Изобретение относится к области микробиологии и молекулярной генетики и может быть использовано в медицинской промышленности при производстве вакцин против Streptococcus pyogenes, стрептококка группы А (СГА).
S. pyogenes (СГА) является основной причиной заболеваемости и смертности от патогенных бактерий во всем мире [Carapetis JR и др. Lancet Infect Dis. 685–694 (2005)]. СГА могут инфицировать как кожу, так и дыхательные пути, и вызывают такие неинвазивные заболевания, как фарингиты, импетиго и скарлатина [Tanz RR, Shulman ST. Pediatr Infect Dis J. 175–176 (2007); Currie BJ. Curr Opin Infect Dis. 132–138 (2006)]. К инвазивным заболеваниям, вызываемым СГА, относятся некротический фасцит, сепсис, токсический шоковый синдром и пневмония. СГА инфекции могут приводить к серьезным осложнениям: острому пострептококковому гломерулонефриту, ревматической лихорадке или ревматическому поражению сердца [Martin JM, Green M. Semin Pediatr Infect Dis. 140–148 (2006)]. Во всем мире ежегодно от заболеваний, вызываемых СГА, умирает около 500000 человек [Carapetis JR и др. Lancet Infect Dis. 685–694 (2005)].
Заболевания, вызываемые стрептококками групп А, рассматриваются как важная социально-экономическая и медицинская проблема во всех странах мира.
Общепризнанным методом лечения бактериальных инфекций, вызванных СГА, является антибиотикотерапия. При лечении СГА инфекций препаратами выбора являются бета-лактамы: бензилпенициллин, ампициллин [Семина Н.А., Сидоренко С.В. и др., Клин. Микробиол. Антимикроб. Химиотер. 6 (4): 306-359 (2004)], ампициллин + сульбактам [Бурбелло А.Т. и др., Соврем. Лекар. Средства, СПб: Нева, 332 (2003)]; макролиды: эритромицин, кларитромицин, рокситромицин, азитромицин; ванкомицин [Зуева Л.П., Поляк М.С. и др., Микробиологический мониторинг, СПб: Медицинский информационно-аналитический центр, 1- 72 (2004)].
В целях профилактики СГА инфекций используют такие антибиотики, как бензатин бензилпенициллин [Бурбелло А.Т. и др., Соврем. Лекар. Средства, СПб: Нева, 323-324 (2003)], эритромицин [Бурбелло А.Т. и др., Соврем. Лекар. Средства, СПб: Нева, 352 (2003)], ампициллин с сульбактамом [Бурбелло А.Т. и др., Соврем. Лекар. Средства, СПб: Нева, 332 (2003)].
Широкое использование антибиотиков в терапии выявило целый ряд недостатков. Установлено, что побочные эффекты применения антибиотиков связаны с изменением биохимических характеристик крови человека, развитием аллергических реакций, дисбактериоза, иммуносупрессии, угнетения кроветворения и рядом других нарушений. Существенным недостатком терапии антибиотиками является также неизбежная селекция антибиотикорезистентных вариантов микроорганизмов, связанная с широким представительством разнообразных мобильных генетических элементов в окружающей микробиоте являющейся практически неистощимым источником генов лекарственной устойчивости. Это приводит к снижению эффективности антибактериальной терапии и необходимости постоянного поиска новых вариантов антибиотиков.
Альтернативой антибиотикотерапии стали разработки вакцинных препаратов, что продолжает оставаться перспективным направлением на протяжении последних десятилетий. В русле этой тенденции проводятся интенсивные исследования по созданию эффективных вакцин против стрептококка группы А.
Вакцины, специфичные в отношении стрептококка группы А, разрабатываются на основе:
– М белка (мультивалентные типоспецифичные вакцины и вакцины на основе консервативного фрагмента М белка);
– группового полисахарида;
– поверхностных бактериальных белков (C5a пептидаза, цистеиновая протеаза (экзотоксин В), фибронектин-связывающий белок (Sfb1), пили (антиген Т), экзотоксин С, сериновая протеаза (SpyCEP), сериновая эстераза (Sse), супероксиддисмутаза, GRAB, MtsA, липопротеины).
Типоспецифичные мультивалентные вакцины на основе М белка состоят из коротких пептидов, гомологичных N-терминальному концу М белка разных emm серотипов, собранных вместе в один большой полипептид. Так созданы 6, 26 и 30 валентные вакцины, включающие пептиды из 6, 26 и 30 серотипов СГА (соответственно) [Kotloff KL и др. JAMA; 292(6), 709–715 (2004); McNeil SA и др. Clin Infect Dis. 41(8). 1114–1122 (2005); Dale JB и др. Vaccine. 29(46). 8175–8178 (2011)]. Основным недостатком таких вакцин является их узкая типовая специфичность, поскольку сегодня существует более 220 серотипов СГА [McMillan D. J. и др. Clinical Microbiology and Infection; 19 (5), 222-229 (2013)].
Консервативный участок М белка также рассматривается некоторыми учеными в качестве компонента вакцины. Так исследуются полноразмерный консервативный фрагмент из C-повторов [Bessen D, Fischetti VA. Infect Immun; 56, 2666–2672 (1988); Bronze MS и др. J Immunol; 148, 888-893 (1992)], маленькие пептиды J8, J14 и p145, гомологичные консервативному участку из С-повторов [ Batzloff M и др. Indian J Med Res; 119, 104-107 (2004)], эпитоп StreptInCor vaccine [Guilherme L и др. Methods; 49 (4), 316-321 (2009)], поливалентный вакцинный кандидат SV1[Bauer MJ и др. Vaccine; 30 (12), 2197-2205 (2012)]. Однако спорным остается вопрос: обладают ли антитела к консервативной области М белка опсонизирующими и защитными свойствами против СГА, так как исследуемая консервативная область белка находится в толще клеточной стенки и недоступна антителам.
Групповой полисахарид СГА также рассматривается в качестве вакцинного кандидата против СГА [Sabharwal H и др. J Infect Dis; 193 (1), 129-135 (2006)]. Однако антитела, синтезируемые к групповому полисахариду могут перекрестно взаимодействовать с тканями человеческого организма [Cunningham MW и др. J Immunol; 141, 2760-2766 (1988); Dudding BA, Ayoub EM. J Exp Med 1968;128:1081-1098], что и является основным недостатком в его использовании в качестве компонента вакцины против СГА.
Аналогами заявляемого изобретения являются вакцинные препараты, полученные на основе рекомбинантных белков. В качестве компонентов вакцины против СГА рассматриваются поверхностные факторы патогенности СГА белковой природы. В качестве вакцинных кандидатов рассмотрены рекомбинантные полипептиды, полученные на основе: C5a пептидаза (ScpA) [Cleary PP и др. J Infect Dis; 188 (6), 809-817 (2003)], цистеиновая протеаза экзотоксин В (Spe B) [Kapur V и др. Microb Pathogen; 16, 443 (1994); Chih-Feng Kuo и др. PLoS One; 10 (1), 1-17 (2015)], фибронектин-связывающий белок (Sfb1) [Kawabata S и др. Infect Immun; 69 (2), 924 -930 (2001)], пили (антиген Т) [Koller T и др. J Med Microbiol; 59, 442-452 (2011)], экзотоксин С (Spe C) [McCormick JK и др. J Immunol; 165 (4), 2306-2312 (2000)], сериновая протеаза (SpyCEP) [Zingaretti C и др. FASEB J; 24 (8), 2839-2848 (2010)], сериновая эстераза (Sse) [Liu M и др. Infect Immun; 75 (7), 3651-3657 (2007)], супероксиддисмутаза [McMillan DJ и др. Indian J Med Res; 119, 121-125 (2004)], GRAB, MtsA [McMillan DJ и др. Vaccine; 22, 2783-2790 (2004)], липопротеины [Lei B и др. J Infect Dis. 189 (1), 79-89 (2004)].
Недостатком вакцинных препаратов полученных на основе одного из перечисленных выше белков является их различная экспрессия на разных фазах роста микроорганизма, а также вариабельность факторов патогенности среди СГА.
В представленном изобретении заявляется химерный белок, состоящий из фрагментов поверхностных белков СГА: Sic, SpySEP, SpeA, ScpA, ScaAB и терминальных доменов флагеллина Salmonella typhiurium FliA и FliC, выполняющих функцию внутреннего адьюванта, являющегося рецептором Toll 5.
СГА секретирует ингибитор комплемент-опосредованного лизиса Sic молекулярной массой 30 кДа [Akesson P и др. J Biol Chem. 271: 1081-1088 (1996)]. Ранее было продемонстрировано, что Sic блокирует комплемент-опосредованный гемолиз, ингибирует активность противобактериальных пептидов и связывается с белками человеческой плазмы (кластерином и гистидин богатым гликопротеином) [Binks M. J. и др. J Biol Chem. 280: 20120–20125 (2005); Fernie-King B.A. и др. Infect Immun. 70: 4908–4916 (2002); Frick I. M. и др. J Biol Chem. 278: 16561–16566 (2003)]. Также было доказано, что Sic связывается с высокомолекулярными и низкомолекулярными кининогенами (ВМК и НМК, соответственно), ингибируя, таким образом, связывание ВМК с клетками эндотелия и уменьшая активацию контактной фазы плазменного протеолиза [Akesson P и др. Microbiology. 156: 3660-3668 (2010)].
Хемокиновая протеаза SpySEP является поверхностно-связанной сериновой протеазой субтилизинового семейства молекулярной массой 170 кДа. Она инактивирует IL8, специфически расщепляя его С-конец [Edwards R. J. и др. J. Infect. Dis. 192: 783-790 (2005)]. Это приводит к снижению привлечения нейтрофилов в очаг инфекции и последующего бактериального клиренса [Zinkernagel A.S. и др. Cell Host Microbe 4: 170-178 (2008); Hidalgo-Grass C. И др. Lancet. 363: 696-703 (2004)]. На мышиной модели было показано, что анти-SpySEP антитела обладают протективной защитой, что доказывает целесообразность его использования в качестве компонента вакцины против СГА [Rodriguez-Ortega M. J. и др. Nat. Biotechnol. 24: 191-197 (2006); Turner C.E. и др. Vaccine. 27: 4923-4929 (2009)].
Стрептококковый пирогенный экзотоксин SpeA – суперантиген молекулярной массой 25,7 кДа, участвующий в развитии клиники скарлатины, а также формировании стрептококкового токсического шокового синдрома, некротического фасцита, острой ревматической лихорадки [Bohach G.A. и др. Crit. Rev. Microbiol. 17: 251–272 (1990); Hauser A.R. и др. J. Clin. Microbiol. 29: 1562–1567 (1991)]. Суперантигены – это семейство белков, характеризующиеся способностью вызывать чрезмерную активацию Т лимфоцитов человека и млекопитающих, что приводит к избыточному высвобождению Т-клеточных медиаторов и провоспалительных цитокинов, приводящих к возникновению токсического шокового синдрома [Thomas Proft и John D. Fraser в книге Ferretti JJ и др. Streptococcus pyogenes: Basic Biology to Clinical Manifestations (2016)]. Вакцинация кроликов SpeA защищала животных от возникновения всех симптомов стрептококкового токсического шокового синдрома, включая некротический фасцит [Schlievert P.M. И др. J. Lab. Clin. Med. 127: 13–22 (1996)]. Мутанты СГА по гену, кодирующему SpeA, стимулировали индукцию протективных антител у кроликов [Roggiani M и др. Infect Immun. 68: 5011-5017 (2000)].
ScpA (С5а пептидаза) – поверхностный белок экспрессируемый всеми серотипами СГА. ScpA – сериновая протеаза молекулярной массой 125 кДа, специфически расщепляющая С5а компонент комплемента человека, обезьяны и коровы, тем самым блокируя сайт связывания С5а с лейкоцитами [Cleary PP и др. Infect Immun. 60: 5219-5223 (1992)]. В результате ScpA ингибирует хемотаксис, задерживая инфильтрацию фагацитов, замедляя клиренс бактерий с подкожных поверхностей и со слизистых оболочек организма хозяина и, таким образом, позволяет СГА создавать очаг инфекции [Ji Y и др. Infect Immun. 64: 503-510 (1996); Ji Y и др. Infect Immun. 66: 5399-5405 (1998)].
Интраназальная иммунизация мышей рекомбинантным ScpA вызывала сильный гуморальный иммунный ответ и приводила к снижению носоглоточной колонизации после заражения различными сиротипами СГА, что свидетельствует о перекрестной защитной реакции [Ji Y и др. Infect Immun. 65: 2080-2087 (1997)]. Также было продемонстрировано, что кроличьи анти-ScpA антитела и человеческая сыворотка против ScpA могут нейтролизовать активность С5а пептидазы как in vitro, так и in vivo у мышей [Wexler DE, Cleary PP. Infect Immun. 50: 757-764 (1985); O’Connor SP, Cleary PP. Infect Dis. 156: 495-504 (1987)].
Белок ScaAB СГА также был выбран в качестве компонента вакцины. ScaAB СГА гомологичен белку ScaAB Streptococcus agalactiae и белку PsaA Streptococcus pnumoniae. Последние являются хорошо изученными белками, с доказанной иммуногенностью и протективностью антител, индуцируемых на их введение [Vorobieva E.I., Meringova L.F. и др. Folia Microbiol.50: 172-176 (2005); Berry AM, Paton JC. Infect Immun. 64: 5255-5262 (1996); Ren B и др. Infect Immun. 71: 75-85 (2003)].
В качестве внутренного адьюванта в состав химерного белка были включены терминальные домены флагеллина Salmonella typhiurium FliA и FliC из-за их способности взаимодействовать с Toll-подобными рецепторами-5 (TLR-5) [Simon R1, Levine MM. Hum Vaccin Immunother. 8: 494-498 (2012); Ramachandran G. и др. PLoS One. 11. (2016)].
Прототипом заявляемого изобретения является 6, 26 и 30 валентные вакцины, состоящие из коротких остатков, гомологичных N-терминальному концу М белка 6, 26 и 30 серотипов СГА, соответственно [Kotloff KL и др. JAMA; 292(6), 709–715 (2004); McNeil SA и др. Clin Infect Dis. 41(8). 1114–1122 (2005); Dale JB и др. Vaccine. 29(46). 8175–8178 (2011)]. Однако такие вакцины обладают протективными свойствами в отношении только тех серотипов СГА, на основе которых они были созданы.
Прототипом заявленного изобретения является рекомбинантный слитый белок SpeAB, состоящий из комбинации инактивированных измененных форм стрептококкового пирогенного экзотоксина А и экзотоксина В СГА [Ulrich RG. J Immune Based Ther Vaccines. (2008); Morefield G. и др. Vaccine. 32: 3810-3815 (2014)]. Авторами на мышиной модели были продемонстрированы протективные свойства анти-SpeAB антител. Однако остается открытым вопрос о стабильности данного слитого белка.
Прототипом заявляемого изобретения является синтетический липопептид, состоящий из В-клеточного эпитопа М белка J14, фибриллобразующего пептида Q11 и липоаминокислотного фрагмента С16 [Azmi F и др. Bioorg Med Chem. 15: 6401-64028 (2014)]. Авторами была доказана иммуногенность липопептида и его стабильность.
Задачей данного изобретения стало получение химерного рекомбинантного белка SGA-CHIM (FliA-FliC-Sic-SpySEP-SpeA-ScpA-ScaAB), состоящего из консервативных и иммуногенных фрагментов аминокислотных последовательностей поверхностных белков: Sic, SpySEP, SpeA, ScpA, ScaAB и терминальных доменов флагеллина Salmonella typhiurium FliA и FliC, выполняющих функцию внутреннего адьюванта; обладающего иммуногенными свойствами за счет синтеза специфических антител, и выраженными протективными свойствами анти-SGA-CHIM антител. Химерный рекомбинантный белок SGA-CHIM используют в качестве вакцинного препарата против Streptococcus pyogenes.
Поставленная задача решалась:
а) получением штамма-продуцента SGA-CHIM;
б) получением химерного рекомбинантного белка SGA-CHIM.
Сущностью предлагаемого изобретения является создание уникального химерного рекомбинантного белка SGA-CHIM, обладающего выраженным иммуногенными и протективным эффектами.
Авторами заявляемого изобретения получены анти-SGA-CHIM антитела при интраназальном и подкожном введении SGA-CHIM мышам.
Авторами заявляемого изобретения изучены:
1) иммуногенность химерного рекомбинантного белка SGA-CHIM;
2) протективные свойства анти-SGA-CHIM антител мыши в отношении СГА in vivo.
Уникальный химерный рекомбинантный белок SGA-CHIM имеет высокую физиологическую стабильность, вызывает синтез анти-SGA-CHIM антител, причем синтезируемые специфические антитела обладают протективными свойствами против Streptococcus pyogenes.
Уникальный химерный рекомбинантный белок SGA-CHIM полезен в качестве вакцинного препарата для профилактики и лечения бактериальных инфекций, вызванных Streptococcus pyogenes.
Результатом заявленного изобретения является создание химерного рекомбинантного белка SGA-CHIM.
Химерный рекомбинантный белок SGA-CHIM получали с использованием генно-инженерных методов. Аминокислотная последовательность химерного рекомбинантного белка SGA-CHIM была сконструирована и транслирована в нуклеотидную последовательность, оптимизированную для экспрессии в E. coli. Синтез нуклеотидной последовательности гена, кодирующей белок SGA-CHIM, осуществляли путем удлинения перекрывающихся олигонуклеотидов, как описано у Majumder K. Gene. 110: 89-94 (1992). Для клонирования на концах гена были добавлены сайты рестрикции для эндонуклеаз EcoRI и XhoI.
Синтетический химерный ген sga-chim был проклонирован в плазмиду pET24b+ (Invitrogen, США). Полученную плазмиду, названную pColdIIsga-chim (6012 п.н.), трансформировали в штамм E. coli BL21 Star F- ompT hsdSB (rB-mB-) gal dcm rne131 (DE3) (Invitrogen, США) с помощью «кальциевого метода». Отбор клонов-трансформантов проводили через 18 часов роста клеток при 37°С на плотной среде с 1% L-агаром (Difco, США), содержащей ампициллин (100мкг/мл). Штамм-продуцент химерного рекомбинантного белка SGA-CHIM назвали E. coli BL21 (DE3) pColdIIsga-chim.
Для получения чистого препарата химерного рекомбинантного белка клетки штамма-продуцентав E. сoli BL21 (DE3) pColdIIsga-chim культивировали на L-бульоне (Difco, США) с добавлением ампициллина (100мкг/мл) до поздней логарифмической фазы роста (ОD600=0,7 ÷ 0,9). Затем продукцию химерного рекомбинантного белка индуцировали добавлением изопропил-бета-D-тиогалактопиранозида (IPTG), и клетки культивировали еще 4,5 часа. После этого клетки осаждали центрифугированием, суспендировали в лизирующем (связывающем) буфере А (20 мМ Na2HPO4, 20 мM NaH2PO4, 500 мM NaCl, 20 мM имидазола, лизоцим (10-3 мг/мл) и PMSF (0,03М), рН=7.4). Суспензию клеток вскрывали ультразвуком (3 раза по 20 секунд с перерывом 40 секунд). Разрушенные клетки подвергали центрифугированию (20000 g в течение 20 минут при температуре 40С). Полученный осадок отделяли от супернатанта и растворяли в связывающем буфере, содержащем мочевину (8 М раствор), материал центрифугировали для отделения лизата от не растворившихся фрагментов. Полученную надосадочную жидкость наносили на Ni-сефарозу.
Очистку рекомбинантного белка проводили с использованием Ni-сефарозы (Amersham, США). Полученный после аффинной хроматографии рекомбинантный белок диализовали против дистиллированной воды рН=8,0. Чистоту и молекулярную массу химерного рекомбинантного полипептида определяли с помощью 14% SDS-PAGE в вертикальном пластинчатом аппарате Mini-PROTEAN II (BioRad, США), сравнивая пробег белка с пробегом белков известной молекулярной массы (Precision Plus Protein standards (№161-0373), Bio-Rad, США). Молекулярная масса SGA-CHIM = (70±0,5) кДа.
Результатом изобретения является химерный рекомбинантный белок SGA-CHIM, состоящий из консервативных и иммуногенных фрагментов аминокислотных последовательностей поверхностных белков: Sic, SpySEP, SpeA, ScpA, ScaAB и терминальных доменов флагеллина Salmonella typhiurium FliA и FliC.
Белок SGA-CHIM обладает следующими свойствами:
а) при интраназальном введении в организм млекопитающим (мышам) вызывает синтез анти-SGA-CHIM антител;
б) синтезируемые специфические антитела по п. (а) обладают протективными свойствами в отношении Streptococcus pyogenes;
Ниже приводятся конкретные примеры, иллюстрирующие некоторые варианты изобретения, но не ограничивающие его.
Пример 1. Выделение и очистка химерного рекомбинантного белка SGA-CHIM.
Клонирование синтетического гена sga-chim осуществляли с использованием экспрессионной плазмиды pET24b+ (Invitrogen, США). При подготовке к клонированию была проведена двойная рестрикция sga-chim (1674 п.н.) и плазмиды pET24b+ (4368 п.н.) ферментами EcoRI и XhoI с образованием липких концов. В ходе клонирования была получена рекомбинантная плазмидная ДНК (обозначенная как pColdIIsga-chim (6042 п.н.). Рекомбинантную плазмидную ДНК pColdIIsga-chim трансформировали в гетерологичную систему E. coli BL21 Star F- ompT hsdSB (rB-mB-) gal dcm rne131 (DE3). Генетическим маркером плазмидной ДНК pET24b+ является ген amp, кодирующий бета-лактамазу, что обеспечивает устойчивость к ампициллину клеток, несущих данную плазмиду. Трансформацию клеток E. coli проводили по Darget (1979). Отбор клонов-трансформантов проводили через 18 часов роста клеток при 37°С на плотной среде с 1% L-агаром (Difco, США), содержащей ампициллин (100мкг/мл). В результате проведенного клонирования были получены рекомбинантные клоны, несущие плазмиду pColdIIsga-chim с синтетическим геном sga-chim. Один из рекомбинантных клонов получил название E. coli BL21 (DE3) pColdIIsga-chim и был использован для дальнейшей работы.
Клон E. coli BL21 (DE3) pColdIIsga-chim проверяли на способность продуцировать белок SGA-CHIM. Для получения чистого препарата SGA-CHIM клетки E. coli BL21 (DE3) pColdIIsga-chim культивировали на L-бульоне (Difco, США) с добавлением ампициллина (100мкг/мл). Продукцию и очистку (с использованием Ni-сефарозы, Amersham, США) SGA-CHIM проводили как описано выше.
Молекулярную массу SGA-CHIM определяли с помощью 14% SDS-PAGE, сравнивая пробег белка SGA-CHIM с пробегом белков известной молекулярной массы (Precision Plus Protein standards (№161-0373), Bio-Rad, США). Молекулярная масса химерного рекомбинантного белка SGA-CHIM оказалась равной (70,0±0,5) кДа.
Аминокислотная последовательность химерного рекомбинантного белка SGA-CHIM представлена в перечне последовательностей под № 1 и состоит из 548 аминокислотных остатков.
Химерный рекомбинантный белок SGA-CHIM содержит аминокислотные последовательности поверхностных белков Sic, SpySEP, SpeA, ScpA, ScaAB и терминальных доменов флагеллина Salmonella typhiurium FliA и FliC, ковалентно связанных с аминокислотными остатками, кодируемыми pET24b+ (Invitrogen, США).
Пример 2. Изучение динамики изменения титра анти-SGA-CHIM антител в ходе интраназальной иммунизации мышей белком SGA-CHIM.
Иммунный ответ приобретенного иммунитета к химерному рекомбинантному белку SGA-CHIM изучали на беспородных белых мышах (самки, возраст – 10 недель, вес 22÷25 г), полученных из питомника лабораторных животных «Рапполово». Иммунизацию проводили двукратно с интервалом в 21 день и интраназально под легким эфирным наркозом. Группе мышей из 50 особей вводили 0,05 мл препарата, содержащего 20 мкг SGA-CHIM. Специфические антитела к SGA-CHIM определяли с 1 по 47 день от начала иммунизации. Специфические антитела определяли с помощью двухслойного метода иммуноферментного анализа (ИФА). В лунки полистеролового планшета с высокой сорбционной емкостью (Costar, США) вносили по 100 мкл SGA-CHIM в концентрации 2 мкг/мл. Сорбцию проводили в 100 мМ бикарбонатном буфере (рН=9,45) в течение 16 ÷ 20 часов при 4°С. После этого содержимое из планшета удаляли и добавляли в лунки по 150 мкл 0,15 М NaCl, рН=7,4 (PBS), содержащего 0,05% Твин–20 (PBST). Инкубацию проводили при 37°С в течение 30 мин. Содержимое планшета удаляли и трижды промывали PBST. Затем в лунки планшета вносили иммунную или нормальную (в качестве отрицательного контроля) сыворотку (с начальным разведением 1:100) с последующим шагом разведения равным двум. Все разведения осуществляли в PBST, и каждую пробу дублировали. Инкубацию проводили в течение 1 часа при 37°С. Далее содержимое планшета удаляли и дважды промывали планшет PBST. После этого в лунки добавляли по 100 мкл А-ПХ конъюгата (белок А стафилококка, ковалентно связанный с пероксидазой хрена (Sigma, США) в концентрации 10-7 моль/л. После часовой инкубации при 37°С содержимое планшета удаляли и трижды отмывали планшет PBST и один раз PBS, чтобы избежать ингибирующего действия Твин-20 на ферментативную активность пероксидазы. Далее для визуализации реакции в лунки планшета вносили по 100 мкл субстратной смеси (0,5 мкг/мл тетраметилбензидина (Sigma, США) в 100 мМ фосфатно–цитратном буфере (рН=5,0), 10 мкл 30% перекиси водорода), которую готовили непосредственно перед применением. Планшет инкубировали в течение 30 мин в темноте при комнатной температуре, и реакцию останавливали внесением в лунки по 30 мкл 50% концентрированной серной кислоты. Реакцию регистрировали при длине волны 450 нм с помощью прибора iMark Microplate Reader, Bio-Rad (США).
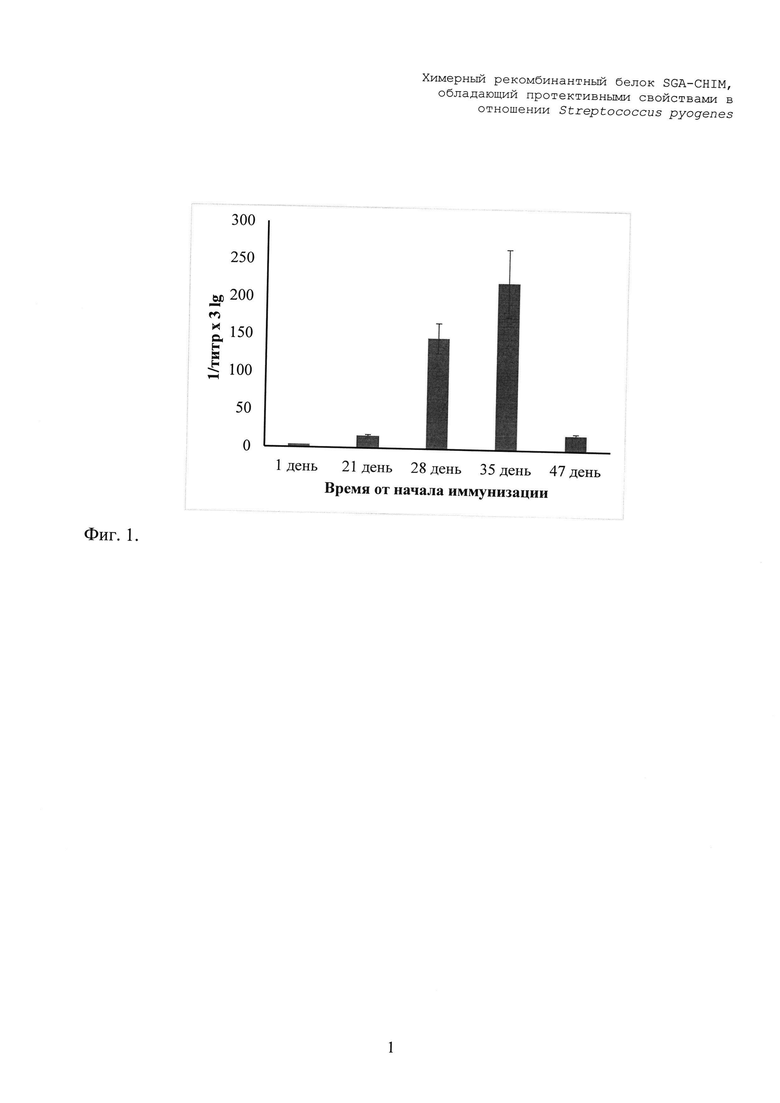
Через три недели от начала иммунизации в крови мышей регистрировали появление специфических антител к SGA-CHIM. На фиг. 2. представлена динамика изменения титра анти-SGA-CHIM антител в ходе интраназальной иммунизации мышей SGA-CHIM.
Максимум иммунного ответа наблюдали на 35 день от начала иммунизации с титром антител равным 222х103. Также отмечали наличие специфических антител через 47 дней с титром 21х103. Результаты, представленные на фиг. 1 демонстрируют, что интраназальное введение SGA-CHIM приводило к выраженной индукции анти-SGA-CHIM в организме мышей.
Пример 3. Изучение протективных свойств анти-SGA-CHIM антител при интраназальном заражении мышей СГА 9189 (серотип М1).
Протективные свойства анти-SGA-CHIM антител изучали на мышах в экспериментах in vivo на модели генерализованной инфекции. Беспородных белых мышей (самок, 22÷25 г) из питомника «Рапполово» в количестве 20 особей сначала иммунизировали по схеме, приведенной в примере 2. Контрольной группе вводили PBS. Затем обеим группам мышей интраназально под легким эфирным наркозом вводили суспензию СГА М1 серотипа(коллекционный номер 9189) в объеме 25 мкл в дозе 10×108 КОЕ на сроке с максимальной выработкой антител к SGA-CHIM (35-й день от начала иммунизации). Streptococcus pyogenes серотипа M1 (9189) был получен из коллекции отдела молекулярной микробиологии ФГБНУ «ИЭМ». Бактерии культивировали в среде THB в течение 24 часов при 37°C в аэробных условиях, отмывали PBS трехкратным центрифугированием при 3500 об/мин в течение 20 минут и концентрировали при необходимости. Полученную суспензию использовали для заражения.
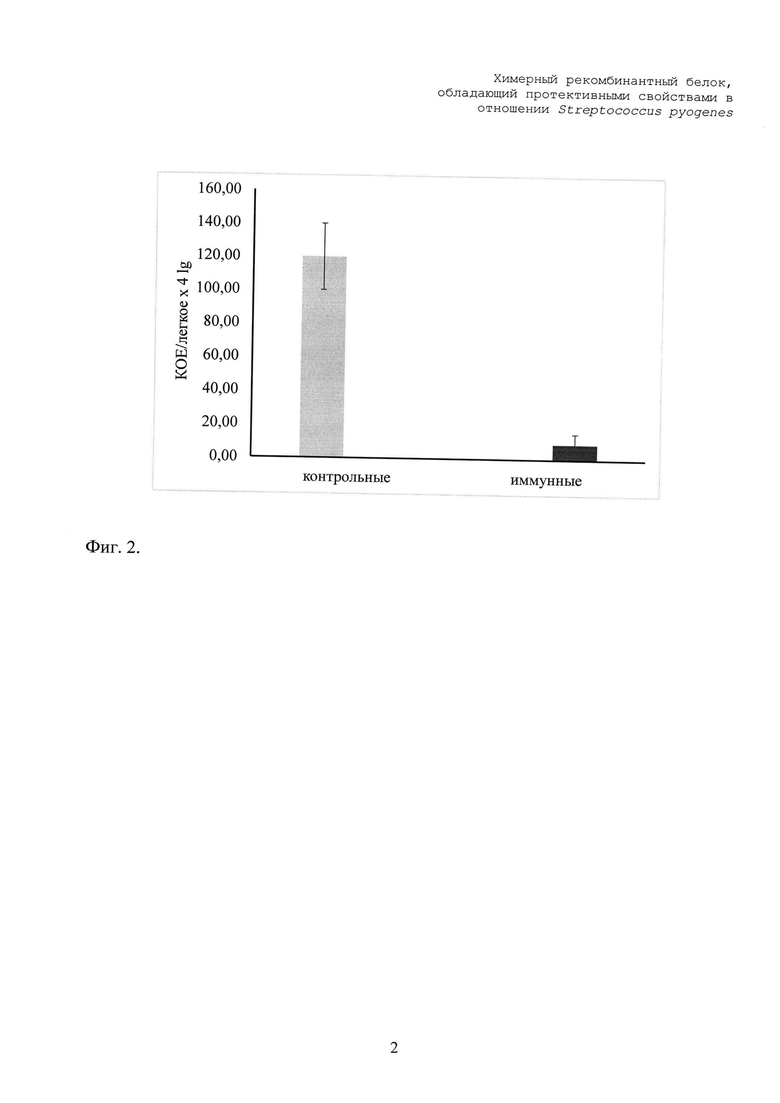
Контроль развития инфекции осуществляли с помощью высева стрептококков из гомогенатов легких мышей на колумбийский агар (Difco, США) с добавлением 5% бараньих эритроцитов через 2 часа после заражения СГА. Ткань легкого измельчали на механическом дезинтеграторе Retsch MM400 в течение 3 минут при интенсивности 29 колебаний в секунду.
Как показано на фиг. 2, через 2 часа после интраназального заражения мышей СГА № 9189 серотипа М1 в легких контрольной группы мышей происходило интенсивное накопление СГА, достигая значения 1,2х106 КОЕ. В противоположность этому через 5 часов после введения бактерий наблюдалась элиминация стрептококков из легких мышей, иммунизированных SGA-CHIM.
Полученные данные убедительно показывают протективность специфических антител, индуцированных введением SGA-CHIM.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ ПОЛУЧЕНИЕ РЕКОМБИНАНТНОГО БЕЛКА PB1, ОБЛАДАЮЩЕГО ПРОТЕКТИВНЫМИ СВОЙСТВАМИ В ОТНОШЕНИИ STREPTOCOCCUS PYOGENES И STREPTOCOCCUS AGALACTIAE | 2008 |
|
RU2378374C2 |
| КОМПЛЕКС РЕКОМБИНАНТНЫХ ПОЛИПЕПТИДОВ, ОБЛАДАЮЩИЙ ПРОТЕКТИВНЫМИ СВОЙСТВАМИ В ОТНОШЕНИИ STREPTOCOCCUS AGALACTIAE И STREPTOCOCCUS PYOGENES | 2014 |
|
RU2587627C2 |
| СПОСОБ ВВЕДЕНИЯ ГЕНОВ ПАТОГЕННЫХ СТРЕПТОКОККОВ В ХРОМОСОМНУЮ ДНК ПРОБИОТИЧЕСКОГО ШТАММА ENTEROCOCCUS FAECIUM L3 ДЛЯ ЭКСПРЕССИИ В ПИЛЯХ | 2015 |
|
RU2640250C2 |
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |
| Живая вакцина на основе штамма пробиотиков ENTEROCOCCUS FAECIUM L3 для профилактики инфекции, вызванной STREPTOCOCCUS PNEUMONIE | 2018 |
|
RU2701733C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pVA891-2[mutR], СПОСОБ ПОЛУЧЕНИЯ МУТАНТНЫХ ПО ГЕНУ mutR ШТАММОВ Streptococcus pyogenes, РЕКОМБИНАНТНЫЕ ШТАММЫ Streptococcus pyogenes (ВАРИАНТЫ) ДЛЯ ПРОФИЛАКТИКИ СТРЕПТОКОККОВЫХ ИНФЕКЦИЙ | 2010 |
|
RU2472856C2 |
| ВАКЦИНА, СПОСОБНАЯ СООБЩАТЬ ХОЗЯИНУ ИММУНИТЕТ ПРОТИВ ИНФЕКЦИИ, ВЫЗВАННОЙ STREPTOCOCCUS ГРУППЫ B, СПОСОБ ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИИ, ВЫЗВАННОЙ STREPTOCOCCUS ГРУППЫ B, И СПОСОБ ОСЛАБЛЕНИЯ ИНФЕКЦИИ, ВЫЗВАННОЙ STREPTOCOCCUS ГРУППЫ B | 1990 |
|
RU2113234C1 |
| Способ получения химерного рекомбинантного белка fliC:pagN | 2015 |
|
RU2627602C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ВАКЦИНЫ | 2010 |
|
RU2444570C1 |
| МУТАНТНЫЕ ФОРМЫ СТРЕПТОЛИЗИНА О | 2008 |
|
RU2498994C2 |


Изобретение относится к области биотехнологии, микробиологии и молекулярной генетики. Заявлен химерный рекомбинантный белок, состоящий из консервативных и иммуногенных фрагментов аминокислотных последовательностей поверхностных белков Streptococcus pyogenes: Sic, SpySEP, SpeA, ScpA, ScaAB и терминальных доменов флагеллина Salmonella typhiurium FliA и FliC, выполняющих функцию внутреннего адъюванта. Заявленный белок вызывает in vivo синтез антител класса G, специфических к указанному химерному белку и обладающих протективными свойствами в отношении Streptococcus pyogenes. Изобретение может быть использовано в медицинской промышленности при производстве вакцин против стрептококка группы А (СГА) Streptococcus pyogenes. 2 ил., 3 пр.
Химерный рекомбинантный белок, состоящий из консервативных и иммуногенных фрагментов аминокислотных последовательностей поверхностных белков Streptococcus pyogenes: Sic, SpySEP, SpeA, ScpA, ScaAB и терминальных доменов флагеллина Salmonella typhiurium FliA и FliC, выполняющих функцию внутреннего адъюванта, аминокислотная последовательность которого представлена под номером SEQ ID NO: 1, вызывающий in vivo синтез антител класса G, специфических к указанному химерному белку и обладающих протективными свойствами в отношении Streptococcus pyogenes.
| GOOD M.F | |||
| et al., Strategic development of the conserved region of the M protein and other candidates as vaccines to prevent infection with group A streptococci, Expert review of vaccines, 2015, Vol.14, No.11, pp.1459-1470 | |||
| RU 2010107878 A, 20.09.2011 | |||
| SONG Y | |||
| et al., Progress in development of group A streptococcus vaccines, Current pharmaceutical biotechnology, 2013, Vol.14, No.11, pp.947-950. |

