ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к рекомбинантному микроорганизму, продуцирующему путресцин или орнитин, и к способу получения путресцина или орнитина с использованием указанного микроорганизма.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Биогенные амины (БА) представляют собой соединения азота, получаемые, главным образом, посредством декарбоксилирования аминокислот или аминирования и трансаминирования альдегидов и кетонов. Эти биогенные амины имеют малую молекулярную массу и синтезируются во время метаболических процессов у микроорганизмов, растений и животных, поэтому они известны как составные элементы, часто обнаруживаемые в их клетках.
Среди них, путресцин обнаруживают у грамотрицательных бактерий или грибов, и он присутствует в высокой концентрации у различных видов, и поэтому полагают, что путресцин играет важную роль в метаболизме микроорганизмов. В целом, путресцин является важным сырьем для синтеза полиамина нейлона-4,6, и его получают преимущественно химическим синтезом. Химический синтез представляет собой 3-хстадийный процесс, включающий реакцию каталитического окисления, реакцию с использованием цианидного соединения и реакцию гидрогенизации с использованием высокого давления водорода. Соответственно, существует потребность в разработке более экологически безопасного и энергоэффективного способа, включающего использование биомассы.
С учетом этих обстоятельств были описаны различные способы получения путресцина в высокой концентрации трансформацией E. coli и микроорганизма рода Corynebacterium (публикация международной заявки на патент № WO 06/005603; публикация международной заявки на патент № WO 09/125924; Qian ZD et al., Biotechnol. Bioeng. 104 (4): 651 - 662, 2009; Schneider et al., Appl. Microbiol. Biotechnol. 88 (4): 859 - 868, 2010; Schneider et al., Appl. Microbiol. Biotechnol. 95: 169 - 178, 2012).
С другой стороны орнитин представляет собой вещество, часто обнаруживаемое у растений, животных и микроорганизмов, и является предшественником для биосинтеза аргинина, пролина и полиамина. Кроме того, орнитин играет важную роль в метаболическом пути образования мочевины из аминокислот или аммиака и ее выведения посредством орнитинового цикла в процессе метаболизма у высших организмов in vivo. Орнитин эффективен при наращивании мышечной массы и уменьшении жировой массы тела, и поэтому его используют в качестве пищевой добавки, а также в качестве фармацевтического лекарственного средства для лечения цирроза печени и печеночной недостаточности. Способы получения орнитина включают способ с использованием казеина молока и пищеварительного фермента и способ с использованием E. coli или микроорганизма рода Corynebacterium (патент Кореи № 10-1372635; T. Gotoh et al., Bioprocess Biosyst. Eng., 33: 773 - 777, 2010).
Биосинтетические пути образования путресцина и орнитина у E. coli и микроорганизма рода Corynebacterium сходны, но также имеют следующие различия. Во-первых, микроорганизм рода Corynebacterium имеет «циклический путь» с превращением глутаминовой кислоты в N-ацетил-L-глутаминовую кислоту и превращением N-ацетил-L-орнитина в L-орнитин посредством argJ (бифункциональной орнитинацетилтрансферазы/N-ацетилглутаматсинтазы, EC 2.3.1.35). В отличие от этого, у E. coli биосинтез путресцина или орнитина проходит по «линейному пути», где функцию argJ микроорганизма рода Corynebacterium выполняют argA (N-ацетилглутаматсинтаза, EC 2.3.1.1) и argE (ацетилорнитиндезацетилаза, EC 3.5.1.16).
Известно, что в ArgJ микроорганизма рода Corynebacterium происходит повторный обмен ацетильной группой между орнитином и глутаминовой кислотой. Однако у E. coli ArgA присоединяет ацетильную группу ацетил-КоА к глутамату с образованием N-ацетилглутамата и ArgE обеспечивает распад N-ацетилорнитина с образованием орнитина и ацетата (Schneider et al., Appl. Microbiol. Biotechnol. 91, 17 - 30., 2011).
В частности, известно, что оперон pta-ackA (pta, фосфотрансацетилаза; ackA, ацетаткиназа) и ацетилкоэнзим-А-синтетаза (acs) являются генами, обеспечивающими синтез ацетил-КоА с использованием ацетата.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Авторы настоящего изобретения приложили значительные усилия для улучшения способности микроорганизма рода Corynebacterium продуцировать орнитин и путресцин и, в результате, обнаружили, что введение argA и argE, имеющих происхождение от E. coli, в микроорганизм рода Corynebacterium может улучшить его способность продуцировать орнитин и путресцин, завершив посредством этого настоящее изобретение.
Техническое решение
Задачей настоящего изобретения является обеспечение рекомбинантного микроорганизма, продуцирующего путресцин или орнитин с высоким выходом.
Другой задачей настоящего изобретения является обеспечение способа получения путресцина или орнитина с использованием указанного выше микроорганизма.
Полезные эффекты изобретения
Была подтверждена возможность увеличения количества путресцина или орнитина, продуцируемого микроорганизмом рода Corynebacterium по настоящему изобретению, продуцирующим путресцин или орнитин, при введении в микроорганизм argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, а также при усилении пути утилизации ацетата. Соответственно, микроорганизм по настоящему изобретению может быть широко использован для получения путресцина или орнитина, а также может быть широко использован в качестве эффективного и желательного, с экономической и экологической точек зрения, средства получения сырья для получения различных полимерных продуктов, где путресцин или орнитин используют в качестве сырья.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На ФИГ. 1 представлена схематическая диаграмма, на которой показан биосинтетический путь (циклический путь) образования путресцина и орнитина у микроорганизма рода Corynebacterium (A) и биосинтетический путь (линейный путь) образования путресцина и орнитина у E. coli (B).
На ФИГ. 2 представлена схематическая диаграмма, на которой показан биосинтетический путь с улучшением способности к продукции путрецина и орнитина посредством введения argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, в микроорганизм рода Corynebacterium, находящийся в состоянии экспрессии argJ.
НАИЛУЧШИЙ ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Согласно одному аспекту настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который введена активность N-ацетилглутаматсинтазы из E. coli и ацетилорнитиндезацетилазы из E. coli.
Согласно типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где N-ацетилглутаматсинтаза, имеющая происхождение от E. coli, состоит из аминокислотной последовательности SEQ ID NO:1.
Согласно другому типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где ацетилорнитиндезацетилаза, имеющая происхождение от E. coli, состоит из аминокислотной последовательности SEQ ID NO:3.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, выбранный из группы, состоящей из Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum и Brevibacterium lactofermentum.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность фосфотрансацетилазного и ацетаткиназного оперона (оперона pta-ackA) дополнительно усилена по сравнению с его эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где фосфотрансацетилазный и ацетаткиназный оперон состоит из аминокислотной последовательности SEQ ID NO:5 или 7.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который дополнительно введена активность ацетил-КоА-синтетазы (acs), имеющей происхождение от E. coli.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где ацетил-КоА-синтетаза (acs), имеющая происхождение от E. coli, состоит из аминокислотной последовательности SEQ ID NO:9.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который дополнительно введена активность орнитиндекарбоксилазы (ODC).
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность (1) орнитинкарбамоилтрансферазы (ArgF), (2) экспортера глутамата или (3) орнитинкарбамоилтрансферазы и экспортера глутамата дополнительно ослаблена по сравнению с их эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность по меньшей мере одного, выбранного из группы, состоящей из ацетил-гамма-глутамилфосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитинаминотрансферазы (ArgD), дополнительно усилена по сравнению с их эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность ацетилтрансферазы дополнительно ослаблена по сравнению с ее эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где ацетилтрансфераза состоит из аминокислотной последовательности SEQ ID NO:30 или 31.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность экспортера путресцина дополнительно усилена по сравнению с его эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где экспортер путресцина состоит из аминокислотной последовательности SEQ ID NO:26 или 28.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина или орнитина, включающий:
(1) культивирование модифицированного микроорганизма рода Corynebacterium, продуцирующего путресцин или орнитин, в среде; и
(2) выделение путресцина или орнитина из культивированного микроорганизма или среды.
В типичном воплощении настоящего изобретения модифицированный микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
Далее настоящее изобретение будет описано подробно.
Согласно одному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который введены активности N-ацетилглутаматсинтазы, имеющей происхождение от E. coli, и ацетилорнитиндезацетилазы, имеющей происхождение от E. coli.
При использовании здесь термин «N-ацетилглутаматсинтаза» относится к ферменту, опосредующему реакцию образования N-ацетилглутамата из глутамата и ацетил-КоА, и образованный таким образом N-ацетилглутамат может быть использован как предшественник орнитина и аргинина.
В настоящем изобретении N-ацетилглутаматсинтаза может включать, например, без ограничения, белок, имеющий аминокислотную последовательность SEQ ID NO:1, и любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что белок обладает существенной N-ацетилглутаматсинтазной активностью.
Кроме того, аминокислотные последовательности белков, демонстрирующих указанную выше активность, могут различаться у разных видов и штаммов микроорганизмов. Соответственно, N-ацетилглутаматсинтаза по настоящему изобретению может представлять собой, например, без ограничения, N-ацетилглутаматсинтазу из E. coli.
Очевидно, что в качестве последовательностей, гомологичных указанной выше последовательности, в объем настоящего изобретения также следует включать аминокислотные последовательности с делецией, модификацией, заменой или добавлением части последовательности, если биологическая активность этих аминокислотных последовательностей идентична или соответствует биологической активности белка по SEQ ID NO:1.
Полинуклеотид, кодирующий N-ацетилглутаматсинтазу по настоящему изобретению, может включать, без ограничения, полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность SEQ ID NO:1, и любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что полинуклеотид обладает активностью, сходной с активностью N-ацетилглутаматсинтазы, и, например, может быть включена полинуклеотидная последовательность SEQ ID NO:2.
При использовании здесь термин «ацетилорнитиндезацетилаза» относится к ферменту, опосредующему реакцию, вовлеченную в образование уксусной кислоты и орнитина, путем опосредования гидролиза ацетилорнитина.
В настоящем изобретении ацетилорнитиндезацетилаза может включать, без ограничения, белок, имеющий аминокислотную последовательность SEQ ID NO:3, и любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что белок обладает существенной активностью по разделению ацетилорнитина на ацетильную группу и орнитин.
Кроме того, аминокислотные последовательности белков, демонстрирующих указанную выше активность, могут различаться у разных видов и штаммов микроорганизмов. Соответственно, ацетилорнитиндезацетилаза по настоящему изобретению может представлять собой, без ограничения, ацетилорнитиндезацетилазу из E. coli. Очевидно, что в качестве гомологичных последовательностей в объем настоящего изобретения также следует включать аминокислотные последовательности с делецией, модификацией, заменой или добавлением части последовательности, если биологическая активность этих аминокислотных последовательностей идентична или соответствует биологической активности белка по SEQ ID NO:3.
Полинуклеотид, кодирующий ацетилорнитиндезацетилазу по настоящему изобретению, может включать, при условии, что полинуклеотид обладает активностью, сходной с активностью белка ацетилорнитиндезацетилазы, белок, имеющий аминокислотную последовательность SEQ ID NO:3, или полинуклеотид, кодирующий белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, например, полинуклеотидную последовательность SEQ ID NO:4.
Кроме того, полинуклеотид, кодирующий N-ацетилглутаматсинтазу или ацетилорнитиндезацетилазу по настоящему изобретению, можно гибридизовать с полинуклеотидной последовательностью SEQ ID NO:2 или SEQ ID NO:4 или зондом, имеющим происхождение от указанной полинуклеотидной последовательности, в жестких условиях, и он может представлять собой модифицированный тип N-ацетилглутаматсинтазы или ацетилорнитиндезацетилазы, обладающий нормальной функцией. Использованный выше термин «жесткие условия» относится к условиям, обеспечивающим специфичность гибридизации полинуклеотидов. Например, конкретные жесткие условия описаны в литературе (например, J. Sambrook et al., supra).
Использованный выше термин «гомология» относится к степени идентичности данной аминокислотной последовательности или полинуклеотидной последовательности и может быть указан в процентах. При использовании здесь гомологичная последовательность, обладающая такой же активностью, что данная полипептидная последовательность или полинуклеотидная последовательность, или сходной активностью, может быть обозначена посредством термина «% гомологии». Например, % гомологии может быть подтвержден с применением стандартного программного обеспечения, то есть BLAST 2.0, для расчета таких параметров, как показатель, идентичность и сходство, или сравнением последовательностей посредством гибридизационных экспериментов по Саузерну, и подходящие условия гибридизации могут быть определены способом, известным специалисту в данной области (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
С другой стороны, микроорганизм по настоящему изобретению может включать как микроорганизмы естественного типа, так и микроорганизмы модифицированного типа, например, микроорганизмы, принадлежащие к роду Escherichia, роду Shigella, роду Citrobacter, роду Salmonella, роду Enterobacter, роду Yersinia, роду Klebsiella, роду Erwinia, роду Corynebacterium, роду Brevibacterium, роду Lactobacillus, роду Selenomanas, роду Vibrio, роду Pseudomonas, роду Streptomyces, роду Arcanobacterium, роду Alcaligenes и так далее. Конкретно, микроорганизм по настоящему изобретению может представлять собой, без ограничения, микроорганизм, принадлежащий к роду Corynebacterium, конкретнее, микроорганизм, выбранный из группы, состоящей из Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum и Brevibacterium lactofermentum, и, конкретнее, Corynebacterium glutamicum.
Конкретно, при использовании здесь термин микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, относится к микроорганизму рода Corynebacterium, продуцирующему путресцин или орнитин в естественном состоянии, или микроорганизму рода Corynebacterium, продуцирующему путресцин или орнитин, полученному приданием способности продуцировать путресцин или орнитин его исходному штамму, неспособному продуцировать путресцин или орнитин.
Микроорганизм, которому придана способность продуцировать путресцин или орнитин, или способный продуцировать путресцин или орнитин, может обладать улучшенной способностью продуцировать орнитин, используемый в качестве исходного вещества для биосинтеза путресцина, благодаря, без ограничения, модификации активности ацетилглутаматсинтазы (превращающей глутамат в N-ацетилглутамат), орнитинацетилтрансферазы (ArgJ, превращающей ацетилорнитин в орнитин), ацетилглутаматкиназы (ArgB, превращающей ацетилглутамат в N-ацетилглутамилфосфат), гамма-глутамилфосфатредуктазы (ArgC, превращающей N-ацетилглутамилфосфат в N-ацетилглутаматполуальдегид) и ацетилорнитинаминотрансферазы (ArgD, превращающей ацетилглутаматполуальдегид в N-ацетилорнитин) в сторону повышения, по сравнению с их эндогенной активностью, для усиления биосинтетического пути от глутамата до орнитина.
Кроме того, микроорганизм может быть модифицирован для ослабления активности орнитинкарбамоилтрансферазы (ArgF, вовлеченной в синтез аргинина из орнитина), белка (белков), вовлеченного в экспорт глутамата, и/или белка (белков), ацетилирующего путресцин, по сравнению с их эндогенной активностью, и/или введения активности орнитиндекарбоксилазы (ODC).
При использовании здесь термин «введение активности» может относиться к активности белка, отсутствующей или слабой у микроорганизма, вводимой впервые или усиленной у соответствующего микроорганизма. Конкретно, он может включать, без ограничения, введение или доставку гена, кодирующего белок, отсутствующий у микроорганизма, в микроорганизм для экспрессии в нем или введение модификации белка для усиления экспрессии белка, не экспрессируемого или почти не экспрессируемого у микроорганизма.
С другой стороны, в настоящем изобретении модификации, такие как введение активности, усиление активности, ослабление активности и так далее, могут быть проведены способом, называемым трансформацией. При использовании здесь термин «трансформация» относится к процессу введения вектора, содержащего полинуклеотид, кодирующий определенный белок, или промоторную последовательность с сильной или слабой активностью и так далее в клетку-хозяина, что обеспечивает экспрессию белка, кодируемого полинуклеотидом, или приводит к модификации хромосомы клетки-хозяина. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид может быть введен в любой форме, при условии, что он может быть введен в клетку-хозяина и обеспечить экспрессию или привести к модификации в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, представляющей собой генную конструкцию, содержащую все обязательные элементы, необходимые для самостоятельной экспрессии. Экспрессионная кассета может обычно содержать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, домен связывания рибосом и сигнал терминации трансляции и может иметь форму вектора экспрессии, способного к самостоятельной репликации. Кроме того, полинуклеотид может, без ограничения, быть введен в клетку-хозяина сам по себе и функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине.
Кроме того, при использовании здесь термин «функционально связанный» относится к функциональной связи промоторной последовательности, инициирующей и опосредующей транскрипцию полинуклеотида, кодирующего определенный белок по настоящему изобретению, с последовательностью гена.
При использовании здесь термин «вектор» относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующего интересующий белок, где интересующий белок функционально связан с подходящей регуляторной последовательностью, таким образом, что интересующий белок может быть экспрессирован у подходящего хозяина. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящий домен связывания рибосом на мРНК, и последовательность для регуляции транскрипции и трансляции. После трансформации в подходящую клетку-хозяина вектор может быть реплицирован или функционировать независимо от генома хозяина, или может быть интегрирован в сам геном хозяина.
Относительно вектора, используемого в настоящем изобретении, может не быть существенных ограничений, при условии, что вектор реплицирован в клетке-хозяине, и может быть использован любой вектор, известный в данной области. Примеры вектора могут включать естественные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, M13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и так далее; и в качестве плазмидного вектора могут быть использованы векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и так далее. Относительно вектора для использования в настоящем изобретении, может не быть существенных ограничений, и может быть использован любой вектор, известный в данной области. Конкретно, могут быть использованы векторы pDZTn, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и так далее.
Как таковой, полинуклеотид, кодирующий белок-мишень, может быть заменен модифицированным полинуклеотидом с использованием вектора для введения в хромосомы бактерий. Введение полинуклеотида в хромосому может быть проведено, без ограничения, любым способом, известным в данной области, например, гомологичной рекомбинацией. Поскольку вектор по настоящему изобретению может быть введен в хромосому гомологичной рекомбинацией, может быть дополнительно включен селективный маркер для подтверждения введения в хромосому. Селективный маркер используют для отбора трансформированных клеток, то есть для подтверждения введения целевого полинуклеотида, и могут быть использованы маркеры, обеспечивающие селектируемые фенотипы, такие как резистентность к лекарственным средствам, потребность в питательных веществах, резистентность к цитотоксическим агентам и экспрессия поверхностных белков. При обработке селективными агентами только клетки, экспрессирующие селективные маркеры, могут выжить или проявлять другие фенотипические признаки, что позволяет легко отбирать трансформированные клетки.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность фосфотрансацетилазного и ацетаткиназного оперона (оперона pta-ackA) дополнительно усилена по сравнению с его эндогенными ферментами.
В настоящем изобретении фосфотрансацетилазный и ацетаткиназный оперон (оперон pta-ackA) представляет собой опероны, содержащие гены, обратно опосредующие метаболический путь, в котором происходит превращение ацетил-КоА, полученного из глюкозы или пирувата, в уксусную кислоту через ацетилфосфат, и метаболический путь противоположного направления.
В настоящем изобретении фосфотрансацетилазный и ацетаткиназный оперон может включать, без ограничения, белки, содержащие аминокислотную последовательность SEQ ID NO:5 или SEQ ID NO:7, или любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, или, наиболее конкретно, на 99% или более гомологичный указанным выше аминокислотным последовательностям, при условии, что белок существенным образом опосредует реакцию образования ацетил-КоА из уксусной кислоты.
Кроме того, поскольку аминокислотные последовательности белков, демонстрирующих указанные активности, могут различаться у разных видов или штаммов данного микроорганизма, фосфотрансацетилазный и ацетаткиназный оперон в настоящем изобретении может не быть ограничен теми источниками, от которых он имеет происхождение. Очевидно, что объем настоящего изобретения может также включать любую аминокислотную последовательность, гомологичную указанным выше последовательностям и обладающую биологической активностью, по существу идентичной или соответствующей белку по SEQ ID NO:5 или SEQ ID NO:7, несмотря на то, что аминокислотная последовательность может иметь делецию, модификацию, замену или добавление части последовательности.
Полинуклеотид, кодирующий фосфотрансацетилазный и ацетаткиназный оперон по настоящему изобретению, может включать полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO:5 или SEQ ID NO:7, или полинуклеотид, кодирующий белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанным выше аминокислотным последовательностям, и, наиболее конкретно, может включать полинуклеотидную последовательность SEQ ID NO:6 или SEQ ID NO:8.
При использовании здесь термин «усиление активности» включает не только оказание эффекта, превосходящего исходную функцию, благодаря введению новой активности или повышению активности самого белка, но также включает повышение его активности посредством повышения активности эндогенного гена, амплификации эндогенного гена под действием внутреннего или внешнего фактора (факторов), устранения регуляторного фактора (факторов), ингибирующего экспрессию гена, увеличения числа копий гена, введения гена извне, модификации последовательности регуляции экспрессии и, конкретно, повышение ферментативной активности, благодаря замене или модификации промотора и мутации гена, и так далее.
Конкретно, в настоящем изобретении повышение активности может быть осуществлено посредством:
1) увеличения числа копий полинуклеотида, кодирующего фермент;
2) модификации последовательности регуляции экспрессии для повышения экспрессии полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосоме для усиления активности фермента; или
4) модификации комбинацией указанного выше;
но способ повышения активности не ограничен указанными вариантами.
Увеличение числа копий полинуклеотида (способ 1) может быть осуществлено в форме функционального связывания полинуклеотида с вектором или введением полинуклеотида в хромосому клетки-хозяина, однако способ не ограничен указанными вариантами. Конкретно, число копий полинуклеотида в хромосоме клетки-хозяина может быть увеличено введением вектора, способного проходить репликацию и функционировать независимо от клетки-хозяина, с которым функционально связан полинуклеотид, кодирующий белок по настоящему изобретению; или оно может быть увеличено введением в вектора, способного интегрировать полинуклеотид в хромосому клетки-хозяина, с которым функционально связан полинуклеотид, в клетку-хозяина.
Далее, модификация последовательности регуляции экспрессии для повышения экспрессии полинуклеотида (способ 2) может быть осуществлена модификацией последовательности посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного усиления активности последовательности регуляции экспрессии или заменой полинуклеотидной последовательности на полинуклеотидную последовательность, обладающую более сильной активностью, однако способ не ограничен указанными вариантами. Последовательность регуляции экспрессии включает, без ограничения, промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосом, и последовательность, регулирующую терминацию транскрипции и трансляции.
Вместо исходного промотора с начальной областью экспрессионной единицы полинуклеотида может быть связан сильный экзогенный промотор. Примерами сильных промоторов могут быть, без ограничения, промотор CJ7, промотор lysCP1, промотор EF-Tu, промотор groEL, промотор aceA или промотор aceB и так далее, и, конкретнее, интенсивность экспрессии может быть улучшена функциональным связыванием с промотором lysCP1 (WO 2009/096689) или промотором CJ7 (патент Кореи № 10-0620092 и WO 2006/065095), имеющими происхождение от Corynebacterium.
Кроме того, модификация полинуклеотидной последовательности в хромосоме (способ 3) может быть осуществлена модификацией последовательности регуляции экспрессии посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного усиления активности полинуклеотидной последовательности или заменой полинуклеотидной последовательности на улучшенную полинуклеотидную последовательность, обладающую более сильной активностью, однако способ не ограничен указанными вариантами.
Конкретно, в настоящем изобретении активность фосфотрансацетилазного и ацетаткиназного оперона (оперона pta-ackA) может быть усилена по сравнению с его эндогенной активностью любым способом, выбранным из группы, состоящей из способа увеличения числа копий оперона в клетке, способа модификации последовательности регуляции экспрессии оперона, способа замены последовательности регуляции экспрессии гена в опероне на последовательность, обладающую более сильной активностью, способа замены генов, кодирующих ферменты в хромосоме, мутантными генами для повышения активности ферментов, составляющих оперон, и способа модификации гена в хромосоме для повышения активности ферментов, составляющих оперон. Конкретно, способ замены последовательности регуляции экспрессии гена в опероне на последовательность, обладающую более сильной активностью, может быть осуществлен, без ограничения, заменой эндогенного промотора ацетилазного и ацетаткиназного оперона на промотор CJ7, промотор lysCP1, промотор EF-Tu, промотор groEL, промотор aceA или промотор aceB и так далее.
При использовании здесь термин «эндогенная активность» относится к активному состоянию фермента в немодифицированном состоянии, исходно присутствующего у микроорганизма, и термин «усиление по сравнению с эндогенной активностью» относится к состоянию повышенной активности фермента, присутствующего у микроорганизма после манипуляции, такой как введение гена, демонстрирующего активность, или увеличение числа копий соответствующего гена, делеция регуляторного фактора, ингибирующего экспрессию гена, или модификация последовательности регуляции экспрессии, например, использование промотора, улучшенного по сравнению с активностью у микроорганизма до манипуляции.
В настоящем изобретении в микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, может быть дополнительно введена активность ацетил-КоА-синтетазы (acs), имеющей происхождение от E. coli.
В настоящем изобретении ацетил-КоА-синтетаза (acs) представляет собой фермент, опосредующий реакцию образования ацетил-КоА из АТФ, уксусной кислоты и КоА.
В настоящем изобретении ацетил-КоА-синтетаза может включать, без ограничения, белки, имеющие аминокислотную последовательность SEQ ID NO:9, или любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что белок обладает существенной активностью по опосредованию синтеза ацетил-КоА.
Кроме того, поскольку аминокислотные последовательности белков, демонстрирующих указанную активность, могут различаться у разных видов или штаммов данного микроорганизма, ацетил-КоА-синтетаза (acs) в настоящем изобретении может не быть ограничена источником, от которого она имеет происхождение, и, например, она может иметь происхождение от E. coli. Очевидно, что объем настоящего изобретения может также включать любую аминокислотную последовательность, гомологичную указанной выше последовательности и обладающую биологической активностью, по существу идентичной или соответствующей белку по SEQ ID NO:9, несмотря на то, что аминокислотная последовательность может иметь делецию, модификацию, замену или добавление части последовательности.
Полинуклеотид, кодирующий ацетил-КоА-синтетазу (acs) по настоящему изобретению, может включать полинуклеотид, кодирующий белок, содержащий аминокислотную последовательность SEQ ID NO:9, или любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, и, наиболее конкретно, он может включать полинуклеотидную последовательность SEQ ID NO:10.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который дополнительно введена активность орнитиндекарбоксилазы (ODC), имеющей происхождение от E. coli.
При использовании здесь термин «орнитиндекарбоксилаза» относится к ферменту, обеспечивающему образование путресцина, опосредуя декарбоксилирование орнитина. Несмотря на то, что у микроорганизма рода Corynebacterium нет фермента биосинтеза путресцина, при введении орнитиндекарбоксилазы (ODC) извне происходит экспорт путресцина из клеток по мере его синтеза. Орнитиндекарбоксилаза, которая может быть введена извне, может быть использована в настоящем изобретении при условии наличия у нее указанной выше активности, независимо от источника, от которого микроорганизм имеет происхождение, и, конкретно, может быть введена орнитиндекарбоксилаза из E. coli.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность (1) орнитинкарбамоилтрансферазы (ArgF), (2) экспортера глутамата или (3) орнитинкарбамоилтрансферазы и экспортера глутамата дополнительно ослаблена по сравнению с их эндогенной активностью. Экспортер глутамата рода Corynebacterium может представлять собой NCgl1221.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность по меньшей мере одного, выбранного из группы, состоящей из ацетил-гамма-глутамилфосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитинаминотрансферазы (ArgD), дополнительно усилена по сравнению с их эндогенной активностью.
Кроме того, микроорганизм рода Corynebacterium может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность ацетилтрансферазы, конкретно, активность NCgl1469, дополнительно ослаблена по сравнению с ее эндогенной активностью.
В заключение, микроорганизм рода Corynebacterium может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность экспортера путресцина, конкретно, активность NCgl2522, дополнительно усилена по сравнению с его эндогенной активностью.
При использовании здесь «ослабление активности» включает не только оказание эффекта, уступающего исходной функции, ввиду снижения или устранения активности самого белка, но также включает снижение его активности посредством снижения активности эндогенного гена, активации регуляторного фактора (факторов), ингибирующего экспрессию гена, уменьшения числа копий гена, модификации последовательности регуляции экспрессии и, конкретно, устранение или снижение ферментативной активности, ввиду замены или модификации промотора и мутации гена, и так далее.
Конкретно, в настоящем изобретении ослабление активности может быть осуществлено посредством:
1) частичного или полного удаления полинуклеотида, кодирующего белок;
2) модификации последовательности регуляции экспрессии для снижения экспрессии полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосомах для ослабления активности белка; и
4) способа, выбранного из комбинации указанного выше;
но способ ослабления активности не ограничен указанными вариантами.
Конкретно, способ частичного или полного удаления полинуклеотида, кодирующего белок, может быть осуществлен заменой полинуклеотида, кодирующего эндогенный целевой белок, в хромосоме на полинуклеотид с частичной делецией полинуклеотидной последовательности или маркерный ген с использованием вектора для введения в хромосомы бактерий. При использовании здесь термин «часть» может варьировать в зависимости от типа полинуклеотида и, конкретно, может относиться к 1-300, конкретнее, 1-100, и, еще конкретнее, 1-50.
Кроме того, способ модификации последовательности регуляции экспрессии может быть осуществлен модификацией последовательности регуляции экспрессии посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного ослабления активности последовательности регуляции экспрессии или заменой полинуклеотидной последовательности на полинуклеотидную последовательность, обладающую более слабой активностью. Последовательность регуляции экспрессии включает промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосом, и последовательность, регулирующую терминацию транскрипции и трансляции.
Кроме того, способ модификации полинуклеотидной последовательности в хромосоме может быть осуществлен модификацией последовательности посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного ослабления активности фермента или заменой полинуклеотидной последовательности на улучшенную полинуклеотидную последовательность, обладающую более сильной активностью.
Кроме того, способ удаления регуляторного фактора, ингибирующего экспрессию полинуклеотида фермента, может быть осуществлен заменой полинуклеотида фактора, ингибирующего экспрессию, на полинуклеотид с частичной делецией полинуклеотидной последовательности или маркерный ген. При использовании здесь термин «часть» может варьировать в зависимости от типа полинуклеотида и, конкретно, может относиться к 1-300, конкретнее, 1-100, и, еще конкретнее, 1-50.
В частности, ацетил-гамма-глутамилфосфатредуктаза (ArgC), ацетилглутаматсинтаза или орнитинацетилтрансфераза (ArgJ), ацетилглутаматкиназа (ArgB), ацетилорнитинаминотрансфераза (ArgD), орнитинкарбамоилтрансфераза (ArgF), белки, вовлеченные в экспорт глутамата, и орнитиндекарбоксилаза (ODC) могут, соответственно, без ограничения, содержать аминокислотную последовательность SEQ ID NO:32, 33, 34, 35, 36, 37 или 38 или, конкретно, любую аминокислотную последовательность, на 70% или более, конкретнее, на 80% или более, и, еще конкретнее, на 90% или более гомологичную указанным выше аминокислотным последовательностям. Кроме того, белок, ацетилирующий путресцин, может, без ограничения, содержать аминокислотную последовательность SEQ ID NO:30 или 31 или, конкретно, любую аминокислотную последовательность, на 70% или более, конкретнее, на 80% или более, и, еще конкретнее, на 90% или более гомологичную указанным выше аминокислотным последовательностям.
Кроме того, в настоящем изобретении экспортер путресцина может содержать аминокислотную последовательность SEQ ID NO:26 или 28 или, конкретно, любую аминокислотную последовательность, на 70% или более, конкретнее, на 80% или более, и, еще конкретнее, на 90% или более гомологичную указанным выше аминокислотным последовательностям.
Среди описанных выше белков, усиление активности ацетил-гамма-глутамилфосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB), ацетилорнитинаминотрансферазы (ArgD), орнитиндекарбоксилазы (ODC) и экспортера путресцина может быть осуществлено, например, способом, выбранным из увеличения числа копий полинуклеотидов, кодирующих белки, модификации последовательности регуляции экспрессии для повышения экспрессии полинуклеотидов, модификации полинуклеотидных последовательностей в хромосоме для усиления активности указанных выше ферментов, удаления регуляторного фактора (факторов), ингибирующего экспрессию полинуклеотидов указанных выше ферментов, или их комбинаций.
Кроме того, ослабление активности орнитинкарбамоилтрансферазы (ArgF), белков, вовлеченных в экспорт глутамата, и белков, ацетилирующих путресцин, может быть осуществлено способом, выбранным из частичного или полного удаления полинуклеотидов, кодирующих белки, модификации последовательности регуляции экспрессии для снижения экспрессии полинуклеотидов, модификации полинуклеотидных последовательностей в хромосоме для ослабления активности белков и их комбинаций.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина или орнитина, включающий:
(1) культивирование микроорганизма рода Corynebacterium, продуцирующего путресцин или орнитин, в среде; и
(2) выделение путресцина или орнитина из культивированного микроорганизма или среды.
В указанном выше способе микроорганизм можно культивировать, без ограничения, в периодической культуре, непрерывной культуре, подпитываемой культуре и так далее, как известно в данной области. В частности, применительно к условиям культивирования, надлежащий pH (то есть оптимальный pH от 5 до 9, конкретно, pH 6-8, и, наиболее конкретно, pH 6,8) можно поддерживать с использованием, без ограничения, основного соединения (например, гидроксид натрия, гидроксид калия или аммиак) или кислого соединения (например, фосфорная кислота или серная кислота). Кроме того, можно поддерживать аэробные условия, добавляя в клеточную культуру кислород или кислород-содержащую смесь газов. В культуре можно поддерживать температуру от 20°C до 45°C и, конкретно, от 25°C до 40°C, и микроорганизм можно культивировать в течение приблизительно 10-160 часов. Путресцин или орнитин, продуцируемый посредством описанного выше культивировании, может секретироваться в культуральную среду или оставаться внутри клеток.
Кроме того, в культуральной среде могут быть использованы источники углерода, такие как, без ограничения, сахара и углеводы (например, глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, крахмал и целлюлоза), масла и жиры (например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например, пальмитиновая кислота, стеариновая кислота и линолевая кислота), спирты (например, глицерин и этанол) и органические кислоты (например, уксусная кислота), по отдельности или в комбинации; источники азота, такие как, без ограничения, азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясной сок, солодовый экстракт, жидкий кукурузный экстракт, соевая мука и мочевина) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония), по отдельности или в комбинации; и источники калия, такие как, без ограничения, дигидрофосфат калия, гидрофосфат калия или соответствующие натрийсодержащие соли, по отдельности или в комбинации. Кроме того, среда может дополнительно содержать, без ограничения, другие необходимые ростостимулирующие вещества, включая соли металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины.
Способ выделения путресцина или орнитина, полученного при культивировании по настоящему изобретению, может быть осуществлен подходящим методом культивирования, известным в данной области, например, таким как периодическая культура, непрерывная культура или подпитываемая культура, и, посредством этого, целевая аминокислота может быть выделена из культуры.
ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие Примеры. Тем не менее, эти Примеры приведены лишь в целях наглядности, и изобретение не следует ограничивать этими Примерами.
Пример 1: Введение argA , имеющего происхождение от E. coli , и argE , имеющего происхождение от E. coli , в штамм, продуцирующий путресцин, и подтверждение способности полученного штамма продуцировать путресцин
1-1. Получение штамма с одновременным введением argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, в ген транспозона штамма, продуцирующего путресцин, на основе ATCC13032
Для подтверждения того, что введение гена argA, имеющего происхождение от E. coli, и гена argE, имеющего происхождение от E. coli, в штамм, продуцирующий путресцин, на основе ATCC13032 может улучшить способность продуцировать путресцин, гены argA и argE вводили в ген транспозона указанного штамма.
В качестве вектора для трансформации, позволяющего осуществлять введение генной области транспозона микроорганизма рода Corynebacterium в хромосому, использовали pDZTn (WO 2009/125992) и в качестве промотора использовали промотор lysCP1 (публикация международной заявки на патент № WO 2009/096689, SEQ ID NO:39).
Конкретно, на основе полинуклеотидной последовательности (SEQ ID NO:2) гена argA, имеющего происхождение от E. coli, кодирующего N-ацетилглутаматсинтазу, получали пару праймеров SEQ ID NO:11 и 12 для получения гомологичных рекомбинантных фрагментов открытой рамки считывания (ORF) argA. Кроме того, на основе полинуклеотидной последовательности (SEQ ID NO:4) гена argE, имеющего происхождение от E. coli, кодирующего ацетилорнитиндезацетилазу, получали пару праймеров SEQ ID NO:15 и 16 для получения гомологичных рекомбинантных фрагментов ORF argE, и на основе полинуклеотидной последовательности (SEQ ID NO:39) lysCP1 получали пару праймеров SEQ ID NO:13 и 14 для получения гомологичных рекомбинантных фрагментов области lysCP1 (Таблица 1).
Таблица 1
(SEQ ID NO:11)
(SEQ ID NO:12)
(SEQ ID NO:13)
(SEQ ID NO:14)
(SEQ ID NO:15)
(SEQ ID NO:16)
Сначала для получения гена argA амплифицировали фрагмент гена размером приблизительно 1,6 тысячи пар оснований (т.п.о.) с использованием хромосомы штамма E. coli W3110 в качестве матрицы с парой праймеров SEQ ID NO:11 и 12. В частности, полимеразную цепную реакцию (ПЦР) проводили, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты и 30 секунд. Полученные таким образом фрагменты подвергали электрофорезу в 0,8%-м агарозном геле и проводили элюирование и очистку полос желаемого размера.
Кроме того, амплифицировали промоторную область lysCP1, проводя ПЦР с использованием хромосомы штамма KCCM10919P (публикация международной заявки на патент № WO 2009/096689) в качестве матрицы с парой праймеров SEQ ID NO:13 и 14, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 30 секунд.
Вектор pDZTn обрабатывали XhoI, после чего каждый из полученных ПЦР-продуктов подвергали клонированию посредством слияния. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech), и полученная таким образом плазмида была названа pDZTn-lysCP1-argA.
Затем получали ПЦР-продукты для получения гена argE, амплифицируя фрагмент гена размером приблизительно 1,4 т.п.о. таким же образом, как описано выше, с использованием хромосомы штамма E. coli W3110 в качестве матрицы с парой праймеров SEQ ID NO:15 и 16, и подвергали их клонированию посредством слияния вместе с промоторной областью lysCP1. Полученная таким образом плазмида была названа pDZTn-lysCP1-argE.
Затем плазмиду pDZTn-lysCP1-argA вводили в штамм KCCM11240P (публикация заявки на патент Кореи № 10-2013-0082478) электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZTn-lysCP1-argA.
Отобранные штаммы культивировали с покачиванием (30°C, 8 часов) в среде CM (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), pH 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма с геном, кодирующим argA, введенным посредством вторичного кроссинговера. Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO:12 и 13, подтвердив введение гена, кодирующего argA, и модифицированный штамм Corynebacterium glutamicum был назван KCCM11240P Tn:lysCP1-argA.
Для введения в штамм с введенным argA, полученный выше, KCCM11240P Tn:lysCP1-argA трансформировали полученной выше pDZTn-lysCP1-argE таким же образом, как описано выше, и введение argE в транспозон отобранного в итоге штамма было подтверждено проведением ПЦР с использованием пары праймеров SEQ ID NO:13 и 16. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE.
1-2. Получение штамма с одновременным введением argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, в ген транспозона штамма, продуцирующего путресцин, на основе ATCC13869
DAB12-a ΔNCgl1469 (публикация заявки на патент Кореи № 10-2013-0082478), представляющий собой штамм, продуцирующий путресцин, на основе Corynebacterium glutamicum ATCC13869, был назван DAB12-b, и в ген его транспозона вводили argA и argE для подтверждения того, что введение генов argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, может быть связано с улучшением способности полученного штамма продуцировать путресцин.
Сначала pDZTn-lysCP1-argA, полученной ранее, трансформировали Corynebacterium glutamicum DAB12-b таким же образом, как в Примере 1-1, и подтверждали введение argA в транспозон. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван DAB12-b Tn:lysCP1-argA.
Затем, для введения argE в штамм с уже введенным argA, DAB12-b Tn:lysCP1-argA трансформировали полученной ранее pDZTn-lysCP1-argE таким же образом, как в Примере 1-1, и подтверждали введение argE в транспозон. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван DAB12-b Tn:lysCP1-argE.
1-3. Оценка способности штамма Corynebacterium, продуцирующего путресцин, с введенным геном argA, имеющим происхождение от E. coli, и геном argE, имеющим происхождение от E. coli, продуцировать путресцин
Проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примерах 1-1 и 1-2, продуцировать путресцин для подтверждения эффекта введения argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, в штамм, продуцирующий путресцин, на продукцию путресцина.
Конкретно, модифицированные штаммы Corynebacterium glutamicum двух разных типов (то есть KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), полученные в Примерах 1-1 и 1-2, и исходные штаммы двух разных типов (то есть KCCM11240P и DAB12-b), соответственно, высевали на твердую среду CM (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, pH 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°C в течение 24 часов.
Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°C и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 2 ниже.
Таблица 2
Как показано в Таблице 2 выше, оба модифицированных штамма Corynebacterium glutamicum с одновременным введением генов argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, продемонстрировали повышение продукции путресцина на 9,8% или более.
Пример 2: Усиление pta-ackA у штамма, продуцирующего путресцин, с введенными argA , имеющим происхождение от E. coli , и argE , имеющим происхождение от E. coli , и подтверждение способности полученного штамма продуцировать путресцин
2-1. Получение штамма с заменой промотора pta-ackA из штамма Corynebacterium, продуцирующего путресцин, на основе ATCC13032
У штамма, продуцирующего путресцин, с введенными генами argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, полученного в Примере 1, проводили дополнительное усиление активности фосфотрансацетилазы и ацетаткиназы (pta-ackA) и оценивали эффект такого усиления на способность штамма продуцировать путресцин.
С этой целью промотор оперона pta-ackA в хромосоме заменяли на промотор, обладающий более сильной активностью, по сравнению с его эндогенным промотором, конкретно перед инициирующим кодоном оперона pta-ackA вводили промотор lysCP1 (публикация международной заявки на патент № WO 2009/096689).
Сначала получали гомологичный рекомбинантный фрагмент, содержащий промотор lysCP1, где оба конца промотора имеют оригинальную хромосомную последовательность pta-ackA. Конкретно, 5’-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum ATCC13032 с парой праймеров SEQ ID NO:17 и 18. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 30 секунд.
Кроме того, промоторную область lysCP1 получали, проводя ПЦР в таких же условиях с использованием пары праймеров SEQ ID NO:14 и 19, и 3’-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum ATCC13032 в качестве матрицы с парой праймеров SEQ ID NO:20 и 21. Праймеры, использованные при получении промотора lysCP1, показаны в Таблице 1 выше и в Таблице 3 ниже.
Таблица 3
(SEQ ID NO:17)
(SEQ ID NO:18)
(SEQ ID NO:19)
(SEQ ID NO:20)
(SEQ ID NO:21)
Каждый из полученных выше ПЦР-продуктов подвергали клонированию посредством слияния с использованием вектора pDZ, обработанного XbaI. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech), и полученная таким образом плазмида была названа pDZ-lysCP1-1’pta-ackA.
Полученную выше плазмиду pDZ-lysCP1-1’pta-ackA вводили, соответственно, в штаммы KCCM11240P и KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-1, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZ-lysCP1-1’pta-ackA.
Отобранные штаммы культивировали с покачиванием (30°C, 8 часов) в среде CM (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), pH 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма, в котором промотор pta-ackA был заменен на промотор lysCP1 посредством вторичного кроссинговера.
Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO:19 и 21, подтвердив введение промотора lysCP1 в хромосому перед инициирующим кодоном pta-ackA. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты.
Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы KCCM11240P lysCP1-1’pta-ackA и KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1’pta-ackA, соответственно.
2-2. Получение штамма с заменой промотора pta-ackA из штамма Corynebacterium, продуцирующего путресцин, на основе ATCC13869
Для подтверждения последовательности гена, кодирующего pta-ackA, имеющего происхождение от Corynebacterium glutamicum ATCC13869, и экспрессированного с него белка, полученного методом, раскрытым в Примере 2-1, проводили ПЦР с использованием геномной ДНК Corynebacterium glutamicum ATCC13869 в качестве матрицы с парой праймеров SEQ ID NO:17 и 22 (Таблицы 3 и 4). В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 3 минут.
Полученные таким образом ПЦР-продукты разделяли электрофорезом и анализировали их последовательности. В результате, было подтверждено, что ген, кодирующий pta-ackA, имеющий происхождение от Corynebacterium glutamicum ATCC13869, содержит полинуклеотидную последовательность, описываемую SEQ ID NO:8, и что белок, кодируемый этим геном, содержит аминокислотную последовательность, описываемую SEQ ID NO:7.
С другой стороны, в результате сравнения аминокислотной последовательности pta-ackA, имеющей происхождение от Corynebacterium glutamicum ATCC13032 (SEQ ID NO:5), и аминокислотной последовательности pta-ackA, имеющей происхождение от Corynebacterium glutamicum ATCC13869, было подтверждено, что эти последовательности гомологичны на 99,4%.
Таблица 4
(SEQ ID NO:22)
(SEQ ID NO:23)
Сначала получали гомологичный рекомбинантный фрагмент, содержащий промотор lysCP1, где оба конца промотора имеют оригинальную хромосомную последовательность pta-ackA. Конкретно, 5’-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum ATCC13869 с парой праймеров SEQ ID NO:17 и 23. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 30 секунд. Кроме того, промоторную область lysCP1 получали, проводя ПЦР в таких же условиях с использованием пары праймеров SEQ ID NO:14 и 19, и 3’-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum ATCC13869 в качестве матрицы с парой праймеров SEQ ID NO:20 и 21. Праймеры, использованные при замене промотора, показаны в Таблицах 1, 3 и 4.
Каждый из полученных ПЦР-продуктов подвергали клонированию посредством слияния с использованием вектора pDZTn, обработанного XhoI. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech), и полученная таким образом плазмида была названа pDZ-lysCP1-2’pta-ackA.
Плазмидой pDZ-lysCP1-2’pta-ackA, полученной выше, трансформировали, соответственно, DAB12-b и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-2, таким же образом, как в Примере 2-1. В результате, было подтверждено, что промотор lysCP1 был введен в хромосому перед инициирующим кодоном pta-ackA. Модифицированные штаммы Corynebacterium glutamicum были названы DAB12-b lysCP1-2’pta-ackA и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-2’pta-ackA, соответственно.
2-3. Оценка способности штамма с усилением pta-ackA продуцировать путресцин
Для подтверждения эффекта усиления pta-ackA у штамма, продуцирующего путресцин, с введенными argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примерах 2-1 и 2-2, продуцировать путресцин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum четырех типов (KCCM11240P lysCP1-1’pta-ackA; KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1’pta-ackA; DAB12-b lysCP1-2’pta-ackA; и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-2’pta-ackA) и исходные штаммы четырех типов (KCCM11240P; KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b; и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду CM (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, pH 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°C в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°C и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 5 ниже.
Таблица 5
Как показано в Таблице 5, при усилении pta-ackA у KCCM11240P и DAB12-b, соответственно, количество продуцируемого путресцина оставалось на том же уровне. Тем не менее, при усилении pta-ackA у модифицированных штаммов Corynebacterium glutamicum двух разных типов с одновременным введением генов argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli (KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, количество продуцируемого путресцина увеличивалось на 14,3% или более по сравнению с исходным штаммом. Кроме того, количество продуцируемого путресцина увеличивалось на 4% или более по сравнению с модифицированными штаммами.
Как таковой, полученный микроорганизм рода Corynebacterium (Corynebacterium glutamicum KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE :lysCP1-1’pta-ackA), обладающий улучшенной способностью продуцировать путресцин, полученный из штамма Corynebacterium glutamicum KCCM11240P, продуцирующего путресцин, введением активности argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli и усилением активности pta-ackA у штамма Corynebacterium glutamicum KCCM11240P, был назван авторами настоящего изобретения CC01-1145 и депонирован в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM) 21 ноября 2014 г. под регистрационным номером KCCM11606P в соответствии с Будапештским соглашением.
Пример 3: Введение acs , имеющего происхождение от E. coli , в штамм, продуцирующий путресцин, с введенными argA , имеющим происхождение от E. coli , и argE , имеющим происхождение от E. coli , и подтверждение способности полученного штамма продуцировать путресцин
3-1. Получение штамма с введением acs, имеющего происхождение от E. coli, в ген транспозона штамма, продуцирующего путресцин, на основе ATCC13032
acs вводили в ген транспозона с использованием промотора lysCP1 для подтверждения того, что введение гена ацетил-КоА-синтетазы (acs), имеющего происхождение от E. coli, в штамм, продуцирующий путресцин, на основе ATCC13032 с уже введенными argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, может улучшить способность продуцировать путресцин.
Конкретно, на основе полинуклеотидной последовательности гена, кодирующего acs, описываемой SEQ ID NO:10, получали пару праймеров SEQ ID NO:24 и 25 для получения гомологичного рекомбинантного фрагмента вокруг ORF-области acs и пару праймеров SEQ ID NO:13 и 14 для получения гомологичного рекомбинантного фрагмента вокруг промоторной области lysCP1, как показано в Таблице 1 выше и в Таблице 6 ниже.
Таблица 6
(SEQ ID NO:24)
(SEQ ID NO:25)
Конкретно, для получения гена acs амплифицировали фрагмент гена размером приблизительно 2 т.п.о. с использованием хромосомы штамма E. coli W3110 в качестве матрицы с парой праймеров SEQ ID NO:24 и 25. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты и 30 секунд. Затем полученные таким образом ПЦР-продукты подвергали электрофорезу в 0,8%-м агарозном геле и проводили элюирование и очистку полос желаемого размера.
Кроме того, получали промоторную область lysCP1, проводя ПЦР с использованием хромосомы штамма KCCM10919P (публикация международной заявки на патент № WO 2009/096689) в качестве матрицы с парой праймеров SEQ ID NO:13 и 14, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 30 секунд.
Вектор pDZ обрабатывали XhoI и каждый из полученных таким образом ПЦР-продуктов подвергали клонированию посредством слияния. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech). Полученная таким образом плазмида была названа pDZTn-lysCP1-acs.
Затем плазмиду pDZTn-lysCP1-acs вводили, соответственно, в KCCM11240P и KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-1, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZTn-lysCP1-acs.
Отобранные штаммы культивировали с покачиванием (30°C, 8 часов) в среде CM (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), pH 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штаммов с геном, кодирующим acs, введенным посредством вторичного кроссинговера. Отобранные в итоге штаммы подвергали ПЦР с использованием пары праймеров SEQ ID NO:13 и 25, подтвердив введение гена, кодирующего acs, и модифицированные штаммы Corynebacterium glutamicum были названы KCCM11240P Tn:lysCP1-acs и KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs, соответственно.
3-2. Получение штамма с введением acs, имеющего происхождение от E. coli, в ген транспозона штамма, продуцирующего путресцин, на основе ATCC13869
Как в Примере 3-1, pDZTn-lysCP1-acs, полученной выше, трансформировали DAB12-b и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-2, соответственно, таким же образом, как в Примере 3-1, подтвердив введение acs в ген транспозона.
Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы DAB12-b Tn:lysCP1-acs и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs, соответственно.
3-3. Оценка способности штамма с введенным acs, имеющим происхождение от E. coli, продуцировать путресцин
Для подтверждения эффекта введения acs в штамм, продуцирующий путресцин, с уже введенными argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примерах 3-1 и 3-2, продуцировать путресцин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum четырех типов (KCCM11240P Tn:lysCP1-acs; KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs; DAB12-b Tn:lysCP1-acs; и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs) и исходные штаммы четырех типов (KCCM11240P; KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b; и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду CM (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, pH 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°C в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4·7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°C и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 7 ниже.
Таблица 7
Как показано в Таблице 7, при введении acs в KCCM11240P и DAB12-b, соответственно, количество продуцируемого путресцина оставалось на том же уровне. Тем не менее, при введении acs в модифицированные штаммы Corynebacterium glutamicum двух разных типов с одновременным введением генов argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli (KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, количество продуцируемого путресцина увеличивалось на 13,5% или более по сравнению с исходным штаммом. Кроме того, количество продуцируемого путресцина увеличивалось на 3,4% или более по сравнению с указанными выше модифицированными штаммами.
Пример 4: Штамм с введением argA , имеющего происхождение от E. coli , и argE , имеющего происхождение от E. coli , и заменой промотора pta-ackA из штамма, продуцирующего путресцин, с улучшенной способностью к экспорту путресцина и способность штамма продуцировать путресцин
4-1. Получение штамма с введением argA и argE, имеющих происхождение от E. coli, и заменой промотора pta-ackA из штамма с улучшенной способностью к экспорту путресцина
На основе штамма KCCM11401P (публикация заявки на патент Кореи № 10-2014-0115244) с улучшенной способностью к экспорту путресцина получали штамм для определения того, может ли введение argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, и усиление активности pta-ackA Corynebacterium улучшить способность продуцировать путресцин.
Конкретно, pDZTn-lysCP1-argA, полученной в Примере 1-1, трансформировали KCCM11401P таким же образом, как в Примере 1-1, и, в результате, введение argA в ген транспозона было подтверждено. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван KCCM11401P Tn:lysCP1-argA.
Кроме того, для введения argE в штамм с уже введенным argA, как в Примере 1-1, pDZTn-lysCP1-argE, полученной в Примере 1-1, трансформировали KCCM11401P Tn:lysCP1-argA таким же образом, как в Примере 1-1, и введение argE в ген транспозона было подтверждено. Отобранный таким образом модифицированный штамм был назван KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE.
Затем pDZ-lysCP1-1’pta-ackA, полученной в Примере 2-1, трансформировали KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE таким же образом, как в Примере 2-1, и было подтверждено, что промотор lysCP1 был введен в хромосому перед инициирующим кодоном pta-ackA. Полученный выше модифицированный штамм Corynebacterium glutamicum был назван KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1’pta-ackA.
4-2. Оценка штамма с введением argA, имеющего происхождение от E. coli, argE, имеющего происхождение от E. coli, и заменой промотора pta-ackA из штамма с улучшенной способностью к экспорту путресцина
Для подтверждения эффекта введения argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, и усиления активности pta-ackA у штамма Corynebacterium glutamicum, продуцирующего путресцин, с улучшенной способностью к экспорту путресцина проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 4-1, продуцировать путресцин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum (KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE, KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1’pta-ackA) и исходный штамм (KCCM11401P), соответственно, высевали на твердую среду CM (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, pH 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°C в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°C и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 8 ниже.
Таблица 8
Как показано в Таблице 8, было подтверждено, что при введении гена argA, имеющего происхождение от E. coli, и гена argE, имеющего происхождение от E. coli, в KCCM11401P с улучшенной способностью к экспорту путресцина количество продуцируемого путресцина увеличивалось на 11,9% по сравнению с исходным штаммом, а при дополнительном усилении штамма посредством pta-ackA количество продуцируемого путресцина увеличивалось на 16,1% по сравнению с исходным штаммом.
Пример 5: Введение argA , имеющего происхождение от E. coli , и argE , имеющего происхождение от E. coli , в штамм, продуцирующий орнитин, и подтверждение способности штамма продуцировать орнитин
5-1. Получение штамма с одновременным введением argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, в ген транспозона штамма, продуцирующего орнитин, на основе KCCM11137P
Для подтверждения того, что введение гена argA, имеющего происхождение от E. coli, и гена argE, имеющего происхождение от E. coli, в штамм KCCM11137P (публикация заявки на патент Кореи № 10-1372635), представляющий собой штамм, продуцирующий орнитин, на основе Corynebacterium glutamicum ATCC13032, может улучшать способность продуцировать орнитин, ген argA и ген argE вводили в ген транспозона указанного штамма с использованием вектора, полученного в Примере 1-1.
Сначала плазмиду pDZTn-lysCP1-argA вводили в штамм KCCM11137P электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор штаммов с введенной плазмидой pDZTn-lysCP1-argA.
Отобранные штаммы культивировали с покачиванием (30°C, 8 часов) в среде CM (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), pH 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма с геном, кодирующим argA, введенным посредством вторичного кроссинговера. Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO:12 и 13, подтвердив введение гена, кодирующего argA, и модифицированный штамм Corynebacterium glutamicum был назван KCCM11137P Tn:lysCP1-argA.
Для введения argE в штамм с уже введенным argA, полученный выше, pDZTn-lysCP1-argE, полученной в Примере 1-1, трансформировали KCCM11137P Tn:lysCP1-argA таким же образом, как в Примере 1-1, и, таким образом, введение argE в ген транспозона было подтверждено.
Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE.
5-2. Оценка способности штамма Corynebacterium, продуцирующего орнитин, с введенным argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, продуцировать орнитин
Для подтверждения эффекта введения argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, на продукцию орнитина штаммом, продуцирующим орнитин, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 5-1, продуцировать орнитин.
Конкретно, модифицированный штамм Corynebacterium glutamicum одного типа (KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE) и исходный штамм одного типа (KCCM11137P), соответственно, высевали на твердую среду CM (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, pH 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°C в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°C и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 9 ниже.
Таблица 9
Как показано в Таблице 9, было подтверждено, что при введение гена argA, имеющего происхождение от E. coli, и гена argE, имеющего происхождение от E. coli, в модифицированный штамм Corynebacterium glutamicum продемонстрировало увеличение количества продуцируемого орнитина на 14,1% по сравнению с исходным штаммом.
Пример 6: Усиление pta-ackA у штамма с введенными argA , имеющим происхождение от E. coli , и argE , имеющим происхождение от E. coli , и подтверждение способности штамма продуцировать орнитин
6-1. Получение штамма с заменой промотора pta-ackA из штамма, продуцирующего орнитин, на основе ATCC13032
Для подтверждения того, что усиление pta-ackA у штамма, продуцирующего орнитин, на основе ATCC13032 с введенными argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, может улучшить способность продуцировать орнитин, перед инициирующим кодоном оперона pta-ackA в хромосому вводили промотор lysCP1 (WO 2009/096689).
Сначала плазмиду pDZ-lysCP1-1’pta-ackA, полученную в Примере 2-1, вводили в штаммы KCCM11137P и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, соответственно, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZ-lysCP1-1’pta-ackA.
Отобранные штаммы культивировали с покачиванием (30°C, 8 часов) в среде CM (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), pH 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма, в котором промотор pta-ackA был заменен на промотор lysCP1 посредством вторичного кроссинговера. Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO:19 и 21, подтвердив введение промотора lysCP1 в хромосому перед инициирующим кодоном оперона pta-ackA. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты.
Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы KCCM11137P lysCP1-1’pta-ackA и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1’pta-ackA, соответственно.
6-2. Оценка способности штамма с усилением активности pta-ackA продуцировать орнитин
Для подтверждения эффекта усиления активности pta-ackA у штамма, продуцирующего орнитин, с введенными argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 6-1, продуцировать орнитин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum двух разных типов (KCCM11137P lysCP1-1’pta-ackA; KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1’pta-ackA) и исходные штаммы двух разных типов (KCCM11137P; KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду CM (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, pH 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°C в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°C и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного орнитина, и результаты показаны в Таблице 10 ниже.
Таблица 10
Как показано в Таблице 10, было подтверждено, что при усилении активности pta-ackA у штамма KCCM11137P количество продуцируемого орнитина не увеличивалось, в то время как в случае штамма KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, представляющего собой модифицированный штамм Corynebacterium glutamicum с одновременным введением гена argA, имеющего происхождение от E. coli, и гена argE, имеющего происхождение от E. coli, количество продуцируемого орнитина увеличивалось на 20,5% по сравнению со штаммом KCCM11137P и на 5,6% по сравнению со штаммом KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE.
Пример 7: Введение acs , имеющего происхождение от E. coli , в штамм с введенными argA , имеющим происхождение от E. coli , и argE , имеющим происхождение от E. coli , и подтверждение способности штамма продуцировать орнитин
7-1. Получение штамма с введением acs, имеющего происхождение от E. coli, в ген транспозона штамма, продуцирующего орнитин, на основе KCCM11137
Для подтверждения того, что введение acs, имеющего происхождение от E. coli, в штамм KCCM11137P (патент Кореи № 10-1372635), представляющий собой штамм, продуцирующий орнитин, на основе Corynebacterium glutamicum ATCC13032, может улучшить способность продуцировать орнитин, acs вводили в ген транспозона с использованием промотора lysCP1.
Сначала плазмиду pDZTn-lysCP1-acs, полученную в Примере 3-1, вводили в штаммы KCCM11137P и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, соответственно, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZTn-lysCP1-acs.
Отобранные штаммы культивировали с покачиванием (30°C, 8 часов) в среде CM (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), pH 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма с геном, кодирующим acs, введенным посредством вторичного кроссинговера.
Отобранные в итоге штаммы подвергали ПЦР с использованием пары праймеров SEQ ID NO:13 и 25, подтвердив введение гена, кодирующего acs. Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы KCCM11137P Tn:lysCP1-acs и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs, соответственно.
7-2. Оценка способности штамма с введенным acs, имеющим происхождение от E. coli, продуцировать орнитин
Для подтверждения эффекта введения acs в штамм, продуцирующий орнитин, с введенными argA, имеющим происхождение от E. coli, и argE, имеющим происхождение от E. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 7-1, продуцировать орнитин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum двух разных типов (KCCM11137P Tn:lysCP1-acs; KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs) и исходные штаммы двух разных типов (KCCM11137P; KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду CM (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, pH 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°C в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°C и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного орнитина, и результаты показаны в Таблице 11 ниже.
Таблица 11
Как показано в Таблице 11, было подтверждено, что при введении acs в штамм KCCM11137P количество продуцируемого орнитина не увеличивалось, в то время как в случае штамма KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, представляющего собой модифицированный штамм Corynebacterium glutamicum с одновременным введением гена argA, имеющего происхождение от E. coli, и гена argE, имеющего происхождение от E. coli, количество продуцируемого орнитина увеличивалось на 17,9% по сравнению со штаммом KCCM11137P и на 3,4% по сравнению со штаммом KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE.
Суммируя вышеизложенное, было подтверждено, что введение argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от E. coli, в штамм Corynebacterium может увеличить количество продуцируемого путресцина и орнитина, и, кроме того, было подтверждено, что усиление активности гена pta-ackA у штамма Corynebacterium или введение acs, имеющего происхождение от E. coli, может дополнительно увеличить количество продуцируемого путресцина и орнитина.
Исходя из изложенного выше, специалисту в области, к которой относится настоящее изобретение, будет ясно, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технической концепции или существенных характеристик настоящего изобретения. В этом отношении, типичные воплощения раскрыты здесь исключительно в целях наглядности, и их не следует толковать как ограничение объема настоящего изобретения. Наоборот, подразумевают, что настоящее изобретение охватывает не только типичные воплощения, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем настоящего изобретения, как определено приложенной формулой изобретения.
--->
<110> CJ CHEILJEDANG CORPORATION
<120> MICROORGANISMS FOR PRODUCING PUTRESCINE OR ORNITHINE AND PROCESS
FOR PRODUCING PUTRESCINE OR ORNITHINE USING THEM
<130> OPA16079
<150> KR10-2015-0102624
<151> 2015-07-20
<160> 39
<170> KopatentIn 2.0
<210> 1
<211> 443
<212> PRT
<213> E.coli N-acetylglutamate synthase
<400> 1
Val Val Lys Glu Arg Lys Thr Glu Leu Val Glu Gly Phe Arg His Ser
1 5 10 15
Val Pro Tyr Ile Asn Thr His Arg Gly Lys Thr Phe Val Ile Met Leu
20 25 30
Gly Gly Glu Ala Ile Glu His Glu Asn Phe Ser Ser Ile Val Asn Asp
35 40 45
Ile Gly Leu Leu His Ser Leu Gly Ile Arg Leu Val Val Val Tyr Gly
50 55 60
Ala Arg Pro Gln Ile Asp Ala Asn Leu Ala Ala His His His Glu Pro
65 70 75 80
Leu Tyr His Lys Asn Ile Arg Val Thr Asp Ala Lys Thr Leu Glu Leu
85 90 95
Val Lys Gln Ala Ala Gly Thr Leu Gln Leu Asp Ile Thr Ala Arg Leu
100 105 110
Ser Met Ser Leu Asn Asn Thr Pro Leu Gln Gly Ala His Ile Asn Val
115 120 125
Val Ser Gly Asn Phe Ile Ile Ala Gln Pro Leu Gly Val Asp Asp Gly
130 135 140
Val Asp Tyr Cys His Ser Gly Arg Ile Arg Arg Ile Asp Glu Asp Ala
145 150 155 160
Ile His Arg Gln Leu Asp Ser Gly Ala Ile Val Leu Met Gly Pro Val
165 170 175
Ala Val Ser Val Thr Gly Glu Ser Phe Asn Leu Thr Ser Glu Glu Ile
180 185 190
Ala Thr Gln Leu Ala Ile Lys Leu Lys Ala Glu Lys Met Ile Gly Phe
195 200 205
Cys Ser Ser Gln Gly Val Thr Asn Asp Asp Gly Asp Ile Val Ser Glu
210 215 220
Leu Phe Pro Asn Glu Ala Gln Ala Arg Val Glu Ala Gln Glu Glu Lys
225 230 235 240
Gly Asp Tyr Asn Ser Gly Thr Val Arg Phe Leu Arg Gly Ala Val Lys
245 250 255
Ala Cys Arg Ser Gly Val Arg Arg Cys His Leu Ile Ser Tyr Gln Glu
260 265 270
Asp Gly Ala Leu Leu Gln Glu Leu Phe Ser Arg Asp Gly Ile Gly Thr
275 280 285
Gln Ile Val Met Glu Ser Ala Glu Gln Ile Arg Arg Ala Thr Ile Asn
290 295 300
Asp Ile Gly Gly Ile Leu Glu Leu Ile Arg Pro Leu Glu Gln Gln Gly
305 310 315 320
Ile Leu Val Arg Arg Ser Arg Glu Gln Leu Glu Met Glu Ile Asp Lys
325 330 335
Phe Thr Ile Ile Gln Arg Asp Asn Thr Thr Ile Ala Cys Ala Ala Leu
340 345 350
Tyr Pro Phe Pro Glu Glu Lys Ile Gly Glu Met Ala Cys Val Ala Val
355 360 365
His Pro Asp Tyr Arg Ser Ser Ser Arg Gly Glu Val Leu Leu Glu Arg
370 375 380
Ile Ala Ala Gln Ala Lys Gln Ser Gly Leu Ser Lys Leu Phe Val Leu
385 390 395 400
Thr Thr Arg Ser Ile His Trp Phe Gln Glu Arg Gly Phe Thr Pro Val
405 410 415
Asp Ile Asp Leu Leu Pro Glu Ser Lys Lys Gln Leu Tyr Asn Tyr Gln
420 425 430
Arg Lys Ser Lys Val Leu Met Ala Asp Leu Gly
435 440
<210> 2
<211> 1332
<212> DNA
<213> E.coli N-acetylglutamate synthase
<400> 2
gtggtaaagg aacgtaaaac cgagttggtc gagggattcc gccattcggt tccctatatc 60
aatacccacc ggggaaaaac gtttgtcatc atgctcggcg gtgaagccat tgagcatgag 120
aatttctcca gtatcgttaa tgatatcggg ttgttgcaca gcctcggcat ccgtctggtg 180
gtggtctatg gcgcacgtcc gcagatcgac gcaaatctgg ctgcgcatca ccacgaaccg 240
ctgtatcaca agaatatacg tgtgaccgac gccaaaacac tggaactggt gaagcaggct 300
gcgggaacat tgcaactgga tattactgct cgcctgtcga tgagtctcaa taacacgccg 360
ctgcagggcg cgcatatcaa cgtcgtcagt ggcaatttta ttattgccca gccgctgggc 420
gtcgatgacg gcgtggatta ctgccatagc gggcgtatcc ggcggattga tgaagacgcg 480
atccatcgtc aactggacag cggtgcaata gtgctaatgg ggccggtcgc tgtttcagtc 540
actggcgaga gctttaacct gacctcggaa gagattgcca ctcaactggc catcaaactg 600
aaagctgaaa agatgattgg tttttgctct tcccagggcg tcactaatga cgacggtgat 660
attgtctccg aacttttccc taacgaagcg caagcgcggg tagaagccca ggaagagaaa 720
ggcgattaca actccggtac ggtgcgcttt ttgcgtggcg cagtgaaagc ctgccgcagc 780
ggcgtgcgtc gctgtcattt aatcagttat caggaagatg gcgcgctgtt gcaagagttg 840
ttctcacgcg acggtatcgg tacgcagatt gtgatggaaa gcgccgagca gattcgtcgc 900
gcaacaatca acgatattgg cggtattctg gagttgattc gcccactgga gcagcaaggt 960
attctggtac gccgttctcg cgagcagctg gagatggaaa tcgacaaatt caccattatt 1020
cagcgcgata acacgactat tgcctgcgcc gcgctctatc cgttcccgga agagaagatt 1080
ggggaaatgg cctgtgtggc agttcacccg gattaccgca gttcatcaag gggtgaagtt 1140
ctgctggaac gcattgccgc tcaggcgaag cagagcggct taagcaaatt gtttgtgctg 1200
accacgcgca gtattcactg gttccaggaa cgtggattta ccccagtgga tattgattta 1260
ctgcccgaga gcaaaaagca gttgtacaac taccagcgta aatccaaagt gttgatggcg 1320
gatttagggt aa 1332
<210> 3
<211> 408
<212> PRT
<213> E.coli Acetylornithine deacetylase
<400> 3
Met Asp Lys Leu Leu Glu Arg Phe Leu Asn Tyr Val Ser Leu Asp Thr
1 5 10 15
Gln Ser Lys Ala Gly Val Arg Gln Val Pro Ser Thr Glu Gly Gln Trp
20 25 30
Lys Leu Leu His Leu Leu Lys Glu Gln Leu Glu Glu Met Gly Leu Ile
35 40 45
Asn Val Thr Leu Ser Glu Lys Gly Thr Leu Met Ala Thr Leu Pro Ala
50 55 60
Asn Val Pro Gly Asp Ile Pro Ala Ile Gly Phe Ile Ser His Val Asp
65 70 75 80
Thr Ser Pro Asp Cys Ser Gly Lys Asn Val Asn Pro Gln Ile Val Glu
85 90 95
Asn Tyr Arg Gly Gly Asp Ile Ala Leu Gly Ile Gly Asp Glu Val Leu
100 105 110
Ser Pro Val Met Phe Pro Val Leu His Gln Leu Leu Gly Gln Thr Leu
115 120 125
Ile Thr Thr Asp Gly Lys Thr Leu Leu Gly Ala Asp Asp Lys Ala Gly
130 135 140
Ile Ala Glu Ile Met Thr Ala Leu Ala Val Leu Gln Gln Lys Lys Ile
145 150 155 160
Pro His Gly Asp Ile Arg Val Ala Phe Thr Pro Asp Glu Glu Val Gly
165 170 175
Lys Gly Ala Lys His Phe Asp Val Asp Ala Phe Asp Ala Arg Trp Ala
180 185 190
Tyr Thr Val Asp Gly Gly Gly Val Gly Glu Leu Glu Phe Glu Asn Phe
195 200 205
Asn Ala Ala Ser Val Asn Ile Lys Ile Val Gly Asn Asn Val His Pro
210 215 220
Gly Thr Ala Lys Gly Val Met Val Asn Ala Leu Ser Leu Ala Ala Arg
225 230 235 240
Ile His Ala Glu Val Pro Ala Asp Glu Ser Pro Glu Met Thr Glu Gly
245 250 255
Tyr Glu Gly Phe Tyr His Leu Ala Ser Met Lys Gly Thr Val Glu Arg
260 265 270
Ala Asp Met His Tyr Ile Ile Arg Asp Phe Asp Arg Lys Gln Phe Glu
275 280 285
Ala Arg Lys Arg Lys Met Met Glu Ile Ala Lys Lys Val Gly Lys Gly
290 295 300
Leu His Pro Asp Cys Tyr Ile Glu Leu Val Ile Glu Asp Ser Tyr Tyr
305 310 315 320
Asn Met Arg Glu Lys Val Val Glu His Pro His Ile Leu Asp Ile Ala
325 330 335
Gln Gln Ala Met Arg Asp Cys Asp Ile Glu Pro Glu Leu Lys Pro Ile
340 345 350
Arg Gly Gly Thr Asp Gly Ala Gln Leu Ser Phe Met Gly Leu Pro Cys
355 360 365
Pro Asn Leu Phe Thr Gly Gly Tyr Asn Tyr His Gly Lys His Glu Phe
370 375 380
Val Thr Leu Glu Gly Met Glu Lys Ala Val Gln Val Ile Val Arg Ile
385 390 395 400
Ala Glu Leu Thr Ala Gln Arg Lys
405
<210> 4
<211> 1227
<212> DNA
<213> E.coli Acetylornithine deacetylase
<400> 4
atggataaac tacttgagcg atttttgaac tacgtgtctc tggataccca atcaaaagca 60
ggggtgagac aggttcccag cacggaaggc caatggaagt tattgcatct gctgaaagag 120
cagctcgaag agatggggct tatcaatgtg accttaagtg agaagggcac tttgatggcg 180
acgttaccgg ctaacgtccc tggcgatatc ccggcgattg gctttatttc tcatgtggat 240
acctcaccgg attgcagcgg caaaaatgtg aatccgcaaa ttgttgaaaa ctatcgcggt 300
ggcgatattg cgctgggtat cggcgatgaa gttttatcac cggttatgtt cccggtgctg 360
catcagctac tgggtcagac gctgattacc accgatggta aaaccttgtt aggtgccgat 420
gacaaagcag gtattgcaga aatcatgacc gcgctggcgg tattgcaaca gaaaaaaatt 480
ccgcatggtg atattcgcgt cgcctttacc ccggatgaag aagtgggcaa aggggcgaaa 540
cattttgatg ttgacgcctt cgatgcccgc tgggcttaca ctgttgatgg tggtggcgta 600
ggcgaactgg agtttgaaaa cttcaacgcc gcgtcggtca atatcaaaat tgtcggtaac 660
aatgttcatc cgggcacggc gaaaggagtg atggtaaatg cgctgtcgct ggcggcacgt 720
attcatgcgg aagttccggc ggatgaaagc ccggaaatga cagaaggcta tgaaggtttc 780
tatcatctgg cgagcatgaa aggcaccgtt gaacgggccg atatgcacta catcatccgt 840
gatttcgacc gtaaacagtt tgaagcgcgt aaacgtaaaa tgatggagat cgccaaaaaa 900
gtgggcaaag ggttacatcc tgattgctac attgaactgg tgattgaaga cagttactac 960
aatatgcgcg agaaagtggt tgagcatccg catattctcg atatcgccca gcaggcgatg 1020
cgcgattgcg atattgaacc ggaactgaaa ccgatccgcg gtggtaccga cggcgcgcag 1080
ttgtcgttta tgggattacc gtgcccgaac ctgttcactg gcggttacaa ctatcatggt 1140
aagcatgagt ttgtgactct ggaaggtatg gaaaaagcgg tgcaggtgat cgtccgtatt 1200
gccgagttaa cggcgcaacg gaagtaa 1227
<210> 5
<211> 858
<212> PRT
<213> Corynebacterium glutamicum ATCC 13032 pta-ackA
<400> 5
Met Ser Asp Thr Pro Thr Ser Ala Leu Ile Thr Thr Val Asn Arg Ser
1 5 10 15
Phe Asp Gly Phe Asp Leu Glu Glu Val Ala Ala Asp Leu Gly Val Arg
20 25 30
Leu Thr Tyr Leu Pro Asp Glu Glu Leu Glu Val Ser Lys Val Leu Ala
35 40 45
Ala Asp Leu Leu Ala Glu Gly Pro Ala Leu Ile Ile Gly Val Gly Asn
50 55 60
Thr Phe Phe Asp Ala Gln Val Ala Ala Ala Leu Gly Val Pro Val Leu
65 70 75 80
Leu Leu Val Asp Lys Gln Gly Lys His Val Ala Leu Ala Arg Thr Gln
85 90 95
Val Asn Asn Ala Gly Ala Val Val Ala Ala Ala Phe Thr Ala Glu Gln
100 105 110
Glu Pro Met Pro Asp Lys Leu Arg Lys Ala Val Arg Asn His Ser Asn
115 120 125
Leu Glu Pro Val Met Ser Ala Glu Leu Phe Glu Asn Trp Leu Leu Lys
130 135 140
Arg Ala Arg Ala Glu His Ser His Ile Val Leu Pro Glu Gly Asp Asp
145 150 155 160
Asp Arg Ile Leu Met Ala Ala His Gln Leu Leu Asp Gln Asp Ile Cys
165 170 175
Asp Ile Thr Ile Leu Gly Asp Pro Val Lys Ile Lys Glu Arg Ala Thr
180 185 190
Glu Leu Gly Leu His Leu Asn Thr Ala Tyr Leu Val Asn Pro Leu Thr
195 200 205
Asp Pro Arg Leu Glu Glu Phe Ala Glu Gln Phe Ala Glu Leu Arg Lys
210 215 220
Ser Lys Ser Val Thr Ile Asp Glu Ala Arg Glu Ile Met Lys Asp Ile
225 230 235 240
Ser Tyr Phe Gly Thr Met Met Val His Asn Gly Asp Ala Asp Gly Met
245 250 255
Val Ser Gly Ala Ala Asn Thr Thr Ala His Thr Ile Lys Pro Ser Phe
260 265 270
Gln Ile Ile Lys Thr Val Pro Glu Ala Ser Val Val Ser Ser Ile Phe
275 280 285
Leu Met Val Leu Arg Gly Arg Leu Trp Ala Phe Gly Asp Cys Ala Val
290 295 300
Asn Pro Asn Pro Thr Ala Glu Gln Leu Gly Glu Ile Ala Val Val Ser
305 310 315 320
Ala Lys Thr Ala Ala Gln Phe Gly Ile Asp Pro Arg Val Ala Ile Leu
325 330 335
Ser Tyr Ser Thr Gly Asn Ser Gly Gly Gly Ser Asp Val Asp Arg Ala
340 345 350
Ile Asp Ala Leu Ala Glu Ala Arg Arg Leu Asn Pro Glu Leu Cys Val
355 360 365
Asp Gly Pro Leu Gln Phe Asp Ala Ala Val Asp Pro Gly Val Ala Arg
370 375 380
Lys Lys Met Pro Asp Ser Asp Val Ala Gly Gln Ala Asn Val Phe Ile
385 390 395 400
Phe Pro Asp Leu Glu Ala Gly Asn Ile Gly Tyr Lys Thr Ala Gln Arg
405 410 415
Thr Gly His Ala Leu Ala Val Gly Pro Ile Leu Gln Gly Leu Asn Lys
420 425 430
Pro Val Asn Asp Leu Ser Arg Gly Ala Thr Val Pro Asp Ile Val Asn
435 440 445
Thr Val Ala Ile Thr Ala Ile Gln Ala Gly Gly Arg Ser Met Ala
450 455 460
Leu Ala Leu Val Leu Asn Ser Gly Ser Ser Ser Ile Lys Phe Gln Leu
465 470 475 480
Val Asn Pro Glu Asn Ser Ala Ile Asp Glu Pro Tyr Val Ser Gly Leu
485 490 495
Val Glu Gln Ile Gly Glu Pro Asn Gly Arg Ile Val Leu Lys Ile Glu
500 505 510
Gly Glu Lys Tyr Thr Leu Glu Thr Pro Ile Ala Asp His Ser Glu Gly
515 520 525
Leu Asn Leu Ala Phe Asp Leu Met Asp Gln His Asn Cys Gly Pro Ser
530 535 540
Gln Leu Glu Ile Thr Ala Val Gly His Arg Val Val His Gly Gly Ile
545 550 555 560
Leu Phe Ser Ala Pro Glu Leu Ile Thr Asp Glu Ile Val Glu Met Ile
565 570 575
Arg Asp Leu Ile Pro Leu Ala Pro Leu His Asn Pro Ala Asn Val Asp
580 585 590
Gly Ile Asp Val Ala Arg Lys Ile Leu Pro Asp Val Pro His Val Ala
595 600 605
Val Phe Asp Thr Gly Phe Phe His Ser Leu Pro Pro Ala Ala Ala Leu
610 615 620
Tyr Ala Ile Asn Lys Asp Val Ala Ala Glu His Gly Ile Arg Arg Tyr
625 630 635 640
Gly Phe His Gly Thr Ser His Glu Phe Val Ser Lys Arg Val Val Glu
645 650 655
Ile Leu Glu Lys Pro Thr Glu Asp Ile Asn Thr Ile Thr Phe His Leu
660 665 670
Gly Asn Gly Ala Ser Met Ala Ala Val Gln Gly Gly Arg Ala Val Asp
675 680 685
Thr Ser Met Gly Met Thr Pro Leu Ala Gly Leu Val Met Gly Thr Arg
690 695 700
Ser Gly Asp Ile Asp Pro Gly Ile Val Phe His Leu Ser Arg Thr Ala
705 710 715 720
Gly Met Ser Ile Asp Glu Ile Asp Asn Leu Leu Asn Lys Lys Ser Gly
725 730 735
Val Lys Gly Leu Ser Gly Val Asn Asp Phe Arg Glu Leu Arg Glu Met
740 745 750
Ile Asp Asn Asn Asp Gln Asp Ala Trp Ser Ala Tyr Asn Ile Tyr Ile
755 760 765
His Gln Leu Arg Arg Tyr Leu Gly Ser Tyr Met Val Ala Leu Gly Arg
770 775 780
Val Asp Thr Ile Val Phe Thr Ala Gly Val Gly Glu Asn Ala Gln Phe
785 790 795 800
Val Arg Glu Asp Ala Leu Ala Gly Leu Glu Met Tyr Gly Ile Glu Ile
805 810 815
Asp Pro Glu Arg Asn Ala Leu Pro Asn Asp Gly Pro Arg Leu Ile Ser
820 825 830
Thr Asp Ala Ser Lys Val Lys Val Phe Val Ile Pro Thr Asn Glu Glu
835 840 845
Leu Ala Ile Ala Arg Tyr Ala Val Lys Phe Ala
850 855
<210> 6
<211> 2579
<212> DNA
<213> Corynebacterium glutamicum ATCC 13032 pta-ackA
<400> 6
atgtctgaca caccgacctc agctctgatc accacggtca accgcagctt cgatggattc 60
gatttggaag aagtagcagc agaccttgga gttcggctca cctacctgcc cgacgaagaa 120
ctagaagtat ccaaagttct cgcggcggac ctcctcgctg aggggccagc tctcatcatc 180
ggtgtaggaa acacgttttt cgacgcccag gtcgccgctg ccctcggcgt cccagtgcta 240
ctgctggtag acaagcaagg caagcacgtt gctcttgctc gcacccaggt aaacaatgcc 300
ggcgcagttg ttgcagcagc atttaccgct gaacaagagc caatgccgga taagctgcgc 360
aaggctgtgc gcaaccacag caacctcgaa ccagtcatga gcgccgaact ctttgaaaac 420
tggctgctca agcgcgcacg cgcagagcac tcccacattg tgctgccaga aggtgacgac 480
gaccgcatct tgatggctgc ccaccagctg cttgatcaag acatctgtga catcacgatc 540
ctgggcgatc cagtaaagat caaggagcgc gctaccgaac ttggcctgca ccttaacact 600
gcatacctgg tcaatccgct gacagatcct cgcctggagg aattcgccga acaattcgcg 660
gagctgcgca agtcaaagag cgtcactatc gatgaagccc gcgaaatcat gaaggatatt 720
tcctacttcg gcaccatgat ggtccacaac ggcgacgccg acggaatggt atccggtgca 780
gcaaacacca ccgcacacac cattaagcca agcttccaga tcatcaaaac tgttccagaa 840
gcatccgtcg tttcttccat cttcctcatg gtgctgcgcg ggcgactgtg ggcattcggc 900
gactgtgctg ttaacccgaa cccaactgct gaacagcttg gtgaaatcgc cgttgtgtca 960
gcaaaaactg cagcacaatt tggcattgat cctcgcgtag ccatcttgtc ctactccact 1020
ggcaactccg gcggaggctc agatgtggat cgcgccatcg acgctcttgc agaagcacgc 1080
cgacttaacc cagaactatg cgtcgatgga ccacttcagt tcgacgccgc cgtcgacccg 1140
ggtgtggcgc gcaagaagat gccagactct gacgtcgctg gccaggcaaa tgtgtttatc 1200
ttccctgacc tggaagccgg aaacatcggc tacaaaactg cacaacgcac cggtcacgcc 1260
ctggcagttg gtccgattct gcagggccta aacaaaccag tcaacgacct ttcccgtggc 1320
gcaacagtcc ctgacatcgt caacacagta gccatcacag caattcaggc aggaggacgc 1380
agctaatggc attggcactt gttttgaact ccggttcatc ttccatcaaa ttccagctgg 1440
tcaaccccga aaactctgcc atcgacgagc catatgtttc tggtcttgtg gagcagattg 1500
gtgagccaaa cggccgcatc gtactcaaaa tagagggtga aaaatatacc ctagagacac 1560
ccatcgcaga tcactccgaa ggcctaaacc tggcgttcga tctcatggac cagcacaact 1620
gtggtccttc ccaactggaa atcaccgcag ttggacaccg cgtggtccac ggcggaatct 1680
tgttctccgc accggaactt atcactgatg aaatcgtgga aatgatccgc gatctcattc 1740
cactcgcacc actgcacaac cctgcaaacg ttgacggcat tgatgttgct cgaaaaattc 1800
tccccgatgt cccacacgta gctgtctttg acaccggttt cttccactca cttccaccag 1860
cagctgcgct gtatgccatc aacaaggatg tcgcagctga acacggaatc aggcgctatg 1920
gtttccacgg cacctcccat gaatttgtgt ccaagcgcgt ggtggaaatt ctggaaaagc 1980
ccaccgaaga catcaacacc atcaccttcc acctgggcaa cggcgcatcc atggctgctg 2040
ttcaaggtgg ccgtgcggta gatacttcca tgggtatgac acctctcgcg ggccttgtca 2100
tgggtacccg aagcggtgac attgatccag gtatcgtctt ccacctttcc cgcaccgctg 2160
gcatgagcat cgatgagatc gataatctgc tgaacaaaaa gtcgggtgta aagggacttt 2220
ccggtgttaa tgatttccgt gaactgcggg aaatgatcga caacaatgat caagatgcct 2280
ggtccgcgta caacatttac atacaccaac tccgccgcta cctcggttcc tacatggtgg 2340
cactgggacg ggtagacacc atcgtgttca ccgccggtgt cggtgaaaat gcccagtttg 2400
tccgtgagga tgccttggca ggtttggaaa tgtacggaat tgagatcgat ccagagcgta 2460
acgcattgcc aaacgatggt cctcgattga tttccaccga tgcctccaag gtgaaggtgt 2520
ttgttattcc aactaatgaa gagttagcta tcgctaggta cgcggtgaag ttcgcttag 2579
<210> 7
<211> 858
<212> PRT
<213> Corynebacterium glutamicum ATCC 13869 pta-ackA
<400> 7
Met Ser Asp Thr Pro Thr Ser Ala Leu Ile Thr Thr Val Asn Arg Ser
1 5 10 15
Phe Asp Gly Phe Asp Leu Glu Glu Val Ala Ala Asp Leu Gly Val Arg
20 25 30
Leu Thr Asp Leu Pro Asp Glu Glu Leu Glu Val Ser Lys Val Leu Ala
35 40 45
Ala Asp Leu Leu Ala Glu Gly Pro Ala Leu Ile Ile Gly Val Gly Asn
50 55 60
Thr Phe Phe Asp Ala Gln Val Ala Ala Ala Leu Gly Val Pro Val Leu
65 70 75 80
Leu Leu Val Asp Lys Gln Gly Lys His Val Ala Leu Ala Arg Thr Gln
85 90 95
Val Asn Asn Ala Gly Ala Val Val Ala Ala Ala Phe Thr Ala Glu Gln
100 105 110
Glu Pro Met Pro Asp Lys Leu Arg Lys Ala Val Arg Asn His Ser Asn
115 120 125
Leu Glu Pro Val Met Ser Ala Glu Leu Phe Glu Asn Trp Leu Leu Lys
130 135 140
Arg Ala Arg Ala Glu His Ser His Ile Val Leu Pro Glu Gly Asp Asp
145 150 155 160
Asp Arg Ile Leu Met Ala Ala His Gln Leu Leu Asp Gln Asp Ile Cys
165 170 175
Asp Ile Thr Ile Leu Gly Asp Pro Val Gln Ile Lys Glu Arg Ala Thr
180 185 190
Glu Leu Gly Leu His Leu Asn Thr Ala Tyr Leu Val Asn Pro Leu Thr
195 200 205
Asp Pro Arg Leu Glu Glu Phe Ala Glu Gln Phe Ala Glu Leu Arg Lys
210 215 220
Ser Lys Ser Val Thr Ile Asp Glu Ala Arg Glu Ile Met Lys Asp Ile
225 230 235 240
Cys Tyr Phe Gly Thr Met Met Val His Asn Gly Asp Ala Asp Gly Met
245 250 255
Val Ser Gly Ala Ala Asn Thr Thr Ala His Thr Ile Lys Pro Ser Phe
260 265 270
Gln Ile Ile Lys Thr Val Pro Glu Ala Ser Val Val Ser Ser Ile Phe
275 280 285
Leu Met Val Leu Arg Gly Arg Leu Trp Ala Phe Gly Asp Cys Ala Val
290 295 300
Asn Pro Asn Pro Thr Ala Glu Gln Leu Gly Glu Ile Ala Val Val Ser
305 310 315 320
Ala Lys Thr Ala Ala Gln Phe Gly Ile Asp Pro Arg Val Ala Ile Leu
325 330 335
Ser Tyr Ser Thr Gly Asn Ser Gly Gly Gly Ser Asp Val Asp Arg Ala
340 345 350
Ile Asp Ala Leu Ala Glu Ala Arg Arg Leu Asn Pro Glu Leu Cys Val
355 360 365
Asp Gly Pro Leu Gln Phe Asp Ala Ala Val Asp Pro Gly Val Ala Arg
370 375 380
Lys Lys Met Pro Asp Ser Asp Val Ala Gly Gln Ala Asn Val Phe Ile
385 390 395 400
Phe Pro Asp Leu Glu Ala Gly Asn Ile Gly Tyr Lys Thr Ala Gln Arg
405 410 415
Thr Gly His Ala Leu Ala Val Gly Pro Ile Leu Gln Gly Leu Asn Lys
420 425 430
Pro Val Asn Asp Leu Ser Arg Gly Ala Thr Val Pro Asp Ile Val Asn
435 440 445
Thr Val Ala Ile Thr Ala Ile Gln Ala Gly Gly Arg Ser Met Ala
450 455 460
Leu Ala Leu Val Leu Asn Ser Gly Ser Ser Ser Ile Lys Phe Gln Leu
465 470 475 480
Val Asn Pro Glu Asn Ser Ala Ile Asp Glu Pro Tyr Val Ser Gly Leu
485 490 495
Val Glu Gln Ile Gly Glu Pro Asn Gly Arg Ile Val Leu Lys Val Glu
500 505 510
Gly Glu Lys Tyr Thr Leu Glu Thr Pro Ile Ala Asp His Ser Glu Gly
515 520 525
Leu Asn Leu Ala Phe Asp Leu Met Asp Gln His Asn Cys Gly Pro Ser
530 535 540
Gln Leu Glu Ile Thr Ala Val Gly His Arg Val Val His Gly Gly Ile
545 550 555 560
Leu Phe Ser Ala Pro Glu Leu Ile Thr Asp Glu Ile Val Glu Met Ile
565 570 575
Arg Asp Leu Ile Pro Leu Ala Pro Leu His Asn Pro Ala Asn Val Asp
580 585 590
Gly Ile Asp Val Ala Arg Lys Ile Leu Pro Asp Val Pro His Val Ala
595 600 605
Val Phe Asp Thr Gly Phe Phe His Ser Leu Pro Pro Ala Ala Ala Leu
610 615 620
Tyr Ala Ile Asn Lys Asp Val Ala Ala Glu His Gly Ile Arg Arg Tyr
625 630 635 640
Gly Phe His Gly Thr Ser His Glu Phe Val Ser Lys Arg Val Val Glu
645 650 655
Ile Leu Glu Lys Pro Thr Glu Asp Ile Asn Thr Ile Thr Phe His Leu
660 665 670
Gly Asn Gly Ala Ser Met Ala Ala Val Gln Gly Gly Arg Ala Val Asp
675 680 685
Thr Ser Met Gly Met Thr Pro Leu Ala Gly Leu Val Met Gly Thr Arg
690 695 700
Ser Gly Asp Ile Asp Pro Gly Val Val Phe His Leu Ser Arg Thr Ala
705 710 715 720
Gly Met Ser Ile Asp Glu Ile Asp Asn Leu Leu Asn Lys Lys Ser Gly
725 730 735
Val Lys Gly Leu Ser Gly Val Asn Asp Phe Arg Glu Leu Arg Glu Met
740 745 750
Ile Asp Asn Asn Asp Gln Asp Ala Trp Ser Ala Tyr Asn Ile Tyr Ile
755 760 765
His Gln Leu Arg Arg Tyr Leu Gly Ser Tyr Met Val Ala Leu Gly Arg
770 775 780
Val Asp Thr Ile Val Phe Thr Ala Gly Val Gly Glu Asn Ala Gln Phe
785 790 795 800
Val Arg Glu Asp Ala Leu Ala Gly Leu Glu Met Tyr Gly Ile Glu Ile
805 810 815
Asp Pro Glu Arg Asn Ala Leu Pro Asn Asp Gly Pro Arg Leu Ile Ser
820 825 830
Thr Asp Ala Ser Lys Val Lys Val Phe Val Ile Pro Thr Asn Glu Glu
835 840 845
Leu Ala Ile Ala Arg Tyr Ala Val Lys Phe Ala
850 855
<210> 8
<211> 2579
<212> DNA
<213> Corynebacterium glutamicum ATCC 13869 pta-ackA
<400> 8
atgtctgaca caccgacctc agctctgatc accacggtca accgcagctt cgatggattc 60
gatttggaag aagtagcagc agaccttgga gttcggctca ccgacctgcc cgacgaagaa 120
ctagaagtat ccaaagttct cgcggcggac ctcctcgctg aggggccagc tctcatcatc 180
ggtgtaggaa acacgttttt cgacgcccag gtcgccgctg ccctcggcgt cccagtgcta 240
ctgctggtag acaagcaagg caagcacgtt gctcttgctc gcacccaggt aaacaatgcc 300
ggcgcagttg ttgcagcagc atttaccgct gaacaagagc caatgccgga taagctgcgc 360
aaggctgtgc gcaaccacag caacctcgaa ccagtcatga gcgccgaact ctttgaaaac 420
tggctgctca agcgcgcacg cgcagagcac tcccacattg tgctgccaga aggtgacgac 480
gaccgcatct tgatggctgc ccaccagctg cttgatcaag acatctgtga catcacgatc 540
ctgggcgatc cagtacagat caaggagcgc gctaccgaac ttggcctgca ccttaacact 600
gcatacctgg tcaatccgct gacagatcct cgcctggagg aattcgccga acaattcgcg 660
gagctgcgca agtcaaagag cgtcactatc gatgaagccc gcgaaatcat gaaggatatt 720
tgctacttcg gcaccatgat ggtccacaac ggcgacgccg acggaatggt atccggtgca 780
gcaaacacca ccgcacacac cattaagcca agcttccaga tcatcaaaac tgttccagaa 840
gcatccgtcg tttcttccat cttcctcatg gtgctgcgcg ggcgactgtg ggcattcggc 900
gactgtgctg ttaacccgaa cccaactgct gaacagcttg gtgaaatcgc cgttgtgtca 960
gcaaaaactg cagcacaatt tggcattgat cctcgcgtag ccatcttgtc ctactccact 1020
ggcaactccg gcggaggctc agatgtggat cgcgccatcg acgctcttgc agaagcacgc 1080
cgactcaacc cagaactatg cgtcgatgga ccacttcagt tcgacgccgc cgtcgacccg 1140
ggtgtggcgc gcaagaagat gccagactct gacgtcgctg gccaggcaaa tgtgtttatc 1200
ttccctgacc tggaagccgg aaacatcggc tacaaaactg cacaacgcac cggtcacgcc 1260
ctggcagttg gtccgattct gcagggcctg aacaaaccag tcaacgacct ttcccgtggc 1320
gcaacagtcc ctgacatcgt caacacagta gccatcaccg caattcaggc aggaggacgc 1380
agctaatgg cattggcact tgttttgaac tccggttcat cttccatcaa attccagctg 1440
gtcaaccccg aaaactctgc catcgacgag ccatatgttt ctggtcttgt ggagcagatt 1500
ggtgagccaa acggccgcat cgtactcaaa gtagagggtg aaaaatacac cctagagaca 1560
cccatcgcag atcactccga aggcctaaac ctggcgttcg atctcatgga ccagcacaac 1620
tgtggtcctt cccaactgga aatcaccgca gttggacacc gcgtggtcca cggtggaatc 1680
ttgttctctg cgccggaact catcactgat gaaatcgttg aaatgatccg cgatctcatt 1740
ccactcgcac cactgcacaa ccctgcaaac gttgacggca ttgatgttgc tcgaaaaatt 1800
ctccccgatg tcccacacgt agctgtcttt gacaccggtt tcttccactc acttccacca 1860
gcagctgcac tgtatgccat caacaaggat gtcgcagctg aacacggaat caggcgctat 1920
ggtttccacg gtacctccca cgaatttgtg tccaagcgcg tggtggaaat tttggaaaag 1980
cccaccgaag acatcaacac catcaccttc cacctgggca acggcgcatc catggctgct 2040
gttcaaggcg gccgtgcggt agatacttcc atgggtatga cacctctcgc gggacttgtc 2100
atgggtaccc gaagcggtga cattgatcca ggtgtcgttt tccatctctc acgcaccgct 2160
ggcatgagca tcgatgagat cgataatctg ctgaacaaaa agtcgggtgt aaagggactt 2220
tccggagtca atgatttccg tgaactgcgg gaaatgatcg acaacaatga tcaagatgcc 2280
tggtccgcgt acaacattta catacaccaa ctccgccgct acctcggttc ctacatggtg 2340
gcactgggac gggtagacac catcgtgttc accgccggtg ttggtgaaaa tgcccagttt 2400
gtccgtgagg atgccttggc aggtttggaa atgtacggca tcgaaatcga tccggagcgc 2460
aacgcactgc caaacgatgg tcctagattg atttccaccg atgcctccaa ggtgaaggtg 2520
tttgttattc caactaatga agagttggct atcgctaggt acgcggtgaa gttcgcttag 2580
2580
<210> 9
<211> 652
<212> PRT
<213> E.coli acetyl-CoA synthetase
<400> 9
Met Ser Gln Ile His Lys His Thr Ile Pro Ala Asn Ile Ala Asp Arg
1 5 10 15
Cys Leu Ile Asn Pro Gln Gln Tyr Glu Ala Met Tyr Gln Gln Ser Ile
20 25 30
Asn Val Pro Asp Thr Phe Trp Gly Glu Gln Gly Lys Ile Leu Asp Trp
35 40 45
Ile Lys Pro Tyr Gln Lys Val Lys Asn Thr Ser Phe Ala Pro Gly Asn
50 55 60
Val Ser Ile Lys Trp Tyr Glu Asp Gly Thr Leu Asn Leu Ala Ala Asn
65 70 75 80
Cys Leu Asp Arg His Leu Gln Glu Asn Gly Asp Arg Thr Ala Ile Ile
85 90 95
Trp Glu Gly Asp Asp Ala Ser Gln Ser Lys His Ile Ser Tyr Lys Glu
100 105 110
Leu His Arg Asp Val Cys Arg Phe Ala Asn Thr Leu Leu Glu Leu Gly
115 120 125
Ile Lys Lys Gly Asp Val Val Ala Ile Tyr Met Pro Met Val Pro Glu
130 135 140
Ala Ala Val Ala Met Leu Ala Cys Ala Arg Ile Gly Ala Val His Ser
145 150 155 160
Val Ile Phe Gly Gly Phe Ser Pro Glu Ala Val Ala Gly Arg Ile Ile
165 170 175
Asp Ser Asn Ser Arg Leu Val Ile Thr Ser Asp Glu Gly Val Arg Ala
180 185 190
Gly Arg Ser Ile Pro Leu Lys Lys Asn Val Asp Asp Ala Leu Lys Asn
195 200 205
Pro Asn Val Thr Ser Val Glu His Val Val Val Leu Lys Arg Thr Gly
210 215 220
Gly Lys Ile Asp Trp Gln Glu Gly Arg Asp Leu Trp Trp His Asp Leu
225 230 235 240
Val Glu Gln Ala Ser Asp Gln His Gln Ala Glu Glu Met Asn Ala Glu
245 250 255
Asp Pro Leu Phe Ile Leu Tyr Thr Ser Gly Ser Thr Gly Lys Pro Lys
260 265 270
Gly Val Leu His Thr Thr Gly Gly Tyr Leu Val Tyr Ala Ala Leu Thr
275 280 285
Phe Lys Tyr Val Phe Asp Tyr His Pro Gly Asp Ile Tyr Trp Cys Thr
290 295 300
Ala Asp Val Gly Trp Val Thr Gly His Ser Tyr Leu Leu Tyr Gly Pro
305 310 315 320
Leu Ala Cys Gly Ala Thr Thr Leu Met Phe Glu Gly Val Pro Asn Trp
325 330 335
Pro Thr Pro Ala Arg Met Ala Gln Val Val Asp Lys His Gln Val Asn
340 345 350
Ile Leu Tyr Thr Ala Pro Thr Ala Ile Arg Ala Leu Met Ala Glu Gly
355 360 365
Asp Lys Ala Ile Glu Gly Thr Asp Arg Ser Ser Leu Arg Ile Leu Gly
370 375 380
Ser Val Gly Glu Pro Ile Asn Pro Glu Ala Trp Glu Trp Tyr Trp Lys
385 390 395 400
Lys Ile Gly Asn Glu Lys Cys Pro Val Val Asp Thr Trp Trp Gln Thr
405 410 415
Glu Thr Gly Gly Phe Met Ile Thr Pro Leu Pro Gly Ala Thr Glu Leu
420 425 430
Lys Ala Gly Ser Ala Thr Arg Pro Phe Phe Gly Val Gln Pro Ala Leu
435 440 445
Val Asp Asn Glu Gly Asn Pro Leu Glu Gly Ala Thr Glu Gly Ser Leu
450 455 460
Val Ile Thr Asp Ser Trp Pro Gly Gln Ala Arg Thr Leu Phe Gly Asp
465 470 475 480
His Glu Arg Phe Glu Gln Thr Tyr Phe Ser Thr Phe Lys Asn Met Tyr
485 490 495
Phe Ser Gly Asp Gly Ala Arg Arg Asp Glu Asp Gly Tyr Tyr Trp Ile
500 505 510
Thr Gly Arg Val Asp Asp Val Leu Asn Val Ser Gly His Arg Leu Gly
515 520 525
Thr Ala Glu Ile Glu Ser Ala Leu Val Ala His Pro Lys Ile Ala Glu
530 535 540
Ala Ala Val Val Gly Ile Pro His Asn Ile Lys Gly Gln Ala Ile Tyr
545 550 555 560
Ala Tyr Val Thr Leu Asn His Gly Glu Glu Pro Ser Pro Glu Leu Tyr
565 570 575
Ala Glu Val Arg Asn Trp Val Arg Lys Glu Ile Gly Pro Leu Ala Thr
580 585 590
Pro Asp Val Leu His Trp Thr Asp Ser Leu Pro Lys Thr Arg Ser Gly
595 600 605
Lys Ile Met Arg Arg Ile Leu Arg Lys Ile Ala Ala Gly Asp Thr Ser
610 615 620
Asn Leu Gly Asp Thr Ser Thr Leu Ala Asp Pro Gly Val Val Glu Lys
625 630 635 640
Leu Leu Glu Glu Lys Gln Ala Ile Ala Met Pro Ser
645 650
<210> 10
<211> 1959
<212> DNA
<213> E.coli acetyl-CoA synthetase
<400> 10
atgagccaaa ttcacaaaca caccattcct gccaacatcg cagaccgttg cctgataaac 60
cctcagcagt acgaggcgat gtatcaacaa tctattaacg tacctgatac cttctggggc 120
gaacagggaa aaattcttga ctggatcaaa ccttaccaga aggtgaaaaa cacctccttt 180
gcccccggta atgtgtccat taaatggtac gaggacggca cgctgaatct ggcggcaaac 240
tgccttgacc gccatctgca agaaaacggc gatcgtaccg ccatcatctg ggaaggcgac 300
gacgccagcc agagcaaaca tatcagctat aaagagctgc accgcgacgt ctgccgcttc 360
gccaataccc tgctcgagct gggcattaaa aaaggtgatg tggtggcgat ttatatgccg 420
atggtgccgg aagccgcggt tgcgatgctg gcctgcgccc gcattggcgc ggtgcattcg 480
gtgattttcg gcggcttctc gccggaagcc gttgccgggc gcattattga ttccaactca 540
cgactggtga tcacttccga cgaaggtgtg cgtgccgggc gcagtattcc gctgaagaaa 600
aacgttgatg acgcgctgaa aaacccgaac gtcaccagcg tagagcatgt ggtggtactg 660
aagcgtactg gcgggaaaat tgactggcag gaagggcgcg acctgtggtg gcacgacctg 720
gttgagcaag cgagcgatca gcaccaggcg gaagagatga acgccgaaga tccgctgttt 780
attctctaca cctccggttc taccggtaag ccaaaaggtg tgctgcatac taccggcggt 840
tatctggtgt acgcggcgct gacctttaaa tatgtctttg attatcatcc gggtgatatc 900
tactggtgca ccgccgatgt gggctgggtg accggacaca gttacttgct gtacggcccg 960
ctggcctgcg gtgcgaccac gctgatgttt gaaggcgtac ccaactggcc gacgcctgcc 1020
cgtatggcgc aggtggtgga caagcatcag gtcaatattc tctataccgc acccacggcg 1080
atccgcgcgc tgatggcgga aggcgataaa gcgatcgaag gcaccgaccg ttcgtcgctg 1140
cgcattctcg gttccgtggg cgagccaatt aacccggaag cgtgggagtg gtactggaaa 1200
aaaatcggca acgagaaatg tccggtggtc gatacctggt ggcagaccga aaccggcggt 1260
ttcatgatca ccccgctgcc tggcgctacc gagctgaaag ccggttcggc aacacgtccg 1320
ttcttcggcg tgcaaccggc gctggtcgat aacgaaggta acccgctgga gggggccacc 1380
gaaggtagcc tggtaatcac cgactcctgg ccgggtcagg cgcgtacgct gtttggcgat 1440
cacgaacgtt ttgaacagac ctacttctcc accttcaaaa atatgtattt cagcggcgac 1500
ggcgcgcgtc gcgatgaaga tggctattac tggataaccg ggcgtgtgga cgacgtgctg 1560
aacgtctccg gtcaccgtct ggggacggca gagattgagt cggcgctggt ggcgcatccg 1620
aagattgccg aagccgccgt agtaggtatt ccgcacaata ttaaaggtca ggcgatctac 1680
gcctacgtca cgcttaatca cggggaggaa ccgtcaccag aactgtacgc agaagtccgc 1740
aactgggtgc gtaaagagat tggcccgctg gcgacgccag acgtgctgca ctggaccgac 1800
tccctgccta aaacccgctc cggcaaaatt atgcgccgta ttctgcgcaa aattgcggcg 1860
ggcgatacca gcaacctggg cgatacctcg acgcttgccg atcctggcgt agtcgagaag 1920
ctgcttgaag agaagcaggc tatcgcgatg ccatcgtaa 1959
<210> 11
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> PlysC-argA-F
<400> 11
gaaaggtgca caaagatggt aaaggaacgt aaaaccg 37
<210> 12
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-argA-RXh
<400> 12
gcccactagt ctcgagcatg cggcgttgat tttg 34
<210> 13
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-PlysC-FXh
<400> 13
gaatgagttc ctcgagccga tgctagggcg aaaa 34
<210> 14
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> PlysC-R
<400> 14
ctttgtgcac ctttcgatct acgtgctgac agttac 36
<210> 15
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PlysC-argE-F
<400> 15
gaaaggtgca caaagatgaa aaacaaatta ccgcc 35
<210> 16
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-argE-RXh
<400> 16
gcccactagt ctcgaggttt gagtcactgt cggtcg 36
<210> 17
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> Pro-pta-FX
<400> 17
ccggggatcc tctagagggg ttctaaaaaa tgtggagt 38
<210> 18
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> pta-PlysC-R
<400> 18
gccgtgcttt tcgccctagc atcggacatc gcctttctaa ttt 43
<210> 19
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> PlysC-F
<400> 19
ccgatgctag ggcgaaaagc acggc 25
<210> 20
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> PlysC-pta-ackA-F
<400> 20
gaaaggtgca caaagatgtc tgacacaccg acctcagctc 40
<210> 21
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Pro-pta-RX
<400> 21
gcaggtcgac tctagattat ccggcattgg ctct 34
<210> 22
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Pta-ackA-R
<400> 22
tgcagtttca ccccttaa 18
<210> 23
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> 13869_pta-PlysC-R
<400> 23
gccgtgcttt tcgccctagc atcggacatc gcctttctag ttt 43
<210> 24
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> PlysC-acs-F
<400> 24
gaaaggtgca caaagatgag ccaaattcac aaa 33
<210> 25
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-acs-RXh
<400> 25
gcccactagt ctcgagaagg cgtttacgcc gcatcc 36
<210> 26
<211> 494
<212> PRT
<213> Corynebacterium glutamicum ATCC 13032 putrescine exporter
<400> 26
Met Thr Ser Glu Thr Leu Gln Ala Gln Ala Pro Thr Lys Thr Gln Arg
1 5 10 15
Trp Ala Phe Leu Ala Val Ile Ser Gly Gly Leu Phe Leu Ile Gly Val
20 25 30
Asp Asn Ser Ile Leu Tyr Thr Ala Leu Pro Leu Leu Arg Glu Gln Leu
35 40 45
Ala Ala Thr Glu Thr Gln Ala Leu Trp Ile Ile Asn Ala Tyr Pro Leu
50 55 60
Leu Met Ala Gly Leu Leu Leu Gly Thr Gly Thr Leu Gly Asp Lys Ile
65 70 75 80
Gly His Arg Arg Met Phe Leu Met Gly Leu Ser Ile Phe Gly Ile Ala
85 90 95
Ser Leu Gly Ala Ala Phe Ala Pro Thr Ala Trp Ala Leu Val Ala Ala
100 105 110
Arg Ala Phe Leu Gly Ile Gly Ala Ala Thr Met Met Pro Ala Thr Leu
115 120 125
Ala Leu Ile Arg Ile Thr Phe Glu Asp Glu Arg Glu Arg Asn Thr Ala
130 135 140
Ile Gly Ile Trp Gly Ser Val Ala Ile Leu Gly Ala Ala Ala Gly Pro
145 150 155 160
Ile Ile Gly Gly Ala Leu Leu Glu Phe Phe Trp Trp Gly Ser Val Phe
165 170 175
Leu Ile Asn Val Pro Val Ala Val Ile Ala Leu Ile Ala Thr Leu Phe
180 185 190
Val Ala Pro Ala Asn Ile Ala Asn Pro Ser Lys His Trp Asp Phe Leu
195 200 205
Ser Ser Phe Tyr Ala Leu Leu Thr Leu Ala Gly Leu Ile Ile Thr Ile
210 215 220
Lys Glu Ser Val Asn Thr Ala Arg His Met Pro Leu Leu Leu Gly Ala
225 230 235 240
Val Ile Met Leu Ile Ile Gly Ala Val Leu Phe Ser Ser Arg Gln Lys
245 250 255
Lys Ile Glu Glu Pro Leu Leu Asp Leu Ser Leu Phe Arg Asn Arg Leu
260 265 270
Phe Leu Gly Gly Val Val Ala Ala Gly Met Ala Met Phe Thr Val Ser
275 280 285
Gly Leu Glu Met Thr Thr Ser Gln Arg Phe Gln Leu Ser Val Gly Phe
290 295 300
Thr Pro Leu Glu Ala Gly Leu Leu Met Ile Pro Ala Ala Leu Gly Ser
305 310 315 320
Phe Pro Met Ser Ile Ile Gly Gly Ala Asn Leu His Arg Trp Gly Phe
325 330 335
Lys Pro Leu Ile Ser Gly Gly Phe Ala Ala Thr Ala Val Gly Ile Ala
340 345 350
Leu Cys Ile Trp Gly Ala Thr His Thr Asp Gly Leu Pro Phe Phe Ile
355 360 365
Ala Gly Leu Phe Phe Met Gly Ala Gly Ala Gly Ser Val Met Ser Val
370 375 380
Ser Ser Thr Ala Ile Ile Gly Ser Ala Pro Val Arg Lys Ala Gly Met
385 390 395 400
Ala Ser Ser Ile Glu Glu Val Ser Tyr Glu Phe Gly Thr Leu Leu Ser
405 410 415
Val Ala Ile Leu Gly Ser Leu Phe Pro Phe Phe Tyr Ser Leu His Ala
420 425 430
Pro Ala Glu Val Ala Asp Asn Phe Ser Ala Gly Val His His Ala Ile
435 440 445
Asp Gly Asp Ala Ala Arg Ala Ser Leu Asp Thr Ala Tyr Ile Asn Val
450 455 460
Leu Ile Ile Ala Leu Val Cys Ala Val Ala Ala Ala Leu Ile Ser Ser
465 470 475 480
Tyr Leu Phe Arg Gly Asn Pro Lys Gly Ala Asn Asn Ala His
485 490
<210> 27
<211> 1485
<212> DNA
<213> Corynebacterium glutamicum ATCC 13032 putrescine exporter
<400> 27
atgacttcag aaaccttaca ggcgcaagcg cctacgaaaa cccaacgttg ggctttcctc 60
gccgttatca gcggtggtct ctttctgatc ggtgtagaca actcgattct ctacaccgca 120
ctccctctgc tgcgtgaaca gctcgcagcc accgaaaccc aagcgttgtg gatcatcaac 180
gcatatcccc tgctcatggc gggccttctt ttgggtaccg gcactttggg tgacaaaatc 240
ggccaccgcc ggatgttcct catgggcttg agcattttcg gaatcgcttc acttggtgct 300
gcgtttgctc caactgcgtg ggctcttgtt gctgcgagag ctttccttgg catcggtgcg 360
gcaacgatga tgcctgcaac cttggctctg atccgcatta cgtttgagga tgagcgtgag 420
cgcaacactg caattggtat ttggggttcc gtggcaattc ttggcgctgc ggcaggcccg 480
atcattggtg gtgcgctgtt ggaattcttc tggtggggtt cggttttcct cattaacgtt 540
ccggtggctg ttatcgcgtt gatcgctacg ctttttgtgg cgccggccaa tatcgcgaat 600
ccgtctaagc attgggattt cttgtcgtcg ttctatgcgc tgctcacact tgctgggttg 660
atcatcacga tcaaggaatc tgtgaatact gcacgccata tgcctcttct tttgggtgca 720
gtcatcatgt tgatcattgg tgcggtgttg tttagcagtc gtcagaagaa gatcgaggag 780
ccacttctag atctgtcgtt gttccgtaat cgccttttct taggcggtgt ggttgctgcg 840
ggcatggcga tgtttactgt gtccggtttg gaaatgacta cctcgcagcg tttccagttg 900
tctgtgggtt tcactccact tgaggctggt ttgctcatga tcccagctgc attgggtagc 960
ttcccgatgt ctattatcgg tggtgcaaac ctgcatcgtt ggggcttcaa accgctgatc 1020
agtggtggtt ttgctgccac tgccgttggc atcgccctgt gtatttgggg cgcgactcat 1080
actgatggtt tgccgttttt catcgcgggt ctattcttca tgggcgcggg tgctggttcg 1140
gtaatgtctg tgtcttccac tgcgattatc ggttccgcgc cggtgcgtaa ggctggcatg 1200
gcgtcgtcga tcgaagaggt ctcttatgag ttcggcacgc tgttgtctgt cgcgattttg 1260
ggtagcttgt tcccattctt ctactcgctg catgccccgg cagaggttgc ggataacttc 1320
tcggcgggtg ttcaccacgc gattgatggc gatgcggcgc gtgcatcttt ggacaccgca 1380
tacattaacg tgttgatcat tgccctagta tgcgcagtag cggctgctct gatcagcagt 1440
taccttttcc gcggaaatcc gaagggagcc aataatgcgc actag 1485
<210> 28
<211> 494
<212> PRT
<213> Corynebacterium glutamicum ATCC 13869 putrescine exporter
<400> 28
Met Ile Ser Glu Thr Leu Gln Ala Gln Ala Pro Thr Lys Thr Gln Arg
1 5 10 15
Trp Ala Phe Leu Ala Val Ile Ser Gly Gly Leu Phe Leu Ile Gly Val
20 25 30
Asp Asn Ser Ile Leu Tyr Thr Ala Leu Pro Leu Leu Arg Glu Gln Leu
35 40 45
Ala Ala Thr Glu Thr Gln Ala Leu Trp Ile Ile Asn Ala Tyr Pro Leu
50 55 60
Leu Met Ala Gly Leu Leu Leu Gly Thr Gly Thr Leu Gly Asp Lys Ile
65 70 75 80
Gly His Arg Arg Met Phe Leu Met Gly Leu Ser Ile Phe Gly Ile Ala
85 90 95
Ser Leu Gly Ala Ala Phe Ala Pro Thr Ala Trp Ala Leu Val Ala Ala
100 105 110
Arg Ala Phe Leu Gly Ile Gly Ala Ala Thr Met Met Pro Ala Thr Leu
115 120 125
Ala Leu Ile Arg Ile Thr Phe Glu Asp Glu Arg Glu Arg Asn Thr Ala
130 135 140
Ile Gly Ile Trp Gly Ser Val Ala Ile Leu Gly Ala Ala Ala Gly Pro
145 150 155 160
Ile Ile Gly Gly Ala Leu Leu Glu Phe Phe Trp Trp Gly Ser Val Phe
165 170 175
Leu Ile Asn Val Pro Val Ala Val Ile Ala Leu Ile Ala Thr Leu Phe
180 185 190
Val Ala Pro Ala Asn Ile Ala Asn Pro Ser Lys His Trp Asp Phe Leu
195 200 205
Ser Ser Phe Tyr Ala Leu Leu Thr Leu Ala Gly Leu Ile Val Thr Ile
210 215 220
Lys Glu Ser Val Asn Thr Ala Arg His Leu Pro Leu Leu Val Gly Ala
225 230 235 240
Ile Ile Leu Leu Ile Ile Gly Ala Val Leu Phe Ser Ser Arg Gln Lys
245 250 255
Lys Ile Glu Glu Pro Leu Leu Asp Leu Ser Leu Phe Arg Asn Arg Leu
260 265 270
Phe Leu Gly Gly Val Val Ala Ala Gly Met Ala Met Phe Thr Val Ser
275 280 285
Gly Leu Glu Met Thr Thr Ser Gln Arg Phe Gln Leu Ser Val Gly Phe
290 295 300
Thr Pro Leu Glu Ala Gly Leu Leu Met Ile Pro Ala Ala Leu Gly Ser
305 310 315 320
Phe Pro Met Ser Ile Ile Gly Gly Ala Asn Leu His Arg Trp Gly Phe
325 330 335
Lys Pro Leu Ile Ser Gly Gly Phe Leu Ala Thr Ala Val Gly Ile Ala
340 345 350
Leu Cys Ile Trp Gly Ala Thr His Thr Asp Gly Leu Pro Phe Phe Ile
355 360 365
Ala Gly Leu Phe Phe Met Gly Ala Gly Ala Gly Ser Val Met Ser Val
370 375 380
Ser Ser Thr Ala Ile Ile Gly Ser Ala Pro Val Arg Lys Ala Gly Met
385 390 395 400
Ala Ser Ser Ile Glu Glu Val Ser Tyr Glu Phe Gly Thr Leu Leu Ser
405 410 415
Val Ala Ile Leu Gly Ser Leu Phe Pro Phe Phe Tyr Ser Leu His Ala
420 425 430
Pro Ala Glu Val Ala Asp Asn Phe Ser Ala Gly Val His His Ala Ile
435 440 445
Tyr Gly Asp Ala Ala Arg Ala Ser Leu Asp Thr Ala Tyr Ile Asn Val
450 455 460
Leu Ile Ile Ala Leu Val Cys Ala Val Ala Ala Ala Leu Ile Ser Ser
465 470 475 480
Tyr Leu Phe Arg Gly Asn Pro Lys Gly Ala Asn Asn Ala His
485 490
<210> 29
<211> 1485
<212> DNA
<213> Corynebacterium glutamicum ATCC 13869 putrescine exporter
<400> 29
atgatttcag aaactttgca ggcgcaagcg cctacgaaaa cccaacgttg ggctttcctc 60
gctgttatca gcggtggtct ctttctgatc ggtgtagaca actcaatcct ctacaccgca 120
ctccccctgc tgcgtgaaca actcgcagcc actgaaaccc aagcgttgtg gatcatcaac 180
gcatatcccc tgctcatggc gggtcttctt ttgggtaccg gcactttggg tgacaaaatc 240
ggccaccgcc ggatgttcct catgggcttg agcattttcg gaatcgcttc acttggcgct 300
gcgtttgctc caactgcgtg ggctcttgtt gctgcgagag ctttccttgg catcggtgcg 360
gcgacgatga tgcccgcaac cttggctctg atccgcatta cgtttgaaga tgaacgcgaa 420
cggaacaccg cgattggcat ttggggttct gtggcaattc ttggcgcggc ggcaggtccg 480
atcattggtg gtgcgctgtt ggaattcttc tggtggggtt cggttttcct cattaacgtt 540
ccggtggctg ttatcgcgtt gatcgctacg ctttttgtgg cgccggccaa tatcgcgaat 600
ccgtccaagc actgggattt cttatcctcg ttctatgcat tgcttaccct tgcaggtttg 660
attgtcacca tcaaagaatc ggtaaacact gcacgtcatc tgccactgct tgtaggtgcc 720
atcatcttgc ttatcattgg tgcggtgttg tttagcagtc gtcagaagaa gatcgaggag 780
ccacttctag atctgtcgtt gttccgtaat cgccttttct taggcggtgt ggttgctgcg 840
ggcatggcga tgtttactgt gtccggtttg gaaatgacta cctcgcagcg tttccagttg 900
tctgtgggtt tcactccact tgaggctggt ttgctcatga tcccagctgc attgggtagc 960
ttcccgatgt ctattatcgg tggtgcaaac ttgcatcgtt ggggcttcaa accgctgatc 1020
agtggtggtt tccttgccac ggcagtcggc atcgccctgt gtatttgggg cgcgactcat 1080
actgatggtt tgccgttttt catcgcgggt ctgttcttca tgggcgcggg tgctggttcg 1140
gtaatgtctg tgtcttccac tgcgattatc ggttccgcgc cggtgcgtaa ggctggcatg 1200
gcgtcgtcga tcgaagaggt ctcttatgag ttcggcacgc tgttgtctgt cgcgattttg 1260
ggtagcttgt tcccattctt ctactcgctg catgccccgg cagaggttgc ggataacttc 1320
tcggcgggtg ttcaccacgc gatttatggc gatgcggcgc gtgcatcttt ggacaccgca 1380
tacattaacg tgttgatcat tgccctagta tgcgcagtag cggctgctct gatcagcagt 1440
taccttttcc gcggaaatcc gaagggagcc aataatgcgc actag 1485
<210> 30
<211> 203
<212> PRT
<213> Corynebacterium glutamicum ATCC 13032 acetyltransferase
<400> 30
Met Ser Pro Thr Val Leu Pro Ala Thr Gln Ala Asp Phe Pro Lys Ile
1 5 10 15
Val Asp Val Leu Val Glu Ala Phe Ala Asn Asp Pro Ala Phe Leu Arg
20 25 30
Trp Ile Pro Gln Pro Asp Pro Gly Ser Ala Lys Leu Arg Ala Leu Phe
35 40 45
Glu Leu Gln Ile Glu Lys Gln Tyr Ala Val Ala Gly Asn Ile Asp Val
50 55 60
Ala Arg Asp Ser Glu Gly Glu Ile Val Gly Val Ala Leu Trp Asp Arg
65 70 75 80
Pro Asp Gly Asn His Ser Ala Lys Asp Gln Ala Ala Met Leu Pro Arg
85 90 95
Leu Val Ser Ile Phe Gly Ile Lys Ala Ala Gln Val Ala Trp Thr Asp
100 105 110
Leu Ser Ser Ala Arg Phe His Pro Lys Phe Pro His Trp Tyr Leu Tyr
115 120 125
Thr Val Ala Thr Ser Ser Ser Ala Arg Gly Thr Gly Val Gly Ser Ala
130 135 140
Leu Leu Asn His Gly Ile Ala Arg Ala Gly Asp Glu Ala Ile Tyr Leu
145 150 155 160
Glu Ala Thr Ser Thr Arg Ala Ala Gln Leu Tyr Asn Arg Leu Gly Phe
165 170 175
Val Pro Leu Gly Tyr Ile Pro Ser Asp Asp Asp Gly Thr Pro Glu Leu
180 185 190
Ala Met Trp Lys Pro Pro Ala Met Pro Thr Val
195 200
<210> 31
<211> 203
<212> PRT
<213> Corynebacterium glutamicum ATCC 13869 acetyltransferase
<400> 31
Met Ser Pro Thr Val Leu Pro Ala Thr Gln Ala Asp Phe Pro Lys Ile
1 5 10 15
Val Asp Val Leu Val Glu Ala Phe Ala Asn Asp Pro Ala Phe Leu Arg
20 25 30
Trp Ile Pro Gln Pro Asp Pro Gly Ser Ala Lys Leu Arg Ala Leu Phe
35 40 45
Glu Leu Gln Ile Glu Lys Gln Tyr Ala Val Ala Gly Asn Ile Asp Val
50 55 60
Ala Arg Asp Ser Glu Gly Glu Ile Val Gly Val Ala Leu Trp Asp Arg
65 70 75 80
Pro Asp Gly Asn His Ser Ala Lys Asp Gln Ala Ala Ile Leu Pro Arg
85 90 95
Leu Val Ser Ile Phe Gly Ile Lys Ala Ala Gln Val Ala Trp Thr Asp
100 105 110
Leu Ser Ser Ala Arg Phe His Pro Lys Phe Pro His Trp Tyr Leu Tyr
115 120 125
Thr Val Ala Thr Ser Ser Ser Ala Arg Gly Thr Gly Val Gly Ser Ala
130 135 140
Leu Leu Asn His Gly Ile Ala Arg Ala Gly Asp Glu Ala Ile Tyr Leu
145 150 155 160
Glu Ala Thr Ser Thr Arg Ala Ala Gln Leu Tyr Asn Arg Leu Gly Phe
165 170 175
Val Pro Leu Gly Tyr Ile Pro Ser Asp Asp Asp Gly Thr Pro Glu Leu
180 185 190
Ala Met Trp Lys Pro Pro Ala Met Pro Thr Val
195 200
<210> 32
<211> 357
<212> PRT
<213> Acetyl gamma glutamyl phosphate reductase (ArgC)
<400> 32
Met Ile Met His Asn Val Tyr Gly Val Thr Met Thr Ile Lys Val Ala
1 5 10 15
Ile Ala Gly Ala Ser Gly Tyr Ala Gly Gly Glu Ile Leu Arg Leu Leu
20 25 30
Leu Gly His Pro Ala Tyr Ala Ser Gly Glu Leu Glu Ile Gly Ala Leu
35 40 45
Thr Ala Ala Ser Thr Ala Gly Ser Thr Leu Gly Glu Leu Met Pro His
50 55 60
Ile Pro Gln Leu Ala Asp Arg Val Ile Gln Asp Thr Thr Ala Glu Thr
65 70 75 80
Leu Ala Gly His Asp Val Val Phe Leu Gly Leu Pro His Gly Phe Ser
85 90 95
Ala Glu Ile Ala Leu Gln Leu Gly Pro Asp Val Thr Val Ile Asp Cys
100 105 110
Ala Ala Asp Phe Arg Leu Gln Asn Ala Ala Asp Trp Glu Lys Phe Tyr
115 120 125
Gly Ser Glu His Gln Gly Thr Trp Pro Tyr Gly Ile Pro Glu Met Pro
130 135 140
Gly His Arg Glu Ala Leu Arg Gly Ala Lys Arg Val Ala Val Pro Gly
145 150 155 160
Cys Phe Pro Thr Gly Ala Thr Leu Ala Leu Leu Pro Ala Val Gln Ala
165 170 175
Gly Leu Ile Glu Pro Asp Val Ser Val Val Ser Ile Thr Gly Val Ser
180 185 190
Gly Ala Gly Lys Lys Ala Ser Val Ala Leu Leu Gly Ser Glu Thr Met
195 200 205
Gly Ser Leu Lys Ala Tyr Asn Thr Ser Gly Lys His Arg His Thr Pro
210 215 220
Glu Ile Ala Gln Asn Leu Gly Glu Val Ser Asp Lys Pro Val Lys Val
225 230 235 240
Ser Phe Thr Pro Val Leu Ala Pro Leu Pro Arg Gly Ile Leu Thr Thr
245 250 255
Ala Thr Ala Pro Leu Lys Glu Gly Val Thr Ala Glu Gln Ala Arg Ala
260 265 270
Val Tyr Glu Glu Phe Tyr Ala Gln Glu Thr Phe Val His Val Leu Pro
275 280 285
Glu Gly Ala Gln Pro Gln Thr Gln Ala Val Leu Gly Ser Asn Met Cys
290 295 300
His Val Gln Val Glu Ile Asp Glu Glu Ala Gly Lys Val Leu Val Thr
305 310 315 320
Ser Ala Ile Asp Asn Leu Thr Lys Gly Thr Ala Gly Ala Ala Val Gln
325 330 335
Cys Met Asn Leu Ser Val Gly Phe Asp Glu Ala Ala Gly Leu Pro Gln
340 345 350
Val Gly Val Ala Pro
355
<210> 33
<211> 388
<212> PRT
<213> Acetyl glutamate synthase or Ornithine acetyl transferase (ArgJ)
<400> 33
Met Ala Glu Lys Gly Ile Thr Ala Pro Lys Gly Phe Val Ala Ser Ala
1 5 10 15
Thr Thr Ala Gly Ile Lys Ala Ser Gly Asn Pro Asp Met Ala Leu Val
20 25 30
Val Asn Gln Gly Pro Glu Phe Ser Ala Ala Ala Val Phe Thr Arg Asn
35 40 45
Arg Val Phe Ala Ala Pro Val Lys Val Ser Arg Glu Asn Val Ala Asp
50 55 60
Gly Gln Ile Arg Ala Val Leu Tyr Asn Ala Gly Asn Ala Asn Ala Cys
65 70 75 80
Asn Gly Leu Gln Gly Glu Lys Asp Ala Arg Glu Ser Val Ser His Leu
85 90 95
Ala Gln Asn Leu Gly Leu Glu Asp Ser Asp Ile Gly Val Cys Ser Thr
100 105 110
Gly Leu Ile Gly Glu Leu Leu Pro Met Asp Lys Leu Asn Ala Gly Ile
115 120 125
Asp Gln Leu Thr Ala Glu Gly Ala Leu Gly Asp Asn Gly Ala Ala Ala
130 135 140
Ala Lys Ala Ile Met Thr Thr Asp Thr Val Asp Lys Glu Thr Val Val
145 150 155 160
Phe Ala Asp Gly Trp Thr Val Gly Gly Met Gly Lys Gly Val Gly Met
165 170 175
Met Ala Pro Ser Leu Ala Thr Met Leu Val Cys Leu Thr Thr Asp Ala
180 185 190
Ser Val Thr Gln Glu Met Ala Gln Ile Ala Leu Ala Asn Ala Thr Ala
195 200 205
Val Thr Phe Asp Thr Leu Asp Ile Asp Gly Ser Thr Ser Thr Asn Asp
210 215 220
Thr Val Phe Leu Leu Ala Ser Gly Ala Ser Gly Ile Thr Pro Thr Gln
225 230 235 240
Asp Glu Leu Asn Asp Ala Val Tyr Ala Ala Cys Ser Asp Ile Ala Ala
245 250 255
Lys Leu Gln Ala Asp Ala Glu Gly Val Thr Lys Arg Val Ala Val Thr
260 265 270
Val Val Gly Thr Thr Asn Asn Glu Gln Ala Ile Asn Ala Ala Arg Thr
275 280 285
Val Ala Arg Asp Asn Leu Phe Lys Cys Ala Met Phe Gly Ser Asp Pro
290 295 300
Asn Trp Gly Arg Val Leu Ala Ala Val Gly Met Ala Asp Ala Asp Met
305 310 315 320
Glu Pro Glu Lys Ile Ser Val Phe Phe Asn Gly Gln Ala Val Cys Leu
325 330 335
Asp Ser Thr Gly Ala Pro Gly Ala Arg Glu Val Asp Leu Ser Gly Ala
340 345 350
Asp Ile Asp Val Arg Ile Asp Leu Gly Thr Ser Gly Glu Gly Gln Ala
355 360 365
Thr Val Arg Thr Thr Asp Leu Ser Phe Ser Tyr Val Glu Ile Asn Ser
370 375 380
Ala Tyr Ser Ser
385
<210> 34
<211> 317
<212> PRT
<213> Acetyl glutamate kinase (ArgB)
<400> 34
Met Asn Asp Leu Ile Lys Asp Leu Gly Ser Glu Val Arg Ala Asn Val
1 5 10 15
Leu Ala Glu Ala Leu Pro Trp Leu Gln His Phe Arg Asp Lys Ile Val
20 25 30
Val Val Lys Tyr Gly Gly Asn Ala Met Val Asp Asp Asp Leu Lys Ala
35 40 45
Ala Phe Ala Ala Asp Met Val Phe Leu Arg Thr Val Gly Ala Lys Pro
50 55 60
Val Val Val His Gly Gly Gly Pro Gln Ile Ser Glu Met Leu Asn Arg
65 70 75 80
Val Gly Leu Gln Gly Glu Phe Lys Gly Gly Phe Arg Val Thr Thr Pro
85 90 95
Glu Val Met Asp Ile Val Arg Met Val Leu Phe Gly Gln Val Gly Arg
100 105 110
Asp Leu Val Gly Leu Ile Asn Ser His Gly Pro Tyr Ala Val Gly Thr
115 120 125
Ser Gly Glu Asp Ala Gly Leu Phe Thr Ala Gln Lys Arg Met Val Asn
130 135 140
Ile Asp Gly Val Pro Thr Asp Ile Gly Leu Val Gly Asp Ile Ile Asn
145 150 155 160
Val Asp Ala Ser Ser Leu Met Asp Ile Ile Glu Ala Gly Arg Ile Pro
165 170 175
Val Val Ser Thr Ile Ala Pro Gly Glu Asp Gly Gln Ile Tyr Asn Ile
180 185 190
Asn Ala Asp Thr Ala Ala Gly Ala Leu Ala Ala Ala Ile Gly Ala Glu
195 200 205
Arg Leu Leu Val Leu Thr Asn Val Glu Gly Leu Tyr Thr Asp Trp Pro
210 215 220
Asp Lys Ser Ser Leu Val Ser Lys Ile Lys Ala Thr Glu Leu Glu Ala
225 230 235 240
Ile Leu Pro Gly Leu Asp Ser Gly Met Ile Pro Lys Met Glu Ser Cys
245 250 255
Leu Asn Ala Val Arg Gly Gly Val Ser Ala Ala His Val Ile Asp Gly
260 265 270
Arg Ile Ala His Ser Val Leu Leu Glu Leu Leu Thr Met Gly Gly Ile
275 280 285
Gly Thr Met Val Leu Pro Asp Val Phe Asp Arg Glu Asn Tyr Pro Glu
290 295 300
Gly Thr Val Phe Arg Lys Asp Asp Lys Asp Gly Glu Leu
305 310 315
<210> 35
<211> 391
<212> PRT
<213> Acetyl ornithine aminotransferase (ArgD)
<400> 35
Met Ser Thr Leu Glu Thr Trp Pro Gln Val Ile Ile Asn Thr Tyr Gly
1 5 10 15
Thr Pro Pro Val Glu Leu Val Ser Gly Lys Gly Ala Thr Val Thr Asp
20 25 30
Asp Gln Gly Asn Val Tyr Ile Asp Leu Leu Ala Gly Ile Ala Val Asn
35 40 45
Ala Leu Gly His Ala His Pro Ala Ile Ile Glu Ala Val Thr Asn Gln
50 55 60
Ile Gly Gln Leu Gly His Val Ser Asn Leu Phe Ala Ser Arg Pro Val
65 70 75 80
Val Glu Val Ala Glu Glu Leu Ile Lys Arg Phe Ser Leu Asp Asp Ala
85 90 95
Thr Leu Ala Ala Gln Thr Arg Val Phe Phe Cys Asn Ser Gly Ala Glu
100 105 110
Ala Asn Glu Ala Ala Phe Lys Ile Ala Arg Leu Thr Gly Arg Ser Arg
115 120 125
Ile Leu Ala Ala Val His Gly Phe His Gly Arg Thr Met Gly Ser Leu
130 135 140
Ala Leu Thr Gly Gln Pro Asp Lys Arg Glu Ala Phe Leu Pro Met Pro
145 150 155 160
Ser Gly Val Glu Phe Tyr Pro Tyr Gly Asp Thr Asp Tyr Leu Arg Lys
165 170 175
Met Val Glu Thr Asn Pro Thr Asp Val Ala Ala Ile Phe Leu Glu Pro
180 185 190
Ile Gln Gly Glu Thr Gly Val Val Pro Ala Pro Glu Gly Phe Leu Lys
195 200 205
Ala Val Arg Glu Leu Cys Asp Glu Tyr Gly Ile Leu Met Ile Thr Asp
210 215 220
Glu Val Gln Thr Gly Val Gly Arg Thr Gly Asp Phe Phe Ala His Gln
225 230 235 240
His Asp Gly Val Val Pro Asp Val Val Thr Met Ala Lys Gly Leu Gly
245 250 255
Gly Gly Leu Pro Ile Gly Ala Cys Leu Ala Thr Gly Arg Ala Ala Glu
260 265 270
Leu Met Thr Pro Gly Lys His Gly Thr Thr Phe Gly Gly Asn Pro Val
275 280 285
Ala Cys Ala Ala Ala Lys Ala Val Leu Ser Val Val Asp Asp Ala Phe
290 295 300
Cys Ala Glu Val Ala Arg Lys Gly Glu Leu Phe Lys Glu Leu Leu Ala
305 310 315 320
Lys Val Asp Gly Val Val Asp Val Arg Gly Arg Gly Leu Met Leu Gly
325 330 335
Val Val Leu Glu Arg Asp Val Ala Lys Gln Ala Val Leu Asp Gly Phe
340 345 350
Lys His Gly Val Ile Leu Asn Ala Pro Ala Asp Asn Ile Ile Arg Leu
355 360 365
Thr Pro Pro Leu Val Ile Thr Asp Glu Glu Ile Ala Asp Ala Val Lys
370 375 380
Ala Ile Ala Glu Thr Ile Ala
385 390
<210> 36
<211> 319
<212> PRT
<213> Ornithine carbamoyl transferase (argF)
<400> 36
Met Thr Ser Gln Pro Gln Val Arg His Phe Leu Ala Asp Asp Asp Leu
1 5 10 15
Thr Pro Ala Glu Gln Ala Glu Val Leu Thr Leu Ala Ala Lys Leu Lys
20 25 30
Ala Ala Pro Phe Ser Glu Arg Pro Leu Glu Gly Pro Lys Ser Val Ala
35 40 45
Val Leu Phe Asp Lys Thr Ser Thr Arg Thr Arg Phe Ser Phe Asp Ala
50 55 60
Gly Ile Ala His Leu Gly Gly His Ala Ile Val Val Asp Ser Gly Ser
65 70 75 80
Ser Gln Met Gly Lys Gly Glu Ser Leu Gln Asp Thr Ala Ala Val Leu
85 90 95
Ser Arg Tyr Val Glu Ala Ile Val Trp Arg Thr Tyr Ala His Ser Asn
100 105 110
Phe His Ala Met Ala Glu Thr Ser Thr Val Pro Leu Val Asn Ser Leu
115 120 125
Ser Asp Asp Leu His Pro Cys Gln Ile Leu Ala Asp Leu Gln Thr Ile
130 135 140
Val Glu Asn Leu Ser Pro Glu Glu Gly Pro Ala Gly Leu Lys Gly Lys
145 150 155 160
Lys Ala Val Tyr Leu Gly Asp Gly Asp Asn Asn Met Ala Asn Ser Tyr
165 170 175
Met Ile Gly Phe Ala Thr Ala Gly Met Asp Ile Ser Ile Ile Ala Pro
180 185 190
Glu Gly Phe Gln Pro Arg Ala Glu Phe Val Glu Arg Ala Glu Lys Arg
195 200 205
Gly Gln Glu Thr Gly Ala Lys Val Val Val Thr Asp Ser Leu Asp Glu
210 215 220
Val Ala Gly Ala Asp Val Val Ile Thr Asp Thr Trp Val Ser Met Gly
225 230 235 240
Met Glu Asn Asp Gly Ile Asp Arg Thr Thr Pro Phe Val Pro Tyr Gln
245 250 255
Val Asn Asp Glu Val Met Ala Lys Ala Asn Asp Gly Ala Ile Phe Leu
260 265 270
His Cys Leu Pro Ala Tyr Arg Gly Lys Glu Val Ala Ala Ser Val Ile
275 280 285
Asp Gly Pro Ala Ser Lys Val Phe Asp Glu Ala Glu Asn Arg Leu His
290 295 300
Ala Gln Lys Ala Leu Leu Val Trp Leu Leu Ala Asn Gln Pro Arg
305 310 315
<210> 37
<211> 533
<212> PRT
<213> Glutamate expoter (NCgl1221)
<400> 37
Met Ile Leu Gly Val Pro Ile Gln Tyr Leu Leu Tyr Ser Leu Trp Asn
1 5 10 15
Trp Ile Val Asp Thr Gly Phe Asp Val Ala Ile Ile Leu Val Leu Ala
20 25 30
Phe Leu Ile Pro Arg Ile Gly Arg Leu Ala Met Arg Ile Ile Lys Arg
35 40 45
Arg Val Glu Ser Ala Ala Asp Ala Asp Thr Thr Lys Asn Gln Leu Ala
50 55 60
Phe Ala Gly Val Gly Val Tyr Ile Ala Gln Ile Val Ala Phe Phe Met
65 70 75 80
Leu Ala Val Ser Ala Met Gln Ala Phe Gly Phe Ser Leu Ala Gly Ala
85 90 95
Ala Ile Pro Ala Thr Ile Ala Ser Ala Ala Ile Gly Leu Gly Ala Gln
100 105 110
Ser Ile Val Ala Asp Phe Leu Ala Gly Phe Phe Ile Leu Thr Glu Lys
115 120 125
Gln Phe Gly Val Gly Asp Trp Val Arg Phe Glu Gly Asn Gly Ile Val
130 135 140
Val Glu Gly Thr Val Ile Glu Ile Thr Met Arg Ala Thr Lys Ile Arg
145 150 155 160
Thr Ile Ala Gln Glu Thr Val Ile Ile Pro Asn Ser Thr Ala Lys Val
165 170 175
Cys Ile Asn Asn Ser Asn Asn Trp Ser Arg Ala Val Val Val Ile Pro
180 185 190
Ile Pro Met Leu Gly Ser Glu Asn Ile Thr Asp Val Ile Ala Arg Ser
195 200 205
Glu Ala Ala Thr Arg Arg Ala Leu Gly Gln Glu Lys Ile Ala Pro Glu
210 215 220
Ile Leu Gly Glu Leu Asp Val His Pro Ala Thr Glu Val Thr Pro Pro
225 230 235 240
Thr Val Val Gly Met Pro Trp Met Val Thr Met Arg Phe Leu Val Gln
245 250 255
Val Thr Ala Gly Asn Gln Trp Leu Val Glu Arg Ala Ile Arg Thr Glu
260 265 270
Ile Ile Ser Glu Phe Trp Glu Glu Tyr Gly Ser Ala Thr Thr Thr Ser
275 280 285
Gly Thr Leu Ile Asp Ser Leu His Val Glu His Glu Glu Pro Lys Thr
290 295 300
Ser Leu Ile Asp Ala Ser Pro Gln Ala Leu Lys Glu Pro Lys Pro Glu
305 310 315 320
Ala Ala Ala Thr Val Ala Ser Leu Ala Ala Ser Ser Asn Asp Asp Ala
325 330 335
Asp Asn Ala Asp Ala Ser Val Ile Asn Ala Gly Asn Pro Glu Lys Glu
340 345 350
Leu Asp Ser Asp Val Leu Glu Gln Glu Leu Ser Ser Glu Glu Pro Glu
355 360 365
Glu Thr Ala Lys Pro Asp His Ser Leu Arg Gly Phe Phe Arg Thr Asp
370 375 380
Tyr Tyr Pro Asn Arg Trp Gln Lys Ile Leu Ser Phe Gly Gly Arg Val
385 390 395 400
Arg Met Ser Thr Ser Leu Leu Leu Gly Ala Leu Leu Leu Leu Ser Leu
405 410 415
Phe Lys Val Met Thr Val Glu Pro Ser Glu Asn Trp Gln Asn Ser Ser
420 425 430
Gly Trp Leu Ser Pro Ser Thr Ala Thr Ser Thr Ala Val Thr Thr Ser
435 440 445
Glu Thr Ser Ala Pro Val Ser Thr Pro Ser Met Thr Val Pro Thr Thr
450 455 460
Val Glu Glu Thr Pro Thr Met Glu Ser Asn Val Glu Thr Gln Gln Glu
465 470 475 480
Thr Ser Thr Pro Ala Thr Ala Thr Pro Gln Arg Ala Asp Thr Ile Glu
485 490 495
Pro Thr Glu Glu Ala Thr Ser Gln Glu Glu Thr Thr Ala Ser Gln Thr
500 505 510
Gln Ser Pro Ala Val Glu Ala Pro Thr Ala Val Gln Glu Thr Val Ala
515 520 525
Pro Thr Ser Thr Pro
530
<210> 38
<211> 711
<212> PRT
<213> Ornithine decarboxylase (ODC)
<400> 38
Met Lys Ser Met Asn Ile Ala Ala Ser Ser Glu Leu Val Ser Arg Leu
1 5 10 15
Ser Ser His Arg Arg Val Val Ala Leu Gly Asp Thr Asp Phe Thr Asp
20 25 30
Val Ala Ala Val Val Ile Thr Ala Ala Asp Ser Arg Ser Gly Ile Leu
35 40 45
Ala Leu Leu Lys Arg Thr Gly Phe His Leu Pro Val Phe Leu Tyr Ser
50 55 60
Glu His Ala Val Glu Leu Pro Ala Gly Val Thr Ala Val Ile Asn Gly
65 70 75 80
Asn Glu Gln Gln Trp Leu Glu Leu Glu Ser Ala Ala Cys Gln Tyr Glu
85 90 95
Glu Asn Leu Leu Pro Pro Phe Tyr Asp Thr Leu Thr Gln Tyr Val Glu
100 105 110
Met Gly Asn Ser Thr Phe Ala Cys Pro Gly His Gln His Gly Ala Phe
115 120 125
Phe Lys Lys His Pro Ala Gly Arg His Phe Tyr Asp Phe Phe Gly Glu
130 135 140
Asn Val Phe Arg Ala Asp Met Cys Asn Ala Asp Val Lys Leu Gly Asp
145 150 155 160
Leu Leu Ile His Glu Gly Ser Ala Lys Asp Ala Gln Lys Phe Ala Ala
165 170 175
Lys Val Phe His Ala Asp Lys Thr Tyr Phe Val Leu Asn Gly Thr Ser
180 185 190
Ala Ala Asn Lys Val Val Thr Asn Ala Leu Leu Thr Arg Gly Asp Leu
195 200 205
Val Leu Phe Asp Arg Asn Asn His Lys Ser Asn His His Gly Ala Leu
210 215 220
Ile Gln Ala Gly Ala Thr Pro Val Tyr Leu Glu Ala Ser Arg Asn Pro
225 230 235 240
Phe Gly Phe Ile Gly Gly Ile Asp Ala His Cys Phe Asn Glu Glu Tyr
245 250 255
Leu Arg Gln Gln Ile Arg Asp Val Ala Pro Glu Lys Ala Asp Leu Pro
260 265 270
Arg Pro Tyr Arg Leu Ala Ile Ile Gln Leu Gly Thr Tyr Asp Gly Thr
275 280 285
Val Tyr Asn Ala Arg Gln Val Ile Asp Thr Val Gly His Leu Cys Asp
290 295 300
Tyr Ile Leu Phe Asp Ser Ala Trp Val Gly Tyr Glu Gln Phe Ile Pro
305 310 315 320
Met Met Ala Asp Ser Ser Pro Leu Leu Leu Glu Leu Asn Glu Asn Asp
325 330 335
Pro Gly Ile Phe Val Thr Gln Ser Val His Lys Gln Gln Ala Gly Phe
340 345 350
Ser Gln Thr Ser Gln Ile His Lys Lys Asp Asn His Ile Arg Gly Gln
355 360 365
Ala Arg Phe Cys Pro His Lys Arg Leu Asn Asn Ala Phe Met Leu His
370 375 380
Ala Ser Thr Ser Pro Phe Tyr Pro Leu Phe Ala Ala Leu Asp Val Asn
385 390 395 400
Ala Lys Ile His Glu Gly Glu Ser Gly Arg Arg Leu Trp Ala Glu Cys
405 410 415
Val Glu Ile Gly Ile Glu Ala Arg Lys Ala Ile Leu Ala Arg Cys Lys
420 425 430
Leu Phe Arg Pro Phe Ile Pro Pro Val Val Asp Gly Lys Leu Trp Gln
435 440 445
Asp Tyr Pro Thr Ser Val Leu Ala Ser Asp Arg Arg Phe Phe Ser Phe
450 455 460
Glu Pro Gly Ala Lys Trp His Gly Phe Glu Gly Tyr Ala Ala Asp Gln
465 470 475 480
Tyr Phe Val Asp Pro Cys Lys Leu Leu Leu Thr Thr Pro Gly Ile Asp
485 490 495
Ala Glu Thr Gly Glu Tyr Ser Asp Phe Gly Val Pro Ala Thr Ile Leu
500 505 510
Ala His Tyr Leu Arg Glu Asn Gly Ile Val Pro Glu Lys Cys Asp Leu
515 520 525
Asn Ser Ile Leu Phe Leu Leu Thr Pro Ala Glu Ser His Glu Lys Leu
530 535 540
Ala Gln Leu Val Ala Met Leu Ala Gln Phe Glu Gln His Ile Glu Asp
545 550 555 560
Asp Ser Pro Leu Val Glu Val Leu Pro Ser Val Tyr Asn Lys Tyr Pro
565 570 575
Val Arg Tyr Arg Asp Tyr Thr Leu Arg Gln Leu Cys Gln Glu Met His
580 585 590
Asp Leu Tyr Val Ser Phe Asp Val Lys Asp Leu Gln Lys Ala Met Phe
595 600 605
Arg Gln Gln Ser Phe Pro Ser Val Val Met Asn Pro Gln Asp Ala His
610 615 620
Ser Ala Tyr Ile Arg Gly Asp Val Glu Leu Val Arg Ile Arg Asp Ala
625 630 635 640
Glu Gly Arg Ile Ala Ala Glu Gly Ala Leu Pro Tyr Pro Pro Gly Val
645 650 655
Leu Cys Val Val Pro Gly Glu Val Trp Gly Gly Ala Val Gln Arg Tyr
660 665 670
Phe Leu Ala Leu Glu Glu Gly Val Asn Leu Leu Pro Gly Phe Ser Pro
675 680 685
Glu Leu Gln Gly Val Tyr Ser Glu Thr Asp Ala Asp Gly Val Lys Arg
690 695 700
Leu Tyr Gly Tyr Val Leu Lys
705 710
<210> 39
<211> 289
<212> DNA
<213> Artificial Sequence
<220>
<223> lysCP1 promoter
<400> 39
ccgatgctag ggcgaaaagc acggcgagca gattgctttg cacttgattc agggtagttg 60
actaaagagt tgctcgcgaa gtagcacctg tcacttttgt ctcaaatatt aaatcgaata 120
tcaatatatg gtctgtttat tggaacgcgt cccagtggct gagacgcatc cgctaaagcc 180
ccaggaaccc tgtgcagaaa gaaaacactc ctctggctag gtagacacag tttattgtgg 240
tagagttgag cgggtaactg tcagcacgta gatcgaaagg tgcacaaag 289
<---
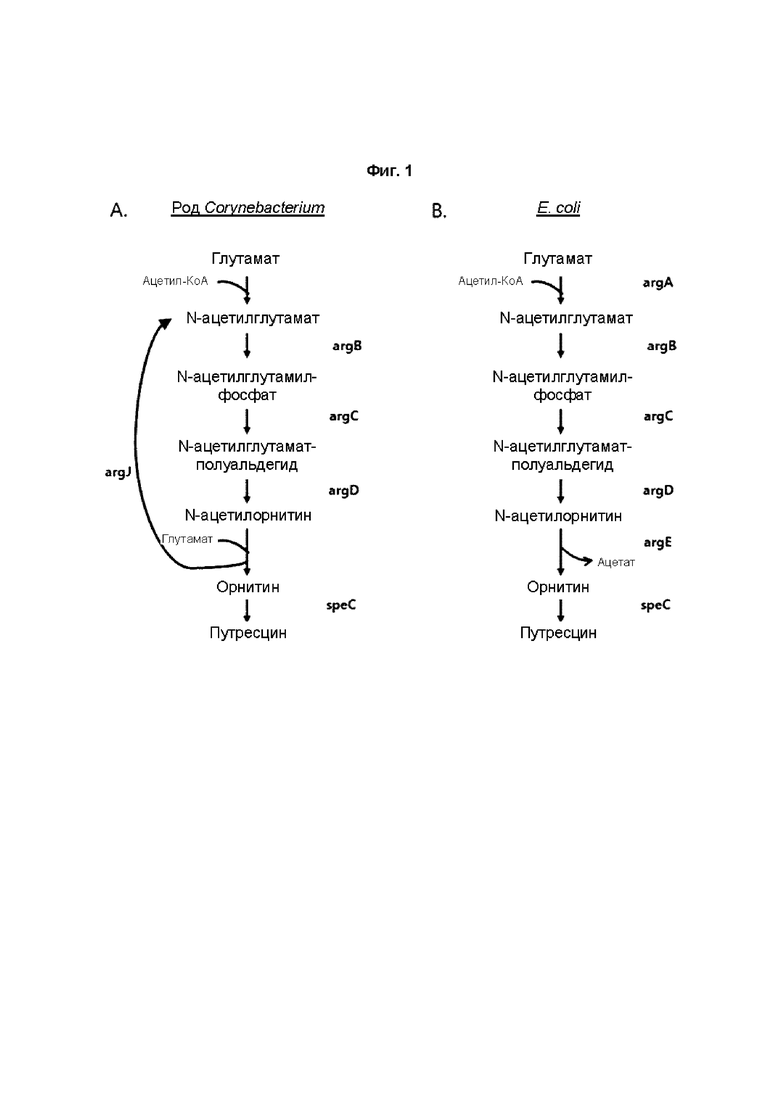
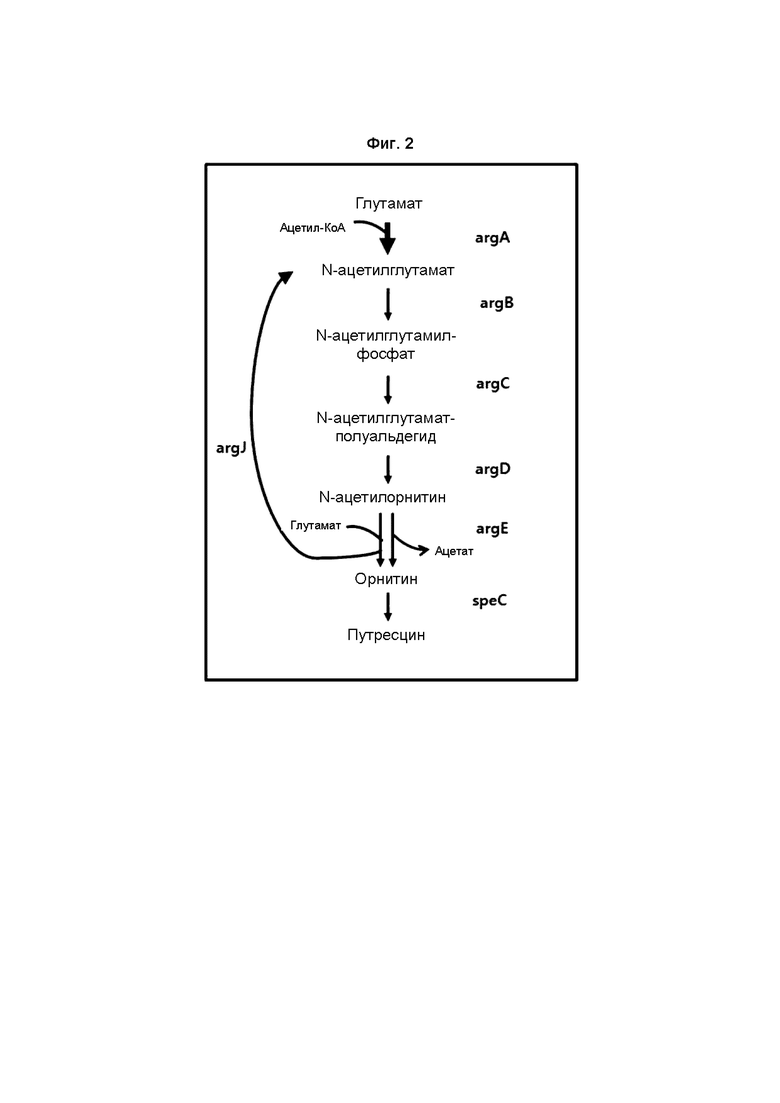
Настоящее изобретение относится к биотехнологии и представляет собой модифицированный микроорганизм рода Corynebacterium, продуцирующий орнитин, где активности N-ацетилглутаматсинтазы из E. coli и ацетилорнитиндезацетилазы из E. coli введены в указанный микроорганизм рода Corynebacterium, обладающий активностью ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ). Изобретение позволяет повысить продукции орнитина. 2 н. и 10 з.п. ф-лы, 2 ил., 11 табл, 7 пр.
1. Модифицированный микроорганизм рода Corynebacterium, продуцирующий орнитин, где активности N-ацетилглутаматсинтазы из E. coli и ацетилорнитиндезацетилазы из E. coli введены в указанный микроорганизм рода Corynebacterium, обладающий активностью ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ).
2. Модифицированный микроорганизм по п. 1, где N-ацетилглутаматсинтаза из E. coli состоит из аминокислотной последовательности SEQ ID NO:1.
3. Модифицированный микроорганизм по п. 1, где ацетилорнитиндезацетилаза из E. coli состоит из аминокислотной последовательности SEQ ID NO:3.
4. Модифицированный микроорганизм по п. 1, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
5. Модифицированный микроорганизм по п. 1, где активность фосфотрансацетилазного и ацетаткиназного оперона (оперона pta-ackA) дополнительно усилена по сравнению с его эндогенной активностью.
6. Модифицированный микроорганизм по п. 5, где фосфотрансацетилазный и ацетаткиназный оперон состоит из аминокислотной последовательности SEQ ID NO:5 или SEQ ID NO:7.
7. Модифицированный микроорганизм по п. 1, в который дополнительно введена активность ацетил-КоА-синтетазы (acs) из E. coli.
8. Модифицированный микроорганизм по п. 7, где ацетил-КоА-синтетаза состоит из аминокислотной последовательности SEQ ID NO:9.
9. Модифицированный микроорганизм по п. 1, где активность (1) орнитинкарбамоилтрансферазы (ArgF), (2) экспортера глутамата или (3) орнитинкарбамоилтрансферазы и экспортера глутамата дополнительно ослаблена по сравнению с их эндогенной активностью.
10. Модифицированный микроорганизм по п. 1, где активность по меньшей мере одного, выбранного из группы, состоящей из ацетил-гамма-глутамилфосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитинаминотрансферазы (ArgD), дополнительно усилена по сравнению с их эндогенной активностью.
11. Способ получения орнитина, включающий:
(1) культивирование модифицированного микроорганизма рода Corynebacterium по любому из пп. 1-10 в среде; и
(2) выделение орнитина из культивированного микроорганизма или среды.
12. Способ по п. 11, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
| KR 101053429 B1, 03.08.2011 | |||
| DE 1642678 A1, 03.06.1971 | |||
| CN 102191291 A, 21.09.2011 | |||
| МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ К ПРОДУКЦИИ ПУТРЕСЦИНА В ВЫСОКОЙ КОНЦЕНТРАЦИИ, СПОСОБ ПОЛУЧЕНИЯ ЭТОГО МИКРООРГАНИЗМА И СПОСОБ ПРОДУКЦИИ ПУТРЕСЦИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2009 |
|
RU2433180C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |

