ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ОТПРАВЛЕННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
[0001] Содержание отправленного в электронном виде перечня
последовательностей в текстовом файле ASCII, имеющем название sequencelisting_PCTascii.txt, созданном 6 ноября 2012 г. и имеющем размер 382 килобайта, поданного вместе с настоящей заявкой, включено в данный документ посредством ссылки во всей своей полноте.
ПРЕДПОСЫЛКИ
Область техники
[0002] Настоящее изобретение относится к способам комбинированной терапии с применением доменов, связывающихся с Psl и PcrV Pseudomonas для применения в предупреждении и лечении инфекции, вызываемой Pseudomonas. Кроме того, настоящее изобретение предусматривает композиции, применимые в таких способах терапии.
Предпосылки настоящего изобретения
[0003] Pseudomonas aeruginosa (P. aeruginosa) представляет собой грамотрицательный условно-патогенный микроорганизм, вызывающий как острые, так и хронические инфекции у ослабленных лиц (Ma et al., Journal of Bacteriology 189 (22): 8353-8356 (2007)). Отчасти это связано с высокой врожденной устойчивостью бактерии к используемым в клинической практике антибиотикам, а отчасти связано с формированием чрезвычайно устойчивых к антибиотикам биопленок (Drenkard E., Microbes Infect 5: 1213-1219 (2003); Hancock & Speert, Drug Resist Update 3: 247-255 (2000)).
[0004] P. aeruginosa является частой причиной внутрибольничных инфекций в западном мире. Это частый возбудитель бактериемии у ожоговых больных и лиц с ослабленным иммунитетом (Lyczak et al, Microbes Infect 2: 1051-1060 (2000)). Он также является наиболее распространенной причиной госпитальной пневмонии, вызываемой грамотрицательными микроорганизмами (Craven et al, Semin Respir Infect 11: 32-53 (1996)), особенно у пациентов, находящихся на искусственной вентиляции легких, и является наиболее распространенным патогеном в легких лиц с муковисцидозом (Pier et al, ASM News 6: 339-347 (1998)).
[0005] Экзополисахарид Psl Pseudomonas, как сообщается, заякорен на поверхности P. aeruginosa и считается важным в облегчении колонизации тканей хозяина и в создании/поддержании формирования биопленки (Jackson, K.D., et al, J Bacteriol 186, 4466-4475 (2004)). Его структура включает богатый маннозой повторяющийся пентасахарид (Byrd, M.S., et al, Mol Microbiol 73, 622-638 (2009)).
[0006] PcrV представляет собой относительно консервативный компонент системы секреции III типа. PcrV, по-видимому, представляет собой интегральный компонент транслокационного аппарата системы секреции III типа, опосредующей доставку секреторных токсинов III типа в эукариотические клетки-мишени (Sawa Т., et al. Nat. Med. 5:392-398 (1999)). Активная и пассивная иммунизация против PcrV уменьшала проявления острого повреждения легких и смертность мышей, инфицированных цитотоксической P. aeruginosa (Sawa et al. 2009). Основной эффект иммунизации против PcrV был связан с блокадой транслокации секреторных токсинов III типа в эукариотические клетки.
[0007] В связи с увеличением устойчивости ко множеству лекарственных средств в данной области техники остается потребность в разработке новых стратегий для идентификации новых профилактических и терапевтических средств, специфичных в отношении Pseudomonas.
КРАТКОЕ ОПИСАНИЕ
[0008] Настоящее изобретение предусматривает связывающую молекулу или ее антиген-связывающий фрагмент, которые специфически связываются с PcrV Pseudomonas, содержащие (a) CDR1 тяжелой цепи, содержащий SYAMN (SEQ ID NO: 218), или его вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; CDR2 тяжелой цепи, содержащий AITISGITAYYTDSVKG (SEQ ID NO: 219), или его вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; и CDR3 тяжелой цепи, содержащий EEFLPGTHYYYGMDV (SEQ ID NO: 220), или его вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; (b) CDR1 легкой цепи, содержащий RASQGIRNDLG (SEQ ID NO: 221), или его вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; CDR2 легкой цепи, содержащий SASTLQS (SEQ ID NO: 222), или его вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; и CDR3 легкой цепи, содержащий LQDYNYPWT (SEQ ID NO: 223), или его вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; или комбинации (a) и (b). В одном варианте осуществления связывающая молекула или ее антиген-связывающий фрагмент специфически связывается с PcrV Pseudomonas и содержит (a) CDR1 тяжелой цепи, содержащий SYAMN (SEQ ID NO: 218), CDR2 тяжелой цепи, содержащий AITISGITAYYTDSVKG (SEQ ID NO: 219), и CDR3 тяжелой цепи, содержащий EEFLPGTHYYYGMDV (SEQ ID NO: 220), и (b) CDR1 легкой цепи, содержащий RASQGIRNDLG (SEQ ID NO: 221), CDR2 легкой цепи, содержащий SASTLQS (SEQ ID NO: 222), и CDR3 легкой цепи, содержащий LQDYNYPWT (SEQ ID NO: 223). В одном варианте осуществления выделенная связывающая молекула или ее антиген-связывающий фрагмент специфически связываются с PcrV Pseudomonas и содержат (a) вариабельную область тяжелой цепи по меньшей мере с 90% идентичностью последовательности по отношению к SEQ ID NO: 216; (b) вариабельную область легкой цепи по меньшей мере с 90% идентичностью последовательности по отношению к SEQ ID NO: 217; или комбинации (a) и (b). В другом варианте осуществления связывающая молекула или ее фрагмент содержат: (a) вариабельную область тяжелой цепи по меньшей мере с 95% идентичностью последовательности по отношению к SEQ ID NO: 216; (b) вариабельную область легкой цепи по меньшей мере с 95% идентичностью последовательности по отношению к SEQ ID NO: 217; или комбинации (a) и (b). В другом варианте осуществления связывающая молекула или ее фрагмент представляют собой V2L2 и содержат: (a) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 216, и (b) вариабельную область легкой цепи, содержащую SEQ ID NO: 217.
[0009] В одном варианте осуществления настоящее изобретение предусматривает выделенную связывающую молекулу или ее антиген-связывающий фрагмент, которые специфически связываются с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-области V2L2. В другом варианте осуществления настоящее изобретение предусматривает выделенную связывающую молекулу или ее антиген-связывающий фрагмент, которые специфически связываются с PcrV Pseudomonas и осуществляют конкурентное ингибирование связывания PcrV Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH и VL V2L2. В одном варианте осуществления связывающая молекула или ее фрагмент представляют собой рекомбинантное антитело. В одном варианте осуществления связывающая молекула или ее фрагмент представляют собой моноклональное антитело. В одном варианте осуществления связывающая молекула или ее фрагмент представляют собой химерное антитело. В одном варианте осуществления связывающая молекула или ее фрагмент представляют собой гуманизированное антитело. В одном варианте осуществления связывающая молекула или ее фрагмент представляют собой антитело человека. В одном варианте осуществления связывающая молекула или ее фрагмент представляют собой биспецифическое антитело.
[0010] В одном варианте осуществления связывающая молекула или ее фрагмент ингибируют доставку секреторных токсинов III типа в клетки-мишени.
[0011] В одном варианте осуществления настоящее изобретение предусматривает биспецифическое антитело, содержащее связывающий домен, который связывается с Psl Pseudomonas, и связывающий домен, который связывается с PcrV Pseudomonas. В одном варианте осуществления домен, связывающийся с Psl, включает scFv-фрагмент, а домен, связывающийся с PcrV, включает интактный иммуноглобулин. В одном варианте осуществления домен, связывающийся с Psl, включает интактный иммуноглобулин, а указанный домен, связывающийся с PcrV, включает scFv-фрагмент. В одном варианте осуществления scFv слит с амино-концом VH-области интактного иммуноглобулина. В одном варианте осуществления scFv слит с карбокси-концом CH3-области интактного иммуноглобулина. В одном варианте осуществления scFv вставлен в шарнирную область интактного иммуноглобулина.
[0012] В одном варианте осуществления домен, связывающийся с Psl, специфически связывается с тем же эпитопом Psl Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), по меньшей мере на 90% идентичные соответствующим областям WapR-004. В одном варианте осуществления домен, связывающийся с Psl, специфически связывается с Psl Pseudomonas и осуществляет конкурентное ингибирование связывания Psl Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH- и VL-области, по меньшей мере на 90% идентичные соответствующим областям WapR-004. В одном варианте осуществления VH и VL WapR-004 содержат SEQ ID NO: 11 и SEQ ID NO: 12, соответственно. В одном варианте осуществления последовательность WapR-004 выбрана из группы, состоящей из SEQ ID NO: 228, SEQ ID NO:229 и SEQ ID NO: 235. В одном варианте осуществления домен, связывающийся с PcrV, специфически связывается с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-области V2L2. В одном варианте осуществления домен, связывающийся с PcrV, специфически связывается с PcrV Pseudomonas и осуществляет конкурентное ингибирование связывания PcrV Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH и VL V2L2. В другом варианте осуществления домен, связывающийся с PcrV, специфически связывается с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-области, по меньшей мере на 90% идентичные соответствующим областям V2L2. В одном варианте осуществления VH и VL V2L2 содержат SEQ ID NO: 216 и SEQ ID NO: 217, соответственно. В одном варианте осуществления VH и VL WapR-004 (SEQ ID NO: 11 и 12, соответственно) и VH и VL V2L2 (SEQ ID NO: 216 и 217, соответственно). В одном варианте осуществления биспецифическое антитело содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 228, SEQ ID NO: 229 и SEQ ID NO: 235.
[0013] В одном варианте осуществления настоящее изобретение предусматривает полипептид, содержащий аминокислотную последовательность SEQ ID NO: 216 или SEQ ID NO: 217. В одном варианте осуществления полипептид представляет собой антитело.
[0014] В одном варианте осуществления настоящее изобретение предусматривает клетку, содержащую или производящую связывающую молекулу или полипептид, раскрытые в данном документе.
[0015] В одном варианте осуществления настоящее изобретение предусматривает выделенную молекулу полинуклеотида, содержащую полинуклеотид, который кодирует связывающую молекулу или полипептид, описанные в данном документе. В одном варианте осуществления молекула полинуклеотида содержит последовательность полинуклеотида, выбранную из группы, состоящей из SEQ ID NO: 238 и SEQ ID NO: 239. В другом варианте осуществления настоящее изобретение предусматривает вектор, содержащий полинуклеотид, описанный в данном документе. В другом варианте осуществления настоящее изобретение предусматривает клетку, содержащую полинуклеотид или вектор.
[0016] В одном варианте осуществления настоящее изобретение предусматривает композицию, содержащую связывающую молекулу, биспецифическое антитело или полипептид, описанные в данном документе, и фармацевтически приемлемый носитель.
[0017] В одном варианте осуществления настоящее изобретение предусматривает композицию, содержащую связывающий домен, который связывается с Psl Pseudomonas, и связывающий домен, который связывается с PcrV Pseudomonas. В одном варианте осуществления домен, связывающийся с Psl, специфически связывается с тем же эпитопом Psl Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), по меньшей мере на 90% идентичные соответствующим областям WapR-004, Cam-003, Cam-004, Cam-005, WapR-001, WapR-002, WapR-003 или WapR-016. В одном варианте осуществления домен, связывающийся с Psl, специфически связывается с Psl Pseudomonas и осуществляет конкурентное ингибирование связывания Psl Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH- и VL-области, по меньшей мере на 90% идентичные соответствующим областям WapR-004, Cam-003, Cam-004, Cam-005, WapR-001, WapR-002, WapR-003 или WapR-016. В одном варианте осуществления VH и VL WapR-004 содержат SEQ ID NO:11 и SEQ ID NO: 12, соответственно, VH и VL Cam-003 содержат SEQ ID NO: 1 и SEQ ID NO: 2, соответственно, VH и VL Cam-004 содержат SEQ ID NO: 3 и SEQ ID NO: 2, соответственно, VH и VL Cam-005 содержат SEQ ID NO:4 и SEQ ID NO: 2, соответственно, VH и VL WapR-001 содержат SEQ ID NO:5 и SEQ ID NO: 6, соответственно, VH и VL WapR-002 содержат SEQ ID NO: 7 и SEQ ID NO: 8, соответственно,VH и VL WapR-003 содержат SEQ ID NO: 9 и SEQ ID NO: 10, соответственно, и VH и VL WapR-016 содержат SEQ ID NO: 15 и SEQ ID NO: 16, соответственно. В одном варианте осуществления домен, связывающийся с PcrV, специфически связывается с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-области V2L2. В одном варианте осуществления домен, связывающийся с PcrV, специфически связывается с PcrV Pseudomonas и осуществляет конкурентное ингибирование связывания PcrV Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH и VL V2L2. В одном варианте осуществления домен, связывающийся с PcrV, специфически связывается с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-области, по меньшей мере на 90% идентичные соответствующим областям V2L2. В одном варианте осуществления VH и VL V2L2 содержат SEQ ID NO: 216 и SEQ ID NO: 217, соответственно. В одном варианте осуществления домен, связывающийся с Psl, содержит VH- и VL-область WapR-004, а указанный домен, связывающийся с PcrV, содержит VH- и VL-область V2L2 или его антиген-связывающие фрагменты.
[0018] В одном варианте осуществления композиция содержит первую связывающую молекулу, содержащую указанный домен, связывающийся с Psl, и вторую связывающую молекулу, содержащую домен, связывающийся с PcrV. В одном варианте осуществления первая связывающая молекула представляет собой антитело или его антиген-связывающий фрагмент, а указанная вторая связывающая молекула представляет собой антитело или его антиген-связывающий фрагмент. В одном варианте осуществления антитела или антиген-связывающие фрагменты независимо выбирают из группы, состоящей из моноклонального антитела, гуманизированного антитела, химерного антитела, антитела человека, Fab-фрагмента, Fab'-фрагмента, F(ab)2-фрагмента и scFv-фрагмента. В одном варианте осуществления связывающие домены, связывающие молекулы или их фрагменты связываются с двумя или более, тремя или более, четырьмя или более или пятью или более различными серотипами P. aeruginosa. В одном варианте осуществления связывающие домены, связывающие молекулы или их фрагменты связываются по меньшей мере с 80%, по меньшей мере с 85%, по меньшей мере с 90% или по меньшей мере с 95% штаммов P. aeruginosa, выделенных от инфицированных пациентов. В одном варианте осуществления штаммы P. aeruginosa выделены из одного или более из легких, мокроты, глаз, гноя, фекалий, мочи, пазух, раны, кожи, крови, кости или жидкости из коленного сустава. В одном варианте осуществления антитело или его антиген-связывающий фрагмент конъюгированы со средством, выбранным из группы, состоящей из противомикробного средства, терапевтического средства, пролекарства, пептида, белка, фермента, липида, модификатора биологического ответа, фармацевтического средства, лимфокина, гетерологичного антитела или его фрагмента, детектируемой метки, полиэтиленгликоля (PEG) и комбинации двух или более из любых указанных средств. В одном варианте осуществления детектируемая метка выбрана из группы, состоящей из фермента, флуоресцентной метки, хемилюминесцентной метки, биолюминесцентной метки, радиоактивной метки или комбинации двух или более из любых указанных детектируемых меток.
[0019] В одном варианте осуществления настоящее изобретение предусматривает способ предупреждения или лечения инфекции, вызываемой Pseudomonas, у субъекта, нуждающегося в этом, включающий введение субъекту эффективного количества композиции, описанной в данном документе, где указанное введение обеспечивает синергический терапевтический эффект в предупреждении или лечении инфекции, вызываемой Pseudomonas, у указанного субъекта, и при этом указанный синергический эффект превышает сумму отдельных эффектов от введения равных молярных количеств отдельных связывающих доменов. В одном варианте осуществления синергический терапевтический эффект приводит к большей процентной выживаемости по сравнению с совокупной процентной выживаемостью субъектов, которым вводили только один из связывающих доменов. В одном варианте осуществления композицию вводят в течение двух или более циклов предупреждения/лечения. В одном варианте осуществления связывающие домены или связывающие молекулы вводят одновременно. В одном варианте осуществления связывающие домены или связывающие молекулы вводят последовательно. В одном варианте осуществления инфекция, вызываемая Pseudomonas, представляет собой инфекцию, вызываемую P. aeruginosa. В одном варианте осуществления субъект является человеком. В одном варианте осуществления инфекция представляет собой инфекцию глаз, инфекцию легких, ожоговую инфекцию, раневую инфекцию, кожную инфекцию, инфекцию крови, инфекцию костей или комбинацию двух или более из указанных инфекций. В одном варианте осуществления субъект страдает от острой пневмонии, ожоговой травмы, инфекции роговицы, муковисцидоза или их комбинации.
[0020] В одном варианте осуществления настоящее изобретение предусматривает способ предупреждения или лечения инфекции, вызываемой Pseudomonas, у субъекта, нуждающегося в этом, включающий введение данному субъекту эффективного количества связывающей молекулы или ее фрагмента, биспецифического антитела, полипептида или композиции, описываемых в данном документе.
[0021] В одном варианте осуществления настоящее изобретение предусматривает набор, содержащий композицию, описываемую в данном документе.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ/ФИГУР
[0022] Фигура 1 (A-F). В фенотипическом скрининге фаговых библиотек антителчеловека с использованием целых клеток идентифицировали функционально активные специфические антитела к P. aeruginosa. (A) Обзор стратегии селекции полного антитела. (B) Блок-схема, описывающая способ выделения генов вариабельных областей от пациентов, недавно подвергавшихся воздействию Р. aeruginosa. (C) Характеристики фаг-дисплейных библиотек scFv, указывающие размер и разнообразие репертуара клонированных антител. (D) Сравнение эффективности селекции по методу фагового дисплея с применением либо библиотеки антител пациентов, либо библиотеки наивных антител при селекции 3064 Δ WapR P. aeruginosa (1) или PAO1 MexAB OprM Δ WapR P. aeruginosa (2) в суспензии. Столбцы обозначают выходные титры (в CFU) в каждом цикле селекции, а кружки обозначают долю дуплицированных последовательностей CDR3 VH, что указывает на обогащение клонами. (E) ELISA-скрининг scFv из фагового дисплея для исследования связывания с несколькими штаммами Р. aeruginosa. Данные ELISA (поглощение при 450 нм) показаны для восьми отдельных гибридов фаг-scFv из этапов селекции и одного нерелевантного гибрида фаг-scFv. (F) FACS-анализ связывания антител, специфичных в отношении P. aeruginosa, с типичными штаммами уникальных серотипов P. aeruginosa. Для каждого тестируемого антитела антитело IgG человека, представляющее собой отрицательный контроль, показано в виде заштрихованного пика.
[0023] Фигура 2 (A-B). Оценивание mAb, способствующих ОРК P. aeruginosa. (A) Анализ опсонофагоцитоза с применением люминесцентного штамма (PAO1.lux) Р. aeruginosa серогруппы 05 с разведением очищенных моноклональных антител, полученных из фагового пэннинга. (В) Анализ опсонофагоцитоза с применением люминесцентного штамма (9882-80.lux) P. aeruginosa серогруппы 011 с разведениями очищенных моноклональных антител WapR-004 и Cam-003, полученных из фагового пэннинга. R347, изотипически сходное моноклональное антитело человека, которое не связывается с антигенами P. aeruginosa, применяли в качестве отрицательного контроля, что показано как на A, так и на B.
[0024] Фигура 3 (A-I). Идентификация Psl P. aeruginosa, экзополисахаридной мишени для антител, полученных из фенотипического скрининга. Реактивность антител определяли посредством непрямого ELISA на планшетах, покрытых указанными штаммами P. aeruginosa: A) PAO1 дикого типа, PAO1ΔwbpL, PAO1ΔrmlC и PAO1ΔgalU; (B) РАО1ΔpslA. Антитело Genway является специфичным в отношении белка наружной мембраны P. aeruginosa, и его применяли в качестве положительного контроля. (C) FACS-анализ связывания Cam-003 с PAO1 и PAO1ΔpslA. Cam-003 обозначается сплошной черной линией и четким пиком; изотипически сходное антитело IgG1 человека, не специфичное в отношении P. aeruginosa, применяли в качестве отрицательного контроля, и оно обозначается серой линией и заштрихованным пиком. (D) LPS, очищенный из PAO1 и PAO1ΔpslA, разделяли с помощью SDS-PAGE и подвергали иммуноблоттингу с антисывороткой, полученной от мышей, вакцинированных PAO1ΔwapRΔalgD, мутантным штаммом с недостаточностью транспорта О-антигена к наружной мембране и выработки альгината. (E) Данные ELISA по связыванию Cam-003 с изогенными мутантами PAO1. Cam-003 способен связываться только со штаммами, экспрессирующими Psl. pPW145 представляет собой вектор экспрессии pUCP, содержащий pslA. (F и G) Анализы опсонофагоцитоза, указывающие, что Cam-003 только опосредует уничтожение штаммов, способных к выработке Psl (PAO1 дикого типа и PAO1ΔpslA, дополненный в транс-положении геном pslA). (H и I) Данные ELISA, указывающие на реактивность антител WaR-001, WapR-004 и WapR-016 к Psl относительно PAO1 ΔwbpLΔalgD и PAO1 ΔwbpLΔalgDΔpslA. R347 применяли в качестве отрицательного контроля во всех экспериментах.
[0025] Фигура 4. MAb к Psl ингибируют прикрепление клеток люминесцентного
штамма PAO1.lux P. aeruginosa к клеткам A549. PAO1.lux в логарифмической фазе роста добавили к конфлюэнтному монослою клеток A549 при значении MOI, составляющем 10, с последующим анализом RLU после повторного промывания для удаления несвязанных P. aeruginosa. Результаты являются репрезентативными для трех независимых экспериментов, выполненных в двух повторностях для каждой концентрации антител.
[0026] Фигура 5 (A-C). In vivo пассированные штаммы P. aeruginosa поддерживают/повышают экспрессию Psl. Антитело Cam-003 показано сплошной черной линией и четким пиком; антитело IgG человека, представляющее собой отрицательный контроль, показано серой линией и заштрихованным пиком. (A) Что касается положительного контроля, Cam-003 анализировали в отношении связывания со штаммами, выращиваемых до логарифмической фазы роста из суточной культуры (~5×108/мл). (B) Инокуляты каждого штамма получали при 5×108 CFU/мл из суточной культуры в чашках с TSA, выращиваемой до образования газона, и тестировали на реактивность относительно Cam-003 с помощью проточной цитометрии. (C) Через четыре часа после интраперитонеального контрольного заражения бактерии собирали от мышей с помощью перитонеального лаважа и анализировали на наличие Psl с Cam-003 с помощью проточной цитометрии.
[0027] Фигура 6 (A-F). Показатели выживаемости животных, обработанных моноклональными антителами Cam-003 или WapR-004 к Psl, в модели острой пневмонии с инфицированием P. aeruginosa. (A-D) Животных обрабатывали с помощью Cam-003 при 45, 15 и 5 мг/кг и R347 при 45 мг/кг или PBS за 24 часа до интраназального инфицирования при помощи (A) PAO1 (1,6×107 CFU), (B) 33356 (3×107CFU), (C) 6294 (7×106 CFU), (D) 6077 (1×106 CFU). (E-F) Животных обрабатывали с помощью WapR-004 при 5 и 1 мг/кг, как указано, с последующим инфицированием при помощи 6077 при (Е) (8×105 CFU) или (F) (6×105 CFU). Тщательно отслеживали выживаемость животных в течение до 72 часов (A-D) или в течение 120 часов (E-F). Во всех экспериментах PBS и R347 служили в качестве отрицательных контролей. Результаты представлены в виде кривых выживаемости Каплана-Мейера; различия в выживаемости рассчитали с помощью логарифмического рангового критерия для Cam-003 в сравнении с R347. (A) Cam-003 (45 мг/кг - P<0,0001; 15 мг/кг - P=0,0003; 5 мг/кг - P=0,0033). (B) Cam-003 (45 мг/кг - P=0,0012; 15 мг/кг - P=0,0012; 5 мг/кг - P=0,0373). (C) Cam-003 (45 мг/кг - P=0,0007; 15 мг/кг - P=0,0019; 5 мг/кг - P=0,0212). (D) Cam-003 (45 мг/кг - P<0,0001; 15 мг/кг - P<0,0001; 5 мг/кг - P=0,0001). Результаты являются репрезентативными для по меньшей мере двух независимых экспериментов. (E) [Cam-003 (5 мг/кг) в сравнении с R347 (5 мг/кг): P=0,02; Cam-003 (1 мг/кг) в сравнении с R347 (5 мг/кг): P=0,4848; WapR-004 (5 мг/кг) в сравнении с R347 (5 мг/кг): P<0,0001; WapR-004 (1 мг/кг) в сравнении с R347 (5 мг/кг): P=0,0886; WapR-004 (5 мг/кг) в сравнении с Cam-003 (5 мг/кг): P=0,0017; WapR-004 (1 мг/кг) в сравнении с Cam-003 (1 мг/кг): P=0,2468; R347 (5 мг/кг) в сравнении с PBS: P=0,6676]; (F) [Cam-003 (5 мг/кг) в сравнении с R347 (5 мг/кг): P=0,0004; Cam-003 (1 мг/кг) в сравнении с R347 (5 мг/кг): P<0,0001; WapR-004 (5 мг/кг) в сравнении с R347 (5 мг/кг): P<0,0001; WapR-004 (1 мг/кг) в сравнении с R347 (5 мг/кг): P<0,0001; WapR-004 (5 мг/кг) в сравнении с Cam-003 (5 мг/кг): P=0,0002; WapR-004 (1 мг/кг) в сравнении с Cam-003 (1 мг/кг): P=0,2628; R347 (5 мг/кг) в сравнении с PBS: P=0,6676]. Результаты являются репрезентативными для пяти независимых экспериментов.
[0028] Фигура 7 (A-F). Моноклональные антитела к Psl, Cam-003 и WapR-004 уменьшают нагрузку органа после индуцирования острой пневмонии. Мышей обрабатывали антителом Cam-003 за 24 часа до инфицирования при помощи (A) PAO1 (1,1×107 CFU), (B) 33356 (1×107 CFU), (C) 6294 (6,25×106 CFU) (D) 6077 (1×106 CFU), и антителом WapR-004 за 24 часа до инфицирования при помощи (Е) 6294 (~1×107 CFU) и (F) 6206 (~1×106 CFU). Через 24 часа после инфицирования животных подвергли эвтаназии с последующим забором органов для идентификации жизнеспособных CFU. Различия в жизнеспособных CFU определяли с использованием U-критерия Манна-Уитни для Cam-003 или WapR-004 в сравнении с R347. (A) Легкое: Cam-003 (45 мг/кг - P=0,0015; 15 мг/кг - P=0,0021; 5 мг/кг - P=0,0015); селезенка: Cam-003 (45 мг/кг - P=0,0120; 15 мг/кг - P=0,0367); почки: Cam-003 (45 мг/кг - P=0,0092; 15 мг/кг - P=0,0056); (B) легкое: Cam-003 (45 мг/кг - P=0,0010; 15 мг/кг - P<0,0001; 5 мг/кг - P=0,0045); (C) легкое: Cam-003 (45 мг/кг - P=0,0003; 15 мг/кг - P=0,0039; 5 мг/кг - P=0,0068); селезенка: Cam-003 (45 мг/кг - P=0,0057; 15 мг/кг - P=0,0230; 5 мг/кг - P=0,0012); (D) легкое: Cam-003 (45 мг/кг - P=0,0005; 15 мг/кг - P=0,0003; 5 мг/кг - P=0,0007); селезенка: Cam-003 (45 мг/кг - P=0,0015; 15 мг/кг - P=0,0089; 5 мг/кг - P=0,0089); почки: Cam-003 (45 мг/кг - P=0,0191; 15 мг/кг - P=0,0355; 5 мг/кг - P=0,0021). (E) Легкое: WapR-004 (15 мг/кг - P=0,0011; 5 мг/кг - P=0,0004; 1 мг/кг - P=0,0002); селезенка: WapR-004 (15 мг/кг - P=0,0001; 5 мг/кг - P=0,0014; 1 мг/кг - P=0,0001); (F) легкое: WapR-004 (15 мг/кг - P<0,0001; 5 мг/кг - P=0,0006; 1 мг/кг - P=0,0079); селезенка: WapR-004 (15 мг/кг - P=0,0059; 5 мг/кг - P=0,0261; 1 мг/кг - P=0,0047); почка: WapR-004 (15 мг/кг - P=0,0208; 5 мг/кг - P=0,0268).
[0029] Фигура 8 (A-G). Моноклональные антитела Cam-003 и WapR-004 к Psl активны в модели кератита и модели термического повреждения с инфицированием P. aeruginosa. Мышей обрабатывали контрольным антителом IgG1 или Cam-003 при 45 мг/кг (А, В) или 15 мг/кг (C, D), или PBS, или контрольным антителом IgG1, или Cam-003 при 45 мг/кг, или WapR-004 при 45 мг/кг, или 15 мг/кг, или 5 мг/кг (F, G) за 24 часа до инфицирования при помощи 6077 (цитотоксичный O11 - 2×106 CFU). Непосредственно перед инфицированием на роговице левого глаза каждого животного были сделаны три царапины на 1 мм с последующим местным нанесением 5 мкл инокулята P. aeruginosa. Через 24 часа после инфицирования были рассчитаны количественные показатели патологии роговицы с последующим удалением глаза для определения жизнеспособных CFU. Различия в количественных показателях патологии и жизнеспособных CFU определяли с помощью U-критерия Манна-Уитни. (A) P=0,0001, (B) P<0,0001, (C) P=0,0003, (D) P=0,0015. (F) и (G) Cam-003 (45 мг/кг) в сравнении с WapR-004 (45 мг/кг): P=0,018; Cam-003 (45 мг/кг) в сравнении с WapR-004 (15 мг/кг): P=0,0025; WapR-004 (45 мг/кг) в сравнении с WapR-004 (15 мг/кг): P=0,1331; R347 (5 мг/кг) в сравнении с контрольным соединением: P<0,0001. Результаты являются репрезентативными для пяти независимых экспериментов. (E) Анализ выживаемости мышей CF-1, обработанных с помощью Cam-003 и R347, в модели термического повреждения с инфицированием P. aeruginosa после инфицирования при помощи 6077 (2×105 CFU) (логарифмический ранговый критерий: R347 в сравнении с 15 мг/кг Cam-003, P=0,0094; R347 в сравнении с 5 мг/кг Cam-003, P=0,0017). Результаты являются репрезентативными по меньшей мере для трех независимых экспериментов, (n) относится к числу животных в каждой группе. Фигура 8 (H). Моноклональные антитела к Psl и к PcrV активны в модели глазного кератита, вызываемого P. aeruginosa, на мышах. Мышам интраперитонеально инъецировали (IP) PBS или контрольное антитело IgG1 (R347) при 45 мг/кг, или WapR-004 (α-Psl) при 5 мг/кг, или V2L2 (α-PcrV) при 5 мг/кг за 16 часов до инфицирования с помощью 6077 (цитотоксичным O11 - 1×106 CFU). Непосредственно перед инфицированием мышей анестезировали с последующим нанесением трех царапин на 1 мм на роговицу и поверхностную строму одного глаза каждой мыши с применением иглы 27 калибра под препаровальной лупой с последующим местным применением штамма 6077 P. aeruginosa в 5 мкл инокулята.
[0030] Фигура 9 (A-C). Антитело Cam-003 с мутацией в Fc, Cam-003-ТМ, обладает уменьшенными ОРК и эффективностью in vivo, но сохраняет активность против прикрепления к клеткам. (A) Анализ ОРК в отношении PAO1.lux с применением Cam-003 и Cam-003-ТМ, обладающих мутациями в Fc-домене, которые препятствуют взаимодействиям Fc с рецепторами Feγ (Oganesyan, V., et al, Acta Crystallogr D Biol Crystallogr 64, 700-704 (2008)). R347 применяли в качестве отрицательного контроля. (B) Анализ прикрепления PAO1 к клеткам с применением Cam-003 и Cam-003-ТМ. (C) В модели острой пневмонии сравнивается эффективность Cam-003 с таковой Cam-003-TM.
[0031] Фигура 10 (A-С). A: Картирование эпитопов и идентификация относительной аффинности связывания моноклональных антител к Psl. Картирование эпитопов выполняли с помощью конкурентного ELISA и подтверждали при помощи проточной системы OCTET® с Psl, полученным из надосадочной жидкости суточной культуры штамма PAO1 P. aeruginosa. Значения относительной аффинности связывания измеряли при помощи прибора FORTEBIO® OCTET® 384. Также показаны концентрации антитела, при которых прикрепление к клеткам максимально ингибировалось, и значения EC50 ОРК для каждого антитела. B, C. Значения относительной аффинности разнообразных мутантных WapR-004, измеренные при помощи прибора FORTEBIO® OCTET® 384. Также показаны значения EC50 ОРК для разнообразных мутантных форм.
[0032] Фигура 11 (A-M). Оценка мутантных клонов WapR-004 (W4) в анализе опсонофагоцитарного уничтожения (ОРК) P. aeruginosa (A-M) с люминесцентным штаммом P. aeruginosa серогруппы 05 (PAO1.lux) с разведениями различных мутантных клонов W4 в формате scFv-Fc. В некоторых случаях IgG1 W4 включали в анализ и обозначали как W4-IgGl. W4-RAD-Cam и W4-RAD-GB представляют одну и ту же последовательность WapR-004RAD, описываемую в данном документе. "W4-RAD" представляет собой краткое название WapR-004RAD, и обозначения W4-RAD-Cam и W4-RAD-GB в панелях D-M представляют два разных препарата WapR-004RAD. (N-Q): Оценка оптимизированных mAb к Psl, полученных в результате оптимизации лидерных молекул (WapR-004) в анализе ОРК P. aeruginosa. (N-O) Анализ ОРК с люминесцентным PAO1.lux с применением разведений очищенных лидерных оптимизированных моноклональных антител. (Р-Q) Повторный анализ ОРК с PAO1.lux с разведениями очищенных mAb для подтверждения результатов. (N-Q): W4-RAD применяли в качестве сравнительного положительного контроля. Во всех экспериментах R347, моноклональное антитело IgGl человека, которое не связывается с антигенами P. aeruginosa, применяли в качестве отрицательного контроля.
[0033] Фигура 12 (A-H). (A) Разнообразие эпитопов PcrV. (B) Анализ процентного ингибирования цитотоксичности для родительского mAb V2L2, mAb166 (положительный контроль) и R347 (отрицательный контроль). (C) Оценивание способности mAb V2L2, mAb166 (положительный контроль) и R347 (отрицательный контроль) предупреждать лизис RBC. (D) Оценивание mAb V2L2, ген которого был изменен с получением гена в конфигурации зародышевой линии (V2L2-GL), и оптимизированных mAb V2L2-GL (V2L2-P4M, V2L2-MFS, V2L2-MD и V2L2-MR) в отношении предупреждения лизиса RBC. (E) Оценивание mAb 1Е6, 1F3, 11А6, 29D2, PCRV02 и V2L7 в отношении предупреждения лизиса RBC. (F) Оценивание mAb V2L2 и 29D2 в предупреждении лизиса RBC. (G-H) Значения относительной аффинности связывания антител V2L2-GL и V2L2-MD.
[0034] Фигура 13 (A-I). Исследование выживаемости in vivo мышей, обработанных антителами к PcrV. (A) Мышей обрабатывали за 24 ч. до инфицирования 1,03×106 CFU 6077 (exoU+) с помощью R347 при 45 мг/кг (отрицательный контроль), mAb 166 при 45 мг/кг, 15,0 мг/кг, 5,0 мг/кг или 1,0 мг/кг (положительный контроль) либо V2L2 при 15 мг/кг, 5,0 мг/кг, 1,0 мг/кг или 0,2 мг/кг. Выживаемость отслеживали в течение 96 часов. (В) Мышей обрабатывали за 24 ч. до инфицирования 2,1×107 CFU 6294 (exoU+) с помощью R347 при 15 мг/кг (отрицательный контроль), mAb166 при 15,0 мг/кг, 5,0 мг/кг или 1,0 мг/кг (положительный контроль) либо V2L2 при 15 мг/кг, 5,0 мг/кг или 1,0 мг/кг. Выживаемость отслеживали в течение 168 часов. Мышей обрабатывали за 24 ч. до инфицирования (C) 6294 (O6) или (D) PA103A с помощью R347 (отрицательный контроль), антитела к PcrV PcrV-02 при 5 мг/кг или V2L2 при 5 мг/кг, 1,0 мг/кг, 0,2 мг/кг или 0,04 мг/кг. Мышей обрабатывали за 24 ч. до инфицирования штаммом 6077 с помощью R347 (отрицательный контроль), антитела к PcrV PcrV-02 при 5 мг/кг, V2L7 (5 мг/кг или 1 мг/кг), 3G5 (5 мг/кг или 1 мг/кг) или 11А6 (5 мг/кг или 1 мг/кг) (E), или V2L7, 1Е6, 1F3, 29D2, R347 при 25 мг/кг или антитела к PcrV PcrV-01 при 1 мг/кг (F), или 21F1, V2L2, 2H3, 4A8, SH3, LE10, R347 при 25 мг/кг или PcrV-02 при 1 мг/кг (G), или 29D2 (1 мг/кг, 3 мг/кг или 10 мг/кг), V2L2 (1 мг/кг, 3 мг/кг или 10 мг/кг), R347 или PcrV-02 при 1 мг/кг (H). Мышей обрабатывали за 24 ч. до инфицирования 6294 (О6) или РА103А с помощью V2L2 (0,04 мг/кг, 0,2 мг/кг, 1 мг/кг или 5 мг/кг), R347 или PcrV-02 при 5 мг/кг. Процентную выживаемость анализировали в модели острой пневмонии.
[0035] Фигура 14. Анализ нагрузки органа у мышей, обработанных V2L2. Мышей обрабатывали за 24 ч. до инфицирования 6206 с помощью (A) R347 (отрицательный контроль), V2L2 при 1 мг/кг, 0,2 мг/кг или 0,07 мг/кг и (B) R347 при 15 мг/кг (отрицательный контроль); mAb166 при 15,0 мг/кг, 5,0 мг/кг или 1,0 мг/кг (положительный контроль) или V2L2 при 5,0 мг/кг, 1,0 мг/кг или 0,2 мг/кг. Колониеобразующие единицы идентифицировали на грамм ткани легкого, селезенки и почки.
[0036] Фигура 15. Анализ нагрузки органа у мышей, обработанных V2L2 и WapR-004 (W4). Мышей обрабатывали за 24 ч. до инфицирования 6206 (O11-ExoU+) с помощью R347 (отрицательный контроль), V2L2 в отдельности или V2L2 (0,1 мг/кг) в комбинации с увеличивающимися концентрациями W4 (0,1, 0,5, 1,0 или 2,0 мг/кг). Колониеобразующие единицы идентифицировали на грамм ткани легкого, селезенки и почки.
[0037] Фигура 16 (A-G). Показатели выживаемости животных, обработанных моноклональным антителом V2L2 к PcrV, в модели острой пневмонии с инфицированием P. aeruginosa. Обозначения V2L2-GL, V2L2-MD, V2L2-PM4, V2L2-A и V2L2-MFS на панелях A-G представляют различные препараты V2L2. (A-C) Животных обрабатывали с помощью V2L2 при 1 мг/кг, 0,5 мг/кг или R347 при 0,5 мг/кг до интраназального инфицирования (A) 6077 (9,75×105 CFU), (B, C) 6077 (9,5×105 CFU). (D-F) Животных обрабатывали с помощью V2L2 при 0,5 мг/кг, 0,1 мг/кг или R347 при 0,5 мг/кг с последующим инфицированием 6077 (D) (1×106 CFU), (Е) (9,5×105 CFU) или F (1,026×106 CFU). Животных обрабатывали с помощью V2L2-MD при 0,04 мг/кг, 0,2 мг/кг, 1 мг/кг или 5 мг/кг, mAb166 (положительный контроль) при 0,2 мг/кг, 1 мг/кг, 5 мг/кг или 15 мг/кг или R347 при 0,5 мг/кг с последующим инфицированием 6206 (2×107 CFU).
[0038] Фигура 17 (A-B). Схематическое представление биспецифических антител к Psl/PcrV (A) Bs1-TNFα/W4, Bs2-TNFα/W4, Bs3-TNFα/W4 и (В) Bs2-V2L2/W4-RAD, Bs3-V2L2/W4-RAD и Bs4-V2L2-W4-RAD. (A) Что касается Bsl-TNFα/W4, scFv W4 слит с амино-концом VL антитела к TNFα с помощью линкера (G4S)2. Что касается Bs2-TNFα/W4, scFv W4 слит с амино-концом VH антитела к TNFα с помощью линкера (G4S)2. Что касается Bs3-TNFα/W4, scFv W4 слит с карбокси-концом CH3 с помощью линкера (G4S)2. (В) Что касается Bs2-V2L2-2C, scFv W4-RAD слит с амино-концом VH V2L2 с помощью линкера (G4S)2. Что касается Bs2-W4-RAD-2С, scFv V2L2 слит с амино-концом VH W4-RAD с помощью линкера (G4S)2. Что касается Bs3-V2L2-2C, scFv W4-RAD слит с карбокси-концом СНЗ с помощью линкера (G4S)2. Что касается Bs4-V2L2-2C, scFv W4-RAD вставлен в шарнирную область, связанную с помощью линкера (G4S)2 на N-концевой и C-концевой части scFv.
[0039] Фигура 18. Оценивание активности scFv WapR-004 (W4) в биспецифических конструктах, изображенных на фигуре 17A. scFv W4 лигировали в два разных биспецифических конструкта (в измененных N-или C-концевых ориентациях), содержащих плечо, связывающееся с TNFα. Каждый биспецифический конструкт W4-TNFα (Bs1-TNFα/W4, Bs2-TNFα/W4 и Bs3-TNFα/W4), так же, как и W4, сохранял способность ингибировать прикрепление клеток при использовании анализа с РАО1.lux (05), что указывает на то, что scFv W4 сохраняет свою активность в биспецифическом формате. R347 применяли в качестве отрицательного контроля.
[0040] Фигура 19 (A-C). Домены, связывающие Psl и PcrV, объединяли в биспецифическом формате путем замены антитела к TNFα из фигуры 17 В на V2L2. Эти конструкты идентичны тем, которые изображены на фигуре 17 В, за исключением применения нестабилизированного W4-scFv вместо стабилизированного scFv W4-RAD. Как W4, так и W4-RAD нацеливаются на идентичные эпитопы и характеризуются одинаковыми функциональными активностями. Процентное ингибирование цитотоксичности анализировали как для BS2-V2L2, так и BS3-V2L2 с применением клеток А549, обработанных с помощью как (А) 6206, так и (В) 6206ΔpslA. (С) BS2-V2L2, BS3-V2L2 и BS4-V2L2 оценивали в отношении их способности предупреждать лизис RBC по сравнению с родительскими контрольными образцами. Все биспецифические конструкты сохраняли активность, направленную против цитотоксичности, подобную таковой родительского антитела V2L2, при использовании клеток, инфицированных 6206 и 6206ΔpslA, и предупреждали лизис RBC подобно родительскому контрольному образцу (V2L2). R347 применяли в качестве отрицательного контроля во всех экспериментах.
[0041] Фигура 20 (A-C). Оценивание биспецифических конструктов антитело к Psl/антитело к PcrV в отношении способствованию ОРК P. aeruginosa. Анализ опсонофагоцитоза был показан с люминесцентным штаммом P. aeruginosa (PAO1.lux) серогруппы O5 с разведениями очищенных биспецифических антител к Psl/TNFα (Bs2-TNFα и Bs3-TNFα); родительских антител W4-RAD или V2L2-IgG1; биспецифических антител к Psl/PcrV Bs2-V2L2, или BS3-V2L2, или Bs2-V2L2-2C, BS3-V2L2-2C, BS4-V2L2-2C, или антитела Bs4-V2L2-2C, содержащего мутацию YTE (Bs4-V2L2-2C-YTE). (А) В то время как антитело Bs2-V2L2 показало уничтожение, сходное с таковым для родительского антитела W4-RAD, уничтожение для антитела Bs3-V2L2 было снижено. (B) В то время как антитела Bs2-V2L2-2C и Bs4-V2L2-2C показали уничтожение, сходное с таковым для родительского антитела W4-RAD, уничтожение для антитела Bs3-V2L2-2C было снижено. C) Обозначения W4-RAD и W4-RAD-YTE представляют различные препараты W4-RAD. Обозначения Bs4-V2L2-2C (старая партия) и Bs4-V2L2-2C (новая партия) представляют различные препараты Bs4-V2L2-2C. Модификация YTE в Bs4-V2L2-2C-YTE представляет собой модификацию, произведенную в отношении антител, которая увеличивают период полувыведения антител. Различные препараты антител Bs4 (старая партия в сравнении с новой партией) показали уничтожение, сходное с таковым для родительского антитела W4-RAD, однако антитела Bs4-V2L2-2C-YTE характеризовались 3-кратным снижением ОРК-активности по сравнению с Bs4-V2L2-2C (см. таблицу EC50). R347 применяли в качестве отрицательного контроля во всех экспериментах.
[0042] Фигура 21 (A-I). Исследование выживаемости in vivo мышей, обработанных биспецифическими антителами Bs2-V2L2, Bs3-V2L2, Bs4-V2L2-2C и Bs4-V2L2-2C-YTE к Psl/PcrV в модельной системе для изучения острой пневмонии с инфицированием 6206. Мышей (n=10) обрабатывали с помощью (A): R347 (отрицательный контроль, 0,2 мг/кг), Bs2-V2L2 (0,28 мг/кг), Bs3-V2L2 (0,28 мг/кг), V2L2 (0,2 мг/кг) или W4-RAD (0,2 мг/кг); (B-C): R347 (отрицательный контроль, 1 мг/кг), Bs2-V2L2 (0,5 мг/кг или 1 мг/кг) или Bs4-V2L2-2C (0,5 мг/кг или 1 мг/кг); (D): R347 (отрицательный контроль, 1 мг/кг), Bs3-V2L2 (0,5 мг/кг или 1 мг/кг) или Bs4-V2L2-2C (0,5 мг/кг или 1 мг/кг); (Е): R347 (отрицательный контроль, 2 мг/кг), комбинации отдельных антител W4 и V2L2 (каждое при 0,5 мг/кг или 1 мг/кг) или Bs4-V2L2-2C (1 мг/кг или 2 мг/кг); (F): R347 (отрицательный контроль, 2 мг/кг), смеси отдельных антител W4 и V2L2 (каждое при 0,5 мг/кг или 1 мг/кг) или Bs4-V2L2-2C (1 мг/кг или 0,5 мг/кг). Через двадцать четыре часа после обработки все мыши были инфицированы ~6,25×105-1×106 CFU/животное 6206 (O11-ExoU+). Всех мышей наблюдали в течение 120 часов. (A) Все контрольные мыши поддались инфекции примерно через 30 часов после инфицирования. Все животные, обработанные с помощью Bs3-V2L2, выжили, наряду с теми, которые получили контрольное V2L2. Примерно 90% животных, иммунизированных с помощью W4-RAD, выжили. В противоположность этому, примерно 50% животных, обработанных с помощью Bs2-V2L2, поддались инфекции через 120 часов. (B-F) Все контрольные мыши поддались инфекции примерно через 48 часов после инфицирования. (B) Bs4-V2L2-2C характеризовался большей активностью по сравнению с Bs2-V2L2 как при 1,0, так и при 0,5 мг/кг. (C) Bs4-V2L2-2C, по-видимому, характеризуется большей активностью по сравнению с Bs2-V2L2 при 1,0 мг/кг (результаты не являются статистически значимыми). (D) Bs4-V2L2-2C характеризовался большей активностью по сравнению с Bs3-V2L2 при 0,5 мг/кг. (E) Bs4-V2L2-2C как при 2 мг/кг, так и при 1 мг/кг характеризовался большей активностью по сравнению со смесью антител как при 1,0, так и при 0,5 мг/кг. (F) Bs4-V2L2 (1 мг/кг) характеризуется сходной активностью как при 1,0, так и при 0,5 мг/кг. (G-H) Как Bs4-V2L2-2C, так и Bs4-V2L2-2C-YTE характеризовались сходной активностью как при 1,0, так и при 0,5 мг/кг. Результаты представлены в виде кривых выживаемости Каплана-Мейера; различия в выживаемости рассчитали с помощью логарифмического рангового критерия для (B) Bs4-V2L2-2C в сравнении с Bs2-V2L2 (1 мг/кг - P=0,034; 0,5 мг/кг - P=0,0002); (D) Bs4-V2L2-2C в сравнении с Bs3-V2L2 (0,5 мг/кг - P<0,0001); (E) Bs4-V2L2-2C (2 мг/кг) в сравнении со смесью антител (каждое при 1 мг/кг) - P=0,0012; Bs4-V2L2-2C (1 мг/кг) в сравнении со смесью антител (каждое при 0,5 мг/кг) - P=0,0002. (G-H) Мышей (n=8) обрабатывали с помощью R347 (отрицательный контроль, 1 мг/кг), Bs4-V2L2-2C (1 и 0,5 мг/кг), и Bs4-V2L2-2C-YTE (1 и 0,5 мг/кг), и 6206 (9e5 CFU). Никакой разницы в выживаемости между животными, обработанными Bs4-V2L2-2C и Bs4-V2L2-2C-YTE в любой дозе, не наблюдалось при использовании логарифмического рангового критерия. (I) Для анализа эффективности каждого конструкта на основе антител мышей обрабатывали при 0,1 мг/кг, 0,2 мг/кг, 0,5 мг/кг, 1 мг/кг, 2 мг/кг, 5 мг/кг, 10 мг/кг или 15 мг/кг и анализировали в отношении выживаемости в модели смертельной пневмонии с инфицированием 6206. Процентная выживаемость указана в таблице с числом животных для каждого сравнения, указанным в скобках.
[0043] Фигура 22. Анализ нагрузки органа животных, обработанных биспецифическими антителами к Psl/PcrV, с применением модели острой пневмонии с инфицированием 6206. Мышей обрабатывали за 24 часа до инфицирования 6206 (O11-ExoU+) с помощью R347 (отрицательный контроль), V2L2 или W4-RAD в отдельности (0,2 мг/кг), Bs2-V2L2 (0,28 мг/кг) или Bs3-V2L2 (0,28 мг/кг). Колониеобразующие единицы идентифицировали на грамм ткани легкого, селезенки и почки. При тестируемой концентрации как Bs2-V2L2, так и Bs3-V2L2 значительно уменьшали нагрузку органа в легком. Тем не менее, ни одному из биспецифических конструктов не удалось значительно повлиять на нагрузку органа в селезенке или почке по сравнению с родительскими антителами.
[0044] Фигура 23 (A-B). Анализ нагрузки органа животных, обработанных
биспецифическими антителами к Psl/PcrV, с применением модельной системы с инфицированием 6294. Мышей обрабатывали за 24 часа до инфицирования 6294 с помощью R347 (отрицательный контроль), V2L2 или W4-RAD в отдельности (0,5 мг/кг), Bs2-V2L2 (0,7 мг/кг) или Bs3-V2L2 (0,7 мг/кг) (A) или V2L2 или W4-RAD в отдельности (0,2 мг/кг), Bs2-V2L2 (0,2 мг/кг), Bs3-V2L2 (0,2 мг/кг) или комбинации отдельных антител W4-RAD и V2L2 (каждое при 0,1 мг/кг) (B). Через двадцать четыре часа после введения антитела всех мышей инфицировали инокулятом, содержащим 2,5×107 CFU 6294 (A) или 1,72×107 CFU 6294 (B). Колониеобразующие единицы идентифицировали на грамм ткани легкого, селезенки и почки. При использовании модельной системы с инфицированием 6294 (A) как Bs2-V2L2, так и Bs3-V2L2 значительно уменьшали нагрузку органа во всех тканях до уровня, сравнимого с таковым для родительского антитела V2L2. Родительское антитело W4-RAD не оказывало эффект на уменьшение нагрузки органа. Bs2-V2L2, Bs3-V2L2 и комбинация W4-RAD+V2L2 значительно уменьшали нагрузку органа во всех тканях до уровня, сравнимого с таковым для родительского антитела V2L2.
[0045] Фигура 24. Исследование выживаемости in vivo мышей, обработанных с помощью Bs2-W4/V2L2 и Bs3-W4/V2L2, в модельной системе с инфицированием 6294. Мышей обрабатывали с помощью R347 (отрицательный контроль, 0,2 мг/кг), Bs2-V2L2 (0,28 мг/кг), Bs3-V2L2 (0,28 мг/кг), V2L2 (0,2 мг/кг) или W4-RAD (0,2 мг/кг). Через двадцать четыре часа после обработки всех мышей инфицировали 6294. Всех мышей наблюдали в течение 120 часов. Все контрольные мыши поддались инфекции примерно через 75 часов после инфицирования. Шестьдесят процентов обработанных с помощью Bs3-V2L2 и 50% обработанных с помощью Bs2-V2L2 животных выжили через 120 часов после инокуляции. Как было видно в исследованиях нагрузки органа, иммунизация с помощью W4-RAD не влияла на выживаемость, при этом все мыши поддались инфекции примерно в то же время, что и представители контрольной группы.
[0046] Фигура 25 (A-D). Анализ нагрузки органа при терапии биспецифическим антителом к Psl/PcrV или комбинацией W4+V2L2 в модельной системе с инфицированием 6206. Применяли субоптимальные концентрации антител (A-C) для обеспечения способности к интерпретации активности антител. (D) Применяли высокие концентрации Bs4. Мышей обрабатывали за 24 часа до инфицирования 6206 с помощью R347 (отрицательный контроль), V2L2 или W4-RAD в отдельности (0,2 мг/кг), Bs2-V2L2 (0,2 мг/кг), Bs3-V2L2 (0,2 мг/кг), Bs4 (15,0, 5,0 и 1,0 мг/кг) или комбинации отдельных антител W4-RAD и V2L2 (каждое при 0,1 мг/кг). Через двадцать четыре часа после введения антитела всех мышей инфицировали инокулятом, содержащим (A), (B) 4,75×105 CFU 6206 (O11-ExoU+), или (C) 7,75×105 CFU 6206 (O11-ExoU+), или (D) 9,5×105 CFU 6206 (O11-ExoU+). Колониеобразующие единицы идентифицировали на грамм ткани легкого, селезенки и почки. При использовании модельной системы с инфицированием 6206 как Bs2-V2L2, так и Bs3-V2L2 уменьшали нагрузку органа в легком, селезенке и почках до уровня, сравнимого с таковым для комбинации W4+V2L2. В легком комбинация Bs2- и Bs3-V2L2 и V2L2 значительно снижала количество бактериальных CFU при использовании критерия Краскела-Уоллиса с апостериорным критерием Данна. Значительных различий в бактериальной нагрузке в селезенке и почке не наблюдали, хотя была отмечена тенденция к снижению. (D) Когда применяли оптимальные концентрации Bs4-V2L2-2C (15,0, 5,0 и 1,0), наблюдался быстрый и эффективный клиренс бактерий из легкого. Кроме того, диссеминация бактерий в селезенку и почки также прекращалась. Звездочки указывают на статистическую значимость по сравнению с контрольным R347 при использовании критерия Краскела-Уоллиса с апостериорным критерием Данна.
[0047] Фигура 26 (A-J). Терапевтическая вспомогательная терапия: Bs4-V2L2-2C+антибиотик. (A)-(B) Мышей обрабатывали за 24 часа до инфицирования 1×106 CFU 6206 с помощью R347 при 0,5 мг/кг (отрицательный контроль) или Bs4-V2L2-2C (0,2 мг/кг или 0,5 мг/кг), или ципрофлоксацина (CIP) (20 мг/кг или 6,7 мг/кг) через 1 час после инфицирования, или комбинации Bs4-V2L2-2C за 24 часа до инфицирования и CIP через 1 час после инфицирования (0,5 мг/кг + 20 мг/кг или 0,5 мг/кг + 6,7 мг/кг или 0,2 мг/кг + 20 мг/кг или 0,2 мг/кг + 6,7 мг/кг, соответственно). (C) Мышей обрабатывали через 1 час после инфицирования 9,5×105 CFU 6206 с помощью R347 при 5 мг/кг, или CIP (20 мг/кг или 6,7 мг/кг), или Bs4-V2L2-2C (1 мг/кг или 5 мг/кг), или комбинации Bs4-V2L2-2C и CIP (5 мг/кг + 20 мг/кг или 5 мг/кг + 6,7 мг/кг или 1 мг/кг + 20 мг/кг или 1 мг/кг + 6,7 мг/кг, соответственно). (D) Мышей обрабатывали через 2 часа после инфицирования 9,5×105 CFU 6206 с помощью R347 при 5 мг/кг, или CIP (20 мг/кг или 6,7 мг/кг), или Bs4-V2L2-2C (1 мг/кг или 5 мг/кг), или комбинации Bs4-V2L2-2C и Cipro (5 мг/кг + 20 мг/кг или 5 мг/кг + 6,7 мг/кг или 1 мг/кг + 20 мг/кг или 1 мг/кг + 6,7 мг/кг, соответственно). (E) Мышей обрабатывали через 2 часа после инфицирования 9,75×105 CFU 6206 с помощью R347 при 5 мг/кг, или Bs4-V2L2-2C (1 мг/кг или 0,5 мг/кг), или CIP (20 мг/кг или 6,7 мг/кг) через 1 час после инфицирования, или комбинации Bs4-V2L2-2C через 2 часа после инфицирования и CIP через 1 час после инфицирования (5 мг/кг + 20 мг/кг или 5 мг/кг + 6,7 мг/кг или 1 мг/кг + 20 мг/кг или 1 мг/кг + 6,7 мг/кг, соответственно). (F) Мышей обрабатывали через 1 час после инфицирования 9,5×105 CFU 6206 с помощью R347 при 5 мг/кг, или меропенема (MEM) (0,75 мг/кг или 2,3 мг/кг), или Bs4-V2L2-2C (1 мг/кг или 5 мг/кг), или комбинации Bs4-V2L2-2C и MEM (5 мг/кг + 2,3 мг/кг или 5 мг/кг + 0,75 мг/кг или 1 мг/кг + 2,3 мг/кг или 1 мг/кг + 0,75 мг/кг, соответственно). (G) Мышей обрабатывали через 2 часа после инфицирования 9,75×105 CFU 6206 с помощью R347 при 5 мг/кг, или Bs4-V2L2-2C (1 мг/кг или 0,5 мг/кг), или MEM (0,75 мг/кг или 2,3 мг/кг) через 1 час после инфицирования, или комбинации Bs4-V2L2-2C через 2 часа после инфицирования и MEM через 1 час после инфицирования (5 мг/кг + 2,3 мг/кг или 5 мг/кг + 0,75 мг/кг или 1 мг/кг + 2,3 мг/кг или 1 мг/кг + 0,75 мг/кг, соответственно). (Н) Мышей обрабатывали через 2 часа после инфицирования 1×106 CFU 6206 с помощью R347 при 5 мг/кг, или Bs4-V2L2-2C (1 мг/кг или 5 мг/кг), или MEM (0,75 мг/кг или 2,3 мг/кг), или комбинации Bs4-V2L2-2C и MEM (5 мг/кг + 2,3 мг/кг или 5 мг/кг + 0,75 мг/кг или 1 мг/кг + 2,3 мг/кг или 1 мг/кг + 0,75 мг/кг, соответственно). (I) Мышей обрабатывали через 4 часа после инфицирования 9,25×105 CFU 6206 с помощью R347 при 5 мг/кг, или CIP (6,7 мг/кг), или Bs4-V2L2-2C (1 мг/кг или 5 мг/кг), или комбинации Bs4-V2L2-2C и CIP (5 мг/кг + 6,7 мг/кг или 1 мг/кг + 6,7 мг/кг, соответственно). (J) Мышей обрабатывали через 4 часа после инфицирования 1,2×106 CFU 6206 с помощью R347 при 5 мг/кг + CIP (6,7 мг/кг), CIP (6,7 мг/кг), или Bs4-V2L2-2C (1 мг/кг или 5 мг/кг), или комбинации Bs4-V2L2-2C и CIP (5 мг/кг + 6,7 мг/кг или 1 мг/кг + 6,7 мг/кг, соответственно). (A-J) Антитело Bs4 в сочетании с CIP или MEM повышает эффективность терапии антибиотиками, что указывает на синергический защитный эффект при сочетании молекул. Кроме того, хотя антибиотик, доставляемый в отдельности или в сочетании с неспецифическим антителом к P. aeruginosa, может снижать или регулировать количество бактериальных CFU в легких, антибиотик в отдельности не защищает мышей от летального исхода в этой планировке. Оптимальная защита в этой планировке требует включения Bs4-V2L2-2C в комбинацию с антибиотиком.
[0048] Фигура 27 (A-C). Разница в функциональной активности биспецифических антител Bs4-WT, Bs4-GL и Bs4-GLO: анализ опсонофагоцитарного уничтожения (A), анализ активности, направленной против прикрепления к клеткам (B), а также анализ активности, направленной против цитотоксичности и лизиса RBC (C).
[0049] Фигура 28 (A-B). Процентная защита от смертельной пневмонии у мышей, подвергнутых контрольному заражению штаммами P. aeruginosa в профилактическом (A) или терапевтическом (B) плане. Процентная выживаемость указана в таблице с числом животных для каждого сравнения, указанным в скобках. Черточки указывают на отсутствие тестирования.
[0050] Фигура 29 (A-B). Показатели выживаемости животных, обработанных биспецифическим антителом Bs4-GLO, в модели смертельной бактериемии с инфицированием P. aeruginosa. (A) Животных обрабатывали с помощью Bs4-GLO при 15 мг/кг, 5 мг/кг, 1 мг/кг или R347 при 15 мг/кг за 24 часа до интраперитонеального инфицирования 6294 (O6) (5,58×107 CFU). (В) Животных обрабатывали с помощью Bs4-GLO при 5 мг/кг, 1 мг/кг, 0,2 мг/кг или R347 при 5 мг/кг за 24 часа до интраперитонеального инфицирования 6206 (O11-ExoU+) (6,48×106 CFU). Результаты представлены в виде кривых выживаемости Каплана-Мейера; различия в выживаемости рассчитали с помощью логарифмического рангового критерия для Bs4-GLO при каждой концентрации в сравнении с R347. (A) Bs4-GLO при всех концентрациях в сравнении с R347 P<0,0001. (B) Bs4-GLO при всех концентрациях в сравнении с R347 P<0,0003. Результаты являются репрезентативными для трех независимых экспериментов.
[0051] Фигура 30 (А-С). Показатели выживаемости животных, обработанных с целью профилактики (предупреждения) с помощью Bs4-GLO, в модели термического повреждения с инфицированием P. aeruginosa. (A) Животных обрабатывали с помощью Bs4-GLO при 15 мг/кг, 5 мг/кг или R347 при 15 мг/кг за 24 часа до индуцирования термического повреждения и подкожного инфицирования штаммом 6077 (O11-ExoU+) P. aeruginosa при 1,4×105 CFU непосредственно под раневую поверхность. (В) Животных обрабатывали с помощью Bs4-GLO при 15 мг/кг или R347 при 15 мг/кг за 24 часа до индуцирования термического повреждения и подкожного инфицирования штаммом 6206 (O11-ExoU+) P. aeruginosa при 4,15×104 CFU непосредственно под раневую поверхность. (C) Животных обрабатывали с помощью Bs4-GLO при 15 мг/кг, 5 мг/кг или R347 при 15 мг/кг за 24 часа до индуцирования термического повреждения и подкожного инфицирования штаммом 6294 (O11-ExoU+) P. aeruginosa при 7,5×101 CFU непосредственно под раневую поверхность. Результаты представлены в виде кривых выживаемости Каплана-Мейера; различия в выживаемости рассчитали с помощью логарифмического рангового критерия для Bs4-GLO при каждой концентрации в сравнении с R347. (A-C) Bs4-GLO при всех концентрациях в сравнении с R347 - P <0,0001. Результаты являются репрезентативными для двух независимых экспериментов с каждым штаммом P. aeruginosa.
[0052] Фигура 31 (A-B). Показатели выживаемости животных, обработанных с целью терапии (лечения) с помощью Bs4-GLO, в модели термического повреждения с инфицированием P. aeruginosa. (A) Животных обрабатывали с помощью Bs4-GLO при 42,6 мг/кг, 15 мг/кг или R347 при 45 мг/кг через 4 часа после индуцирования термического повреждения и подкожного инфицирования штаммом 6077 (O11-ExoU+) P. aeruginosa при 1,6×105 CFU непосредственно под раневую поверхность. (B) Животных обрабатывали с помощью Bs4-GLO при 15 мг/кг, 5 мг/кг или R347 при 15 мг/кг через 12 часов после индуцирования термического повреждения и подкожного инфицирования штаммом 6077 (O11-ExoU+) P. aeruginosa при 1,0×105 CFU непосредственно под раневую поверхность. Результаты представлены в виде кривых выживаемости Каплана-Мейера; различия в выживаемости рассчитали с помощью логарифмического рангового критерия для Bs4-GLO при каждой концентрации в сравнении с R347. (A) Bs4-GLO при обеих концентрациях в сравнении с R347 - P<0,0004. (B) Bs4-GLO при 5 мг/кг в сравнении с R347 - P<0,048. Результаты являются репрезентативными для двух независимых экспериментов.
[0053] Фигура 32 (A-B). Терапевтическая вспомогательная терапия: Bs4GLO + ципрофлоксацин (CIP). (A) Мышей обрабатывали через 4 часа после инфицирования 9,5×105 CFU 6206 с помощью R347 при 5 мг/кг + CIP (6,7 мг/кг), или Bs4-WT (1 мг/кг или 5 мг/кг), или комбинации Bs4-WT и CIP (5 мг/кг + 6,7 мг/кг или 1 мг/кг + 6,7 мг/кг, соответственно). (B) Мышей обрабатывали через 4 часа после инфицирования 9,5×105 CFU 6206 с помощью R347 при 5 мг/кг + CIP (6,7 мг/кг), или Bs4-GLO (1 мг/кг или 5 мг/кг), или комбинации Bs4-GLO и CIP (5 мг/кг + 6,7 мг/кг или 1 мг/кг + 6,7 мг/кг, соответственно).
[0054] Фигура 33 (A-B). Терапевтическая вспомогательная терапия: Bs4-GLO + меропенем (MEM). (A) Мышей обрабатывали через 4 часа после инфицирования 9,5×105 CFU 6206 с помощью R347 при 5 мг/кг + MEM (0,75 мг/кг), или Bs4-WT (1 мг/кг или 5 мг/кг), или комбинации Bs4-WT и MEM (5 мг/кг + 0,75 мг/кг или 1 мг/кг + 0,75 мг/кг, соответственно). (B) Мышей обрабатывали через 4 часа после инфицирования 9,5×105 CFU 6206 с помощью R347 при 5 мг/кг + MEM (0,75 мг/кг), или Bs4-GLO (1 мг/кг или 5 мг/кг), или комбинации Bs4-GLO и MEM (5 мг/кг + 0,75 мг/кг или 1 мг/кг + 0,75 мг/кг, соответственно).
[0055] Фигура 34 (A-C). Терапевтическая вспомогательная терапия: Bs4-GLO + антибиотик в модели смертельной бактериемии. Мышей обрабатывали за 24 часа до интраперитонеального инфицирования штаммом 6294 (O6) P. aeruginosa при 9,3×107 с помощью Bs4-GLO (при 0,25 мг/кг или 0,5 мг/кг) или R347 (отрицательный контроль). Через час после инфицирования мышей обрабатывали подкожно с помощью (A) 1 мг/кг CIP, (B) 2,5 мг/кг MEM или (C) 2,5 мг/кг ТОВ. Результаты представлены в виде кривых выживаемости Каплана-Мейера; различия в выживаемости рассчитали с помощью логарифмического рангового критерия для Bs4-GLO при каждой концентрации в сравнении с R347.
[0056] Фигура 35 (A-B). Схематическое представление альтернативных форматов для конструктов Bs4. (A) Вариабельные области антител к PcrV присутствуют отдельно на тяжелых и легких цепях, в то время как вариабельные области антител к Psl присутствуют в качестве scFv в шарнирной области тяжелой цепи, и (B) вариабельные области антител к Psl присутствуют отдельно на тяжелых и легких цепях, в то время как вариабельные области антител к PcrV присутствуют в качестве scFv в шарнирной области тяжелой цепи.
ПОДРОБНОЕ ОПИСАНИЕ
I. ОПРЕДЕЛЕНИЯ
[0057] Следует отметить, что формы единственного числа объектов относятся к одному или нескольким из этих объектов, например, выражение "связывающая молекула, которая специфически связывается с Psl и/или PcrV Pseudomonas" понимают как представляющее одну или несколько связывающих молекул, которые специфически связываются с Psl и/или PcrV Pseudomonas. В силу этого, формы единственного числа, выражения "один или несколько" и "по меньшей мере один" можно применять в данном документе взаимозаменяемо.
[0058] Применяемое в данном документе выражение "полипептид" предназначено охватывать "полипептид" в единственном числе, а также "полипептиды" во множественном числе и относится к молекуле, состоящей из мономеров (аминокислот), линейно соединенных амидными связями (также известными как пептидные связи). Выражение "полипептид" относится к любой цепи или цепям из двух или более аминокислот и не относится к конкретной длине продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептиды, "белок", "цепь аминокислот" или любое другое выражение, применяемое для обозначения цепи или цепей из двух или более аминокислот, включены в определение "полипептид", и выражение "полипептид" можно применять вместо любого из указанных выражений или взаимозаменяемо с любым из них. Выражение "полипептид" также предназначено для обозначения продуктов постэкспрессионных модификаций полипептида, включая, без ограничений, гликозилирование, ацетилирование, фосфорилирование, амидирование, получение производных с помощью известных защитных/блокирующих групп, протеолитическое расщепление или модификацию с помощью аминокислот, не встречающихся в природе. Полипептид может быть получен из природного биологического источника или создан с помощью рекомбинантной технологии, но не обязательно, чтобы он был транслирован из определенной последовательности нуклеиновой кислоты. Он может быть получен любым способом, в том числе путем химического синтеза.
[0059] Полипептид, раскрытый в данном документе, может иметь размер около 3 или более, 5 или более, 10 или более, 20 или более, 25 или более, 50 или более, 75 или более, 100 или более, 200 или более, 500 или более, 1000 или более или 2000 или более аминокислот.Полипептиды могут иметь определенную трехмерную структуру, хотя они не обязательно имеют такую структуру. Полипептиды с определенной трехмерной структурой называются свернутыми, а полипептиды, не обладающие определенной трехмерной структурой, а скорее способные принимать большое число различных конформаций, называются несвернутыми. Применяемое в данном документе выражение "гликопротеин" относится к белку, связанному по меньшей мере с одним углеводным фрагментом, присоединенным к белку посредством кислородсодержащей или азотсодержащей боковой цепи аминокислотного остатка, например, серинового остатка или аспарагинового остатка.
[0060] Под "выделенным" полипептидом или его фрагментом, вариантом или производным подразумевается полипептид, не находящийся в своем естественном окружении. Не требуется конкретный уровень очистки. Например, выделенный полипептид может быть удален из его нативной или естественной окружающей среды. Полученные рекомбинантным путем полипептиды и белки, экспрессирующиеся в клетках-хозяевах, считаются выделенными, как раскрыто в данном документе, поскольку являются нативными или рекомбинантными полипептидами, разделенными, фракционированными или частично или в значительной степени очищенными с помощью любой подходящей методики.
[0061] Другие полипептиды, раскрытые в данном документе, представляют собой фрагменты, производные, аналоги или варианты вышеупомянутых полипептидов и любую их комбинацию. Выражения "фрагмент", "вариант", "производное" и "аналог", обозначающие связывающую молекулу, такую как антитело, которая специфически связывается с Psl и/или PcrV Pseudomonas, как раскрыто в данном документе, включают любые полипептиды, которые сохраняют по меньшей мере некоторые свойства связывания с антигеном соответствующего нативного антитела или полипептида. Фрагменты полипептидов включают, например, протеолитические фрагменты, а также делеционные фрагменты, в дополнение к фрагментам специфических антител, рассматриваемым где-либо в данном документе. Варианты связывающей молекулы, например, антитела, которое специфически связывается с Psl и/или PcrV Pseudomonas, как раскрыто в данном документе, включают фрагменты, описанные выше, а также полипептиды с аминокислотными последовательностями, измененными в связи с аминокислотными заменами, делециями или вставками. Варианты могут быть встречающимися в природе или не встречающимися в природе. Не встречающиеся в природе варианты можно получить при помощи методик мутагенеза, известных в данной области техники. Варианты полипептидов могут включать консервативные или неконсервативные аминокислотные замены, делеции или добавления. Производные связывающей молекулы, например, антитела, которое специфически связывается с Psl и/или PcrV Pseudomonas, как раскрыто в данном документе, являются полипептидами, измененными таким образом, чтобы они проявляли дополнительные признаки, не обнаруживаемые в нативном полипептиде. Примеры включают белки слияния. Варианты полипептидов также могут называться в данном документе "аналогами полипептидов". Применяемое в данном документе выражение "производное" связывающей молекулы, например, антитела, которое специфически связывается с Psl и/или PcrV Pseudomonas, относится к объекту-полипептиду, имеющему один или несколько остатков, химически дериватизированных с помощью реакции функциональной боковой группы. Также в качестве "производных" включены те пептиды, которые содержат одну или несколько встречающихся в природе аминокислотных производных двадцати стандартных аминокислот. Например, 4-гидроксипролин может быть замещен на пролин; 5-гидроксилизин может быть замещен на лизин; 3-метилгистидин может быть замещен на гистидин; гомосерин может быть замещен на серии; и орнитин может быть замещен на лизин.
[0062] Выражение "полинуклеотид" предназначено охватывать нуклеиновую кислоту в единственном числе, а также нуклеиновые кислоты во множественном числе и относится к молекуле или конструкту выделенной нуклеиновой кислоты, например, матричной РНК (мРНК) или плазмидной ДНК (пДНК). Полинуклеотид может содержать традиционную фосфодиэфирную связь или нетрадиционную связь (например, амидную связь, такую как обнаруживаемую в пептидных нуклеиновых кислотах (PNA)). Выражение "нуклеиновая кислота" относится к какому-либо одному или нескольким сегментам нуклеиновой кислоты, например, фрагментам ДНК или РНК, имеющимся в полинуклеотиде. Под "выделенными" нуклеиновой кислотой или полинуклеотидом подразумевается молекула нуклеиновой кислоты, ДНК или РНК, которая была удалена из ее нативной окружающей среды. Например, рекомбинантный полинуклеотид, кодирующий связывающую молекулу, например, антитело, которое специфически связывается с Psl и/или PcrV Pseudomonas, содержащийся в векторе, считается выделенным, как раскрыто в данном документе. Дополнительные примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, содержащиеся в гетерологичных клетках-хозяевах, или очищенные (частично или в значительной степени) полинуклеотиды в растворе. Выделенные молекулы РНК включают in vivo или in vitro РНК-транскрипты полинуклеотидов. Выделенные полинуклеотиды или нуклеиновые кислоты дополнительно включают такие молекулы, полученные синтетически. В дополнение, полинуклеотид или нуклеиновая кислота могут представлять собой или могут включать регуляторный элемент, такой как промотор, сайт связывания рибосомы или терминатор транскрипции.
[0063] Применяемое в данном документе выражение "кодирующая область" представляет собой часть нуклеиновой кислоты, состоящую из кодонов, транслируемых в аминокислоты. Хотя "стоп-кодон" (TAG, TGA или TAA) не транслируется в аминокислоту, он может считаться частью кодирующей области, однако любые фланкирующие последовательности, например, промоторы, сайты связывания рибосомы, терминаторы транскрипции, интроны и т.п., не являются частью кодирующей области. В одном полинуклеотидном конструкте, например, в одном векторе, или в отдельных полинуклеотидных конструктах, например, в отдельных (различных) векторах, могут иметься две или более кодирующих области. Кроме того, любой вектор может содержать одну кодирующую область или может содержать две или более кодирующие области, например, один вектор может по отдельности кодировать вариабельную область тяжелой цепи иммуноглобулина и вариабельную область легкой цепи иммуноглобулина. В дополнение, вектор, полинуклеотид или нуклеиновая кислота могут кодировать гетерологичные кодирующие области, либо слитые, либо не слитые с нуклеиновой кислотой, кодирующей связывающую молекулу, которая специфически связывается с Psl и/или PcrV Pseudomonas, например, антитело или его антиген-связывающий фрагмент, вариант или производное. Гетерологичные кодирующие области включают, без ограничений, специализированные элементы или мотивы, такие как кодирующие секреторный сигнальный пептид или гетерологичный функциональный домен.
[0064] В определенных вариантах осуществления полинуклеотид или нуклеиновая кислота представляют собой ДНК. В случае ДНК полинуклеотид, содержащий нуклеиновую кислоту, которая кодирует полипептид, обычно может включать промотор и/или другие элементы, осуществляющие контроль транскрипции или трансляции, функционально связанные с одной или несколькими кодирующими областями. Функциональная связь наблюдается тогда, когда кодирующая область для продукта гена, например, полипептида, связана с одной или несколькими регуляторными последовательностями таким образом, чтобы экспрессия продукта гена была помещена под влияние или контроль регуляторной последовательности (регуляторных последовательностей). Два фрагмента ДНК (такие как кодирующая область для полипептида и промотор, связанный с ней) "функционально связаны", если индуцирование функции промотора приводит в результате к транскрипции мРНК, кодирующей желаемый продукт гена, и если природа связи между двумя фрагментами ДНК не препятствует способности регуляторных последовательностей, отвечающих за экспрессию, управлять экспрессией продукта гена или не препятствует способности ДНК-матрицы транскрибироваться. Таким образом, промоторная область может быть функционально связанной с нуклеиновой кислотой, кодирующей полипептид, если промотор способен воздействовать на транскрипцию этой нуклеиновой кислоты. Промотор может быть промотором, специфичным в отношении клеток, управляющим существенной транскрипцией ДНК только в предопределенных клетках. Другие элементы, осуществляющие контроль транскрипции, помимо промотора, например, энхансеры, операторы, репрессоры и сигналы терминации транскрипции, могут быть функционально связанными с полинуклеотидом для управления транскрипцией, специфичной в отношении клеток. Подходящие промоторы и другие области, осуществляющие контроль транскрипции, раскрыты в данном документе.
[0065] Разнообразные области, осуществляющие контроль транскрипции, известны специалистам в данной области. Они включают, без ограничений, области, осуществляющие контроль транскрипции, функционирующие в клетках позвоночных, такие как, без ограничений, промоторные и энхансерные сегменты цитомегаловирусов (немедленно-ранний промотор вместе с интроном-A), вируса обезьян 40 (ранний промотор) и ретровирусов (такой как вирус саркомы Рауса). Другие области, осуществляющие контроль транскрипции, включают таковые, полученные из генов позвоночных, таких как гены актина, белка теплового шока, гормона роста крупного рогатого скота и β-глобина кролика, а также другие последовательности, способные осуществлять контроль экспрессии генов в эукариотических клетках. Дополнительные подходящие области, осуществляющие контроль транскрипции, включают тканеспецифичные промоторы и энхансеры, а также лимфокин-индуцируемые промоторы (например, промоторы, индуцируемые интерферонами или интерлейкинами).
[0066] Аналогично, разнообразные элементы, осуществляющие контроль трансляции, известны специалистам в данной области техники. Они включают, без ограничений, сайты связывания рибосомы, кодоны, инициирующие и терминирующие трансляцию, и элементы, полученные из пикорнавирусов (в частности, внутренний сайт связывания рибосомы или IRES, также называемый CITE-последовательностью).
[0067] В других вариантах осуществления полинуклеотид может представлять собой РНК, например, в виде матричной РНК (мРНК).
[0068] Кодирующие области полинуклеотидов и нуклеиновых кислот могут быть связаны с дополнительными кодирующими областями, которые кодируют секреторные или сигнальные пептиды, которые управляют секрецией полипептида, кодируемого полинуклеотидом, раскрытым в данном документе, например, полинуклеотидом, кодирующим связывающую молекулу, которая специфически связывается с Psl и/или PcrV Pseudomonas, например, антитело или его антиген-связывающий фрагмент, вариант или производное. Согласно гипотезе сигнальной последовательности, белки, секретируемые в клетках млекопитающих, имеют сигнальный пептид или секреторную лидерную последовательность, отщепляющуюся от зрелого белка, как только инициируется экспорт растущей белковой цепи через гранулярный эндоплазматический ретикулум. Специалистам в данной области известно, что полипептиды, секретируемые в клетках позвоночных, обычно имеют сигнальный пептид, слитый с N-концом полипептида, который отщепляется от полного полипептида или такового "полной длины" с получением секретированной или "зрелой" формы полипептида. В некоторых вариантах осуществления применяют нативный сигнальный пептид, например, сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина, или функциональное производное этой последовательности, сохраняющее способность управлять секрецией полипептида, функционально связанного с ним. Альтернативно, можно применять гетерологичный сигнальный пептид млекопитающих или его функциональное производное. Например, лидерная последовательность дикого типа может быть заменена на лидерную последовательность тканевого активатора плазминогена (TPA) человека или β-глюкуронидазы мыши.
[0069] В данном документе раскрыты определенные связывающие молекулы или их антиген-связывающие фрагменты, варианты или производные. Если конкретно не указана ссылка на полноразмерные антитела, такие как встречающиеся в природе антитела, выражение "связывающая молекула" охватывает полноразмерные антитела, а также антиген-связывающие фрагменты, варианты, аналоги или производные таких антител, например, молекулы встречающихся в природе антител или иммуноглобулинов или сконструированные молекулы или фрагменты антител, связывающиеся с антигеном подобно молекулам антител.
[0070] Применяемое в данном документе выражение "связывающая молекула" относится в своем самом широком смысле к молекуле, которая специфически связывается с антигенной детерминантой. Как описано дополнительно в данном документе, связывающая молекула может содержать один или несколько связывающих доменов, раскрытых в данном документе. Применяемое в данном документе выражение "связывающий домен" включает в себя участок, который специфически связывается с антигенной детерминантой. Неограничивающим примером антиген-связывающей молекулы является антитело или его фрагмент, сохраняющие антиген-специфическое связывание.
[0071] Выражения "антитело" и "иммуноглобулин" можно применять в данном документе взаимозаменяемо. Антитело (или его фрагмент, вариант или производное, раскрытые в данном документе) содержит по меньшей мере вариабельный домен тяжелой цепи и по меньшей мере вариабельные домены тяжелой цепи и легкой цепи. Основные структуры иммуноглобулинов изучены довольно хорошо в системах позвоночных. См., например, Harlow et al, Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988).
[0072] Как будет более подробно рассматриваться ниже, выражение "иммуноглобулин" включает разнообразные широкие классы полипептидов, между которыми можно провести различие с помощью методов биохимии. Специалисты в данной области примут во внимание, что тяжелые цепи классифицируют как гамма, мю, альфа, дельта или эпсилон (γ, μ, α, δ, ε), при этом среди них существует несколько подклассов (например, γ1-γ4). Именно природа этой цепи определяет "класс" антитела как IgG, IgM, IgA, IgG или IgE, соответственно. Подклассы иммуноглобулинов (изотипы), например, IgG1 IgG2, IgG3, IgG4, IgA1 и т.д. хорошо охарактеризованы и, как известно, придают функциональную специализацию. Модифицированные версии каждого из этих классов и изотипов легко различимы для специалиста в данной области ввиду настоящего изобретения и, соответственно, находятся в пределах объема настоящего изобретения.
[0073] Легкие цепи классифицируют как каппа либо лямбда (κ, λ). Тяжелая цепь каждого класса может быть связана с легкой цепью либо каппа-, либо лямбда-типа. В большинстве случаев легкие и тяжелые цепи ковалентно связаны друг с другом, а "хвостовые" части двух тяжелых цепей связаны друг с другом с помощью ковалентных дисульфидных связей или нековалентных связей, когда иммуноглобулины образуются гибридомами, В-клетками либо клетками-хозяевами, созданными с помощью методов генной инженерии. В тяжелой цепи аминокислотные последовательности проходят от N-конца на раздвоенных концах Y-конфигурации до С-конца на нижней части каждой цепи.
[0074] Как легкая, так и тяжелая цепи разделены на области структурной и функциональной гомологии. Выражения "константный" и "вариабельный" применяют в функциональном отношении. В связи с этим следует принять во внимание, что вариабельные домены частей как легкой (VL), так и тяжелой (VH) цепи определяют распознавание антигена и специфичность в его отношении. Наоборот, константные домены легкой цепи (CL) и тяжелой цепи (CH1, CH2 или CH3) придают важные биологические свойства, такие как секреция, трансплацентарная подвижность, связывание с Fc-рецепторами, связывание комплемента и т.п. Принято, что нумерация доменов константных областей возрастает по мере того, как они становятся более отдаленными от антиген-связывающего участка или амино-конца антитела. N-концевая часть представляет собой вариабельную область, а в C-концевой части находится константная область; CH3- и CL-домены фактически включают карбокси-конец тяжелой и легкой цепи, соответственно.
[0075] Как указано выше, вариабельная область позволяет связывающей молекуле избирательно распознавать эпитопы антигенов и специфически связываться с ними. Это значит, что VL-домен и VH-домен или подмножество гипервариабельных участков (CDR) связывающей молекулы, например, антитела, объединяются с образованием вариабельной области, которая определяет трехмерный антиген-связывающий участок. Эта четвертичная структура связывающей молекулы образует антиген-связывающий участок, находящийся в конце каждого плеча Y. В частности, антиген-связывающий участок определяется тремя CDR на каждой из цепей VH и VL.
[0076] Во встречающихся в природе антителах шесть "гипервариабельных участков" или "CDR", имеющихся в каждом антиген-связывающем домене, являются короткими, несмежными последовательностями аминокислот, которые расположены определенным образом для образования антиген-связывающего домена, поскольку антитело приобретает свою трехмерную конфигурацию в водной окружающей среде. Остальные аминокислоты антиген-связывающих доменов, называемые "каркасными" областями, показывают меньшую межмолекулярную изменчивость. Каркасные области по большей части принимают конформацию β-листа, и CDR образуют петли, которые соединяются со структурой β-листа и в некоторых случаях образуют ее часть. Таким образом, каркасные области действуют с формированием остова, который обеспечивает расположение CDR в правильной ориентации с помощью нековалентных взаимодействий между цепями. Антиген-связывающий домен, образованный расположенными CDR, определяет поверхность, комплементарную эпитопу иммунореактивного антигена. Эта комплементарная поверхность способствует нековалентному связыванию антитела с его когнатным эпитопом. По аминокислотам, включающим CDR и каркасные области, соответственно, специалист в данной области может без труда идентифицировать любую данную вариабельную область тяжелой или легкой цепи, поскольку они были точно определены (см. "Sequences of Proteins of Immunological Interest," Kabat, E., et al, U.S. Department of Health и Human Services, (1983); и Chothia and Lesk, J. Mol. Biol, 196: 901-917 (1987), которые включены в данный документ посредством ссылки во всей своей полноте).
[0077] В случае, когда существует два или более определения выражения, применяемого и/или принятого в данной области техники, определение выражения, применяемого в данном документе, предназначено включать все такие значения, если прямо не указано обратное. Конкретным примером является применение выражения "гипервариабельный участок" ("CDR") для описания несмежных антиген-связывающих участков, обнаруживаемых в вариабельной области как тяжелой, так и легкой цепей полипептидов. Эта конкретная область была описана в Kabat et al., U.S. Dept. of Health and Human Services, "Sequences of Proteins of Immunological Interest" (1983), и в Chothia et al., J. Mol. Biol. 196: 901-917 (1987), которая включена в данный документ посредством ссылки, где определения включают перекрывания или подмножества аминокислотных остатков при сравнении друг с другом. Тем не менее, применение каждого определения для обозначения CDR антитела или его вариантов предназначено находиться в пределах объема выражения, как определяется и применяется в данном документе. Соответствующие аминокислотные остатки, охватываемые CDR, как определено в каждой из вышеупомянутых ссылок, изложены ниже в таблице I для сравнения. Точное число остатков, охватываемых конкретным CDR, будет варьировать в зависимости от последовательности и размера CDR. Специалисты в данной области могут обычным образом определить, какие остатки включаются в конкретный CDR, с учетом аминокислотной последовательности вариабельной области антитела.

[0078] Kabat и соавт.также определили систему нумерации последовательностей вариабельных доменов, применимую к любому антителу. Специалист в данной области может однозначно установить данную систему "нумерации по Kabat" для любой последовательности вариабельного домена, не опираясь ни на какие экспериментальные данные за рамками самой последовательности. Применяемое в данном документе выражение "нумерация по Kabat" относится к системе нумерации, изложенной в Kabat et al, U.S. Dept. of Health and Human Services, "Sequence of Proteins of Immunological Interest" (1983). Если не указано иное, ссылки на нумерацию конкретных положений аминокислотных остатков в связывающей молекуле, которая специфически связывается с Psl и/или PcrV Pseudomonas, например, в антителе или его антиген-связывающем фрагменте, варианте или производном, раскрытой в данном документе, соответствуют системе нумерации по Kabat.
[0079] Связывающие молекулы, например, антитела или их антиген-связывающие фрагменты, варианты или производные, включают, без ограничений, поликлональные, моноклональные, человеческие, гуманизированные или химерные антитела, одноцепочечные антитела, эпитоп-связывающие фрагменты, например, Fab, Fab' и F(ab')2, Fd, Fv, одноцепочечные Fv (scFv), одноцепочечные антитела, Fv, образованные за счет дисульфидньгх связей (sdFv), фрагменты, содержащие либо VL-, либо VH-домен, фрагменты, получаемые из экспрессионной библиотеки Fab. Молекулы scFv известны в данной области техники и описаны, например, в патенте США №5892019. Молекулы иммуноглобулинов или антител, охватываемые настоящим изобретением, могут принадлежать к любому типу (например, IgG, IgE, IgM, IgD, IgA и IgY), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу молекул иммуноглобулинов.
[0080] Под выражением "специфически связывается" обычно подразумевают, что связывающая молекула, например, антитело или его фрагмент, вариант или производное, связывается с эпитопом посредством своего антиген-связывающего домена, и что связывание предполагает некоторую комплементарность между антиген-связывающим доменом и эпитопом. Согласно данному определению говорят, что связывающая молекула "специфически связывается" с эпитопом, когда она связывается с этим эпитопом посредством своего антиген-связывающего домена более легко, чем если бы она связывалась со случайным, неродственным эпитопом. Выражение "специфичность" применяется в данном документе для определения относительной аффинности, с которой определенная связывающая молекула связывается с определенным эпитопом. Например, могут полагать, что связывающая молекула "A" имеет более высокую специфичность в отношении данного эпитопа, чем связывающая молекула "B", или могут говорить, что связывающая молекула "A" связывается с эпитопом "C" с более высокой специфичностью, чем специфичность, которой она характеризуется в отношении родственного эпитопа "D".
[0081] Под выражением "преимущественно связывается" подразумевают, что антитело специфически связывается с эпитопом более легко, чем если бы оно связывалось с родственным, сходным, гомологичным или аналогичным эпитопом. Таким образом, антитело, которое " преимущественно связывается" с данным эпитопом, будет с большей вероятностью связываться с этим эпитопом, чем с родственным эпитопом, несмотря на то, что такое антитело может вступать в перекрестную реакцию с родственным эпитопом.
[0082] В качестве неограничивающего примера связывающая молекула, например, антитело, может считаться преимущественно связывающейся с первым эпитопом, если она связывается с указанным первым эпитопом при константе диссоциации (KD), меньшей, чем KD антитела для второго эпитопа. В другом неограничивающем примере связывающая молекула, такая как антитело, может считаться преимущественно связывающейся с первым антигеном, если она связывается с первым эпитопом с аффинностью, величина которой по меньшей мере на один порядок меньше, чем KD антитела для второго эпитопа. В другом неограничивающем примере связывающая молекула может считаться преимущественно связывающейся с первым эпитопом, если она связывается с первым эпитопом с аффинностью, величина которой по меньшей мере на два порядка меньше, чем KD антитела для второго эпитопа.
[0083] В другом неограничивающем примере связывающая молекула, например, антитело или его фрагмент, вариант или производное, может считаться преимущественно связывающейся с первым эпитопом, если она связывается с первым эпитопом при скорости диссоциации (k(off)), меньшей, чем k(off) антитела для второго эпитопа. В другом неограничивающем примере связывающая молекула может считаться преимущественно связывающейся с первым эпитопом, если она связывается с первым эпитопом с аффинностью, величина которой по меньшей мере на один порядок меньше, чем k(off) антитела для второго эпитопа. В другом неограничивающем примере связывающая молекула может считаться преимущественно связывающейся с первым эпитопом, если она связывается с первым эпитопом с аффинностью, величина которой по меньшей мере на два порядка меньше, чем k(off) антитела для второго эпитопа.
[0084] Можно говорить, что связывающая молекула, например, антитело или его фрагмент, вариант или производное, раскрытая в данном документе, связывается с антигеном-мишенью, например, с полисахаридом, раскрытым в данном документе, или с его фрагментом или вариантом, при скорости диссоциации (k(off)), меньшей или равной 5×10-2 с-1, 10-2 с-1, 5×10-3 с-1 или 10-3 с-1. Можно говорить, что связывающая молекула, раскрытая в данном документе, связывается с антигеном-мишенью, например, с полисахаридом, при скорости диссоциации (k(off)), меньшей или равной 5×10-4 с-1, 10-4 с-1, 5×10-5 с-1 или 10-5 с-1, 5×10-6 с-1, 10-6 с-1, 5×10-7 с-1 или 10-7 с-1.
[0085] Можно говорить, что связывающая молекула, например, антитело или антиген-связывающий фрагмент, вариант или производное, раскрытая в данном документе, связывается с антигеном-мишенью, например, с полисахаридом, при скорости ассоциации (k(on)), большей или равной 103 М-1 с-1, 5×103 М-1 с-1, 104 М-1 с-1 или 5×104 М-1 с-1. Можно говорить, что связывающая молекула, например, антитело или антиген-связывающий фрагмент, вариант или производное, раскрытая в данном документе, связывается с антигеном-мишенью, например, с полисахаридом, при скорости ассоциации (k(on)), большей или равной 105 М-1 с-1, 5×105 М-1 с-1, 106 М-1 с-1, или 5×106 M-1 c-1, или 107 М-1 с-1.
[0086] Говорят, что связывающая молекула, например, антитело или его фрагмент, вариант или производное, осуществляет конкурентное ингибирование связывания эталонного антитела или антиген-связывающего фрагмента с данным эпитопом, если она преимущественно связывается с этим эпитопом до такой степени, что в какой-то мере блокирует связывание эталонного антитела или антиген-связывающего фрагмента с эпитопом. Конкурентное ингибирование можно определить с помощью любого способа, известного в данной области техники, например, с помощью конкурентных ELISA-анализов. Можно говорить, что связывающая молекула осуществляет конкурентное ингибирование связывания эталонного антитела или антиген-связывающего фрагмента с данным эпитопом по меньшей мере на 90%, по меньшей мере на 80%), по меньшей мере на 70%, по меньшей мере на 60% или по меньшей мере на 50%.
[0087] Применяемое в данном документе выражение "аффинность" относится к мере силы связывания отдельного эпитопа с CDR молекулы иммуноглобулина. См., например, Harlow et al, Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988), страницы 27-28. Применяемое в данном документе выражение "авидность" относится к общей стабильности комплекса между популяцией иммуноглобулинов и антигеном, то есть к функциональной прочности связывания смеси иммуноглобулинов с антигеном. См., например, Harlow, страницы 29-34. Авидность относится как к аффинности отдельных молекул иммуноглобулинов в популяции в отношении конкретных эпитопов, так и также к валентностям иммуноглобулинов и антигена. Например, взаимодействие между бивалентным моноклональным антителом и антигеном с высокоповторяющейся структурой эпитопа, таким как полимер, может характеризоваться высокой авидностью.
[0088] Связывающие молекулы или их антиген-связывающие фрагменты, варианты или производные, раскрытые в данном документе, также могут быть описаны или определены относительно их перекрестной реактивности. Применяемое в данном документе выражение "перекрестная реактивность" относится к способности связывающей молекулы, например, антитела или его фрагмента, варианта или производного, специфичной в отношении одного антигена, реагировать со вторым антигеном; к мере родства между двумя различными антигенными веществами. Таким образом, связывающая молекула является перекрестно-реактивной, если она связывается с эпитопом, отличным от эпитопа, индуцировавшего ее образование. Перекрестно-реактивный эпитоп, как правило, содержит много таких же комплементарных структурных признаков, что и индуцирующий эпитоп, и в некоторых случаях может фактически быть более подходящим, чем исходный.
[0089] Связывающая молекула, например, антитело или его фрагмент, вариант или производное, также может быть описана или определена относительно ее аффинности связывания с антигеном. Например, связывающая молекула может связываться с антигеном при константе диссоциации KD, не превышающей 5×10-2 М, 10-2 М, 5×10-3 М, 10-3 М, 5×10-4 М, 10-4 М, 5×10-5 М, 10-5 М, 5×10-6 М, 10-6 М, 5×10-7 М, 10-7 М, 5×10-8 М, 10-8 М, 5×10-9 М, 10-9 М, 5×10-10 М, 10-10 М, 5×10-1 М, 10-1 М, 5×10-12 М, 10-12 М, 5×10-13 М, 10-13 М, 5×10-14 М, 10-14 М, 5×10-15 Мили 10-15 М.
[0090] Фрагменты антител, в том числе одноцепочечные антитела, могут содержать вариабельную область (вариабельные области) отдельно или в комбинации со всем или частью из следующего: шарнирная область, CH1-, CH2- и CH3-домены. Также включены антиген-связывающие фрагменты, также содержащие любую комбинацию вариабельной области (вариабельных областей) с шарнирной областью, CH1-, CH2- и CH3-доменами. Связывающие молекулы, например, антитела или их антиген-связывающие фрагменты, раскрытые в данном документе, могут происходить из любых животных, включая птиц и млекопитающих. Антитела могут быть антителами человека, мыши, осла, кролика, козы, морской свинки, верблюда, ламы, лошади или курицы. В другом варианте осуществления вариабельная область может происходить из хрящевых рыб (например, из акул). Применяемое в данном документе выражение "антитела человека" включает антитела, имеющие аминокислотную последовательность иммуноглобулина человека, и включает антитела, выделенные из библиотек иммуноглобулинов человека или из животных, трансгенных по одному или нескольким иммуноглобулинам человека и не экспрессирующих эндогенные иммуноглобулины, как описано ниже и, например, в патенте США №5939598 Kucherlapati et al.
[0091] Применяемое в данном документе выражение "часть тяжелой цепи" включает аминокислотные последовательности, полученные из тяжелой цепи иммуноглобулина. Связывающая молекула, например, антитело, содержащая часть тяжелой цепи, содержит по меньшей мере одно из следующего: CH1-домен, шарнирный (например, верхняя, средняя и/или нижняя шарнирная область) домен, CH2-домен, CH3-домен или их вариант или фрагмент.Например, связывающая молекула, например, антитело или его фрагмент, вариант или производное, может содержать полипептидную цепь, содержащую CH1-домен; полипептидную цепь, содержащую CH1-домен, по меньшей мере часть шарнирного домена и CH2-домен; полипептидную цепь, содержащую CHI-домен и CH3-домен; полипептидную цепь, содержащую CH1-домен, по меньшей мере часть шарнирного домена и CH3-домен, или полипептидную цепь, содержащую CH1-домен, по меньшей мере часть шарнирного домена, CH2-домен и CH3-домен. В другом варианте осуществления связывающая молекула, например, антитело или его фрагмент, вариант или производное, содержит полипептидную цепь, содержащую CH3-домен. Дополнительно, в связывающей молекуле для применения в настоящем изобретении может недоставать по меньшей мере части CH2-домена (например, всего или части CH2-домена). Как изложено выше, специалисту в данной области будет понятно, что данные домены (например, части тяжелой цепи) можно модифицировать таким образом, чтобы они отличались по аминокислотной последовательности от встречающейся в природе молекулы иммуноглобулина.
[0092] Части тяжелой цепи связывающей молекулы, например, антитела, раскрытого в данном документе, могут быть получены из различных молекул иммуноглобулинов. Например, часть тяжелой цепи полипептида может содержать CH1-домен, полученный из молекулы IgG1, и шарнирную область, полученную из молекулы IgG3. В другом примере часть тяжелой цепи может содержать шарнирную область, полученную частично из молекулы IgG1 и частично из молекулы IgG3. В другом примере часть тяжелой цепи может содержать химерную шарнирную область, полученную частично из молекулы IgG1 и частично из молекулы IgG4.
[0093] Применяемое в данном документе выражение "часть легкой цепи" включает аминокислотные последовательности, полученные из легкой цепи иммуноглобулина. Часть легкой цепи содержит по меньшей мере одно из VL- или CL-домена.
[0094] Связывающие молекулы, например, антитела или их антиген-связывающие фрагменты, варианты или производные, раскрытые в данном документе, могут быть описаны или определены относительно эпитопа (эпитопов) или части (частей) антигена, например, полисахарида-мишени, который они распознают или с которым специфически связываются. Часть полисахарида-мишени, которая специфически взаимодействует с антиген-связывающим доменом антитела, представляет собой "эпитоп" или "антигенную детерминанту". Антиген-мишень, например, полисахарид, может содержать один эпитоп, но обычно содержит по меньшей мере два эпитопа и может включать любое число эпитопов в зависимости от размера, конформации и типа антигена.
[0095] Как указывалось ранее, структуры субъединиц и трехмерная конфигурация константных областей иммуноглобулинов различных классов хорошо известны. Применяемое в данном документе выражение "VH-домен" включает амино-концевой вариабельный домен тяжелой цепи иммуноглобулина, и выражение "CH1-домен" включает первый (наиболее близкий к амино-концу) домен константной области тяжелой цепи иммуноглобулина. CH1-домен прилегает к VH-домену и является амино-концевым по отношению к шарнирной области тяжелой цепи молекулы иммуноглобулина.
[0096] Применяемое в данном документе выражение "CH2-домен" включает часть молекулы тяжелой цепи, продолжающуюся, например, от примерно остатка 244 до остатка 360 антитела согласно традиционным схемам нумерации (остатки 244-360 согласно системе нумерации по Kabat и остатки 231-340 согласно EU-системе нумерации; см. Kabat EA et al. в цитируемой работе). CH2-домен уникален в том, что он не соединен тесно с другим доменом. Наоборот, две разветвленных углеводных цепи с N-связями размещены между двумя CH2-доменами интактной нативной молекулы IgG. Также хорошо подтверждено документально, что CH3 домен продолжается от CH2-домена до С-конца молекулы IgG и содержит приблизительно 108 остатков.
[0097] Применяемое в данном документе выражение "шарнирная область" включает часть тяжелой цепи молекулы, которая присоединяет CH1-домен к CH2-домену. Эта шарнирная область содержит приблизительно 25 остатков и является гибкой, что, таким образом, позволяет двум N-концевым антиген-связывающим областям перемещаться независимо. Шарнирные области могут быть подразделены на три различных домена: верхний, средний и нижний шарнирные домены (Roux et al, J. Immunol. 161: 4083 (1998)).
[0098] Применяемое в данном документе выражение "дисульфидная связь" включает ковалентную связь, образованную между двумя атомами серы. Аминокислота цистеин содержит тиоловую группу, которая может образовывать дисульфидную связь или мостик со второй тиоловой группой. В большинстве встречающихся в природе молекул IgG CH1- и CL-области связаны дисульфидной связью, а две тяжелые цепи связаны двумя дисульфидными связями в положениях, соответствующих 239 и 242 согласно системе нумерации по Kabat (положение 226 или 229 согласно EU-системе нумерации).
[0099] Применяемое в данном документе выражение "химерное антитело" будет означать любое антитело, где иммунореактивная область или участок получены или происходят из одного вида, а константная область (которая может быть интактной, неполной или модифицированной) получена из второго вида. В некоторых вариантах осуществления область или участок, связывающиеся с мишенью, будут происходить из источника, не являющегося человеком (например, из мыши или примата), а константная область - из человека.
[0100] Применяемое в данном документе выражение "биспецифическое антитело" относится к антителу, которое содержит участки связывания для двух различных антигенов в пределах одной молекулы антитела. Следует понимать, что другие молекулы, в дополнение к структуре канонического антитела, могут быть сконструированы с двумя специфичностями связывания. Дополнительно следует понимать, что связывание антигена с биспецифическими антителами может быть одновременным или последовательным. Триомы и гибридные гибридомы представляют собой два примера клеточных линий, которые могут секретировать биспецифические антитела. Биспецифические антитела также могут быть сконструированы с помощью способов рекомбинации. (Strohlein and Heiss, Future Oncol. 6: 1387-94 (2010); Mabry and Snavely, IDrugs. 73: 543-9 (2010)).
[0101] Применяемое в данном документе выражение "сконструированное антитело" относится к антителу, в котором вариабельный домен в тяжелой или в легкой цепи либо в них обеих изменен путем по меньшей мере частичного замещения одного или нескольких CDR из антитела с известной специфичностью и, при необходимости, путем частичного замещения каркасной области и изменения последовательностей. Хотя CDR могут быть получены из антитела того же класса или даже подкласса, что и антитело, из которого получены каркасные области, предусматривается, что CDR будут получены из антитела другого класса и предпочтительно из антитела от другого вида. Сконструированное антитело, в котором один или несколько "донорных" CDR из антитела, не являющегося антителом человека, с известной специфичностью пересаживают в каркасную область тяжелой или легкой цепи человека, называется в данном документе "гуманизированным антителом". Для передачи антиген-связывающей способности одного вариабельного домена другому может не быть необходимым замещение всех CDR полными CDR из донорной вариабельной области. Наоборот, может быть необходимым только перенос тех остатков, которые являются необходимыми для поддерживания активности участка, связывающегося с мишенью. С учетом пояснений, изложенных, например, в патентах США №5585089, 5693761, 5693762 и 6180370, получение функционального сконструированного или гуманизированного антитела путем проведения стандартных экспериментов либо путем тестирования по методу проб и ошибок будет находиться вполне в пределах компетенции специалиста в данной области техники.
[0102] Применяемое в данном документе выражение "правильно свернутый полипептид" включает полипептиды (например, антитела к Psl и PcrV Pseudomonas), в которых все функциональные домены, содержащие полипептид, являются явно активными. Применяемое в данном документе выражение "неправильно свернутый полипептид" включает полипептиды, в которых по меньшей мере один из функциональных доменов полипептида не является активным. В одном варианте осуществления правильно свернутый полипептид содержит полипептидные цепи, связанные по меньшей мере одной дисульфидной связью, и напротив, неправильно свернутый полипептид содержит полипептидные цепи, не связанные по меньшей мере одной дисульфидной связью.
[0103] Применяемое в данном документе выражение "сконструированный" включает воздействие на молекулы нуклеиновых кислот или полипептидов с помощью способов синтеза (например, с помощью рекомбинантных методик, синтеза пептидов in vitro, с помощью ферментативного или химического сочетания пептидов или некоторых комбинаций данных методик).
[0104] Применяемые в данном документе выражения "связанный", "слитый" или "слияние" применяют взаимозаменяемо. Эти выражения относятся к соединению двух или более элементов или компонентов с помощью любых способов, включая способы химической конъюгации или рекомбинации. "Внутрирамочное слияние" относится к соединению двух или более открытых рамок считывания полинуклеотидов (ORF) с образованием непрерывной более длинной ORF таким образом, чтобы поддерживать правильную трансляционную рамку считывания исходных ORF. Таким образом, рекомбинантный белок слияния представляет собой один белок, содержащий два или более сегмента, которые соответствуют полипептидам, кодируемым исходными ORF (такие сегменты обычно не соединены подобным образом в природе). Хотя рамка считывания делается, таким образом, непрерывной во всех слитых сегментах, сегменты могут быть физически или пространственно разделены с помощью, например, внутрирамочной линкерной последовательности. Например, полинуклеотиды, кодирующие CDR вариабельной области иммуноглобулина, могут быть слитыми, внутрирамочными, но разделенными полинуклеотидом, кодирующим по меньшей мере одну каркасную область иммуноглобулина или дополнительные области CDR, поскольку "слитые" CDR подвергаются котрансляции как часть непрерывного полипептида.
[0105] В отношении полипептидов "линейная последовательность" или "последовательность" представляет собой порядок аминокислот в полипептиде в направлении от амино-конца до карбокси-конца, при котором остатки, являющиеся соседними относительно друг друга в последовательности, являются смежными в первичной структуре полипептида.
[0106] Выражение "экспрессия", применяемое в данном документе, относится к способу, с помощью которого ген вырабатывает биохимическое вещество, например, полипептид. Способ включает любое проявление функционального присутствия гена в клетке, включая, без ограничения, нокдаун гена, а также как транзиентную экспрессию, так и стабильную экспрессию. Он включает, без ограничений, транскрипцию гена в матричную РНК (мРНК) и трансляцию таких мРНК в полипептид (полипептиды). Если конечный желаемый продукт представляет собой биохимическое вещество, экспрессия включает создание таких биохимических веществ и любых предшественников. При экспрессии гена вырабатывается "продукт гена". Как применяется в данном документе, продуктом гена может быть либо нуклеиновая кислота, например, матричная РНК, вырабатываемая путем транскрипции гена, либо полипептид, транслируемый из транскрипта. Продукты генов, описываемые в данном документе, дополнительно включают нуклеиновые кислоты с посттранскрипционными модификациями, например, полиаденилированием, или полипептиды с посттрансляционными модификациями, например, метилированием, гликозилированием, добавлением липидов, ассоциацией с другими белковыми субъединицами, протеолитическим расщеплением и т.п.
[0107] Применяемые в данном документе выражения "лечить" или "лечение" относятся как к терапевтическому лечению, так и к мерам профилактики или предупреждения, где целью является предупреждение или замедление (облегчение) нежелательного физиологического изменения, инфекции или нарушения. Благоприятные или желаемые клинические результаты включают, без ограничений, смягчение симптомов, уменьшение степени тяжести заболевания, стабилизацию (т.е. отсутствие ухудшения) состояния заболевания, устранение или ослабление возбудителя инфекции, такого как P. aeruginosa, у субъекта, задержку или замедление прогрессирования заболевания, улучшение или временное облегчение состояния заболевания и ремиссию (частичную либо полную), поддающиеся или не поддающиеся выявлению. "Лечение" может также означать увеличение выживаемости по сравнению с ожидаемой выживаемостью в том случае, если лечение не было получено. Нуждающиеся в лечении включают таковых, уже имеющих инфекцию, состояние или нарушение, а также таковых, имеющих предрасположенность к состоянию или нарушению, или таковых, у которых состояние или нарушение нужно предупреждать, например, у ожоговых пациентов или пациентов с ослабленным иммунитетом, восприимчивых к инфекции, вызываемой P. aeruginosa.
[0108] Под "субъектом", или "лицом", или "животным", или "пациентом", или "млекопитающим" подразумевается любой субъект, в частности, субъект-млекопитающее, для которого желаемыми являются диагностика, прогнозирование или терапия. Субъекты-млекопитающие включают людей, домашних животных, сельскохозяйственных животных и зоопарковых, используемых в спорте или комнатных животных, таких как собаки, кошки, морские свинки, кролики, крысы, мыши, лошади, крупный рогатый скот, коровы, медведи и т.д.
[0109] Применяемые в данном документе фразы, такие как "субъект, который может получить пользу от введения доменов или молекул, связывающихся с Psl и PcrV Pseudomonas" и "животное, нуждающееся в лечении" включают субъектов, таких как субъекты-млекопитающие, которые могут получить пользу от введения доменов, связывающихся с Psl и PcrV Pseudomonas, или молекулы, связывающейся с таковыми, такой как антитело, содержащее один или несколько связывающих доменов. Такие связывающие домены или связывающие молекулы могут применяться, например, для выявления Psl и PcrV Pseudomonas (например, для диагностической процедуры) и/или для лечения, т.е. временного облегчения или предупреждения заболевания, с помощью молекул, связывающихся с Psl и PcrV Pseudomonas. Как более подробно описано в данном документе, молекулы, связывающиеся с Psl и PcrV Pseudomonas, могут применяться в неконъюгированной форме или могут быть конъюгированными, например, с лекарственным средством, пролекарством или радиоактивным изотопом.
[0110] Применяемое в данном документе выражение "синергический эффект" относится к большему, чем аддитивный, терапевтическому эффекту, производимому комбинацией соединений, причем терапевтический эффект, полученный с помощью комбинации, превышает аддитивные эффекты, которые могли бы возникать в других случаях в результате независимого введения соединений по отдельности. Определенные варианты осуществления включают способы получения синергического эффекта при лечении инфекций, вызываемых Pseudomonas, причем указанный эффект по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 100%, по меньшей мере на 200%, по меньшей мере на 500%) или по меньшей мере на 1000% превышает соответствующий аддитивный эффект.
[0111] "Совместное введение" относится к введению различных соединений, таких как домен, связывающийся с Psl и PcrV, или связывающая молекула, содержащая один или оба домена, связывающихся с Psl и PcrV, таким образом, что соединения вызывают синергический эффект в отношении иммунитета к P. aeruginosa. Соединения могут быть введены в одной и той же или в разных композициях, которые в случае введения по отдельности вводят через короткий временной промежуток между каждыми двумя введениями, как правило, в течение 24 часов между каждыми двумя введениями, и более типично в пределах приблизительно 1-8 часов между между каждыми двумя введениями, и еще более типично в пределах 1-4 часов между между каждыми двумя введениями или в режиме, близком к одновременному введению. Относительные количества представляют собой дозировки, при которых достигается желаемый синергизм.
II. СВЯЗЫВАЮЩИЕ ДОМЕНЫ И СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ
[0112] Антитела, которые связываются с Psl, и форматы для применения этих антител были описаны в данной области техники. См., например, Международную заявку на патент №PCT/US2012/041538, поданную 8 июня 2012 г., и PCT/US2012/63639, поданную 6 ноября 2012 г. (номер дела патентного поверенного AEMS-115W01, под названием "ПОЛИСПЕЦИФИЧЕСКИЕ И ПОЛИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ"), которые включены в данный документ посредством ссылки во всей своей полноте.
[0113] Один вариант осуществления направлен на связывающие домены, которые специфически связываются с PcrV Pseudomonas, причем связывание может нарушить активность системы секреции токсинов III типа. В некоторых вариантах осуществления связывающие домены характеризуются такой же специфичностью связывания Pseudomonas, что и антитело V2L2.
[0114] Другой вариант осуществления направлен на связывающие домены, которые специфически связываются с Psl или PcrV Pseudomonas, причем введение обоих связывающих доменов приводит к синергическим эффектам, противодействующим инфекциям, вызываемым Pseudomonas, посредством (a) ингибирования прикрепления P. aeruginosa к эпителиальным клеткам, (b) способствования, опосредования или усиления опсонофагоцитарного уничтожения (ОРК) Р. aeruginosa, (c) ингибирования прикрепления P. aeruginosa к эпителиальным клеткам или (d) нарушения работы системы секреции токсинов III типа. В определенных вариантах осуществления связывающие домены характеризуются такой же специфичностью связывания Pseudomonas, что и антитела Cam-003, WapR-004, V2L2 или 29D2.
[0115] Другие варианты осуществления направлены на выделенную(ые) связывающую(ие) молекулу(ы), содержащую один или оба связывающих домена, которые специфически связываются с Psl и/или PcrV Pseudomonas, причем введение связывающих молекул приводит к синергическим эффектам, противодействующим инфекциям, вызываемым Pseudomonas. В определенных вариантах осуществления связывающая молекула может содержать связывающий домен из антител или их фрагментов, которые включают, без ограничения, Cam-003, WapR-004, V2L2 или 29D22.
[0116] Применяемые в данном документе выражения "связывающий домен" или "антиген-связывающий домен" включают участок, который специфически связывается с эпитопом антигена (например, с эпитопом Psl или PcrV Pseudomonas). Антиген-связывающий домен антитела обычно включает по меньшей мере часть вариабельной области тяжелой цепи иммуноглобулина и по меньшей мере часть вариабельной области легкой цепи иммуноглобулина. Участок связывания, образованный этими вариабельными областями, определяет специфичность антитела.
[0117] Настоящее изобретение более конкретно направлено на композицию, содержащую по меньшей мере два домена, связывающихся с Pseudomonas, причем один связывающий домен специфически связывается с Psl, а другой связывающий домен специфически связывается с PcrV. В одном варианте осуществления композиция содержит один связывающий домен, который специфически связывается с тем же эпитопом Psl Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL) WapR-004, Cam-003, Cam-004, Cam-005, WapR-001, WapR-002, WapR-003 или WapR-016. В определенных вариантах осуществления второй связывающий домен специфически связывается с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL) V2L2 или 29D2.
[0118] В одном варианте осуществления композиция содержит один связывающий домен, который специфически связывается с Psl Pseudomonas и/или осуществляет конкурентное ингибирование связывания Psl Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH- и VL-области WapR-004, Cam-003, Cam-004, Cam-005, WapR-001, WapR-002, WapR-003 или WapR-016. В определенных вариантах осуществления второй связывающий домен специфически связывается с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL) V2L2 или 29D2, и/или осуществляет конкурентное ингибирование связывания PcrV Pseudomonas с таковыми.
[0119] Другой вариант осуществления направлен на выделенную связывающую молекулу, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с тем же эпитопом PcrV Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-область V2L2 или 29D2.
[0120] Также включена выделенная связывающая молекула, например, антитело или его фрагмент, которая специфически связывается с PcrV Pseudomonas и осуществляет конкурентное ингибирование связывания PcrV Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH и VL V2L2 или 29D2.
[0121] Один вариант осуществления направлен на выделенную связывающую молекулу, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с тем же эпитопом Psl Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-область WapR-001, WapR-002 или WapR-003.
[0122] Также включена выделенная связывающая молекула, например, антитело или его фрагмент, которая специфически связывается с Psl Pseudomonas и осуществляет конкурентное ингибирование связывания Psl Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH и VL WapR-001, WapR-002 или WapR-003.
[0123] Дополнительно включена выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с тем же эпитопом Psl Pseudomonas, что и антитело или его антиген-связывающий фрагмент, содержащие VH- и VL-области WapR-016.
[0124] Также включена выделенная связывающая молекула, например, антитело или его фрагмент, которая специфически связывается с Psl Pseudomonas и осуществляет конкурентное ингибирование связывания Psl Pseudomonas с антителом или его антиген-связывающим фрагментом, содержащими VH и VL WapR-016.
[0125] Способы создания антител хорошо известны в данной области техники и описаны в данном документе. Как только получены антитела к различным фрагментам Psl или PcrV Pseudomonas или к таковым полной длины без сигнальной последовательности, можно определить то, какими именно являются аминокислоты или эпитоп Psl или PcrV Pseudomonas, с которыми связывается антитело или антиген-связывающий фрагмент, с помощью протоколов картирования эпитопов, описываемых в данном документе, а также способов, известных в данной области техники (например, сэндвич-ELISA двойных антител, описанный в "Chapter 11 -Immunology," Current Protocols in Molecular Biology, Ed. Ausubel et al, v. 2, John Wiley & Sons, Inc. (1996)). Дополнительные протоколы картирования эпитопов можно найти в Morris, G. Epitope Mapping Protocols, New Jersey: Humana Press (1996), оба из которых включены в данный документ посредством ссылки во всей своей полноте. Картирование эпитопов можно также выполнять с помощью коммерчески доступных средств (т.е. ProtoPROBE, Inc. (Милуоки, Висконсин)).
[0126] В определенных аспектах настоящее изобретение направлено на связывающую молекулу, например, антитело или его фрагмент, вариант или производное, которая специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся константой диссоциации (KD), меньшей, чем KD для указанного эталонного моноклонального антитела.
[0127] В определенных вариантах осуществления молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его антиген-связывающий фрагмент, вариант или производное, раскрытая в данном документе, специфически связывается по меньшей мере с одним эпитопом Psl или PcrV, т.е. связывается с таким эпитопом более легко, чем если бы она связывалась с неродственным или случайным эпитопом; преимущественно связывается по меньшей мере с одним эпитопом Psl или PcrV, т.е. связывается с таким эпитопом более легко, чем если бы она связывалась с родственным, сходным, гомологичным или аналогичным эпитопом; осуществляет конкурентное ингибирование связывания эталонного антитела, которое само специфически или преимущественно связывается с определенным эпитопом Psl или PcrV; или связывается по меньшей мере с одним эпитопом Psl или PcrV с аффинностью, характеризующейся константой диссоциации KD, меньшей, чем около 5×10-2 М, около 102 М, около 5×10-3 М, около 10-3 М, около 5×10-4 М, около 10-4 М, около 5×10-5 М, около 10-5 М, около 5×10-6 М, около 10-6 М, около 5×10-7 М, около 10-7 М, около 5×10-8 М, около 10-8 М, около 5×10-9 М, около 10-9 М, около 5×10-10 М, около 10-10 М, около 5×10-11 М, около 10-1 М, около 5×10-12 М, около 10-12 М, около 5×10-13 М, около 10-13 М, около 5×10-14 М, около 10-14 М, около 5×10-15 М или около 10-15 М.
[0128] Применяемое в отношении констант диссоциации, характеризующих связывание, выражение "около" предусматривает степень изменчивости, характерную для способов, используемых для измерения аффинности антител. Например, в зависимости от уровня точности применяемых измерительных приборов, среднеквадратической ошибки на основе числа измеренных образцов и ошибки округления выражение "около 10-2 М" может включать, например, от 0,05 М до 0,005 М.
[0129] В конкретных вариантах осуществления связывающая молекула, например, антитело или его антиген-связывающий фрагмент, вариант или производное, связывается с Psl и/или PcrV Pseudomonas при скорости диссоциации (k(off)), меньшей или равной 5×10-2 с-1, 10-2 с-1, 5×10-3 с-1 или 10-3 с-1. Альтернативно, антитело или его антиген-связывающий фрагмент, вариант или производное связывается с Psl и/или PcrV Pseudomonas при скорости диссоциации (k(off)), меньшей или равной 5×10-4 с-1, 10-4 с-1, 5×10-5 с-1 или 10-5 с-1, 5×10-6 с-1, 10-6 с-1, 5×10-7 с-1 или 10-7 с-1.
[0130] В других вариантах осуществления связывающая молекула, например, антитело или его антиген-связывающий фрагмент, вариант или производное, раскрытые в данном документе, связывается с Psl и/или PcrV Pseudomonas при скорости ассоциации (k(on)), большей или равной 103 М-1 с-1, 5×103 М-1 с-1, 104 М-1 с-1 или 5×104 М-1 с-1. Альтернативно, связывающая молекула, например, антитело или его антиген-связывающий фрагмент, вариант или производное, раскрытые в данном документе, связывается с Psl и/или PcrV Pseudomonas при скорости ассоциации (k(on)), большей или равной 105 М-1 с-1, 5×105 М-1 с-1, 106 М-1 с-1, или 5×106 М-1 с-1, или 107 М-1 с-1.
[0131] В различных вариантах осуществления молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его антиген-связывающий фрагмент, вариант или производное, описываемые в данном документе, способствует опсонофагоцитарному уничтожению Pseudomonas или ингибирует связывание Pseudomonas с эпителиальными клетками. В определенных вариантах осуществления, описываемых в данном документе, Psl- или PcrV-мишень Pseudomonas представляет собой Psl или PcrV Pseudomonas aeruginosa. В других вариантах осуществления определенные связывающие молекулы, описываемые в данном документе, могут связываться с молекулами родственных по структуре полисахаридов независимо от их источника. Такие Psl-подобные молекулы, как следует ожидать, являются идентичными или обладают достаточным структурным родством по отношению к Psl P. aeruginosa, что позволяет осуществлять специфическое распознавание одной или несколькими раскрытыми связывающими молекулами. В других вариантах осуществления определенные связывающие молекулы, описываемые в данном документе, могут связываться с молекулами родственных по структуре полипептидов независимо от их источника. Такие PcrV-подобные молекулы, как следует ожидать, являются идентичными или обладают достаточным структурным родством по отношению к PcrV P. aeruginosa, что позволяет осуществлять специфическое распознавание одной или несколькими раскрытыми связывающими молекулами. Таким образом, например, определенные связывающие молекулы, описываемые в данном документе, могут связываться с Psl-подобными и/или PcrV-подобными молекулами, вырабатываемыми другим видом бактерий, например, с Psl-подобными или PcrV-подобными молекулами, вырабатываемыми другим видом Pseudomonas, например, Pseudomonas fluorescens, Pseudomonas putida или Pseudomonas alcaligenes. Альтернативно, определенные связывающие молекулы, описываемые в данном документе, могут связываться с Psl-подобными и/или PcrV-подобными молекулами, вырабатываемыми синтетически или клетками-хозяевами, генетически модифицированными с целью получения Psl-подобных или PcrV-подобных молекул.
[0132] Если конкретно не отмечено, применяемое в данном документе выражение "его фрагмент" в отношении связывающей молекулы, например, антитела, относится к антиген-связывающему фрагменту, т.е. части антитела, которая специфически связывается с антигеном.
[0133] Молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их антиген-связывающие фрагменты, варианты или производные, могут содержать константную область, опосредующую одну или несколько эффекторных функций. Например, связывание C1-компонента системы комплемента с константной областью антитела может активировать систему комплемента. Активация комплемента важна в опсонизации и лизисе патогенов. Активация комплемента также стимулирует воспалительный ответ и может также быть вовлечена в аутоиммунные реакции гиперчувствительности. Дополнительно, антитела связываются с рецепторами различных клеток посредством Fc-области, при этом участок связывания Fc-рецептора Fc-области антитела связывается с Fc-рецептором (FcR) клетки. Существует ряд Fc-рецепторов, являющихся специфичными в отношении антител различных классов, включая IgG (гамма-рецепторы), IgE (эпсилон-рецепторы), IgA (альфа-рецепторы) и IgM (мю-рецепторы). Связывание антитела с Fc-рецепторами на поверхностях клеток запускает ряд важных и разнообразных биологических ответов, включая поглощение и разрушение нагруженных антителами частиц, клиренс иммунных комплексов, лизис нагруженных антителами клеток-мишеней клетками-киллерами (называемый антителозависимой клеточно-опосредованной цитотоксичностью или ADCC), высвобождение медиаторов воспаления, плацентарный перенос и регуляцию выработки иммуноглобулинов.
[0134] Соответственно, определенные варианты осуществления, раскрытые в данном документе, включают молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, например, антитело или его антиген-связывающий фрагмент, вариант или производное, в которой по меньшей мере доля одного или нескольких доменов константной области была удалена или иным образом изменена так, чтобы обеспечивать желаемые биохимические характеристики, такие как сниженные эффекторные функции, способность к димеризации за счет нековалентных связей, повышенная способность к локализации в месте опухоли, сниженный период полувыведения из сыворотки или повышенный период полувыведения из сыворотки по сравнению с полным, неизмененным антителом, обладающим приблизительно такой же иммуногенностью. Например, определенные связывающие молекулы, описываемые в данном документе, являются антителами с удаленными доменами, которые содержат полипептидную цепь, подобную тяжелой цепи иммуноглобулина, но в которых недостает по меньшей мере части одного или нескольких доменов тяжелой цепи. Например, в определенных антителах будет удален один целый домен константной области модифицированного антитела, например, будет удален весь CH2-домен или его часть.
[0135] Модифицированные формы молекул, связывающихся с Psl и/или PcrV Pseudomonas, например, антитела или их антиген-связывающие фрагменты, варианты или производные, могут быть созданы из полных антител-предшественников или родительских антител с применением методик, известных в данной области техники. Иллюстративные методики рассматриваются в другом месте в данном документе.
[0136] В определенных вариантах осуществления как вариабельные, так и константные области молекул, связывающихся с Psl и/или PcrV Pseudomonas, например, антител или антиген-связывающих фрагментов, являются полностью человеческими. Полностью человеческие антитела могут быть созданы С помощью методик, известных в данной области техники и описываемых в данном документе. Например, полностью человеческие антитела к конкретному антигену можно получить путем введения антигена трансгенному животному, модифицированному с целью получения таких антител в ответ на антигенную стимуляцию, но эндогенные локусы которого были деактивированы. Иллюстративные методики, которые можно применять для создания таких антител, описаны в патентах США 6150584; 6458592; 6420140. Другие методики известны в данной области техники. Полностью человеческие антитела можно получать подобным образом с помощью различных технологий дисплея, например, фагового дисплея или других систем вирусного дисплея, описываемых более подробно в другом месте в данном документе.
[0137] Молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их антиген-связывающие фрагменты, варианты или производные, раскрытые в данном документе, могут быть созданы или произведены с помощью методик, известных в данной области техники. В определенных вариантах осуществления связывающие молекулы или их фрагменты являются "полученными рекомбинантным путем", т.е. получены с помощью технологии рекомбинантных ДНК. Иллюстративные методики создания молекул антител или их фрагментов рассматриваются более подробно в другом месте в данном документе.
[0138] В определенных молекулах, связывающихся с Psl и/или PcrV Pseudomonas, например, антителах или их антиген-связывающих фрагментах, вариантах или производных, описываемых в данном документе, Fc-часть можно подвергнуть мутации для уменьшения эффекторной функции с помощью методик, известных в данной области техники. Например, делеция или инактивация (с помощью точечных мутаций или других средств) домена константной области может уменьшать связывание циркулирующего модифицированного антитела с Fc-рецепторами, таким образом повышая локализацию в опухоли. В других случаях может быть так, что модификации константной области умеряют связывание комплемента и таким образом уменьшают период полувыведения из сыворотки и неспецифическую ассоциацию конъюгированного цитотоксина. Еще несколько модификаций константной области можно применять для модификации дисульфидных связей или олигосахаридных фрагментов, что дает возможность увеличения локализации в связи с повышенной специфичностью по отношению к антигену или гибкостью антитела. Полученные в результате физиологический профиль, биодоступность и другие биохимические эффекты модификаций, такие как локализация, биораспределение и период полувыведения из сыворотки, можно легко измерить и оценить количественно с применением хорошо известных иммунологических методик без ненужных экспериментов.
[0139] В определенных вариантах осуществления молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их антиген-связывающие фрагменты, варианты или производные, не будут вызывать вредный иммунный ответ у животного, подлежащего лечению, например, у человека. В одном варианте осуществления молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их антиген-связывающие фрагменты, варианты или производные, модифицированы с целью уменьшения их иммуногенности с помощью методик, принятых в данной области техники. Например, антитела могут быть гуманизированными, деиммунизированными, или могут быть созданы химерные антитела. Эти типы антител получены из антитела, не являющегося антителом человека, обычно из антитела мыши или примата, которое сохраняет или в значительной степени сохраняет антиген-связывающие свойства родительского антитела, но которое является менее иммуногенным у людей. Этого можно достичь разными способами, включая (a) пересадку целых вариабельных доменов, полученных не от человека, в константные области, полученные от человека, с образованием химерных антител; (b) пересадку по меньшей мере части одного или нескольких гипервариабельных участков (CDR), полученных не от человека, в каркасные и константные области, полученные от человека, с сохранением критически важных каркасных остатков или без такового или (c) трансплантацию целых вариабельных доменов, полученных не от человека, однако, с "маскировкой" их участком, подобным полученному от человека, путем замещения поверхностных остатков. Такие способы раскрыты в Morrison et al., Proc. Natl. Acad. Set #7: 6851-6855 (1984); Morrison et al, Adv. Immunol 44: 65-92 (1988); Verhoeyen et al, Science 239: 1534-1536 (1988); Padlan, Molec. Immun. 25: 489-498 (1991); Padlan, Molec. Immun. 31: 169-217 (1994) и в патентах США №№5585089, 5693761, 5693762 и 6190370, все из которых включены в данный документ посредством ссылки во всей своей полноте.
[0140] Деиимунизацию также можно применять для уменьшения иммуногенности антитела. Применяемое в данном документе выражение "деиммунизация" включает изменение антитела с целью модификации T-клеточных эпитопов (см., например, WO 9852976 A1, WO 0034317 A2). Например, анализируют последовательности VH и VL исходного антитела и составляют "карту" T-клеточных эпитопов человека из каждой V-области, показывающую расположение эпитопов по отношению к гипервариабельным участкам (CDR) и другим ключевым остаткам в последовательности. Отдельные T-клеточные эпитопы из карты T-клеточных эпитопов анализируют в целях идентификации альтернативных аминокислотных замен с низким риском изменения активности конечного антитела. Ряд альтернативных последовательностей VH и VL разработаны таким образом, что они содержат комбинации аминокислотных замен, и эти последовательности впоследствии включают в ряд связывающих полипептидов, например, антител или их антиген-связывающих фрагментов, специфичных в отношении Psl и/или PcrV Pseudomonas, раскрытых в данном документе, которые затем тестируют в отношении функции. Гены полной тяжелой и легкой цепей, содержащие последовательности, кодирующие модифицированную V-область и С-область человека, затем клонируют в векторы экспрессии, и полученные в результате плазмиды вводят в линии клеток для получения полного антитела. Антитела затем сравнивают в соответствующих биохимических и биологических анализах, и идентифицируют оптимальный вариант.
[0141] Молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их антиген-связывающие фрагменты, варианты или производные, можно образовать с помощью любого подходящего способа, известного в данной области техники. Поликлональные антитела к антигену, представляющему интерес, можно получить с помощью различных методов, хорошо известных в данной области техники. Например, антитело к Psl и/или PcrV Pseudomonas или его антиген-связывающий фрагмент можно вводить различным животным-хозяевам, включая, без ограничений, кроликов, мышей, крыс, кур, хомяков, коз, ослов и т.д., с индуцированием выработки сыворотки, содержащей поликлональные антитела, специфичные в отношении антигена. Для усиления иммунного ответа можно применять различные адъюванты в зависимости от вида-хозяина, и они включают, без ограничений, адъювант Фрейнда (полный и неполный), минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, полиолы-плюроники, полианионы, пептиды, масляные эмульсии, гемоцианин лимфы улитки, динитрофенол и потенциально применимые адьюванты для человека, такие как BCG (бацилла Кальметта-Герена) и Corynebacterium parvum. Такие адъюванты также хорошо известны в данной области техники.
[0142] Моноклональные антитела можно получать при помощи широкого разнообразия методик, известных в данной области техники, включая применение гибридомной, рекомбинантной технологий и технологии фагового дисплея или их комбинации. Например, моноклональные антитела можно получать при помощи гибридомных методик, включая таковые, известные в данной области техники и изложенные, например, в Harlow et al, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 2nd ed. (1988).
[0143] ДНК, кодирующую антитела или фрагменты антител (например, антиген-связывающие участки), также можно получить из библиотек антител, таких как фаг-дисплейные библиотеки. В частности, такой фаг можно использовать для отображения антиген-связывающих доменов, экспрессирующихся в репертуаре или комбинаторной библиотеке антител (например, человека или мыши). Фаг, экспрессирующий антиген-связывающий домен, который связывается с антигеном, представляющим интерес, можно отобрать или идентифицировать при помощи антигена, например, применяя меченый антиген или антиген, связанный с твердой поверхностью или гранулой или захваченный на них. Фаг, применяемый в этих способах, обычно является нитевидным фагом, включающим связывающие домены fd и M13, экспрессируемые фагом, при этом домены scFv, Fab, Fv OE DAB (отдельная Fv-область легкой или тяжелой цепи) или стабилизированные дисульфидными связями Fv-домены антитела слиты рекомбинантным путем с белком, кодируемым либо геном III, либо геном VIII фага. Иллюстративные способы изложены, например, в EP 368684 B1; патенте США 5969108, Hoogenboom, H.R. and Chames, Immunol. Today 21: 371 (2000); Nagy et al. Nat. Med. 5: 801 (2002); Huie et al, Proc. Natl. Acad. Sci. USA 982682 (2001); Lui et al, J. Mol Biol 3151063 (2002), каждый из которых включен в данный документ посредством ссылки. В некоторых публикациях (например, Marks et al, Bio/Technology 10: 119-783 (1992)) было описано получение высокоаффинных антител человека с помощью шаффлинга цепей, а также комбинаторного инфицирования и in vivo рекомбинации в качестве стратегии конструирования больших фаговых библиотек. В другом варианте осуществления можно применять рибосомный дисплей для замены бактериофага в качестве платформы для дисплея (см., например, Hanes et al., Nat. Biotechnol. 75: 1287 (2000); Wilson et al, Proc. Natl. Acad. Sci. USA 95:3750 (2001); или Irving et al., J. Immunol. Methods 245:31 (2001)). В еще одном варианте осуществления библиотеки на клеточных поверхностях могут быть подвергнуты скринингу на наличие антител (Boder et al., Proc. Natl. Acad. Sci. USA 97: 10701 (2000); Daugherty et al., J. Immunol. Methods 243: 211 (2000)). Такие методы обеспечивают альтернативы традиционным гибридомным методикам выделения и последующего клонирования моноклональных антител.
[0144] В способах фагового дисплея функциональные домены антител отображены на поверхности фаговых частиц, несущих последовательности полинуклеотидов, кодирующие их. Например, последовательности ДНК, кодирующие VH- и VL-области, амплифицируют из библиотек кДНК животных (например, из библиотек кДНК лимфоидной ткани человека или мыши) или библиотек синтетических кДНК. В некоторых вариантах осуществления ДНК, кодирующие VH- и VL-области, соединяют с помощью scFv-линкера путем ПЦР и клонируют в вектор-фагмиду (например, pCANTAB 6 или pComb 3 HSS). Вектор вводят в Е. coli путем электропорации, и Е. coli инфицируют фагом-помощником. Фаг, применяемый в этих способах, обычно является нитевидным фагом, в том числе fd и M13, и VH-или VL-области, как правило, слиты рекомбинантным путем либо с геном III, либо с геном VIII фага. Фаг, экспрессирующий антиген-связывающий домен, который связывается с антигеном, представляющим интерес (т.е. Psl или PcrV Pseudomonas), можно отобрать или идентифицировать при помощи антигена, например, применяя меченый антиген или антиген, связанный с твердой поверхностью или гранулой или захваченный на них.
[0145] Дополнительные примеры способов фагового дисплея, которые можно применять для создания антител, включают раскрытые в Brinkman et al., J. Immunol. Methods 752: 41-50 (1995); Ames et al, J. Immunol. Methods 754: 177-186 (1995); Kettleborough et al, Eur. J. Immunol. 24: 952-958 (1994); Persic et al., Gene 187: 9-18 (1997); Burton et al, Advances in Immunology 57: 191-280 (1994); заявке по PCT № PCT/GB91/01134; публикациях no PCT WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401 и патентах США №№5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698;5427908; 5516637; 5780225; 5658727; 5733743 и 5969108; все из которых включены в данный документ посредством ссылки во всей своей полноте.
[0146] Как описано в вышеприведенных источниках и нижеследующих примерах, после отбора фага можно выделять области, кодирующие антитела, из фага и применять для образования полного антитела, включая антитела человека или любой другой желаемый антиген-связывающий фрагмент, и экспрессировать в любом желаемом хозяине, включая клетки млекопитающих, клетки насекомых, клетки растений, дрожжи и бактерии. Например, также можно использовать методики рекомбинантного получения Fab-, Fab'- и P(ab')2-фрагментов, применяя способы, известные в данной области техники, такие как раскрытые в публикации по PCT WO 92/22324; Mullinax et al, BioTechniques 12 (6): 864-869 (1992); и Sawai et al, AJRI 54: 26-34 (1995); и Better et al, Science 240: 1041-1043 (1988) (указанные источники включены посредством ссылки во всей своей полноте).
[0147] Примеры методик, которые можно применять для получения одноцепочечных Fv и антител, включают методики, описанные в патентах США №№4946778 и 5258498; Huston et al., Methods in Enzymology 203: 46-88 (1991); Shu et al., PNAS 90: 7995-7999 (1993); и Skerra et al, Science 240: 1038-1040 (1988). В определенных вариантах осуществления, таких как терапевтическое введение, можно применять химерные, гуманизированные антитела или антитела человека. Химерное антитело представляет собой молекулу, в которой различные части антитела получены от различных видов животных, как, например, антитела, имеющие вариабельную область, полученную из моноклонального антитела мыши, и константную область из иммуноглобулина человека. Способы получения химерных антител известны в данной области техники. См., например, Morrison, Science 229: 1202 (1985); Oi et al, BioTechniques 4: 214 (1986); Gillies et al, J. Immunol. Methods 125: 191-202 (1989); патенты США №№5807715; 4816567 и 4816397, которые включены в данный документ посредством ссылки во всей своей полноте. Гуманизированные антитела представляют собой молекулы антител, полученные из антитела от вида, отличного от человека, которое связывается с желаемым антигеном, имеющие один или несколько гипервариабельных участков (CDR) от вида, отличного от человека, и каркасные области из молекулы иммуноглобулина человека. Часто каркасные остатки в каркасных областях от человека замещены соответствующими остатками из антитела, являющегося донором CDR, для изменения, предпочтительно улучшения, связывания с антигеном. Эти замены в каркасных областях идентифицируют с помощью способов, хорошо известных в области техники, например, путем моделирования взаимодействий CDR и каркасных остатков для идентификации каркасных остатков, важных для связывания с антигеном, и сравнения последовательностей для идентификации необычных каркасных остатков в конкретных положениях (см., например, Queen et al, патент США №5585089; Riechmann et al, Nature 332: 323 (1988), которые включены в данный документ посредством ссылки во всей своей полноте). Антитела можно гуманизировать при помощи разнообразных методик, известных в данной области техники, в том числе, например, пересадки CDR (EP 239400; публикация по PCT WO 91/09967; патенты США №№5225539; 5530101 и 5585089), гиперхимеризации или изменения поверхности (EP 592106; EP 519596; Padlan, Molecular Immunology 28 (4/5): 489-498 (1991); Studnicka et al., Protein Engineering 7 (6): 805-814 (1994); Roguska et al., PNAS 91: 969-973 (1994)) и шаффлинга цепей (патент США №5565332).
[0148] Полностью человеческие антитела являются особенно желательными для терапевтического лечения пациентов-людей. Антитела человека можно создать с помощью разнообразных способов, известных в данной области техники, в том числе способов фагового дисплея, описанных выше, применяя библиотеки антител, полученные из последовательностей иммуноглобулина человека. См. также патенты США №4444887 и 4716111 и публикации по PCT WO 98/46645, WO 98/50433, WO 98/24893, WO 98/16654, WO 96/34096, WO 96/33735 и WO 91/10741; все из которых включены в данный документ посредством ссылки во всей своей полноте.
[0149] Антитела человека также можно получать, применяя трансгенных мышей, не способных к экспрессии функциональных эндогенных иммуноглобулинов, но в которых может происходить экспрессия генов иммуноглобулинов человека. Например, генные комплексы тяжелых и легких цепей иммуноглобулинов человека можно вводить случайно или с помощью гомологичной рекомбинации в эмбриональные стволовые клетки мышей. В дополнение, различные компании могут заниматься обеспечением антител человека, получаемых в трансгенных мышах, направленных против выбранного антигена, с применением технологии, подобной описанной выше.
[0150] Полностью человеческие антитела, распознающие выбранный эпитоп, можно образовывать с помощью методики, называемой "управляемой селекцией". В этом подходе выбранное моноклональное антитело, не являющееся антителом человека, например, антитело мыши, применяют для управления селекцией полностью человеческого антитела, распознающего тот же эпитоп (Jespers et al., Bio/Technology 12: 899-903 (1988). См. также патент США №5565332).
[0151] В другом варианте осуществления ДНК, кодирующую желаемые моноклональные антитела, можно без труда выделить и секвенировать с помощью традиционных методов (например, с помощью применения олигонуклеотидных зондов, способных к специфическому связыванию с генами, кодирующими тяжелые и легкие цепи антител мыши). Выделенные и субклонированные клетки гибридомы или выделенный фаг, например, могут служить источником такой ДНК. После выделения ДНК можно поместить в векторы экспрессии, которые затем вводят путем трансфекции в прокариотические или эукариотические клетки-хозяева, такие как клетки E. coli, клетки COS обезьяны, клетки яичников китайского хомячка (CHO) или клетки миеломы, в которых не вырабатываются иммуноглобулины иным путем. Более конкретно, выделенную ДНК (которая может быть синтетической, как описано в данном документе) можно применять для клонирования последовательностей константных и вариабельных областей для производства антител, как описано в патенте США №5658570 Newman et al, заявка на который была подана 25 января 1995 г., включенном в данный документ посредством ссылки. Трансформированные клетки, экспрессирующие желаемое антитело, можно выращивать в относительно больших количествах, обеспечивая поставки иммуноглобулина для клинических и коммерческих нужд.
[0152] В одном варианте осуществления выделенная связывающая молекула, например, антитело, содержит по меньшей мере один CDR тяжелой или легкой цепи молекулы антитела. В другом варианте осуществления выделенная связывающая молекула содержит по меньшей мере два CDR из одной или нескольких молекул антител. В другом варианте осуществления выделенная связывающая молекула содержит по меньшей мере три CDR из одной или нескольких молекул антител. В другом варианте осуществления выделенная связывающая молекула содержит по меньшей мере четыре CDR из одной или нескольких молекул антител. В другом варианте осуществления выделенная связывающая молекула содержит по меньшей мере пять CDR из одной или нескольких молекул антител. В другом варианте осуществления выделенная связывающая молекула согласно настоящему описанию включает по меньшей мере шесть CDR из одной или нескольких молекул антител.
[0153] В конкретном варианте осуществления аминокислотную последовательность вариабельных доменов тяжелых и/или легких цепей можно изучить с целью идентификации последовательностей гипервариабельных участков (CDR) с помощью способов, хорошо известных в данной области техники, например, с помощью сравнения с известными аминокислотными последовательностями других вариабельных областей тяжелой и легкой цепей для определения областей гипервариабельности последовательностей. Применяя стандартные методики рекомбинантных ДНК, можно вставить один или несколько CDR в каркасные области, например, в каркасные области от человека, для гуманизации антитела, не являющегося антителом человека. Каркасные области могут быть встречающимися в природе или консенсусными каркасными областями и предпочтительно каркасными областями от человека (см., например, Chothia et al, J. Mol. Biol. 278-457-479 (1998) в отношении перечня каркасных областей от человека). Полинуклеотид, образованный комбинацией каркасных областей и CDR, кодирует антитело, которое специфически связывается по меньшей мере с одним эпитопом желаемого антигена, например, Psl или PcrV. В каркасных областях могут быть произведены одна или несколько аминокислотных замен, и аминокислотные замены улучшают связывание антитела с его антигеном. Дополнительно, такие способы можно применять для произведения аминокислотных замен или делеций одного или нескольких цистеиновых остатков в вариабельных областях, принимающих участие в образовании дисульфидной связи внутри цепи, с образованием молекул антител, в которых недостает одной или нескольких дисульфидных связей внутри цепи. Другие изменения полинуклеотида охвачены настоящим изобретением и находятся в пределах квалификации специалиста в данной области.
[0154] Также обеспечиваются связывающие молекулы, содержащие таковые, состоящие главным образом из или состоящие из вариантов (включая производные) молекул антител (например, VH-области и/или VL-области), описываемых в данном документе, и эти связывающие молекулы или их фрагменты специфически связываются с Psl или PcrV Pseudomonas. Для внедрения мутаций в нуклеотидную последовательность, кодирующую связывающую молекулу или ее фрагмент, которые специфически связываются с Psl и/или PcrV Pseudomonas, можно применять стандартные методики, известные специалистам в данной области, включая, без ограничений, сайт-направленный мутагенез и ПЦР-опосредованный мутагенез, который приводит в результате к аминокислотным заменам. Варианты (включая производные) кодируют полипептиды, содержащие менее чем 50 аминокислотных замен, менее чем 40 аминокислотных замен, менее чем 30 аминокислотных замен, менее чем 25 аминокислотных замен, менее чем 20 аминокислотных замен, менее чем 15 аминокислотных замен, менее чем 10 аминокислотных замен, менее чем 5 аминокислотных замен, менее чем 4 аминокислотных замены, менее чем 3 аминокислотных замены или менее чем 2 аминокислотных замены по сравнению с эталонной VH-областью, VHCDR1, VHCDR2, VHCDR3, VL-областью, VLCDR1, VLCDR2 или VLCDR3. "Консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток замещается аминокислотным остатком, имеющим боковую цепь с одноименным зарядом. Семейства аминокислотных остатков, имеющих боковые цепи с одноименными зарядами, определены в данной области техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Альтернативно, мутации можно внедрять случайно по всей кодирующей последовательности или в ее части, как, например, путем насыщающего мутагенеза, и полученных в результате мутантов можно подвергнуть скринингу в отношении биологической активности для идентификации мутантов, сохраняющих активность (например, способность к связыванию с Psl или PcrV Pseudomonas).
[0155] Например, возможно внедрять мутации только в каркасные области или только в CDR-области молекулы антитела. Внедренные мутации могут быть молчащими или нейтральными миссенс-мутациями, т.е. не оказывающими или оказывающими небольшой эффект на способность антитела к связыванию с антигеном. Эти типы мутаций могут быть применимыми для оптимизации частоты использования код она или улучшения выработки антител гибридомой. Альтернативно, ненейтральные миссенс-мутации могут изменять способность антитела к связыванию с антигеном. Большинство молчащих и нейтральных миссенс-мутаций, вероятно, локализованы в каркасных областях, тогда как большинство ненейтральных миссенс-мутаций, вероятно, локализованы в CDR, хотя это не является обязательным требованием. Специалист в данной области техники будет способен разработать и протестировать мутантные молекулы с желаемыми свойствами, такими как отсутствие изменения активности связывания с антигеном или изменение активности связывания (например, улучшения активности связывания с антигеном или изменение специфичности антитела). После мутагенеза кодируемый белок может экспрессироваться обычным образом, и функциональную и/или биологическую активность кодируемого белка (например, способность к связыванию по меньшей мере с одним эпитопом Psl или PcrV Pseudomonas) можно определять при помощи методик, описываемых в данном документе, или при помощи методик стандартных модификаций, известных в данной области техники.
[0156] Один вариант осуществления предусматривает биспецифическое антитело, содержащее домен, связывающийся с Psl и PcrV Pseudomonas, раскрытый в данном документе. В некоторых вариантах осуществления биспецифическое антитело содержит первый домен, связывающийся с Psl, и второй домен, связывающийся с PcrV. Предполагаются биспецифические антитела с более чем двумя валентностями. Например, триспецифические антитела могут быть также получены с применением способов, описанных в данном документе (Tutt et al., J. Immunol, 147: 60 (1991)).
[0157] Один вариант осуществления предусматривает способ получения биспецифического антитела, в котором используется одна легкая цепь, которая может образовывать пару с обоими вариабельными доменами тяжелой цепи, присутствующими в биспецифической молекуле. Для идентификации этой легкой цепи могут применяться различные стратегии. В одном варианте осуществления идентифицируют ряд моноклональных антител для каждого антигена, на который может быть нацелено биспецифическое антитело, с последующим определением того, какая из легких цепей, используемых в этих антителах, способна функционировать в паре с тяжелой цепью любого из идентифицированных антител ко второй мишени. Таким образом может быть идентифицирована легкая цепь, которая может функционировать с двумя тяжелыми цепями для обеспечения связывания с обоими антигенами. В другом варианте осуществления дисплейные методики, такие как фаговый дисплей, могут обеспечить идентификацию легкой цепи, которая может функционировать с двумя или более тяжелыми цепями. В одном варианте осуществления конструируют фаговую библиотеку, которая содержит разнообразный репертуар вариабельных доменов тяжелой цепи и один вариабельный домен легкой цепи. Эта библиотека может быть дополнительно использована для идентификации антител, которые связываются с различными антигенами, представляющими интерес. Таким образом, в определенных вариантах осуществления идентифицированные антитела будут иметь общую легкую цепь.
[0158] В определенных вариантах осуществления биспецифическое антитело содержит по меньшей мере один одноцепочечный Fv (scFv). В определенных вариантах осуществления биспецифическое антитело содержит два scFv. Например, scFv может быть слит с одним или обоими содержащими CH3-домен полипептидами, содержащимися в антителе. Некоторые способы включают получение биспецифической молекулы, где одна или обе константные области тяжелой цепи, содержащие по меньшей мере CH3-домен, используются в сочетании с одноцепочечным Fv-доменом для обеспечения связывания антигена.
III. ПОЛИПЕПТИДЫ-АНТИТЕЛА
[0159] Настоящее изобретение дополнительно направлено на выделенные полипептиды, образующие связывающие молекулы, например, антитела или их антиген-связывающие фрагменты, которые специфически связываются с Psl и/или PcrV Pseudomonas, и на полинуклеотиды, кодирующие такие полипептиды. Связывающие молекулы, например, антитела или их фрагменты, раскрытые в данном документе, содержат полипептиды, например, аминокислотные последовательности, кодирующие, например, антиген-связывающие области, специфичные в отношении Psl и/или специфичные в отношении PcrV, полученные из молекул иммуноглобулинов. Полипептид или аминокислотная последовательность, "полученные из" указанного белка, относятся к происхождению полипептида. В определенных случаях полипептид или аминокислотная последовательность, полученные из конкретного исходного полипептида или аминокислотной последовательности, имеют аминокислотную последовательность, главным образом идентичную таковой в исходной последовательности или ее части, где часть состоит по меньшей мере из 10-20 аминокислот, по меньшей мере из 20-30 аминокислот, по меньшей мере из 30-50 аминокислот, или которая может быть иным способом идентифицирована специалистом в данной области техники как происходящая из исходной последовательности.
[0160] Также раскрыта выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая аминокислотную последовательность вариабельной области тяжелой цепи иммуноглобулина (VH), по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичную по отношению к одной или нескольким из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15 или SEQ ID NO: 74, показанным в таблице 2.
[0161] Дополнительно раскрыта выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая аминокислотную последовательность вариабельной области тяжелой цепи иммуноглобулина (VH), идентичную или идентичную за исключением одной, двух, трех, четырех, пяти или более аминокислотных замен по отношению к одной или нескольким из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15 или SEQ ID NO: 74, показанным в таблице 2.
[0162] Некоторые варианты осуществления включают выделенную связывающую молекулу, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащую VH, где одна или несколько VHCDR1-, VHCDR2- или VHCDR3-областей VH по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичны по отношению к одной или нескольким эталонным аминокислотным последовательностям VHCDR1, VHCDR2 или VHCDR3 тяжелой цепи с одним или несколькими из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15 или SEQ ID NO: 74, показанным в таблице 2.
[0163] Дополнительно раскрыта выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая VH, где одна или несколько VHCDR1-, VHCDR2- или VHCDR3-областей VH идентичны или идентичны за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким эталонным аминокислотным последовательностям VHCDR1, VHCDR2 и/или VHCDR3 тяжелой цепи с одним или несколькими из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15 или SEQ ID NO: 74, показанным в таблице 2. Таким образом, согласно данному варианту осуществления VH содержит одну или несколько из VHCDR1, VHCDR2 или VHCDR3, идентичных или идентичных за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким аминокислотным последовательностям VHCDR1, VHCDR2 или VHCDR3, показанным в таблице 3.
[0164] Также раскрыта выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая аминокислотную последовательность вариабельной области легкой цепи иммуноглобулина (VL), по меньшей мере на 80%, 85%, 90%, 95% или 100%) идентичную по отношению к одной или нескольким из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14 или SEQ ID NO: 16, показанным в таблице 2.
[0165] В некоторых вариантах осуществления раскрыта выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая аминокислотную последовательность VL, идентичную или идентичную за исключением одной, двух, трех, четырех, пяти или более аминокислотных замен по отношению к одной или нескольким из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14 или SEQ ID NO: 16, показанным в таблице 2.
[0166] Также обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая VL, где одна или несколько VLCDR1-, VLCDR2- или VLCDR3-областей VL по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичны по отношению к одной или нескольким эталонным аминокислотным последовательностям VLCDR1, VLCDR2 или VLCDR3 легкой цепи с одним или несколькими из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14 или SEQ ID NO: 16, показанным в таблице 2.
[0167] Дополнительно обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая VL, где одна или несколько VLCDR1-, VLCDR2- или VLCDR3-областей VL идентичны или идентичны за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким эталонным аминокислотным последовательностям VLCDR1, VLCDR2 и/или VLCDR3 легкой цепи с одним или несколькими из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14 или SEQ ID NO: 16, показанным в таблице 2. Таким образом, согласно данному варианту осуществления VL содержит одну или несколько из VLCDR1, VLCDR2 или VLCDR3, идентичных или идентичных за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким аминокислотным последовательностям VLCDR1, VLCDR2 или VLCDR3, показанным в таблице 3.
[0168] В других вариантах осуществления выделенное антитело или его антиген-связывающий фрагмент, которые специфически связываются с Psl Pseudomonas, содержат таковые, состоят главным образом из или состоят из аминокислотных последовательностей VH и VL, по меньшей мере на 80%, 85%, 90%), 95% или 100% идентичных по отношению к (a) SEQ ID NO: 1 и SEQ ID NO: 2, соответственно; (b) SEQ ID NO: 3 и SEQ ID NO:2, соответственно, (c) SEQ ID NO: 4 и SEQ ID NO: 2, соответственно; (d) SEQ ID NO: 5 и SEQ ID NO: 6, соответственно; (e) SEQ ID NO: 7 и SEQ ID NO: 8, соответственно; (f) SEQ ID NO: 9 и SEQ ID NO: 10, соответственно; (g) SEQ ID NO: 11 и SEQ ID NO: 12, соответственно; (h) SEQ ID NO: 13 и SEQ ID NO: 14, соответственно; (i) SEQ ID NO: 15 и SEQ ID NO: 16, соответственно; или (j) SEQ ID NO: 74 и SEQ ID NO: 12, соответственно. В определенных вариантах осуществления вышеописанное антитело или его антиген-связывающий фрагмент содержат VH с аминокислотной последовательностью SEQ ID NO: 11 и VL с аминокислотной последовательностью SEQ ID NO: 12. В некоторых вариантах осуществления вышеописанное антитело или его антиген-связывающий фрагмент содержат VH с аминокислотной последовательностью SEQ ID NO: 1 и VL с аминокислотной последовательностью SEQ ID NO: 2. В других вариантах осуществления вышеописанное антитело или его антиген-связывающий фрагмент содержат VH с аминокислотной последовательностью SEQ ID NO: 11 и VL с аминокислотной последовательностью SEQ ID NO: 12.
[0169] В некоторых вариантах осуществления обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая VH иммуноглобулина и VL иммуноглобулина, каждая из которых содержит гипервариабельный участок 1 (CDR1), CDR2 и CDR3, причем CDR1 VH представляет собой PYYWT (SEQ ID NO: 47), CDR2 VH представляет собой YIHSSGYTDYNPSLKS (SEQ ID NO: 48), CDR3 VH выбран из группы, состоящей  представляет собой RASQSIRSHLN (SEQ ID NO: 50), CDR2 VL представляет собой GASNLQS (SEQ ID NO: 51), a CDR3 VL выбран из группы, состоящей из
представляет собой RASQSIRSHLN (SEQ ID NO: 50), CDR2 VL представляет собой GASNLQS (SEQ ID NO: 51), a CDR3 VL выбран из группы, состоящей из  NO:287), причем положения CDR VH и VL соответствуют системе нумерации по Kabat.
NO:287), причем положения CDR VH и VL соответствуют системе нумерации по Kabat.
[0170] В некоторых вариантах осуществления обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая VH иммуноглобулина и VL иммуноглобулина, каждая из которых содержит гипервариабельный участок 1 (CDR1), CDR2 и CDR3, причем CDR1 VH представляет собой PYYWT (SEQ ID NO: 47), CDR2 VH представляет собой YIHSSGYTDYNPSLKS (SEQ ID NO: 48), CDR1 VL представляет собой RASQSIRSHLN (SEQ ID NO: 50), CDR2 VL представляет собой GASNLQS (SEQ ID NO: 51), a CDR3 VH и CDR3 VL содержат, соответственно, ADWDRLRALDI 

[0171] В некоторых вариантах осуществления обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащая VH иммуноглобулина и VL иммуноглобулина, причем VH содержит 


[0172] Кроме того, раскрыт одноцепочечный Fv-фрагмент (scFv) выделенного антитела, который специфически связывается с Psl Pseudomonas ("scFv, связывающийся с Psl"), содержащий формулу VH-L-VL или, альтернативно, VL-L-VH, где L представляет собой линкерную последовательность. В определенных аспектах линкер может содержать (a) [GGGGS]n, где n равно 0, 1, 2, 3, 4 или 5, (b) [GGGG]n, где n равно 0, 1, 2, 3, 4 или 5, или комбинацию (a) и (b). Например, иллюстративный линкер содержит GGGGSGGGGSGGGGSGGGGSGGGGS (SEQ ID NO: 326). В определенных вариантах осуществления линкер дополнительно содержит аминокислоты ala-leu на С-конце линкера. В определенных вариантах осуществления scFv, связывающийся с Psl, содержит аминокислотную последовательность SEQ ID NO: 240, SEQ ID NO: 241, SEQ ID NO: 242, SEQ ID NO: 243, SEQ ID NO: 244, SEQ ID NO: 245, SEQ ID NO: 246, SEQ ID NO: 247, SEQ ID NO: 248, SEQ ID NO: 249, SEQ ID NO: 250, SEQ ID NO: 251, SEQ ID NO: 252, SEQ ID NO: 253, SEQ ID NO: 254 или SEQ ID NO: 262.
[0173] Кроме того, раскрыт одноцепочечный Fv-фрагмент (scFv) выделенного антитела, который специфически связывается с PcrV Pseudomonas ("scFv, связывающийся с PcrV"), содержащий формулу VH-L-VL или, альтернативно, VL-L-VH, где L представляет собой линкерную последовательность. В определенных аспектах линкер может содержать (a) [GGGGS]n, где n равно 0, 1, 2, 3, 4 или 5, (b) [GGGG]n, где n равно 0, 1, 2, 3, 4 или 5, или комбинацию (a) и (b). Например, иллюстративный линкер содержит GGGGSGGGGSGGGGSGGGGSGGGGS (SEQ ID NO:326). В определенных вариантах осуществления линкер дополнительно содержит аминокислоты ala-leu на C-конце линкера.
[0174] Также раскрыта выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с PcrV Pseudomonas, содержащая аминокислотную последовательность вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL) иммуноглобулина, по меньшей мере на 80%, 85%), 90%, 95% или 100% идентичную по отношению к SEQ ID NO: 216 или SEQ ID NO: 217.
[0175] Дополнительно раскрыта выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с PcrV Pseudomonas, содержащая VH, где одна или несколько VHCDR1-, VHCDR2- или VHCDR3-областей VH идентичны или идентичны за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким эталонным аминокислотным последовательностям VHCDR1, VHCDR2 и/или VHCDR3 тяжелой цепи с одним или несколькими из SEQ ID NO: 218-220, показанным в таблице 3. Таким образом, согласно данному варианту осуществления VH содержит одну или несколько из VHCDR1, VHCDR2 или VHCDR3, идентичных или идентичных за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким аминокислотным последовательностям VHCDR1, VHCDR2 или VHCDR3, показанным в таблице 3.
[0176] Дополнительно обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с PcrV Pseudomonas, содержащая VL, где одна или несколько VLCDR1-, VLCDR2- или VLCDR3-областей VL идентичны или идентичны за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким эталонным аминокислотным последовательностям VLCDR1, VLCDR2 и/или VLCDR3 легкой цепи с одним или несколькими из SEQ ID NO: 221-223, показанным в таблице 3. Таким образом, согласно данному варианту осуществления VL содержит одну или несколько из VLCDR1, VLCDR2 или VLCDR3, идентичных или идентичных за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким аминокислотным последовательностям VLCDR1, VLCDR2 или VLCDR3, показанным в таблице 3.
[0177] Также обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с PcrV Pseudomonas, содержащая VH и VL, причем VH содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 255 и SEQ ID NO: 257, и при этом VL содержит аминокислотную последовательность SEQ ID NO: 256.
[0178] Дополнительно обеспечивается выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с PcrV Pseudomonas, содержащая VH и VL, каждая из которых содержит CDR1, CDR2 и CDR3, причем (a) CDR1 VH представляет собой SYAMS (SEQ ID NO: 311) или ее вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены, CDR2 VH представляет собой AISGSGYSTYYADSVKG (SEQ ID NO: 312) или ее вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; и CDR3 VH представляет собой EYSISSNYYYGMDV (SEQ ID NO: 313) или ее вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; или (b) при этом CDR1 VL представляет собой WASQGISSYLA (SEQ ID NO: 314) или ее вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены, CDR2 VL представляет собой AASTLQS (SEQ ID NO: 315) или ее вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены, и CDR3 VL представляет собой QQLNSSPLT (SEQ ID NO: 316) или ее вариант, содержащий 1, 2, 3 или 4 консервативные аминокислотные замены; или (с) имеет место комбинация (a) и (b); причем положения CDR VH и VL соответствуют системе нумерации по Kabat. В определенных аспектах данного варианта осуществления (a) VH содержит аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную по отношению к SEQ ID NO: 317, (b) VL содержит аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную по отношению к SEQ ID NO:318; или (c) имеет место комбинация (a) и (b).
[0179] Также раскрыта выделенная биспецифическая связывающая молекула, например, биспецифическое антитело или его антиген-связывающий фрагмент, которая специфически связывается как с Psl Pseudomonas, так и с PcrV Pseudomonas, содержащая аминокислотную последовательность вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL) иммуноглобулина, по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичную по отношению к SEQ ID NO: 228, SEQ ID NO: 229 или SEQ ID NO: 235.
[0180] В определенных вариантах осуществления биспецифическое антитело, раскрытое в данном документе, имеет структуру Bs1, Bs2, Bs3 или Bs4, все из которых показаны на фиг. 17. В определенных биспецифических антителах, раскрытых в данном документе, связывающий домен, который специфически связывается с Psl Pseudomonas, включает в себя молекулу scFv, связывающуюся с Psl. В других аспектах связывающий домен, который специфически связывается с Psl Pseudomonas, содержит обычную тяжелую цепь и легкую цепь. Аналогично, в определенных биспецифических антителах, раскрытых в данном документе, связывающий домен, который специфически связывается с PcrV Pseudomonas, включает в себя молекулу scFv, связывающуюся с PcrV. В других аспектах связывающий домен, который специфически связывается с PcrV Pseudomonas, содержит обычную тяжелую цепь и легкую цепь.
[0181] В определенных аспектах биспецифическое антитело, раскрытое в данном документе, имеет структуру Bs4, подробно раскрытую в предварительной заявке на патент США №61/624651, поданной 16 апреля 2012 г., и в международной заявке № PCT/US2012/63639, поданной 6 ноября 2012 г. (номер дела патентного поверенного AEMS-115W01, под названием "ПОЛИСПЕЦИФИЧЕСКИЕ И ПОЛИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ"), которые включены в данный документ посредством ссылки во всей своей полноте. Например, настоящее изобретение обеспечивает биспецифическое антитело, в котором молекулу scFv, связывающуюся с Psl, вставляют в шарнирную область каждой тяжелой цепи антитела к PcrV или его фрагмента.
[0182] Настоящее изобретение обеспечивает выделенную связывающую молекулу, например, биспецифическое антитело, содержащую тяжелую цепь антитела и легкую цепь антитела, причем тяжелая цепь антитела содержит формулу VH-CH1-H1-L1-S-L2-H2-CH2-CH3, причем CH1 представляет собой домен-1 константной области тяжелой цепи, H1 представляет собой первый фрагмент шарнирной области тяжелой цепи, L1 представляет собой первый линкер, S представляет собой молекулу scFv, связывающуюся с PcrV, L2 представляет собой второй линкер, H2 представляет собой второй фрагмент шарнирной области тяжелой цепи, CH2 представляет собой домен-2 константной области тяжелой цепи, а CH3 представляет собой домен-3 константной области тяжелой цепи. В определенных аспектах VH содержит аминокислотную последовательность SEQ ID NO: 255, SEQ ID NO: 257 или SEQ ID NO: 317. В определенных аспектах L1 и L2 являются одинаковыми или различными и независимо содержат (a) [GGGGS]n, где n равно 0, 1, 2, 3, 4 или 5, (b) [GGGG]n, где n равно 0, 1, 2, 3, 4 или 5, или комбинацию (a) и (b). В некоторых вариантах осуществления H1 содержит EPKSC (SEQ ID NO: 320), а H2 содержит DKTHTCPPCP (SEQ ID NO: 321).
[0183] В определенных аспектах S включает в себя молекулу scFv, связывающуюся с Psl, имеющую аминокислотную последовательность SEQ ID NO: 240, SEQ ID NO: 241, SEQ ID NO: 242, SEQ ID NO: 243, SEQ ID NO: 244, SEQ ID NO: 245, SEQ ID NO: 246, SEQ ID NO: 247, SEQ ID NO: 248, SEQ ID NO: 249, SEQ ID NO: 250, SEQ ID NO: 251, SEQ ID NO: 252, SEQ ID NO: 253, SEQ ID NO: 254, или SEQ ID NO: 262, или любую комбинацию двух или более этих аминокислотных последовательностей.
[0184] В дополнительных аспектах CH2-CH3 содержит (SEQ ID NO: 322), причем X1 представляет собой М или Y, X2 представляет собой S или T, а X3 представляет собой T или E. В дополнительных аспектах легкая цепь антитела содержит VL-CL, причем CL представляет собой константную область легкой цепи каппа-типа антитела или константную область легкой цепи лямбда-типа антитела. В дополнительных аспектах VL содержит аминокислотную последовательность SEQ ID NO:256 или SEQ ID NO: 318. CL может содержать, например, аминокислотную последовательность SEQ ID NO: 323.
[0185] Дополнительно обеспечивается выделенная связывающая молекула, например, биспецифическое антитело, которая специфически связывается как с Psl Pseudomonas, так и с PcrV Pseudomonas, содержащая VH, содержащую аминокислотную последовательность SEQ ID NO: 264, и VL, содержащую аминокислотную последовательность SEQ ID NO: 263.
[0186] В некоторых вариантах осуществления биспецифические антитела по настоящему изобретению могут представлять собой тандемы одноцепочечных (sc) Fv-фрагментов, которые содержат два различных scFv-фрагмента (т.е. V2L2 и W4), ковалентно связанных вместе с помощью линкера (например, полипептидного линкера). (Ren-Heidenreich et al Cancer 700: 1095-1103 (2004); Korn et al. J Gene Med 6:642-651 (2004)). В некоторых вариантах осуществления линкер может содержать или представлять собой всю константную область тяжелой цепи полипептида или ее часть, такую как CH1-домен. В некоторых вариантах осуществления два фрагмента антител могут быть ковалентно связаны вместе с помощью линкера полиглицин-серин или полисерин-глицин, как описано, например, в патентах США №№7112324 и 5525491, соответственно. Способы создания биспецифических антител с тандемными scFv, описаны, например, в Maletz et al. Int J Cancer 93: 409-416 (2001); и Honemann et al Leukemia 75: 636-644 (2004). Альтернативно, антитела могут представлять собой "линейные антитела", описанные, например, в Zapata et al. Protein Eng. 5: 1057-1062 (1995). Вкратце, эти антитела содержат пару тандемных Fd-сегментов (VH-CH1-VH-CH1), которые образуют пару антиген-связывающих участков.
[0187] Настоящее изобретение также охватывает различные формы таких биспецифических антител, как молекулы тетравалентного иммуноглобулина с двойным вариабельным доменом (DVD-Ig), описанные в Wu et al. (2007) Nat Biotechnol 25 (11): 1290-1297. Молекулы DVD-Ig разработаны так, что два разных вариабельных домена легкой цепи (VL) из двух разных родительских антител связаны в тандем, за которым расположен константный домен легкой цепи, непосредственно или через короткий линкер с помощью методик рекомбинантной ДНК. Например, легкая цепь полипептида DVD-Ig может содержать в тандеме (a) VL от V2L2 и (b) VL от WapR-004. Аналогично, тяжелая цепь содержит два различных вариабельных домена тяжелой цепи (VH), связанные в тандем, за которым расположены константный CH1-домен и Fc-область. Например, тяжелая цепь полипептида DVD-Ig может содержать в тандеме (a) VH от V2L2 и (b) VH от WapR-004. В этом случае экспрессия двух цепей в клетке приводит к образованию гетеротетрамера, содержащего четыре антиген-связывающих участка, два из которых специфически связываются с V2L2 и два из которых специфически связываются с Psl. Способы создания молекул DVD-Ig из двух родительских антител дополнительно описаны, например, в публикации по PCT WO 2008/024188 и WO 2007/024715.
[0188] В определенных вариантах осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, описываемые в данном документе, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся константой диссоциации (KD), не превышающей 5×10-2 М, 10-2 М, 5×10-3 М, 10-3 М, 5×10-4 М, 10-4 М, 5×10-5 М, 10-5 М, 5×10-6 М, 10-6 М, 5×10-7 М, 10-7 М, 5×10-8 М, 10-8 М, 5×10-9 М, 10-9 М, 5×10-10 М, 10-10 М, 5×10-11 М, 10-1 М, 5×10-12 М, 10-12 М, 5×10-13 М, 10-13 М, 5×10-14 М, 10-14 М, 5×10-15 М или 10-15 М.
[0189] В конкретных вариантах осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, описываемые в данном документе, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся константой диссоциации (KD) в диапазоне от около 1×10-10 до около 1×10-6 М. В одном варианте осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, описываемые в данном документе, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся KD, равной около 1,18×10-7 М, как определено в анализе связывания при помощи OCTET®, описываемом в данном документе. В другом варианте осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, описываемые в данном документе, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся KD, равной около 1,44×10-7 М, как определено в анализе связывания при помощи OCTET®, описываемом в данном документе.
[0190] Некоторые варианты осуществления включают выделенные связывающие молекулы, например, антитело или его фрагмент, описываемые выше, которые (a) могут ингибировать прикрепление Pseudomonas aeruginosa к эпителиальным клеткам, (b) могут способствовать ОРК P. aeruginosa или (c) могут ингибировать прикрепление P. aeruginosa к эпителиальным клеткам и могут способствовать ОРК P. aeruginosa.
[0191] В некоторых вариантах осуществления включена выделенная связывающая молекула, например, антитело или его фрагмент, описываемые выше, где максимальное ингибирование прикрепления P. aeruginosa к эпителиальным клеткам достигается при концентрации антитела, равной около 50 мкг/мл или менее, 5,0 мкг/мл или менее или около 0,5 мкг/мл или менее, или при концентрации антитела, находящейся в диапазоне от около 30 мкг/мл до около 0,3 мкг/мл, или при концентрации антитела, равной около 1 мкг/мл, или при концентрации антитела, равной около 0,3 мкг/мл.
[0192] Определенные варианты осуществления включают выделенную связывающую молекулу, например, антитело или его фрагмент, описываемые выше, где EC50 ОРК составляет менее чем около 0,5 мкг/мл, менее чем около 0,05 мкг/мл, или менее чем около 0,005 мкг/мл, или где EC50 ОРК находится в диапазоне от приблизительно 0,001 мкг/мл до около 0,5 мкг/мл, или где EC50 ОРК находится в диапазоне от около 0,02 мкг/мл до около 0,08 мкг/мл, или где EC50 ОРК находится в диапазоне от около 0,002 мкг/мл до около 0,01 мкг/мл, или где EC50 ОРК составляет менее чем около 0,2 мкг/мл, или где EC50 ОРК составляет менее чем около 0,02 мкг/мл. В определенных вариантах осуществления молекула, связывающаяся с Psl Pseudomonas, например, антитело или его фрагмент, вариант или производное, описываемые в данном документе, специфически связывается с тем же эпитопом Psl, что и моноклональное антитело WapR-004, WapR-004RAD, Cam-003, Cam-004 или Cam-005, или будет осуществлять конкурентное ингибирование связывания такого моноклонального антитела с Psl Pseudomonas.
WapR-004RAD является идентичным по отношению к WapR-004 за исключением аминокислотной замены G98A в аминокислотной последовательности SEQ ID NO: 11 VH.
[0193] Некоторые варианты осуществления включают мутантные WapR-004 (W4), содержащие аминокислотную последовательность молекулы scFv-Fc, идентичную или идентичную за исключением одной, двух, трех, четырех, пяти или более аминокислотных замен по отношению к одной или нескольким из SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93, SEQ ID NO: 94, SEQ ID NO: 95, SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103, SEQ ID NO: 104, SEQ ID NO: 105, SEQ ID NO: 106, SEQ ID NO: 107, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ID NO: 110, SEQ ID NO: 111, SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, SEQ ID NO: 145 или SEQ ID NO: 146.
[0194] Другие варианты осуществления включают мутантные WapR-004 (W4), содержащие аминокислотную последовательность молекулы scFv-Fc, по меньшей мере на 80%, 85%), 90%, 95% или 100%» идентичную по отношению к одной или нескольким из SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93, SEQ ID NO: 94, SEQ ID NO: 95, SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103, SEQ ID NO: 104, SEQ ID NO: 105, SEQ ID NO: 106, SEQ ID NO: 107, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ID NO: 110, SEQ ID NO: 111, SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, SEQ ID NO: 145 или SEQ ID NO: 146.
[0195] В некоторых вариантах осуществления молекула, связывающаяся с Psl Pseudomonas, например, антитело или его фрагмент, вариант или производное, описываемые в данном документе, специфически связывается с тем же эпитопом, что и моноклональное антитело WapR-001, WapR-002 или WapR-003, или будет осуществлять конкурентное ингибирование связывания такого моноклонального антитела с Psl Pseudomonas.
[0196] В определенных вариантах осуществления молекула, связывающаяся с Psl Pseudomonas, например, антитело или его фрагмент, вариант или производное, описываемые в данном документе, специфически связывается с тем же эпитопом, что и моноклональное антитело WapR-016, или будет осуществлять конкурентное ингибирование связывания такого моноклонального антитела с Psl Pseudomonas.





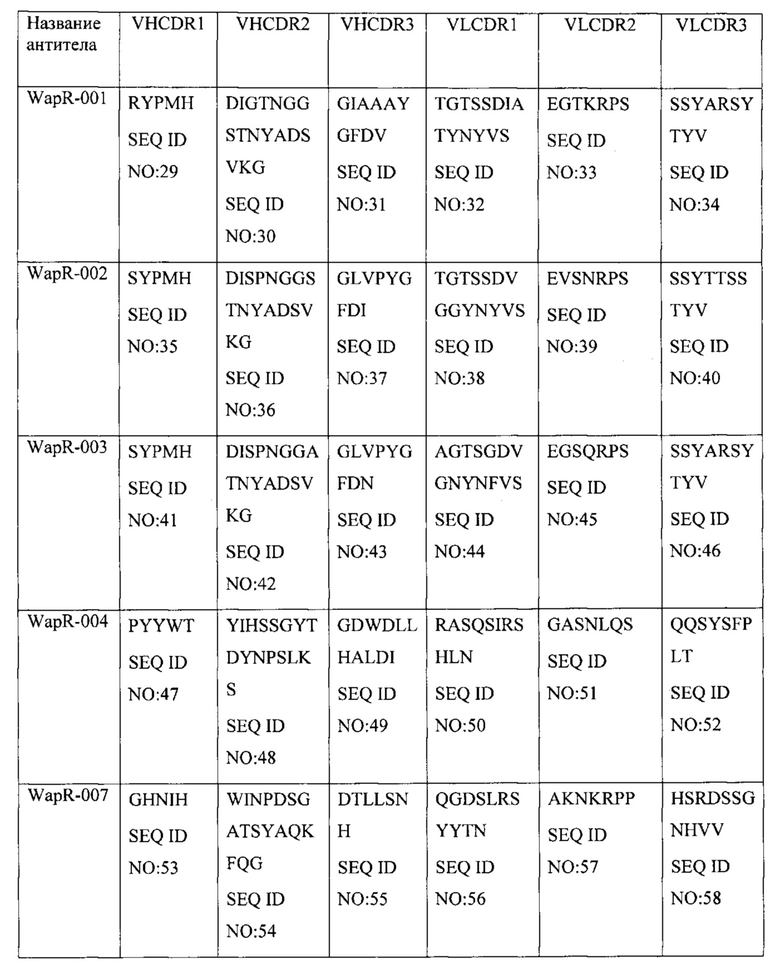
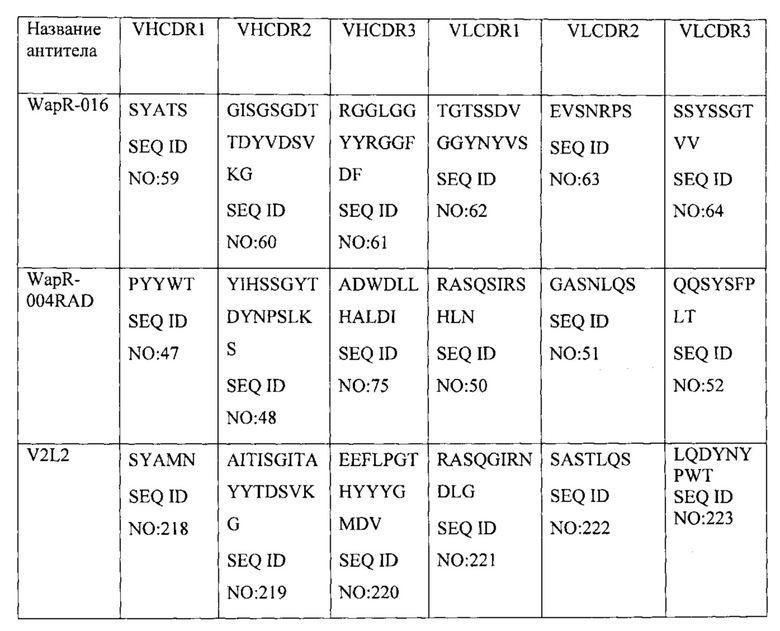
[0197] В определенных вариантах осуществления молекула, связывающаяся с PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, описываемые в данном документе, специфически связывается с тем же эпитопом PcrV, что и моноклональное антитело V2L2, и/или будет осуществлять конкурентное ингибирование связывания такого моноклонального антитела с PcrV Pseudomonas.
[0198] Например, в определенных аспектах молекула, связывающаяся с PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, включает в себя V2L2-GL и/или V2L2-MD.
[0199] В определенных вариантах осуществления молекула, связывающаяся с PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, описываемые в данном документе, специфически связывается с тем же эпитопом PcrV, что и моноклональное антитело 29D2, и/или будет осуществлять конкурентное ингибирование связывания такого моноклонального антитела с PcrV Pseudomonas.
[0200] Любые молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, описываемые в данном документе, могут дополнительно содержать дополнительные полипептиды, например, сигнальный пептид, для управления секрецией кодируемого полипептида, константные области антител, описываемые в данном документе, или другие гетерологичные полипептиды, описываемые в данном документе. Дополнительно, связывающие молекулы или их фрагменты согласно настоящему описанию содержат фрагменты полипептидов, описываемые в другом месте. Дополнительно, молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, описываемые в данном документе, могут представлять собой полипептиды слияния, Fab-фрагменты, scFv или другие производные, описываемые в данном документе.
[0201] Также, как более подробно рассматривается в другом месте в данном документе, настоящее изобретение включает композиции, содержащие молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, описываемые в данном документе.
[0202] Специалисту в данной области также будет понятно, что молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, описываемые в данном документе, можно модифицировать таким образом, чтобы они отличались по аминокислотной последовательности от встречающихся в природе связывающих полипептидов, из которых они были получены. Например, последовательность полипептида или аминокислотная последовательность, полученные от указанного белка, могут быть подобными, например, обладать определенной процентной идентичностью, по отношению к исходной последовательности, например, они могут быть на 60%, 70%, 75%, 80%), 85%), 90% или 95%) идентичными по отношению к исходной последовательности.
[0203] Как известно в данной области техники, "идентичность последовательностей" между двумя полипептидами определяют путем сравнения аминокислотной последовательности одного полипептида с последовательностью второго полипептида. При помощи способов и компьютерных программ/программного обеспечения, известных в данной области техники, таких как, без ограничений, программа BESTFIT (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Мэдисон, Висконсин 53711) можно определить, является ли любой конкретный полипептид по меньшей мере на около 70%, 75%, 80%), 85%, 90% или 95% идентичным по отношению к другому полипептиду, если это обсуждается в данном документе. В BESTFIT применяется алгоритм поиска локальной гомологии Смита-Уотермана, приведенный в Advances in Applied Mathematics 2: 482-489 (1981), для поиска наилучшего сегмента гомологии между двумя последовательностями. При применении BESTFIT или любой другой программы для выравнивания последовательностей для определения того, является ли конкретная последовательность, например, на 95% идентичной по отношению к эталонной последовательности, параметры устанавливают, разумеется, таким образом, чтобы процентное значение идентичности рассчитывалось по всей длине эталонной последовательности полипептида, и чтобы допускалось до 5% гэпов в гомологии от общего числа аминокислот в эталонной последовательности.
[0204] Процент "идентичности последовательностей" также можно определить путем сравнения двух последовательностей, подвергнутых оптимальному выравниванию, в окне сравнения. Чтобы произвести оптимальное выравнивание последовательности для сравнения, часть последовательности полинуклеотида или полипептида в окне сравнения может содержать добавления или делеции, называемые гэпами, в то время как эталонная последовательность сохраняется неизменной. Оптимальное выравнивание представляет собой такое выравнивание, в котором даже при наличии гэпов образуется наибольшее возможное число "идентичных" положений между эталонной и сравниваемой последовательностями. Процент "идентичности последовательностей" между двумя последовательностями можно определить при помощи версии программы "BLAST 2 Sequences", которая была доступна из Национального центра биотехнологической информации по состоянию на 1 сентября 2004 г., каковая программа включает в себя программы BLASTN (для сравнения нуклеотидных последовательностей) и BLASTP (для сравнения полипептидных последовательностей), каковые программы основаны на алгоритме Карлина-Альтшуля (Proc. Natl. Acad. Sci. USA 90 (12): 5873-5877, 1993). При использовании "BLAST 2 Sequences" для длины слова (3), штрафа за открытие гэпа (11), штрафа за продолжение гэпа (1), величины максимального продления с использованием гэпов (50), ожидаемого значения (10) и каких-либо других необходимых параметров, включая, без ограничения, опцию "matrix", можно применять параметры, которые были параметрами по умолчанию по состоянию на 1 сентября 2004 г.
[0205] Кроме того, можно произвести нуклеотидные или аминокислотные замены, делеции или вставки, приводящие к консервативным заменам или изменениям в областях "несущественных" аминокислот. Например, последовательность полипептида или аминокислотная последовательность, полученные из указанного белка, могут быть идентичными по отношению к исходной последовательности за исключением одной или нескольких отдельных аминокислотных замен, вставок или делеций, например, одной, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, пятнадцати, двадцати или более отдельных аминокислотных замен, вставок или делеций. В некоторых вариантах осуществления в последовательности полипептида или аминокислотной последовательности, полученных из указанного белка, имеется от одной до пяти, от одной до десяти, от одной до пятнадцати или от одной до двадцати отдельных аминокислотных замен, вставок или делеций по сравнению с исходной последовательностью.
[0206] Молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, описываемые в данном документе, может содержать белок слияния, состоять главным образом из или состоять из такового. Белки слияния являются химерными молекулами, которые содержат, например, антиген-связывающий домен иммуноглобулина по меньшей мере с одним целевым участком связывания и по меньшей мере с одной гетерологичной частью, т.е. частью, с которой он не связан естественным образом в природе. Аминокислотные последовательности могут обычно существовать в отдельных белках, которые сближены в полипептиде слияния, или они могут обычно существовать в одном и том же белке, но размещаться в полипептиде слияния в новом расположении. Белки слияния можно создать, например, путем химического синтеза или путем создания и трансляции полинуклеотида, в котором пептидные области кодируются в желаемом взаиморасположении.
[0207] Выражение "гетерологичный" применительно к полинуклеотиду, полипептиду или другому фрагменту означает, что полинуклеотид, полипептид или другой фрагмент получены из объекта, отличного от остальной части объекта, с которым их сравнивают. В неограничивающем примере "гетерологичный полипептид", подлежащий слиянию со связывающей молекулой, например, с антителом или его антиген-связывающим фрагментом, вариантом или производным, получен из полипептида, не являющегося иммуноглобулином, того же вида, или полипептида, являющегося иммуноглобулином или не являющегося иммуноглобулином, другого вида.
IV. БЕЛКИ СЛИЯНИЯ И КОНЪЮГАТЫ АНТИТЕЛ
[0208] В некоторых вариантах осуществления молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, можно многократно вводить в конъюгированной форме. В еще одном варианте осуществления молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, можно вводить в неконъюгированной форме, а затем в конъюгированной форме, или наоборот.
[0209] В конкретных вариантах осуществления молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, могут быть конъюгированы с одним или несколькими противомикробными средствами, например, с полимиксином B (PMB). PMB представляет собой небольшой липопептидный антибиотик, одобренный для лечения инфекций, вызываемых грамотрицательными микроорганизмами с множественной лекарственной устойчивостью. В дополнение к его бактерицидной активности, РМВ связывается с липополисахаридом (LPS) и нейтрализует его провоспалительные эффекты. (Dixon, R.A. & Chopra, I.J Antimicrob Chemother 18, 557-563 (1986)). Считается, что LPS в значительной мере содействует воспалению и началу сепсиса, вызываемого грамотрицательными микроорганизмами (Guidet, B., et al, Chest 106, 1194-1201 (1994)). Как было показано, конъюгаты PMB с молекулами носителя нейтрализуют LPS и опосредуют защиту в моделях эндотоксикоза и инфекции на животных (Drabick, J.J., et al. Antimicrob Agents Chemother 42, 583-588 (1998)). Также раскрыт способ присоединения одной или нескольких молекул РМВ к цистеиновым остаткам, внедренным в Fc-область моноклональных антител (mAb) согласно настоящему изобретению. Например, конъюгаты Cam-003-PMB сохраняли специфичное mAb-опосредованное связывание с P. aeruginosa и также сохраняли ОРК-активность. Кроме того, конъюгаты mAb-PMB связывались с LPS и нейтрализовали его in vitro. В некоторых вариантах осуществления молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, могут быть объединены с антибиотиками (например, ципрофлоксацином, меропенемом, тобрамицином, азтреонамом).
[0210] В некоторых вариантах осуществления молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, описываемые в данном документе, может содержать гетерологичную аминокислотную последовательность или один или несколько других фрагментов, обычно не связанных с антителом (например, противомикробное средство, терапевтическое средство, пролекарство, пептид, белок, фермент, липид, модификатор биологического ответа, фармацевтическое средство, лимфокин, гетерологичное антитело или его фрагмент, детектируемую метку, полиэтиленгликоль (PEG) и комбинацию любых двух или более из указанных средств). В дополнительных вариантах осуществления молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, может содержать детектируемую метку, выбранную из группы, состоящей из фермента, флуоресцентной метки, хемилюминесцентной метки, биолюминесцентной метки, радиоактивной метки или комбинации любых двух или более из указанных детектируемых меток.
V. ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ
[0211] В данном документе также обеспечиваются молекулы нуклеиновых кислот, кодирующие молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, описываемые в данном документе.
[0212] В одном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей аминокислотную последовательность вариабельной области тяжелой цепи (VH) иммуноглобулина, по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичную по отношению к одной или нескольким из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ IS NO: 74 или SEQ ID NO:216, показанным в таблице 2.
[0213] В одном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность SEQ ID NO: 257 или SEQ ID NO: 259 вариабельной области тяжелой цепи (VH) иммуноглобулина, состоящий главным образом из или состоящий из таковой. Например, последовательности нуклеиновых кислот SEQ ID NO: 261 и SEQ ID NO: 259, соответственно.
[0214] В другом варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей аминокислотную последовательность VH, идентичную или идентичную за исключением одной, двух, трех, четырех, пяти или более аминокислотных замен по отношению к одной или нескольким из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 74 или SEQ ID NO: 216, показанным в таблице 2.
[0215] В дополнительном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий нуклеиновую кислоту, кодирующую VH, состоящий главным образом из или состоящий из таковой, где одна или несколько VHCDR1-, VHCDR2- или VHCDR3-областей VH идентичны или идентичны за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким эталонным аминокислотным последовательностям VHCDR1, VHCDR2 и/или VHCDR3 тяжелой цепи с одним или несколькими из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 74 или SEQ ID NO: 216, показанным в таблице 2.
[0216] В другом варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей выделенную связывающую молекулу, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащую VH, где одна или несколько VHCDR1-, VHCDR2- или VHCDR3-областей VH идентичны или идентичны за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким эталонным аминокислотным последовательностям VHCDR1, VHCDR2 и/или VHCDR3 тяжелой цепи с одним или несколькими из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15 или SEQ ID NO: 74, показанным в таблице 2.
[0217] В дополнительном варианте осуществления обеспечивается выделенная связывающая молекула, например, антитело или антиген-связывающий фрагмент, содержащая VH, кодируемая полинуклеотидом, которая специфически или преимущественно связывается с Psl и/или PcrV Pseudomonas.
[0218] В другом варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей аминокислотную последовательность вариабельной области легкой цепи (VL) иммуноглобулина, по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичную по отношению к одной или нескольким из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16 или SEQ ID NO: 217, показанным в таблице 2.
[0219] В другом варианте осуществления обеспечивается выделенный полинуклеотид, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность SEQ ID NO: 256 вариабельной области легкой цепи (VL) иммуноглобулина, т.е. последовательность нуклеиновой кислоты SEQ ID NO: 260, состоящий главным образом из или состоящий из таковой.
[0220] В дополнительном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей аминокислотную последовательность VL, идентичную или идентичную за исключением одной, двух, трех, четырех, пяти или более аминокислотных замен по отношению к одной или нескольким из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16 или SEQ ID NO: 217, показанным в таблице 2.
[0221] В другом варианте осуществления обеспечивается выделенный полинуклеотид, содержащий нуклеиновую кислоту, кодирующую VL, состоящий главным образом из или состоящий из таковой, где одна или несколько VLCDR1-, VLCDR2- или VLCDR3-областей VL по меньшей мере на 80%, 85%, 90%, 95% или 100%) идентичны по отношению к одной или нескольким эталонным аминокислотным последовательностям VLCDR1, VLCDR2 или VLCDR3 легкой цепи с одним или несколькими из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14 или SEQ ID NO: 16 или SEQ ID NO:217, показанным в таблице 2.
[0222] В дополнительном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей выделенную связывающую молекулу, например, антитело или его антиген-связывающий фрагмент, которая специфически связывается с Psl Pseudomonas, содержащую VL, где одна или несколько VLCDR1-, VLCDR2- или VLCDR3-областей VL идентичны или идентичны за исключением четырех, трех, двух или одной аминокислотной замены по отношению к одной или нескольким эталонным аминокислотным последовательностям VLCDR1, VLCDR2 и/или VLCDR3 легкой цепи с одним или несколькими из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16 или SEQ ID NO: 217, показанным в таблице 2.
[0223] В другом варианте осуществления обеспечивается выделенная связывающая молекула, например, антитело или антиген-связывающий фрагмент, содержащая VL, кодируемая полинуклеотидом, которая специфически или преимущественно связывается с Psl и/или PcrV Pseudomonas.
[0224] В одном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей молекулу scFv, содержащую VH и VL, где scFv по меньшей мере на 80%, 85%, 90%, 95% или 100% идентична по отношению к одной или нескольким из SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69 или SEQ ID NO: 70, показанным в таблице 4.


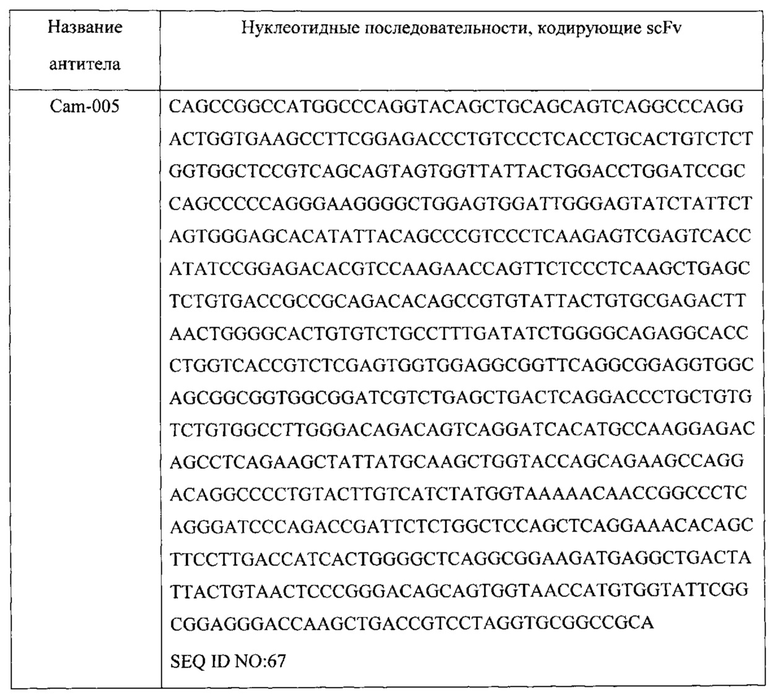

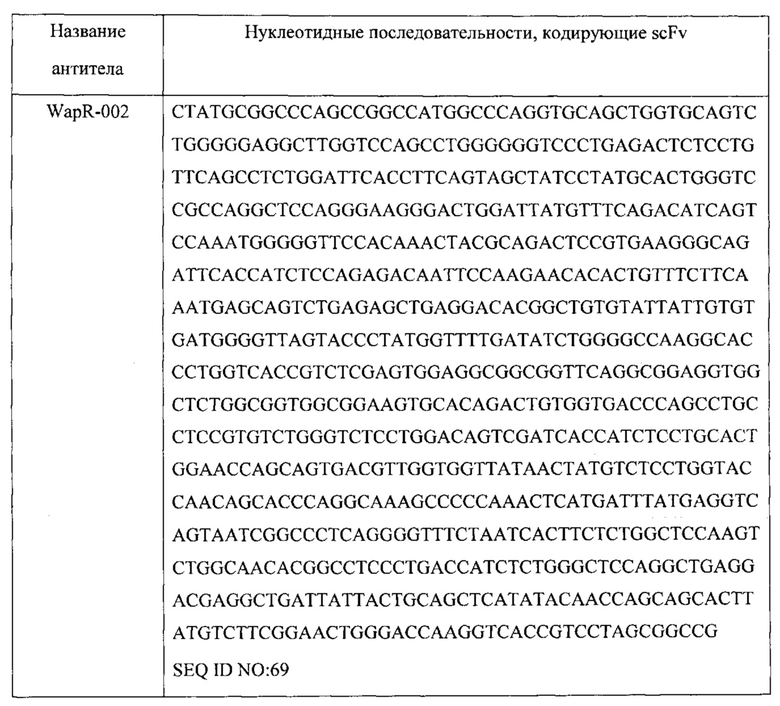





[0225] В некоторых вариантах осуществления выделенное антитело или его антиген-связывающий фрагмент, кодируемые одним или несколькими вышеописанными полинуклеотидами, которые специфически связываются с Psl и/или PcrV Pseudomonas, содержат таковые, состоят главным образом из или состоят из аминокислотных последовательностей VH и VL, по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичных по отношению к
(a) SEQ ID NO: 1 и SEQ ID NO: 2, соответственно; (b) SEQ ID NO: 3 и SEQ ID NO: 2, соответственно; (c) SEQ ID NO: 4 и SEQ ID NO: 2, соответственно; (d) SEQ ID NO: 5 и SEQ ID NO: 6, соответственно; (e) SEQ ID NO: 7 и SEQ ID NO: 8, соответственно; (f) SEQ ID NO: 9 и SEQ ID NO: 10, соответственно; (g) SEQ ID NO: 11 и SEQ ID NO: 12, соответственно; (h) SEQ ID NO: 13 и SEQ ID NO: 14, соответственно; (i) SEQ ID NO: 15 и SEQ ID NO: 16, соответственно; или (j) SEQ ID NO: 74 и SEQ ID NO: 12, соответственно.
[0226] В определенных вариантах осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, кодируемая одним или несколькими полинуклеотидами, описанными выше, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся константой диссоциации (KD), не превышающей 5×10-2 М, 10-2 М, 5×10-3 М, 10-3 М, 5×10-4 М, 10-4 М, 5×10-5 М, 10-5 М, 5×10-6 М, 10-6 М, 5×10-7 М, 10-7 М, 5×10-8 М, 10-8 М, 5×10-9 М, 10-9 М, 5×10-10 М, 10-10 М, 5×10-11 М, 10-11 М, 5×10-12 М, 10-12 М, 5×10-13 М, 10-13 М, 5×10-14 М, 10-14 М, 5×10-15 М или 10-15 М.
[0227] В конкретных вариантах осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, кодируемая одним или несколькими полинуклеотидами, описанными выше, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся константой диссоциации (KD) в диапазоне от около 1×10-10 до около 1×10-6 М. В одном варианте осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, кодируемая одним или несколькими полинуклеотидами, описанными выше, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся KD, равной около 1,18×10-7 М, как определено в анализе связывания при помощи OCTET®, описываемом в данном документе. В другом варианте осуществления выделенная связывающая молекула, например, антитело или его антиген-связывающий фрагмент, кодируемая одним или несколькими полинуклеотидами, описанными выше, специфически связывается с Psl и/или PcrV Pseudomonas с аффинностью, характеризующейся KD, равной около 1,44×10-7 М, как определено в анализе связывания при помощи OCTET®, описываемом в данном документе.
[0228] В определенных вариантах осуществления молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, кодируемая одним или несколькими полинуклеотидами, описанными выше, специфически связывается с тем же эпитопом Psl, что и моноклональные антитела WapR-004, WapR-004RAD, Cam-003, Cam-004 или Cam-005, или будет осуществлять конкурентное ингибирование связывания таких моноклональных антител с Psl Pseudomonas; и/или специфически связывается с тем же эпитопом PcrV, что и моноклональное антитело V2L2, или будет осуществлять конкурентное ингибирование связывания такого моноклонального антитела с PcrV Pseudomonas. WapR-004RAD является идентичным по отношению к WapR-004 за исключением замены нуклеиновой кислоты G293C в последовательности нуклеиновой кислоты, соответствующей VH, кодирующей аминокислотную последовательность VH с SEQ ID NO: 11 (замена нуклеотида в части SEQ ID NO: 71, кодирующей VH, в положении 317). Последовательность нуклеиновой кислоты, кодирующая VH WapR-004RAD, представлена как SEQ ID NO: 76.
[0229] В некоторых вариантах осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей аминокислотную последовательность молекулы W4 с мутацией в scFv-Fc, идентичную или идентичную за исключением одной, двух, трех, четырех, пяти или более аминокислотных замен по отношению к одной или нескольким из SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93, SEQ ID NO: 94, SEQ ID NO: 95, SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103, SEQ ID NO: 104, SEQ ID NO: 105, SEQ ID NO: 106, SEQ ID NO: 107, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ID NO: 110, SEQ ID NO: 111, SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, SEQ ID NO: 145 или SEQ ID NO: 146.
[0230] В других вариантах осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, кодирующей аминокислотную последовательность молекулы W4 с мутацией в scFv-Fc, по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичную по отношению к одной или нескольким из SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93, SEQ ID NO: 94, SEQ ID NO: 95, SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103, SEQ ID NO: 104, SEQ ID NO: 105, SEQ ID NO: 106, SEQ ID NO: 107, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ID NO: 110, SEQ ID NO: 111, SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, SEQ ID NO: 145 или SEQ ID NO: 146.
[0231] В одном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, которая кодирует молекулу W4 с мутацией в scFv-Fc, где нуклеиновая кислота по меньшей мере на 80%, 85%, 90%, 95% или 100%) идентична по отношению к одной или нескольким из SEQ ID NO: 147, SEQ ID NO: 148, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151 или SEQ ID NO: 152, SEQ IS NO: 153, SEQ ID NO: 154, SEQ ID NO: 155, SEQ ID NO: 156, SEQ ID NO: 157, SEQ ID NO: 158, SEQ ID NO: 159, SEQ ID NO: 160, SEQ ID NO: 161, SEQ ID NO: 162, SEQ ID NO: 163, SEQ ID NO: 164, SEQ ID NO: 165, SEQ ID NO: 166, SEQ ID NO: 167, SEQ ID NO: 168, SEQ ID NO: 169, SEQ ID NO: 170, SEQ ID NO: 171, SEQ ID NO: 172, SEQ ID NO: 173, SEQ ID NO: 174, SEQ ID NO: 175, SEQ ID NO: 176, SEQ ID NO: 177, SEQ ID NO: 178, SEQ ID NO: 179, SEQ ID NO: 180, SEQ ID NO: 181, SEQ ID NO: 182, SEQ ID NO: 183, SEQ ID NO: 184, SEQ ID NO: 185, SEQ ID NO: 186, SEQ ID NO: 187, SEQ ID NO: 188, SEQ ID NO: 189, SEQ ID NO: 190, SEQ ID NO: 191, SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194, SEQ ID NO: 195, SEQ ID NO: 196, SEQ ID NO: 197, SEQ ID NO: 198, SEQ ID NO: 199, SEQ ID NO: 200, SEQ ID NO: 201, SEQ ID NO: 202, SEQ ID NO: 203, SEQ ID NO: 204, SEQ ID NO: 205, SEQ ID NO: 206, SEQ ID NO: 207, SEQ ID NO: 208, SEQ ID NO: 209, SEQ ID NO: 210, SEQ ID NO: 211, SEQ ID NO: 212, SEQ ID NO: 213, SEQ ID NO: 214 или SEQ ID NO: 215.
[0232] В одном варианте осуществления обеспечивается выделенный полинуклеотид, содержащий таковую, состоящий главным образом из или состоящий из нуклеиновой кислоты, которая кодирует полипептид V2L2, где нуклеиновая кислота по меньшей мере на 80%, 85%, 90%, 95% или 100% идентична по отношению к одной или нескольким из SEQ ID NO: 238 или SEQ ID NO: 239.
[0233] В других вариантах осуществления молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, кодируемая одним или несколькими полинуклеотидами, описанными выше, специфически связывается с тем же эпитопом, что и моноклональные антитела WapR-001, WapR-002 или WapR-003, или будет осуществлять конкурентное ингибирование связывания таких моноклональных антител с Psl Pseudomonas.
[0234] В определенных вариантах осуществления молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, кодируемая одним или несколькими полинуклеотидами, описанными выше, специфически связывается с тем же эпитопом, что и моноклональное антитело WapR-016, или будет осуществлять конкурентное ингибирование связывания такого моноклонального антитела с Psl Pseudomonas.
[0235] Настоящее изобретение также включает фрагменты полинуклеотидов, описанных в другом месте в данном документе. Дополнительно также обеспечиваются полинуклеотиды, которые кодируют полипептиды слияния, Fab-фрагменты и другие производные, описанные в данном документе.
[0236] Полинуклеотиды можно получать или производить с помощью любого способа, известного в данной области техники. Например, если известна нуклеотидная последовательность для антитела, полинуклеотид, кодирующий антитело, можно собрать из химически синтезированных олигонуклеотидов (например, как описано в Kutmeier et al., BioTechniques 77: 242 (1994)), что, вкратце, включает синтез перекрывающихся олигонуклеотидов, содержащих части последовательности, кодирующей антитело, отжиг и лигирование этих олигонуклеотидов и затем амплификацию лигированных олигонуклеотидов с помощью ПЦР.
[0237] Альтернативно, полинуклеотид, кодирующий молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, можно создать из нуклеиновой кислоты из подходящего источника. Если клон, содержащий нуклеиновую кислоту, кодирующую конкретное антитело, недоступен, а последовательность, кодирующая молекулу антитела, известна, то нуклеиновую кислоту, кодирующую антитело, можно синтезировать химически или получить из подходящего источника (которым является, например, библиотека кДНК, кодирующих антитела, или библиотека кДНК, образованная из любой ткани или любых клеток, экспрессирующих антитела, или, например, гибридомных клеток, выбранных для экспрессии антитела, или нуклеиновая кислота, предпочтительно РНК с поли-A, выделенная из таковых) посредством ПЦР-амплификации с помощью синтетических праймеров, которые гибридизируются с 3'- и 5'-концами последовательности, или посредством клонирования с помощью олигонуклеотидного зонда, специфичного для конкретной последовательности гена, для идентификации, например, кДНК-клона из библиотеки кДНК, который кодирует антитело. Амплифицированные нуклеиновые кислоты, образованные с помощью ПЦР, можно клонировать в реплицируемые клонирующие векторы с помощью любого способа, хорошо известного в данной области техники.
[0238] Когда нуклеотидная последовательность и соответствующая аминокислотная последовательность молекулы, связывающейся с Psl и/или PcrV Pseudomonas, например, антитела или его фрагмента, варианта или производного, определена, на эту нуклеотидную последовательность можно воздействовать с помощью хорошо известных в данной области техники способов воздействия на нуклеотидные последовательности, например, методик рекомбинантных ДНК, сайт-направленного мутагенеза, ПЦР и т.д. (см., например, методики, описанные в Sambrook et al., Molecular Cloning, A Laboratory Manual, 2d Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1990) и Ausubel et al., eds., Current Protocols in Molecular Biology, John Wiley & Sons, NY (1998), оба из которых включены в данный документ посредством ссылки во всей своей полноте), чтобы образовать антитела, имеющие другую аминокислотную последовательность, например, чтобы создать аминокислотные замены, делеции и/или вставки.
[0239] Полинуклеотид, кодирующий молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, можно составить из любого полирибонуклеотида или полидезоксирибонуклеотида, который может представлять собой немодифицированную РНК или ДНК или модифицированную РНК или ДНК. Например, полинуклеотид, кодирующий молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, можно составить из одно- и двухцепочечной ДНК, ДНК, которая является смесью одно- и двухцепочечных областей, одно- и двухцепочечной РНК и РНК, которая является смесью одно- и двухцепочечных областей, гибридных молекул, содержащих ДНК и РНК, которые могут быть одноцепочечными или, более типично, двухцепочечными, или смесью одно- и двухцепочечных областей. Кроме того, полинуклеотид, кодирующий молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, можно составить из РНК, или ДНК, или как РНК, так и ДНК, содержащих трехцепочечные области. Полинуклеотид, кодирующий молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, может также содержать одно или несколько модифицированных оснований или скелеты ДНК или РНК, модифицированные для стабильности или в других целях. "Модифицированные" основания включают, например, тритилированные основания и нетипичные основания, такие как инозин. Можно осуществлять разнообразные модификации по отношению к ДНК и РНК; таким образом, "полинуклеотид" охватывает химически, ферментативно или метаболически модифицированные формы.
[0240] Выделенный полинуклеотид, кодирующий неприродный вариант полипептида, полученного из иммуноглобулина (например, части тяжелой цепи или части легкой цепи иммуноглобулина), можно создать путем внедрения одной или нескольких нуклеотидных замен, добавлений или делеций в нуклеотидную последовательность, кодирующую иммуноглобулин, таким образом, чтобы одна или несколько аминокислотных замен, добавлений или делеций были внедрены в кодируемый белок. Мутации можно внедрять посредством стандартных методик, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Консервативные аминокислотные замены осуществляют по одному или нескольким остаткам несущественных аминокислот.
VI. ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ-АНТИТЕЛ
[0241] Как хорошо известно, РНК можно выделять из клеток исходной гибридомы или из других трансформированных клеток посредством стандартных методик, таких как извлечение с помощью гуанидинизотиоцианата и осаждение с последующим центрифугированием или хроматографией. Если необходимо, мРНК можно выделять из общей РНК посредством стандартных методик, таких как хроматография на олиго-dT-целлюлозе. Подходящие методики известны в данной области техники.
[0242] В одном варианте осуществления кДНК, которые кодируют легкую и тяжелую цепи молекулы, связывающейся с Psl и/или PcrV Pseudomonas, например, антитела или его фрагмента, варианта или производного, можно создать либо одновременно, либо по отдельности с помощью обратной транскриптазы и ДНК-полимеразы в соответствии с хорошо известными способами. ПЦР можно запускать с помощью консенсусных праймеров для константной области или с помощью более специфичных праймеров на основе опубликованных последовательностей ДНК и аминокислот, соответствующих тяжелым и легким цепям. Как рассматривалось выше, ПЦР также можно применять для выделения ДНК-клонов, кодирующих легкую и тяжелую цепи антител. В этом случае можно осуществлять скрининг библиотек с помощью консенсусных праймеров или больших гомологичных зондов, таких как зонды для константных областей мыши.
[0243] ДНК, обычно плазмидную ДНК, можно выделять из клеток посредством методик, известных в данной области техники, подвергать рестрикционному картированию и секвенировать в соответствии со стандартными, хорошо известными методиками, подробно изложенными, например, в вышеприведенных источниках литературы, касающихся методик рекомбинантных ДНК. Разумеется, ДНК может быть синтетической согласно настоящему изобретению в любой момент времени в ходе способа выделения или последующего анализа.
[0244] После воздействия на выделенный генетический материал с получением молекулы, связывающейся с Psl и/или PcrV Pseudomonas, например, антитела или его фрагмента, варианта или производного, по настоящему изобретению, полинуклеотиды, кодирующие молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, обычно вставляют в вектор экспрессии для введения в клетки-хозяева, которые можно применять для получения желаемого количества молекул, связывающихся с Psl и/или PcrV Pseudomonas.
[0245] Рекомбинантная экспрессия антитела или его фрагмента, производного или аналога, например, тяжелой или легкой цепи антитела, которое связывается с молекулой-мишенью, описанной в данном документе, например, Psl и/или PcrV, требует конструирования вектора экспрессии, содержащего полинуклеотид, который кодирует антитело. Когда полинуклеотид, кодирующий молекулу антитела, или тяжелую или легкую цепь антитела, или его часть (содержащую вариабельный домен тяжелой или легкой цепи) по настоящему изобретению, получен, можно получить вектор для выработки молекулы антитела посредством технологии рекомбинантных ДНК с помощью методик, хорошо известных в данной области техники. Таким образом, способы получения белка посредством экспрессии полинуклеотида, содержащего нуклеотидную последовательность, кодирующую антитело, описаны в данном документе. Способы, которые хорошо известны специалистам в данной области, можно применять для конструирования векторов экспрессии, содержащих последовательности, кодирующие антитела, и соответствующие сигналы регуляции транскрипции и трансляции. Эти способы включают, например, in vitro методики рекомбинантных ДНК, методики синтеза и in vivo генетическую рекомбинацию. Настоящее изобретение, таким образом, обеспечивает реплицируемые векторы, содержащие нуклеотидную последовательность, кодирующую молекулу антитела по настоящему изобретению, или его тяжелую или легкую цепь, или вариабельный домен тяжелой или легкой цепи, функционально связанную с промотором. Такие векторы могут содержать нуклеотидную последовательность, кодирующую константную область молекулы антитела (см., например, публикацию по PCT WO 86/05807; публикацию по РСТ WO 89/01036 и патент США №5122464), а вариабельный домен антитела можно клонировать в такой вектор для экспрессии всей тяжелой или легкой цепи.
[0246] Выражение "вектор" или "вектор экспрессии" применяется в данном документе для обозначения векторов, применяемых в соответствии с настоящим изобретением в качестве носителей для введения в клетку-хозяина и экспрессии желаемого гена в ней. Как известно специалистам в данной области, такие векторы можно без труда выбрать из группы, состоящей из плазмид, фагов, вирусов и ретровирусов. Векторы, совместимые с настоящим изобретением, как правило, будут содержать селектируемый маркер, соответствующие сайты рестрикции для облегчения клонирования желаемого гена и обладать способностью проникать в эукариотические или прокариотические клетки и/или реплицироваться в них.
[0247] Применительно к целям настоящего изобретения можно использовать множество векторных систем экспрессии. Например, в одном классе векторов используются элементы ДНК, которые получают из вирусов животных, таких как вирус папилломы крупного рогатого скота, вирус полиомы, аденовирус, вирус коровьей оспы, бакуловирус, ретровирусы (RSV, MMTV или MOMLV) или вирус SV40. Другие включают применение полицистронных систем с внутренними сайтами связывания рибосомы. Дополнительно можно осуществлять селекцию клеток, в хромосомы которых интегрирована ДНК, путем введения одного или нескольких маркеров, которые обеспечивают возможность селекции трансфицированных клеток-хозяев. Маркер может обеспечивать прототрофность ауксотрофного хозяина, устойчивость к биоцидам (например, к антибиотикам) или устойчивость к тяжелым металлам, таким как медь. Селектируемый маркерный ген может быть либо непосредственно связанным с последовательностями ДНК, которые необходимо экспрессировать, либо введенным в ту же клетку путем котрансформации. Для оптимального синтеза мРНК могут также быть необходимыми дополнительные элементы. Эти элементы могут включать сигнальные последовательности, сигналы для сплайсинга, а также промоторы, участвующие в транскрипции, энхансеры и сигналы терминации.
[0248] В некоторых вариантах осуществления клонируемые гены вариабельных областей вставляют в вектор экспрессии наряду с синтетическими генами константной области тяжелой и легкой цепи (например, человека), обсуждаемыми выше. Разумеется, любой вектор экспрессии, который может вызывать экспрессию в эукариотических клетках, можно применять в настоящем изобретении. Примеры подходящих векторов включают, без ограничений, плазмиды pcDNA3, pHCMV/Zeo, pCR3.1, pEF1/His, pIND/GS, pRc/HCMV2, pSV40/Zeo2, pTRACER-HCMV, pUB6/V5-His, pVAX1 и pZeoSV2 (доступны от Invitrogen, Сан-Диего, Калифорния) и плазмиду pCI (доступна от Promega, Мэдисон, Висконсин). В целом, скрининг больших количеств трансформированных клеток для выявления тех, которые экспрессируют тяжелые и легкие цепи иммуноглобулина на достаточно высоких уровнях, является стандартной экспериментальной работой, которую можно проводить, например, с помощью роботизированных систем.
[0249] В более общем смысле, когда вектор или последовательность ДНК, кодирующие мономерную субъединицу молекулы, связывающейся с Psl и/или PcrV Pseudomonas, например, антитела или его фрагмента, варианта или производного по настоящему изобретению, получены, вектор экспрессии можно вводить в подходящую клетку-хозяина. Введение плазмиды в клетку-хозяина можно осуществлять с помощью различных методик, хорошо известных специалистам в данной области. Они включают, без ограничений, трансфекцию (в том числе электрофорез и электропорацию), слияние протопластов, осаждение фосфатом кальция, слияние клеток с ДНК в оболочке, микроинъекцию и инфицирование интактным вирусом. См. Ridgway, A.A.G. "Mammalian Expression Vectors" Vectors, Rodriguez and Denhardt, Eds., Butterworths, Boston, Mass., Chapter 24.2, pp. 470-472 (1988). Как правило, введение плазмиды в хозяина осуществляют посредством электропорации. Клетки-хозяева, содержащие конструкт для экспрессии, выращивают в условиях, подходящих для выработки легких цепей и тяжелых цепей, и анализируют в отношении белкового синтеза тяжелой и/или легкой цепи. Иллюстративные методики анализа включают твердофазный иммуноферментный анализ (ELISA), радиоиммунологический анализ (RTA) или анализ с помощью сортировки клеток, активированной флуоресценцией (FACS), иммуногистохимический анализ и т.п.
[0250] Вектор экспрессии переносят в клетку-хозяина с помощью стандартных методик, и трансфицированные клетки затем культивируют с помощью стандартных методик с получением антитела для применения в способах, описываемых в данном документе. Таким образом, настоящее изобретение включает клетки-хозяева, содержащие полинуклеотид, кодирующий молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, например, антитело, или его фрагмент, вариант или производное, или его тяжелую или легкую цепь, функционально связанный с гетерологичным промотором. В некоторых вариантах осуществления для экспрессии двухцепочечных антител векторы, кодирующие как тяжелую, так и легкую цепи, могут коэкспрессироваться в клетке-хозяине для экспрессии целой молекулы иммуноглобулина, как подробно изложено ниже.
[0251] Определенные варианты осуществления включают выделенный полинуклеотид, содержащий нуклеиновую кислоту, которая кодирует вышеописанные VH и VL, где связывающая молекула или ее антиген-связывающий фрагмент, экспрессируемые полинуклеотидом, специфически связываются с Psl и/или PcrV Pseudomonas. В некоторых вариантах осуществления описанный полинуклеотид, кодирующий молекулу scFv, содержащую VH и VL, по меньшей мере на 80%, 85%, 90%, 95% или 100% идентичен по отношению к одной или нескольким из SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69 или SEQ ID NO: 70, показанным в таблице 4.
[0252] Некоторые варианты осуществления включают векторы, содержащие вышеописанные полинуклеотиды. В дополнительных вариантах осуществления полинуклеотиды функционально связаны с промотором. В дополнительных вариантах осуществления настоящее изобретение обеспечивает клетки-хозяева, содержащие такие векторы. В дополнительных вариантах осуществления настоящее изобретение обеспечивает векторы, в которых полинуклеотид функционально связан с промотором, где векторы могут экспрессировать связывающую молекулу, которая специфически связывается с Psl и/или PcrV Pseudomonas в подходящей клетке-хозяине.
[0253] Также обеспечивается способ получения связывающей молекулы или ее фрагмента, которые специфически связываются с Psl и/или PcrV Pseudomonas, включающий культивирование клетки-хозяина, содержащей вектор, содержащий вышеописанные полинуклеотиды, и извлечение указанного антитела или его фрагмента. В дополнительных вариантах осуществления настоящее изобретение обеспечивает выделенную связывающую молекулу или ее фрагмент, полученные с помощью вышеописанного способа.
[0254] Применяемое в данном документе выражение "клетки-хозяева" относится к клеткам, которые содержат векторы, сконструированные с помощью методик рекомбинантных ДНК и кодирующие по меньшей мере один гетерологичный ген. В описаниях способов выделения антител из рекомбинантных хозяев выражения "клетка" и "культура клеток" применяют взаимозаменяемо для обозначения источника антитела, если явно не указано иное. Другими словами, извлечение полипептида из "клеток" может означать либо таковое из осажденных центрифугированием целых клеток, либо таковое из культуры клеток, содержащей как среду, так и суспендированные клетки.
[0255] Для экспрессии молекул антител для применения в способах, описываемых в данном документе, можно использовать множество векторных систем экспрессии в хозяине. Такие системы экспрессии в хозяине представляют носители, с помощью которых кодирующие последовательности, представляющие интерес, можно получать и впоследствии очищать, но также представляют клетки, которые могут, будучи трансформированными или трансфицированными с помощью подходящих нуклеотидных кодирующих последовательностей, экспрессировать молекулу антитела по настоящему изобретению in situ. Они включают, без ограничений, микроорганизмы, такие как бактерии (например, E. coli, B. subtilis), трансформированные рекомбинантными векторами экспрессии на основе ДНК бактериофага, плазмидной ДНК или космидной ДНК, содержащими последовательности, кодирующие антитела; дрожжи (например, Saccharomyces, Pichia), трансформированные рекомбинантными векторами экспрессии в дрожжах, содержащими последовательности, кодирующие антитела; системы клеток насекомых, инфицированные рекомбинантными векторами экспрессии на основе вируса (например, бакуловируса), содержащими последовательности, кодирующие антитела; системы растительных клеток, инфицированные рекомбинантными векторами экспрессии на основе вируса (например, вируса мозаики цветной капусты, CaMV; вируса табачной мозаики, TMV) или трансформированные рекомбинантными векторами экспрессии на основе плазмиды (например, Ti-плазмиды), содержащими последовательности, кодирующие антитела; или системы клеток млекопитающих (например, клетки COS, CHO, BLK, 293, 3T3), содержащие рекомбинантные конструкты для экспрессии, содержащие промоторы, полученные из генома клеток млекопитающих (например, промотор гена металлотионеина) или из вирусов млекопитающих (например, поздний промотор аденовируса; промотор вируса коровьей оспы 7.5K). Бактериальные клетки, такие как клетки Escherichia coli, или эукариотические клетки, в частности, для экспрессии полной молекулы рекомбинантного антитела, применяют для экспрессии молекулы рекомбинантного антитела. Например, клетки млекопитающих, такие как клетки яичников китайского хомячка (CHO), совместно с вектором, таким как главный средне-ранний промоторный элемент гена из цитомегаловируса человека, являются эффективной системой экспрессии антител (Foecking et al, Gene 45:101 (1986); Cockett et al, Bio/Technology 8: 2 (1990)).
[0256] Линия клеток-хозяев, применяемых для экспрессии белка, зачастую происходит из млекопитающих; специалисты в данной области наделены способностью определять конкретные линии клеток-хозяев, которые наилучшим образом подходят для экспрессии в них желаемого продукта гена. Иллюстративные линии клеток-хозяев включают, без ограничений, CHO (клетки яичников китайского хомячка), DG44 и DUXB11 (линии клеток яичников китайского хомячка, DHFR-отрицательные), HELA (клетки карциномы шейки матки человека), CVI (линия клеток почки обезьяны), COS (производное CVI с T-антигеном SV40), VERY, ВНК (клетки почки детеныша хомячка), MDCK, 293, WI38, R1610 (фибробласты китайского хомячка), BALBC/3T3 (фибробласты мыши), HAK (линия клеток почки хомячка), SP2/0 (клетки миеломы мыши), P3×63-Ag3.653 (клетки миеломы мыши), BFA-lclBPT (эндотелиальные клетки крупного рогатого скота), RAJI (лимфоциты человека) и 293 (клетки почки человека). Линии клеток-хозяев обычно доступны из коммерческих служб, Американской коллекции тканевых культур или из опубликованной литературы.
[0257] Кроме того, можно выбрать такой штамм клеток-хозяев, который модулирует экспрессию вставленных последовательностей или модифицирует и осуществляет процессинг продукта гена определенным желаемым способом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важными для функции белка. Разные клетки-хозяева имеют характеристики и определенные механизмы для посттрансляционного процессинга и модификации белков и продуктов генов. Можно выбирать подходящие линии клеток или системы экспрессии в хозяине для обеспечения правильной модификации и процессинга экспрессируемого чужеродного белка. В связи с этим можно применять эукариотические клетки-хозяева, которые обладают клеточным аппаратом для надлежащего процессинга первичного транскрипта, гликозилирования и фосфорилирования продукта гена.
[0258] Для длительной выработки рекомбинантных белков с высоким выходом стабильная экспрессия является предпочтительной. Например, можно сконструировать линии клеток, в которых происходит стабильная экспрессия молекул антител. Вместо применения векторов экспрессии, которые содержат вирусные точки начала репликации, клетки-хозяева можно трансформировать с помощью ДНК, регулируемой подходящими элементами регуляции экспрессии (например, промотором, энхансером, последовательностями, терминаторами транскрипции, сайтами полиаденилирования и т.д.), и селектируемого маркера. После введения чужеродной ДНК сконструированным клеткам можно позволить расти в течение 1-2 суток в обогащенной среде, а затем их переносят в селективную среду. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость к селекции и позволяет клеткам стабильно интегрировать плазмиду в их хромосомы и расти с формированием очагов, которые, в свою очередь, можно клонировать, и количество которых можно увеличить до линии клеток. Этот способ можно преимущественно применять для конструирования линий клеток, в которых происходит стабильная экспрессия молекулы антитела.
[0259] Можно применять ряд систем для селекции, в том числе, без ограничений, можно использовать гены тимидинкиназы вируса простого герпеса (Wigler et al., Cell 11: 223 (1977)), гипоксантин-гуанинфосфорибозилтрансферазы (Szybalska & Szybalski, Proc. Natl. Acad. Sci. USA 48: 202 (1992)) и аденинфосфорибозилтрансферазы (Lowy et al., Cell 22: 817 1980) в клетках tk-, hgprt- или aprt-, соответственно. Также можно использовать устойчивость к антиметаболитам в качестве основы для селекции по следующим генам: dhfr, который придает устойчивость к метотрексату (Wigler et al, Natl. Acad. Sci. USA 77: 351 (1980); O'Hare et al., Proc. Natl. Acad. Sci. USA 78: 1527 (1981)); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan & Berg, Proc. Natl. Acad. Sci. USA 78: 2072 (1981)); neo, который придает устойчивость к аминогликозиду G-418, Clinical Pharmacy 12: 488-505; Wu and Wu, Biotherapy 3: 87-95 (1991); Tolstoshev, Ann. Rev. Pharmacol. Toxicol. 52: 573-596 (1993); Mulligan, Science 260: 926-932 (1993); и Morgan and Anderson, Ann. Rev. Biochem. 62: 191-217 (1993); TIB TECH 11 (5): 155-215 (May, 1993); и hygro, который придает устойчивость к гигромицину (Santerre et al., Gene 30: 147 (1984). Способы, широко известные в области техники, к которой относится технология рекомбинантных ДНК, которые можно применять, описаны в Ausubel et al. (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, NY (1993); Kriegler, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY (1990); и в Chapters 12 and 13, Dracopoli et al. (eds), Current Protocols in Human Genetics, John Wiley & Sons, NY (1994); Colberre-Garapin et al., J. Mol. Biol. 150: 1 (1981), которые включены в данный документ посредством ссылки во всей своей полноте.
[0260] Уровни экспрессии молекулы антитела можно повышать посредством амплификации вектора (в отношении обзора см. Bebbington and Hentschel, The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells in DNA cloning, Academic Press, New York, Vol. 3. (1987)). Если маркер в векторной системе, экспрессирующей антитело, является амплифицируемым, то при повышении уровня ингибитора, присутствующего в культуре клеток-хозяев, будет повышаться число копий маркерного гена. Поскольку амплифицируемая область связана с геном антитела, выработка антитела будет также повышаться (Crouse etal, Mol. Cell. Biol. 3: 257 (1983)).
[0261] Выработка in vitro позволяет увеличить масштаб с получением больших количеств желаемых полипептидов. Методики культивирования клеток млекопитающих в условиях тканевой культуры известны в данной области техники и включают гомогенную суспензионную культуру, например, в аэролифтном реакторе или в реакторе с непрерывным перемешиванием, или иммобилизованную или фиксированную культуру клеток, например, в полых волокнах, микрокапсулах, на агарозньгх микрогранулах или в керамических картриджах. Если необходимо и/или желательно, растворы полипептидов можно очищать с помощью общепринятых способов хроматографии, например, гель-фильтрационной, ионнообменной хроматографии, хроматографии на DEAE-целлюлозе или (иммуно-)аффинной хроматографии, например, после предпочтительного биосинтеза синтетической шарнирной области полипептида или перед или вслед за этапом HIC-хроматографии, описанным в данном документе.
[0262] Конструкты, кодирующие молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, раскрытые в данном документе, могут также экспрессироваться клетками, не являющимися клетками млекопитающих, такими как клетки бактерий, или дрожжей, или растений. Бактерии, которые легко принимают нуклеиновые кислоты, включают представителей семейства Enterobacteriaceae, таких как штаммы Escherichia coli или Salmonella; Bacillaceae, таких как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Дополнительно следует иметь в виду, что если гетерологичные полипептиды экспрессируются в бактериях, они, как правило, входят в состав телец-включений. Гетерологичные полипептиды нужно выделять, очищать и затем собирать в функциональные молекулы. Если желательны тетравалентные формы антител, субъединицы затем будут самостоятельно собираться в тетравалентные антитела (WO 02/096948 A2).
[0263] В бактериальных системах количество векторов экспрессии можно преимущественно выбирать в зависимости от предполагаемого применения экспрессируемой молекулы антитела. Например, если необходимо получить большое количество такого белка для создания фармацевтических композиций на основе молекулы антитела, могут быть желательными векторы, которые управляют экспрессией высоких уровней продуктов на основе белка слияния, которые легко очищаются. Такие векторы включают, без ограничений, вектор экспрессии pUR278 в E. coli (Ruther et al., EMBO J. 2: 1791 (1983)), в котором последовательность, кодирующая антитело, может быть отдельно лигирована в вектор внутрирамочно с областью, кодирующей lacZ, таким образом, что вырабатывается белок слияния; векторы pIN (Inouye & Inouye, Nucleic Acids Res. 13: 3101-3109 (1985); Van Heeke & Schuster, J. Biol. Chem. 24: 5503-5509 (1989)) и т.п. Также можно применять векторы pGEX для экспрессии чужеродных полипептидов в виде белков слияния с глутатион-8-трансферазой (GST). В целом, такие белки слияния являются растворимыми, и их можно легко очищать из лизированных клеток посредством адсорбции и связывания с матрицей на основе глутатион-агарозных гранул с последующим элюированием в присутствии свободного глутатиона. Векторы pGEX разработаны таким образом, что они включают сайты расщепления протеазами тромбином или фактором Ха, так что клонируемый целевой продукт гена можно высвобождать из GST-фрагмента.
[0264] В дополнение к прокариотам можно применять эукариотические микробы. Saccharomyces cerevisiae, или обыкновенные пекарские дрожжи, являются наиболее часто применяемыми среди эукариотических микроорганизмов, хотя широко доступен ряд других штаммов, например, Pichia pastoris.
[0265] Для экспрессии в Saccharomyces, например, широко применяется плазмида YRp7 (Stinchcomb et al, Nature 282: 39 (1979); Kingsman et al, Gene 7: 141 (1979); Tschemper et al, Gene 70:157 (1980)). Эта плазмида уже содержит ген TRP1, который обеспечивает селектируемый маркер для мутантного штамма дрожжей, у которого отсутствует способность расти в присутствии триптофана, например, №44076 в ATCC или PEP4-1 (Jones, Genetics 85: 12 (1977)). Наличие повреждения trpI в качестве характеристики генома дрожжевых клеток-хозяев в этом случае обеспечивает эффективную среду для выявления трансформации с помощью роста в отсутствие триптофана.
[0266] В системе на основе клеток насекомых, как правило, в качестве вектора для экспрессии чужеродных генов применяют вирус ядерного полиэдроза Autographa californica (AcNPV). Вирус растет в клетках Spodoptera frugiperda. Последовательность, кодирующую антитело, можно отдельно клонировать в несущественные области вируса (например, ген полиэдрина) и помещать под контроль промотора AcNPV (например, промотора гена полиэдрина).
[0267] Когда молекула, связывающаяся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, раскрытые в данном документе, экспрессирована рекомбинантным путем, ее можно очищать с помощью любого известного в области техники способа очистки молекулы иммуноглобулина, например, с помощью хроматографии (например, ионнообменной, аффинной хроматографии, в частности, с аффинностью в отношении специфического антигена после добавления белка A, и колоночной хроматографии с разделением по размеру), центрифугирования, дифференциальной растворимости или с помощью любой другой стандартной методики очистки белков. Другой способ повышения аффинности антител по настоящему изобретению раскрыт в US 2002 0123057 A1.
VII. ИДЕНТИФИКАЦИЯ СВЯЗЫВАЮЩИХ МОЛЕКУЛ, ИНДИФФЕРЕНТНЫХ В ОТНОШЕНИИ СЕРОТИПА
[0268] Настоящее изобретение охватывает индифферентный в отношении мишени подход с использованием целых клеток для идентификации серотип-независимых терапевтических связывающих молекул, например, антител или их фрагментов, с превосходными или желаемыми видами терапевтической активности. Способ можно использовать для идентификации связывающих молекул, которые могут противодействовать нежелательной активности возбудителя инфекции, например, бактериального патогена, нейтрализовать, устранять или блокировать ее. Как известно в данной области техники, у многих возбудителей инфекции проявляется значительное разнообразие их преобладающих поверхностных антигенов, что позволяет им избегать иммунологического надзора. С помощью способа идентификации, описанного в данном документе, можно идентифицировать связывающие молекулы, которые нацеливаются на антигены, являющиеся общими для многих различных видов Pseudomonas или других грамотрицательных патогенов, таким образом обеспечивая терапевтическое средство, которое может нацеливаться на многочисленные патогены из многочисленных видов. Например, способ был использован для идентификации группы связывающих молекул, которые связываются с поверхностью клеток P. aeruginosa серотип-независимым образом и, будучи связанными с бактериальными патогенами, опосредуют опсонофагоцитарную активность (OPK), способствуют таковой или усиливают таковую в отношении бактериальных клеток, таких как бактериальные патогены, например, условно-патогенные виды Pseudomonas (например, Pseudomonas aeruginosa, Pseudomonas fluorescens, Pseudomonas putida и Pseudomonas alcaligenes), и/или ингибируют прикрепление таких бактериальных клеток к эпителиальным клеткам.
[0269] В определенных вариантах осуществления раскрывается способ идентификации связывающих молекул, индифферентных в отношении серотипа, включающий (a) получение библиотек наивных антител и/или антител фазы выздоровления в фаге, (b) удаление серотип-специфических антител из библиотеки посредством элиминирующего пэннинга, (c) скрининг библиотеки в отношении антител, которые специфически связываются с целыми клетками независимо от серотипа, и (d) скрининг оставшихся антител в отношении желаемых функциональных свойств.
[0270] В некоторых вариантах осуществления обеспечивается подход по методу фенотипического скрининга с использованием целых клеток, раскрытый в данном документе, с фаговыми библиотеками антител, полученных либо от "наивных", либо от выздоравливающих пациентов, инфицированных P. aeruginosa. С помощью метода пэннинга, при котором изначально осуществлялась селекция против серотип-специфической реактивности, были выделены различные клоны, которые связываются с целыми клетками P. aeruginosa. Выбранные клоны были преобразованы в антитела IgG1 человека, и было подтверждено, что они реагируют с клиническими изолятами P. aeruginosa вне зависимости от группы серотипической классификации или участка изъятия ткани (см. примеры). Этапы скрининга в отношении функциональной активности, описанные в данном документе, указывали, что антитела были эффективными при предупреждении прикрепления P. aeruginosa к клеткам млекопитающих и опосредовали опсонофагоцитарное уничтожение (OPK) зависимым от концентрации и независимым от серотипа способом.
[0271] В дополнительных вариантах осуществления вышеописанные связывающие молекулы или их фрагменты, антитела или их фрагменты или композиции связываются с двумя или более, тремя или более, четырьмя или более или пятью или более различными серотипами P. aeruginosa или по меньшей мере с 80%, по меньшей мере с 85%, по меньшей мере с 90% или по меньшей мере с 95% штаммов P. aeruginosa, выделенных от инфицированных пациентов. В дополнительных вариантах осуществления штаммы P. aeruginosa выделены из одного или более из легких, мокроты, глаз, гноя, фекалий, мочи, пазух, раны, кожи, крови, кости или жидкости из коленного сустава.
VIII. ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С PSL И/ИЛИ PCRV PSEUDOMONAS
[0272] Фармацевтические композиции, применяемые в настоящем изобретении, содержат фармацевтически приемлемые носители, хорошо известные специалистам в данной области. Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Определенные фармацевтические композиции, раскрытые в данном документе, можно вводить перорально в виде приемлемой лекарственной формы, включая, например, капсулы, таблетки, водные суспензии или растворы. Определенные фармацевтические композиции можно вводить в составе назального аэрозоля или путем ингаляции. Могут также присутствовать консерванты и другие добавки, такие как, например, противомикробные средства, антиоксиданты, хелатирующие средства и инертные газы и т.п. Подходящие составы для применения в терапевтических способах, раскрытых в данном документе, описаны в Remington's Pharmaceutical Sciences, Mack Publishing Co., 16th ed. (1980). [0273] Количество молекулы, связывающейся с Psl и/или PcrV Pseudomonas, например, антитела или его фрагмента, варианта или производного, которую можно объединять с материалами-носителями с получением единичной лекарственной формы, будет варьировать в зависимости от обрабатываемого хозяина и конкретного способа введения. Схему дозирования также можно установить таким образом, чтобы обеспечивать оптимальный желаемый эффект (например, терапевтический или профилактический эффект). Композиции могут также содержать молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, диспергированные в биологически совместимом материале-носителе, который выполняет функцию подходящей системы доставки или скрепления соединений.
IX. СПОСОБЫ ЛЕЧЕНИЯ С ПОМОЩЬЮ ТЕРАПЕВТИЧЕСКИХ СВЯЗЫВАЮЩИХ МОЛЕКУЛ
[0274] Способы получения и введения молекул, связывающихся с Psl и/или PcrV Pseudomonas, например, антитела или его фрагмента, варианта или производного, раскрытых в данном документе, субъекту, нуждающемуся в этом, хорошо известны специалистам в данной области или могут быть без труда определены ими. Путь введения молекул, связывающихся с Psl и/или PcrV Pseudomonas, например, антитела или его фрагмента, варианта или производного, может быть, например, пероральным, парентеральным, ингаляционным или местным. Выражение "парентеральное", применяемое в данном документе, включает, например, внутривенное, внутриартериальное, интраперитонеальное, внутримышечное или подкожное введение. Подходящей формой для введения будет раствор для инъекций, в частности, для внутривенных или внутриартериальных инъекций или капельного введения. Однако в других способах, совместимых с идеями, изложенными в данном документе, молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитело или его фрагмент, вариант или производное, раскрытые в данном документе, могут быть доставлены непосредственно к участку с нежелательной клеточной популяцией, например, к очагу инфекции, что, таким образом, увеличивает воздействие терапевтического средства на пораженную ткань. Например, молекулу, связывающуюся с Psl и/или PcrV Pseudomonas, можно непосредственно вводить в ткань глаза, область ожоговой травмы или ткань легких.
[0275] Молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, раскрытые в данном документе, можно вводить в фармацевтически эффективном количестве для лечения in vivo инфекции, вызываемой Pseudomonas. В этом отношении следует иметь в виду, что раскрытые связывающие молекулы будут составлять таким образом, чтобы облегчать введение и способствовать стабильности активного средства. Для целей настоящей заявки фармацевтически эффективное количество будет означать количество, достаточное для достижения эффективного связывания с мишенью и получения пользы, например, лечения, улучшения, облегчения, устранения или предупреждения инфекции, вызываемой Pseudomonas.
[0276] Некоторые варианты осуществления направлены на способ предупреждения или лечения инфекции, вызываемой Pseudomonas, у субъекта, нуждающегося в этом, включающий введение данному субъекту эффективного количества связывающей молекулы или ее фрагмента, антитела или его фрагмента, композиции, полинуклеотида, вектора или клетки-хозяина, описываемых в данном документе. В дополнительных вариантах осуществления инфекция, вызываемая Pseudomonas, представляет собой инфекцию, вызываемую P. aeruginosa. В некоторых вариантах осуществления субъектом является человек. В некоторых вариантах осуществления инфекция представляет собой инфекцию глаз, инфекцию легких, ожоговую инфекцию, раневую инфекцию, кожную инфекцию, инфекцию крови, инфекцию костей или комбинацию двух или более из указанных инфекций. В дополнительных вариантах осуществления субъект страдает от острой пневмонии, ожоговой травмы, инфекции роговицы, муковисцидоза или их комбинации.
[0277] Определенные варианты осуществления направлены на способ блокирования или предупреждения прикрепления P. aeruginosa к эпителиальным клеткам, включающий приведение смеси эпителиальных клеток и P. aeruginosa в контакт со связывающей молекулой или ее фрагментом, антителом или его фрагментом, композицией, полинуклеотидом, вектором или клеткой-хозяином, описываемыми в данном документе.
[0278] Также раскрывается способ усиления ОРК P. aeruginosa, включающий приведение смеси фагоцитарных клеток и P. aeruginosa в контакт со связывающей молекулой или ее фрагментом, антителом или его фрагментом, композицией, полинуклеотидом, вектором или клеткой-хозяином, описываемыми в данном документе. В дополнительных вариантах осуществления фагоцитарные клетки являются дифференцированными клетками HL-60 или полиморфноядерными лейкоцитами человека (PMN).
[0279] В соответствии с объемом настоящего изобретения молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, можно вводить человеку или другому животному в соответствии с вышеуказанными способами лечения в количестве, достаточном для получения терапевтического эффекта. Молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, раскрытые в данном документе, можно вводить такому человеку или другому животному в традиционной лекарственной форме, полученной путем объединения антитела по настоящему изобретению с традиционным фармацевтически приемлемым носителем или разбавителем в соответствии с известными методиками.
[0280] Эффективные дозы композиций по настоящему изобретению для лечения инфекции, вызываемой Pseudomonas, варьируют в зависимости от множества различных факторов, включая способ введения, целевой участок, физиологическое состояние пациента, того, является ли пациент человеком или животным, других вводимых лекарственных препаратов и того, является ли лечение профилактическим или терапевтическим. Как правило, пациент является человеком, но также можно лечить млекопитающих, отличных от человека, включая трансгенных млекопитающих. Дозировки для лечения можно титровать с помощью обычных способов, известных специалистам в данной области, для получения оптимальных безопасности и эффективности.
[0281] Молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, можно вводить многократно с различной частотой, зависящей от различных факторов, известных специалистам в данной области. Альтернативно, молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, можно вводить в виде состава с замедленным высвобождением, причем в этом случае требуется менее частое введение. Дозировка и частота варьируют в зависимости от периода полувыведения антитела из организма пациента.
[0282] Композиции по настоящему раскрытию можно вводить любым подходящим способом, например, парентерально, интравентрикулярно, перорально, в составе ингаляционного спрея, местно, ректально, назально, трансбуккально, вагинально или через имплантируемый резервуар. Выражение "парентеральное", используемое в данном документе, включает методики подкожных, внутривенных, внутримышечных, внутрисуставных, интрасиновиальных, интрастернальных, интратекальных, внутрипеченочных, внутриочаговьгх и интракраниальных инъекции или инфузии.
X. СИНЕРГИЗМ
[0283] Chou и Talalay (Adv. Enzyme Regul, 22: 27-55 (1984)) разработали математический способ описания экспериментальных данных в отношении комбинированных эффектов лекарственных средств качественным и количественным способом. Для несовместимых лекарственных средств они показали, что обобщенное уравнение изоболы применяется для любой степени эффекта (см. стр. 52 в Chou and Talalay). Изобола или изоболограмма представляет собой графическое представление всех комбинаций доз двух лекарственных средств, которые имеют одинаковую степень эффекта. В изоболограммах прямая линия указывает на аддитивные эффекты, вогнутая кривая (кривая ниже прямой линии) представляет синергические эффекты, а выпуклая кривая (кривая выше прямой линии) представляет антагонистические эффекты. Эти кривые также показывают, что комбинация двух несовместимых лекарственных средств, является ли эта комбинация аддитивной, синергической или антагонистической, будет демонстрировать одинаковый тип эффекта во всем диапазоне концентраций. Большинство комбинаций лекарственных средств демонстрируют аддитивный эффект. В некоторых случаях, однако, комбинации демонстрируют эффект, больший или меньший, чем аддитивный. Эти комбинации называют, соответственно, антагонистическими или синергическими. Комбинация проявляет терапевтический синергизм, если ее эффект терапевтически превосходит эффект одной или другой из ее составляющих, применяемых в оптимальной дозе. См., T.H. Corbett et al., Cancer Treatment Reports, 66, 1187 (1982). Tallarida RJ (J Pharmacol Exp Ther. 2001 Sep; 298 (3): 865-722) также отмечает, что "два лекарственных средства, которые производят явно сходные эффекты, будут иногда производить усиленные или ослабленные эффекты при одновременном применении. Для того, чтобы различить эти случаи от попросту аддитивного действия, необходима количественная оценка".
[0284] Синергический эффект может быть измерен при помощи способа по Chou-Talalay с использованием показателя аддитивности (CI) (см. Chang et al., Cancer Res. 45: 2434-2439, (1985)), который основан на принципе медианного эффекта. С помощью этого способа рассчитывают степень синергизма, аддитивности или антагонизма двух лекарственных средств при различных уровнях цитотоксичности. Если значение CI меньше 1, то наблюдается синергизм двух лекарственных средств. Если значение CI равно 1, то наблюдается аддитивный эффект, но не наблюдается синергический эффект.Значения CI больше 1 указывают на антагонизм. Чем меньше значение CI, тем больше синергический эффект. В другом варианте осуществления синергический эффект определяют с применением фракционной ингибирующей концентрации (FIC). Это фракционное значение определяют путем выражения IC50 лекарственного средства, действующего в комбинации, в зависимости от IC50 лекарственного средства, действующего отдельно. Для двух взаимодействующих лекарственных средств сумма значений FIC каждого лекарственного средства представляет меру синергического взаимодействия. Если значение FIC меньше 1, то наблюдается синергизм двух лекарственных средств. Значение FIC, равное 1, указывает на аддитивный эффект. Чем меньше значение FIC, тем существеннее синергическое взаимодействие.
[0285] В некоторых вариантах осуществления синергический эффект достигается в лечении инфекции, вызываемой Pseudomonas, где одно или несколько связывающих средств вводят в "низкой дозе" (т.е, применяя дозу или дозы, которые рассматривались бы нетерапевтическими при введении в отдельности), причем введение связывающего средства в низкой дозе в комбинации с другими связывающими средствами (вводимыми либо в низкой, либо в терапевтической дозе) приводит к синергическому эффекту, который превышает аддитивные эффекты, которые могли бы возникать в других случаях в результате независимого введения связывающих средств по отдельности. В некоторых вариантах осуществления синергический эффект достигается посредством введения одного или нескольких связывающих средств, вводимых в "низкой дозе", причем низкая доза предусматривается для снижения или во избежание токсичности или других нежелательных побочных эффектов.
XI. ИММУНОЛОГИЧЕСКИЕ АНАЛИЗЫ
[0286] Молекулы, связывающиеся с Psl и/или PcrV Pseudomonas, например, антитела или их фрагменты, варианты или производные, можно анализировать в отношении иммуноспецифического связывания любым способом, известным в данной области техники. Иммунологические анализы, которые можно применять, включают, без ограничений, системы конкурентного и неконкурентного анализа с использованием методик, таких как вестерн-блоттинг, радиоиммунологические анализы, ELISA (твердофазный иммуноферментный анализ), иммунологические "сэндвич"-анализы, анализы на основе иммунопреципитации, реакции преципитации, реакции диффузионной преципитации в геле, анализы на основе иммунодиффузии, анализы на основе агглютинации, анализы на основе фиксации комплемента, иммунорадиометрические анализы, флуоресцентные иммунологические анализы, иммунологические анализы с белком A, и это лишь некоторые из них. Такие анализы являются стандартными и хорошо известны в данной области техники (см., например, Ausubel et al., eds, Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, Vol. 1 (1994), который включен в данный документ посредством ссылки во всей своей полноте).
Иллюстративные иммунологические анализы вкратце описаны ниже (но не подразумеваются как ограничивающие).
[0287] Существуют разнообразные способы, доступные для измерения аффинности взаимодействия антитело-антиген, но относительно немного для определения констант скорости. Большинство способов основаны на мечении антитела либо антигена, что неизбежно усложняет стандартные измерения и вносит неточности в измеряемые значения количества. Аффинность антитела можно измерять с помощью ряда способов, включая OCTET®, BIACORE®, ELISA и FACS.
[0288] В системе OCTET® используются биосенсоры в формате 96-луночного планшета для представления результатов кинетического анализа. Факты связывания белка и диссоциации можно отслеживать путем измерения связывания одного белка в растворе со вторым белком, иммобилизованным на биосенсоре ForteBio. В случае измерения связывания антител к Psl или PcrV с Psl или PcrV Psl или PcrV иммобилизуют на головках OCTET® с последующим анализом связывания антитела, которое находится в растворе. Ассоциацию и диссоциацию антитела с иммобилизованными Psl или PcrV затем выявляют с помощью сенсорного прибора. Данные затем собирают и экспортируют в GraphPad Prism для построения кривой аффинности.
[0289] Поверхностный плазмонный резонанс (SPR), осуществляемый на BIACORE®, дает ряд преимуществ над традиционными способами измерения аффинности взаимодействий антитело-антиген: (i) нет необходимости в мечении антитела либо антигена; (ii) антитела не нужно предварительно очищать, можно непосредственно применять надосадочную жидкость культуры клеток; (iii) обеспечивается возможность измерений в реальном времени, дающих возможность осуществления быстрого полуколичественного сравнения различных взаимодействий с моноклональным антителом, и эти измерения являются достаточными для многих целей оценивания; (iv) биоспецифическую поверхность можно регенерировать таким образом, что группу различных моноклональных антител можно легко сравнивать в идентичных условиях; (v) аналитические методы полностью автоматизированы, и можно осуществлять обширные серии измерений без вмешательства пользователя. BIAapplications Handbook, версия AB (переиздание 1998 г. ), BIACORE®, кодовый № BR-1001-86; BIAtechnology Handbook, версия AB (переиздание 1998 г. ), BIACORE®, кодовый № BR-1001-84.
[0290] Исследования связывания на основе SPR требуют, чтобы один участник пары связывания был иммобилизован на поверхности сенсора. Иммобилизованный партнер связывания называют лигандом. Партнер связывания в растворе называют аналитом. В некоторых случаях лиганд прикреплен к поверхности опосредованно путем связывания с другой иммобилизованной молекулой, которую называют молекулой захвата. Отклик SPR отражает изменение массовой концентрации на поверхности детектора по мере того, как происходит связывание аналитов или диссоциация.
[0291] На основе SPR с помощью измерений в реальном времени на BIACORE® отслеживают взаимодействия, как только они происходят.Методика подходит для определения кинетических параметров. Чрезвычайно просто осуществлять сравнительное ранжирование значений аффинности, и из данных сенсограммы можно получить как кинетические константы, так и константы аффинности.
[0292] Если аналит вносят дискретно пульсовым методом на всю поверхность с лигандом, полученную сенсограмму можно разделить на три основные фазы: (i) ассоциация аналита с лигандом во время внесения образца; (ii) состояние равновесия или стационарное состояние во время внесения образца, при котором скорость связывания аналита уравновешена диссоциацией из комплекса; (iii) диссоциация аналита из поверхности во время протекания буфера.
[0293] Фазы ассоциации и диссоциации обеспечивают информацию о кинетике взаимодействия аналита с лигандом (ka и kd, скорости образования комплексов и диссоциации, kd/ka=KD). Фаза равновесия обеспечивает информацию об аффинности взаимодействия аналита с лигандом (KD).
[0294] Программное обеспечение BIAevaluation обеспечивает обширные возможности для построения кривых при помощи как численного интегрирования, так и глобальных алгоритмов подбора. При помощи подходящего анализа данных можно получить отдельные константы скорости и аффинности взаимодействия на основании простых исследований с помощью BIACORE®. Значения аффинности, которые могут быть измерены с помощью этой методики, находятся в очень широком диапазоне, варьируя в диапазоне от мМ до пМ.
[0295] Специфичность в отношении эпитопа является важной характеристикой моноклонального антитела. Картирование эпитопов с помощью BIACORE®, в отличие от традиционных методик с применением радиоиммунологического анализа, ELISA или других способов на основе адсорбции на поверхности, не требует мечения или очищения антител и позволяет осуществлять тесты на специфичность в отношении нескольких сайтов с применением последовательности нескольких моноклональных антител. Дополнительно, большое число анализов можно производить автоматически.
[0296] В парных экспериментах по связыванию тестируется способность двух mAb к одновременному связыванию с одним и тем же антигеном. MAb, направленные против отдельных эпитопов, будут связываться независимо, тогда как mAb, направленные против идентичных или близкородственных эпитопов, будут препятствовать друг другу в связывании. Эти эксперименты по связыванию, осуществляемые с помощью BIACORE®, не вызывают затруднений при проведении.
[0297] Например, можно применять молекулу захвата для связывания с первым mAb с последующим последовательным добавлением антигена и второго mAb. На сенсограммах будет выявлено: 1) какое количество антигена связывается с первым mAb, 2) до какой степени второе mAb связывается с антигеном, прикрепленным к поверхности, 3) если второе mAb не связывается, изменяет ли результаты изменение порядка проведения парного теста на обратный.
[0298] Пептидное ингибирование является другой методикой, применяемой для картирования эпитопов. Этот способ может дополнять парные исследования связывания антител и может соотносить функциональные эпитопы со структурными признаками, когда известна первичная последовательность антигена. Пептиды или фрагменты антигена тестируют в отношении ингибирования связывания различных mAb с иммобилизованным антигеном. Полагают, что пептиды, препятствующие связыванию данного mAb, являются родственными по структуре эпитопу, определенному с помощью этого mAb.
XII. ВВЕДЕНИЕ
[0299] Композицию, содержащую домен, связывающийся с Psl, или домен, связывающийся с PcrV, либо композицию, содержащую как домен, связывающийся с Psl, так и домен, связывающийся с PcrV, вводят таким образом, чтобы они обеспечивали синергический эффект в лечении инфекции, вызываемой Pseudomonas, у пациента. Введение может осуществляться любыми подходящими способами, при условии, что введение обеспечивает необходимый терапевтический эффект, т.е. синергизм. В определенных вариантах осуществления антитела вводят в ходе одного и того же цикла терапии, например, оба антитела вводят субъекту в ходе одного цикла терапии в течение установленного периода времени. В некоторых вариантах осуществления введение антител может осуществляться в ходе последовательного введения в отдельных циклах терапии, например, в первом цикле терапии, включающем введение антитела к Psl, и во втором цикле терапии, включающем введение антитела к PcrV. Дозировка связывающих доменов, вводимых пациенту, будет также зависеть от частоты введения и может быть без труда определена специалистом в данной области.
[0300] В других вариантах осуществления связывающие домены вводят больше одного раза в ходе цикла лечения. Например, в некоторых вариантах осуществления связывающие домены вводят один раз в неделю в течение трех недель подряд в трех- или четырехнедельном цикле лечения.
[0301] Введение композиции, содержащей один или несколько связывающих доменов, может осуществляться в один день или в различные дни при условии, что введение обеспечивает необходимый терапевтический эффект.
[0302] Специалистам в данной области будет очевидно, что другие дозы или частоты введения, которые обеспечивают желаемый терапевтический эффект, являются подходящими для применения в настоящем изобретении.
XII. НАБОРЫ
[0303] В других вариантах осуществления настоящее изобретение обеспечивает наборы, которые можно применять для выполнения способов, описанных в настоящем документе. В определенных вариантах осуществления набор содержит связывающую молекулу, раскрытую в данном документе, в одном или нескольких контейнерах. Специалист в данной области без труда признает, что раскрытые связывающие домены, полипептиды и антитела по настоящему изобретению могут быть легко включены в один из установленных форматов набора, которые хорошо известны в данной области техники.
***
[0304] В практических аспектах настоящего изобретения будут использоваться, если не указано иное, традиционные методики клеточной биологии, культивирования клеток, молекулярной биологии, биологии трансгенных организмов, микробиологии, рекомбинантных ДНК и иммунологии, которые находятся в пределах компетентности специалиста в данной области. Такие методики в полном объеме объясняются в литературе. См., например, Molecular Cloning A Laboratory Manual, 2nd Ed., Sambrook et al, ed., Cold Spring Harbor Laboratory Press: (1989); Molecular Cloning: A Laboratory Manual, Sambrook et al, ed., Cold Springs Harbor Laboratory, New York (1992), DNA Cloning, D.N. Glover ed., Volumes I and II (1985); Oligonucleotide Synthesis, M.J. Gait ed., (1984); Mullis et al, патент США №4,683,195; Nucleic Acid Hybridization, B.D. Hames & S.J. Higgins eds. (1984); Transcription And Translation, B.D. Hames & S.J. Higgins eds. (1984); Culture Of Animal Cells, R.I. Freshney, Alan R. Liss, Inc., (1987); Immobilized Cells And Enzymes, IRL Press, (1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); научный труд Methods In Enzymology, Academic Press, Inc., N.Y.; Gene Transfer Vectors For Mammalian Cells, J.H. Miller and M.P. Calos eds., Cold Spring Harbor Laboratory (1987); Methods In Enzymology, Vols. 154 and 155 (Wu et al eds.); Immunochemical Methods In Cell And Molecular Biology, Mayer and Walker, eds., Academic Press, London (1987); Handbook Of Experimental Immunology, Volumes I-IV, D.M. Weir and C.C. Blackwell, eds., (1986); Manipulating the Mouse Embryo, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., (1986); и в Ausubel et al, Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1989).
[0305] Общие принципы конструирования антител изложены в Antibody Engineering, 2nd edition, C.A.K. Borrebaeck, Ed., Oxford Univ. Press (1995). Общие принципы конструирования белков изложены в Protein Engineering, A Practical Approach, Rickwood, D., et al., Eds., IRL Press at Oxford Univ. Press, Oxford, Eng. (1995). Общие принципы относительно антител и связывания антител с гаптенами изложены в Nisonoff, A., Molecular Immunology, 2nd ed., Sinauer Associates, Sunderland, MA (1984); и Steward, M.W., Antibodies, Their Structure and Function, Chapman and Hall, New York, NY (1984). Кроме того, стандартные способы иммунологии, известные в данной области техники и не описанные конкретно, как правило, соответствуют таковым в Current Protocols in Immunology, John Wiley & Sons, New York; Stites et al. (eds), Basic and Clinical-Immunology (8th ed.), Appleton & Lange, Norwalk, CT (1994), и Mishell and Shiigi (eds), Selected Methods in Cellular Immunology, W.H. Freeman and Co., New York (1980).
[0306] Авторитетные справочные издания, в которых изложены общие принципы иммунологии, включают Current Protocols in Immunology, John Wiley & Sons, New York; Klein, J., Immunology: The Science of Self-Nonself Discrimination, John Wiley & Sons, New York (1982); Kennett, R., et al., eds., Monoclonal Antibodies, Hybridoma: A New Dimension in Biological Analyses, Plenum Press, New York (1980); Campbell, A., "Monoclonal Antibody Technology" в Burden, R., et al, eds., Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 13, Elsevere, Amsterdam (1984), Kuby Immunology 4th ed. Ed. Richard A. Goldsby, Thomas J. Kindt and Barbara A. Osborne, H. Freemand & Co. (2000); Roitt, I., Brostoff, J. and Male D., Immunology 6th ed. London: Mosby (2001); Abbas A., Abul, A. and Lichtman, A., Cellular and Molecular Immunology Ed. 5, Elsevier Health Sciences Division (2005); Kontermann and Dubel, Antibody Engineering, Springer Verlan (2001); Sambrook and Russell, Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Press (2001); Lewin, Genes VIII, Prentice Hall (2003); Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Press (1988); Dieffenbach and Dveksler, PCR Primer Cold Spring Harbor Press (2003).
ПРИМЕРЫ
Пример 1. Конструирование и скрининг фаг-дисплейных библиотек антител человека
[0307] В этом примере описывается подход по методу пэннинга, индифферентного в отношении мишени, с помощью целых клеток, в котором применяют фаговые библиотеки антител человека, полученные как от "наивных", так и от выздоравливающих пациентов, инфицированных P. aeruginosa, для идентификации новых антигенов, защищающих от инфекции, вызываемой Pseudomonas (фигура 1A). Анализы, включенные в этапы функционального скрининга in vitro, включают анализы опсонофагоцитарного уничтожения (ОРК) и анализы прикрепления к клеткам с применением эпителиальных клеток линии A549. Кандидатов-лидеров, определенных на основе превосходящей активности in vitro, тестировали в моделях острой пневмонии, кератита и ожоговой инфекции с инфицированием Р. aeruginosa.
[0308] На фигуре 1B показано конструирование фаг-дисплейной библиотеки антител пациента. Через 7-10 дней после постановки диагноза смешивали цельную кровь от 6 выздоравливающих пациентов с последующей экстракцией РНК и конструированием фаговых библиотек согласно описанному ранее (Vaughan, T.J., et al, Nat Biotechnol 14, 309-314 (1996); Wrammert, J., et al, Nature 453, 667-671 (2008)). На фигуре 1C показано, что конечная библиотека клонированных scFv содержит 5,4×108 трансформантов, и секвенирование выявило, что 79% генов scFv были полной длины и внутрирамочными. Петли CDR3 VH, зачастую важные для определения специфичности в отношении эпитопа, перед селекцией в библиотеке различались на 84% на аминокислотном уровне.
[0309] В дополнение к библиотеке от пациентов, для выделения антител (Vaughan, T.J., et al, Nat Biotechnol 14, 309-314 (1996)) применяли фаг-дисплейную библиотеку наивных scFv человека, содержащую до 1×1011 участников связывания (Lloyd, C, et al, Protein Eng Des Sel 22, 159-168 (2009)). Убитую нагреванием P. aeruginosa (1×109) иммобилизировали в пробирках IMMUNO™ (Nunc; MAXISORP™) с последующими циклами селекции по методу фагового дисплея согласно описанному (Vaughan, T.J., et al, Nat Biotechnol 14, 309-314 (1996)), за исключением того, что в качестве элюирующего буфера применяли триэтаноламин (100 нМ). Для селекции P. aeruginosa в суспензии убитые нагреванием клетки блокировали с последующим добавлением блокированного фага к клеткам. После промывания применяли элюированный фаг для инфицирования клеток E. coli согласно описанному (Vaughan, 1996). Освобождение фага из E. coli и связывание с убитой нагреванием P. aeruginosa выполняли с помощью ELISA согласно описанному (Vaughan, 1996).
[0310] После разработки и валидации методики селекции в отношении аффинности с использованием целых клеток как новую библиотеку от выздоравливающего пациента, так и ранее сконструированную наивную библиотеку (Vaughan, T.J., et al, Nat Biotechnol 14, 309-314 (1996)) подвергали селекции в отношении аффинности в суспензиях штамма 3064 P. aeruginosa, имеющего полный O-антиген, а также штамма с изогенной мутацией в wapR, у которого отсутствует поверхностная экспрессия O-антигена. На фигуре 1D показано, что выходные титры в последовательных циклах селекции в библиотеках от пациентов, как было обнаружено, увеличиваются с большей скоростью применительно к библиотеке от пациента, чем применительно к наивной библиотеке (1×107 в сравнении с 3×105 в цикле 3, соответственно). В дополнение, также было обнаружено, что уровень дупликации последовательностей петель CDR3 VH в библиотеках (мера обогащения клонами в ходе селекции) является более высоким в библиотеке от пациента, достигая 88-92%, по сравнению с 15-25% в наивной библиотеке в цикле 3 (фигура 1D). Отдельные фаги, экспрессирующие scFv, из циклов селекции в отношении аффинности затем подвергали скринингу с помощью ELISA на реактивность в отношении штаммов гетерологичных серотипов P. aeruginosa (фигура 1E). Планшеты для ELISA (Nunc; MAXISORP™) покрывали штаммами P. aeruginosa из суточных культур согласно описанному (DiGiandomenico, A., et al, Infect Immun 72, 7012-7021 (2004)). Разведенные антитела добавляли в блокированные планшеты на 1 час, промывали и обрабатывали вторичными антителами к иммуноглобулинам человека, конъюгированными с HRP, в течение 1 часа с последующими проявлением и анализом согласно описанному (Ulbrandt, N.D., et al, J Virol 80, 7799-7806 (2006)). Преобладающий вид фага, полученный из циклов селекции с использованием целых клеток в обеих библиотеках, давал серотип-специфическую реактивность (данные не показаны). Клоны, проявляющие связывание, независимое от серотипа, в отсутствие неспецифического связывания с E. coli или бычьим сывороточным альбумином, отбирали для дополнительного оценивания.
[0311] Что касается экспрессии IgG, VH- и VL-цепи выбранных антител клонировали в векторы экспрессии IgGl человека, подвергали коэкспрессии в клетках HEK293 и очищали с помощью аффинной хроматографии с белком А согласно описанному (Persic, L., et al, Gene 187, 9-18 (1997)). Для антител IgGl человека, созданных с вариабельными областями от этих выбранных фагов, действующих независимо от серотипа, была подтверждена специфичность в отношении P. aeruginosa, и им отдавали предпочтение при последующем анализе путем связывания с использованием целых клеток с преобладающими клинически значимыми серотипами с помощью FACS-анализа (фигура 1F), поскольку этот способ является более строгим, чем ELISA. Что касается анализов связывания на основе проточной цитометрии, штаммы P. aeruginosa в середине фазы логарифмического роста концентрировали в PBS до OD650, составляющей 2,0. После инкубирования антитела (10 мкг/мл) и бактерий (~1×107 клеток) в течение 1 часа при 4°C со встряхиванием промытые клетки инкубировали с антителом козы к IgG человека, конъюгированным с ALEXA FLUOR 647® (Invitrogen, Карлсбад, Калифорния), в течение 0,5 часа при 4°C. Промытые клетки окрашивали зеленым красителем для бактерий BACLIGHT™ согласно рекомендациям (Invitrogen, Карлсбад, Калифорния). Образцы прогоняли на проточном цитометре LSR II (BD Biosciences) и анализировали при помощи BD FacsDiva (версия 6.1.3) и Flow Jo (версия 9.2; TreeStar). Антителам, проявляющим связывание, выявляемое при помощи FACS, дополнительно отдавали предпочтение при тестировании функциональной активности в анализе опсонофагоцитарного уничтожения (OPK).
Пример 2. Оценивание mAb, способствующих ОРК P. aeruginosa.
[0312] В этом примере описывается оценивание антител IgG1 человека, которым отдали предпочтение, в отношении способствования ОРК P. aeruginosa. На фигуре 2A показано, что за исключением WapR-007 и антитела R347, представляющего собой отрицательный контроль, все антитела опосредовали уничтожение люминесцентного штамма (РАО1.lux) P. aeruginosa серогруппы 05, зависимое от концентрации. WapR-004 и Cam-003 проявляли превосходящую OPK-активность. Анализы ОРК выполняли согласно описанному в (DiGiandomenico, A., et al, Infect Immun 72, 7012-7021 (2004)), с модификациями. Вкратце, анализы выполняли в 96-луночных планшетах с применением по 0,025 мл каждого из компонентов ОРК: штаммов P. aeruginosa; разведенной сыворотки крольчонка; дифференцированных клеток HL-60 и моноклонального антитела. В некоторых анализах ОРК применяли люминесцентные штаммы P. aeruginosa, сконструированные согласно описанному (Choi, K.H., et al, Nat Methods 2, 443-448 (2005)). Люминесцентные анализы ОРК выполняли согласно описанному выше, но с определением относительных единиц активности люциферазы (RLU) при помощи многометочного планшет-ридера Perkin Elmer ENVISION (Perkin Elmer).
[0313] Оценивали способность антител WapR-004 и Cam-003 опосредовать OPK-активность против другого штамма клинически значимого серотипа, имеющего O-антиген, 9882-80.lux. На фигуре 2B показано, что усиленная OPK-активность WapR-004 и Сат-003 распространяется на штамм 9882-80 (O11).
[0314] В дополнение, в этом примере описывается оценивание мутантных WapR-004 (W4) в формате scFv-Fc в отношении способствования ОРК P. aeruginosa. Один мутант, Wap-004RAD (W4-RAD), был особым образом создан посредством сайт-направленного мутагенеза с удалением RGD-мотива в VH. Другие мутантные W4 получали, как указано ниже. Гнездовую ПЦР выполняли согласно описанному (Roux, K.H., PCR Methods Appl 4, S185-194 (1995)) с амплификацией W4-вариантов (полученных в результате соматической гипермутации) из библиотеки scFv, полученной от выздоравливающих пациентов, инфицированных P. aeruginosa, для анализа. Именно из этой библиотеки получали WapR-004. Фрагменты вариантов W4 субклонировали и секвенировали при помощи стандартных методов, известных в данной области техники. Мутантные легкие цепи (LC) W4 рекомбинировали с тяжелой цепью (HC) WapR-004 с получением мутантных W4 в формате scFv-Fc. В дополнение, мутантные тяжелые цепи (HC) WapR-004 RAD рекомбинировали с родительскими LC M7 и M8 в формате scFv-Fc. Конструкты получали при помощи стандартных методов, известных в данной области техники. На фигуре 11 (A-M) показано, что, за исключением антитела R347, представляющего собой отрицательный контроль, все мутантные WapR-004 (W4) опосредовали уничтожение люминесцентного штамма (PAO1.lux) P. aeruginosa серогруппы 05, зависимое от концентрации.
[0315] Ген вариабельной области WapR-004-RAD изменяли с получением гена в конфигурации зародышевой линии для снижения потенциальной иммуногенности, производя WapR-004, кодируемое геном в конфигурации зародышевой линии ("WapR-004-GL"), которое подвергали оптимизации лидерных молекул посредством сайт-направленного мутагенеза. Клоны с улучшенной аффинностью в отношении Psl отбирали на этапах конкурентного скрининга. Лучшие клоны ранжировали по улучшению аффинности и анализировали в функциональном анализе in vitro. 14 клонов, подвергнутых оптимизации лидерных молекул, представляют собой Psl0096, Psl0170, Psl0225, Psl0304, Psl0337, Psl348, Psl0567, Psl0573, Psl0574, Psl0582, Psl0584, Psl0585, Psl0588 и Psl0589.
Пример 3. Антитела к P. aeruginosa, действующие независимо от серотипа, нацеливаются на экзополисахарид Psl
[0316] В этом примере описывается идентификация мишени для антител к P. aeruginosa, полученных из фенотипического скрининга. Анализ мишени проводили для тестирования того, нацеливались ли антитела, действующие независимо от серотипа, на белковые или углеводные антигены. В ELISA в отношении экстрактов из целых клеток PAO1, подвергнутых полному расщеплению с помощью протеиназы K, не наблюдали снижения связывания, что позволяет предположить, что реактивность была нацелена на доступные поверхностные углеводные остатки (данные не показаны). Конструировали изогенных мутантов по генам, отвечающим за биосинтез O-антигена, альгината и центрального олигосахарида LPS: wbpL (без O-антигена); wbpL/algD (без O-антигена и альгината); rmlC (без O-антигена и с усеченной внешней частью центрального олигосахарида) и galU (без O-антигена и с усеченной внутренней частью центрального олигосахарида). Мутантных P. aeruginosa конструировали на основе стратегии замещения аллелей, описанной Schweizer (Schweizer, H.P., Mol Microbiol 6, 1195-1204 (1992); Schweizer, H.D., Biotechniques 15, 831-834 (1993)). Векторы мобилизовали из штамма S17.1 E. coli в штамм PAO1 P. aeruginosa; рекомбинантов выделяли согласно описанному (Hoang, T.T., et al, Gene 212, 77-86 (1998)). Делецию в генах подтверждали с помощью ПЦР. Мутантных P. aeruginosa дополняли конструктами на основе pUCP30T, имеющими гены дикого типа. Реактивность антител определяли посредством непрямого ELISA на планшетах, покрытых вышеуказанными штаммами Р. aeruginosa: На фигуре 3A показано, что связывание Cam-003 с мутантом по wbpL или двойным мутантом по wbpL/algD не подвергалось воздействию, однако связывание с мутантами по rmlC и galU прекращалось. Несмотря на то, что эти результаты согласовывались со связыванием с центральным олигосахаридом LPS, реактивность в отношении LPS, очищенного из PAO1, не наблюдалась. Недавно было показано, что гены rmlC и galU требуются для биосинтеза экзополисахарида Psl, повторяющегося пентасахаридного полимера, состоящего из D-маннозы, L рамнозы и D-глюкозы. Тестировали связывание Cam-003 с изогенным FAO1ΔpslA, характеризующимся нокаутом pslA, поскольку pslA требуется для биосинтеза Psl (Byrd, M.S., et al, Mol Microbiol 73, 622-638 (2009)). Связывание Cam-003 с PAO1ΔpslA прекращалось при тестировании с помощью ELISA (фигура 3B) и FACS (фигура 3C), несмотря на то, что молекула LPS этого мутанта не подвергалась воздействию (фигура 3D). Связывание с Cam-003 восстанавливалось у тройного мутанта PAO1×wbpL/algD/pslA, дополненного pslA (фигура 3Е), так же, как и способность Cam-003 опосредовать опсоническое уничтожение дополненного PAO1ΔpslA, в отличие от мутанта (фигуры 3F и 3G). Связывание антитела Cam-003 с экзополисахаридом Pel мутантного штамма также не подвергалось воздействию, что дополнительно подтверждает, что Psl является мишенью для антитела (фигура 3Е). Анализы связывания подтверждали, что остальные антитела также связываются с Psl (фигуры 3H и 31).
Пример 4. MAb к Psl блокируют прикрепление P. aeruginosa к культивируемым эпителиальным клеткам
[0317] В этом примере показано, что антитела к Psl блокируют присоединение P. aeruginosa к эпителиальным клеткам. Антитела к Psl добавляли к конфлюэнтному монослою клеток A549 (линии клеток аденокарциномы из базального эпителия альвеол человека), выращиваемых в непрозрачных 96-луночных планшетах (Nunc Nunclon Delta). Люминесцентный штамм PAO1 P. aeruginosa (PAO1.lux) в логарифмической фазе роста добавляли при значении MOI, составляющем 10. После инкубирования PAO1.lux с клетками А549 при 37°C в течение 1 часа клетки A549 промывали с последующим добавлением LB+0,5% глюкозы. Бактерии оценивали количественно после непродолжительного инкубирования при 37°C, как выполнялось в анализе ОРК, описанном в примере 2. Измерения от лунок без клеток A549 применяли для введения поправки на неспецифическое связывание. На фигуре 4 показано, что, за исключением Cam-005 и WapR-007, все антитела уменьшали присоединение PAO1.lux к клеткам A549 в зависимости от дозы. MAb, показавшие наилучшие результаты в анализах ОРК, WapR-004 и Cam-003 (см. фигуры 2A-B и пример 2), были также наиболее активными в ингибировании прикрепления клеток P. aeruginosa к клеткам легочного эпителия A549, обеспечивая уменьшение на до ~80% по сравнению с отрицательным контролем. WapR-016 было третьим наиболее активным антителом, показывающим ингибирующую активность, сходную с таковой WapR-004 и Cam-003, но при концентрации антитела, в 10 раз более высокой.
Пример 5. In vivo пассированные штаммы P. aeruginosa поддерживают/повышают экспрессию Psl
[0318] Для тестирования того, поддерживается ли экспрессия Psl in vivo, мышам интраперитонеально инъецировали изоляты P. aeruginosa с последующим сбором бактерий с помощью перитонеального лаважа через четыре часа после инфицирования. Наличие Psl анализировали с контрольным антителом и Cam-003 с помощью проточной цитометрии, поскольку условия для связывания антител являются более жесткими и позволяют проводить количественную оценку клеток, в которых имеет место или отсутствует экспрессия Psl. Что касается связывания ex vivo, бактериальный инокулят (0,1 мл) получали из суточной культуры в чашке с TSA и доставляли интраперитонеально мышам BALB/c. Через 4 часа после контрольного заражения бактерии собирали, RBC лизировали, подвергали действию ультразвука и ресуспендировали в PBS с добавлением 0,1% Tween-20 и 1% BSA. Образцы окрашивали и анализировали согласно описанному ранее в примере 1. На фигуре 5 показано, что бактерии, собранные после перитонеального лаважа с тремя штаммами P. aeruginosa дикого типа, показывали сильное окрашивание с помощью Cam-003, которое было сравнимым с таковым у культивируемых бактерий в логарифмической фазе роста (сравни фигуры 5A и 5C). Пассированные in vivo бактерии дикого типа проявляли усиленное окрашивание по сравнению с инокулятом (сравни фигуры 5B и 5C). Psl не был выявлен для штамма 6077 и был выявлен на минимальном уровне для штаммов PAO1 (O5) и 6206 (O11, цитотоксичный) в инокуляте. Применительно к инокуляту связывание Cam-003 с бактериями увеличивалось, что указывает на то, что экспрессия Psl поддерживается или повышается in vivo. Штаммы 6077, PAO1 и 6206 дикого типа экспрессируют Psl после пассирования in vivo, однако штамм PAO1, имеющий делецию в pslA (PAO1ΔpslA), не способен реагировать с Cam-003. По этим результатам дополнительно делается акцент на Psl в качестве мишени для моноклональных антител.
Пример 6. Показатели выживаемости животных, обработанных моноклональными антителами Cam-003 и WapR-004 к Psl, в модели острой пневмонии с инфицированием Р. aeruginosa
[0319] Антитела или PBS вводили за 24 часа до инфицирования в каждой модели.
Модели острой пневмонии, кератита и термического повреждения с инфицированием P. aeruginosa функционировали согласно описанному (DiGiandomenico, A., et al, Proc Natl Acad Sci USA 104, 4624-4629 (2007)) с модификациями. В модели острой пневмонии мышей BALB/c (The Jackson Laboratory) инфицировали штаммами P. aeruginosa, суспендированными в 0,05 мл инокулята. В модели термического повреждения мыши CF-1 (Charles River) получали ожог 10% общей площади поверхности тела с помощью металлического клейма, нагретого до 92°C, в течение 10 секунд. Животных инфицировали подкожно штаммом 6077 P. aeruginosa в указанной дозе. Что касается экспериментов, связанных с нагрузкой органа, у мышей индуцировали острую пневмонию с последующим сбором легких, селезенок и почек через 24 часа после инфицирования для определения CFU.
[0320] В модели острой смертельной пневмонии оценивали действие моноклональных антител Cam-003 и WapR-004 против штаммов P. aeruginosa, представляющим наиболее часто встречающиеся серотипы, ассоциированные с заболеванием с клиническими проявлениями. На фигурах 6A и 6C показана значительная зависимая от концентрации выживаемость обработанных с помощью Cam-003 мышей, инфицированных штаммами PAO1 и 6294, по сравнению с представителями контрольной группы. На фигурах 6B и 6D показано, что полную защиту от контрольного заражения 33356 и цитотоксичным штаммом 6077 предоставляло Cam-003 при 45 и 15 мг/кг, тогда как при 5 мг/кг наблюдали 80 и 90% выживаемость в отношении 33356 и 6077, соответственно. На фигурах 6E и 6F показана значительная зависимая от концентрации выживаемость мышей, обработанных с помощью WapR-004, в модели острой пневмонии со штаммами 6077 (O11) (8×105 CFU) (фигура 6Е) или 6077 (O11) (6×105 CFU) (фигура 6F).
[0321] Cam-003 и WapR-004 затем изучали в отношении их способности снижать нагрузку органа, вызываемую P. aeruginosa, в легких и распространение в отдаленные органы, и позже животных обрабатывали с помощью WapR-004, Cam-003 в различных концентрациях или контрольными антителами в нескольких различных концентрациях. Cam-003 было эффективным при снижении нагрузки на легкие, вызываемой P. aeruginosa, в отношении всех четырех тестируемых штаммов. Cam-003 было наиболее эффективным против высокопатогенного цитотоксичного штамма 6077, где низкая доза была такой же эффективной, как и более высокая доза (фигура 7D). Cam-003 также обладало выраженным эффектом в отношении снижения диссеминации в селезенку и почки у мышей, инфицированных PAO1 (фигура 7A), 6294 (фигура 7C) и 6077 (фигура 7D), тогда как у мышей, инфицированных 33356, не наблюдалась диссеминация в эти органы (фигура 7B). На фигурах 7E и 7F показано, что, аналогично, WapR-004 снижало нагрузку органа после индуцирования острой пневмонии с помощью 6294 (O6) и 6206 (O11). В частности, WapR-004 было эффективным при снижении диссеминации P. aeruginosa в селезенку и почки у инфицированных мышей.
Пример 7. Конструирование моноклонального антитела V2L2 к PcrV
[0322] Мышей VelocImmune® (Regeneron Pharmaceuticals) иммунизировали с помощью способа ультракороткой иммунизации с r-PcrV, и за определением сывороточных титров следовало изучение связывания с PcrV и нейтрализации гемолитической активности живых P. aeruginosa. Мышей, в сыворотке которых демонстрировалась антигемолитическая активность, умерщвляли, и собирали селезенку и лимфатические узлы (аксиальные, паховые и подколенные). Популяции клеток из этих органов подвергали пэннингу с биотинилированными r-Pcrv для селекции В-клеток, специфичных к PcrV. Выбранные клетки затем сливали с миеломными клетками-партнерами мыши P3X63-Ag8 и высевали в количестве 25000 клеток/лунка в среду для селекции гибридом. Через 10 дней среду из лунок с гибридомами полностью меняли на свежую среду, а еще через 3-4 дня надосадочную жидкость гибридом анализировали в отношении антигемолитической активности. Колонии, демонстрирующие антигемолитическую активность, клонировали по методу серийных разведений при 0,2 клетки/лунка в 96-луночном планшете, и анализ антигемолитической активности повторяли. Клоны, демонстрирующие антигемолитическую активность, приспосабливали к среде для культивирования гибридомы, содержащей IgG на сверхнизком уровне. IgG из кондиционированной среды очищали и анализировали в отношении антигемолитической активности in vitro и в отношении защиты от инфекции, вызываемой P. aeruginosa, in vivo. Антитела также распределяли посредством конкурентного анализа в различные группы. Вариабельные (V) домены антител, представляющих интерес, субклонировали из кДНК, полученных из их различных соответствующих клонов. Субклонированные V-сегменты сливали внутрирамочно с кДНК для соответствующего константного домена в плазмиде для экспрессии у млекопитающих. Рекомбинантные IgG экспрессировали и очищали из клеток HEK293. В случаях, когда из определенного клона получали более чем одну последовательность кДНК, кодирующую V, для идентификации функциональных IgG экспрессировали и характеризовали все комбинации вариабельных тяжелых и легких цепей.
Пример 8. Уровни выживаемости животных, обработанных моноклональными антителами Cam-003, WapR-004 к Psl и моноклональным антителом V2L2 к PcrV, в модели инфекции роговицы, вызываемой P. aeruginosa
[0323] Эффективность Cam-003 и WapR-004 затем оценивали в модели инфекции роговицы, вызываемой P. aeruginosa, в которой делается акцент на способность патогенов прикрепляться к поврежденной ткани и колонизировать ее. На фигурах 8 A-D и 8 F-G показано, что у мышей, получавших Cam-003 и WapR-004, была в значительно меньшей степени выражена патология и они имели уменьшенное количество бактерий в общих гомогенатах глаза по сравнению с тем, что наблюдалось у животных, обработанных отрицательным контролем. На фигуре 8E показано, что Cam-003 было также эффективным при тестировании в модели термического повреждения, обеспечивая значительную защиту при 15 и 5 мг/кг по сравнению с обработкой контрольным антителом. Фигура 8 (Н). Активность моноклональных антител V2L2 к Psl и PcrV тестировали в модели глазного кератита с инфицированием P. aeruginosa на мышах. Мышам C3H/HeN интраперитонеально (IP) инъецировали PBS или контрольное антитело IgGl (R347) при 45 мг/кг, или WapR-004 (α-Psl) при 5 мг/кг, или V2L2 (α-PcrV) при 5 мг/кг за 16 часов до инфицирования 6077 (цитотоксичным O11-1×106 CFU). Непосредственно перед инфицированием мышей анестезировали с последующим нанесением трех царапин на 1 мм на роговицу и поверхностную строму одного глаза каждой мыши с применением иглы 27 калибра под препаровальной лупой с последующим местным применением штамма 6077 P. aeruginosa в 5 мкл инокулята. Глаза фотографировали через 48 часов после инфицирования с последующей классификацией инфекции роговицы путем визуализации глаз под препаровальной лупой. Классификацию инфекции роговицы выполняли согласно описанному ранее в Preston et al. (Preston, MJ., 1995, Infect. Immun. 63: 3497). Вкратце, инфицированные глаза классифицировал через 48 часов после инфицирования штаммом 6077 исследователь, который не был осведомлен о способах лечения животных. Использовалась следующая схема классификации: степень 0, глаз макроскопически идентичен неинфицированному глазу; степень 1, незначительное помутнение, частично закрывающее зрачок; степень 2, плотное помутнение, закрывающее зрачок; степень 3, плотное помутнение, полностью закрывающее зрачок; степень 4, перфорация роговицы (сморщивание глазного яблока). Мыши, систематически получавшие дозы (IP) Cam-003 или WapR-004RAD, показали в значительно меньшей степени выраженную патологию и уменьшенное количество бактериальных колониеобразующих единиц (CFU) в общих гомогенатах глаза по сравнению с тем, что наблюдалось у животных, обработанных контрольным mAb R347. Сходные результаты наблюдались у животных, обработанных V2L2, в сравнении с представителями контрольной группы, обработанными R347.
Пример 9. Антитело Cam-003 с мутацией в Fc, Cam-003-TM, обладает уменьшенными ОРК и эффективностью in vivo, но сохраняет активность против прикрепления к клеткам
[0324] С учетом возможного двойного механизма действия было создано Cam-003 с мутацией в Fc, Cam-003-ТМ, которое имеет мутации в Fc-домене, ослабляющие его взаимодействие с рецепторами Feγ (Oganesyan, V., et al., Acta Crystallogr D Biol Crystallogr 64, 700-704 (2008)), для идентификации того, коррелирует ли защита в большей степени с активностью против прикрепления к клеткам или с ОРК-активностью. Мутантных P. aeruginosa конструировали на основе стратегии замещения аллелей, описанной Schweizer (Schweizer, H.P., Mol Microbiol 6, 1195-1204 (1992); Schweizer, H.D., Biotechniques 15, 831-834 (1993)). Векторы мобилизовали из штамма SI7.1 E. coli в штамм PAO1 P. aeruginosa; рекомбинантов выделяли согласно описанному (Hoang, T.T., et al., Gene 212, 77-86 (1998)). Делецию в генах подтверждали с помощью ПЦР. Мутантных P. aeruginosa дополняли конструктами на основе pUCP30T, имеющими гены дикого типа. На фигуре 9A показано, что Cam-003-TM проявляло 4-кратное снижение OPK-активности по сравнению с Cam-003 (значение EC50 составляло 0,24 и 0,06, соответственно), но было таким же эффективным в анализе прикрепления к клеткам (фигура 9B). На фигуре 9C показано, что Cam-003-TM также было менее эффективным против пневмонии, что позволяет предположить, что для оптимальной защиты необходима оптимальная OPK-активность. Анализы OPK и прикрепления к клеткам выполняли согласно описанному ранее в примерах 2 и 4, соответственно.
Пример 10. Картирование эпитопов и сравнительная аффинность антител к Psl
[0325] Картирование эпитопов выполняли с помощью конкурентного ELISA и подтверждали при помощи проточной системы OCTET® с Psl, полученным из надосадочной жидкости суточной культуры штамма PAO1 P. aeruginosa. Для конкурентного ELISA антитела биотинилировали при помощи сульфо-NHS-биотина EZ-Link и набора для биотинилирования (Thermo Scientific). Планшеты, покрытые антигеном, обрабатывали биотинилированными антителами при EC50, совместно инкубированными с немечеными антителами. После инкубирования со стрептавидином, конъюгированным с HRP (Thermo Scientific), в планшетах производили проявление согласно описанному выше. В конкурентных экспериментах с mAb к Psl определили, что антитела нацеливаются по меньшей мере на три уникальных эпитопа, и их назвали антителами к эпитопам класса 1, 2 и 3 (фигура 10A). Антитела к эпитопам класса 1 и 2 не конкурируют за связывание, однако антитело к эпитопу класса 3, WapR-016, частично ингибирует связывание антител к эпитопам классов 1 и 2.
[0326] Аффинность антител определяли в анализах связывания при помощи OCTET®, применяя Psl, полученный из надосадочной жидкости суточных культур PAO1. KD определяли путем усреднения данных о кинетике связывания каждого антитела в семи концентрациях. Измерения аффинности производили при помощи прибора FORTEBIO® OCTET® 384, применяя планшеты на 384 лунки со скошенным дном. Надосадочную жидкость суточных культур PAO1 ± ген pslA применяли в качестве источника Psl. Образцы загружали на сенсоры OCTET на основе аминопропилсилана (гидратированного в PBS) и блокировали с последующим измерением связывания mAb к Psl при нескольких концентрациях и диссоциацией в PBS+1% BSA. Все методы выполняли согласно описанному (Wang, X., et al, J Immunol Methods 362, 151-160). На основании необработанных данных об ассоциации и диссоциации в виде ДнМ построили кривую с помощью GraphPad Prism. На фигуре 10A показаны значения относительной аффинности связывания антител к Psl, охарактеризованных выше. Антитела к эпитопам класса 2 обладали наиболее высокой аффинностью среди всех антител к Psl. На фигуре 10A также показан перечень данных из экспериментов в отношении прикрепления к клеткам и OPK. На фигуре 10 В показаны значения относительной аффинности связывания и значения EC50 OPK мутантного Wap-004RAD (W4RAD), а также других мутантных W4, подвергнутых оптимизации лидерных молекул посредством сайт-направленного мутагенеза согласно описанному в примере 2. На фигуре 10C показаны значения относительной аффинности связывания Wap-004RAD (W4RAD), WapR-004, кодируемого геном в конфигурации зародышевой линии (W4RAD-GL), а также моноклональных антител к Psl, подвергнутых оптимизации лидерных молекул (Psl0096, Psl0170, Psl0225, Psl0304, Psl0337, Psl348, Psl0567, Psl0573, Psl0574, Psl0582, Psl0584, Psl0585, Psl0588 и Psl0589). Особо отмеченные клоны Psl0096, Psl0225, Psl0337, Psl0567 и Psl0588 выбирали, исходя из их усиленной OPK-активности, как показано в примере 10 ниже.
Пример 11. Оценивание мутантных клонов WapR-004 (W4), подвергнутых оптимизации лидерных молекул, и моноклональных антител к Psl, подвергнутых оптимизации лидерных молекул, в анализе опсонофагоцитарного уничтожения (OPK) P. aeruginosa [0327] В данном примере описывается оценивание мутантных клонов WapR-004 (W4), подвергнутых оптимизации лидерных молекул, и моноклональных антител к Psl, подвергнутых оптимизации лидерных молекул, в способствовании ОРК P. aeruginosa с использованием способа, описанного в примере 2. На фигурах 11A-Q показано, что, за исключением антитела R347, представляющего собой отрицательный контроль, все антитела опосредовали уничтожение люминесцентного штамма (PAO1.lux) P. aeruginosa серогруппы 05, зависимое от концентрации.
Пример 12. Моноклональное антитело V2L2 к PcrV снижает смертность от острой пневмонии, вызываемой несколькими штаммами
[0328] Разнообразие эпитопов PcrV анализировали с использованием трех подходов: способа проточной цитометрии на основе гранул, конкурентного ELISA и вестерн-блоттинга фрагментированных rPcrV. В конкурентных экспериментах с mAb к PcrV определили, что антитела нацеливаются по меньшей мере на шесть уникальных эпитопов, и их назвали антителами к эпитопам класса 1, 2, 3, 4, 5 и 6 (фигура 12A). Антитела к эпитопам класса 2 и 3 частично конкурируют за связывание. MAb, представляющие дополнительные классы эпитопов, класс 1 (V2L7, 3G5, 4C3 и 11A6), класс 2 (1E6 и 1F3), класс 3 (29D2, 4A8 и 2H3), класс 4 (V2L2) и класс 5 (21F1, LE10 и SH3), тестировали в отношении защиты in vivo согласно описанному ниже.
[0329] Новые mAb к PcrV выделяли с использованием технологии гибридом, и наиболее действенные ингибиторы T3SS выбирали с использованием анализа ингибирования лизиса эритроцитов кролика. Анализ процентного ингибирования цитотоксичности проводили для родительского mAb V2L2, mAb 166 (положительного контроля) и R347 (отрицательного контроля), где антитела добавляли к линии бронхоэпителиальных клеток A549, культивируемых в сочетании со штаммом 6077 P. aeruginosa (exoU+) в логарифмической фазе роста при значении MOI, составляющем приблизительно 10. Лизис A549 анализировали по измерению активности высвобожденной лактатдегидрогеназы (LDH), и лизис в присутствии mAb сравнивали с таковым в лунках без mAb для определения процентного ингибирования. MAb V2L2, mAb 166 (положительный контроль) и R347 (отрицательный контроль) оценивали в отношении их способности предупреждать лизис RBC, где антитела смешивали с 6077 P. aeruginosa (exoU+) в логарифмической фазе роста и отмытыми эритроцитами кролика (RBC) и инкубировали в течение 2 часов при 37°. Интактные RBC осаждали, и степень лизиса определяли путем измерения OD405 надосадочной жидкости клеток. Лизис в присутствии mAb к PcrV сравнивали с таковым в лунках без mAb для определения процентного ингибирования. Антитело, представляющее собой положительный контроль, mAb 166, является ранее охарактеризованным антителом к PcrV (J Infect Dis. 186: 64-73 (2002), Crit Care Med. 40: 2320-2326 (2012)).(B) Родительское mAb V2L2 демонстрировало ингибирование цитотоксичности при IC50, составляющей 0,10 мкг/мл, и характеризовалось концентрацией IC50, в 28 раз более низкой, чем таковая для mAb166 (IC50, составляющая 2,8 мкг/мл). (C) V2L2 также демонстрировало предупреждение лизиса RBC при IC50, составляющей 0,37 мкг/мл, и характеризовалось концентрацией IC50, в 10 раз более низкой, чем таковая для mAb166 (IC50, составляющая 3,7 мкг/мл).
[0330] Ген вариабельной области V2L2 полностью изменяли с получением гена в конфигурации зародышевой линии для снижения потенциальной иммуногенности. V2L2 подвергали созреванию аффинности с использованием подхода на основе экономного мутагенеза для рандомизации в каждом положении с участием 20 аминокислот во всех шести CDR, идентифицируя отдельные мутации, улучшающие аффинность. Затем применяли комбинаторную библиотеку, кодирующую все возможные комбинации отдельных мутаций с улучшенной аффинностью. Клоны с улучшенной аффинностью к PcrV отбирали с использованием анализа связывания ELISA в формате IgG. Лучшие клоны ранжировали по улучшению аффинности и анализировали в функциональном анализе in vitro. CDR V2L2 подвергались систематическому мутагенезу, и клоны с улучшенной аффинностью к PcrV отбирали на этапах конкурентного скрининга. Клоны ранжировали по увеличению аффинности и анализировали в функциональном анализе. Как показано на фигуре 12D, лизис RBC анализировали для mAb V2L2, ген которого полностью изменяли с получением гена в конфигурации зародышевой линии (V2L2-GL), оптимизированных mAb V2L2-GL (V2L2-P4M, V2L2-MFS, V2L2-MD и V2L2-MR) и антитела R347, представляющего собой отрицательный контроль, с использованием клеток А549, инфицированных штаммом 6077 Pseudomonas. V2L2-GL, V2L2-P4M, V2L2-MFS, V2L2-MD и V2L2-MR демонстрировали предупреждение лизиса RBC. Как показано на фигуре 12Е, mAb 1Е6, 1F3, 11A6, 29D2, PCRV02 и V2L7 демонстрировали предупреждение лизиса RBC. Как показано на фигуре 12F, V2L2 было более действенным в предупреждении лизиса RBC, чем 29D2.
[0331] Показатели кинетики связывания V2L2-GL и V2L2-MD измеряли с использованием прибора Bio-Rad ProteOn™ XPR36. Антитела захватывали на биосенсорном чипе GLC с использованием реагентов, представляющих собой антитела к IgG человека. Белок rPcrV вводили в нескольких концентрациях, и фаза диссоциации продолжалась в течение 600 секунд. Данные собирали и анализировали с использованием программного обеспечения ProteOn Manager. На фигуре (G-H) показаны значения относительной аффинности связывания антител (G) V2L2-GL и (Н) V2L2-MD. Клон V2L2-MD характеризовался Kd, увеличенной в 2-3 раза по сравнению с таковой V2L2-GL.
[0332] Эффект in vivo введения антител к PcrV исследовали на мышах с использованием модели острой пневмонии. Группы мышей обрабатывали антителом V2L2 при увеличивающихся концентрациях, антителом к PcrV, представляющим собой положительный контроль (mAb166), либо отрицательным контролем (R347), как показано на фигуре 13 (A-B). Группы мышей также обрабатывали антителом V2L2 при увеличивающихся концентрациях, антителом PcrV-02 к PcrV либо отрицательным контролем (R347), как показано на фигуре 13 (C-D). Через двадцать четыре часа после обработки всех мышей инфицировали 5×107 CFU (C) 6294 (О6) или (D) PA103A (O11) Pseudomonas aeruginosa. Как показано на фигуре 13, почти все животные, обработанные контрольным соединением, поддались инфекции через 48 часов после инфицирования. Однако, V2L2 показывал дозозависимый эффект, выражающийся в улучшении выживаемости, даже к 168 часам после инфицирования. Дополнительно, V2L2 обеспечивало значительно более действенную защиту, чем mA166, в тех же дозах (P=0,025, 5 мг/кг для штамма 6077; P<0,0001, 1 мг/кг для штамма 6294).
[0333] Группы мышей обрабатывали 11A6, 3G5 или V2L7 при увеличивающихся концентрациях, 29D2, 1F3, 1E6, V2L2, LE10, SH3, 4A8, 2H. или 21F1 при одинаковых концентрациях, 29D2 при увеличивающихся концентрациях, V2L2 при увеличивающихся концентрациях, антителом PcrV-02 к PcrV либо отрицательным контролем (R347), как показано на фигуре 13 (E-H). Мышам инъецировали интраперитонеально (IP) mAb за 24 часа до интраназального инфицирования штаммом 6077 Pseudomonas (1×106 CFU/животное). Как показано на фигуре 13E, mAb 11A6, 3G5 и V2L7 не обеспечивали защиту in vivo. Как показано на фигуре 13F, mAb 29D2 обеспечивает защиту in vivo. Как показано на фигуре 13G, mAb V2L2 также обеспечивает защиту in vivo. На фигуре 13H показано сравнение in vivo 29D2 и V2L2. На фигуре 131 показано, что mAb V2L2 защищает от дополнительных штаммов Pseudomonas (т.е. 6294 и PA103A).
[0334] Нагрузку органа мышей, инфицированных Pseudomonas, также исследовали в ответ на введение V2L2. На фигуре 14 (A) мышей обрабатывали с помощью R347 (контроль) при 1 мг/кг либо V2L2 при 1 мг/кг, 0,2 мг/кг или 0,07 мг/кг и затем инфицировали интраназально 1,2×106 CFU 6206 Pseudomonas. На фигуре 14 (B) мышей также обрабатывали с помощью R347 (отрицательный контроль) при 15 мг/кг; mAb166 (положительный контроль) при 15,0 мг/кг, 5,0 мг/кг или 1,0 мг/кг либо V2L2 при 5,0 мг/кг, 1,0 мг/кг или 0,2 мг/кг и затем инфицировали интраназально 5,5×106 CFU 6206 Pseudomonas. Как показано на фигуре 14 (A-B), в то время как V2L2 оказывало небольшой эффект на почечный клиренс, оно значительно снижало диссеминацию как в легкие, так и в селезенку дозозависимым образом. Кроме того, V2L2 обеспечивало значительно большее снижение CFU в органе, чем mA166 в тех же дозах (P<0,0001, 1 мг/кг, легкое).
Пример 13. Активность in vivo комбинированной терапии с применением антител WapR-004 (к Psl) и V2L2 (к PcrV)
[0335] Эффект in vivo комбинированного введения доменов, связывающихся с Psl и PcrV, дополнительно исследовали на мышах с применением антител V2L2 и WapR-004 (RAD). Группы мышей обрабатывали с помощью R347 (2,1 мг/кг - отрицательный контроль), V2L2 (0,1 мг/кг), W4-RAD (0,5 мг/кг) или комбинации V2L2/W4 (каждое при 0,1, 0,5, 1,0 либо 2,0 мг/кг). Через двадцать четыре часа после введения антитела всех мышей инфицировали инокулятом, содержащим 5,25×105 CFU 6206 (O11-ExoU+). Через двадцать четыре часа после инфицирования легкие, селезенки и почки собирали, гомогенизировали и высевали для определения количества колониеобразующих единиц (CFU) на грамм ткани. Как показано на фигуре 15, как V2L2, так и W4 при тестируемых концентрациях были эффективными в снижении нагрузки органа, а комбинация V2L2/W4 продемонстрировала дополнительный эффект в отношении тканевого клиренса. Гистологическое исследование ткани легкого выявило меньшее кровоизлияние, меньший отек и меньший воспалительный инфильтрат по сравнению с мышами, получавшими V2L2 или WapR-004 в отдельности (таблица 5).
[0336] Животных, иммунизированных сходным образом, также оценивали в отношении выживаемости при острой инфекционной пневмонии.
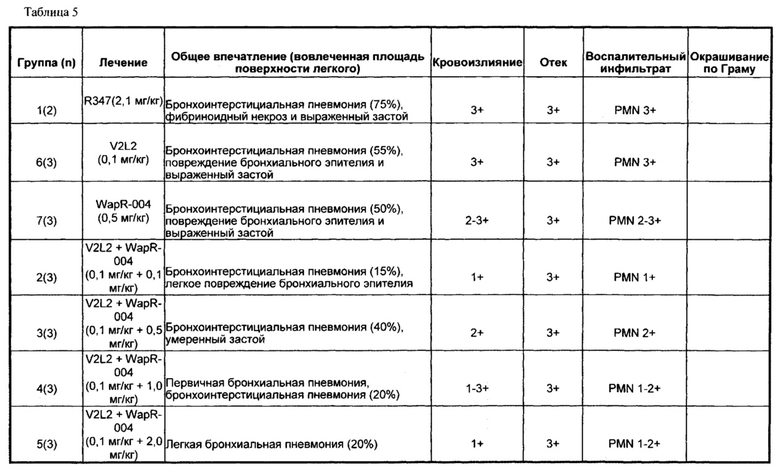
Пример 14. Показатели выживаемости животных, обработанных моноклональным антителом V2L2 к PcrV, в модели острой пневмонии с инфицированием P. aeruginosa
[0337] Оценивали действие моноклональных антител V2L2-GL, V2L2-MD, V2L2-A, V2L2-C, V2L2-PM4 и V2L2-MFS против штамма 6077 P. aeruginosa в модели острой смертельной пневмонии, как ранее описано в примере 11. На фигурах 16 (A-F) показана выживаемость всех мышей, обработанных с помощью V2L2, инфицированных штаммом 6077, по сравнению с представителями контрольной группы. Однако не наблюдалось значительных различий в выживаемости между группами, обработанными антителами V2L2 в дозах 0,5 мг/кг и 1 мг/кг (A-C) либо 0,5 мг/кг и 0,1 мг/кг (D-F). На фигурах 16 (G-I) показана выживаемость всех мышей, обработанных с помощью V2L2, инфицированных штаммом 6077, по сравнению с представителями контрольной группы. Не наблюдалось значительных различий в выживаемости между группами, обработанными антителами V2L2 в дозах 0,5 мг/кг и 1 мг/кг (G-I). (A-H)
[0338] Все контрольные мыши поддались инфекции примерно через 48 часов после инфицирования.
Пример 15. Конструирование биспецифических антител WapR-004/V2L2
[0339] На фигуре 17A показаны модели биспецифических конструктов на основе антитела к TNFα. Что касается Bsl-TNFα/W4, scFv W4 слит с амино-концом VH антитела к TNFα с помощью линкера (G4S)2. Что касается Bs2-TNFα/W4, scFv W4 слит с амино-концом VH антитела к TNFα с помощью линкера (G4S)2. Что касается Bs3-TNFα/W4, scFv W4 слит с карбокси-концом CH3 с помощью линкера (G4S)2.
[0340] Поскольку комбинация WapR-004+V2L2 обеспечивает защиту при контрольном заражении Pseudomonas, получали биспецифические конструкты, содержащие scFv WapR-004 (W4-RAD) и IgG V2L2 (фигура 17 В). Для образования Bs2-V2L2-2C scFv W4-RAD сливали с N-концом VH V2L2 с помощью линкера (G4S)2. Для образования Bs3-V2L2-2C scFv W4-RAD сливали с С-концом СНЗ с помощью линкера (G4S)2. Для образования Bs4-V2L2-2C scFv W4-RAD вставляли в шарнирную область, связанную с помощью линкера (G4S)2 на N-концевой и С-концевой части scFv. Для образования Bs2-W4-RAD-2C, scFv V2L2 сливали с амино-концом VH W4-RAD с помощью линкера (G4S)2.
[0341] Для образования scFv W4-RAD для конструкта Bs3 VH и VL W4-RAD амплифицировали с помощью ПЦР. Праймерами, применяемыми для амплификации VH W4-RAD, были прямой праймер для VH W4-RAD, который охватывает линкер (G4S)2 и 22 п.о. N-концевой последовательности VH (GTAAAGGCGGAGGGGGATCCGGCGGAGGGGGCTCTGAGGTGCAGCTGTTGG AGTCGG (SEQ ID NO: 224)); и обратный праймер для VH W4-RAD, который охватывает часть линкера (G4S)4 и 22 п.о. C-концевой последовательности VH (GATCCTCCGCCGCCGCTGCCCCCTCCCCCAGAGCCCCCTCCGCCACTCGAGA CGGTGACCAGGGTC (SEQ ID NO: 225). Аналогично, VL W4-RAD амплифицировали с помощью ПЦР с использованием следующих прайм еров: прямого праймера для VL W4-RAD, который охватывает часть линкера (G4S)2 и 22 п.о. N-концевой последовательности VL (AGGGGGCAGCGGCGGCGGAGGATCTGGGGGAGGGGGCAGCGAAATTGTGTT GACACAGTCTC (SEQ ID NO: 226)); и обратного праймера для VL W4-RAD, который охватывает часть векторной последовательности и 22 п.о. C-концевой последовательности VL (CAATGAATTCGCGGCCGCTCATTTGATCTCCAGCTTGGTCCCAC (SEQ ID NO: 227). Перекрывающиеся фрагменты затем сливали вместе с образованием scFv W4-RAD.
Последовательность scFv W4-RAD в векторе Bs3: подчеркнутые последовательности представляют собой линкер G4S

[0342] После амплификации фрагмента scFv W4-RAD его затем очищали из геля и лигировали в вектор Bs3, который расщепляли с помощью BamHI/Notl. Лигирование осуществляли с использованием системы In-Fusion с последующим введением в компетентные клетки Stellar путем трансформации. Колонии секвенировали для подтверждения наличия правильной вставки scFv W4-RAD.
[0343] Для образования Bs3-V2L2-2C, часть IgG в векторе Bs3 заменяли на IgG V2L2. Вкратце, вектор Bs3, который содержит scFv W4-RAD, расщепляли с помощью BssHII/SalI, и полученную полосу вектора очищали из геля. Подобным образом, вектор, содержащий вектор V2L2, расщепляли с помощью BssHII/SalI, и вставку V2L2 очищали из геля. Вставку V2L2 затем лигировали в вектор Bs3 с scFv W4-RAD, и колонии секвенировали для подтверждения наличия правильной вставки IgG V2L2.
[0344] Подобный подход применяли для образования Bs2-V2L2-2C.
Последовательности scFv W4-RAD-VH V2L2 в векторе Bs2: подчеркнутые последовательности представляют собой линкер G4S

[0345] Для амплификации scFv W4-RAD применяли следующие праймеры.
Праймер для VH (прямой) и праймер для VL (обратный): прямой праймер для VH W4-RAD для вектора Bs2, который охватывает некоторый интрон, 3'-концевой сигнальный пептид и 22 п.о. N-концевой последовательности VH W4-RAD (TTCTCTCCACAGGTGTACACTCCGAGGTGCAGCTGTTGGAGTCGG (SEQ ID NO: 230)) и обратный праймер для VL W4-RAD для вектора Bs2, который охватывает линкер (G4S)2 и 32 п.о. C-концевой последовательности VL (CCCCCTCCGCCGGATCCCCCTCCGCCTTTGATCTCCAGCTTGGTCCCACAGCC GAAAG (SEQ ID NO: 231)).
[0346] Для амплификации области VH V2L2 применяли следующие праймеры: прямой праймер для VH V2L2, который охватывает линкер (G4S)2 и 22 п.о. N-концевой последовательности VH V2L2 (GGCGGAGGGGGATCCGGCGGAGGGGGCTCTGAGATGCAGCTGTTGGAGTCT GG (SEQ ID NO:232)), и обратный праймер для VH V2L2, который охватывает некоторые N-концевые последовательности CH1 и 22 п.о. C-концевой последовательности VH V2L2 (ATGGGCCCTTGGTCGACGCTGAGGAGACGGTGACCGTGGTC (SEQ ID NO: 233)).
[0347] Эти праймеры затем применяли для амплификации VH V2L2, которую затем соединяли путем перекрывания scFv W4-RAD и VH V2L2 с получением scFv W4-RAD-VH V2L2. scFv W4-RAD-VH V2L2 затем лигировали в вектор Bs2 путем очищения scFv W4-RAD-VH V2L2 из геля (после ПЦР с перекрывающимися праймерами); вектор Bs2 расщепляли с помощью BsrGI/SalI, и полосу вектора очищали из геля. scFv W4-RAD-VH V2L2 затем лигировали в вектор Bs2 с помощью системы In-Fusion и вводили в компетентные клетки Stellar путем трансформации, а в колониях подтверждали наличие правильной вставки scFv W4-RAD-VH V2L2. Для замены VL в векторе Bs2 на VL V2L2 вектор Bs2, который содержит scFv W4-RAD-VH V2L2, расщепляли с помощью BssHII/BsiWI, и полосу вектора очищали из геля. Вектор pOE-V2L2 затем расщепляли с помощью BssHII/BsiWI, и вставку VL V2L2 очищали из геля. Вставку VL V2L2 затем лигировали в вектор Bs2 с scFv W4-RAD-VH V2L2, и колонии секвенировали для подтверждения наличия правильной вставки IgG V2L2.
[0348] В заключение применяли сходный подход на основе ПЦР для образования конструкта Bs4-V2L2-2C. Шарнирная область и линкерная последовательность показаны ниже.
Шарнирная область и линкерная последовательность

Последовательности scFv W4-RAD в векторе Bs4: scFv W4-RAD выделены жирным курсивом, при этом линкеры G4S подчеркнуты и выделены жирным курсивом; шарнирные области подчеркнуты двойной чертой

scFv W4-RAD показаны жирным курсивом, при этом линкеры G4S подчеркнуты и выделены жирным курсивом

[0349] ScFv W4-RAD образовывали при помощи ПЦР и следующих праймеров: прямой праймер для VH W4-RAD для вектора Bs4, который охватывает некоторые линкерные последовательности и 24 п. о. N-концевой последовательности VH W4-RAD (GAGGTGCAGCTGTTGGAGTCGGGC (SEQ ID NO:236)); и обратный праймер для VL W4-RAD для вектора Bs4, который охватывает некоторую шарнирную последовательность, линкер и 21 п.о. C-концевой последовательности VL W4-RAD (GTGTGAGTTTTGTCggatccCCCTCCGCCAGAGCCACCTCCGCCTTTGATCTCCA GCTTGGTCCC (SEQ ID NO: 237)).
[0350] ScFv W4-RAD затем лигировали в вектор Bs4 с получением Bs4-V2L2-2C путем очищения scFv W4-RAD из геля (после ПЦР); вектор Bs4-V2L2 расщепляли с помощью BamHI, и полосу вектора очищали из геля. ScFv W4-RAD лигировали в вектор Bs4 с помощью системы In-Fusion, и вектор вводили в компетентные клетки Stellar путем трансформации. Колонии секвенировали для подтверждения наличия правильной вставки scFv W4-RAD.
[0351] Последовательности легкой цепи и тяжелой цепи конструкта Bs4-V2L2-2C приведены под SEQ ID NO 327 и 328, соответственно.
Пример 16. Биспецифическое антитело к Psl/PcrV способствует выживаемости в моделях пневмонии
[0352] В качестве исходной задачи тестировали биспецифические антитела Bs2 и Bs3 для изучения того, сохраняют ли они свою активность W4 или V2L2 в биспецифическом формате. Для scFv родительского W4 образовывали биспецифическое антитело, содержащее W4 и плечо, связывающееся с TNF-альфа. Анализ прикрепления к клеткам проводили, как описано выше, с применением люминесцентного штамма PAO1.lux P. aeruginosa. Как показано на фигуре 18, все биспецифические конструкты проявляли свойства, аналогичные таковым родительского конструкта W4-IgG1.
[0353] Как показано на фигуре 19 (A-C), процентное ингибирование цитотоксичности анализировали как для Bs2-V2L2, так и Bs3-V2L2 с применением клеток, инфицированных как (A) 6206, так и (B) 6206ΔpslA, и (C) процентное ингибирование лизиса RBC анализировали для Bs2-V2L2-2C, Bs3-V2L2-2C и Bs4-V2L2-2C с применением клеток, инфицированных 6206. Как показано на фигуре 19 (A-C), все биспецифические антитела сохраняли активность, направленную против цитотоксичности, и ингибировали лизис RBC на уровнях, сходных с таковыми родительского антитела V2L2, при использовании клеток, инфицированных 6206 и 6206ΔpslA.
[0354] Способность биспецифических антител Bs2 и Bs3 опосредовать OPK P. aeruginosa оценивали при помощи способа, описанного в примере 2. (A) В то время как антитело Bs2-V2L2 показало уничтожение, сходное с таковым для родительского антитела W4-RAD, уничтожение для антитела Bs3-V2L2 было снижено (фигура 20A). (B) В то время как антитела Bs2-V2L2-2C и Bs4-V2L2-2C показали уничтожение, сходное с таковым для родительского антитела W4-RAD, уничтожение для антитела Bs3-V2L2-2C было снижено (фигура 20B). На фигуре 20С показано, что различные препараты антител Bs4 (старая партия в сравнении с новой партией) показали уничтожение, сходное с таковым для родительского антитела W4-RAD, однако антитела Bs4-V2L2-2C-YTE характеризовались 3-кратным снижением ОРК-активности по сравнению с Bs4-V2L2-2C. YTE-мутант содержит комбинацию трех "мутаций YTE": M252Y, S254T и T256E, внедренных в тяжелую цепь IgG, причем нумерация соответствует EU-индексу, приведенному у Kabat. См. патент США №7658921, включенный в данный документ посредством ссылки. Было показано, что мутация YTE увеличивает период полувыведения антител из сыворотки приблизительно в четыре раза по сравнению с таковым у вариантов дикого типа того же антитела. См., например, Dall'Acqua et al., J. Biol. Chem. 281: 23514-24 (2006) и патент США №7083784, которые настоящим включены посредством ссылки во всей своей полноте.
[0355] После подтверждения того, что как W4, так и V2L2 сохраняли активность в биспецифическом формате, конструкты Bs2-V2L2, Bs3-V2L2 и Bs4-V2L2 оценивали в отношении выживаемости при острой инфекционной пневмонии. Как показано на фигуре 21A, все контрольные мыши поддались инфекции примерно через 30 часов после инфицирования. Все животные, обработанные с помощью Bs3-V2L2, выжили, наряду с теми, которые получили контрольное V2L2. Примерно 90% животных, иммунизированных с помощью W4-RAD, выжили. В противоположность этому, на фигурах B-F показано, что примерно 50% животных, обработанных с помощью Bs2-V2L2, поддались инфекции через 120 часов. Все контрольные мыши поддались инфекции примерно через 48 часов после инфицирования. На фигурах G-H не показаны различия в выживаемости между мышами, обработанными с помощью Bs4-V2L2-2C и Bs4-V2L2-2C-YTE в любой дозе. Эти результаты позволяют предположить, что в модели острой пневмонии с инфицированием 6206 оба антитела функционируют эквивалентно. На фигуре 21I показано, что Bs2-V2L2, Bs4-V2L2-2C и смесь антител W4-RAD+V2L2 являются наиболее эффективными в защите от смертельной пневмонии у мышей, подвергнутых контрольному заражению штаммом 6206 P. aeruginosa (ExoU+).
[0356] Нагрузку органа также оценивали для мышей, иммунизированных сходным образом, как описано выше. После иммунизации, описанной выше, мышей подвергали контрольному заражению 2,75×105 CFU 6206. Как показано на фигуре 22, при тестируемой концентрации как Bs2-V2L2, так и Bs3-V2L2 значительно уменьшали нагрузку органа в легком. Тем не менее, ни одному из биспецифических конструктов не удалось значительно повлиять на нагрузку органа в селезенке или почке по сравнению с родительскими антителами, что было обусловлено применением субоптимальных концентраций биспецифических конструктов. Применяли субоптимальные концентрации для обеспечения способности к интерпретации активности антител.
[0357] Эффекты биспецифических антител в отношении выживаемости и нагрузки органа также рассматривали с применением штамма 6294. При использовании модельной системы с инфицированием 6294 как Bs2-V2L2, так и Bs3-V2L2 значительно уменьшали нагрузку органа во всех тканях до уровня, сравнимого с таковым для родительского антитела V2L2. Родительское антитело W4-RAD не оказывало эффект на уменьшение нагрузки органа (фигура 23A). Как показано на фигуре 23 В, Bs2-V2L2, Bs3-V2L2 и комбинация W4-RAD+V2L2 значительно уменьшали нагрузку органа во всех тканях до уровня, сравнимого с таковым для родительского антитела V2L2.
[0358] Данные о выживаемости для иммунизированных мышей были аналогичны данным для мышей, подвергнутых контрольному заражению 6294, описанным ранее. Как показано на фигуре 24, Bs3-V2L2 показало аналогичную активность в отношении выживаемости у мышей, обработанных с помощью V2L2 в отдельности, тогда как для мышей, обработанных с помощью Bs2-V2L2 в отдельности, был показан немного более низкий уровень защиты при контрольном заражении.
[0359] Нагрузку органа также оценивали у животных, обработанных биспецифическими антителами, в сравнении с животными, обработанными комбинацией, как описано выше. Как показано на фигуре 25 (A-C), как Bs2-V2L2, так и Bs3-V2L2 уменьшали нагрузку органа в легком, селезенке и почках до уровня, сравнимого с таковым для комбинации W4+V2L2. В легком комбинация Bs2- и Bs3-V2L2 и V2L2 значительно снижала количество бактериальных CFU при использовании критерия Краскела-Уоллиса с апостериорным критерием Данна. Значительных различий в бактериальной нагрузке в селезенке и почке не наблюдали, хотя была отмечена тенденция к снижению. Исследование нагрузки органа также проводили с Bs4-GLO с применением 6206 в модели пневмонии. Как показано на фигуре 25 (D), при применении более высоких концентраций антитела у мышей с целью профилактики наблюдали значительный (критерий Краскела-Уоллиса с апостериорным критерием Данна) уровень снижения бактериальной нагрузки в легких. Значительное снижение диссеминации бактерий в селезенку и почки также наблюдали при применении более высоких концентраций Bs4-GLO в этой модели.
[0360] Эти результаты подтверждали в гистологическом исследовании ткани легкого иммунизированных мышей BALB/c, подвергнутых контрольному заражению с применением 1,33×107 CFU штамма 6294 P. aeruginosa (таблица 6A), с применением 1,7×107 CFU штамма 6294 P. aeruginosa (таблица 6B) и с применением 9,25×105 CFU штамма 6206 P. aeruginosa (таблица 7).
Пример 17. Терапевтическая вспомогательная терапия: Bs4-V2L2-2C+антибиотик
[0361] Эффект биспецифического антитела Bs4 и антибиотика как средства вспомогательной терапии, действующих против штамма 6206 P. aeruginosa, в отношении выживаемости оценивали в модели острой смертельной пневмонии, как ранее описано в примере 6 (фигура 26 (A-J)). (A-B) Мышей обрабатывали с помощью R347 (отрицательный контроль) или Bs4-V2L2-2C за 24 часа до инфицирования 6206, или ципрофлоксацина (CIP) через 1 час после инфицирования, или комбинации Bs4-V2L2-2C за 24 часа до инфицирования и Cipro через 1 час после инфицирования. (C) Мышей обрабатывали через 1 час после инфицирования 6206 с помощью R347, или CIP, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и CIP. (D) Мышей обрабатывали через 2 часа после инфицирования 6206 с помощью R347, или CIP, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и CIP. (E) Мышей обрабатывали с помощью R347 или Bs4-V2L2-2C через 2 часа после инфицирования 6206, или CIP через 1 час после инфицирования, или комбинации Bs4-V2L2-2C через 2 часа после инфицирования и CIP через 1 час после инфицирования. (F) Мышей обрабатывали через 1 час после инфицирования 6206 с помощью R347, или меропенема (MEM), или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и MEM. (G) Мышей обрабатывали с помощью R347 или Bs4-V2L2-2C через 2 часа после инфицирования 6206, или MEM через 1 час после инфицирования, или комбинации Bs4-V2L2-2C через 2 часа после инфицирования и MEM через 1 час после инфицирования. (H) Мышей обрабатывали через 2 часа после инфицирования 6206 с помощью R347, или Bs4-V2L2-2C, или MEM, или комбинации Bs4-V2L2-2C и MEM. (I) Мышей обрабатывали через 4 часа после инфицирования 6206 с помощью R347, или Cipro, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и Cipro. Все контрольные мыши поддались инфекции примерно через 24 часа после инфицирования. Как показано на фигурах 26 (A-I), антитело Bs4 в сочетании с CIP или MEM повышает эффективность терапии антибиотиками, что указывает на синергический защитный эффект при сочетании молекул. В дополнительных исследованиях внимание сосредоточили на уровне бактериальной нагрузки у мышей, обработанных с помощью Bs4 или CIP в отдельности или в комбинации (Bs4+CIP). Как показано на фигуре 26 (J), уровень бактериальной нагрузки во всех органах (легкое, селезенка и почки) был сходным при обработке 347+CIP и Bs4+CIP, однако только мыши, для которых Bs4 включили в комбинацию с CIP, выживают после инфицирования (фигуры 26 (A-E, I)). Вместе эти данные показывают, что антибиотики являются важными для снижения бактериальной нагрузки в этой планировке модели на животных, однако специфическое антитело необходимо для снижения бактериальной патогенности, что, таким образом, защищает нормальный иммунитет хозяина.
[0362] Эффект биспецифического антитела Bs4 и антибиотика тобрамицина как средства вспомогательной терапии, действующих против штамма 6206 P. aeruginosa, в отношении выживаемости оценивали в модели острой смертельной пневмонии, как ранее описано в примере 6. Мышей обрабатывали с помощью R347 (отрицательный контроль) или Bs4-V2L2-2C за 24 часа до инфицирования 6206, или тобрамицина через 1 час после инфицирования, или комбинации Bs4-V2L2-2C за 24 часа до инфицирования и тобрамицина через 1 час после инфицирования. Мышей также обрабатывали через 1 час после инфицирования 6206 с помощью R347, или тобрамицина, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и тобрамицина. Кроме того, мышей также обрабатывали через 2 часа после инфицирования 6206 с помощью R347, или тобрамицина, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и тобрамицина. Дополнительно, мышей обрабатывали с помощью R347 или Bs4-V2L2-2C через 2 часа после инфицирования 6206, или тобрамицина через 1 час после инфицирования, или комбинации Bs4-V2L2-2C через 2 часа после инфицирования и тобрамицина через 1 час после инфицирования. Мышей также обрабатывали через 4 часа после инфицирования 6206 с помощью R347, или тобрамицина, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и тобрамицина.
[0363] Эффект биспецифического антитела Bs4 и антибиотика азтреонама как средства вспомогательной терапии, действующих против штамма 6206 P. aeruginosa, в отношении выживаемости оценивали в модели острой смертельной пневмонии, как ранее описано в примере 6. Мышей обрабатывали с помощью R347 (отрицательный контроль) или Bs4-V2L2-2C за 24 часа до инфицирования 6206, или азтреонама через 1 час после инфицирования, или комбинации Bs4-V2L2-2C за 24 часа до инфицирования и азтреонама через 1 час после инфицирования. Мышей также обрабатывали через 1 час после инфицирования 6206 с помощью R347, или азтреонама, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и азтреонама. Кроме того, мышей также обрабатывали через 2 часа после инфицирования 6206 с помощью R347, или азтреонама, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и азтреонама. Дополнительно, мышей обрабатывали с помощью R347 или Bs4-V2L2-2C через 2 часа после инфицирования 6206, или азтреонама через 1 час после инфицирования, или комбинации Bs4-V2L2-2C через 2 часа после инфицирования и азтреонама через 1 час после инфицирования. Мышей обрабатывали через 4 часа после инфицирования 6206 с помощью R347, или азтреонама, или Bs4-V2L2-2C, или комбинации Bs4-V2L2-2C и азтреонама.
Пример 18. Конструирование биспецифического антитела BS4-GLO
[0364] Образовывали биспецифический конструкт Bs4-GLO (ген которого изменили с получением гена в конфигурации зародышевой линии, а также подвергнутый оптимизации лидерных молекул), содержащий scFv, связывающийся с Psl (scFv Psl0096) и V2L2-MD (VH+VL), как показано на фигуре 35A. Легкая цепь Bs4-GLO содержит вариабельную область легкой цепи антитела к PcrV, ген которого изменили с получением гена в конфигурации зародышевой линии, а также подвергнутого оптимизации лидерных молекул (т.е. V2L2-MD). Тяжелая цепь Bs4-GLO содержит формулу VH-CH1-H1-L1-S-L2-H2-CH2-CH3, причем CH1 представляет собой домен-1 константной области тяжелой цепи, H1 представляет собой первый фрагмент шарнирной области тяжелой цепи, L1 представляет собой первый линкер, S представляет собой молекулу scFv, связывающуюся с PcrV, L2 представляет собой второй линкер, H2 представляет собой второй фрагмент шарнирной области тяжелой цепи, CH2 представляет собой домен-2 константной области тяжелой цепи, а CH3 представляет собой домен-3 константной области тяжелой цепи.
Легкая цепь Bs4-GLO

Вариабельная область легкой цепи V2L2 (т.е. V2L2-MD) GLO (ген которого изменили с получением гена в конфигурации зародышевой линии, а также подвергнутого оптимизации лидерных молекул) подчеркнута.
Тяжелая цепь Bs4-GLO

KSLSLSPGK Вариабельная область тяжелой цепи V2L2 (т.е. V2L2-MD) GLO (ген которого изменили с получением гена в конфигурации зародышевой линии, а также подвергнутого оптимизации лидерных молекул) подчеркнута; CH1 заключен в скобки []; scFv W4-RAD (т.е. Psl0096) GLO (ген которого изменили с получением гена в конфигурации зародышевой линии, а также подвергнутого оптимизации лидерных молекул) выделен жирным курсивом, при этом линкеры G4S подчеркнуты и выделены жирным курсивом; шарнирные области подчеркнуты двойной чертой.
[0365] Альтернативный биспецифический конструкт Bs4-GLO, содержащий scFv, связывающийся с PcrV, и VH+VL антитела к Psl, показан на фигуре 35 В и образован сходным образом.
Пример 19. Оценивание функциональной активности и эффективности биспецифического антитела Bs4-GLO
[0366] Биспецифические антитела Bs4-WT (также называемые в данном документе Bs4-V2L2-2C), Bs4-GL (содержащие вариабельные области антител к PcrV и Psl, гены которых изменили с получением генов в конфигурации зародышевой линии) и Bs4-GLO, полученные согласно описанному в примере 18, тестировали в отношении различий в функциональной активности в анализе опсонофагоцитарного уничтожения (фигура 27A), описанном ранее в примере 2, в анализе активности, направленной против прикрепления к клеткам (фигура 27 В), описанном ранее в примере 4, и в анализе активности, направленной против цитотоксичности и лизиса RBC (фигура 27C), описанном ранее в примере 12. Различий в функциональной активности антител in vitro не наблюдали.
[0367] In vivo эффективность Bs4-GLO исследовали следующим образом. Для оценивания профилактических свойств мышей обрабатывали с целью профилактики с помощью Bs4-GLO в нескольких концентрациях (т.е., 0,007 мг/кг, 0,02 мг/кг, 0,07 мг/кг, 0,2 мг/кг, 0,5 мг/кг, 1 мг/кг, 3 мг/кг, 5 мг/кг, 10 мг/кг или 15 мг/кг) (фигура 28A) за 24 часа до инфицирования следующими штаммами Р. aeruginosa: 6206 (1,0×106), 6077 (1,0×106), 6294 (2,0×107) или PA103 (1,0×106). Для оценивания терапевтических свойств мышей обрабатывали с целью терапии с помощью Bs4-GLO в нескольких концентрациях (т.е., 0,03 мг/кг, 0,3 мг/кг, 0,5 мг/кг, 1 мг/кг, 2 мг/кг, 5 мг/кг, 10 мг/кг, 15 мг/кг или 45 мг/кг) (фигура 28B) через час после инфицирования следующими штаммами P. aeruginosa: 6206 (1,0×106), 6077 (1,0×106), 6294 (2,0×107) или PA103 (1,0×106).
[0368] Эффект биспецифического антитела Bs4-GLO, действующего против различных штаммов P. aeruginosa, в отношении выживаемости оценивали в модели острой смертельной пневмонии, как ранее описано в примере 6. На фигуре 29 представлены показатели выживаемости животных, обработанных с помощью Bs4-GLO, в модели смертельной бактериемии с инфицированием P. aeruginosa. Аспекты модели бактериемии подробно раскрыты в предварительной заявке на патент США №61/723128, поданной 6 ноября 2012 г. (номер дела патентного поверенного ATOX-500P1, под названием "СПОСОБЫ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, АССОЦИИРОВАННЫХ С S. AUREUS"), которая включена в данный документ посредством ссылки во всей своей полноте.
[0369] Животных обрабатывали с помощью Bs4-GLO или R347 за 24 часа до интраперитонеального инфицирования (A) 6294 (O6) или (B) 6206. Bs4-GLO при всех тестируемых концентрациях является эффективным в защите мышей, подвергнутых контрольному заражению штаммами (A) 6294 и (B) 6206 P. aeruginosa, от смертельной пневмонии.
[0370] Эффект биспецифического антитела Bs4-GLO, действующего против различных штаммов P. aeruginosa, в отношении выживаемости оценивали в модели термического повреждения с инфицированием P. aeruginosa. На фигуре 30 представлены показатели выживаемости животных, обработанных с целью профилактики с помощью Bs4-GLO, в модели термического повреждения с инфицированием P. aeruginosa. Животных обрабатывали с помощью Bs4-GLO или R347 за несколько часов до индуцирования термического повреждения и подкожного инфицирования штаммом (A) 6077 (O11-ExoU+), или (B) 6206 (O11-ExoU+), или (C) 6294 (O6) P. aeruginosa непосредственно под раневую поверхность. Bs4-GLO при всех тестируемых концентрациях является эффективным в предупреждении в модели термического повреждения с инфицированием P. aeruginosa на мышах, подвергнутых контрольному заражению штаммами (A) 6077, (B) 6206 и (C) 6294 P. aeruginosa.
[0371] На фигуре 31 представлены показатели выживаемости животных, обработанных с целью терапии с помощью Bs4-GLO, в модели термического повреждения с инфицированием P. aeruginosa. (A) Животных обрабатывали с помощью Bs4-GLO или R347 (A) через 4 часа или (B) через 12 часов после индуцирования термического повреждения и подкожного инфицирования штаммом 6077 (O11-ExoU+) P. aeruginosa непосредственно под раневую поверхность. Bs4-GLO при всех тестируемых концентрациях является эффективным в лечении в модели термического повреждения с инфицированием P. aeruginosa на мышах, обработанных с помощью Bs4-GLO (A) через 4 часа или (B) через 12 часов после индуцирования термического повреждения и подкожного инфицирования штаммом 6077 P. aeruginosa.
Пример 20. Терапевтическая вспомогательная терапия: Bs4-GLO+антибиотик
[0372] Эффект биспецифического антитела Bs4-GLO и антибиотика как средства вспомогательной терапии, действующих против штамма 6206 P. aeruginosa, в отношении выживаемости оценивали в модели острой смертельной пневмонии, как ранее описано в примере 6.
[0373] На фигуре 32 показана терапевтическая вспомогательная терапия при помощи ципрофлоксацина (CIP). (A) Мышей обрабатывали через 4 часа после инфицирования штаммом 6206 P. aeruginosa с помощью R347+CIP, или Bs4-WT, или комбинации Bs4-WT и CIP. (B) Мышей обрабатывали через 4 часа после инфицирования штаммом 6206 P. aeruginosa с помощью R347+CIP, или Bs4-GLO, или комбинации Bs4-GLO и CIP. (A-B) Антитело Bs4-WT или Bs4-GLO в сочетании с CIP повышало эффективность терапии антибиотиками.
[0374] На фигуре 33 показана терапевтическая вспомогательная терапия при помощи меропенема (MEM). (A) Мышей обрабатывали через 4 часа после инфицирования штаммом 6206 P. aeruginosa с помощью R347+MEM, или Bs4-WT, или комбинации Bs4-WT и MEM. (B) Мышей обрабатывали через 4 часа после инфицирования штаммом 6206 P. aeruginosa с помощью R347+MEM, или Bs4, или комбинации Bs4-GLO и MEM. (A-B) Антитело Bs4-WT или Bs4-GLO в сочетании с MEM повышало эффективность терапии антибиотиками.
[0375] На фигуре 34 показана терапевтическая вспомогательная терапия: Bs4-GLO с антибиотиком в модели смертельной бактериемии. Мышей обрабатывали за 24 часа до интраперитонеального инфицирования штаммом 6294 P. aeruginosa с помощью Bs4-GLO в указанных концентрациях, которые, как ранее определили, являются субтерапевтическими защитными дозами в этой модели, и R347 (отрицательный контроль). Через час после инфицирования мышей обрабатывали подкожно с помощью антибиотиков (A) ципрофлоксацина (CIP), (B) меропенема (MEM) или (C) тобрамицина (ТОВ) в указанных концентрациях, которые, как ранее определили, являются субтерапевтическими защитными дозами. Тщательно отслеживали выживаемость животных в течение до 72 часов после инфицирования.
Антитело Bs4-GLO в сочетании с CIP, MEM либо TOB в субтерапевтических защитных дозах повышает эффективность терапии антибиотиками.
***
[0376] Настоящее изобретение не подлежит ограничению по объему конкретными описанными вариантами осуществления, которые, как предполагается, являются единичными иллюстрациями отдельных аспектов настоящего изобретения, и любые композиции или способы, являющиеся функционально эквивалентными, находятся в пределах объема настоящего изобретения. В действительности, различные модификации настоящего изобретения, в дополнение к показанным и описанным в данном документе, станут очевидными специалистам в данной области из предшествующего описания и сопроводительных графических материалов. Предполагается, что такие модификации находятся в пределах объема прилагаемой формулы изобретения.
[0377] Все публикации и заявки на патенты, упоминаемые в настоящем описании, включены в данный документ посредством ссылки в такой же степени, как если бы каждая отдельная публикация или заявка на патент была конкретно и отдельно указана как включенная посредством ссылки. Кроме того, предварительные заявки на патенты США №№61/556645, поданная 7 ноября 2011 г.; 61/624651, поданная 16 апреля 2012 г.; 61/625299, поданная 17 апреля 2012 г.; 61/697585, поданная 6 сентября 2012 г., и международная заявка на патент №: PCT/US2012/63639, поданная 6 ноября 2012 г. (номер дела патентного поверенного AEMS-115WO1, под названием "ПОЛИСПЕЦИФИЧЕСКИЕ И ПОЛИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ") включены посредством ссылки во всей своей полноте во всех отношениях.
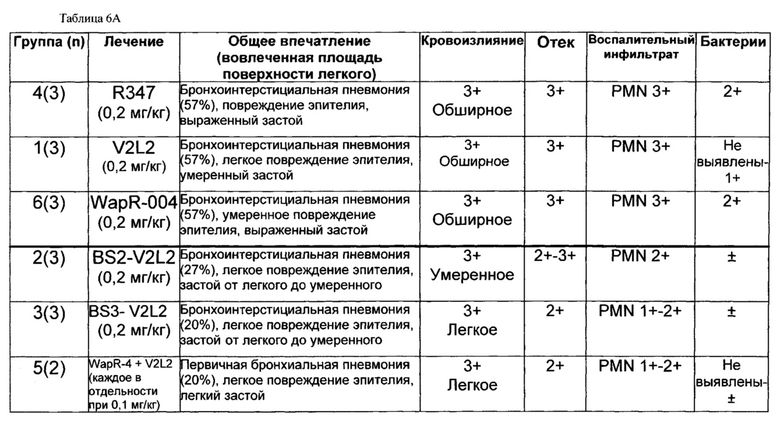
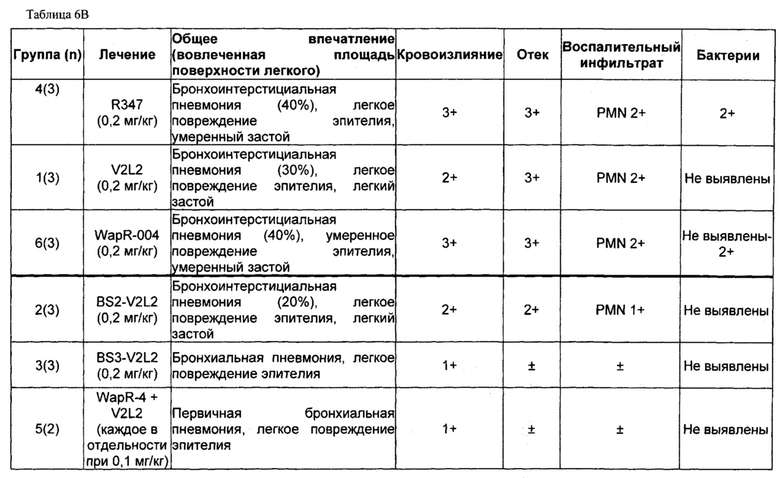
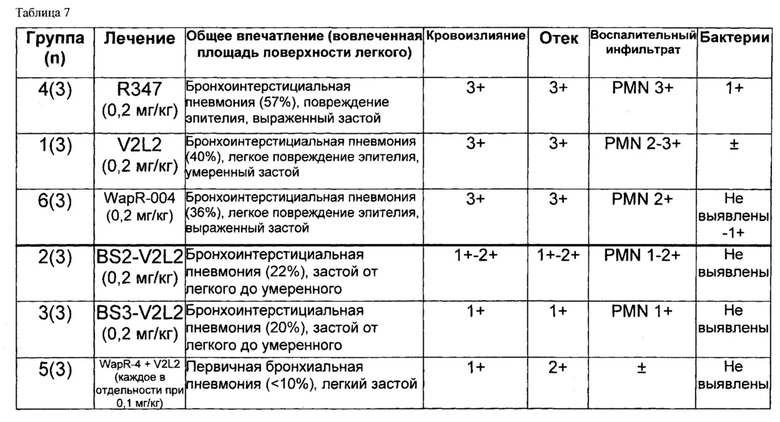
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С PSL PSEUDOMONAS, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2708977C2 |
| АНТИТЕЛА, КОТОРЫЕ СПЕЦИФИЧНО СВЯЗЫВАЮТ АЛЬФА-ТОКСИН STAPHYLOCOCCUS AUREUS, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620065C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, АССОЦИИРОВАННЫХ С S. AUREUS | 2013 |
|
RU2661406C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ | 2018 |
|
RU2791256C2 |
| НЕЙТРАЛИЗУЮЩИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, НАПРАВЛЕННЫЕ ПРОТИВ ВИРУСА ГРИППА, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2721706C2 |
| АНТИТЕЛА К ПОВЕРХНОСТНЫМ ДЕТЕРМИНАНТАМ S. AUREUS | 2013 |
|
RU2698131C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ASCT2, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2710194C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ ПО ОТНОШЕНИЮ К HER3, И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620068C2 |
| АНТИТЕЛА К ПОВЕРХНОСТНЫМ ДЕТЕРМИНАНТАМ S. AUREUS | 2013 |
|
RU2808018C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К OX40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2709742C2 |



























































































































































































































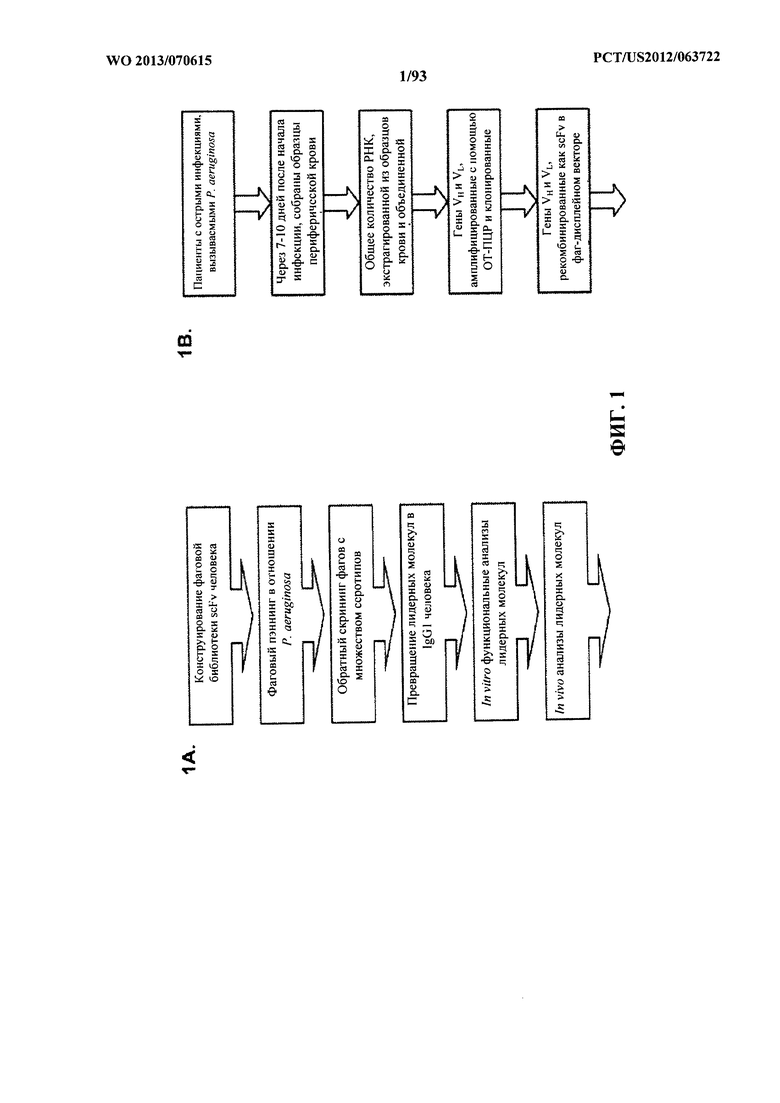
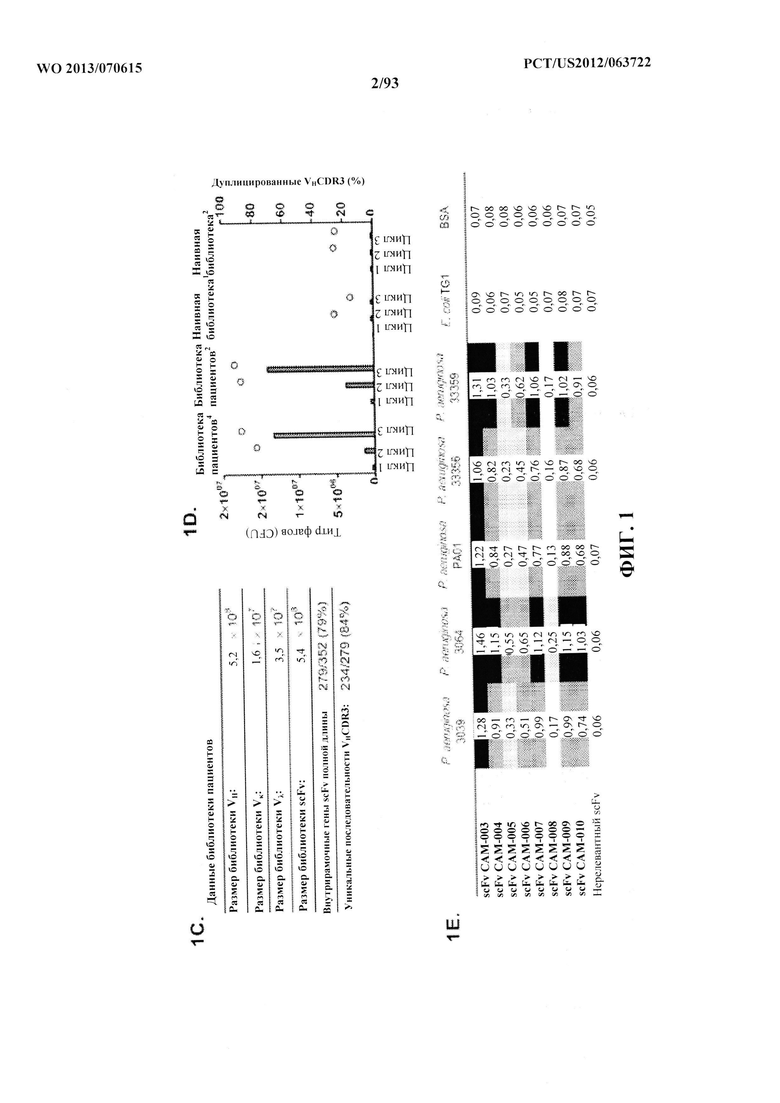
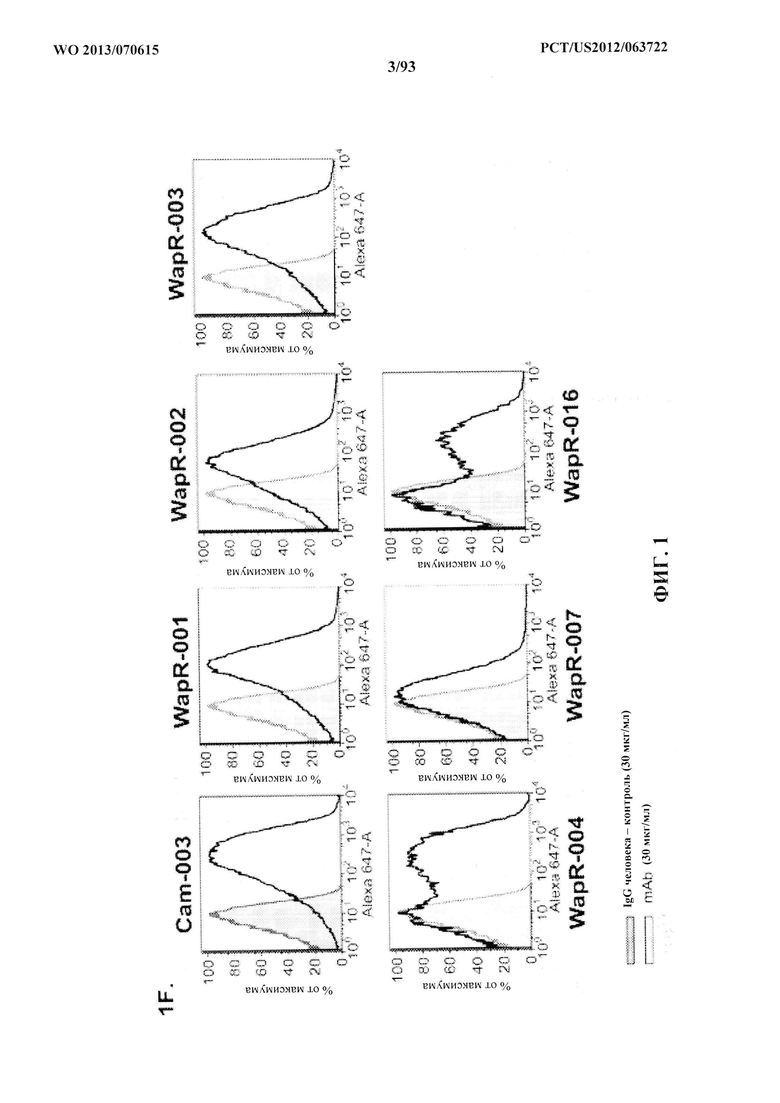
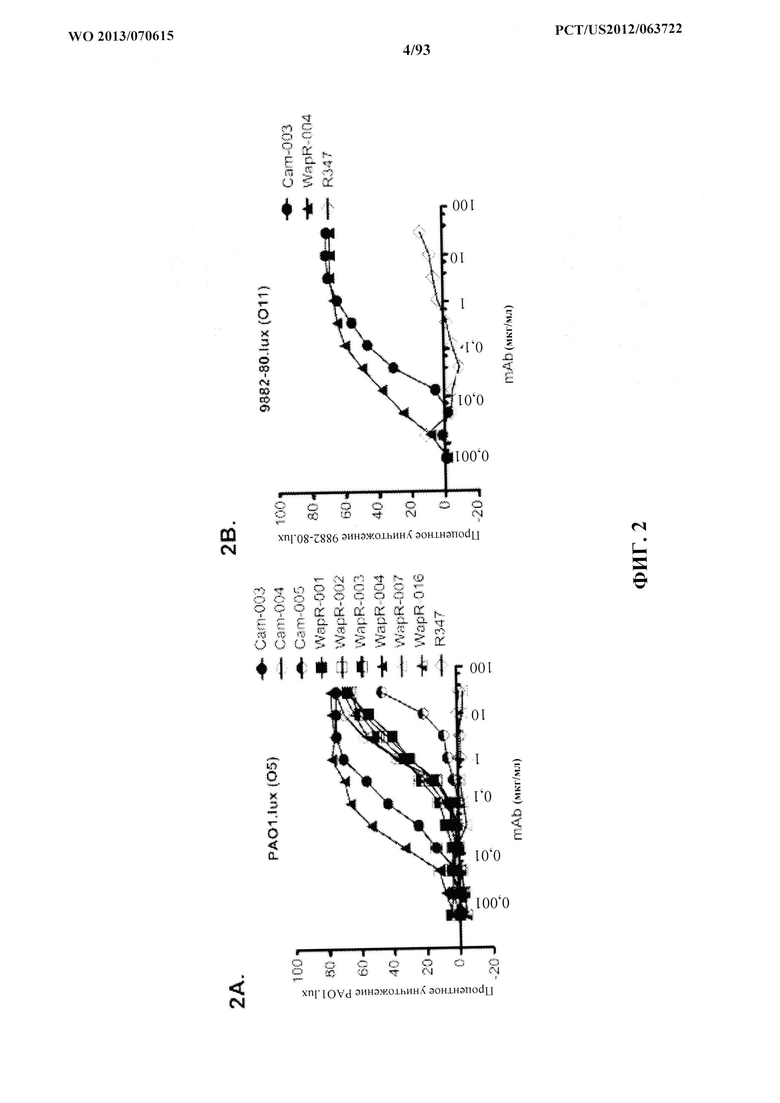
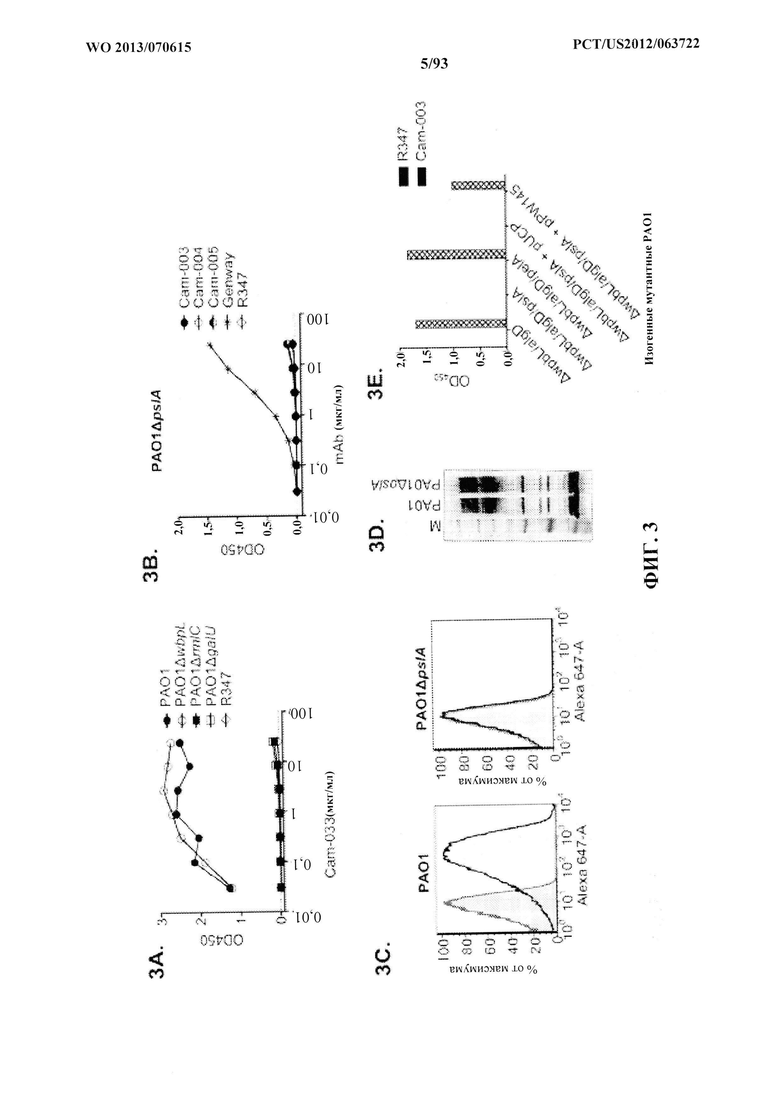
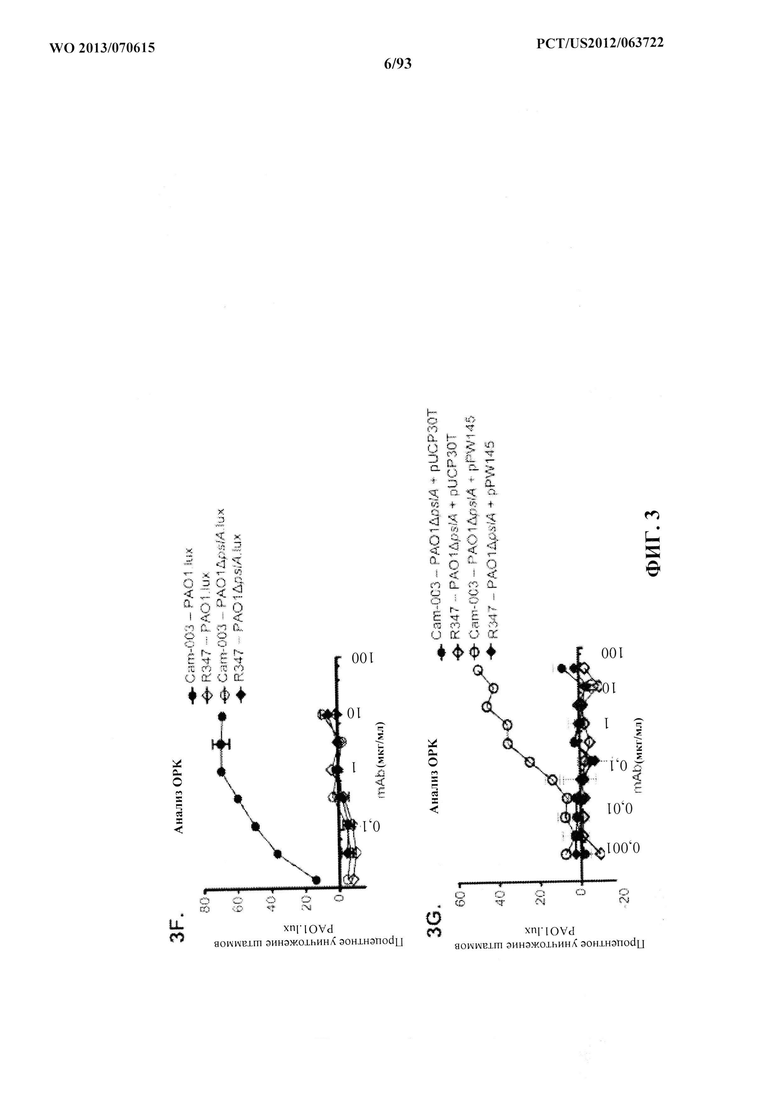
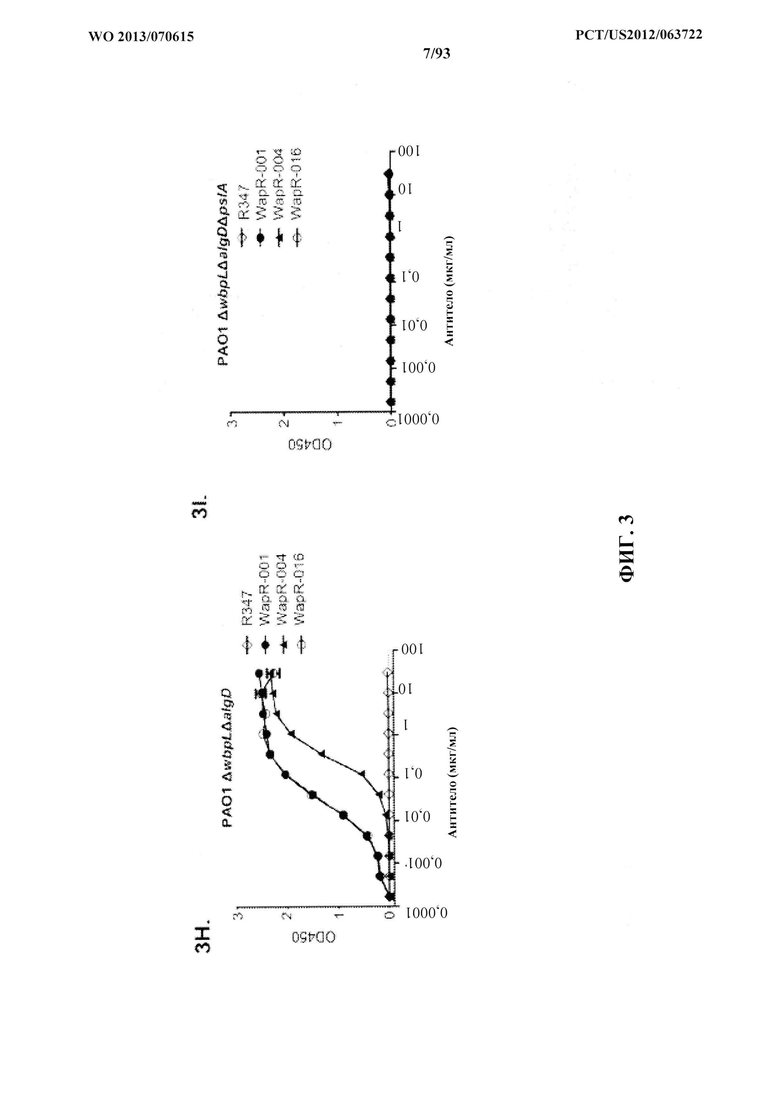
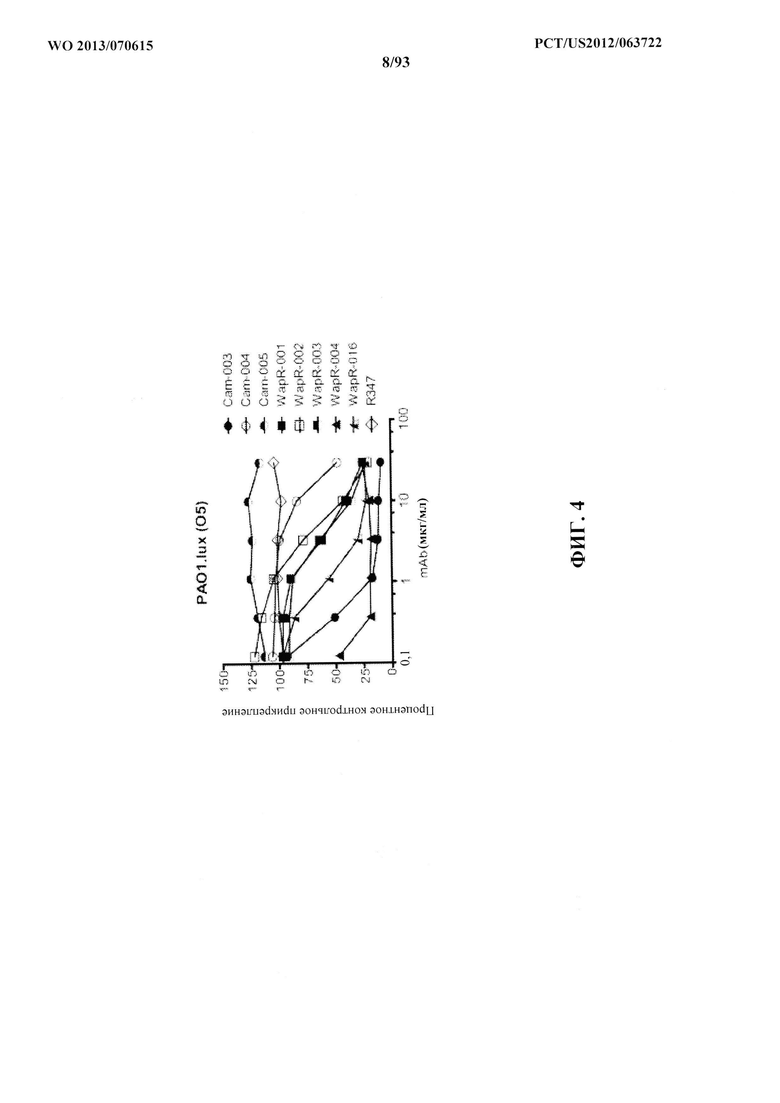
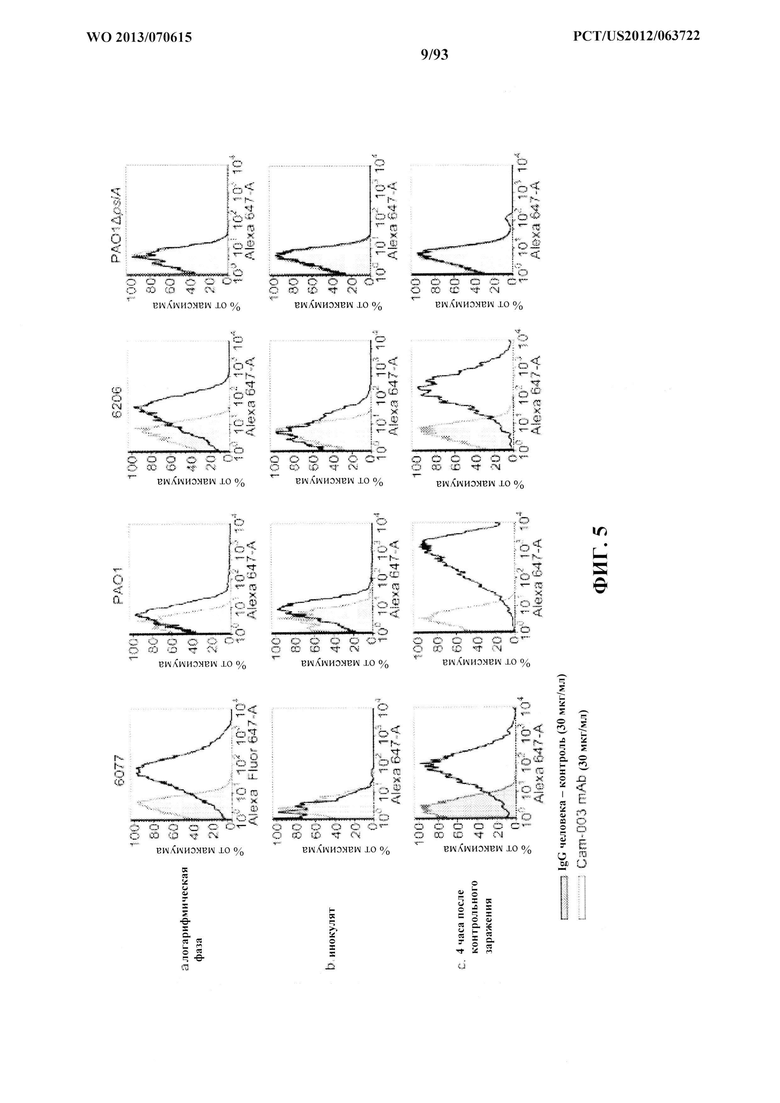
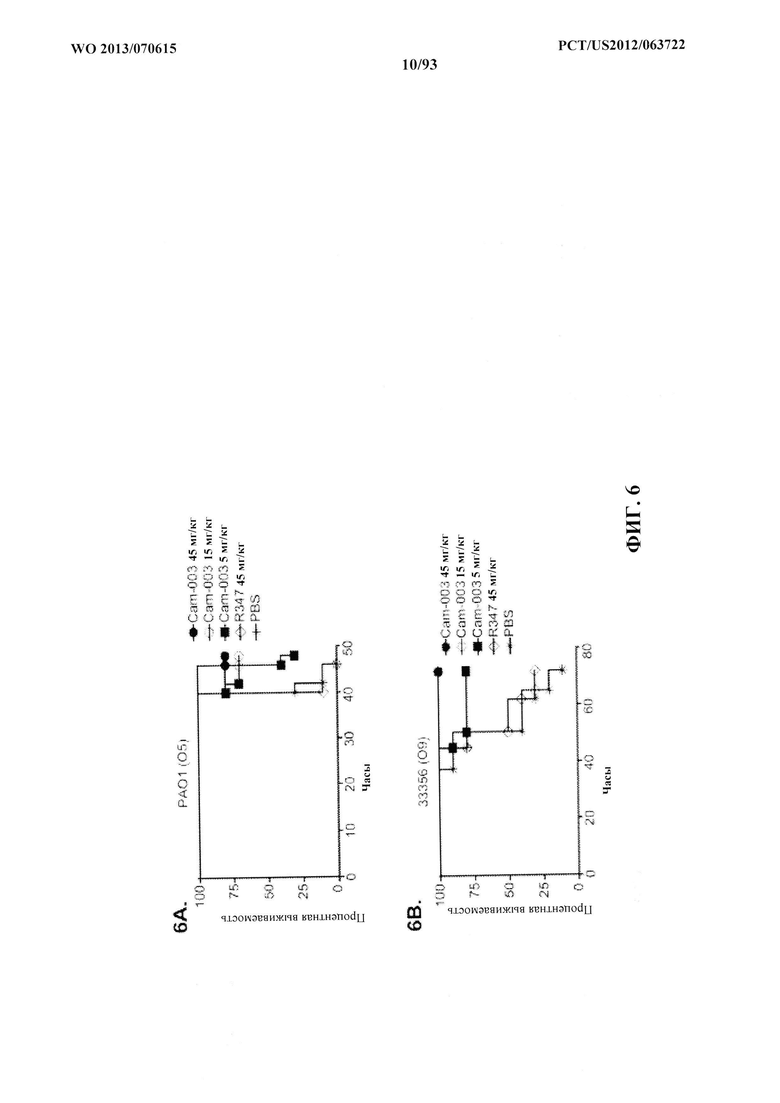
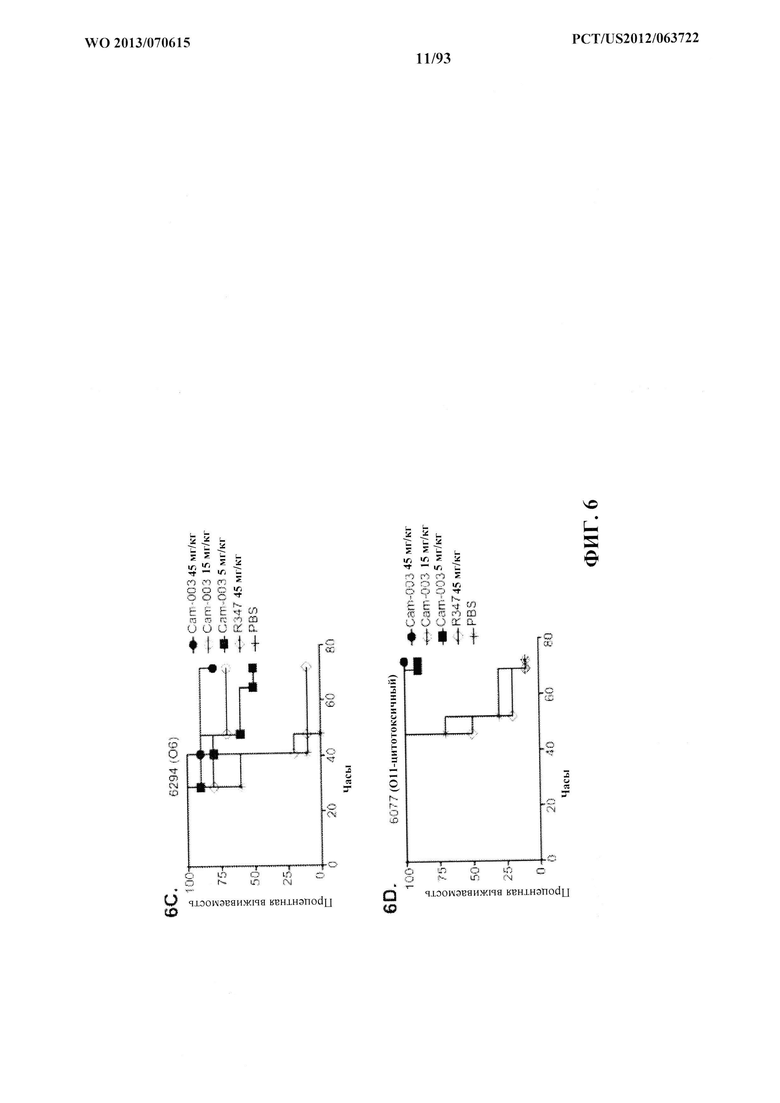
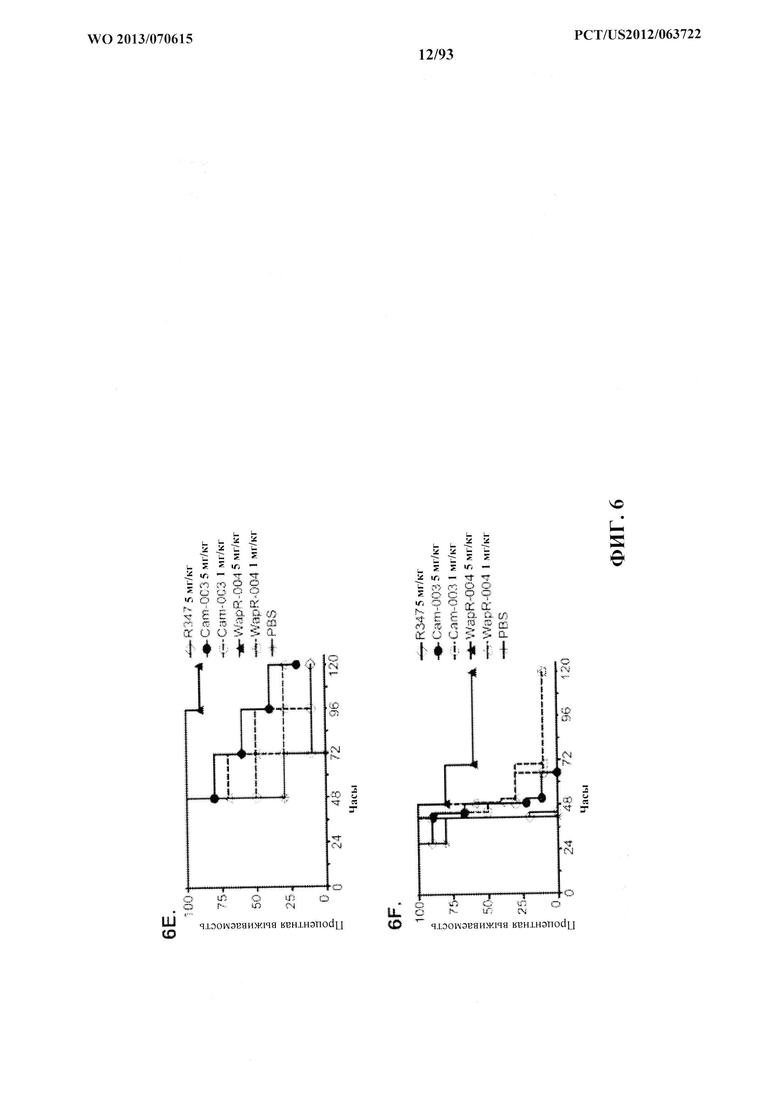
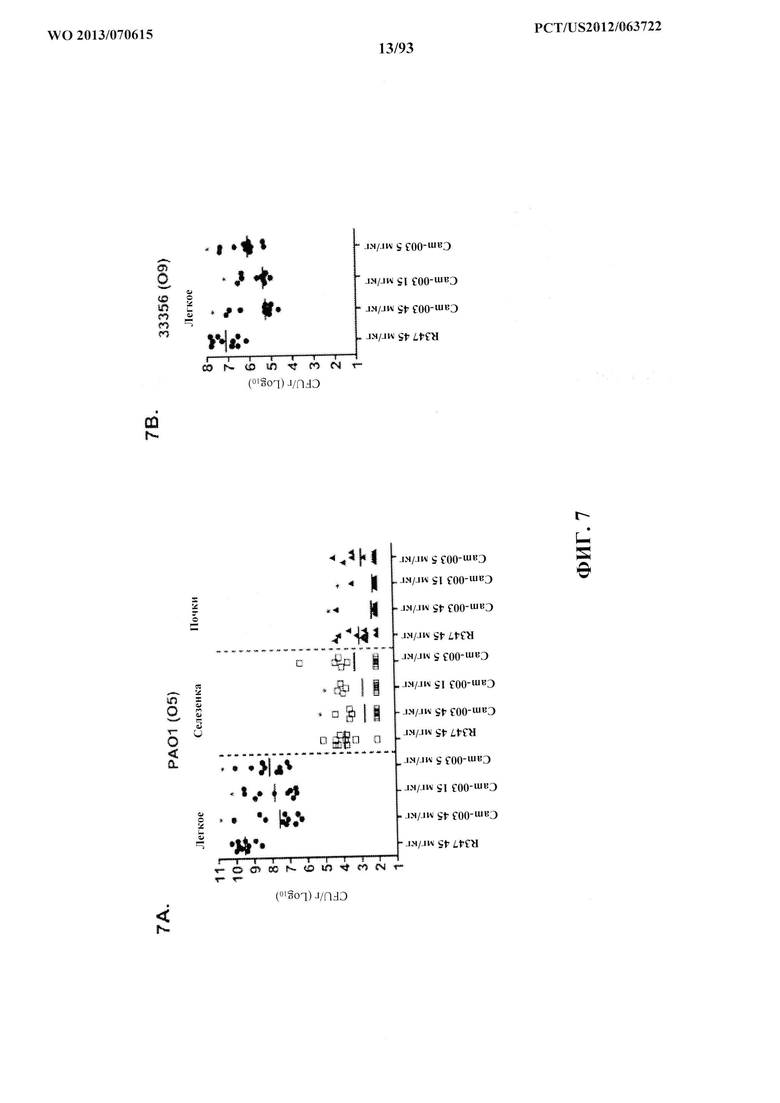
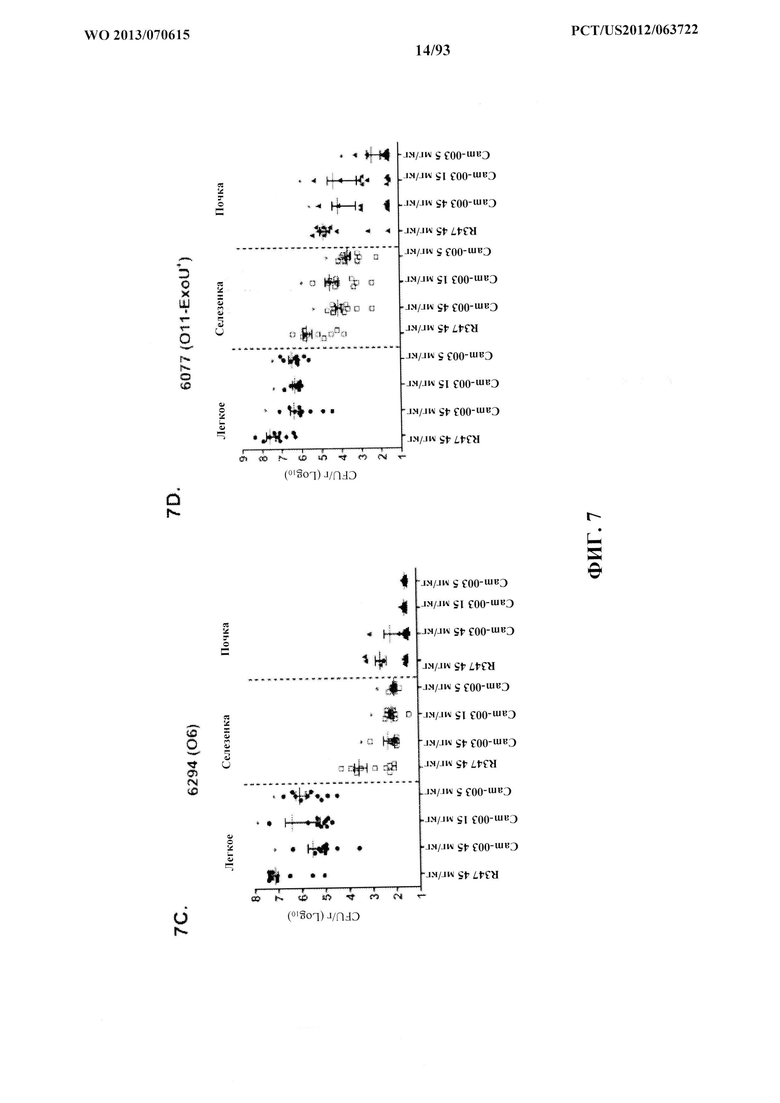
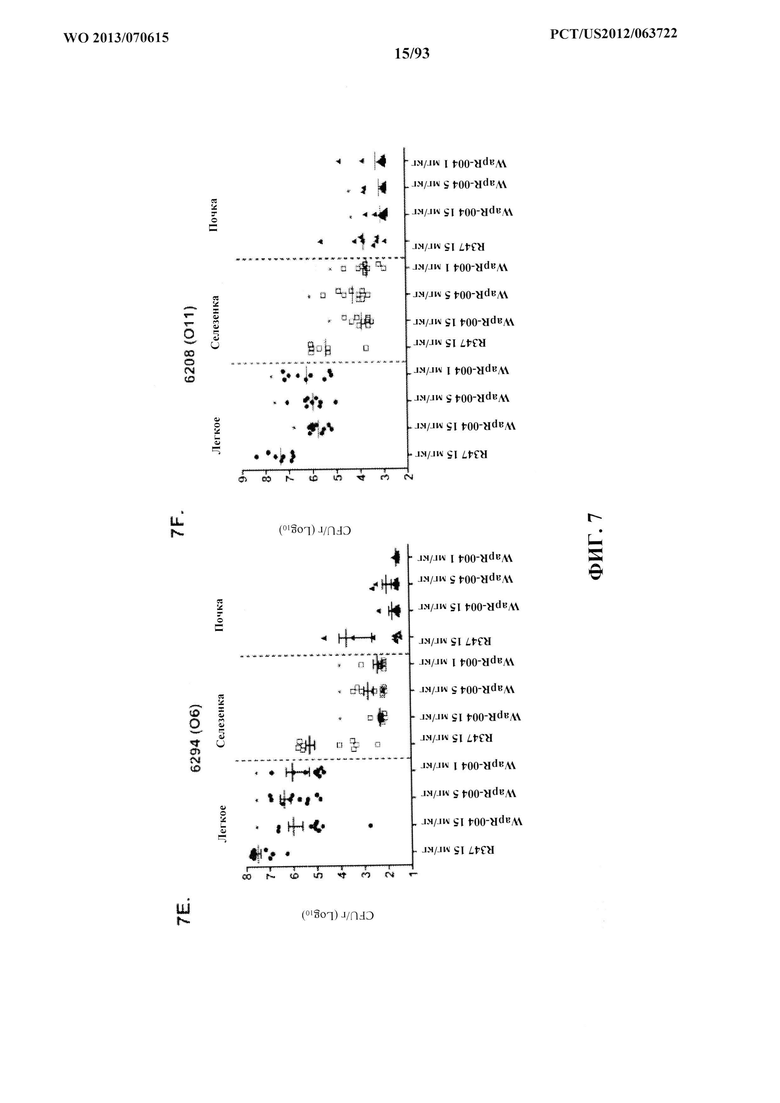
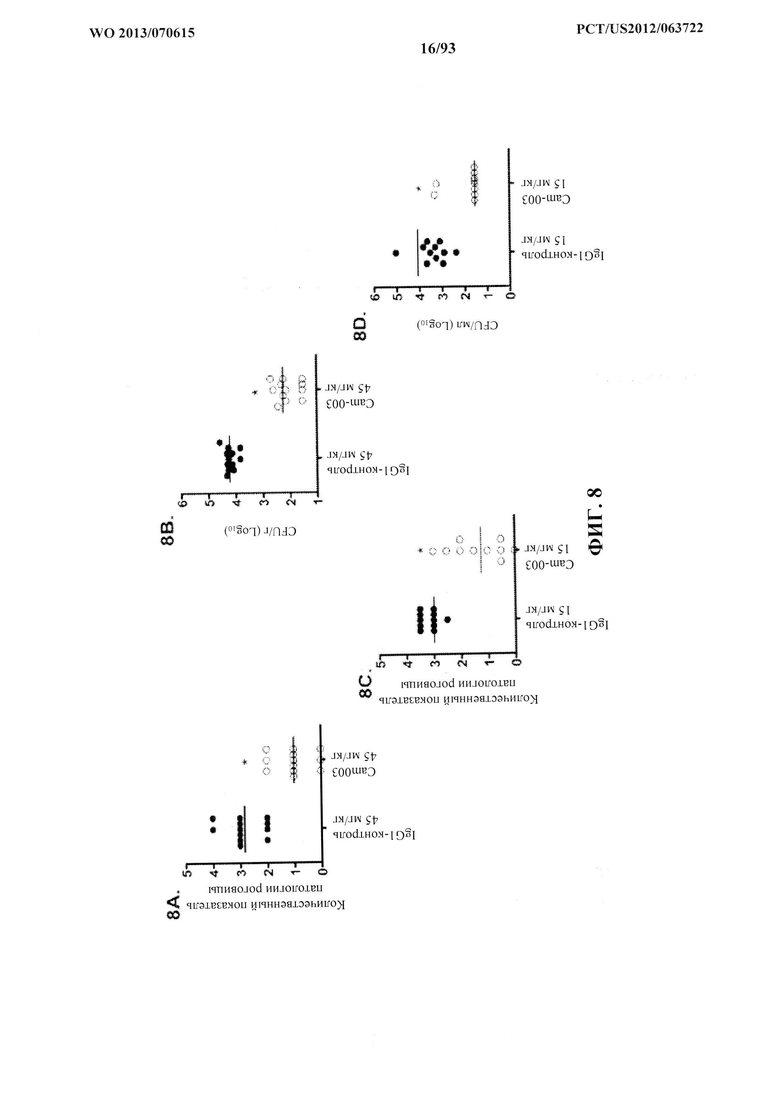
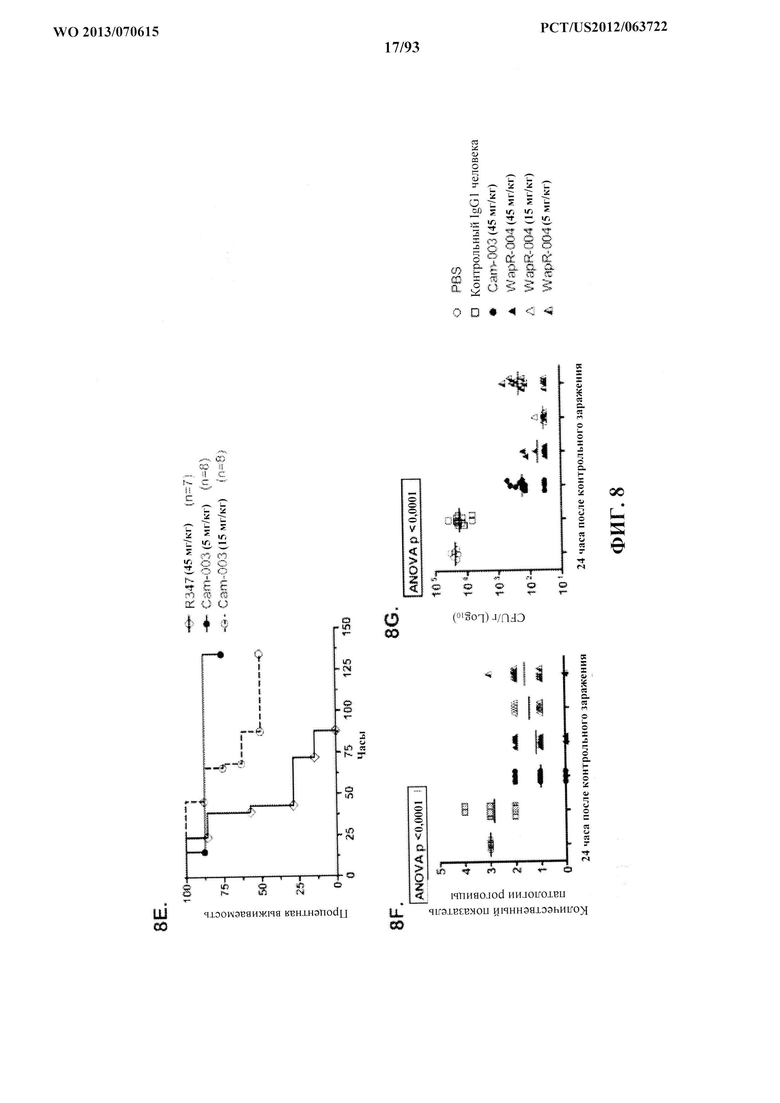
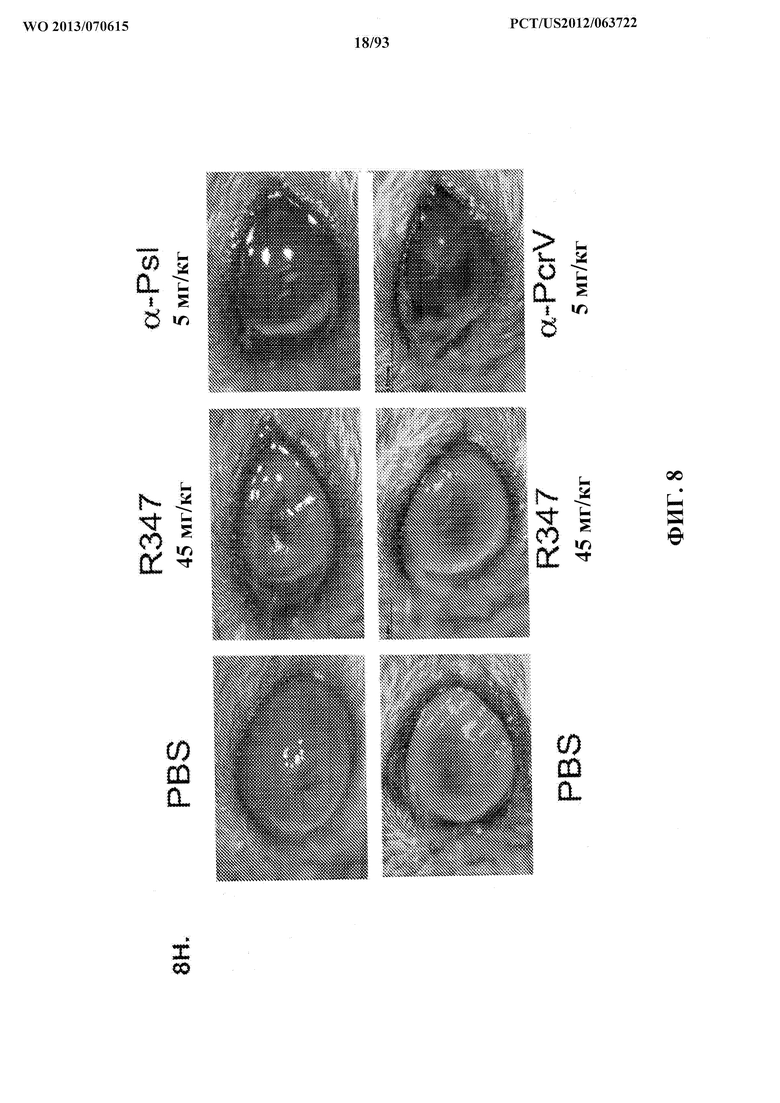
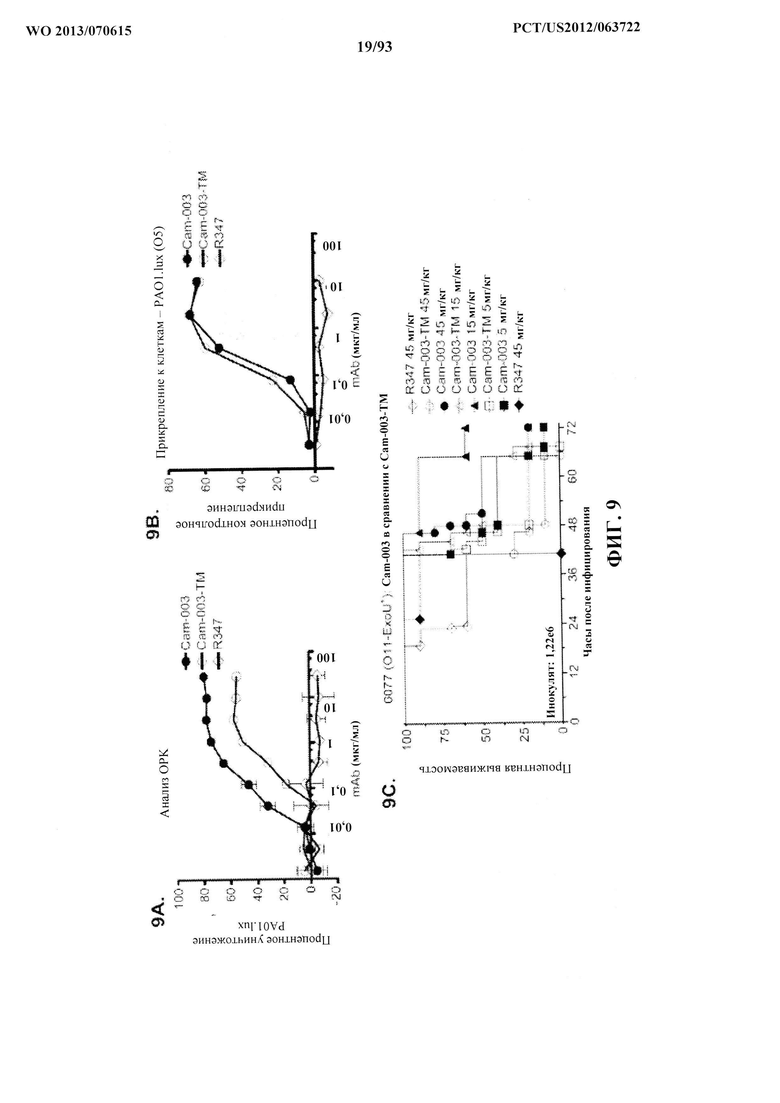
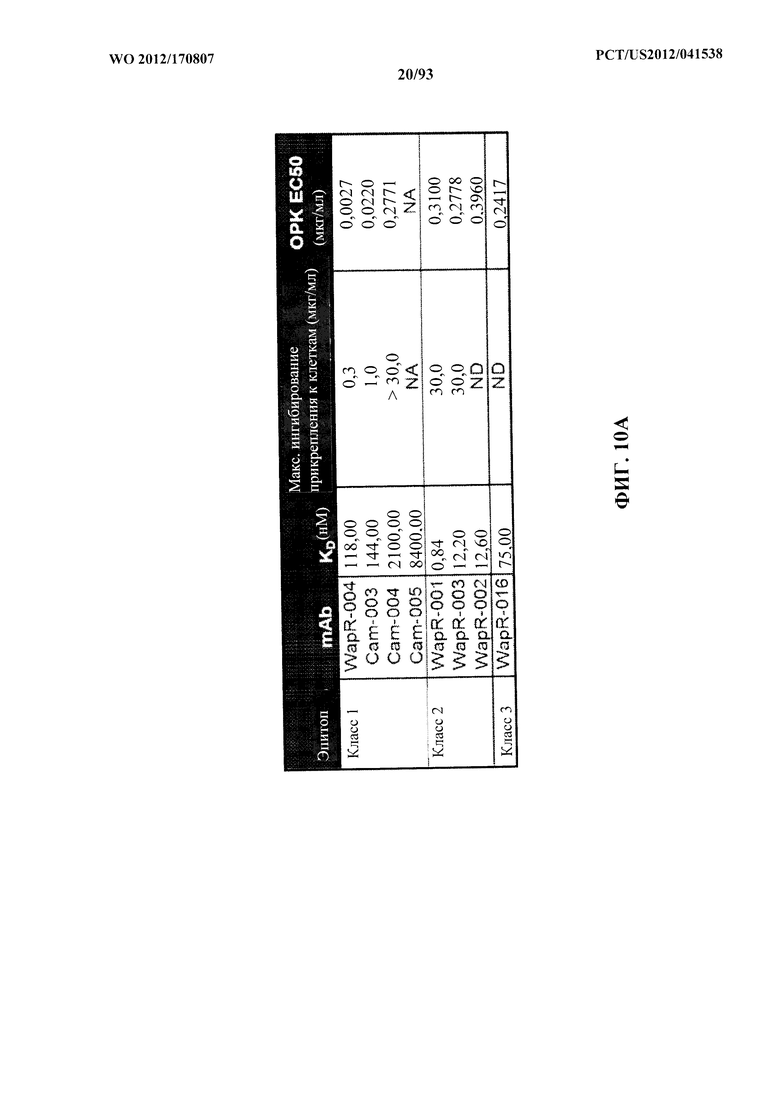
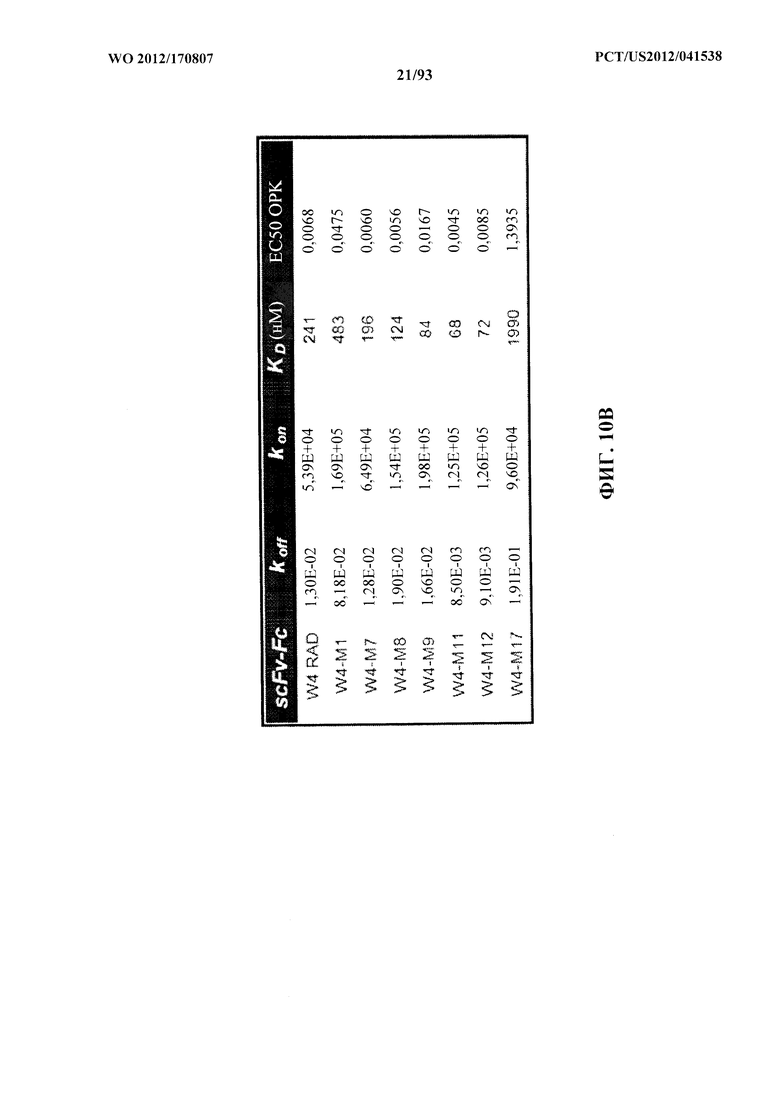
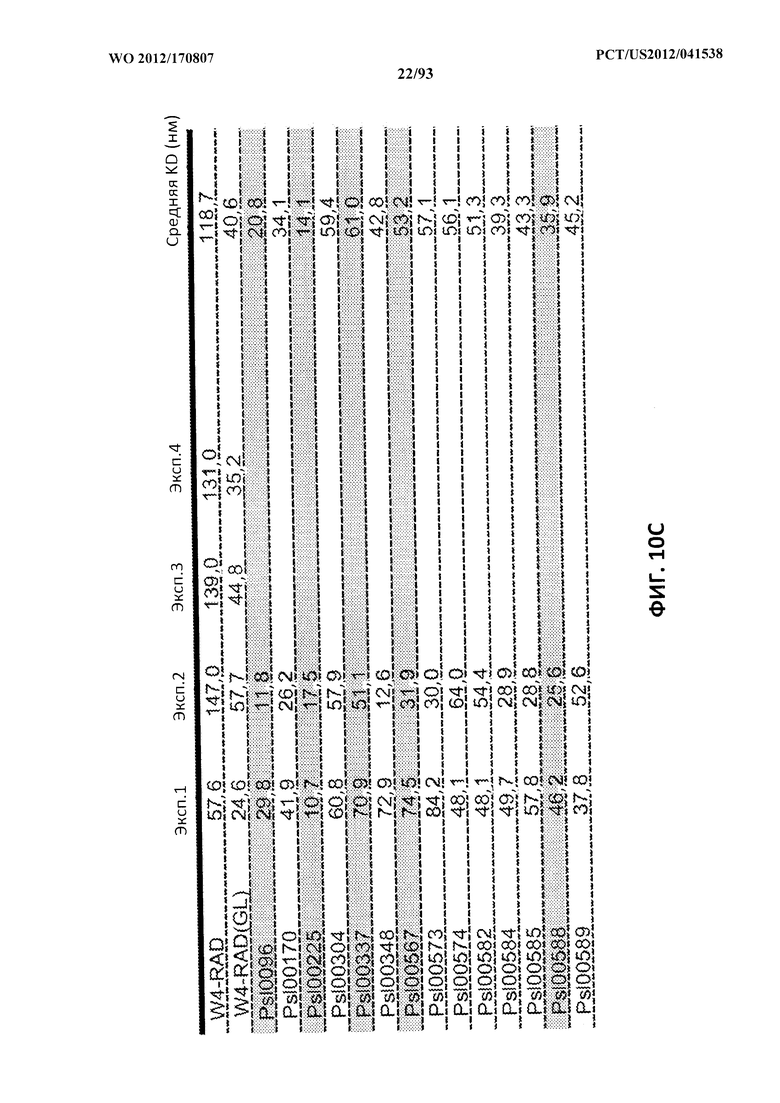
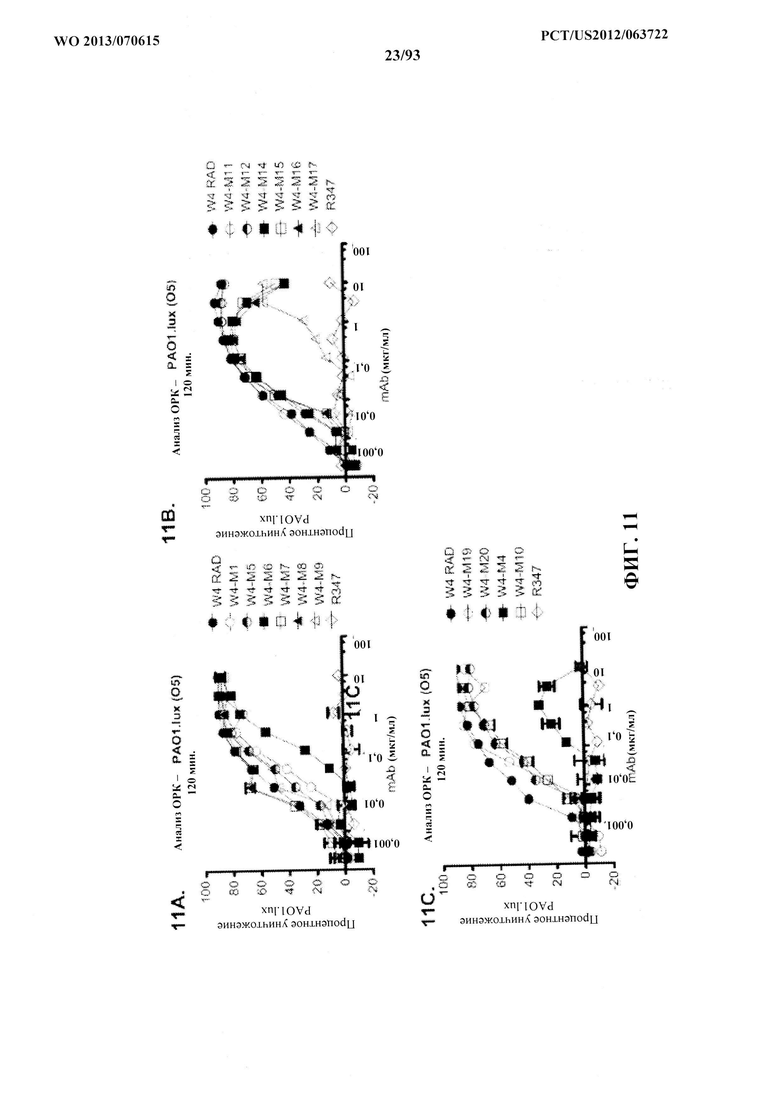
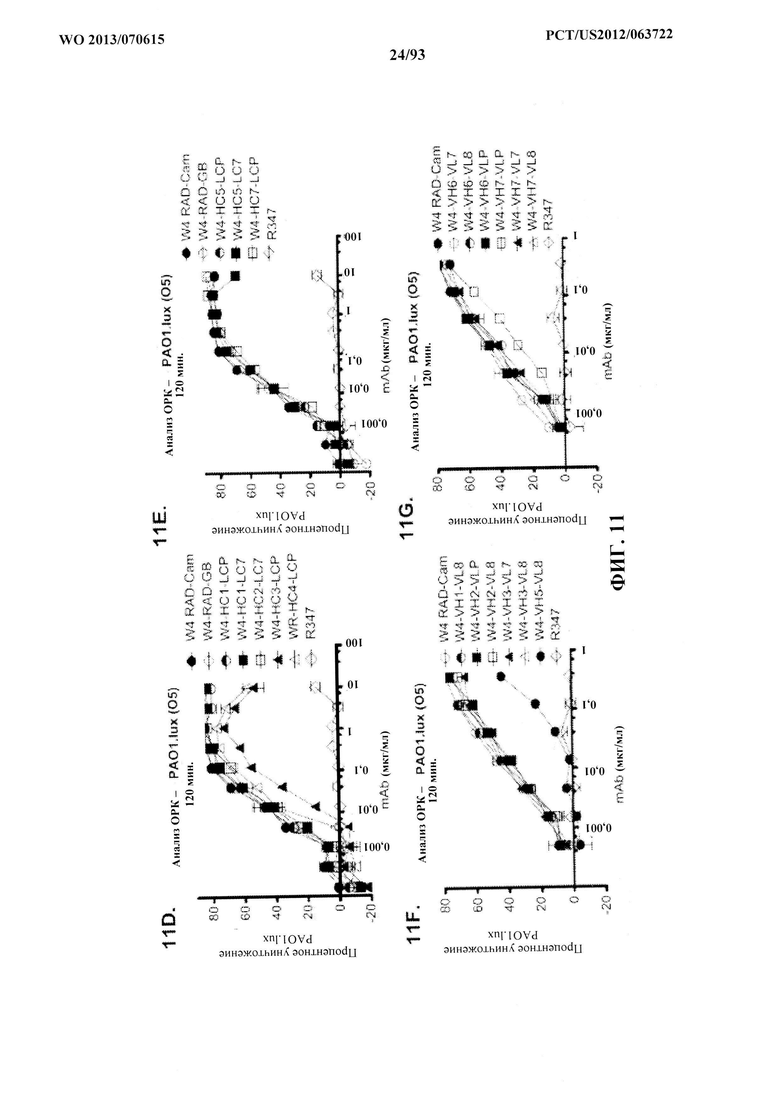
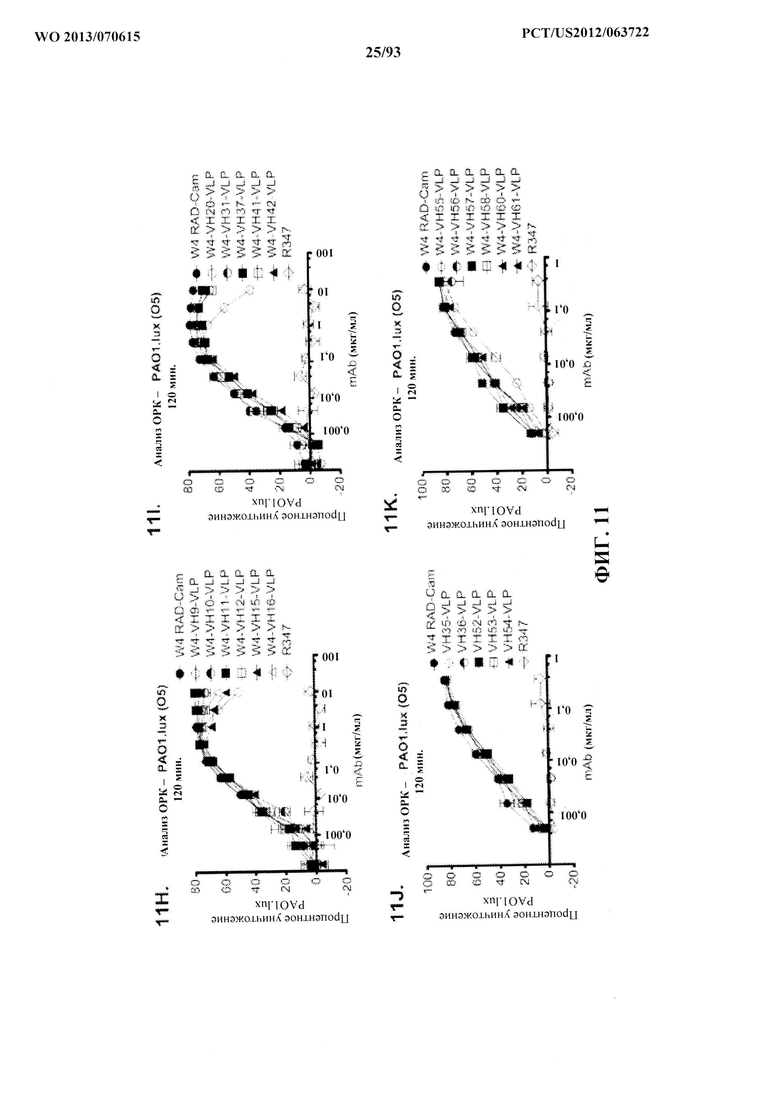
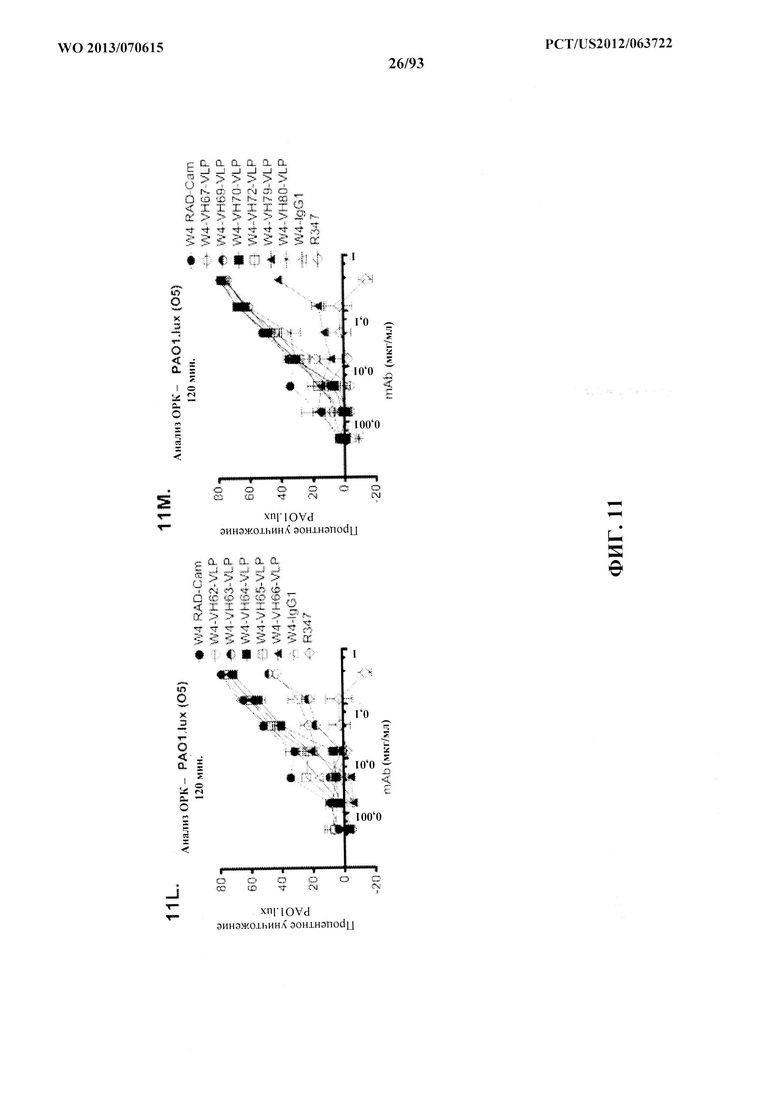
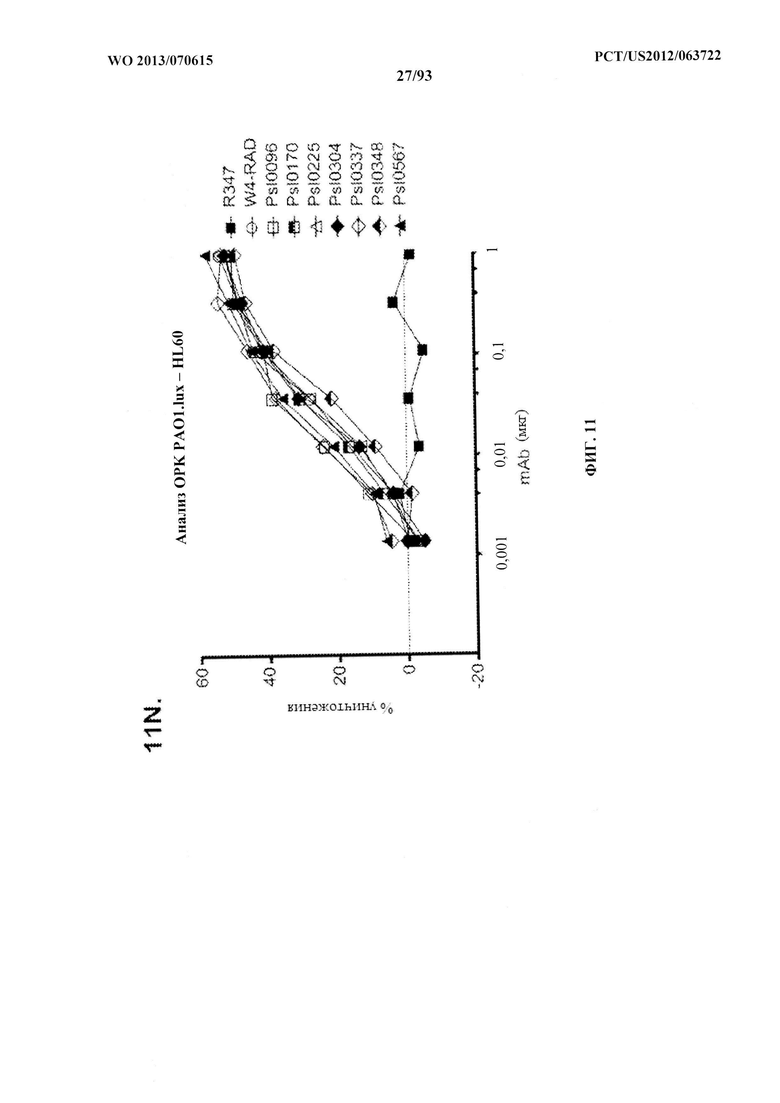
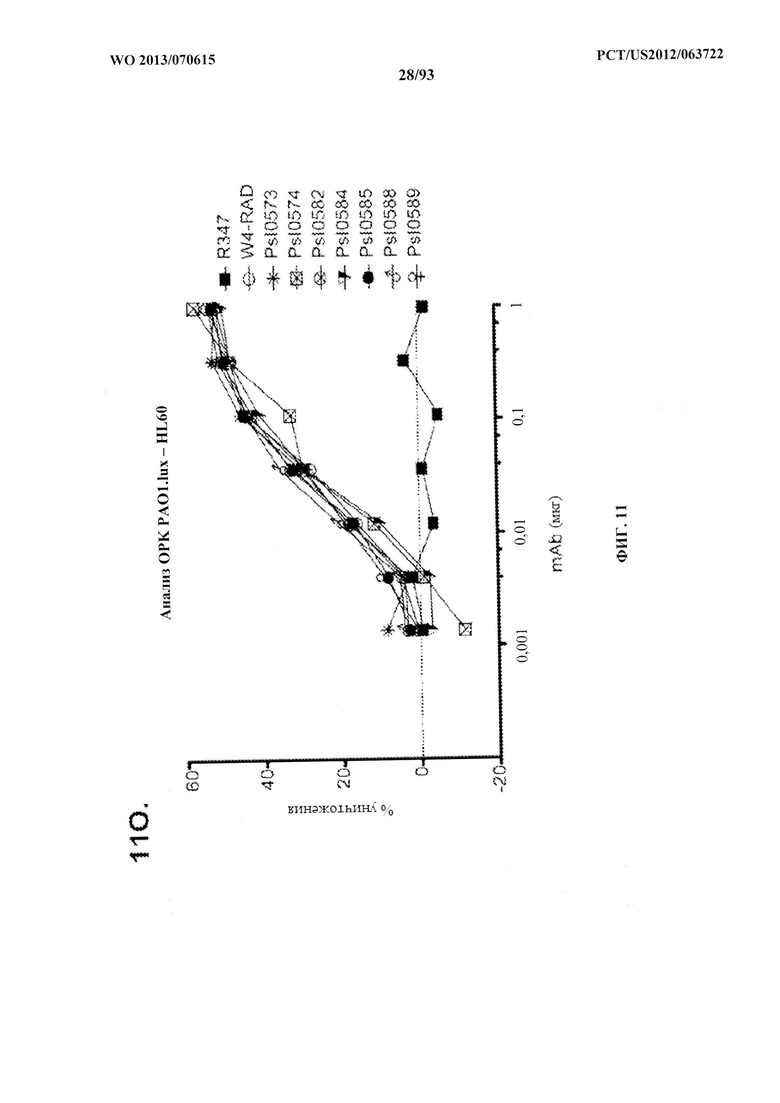
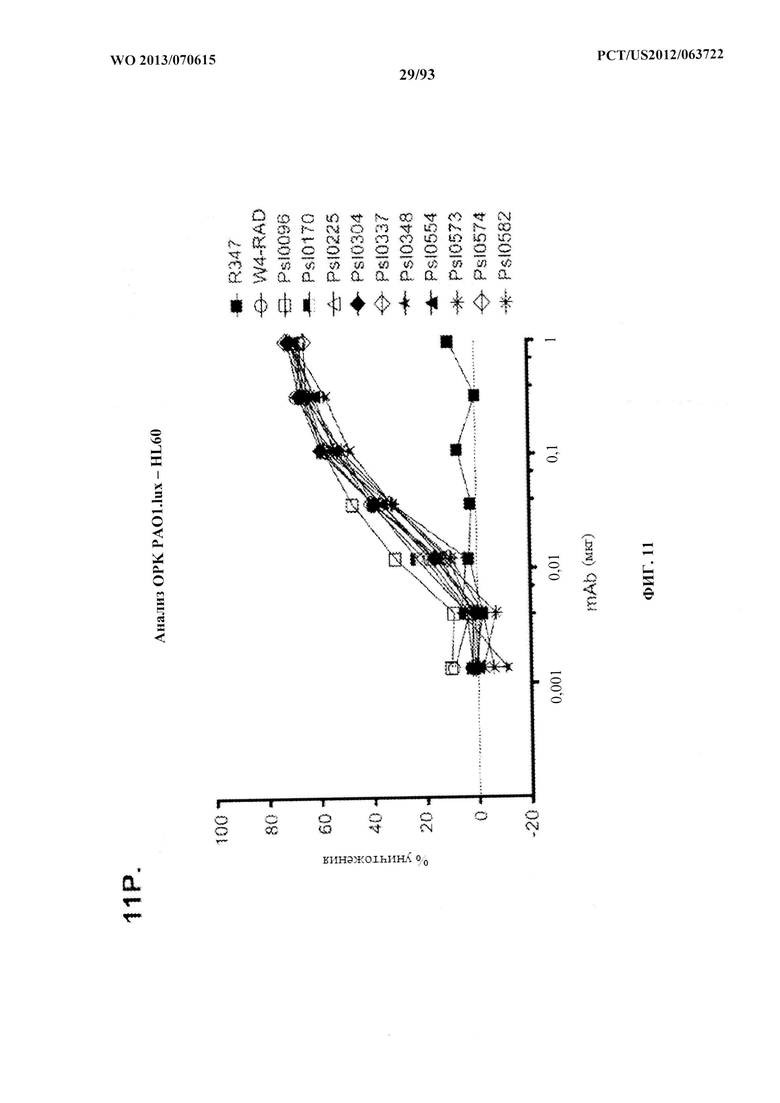
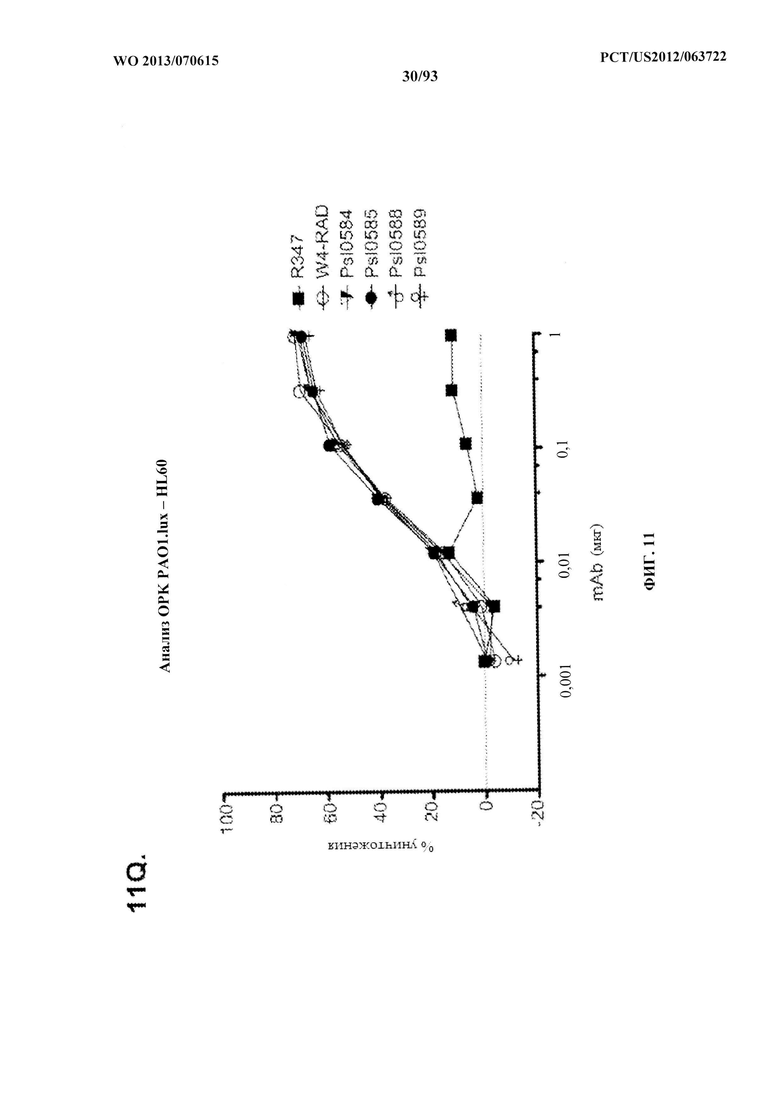
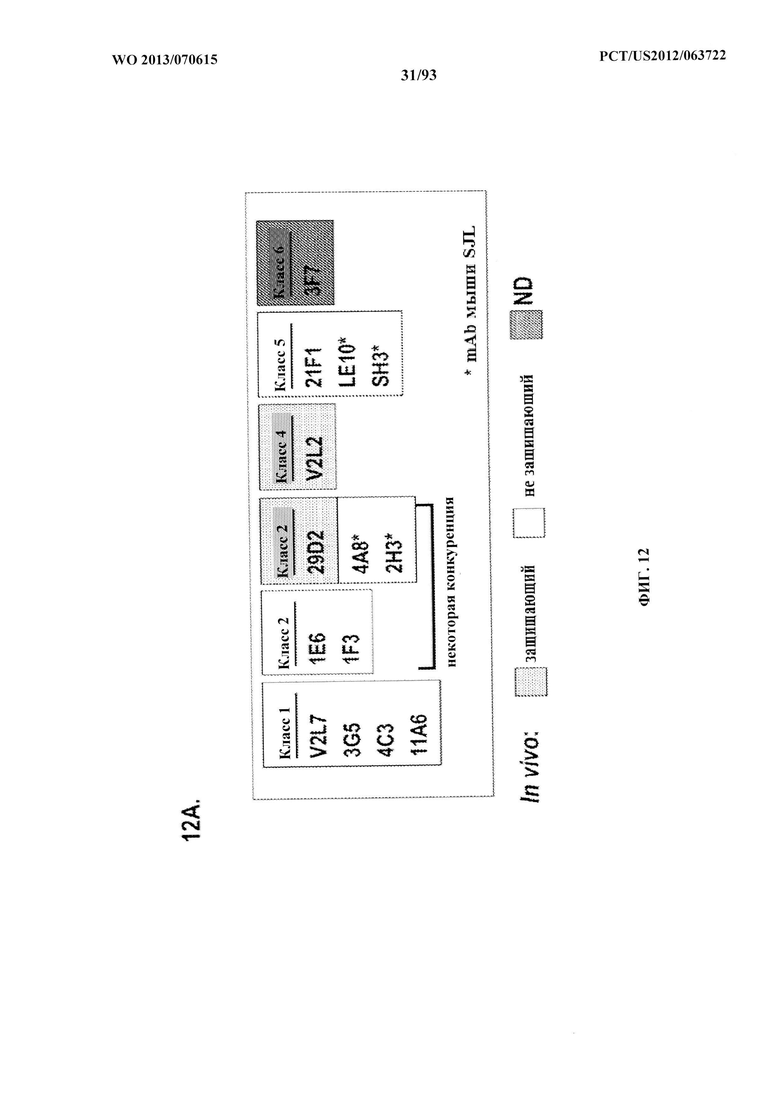
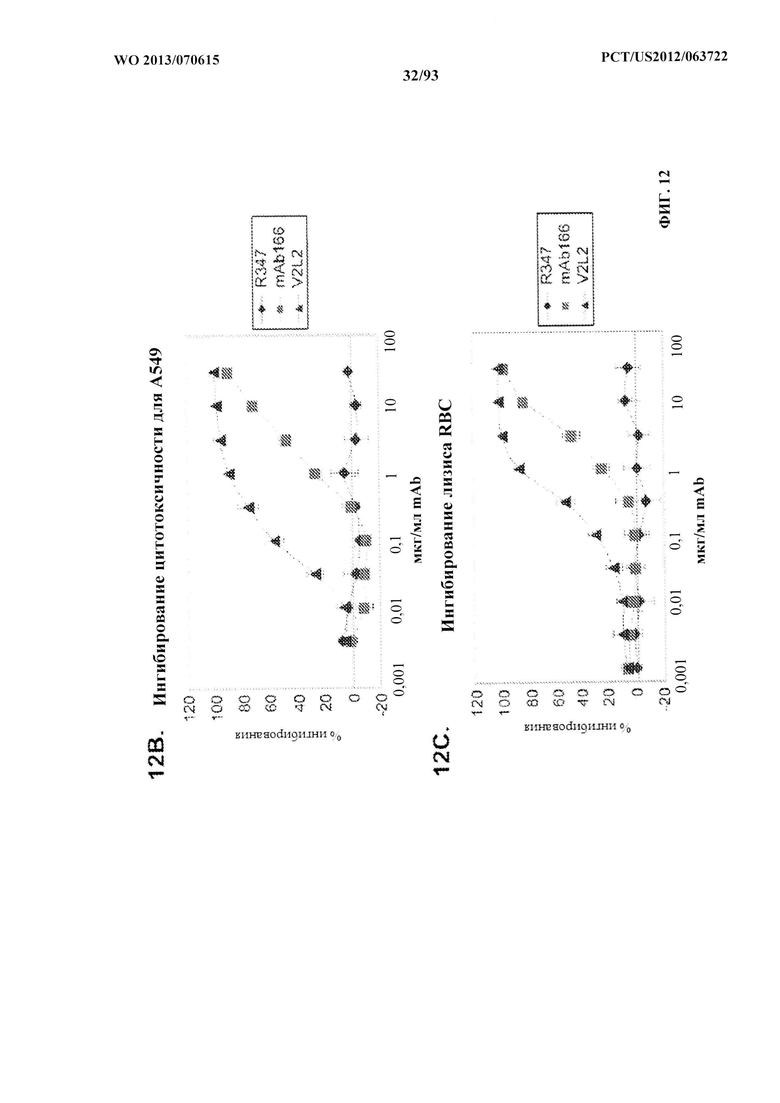
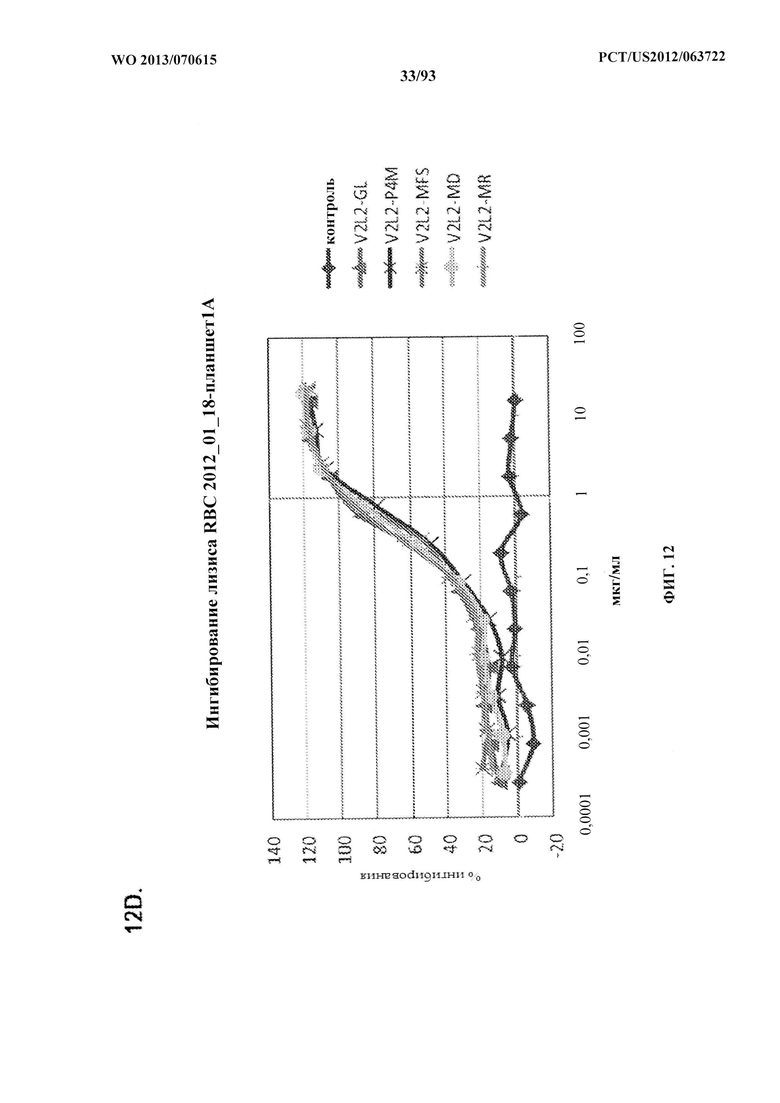
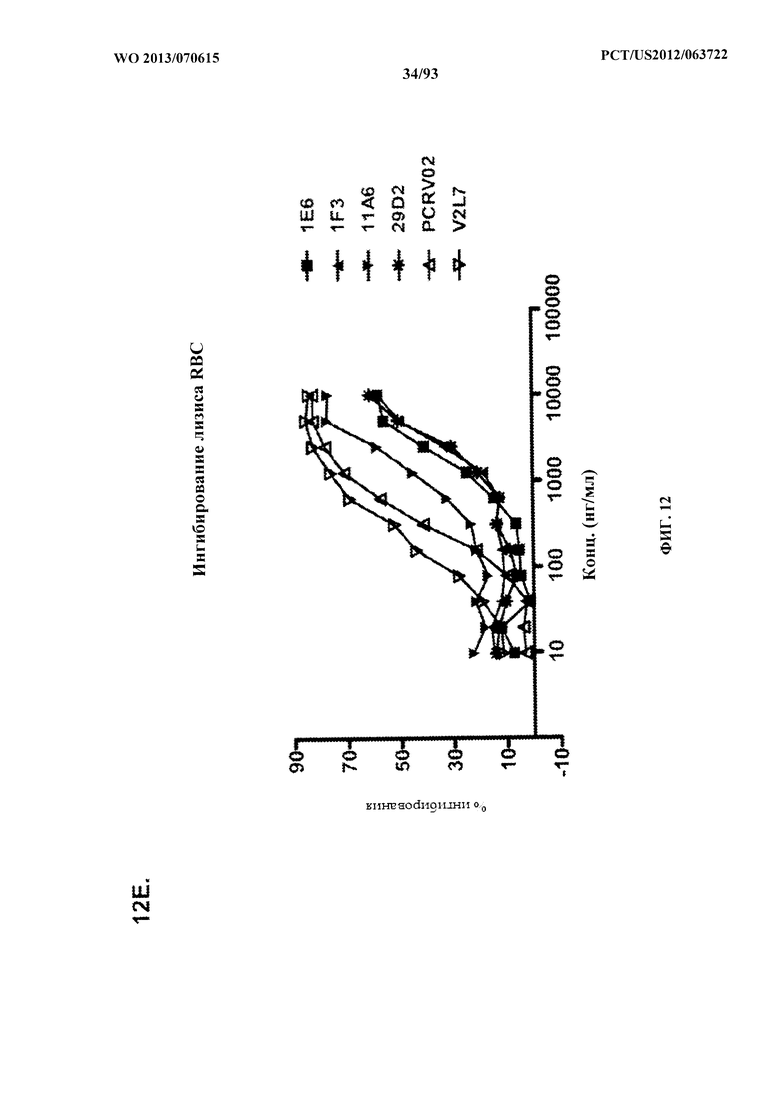
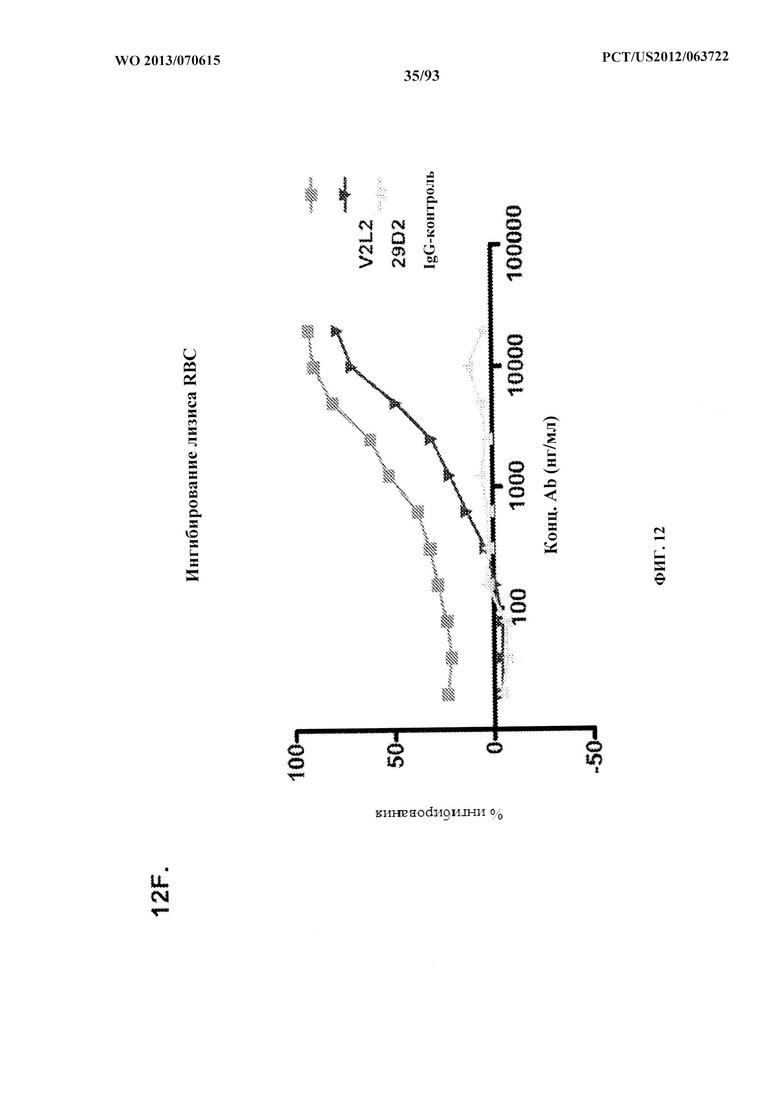
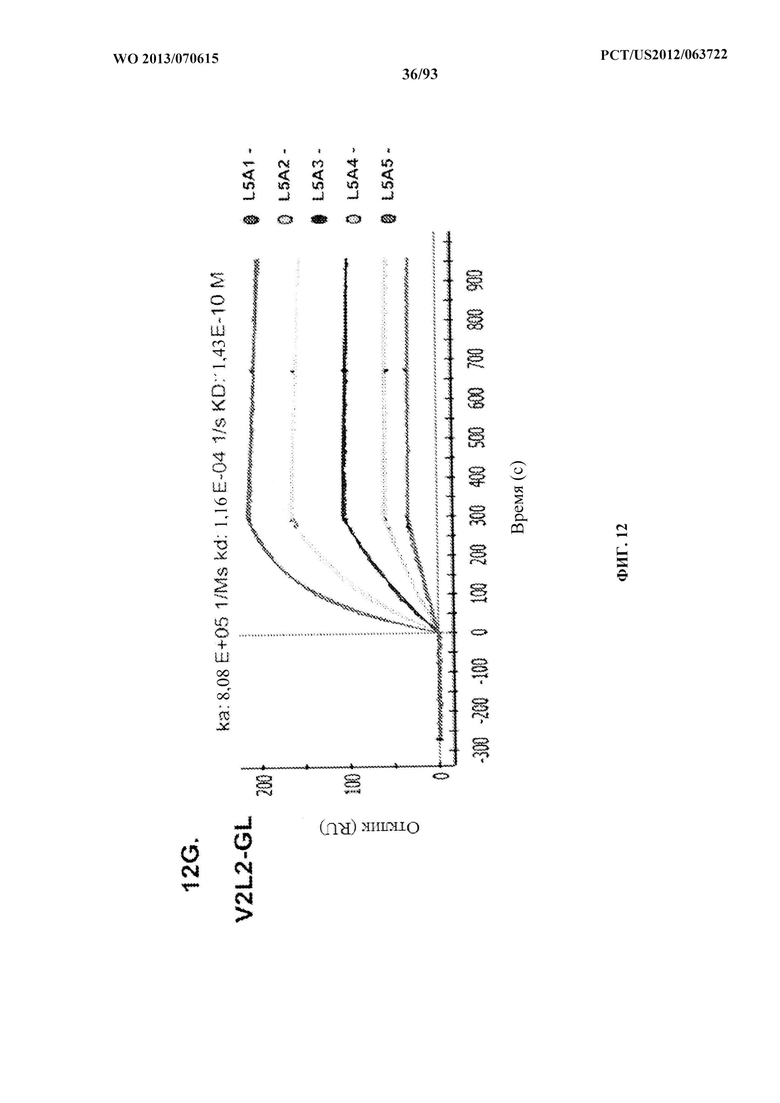
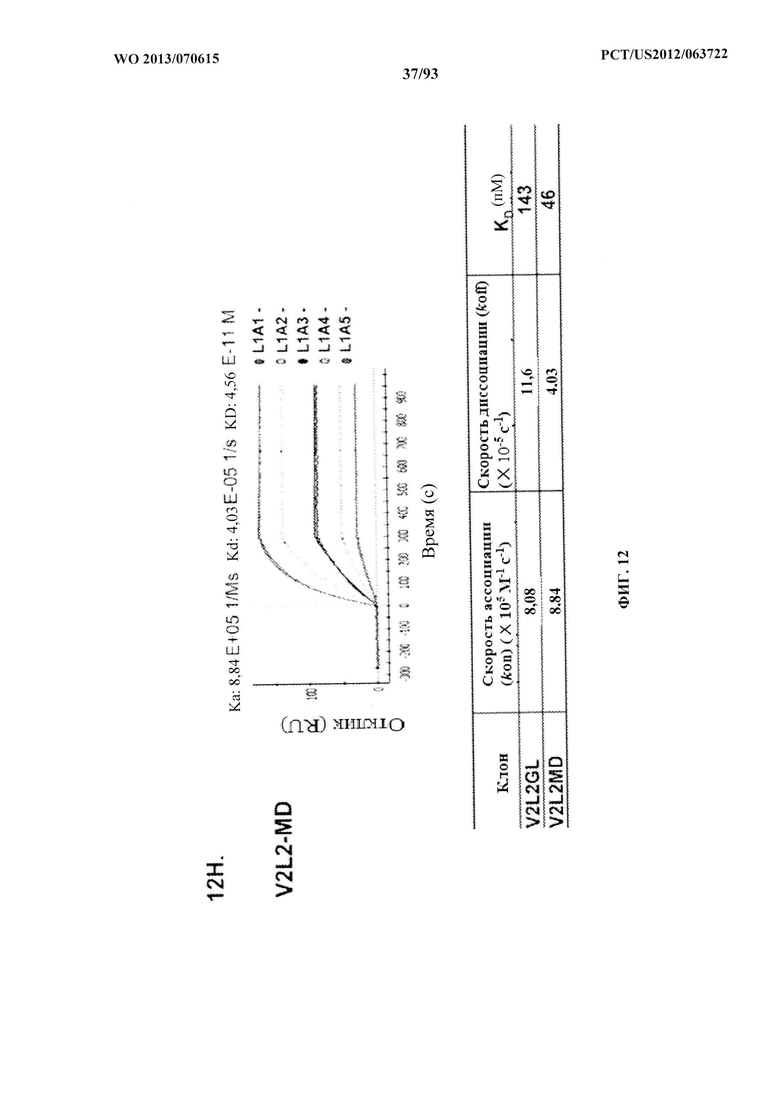
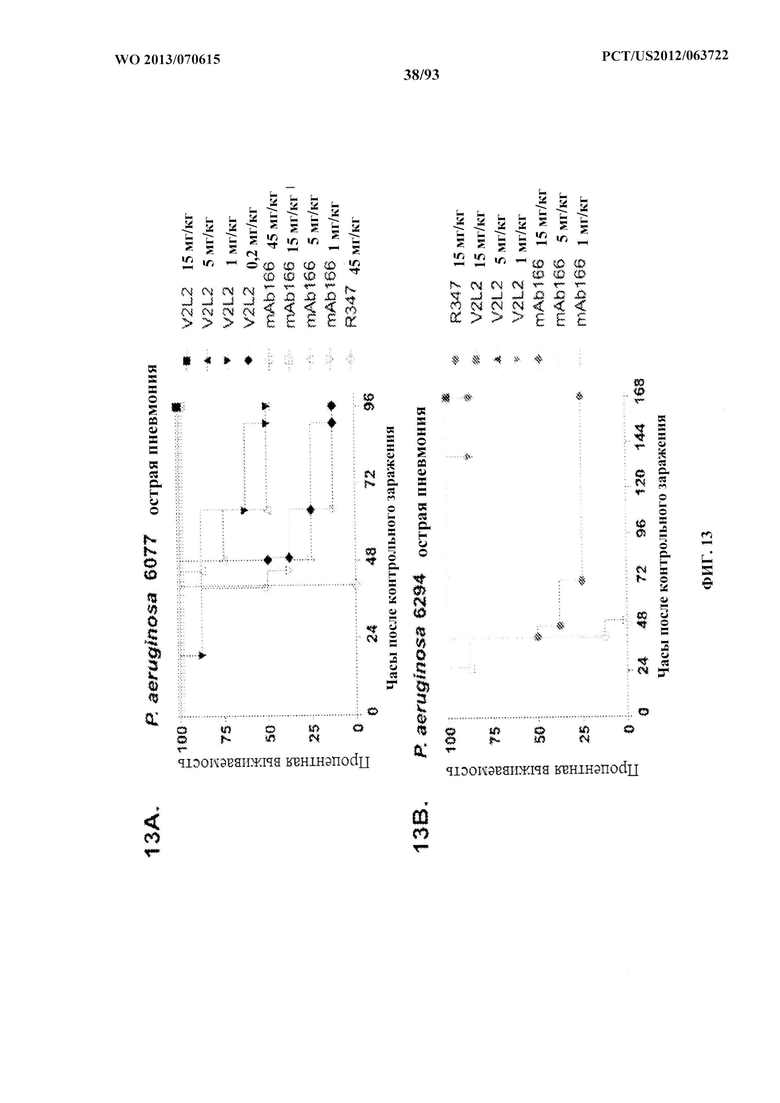
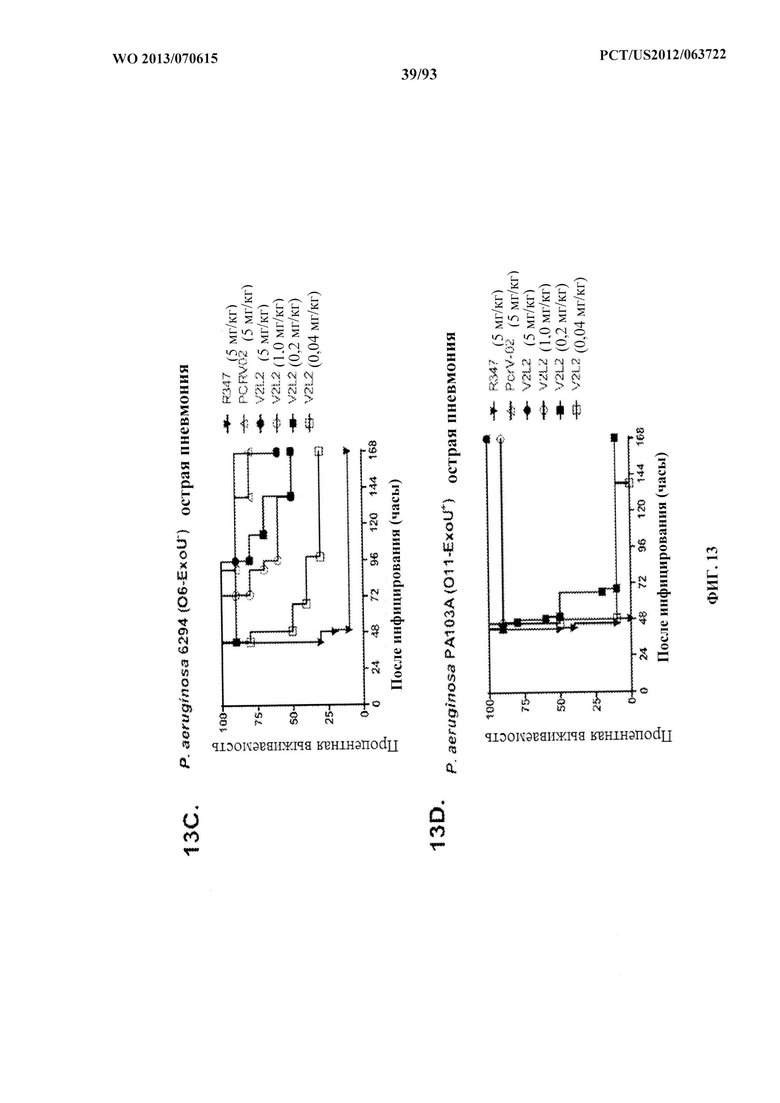
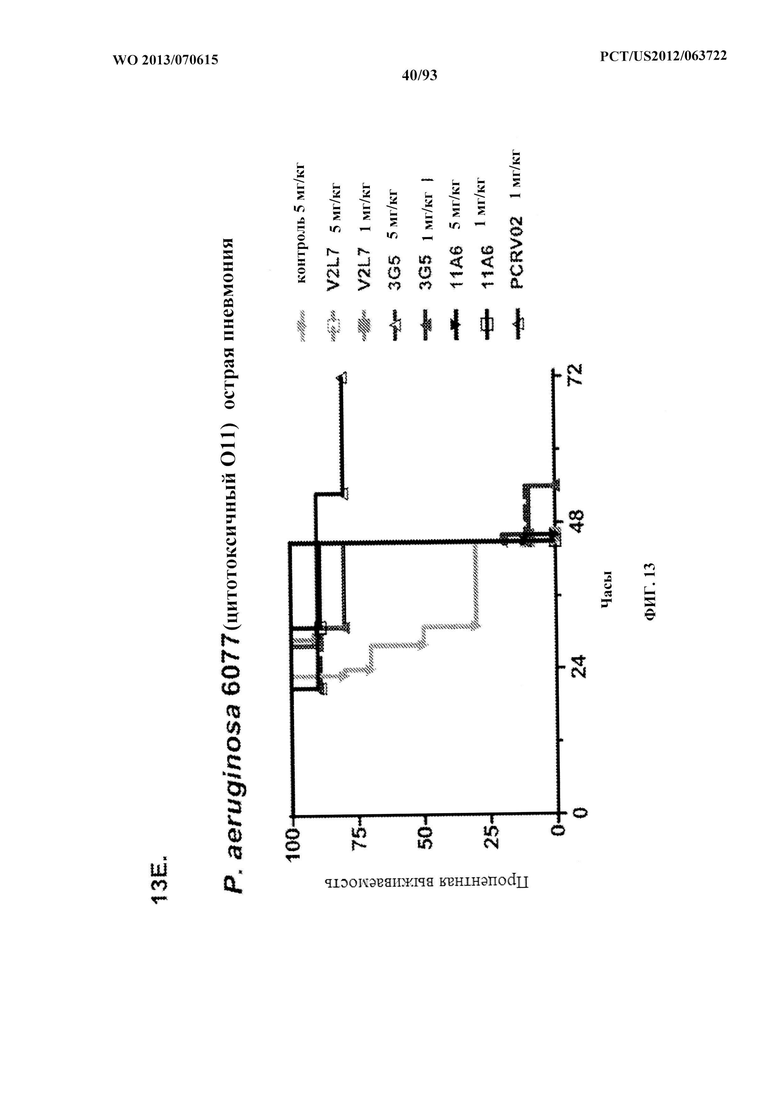
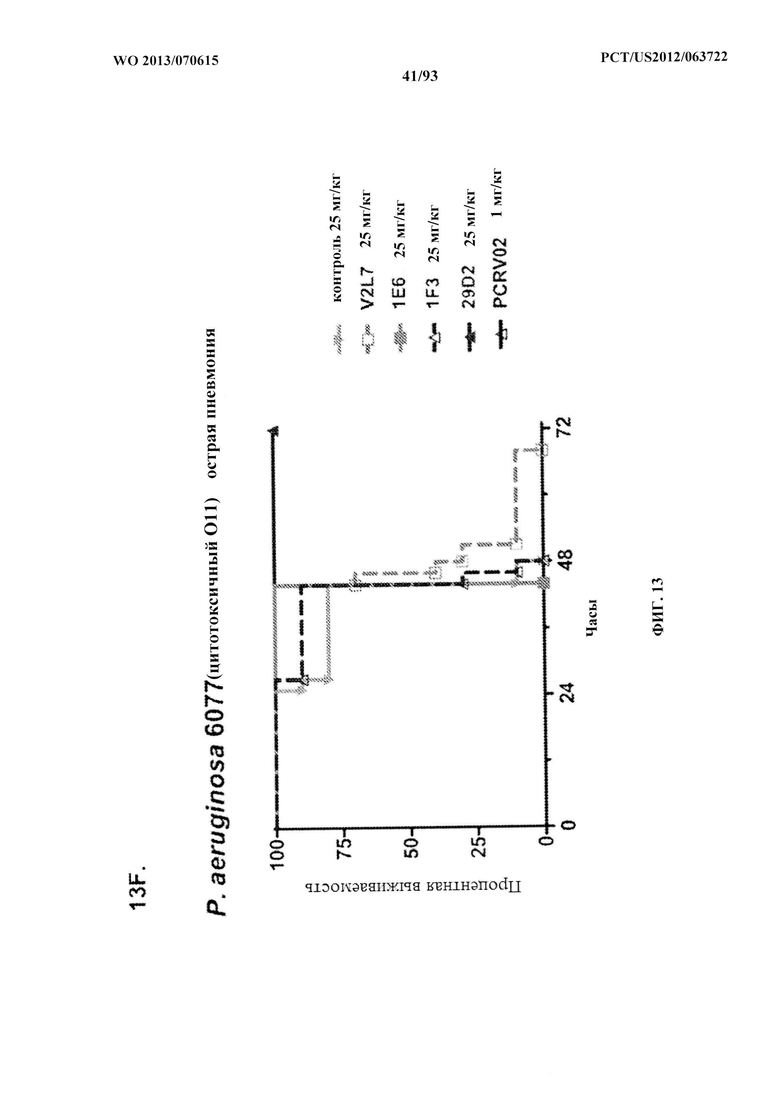
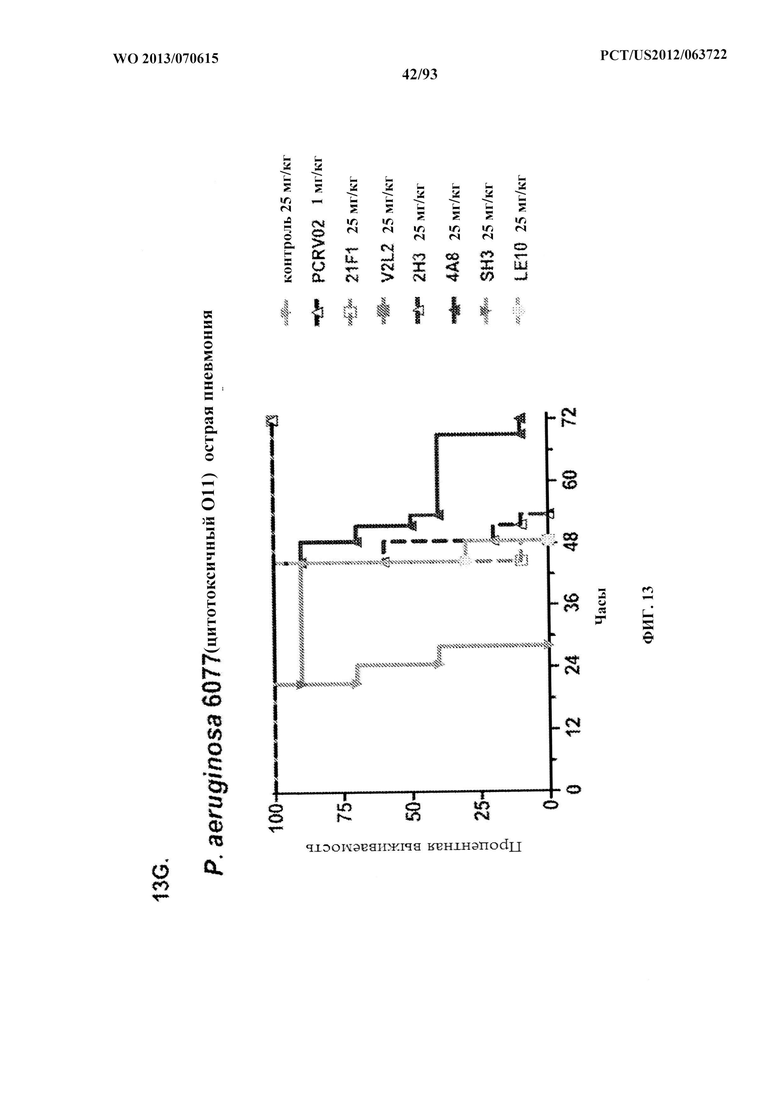
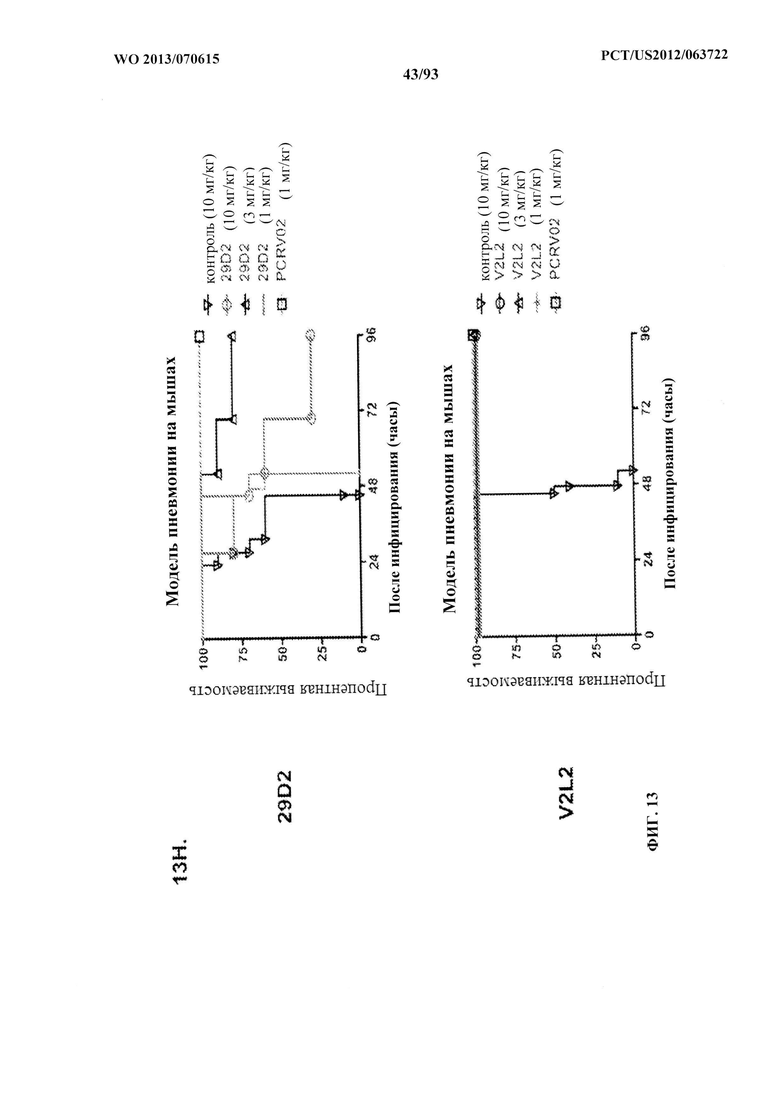
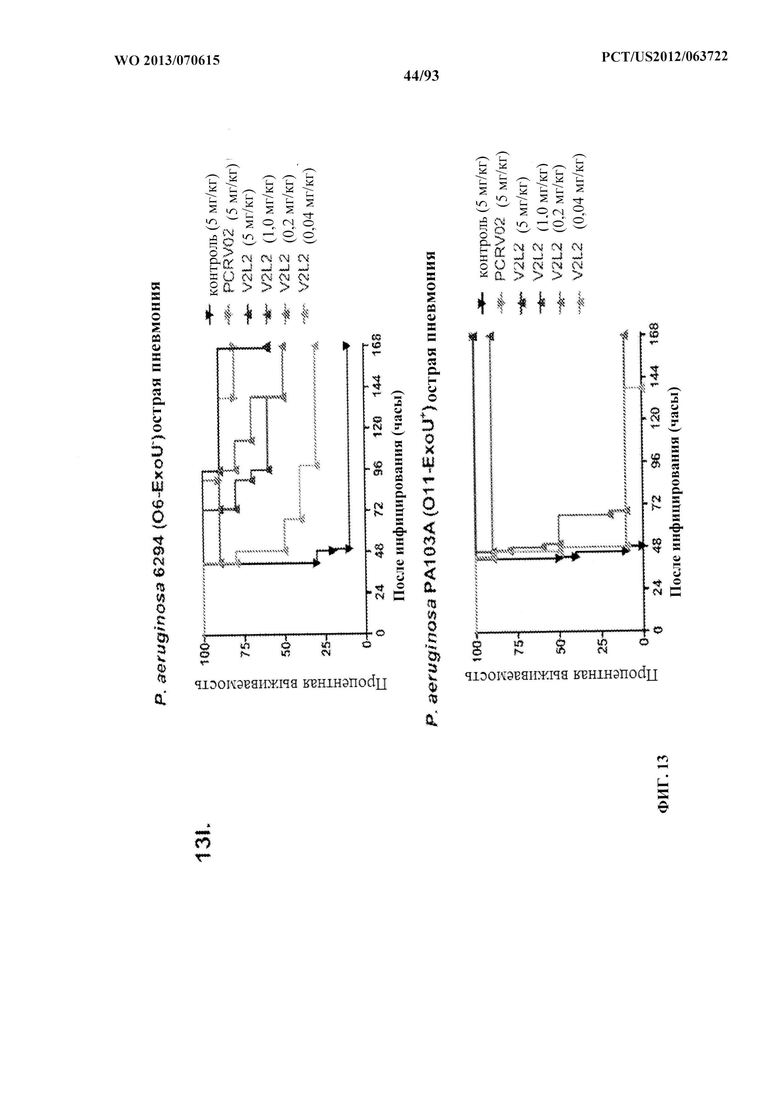
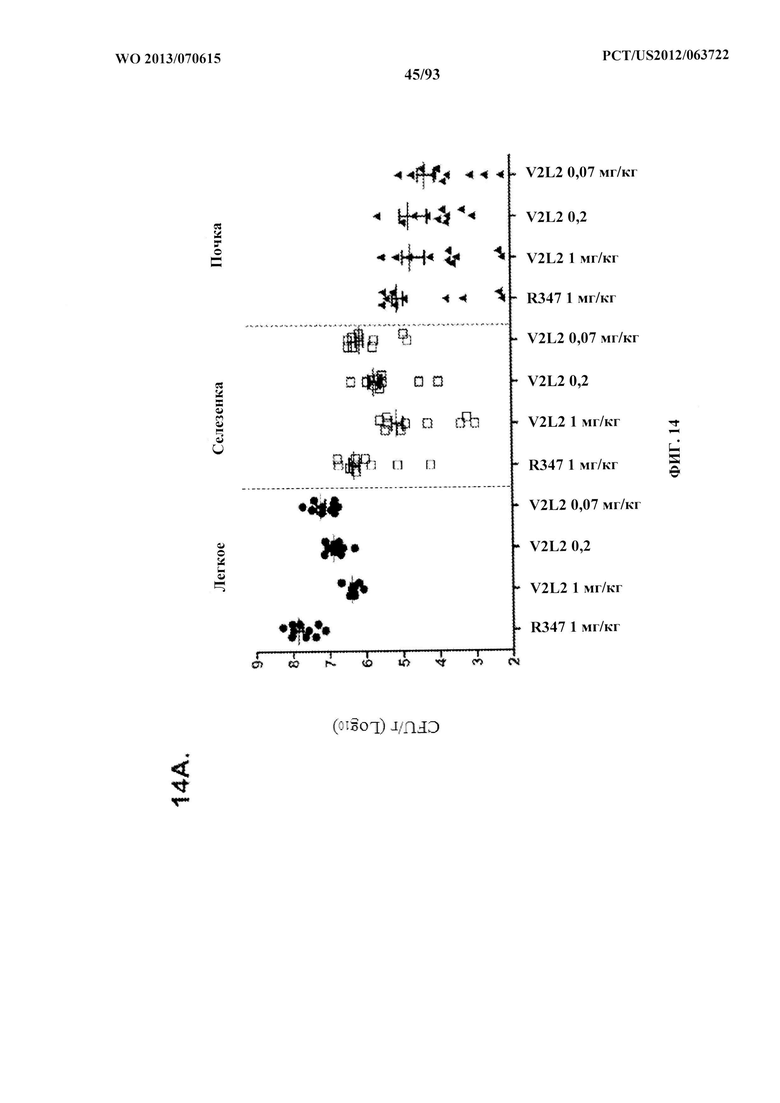
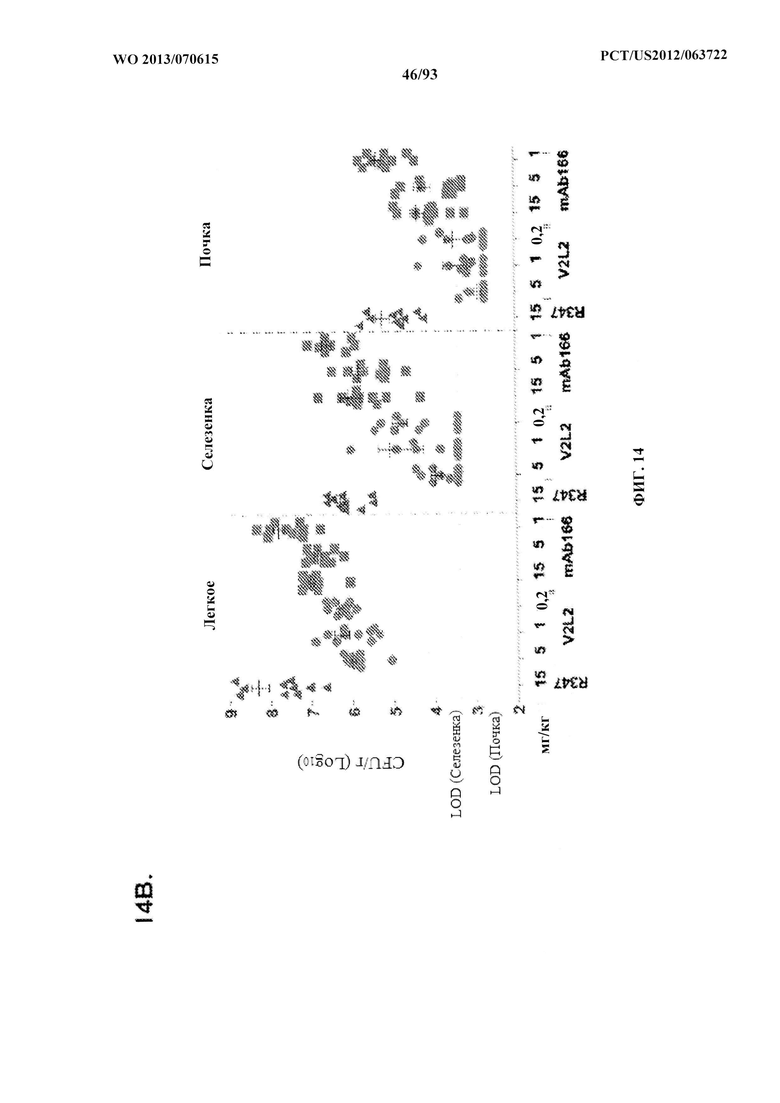
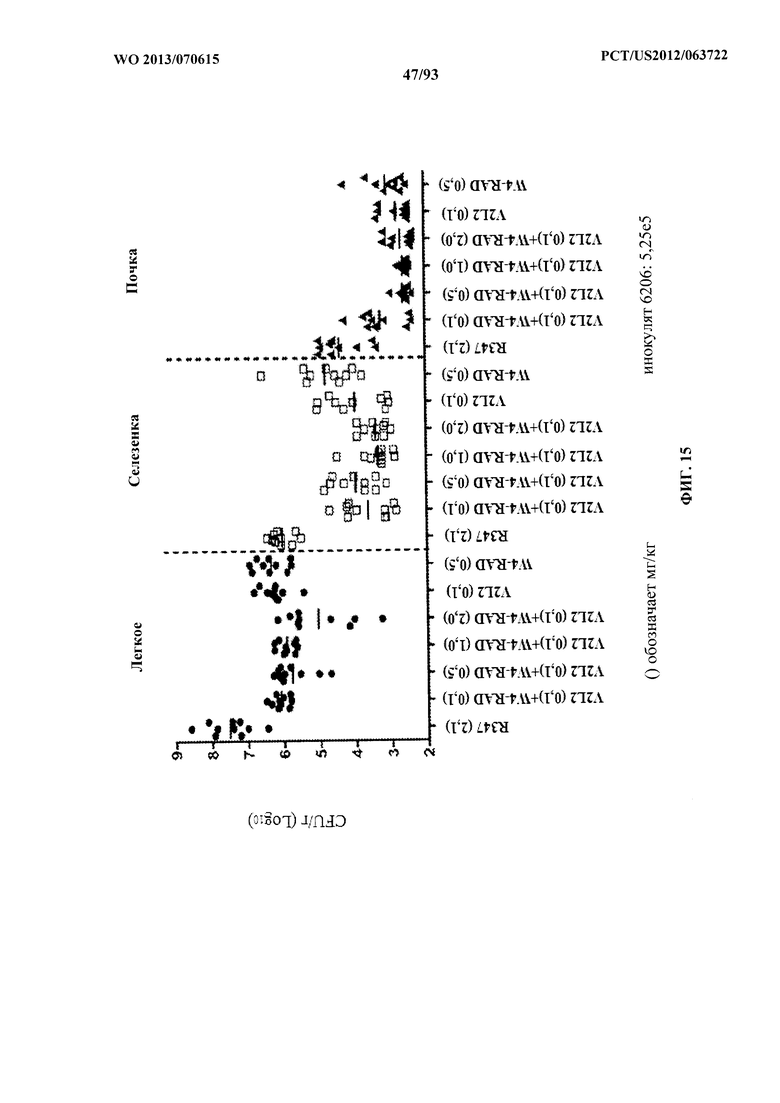
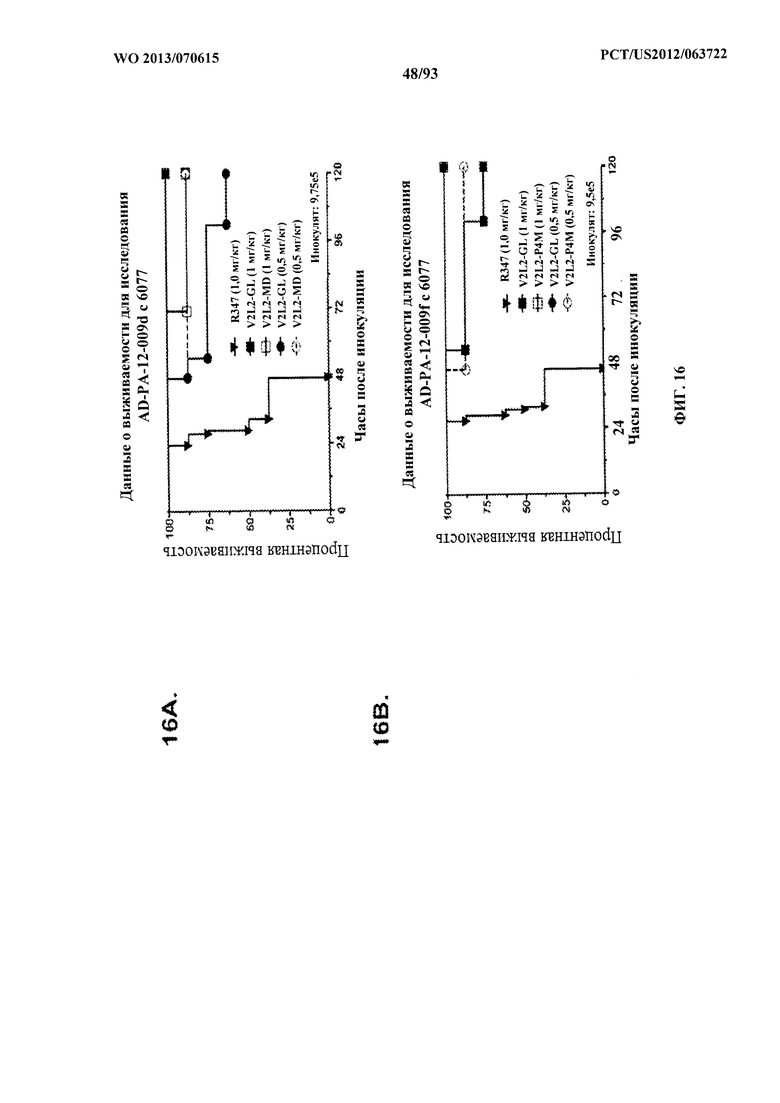
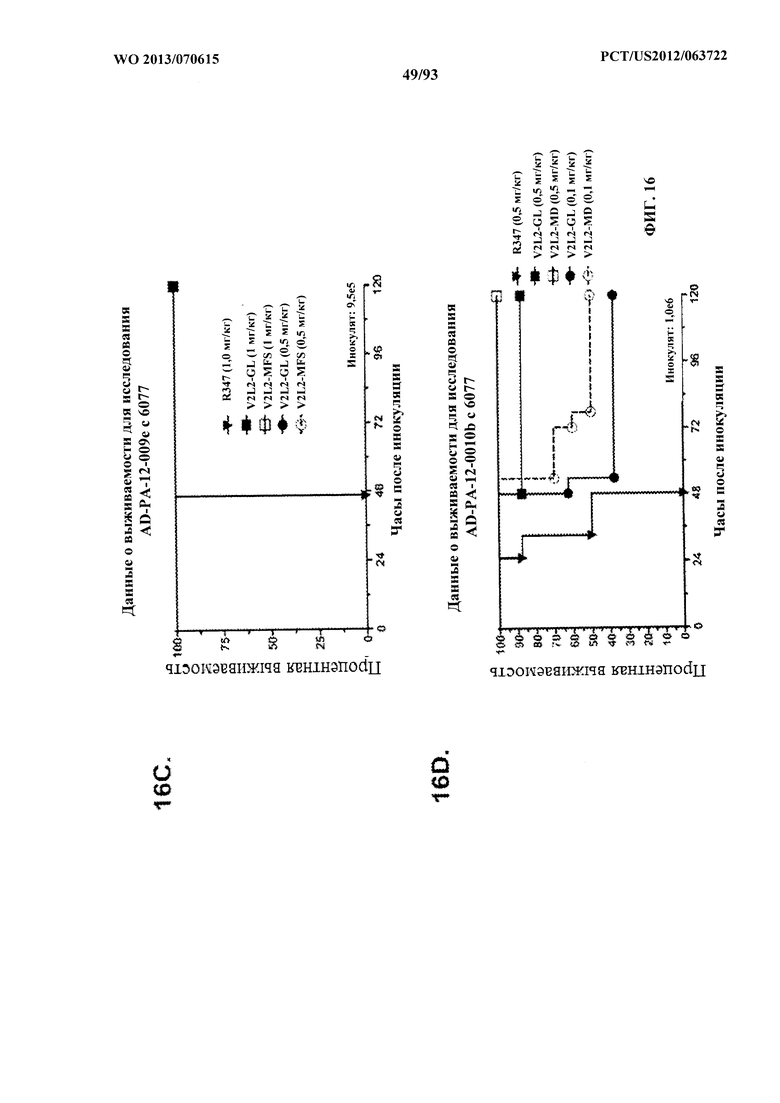
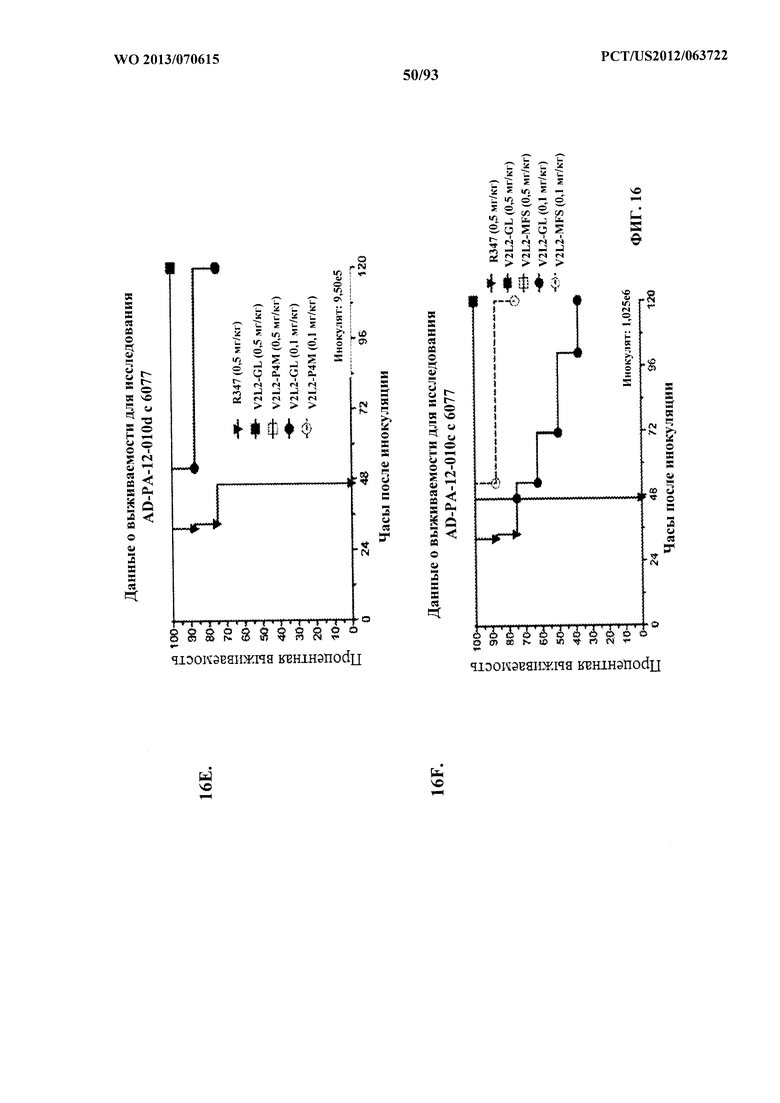
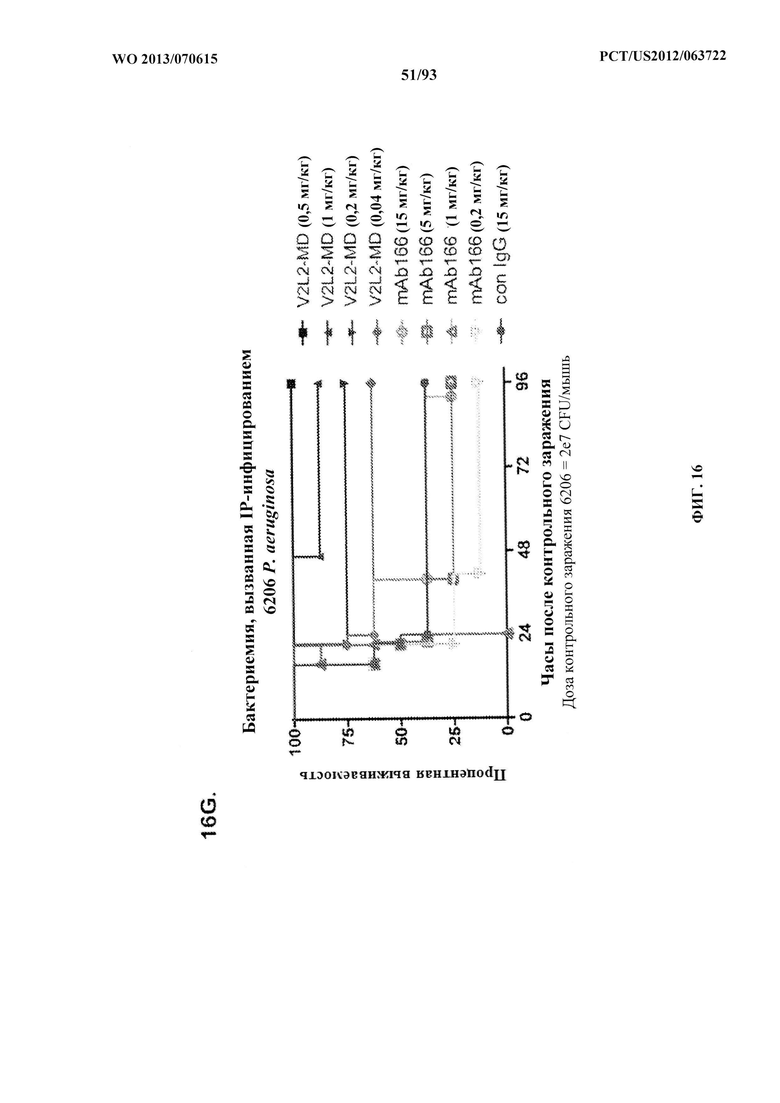
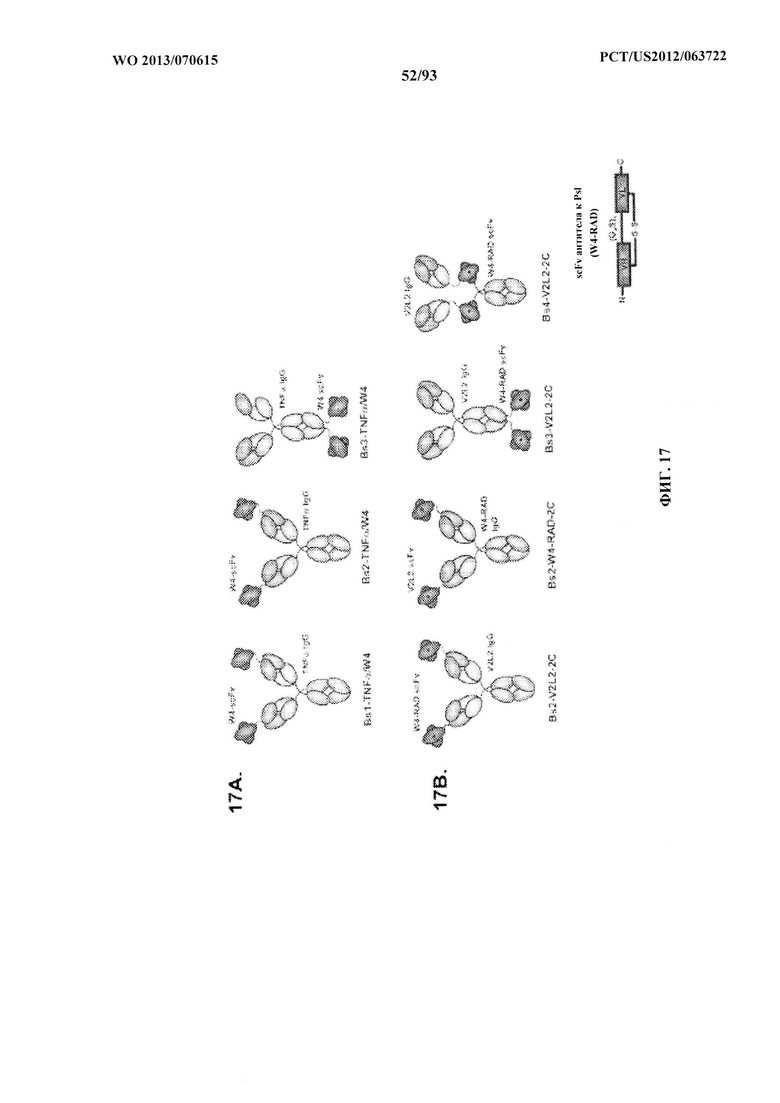
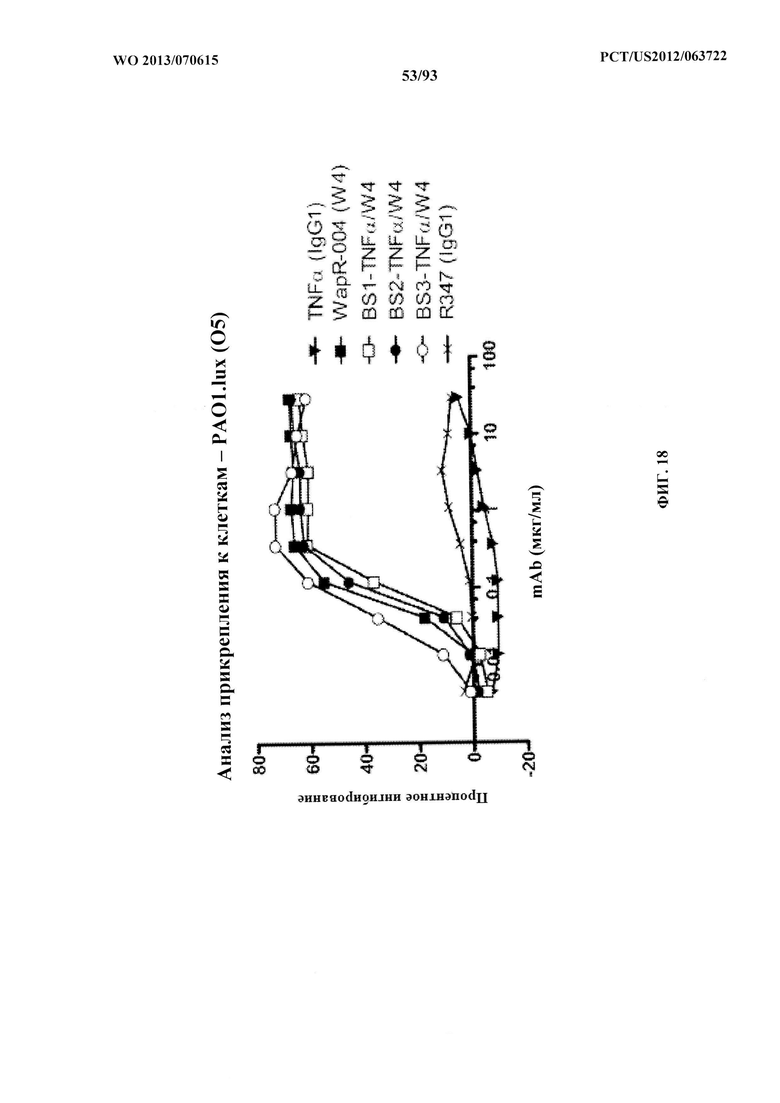
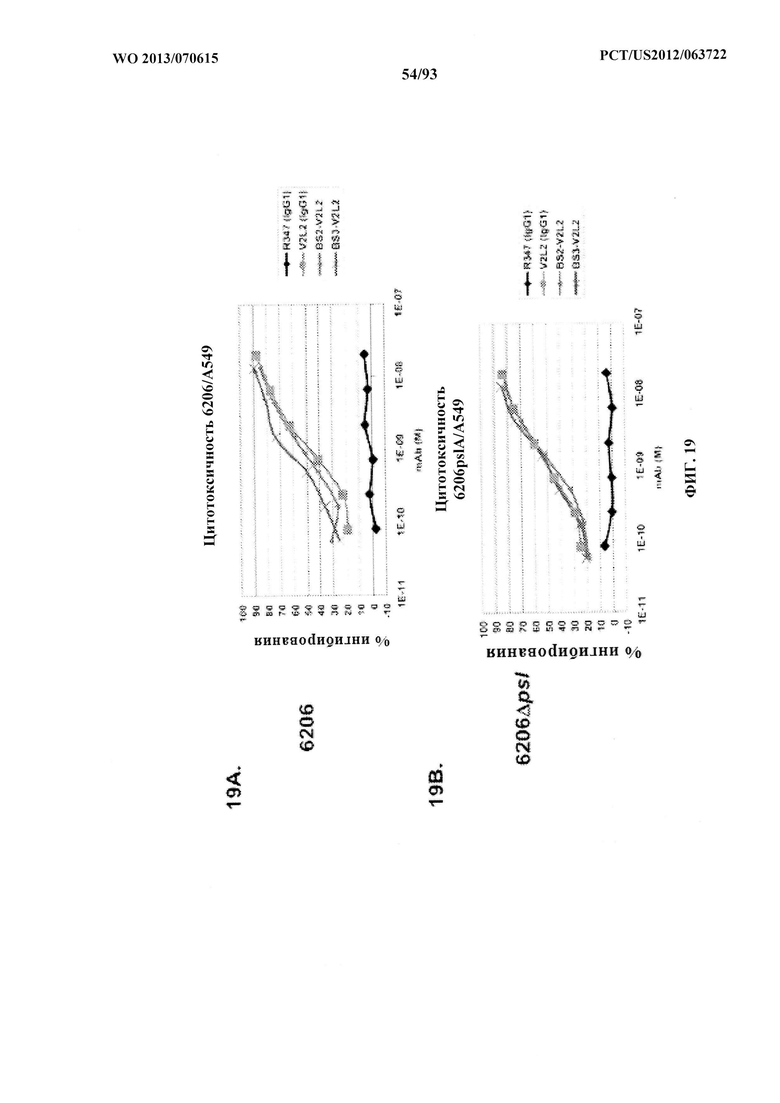
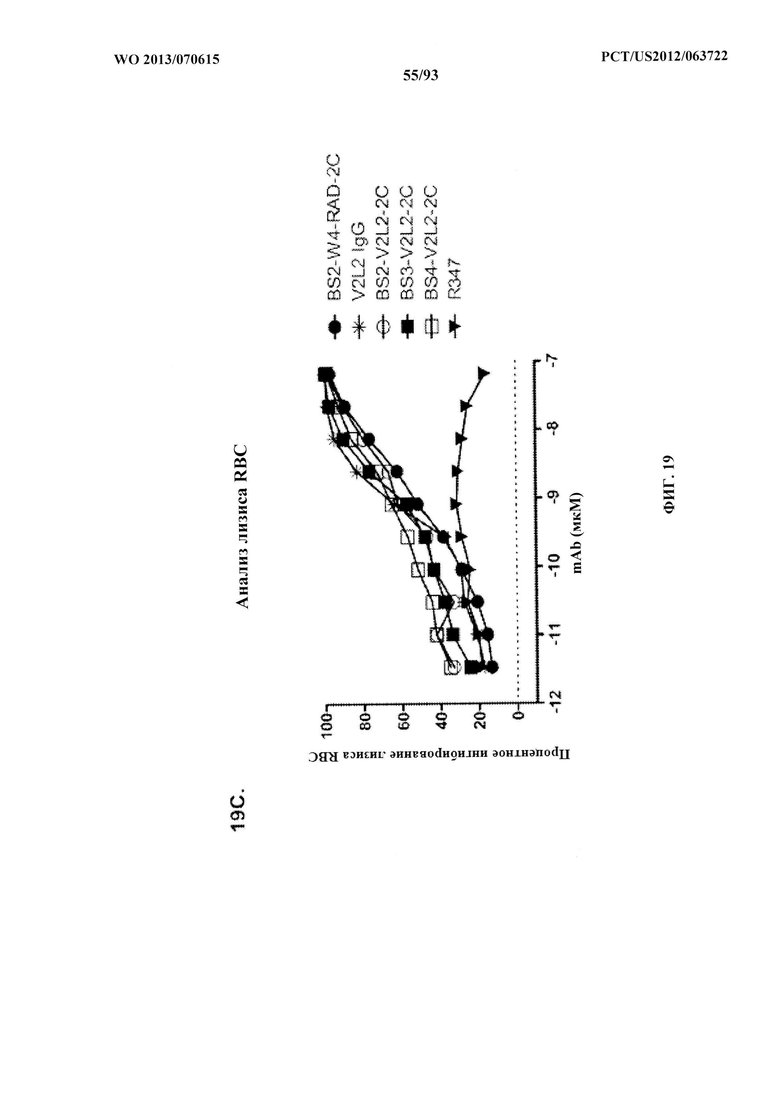
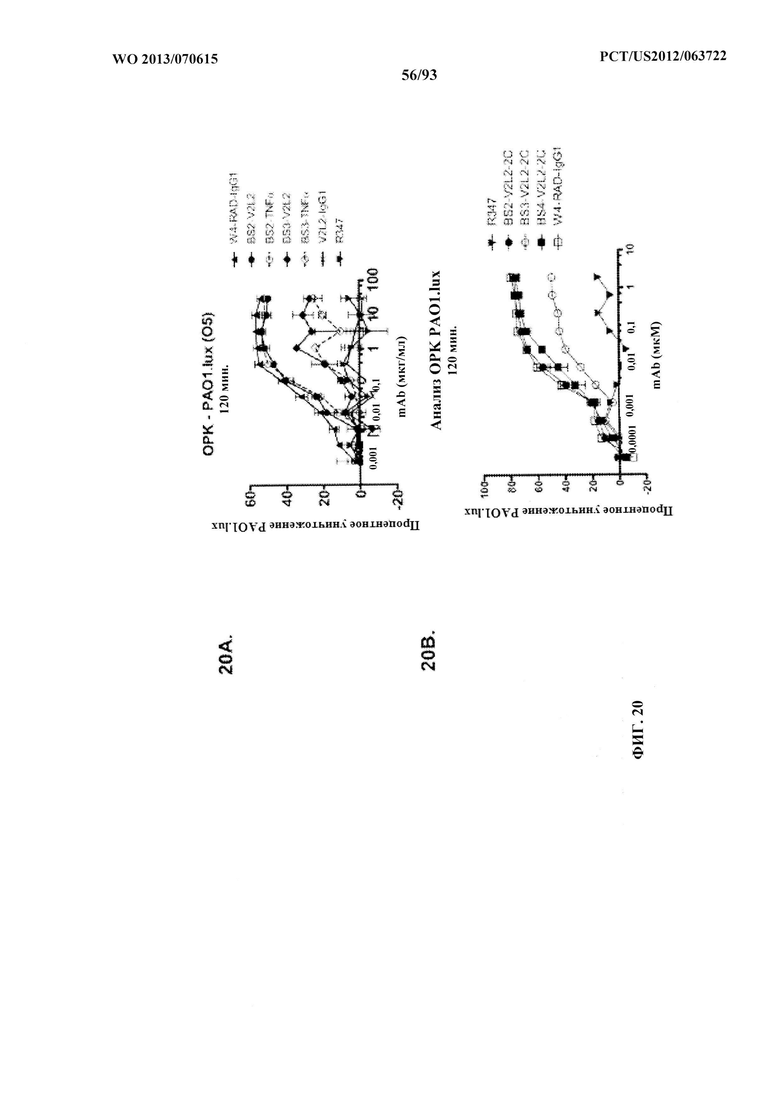
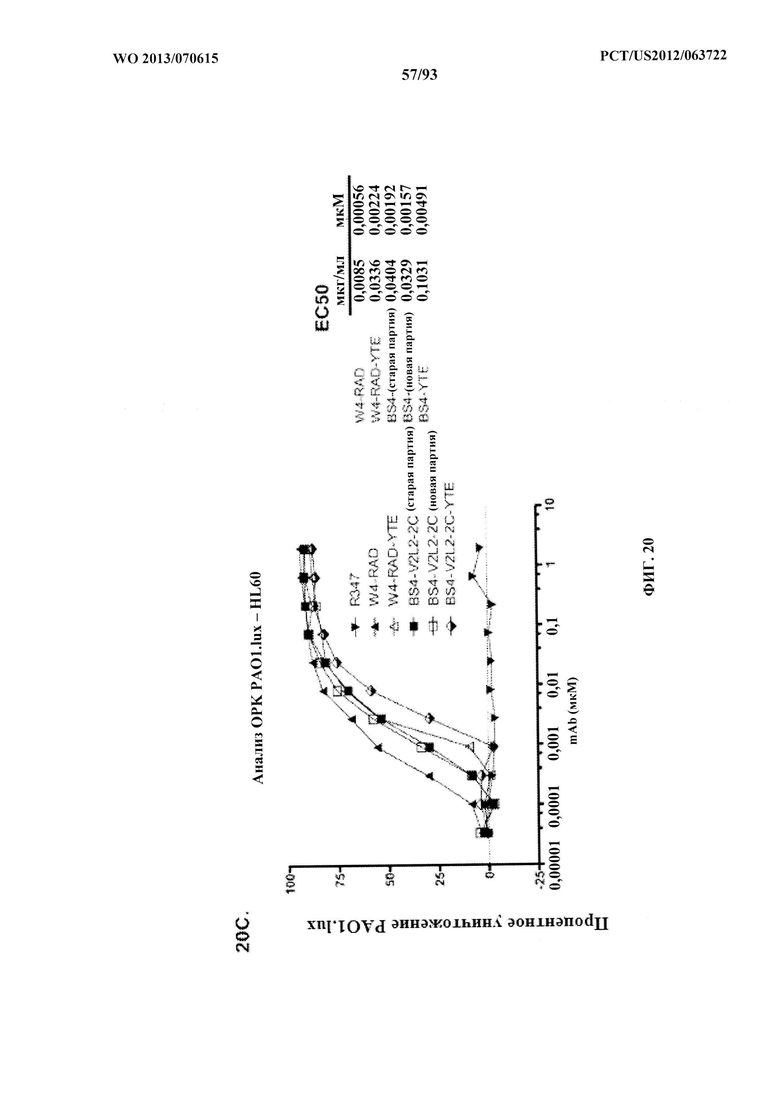
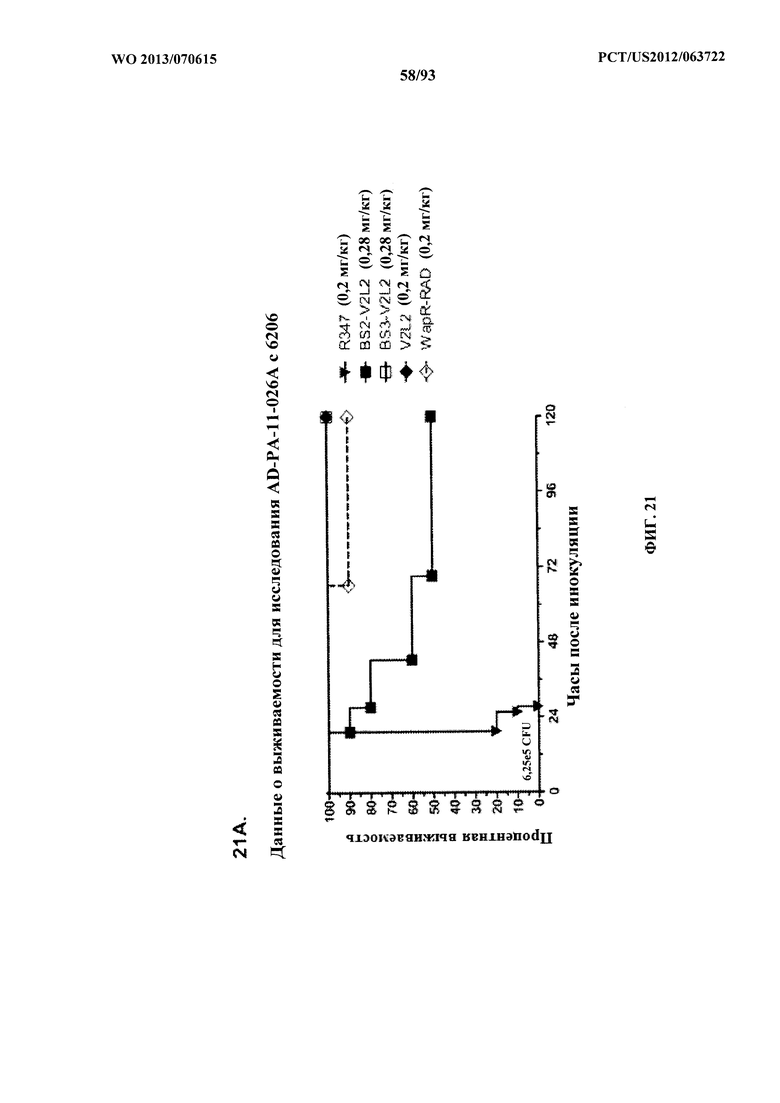
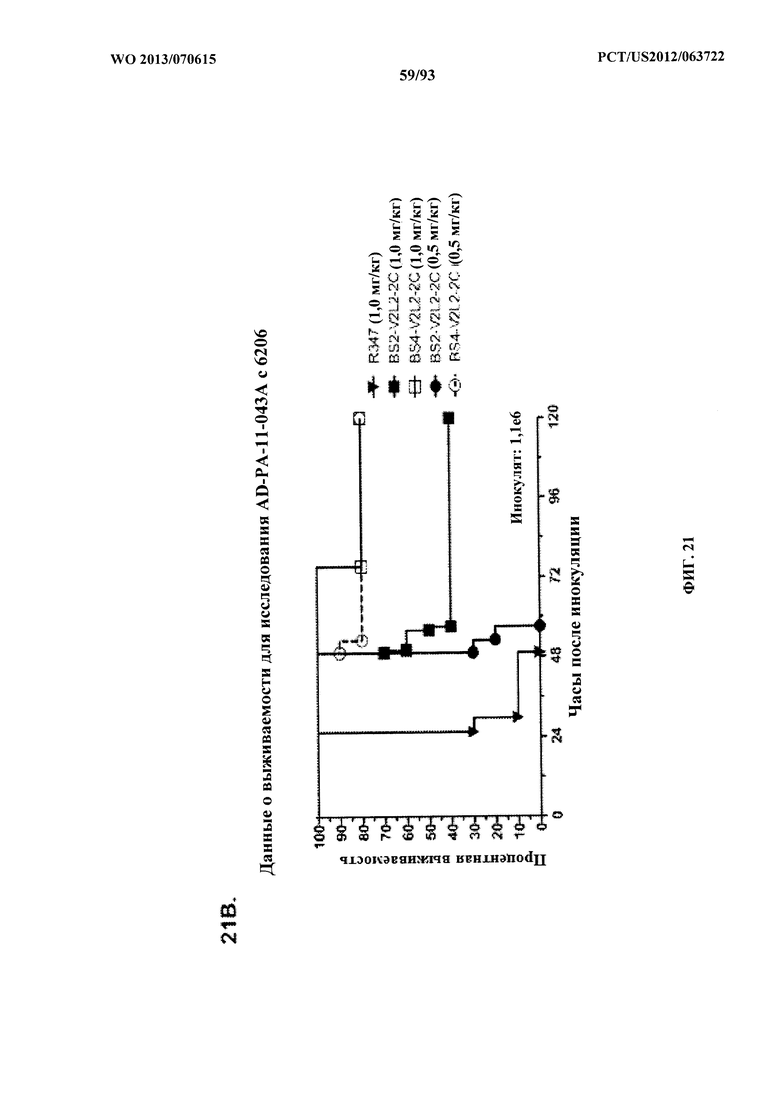
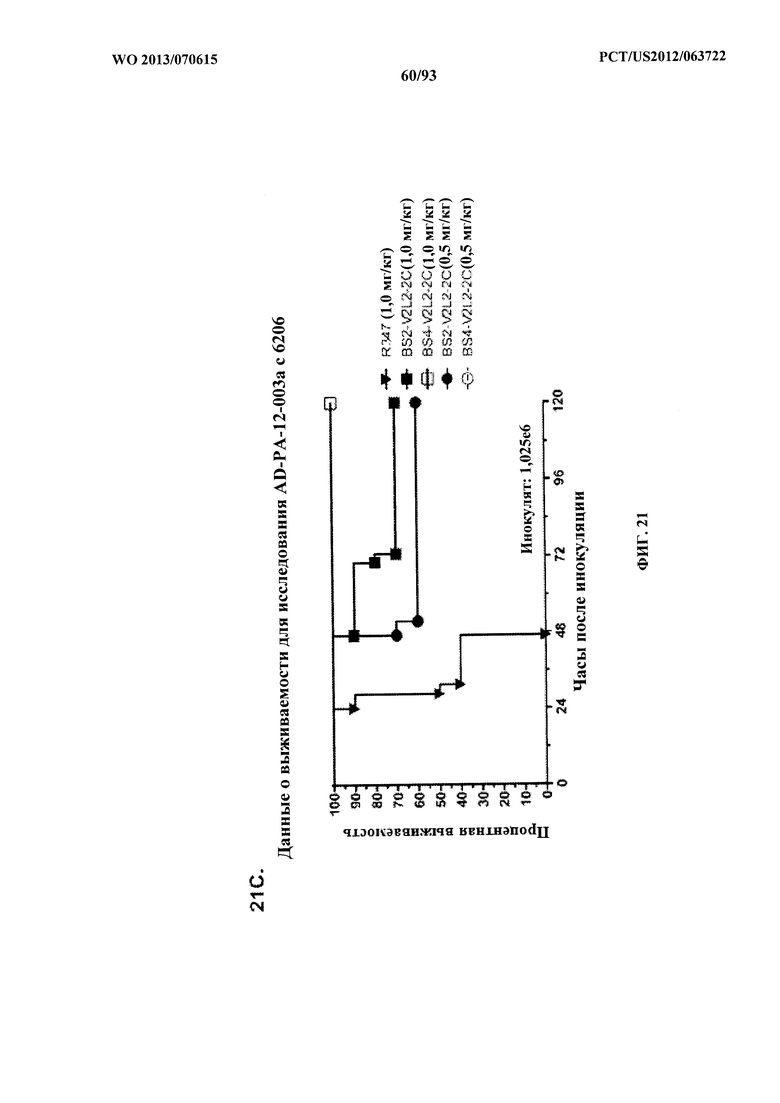
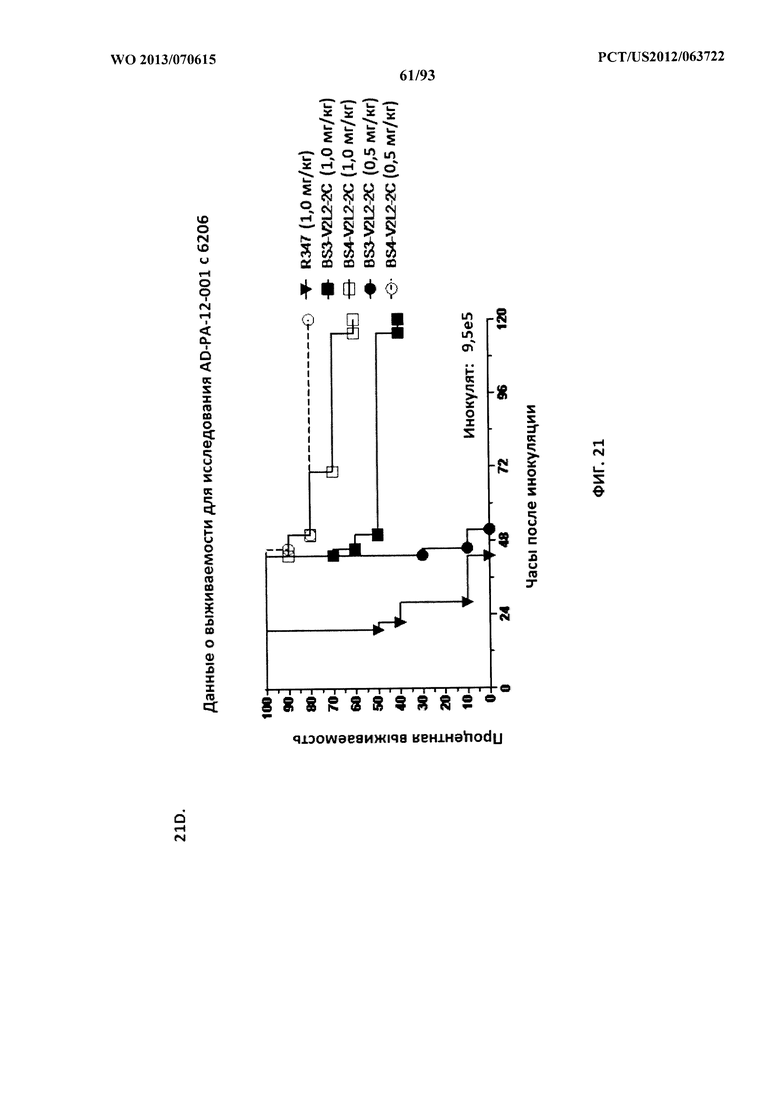
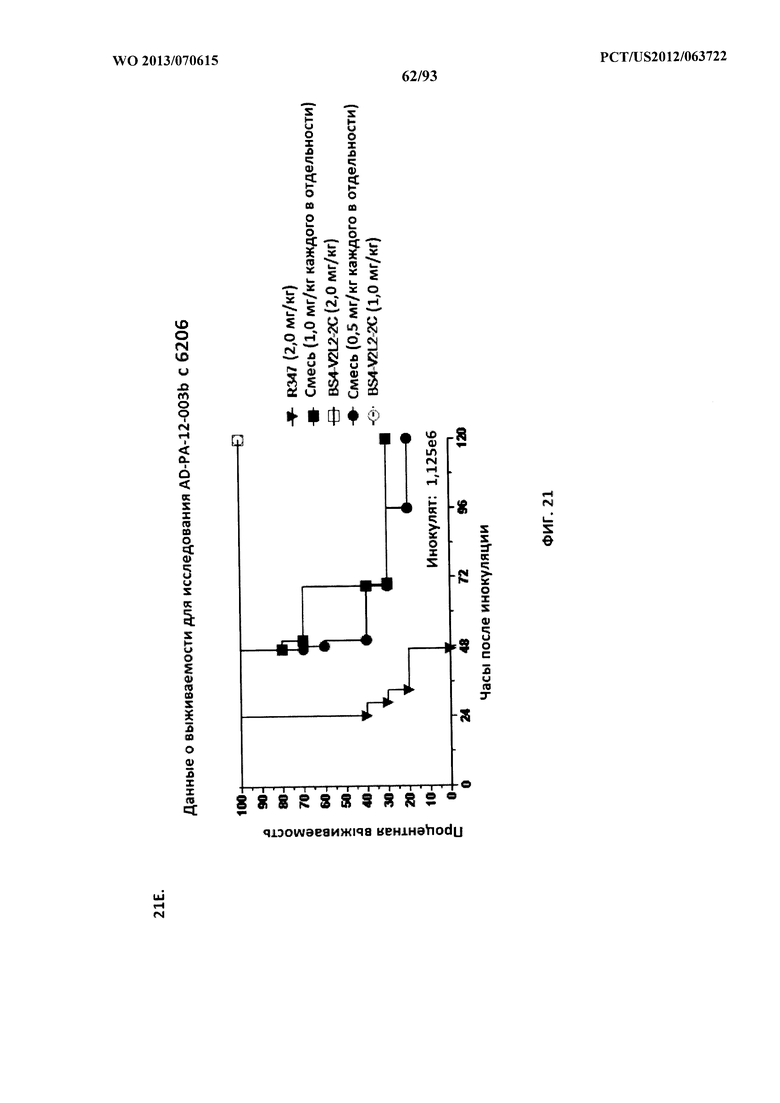
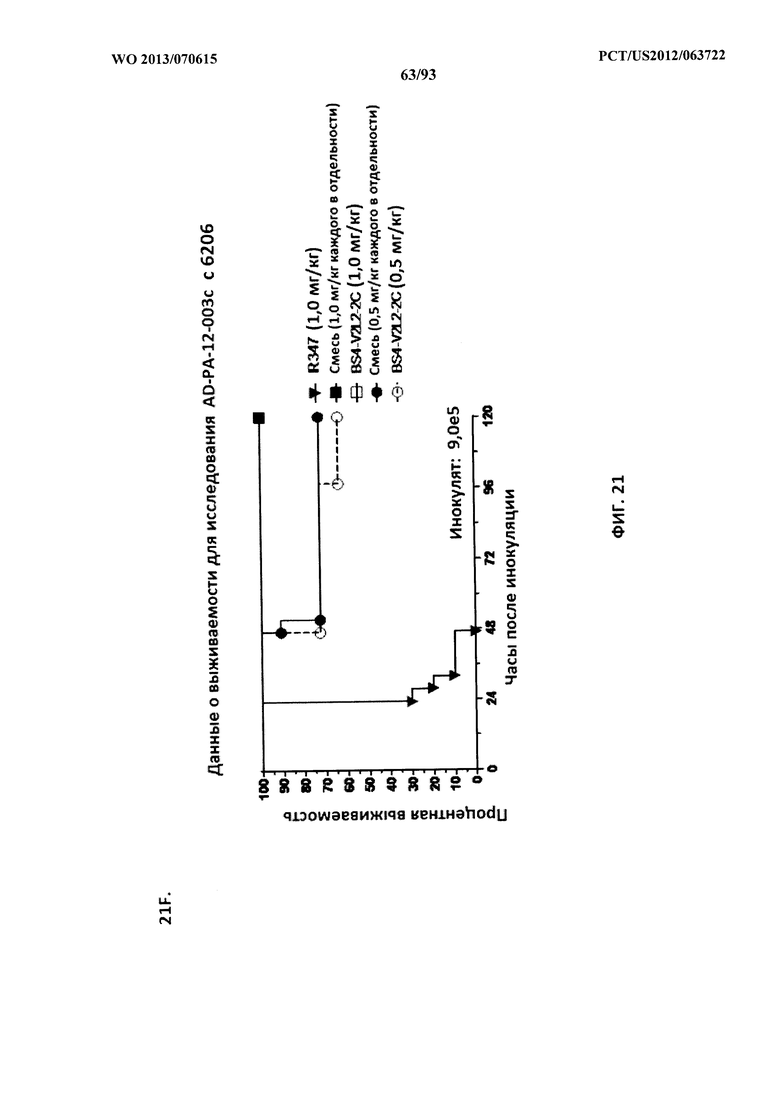
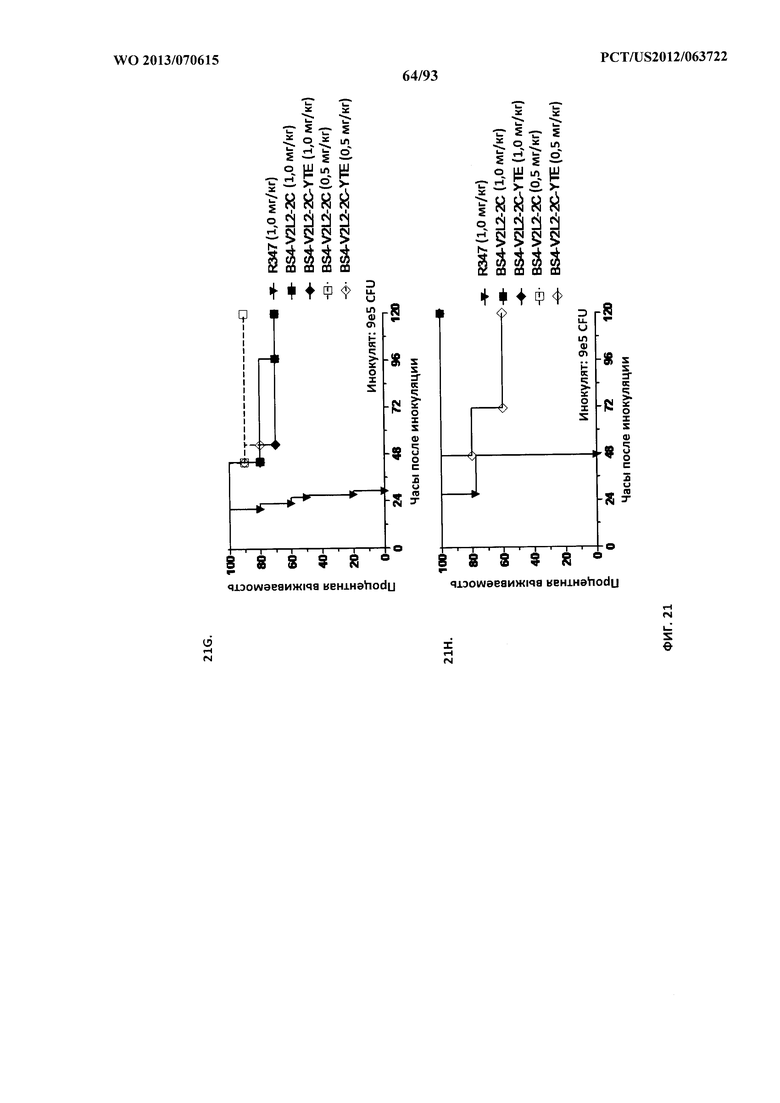

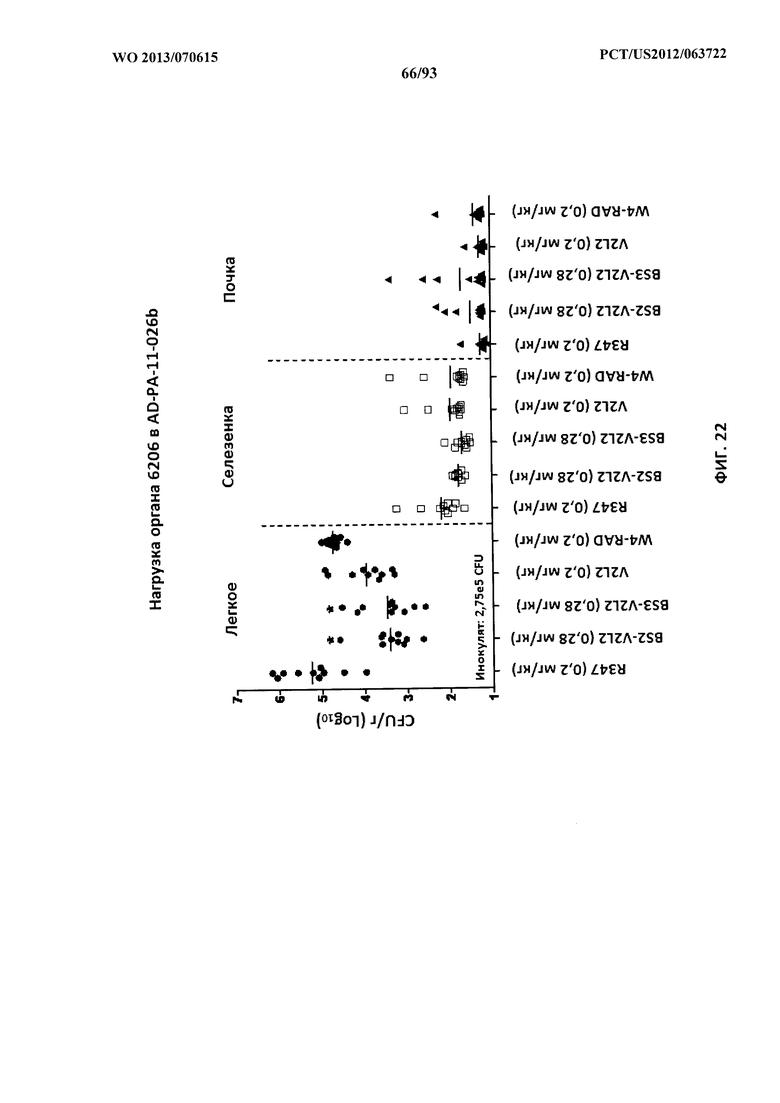
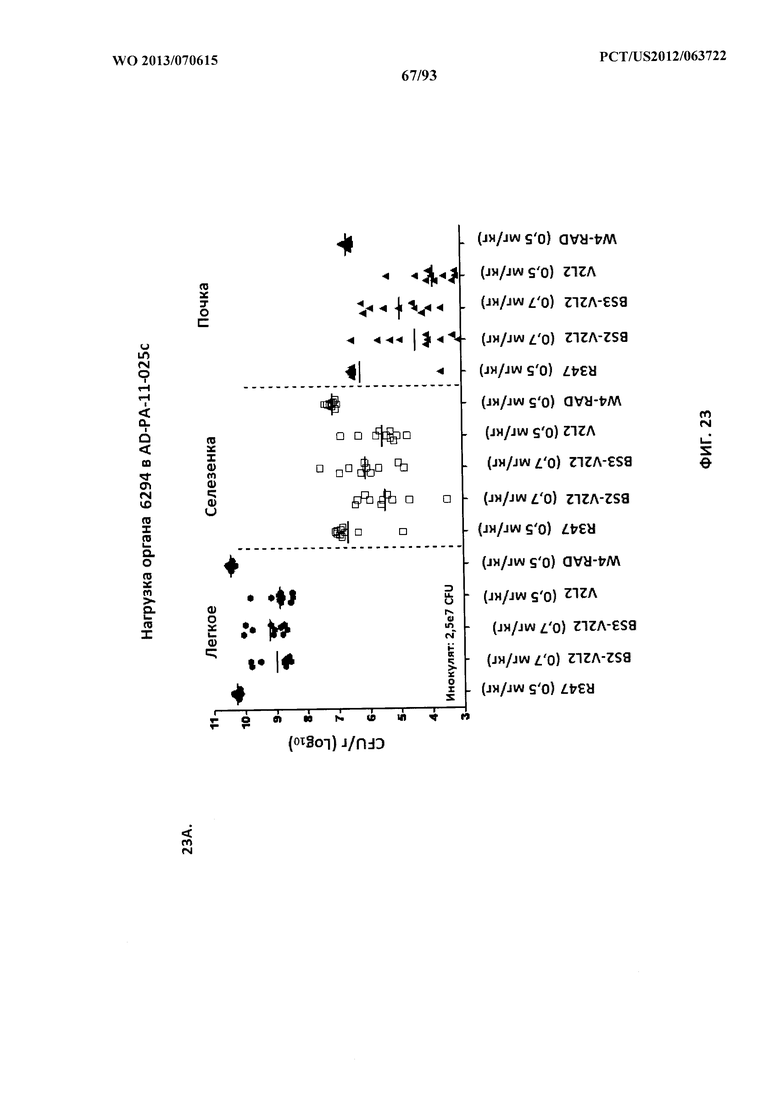
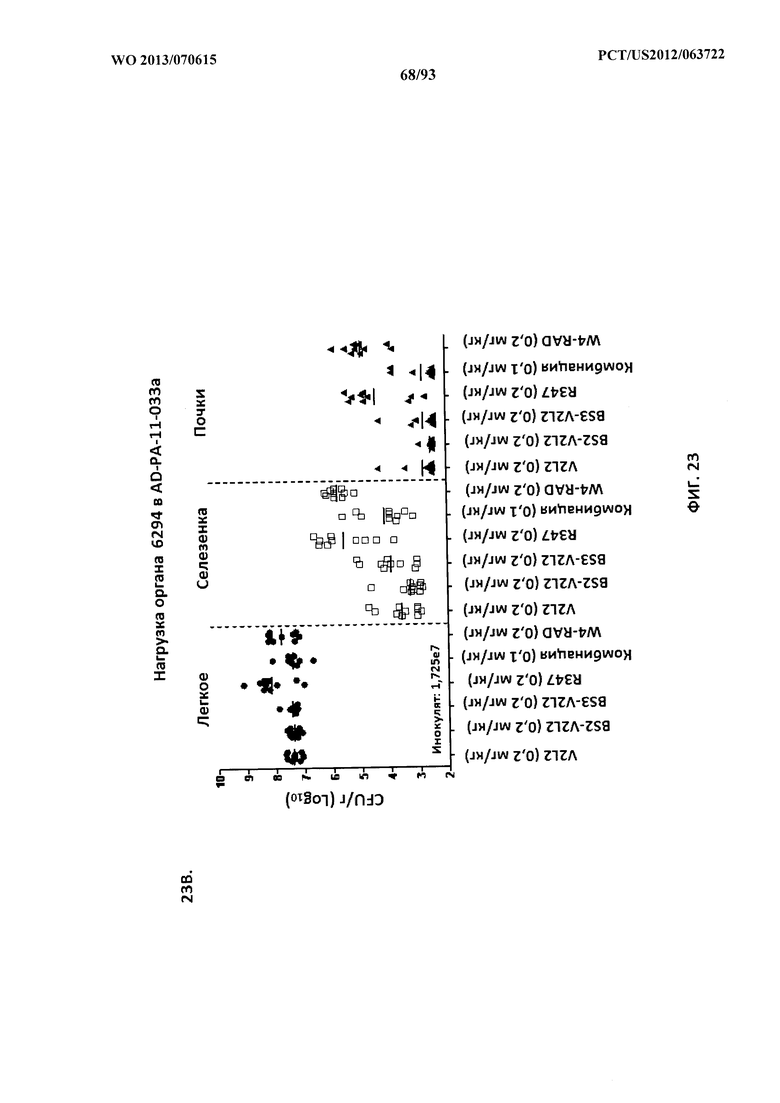
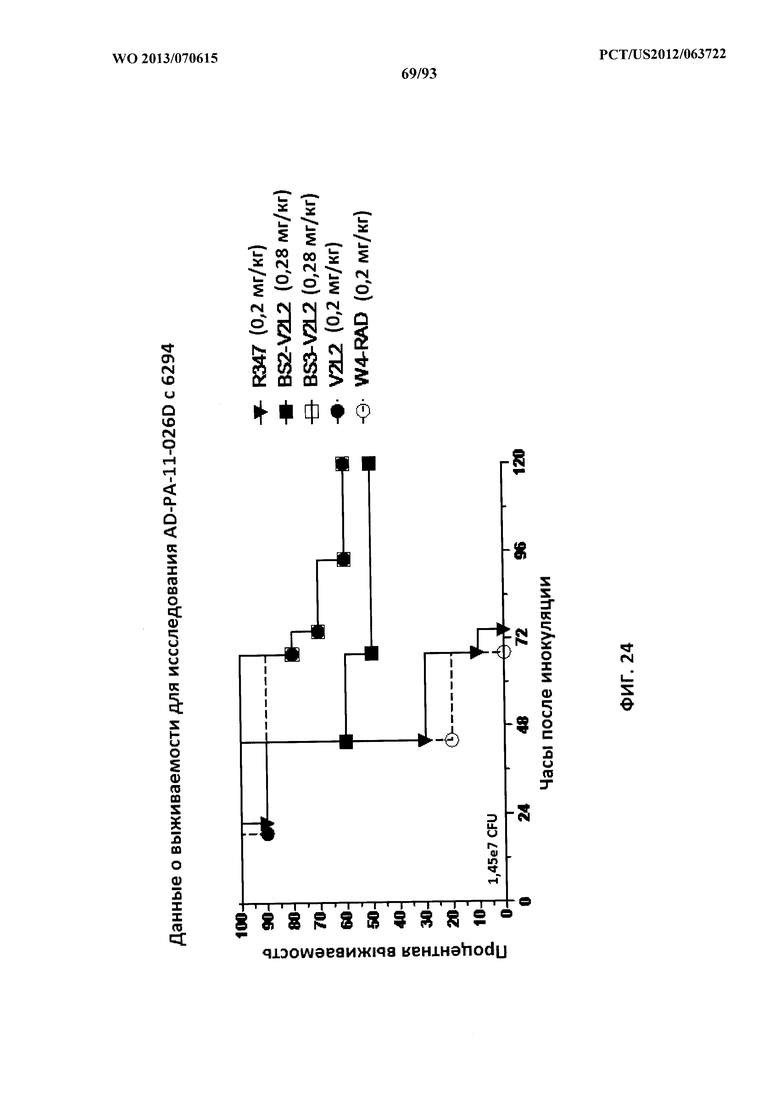
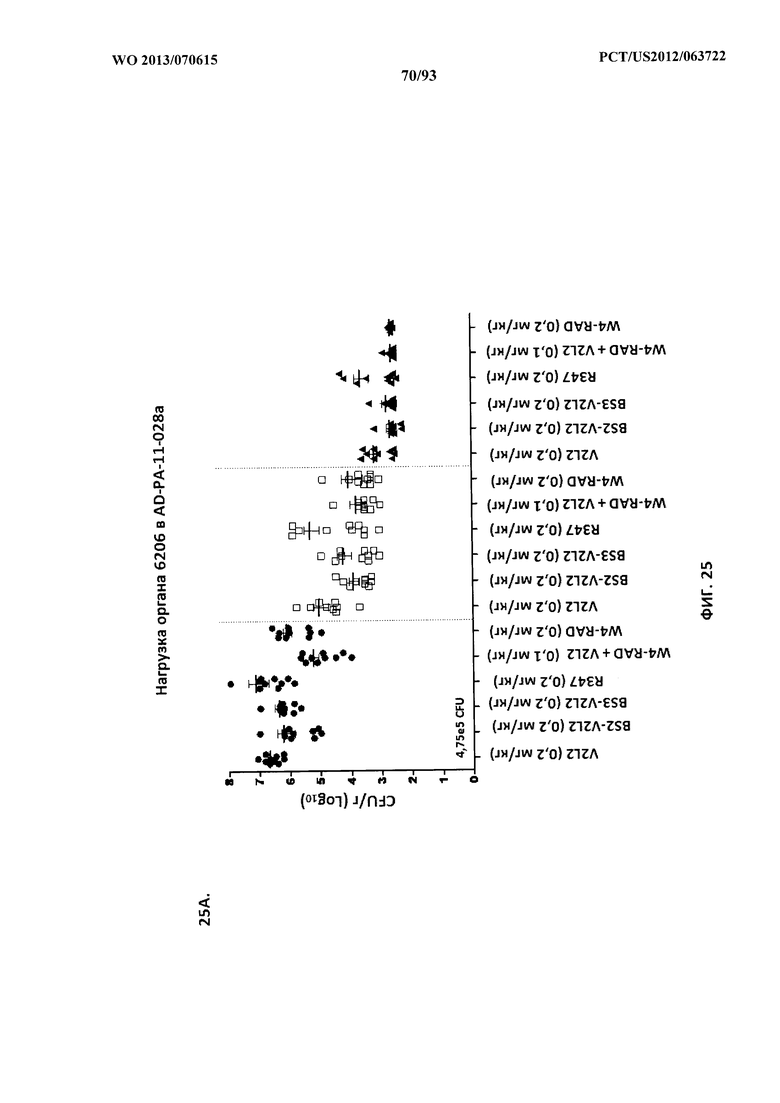
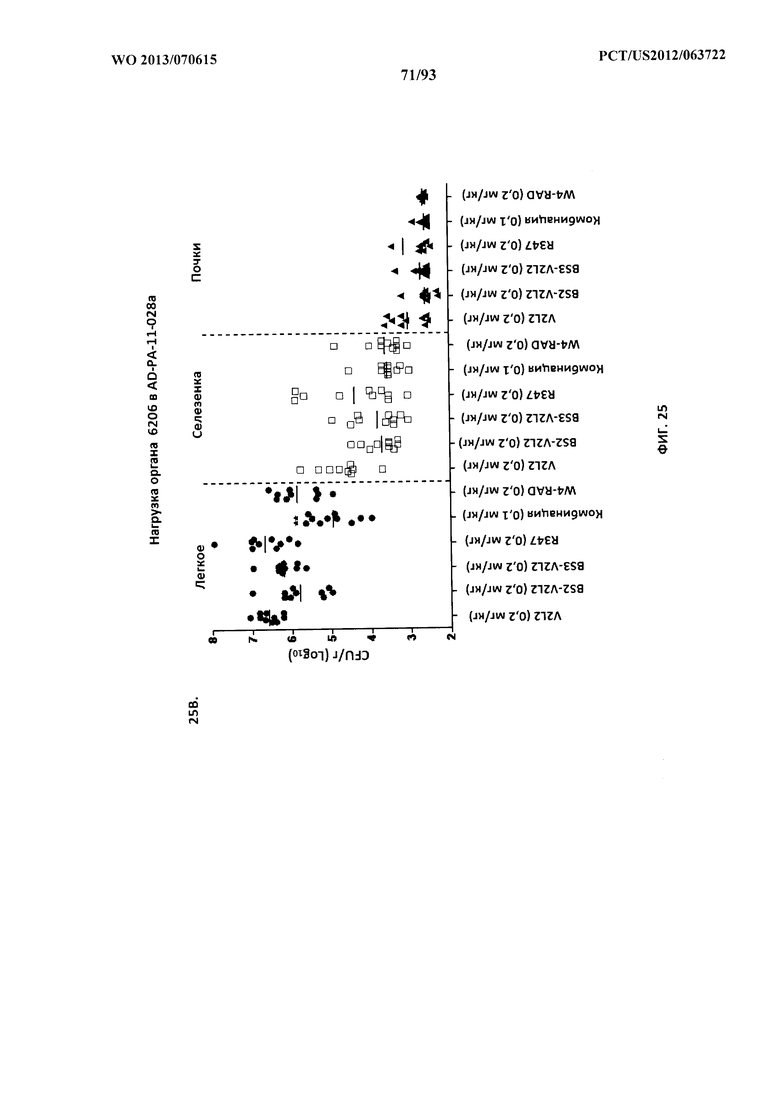

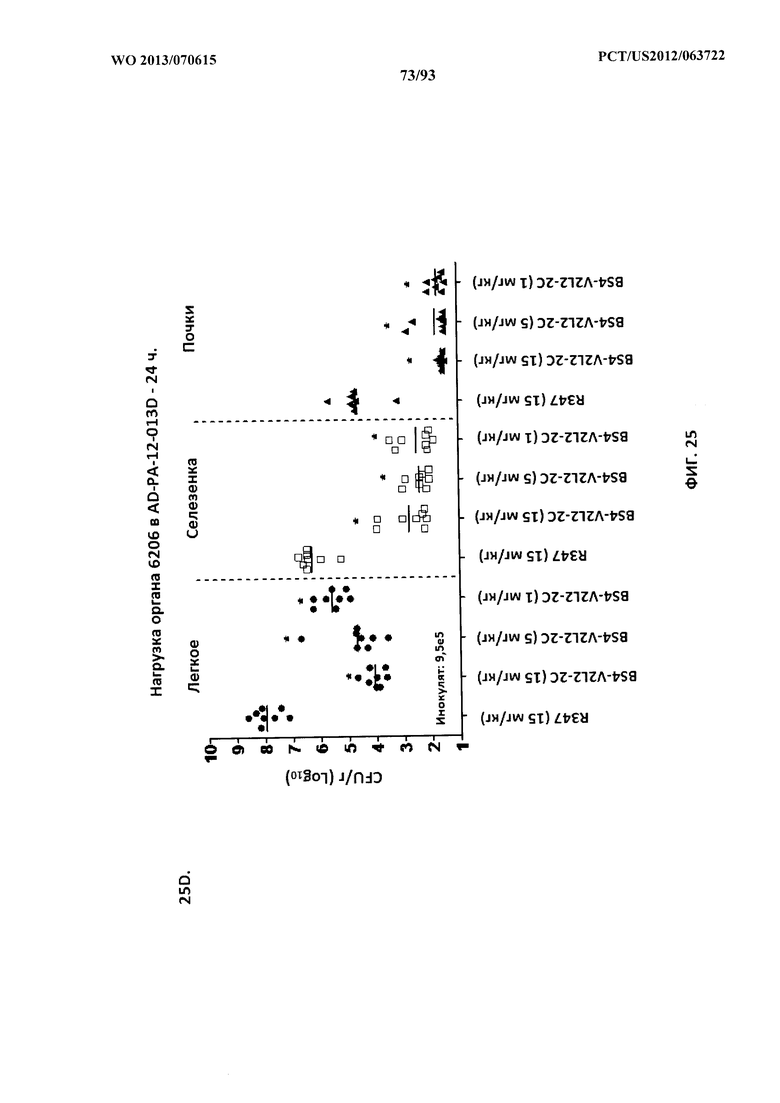
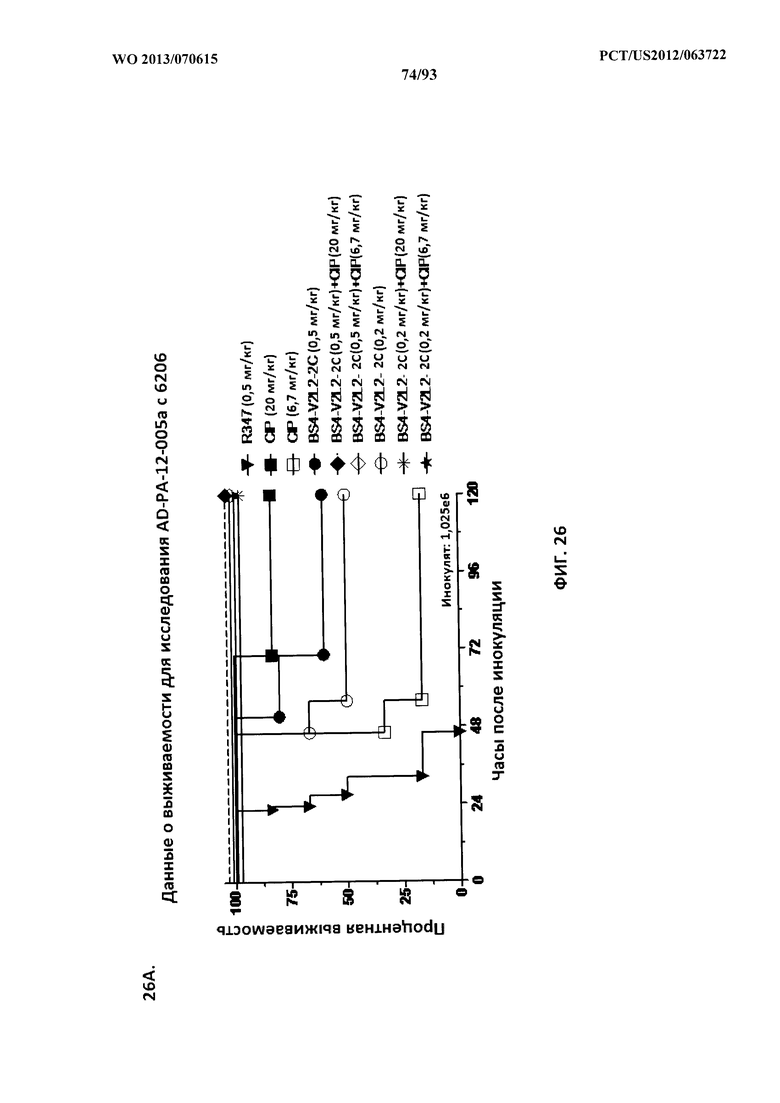
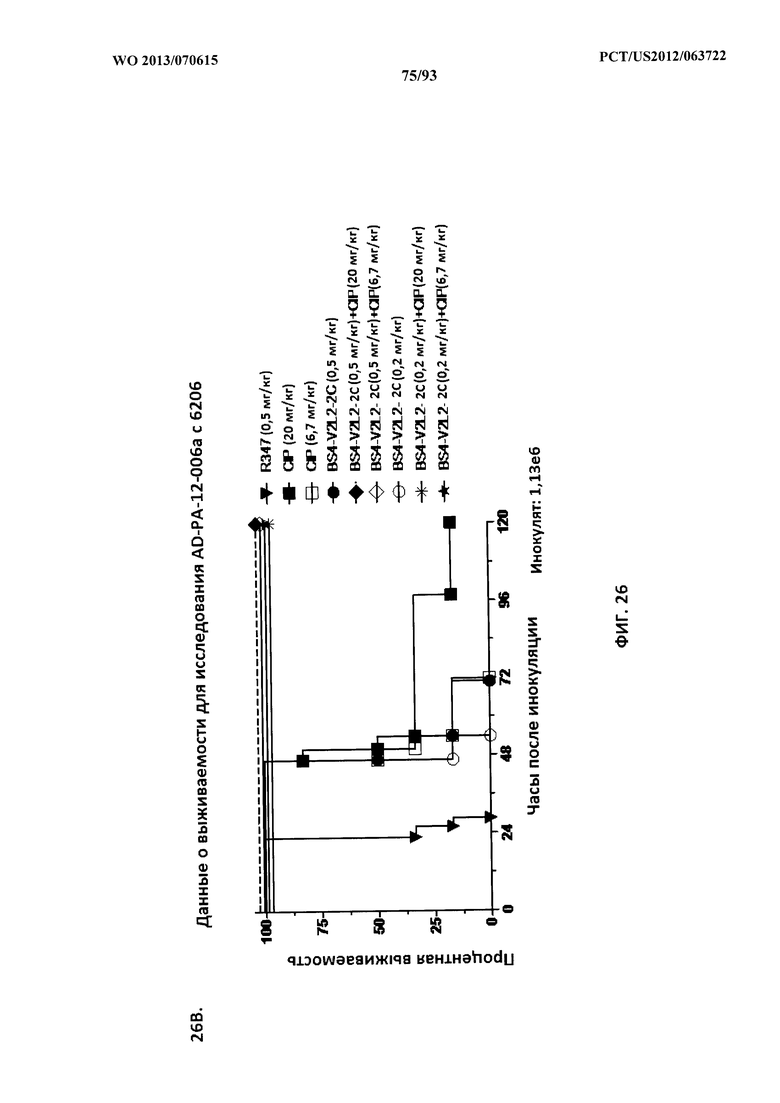
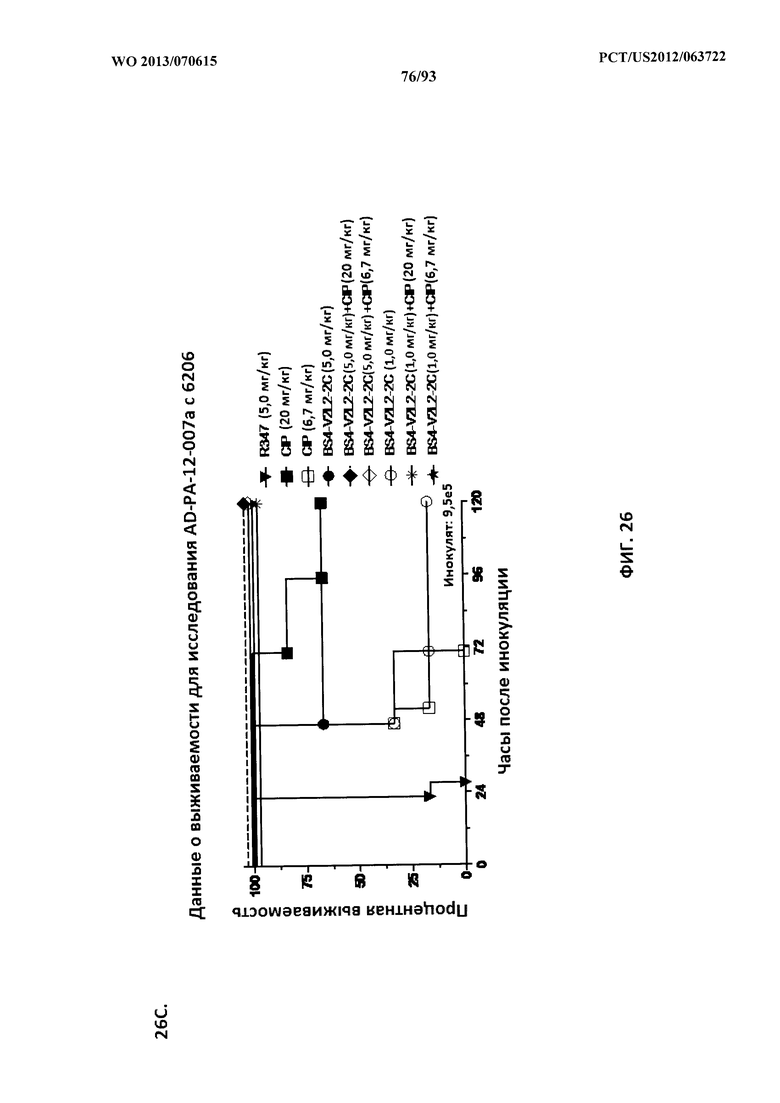
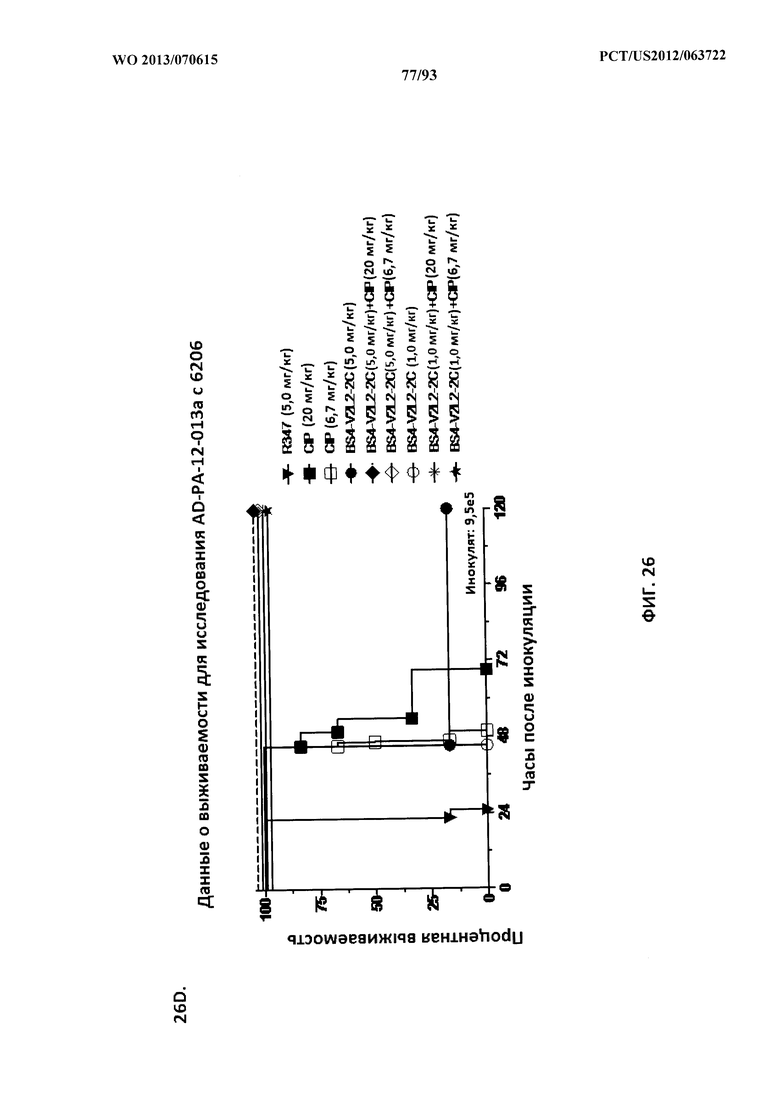
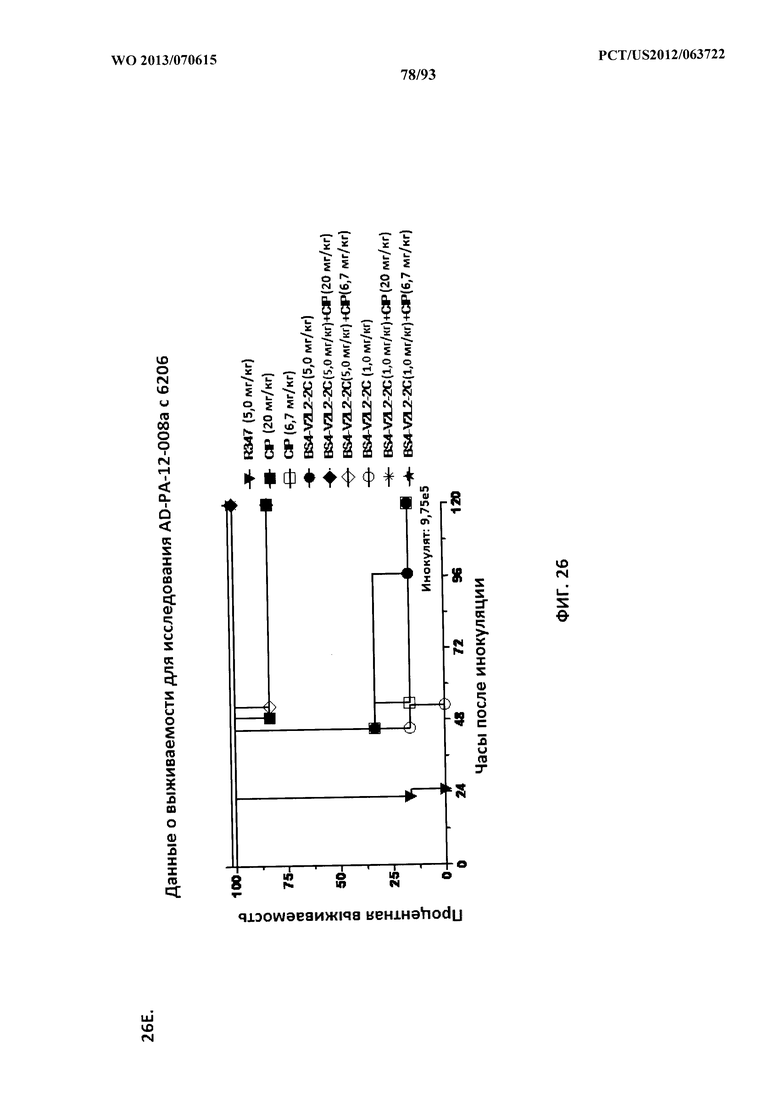
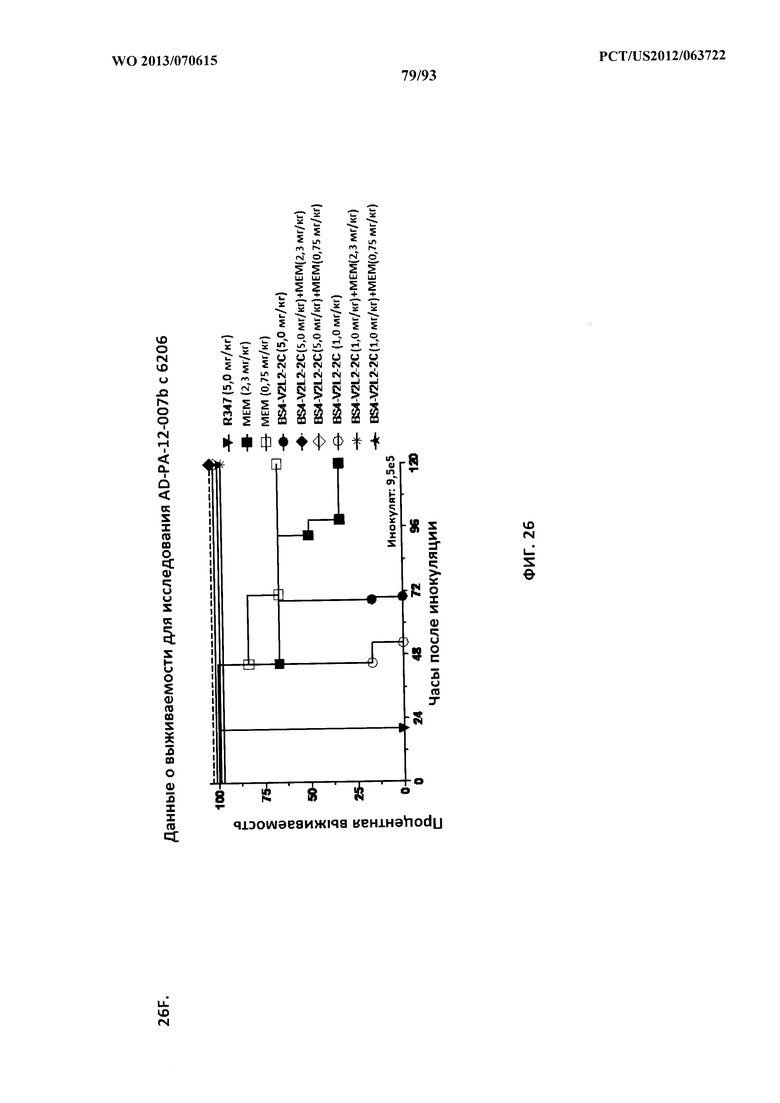
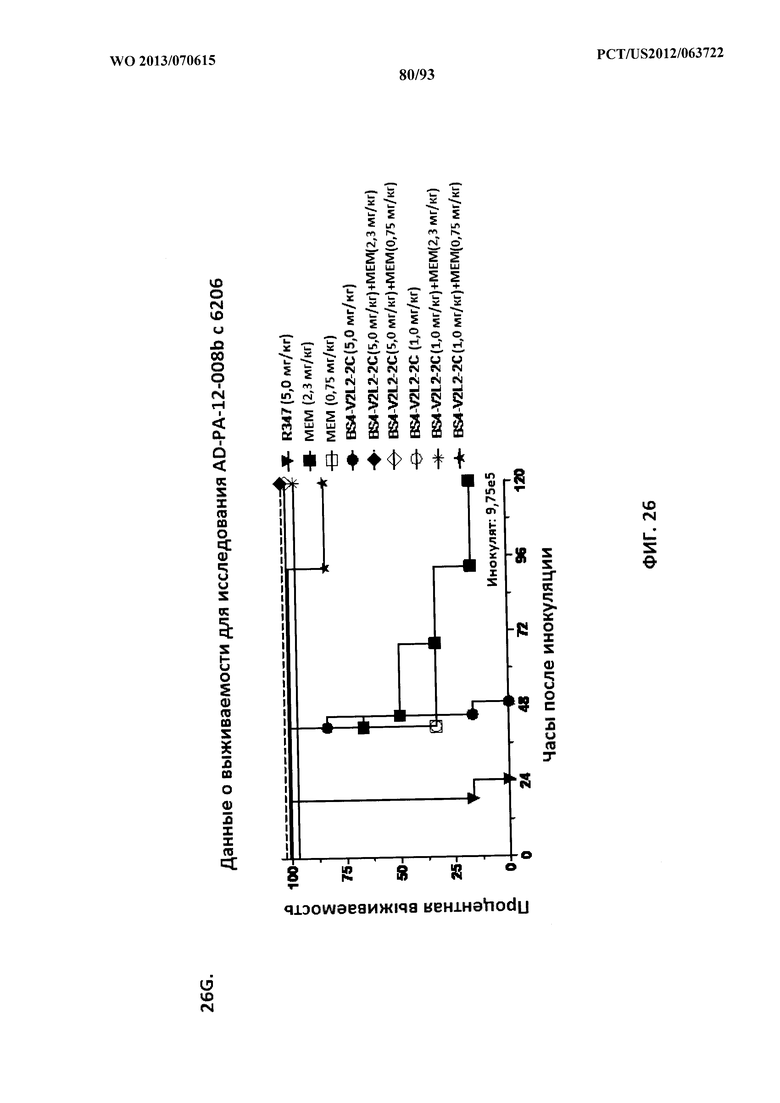
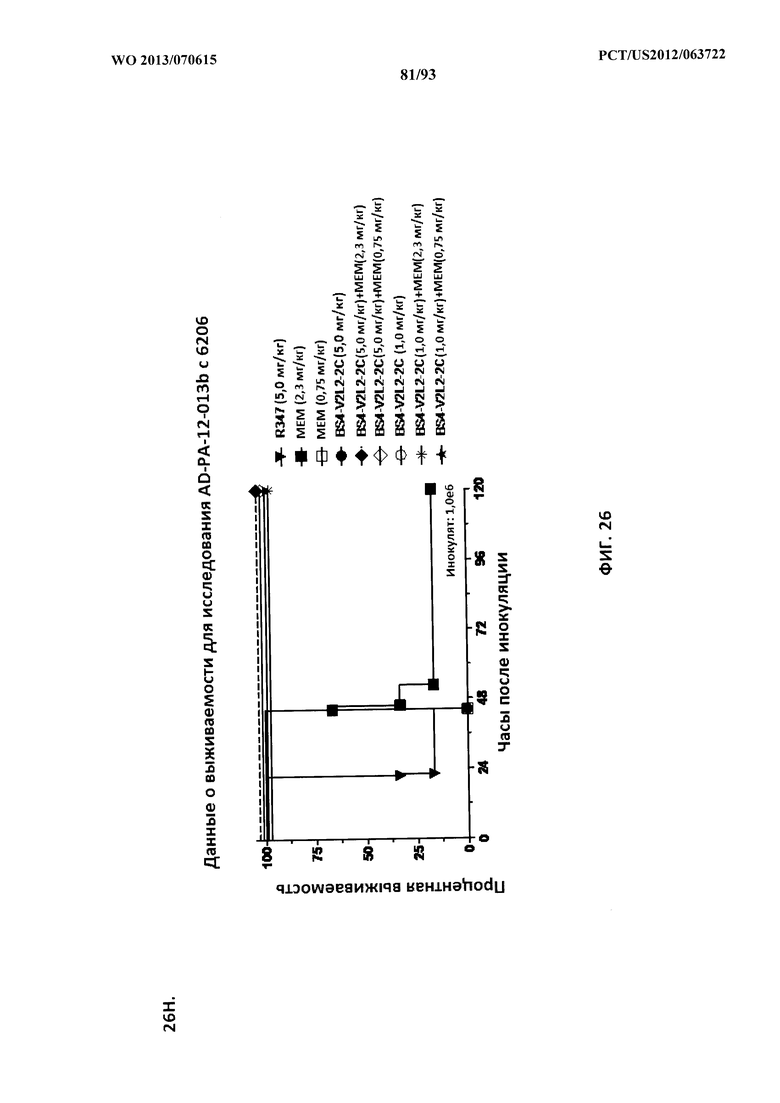
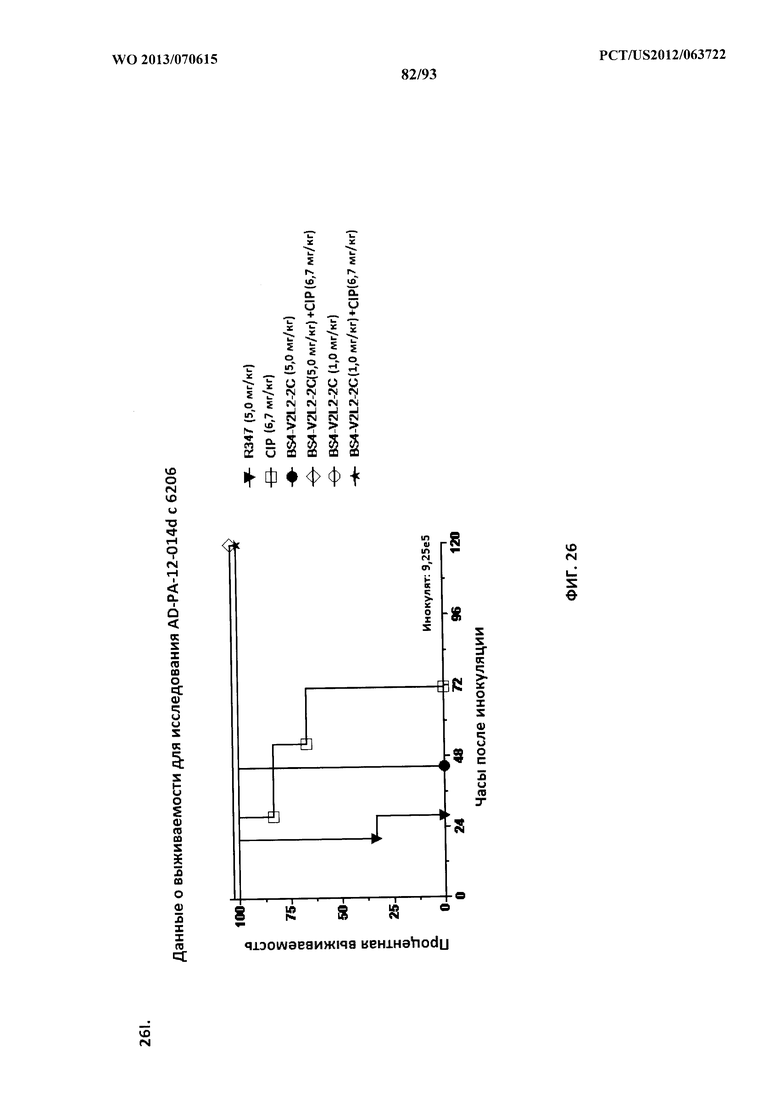

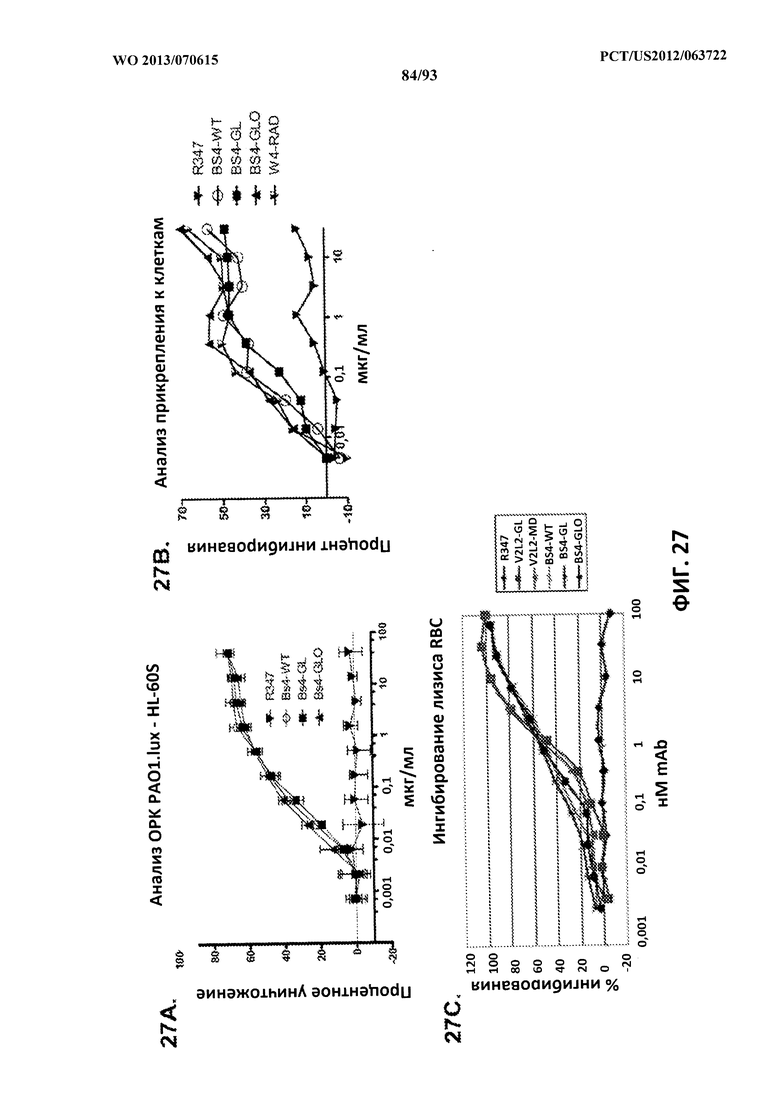
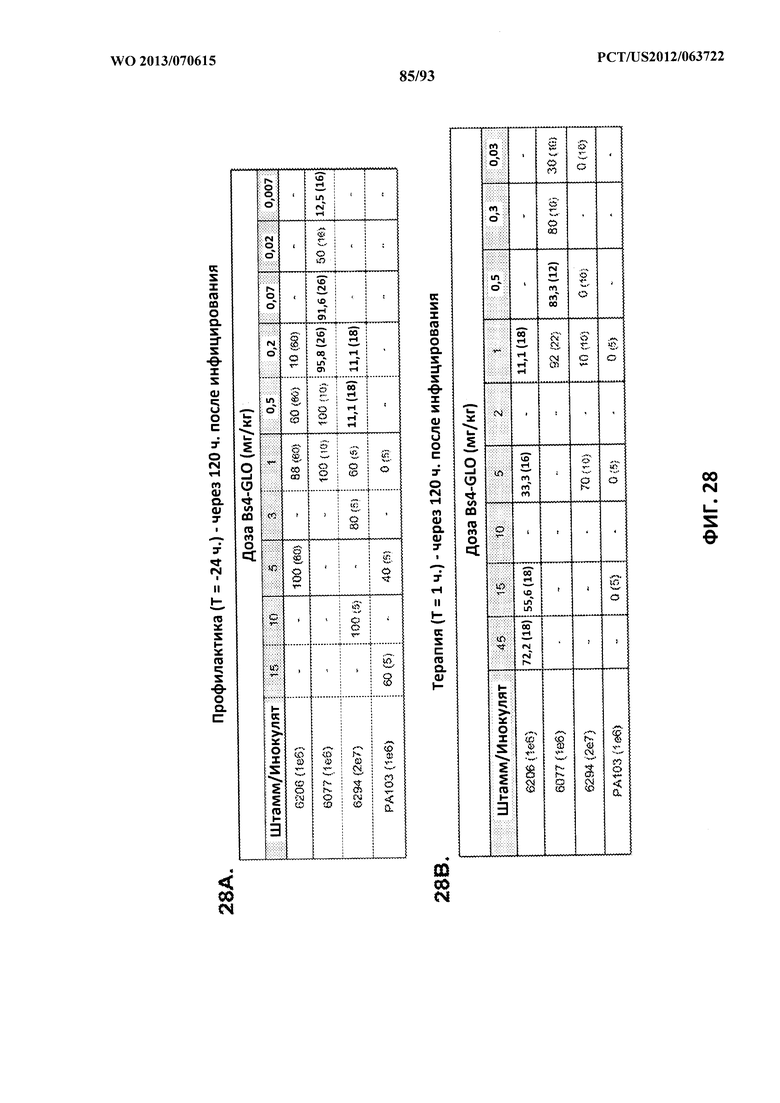
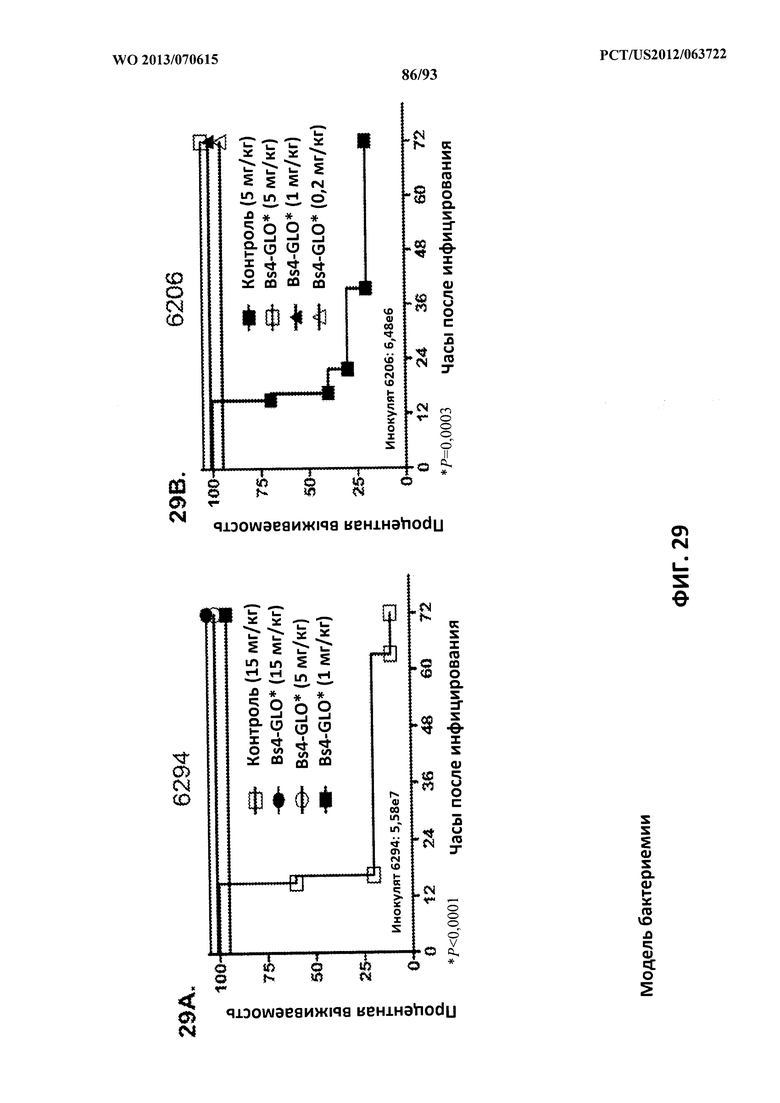
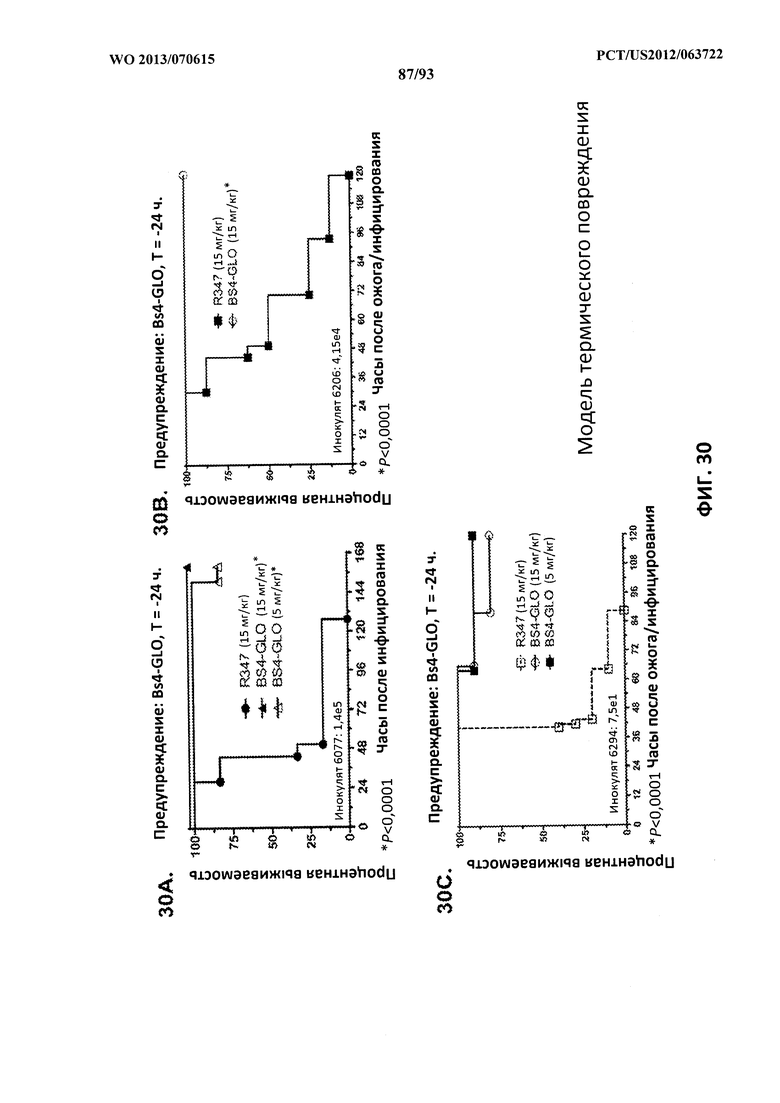
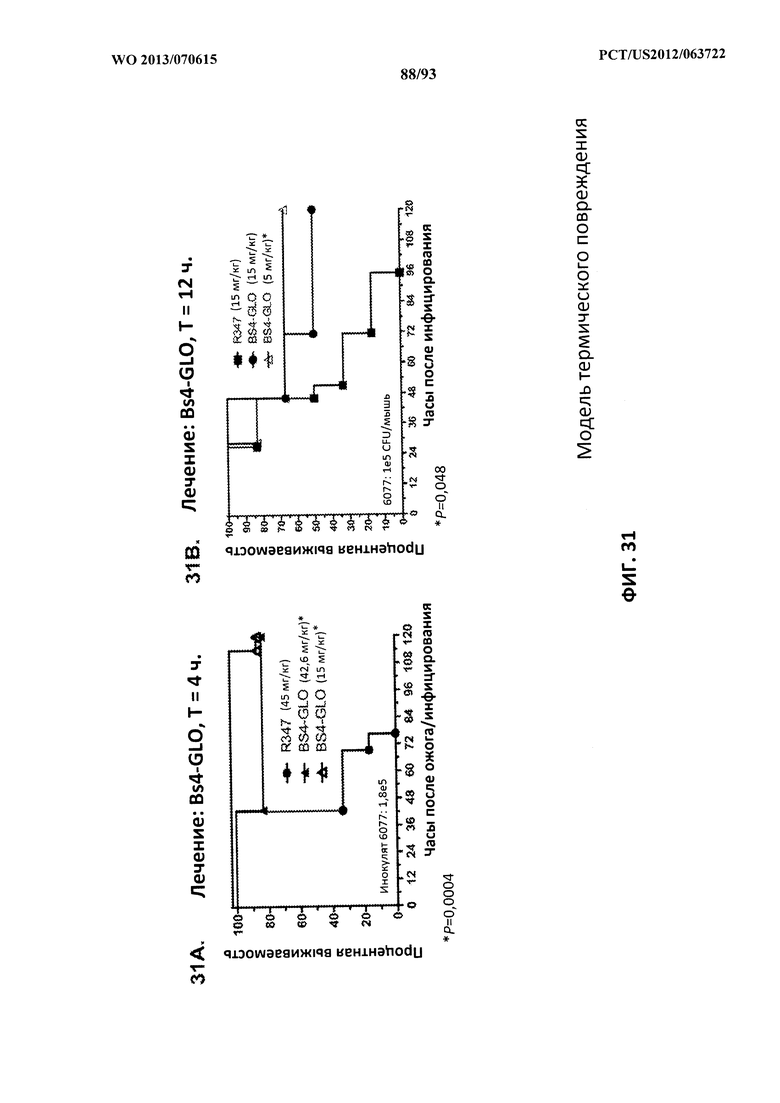
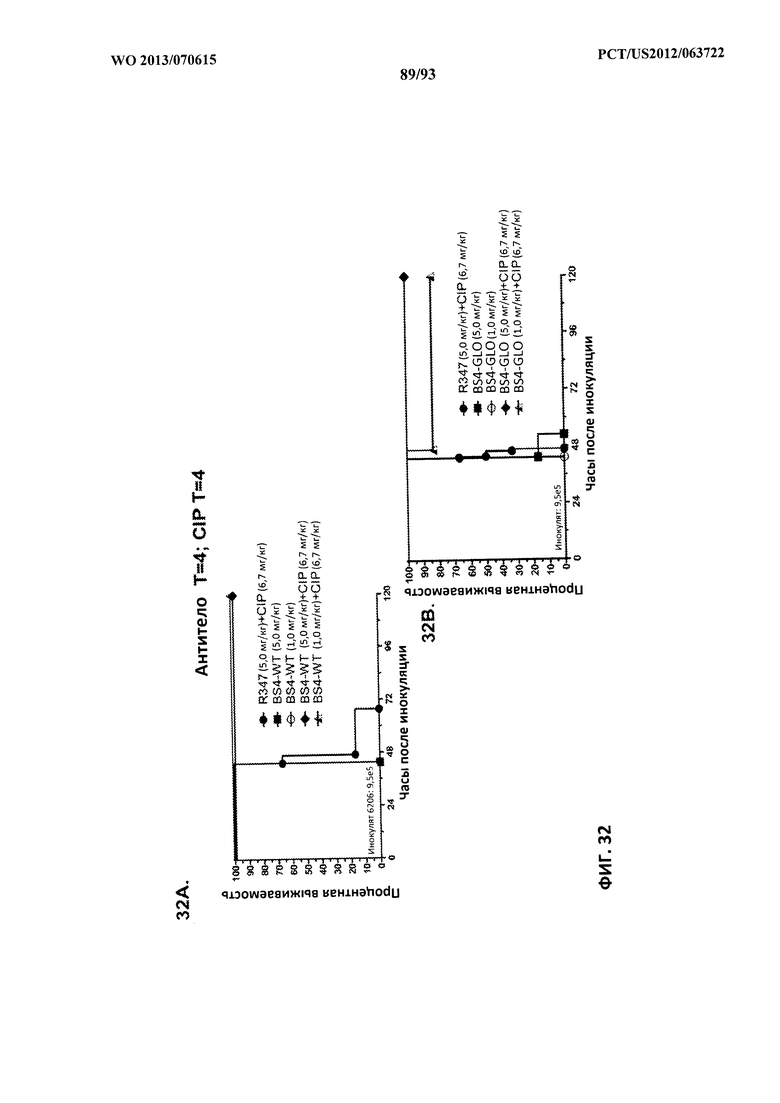
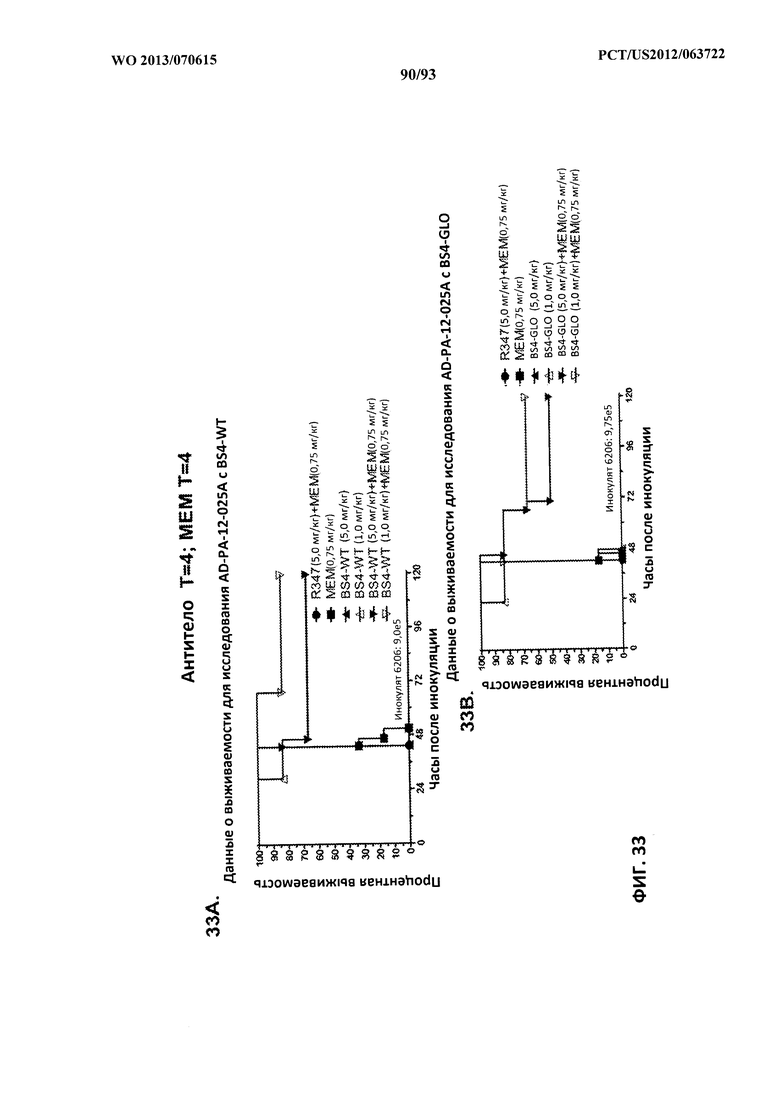
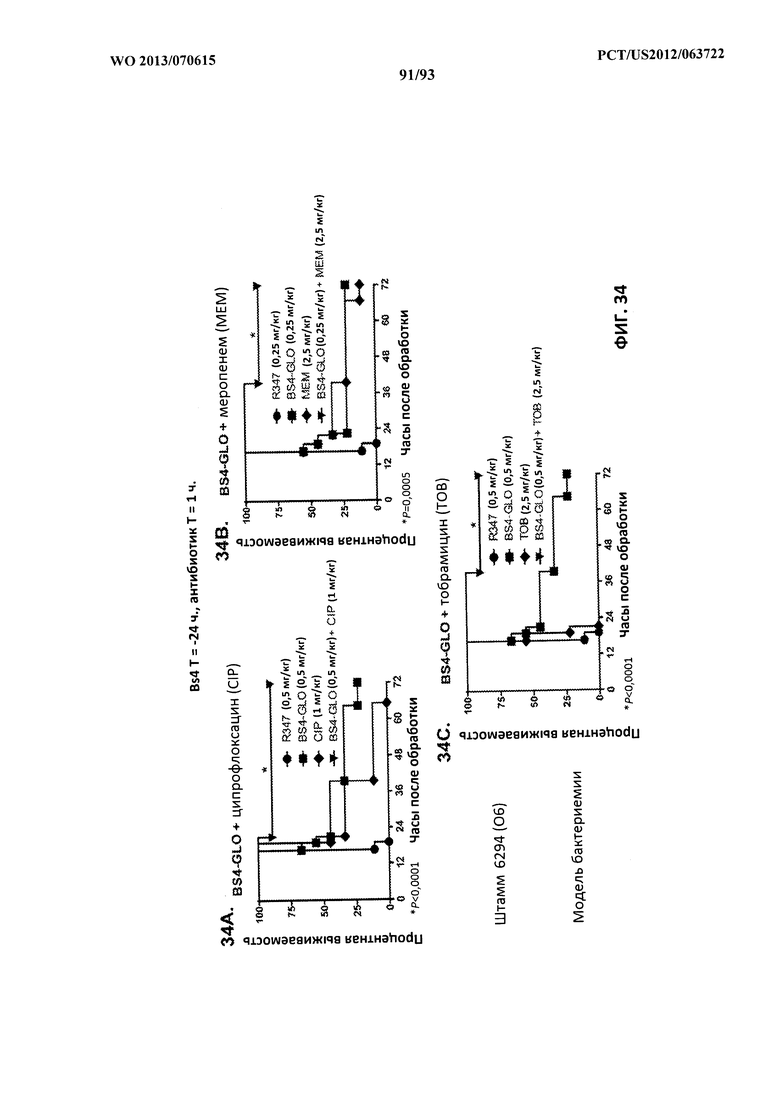


Настоящее изобретение относится к иммунологии. Предложено антитело и его антигенсвязывающий фрагмент, которые специфически связываются с PcrV Pseudomonas, а также биспецифическое антитело, содержащее домен, который связывается с Psl Pseudomonas, и домен, который связывается с PcrV Pseudomonas. Кроме того, описаны: молекула полинуклеотида, кодирующая антитело по изобретению; композиция, содержащая антитело по изобретению, и способ профилактики или лечения инфекции, вызванной Pseudomonas. Данное изобретение может найти применение в терапии. 5 н. и 10 з.п. ф-лы, 35 ил., 7 табл., 20 пр.
1. Антитело или его антигенсвязывающий фрагмент, которые специфически связываются с PcrV Pseudomonas и содержат набор CDR: HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, в котором:
HCDR1 содержит аминокислотную последовательность SYAMN (SEQ ID NO:218);
HCDR2 содержит аминокислотную последовательность AITMSGITAYYTDDVKG (аминокислоты 50-66 SEQ ID NO:264);
HCDR3 содержит аминокислотную последовательность EEFLPGTHYYYGMDV (SEQ ID NO:220);
LCDR1 содержит аминокислотную последовательность RASQGIRNDLG (SEQ ID NO:221);
LCDR2 содержит аминокислотную последовательность SASTLQS (SEQ ID NO:222); и
LCDR3 содержит аминокислотную последовательность LQDYNYPWT (SEQ ID NO:223).
2. Антитело по п.1, содержащее:
(а) вариабельную область тяжелой цепи, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:216; и
(b) вариабельную область легкой цепи, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:217.
3. Антитело по п.1, которое представляет собой рекомбинантное антитело, моноклональное антитело, химерное антитело, гуманизированное антитело, полностью человеческое антитело, биспецифическое антитело или любую их комбинацию.
4. Антитело по п.1, содержащее:
(а) вариабельную область тяжелой цепи, содержащую SEQ ID NO:216 или аминокислоты 1-124 SEQ ID NO:264; и
(b) вариабельную область легкой цепи, содержащую SEQ ID NO:217 или аминокислоты 1-107 SEQ ID NO:263.
5. Биспецифическое антитело, содержащее связывающий домен, который связывается с Psl Pseudomonas, и связывающий домен, который связывается с PcrV Pseudomonas, где домен, связывающий PcrV Pseudomonas, содержит:
CDR1 тяжелой цепи, содержащий аминокислоты SYAMN (SEQ ID NO:218);
CDR2 тяжелой цепи, содержащий аминокислоты AITMSGITAYYTDDVKG (аминокислоты 50-66 SEQ ID NO:264);
CDR3 тяжелой цепи, содержащий аминокислоты EEFLPGTHYYYGMDV (SEQ ID NO:220);
CDR1 легкой цепи, содержащий аминокислоты RASQGIRNDLG (SEQ ID NO:221);
CDR2 легкой цепи, содержащий аминокислоты SASTLQS (SEQ ID NO:222); и
CDR3 легкой цепи, содержащий аминокислоты LQDYNYPWT (SEQ ID NO:223).
6. Биспецифическое антитело по п.5, где
(а) домен, связывающийся с Psl, содержит scFv-фрагмент, а домен, связывающийся с PcrV, содержит интактный иммуноглобулин, или
(b) домен, связывающийся с Psl, содержит интактный иммуноглобулин, а домен, связывающийся с PcrV, содержит scFv-фрагмент.
7. Биспецифическое антитело по п.6, где
(а) scFv слит с аминоконцом VH-области интактного иммуноглобулина;
(b) scFv слит с карбоксиконцом CH3-области интактного иммуноглобулина, или
(с) scFv встроен в шарнирную область интактного иммуноглобулина.
8. Биспецифическое антитело по п.5, где домен, связывающийся с Psl, содержит:
CDR1 тяжелой цепи, содержащий аминокислоты PYYWT (SEQ ID NO:47);
CDR2 тяжелой цепи, содержащий аминокислоты YIHSSGYTDYNPSLKS (SEQ ID NO:48);
CDR3 тяжелой цепи, содержащий аминокислоты ADWDRLRALDI (SEQ ID NO:258);
CDR1 легкой цепи, содержащий аминокислоты RASQSIRSHLN (SEQ ID NO:50);
CDR2 легкой цепи, содержащий аминокислоты GASNLQS (SEQ IDNO:51); и
CDR3 легкой цепи, содержащий аминокислоты QQSTGAWNW (SEQ ID NO:280).
9. Биспецифическое антитело по п.5, где домен, связывающийся с PcrV, содержит
VH-область, которая на 90% идентична SEQ ID NO:216, и
VL-область, которая на 90% идентична SEQ ID NO:217.
10. Биспецифическое антитело по п.5, содержащее тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:264.
11. Биспецифическое антитело по п.5, содержащее легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:263.
12. Выделенная молекула полинуклеотида c последовательностью нуклеиновой кислоты, которая кодирует антитело по п.1 или биспецифическое антитело по п.5.
13. Композиция для профилактики или лечения инфекции, вызываемой Pseudomonas, у индивида, содержащая фармацевтически эффективное количество антитела по п.1 или биспецифического антитела по п.5 и фармацевтически приемлемый носитель.
14. Способ профилактики или лечения инфекции, вызванной Pseudomonas, у индивида, включающий введение индивиду эффективного количества композиции по п.13.
15. Способ по п.14, дополнительно включающий введение индивиду антибиотика.
| Колосоуборка | 1923 |
|
SU2009A1 |
| РАСПЫЛИТЕЛЬНАЯ ГОЛОВКА С СОПЛАМИ, ВЫПОЛНЕННЫМИ СВЕРЛЕНИЕМ | 2000 |
|
RU2248826C2 |
| IMAMURA Y | |||
| et al | |||
| "Effect of anti-PcrV antibody in a murine chronic airway Pseudomonas aeruginosa infection model", European Respiratory Journal, 2007, 29(5): 965-968 | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| ОБОДОВА М | |||
| А | |||
| и др | |||
| "Получение моноклональных антител к экзотоксину а синегнойной палочки и оценка их протективных свойств", Вестник ВолГМУ, 2004, 12:14-16. | |||

