ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Эта заявка является частичным продолжением Заявки США №12/922,441, поданной 13 сентября 2010 г., которая является заявкой США в национальной фазе на основании заявки PCT/EP2009/001733, поданной 11 марта 2009 г., которая заявляет приоритет Европейской заявки №08360005.6, поданной 14 марта 2008 г., и приоритет предварительной заявки США №61/043,884, поданной 10 апреля 2008 г.
Список последовательностей
Настоящая заявка содержит Список последовательностей, представленный через EFS-Web и включенный в это описание путем ссылки в полном объеме. Указанная ASCII-копия, созданная 30 марта 2011 г., имеет название 13026944.txt и размер 56764 байт.
Описание
КСФ-1 (Колониестимулирующий фактор-1) является цитокином, который, в частности, экспрессируется различными типами клеток. Он является фактором дифференциации, роста и выживания для клеток линии мононуклеарных фагоцитов, которые экспрессируют рецептор для КСФ-1 (КСФ-1R) (SHERR. Colony-stimulating factor-1 receptor, blood. 1990, т.75, №1, стр.1-12). КСФ-1R является рецептором тирозинкиназы, который кодируется протоонкогеном c-fms, который содержит внутриклеточный домен киназы и лиганд-связывающий внеклеточный участок, организованный в пять иммуноглобулиноподобных субдоменов. Реакция на КСФ-1 ведет к увеличению выживаемости, роста, дифференциации и обратимых изменений в функциях. Сам ген c-fms является маркером дифференциации макрофагов. Степень экспрессии c-fms выше по сравнению с другими специфическими к макрофагам генами, включая лизоцим и специфический к макрофагам белок тирозинфосфатазу (HUME, et al. Regulation of CSF-1 receptor expression. Molecular reproduction и development. 1997, т.46, №1, стр.46-52).
Помимо клеток линии мононуклеарных фагоцитов, КСФ-1R также экспрессируется многими типами опухолей человека. В случае рака молочной железы экспрессия КСФ-1R связана с большими размерами опухолей и сниженной выживаемостью (KLUGER, et al. Macrophage colony-stimulating factor-1 receptor expression is associated with poor outcome in breast cancer by large cohort tissue microarray analysis. Clinical cancer research. 2004, т.10, №1, стр.173-7; SCHOLL, et al. Anti-colony-stimulating factor-1 antibody staining in primary breast adenocarcinomas correlates with marked inflammatory cell infiltrates and prognosis. Journal of the National Cancer Institute. 1994, т.86, №2, стр.120-6). При эпителиальном раке яичников большинство первичных опухолей и метастазов сильно экспрессируют КСФ-1R, и метастазы часто коэкспрессируют КСФ-1 и КСФ-1R. КСФ-1R также экспрессируется макрофагами, которые инфильтруют в опухоли (CHAMBERS, et al. Overexpression of epithelial macrophage colony-stimulating factor (CSF-1) и CSF-1 receptor: a poor prognostic factor in epithelial ovarian cancer, contrasted with a protective effect of stromal CSF-1. Clinical Cancer Research. 1997, т.3, №6, стр.999-1007). При раке яичников и эндометрия нозерн-блоттинг показывает, что подавляющее большинство опухолей коэкспрессирует КСФ-1 и КСФ-1R, в то время, как экспрессия КСФ-1R лишь едва заметно обнаруживается в образцах нормальной ткани эндометрия (BAÏOCCHI, et al. Expression of the macrophage colony-stimulating factor и its receptor in gynecologic malignancies. Cancer. 1991, т.67, №4, стр.990-6). При карциноме шейки матки экспрессия КСФ-1R подвергается повышающей регуляции как в строме опухолей, так и в эпителии опухолей, по сравнению с нормальным эндометрием (KIRMA, et al. Elevated expression of the oncogene c-fms and its ligand, the macrophage colony-stimulating factor-1, in cervical cancer and the role of transforming growth factor-betal in inducing c-fms expression. Cancer res.. 2007, т.67, №5, стр.1918-26). При раке почек инфильтрация опухолево-ассоциированных макрофагов с высоким уровнем экспрессии КСФ-1R связана с прогрессированием опухоли (HEMMERLEIN, et al. Expression of acute и late-stage inflammatory antigens, c-fms, CSF-1, and human monocytic serine esterase 1, in tumor-associated macrophages of renal cell carcinomas. Cancer immunology, immunotherapy. 2000, т.49, №9, стр.485-92). КСФ-1R экспрессируется почти 100%-ми образцов внутриэпителиальной неоплазии предстательной железы или рака (IDE, et al. Expression of colony-stimulating factor 1 receptor during prostate development and prostate cancer progression. Proc. Natl. Acad. Sci. U.S.A.. 2002, т.99, №22, стр.14404-9). Экспрессия КСФ-1R также была обнаружена при остром миелобластном лейкозе и В-клеточном хроническом лимфоцитарном лейкозе (RAMBALDI, et al. Expression of the macrophage colony-stimulating factor and c-fms genes in human acute myeloblastic leukemia cells. Journal of Clinical Investigation. 1988, т.81, №4, стр.1030-5).
Работа, проведенная в области иммуногистохимии и in situ гибридизации, продемонстрировала специфичность экспрессии КСФ-1 в клетках инвазивного рака молочной железы, в то время, как подобное вырабатывание в клетках интраканальных или неинвазивных опухолей не наблюдалось (SCHOLL, et al. Anti-colony-stimulating factor-1 antibody staining in primary breast adenocarcinomas correlates with marked inflammatory cell infiltrates and prognosis. Journal of the National Cancer Institute. 1994, т.86, №2, стр.120-6; TANG, et al. M-CSF (monocyte colony stimulating factor) and M-CSF receptor expression by breast tumor cells: M-CSF mediated recruitment of tumor infiltrating monocytes? Journal of cellular biochemistry. 1992, т.50, №4, стр.350-6). Вырабатывание КСФ-1 клетками инвазивных опухолей коррелирует с увеличением его концентрации в плазме пациентов, где она может превышать 1000 пг/мл, по сравнению с показателем, меньшим, чем 300 пг/мл, у нормальных субъектов. Высокая концентрация в сыворотке коррелирует с запущенными стадиями болезни и неблагоприятным краткосрочным прогнозом (SCHOLL, et al. Circulating levels of colony-stimulating factor 1 as a prognostic indicator in 82 patients with epithelial ovarian cancer. British journal of cancer. 1994, т.69, №2, стр.342-6; SCHOLL. Circulating levels of the macrophage colony stimulating factor CSF-1 in primary и metastatic breast cancer patients. A pilot study. Breast cancer research and treatment. 1996, т.39, №3, стр.275-83). Кроме того, было продемонстрировано, что КСФ-1 стимулирует подвижность и инвазивность клеток опухолей (DORSCH, et al. Macrophage colony-stimulating factor gene transfer into tumor cells induces macrophage infiltration but not tumor suppression. European journal of immunology. 1993, т.23, №1, стр.186-90; WANG, et al. Induction of monocyte migration by recombinant macrophage colony-stimulating factor. Journal of immunology. 1988, т.141, №2, стр.575-9; FILDERMAN, et al. Macrophage colony-stimulating factor (CSF-1) enhances invasiveness in CSF-1 receptor-positive carcinoma cell lines. Cancer res.. 1992, т.52, №13, стр.3661-6).
КСФ-1 также имеет хемотаксическое влияние на предшественники миелоидной линии, что способствует инфильтрации моноцитов в опухоль. Однако присутствие этих моноцитов является недостаточным для наблюдения разрушения опухоли иммунной системой (DORSCH, et al. Macrophage colony-stimulating factor gene transfer into tumor cells induces macrophage infiltration but not tumor suppression. European journal of immunology. 1993, т.23, №1, стр.186-90). Обнаруживается, что при высоком содержании в сыворотке, которое обычно наблюдается у пациентов, страдающих от опухолей молочной железы, яичников или поджелудочной железы, КСФ-1 ориентирует дифференциацию этих моноцитов в макрофаги, а не в дендритные клетки, способные вырабатывать опухолевые антигены, и, следовательно, запускать эффективную цитотоксическую иммунную реакцию, направленную против клеток опухолей (SCHOLL. Circulating levels of the macrophage colony stimulating factor CSF-1 in primary и metastatic breast cancer patients. A pilot study. Breast cancer research and treatment. 1996, т.39, №3, стр.275-83; BARON, et al. Modulation of MHC class II transport and lysosome distribution by macrophage-colony stimulating factor in human dendritic cells derived from monocytes. Journal of cell science. 2001, т.114, ч. 5, стр.999-1010).
КСФ-1 также является существенным для пролиферации и дифференциации остеокластов (CECCHINI, et al. Role of CSF-1 in bone and bone marrow development. Molecular reproduction и development. 1997, т.46, №1, стр.75-83). Остеокласты являются многоядерными клетками, которые экспрессируют КСФ-1R из кроветворных клеток-посредников, которые в первую очередь отвечают за распад минерализированных костей во время развития, гомеостаза и восстановления костей. При различных нарушениях скелетных костей, таких, как остеопороз, гиперкальциемия или злокачественность, ревматоидный артрит, метастазы опухолей и болезнь Педжета, резорбция костей остеокластами превышает формирование костей остеобластами, что ведет к снижению костной массы, ломкости скелетных костей и переломов костей (BRUZZANITI, et al. Molecular regulation of osteoclast activity. Reviews in endocrine. 2006, т.7, №1-2, стр.123-39). Например, у пациентов с запущенным раком молочной железы часто развиваются метастазы в кости. Метастаз в кость ведет к хронической боли и высокому риску переломов из-за вызванной опухолью потери костной массы (остеолиза), которая вызывается повышенной активностью остеокластов (CICEK, et al. Breast cancer bone metastasis and current small therapeutics. Cancer metastasis reviews. 2006, т.25, №4, стр.635-44). Было продемонстрировано, что остеолиз связан с высоким уровнем циркулирующего КСФ-1 (KITAURA, et al. The journal of clinical investigation. M-CSF mediates TNF-induced inflammatory osteolysis. 2005, т.115, №12, стр.3418-27).
Путь КСФ-1 также является посредником кишечного воспаления при таких болезнях, как воспалительная болезнь кишечника (MARSHALL, et al. Blockade of colony stimulating factor-1 (CSF-I) leads to inhibition of DSS-induced colitis. Inflammatory bowel diseases. 2007, т.13, №2, стр.219-24), пролиферация макрофагов во время острого отторжения аллотрансплантатов (JOSE, et al. Blockade of macrophage colony-stimulating factor reduces macrophage proliferation and accumulation in renal allograft rejection. American journal of transplantation. 2003, т.3, №3, стр.294-300) и при репликации ВИЧ-1 в инфицированных макрофагах (KUTZA, et al. Macrophage colony-stimulating factor antagonists inhibit replication of HIV-1 in human macrophages. Journal of immunology. 2000, №164, стр.4955-4960).
Поэтому предлагается ингибирование активности КСФ-1 различными соединениями для лечения раки и распада костей.
Уровень техники
Документ WO 01/30381 касается применения ингибиторов активности КСФ-1 при производстве медикаментов для лечения опухолевых болезней. Два предложенных подхода к ингибированию активности КСФ-1 состоят в подавлении активности самого КСФ-1 и подавлении активности КСФ-1R. Нейтрализации антител против КСФ-1 или его рецептора отдают предпочтение для ингибирования активности КСФ-1.
В документе WO 03/059395 описываются комбинированные продукты, включающие вещество, способное ингибировать активность КСФ-1, и вещество, обладающее цитотоксической активностью, для лечения рака.
В документе WO 2005/068503 описывается способ профилактики и лечения остеолиза, метастазов рака и потери костной массы, связанной с метастазами рака, путем введения субъекту антитела против КСФ-1.
Документ EP 1488792 А касается применения соединений моно- и/или бициклического арил- или гетероарилхиназолина, которые демонстрируют избирательное ингибирование дифференциации, пролиферации или высвобождения медиатора путем эффективного ингибирования тирозинкиназной активности КСФ-1R. Эта заявка также касается применения таких соединений для производства медикамента для ингибирования избыточной пролиферации клеток.
Документ US 2005059113 касается антител и их антиген-связывающих частей, которые специфически связываются с аКСФ-1. Изобретение также касается человеческих антител против КСФ-1 и их антиген-связывающих частей. Описанное в этой заявке изобретение также обеспечивает способы генной терапии с применением молекул нуклеиновых кислот, которые кодируют молекулы тяжелого и/или легкого иммуноглобулина, которые включают человеческие антитела против КСФ-1.
В публикациях Roussel и Sherr, 1989, PNAS, 86, 7924-7927 и Ashmun et al., 1989, Blood, 73, 827-837, описываются моноклональные антитела к человеческому рецептору КСФ-1 (например, 12-3A3 и 2-4А5), которые специфически блокируют связывание КСФ-1 с человеческим рецептором, таким образом, ингибируя лиганд-зависимый рост. Распознанный эпитоп находился между позициями аминокислот 349-512.
Документ WO 2009/026303 предлагает антиген-связывающие белки, способные конкурировать с КСФ-1, и, следовательно, предотвращать связывание КСФ-1 с его рецептором и в некоторых вариантах осуществления ингибировать связывание между IL-34 и КСФ-1R. Кроме того, в экспериментальном разделе документа WO 2009/026303 указывается, что полученные авторами изобретения антитела являются связывающими эпитопами, которые находятся, главным образом, на N-конце между аминокислотами с 20 по 223 последовательности SEQ ID NO 29 (что соответствует Ig-подобной петле 1 и Ig-подобной петле 2 в WO 2009/026303) КСФ-1R и требуют присутствия участков Ig-подобной петли 1 и Ig-подобной петли 2.
Описание изобретения
Настоящее изобретение касается антитела, специфически связывающегося с КСФ-1R, более конкретно - с человеческим КСФ-1R.
По всему тексту заявки термины в форме единственного числа применяются в значении "как минимум один", "как минимум первый", "один или несколько" или "несколько" в отношении указанных компонентов или этапов, если контекстом не предусматривается иного. Например, термин "клетка" охватывает множество клеток, включая их смеси.
Термин "и/или" по всему тексту данного документа охватывает значения "и", "или" и "все или любая другая комбинация элементов, объединенных указанным термином".
Термин "около" или "приблизительно" в контексте данного описания означает в пределах 20%, в предпочтительном варианте - в пределах 10%, в более предпочтительном варианте - в пределах 5% от указанного значения или диапазона.
В контексте данного описания термины "включая" и "включать" означают, что наборы частей, продуктов, композиций и способов включают указанные компоненты или этапы, но не исключают других. Термин "состоит главным образом из…", применяемый для определения продуктов, композиций и способов, означает исключение других компонентов или этапов любого существенного значения. Таким образом, композиция, состоящая главным образом из указанных компонентов, не исключает микропримесей и фармацевтически приемлемых носителей. "Состоит из…" означает исключение других компонентов в количестве, выходящем за пределы микропримесей, или других этапов.
В контексте данного описания термин "специфически связывается с…" касается реакции связывания, являющейся определяющей для присутствия белка-мишени в присутствии гетерогенной популяции белков и других высокомолекулярных соединений. Таким образом, в указанных аналитических условиях антитело согласно изобретению предпочтительно связывается с как минимум частью КСФ-1R и не связывается в существенном количестве с другими компонентами, присутствующими в испытуемом образце. Специфическое связывание между антителом согласно изобретению и мишенью КСФ-1R означает, что аффинность связывания составляет как минимум 103 М-1, в предпочтительном варианте - 105 М-1, 106 М-1, 107 М-1, 108 М-1, 109 М-1 или 1010 М-1. В особенно предпочтительном варианте осуществления аффинность связывания составляет как минимум 109 М-1 или 1010 М-1.
В контексте данного описания термин "КСФ-1R" касается человеческого рецептора КСФ1. Человеческий рецептор КСФ-1 был секвенован, и его аминокислотная последовательность показана у SEQ ID NO: 29.
В контексте данного описания термин "антитело" или "Ab" употребляется в самом широком значении. Таким образом, "антитело" или "Ab" может быть естественным или искусственным, таким, как моноклональные антитела (mAbs), полученные с применением традиционных гибридомных технологий, рекомбинантных технологий, и/или их функциональные фрагменты. Антитела согласно настоящему изобретению охватывают как интактные молекулы иммуноглобулина, например, поликлональное антитело, моноклональное антитело (mAb), моноспецифическое антитело, биспецифическое антитело, полиспецифическое антитело, человеческое антитело, животное антитело (например, антитело камелидов), химерные антитела, а также части, фрагменты, участки, пептиды и их производные (обеспечиваемые любым известным способом, включая, помимо прочих, ферментативное расщепление, пептидный синтез или рекомбинантные технологии), например, иммуноглобулин, лишенных легких цепей (см., например, документ US 6,005,079), Fab, Fab', F(ab')2, Fv, scFv, фрагмент антитела, диатело, Fd, CDR-участки или любая часть или пептидная последовательность антитела, способная связываться с антигеном или эпитопом. Антитело считается "способным к связыванию" с молекулой, если оно способно специфически реагировать с молекулой для связывания таким образом молекулы с антителом. Фрагменты или части антител могут не иметь Fc-фрагмента интактного антитела, быстрее выводиться из системы кровообращения и могут обладать меньшим неспецифическим тканевым связыванием по сравнению с интактным антителом. Примеры антител могут быть получены из интактных антител с применением способов, общеизвестных среди специалистов в данной области, например, путем протеолитического расщепления ферментами, такими, как папаин (для получения фрагментов Fab) или пепсин (для получения фрагментов F(ab')2 (см., например, Wahl et al., 24 J. Nucl. Med. 316-25 (1983). Части антител могут быть получены любым из вышеуказанных способов или путем экспрессии части рекомбинантной молекулы. Например, CDR-участок (участки) рекомбинантного антитела выделяют и субклонируют в соответствующий вектор экспрессии.
В контексте данного описания термин "вариабельный участок" касается вариабельного участка или домена легкой цепи (VL) или тяжелой цепи (VH), которые содержат детерминанты для распознавания специфичности связывания. Вариабельные домены участвуют в распознавании антигенов и образуют антиген-связывающий центр. В контексте данного описания термин "каркасный участок" означает части вариабельных участков легкой и тяжелой цепи, которые являются как минимум на 85% гомологичными (то есть, отличными от CDR) среди различных антител в одном виде. В контексте данного описания термин "гомологичный" означает сравнение аминокислот двух полипептидов, которые при выравнивании с применением алгоритма Смита-Уотермана (SMITH, et al. Identification of common molecular subsequences. Journal of Molecular Biology. 1981, №147, стр.195-7.) имеют приблизительно указанный процент одинаковых аминокислот. Например, "на 85% гомологичный" означает сравнение аминокислот двух полипептидов, которые при оптимальному выравнивании имеют 85% идентичности аминокислот. Вариабельный участок тяжелой и легкой цепи разделяется на сегменты, которые включают четыре каркасных субучастка (FR1, FR2, FR3 и FR4), которые прерываются тремя отрезками гипервариабельных последовательностей или участков определения комплементарности (CDR), как определяется в базе данных Kabat (Kabat et al., op.cit.), причем CDR1 находится между FR1 и FR2, CDR2 - между FR2 и FR3, и CDR3 - между FR3 и FR4. Без указания конкретных субучастков как FR1, FR2, FR3 или FR4, каркасный участок, относительно других, представляет комбинированные FR в пределах вариабельного участка одной естественной иммуноглобулиновой цепи. В контексте данного описания FR представляет один из четырех субучастков, и FR представляет два или более из четырех субучастков, которые составляют каркасный участок. Последовательности каркасных участков разных легких или тяжелых цепей являются относительно консервативными в пределах вида. Каркасный участок антитела представляет собой комбинированные каркасные участки составляющих легких и тяжелых цепей и служит для позиционирования и выравнивания CDR. CDR в первую очередь отвечают за создание сайта связывания антитела, обеспечение специфичности связывания и аффинности к эпитопу антигена. В пределах вариабельных участков тяжелых или легких цепей, которые обеспечивают связывание антигена, участки являются меньшими последовательностями, которые называются "гипервариабельными" из-за их чрезвычайной вариабельности между антителами различной специфичности. Такие гипервариабельные участки также называются "участками определения комплементарности" или "CDR-участками". Эти CDR-участки отвечают за основную специфичность антитела к определенной структуре антигенной детерминанты. CDR представляют прерывистые отрезки аминокислот в пределах вариабельных участков, однако, независимо от вида, было обнаружено, что позиционное расположение этих критичных аминокислотных последовательностей в пределах вариабельных участков тяжелой и легкой цепей является подобным в пределах аминокислотных последовательностей вариабельных цепей. Каждая из вариабельных тяжелых и легких цепей всех антител имеет 3 CDR-участка, каждый из которых прерывается другими (которые называются L1, L2, L3, H1, Н2, Н3) для соответствующих легких (L) и тяжелых (Н) цепей. Акцептированные CDR-участки были описаны у публикации Kabat et al, 252 J. Biol. Chem. 6609-16 (1977), и CDR-петли могут распознаваться путем применения этих правил во время исследования линейной аминокислотной последовательности. Однако правила определения CDR-Н3-петли могут быть разными (см. Раздел 4, Antibody Engineering: Methods & Protocols, (Lo, ed. Humana Press, Totowa, NJ, 2004)), и фактические пределы некоторых CDR-H3-петель могут не поддаваться распознанию без экспериментов, таких, как круговой дихроизм, ядерный магнитный резонанс или рентгеновская кристаллография. У всех видов млекопитающих пептиды антител содержат постоянные (то есть высококонсервативные) и вариабельные участки, и в пределах последних существуют CDR и так называемые "каркасные участки", которые состоят из аминокислотных последовательностей в пределах вариабельного участка тяжелой или легкой цепи, но за пределами CDR. CDR-участки также могут определяться с применением номенклатуры Chothia (CHOTHIA and LESK. Canonical structures for the hypervariable regions of immunoglobulins (1987) J Mol Biol. 1987 Aug 20; 196 (4): 901-17). Таким образом, в некоторых вариантах осуществления CDR представляют собой CDR согласно определению Kabat, а в других вариантах осуществления CDR представляют собой CDR согласно определению Chothia. Относительно антигенной детерминанты, распознаваемой CDR-участками антитела, это также называется "эпитопом". Другими словами, эпитоп означает часть любой молекулы, которая может распознаваться или связываться антителом (соответствующий антитело-связывающий участок может называться паратопом). В целом эпитопы состоят из химически активных поверхностных групп молекул, например, аминокислот или сахарных боковых цепей, и обладают конкретными трехмерными структурными характеристиками, а также конкретными зарядными характеристиками.
Термин "моноклональное антитело" или "mAb" в контексте данного описания означает антитело, происходящее из одного клона. Моноклональные антитела могут быть получены с применением гибридомных технологий, таких, как описываемые в публикациях HARLOW. Antibodies: A Laboratory manual. 2nd edition. Cold Spring Harbor: Laboratory press, 1988, и HAMMERLING, et al. Monoclonal Antibodies и T Cell Hybridomas. New York: Elsevier, 1981. p.563-681.
В контексте данного описания термин "человеческое антитело" означает антитело, имеющее вариабельные и постоянные участки, которые являются взятыми из последовательностей иммуноглобулина зародышевой линии человека или являются их близкими аналогами. Человеческое антитело согласно изобретению может включать аминокислотные остатки, которые не кодируются последовательностями иммуноглобулина зародышевой линии человека (например, мутации, включенные путем случайного или сайт-специфического мутагенеза in vitro или путем соматической мутации in vivo). Таким образом, в контексте данного описания термин "человеческое антитело" означает антитело, в котором практически каждая часть белка является по сути подобной антителу зародышевой линии человека. "По сути подобное" означает антитело, имеющее нуклеиновокислотную последовательность, которая является как минимум на 80, в предпочтительном варианте - на 85, в более предпочтительном варианте - на 90, и в более предпочтительном варианте - на 95% гомологичной нуклеиновокислотной последовательности антитела зародышевой линии человека.
В контексте данного описания термин "Fab" означает участки молекул антитела, которые включают вариабельный участок тяжелой цепи и легкой цепи, и которые демонстрируют связывающую активность. "Fab" включает агрегаты одной тяжелой и одной легкой цепи (общеизвестные как Fab), независимо от того, являются ли какие-либо из вышеуказанных ковалентно или нековалентно агрегированными, при условии, что агрегация способна избирательно реагировать с конкретным антигеном или семейством антигенов. Фрагмент Fab является гетеродимером, который включает VL и второй полипептид, который включает домены VH и СН1. В предпочтительном варианте осуществления антитело представляет собой Fab' фрагмент. Fab' фрагменты отличаются от Fab фрагментов тем, что Fab' фрагмент содержит несколько остатков на карбокси-конце тяжелоцепочечного СН1-домена, включая один или несколько цистеинов из "шарнирного участка" антитела.
"F(ab')2" означает фрагмент антитела, полученного путем пепсиновой обработки антитела, или равноценный белок, полученный другими способами, такими, как рекомбинантные технологии. F(ab')2 фрагмент имеет два антиген-связывающих центра и сохраняет способность к сшиванию антигена.
"Fv" является минимальным фрагментом антитела, который содержит полный сайт распознавания и связывания антигена. Этот участок состоит из димера одного тяжело- и одного легкоцепочечного вариабельного домена в тесной нековалентной связи. Именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют для определения антиген-связывающего центра на поверхности VH-VL-димера. Вместе шесть CDR обеспечивают антиген-связывающую специфичность антитела. Однако даже один вариабельный домен (или половина Fv, включающая только три CDR, специфических к антигену) обладает способностью к распознаванию и связыванию антигена, хотя и с более низкой аффинностью по сравнению с полным сайтом связывания.
"Одноцепочечный Fv" или "scFv" означает VH- и VL-домены антитела, причем эти домены присутствуют в одной полипептидной цепи. В предпочтительном варианте scFv также включает полипептидный линкер между VH- и VL-доменами, который позволяет scFv образовывать нужную структуру для связывания антигена (LENNARD. Standard protocols for the construction of scFv libraries. Methods in molecular biology. 2002, №178, стр.59-71).
В контексте данного описания термин "фрагмент антитела" означает один или несколько фрагментов антитела, которые сохраняют способность к специфическому связыванию с КСФ-1R.
Термин "диатела" означает малые фрагменты антитела с двумя антиген-связывающими центрами, и эти фрагменты включают вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH VL). При применении линкера, который является слишком коротким для обеспечения создания пар между двумя доменами на одной цепи, домены вынуждены образовывать пары с комплементарными доменами другого цепи и создавать два антиген-связывающие центры. Диатела могут связываться с одним или несколькими эпитопами. Диатела подробнее описываются в публикациях POLJAK. Production и structure of diabodies. Structure. 1994, т.2, №12, стр.1121-3, HUDSON, et al. High avidity scFv multimers; diabodies и triabodies. Journal of immunological methods. 1999, т.231, №1-2, стр.177-89, и KIPRIYANOV. Generation of bispecific и tandem diabodies. Methods in molecular biology. 2002, №178, стр.317-31.
Были разработаны разные технологии получения фрагментов антител. Традиционно эти фрагменты получали путем протеолитического расщепления интактных антител. Однако эти фрагменты теперь могут образовываться непосредственно рекомбинантными клетками-хозяевами. Фрагменты антител Fab, Fv и scFv могут быть экспрессированы и секретированы из Е. coli, что обеспечивает возможность выработки большого количества этих фрагментов. Технологии выработки фрагментов антител известны специалистам в данной области. В других вариантах осуществления нужным антителом является одноцепочечный Fv фрагмент (scFv).
В контексте данного описания "доменные антитела" (dAbs) состоят из наименьших функциональных связывающих единиц антител, которые отвечают вариабельным участкам тяжелых (VH) или легких (VL) цепей антител. Доменные антитела имеют молекулярную массу приблизительно 13 кДа или менее одной десятой размера полного антитела.
В контексте данного описания "Fd" означает фрагмент антитела, который состоит из доменов VH и CH1.
Термин "антитело" или "Ab" также касается других фрагментов антител, общеизвестных среди специалистов в данной области, например, описанных в публикациях HOLLIGER, et al. Engineered antibody fragments и the rise of single domains. Nature biotechnology. 2005, т.23, №9, стр.1126-36, и HOOGENBOOM, et al. Natural и designer binding sites made by phage display technology. Immunology today. 2000, т.21, №8, стр.371-8.
Согласно одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает:
(i) как минимум один CDR, причем вышеупомянутый CDR включает как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2, - последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, или последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
или,
(ii) как минимум один CDR, причем вышеупомянутый CDR включает как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4, последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4 или последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, причем вышеупомянутые CDR являются, независимо один от другого, выбранными из группы CDR, которые включают как минимум пять последовательных аминокислот:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4,
- или последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4 или 5, в более предпочтительном варианте - 6 CDR, причем вышеупомянутые CDR являются, независимо один от другого, выбранными из группы CDR, которые включают как минимум пять последовательных аминокислот:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4,
- или последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает:
(i) 2, в более предпочтительном варианте - 3 CDR, причем вышеупомянутые CDR являются, независимо один от другого, выбранными из группы CDR, которые включают как минимум пять последовательных аминокислот:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2, или
(ii) 2, в более предпочтительном варианте - 3 CDR, причем вышеупомянутые CDR являются, независимо один от другого, выбранными из группы CDR, которые включают как минимум пять последовательных аминокислот:
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4
- или последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает:
(i) как минимум один CDR, выбранный, независимо от других, из группы CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
или
(ii) как минимум один CDR, выбранный, независимо от других, из группы CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, выбранный, независимо от других, из группы CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO:2,
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4 или 5, в более предпочтительном варианте - 6 CDR, выбранных, независимо один от другого, из группы CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает:
(i) 2, в более предпочтительном варианте - 3 CDR, причем вышеупомянутые CDR являются, независимо один от другого, выбранными из группы CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO:2,
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2,
или
(ii) 2, в более предпочтительном варианте - 3 CDR, причем вышеупомянутые CDR являются, независимо один от другого, выбранными из группы CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает (i) как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 11, 12 или 13; или (ii) как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 14, 15 или 16.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 11, 12, 13, 14, 15 или 16.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4, 5 в более предпочтительном варианте - 6 CDR, которые включают аминокислотная последовательность, изложенную в любой из последовательностей SEQ ID NO: 11, 12, 13, 14, 15 или 16.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает (i) как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 11, 12 или 13; или (ii) как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 14, 15 или 16.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 11, 12, 13, 14, 15 или 16.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4, 5 в более предпочтительном варианте - 6 CDR, как изложено в любой из последовательностей SEQ ID NO: 11, 12, 13, 14, 15 или 16.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает (i) как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 17, 18 или 19; или (ii) как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 20, 21 или 22.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 17, 18, 19, 20, 21 или 22.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4, 5 в более предпочтительном варианте - 6 CDR, которые включают аминокислотная последовательность, изложенную в любой из последовательностей SEQ ID NO: 17, 18, 19, 20, 21 или 22.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает (i) как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 17, 18 или 19; или (ii) как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 20, 21 или 22.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 17, 18, 19, 20, 21 или 22.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4, 5 в более предпочтительном варианте - 6 CDR, как изложено в любой из последовательностей SEQ ID NO: 17, 18, 19, 20, 21 или 22.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает (i) как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 23, 24 или 25; или (ii) как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 26, 27 или 28.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, который включает аминокислотную последовательность, изложенную в любой из последовательностей SEQ ID NO: 23, 24, 25, 26, 27 или 28.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4, 5 в более предпочтительном варианте - 6 CDR, которые включают аминокислотная последовательность, изложенную в любой из последовательностей SEQ ID NO: 23, 24, 25, 26, 27 или 28.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает (i) как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 23, 24 или 25; или (ii) как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 26, 27 или 28.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает как минимум один CDR, как изложено в любой из последовательностей SEQ ID NO: 23, 24, 25, 26, 27 или 28.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает 2, 3, 4, 5 в более предпочтительном варианте - 6 CDR, как изложено в любой из последовательностей SEQ ID NO: 23, 24, 25, 26, 27 или 28.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает три CDR, как изложено в последовательностях SEQ ID NO: 11, 12 и 13.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает три CDR, как изложено в последовательностях SEQ ID NO: 14, 15 и 16.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает три CDR, как изложено в последовательностях SEQ ID NO: 17, 18 и 19.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает три CDR, как изложено в последовательностях SEQ ID NO: 20, 21 и 22.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает три CDR, как изложено в последовательностях SEQ ID NO: 23, 24 и 25.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R, более конкретно - с человеческим КСФ-1R, и включает вариабельный участок, причем вышеупомянутый вариабельный участок включает три CDR, как изложено в последовательностях SEQ ID NO: 26, 27 и 28.
В оптимальных вариантах осуществления вышеупомянутый вариабельный участок также включает один, в более предпочтительном варианте - два, в более предпочтительном варианте - три, и в наиболее предпочтительном варианте - четыре каркасных участка, в более предпочтительном варианте - человеческие FR. В контексте данного описания "человеческий FR" является каркасным участком, который является как минимум на 75% гомологичным каркасному участку естественного человеческого антитела.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает вариабельный участок, причем вариабельный участок включает аминокислотную последовательность, изложенную в SEQ ID NO: 6.
В более предпочтительном варианте осуществления антитело согласно изобретению специфически связывается с КСФ-1R и включает вариабельный участок, причем вариабельный участок представлен в SEQ ID NO: 6.
В другом предпочтительном варианте осуществления антитело согласно изобретению специфически связывается с КСФ-1R и включает вариабельный участок, причем вариабельный участок включает аминокислотную последовательность, изложенную в SEQ ID NO: 9.
В другом более предпочтительном варианте осуществления антитело согласно изобретению специфически связывается с КСФ-1R и включает вариабельный участок, причем вариабельный участок представлен в SEQ ID NO: 9.
В другом варианте осуществления антитело согласно изобретению специфически связывается с КСФ-1R и включает два вариабельных участка, причем вариабельные участки являются, независимо один от другого, выбранными из группы, к которой относятся
(i) вариабельные участки, которые включают CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
(ii) вариабельные участки, которые включают CDR, как изложено в следующих последовательностях:
- последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4;
(iii) вариабельные участки, которые включают три CDR, как изложено в последовательностях SEQ ID NO: 11, 12 и 13;
(iv) вариабельные участки, которые включают три CDR, как изложено в последовательностях SEQ ID NO: 14, 15 и 16;
(v) вариабельные участки, которые включают три CDR, как изложено в последовательностях SEQ ID NO: 17, 18 и 19;
(vi) вариабельные участки, которые включают три CDR, как изложено в последовательностях SEQ ID NO: 20, 21 и 22;
(vii) вариабельные участки, которые включают три CDR, как изложено в последовательностях SEQ ID NO: 23, 24 и 25,
и
(viii) вариабельные участки, которые включают три CDR, как изложено в последовательностях SEQ ID NO: 26, 27 и 28.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает:
(i) первый вариабельный участок, причем вышеупомянутый вариабельный участок включает:
- CDR, представленный в - последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- CDR, представленный в - последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- CDR, представленный в - последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
и
(ii) второй вариабельный участок, причем вышеупомянутый вариабельный участок включает:
- CDR, представленный в - последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- CDR, представленный в - последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- CDR, представленный в - последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает:
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 11, 12 и 13, и
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 14, 15 и 16.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает:
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 17, 18 и 19, и
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 20, 21 и 22.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает:
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 23, 24 и 25, и
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 26, 27 и 28.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает:
- вариабельный участок, изложенный в SEQ ID NO: 6, и
- вариабельный участок, изложенный в SEQ ID NO: 9.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает:
(i) вариабельный участок тяжелой цепи, который включает:
- CDR, представленный в последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- CDR, представленный в последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- CDR, представленный в последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
и
(ii) вариабельный участок легкой цепи, который включает:
- CDR, представленный в последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- CDR, представленный в последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- CDR, представленный в последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает (i) вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 11, 12 и 13, и (ii) вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 14, 15 и 16.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает (i) вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 17, 18 и 19, и (ii) вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 20,21 и 22.
Согласно еще одному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает (i) вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 23, 24 и 25, и (ii) вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 26,27 и 28.
Согласно предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает (i) вариабельный участок тяжелой цепи, как изложено в последовательности SEQ ID NO: 6, и (ii) вариабельный участок легкой цепи, как изложено в последовательности SEQ ID NO: 9.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
(i) вариабельные участки, которые включают:
- CDR, представленный в последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- CDR, представленный в последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- CDR, представленный в последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
и
(ii) вариабельный участок, который включает:
- CDR, представленный в последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- CDR, представленный в последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- CDR, представленный в последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 11, 12 и 13, и
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 14, 15 и 16.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 17, 18 и 19, и
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 20, 21 и 22.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 23, 24 и 25, и
- вариабельный участок, который включает три CDR, как изложено в последовательностях SEQ ID NO: 26, 27 и 28.
Согласно более предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок, изложенный в SEQ ID NO: 6, и
- вариабельный участок, изложенный в SEQ ID NO: 9.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
(i) вариабельный участок тяжелой цепи, который включает:
- CDR, представленный в последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- CDR, представленный в последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- CDR, представленный в последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2, и
(ii) вариабельный участок легкой цепи, который включает:
- CDR, представленный в последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- CDR, представленный в последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- CDR, представленный в последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 11, 12 и 13, и
- вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 14, 15 и 16.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 17, 18 и 19, и
- вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 20, 21 и 22.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 23, 24 и 25, и
- вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 26, 27 и 28.
Согласно более предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и представляет собой scFv, причем вышеупомянутый scFv включает:
- вариабельный участок тяжелой цепи, изложенный в SEQ ID NO: 6, и
- вариабельный участок легкой цепи, изложенный в SEQ ID NO: 9.
в более предпочтительном варианте осуществления предлагается scFv, в котором как минимум одна аминокислота является замещенной (согласно Таблице 1 и Таблице 2) в пределах аминокислотной последовательности, изложенной в SEQ ID NO 6 и 9. В особенно предпочтительном варианте осуществления предлагается человеческое антитело, в котором все аминокислоты, показанные в Таблице 1 и Таблице 2, являются замещенными (согласно Таблице 1 и Таблице 2) в пределах аминокислотной последовательности, изложенной в SEQ ID NO 6 и 9.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает тяжелую цепь, изложенную в SEQ ID NO: 2.
Согласно еще одному предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает легкую цепь, изложенную в SEQ ID NO: 4.
Согласно более предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает тяжелую цепь, изложенную в SEQ ID NO: 2 и легкую цепь, изложенную в SEQ ID NO: 4.
Согласно более предпочтительному варианту осуществления, антитело согласно изобретению специфически связывается с КСФ-1R и включает две тяжелых цепи, изложенных в SEQ ID NO: 2, и две легких цепи, изложенных в SEQ ID NO: 4. Это конкретное антитело по всему тексту данной заявки указывается как CXIIG6.
Согласно еще одному предпочтительному варианту осуществления, настоящее изобретение касается человеческого антитела, которое специфически связывается с КСФ-1R и включает:
(i) вариабельный участок тяжелой цепи, который включает:
- CDR, представленный в последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2,
- CDR, представленный в - последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2, и
- CDR, представленный в - последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
и
(ii) вариабельный участок легкой цепи, который включает:
- CDR, представленный в - последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4,
- CDR, представленный в - последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4, и
- CDR, представленный в - последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, настоящее изобретение касается человеческого антитела, которое специфически связывается с КСФ-1R и включает:
- вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 11, 12 и 13, и
- вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 14, 15 и 16.
Согласно еще одному варианту осуществления, настоящее изобретение касается человеческого антитела, которое специфически связывается с КСФ-1R и включает:
- вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 17, 18 и 19, и
- вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 20, 21 и 22.
Согласно еще одному варианту осуществления, настоящее изобретение касается человеческого антитела, которое специфически связывается с КСФ-1R и включает:
- вариабельный участок тяжелой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 23, 24 и 25, и
- вариабельный участок легкой цепи, который включает три CDR, как изложено в последовательностях SEQ ID NO: 26, 27 и 28.
Согласно предпочтительному варианту осуществления, настоящее изобретение касается человеческого антитела, которое специфически связывается с КСФ-1R и включает:
- вариабельный участок тяжелой цепи, изложенный в SEQ ID NO: 6, и
- вариабельный участок легкой цепи, изложенную в SEQ ID NO: 9.
В более предпочтительном варианте осуществления предлагается человеческое антитело, в котором как минимум одна аминокислота является замещенной (согласно Таблице 1 и Таблице 2) в пределах аминокислотной последовательности, изложенной в SEQ ID NO: 6 и 9. В более предпочтительном варианте осуществления предлагается человеческое антитело, в котором все аминокислоты, показанные в Таблице 1 и Таблице 2, являются замещенными (согласно Таблице 1 и Таблице 2) в пределах аминокислотной последовательности, изложенной в SEQ ID NO: 6 и 9.
Согласно еще одному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает:
(а) первый вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 имеет как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 45 и заканчивается в позиции 54 SEQ ID NO: 2;
CDR2 имеет как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 66 и заканчивается в позиции 87 SEQ ID NO: 2; и
CDR3 имеет как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 117 и заканчивается в позиции 126 SEQ ID NO: 2;
и
(b) второй вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 имеет как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 44 и заканчивается в позиции 56 SEQ ID NO: 4;
CDR2 имеет как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 66 и заканчивается в позиции 76 SEQ ID NO: 4; и
CDR3 имеет как минимум пять последовательных аминокислот последовательности, которая начинается с позиции 109 и заканчивается в позиции 117 SEQ ID NO: 4.
Согласно еще одному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R и включает:
(а) первый вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 имеет аминокислотную последовательность, выбранную из группы, к которой относятся: SEQ ID NO: 11, 17 и 23;
CDR2 имеет аминокислотную последовательность, выбранную из группы, к которой относятся: SEQ ID NO: 12, 18 и 24; и
CDR3 имеет аминокислотную последовательность, выбранную из группы, к которой относятся: SEQ ID NO: 13, 19 и 25;
и
(b) второй вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 имеет аминокислотную последовательность, выбранную из группы, к которой относятся: SEQ ID NO: 14, 20 и 26;
CDR2 имеет аминокислотную последовательность, выбранную из группы, к которой относятся: SEQ ID NO: 15, 21 и 27; и
CDR3 имеет аминокислотную последовательность, выбранную из группы, к которой относятся: SEQ ID NO: 16, 22 и 28.
Согласно еще одному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает любой из компонентов (i), (ii) или (iii):
(i)
(a) первый вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 является изложенным в SEQ ID NO: 11;
CDR2 является изложенным в SEQ ID NO: 12; и
CDR3 является изложенным в SEQ ID NO: 13;
и
(b) второй вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 является изложенным в SEQ ID NO: 14;
CDR2 является изложенным в SEQ ID NO: 15; и
CDR3 является изложенным в SEQ ID NO: 16; или
(ii) (а) первый вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 является изложенным в SEQ ID NO: 17;
CDR2 является изложенным в SEQ ID NO: 18; и
CDR3 является изложенным в SEQ ID NO: 19;
и
(b) второй вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 является изложенным в SEQ ID NO: 20;
CDR2 является изложенным в SEQ ID NO: 21; и
CDR3 является изложенным в SEQ ID NO: 22;
или
(iii) (а) первый вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности; где:
CDR1 является изложенным в SEQ ID NO: 23;
CDR2 является изложенным в SEQ ID NO: 24; и
CDR3 является изложенным в SEQ ID NO: 25;
и
(b) второй вариабельный участок, который определяется следующей формулой
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
где:
FR1, FR2, FR3 и FR4 являются каркасными участками;
CDR1, CDR2 и CDR3 являются участками определения комплементарности;
где:
CDR1 является изложенным в SEQ ID NO: 26;
CDR2 является изложенным в SEQ ID NO: 27; и
CDR3 является изложенным в SEQ ID NO: 28.
Согласно еще одному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает:
- первый вариабельный участок, который включает аминокислотную последовательность SEQ ID NO: 6; и
- второй вариабельный участок, который включает аминокислотную последовательность SEQ ID NO: 9.
Согласно еще одному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает:
- первый вариабельный участок, который включает аминокислотную последовательность SEQ ID NO: 2; и
- второй вариабельный участок, который включает аминокислотную последовательность SEQ ID NO: 4.
Согласно еще одному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает:
- тяжелую цепь, выбранную из группы, к которой относятся SEQ ID NO: 37 (см. Фиг.32) и SEQ ID NO: 38, и
- легкую цепь, выбранную из группы, к которой относятся SEQ ID NO: 39, SEQ ID NO: 40 и SEQ ID NO: 41.
Согласно еще одному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает:
- первый вариабельный участок, выбранный из группы, к которой относятся SEQ ID NO: 42 и SEQ ID NO: 43; и
- второй вариабельный участок, выбранный из группы, к которой относятся SEQ ID NO: 44, SEQ ID NO: 45 и SEQ ID NO: 46.
Согласно предпочтительному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает (а) тяжелую цепь, которая состоит из SEQ ID NO: 37, и (b) легкую цепь, которая состоит из SEQ ID NO: 39.
Согласно еще одному предпочтительному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает (а) тяжелую цепь, которая состоит из SEQ ID NO: 38, и (b) легкую цепь, которая состоит из SEQ ID NO: 40.
Согласно одному предпочтительному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает (а) тяжелую цепь, которая состоит из SEQ ID NO: 37, и (b) легкую цепь, которая состоит из SEQ ID NO:41.
Согласно предпочтительному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает (а) первый вариабельный участок, который состоит из SEQ ID NO: 42, и (b) второй вариабельный участок, который состоит из SEQ ID NO: 44.
Согласно еще одному предпочтительному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает (а) первый вариабельный участок, который состоит из SEQ ID NO: 43, и (b) второй вариабельный участок, который состоит из SEQ ID NO:45.
Согласно одному предпочтительному варианту осуществления, настоящее изобретение касается выделенного, рекомбинантного или очищенного антитела, которое специфически связывается с КСФ-1R, в более предпочтительном варианте - человеческого КСФ-1R, которое включает (а) первый вариабельный участок, который состоит из SEQ ID NO: 42, и (b) второй вариабельный участок, который состоит из SEQ ID NO: 46.
Антитело, более конкретно - человеческое антитело согласно изобретению, может относиться к разным изотипам, таким, как IgG, IgA, IgM или IgE. В предпочтительном варианте осуществления антитело, более конкретно - человеческое антитело согласно изобретению представляет собой IgG.
В соответствующем варианте осуществления человеческое антитело включает модифицированный или немодифицированный постоянный участок человеческого IgG1, IgG2, IgG3 или IgG4. В предпочтительном варианте осуществления постоянным участком является человеческий IgG1 или IgG4, который необязательно может быть модифицирован для усиления или ослабления определенных свойств.
В случае IgG1 модификации постоянного участка, в частности, шарнирного или СН2-участка, могут повышать или снижать эффекторную функцию, включая активность ADCC и/или CDC. В других вариантах осуществления IgG2 постоянный участок модифицируют для уменьшения образования агрегатов антитело-антиген. В случае IgG4 модификации постоянного участка, в частности, шарнирного участка, могут уменьшать образование полуантител.
Необходимая аффинность связывания может сохраняться, даже несмотря на мутацию одной или нескольких аминокислот в антителе. Эти варианты имеют как минимум одну аминокислоту в антителе, замененную на другой остаток. Согласно еще одному варианту осуществления, настоящее изобретение предлагает антитело, которое специфически связывается с КСФ1, как было описано выше, в котором как минимум одна из аминокислот, включенных в CDR, является консервативно замещенной. Консервативные замещения показаны в Таблице 3.
Настоящее изобретение также касается способа модификации антитела согласно изобретению через созревание аффинности.
В контексте данного описания "созревание аффинности" означает замещение одной или нескольких аминокислот, которые содержатся в одном или нескольких CDR, причем вышеупомянутое замещение в результате улучшает аффинность антитела к КСФ-1R по сравнению с исходным антителом, не имеющим таких замещений. Процессы созревания аффинности известны специалистам в данной области. См., например, способы, описанные в публикациях MARKS, et al. By-passing immunization: building high affinity human antibodies by chain shuffling. Biotechnology. 1992, т.10, №7, стр.779-83; BARBAS, et al. In vitro evolution of a neutralizing human antibody to human immunodeficiency virus type 1 to enhance affinity and broaden strain cross-reactivity. Proceedings of the National Academy of Sciences of the United States of America. 1994, т.91, №9, стр.3809-13; SCHIER. Identification of functional и structural amino-acid residues by parsimonious mutagenesis. Gene. 1996, т.169, №2, стр.147-55; YELTON. Affinity maturation of the BR96 anti-carcinoma antibody by codon-based mutagenesis. J. immunol. 1995, т.155, №4, стр.1994-2004; JACKSON, et al. In vitro antibody maturation. Improvement of a high affinity, neutralizing antibody against IL-1 beta. J. immunol. 1995, т.154, №7, стр.3310-9. и HAWKINS, et al. Selection of phage antibodies by binding affinity. Mimicking affinity maturation. Journal of molecular biology. 1992, т.226, №3, стр.889-96.
Настоящее изобретение также касается антитела, которое специфически связывается с КСФ-1R, полученного путем созревания аффинности, как описано выше.
В другом варианте осуществления настоящее изобретение обеспечивает варианты описанного выше антитела, которые имеют аминокислотная последовательность, которая является как минимум на 80%, в предпочтительном варианте - как минимум на 85%, в более предпочтительном варианте - как минимум на 90%, в более предпочтительном варианте - как минимум на 98% гомологичной аминокислотной последовательности вышеописанного антитела.
В другом варианте осуществления антитело согласно изобретению специфически связывается с более, чем одним эпитопом. Например, антитело согласно изобретению может связываться с двумя разными эпитопами КСФ-1R. В альтернативном варианте антитело согласно изобретению может быть способным к связыванию с КСФ-1R и другой молекулой. В контексте данного описания антитела, которые специфически связываются более, чем с одним эпитопом, могут быть сшитыми антителами. Например, антитело может быть соединено с авидином, а другое - с биотином. Сшитые антитела могут быть получены с применением любых удобных способов сшивания, общеизвестных среди специалистов в данной области. Способы образования биспецифических антител из фрагментов антител были описаны, например, в публикациях BRENNAN, et al. Preparation of bispecific antibodies by chemical recombination of monoclonal immunoglobulin G1 fragments. Science. 1985, т.229, №4708, стр.81-3, и SHALABY, et al. Development of humanized bispecific antibodies reactive with cytotoxic lymphocytes and tumor cells overexpressing the HER2 protooncogene. The Journal of experimental medicine. 1992, т.175, №1, стр.217-25; KOSTELNY, et al. Formation of a bispecific antibody by the use of leucine zippers. J. immunol. 1992, т.148, №5, стр.1547-33.
Согласно предпочтительному варианту осуществления, антитело, которое специфически связывается более, чем с одним эпитопом согласно изобретению, является диателом.
Согласно еще одному предпочтительному варианту осуществления, антитело, которое специфически связывается более, чем с одним эпитопом согласно изобретению, является линейным антителом, как описано в публикации ZAPATA, et al. Engineering linear F(ab')2 fragments for efficient production in Escherichia coli and enhanced antiproliferative activity. Protein engineering. 1995, т.8, №10, стр.1057-62.
В предпочтительном варианте осуществления антитело согласно изобретению специфически связывается с как минимум одним эпитопом, расположенным между позициями аминокислот с 20 по 41 SEQ ID N0: 29 (то есть, в N-конечной части человеческого домена D1). В предпочтительном варианте осуществления антитело согласно изобретению связывается с одним эпитопом, расположенным между позициями аминокислот с 20 по 41 SEQ ID N0: 29 (то есть, в N-конечной части человеческого домена D1) и не связывается ни с одним эпитопом, расположенным между позициями аминокислот с 42 по 90 и/или между позициями аминокислот с 91 по 104 и/или между позициями аминокислот с 105 по 199 и/или между позициями аминокислот с 200 по 298 SEQ ID N0: 29. Согласно предпочтительному варианту осуществления, антитело согласно настоящему изобретению способно к распознаванию минимального эпитопа, расположенного между позициями аминокислот с 20 по 41 SEQ ID N0: 29 (то есть, в N-конечной части человеческого домена D1). Согласно предпочтительному варианту осуществления, антитело согласно настоящему изобретению способно к распознаванию и связыванию с последовательностью pTG18016 (см. Фиг.19 и соответствующий пример).
В предпочтительном варианте осуществления антитело согласно изобретению не конкурирует с лигандом IL-34 за связывание с рецептором КСФ-1R. Термин "не конкурирует с лигандом IL-34" в контексте данного описания означает отсутствие ингибирования связывания лиганда IL34 с его рецептором КСФ-1R.
В предпочтительном варианте осуществления антитело согласно изобретению частично конкурирует с лигандом КСФ-1 за связывание с рецептором КСФ-1R. Термин "частично конкурирует с лигандом КСФ-1" в контексте данного описания касается ингибирования связывания лиганда КСФ-1 с его рецептором КСФ-1R, которое является меньшим, чем 100%, в предпочтительном варианте - меньшим, чем 50%, в более предпочтительном варианте - меньшим, чем 20%, в наиболее предпочтительном варианте - меньшим, чем 10%. Этот частичный ингибитор только уменьшает, но полностью не исключает лигандное связывание, и ингибирование называется частичным ингибированием.
В предпочтительном варианте осуществления антитело согласно изобретению может частично предотвращать связывание КСФ1 с его рецептором КСФ-1R и не может полностью ингибировать вышеупомянутое связывание. То есть, антитела согласно изобретению могут уменьшать связывание КСФ-1 с КСФ-1R приблизительно на 5-10%. Согласно отдельному варианту осуществления, вышеупомянутое уменьшение связывания измеряется, как описано в представленном разделе "Эксперименты", путем измерения связывания КСФ-1 (который имеет аминокислотную последовательность, которая проходит между позициями с 1 по 444 SEQ ID: 47 - см. Фиг.21) с рецептором КСФ-1R, который имеет аминокислотную последовательность, которая проходит от Ile 20 до Glu 512 у SEQ ID NO: 29.
В предпочтительном варианте осуществления антитела согласно изобретению характеризуются высокоаффинным связыванием с КСФ1-К. То есть, антитела согласно изобретению имеют показатель Ki менее 1 нМ, в предпочтительном варианте - менее 0,8 нМ, в более предпочтительном варианте - менее 0,6 нМ. Благодаря такой неожиданно высокой аффинности, антитела согласно изобретению могут вводиться в меньшем количестве, и, следовательно, исключать возможные побочные эффекты.
В конкретных вариантах осуществления антитело согласно настоящему изобретению является антагонистическим антителом, которое частично или полностью блокирует или ингибирует биологическую активность полипептидов или клеток, с которыми оно специфически или предпочтительно связывается, то есть, экспрессирующих КСФ1-R клеток, в более предпочтительном варианте - экспрессирующих КСФ1-R1 человеческих клеток, в наиболее предпочтительном варианте - экспрессирующих КСФ1-R человеческих раковых клеток.
Согласно конкретному варианту осуществления настоящего изобретения, антитело согласно настоящему изобретению обладает как минимум одним из нижеуказанных благоприятных свойств:
- In vitro испытание: антитело согласно изобретению не конкурирует с лигандом IL-34 за связывание с рецептором КСФ-1R;
- In vitro испытание: антитело согласно изобретению частично конкурирует с лигандом КСФ-1 за связывание с рецептором КСФ-1R;
- ADCC-испытание: в присутствии нормальных мононуклеарных клеток периферической крови человека антитело согласно настоящему изобретению обеспечивает антителозависимую клеточную цитотоксичность (ADCC) против экспрессирующих КСФ1-R клеток, в частности, экспрессирующих КСФ1-R раковых клеток человека;
- In vivo испытание: антитело согласно настоящему изобретению обеспечивает противоопухолевый эффект против отличных от человека животных, которые являются носителями экспрессирующих КСФ1-R раковых клеток человека;
- In vivo испытание: антитело согласно настоящему изобретению обеспечивает противоопухолевый эффект против животных, включая человека, которые являются носителями экспрессирующих КСФ1-R раковых клеток человека,
- In vivo испытание: антитело согласно настоящему изобретению ингибирует зависящую от КСФ-1 пролиферацию клеток AML5.
- In vivo испытание: антитело согласно настоящему изобретению частично ингибирует зависящее от КСФ1-R фосфорилирование КСФ-1R;
- In vivo испытание: антитело согласно настоящему изобретению не имеет прямого антагонистического воздействия на КСФ1-1R.
Антитело согласно изобретению может быть гликозилированным или негликозилированным.
В контексте данного описания термин "гликозилирование" означает присутствие углеводных единиц, которые являются ковалентно присоединенными к антителу.
В другом варианте осуществления антитело согласно изобретению является конъюгированным с радиосенсибилизирующим агентом, рецептором и/или цитотоксичным агентом.
В контексте данного описания термин "радиосенсибилизатор" означает молекулу, которая делает клетки более чувствительными к лучевой терапии. К радиосенсибилизаторам, помимо прочих, относятся метронидазол, мизонидазол, десметилмизонидазол, пимонидазол, етанидазол, ниморазол, митомицин С, RSU 1069, SR 4233, Е09, RB 6145, никотинамид, 5-бромодезоксиуридин (BUdR), 5-йододезоксиуридин (IUdR), бромодезоксицитидин, фтородезоксиуридин (FUdR), гидроксимочевина и цисплатин.
В контексте данного описания термин "рецептор" означает соединение, способное специфически связываться с лигандом. Согласно предпочтительному варианту осуществления изобретения, рецептором является биотин.
В контексте данного описания термин "цитотоксичный агент" означает соединение, являющееся прямо токсичным для клеток, препятствуя их воспроизводству или росту. Согласно предпочтительному варианту осуществления, цитотоксичный агент, применяемый в контексте настоящего изобретения, выбирают из группы, к которой относятся противораковое терапевтическое средство, токсин (например, ферментно активный токсин бактериального, грибкового, растительного или животного происхождения, или его фрагменты) или радиоактивный изотоп.
В другом варианте осуществления антитело согласно изобретению является конъюгированным с маркирующим агентом.
В контексте данного описания "маркирующий агент" означает обнаружимое соединение. Маркирующий агент может обнаруживаться сам по себе (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментной метки, может катализировать химическую модификацию обнаружимого субстратного соединения.
В контексте данного описания термин "конъюгированный" означает, что антитело согласно изобретению и маркирующий агент являются ковалентно или нековалентно связанными.
"Ковалентная связь" означает соединение через химически активные функциональные группы, необязательно с промежуточным использованием кросс-линкера или другого активирующего агента (см., например HERMANSON. Bioconjugate techniques. Academic press, 1996). Антитело согласно изобретению и/или конъюгированный агент могут быть модифицированы для обеспечения возможности их связывания, например, путем замещения на активированной карбонильной группе (включая активированные in situ) или на имидоэфире, путем добавления на ненасыщенной карбонильной группе, путем восстановительного аминирования, нуклеофильного замещения на насыщенном атоме углерода или на гетероатоме, путем реакции на ароматических циклах. В частности, соединение может осуществляться с использованием гомобифункциональных или гетеробифункциональных сшивающих реагентов. Гомобифункциональные кросс-линкеры, включая глутаральдегид, янтарную кислоту и бис-имидоэфир, такой, как DMS (диметил-суберимидат), могут применяться для соединения аминных групп, которые могут присутствовать на разных компонентах. Многочисленные примеры приводятся в публикации HERMANSON. Bioconjugate techniques. Academic press, 1996. стр.118-228 и хорошо известны специалистам в данной области. К гетеробифункциональным кросс-линкерам относятся те, которые имеют как аминные, так и сульфгидрильные реакционноспособные группы, карбонильные реакционноспособные и сульфгидрильные реакционноспособные группы и сульфгидрильные реакционноспособные группы и фотореактивные линкеры. Подходящие гетеробифункциональные кросс-линкеры описываются, например, в публикации HERMANSON. Bioconjugate techniques. Academic press, 1996. стр.229-285. Примерами являются, например, SPDP (N-сукцинимидил 3-(2-пиридилдитио)пропионат), SMBP (сукцинимидил-4-(р-малеимидофенил)бутират), SMPT (сукцинимидилоксикарбонил-α-метил-α-(2-пиридилдитио)толуол), MBS (m-малеимидобензоил-N-гидроксисукцинимидный эфир), SIAB (N-сукцинимидил (4 йодоацетил)аминобензоат), GMBS (γ-малеимидобутирилокси)сукцинимидный эфир), SIAX (сукцинимидил-6- йодоацетиламиногексонат, SIAC (сукцинимидил-4-йодоацетиламинометил), NPIA (р-нитрофенилйодоацетат). Другие примеры могут применяться для соединения углеводсодержащих молекул (например, env гликопротеины, антитела) с сульфгидрильными реакционноспособными группами. Примерами могут быть МРВН (4-(4-N малеимидофенил) масляной кислоты гидразид) и PDPH (4-(N-малеимидометил) циклогексан-1-карбоксил-гидразид (М2С2Н и 3-2(2-пиридилдитио)пропионил гидразид).
Согласно еще одному варианту осуществления, настоящее изобретение касается нуклеиновокислотной последовательности, кодирующей антитело согласно изобретению.
Термин "нуклеиновокислотная последовательность" касается линейной последовательности нуклеотидов. Нуклеотиды представляют собой линейную последовательность полирибонуклеотидов или полидезоксирибонуклеотидов, или их смесь. Примерами полинуклеотидов в контексте настоящего изобретения могут быть одно- и двухцепочечные ДНК, одно- и двухцепочечные РНК и гибридные молекулы, включающие смеси одно- и двухцепочечных ДНК и РНК. Кроме того, полинуклеотиды согласно настоящему изобретению могут включать один или несколько модифицированных нуклеотидов.
Согласно предпочтительному варианту осуществления изобретения, нуклеиновокислотная последовательность согласно изобретению включается в вектор.
Вектор может иметь плазмидное или вирусное происхождение и в соответствующих случаях может комбинироваться с одним или несколькими веществами, улучшающими эффективность и/или устойчивость трансфекции вектора. Эти вещества широко задокументированы в литературе, доступной специалистам в данной области (см., например, FELGNER, et al. Cationic liposome mediated transfection. Proceedings of the Western Pharmacology Society. 1989, т.32, стр.115-21; HODGSON, et al. Virosomes: cationic liposomes enhance retroviral transduction. Nature biotechnology. 1996, т.14, №3, стр.339-42; REMY, et al. Gene transfer with a series of lipophilic DNA-binding molecules. Bioconjugate chemistry. 1994, т.5, №6, стр.647-54). В качестве неограничивающей иллюстрации вещества могут представлять собой полимеры, липиды, в частности, катионные липиды, липосомы, ядерные белки или нейтральные липиды. Эти вещества могут использоваться отдельно или в комбинации. Может быть предусмотрена комбинация рекомбинантного плазмидного вектора, который комбинируется с катионными липидами (DOGS, DC-CHOL, спермин-chol, спермидин-chol и т.п.), лизофосфолипидами (например, гексадецилфосфохолином) и нейтральными липидами (DOPE).
Согласно предпочтительному варианту осуществления, катионными липидами, которые могут применяться согласно настоящему изобретению, являются катионные липиды, описанные в документе EP 901463, в более предпочтительном варианте - pcTG90.
Выбор плазмид, которые могут применяться в контексте настоящего изобретения, огромен. Они могут представлять собой векторы клонирования и/или векторы экспрессии. В целом они известны специалистам в данной области, и, хотя многие из них могут быть приобретены в торговой сети, их также можно построить или модифицировать с применением технологий генетической манипуляции. В качестве примеров могут служить плазмиды, взятые из pBR322 (Gibco BRL), pUC (Gibco BRL), pBluescript (Stratagene), pREP4, pCEP4 (Invitrogen) или p Poly (LATHE, et al. Plasmid and bacteriophage vectors for excision of intact inserts. Gene. 1987, т.57, №2-3, стр.193-201). В предпочтительном варианте, плазмида, которая может применяться в контексте настоящего изобретения, содержит источник репликации, который гарантирует запуск репликации в клетке-продуценте и/или клетке-хозяине (например, источник ColE1 выбирают для плазмиды, предназначенной для вырабатывания в Е. coli, а систему oriP/EBNA1 выбирают в случае необходимости саморепликации плазмиды в клетке-хозяине млекопитающего; см. LUPTON, et al. Mapping genetic elements of Epstein-Barr virus that facilitate extrachromosomal persistence of Epstein-Barr virus-derived plasmids in human cells. Molecular and cellular biology. 1985, т.5, №10, стр.2533-42; YATES, et al. Stable replication of plasmids derived from Epstein-Barr virus in various mammalian cells. Nature. 1985, т.313, №6005, стр.812-5). Плазмида дополнительно может включать селекционный ген, позволяющий отбирать или распознавать трансфицированные клетки (комплементация ауксотрофной мутации, ген, кодирующий резистентность к антибиотику, и т.п.). Естественно, плазмида может содержать дополнительные элементы, улучшающие ее сохранение и/или ее устойчивость в данной клетке (последовательность, способствующую сохранению плазмиды в мономерной форме (SUMMERS, et al. Multimerization of high copy number plasmids causes instability: CoIE1 encodes a determinant essential for plasmid monomerization and stability. Cell. 1984, т.36, №4, стр.1097-103, последовательности для включения в клеточный геном).
В отношении вирусного вектора может быть предусмотрен вектор, взятый из поксвируса (вирус вакцины, в частности, MVA, поксвирус канареек и т.п.), из аденовируса, из ретровируса, из вируса герпеса, из альфа-вируса, из вируса пенистости или из аденовирус-ассоциированного вируса. Возможно применение компетентных к репликации или дефицитных по репликации вирусных векторов. Предпочтение отдают применению вектора, который не интегрируется. В этом отношении аденовирусные векторы и векторы, взятые из поксвируса, в более предпочтительном варианте - вирус вакцины и MVA, являются особенно подходящими для осуществления настоящего изобретения.
Согласно предпочтительному варианту осуществления, вирусный вектор согласно изобретению взят из модифицированного вируса коровьей оспы Ankara (MVA). Векторы MVA и способы получения таких векторов полностью описываются в европейских патентах EP 83286 и EP 206920, а также в публикации SUTTER, et al. Nonreplicating vaccinia vector efficiently expresses recombinant genes. Proc. Natl. Acad. Sci. U.S.A. 1992, т.89, №22, стр.10847-51. Согласно более предпочтительному варианту осуществления, нуклеиновокислотная последовательность согласно изобретению может быть вставлена в делецию I, II, III, IV, V и VI вектора MVA, в более предпочтительном варианте - в делецию III (MEYER, et al. Mapping of deletions in the genome of the highly attenuated vaccinia virus MVA and their influence on virulence. The Journal of general virology. 1991, т.72, ч. 5, стр.1031-8.; SUTTER, et al. A recombinant vector derived from the host range-restricted and highly attenuated MVA strain of vaccinia virus stimulates protective immunity in mice to influenza virus. Vaccine. 1994, т.12, №11, стр.1032-40).
Ретровирусы обладают свойством инфицирования и, в большинстве случаев, интегрирования в делящиеся клетки и в этом отношении являются особенно подходящими для применения в лечении рака. Рекомбинантный ретровирус согласно изобретению, как правило, содержит последовательность LTR, участок инкапсулирования и нуклеотидную последовательность согласно изобретению, которая находится под контролем ретровирусной LTR или внутреннего промотора, такого, как описываемые ниже. Рекомбинантный ретровирус может происходить от ретровируса любого происхождения (мышей, приматов, кошачьих, человека и т.п.), в частности, от MoMuLV (вируса мышиного лейкоза Молони), MVS (вируса саркомы мышей) или Мышиного ретровируса Фрейда (Fb29). Он распространяется в линии инкапсулирующих клеток, способной обеспечивать in trans вирусные полипептиды gag, pol и/или env, которые требуются для составления вирусной частицы. Такие линии клеток описываются в литературе (РА 317, Psi CRIP GP+Am-12 etc). Ретровирусный вектор согласно изобретению может содержать модификации, в частности, в LTR (замена промоторного участка эукариотным промотором) или участке инкапсулирования (замена гетерологичным участком инкапсулирования, например, VL3O-типа) (см. документ US 5747323).
Также следует указать на применение аденовирусного вектора, в котором полностью или частично отсутствует как минимум один участок, который является существенным для репликации и выбирается из участков Е1, Е2, Е4 и L1-L5 с целью избежания распространения вектора в организме-хозяине или окружающей среде. Предпочтение отдается делеции участка Е1. Однако возможна комбинация с другими модификациями / делециями, влияющими, в частности, на все или на часть участков Е2, Е4 и/или L1-L5, настолько, чтобы недостающие существенные функции дополнялись in trans путем дополнения линии клеток и/или хелперного вируса. В этом отношении возможно применение векторов второго поколения существующего уровня техники (см., например, международные заявки WO 94/28152 и WO 97/04119). В качестве иллюстрации, делеция большей части участка Е1 и единицы транскрипции Е4 является особенно предпочтительной. С целью повышения способности к клонированию аденовирусный вектор дополнительно может быть полностью или частично лишен несущественного участка Е3. Согласно еще одному альтернативному варианту, возможно применение минимального аденовирусного вектора, который сохраняет последовательности, существенные для инкапсуляции, то есть, 5' и 3' ITR (обращенного концевого повтора), и участка инкапсулирования. Различные аденовирусные векторы и способы их получения известны специалистам (см., например, GRAHAM, et al. Methods in molecular biology. Edited by MURREY. The human press inc., 1991. p.109-128).
Кроме того, происхождение аденовирусного вектора согласно изобретению может быть разным, как с точки зрения вида, так и с точки зрения серотипа. Вектор может быть взят из генома аденовируса человеческого или животного (собак, птиц, крупного рогатого скота, мышей, овец, свиней и т.п.) происхождения или из гибрида, который включает фрагменты аденовирусного генома как минимум из двух различных источников. В качестве более конкретного примера можно привести аденовирусы CAV-1 или CAV-2 собаки, аденовирус DAV птиц или аденовирус Bad 3-го типа крупного рогатого скота (ZAKHARCHUK, et al. Physical mapping and homology studies of egg drop syndrome (EDS-76) adenovirus DNA. Archives of virology. 1993, т.128, №1-2, стр.171-6; SPIBEY, et al. Molecular cloning and restriction endonuclease mapping of two strains of canine adenovirus type 2. The Journal of general virology. 1989, т.70, ч. 1, стр. 165-72; JOUVENNE, et al. Cloning, physical mapping and cross-hybridization of the canine adenovirus types 1 and 2 genomes. Gene. 1987, т.60, №1, стр.21-8.; MITTAL, et al. Development of a bovine adenovirus type 3-based expression vector. The Journal of general virology. 1995, т.76, ч.1, стр.93-102). Однако предпочтение отдается аденовирусному вектору человеческого происхождения, который в предпочтительном варианте является взятым из аденовируса серотипа C, в частности, аденовируса 2-го или 5-го типа серотипа C.
Термин "компетентный к репликации" в контексте данного описания относится к вирусному вектору, способному к репликации в клетке-хозяине в отсутствие какой-либо транс-комплементации.
Согласно предпочтительному варианту осуществления изобретения, компетентный к репликации вектор является компетентным к репликации аденовирусным вектором. Эти компетентные к репликации аденовирусные векторы хорошо известны специалистам в данной области. Среди них особое предпочтение отдается аденовирусным векторам, делетированным на участке E1b, кодирующем 55 кДа ингибитор Р53, как в вирусе ONYX-015 (BISCHOFF, et al. An adenovirus mutant that replicates selectively in p53-deficient human tumor cells. Science. 1996, т.274, №5286, стр.373-6; He HEISE, et al. An adenovirus E1A mutant that demonstrates potent and selective systemic anti-tumoral efficacy. Nature Medicine. 2000, т.6, №10, стр.1134-9; документ WO 94/18992). Соответственно, этот вирус может использоваться для избирательного инфицирования и уничтожения опухолевых клеток с дефицитом р53. Специалист в данной области также может выполнить мутацию и разрушение ингибиторного гена р53 в аденовирусе 5 или других вирусах в соответствии с общепринятыми технологиями. Аденовирусные векторы, делетированные в участке связывания Rb E1A, также могут использоваться согласно настоящему изобретению. Например, вирус Delta24, являющийся мутантным аденовирусом с делецией 24 пар оснований в участке E1A (FUEYO, et al. A mutant oncolytic adenovirus targeting the Rb pathway produces anti-glioma effect in vivo. Oncogene. 2000, т.19, №1, стр.2-12.). Delta24 имеет делецию в участке связывания Rb и не связывается с Rb. Таким образом, репликация мутантного вируса ингибируется Rb в нормальной клетке. Однако, если Rb инактивируется, и клетка становится неопластической, Delta24 перестает ингибироваться. В то же время, мутантный вирус эффективно реплицируется и подвергается лизису в клетках с дефицитом Rb.
Аденовирусный вектор согласно настоящему изобретению может вырабатываться in vitro в Escherichia coli (Е. coli) путем сшивания или гомологичной рекомбинации (см., например, международную заявку WO 96/17070) или путем рекомбинации в дополняющей линии клеток.
Согласно предпочтительному варианту осуществления изобретения, вектор также включает элементы, необходимые для экспрессии антитела согласно изобретению.
Элементы, необходимые для экспрессии, состоят из всех элементов, которые обеспечивают возможность транскрипции нуклеиновокислотной последовательности в РНК и мРНК в полипептид. Эти элементы, в частности, включают промотор, который может быть регулируемым или конститутивным. Естественно, промотор подходит к выбранному вектору и клетке-хозяину. Примерами могут быть эукариотные гены-промоторы PGK (фосфоглицераткиназа), МТ (металлотионеин; MCIVOR. Human purine nucleoside phosphorylase and adenosine deaminase: gene transfer into cultured cells and murine hematopoietic stem cells by using recombinant amphotropic retroviruses. Molecular and cellular biology. 1987, т.7, №2, стр.838-46), α-1 антитрипсин, CFTR, поверхностно-активное вещество, иммуноглобулин, актин (TABIN, et al. Adaptation of a retrovirus as a eukaryotic vector transmitting the herpes simplex virus thymidine kinase gene. Molecular and cellular biology. 1982, т.2, №4, стр.426-36) и SRα (TAKEBE, et al. SR alpha promoter: an efficient and versatile mammalian cDNA expression system composed of the simian virus 40 early promoter and the R-U5 segment of human T-cell leukemia virus type 1 long terminal repeat. Molecular and cellular biology. 1988, т.8, №1, стр.466-72), ранний промотор вируса SV40 (вирус обезьян), LTR вируса RSV (вируса саркомы Рауса), промотор HSV-1 ТК, ранний промотор вируса CMV (Цитомегаловирус), промоторы р7.5К pH5R, pK1L, р28 и p11 вируса вакцины, химерные промоторы, такие, как p11K7.5 и E1A и аденовирусные промоторы MLP. Промотор также может быть промотором, который стимулирует экспрессию в опухолевых или раковых клетках. Отдельно можно упомянуть промоторы гена MUC-1, который сверхэкспрессируется при раке молочной железы и предстательной железы (CHEN, et al. Breast cancer selective gene expression and therapy mediated by recombinant adenoviruses containing the DF3/MUC1 promoter. The Journal of clinical investigation. 1995, т.96, №6, стр.2775-82), гена CEA (эмбрионального антигена карциномы), который сверхэкспрессируется при раке толстой кишки (SCHREWE, et al. Cloning of the complete gene for carcinoembryonic antigen: analysis of its promoter indicates a region conveying cell type-specific expression. Molecular and cellular biology. 1990, т.10, №6, стр.2738-48), гена тирозиназы, который сверхэкспрессируется при меланомах (VILE, et al. Use of tissue-specific expression of the herpes simplex virus thymidine kinase gene to inhibit growth of established murine melanomas following direct intratumoral injection of DNA. Cancer res. 1993, т.53, №17, стр.3860-4), гена ERBB-2, который сверхэкспрессируется при раке молочной железы и поджелудочной железы (HARRIS, et al. Gene therapy for cancer using tumor-specific prodrug activation. Gene therapy. 1994, т.1, №3, стр.170-5) и гена α-фетопротеина, который сверхэкспрессируется при раке печени (KANAI, et al. In vivo gene therapy for alpha-fetoprotein-producing hepatocellular carcinoma by adenovirus-mediated transfer of cytosine deaminase gene. Cancer res. 1997, т.57, №3, стр.461-5). Особое предпочтение отдается раннему промотору цитомегаловируса (CMV).
Однако при применении вектора, взятого из вируса вакцины (например, вектора MVA) особое предпочтение отдается промотору гена 7.5К тимидинкиназы и промотору pH5R.
Кроме того, необходимые элементы включают дополнительные элементы, улучшающие экспрессию нуклеиновокислотной последовательности согласно изобретению или ее сохранение в клетке-хозяине. В частности, можно упомянуть интронные последовательности, сигнальные последовательности секреции, последовательность локализации нуклеаров, внутренние центры реинициации трансляции IRES-типа, поли(А)-последовательности терминации транскрипции, трехкомпонентные лидерные последовательности и источники репликации. Эти элементы известны специалистам в данной области. Среди сигнальных последовательностей секреции особое предпочтение отдается последовательностям, кодирующим полипептиды, как изложено в SEQ ID NO 5 и/или 8.
Рекомбинантный вектор согласно изобретению также может включать один или несколько дополнительных нужных генов, причем эти гены могут пребывать под контролем одних и тех же регулирующих элементов (полицистронной кассеты) или независимых элементов. Отдельно можно упомянуть гены, кодирующие интерлейкины IL-2, IL-4, IL-7, IL-10, IL-12, IL-15, IL-18, хемокины, такие, как CCL19, CCL20, CCL21, CXCL-14, интерфероны, фактор некроза опухолей (TNF) и факторы, воздействующие на врожденный иммунитет и ангиогенез (например PAl-1, означающий ингибитор активатора плазминогена). В одном конкретном варианте осуществления рекомбинантный вектор согласно изобретению включает нужный ген, кодирующий IL-2.
Настоящее изобретение также касается клетки, включающей нуклеиновокислотную последовательность согласно изобретению. В предпочтительном варианте осуществления, клетка согласно изобретению является эукариотной клеткой, в более предпочтительном варианте - клеткой млекопитающего. Клетки млекопитающих, используемые в качестве хозяев для экспрессии, хорошо известны среди специалистов в данной области, и к ним относятся многие иммортализованные линии клеток, к которым, помимо прочих, относятся клетки яичника китайского хомячка (СНО), клетки почки новорожденного хомяка (ВНК) и многие другие. К подходящим эукариотным клеткам относятся дрожжи и другие грибки.
Настоящее изобретение также касается способа получения антитела согласно изобретению, включающего культивирование клеток согласно изобретению в условиях, обеспечивающих возможность экспрессии антитела и очистку антитела от клеток или среды, окружающей клетки.
В другом варианте осуществления настоящее изобретение касается фармацевтической композиции, включающей антитело, нуклеиновокислотную последовательность или вектор согласно изобретению и фармацевтически приемлемый носитель. В предпочтительном варианте осуществления фармацевтическая композиция также включает нужное соединение.
Фармацевтически приемлемый носитель в предпочтительном варианте является изотоническим, гипотоническим или слабогипертоническим и обладает относительно малой ионной силой, каковым является, например, раствор сахарозы. Кроме того, такой носитель может содержать любой растворитель или водную или частично водную жидкость, такую, как непирогенная стерильная вода. Кроме того, уровень pH фармацевтической композиции регулируют и буферируют таким образом, чтобы он соответствовал требованиям к применению in vivo. Фармацевтическая композиция также может включать фармацевтически приемлемый разбавитель, адъювант или формообразующее, а также солюбилизаторы, стабилизаторы и консерванты. для инъекционного введения предпочтение отдается водному, неводному или изотоническому раствору. Она может обеспечиваться в однократной дозе или в нескольких дозах в жидкой или сухой (порошок, лиофилизат и т.п.) форме, влагосодержание которой может быть восстановлено во время применения при помощи соответствующего разбавителя.
Настоящее изобретение также касается комплекта компонентов, включающего (i) фармацевтическую композицию, антитело, нуклеиновокислотную последовательность или вектор согласно изобретению и (ii) нужное соединение.
В контексте данного описания термин "нужное соединение" касается терапевтического соединения, в предпочтительном варианте - противоракового терапевтического средства или соединения, применимого для лечения снижения костной массы.
Согласно предпочтительному варианту осуществления, противораковое терапевтическое средство выбирают из группы, к которой относятся Abraxane (наночастичная композиция альбумин-стабилизированного паклитаксела), Adriamycin (гидрохлорид доксорубицина), Adrucil (фтороурацил), Aldara (имиквимод), Алемтузумаб, Alimta (пеметрексед динатрий), аминолевулиновая кислота, Анастрозол, Апрепитант, Arimidex (анастрозол), Aromasin (эксеместан), Arranon (неларабин), триоксид мышьяка, Avastin (Bevacizumab), Азацитидин, Bevacizumab, Бексаротен, Бортезомиб, Campath (алемтузумаб), Camptosar (гидрохлорид иринотекана), Капецитабин, Карбоплатин, Цетуксимаб, Цисплатин, Clafen (циклофосфамид), Клофарабин, Clofarex (клофарабин), Clolar (клофарабин), Циклофосфамид, Цитарабин, Cytosar-U (цитарабин), Cytoxan (циклофосфамид), Dacogen (децитабин), Дазатиниб, Децитабин, DepoCyt (липосомальный цитарабин), DepoFoam (липосомальный цитарабин), Дексразоксан гидрохлорид, Доцетаксел, Doxil (гидрохлорид доксорубицина, липосомальная форма), Гидрохлорид доксорубицина, Гидрохлорид доксорубицина, липосомальная форма, Dox-SL (гидрохлорид доксорубицина, липосомальная форма), Efudex (Фтороурацил), Ellence (гидрохлорид эпирубицина), Eloxatin (оксалиплатин), Emend (апрепитант), Гидрохлорид эпирубицина, Erbitux (цетуксимаб), Гидрохлорид эрлотиниба, Evacet (гидрохлорид доксорубицина, липосомальная форма), Evista (гидрохлорид ралоксифена), Эксеместан, Faslodex (фулвестрант), Femara (Летрозол), Fluoroplex (фтороурацил), Фтороурацил, Фулвестрант, Гефинитиб, Гидрохлорид гемцитабина, Гемтузумаб озогамицин, Gemzar (гидрохлорид гемцитабина), Gleevec (иматиниб мезилат), Herceptin (трастузумаб), Hycamtin (гидрохлорид топотекана), Иматиниб мезилат, Имиквимод, Iressa (гефинитиб), Гидрохлорид иринотекана, Иксабепилон, Ixempra (иксабепилон), Keoxifene (гидрохлорид ралоксифена), Kepivance (палифермин), Дитозилат лапатиниба, Леналидомид, Летрозол, Levulan (аминолевулиновая кислота), LipoDox (гидрохлорид доксорубицина, липосомальная форма), липосомальный цитарабин, Methazolastone (темозоломид), Methotrexate, Mylosar (азацитидин), Mylotarg (гемтузумаб озогамицин), наночастичный паклитаксел (наночастичная композиция альбумин-стабилизированного паклитаксела), Неларабин, Neosar (циклофосфамид), Nexavar (тозилат сорафениба), Нилотиниб, Nolvadex (цитрат тамоксифена), Oncaspar (пэгаспаргаза), Оксалиплатин, Паклитаксел, наночастичная композиция альбумин-стабилизированного паклитаксела, Палифермин, Панитумумаб, Paraplat (карбоплатин), Paraplatin (карбоплатин), Пэгаспаргаза, Пеметрексед динатрий, Platinol-AQ (цисплатин), Platinol (цисплатин), гидрохлорид ралоксифена, Revlimid (Леналидомид), Rituxan (ритуксимаб), Ритуксимаб, Sclerosol Intrapleural Aerosol (тальк), тозилат сорафениба, Sprycel (дазатиниб), стерильный порошок талька (тальк), Steritalc (тальк), малат сунитиниба, Sutent (малат сунитиниба), Synovir (талидомид), Тальк, цитрат тамоксифена, Tarabine PFS (цитарабин), Tarceva (гидрохлорид эрлотиниба), Targretin (бексаротен), Tasigna (нилотиниб), Taxol (паклитаксел), Taxotere (доцетаксел), Temodar (темозоломид), Темозоломид, Темсиролимус, Thalomid (талидомид), Талидомид, Totect (дексразоксан гидрохлорид), Гидрохлорид топотекана, Torisel (темсиролимус), Трастузумаб, Trisenox (триоксид мышьяка), Tykerb (дитозилат лапатиниба), Vectibix (панитумумаб), Velcade (бортезомиб), Vidaza (азацитидин), Вориностат, Xeloda (капецитабин), Zinecard (дексразоксан гидрохлорид), Золедроновая кислота, Zolinza (Вориностат) и Zometa (золедроновая кислота).
Согласно предпочтительному варианту осуществления изобретения, соединением, применимым для лечения снижения костной массы, является бифосфонат, селективные модуляторы рецепторов эстрогена (SERM), паратиреоидный гормон (РТН) (например, терипаратид (Forteo)), ранелат стронция, Denosumab или кальцитонин, или их комбинация. Согласно более предпочтительному варианту осуществления, бифосфонат выбирают из группы, к которой относятся Alendronate (Fosamax, Fosamax Plus D), Etidronate (Didronel), Ibandronate (Boniva), Pamidronate (Aredia), Risedronate (Actonel, Actonel W/Calcium), Tiludronate (Skelid) и золедроновая кислота (Reclast, Zometa). Согласно более предпочтительному варианту осуществления, SERM выбирают из группы, к которой относятся ралоксифен (Evista), базедоксифен / премарин (Aprelal) и тамоксифен.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению для лечения болезней, связанных с повышенной активностью остеокластов. К таким болезням, помимо прочих, относятся эндокринопатии (повышенный уровень кортизона, гипогенитализм, первичный или вторичный гиперпаратиреоидизм, гипертиреоидизм), гиперкальциемия, дефицитные состояния (рахит/остеомаляция, цинга, неполноценное питание), хронические болезни (синдромы мальабсорбции, хроническая почечная недостаточность (нефрогенная остеодистрофия), хроническая печеночная недостаточность (гепатогенная остеодистрофия), лекарственные препараты (глюкокортикоиды (глюкокортикоидный остеопороз), антиандрогенная терапия, терапия с применением ингибитора ароматазы, гепарин, алкоголь) и наследственные заболевания (незавершенный остеогенез, гомоцистинурия), остеопороз, остеопетроз, воспаление костей, связанное с артритом и ревматоидным артритом, пародонтоз, фиброзная дисплазия и/или болезнь Педжета.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению для лечения болезней, связанных с воспалением и/или аутоиммунностью. К таким болезням, помимо прочих, относятся серонегативная спондилоартропатия (псориатический артрит, анкилозирующий спондилит, синдром Рейтера, спондилоартропатия, связанная с воспалительной болезнью кишечника), расшатывание протезных суставов, болезни соединительной ткани (ювенильный ревматоидный артрит, ревматоидный артрит, системная красная волчанка (СКВ) и волчаночный нефрит, склеродерма, синдром Шегрена, смешанное заболевание соединительной ткани, полимиозит, дерматомиозит), воспалительная болезнь кишечника (например, болезнь Крона; язвенный колит), болезнь Уиппла, артрит, связанный с гранулематозным илеоколитом, воспалительные кожные состояния (аутоиммунный буллезный пемфигоид, аутоиммунная обыкновенная пузырчатка, экзема, дерматит), воспаление легких (альвеолит, легочный фиброз, саркоидоз, астма, бронхит, облитерирующий бронхиолит), воспалительное заболевание почек (гломерулонефрит, отторжение почечного аллотрансплантата, воспаление почечных канальцев), атеросклероз, системный васкулит (височный артерии/гигантоклеточный артериит, артериит Такаясу, нодозный полиартериит, болезнь Кавасаки, гранулематоз Вегенера, синдром Черджа-Стросс, микроскопическая ангиопатия, некротизирующий гломерулонефрит, пурпура Шенлейна-Геноха, эссенциальный криоглобулинемический васкулит и другие васкулиты мелких сосудов, болезнь Бехчета), болезни, связанные с активацией макрофагов (синдром активации макрофагов (MAS), приобретенная болезнь Стилла, гематофагоцитарный синдром), ревматическая полимиалгия, первичный склероз желчных путей, склерозирующий холангит, аутоиммунный гепатит, сахарный диабет первого типа, тиреоидит Хашимото, базедова болезнь, рассеянный склероз (MS), синдром Гийена-Барре, болезнь Аддисона и/или феномен Рейно, синдром Гудпасчера.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению для лечения рака.
В контексте данного описания термин "рак" означает, помимо прочих, аденокарциному, ацинарноклеточную аденокарциному, рак коры надпочечников, рак альвеолярных клеток, недифференцированный мелкоклеточный рак, базалоидная карцинома, базальноклеточная карцинома, бронхиолярная карцинома, бронхогенная карцинома, почечная карцинома, эмбриональный рак, анометроидная карцинома, фиброламеллярная печеночно-клеточная карцинома, фолликулярные карциномы, гигантоклеточные карциномы, гепатоцеллюлярная карцинома, интраэпидермальный рак, интраэпителиальный рак, лептоманигиальная (leptomanigio) карцинома, медуллярная карцинома, меланотическая карцинома, менингиальная карцинома, мезометанефрома, овсяно-клеточная карцинома, плоскоклеточная карцинома, карцинома потовых желез, переходно-клеточная карцинома, тубулярная карцинома, амелобластосаркома, псаммома, ботриоидная саркома, эндометриальная стромальная саркома, саркома Юинга, веретеноклеточная саркома, гигантоклеточная саркома, гранулезная саркома, иммунобластная саркома, юкстакортикальная остеогенная саркома, саркома coppices, лейкоцитарная саркома (лейкоз), лимфатическая саркома (лимфосаркома), медуллярная саркома, миелоидная саркома (гранулоцитарная саркома), остеогенная саркома, периостальная саркома, ретикулосаркома (гистиоцитарная лимфома), круглоклеточная саркома, веретеноклеточная саркома, синовиальная саркома, телеангиэктатическая аудиогенная саркома, лимфома Беркитта, узелковая недифференцированная лимфома (NPDL), узелковая смешанная лимфома (NML), неходжкинская лимфома (NH) и диффузные лимфомы. Согласно предпочтительному варианту осуществления, способ согласно изобретению направлен на лечение метастазов рака в кости, причем метастатический рак представляет собой рак молочной железы, легких, почек, множественную миелому, рак щитовидной железы, предстательной железы, аденокарциному, злокачественность кровяных клеток, включая лейкоз и лимфому; раковые образования головы и шеи; рак желудочно-кишечного тракта, включая рак пищевода, рак желудка, рак толстой кишки, рак кишечника, рак ободочной и прямой кишки, рак прямой кишки, рак поджелудочной железы, рак печени, рак желчных протоков или желчного пузыря; злокачественности женских половых путей, включая карциному яичника, рак эндометрия, вагинальный рак и рак шейки матки; рак мочевого пузыря; рак головного мозга, включая нейробластому; саркому, остеосаркому; и рак кожи, включая злокачественную меланому или плоскоклеточный рак.
Настоящее изобретение также касается способа усовершенствования лечения страдающего от рака пациента, подвергаемого химиотерапевтическому лечению с применением противоракового терапевтического средства, включая комбинированное лечение вышеупомянутого пациента с применением вышеописанного способа.
Настоящее изобретение также касается способа повышения цитотоксичной эффективности цитотоксичных медикаментов или лучевой терапии, включая комбинированное лечение пациента, нуждающегося в таком лечении, с применением вышеописанного способа.
Настоящее изобретение также касается способа усовершенствования лечения пациента с болезнью, связанной с повышенной активностью остеокластов, который подвергается лечению с применением бифосфоната, селективных модуляторов рецепторов эстрогена (SERM), паратиреоидного гормона (РТН) (например, терипаратида (Forteo)), ранелата стронция, Denosumab или кальцитонина, или их комбинации, включая комбинированное лечение вышеупомянутого пациента с применением вышеописанного способа.
В другом варианте осуществления использование антитела согласно изобретению предусмотрено при производстве медикамента для профилактики или лечения метастазов рака в кости у пациента, страдающего от метастатического рака. В соответствующем варианте осуществления метастатический рак представляет собой рак молочной железы, легких, почек, множественную миелому, рак щитовидной железы, предстательной железы, аденокарцинома, злокачественность кровяных клеток, включая лейкоз или лимфому; раковые образования головы и шеи; рак желудочно-кишечного тракта, включая рак пищевода, рак желудка, рак толстой кишки, рак кишечника, рак ободочной и прямой кишки, рак прямой кишки, рак поджелудочной железы, рак печени, рак желчных протоков или желчного пузыря; злокачественности женских половых путей, включая карциному яичника, рак эндометрия, вагинальный рак, или рак шейки матки; рак мочевого пузыря; рак головного мозга, включая нейробластому; саркому, остеосаркому; или рак кожи, включая злокачественную меланому или плоскоклеточный рак.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению при производстве медикамента.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению при производстве медикамента для лечения страдающего от рака пациента.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению при производстве медикамента для лечения пациента с болезнью, связанной с повышенной активностью остеокластов.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению при производстве медикамента для лечения пациента с воспалительной болезнью, более конкретно - с воспалительной болезнью кишечника.
Согласно еще одному варианту осуществления, настоящее изобретение касается применения антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению при производстве медикамента для лечения пациента, страдающего от ревматоидного артрита.
Введение антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению может осуществляться известными специалистам в данной области способами. Предпочтительными путями введения, помимо прочих, являются внутрикожный, подкожный, пероральный, парентеральный, внутримышечный, интраназальный, сублингвальный, внутритрахеальный, ингаляционный, глазной, вагинальный и ректальный. Согласно предпочтительному варианту осуществления, антитело, нуклеиновокислотную последовательность, вектор, фармацевтическую композицию или комплект компонентов согласно изобретению вводят системно.
Введение может осуществляться однократной дозой или повторным введением доз один или несколько раз с определенными интервалами времени. Желательно введение антитела, нуклеиновокислотной последовательности, вектора, фармацевтической композиции или комплекта компонентов согласно изобретению от 1 до 10 раз с недельными интервалами.
Как правило, приемлемая доза антитела составляет приблизительно от 2 мг/кг до 30 мг/кг, от 0,1 мг/кг до 30 мг/кг или от 0,1 мг/кг до 10 мг/кг массы тела. Приемлемая доза вектора согласно изобретению колеблется приблизительно от 104 до 1010 БОЕ (бляшкообразующих единиц), желательно - приблизительно от 105 до 108 БОЕ для вектора MVA, в то время, как для вектора на основе аденовируса она колеблется приблизительно от 105 до 1013 ИЕ (инфекционных единиц), желательно - приблизительно от 107 до 1012 ИЕ. Композиция на основе векторных плазмид может вводиться в дозах от 10 мкг до 20 мг, предпочтительно - от 100 мкг до 2 мг.
Если применение или способ согласно изобретению предназначаются для лечения рака, способ или применение согласно изобретению могут осуществляться в связи с одним или несколькими традиционными режимами лечебного воздействия (например, лучевой терапии, химиотерапии и/или хирургии). Применение нескольких терапевтических подходов обеспечивает для пациента более универсальное вмешательство. Во одном варианте осуществления способу согласно изобретению может предшествовать, или после него может следовать, хирургическое вмешательство. В другом варианте осуществления способу может предшествовать, или после него может следовать, лучевая терапия (например, гамма-излучение). Специалисты в данной области смогут легко определить надлежащие протоколы лучевой терапии и параметры, которые должны применяться (см., например, PEREZ. Principles and practice of radiation oncology. 2nd edition. LIPPINCOTT, 1992).
Краткое описание фигур
Фигура 1 показывает специфическое окрашивание трансфицированных КСФ-1R клеток NIH/3T3 с применением mAb CXIIG6.
Фигура 2 показывает ингибирование связывания КСФ-1 с КСФ-1R поверхности клеток в присутствии mAb CXIIG6.
Фигура 3 показывает специфическое блокирование растворимого человеческого КСФ-1R с применением mAb CXIIG6 (ctrl' означает контроль; 'neg SN' означает отрицательный контрольный супернатант гибридомы).
Фигура 4 показывает ингибирование дифференциации остеокластов человека и секреции матриксной металлопротеиназы-9 (ММР-9) в присутствии mAb CXIIG6 ('ctrl' означает контроль; 'CXIIG6 SN' означает супернатант гибридомы CXIIG6; 'neg SN' означает отрицательный контрольный супернатант гибридомы).
Фигура 5 показывает отсутствие перекрестной реактивности mAb CXIIG6 с другими рецепторами тирозинкиназы, имеющими гомологию с КСФ-1R ('SN' означает супернатант гибридомы).
Фигура 6 показывает нуклеиновокислотную последовательность (SEQ ID NO: 1) и предсказанную аминокислотную последовательность (SEQ ID NO: 2) тяжелой цепи CXIIG6. Праймерные последовательности, включающие сайты рестрикции для клонирования, добавленные к нуклеотидным последовательностям, подчеркнуты. Сайты рестрикции показаны подчеркнутым курсивом. Аминокислотные последовательности V-доменов выделены жирным шрифтом.
Фигура 7 показывает нуклеиновокислотную последовательность (SEQ ID NO: 3) и предсказанную аминокислотную последовательность (SEQ ID NO: 4) легкой цепи CXIIG6. Праймерные последовательности, включающие сайты рестрикции для клонирования, добавленные к нуклеотидным последовательностям, подчеркнуты. Сайты рестрикции показаны подчеркнутым курсивом. Аминокислотные последовательности V-доменов выделены жирным шрифтом.
Фигура 8 показывает плазмидную последовательность pTG17753.
Фигура 9 показывает плазмидную последовательность pTG17727.
Фигура 10 показывает плазмидную последовательность pOptiVEC™.
Фигура 11 показывает плазмидную последовательность pTG17895.
Фигура 12 показывает плазмидную последовательность pTG17812.
Фигура 13 показывает плазмидную последовательность pTG17868.
Фигура 14 показывает плазмидную последовательность pTG17869.
Фигура 15 показывает гуманизированные варианты легкой цепи CXIIG6.
Фигура 16 показывает гуманизированные варианты IgG1 тяжелой цепи CXIIG6.
Фигура 17 показывает специфическое блокирование растворимого человеческого КСФ-1R рекомбинантным мышиным CXIIG6 и химерным CXIIG6 IgG1.
Фигура 18 показывает ингибирование дифференциации остеокластов человека и секреции матриксной металлопротеиназы-9 (ММР-9) в присутствии рекомбинантного мышиного CXIIG6 и химерного CXIIG6 IgG1.
Фигура 19 показывает различные последовательности КСФ-1R, применяемые для картирования эпитопа моноклональных антител согласно изобретению относительно эпитопов реализуемых на рынке антител против КСФ-1R. Человеческая последовательность представлена в виде незакрашенного блока, а мышиная последовательность - в виде закрашенного блока. Антитела с зачеркнутыми названиями указывают, что они не связываются с последовательностью.
Фигура 20 показывает пределы Ig-подобных доменов различных последовательностей КСФ-1R, применяемых для картирования эпитопа.
Фигура 21: Кривая конкуренции между антителом 125I-H27K15 и немеченным H27K15 на EL4-КСФ-1R.
Фигура 22: Кривая конкуренции между антителом 125I-H27K15 и различными антителами на EL4-КСФ-1R.
Фигура 23: Кривые конкуренции после инкубации: КСФ-1/mAb.
Фигура 24: Кривые конкуренции до инкубации: mAb/КСФ-1.
Фигура 25: Кривые конкуренции при совместной инкубации: mAB+КСФ-1.
Фигура 26: Кривые конкуренции при совместной инкубации: mAB+КСФ-1R.
Фигура 27: Отсутствие перекрестной реактивности вариантов mAB.
Фигура 28: Блокирование растворимого КСФ-1R. Три варианта hCXIIG6 и chCXIIG6 блокируют растворимый человеческий CD115 и восстанавливают зависящий от КСФ-1 рост клеток M-NFS-60. Результаты выражены как средние значения+/-стандартная погрешность среднего лунок в четырех экземплярах. Значения ND50 вычисляли, применяя пятипараметрическое логистическое уравнение GraphPadPrism.
Фигуры 29: A/B/C/D: Ингибирование КСФ-1-зависимой пролиферации клеток AML3. Как показано на Фигуре 29А, клетки AML5 окрашивали CD115 mAN 3291 против человека (левый график) или chCXIIG6 (правый график). Для каждого графика левая гистограмма соответствует клеткам AML5, окрашенным контрольными изотопами (соответственно, мышиным IgG1 и ритуксимабом). Фигура 29В показывает, что рост клеток AML5 стимулируется КСФ-1 в зависимости от дозы. Фигура 29С показывает, что варианты hCXIIG6 и chCXIIG6 ингибируют зависящую от КСФ-1 пролиферацию клеток AML5, демонстрируя результаты трех независимых экспериментов. Фигура 29D включает значения квадратов ЕС50 и R, вычисленные при помощи GraphPad Prism по результатам, показанным на Фигуре 29С.
Фигуры 30А/В: активность ADCC на клетках-мишенях EL4-КСФ-R. На Фиг.30А РВМС донора крови #1 использовали в качестве эффекторных клеток для измерения цитотоксичной активности H27K5, H27K15 и H19K12 на клетках-мишенях EL4-CD115 при соотношении Е:Т 25. Как показано на 30В, РВМС донора #2 испытывали в таком же анализе, как на Фигуре 30А при указанных соотношениях Е:Т. Звездочка (*) указывает p<0,05 по сравнению с ритуксимабом, а символ тэта (Ф) указывает p<0,05 по сравнению с chCXIIG6.
Фигуры 31А/В: Терапевтический эффект в модели опухоли хориокарциномы BeWo.
Фигура 32: Список последовательностей с SEQ ID NO: 37 по SEQ ID NO: 47.
ПРИМЕРЫ
Специфическое окрашивание трансфицированных клеток КСФ-1R NIH/3T3 с применением mAb CXIIG6
Линию клеток В4-800-5 получали путем стабильной трансфекции клеток NIH/3T3 плазмидой экспрессии, кодирующей человеческий КСФ-1 R полной длины. Экспрессию КСФ-1 R поверхности клеток на клетках В4-800-5 проверяли путем непрямого иммунного окрашивания с использованием mAbs 61701 против человеческого КСФ-1 R (мышиный IgG1, R&D Systems) или 2-4А5-4 (крысиный IgG1,k, GeneTex), сравнивали с контрольными изотипами (Фигура 1, верхние и средние графики). Супернатанты культур из гибридомы CXIIG6 или из отрицательной контрольной гибридомы использовали для иммунного окрашивания клеток В4-800-5 или родительских клеток NIH/3T3 (Фигура 1, нижние графики).
Анализ проточной цитометрии показал, что супернатант культуры из гибридомы CXIIG6 избирательно окрашивал клетки В4-800-5, демонстрируя специфичность mAb к КСФ-1 R поверхности клеток.
Частичное ингибирование связывания КСФ-1 с КСФ-1R поверхности клеток
3×105 клеток ТНР-1 (линия КСФ-1 R-положительных клеток моноцитарного лейкоза человека) инкубировали в течение 30 мин при 4°C в присутствии супернатантов культур гибридомы, сыворотки интактных или иммунизированных против КСФ-1R мышей (разведение 1:1000), mAb против КСФ-1R 2-4А5-4 (GeneTex) или контрольного крысиного IgG1 (10 мкг/мл), или без реагента. После двух промываний холодным PBS клетки инкубировали с 1 мкг/мл биотинилированного рекомбинантного человеческого КСФ-1 в течение 30 мин. Клетки дважды промывали и дополнительно инкубировали в течение 30 мин при 4°C с использованием 10 мкг/мл стрептавидин-Alexa Fluor 488 (Invitrogen). После промывания PBS и фиксации 4% параформальдегидом окрашивание клеток анализировали при помощи проточной цитометрии.
Сниженная интенсивность флуоресценции по сравнению с контрольными образцами отражает ингибирование связывания КСФ-1 с КСФ-1R поверхности клеток. Сыворотка иммунизированной КСФ-1R мыши блокирует связывание КСФ-1 с клетками ТНР-1 (Фигура 2). Хотя отрицательный контрольный супернатант гибридомы или нейтральное mAb эффекта не демонстрируют, супернатант культуры из гибридомы CXIIG6, по-видимому, ингибирует, как минимум частично, связывание КСФ-1 с клетками ТНР-1 (Фигура 2, правый нижний график).
Первая локализация сайта связывание mAb CXIIG6.
Для определения сайта связывания mAb CXIIG6 на КСФ-1R выполняли вестерн-блоттинг с использованием растворимых форм человеческого КСФ-1R, включавших либо пять внеклеточных иммуноглобулин-подобных доменов (от Met 1 до Glu 512, R&D Systems) или только три N-конечных иммуноглобулин-подобных домена внеклеточного участка КСФ-1R (от Met 1 до Ser 290), причем и те, и другие были слиты на их С-концах с Fc-участком человеческого IgG1. Растворимую форму EGFR, слитую с Fc-участком человеческого IgG1 (R&D Systems), использовали в качестве отрицательного контроля.
Сто нанограммов каждого растворимого рецептора подвергали электрофорезу в природных условиях перед переносом на нитроцеллюлозный лист и зондированием супернатантами гибридомы, кроличьим pAb c-fms/ КСФ-1R Н300 (Santa Cruz Biotechnology), мышиным mAb 61701 (R&D Systems) или сывороткой интактных или КСФ-1R-иммунизированных мышей.
Обе растворимые формы КСФ-1R обнаруживались как широкие полосы при зондировании pAb c-fins/ КСФ-1R Н300, mAb 61701 или сывороткой иммунизированных мышей. Не наблюдалось обнаружимых сигналов при сыворотке интактных мышей или отрицательном контрольном супернатанте гибридомы. Супернатант гибридомы CXIIG6 распознавал КСФ-1R1-290:Fc, а также КСФ-1R1-512:Fc, но не распознавал EGFR:Fc, что указывало на то, что CXIIG6 специфически связывается с эпитопом, находящимся в пределах трех N-конечных иммуноглобулин-подобных доменов (между остатками с 1 по 290) человеческого КСФ-1R.
Специфическое блокирование растворимого человеческого КСФ-1R с применением mAb CXIIG6
КСФ-1-зависимую линию клеток M-NFS-60 миелоидного лейкоза мышей (# CRL-1838, АТСС) использовали для оценки блокирующей активности супернатанта гибридомы CXIIG6 на человеческом и мышином КСФ-1R. Пять нанограммов растворимого человеческого КСФ-1R (КСФ-1R1-512:Fc от R&D Systems) предварительно инкубировали в белых 96-луночных микропланшетах с последовательно разводимыми супернатантами гибридомы, mAb 61701 (R&D Systems) или мышиным изотипическим контрольным mAb. Клетки 10Е4 M-NFS-60, которые культивировали в течение суток в отсутствие КСФ-1, затем собирали в лунки для культуры вместе с 0,1 нг человеческого КСФ-1 в конечном аналитическом объеме 100 мкл. Культуры инкубировали в течение 48 ч при 37°С и пролиферацию количественно определяли путем включения BrdU с применением ELISA-анализа пролиферации клеток (Roche).
Растворимый человеческий КСФ-1R полностью ингибировал пролиферацию клеток M-NFS-60, опосредованную человеческим КСФ-1, которая обнаруживалась в присутствии отрицательного супернатанта гибридомы, содержащего или не содержащего отрицательный контрольный IgG1 (Фигура 3; средн. +/- стандартная погрешность среднего 3 лунок). В отличие от него, супернатант гибридомы CXIIG6 и положительное контрольное mAb 61701 были способны возобновлять пролиферацию клеток в зависимости от дозы, продемонстрировав, что они способны нейтрализовать растворимый человеческий КСФ-1R.
В этом анализе активная пролиферация клеток M-NFS-60 в присутствии слабо разведенного супернатанта гибридомы CXIIG6 продемонстрировала, что mAb CXIIG6 не способно блокировать мышиный КСФ-1R, экспрессируемый клетками M-NFS-60. Кроме того, в анализе поддерживаемой КСФ-1 пролиферации M-NFS-60, выполнявшемся в отсутствие растворимого КСФ-1R, обработка mAb AFS98 против мышиного КСФ-1R (eBioscience) в результате обеспечивала заметное зависимое от концентрации снижение роста клеток (данные не показаны). Супернатант гибридомы CXIIG6, как и отрицательные контрольные антитела и отрицательный супернатант гибридомы, не вызывал снижения пролиферации клеток. Эти результаты демонстрируют, что mAb CXIIG6 специфически направлено на человеческий КСФ-1R.
Ингибирование дифференциации остеокластов человека и секреции матриксной металлопротеиназа-9 (ММР-9)
Остеокласты получали из человеческих моноцитов, получаемых путем декантирования РВМС здорового донора крови. В общих чертах, моноциты высевали в количестве 2×10Е4 клеток на лунку в 96-луночные планшеты и обрабатывали в течение 45 мин супернатантами культур гибридомы, mAbs против человеческого КСФ-1R 61701 (R&D Systems) или 2-4А5-4 (GeneTex), mAb против человеческого КСФ-1 26730 (R&D Systems), контролями мышиного или крысиного изотипа или сыворотками интактных и КСФ-1 R-иммунизированных мышей, разведенными в культуральной среде гибридомы. Полную среду а-МЕМ добавляли в лунки с культурой с содержание или без содержания человеческого КСФ-1 и RANKL (PeproTech, 25 и 40 нг/мл, соответственно). Супернатанты гибридомы, mAbs и/или среду с содержанием или без содержания цитокинов пополняли раз в 3 дня в течение 9 дней. Кондиционированные супернатанты культур собирали на 9-й день и подвергали анализу на общий человеческий ММР-9 с применением ELISA-анализа (R&D Systems). Образование остеокластов оценивали путем окрашивания устойчивой к тартрату кислой фосфатазы (TRAP) с применением комплекта кислой фосфатазы лейкоцитов от Sigma-Aldrich.
КСФ-1 + RANKL вызывали дифференциацию моноцитов в остеокласты, определяемые как большие многоядерные TRAP-положительные клетки, в то время, как в отсутствие цитокинов TRAP-положительные остеокласты не образовывались. Добавление 0,5 мкг/мл mAb 26730 против КСФ-1 полностью исключало дифференциацию остеокластов, на что указывает отсутствие секреции ММР-9. MAbs 61701 против КСФ-1R или 2-4А5-4 в одинаковой концентрации и сыворотка иммунизированных мышей (разведение 1:1000) ингибировали образование остеокластов лишь частично (Фигура 4; с цитокинами (+) или без них (-); средн. +/- стандартная погрешность среднего 3 лунок; *: средний показатель 2 лунок). Обработка супернатантом культуры гибридомы CXIIG6, разведенным до 1:20 или 1:100, значительно снижала уровень выработки ММР-9 по сравнению с двумя отрицательными контрольными супернатантами гибридомы (А, В). Эти результаты демонстрируют, что mAb CXIIG6 ингибирует дифференциацию остеокластов из человеческих моноцитов путем блокирования функции КСФ-1R поверхности клеток.
Ингибирование зависящего от КСФ-1 фосфорилирования КСФ-1R
Линию клеток В4-800-5, полученная путем стабильной трансфекции клеток NIH/3T3 плазмидой, экспрессирующей человеческий КСФ-1R, использовали для исследования влияния супернатанта гибридомы CXIIG6 на зависящее от КСФ-1 фосфорилирование КСФ-1R. Клетки высевали в количестве 2×10Е5 клеток на 60 мм чашку Петри и культивировали в течение 48-72 ч. После депривации сыворотки в течение 1 ч при 37°C клетки обрабатывали в течение 1 ч при 37°C культуральной средой, содержащей супернатант гибридомы CXIIG6, mAb 2-4А5-4 (NeoMarkers) или изотипические контрольные mAb (разведенные в отрицательном супернатанте гибридомы), а затем стимулировали 100 нг/мл hКСФ-1 или оставляли нестимулированными на 5 мин при 37°C. Слои клеток затем подвергали лизису и извлекали общие белки. 10 мкг белков анализировали путем зондирования вестерн-блоттингов кроличьими pAb с-fms/КСФ-1R Н300 или кроличьими pAb р-с-fms/КСФ-1R (Tyr708)-R (Santa Cruz Biotechnology), а затем иммуноглобулинами козы против кроликаHRP.
В отсутствие КСФ-1 ни супернатант гибридомы CXIIG6, ни mAb 2-4А5-4 не вызывали фосфорилирования рецептора, которое наблюдалось в случае антитела, специфического к КСФ-1R, фосфорилированного в позиции 708, демонстрируя, что только mAb CXIIG6 не оказывает агонистического влияния. После стимулирования с использованием КСФ-1 КСФ-1R фосфорилировали на Tyr708 и количество КСФ-1R в изотипических обработанных контролем клетках снижалось по сравнению с нестимулированными клетками, что свидетельствовало о распаде рецептора (данные не показаны). Предварительная обработка супернатантом гибридомы CXIIG6 или mAb 2-4А5-4 не способствовала исчезновению КСФ-1R. Фосфорилирование КСФ-1R уменьшалось после обработки супернатантом гибридомы CXIIG6 или mAb 2-4А5-4. Эти результаты показывают, что mAb CXIIG6 может блокировать зависящее от КСФ-1 фосфорилирование КСФ-1R.
Перекрестная реактивность mAb CXIIG6
Перекрестная реактивность mAb CXIIG6 испытывали путем ELISA-анализа на ряде очищенных растворимых рецепторов, относящихся к III типу подсемейства рецепторов тирозинкиназы и демонстрирующих гомологию к КСФ-1R в их внеклеточных Ig-подобных доменах: растворимом VEGFR-1, VEGFR-2, Flt-3 и PDGFRβ (все четыре экспрессировались как слитые белки Fc), а также PDGFRα и SCFR (c-kit) получали от R&D Systems и использовали для покрытия ELISA-планшетов. Растворимый EGFR (R&D Systems) из подсемейства EGFR рецепторов тирозинкиназы использовали в качестве отрицательного контроля.
Супернатанты культур из гибридомы CXIIG6 (CXIIG6 SN) или отрицательной контрольной гибридомы или мышиного IgG1 61701 против КСФ-1R (R&D Systems) инкубировали на покрытом ELISA-планшете при концентрации антитела 500 нг/мл. После промывания ELISA-планшета связанные антитела обнаруживали с использованием конъюгированного с пероксидазой Ig козы против мыши (Sigma) и измеряли OD(450-540 нм). Результаты показанные на Фигуре 5, показывают, что, подобно mAb 61701, CXIIG6 сильно связывает КСФ-1R, в то время, как на любом другом рецепторе тирозинкиназы специфических сигналов не обнаруживалось. Это демонстрирует, что среди различных испытанных рецепторов тирозинкиназы III типа CXIIG6 является специфическим к КСФ-1R.
Построение векторов экспрессии для mAb CXIIG6
Комплект для экспрессии антител OptiCHOтм (Invitrogen, Catalog №12762-019) использовали для клонирования генов, кодирующих тяжелые и легкие цепи CXIIG6 с целью получения mAb CXIIG6 в линии клеток млекопитающих DG44. Комплект для экспрессии антител OptiCHOтм включает: (1) Вектор pOptiVECтм - бицистронную плазмиду, позволяющую клонировать нужный ген после промотора CMV. Транскрипция нужного гена отделяется от ауксотрофного маркера отбора дигидрофолатредуктазы (DHFR) участком внутренней посадки рибосомы (IRES), что обеспечивает возможность транскрипции нужного гена и маркера отбора на одной мРНК; (2) Вектор pcDNAтм3.3, позволяющий клонировать нужный ген после промотора CMV. PcDNAтм3.3 содержит ген резистентности к неомицину, обеспечивающий возможность отбора с применением Geneticin®. Векторы pOptiVECтм и pcDNAтм3.3 содержат ТК поли-А последовательность, направляющую надлежащую обработку 3'-конца мРНК нужного гена.
Специфические праймеры (см. Таблицу 4) синтезировали и использовали для ПЦР-амплификации и клонирования полных генов CXIIG6 тяжелой цепи и легкой цепи (соответственно SEQ ID NO: 1 и SEQ ID NO: 3; см., соответственно, Фигуру 6 и Фигуру 7). Обратные праймеры включали консенсусную последовательность Козака для эффективной эукариотной трансляции (KOZAK М. An analysis of 5'-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Res. 1987, 15 (20): 8125-8148).
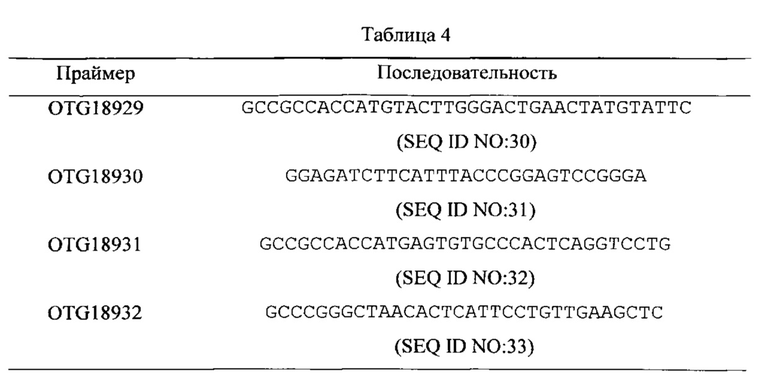
CXIIG6 тяжелой цепи подвергали ПЦР-амплификации путем использования OTG18929 и OTG18930 с плазмидой pTG17753 (Фигура 8) в качестве матрицы и клонировали в вектор pOptiVEC™-TOPO® (pOptiVEC™-TOPO® ТА Cloning® Kit, Invitrogen, Catalog №12744-017-01) и векторы pcDNAтм3.3-TOPO® (pcDNA™3.3-TOPO® ТА Cloning® Kit, Invitrogen, Catalog №K8300-01) для получения, соответственно, pTG17786 и pTG17789.
Легкую цепь CXIIG6 подвергали ПЦР-амплификации путем использования OTG18931 и OTG18932 с плазмидой pTG17727 (Фигура 9) в качестве матрицы и клонировали в вектор pOptiVEC™-TOPO® (pOptiVEC™-TOPO® ТА Cloning® Kit, Invitrogen, Catalog №12744-017-01) и векторы pcDNA™3.3-TOPO® (pcDNA™3.3-TOPO® ТА Cloning® Kit, Invitrogen, Catalog №K8300-01) для получения, соответственно, pTG17788 и pTG17787.
Нуклеотидную последовательность полной экспрессионной кассеты, включающей CMV промотор и ТК поли-А-сигнал pTG17786, pTG17787, pTG17788 и pTG17789, секвенировали и обнаруживали соответствие их теоретическим последовательностям.
Образование химерных антител из mAb CXIIG6
Вариабельные домены mAb CXIIG6 комбинировали с человеческими постоянными участками.
Для образования химерной легкой цепи (называемой химерной CXIIG6 Igκ-цепью) составляли теоретическую последовательность путем соединения последовательности, кодирующей VK домен CXIIG6 (из SEQ ID NO: 3) с последовательностью, кодирующей человеческий участок IGKC (учетный номер GenBank: J00241). Этот фрагмент ДНК XbaI NotI сохранял такую же нетранслированную последовательность на 5'-конце, включая последовательность Козака, как та, что использовалась в мышиной версии (как в описанных выше pTG17787 и pTG17788). Последовательность легкой цепи химерного CXIIG6 подвергали кодон-оптимизации для экспрессии в СНО, собирали из синтетических олигонуклеотидов и субклонировали в pOptiVEC™ (Фигура 10) через XbaI NotI от GeneArt AG. Кодон-оптимизированная нуклеиновокислотная последовательность полученной легкой цепи химерного CXIIG6 (вариабельные и постоянные участки) изложена в SEQ ID NO: 34. Полученная плазмида называлась pTG17895 (Фигура 11).
Для образования химерных тяжелых цепей IgG1 и IgG4 (соответственно называемых химерной цепью CXIIG6 IgG1 и химерной цепью CXIIG6 IgG4) составляли теоретические последовательности путем соединения последовательности, кодирующей VH домен CXIIG6 (из SEQ ID NO: 1), с последовательностями, кодирующими человеческий участок IGHG1C (учетный номер GenBank: J00228) или человеческий участок IGHG4C (учетный номер GenBank: K01316). Эти фрагменты ДНК XbaI NotI сохраняли такую же нетранслированную последовательность на 5-конце, включая последовательность Козака, как та, что использовалась в мышиной версии (как в pTG17786 и pTG17789, как описывается выше). Тяжелые цепи химерного CXIIG6 затем подвергали кодон-оптимизации для экспрессии в СНО, синтезировали и клонировали в pTG17812 (Фигура 12) через XbaI NotI от GeneArt AG. Кодон-оптимизированная нуклеиновокислотная последовательность полученной тяжелой цепи химерного CXIIG6 IgG1 (вариабельные и постоянные участки) изложена в SEQ ID NO: 35, и полученные плазмиды назывались pTG17868 (Фигура 13). Кодон-оптимизированная нуклеиновокислотная последовательность полученной тяжелой цепи химерного CXIIG6 IgG4 (вариабельные и постоянные участки) изложена в SEQ ID NO: 36, и полученные плазмиды назывались pTG17869 (Фигура 14).
Образование гуманизированных антител из mAb CXIIG6
Для образования гуманизированных легкоцепочечных вариантов выполняли аминокислотные замещения согласно Таблице 2 в пределах вариабельного участка легкой цепи, изложенного в SEQ ID NO: 9.
Последовательности ДНК составляли путем соединения модифицированных последовательностей, имеющих замещения CXIIG6 VK домена (из SEQ ID NO: 3) с последовательностью, кодирующей человеческий участок IGKC (учетный номер GenBank: J00241). Этот фрагмент ДНК XbaI NotI сохранял такую же нетранслированную последовательность на 5'-конце, включая последовательность Козака, как та, что использовалась в мышиной версии (как в pTG17787 и pTG17788, как описывается выше). Последовательности легкой цепи гуманизированного CXIIG6 затем подвергали кодон-оптимизации для экспрессии в СНО, собирали из синтетических олигонуклеотидов и клонировали в pOptiVEC™ (Фигура 10) через XbaI NotI от GeneArt AG. Полученные гуманизированные варианты легкой цепи CXIIG6 и плазмиды перечислены на Фигуре 15.
Для образования гуманизированных вариантов тяжелой цепи выполняли аминокислотные замещения согласно Таблице 1 в пределах вариабельного участка тяжелой цепи, изложенного в SEQ ID NO:6.
Последовательности ДНК составляли путем соединения модифицированных последовательностей, имеющих замещения CXIIG6 VH домена (из SEQ ID NO:1) с последовательностями, кодирующими человеческий участок IGHG1C (учетный номер GenBank: J00228). Фрагменты ДНК XbaI ApaI сохраняли такую же нетранслированную последовательность на 5-конце, включая последовательность Козака, как та, что использовалась в мышиной версии (как в pTG17786 и pTG17789, как описывается выше). Последовательности ДНК затем подвергали кодон-оптимизации для экспрессии в СНО, синтезировали и клонировали в pTG17812 (Фигура 12) через XbaI ApaI от GeneArt AG. Полученные гуманизированные варианты IgG1 тяжелой цепи CXIIG6 и плазмиды перечислены на Фигуре 16.
In vitro ингибиторная активность рекомбинантного мышиного CXIIG6 и химерного CXIIG6 IgG1
Для того, чтобы определить, способны ли очищенное рекомбинантное мышиное CXIIG6 (як описано выше) и его химерный вариант IgG1 (химерный CXIIG6 IgG1, как описано выше) блокировать растворимый человеческий КСФ-1R, выполняли исследования влияния величины дозы в моделях пролиферации клеток M-NFS-60 и дифференциации остеокластов (як описано выше). Очищенный поликлональный мышиный IgG2a от Rockland (Rockland, 010-0141) и химерный IgG1, полученный заявителем, испытывали параллельно как контрольные антитела. Блокирующее воздействие оценивали путем подвергания клеток действию ряда концентраций активных антител против КСФ-1R, измеряемому через связывание антигена при SPR-анализе с использованием биодатчика. Сравнение между mAb CXIIG6 и их соответствующим контрольным mAb осуществляли путем загрузки одинакового количества общего антитела (SPR-анализ с использованием биодатчика путем Fc-связывания).
Биоанализ M-NFS-60: при биоанализе M-NFS-60 клетки обрабатывали активными mAb CXIIG6 в количестве от 0,23 нг/мл до 0,5 мкг/мл (рекомбинантное мышиное CXIIG6; химерный CXIIG6 IgG1) или соответствующими концентрациями контрольных mAb в присутствии 50 нг/мл человеческого растворимого КСФ-1R и 1 нг/мл человеческого КСФ-1 в течение 48 ч. Результаты, показанные на Фигуре 17. показывают, что рост клеток M-NFS-60 повышался в ответ на увеличение концентрации обоих mAb CXIIG6 (рекомбинантного мышиного CXIIG6; химерного CXIIG6 IgG1), демонстрируя, что они антагонизируют связывание растворимого КСФ-1R с КСФ-1 (средн. +/- стандартная погрешность среднего трех лунок). Химерный CXIIG6 IgG1 был так же эффективен, как рекомбинантный мышиный CXIIG6, при возобновлении пролиферации клеток. Контрольный мышиный IgG2a и химерный IgG1 не имели воздействия на нейтрализацию КСФ-1 растворимым КСФ-1R в их соответствующим диапазоне концентраций. Эти результаты показывают, что очищенное рекомбинантное мышиное CXIIG6 и химерный CXIIG6 IgG1 ингибируют растворимый человеческий КСФ-1R.
Биоанализ остеокластов: при биоанализе остеокластов, декантированные человеческие моноциты инкубировали в течение 8 дней с активными mAb CXIIG6 в количестве от 0,85 нг/мл до 0,62 мкг/мл (рекомбинантное мышиное CXIIG6; химерный CXIIG6 IgG1) в присутствии 25 нг/мл КСФ-1 (ImmunoTools) и 40 нг/мл RANKL. Среду и все добавленные агенты пополняли на 4-й и 6-й дни, и общий ММР-9 измеряли в культуральной среде, которую кондиционировали с 6-го по 8-й дни. Результаты, показанные на Фигуре 18, показывают, что, по сравнению с контрольными антителами, рекомбинантное мышиное CXIIG6 и его химерный вариант (химерный CXIIG6 IgG1) значительно снижали образование ММР-9 параллельно с дифференциацией остеокластов, указывая на то, что произошло замедление роста (Фигура 18: средн. +/- стандартная погрешность среднего трех лунок).
Эти результаты дополнительно демонстрируют, что очищенное рекомбинантное мышиное CXIIG6 и химерный CXIIG6 IgG1 ингибируют функцию КСФ-1R поверхности клеток.
Картирование эпитопа антитела согласно изобретению по сравнению с серийно выпускаемыми антителами
Эти эксперименты разрабатывались с целью определения локализации эпитопа антител согласно настоящему изобретению и более точного определения, связываются ли они с идентичными или с различными эпитопами по сравнению с многими антителами против КСФ-1R серийного производства.
Химерное CXIIG6 (chCXIIG6) согласно изобретению, mAb 3291 (мышиный IgG1, клон 61701, R&D Systems МАВ3291) и mAb JF14 (мышиный IgG1, Santa Cruz sc-80174) связывают человеческий КСФ-1R, в отличие от мышиного. Моноклональные антитела 3291 и JF14 были выбраны, поскольку они были выработаны против N-конечного компонента человеческого КСФ-1R. С целью картирования сайтов связывания mAb на КСФ-1R строили усеченные мутанты человеческого КСФ-1R и химеры между человеческим и мышиным D1-D3 КСФ-1R, слитые с гистидиновыми тагами (детали см. на Фигурах 19 и 20). Эти последовательности экспрессировали как секретированные белки с использованием клеток СНО. Экспрессию последовательностей проверяли путем иммуноблот-анализа с использованием антитела против гистидинового тага для обнаружения. Все последовательности захватывали на ELISA-планшете путем инкубации супернатанта культуры СНО в лунках, покрытых антителом против гистидинового тага. Антитела биотинилировали, используя Biotin-NHS (Sigma ref B3295-10MG). Биотинилированные антитела добавляли в каждую лунку и связывание обнаруживали при помощи стрептавидина-HRP. Биотинилированный ритуксимаб (Du et al., 2007, J. Biol. Chem. 282 (20), 15073-15080, NCBI Access Numbers 20SLH & 20SLL) использовали в качестве отрицательного изотипического контроля (не связывающегося с КСФ-1R). Результаты показаны на Фигуре 19.
Результаты: Как ожидалось, ритуксимаб не связывается ни с одной из испытываемых последовательностей. Химерное CXIIG6, mAb 3291 и mAb JF14 способны связываться с выделенным доменом 1 (D1) человеческого КСФ-1R (см. последовательность pTG18038; Фигура 19). Кроме того, замена человеческого D1 на мышиный D1 полностью исключает связывание трех mAb (см. последовательность pTG 18003; Фигура 19). Результаты, полученные с использованием последовательностей pTG18015 и pTG 18000 с сочетанием человеческого и мышиного субдоменов D1 показывают, что связывание mAbs 3291 и JF14 требует как N-конечной, так и центральной части человеческого домена D1 КСФ-1R, в то время, как связывание моноклонального антитела согласно изобретению требует только N-конечной части человеческого домена D1 (см. последовательность pTG18016; Фигура 19).
Эти данные показывают, что эпитоп моноклонального антитела согласно изобретению связывается с человеческим КСФ-1R на другом эпитопе по сравнению с моноклональными антителами 3291 и JF14. Интересно, что N-конечные части человеческого и мышиного КСФ-1R отличаются лишь четырьмя остатками: I20А, V27G, K33E и А36Е (начальный метионин представляет остаток 1).
Моноклональные антитела H19K12, H27K5 и H27K15
Образование гибридомы CXIIG6, вырабатывающей мышиный IgG2a с блокирующей активностью в отношении функции человеческого КСФ-1R, описывалось выше. Тяжелые и легкие цепи CXIIG6 IgG2a клонировали и секвенировали (см. Фигуры 6 и 7). Химерный вариант моноклонального антитела (chCXIIG6) строили, комбинируя вариабельные домены VH и VL mAb CXIIG6 с постоянными участками человеческого IgG1 (см. выше). Для повышения гомологии с последовательностями человеческих антител и уменьшения потенциальной иммуногенности моноклональных антител образовывали гуманизированные варианты путем введения мутаций в VH- и VL-домены (см. Фигуры 15 и 16).
262 варианта гуманизированного моноклонального антитела согласно изобретению временно экспрессировали в клетках яичника китайского хомячка (СНО) и испытывали на их способность связывания с КСФ-1R. Из этого первого отбора выбирали тридцать один вариант гуманизированного CXIIG6 (hCXIIG6) на основе (i) аффинности к человеческому КСФ-1R, (ii) наивысшей гомологии с последовательностями человеческой зародышевой линии (в предпочтительном варианте вышеупомянутая гомология составляет как минимум 76%, в более предпочтительном варианте - как минимум 85%) и (iii) наинизшей in silico иммуногенности. Исследования аффинности при помощи микровесов на кристалле кварца (Attana) показали, что все выбранные варианты hCXIIG6 обладают аффинностью к рекомбинантному человеческому КСФ-1R, в пределах от 10-9 до 10-10 М, подобной показателю родительского mAb chCXIIG6. Три из 30 гуманизированных вариантов, характеризуемые высокой аффинностью к КСФ-1R, наивысшей степенью человеческой гомологии и наинизшей in silico иммуногенностью, были отобраны для дальнейших исследований: H19K12, H27K5 и H27K15.
Биохимическая характеризация моноклонального антитела H27K15 согласно изобретению
А-радиоактивное мечение антитела H27K15 йодом-125
Материалы
Моноклональное антитело H27K15 согласно изобретению обеспечивалось в виде водного раствора (PBS, pH 7,5) в концентрации 2,1 мг/мл.
Радионуклид йода-125 приобретали у Perkin Elmer в форме йодида натрия в 10-5 N гидроксида натрия (удельная активность: 643,8 ГБк/мг - чистота радионуклида: 99,95%).
Хлорамин-Т (N-хлоро-р-толуолсульфонамид, PM: 227,6 г.моль-1), метабисульфит натрия (PM=190,1 г.моль-1), альбумин сыворотки крупного рогатого скота (BSA) и трихлоруксусную кислоту приобретали у Sigma.
Фосфатно-буферный раствор 0,1М, pH 7,2 приготавливали в нашей лаборатории.
Способ
К 100 мкг антитела добавляли 1,2 мКи раствора Na125I (44,4 МБк) и 7,6 мкл свежеприготовленного раствора хлорамина-Т (1 мг/мл в фосфатном буфере). После 2 мин при комнатной температуре реакцию останавливали путем добавления 12,7 мкл метабисульфита натрия (1 мг/мл в фосфатном буфере).
Невключенный йод-125 удаляли путем гель-фильтрации на колонке Sephadex G-25 (PD-10, Pharmacia), предварительно насыщенной элюирующим буфером (фосфатный буфер, 0,1 M, pH 7,2 / 0,5% BSA). Колонку элюировали в 40 аликвотах 0,5 мл. Радиоактивность в каждой фракции измеряли в автоматическом гамма-счетчике, калиброванном для радионуклида йода-125 (автоматический гамма-счетчик Wallace Wizard 2470, калиброванный для радионуклида йода-125 - Perkin Elmer) и собирали фракции, содержащие нужный меченный радиоактивным йодом продукт.
Радиохимическую чистоту радиоактивно меченного соединения определяли путем ITLC-анализа, осуществляемого на предварительно покрытых силикагелевых планшетах Gelman Sciences, используя 10% ТСА в качестве элюента.
Результаты и характеристики растворов радиоактивно меченных антител
Перед очищением, радиохимический выход, определяемый с помощью ITLC, составлял 97,8%.
Характеристики раствора радиоактивно меченного антитела показаны ниже:
Чистоту и целостность радиоактивно меченного антитела определяли при помощи электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) в восстановительных и невосстановительных условиях. Для калибровки геля использовали широкодиапазонный стандарт молекулярной массы (Biorad Laboratories), состоящий их 10 маркеров молекулярной массы от 10 до 250 кДа. Гели окрашивали Кумасси голубым. На втором этапе гели высушивали и выкладывали на экран для авторадиографии с помощью прибора Phosphorlmager 445 SI (Amersham Biosciences). Для анализа данных использовали программу ImageJ (Molecular Dynamics).
Электрофоретические спектры, полученные после окрашивания Кумасси голубым и авторадиографии, оказались идентичными. В невосстановительных условиях визуализируется одна основная полоса при 150 кДа, соответствующая полной молекуле IgG. В восстановительных условиях обнаруживаются полосы 50 кДа и 25 кДа, соответствующие тяжелым и легким цепям антитела, соответственно. Эти результаты электрофореза подтверждают, что способ с применением хлорамина-Т не влияет на целостность антитела.
В- Анализы связывания радиоактивно меченного антитела H27K15 на клетках EL4-KCФ-1R
b1 Измерение аффинного связывания 125I-H27K15
Равновесную константу диссоциации (KD) использовали в качестве меры аффинности связывания. KD для антитела 125I-H27K15 определяли при помощи анализа насыщения, при котором увеличивающуюся концентрацию радиоактивно меченного антитела добавляли к неизменному количеству клеток.
Материалы
Раствор радиоактивно меченного антитела в PBS, содержащий 0,5% BSA
Растворы радиоактивно меченного антитела с 100-кратным молярным избытком немеченного антитела (неспецифическое связывание): приготавливали раствор, содержащий 45 мкг/мл немеченного антитела и 0,45 мкг/мл радиоактивно меченного антитела, а затем выполняли серию 4-кратных разведений в PBS/BSA 0,5%.
Клетки: приготавливали суспензию клеток EL4-KCФ-1R в концентрации 40×106 клеток/мл в PBS с содержанием 0,5% BSA.
Способ
Клетки EL4-KCФ-1R инкубировали (2×106 в 50 мкл PBS/BSA 0,5%) с увеличивающимися концентрациями радиоактивно меченного антитела в окончательном объеме 150 мкл в течение 1 ч на льду при перемешивании.
Неспецифическое связывание определяли путем совместной инкубации избыточного количества немеченного антитела (с 100-кратным молярным избытком).
После инкубации реакционные смеси помещали на 200 мкл дибутилфталатной масляной ванны и центрифугировали в микроцентрифужных пробирках при 12000 об/мин в течение 3 мин. Пробирки замораживали в жидком азоте и концы пробирок, содержащие клеточный дебрис, срезали для определения радиоактивности, используя автоматический гамма-счетчик Wallace Wizard 2470, калиброванный для радионуклида йода-125 (эффективность: 63%).
Неспецифическое связывание определяли путем измерения связанной радиоактивности четырехточечной концентрации радиоактивно меченного антитела, содержащего 100-кратный молярный избыток немеченного антитела. Строили график сравнения связанной фракции со свободной фракцией, и линейную регрессию использовали с целью определения наклона прямой линии. Специфическое связывание определяли путем вычитания связывания, наблюдаемого в присутствии (неспецифическое связывание) от наблюдаемого в отсутствие (общее связывание) избыточного немеченного антитела.
Результаты
Константу аффинности гуманизированного антитела 125I-H27K15 для клеток EL4-KСФ-1R и максимальное количество сайтов связывания на клетку определяли при помощи программы Prism.
Характеристики связывания антитела 125I-H27K15 для клеток EL4-KCФ-1R показаны ниже:
b2 Измерение константы ингибирования немеченного H27K15
Цель этого исследования состояла в то, чтобы определить, является ли аффинность иммунологически радиоактивно меченного антитела такой же, как у немеченного антитела. Выполняли анализ связывания с радиоактивно меченным смещением при неизменной концентрации радиоактивно меченного антитела и последовательных разведениях немеченного антитела.
Материал
Растворы немеченного антитела: приготавливали раствор при 450 нМ (67,5 мкг/мл), а затем выполняли серию 2,5-кратных разведений в PBS/BSA 0,5%. Диапазон концентрации: от 450 нМ до 7,56 пМ.
Клетки: приготавливали суспензию клеток EL4-KCФ-1R в концентрации 40×106 клеток/мл в PBS, содержащем 0,5% BSA.
Способ: клетки инкубировали (2×106 в 50 мкл PBS/BSA 0,5%) с 50 мкл радиоактивно меченного антитела при 1,5 нМ в отсутствие (общее связывание: контрольная лунка) и в присутствии различных концентраций (как указано выше) в 50 мкл немеченного антитела в течение 1 ч на льду при перемешивании.
После инкубации реакционные смеси помещали на 200 мкл дибутилфталатной масляной ванны и центрифугировали в микроцентрифужных пробирках при 12000 об/мин в течение 3 мин. Пробирки замораживали в жидком азоте и концы пробирок, содержащие клеточный дебрис, срезали для определения радиоактивности с применением гамма-счетчика.
Процент относительного связывания радиоактивно меченного антитела вычисляли следующим образом:
Радиоактивность в контрольной лунке
Радиоактивность в тест-лунке ×100
(Контрольная лунка не содержит немеченного антитела)
Результаты
Данные связывания представлены на Фигуре 21.
Значение IC50, полученное на основе кривой конкуренции, определяли при помощи программы Prism и переводили в абсолютную константу ингибирования Ki, применяя уравнение Ченга-Прусоффа (Biochem. Pharmacol., 22, 3099-3108, 1973):
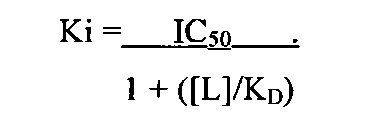
[L] - концентрация радиоактивно меченного антитела, применяемого при анализе
IC50 - концентрация немеченного антитела, необходимая для достижения 50% ингибирования радиоактивно меченного антитела
Значение IC50 и константа ингибирования показаны ниже:
b3 Исследование конкуренции между 125I-H27K15 и различными антителами
Материалы
Раствор радиоактивно меченного антитела H27K15 в PBS, содержащем 0,5% BSA: см. выше
Растворы немеченного антитела
Различные антитела представлены в таблице:
Для каждого антитела приготавливали раствор в количестве 450 нМ (67,5 мкг/мл), а затем выполняли серию 2,5-кратных разведений в PBS/BSA 0,5%. Диапазон концентрации: от 450 нМ до 7,56 пМ.
Клетки: приготавливали суспензию клеток EL4-KCФ-1R в концентрации 40×106 клеток/мл в PBS, содержащем 0,5% BSA.
Способ
Клетки инкубировали (2×106 в 50 мкл PBS/BSA 0,5%) с 50 мкл антитела 125I-H27K15 при 1,5 нМ в отсутствие (общее связывание: контрольная лунка) и в присутствии различных концентраций (как указано выше) в 50 мкл немеченного конкурирующего антитела в течение 1 ч на льду при перемешивании.
После инкубации реакционные смеси помещали на 200 мкл дибутилфталатной масляной ванны и центрифугировали в микроцентрифужных пробирках при 12000 об/мин в течение 3 мин. Пробирки замораживали в жидком азоте и концы пробирок, содержащие клеточный дебрис, срезали для определения радиоактивности с применением гамма-счетчика.
Процент относительного связывания радиоактивно меченного антитела вычисляли следующим образом:
Радиоактивность в тест-лунке×100
Радиоактивность в контрольной лунке
Контрольная лунка не содержала конкурирующего антитела.
Результаты
Данные связывания представлены на Фигуре 22
Значение IC50, полученное на основе кривой конкуренции, определяли при помощи программы Prism и переводили в абсолютную константу ингибирования Ki, применяя уравнение Ченга-Прусоффа:
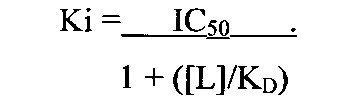
[L] - концентрация антитела 125I-H27K15, применяемого при анализе
IC50 - концентрация конкурирующего антитела, необходимая для достижения 50% ингибирования антитела 125I-H27K15
KD - константа аффинности антитела 125I-H27K15.
Для каждого конкурирующего антитела значения IC50 и константы Ki показаны ниже:
Аффинность моноклональных антител согласно изобретению (H27K15, chCXIIG6, H19K12 и H27K5) вполне подобна значению Ki от 0,5 до 0,8 нМ.
Значения Ki моноклональных антител серийного производства 3291 и JF14 превышают 1 нМ. Таким образом, эти антитела обладают меньшей аффинностью к антигену КСФ-1R по сравнению с моноклональными антителами согласно изобретению.
С - Анализы конкурентного связывания человеческого КСФ-1 с человеческим КСФ-1R D1-D5
КСФ-1 (1-444) биотинилировали, используя Биотин-NHS (Sigma ref. В3295-10MG). Биотинилированный КСФ-1 (100 мкл/лунку при 0,012 мкг/мл) добавляли в лунки, покрытые человеческим КСФ-1R FC-D1-D5 (100 мкл/лунку при 1 мкг/мл) (R&D systems ref. 329-MR-100) и связанные молекулы обнаруживали, используя стрептавидин-HRP. Для экспериментов с конкуренцией увеличивающееся количество моноклональных антител (100 мкл/лунку в концентрации от 0,030 мкг/мл до 200 мкг/мл) инкубировали до, после или совместно с биотинилированным КСФ-1.
Авторами было испытано серийно выпускаемое моноклональное антитело 3291 (R&D Systems ref. МАВ3291), одно моноклональное антитело согласно изобретению H27K15, моноклональное антитело X (WO 2009/026303) в качестве положительного контроля, о котором известно, что оно конкурирует за связывание КСФ-1 с его рецептором, и Ритуксимаб в качестве отрицательного контроля.
В случае предварительной инкубации моноклональные антитела инкубировали с покрытым человеческим КСФ-1R за один час до добавления биотинилированного КСФ-1.
В случае последующей инкубации биотинилированное КСФ-1 инкубировали с покрытым человеческим КСФ-1R за один час до добавления моноклональных антител.
В случае совместной инкубации биотинилированное КСФ-1 и моноклональные антитела инкубировали вместе с покрытым человеческим КСФ-1R.
Результаты: ранее было продемонстрировано (см. выше), что антитела 3291 и H27K15 распознают один эпитоп (т.е., Ig-подобный домен 1), отличный от распознаваемого положительным контрольным антителом X, которое, как известно, связывается с тем же сайтом связывания, что и КСФ-1 (т.е., в Ig-подобных доменах 2-3). В данном случае показывается, что, в отличие от положительного контроля, антитела 3291 и H27K15 являются частичными конкурентами за связывание КСФ-1, даже при высокой дозе антитела (то есть, при ~100-кратном IC50). При любых условиях (предварительной, совместной или последующей инкубации) и при насыщенных дозах Н27К15 может уменьшать связывание КСФ-1 с КСФ-1R приблизительно на 10-20% (см. Фигуры 23, 24 и 25), тогда, как антитело 3291 может уменьшать такое же связывание приблизительно на 30-40%, в зависимости от условий эксперимента.
Соответственно, моноклональные антитела согласно настоящему изобретению способны частично предотвращать связывание КСФ1 с его рецептором КСФ-1R.
Это было подтверждено в следующем эксперименте.
ELISA-микропланшеты покрывали 0,1 мкг (то есть, 100 мкл при 1 мкг/мл) человеческого КСФ1 1-444 (Geneart - см. SEQ ID NO: 47, Фигура 32). 100 мкл человеческого КСФ-1R Fc-D1-D5 (R&D systems ref 329-MR-100) при 0,125 мкг/мл совместно инкубировали в присутствии увеличивающейся концентрации антитела. Человеческий КСФ-1R Fc-D1-D5, связанный с КСФ1, обнаруживали, используя антитело против IgG-Fc-HRP человека (Bethyl А80-204Р). Поскольку антитело, используемое для обнаружения, связывается с человеческим Fc, для конкурентного связывания использовали мышиное антитело CXIIG6 вместо антитела H27K15. Конкурирующее mAb X (WO 2009/026303) использовали в качестве положительного контроля, поскольку известно, что оно связывается с человеческим КСФ-1R в том же месте, что и человеческий КСФ-1. IgG2a крысы использовали в качестве отрицательного контроля (иррелевантное изотипическое антитело, которое не связывается с hCD115).
Результаты: mAb X, как указывается в WO 2009/026303, полностью ингибирует связывание КСФ1 (1-444) с человеческим КСФ-1R Fc-D1-D5, как ожидалось для конкурирующего антитела. И 3291, и mCXIIG6 лишь частично уменьшают связывание человеческого КСФ-1R Fc-D1-D5 с КСФ1 даже при высокой дозе антитела (то есть, 100 мкг/мл). MCXIIG6 и 3291 уменьшали связывание КСФ1 с Fc-hCD115 приблизительно на 5-10% и 10-20%, соответственно (см. Фиг.26). Эти результаты согласуются с вышеописанными, демонстрируя частичное ингибирование связывания КСФ1 с человеческим КСФ-1R Fc-D1-D5 со стороны H27K15 или mAb 3291.
Фигура 26 показывает частичное уменьшение связывания рекомбинантного человеческого Fc - человеческого D1-D5 CD115 с человеческим КСФ1 различными mAb (эксперимент с совместной инкубацией). Человеческий КСФ1 (0,1 мкг/мл) наносили на 96-луночный планшет и инкубировали в течение 1 ч 45 мин со смесью (эксперимент с совместной инкубацией) рекомбинантного человеческого КСФ-1R Fc-D1-D5 (0,125 мкг/мл) и увеличивающимися концентрациями mAb. CD115, связывающийся с hКСФ-1, обнаруживали, используя Fc-IgG против человека, конъюгированное с HRP.
D- Исследование конкуренции между 125I-H27K15 и IL-34, известными как природные лиганды рецептора КСФ-1R.
Материалы
Растворы радиоактивно меченного антитела в PBS, содержащем 0,5% BSA 125I-H27K15 (см. выше)
Растворы лигандов: рекомбинантный человеческий IL-34 (26,1 кДа) приобретали у R&D Systems в концентрации 0,422 мг/мл. Приготавливали раствор при 450 нМ, а затем выполняли серию 2,5-кратных разведений в PBS/BSA 0,5%. Диапазон концентрации: от 450 нМ до 7,56 пМ.
Клетки: приготавливали суспензию клеток EL4-КСФ-1R в концентрации 40×106 клеток/мл в PBS, содержащем 0,5% BSA.
Способ
Во время этого конкурентного исследования выполняли два разных протокола с природными лигандами.
Согласно первому, клетки совместно инкубировали (2×106 в 50 мкл PBS/BSA 0,5%) с 50 мкл антитела 125I-H27K15 при 1,5 нМ в отсутствие (общее связывание: контрольная лунка) и в присутствии различных концентраций (как указано выше) в 50 мкл немеченного конкурирующего антитела в течение 1 ч на льду при перемешивании.
Согласно второму протоколу, инкубировали клетки EL4-КСФ-1R с увеличивающимися концентрациями лиганда в течение 30 мин на льду перед добавлением радиоактивно меченного антитела H27K15. Затем вторую инкубацию также выполняли на льду в течение 1 часа при перемешивании.
Для двух экспериментов, реакционные смеси помещали на 200 мкл дибутилфталатной масляной ванны и центрифугировали в микроцентрифужных пробирках при 12000 об/мин в течение 3 мин. Пробирки замораживали в жидком азоте и концы пробирок, содержащие клеточный дебрис, срезали для определения радиоактивности с применением гамма-счетчика.
Процент относительного связывания радиоактивно меченного антитела вычисляли следующим образом:
Радиоактивность в тест-лунке×100
Радиоактивность в контрольной лунке
(Контрольная лунка не содержит лиганда)
Результаты: смещения связывания антитела 125I-H27K15 не наблюдалось. Следовательно, Антитело H27K15 и лиганд IL-34 распознают разные эпитопы антигена КСФ-1R.
Кроме того, выполняли исследование конкурентного связывания между 125I-IL-34 и гуманизированным антителом H27K15, и было продемонстрировано, что связывание 125I-IL-34 не смещалось с повышением концентрации антитела H27K15, показывая, что антитело согласно изобретению не конкурирует с IL-34 за связывание с рецептором КСФ-1R.
Биологическая характеризация вариантов hCXIIG6 H27K5, H27K15 и H19K12.
A. Экспрессия и очищение моноклональных антител
Варианты H19K12, H27K5, H27K15 химерных CXIIG6, hCXIIG6 и ритуксимаб экспрессировались либо путем временной трансфекции адгезивных клеток СНО-К1 или CHO-DG44, либо через поликлональные пулы стабильных трансфектантов СНО-DG44.
B. Специфичность H27K5, H27K15 и H19K12 к КСФ-1R среди других рецепторов тирозинкиназы
Способ
На микротитровальные планшеты (Maxisorp, Nunc) наносили 300 нг на лунку каждого из следующих растворимых рецепторов, приобретенных у R&D Systems: КСФ-1R20-512-Fc (cat. 329-MR/CF), (EGFR)2-Fc-6xhis (cat. 1129-ER), Flt-3-Fc-6xhis (cat. 368-ST/CF), PDGFRp-Fc-6xhis (cat. 385-PR/CF), рецептора фактора роста эндотелия сосудов (VEGFR)2-Fc-6xhis (cat. 357-KD/CF), VEGFRl-Fc-6xhis (cat. 321-FL-050), человеческого SCFR (cat. 332-SR/CF) и PDGFRα ECDs (cat. 322-PR/CF). В лунки добавляли по 50 мкл H27K5, H27K15, H19K12 mAbs или ритуксимаба (500 нг/мл в PBS) и планшеты инкубировали в течение 1 ч при 37°C. Для контролирования эффективного покрытия лунок IgG против человека (IgG козы против человека, Sigma cat. 13382) также инкубировали с лунками, покрытыми слитыми белками рецептора-человеческого Fc Flt-3-Fc-, PDGFRβ-Fc, VEGFR2-Fc, VEGFR1-Fc, в то время, как покрытые PDGFRα- и SCFR-лунки инкубировали, соответственно, с антителом против PDGF-Ra (мышиный IgG1, R&DSystems cat. МАВ322) и IgG козы против SCF-R (R&DSystems cat. AF332). Связывание антитела обнаруживали при помощи конъюгированного с пероксидазой хрена (HRP) Abs, направленного против каппа-легкой цепи человеческого Ig (Bethyl Laboratories, cat. A80-115P), IgG козы (Santa Cruz cat. sc2033) или мышиного Ig (Sigma cat. A 0412). После обнаружения тетраметилбензидином (Sigma, cat. T8665) (0,1 мг/мл в 0,05 M ацетата натрия, 0,05 М лимонной кислоты, Н2О2 1/7000), измеряли оптическую плотность при 450 нМ и корректировали путем вычитания оптической плотности при 540 нМ при помощи спектрофотометра (Berthold, Tristar LB941).
Результаты
Как и chCXIIG6, ни один из гуманизированных вариантов не демонстрировал иммунологической реактивности ни с одним из других рецепторов ECD, в то время, как они сильно связываются с иммобилизованным КСФ-1R ECD (Фигура 27). Этот результат показывает, что гуманизация не меняет специфичности mAb H19K12, H27K5 и H27K15 к КСФ-1R среди других рецепторов тирозинкиназы.
С.КСФ-1R-блокирующая активность вариантов hCXIIG6
С.1 Блокирование растворимого КСФ-1R-Fc, испытываемое в анализе пролиферации клеток M-NFS-60
Способ
Клетки M-NFS-60 дважды промывали полной средой RPMI-1640 без КСФ-1 и инкубировали в течение суток в той же среде для депривации КСФ-1. Для анализа нейтрализации растворимого человеческого КСФ-1R 5 нг человеческого КСФ-1R20-512-Fc инкубировали в течение 30 мин в белых 96-луночных микропланшетах (ViewPlateTM-96, Packard) с последовательными разведениями антителами (mAb H27K5, H27K15, H19K12, chCXIIG6 или ритуксимабом в полной среде RPMI-1640. 104 КСФ-1-депривированные клетки затем добавляли в лунки с культурой вместе с 0,1 нг человеческого КСФ-1 в конечном аналитическом объеме 100 мкл. Клетки инкубировали в течение 48 ч при 37°C и пролиферацию количественно определяли после включения 5-бромо-2'-дезоксиуридина (BrdU) в течение 2 ч с применением ELISA-анализа пролиферации клеток от Roche (cat. #11647223001) согласно протоколу производителя. Оптическую плотность измеряли в спектрофотометре (Berthold Tristar LB941).
Результаты
Линия клеток миелогенного лейкоза мышей M-NFS-60 зависит от КСФ-1 в своем росте и ранее применялась для демонстрации КСФ-1R-блокирующей активности мышиного mAb CXIIG6. В этом анализе клетки M-NFS-60 культивируют с человеческим КСФ-1 и человеческим растворимым гомодимерным дисульфидно-связанным КСФ-1 R20-512-Fc. Захват КСФ-1 растворимым КСФ-1R ведет к ингибированию пролиферации клеток. Добавление антител, нейтрализующих человеческий КСФ-1R, препятствует захвату КСФ-1 растворимым КСФ-1R и восстанавливает рост клеток. В присутствии отрицательного контрольного ритуксимаба растворимый КСФ-1R полностью ингибировал пролиферацию клеток М-NFS-60, опосредованную КСФ-1 (Фигура 28). H19K12, H27K5 и H27K15 обеспечивали зависящую от дозы нейтрализацию растворимого КСФ-1 R20-512-Fc и восстанавливали пролиферацию клеток, как и chCXIIG6. Показатели ND50 (нейтрализующая доза, обеспечивающая 50% от максимального КСФ-1 R-блокирующего воздействия), рассчитанные при помощи программы GraphPad Prism с применением 5-параметрического логистического уравнения, были подобными для 3-х испытываемых вариантов hCXIIG6 (29,8, 30,7 и 29,1 нг/мл, соответственно, для H19K12, H27K5 и H27K15) и существенно не отличались от показателя родительского mAb chCXIIG6 (18,1 нг/мл).
С.2 Блокирование КСФ-1R поверхности клеток, испытываемое при анализе на основе клеток AML5
Способы
Иммуноцитохимический анализ и анализ проточной цитометрии клеток AML5 Клетки AML5 (OCI-AML5, DSMZ cat. АСС-247) анализировали путем иммуноцитохимического анализа и анализа проточной цитометрии. 5×105 клеток инкубировали в течение 30 мин на льду с mAb 3291 против человеческого КСФ-1R (мышиный IgG1, клон 61701, 10 мкг/мл) (R&D Systems cat. #МАВ3291), изотипическим контрольным мышиным IgG1 (R&D Systems cat. #МАВ002, 10 мкг/мл), chCXIIG6 (10 мкг/мл) или ритуксимабом в 100 мкл PBS. Затем клетки метили в течение 30-45 мин на льду с конъюгированным с фикоэритрином Ig козы против мыши (BD Pharmingen cat. #550589) или конъюгированным с флуоресцеинизотиоцианатом IgG против человека F(ab')2 (Millipore cat. #AQ112F).
Влияние КСФ-1 на пролиферацию клеток AML5
Клетки AML5 помещали в количестве 2,5 Е4 клеток /100 мкл/ лунку в 96-луночные планшеты с плоским дном (в экспериментах 1 и 2, ТРР cat. #92096) или с круглым дном (в эксперименте 3, Falcon cat. #3077). В каждую лунку добавляли 100 мкл среды, содержащей человеческий КСФ-1. Клетки культивировали в течение 48 ч и их пролиферацию измеряли после 4-часового включения BrdU, применяя ELISA-анализ пролиферации клеток от Roche (cat. #11647223001) согласно протоколу производителя. Оптическую плотность измеряли в спектрофотометре (Berthold, Tristar LB941). Среднее значение оптической плотности +/- стандартная погрешность среднего вычисляли для лунок в четырех экземплярах и строили log-график [КСФ-1] (нг/мл), применяя GraphPad Prism для подгонки кривой нелинейной регрессии с применением 3-параметрического уравнения.
Влияние вариантов hCXIIG6 на пролиферацию клеток AML5
Клетки культивировали и помещали в 96-луночные планшеты, как описывается выше. Дифференцированное количество chCXIIG6, H27K15, H27K5, H19K12 или отрицательного контрольного ритуксимаба добавляли в 50 мкл среды. После 30-минутной инкубации при 37°C в каждую лунку добавляли 50 мкл среды, содержащей 40 нг/мл hКСФ-1. Каждый эксперимент выполняли на 4 идентичных 96-луночных планшетах, каждый из которых содержал по два образца каждой концентрации mAb. Клетки культивировали в течение 48 ч и их пролиферацию измеряли после 4-часового включения BrdU, как описывалось выше. Результат для каждого планшета сначала анализировали, применяя GraphPad Prism для подгонки кривой нелинейной регрессии с 5-параметрическим уравнением. Планшеты, обеспечивавшие результаты, которые подгонялись при помощи GraphPad Prism как минимум для 3-х из 4-х испытанных mAb, выбирали для вычисления средних значений +/- стандартная погрешность среднего. Таким образом, эксперименты 1 и 2 анализировали на основе результатов 2-х из 4-х планшетов (средние значения, рассчитанные по показателям оптической плотности четырех образцов), а эксперимент 3 - на основе результатов 3-х из 4-х планшетов (средние значения, рассчитанные по 6 показателям оптической плотности). Квадраты ЕС50 и R вычисляли при помощи GraphPad Prism для каждого испытанного mAb.
Результаты
Известно, что колониестимулирующий фактор-1 стимулирует рост линии клеток острого миелоидного лейкоза человека OCI-AML5 (AML5) (Drexler HG, Zaborski М, Quentmeier Н., 2007. Cytokine response profile of human myeloid factor-dependent leukaemia cell lines. Leukemia 11: 701-8). Клетки AML5 анализировали, применяя иммуноцитохимический анализ и анализ проточной цитометрии. Окрашивание было положительным для серийно выпускаемого мышиного IgG1 3291 против hКСФ-1R (по сравнению с изотипическим контрольным мышиным IgG1) и chCXIIG6 (по сравнению с ритуксимабом), демонстрируя, что клетки AML5 экспрессируют поверхностный КСФ-1R (Фигура 29А).
Для создания модели для исследования влияния блокирования КСФ-1R сначала было необходимо проверить. Вызывает ли КСФ-1 зависящее от дозы повышение пролиферации клеток AML5. Фигура 29В показывает, что хотя AML5 способны расти в отсутствие экзогенного КСФ-1, добавление фактора роста в культуральную среду повышает пролиферацию клеток в зависимости от дозы. Таким образом, рост клеток AML5 не имеет строгой зависимости от КСФ-1, однако они являются КСФ-1-чувствительными.
Затем испытывали влияние вариантов hCXIIG6 и chCXIIG6 на клетки AML5, культивируемые с 10 нг/мл КСФ-1, при концентрации, вызывающей приближенную к максимальной пролиферацию (Фигура 29 В). Фигура 29С показывает результаты 3-х независимых экспериментов. Гуманизированные варианты H27K5, H27K15 и H19K12, а также chCXIIG6 вызывали зависящее от дозы снижение пролиферации клеток AML5. Наблюдалась изменчивость показателей между экспериментами, отражаемая в различиях ЕС50 для каждого mAb и довольно низких квадратах R, которые рассчитывали при помощи GraphPad Prism, в частности, в эксперименте 1 (Фигура 29D). Таким образом, варианты hCXIIG6 не могут быть достоверно классифицированы по их ЕС50 в этом анализе. Все 3 варианта hCXIIG6 демонстрировали одинаковую эффективность в ингибировании зависящей от КСФ-1 пролиферации клеток AML5.
С.3 Неагонистическое влияние на сигнал КСФ-1R и ингибиторное влияние на зависящее от КСФ-1 фосфорилирование
Способы
В4-800-5 представляет собой линию рекомбинантных клеток, полученных путем стабильной трансфекции клеток мышиного NIH/3T3 с использованием pTG17366, плазмиды экспрессии, кодирующей человеческий КСФ-1R. Клетки В4-800-5 высевали в количестве 2×105 клеток на 60 мм чашку Петри и культивировали в течение 72 ч. Клетки подвергали депривации сыворотки культурой в 1 мл среды DMEM, содержащей 1% FCS, при 37°C в течение 1 ч перед экспериментами.
Для исследования блокирования КСФ-1R или влияния антител в отсутствие сшивания клетки обрабатывали 1 мл DMEM-1% FCS, содержащим chCXIIG6, H19K12, H27K5, H27K15 или изотипический контрольный человеческий IgG1 ритуксимаб в течение 1 ч при 37°C. В культуру клеток добавляли 100 нг/мл hКСФ-1 (ImmunoTools cat. 11343115) в течение 5 мин при 37°C или клетки оставляли нестимулированными.
В эксперименте со сшиванием антитела клетки обрабатывали 1 мл DMEM-1% FCS, содержащим mAb chCXIIG6, H19K12, H27K5, H27K15, изотипический контрольный человеческий IgG1 ритуксимаб, рекомбинантный мышиный IgG2a CXIIG6 или изотипический контрольный мышиный IgG2a (R&D Systems cat. МАВ003) в течение 1 ч на льду. Клетки промывали PBS ледяной температуры. К клеткам добавляли мкг/мл поликлонального IgG козы против мыши (R&D Systems cat. AF007), 20 мкг/мл поликлонального IgG козы против человека FcHRP (Jackson cat. 109-035-098) или 100 нг/мл hКСФ-1 в течение 10 мин при 37°C или клетки оставляли необработанными. Среду удаляли и слои клеток подвергали лизису путем добавления экстракционного буфера (62 мМ Tris, 10% глицерина, 2% SDS, 100 мМ DTT, рН 6,8) в чашках Петри. Экстракты клеток анализировали путем вестерн-блоттинга и зондировали следующими антителами: поликлональным кроличьим против фосфо-КСФ-1RTyr708 (Santa Cruz Biotechnology cat sc-33358-R), поликлональным кроличьим против КСФ-1R (Н300, Santa Cruz Biotechnology cat. sc-13949, разведение 1/200) и моноклональным мышиным против β-актина (Sigma-Aldrich cat. А2228, разведение 1/2000) для обнаружения тирозин 708-фосфорилированного КСФ-1R, полного КСФ-1R и β-актина, соответственно. Кроличьи и мышиные первичные антитела обнаруживали при помощи поликлонального козьего против кролика или кроличьего IgHRP против мыши (DakoCytomation cat. Р0448, Р0260), соответственно.
Результаты
Авторами исследовалось влияние вариантов hCXIIG6 H19K12, H27K5 и H27K15 и chCXIIG6 на зависящее от КСФ-1 фосфорилирование тирозина708 и отсутствие их агонистического влияния. В отсутствие КСФ-1 ни одно mAb не вызывало фосфорилирования рецептора при испытании в количестве 0,1, 1 или 10 мкг/мл, как наблюдалось в случае антитела, специфического к КСФ-1R, фосфорилированному на тирозине708. Таким образом, было продемонстрировано, что ни одна из производных CXIIG6-mAb, в химерных или гуманизированных формах, не оказывала никакого агонистического влияния при применении в отдельности к клеткам В4-800-5.
После стимулирования КСФ-1 полоса ~150 кДа, соответствующая КСФ-1R полной длины, фосфорилировалась, и общее клеточное содержание КСФ-1R снижалось в клетках, культивируемых с изотипическим контрольным ритуксимабом или без антитела. Три других полосы низкой молекулярной массы обнаруживались при помощи антитела против фосфо-КСФ-1RTyr 708. При инкубации клеток В4-800-5 при 37°C с chCXIIG6 или вариантами hCXIIG6 перед стимуляцией КСФ-1 интенсивность 4-х полос, распознаваемых mAb против фосфо-КСФ-1RTyr 708, снижалась по сравнению с показателями, наблюдаемыми в обработанных ритуксимабом клетках. Снижение интенсивности полосы 150 кДа было сравнимым с наблюдаемым ранее для CXIIG6 гибридомного происхождения в подобных условиях. Снижение интенсивности двух нижних полос оказалось менее резким. Ингибиторное влияние вариантов chCXIIG6 и hCXIIG6 на фосфорилирование КСФ-1RTyr 708 наблюдалось при 0,1, 1 и 10 мкг/мл.
Для прогнозирования возможного развития гуморального ответа на hCXIIG6 в mAb подвергнутых лечению пациентов (ответа человеческого антитела против иммуноглобулина человека) далее исследовали влияние сшивания mAb вторичными антителами против IgG на фосфорилирование рецептора КСФ-1R. Сшивание мышиного CXIIG6 создавало лишь слабую полосу 150 кДа, обнаруживаемую антителом против фосфо-КСФ-1RTyr708, как ранее наблюдалось при CXIIG6 гибридомного происхождения, и обнаруживалась легкая понижающая регуляция общего КСФ-1R, возможно, отражающая слабую активацию КСФ-1R. После сшивания варианты hCXIIG6 и chCXIIG6 также создавали полосу 150 кДа с низкой интенсивностью. Интенсивность полосы 150 кДа фосфорилированного КСФ-1R была чрезвычайно слабой по сравнению с наблюдаемой после стимулирования 100 нг/мл КСФ-1. Полосы нижнего MW, образованные КСФ-1, не обнаруживались антителом против КСФ-1RTyr 708 после сшивания каких бы то ни было производных mAb против КСФ-1R, ни одного из вариантов chCXIIG6 или hCXIIG6.
Эти эксперименты демонстрируют, что варианты hCXIIG6 H19K12, H27K5 и H27K15, как и chCXIIG6, способны частично ингибировать зависящее от КСФ-1 фосфорилирование КСФ-1R и не обладают прямой агонистической активностью в отношении КСФ-1R. Сшивание mAb на поверхности клеток имеет лишь минимальное влияние на фосфорилирование КСФ-1R.
D- Цитотоксичность для клеток-мишеней EL4-KCФ-1R
Способ
Линию рекомбинантных клеток EL4-KCФ-1R образовывали путем стабильной трансфекции образованной от лимфомы мыши линии Т-клеток EL4 (АТСС cat. TIB-39) лентивирусным вектором, кодирующим человеческий КСФ-1R полной длины. Поверхностную экспрессию КСФ-1R проверяли путем иммунного окрашивания mAb CXIIG6 (мышиный IgG2a) или 3291 и анализа проточной цитометрии (данные не показаны).
Клетки EL4-КСФ-R1 промывали в полной среде DMEM, ресуспендировали в той же среде и высевали в 96-луночные планшеты в количестве 2×104 клеток на лунку в 50 мкл. Клетки опсонизировали в течение 45-60 мин на льду с 50 мкл антител, разведенных в культуральной среда. Затем добавляли 50 мкл мононуклеарных клеток периферической крови человека (РВМС) в различных соотношениях эффектора и мишени (Е:Т). Гуманизированные варианты CXIIG6, chCXIIG6 и отрицательный контрольный ритуксимаб испытывали в количестве 10 нг/мл, 0,3 и 10 мкг/мл. Параллельно испытывали контрольные лунки, содержащие 150 мкл культуральной среды, клетки-мишени или только РВМС, для измерения (i) фона культуральной среды (CM), (ii) фона культуральной среды + лизисного раствора, (iii) спонтанного высвобождения (SR) клеток-мишеней, (iv) спонтанное высвобождение эффекторных клеток при каждой концентрации РВМС и (v) максимального высвобождения (MR) клеток-мишеней в присутствии лизисного раствора. Планшеты центрифугировали при 250×g в течение 4 мин и инкубировали в течение суток при 37°C. На следующий день в контрольные лунки, содержащие культуральную среду или только клетки-мишени, добавляли 15 мкл лизисного раствора 10 X (Promega cat. G182A) и планшеты дополнительно инкубировали в течение 45 мин при 37°C. Лактатдегидрогеназу (LDH) количественно определяли в супернатантах культур, используя нерадиоактивную пробу на цитотоксичность CytoTox 96® (Promega cat. G1780) в соответствии с инструкциями производителя. Оптическую плотность записывали при 490 нМ при помощи считывающего устройства TriStar LB 941 и программы MikroWin 2000 от Berthold Technologies.
Средний фон СМ вычитали от значений экспериментального значения, SR мишени и SR эффектора. Среднее значение СМ + фон лизисного раствора вычитали от значений MR мишеней. Статистический анализ осуществляли, используя значения оптической плотности, после вычитания средних значений контрольных СМ и SR, с применением программы GraphPad Prism и непараметрического одностороннего критерия Манна-Уитни (для сравнения chCXIIG6 или каждого варианта hCXIIG6 с ритуксимабом) или двустороннего t-критерия (для сравнения каждого варианта hCXIIG6 с chCXIIG6). Процент лизиса вычисляли по следующей формуле:

Результаты
ADCC-активность chCXIIG6 и вариантов hCXIIG6 (человеческого IgG1) испытывали на клетках-мишенях EL4-KCФ-1R, используя РВМС разных доноров крови.
Эффекторные клетки донора #1 использовали в соотношении Е:Т 25:1 (Фигура 30А). В этом эксперименте свыше 10% прямой лизис наблюдался в отсутствие mAb, что может отражать активность клеток NK, содержащихся в РВМС. Значения, полученные с отрицательным контрольным ритуксимабом, пребывали в том же диапазоне. Путем анализа проточной цитометрии было подтверждено, что клетки EL4-КСФ-1R не связывают ритуксимаб (данные не показаны). При 10 нг/мл mAb специфический лизис (превышающий неспецифический лизис, наблюдаемый в присутствии ритуксимаба) не обнаруживался для mAb против КСФ-1R. При 0,3 мкг/мл mAb chCXIIG6 вызывал цитотоксичность свыше 20%. Лизис со стороны 3-х вариантов hCXIIG6 был в одинаковом порядке величин. При высоких концентрациях mAb (10 мкг/мл) все 3 варианта hCXIIG6 обеспечивали приближенную к 35% цитотоксичность, в то время, как лизис со стороны chCXIIG6 оставался меньшим, чем 20%.
Другой эксперимент выполняли, используя эффекторные клетки донора #2 при соотношении Е:Т 25:1, 50:1 или 100:1 (Фигура 30В). Прямой лизис не обнаруживался, в отличие от первого эксперимента. Фоновая цитотоксичность была ниже 10% для ритуксимаба при всех испытываемых соотношениях Е:Т. Результаты хорошо коррелировали с результатами предыдущих экспериментов: не наблюдалось специфического лизиса при низких концентрациях mAb (10 нг/мл), однако при 0,3 и 10 мкг/мл chCXIIG6 и 3 варианта hCXIIG6 вызывали значительный специфический лизис клеток EL4-КСФ-1R. При обеих этих концентрациях mAb и всех испытываемых соотношениях Е:Т лизис клеток-мишеней при H27K5, H27K15 и H19K12 также был выше, чем при chCXIIG6 (р=0,02).
Эти результаты демонстрируют, что chCXIIG6 и варианты hCXIIG6 обладают потенциалом уничтожения клеток-мишеней, экспрессирующих поверхностный КСФ-1R.
Терапевтический эффект химерных и гуманизированных mAb против КСФ-1R в модели опухоли BeWo
Поскольку chCXIIG6 и варианты hCXIIG6 являются специфическими к человеческому КСФ-1R и не распознают мышиный гомолог, их влияние in vivo может исследоваться только в мышиных моделях с применением положительных в отношении человеческого КСФ-1R опухолях. Однако в отсутствие человеческого КСФ-1, человеческий КСФ-1R остается неактивным, и блокирование его функции не может быть исследовано. Кроме того, блокирование КСФ-1 R-положительных клеток-хозяев (опухолево-ассоциированных макрофагов, остеокластов), которое должно быть терапевтически благоприятным, в системе этой модели неосуществимо. В последующих экспериментах с применением положительных в отношении человеческого КСФ-1R опухолях BeWo только цитотоксичное влияние mAb на клетки опухолей в результате могут обеспечивать терапевтическую эффективность.
Клетки хориокарииномы человека BeWo экспрессируют поверхностный КСФ-1R
Клетки BeWo, культивированные in vitro, подвергали иммунному окрашиванию mAb H27K15 (гуманизированным антителом против КСФ-1R) и флуоресценцию анализировали при помощи конфокальной микроскопии. Специфическое окрашивание, наблюдаемое при H27K15, по сравнению с отрицательным контрольным ритуксимабом, показывает, что клетки BeWo экспрессируют человеческий КСФ-1R на их поверхности. Клетки BeWo не секретируют КСФ-1, как было обнаружено путем ELISA-титрования супернатантов культур (результат не показан).
Твердую опухоль, развившуюся из клеток BeWo, подкожно вживленную мышам линии NMR1, лишенным вилочковой железы, включали в ОСТ для иммуногистохимического анализа. Замороженные срезы ткани окрашивали мышиным вариантом CXIIG6 или изотипическим контролем. Сильное специфическое окрашивание наблюдалось по всей опухоли, что отражало экспрессию на поверхности клеток и в цитоплазме hКСФ-1R в клетках опухоли BeWo in vivo.
Эксперимент 1: Терапевтический эффект химерного CXIIG6 в модели опухоли хориокарииномы BeWo
Четыре миллиона клеток BeWo подкожно вживляли в бок мышей линии NMR1, лишенных вилочковой железы. Группе из 11 мышей вводили 3 инъекции химерного CXIIG6 (chCXIIG6, 50 мг/кг в PBS, внутрибрюшинно, с введением в 1, 3 и 7-й дни), в то время, как другая группа из 11 мышей получала изотипический контрольный ритуксимаб на основе той же схемы. Ингибирование роста клеток (Фигура 31А) и продление выживаемости мышей (Фигура 31В) наблюдали у мышей, получавших chCXIIG6, что демонстрировало наличие терапевтического эффекта воздействия mAb на положительные в отношении человеческого КСФ-1R опухоли BeWo.
Эксперимент 2: Терапевтический эффект химерного CXIIG6 и гуманизированного H27K15 в опухолевой модели хориокарииномы BeWo
Повторяли вышеизложенный протокол с двумя модификациями:
- испытывали mAb chCXIIG6 или гуманизированное H27K15 (10 мышей на группу) и сравнивали с изотипическим контрольным ритуксимабом
- mAb вводили путем инъекции 3 раза в неделю в течение 3 недель, а не один раз в неделю
Продление лечения с применением chCXIIG6 или H27K15 не улучшало результаты в отношении ингибирования роста опухолей или выживание мышей по сравнению с первым экспериментом. Однако оба mAb ингибировали рост опухоли, в отличие от группы, получавшей изотипический контрольный ритуксимаб. В случае H27K15 уменьшение объема опухоли было статистически значимым на 14-й день согласно критерию Манна-Уитни и приближенным к значимому в более поздние моменты времени (17 и 21-й дни). В случае chCXIIG6 уменьшение объема опухоли было приближенным к статистически значимому.
Список источников
- WO 01/30381.
- WO 03/059395.
- WO 2005/068503.
- ЕР 1488792 A.
- US 2005059113.
- ЕР 901463 В.
- ЕР 83286 А.
- ЕР 206920 А.
- US 5747323.
- WO 94/28152.
- WO 97/04119.
- WO 94/18992.
- WO 96/17070.
- WO 2009/026303.
- SHERR. Colony-stimulating factor-1 receptor, blood. 1990, vol.75, no.1, p.1 -12.
- HUME, et al. Regulation of CSF-1 receptor expression. Molecular reproduction and development. 1997, vol.46, no.1, p.46-52.
- Roussel AND SHERR, 1989, PNAS, 86, 7924-7927.
- ASHMUN et al., 1989, Blood, 73, 827-837.
- KLUGER, et al. Macrophage colony-stimulating factor-1 receptor expression is associated with poor outcome in breast cancer by large cohort tissue microarray analysis. Clinical cancer research. 2004, vol.10, no.1, p.173-7.
- SCHOLL, et al. Anti-colony-stimulating factor-1 antibody staining in primary breast adenocarcinomas correlates with marked inflammatory cell infiltrates and prognosis. Journal of the National Cancer Institute. 1994, vol.86, no.2, p.120-6.
- CHAMBERS, et al. Overexpression of epithelial macrophage colony-stimulating factor (CSF-1) and CSF-1 receptor: a poor prognostic factor in epithelial ovarian cancer, contrasted with a protective effect of stromal CSF-1. Clinical Cancer Research. 1997, vol.3, no.6, p.999-1007.
- BAÏOCCHI, et al. Expression of the macrophage colony-stimulating factor and its receptor in gynecologic malignancies. Cancer. 1991, vol.67, no.4, p.990-6.
- KIPMA, et al. Elevated expression of the oncogene c-fms and its ligand, the macrophage colony-stimulating factor-1, in cervical cancer and the role of transforming growth factor-betal in inducing c-fms expression. Cancer res.. 2007, vol.67, no.5, p.1918-26.
- HEMMERLEIN, et al. Expression of acute and late-stage inflammatory antigens, c-fms, CSF-1, and human monocytic serine esterase 1, in tumor-associated macrophages of renal cell carcinomas. Cancer immunology, immunotherapy. 2000, vol.49, no.9, p.485-92.
- IDE, et al. Expression of colony-stimulating factor 1 receptor during prostate development and prostate cancer progression. Proc. Natl. Acad. Sci. U.S.A.. 2002, vol.99, no.22, p.14404-9.
- RAMBALDI, et al. Expression of the macrophage colony-stimulating factor and c-fms genes in human acute myeloblastic leukemia cells. Journal of Clinical Investigation. 1988, vol.81, no.4, p.1030-5.
- SCHOLL, et al. Anti-colony-stimulating factor-1 antibody staining in primary breast adenocarcinomas correlates with marked inflammatory cell infiltrates and prognosis. Journal of the National Cancer Institute. 1994, vol.86, no.2, p.120-6.
- TANG, et al. M-CSF (monocyte colony stimulating factor) and M-CSF receptor expression by breast tumor cells: M-CSF mediated recruitment of tumor infiltrating monocytes?. Journal of cellular biochemistry. 1992, vol.50, no.4, p.350-6.
- SCHOLL, et al. Circulating levels of colony-stimulating factor 1 as a prognostic indicator in 82 patients with epithelial ovarian cancer. British journal of cancer. 1994, vol.69, no.2, p.342-6.
- SCHOLL. Circulating levels of the macrophage colony stimulating factor CSF-1 in primary and metastatic breast cancer patients. A pilot study. Breast cancer research and treatment. 1996, vol.39, no.3, p.275-83.
- DORSCH, et al. Macrophage colony-stimulating factor gene transfer into tumor cells induces macrophage infiltration but not tumor suppression. European journal of immunology. 1993, vol.23, no.1, p.186-90.
- WANG, et al. Induction of monocyte migration by recombinant macrophage colony-stimulating factor. Journal of immunology. 1988, vol.141, no.2, p.575-9.
- FILDERMAN, et al. Macrophage colony-stimulating factor (CSF-1) enhances invasiveness in CSF-1 receptor-positive carcinoma cell lines. Cancer res.. 1992, vol.52, no.13, p.3661-6.
- DORSCH, et al. Macrophage colony-stimulating factor gene transfer into tumor cells induces macrophage infiltration but not tumor suppression. European journal of immunology. 1993, vol.23, no.1, p.186-90.
- SCHOLL. Circulating levels of the macrophage colony stimulating factor CSF-1 in primary and metastatic breast cancer patients. A pilot study. Breast cancer research and treatment. 1996, vol.39, no.3, p.275-83.
- BARON, et al. Modulation of MHC class II transport and lysosome distribution by macrophage-colony stimulating factor in human dendritic cells derived from monocytes. Journal of cell science. 2001, vol.114, no.pt5, p.999-1010.
- CECCHINI, et al. Role of CSF-1 in bone and bone marrow development. Molecular reproduction and development. 1997, vol.46, no.1, p.75-83.
- BRUZZANITI, et al. Molecular regulation of osteoclast activity. Reviews in endocrine. 2006, vol.7, no.1-2, p.123-39.
- CICEK, et al. Breast cancer bone metastasis and current small therapeutics. Cancer metastasis reviews. 2006, vol.25, no.4, p.635-44.
- KITAURA, et al. The journal of clinical investigation. M-CSF mediates TNF-induced inflammatory osteolysis. 2005, vol.115, no.12, p.3418-27.
- MARSHALL, et al. Blockade of colony stimulating factor-1 (CSF-I) leads to inhibition of DSS-induced colitis. Inflammatory bowel diseases. 2007, vol.13, no.2, p.219-24.
- JOSE, et al. Blockade of macrophage colony-stimulating factor reduces macrophage proliferation and accumulation in renal allograft rejection. American journal of transplantation. 2003, vol.3, no.3, p.294-300.
- KUTZA, et al. Macrophage colony-stimulating factor antagonists inhibit replication of HIV-1 in human macrophages. Journal of immunology. 2000, no.164, p.4955-4960.
- SMITH, et al. Identification of common molecular subsequences. Journal of Molecular Biology. 1981, no. 147, p.195-7.
- HARLOW. Antibodies: A Laboratory manual. 2nd edition. Cold Spring Harbor: Laboratory press, 1988.
- HAMMERLING, et al. Monoclonal Antibodies and T Cell Hybridomas. New York: Elsevier, 1981. p.563-681.
- SMITH, et al. Identification of common molecular subsequences. Journal of Molecular Biology. 1981, no.147, p.195-7.
- LENNARD. Standard protocols for the construction of scFv libraries. Methods in molecular biology. 2002, no.178, p.59-71.
- POLJAK. Production and structure of diabodies. Structure. 1994, vol.2, no. 12, p.1121-3.
- HUDSON, et al. High avidity scFv multimers; diabodies and triabodies. Journal of immunological methods. 1999, vol.231, no.1-2, p.177-89.
- KIPRIYANOV. Generation of bispecific and tandem diabodies. Methods in molecular biology. 2002, no.178, p.317-31.
- HOLLIGER, et al. Engineered antibody fragments and the rise of single domains. Nature biotechnology. 2005, vol.23, no.9, p.1126-36.
- HOOGENBOOM, et al. Natural and designer binding sites made by phage display technology. Immunology today. 2000, vol.21, no.8, p.371-8.
- MARKS, et al. By-passing immunization: building high affinity human antibodies by chain shuffling. Biotechnology. 1992, vol.10, no.7, p.779-83.
- BARBAS, et al. In vitro evolution of a neutralizing human antibody to human immunodeficiency virus type 1 to enhance affinity and broaden strain cross-reactivity. Proceedings of the National Academy of Sciences of the United States of America. 1994, vol.91, no.9, p.3809-13.
- SCHIER. Identification of functional and structural amino-acid residues by parsimonious mutagenesis. Gene. 1996, vol.169, no.2, p.147-55.
- YELTON. Affinity maturation of the BR96 anti-carcinoma antibody by codon-based mutagenesis. J. Immunol.. 1995, vol.155, no.4, p.1994-2004.
- JACKSON, et al. In vitro antibody maturation. Improvement of a high affinity, neutralizing antibody against IL-1 beta. J. immunol.. 1995, vol.154, no.7, p.3310-9.
- HAWKINS, et al. Selection of phage antibodies by binding affinity. Mimicking affinity maturation. Journal of molecular biology. 1992, vol.226, no.3, p.889-96.
- SMITH, et al. Identification of common molecular subsequences. Journal of Molecular Biology. 1981, no. 147, p.195-7.
- BRENNAN, et al. Preparation of bispecific antibodies by chemical recombination of monoclonal immunoglobulin G1 fragments. Science. 1985, vol.229, no.4708, p.81-3.
- SHALABY, et al. Development of humanized bispecific antibodies reactive with cytotoxic lymphocytes and tumor cells overexpressing the HER2 protooncogene. The Journal of experimental medicine. 1992, vol.175, no.1, p.217-25.
- KOSTELNY, et al. Formation of a bispecific antibody by the use of leucine zippers. J. immunol.. 1992, vol.148, no.5, p.1547-33.
- ZAPATA, et al. Engineering linear F (ab')2 fragments for efficient production in Escherichia coli and enhanced antiproliferative activity. Protein engineering. 1995, vol.8, no. 10, p.1057-62.
- HERMANSON. Bioconjugate techniques. Academic press, 1996.
- HERMANSON. Bioconjugate techniques. Academic press, 1996. p.118-228.
- HERMANSON. Bioconjugate techniques. Academic press, 1996. p.229-285.
- FELGNER, et al. Cationic liposome mediated transfection. Proceedings of the Western Pharmacology Society. 1989, vol.32, p.115-21.
- HODGSON, et al. Virosomes: cationic liposomes enhance retroviral transduction. Nature biotechnology. 1996, vol.14, no.3, p.339-42.
- REMY, et al. Gene transfer with a series of lipophilic DNA-binding molecules. Bioconjugate chemistry. 1994, vol.5, no.6, p.647-54.
- LATHE, et al. Plasmid and bacteriophage vectors for excision of intact inserts. Gene. 1987, vol.57, no.2-3, p.193-201.
- LUPTON, et al. Mapping genetic elements of Epstein-Barr virus that facilitate extrachromosomal persistence of Epstein-Barr virus-derived plasmids in human cells. Molecular and cellular biology. 1985, vol.5, no. 10, p.2533-42.
- YATES, et al. Stable replication of plasmids derived from Epstein-Barr virus in various mammalian cells. Nature. 1985, vol.313, no.6005, p.812-5.
- SUMMERS, et al. Multimerization of high copy number plasmids causes instability: CoIE1 encodes a determinant essential for plasmid monomerization and stability. Cell. 1984, vol.36, no.4, p.1097-103.
- SUTTER, et al. Nonreplicating vaccinia vector efficiently expresses recombinant genes. Proc. Natl. Acad. Sci. U.S.A.. 1992, vol.89, no.22, p.10847-51.
- MEYER, et al. Mapping of deletions in the genome of the highly attenuated vaccinia virus MVA and their influence on virulence. The Journal of general virology. 1991, vol.72, no.Pt5, p.1031-8.
- SUTTER, et al. A recombinant vector derived from the host range-restricted and highly attenuated MVA strain of vaccinia virus stimulates protective immunity in mice to influenza virus. Vaccine. 1994, vol.12, no. 11, p.1032-40.
- GRAHAM, et al. Methods in molecular biology. Edited by MURREY. The human press inc, 1991. p.109-128.
- ZAKHARCHUK, et al. Physical mapping and homology studies of egg drop syndrome (EDS-76) adenovirus DNA. Archives of virology. 1993, vol.128, no.1-2, p.171-6.
- SPIBEY, et al. Molecular cloning and restriction endonuclease mapping of two strains of canine adenovirus type 2. The Journal of general virology. 1989, vol.70, no. Pt 1, p.165-72.
- JOUVENNE, et al. Cloning, physical mapping and cross-hybridization of the canine adenovirus types 1 and 2 genomes. Gene. 1987, vol.60, no.1, p.21-8.
- MITTAL, et al. Development of a bovine adenovirus type 3-based expression vector. The Journal of general virology. 1995, vol.76, no. Pt 1, p.93-102.
- BISCHOFF, et al. An adenovirus mutant that replicates selectively in p53-deficient human tumor cells. Science. 1996, vol.274, no.5286, p.373-6.
- HEISE, et al. An adenovirus E1A mutant that demonstrates potent and selective systemic anti-tumoral efficacy. Nature Medicine. 2000, vol.6, no.10, p.1134-9.
- FUEYO, et al. A mutant oncolytic adenovirus targeting the Rb pathway produces anti-glioma effect in vivo. Oncogene. 2000, vol.19, no.1, p.2-12.
- MCIVOR. Human purine nucleoside phosphorylase and adenosine deaminase: gene transfer into cultured cells and murine hematopoietic stem cells by using recombinant amphotropic retroviruses. Molecular and cellular biology. 1987, vol.7, no.2, p.838-46.
- TABIN, et al. Adaptation of a retrovirus as a eukaryotic vector transmitting the herpes simplex virus thymidine kinase gene. Molecular and cellular biology. 1982, vol.2, no.4, p.426-36.
- TAKEBE, et al. SR alpha promoter: an efficient and versatile mammalian cDNA expression system composed of the simian virus 40 early promoter and the R-U5 segment of human T-cell leukemia virus type 1 long terminal repeat. Molecular and cellular biology. 1988, vol.8, no.1, p.466-72.
- CHEN, et al. Breast cancer selective gene expression and therapy mediated by recombinant adenoviruses containing the DF3/MUC1 promoter. The Journal of clinical investigation. 1995, vol.96, no.6, p.2775-82.
- SCHREWE, et al. Cloning of the complete gene for carcinoembryonic antigen: analysis of its promoter indicates a region conveying cell type-specific expression. Molecular and cellular biology. 1990, vol.10, no.6, p.2738-48.
- VILE, et al. Use of tissue-specific expression of the herpes simplex virus thymidine kinase gene to inhibit growth of established murine melanomas following direct intratumoral injection of DNA. Cancer res.. 1993, vol.53, no.17, p.3860-4.
- HARRIS, et al. Gene therapy for cancer using tumor-specific prodrug activation. Gene therapy. 1994, vol.1, no.3, p.170-5.
- KANAI, et al. In vivo gene therapy for alpha-fetoprotein-producing hepatocellular carcinoma by adenovirus-mediated transfer of cytosine deaminase gene. Cancer res.. 1997, vol.57, no.3, p.461-5.
- PEREZ. Principles and practice of radiation oncology. 2nd edition. LIPPINCOTT, 1992.
- KOZAK M. An analysis of 5'-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Res. 1987, 15 (20): 8125-8148.
- CHOTHIA and LESK. Canonical structures for the hypervariable regions of immunoglobulins (1987) J Mol Biol. 1987 Aug 20; 196 (4): 901-17.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ CSF-1R | 2009 |
|
RU2547586C2 |
| МОДУЛЯЦИЯ АКТИВАЦИИ МАКРОФАГОВ | 2012 |
|
RU2639553C2 |
| АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CSF-1R И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2658603C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2761663C2 |
| АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО VISTA И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2746994C2 |
| ПОЛНОСТЬЮ ЧЕЛОВЕЧЕСКОЕ АНТИТЕЛО, СПЕЦИФИЧНОЕ K ИНТЕРЛЕЙКИНУ 1-АЛЬФА | 2021 |
|
RU2838383C1 |
| ТРИСПЕЦИФИЧНЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, КОТОРЫЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАЮТ АНТИГЕНЫ МНОЖЕСТВА ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2718692C2 |
| КОНЪЮГАТЫ АНТИ-РТК7 АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2015 |
|
RU2708075C2 |
| ВЕЩЕСТВА И СПОСОБЫ ДЛЯ ПРИМЕНЕНИЯ ПРИ ПРЕДУПРЕЖДЕНИИ И/ИЛИ ЛЕЧЕНИИ БОЛЕЗНИ ГЕНТИНГТОНА | 2015 |
|
RU2742493C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ ПРЕИМУЩЕСТВЕННО С ВНЕКЛЕТОЧНЫМ ДОМЕНОМ 4 ЧЕЛОВЕЧЕСКОГО CSF-1R, И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2565541C2 |

































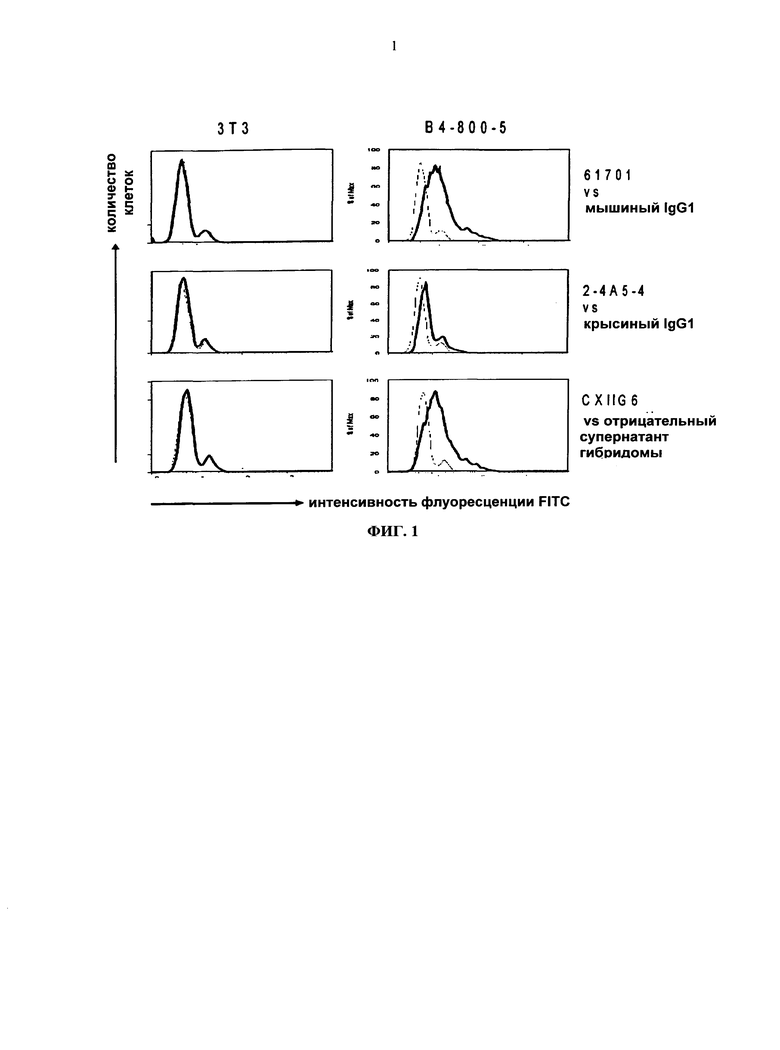
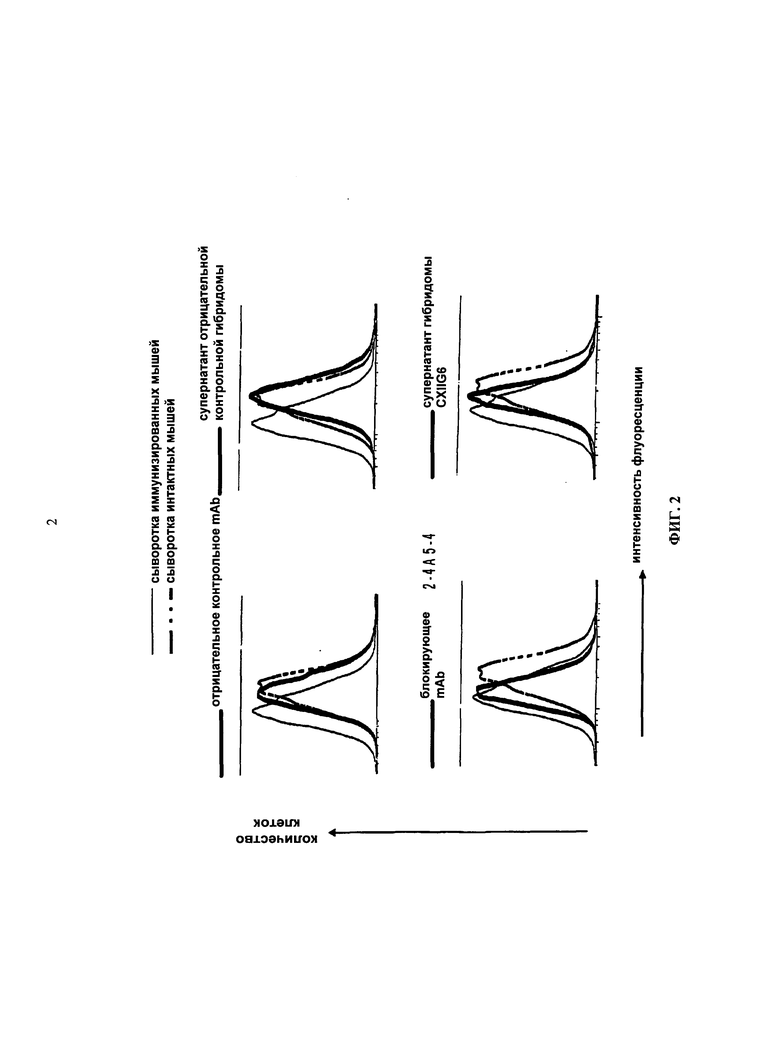
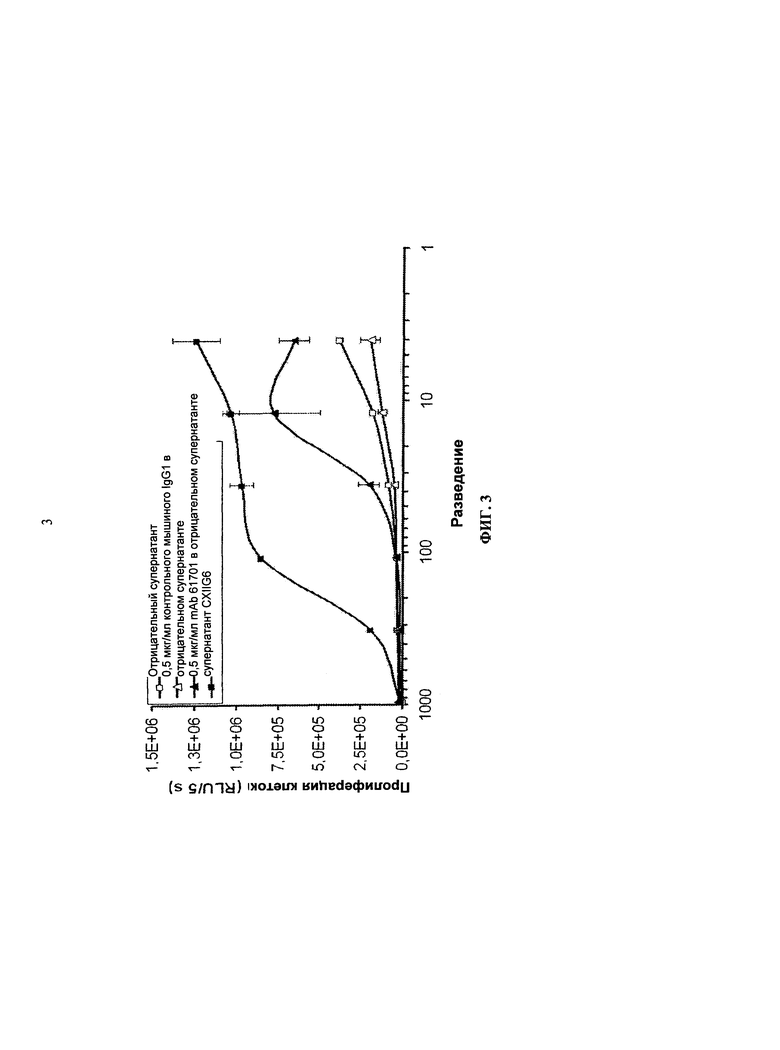
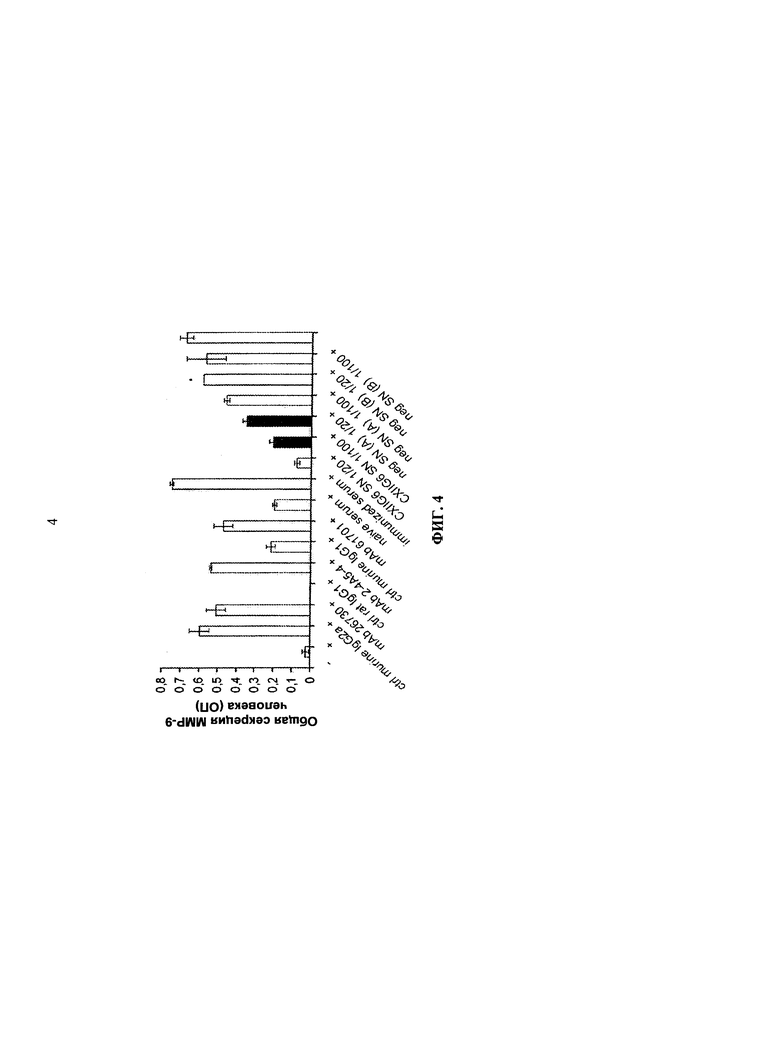
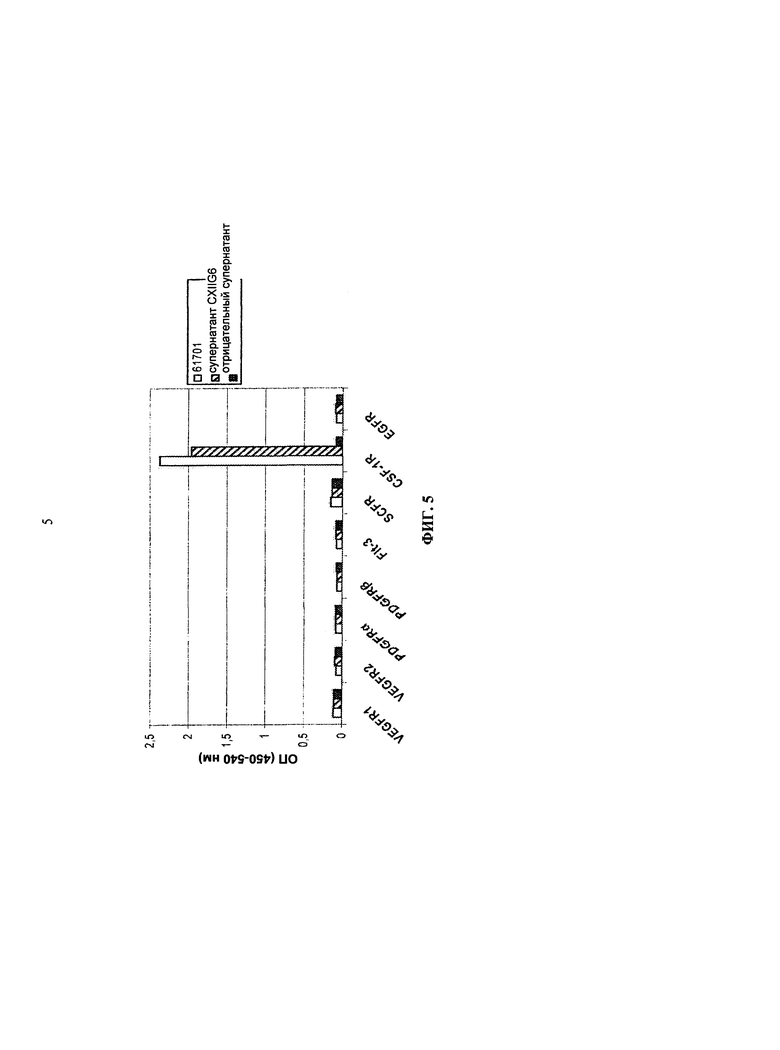


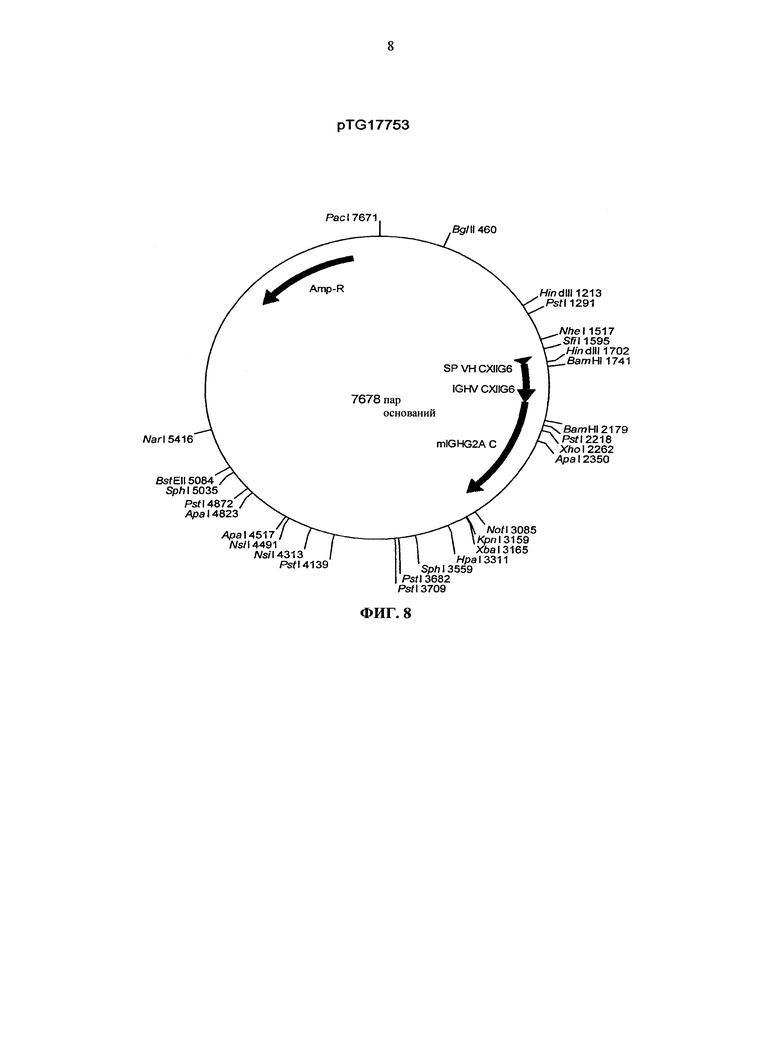
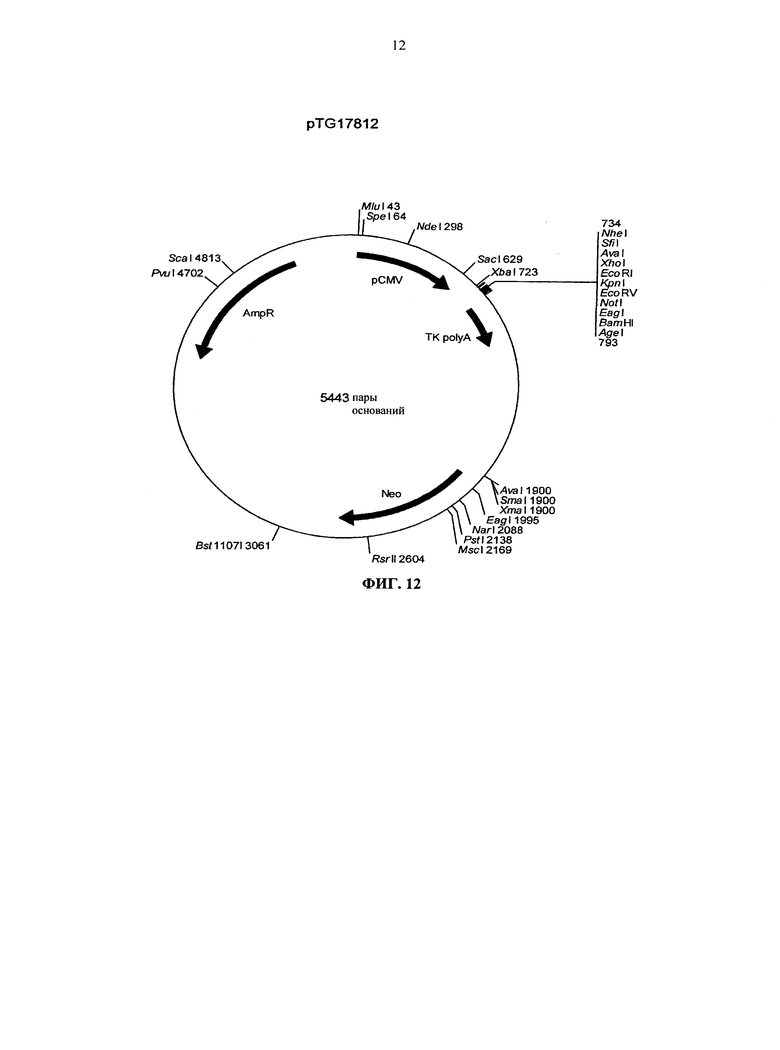
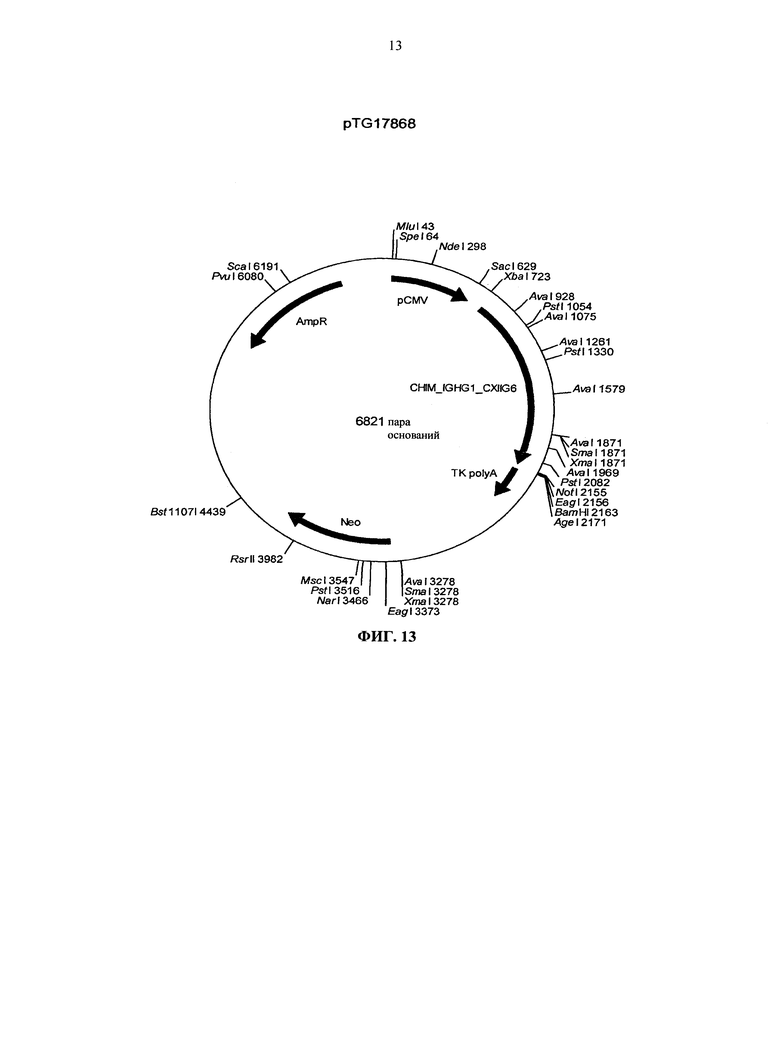
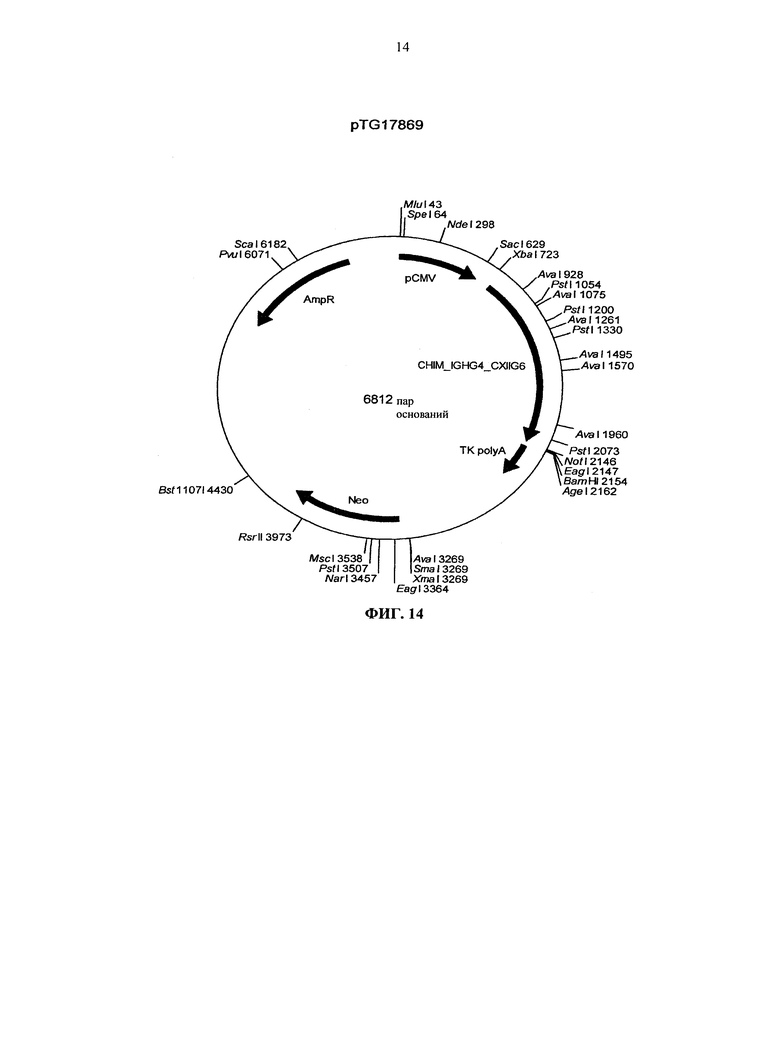
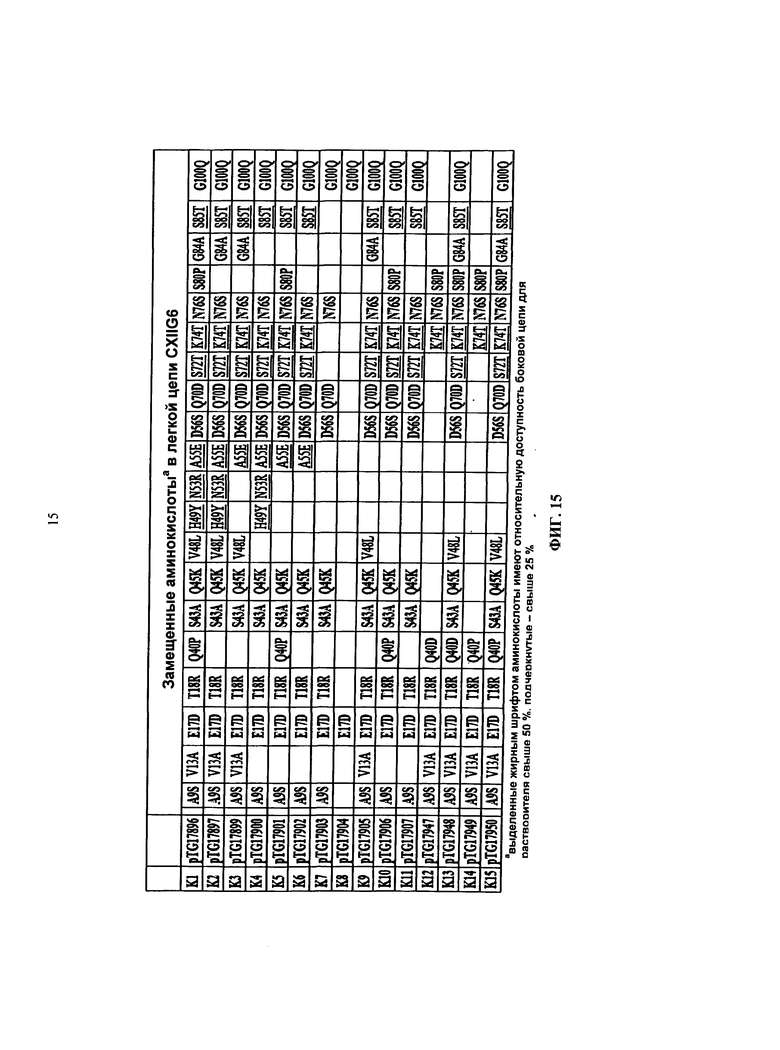
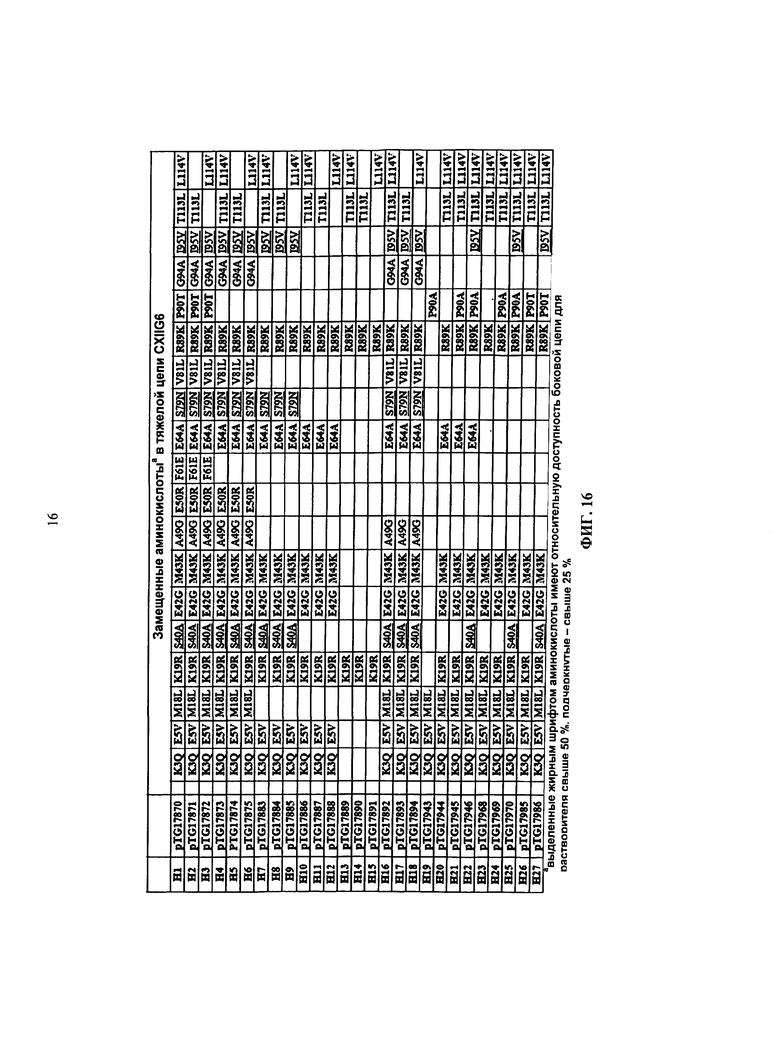
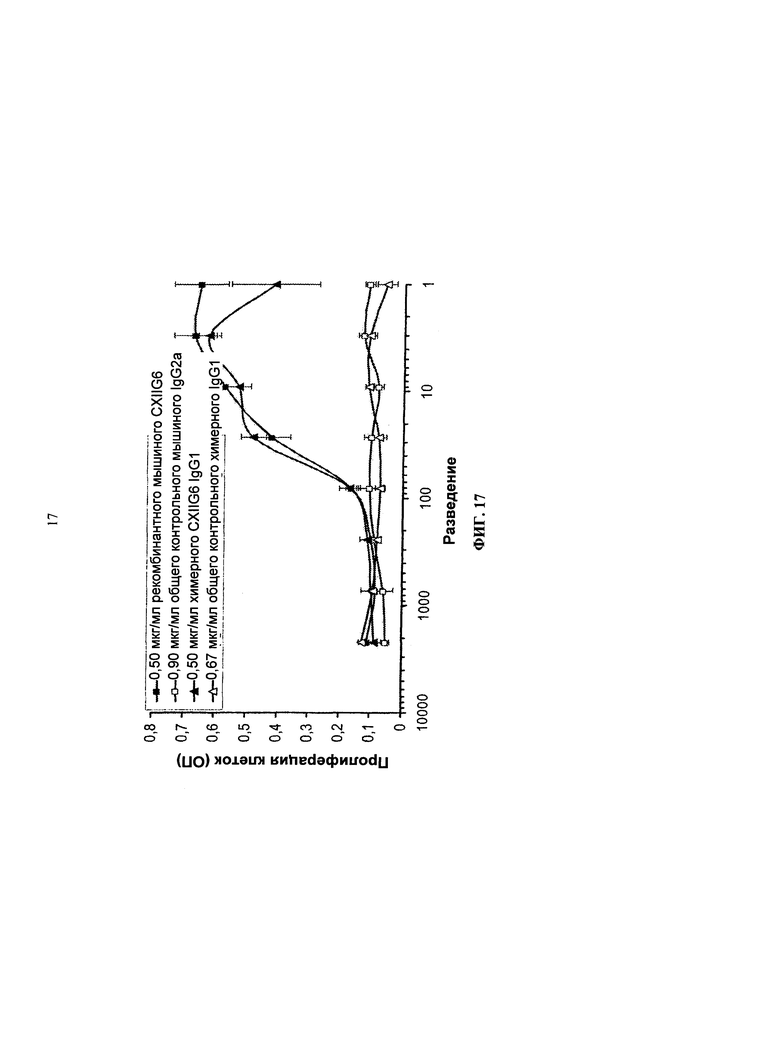
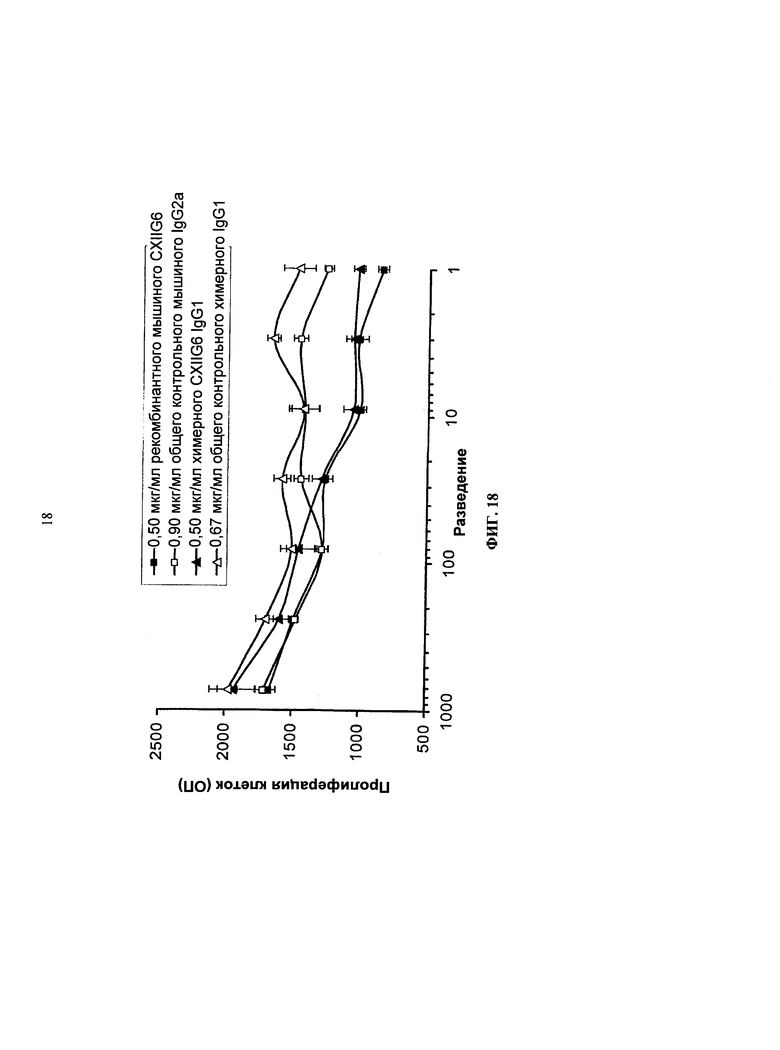


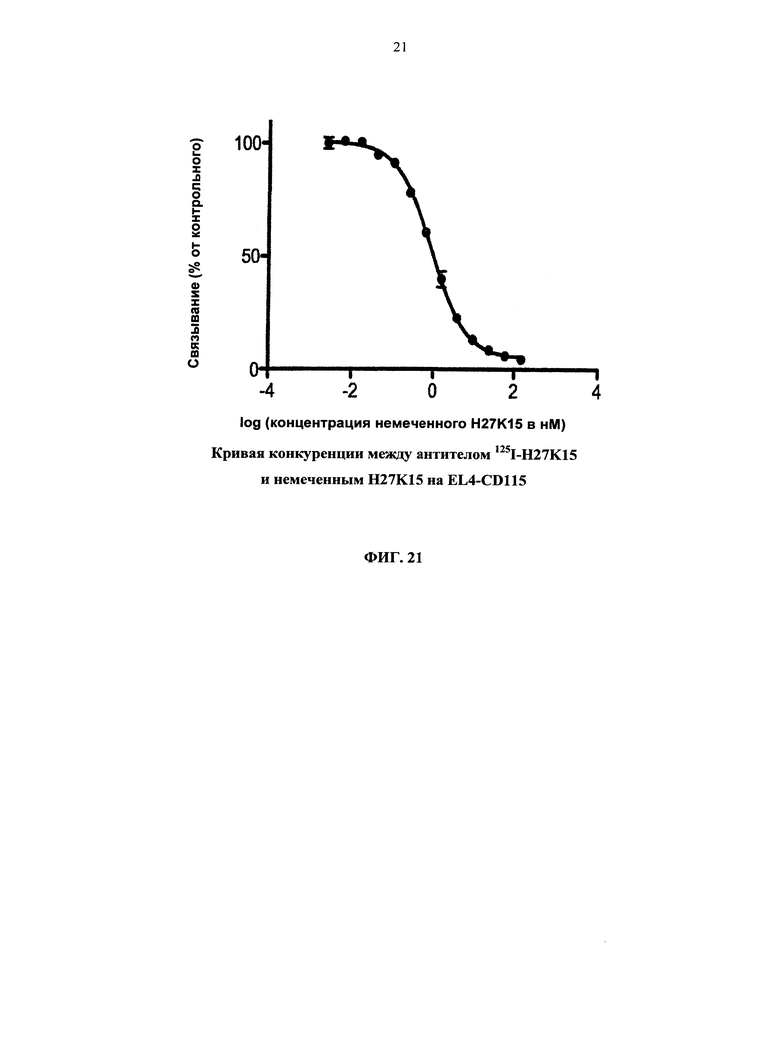

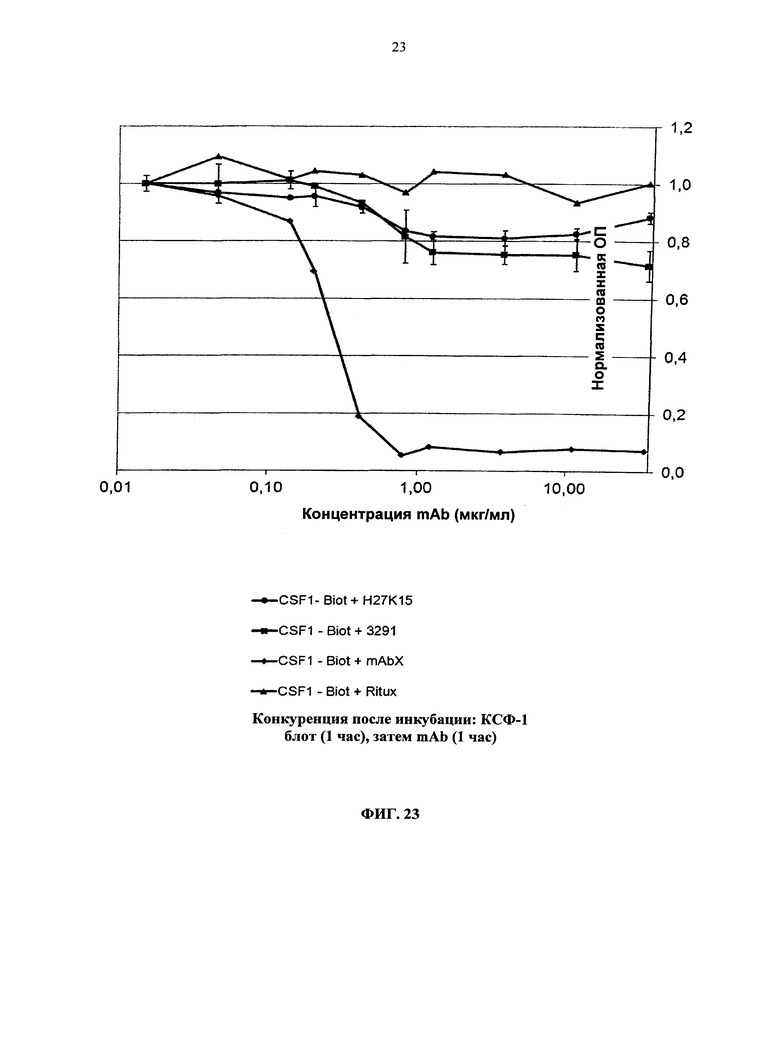
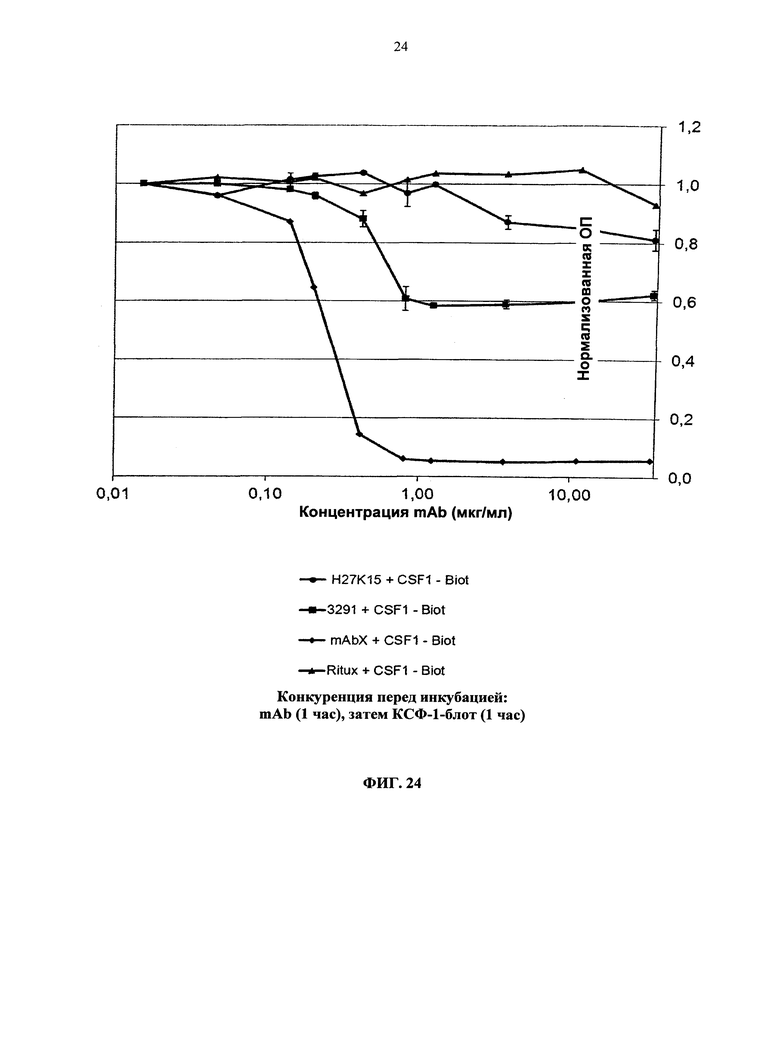
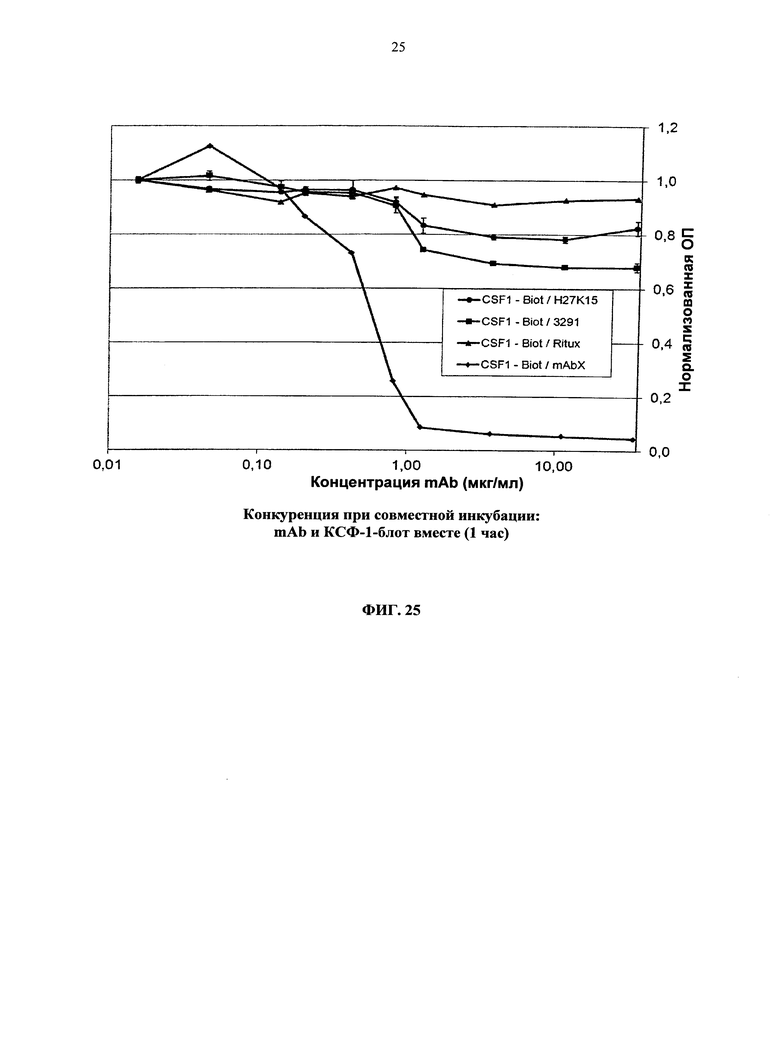
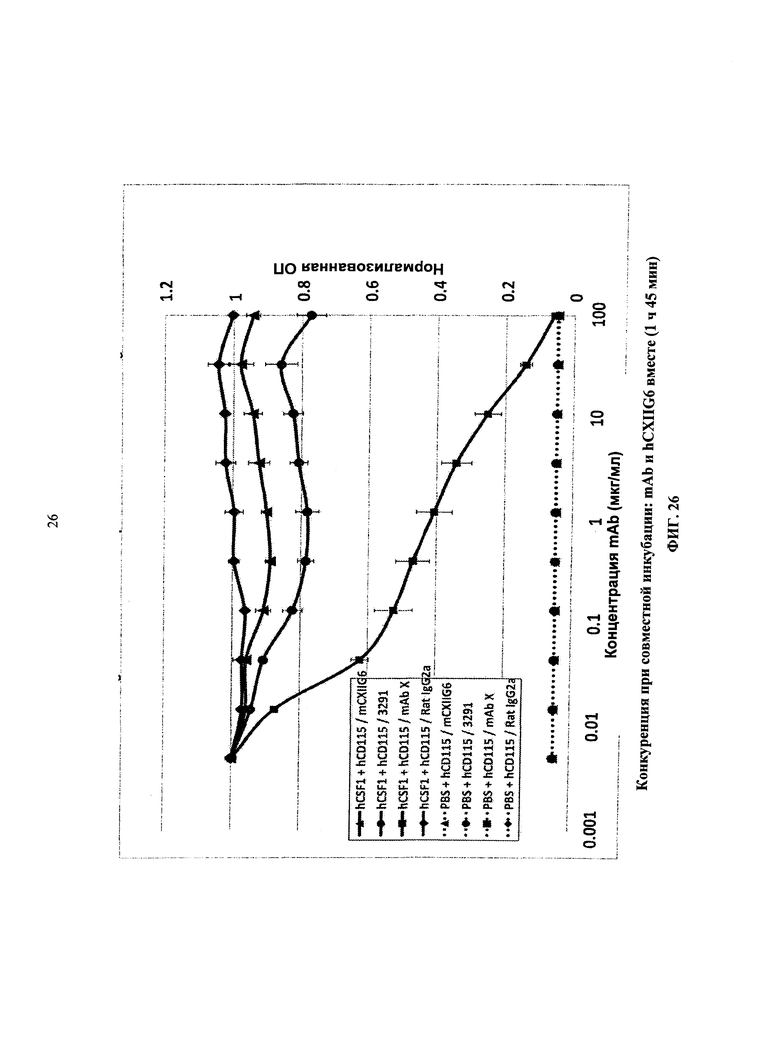
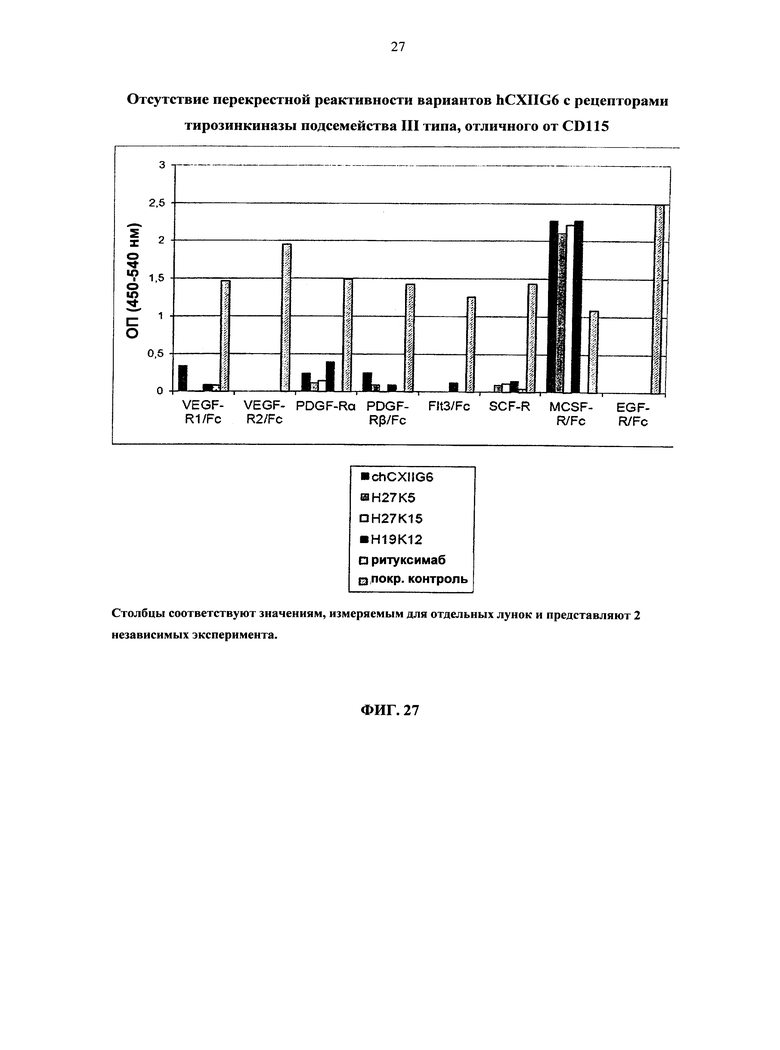
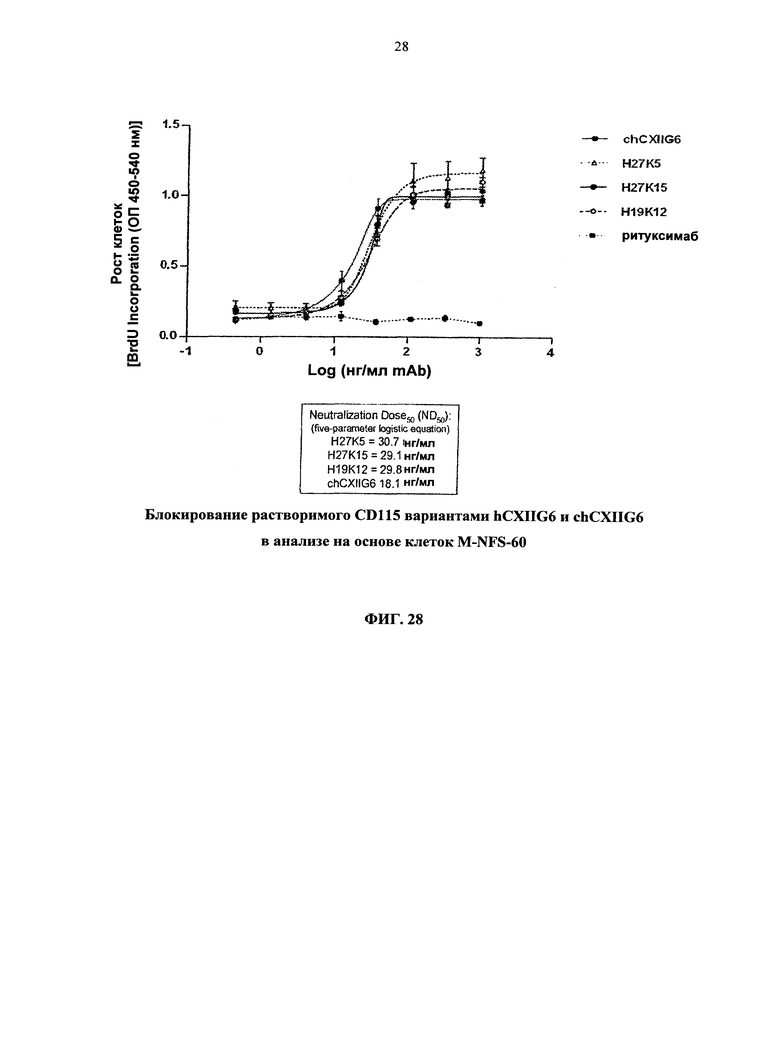
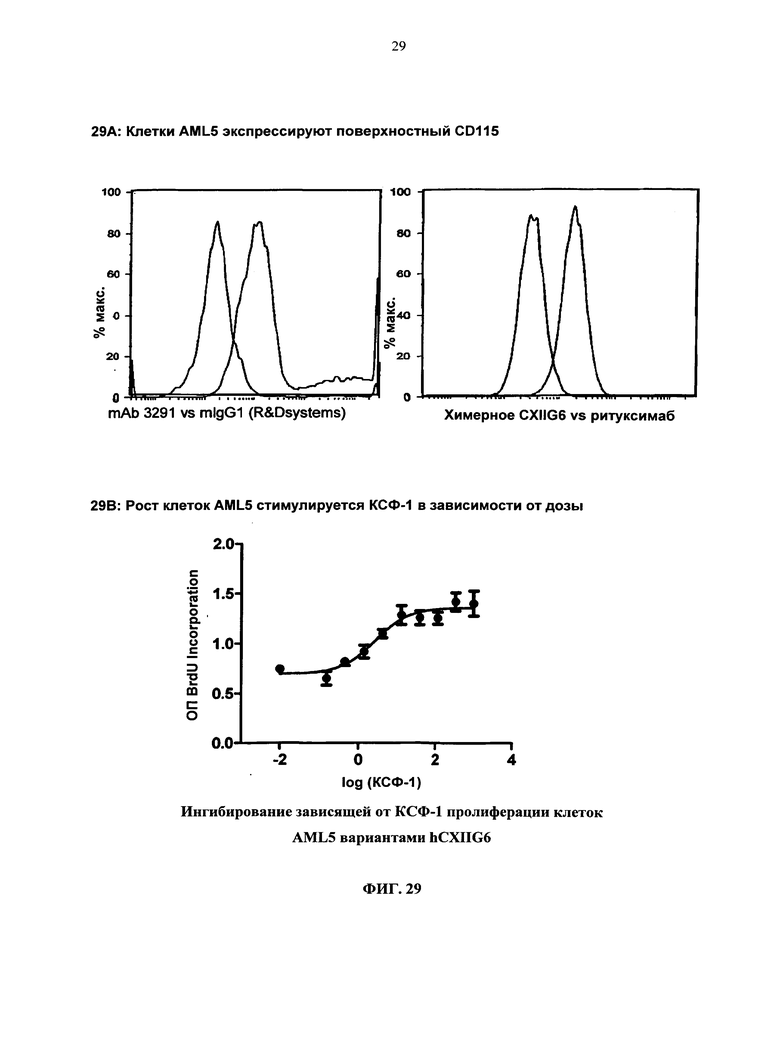
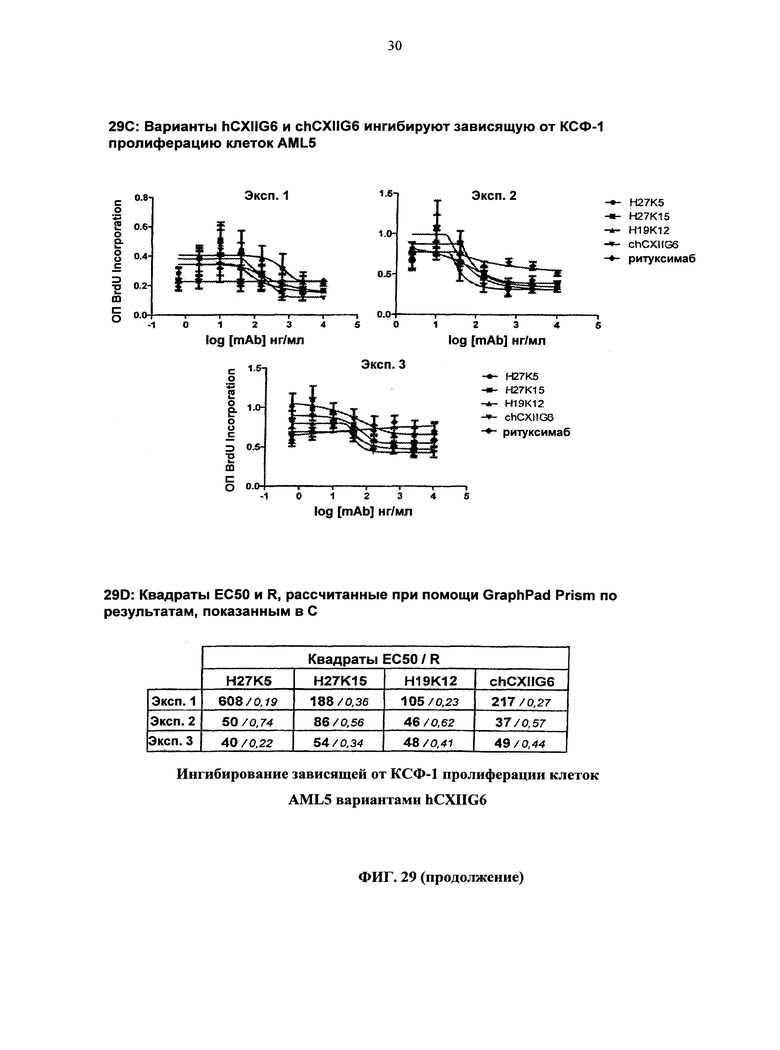
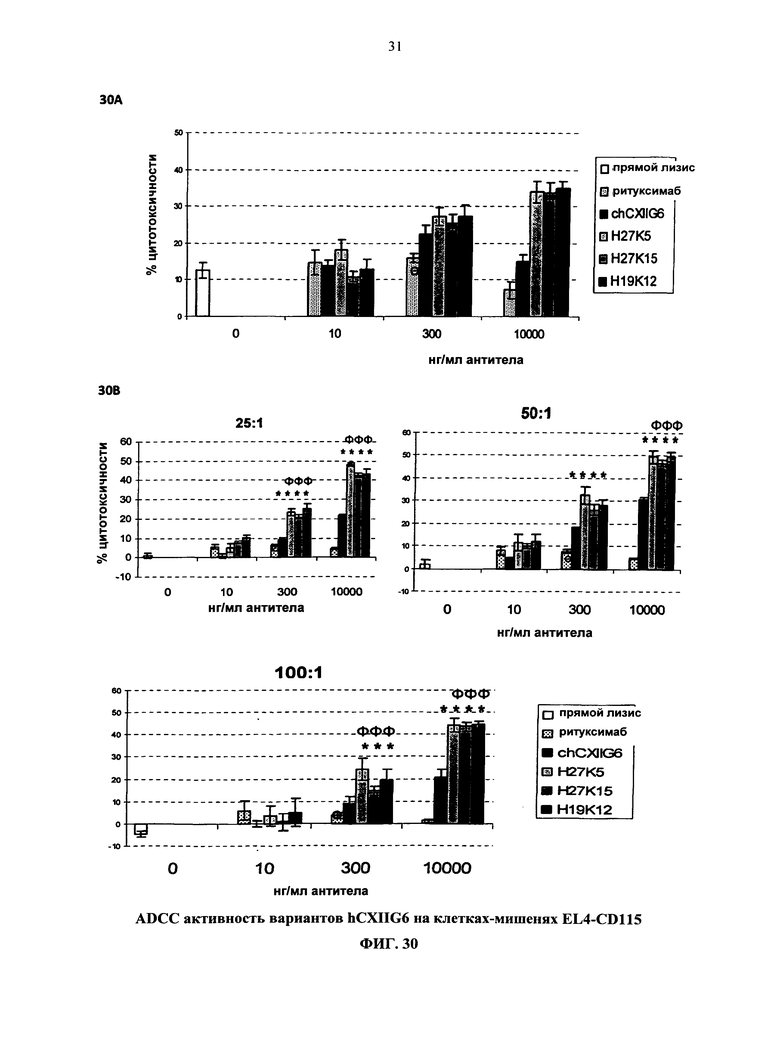
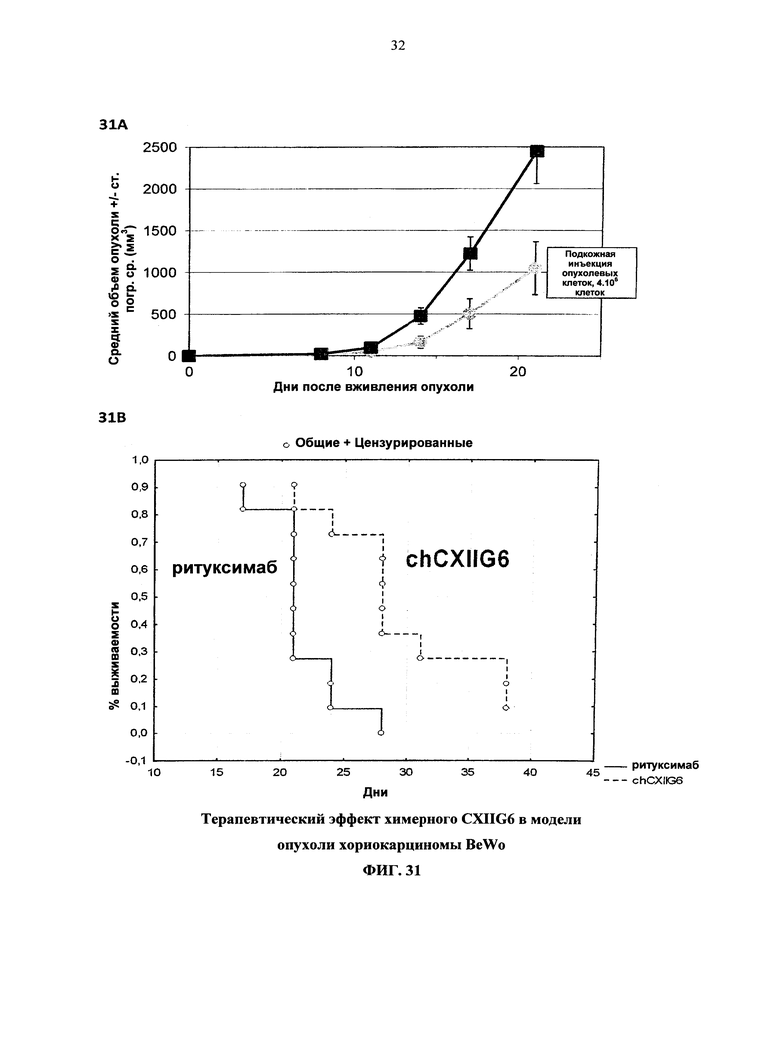








Настоящее изобретение относится к области биотехнологии. Предложено выделенное рекомбинантное или очищенное антитело, которое специфически связывается с рецептором колониестимулирующего фактора-1 (КСФ-1R, CSF-1R), охарактеризованное аминокислотными последовательностями вариабельных доменов. Также предложены нуклеиновая кислота, кодирующая антитело по изобретению, вектор, клетка и способ получения антитела. Кроме того, описана фармацевтическая композиция, а также применение антитела и фармацевтической композиции по изобретению в качестве лекарственного средства для лечения рака, болезни, связанной с повышенной активностью остеокластов, воспалительной болезни и ревматоидного артрита. Данное изобретение может найти дальнейшее применение в терапии заболеваний, ассоциированных с CSF-1R. 9 н. и 10 з.п. ф-лы, 32 ил., 4 табл.
1. Выделенное рекомбинантное или очищенное антитело, которое специфически связывается с КСФ-1R, в более предпочтительном варианте с человеческим КСФ-1R, характеризующееся тем, что указанное антитело включает:
(i) (а) первый вариабельный участок, состоящий из SEQ ID NO: 42, и (b) второй вариабельный участок, состоящий из SEQ ID NO: 44; или
(ii) (а) первый вариабельный участок, состоящий из SEQ ID NO: 43, и (b) второй вариабельный участок, состоящий из SEQ ID NO: 45; или
(iii) (а) первый вариабельный участок, состоящий из SEQ ID NO: 42, и (b) второй вариабельный участок, состоящий из SEQ ID NO: 46.
2. Антитело по п.1 (i), отличающееся тем, что включает (а) тяжелую цепь, которая состоит из SEQ ID NO: 37, и (b) легкую цепь, которая состоит из SEQ ID NO: 39.
3. Антитело по п.1 (ii), отличающееся тем, что включает (а) тяжелую цепь, которая состоит из SEQ ID NO: 38, и (b) легкую цепь, которая состоит из SEQ ID NO: 40.
4. Антитело по п.1 (iii), отличающееся тем, что включает (а) тяжелую цепь, которая состоит из SEQ ID NO: 37, и (b) легкую цепь, которая состоит из SEQ ID NO: 41.
5. Антитело по п.1, которое является поликлональным антителом, моноклональным антителом, Fab, Fab', F(ab')2, Fv, scFv, антигенсвязывающим фрагментом или диабоди.
6. Нуклеиновая кислота, кодирующая антитело по любому из пп.1-5.
7. Вектор для обеспечения экспрессии антитела по любому из пп.1-5, содержащий нуклеиновую кислоту по п.6.
8. Вектор по п.7, который имеет плазмидное или вирусное происхождение.
9. Вектор по п.8, который имеет вирусное происхождение и который происходит из поксвируса, аденовируса, ретровируса, вируса герпеса, альфа-вируса, пенистого вируса или аденоассоциированного вируса.
10. Вектор по п.9, где указанный поксвирус является вирусом осповакцины или поксвирусом канареек.
11. Вектор по п.10, где указанный вирус осповакцины является MVA.
12. Клетка для получения антитела по пп.1-5, включающая нуклеиновую кислоту по п.6.
13. Клетка по п.12, которая является эукариотической клеткой.
14. Клетка по п.13, где указанная клетка является клеткой млекопитающего, предпочтительно СНО или ВНК клеткой.
15. Способ получения антитела по любому из пп.1-5, включающий культивирование клетки по любому из пп.12-14 в условиях, обеспечивающих экспрессию антитела, и выделение антитела из клетки или из окружающей клетку среды.
16. Фармацевтическая композиция для лечения болезней, выбранных из группы, состоящей из рака, болезни, связанной с повышенной активностью остеокластов, воспалительной болезни и ревматоидного артрита, включающая антитело по любому из пп.1-5 в терапевтически эффективном количестве и фармацевтически приемлемый носитель.
17. Применение антитела по любому из пп.1-5 в качестве лекарственного средства для лечения болезней, выбранных из группы, состоящей из рака, болезни, связанной с повышенной активностью остеокластов, воспалительной болезни и ревматоидного артрита.
18. Применение фармацевтической композиции по п.16 в качестве лекарственного средства для лечения болезней, выбранных из группы, состоящей из рака, болезни, связанной с повышенной активностью остеокластов, воспалительной болезни и ревматоидного артрита.
19. Применение антитела по любому из пп.1-5 для лечения болезней, выбранных из группы, состоящей из рака, болезни, связанной с повышенной активностью остеокластов, воспалительной болезни и ревматоидного артрита.
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| ASHMUN R.A | |||
| et al., "Monoclonal antibodies to the human CSF-1 receptor (c-fms proto-oncogene product) detect epitopes on normal mononuclear phagocytes and on human myeloid leukemic blast cells." Blood, 1989; 73(3):827-37 | |||
| ПИЛИПЕНКО И.В | |||
| и др., "Ассоциативный анализ полиморфизмов генов р53 и c-fms с развитием рака молочной железы у русских женщин", Сибирский онкологический журнал | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

