ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка претендует на дату приоритета временной патентной заявки США № 61/331550, поданной 5 мая 2010 г., раскрытие которой настоящим включено сюда в качестве ссылки.
ПЕРЕЧНИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Данная заявка содержит перечни последовательностей, которые были направлены на рассмотрение с помощью системы EFS-Web и настоящим целиком включены сюда в качестве ссылок. Указанная копия в формате ASCII, созданная 2 мая 2011 г., названа Sequence Listing_Staphylococcus Aureus Leucocidins_ST25.txt и имеет размер 102 килобайта.
ИЗВЕСТНЫЙ УРОВЕНЬ ТЕХНИКИ
Бактерии Staphylococcus aureus или стафилококки ("staph") обычно присутствуют на коже или в носовой полости людей и животных. Стафилококковые бактерии обычно являются безвредными, если только они не попадают в организм через порез или другую рану. Как правило, стафилококковые инфекции создают незначительные кожные проблемы для здоровых людей. Исторически, стафилококковые инфекции лечат антибиотиками широкого спектра действия, такими как метициллин. Однако в настоящее время появились определенные штаммы стафилококков, резистентные к метициллину и другим β-лактамовым антибиотикам, таким как пенициллин и цефалоспорины. Они называются метициллин-резистентным Staphylococcus aureus (также известным как Staphylococcus aureus с множественной лекарственной резистентностью, или "MRSA").
Стафилококковые инфекции, включая MRSA, обычно начинаются с появления маленьких красных бугорков, напоминающих прыщи, нарывы или укусы пауков. Такие бугорки или пятна могут быстро превращаться в глубокие, болезненные абсцессы, требующие хирургического дренирования. Иногда бактерии остаются в пределах кожи. В некоторых случаях, они могут проникать далеко вглубь тела, вызывая потенциально опасные для жизни инфекции в широком спектре человеческих тканей, включая кожу, мягкие ткани, кости, суставы, хирургические раны, кровоток, сердечные клапаны, легкие или другие органы. Таким образом, инфекции S. aureus могут приводить к потенциально смертельным болезням, таким как некротизирующий фасцит, пневмония, эндокардит, сепсис, токсический шок, и различные формы пневмонии. Инфекция MRSA создает особенно много проблем в условиях больницы или медицинских учреждений по уходу за инвалидами и престарелыми, где пациенты часто имеют открытые раны, подвергаются инвазивным процедурам с помощью разных приспособлений, и имеют ослабленные иммунные системы и, таким образом, подвержены большему риску инфекции, чем население в целом. Сотрудники, не придерживающиеся надлежащих санитарных процедур, могут переносить бактерии MRSA от одного пациента к другому.
S. aureus продуцирует широкий спектр вирулентных факторов и токсинов, позволяющих этой бактерии нейтрализовать и противостоять атакам разных видов иммунных клеток, конкретнее, субпопуляций белых кровяных клеток, образующих первичную защитную систему организма. Продуцирование таких вирулентных факторов и токсинов позволяет S. aureus поддерживать инфекционное состояние. См. Nizet, J. Allergy Clin. Immunol. 120:13-22 (2007). Наряду с этими вирулентными факторами, S. aureus продуцирует несколько бикомпонентных лейкотоксинов, которые повреждают мембраны защитных клеток хозяина и эритроцитов в результате синергичного действия двух неассоциированных белков или субъединиц. См. Supersac, et al., Infect. Immun. 61:580-7 (1993). Из таких бикомпонентных лейкотоксинов лучше всего исследованы гамма-гемолизин (HlgAB и HlgCB) и лейкоцидин Пантона-Валентайна (Pantone-Valentine Leukocidin, PVL).
Токсичность лейкоцидинов по отношению к клеткам млекопитающих связана с действием двух компонентов. Первая субъединица называется субъединицей класса S (т.е. "медленно элюирующаяся"), и вторая субъединица называется субъединицей класса F (т.е. "быстро элюирующаяся"). S- и F-субъединицы действуют синергично, образуя поры в белых кровяных клетках, включая моноциты, макрофаги, дендритные клетки и нейтрофилы (коллективно называемые фагоцитами). См. Menestrina, et al., Toxicol. 39:1661-1672 (2001). Механизм, по которому бикомпонентные токсины образуют поры в мембранах клетки-мишени, до конца не выяснен. Предложенный механизм действия этих токсинов включает связывание S-субъединицы с мембраной клетки-мишени, наиболее вероятно, через рецептор, с последующим связыванием F-субъединицы с S-субъединицей, с образованием при этом олигомера, который в свою очередь образует предпору (pre-pore), проникающую в мембрану клетки-мишени (Jayasinghe, et al., Protein. Sci. 14:2550-2561 (2005)). Поры, образованные бикомпонентными лейкотоксинами, типично являются катионселективными. Образование пор вызывает гибель клетки вследствие лизиса, который в тех случаях, когда мишенями являются белые кровяные клетки, как сообщалось, вызван нарушением осмотического баланса вследствие притока катионов (Miles, et al., Biochemistry 40:8514-8522 (2001)).
Известно, что в дополнение к PVL (также известным как лейкоцидин S/F-PV или LukSF-PV) и гамма-гемолизину (HlgAB и HlgCB), репертуар бикомпонентных лейкотоксинов, продуцируемых S. aureus, включает лейкоцидин E/D (LukED) и лейкоцидин M/F' (LukMF'). Таким образом, субъединицы S-класса таких бикомпонентных лейкоцидинов включают HlgA, HlgC, LukE, LukS-PV и LukM, и субъединицы F-класса включают HlgB, LukD, LukF-PV и LukF'-PV (Menestrina, et al., supra.). S- и F-субъединицы S. aureus не являются лейкоцидин-специфичными. Это означает, что они являются взаимозаменяемыми таким образом, что другие бикомпонентные комбинации могут создавать функциональные поры в белой кровяной клетке, значительно расширяя репертуар лейкотоксинов (Meyer, et al., Infect. Immun. 77:266-273 (2009)).
Разработка эффективной терапии для лечения инфекции MRSA является особенно сложной проблемой. Было обнаружено, что в дополнение к вышеупомянутой резистентности к метициллину и родственным антибиотикам, MRSA также проявляет значительные уровни резистентности к макролидам (например, эритромицину), комбинациям ингибитора бета-лактамазы (например, уназин, аугментин) и фторхинолонам (например, ципрофлоксацин), а также к клиндамицину, триметоприму/сульфаметоксизолу (Bactrim) и рифампину. В случае серьезной инфекции S. aureus, клинические врачи прибегают к использованию внутривенно ванкомицина. Однако имеются сообщения о резистентности S. aureus к ванкомицину. Таким образом, существует потребность в разработке новых препаратов антибиотиков, эффективно борющихся с инфекцией S. aureus.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Заявители обнаружили и охарактеризовали другой бикомпонентный член защитной системы нативного Staphylococcus aureus. Новый охарактеризованный полипептидный компонент нативной S-субъединицы, обозначенный здесь "LukA", охватывает нативные полипептиды и их аналоги, имеющие степень подобия последовательностей, по меньшей мере, 70% с последовательностями нативных полипептидов. Таким образом, один аспект настоящего изобретения касается изолированного и/или очищенного LukA. Другой аспект настоящего изобретения касается изолированного и/или очищенного полинуклеотида, кодирующего LukA, трансформированного хозяина (например, клетки), содержащего полинуклеотид, и способов получения рекомбинантного LukA путем экспрессии полинуклеотида в трансформированном хозяине.
Новый охарактеризованный полипептидный компонент F-субъединицы, обозначенный здесь "LukB", охватывает нативные полипептиды и их аналоги, имеющие степень подобия последовательностей, составляющую, по меньшей мере, 70% с последовательностями нативных полипептидов. Таким образом, другой аспект настоящего изобретения касается изолированного и/или очищенного LukB. Другой аспект настоящего изобретения касается изолированного и/или очищенного полинуклеотида, кодирующего LukB, трансформированного хозяина (например, клетки), содержащего полинуклеотид, и способов получения рекомбинантного LukB путем экспрессии полинуклеотида в трансформированном хозяине.
Еще один аспект данной заявки касается терапевтических композиций, пригодных для ингибирования начальных проявлений или лечения инфекции Staphyloccocus aureus, содержащих терапевтически эффективные количества LukA и/или LukB в композиции с фармацевтически приемлемым носителем. Таким образом, в одном варианте исполнения, терапевтическая композиция содержит терапевтически эффективное количество LukA. В другом варианте исполнения, терапевтическая композиция содержит терапевтически эффективное количество LukB. В еще одном варианте исполнения, терапевтическая композиция содержит терапевтически эффективные количества обоих LukA и LukB. В еще одних вариантах исполнения, композиция содержит аналог LukA с отсутствующими 10 C-концевыми остатками, являющийся нетоксичным (обозначается здесь LukAΔ10C или rLukAΔ10C). Такие композиции пригодны для комплексного терапевтического применения. В некоторых вариантах исполнения, композиции называются противовоспалительными композициями и могут быть использованы для лечения острых воспалительных состояний или расстройств, в частности, локализованных острых воспалительных состояний.
Такое применение использует сделанные заявителями дополнительные открытия того, что в физиологических условиях (т.е. LukAB продуцируется непосредственно S. aureus), комплекс LukAB обладает исключительной специфичностью к фагоцитам, но не к другим ядросодержащим клеткам, таким как эпителиальные клетки и эндотелиальные клетки. Это означает, что комплекс образует поры в мембранах таких видов клеток, тем самым вызывая гибель клетки, что называется здесь "LukAB-опосредованной цитотоксичностью". С другой стороны, LukAB обладает относительно небольшой или пренебрежимо малой специфичностью по отношению к другим ядросодержащим клеткам млекопитающих. Таким образом, противовоспалительные композиции по настоящему изобретению используют специфичность LukAB к человеческим фагоцитам в целях лечения острых воспалительных состояний, характеризующихся массивной инфильтрацией фагоцитов к месту воспаления.
В других вариантах исполнения, терапевтические композиции могут быть названы композицией (активной) вакцины. Композиции могут быть использованы для индукции продуцирования нейтрализующих антител к LukA и к LukB у субъекта с риском инфекции S. aureus или субъекта с диагностированной инфекцией S. aureus, такой как MRSA.
Другие аспекты настоящего изобретения касаются антител, которые специфически связывают LukA, антител, которые специфически связывают LukB, терапевтических композиций, содержащих антитела LukA и/или LukB, и их применения для лечения инфекционных состояний, связанных с S. aureus. Такие терапевтические композиции могут быть названы пассивными композициями вакцины. Таким образом, в одном варианте исполнения, терапевтическая композиция содержит терапевтически эффективное количество антител к LukA. В другом варианте исполнения, терапевтическая композиция содержит терапевтически эффективное количество антител к LukB. В еще одном варианте исполнения, терапевтическая композиция содержит терапевтически эффективные количества как антител к LukA, так и антител к LukB.
Пассивные и активные композиции вакцин по настоящему изобретению используют еще одно сделанное заявителями открытие - что инфекционные вирулентные штаммы S. aureus, такие как MRSA, экспрессируют LukA и LukB. Неизменность LukA и LukB для широкого спектра штаммов S. aureus позволяет вакцинам по настоящему изобретению обеспечивать полный спектр терапевтической эффективности. LukA, LukB, антитела к LukA и антитела к LukB также называются здесь активными агентами.
Другой аспект настоящего изобретения касается способов применения LukAB, LukA и/или LukB для идентификации потенциальных ингибиторов LukAB-опосредованной цитотоксичности. Такие способы могут использовать комплекс LukAB, per se, в комбинации с фагоцитом, или его часть, связывающуюся с мембраной фагоцита. Идентифицированные таким образом ингибиторы могут быть кандидатами для терапии с целью лечения инфекции S. aureus.
Еще один аспект настоящего изобретения касается способа прогнозирования или оценки тяжести инфекции S. aureus, предусматривающего детектирование присутствия или количества LukA и/или LukB, или детектирование соответствующих генов LukA и/или LukB, в биологическом образце, полученном от инфицированного субъекта. Этот аспект настоящего изобретения основан на еще одном сделанном заявителями открытии - того, что из многих цитотоксинов, продуцируемых S. aureus, LukAB проявляет сильную токсичность по отношению к человеческим фагоцитам. Таким образом, детектирование присутствия или относительно высоких количеств LukA и/или LukB, или их соответствующих генов (например, демонстрируемое штаммами S. aureus Newman, 4645 и штаммами MRSA USA300 и USA500) по сравнению с контролем (например, штаммами S. aureus USA100 и USA400), который продуцирует небольшие или недектируемые количества LukA и/или LukB, указывает на тяжелую инфекцию S. aureus.
Эти и другие аспекты настоящего изобретения более полно описаны ниже.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
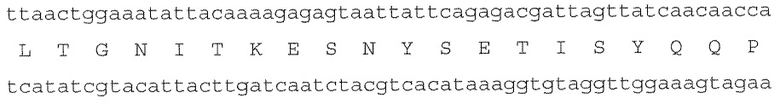
Фигура 1 представляет собой выравнивание, содержащее аминокислотную последовательность мажоритарной (majority) последовательности LukA (обозначенной как SEQ ID NO:1) и полипептидов LukA от тринадцати (13) разных штаммов S. aureus, которым она соответствует (обозначенным как SEQ ID NO:2-14).
Фигура 2 представляет собой выравнивание, содержащее аминокислотную последовательность мажоритарной последовательности LukB (обозначенной как SEQ ID NO:15) и полипептидов LukB от двенадцати (12) разных штаммов S. aureus, которым она соответствует (обозначенным как SEQ ID NO:16-27).
Фигура 3. LukAB является сильным стафилококковым цитотоксином, который нацелен на и уничтожает первичные человеческие фагоциты. (a) Интоксикация первичных человеческих мононуклеарных клеток периферической крови (PBMC) фильтратом культуры (2,5% об./об.) S. aureus, штамм Newman (дикого типа, WT) и указанных изогенных мутантных штаммов. Жизнеспособность клеток контролируют с помощью CellTiter, где клетки, обработанные средой, принимались за 100%. Результаты представляют средние значения для трех параллельных образцов + стандартное отклонение (S.D.). (b) Интоксикация первичных человеческих моноцитов, макрофагов и дендритных клеток (DC) фильтратом культуры (2,5% об./об.) S. aureus, штамм Newman (WT) и указанных изогенных мутантных штаммов. Жизнеспособность клеток контролируют, как описано выше. Результаты представляют среднее для двух доноров, где клетки от каждого донора интоксицируют тремя независимыми препаратами экзопротеина, + S.E.M. (стандартная ошибка средней). (c) Интоксикация первичных человеческих PMN (полиморфоядерных нейтрофилов) различными разбавлениями фильтратов культур S. aureus штамм Newman (WT) и указанных изогенных мутантных штаммов. Жизнеспособность клеток контролируют, как описано выше. Результаты представляют среднее для PMN, выделенных у четырех доноров ±S.E.M. (d) Интоксикация первичных человеческих PMN очищенными rLukA, rLukB, или комбинацией rLukA и rLukB в указанных концентрациях. * указывает статистическую значимость при сравнении с вместе взятыми rLukA и rLukB, P<0,05. Для панелей (a-c) * указывает статистическую значимость при сравнении с WT, ** указывает статистическую значимость при сравнении с ΔLukAB/p, P<0,05 (t-критерий Стьюдента p<0,05).
Фигура 4. LukAB предпочтительно нацелен на человеческие фагоцитарные клетки. Интоксикация (a, c и d) PMN-HL60 или (b) клеток THP1 различными разведениями фильтрата культуры S. aureus WT штамм Newman, изогенных мутантных штаммов, не содержащих указанных генов/токсинов, (c) фильтратом культуры S. aureus WT, содержащей пустую плазмиду (WT/p), штамма, не содержащего LukAB, с пустой плазмидой (ΔLukAB/p) и штамма, не содержащего LukAB, с LukAB-комплементирующей плазмидой (ΔLukAB/pLukAB), или (d) очищенным рекомбинантным LukA (rLukA), LukB (rLukB) или комбинаций rLukA и rLukB (rLukA+rLukB) в указанных концентрациях. Для интоксикаций вместе взятыми rLukA и rLukB, общая концентрация белка складывается из равных количеств rLukA и rLukB (например, 2,8 мкг общего белка соответствует 1,4 мкг rLukA и 1,4 мкг rLukB). (e) Интоксикация указанных человеческих клеточных линий 10 мкг/мл rLukAB. Жизнеспособность клеток контролируют с помощью CellTiter, принимая клетки, обрабатываемые средой, за 100%. Результаты представляют среднее для трех параллельных образцов ±S.D. Символ звездочки (*) обозначает статистически значимое различие по сравнению с WT (однофакторный дисперсионный анализ).
Фигура 5. LukAB является важным токсином разных стафилококковых штаммов. (A) Экспрессия LukB различными штаммами S. aureus, определенная методом вестерн-блоттинга с использованием анти-LukB поликлональных сывороток. (B) Интоксикация PMN-HL60 разбавлениями экзопротеинов от разных штаммов S. aureus. Жизнеспособность клеток контролируют с помощью CellTiter, принимая клетки, обрабатываемые средой, за 100% жизнеспособности. (C) Экспрессия LukB и α-токсина WT и LukAB-изогенными штаммами, определенная методом вестерн-блоттинга с использованием токсин-специфичных сывороток. (D) Интоксикация PMN-HL60 экзопротеинами WT-штаммов Newman (New.) и 4645 и LukAB-изогенных штаммов. Жизнеспособность клеток контролируют, как на панели B. Результаты представляют среднее для трех параллельных образцов + S.D. * обозначает статистически значимое различие по сравнению с Newman (C) или с WT (E) (t-критерий Стьюдента p<0,05).
Фигура 6. LukAB разрушает плазматические мембраны клеток-мишеней. (a) Полученные методом световой микроскопии изображения клеток PMN-HL60, интоксицированных фильтратом культуры S. aureus, штамма дикого типа (WT), и изогенного штамма, не содержащего LukAB (ΔLukAB). (b-c) Интоксикация клеток PMN-HL60 фильтратами культур штамма WT (WT/p), изогенного штамма, не содержащего LukAB (ΔLukAB/p), комплементированного штамма (ΔLukAB/pLukAB), или псевдо-интоксикация средой. Клетки с нарушенными мембранами окрашивают SYTOX Green, визуализируют методом флуоресцентной микроскопии (c) и измеряют интенсивность зеленой флуоресценции (b). (d) PMN инфицировали ex vivo S. aureus, штамм Newman (MSSA) или штамм USA300 LAC (MRSA), и указанными изогенными мутантами с различными кратностями инфекции (MOI). Повреждение мембраны контролируют с помощью красителя SYTOX green. Результаты представляют среднее для трех параллельных образцов ±S.D. Символы звездочки (*) обозначают статистически значимое различие по сравнению с WT (t-критерий Стьюдента p<0,05).
Фигура 7. LukAB защищает S. aureus от медиируемой хозяином гибели путем нацеливания на фагоциты и их уничтожения. (a) Инфекция клеток PMN-HL60 S. aureus WT, штаммом, не содержащим IukAB, и штаммом, не содержащим IukAB, с IukAB-комплементирующей плазмидой (ΔlukAB/plukAB), при различных кратностях инфекции (MOI). Клетки млекопитающих с нарушенными мембранами контролируют с помощью SYTOX Green, как описано на Фигуре 6. Результаты представляют среднее для трех параллельных образцов + S.D. (b) Жизнеспособность указанных штаммов S. aureus при ex vivo инфицировании человеческой цельной крови. Результаты представляют среднее для цельной крови, полученной от 12 доноров + S.E.M. (c) Жизнеспособность указанных S. aureus штаммов Newman при инфицировании первичных человеческих нейтрофилов (PMN). Результаты представляют среднее для PMN, полученных от 12 доноров + S.E.M. (d) Интоксикация первичных человеческих PMN различными разбавлениями фильтрата культуры штаммов WT/p, ΔlukAB/p и ΔlukAB/plukAB. В качестве индикатора клеточного лизиса измеряют высвобождение LDH (лактатдегидрогеназы). Результаты представляют среднее для PMN, полученных от 6 доноров + S.E.M. Символы звездочки (*) обозначают статистически значимое различие по сравнению с WT штаммом Newman (t-критерий Стьюдента p<0,05).
Фигура 8. LukAB важен для патогенеза S. aureus in vivo, (a) Биолюминесцентные изображения почек мышей, инфицированных WT штаммом LAC S. aureus, содержащим pXen1 или pLukAB.Xen1. Показаны почки двух типичных мышей в каждой группе. (b) Бактериальная нагрузка, выделенная из почек мышей, инфицированных ретроорбитально указанными штаммами S. aureus LAC. Каждая точка данных представляет число бактерий (CFU, колониеобразующих единиц) на миллилитр гомогената ткани для отдельного животного. Пунктирная линия показывает предел детектирования. Для панелей (A-C и E) * указывает статистическую значимость при сравнении с WT, ** указывает статистическую значимость при сравнении с ΔLukAB/p, P<0,05.
Фигура 9. LukAB уничтожает человеческие фагоциты путем образования пор в клеточных мембранах. (a) Клетки PMN-HL60 интоксицируют rLukA+rLukB и связывание токсина контролируют методами ДСН-ПААГ (электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия) и иммуноблоттинга с использованием антител, специфичных к LukA или LukB. (b) PMN-HL60 инкубируют с rLukAB и связывание токсина определяют методом FACS (сортировки клеток с активируемой флуоресценцией) с использованием кроличьего анти-His антитела, (c) Клетки PMN-HL60 интоксицируют rLukAB и образование олигомеров LukAB в плазматической мембране определяют методом ДСН-ПААГ и иммуноблоттинга с использованием антитела к LukB, (d) PMN-HL60 интоксицируют rLukAB или обрабатывают сапонином в присутствии или в отсутствие ПЭГ-400. Поры LukAB детектируют бромидом этидия. (e) Жизнеспособность PMN-HL60, обработанных как указано на панели, определяют с помощью CellTiter, принимая клетки, обрабатываемые средой, за 100%. Результаты на панелях (d) и (e) представляют среднее для трех параллельных образцов ±SEM. * обозначает статистически значимое различие с ПЭГ (панели d-e) (t-критерий Стьюдента p<0,05).
Фигура 10. Цитотоксичность LukAB может быть нейтрализована с помощью α-LukA поликлонального антитела. PMN-HL60, интоксицированные 5% (об./об.) фильтратом культуры S. aureus штамм Newman, инкубируют с указанными количествами α-LukA поликлональных антител или неиммунной сывороткой, полученной из разных порций крови (production bleeds) двух разных кроликов. Жизнеспособность клеток контролируют с помощью CellTiter, принимая клетки, обрабатываемые средой, за 100%. Результаты представляют среднее для трех параллельных образцов ± стандартное отклонение (S.D.).
Фигура 11. C-концевой выступающий участок LukA необходим для цитотоксичного эффекта LukAB, но не влияет на распознавание α-LukA поликлональным антителом. (a) Выравнивание аминокислотных последовательностей различных S-субъединиц лейкотоксина S. aureus (обозначенных как SEQ ID NO:44-49), проведенное с использованием метода MegAlign Clustal W прикладной программы Lasergene. N- и C-концевые выступающие участки, присутствующие только в последовательности LukA, обведены рамкой. (b) Окрашивание кумасси голубым 2 мкг рекомбинантного LukA (rLukA), LukB (rLukB), LukA, не имеющего C-концевого выступающего участка (rLukAΔ10C) и LukA, не имеющего N-концевого выступающего участка (rΔ33NLukA), выделенного из E. coli и разделенного методом ДСН-ПААГ, сопровождаемое интоксикацией PMN-HL60 различными количествами rLukA, rLukAΔ10C и rΔ33NLukA в комбинации с rLukB. Конечная концентрация белка состоит из равных количеств rLukA, rLukAΔ10C или rΔ33NLukA и rLukB. Результаты представляют среднее для трех параллельных образцов ±S.D. (c) Иммуноблот, демонстрирующий эквивалентность распознавания 6×His-меченого rLukAΔ10C как α-LukA, так и α-His поликлональными антителами.
ДЕТАЛЬНОЕ ОПИСАНИЕ
Приведенное далее раскрытие касается, в последовательном порядке, полипептидов LukA, полипептидов LukB, полинуклеотидов LukA и LukB, антител к LukA и к LukB, терапевтических композиций, содержащих LukA и/или LukB, или антитела к LukA и/или к LukB, способов применения терапевтических композиций, способов идентификации ингибиторов LukAB-опосредованной цитотоксичности и способов прогнозирования или оценки тяжести инфекции S. aureus.
Полипептиды LukA
Полипептиды, нативные для Staphylococcus aureus, были изолированы и идентифицированы заявителями как проявляющие профиль активности известной S-субъединицы лейкоцидинов (например, LukS-PVL, LukE и HlgC). Такие полипептиды, обозначенные здесь коллективно как LukA, специфически нацелены на и связывают человеческие фагоциты (но не человеческие эпителиальные или человеческие эндотелиальные клетки, или мышиные клетки), и после связывания с мембраной фагоцита, LukA олигомеризуется с F-субъединицей лейкоцидина S. aureus (например, LukF-PVL, LukD и HlgB и LukB, раскрытые здесь), а после олигомеризации образуют трансмембранную пору (что обобщенно называется LukA-активностью). Выравнивание, приведенное на Фиг. 1, содержит аминокислотные последовательности мажоритарной (majority) последовательности LukA (обозначенной здесь SEQ ID NO:1) и полипептидов LukA от 13 разных штаммов S. aureus, которым они соответствуют (обозначены здесь SEQ ID NO:2-14).
N-концевые 27 аминокислотных остатков каждой из SEQ ID NO:1-14 представляют нативную секреторную/сигнальную последовательность. Таким образом, зрелая секретируемая форма LukA, представленная аминокислотными остатками 28-351 в каждой из SEQ ID NO:1-14, может быть обозначена здесь "LukA(28-351)" или "зрелый LukA". Соответственно, незрелая форма LukA может быть обозначена здесь "LukA(1-351)".
Консенсусная последовательность LukA, определенная на основании SEQ ID NO:2-14 (которые не являются исчерпывающими по отношению к нативному LukA S. aureus) будет, таким образом, включать различающиеся варианты минимум в 64 положениях LukA (где последовательно расположенные положения изменчивости обозначены X1-X64), определяемые следующим образом: 8 (X1=L или F), 16 (X2=A или V), 17 (X3=I или L), 24 (X4=T или Ν), 26 (X5=Q или E), 31 (X6=H или N), 38 (X7=N или T), 46 (X8=S или A), 50 (X9=E или D), 55 (X10=T или N), 56 (X11=N или D), 61 (X12=S или T), 62 (X13=T или P), 63 (X14=A, G или V), 73 (X15=I или V), 78 (X16=E или V), 77 (X17=T или S), 80 (X18=V или E), 83 (X19=E или K), 84 (X20=E или K), 105 (X21=V или I), 124 (X22=K или R), 125 (X23=E или N), 127 (X24=K, T или N), 129 (X25=S или A), 130 (X26=N или S), 135 (X27=K или Q), 146 (X28=R или S), 148 (X29=R или P), 173 (X30=S или N), 174 (X31=S или L), 181 (X32=T или V), 184 (X33=I или V), 195 (X34=T или S), 202 (X35=N или K), 208 (X36=S или I), 214 (X37=W или R), 221 (X38=I или V), 229 (X39=G или N), 231 (X40=V или I), 237 (X41=E или D), 239 (X42=L или F), 243 (X43=N или T), 246 (X44=I или L), 247 (X45=A или S), 278 (X46=L или I), 283 (X47=S или T), 285 (X48=E или D), 288 (X49=Q или R), 299 (X50=I или V), 303 (X51=R или K), 309 (X52=A или G), 310 (X53=P или Q), 315 (X54=K или Q), 318 (X55=D или E), 322 (X56=L или F), 325 (X57=T или V), 338 (X58=V или I), 339 (X59=D или E), 342 (X60=S или T), 344 (X61=D, E или Q), 347 (X62=P или S), 348 (X63=Y или F) и 349 (X64=K или R).
Полипептиды LukB
Полипептиды, нативные для Staphylococcus aureus, были идентифицированы заявителями как проявляющие профиль активности известной F-субъединицы лейкоцидинов (например, LukF-PVL, LukD и HlgB). Такие полипептиды, обозначенные здесь коллективно как LukB, специфически олигомеризуются с S-субъединицей лейкоцидина S. aureus (например, LukS-PVL, LukE и HlgC и LukA, раскрытыми здесь), связанной с человеческим фагоцитом; и после олигомеризации образуют трансмембранную пору в фагоците (что обобщенно называется LukB-активностью). Выравнивание, приведенное на Фиг. 2, содержит аминокислотные последовательности мажоритарной последовательности LukB (обозначенной здесь SEQ ID NO:15) и полипептидов LukB от 12 разных штаммов S. aureus, которым они соответствуют (обозначены здесь SEQ ID NO:16-27).
N-концевые 29 аминокислотных остатков каждой из SEQ ID NO:15-27 представляют собой секреторную/сигнальную последовательность. Таким образом, зрелая секретируемая форма LukB, представленная аминокислотными остатками 30-339 в каждой из SEQ ID NO:16-27, может быть обозначена здесь "LukB(30-339)" или "зрелая LukB". Соответственно, незрелая форма LukB может быть обозначена здесь "LukA(1-339)".
Консенсусная последовательность LukB, определенная на основании SEQ ID NO:15-28 (которые не являются исчерпывающими по отношению к нативному LukB S. aureus) будет, таким образом, включать различающиеся варианты минимум в 49 положениях LukB (где последовательно расположенные положения изменчивости обозначены X1-X49), определяемые следующим образом: 5 (X1=L или V), 6 (X2=C или Y), 13 (X3=S или T), 15 (X4=A или T), 16 (X5=L или I), 19 (X6=A или T), 20 (X7=L или F), 23 (X8=F или L), 26 (X9=S или T), 28 (X10=Y или F), 34 (X11=E или K), 36 (X12=K или T), 37 (X13=Q, T или A), 46 (X14=D или E), 59 (X15=S или T), 60 (X16=Q или E), 62 (X17=N или K), 64 (X18=T или S), 75 (X19=P или K), 95 (X20=K или R), 98 (X21=N, d или E), 126 (X22=S или делеция), 159 (X23=R или Q), 163 (X24=T или P), 170 (X25=S или K), 187 (X26=L или I), 190 (X27=S или P), 192 (X28=S или T), 193 (X29=S или T), 193 (X29=H или N), 197 (X30=G или A), 204 (X31=S или L), 222 (X32=D или N), 224 (X33=T или V), 247 (X34=N или D), 270 (X35=N или K), 272 (X36=K или E), 276 (X37=R, Q или K), 287 (X38=D или E), 290 (X39=L или I), 294 (X40=K или R), 309 (X41=Q или K0, 327 (X42=D или N), 329 (X43=L или F), 330 (X44=I или V), 332 (X45=t или V), 333 (X46=f, I или L), 336 (X47=K или N) и 338 (X48=K или Q).
Лейкоцидины LukA и LukB могут отличаться от нативных полипептидов, обозначенных как SEQ ID NO:2-14 и 16-27, соответственно, вследствие одной или нескольких дополнительных инсерций, замещений или делеций аминокислот, например, один или несколько аминокислотных остатков в SEQ ID NO:2-14 или 16-27 могут быть замещены на другую аминокислоту схожей полярности, которая выступает в роли функционального эквивалента, приводя к молчащему изменению. Это означает, что изменение по сравнению с нативной последовательностью существенно не ослабит основные свойства нативных LukA и LukB. Примеры включают SEQ ID NO:1 и 15. Любой такой аналог LukA или LukB может быть подвергнут скринингу в соответствии с протоколами, раскрытыми здесь (например, анализ клеточной токсичности и анализ повреждения мембраны) для определения того, сохраняет ли он активность нативного LukA или LukB. Замещения в таких лейкоцидинах могут быть выбраны из других членов класса, к которому принадлежит данная аминокислота. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин. Полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин. Положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин. Отрицательно заряженные (кислотные) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту.
В других вариантах исполнения, неконсервативные изменения (например, одно или аминокислотных замещений, делеций и/или аддиций) могут быть проведены с целью инактивации или детоксификации LukA и LukB. В одном варианте исполнения, нетоксичный аналог LukA отличается от нативных полипептидов тем, что C-концевые аминокислоты в положениях 342-351 подвергаются делеции. За исключением SEQ ID NO:4-6 (которые содержат 9 аминокислот в этих положениях), в аналоге отсутствуют 10 C-концевых аминокислотных остатков. Коллективно, такие аналоги обозначаются LukAΔ10C. Детоксифицированные LukA и LukB могут быть использованы в композициях активных вакцин, описанных здесь. Молекулярные изменения могут быть проведены способами, хорошо известными специалистам в данной области техники, включая метод удлинения праймера на плазмидной матрице с использованием одноцепочечных матриц (Kunkel, Proc. Acad. Sci., USA 82:488-492 (1985)), двухцепочечных ДНК-матриц (Papworth, et al., Strategies 9(3):3-4 (1996)), и ПЦР-клонирование (Braman, J. (ed.), IN VITRO MUTAGENESIS PROTOCOLS, 2nd ed. Humana Press, Totowa, N.J. (2002). Способы определения того, будет ли данное молекулярное изменение LukA или LukB уменьшать цитотоксичность LukAB, описаны здесь.
Таким образом, учитывая приведенное выше описание и в целях настоящего изобретения, LukA может быть более широко описан как любая из последовательностей SEQ ID NO:1-14 (например, SEQ ID NO:2, которая представляет собой полипептид LukA, нативный для штамма Newman S. aureus), или (нативный или ненативный) полипептид, имеющий, по меньшей мере, примерно 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% подобия с этими последовательностями.
Аналогично, с учетом приведенного выше описания и в целях настоящего изобретения, LukB может быть более широко описан как любая из последовательностей SEQ ID NO:15-27 (например, SEQ ID NO:27, которая представляет собой полипептид LukB, нативный для штамма Newman S. aureus), или (нативный или ненативный) полипептид, имеющий, по меньшей мере, примерно 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% подобия с этими последовательностями.
Таким образом, если не будет указано иное, как незрелая, так и зрелая формы нативных LukA и LukB, и последовательности, имеющие менее 100% подобия с нативной LukA (т.е. нативные последовательности и аналоги в равной степени, коллективно обозначаемые здесь "LukA" и "LukB"), могут быть использованы в композициях и способах, и для приготовления антител к LukA и к LukB по настоящему изобретению.
Полинуклеотиды, кодирующие LukA и LukB, и способы синтеза или изоляции LukA и LukB
Лейкоцидины LukA и LukB могут быть синтезированы методами рекомбинантных ДНК, которые хорошо известны специалистам в данной области техники. Например, нуклеотидная последовательность (обозначенная SEQ ID NO:28), кодирующая полипептид LukA S. aureus (Newman) (SEQ ID NO:2), приведена ниже. Вырожденные последовательности (например, такие, которые могут быть полезны с учетом предпочтительно используемых кодонов у хозяев, выбранных для рекомбинантной экспрессии), кодирующие этот полипептид, и полинуклеотиды, кодирующие другие полипептиды LukA, известны в данной области техники или могут быть сконструированы квалифицированными специалистами в данной области техники.





Нуклеотидная последовательность (обозначенная здесь SEQ ID NO:29), кодирующая полипептид LukB S. aureus (Newman), (SEQ ID NO:27), приведена ниже. Вырожденные последовательности (например, такие, которые могут быть полезны с учетом предпочтительно используемых кодонов у хозяев, выбранных для рекомбинантной экспрессии), кодирующие этот полипептид, и полинуклеотиды, кодирующие другие полипептиды LukB, известны в данной области техники или могут быть сконструированы квалифицированными специалистами в данной области техники.




Полинуклеотиды, кодирующие LukA и LukB, могут быть экспрессированы в хозяине, таком как бактерии (E. coli), растения и/или дрожжи, и затем выделены и очищены. Альтернативно, лейкоцидины LukA и LukB могут быть выделены из бактерий S. aureus (например, штамм Newman) в соответствии со стандартными методиками. Таким образом, такие лейкоцидины могут быть выделены (из нативной или ненативной среды). Они также могут быть очищены так, чтобы они по существу не содержали других белков и клеточных компонентов, с которыми LukA и LukB S. aureus ассоциированы в своем нативном состоянии (т.е. белков и клеточных компонентов, присутствующих в клетках S. aureus) или ненативном состоянии (т.е. белков и клеточных компонентов рекомбинантной клетки-хозяина). Пригодные схемы очистки, которые типично предусматривают комбинацию, по меньшей мере, двух последовательных процедур, известны специалистам в данной области техники. См. Deutscher, Methods in Enzymology, 182 (1990); и Scopes, Protein Purification: Principles and Practice, Springer-Verlag, N.Y. (1982), использующие одну или комбинацию двух или больше стандартных методик, таких как аффинная хроматография на колонке и катионообменная жидкостная хроматография.
Антитела к LukA и антитела к LukB
Аспекты настоящего изобретения касаются антител к LukA, которые специфически связываются с LukA, и антител к LukB, которые специфически связываются с LukB, терапевтических композиций, содержащих антитела, и способов их применения. В целях настоящего изобретения, термин "антитело" включает моноклональные антитела, поликлональные антитела, фрагменты антител и генетически модифицированные формы антител и их комбинации. Более конкретно, термин "антитело", который используется взаимозаменяемо с термином "иммуноглобулин", включает полноразмерные (т.е. встречающиеся в природе или полученные с помощью процессов рекомбинации фрагментов нормального гена иммуноглобулина) иммуноглобулиновые молекулы (например, IgG-антитело) и их иммунологически активные фрагменты (т.е. включая специфический связывающий участок полноразмерной молекулы иммуноглобулина), которые также могут быть природными или синтетическими по характеру. Соответственно, термин "фрагмент антитела" включает участок антитела, такой как F(ab')2, F(ab)2, Fab', Fab, Fv, scFv и т.п. Независимо от структуры, фрагмент антитела связывается с тем же самым антигеном, который распознается полноразмерным антителом и, в контексте настоящего изобретения, специфически связывается с LukA, LukB или комплексом LukAB. Способы получения и скрининга фрагментов антител хорошо известны специалистам в данной области техники.
В некоторых вариантах исполнения, антитела к LukA по настоящему изобретению могут обладать некоторой степенью перекрестной реактивности с другими S-субъединицами лейкоцидина Staphylococcus, такими как HlgC, LukS-PVL, HlgA, LukS-I, LukE, LukEv и LukM. Аналогично, в некоторых вариантах исполнения, антитела к LukB по настоящему изобретению могут обладать некоторой степенью перекрестной реактивности с другими F-субъединицами лейкоцидина Staphylococcus, такими как LukF'-PV, LukF-PV, LukDv, LukD, LukF-I и HlgB. Антитела к LukA и/или к LukB могут ингибировать или понижать активность LukA и активность LukB, соответственно. В некоторых вариантах исполнения, антитела к LukA и/или к LukB нейтрализуют (например, по существу устраняют) активность LukA и LukB, соответственно.
Природные антитела типично имеют две идентичные тяжелые цепи и две идентичные легкие цепи, причем каждая легкая цепь ковалентно связана с тяжелой цепью межцепной дисульфидной связью, и множество дисульфидных связей дополнительно связывают две тяжелые цепи друг с другом. Индивидуальные цепи могут укладываться в домены, имеющие близкие размеры (110-125 аминокислот) и структуры, но разные функции. Легкая цепь может включать один вариабельный домен (VL) и/или один константный домен (CL). Тяжелая цепь также может содержать один вариабельный домен (VH) и/или, в зависимости от класса или изотипа антитела, три или четыре константных домена (CHI, CH2, CH3 и CH4). У людей встречаются изотипы IgA, IgD, IgE, IgG и IgM, причем IgA и IgG дополнительно делятся на подклассы или субтипы (IgA1-2 и IgG1-4).
В общем, вариабельные домены демонстрируют значительную изменчивость аминокислотных последовательностей разных антител, особенно на участке антигенсвязывающего сайта. В каждом VL и VH присутствуют три области, называемые гипервариабельными участками или участками, определяющими комплементарность (CDR), которые поддерживаются менее вариабельными участками, называемыми каркасными вариабельными областями. Антитела по изобретению включают IgG моноклональные антитела, но антитела по настоящему изобретению также включают фрагменты антител или генно-модифицированные формы. Они представляют собой, например, фрагменты или белки Fv, в которых CDR и/или вариабельные домены приведенных в качестве примеров антител сконструированы в виде одноцепочечных антигенсвязывающих белков генно-инженерными методами.
Часть антитела, состоящая из доменов VL и VH, обозначается Fv (вариабельный фрагмент) и образует антигенсвязывающий сайт. Одноцепочечный Fv (scFv или SCA) представляет собой фрагмент антитела, содержащий VL-домен и VH-домен в одной полипептидной цепи, причем N-конец одного домена и C-конец другого домена соединены гибким линкером. Пептидные линкеры, используемые для получения одноцепочечного антитела, типично являются гибкими пептидами, выбранными таким образом, чтобы обеспечить надлежащую трехмерную укладку доменов VL и VH. Линкер обычно содержит от 10 до 50 аминокислотных остатков и, в некоторых случаях, является более коротким, например, примерно от 10 до 30 аминокислотных остатков, или от 12 до 30 аминокислотных остатков, или даже от 15 до 25 аминокислотных остатков. Пример таких линкерных пептидов включает повторы четырех глициновых остатков с последующим сериновым остатком.
Одноцепочечные антитела не содержат некоторой части или всех константных доменов цельных антител, из которых их получают. Благодаря этому, они позволяют решить некоторые проблемы, ассоциированные с использованием цельных антител. Например, одноцепочечные антитела имеют тенденцию к отсутствию некоторых нежелательных взаимодействий между константными участками тяжелой цепи и другими биологическими молекулами. Дополнительно, одноцепочечные антитела значительно меньше цельных антител и могут обладать большей проницаемостью, чем цельные антитела, что позволяет одноцепочечным антителам более эффективно локализоваться и связывать антигенсвязывающие сайты-мишени. Кроме того, вследствие относительно малого размера одноцепочечные антитела будут с меньшей вероятностью вызывать нежелательный иммунный ответ у реципиента, чем цельные антитела.
Fab (фрагмент, антигенсвязывающий) относится к фрагментам антител, состоящим из доменов VL, CL, VH и CH1. Фрагменты, образующиеся после гидролиза папаином, называются просто Fab и не сохраняют шарнирный участок тяжелой цепи. После гидролиза пепсином образуются различные Fab, сохраняющие шарнирный участок тяжелой цепи. Такие фрагменты с интактными межцепными дисульфидными связями называются F(ab')2, а в тех случаях, когда дисульфидные связи не сохраняются, образуется одиночный Fab'. Фрагменты F(ab')2 обладают более высокой авидностью к антигену, чем моновалентные фрагменты Fab.
Fc (фрагмент, кристаллизующийся) используется для обозначения части или фрагмента антитела, включающего спаренные константные домены тяжелой цепи. В IgG-антителе, например, Fc содержит домены CH2 и CH3. Fc IgA- или IgM-антитела дополнительно включает домен CH4. Fc ассоциирован со связыванием Fc-рецептора, активацией комплемент-опосредованной цитотоксичности и антитело-зависимой клеточной цитотоксичностью (ADCC). Для таких антител, как IgA и IgM, которые представляют собой комплексы множества IgG-подобных белков, для образования комплекса требуются константные домены Fc.
Наконец, шарнирный участок разделяет Fab- и Fc-участки антитела, обеспечивая подвижность Fab по отношению друг к другу и по отношению к Fc, а также включает множество дисульфидных связей для ковалентного связывания двух тяжелых цепей.
"Специфичность" антитела относится к селективному распознаванию антителом определенного эпитопа антигена. Термин "эпитоп" включает любую белковую детерминанту, способную специфически связываться с иммуноглобулином или T-клеточным рецептором, или иным образом взаимодействовать с молекулой. Эпитопные детерминанты обычно состоят из химически активных поверхностных группировок молекул, таких как аминокислоты или углеводы, или боковые цепи сахаров, и обычно обладают специфическими трехмерными структурными характеристиками, а также специфическими характеристиками заряда. Эпитоп может быть "линейным" или "конформационным". В линейном эпитопе все точки взаимодействия между белком и взаимодействующей молекулой (такой как антитело) расположены линейно вдоль первичной аминокислотной последовательности белка. В конформационном эпитопе точки взаимодействия распределены по аминокислотным остаткам белка, расположенным на некотором расстоянии друг от друга, т.е. несмежным аминокислотам, сближающимся в результате укладки третичной структуры белка. Эпитопы, образованные смежными аминокислотами, типично сохраняются после воздействия денатурирующих растворителей, в то время как эпитопы, образующиеся при третичной укладке, типично исчезают после обработки денатурирующими растворителями. Эпитоп типично включает, по меньшей мере, 3 и, как правило, по меньшей мере, 5 или 8-10 аминокислот с уникальной пространственной конформацией. Антитела, распознающие один и тот же эпитоп, могут быть определены с помощью одного иммуноанализа, демонстрирующего способность одного антитела блокировать связывание другого антитела с антигеном-мишенью.
Моноклональные антитела по настоящему изобретению могут быть мышиными, человеческими, гуманизированными или химерными. Гуманизированное антитело представляет собой рекомбинантный белок, в котором CDR антитела одного вида; например, антитела грызуна, кролика, собаки, козы, лошади или курицы (или любого другого пригодного антитела животного), переносят из тяжелых и легких вариабельных цепей антитела грызуна в тяжелые и легкие вариабельные домены человека. Константные домены молекулы антитела берут из человеческого антитела. Способы получения гуманизированных антител хорошо известны специалистам в данной области техники. Химерные антитела предпочтительно имеют константные участки, полученные по существу или исключительно из константных участков человеческого антитела, и вариабельные участки, полученные по существу или исключительно из последовательности вариабельного участка млекопитающего, отличного от человека. Эффективность процесса химеризации можно также увеличить путем замены вариабельных участков - кроме гипервариабельных участков или участков, определяющих комплементарность (CDR), мышиного (или другого не являющегося человеком млекопитающего) антитела на соответствующие человеческие последовательности. Вариабельные участки, отличные от CDR, также известны как вариабельные каркасные участки (FR). Еще одни моноклональные антитела по настоящему изобретению являются биспецифичными, т.е. они обладают специфичностью по отношению к обоим LukA и LukB. Биспецифичные антитела предпочтительно являются человеческими или гуманизированными.
Вышеописанные антитела могут быть получены в соответствии со стандартными методиками. Например, LukA, LukB (в значении, в котором эти термины используются здесь, включают их нетоксичные аналоги, такие как LukAΔ10C) или иммунологически активный фрагмент LukA или LukB могут быть введены субъекту (например, млекопитающему, такому как человек или мышь). Лейкоцидины могут быть использованы сами по себе в качестве иммуногенов или они могут быть присоединены к белку-носителю или другим объектам, таким как бусины, например, сефарозные бусины. После продуцирования млекопитающим антител выделяют смесь антитело-продуцирующих клеток, таких как спленоциты, из которых могут быть получены поликлональные антитела. Моноклональные антитела могут продуцироваться путем изоляции индивидуальных антитело-продуцирующих клеток из смеси и их иммортализации, например, путем слияния с опухолевыми клетками, такими как клетки миеломы. Полученные гибридомы сохраняют в культуре и моноклональные антитела собирают из культуральной среды.
Методики приготовления рекомбинантных моноклональных антител хорошо известны специалистам в данной области техники. Рекомбинантные поликлональные антитела могут быть получены способами, аналогичными описанным в публикации патентной заявки США 2002/0009453, с использованием LukA, LukB или LukAB в качестве иммуногена (иммуногенов).
Терапевтические композиции
LukA и LukB могут быть включены в терапевтическую композицию, предназначенную для использования в качестве противовоспалительного агента при лечении острых воспалительных состояний, включая локализованные острые воспалительные состояния. LukA и LukB также могут быть включены в терапевтическую композицию для использования в качестве активной вакцины. Антитела к LukA и к LukB могут быть включены в терапевтическую композицию для использования в качестве пассивной вакцины. Пассивные и активные вакцины могут быть использованы профилактически для ингибирования начальных проявлений инфекции S. aureus, или терапевтически для лечения инфекций S. aureus, в частности, таких инфекций S. aureus, как MRSA, известная как резистентная, или в случае, когда определенный субъект окажется невосприимчивым к лечению другими обычными методами терапии антибиотиками.
В вариантах исполнения, в которых терапевтическая композиция предназначена для использования в качестве активной вакцины, LukA и/или LukB могут быть изменены таким образом, чтобы они проявляли пониженную токсичность. Молекулярные изменения описаны выше. Таким образом, могут быть использованы их нетоксичные аналоги, такие как LukAΔ10C. Заявители считают, что антитела, продуцируемые в ответ на нетоксичный иммуноген, будут нейтрализовать токсичные нативные LukA или LukAB. Другие изменения в целях снижения токсичности LukA и LukB включают химическую обработку (например, модифицирование определенных аминокислотных остатков) или конъюгирование с другим фрагментом (например, с другим бактериальным антигеном, таким как бактериальный полисахарид или бактериальный гликопротеин). Известны химические изменения других токсинов S. aureus в целях инактивации или детоксификации (или снижения токсичности). Способы определения того, будет ли данное изменение снижать токсичность LukA или LukB, известны специалистам в данной области техники и/или описаны здесь.
Терапевтические композиции по настоящему изобретению готовят путем составления композиций LukA и LukB, или антител к LukA и к LukB, с фармацевтически приемлемым носителем и, необязательно, фармацевтически приемлемым эксципиентом. В используемом здесь значении термины "фармацевтически приемлемый носитель" и "фармацевтически приемлемый эксципиент" (например, добавки, такие как разбавители, иммуностимуляторы, адъюванты, антиоксиданты, консерванты и солюбилизирующие агенты) являются нетоксичными для клетки или млекопитающего, которые подвергаются их воздействию в используемых дозировках и концентрациях. Примеры фармацевтически приемлемых носителей включают воду, например, забуференную фосфатом, цитратом и другой органической кислотой. Типичные примеры фармацевтически приемлемых эксципиентов, которые могут быть пригодны для использования по настоящему изобретению, включают антиоксиданты, такие как аскорбиновая кислота; низкомолекулярные (менее примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; адъюванты (выбранные так, чтобы избежать адъювант-индуцируемой токсичности, такие как β-глюкан, как описано в патенте США 6355625, или гранулоцитарный колониестимулирующий фактор (GCSF)); гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахароспирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEEN®, полиэтиленгликоль (ПЭГ) и PLURONICS®.
Как указано в другом месте данного описания, терапевтические композиции по настоящему изобретению могут дополнительно содержать, по меньшей мере, один дополнительный активный агент.
Терапевтические композиции по настоящему изобретению могут быть подготовлены для хранения путем смешения активного ингредиента (ингредиентов), имеющих желательную степень чистоты, с фармацевтически приемлемым носителем и, необязательно, эксципиентом и/или дополнительным активным агентом, в форме лиофилизированных композиций или водных растворов.
Применение терапевтических композиций -- Показания
Острые воспалительные состояния
Под воспалением обычно понимают защитную биологическую реакцию, направленную на удаление вредных инвазивных стимулов, таких как патогены (например, бактерии и вирусы), поврежденные клетки и раздражители, и на инициирование исцеления. Воспаление более конкретно понимают как реакцию васкуляризованной живой ткани на повреждение. По существу, воспаление представляет собой фундаментальный стереотипный комплекс цитологических и химических реакций пораженных кровеносных сосудов и прилегающих тканей в ответ на повреждение или аномальное раздражение, вызванное физическим, химическим или биологическим агентом. Воспаление обычно приводит к скоплению жидкости и кровяных клеток на месте повреждения и обычно является процессом заживления. Без воспалительного процесса раны и инфекции не излечивались бы и прогрессирующее разрушение ткани становилось бы угрожающим для жизни. Острое воспаление относится к начальной реакции организма на инвазивные стимулы и включает рекрутмент плазмы и белых кровяных клеток (лейкоцитов) к поврежденным или инфицированным тканям. Пролонгированное воспаление, также называемое хроническим воспалением, связано с постепенным изменением типа иммунных клеток, присутствующих на месте воспаления, и характеризуется одновременным разрушением и исцелением ткани от воспалительного процесса.
Однако воспаление иногда причиняет вред, обычно в результате нарушения нормального развития воспаления. Воспалительными являются болезни, относящиеся к, характеризующиеся, вызывающие, вызванные или подверженные влиянию воспаления. "Острые воспалительные состояния" как термин, используемый здесь, в соответствии с нормальной медицинской лексикой, относится к воспалительным состояниям с быстрым началом и тяжелыми симптомами. Длительность начального периода, от нормального состояния пациента до серьезных проявлений симптомов воспаления, обычно не превышает примерно 72 часов. Острые воспалительные состояния следует отличать от хронических воспалительных состояний, которые представляют собой воспалительные состояния большой продолжительности, характеризующиеся незначительными изменениями или медленным развитием болезни. Различие между острыми и хроническими состояниями хорошо известны специалистам в области медицины.
Основные иммунные клетки, принимающие участие в острой стадии воспаления, а также в острых воспалительных расстройствах, включают мононуклеарные клетки (например, моноциты, которые в ответ на воспаление дифференцируются в макрофаги), дендритные клетки и нейтрофилы (которые мигрируют к месту воспаления). Эти иммунные клетки способствуют проявлению воспалительного ответа путем высвобождения воспалительных медиаторов, таких как гистамин, интерферон-гамма, интерлейкин-8, лейкотриен B4, окись азота и т.д., и путем поглощения бактерий, вирусов и клеточных обломков (процесс, известный как фагоцитоз). Такие клетки известны специалистам в данной области техники коллективно как фагоциты.
Заявители обнаружили, что LukAB нацелен на и уничтожает человеческие фагоциты, и что такая LukAB-опосредованная цитотоксичность является по существу специфической по отношению к этим клеткам, но не к другим ядросодержащим клеткам млекопитающих. Без намерения ограничиваться какой-либо конкретной теорией действия, заявители считают, что комплекс LukA/LukB образует поры в плазматической мембране инфильтрующих фагоцитов, вызывая гибель клеток и тем самым уменьшая воспаление. Таким образом, противовоспалительные композиции по настоящему изобретению могут быть пригодны для лечения острых воспалительных состояний у млекопитающих, таких как человек, независимо от их причины, например, любой бактериальной или вирусной инфекции и, в предпочтительных вариантах исполнения, локализованных острых воспалительных состояний. Другие примеры таких состояний включают аллергический контактный дерматит, острую гиперчувствительность, острое неврологическое воспалительное поражение (например, вызванное острой инфекцией), острый инфаркт миокарда, острое нейрональное поражение, вызванное хирургией с использованием искусственного кровообращения, и острые локализованные противовоспалительные состояния, вызванные бактериальной или вирусной инфекцией.
В предпочтительных вариантах исполнения, острое воспалительное состояние представляет собой инфицированную рану на коже или в мягкой ткани. Раны, поддающиеся лечению по изобретению, могут иметь форму проколов, разрезов или разрывов живых тканей. Раны кожи могут проникать через эпидермис, дерму или, в случае глубоких ран, в подкожные ткани. Таким образом, раны, поддающиеся лечению терапевтическими композициями по настоящему изобретению, включают глубокие грудинные раны, например, после хирургии на открытом сердце, и послеоперационные раны после брюшной и любых других типов хирургии. Другие раны включают раны, причиненные травмами, такими как выстрелами из огнестрельного оружия, ножами или любыми другими предметами, способными причинять разрез или разрыв кожи. Раны, возникающие как побочные эффекты медикаментозного лечения или как симптомы различных патологий (например, язвы, ассоциированные с саркомой Капоши), а также внутренние раны (например, трещины заднего прохода и раны или поражения желудочно-кишечного тракта, такие как язвы желудка или кишечника) также могут поддаваться лечению с помощью настоящего изобретения.
Еще одни острые воспалительные состояния, которые могут поддаваться лечению терапевтическими композициями по настоящему изобретению, включают конъюнктивит, воспаление радужной оболочки глаза, увеит, центральный ретинит, наружный отит, средний гнойный острый отит, мастоидит, лабиринтит, хронический ринит, острый ринит, синусит, фарингит, тонзиллит (tonsillitio), контактный дерматит, дермонекроз, диабетический полиневрит, полимиозит, оссифицирующий миозит, дегенеративный артрит, ревматоидный артрит, периартрит плечевого сустава и деформирующий остит.
Инфекции S. aureus
Настоящее изобретение также предусматривает способ ингибирования начальных проявлений или лечения инфекции S. aureus путем введения композиций антитела субъекту-млекопитающему, нуждающемуся в этом. В целях настоящего изобретения, целевая популяция субъектов включает млекопитающих, таких как людей, инфицированных, или с риском инфицирования S. aureus. В некоторых вариантах исполнения, субъект, лечение которого должно проводиться, инфицирован S. aureus, включая MRSA, и/или уже получал лечение антибиотиками или другими терапевтическими агентами, но лечение оказалось безуспешным.
Терапевтически эффективные количества
В контексте лечения острых воспалительных состояний, количества LukA и LukB являются терапевтически эффективными в том смысле, что лечение позволяет достичь любой одной или нескольких целей из уменьшения числа симптомов, снижения тяжести, по меньшей мере, одного симптома или задержки дальнейшего развития, по меньшей мере, одного симптома, или даже полного облегчения острого воспалительного состояния.
В контексте использования терапевтических композиций в качестве пассивных или активных вакцин в связи с инфекцией S. aureus, терапевтически эффективные количества LukA и LukB, или антител к LukA и к LukB, также являются профилактически эффективными в том смысле, что введение композиции позволяет достичь любой одной или нескольких целей из ингибирования или профилактики инфекции S. aureus у субъектов, подверженных риску, и в случае субъектов-млекопитающих, инфицированных S. aureus, уменьшения числа симптомов, снижения тяжести, по меньшей мере, одного симптома или задержки дальнейшего развития, по меньшей мере, одного симптома, или даже полного облегчения инфекции.
В общем, терапевтически эффективные количества LukA, LukB и антител к LukA и к LukB могут быть определены в соответствии со стандартными процедурами, которые принимают во внимание многочисленные факторы, включая, например, концентрации этих активных агентов в композиции, способ и частоту введения, тяжесть острого воспалительного состояния или инфекции S. aureus, лечение которой должно проводиться (или которая должна быть предотвращена) и характеристики субъекта, такие как возраст, вес и общее состояние здоровья и иммунной системы. Общие рекомендации можно найти, например, в публикациях Международной конференции по гармонизации (International Conference on Harmonization) и REMINGTON'S PHARMACEUTICAL SCIENCES (Mack Publishing Company 1990). Клинический врач может вводить LukA и LukB или антитела к LukA и к LukB до достижения дозировки, обеспечивающей желательный или требуемый профилактический или терапевтический эффект. Ход такого лечения можно легко контролировать с помощью обычных анализов.
Терапевтически эффективные количества LukA и LukB типично составляют 1-400 мкг каждого из LukA и LukB, на дозу или в сутки. Предпочтительно, количества LukA и LukB являются по существу одинаковыми. Терапевтически эффективные количества композиций антител типично составляют, по меньшей мере, 50 мг композиции на килограмм веса тела (мг/кг), включая по меньшей мере 100 мг/кг, по меньшей мере 150 мг/кг, по меньшей мере 200 мг/кг, по меньшей мере 250 мг/кг, по меньшей мере 500 мг/кг, по меньшей мере 750 мг/кг и по меньшей мере 1000 мг/кг, на дозу или в сутки. Дозировки композиций моноклональных антител могут быть ниже, такими как примерно одна десятая от композиций не-моноклональных антител, такими как, по меньшей мере, примерно 5 мг/кг, по меньшей мере, примерно 10 мг/кг, по меньшей мере, примерно 15 мг/кг, по меньшей мере, примерно 20 мг/кг или, по меньшей мере, примерно 25 мг/кг.
Способы введения
Перед введением терапевтические композиции по настоящему изобретению могут быть стерилизованы, что можно легко осуществить путем фильтрации через мембраны для стерильной фильтрации, перед или после лиофилизации и восстановления. Терапевтические композиции могут быть помещены в контейнер, имеющий стерильное входное отверстие, например, пакетик с раствором для внутривенного введения или флакон, имеющий пробку, которую прокалывают иглой для подкожных инъекций.
Противовоспалительная композиция может быть введена любым числом путей в соответствии с принятой медицинской практикой. Предпочтительные способы включают внутривенное, внутримышечное, подкожное и чрескожное введение, с использованием методик, известных специалистам в данной области техники. Могут быть предусмотрены другие пути введения. В случае лечения острых воспалительных состояний, которые являются локализованными, несистемное введение может быть предпочтительным, и в этом случае введение терапевтической композиции осуществляется в месте острого воспаления или вокруг него.
Комбинированная терапия
В некоторых вариантах исполнения, терапевтическую композицию вводят как часть комбинированной терапии в сочетании с другим активным агентом, в зависимости от характера острого воспалительного состояния или инфекции S. aureus, лечение которой проводится. Такие дополнительные активные агенты включают антиинфекционные агенты, антибиотические агенты и антимикробные агенты. Типичные примеры антиинфекционных агентов, которые могут быть пригодными для использования по настоящему изобретению, включают ванкомицин и лизостафин. Типичные примеры антибиотических агентов и антимикробных агентов, которые могут быть пригодными для использования по настоящему изобретению, включают пенициллиназа-резистентные пенициллины, цефалоспорины и карбапенемы, включая ванкомицин, лизостафин, пенициллин G, ампициллин, оксациллин, нафциллин, клоксациллин, диклоксациллин, цефалотин, цефазолин, цефалексин, цефадрин, цефамандол, цефокситин, имипенем, меропенем, гентамицин, тейкопланин, линкомицин и клиндамицин. Дозировки этих антибиотиков хорошо известны специалистам в данной области техники. См. например, MERCK MANUAL OF DIAGNOSIS AND THERAPY, Section 13, Ch. 157, 100th Ed. (Beers & Berkow, eds., 2004). Противовоспалительные, антиинфекционные, антибиотические и/или антимикробные агенты могут быть скомбинированы перед введением, или могут вводиться одновременно (как часть одной и той же композиции или с помощью разных композиций), или последовательно с терапевтическими композициями по настоящему изобретению.
В некоторых вариантах исполнения, композиция антитела к LukA и/или антитела к LukB является мультивалентной, поскольку также содержит антитело, которое специфически связывает другой бактериальный антиген (и которое необязательно нейтрализует другой бактериальный антиген). Антитела могут специфически связывать любые из антигенов, описанных здесь в контексте композиций вакцины. Таким образом, например, другое антитело может специфически связывать полисахариды или гликопротеины, включая S. aureus типа 5, S. aureus типа 8, S. aureus 336, компоненты лейкоцидинов, такие как PVL (включая индивидуальные субъединицы PVL, LukS-PV и LukF-PV), субъединицы гамма-гемолизина (HlgA, HlgB и HlgC), LukE или LukD S. aureus, LukM или LukF'-PV S. aureus, липотейхоевую кислоту (LTA) и белковые микробные поверхностные компоненты, распознающие адгезивные молекулы матрикса (MSCRAMM).
Схемы лечения
Терапевтические композиции по настоящему изобретению могут быть введены в виде разовой дозы или в соответствии с протоколом введения многократных доз. Например, вводят относительно небольшое число доз терапевтической композиции, такое как одна или две дозы. В вариантах исполнения, включающих обычную терапию антибиотиками, которая обычно предусматривает введение множества доз на протяжении нескольких дней или недель, антибиотики могут вводиться один, два или три или больше раз в день на протяжении периода времени, такого как, по меньшей мере, 5 дней, 10 дней или даже 14 или больше дней, в то время как композиция антитела обычно вводится всего один или два раза. Однако разные дозировки, время введения доз и относительные количества терапевтической композиции и антибиотиков могут быть выбраны и откорректированы рядовым специалистом в данной области техники.
Способы идентификации ингибиторов LukAB-опосредованной цитотоксичности и измененные формы LukA и LukB, обладающие меньшей токсичностью
Антитела к LukA и к LukB и их фрагменты, а также другие потенциальные терапевтические частицы (например, малые органические молекулы) могут быть использованы в различных способах (включая форматы анализов или скрининг) для оценки их способности ингибировать LukAB-опосредованную цитотоксичность. Как описано ниже, разработаны такие способы идентификации агентов, которые ингибируют определенный аспект каскада событий, приводящего к LukAB-опосредованной цитотоксичности и лизису человеческих фагоцитов. Также разработаны способы идентификации измененных форм LukA и LukB, обладающих пониженной токсичностью по сравнению с их нативными аналогами. События-мишени, являющиеся частью каскада, включают, например, связывание LukA с мембранами фагоцитов, связывание LukB с LukA (олигомеризация LukAB) и блокировка мембранной поры, образованной олигомером LukAB. Форматы анализов обычно требуют использования человеческих фагоцитов (или их LukAB мембранно-связывающего участка), пригодной культуральной среды и очищенного LukA или очищенных LukA и LukB.
Квалифицированному специалисту будет понятно, что приведенные далее протоколы являются только иллюстративными, и что различные рабочие параметры, такие как условия реакции, выбор детектируемой метки и аппаратура (например, инструменты для детектирования и количественного определения), могут меняться в зависимости от ситуации.
Приведенные далее способы обычно предназначены для идентификации агентов, ингибирующих LukAB-цитотоксичность, не обязательно позволяя точно определить задействованное событие каскада.
Для идентификации ингибиторов LukAB-цитотоксичности, человеческие фагоциты (например, клетки PMN-HL60) могут быть высеяны на 384-луночный черный обработанный планшет для тканевых культур с прозрачным дном (Corning) при 5×103 клеток/лунку в конечном объеме 50 мкл среды RPMI (Gibco) с добавкой 10% термоинактивированной сыворотки плода коровы (FBS). Клетки затем могут быть введены в контакт/смешаны с/введены в реакцию/обработаны тестируемым соединением/молекулой (около 5 мкл/разные концентрации) и затем интоксицированы LukA и LukB, которые в предпочтительных вариантах исполнения являются по существу очищенными (5 мкл около 0,001-2 мкМ раствора), предпочтительно добавляются вместе, в условиях культивации, позволяющих провести интоксикацию фагоцитов с помощью LukA и LukB, например, в течение 1 ч при 37°C, 5% CO2. В качестве контроля, клетки могут быть обработаны культуральной средой (100% жизнеспособные) и 0,1% об./об. Triton X100 (100% гибель).
В этих вариантах исполнения, клетки, обработанные как описано выше, могут затем инкубироваться с красителем для контроля жизнеспособности клеток, например, с помощью набора CellTiter (Promega) (который позволяет определять жизнеспособность клеток по оптической плотности путем измерения числа жизнеспособных клеток в культуре посредством количественного определения метаболической активности клеток), и инкубироваться в течение дополнительного периода времени (например, примерно 2 ч при 37°C, 5% CO2). Жизнеспособность клеток может быть затем определена, например, путем измерения колориметрической реакции при 492 нм с помощью планшет-ридера, например, Envision 2103 Multi-label Reader (Perkin-Elmer). Процент жизнеспособных клеток может быть рассчитан, например, с помощью приведенного ниже уравнения: % жизнеспособности=100×[(Аb492Образец-Ab492Triton X)/(Ab492Среда для тканевых культур). Увеличение 100% жизнеспособности позволяет предположить наличие ингибирования LukAB цитотоксичности.
Вариант этого анализа называется анализом повреждения мембраны. В таких вариантах исполнения, клетки, обработанные как описано выше (например, вплоть до и включая стадию обработки клеток тестируемым соединением/молекулой и затем интоксицирования клеток очищенным LukA, могут затем инкубироваться с не проникающим в клетку флуоресцентным красителем, таким как SYTOX green (зеленый) (0,1 мкМ; Invitrogen) (в соответствии с инструкциями производителя), и инкубироваться, например, в течение дополнительных 15 минут при комнатной температуре в темноте. Флуоресценция, как индикатор повреждения мембраны, может быть затем измерена с помощью планшет-ридера, такого как Envision 2103 Multilabel Reader (Perkin-Elmer) при длине волны возбуждения 485 нм, длине волны излучения 535 нм. Уменьшение флуоресценции позволяет предположить ингибирование LukAB-цитотоксичности.
Взятые вместе, эти анализы способствуют идентификации соединений, которые ингибируют или ослабляют цитотоксичные эффекты LukAB по отношению к клеткам человеческих фагоцитов.
Дополнительные способы могут быть использованы независимо или в сочетании со способами, описанными выше; в частности, если вышеописанные способы выявляют ингибирующую активность, то это позволит квалифицированному специалисту в данной области техники более точно определить, на какое именно событие биохимического каскада влияет или нацелен агент. Такие события включают связывание LukA с мембранами фагоцитов, связывание LukB с LukA (олигомеризация LukAB) и блокировку мембранных пор, образованных олигомером LukAB.
Скрининг связывания ингибиторов LukA с клетками-мишенями
Для скрининга ингибиторов, блокирующих или ослабляющих связывание LukA с клетками-мишенями, которое считается первой стадией процесса интоксикации, человеческие фагоциты (например, PMN-HL60 клетки) могут быть высеяны в 384-луночные плоскодонные обработанные планшеты для тканевых культур (Corning) в количестве 2,5×103 клеток/лунку в конечном объеме 50 мкл среды RPMI (Gibco) с добавкой 10% термоинактивированной сыворотки плода коровы (FBS). Клетки могут быть затем обработаны тестируемым соединением/молекулой (около 5 мкл/разные концентрации) и интоксицированы очищенным флуоресцентно меченым LukA (например, FITC, Cy3, Cy5, APC, PE) 5 мкл около 0,01-2 мкМ раствора в течение 1 ч при 37°C, 5% CO2. Для оценки эффективности тестируемых соединений/молекул, связанная с клетками флуоресценция может быть измерена в качестве индикатора связывания LukA с клетками, например, с помощью автоматической флуоресцентной системы микроскопической визуализации, предназначенной для многопараметрического скрининга и многопараметрического анализа (например, Cellomics ArrayScan HCS Reader (Thermo Scientific) (возбуждение 485 нм, излучение 535 нм)).
Скрининг ингибиторов олигомеризации/взаимодействия LukA-LukB
Для скрининга ингибиторов, блокирующих или ослабляющих взаимодействие LukA/LukB, которое считается второй стадией процесса интоксикации, человеческие фагоциты (например, клетки PMN-HL60) могут быть высеяны на 384-луночные плоскодонные обработанные планшеты для тканевых культур (Corning) в количестве 2,5×103 клеток/лунку в конечном объеме 50 мкл среды RPMI (Gibco) с добавкой 10% термоинактивированной сыворотки плода коровы (FBS). Клетки могут быть затем обработаны тестируемым соединением/молекулой и затем интоксицированы смесью очищенного LukA и очищенного LukB, где LukB является флуоресцентно-меченым флуоресцентной молекулой, такой как FITC, Cy3, Cy5, APC и PE, и оставлены до завершения процесса интоксикации (например, в течение 1 ч при 37°C, 5% CO2). Для оценки эффективности тестируемых соединений/молекул, связанная с клетками LukB-FITC флуоресценция может быть измерена в качестве индикатора взаимодействия LukA/LukB-FITC, с использованием, например, автоматической флуоресцентной системы микроскопической визуализации, предназначенной для многопараметрического скрининга и многопараметрического анализа (например, Cellomics ArrayScan HCS Reader (Thermo Scientific) (возбуждение 485 нм, излучение 535 нм)).
Скрининг ингибиторов образования пор LukAB
Для скрининга ингибиторов, блокирующих или ингибирующих образование пор LukAB, эффекторной молекулы, приводящей к клеточному лизису, человеческие фагоциты (например, клетки PMN-HL60) могут быть высеяны на 384-луночный черный обработанный планшет для тканевых культур с прозрачным дном (Corning) в количестве 2,5×103 клеток/лунку в конечном объеме 50 мкл среды RPMI (Gibco) с добавкой 10% термоинактивированной сыворотки плода коровы (FBS). Клетки могут быть затем обработаны тестируемым соединением/молекулой (около 5 мкл, с разными концентрациями) и затем интоксицированы очищенным LukAB (около 0,001-2 мкМ) в течение 10 минут при 37°C, 5% CO2. В качестве контроля, клетки PMN-HL60 могут быть обработаны культуральной средой (отрицательный контроль) и 0,1% об./об. Triton X100 (положительный контроль).
Для прямой оценки пор LukAB на поверхности клетки-хозяина может быть использован анализ притока бромида этидия (EB). EB представляет собой катионный краситель малого размера, не проникающий в здоровые клетки-хозяева. После образования катионных пор LukAB, EB поступает в клетки и связывается с ДНК, что приводит к флуоресценции. Клетка, обработанная таким образом, может быть затем инкубирована с EB (5 мкМ) в течение еще 5 минут при комнатной температуре в темноте. Для оценки эффективности тестируемых соединений/молекул при ингибировании образования LukAB-пор, флуоресценция может быть измерена в качестве индикатора порообразования, с использованием планшет-ридера, такого как Envision 2103 Multilabel Reader (Perkin-Elmer) при возбуждении 530 нм, излучении 590 нм. Этот анализ способствует идентификации молекул, которые могут блокировать или ингибировать LukAB-поры, снижая тем самым LukAB-опосредованную токсичность.
Способ определения продуцирования LukAB клиническими изолятами S. aureus для прогнозирования тяжести инфекции
Еще один аспект настоящего изобретения касается способа прогнозирования или оценки тяжести инфекции S. aureus, предусматривающего детектирование присутствия или количества LukA и/или LukB, или их соответствующих генов в биологическом образце, полученном от инфицированного субъекта. Таким образом, детектирование присутствия или относительно высоких количеств LukA и/или LukB, или детектирование их соответствующих генов (например, наблюдаемое для S. aureus штамм Newman, 4645 и штаммов MRSA USA300 и USA500) по сравнению с контролем (например, штаммы S. aureus USA100 и USA400), продуцирующим небольшие или недектируемые количества LukA и/или LukB, является показателем тяжелой инфекции. Что касается детектирования или присутствия относительных количеств LukA и/или LukB, можно сослаться на иллюстрации, приведенные на Фиг. 4A. Типичные примеры вариантов исполнения способа описаны ниже.
Иммуноблот-анализ для определения уровней LukA и LukB
Для определения уровней LukAB (т.е. продуцирования LukAB), биологический образец, например, жидкость (например, кровь) или образец ткани, получают от инфицированного субъекта, с последующим помещением культуры в пригодные условия культивирования для обеспечения роста S. aureus, получают супернатант культуры, выделяют из него бактериальные белки, идентифицируют LukA и/или LukB, и затем количественно определяют LukA и/или LukB. Более конкретно, в одном варианте исполнения, штаммы клинического изолята могут быть выбраны и выращены в пригодной культуральной среде, например, в культуральной среде Royal Park Memorial Institute 1640 (RPMI; Invitrogen) с добавкой 1% казаминовых кислот (RPMI+CAS) в пригодных условиях культивирования, например, в течение 12-18 часов при 37°C со встряхиванием при 180 об/мин. Бактерии могут быть затем осаждены путем центрифугирования, и собирают супернатанты культур. Супернатанты культур (около 30 мкл) могут быть затем смешаны с 10 мкл буфера ДСН-Лэммли и нагреты до 95°C в течение 10 минут. Белки могут быть затем разделены, например, с помощью 15% гелей ДСН-ПААГ и затем перенесены на твердую подложку, например, нитроцеллюлозную мембрану. Мембраны можно затем инкубировать с антителами, направленными против LukA или LukB (например, кроличьими поликлональными антителами), и присутствие LukA или LukB можно визуализировать путем детектирования комплексов антитело-LukA/антитело-LukB со вторичным антителом, конъюгированным с флуорофором (например, анти-кроличье антитело, конъюгированное с AlexaFluor-680; Invitrogen). Мембраны можно затем высушить и отсканировать, например, с помощью системы инфракрасной визуализации Odyssey (LI-COR Biosciences) для определения количества LukA и LukB.
Полимеразная цепная реакция (ПЦР) для определения присутствия генов LukA и/или LukB
Для определения присутствия генов, кодирующих LukAB, получают от инфицированного субъекта биологический образец с последующим помещением культуры в пригодные условия культивирования для обеспечения роста S. aureus, экстрагируют нуклеиновую кислоту из культуры S. aureus и затем проводят, по меньшей мере, один цикл амплификации нуклеиновой кислоты с использованием ПЦР или другого пригодного протокола амплификации, используя LukA и/или LukB-специфичные праймеры и детектируя LukA и/или LukB. Таким образом, в одном типичном примере варианта исполнения, после подготовки исходного образца, штаммы клинического изолята могут выращиваться на твердой среде, например, триптиказо-соевом бульоне (TSB), отвержденном 1,5% агара, при 37°C. Колонии S. aureus могут быть затем выбраны и подвергнуты ферментативному гидролизу, например, с помощью 2 мг/мл лизостафина (AMBI PRODUCTS LLC) в TSM-буфере (100 мМ TRIS pH 7, 500 мМ сахарозы, 10 мМ MgCl2) в течение 10 минут при 37°C. Образцы затем можно центрифугировать, супернатант отбрасывают и осадок ресуспендируют в 100 мкл стерильной воды, с последующим кипячением в течение пяти минут при 100°C и центрифугированием. Супернатант обеспечивает исходный материал и ДНК-матрицу для реакции амплификации, такой как ПЦР, с использованием LukA и/или LukB-специфичных праймеров.
Рабочие примеры
Изобретение будет далее описано со ссылкой на приведенные ниже неограничивающие примеры. Если не указано иное, все доли определяются по весу.
Пример 1: Экспрессия и очистка рекомбинантного LukA и LukB в нативных условиях: система экспрессии pMAL
Гены LukA и LukB амплифицируют из ДНК S. aureus с помощью Taq-полимеразы в стандартных условиях ПЦР при температуре отжига 55°C с использованием указанных далее праймеров: 5'-ccc-GTCGAC-tta-TCCTTCTTTATAAGGTTTATTGTC-3' (SEQ ID NO:30) и 5'-ccc-GAAGGATTTCACATCATCATCATCATCACAATTCAGCTCATAAAGACTCTC-3' (SEQ ID NO:31) для LukA и 5'-CCCCGAAGGATTTCaCATCATCATCATCATCACAAGATTAATTCTGAAATCAAACAAG-3' (SEQ ID NO:32) и 5'-ccc-GTCGAC-tta-TTTCTTTTCATTATCATTAAGTACTT-3' (SEQ ID NO:33) для LukB. Генные продукты LukA и LukB гидролизуют Nde1 и Sal1 (New England BioLabs) и лигируют в вектор pMAL-c4X (New England BioLabs). Конструкты трансформируют в E. coli штамм DH5α и плазмидные вставки подтверждают путем секвенирования. Трансформанты выращивают в среде Terrific Broth со 100 мкг/мл ампициллина и 0,2% глюкозы при 37°C до достижения культурами значения A600, равного приблизительно 0,5. Экспрессию 6-his-меченого MBP-LukA или 6-his-меченого MBP-LukB индуцируют с помощью 0,3 мМ изопропил-β-D-1-тиогалактопиранозида (IPTG) при 16°C, в течение ночи, со встряхиванием при 180 об/мин.
После индуцирования клетки собирают центрифугированием при 4000 об/мин при 4°C в течение 20 мин и ресуспендируют в охлажденном на льду буфере для колонок (20 мМ Tris-HCl, 200 мМ NaCl и 1 мМ ЭДТА) с добавкой не содержащего ЭДТА ингибитора протеазы (Roche). Бактериальные клетки озвучивают на льду в течение 1 мин (импульсы 10 с). Образцы центрифугируют при 10000 об/мин при 4°C в течение 30 мин и супернатант собирают и наносят на колонку с амилозной смолой. Колонки промывают дважды буфером для колонок и очищенный 6-his-меченый MBP-LukA или 6-his-меченый MBP-LukB элюируют в 10 фракциях буфером для колонок с добавкой 10 мМ мальтозы.
Пример 2: Экспрессия и очистка рекомбинантных активных токсинов LukA, LukAΔ10C, Δ33NLukA и LukB: система экспрессии pET14b
Гены LukA, LukAΔ10C, Δ33NLukA и LukB амплифицируют из ДНК S. aureus с помощью полимеразы Vent (New England BioLabs) в стандартных условиях ПЦР при температуре отжига 55°C с использованием приведенных далее праймеров: 5'-cccc-CTCGAG-AATTCAGCTCATAAAGACTCTCAAG-3' (SEQ ID NO:34) и 5'-cccc-GGATCC-tta-TCCTTCTTTATAAGGTTTATTGTC-3' (SEQ ID NO:35) для LukA; 5'-cccc-CTCGAG-AATTCAGCTCATAAAGACTCTCAAG (SEQ ID NO:34) и 5'-cccc-GGATCC-tta-ATATTTATCAACGACTTTAACTG (SEQ ID NO:36) для LukAΔ10C; 5'-cccc-CTCGAG-TCAACAGCACCGGATGATATTG (SEQ ID NO:37) и 5'-cccc-GGATCC-tta-TCCTTCTTTATAAGGTTTATTGTC (SEQ ID NO:35) для Δ33NLukA; 5'-cccc-CTCGAG-AAGATTAATTCTGAAATCAAACAAG-3' (SEQ ID NO:38) и 5'-cccc-GGATCC-tta-TTTCTTTTCATTATCATTAAGTACTTT-3' (SEQ ID NO:39) для LukB. Генные продукты гидролизуют Xho1 и BamH1 (New England BioLabs) и лигируют в вектор pET14b (Novagen) со слиянием кодирующей последовательности гистидиновой метки с 5'-областью генов. Экспрессионные плазмиды трансформируют в E. coli штамм DH5α и плазмидные вставки подтверждают путем секвенирования. Плазмиды очищают и трансформируют в экспрессионный штамм E. coli T7 lysY/lq (New England BioLabs).
Для очистки в денатурирующих условиях трансформанты выращивают в среде Terrific Broth со 100 мкг/мл ампициллина при 37°C до достижения культурами величины A600, равной приблизительно 0,5. Экспрессию 6-his-меченого LukA или 6-hisLukB индуцируют с помощью 0,4 мМ IPTG при 37°C, в течение 3 ч, со встряхиванием при 180 об/мин. После индукции клетки собирают путем центрифугирования при 4000 об/мин при 4°C в течение 15 мин и затем ресуспендируют в 1× TBS (50 мМ Tris, 150 мМ NaCl, pH 7,5). Бактериальные клетки озвучивают на льду в течение 2 мин (импульсы по 10 с). Озвученные бактерии подвергают ультрацентрифугированию в течение 30 мин при 50000 об/мин. Осадки ресуспендируют в буфере для лизиса (100 мМ NaH2PO4, 10 мМ Tris, 8M мочевины, pH 8,0) и инкубируют при комнатной температуре в течение 30 мин на ротаторе (rotisserie). Образцы центрифугируют при 13000 об/мин в течение 30 мин и супернатанты наносят на колонку, содержащую смолу Ni-NTA (Qiagen). Колонку дважды промывают буфером для промывки (100 мМ NaH2PO4, 10 мМ Tris, 8M мочевины, pH 6,3) и белок элюируют из колонки с помощью буфера для элюирования (100 мМ NaH2PO4, 10 мМ Tris, 8M мочевины) при pH 5,9 и при pH 4,5. Фракции, содержащие очищенный белок, определенные методом ДСН-ПААГ, объединяют, разводят 1:1 забуференным tris солевым раствором (TBS; 500 мМ Tris, 1,5 M NaCl, pH 7,5) и подвергают диализу в TBS при 4°C в течение ночи для удаления мочевины и обеспечения возможности повторной укладки. Очищенные 6-his-меченые LukA и LukB количественно определяют с помощью набора для анализа белков Thermo Scientific Pierce BCA Protein Assay Kit.
Для очистки в нативных условиях трансформанты выращивают в бульоне Луриа-Бертани со 100 мкг/мл ампициллина при 37°C до достижения культурами величины A600, равной приблизительно 0,5. Экспрессию 6-his-меченого LukA, 6-his-меченого LukAΔ10C, 6-his-меченого Δ33NLukA или 6-hisLukB индуцируют с помощью 0,05-0,1 мМ IPTG при 25-30°C, в течение 3 ч, со встряхиванием при 220 об/мин. После индукции клетки собирают центрифугированием при 4000 об/мин при 4°C в течение 15 мин и затем ресуспендируют в 1× TBS (50 мМ Tris, 600 мМ NaCl, pH 7,5) с 10 мМ имидазола и коктейля HALT ингибиторов протеазы, не содержащего ЭДТА (Thermo Scientific). Бактериальные клетки озвучивают на льду. Озвученные бактерии центрифугируют в течение 20 мин при 20000 об/мин. Супернатанты инкубируют со смолой Ni-NTA (Qiagen) в течение 1 ч при 4°C на ротаторе. Образцы наносят на колонку и колонку промывают буфером для промывки 1× TBS (50 мМ Tris, 600 мМ NaCl, pH 7,5) с 25 мМ имидазола. Белок элюируют из колонки с помощью 50-500 мМ имидазола в буфере для элюирования 1× TBS (50 мМ Tris, 600 мМ NaCl, pH 7,5). Фракции, содержащие очищенный белок, определенные методом ДСН-ПААГ, объединяют, разводят 1:1 в 1× TBS (50 мМ Tris, 150 мМ NaCl, pH 7,5) и диализируют в 1× TBS при 4°C в течение ночи. Очищенные 6-his-меченые LukA и LukB количественно определяют с помощью набора для анализа белков Thermo Scientific Pierce BCA Protein Assay Kit.
Пример 3: Экспрессия и очистка денатурированных рекомбинантных LukA и LukB
Гены LukA и LukB амплифицируют из ДНК S. aureus с помощью полимеразы Vent (New England BioLabs) в стандартных условиях ПЦР при температуре отжига 55°C с использованием следующих праймеров: 5'-ggg-CATATG- AATTCAGCTCATAAAGACTCTCAA-3' (SEQ ID NO:40) и 5'-ccc-GTCGAC- TCCTTCTTTATAAGGTTTATTGTC-3' (SEQ ID NO:41) для LukA, и 5'-ggg-CATATG-AAGATTAATTCTGAAATCAAACAAG-3' (SEQ ID NO:42) и 5'-ccc-GTCGAC-TTTCTTTTCATTATCATTAAGTACTT-3' (SEQ ID NO:43) для LukB. Генные продукты LukA и LukB гидролизуют Nde1 и Sal1 (New England BioLabs) и лигируют в вектор pET41b (Novagen). Конструкты сначала трансформируют в клетки DH5α и затем трансформируют в экспрессионный штамм E. coli ER2566 (New England BioLabs). Трансформанты выращивают в среде Terrific Broth с канамицином, 25 мкг/мл, в течение 2,5 ч при 37°C, и экспрессию LukA и LukB индуцируют с помощью 0,3 мМ IPTG при 37°C в течение 2 ч со встряхиванием при 180 об/мин. Клетки осаждают и ресуспендируют в 1× TBS (500 мМ Tris, 1,5 M NaCl, pH 7,5) и озвучивают на льду в течение 1 мин (импульсы по 10 с). Озвученные бактерии подвергают ультрацентрифугированию в течение 30 мин при 50000 об/мин.
Для очистки C-концевых 6-his-меченых LukA и LukB в денатурирующих условиях, осадки ресуспендируют в буфере для лизиса (100 мМ NaH2PO4, 10 мМ Tris, 8 M мочевины, pH 8,0) и инкубируют при комнатной температуре в течение 30 мин на ротаторе. Образцы центрифугируют при 13000 об/мин в течение 30 мин и супернатанты наносят на колонку Ni-NTA. Колонки промывают дважды буфером для промывки (100 мМ NaH2PO4, 10 мМ Tris, 8 M мочевины, pH 6,3) и LukA и LukB элюируют из колонки с помощью буфера для элюирования (100 мМ NaH2PO4, 10 мМ Tris, 8 M мочевины) при pH 5,9 и при pH 4,5. Очищенные 6-his-меченые LukA и LukB количественно определяют с помощью набора для анализа белков BioRad DC Protein Assay.
Пример 4: Продуцирование поликлональных антител к LukA и к LukB
Денатурированный рекомбинантный LukA (250 мкг), эмульгированный в полном адъюванте Фрейнда (FCA), вводят инъекцией новозеландским белым кроликам. Животным вводят бустер-дозу рекомбинантного LukA (125 мкг), эмульгированного в неполном адъюванте Фрейнда (FCA), в день двадцать один (21) и в день сорок девять (49).
Денатурированный рекомбинантный LukB (250 мкг), эмульгированный в полном адъюванте Фрейнда (FCA), вводят инъекцией новозеландским белым кроликам. Животным вводят бустер-дозу рекомбинантного LukB (125 мкг), эмульгированного в неполном адъюванте Фрейнда (FCA), в день двадцать один (21) и в день сорок девять (49).
Пример 5: Лейкоцидин A/B несет основную ответственность за цитотоксин-опосредованную гибель человеческих фагоцитов вследствие разрыва мембраны
Использованные клеточные линии
В качестве модели для изучения, каким образом LukAB выбирает мишень и уничтожает клетки человеческих фагоцитов, была использована HL-60 (ATCC CCL-240, человеческая промиелоцитарная клеточная линия). Клетки HL-60 выращивают в среде RPMI (Gibco) с добавкой 10% термоинактивированной сыворотки плода коровы (FBS) при 37°C с 5% CO2. Для дифференцировки HL-60 в нейтрофилоподобные клетки (PMN-HL60), в культуры добавляют 1,5% (об./об.) диметилсульфоксида (ДМСО) и выращивают в течение 4 дней.
Использованные способы/анализы
Анализ клеточной токсичности
Для оценки жизнеспособности клеток млекопитающих после интоксикации лейкоцидином AB Staphylococcus aureus (LukAB), клетки PMN-HL60 высеивают на 96-луночные плоскодонные обработанные планшеты для тканевых культур (Corning) в количестве 1×105 клеток/лунку в конечном объеме 100 мкл RPMI (Gibco) с добавкой 10% термоинактивированной сыворотки плода коровы (FBS). Клетки интоксицируют в течение 2 ч при 37°C, 5% CO2 с серийными 2-кратными разбавлениями фильтрата культуры Staphylococcus aureus штамм Newman в интервале значений от 20% до 0,16% об./об. с тремя параллельными опытами. Эксперименты проводят с использованием экзопротеинов штамма дикого типа и экзопротеинов штамма, не содержащего LukAB (мутантный штамм). Контроли для 100% жизнеспособности включали 20% об./об. среды для тканевых культур (RPMI+10% термоинактивированной сыворотки плода коровы) и 20% об./об. питательной среды. S. aureus (RPMI+казаминовые кислоты). 0,1% об./об. Triton X100 был использован в качестве контроля для 100% гибели клеток. После интоксикации, 10 мкл CellTiter (Promega) добавляют в каждую лунку и клетки инкубируют дополнительно еще 3 ч при 37°C, 5% CO2. CellTiter контролирует метаболически активные клетки (изменение окраски), т.е. способность, которую утрачивают погибшие клетки. Колориметрическую реакцию измеряют при 492 нм с помощью прибора Perkin Elmer Envision 2103 Multilabel Reader. Процент жизнеспособных клеток рассчитывают по приведенному ниже уравнению: % жизнеспособности=100×[(Ab492Образец-Ab492Triton X)/(Ab492Среда для тканевых культур)].
Анализ повреждения мембраны
Альтернативным анализом с целью измерения LukAB-опосредованной цитотоксичности является оценка целостности мембраны клетки-хозяина. Для этого используют анализ проницаемости красителя SYTOX green (Invitrogen). Здоровые клетки являются непроницаемыми для SYTOX green, но становятся проницаемыми для красителя при нарушении целостности клеточной мембраны. Внутри клетки SYTOX green связывается с ДНК и проявляет сильную флуоресценцию.
Для оценки целостности мембраны клетки-хозяина после интоксикации LukAB или ex vivo инфекции штаммами S. aureus, клетки PMN-HL60 высеивают на 96-луночные плоскодонные обработанные планшеты для тканевых культур (Corning) в количестве 1×105 клеток/лунку в конечном объеме 100 мкл RPMI (Gibco) с добавкой 10% термоинактивированной сыворотки плода коровы (FBS). Клетки интоксицируют разбавлениями фильтрата культуры S. aureus штамм Newman в интервале значений от 20% до 0,16% об./об., или инфицируют с MOI (кратностями инфекции) в интервале значений 1-100 с тремя параллельными опытами в течение 2 ч при 37°C, 5% CO2. Эксперименты проводят с использованием штамма дикого типа и штамма, не содержащего LukAB (мутантный штамм). Контроли для определения фонового уровня флуоресценции включают 20% об./об. среды для тканевых культур (RPMI+10% термоинактивированной сыворотки плода коровы) и 20% об./об. питательной среды S. aureus (RPMI+казаминовые кислоты). После интоксикация или инфекции клетки переносят на 96-луночный планшет с v-образным дном лунок (Corning) и центрифугируют при 1500 об/мин в течение 5 мин. Супернатант отбрасывают и осадки ресуспендируют в 100 мкл PBS + SYTOX green (0,1 мкМ; Invitrogen). Клетки затем переносят на 96-луночный черный обработанный планшет для тканевых культур с прозрачным дном (Corning) и инкубируют при комнатной температуре в темноте в течение 10 мин. Флуоресценцию измеряют с помощью прибора Perkin Elmer Envision 2103 Multilabel Reader (возбуждение 485 нм, излучение 535 нм).
Результаты
LukAB нацелен на человеческие первичные фагоциты и уничтожает их
Интоксикация первичных человеческих периферических мононуклеарных клеток (PBMC) (Фиг. 3a), моноцитов, макрофагов, дендритных клеток (Фиг. 3b) и полиморфноядерных клеток (PMN) (Фиг. 3c) фильтрованными супернатантами культур S. aureus штамм Newman приводит к сильной гибели клеток, по результатам определения с помощью анализа клеточной токсичности (Фиг. 3a-c). Интоксикация этих клеток фильтрованными супернатантами культур S. aureus штамм Newman, не содержащих гемолизина (hla), γ-гемолизина (hlg), лейкоцидина E/D (LukED) или лейкоцидина A/B (LukAB), продемонстрировала, что экзопротеины hla-, hlg- и LukED-негативных штаммов являются такими же цитотоксичными, как и штамм Newman дикого типа (WT) (при определении с помощью анализа клеточной токсичности), указывая на то, что ни один из вышеуказанных лейкотоксинов, продуцируемых Newman, не участвует в цитотоксин-медиируемой гибели этих клеток (Фиг. 3a-c). В отличие от них, очень низкая гибель клеток наблюдалась в тех случаях, когда клетки интоксицируют экзопротеинами штамма Newman, не содержащего LukAB (ΔlukAB). Отсутствие цитотоксичной активности у штамма Newman, не содержащего LukAB, устраняется путем введения генов IukAB в трансформант (trans) в плазмиде (ΔlukAB/pLukAB) (Фиг. 3c). Важно отметить, что этот фенотип является полностью зависимым от LukAB, по результатам определения путем интоксицирования PMN очищенными рекомбинантными LukA и LukB. Индивидуальные субъединицы не демонстрируют детектируемой цитотоксичности по отношению к PMN (Фиг. 3d). В отличие от них, комбинация обеих субъединиц приводит к сильной цитотоксичности по отношению к таким клеткам доза-зависимым образом (Фиг. 3d). В общем, эти результаты демонстрируют, что LukAB является ответственным за способность S. aureus нацеливаться на и уничтожать первичные человеческие фагоциты, ключевые иммунные клетки, необходимые для защиты хозяина от инфекционных агентов.
LukAB предпочтительно уничтожает человеческие фагоцитарные клетки
Интоксикация нейтрофилоподобной клеточной линии (PMN-HL60) и макрофагоподобной клеточной линии (THP1+PMA) фильтрованными супернатантами культур S. aureus штамм Newman приводила к сильной гибели клеток, по результатам определения с помощью анализа клеточной токсичности (Фиг. 3). Интоксикация этих клеток фильтрованными супернатантами культур S. aureus WT и мутантных штаммов с изогенными цитотоксинами (hla, hlgABC, LukED и LukAB) продемонстрировала, что LukAB отвечает за способность S. aureus уничтожать эти клетки, по результатам определения с помощью анализа клеточной токсичности (Фиг. 4a и 4b). Отсутствие цитотоксичной активности штамма Newman, не содержащего LukAB, устраняется путем трансформации штамма Newman, не содержащего LukAB, плазмидой, экспрессирующей LukAB (ΔlukAB/pLukAB) (Фиг. 4c). Экзопротеины этого штамма были чрезвычайно цитотоксичными по отношению как к клеткам PMN-HL60, так и клеткам THP-1+PMA, что является сильным доказательством того, что LukAB является сильным стафилококковым токсином, нацеленным на человеческие клетки и уничтожающим их.
Для того чтобы дополнительно исключить влияние других факторов, присутствующих в супернатантах культур S. aureus, клетки PMN-HL60 интоксицируют очищенными рекомбинантными LukA или LukB. Индивидуальные субъединицы не проявляют детектируемой цитотоксичности по отношению к PMN-HL60 (Фиг. 3d). В отличие от них, комбинация обеих субъединиц приводит к сильной цитотоксичности по отношению к этим клеткам доза-зависимого характера (Фиг. 3d). В дополнение к клеткам PMN-HL60 и THP-1+PMA, несколько других человеческих клеточных линий, включая миелоидного предшественника, из которого дифференцируются PMN-HL60 (HL60), моноцитарного предшественника, из которого дифференцируются THP-1+PMA (THP-1), лимфоциты (HuT и клетки Jurkat) и эпителиальные клетки (293T и HepG2) также интоксицируют рекомбинантным LukAB (Фиг. 4e). Эти результаты демонстрируют, что LukAB предпочтительно нацелен на и уничтожает человеческие фагоцитарные клетки, и не оказывает никакого влияния на человеческие лимфоциты или эпителиальные клетки. Взятые вместе эти результаты демонстрируют, что LukAB играет значительную роль в S. aureus-медиируемой гибели фагоцитов.
LukAB продуцируется клинически релевантными штаммами S. aureus
Иммуноблот-анализы с поликлональным антителом, индуцированным против LukB, показали, что LukB продуцируется рядом стафилококковых штаммов, включая штаммы MRSA, ассоциированные с госпитальными и внегоспитальными инфекциями (USA300, 400 и 500; Фиг. 5a). Важно, что уровни LukB ассоциированы с цитотоксичным фенотипом этих штаммов (Фиг. 5a и 5b). Штаммы, продуцирующие высокие уровни LukB (например, Newman, 4645, USA500 и USA300), были более цитотоксичными по отношению к клеткам PMN-HL60, чем штаммы, продуцирующие низкие или недектируемые уровни LukB (например, USA100 и USA400) (Фиг. 5b). Для изучения роли LukAB в штаммах MRSA был создан изогенный мутант LukAB в клиническом изоляте клона USA тип 300 LA (Фиг. 5c). Как и в случае штамма Newman, экзопротеины штамма USA300, не содержащего LukAB, были заметно менее цитотоксичными, чем экзопротеины родительского штамма (Фиг. d). Эти данные демонстрируют, что LukAB является важным цитотоксином, продуцируемым штаммами MRSA.
LukAB повреждает мембраны человеческих фагоцитов
Интоксикация PMN-HL60 экзопротеинами S. aureus приводит к округлению формы клетки и набуханию ядра, по фенотипу, зависимому от LukAB (Фиг. 6a). Такой фенотип округления клетки и набухания был ассоциирован с повышенной проницаемостью мембраны, определенной с помощью анализа повреждения мембран (Фиг. 6b и 6c). Важно, что экзопротеины LukAB-отрицательного штамма демонстрировали незначительный или отсутствие эффекта на проницаемость мембраны, по фенотипу, который можно было устранить путем продуцирования LukAB из плазмиды (Фиг. 6b), причем рекомбинантный LukAB, но не индивидуальные субъединицы токсина, вызывали доза-зависимое повреждение мембраны (Фиг. 6c). Кроме того, инфекция первичных человеческих PMN штаммом метициллин-чувствительного S. aureus (MSSA) и штаммом метициллин-резистентного S. aureus (MRSA) USA300 приводит к LukAB-зависимому повреждению мембраны (Фиг. 6d). Эти результаты демонстрируют, что LukAB повреждает плазматическую мембрану клетки-хозяина при ex vivo инфекции.
LukAB защищает S. aureus от хозяин-опосредованной гибели путем нацеливания на фагоциты и их уничтожения
Инфекция клеток PMN-HL60 S. aureus WT, ΔlukAB и ΔlukAB, содержащими экспрессионную плазмиду IukAB (ΔlukAB/pLukAB), показала, что LukAB является необходимым для обеспечения способности S. aureus разрывать мембрану фагоцитов в процессе взаимодействия стафилококк-фагоцит (Фиг. 7a), определяемой с помощью анализа повреждения мембраны. Важно, что S. aureus с перепродуцированием LukAB (ΔlukAB/plukAB) продемонстрировал более сильное повреждение мембраны, чем штамм дикого типа (WT) (Фиг. 7a), указывая на то, что LukAB сильно повреждает мембраны клетки-хозяина. Инфекция человеческой цельной крови (Фиг. 7b) и очищенных первичных человеческих нейтрофилов (PMN; Фиг. 7c) показала, что IukAB-отрицательный стафилококк уничтожался более эффективно по сравнению с WT-штаммом (Фиг. 7b и 7c). Важно, что ослабленный фенотип, проявляемый ΔlukAB-отрицательными стафилококками, можно было устранить с помощью экспрессионной плазмиды IukAB (Фиг. 7b и 7c). Интоксикация первичных человеческих PMN фильтратом культуры S. aureus WT, ΔlukAB и мутантным штаммом ΔlukAB, содержащим экспрессионную плазмиду IukAB, показала, что LukAB нацелен на и уничтожает первичные человеческие PMN (Фиг. 7d). Эти данные являются сильным свидетельством того, что LukAB представляет собой сильный стафилококковый цитотоксин, который нацелен на и уничтожает PMN путем разрыва мембраны, тем самым защищая S. aureus от PMN-медиируемой гибели.
LukAB способствует патогенезу S. aureus in vivo
Мыши, инфицированные ретроорбитально S. aureus, содержащим конструкт репортера люциферазы со слитым с ним промотором LukAB (pLukAB-Xen1), продемонстрировали активность промотора LukAB при почечных абсцессах, в то время как не содержащий промотора репортер (pXen1) не проявлял активности (Фиг. 8a). Эти данные демонстрируют, что LukAB экспрессируется in vivo в модели инфекции почечного абсцесса. В дополнение к этому, мыши, инфицированные ретроорбитально S. aureus WT, но не штаммом S. aureus, не содержащим LukAB, демонстрируют обширную колонизацию почек. Недостаточная колонизация, наблюдаемая для LukAB-отрицательного штамма, восстанавливается до уровней WT путем обеспечения LukAB у трансформантов (trans) (Фиг. 8b). Взятые вместе эти данные демонстрируют, что LukAB является важным стафилококковым цитотоксином, способствующим патогенезу бактерии.
LukAB образует поры в мембране клетки-мишени, которые могут быть заблокированы полиэтиленгликолем
Интоксикация клеток PMN-HL60 рекомбинантным LukAB (rLukAB) показала, что как rLukA, так и rLukB связываются с клетками-мишенями, при определении методом иммуноблоттинга с LukA и LukB специфичными антителами (Фиг. 9a). Связывание 6His-меченого rLukAB с PMN-HL60 было также подтверждено путем сортировки клеток с возбуждением флуоресценции (FACS) и с помощью His-специфичного антитела (Фиг. 9b). Олигомеры rLukAB также детектируются методом иммуноблоттинга на мембранах PMN-HL60 после интоксикации рекомбинантным токсином, что указывает на образование структур LukAB более высокого порядка на мембранах клетки-мишени (Фиг. 9c). Важно отметить, что интоксикация PMN-HL60 rLukAB продемонстрировала, что LukAB образует на мембранах клеток-мишеней проницаемые для бромида этидия поры, которые могут быть заблокированы с помощью молекул полиэтиленгликоля (ПЭГ) (Фиг. 9d), и что блокировка LukAB-пор повышает жизнеспособность клеток (Фиг. 9E). Кроме того, ПЭГ специфически блокирует LukAB-поры, поскольку поры, образованные в мембранах PMN-HL60 сапонином, не блокировались ПЭГ и, в результате, эти клетки не были защищены от опосредованной порами гибели. Эти данные демонстрируют, что LukAB-поры могут быть заблокированы малыми молекулами, и блокирование LukAB-пор защищает клетки от LukAB-опосредованной гибели.
Нейтрализация цитотоксичности фильтрата культур S. aureus α-LukA поликлональным антителом
Интоксикация PMN-HL60 фильтратом культуры S. aureus, предварительно инкубированного с различными количествами α-LukA поликлональных антител, индуцированных у двух разных кроликов, приводит к доза-зависимому снижению токсичности фильтрата культуры (Фиг. 10). Этот нейтрализующий эффект не наблюдался, когда фильтрат культуры предварительно инкубировался с неиммунной сывороткой. Важно, что антитела, вырабатываемые двумя разными кроликами, вели себя очень похоже, и нейтрализующая способность антител возрастала с увеличением их степени зрелости, что видно при сравнении нейтрализующего эффекта антител из проб крови, взятых в более поздние моменты времени, с нейтрализующим эффектом антител из крови, взятой раньше (Фиг. 10). Эти данные показывают, что цитотоксичность, наблюдающаяся для фильтрата культуры S. aureus, может быть нейтрализована с помощью α-LukA поликлональных антител.
Идентификация нецитотоксичного LukA усеченного мутанта, который бы еще распознавался α-LukA поликлональным антителом
LukA отличается от других S-субъединиц стафилококкового лейкотоксина тем, что он имеет выступающие участки на обоих N- и C-концах. Эти выступы состоят из 33 аминокислот на N-конце и 10 аминокислот на C-конце (Фиг. 11a). Интоксикация PMN-HL60 очищенным рекомбинантным LukA, не имеющим N-концевого выступа (rΔ33NLukA) в комбинации с очищенным rLukB, приводит к сильной цитотоксичности по отношению к клеткам, сравнимой с показателями для очищенного rLukA+rLukB (Фиг. 11b). Однако интоксикация PMN-HL60 очищенным рекомбинантным LukA, не имеющим C-концевого выступающего участка (rLukAΔ10C) в комбинации с rLukB не вызывает цитотоксичного эффекта (Фиг. 11b). Эти данные демонстрируют, что N-концевой выступающий участок является несущественным для цитотоксичного эффекта LukA, но C-концевой выступающий участок необходим для токсичности. Важно, что α-LukA поликлональное антитело, нейтрализующее эффект LukAB (Фиг. 10), продолжает распознавать 6×His-меченый нецитотоксичный rLukAΔ10C мутант, так же как и α-His поликлональное антитело (Фиг. 11c). Эти данные позволяют предположить, что rLukAΔ10C могут быть использованы для in vivo получения α-LukA поликлональных антител, представляющих собой нейтрализующие антитела. Таким образом, rLukAΔ10C могут быть использованы в композиции активной вакцины.
Все патентные публикации и непатентные публикации указывают на уровень знаний квалифицированных специалистов в области техники, к которой относится данное изобретение. Все эти публикации настоящим включены сюда в качестве ссылок в такой же степени, как если бы для каждой индивидуальной публикации было конкретно и индивидуально указано, что она включается в качестве ссылки.
Хотя изобретение было описано здесь со ссылками на конкретные варианты исполнения, следует понимать, что эти варианты исполнения являются просто иллюстрациями принципов и применений настоящего изобретения. Следует понимать поэтому, что могут быть выполнены многочисленные модификации иллюстративных вариантов исполнения, и что могут быть разработаны другие схемы, не выходящие за пределы сущности и объема настоящего изобретения, определяемые приложенной формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЙКОЦИДИНЫ STAPHYLOCOCCUS AUREUS, ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2795545C2 |
| ЛЕЙКОЦИДИНЫ STAPHYLOCOCCUS AUREUS, ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2644237C2 |
| СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ СТАФИЛОКОККА ЗОЛОТИСТОГО И СВЯЗАННЫХ С НИМИ НАРУШЕНИЙ | 2012 |
|
RU2635462C2 |
| СПОСОБ ИНГИБИРОВАНИЯ ТОКСИЧНОСТИ ЛЕЙКОЦИДИНА ED STAPHYLOCOCCUS AUREUS У СУБЪЕКТА | 2012 |
|
RU2609650C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ, ВЫЗВАННЫХ Staphylococcus aureus | 2015 |
|
RU2636780C1 |
| АНТИТЕЛА ПРОТИВ СТЕНОЧНОЙ ТЕЙХОЕВОЙ КИСЛОТЫ И ИХ КОНЪЮГАТЫ | 2014 |
|
RU2687044C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ЛЕЙКОТОКСИНОВ STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2805969C2 |
| АНТИТЕЛА, КОТОРЫЕ СПЕЦИФИЧНО СВЯЗЫВАЮТ АЛЬФА-ТОКСИН STAPHYLOCOCCUS AUREUS, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620065C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ГРИБКОВЫХ И БАКТЕРИАЛЬНЫХ ПАТОГЕНОВ | 2014 |
|
RU2717306C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ АНТИГЕНОВ STAPHYLOCOCCUS AUREUS | 2010 |
|
RU2536981C9 |






























































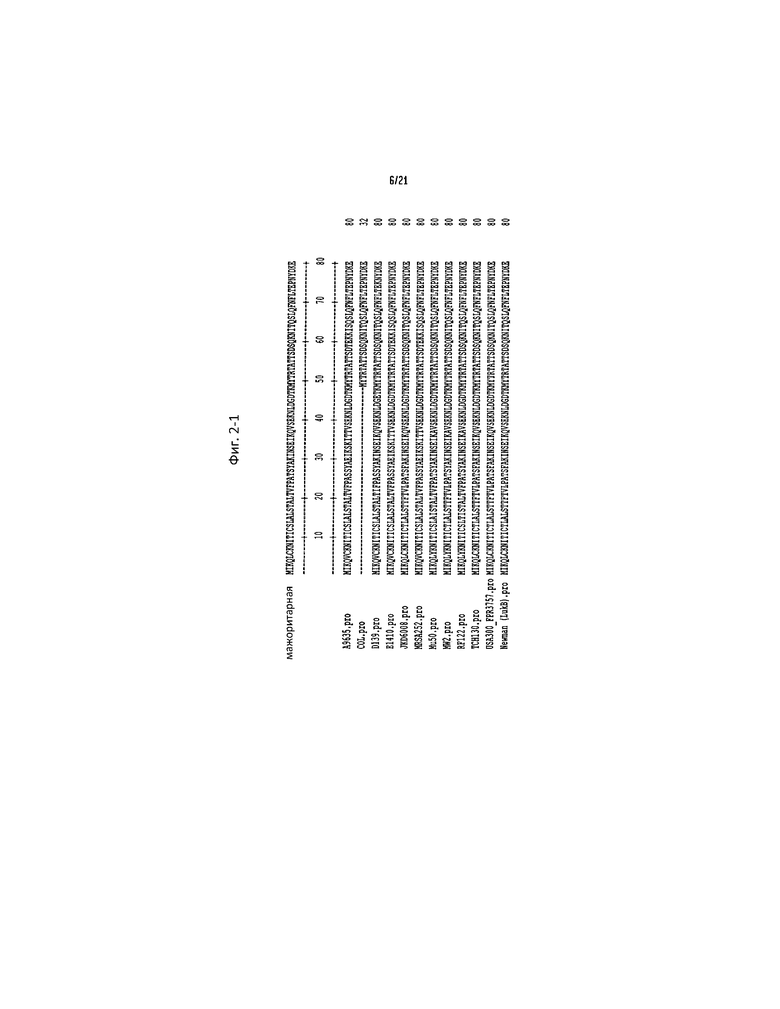
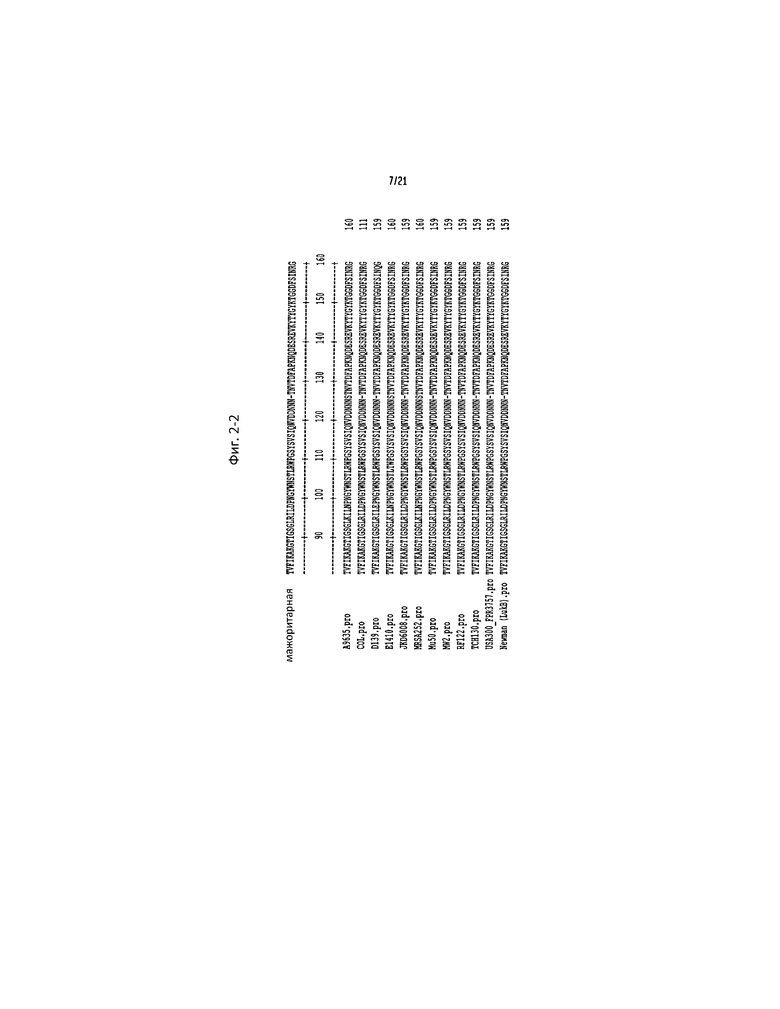




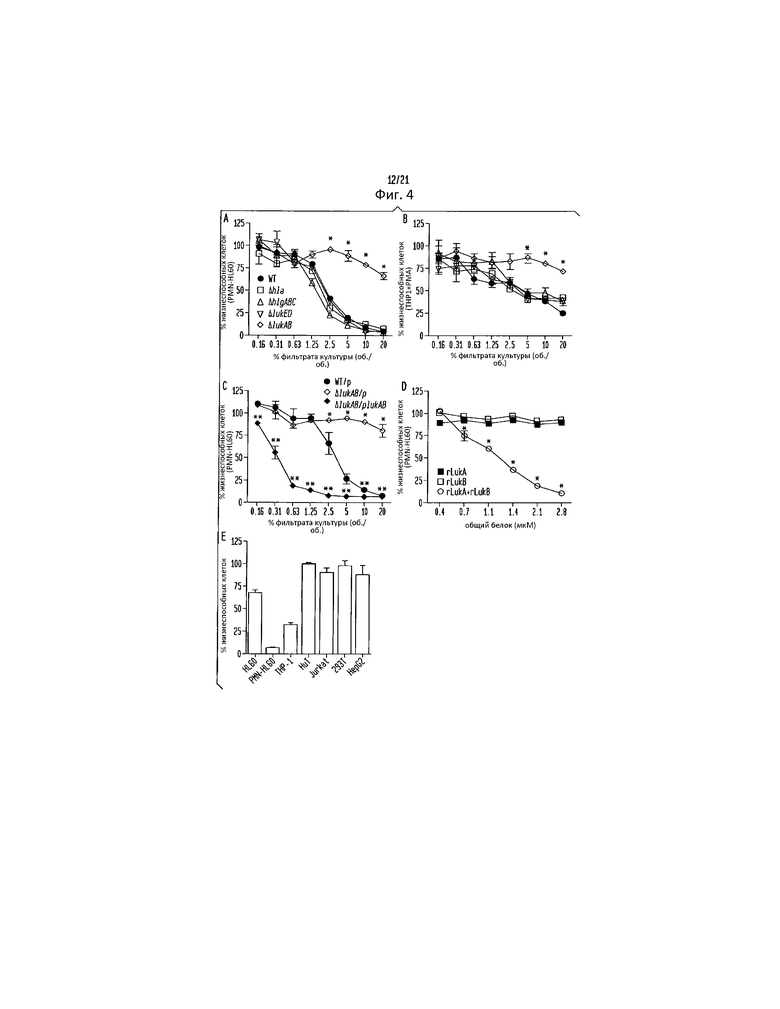



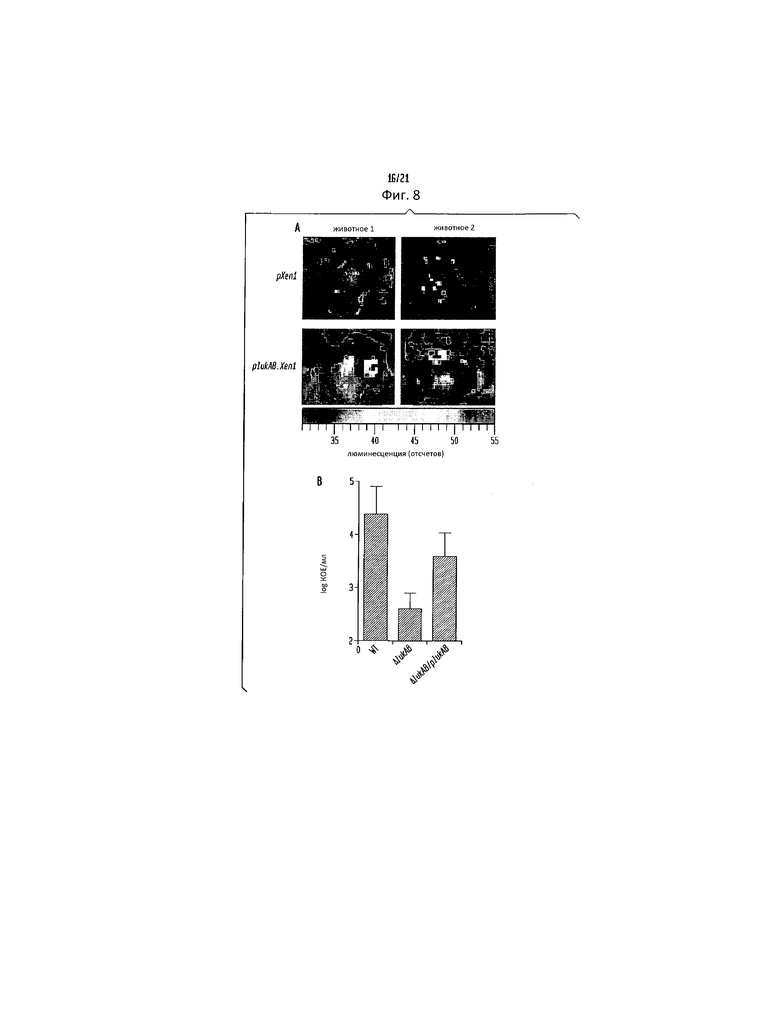

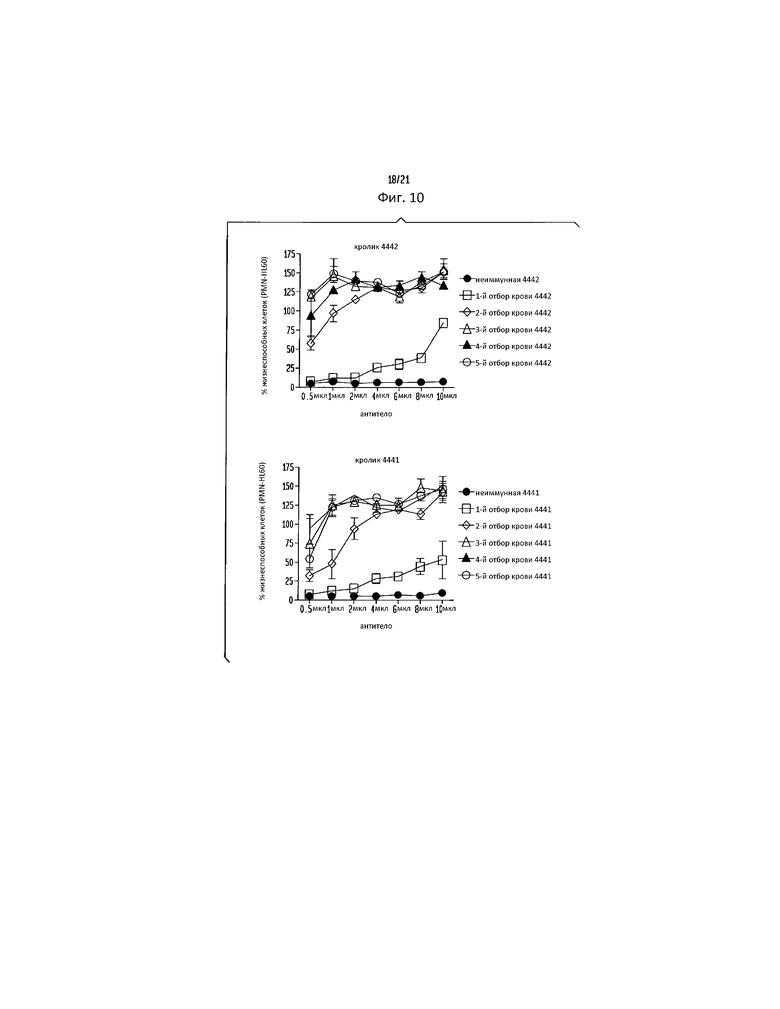

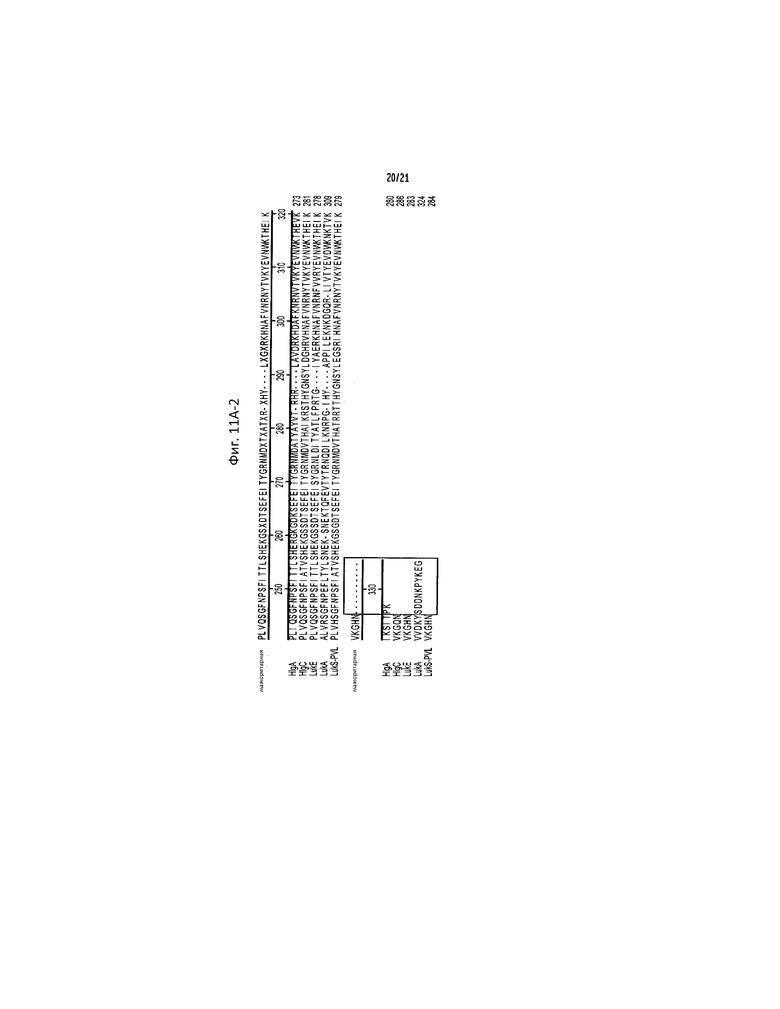
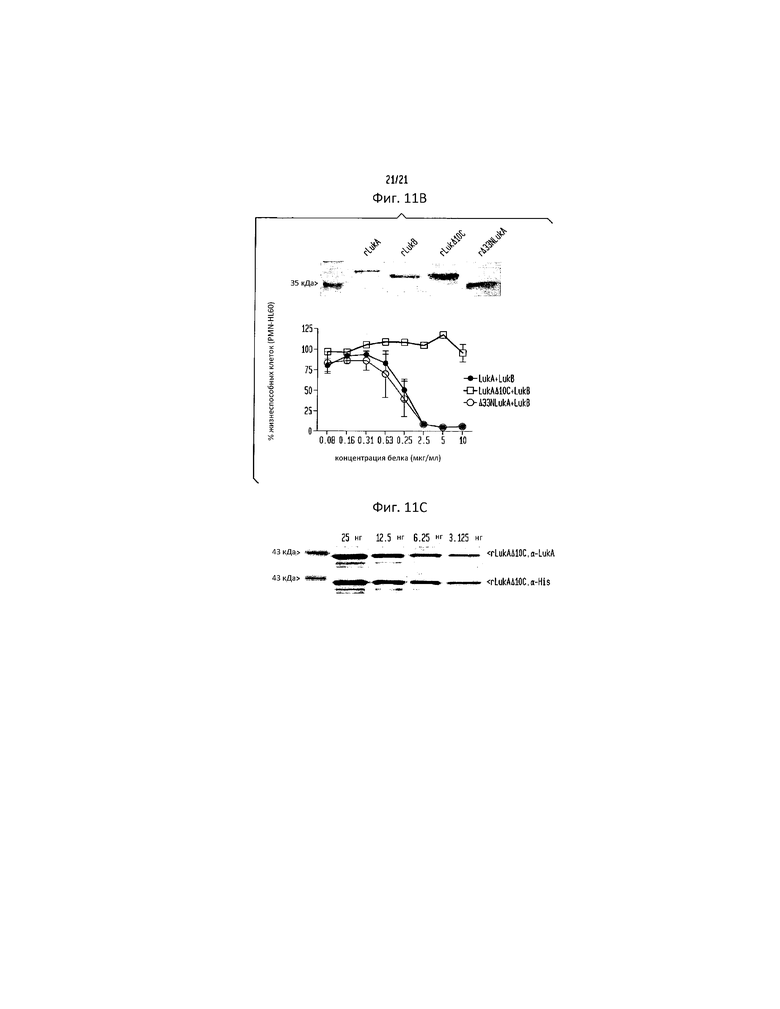
Настоящее изобретение относится к биотехнологии, в частности к композиции и способу ингибирования начала инфекции Staphylococcus aureus с применением такой композиции. Указанная композиция содержит терапевтически эффективное количество аналога полипептида лейкоцидина А S. Aureus. Настоящее изобретение позволяет повысить эффективность лечения инфекции S. aureus благодаря получению раскрытой вакцинной композиции. 2 н. и 8 з.п. ф-лы, 21 ил., 5 пр.
1. Композиция, пригодная для ингибирования начала инфекции Staphyloccocus aureus, причем указанная композиция содержит:
терапевтически эффективное количество аналога полипептида лейкоцидина A Staphyloccocus aureus, содержащего аминокислотную последовательность аминокислотных остатков 28-351 SEQ ID NO:10 или аминокислотную последовательность, имеющую по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью аминокислотных остатков 28-351 SEQ ID NO: 10, где указанный аналог полипептида лейкоцидина А содержит одну или более неконсервативных аминокислотных замен или делеций в аминокислотных остатках 342-351 SEQ ID NO: 10, которые делают указанный аналог полипептида лейкоцидина А нецитотоксичным при комбинировании с полипептидом лейкоцидина B S. aureus.
2. Композиция по п. 1, где указанный аналог полипептида лейкоцидина A содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью аминокислотных остатков 28-351 SEQ ID NO:10.
3. Композиция по п. 1, где указанный аналог полипептида лейкоцидина A не содержит аминокислотную последовательность аминокислотных остатков 342-351 SEQ ID NO:10.
4. Композиция по п. 1, дополнительно содержащая терапевтически эффективное количество полипептида лейкоцидина B S. aureus, содержащего любую аминокислотную последовательность из SEQ ID NO: 15-27, или последовательность, имеющую по меньшей мере 90% сходства последовательности с любой из последовательностей SEQ ID NO: 15-27.
5. Композиция по п. 1, дополнительно содержащая терапевтически эффективное количество полипептида лейкоцидина B S. aureus, содержащего аминокислотную последовательность аминокислотных остатков 30-339 SEQ ID NO: 16 или аминокислотную последовательность, имеющую по меньшей мере 90% сходство последовательности с аминокислотной последовательностью аминокислотных остатков 30-339 SEQ ID NO: 16.
6. Композиция по п. 5, где полипептид лейкоцидина B содержит аминокислотную последовательность, имеющую по меньшей мере 95% сходство последовательности с аминокислотной последовательностью аминокислотных остатков 30-339 SEQ ID NO: 16.
7. Композиция по п. 1, дополнительно содержащая фармацевтически приемлемый носитель.
8. Композиция по п. 5, дополнительно содержащая адъювант.
9. Способ ингибирования начала инфекции Staphyloccocus aureus, включающий введение нуждающемуся в этом субъекту-млекопитающему композиции по п. 1.
10. Способ по п. 9, где инфекция S. aureus является инфекцией метициллин-резистентного S. aureus.
| WO 2009111177 A2, 11.09.2009 | |||
| WO 2008068533 A2, 12.06.2008 | |||
| RAINARD P | |||
| et al., Leucotoxic activities of Staphylococcus aureus strain isolated from cows, ewes and goats with mastitis: importance of LukM/LukF´-PV leucotoxin, Clinical and diagnostic laboratory immunology, 2003, Vol.10, No.2, pp.272-277 | |||
| БЕСКЛЕТОЧНАЯ АНТИСТАФИЛОКОККОВАЯ ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ ХРОНИЧЕСКОЙ СТАФИЛОКОККОВОЙ ИНФЕКЦИИ | 1995 |
|
RU2122862C1 |

