Изобретение относится к области микробиологии и вакцинной технологии и направлено на разработку вакцины, способной вырабатывать иммунитет против инфекции, вызванной бактериями группы B Streptococcus.
Бактерии рода Streptococcus являются возбудителями многих заболеваний человека и животных. Стрептококки подразделяются на иммунологические группы на основе присутствия специфических углеводных антигенов на их клеточных поверхностях. В настоящее время известны трептококки групп A- O (Davis B.D. и др., Microbilogy, 3-е изд., с. 609 (Harper & Rom, 1980).
Стрептококки являются одними из наиболее известных и серьезных возбудителей заболеваний человека.
Хотя стрептококки группы B ассоциируются с болезнями животных (например, такими, как маститы у рогатого скота), однако бактерия Streptococcus agalactiae (стрептококк группы B) является основной причиной возникновения неонатального сепсиса у человека в США, который по установленным данным уносит свыше 6000 жизней ежегодно Hill H.P. и др., Sexually Transmitted Diseases, McGraw Hill, с. 397-407). Стрептококки группы B являются также основными патогенами затяжных менингитов у детей, послеродовых эндометритов, и нарушений иммунологической реактивности у взрослых людей (Patt erson, M.J. и др., Bact. Rev. 40:774-792(1976)). Хотя человеческий организм и восприимчив к действию антибиотиков, однако, острое начало заболевания и быстрое развитие сепсиса у новорожденных и менингитов у детей приводит к высокому проценту заболеваемости (50%) и смертности (20%) Baker C.J. и др., New Eng. J. Med. (Editorial) 314(26): 1702-1704 (1986); Baker, C.J. и др., J. Infect.Dis. 136:137-152 (1977)).
Streptococcus группы B является основным компонентом нормальной микрофлоры влагалища и прямой кишки человека. Хотя в большинстве случаев инфецирование новорожденных происходит интернатально в результате вагинальной колонизации, однако, имеются случаи также внутрибольничного заражения новорожденных, которые описаны, например, Patterson M.J. и др., Bact. Rev. 40: 774-792 (1976)). Однако лишь небольшой процент инфекций у детей, вызванных стрептококком группы B, развиваются в серьезные заболевания. При этом следует отметить, что роль хозяйских факторов и детерминат бактериальной вирулентности в механизме перехода от колонизации до инфекции не совсем ясна.
Очевидно, за вирулентность и иммунитет ответственны некоторые белки стрептококков группы B (Ferrieri P., Rev. Infect. Dis. 10:S363, 1988). В 1975 г. Lancefield обнаружил способность белков C стрептококков группы B вырабатывать защитный иммунитет (Lancefield, R. C. и др., J. Exp.Med. 142: 165-179 (1975)). По всей вероятности указанная группа белков содержит несколько различных полипептидов и антигенных детерминант. С точки зрения этих исследователей, усилия по предупреждению инфекций, вызываемых Streptococcus группы B, должны быть направлены на профилактическое введение антибиотиков и на разработку вакцины против стрептококков группы B (Baker C.J. и др., Rev. of Infect. Dis. 7:458-467 (1985)), Baker C.J. и др., New Eng. J.Med.(Editor al) 314(26): 1072-1704 (1986)). Полисахаридные вакцины против стрептококков группы B описаны Kasper D. L. (патенты США 4 207 414 и заменяющий патент PE3172, а также патент США 4 324 887, 4 356 263, 4 367 221, 4 367 221, 4 367 222 и 4 367 223), Carlo D. L. (патент США 4 413 057, публ. Европатента 38265), и Yavordios D. и др. (публ. Европатент 71515); все указанные работы вводятся в настоящее описание посредством ссылки.
За исключением небольшой группы детей с установленным риском материнской колонизации стрептококками группы B и другими факторами риска в перинатальном периоде, в большинстве случаев использование антибиотиков в профилактических целях либо не проводилось, либо не давало эффективных результатов (Boyer K. M. , и др., New Eng. J. Med. 314(26):1665-1669(1986)). Интернатальная химиопрофилактика также не получила широкого распространения по следующим причинам: практически невозможно установить материнскую колонизацию Streptococcus группы B быстрым, надежным и экономически эффективным способом; около 40% новорожденных появляется на свет в обстановке пониженного риска; практически трудно выявить и/или обработать всех матерей или детей с потенциально высоким риском; и фактически невозможно осуществить профилактическую обработку антибиотиками для предупреждения затяжного менингита (7200 случаев в год в США) или послеродового эндометрита (45000 случаев в год) (Baker C.J. и др., New Eng. J. Med. (Editorial) 314:1702-1704, (1986).
Хранение микроорганизмов.
Плазмиды pJMSI и pJMS23 происходят от плазмиды pUX12, которая содержит ДНК, способную кодировать антигенные белки Streptococcus, которые могут быть использованы в соответствии с настоящим изобретением. Плазмида pUX12 происходит от плазмиды pUC12. Плазмиды pJMSI и pJM223 были депонированы 15.099.1989 в Американской коллекции типовых культур, Роквил, M.D. под входящими номерами АТСС 40659 и АТСС 40660 соответственно.
Streptococcus agalactiae является наиболее известным патогеном, вызывающим сепсис новорожденных в США, и уносящим от 6000 до 10000 жизней в год. Хотя группоспецифический капсульный полисахарид стрептококка группы B является иммуногенным и имеет важные защитные антигены, однако, клинические испытания полисахаридной вакцины показали недостаточную реактивность (Baker C. J. и др., New Engl. J. Med. 319:1180 (1980); Insel R.A. и др. New Eng. J. Med (Editorial) 319:(18):1219-1220 (1988)).
Настоящее изобретение направлено на разработку конъюгатной вакцины к стрептококкам группы B (т. е. Streptococcus agalactiae), где используется протективный белковый антиген, полученный в результате экспрессии гена, клонированного из Streptococcus группы B. Эта новая конъюгатная вакцина имеет то преимущество, что она вырабатывает Т-независимый иммунитет посредством адъювантного действия белка-носителя, а также вносит дополнительные протективные эпитопы, которые присутствуют на клонированном белке Streptococcus группы B (Insel R. A. и др., New Eng, J. Med (Editorial) 319(18): 1219-1220, (1988)); Baker C. J. и др. , Rev. of Invec. Dis. 7: 458-467 (1985)).
В частности, настоящее изобретение относится к конъюгатной вакцине, способной сообщать иммунитет хозяина против инфекции, вызываемой стрептококками группы B и содержащей (a) полисахарид, конъюгированный с (b) белком, причем как полисахарид, так и белок являются характерными молекулами Streptococcus группы B.
Изобретение также относится к способу предупреждения или аттенуации инфекции, вызываемой Streptococcus группы B, который заключается в том, что пациенту с предполагаемым риском заражения вводят эффективное количество конъюгатной вакцины, способной сообщать иммунитет хозяина против указанной инфекции, и содержащей: (a) полисахарид, конъюгированный с (b) белком; причем как полисахарид, так и белок являются характерными молекулами Streptococcus группы B.
Кроме того, изобретение относится к способу предупреждения или ослабления инфекции, вызываемой Streptococcus группы B, который заключается в том, что беременным женщинам вводят эффективное количество конъюгироватной вакцины, способной сообщать иммунитет еще неродившемуся ребенку против указанной инфекции и содержащей: (a) полисахарид, конъюгированный с (b) белком; причем как полисахарид, так и белок являются характерными молекулами Streptococcus группы B.
Изобретение также относится к способу предупреждения или ослабления инфекции, вызванной Streptococcus группы B, который заключается в том, пациенту с предполагаемым риском заражения указанной инфекцией вводят эффективное количество антисыворотки, взятой от другого индивидуума, которому была введена конъюгатная вакцина, способная сообщать иммунитет против инфекции и содержащая: a) полисахарид, конъюгированный с b) белком; причем как полисахарид, так и белок являются характерными молекулами Streptococcus группы B.
Фиг. 1, 2, 3 иллюстрируют модификацию плазмиды puC12, осуществленную для получения плазмиды puX12; на фиг. 2 представлены рестрикционные карты вставок стрептококков группы B в гибридных плазмидах: (a) pPHC10. Фрагменты субклонировали из рекомбинантных фагов λα+ и λβ+ соответственно, в соответствующие сайты полилинкера p C18. Рестрикционные сайты обозначены следующими аббревиатурами: E - EcoRI; BL - BalI; EV - EcoRV; Hp - HpaI; P - PstI; H - Hind III; Bg lII. ▪ - омега, инсерции фрагмента; ←_ направление транскрипции; kb - тысяча пар оснований ДНК; на фиг. 3 представлена рестрикционная карта ДНК-вставки GBC в плазмиде pJMS23 и Hind III - фрагмент 5,5 kb. Рестрикционные эндонуклеазные сайты показаны на нижней линии, а расположение TnS - инерций - на верхней линии и обозначены черточками и кружками. Результаты Вестерн-блоттинга, представляющие экспрессию предполагаемых клонированных генов, проиллюстрированы кружочками: • - полноразмерные генные продукты,  - частичные генные продукты; ° - генные продукты не обнаружены. Сайты расщепления рестрикционных эндонуклеаз обозначены следующими аббревиатурами: B - Bam HI; Hc - Hin cIII; Hd - Hind III; N - NheI; S - StyI; на фиг. 4 представлена рестрикционная и транскрипционная карта плазмиды pUX12; на фиг. 5 - модификации, внесенные в плазмиду pUX12 для получения плазмиды pUX12+1 с рамкой считывания +1 (A), и плазмиды pUX12-1 с рамкой считывания - 1 (C). (B) представляет собой конструкцию, которая способна дополнительно продуцировать плазмиду с рамкой считывания -1; на фиг. 6 - исследования иммунитета мышей с использованием кроличьей антисыворотки против S1 и S23. Иммунитет наблюдали у мышей, инокулированных антисывороткой против S1 (p < 0,002) или антисывороткой против S23 (p < 0,022). Из-за размера исследуемых проб, различия между наблюдаемыми статистическими значениями в экспериментах S1 и S23 являются незначительными. На этой фигуре вверху над каждой полосой указано количество мышей, выживших после испытания.
- частичные генные продукты; ° - генные продукты не обнаружены. Сайты расщепления рестрикционных эндонуклеаз обозначены следующими аббревиатурами: B - Bam HI; Hc - Hin cIII; Hd - Hind III; N - NheI; S - StyI; на фиг. 4 представлена рестрикционная и транскрипционная карта плазмиды pUX12; на фиг. 5 - модификации, внесенные в плазмиду pUX12 для получения плазмиды pUX12+1 с рамкой считывания +1 (A), и плазмиды pUX12-1 с рамкой считывания - 1 (C). (B) представляет собой конструкцию, которая способна дополнительно продуцировать плазмиду с рамкой считывания -1; на фиг. 6 - исследования иммунитета мышей с использованием кроличьей антисыворотки против S1 и S23. Иммунитет наблюдали у мышей, инокулированных антисывороткой против S1 (p < 0,002) или антисывороткой против S23 (p < 0,022). Из-за размера исследуемых проб, различия между наблюдаемыми статистическими значениями в экспериментах S1 и S23 являются незначительными. На этой фигуре вверху над каждой полосой указано количество мышей, выживших после испытания.
Описание предпочтительного осуществления изобретения.
Иммунопрофилактика матерей с использованием вакцины против Streptococcus группы B была предложена в целях защиты матери и ребенка от заражения, осуществляемой посредством перинатального переноса антител (Baker C.J. и др., New Eng. J. Med. (EditoriaI) 314(26): 1702-1704 (1986): Baker C.J. и др., New Eng. J. Med. 319:1180 (1988); Baker C.J. и др., J. Infect. Dic. 7:458 (1985)). Как и в случае других инкапсулированных бактерий, восприимчивость к инфекции коррелирует с отсутствием группоспецифических антител (Kasper D.L., и др. , J. Clin. Invest. 72:260-269 (1983); Kasper D.L. и др., Anti biot. Chemother. 35: 90-100 (1985)). Отсутствие "опсонически" активных группоспецифических антикапсулярных антител к Streptococcus группы B является фактором риска для развития заболевания в результате колонизации стрептококками группы B (Kasper D.L. и др., J.Infect.Dis. 153: 407-415, 1986).
Один из способов вакцинирования предусматривает использование очищенных группоспецифических капсулярных полисахаридов.
Способы получения таких вакцин и их использование для иммунизации против Streptococcus группы B были описаны Kasper D.L. (патент CША 4 207 414 и замена патента США PE31672, а также патенты США NN 4 324 887, 4 356 263, 4 367 221, 4 367 222 и 4 367 223), Carlo D.J. (патент США 4 413 057, публикация Европатента 38265) и Yavordios D. и др., (публикация Европатента 71515) (все указанные работы вводятся в настоящее описание посредством ссылки).
Хотя полисахаридная капсула стрептококков группы B хорошо изучена, и уже известно, что она играет важную роль в индуцировании как вирулентности, так и иммунитета (Kasper D.L. J. Infect. Dis. 153:407 (1986)), однако, было обнаружено, что ее иммуногенность варьируется в зависимости от вида капсулы и факторов иммунной системы хозяина (Baker C.J., и др.; Rev. Infect. Dis. 7: 458-467 (1985)). Недавно проведенные клинические исследования по оценке вакцины, содержащей капсульный полисахарид стрептококков группы B, показали иммунный ответ в целом 63%, что свидетельствует о том, что указанная вакцина не обладает оптимальной иммуногенностью (Baker C.J. и др., New Eng. J. Med 319(18):1180-1185 (1988)).
Различие в иммуногенности наблюдалось также и при испытаниях капсульных полисахаридов других бактерий. Например, вакцина против менингококка типа C является высокоактивной, тогда как полисахаридная вакцина против менингококка типа B является неиммуногенной (Kasper D.L. и др. J. Infect. Dis. 153: 407-415 (1986)). Т-независимые функции иммунной системы хозяина часто бывают необходимы для повышения иммунного ответа к полисахаридным антигенам. Отсутствие Т-независимого иммунного ответа к полисахаридным антигенам может быть причиной низкого уровня антител против Streptococcus группы B у матерей, дети которых впоследствии заражаются стрептококками группы В. Кроме того, дети в возрасте от 18 до 24 месяцев обычно плохо вырабатывают иммунный ответ к Т-независимым антигенам.
Детерминанты вирулентности и иммунитета у Streptococcus группы B.
Существуют пять серотипов стрептококков группы B, которые имеют общий группоспецифический полисахаридный антиген. Однако антитела этой группы антигена не являются протективными в живых моделях. Первоначально с помощью техники, использующей преципитин, Lancefield подразделял стрептококки группы B на четыре серотипа (Ia, Ib, II и III). Состав и структура уникальных группоспецифических капсульных полисахаридов для каждого серотипа были затем определены Jennings H.J. и др., Biochem. 22:1258-1264 (1983), Kasper D.L. и др. J. Infect. Dis. 153: 407-415(1986), Wessels M.P., и др. Trans. Assoc. Amer. Phys. 98: 384-391 (1985). Пятый серотип Ic был определен Wilkinson путем идентификации белкового антигена (первоначального названного белком Ibc), присутствующего во всех штаммах серотипа Ib и в некоторых штаммах с капсулой типа Ia (Wilkinson H.W., и др. J. Bacteriol. 97:629-634 (1969), Wilkinson H. W., и др. Infec. and Immun. 4:596-604 (1971). Затем было установлено, что этот белок по своей распространенности варьируется между различными серотипами стрептококков группы B, и отсутствует в серотипе Ia (Johnson D.R. и др. J. Clin. Microbiol. 19:506-510 (1984)).
Недавно номенклатура была изменена и классификация серотипов Streptococcus группы B проводилась лишь на основании капсульных группоспецифических полисахаридов; при этом был также описан пятый капсульный тип (тип IV) (Pritchard D.G. и др., Rev. Infec. Dis. 10(8):5367-5371, 1988). Поэтому при типировании штаммов стрептококков группы B больше не принимался во внимание антигенный Ibc-белок, который теперь уже именуется белком C. Штамм типа Ic был переклассифицирован в серопит Ia на основании состава его капсульного полисахарида, учитывая также дополнительную информацию относительно белка C.
Иммунологические, эпидемиологические и генетические данные позволяют предположить, что группоспецифическая капсула играет важную роль при выработке иммунитета к инфекциям, вызываемым стрептококками группы B. В последнее время состав и структура группоспецифических капсульных полисахаридов и их роль в вирулентности и иммунитете являются объектом интенсивных исследований (Ferri eri P. и др., Infec/ Immun. 27:1023-1032 (1980), Kranse R.M. и др. , J. Exp. Med. 142:165-179 (1975), Levy N.J. и др., J. Infec. Dis. 149: 851-860 (1984), Wagner B., и др., J. Gen. Microboil. 118:95-105 (1980), Wessel M.R. и др., Trans. Assoc, Amer. Phys. 98:384-391 (1985)).
Специалисты придерживаются различных мнений относительно структурного расположения группоспецифических и стрептококковых полисахаридов группы B на поверхности клетки относительно иммунологически важных факторов, связанных с группоспецифическими полисахаридами, а также относительно механизмов капсуло-детерминированной вирулентности стрептококков группы B (Kasper D.L. и др. , J. Infec. Dis. 153:407-415 (1986)). Для изучения роли капсулы в вирулентности Rubens и др. использовали метод транспозонного мутагенеза в целях получения изогенной линии Streptococcus группы B типа III, которая является неинкапсулированной (Rubens C. E. и др., Proc. Nat I. Acad. Sci. США, 84: 7208-7212 (1987)). Этот эксперимент показал, что, например, в модели новорожденной крысы потеря капсулы приводит к значительному снижению вирулентности. Однако вирулентность клинических изолятов с одинаковым строением капсулы также широко варьировалась. Это дает основание предположить, что помимо капсулы, в патогенезе стрептококков группы B играют роль и другие бактериальные факторы.
Были также описаны некоторые белки и другие бактериальные продукты, которые входят в состав стрептококков группы B и роль которых в вирулентности и иммунитете еще не установлена, например, такие, как CAMP (Кристин-Аткинс-Мунх-Петерсона) фактор, пигмент (вероятно каротиноид), R-антиген, X-антиген, антифагоцитные факторы, и недостаточно определенные "легочные токсины (Ferrieri P. и др., J. Exp. Med. 151: 56-68 (1980), Ferrieri P. и др., Rev. Inf. Dis. 10(2): 1004-1071 (1988), Hill H.R. и др., Sexually Transmitted Diseases Mc Graw Hill, стр. 397-407). Белок C обсуждается ниже.
Изогенные линии стрептококков группы B, не содержащие гемолизина, не показывают уменьшения вирулентности в моделях новорожденных крыс (Weiser J.N. и др. , Infec. and Immun. 55:2314-2316 (1987)). Однако в клинических изолятах, ассоциированных с инфекцией, гемолизин и нейраминидаз присутствуют не всегда. CAMP-фактор является внеклеточным белком стрептококка группы B с молекулярной массой 23500 Да, который в присутствии стафилококкового бета-токсина (сфингомиелиназа) вызывает лизис мембран эритроцитов. Ген, кодирующий CAMP-фактор в стрептококке группы B, недавно удалось клонировать и экспрессировать в E.coli (Schneewind O., и др., Infec. and Immun. 56:2174-2179 (1988)). Роль (если она вообще имеет место) CAMP-фактора, X- и R-антигенов, и других факторов, перечисленных выше, в патогенезе стрептококков группы B не раскрывается в прототипах (Fehrenbach F., и др., In : Bacterial Protein Toxins, Gustav Fischer VerIag, Штуттгарт (1988); Hill H.P. и др., Sexyally Transmitted Diseases, McGraw Hill, W.Y, стр. 297-407 (1984)).
Белки C относятся к группе белковых антигенов, ассоциирующихся с клеточной поверхностью стрептококков группы B, и первоначально были экстрагированы из стрептококков группы B (WiIkinson H.W., и др., J. Bacteri 01.97:629-634 (1969), Wilkinson H. W. и др., Infec and Immun 4:597-604 (1971)). Эти исследователи использовали нагретую соляную кислоту (HCl) для экстракции клеточной стенки и трихлоруксусную кислоту (TCA) для осаждения белковых антигенов. Были описаны две популяции белков C, отличающихся по своей антигенности, а именно: (1) группа белков, подверженных разложению пепсином, но не трипсином, и именуемых либо TR - (трипсин-резистентными), либо α -белками, (2) другая группа белков стрептококков группы B, подверженных разложению как пепсином, так и трипсином, и обозначаемых либо TS - (трипсин-восприимчивыми) или β -белками (Bevanger L. и др., Acta Path. Microbio. Scand Sect. B. 87: 51-54 (1979), Bevanger L. и др., Acta Path. Microbio. Scand. Sect. B 89: 205-209 (1981), Bevanger L. и др., Acta Path. Microbio. Scand. Sect. B 91: 231-234 (1983), Bevanger L. и др., Acta Path. Microbio. Scand. Sect. B. 93: 113-119 (1985), Bevanger L. и др., Asta Path. Microbi ol. Immunol. Scand. Sect. B. 93: 121-124 (1985), Johnson D.R., J. Clin. Microbi ol. 19:506-510 (1984), Russell-J ones G.J., и др., J. EXP. Med. 160:1476-1484 (1984).
В 1975 г. Lancefield и др. использовали исследования мышиного иммунитета, проводимые с помощью антисыворотки, взятой у кроликов, для определения функциональной способности белков C к продуцированию защитного иммунитета против штаммов стрептококков группы B, несущих аналогичные белковые антигены (Lancefi eld R. C. и др., J. Exp. Med. 142:165-179 (1975)). Многими исследователями были получены неочищенные препараты антигенных белков, происходящих от стрептококков группы B и называемых белками C, путем химического экстрагирования из клеточной стенки с использованием либо HCl, либо детергентов (Bevanger L. , и др., Acta Path. Microbio. Scand. Sect. B. 89: 205-209 (1981), Bevanger L. и др., Acta Path. Microbio. Scand. Sect. B. 93: 113-119 (1985), Russell-Jones G.J., и др., J. Exp. Meg. 160:1476-1484 (1984), Valtonen M. V. и др., Microb. Path. 1:191-204 (1986), Wilkinson H.W., и др., Infec. and Immun 4:596-604 (1971). Полученные значения размеров этих антигенов варьируются между 10 и 90 кДа, но ни один вид этих белков не был выделен или охарактеризован (Ferri eri P. и др. , Rev. Inf. Dis. 10(2): 1004-1071 (1988)).
Посредством скрининга с использованием протективной антисыворотки белки C могут быть обнаружены приблизительно в 60% клинических изолятов стрептококков группы B и были обнаружены во всех серотипах, но с различной частотой встречаемости (Johnson D. R., и др., J Clin. MicrobioI. 19:506-510 (1984)) Отдельные изоляты стрептококков группы B ( далее сокращенно - стрептококки B) могут иметь как TR-, так и TS -антигены, или либо один, либо другой из этих антигенов. За исключением способности частично очищенных антигенов вырабатывать защитный иммунитет, роль этих антигенов в патогенезе не исследовалась in vitro. In vivo - исследования штаммов стрептококков В,имеющих белки C, позволяют предположить, что белки C могут быть ответственными за резистентность к опсонизации (Payne N. R. и др., J.Infec. Dis 151:672-681 (1982)), и могут ингибировать внутриклеточный киллинг стрептококков B после фагоцитоза (Payne N. R. и др., Infec. and Immun 55:1243-1251 (1987). Было установлено, что линии типа II стрептококков B, несущие белки C, являются более вирулентными в модели сепсиса новорожденных крыс (Ferrieri P. и др., Immun. 2751023-1032 (1980), Ferrieri P. и др., Rev. Inf. Dis. 102:1004-1071 (1988). Поскольку генетические данные о белках C отсутствуют, то изогенные линии, не содержащие белков C, ранее не были исследованы. Очевидно, что один из TS, или β - белков C связываются с IgA (Russell-Jones G.J. и др., L. Exp. Med. 160: 1476-1484 (1984)). Однако влияние связывания IgA белками C на вирулентность, если оно вообще имеет место, не раскрывается.
В 1986 г. Вальтонен и др. выделили белки стрептококков группы B из супернатантов клеточных культур, которые вырабатывали иммунитет в мышиных моделях (Waltonen M.V. и др., Microb. Path. 1:191-204 (1986))/ Они идентифицировали и частично очистили трипсин-резистентные белки стрептококков B с молекулярной массой 14 000 Да. Антисыворотка с этими белками, полученная от кроликов, индуцировала у мышей иммунитет против их последующего заражения стрептококками группы B типа lb (иммунитет 89%).Этот белок по определению Лансфилда является белком C. Однако при использовании антисыворотки против этого белка для экстрактов иммунопреципитата антигенов стрептококков B был обнаружен ряд реактивных белков с более высоким молекулярным весом. Это позволяет предположить, что белок с молекулярной массой 14 000 может представлять собой общий эпитоп для нескольких белков стрептококков B, либо он представляет собой продукт разложения, обнаруженный в супернатантах культур стрептококков. B. Разница в размерах белков C, выделенных из бактериальных клеток и из супернатантов, дает основание предположить, что белки C могут представлять семейство генов и поддерживать антигенное многообразие как механизм защиты против иммунной системы.
Разнообразие полученных молекулярных масс и трудности в очистке отдельных белков C влекут за собой проблемы, аналогичные тем, с которыми сталкивались многие исследователи при выделении белка M стрептококков группы A (Dale J. B., и др., Infec. and Immun 46(1)267-269 (1984), Fis chett: V.A и др., J, Exp. Meo. 144:32-53 (1976), Fischetti V.A. и др., J.Exp. Med. 146:1108-1123 (1977). Ген, кодирующий белок M, выделен и клонирован, в результате чего было установлено, что он содержит ряд повторяющихся ДНК-последовательностей (Hollin gshea S.K., т др., J. Biol. Chem. 261:1677-1686 (1986), Scott J. R., и др. , Prolc. Natl, Acad. Sci. США 82:1822-1826 (1986), Scott J. R. и др., Infec. fnd Immun. 52:609-612 (1986)). Эти повторяющиеся последовательности могут быть ответственными за посттранскрипционный процессинг, результатом которого является разнообразие размеров продуцируемых белков M. Механизм указанного эффекта не установлен. Различие молекулярных масс, полученных для белков C стрептококков B, может быть результатом аналогичного процесса.
Cleat и др. попытались клонировать белки C, используя препараты антисыворотки против стрептококков B, полученной от Bevanger (α и β) , для скринирования библиотеки ДНК стрептококков B в E.coli (Bevanger L. и др., Acta Path. Microbiol. Immunol. Scand. Sect. B. 93:113-119 (1985), Claet P.H. и др., In f ec. and Immun., 55(5):1151-1155 (1987); все указанные работы вводятся в настоящее описание посредством ссылки). Эти исследователи описали два клона, которые продуцировали белки, связывающие с антистрептококковыми антителамии. Однако им не удалось установить, обладают ли клонированные белки способностью вырабатывать защитные антитела, либо преобладание этих генов связано с линиями стрептококков B, которые, как известно, содержат белки C. Роль последовательностей клонированных генов в вирулентности стрептококков B не исследована. После того, как было установлено, что белки C обладают способностью вырабатывать защитные антитела, этим исследователям все же не удалось получить доказательство того, что любой из этих клонов кодирует белок C.
Конъюгантная вакцина изобретения.
Изобретение позволяет устранить описанные выше недостатки уже имеющихся вакцин против стрептококков B посредством разработки конъюгатной вакцины, в которой капсульные полисахариды ковалентно связаны с остовом белка. Этот способ направлен на продуцирование T-зависимосго гуморального иммунного ответа на капсульные полисахаридные антигены и позволяет "обойти" требования T-независимости для продуцирования антитела (Baker C. и др., Rev. Infec. Dic. 7:458-467 (1985), Kasper D.L. и др., J. Infes. Dis. 153:407-415 (1986), все указанные работы вводятся в настоящее описание в виде ссылки).
В конъюгантной вакцине антигенная молекула такая, как капсульный полисахарид стрептококка B (обсуждается ниже), ковалентно связана с белком-носителем или полипептидом. Эта связь служит для повышения антигенности конъюгантной молекулы. Способы получения конъюгантных вакцин из антигенной молекулы и белка-носителя или полипептида известны специалистам (Jacob C.O. и др., Eur. J. Immunol. 16:1057-1062 (1986); Parker. M. P. и др., Modern Approaches to Vaccines, Chanock R.M., и др., Eds., стр.133-138, Cold Spring Harbor Laboratory, Cold Spring Habor, Ny (1983); Zurawsk: V.P., и др., J. Immun ol. 121: -122-129 (1978); Kl: pstein F.A., и др., Infect. Immun. 37:550-557 (1982); Bessler W. G. Immunobiol. 170:239-244 (1985); Posnett D.N. и др., J. Biol. Chem. 263: 1719-1725 (1988); Chose A.C. и др., Molec, Immunol. 25:223-230 (1988); все указанные работы вводятся в настоящее описание в виде ссылки).
Ранее разработанная модель конъюгатной вакцины была направлена против Hemophilus influenzae (Anderson P. , Infec. and Immun. 39:322-238 (1983); Chu, C. и др. , Infec. Immun. 40:245-256 (1983); Lepow M. Pediat. Infect. Dis. J. 6:804-807 (1987); все указанные работы вводятся в настоящее описание посредством ссылки), и эта модель может быть использована при конструировании новых вакцин изобретения. Другие способы получения таких конъюгатных вакцин раскрываются Anderson P. W. и др. (публикация Европатента 245045; Anderson P.W. и др., патент США NN 5673574 и 4761283; Prank P. и др., патент США 4789735; публикация Европатента 206852; Gordon L. K. патент 4619828; и Beachey E. H. патент США N 4284537; все указанные работы вводятся в настоящее описание посредством ссылки).
В конъюгатных вакцинах против Hemophilus influenzae для остовов белков использовали белки, не имеющие общих антигенных свойств с целевым организмом, из которого были получены бактериальные капсульные полисахариды (Ward J. и др., Vaccines, Plotkin S.A. и др., Eds., Saunder, Филадельфия, с. 300 (1988)).
В противоположность этому в конъюгатной вакцине изобретения в качестве остова используются иммуногенные белки стрептококков B. Как было установлено, такой способ позволяет получить более эффективные вакцины (Insel R.A. и др., New Eng. J. Med. (Editorial) 319(18): 1219-1220 (1988)). Конъюгатная вакцина изобретения, состоящая из белка и полисахарида, является первой вакциной, в которой могут быть использованы белки стрептококков B. В конъюгатных вакцинах изобретения в качестве белков могут быть использованы любые белки, которые являются характерными для стрептококков группы B. Однако для целей изобретения предпочтительно использовать белок C стрептококков группы B. Как будет показано ниже, плазмиды pjMS1 и pJMS23 содержат ДНК, кодирующую стрептококковый белок C. Наиболее предпочтительными белками C являются белки, полученные путем экспрессии указанных ДНК в бактериях.
Как указывалось выше, изобретение относится к клонированию и экспрессии генов, кодирующих протективные белковые антигены стрептококков B. Указанные белки предпочтительно используются в качестве белковых остовов, с которыми связываются капсульные полисахариды стрептококков B с образованием конъюгатной вакцины против указанных бактерий. Кроме того, роль этих белков в вирулентности и иммунности стрептококков группы B может быть использована для разработки дополнительных средств терапии против инфекции, вызываемой стрептококками B. Выделение и характеризация этих генов бактериального происхождения позволит осуществлять обработку генных продуктов в целях оптимизации адъювантных и антигенных свойств полипептидного остова/носителя конъгатной вакцины.
Генетические исследования белков C.
Таким образом, изобретение относится к клонированию белков C стрептококков группы B, их роли и вирулентности и иммунитете, и их способности служить в качестве иммуногена для конъюгатной вакцины против стрептококков группы B.
Несмотря на большое количество литературы, посвященной клонированию во многих группах Streptococcous, лишь ограниченное количество работ раскрывает манипуляции, осуществленные в Steptococcus группы B (Macrina F.L., Ann. Rev. MicrobioI. 38: 193-219 (1984), Waneger A.R. и др., Infec. and Immun. 55: 1170-1175 (1987)). Наиболее широко используемая техника манипулярования в Streptococcus группы B относится к разработке Tn 916 и его использование в транспозонном мутагенезе (Rubens C.E. и др., Proc. Natl. Acad. Sci. США 85: 7208-7212 (1987), Wanger A. R., и др., Res. Vet. Sci. 38:202-208 (1985)). Однако, поскольку очевидно, что имеется более чем один ген, кодирующий белки C, и иммунная антисыворотка связывается с несколькими белками, то скринирование на гены для белка C путем траспозонного мутагенеза является практически неэффективным.
В изобретении клонирование белков C и любых других белков, оказывающих влияние на вирулентность стрептококков группы B или на иммунитет против стрептококков группы B, осуществляется с использованием нового плазмидного вектора. Для этих целей желательно использовать такой вектор клонирования, который можно было бы быстро скринировать на экспрессию белков, способных связываться с естественно продуцируемым антителам к стрептококкам группы B. Поскольку эти антитела являются гетерологичными поликлональными антителами, а не моноклональными антителами, то необходимо, чтобы используемый вектор можно было бы легко скринировать посредством многих позитивных клонов для идентификации нужных генов.
Для скринирования клонов на экспрессию антигенов, связывающихся со специфической антисывороткой, могут быть использованы стандартные способы (Aruffo A. и др., Proc. Natl. Acad. Sci. США 84:8573-8577 (1977). Наиболее широко используемая система GT11, была разработана Young и Davis (Huynh T.V. и др. , DNA CIOning, A Practi cal Approach vol.1. (Glover D.M. ed.) IRL Press, Вашингтон, стр. 49-78); Wong W. W. и др., J. Immunol. Methods. 82: 303-313 (1985); все указанные работы вводятся в настоящее описание в виде ссылки). Эти способы позволяют проводить быстрое скринирование клонов, экспрессированных в лизоненном фаге, продукты которого выделяются посредством фагового лизиса. При использовании этой системы обычно сталкиваются с проблемами, связанными с необходимостью субклонирования ДНК-фрагментов в плазмидные векторы для более детального рестрикционного картирования, получения зондов и ДНК-секвенирования. Кроме того, получение ДНК из фаговых культур является довольно трудоемкой процедурой и ограничивает число потенциально положительных клонов, которые могли бы быть эффективно исследованы. И наконец, получение сырых экстрактов белка из клонированных генов в фаговых векторах-хозяевах является проблематичным.
Для решения указанных проблем в изобретении используется плазмидный вектор, который была разработан в целях скрининга клонированной бактериальной хромосомной ДНК для экспрессии белков, причастных к вирулентности и/или иммунитету. Кроме того, изобретение относится к разработке и использованию эффективного вектора клонирования, который может быть быстро отобран для экспрессии белков, связывающихся с естественно продуцированными антителами к стрептококкам группы B. Этот вектор получали путем модификации обычно используемого плазмидного вектора клонирования, puC12 (Messing J. и др., Gene 19: 269-276 (1982); Norrander J. и др., Gene 26:101-106 1983; Vieira J. и др., Gene 19:259-268 (1982)); все указанные работы вводятся в настоящее описание в виде ссылки). Изобретение относится к описанному ниже вектору и его функциональным эквивалентам.
Используя эту систему, можно легко обрабатывать плазмидные клоны, картировать с помощью рестрикционных эндонуклеаз, и исследовать последовательности ДНК-вставок, полученные зонды и генные продукты, не прибегая к обязательному субклонированию. puC12 представляет собой плазмиду в 2,73 кв с высоким числом копий, несущую ColEI-начало репликации, резистентность к ампициллину и полилинкер в IacZ-гене (Ausub el F.M. и др., Current Topics in Molecular BioIOOy; Greene PubI. Assn./ Wiley Interscience, NY, (1987)); эта работа вводится в настоящее описание посредством ссылки).
Некоторые модификации были осуществлены в полилинкере puC12 (Aruffo A., и др., Proc. NatI. Acad. Sci. США 84:8573-8577 (1987), эта работа вводится в настоящее описание посредством ссылки). В целом схема изменения puC12 сводится к модификации полилинкера для получения идентичных, но не "липких" BstXI-сайтов для клонирования; к добавлению "stuffer" - фрагмента для легкого отделения линейной хозяйской плазмиды, и к обеспечению экспрессии от Iac-промтора во всех трех трансляционных рамках считывания.
Для получения сайта для инсерции чужеродной ДНК с высокой степенью эффективности и минимизации возможности самолигирования плазмиды к полилинкеру добавляют BstXI с инвертированными не "липкими" концами. Как показано на фиг. 1, puC12 сначала разрезают BamHI (Стадия 1) и плазмиду с двумя синтетическими олигонуклеотидными адапторами, которые являются частично комплементарными: с 15-мером (GATCCATTGTGCTGG) и 11-мером (GTAACACGACC) (Стадия 2). При лигировании этих адапторов в puX12, образуются два новых BstXI-сайта, но при этом также сохраняются первоначальные BamHI - сайты (Стадия 3). Затем плазмиду обрабатывают полинуклеотид-киназой и лигируют с образованием замкнутой кольцевой плазмиды (Стадия 4). При обработке этой плазмиды BstXI полученные концы были идентичными и не "липкими" (обе имеют GTGT-выступа) (Стадия 5).
Вторую модификацию в полилинкере осуществляли для очистки плазмиды в целях клонирования без загрязнения из частично разрезанной плазмиды, способной к самолигированию. Fnu 2 фрагмент в 365 п. о. и с тупым концом получали из плазмиды pCbM. Эта кассета или фрагмент - "stuffer", который не содержал BstXI-сайт, был лигирован тупым концом к двум синтетическим олигонуклеотидам, которые являются частично комплементарными: 12-мер (ACACGAGATTTC) и 8-мер (CTCTAAAG) (Стадия 6). Полученный фрагмент с адапторами имел "выступы" в 4 п. о. (ACAC), которые являются комплементарными концами модифицированной плазмиды puC12, показанной в Стадии 5. Модифицированную плазмиду рuC12 лигировали к pCDM-вставке с адапторами, и полученную конструкцию обозначали puX12 (фиг. 4). Плазмида puX12 может быть снова получена из плазмид pJMS1 или pJMS23 путем вырезания введенных ДНК-последовательностей Streptococcus. Альтернативно она может быть образована с помощью рекомбинантной техники (или путем гомологичной рекомбинации) с использованием плазмиды puC12.
Поскольку puX12 используется в качестве экспрессирующего вектора, то последующую модификацию полилинкера предпочтительно осуществлять таким образом, чтобы он содержал все три возможные рамки считывания для Iac-промотора. Эти модификации осуществляют с учетом правильной трансляционной рамки считывания для фрагментов клонированного гена с частотой повторяемости один к шести. Например, клонированный фрагмент может быть введен в вектор в одной из двух ориентаций и в одной из трех рамок считывания. Для конструирования рамки считывания +1 плазмиду puX12 разрезали рестриктазой EcoRI в уникальном сайте полилинкера. Однонитевые липкие 5' - концы затупляли, используя 5' - 3'- полимеразную активность ДНК-полимеразы T4, т два тупых конца лигировали. В результате этих манипуляций теряли EcoRI-сайт и создавали новый XmnI-сайт (фиг. 5). Эту конструкцию подтверждали путем проверки на отсутствие EcoRI-сайта и присутствие нового XmnI-сайта в полилинкере. Помимо этого, осуществляли секвенирование двухнитевой ДНК на +1-модифицированной плазмиде рuX12 с использованием стандартных праймеров секвенирования (Ausu b el P. M., и д. р. , Current Topic in Molecu lar Biology; Greene Publ. Assh/Wiley Interscience, NY (1987)). Секвенирование ДНК-последовательности показало добавление 4 п. о. к полилинкеру и подтвердило модификацию puX12 в +1 рамку считывания. Эту плазмиду обозначали puX12+1.
Для конструирования рамки считывания -1 вектор puX12 разрезали рестриктирующим ферментом Sac1 в уникальном сайте полилинкера puX12. Однонитевые липкие 3'-концы обрезали для получения тупых концов с использованием эндонуклеазной активности 3'- 5' полимеразы T4 и полученные тупые концы лигировали. В полученной последовательности должен отсутствовать Sacl-сайт и присутствовать новый Fnu 2-сайт (фиг. 5). Однако рестрикционное картирование плазмы puX12-1 показало, что там, где отсутствовал Sacl-сайт, не обнаруживалось наличия и FnuD 2-сайта. Кроме того, на полилинкере уже не присутствовали Smal/Xmal-сайты. Несколько возможных конструкций puX12-1 секвенировали из мини-препа, двухнитевой ДНК. Из 6 модифицированных плазмид, которые были секвенированы, была обнаружена одна, где отсутствовали 10 нуклеотидов, в результате чего образовалась рамка считывания - 1 (фиг. 5). В данном случае, очевидно, ДНК-полимераза T4 имела дополнительную экзонуклеазную активность и отрезала дополнительную двухнитевую часть полилинкера. Тем не менее, полученная плазмида имела рамку считывания -1. Эту плазмиду обозначали puX12-1.
Использование векторов puX-12 в клонировании антигенных белков Streptococcus группы B подробно обсуждается в приведенных ниже примерах. Короче говоря, ДНК, происходящую из стрептококков B, или ДНК, комплементарную такой ДНК, вводят в векторы puX12, puX12+1 или puX12-1 и трансформируют в E.coli. Клонированную ДНК экспрессируют в E.coli, и клеточный лизат тестировали на содержание любого белка, способного связываться с антисывороткой к стерптококкам группы B.
Может быть осуществлено несколько представляющих интерес модификаций puX12, которые позволят увеличить ее эффективность. Например, Iac-промотор может быть заменен другим промотором, начало реапликации может быть модифицировано с образованием вектора с низким числом копий, а также может быть заменен маркер резистентности к лекарственному средству.
Изобретение относится к вакцине, содержащей полисахарид (такой, как капсульный полисахарид), который является характерным полисахаридом стрептококков B, конъюгированным с белком, который также является характерным белком стрептококков группы B. Полисахарид и белок указанной конъюгированной вакцины могут быть идентичными молекуле, являющейся характерной для стрептококков B, либо они могут быть функциональными производными таких молекул. Примерами функциональных производных являются фрагменты натуральных белков и/или варианты натуральных белков (таких, как белки, имеющие модифицированную аминокислотную последовательность, но сохраняющие при этом в основном те же иммуногенные, вирулентные или антигенные свойства, что и натуральные молекулы). В целях изобретения может быть использован любой полисахарид, который при введении млекопитающему (человеку или животному) вырабатывает антитела, способные реагировать со стрептококками группы B. Примерами предпочтительных полисахаридов изобретения являются капсульный полисахарид стрептококка B или его эквиваленты. В целях изобретения может быть использован любой белок, который при введении млекопитающему (человеку или животному) либо вырабатывает антитела, способные реагировать с белком, экспрессированным стрептококком группы B, либо способствует повышению иммуногенности полисахарида для продуцирования антител к полисахариду стрептококка B. Примерами предпочтительных белков изобретения являются белки C стрептококков группы B или их эквиваленты.
Используемые в изобретении полисахарид или белок называют характерными для бактерий, если они в основном аналогичны по своей структуре или последовательности молекуле, естественно ассоциированной с бактерией. Этот термин также относится к молекулам, которые являются специфическими для данного организма, и к молекулам, которые, хотя и присутствуют в других организмах, но при этом являются причастными к вирулентности или антигенности бактерий в хозяйском организме человека или животного.
Вакцина изобретения может сообщать резистентность к стрептококкам группы B посредством либо пассивной иммунизации, либо активной иммунизации. В одном из вариантов пассивной иммунизации вакцину вводят здоровому хозяину (человеку или животному), после чего выделяют полученную антисыворотку и вводят ее непосредственно реципиенту, предположительно инфецированному стрептококками группы B.
При осуществлении пассивной иммунизации этого типа возможность мечения антител или фрагментов антител токсинными метками дает дополнительные средства обработки инфекций, вызываемых стрептококками B. В рассматриваемом варианте антитела или фрагменты антител, способные распознавать антигены Streptococus группы B, метят молекулами токсина перед их введением пациенту. При связывании такой токсин-дериватизированной молекулы с клеткой стрептококка B токсинная часть указанной молекулы может способствовать уничтожению этой клетки.
Во втором варианте осуществления иммунизации вакцину вводят женщинам (во время или до беременности или родов) в течение времени или в количестве, достаточном для продуцирования антисыворотки, являющейся протективной как для женщины, так и для плода или новорожденного (посредством пассивного введения антител через планцету).
Таким образом, изобретение позволяет получить средство предупреждения или ослабления инфекции, вызываемой стрептококками B или организмами, имеющими антигены, которые могут быть распознаваемыми и связываемыми антисывороткой к полисахариду и/или белку конъюгированной вакцины. Вакцина в соответствии с изобретением является пригодной для предупреждения или ослабления заболевания, если ее введение пациенту приводит либо к полному или частичному ослаблению (т. е. суппрессии) симптомов или течения болезни, либо к выработке полного или частичного иммунитета к указанной болезни.
Введение вакцины или сыворотки, которую она продуцирует, может быть осуществлено либо в профилактических, либо в терапевтических целях. Профилактическое введение обычно проводят до появления каких-либо симптомов заболевания, вызываемого стрептококками группы B. Профилактическое введение служит целям предупреждения или ослабления любой последующей инфекции. Терапевтическое введение лекарственного средства обычно осуществляют при наличии симптомов, характерных для данного заболевания. Терапевтическое введение служит целям подавления любой имеющейся инфекции.
Таким образом, противовоспалительные средства изобретения могут быть введены либо до начала заражения (т.е. для предупреждения или подавления ранних признаков заболевания) или после начала уже установленного заболевания.
Композиция изобретения является фармакологически приемлемой, если ее введение не оказывает неблагоприятного воздействия на пациента-реципиента. Указанное средство вводится пациенту в терапевтически эффективном количестве, т.е., в таком количестве, которое оказывает физиологически ощутимое воздействие на пациента. Это физиологически ощутимое воздействие выражается в том, что данное лекарственное средство вызывает физиологические обнаружимые изменения у пациента.
Для каждого специалиста очевидно, что вакцина изобретения, предназначенная для введения пациенту, может содержать в своем составе соли, буферы, адъюванты, или другие вещества, необходимые для повышения эффективности композиции. Адъювантами являются вещества, которые могут быть использованы специально для повышения специфического иммунного ответа. Обычно адъювант и композицию смешивают до введения в иммунную систему, либо их вводят отдельно, но в то же самое место иммунизируемого животного. Приблизительно адъюванты могут быть отнесены к нескольким группам по их составу. Такими группами являются масляные адъюванты (например, полный и неполный адъювант Фрейнда), минеральные соли (например, AlK(SO4)2, AlNa(SO4)2, AlNH4(SO4), кремнозем, каолин, и уголь полинуклеотиды (например, поли IC и поли AU-кислоты), и некоторые натуральные вещества (например, воск D из Mycobacterium tuberculosis, а также вещества, обнаруженные в Coryheba cterium parvum, или Bordetella pertussis, и членах рода Brucella. Из перечисленных веществ особенно предпочтительными адъювантами являются сапонины, например, такие, как Quil A (Superfos A/S, Дания).
Примеры материалов, подходящих для использования в вакционной композиции, приводятся в работе Remington Pharmaceuti-cal Sciences (Osol A., Ed., Mack Publishing Co. , Easton, PA, с. 1324-1341 (1980), которая вводится в настоящее описание посредством ссылки).
Терапевтические композиции изобретения могут быть введены парентерально путем инъекции, быстрой инфузии, назофарингеальной абсорбции (через носоглотку), чрезкожно, или перорально. Альтернативно эти композиции могут быть введены внутримышечно или внутривенно. Композиции для парентерального введения могут быть изготовлены в виде стерильных или безводных растворов, суспензий и эмульсий. Примерами безводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, например оливковое масло, и инъецируемые органические сложные эфиры, такие, как этилолеат. Для повышения проницаемости кожи или абсорбции антигена могут быть использованы носители или окклюзионные повязки. Дозы жидких форм для перорального введения могут состоять из липосомного раствора, содержащего жидкую лекарственную форму. Для суспендирования липосом могут служить эмульсии, суспензии, растворы, сиропы, и эликсиры, содержащие инертные разбавители, обычно используемые в этих целях, такие, как очищенная вода. Помимо инертных разбавителей, указанные композиции могут содержать также адъюванты, смачивающие агенты, эмульгирующие и суспендирующие агенты, или подслащивающие, ароматизирующие или отдушивающие агенты.
При режиме многократного введения вакцины могут быть использованы различные способы хронометража иммунизации. Например, композиции изобретения можно использовать более одного раза для повышения уровней и разнообразия иммуноглобулинов, экспрессируемых иммунизованным животным. Обычно при многократной иммунизации вакцину вводят через один-два месяца.
Согласно изобретению, эффективным количеством терапевтической композиции является количество, достаточное для достижения желаемого биологического действия. Как правило, эффективное количество композиции может варьироваться в зависимости от возраста человека или животного, его состояния, пола, и степени заболевания, если оно имеется, а также других факторов, которые могут быть установлены самим специалистом.
Антигенные препараты изобретения могут быть введены в виде одноразовых или в многократных дозах. Эффективные количества композиций изобретения могут варьироваться от 0,01-1,000 мкг/мл на дозу, более предпочтительно от 0,1 до 500 мкг/мл, а наиболее предпочтительно от 10 до 300 мкг/мл на дозу.
Для лучшего понимания изобретения ниже приводятся конкретные примеры, иллюстрирующие некоторые варианты изобретения, но не ограничивающие его.
Пример 1. Эффективность клонирования векторов puX12.
Было проведено несколько тестов на эффективность клонирования векторов puX12 и правильность транскрибирования модифицированной рамки считывания. Результаты проведенных экспериментов кратко систематизированы ниже:
1. Для клонирования ДНК-фрагмента в puX12 три конструкции puX12 (исходная конструкция с рамкой считывания "O"), puX12+1 и puX12-1 смешивали в эквимолярных концентрациях. Затем плазмиды разрезали BstXl для расщепления "Stuffer"-фрагмента в полилинкере. Этот фрагмент выделяли из плазмиды с использованием либо агарозы с низкой температурой плавления, либо градиент ацетата калия (Arufffo A. и др. , Proc. Nat I. Acad. Sci. США 84:8573-8577 (1987), Ausubel F. M. , и др., Current Topics in Molecular Biology; Greene Publ. Assun/Wiley Intersc ience, Ny (1987)). Клонированную ДНК разрезали рестриктазой для получения тупых концов (при этом может быть использована любая рестриктаза). Если необходимо, то для получения тупых концов может быть модифицирована двухнитевая ДНК с однонитевыми концами. Тупые концы ДНК-фрагментов смешивали с двумя синтетическими олигонуклеотидными адапторами. При этом использовали те же 12-мер и 8-мер, что при получении "Stuffer" - фрагмента. Модифицированные ДНК-фрагменты выделяли из невключенных синтетических олигонуклеотидов с использованием градиента ацетата калия. Эти фрагменты затем лигировали в семейство линейных puX12-плазмид и использовали для трансформации E.coli.
Для подтверждения самолигирования puX12-векторов при низкой частоте в условиях оптимизации клонирования вставок с адапторами, второй маркер устойчивости к лекарственному средству клонировали в puX12. Как показано на фиг. 1, puX12 имеет ген β -лактамазы и несет устойчивость к ампициллину (ampR). Клонирование второго маркера проводят для того, чтобы оценить количество клонов, содержащих оба маркера устойчивости к лекарственному средству, по отношению к клонам плазмид puX12, которые самолигировались в обычных условиях клонирования, в результате чего экспрессировалась лишь устойчивость к ампициллину. Ген устойчивости к тетрациклину tetR от плазмиды PBP322 клонировали в полилинкер puX12 с адапторами, описанными выше. Затем проводили ряд проверочных лигирований для установления оптимальной концентрации олигонуклеотидного адаптора на концах фрагмента, и относительного количества модифицированной вставки в линейную плазмиду puX12 для лигирования и трансформации. Используя в качестве маркера ген tetR, можно определить параметры клонирования так, чтобы более чем 99% трансформантов, выбранных на ампицилинсодержащих чашках, также содержали tetR-маркер. Таким образом, частота самолегирования в такой системе является крайне низкой и поэтому нет необходимости проводить скринирование на присутствие вставки в полилинкере до проведения скрининга библиотеки в puX12.
2. Для подтверждения положения трансляционной рамки считывания в полилинкер puX12 клонировали структурный ген, продукт и последовательность которого известны, и в котором отсутствовал его собственный промотор. Для этих целей выбирали мутант tox-структурного гена, содержащегося в плазмиде (Costa T. T. и др., T. Bacteriol. 148(1): 124-130 (1981), Michel T.L., и др., T.Virol. 42: 510-518 (1982); (эти работы вводятся в настоящее описание в виде ссылки)). Эту плазмиду pABC402 обрабатывали рестрикционными эндонуклеазами ApAI и HindIII (Bishai W. R. и др., J. Bacteriol. 169:1554-1563 (1987), Bishai W. R и др., J. Bacteriol. 169:5140-5151 (1987) указанные работы вводятся в настоящее описание в виде ссылки). Apal-сайт структурного гена расположен близ N-конца, а Hind III сайт расположен как раз за пределами C-конца tox-гена. Этот рестрикционный фрагмент (1,2 кв) выделяли из оставшихся 4,1 кв вектора pABC402, используя низкоплавкую агарозу.
Чтобы получить тупые концы для лигирования, tox-фрагмент обрабатывали ДНК-полимеразой T4. С помощью эндонуклеазной активности полимеразы отрезали Apal 3 - концы, а с помощью полимеразной активности затупляли 5' - выступ у Hind III-сайта (Maniatis T. и др., Molecular Cloning. A Laboratory Manual, Cold Spring Habor Press, Cold Spring Harbor, NY (1982)). Этот очищенный фрагмент с тупыми концами лигировали в смесь puX12, которые содержали все три рамки считывания. Затем произвольно отбирали отдельные трансформанты и скринировали путем рестрикционного картирования для определения ориентации и наличия рамок считывания вставок. Кроме того, определяли нуклеотидные последовательности областей полилинкера (адаптора) вставки. Затем идентифицировали все шесть возможных ориентаций и комбинаций рамок считывания. И наконец, экстракты из этих клонов скринировали с использованием вестерн-блот-зондов с антисывороткой к дифтерийному токсину (Blake M.S. и др. Analyt. Biochem. 136: 175-179 (1984), Murphy J.P. и др., Curr. Topics Microbiol. and Immun. 118:235-251 (1985)).
Реактивные белки, родственные токсину, были обнаружены лишь в клонах, которые содержали структурный ген в правильной ориентации и рамку считывания. Эту плазмиду обозначали puDTAH. ДНК-последовательность полилинкера и начало структурного гена tox показаны в табл.1. Изображенная последовательность представляет собой ДНК-последовательность начала структурного гена tox' в puDTAH-1. ATG является инициирующим сигналом для транскрипта (IаcZ)', Gat является началом модифицированного полилинкера puX12 и GCC является началом правильной трансляционной рамки считывания для tox' - гена.
Пример 2. Очистка хромосомной ДНК от стрептококка группы B.
Для осуществления очистки хромосомную ДНК, происходящую от стрептококка группы B, выделяли из A909-штамма стрептококков группы B (Lance field R.C., и др., J. exp. Med. 1: 165-179 (1975) используя метод Hull R.A. и др., Infect. and Immun. 33:933-938 (1981)), модифицированный Рубенсом и др. (Rubenc C. E. и др., Prooc. NatI. Acad. SCi. США 84:7202-7212 (1987); обе эти работы вводятся в настоящее описание посредством ссылки). Вкратце, для превращения штамма A909 (Ia/с) стрептококков группы B в протопласты использовали мутанолизин. Было установлено, что полученный поверхностный экстракт содержал большое количество белков, иммунореагирующих с протективной антисывороткой, полученной против полной бактерии. Нерастворимую белковую фракцию частично очищали с использованием стандартной колоночной хроматографии. Две фракции, одна из которых являлась высококонцентрированной лишь для видов 14 кДа, использовали для иммунизации кроликов. Было установлено, что антисыворотка, продуцированная против этих частично очищенных белков стрептококков B, способна сообщать пассивный иммунитет у мышей при анализе на вирулентность у мышей при анализе на вирулентность против гетерологических капсул стрептококков группы B, несущих белки C.
ДНК стрептококка B очищали путем центрифугирования в градиенте плотности хлорида цезия (CsCl) и хромосомную ДНК исчерпывающе диализовали против TAE-буфера, pH 8,0 (Маниатис T, и др., Molecular Clioning, A Laboratory Manual, Cold Spring Harbor Press, Cold Spring Harbor, NY (1982)). Штамм A909 стрептококков группы B имеет капсулу типа 1 и экспрессирует белки C, и ранее использовался для изучения белков C (Valtonen M. V. и др., Microb. Path. 1: 191-204 (1986)). Этот штамм стрептококков B также использовали при получении протективной антисыворотки для скрининга.
Выход хромосомной ДНК стрептококков группы B составляет в среднем 3-5 мг на каждые 500 мл ночной культуры стрептококков B. Очищенную ДНК отдельно переваривали 24 обычно используемыми рестрикционными эндонуклеазами, и полученные фрагменты помещали на 1,0%-ный агарозный гель. Ферменты выбирали в широком диапазоне, включая и те, которые имели уникальные сайты на полилинкерах, обычно используемые при клонировании векторов. Окрашивание геля этидийбромидом (EtBr) показало, что все рестрикционные ферменты давали распределение по размерам отдельных фрагментов ДНК стрептококков B. Это позволяет предположить, что ДНК стрептококков группы B не является модифицированной для любого из тестированных рестрикционных ферментов.
Для испытания на присутствие каких-либо ингибиторов лигирования ДНК вышеописанные эндонуклеазные перевары осаждали этанолом, помещали в буфер для лигирования и инкубировали с ДНК-лигазой при 14oC в течение ночи. Затем эти образцы снова наносили на 0,1%-ный агарозный гель и окрашивали бромистым этидием. Полученная рестрикционная картина показала более высокомолекулярное распределение. Следовательно, ингибирование лигированных ДНК стрептококков B отсутствовало.
Пример 3. Получение библиотеки хромосомных ДНК стрептококков группы B.
Получение библиотеки хромосомной ДНК стрептококков группы B в puX12 и ее трансформацию в E. coli осуществляли следующим образом. В целях гидролиза хромосомной ДНК стрептококков группы B для клонирования выбирали 4 реастрикционных фермента, которые дают широкий спектр распределения по размерам рестрикционных фрагментов. puX12-вектор и адапторы являются наиболее эффективными, если клонировать фрагменты с тупыми концами. Выбранные ферменты распознают сайты 4 п.о. и оставляют тупыми концы. ДНК стрептококков B отдельно переваривают Alu 1, Fund 2, Hae III и Rsal.
Полученные фрагменты смешивали, очищали фенол/хлороформом, этанол осаждали и ресуспендировали в буфере для лигирования (Maniatis, T. и др., Molecular Cloning. A Laboratory Manual, Cold Spring Harbor Press, Cold Spring Harbor, NY (1982)). 1 мкг ДНК-фрагментов от стрептококка B смешивали с 3 мкг 12-мерного и 2 мкг 8-мерного олигонуклеотидных адапторов. Добавляли 3 мкл ДНК-лигазы T4 (600 ед., New England Biotab), и реакционную смесь выдерживали в течение ночи при температуре 14oC. Затем из ДНК-фрагментов Strertococcus группы B (Str. B) выделяли три свободных линкера с использованием этилацетата с градиентом скорости (Aruffo A. и др., Proc. Natl. Acad. Sci. США, 84: 8573-8577 (1978)).
Плазмиду puX12, содержащую все три трансляционные рамки считывания, переваривали DstX1, и удаляли "stuffer" - фрагмент с использованием низкоплавкого агарозного геля. Библиотеку Ctr. B получали путем смешивания 10 нг адаптированных фрагментов Str. B с 100 нг линейного puX12-вектора в 100 мкл буфера для лигирования, к которому была добавлена 0,1%-ная ДНК-лигаза T4. Реакцию лигирования проводили в течение ночи при 14oC, а затем использовали для трансформации штамма MC1061 E. coli на чашках, содержащих ампициллин (Ausubel F.M., и др., Current Topics Molecuiar Biology (1987)).
16 полученных трансформантов выделяли, культивировали в течение ночи в LB-среде и плазмидную ДНК выделяли с использованием мини-препов, плазмидную ДНК переваривали BamHI и наносили на 1,0%-ный агарозный гель. Все скринированные плазмиды содержали вставки в puX12-вектора и средний размер вставки составлял 1,4 kв. До сих пор скринирование плазмидных ДНК, полученных из более чем 200 клонов, обнаружило лишь один клон, где отсутствовала вставка в полилинкере.
Пример 4. Характеризация протективной антисыворотки, используемой в скрининге библиотеки.
Как обсуждалось ранее, рядом исследователей с использованием различной техники проводилась частичная очистка белков C и протективной антисыворотки, полученной рядом исследователей (Bevanger L. и др., Acta Path. Microbio Scand. Sect. B. 93: 113 - 119 (1985), Russell-I ones G.J. и др., J. Exp. Med. 160:1476 - 1484 (1984), Wilkinson H.W. и др., Infect. and Immum. 4:596 - 604 (1971)).
Серию экспериментов проводили в целях дублирования работ Valtonen, Kasper и Levy, которые выделяли белок с Mw 14000 из супернатантов Str.B,которые вырабатывали протективную антисыворотку (Valtonen M.V. и др., Microb. Path. 1: 191 - 204 (1982)); эта работа вводится в настоящее описание посредством ссылки). Этот эксперимент показал, что если перед концентрированием и очисткой белков C (Wong W.W. и др., J. Immunol. Methods. 82:303 - 313 (1985)) к супернатантам культур Str. B добавить протеолитические ингибиторы, то белок 14000 больше не обнаруживается в супернатанте. Это говорит о том, что указанный белок является продуктом протеолиза белков C с большей молекулярной массой, присутствующих в супернатантах культур Str. B.
Пример 5. Оптимизация условий скрининга для экспрессии в плазмидном векторе.
Как обсуждалось выше, наиболее часто используемые векторы для обнаружения экспрессии основаны на λ GTII (Young R.A. и др., Proc. Natl. Acad. Sci. США 80:1194 - 1198 (1983)).
Повысить чувствительность обнаружения экспрессии из плазмидного вектора puX12 можно путем объединения двух описанных ранее процедур для серологического теста бактериальных колоний. Трансформанты из библиотеки засевали на чашки на ночь и полученные колонии переносили на нитроцеллюлозные фильтры (Bio-Rad). Эти колонии подвергали лизису путем помещения фильтров в атмосферу, насыщенную хлороформом (CHCL3), в закрытый контейнер в течение 30 мин. Затем фильтры помещали в буфер для лизиса и инкубировали в течение ночи в соответствии с описанием Хельфмана (Helfman. M и др., Proc. Nat I. Acad. США 80:31-35 (1983)). Серологическое тестирование проводили с использованием коммерческого лизата (отношение 1: 200) и очищенных антикроличьих IoG козы (отношение 1:3000), конъюгированные с пероксидазой хрена в наборе для экспресс-блот-анализа, полученном Bio-Rad Lab. путем предварительной обработки колоний хлороформом и инкубирования с ДНКазой и лизоцимом, описанного выше, можно снизить требуемое относительное количество антител от 1:500 до 1:5000.
Пример 6. Начальный анализ политивных клонов и их белковых продуктов.
Библиотеку хромосомных ДНК стрептококков группы B в векторе puX12 скринировали с использованием описанной выше протективной антисыворотки против белков C. Библиотека Str. B имела средний фрагмент размером 1,4 kв. Трансформанты скринировали в соответствии с приведенным выше описанием, а затем субклонировали, и снова скринировали три раза с использованием антисыворотки. Из скринированных 20000 клонов было независимо выделено 35 клонов, реагировавших с протективной антисывороткой. Эти клоны обозначали SI - S35, а плазмиды, содержащие эти клоны, обозначали pJMSI-pJMS35. Размеры клонов варьировались от 0,9 до 13,7 kв, а средний размер составлял 4,5 kв.
Плазмидную ДНК выделяли из клонов посредством электрофореза в мини-препаратах, а вставки исследовали с использованием 4 рестрикционных эндонуклеаз. 14 клонов можно разделить на три группы по идентичности размеров вставок и по общей картине рестрикционно-эндонуклеазного картирования внутри каждой группы. Как обсуждается ниже, клоны S1 и S23 являются членами различных групп.
Последующее сравнение рестрикционных картин отдельных клонов позволило идентифицировать 24 клона, которые имели общие рестрикционные фрагменты. Клоны S1 и S23 не обнаруживали каких-либо общих рестрикционных фрагментов.
Получали экстракты клонов, наносили на смесь для вестерн-блоттинга и зондировали с антисывороткой, использованной при скрининге библиотеки. Было идентифицировано 6 классов (по размеру) белковых антигенов (A-F). Совместное использование данных рестрикционно-эндонуклеазного картирования и вестерн-блоттинга позволило классифицировать 24 из 35 клонов по 6 различным картинам белковых антигенов (табл. 2). Эта начальная классификация давала только грубый обзор возможного числа имеющихся генов. Было установлено, что S1 имеет размер 3,5 kв и принадлежит к антигенному белку класса A. S23 имеет размер 13,7 kв и принадлежит к антигенному белку класса D.
Когда вестерн-блотты экстратов клонов зондировали с антисывороткой против Str. B-штамма, не экспрессирующего белки C, то наблюдалась только одна положительная группа клонов (профиль белка B). Это говорит о том, что большинство положительных клонов экспрессируют белки, которые являются уникальными для штаммов, несущих белки C; эти белки не являются общими для всех штаммов стрептококков группы B.
Пример 7. Характеризация клонированных генных последовательностей.
Фактическое число белков C, экспрессируемых стрептококками группы B, не определялось. Недавние иммунологические исследования Брэди и др. по характеризации белков C с помощью типоспецифических антисывороток, полученных из СДС, позволили идентифицировать 4 отдельных антигена (Brady L.J. и др. J. Infect. Dis.
158(5): 965 - 972 (1988)). Предварительная генетическая и иммунологическая характеризация предлагаемых клонов белка C Str. B позволяет сделать вывод о том, что 4 или 5 генов кодируют белки, присутствующие в штаммах стрептококков B, известных как несущие белки C. Для определения, являются ли клонированные генные продукты белками C, были проведены две серии испытаний.
Как указывалось выше, исследование белков C выявило два фенотипа: одна группа белков является восприимчивой к разложению пепсином, но не трипсином (обозначается ТР или α ), а другая группа белков является восприимчивой к разложению как пепсином, так и трипсином (называемая TS или β ) (Johnson D. R. и др., J. Clin Microbiol 19:506 - 510 (1984), Russell-Jones G.J., и др., J. Exp. Med. 160:1476 - 1484 (1984)).
Для скрининга клонированных генных продуктов на вестерн-блоттах использовали типоспецифические антисыворотки α и β (Bevanger L. и др., Acta Path. Microbio. Scand. Sect. B. 87: 51 - 54 (1979); Bevanger L. и др., Acta. Path. Microbio. Scand Sect. B. 89;205 - 209 (1981); Bevanger L. и др., Acta. Path. Microbio, Scand. Sec. B. 91: 231 - 234 (1983); Bevanger L. и др., Acta. Path. Microbio. Scand. Sect. B. 93: 113 - 119 (1985); Bevanger L. и др., Act a. Path, Microbiol, Immunol. Scand. Sec. B. 93:121 - 124 (1985); указанные работы выводятся в настоящее описание в виде ссылки).
Серологическое типирование с использованием α -типоспецифической антисыворотки идентифицировало профиль белка D, а с использованием β -типоспецифической антисыворотки - профиль белка A. Эти белки подвергали перевариванию пепсином и трипсином. Профиль белка D показал чувствительность к пепсину, но не к трипсину, а профиль белка A оказался восприимчивым как к пепсину, так и к трипсину. Эти результаты полностью совпадают с предыдущими исследованиями и подтверждают, что по крайней мере, два из генов белков C были клонированы.
Наиболее важным и характерным свойством белков C является их способность продуцировать протективные антитела против штаммов Str. B, экспрессирующих белки C. Для получения антисыворотки против клонированных генных продуктов могут быть использованы несколько способов. Например, лизаты E. coli - клонов могут быть непосредственно инъецированы кроликам, чтобы определить, содержат ли эти лизаты белки, способные вырабатывать антитела к любым введенным белкам E. coli или Streptococcus B. Полученные антисыворотки могут быть предварительно абсорбированы с лизатом E. coli перед тестированием для снижения уровня перекрестно-реагирующих антител в обоих блотах колоний, используемых для скринирования клонов на экспрессию, и в вестерн-блоттах, используемых для анализа обоих клеточных экстрактов белков Str. B и частично очищенных белков Str. B.
Наиболее характерные клоны из белковых профилей A, B и D обрабатывали ультразвуком и инъецировали кроликам для продуцирования антисыворотки против клонированных белковых антигенов стрептококков B (Lancefield R.C. и др.,, J. Exp. Med. 142: 165 - 179 (1975), Valtonen M.V. и др., Microb. Path. I: 191 - 204 (1986). Контрольным кроликам инъецировали E. coli, несущие puX12 без вставки в полилинкере. Антисыворотку предварительно адсорбировали лизатом E. coli и скринировали сначала на вестерн-блоттах против экстрактов клонов в библиотеке. Это позволяет определить, имеются ли перекрестно-реагирующие эпитопы между клонами, и подтвердить, что эти антисыворотки направлены против клонированных белков, идентифицированных в предыдущем туре скринирования.
Альтернативно предварительно адсорбированная антисыворотка может быть тестирована в мышиной модели иммунитета. При использовании этой классической модели мышам внутрибрюшинно инъецировали кроличью антисыворотку (LancefieId R.C., и др., J. Exp. Med. 142:165-179 (1975)). На следующий день мышам снова внутрибрюшинно инъецировали с LD90 жизнеспособные стрептококки группы B, известные как несущие белки C. Затем подсчитывали число мышей, погибших в течение 48 ч.
Для оценки иммуногенности белков, экспрессированных при клонировании генных последовательностей, культивировали клетки E.coIi, содержащие pJMSI и pJMS23, и использовали их для получения клеточных экстрактов. Затем эти экстракты использовали для иммунизации кроликов. Антисыворотку, продуцированную в ответ на иммунизацию экстрактами S1 и S23, тестировали с использованием мышиной иммунной модели.
При осуществлении эксперимента на мышиной иммунной модели было установлено, что все антисыворотки, продуцированные от клонов, представляющих профили белков A и D (S1 и S23 соответственно), являются протективными. Антисыворотка от клона, представляющего белковый профиль C, а также контрольная антисыворотка не обнаруживали протективности. Антисыворотка, продуцированная против клонов, экспрессирующих белковый профиль C, также связывается с белками, экстрагированными из штаммов стрептококков группы B, которые не несут белок C. Следовательно, эта группа клонов не кодирует белки C. Таким образом, пять из шести групп клонов не кодирует белки, которые являются уникальными для штаммов стрептококков B, экспрессирующих белки C.
Предварительные биохимический, иммунологический и функциональный анализы двух групп клонов показали, что могут быть успешно клонированы, по крайней мере, два гена белков C (S1 и S23). Этот эксперимент является первым свидетельством того, что однополипептидный генный продукт, клонированный из Streptococcus группы C, может вырабатывать протективный иммунитет. Было установлено, что антитела к S1 способны связываться с двумя полосами экстракта A909 с 50 и 60 кДа, а антитела к S23 способны связываться с регулярно повторяющимся элементом полос на поверхности экстракта Str/ B, в пределах Mw от > 180 до 40 кДа. Моноклональное антитело, происходящее от экстракта A909, показало такую же повторяющуюся картину иммунореактивности. Это указывает на то, что в белках разной молекулярной массы распознается один эпитоп, что позволяет предположить, что эти белки имеют регулярно повторяющуюся структуру. Белки, распознаваемые антисывороткой к S1, чувствительны к пепсину и трипсину, а белки, распознаваемые антисывороткой к S23, чувствительны к пепсину, но не к трипсину. Этот эксперимент свидетельствует о том, что эти белки, частично очищенные из Str. В и экспрессированные при клонировании генов Str. B, являются α - и β - антигенами белка C Streptococcus группы B.
35 возможных клонов белка C, описанного выше, могут быть оценены как генетически, так и иммунологически для определения числа присутствующих генов. Кроме того, выделение этих клонов позволило бы идентифицировать гены, несущие протективный иммунитет к инфекции, вызываемой стрептококками группы B. Вероятно, что протективная антисыворотка, используемая для получения исходных клонов, также может обнаруживать и другие белки, не являющиеся белками C. Использование таких других белков в терапии против инфекции, вызываемой стрептококками B, также предусматривается изобретением. Поскольку целью изобретения является выделение и идентификация белков, причастных к иммунитету, то антисыворотка, полученная против белков, экспрессированных этими клонами, может быть также исследована в модели мышиного иммунитета. Гены, экспрессирующие протективные белки, являются предпочтительными для конъюгатной вакцины.
Как обсуждалось выше, предварительное скринирование хромосомной ДНК Str. B в E.coIi/puX12-векторной библиотеке с протективной антисывороткой дало 35 независимо выделенных клонов. Совместное использование данных рестрикционно-эндонуклеазного картирования клонированных фрагментов и вестерн-блотт-анализ белковых экстрактов из клонов позволило предварительно классифицировать 24 и 35 клонов по 6 различным группам белковых антигенов (табл. 2). Это исследование позволяет определить возможное число выделенных генов.
Для дополнительной характеристики указанных клонов использовали предпочтительно блоты колоний для того, чтобы определить, которые из клонов имеют общие ДНК-последовательности. Для этого одну колонию от каждого клона помещали в лунки планшеты для микротитрования, содержащие питательную среду LB, и культивировали при 37oC в течение ночи. Контрольные колонии содержали хозяйский штамм E.coIi и штамм E.coIi, несущий puX12. Ночные культуры переносили на нитроцеллюлозный фильтр на агаровых чашках, содержащих ту же самую культуральную среду. Эти чашки культивировали более 8 ч при 37oC и получали нитроцеллюлозный фильтр, содержащий свежекультивированные колонии, которые могут быть скринированы для ДНК-ДКН-гибридизации. Зонды получали из ДНК-вставок Str. B в библиотеке puX12. Для получения плазмидной ДНК из клонов использовали мини-препараты. Полилинкер в puX12 имел BamHI- и BstXI-сайты на каждой стороне вставки, поэтому вставку Str. B вырезали с использованием либо BamHI, либо BstHI. Хромосомная ДНК Str. B содержит несколько BamHI - сайтов и многие вставки удаляли из вектора в одном фрагменте, полученном в результате переваривания BamHI. Для отделения плазмидного вектора от вставок использовали легкоплавкую агарозу. Эти вставки могут быть вырезаны из агарозного геля и подвергнуты непосредственному рандомизированному праймерному мечению. В результате этого можно идентифицировать клоны, которые имеют общие ДНК-последовательности.
Таким образом, на основании информации, полученной в результате анализа блотов колоний, 35 клонов разделяли на группы по наличию общих ДНК-последовательностей. Эти группы картировали с использованием нескольких рестрикционных эндонуклеаз для определения соотношения каждого клона с другими внутри данной области ДНК. Поскольку хозяйская плазмида puX12 содержит много уникальных сайтов, которые присутствуют только в полилинкере, то в основном рестрикционное картирование может быть проведено с использованием мини-препаратов плазмидной ДНК, которое не требует отдельной очистки вставок. Совместное использование данных блот-анализа колоний с детальным рестрикционным картированием позволяет получить удовлетворительную оценку числа имеющихся генных локусов. Если некоторые группы клонов не представляют нужные гены целиком, то эти клоны могут быть использованы для выделения других более полных копий генов из хромосомной библиотеки. Однако, если изначально выделенные 35 клонов имеют широкое распределение по средним размерам,то вероятно, некоторые из них могут представлять полную открытую рамку считывания.
Перед тем, как приступить в генетическому анализу, предпочтительно получить антисыворотку против клонированных генных продуктов и использовать ее в мышиной иммунной модели для определения протективной способности данной сыворотки против инфекции, вызванной стрептококками группы B (LancefieId R.C. и др., J.Exp.Mrd. 142:165-179 (1975), VaItonen M.V. и др., Microb. Path. 1: 191-204 (1986).
Клон, экспрессированные белки которого способны вырабатывать протективные антитела, является предпочтительным кандидатом для использования в конъюгатной вакцине. Клоны, экспрессированные белки которых не вырабатывают защитных антител, могут быть дополнительно проанализированы на возможность их использования так же, как кандидатов для вакцины. Поскольку белки C являются мембранно-ассоциированными, то неспособность белка, экспрессированного клоном, вырабатывать защитные антитела, очевидно объясняется тем фактом, что этот белок является нестабильным в E.coIi и в высококопийном векторе. С аналогичной проблемой сталкиваются и при клонировании других мембранных белков от Streptococcus группы A и группы B (Kehoe M. и др. Infec. and Immun. 43: 804-810 (1984), SchneewindO, и др. , Infect. and Immun. 56:2174-2179 (1988). Некоторые из 35 клонов, выделенных в предварительном анализе, показали небольшую морфологию колоний. Кроме того, некоторые их этих клонов являются нестабильными и в них обнаруживалось отсутствие в полилинкере puX12 части ДНК-вставки Str. B. Для стабилизации этих клонов может быть использовано несколько способов, например, клонирование в низкокопийный вектор или позади промотора, который может быть негативно регулируемым; культивирование клонов при 30oC вместо 37oC; клонирование в вектор, который был адаптирован для аккумулирования мембранных белков. Кроме того, можно трансформировать плазмиды в E. coIi - хозяина, pcnB, который ограничивает число копий pBR322-присходящих плазмид, подобных puX12 (LopiIato J. и др., MoI, Gen. Genet. 205:285-290 (1986); эта работа вводится в настоящее описание посредством ссылки).
Неспособность клона экспрессировать белок, вырабатывающий защитные антитела, может также свидетельствовать о том, что в экспрессированном белке отсутствует эпитоп, ответственный за иммунитет. Это может быть в том случае, если полный ген не был клонирован, или не может быть экспрессирован в E. coIi. Эта проблема может также возникнуть, если имеет место пост-транскрипционный процессинг белков C в стрептококках группы B, но отсутствует для клонированных генов белка C в E. coIi. В данном случае необходимо либо субклонировать из полного гена и/или перенести его в альтернативный хозяйский фон, где он может быть экспрессирован.
Неспособность клона экспрессировать белок, вырабатывающий защитные антитела, может указывать на то, что антитела, полученные при антигенах, продуцированных в E.coIi, могут отличаться от антител, полученных от животного посредством нативных белков C в Streptococcus группы B. Кроме того, лизированные бактериальные экстракты, используемые для иммунизации кроликов, содержат некоторое количество антигенов белка E.coIi. Поэтому иногда необходимо при получении антисыворотки для тестирования в модели животного получать ее не от полного организма, а от частично очищенных генных продуктов.
Любые клонированные белки стрептококков группы B, которые способны вырабатывать защитные антитела, могут быть названы белками C. Антисыворотка, полученная для этой серии экспериментов, может быть также использована для определения локализации этих белков.
Пример 8. Картирование, характеризация и секвенирование генов белка C.
Для дальнейшего определения характеристик генов белка C может быть получена детальная структурно-генетическая карта описанных выше клонов генов белка C, и определена их ДНК последовательность. Такое картирование предпочтительно проводить с использованием геномного саузен-блоттирования. При определении ДНК-последовательностей генов белка C может получить информацию о структуре н генов, включая их сайты связывания с рибосомой, потенциальные промоторы, сигнальные последовательности, и любые нетипично повторяющиеся последовательности. ДНК-последовательности предпочтительно сравнить с библиотекой известных ДНК-последовательностей для того, чтобы определить, имеется ли гомология с другими генами, которые уже были охарактеризованы. Кроме того, последовательности белков C могут быть определены из ДНК-последовательностей их генов. Иногда можно сделать некоторые предположения относительно структуры, функции и клеточной локализации белка на основании анализа его последовательности.
Геномные саузерн-блотты, например, предпочтительно использовать для определения того, являются ли гены сцепленными. Для осуществления этой техники хромосомную ДНК стрептококков B переваривали отдельно несколькими рестрикционными эндонуклеазами, которые распознают последовательности, содержащие 6 или более пар оснований. Целью это процедуры является получить более крупные сегменты хромосомной ДНК, которая может нести более чем один ген. Отдельные эндонуклеазные перевары затем удаляли из агарозного геля и переносили на нитроцеллюлозу. После этого саузерн-блотты зондировали меченными вставками, происходящими из вышеописанной библиотеки. Если обнаружится, что два клона не соответствуют блотам колонии или эндонуклеазное картирование указывает на аналогичные хромосомные полосы, то это свидетельствует о том, что либо они представляют собой часть одного и того же гена, любо они представляют собой два гена, тесно сцепленных на хромосоме. В любом случае существует несколько способов клонирования из этих более крупных генных сегментов для дальнейшего исследования. Одним способом является получение космидной библиотеки Streptococcus группы B и скрининг на гибридизацию с одним из нужных зондов. Если при этом получают клон, который содержит два или несколько представляющих интерес гена, то он может быть картирован с использованием эндокуклеах и исследован на экспрессию протективных антигенов, как указано выше для описанных ранее клонов.
Идентификация вышеуказанных клонов позволяет определить их ДНК-последовательности. Если эти клоны находятся в плазмиде puX12, то можно использовать секвенирование двухнитевой ДНК с обратной транскриптазой для последовательности от олигонуклеотидных праймеров, полученных для полилинкера. Эта техника была использована ранее для характеризации плазмиды puX12 и является быстрым способом секвенирования множества плазмиды puX12, и является быстрым способом секвенирования множества дополнительных олигонуклеидных праймеров для определения последовательности гена, имеющего более 600 п. о. Поэтому ДНК-секвенирование для генов белка C предпочтительно осуществлять путем субклонирования в М13, однонитевую ДНК-секвенирующую систему (AUSUbel F.M. и др., Current Topicin Molecular Biology (1987)).
Выявление ДНК-последовательностей белков C дает важную информацию о структуре, функции и регуляции генов и их белковых продуктов. Как обсуждалось ранее, гетерогенность размеров белков C, выделенных многими исследователями, и их заметное различие в антигенности дают основание предположить либо наличие семейства генов, либо существование пост-транскрипционного механизма для модификации продуктов гена белков C (Ferrieri P., и др., Infect. Immun. 27-1023-1032 (1980)). Белок M стрептококков группы A обсуждался ранее как пример этого явления (Scott J. R. и др., Proc, Natl. Acad. Sci. США 82: 1822-1826 (1985)). Хотя ДНК последовательности белков M не обнаруживает гомологии с хромосомной ДНК Str. В при гибридизации, однако, могут существовать структурные гомологии между их ДНК-последовательностями (Holliugshead S. K. и др., J, Biol. Chem. 261-1677-1686 (1986), Scott J. R. и др., Proc. Natl. Acad Sci США 82:1822-1826 (1985), Scott J. R. и др., Intect and Immun. 52: 609-612 (1986)). ДНК-последовательности белков C предпочтительно сравнивают с библиотекой известных ДНК-последовательностей. Кроме того, аминокислотные последовательности, происходящие от ДНК-последовательностей, сравнивают с библиотекой известных аминокислотных последовательностей.
Пример 9. Распространенность генов белков C. Для определения распространенности генов белков C хромосомную ДНК, полученную из клинических и лабораторных изолятов различных серотипов стрептококков группы B, зондировали на Саузерн-блотах с генами белка C. Кроме того, для получения информации регуляции экспрессии генов белка C проводили сравнение фенотипической экспрессии, определенной методом, использующим преципитин, с генетической структурой, установленной методом ДНК-ДНК-гибридизации. Зонды генов белков C использовали для скининга хромосомной ДНК от других типов стрептококков и других бактериальных патогенов.
Получали и метили зонды от генов белков B, происходящих из изоляторов стрептококков группы B, которыми являются большинство исходных типовых штаммов, использованных Lancefi eld (Lancefi eld R.C., и др., J. Exp. Med. 142:165-179 (1975)). Блоты колоний из 24 клинических и лабораторных изоляторов Str. В скринировали, используя технику микротитрирования, описанную выше. Способность различных штаммов к гибридизации с генами белков затем сравнивали с фенотипическими характеристиками этих организмов при связывании в целях типирования антисыворотки, направленной против белков C. Таким способом можно определить, какие штаммы несут тот или иной, либо все гены белка C, и несут ли некоторые штаммы молчащие или криптические копии этих генов.
Те штаммы, которые гебридизировались с зондами генов белка C на блотах колоний, затем скринировали с использованием геномных саузерн-блоттов для определения размера, структуры и расположения генов белка C. Хромосомную ДНК выделяли из штаммов стрептококков группы B, которые показали связывание на блотах колоний, переваривали рестриктирующими эндонуклеазами, помещали на агарозный гель и блотировали на нитроцеллюлозу. Эти саузерн-блотты зондировали с использованием зондов генов белков C. Таким способом можно определить, имеются ли различия в расположении и размерах этих генов для различных серотипов стрептококков группы B и сравнить клинические изоляторы (т.е., потенциально вирулентные) с лабораторными штаммами (и с теми, которые заносятся клинически, но не связаны с инфекцией).
Зонды генов белка C предпочтительно также использовать для скрининга других стрептококковых штаммов и ряда патогенных бактерий. Известно, что штаммы стрептококков имеют и другие общие для них белки, ассоциирующие с вирулентностью, например белки M и G (Fahnestock S.R. и др., J. Bact. 167 (3): 870-880 (1986), Heath. G и др., Infec. and Immun/ 55:1233-1238 (1987), Scott J.R. и др. Infect. And Immun 52:609-612 (1986), Walker J. A. и др., Inf ect. and Immun. 55-1184-1189 (1987); все указанные работы вводятся в настоящее описание посредством ссылки). Испытуемые штаммы сначала скринировали с использованием блотов колоний, чтобы определить, имеют ли она какие-либо гомологичные последовательности с зондами генов белков C. Затем получали саузерн-блотты с использованием хромосомной ДНК от бактериальных штаммов, которые показали положительную реакцию на блотах колоний. Затем эти блоты зондировали генами белка C для определения локализации и областей гомологии, таких, как область белка C, служащая "мембранным якорем" (т. е. мембранно-связывающим фрагментом), связывающая с Fc-фрагментом иммуноглобулинов, или имеющая общие участки гомологии с другими генами с аналогичными функциями в других бактериях.
Пример 10. Модификация генов белка C в Streptococcus группы B.
Многие исследователи приписывали белкам C ряд возможных свойств, ассоциированных с вирулентностью, включая резистентность к опсонизации и ингибирование внутриклеточного киллинга после фагоцитоза (Payne N.R. и др., J. Inf ect. Dis 151:672-681 (1985), Payne N.R. и др., Infect. and Immun. 55: 1243-1251 (1987)). Для лучшего понимания роли белков C в вирулентности бактерии конструировали изогенные штаммы, в которых гены белка C являются отдельно мутированными. Эти штаммы могут быть тестированы на вирулентность в модели новорожденных крыс (Zeligs B.J. и др., Infect. and Immun. 37:255-263 (1982)). Для создания изогенных штаммов в целях выяснения роли белков C в вирулентности стрептококков группы B могут быть использованы два метода. Предпочтительным способом является транспозонный мутагенез с самоконъюгирующимся транспозоном Tn 916. Альтернативно может быть использован сайт-специфический мутагенез. Отсутствие эффективных методов для генетических манипуляций в стрептококках группы B приводит к необходимости разрабатывать новые генетические способы модификации генов в стрептококках группы B и создавать изогенные штаммы для исследования вирулентности (Lopilato J. и др. , MOL, Gen, Gend. 205:285-290 (1986); эта работа вводится в настоящее описание в виде ссылки).
Транспозонный инсерционный мутагенез обычно используют для конструирования изогенных штаммов, которые различаются в своей экспрессии антигенов, ассоциирующихся с вирулентностью, и использование этого метода для стрептококков группы B хорошо описаны в литературе (Caparon M.G. и др., Proc. Natl, Acad. Sci. США 84: 8677-8681 (1987), Rubens C.E. и др., Proc. Natl, Acad. Sci. США 84:7208-7217 (1987), Wan ger F.R. Re S. Vet Sci. 38:202-208 (1985), Wepser J. N. Traus. Assoc. Am er. Phys. 98:384-391 (1985); все эти работы вводятся в настоящее описание в виде ссылки). Rubens и др. описывают использование Th 916 для изучения капсулы стрептококков группы B (Rubens C.E., Proc. Natl. Acad. Sci. США 84:7208-7212 (1987)). Как было описано раннее, самоконъюгирующийся транспозон Tn916 может быть изготовлен из Streptococcus faecalis и введен в Streptococcus группы B (Wander A.R., Res. Vet. Sci. 38-202-208 (1985); эта работа вводится в настоящее описание в виде ссылки). Штаммы отбирали на присутствие маркера устойчивости к антибиотику и скринировали на блотах колоний на отсутствие экспрессии белков C, обнаруживаемых посредством специфической антисыворотки, полученной в соответствии с приведенным выше описанием. Изоляты, не обнаружившие экспрессии белков C, могут быть затем картированы, с использованием геномных с Саузерн-блотов для определения локализации инсерциии внутри генов белка C. Первоначальный Tn916-штамм содержал tetR (устойчивость к тетрациклину); однако, недавно в Tn916 был клонирован маркер устойчивости к эритромицину (Rubens C.E. и др., Plasmid, 20: 137-142 (1988)). Необходимо отметить, что в соответствии с Th916-мутагенезом мутантный штамм несет лишь одну копию транспозона и он локализируется внутри гена белка C.
Этот способ может быть использован непосредственно для делетирования генов белка C в стрептококках группы B, если только ген белка C не является главным для выживания стрептококков группы B. Однако ранее указывалось, что штаммы стрептококков группы B страдают от недостатка обнаруживаемого белка C, и исходя из вирулентности бактерии, было бы странным, если этот ген играл главную роль для выживания in vitro. Дополнительное использование Tn916 позволило бы идентифицировать возможные регуляторные элементы генов белка C.
В случае, когда желательно получить конкретно определенные мутации, или белок C является главным для выживаемости стрептококков группы C, то может быть использована техника сайт-направленного мутагенеза (например, для продуцирования условных мутантов). Так, например, сайт-специфический мутагенез может быть использован для генетического анализа белков стрептококков группы B. Проблема, препятствующая дальнейшей разработке указанной техники манипуляции в Streptococcus группы B, заключается в трудности трансформации стрептококков группы B. С помощью электропорации можно ввести ДНК в бактерию, которая трудно поддается трансформации другими способами (Shigekawa R. и др. , Biotech 6:742-751 (1988), эта работа вводится в настоящее описание посредством ссылки). Для преодоления вышеуказанных трудностей может быть проведена трансформация стрептококков группы B с помощью электропорации в определенных условиях. Так, например, можно использовать сайт-направленный мутагенез для оценки комплементации и для введения генов белка C в штаммы стрептококков группы B, которые не экспрессируют белки C. Для вставки нативных или мутированных генов белка C в штаммы стрептококков группы B могут быть использованы несколько способов. Например, в клоны гена белка C в puX12 может быть введен маркер устойчивости к лекарственному средству. Этот маркер может быть экспрессирован в Streptococcus B, но желательно, там, где он обычно отсутствует. Этот модифицированный клон белка puX12 трансформируют в Streptococcus B с использованием электропорации (Shigekawa K. и др., Biotec 6: 742-751 (1988); эта работа вводится в настоящее описание в виде ссылки). Поскольку плазмида puX12 не может реплицировать в стрептококках группы B, то в тех штаммах, которые приобрели фенотип устойчивости к лекарственному средству, это было бы возможным посредством гомологичной рекомбинации между геном белка C на хромосоме хозяина GB и мутантным белком C на плазмиде puX12. Мутанты скринировали согласно с приведенным выше описанием. Если в штамме-реципиенте отсутствуют гомологичные последовательности, то можно сконструировать вектор, содержащий ген белка C, введенный в известный стрептококковый ген, т. е. нативный маркерный ген устойчивости к лекарственному средству от Streptococcus группы B. После электропорации такая плазмидная конструкция может быть интегрирована в хромосому посредством гомологичной рекомбинации.
Альтернативно модифицированные гены белка C могут быть введены в хромосому стрептококка группы B путем инсерции генов в самоконъюгирующийся транспозон Tn916 и вставки модифицированных транспозонов посредством спаривания от Streptococcus faecalis. Этот способ с успехом используют для модификации Tn916 геном эритромицина и введения этого гена в хромосому стрептококка группы B (Rubens C.E. и др., Plasmid 20: 137-142 (1988)). Необходимо отметить, что после Tn916-мутагенеза мутанный штамм несет лишь одну копию транспозона и этот транспозон локализуется внутри гена белка C.
Пример 11. Оценка роли белков C в вирулентности стрептококков группы B.
В предыдущих исследованиях по сравнению штаммов стрептококков группы B, содержащих и не содержащих белки C, использованы изоляты, которые, как известно, не были изогенными (Ferrieri P.и др., Rev. Inf. Dis. 10(2): 1004-1071 (1988)). Поэтому определить, обусловлена ли наблюдаемая разница в вирулентности белками C или какими-либо другими факторами, не представлялось возможным. Конструирование изогенных штаммов, имеющих либо интактные гены белка C, либо делеции генов белка C, позволяет определить роль белка C в вирулентности бактерии. Эти штаммы испытывали на вирулентность в моделях новорожденных крыс, а на иммуногенные свойства - в иммунной мышиной модели. Второй важный тест на вирулентность заключается в проведении испытания на способность гена сохранять вирулентность посредством аллельного замещения в мутантном штамме. Путем инсерции генов белка C в штаммы стрептококков группы B, которые либо не несут этот ген, либо несут инактивированные гены белка C, можно оценить действие белка C посредством исследования вирулентности полученной конструкции в вышеуказанных живых моделях.
Изогенные штаммы стрептококков группы B, в которых гены белка C являются отдельно мутированными, могут быть получены либо путем транспозонного мутагенеза, либо сайт-специфического матегенеза. Эти штаммы предпочтительно оценивались на геномных Сайзерн-блотах, чтобы убедиться, что только одна инсерция присутствует в хромосоме. Расположение этих инсерций может быть определено с использованием техники тонкого структурно-генетического картирования, описанного выше. Затем изогенные штаммы тестируют на вирулентность в модели новорожденных крыс (Zeli gs B.J. и др., Infect and Immun 37: 255-263 (1982)).
Транспозонный мутагенез позволяет идентифицировать гены, участвующие в регуляции белков C. Например, штаммы, несущие гены белка C дикого типа, которые после транспозонного мутагенеза не обнаруживают больше экспрессии белка C и в которых этот транспозон не находится внутри структурного гена белка C, имеют мутации в последовательностях, участвующих в регуляции экспрессии генов белка C. Этот метод успешно использовался при характеризации mry-локуса в стрептококках группы C, который участвует в регуляции белка M (Caparen M.G. и др. , Proc. Natl. Acad. Sci, США 84:8677-8681 (1987), Robbins J.C. и др., J.Bacteriol. 169: 5633-5640 (1987); эти работы вводятся в настоящее описание посредством ссылки). Указанные методы могут быть также использованы для продуцирования штаммов, способных к сверхсинтезу белков C, или штаммов, которые продуцируют белки C с измененными вирулентностью или иммунитетом.
Пример 12. Локализация белков C в стрептококках группы B и оценка их способности к связыванию с иммуноглобулинами.
Лансефилд и др. показали, что антитело к белкам C связывается с внешней мембраной стрептококков группы B (Lancefield R.C. и др., J. Exp. Med. 142: 165-179 (1975), Wagner B. и др., J. Gem. Microbiol 118: 95-105 (1980)). Это позволяет предположить, что белок C является внешнемембранным белком. Белки C могут быть также выделены из супернатантов культур стрептококков группы B, что говорит о том, что эти белки могут быть либо секретированы стрептококками группы B, либо "потеряны" ими с клеточной поверхности при высокой скорости. ДНК - и белковые последовательности, происходящие от генов белка C, являются ценным материалом при определении структуры и функции белков C. Одним, возможно, из наиболее важных факторов, обычно приписываемых белкам C, является их способность связываться с иммуноглобулином IgA (Ferrieri P. и др. , Rev. Inf. Dis. 10(2): 1004-1071 (1988), Russell-Jores G.J. и др., J. Exp. Med, 160: 1467-1475 (1984)).
Для определения локализации поверхностных детерминант клеток, распознаваемых специфическим антителом, может быть использована иммуноэлектронная микроскопия. Антисыворотку, продуцированную против клонов белка с Str.B, инкулировали со штаммами Str.B, несущими белок C. Как было описано ранее, для обнаружения антигена на клеточной поверхности использовали ферритин-конъюгированный козий антикроличий IgG (Rubens C.E. и др., Proc.Natl. Acad. Sci. США 84: 7208-7212 1987, Wagner B. и др., J. Gen.Mierobiol. 118:95-105 (1980)).
Несложный способ определения способности белков C к связыванию иммуноглобулинов может быть осуществлен с использованием вестерн-блоттинга. В этих целях клеточные экстракты клонов E.coli, содержащих гены белка C, и штаммов Str.B, несущих белки C, подвергают ДСН-электрофорезу в ПААГ и переносят на нитроцеллюлозу. В качестве контроля используют экстракты E.coli, несущие плазму puX12 дикого типа; штаммы стрептококков группы B, которые не содержали генов белка C; и изогенные штаммы стрептококков группы B, в которых гены белка C были инактивированы. Вестерн-блотты могут быть отдельно зондированы меченными иммуноглобулинами, например, такими, как IgG, IgM, IgA, и их компонентами, например, Fc или F(ab)2-фрагментами (Heath D.G., и др. Infect. and Immun. 55:1233-1238 1987, Russell-Jones G.J. и др., L.Exp.Meo. 160: 1467-1475 (1984)). Иммуноглобулины предпочтительно йодируют с использованием йодогена или хлорамина T.
Более специфический способ измерения способности белков C связываться с имунноглоблулинами и их компонентами заключается в очистке белков C и их непосредственного использования в анализе на связывание (Fahnestock S.R. и др. , J. Bact. 167(3): 870-880 (1986), Heath D.G и др., Infect and Immun 55: 1233-1238 (1987)). Очистку белка C можно проводить с использованием последовательности белка. Кроме того, поскольку гены белка C можно экспрессировать в E. coli, то можно сконструировать штаммы E.coli, способные к сверхпродуцированию белка C, и тем самым получать большие количества белков C для очистки.
Хотя настоящее изобретение описано выше на предпочтительных примерах его осуществления, однако, эти примеры не следует рассматривать как некое ограничение, и каждый специалист может внести различные изменения, не выходящие при этом за рамки существа и объема настоящего изобретения.
Пример 13. Использование антигенов клонированного белка C стрептококков группы B в конъюгатной вакцине.
Описанные выше антигены протективного белка C стрептококков группы B тестировали на возможность использования их в конъюгантной вакцине. Для оценки такой возможности получали клеточные экстракты E.coli, содержащих pJMSI или pJMS23, в соответствии с описанным выше процедурами, и использовали их для иммунизации кроликов. Полученную антисыворотку испытали в машиной модели на летательность для того, чтобы оценить способность данной антисыворотки продуцировать у мышей иммунитет против инфекции, вызываемой штаммом H36B стрептококков группы B. Штамм H36B содержит белок C Str.B. Для контроля определяли протективную способность антисыворотки на мышах против штамма 515 стрептококков (который не содержит белка C).
Результаты эксперимента показаны на фиг. 6.
Хотя изобретение описано выше на предпочтительных примерах его осуществления, однако эти примеры не следует рассматривать как некое ограничение и в него могут быть внесены различные изменения, не выходящие за рамки существа и объема настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ МОЛЕКУЛА ДНК, КОДИРУЮЩАЯ БЕЛОК АЛЬФА-АНТИГЕНА STREPTOCOCCUS ГРУППЫ В, ПЛАЗМИДНЫЙ ВЕКТОР | 1993 |
|
RU2209247C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ОТНОСЯЩИЕСЯ К СИНТЕТИЧЕСКИМ БЕТА-1,6-ГЛЮКОЗАМИНОЛИГОСАХАРИДАМ | 2009 |
|
RU2532911C2 |
| Живая вакцина на основе штамма пробиотиков ENTEROCOCCUS FAECIUM L3 для профилактики инфекции, вызванной STREPTOCOCCUS PNEUMONIE | 2018 |
|
RU2701733C1 |
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |
| КОМПЛЕКС РЕКОМБИНАНТНЫХ ПОЛИПЕПТИДОВ, ОБЛАДАЮЩИЙ ПРОТЕКТИВНЫМИ СВОЙСТВАМИ В ОТНОШЕНИИ STREPTOCOCCUS AGALACTIAE И STREPTOCOCCUS PYOGENES | 2014 |
|
RU2587627C2 |
| БЕЛКОВЫЙ ХИМЕРНЫЙ РЕЦЕПТОР, ДНК, КОДИРУЮЩАЯ БЕЛКОВЫЙ ХИМЕРНЫЙ РЕЦЕПТОР | 1994 |
|
RU2158305C2 |
| СПОСОБ ВВЕДЕНИЯ ГЕНОВ ПАТОГЕННЫХ СТРЕПТОКОККОВ В ХРОМОСОМНУЮ ДНК ПРОБИОТИЧЕСКОГО ШТАММА ENTEROCOCCUS FAECIUM L3 ДЛЯ ЭКСПРЕССИИ В ПИЛЯХ | 2015 |
|
RU2640250C2 |
| ВАКЦИНЫ ПРОТИВ ВИРУСА ПРОСТОГО ГЕРПЕСА | 2010 |
|
RU2575067C2 |
| СПОСОБ НАПРАВЛЕНИЯ (ИНДУКЦИИ) ИММУННОГО ОТВЕТА, ДНК, ВЕКТОР, СПОСОБ ЛЕЧЕНИЯ | 1992 |
|
RU2161044C2 |
| ВАКЦИНЫ С ИСПОЛЬЗОВАНИЕМ ВЕЗИКУЛ НА ОСНОВЕ GNA 1870 ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ДЛЯ ПРОФИЛАКТИКИ ЗАБОЛЕВАНИЙ, ВЫЗЫВАЕМЫХ NEISSERIA MENINGITIDIS | 2006 |
|
RU2432962C2 |
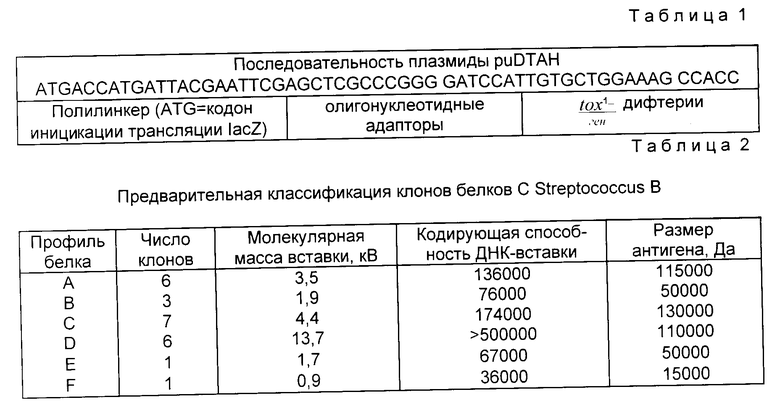
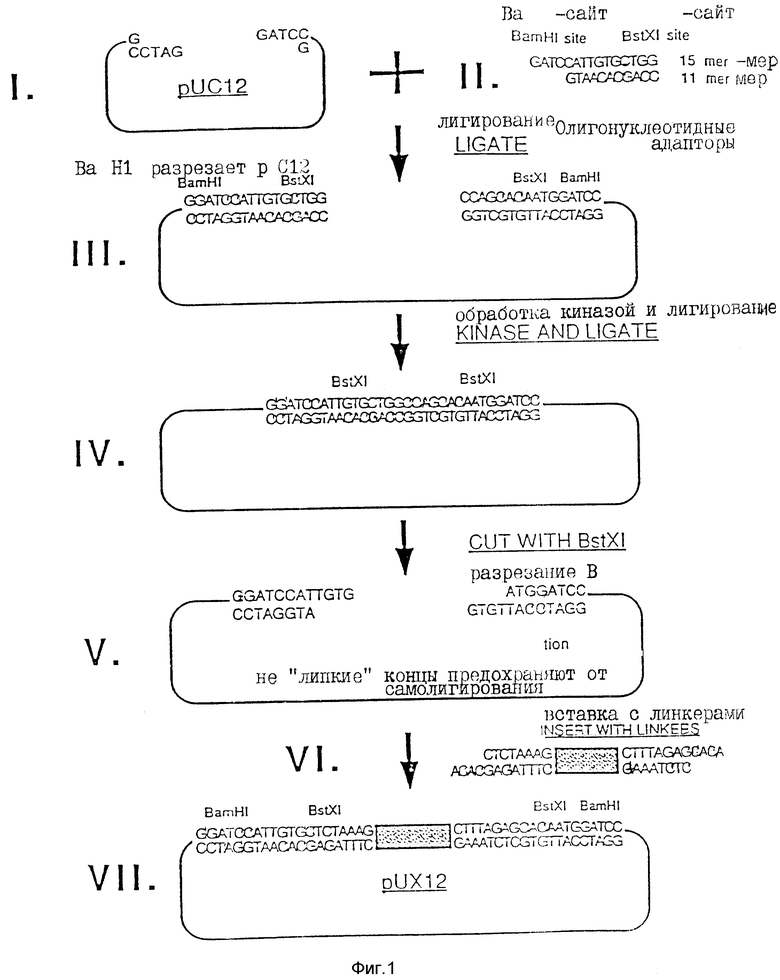
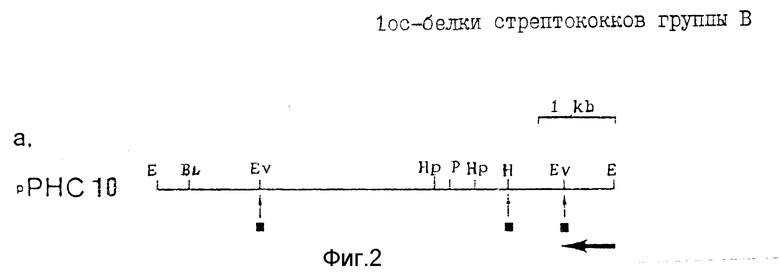
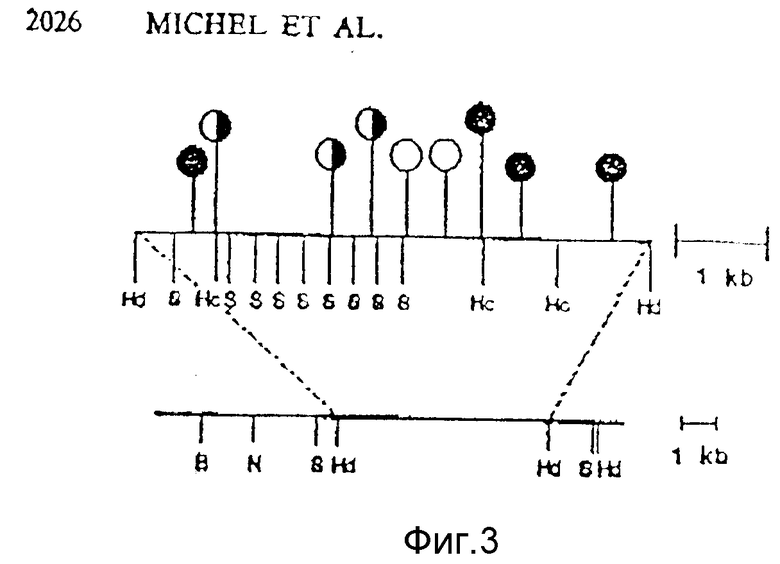

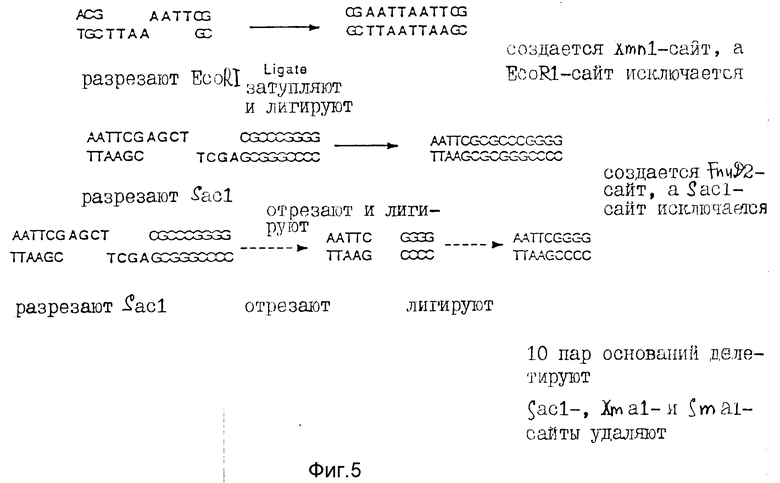
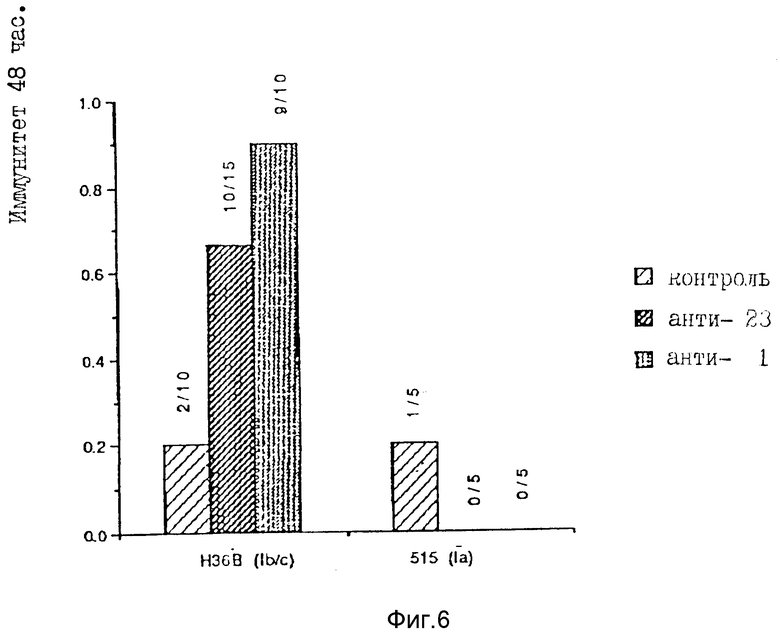
Изобретение относится к вакцине, способной вырабатывать иммунитет у реципиента против инфекции, вызванной стрептококками группы B. Вакцина включает в себя полисахарид, конъюгированный с белком. При этом указанные белок и полисахарид являются характерными молекулами стрептококков группы B. Используемый белок является рекомбинантным белком S1 и кодируется плазмидой pJMSI АТСС N 40659. Вакцина индуцирует T-независимый иммунитет против инфекции, вызываемой стрептококками группы B. 3 с. и 5 з.п. ф-лы, 6 ил., 2 табл.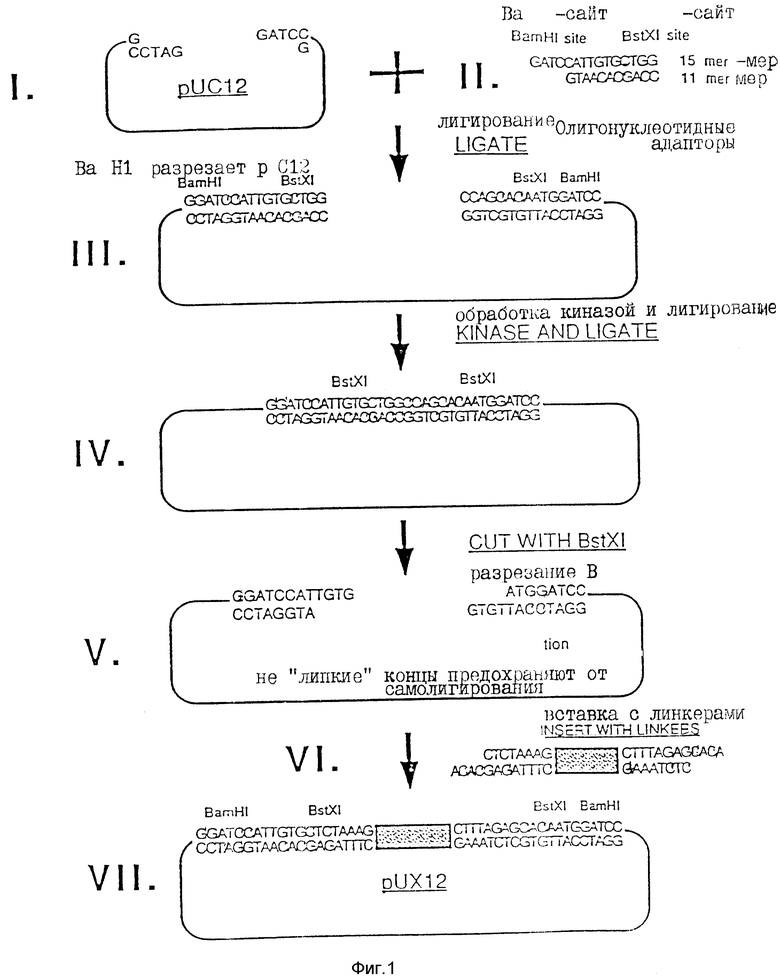
| US, патент, 4367223, A 61 K 39/09, 1983 | |||
| Rev.Jnfec | |||
| Disiases, 1985, N 4, v | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Приспособление для отопления печей нефтью | 1922 |
|
SU458A1 |

