее хорошо изученными миристоилированными белками.
Миристоилирование белков катализируют посредством N-миристоилтрансферазы (NMT). NMT отвесттвенна за данную активность в эукариотических клетках и работает за счет модификации своего полипептидного субстрата после удаления остатка инициаторного метионина метиониламинопептидазой. Данная модификация в основном проитекает в качестве котрансляционного процесса, хотя миристоилирование также может происходить посттрансляцонно после протеолитического расщепления белков, обычно во время апоптоза. Два изомера фермента NMT млекопитающих были клонированы и названы NMT1 и NMT2. NMT необходимы для выживания клеток. Две NMT присутствуют во всех нормальных клетках.
Все еще существует потребность в соединениях, композиции и способе лечения рака.
Данная дополнительная информация приведена с целью сделать известной информацию, согласно данной заявке предположительно относящуюся к настоящему изобретению. Признание не обязательно предполагается, и не подразумевается, что какая-либо предшествующая информация составляет уровень техники, противоречащий настоящему изобретению.
Краткое описание изобретения
Согласно одному аспекту настоящего изобретения предложены соединения и композиции для лечения субъекта, больного раком. Также предложены способы идентификации субъекта, больного раком, которые приемлемы для лечения при помощи соединений, композиции и способов, описанных в данном документе.
Согласно одному аспекту предложен способ лечения субъекта, имеющего рак, с недостатком NMT2, включающий: введение упомянутому субъекту ингибитора NMT.
В конкретном аспекте упомянутый ингибитор NMT включает ингибитор NMT1. В конкретном аспекте, упомянутый ингибитор NMT1 включает небольшую молекулу, антитело, пептидный фрагмент, нуклеиновую кислоту или их комбинации.
В конкретном аспекте, упомянутая небольшая молекула включает Tris-DBA, НМД, или DDD85646, DDD86481 или их производные.
В конкретном аспекте упомянутое антитело представляет собой моноклональное антитело или поликлональное антитело.
В конкретном аспекте упомянутая нуклеиновая кислота включает молекулу дцРНК (двухцепочечная РНК), молекулу иРНК, молекулу микроРНК, рибозим, молекулу кшРНК или миРНК.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
Согласно другому аспекту предложен способ лечения субъекта, больного раком, или предположительно больного раком, включающий: тест, обеспечивающий результаты анализа на определение экспрессирует ли образец от субъекта NMT2, и введение ингибитора NMT1 субъекту, если образец с недостатком NMT2.
Согласно другому аспекту предложен способ, включающий: получение образца от субъекта, больного раком, или предположительно больного раком; обработку упомянутого образца; выполнение анализа связывания, включающего контактирование обработанного образца с антителом к NMT2 с образованием комплекса между антителом и белком NMT2, присутствующем в обработанном образце, упомянутый анализ связывания, дающий по меньшей мере один результат, свидетельствующий об упомянутом комплексе; где введение ингибитора NMT1 упомянутому субъекту назначают в случае, если количество упомянутого образца белка NMT2 является низким или отсутствует, возможно по сравнению с контролем.
Согласно другому аспекту предложен способ, включающий: получение образца от субъекта, больного раком или предположительно больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с антителом к белку NMT2 с образованием комплекса между антителом и белком NMT2, присутствующим в упомянутом образце, упомянутый анализ связывания, дающий по меньшей мере один результат, свидетельствующий об упомянутом комплексе; и введение ингибитора NMT1 упомянутому субъекту, в случае когда количество белка NMT2 в упомянутом образце является низким или отсутствует, возможно по сравнению с контролем.
В конкретном аспекте упомянутый анализ для определения экспрессирует ли упомянутый образец от субъекта NMT2, включает проведение анализа связывания обработанного образца с антителом к NMT2 с образованием комплекса между антителом и NMT2, присутствующим в обработанном образце, упомянутого анализа связывания, дающего по меньшей мере один результат, свидетельствующий об упомянутом комплексе.
В конкретном аспекте упомянутый анализ связывания включает сортировку клеток с активацией флуоресценции, твердофазный иммуноферментный анализ, иммуногистохимию, количественную иммуногистохимию, резонансный перенос энергии флуоресценции, Ферстеровский резонансный перенос энергии, биомолекулярную комплементацию флуоресценции, масс-спектрометрию, иммуноблотинг или коиммунопреципетацию.
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса, образованного между упомянутым антителом и упомянутым NMT2 в упомянутом образце.
В конкретном аспекте упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложен способ, включающий: получение образца от субъекта, больного раком или предположительно больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с детектируемой меткой, связывающейся с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, упомянутый анализ связывания, дающий по меньшей мере один результат, свидетельствующий об упомянутом комплексе; где введение ингибитора NMT1 упомянутому субъекту показано, в случае когда количество нуклеиновой кислоты NMT2 в упомянутом образце является низким или отсутствует, возможно по сравнению с контролем.
В другом аспекте предложен способ, включающий: получение упомянутого образца от субъекта, больного раком или предположительно больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, упомянутый анализ связывания, дающий по меньшей мере один результат, означающий упомянутый комплекс; и введение ингибитора NMT1 упомянутому субъекту показано в случае, когда количество нуклеиновой кислоты NMT2 в упомянутом образце является низким или отсутствует, возможно по сравнению с контролем.
В конкретном аспекте упомянутый анализ для определения экспрессирует ли упомянутый образец от субъекта NMT2, включает проведение анализа связывания, включающего контактирование обработанного образца с детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, упомянутый анализ связывания, дающий по меньшей мере один результат, свидетельствующий об упомянутом комплексе.
В конкретном аспекте упомянутый анализ связывания включает гибридизационный анализ при помощи детектируемо меченных проб ДНК или РНК.
В конкретном аспекте упомянутый гибридизационный анализ является количественным или полуколичественным.
В конкретном аспекте упомянутый гибридизационный анализ представляет собой РВ-ПЦР, гибридизацию in situ, РНК-РНК гибридизация в растворе (RNA protection assay, RPA), микроэррей кДНК и олигонуклеотида, репрезентативный разностный анализ ("RDA"), дифференциальный дисплей, анализ маркерных экспрессируемых последовательснотей (EST), серийный анализ экспрессии генов ("SAGE"), и мульиплексную амплификацию, опосредованную лигированием, при помощи Luminex FlexMAP ("LMF").
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце.
В конкретном аспекте упомянутое оборудование представляет собой спектрофтометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложен способ, включающий получение образца от субъекта, больного раком или предположительно больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с антителом, к которому присоединяется миристоилированный белок, или азидо-биотин меченными миристоилированными белками, внутри образца с образованием комплекса между детектируемой меткой и миристоилированным белком, присутствующим в образце, упомянутый анализ связывания, дающий по меньшей мере один профиль миристоилирования, свидетельствующий об упомянутом комплексе, где введение ингибитора NMT1 упомянутому субъекту показано в случае, когда упомянутый профиль миристоилирования означает рак с недостатком NMT2, возможно по сравнению с контролем.
В другом аспекте предложен способ, включающий получение образца от субъекта, больного раком или предположительно больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с антителом, которое связывается с миристоилированным белком или азидо-биотин меченными миристоилированными белками, внутри образца с образованием комплекса между детектируемой меткой и миристоилированным белком, присутствующим в образце, упомянутый анализ связывания, дающий по меньшей мере один профиль миристоилирования, свидетельствующий об упомянутом комплексе; и введение ингибитора NMT1 упомянутому субъекту показано в случае, когда упомянутый профиль миристоилирования означает, что упомянутый рак с недостатком NMT2, возможно по сравнению с контролем.
В конкретном аспекте упомянутая обработка включает обработку упомянутого образца алкинил-миристатом и детиобиотин азидо-ПЭГ-биотином.
В конкретном аспекте упомянутое связывание включает сортировку клеток с акивацией флуоресценции, твердофазный иммуноферментный анализ, иммуногистохимию, количественную иммуногистохимию, резонансный перенос энергии флуоресценции, Ферстеровский резонансный перенос энергии, биомолекулярную комплементацию флуоресцениции, масс-спектрометрию, иммуноблотинг или коиммунопреципитацию.
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса, образованного между упомянутым антителом и упомянутым NMT2 в упомянутом образце.
В конкретном аспекте упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложено применение ингибитора NMT для лечения субъекта, больного раком с недостатком NMT2.
В конкретном аспекте упомянутый ингибитор NMT включает ингибитор NMT1.
В конкретном аспекте упомянутый ингибитор NMT1 включает небольшую молекулу, антитело, пептидный фрагмент, нуклеиновую кислоту или их комбинацию.
В конкретном аспекте упомянутая небольшая молекула включает Tris-DBA, НМА, или DDD85646, DDD86481 или их производные.
В конкретном аспекте упомянутая небольшая молекула включает DDD86481.
В конкретном аспекте упомянутое антитело является моноклональным антителом или поликлональным антителом.
В конкретном аспекте упомянутая нуклеиновая кислота включает молекулу дцРНК, молекулу иРНК, молекулу микроРНК, рибозим, молекулу кшРНК или молекулу миРНК.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложено применение ингибитора NMT1 для лечения субъекта, больного раком или предположительно больного раком, где упомянутый ингибитор NMT1 показан к применению в случае, когда количество белка NMT2 в образце от упомянутого субъекта является низким или отсутствует, возможно по сравнению с контролем, где анализ связывания, включающий контактирование обработанного образца от упомянутого субъекта с антителом к NMT2 с образованием комплекса между антителом и белком NMT2, присутствующим в обработанном образце, дает по меньшей мере один резкультат анализа, свидетельствующий об упомянутом комплексе; где упомянутый результат анализа означает упомянутое количество белка NMT2 в образце.
В конкретном аспекте упомянутый анализ для определения экспрессирует ли упомянутый образец от субъекта NMT2, включает проведение анализа связывания, включающего контактирование обработанного образца с антителом к NMT2 с образованием комплекса между антителом и NMT2, присутствующим в обработанном образце, упомянутый анализ связывания, дающий по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе.
В конкретном аспекте упомянутый анализ связывания включает сортировку клеток с активацией флуоресценции, твердофазный иммуноферментный анализ, иммуногистохимию, количественную иммуногистохимию, резонансный перенос энергии флуоресценции, Ферстеровский резонансный перенос энергии, биомолекулярную комплементацию флуоресценции, масс-спектрометрию, иммуноблотинг или коиммунопреципитацию.
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса, образованного между упомянутым антителом и упомянутым NMT2 в упомянутом образце.
В конкретном аспекте упомянутое оборуование представляет собой спектрофотметр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложено применение ингибитора NMT1 для лечения субъекта, больного раком или предположительно больного раком, где упомянутый ингибитор NMT1 показан к применению в случае, когда количество нуклеиновой кислоты NMT2 в образце от упомянутого субъекта является низким или отсутствует, возможно по сравнению с контролем, где анализ связывания, включающий контактирование обработанного образца от упомянутого субъекта с детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2 в упомянутом образце, упомянутый анализ связывания, дающий по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе.
В конкретном аспекте упомянутый анализ для определения экспрессирует ли упомянутый образец от субъекта NMT2, включает проведение анализа связывания, включающего контактирование обработанного образца с детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, упомянутый анализ связывания, дающий по меньшей мере один результат анализа, означающий упомянутый комплекс.
В конкретном аспекте упомянутый анализ связывания включает гибридизационный анализ при помощи меченных проб ДНК или РНК.
В конкретном аспекте упомянутый гибридизационный анализ является количественным или полуколичественным.
В конкретном аспекте упомянутый гибридизационный анализ представляет собой РВ-ПЦР, гибридизацию in situ, РНК-РНК гибридизация в растворе (RNA protection assay, RPA), микроэррей кДНК и олигонуклеотида, репрезентативный разностный анализ ("RDA"), дифференциальный дисплей, анализ маркерных экспрессируемых последовательснотей (EST), серийный анализ экспрессии генов ("SAGE"), и мульиплексную амплификацию, опосредованную лигированием, при помощи Luminex FlexMAP ("LMF").
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса между детектируемой меткой и нуклеиновой кислотой NMT2.
В конкретном аспекте упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическе устройство ли элктрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В одном аспекте предложено применение ингибитора NMT1 для лечения субъекта, больного раком, где упомянутое применение ингибитора NMT1 показано в случае, когда профиль миристоилирования образца от субъекта свидетельствует о раке с недостатком NMT2, возможно по сравнению с контролем, где проведение анализа связывания, включающего контактирование обработанного образца с антителом, которое связывается с миристоилированным белком, или азидо-биотин меченными миристоилированными белками, внутри образца с образованием комплекса между детектируемой меткой и миристоилированным белком, присутствующем в образце, упомянутый анализ связывания дающий по меньшей мере один профиль миристоилирования, свидетельствующий об упомянутом комплексе.
В конкретном аспекте упомянутая обработка включает обработку упомянутого образца алкинил-миристатом и детиобиотин азидо-ПЭГ-биотином.
В конкретном аспекте упомянутый анализ связывания включает сортировку клеток с активированной флуоресценции, твердофазный иммуноферментный анализ, иммуногистохимию, количественную иммуногистохимию, резонансный перенос энергии флуоресценции, Ферстеровский резонансный перенос энергии, биомолекулярную комплементацию флуоресценции, масс-спектрометрию, иммуноблотинг или коиммунопреципитацию.
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса, образованного между упомянутым антителом и упомянутым NMT2 в образце.
В конкретном аспекте упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложен способ идентификации субъекта, приемлемого для лечения при помощи ингибитора NMT1, включающий получение образца от упомянутого субъекта, больного раком или предположительно больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с антителом к NMT2 с образованием комплекса между антителом и белком NMT2, присутствующем в обработанном образце, упомянутый анализ связывания, дающий по меньшей мере один результат анализа, означающий упомянутый комплекс; где лечение упомянутым ингибитором NMT1 показано в случае, когда количество белка NMT2 в упомянутом образце является низким или отсутствует, возможно по сравнению с контролем.
В конкретном аспекте упомянутый анализ для определения экспрессирует ли упомянутый образец от субъекта NMT2, включает проведение анализа связывания, включающего контактирование обработанного образца с антителом к NMT2 с образованием комплекса между антителом и NMT2, присутствующим в обработанном образце, упомянутый анализ связывания, дающий по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе.
В конкретном аспекте упомянутый анализ связывания включает сортировку клеток с активированной флуоресценции, твердофазный иммуноферментный анализ, иммуногистохимию, количественную иммуногистохимию, резонансный перенос энергии флуоресценции, Ферстеровский резонансный перенос энергии, биомолекулярную комплементацию флуоресценции, масс-спектрометрию, иммуноблотинг или коиммунопреципитацию.
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса, образованного между упомянутый антителом и упомянутым NMT2 в упомянутом образце.
В конкретном аспекте упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложен способ идентификации субъекта, приемлемого для лечения ингибитором NMT1, включающий получение образца от субъекта, больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, упомянутый анализ связывания, дающий по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе; где введение ингибитора NMT1 упомянутому субъекту показано в случае, когда количество нуклеиновой кислоты NMT2 в упомянутом образце является низким или отсутствует, возможно по сравнению с контролем.
В конкретном аспекте упомянутый анализ для определения экспрессирует ли упомянутый образец от субъекта NMT2, включает проведение анализа связывания, включающего контактирование обработанного образца с детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, упомянутый анализ связывания дающий по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе.
В конкретном аспекте упомянутый анализ включает гибридизационный анализ при помощи детектируемо меченных проб ДНК или РНК.
В конкретном аспекте упомянутый гибридизационный анализ является количественным или полуколичественным.
В конкретном аспекте упомянутый гибридизационный анализ представляет собой РВ-ПЦР, гибридизацию in situ, РНК-РНК гибридизация в растворе (RNA protection assay, RPA), микроэррей кДНК и олигонуклеотида, репрезентативный разностный анализ ("RDA"), дифференциальный дисплей, анализ маркерных экспрессируемых последовательснотей (EST), серийный анализ экспрессии генов ("SAGE"), и мульиплексную амплификацию, опосредованную лигированием, при помощи Luminex FlexMAP ("LMF").
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для определения комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце.
В конкретном аспекте упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте упомянутый субъект является человеком.
В другом аспекте предложен способ идентификации субъекта, приемлемого для лечения при помощи ингибитора NMT1, включающий получение образца от субъекта, больного раком или предположительно больного раком; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с антителом к миристоилированному белку или азидо-биотин меченными миристоилированными белками, внутри образца с образованием комплекса между детектируемой меткой и миристоилированным белком, присутствующем в образце, упомянутый анализ связывания, дающий по меньшей мере один профиль миристоилирования, свидетельствующий об упомянутом комплексе; где введение ингибитора NMT1 упомянутому субъекту показано в случае, когда упомянутый профиль миристоилирования означает упомянутый рак с недостатком NMT2, возможно по сравнению с контролем.
В конкретном аспекте упомянутая обработка включает обработку упомянутого образца алкинил-миристатом и детиобиотин азидо-ПЭГ-биотином.
В конкретном аспекте упомянутый анализ связывания включает сортировку клеток с активацией флуоресценции, твердофазный иммуноферментный анализ, иммуногистохимию, количественную иммуногистохимию, резонансный перенос энергии флуоресценции, Ферстеровский резонансный перенос энергии, биомолекулярную комплементацию флуоресценции, масс-спектрометрию, иммуноблоттинг или коиммунопреципитацию.
В конкретном аспекте для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для обнаружения комплекса между упомянутый антителом и упомянутым NMT2 в упомянутом образце.
В конкретном аспекте упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
В конкретном аспекте упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
В конкретном аспекте, где упомянутый субъект является человеком.
В другом аспекте предложен набор для идентификации субъекта приемлемого для лечения ингибитором NMT1, включающий антитело к NMT2; инструкции для идентификации субъекта согласно способу по любому из п.п. 68-74.
В конкретном аспекте набор дополнительно включает контроль.
В другом аспекте предложен набор для идентификации субъекта, приемлемого для лечения ингибитором NMT1, включающий нуклеиновую кислоту для связывания с NMT2; инструкции для идентификации субъекта согласно способу по любому из п.п. 75-83.
В конкретном аспекте набор дополнительно включает контроль.
Набор по п. 93, где упомянутая нуклеиновая кислота представляет собой РНК или ДНК.
В другом аспекте предложен набор для идентификации субъекта, приемлемого для лечения ингибитором NMT1, включающий NeutrAvidinTM-HRP; и инструкции для идентификации упомянутого субъекта по любому из п.п. 84-90.
В конкретном аспекте набор дополнительно включает контроль.
В конкретном аспекте набор дополнительно включает алкинил-миристат или детиобиотин азидо-ПЭГ-биотин.
Краткое описание графических материалов
Воплощения настоящего изобретения будут сейчас описаны посредством только примеров со ссылкой на прикрепленные фигуры, где:
На фигуре 1 изображен иммуноблот экспрессии NMT1 и NMT2 в одном типе нормальных В-клеток (L0) и различных В-клеточных лимфомах и Т-клеточных лейкемиях;
Фигура 2 представляет собой графическую иллюстрацию чувствительности различных нормальных клеток и различных В-клеточных лимфом и Т-клеточных лейкемий к ингибиторам NMT трис-дебензилиденацетон-дипалладий (Tris-DBA);
Фигура 3 представляет собой диаграмму, иллюстрирующую ингибирование N-миристоилтрансферазы (NMT) посредством трис-дебензилиденацетон-дипалладия (Tris-DBA); и
Фигура 4 представляет собой иммуноблотты, изображающие клеточные линии лимфом, изученные с антителами к NMT1 и NMT2.
Фигура 5 представляет собой линейный график, показывающий чувствительность ингибиторов NMT на клеточной линии лимфомы Беркитта в сравнении с иммортализованной нормальной клеточной линией В-лимфоцитов;
Фигура 6 отображает результаты трансфекции В-клеток лимфомы Рамоса при помощи pcDNA3.1-V5-NMT2, показывающие увеличенную устойчивость к Tris-DBA (5 мкг/мл) 2,5 кратно vs клеток контроля, трансфецированных пустым плазмидным вектором (панель А), показывающих жизнеспособность клеток и (панель В) иммуноблот;
Фигура 7 отображает различия в уровнях белка NMT2, присутствующих в различных клеточных линиях лимфоцитов и твердых лимфоидных опухолях;
Фигуры 8А и Б отображают различия в уровнях белка NMT2, присутствующего в различных твердых лимфоидных опухолях, пронанализированных посредством иммуногистохимии: NMT иммуногистохимическое окрашивание нормальных лимфотических узлов, лимфомы Беркитта (BL) и диффузной В-крупноклеточной лимфомы (DLBCL), в панели D log2(интенсивность флуоресценции микроаррей NMT) для NMT1 и NMT2 приведена для всех клеточных линий базы данных CCLE, в панели D log2(интенсивность флуоресценции микроаррей NMT) для NMT1 и NMT2 приведена для всех 100 клеточных линий базы данных CCLE с наименьшим уровнем экспрессии NMT2, (панель Е) экспрессию NMT2 в лимфоме Беркитта, диффузной В-крупноклеточной лимфоме и фолликулярной лимфоме анализировали посредством иммуногистохимии и окрашивание пероксидазой оценивали при помощи Image J. Содержание NMT2 в нормльном лимфотическом узле составляет 0,392 +/- 0,3 (относительная единица, данные не приведены).
Фигура 9 отображает остаточную жизнеспособность различных клеточных линий В-лимфоцитов, обработанных DDD85646 (панель A), DDD86481 (панель В), и DDD73226 (панель С) в течение 72 часов, панель D изображает подтверждение ингибирования миристоилирования посредством DDD86481 в IM9 (i) и BL2 (ii) лимфоцитах, панель Ε изображает минимальную дозу DDD86481, необходимую для ингибирования миристоилирования в клеточных линиях ΙΜ-9, BL2 и Рамоса;
Фигура 10 (панель А) отображает чувствительность различных иммортализованных нормальных L0 В-лимфоцитов, В-клеток злокачественной лимфомы (Рамоса и BL2), и Т-лейкемических клеток (СЕМ) к ингибитору NMT Tris-DBA в течение 24 часов, (панель В) показывает, что трансфекция В-клеток лимфомы Рамоса при помощи pcDNA3.1-V5-NMT2 повышала устойчивость к Tris-DBA (5 мкг/мл) 2,5 кратно vs контрольных клеток, трансфецированных пустым плазмидным вектором.
Фигура 11 отображает экспрессионные уровни NMT в 967 клеточных линиях рака базы данных (CCLE) (панель A), NMT2 экспрессию в выбранном раке при помощи диаграммы «ящик с усами» (панель Б), 50 CCLE клеточных линий с самой низкой экспрессией NMT2 приведены и сортированы по типу рака;
Фигура 12 показывает, что NMT расщепляются во время апоптоза, но остаются активными;
Фигура 13 изображает очистку рекомбинантной GST- и His6-меченных hNMT1 и hNMT2;
Фигура 14 отображает сравнение ферментативных активностей рекомбинантного полноразмерного His6-NMT1 и рекомбинантного "каспазо-процессированного" ct-His6-NMT1;
Фигура 15 отображает сравнение ЕС50 и IC50 различных ингибиторов NMT и различных клеточных линий;
Фигура 16 отображает индукцию апоптоза DDD86481;
Фигура 17 отображает (панель А) иммуноблот с видимыми лимфоидными кетками, изученными на присутствие NMT2, и (панель В) экпрессионный уровень NMT2 мРНК, показанный в виде log2(микроэррей интенсивности флуоресценции NMT2) для указанных клеточных линий базы данных CCLE;
Фигура 18 отображает применение детиобиотин-ПЭГ-азида для посттрансляционных ω-алкинил-миристоилированных белков (pull-down) в лейкемических Т-клетках Jurkat при помощи стрептавидин-сефарозных гранул;
Фигура 19 отображает повышенное применение детиобиотин-ПЭГ-азидной пробы в отношении посттрансляционных ω-алкинил-миристоилированных белков (pull-down) в лейкемических Т-клетках Jurkat при помощи стрептавидин-магнитных гранул;
Фигура 20 отображает профили миристоилирования «нормальных» иммортализированных В-клеток (IM9) и BL клеток (BL2 и Рамоса), меченных алкинил-миристатом;
Фигура 21 отображает графики время и дозозависимой цитотоксичности комбинации DDD86481 и доксорубиксина.
Фигура 22 отображает иммуноблоттинг, проведенный с клетками, инкубированными с ДМСО, стауроспорином, α FAS или только носителем;
Фигура 23 показывает, что каспазо-процессированная NMT2 является в 3-4 раза более активной, чем полноразмерная NMT2;
Фигура 24 отображает иммуноблоты «нормальных» В-клеток (IM9) и злокачественных BL клеток (Рамос, BL2), обработанных 1 мкМ SAHA (HDAC класс I/II ингибитор) в течение 24 часов;
Фигура 25 отображает, что уровни белка NMT2 снижены в различных В-клеточных линиях;
Фигура 26 отображает, что протеасомная деградация не является причиной элиминации NMT2 в BL клетках;
Фигура 27 отображает, что каспаза-8 расщепляет NMT1, но не NMT2;
Фигура 28 отображает, что как NMT1, так и NMT2 расщепляются каспазой-3;
Фигура 29 отображает сайты расщепления каспазы NMT1 и NMT2, как идентифицировано посредством расщепления по Эдману, приведены жирным шрифтом и положительно заряженный бокс лизина (К) выделен светлым на аминокислотных последовательностях NMT1 и NMT2 (аминокислоты 1-80);
Фигура 30 отображает подтверждение сайтов расщепления NMT посредством сайт-направленного мутагенеза;
Фигура 31 отображает сайты расщепления каспазы NMT1 и NMT2, как идентифицировано посредством расщепления по Эдману, выделенные жирным шрифтом, и положительно заряженный бокс лизина (К), выделенный светлым на аминокислотных последовательностях NMT1 и NMT2 (аминокислоты 1-80);
Фигура 32 отображает изменения уровней NMT в результате апоптоза клеток;
Фигура 33 отображает начальную активность NMT в лизатах временно трансфецированных клеток COS7;
Фигура 34 отображает начальную активность NMT в лизатах временно трансфецированных клеток COS7.
Фигура 35 отображает активность NMT в клетках COS7, временно экспрессирующих V5-NMT1 и V5-NMT2, инкубированных со стауроспорином (2,5 мкМ) и циклогексимидом (5 мкг/мл);
Фигура 36 отображает очистку рекомбинантной гексагистидин(Н1з)-меченной полноразмерной и каспазо-процессированной ηΝΜΤΊ;
Фигура 37 отображает очистку рекомбинантной гексагистидин(Н1в)-меченной полноразмерой и каспазо-процессированной hNMT2;
Фигура 38 отображает активность NMT очищенных полноразмерой и каспазо-процессированой гексагистидин(His)-NMT, анализированной при помощи анализа миристоилирования пептидов;
Фигура 39 отображает субклеточное фракционирование эндогенных NMTs в клетках HeLa во время апоптоза;
Фигура 40 отображает определение количества NMT в различных фракциях после субклеточного фракционирования эндогенных NMT в клетках HeLa в течение апоптоза; и
Фигура 41 отображает субклеточное фракционирование клеток HeLa преерпевших апоптоз и меченных алкинил-миристатом; и
Фигура 42 отображает влияние 2-гидроксимиристиновой кислоты (НМА) на индукцию апоптоза. Т-клетки Jurkat обрабатывали или не обрабатывали НМА (1 мм) и апоптоз индуцировали при помощи анти-Fas (150 нг/мл) и циклогексимида (5 мкг/мл).
В последующем подробном описании числа жирным шрифтом служат для идентификации составляющих частей, которые описаны и относятся к иллюстрациям, изображающим различные воплощения изобретения. Следует отметить, что в описании различных воплощений настоящего изобретения одни и те же ссылочные номера применяют для обозначения одинаковых из сходных элементов. Более того, для простоты, части не указаны на некоторых фигурах графических материалов.
Подробное описание изобретения
Как описано в деталях ниже, в данном документе описаны соединения, композиция и способы лечения субъекта, больного раком. Также в данном документе описаны способы идентификации субъекта, имеющего рак, приемлемые для лечения соединениями, композицией и способами, описанными в данном документе. Также в данном документе описаны способы идентификации субъекта, больного раком.
Настоящая заявка предлагает способы и композиции для лечения у субъекта рака с недостатком NMT. NMT-недостаточный рак включает рак с недостатком NMT2 или NMT1. В конкретном примере рак с недостатком NMT, представляет собой рак с недостатком NMT2.
Термин «рак», как употреблено в данном документе, относится к ряду условий, вызываемых ненормальным, неконтролируемым ростом клеток. Клетки способные вызывать рак относятся к «раковым клеткам», обладают характерными свойствами, такими как неконтролируемая пролиферация, бессмертие, метаститический потенциал, быстрая скорость роста и пролиферации и/или определенные типичные морфологические черты. Раковые клетки могут быть в виде опухоли, но такие клетки также могут существовать по отдельности внутри субъекта, или могут не быть опухолегенными раковыми клетками. Рак может быть детектирован любым числом способов, включая, но не ограничено, детектирование присутствия опухоли или опухолей (например, посредством клинических или радиологических способов), исследования клеток внутри опухоли или от другого биологического образца (например, тканевого биоптата), измерения маркеров крови, указывающих на рак, и детектирование генотипа, указывающего на рак. Однако отрицательный результат одного или более из упомянутых способов детекции не обязательно означает отсутствие рака, например, пациент, проявляющий полный ответ на лечения рака, все еще может иметь рак, что подтверждается последующим рецидивом.
В конкретном примере настоящего раскрытия рак представляет собой лимфому.
Термин «лимфома», как употреблено в данном документе относится к злокачественному росту В- или Т-клеток в лимфатической системе. «Лимфома» включает различные типы злокачественного роста, включая Лимфому Ходжкина, и неходжкинскую лимфому. Термин «неходжкинекая лимфома», как употребелено в данном документе относится к злокачественному росту В- или Т-клеток в лимфатической системе, который не является лимфомой Ходжкина (которая характеризуется, например, посредством присутствия клеток Рид-Штернберга в раковой области). Неходжкинская лимфома охватывает 29 типов лимфом, различия между которыми основаны на типе раковых клеток.
В более конкретном примере настоящего раскрытия рак представляет собой В-лимфому.
Таким образом, в одном воплощении соединения, композиции и способы являются приемлемыми для лечения субъекта, больного В-клеточной лимфомой.
Примеры В-клеточных лимфом включают, но не ограничены, например, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийно-клеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема и лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому. Также рассматривают лечение детских лимфом, таких как лимфомы Беркитта, диффузной В-крупноклеточной лимфомы, фолликулярной лимфомы, В-лимфобластной лимфомы из клеток предшественников, Т-лимфобластной лимфомы из клеток предшественников, и анапластической крупноклеточной лимфомы.
В другом воплощении рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легких, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
Термин «субъект», употребляемый в данном документе, относится к животному и может включать, например, одомашненных животных, таких как коты, собаки и т.д., домашний скот (например, крупный рогатый скот, лошади, свиньи, овцы, козы и т.д.), лабораторных животных (например, мыши, кролики, крысы, морские свинки и т.д.), млекопитающих, млекопитающих, не являющихся человеком, приматов, приматов, не являющихся человеком, грызунов, птиц, рептилий, земноводных, рыб и любых других животных. В конкретном примере субъект является человеком.
Термин «лечение» или «лечить», как употреблено в данном документе, относится к получению благоприятных или желаемых результатов, включая клинические результаты. Благоприятные или желаемые клинические результаты могут включать, но не ограничены, облегчение или улучшение одного или более симптомов или состояний, уменьшение степени заболевания, стабилизированное (т.е. не ухудшающееся) состояние заболевания, предупреждение распространения заболевания, приостановку или замедление прогресса заболевания, улучшение или временное облегчение состояния заболевания, уменьшение повторения заболевания и ремиссию (частичную или полную), детектируемую или недетектируемую. «Лечение» или «лечение» также означает продление выживания по сравнению с ожидаемым выживанием при отсутствии лечения. «Лечение» и «лечение», как употреблено в данном документе, также включает профилактическое лечение. Например, субъект с ранней стадией рака, например, с ранней стадией лимфомы, может получать лечение для предупреждения прогресса или альтернативно субъект в ремиссии может получать лечение соединением или композицией, описанной в данном документе, для предупреждения повторения.
В данном документе показано, что клетки В-клеточной лимфомы экспрессируют NMT1, но не NMT2. Это сильно отличается от лейкемических и других клеток, экпрессирующих как NMT1, так и NMT2 (как показано на фигурах 1 и 4).
Как показано в данном документе, клетки В-лимфомы чувствительны к ингибированию клеточной жизнеспособности ингибиторами NMT.
В одном примере NMT ингибитор представляет собой трис-дибензилиденацетон-дипалладий (Tris-DBA) (фигура 2).
В других примерах ингибитор NMT 2-гидроксимиристат (НМА) применяют для ингибирования клеток В-лимфомы.
В еще одном примере для ингибирования клеток применяют В-лимфомы ингибитор NMT пиразол сульфонамид T. brucie (DDD85646) [J.A. Frearson et al (2010) Nature. 464.728-723)] (фигура 5).
В другом примере ингибитор представляет собой DDD86481.
В конкретном примере лечение субъекта, больного В-лимфомой, включает введение упомянутому субъекту ингибитора NMT.
Соединения ингибитора NMT или производные могут быть применены в настоящем изобретении для лечения рака с недостатком NMT2.
Термин «с недостатком», как употреблено в данном документе, относится в широком смысле к ингибированию, уменьшению или элиминации (по сравнению с диким типом или контрольными образцами), например, синтеза, уровней, активности или функции NMT, а также ингибированию индукции или стимуляции синтеза, уровней, активности или функции белка (например, NMT1 или NMT2). Термин также относится к любому метаболическому или регуляторному пути, который может регулировать синтез, уровни, активность или функцию NMT. Термин также включает ингибирование, снижение или элиминацию в результате связывания с другими молекулами и образования комплекса. Поэтому, термин " с недостатком NMT" относится к таковому, который приводит к ингибированию, снижению или элиминации функции белка или функции белкового пути. Однако термин не подразумевает, что каждая или все эти функции должны быть ингибированы в одно и тоже время.
В некоторых примерах, рак может быть идентифицирован в качестве с недостатком NMT посредством определения присутствия мутации в гене NMT. Такие способы детекции нуклеиновой кислоты и амплификации хорошо известны специалистам в области техники.
Например, амплифицируемая нуклеиновая кислота может быть получена из биологического образца. Различные способы (такие как экстрация фенол-хлороформом) экстракции приемлемы для выделения ДНК или РНК. Нуклеиновая кислота, выделенная из образца, может быть амплифицирована при помощи способов амплификации нуклеиновых кислот, хорошо известны в области техники. Неограничивающие примеры включают полимеразную цепную реакцию (ПЦР), полимеразуню цепную реакцию с обратной транскрипцией (ОТ-ПЦР), гнездовую ПЦР, лигазную цепную реакцию, амплифицируемые РНК-репортеры, Q-бета репликацию, амплификацию на основе транскрипции, бумеранг-ДНК-амплификацию, амплификацию замещением цепей, технологию циклического зонда, изотермической амплификации, основанной на последовательности нуклеиновых кислот (NASBA), или могут быть применены другие репликационные анализы последовательностей или способы сигнальной амплификации.
Способы амплификации хорошо известны в области техники. Некоторые способы применяют обратную транскрипцию РНК в кДНК.
В одном примере ПЦР применяют для амплификации интересующей мишенной последовательности, например, последовательности NMT2.
Нуклеиновые кислоты могут быть амплифицированы перед детекцией или могут быть детектированы непосредственно во время этапа амплификации, например, способами «в реальном времени». В некоторых воплощениях мишенную последовательность амплифицируют при помощи меченного праймера такого, что полученный ампликон является детектируемо меченным. В некоторых воплощениях праймер является флуоресцентномеченным. В некоторых воплощениях мишенную последовательность амплифицируют, и полученный ампликон детектируют при помощи электрофореза.
Уровень экспрессии гена может быть определен посредством оценки количества мРНК NMT2 в образце. Способы измерения мРНК в образцах известны в области техники. Для измерения уровней мРНК клетки в образцах могут быть лизированы, и уровни мРНК в лизатах или очищенной РНК или полуочищенной РНК, могут быть измерены посредством различных способов, известных специалистам в области техники. Такие способы включают, без ограничения, гибридизационные исследования при помощи детектируемо меченных ДНК или РНК проб, например, нозерн-блотта, или колличественных или полуколличественных РВ-ПЦР способов при помощи соответствующих олигонуклеотидных праймеров. Альтернативно, колличественный или полуколичественный гибридизационный анализ in situ может быть выполнен при помощи, например, кусочков ткани, или нелизированных клеточных суспензий, и детектируемо меченных, например, флуоресцентно или ферментно-меченных ДНК или РНК проб. Дополнительные способы для колличественной оценки мРНК включают РНК-РНК гибридизация в растворе (RNA protection assay, RPA), микроэррей кДНК и олигонуклеотида, репрезентативный разностный анализ ("RDA"), дифференциальный дисплей, анализ маркерных экспрессируемых последовательснотей (EST), серийный анализ экспрессии генов ("SAGE"), и мульиплексную амплификацию, опосредованную лигированием, при помощи Luminex FlexMAP ("LMF").
Амплификация также может быть мониторирована при помощи способов «в реальном времени». ПЦР в реальном времени позволяет детектировать и колличественно оценивать мишенную нуклеиновую кислоту. Обычно данный подход к количественной ПЦР применяет флуоресцентный краситель, который может специальным красителем для двуцепочечной ДНК, таким как SYBR Green. RTM. I. Альтернативно другие флуоресцентные красители, например, FAM или HEX, могут быть конъюгированы с олигонуклеотидной пробой или праймером. Различное оборудование для проведения ПЦР в реальном времени известно в области техники. Флуоресцентный сигнал, генерируемый в каждом цикле ПЦР, пропорционален количеству ПЦР-продукта. Единицу флуоресценции против номера цикла применяют для описания кинетики амплификации, и пороговое значение флуоресценции применяют для определения дробного номера цикла, связанного с изначальной концентрации матрицы. В случае, когда амплификацию проводят и детектируют на оборудовании, способном считывать флуоресценцию в течение термоциклирования, предполагаемый продукт ПЦР может быть отдифференцирован от неспецифичных продуктов при помощи анализа плавления. За счет измерения изменения флуоресценции во время градуального повышения температуры реакции после амплификации и генерации сигнала, становиться возможным определение (Act) предполагаемого продукта(тов), также как и неспецифического продукта.
Способы могут включать амплификацию множества нуклеиновых кислот в образце, также известную как «мультиплексная детекция» или «мультиплексирование». Как употреблено в данном документе термин «мультиплексная ПЦР» относится к ПЦР, которая вовлекает добавление более чем одного набора ПЦР праймеров в реакцию с целью детектировать и количественно оценить множество нуклеиновых кислот, включая нуклеиновые кислоты из одного или более мишенных генных маркеров. Более того, мультиплексирование с внутренним контролем, например, геном 18s рРНК, GADPH или бета-актина) обеспечивает внутренний контроль для ПЦР без реакции.
В некоторых примерах рак может быть идентифицирован в качестве с недостатком NMT посредством определения эпигенетической инактивации гена NMT.
В некоторых примерах, рак может быть идентифицирован в качестве с недостатком NMT за счет определения активности NMT (включая NMT1 или NMT2) в образце клеток от субъекта. Активность может быть определена относительно контроля, например, в случае дефектов в раковых клетках, относительно нераковых клеток, предпочтительно из той же ткани. Таким образом, рак с недостатком NMT, может иметь пониженную или элиминированную активность и/или экспрессию NMT. Активность NMT может быть определена за счет применения способов, хорошо известных в области техники, и/или описанных в данном документе. В данных примерах, рак с недостатком NMT, обладает пониженной или элиминированной активностью.
В некоторых примерах рак может быть идентифицирован в качестве с недостатком NMT (например, NMT1, NMT2 или обоих), за счет определения количества, концентрации и/или уровней NMT белка(в).
В некоторых примерах рак может быть идентифицирован в качестве с недостатком NMT, за счет определения количества миристоилированных белков в биологическом образце от субъекта, больного раком, или предположительно больного раком. В данном примере присутствие, отсутствие или количество миристоилированного белка может быть определено, например, при помощи клик-химии при помощи соответствующих жирнокислотных аналогов. Неограничивающие способы описаны в данном документе. Альтернативные способы определения присутствия, отсутствия или количества миристоилированных белков хорошо известны специалистам в области техники. Образец, обладающий пониженным количеством миристоилированного белка в образце (возможно по сравнению с контролем) является показателем образца с недостатком NMT, или рака с недостатком NMT. В некоторых примерах, образец, обладающий пониженным количеством миристоилированного белка в образце, является показателем образца с недостатком NMT2, или рака с недостатком NMT2.
В некоторых примерах, рак может быть идентифицирован в качестве с недостатком NMT за счет определения количества ацилирования белков в биологическом образце от субъекта, больного раком или предположительно больного раком. В данном примере присутствие, может быть определено отсутствие или дана количественная оценка ацилирования белков. Такие способы будут известны специалистам в области техники. Образец, обладающий пониженным количеством ацилирования белков в образце (возможно по сравнению с контролем) является показателем образца с недостатком NMT, или рака с недостатком NMT. В некоторых примерах образец, обладающий пониженным количеством ацилирования белков в образце, является показателем образца с недостатком NMT2, или рака с недостатком NMT2.
В некоторых примерах рак может быть идентифицирован в качестве с недостатком NMT, за счет определения присутствия одной или более вариаций последовательностей, таких как мутации, и полиморфизмы могут включать делецию, инсерцию или замену одного или более нуклеотидов, относительно дикого типа нуклеотидной последовательности. Одна или более вариация может быть в кодирующем или некодирующем участке последовательности нуклеиновой кислоты, и может снижать или полностью прекращать экспрессию или функцию NMT. Таким образом, вариантная нуклеотидная последовательность может кодировать вариантный полипептид, который обладает пониженной или полностью отсутствующей активностью, или может кодировать дикий тип полипептида, экспрессирующийся немного или не экспрессирующийся внутри клетки, например, в результате измененной активности регуляторного элемента.
В некотором примере, рак может быть идентифицирован в качестве с недостатком NMT, за счет определения гена(ов), которые влияют или отрицательно регулируют экспрессию.
Ряд способов может быть применен для определения присутствия или отсутствия отдельной нуклеокислотной последовательности в образце, полученном от субъекта.
В некоторых примерах, рак может быть идентифицирован в качестве с недостатком NMT за счет оценки уровня экспрессии или активности положительного или отрицательного регулятора компонента NMT NMT-пути. Уровни экспрессии могут быть определены, например, посредством иммунологических исследований, таких как иммуноблот и ELISA, и способов детекции нуклеиновых кислот.Таких как ОТ-ПЦР, нанострунного способа, секвенирования РНК, гибридизации нуклеиновых кислот или кариотипного анализа.
В некоторых примерах рак может быть идентифицирован в качестве с недостатком NMT1 и/или NMT2 за счет определения присутствия в клеточном образце от индивидуума одной или более вариаций, например, полиморфизмов или мутаций в NMT1 и/или NMT2.
Мутации и полиморфизмы, ассоциированные с раком, также могут быть детектированы по уровню белка за счет детекции присутствия варианта (т.е. мутантного или аллельного варианта) полипептида.
В другом примере предложен способ лечения субъекта, больного раком, где упомянутый рак включает раковые клетки с недостатком NMT2, включая введение упомянутому субъекту ингибитора NMT и/или ингибитора NMT1.
Термин «ингибировать» или «ингибитор», как употреблено в данном документе, относится к способу или технике, при которых ингибируют синтез, уровни, активность или функцию интересующего белка, например NMT2. Термин также относится к любому метаболическому или регуляторному пути, который может регулировать синтез, уровни, активность или функцию интересующего белка. Термин включает связывание с другими молекулами и образование комплекса. Более того, термин «ингибитор» относится к любому агенту или соединению, применение которого приводит к ингибированию функции белка или функции белкового пути. Однако термин не подразумевает, что каждая или все из этих функций должны быть ингибированы в одно и тоже время.
В другом примере предложен способ лечения субъекта, больного раком, где упомянутый рак включает раковые клетки с недостатком NMT1, включающий введение упомянутому субъекту ингибитора NMT и/или ингибитора NMT2.
В некоторых примерах способы лечения включают введение субъекту терапевтически эффективного количества соединений, описанного в данном документе, и возможно состоят из однократного введения или применения, или альтернативно включают серию введений или применений. В конкретном примере упомянутое соединение представляет собой ингибитор NMT, ингибитор NMT1 и/или ингибитор NMT2.
В более конкретном примере ингибитор NMT представляет собой Tris-DBA, НМА, DDD85646, DDD86481 или их производные.
В других примерах соединения и/или композиции предложены в фармацевтически эффективном количестве, приемлемом для введения субъекту.
Термин «фармацевтически эффективное количество», как употреблено в данном документе, относится к количеству лекарственного средства или фармацевтического агента, которое будет вызывать биологический или медицинский ответ ткани, системы, животного или человека, представляющих интерес для исследователя или клинициста. Данное количество может представлять собой терапевтически эффективное количество.
Соединения и композиции предложены в фармацевтически приемлемой форме.
Термин «фармацевтически приемлемый», как употреблено в данном документе» включает соединения, материалы, композиции и/или дозированные формы (такие как единичные дозированные формы), которые приемлемы для применения в контакте с тканями субъекта без излишней токсичности, раздражения, аллергической реакции или других проблем или осложнений, в соответствии с соотношением риск/польза. Каждый носитель, эксципиент и т.д. также является «приемлемым» в смысле совместимости с другими ингредиентами препарата.
Действительное вводимое количество и скорость и продолжительность введения будут зависеть от природы и серьезности того, что подлежит лечению. Предписание лечения, решения по дозированию и т.д. находится в компетенции обычных врачей общей практики и других врачей, и обычно учитывает нарушение, подлежещее лечению, состояние отдельного пациента, сайт доставки, способ введения и другие факторы, известные врачам общей практики.
Соединение или композиция могут быть введены по отдельности или в комбинации с другими лечениями, одновременно или последовательно, в зависимости от состояния, подлежащего лечению.
Препараты могут для удобства могут быть представлены в единичных дозированных формах и могут быть получены посредством любых способов, хорошо известных в области фармакологии. Такие способы включают этап доставки активного соединения в ассоциации с носителем, который может состоять из одного или более ингредиентов. Обычно, препараты получают посредством однородной и равномерной доставки в ассоциации активного соединения с жидкими носителями или окончательно разделенными твердыми носителями или с обоими, и затем если необходимо формование продукта.
Соединения и композиции могут быть введены субъекту посредством обычного пути введения, или системно/переферически или в сайте желаемого воздейтсвия, включая, но не ограничено, оральный (например, проглатывание); местный (включая чрескожный, интраназальный, окулярный, буккальный и сублингвальный); легочный (например, посредством ингаляции или инсуфляции при помощи, например, аэрозоля, например, через рот или нос); ректальный; вагинальный; парентеральный, например, посредством инъекции, включая подкожный, внутрикожный, внутривенный, внутриартериальный, интракардиальный, интратекальный, интраспинальный, интракапсулярный, субкапсулярный, интраорбитальный, интраперитонеальный, интратрахеальный, субкутикулярный, внутрисуставной, субарахноидальный и интрастернальный пути; посредством имплантации депо / например, подкожно или внутримышечно.
Препараты, приемлемые для орального применения (например, посредством проглатывания) могут быть представлены в виде дискретных единиц, таких как капсулы, саше или таблетки, каждые включающие определенное количество активного соединения; в виде порошка или гранул; в виде раствора или суспензии в водной или неводной жидкости; или в виде жидкой эмульсии типа масло в воде или вода в масле; в виде болюсов; в виде электуария или в виде пасты.
Препараты, приемлемые для парентерального введения (например, посредством инъекции, включая кожные, подкожные, внутримышечные, внутривенные и внутрикожные), включают водные и неводные изотонические апирогенные стерильные инъекционные растворы, которые могут включать антиоксиданты, буферы, консерванты, стабилизаторы, бактериостатики растворы, которые делают препарат изотоническим с кровью предназначенного реципиента; и водные и неводные стерильные суспензии, которые могут включать суспендирующие агенты и загустители, и липосомы или другие системы микрочастиц, которые разработаны для доставки соединения к компонентам крови или одному или более органам. Примеры приемлемых изотонических носителей для применения в таких препаратах включают инъекции хлорида натрия, раствора Рингера, раствора Рингера с лактатом.
Препараты могут быть представлены в однодозовых или многодозовых запаянных контейнерах, например, ампулах или сосудах. И могут храниться в высушенном замораживанием (лиофилизированном) состоянии, требуя только добавления стерильного жидкого носителя, например воды для инъекций, непосредственно перед применением. Приготовленные для немедленного приема инъекционные растворы и суспензии могут быть получены из стерильных порошков, гранул и таблеток. Препараты могут быть в виде липосом или других систем микрочастиц, которые разработаны для доставки активного соединения к компонентам крови или одному или более органам.
Композиции, содержащие соединения, раскрытые в данном документе, могут быть применены в способах, описанных в данном документе в комбинации со стандартными хемотерапевтическими режимами или в сочетании с лучевой терапией.
В случае лимфомы у пациента известные способы лечения зависят от субъекта, подлежащего лечению, типа заболевания и его стадии. Существующие способы лечения лимфомы известны квалифицированным специалистам. Соответственно, известные способы лечения могут быть применены вместе с ингибиторами NMT, раскрытыми в данном документе.
Распространенные комбинации лекарственных средств для применения в лечении лимфом включают, но не ограничены, CHOP (т.е. циклофосфамид, доксорубицин, винкристин и преднизон), GAP-ВОР (т.е. циклофосфамид, доксорубицин, прокарбазин, блеомицин, винкристин и преднизон), m-BACOD (т.е. метатрексат, блеомицин, доксорубицин, циклофосфамид, винкристин, дексаметазон и лейковорин), ProMACE-МОРР (т.е. преднизон, метотрексат, доксорубицин, циклофосфамид, этопозид, лейковорин со стандартным МОРР), ProMACE-CytaBOM (преднизон, доксорубицин, циклофосфамид, этопозид, цитарабин, блеомицин. Винкристин, метатрексат и лейковорин) и МАСОР-В (метотрексат, доксорубицин, циклофосфамид, винкристин, преднизон, блеомицин и лейковорин). Для рецидивирующей агрессивной неходжкинской лимфомы могут быть применены следующие хемотерапевтические комбинации лекарственных средств с соединениями и композициями, описанными в данном документе: IMVP-16 (т.е. ифосфамид, метотрексат и этопозид), MIME (т.е. метил-gag, ифосфамид, метотрексат и этопозид), DHAP (т.е. дексаметазон, - 16 высоко дозированный цитарабин и цисплатин), ESHAP (т.е. этопозид, метилпреднизон, высокодозированный цитарабин и цисплатин), CEFF(B) (т.е. циклофосфамид, этопозид, прокарбазин, преднизон и блеомицин), и CAMP (т.е. ломустин, митоксантрон, цитарабин и преднизон).
Лечение при резервной химиотерапии, применяемое для определенных лимфом, таких как рецидивирующая устойчивая ходжкинская лимфома, включают но не ограничены VABCD (т.е. винбластин, доксорубицин, дакарбазин, ломустин и блеомицин), ABDIC (т.е. доксорубицин, блеомицин, дакарбазин, ломустин и преднизоне), CBVD (т.е. ломустин, блеомицин, винбластин, дексаметазон), PCVP (т.е. винбластин, прокарбазин, циклофосфамид и преднизон), СЕР (т.е. ломустин, этопозид и преднизон), EVA (т.е. этопозид, винбластин и доксорубицин), MOPLACE (т.е. циклофосфамид, этопозид, преднизон, метотрексат, цитаравин и винкристин), MIME (т.е. метил-gag, ифосфамид, метотрексат и этопозид), MINE (т.е. митоквазон, ифосфамид, винорелбин и этопозид), МТХ-СНОР (т.е. метотрексат и CHOP), СЕМ (т.е. ломустин, этопозид и метотрексат), CEVD (т.е. ломустин, этопозид, виндезин и дексаметазон), CAVP (т.е. ломустин, мельфалан, этопозид и преднизон), EVAP (т.е. этопозид, винбластин, цитарабин и цисплатин), и EPOCH (т.е. этопозид, винкристин; доксорубицин, циклофосфамид и преднизон).
Следует понимать, что альтернативные способы ингибирования NMT1 или NMT2 могут быть применены в стратегии синтетической летальности для лечения рака и в частности лечения В-клеточной лимфомы. Например, экспрессия NMT1 или NMT2 может быть ингибирована при помощи технологии антисмысловой или иРНК. Применение данных подходов для пониженной регуляции экспрессии активности гена и/или белка известно специалистам в области техники.
В другом воплощении настоящего раскрытия предложен способ определения пользы лечения пациента ингибитором NMT2 и/или ингибитором.
В одном примере способ настоящего раскрытия включает качественное или количественное определение, анализ или измерение образца от субъекта, больного раком или предположительного больного раком, на присутствие или отсутствие, или количество или концентрацию NMT1 и/или NMT2.
В другом примере способ настоящего раскрытия включает качественное или количественное определение, анализ или измерение образца от субъекта, больного раком или предположительно больного раком, на присутствие или отсутствие или количества или концентрацию миристоилированных белков.
В другом примере способ настоящего раскрытия включает качественное или количественное определение, анализ или измерение образца от субъекта, больного раком или предположительно больного раком, на присутствие или отсутствие или количество или концентрацию ацилированных белков.
Термин «образец», как употреблено в данном документе относится к любому образцу от субъекта, включая, но не ограничено, жидкий, клеточный или тканевый образец, который включает раковые клетки или который предположительно включает раковые клетки, который может быть оценен в отношении уровней экспрессии, уровней белка, уровней ферментативной активности и т.п. Образец может включать, например, образец крови, образец костного мозга, биопсию, замороженный тканевой образец, свежий тканевой образец, клеточный образец и/или парафиновый срез, материал из которого может быть выделена РНК в достаточном количестве и с соответствующем качеством, что позволит измерить относительные уровни мРНК, или материал, из которого могут быть выделены полипептиды в достаточных количествах для измерения относительных уровней полипептидов.
Определение, анализ или измерение NMT1 или NMT2, или присутствие или отсутствие NMT1 и/или NMT2 может быть скоррелировано у пациента с пользой лечения рака ингибитором NMT1 или NMT2.
Определение, анализ или измерение миристоилированных белков или присутствие или отсутствие миристоилированных белков может быть скоррелировано с пользой лечения рака у пациента ингибитором NMT1 или NMT2.
Определение, анализ или измерение ацилированных белков или присутствие или отсутствие миристоилированных белков может быть скоррелировано с пользой лечения рака у пациента ингибитором NMT1 или ингибитором NMT2.
В конкретном примере антитела по настоящему изобретению являются иммунореактивными или иммуноспецифичными и поэтому специфично и селективно связывают интересующий белок, например, белок NMT1 или NMT2. В одном примере, могут быть применены антитела, которые являются иммунореактивными и иммуноспецифичными для NMT1 или NMT2 человека. Антитела к NMT1 или NMT2 человека являются предпочтительно иммуноспецифичными. Термин «антитело» и «антитела» включает, но не ограничен, моноклональные и поликлональные антитела.
В другом примере антитела по настоящему изобретению являются иммунореактивными или иммуноспецифичными к, и поэтому специфично и селективно связывают как белок NMT1, так и NMT2. В данном примере применяют антитела, которые являются иммунореактивными и иммуноспецифичными к NMT1 или NMT2 человека. Антитела к NMT1 или NMT2 человека предпочтительно являются иммуноспецифичными. В данном примере и в связи с различной молекулярной массой NMT1 и NMT2, возможна идентификация присутствия или отсутствия обоих белков при помощи, например, SDS-PAGE и иммуноблотинга. Термин «антитело» и «антитела» включает, но не ограничен, моноклональные и поликлональные антитела.
Термин «связывают специфично» относится к высоко авидному и/или высоко афинному связыванию антитела со специфичным полипептидом, например, эпитопом NMT1 или NMT2. Связывания антитела с его эпитопом на данном специфичном полипептиде является более сильным, чем связывание этого же антитела с любым другим эпитопом, в частности такими, которые могут быть представлены на молекулах в ассоциации с или в том же образце, в качестве интересующего специфичного полипептида. Антитела, которые специфично связываются с интересующим полипептидом могут быть способны к связыванию другими полипептидами на слабом, еще детектируемом уровне. Такое слабое связывание или фоновое связывание, быстро отличимо от специфичного связывания антитела с интересующим соединением или полипептидом, например, посредством применения соответствующих контролей, как будет известно специалистам в области техники.
В одном примере образец, содержащий раковые клетки или предположительно содержащий раковые клетки, получают от субъекта, больного раком. Взятие такого образца хорошо известно квалифицированным специалистам. В конкретном примере образец представляет собой образец крови. Способы получения образца, обработки и/или хранения такого образца также хорошо известны квалифицированным специалистам.
В конкретном примере детекцию, анализ или измерение белка NMT1 или NMT2 внутри образца проводят при помощи иммуногистохимии. В более конкретном примере детекцию, анализ или измерение NMT2 внутри образца проводят при помощи иммуногистохимии. Специалистам в области техники будет понятно, что другие иммунологические исследования, как качественные, так и количественные, могут быть применены в настоящем изобретении.
В дополнительных примерах, иммуногистохимия (IHC) может быть выполнена посредством любого приемлемого способа или системы иммуногистохимии. Неограничивающие примеры включают автоматизированные системы, количественную ИГХ, полуколичественную ИГХ и ручные способы.
Термин «качественная» иммуногистохимия относится к автоматизированному способу сканирования и подсчета образцов, которые подвергают иммуногистохимии, для идентификации и количественного определения присутствия специфичных биомаркеров, таких как антиген или другой белок. Например, количественное определение NMT1 и/или NMT2. Подсчет, сделанный для образца, является численным представлением интенсивности иммуногистохимического окрашивания образца, и представляет количество целевого биомаркера (такого как NMT1 или NMT2) присутствующего в образце. Как определено в данном документе оптическая плотность (ОП) является численной оценкой, которая представляет интенсивность окрашивания, а также процент окрашенных клеток. Как употреблено в данном документе, полуколичественная иммуногистохимия относится к подсчету иммуногистохимических результатов человеческим глазом, где опытный рабочий определяет результаты численно (например, 0 [слабое окрашивание или отсутствие окрашивания], 1 или 2 [сильное окрашивание]).
Автоматизированная обработка, сканирование и системы анализа, хорошо известны в области техники и могут быть применены с настоящим изобретением. Такие системы могут включать автоматизированное сканирование и микроскопическое сканирование, анализ компьютерного изображения, сравнение серийных срезов (с контролем для вариации в ориентации и размере образца), цифровая подготовка отчета, и архивирование и трэкинг образцов (такие как стекла на которые помещают тканевые срезы). Коммерчески доступны системы клеточной визуализации, которые сочетают обычные световые микроскопы с системами цифровой обработки изображения для выполнения количественного анализа на клетках и тканях, включая иммуноокрашенные образцы.
На практике в примере, в котором образец от пациента определяют как имеющий низкое (например, слабое или отсутствующее) окрашивание NMT2 опухоли, пациента рассматривают как хорошего кандидата для лечения ингибитором NMT1. В другом конкретном примере пациента, определенного как обладающего высоким (например, сильным) окрашиванием опухоли NMT2 рассматривают в качестве плохого кандидата для лечения ингибитором NMT1.
Следует понимать, что точка отсечки для ИГХ отрицательного vs положительного определения является полуколичественным определением и определяется опытным патоморфологом при помощи полуколичественных способов и световой микроскопии.
Например, ИГХ может быть оценена любым из следующих способов: 1. Любое окрашивание vs неокрашивания; 2. сильное vs слабого окрашивания; 3. Отсутствие vs слабое vs сильное; 4. H Score, состоящий из формулы (% отсутствия × 0) + (% слабое × 100); + (% среднее × 200) + (% сильное × 300); 5. Программное обеспечение для анализа компьютерных изображений для количественной оценки.
Точки отсечки изначально определяют посредством переменной лекарственной чувствительности in vitro. Проведенные ранее исследования лекарственных средств, точка отсечки чувствительности vs нечувствительности будет в дальнейшем уточнена и проверена.
В одном примере при определении будет ли окрашивание NMT2 опухоли высоким (например, сильным) или низким (например, слабым или отсутствующим) образец от пациента может быть сравнен с одним или более контрольным образцом, имеющим известный и/или установленный уровень окрашивания NMT2. В одном примере контрольный образец представляет собой образец от пациента, который имеет известные и/или установленные уровни окрашивания NMT2 опухоли и/или известный клинический исход. В одном примере контроль представляет собой клеточную линию, имеющую известное количество NMT2 окрашивания.
Продолжительные способы лечения пациентов, рассматриваемых в качестве плохих кандидатов для лечения ингибитором NMT1, известны специалистам в области техники.
Следует понимать, что при некоторых условиях пациент, который изначально отвечает на лечение ингибитором NTM1, может иметь рецидив. Такой рецидив может манифестировать несколькими путями, включая, но не ограничено, повторение первичной опухоли и развитие метастаз. Кроме того или альтернативно может возникнуть другая дополнительная опухоль.
В соответствии с одним аспектом настоящего изобретения, предложен способ, включающий: а) получение образца от субъекта, больного или предположительно больного раком; b) контактирование образца с антителом к NMT2 с образованием комплекса между антителом и NMT2, присутствующим в образце; с) измерение комплекса, образованного для определения количества или концентрации NMT2 в образце; и d) определение пользы лечения ингибитором NMT1 упомянутого рака в упомянутом субъекте. Где определение пользы лечения ингибитором NMT1 определяют посредством уровня NMT2 в упомянутом образце.
В конкретном аспекте введение ингибитора NMT1 упомянутому субъекту назначают в случае, когда количество NMT2 в упомянутом образце является низким или отсутствует, возможно по сравнению с контролем.
В соответствии с одним аспектом настоящего изобретения предложен способ, включающий получение образца от субъекта; обработку упомянутого образца; проведение анализа связывания, включающего контактирование обработанного образца с антителом к NMT2 с образованием комплекса между антителом и NMT2, присутствующим в образце, упомянутый анализ связывания, дающий по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе; и введение ингибитора NMT1 упомянутому субъекту в случае, когда количество NMT2 в упомянутом образце является низким или отсутствует, возможно по сравнению с контролем.
В некоторых аспектах для определения количества комплекса в образце применяют оборудование, имеющее детектор для обнаружения комплекса, образованного между антителом и NMT2 в упомянутом образце. В некоторых примерах, оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство. В некоторых примерах антитело к NMT2 является моноклональным или поликлональным антителом.
Другие способы, которые могут быть применены при детекции, анализе или измерении NMT1 или NMT2 включают, но не ограничены иммуноблот, ELISA, непрямую иммунофлуоресценцию, технология мультиплексирования на частицах, иммунопреципетацию и масс-спектрометрию образца, полученного от субъекта. На практике в примере, в котором образец от пациента определяют в качестве имеющий низкое или отсутствующее окрашивание NMT2, субъект рассматривают в качестве хорошего кандидата для терапии ингибитором NMT.
В одном примере образец анализируют посредством световой микроскопии при помощи прямого исследования или за счет захвата изображения и анализа, или применения флуоресцентной микроскопии при помощи прямого исследования захвата и анализа изображения.
В другом примере способ по настоящему раскрытию включает качественное и количественное определение, анализ или измерение активности белка NMT1 и/или NMT2 в биологическом образце от субъекта, больного раком на присутствие или отсутствие или количество активности NMT1 и/или NMT2. В данном примере применения субстратов (натуральных или синтетических) NMT1 или NMT2 применяют для идентификации образца, в котором активность NMT1 или NMT2 присутствует, отсутствует или ее количества.
На практике в примере, в котором образец от субъекта, определяют как с недостатком NMT2, субъекта рассматривают в качестве хорошего кандидата для введения ингибитора NMT1.
На практике в примере, в котором образец от субъекта определяют в качестве имеющего низкие или отсутствующие уровни белка NMT2, субъект рассматривают в качестве хорошего кандидата для введения ингибитора NMT1.
На практике в примере, в котором образец от субъекта определяют в качестве имеющего низкую или отсутствующую активность NMT2, субъект рассматривают в качестве хорошего кандидата для введения ингибитора NMT1.
На практике в примере, в котором образец от субъекта определяют в качестве имеющего низкое или отсутствующее количество миристоилированного белка, субъект рассматривают в качестве хорошего кандидата для введения ингибитора NMT1.
На практике в примере, в котором образец от субъекта определяют в качестве имеющего низкое или отсутствующее количество ацилированного белка, субъекта рассматривают в качестве хорошего кандидата для введения ингибитора NMT2.
В другом примере способ по настоящему раскрытию включает идентификацию мутации, делеции и т.п. в гене NMT1 или NMT2 в образце от субъекта, больного раком или предположительно больного раком. При этом упомянутая мутация, делеция и т.п. в гене NMT1 или NMT2 приводит к потере снижения активности белка NMT1 или NMT2 в раковых клетках внутри образца. Способы идентификации упомянутых мутаций, делеций и т.п. в NMT1 или NMT2 известны специалистам в области техники. И включают, но не ограничены RFLP, ОТ-ПЦР, микроаррей и/или любой тип секвенирования ДНК. На практике в одном примере, в котором образец от пациента определяют в качестве имеющего мутацию, делецию или т.п. в NMT2, которая приводит к низкой или отсутствующей активности белка NMT2, субъект рассматривают в качестве хорошего кандидата для терапии ингибитором NMT.
В другом примере способ по настоящему раскрытию включает идентификацию мутации, делеции или т.п. в мРНК NMT1 или NMT2 в образце от субъекта, больного раком или предположительно больного раком. При этом упомянутая мутация, делеция или т.п. в мРНК NMT1 или NMT2 приводит к потере снижения активности белка NMT1 или NMT2 в раковых клетках внутри упомянутого образца. Способы идентификации таких мутаций, делеций или т.п.в мРНК NMT1 или NMT2 известны специалистам в области техники, и включают но не ограничены, Нозерн-блот, ОТ-ПЦР, микроаррей и/или любой приемлемый тип секвенирования мРНК. На практике в примере в котором образец от пациента определяют в качестве имеющего мутацию, делецию или т.п. в мРНК NMT2, которая приводит к низкой или отсутствующей активности белка NMT2, субъект рассматривают в качестве хорошего кандидата для терапии ингибитором NMT.
В другом примере способа по настоящему раскрытию, предложен способ лечения субъекта, больного раком, ассоциированный с дефектом в NMT1 или NMT2, включающий введение упомянутому субъекту ингибитора NMT. В конкретном примере рак ассоциирован с дефектом в NMT2, и ингибитор представляет собой ингибитор NMT1.
В другом примере предложено применение ингибитора NMT1 для лечения рака с недостатком NMT2.
В другом примере предложено применение ингибитора NМТ1 для лечения рака, где упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, лимфому Беркитта, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легкх, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному. В более конкретном примере упомянутый рак определяют в качестве рака с недостатком NMT2.
Примеры ингибиторов включают, но не ограничены небольшие молекулы, антитела, пептидные фрагменты и/или молекулы нуклеиновых кислот.
Конкретные примеры небольших молекул включают Tris-DBA, НМА, DDD85646, DDD86481 и их производные. Термин «производные», как определено в данном документе включает, но не ограничен, соли, координационные комплексы, сложные эфиры, такие как in vivo гидролизуемые сложные эфиры, свободные кислоты или основания, гидраты, пролекарственные средства или липиды, агенты слияния.
Пептидные фрагменты могут быть получены целиком или частично посредством химического синтеза, который активирует сайт NMT1. Пептидные фрагменты могут быть получены согласно установленным, стандартным жидко или твердо-фазным способам пептидного синтеза, хорошо известным специалистам в области техники.
Ингибиторы нуклеиновых кислот или их комплементы, ингибируют активность или функцию посредством понижающей регуляции продукции активного полипептида. Это может быть мониторировано при помощи обычных способов, хорошо известных в области техники, например, посредством скрининга при помощи ПЦР в реальном времени, как описано в примерах.
Примеры ингибиторов нуклеиновых кислот включают технологию асРНК или иРНК, применение которых для пониженной регуляции экспрессии гена хорошо известно в области техники. Анти-смысловые олигонуклеотиды могут быть разработаны для гибридизации с комплементарной последовательностью нуклеиновой кислоты, пре-мРНК или зрелой мРНК, интерферирующими с продукцией компонента пути эксцизионной репарации оснований так, что его экспрессия понижена или фактически полностью предотвращена. В дополнение к мишенной кодирующей последовательности, могут быть применены антисмысловые техники для нацеливания контрольных последовательностей гена, например, в 5'-фланкирующей последовательности, таким образом, антисмысловые олигонуклеотиды могут интерферировать с экспрессией контрольных последовательностей.
Альтернативно антисмысловым технологиям, применяют копию всего или части мишенного гена, встроенную в смысловую, в той же ориентации, как в мишенном гене, для достижения снижения экспрессии мишенного гена посредством соэкспрессии.
Кроме того, может быть применен сайленсинг дцРНК. Сайленсинг опосредованный дцРНК является геноспецифичным и часто называется РНК интерференцией (РНКи).
В другом примере применяют нуклеиновую кислоту, которая при транскрипции дает рибозим, способный разрезать нуклеиновую кислоту в конкретном сайте и таким образом также полезна при влиянии на NMT.
В еще одном примере могут быть применены небольшие молекулы РНК для регуляции экспрессии гена. Они включают нацеленную деградацию мРНК малыми интерферирующими РНК (миРНК), посттранскрипционный сайленсинг генов (PTGs), соответствующе регулируемую трансляционную репрессию специфичной последовательности мРНК посредством микро-РНК (микроРНК) и нацеленный транскрипционный сайленсинг генов.
В еще одном примере может быть применена экспрессия молекул коротких шпилечных РНК (кшРНК) в клетке. кшРНК состоит из коротких инвертированных повторов, разделенных небольшой петлевой последовательностью. Один инвертированный повтор комплементарен мишенному гену. В клетке DICER процессирует кшРНК в миРНК, которая деградирует мишенный ген мРНК NMT и подавляет экспрессию. В предпочтительном воплощении кшРНК продуцируют эндогенно (внутри клетки) посредством транскрипции с вектора.
Дефект в NMT1 или NMT2, представляет собой недостаток NMT1 или NMT2, соответственно фенотип, который может быть дефицитным по компоненту пути, опосредованному NMT1 или NMT2, т.е. экспрессия активности компонента пути может быть снижена или утрачена в раковой клетке относительно контрольных клеток. В некоторых воплощениях раковая клетка может быть с недостатком NMT1 или NMT2, т.е. экспрессия активности NMT1 или NMT2 может быть снижена или утрачена в раковой клетке относительно контрольных клеток.
Соответственно, предложено применение NMT2 в качестве маркера для одного или более диагнозов, прогнозов, классификации или мониторинга рака у субъекта. В некоторых примерах NMT2 измеряют при помощи исследования, выбранного из иммунологических исследований или детекции нуклеиновых кислот или активности белка.
Термин «прогноз», как употреблено в данном документе относится к предсказанию вероятности смерти вследствие рака или прогресса, включая рецидив, распространение метастаз и лекарственную устойчивость, неопластического заболевания, такого как рак молочных желез.
Термин «прогностический маркер», как употреблено в данном документе относится к маркеру, который информирует о результате для пациента в отсутствии системной терапии или означает результат отличный от такового пациентов без маркера, несмотря на эмпирическую (не нацеленную на маркер) системную терапию.
Термин «прогностический маркер», как употреблено в данном документе, относится к маркеру, который предсказывает дифференцированную эффективность (пользу) конкретной терапии на основании статуса маркера.
Термин «диагноз», как употреблено в данном документе, относится к идентификации молекулярного и/или патологического состояния, заболевания или состояния такого, как идентификация рака молочных желез или другого типа рака.
Также предложено применение миристоилирования белка в качестве маркера для одного или более диагноза, прогноза, классификации или мониторинга рака у субъекта.
Также предложено применение ацилирования белка в качестве маркера для одного или более диагноза, прогноза, классификации или мониторинга рака у субъекта.
В некоторых примерах упомянутый рак представляет собой лимфому. В более конкретных примерах упомянутая лимфома представляет собой В-клеточную лимфому. В более конкретных примерах В-клеточная лимфома представляет собйо фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийно-клеточную лимфому, В-ХЛЛ-ЛЛ, иммуноцитому/болезнь Вальдентсрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому или анапластическую крупноклеточную лимфому.
В некоторых примерах рак представляет собой острую миелоидную лейкемию, В-клеточную лимфому, хроническую миелоидную бластную лейкемию, лимфому Беркитта, диффузную В-крупноклеточную лимфому, плазмаклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному молочных желез, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути.
В некоторых примерах рак представляет собой В-клеточную лимфому, лимфому беркитта, диффузную крупноклеточную лимфому, острую миелоидную лейкемию, миелому, светлоклеточную карциному яичника, переходно-клеточную карциному (рак мочевыводящего пути и мочевого пузыря), хроническую миелогенную лейкеимию (ХМЛ), лимфому-ХЛЛ, мелкоклеточную карциному легких, карциному молочных желез, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легких, остеосаркому, меланому, аденокарциному желудка, аденокарциному эндометрия, эзофагеальную сквамозную аденокарциному.
Способы по изобретению обычно практикуют посредством обеспечения соединений и/или композиций, применяемых в таком способе в виде набора. Такой набор предпочтительно включает композицию. Такой набор предпочтительно включает инструкции для его применения.
Для лучшего понимания изобретения, описанного в данном документе, приведены следующие примеры. Следует понимать, что данные примеры приведены только в целях иллюстрации. Поэтому, они не должны ограничивать объем данного изобретения никоим образом.
Примеры
В следующих примерах применяют стандартные способы, которые будут оценены специалистами в области техники. Материалы и методы Антитела и реагенты
Трис дибутилбензинилиден ацетон палладий (Tris-DBA) был любезно предоставлен доктором Arbiser (U. Alabama). DDD85646 был синтезирован как описано [J.A. Frearson et al (2010) Nature. 464.728-723)] и/или DDD86481 был получен от доктора David Gray и Paul Wyatt, Dundee University).
Мышиные антитела κ NMT1 (клон 14; 1:1000) и мышиные антитела к NMT2 (клон 30; 1:2000) были получены от BD Biosciences, Сан-Хосе, калифорния, США. Кроличьи антитела к NMT1 (поликлональные, 1:3000) закуплены у Proteintech, Чикаго, Иллинойс, США. Кроличьи антитела к GFP (1:20,000), к PARP-1 (1:5000), к GAPDH (1:5000) и антитела к c-terminal РАК2 (1:2000) были получены от Eusera (www.eusera.com), Эдмонтон, Альберта, Канада. Мышиные антитела к α-тубулину (1:15,000) и кроличьи антитела к V5 (1:10,000) были закуплены у Sigma Aldrich, Сент-Луис, Миссури, США. Мышиные антитела к His (1:2000) получены от Qiagen, Германия. Кроличьи антитела к расщепленной каспазе 8 (1:1000) и расщепленной каспазе 3 (1:1000) были закуплены у Cell Signaling, Данверс, Массачусетс, США. Киты для детекции методом Вестерн-блот Enhanced chemiluminesce (ECL) Plus и ECL Prime были закуплены y GE Healthcare, Питсбург, Пенсельвания, США. Если не указано иное все примененнеы химические реагенты были закуплены у Sigma-Aldrich (Сент-Луис, Миссури, США) и были самого высокого качества.
ДНК конструкты. Инжинирование V5- и His-vtxtyys[NMT1 и NMT2 конструктов.
NMT1 и NMT2 исходные вектора, предназначенные для работы с клонирующей системой Gateway (Life Technologies, Гранд-Айленд, Нью-Йорк, США) были закуплены у Genecopoeia (Роквиль, Мэриленд, США). Гены NMT1 и NMT2 инкорпорировали в вектор доставки pcDNA3.1/nV5 DEST (Life Technologies) при помощи фермента для клонирования LR (Life Technologies) согласно инструкциям производителя для получения плазмид N-терминально меченных NMT (His-NMT1, His-NMT2, V5-NMT1 и V5-NMT2). V5-меченные NMT конструкты применяли для экспрессии в клетках млекопитающих, при этом His-NMT конструкты применяли для бактериальной экспрессии. Продукты клонирования подтверждали посредством секвенирования ДНК (Eurofins MWG Operan, Хантсвилл, Алабама, США).
Клеточная культура
В-клетки были получены от доктора Jim Stone или из АТСС. Все реагенты для работы с клеточной культурой были закуплены у Invitrogen. В-клетки культивировали при 37°С и 5% CO2 во влажной камере и поддерживали в среде RPMI, дополненной 10% фетальной бычьей сыворотки, 100 ед/мл пенициллина и 0,1 мг/мл стрептомицина.
Клеточный лизис.Клетки промывали в холодном PBS, лизировали в 0,1% буфере SDS-RIPA [50 мМ Трис, 150 мМ NaCl, 1% Igepal СА-630, 0,5% NaDC, 2 мМ MgCl2, и 1х ингибитор протеазы (Roche Diagnostics); рН 8,0] и покачивали в течение 15 минут при 4°С. Супернатант клеточных лизатов получали после 15 мин центрифугирования при 16000 об/мин при 4°С.
Индукция апоптоза.
Если не указано иное апоптоз индуцировали при помощи 2,5 мкМ стауроспорина (STS) (Sigma Aldrich, Сент-Луис, Миссури, США) и 5 мкг/мл циклогексимида (ICN Biochemicals Inc. Аврора, Огайо, США) с целью ингибирования трансляции белка и усиления индукции апоптоза. Инкубация с ингибиторами NMT.
Трис дибутилбензинилиден ацетон палладий (Tris-DBA) был любезно предоставлен доктором Arbiser. Клетки инкубировали при повышающихся концентрациях в течение 24 часов с Tris-DBA (или ДМСО для контроля) или в течение 24 часов и 48 часов с DDD85646.
В-клеточная трансфекция.
В-клетки трансфецировали при помощи системы трансфекции Neon® (Life technologies) согласно инструкциям производителя и оптимизированному протоколу для трансфекции В-клеток Рамоса (импульсное напряжение 1300 В; длительность импульса 20 мсек, 2 пульса и 7,7×106 клеток/мл), адаптированному для 100 мкл наконечников. Классически проводили две трансфекции для получения достаточного количества живых клеток для проведения анализа жизнеспособности.
Анализ жизнеспособности клеток.
В и Τ клеточную жизнеспособность измеряли при помощи способа вытеснения трипанового синего. Клетки выращивали в конфлюентных состояниях (2×106 клеток/мл максимум). Обеспечивающих минимальный базовый апоптоз. После инкубации с ингибиторами приблизительно 20000 клеток (10 мкл) инкубировали с 10 мкл ТС10™ красителя трипанового синего (Biorad) в течение 15 минут. Жизнеспособность клеток оценивали при помощи ТС10™ автоматического цитометра (Biorad).
In vitro исследование активности NMT.
Протокол исследования активности N-миристоилтрансферазы адаптировали от Raju, R.V., and Sharma, R.К. (1999) Preparation and assay of myristoyl-CoA:protein N-myristoyltransferase. Methods Mol Biol 116, 193-211. [3H] миристоил-СоА синтезировали для каждого эксперимента, как описано ранее Towler, D., and Glaser, L. (1986) Protein fatty acid acylation: enzymatic synthesis of an N-myristoylglycyl peptide. Proc Natl Acad Sci U S A 83, 2812-2816. Вкратце, клетки ресуспендировали в 0,25 M сахарозном буфере (50 мМ NaH2PO4, рН 7,4) и подвергали 2 раундам обработки ультразвуком при уровне 6,0 на соникаторе Брансона. Реакционная смесь состояла из 10 мкл клеточного экстракта (приблизительно 20 мкг белков), инкубированного в буфере для активности NMT (0,26 M Трис-HCl, 3,25 мМ EGTA, 2,92 мМ ЭДТА и 29,25 мМ 2-меркаптоэтанол, 1% Тритон Х-100, рН 7,4) и миристоилированный или немиристоилированный декапептид, соответствующий N-терминальной последовательности усеченного-Bid (0,1 мМ растворенный в воде). Реакцию запускали добавлением 7,4 мкл (≈10 пкмоль) свеже синтезированного [3Н] миристоил-СоА (объем конечной смеси = 25 мкл) и инкубировали в течение 15 мин при 30°С. Реакцию останавливали посредством нанесения 15 мкл реакционной смеси на Р81 фосфоцеллюлозный бумажный диск (Whatman, Кент, Великобритания) и высушивали в течение 30 секунд. Диски промывали (промывочный буфер: 25 мМ Трис-буфер, рН 7,4) для удаления остаточной радиоактивности ([3Н]-миристат и d [3Н]-иристоил-СоА) в то время как [3Н]-иристоилпептид сохранялся на фосфоцеллюлозной бумаге. Радиоактивность оценивали посредством жидкостно-сцинтилляционного измерения и переводили в пикомоли миристоилированного пептида (Raju, R.V., and Sharma, R.К. (1999) Preparation and assay of myristoyi-CoA:protein N-myristoyltransferase. Methods Mol Biol 116, 193-211.
РВ-ПЦР
РВ-ПЦР проводили с пробами Taqman NMT1 и NMT2 при помощи 18S пробы в качестве контроля. Различия в номере времени цикла (Δct) вычисляли посредством вычитания времени цикла (ct), при котором мы видим экспоненциальное повышение в экспрессии 18S внутреннего контроля для каждого типа клеток из времени цикла NMT, снова в точке, где видно экспоненциальное увеличение сигнала.
Мутация сайта расщепления каспазой в V5-NMT посредством сайт-направленного мутагенеза
Сайты расщепления каспазой NMT1 и NMT2 идентифицировали посредством расщепления по Эдману (Alphalyse), мутировали посредством сайт-направленного мутагенеза. Поэтому мы применяли ранее клонированный V5-NMT1 и V5-NMT2 вектора gateway для мутирования идентифицированных сайтов расщепления каспазой. Остаток Asp-72 V5-NMT1 и Asp-25, Asp-67 и обоих Asp-25,67 (двойной мутант) остатков V5-NMT2 мутировали при помощи кита Quickchange® для сайт-направленного мутагенеза (Agilent Technologies) согласно инструкциям производителя.
Вкратце, сайт-направленный мутагенез проводили при помощи 5-50 нг дцДНК (вектор), 5 мкл 10х реакционного буфера, 125 нг праймеров, растворенных в безнуклеазной воде и доводили до конечного объема реакции 50 мкл безнуклеазной водой. 2,5 ед ДНК-полимеразы PfuTurbo (Agilent Technologies) добавляли к смеси перед началом реакции, которую проводили в течение 18 циклов в амплификатор Eppendorf Mastercycler 1. Следовательно, исходную дцДНК разрушали посредством добавления 10 ед рестриктазы Dpn I к каждой реакции и инкубирования в течение 1 ч при 37°С. Дополнительно праймеры для сайт-направленного мутагенеза разрабатывали при помощи программы PrimerX (http://www.bioinformatics.org/primerx/) (указаны в таблице 2,6). Важно, остатки аспартата (D) сайтов расщепления каспазой мутировали в остатки глутамата (Е) для получения следующих векторов; V5-NMT1 (D72E), V5-NMT2 (D25E), V5-NMT2 (D67E) и V5-NMT2 (D25.67E). Мутации подтверждали посредством автоматического секвенирования ДНК (Eurofins MWG Operan).
Получение His-меченных каспазо-расщепленных, усеченных NMT векторов
Следующие каспазо-расщепленные усеченные NMT получали посредством молекулярного клонирования; His-73NMT1, His-26NMT2 и His-68NMT2. Усеченные последовательности ДНК клонировали в вектор pET-19b (Novagen®), который имел N-терминальную гексагистидин (His)-меченную последовательность.
Ранее клонированные вектора gateway, GFP-NMT1 и GFP-NMT2 применяли в качестве матриц для данных ПЦР реакций. ПЦР реакции проводили при помощи полимеразы Platinum Pfx® (Life Technologies), и ПЦР реакции проводили согласно руководству производителя. Каждую ПЦР реакцию готовили следующим образом; 200 нг матрицы ДНК, 5 мкл 10Х буфера для амплификации Pfx, 5 мкл 10Х раствора усиливающего ПЦР, 1 мМ MgSO4, 0,3 мМ смеси dNTP, 0,5 мкМ каждого из прямого и обратного праймеров (указаны в таблице 2,6), 1 ед ДНК полимеразы Platinum Pfx® и безнуклеазной воды до 50 мкл. Реакцию проводили в амплификаторе Eppendorf Mastercycler 1, согласно руководтсву, прилагаемому к киту ДНК полимеразы Platinum Pfx®, и реакцию продолжали 30 циклов.
Ингибирование каспаз
Клетки предварительно обрабатывали 10 мкМ установленными ингибиторами каспазы, закупленными у EMD Chemicals, в течение 1 часа для индукции апоптоза. Примененные ингибиторы каспаз представляли собой обычный ингибитор каспазы (z-VAD-FMK), каспазу-3 (z-DEVD-FMK), каспазу-8 (z-IETD-FMK), каспазу-9 (z-LEHD-FMK) и отрицательный контроль (z-FA-FMK). Контрольные клетки обрабатывали ДМСО (1:1000).
Обработка клеток MG-132
Клетки IM9, КМН2, BL2 и клетки Рамос помещали в планшет (3×106 клеток на лунку в 6-луночных планшетах) и обрабатывали 10 мкМ MG-132 в течение 5 часов. Эквивалентное количество ДМСО добавляли к контрольным образцам. Затем клетки лизировали и подвергали анализу при помощи SDS-PAGE/Вестерн-блота.
Обработка клеток субероиланилид гидроксамовой кислоты (SAHA)
Клетки IM9, BL2 и клетки Рамоса помещали в планшет с 3×106 клеток на лунку в 6-луночных планшетах и обрабатывали в течение 24 часов 1 мкМ субероиланилид гидроксамовой кислотой (SAHA), которая является обратимодействующим ингибитором класса I и класса II гистон деацетилаз (HDACs). Эквивалентное количество ДМСО добавляли к клеткам в качестве контроля. Клетки лизировали и подвергали анализу при помощи SDS-PAGE/Вестерн-блота.
Индукция апоптоза
150 или 300 нг/мл мышиных антител к Fas применяли для стимуляции апоптоза через внешний путь и 2,5 мкМ стауроспорина (STS) применяли для стимуляции апоптоза через внутренний путь. Где указано циклогексимид (5 мкг/мл) применяли для ингибирования трансляции белка и в дальнейшем обеспечения клеточной гибели.
Обработка клеток ингибитором NMT, 2-гидроксимиристиновой кислотой (НМА)
НМА омыливали и конъюгировали с бычьим сывороточным альбумином (BSA) перед добавлением к клеткам для усиления их клеточного захвата, как описано ранее (Yap et al., 2010). Вкратце, НМА инкубировали с 20% молярным избытком гидроксида калия (КОН) при 65°С в течение 15 мин. Впоследствии, 20Х раствор готовили при помощи растворения КОН омыленной НМА в не содержащей сыворотку среде RPMI, содержащей 20% свободный от жирных кислот BSA при 37°С, с последующей дополнительной инкубацией в течение 15 мин при 37°С. Учитывая, что миристат натрия применяли в качестве контроля, его также инкубировали в несодержащей сыворотке среде RPMI, содержащей 20% свободный от жирных кислот BSA при 37°С перед добавлением к клеткам. Впоследствии Т-клетки Jurkat (1×107 клетки) инкубировали или с 1 мМ НМА или 1 мМ миристатом натрия конъюгировали со свободным от жирных кислот BSA (конечная концентрация в клеточной культуре 1%) в течение 1 ч при 37°С в среде RPMI (без FBS, добавок или антибиотиков). После инкубации апоптоз индуцировали при помощи антител к Fas (150 нг/мл) или STS (2,5 мкМ) с циклогексимидом (5 мкг/мл) или только носителем (ДМСО). Образцы собирали каждые 2 ч в течение периода 8 ч, промывали холодным PBS и лизировали 0,3 мл буфера 1% додецилсульфата натрия для анализа (SDS)-радио-иммунопреципитации (RIPA) и подвергали 2 раундам обработки ультразвуком в течение 15 секунд при выходном напряжении 5,5-6,5 на ультразвуковом дезинтеграторе Бронсона и помещали на лед на 2 минуты между каждым циклом.
Обработка клеточных линий лимфоцитов ингибитором NMT Tris-DBA
2×106 клеток (СЕМ, L0, BL2 и Ramos) выращивали в 6 луночных планшетах и инкубировали с повышающимися количествами (0, 1, 2 и 5 мкг/мл) Трис (дибензилиденацетон) дипалладия (Tris-DBA) (Bhandarkar et al., 2008). Жизнеспособность клеток, обработанных Tris-DBA измеряли при помощи анализа вытеснения трипанового синего (Hudson, 1976).
Обработка клеточных линий лимфоцитов ингибиторами NMT, DDD85646 и DDD73226
1×105 клеток [КМН2 (лимфома Ходжкина), IM9 ("нормальные" EBV иммортализованные В-клетки), BL2, Рамос] (100 мкл/лунку) помещали в 96-луночные планшеты и обрабатывали при помощи повышающихся количеств ингибитора NMT на основе пиразол сульфонамида DDD85646 [молекулярная масса (MW) 495,43] (Frearson et al., 2010) и наименее потентного аналога на основе пиразол сульфонамида DDD73228 (MW=362,5) в течение 24, 48 и 72 часов. Лекарственные средства растворяли в ДМСО для получения 100Х стоковых растворов для каждой тестируемой концентрации, так, что конечный объем ДМСО в каждой лунке составил 1%. Жизнеспособность клеток, обработанных данными ингибиторами, измеряли при помощи исследования вытеснения трипанового синего (Hudson, 1976).
Обработка клеточных линий лимфоцитов ингибитором NMT, DDD86481 1×105 клеток [КМН2 (лимфома Ходжкина), IM9 ("нормальные" EBV иммортализованные В-клетки), BL2, Рамос] (100 мкл/лунку) помещали в 96-луночный планшет и обрабатывали повышающимися количествами DDD86481 [MW 610.5]. Лекарственные средства растворяли в ДМСО для получения 100Х стоковых растворов для каждой тестируемой концентрации так, чтобы конечный объем ДМСО в каждой лунке составил 1%. MTS [(3-(4,5-диметилтиазол-2-ил)-5-(3-карбоксиметоксифенил)-2-(4-сульфофенил)-2Н-тетразолиум)] анализа применяли для измерения цитотоксичности лекарственного средства (Cory et al., 1991).
Лизис клеток
Обычно, клетки промывали холодным PBS, собирали, лизировали в 0,1% буфере SDS-RIPA или 0,1% буфере SDS-RIPA-HEPES (в случае, когда клетки метили алкин-жирными кислотами), дополненным 1Х полным ингибитором протеазы. Суспензию клеток покачивали в течение 15 мин при 4°С. затем лизаты клеток центрифугировали при 16000 об/мин в течение 10 мин при 4°С и собирали постядерный супернатант.Концентрации белка измеряли при помощи PierceTM bicinchoninic acid (ВСА) protein assay kit (Thermo Fisher Scientific, валтам, Массачусетс).
Лизис тканевых образцов лимфомы
Ткани диффузной В-крупноклеточной лимфомы человека (DLBCL) и фолликулярной лимфомы (FL) были любезно предоставлены доктором Raymond Lai (Cross Cancer Institute, Альберта, Канада). Замороженные опухолевые ткани разрезали на небольшие (~1 мм3) кусочки и смешивали с 1% SDS-RIPA с 1Х полным ингибитором протеазы. Образцы гомогенизировали при помощи небольшого гомогенизатора Даунсона и затем повторно обрабатывали ультразвуком в течение 2 мин с интервалами 1 мин (на льду) при выходном напряжении 6.0 (ультразвуковой дезинтегратор Брансона 450) до расстворения опухолевых тканей в лизирующем буфере. Затем образцы центрифугировали при 16000 об/мин в течение 10 мин при 4°С и собирали постядерный супернатант для анализа Вестерн-блотом.
Вестерн-блот при помощи сканера Odyssey
Данный способ применяли только для приведенного блота расщепленной каспазы-3, все другие Вестерн-блоты делали при помощи стандартного способа. После электрофореза гели переносили на нитроцеллюлозную мембрану и блокировали в 5% NFM в PBS с 0,1% Tween 20 (PBS-T) в течение 1 часа. Первичное антитело (кроличье к расщепленной каспазе-3; клеточный сигналинг) также разводили 1:2000 в 5%NFM в PBS-T, добавленном к мембране и инкубировали в течение 24 часов. Затем мембраны подвергали 6 циклам промывки по 5 мин каждый; 2 промывки в PBS, 2 промывки в PBS-T, и наконец 2 промывки в PBS. Вторичное антитело (Alex Fluor 680 goat-anti-rabbit; Life Technologies) добавляли после разведения в PBS-T (1:5000) и блоты оборачивали алюминиевой фольгой и инкубировали со вторичным антителом в течение 1 часа и подвергали таким же 6Х циклам промывки с PBS и PBS-T как раньше. Блоты сканировали на системе для флуоресцентного изображения Odyssey ® от LI-COR Inc. с сканирующим разрешением 84 мкм (Интенсивность: 5-7) на канале 700 (для кролика).
Субклеточное фракционирование
Клетки HeLa выращивали до конфлюентности в 150 мм чашках и индуцировали для апоптоза с антителами к Fas (300 нг/мл) (внешний) или STS (внутренний) в течение периода 5 ч. Клетки затем метаболически метили при помощи алкин-С14 (описано в деталях в разделе ниже) (Yap et al., 2010) и подвергали гипотоническому лизису.
Вкратце клетки промывали холодным буфером PBS, соскабливали с чашек при помощи скребка для клеток и собирали. Затем образцы центрифугировали при 2000 об/мин в течение 5 мин, супернатант аспирировали и клетки ресуспендировлаи в 600 мкл гипотонического буфера, что вызывало набухание клеток (Мд ресуспендирующий буфер), и инкубировали на льду в течение 20 мин. Клеточную суспензию переносили в гомогенизатор Даунса и гомогенизировали при помощи 45 ударов при помощи плотного пестика. Затем 400 мкл свободного от ЭДТА 2,5Х буфера для гомогенизации (НВ) добавляли к гомогенату и клетки гомогенизировали при помощи 15 ударов в гомогенизаторе Даунса при помощи широкого пестика. Затем гомогенат центрифугировали при 1000 об/мин в течение 5 мин и собирали постядерный супернатант (PNS). Затем 200 мкл PNS сохраняли, и остаток (~750 мкл) центрифугировали при 100000 об/мин в течение 45 мин в роторе Beckman TLA 120.2, что приводило к цитоплазматическому фракционированию (S100) и светлому мембранному осадку (Р100).
Осадок Р100 промывали один раз 1Х НВ и Р100 фракции доводили до сходного объема S100 фракции при помощи 1Х НВ. После этого полученный постядерный осадок (PNS), цитоплазматические фракции (S100) и мембранные фракции (Р100) доводили до содержания 1% SDS. Затем такой же объем фракции подвергали анализу Вестер-блот, и количественно оценивали уровни белка при помощи программного обеспечения Image J software (http://rsbweb.nih.gov/ij/).
Метаболическое мечение клеток при помощи био-ортогональной ω-алкинил-миристиновой кислоты и клик-химии
Метаболическое мечение клеток при помощи ω-алкинил-миристиновой кислоты
Клетки метили при помощи 100-25 мкМ ω-алкинилмиристиновой кислоты 30 мин перед сбором клеток и лизировали 0,1% буфером SDS-RIPA-HEPES. Белки (50-30 мкг) из полученных клеточных лизатов реагировали с 100 мкМ азидо-биотином при помощи клик-химии и обрабатывали, как описано ранее (Yap et al., 2010). Вкратце клетки лишали жирных кислот посредством инкубирования в среде (RPMI или DMEM), дополненной 5% фетальной бычьей сыворотки, адсорбированной на системе декстран-древесный уголь (DCC-FBS) в течение 1 ч перед мечением. Затем, ω-алкинилмиристиновую кислоту растворяли в ДМСО для получения 25 или 100 мМ стоковых растворов. Обычно, клеточный захват ω-алкинилмиристиновой кислоты улучшали при помощи сапонификации и конъюгирования с BSA. Таким образом, перед мечением ω-алкинилмиристиновую кислоту сапонифицировали с 20% молярным избытком гидроксида калия при 65°С в течение 15 мин. Затем, свободную от сыворотки среду для культивирования (предварительно нагретую), содержащую 20% свободный от жирных кислот BSA при 37°С добавляли к сапонифицированной ω-алкинилмиристиновой кислоте и инкубировали в течение дополнительных 15 мин при 37°С.
Затем клетки, лишенные жирных кислот, промывали один раз теплым PBS и инубировали в свежей RPMI или DMEM без добавок. Затем соответствующий объем 20Х конъюгата жирная кислота-BSA в свободной от сыворотке среде добавляли к клеткам, так чтобы конечная концентрация BSA составляла 1%, и ω - алкинилмиристиновая кислота была в индикаторной концентрации (25 мкМ или 100 мкМ) для каждого соответствующего эксперимента. ДМСО или немеченные жирные кислоты, конъюгированные с BSA, применяли в качестве контролей для выполнения экспериментов. Клетки метили при помощи ω-алкинилмиристиновой кислоты в течение 30 мин при 37°С в 5% CO2-инкубаторе перед сбором. Клетки собирали в 0,1% буфере SDS-RIPA-HEPES, дополненном 1Х полным ингибитором протеазы (ЭДТА-свободный).
Клик-химия
После мечения клеток ω-алкинилмиристиновой кислотой полученные лизаты подвергали клик-химии с азидо-биотином, позволяющей детектировать миристоилированные белки посредством ECL, как описано ранее (Yap et al., 2010). Обычно, лизаты клеток (30-50 мкг белка) доводили до содержания 1% SDS и инкубировали с 100 мкМ Трис-(бензилтриазолилметил)амином (ТВТА), 1 мМ CuS04, 1 мМ трис-карбоксиэтилфосфином (ТСЕР), и 100 мкМ азидо-биотином при 37°С в течение 30 мин в темноте, с целью продолжения клик-реакции. Затем, 10 объемов охлажденного на льду ацетона добавляли для остановки клик-реакции, и белки осаждали при -20°С в течение ночи. Затем, белки, осажденные ацетоном, центрифугировали при 16000 об/мин в течение 10 мин, ресуспендировлаи в 1Х SDS-PAGE буфере для образцов с 20 мМ DTT. Затем образцы нагревали при 95°С в течение 5 мин и подвергали SDS-PAGE, и переносили на PVDF мембраны для анализа Вестерн-блотом.
Обработка PVDF мембран нейтральным Трис-HCl или КОН
Образцы белков наносили в дупликатах в гели для SDS-PAGE для экспериментов, при которых мембраны PVDF обрабатывали КОН или нейтральным Трис-HCl. После электрофореза, мембраны PVDF, обработанные КОН, инкубировали в 100 мл 0,1 N КОН в метаноле [1 N КОН в H2O: метанол 1:9 (v/v)], при этом другую инкубировали в 0,1 N Трис-HCl рН 7,0 в метаноле [1 N Трис-HCl, рН 7,0:метанол 1:9 (о/о)] при комнатной температуре в течение 45 мин с бережным покачиванием. Затем обработанные мембраны хорошо промывали PBS, исследовали при помощи стрептавилин-HRP или нейтравидин-HRP, и детектировали при помощи ECL.
Радиоактивное исследование активности ΝΜΤ в клетках претерпевших апоптоз
Активность ΝΜΤ измеряли посредством адаптирования протокола, разработанного Sharma laboratory (King and Sharma, 1991; Raju and Sharma, 1999). Стоковый раствор [3Н]-миристоил-СоА готовили свежим и синтезировали как описано ранее (Towler and Glaser, 1986). Для получения 200 мкл стокового раствора [3Н]-миристоил-СоА, 97 мкл миристоил-СоА-генерирующим буфером комбинировали с 20 мкл 50 мМ АТР, 10 мкл 20 мМ LiCoA, 60 мкл ацил-СоА синтазы Pseudomonas и 13 мкл [9,10 - 3Н]-миристиновой кислоты. Раствор смешивают осторожно и инкубируют при 37°С в течение 30 мин.
Для данного исследования COS-7 клетки временно трансфецированные плазмидами, кодирующими V5-NMT1 и V5-NMT2, инкубировали для подвергания апоптозу с STS (2,5 мкМ) и циклогексимидом (5 мкг/мл) или нет, и собирали во временных точках 0, 1, 2, 4 и 8 ч. Клетки обрабатывали ультразвуком при помощи ультразвукового дезинтегратора Брансона 450 и ~20 мкг лизат (лизированный в 50 мМ NaH2P04 рН 7,4, 0,25 M сахарозный буфер) применяли для каждой реакции. Для каждой реакции исследования NMT 3,85 мкл буфера для исследования NMT, 1,25 мкл 20% Triton Х-100 и 2,5 мкл (0,1 мМ) миристоилированного (BID_G: GNRSSHSRLG) или немиристоилированного (BID_A: ANRSSHSRLG) декапептида (закупленного у Peptide 2.0 Inc.) соответствующего N-терминальной последовательности ct-Bid (1 мМ стока этанола) добавляли в микроцентрифужную пробирку и хранили на льду до начала реакции. В конце реакции 7,4 мкл (10 пикомоль) свежесинтезированного [3Н] миристоил-СоА добавляли в каждую микроцентрифужную пробирку. Содержащую смесь для исследования NMT с 15 секундными интервалами, и 25 мкл реакций инкубировали в течение 15 мин при 30°С. Реакцию останавливали посредством нанесения 15 мкл реакционной смеси на Р81 фосфоцеллюлозный бумажный диск (Whatman) с 15 секундными интервалами и высушивали феном для волос в течение 30 секунд.
Затем, р81 диски фосфоцеллюлозной бумаги переносили на устройство промывки и промывали 3 раза, 30 мин каждый, с промывочным буфером для исследования NMT в течение периода 90 мин. Затем радиоактивность, оставшуюся на фосфоцеллюлозе (соответствующую миристоилированному пептиду) количественно определяли в 5 мл жидкостно-сцинтилляционной смеси при помощи сцинтилляционного счетчика Beckman Coulter LS6500. Активность NMT вычисляли в виде фемтомоль/мин/мкг белка.
Применение радиоактивного исследования для измерения активности NMT в очищенных His-меченных
Активность NMT полноразмерной и усеченной His-меченных NMT измеряли при помощи такого же протокола, как описано ранее. Однако, только 1 мкг очищенного белка (объем доводили до 10 мкл в буфере для исследования NMT) применяли в данном исследовании.
In vitro исследование расщепления NMT
Исследование расщепления NMT in vitro проводили посредством инкубирования 10 мкг очищенной His-NMT1 и HiS-NMT2 с 1 мкг рекомбинантной активной каспазой-3 или -8 человека в буфере для расщепления каспазой в 100 мкл реакций в течение 1 часа при 37°С. Реакцию останавливали добавлением 10 мкМ обычного ингибитора каспазы z-VAD-FMK и затем добавляли 5Х буфер для нанесения образцов. Образцы после реакции разделяли в геле 10% SDS-PAGE и переносили на мембрану PVDF. Бэнды визуализировлаи при помощи окрашивания Кумасси голубым, и фрагменты расщепления вырезали из мембраны и отправляли на расщепление по Эдману в Alphalyse в Palo Alto, Калифорния.
Очистка белка рекомбинантных His-NMT
Lemo21(DE3) pLysS компетентные клетки (New England Biolabs) трансформировали с векторами His-NMT1 и His-NMT2 согласно протоколу производителя. Белки NMT1 и NMT2 готовили следующим образом: 3 мл стартовой культуры выращивали в LB бульоне (1% триптон, 0,5% дрожжевой экстракт, 0,5% NaCl, 100 мкг/мл ампициллина и 34 мкг/мл хлорамфеникола) в течение 4 ч при 37°С, при этом перемешивая при 225 об/мин. Целую культуру применяли для инокуляции 50 мл культуры, которую выращивали при 37°С в течение 16 ч. Затем бактериальные клетки осаждали центрифугированием при 6000 × об/мин в течение 10 мин при комнатной температуре. Бактериальный осадок ресуспендировали в 10 мл LB и 5 мл ресуспензии применяли для инокуляции 500 мл LB, который инкубировали при 37°С с активным покачиванием (225 об/мин) до достижения ОП600 0,5-0,6.
Затем экспрессию белка индуцировали посредством добавления 0,5 мМ изопропил-β-D-1-тиогалактопиранозида (IPTG) при 30°С с активным покачиванием (225 об/мин) в течение 4 ч. Суспензию центрифугировали при 6000 × об/мин при 4°С в течение 20 мин и бактериальный осадок замораживали в течение ночи. Затем, размороженный бактериальный осадок ресуспендировали в 25 мл буфера для лизиса бактерий, дополненного полным ингибитором протеазы (CPI) от Roche и инкубировали на льду в течение 30 мин. Затем образцы обрабатывали ультразвуком 3 раза в течение 1 мин с интервалами 1 мин при выходном напряжении 5.0 (ультразвуковой дезинтегратор Брансона 450) и инкубировали при 37°С в течение 30 мин перед центрифугированием при 15,000 × об/мин в течение 1 ч при 4°С.
При этом, Ni-NTA агарозные гранулы (His-Pure Ni-NTA смола от Thermo Fisher Scientific) упаковывали в колонки согласно инструкциям производителя и промывали при помощи буфера для колонки три раза. Прозрачный надосадок, полученный после предыдущего центрифугирования затем наносили на колонку и инкубировали с бережным покачиванием при 4°С в течение 1 ч. Затем образец элюировали с колонки за счет гравитационного тока и колонку промывали дважды при помощи 5 мл промывочным буфером (буфер для колонки с 20 мМ имидазолом, рН 8,0). Затем 2×5 мл элюирующего буфера 1 (буфер для колонки с 75 мМ имидазолом, рН 8,0) применяли для элюции первых двух фракций и 2×5 мл элюирующего буфера 2 (буфер для колонки с 150 мМ имидазолом, рН 8,0) применяли для элюции следующих двух фракций. Последнюю фракцию элюировали элюирующим буфером 3 (буфер для колонки с 250 мМ имидазола, рН 8,0). Наконец 20% глицерола добавляли к каждой из фракций, собранных перед замораживанием при -80°С. Образцы с каждого этапа способа очистки белка собирали, подвергали 12,5% SDS-PAGE электрофорезу в геле и окрашивали Кумасси голубым с целью определения какая фракция(и) обладает наиболее высокой концентрацией белка.
Поскольку образцы от первых четырех элюций обладали наивысшей концентрацией белка, их объединяли и диализировали для удаления любого присутствия имидазола. Поэтому, объединенные образцы наносили на диализную трубку Spectra/Por 2 (Spectrum Laboratories) с границей отсечки по молекулярной массе (MWCO) 12-14 кДа для удаления любых небольших деградированных фрагментов белка. Образцы белка затем диализовали в 4 л охлажденного диализного буфера в течение 2 ч при 4°С с бережным перемешиванием, с последующим диализом в течение ночи с тем же буфером при 4°С с другими 4 л свежего охлажденного буфера. Диализованные образцы белков концентрировали посредством центрифугирования в Amicon Ultra-15 ценитрифужным фильтром с MWCO 30 кДа (Millipore) при 5000 об/мин, 4°С до достижения желаемых объемов. РВ-ПЦР клеточных линий В-клеточной лимфомы
РНК изолировали из клеток IM9, КМН2, Рамоса и BL2 при помощи реагента TRIzol® согласно протоколам производителя. Затем, кит для обратной транскрипции High Capacity cDNA Reverse Transcription kit с рандомными праймерами от Applied Biosciences применяли для синтеза кДНК из изолированной РНК согласно инструкциям производителя. Колличественные РВ-ПЦР реакции проводили при помощи TaqMan® Universal Master Mix II и NMT1, NMT2 и 18 Taqman® проб, закупленных у Life Technologies (карлсбад, Калифорния) и три репликата каждой реакции проводили согласно руководству поставщика. РВ-ПЦР проводили при помощи Mastercycler® ер realplex thermocycler (Eppendorf) и результаты анализировали при помощи программного обеспечения Realplex (Eppendorf).
Исследование вытеснением трипанового синего
Жизнеспособность клеток, обработанных Tris-DBA, DDD73228, и DDD85646 измеряли посредством инкубирования клеток с красителем ТС10ТМ Trypan Blue Dye (Biorad) (Hudson, 1976), согласно инструкциям производителя. Жизнеспособность клеток затем количественно оценивали при помощи ТС10ТМ автоматического цитометра (Biorad).
Исследование MTS [(3-(4,5-диметилтиазол-2-ил)-5-(3-карбоксиметоксифенил)-2-(4-сульфофенил)-2Н-тетразолиий)]
Жизнеспособность клеток, обработанных DDD86481, измеряли посредством нерадиоактивного анализа клеточной пролиферации при помощи CellTiter 96 AQueous (MTS) (Cory et al., 1991) от Promega согласно инструкциям производителя.
Иммуногистохимия
Опухоли, В-клеточные лимфомы, фиксировали в формалине и заключали в парафин и нарезали на микротоме на желаемую толщину (~5 микрон или мкм) и прикрепляли к положительно заряженным предметным стеклам Superfrost® plus (Fisher Scientific). До окрашивания стекла с опухолевыми тканями депарафинизировали посредством опускания в ксилол 3 раза в течение 10 мин каждый, серии промывок в этаноле [20 опусканий в 100% этанол (повторы 4 раза), 20 опусканий в 80% этанол, и 20 опусканий в 50% этанол] и окончательной промывки под проточной холодной водой в течение 10 мин.
Для демаскирования антигена стекла помещали в штатив-рамку для окраски микропрепаратов и помещали в ультразвуковом автоклаве Nordicware®, и добавляли к нему 800 мл 10 мМ цитратного буфера рН 6,0. автоклав плотно закрывали и обрабатывали ультразвуком на высоком режиме в течение 20 мин. Затем, стекла промывали в холодной проточной воде в течение 10 мин. Блокирование пероксидазой проводили посредством замачивания стекол в 3% H2O2 в метаноле в течение 10 мин и промывки теплой проточной водой в течение 10 мин до промывки в PBS в течение 3 мин.
Затем, избыток PBS удаляли и вокруг образца рисовали гидрофобное кольцо при помощи гидрофобного карандаша (PAP pen, Sigma-Aldrich). Кроличьи антитела к NMT1 (Proteintech) или к NMT2 (Origene) разводили буфером для разведения антител Dako (Dako, Agilent Technologies) в разведении 1:50 и добавляли при помощи Пастеровской пипетки (~400 мкл на стекло) и инкубировали во влажной камере в течение ночи при 4°С. Затем слайды промывали в PBS дважды в течение 5 мин каждый, к каждому стеклу добавляли ~4 капли DAKO EnVision+System-HRP-меченного полимера (к кролику) (Dako, Agilent Technologies) и инкубировали при комнатной температуре в течение 30 мин. Стекла промывали снова в PBS дважды в течение 5 мин каждый, 4 капли жидкого DAB (диаминобензидина) + субстрат хромогена (готовили согласно инструкциям производителя; Dako, Agilent Technologies) добавляли, выдерживали в течение 5 мин и промывали под проточной холодной водой в течение 10 мин.
Затем стекла замачивали в 1% CuSO4 в течение 5 мин, промывали быстро под проточной холодной водой, окрашивали при помощи гематоксилина в течение 60 с и промывали проточной холодной водой до тех пор пока вода не станет прозрачной. Затем стекла опускали в карбонат лития 3 раза и быстро промывали проточной водой из под крана. Стекла дегидрировали в серии этапов; 20 опусканий в 50% этанол, 20 опусканий в 80% этанол и 20 опусканий в 100% этанол (повтор 4 раза) и наконец в ксилол (3 раза по 10 мин для каждого). Затем покровные стекла добавляли к стеклам и проводили микроскопию образцов опухоли при помощи Nikon eclipse 80i microscop. Изображения создавали при помощи научной камере Qlmaging (Qimaging).
Пример 1
Фигура 1 изображает анализ экспрессии NMT1 и NMT2 в нормальных клетках и различных В-клеточных лимфомах и Т-клеточных лейкемиях. Данная фигура показывает почти полное отсутствие экспрессии NMT2 в В-лимфотических клеточных линиях (BL-2, Рамос), которые экспрессируют только NMT1 по сравнению с нормальными В-клетками (EBV трансформированные В-лимфоциты человека, L0) и Т-клеточные линии лейкемии человека (Jurkat, MOLT-4, СЕМ).
Без связи с какой-либо теорией, эти клетки экспрессируют только один изофермент NMT, например, клетки лимфомы беркитта, которые показывают почти полное отсутствие NMT2, вероятно обладают измененными профилями миристоилирования.
Образец, обладающий пониженным количеством миристоилированного белка в образце (возможно по сравнению с контролем) является показателем образца с недостатком NMT, или рака с недостатком NMT. Такой рак с недостатком NMT приемлем для лечения ингибитором NMT1.
Пример 2
Фигура 2 изображает чувствительность различных В-клеточных лимфом и Т-клеточных лейкемий к ингибитору NMT трис-дибензилиденацетон-дипалладию (Tris-DBA). Различные В- и Т-клетки инкубировали в течение 24 ч с повышающейся концентрацией Tris-DBA. Жизнеспособность клеток измеряли при помощи способа вытеснения трипанового синего и подводили до 100% контроля. Кривые выживаемости клеток, измеренной посредством вытеснения трипанового синего, показывают, что В-клеточные лимфомы более чувствительны к ингибитору NMT трис-дибензилиденацетон-дипалладию (Tris-DBA).
Пример 3
Фигура 3 изображает ингибирование N-миристоилтрансферазы (NMT) посредством трис-дибензилиденацетон-дипалладия (Tris-DBA).
Активность NMT оценивали при помощи исследования миристоилирования пептида с очищенными рекомбинантными NMT1 и NMT2. NMT активность вычисляли исходя из количества радиомеченного миристоилпептида, полученного и детектированного на фосфоцеллюлозной бумаге (адаптировано от King et al. 1991, Anal Biochem.).
Данная фигура показывает, что трис-дибензилиденацетон-дипалладий (Tris-DBA) ингибирует NMT in vitro при помощи очищенных рекомбинантных ферментов NMT.
Пример 4
Фигура 4 изображает результаты иммуноблота, в котором клеточные линии лимфомы изучены с антителами к NMT1 (панель А) и NMT2 (панель В). Легенда на фигуре 4 соответствует следующему: IM9: В-лимфобласт; BL2: лимфома Беркитта; СЕМ: Т-клеточная лейкемия; Karpas 299: Т-клеточная лейкемия; Sup-M2: ALCL; UCONN: ALCL (ALCL: анапластическая крупноклеточная лимфома); DAUDI: лимфома Беркитта; Ramos: лимфома Беркитта BJAB: лимфома Беркитта; HD-MYZ: лимфома Ходжкина; КМ-Н2: лимфома Ходжкина; L428: лимфома Ходжкина; Jurkat: Т-клеточная лейкемия.
Пример 5
Фигура 5 изображает эффективность ингибиторов NMT в отношении клеточной линии лимфомы Беркитта Ramos по сравнению с иммортализованной нормальной клеточной линией В-лимфоцитов (IM9) после 48 часов при различных концентрациях.
Пример 6
В данном примере трансфекция В-лимфотических клеток линии Ramos (которая, как показано в данном документе, экспрессирует NMT1) с pcDNA3.1-V5-NMT2 повышенной устойчивостью к Tris-DBA (5 мкг/мл) 2,5 кратно vs контрольных клеток, трансфецированных пустым плазмидным вектором. На фигуре 6,20×106 Ramos В-лимфотические клетки трансфецировали 32 мкг ДНК (pcDNA3.1-V5-nycnrà или pcDNA3.1-V5-NMT2) при помощи трасфецирующей системы Neon (Invitrogen) согласно рекомендованному протоколу для клеточной линии Ramos (1,350 Вольт, 30 мсек). Трансфецированные клетки центрифугировали в течение 5 минут при 1200 об/мин для удаления умерших клеток и клеточного осадка. Клетки в надосадке оставляли восстанавливаться в течение 6 часов в полной RPMI. После промывки PBS, клетки ресуспендировали и выращивали в RPMI, содержащей Tris-DBA (5 мкг/мл) в течение 24 часов затем подсчитывали при помощи способа вытеснения трипанового синего (панель А). Клетки лизировали и проводили Вестерн-блот (ECL) для подтверждения экспрессии NMT2 при помощи антител к NMT2, и GAPDH для нагрузки контроля (панель В).
Пример 7
В данном примере РВ-ПЦР выполняли при помощи проб Taqman к NMT1 и NMT2 при помощи 18S пробы в качестве внутреннего контроля. Различия в номере времени цикла (∆ct) вычисляли посредством вычитания времени цикла (et), при котором мы видим экспоненциальное увеличение экспрессии 18S внутреннего контроля для каждого типа клеток из времени цикла NMT, против точки, где видно экспоненциальное увеличение сигнала. Как показано в таблице 1, ниже, отношение экспрессии NMT2 к NMT1 понижено (до 60 кратно) в клеточных линиях В-лимфомы. Без связи с какой-либо теорией, данные результаты могут подтвердить, что снижение в мРНК, кодирующей NMT2 может быть ответственным за снижение уровней белка NMT2, оцененных посредством Вестерн-блота.
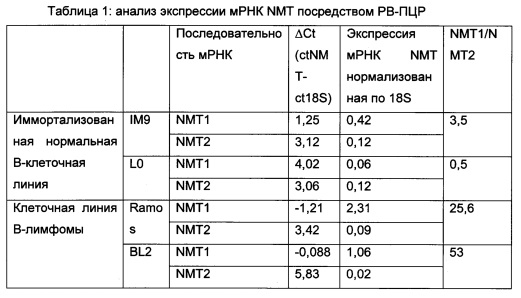
Пример 8
В данном примере на фигуре 7 приведены различия в уровнях белка NMT2 присутствующего в различных лимфотических клеточных линиях и твердых лимфоидных опухолях. (А) Уровни NMT1 и NMT2 оценивали посредством Вестерн-блота в лизатах различных типов твердой лимфомы человека (диффузной В-крупнокпеточной лимфомы (DLBCL) и фолликулярной лимфомы (FL). NMT детектировали посредством Вестерн-блота при помощи одинаковых концентраций антител для обеих панелей: кроличьи антитела к NMT1 (1:2500), мышиные монокпональные антитела к NMT2 (1:2500). Показано, что многочисленные образцы опухолей каждого типа подпадали под средний уровень экспрессии NMT2 (0,392 +/-0,30).
В панеле А, показано, что клеточные линии BL-2 лимфомы Беркитта, Daudi, Ramos и BJAB с недостатком NMT2.
В панеле В, показано, что 4 из 5 лизатов DLBCL и 1 из 6 лизатов опухоли FL человека имели отмеченное снижение в NMT2.
Пример 9
В данном примере в панелях А и В на фигуре 8 определяли различия в уровнях белка NMT2, присутствующего в различных твердых лимфотических опухолях, посредством иммуногистохимии.
В-клеточные лимфотические опухоли фиксировали в формалине, помещали в парафин, нарезали на микротоме на желаемую толщину (~5 микрон или мкм) и закрепляли на положительно заряженном стекле Superfrost® plus (Fisher Scientific). До окрашивания стекла содержащие опухолевые ткани депарафинизировали посредством погружения в ксилол 3 раза в течение 10 мин каждый, серии промываний в этаноле [20 погружений в 100% этанол (4 повтора), 20 погружений в 80% этанол и 20 погружений в 50% этанол] и конечной промывки в проточной холодной воде в течение 10 мин.
Для демаскирования, стекла помещали в штатив-рамку для окраски микропрепаратов и помещали в ультразвуковой автоклав Nordicware® и к ним добавляли 800 мл 10 мМ цитратного буфера рН 6,0. Автоклав плотно закрывали, помещали в обрабатывали ультразвуком в течение 20 мин. Затем, стекла промывали в холодной проточной воде в течение 10 мин. Блокирование пероксидазой выполняли посредством замачивания стекол в 3% H2O2 в метаноле в течение 10 мин и промывки теплой проточной водой в течение 10 мин перед промывкой в PBS в течение 3 мин.
Затем, избыток PBS удаляли, и вокруг образца рисовали гидрофобное кольцо при помощи гидрофобного карандаша (PAP pen, Sigma-Aldrich). Кроличьи антитела к NMT1 (Proteintech) или кроличьи антитела к NMT2 (Oxigene) разводили при помощи буфера для разведения антител Dako (Dako, Agilent Technologies) при разведении 1:50 и добавляли пастеровской пипеткой (~400 мкл на слайд) и инкубировали во влажной камере в течение ночи при 4°С. Затем стекла дважды промывали в PBS в течение 5 мин каждый раз и ~4 капли DAKO EnVision+System-HRP-меченного полимера (к кролику) (Dako, Agilent Technologies) добавляли к каждому стеклу и инкубировали при комнатной температуре в течение 30 мин. Стекла снова дважды промывали в PBS в течение 5 мин каждый, 4 капли жидкого DAB (диаминобензидин) + субстрат хромоген (приготовленного согласно инструкциям производителя; Dako, Agilent Technologies) добавляли, выжидали в течении 5 мин и промывали под проточной холодной водой в течении 10 мин.
Затем стекла замачивали в 1% CuSO4 в течение 5 мин, промывали быстро проточной холодной водой, окрашивали для подсчета при помощи гематоксилина в течение 60 с и промывали проточной холодной водой до прозрачности воды. Затем стекла погружали в карбонат лития 3 раза и быстро промывали водой из под крана. Затем стекла дегидрировали в серии этапов; 20 погружений в 50% этанол, 20 погружений в 80% этанол и 20 погружений в 100% этанол (повтор 4 раза) и наконец в ксилол (3 раза по 10 мин каждый). На стекла накладывали покровные стекла и проводили микроскопию опухолевых образцов при помощи микроскопа Nikon eclipse 80i. Изображения получали при помощи научной камеры Qlmaging (Qimaging).
NMT иммуногистохимическое окрашивание нормальных лимфотических узлов, лимфомы Беркитта (BL диффузной В-крупноклеточной лимфомы (DLBCL или LCL). A) NMT1 окрашивание: как нормальные лимфатические узлы, так и каждый из трех независимых случаев нелеченной лимфомы Беркитта (BL1-3) и DLBCL (LCL1-3) окрашивались однородно и сильно положительно (коричневая окраска продукта пероксидазной реакции [проявляющаяся как темно серая]) для белка NMT1. Различия не наблюдались. Отрицательный контроль получали посредством исключения первичного антитела. Верхний ряд: N1, N2 нормальные лимфотические узлы, Neg отрицательный контроль. Средний ряд: три случая лимфомы Беркитта (BL1-3). Нижний ряд: три случая DLBCL (LCL1-3). Как нормальные лимфотические узлы, так и лимфома проявляли сильное окрашивание. В) NMT2 окрашивание: нормальные лимфотические узлы окрашивались как однородно, так и сильно положительно (коричневое окрашивание, проявляющееся как темно серое) для белка NMT2. Резкое отличие, каждый из трех независимых случаев нелеченной лимфомы Беркитта и DLBCL проявляют очень слабое окрашивание для NMT2 (показано как светло-голубое или светло-серое). Отрицательный контроль получали за счет исключения первичного антитела. Верхний ряд: N1, N2 нормальный лимфатический узел, Neg отрицательный контроль. Средний ряд: три лимфомы Беркитта (BL1-3); нижний ряд: три DLBCL(LCL1-3).
Фигура 8 панель С и панель D демонстрируют, что экспрессия NMT1 не существенно повышена в клеточных линиях, утративших экспрессию NMT2. В панеле С log2(микроэррей интенсивность флуоресценции NMT) для NMT1 и NMT2 нанесена для всех трех клеточных линий базы данных CCLE. В панеле D log2(микроэррей интенсивность флуоресценции NMT) для NMT1 и NMT2 нанесена для 100 клеточных линий базы данных CCLE с наименьшим экспрессионным уровнем NMT2.
Фигура 8 панель Ε изображает пример, в котором определения ΝΜΤ2 колличественно оценивали при помощи компьютерной световой микроскопии, дополненной денситометрическими измерениями. Визуально, данные числа, связанные с силой окрашивания злокачественных лимфоцитов. Выбранные границы представляли собой сильное vs отсутствие/слабое окрашивание.
Пример 10
В данном примере на фигуре 9 приведена остаточная жизнеспособность различных В-лимфотических клеточных линий, обработанных DDD85646 в течение 72 часов (фигура 9А). Кривые на фигуре 9А нанесены согласно 4 параметровому выравниванию, как описано в Leatherbarrow, R.J. (2009) GraFit Version 6, Erithacus Software Ltd., Horley, U.K. для анализа GraFit при помощи формулы:
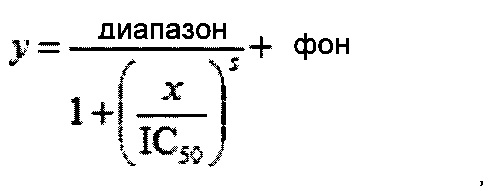
где диапазон представляет собой соответствующее неингибированное значение минус фон, и s представляет собой коэффициент уклрна. Формула предполагает, что у согласуется с повышением х. А и В. Сводные графики (n=4).
На фигуре 9 панель А, повышающиеся концентрации DDD85646 применяли для лечения клеточных линий BL (BL-2 nd Ramos) наряду с относительными контролями [IM9 ("нормальный" В-лимфоцит) и КМН2 (HL клеточная линия), которая экспрессирует обе NMTs]. Исследование трипановым синем применяли для измерения цитотоксичности DDD85646 при 24, 48 и 72 часов. Хотя данные собирали во временных точках 24 ч и 48 ч (данные не приведены), наибольшее заметное воздействие DDD85646 наблюдалось во временной точке 72 ч.
Коэффициенты выживаемости были понижены в примененных В-лимфотических клеточных линиях (Ramos: DDD85646 ЕС50=0,37+/-0,05 мкМ и BL2: DDD85646 ЕС50=0,43+/-0,2 мкМ) зависимым от концентрации DDD85646 образом. Коэффициенты выживаемости контрольных клеточных линий, IM-9 (DDD85646 ЕС50=7,08+/-2,32 мкМ) и КМН2 (DDD85646 ЕС50=29,6+/-10,6 мкМ) были высокими. Данные ЕС50 суммированы следующим образом.
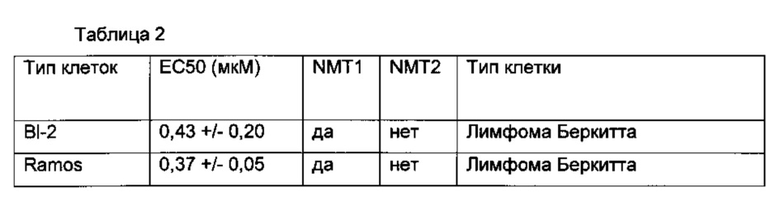
DDD85646 проявляли низкий ЕС50 (концентрация, которая убивала 50% раковых клеток) в клетках BL, и проявляли более высокий ЕС50 в клетках HL (КМН2). Таким образом, DDD85646 проявляли высокий индекс селективной гибели для раковых клеток. Как употреблено в данном документе, мы определяем индекс селективной гибели как отношение ЕС50 «нормальных» иммортализованных В-клеток/ЕС50 злокачественных клеток. Индекс селективной гибели для DDD85646 составил 16,4 и 19,1 для клеток BL-2 и Ramos соответственно.
На фигуре 9, панель В DDD86481 применяли для обработки В-клеток и контрольных клеточных линий [IM9 ("нормальный" В-лимфоцит) и КМН2 (HL клеточная линия), которая экспрессирует обе NMTs]. Жизнеспособность клеток измеряли при помощи MTS-анализа при 48 ч и 72 ч, хотя мы не наблюдали существеенного изменения клеточной жизнеспособности при 48 ч (данные не приведены), мы обнаружили, что коэффициент выживаемости исследованных клеточных линий BL, снижался существенно во временной точке 72 ч (Ramos: DDD86481 ЕС50=42 нМ и BL2: DDD86481 ЕС50=58 нМ) зависимым от концентрации DDD86481 образом. Коэффициенты выживаемости контрольных клеточных линий, IM-9 (DDD86481 ЕС50=2,2 мкМ) и КМН2 (DDD86481 ЕС50=12,6 мкМ) были выше. Индекс селективности, вычисленный для DDD86481, составил 37,9 и 52,3 для клеток BL-2 и Ramos, соответственно. Данные ЕС50 суммированы далее.
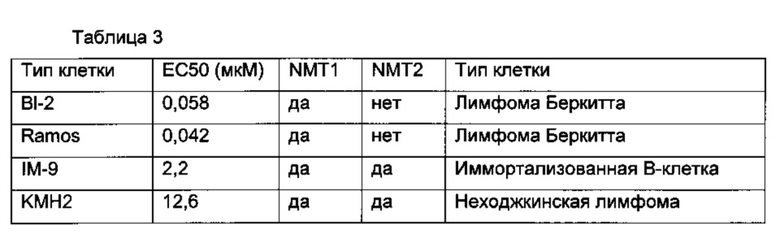
Согласно таблице, представляющей описание клеточного типа и EC50's для DDD85646 и DDD864841, В-клеточные клетки (BL-2 и Ramos), экспрессирующие только одну NMT (NMT1) являются в ~15-72 раз более чувствительными к DDD85646 или DDD86481, чем иммортализованные «нормальные» В-клетки или клеточные линии неходжкинской лимфомы, экспрессирующие обе NMT.
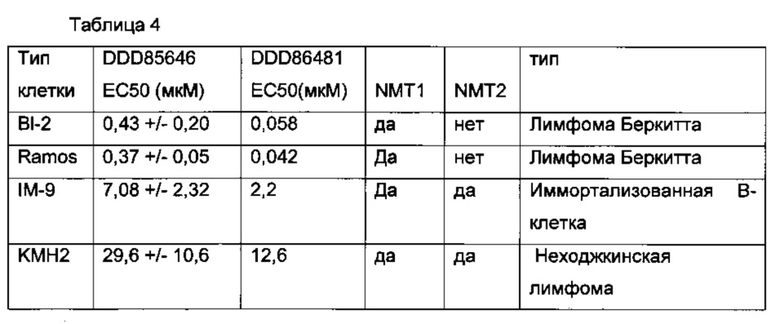
В следующей таблице приведены суммарные данные фармакологических EC50'S и EC90'S для DDD86481 в клеточных линиях лимфомы Беркитта (BL-2 и Ramos) и «нормальных» иммортализованных В-клетках (данные, вычисленные исходя из фиг. 9В).
Индексы селективности показывают отношения EC50'S и EC90'S, которые указывают, что DDD86481 преимущественно убивает раковые клетки посредством фактора >300 кратного (поскольку концентрация необходимая для убийства нормальной В-клеточной линии является намного более высокой).
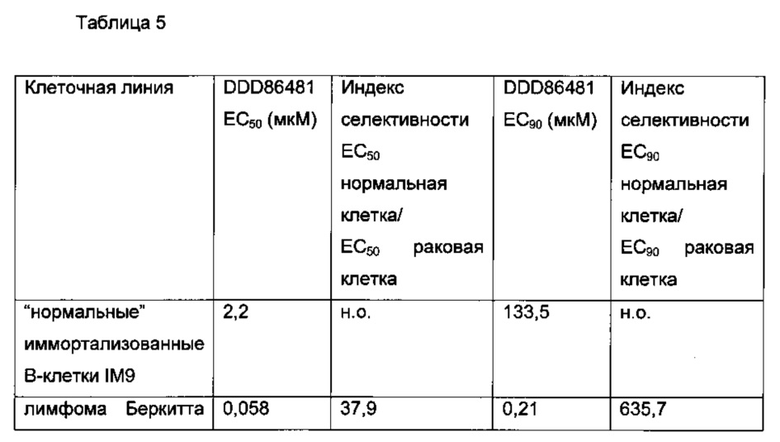

На фигуре 9C, DDD73226 применяли для обработки клеточных линий BL (BL-2 и Ramos) наряду с соответствующими контролями [IM9 ("нормальный" В-лимфоцит) и КМН2 (HL клеточная линия), экспрессирующая обе NMT)]. Цитотоксичность DDD73226 измеряли при помощи испытания вытеснением трипанового синего при 72 ч. Не наблюдалось существенных изменений в жизнеспособности клеток на 24 и 48 ч (данные не приведены). Также не было существенного изменения в жизнеспособности клеток IM9, КМН2 и Ramos в случае, когда клетки обрабатывали DDD73226 при концентрациях до 100 мкМ во временной точке 72 ч. Однако было незначительное снижение жизнеспособности клеток, наблюдаемое в клетках BL2, обработанных 100 мкМ DDD73226 на 72 ч, при сравнении с другими клетками. Без связи с какой-либо теорией это может быть обусловлено тем, что в дополнение к обладанию существенно сниженными уровнями NMT2, BL2 также имеют более низкие уровни по сравнению с клеточными линиями Ramos и другими и поэтому могут быть чувствительными к действию даже менее эффективных ингибиторов NMT.
На фигуре 9D (панели i и ii), представлены эксперименты по определению зависимости от времени, где клетки IM-9 и BL2 метили при помощи 100 мкМ ω-алкинил-миристата в течение 1 ч после обработки DDD86481 (0,1 и 10 мкМ) для 0, 1 или 4 ч. Проводили реакцию образцов белка с азидо-биотином при помощи клик-химии и визуализировали посредством вестерн-блота с NeutrAvidin™-HRP. Ингибиторное действие DDD86481 было быстрым, с почти полным ингибированием миристоилирования в клетках (как IM-9, так и BL2), обработанных 1 мкМ DDD86481 после одного часа обработки.
На фигуре 9Е, определяли минимальную дозу DDD86481, необходимую для ингибирования миристоилирования в клеточных линиях IM-9, BL2 и Ramos. Клетки обрабатывали DDD86481 при 0, 10, 50, 100, 500 и 1000 нМ концентрациях в течение 2 ч с метаболическим мечением клеток при помощи алкинил-миристата на последнем 1 ч обработки. DDD86481 ингибировал миристоилирование зависимым от концентрации образом во всех исследуемых клеточных линиях. Клеточные линии BL-2 и Ramos были более чувствительны к ингибиторному действию DDD86481, поскольку снижение во включении метки алкин-миристата в миристоилированные белки было очевидным, начиная с 50 нМ для обеих клеточных линий BL при сравнении с IM-9, где включение метки алкин-миристата снижалось начиная с концентрации 100 нМ.
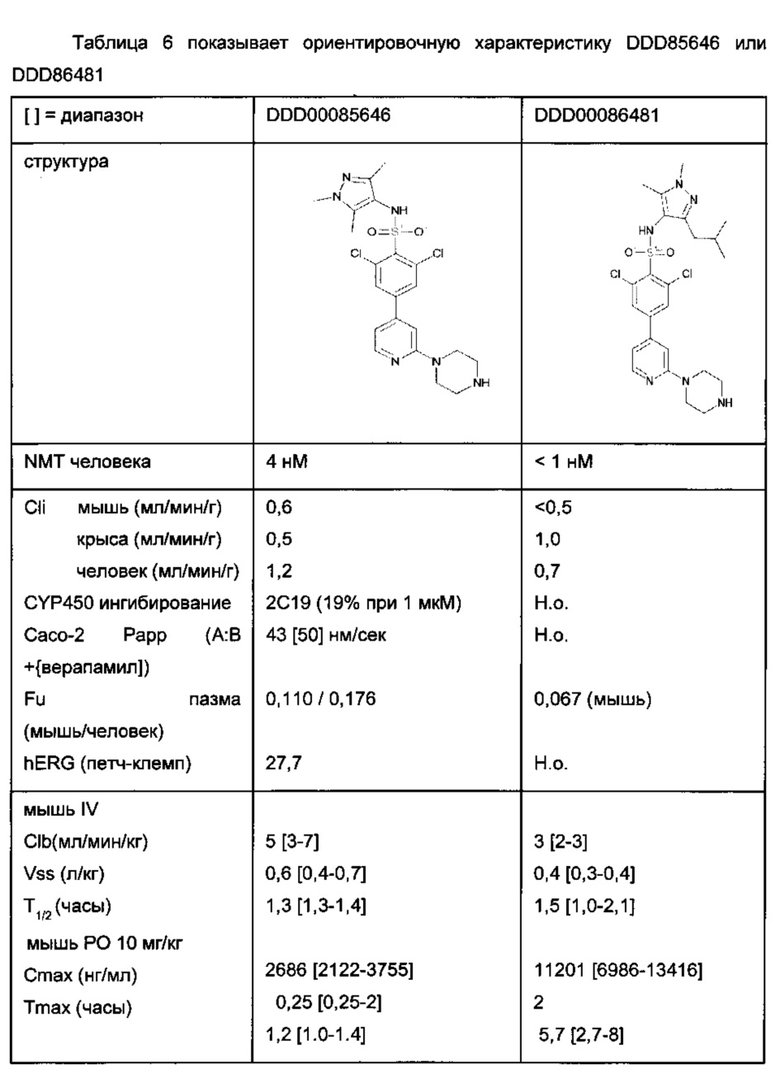

Пример 11
В данном примере на фигуре 10 показана чувствительность различных иммортализованных нормальных L0 В-лимфоцитов (полученных от доктора Riabowol, U. of Calgary), злокачественных В-лимфотических клеток (Ramos и BL2), и, Т-клеточной лейкемии (СЕМ) к ингибитору NMT Tris-DBA в течение 24 часов (А) остаточная жизнеспособность клеточных линий, обработанных Tris-DBA в течение 24 часов (n=9) (В) эффект Tris-DBA на активность N-миристоилтрансферазы в клетках COS7, временно экспрессирующих NMT1 или NMT2, инкубированных в течение 24 ч с Tris-DBA. In vitro IC50 для Tris-DBA составила приблизительно 2 мкМ для NMT1 и 5 мкМ для NMT2. Клеточные линии лимфомы Беркитта Ramos и BL-2 более чувствительны к ингибитору NMT Tris-DBA.
Пример 12
В данном примере на фигуре 11, определяли уровни экспрессии NMT в 967 клеточных линиях рака базы данных (CCLE). Панель А) базу данных CCLE изучали в отношении экспрессии NMT1 и NMT2 и соответствующие уровни NMT наносили. NMT2 показывает большой диапазон экспрессии (~3-11) по сравнению с NMT1 (~6-9). Панель В) Диаграммы вида «ящик с усами» раковых клеточных линий из нескольких подтипов рака нанесены и сравнены с диаграммой всех 967 клеточных линий рака. Т-тест Стьюдента проводили сравнивая каждую группу со всеми опухолями. ** означает Р уровни <0,001. Панель С) база данных CCLE изучали в отношении 50 клеточных линий, которые экспрессировали самые низкие уровни NMT2 и приводили в формате таблицы. Опухолевые клеточные линии, полученные из различных гематопоэтических и лимфоидных тканей, представляют 76% (38 из 50) клеточных линий с самой низкой экспрессией NMT2 и отмечены рахными цветами.
Пример 13
В данном примере на фигуре 12 NMT расщепляются в течение апоптоза, но сохраняют активность. Панель А и В. Лизаты клеток Jurkat, индуцированные к апоптозу, опосредованному антителами к Fas или стауроспорином, изучали при помощи антител к NMT1 и антител к NMT2 посредством Вестерн-блота, и показано время зависимое расщепление обоих ферментов. Вестерн-блот проводили на одинаковых образцах при помощи антител к РАК2 и GAPDH. Панель С и D. Включение ω-алкинил-миристата в клетки Jurkat, индуцированные к апоптозу, опосредованному антителами к Fas или стауроспорином, иллюстрирует изменение в профилях миристоилирования, поскольку клетки начинают окрашиваться и подтверждает, что несмотря на расщепление NMT, они все еще активны. Клетки Jurkat метаболически метили при помощи 25 мкМ-алкинил-миристата после индукции апоптоза при помощи антител к FAS (100 нг/мл) и циклогексимида (5 мкг/мл) (С) или стауроспорина (2,5 мкМ) и циклогексимида (5 мкг/мл) (D). Образцы белка реагировали с азидо-биотином при помощи клик-химии и визуализировали посредством Вестерн-блота при помощи NeutrAvidinTM-HRP. Перед оценкой включения метки посредством Вестерн-блота мембраны инкубировали в 0,1 M КОН (В и D). Панель (Е) при различном указанном времени, активность NMT оценивали при помощи ct-Bid N-терминального декапептида и [3Н]-миристоил-СоА. Несмотря на то, что оба фермента NMT расщеплялись, оба сохраняли каталитическую активность, и только NMT1 проявляла существенную потерю активности. (F) Воспроизведение расщепления NMT1 каспазой-8 и каспазой-3, и расщепление NMT2 каспазой-3 in vitro (гель окрашенный Кумасси). Очищенный His-ΝΜΤΊ и His-NMT2 (5 мкг каждого) инкубировали с активной каспазой 3 и 8 (500 нг кажого) в течение 1 часа при 37°С. Общий ингибитор каспаз (z-VAD-fmk) добавляли для остановки реакции в конце инкубации. Образцы немедленно кипятили с 5х буфером для нанесения образцов с бета-меркаптоэтанолом и наносили на 10% акриламидный гель и переносили на PVDF мембрану. Затем мембрану окрашивали при помощи Кумасси голубого для визуализации белков. Расщепленные фрагменты, которые отправляли на расщепление по Эдману, были в боксе и делали возможной идентификацию сайтов расщепления каспазой в NMT1 после D72 и NMT2 после D25 и D67.
Пример 14
В данном примере на фигуре 13 приведена очистка рекомбинантных GST- и His6-меченных hNMT1 и hNMT2. Панель (A) GST-NMT1 и (В) GST-NMT2 очистка на глутатионагарозе, (С) очистка His6-NMT1 и (D) His6-NMT2 посредством Ni-хелатирующей хроматографии и ионнообменной хроматографии на Resource S. На фигуре 13, панели Си D, дорожки представляют собой следующее. Панель (С) гель А: очистка NMT1: 1) голубой маркер, 2) Ni-NTA пул колонки, 3) пул гель-фильтрации, 4) FT Resource S, 5) фракция 8 (f8), 6) f16, 7) f18, 8) f19, 9) f20, 10) f21, 11) f22, 12) f41, 13) f42, 14) f66, 15) f67. Панель (D) гель В: очистка NMT2: 1) голубой маркер, 2) Ni-NTA пул колонки, 3) пул гель-фильтрации, 4) FT Resource S, 5) фракция 21 (f21), 6) f28, 7) f38, 8) f39, 9) f40, 10) f41, 11) f42, 12) f44.
Пример 15
В данном примере на фигуре 14 приведено сравнение ферментативных активностей между очищенной рекомбинантной полноразмерной His6-NMT1 и рекомбинантной "каспазо-отщепленной" ct-His6-NMT1. Полноразмерная His6-NMT1 (аа 73-496). Активность NMT оценивали при помощи исследования миристоилирования пептида, адаптированного от King et al. 1999, Anal Biochem. Эксперименты в дупликате.
Пример 16
В данном примере на фигуре 15 показано сравнение ЕС50 и IC50 различных ингибиторов NMT и различных клеточных линий. График показывает корреляцию между биохимической активностью 8 ингибиторов NMT в биохимическом ферментативном анализе (IC50) и их активность в исследовании жизнеспособности клеток с аламаровым синем при помощи 7 клеточных линий (ЕС50). Регрессионный анализ проводили на каждой клеточной линии и получали среднее значение R2 0,9 (диапазон 0,82-0,96). Данный высокий уровень корреляции между различными IC50 и ЕС50 решительно подтверждает, что активность молекул в исследовании жизнеспособности клеток обусловлена их ингибирующей активностью при NMT. Необходимо отметить, что 2 пары молекул обладают сходными активностями в биохимическом исследовании.
Пример 17
В данном примере на фигуре 16 показана DDD86481 индукция апоптоза в клетках BL.
"Нормальные" иммортализованные В-лимфоциты (IM9), клетки КМН2 (Ходжкинская лимфома) и BL (BL2 и Ramos) инкубировали с повышающимися концентрациями (нМ) DDD86481 и мониторировали расщепление поли-ADP-рибозополимеразы-1 (PARP-1) и каспазы-3 после временной точки 72 ч. Вестерн-блот проводили на клеточных лизатах для мониторинга расщепления или PARP-1 и каспазы-3, а также присутствия GAPDH в качестве контроля нанесения (композитные гели). Как PARP-1, так и каспазо-3 расщепление происходило дозо-зависимым образом в случае обработки клеточных линий BL ингибитором, показывая, что данные клетки претерпели апоптоз. Не наблюдалось расщепления ни PARP-1 ни каспазой-3 в "нормальных" клетках IM9, обработанных ингибитором. PARP-1 расщепление клеток КМН2 (HL) наблюдали при более высоких концентрациях, хотя оно было минимальным по сравнению с расщеплением PARP-1, наблюдаемым в клетках BL. Таким образом, мы обнаружили, что клетки BL более склонны к апоптозу, чем «нормальные» В-клетки при обработке DDD86481.
Пример 18
В данном примере на фигуре 17 предложена оценка порогового значения для утраты NMT2 в опухолях человека. В таблице 7 показан уровень экспрессии мРНК NMT2 в виде log2 (микроэррей интенсивность флуоресценции NMT2) для указанных клеточных линий базы CCLE, где доступно.
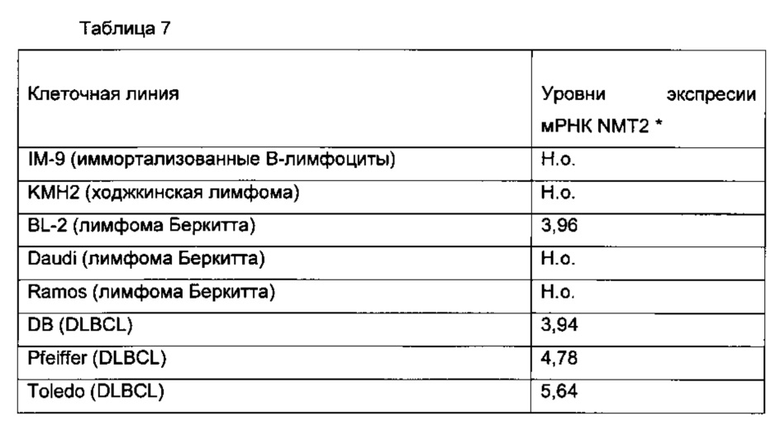
На фигуре 17 панель А, указанные лимфоидные клеточные линии изучали на присутствие NMT2 посредством Вестерн-блота. На панеле В фигуры 17, посредством сравнения панели А и таблицы 7, мы демонстрируем, что при 1од2(микроэррей интенсивность флуоресценции NMT2) = 5,64, наблюдаемой в клетках Toledo, NMT2 не детектируется посредством Вестерн-блота. Хотя действительное пороговое значение для утраты экспрессии NMT2 может быть более 5,64, мы, в данном примере, используем данное значение в качестве порогового значения для установления приблизительного распространения утраты NMT2 в человеческой популяции (панель В). Данные показывают, что утрата NMT2 распространена не только в лимфоме Беркитта и диффузной В-крупноклеточной лимфоме, но также в широком спектре других типов опухолей (см. также Фигуру 11С для списка из 50 клеточных линий, экспрессирующих по меньшей мере NMT2, и таблицу для распространенности утраты NMT2 в различных типах рака).
Соответственно в данном документе предложен способ идентификации таких типов рака с недостатком NMT2, приемлемых для лечения одним или более ингибиторами NMT1. В одном примере образец от пациента с уровнями или концентрацией мРНК ниже приблизительного порогового значения, указывает, что упомянутый пациент является хорошим кандидатом для лечения ингибитором NMT1. В некоторых примерах, уровень мРНК ниже приблизительного порогового значения рассматривают в качестве низкой экспрессии. В одном примере образец от пациента с уровнями или концентрацией мРНК ниже порогового значения указывает, что упомянутый пациент является плохим кандидатом для лечения ингибитором NMT1. В некоторых примерах уровень мРНК выше приблизительного порогового значения рассматривают в качестве высокой экспрессии.
Следует понимать, что в дополнение к или вместо пороговое значение, установленное в отношении клеток Toledo, альтернативный источник может служить в качестве приемлемх пороговых значений. В некотором примере источник или контроль, примененный для получения порогового значения, представляет собой тот же тип клеток, что и рак подлежащий тестированию. В некоторых примерах пороговые значения источника или контроля хранят или находят в базе данных.
Пример 19
В данном примере на фигуре 18 показано применение детиобиотин-ПЭГ-азидной пробы в отношении посттрансляционно со-алкинил-миристоилированным белкам (pull-down) в лейкемических Т-клетках при помощи стрептавидин-сефарозных гранул.
Детиобиотин (показан в I) является аналогом биотина, который обратимо связывает авидин и авидин-подобные белки. Мы разработали и заказали синтез детиобиотин-ПЭГ-азида (I.) и биотин-ПЭГ-азида (II.).
В (В) Т-клетки Jurkat выращивали в присутствии ω-алкинил-миристата и вводили для подвергания апоптозу, опосредованному антителами к Fas в присутствии циклогексимида. В результате клеточного лизиса, лизаты реагировали ил не реагировали с детиобиотин-ПЭГ-азидом при помощи клик-химии, смешивали с нейтравидин-сефарозными гранулами, промывали и элюировали 10 мМ биотином. Панель В (i) показывает, что детиобиотинилированный посттранскрипционно миристоилированный белок может быть «опущен» и эффективно восстановлен из линий нейтравидиновых гранул 5,6,7 и В (Ni). В панеле В (¡i), показан контроль, в котором исключен детиобиотин-ПЭГ-азид из реакции клик-химии. В этом случае очень мало белков связано с нейтравидиновыми гранулами и очень мало белков элюируется (только полоса небольшой интенсивности может быть увидена на уровне 75 кДа). Данные результаты показывают рзвитие способа, делающего возможным протеомный анализ и оценку клеточных содержаний ко- и посттрансляционно миристоилированных белков или миристоиломов.
Пример 20
В данном примере фигура 19 изображает масштабированное применение пробы детиобиотин-ПЭГ-азида в отношении посттрансляционно со-алкинил-миристоилированных белков (pull-down) в лейкемических Т-клетках Jurkat при помощи стрептавидин-магнитных гранул. Клетки Jurkat индуцировали к апоптозу 2,5 мкМ стауроспорином в течение 3 ч, затем метили 100 мкМ миристатом (С14) (панель А) или алкинил-миристатом (Alk С14) (Панель В) обоими конъюгированными с БСА в течение 1 ч при 37°С. Клетки собирали, лизировали и реагировали с азидо-детиобиотином при помощи клик-химии. Детиобиотин-алкинил-миристоилированные белки связывали со стрептавидин-магнитными гранулами, промывали и элюировали 50 мМ биотином с 2% SDS при 37°С в течение 15 мин. Элюция 1 (В панель) содержит большинство детиобиотин-миристоилированных белков, в тоже время мало обнаружено в элюции 1 из клеток, меченных миристатом (А панель). Время: 5 секунд.
Пример 21
В данном примере на фигуре 20 показаны миристоилированные профили «нормальных» иммортализованных В-клеток (IM9) и BL клеток (BL2 и Ramos), меченных алкинил-миристатом. Клетки метаболически метили 100 мкМ алкинил-миристатом в течение 1 часа перед сбором. Образцы белка реагировали с азидо-биотином при помощи клик-химии, 25 мкг белка из каждого клеточного лизата, отделяли посредством SDS-PAGE и визуализировали при помощи Вестерн-блота, используя NeutrAvidinTM-HRP. Стрелки указывают на биотинилированные-алкинил миристоилированные белки, которые присутствуют в «нормальном» иммортализованном В-лимфоците IM9, но не видны в клетках BL (BL2 и Ramos).
Пример 22
В данном примере на фигуре 21 показаны время и дозозависимые графики цитотоксичности от комбинации DDD86481 и доксорубицина. Клеточные линии В-лимфоцитов IM-9 (А) и BL-2 (В) обрабатывали повышающимися концентрациями DDD86481 (0, 0,001, 0,01, 0,1, 1,0, 10 и 100 мкМ) при 0 (голубой), 17 (красный) и 86 M (серый) гидроксидоксорубицина для 24, 48 или 72 часов (сверху вниз). MTS исследования демонстрируют клеточную гибель, индуцированную концентрациями доксорубицина и DDD86481, которые по отдельноти являются намного менее токсичными; данный эффект соответствует синергичному взаимодействию. Это характеризуется посредством 6-кратного снижения EC50's в случае, когда оба содинения применяют вместе в сравнении с EC50's для DDD86481, применяемого отдельно во временных точках 24 и 48 часов.
Пример 23
В данном примере в таблице 8 предложена оценка приблизительной распространенности утраты NMT2 во множестве раков человека в Северной Америке. Мы установили минимальное пороговое значение 5,64 для 1од2(микроэррей интенсивность флуоресценции), которое соответствует клеткам без NMT2 (Toledo, см. пример 18) и обнаружили, что 114 из 967 клеточных линий соответствуют данному пороговому значению (~12%). Коэффициент заболеваемости в Северной Америке для каждого типа рака, число пациентов, которые выказывали пользу от терапии ингибитором NMT (заболеваемость X % ниже порогового значения) и основание для медицинских нужд выбранных раков также показано.
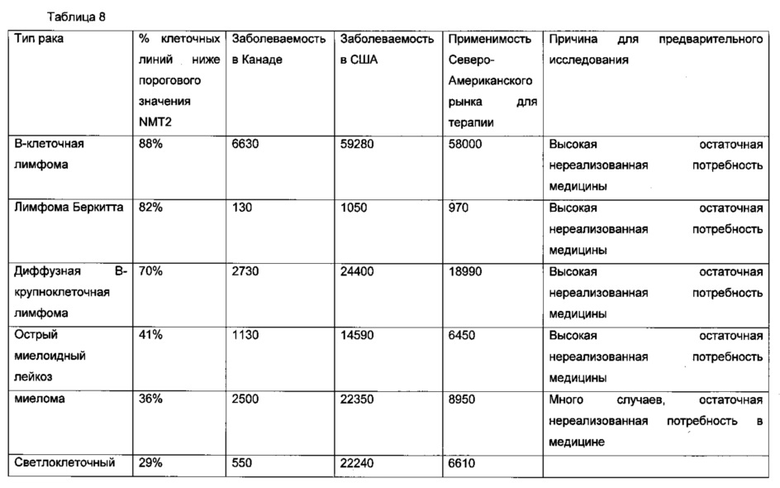
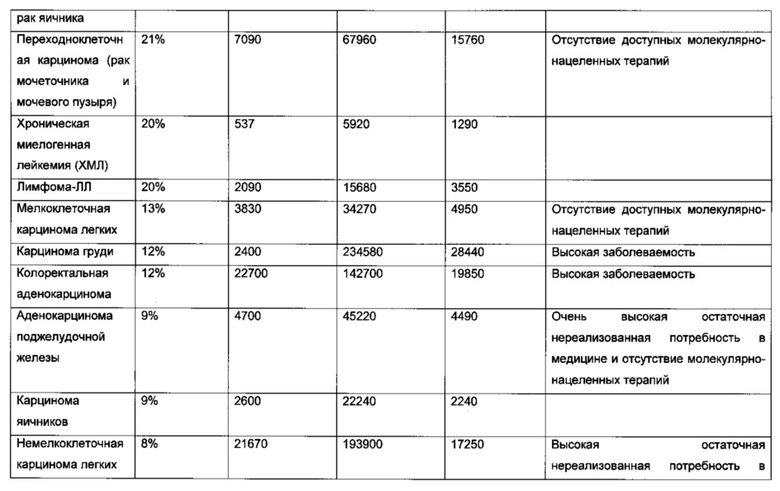
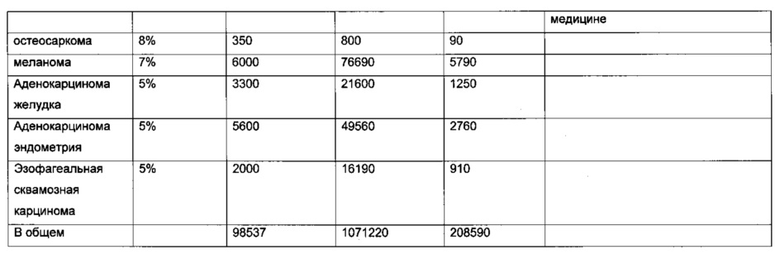
Пример 24
В данном примере на фигуре 22 приведен иммуноблот с клетками, инкубированными с ДМСО, стауроспорином, aFAS или только носителем. Полученные иммуноблоты изучали с применением антител к NMT1, антител к NMT2, антител к PARP-1 и антител к GAPDH. Эти данные показывают, что NMT расщепляется при индукции апоптоза.
В частности в данном примере на фигуре 22А исследовали роль(и) NMT в живых и мертвых клетках. Т-клетки Jurkat, индуцированные к программированной клеточной гибели при помощи антител к Fas или STS анализировали на содержание в них NMT. Т-клетки Jurkat обрабатывали ДМСО, стауроспорином (2,5 мкМ) или антителом к Fas (300 нг/мл) с циклогексемидом (5 мкг//мл) и образцы собирали во временных точках 0, 2, 4, 6 и 8 ч. Клетки лизировали, и присутствие NMT1, NMT2, PARP-1 и GAPDH оценивали посредством Вестерн-блота при помощи ECL. Обработка Т-клеток Jurkat или анти-Fas или STS приводила к время зависимому расщеплению как NMT1, так и NMT2 (фигура 22А). Расщепление NMT1 начиналось спуся 2 ч после индукции клеточной смерти, в то время как NMT2 детектировался лишь после 4 ч апоптоза.
Расщепление NMT1, мигрирующего в виде различимого 67 кДа белка (предпологаемая M.W. = 56,8 кДа) приводило к различимому ~46 кДа фрагменту (фиг. 3.1). Расщепление NMT2, мигрирующего в виде различимого 65 кДа белка (предполагаемая M.W. = 56,9), давало различимый ~55 кДа фрагмент в ранних временных точках, который превращался в более короткий фрагмент, мигрирующий на уровне или ниже ~46 кДа в более поздних временных точках (4 и 8 ч). Этот более короткий фрагмент NMT2 был виден с наложившимся неспецифичным белковым бендом. Однако он был более заметен в клеточных лизатах Jurkat, обработанных Fas, на фигурах 3.2 (8 ч) и 3.4 А (4 ч и 8 ч). Таким бразом, расщепление NMT1 и NMT2 приводит первично и изначально к потере ~11 кДа и ~10 кДа фрагментов, соответственно. Причина несоотвествия между видимыми миграциями и предполагаемыми молекулярными массами NMT не известна. Время расщепления NMT соответствовало апоптотическому маркеру PARP-1, подтверждая, что расщепление происходит под действием каспаз.
NMT1 представляет собой субстрат каспазы-3 или -8, и NMT2 представляет собой субстрат каспазы-3.
В эксперименте на фигуре 22 панель В, для исследования того, будут ли каспазы вовлечены в расщепление NMT, мы индуцировали апоптоз, опосредованный анти-Fas, в Т-клетках Jurkat в присуствии или отсутствии хорошо охарактеризованных необратимых ингибиторов каспаз-3, -8, и -9 наряду с обычным ингибитором каспаз, и анализировали содержания в клетках обеих NMT. Т-клетки Jurkat обрабатывали 10 мкМ ингибиторов каспаз в течение одного часа и затем обрабатывали анти-Fas (300 нг/мл) и циклогексимидом (5 мкг/мл) для индукции апоптоза. Образцы собирали во временных точках 0, 4 и 8 ч. Клетки лизировали, и присутствие NMT1, NMT2 и PARP-1 оценивали посредством Вестерн-блота при помощи ECL. Затем блоты разрезали и повторно исследовали с анти-тубулином (контроль нанесения) (композитные гели).
Расщепление NMT1 отменяли посредством обычных или каспазо-8-специфичных ингибиторов, тогда как оно только частично блокировалось каспазо-3-специфичным ингибитором (фигура 22 В), это подтверждает, что NMT1 является субстратом каспазы-3 или -8. Для сравнения расщепление NMT2 заметно ингибировалось всеми примененными ингибиторами каспаз (фигура 22В). Это подтверждает, что NMT2 вероятно представляет собой субстрат каспазы-3, поскольку ингибирование инициатора каспаз-8 или -9, будет напротив ингибировать расщепление и активацию эффектора каспазы-3. Развитие апоптоза, индуцированного анти-Fas, проверяли посредством Вестерн-блота при помощи антитела к PARP-1 (фигура 22В). Степень PARP-1 расщепления соответствовала и была соизмерима с таковой NMT1 и NMT2.
Пример 25
В данном примере на фигуре 23 показано, что каспазо отщепленная NMT2 является в 3-4 раза более активной, чем полноразмерная NMT2. NMT активность очищенных плноразмерных и каспазо-расщепленных гексагистидин(His)-NМТ оценивали при помощи исследования миристоилирования пептида. N-миристоилтрансферазную активность вычисляли исходя из количества радиоактивномеченного миристоилпептида, продуцированного и детектированного на фосфоцеллюлозной бумаге (адаптировано от King et al. 1991, Anal Biochem.). Каждое NMT-исследование выполняли в трипликатах. Примененный контроль представлял собой элюцию 5 из очистки His-ΝΜT1. Каспазо-отщепленная NMT2 являлась в 3-4 раза более активной, чем полноразмерная NMT2.
Пример 26
В данном примере на фигуре 24 показано, что в снижение уровней NMT2 в клетках BL вовлечено действие гистон деацитилаз.
В экспериментах на фигуре 24 мы хотели оценить возможное вовлечение сайленсинга генов в локусе NMT2. Ацетилирование ε-аминогруппы остатков лизина на гистонах посредством гистонацетилаз (HATs) приводит к снижению положиельно заряженных гистонов, которые ослабляют конформацию хроматина и обеспечивают транскрипционному аппарату лучший доступ к ДНК (Barneda-Zahonero, В., and M. Parra. 2012. Histone deacetylases and cancer. Mol Oncol. 6:579-589.). Поэтому, ацетилирование гистонов обычно ассоциировано с активацией генов. Напротив, удаление ацетильных групп из гистонов посредством гистондеацетилаз (HDACs) индуцирует конденсацию хроматина и приводит к транскрипционной репрессии или сайленсингу генов.
Для проверки того была ли редукция в мРНК NMT2 результатом сайленсинга генов, мы обработали «нормальные» В-клетки (IM9) и злокачественные клетки BL субероиланилид гидроксамовой кислотой (SAHA) (также называемой вориностат), ингибитором гистон ацетилазы класса I и II, в течение 24 часов и мониторировали уровни NMT1 и NMT2 посредством Вестерн-блота. В частности, «нормальные» В-клетки (IM9) и злокачественные клетки BL (Ramos, BL2) обрабатывали 1 мкМ SAHA (HDAC класс I/II ингибитор) в течение 24 часов. Затем клетки лизировали, подвергали SDS-PAGE, и проводили Вестерн-блот с антителами к NMT1, NMT2, p21/WAF1 и GAPDH.
Мы обнаружили, что применение SAHA повышало уровни NMT2 в клетках BL, где уровни NMT2 обычно отсутствуют, в тоже время уровни NMT1 оставались относительно неизмененными. p21/WAF1, белок связывался и ингибировал активность комплексов циклин зависимой киназы 1 (СDК1) или CDK2, мы применяли в качестве контроля для верификации эффективности SAHA, которая известна повышением генной экспрессии p21/WAF1 в клетках. Все клетки, обработанные SAHA, содержали повышенные уровни p21/WAF1. Без связи с какой-либо теорией эти данные подтверждают, что HDAC класса I или II подавляет ген NMT2 за счет деацетилирования гистонов и таким образом компактизации локуса NMT2 в клетках BL.
В одном примере ингибитор HDAC применяют для лечения рака с недостатком NTM2.
Пример 27
В данном примере на фигуре 25 показано, что уровни белка NMT2 снижены в различных клеточных линиях. Лизаты из различных клеточных линий лимфоцитов, собирали от отдельных исследователей и анализировали в отношении их содержания NMT. Результаты демонстрируют, что уровни белка NMT2 были снижены во всех изученных клеточных линиях лимфомы Беркитта (BL2, Daudi, Ramos и BJAB), по сравнению с «нормальными» иммортализованными клеточными линиями лимфобластов, IM9, L0 и VDS.
Пример 28
В данном примере на фигуре 26 показано, что протеасомная деградация не является причиной истощения NMT2 в клетках BL. В данных экспериментах, «нормальные» иммортализованные В-лимфоциты (IM9) и злокачественные В-клетки лимфомы [лимфома Ходжкина (КМН2) и BL (Ramos, BL2)] обрабатывали 10 мкМ протеасомным ингибитором (MG-132) в течение 5 ч. Вестерн-блот проводили на клеточных лизатах для мониторинга уровней NMT1, NMT2, МсИ и GAPDH.
Для исследования будет ли деградация NMT2 увеличиваться в результате дестабилизирующей мутации или присутствия неизвестного фактора, который может дестабилизировать NMT2 в раковых клетках, мы обработали нормальные и злокачественные лимфоциты протеасомным ингибитором деградации MG-132 в течение 5 ч. Клеточные лизаты анализировали Вестер-блотом на присутсвие NMT1, NMT2 и белка миелоидного лейкоза-1 (Mcl-1), который применяли в качестве контроля для проверки эффективности MG-132, поскольку он подвержен протеасомной деградации. Результаты показывают, что уровни NMT2 не повышаются в клетках BL при обработке MG-132, подтверждая, что отсутствие дестабилизирующей мутации или дестабилизирующего фактора, присутствующих в злокачественных лимфоцитах, приводядя к деградации NMT2. Обработка клеток MG-132 приводила к повышенным уровням Mcl-1 во всех типах клеток.
Пример 29
В данном примере на фигуре 27, показано, что каспаза-8 расщепляет NMT1, но не NMT2. Jurkat Т-клетки (А) и Т-клетки Jurkat, экспрессирующие каспазо-8 доминантный негативный мутант (В) обрабатывали ДМСО или антителами к Fas (150 нг/мл) и образцы собирали в точах 0, 4 и 8 ч. Клетки лизировали, и присутствие NMT1, PARP-1 и расщепленную каспазу-8 оценивали при помощи Вестерн-блота при помощи ECL. * означает неспецифичный бенд.
Рсщепление NMT1 исследовали на Т-клетках Jurkat дикого типа и экспрессирующих каспазо-8 доминантный негативный мутант (C8DN) (Juo et al., 1998). Экспрессия C8DN в апоптотических Т-клетках Jurkat отмняла расщепление NMT1 по сравнению с уровнями, обнаруженными в Т-клетках Jurkat дикого типа (фиг. 27А и В). Минимальное количество расщепления NMT2 наблюдали в клетках Jurkat-C8DN после 8 ч индукции апоптоза (Fig. 27В). Расщепление PARP-1 и каспазы-8 также ингибировано в клетках T-C8DN.
Пример 30
В данном примере на фигуре 28 как NMT1, так и NMT2 расщепляли каспазой-3. MCF7 (А) и MCF7, экспрессирующие каспазу 3 (МСР7/каспаза 3) (В) обрабатывали ДМСО или STS (2,5 мкМ) циклогексимидом (5 мкг/мл) и образцы собирали в точке 0, 4 и 8 ч. Клетки лизировали, и присутствие NMT2, PARP-1 и расщепленной каспазы-3 оценивали при помощи Вестерн-блота при помощи ECL.
Было показано, что ни ΝΜΤ1, ни NMT2 не были расщеплены в апоптотических клетках MCF-7 во временной точке 8 ч (фиг. 28А). Для сравнения оба фермента быстро расщепляли в апоптотических клетках MCF-7, стабильно экспрессирующих каспазу-3 (Kagawa et al., 2001) (фиг. 28В). Расщепление известного субстрата каспазы-3/-7 PARP-1 также подтверждали в обеих клеточных линиях MCF-7 в случае индукции апоптоза (фиг. 27А и В) (Walsh et al., 2008). Поэтому, обе NMT оказались субстратом для каспазы-3.
Пример 31
В данном примере на фигуре 29 показаны жирным шрифтом сайты расщепления каспазы NMT1 и NMT2, как идентифицировано при помощи расщепления по Эдману и бокс положительно заряженных лизинов (К) выделен на NMT1 и NMT2 аминокислотных последовательностях (аминокислоты 1-80).
N-терминальное секвенирование выявило, что NMT2 расщеплялась по обоим сайтам D-25 и D-67 (фиг. 29). Сайты расщепления для обеих NMT1 и NMT2 локализованы в N-конце ферментов, а не в С-терминальном каталитическом домене, подтверждая, что расщепленные ферменты могут все еще быть активными в течение апоптоза.
Пример 32
В данном примере на фигуре 30, изображено подтверждение сайтов расщепления NMT посредством сайт-направленного мутагенеза. HeLa клетки временно экспрессирующие дикий тип, и мутантный конструкт V5-NMT инкубировали с STS (2,5 мкМ). Клети трансфецированные V5-NMT1 или V5-NMT2 конструктами лизировали во временных точках 4 ч и 5 ч, соответственно. Вестерн-блот проводили на образцах при помощи антител к V5 и α-тубулину (контроль нанесения).
Для подтверждения сайтов расщепления NMT, выявленных посредством N-терминального секвенирования мы сконструировали N-терминально меченные V5-NMT вектора и применяли кит Quickchange® для сайт-направленного мутагенеза (Stratagene) для создания точечных мутаций в сайтах расщепления NMT. Инжинированная мутация D72E в V5-NMT1 отменяла расщепление каспазой V5-NMT1 в соответствующе трансфецированных клетках HeLa, индуцированных к апапоптозу при помощи STS в течение 4 ч в то время как уровни WT V5-NMT1 были существенно снижены в данных клетках (Фиг. 30). Это подвердило, что D72 является сайтом расщепления каспазой в NMT1.
Поскольку N-терминальное секвенирование выявило два сайта расщепления каспазами (D25 и D67) для NMT2 (фиг. 30), мы мутировали оба остатка D25 и D67 в остатки глутамата (Е) независимо [V5-NMT2 (D25E), V5- NMT2 (D67E)] и вместе [V5-NMT2 (D25.67E)]. Мы наблюдали, что расщепление V5-NMT2 (D25.67E) двойного мутанта было существенно отменена в соответствующе трансфецированных апоптотических клетках HeLa, в тоже время мутанты по одной позиции [V5-NMT2 (D25E), V5-NMT2 (D67E)] имели различные эффекты (Фиг. 30) и дикий тип V5-NMT2 в основном расщеплялся спустя 5 ч после индукции апоптоза (Фиг. 3.8). При сравнении целостности отдельных мутантов в течение апоптоза, мы наблюдали, что V5-NMT2 (D25E) был воспроизводимо чуть менее расщеплен, чем V5-NMT2 (D67E) (фиг. 30), подтверждая, что D25 может быть первичым сайтом расщепления NMT2, в то же время D67 может быть вторичным сайтом расщепления.
Прмер 33
В данном примере на фигуре 31 изображены изменения в профиле миристоилирования в результате апоптоза клеток. Клетки Jurkat метаболически метили 25 мкМ алкинил-миристатом после индукции апоптоза антителами к Fas (150 нг/мл) и циклогексимидом (5 мкг/мл). Образцы белка реагировали с азидо-биотином при помощи клик-химии и визуализировали посредством Вестерн-блота при помощи NeutrAvidinTM-HRP. Перед оценкой включения метки посредством Вестерн-блота, мембраны инкубировали в 0,1 M Трис-HCl или 0,1 M КОН.
Клетки Jurkat индуцировали к апоптозу при помощи антител к Fas и метаболически метили в течение 30 мин ω-алкинил-миристимовой кислотой перед сбором клеток в различных временных точках (0, 1, 2, 4 и 8 ч). Циклогексимид также добавляли к клеткам для ингибирования трансляции белка как часть апоптатического стимула, таким образом, блокируя котрансляционное миристоилирование во время апоптоза. Клеточные лизаты реагировали с азидо-биотином при помощи клик-химии и биотинилированные-миристоилированные белки визуализировали посредством Вестерн-блот анализа при помощи NeutrAvidinTM-HRP, как описано ранее (Yap et al., 2010).
Профиль миристоилирования в точке 0 ч коррелировал с котрансляционным паттерном миристолирования, наблюдаемым ранее в неапоптатических клетках (Фиг. 31) (Martin et al., 2008; Yap et al., 2010). Преднамеренно низкая экспозиция показывает только несколько миристоилированных белков в клеточных лизатах. Обработка мембран 0,1 M КОН подтвердила, что ω-алкинил-миристиновая кислота инкорпорирует в белки за счет щелочеустойчивой амидной связи, поскольку щелочная обработка удаляла метку только из нескольких белковых бендов. Даная щелочная обработка гидролизует тиоэфирные связи, образуемые в пальмитоилированных белках (Arman and Mensa-Wilmot, 1999; Zhao et al., 2000; Vilas et al., 2006). После индукции апоптоза при помощи антител к Fas и циклогексимиду в течение 1 ч, котрансляционное миристоилирование было снижено, предположительно в результате ингибирования трансляции белка циклогексимидом. Это влекло за собой изменение в клеточном содержании миристолированных белков, как проиллюстрировано посредством основных различий в электрофоретических миристоилированных профилях белков, начиная спустя 2 ч после индукции апоптоза и постоянно в течение эксперимента (фиг. 31).
Пример 34
В данном примере на фигуре 32 изображены изменения уровней NMT в результате того, что клетки были подвергнуты апоптозу. Клетки Jurkat метаболически метили 25 мкМ алкинил-миристатом после индукции апоптоза антителами к Fas (150 нг/мл) и циклогексимидом (5 мкг/мл). Вестерн-блот проводили на тех же образцах, что на фигуре 31 при помощи антител к NMT1, NMT2 и GAPDH. (*) изображает неспецифические бенды.
Данные результаты показывают, что хотя NMT расщепляются в течение апоптоза в различной степени, активность миристоилирования сохраняется в клетках до 8 ч после индукции апоптоза.
Пример 35
В данном примере на фигуре 33 изображена индукция к апоптозу клеток COS7, временно экспрессирующих V5-NMT1 и V5-NMT2, при помощи стауроспорина и циклогексимида. NMT активность оценивали при помощи исследования миристоилирования пептидов и вестерн-блот проводили на образцах с применением антител к v5 и альфа-тубулину (контроль нанесения).
Фигура 34 изображает начальную активность NMT в лизатах временно трансфецированных клеток COS7. Клетки COS7, временно экспрессирующие V5-NMT1 и V5-NMT2 инкубировали с STS (2,5 мкМ) и циклогексимидом (5 мкг/мл). NMT активность оценивали посредством исследования миристоилирования пептидов, как описано в материалах и методах. N-миристоилтрансферазную активность вычисляли исходя из количества продуцируемого радиоактивномеченного миристоилпептида и детектировали на фосфоцеллюлозной бумаге (адаптировано King et al.1991, Anal Biochem.). Уровни активности нормализовали к активности NMT1 в t=0 ч. NMT1 представляет среднее трех независимых экспериментов, выполненных в дупликатах. NMT2 представляет среднее четырех независимых экспериментов, выполненных в дупликатах.
Фигура 35 изображает активность NMT в клетках COS7, временно экспрессирующих V5- NMT1 и V5-NMT2, инкубированных со стауроспорином (2,5 мкМ) и циклогексимидом (5 мкг/мл). NMT активность оценивали при помощи исследования миристоилирования пептидов, как описано в материалах и методах. N-миристоилтрансферазную активность вычисляли исходя из количества продуцируемого радиоактивномеченного миристоилпептида и детектировали на фосфоцеллюлозной бумаге (адаптировано от King et al.1991, Anal Biochem.). NMT активность нормализовали к 100% в t=0 ч для каждой NMT. NMT1 представляет среднее трех независимых экспериментов, выполненных в дупликатах. Различия обозначены посредством (*), и показывают статистическую значимость (*<р=0,05, **<р=0,005) при сравнении с временной точкой 0 ч для обеих NMT.
Для оценки будут ли расщепленные NMT все еще каталитически активными, мы измеряли V5-NMT ферментативную активность временно трансфецированных клеток, экспрессирующих обе V5-NMT при помощи фильтоснованного анализа пептидов (King and Sharma, 1991; Raju and Sharma, 1999) в течение наступления апоптоза. COS-7 клетки, временно трансфецированные V5-NMT1, V5-NMT2 или пустым вектором индуцировали к STS/циклогексимид индуцированному апоптозу и применяли в качестве источника фермента. NMT активность измеряли в различное время апоптоза при помощи [3Н]-миристоил-СоА и миристоилируемые или немиристоилируемые (G->A) усеченные Bid декапептиды применялив в качестве субстратов (King and Sharma, 1991; Raju and Sharma, 1999). Активность NMT вычисляли исходя из количества радиоактивномеченного пептида, который оставался свзанным на фосфоцеллюлозной бумаге, и детектировали посредством сцинтилляционного измерения.
Хотя трансфецированные клетки COS-7 экспрессировали сходные уровни химерных NMT (фиг. 32), такие экспрессирующие V5-NMT1 проявляли почти в 5 раз более высокую активность NMT, чем таковые экспрессирующие V5-NMT2 в t=0 ч (фиг. 34). Количество интактных V5-NMT, обнаруженных в клеточных лизатах, снижалось с течением времени индукции апоптоза (фиг. 33) и имело сходную тенденцию с эндогенными NMT (фиг. 32), хотя почти все из повышенно экспрессируемых NMT1 были расщеплены после 4 ч и 8 ч апоптоза и все из повышенно экспрессируемых NMT2 после 8 ч (фиг. 3.33). Хотя более чем 90% V5-NMT1 расщеплено (фиг. 3.33) в 4 ч после индукции апоптоза, активность NMT в таких клетках оставалась относительно неизмененной до 8 ч после инициации клеточной гибели. Было небольшая тенденция к повышению (хотя не существенное) в NMT каталитической активности в t=2 ч и 4 ч, в лизатах клеток COS-7, повышенно экспрессирующих NMT1, по сравнению с активностью в t=0 ч (фиг. 3.35). Это подтверждает, что расщепленная V5-NMT1 является каталитически активной во время апоптоза, в случае инициации посттрансляционного миристоилирования. Однако мы наблюдали существенно снижение активности NMT (20%, р<0,05) в 8 ч после индукции апоптоза при сравнении с временной точкой 0 ч в клетках, трансфецированных V5-NMT1 (фиг. 35).
Активность NMT клеток, экспрессирующих V5-NMT2 не была существенно изменена с 0 ч до 4 ч, до тех пор, пока расщепление V5-NMT2 каспазой приводило к статистически значимому снижению (р<0,005) (33% снижение по сравнению с активностью в t=0 ч) в ферментативной активности после 8 ч индукции апоптоза (фиг. 35). Интересно, что хотя уровни NMT2 существенно понижены из-за расщепления каспазой во время апоптоза (фиг. 33), 66% активности NMT2 все еще сохранялось после 8 ч индукции апоптоза (фиг. 33), означая, что NMT2 также играет роль в посттрансляционном миристоилировании белков в течение апоптоза.
Пример 36
В данном примере на фигуре 36 изображена очистка рекомбинантной гексагистидин(Н1в)-меченной полноразмерной и каспазорасщепленной hNMT1. Очистку проводили при помощи Ni-NTA хроматографии (см. материалы и методы). Очищенные белки визуализировлаи при помощи окрашивания геля кумасси голубым. (FT: проточный, W: промывка и Е: элюированные фракции.
Фигура 37 изображает очистку рекомбинантной гексагистидин(Н|з)-меченной полноразмерной и каспазорасщепленной hNMT2. Очистку выполняли при помощи Ni-NTA хроматографии (см. материалы и методы). Очищенные белки визуализировали посредством окрашивания гелей кумасси голубым. (FT: проточный, W: промывка и Е: элюированные фракции).
Фигура 38 изображает активность NMT очищенных полноразмерных и каспазорасщепленных гексагистидин(Нis)-NМТ, оцененную при помощи исследования миристоилирования пептидов. N-миристоилтрансферазную активнсоть вычисляли исходя из количества радиоактивномеченного миристоилпептида, продуцированного и детектированного на фосфоцеллюлозной бумаге (адаптировано от King et al. 1991, Anal Biochem.). Каждое исследование NMT выполняли в трипликатах. Примененный контроль представлял собой элюцию 5 из His-NMT1 очистки.
Мы получили His-меченный полноразмерный (His-NMT1 и His-NMT2) и каспазорасщепленный (His-73NMT1, His-26NMT2 и His-68NMT2) человеческие NMT вектора, и применяли данные вектора для бактериальной экспрессии и очистки белка (фиг. 36 и 337). При помощи фильтр-основанного анализа NMT исследования (King and Sharma, 1991;. Raju and Sharma, 1999), мы обнаружили, что обе и полноразмерная и каспазорасщепленная NMT1 и NMT2 были каталитически активны при сравнении с контролем во всех исследованиях in vitro (фиг. 37). Расщепление NMT2 повышало ее активность, поскольку каспазо расщепленная NMT2 обладала в ~3 (His-26NMT2) и ~4 раза (His-68NMT2) большей активностью при сравнении с полноразмерной NMT2 (фиг 38).
Пример 37
В данном примере на фигуре 39 изображено субклеточное фракционирование эндогенных NMTs в клетках HeLa в течение апоптоза. Клетки HeLa обрабатывали ДМСО, STS (2,5 мкМ) или анти-Fas (300 нг/мл) циклогексимидом (5 мкг/мл) и образцы собирали во временной точке 5 ч и подвергали субклеточному фракционированию как описано в материалах и методах. Такой же объем фракций Р100 и S100 подвергали Вестерн-Блоту с антителами к NMT1 и NMT2.
Фигура 40 изображает колличественную оценку NMT в различных фракциях после субклеточного фракционирования эндогенных NMT в клетках HeLa в течение апоптоза. Клетки HeLa обрабатывали ДМСО, STS (2,5 мкМ) или анти-Fas (300 нг/мл) циклогексимидом (5 мкг/мл) и образцы собирали во временной точке 5 ч и подвергали субклеточному фракционированию (фиг. 39). Уровни полноразмерной (полная) и расщепленной NMT1 (А) и NMT2 (В) между Р100 (Р) и S100 (S) фракциями колличественно оценивали при помощи Image J (http://rsbweb.nih.gov/ij/). Показанные процентные отношения вычисляли в качестве уровней каждого бенда всех двух фракций (P100+S100). Графики представляют среднее трех независимых экспериментов.
Фигура 4 изображает субклеточное фракционирование клеток HeLa, подвергающихся апоптозу, меченных алкинил-миристатом. Перед субклеточным фракционированием (фиг. 39 и 40), клетки HeLa метаболически метили при помощи 25 мкм алкинил-миристат после индукции апоптоза. После фракционирования обазцы белков реагировали с азидо-биотином при помощи клик-химии, и визуализировали посредством вестерн-блота при помощи NeutrAvidinTM-HRP. Перед оценкой включения метки посредством Вестерн-блота мембраны инкубировали в 0,1 M нейтарльном Трис-HCl (А) или 0,1 M КОН (В).
Каспазное расщепление многих белков части приводит к изменению в клеточной локализации (Enari et al., 1998; Zha et al., 2000; Jakobi, 2004; Vilas et al., 2006); поэтому для определения локалзации расщепленных NMT в течение апоптоза мы провели эксперименты по субклеточному фракционированию (фиг. 39) в нормальных и апоптатических клетках. Мы обнаружили, что NMT1 первично локализовалась во фракции рибосомы/мембрана в необработанных клетках HeLa (63,9% в осадке) (фиг. 40). Было отмечено видимое увеличение расщепленной каспазоусеченной NMT1 в цитоплазматических фракциях клеток HeLa, индуцированных для подвергания апоптозу при помощи STS или анти-Fas вместе с циклогексимидом (54% в цитоплазме в анти-Fas обработанных клетках и 60% в цитозоле в STS обработанных клетках) (фиг. 39 и 40). Напротив, большинство из NMT2 (61,7%) локализованы в цитоплазме перед индукцией апоптоза (фиг. 39 и 40). Однако больший фрагмент каспазорасщепленной NMT2 (~55 кДа) в основном локализовался в мембранном осадке (94,7% в Fas-обработанных и 80% в STS-обработанных) в апоптатических клетках (фиг. 39 и 40). Мы не наблюдали присутствия меньшего фрагмента расщепления NMT2 (~46 кДа) в течение нашего фракционирования, возможно, потому что клетки были индуцированы к апоптозу в течение максимум 5 ч.
В случае метаболического мечения клеток алкинил-миристатом перед индукцией апоптоза или нет и субклеточным фракционированием мы наблюдали изменение в профиле миристоилирования после индукции апоптоза как видно на фигуре 31 и обнаружили, что посттрансляционные миристоилированные белки в основном локализованы в мембранах (Р100) по сравнению с цитоплазмой (S100) фиг. 40). Это подтверждает, что добавление группы миристоила к данным посттрансляционно миристоилированным белкам кажется достаточным для обеспечения стабильного заякоривания в мембране.
Пример 38
В данном примере фигура 42 изображает влияние 2-гидроксимиристиновой кислоты (НМА) на индукцию апоптоза. Т-клетки Jurkat обрабатывали или не обрабатывали НМА (1 мМ), и апоптоз индуцировали анти-Fas (150 нг/мл) и циклогексимидом (5 мкг/мл). Контрольные клетки обрабатывали ДМСО. Образцы собирали во временных точках 0, 2, 4, 6 и 8 ч. Клетки лизировали, и образцы отделяли посредством SDS-PAGE и иммуноблотировали с антителами к PARP-1, РАК2, NMT1 и NMT2 (композитные гели).
Расщепление PARP-1 происходило через 2 ч в клетках обработанных 1 мМ НМА и анти-Fas по сравнению с клетками, обработанными 1 мм миристатом натрия и анти-Fas. Сходная тенденция была выявлена также в клетках, обработанных HMA/STS, но в меньшей степени, чем таковая с анти-Fas, в присутствии HMA/STS клетки, которые проявляли более PARP-1 и РАК2 расщепление в 2 ч после инкубации апоптоза чем клетки, обработанные STS и 1 мМ миристатом натрия (фиг. 42). Также наблюдалась сходная стимуляция расщепления NMT1 NMT2. Это показывает, что ингибировнаие NMT усиливает индукцию апоптоза в клетках. Поскольку ингибирование NMT усиливало начало апоптоза кажется, что NMT играют общую, способствующую выживанию роль в клетках.
Все публикации, патенты и заявки на патент, упомянутые в данном описании, являются индикатором уровня техники для специалистов в области техники, к которой относится данное изобретение, и включены в данный документ посредством ссылки в той же степени, как если бы каждая отдельная патентная публикация или заявка на патент, была конкретно и отдельно включена посредством ссылки.
Таким образом, изобретение описано, будет очевидным, что оно может быть варьировано многими путями. Такие вариации не рассматриваются в качестве отклонения от сущности и объема изобретения, и все такие модификации, которые будут очевидны специалистам в области техники, предназначены быть включенными в объем последующей формулы изобретения.
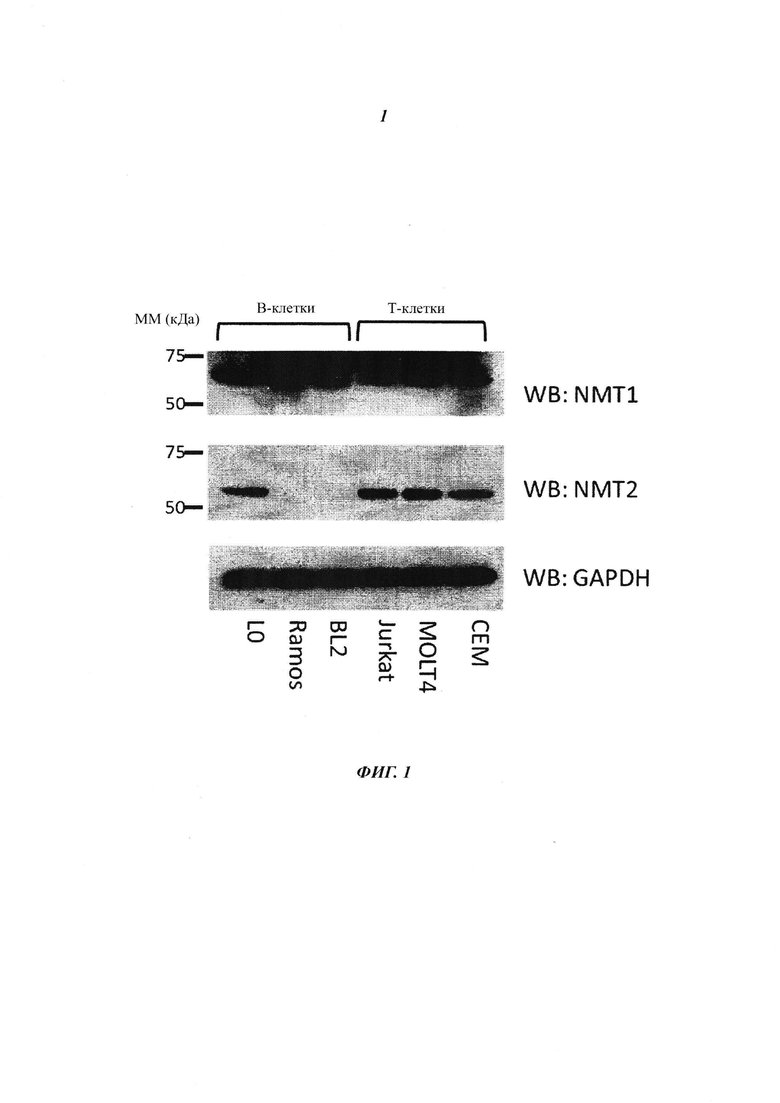
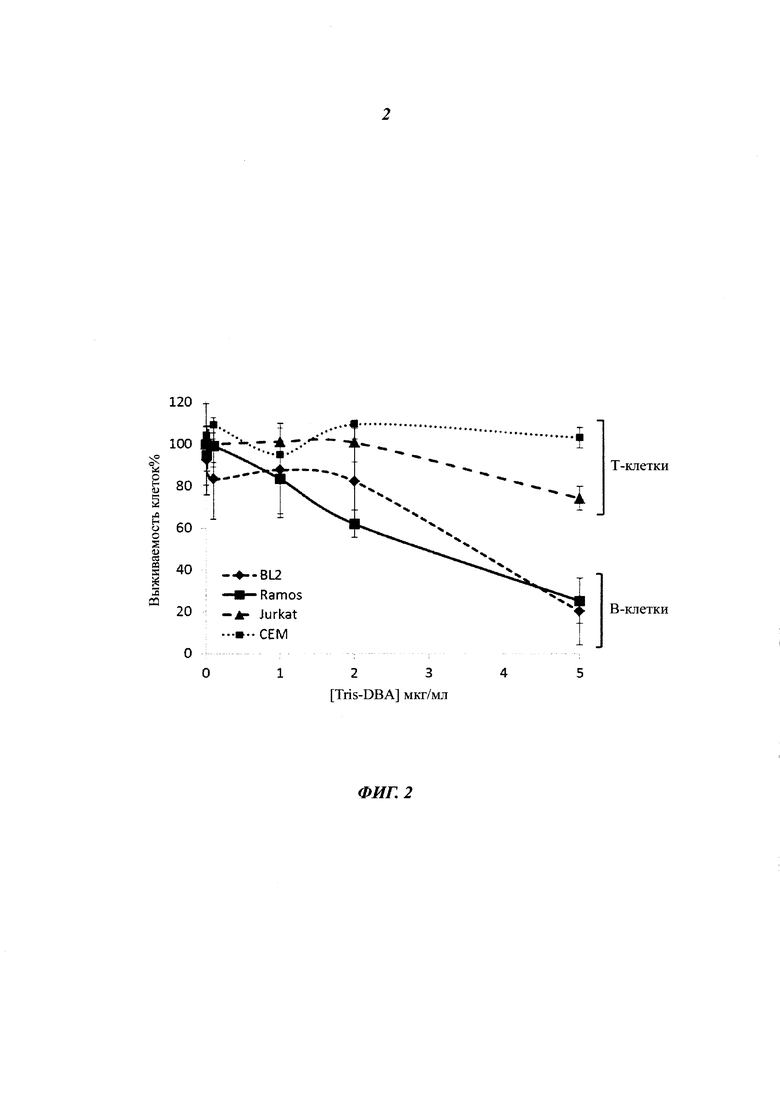
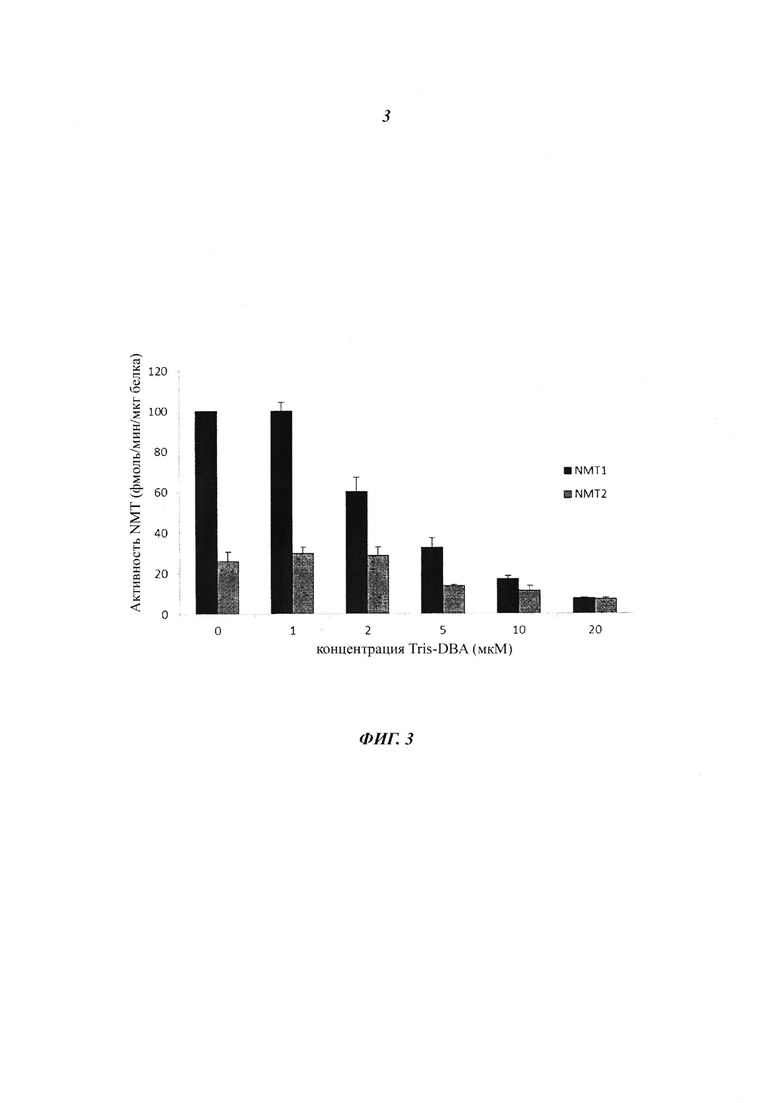
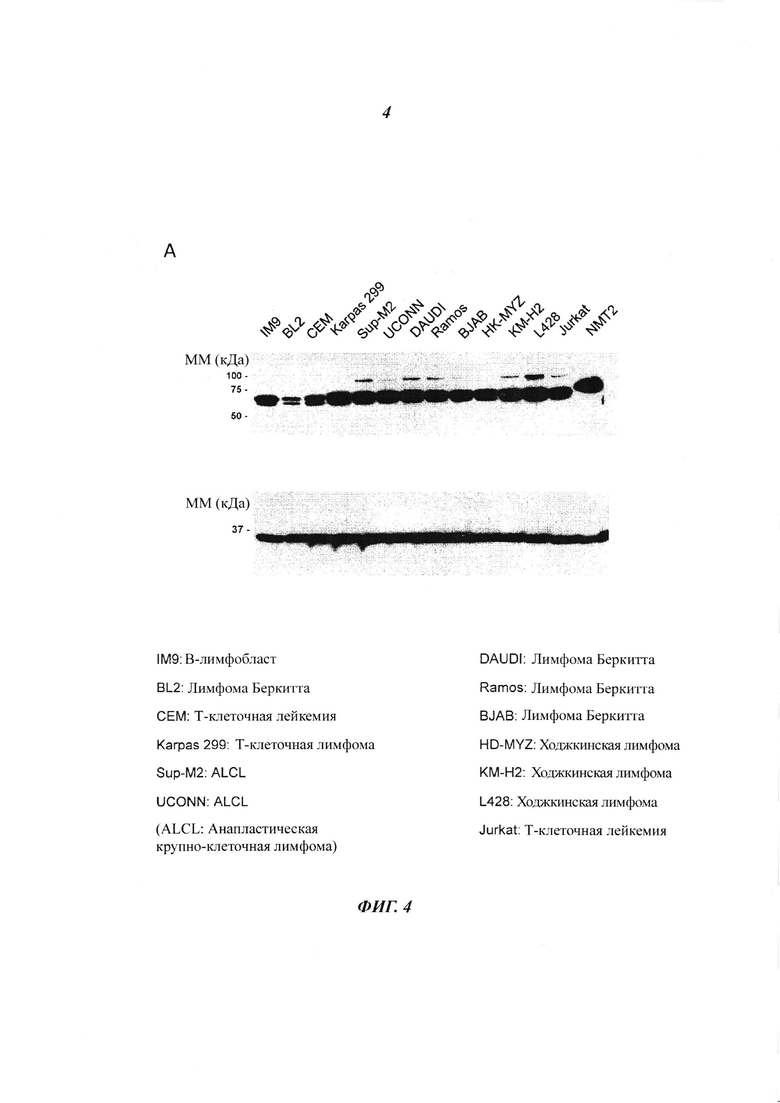
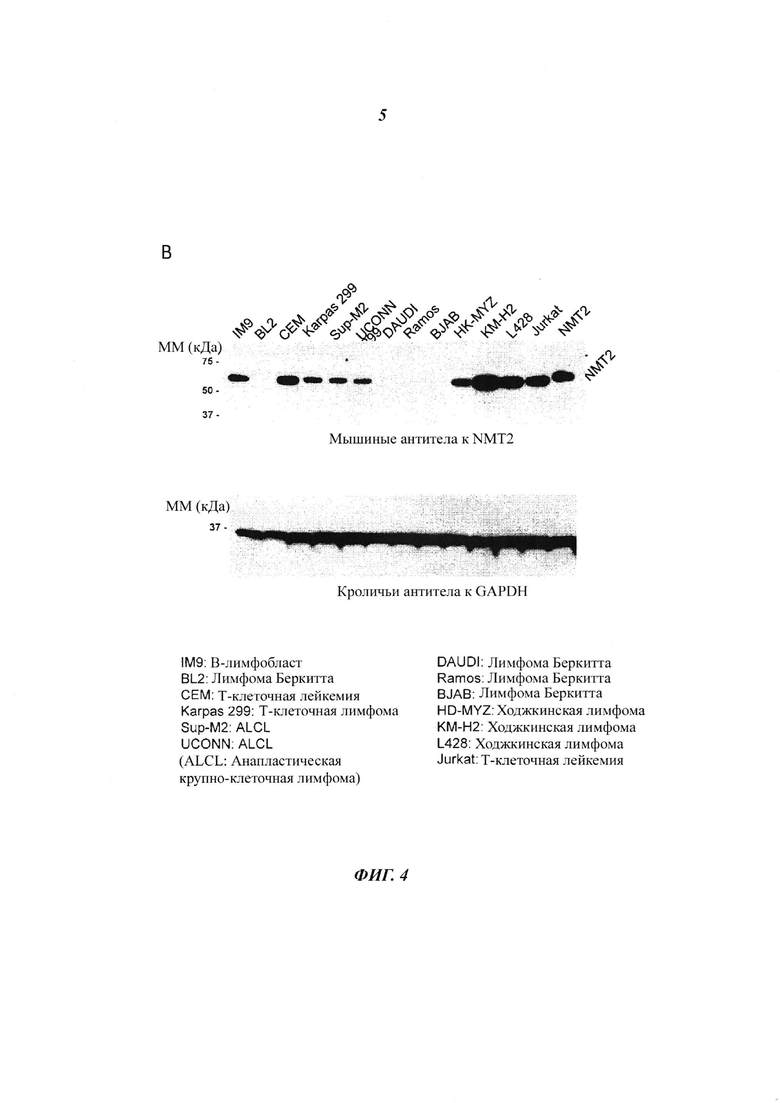
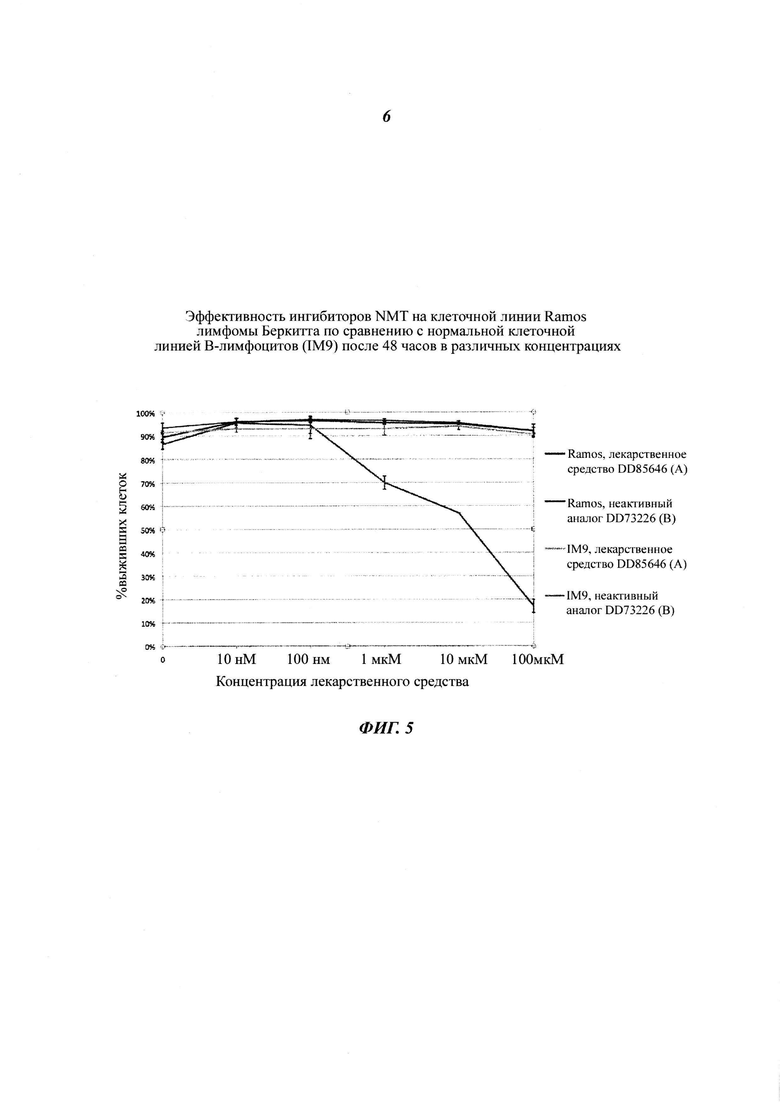
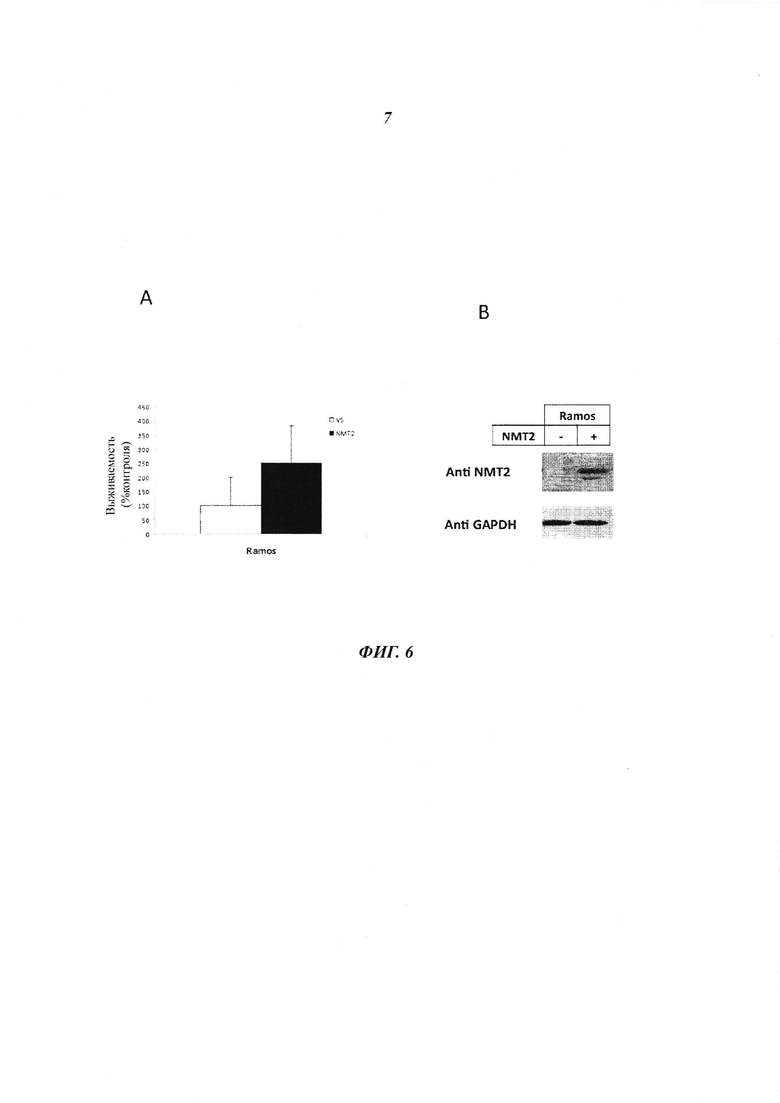
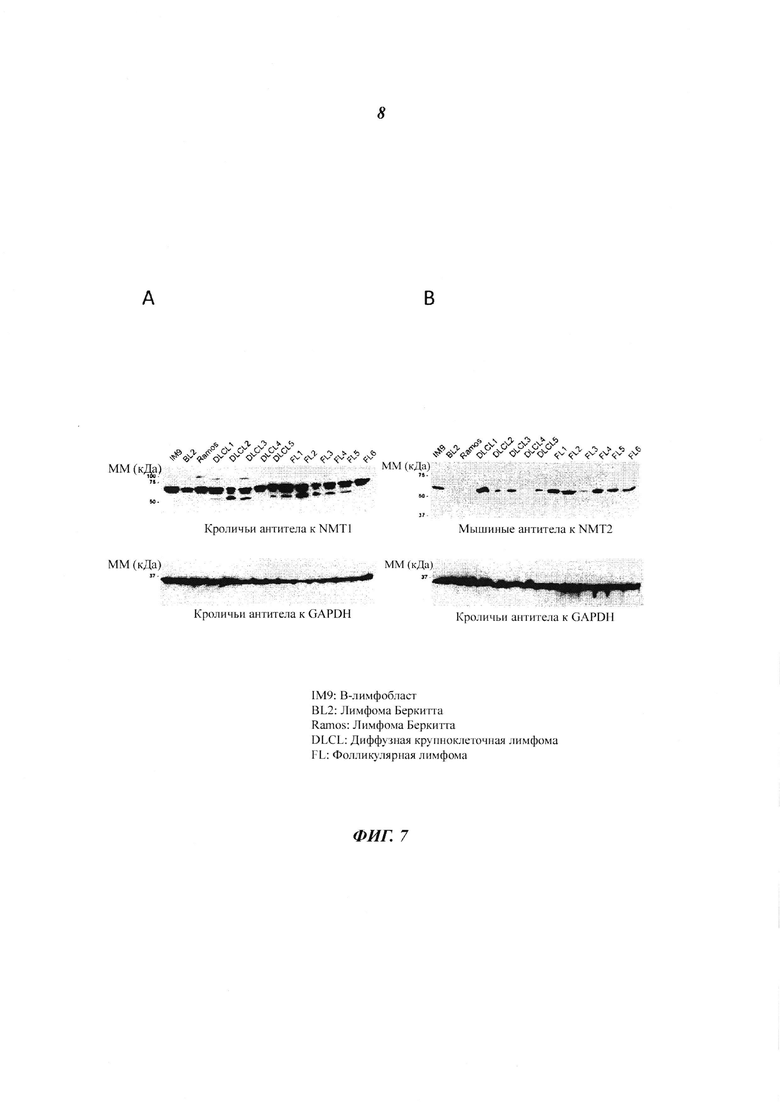

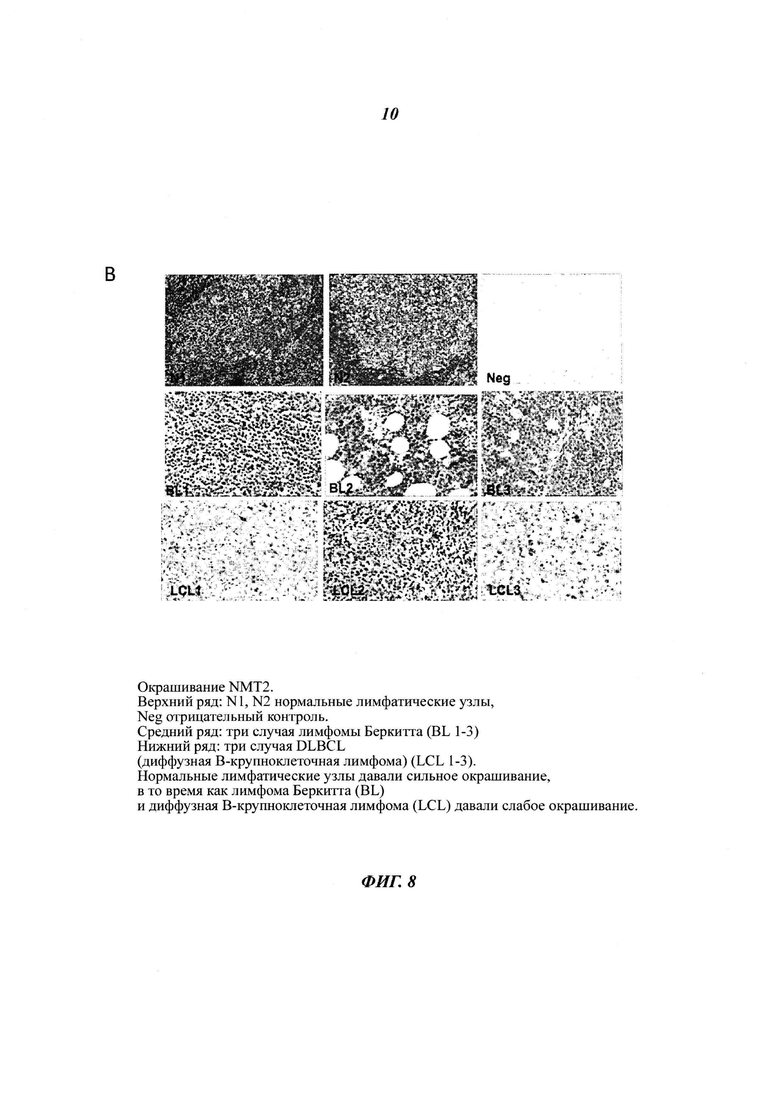
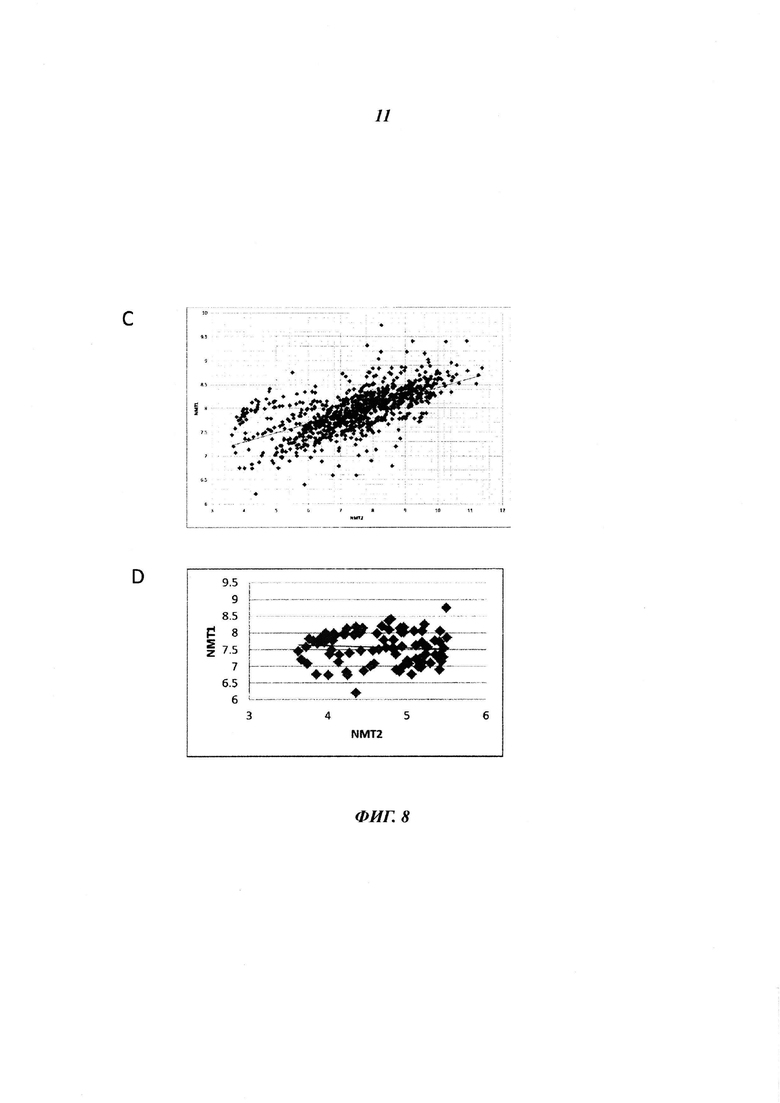
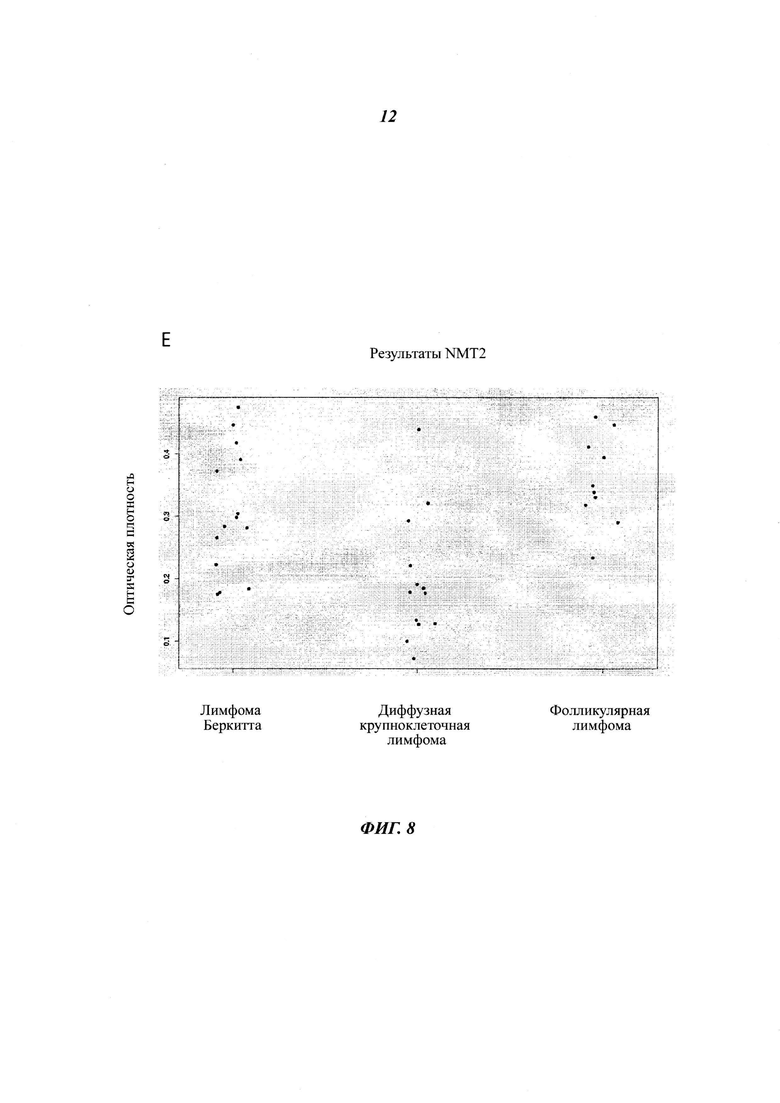
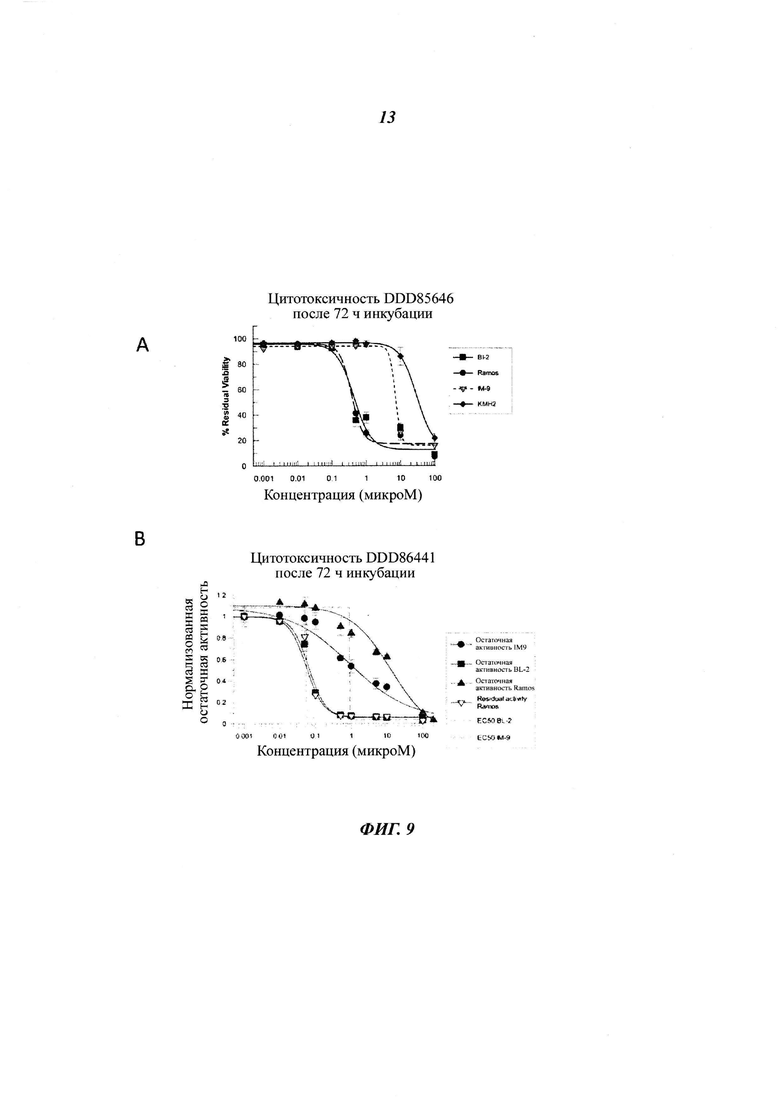

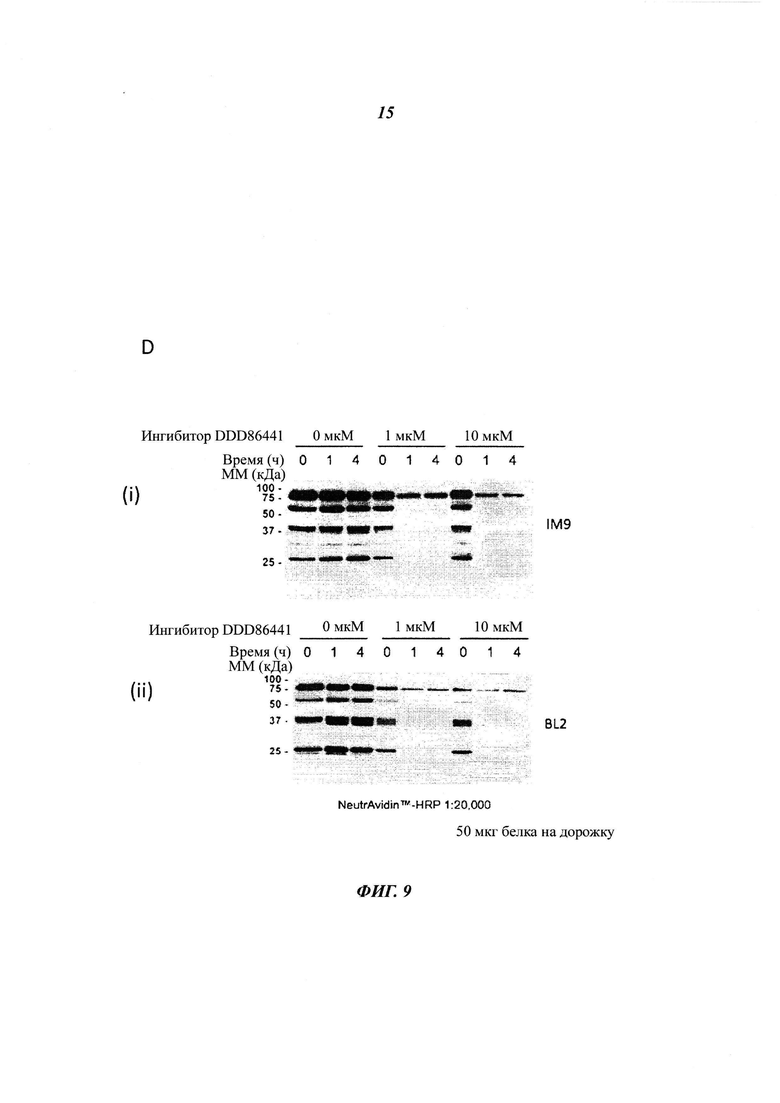
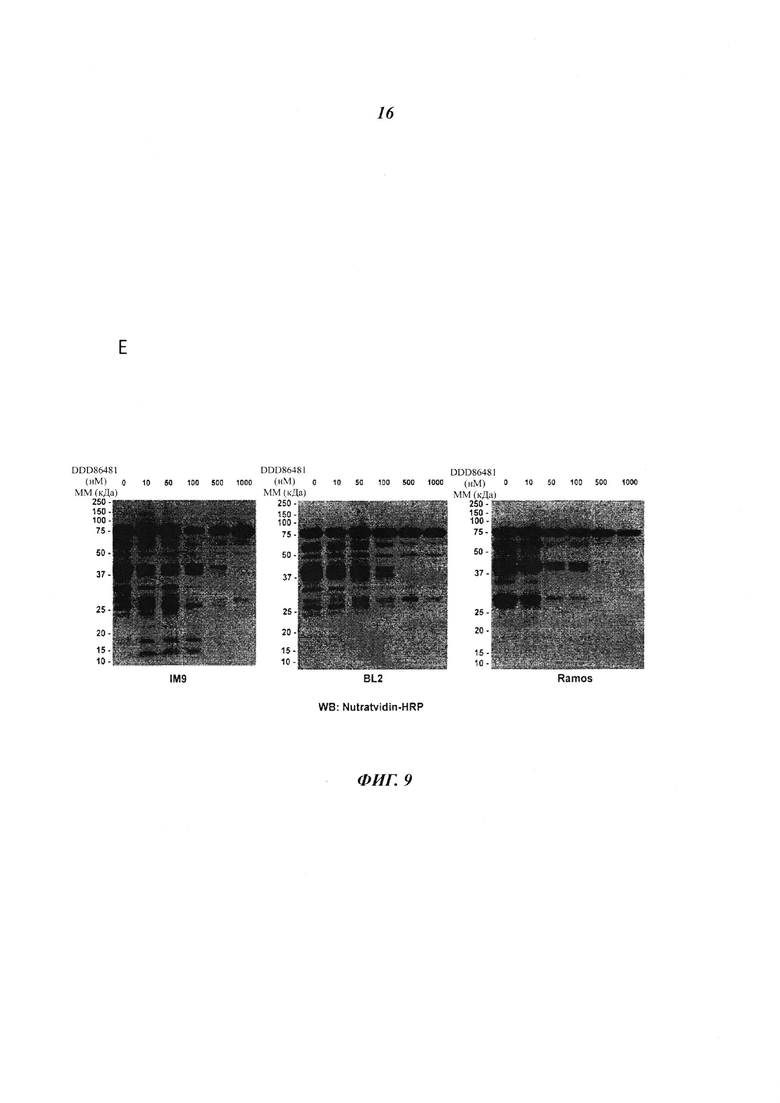
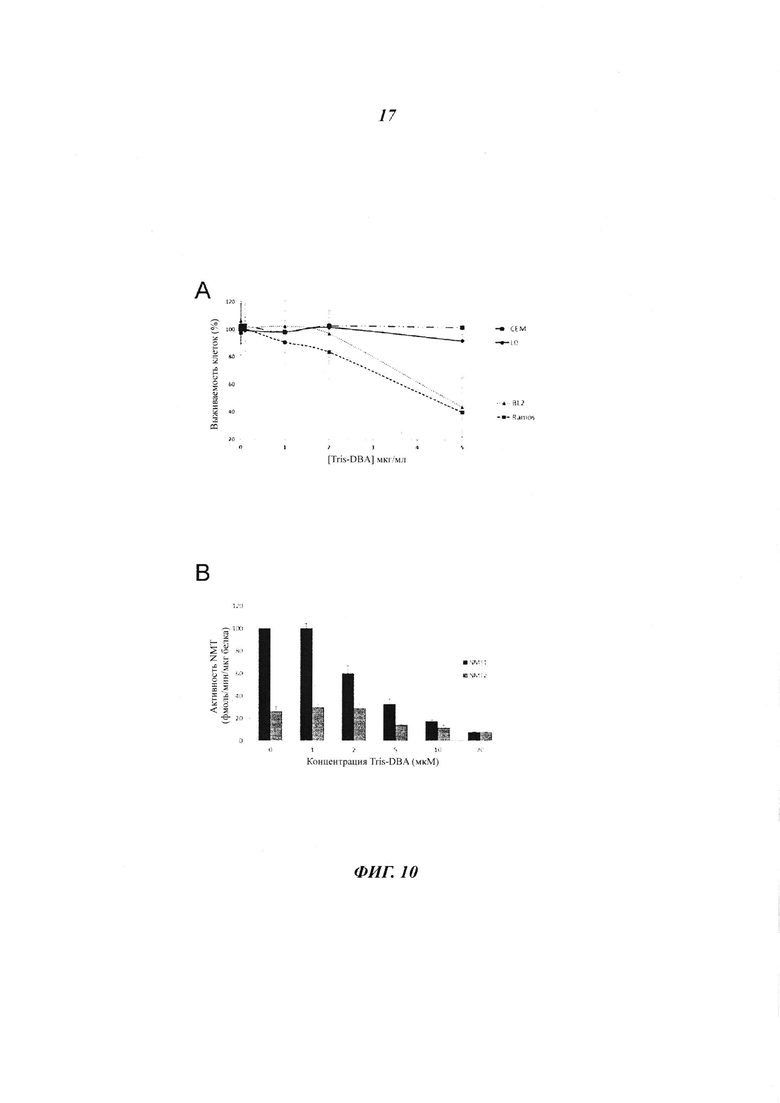
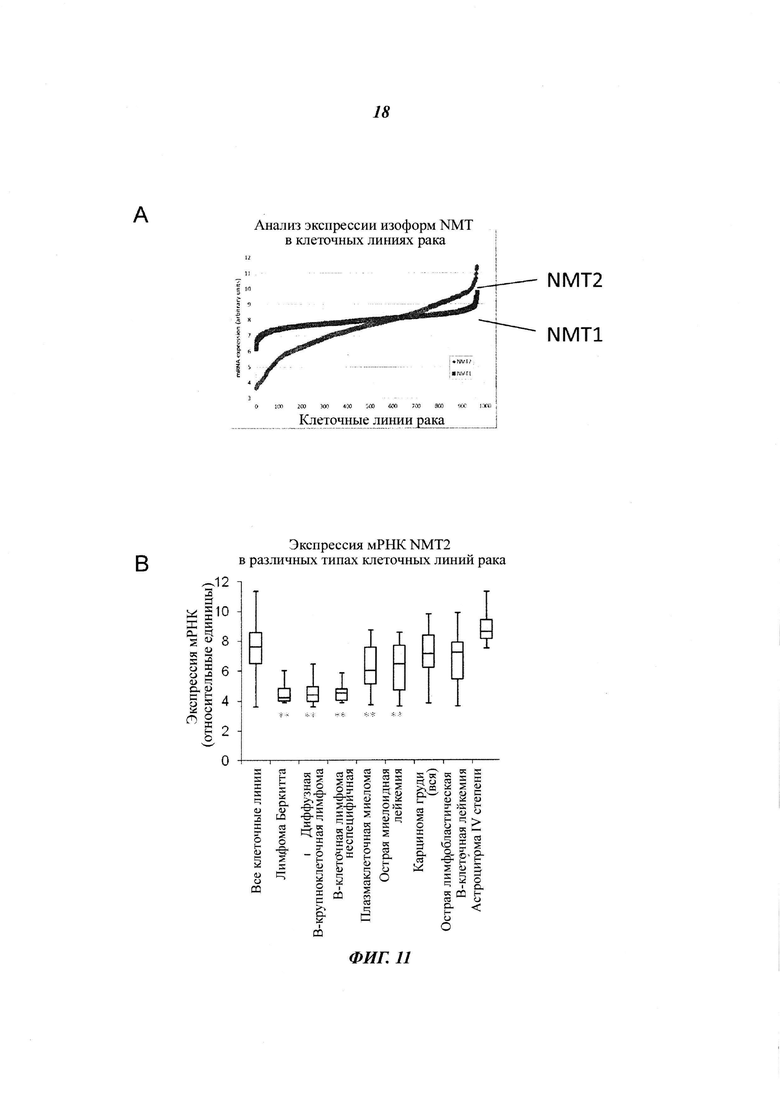

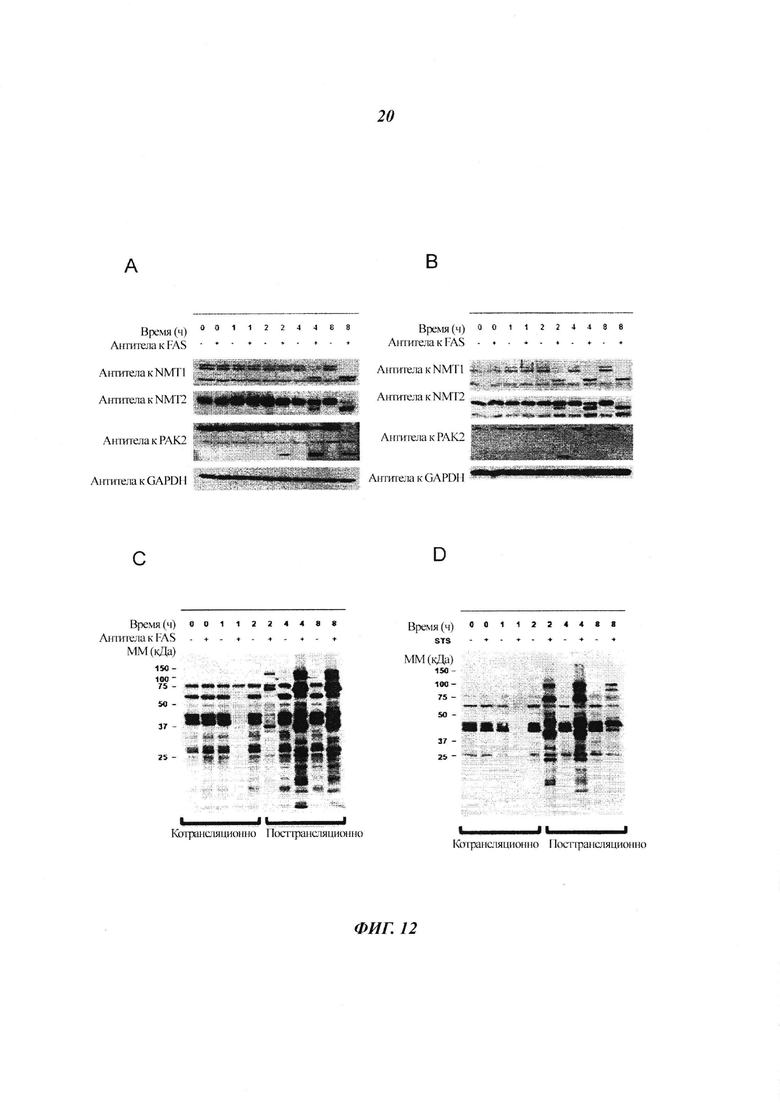
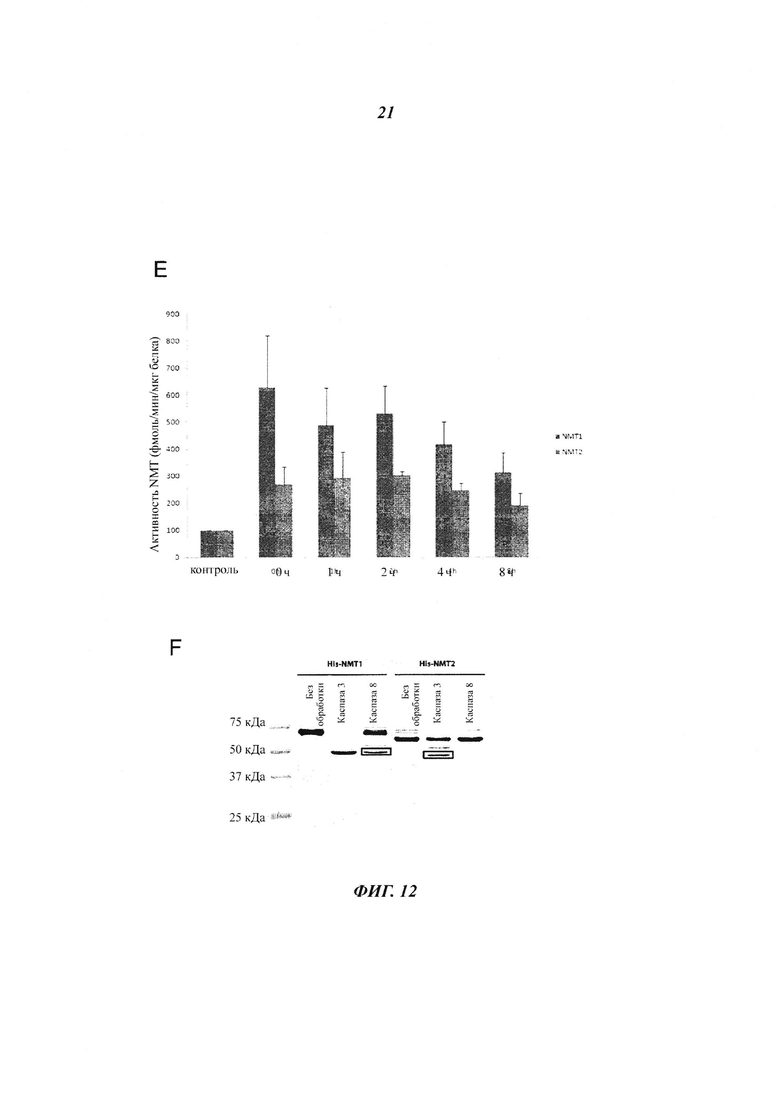
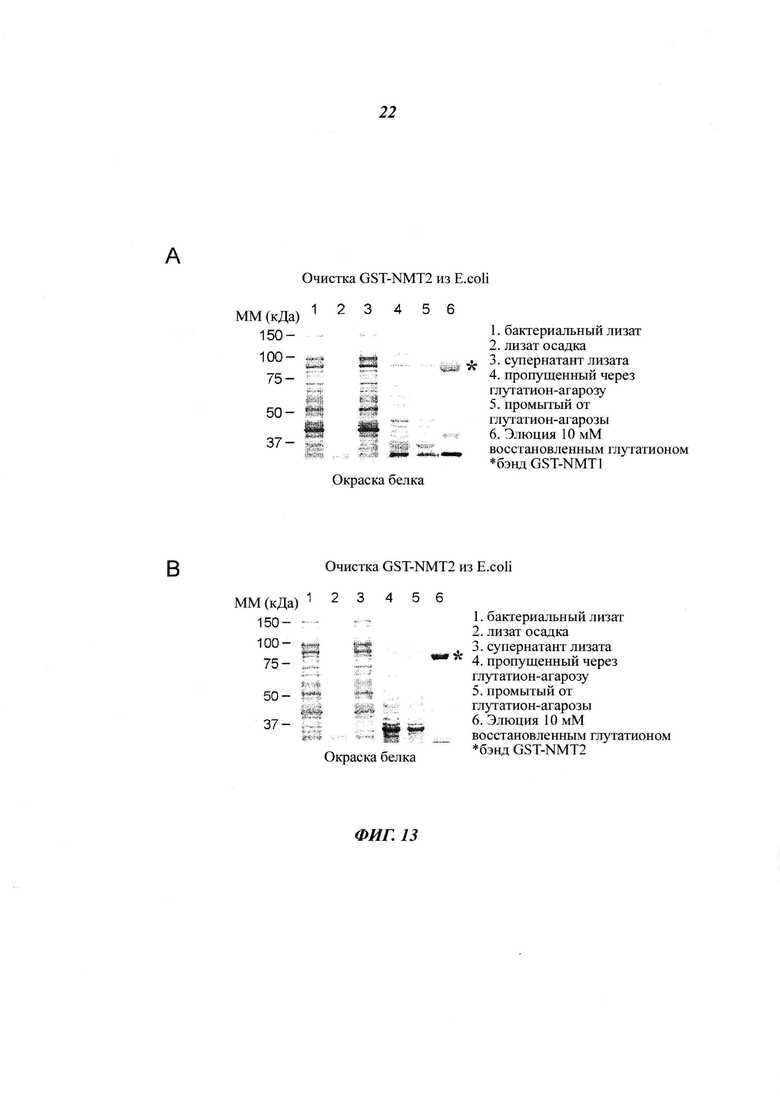
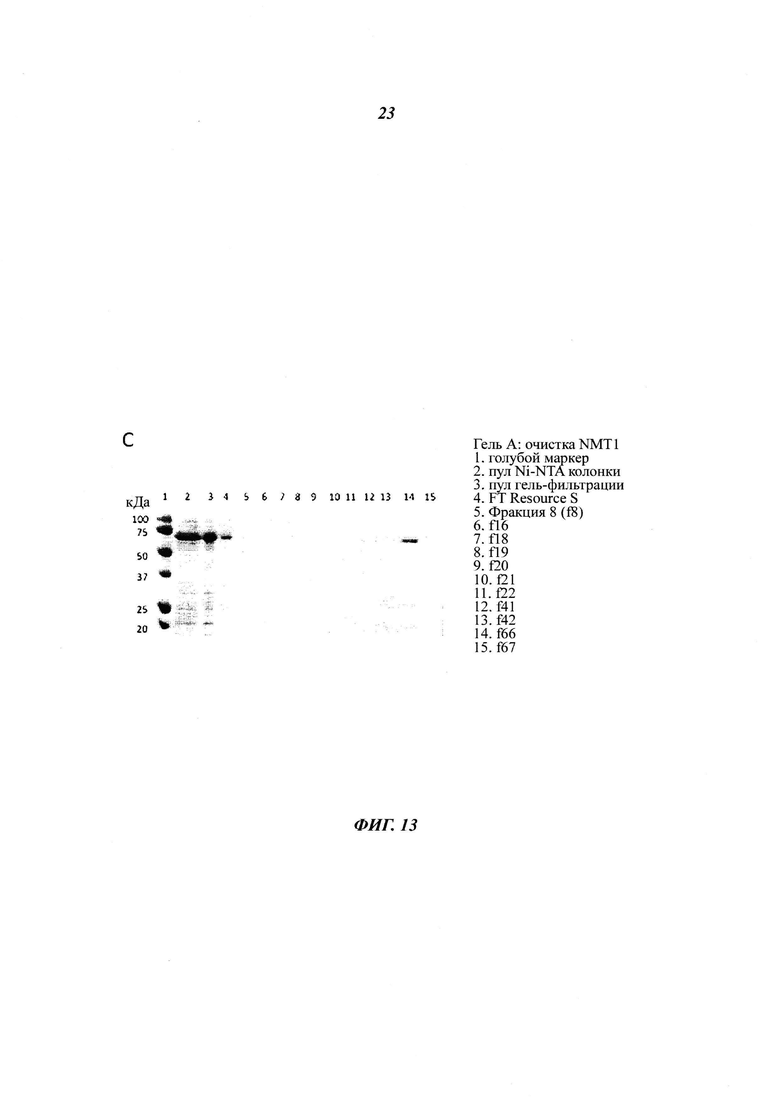

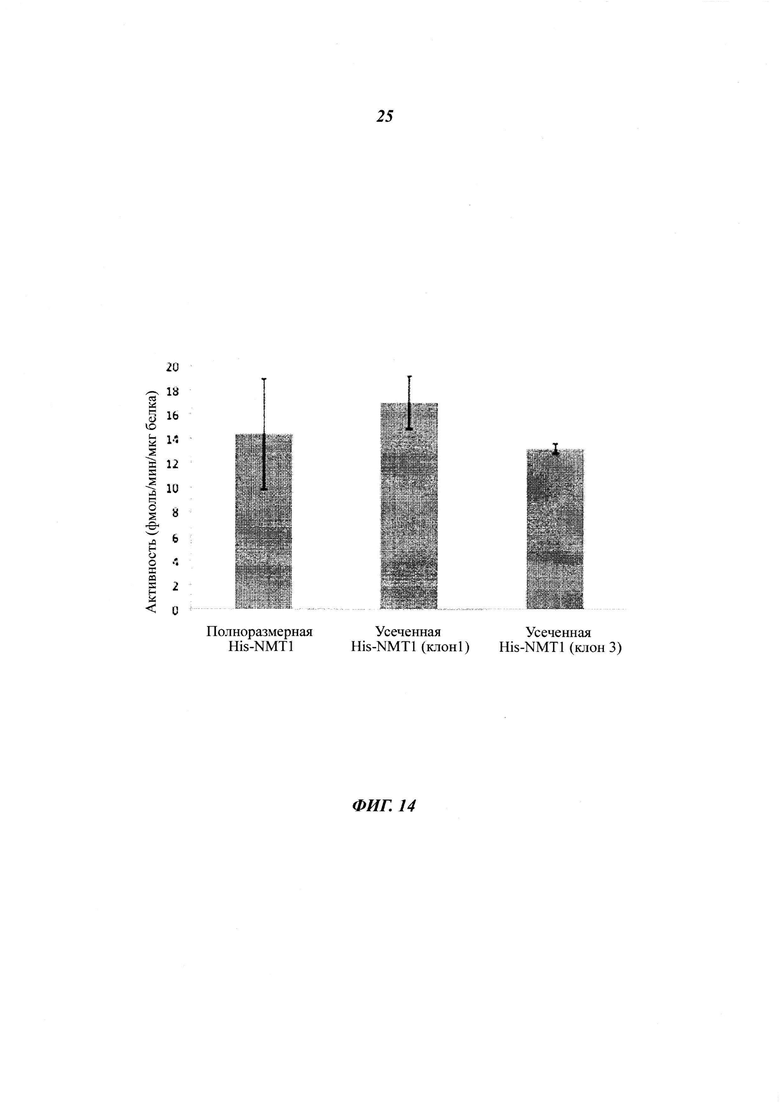

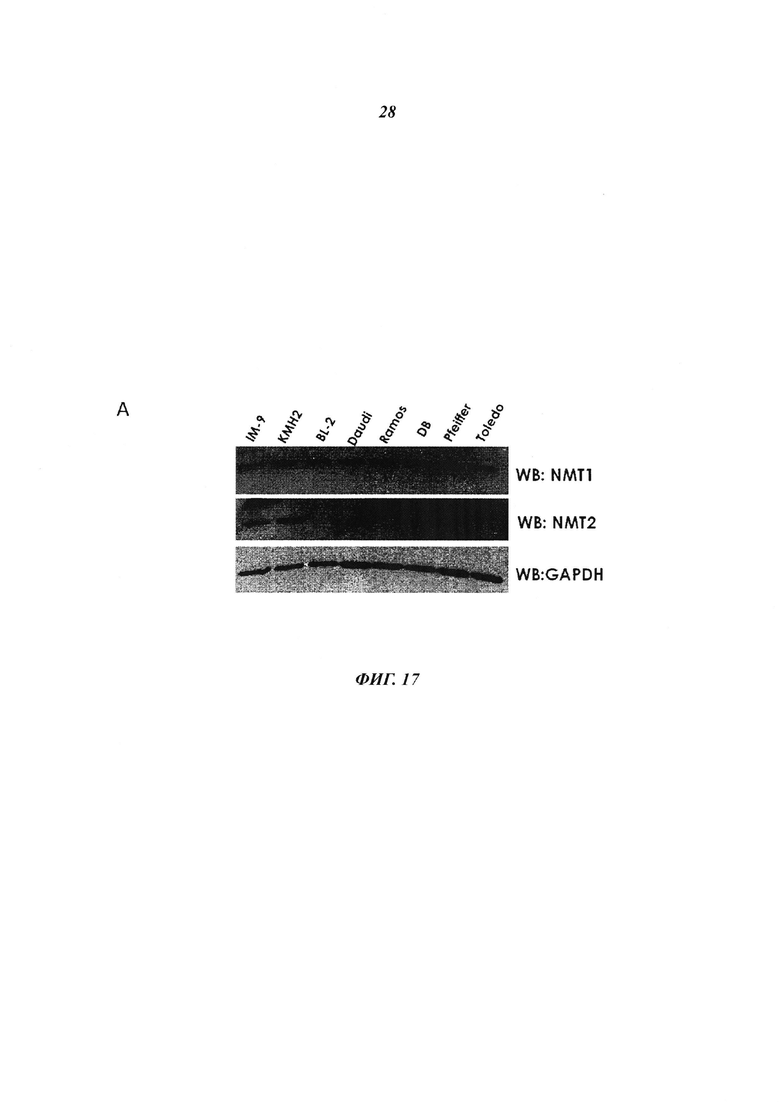
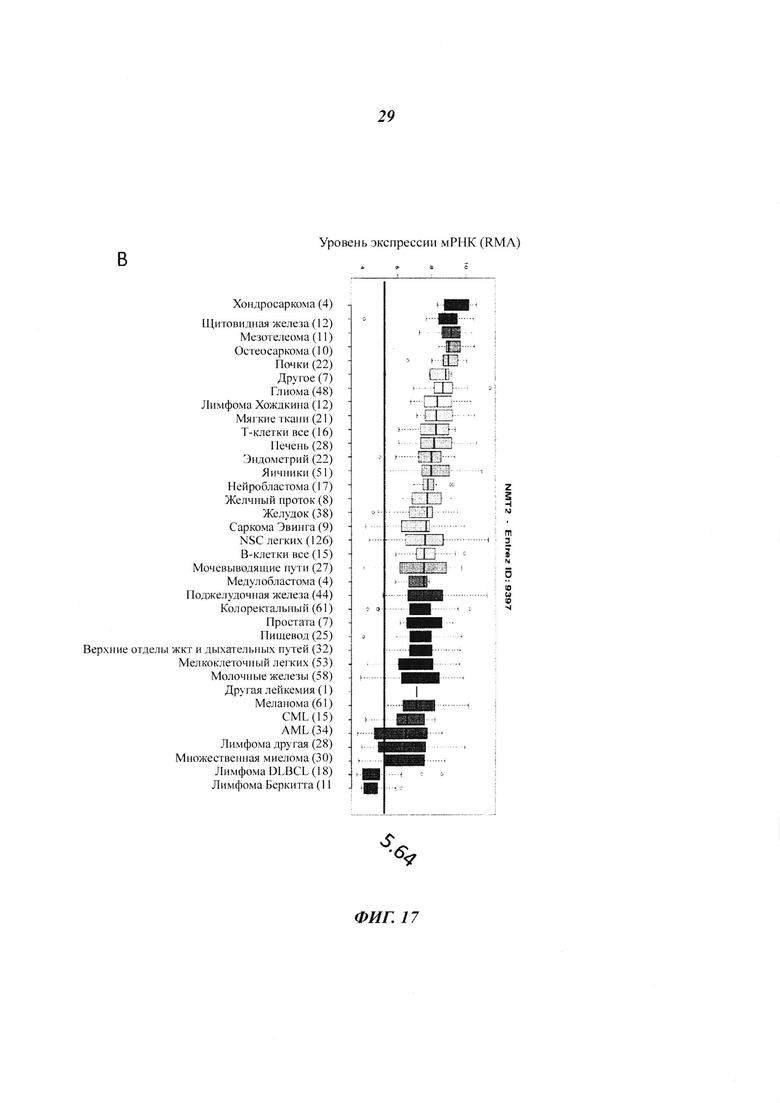
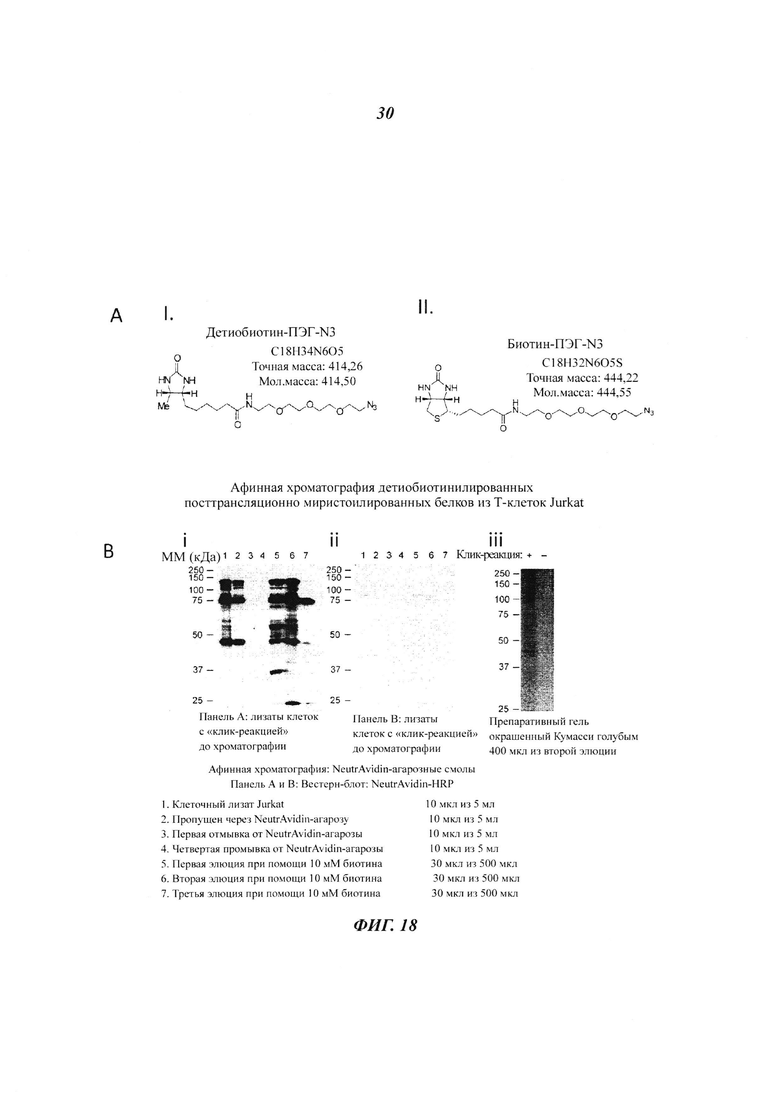
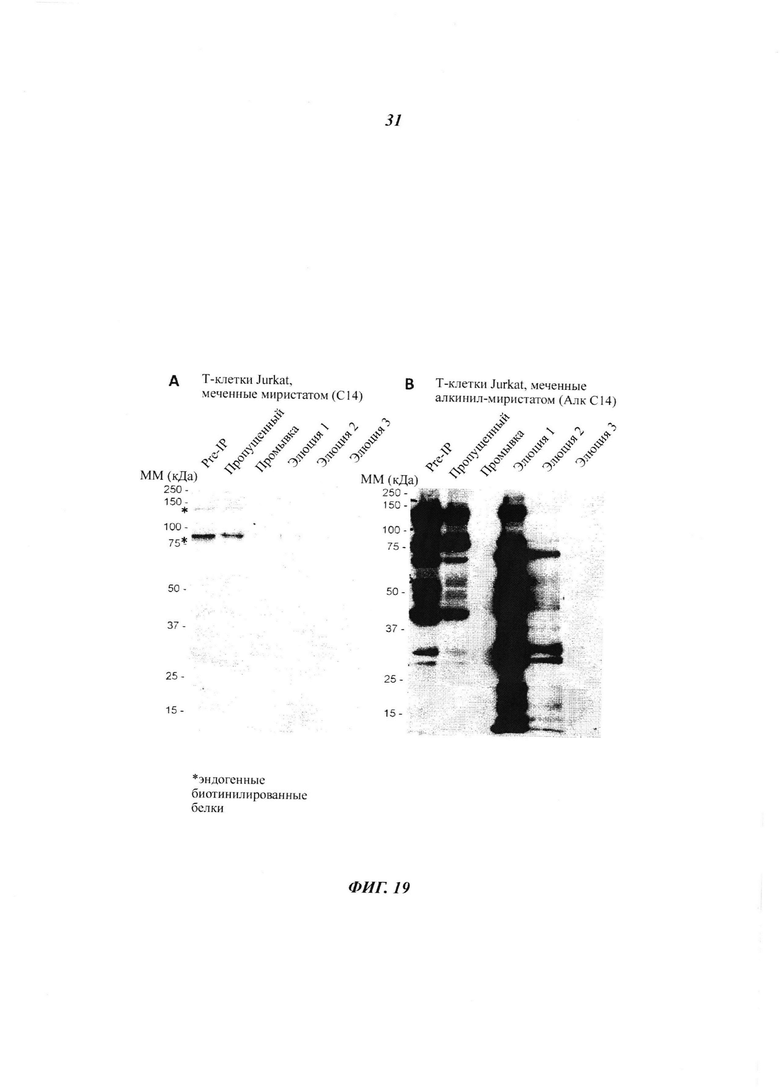
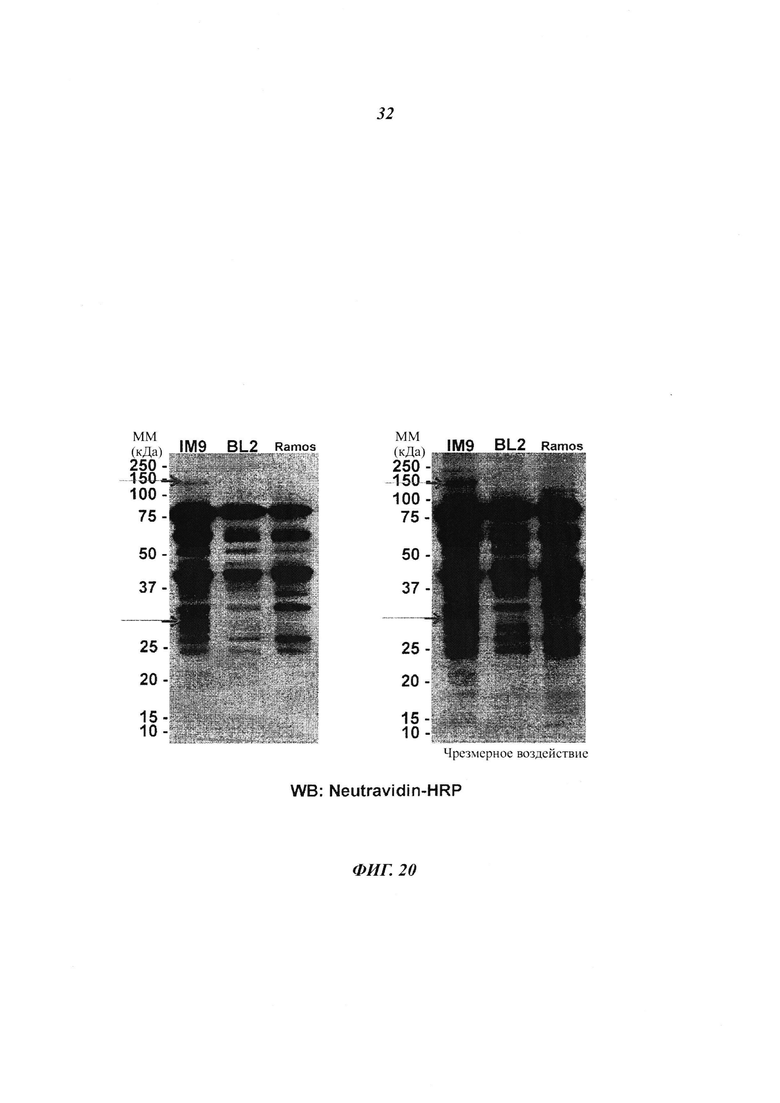
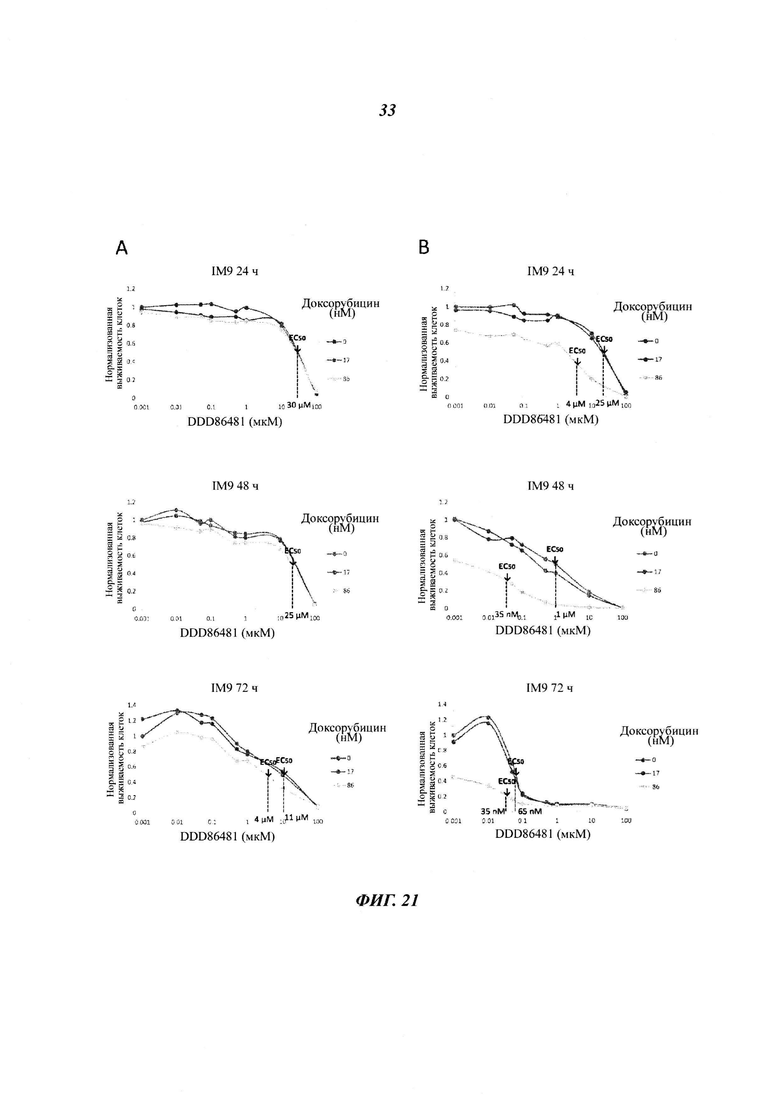
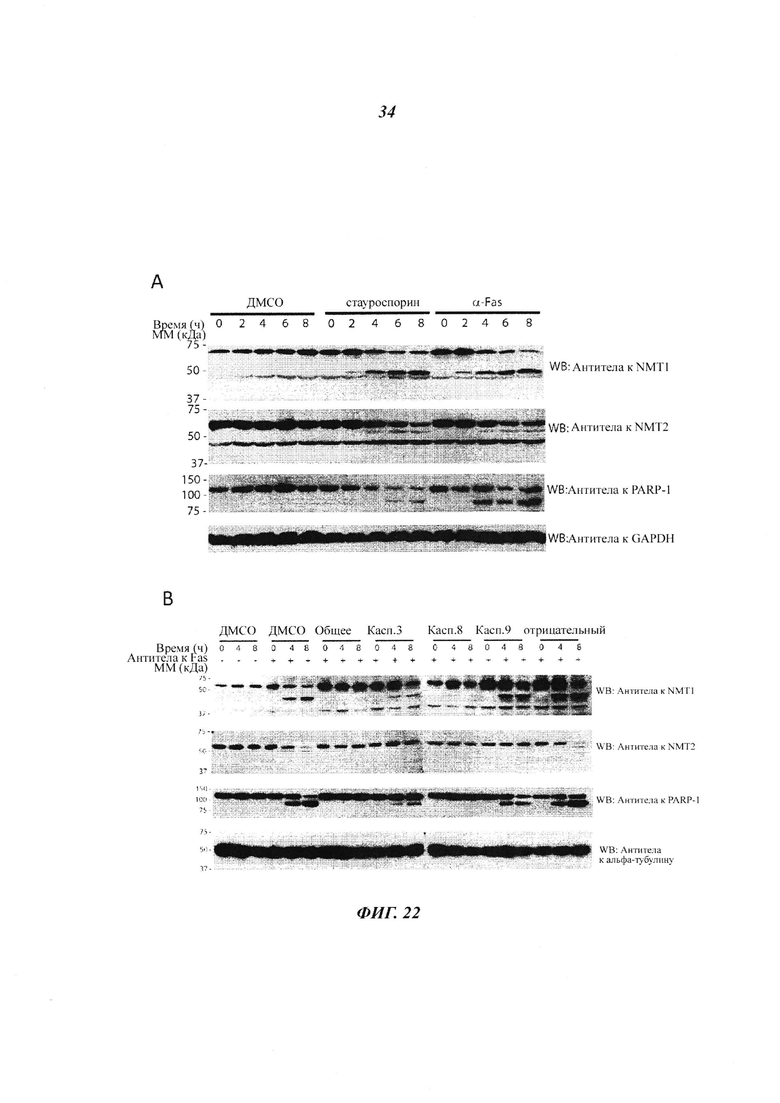
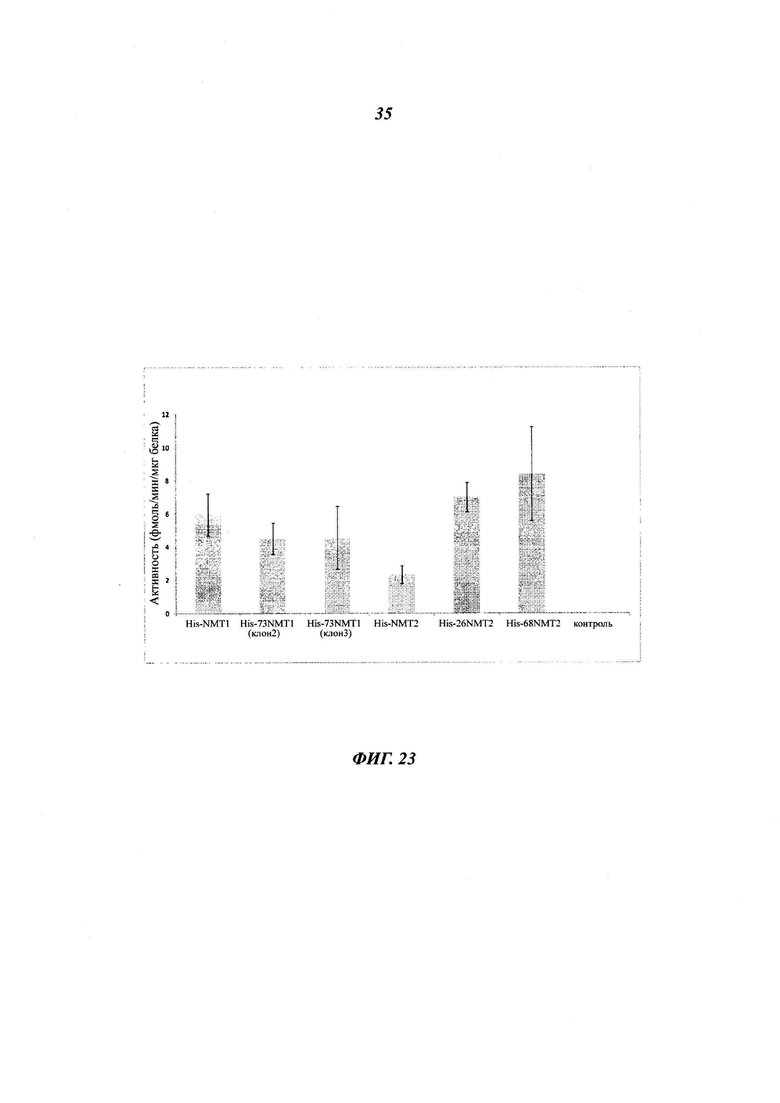
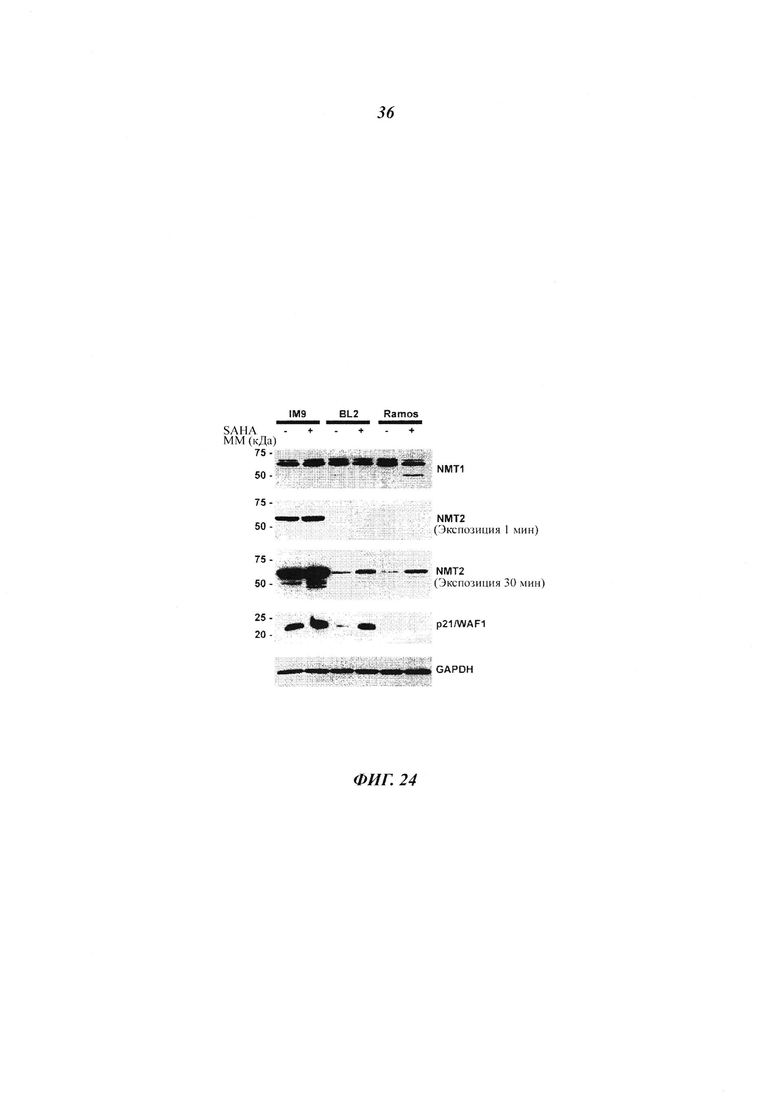
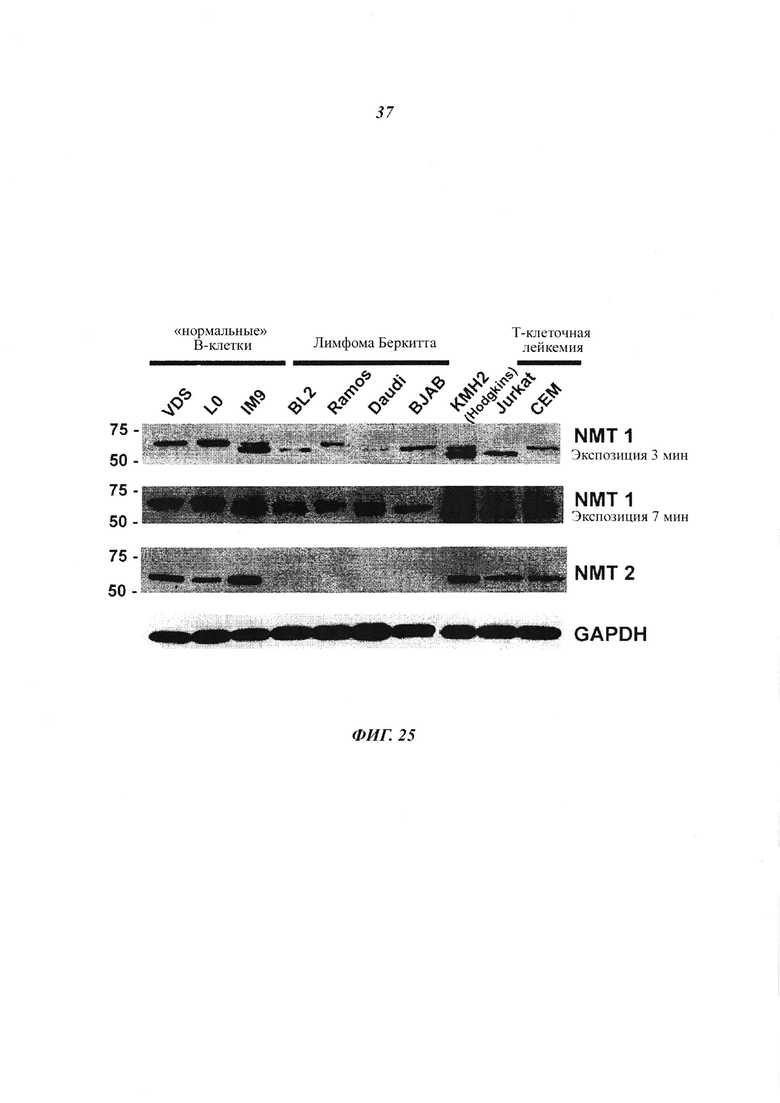
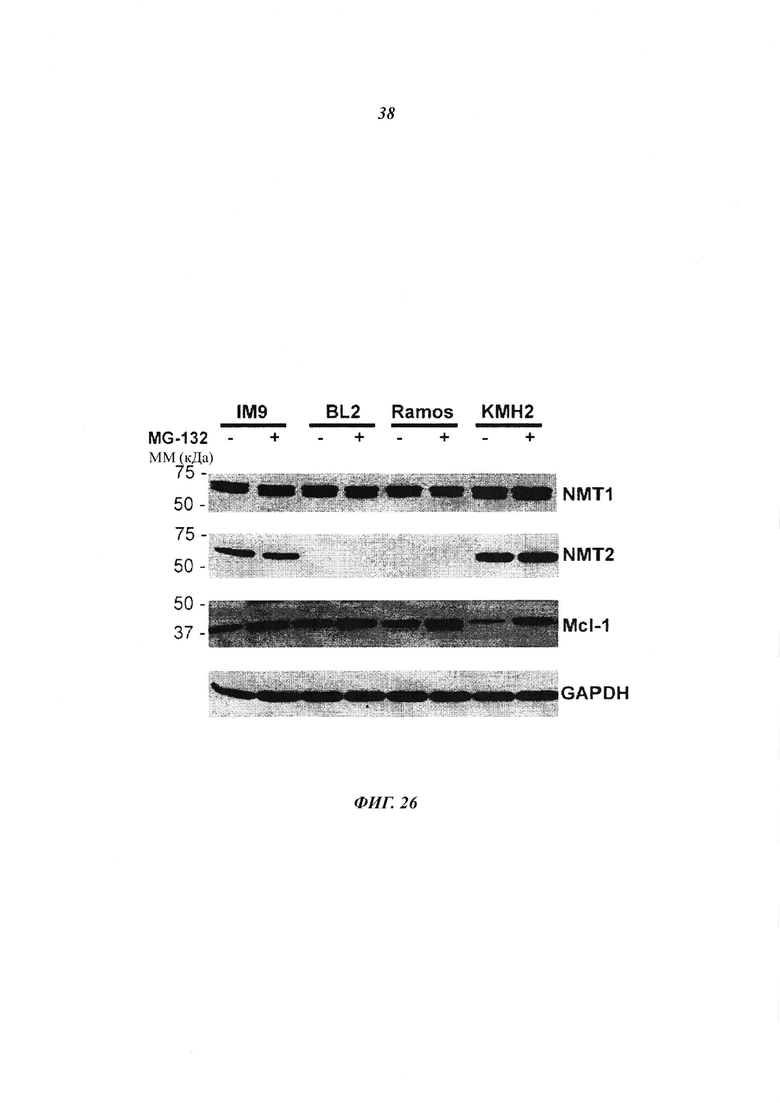
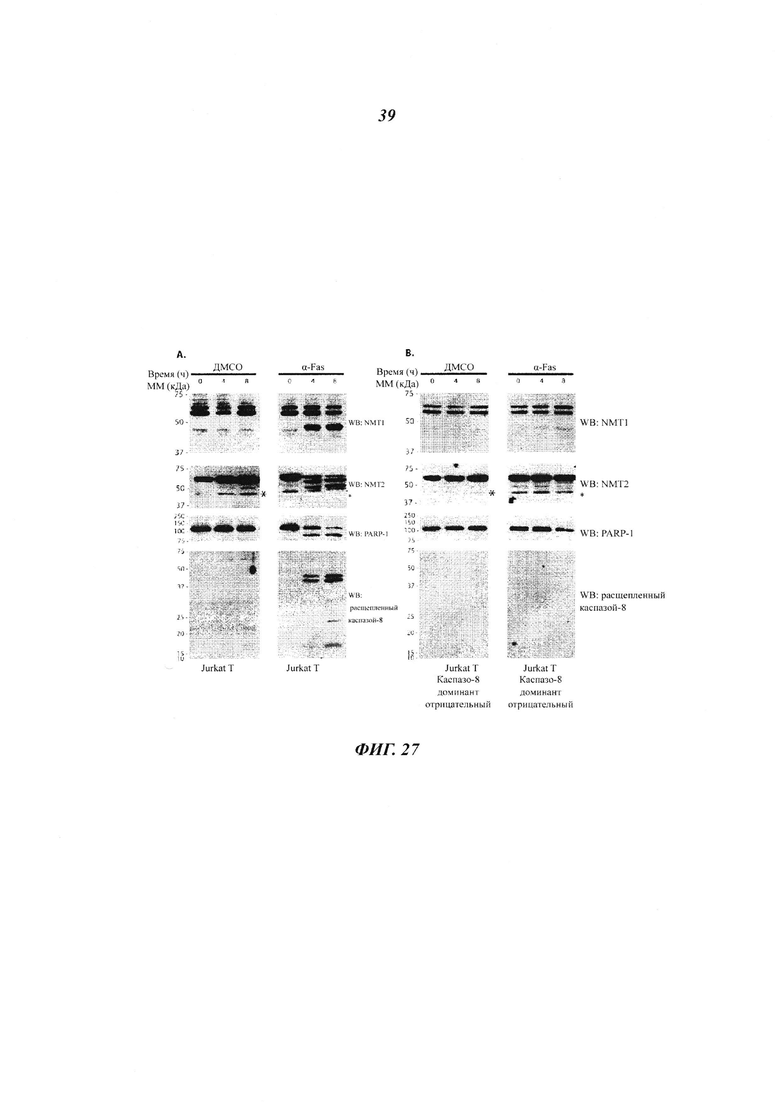
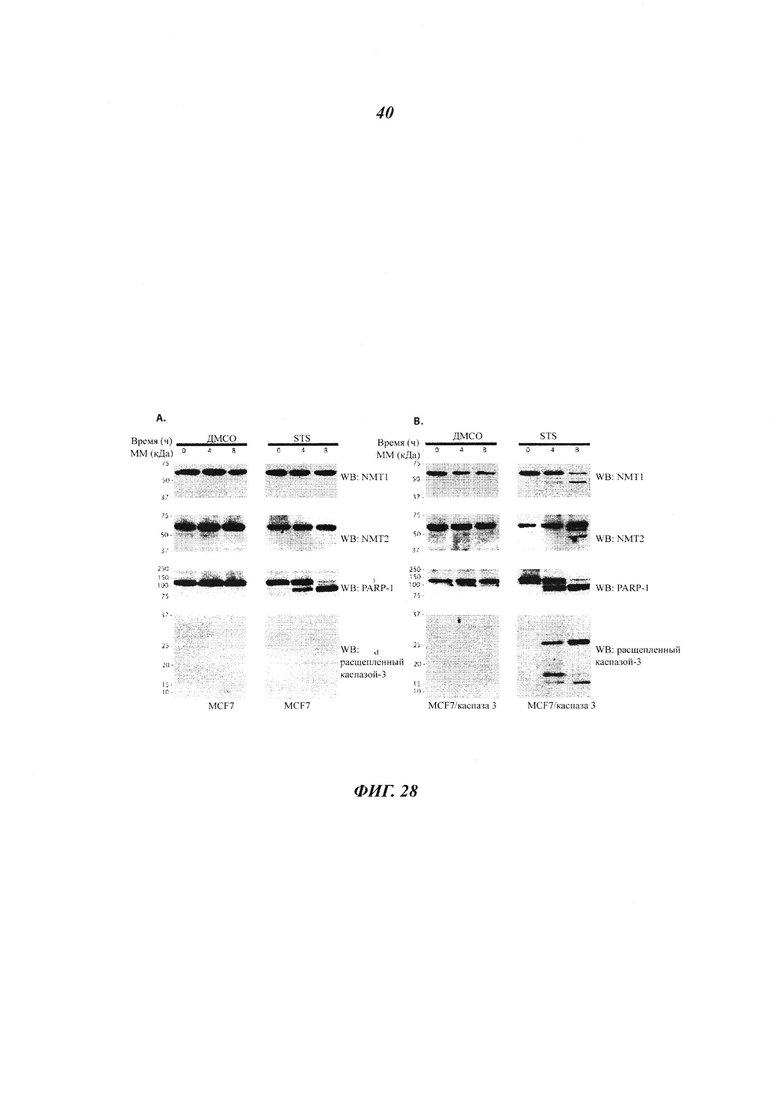

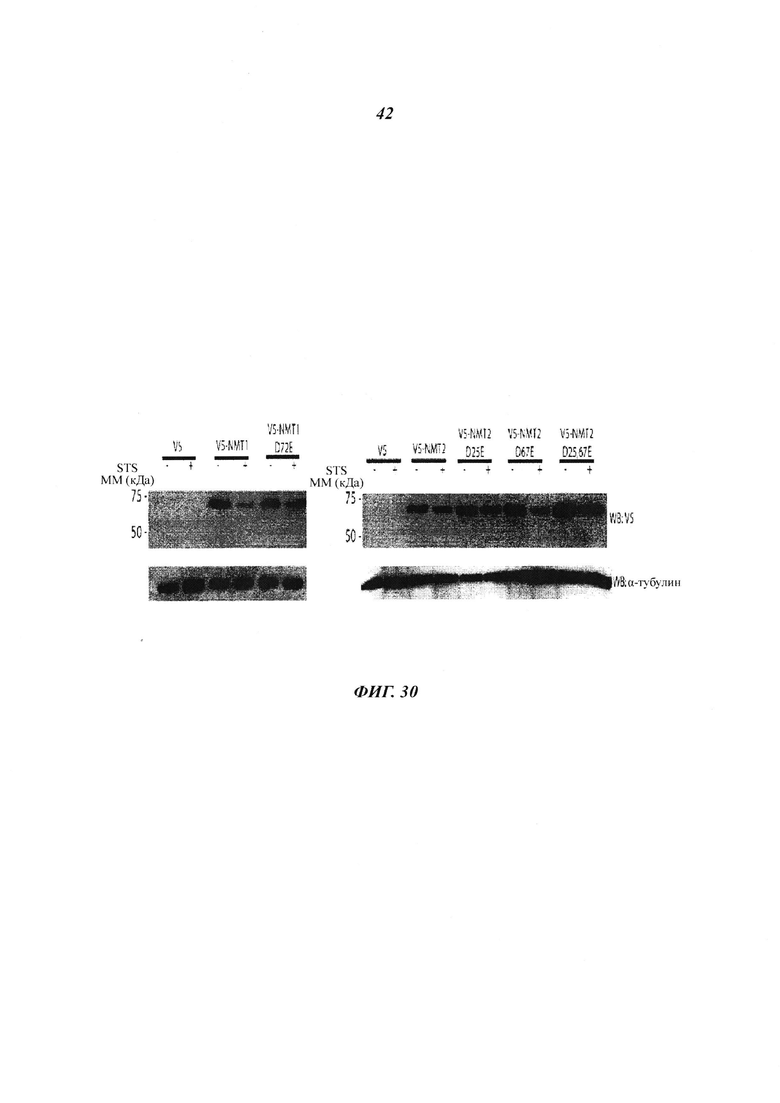
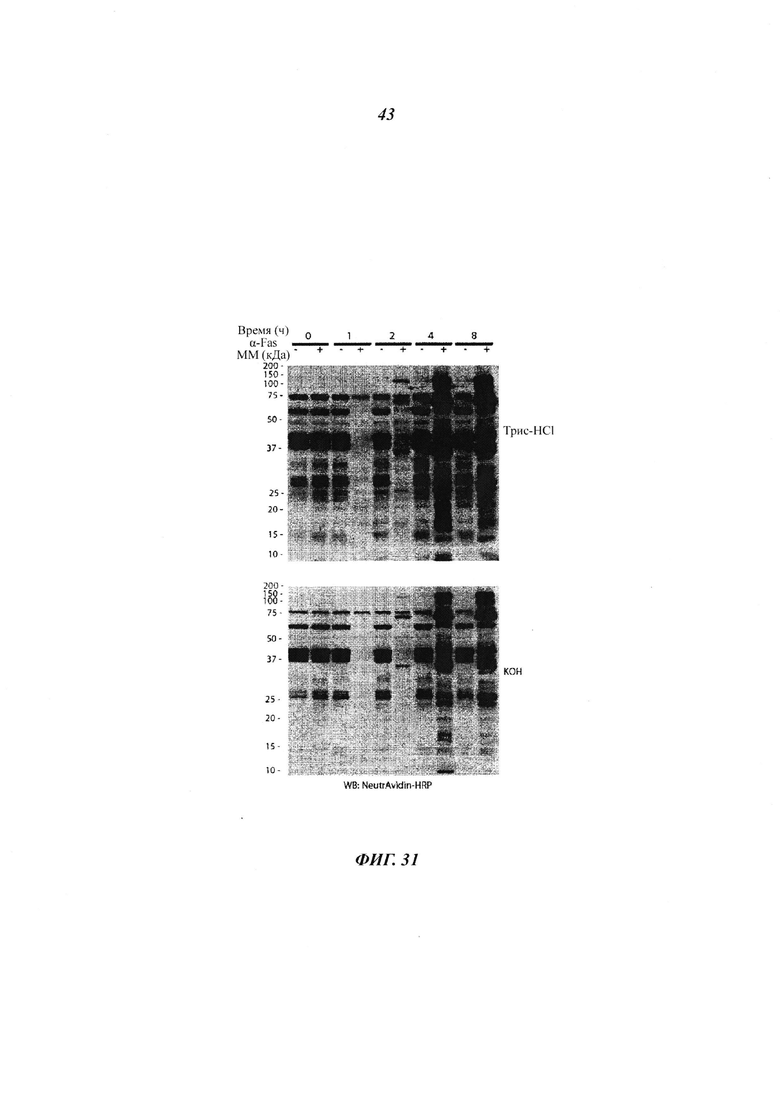


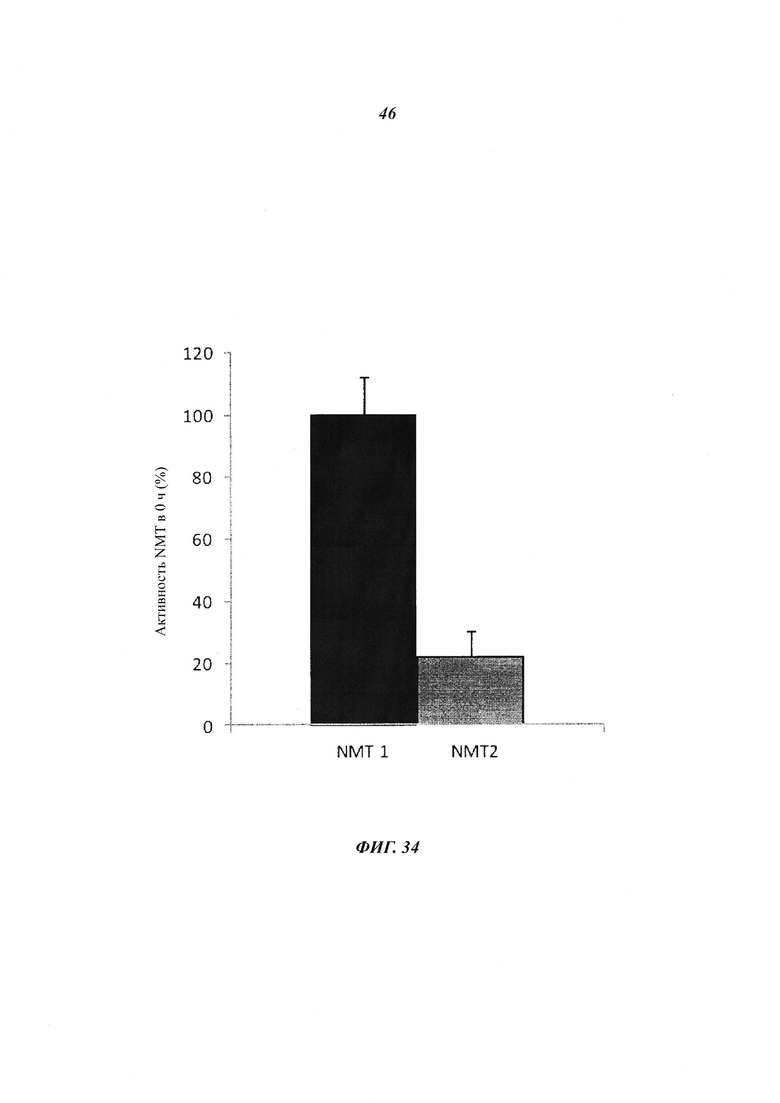
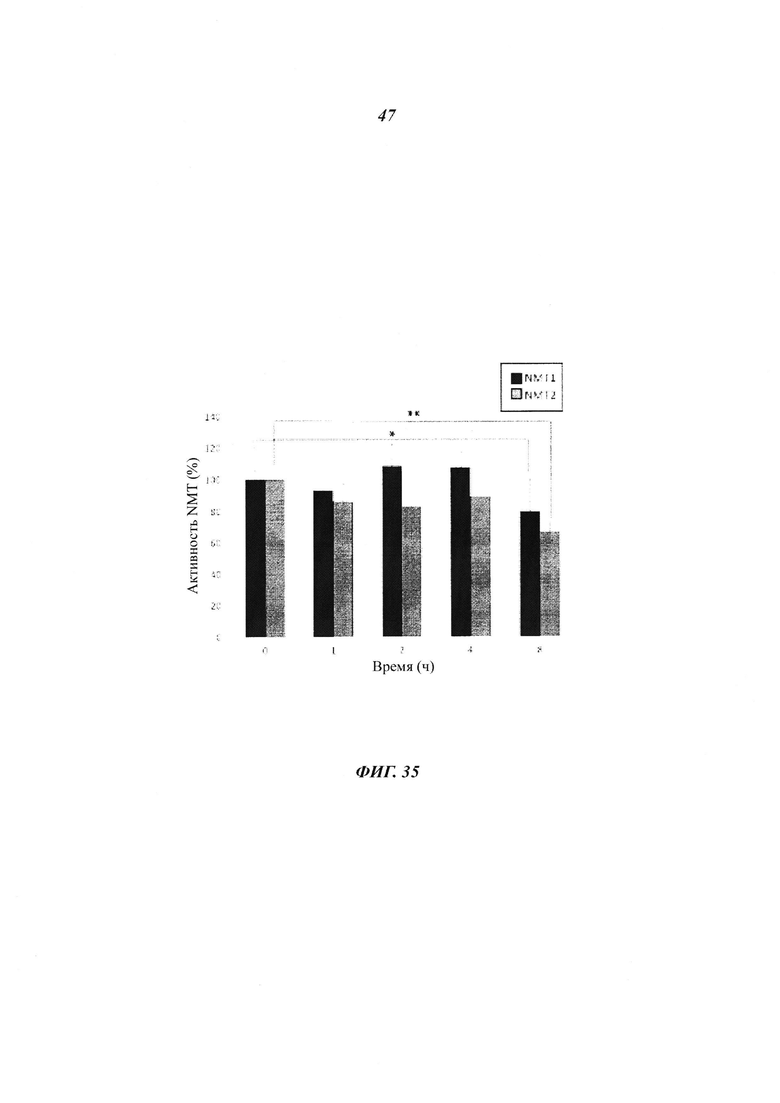

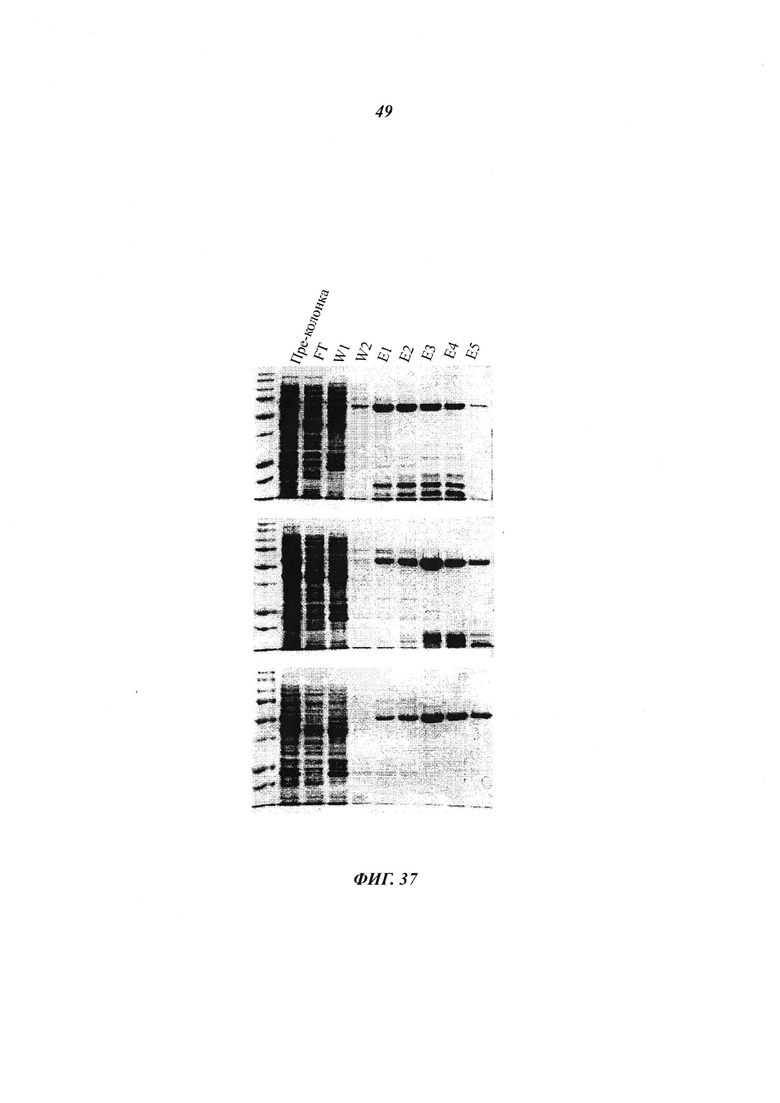
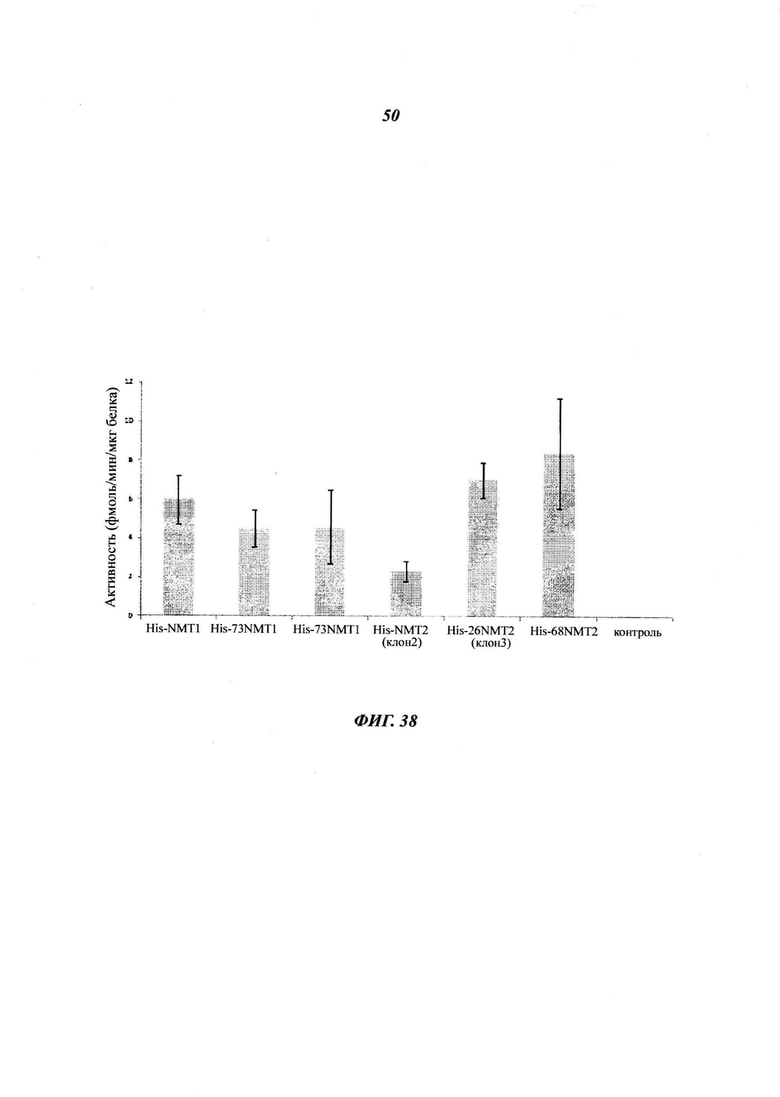
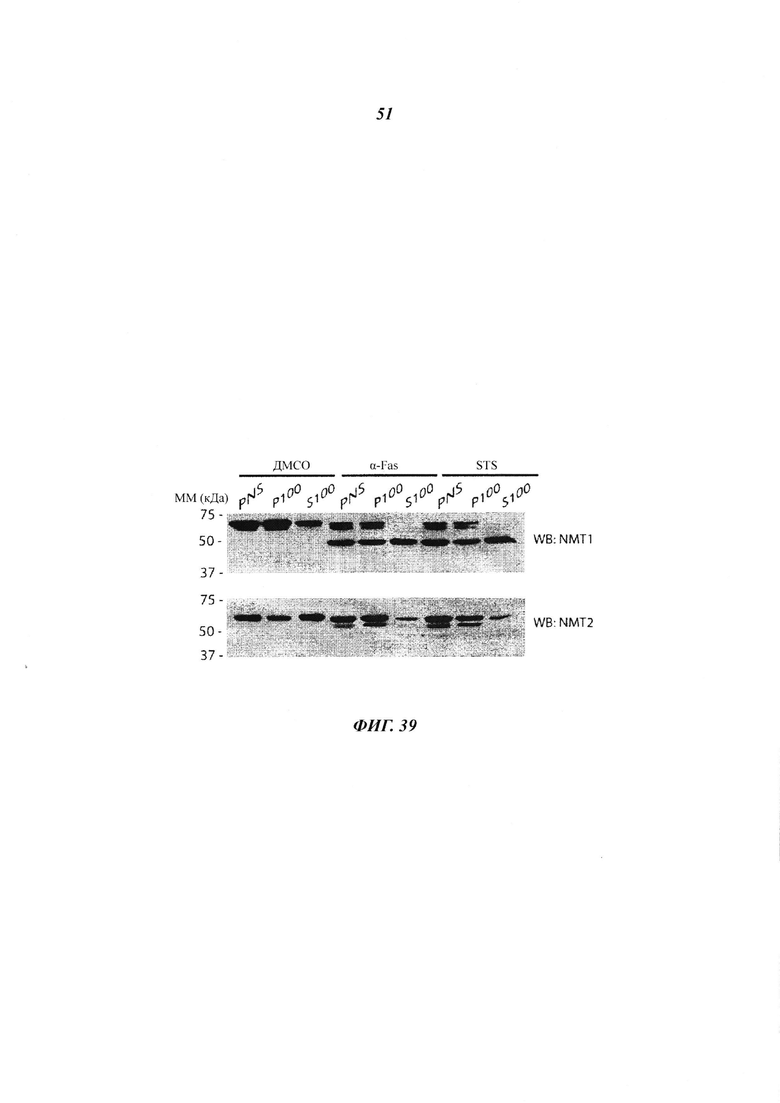
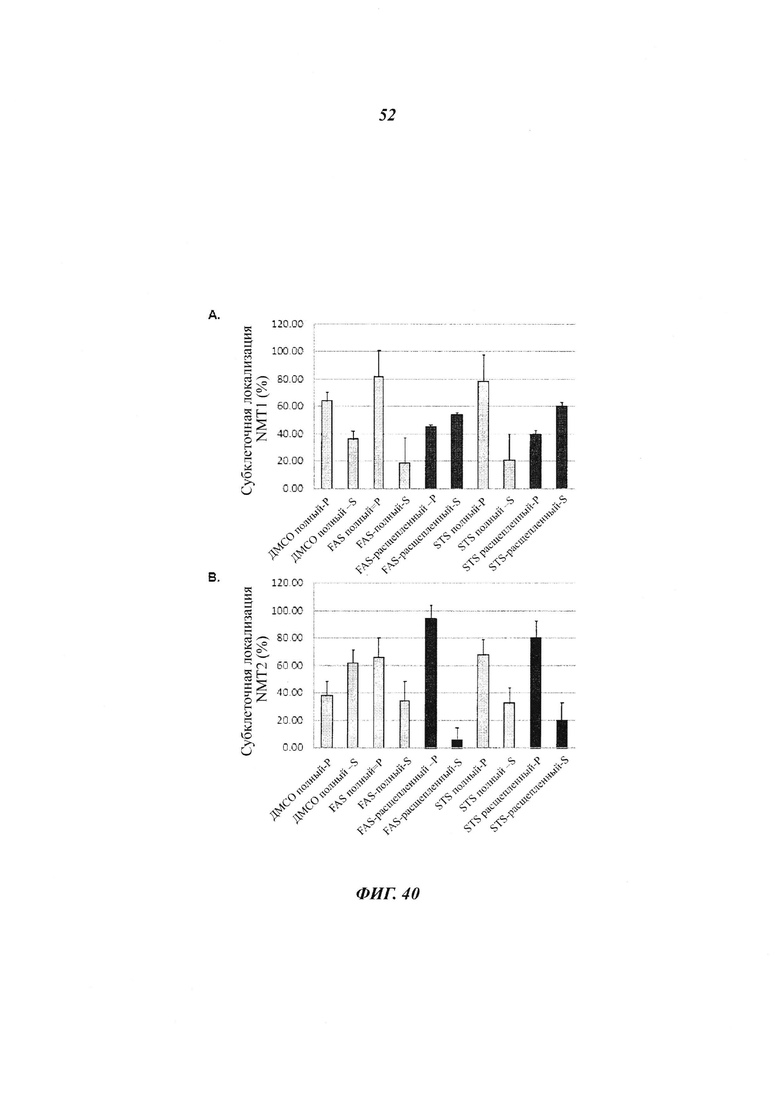
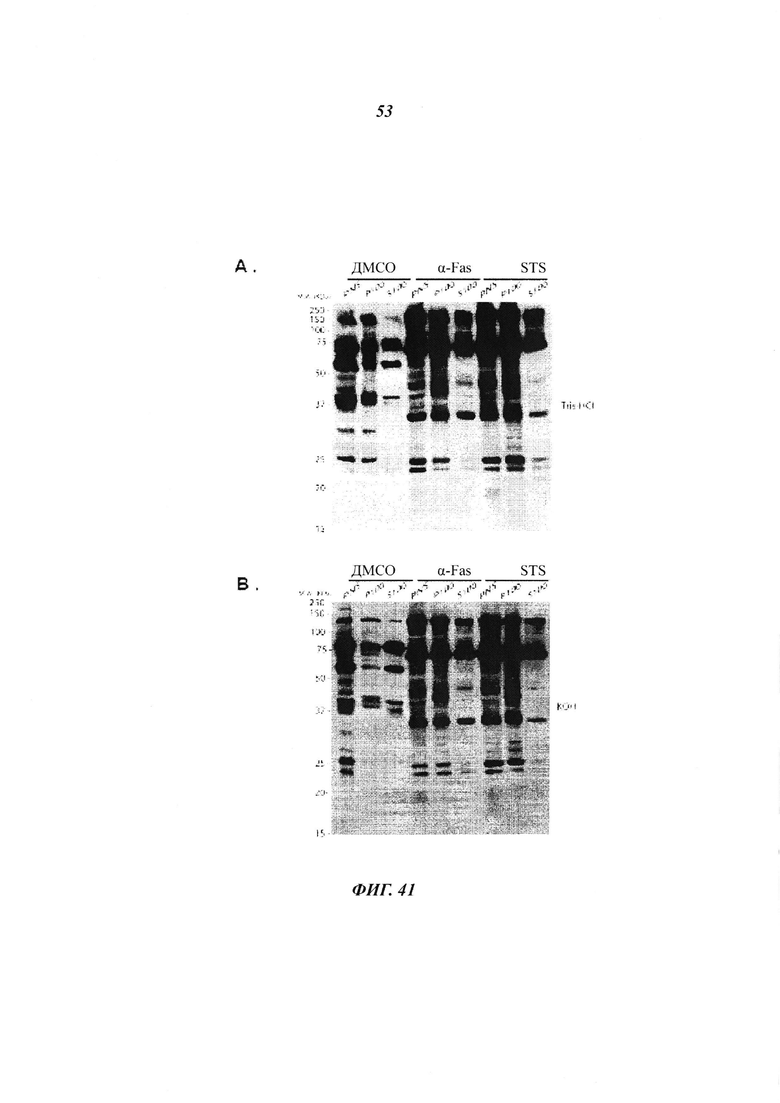
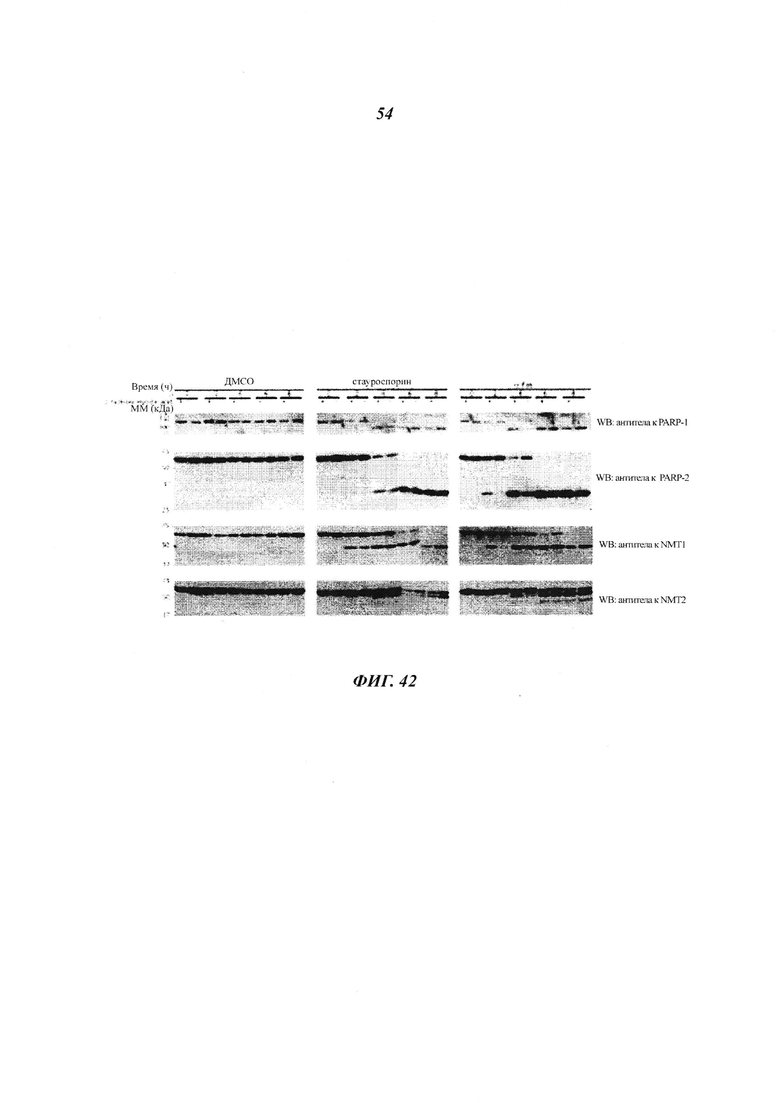
Настоящая группа изобретений относится к медицине, а именно к онкологии, и касается лечения рака. Для этого идентифицируют субъектов, больных раком, дефицитных по изомеру фермента N-миристоилтрансферазы 2 (NMT2). Лечение этой группы больных осуществляют путем введения ингибитора NMT или изофермента NMT1. Это обеспечивает повышение эффективности лечения данной группы больных, поскольку рак, дефицитный по NMT2 наиболее чувствителен к лечению ингибиторами NMT. 3 н. и 13 з.п. ф-лы, 8 табл., 38 пр., 42 ил.
1. Способ идентификации субъекта, приемлемого для лечения ингибитором N-миристоилтрансферазы (NMT) или изофермента NMT1, включающий: проведение анализа на образце, полученном из субъекта, больного раком или предположительно больного раком, для определения того, экспрессирует ли упомянутый образец от субъекта NMT2; где лечение упомянутым ингибитором NMT или упомянутым ингибитором NMT1 назначают в случае, если количество белка NMT2 или его нуклеиновой кислоты в упомянутом образце является низким или отсутствует, или в случае, если упомянутый рак имеет недостаток NMT2, где упомянутый анализ включает проведение анализа связывания, включающего контактирование обработанного образца с:
а) антителом к NMT2 с образованием комплекса между антителом и NMT2, присутствующим в обработанном образце, где упомянутый анализ связывания дает по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе, где упомянутый анализ связывания включает иммуногистохимию, где в случае, если упомянутый по меньшей мере один результат анализа численно определен как 0, то количество упомянутого белка NMT2 в упомянутом образце считается низким или отсутствующим, где в случае, если упомянутый по меньшей мере один результат анализа численно определен как 1 или 2, то количество упомянутого белка NMT2 в упомянутом образце считается высоким или сильным; или
б) детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, при этом упомянутый анализ связывания дает по меньшей мере один результат анализа, означающий упомянутый комплекс, где упомянутый анализ связывания включает гибридизационный анализ при помощи детектируемо меченных проб ДНК или РНК, где упомянутый гибридизационный анализ является количественным или полуколичественным, где упомянутый гибридизационный анализ представляет собой РВ-ПЦР, гибридизацию in situ, РНК-РНК гибридизацию в растворе (RPA), микроэррей кДНК и олигонуклеотида, репрезентативный разностный анализ (RDA), дифференциальный дисплей, анализ маркерных экспрессируемых последовательностей (EST), серийный анализ экспрессии генов (SAGE), и мультиплексную амплификацию, опосредованную лигированием, при помощи Luminex FlexMAP (LMF), где определяют уровень или концентрацию мРНК NMT2 и, если указанный уровень или концентрация ниже порогового значения, то экспрессия NMT2 считается низкой или отсутствующей; или в) антителом, которое связывается с миристоилированным белком или азидобиотин-меченными миристоилированными белками, внутри образца с образованием комплекса между детектируемой меткой и миристоилированным белком, присутствующим в образце, где упомянутый анализ связывания дает по меньшей мере один профиль миристоилирования, свидетельствующий об упомянутом комплексе, где упомянутая обработка включает обработку упомянутого образца алкинил-миристатом и детиобиотин азидо-ПЭГ-биотином.
2. Способ по п. 1(a), где лечение упомянутым ингибитором NMT или упомянутым ингибитором NMT1 назначают в случае, если количество белка NMT2 или его нуклеиновой кислоты в упомянутом образце является низким или отсутствует или если упомянутый рак имеет недостаток NMT2 по сравнению с контролем.
3. Способ по п. 1(б), где указанное пороговое значение равно 5,64 log2 (микроэррей интенсивность флуоресценции NMT2).
4. Способ по п. 1(в), где упомянутый анализ связывания включает клеточный сортинг с активацией флуоресценции, твердофазный иммуноферментный анализ, иммуногистохимию, количественную имуногистохимию, резонансный перенос энергии флуоресценции, Ферстеровский резонансный перенос энергии, биомолекулярную флуоресцентную комплементацию, масс-спектрометрию, иммуноблоттинг или коиммунопреципитацию.
5. Способ по любому из пп. 1-4, где для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для выявления комплекса, образованного между упомянутым антителом и упомянутым NMT2 в упомянутом образце или между упомянутой детектируемой меткой и нуклеиновой кислотой NMT2 в упомянутом образце, предпочтительно где упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
6. Способ по п. 1, где упомянутый рак представляет собой лимфому, В-клеточную лимфому, фолликулярную лимфому, диффузную В-крупноклеточную лимфому, мантийноклеточную лимфому, В-ХЛЛ/ЛЛ, иммуноцитому/болезнь Вальденстрема, лимфому слизистых оболочек (MALT-тип)/моноцитоидную В-клеточную лимфому, лимфому Беркитта, детскую лимфому, анапластическую крупноклеточную лимфому, острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легких, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
7. Способ идентификации субъекта, приемлемого для лечения ингибитором NMT или ингибитором NMT1, включающий: проведение анализа на образце, полученным из субъекта, больного раком или предположительно больного раком, для определения того, экспрессирует ли упомянутый образец от субъекта NMT2; где лечение упомянутым ингибитором NMT или упомянутым ингибитором NMT1 назначают в случае, если количество белка NMT2 или его нуклеиновой кислоты в упомянутом образце является низким или отсутствует, или в случае, если упомянутый рак имеет недостаток NMT2, где упомянутый рак представляет собой острую миелоидную лейкемию, бластный криз хронического миелоидного лейкоза, плазмоклеточную миелому, аденокарциному кишечника, смешанную аденосквамозную карциному легких, мелкоклеточную карциному легких, рак легких, сквамозную карциному пищевода, рак кости, дуктальную карциному груди, диффузную аденокарциному желудка, медуллярную карциному щитовидной железы, переходно-клеточную карциному мочевыводящего пути, миелому, светлоклеточный рак яичников, переходно-клеточную карциному (рак мочеточника и мочевого пузыря), хронический миелогенный лейкоз (CML), лимфому-ХЛЛ, карциному груди, колоректальную аденокарциному, аденокарциному поджелудочной железы, карциному яичника, немелкоклеточную карциному легких, остеосаркому, меланому, аденокарциному желудка, эндометриальную аденокарциному, эзофагеальную сквамозную карциному.
8. Способ по п. 7, где лечение упомянутым ингибитором NMT или упомянутым ингибитором NMT1 назначают в случае, если количество белка NMT2 или его нуклеиновой кислоты в упомянутом образце является низким или отсутствует или если упомянутый рак имеет недостаток NMT2 по сравнению с контролем.
9. Способ по п. 7, где упомянутый анализ включает проведение анализа связывания, включающего контактирование обработанного образца с:
а) антителом к NMT2 с образованием комплекса между антителом и NMT2, присутствующим в обработанном образце, где упомянутый анализ связывания дает по меньшей мере один результат анализа, свидетельствующий об упомянутом комплексе; или
б) детектируемой меткой, которая связывается с нуклеиновой кислотой NMT2 с образованием комплекса между детектируемой меткой и нуклеиновой кислотой NMT2, присутствующей в образце, при этом упомянутый анализ связывания дает по меньшей мере один результат анализа, означающий упомянутый комплекс; или антителом, связывающимся с миристоилированным белком; или
в) антителом, которое связывается с миристоилированным белком или азидобиотин-меченными миристоилированными белками, внутри образца с образованием комплекса между детектируемой меткой и миристоилированным белком, присутствующим в образце, где упомянутый анализ связывания дает по меньшей мере один профиль миристоилирования, свидетельствующий об упомянутом комплексе.
10. Способ по п. 9(б), где упомянутый анализ связывания включает гибридизационный анализ при помощи детектируемо меченных проб ДНК или РНК, предпочтительно где упомянутый гибридизационный анализ является количественным или полуколичественным.
11. Способ по п. 10, где упомянутый гибридизационный анализ представляет собой РВ-ПЦР, гибридизацию in situ, РНК-РНК гибридизацию в растворе (RPA), микроэррей кДНК и олигонуклеотида, репрезентативный разностный анализ (RDA), дифференциальный дисплей, анализ маркерных экспрессируемых последовательностей (EST), серийный анализ экспрессии генов (SAGE), и мультиплексную амплификацию, опосредованную лигированием, при помощи Luminex FlexMAP (LMF).
12. Способ по п. 9(в), где упомянутая обработка включает обработку упомянутого образца алкинил-миристатом и детиобиотин азидо-ПЭГ-биотином.
13. Способ по любому из пп. 9-12, где для определения количества комплекса в упомянутом образце применяют оборудование, имеющее детектор для выявления комплекса, образованного между упомянутым антителом и упомянутым NMT2 в упомянутом образце или между упомянутой детектируемой меткой и нуклеиновой кислотой NMT2 в упомянутом образце, предпочтительно где упомянутое оборудование представляет собой спектрофотометр, спектрофлуориметр, оптическое устройство или электрохимическое устройство.
14. Способ по п. 9, где упомянутый субъект представляет собой человека.
15. Набор для идентификации субъекта, приемлемого для лечения ингибитором NMT или ингибитором NMT1, включающий:
а) антитело к NMT2, или
б) нуклеиновую кислоту для связывания с NMT2, предпочтительно где упомянутая нуклеиновая кислота представляет собой РНК или ДНК, и NeutrAvidinTM-HRP, ω-алкинил-миристат или детиобиотин азидо-ПЭГ-биотин; или
в) антитело, которое связывается с миристоилированным белком или азидобиотин-меченными миристоилированными белками, ω-алкинил-миристат или детиобиотин азидо-ПЭГ-биотин;
и инструкции по идентификации субъекта согласно способу по любому из пп. 1-14.
16. Набор по п. 15, дополнительно включающий контроль.
| ИНГИБИТОРЫ АНГИОПОЭТИНПОДОБНОГО БЕЛКА 4, ИХ КОМБИНАЦИИ И ПРИМЕНЕНИЕ | 2005 |
|
RU2392966C2 |
| Способ флегматизации высокоплотного двухосновного сферического пороха | 2017 |
|
RU2670837C1 |
| BHANDARKAR S.S | |||
| et al | |||
| "Tris(Dibenzylideneacetone) Dipalladium, a N-Myristoyltransferase-1 inhibitor, is effective against melanoma growth on vitro and vivi" | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| KUMAR S | |||
| et al | |||
| "The potential use of N-myristoyltransderase as a biomarker in the early diagnosis of colon cancer" | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |

