Изобретение относится к биотехнологии, конкретно к биофармакологии, к областям получения и производства иммунобиологических препаратов, медицинской микробиологии, созданию фармакологических и вакцинных композиций, биотехнологиям продукции рекомбинантных белков, и касается разработки нового способа получения рекомбинантного химерного белка на основе белков fliC и pagN Salmonella typhimurium (fliC:pagN), обладающего высокой иммуногенностью и адъювантными свойствами, применимого для создания высокоэффективной конъюгированной вакцины в целях профилактики сальмонеллезной инфекции.
Основной сферой применения предлагаемого способа получения рекомбинантного химерного fliC:pagN являются биомедицина, создание медицинских профилактических препаратов, лабораторные биологические и медицинские исследования, создание субъединичных и конъюгированных вакцин и вакцинных штаммов для профилактики кишечных инфекций, вызванных бактериями рода Salmonellae.
Белок fliC, наряду с fliB, является флагеллином (основным компонентом флагелл - органелл бактерии, обеспечивающих ее подвижность) Salmonella sp. (Bonifield H.R., Hughes K.T. // J. Bacteriol. - 2003. - V. 185. - P. 3567-3574). Вместе с этим, FliC, как и некоторые другие флагеллины, в силу его экспозиции на поверхности бактерий, является классическим «молекулярным паттерном, ассоциированным с повреждениями» (DAMP, damage-associated molecular pattern), и распознается одним из рецепторов «примитивной» системы врожденного иммунитета, а именно толл-подобным рецептором 5 (TLR5, Yoon S.I., Kurnasov О., Natarajan V., Hong М., Gudkov А.V., Osterman A.L., Wilson I.A. // Science. - 2012. - V. 335. - P. 859-64). Рецепторы этого типа, в свою очередь, обозначаются, как паттерн-распознающие рецепторы (PRR), которые узнают паттерны, ассоциированные с патогенами (PAMP, Kumar S. // Crit. Rev. Microbiol. - 2013. - V. 39. - P. 229-246). Необходимо отметить, что многие из PRR рассматриваются в качестве важнейших мишеней для молекулярных вакцинных адъювантов (Olive С. // Expert Rev. Vaccines. - 2012. - V. 11. - P. 237-256), в том числе, и система TLR5-флагеллин (Mizel S.B. // J. Immunol. - 2010. - V. 185. - Р. 5677-5682). Также существенно, что для индукции выработки антител флагеллин использует дополнительные механизмы, не связанные напрямую с активацией системы врожденного иммунитета (Sanders C.J. // Eur. J. Immunol. - 2009. - V. 39. - P. 359-371), что можно рассматривать как преимущество с точки зрения эффективности протективного гуморального иммунного ответа.
Белок pagN является мембранным белком сальмонелл с рядом доменов, экспонированных во внешнюю среду. Функцией этого белка является адгезия к клеткам организма-хозяина и участие в инвазии патогена в клетку (Lambert М.А., Smith S.G. // ВМС Microbiol. - 2008. - V. 8. - P. 142). Таким образом, pagN является важнейшим участником процесса инфекции сальмонелл, и его связывание, в частности, антителами, может блокировать процесс инвазии. Недавние исследования показали, что pagN является достаточно эффективным протективным антигеном для профилактики сальмонеллезной инфекции (Yang Y., Wan С., Xu Н., Aguilar Z.P., Tan Q., Xu F., Lai W., Xiong Y., Wei H. // Microbes Infect. - 2013. - V. 15. - P. 388-398). Вместе с тем, условия, при которых pagN обеспечивал бы полную защиту от инфекции, найдены пока не были. Уместно отметить, что наиболее эффективные с точки зрения протективного эффекта антигены сальмонелл не являются иммунодоминантными (Barat S., Wilier Y., Rizos K., Claudi В., 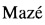 A., Schemmer A.K., Kirchhoff D., Schmidt A., Burton N., Bumann D. // PLoS Pathog. - 2012. - V. 8. - P.e1002966), а, следовательно, иммунный ответ к ним может быть усилен за счет адъювантов, в том числе и молекулярных.
A., Schemmer A.K., Kirchhoff D., Schmidt A., Burton N., Bumann D. // PLoS Pathog. - 2012. - V. 8. - P.e1002966), а, следовательно, иммунный ответ к ним может быть усилен за счет адъювантов, в том числе и молекулярных.
Конъюгированные сальмонеллезные вакцины используют в качестве активатора иммунного ответа экзополисахарид сальмонелл, в частности, экзополисахарид S. typhi, как наиболее изученный на данный момент антиген (Robbins J.D., Robbins J.B. // J. Infect. Dis. - 1984. - V. 150. - P. 436-449). Данный экзополисахарид известен под названием Vi-антигена и использовался как в качестве самостоятельного антигена, так и в качестве конъюгированной вакцины (Anwar Е., Goldberg Е., Fraser A., Acosta C.J., Paul М., Leibovici L. // Cochrane Database Syst. Rev. - 2014. - V. 1. - P.:CD001261), однако даже конъюгированные с известными антигенами-партнерами препараты Vi-антигена до сих пор не одобрены в качестве действующих вакцин и испытывают определенные проблемы с эффективностью (Pier G.B. // Hum. Vaccin. - 2007. - V. 3. - Р. 39-40). В связи с этим постоянно ведется поиск новых антигенов, которые можно было бы использовать в качестве партнеров для конъюгации. Немаловажной является и проблема иммунной интерференции (Borrow R., Dagan R., Zepp F., Hallander H., Poolman J. // Expert Rev. Vaccines. - 2011. - V. 10. - P. 1621-1631), поскольку брюшнотифозная вакцина не является календарной, а вакцины для профилактики вспышек кишечных инфекций, вызываемых Salmonella sp, могут применяться в разное время для разных групп населения. В силу этого вероятность интерференции иммунного ответа к сальмонеллезным вакцинам, конъюгированным с традиционными «календарными» антигенами, такими как дифтерийный и столбнячный токсоиды, возрастает (Dagan R., Poolman J., Siegrist C.A. // Vaccine. - 2010. - V. 28. - P. 5513-5523). Таким образом, идентификация альтернативных партнеров для конъюгации с полисахаридными антигенами сальмонелл, в первую очередь, с Vi-антигеном, представляется важной для создания эффективных вакцинных препаратов.
Происходящий в настоящее время пересмотр парадигмы механизма участия белкового партнера в активации и модификации иммунного ответа к конъюгированному полисахаридному антигену диктует новые стратегии создания конъюгированных вакцин, которые учитывали бы новые данные о механизме активации Т-клеточного иммунитета к конъюгированным антигенам (Avci F.Y. // Curr. Top. Med. Chem. - 2013. - V. 13. - P. 2535-2540; Berti F., Adamo R. // ACS Chem. Biol. - 2013. - V. 8. - P. 1653-1663; Колесников A.B., Козырь A.B., Шемякин И.Г., Дятлов И.А. // Журн. Микробиол. Эпидемиол. Иммунобиол. - 2015. - №3. - С. 97-106). В этой связи особое значение приобретают партнеры для конъюгации, способные повысить эффективность презентации фрагментов вакцинных конъюгатов антиген-презентирующими клетками (АПК) (Avci F.Y., Li X., Tsuji М., Kasper D.L. // Nat. Med. - 2011. - V. 17. - P. 1602-1609). Одним из возможных путей достижения данной цели является включение в состав белкового партнера для конъюгации антигенов, специфически распознаваемых и быстро процессируемых клетками иммунной системы, в частности, за счет рецептор-зависимого эндоцитоза в АПК (Bates J.T., Graff А.Н., Phipps J.P., Grayson J.M., Mizel S.B. // J. Immunol. - 2011. - V. 186. - P. 6255-6262).
На объединении свойств специфического антигена и белка, способного усилить процессинг и презентацию АПК фрагментов конъюгированной вакцины, построена ранее не применявшаяся концепция использования слитого белка fliC:pagN в качестве антигена-партнера для конъюгированных сальмонеллезных вакцин, в первую очередь, вакцин, предназначенных для профилактики брюшнотифозной инфекции.
Известен рекомбинантный секретируемый антиген сальмонеллы SseB, который индуцирует сильный Т-клеточный иммунный ответ и может рассматриваться как гомологичный партнер для конъюгированной сальмонеллезной вакцины (Reynolds C.J., Jones С., Blohmke C.J., Darton Т.С., Goudet A., Sergeant R., Maillere В., Pollard A.J., Altmann D.M., Boyton R.J. // Immunology. - 2014. - V. 143. - P. 438-446). Однако SseB это белок, который секретируется уже эндоцитированной сальмонеллой, поэтому, несмотря на обнаружение сильного Т-клеточного ответа к очищенному белку нет гарантий отсутствия у сальмонелл механизма противодействия распознаванию данного антигена, поскольку указанный антиген активно секретируется патогеном и экспонируется в контексте МНС на поверхности клеток в процессе инфекции, не вызывая быстрого и эффективного иммунного ответа на патоген. Кроме того, поздняя секреция SseB не обеспечивает продукцию антител, способных к блокировке инвазии патогена в макрофаги хозяина как наиболее важного аспекта сальмонеллезной инфекции.
Известен способ получения рекомбинантных антигенов для создания конъюгированной брюшнотифозной вакцины на основе генетически детоксифицированного экзотоксина P. aeruginosa (Szu S.C., Taylor D.N., Trofa A.C., Clements J.D., Shiloach J., Sadoff J.C., Bryla D.A., Robbins J.B. // Infect. Immun. - 1994. - V. 62. - P. 4440-4444). Однако, несмотря на многолетние исследования, эффективность данного носителя в качестве модификатора протективного иммунного ответа к Vi-антигену вызывает сомнения (Pier G.B. // Hum. Vaccin. - 2007. - V. 3. - Р. 39-40).
Известен способ получения рекомбинантных антигенов для создания конъюгированной брюшнотифозной вакцины на основе генетически детоксифицированного дифтерийного токсина CRM197 (Micoli F., Rondini S., Pisoni I., Proietti D., Berti F., Costantino P., Rappuoli R., Szu S., Saul A., Martin L.B. // Vaccine. - 2011. - V. 29. - P. 712-720). Несмотря на успех конъюгатов с использованием CRM197 в случае других конъюгированных вакцин и хорошо известные корреляты защиты для таких вакцин, вакцина на основе Vi-CRM до сих пор не прошла клинические испытания.
Известен способ получения рекомбинантного белка fliC продукцией в Е. coli (Патент US 8449891), однако, помимо того, что белок не содержит второй задействованный в настоящем изобретении белок - активатор иммунного ответа pagN, получаемый по данному способу fliC находится преимущественно в нерастворимой фракции клеточного лизата (тельцах включения), таким образом, получение растворимого белка fliC в данном случае требует процедуры рефолдинга.
Известен по существу наиболее близкий к заявленному изобретению способ получения рекомбинантного антигена fliC для последующего создания конъюгированной сальмонеллезной вакцины (Simon R., Wang J.Y., Boyd М.А., Tulapurkar M.E., Ramachandran G., Tennant S.M., Pasetti M., Galen J.E., Levine M.M. // PLoS One. - 2013. - V. 8. - P.e64680). Однако этот способ не использует в полной мере свойства fliC как молекулярного адъюванта с целью индукции антител, физически блокирующих инвазию сальмонелл в макрофаги организма-хозяина, поскольку не формируется слитый белок с другими потенциально нейтрализующими антигенами. Кроме того, антитела к самому fliC, которые могут формироваться в процессе иммунного ответа, не способны к блокировке инвазии патогена в макрофаги организма-хозяина.
Изобретение решает задачу создания способа получения растворимого рекомбинантного химерного белка fliC:pagN, который может быть использован в виде активатора иммунного ответа при разработке и производстве конъюгированных вакцин против брюшнотифозной инфекции и других вакцинных препаратов.
Поставленная задача решается путем создания способа получения рекомбинантного химерного белка fliC:pagN, включающего конструирование рекомбинантной плазмиды pETfliCpagN длиной 7532 п.н., несущей последовательности, кодирующие белки fliC (включая лидерную последовательность данного белка) и pagN S. typhimurium, разделенные линкерной последовательностью и слитые на 3'-конце с синтетической последовательностью, кодирующей шесть гистидинов, трансформацию клеток Е. coli BL21(DE3) экспрессионной плазмидной ДНК pETfliCpagN, кодирующей рекомбинантный химерный белок fliC:pagN, культивирование полученного штамма BL-fliCpagN при температуре 37°С и продукцию химерного белка fliC:pagN в Е. coli в течение 10 часов при температуре 20°С, получение периплазматической фракции бактерий-продуцентов обработкой клеток лизоцимом в 20% растворе сахарозы с последующим центрифугированием, очистку химерного белка fliC:pagN из бактериальной периплазмы металл-хелатной, анионообменной и гель-фильтрационной хроматографией с получением гомогенного конечного препарата белка fliC:pagN с выходом не менее 0,3 г белка с литра бактериальной культуры, обладающего 98% чистоты по данным электрофореза и денситометрии и содержащего бактериальный эндотоксин в количестве не более 5 IU/мг.
Техническим результатом изобретения является способ продукции и очистки из периплазмы бактерий гомогенного рекомбинантного химерного белка fliC:pagN с чистотой 98%, содержанием бактериального эндотоксина не более 5 IU/мг и выходом 0,3 г с литра бактериальной культуры, применимый для получения больших количеств данного белка - активатора иммунного ответа, пригодного для использования в биотехнологии и фармакологии при разработке и производстве вакцинных препаратов.
Технический результат изобретения достигается за счет получения последовательностей ДНК, определяющих синтез белков fliC и pagN, при помощи ПЦР с геномной ДНК S. typhimurium с использованием специфических праймеров, кодирующих сайты расщепления эндонуклеазами рестрикции NdeI, SalI, XhoI и линкерную последовательность, объединяющую данные белки в составе химерного белка.
Также технический результат изобретения достигается за счет конструирования векторной плазмиды pETfliCpagN, содержащей промоторную последовательность ДНК бактериофага Т7 и последовательности ДНК, кодирующие белки fliC и pagN S. typhimurium, линкерную последовательность, объединяющую данные белки в составе химерного белка, и синтетическую последовательность шесть гистидинов для очистки белка металл-хелатной хроматографией.
Также технический результат изобретения достигается за счет получения штамма BL-fliCpagN на основе штамма E. coli BL21(DE3), трансформированного плазмидной ДНК pETfliCpagN, являющегося продуцентом химерного белка fliC:pagN.
Также технический результат изобретения достигается за счет продукции рекомбинантного химерного белка fliC:pagN в бактериях Е. coli штамма BL-fliCpagN в течение 10 часов при температуре 20°С, что обеспечивает накопление рекомбинантного белка в периплазме бактерий преимущественно в растворенном состоянии.
Также технический результат изобретения достигается получением периплазматической фракции бактериальных белков за счет процедуры обработки бактерий лизоцимом в присутствии 20% раствора сахарозы, что приводит к разрушению клеточной стенки и разделению периплазмы и сферопластов бактерий.
Также технический результат изобретения достигается благодаря схеме хроматографической очистки химерного белка fliC:pagN, включающей этапы металл-хелатной хроматографии на колонке с иммобилизованным бивалентным металлом, анионообменной и гель-фильтрационной хроматографии, позволяющей получить гомогенный белок, характеризующийся 98% чистотой по данным электрофореза и денситометрии и содержанием бактериального эндотоксина не более 5 IU/мг белка.
Отличием предлагаемого способа является высокий выход растворимого рекомбинантного белка fliC:pagN (0,3 г с литра бактериальной культуры), достигаемый за счет особенностей конструирования продуцента BL-fliCpagN и понижения температуры его культивации при синтезе химерного белка fliC:pagN. Необходимо отметить, что в большинстве случаев при продукции белка fliC в Е. coli происходит его накопление в цитоплазме бактерий в виде телец включения (Патенты WO 2007125535, US 8449891).
В предлагаемом техническом решении химерный белок fliC:pagN синтезируется бактериями в растворимой форме с нативной N-концевой последовательностью флагеллина, что способствует корректному фолдингу данного домена химерного белка и оказывается возможным благодаря направленному накоплению fliC:pagN в периплазме бактерий, сопровождающемуся отщеплением лидерной последовательности fliC S. typhimurium.
Предлагаемое техническое решение предусматривает проведение трех этапов хроматографической очистки, включая металл-хелатную хроматографию на смоле с иммобилизованными ионами кобальта, а также анионообменную и гель-фильтрационную хроматографии, обеспечивающих получение гомогенного химерного белка fliC:pagN с высокой степенью чистоты (до 98% по данным денситометрии).
Существенным достоинством предлагаемого способа получения рекомбинантного химерного белка fliC:pagN является низкий уровень содержания бактериального эндотоксина в конечном препарате (5 IU/мг белка), допустимый для применения в инъекциях как по российским, так и по международным стандартам (XII издание Государственной фармакопеи Российской Федерации; Международная фармакопея ВОЗ, документ QAS/11.452 FINAL, июль 2012 г.). Таким образом, полученный по данному способу рекомбинантный белок fliC:pagN пригоден для разработки и производства вакцин.
Изобретение осуществляют следующим образом.
Конструируют рекомбинантную плазмидную ДНК pETfliCpagN на основе коммерчески доступной плазмиды pET22b(+) (Novagen, США). Фрагменты ДНК, кодирующие флагеллин fliC и мембранный белок pagN, получают прямым ПЦР с генома S. typhimurium с использованием праймеров FliC_FNde/FliC_RSal и PagN_FSal/PagN_RXho, представленных на Фиг. 1. В составе последовательности ДНК, кодирующей флагеллин, амплифицируют лидерный пептид, определяющий транслокацию предшественника флагеллина в периплазму бактерий. В состав праймеров, примененных для амплификации, вводят кодоны, определяющие синтез линкерного пептида GSVDSSSGGN. Также в состав праймеров вводят сайты эндонуклеаз рестрикции NdeI (5'-конец фрагмента ДНК, кодирующего ген fliC, праймер FliC_FNde), SalI (3'-конец фрагмента ДНК, кодирующего ген fliC - FliC_RSal, 5'-конец фрагмента ДНК, кодирующего ген pagN - PagN_FSal) и XhoI (3'-конец фрагмента ДНК, кодирующего ген pagN - PagN_RXho).
Полученные фрагменты ДНК расщепляют соответствующими эндонуклеазами рестрикции (NdeI и SalI - фрагмент fliC; SalI и XhoI - фрагмент pagN) и затем последовательно клонируют в плазмиду pET22b(+). Единичные клоны, содержащие инсерты fliC (1500 п.н.) и pagN (700 п.н.), идентифицируют при помощи ПЦР с праймеров T7forward и T7reverse (Sigma-Aldrich, США). Выделяют плазмидную ДНК и верифицируют корректность встроенных последовательностей ДНК методом капиллярного секвенирования. Нуклеотидная и аминокислотная последовательности экспрессионной конструкции, кодирующей продукцию химерного белка fliC:pagN, представлены на Фиг. 2. Структура экспрессионной конструкции для продукции химерного белка fliC:pagN представлены на Фиг. 3.
Плазмидой pETfliCpagN трансформируют клетки штамма Е. coli BL21(DE3) и получают единичные колонии продуцента белка fliC:pagN (BL-fliCpagN). На основании аналитической экспрессии и определения уровня содержания рекомбинантного белка в периплазматической фракции выделяют клоны-суперпродуценты белка. Проводят препаративную экспрессию белка клонов-суперпродуцентов. Для этого бактериальные клоны выращивают в течение ночи в среде 2xYT, содержащей ампициллин и 1% глюкозы, 10 мл ночной культуры помещают в 1 литр свежей среды, содержащей 0,1% глюкозы и ампициллин, и наращивают культуру до достижения 1 единицы оптической плотности. Культуру охлаждают до 20°С и индуцируют синтез белка добавлением 0,2 мМ ИПТГ. Экспрессию белка fliC:pagN проводят при 20°С в течение 10 часов. Бактериальную биомассу собирают центрифугированием, анализируют продукцию белка электрофоретически (Фиг. 4) и хранят при -70°С.
Выделение белка fliC:pagN проводят при температуре +4°С из периплазматической фракции бактерий. Для получения бактериальной периплазмы 50 г бактериальной биомассы суспендируют в 100 мл раствора, содержащего 20% сахарозы, 30 мМ трис-HCl рН 8, 100 мМ хлорида натрия, ингибитор протеаз Complete (Roche Life Sciences, США) и 100 мкг/мл лизоцима, и выдерживают 30 минут во льду с периодическим встряхиванием. Сферопласты отделяют от периплазматической фракции центрифугированием в течение 30 минут при 10000 g.
Белок fliC:pagN выделяют из периплазматической фракции металл-хелатной хроматографией на колонке Talon (Clontech, США) в буфере, содержащем 20 мМ трис-HCl рН 8 и 100 мМ хлорида натрия. Элюцию белка с колонки проводят буфером, содержащим 20 мМ трис-HCl рН 8, 100 мМ хлорида натрия, 250 мМ имидазола.
Очистку белка проводят анионообменной хроматографией на колонке Q-Sepharose (GE Healthcare, США). Белок разводят в 10 раз на буфер, содержащий 20 мМ трис-HCl рН 8, 20 мМ хлорида натрия, наносят на колонку, уравновешенную тем же буфером, и элюируют градиентом хлорида натрия от 20 до 500 мМ в течение 60 минут. Фракции, содержащие белок fliC:pagN, идентифицируют электрофорезом в 10% ПААГ, концентрируют при помощи центрифужных концентраторов Amicon Ultra-15 (Millipore, США) и наносят на гель-фильтрационную колонку Superdex 200 10/300 GL, уравновешенную буфером, содержащим 20 мМ трис-HCl рН 7,5, 150 мМ хлорида натрия. Фракции, содержащие целевой белок, определяют электрофоретически, объединяют и концентрируют. Препарат белка fliC:pagN хранят в аликвотах при температуре -70°С.
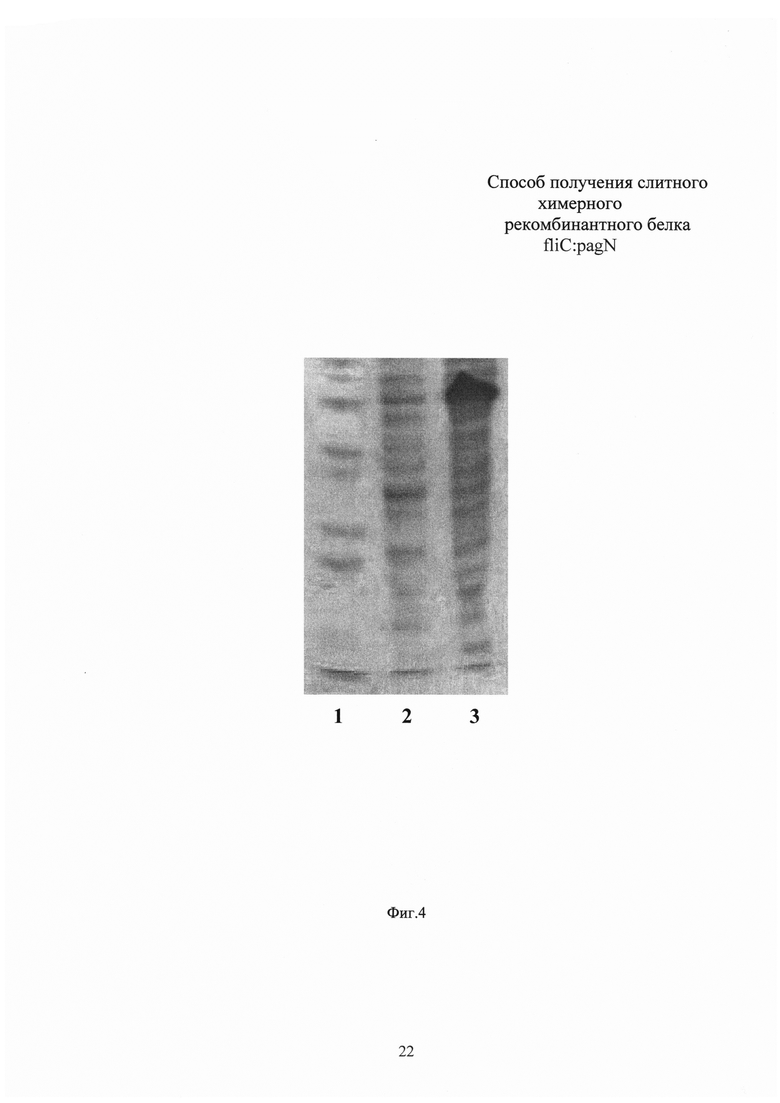
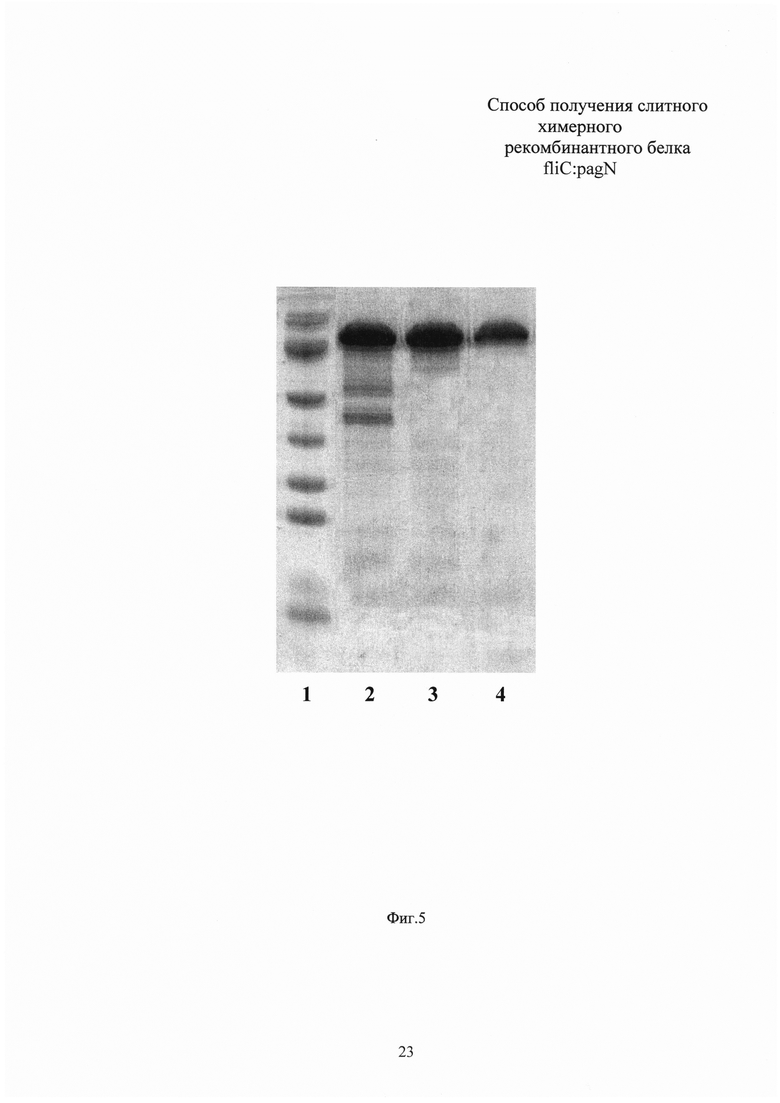
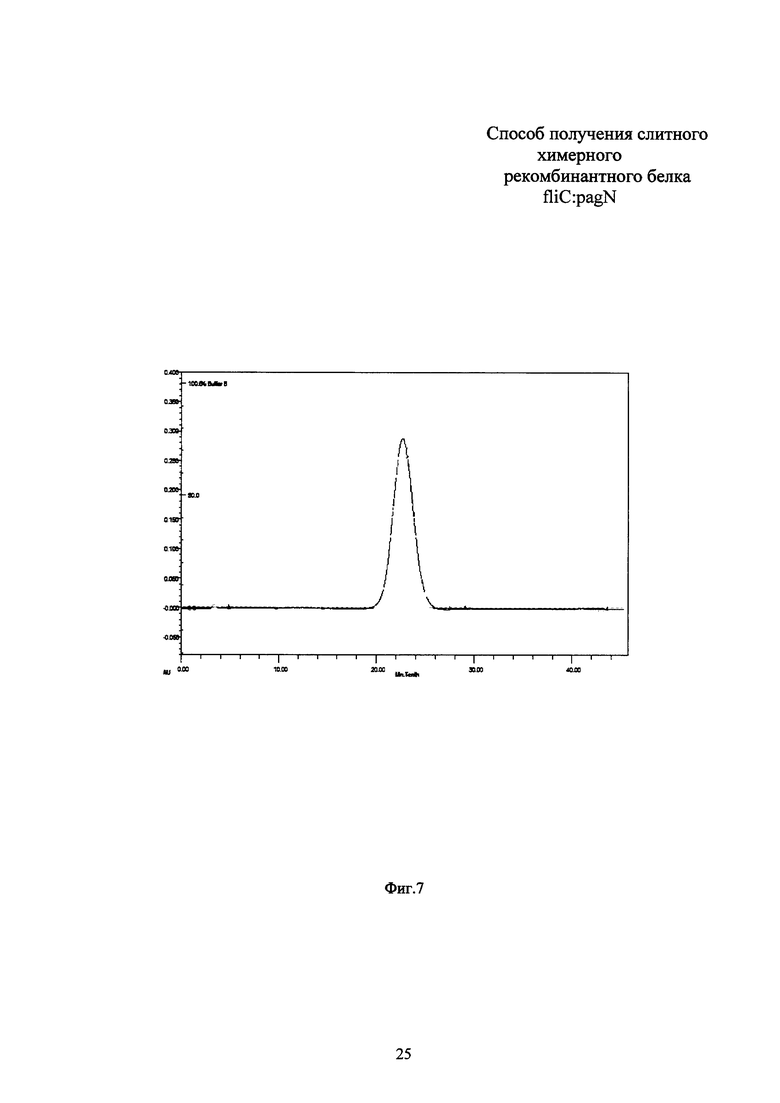
Измерение концентрации белка в полученном препарате проводят по методу Брэдфорд. Выход рекомбинантного белка fliC:pagN составляет 0,3 г с литра бактериальной культуры. Чистоту полученного белкового препарата анализируют электрофорезом в 10% ПААГ с денситометрией, гомогенность препарата оценивают при помощи HPLC-SEC (Фиг. 5, 6). Чистота полученного препарата fliC:pagN по данным денситометрии составляет 98%, препарат гомогенен по данным HPLC-SEC. Иммунологическую идентичность подтверждают Вестерн-блотом с окрашиванием моноклональными антителами к флагеллину S. typhimurium (Фиг. 7). Содержание эндотоксина в конечном препарате белка fliC:pagN составляет менее 5 IU/мг. N-концевым секвенированием верифицируют отсутствие лидерного пептида на N-конце рекомбинантного белка.
Изобретение иллюстрируют следующие графические материалы.
Фиг. 1. Последовательности праймеров, использованных при получении экспрессионной конструкции химерного рекомбинантного белка fliC:pagN для продукции в клетках Е. coli.
Фиг. 2. Нуклеотидная последовательность, кодирующая продукцию белка fliC:pagN и аминокислотная последовательность белка fliC:pagN. А - Нуклеотидная последовательность, кодирующая продукцию белка fliC:pagN; В - аминокислотная последовательность белка fliC:pagN; С - полная последовательность экспрессионной плазмиды pETfliCpagN.
Фиг. 3. Карта экспрессионной конструкции, использованной для получения химерного рекомбинантного белка fliC:pagN в клетках Е. coli.
Фиг. 4. Продукция слитого химерного рекомбинантного белка fliC:pagN в клетках Е. coli. Дорожка 1 - маркер молекулярной массы (Spectra Multicolor Broad Range Protein Ladder, Thermo Scientific, USA), дорожка 2 - периплазматическая фракция клеток BL-fliCpagN
Фиг. 5. Результаты выделения и очистки химерного рекомбинантного белка fliC:pagN. Дорожка 1 - маркер молекулярной массы SM0671 (Thermo Scientific, США), дорожка 2 - белок fliC:pagN, очищенный из периплазматической фракции металл-хелатной хроматографией на колонке Talon (Clontech, США), дорожка 3 - белок fliC:pagN, очищенный анионообменной хроматографией на колонке Q-Sepharose (GE Healthcare, США), дорожка 4 - белок fliC:pagN, очищенный гель-фильтрационной хроматографией на колонке Superdex 200 (GE Healthcare, США).
Фиг. 6. Вестерн-блот по окрашиванию полученного рекомбинантного белка fliC:pagN моноклональными антителами к fliC. Дорожка 1 - маркер молекулярной массы SM0671 (Thermo Scientific, США), дорожка 2 - белок fliC:pagN, окрашенный моноклональными антителами Anti-Flagellin FliC antibody (Invivogen, США).
Фиг. 7. Результаты HPLC-SEC очищенного препарата рекомбинантного белка fliC:pagN.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Создание экспрессионной конструкции для продукции химерного рекомбинантного белка fliC:pagN.
Для получения геномной ДНК бактерии S. typhimurium культивируют в течение ночи в 10 мл среды 2xYT при 37°С и собирают центрифугированием. Бактериальную биомассу замораживают в жидком азоте, размораживают и суспендируют в буфере, содержащем 20 мМ трис-HCl, 50 мМ хлорида натрия, 70 мМ глюкозы, 10 мМ ЭДТА, 20 мкг/мл лизоцима. Обработку лизоцимом проводят в течение 15 минут при +4°С, после чего к суспензии добавляют двухкратный объем раствора, содержащего 1% додецилсульфата натрия и 0,2 М NaOH и инкубируют при +4°С в течение 10 минут до появления вязкости. Полученный лизат троекратно экстрагируют смесью фенол/хлороформ (1:1 v/v) с промежуточным центрифугированием при 20000 g и отбором водной фазы. К водной фазе после третьего раунда экстракции добавляют 1/10 объема ацетата натрия рН 5,2 и 3 объема изопропанола, инкубируют смесь при +4°С в течение 30 минут и отделяют осадок центрифугированием при 20000 g. Полученный осадок промывают 80% этанолом, подсушивают и растворяют в 100 мкл бидистиллированной воды.
Фрагменты ДНК, кодирующие флагеллин fliC и мембранный белок pagN, получают ПЦР-амплификацией с геномной ДНК S. typhimurium с использованием пар праймеров FliC_FNde/FliC_RSal и PagN_FSal/PagN_RXho. ПЦР-амплификацию проводят с понижением температуры отжига праймеров с 65°С до 54°С (1°С на каждом цикле), после чего проводят дополнительно 20 циклов амплификации при температуре отжига праймеров 60°С. В состав праймеров введены сайты эндонуклеаз рестрикции NdeI (5'-конец фрагмента ДНК, кодирующего ген fliC, праймер FliC_FNde), SalI (3'-конец фрагмента ДНК, кодирующего ген fliC - FliC_RSal, 5'-конец фрагмента ДНК, кодирующего ген pagN - PagN_FSal) и XhoI (3'-конец фрагмента ДНК, кодирующего ген pagN - PagN_RXho). В ПЦР-реакции содержится 20 рМ каждого праймера и 1 мкг геномной ДНК. Полученные продукты реакции анализируют электрофоретически в агарозном геле на наличие фрагментов ДНК длиной 1500 п.н. (fliC) и 700 п.н. (pagN).
Полученные с помощью ПЦР фрагменты ДНК обрабатывают эндонуклеазами рестрикции (NdeI и SalI - фрагмент fliC; SalI и XhoI - фрагмент pagN), разделяют в агарозном геле, вырезают из геля и очищают от контаминаций при помощи набора QIAquick Gel Extraction Kit (Qiagen, UK). На первом этапе проводят клонирование фрагмента fliC в вектор pET22b(+), расщепленный по сайтам эндонуклеаз рестрикции NdeI и SalI. Единичные клоны, содержащие плазмиду pET22b(+) со встроенным фрагментом флагеллина S. typhimurium получают электротрансформацией клеток штамма Е. coli DH12S (Thermo Fisher Scientific, США). Клоны, содержащие инсерт длиной 1500 п.н., идентифицируют при помощи ПЦР с праймеров T7forward и T7reverse (Sigma-Aldrich, США). Выделяют плазмидную ДНК pETfliC и верифицируют корректность встроенной последовательности гена fliC методом капиллярного секвенирования.
Полученную плазмиду pETfliC расщепляют по сайтам SalI и XhoI и лигируют с полученным ПЦР-фрагментом pagN, расщепленным по концевым сайтам соответствующих эндонуклеаз рестрикции. Электротрансформацией получают единичные клоны DH12S-pETfliCpagN и анализируют наличие встроенного фрагмента pagN с проверкой его корректной ориентации в конструкции по наличию инсерта 700 п.н. при помощи ПЦР с праймеров T7forward/PagN_RXho.
Праймеры, используемые для ПЦР-амплификации, содержат сайты эндонуклеаз рестрикции NdeI (N-конец гена, праймер MBP_FNde), NheI и XhoI (С-конец гена, праймер MBP_RNheXho). Выделяют плазмидную ДНК pETfliCpagN и верифицируют корректность встроенной последовательности гена pagN методом капиллярного секвенирования.
Пример 2. Получение продуцентов и продукция химерного рекомбинантного белка fliC:pagN в Е. coli.
Штамм-продуцент Е. coli BL-fliCpagN получают электротрансформацией компетентных клеток Е. coli BL21(DE3) плазмидой pETfliCpagN. Проводят аналитическую экспрессию единичных клонов BL21(DE3), несущих плазмиду pETfliCpagN. Для этого едничные колонии Е. coli BL-fliCpagN выращивают в течение ночи при 37°С в 5 мл среды 2xYT с добавлением 1% глюкозы и 50 мкг/мл ампициллина, засевают 100 мкл ночной культуры в 10 мл среды 2xYT, содержащей 0.1% глюкозы и 50 мкг/мл ампициллина, и выращивают при 37°С в течение 3 часов. Выросшие культуры охлаждают до 20°С, добавляют ИПТГ до 0,2 мМ и проводят экспрессию 10 часов при температуре 20°С. Бактериальные клетки с 5 мл культуры продуцента собирают центрифугированием при 5000 об/мин 10 минут и выделяют периплазматическую фракцию. Для выделения периплазматической фракции бактериальную биомассу суспендируют в 1 мл раствора, содержащего 30 мМ трис-HCl рН 8, 100 мМ хлорида натрия, ингибитор протеаз Complete (Roche Life Sciences, США) и 100 мкг/мл лизоцима, и выдерживают 30 минут во льду с периодическим встряхиванием. Сферопласты отделяют от периплазматической фракции центрифугированием в течение 30 минут при 12000 об/мин, и отбирают равные аликвоты на электрофорез. Анализируют уровень содержания белка fliC:pagN в периплазматической фракции денатурирующим электрофорезом в 12% полиакриламидном геле. После окончания электрофореза гель окрашивают кумасси R-250 по стандартной методике и сканируют с помощью денситометра Typhoon FLA9500 (GE Healthcare, США). Клоны, обнаруживающие наибольший выход белка (образование на геле яркой полосы с молекулярной массой 75 кДа), являются суперпродуцентами, их выращивают на богатой среде (2xYT) в течение ночи, добавляют глицерин до 15% и замораживают при -70°С для хранения.
Для препаративной экспрессии штаммы-суперпродуценты Е. coli BL-fliCpagN выращивают в течение ночи при 37°С с добавлением 1% глюкозы и 50 мкг/мл ампициллина на среде 2xYT, помещают 10 мл ночной культуры в 1000 мл среды 2xYT, содержащей 0.1% глюкозы и 50 мкг/мл ампициллина, и выращивают при 37°С до достижения 1 единицы оптической плотности. Выросшую культуру охлаждают до 20°С, добавляют ИПТГ до 0,2 мМ и проводят экспрессию 10 часов при 20°С. Далее клетки собирают центрифугированием при 5000 об/мин 10 минут и хранят в виде замороженных осадков при температуре -70°С.
Пример 3. Выделение и очистка химерного рекомбинантного белка fliC:pagN из бактериальной биомассы.
Выделение белка fliC:pagN и хроматографические процедуры проводят при температуре +4°С. Выделение и очистку белка осуществляют из периплазматической фракции Е. coli. Для получения периплазматической фракции 50 г бактериальной биомассы суспендируют в 100 мл раствора, содержащего 20% сахарозы, 30 мМ трис-HCl рН 8, 100 мМ хлорида натрия, ингибитор протеаз Complete EDTA-free (Roche Life Sciences, США) и 100 мкг/мл лизоцима, и инкубируют 30 минут во льду с периодическим встряхиванием. Сферопласты отделяют от периплазматической фракции центрифугированием в течение 30 минут при 10000 g.
Раствор белков периплазмы наносят на металл-хелатную колонку Talon (Clontech, США) в буфере, содержащем 20 мМ трис-HCl рН 8 и 100 мМ хлорида натрия. Промывают колонку 10 объемами того же буфера. Элюцию белка с колонки проводят буфером, содержащим 20 мМ трис-HCl рН 8, 100 мМ хлорида натрия, 250 мМ имидазола, и с помощью электрофореза выявляют фракции, содержащие очищенный белок fliC:pagN.
Белок, элюированный с металл-хелатного сорбента, разводят в 10 раз на буфер, содержащий 20 мМ трис-HCl рН 8, 20 мМ хлорида натрия и наносят на анионообменную колонку Q-Sepharose (GE Healthcare, США), уравновешенную тем же буфером. Элюируют белок градиентом хлорида натрия от 20 до 500 мМ на том же буфере в течение 60 минут. Фракции, содержащие очищенный белок fliC:pagN, определяют электрофорезом в 10% ПААГ, собирают и концентрируют при помощи центрифужных концентраторов Amicon Ultra-15 (Millipore, США).
Сконцентрированный белок наносят на гель-фильтрационную колонку Superdex 200 10/300 GL, уравновешенную буфером, содержащим 20 мМ трис-HCl рН 7,5, 150 мМ хлорида натрия. Фракции, содержащие целевой белок, определяют электрофоретически, объединяют и концентрируют. Препарат белка fliC:pagN хранят в аликвотах при температуре -70°С.
Пример 4. Валидация препарата химерного рекомбинантного белка fliC:pagN для применения при разработке вакцин.
Измерение концентрации белка в полученном препарате проводят по методу Брэдфорд с использованием набора Quick Start™ Bradford Protein Assay (Bio-Rad, США). Выход рекомбинантного белка fliC:pagN составляет 0,3 г с литра бактериальной культуры.
Чистоту полученного белкового препарата анализируют электрофорезом в 10% денатурирующем ПААГ. После прохождения электрофореза гель окрашивают кумасси R-250 по стандартной методике и сканируют с помощью денситометра Typhoon FLA9500 (GE Healthcare, США). Чистота полученного препарата fliC:pagN по данным денситометрии составляет 98%.
Гомогенность препарата белка fliC:pagN тестируют при помощи хроматографии высокого давления HPLC-SEC. HPLC-SEC проводят с использованием колонки Waters BioSuite High Resolution. 100 мкг белка наносят на колонку в буфере, содержащем 100 мМ фосфата натрия, рН 6.5. Единственный пик на хроматограмме свидетельствует о гомогенности препарата белка fliC:pagN и отсутствии контаминаций.
Иммунологическую идентичность подтверждают Вестерн-блотом с окрашиванием моноклональными антителами к флагеллину S. typhimurium. Белок после электрофореза в денатурирующем геле полусухим способом в буфере, содержащем 25 мМ трис-HCl, 190 мМ глицина, 20% метанола и 0,1% додецилсульфата натрия, переносят в течение 1 часа на мембрану Hybond-C Extra (GE Healthcare Life Sciences, США) при силе тока 1 мА/см. Мембрану в течение 30 минут блокируют фосфатным буфером, содержащим 0,05% Tween-20 и 3% бычьего сывороточного альбумина. Проводят окрашивание мембраны моноклональными антителами Anti-Flagellin FliC antibody (Invivogen, США), трижды промывают фосфатным буфером, содержащим 0,01% Tween-20, и проявляют с использованием набора WesternBreeze Chromogenic Kit, anti-mouse (Thermo Fisher Scientific, США). Наличие на мембране полосы с молекулярной массой 75 кДа свидетельствует о иммунологической идентичности рекомбинантного флагеллина в составе фьюжн-белка fliC:pagN нативному флагеллину S. typhimurium.
Содержание эндотоксина в конечном препарате белка fliC:pagN определяют при помощи ЛАЛ-теста (Charles River Endosafe, США), оно составляет менее 5 IU/мг.
Методом N-концевого секвенирования определяют первые 8 аминокислот очищенного рекомбинантного белка. Согласно результатам N-концевого секвенирования первые 8 аминокислот белка имеют последовательность LGTAIERL.
| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантная плазмида pFM-FliC_syn, обеспечивающая экспрессию рекомбинантного белка флагеллина Salmonella (FliC), моноплазмидный штамм бактерий Escherichia coli FM-FliC_syn и способ получения рекомбинантного белка флагеллина Salmonella (FliC) | 2023 |
|
RU2831404C1 |
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |
| Химерный рекомбинантный белок, обладающий протективными свойствами в отношении Streptococcus pyogenes | 2016 |
|
RU2685957C2 |
| Рекомбинантная плазмидная ДНК pPA-FlicC, кодирующая синтез рекомбинантного флагеллина-С Pseudomonas aeruginosa, штамм Escherichia coli PA-FlicC - продуцент гибридного рекомбинантного белка и способ получения указанного белка | 2022 |
|
RU2793753C1 |
| ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ФЛАГЕЛЛИНА | 2012 |
|
RU2524133C2 |
| Способ получения рекомбинантного бруцеллезного белка rLptD | 2023 |
|
RU2815099C1 |
| Гибридный белок, ДНК, генетическая конструкция, рекомбинантная клетка, вакцина на основе гибридного белка для профилактики и лечения туберкулеза (варианты) | 2015 |
|
RU2615440C2 |
| Способ получения рекомбинантного экзопротеина А Pseudomonas aeruginosa | 2016 |
|
RU2636346C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PSAV27, КОДИРУЮЩАЯ СИНТЕЗ РАСТВОРИМОГО СТРЕПТАВИДИНА ИЗ STREPTOMYCES AVIDINII, И БАКТЕРИАЛЬНЫЙ ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ РАСТВОРИМОГО СТРЕПТАВИДИНА ИЗ STREPTOMYCES AVIDINII | 1999 |
|
RU2153535C1 |
| СПОСОБ ЭКСПРЕССИИ БЕЛКА CRM197 | 2019 |
|
RU2803949C1 |







Настоящее изобретение относится к области биохимии, в частности к способу получения слитого химерного рекомбинантного белка fliC:pagN. Для осуществления способа конструируют рекомбинантную плазмиду pETfliCpagN, затем трансформируют клетки Escherichia coli BL21(DE3) указанной плазмидой и культивируют полученный штамм BL-fliCpagN при температуре 37°С. Затем получают периплазматическую фракцию бактерий-продуцентов обработкой клеток лизоцимом в 20% растворе сахарозы с последующим центрифугированием, проводят очистку химерного белка fliC:pagN металл-хелатной, анионообменной и гель-фильтрационной хроматографией с получением гомогенного конечного препарата белка fliC:pagN. Рекомбинантная плазмида pETfliCpagN содержит промотор бактериофага Т7 и несёт последовательности, кодирующие белки fliC, включая лидерную последовательность данного белка, и pagN S.typhimurium, объединенные короткой линкерной последовательностью GSVDSSSGGN, и слитые на 3'-конце с синтетической последовательностью, кодирующей шесть гистидинов. Заявленный способ получения fliC:pagN позволяет синтезировать данный белок в растворимой форме с нативной N-концевой аминокислотной последовательностью, а также с выходом не менее 0,3 г белка с литра бактериальной культуры, обладающего 98% чистоты. 7 ил., 4 пр.
Способ получения слитого химерного рекомбинантного белка fliC:pagN (SEQ ID NO: 2), включающий конструирование рекомбинантной плазмиды pETfliCpagN (SEQ ID NO: 3) длиной 7532 п.н., содержащей промотор бактериофага Т7 и несущей последовательности, кодирующие белки fliC, включая лидерную последовательность данного белка, и pagN S. typhimurium, объединенные короткой линкерной последовательностью GSVDSSSGGN, и слитые на 3'-конце с синтетической последовательностью, кодирующей шесть гистидинов, трансформацию клеток Escherichia coli BL21(DE3) экспрессионной плазмидной ДНК pETfliCpagN, кодирующей рекомбинантный химерный белок fliC:pagN, культивирование полученного штамма BL-fliCpagN при температуре 37°С и продукцию химерного белка fliC:pagN в Escherichia coli при температуре 20°С в течение 10 часов, получение периплазматической фракции бактерий-продуцентов обработкой клеток лизоцимом в 20% растворе сахарозы с последующим центрифугированием, очистку химерного белка fliC:pagN из бактериальной периплазмы металл-хелатной, анионообменной и гель-фильтрационной хроматографией с получением гомогенного конечного препарата белка fliC:pagN с выходом не менее 0,3 г белка с литра бактериальной культуры, обладающего 98% чистоты по данным электрофореза и денситометрии и содержащего бактериальный эндотоксин в количестве не более 5 IU/мг.
| SIMON R., et al., Sustained Protection in Mice Immunized with Fractional Doses of Salmonella Enteritidis Core and O Polysaccharide-Flagellin Glycoconjugates, PLOS ONE, 2013, Vol.8, Issue 5, e64680 | |||
| MILLER C., et al., PanG, a New Ketopantoate Reductase Involved in PantothenateSynthesis, Journal of Bacteriology, 2013, Vol.195, No.5, pp.965-976 | |||
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |

