Изобретение относится к области биотехнологии, в частности к антителам, связывающим производные морфина и нейтрализующим их биологическую активность.
В настоящее время, по данным федерального статистического наблюдения Минздрава России, среди зарегистрированных больных наркоманией подавляющее большинство составляют больные с зависимостью от опиатов (78,4%) [Киржанова В.В., Григорова Н.И. О заболевании наркологическими расстройствами в 2014 г. //Вопросы наркологии, 2015, 4:19- 28]. В этой связи в нашей стране проводится активный поиск эффективных путей лечения и профилактики наркомании. Изучается возможность создания антиидиотипической вакцины для иммунотерапии опийной зависимости. Проводятся исследования в области усовершенствования иммунохимических методов детекции психоактивных веществ [Любавина И.А., Зинченко А.А., Лапенков М.И., Николаева Т.Л. Экспресс-метод выявления морфина в водных образцах с помощью иммунохроматографии с использованием моноклональных антител, меченных коллоидным золотом // Биоорганическая химия, 2005, 31:108-112].
Лечение и профилактика наркотической зависимости тесно связаны с разработкой новых эффективных методов иммунодиагностики и иммунотерапии с применением высокоспецифичных антител к психоактивным веществам (ПАВ). Создание иммунодиагностических и лечебных препаратов на основе поли- и моноклональных антител против ПАВ может лечь в основу высокоэффективного биотехнологического производства.
Поскольку основными активными метаболитами героина у человека являются 6-моноацетилморфин, морфин и морфин-6-глюкуронид [Берзина А.Г., Гамалея Н.Б., Ульянова Л.И, Шестаков К.А., Ульянова М.А., Капанадзе К.Д., Станкова Н.В., Ревякин А.О., Фокин Ю.В., Кротов Г.И., Родченков Г.М. Методологические подходы к разработке вакцины для лечения зависимости от опиатов // Наркология, 2015, 11 (167):25-31], для лечения и профилактики зависимости от этих опиатов, а также для разработки методов специфической диагностики, необходимо, прежде всего, получить антитела, реагирующие как с этими наркотиками, так и с их активными производными. Такие антитела могут быть получены при иммунизации лабораторных животных конъюгированными с белками производными морфина по гидроксильным остаткам при атомах углерода в 6 и 3 положениях фенантренового кольца молекулы морфина, например 6-гемисукцинильным эфиром морфина (ГСМ) и 3-О-карбоксиметильным эфиром морфина (КММ) [Берзина А.Г., Ульянова Л.И., Гамалея Н.Б., Станкова Н.В., Капанадзе Г.Д. Изучение иммунного ответа у мини-свиней светлогорской популяции к двум производным морфина // Биомедицина, 2018, 2:15-21].
Наиболее близкими по технической сущности к заявляемому изобретению являются антитела 9 В1 к морфину [Pozharski Е., Wilson М.А., Hewagama A., Shanafelt А.В., Petsko G., Ringe D.. Anchoring a Cationic Ligand: The Structure of the Fab Fragment of the Anti-morphine Antibody 9B1 and its Complex with Morphine, J. Mol. Biol, 2004, 337:691-697], для которых был определен морфин-связывающий паратоп этого антитела и показано, что специфическое взаимодействие 9 В1 с морфином в основном определяется топологической и электростатической комплементарностью объектов, а также гидрофобными взаимодействиями. Сравнительный анализ последовательности гипервариабельных участков (CDR) тяжелой и легкой цепей показал отсутствие гомологий между антителами 9 В1 и 6G1.
Технической задачей, решаемой авторами, являлось расширение круга антител, способных специфически связываться с производными морфина и ингибировать биологическую активность наркотика. При этом было важно, чтобы антитела не затрагивали эндогенные опиоидные пептиды, которые выполняют важную роль в физиологии организма.
Технический результат достигался созданием моноклонального антитела (MAT) 6G1, которое специфично и с высокой аффинностью связывается с молекулами производных морфина и не связывается с эндогенными опиоидными пептидами человека, такими как эндорфин, энкефалин и др. Оно распознает 6-гемисукцинильный эфир морфина, конъюгированный как с бычьим сывороточным альбумином (БСА), так и с лизоцимом, не связывается с КММ и 2-р-карбокси-фенилазометильным производным морфина (ФАМ) и блокирует биологическую активность морфина.
Моноклональное антитело 6G1 содержит вариабельную область тяжелой цепи по SEQ ID NO: 3 и вариабельную область легкой цепи по SEQ ID NO: 4, а также гипервариабельные участки (CDR) вариабельной области тяжелой цепи по SEQ ID NO: 5-7 и гипервариабельные участки вариабельной области легкой цепи по SEQ ID NO: 8-10. Сравнительный анализ последовательности CDR тяжелой и легкой цепей показал отсутствие гомологий между антителом ближайшего аналога и антителом 6G1.
Технология получения антитела включала в себя иммунизацию животных антигеном, представляющим собой производные морфина: ГСМ и КММ, конъюгированные с бычьим сывороточным альбумином (БСА) с помощью бифункционального реагента 1-этил-3-(3-диметиламинопропил) карбодиимида гидрохлорида, которые были получены по методикам, описанным в статье [Берзина А.Г., Гамалея Н.Б., Ульянова Л.И, Шестаков К.А., Ульянова М.А., Капанадзе К.Д., Станкова Н.В., Ревякин А.О., Фокин Ю.В., Кротов Г.И., Родченков Г.М. Методологические подходы к разработке вакцины для лечения зависимости от опиатов // Наркология, 2015, 11(167):25-31]. В качестве антигенов для отбора позитивных продуцентов моноклональных антител при постановке ИФА использовали конъюгаты ГСМ и КММ с лизоцимом, а также конъюгат другого производного морфина - 2-р-карбокси-фенилазометильного (ФАМ).
Полученное мышиное моноклональное антитело (МАТ) распознает ГСМ, конъюгированный как с БСА, так и с лизоцимом, не связывается с КММ и ФАМ и блокирует биологическую активность морфина.
Свойства и структура антитела иллюстрируются следующими графическими материалами.
На Фиг. 1 показаны результаты электрофореза в полиакриламидном геле мышиного MAT 6G1 (4-20% полиакриламидный гель):
а - невосстанавливающие условия;
б - маркеры молекулярных весов (кД);
в - восстанавливающие условия;
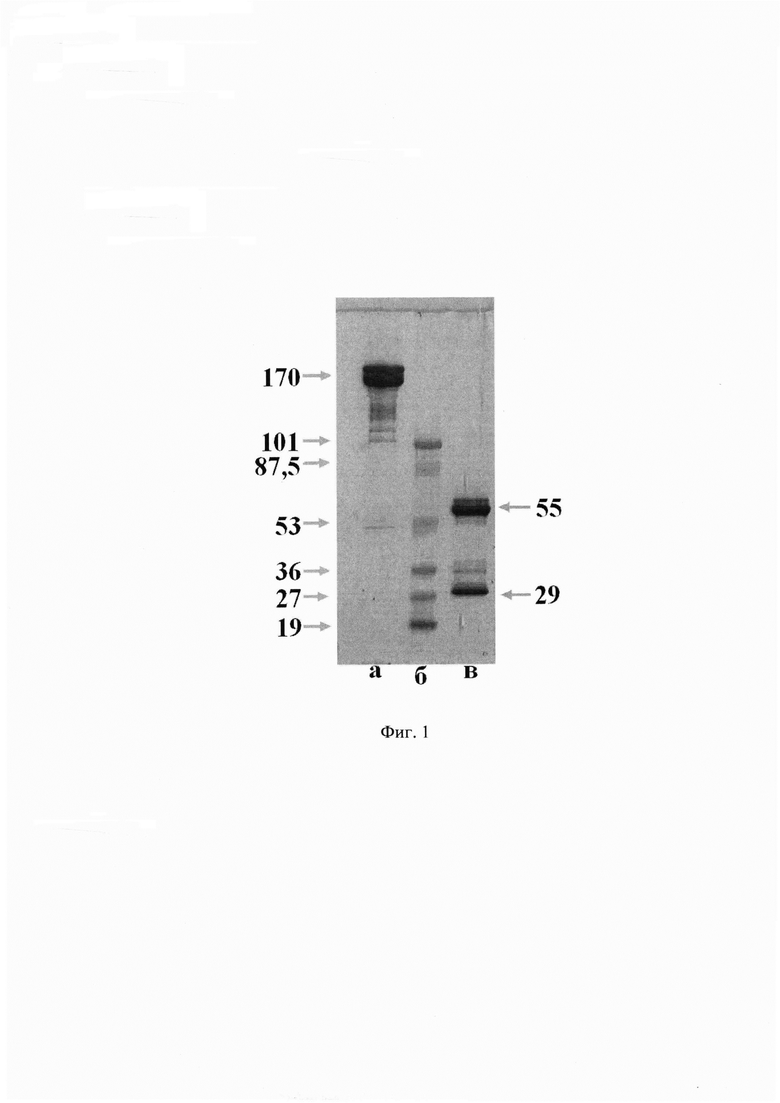
На Фиг. 2 демонстрируется отсутствие конкуренции MAT 6G1 с эндогенными опиоидными пептидами.
Сущность и промышленная применимость изобретения поясняются следующими примерами:
Пример 1. Получение антигенов для иммунизации и скрининга гибри-дом, иммунизация мышей и отбор гибридомы sG1.
Пример 1.1. Получение антигенов. Для иммунизации использовали ГСМ, конъюгированный с БСА с помощью бифункционального реагента 1-этил-3-(3-диметиламинопропил) карбодиимида гидрохлорида по методике, описанной в статье [Берзина А.Г., Гамалея Н.Б. и др. Методологические подходы к разработке вакцины для лечения зависимости от опиатов // Наркология, 2015, 11(167):25-31]. В качестве антигенов для адсорбции на твердой фазе при постановке ИФА использовали ГСМ, КММ и ФАМ, конъюгированные с лизоцимом по описанным методикам [там же]. Приготовленные конъюгаты производных морфина аликвотировали и хранили при -80°С.
Пример 1.2. Иммунизация мышей и скрининг гибридомы. Мышей линии Balb/c иммунизировали полученным, как описано в примере 1.1., препаратом ГСМ-БСА, эмульгированным в полном адъюванте Фрейнда (ПАФ), в дозе 10 мкг на мышь в подошвенный апоневроз задних конечностей. Через 4 недели животным вводили внутривенно 5 мкг антигена в физиологическом растворе. На четвертый день после инъекции животных умерщвляли цервикальной дислокацией и выделяли лимфоциты паховых и брюшных лимфоузлов. Полученные лимфоциты смешивали с клетками миеломы SP 2/0 в соотношении 2:1. Гибридизацию проводили 50% раствором полиэтиленгликоля (ПЭГ) с молекулярной массой 1500 Да в течение 1,5 мин, затем 4-кратно добавляли с интервалом в 1 мин среду RPMI-1640 в объеме, равном объему раствора ПЭГ. После слияния клетки дважды отмывали культуральной средой и высевали в 96-луночные культуральные планшеты с суточными перитонеальными макрофагами (5-10 клеток на лунку) из расчета 5⋅104 клеток миеломы на лунку. Селекцию гибридом проводили в среде RPMI-1640 с добавлением 10% бычьей фетальной сыворотки, содержащей 10-4 М гипоксантина, 4⋅10-7 М аминоптерина и 1,6⋅10-5 М тимидина.
Скрининг антител проводили с помощью непрямого метода ИФА, используя в качестве антигенов для адсорбции на твердой фазе конъюгаты производных морфина с лизоцимом. При постановке теста использовали антивидовые иммунопероксидазные конъюгаты фирмы Sigma-Aldrich.
Первичный отбор позитивных клонов проводили по связыванию моноклональных антител, секретируемых гибридомами, с антигенами в ИФА. Для этого в лунки полистирольных планшетов Corning (Costar) адсорбировали 5 мкг/мл ГСМ-лизоцима в 20 мМ боратном буфере с рН 8,0, содержащем 0,15 М NaCl, в течение 20 часов во влажной камере. По окончании адсорбции планшеты отмывали промывочным буфером (20 мМ боратный буфер с рН 8,0, содержащий 0,15 М NaCl и 0,05% Tween-20). Затем в каждую лунку вносили по 100 мкл промывочного буфера, содержащего 1 мг/мл БСА, и по 50 мкл культуральных сред, содержащих МАТ. Планшеты инкубировали 1 час при 37°С и перемешивании. По окончании инкубации проводили отмывку несвязавшихся с антигеном антител промывочным буфером и вносили в лунки раствор конъюгированных с пероксидазой хрена иммуноглобулинов козы к иммуноглобулинам мыши (Sigma-Aldrich, США) согласно приложенной инструкции. Планшеты инкубировали в течение 1 часа при 37°С и перемешивании, после чего тщательно отмывали и проводили окрашивание с помощью раствора субстрата тетраметилбензидина (Хема, Россия). Культуральные среды клонов, показавшие специфическое связывание с антигенами, повторно проверялись в аналогичном эксперименте на КММ-лизоцим, ГСМ-лизоцим, ФАМ-лизоцим и немодифицированный лизоцим.
На основе проведенного скрининга был отобран клон sG1, продуцирующий специфические антитела к производному морфина ГСМ-лизоцим, не распознающие при этом КММ- и ФАМ-лизоцим. Свойства моноклональных антител 6G1 приведены в Таблице 1.

В результате последующего внутрибрюшинного размножения, криоконсервирования и контроля чистоты клеток был получен штамм гибридомы sG1, продуцент моноклонального антитела 6G1.
Пример 2. Исследование свойств моноклонального антитела 6G1.
Пример 2.1. Очистка MAT 6G1. Антитело 6G1 выделяли из асцитной жидкости мышей линии Balb/c с внутрибрюшинно привитой гибридомой sG1 с помощью аффинной хроматографии на сорбенте MabSelect-сефарозе (General Electrics, США) согласно инструкции.
Пример 2.2. Изучение молекулярных свойств антитела 6G1. По результатам электрофореза в 4-20% полиакриламидном геле с додецилсульфатом натрия молекулярный вес антитела 6G1 в невосстанавливающих условиях соответствует ожидаемому для нативного IgG мыши 170 кДа (Фиг. 1а), в восстанавливающих условиях антитело диссоциирует на тяжелую (55 кДа) и легкую (29 кДа) цепи (Фиг. 1в). Изотип антитела 6G1, установленный с помощью набора «Mouse monoclonal antibody isotyp-ing reagents» (Sigma, США), - IgG2a. Препараты очищенного антитела 6G1 использовали для изучения его биологических свойств.
Пример 2.3. Исследование биологических свойств MAT 6G1.
Пример 2.3.1. Изучение биологической активности MAT 6G1 прово-дили на модели пролиферативной реакции культур лимфоцитов in vitro, выделенных из периферической крови 6 здоровых добровольцев на градиенте плотности фиколлаверографина (плотность 1,077 г/см). В Таблице 2 представлены результаты оценки нейтрализующего действия налоксона и MAT 6G1 на стимуляцию морфином синтеза ДНК в культуре лимфоцитов при их совместном введении с морфином.
Анализ данных, приведенных в Таблице 2, показал наличие воздействия MAT 6G1 на специфическую активность морфина. Как видно из полученных данных, MAT 6G1 обладают способностью полностью блокировать усиление морфином синтеза ДНК в культурах лимфоцитов крови человека. Этот эффект сходен с действием налоксона, который при совместном введении с морфином полностью устраняет стимулирующее влияние опиата на лимфоциты. Следует отметить, однако, что механизм нейтрализующего действия налоксона отличается от механизма действия первичных MAT 6G1. Блокада налоксоном эффекта морфина осуществляется на уровне μ-опиоидных рецепторов, присутствующих на культивируемых лимфоцитах человека. Блокада первичными MAT 6G1 не связана с их действием на рецепторы лимфоцитов, поскольку сами по себе такие антитела эффекта на пролиферацию не оказывают. Блокада в данном случае обусловлена связыванием наркотика с антителами в реакционной смеси, в результате чего действие морфина на μ-опиоидные рецепторы лимфоцитов снижается.
Примечание: Сравнение по одному признаку со спонтанной пролиферацией - СП (РСП) и морфином (РМ) проведено с использованием непараметрического критерия Вилкоксона (W) для двух зависимых групп без учета характера распределения вариант [Гланц С. Медико-биологическая статистика, Практика. 1999, 459 с.]. В скобках приведен процент изменения от средних значений СП и М (Δ % и Δм % соответственно).
Пример 2.3.2. Изучение перекрестного связывания MAT 6G1 с некоторыми эндогеннными опиоидными пептидами. Для проверки перекрестного взаимодействия MAT 6G1 с эндогенными опиоидными пептидами провели конкурентный твердофазный иммуноферментный анализ. В качестве конкурента использовали β-эндорфин, Leu- и Met-энкефалины. Для этого в лунки полистирольных планшетов Corning (Costar) вносили 5 мкг/мл ГСМ-БСА в 20 мМ боратном буфере с рН 8,0, содержащем 0,15 М NaCl, и выдерживали в течение 20 часов во влажной камере. По окончании адсорбции планшеты отмывали промывочным буфером (20 мМ боратный буфер с рН 8,0, содержащий 0,15 М NaCl и 0,05% Tween-20). Затем в каждую лунку вносили по 100 мкл промывочного буфера, содержащего 1 мг/мл БСА, 100 мкг/мл конкурента и MAT 6G1 в различной концентрации. Планшеты инкубировали 1 час при 37°С и перемешивании. По окончании инкубации проводили отмывку несвязавшихся с антигеном антител промывочным буфером и вносили в лунки раствор конъюгированных с пероксидазой хрена иммуноглобулинов козы к иммуноглобулинам мыши (Sigma, США) согласно приложенной инструкции. Планшеты инкубировали в течение 1 часа при 37°С и перемешивании, после чего тщательно отмывали и проводили окрашивание с помощью раствора субстрата тетраметилбензидина (Хема, Россия). Результаты конкурентного анализа с эндогенными опиоидными пептидами представлены на Фиг. 2.
Как видно из представленных данных, моноклональные антитела 6G1 не конкурируют с отобранными опиоидными пептидами даже при очень большом молярном избытке конкурента.
Пример 3. Синтез и секвенирование ДНК, кодирующей вариабельные части легкой и тяжелой цепей моноклонального антитела 6G1.
Из клеток гибридомы sG1 выделяли РНК, на матрице которой с помощью наборов синтетических праймеров (Immunogenetics Information System http://www.imgt.org) были амплифицированы фрагменты ДНК, кодирующие вариабельные области тяжелой (VH) и легкой (VL) цепей антитела. Полученные фрагменты ДНК были клонированы в вектор pALTA (Евроген, Россия) и секвенированы с внешних праймеров (М13). Последовательности фрагментов ДНК, кодирующих VH и VL, представлены на SEQ ID NO: 1 и SEQ ID NO: 2, вычисленные аминокислотные последовательности VH и VL представлены на SEQ ID NO: 3 и SEQ ID NO: 4. Анализ последовательностей аминокислот вариабельных областей тяжелой и легкой цепей моноклонального антитела 6G1 производили по Кэботу (Kabat Е.А., Wu Т.Т., Perry Н., Gottesman K. and Foeller С.(1991) SEQuences of Proteins of Immunological Interest, Fifth Edition. NIH Publication No 91:3242), что позволило выделить участки CDR, определяющие комплементарность антитела антигену.
Сравнение последовательностей участков CDR с последовательностями известных антител к компонентам морфина показало отсутствие гомологии с аналогичными участками известных антител.
Моноклональное антитело 6G1 по настоящему изобретению может служить основой для конструирования химерных и гуманизированных антител, пригодных для создания лекарственных препаратов, блокирующих биологическую активность морфина.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО 3К11 К ПРОИЗВОДНЫМ МОРФИНА | 2018 |
|
RU2702002C1 |
| Моноклональное антиидиотипическое антитело АИ-К11В, обладающее антигенными свойствами морфина | 2020 |
|
RU2745374C1 |
| Моноклональное антиидиотипическое антитело АИ-G1, обладающее антигенными свойствами морфина | 2019 |
|
RU2717989C1 |
| ИММУНОГЕН ДЛЯ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ЗАВИСИМОСТИ ОТ ОПИАТОВ | 2013 |
|
RU2548802C1 |
| Антитела против рецептора фактора роста эндотелия сосудов 1 (VEGFR1) человека | 2023 |
|
RU2838680C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2432362C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К С-КОНЦЕВОМУ ФРАГМЕНТУ АНТИМЮЛЛЕРОВА ГОРМОНА | 2018 |
|
RU2765689C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО СС3-4 К КОНФОРМАЦИОННОМУ ЭПИТОПУ С3 ЧЕЛОВЕКА, ШТАММ ГИБРИДНОЙ ДНК МЫШИ РККК(П)764Д - ПРОДУЦЕНТ МОНОКЛОНАЛЬНОГО АНТИТЕЛА СС3-4 | 2015 |
|
RU2584582C1 |
| Антитела против рецептора стабилин-1 (stabilin-1) человека | 2023 |
|
RU2823309C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2694412C2 |








Изобретение относится к области биохимии, в частности к моноклональному антителу к 6- гемисукцинильному эфиру морфина, способное блокировать биологическую активность морфина. Изобретение позволяет эффективно связываться с производным морфина и ингибировать биологическую активность наркотика. 2 з.п. ф-лы, 2 ил., 2 табл., 3 пр.
1. Моноклональное антитело 6G1 к 6-гемисукцинильному эфиру морфина, способное блокировать биологическую активность морфина, содержащее гипервариабельные участки тяжелой цепи с последовательностями SEQ ID NO: 5-7 и гипервариабельные участки легкой цепи с последовательностями SEQ ID NO: 8-10.
2. Моноклональное антитело по п. 1, имеющее последовательность вариабельной области тяжелой цепи по SEQ ID NO: 3 и последовательность вариабельной области легкой цепи по SEQ ID NO: 4.
3. Моноклональное антитело по п. 2, кодируемое ДНК, содержащей последовательности нуклеотидов по SEQ ID NO: 1 и SEQ ID NO: 2.
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2432362C2 |
| СИНТЕТИЧЕСКИЙ ИММУНОГЕН ДЛЯ ЗАЩИТЫ И ЛЕЧЕНИЯ ОТ ЗАВИСИМОСТИ ОТ ПСИХОАКТИВНЫХ ВЕЩЕСТВ | 2016 |
|
RU2643329C1 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ НАРКОТИЧЕСКОЙ ЗАВИСИМОСТИ | 2000 |
|
RU2182492C1 |
| WO 2006004390 A3, 12.01.2006 | |||
| ADACHI Y | |||
| et al., Distinct germinal center selection at local sites shapes memory B cell response to viral escape, J | |||
| Exp | |||
| Med., 2015, v.212, n.10, pp.1709-1723. | |||

