УРОВЕНЬ ТЕХНИКИ
Область техники, к которой относится изобретение
Настоящее изобретение относится к моноклональным антителам (например, 8F5 и 8C5), которые можно применять, например, для профилактики, лечения и диагностики болезни Альцгеймера или других нейродегенеративных расстройств.
Известный уровень техники
Болезнь Альцгеймера (AD) является нейродегенеративным заболеванием, характеризуемым прогрессирующей потерей когнитивных способностей и характерными нейропатологическими признаками, включая отложения амилоида, нейрофибриллярные клубки и утрату нейронов в некоторых областях головного мозга, см. Hardy and Selkoe (Science 297, 353 (2002); Mattson (Nature 431, 7004 (2004)). Основными составляющими амилоидных отложений являются амилоидные бета-пептиды (Aβ), при этом наиболее известным является тип пептида длиной в 42 аминокислоты (Aβ1-42).
В частности, амилоидный белок β(1-42) является полипептидом, имеющим 42 аминокислоты, который происходит из белка-предшественника амилоида (APP) в результате протеолитического процессинга. Указанный белок, кроме человеческих вариантов, включает также изоформы амилоидного белка β(1-42), присутствующего в других организмах, отличных от человека, в частности других млекопитающих, в частности в организме крысы. Указанный белок, который склонен к полимеризации в водной среде, может присутствовать в самых разных молекулярных формах.
Простая корреляция отложения нерастворимого белка с наличием или прогрессированием связанных с деменцией расстройств, таких, например, как болезнь Альцгеймера, оказалась не доказанной (Terry et al., Ann. Neurol. 30. 572-580 (1991); Dickson et al., Neurobiol. Aging 16, 285-298 (1995)). Напротив, утрата синапсов и когнитивного восприятия, по-видимому, лучше коррелирует с растворимыми формами Aβ(1-42) (Lue et al., Am. J. Pathol. 155, 853-862 (1999); McLean et al., Ann. Neurol. 46, 860-866 (1999)).
Хотя ранее были получены поликлональные и моноклональные антитела против Aβ(1-42), не одно из них, как было доказано, не дает требуемого терапевтического эффекта, не вызывая также серьезных побочных эффектов у животных и/или человека. Например, результаты пассивной иммунизации, полученные в доклинических исследованиях на очень старых мышах APP23, которые получали направленное на N-конце анти-Aβ(1-42)-антитело один раз в неделю в течение 5 месяцев, показывают терапевтически значимый побочный эффект. В частности, у таких мышей наблюдали увеличение количества и тяжести микрогеморрагий по сравнению с мышами, которых лечили физиологическим раствором (Pfeifer et al., Science 2002 298: 1379). Недавно описано также сходное увеличение геморрагии у очень старых (>24 месяцев) мышей Tg2576 и PDAPP (Wilcock et al., J. Neuroscience 2003, 23: 3745-51; Racke et al., J. Neuroscience 2005, 25: 629-636). У обеих линий инъекция анти-Aβ(1-42) приводила к значительному увеличению микрогеморрагий. Таким образом, существует огромная терапевтическая необходимость в разработке биологических средств, которые предотвращают или замедляют прогрессирование заболевания, не проявляя негативных и потенциально летальных эффектов в организме человека. Такая необходимость особенно очевидна в виду увеличения продолжительности жизни в общей популяции и связанного с таким увеличением возрастания количества пациентов, у которых ежегодно диагностируют болезнь Альцгеймера. Кроме того, такие антитела позволят правильно диагностировать болезнь Альцгеймера у пациентов, испытывающих симптомы такого заболевания, диагноз которого в настоящее время может быть подтвержден только при аутопсии. Кроме того, антитела позволят выяснять биологические свойства белков и других биологических факторов, ответственных за такое истощающее заболевание.
Все патенты и публикации, упоминаемые в данном описании, таким образом, включены в виде ссылки в полном объеме.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к изолированному антителу, которое связывается с глобуломером амилоидного бета-(Aβ)-белка с более высокой специфичностью, чем с мономером амилоидного бета-белка. Таким образом, наблюдается предпочтительное связывание. Антитело может представлять собой, например, моноклональное антитело, такое как 8F5 или 8C5. Отношение специфичности связывания с глобуломером по сравнению с мономером составляет по меньшей мере 1,4. В частности, отношение предпочтительно составляет по меньшей мере от примерно 1,4 до меньшей мере примерно 16,9. (Отношение 1,0-17,5, включая конечные точки) также входит в объем настоящего изобретения, так же как их десятые доли. Например, 1,1, 1,2, 1,3, .., 2,0, 2,1, 2,2.., 17,1, 17,2, 17,3, 17,4, 17,5, а также все целые числа меду ними и их доли входят в объем настоящего изобретения.) Мономер бета-амилоидного белка может представлять собой, например, мономер Aβ(1-42) или мономер Aβ(1-40).
Кроме того, настоящее изобретение также охватывает моноклональное антитело (называемое в данном описании «8F5»), продуцируемое гибридомой, имеющей номер PTA-7238 в Американской коллекции типовых культур, а также гибридому, которая продуцирует такое моноклональное антитело (т.е. 8F5). Также настоящее изобретение относится к моноклональному антителу (называемому в данном описании «8C5»), продуцируемому гибридомой, имеющей номер A-7407 в Американской коллекции типовых культур PT, а также к гибридоме, которая продуцирует такое моноклональное антитело (т.е. 8C5).
Кроме того, настоящее изобретение относится к моноклональному антителу, содержащему вариабельную тяжелую цепь, кодируемую последовательностью SEQ ID NO: 1. Такое антитело может быть мышиным, человеческим или гуманизированным.
Кроме того, настоящее изобретение относится к моноклональному антителу, содержащему вариабельную легкую цепь, кодируемую последовательностью SEQ ID NO: 2. Такое антитело также может быть мышиным, человеческим или гуманизированным. Антитело может дополнительно содержать вариабельную тяжелую цепь, кодируемую последовательностью SEQ ID NO: 1, и может быть человеческим или гуманизированным.
Кроме того, настоящее изобретение относится к моноклональному антителу, содержащему последовательность SEQ ID NO: 3. Антитело может быть мышиным, человеческим или гуманизированным.
Кроме того, настоящее изобретение охватывает моноклональное антитело, содержащее последовательность SEQ ID NO: 4. Такое антитело может быть мышиным, человеческим или гуманизированным. Такое антитело дополнительно может содержать последовательность SEQ ID NO: 3 и может быть мышиным, человеческим или гуманизированным.
Дополнительно, настоящее изобретение относится к моноклональному антителу, содержащему вариабельную тяжелую цепь, кодируемую последовательностью SEQ ID NO: 11. Такое антитело может быть мышиным, человеческим или гуманизированным.
Кроме того, настоящее изобретение относится к моноклональному антителу, содержащему вариабельную легкую цепь, кодируемую последовательностью SEQ ID NO: 12. Такое антитело также может быть мышиным, человеческим или гуманизированным. Антитело дополнительно может содержать вариабельную тяжелую цепь, кодируемую последовательностью SEQ ID NO: 11 и может быть человеческим или гуманизированным.
Кроме того, настоящее изобретение относится к моноклональному антителу, содержащему последовательность SEQ ID NO: 19. Антитело может быть мышиным, человеческим или гуманизированным.
Кроме того, настоящее изобретение охватывает моноклональное антитело, содержащее последовательность SEQ ID NO: 20. Такое антитело может быть мышиным, человеческим или гуманизированным. Такое антитело дополнительно может содержать последовательность SEQ ID NO: 19 и может быть мышиным, человеческим или гуманизированным.
Настоящее изобретение также относится к изолированному антителу, которое связывается с глобуломером бета-амилоидного белка с более высокой специфичностью, чем с фибриллой бета-амилоидного белка. Такое антитело может быть, например, моноклональным и может быть моноклональным антителом, продуцируемым гибридомой, имеющей номер PTA-7243 в Американской коллекции типовых культур, или гибридомой, имеющей номер PTA-7407 в Американской коллекции типовых культур. Гибридомы, продуцирующие такие моноклональные антитела, также входят в объем настоящего изобретения.
Кроме того, настоящее изобретение относится к антителу, в котором по меньшей мере одна из областей, определяющих комплементарность (CDR), вариабельной тяжелой цепи выбрана из группы, состоящей из последовательностей SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7.
Кроме того, настоящее изобретение также относится к антителу, в котором по меньшей мере одна из CDR вариабельной легкой цепи выбрана из группы, состоящей из SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10. Такое антитело дополнительно может содержать по меньшей мере одну CDR вариабельной тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7.
Настоящее изобретение также относится к антителу, в котором по меньшей мере одна из CDR вариабельной тяжелой цепи выбрана из группы, состоящей из SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15.
Кроме того, настоящее изобретение также охватывает антитело, в котором по меньшей мере одна из CDR вариабельной легкой цепи выбрана из группы, состоящей из SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18. Такое антитело дополнительно может содержать по меньшей мере одну CDR вариабельной тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15.
Кроме того, настоящее изобретение охватывает способ лечения или профилактики болезни Альцгеймера у пациента, нуждающегося в таком лечении или профилактике. Такой способ включает в себя введение любого одного или нескольких изолированных антител, описанных выше, пациенту в количестве, достаточном для осуществления лечения или профилактики.
Изолированное антитело может быть введено, например, путем, выбранным из группы, состоящей из внутримышечного введения, внутривенного введения и подкожного введения.
Настоящее изобретение также относится к способу диагностирования болезни Альцгеймера у пациента, у которого предполагается наличие такого заболевания. Указанный способ включает в себя стадии: 1) выделения биологического образца из организма пациента; 2) осуществления контакта биологического образца по меньшей мере с одним из антител, описанных выше, в течение периода и в условиях, достаточных для образования комплексов антиген/антитело; и 3) выявления наличия комплексов антиген/антитело в указанном образце, при этом присутствие комплексов свидетельствует о диагнозе болезни Альцгеймера у пациента. Антигеном может быть, например, глобуломер, или его часть, или фрагмент, которые обладают такими же функциональными свойствами, что и полный глобуломер (например, активностью связывания).
Кроме того, настоящее изобретение относится к другому способу диагностирования болезни Альцгеймера у пациента, у которого предполагается наличие такого заболевания. Указанный способ включает в себя стадии: 1) выделения биологического образца из организма пациента; 2) осуществления контакта биологического образца с антигеном в течение периода и в условиях, достаточных для образования комплексов антитело/антиген; 3) добавления конъюгата к полученным в результате комплексам антитело/антиген в течение периода и в условиях, достаточных для того, чтобы обеспечить возможность для связывания конъюгата со связанным антителом, при этом конъюгат содержит одно из антител, описанных выше, связанное с генерирующим сигнал соединением, способным генерировать регистрируемый сигнал; и 4) выявления наличия антитела, которое может присутствовать в биологическом образце, посредством регистрации сигнала, создаваемого генерирующим сигнал соединением, при этом сигнал свидетельствует о диагнозе болезни Альцгеймера у пациента. Антигеном может быть глобуломер, или его часть, или фрагмент, обладающие такими же функциональными свойствами, как и полный глобуломер (например, активностью связывания).
Настоящее изобретение относится к дополнительному способу диагностирования болезни Альцгеймера у пациента, у которого предполагается наличие болезни Альцгеймера. Такой способ включает в себя стадии: 1) выделения биологического образца из организма пациента; 2) осуществление контакта биологического образца с анти-антителом, при анти-антитело специфично по отношению к одному из антител, описанных выше, в течение периода и в условиях, достаточных для обеспечения образования комплексов анти-антитело/антитело, при этом комплексы содержат антитело, присутствующее в биологическом образце; 2) добавления конъюгата к полученным в результате комплексам анти-антитело/антитело в течение периода и в условиях, достаточных для обеспечения связывания конъюгата со связанным антителом, при этом конъюгат содержит антиген, который связывается с генерирующим сигнал соединением, способным генерировать регистрируемый сигнал; и 3) выявления сигнала, создаваемого генерирующим сигнал соединением, при этом сигнал свидетельствует о диагнозе болезни Альцгеймера у пациента.
Кроме того, настоящее изобретение относится к композиции, содержащей любое одно или несколько антител, описанных выше (например, 8F5 и 8C5).
Настоящее изобретение относится к другому способу профилактики или лечения болезни Альцгеймера у пациента, нуждающегося в такой профилактике или лечении. Такой способ включает в себя стадию введения пациенту композиции, описанной непосредственно выше, в количестве, достаточном для осуществления профилактики или лечения.
Кроме того, настоящее изобретение охватывает вакцину, содержащую по меньшей мере одно из антител, описанных выше, и фармацевтически приемлемый адъювант.
Кроме того, настоящее изобретение относится к следующему способу профилактики или лечения болезни Альцгеймера у пациента, нуждающегося в такой профилактике или лечении. Такой способ включает в себя стадию введения пациенту вакцины, указанной выше, в количестве, достаточном для осуществления профилактики или лечения.
Кроме того, настоящее изобретение охватывает способ идентификации соединений, подходящих для активной иммунизации пациента, у которого предполагается развитие болезни Альцгеймера. Такой способ включает в себя: 1) экспонирование одного или нескольких представляющих интерес соединений с одним или несколькими антителами, описанными выше, в течение периода и в условиях, достаточных для связывания одного или нескольких соединений с антителом или антителами; 2) идентификацию таких соединений, которые связываются с антителом или антителами, идентифицированные соединения могут быть использованы для активной иммунизации пациента, у которого предполагается развитие болезни Альцгеймера.
Также настоящее изобретение относится к набору, содержащему: a) по меньшей мере одно из изолированных антител, описанных выше, и b) конъюгат, содержащий антитело, связанное с генерирующим сигнал соединением, при этом антитело в конъюгате отличается от изолированного антитела. Набор также может содержать вкладыш в упаковку с инструкциями о том, как использовать компоненты набора.
Настоящее изобретение также охватывает набор, содержащий: a) анти-антитело к одному из антител, описанных выше, и b) конъюгат, содержащий антиген, связанный с генерирующим сигнал соединением. Антиген может представлять собой глобуломер, или его фрагмент, или часть, обладающие такими же функциональными свойствами, как и глобуломер (например, активность связывания). И, кроме того, набор также может содержать вкладыш в упаковку с инструкциями о том, как использовать компоненты набора.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 иллюстрирует избирательность 8F5 в отношении глобуломеров по сравнению с мономерами Aβ(1-42), Aβ(1-40) и sAPP. Коэффициенты избирательности для 8F5 могут быть рассчитаны в виде отношений между значениями EC50 (по сравнению с мономером Aβ(1-42) в HFIP: 555,8/90,74 = 6,1; по сравнению с мономером Aβ(1-42) в NH4OH: 1007/90,74 = 11,1; по сравнению с мономером Aβ(1-40): 667,8/90,74 = 7,4 по сравнению с sAPP: >100).
Фигура 2 иллюстрирует анализ в SDS-ПААГ связанных с фибриллами тяжелых и легких цепей антител (дорожки 4, 6, 8) и соответствующих несвязанных свободных фракций (дорожки 3, 5, 7) в надосадках.
Фигура 3 иллюстрирует содержание Aβ42 и Aβ40 в образцах спинномозговой жидкости (СМЖ) пациентов с умеренными когнитивными нарушениями (MCI, слева) или болезнью Альцгеймера (AD, справа). В обеих группах можно видеть, что 8F5 улавливает большую часть Aβ(1-42) и меньшее или равное количество Aβ(1-40) по сравнению со стандартным антителом 6E10 или по сравнению с прямым анализом образца в тех же самых анализах ELISA.
Фигура 4 иллюстрирует индекс распознавания новых объектов, основанный на затраченном времени, с использованием неизвестного объекта по сравнению с известным объектом в трех группах трансгенных мышей APP (т.е. 6G1, 8F5, PBS) и одной группе нетрансгенного потомства одного помета (дикий тип). Животных (количество указано под столбиками) иммунизировали моноклональными антителами 6G1 или 8F5 или обрабатывали наполнителем (т.е. фосфатно-солевым буфером; PBS, и дикий тип) один раз в неделю посредством внутрибрюшинной инъекции в течение трех недель. В день последней инъекции выполняли задачу распознавания нового объекта. Различие между группами «PBS» и «дикий тип» свидетельствует о нарушении когнитивной способности у трансгенных мышей APP в данной модели. Мыши, которым инъецировали PBS, выполняли задачу на уровне случайности (т.е. не значимое отличие от 50), тогда как все другие мыши показали распознавание объектов (t-критерий; звездочки). В том случае, когда выполнение задачи трансгенными мышами APP, обработанными антителами, сравнивали с контрольными группами, обнаружили значимое отличие по сравнению с мышами, обработанными PBS, но не по сравнению с мышами дикого типа (ANOVA с апостериорным t-критерием; кружки), что свидетельствует о том, что обработка антителами восстанавливала нарушенную когнитивную способность у указанных трансгенных мышей APP.
Фигура 5(A) иллюстрирует последовательность ДНК (SEQ ID NO: 1), кодирующей вариабельную тяжелую цепь моноклонального антитела, называемого в данном описании «8F5», и фигура 5(B) иллюстрирует последовательность ДНК (SEQ ID NO: 2), кодирующую вариабельную легкую цепь моноклонального антитела 8F5. (Определяющие комплементарность области (CDR) подчеркнуты в каждой последовательности; см. также фигуру 6.)
Фигура 6(A) иллюстрирует аминокислотную последовательность (SEQ ID NO: 3) вариабельной тяжелой цепи моноклонального антитела 8F5, и Фигура 6(B) иллюстрирует аминокислотную последовательность (SEQ ID NO: 4) вариабельной легкой цепи моноклонального антитела 8F5. Одна CDR вариабельной тяжелой цепи представлена аминокислотной последовательностью SYGMS (SEQ ID NO: 5). Другая CDR вариабельной тяжелой цепи представлена аминокислотной последовательностью ASINSNGGSTYYPDSVKG (SEQ ID NO: 6), и еще одна CDR вариабельной тяжелой цепи представлена аминокислотной последовательностью SGDY (SEQ ID NO: 7). Одна CDR вариабельной легкой цепи представлена аминокислотной последовательностью RSSQSLVYSNGDTYLH (SEQ ID NO: 8). Другая CDR вариабельной легкой цепи представлена аминокислотной последовательностью KVSNRFS (SEQ ID NO: 9), и еще одна CDR вариабельной легкой цепи представлена аминокислотной последовательностью SQSTHVPWT (SEQ ID NO: 10). Все описанные выше CDR подчеркнуты на фигуре 6(A) и 6(B).
На фигуре 7 показано связывание антител в различных концентрациях с поперечными срезами неокортекса головного мозга у пациентов с болезнью Альцгеймера (AD) или у старых трансгенных мышей APP. В частности, фигура 7(A) иллюстрирует исследование амилоидных отложений посредством окрашивания Конго красным в виде бляшек в ткани головного мозга и в виде церебральной амилоидной ангиопатии (CAA) в сосудах головного мозга у трансгенных мышей APP линии Tg2576 и у пациентов с AD (RZ55). Фигура 7(B) иллюстрирует, что окрашивание паренхимных отложений Aβ (амилоидные бляшки) у пациентов с AD (RZ16) происходит только в случае 6G1 и коммерчески доступного антитела 6E10, тогда как 8F5 и 8C5 дают значительно более слабое окрашивание. Фигура 7(C) иллюстрирует, что сильное окрашивание паренхимных отложений Aβ (амилоидные бляшки) у мышей TG2576 происходит только в случае 6G1 и коммерчески доступного антитела 6E10, тогда как 8F5 и 8C5 дают значительно более слабое окрашивание. Фигуры 7 (D)-(G) иллюстрируют количественный анализ окрашивания бляшек Aβ на гистологических изображениях с использованием анализа изображений. Значения оптической плотности (0% = нет окрашивания) рассчитывали на основании значений серой шкалы для бляшек, из которых вычитали значения серой шкалы для фоновой ткани. (Фиг. (D)= связывание 0,7 мкг/мл антитела у мышей Tg2576; фиг. (E)= связывание 0,07-0,7 мкг/мл антитела у мышей APP/L; фиг. (F)= связывание 0,7 мкг/мл антитела у пациента с AD (RZ55); и фиг. (G)= связывание 0,07-0,7 мкг/мл антитела у пациента с AD (RZ16).) Различия между окрашиванием коммерчески доступных антител 6E10 (звездочки) и 4G8 (кружочки) и антител 6G1, 8C5 и 8F5 (одна звездочка/кружочек: p <0,05, две звездочки/кружочка: p <0,01, и три звездочки/кружочка: p <0,001 по сравнению с контролем; апостериорный t-критерий Бонферрони после ANOVA с p < 0,001) были оценены статистически (фиг. (D) и фиг. (E)). На фиг. (E) и (G) антитела 8C5 и 8F5 всегда давали значимо менее интенсивное окрашивание, чем коммерчески доступные антитела 6E10 и 4G8 (p <0,05 на основании оценки апостериорного t-критерия после p <0,001 в ANOVA). Фигура (H) иллюстрирует, что интенсивное окрашивание сосудистых отложений Aβ (стрелки) происходит только в случае 6G1 и коммерчески доступного антитела 6E10, тогда как окрашивание 8F5 или 8C5 было намного слабее. Количественно сходная ситуация обнаружена у мышей Tg2576 (не показано).
Фигура 8 иллюстрирует избирательность 8C5 по отношению к глобуломерам по сравнению с мономерами Aβ(1-42), Aβ(1-40) и sAPP. Коэффициенты избирательности для 8C5 можно рассчитать в виде отношений между значениями EC50 (по сравнению с мономером Aβ(1-42) в HFIP: 2346/568,2 = 4,1; по сравнению с мономером Aβ(1-42) в NH4OH: >100; по сравнению с мономером Aβ(1-40): >100; по сравнению с sAPP: >100).
Фигура 9(A) иллюстрирует нуклеотидную последовательность (SEQ ID NO: 11), кодирующую тяжелую цепь 8C5, и фигура 9(B) иллюстрирует нуклеотидную последовательность (SEQ ID NO: 12), кодирующую легкую цепь 8C5. Нуклеотидные последовательности, кодирующие соответствующие CDR, указанные на фигуре 10(A) и 10(B), подчеркнуты.
Фигура 10(B) иллюстрирует аминокислотную последовательность (SEQ ID NO: 19) вариабельной тяжелой цепи моноклонального антитела 8C5, и Фигура 10(B) иллюстрирует аминокислотную последовательность (SEQ ID NO: 20) вариабельной легкой цепи моноклонального антитела 8F5. Одна CDR вариабельной тяжелой цепи представлена аминокислотной последовательностью SYGMS (SEQ ID NO: 13). Другая CDR вариабельной тяжелой цепи представлена аминокислотной последовательностью SIKNNGGSTYYPDSLKG (SEQ ID NO: 14), и еще одна CDR вариабельной тяжелой цепи представлена аминокислотной последовательностью SGDY (SEQ ID NO: 15). Одна CDR вариабельной легкой цепи представлена аминокислотной последовательностью RSSQSLVHSNGDTFLH (SEQ ID NO: 16). Другая CDR вариабельной легкой цепи представлена аминокислотной последовательностью KVSNRFS (SEQ ID NO: 17), и еще одна CDR вариабельной легкой цепи представлена аминокислотной последовательностью SQSIHVPWT (SEQ ID NO: 18). Все описанные выше CDR подчеркнуты на фигуре 10(A) и 10(B).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к моноклональному антителу, называемому в данном описании «8F5», а также к другим родственным антителам (например, 8C5). Такие антитела могут быть использованы, например, для диагностики, профилактики и лечения болезни Альцгеймера и других нейродегенеративных расстройств.
Моноклональное антитело 8F5, а также моноклональное антитело 8C5 обладают множеством интересных свойств, которые делают их очень интересными кандидатами для использования в терапии, а также очень полезными кандидатами для использования в диагностике. Например, моноклональные антитела 8F5 и 8C5 предпочтительно связываются с глобуломерами Aβ(1-42) по сравнению с мономерами или фибрилами.
Термин «Aβ(X-Y)» в данном описании относится к аминокислотной последовательности от положения аминокислоты X до положения аминокислоты Y белка β-амилоида человека, включая X и Y, и, в частности, относится к аминокислотной последовательности от положения аминокислоты X до положения аминокислоты Y аминокислотной последовательности DAEFRHDSGY EVHHQKLVFF AEDVGSNKGA IIGLMVGGW IA или любого из встречающихся в природе вариантов, в частности вариантов по меньшей мере с одной мутацией, выбранной из группы, состоящей из A2T, H6R, D7N, A21G («Фламандская»), E22G («Арктическая»), E22Q («Голландская»), E22K («Итальянская»), D23N («Айова»), A42T и A42V, где номера указаны относительно стартового положения пептида Aβ, включая положение X и положение Y, или последовательности, имеющей до трех дополнительных аминокислотных замен, ни одна из которых не может предотвращать образование глобуломера. «Дополнительную» аминокислотную замену определяют в данном описании как любое отклонение от канонической последовательности, которое не встречается в природе.
Более конкретно, термин «Aβ(1-42)» в данном описании относится к аминокислотной последовательности от положения аминокислоты 1 до положения аминокислоты 42 белка β-амилоида человека, включая аминокислоты 1 и 42, и, в частности, относится к аминокислотной последовательности от положения аминокислоты 1 до положения аминокислоты 42 аминокислотной последовательности DAEFRHDSGY EVHHQKLVFF AEDVGSNKGA IIGLMVGGW IA (соответствующей положениям аминокислот 1-42) или любому из ее встречающихся в природе вариантов. Такие варианты могут представлять собой, например, варианты по меньшей мере с одной мутацией, выбранной из группы, состоящей из A2T, H6R, D7N, A21G («Фламандская»), E22G («Арктическая»), E22Q («Голландская»), E22K («Итальянская»), D23N («Айова»), A42T и A42V, где номера указаны относительно начала пептида Aβ, включая 1 и 42, или последовательность, имеющую до трех дополнительных аминокислотных замен, ни одна из которых не может предотвращать образование глобуломера. Подобным образом, термин «Aβ(1-40)» в данном описании относится к аминокислотной последовательности от положения аминокислоты 1 до положения аминокислоты 40 белка β-амилоида человека, включая 1 и 40, и, в частности, относится к аминокислотной последовательности от положения аминокислоты 1 до положения аминокислоты 40 аминокислотной последовательности DAEFRHDSGY EVHHQKLVFF AEDVGSNKGA IIGLMVGGW или ее любого встречающегося в природе варианта. Такие варианты включают, например, варианты по меньшей мере с одной мутацией, выбранной из группы, состоящей из A2T, H6R, D7N, A21G («Фламандская»), E22G («Арктическая»), E22Q («Голландская»), E22K («Итальянская») и D23N («Айова»), где номера указаны относительно стартового положения пептида Aβ, включая 1 и 40, или последовательность, содержащую до трех дополнительных аминокислотных замен, ни одна из которых не может предотвращать образование глобуломера.
Термин «глобуломер Aβ(X-Y)» (также известный как глобулярный олигомер «Aβ(X-Y)») в данном описании относится к растворимой, глобулярной, нековалентной ассоциации пептидов Aβ(X-Y), которые определены выше, обладающей гомогенностью и особыми физическими свойствами. Глобуломеры Aβ(X-Y) являются стабильными, нефибриллярными, олигомерными сборными структурами пептидов Aβ(X-Y), которые можно получить при инкубации с анионными детергентами. В отличие от мономера и фибрилл такие глобуломеры характеризуются определенным количеством субъединиц в сборке (например, формы ранней сборки, n = 3-6, олигомеры «олигомеры A», и формы поздней сборки, n = 12-14, «олигомеры B», которые описаны в публикации международной заявки PCT No. WO 04/067561). Глобуломеры имеют 3-мерную структуру глобулярного типа («расплавленная глобула», см. Barghorn et al., 2005, J. Neurochem., 95, 834-847). Они могут быть охарактеризованы одним или несколькими из следующих признаков:
- возможность отщепления N-концевых аминокислот X-23 протеазами, которые делают случайный выбор (такими как термолизин или эндопротеиназа GluC), с получением укороченных форм глобуломеров Aβ(X-Y);
- недоступность C-концевых аминокислот 24-Y для делающих случайный выбор протеаз и антител; и
- укороченные формы таких глобуломеров Aβ(X-Y) сохраняют 3-мерную структуру кора глобуломеров с лучшей доступностью эпитопа кора Aβ(20-Y) в конформации глобуломера.
Согласно изобретению и, в частности, в целях оценки аффинностей связывания антител согласно настоящему изобретению термин «глобуломер Aβ(X-Y)» в данном описании относится к продукту, который может быть получен способом, описанным в публикации международной заявки No. WO 04/067561, которая включена в данное описание в виде ссылки в полном объеме. Способ включает в себя разворачивание природного, рекомбинантного или синтетического пептида Aβ(X-Y) или его производного; воздействие на по меньшей мере частично развернутый пептид Aβ(X-Y) или его производное детергентом, уменьшение действия детергента и продолжение инкубации.
В целях разворачивания пептида можно обеспечить воздействие на белок агентов, разрушающих водородные связи, таких, например, как гексафторизопропанол (HFIP). Время действия в течение нескольких минут, например приблизительно от 10 до 60 минут, достаточно, когда температура действия составляет примерно от 20 до 50ºC и, в частности, примерно от 35 до 40ºC. Последующее растворение остатка, полученного после упаривания досуха, предпочтительно в концентрированной форме в подходящих органических растворителях, смешиваемых с водными буферами, такими как, например, диметилсульфоксид (ДМСО), дает суспензию по меньшей мере частично развернутого пептида или его производного, которую можно использовать позже. При необходимости исходную суспензию можно хранить при низкой температуре, например при температуре около -20ºC в течение необходимого периода времени.
Альтернативно пептид или его производное может находиться в слегка подкисленном, предпочтительно водном растворе, например в водном растворе примерно 10 мМ HCl. После инкубационного периода времени, составляющего приблизительно несколько минут, нерастворимые компоненты удаляют центрифугированием. Подходящим является центрифугирование в течение нескольких минут при 10000. Указанные стадии способа предпочтительно осуществляют при комнатной температуре, т.е. при температуре в диапазоне от 20 до 30ºC. Надосадок, полученный после центрифугирования, содержит пептид Aβ(X-Y) или его производное, и его можно хранить при низкой температуре, например при температуре около -20ºC в течение необходимого периода времени.
Следующая экспозиция с детергентом имеет отношение к олигомеризации пептида или его производного с получением промежуточного типа олигомеров (в публикации международной заявки No. WO 04/067561 называемых олигомерами A). В указанных целях обеспечивают возможность воздействия детергента на необязательно по меньшей мере частично развернутый пептид или его производное вплоть до получения достаточного количества промежуточного олигомера. Предпочтительно применение ионных детергентов, в частности анионных детергентов.
Согласно конкретному варианту используют детергент формулы (I):
R-X,
в которой радикал «R» означает неразветвленный или разветвленный алкил, имеющий от 6 до 20 и предпочтительно от 10 до 14 атомов углерода, или неразветвленный или разветвленный алкенил, имеющий от 6 до 20 и предпочтительно от 10 до 14 атомов углерода, а радикал «X» означает кислотную группу или ее соль, при этом X предпочтительно выбран из -COO-M+, -SO3 -M+ и наиболее предпочтительно -OSO3 -M+, и M+ означает катион водорода или неорганический или органический катион, предпочтительно выбранный из катионов щелочных металлов, катионов щелочноземельных металлов и катионов аммония. Наиболее предпочтительными являются детергенты формулы (I), в которых R означает неразветвленный алкил, из которых, в частности, следует упомянуть алк-1-ильные радикалы. Особенно предпочтительным является додецилсульфат натрия (SDS). Также предпочтительно могут быть использованы лауриновая кислота и олеиновая кислота. Также особенно предпочтительной является натриевая соль детергента лауроилсаркозина (также известная как саркозил-NL-30 или Gardol®).
Время действия детергента, в частности, зависит от того, развернут ли, и если развернут, то в какой степени, пептид или его производное, подвергаемое олигомеризации. Если в соответствии с протоколом стадии разворачивания пептид или его производное обработали заранее агентом, разрывающим водородные связи (т.е., в частности, гексафторизопропанолом), то достаточно время действия в диапазоне нескольких часов, преимущественно примерно от 1 до 20 и, в частности, примерно от 2 до 10 часов, когда температура во время действия составляет примерно от 20 до 50ºC и, в частности, примерно от 35 до 40ºC. Если исходной точкой является менее развернутый или по существу неразвернутый пептид или его производное, то, соответственно, необходимы более длительные периоды времени действия. Если пептид или его производное были предварительно обработаны, например, согласно способу, указанному выше в качестве альтернативы обработке HFIP, или указанный пептид или его производное непосредственно подвергают олигомеризации, то достаточным является время действия в диапазоне примерно от 5 до 30 часов и, в частности, примерно от 10 до 20 часов, когда температура во время действия составляет примерно от 20 до 50ºC и, в частности, примерно от 35 до 40ºС. После инкубации нерастворимые компоненты преимущественно удаляют центрифугированием. Достаточно центрифугирование в течение нескольких минут при 10000 g.
Выбираемая концентрация детергента зависит от используемого детергента. Если используют SDS, то подходящей оказывается концентрация в диапазоне от 0,01 до 1% мас., предпочтительно от 0,05 до 0,5% мас., например около 0,2% мас. Если используют лауриновую кислоту или олеиновую кислоту, то необходимы несколько более высокие концентрации, например в диапазоне от 0,05 до 2% мас., предпочтительно от 0,1 до 0,5% мас., например около 0,5% мас. Действие детергента должно происходить при концентрации соли примерно в физиологическом диапазоне. Таким образом, подходящими являются, в частности, концентрации NaCl в диапазоне от 50 до 500 мМ, предпочтительно от 100 до 200 мМ и более конкретно примерно 140 мМ.
Последующее уменьшение действия детергента и продолжение инкубации связано с последующей олигомеризацией с получением глобуломера Aβ(X-Y) согласно изобретению (называемого в публикации международной заявки No. WO 04/067561 олигомером B). Так как композиция, получаемая на предшествующей стадии, обычно содержит детергент и концентрацию соли в физиологическом диапазоне, то затем требуется уменьшить действие детергента, а также предпочтительно уменьшить концентрацию соли. Это можно осуществить посредством уменьшения концентрации детергента и соли, например, соответствующим образом разбавляя водой или буфером с низкой концентрацией соли, например трис-HCl, pH 7,3. Показано, что подходящими являются коэффициенты разбавления в диапазоне примерно от 2 до 10, преимущественно в диапазоне примерно от 3 до 8 и, в частности, примерно 4. Уменьшения действия детергента также можно достичь добавлением веществ, которые могут нейтрализовать такое действие детергента. Примеры веществ включают такие вещества, которые способны к образованию комплексов с детергентами, подобные веществам, способным стабилизировать клетки в процессе очистки и экстракции, в частности, например блок-сополимеры EO/PO, в частности блок-сополимер, продаваемый под торговой маркой Pluronic® F 68. В равной мере могут быть использованы алкоксилированные и, в частности, этоксилированные алкилфенолы, такие как этоксилированные t-оксилфенолы серии Triton® X, в частности тритон X100, 3-(3-холамидопропилдиметиламмонио)-1-пропансульфонат (CHAPS®), или алкоксилированные и, в частности, этоксилированные сложные эфиры жирной кислоты и сорбитана, такие как эфиры серии Tween®, в частности твин 20, в диапазоне концентраций около или выше конкретной критической концентрации мицеллообразования.
Затем раствор инкубируют вплоть до образования достаточного количества глобуломера Aβ(X-Y). Достаточным является время действия в диапазоне нескольких часов, предпочтительно в диапазоне примерно от 10 до 30 часов и, в частности, в диапазоне примерно от 15 до 25 часов, когда температура во время действия составляет примерно от 20 до 50ºC и, в частности, примерно от 35 до 40ºC. Затем раствор можно концентрировать, и возможный остаток может быть удален центрифугированием. Также достаточно центрифугирование в течение нескольких минут при 10000 g. Надосадок, полученный после центрифугирования, содержит глобуломер Aβ(X-Y), который описан выше.
Глобуломер Aβ(X-Y) в конце концов может быть извлечен, например, ультрацентрифугированием, диализом, преципитацией или центрифугированием. Кроме того, предпочтительно, если электрофоретическое разделение глобуломеров Aβ(X-Y) в денатурирующих условиях, например в SDS-ПААГ, дает двойную полосу (например, с кажущейся молекулярной массой 38/48 кД в случае Aβ(1-42)), и особенно предпочтительно, если после обработки олигомеров глутаральдегидом перед разделением указанные две полосы сливаются в одну. Также предпочтительно, если эксклюзионная хроматография по размеру глобуломеров дает один пик (например, соответствующий молекулярной массе примерно 60 кД в случае Aβ(1-42)). Способ, в частности, подходит для получения глобуломеров Aβ(1-42), исходя из пептида Aβ(1-42).
Предпочтительно глобуломер проявляет аффинность к нейронным клеткам, а также оказывает нейромодулирующее действие. «Нейромодулирующее действие» определяют как длительное ингибирующее нейрон действие, приводящее к дисфункции нейрона в отношении пластичности нейронов.
Согласно другому аспекту изобретения термин «глобуломер Aβ(X-Y)» в данном описании относится к глобуломеру, состоящему по существу из субъединиц Aβ(X-Y), при этом предпочтительно в среднем по меньшей мере 11 из 12 субъединиц являются субъединицами Aβ(X-Y)-типа, более предпочтительно меньше 10% глобуломеров содержат какие-либо пептиды, не являющиеся пептидами Aβ(X-Y), и наиболее предпочтительное содержание пептидов, не являющихся пептидами Aβ(X-Y), в препарате ниже порога регистрации. Более конкретно, термин «глобуломер Aβ(1-42)» в данном описании относится к глобуломеру, содержащему единицы Aβ(1-42), которые описаны выше; термин «глобуломер Aβ(12-42)» в данном описании относится к глобуломеру, содержащему единицы Aβ(12-42), которые описаны выше; и термин «глобуломер Aβ(20-42)» в данном описании относится к глобуломеру, содержащему единицы Aβ(20-42), которые определены выше.
Термин «поперечно сшитый глобуломер Aβ(X-Y)» в данном описании относится к молекуле, получаемой из глобуломера Aβ(X-Y), который описан выше, поперечным сшиванием, предпочтительно поперечным сшиванием химическим способом, более предпочтительно поперечным сшиванием альдегидом и наиболее предпочтительно поперечным сшиванием глутаральдегидом единиц, составляющих глобуломер. В другом аспекте изобретения поперечно сшитый глобуломер по существу представляет собой глобуломер, в котором единицы по меньшей мере частично связаны ковалентными связями, а не удерживаются вместе только нековалентными взаимодействиями.
Термин «производное глобуломера Aβ(X-Y)» в данном описании, в частности, относится к глобуломеру, который метят ковалентным связыванием с группой, которая облегчает регистрацию, предпочтительно с флуорофором, например изотиоцианатом флуоресцеина, фикоэритрином, флуоресцирующим белком Aequorea victoria, флуоресцирующим белком Dictyosoma или любой их комбинацией или их флуоресцентно-активными производными; хромофором; хемилюминофором, например люциферазой, предпочтительно люциферазой Photinus pyralis, люциферазой Vibrio fischeri или любой их комбинацией или хемилюминесцентно-активными производными; ферментативно активной группой, например пероксидазой, такой как пероксидаза хрена, или ее ферментативно активным производным; электроноплотной группой, например группой, содержащей тяжелый металл, такой как группа, содержащая золото; гаптеном, например полученным на основе фенола гаптеном; сильной антигенной структурой, например пептидной последовательностью, которая по расчетам является антигенной, например согласно алгоритму Kolaskar и Tongaonkar; аптамером для другой молекулы; хелатирующей группой, например гексагистидинилом; природной или полученной из природной белковой структурой, опосредующей дополнительные специфичные белок-белковые взаимодействия, например представителем пары fos/jun; магнитной группой, например ферромагнитной группой; или радиоактивной группой, такой как группа, содержащая 1H, 14C, 32P, 35S или 125I или любую их комбинацию; или к глобуломеру, маркированному ковалентным или нековалентным связыванием благодаря высокоаффинному взаимодействию, предпочтительно ковалентным связыванием с группой, которая способствует инактивации, удалению, распаду и/или преципитации, предпочтительно, маркированному группой, которая стимулирует распад in vivo, более предпочтительно убиквитином, при этом особенно предпочтительна сборка такого маркированного олигомера in vivo; или к глобуломеру, модифицированному любой комбинацией указанного выше. Такие группы для мечения и маркировки и способы их связывания с белками известны в данной области. Мечение и/или маркировку можно осуществлять до, во время или после глобуломеризации. В другом аспекте изобретения производное глобуломера представляет собой молекулу, получаемую из глобуломера в результате реакции мечения и/или маркировки. Соответственно, термин «производное мономера Aβ(X-Y)» в данном описании, в частности, относится к мономеру Aβ, который метят или маркируют, как описано для глобуломера.
Термин «более высокая аффинность» в данном описании относится к степени взаимодействия, при котором равновесие между несвязанным антителом и несвязанным глобуломером с одной стороны и комплексом антитело-глобуломер с другой стороны сдвинуто в пользу комплекса антитело-глобуломер. Подобным образом, термин «более низкая аффинность» в данном описании относится к степени взаимодействия, при котором равновесие между несвязанным антителом и несвязанным глобуломером с одной стороны и комплексом антитело-глобуломер с другой стороны сдвинуто в пользу несвязанного антитела и несвязанного глобуломера.
Термин «мономер Aβ(X-Y)» в данном описании относится к изолированной форме пептида Aβ(X-Y), предпочтительно к форме пептида Aβ(X-Y), которая по существу не подвергается нековалентным взаимодействиям с другими пептидами Aβ. На практике мономер Aβ(X-Y) обычно получают в форме водного раствора. Предпочтительно водный раствор мономера содержит от 0,05 до 0,2%, более предпочтительно примерно 0,1% NaOH, например при использовании для определения аффинности связывания антитела согласно настоящему изобретению. В другом предпочтительном случае водный раствор мономера содержит от 0,05 до 0,2%, более предпочтительно примерно 0,1% NaOH. При использовании может требоваться разбавление раствора подходящим образом. Кроме того, обычно рекомендуется использование раствора в течение 2 часов, в частности в течение 1 часа, и в основном в течение 30 минут после его приготовления.
Термин «фибрилла» в данном описании относится к молекулярной структуре, которая содержит совокупность нековалентно связанных отдельных пептидов Aβ(X-Y), которые имеют фибриллярную структуру под электронным микроскопом, которая связывает Конго красный, дает двойное лучепреломление в поляризованном свете и картина дифракции рентгеновских лучей которой представляет поперечную β-структуру. Фибриллу также можно определить как молекулярную структуру, получаемую способом, который включает в себя самоиндуцируемую агрегацию в виде полимера подходящего пептида Aβ в отсутствие детергентов, например в 0,1 М HCl, приводящую к образованию агрегатов из более чем 24, предпочтительно более чем 100 единиц. Такой способ хорошо известен в данной области. Соответственно, фибриллу Aβ(X-Y) используют в форме водного раствора. В особенно предпочтительном варианте осуществления изобретения водный раствор фибрилл получают растворением пептида Aβ в 0,1% NH4OH, его разбавлением 1:4 с использованием 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, с последующем доведением pH снова до 7,4, инкубацией раствора при 37ºC в течение 20 ч с последующим центрифугированием при 10000 g в течение 10 мин и ресуспендированием в 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4.
Термин «фибрилла Aβ(X-Y)» в данном описании относится к фибрилле, содержащей субъединицы Aβ(X-Y), при этом предпочтительно в среднем по меньшей мере 90% субъединиц являются субъединицами Aβ(X-Y)-типа, более предпочтительно по меньшей мере 98% субъединиц являются субъединицами Aβ(X-Y)-типа, и наиболее предпочтительно содержание пептидов, не являющихся пептидами Aβ(X-Y), ниже порога регистрации.
Снова возвращаясь к 8F5, как показано на фигуре 1, а также 8C5 (фигура 8), моноклональные антитела, специфичные по отношению к глобуломеру Aβ(1-42), 8F5 и 8C5, преимущественно узнают формы глобуломера Aβ(1-42), а не стандартные препараты мономеров Aβ(1-40) или Aβ(1-42), включая агрегированные Aβ(1-42), в отличие от неспецифичных антител 6G1 и 6E10. В частности, 8F5 выявляет глобуломеры Aβ(1-42) только при анализе в нативном ПААГ-Вестерн-блоте, но не при анализе в SDS-ПААГ-Вестерн-блоте, что свидетельствует о связывании с более сложным диссоциируемым детергентом межсубъединичным эпитопом в коровой структуре глобуломера Aβ(1-42). Межсубъединичный эпитоп определяют как сложный нелинейный протяженный в пространстве эпитоп, локализованный по меньшей мере на двух субъединицах. Более конкретно, дот-блот-анализ против разных стандартных препаратов Aβ(1-42) и Aβ(1-40) показал значимые различия в распознавании глобуломера Aβ(1-42) по сравнению с неглобуломерными формами Aβ (стандартный препарат мономера Aβ(1-40)/(1-42), агрегированный Aβ(1-42)) в случае специфичных 8F5 и 8C5, но не в случае изоформ неспецифичных антител 6G1 и 6E10. Специфичность по отношению к глобуломеру антител 8F5 и 8C5, но не 6G1 и 6E10 подтвердили количественным анализом связывания глобуломера Aβ(1-42), мономера Aβ(1-42), мономера Aβ(1-40) и растворимого белка предшественника амилоида альфа в ELISA типа «сэндвич». Кроме того, так как указанные антитела достигают глобуломера после нативного, но не после SDS-Вестерн-блоттинга, то вероятно, что каждое антитело узнает структурный нелинейный эпитоп между субъединицами в области от 20 до 30 аминокислоты Aβ(1-42). Такая специфичность по отношению к глобуломерам является важной, поскольку специфичное целенаправленное воздействие на форму глобуломера Aβ антителом, предпочтительно действующим на глобуломер, таким как, например, 8F5 или 8C5, позволит: 1) избегать использования в качестве мишени нерастворимых амилоидных отложений, связывание с которыми может являться причиной воспалительных побочных эффектов, наблюдаемых во время иммунизации нерастворимым Aβ; 2) не затрагивать мономеры Aβ и APP, которые, как сообщается, обладают прогнозируемыми физиологическими функциями (Plan et al., J. of Neuroscience 23: 5531-5535 (2003); и 3) увеличивать биодоступность антитела, так как оно не может быть скрыто или сделано недоступным в результате пространственного связывания с нерастворимыми отложениями.
Настоящее изобретение также относится к изолированным нуклеотидным последовательностям (или их фрагментам), кодирующим вариабельные легкие и тяжелые цепи моноклонального антитела 8F5 и 8CD, а также к таким нуклеотидным последовательностям (или их фрагментам), которые имеют последовательности, содержащие, соответствующие, идентичные, гибридизующиеся или комплементарные последовательностям по меньшей мере примерно на 70% (например, 70 71, 72, 73, 74, 75, 76, 77, 78 или 79%), предпочтительно по меньшей мере примерно на 80% (например, 80, 81, 82, 83, 84, 85, 86, 87, 88 или 89%) и более предпочтительно по меньшей мере примерно на 90% (например, 91, 92, 93, 94, 95, 96, 97, 98 или 99%) идентичным таким кодирующим нуклеотидным последовательностям. (Все целые числа (и их части) в диапазоне и включая 70 и 100% считаются входящими в объем настоящего изобретения при упоминании в отношении идентичности в процентах.) Такие последовательности могут быть получены из любого источника (например, выделены из природного источника, получены полусинтетическим способом или синтезированы de novo). В частности, такие последовательности могут быть выделены или получены из других источников, отличных от источников, описанных в примерах (например, бактерий, грибов, водорослей, мыши или человека).
Кроме нуклеотидных последовательностей, описанных выше, настоящее изобретение также относится к аминокислотным последовательностям вариабельных легких и тяжелых цепей моноклонального антитела 8F5 и моноклонального антитела 8C5 (или фрагментам таких аминокислотных последовательностей). Кроме того, настоящее изобретение также относится к аминокислотным последовательностям (или их фрагментам), содержащим, соответствующим, идентичным или комплементарным последовательностям по меньшей мере примерно на 70%, предпочтительно по меньшей мере примерно на 80% и более предпочтительно по меньшей мере примерно на 90% идентичным аминокислотным последовательностям белков согласно настоящему изобретению. (И при этом также все целые числа (и их части) в диапазоне и включая 70 и 100% (которые указаны в связи с идентичностью нуклеотидных последовательностей, описанной выше) также считают входящими в объем настоящего изобретения при упоминании в отношении идентичности в процентах.)
В целях настоящего изобретения «фрагмент» нуклеотидной последовательности определяют как непрерывную последовательность, состоящую примерно по меньшей мере из 6, предпочтительно по меньшей мере примерно из 8, более предпочтительно по меньшей мере примерно из 10 нуклеотидов и еще более предпочтительно по меньшей мере примерно из 15 нуклеотидов, соответствующих области конкретной нуклеотидной последовательности.
Термин «идентичность» относится к соотнесению двух последовательностей нуклеотид за нуклеотидом на протяжении определенного окна сравнения или участка. Таким образом, идентичность определяют как степень сходства, соответствия или эквивалентности между одинаковыми нитями (либо смысловыми, либо антисмысловыми) двух участков ДНК (или двух аминокислотных последовательностей). «Идентичность последовательностей в процентах» рассчитывают посредством сравнения двух оптимально выровненных последовательностей на протяжении конкретной области, определения количества положений, в которых встречаются идентичные основания или аминокислоты в обеих последовательностях, чтобы получить количество совпадающих положений, деления количества таких положений на общее количество положений в сравниваемом участке и умножения результата на 100. Оптимальное выравнивание последовательностей можно осуществить с использованием алгоритма Smith and Waterman, Appl. Math. 2: 482 (1981), алгоритма Needleman and Wunsch, J. MoI. Biol. 48: 443 (1970), способом Pearson and Lipman, Proc. Natl. Acad. Sci. (USA) 85: 2444 (1988) и с помощью компьютерных программ, которые основаны на соответствующих алгоритмах (например, Clustal Macaw Pileup (http://cmgm.Stanford.edu/biochem218/11Multiple.pdf; Higgins et al., CABIOS. 5L151-153 (1989)), FASTDB (Intelligenetics), BLAST (National Center for Biomedical Information; Altschul et al., Nucleic Acids Research 25: 3389-3402 (1997)), PILEUP (Genetics Computer Group, Madison, WI) или GAP, BESTFIT, FASTA и TFASTA (Wisconsin Genetics Software Package Release 7.0, Genetics Computer Group, Madison, WI). (См. патент США No. 5912120.)
В целях настоящего изобретения «комплементарность» определяют как степень родства между двумя участками ДНК. Комплементарность определяют измерением способности смысловой нити одного участка ДНК гибридизоваться с антисмысловой нитью другого участка ДНК в подходящих условиях с образованием двойной спирали. «Комплемент» определяют как последовательность, которая спаривается с данной последовательностью на основании канонических правил спаривания оснований. Например, последовательность A-G-T в одной нуклеотидной нити «комплементарная» T-C-A в другой нити.
В двойной спирали аденин находится в одной нити, тимин находится в другой нити. Подобным образом, где в одной нити находится гуанин, там в другой нити находится цитозин. Чем ближе родство между нуклеотидными последовательностями двух участков ДНК, тем больше способность образовывать гибридные дуплексы между нитями двух участков ДНК.
«Сходство» между двумя аминокислотными последовательностями определяют как наличие серии идентичных, а также консервативных аминокислотных остатков в обеих последовательностях. Чем выше степень сходства между двумя аминокислотными последовательностями, тем выше соответствие, сходство или эквивалентность двух последовательностей. («Идентичность" между двумя аминокислотными последовательностями определяют как наличие серии в точности одинаковых или инвариантных аминокислотных остатков в обеих последовательностях.) Определения «комплементарность», «идентичность» и «сходство» хорошо известны специалистам в данной области.
«Кодируемый» относится к последовательности нуклеиновой кислоты, которая кодирует последовательность полипептида, при этом последовательность полипептида или ее часть содержит аминокислотную последовательность по меньшей мере из 3 аминокислот, более предпочтительно по меньшей мере из 8 аминокислот и еще более предпочтительно по меньшей мере из 15 аминокислот из полипептида, кодируемого последовательностью нуклеиновой кислоты.
Кроме того, молекула нуклеиновой кислоты может «гибридизоваться» с другой молекулой нуклеиновой кислоты, когда однонитевая форма молекулы нуклеиновой кислоты может отжигаться с другой молекулой нуклеиновой кислоты в подходящих условиях температуры и ионной силы (см. Sambrook et al., "Molecular Cloning: A Laboratory Manual, Second Edition (1989), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York). Условия температуры и ионной силы определяют «жесткость» гибридизации.
Термин «гибридизация» в используемом в данном описании смысле обычно используют для обозначения гибридизации нуклеиновых кислот в подходящих условиях жесткости, которые могут быть легко определены специалистами в данной области в зависимости от природы последовательности зонда и последовательностей мишени. Условия гибридизации и промывки хорошо известны в данной области, и корректировка условий в зависимости от требуемой жесткости в отношении разного времени инкубации, температуры и/или ионной силы раствора легко осуществима. См., например, публикацию Sambrook, J. et al., Molecular Cloning: A Laboratory Manual, 2nd edition, Cold spring harbor Press, Cold Spring harbor, N. Y., 1989, которая указана выше и включена в данное описание в виде ссылки. (См. также Short Protocols in Molecular Biology, ed. Ausubel et al. and Tijssen, Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, "Overview of principles of hybridization and the strategy of nucleic acid assays" (1993), обе публикации включены в данное описание в виде ссылки.) В частности, выбор условий определяется длиной последовательностей, подвергаемых гибридизации, в частности длиной последовательности зонда, относительным содержанием G-C в нуклеиновых кислотах и количеством разрешенных ошибочных спариваний. Условия низкой жесткости предпочтительны, когда требуется частичная гибридизация между нитями, которые имеют более низкую степень комплементарности. В том случае, когда требуется абсолютная или почти абсолютная комплементарность, предпочтительны условия высокой жесткости. В случае типичных условий высокой жесткости раствор для гибридизации содержит 6 X S.S.C, 0,01 М EDTA, 1 x раствор Денхардта и 0,5% SDS. Гибридизацию осуществляют при 68 градусах по Цельсию в течение примерно 3-4 часов в случае фрагментов клонированной ДНК и в течение примерно 12-16 часов в случае суммарной эукариотической ДНК. Для умеренной жесткости можно использовать предварительную гибридизацию на фильтре и гибридизацию с использованием раствора 3 X хлорида натрия, цитрата натрия (SSC), 50% формамида (0,1 М такого буфера с pH 7,5) и 5 X раствора Денхардта. Затем может быть осуществлена предварительная гибридизация при 37 градусах по Цельсию в течение 4 часов с последующей гибридизацией при 37 градусах по Цельсию с использованием количества меченого зонда, равного 3000000 имп./мин в течение 16 часов с последующей промывкой в растворе 2 X SSC и 0,1% SDS, при этом промывку проводят 4 раза по 1 минуте каждую при комнатной температуре и 4 раза при 60 градусах по Цельсию в течение 30 минут каждую. После сушки экспонируют с пленкой. Для более низкой жесткости температуру гибридизация снижают примерно до температуры на 12 градусов по Цельсию ниже температуры плавления (Тm) дуплекса. Известно, что Tm является функцией содержания G-C и длины дуплекса, а также ионной силы раствора.
«Гибридизация» требует, чтобы две нуклеиновых кислоты содержали комплементарные последовательности. Однако в зависимости от жесткости гибридизации могут происходить ошибочные спаривания оснований. Как указано выше, подходящая жесткость для гибридизации нуклеиновых кислот зависит от длины нуклеиновых кислот и степени комплементарности. Такие переменные хорошо известны в данной области. Более конкретно, чем больше сходство или гомология двух нуклеотидных последовательностей, тем выше значение Tm для гибридов нуклеиновых кисло, имеющих такие последовательности. Получены уравнения расчета Tm для гибридов длиной более 100 нуклеотидов (см. Sambrook et al., выше). В случае гибридизации более коротких нуклеиновых кислот более важным становится положение ошибочных спариваний, и длина олигонуклеотида определяет его специфичность (см. Sambrook et al., выше).
В используемом в данном описании смысле «изолированный фрагмент или последовательность нуклеиновой кислоты» представляет собой полимер РНК или ДНК, который является однонитевым или двунитевым, необязательно содержащим синтетические, неприродные или измененные нуклеотидные основания. Изолированный фрагмент нуклеиновой кислоты в форме полимера ДНК может состоять из одного или нескольких участков кДНК, геномной ДНК или синтетической ДНК. («Фрагмент» конкретного полинуклеотида относится к полинуклеотидной последовательности, которая содержит непрерывную последовательность, состоящую по меньшей мере примерно из 6 нуклеотидов, предпочтительно по меньшей мере примерно из 8 нуклеотидов, более предпочтительно по меньшей мере примерно из 10 нуклеотидов, еще более предпочтительно по меньшей мере из 15 нуклеотидов и наиболее предпочтительно по меньшей мере примерно из 25 нуклеотидов, идентичных или комплементарных области конкретной нуклеотидной последовательности). Нуклеотиды (обычно находящиеся в форме 5'-монофосфата) называют, используя их однобуквенные обозначения: «A» для аденилата или дезоксиаденилата (для РНК или ДНК соответственно), «C» для цитидилата или дезоксицитидилата, «G» для гуанилата или дезоксигуанлата, «U» для уридилата, «T» для дезокситимидилата, «R» для пуринов (A или G), «Y» для пиримидинов (C или T), «K» для G или T, «H» для A или C или T, «I» для инозина и «N» для любого нуклеотида.
Термины «фрагмент или субфрагмент, который является функционально эквивалентным» и «функционально эквивалентный фрагмент или субфрагмент» используют в данном описании взаимозаменяемо. Такие термины относятся к части или подпоследовательности изолированного фрагмента нуклеиновой кислоты, который сохраняет способность изменять экспрессию гена или давать определенный фенотип, независимо от того, кодирует ли фрагмент или субфрагмент активный фермент или не кодирует. Например, фрагмент или субфрагмент можно использовать для создания химерных конструкций, для того чтобы получить требуемый фенотип у трансформированного растения. Химерные конструкции могут быть созданы для применения в случае косупрессии или антисмыслового действия посредством связывания фрагмента нуклеиновой кислоты или его субфрагмента, независимо от того, кодирует ли он активный фермент или нет, в подходящей ориентации относительно последовательности промотора растений.
Термины «гомология», «гомологичный», «по существу сходный» и «по существу соответствующий» используют в данном описании взаимозаменяемо. Термины относятся к фрагментам нуклеиновой кислоты, в которых изменения в одном или нескольких нуклеотидных основаниях не влияют на способность фрагмента нуклеиновой кислоты опосредовать экспрессию гена или вызывать определенный фенотип. Указанные термины также относятся к модификациям фрагментов нуклеиновой кислоты согласно настоящему изобретению, таким как делеция или инсерция одного или нескольких нуклеотидов, которые по существу не изменяют функциональные свойства полученного в результате фрагмента нуклеиновой кислоты по сравнению с исходным немодифицированным фрагментом. Поэтому ясно и понятно специалистам в данной области, что объем изобретения охватывает больше, чем конкретные приведенные в качестве примера последовательности.
«Ген» относится к фрагменту нуклеиновой кислоты, который экспрессирует конкретный белок, включая регуляторные последовательности, предшествующие (5'-некодирующие последовательности) и следующие (3'-некодирующие последовательности) за кодирующей последовательностью.
«Нативный ген» относится к гену, который встречается в природе со своими собственными регуляторными последовательностями. Напротив, «химерная конструкция» относится к комбинации фрагментов нуклеиновых кислот, которые обычно не встречаются вместе в природе. Соответственно, химерная конструкция может содержать регуляторные последовательности и кодирующие последовательности, которые получены из разных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные в другом порядке, отличном от обычного расположения, встречающегося в природе. (Термин «изолированная» означает, что последовательность изъята из ее природного окружения.)
«Чужеродный» ген относится к гену, в норме не встречающемуся в организме хозяина, но который введен в организм хозяина посредством переноса гена. Чужеродные гены могут содержать нативные гены, введенные в ненативный организм, или химерные конструкции. «Трансген» означает ген, который был введен в геном способом трансформации.
«Кодирующая последовательность» относится к последовательности ДНК, которая кодирует специфичную аминокислотную последовательность. Термин «регуляторные последовательности» относится к нуклеотидным последовательностям, расположенным выше (5'-некодирующие последовательности), в пределах или ниже (3'-некодирующие последовательности) кодирующей последовательности и которые влияют на транскрипцию, процессинг и стабильность РНК или трансляцию связанной кодирующей последовательности. Регуляторные последовательности могут включать в себя без ограничения промоторы, лидерные последовательности трансляции, интроны и узнаваемые последовательности полиаденилирования.
«Промотор» или «последовательность регуляции гена» относятся к последовательности ДНК, способной контролировать экспрессию кодирующей последовательности или функциональной РНК. Последовательность состоит из проксимальных и более удаленных выше расположенных элементов, последние элементы часто называют энхансерами. Соответственно, «энхансер» означает последовательность ДНК, которая может стимулировать активность промотора или последовательности регуляции гена и может представлять собой естественный элемент промотора или гетерологичный элемент, встроенный для повышения уровня или тканевой специфичности промотора. Промоторные последовательности также могут быть расположены в транскрибируемых частях генов и/или ниже транскрибируемых последовательностей. Промоторы могут быть получены как целое из нативного гена или могут состоять из разных элементов, полученных из разных промоторов, встречающихся в природе, или даже содержать участки синтетической ДНК. Специалистам в данной области понятно, что разные промоторы могут управлять экспрессией гена в разных тканях и типах клеток или на разных стадиях развития или в ответ на разные условия окружающей среды. Промоторы, которые вызывают экспрессию гена в большинстве типов клеток-хозяев, в большинстве случаев обычно называют «конститутивными промоторами». Постоянно открывают новые промоторы различных типов, используемые в клетках растений; многочисленные примеры можно найти в сборнике Okamuro and Goldberg, Biochemistry of Plants 15: 1-82 (1989). Кроме того, известно, что в связи с тем, что в большинстве случаев точные границы регуляторных последовательностей полностью не определены, фрагменты ДНК с некоторыми изменениями могут обладать идентичной промоторной активностью.
«Интрон» представляет собой промежуточную последовательность в гене, которая не кодирует часть последовательности белка. Таким образом, такие последовательности транскрибируются в РНК, но затем вырезаются и не транслируются. Термин также используют по отношению к вырезанным последовательностям РНК. «Экзон» представляет собой часть генной последовательности, которая транскрибируется и находится в зрелой матричной РНК, полученной с данного гена, но не обязательно часть последовательности, которая кодирует конечный генный продукт.
«Лидерная последовательность трансляции» относится к последовательности ДНК, расположенной между последовательностью промотора гена и кодирующей последовательностью. Лидерная последовательность трансляции присутствует в полностью процессированной мРНК выше стартовой последовательности трансляции. Лидерная последовательность трансляции может влиять на процессинг первичного транскрипта мРНК, стабильность мРНК или эффективность трансляции. Примеры лидерных последовательностей трансляции описаны (Turner, R. and Foster, G. D. (1995) Molecular Biotechnology 3: 225).
Термин «3'-некодирующие последовательности» относится к последовательностям ДНК, расположенным ниже кодирующей последовательности, которые включают в себя узнаваемые последовательности полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, способные влиять на процессинг мРНК или экспрессию генов. Сигнал полиаденилирования обычно характеризуют по влиянию добавления участков полиадениловой кислоты к 3'-концу предшественника мРНК. Примеры использования разных 3'-некодирующих последовательностей приведены в Ingelbrecht et al., Plant Cell 1:671-680 (1989).
«РНК-транскрипт» относится к продукту, получаемому в результате катализируемой РНК-полимеразой транскрипции последовательности ДНК. В том случае, когда РНК-транскрипт является точной комплементарной копией последовательности ДНК, его называют первичным транскриптом, или РНК-транскрипт может представлять собой последовательность РНК, полученную в результате посттранскрипционного процессинга первичного транскрипта, и его называют зрелой РНК. «Матричная РНК (мРНК)» относится к РНК, которая не имеет интронов и которая может быть транслирована клеткой с образованием белка. «кДНК» относится к ДНК, которая комплементарна и синтезируется на матрице мРНК с использованием обратной транскриптазы. кДНК может быть однонитевой или может быть превращена в двунитевую форму с использованием фрагмента Кленова ДНК-полимеразы I. «Смысловая» РНК относится к РНК-транскрипту, который включает мРНК и может быть транслирована с образованием белка в клетке или in vitro. «Антисмысловая РНК» относится к РНК-транскрипту, который комплементарен полному или части первичного транскрипта мишени или мРНК и который блокирует экспрессию гена-мишени (патент США No. 5107065). Комплементарность антисмысловой РНК может быть с любой частью транскрипта конкретного гена, т.е. с 5'-некодирующей последовательностью, 3'-некодирующей последовательностью, интронами или кодирующей последовательностью. «Функциональная РНК» относится к антисмысловой РНК, рибосомной РНК или другой РНК, которая может быть нетранслируемой, но при этом влиять на клеточные процессы. Термины «комплемент» и «обратный комплемент» используют в данном описании взаимозаменяемо по отношению к транскриптам мРНК и предназначены для определения РНК антисмысловой по отношению к матричной РНК.
Термин «эндогенная РНК» относится к любой РНК, которая кодируется любой последовательностью нуклеиновой кислоты, присутствующей в геноме хозяина до трансформации рекомбинантной конструкцией согласно настоящему изобретению, независимо от того, встречается ли она в природе или не встречается в природе, т.е. введена рекомбинантными способами, в результате мутагенеза и т.д.
Термин «не встречающаяся в природе» означает искусственную, не соответствующую тому, что обычно встречается в природе.
Термин «функционально связанный» относится к ассоциации последовательностей нуклеиновых кислот в одном фрагменте нуклеиновой кислоты, так что функция одной последовательности регулируется другой. Например, промотор функционально связан с кодирующей последовательностью, когда он способен регулировать экспрессию данной кодирующей последовательности (т.е. такая кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации. В другом примере комплементарные области РНК согласно изобретению могут быть функционально связаны, либо прямо, либо опосредованно, с 5'-стороны по отношению к мРНК-мишени, или с 3'-стороны по отношению к мРНК-мишени, или в пределах мРНК-мишени, или первая комплементарная область находится в 5'-стороны, а ее комплемент с 3'-стороны по отношению к мРНК-мишени.
Термин «экспрессия» в используемом в данном описании смысле относится к продукции функционального конечного продукта. Экспрессия гена заключается в транскрипции гена и трансляции мРНК с образованием предшественника или зрелого белка. «Антисмысловое ингибирование» относится к продукции антисмысловых транскриптов РНК, способных подавлять экспрессию белка-мишени. «Косупрессия» относится к продукции смысловых РНК-транскриптов, способных подавлять экспрессию идентичных или по существу сходных чужеродных или эндогенных генов (патент США No. 5231020).
«Зрелый» белок относится к посттранскрипционно процессированному полипептиду; т.е. полипептиду, из которого были удалены любые пре- или пропептиды, имеющиеся в первичном продукте трансляции. Белок-«предшественник» относится к первичному продукту трансляции мРНК; т.е. продукту, в котором еще присутствуют пре- и пропептиды. Пре- и пропептиды могут представлять собой без ограничения сигналы внутриклеточной локализации.
«Стабильная трансформация» относится к переносу фрагмента нуклеиновой кислоты в геном организма-хозяина, результатом которого является генетически стабильное наследование. Напротив, «временная трансформация» относится к переносу фрагмента нуклеиновой кислоты в ядро или ДНК-содержащую органеллу организма-хозяина, результатом которого является экспрессия генов без интеграции или стабильного наследования. Организмы-хозяева, содержащие трансформированные фрагменты нуклеиновой кислоты, называют «трансгенными» организмами. Термин «трансформация» в используемом в данном описании смысле относится как к стабильной трансформации, так и к временной трансформации.
Стандартные способы технологии рекомбинантной ДНК и молекулярного клонирования, используемые в данном случае, хорошо известны в данной области и более подробно описаны в Sambrook, J., Fritsch, E. F. и Maniatis, T., Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, 1989 (в дальнейшем «Sambrook»).
Термин «рекомбинант» относится к искусственной комбинации двух в других обстоятельствах отделенных друг от друга участков последовательности, например посредством химического синтеза или в результате обработки изолированных фрагментов нуклеиновых кислот способами генетической инженерии.
«ПЦР» или «полимеразная цепная реакция» представляет собой способ синтеза больших количеств специфичных фрагментов ДНК, состоит из серии повторяющихся циклов (Perkin Elmer Cetus Instruments, Norwalk, CT). Обычно двунитевую ДНК денатурируют при нагревании, два праймера, комплементарных 3'-границам фрагмента-мишени, отжигают при низкой температуре и затем удлиняют при средней температуре. Одну группу указанных трех последовательных стадий называют циклом.
Полимеразная цепная реакция («ПЦР») является эффективным способом, применяемым для амплификации ДНК в миллионы раз посредством многократной репликации матрицы в течение короткого периода времени (Mullis et al., Cold Spring Harbor Symp. Quant. Biol. 51: 263-273 (1986); Erlich et al., заявка на выдачу европейского патента No. 50424; заявка на выдачу европейского патента No. 84796; заявка на выдачу европейского патента No. 258017; заявка на выдачу европейского патента No. 237362; Mullis, заявка на выдачу европейского патента No. 201184; Mullis et al., патент США No. 4683202; Erlich, патент США No. 4582788; и Saiki et al., патент США No. 4683194). В способе используют наборы специфичных синтезированных in vitro олигонуклеотидов, служащих затравкой для синтеза ДНК. Конструирование праймеров зависит от последовательностей ДНК, которые необходимо анализировать. Способ осуществляют в несколько циклов (обычно 20-50), проводя плавление матрицы при высокой температуре, обеспечивая возможность отжига праймеров с комплементарными последовательностями матрицы и затем реплицируя матрицу с помощью ДНК-полимеразы.
Продукты ПЦР-реакций анализируют разделением в агарозных гелях с последующим окрашиванием бромидом этидия и визуализацией при просвечивании УФ. Альтернативно к ПЦР можно добавить радиоактивные dNTP, чтобы включить метку в продукты. В данном случае продукты ПЦР визуализируют экспонированием геля с рентгеновской пленкой. Дополнительное преимущество радиоактивного мечения продуктов ПЦР заключается в том, что можно количественно определить уровни отдельных продуктов амплификации.
Термины «рекомбинантная конструкция», «экспрессирующая конструкция» и «рекомбинантная экспрессирующая конструкция» используют в данном описании взаимозаменяемо. Указанные термины относятся к функциональной единице генетического материала, которая может быть встроена в геном клетки с использованием стандартной методики, хорошо известной специалисту в данной области. Такая конструкция может быть отдельной единицей или может быть использована в соединении с вектором. Если используют вектор, то выбор вектора зависит от способа, который будет использован для трансформации растений-хозяев, которые хорошо известны специалистам в данной области. Например, может быть использована плазмида. Специалисту известны генетические элементы, которые должны присутствовать в векторе, чтобы успешно трансформировать, отбирать и размножать клетки-хозяева, содержащие любой изолированный фрагмент нуклеиновой кислоты согласно изобретению. Специалисту также будет известно, что разные независимые события трансформации приведут к разным уровням и картинам экспрессии (Jones et al., (1985) EMBO J. 4: 2411-2418; De Almeida et al., (1989) Mol. Gen. Genetics 218: 78-86) и что, следовательно, необходимо проводить скрининг множества событий, чтобы получить линии, имеющие требуемый уровень и картину экспрессии. Такой скрининг может быть осуществлен посредством Саузерн-анализа ДНК, Нозерн-анализа экспрессии мРНК, Вестерн-анализа экспрессии белка или фенотипического анализа.
Подразумевается, что термин «моноклональное антитело» в используемом в данном описании смысле относится к одному из препаратов молекул антител, содержащему антитела, которые имеют общую аминокислотную последовательность тяжелой цепи и общую аминокислотную последовательность легкой цепи, в отличие от антитела из препарата «поликлональных» антител, который содержит смесь разных антител. Моноклональное антитела могут быть созданы несколькими новыми способами, подобными фаговому, бактериальному, дрожжевому или рибосомному дисплею, а также классическими способами, например полученные из гибридом антитела (например, антитело, секретируемое гибридомой, полученной с использованием методики гибридом, например стандартной методики гибридом Kohler and Milstein ((1975) Nature 256: 495-497). Таким образом, полученное не из гибридом агонистическое антитело согласно изобретению все еще называют моноклональным антителом, хотя оно может быть получено с использованием неклассической методики.
Подразумевается, что «изолированное антитело» в используемом в данном описании смысле относится к антителу, которое, по существу, не содержит других антител, обладающих другими антигенными специфичностями (например, изолированное антитело, которое специфично связывается с глобуломером, по существу не содержит антител, которые специфично связывают другие антигены, отличные от глобуломера). Однако изолированное антитело, которое специфично связывает глобуломер, может перекрестно взаимодействовать с другими антигенами. Кроме того, изолированное антитело по существу может не содержать другого клеточного материала и/или химических веществ.
Термин «антигенсвязывающая часть» антитела (или просто «часть антитела») в используемом в данном описании смысле относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфично связываться с антигеном. Было показано, что антигенсвязывающая функция антитела может быть осуществлена фрагментами полноразмерного антитела. Такие варианты антител также могут иметь биспецифичную форму, форму с двойной специфичностью или форму с множественной специфичностью; могут специфично связываться с двумя или несколькими разными антигенами. Примеры связывающих фрагментов, охватываемые термином «антигенсвязывающая часть» антитела, включают (i) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, Ch и CH1; (ii) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в области шарнира; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341: 544-546), который содержит один вариабельный домен; и (vi) изолированная область определения комплементарности (CDR). Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть связаны с использованием рекомбинантных способов, синтетическим линкером, который позволяет получать их в виде одной белковой цепи, в которой области VL и VH спарены с образованием формы моновалентных молекул (известных как одноцепочечный Fv (scFv); см., например Bird et al. (1988) Science 242: 423- 426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA, 85: 5879-5883). Подразумевается, что такие одноцепочечные антитела также охватываются термином «антигенсвязывающая часть» антитела. Термин также охватывает другие формы одноцепочечных антител, такие как диантитела. Диантитела являются бивалентными, биспецифичными антителами, в которых домены VH и VL экспрессированы в виде одной полипептидной цепи, но с использованием линкера, который слишком короток, чтобы позволить спариваться двум доменам на одной и той же цепи, тем самым вынуждая домены спариваться с комплементарными доменами другой цепи с образованием при этом двух антигенсвязывающих сайтов (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90: 6444-6448; Poljak, R.J., et al. (1994) Structure 2: 1121-1123). Такие связывающие части антител известны в данной области (Kontermann and Dubel eds., Antibody Engineering (2001) Springer-Verlag. New York. 790 pp. (ISBN 3-540-41354-5).
Кроме того, антитело или его антигенсвязывающая часть может быть частью более крупных иммуноадгезионных молекул, образованных ковалентным или нековалентным связыванием антитела или части антитела с одним или несколькими другими белками или пептидами. Примеры таких иммуноадгезионных молекул включают использование коровой области стрептавидина для получения тетрамерной молекулы scFv (Kipriyanov, S.M., et al. (1995) Human Antibodies and Hybridomas 6: 93-101) и использование остатка цистеина, маркерного пептида и C-концевой полигистидиновой метки для получения бивалентных и биотинилированных молекул scFv (Kipriyanov, S. M., et al. (1994) Mol. Immunol. 31: 1047-1058). Части антител, такие как Fab- и F(ab')2-фрагменты, могут быть получены из целых антител с применением обычных способов, таких как расщепление целых антител папаином или пепсином соответственно. Кроме того, антитела, части антител и иммуноадгезионные молекулы могут быть получены с применением стандартной методики рекомбинантной ДНК, которая описана в данной публикации.
Подразумевается, что термин «рекомбинантное антитело человека» в используемом в данном описании смысле включает все антитела человека, которые получены, экспрессированы, созданы или изолированы способами рекомбинации, такие как антитела, экспрессированные с использованием рекомбинантного экспрессирующего вектора, трансфицированного в клетку-хозяина, антитела, выделенные из рекомбинантной комбинаторной библиотеки антител человека (Hoogenboom H. R., (1997) TIB Tech. 15:62-70; Azzazy H., and Highsmith W. E., (2002) Clin. Biochem. 35: 425-445; Gavilondo J.V., and Larrick J. W. (2002) BioTechniques 29: 128-145; Hoogenboom H., and Chames P. (2000) Immunology Today 21: 371-378), антитела, выделенные из организма животного (например, мыши), который является трансгенным по генам иммуноглобулина человека (см., например Taylor, L. D., et al. (1992) Nucl. Acids Res. 20: 6287-6295; Kellermann S-A., and Green L. L. (2002) Current Opinion in Biotechnology 13: 593-597; Little M. et al. (2000) Immunology Today 21: 364-370), или антитела, полученные, экспрессированные, созданные или изолированные любыми другими способами, которые заключаются в сплайсинге последовательностей гена иммуноглобулина человека в другие последовательности ДНК. Такие рекомбинантные антитела человека имеют вариабельные и константные области, полученные из последовательностей иммуноглобулинов зародышевой линии. Однако в некоторых вариантах такие рекомбинантные антитела человека подвергают мутагенезу in vitro (или когда используют животное, трансгенное по последовательностям Ig человека, соматическому мутагенезу in vivo), и, таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител представляют собой последовательности, которые, несмотря на то что получены и являются родственными последовательностям VH и VL человека зародышевой линии, могут не существовать в природе в репертуаре антител человека зародышевой линии in vivo (см. также Kabat et al. Sequences Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242, 1991). Однако антитела человека согласно настоящему изобретению могут содержать аминокислотные остатки, не кодируемые последовательностями иммуноглобулинов человека зародышевой линии (например, мутации, введенные посредством случайного или сайт-специфичного мутагенеза in vitro или посредством соматических мутаций in vivo). (См. также Harlow and Lane, Antibodies: A Laboratory Manual, New York: Gold Spring Harbor Press, 1990.)
Термин «химерное антитело» относится к антителам, которые содержат последовательности вариабельных областей тяжелой и легкой цепи от одного вида и последовательности константой области из других видов, таким как антитела, имеющие вариабельные области тяжелой и легкой цепей мышей, связанные с константными областями человека.
Термин «CDR-привитое антитело» относится к антителам, которые содержат последовательности вариабельных областей тяжелой и легкой цепей одного вида, но в которых последовательности одной или нескольких CDR-областей VH и/или VL заменены последовательностями CDR другого вида, таким как антитела, имеющие вариабельные области тяжелой и легкой цепи мыши, в которых одна или несколько CDR мыши (например, CDR3) были заменены последовательностями CDR человека.
Рекомбинантные антитела человека согласно настоящему изобретению имеют вариабельные области и также могут содержать константные области, полученные из последовательностей иммуноглобулинов человека зародышевой линии (см. Kabat et al. (1991), выше). Однако в некоторых вариантах такие рекомбинантные антитела человека подвергают мутагенезу in vitro (или, когда используют животное, трансгенное по последовательностям Ig человека, соматическому мутагенезу in vivo), и, таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител представляют собой последовательности, которые, несмотря на то, что они получены и являются родственными последовательностям VH и VL человека зародышевой линии, могут не существовать в природе в репертуаре антител человека зародышевой линии in vivo. Однако в некоторых вариантах такие рекомбинантные антитела являются результатом избирательного мутагенеза, или обратного мутирования, или и того и другого.
Термин «обратное мутирование» относится к способу, при котором некоторые или все соматически мутированные аминокислоты антитела человека заменены соответствующими остатками зародышевого типа из гомологичной последовательности антитела зародышевой линии. Последовательности тяжелой и легкой цепи антитела человека согласно изобретению выравнивают по отдельности с последовательностями зародышевой линии в базе данных VBASE, чтобы идентифицировать последовательности с наиболее высокой гомологией. VBASE представляет собой полный справочник всех последовательностей вариабельных областей зародышевой линии человека, собранных на основании опубликованной информации о последовательностях, включая текущие выпуски библиотек данных GenBank и EMBL. База данных разработана в MRC Centre for Protein Engineering (Cambridge, UK) в виде депозитария секвенированных генов антител человека (страница Интернета: http://www.mrc-cpe.cam.ac.uk/vbase-intro.php?menu=901). Отличия, имеющиеся в антителе человека согласно изобретению, возвращают к последовательности зародышевого типа путем мутирования нуклеотидов в определенных положениях, кодирующих такие отличающиеся аминокислоты. Роль каждой аминокислоты, идентифицированной таким образом в качестве кандидата для обратного мутирования, необходимо исследовать в отношении прямой или опосредованной роли в связывании антигена, и любую аминокислоту, которая, как обнаружено, после мутирования влияет на какое-либо требуемое свойство антитела человека, необходимо включить в конечное антитело человека. Чтобы минимизировать количество аминокислот, подвергаемых обратному мутированию, аминокислоты в тех положениях, в которых они, как было обнаружено, отличаются от наиболее близкой последовательности зародышевой линии, но идентичны соответствующей аминокислоте во второй последовательности зародышевой линии, могут быть оставлены при условии, что вторая последовательность зародышевой линии идентична и коллинеарна последовательности антитела человека согласно изобретению на протяжении, по меньшей мере 10, предпочтительно 12 аминокислот с обеих сторон рассматриваемой аминокислоты. Обратное мутирование может осуществляться на любой стадии оптимизации антител.
«Меченый связывающий белок» представляет собой белок, в котором антитело или часть антитела согласно изобретению дериватизованы или связаны с другой функциональной молекулой (например, другим пептидом или белком). Например, меченый связывающий белок согласно изобретению может быть дериватизован функциональным связыванием антитела или части антитела согласно изобретению (посредством химического связывания, генетического слияния, нековалентного связывания или иным образом) с одной или несколькими другими компонентами молекулы, такими как другое антитело (например, биспецифичное антитело или диантитело), регистрируемый агет, цитотоксический агент, фармацевтическое средство и/или белок или пептид, который может опосредовать ассоциацию антитела или части антитела с другой молекулой (такой как область кора стрептавидина или полигистидиновая метка).
В целях настоящего изобретения «гликозилированный связывающий белок» относится к белку в том случае, когда антитело или его антигенсвязывающая часть содержит один или несколько углеводных остатков. Продуцированный in vivo белок может подвергаться дальнейшему процессингу, называемому посттрансляционной модификацией. В частности, остатки сахаров (гликозильные остатки) могут быть добавлены ферментативно, способом, называемым гликозилированием. Полученные в результате белки, несущие ковалентно связанные боковые олигосахаридные цепи, называют гликозилированными белками или гликобелками. Антитела являются гликобелками с одним или несколькими углеводными остатками в Fc-домене, а также в вариабельном домене. Углеводные остатки в Fc-домене оказывают важное влияние на эффекторную функцию Fc-домена, с минимальным влиянием на связывание антигена или время полужизни антитела (R. Jefferis, Biotechnol. Prog. 21 (2005), pp. 11-16). Напротив, гликозилирование вариабельного домена может оказывать влияние на антигенсвязывающую активность антитела. Гликозилирование вариабельного домена может оказывать негативное влияние на аффинность связывания антитела, вероятно, вследствие стерических препятствий (Co, M.S., et al., Mol. Immunol. (1993) 30: 1361- 1367) или приводит к повышенной аффинности по отношению к антигену (Wallick, S. C, et al., Exp. Med. (1988) 168: 1099-1109; Wright, A., et al., EMBO J. (1991) 10: 2717 2723). Кроме того, могут быть получены мутанты по сайту гликозилирования, в которых мутирован сайт O- или N-связанного гликозилирования связывающего белка. Специалист в данной области может создать такие мутанты, используя стандартные хорошо известные способы. Также предполагаются мутанты по сайту гликозилирования, которые сохраняют биологическую активность, но имеют повышенную или пониженную активность связывания.
Кроме того, гликозилирование антитела или антигенсвязывающей части согласно изобретению может быть модифицировано. Например, может быть получено агликозилированное антитело (т.е. антитело, без гликозилирования). Гликозилирование может быть изменено, например, чтобы повысить аффинность антитела по отношению к антигену. Такие углеводные модификации могут быть осуществлены, например, посредством изменения одного или нескольких сайтов гликозилирования в последовательности антитела. Например, может быть получена одна или несколько аминокислотных замен, которые приводят к элиминации одного или нескольких сайтов гликозилирования вариабельной области, чтобы исключить гликозилирование в данном сайте. Такое гликозилирование может увеличивать аффинность антитела по отношению к антигену. Такой способ описан более подробно в публикации международной заявки на выдачу патента No. WO 03/016466A2
и патентах США No. 5714350 и 6350861, которые включены в данное описание в виде ссылки в полном объеме.
Дополнительно или альтернативно может быть получено модифицированное антитело, которое имеет измененный тип гликозилирования, например гипофукозилированное антитело, имеющее меньшее количество остатков фукозила, или антитело, имеющее повышенное количество разветвляющихся структур GlcMAc. Показано, что такие измененные картины гликозилирования увеличивают ADCC-способность антитела. Такие углеводные модификации могут быть осуществлены, например, посредством экспрессии антитела в клетке-хозяине с измененным аппаратом гликозилирования. Клетки с измененным аппаратом гликозилирования, описанные в данной области, могут быть использованы в качестве клеток-хозяев, чтобы экспрессировать в них рекомбинантные антитела согласно изобретению и таким образом получить антитело с измененным гликозилированием (см., например, Shields, R. L. et al. (2002) J. Biol. Chem. 277: 26733-26740; Umana et al. (1999) Nat. Biotech. 17: 176-1, а также в Европейском патенте No: EP 1176195; публикациях международных заявок на выдачу патента WO 03/035835 и WO 99/5434280, которые включены в данное описание в виде ссылки в полном объеме).
Гликозилирование белка зависит от аминокислотной последовательности представляющего интерес белка, а также клетки-хозяина, в которой белок экспрессируется. Разные организмы могут продуцировать разные ферменты гликозилирования (например, гликозилтрансферазы и гликозидазы) и имеют разные доступные субстраты (сахара нуклеотидов). Вследствие таких факторов картина гликозилирования белков и состав остатков гликозила могут различаться в зависимости от системы хозяина, в которой экспрессируется конкретный белок. Гликозильные остатки, применимые в изобретении, могут включать без ограничения глюкозу, галактозу, маннозу, фукозу, N-ацетилглюкозамин и сиаловую кислоту. Предпочтительно гликозилированный связывающий белок содержит такие гликозильные остатки, чтобы картина гликозилирования представляла собой картину гликозилирования у человека.
Специалистам в данной области известно, что разное гликозилирование белков может приводить к разным свойствам белков. Например, эффективность терапевтического белка, продуцируемого в хозяине-микроорганизме, таком как дрожжи, и гликозилированного с использованием эндогенного пути дрожжей, может быть снижена по сравнению с эффективностью такого же белка, экспрессированного в клетках млекопитающих, таких как линия клеток CHO. Такие гликобелки также могут быть иммуногенными в организме человека и иметь уменьшенное время полужизни in vivo после введения. Специфичные рецепторы человека и других животных могут узнавать специфичные гликозильные остатки и стимулировать быстрый клиренс белка из кровообращения. Другие неблагоприятные эффекты могут включать изменения фолдинга белка, растворимости, чувствительности к протеазам, передвижения, транспорта, компартментализации, секреции, узнавания другими белками или факторами, антигенности или аллергенности. Соответственно, практик может предпочесть терапевтический белок со специфичным составом и картиной гликозилирования, например составом и картиной гликозилирования, идентичными или по меньшей мере сходными с составом и картиной гликозилирования, возникающими в клетках человека или в видоспецифичных клетках конкретного животного.
Экспрессия гликозилированных белков, отличных от белков клетки-хозяина, может быть осуществлена посредством генетической модификации клетки-хозяина так, чтобы она экспрессировала гетерологичные ферменты гликозилирования. Применяя способы, известные в данной области, специалист может создать антитела или их антигенсвязывающие части, имеющие гликозилирование белка как в организме человека. Например, штаммы дрожжей были генетически модифицированы для экспрессии не встречающихся в природе ферментов гликозилирования, так что гликозилированные белки (гликобелки), продуцируемые в таких штаммах дрожжей, имеют гликозилирование белка, идентичное гликозилированию в клетках животных, в частности в клетках человека (публикации заявок на выдачу патента США No. 20040018590 и 20020137134 и публикации международной заявки No. WO 05/100584 A2).
Кроме того, специалисту в данной области будет понятно, что представляющий интерес белок может быть экспрессирован с использованием библиотеки клеток-хозяев, генетически сконструированных для экспрессии различных ферментов гликозилирования, так что представители клеток-хозяев в библиотеке продуцируют представляющий интерес белок с разными вариантами гликозилирования. Затем специалист может отобрать и выделить представляющий интерес белок с конкретными новыми картинами гликозилирования. Предпочтительно белок, имеющий особую выбранную новую картину гликозилирования, обладает улучшенными или измененными биологическими свойствами.
Изобретение также относится к способу получения моноклональных антител согласно изобретению из организма животных, отличных от человека и отличных от мыши, с использованием иммунизации отличных от человека трансгенных животных, которые имеют локусы иммуноглобулина человека. Таких животных можно получить, используя способы, известные в данной области. В предпочтительном варианте отличным от человека животным могут быть крысы, овцы, свиньи, козы, крупный рогатый скот или лошади. Продуцирующие антитела иммортализованные гибридомы могут быть получены от иммунизированного животного. После иммунизации животного умерщвляют и B-клетки селезенки сливают с иммортализованными клетками миеломы, как хорошо известно в данной области. См., например, Harlow and Lane, выше. В предпочтительном варианте клетки миеломы не секретируют полипептиды иммуноглобулина (не секретирующая линия клеток). После слияния и селекции с использованием антибиотиков проводят скрининг гибридом, используя антиген (например, глобуломер), или его часть, или клетку, экспрессирующую представляющий интерес антиген. В предпочтительном варианте начальный скрининг осуществляют, используя твердофазный иммуноферментный анализ (ELISA) или радиоиммуноанализ (РИА), предпочтительно ELISA. Пример скрининга с помощью ELISA предлагается в публикации международной заявки No. WO 00/37504, включенной в данное описание в виде ссылки.
Продуцирующие антитела гибридомы отбирают, клонируют и проводят дополнительный скрининг в отношении требуемых характеристик, включая устойчивый рост гибридом, высокую продукцию антител и требуемые свойства антител, которые осуждаются ниже. Гибридомы можно культивировать и размножить in vivo в сингенных животных, в животных, у которых отсутствует иммунная система, например мышах nude, или в культуре клеток in vitro. Способы селекции, клонирования и размножения гибридом хорошо известны специалистам в данной области. Предпочтительно иммунизированным животным является животное, отличное от человека, которое экспрессирует гены иммуноглобулина человека, и B-клетки селезенки сливают с миеломой, полученной из того же вида животного, отличного от человека.
В одном аспекте изобретение относится к гибридомам, которые продуцируют моноклональные антитела, используемые для лечения, диагностики и профилактики болезни Альцгеймера. В предпочтительном варианте гибридомы являются мышиными гибридомам. В другом предпочтительном варианте гибридомы получают, используя вид, отличный от человека и отличный от мыши, такой как крысы, овцы, свиньи, козы, крупный рогатый скот или лошади. В другом варианте гибридомы являются гибридомами человека, в которых не секретирующая миелома человека слита с клеткой человека, экспрессирующей антитело против глобуломера.
Рекомбинантные антитела могут быть получены из одиночных изолированных лимфоцитов с использованием способа, называемого в данной области способом получения антител из селектируемых лимфоцитов (SLAM), который описан в патенте США No. 5627052, публикации международной заявки на выдачу патента No. WO 92/02551 и в Babcock, J. S. et al. (1996) Proc. Natl. Acad. Sci. USA 93: 7843-7848. В указанном способе отдельные клетки, секретирующие представляющие интерес антитела (например, лимфоциты, полученные от иммунизированного животного), подвергают скринингу, используя анализ антигенспецифичных гемолитических бляшек, в котором антиген (например, глобуломер) или его фрагмент связывают с эритроцитами барана, используя линкер, такой как биотин, и применяют для идентификации отдельных клеток, которые секретируют антитела, специфичные по отношению к антигену. После идентификации представляющих интерес секретирующих антитела клеток кДНК вариабельных областей тяжелой и легкой цепи «спасают» из клеток посредством ПЦР с обратной транскриптазой, и указанные вариабельные области затем могут быть экспрессированы в контексте подходящих константных областей иммуноглобулина (например, константных областей человека) в клетках-хозяевах млекопитающих, таких как клетки COS или CHO. Клетки-хозяева, трансфицированные амплифицированными последовательностями иммуноглобулина, полученными из отобранных in vivo лимфоцитов, затем могут быть подвергнуты дополнительному анализу и селекции in vitro, например пэннингом трансфицированных клеток, чтобы выделить клетки, экспрессирующие антитела к IL-18. Амплифицированные последовательности иммуноглобулина затем могут быть подвергнуты обработке in vitro, например, способами обеспечения созревания аффинности in vitro, такими как способы, описанные в публикации международной заявки No. WO 97/29131 и публикации международной заявки No. WO 00/56772.
Термин «химерное антитело» относится к антителам, которые содержат последовательности вариабельных областей тяжелой и легкой цепи одного вида и последовательности константных областей другого вида, таким как антитела, имеющие вариабельные области тяжелой и легкой цепи мыши, связанные с константными областями человека.
Термин «CDR-привитое антитело» относится к антителам, которые содержат последовательности вариабельных областей тяжелой и легкой цепи одного вида, но в которых последовательности одной или нескольких CDR-областей VH и/или VL заменены последовательностями CDR другого вида, таким как антитела, имеющие вариабельные области тяжелой и легкой цепи мыши, в которых одна или несколько CDR мыши (например, CDR3) были заменены последовательностями CDR человека.
Термин «гуманизированное антитело» относится к антителам, которые содержат последовательности вариабельных областей тяжелой и легкой цепи вида, отличного от человека (например, мыши), но в которых по меньшей мере часть последовательности VH и/или VL была изменена так, чтобы она была более «подобной человеческой», т.е. более сходной с вариабельными последовательностями человека зародышевой линии. Одним из типов гуманизированного антитела является CDR-привитое антитело, в котором последовательности CDR человека введены в последовательности VH и VL вида, отличного от человека, чтобы заменить соответствующие последовательности CDR вида, отличного от человека. В частности, термин «гуманизированное антитело» означает антитело или его вариант, производное, аналог или фрагмент, который иммуноспецифично связывается с представляющим интерес антигеном и который содержит каркасную область (FR), по существу имеющую аминокислотную последовательность антитела человека, и определяющую комплементарность область (CDR), по существу имеющую аминокислотную последовательность антитела отличного от человека вида. В используемом в данном описании смысле термин «по существу» в контексте CDR относится к CDR, имеющей аминокислотную последовательность по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% идентичную аминокислотной последовательности CDR антитела вида, отличного от человека. Гуманизированное антитело содержит по меньшей мере один и обычно два вариабельных домена (по существу любые из Fab, Fab', F(ab')2, FabC, Fv), в которых все или по существу все области CDR соответствуют CDR-областям иммуноглобулина отличного от человека вида (т.е. донорного антитела), и все или по существу все каркасные области являются каркасными областями консенсусной последовательности иммуноглобулина человека. Предпочтительно гуманизированное антитело также содержит по меньшей мере часть константной области иммуноглобулина (Fc), обычно Fc иммуноглобулина человека. В некоторых вариантах гуманизированное антитело содержит как легкую цепь, так и по меньшей мере вариабельный домен тяжелой цепи. Антитело также может содержать области CH1, шарнира, CH2, CH3 и CH4 тяжелой цепи. В некоторых вариантах гуманизированное антитело содержит только гуманизированную легкую цепь. В других вариантах гуманизированное антитело содержит только гуманизированную тяжелую цепь. В конкретных вариантах гуманизированное антитело содержит только гуманизированный вариабельный домен легкой цепи и/или гуманизированную тяжелую цепь.
Гуманизированное антитело может быть выбрано из любого класса иммуноглобулинов, включая IgM, IgG, IgD, IgA и IgE, и любого изотипа, включая без ограничения IgG1, IgG2, IgG3 и IgG4. Гуманизированное антитело может содержать последовательности более чем из одного класса или изотипа, и могут быть выбраны конкретные константные домены, чтобы оптимизировать требуемые эффекторные функции, с использованием способов, хорошо известных в данной области.
Каркасные области и области CDR гуманизированного антитела не должны точно соответствовать исходным последовательностям, например, CDR донорного антитела или консенсусный каркас могут быть подвергнуты мутагенезу путем замены, инсерции и/или делеции по меньшей мере одного аминокислотного остатка, так что остаток CDR или каркаса в данном сайте не соответствует донорному антителу или консенсусному каркасу. Однако в предпочтительном варианте такие мутации не будут обширными. Обычно по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% остатков гуманизированного антитела будут соответствовать остаткам исходных последовательностей FR и CDR. В используемом в данном описании смысле термин «консенсусный каркас» относится к каркасной области в консенсусной последовательности иммуноглобулина. Кроме того, в используемом в данном описании смысле термин «консенсусная последовательность иммуноглобулина» относится к последовательности, образованной из наиболее часто встречающихся аминокислот (или нуклеотидов) в семействе родственных последовательностей иммуноглобулина (см., например, Winnaker, From Genes to Clones (Verlagsgesellschaft, Weinheim, Germany 1987). В семействе иммуноглобулинов каждое положение в консенсусной последовательности занято аминокислотой, встречающейся наиболее часто в данном положении в семействе. Если две аминокислоты встречаются одинаково часто, то любая из них может быть включена в консенсусную последовательность.
Термин «активность» включает такие активности, как специфичность/аффинность связывания антитела по отношению к антигену.
Термин «эпитоп» включает любую полипептидную детерминанту, способную к специфичному связыванию с иммуноглобулином или T-клеточным рецептором. В некоторых вариантах эпитопные детерминанты включают химически активные поверхностные группировки молекул, таких как аминокислоты, боковые цепи сахаров, фосфорил или сульфонил, и в некоторых вариантах могут специфичные трехмерные структурные параметры и/или специфичные параметры заряда. Эпитоп представляет собой область антигена, которая связывается антителом. В некоторых вариантах говорят, что антитело специфично связывает антиген, когда оно предпочтительно узнает свой антиген-мишень в сложной смеси белков и/или макромолекул.
Термин «резонанс поверхностного плазмона» в используемом в данном описании смысле относится к оптическому явлению, которое позволяет проводить анализ биоспецифичных взаимодействий в режиме реального времени посредством выявления изменений в концентрациях белков в биосенсорной матрице, например, используя систему BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, NJ). Дополнительные описания см. в Jonsson, U., et al. (1993) Ann. Biol. Clin. 51: 19-26; Jonsson, U., et al. (1991) Biotechnigues 11: 620-627; Johnsson, B., et al. (1995) J. Mol. Recognit. 8: 125-131; и Johnnson, B., et al. (1991) Anal. Biochem. 198: 268-277.
Подразумевается, что термин «Kon» в используемом в данном описании смысле относится к константе скорости ассоциации антитела с антигеном с образованием комплекса антитело/антиген, которая известна в данной области.
Подразумевается, что термин «Koff» в используемом в данном описании смысле относится к константе скорости диссоциации антитела из комплекса антитело/антиген, которая известна в данной данной области.
Подразумевается, что термин «Kd» в используемом в данном описании смысле относится к «константе диссоциации» конкретного взаимодействия антитело-антиген, которая известна в данной области.
Термин «меченый связывающий белок» в используемом в данном описании смысле относится к белку с введенной меткой, которая обеспечивает идентификацию связывающего белка. Предпочтительно метка представляет собой регистрируемый маркер, например введение радиоактивно меченой аминокислоты или связывание с полипептидом остатков биотинила, которые могут быть выявлены с использованием меченого авидина (например, стрептавидин, содержащий флуоресцирующий маркер или ферментативную активность, которые могут быть выявлены оптическими или колориметрическими способами). Примеры меток полипептидов включают без ограничения следующие метки: радиоизотопы или радионуклиды (например, 3H, 14C, 35S, 90Y, 99Tc, 111In, 125I, 131I, 177Lu, 166Ho или 153Sm); флуоресцирующие метки (например, ФИТЦ, родамин или лантанидные люминоформы), ферментативные метки (например, пероксидаза хрена, люцифераза или щелочная фосфатаза); хемилюминесцентные маркеры; группы биотинила; предварительно определяемые полипептидные эпитопы, узнаваемые вторым репортером (например, парные последовательности лейциновой молнии, сайты связывания второго антитела, связывающие металлы домены или эпитопные метки); и магнитные средства, такие как хелаты гадолиния.
Термин «конъюгат антитела» относится к связывающему белку, такому как антитело, химически связанному со вторым химическим остатком, таким как терапевтическое или цитотоксическое средство. Термин «средство» используют в данном описании для обозначения химического соединения, смеси химических соединений, биологической макромолекуле или экстракту, полученному из биологических материалов. Предпочтительно терапевтическое или цитотоксическое средство включает без ограничения коклюшный токсин, таксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, а также аналоги и гомологи указанных средств.
Термины «кристалл» и «кристаллизованный» в используемом в данном описании смысле относится к антителу или его антигенсвязывающей части, которые существуют в форме кристалла. Кристаллы представляют собой одну из форм твердого состояния вещества.
Термин «иммунизировать» относится в данном описании к способу презентирования антигена иммунному репертуару, при этом такой репертуар существует в природном генетически неизмененном организме или трансгенном организме, модифицированном так, чтобы иметь искусственный иммунный репертуар человека. Подобным образом «иммуногенный препарат» представляет собой препарат антигена, который содержит адъюванты или другие добавки, которые могут усиливать иммуногенность антигена. Примером может быть совместная инъекция очищенной формы рецептора GLP-1 с полным адъювантом Фрейнда мыши. «Гипериммунизация», как определено в данном описании, представляет собой осуществление серии многократных презентаций антигена в иммуногенном препарате организму животного-хозяина с целью развития сильного иммунного ответа.
Одним из способов измерения кинетики связывания антитела является резонанс поверхностного плазмона. Термин «резонанс поверхностного плазмона» в используемом в данном описании смысле относится к оптическому явлению, которое позволяет проводить анализ биоспецифичных взаимодействий в режиме реального времени посредством выявления изменений в концентрациях белков в биосенсорной матрице, например, используя систему BIAcore (Biacore International, Uppsala, Sweden and Piscataway, NJ). Дополнительные описания см. в Jonsson, U., et al. (1993) Annales de Biologie Clinique (Paris) 51: 19-26; Jonsson et al. (1991) Biotechniques 11: 620-627; Johnsson et al. (1995) Journal of Molecular Recognition 8: 125-131; и Johnnson et al. (1991) Analytical Biochemistry 198: 268-277.
«Фармацевтически приемлемый носитель» включает любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, средства для изотоничности и средства, замедляющие всасывание, и тому подобные, которые являются физиологически совместимыми. Примеры фармацевтически приемлемых носителей включают один или несколько из следующих носителей: воду, физиологический раствор, фосфатно-солевой буфер, декстрозу, глицерин, этанол и тому подобные, а также их комбинации. Во многих случаях предпочтительно введение средств для изотоничности, например сахаров, полиспиртов, таких как маннит, сорбит, или хлорида натрия в композиции. Фармацевтически приемлемые носители дополнительно могут содержать минорные количества вспомогательных веществ, таких как увлажняющие или эмульгирующие средства, консерванты или буферы, которые увеличивают срок хранения или эффективность антитела или части антитела.
Фармацевтические композиции согласно изобретению могут содержать «терапевтически эффективное количество» или «профилактически эффективное количество» антитела или части антитела согласно изобретению. «Терапевтически эффективное количество» относится к количеству, которое в необходимых дозах в течение необходимого периода времени эффективно для получения требуемого терапевтического результата. Терапевтически эффективное количество антитела или части антитела может быть определено специалистом в данной области и может варьировать в зависимости от таких факторов, как состояние болезни, возраст, пол и масса пациента и способность антитела или части антитела вызывать требуемый ответ у пациента. Терапевтически эффективное количество также представляет собой такое количество, при котором любые токсические или вредные эффекты антитела или части антитела перевешиваются терапевтически полезными эффектами. «Профилактически эффективное количество» относится к количеству, которое в необходимых дозах и в течение необходимых периодов времени эффективно для достижения требуемого профилактического результата. Обычно в связи с тем, что профилактическую дозу используют для пациентов до заболевания или на более ранней стадии заболевания, профилактически эффективное количество будет меньше, чем терапевтически эффективное количество.
Антитела и части антител согласно изобретению могут быть введены в фармацевтическую композицию, подходящую, например, для парентерального введения. Предпочтительно антитело или части антител будут приготовлены в виде инъекционного раствора, содержащего 0,1-250 мг/мл антитела. Инъекционный раствор может быть представлен либо в виде жидкой, либо в виде лиофилизованной формы во флаконе из бесцветного или желтого стекла, ампуле или предварительно заполненном шприце. Буфер может содержать L-гистидин (1-50 мМ), оптимально 5-10 мМ, pH 5,0-7,0 (оптимально pH 6,0). Другие подходящие буферы включают без ограничения сукцинат натрия, цитрат натрия, фосфат натрия или фосфат калия. Можно использовать хлорид натрия, чтобы модифицировать токсичность раствора, в концентрации 0-300 мМ (оптимально 150 мМ в случае жидкой дозированной формы). В случае лиофилизированной лекарственной формы могут быть включены криопротекторы, в большинстве случаев 0-10% сахарозы (оптимально 0,5-1,0%). Другие подходящие криопротекторы включают трегалозу и лактозу. В случае лиофилизированной лекарственной формы могут быть включены наполнители, в большинстве случаев 1-10% маннита (оптимально 2-4%). Стабилизаторы могут быть использованы как в жидких, так и в лиофилизированных лекарственных формах, в большинстве случаев 1-50 мМ L-метионин (оптимально 5-10 мМ). Другие подходящие наполнители включают глицин, аргинин, могут быть включены в виде 0-0,05% полисорбата-80 (оптимально 0,005-0,01%). Дополнительные поверхностно-активные вещества включают без ограничения полисорбат 20 и поверхностно-активные вещества BRIJ.
Композиции согласно настоящему изобретению могут быть в различных формах. Формы включают, например, жидкие, полутвердые и твердые дозированные формы, например жидкие растворы (например, инъекционные растворы и растворы для инфузии), дисперсии или суспензии, таблетки, пилюли, порошки, липосомы и суппозитории. Предпочтительная форма зависит от предполагаемого способа введения и терапевтического применения. Типичные предпочтительные композиции имеют форму инъекционных растворов или растворов для инфузии, таких как композиции, сходные с композициями, применяемыми для пассивной иммунизации человека другими антителами. Предпочтительным способом введения является парентеральный (например, внутривенный, подкожный, внутрибрюшинный, внутримышечный). В предпочтительном варианте антитело вводят посредством внутривенной инфузии или инъекции. В другом предпочтительном варианте антитело вводят посредством внутримышечной или подкожной инъекции.
Терапевтические композиции обычно должны быть стерильными и стабильными в условиях производства и хранения. Композиция может быть приготовлена в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Стерильные инъекционные растворы могут быть приготовлены добавлением активного соединения (т.е. антитела или части антитела) в требуемом количестве в подходящий растворитель с одним или с комбинацией необходимых ингредиентов, перечисленных выше, с последующей стерилизацией фильтрованием. В общем, дисперсии готовят добавлением активного соединения в стерильный наполнитель, который содержит основную дисперсионную среду и необходимые другие ингредиенты из тех ингредиентов, которые перечислены выше. В случае стерильных лиофилизированных порошков для приготовления стерильных инъекционных растворов предпочтительными способами приготовления являются вакуумная сушка и сушка распылением, которые дают порошок активного ингредиента плюс любой требуемый дополнительный ингредиент из его предварительно стерильно профильтрованного раствора. Нужную текучесть раствора можно поддерживать, например, используя покрытие, такое как лецитин, поддерживая требуемый размер частиц в случае дисперсии и используя поверхностно-активные вещества. Инъекционные композиции длительного всасывания могут быть получены включением в композицию средства, которое замедляет всасывание, например моностеаратных солей и желатина.
Антитела и части антител согласно настоящему изобретению могут быть введены различными способами, известными в данной области, хотя в случае многих терапевтических применений предпочтительным путем/способом введения является подкожная инъекция, внутривенная инъекция или инфузия. Как будет понятно специалисту в данной области, путь и/или способ введения будет варьировать в зависимости от требуемых результатов. В некоторых вариантах активное соединение может быть приготовлено с носителем, который будет защищать соединение от быстрого высвобождения, например в виде препарата контролируемого высвобождения, включая имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Можно использовать биодеградируемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Многие способы приготовления таких препаратов запатентованы или, в общем, известны специалистам в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
В некоторых вариантах антитело или часть антитела согласно изобретению могут быть введены перорально, например, с инертным разбавителем или усвояемым съедобным носителем. Соединение (и другие ингредиенты, если они необходимы) также могут быть заключены в желатиновую капсулу с твердой или мягкой оболочкой, спрессованы в таблетки или непосредственно введены в пищу пациента. В случае перорального терапевтического введения соединения могут быть включены с эксципиентами и использованы в форме принимаемых внутрь таблеток, буккальных таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, облаток и тому подобного. Чтобы ввести соединение согласно изобретению другим путем, отличным от парентерального введения, может быть необходимо покрывание соединения или совместное введение соединения с веществом, которое предотвращает его инактивацию.
В композицию также могут быть введены дополнительные активные соединения. В некоторых вариантах антитело или часть антитела согласно изобретению готовят вместе в одном препарате и/или вводят совместно с одним или несколькими дополнительными терапевтическими средствами, которые применимы для лечения болезни Альцгеймера или родственных заболеваний или состояний. Например, одно из антител согласно настоящему изобретению или часть такого антитела можно приготовить в одном препарате и/или ввести вместе с одним или несколькими дополнительными антителами, которые связывают другие мишени.
В некоторых вариантах моноклональное антитело согласно настоящему изобретению или его фрагмент могут быть связаны со средством, увеличивающим время полужизни, известным в данной области. Такие средства включают без ограничения Fc-домен, полиэтиленгликоль и декстран. Такие средства описаны, например, в заявке на выдачу патента США с регистрационным No. 09/428082 и в опубликованной заявке PCT No. WO 99/25044, которые включены в данное описание в виде ссылки для любых целей.
Кроме обсуждаемых выше способов специалистам известны стандартные источники, в которых описаны конкретные условия и способы конструирования, обработки и выделения макромолекул (например, молекул ДНК, плазмид, и т.д.), создания рекомбинантных организмов и скрининга и выделения клонов (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press (1989); Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor Press (1995); Birren et al., Genome Analysis: Detecting Genes, 1, Cold Spring Harbor, New York (1998); Birren et al., Genome Analysis: Analyzing DNA, 2, Cold Spring Harbor, New York (1998); Plant Molecular Biology: A Laboratory Manual, eds. Clark, Springer, New York (1997)).
Применения моноклонального антитела
Моноклональные антитела согласно настоящему изобретению (например, 8F5 и 8CF) имеют множество интересных применений. Например, моноклональные антитела могут быть использованы для профилактики, лечения и диагностики болезни Альцгеймера, как описано выше. Кроме того, антитела могут быть использованы для разработки анти-антител. Кроме того, гибридома, продуцирующая соответствующее антитело, обеспечивает возможность стабильного источника для непрерывного получения идентичных моноклональных антител, т.е. реагентов), гарантируя при этом идентичность антител в разных экспериментах, а также при терапевтических применениях.
Также способы согласно настоящему изобретению позволяют специалисту получать соответствующие количества исходного материала для применения при получении дополнительных материалов, которые, в свою очередь, могут быть использованы для получения моноклональных антител (или других антител) для лечения болезни Альцгеймера. Как указано выше, антитела также могут быть использованы для пассивной иммунизации, чтобы предотвратить болезнь Альцгеймера или другие родственные неврологические состояния, которые характеризуются такими же симптомами, как и болезнь Альцгеймера, например нарушение когнитивной функции.
В одном варианте диагностики согласно настоящему изобретению антитело, предлагаемое в настоящем изобретении (например, 8F5), или его часть наносят в виде покрытия на твердую фазу (или оно присутствует в жидкой фазе). Тестируемый или биологический образец (например, цельную кровь, спинномозговую жидкость, сыворотку и т.д.) затем подвергают контакту с твердой фазой. Если антиген (например, глобуломер) присутствует в образце, такие антигены связываются с антителами на твердой фазе и затем их выявляют либо прямым, либо непрямым способом. Прямой способ заключается в простом выявлении наличия комплекса как такового и, следовательно, наличия антигенов. В непрямом способе конъюгат добавляют к связанному антигену. Конъюгат содержит второе антитело, которое связывается со связанным антигеном, который связан с генерирующим сигнал соединением или меткой. Если второе антитело связывается со связанным антигеном, то генерирующее сигнал соединение дает измеряемый сигнал. Затем такой сигнал свидетельствует о присутствии антигена в тестируемом образце.
Примеры твердых фаз, используемых в диагностических иммуноанализах, представляют собой пористые и непористые материалы, частицы латекса, магнитные частицы, микрочастицы (см., например, патент США No. 5705330), шарики, мембраны, лунки для микротитрования и пластиковые пробирки. Выбор материала твердой фазы и способа мечения антигена или антитела, присутствующего в конъюгате, если это требуется, делают на основе рабочих характеристик требуемой формы анализа.
Как указано выше, конъюгат (или индикаторный реагент) будет содержать антитело (или возможно анти-антитело, в зависимости от анализа), связанное с генерирующим сигнал соединением или меткой. Такое генерирующее сигнал соединение или «метка» могут быть выявлены как таковые или могут быть подвергнуты взаимодействию с одним или несколькими дополнительными соединениями для образования регистрируемого продукта. Примеры генерирующих сигнал соединений включают хромогены, радиоизотопы (например, 125I, 131I, 32P, 3H, 35S и 14C), хемилюминесцентные соединения (например, акридиний), частицы (видимые или флуоресцирующие), нуклеиновые кислоты, комплексообразующие агенты или катализаторы, такие как ферменты (например, щелочная фосфатаза, кислая фосфатаза, пероксидаза хрена, бета-галактозидаза и рибонуклеаза). В случае использования ферментов (например, щелочной фосфатазы или пероксидазы хрена) добавление хромогенного, флуоресцирующего или люминесцирующего субстрата приводит к генерированию регистрируемого сигнала. Также применимы другие системы регистрации, такие как флуоресценция с разрешением во времени, флуоресценция, основанная на внутреннем отражении, амплификация (например, полимеразная цепная реакция) и спектроскопия комбинационного рассеяния.
Примеры биологических жидкостей, которые можно тестировать указанными выше способами иммуноанализа, включают плазму, цельную кровь, высушенную цельную кровь, сыворотку, спинномозговую жидкость или водные или водно-органические экстракты тканей и клеток.
Настоящее изобретение также охватывает способ выявления наличия антител в тестируемом образце. Указанный способ включает в себя стадии: (a) осуществления контакта тестируемого образца, предположительно содержащего антитела, с использованием анти-антитела, специфичного по отношению к антителам в образце пациента, в течение периода и в условиях, достаточных для обеспечения возможности образования комплексов анти-антитело/антитело, при этом анти-антитело представляет собой антитело согласно настоящему изобретению, которое связывается с антителом в образце пациента; (b) добавления конъюгата к полученном в результате комплексам анти-антитело/антитело, при этом конъюгат содержит антиген (который связывается с анти-антителом), связанный с генерирующим сигнал соединением, позволяющим выявлять регистрируемый сигнал; и (d) выявления наличия антител, которые могут присутствовать в тестируемом образце, посредством регистрации сигнала, создаваемого генерирующим сигнал соединением. Можно использовать контроль или калибровочный стандарт, который содержит антитело к анти-антителу.
Настоящее изобретение также относится к вакцине, содержащей одно из антител, описанных в данной публикации, или его часть и фармацевтически приемлемый адъювант (например, адъювант Фрейнда или фосфатно-солевой буфер).
Наборы также включены в объем настоящего изобретения. Более конкретно, настоящее изобретение относится к наборам для определения наличия антигенов (например, глобуломеров) у пациента, у которого предполагается наличие болезни Альцгеймера или другого состояния, характеризуемого когнитивным нарушением. В частности, набор для определения наличия антигенов в тестируемом образце содержит a) антитело, которое определено в данном описании, или его часть; и b) конъюгат, содержащий второе антитело (обладающее специфичностью по отношению к антигену), связанный с генерирующим сигнал соединением, способным генерировать регистрируемый сигнал. Набор также может содержать контроль или калибровочный стандарт, который включает в себя реагент, который связывается с антигеном, а также инструкцию, в которой подробно указано, как следует использовать набор и компоненты набора.
Настоящее изобретение также относится к набору для выявления антител в тестируемом образце. Набор может содержать a) анти-антитело, специфичное (например, одно из антител согласно настоящему изобретению) по отношению к представляющему интерес антителу, и b) антиген или его часть, которые определены выше. Также может быть включен контроль или калибровочный стандарт, содержащий реагент, который связывается с антигеном. Более конкретно набор может содержать a) анти-антитело (такое как одно из антител согласно настоящему изобретению), специфичное по отношению к антителу, и b) конъюгат, содержащий антиген (например, глобуломер), связанный с генерирующим сигнал соединением, способным генерировать регистрируемый сигнал. И также набор может содержать контроль или калибровочный стандарт, содержащий реагент, которые связывается с антигеном, и также может содержать инструкцию или вкладыш в упаковку, в котором описано, как следует использовать набор, и компоненты набора.
Набор также может содержать одну емкость, такую как флакон, колбы или полоски, при этом каждая емкость содержит заданную твердую фазу, а другие емкости содержат соответствующие конъюгаты. Такие наборы также могут содержать флаконы или емкости с другими реагентами, необходимыми для проведения анализа, такими как реагенты для промывки, обработки и регистрации.
Также следует отметить, что настоящее изобретение не только содержит полноразмерные антитела, описанные выше, но также их части или фрагменты, например, Fab-часть. Кроме того, настоящее изобретение охватывает любое антитело, имеющее такие же свойства, как предлагаемые антитела, касающиеся, например, специфичности связывания, структуры, и т.д.
Информация о депозитах: Гибридома (ML5-8F5.1F2.2A2), которая продуцирует моноклональное антитело 8F5, депонирована в Американской коллекции типов культур, 10801 University Boulevard, Manassas, Virginia 20110 1 декабря 2005 по условиям Будапештского договора с присвоением номера ATCC No. PTA-7238.
Гибридома (ML5-8C5.2C1.8E6.2D5), которая продуцирует моноклональное антитело 8C5, депонирована в Американской коллекции типов культур, 10801 University Boulevard, Manassas, Virginia 20110 on February 28, 2006 по условиям Будапештского договора с присвоением номера ATCC No. PTA-7407.
Настоящее изобретение может быть проиллюстрировано с помощью следующих неограничивающих примеров:
ПРИМЕР I(a)
ПОЛУЧЕНИЕ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ 8F5 И 8C5
Мышей Balb/c иммунизировали подкожно 50 микрограммами глобуломера Aβ(1-42), как описано в Barghorn et al., 2005, J. Neurochem, 95, 834-847, в CFA (Sigma), и дважды подвергали бустер-иммунизации с интервалами в один месяц. Собирали селезенки и клетки селезенки сливали с клетками миеломы мыши SP2/0 в отношении 5:1, используя основанный на ПЭГ способ. Слитые клетки высевали в 96-луночные планшеты в среду для селекции с азасерином/гипоксантином в концентрации 2×105 клеток/мл по 200 мкл на лунку. Клеткам давали возможность расти, чтобы образовались видимые колонии, и надосадки анализировали в отношении реактивности по отношению к олигомеру A-бета прямым анализом ELISA. Гибридомы, секретирующие антитела к олигомерам A-бета, субклонировали посредством лимитирующего разведения до тех пор, пока не выявляли стабильную экспрессию антител.
ПРИМЕР II
ПРЕИМУЩЕСТВЕННОЕ СВЯЗЫВАНИЕ ГЛОБУЛОМЕРА АНТИТЕЛАМИ 8F5 и 8C5 ПО СРАВНЕНИЮ С МОНОМЕРНЫМИ ПРЕПАРАТАМИ AB(1-40) и AB (1-42)
Чтобы тестировать избирательность 8F5, использовали два различным образом растворенных препарата мономера Aβ(1-42), а также свежеприготовленный Aβ(1-40) в качестве заменителей мономеров. Проводили два типа экспериментов. В первом эксперименте 8F5 тестировали в отношении избирательности по отношению к глобуломеру Aβ с помощью ELISA типа «сэндвич» с использованием полученного на основе глобуломера, но не специфичного для конформера мАт 6G1 (см. S. Barghorn et al. J. Neurochemistry, 95: 834 (2005)), в качестве улавливающего антитела. Биотинилированный 8F5 использовали в качестве второго и избирательного по отношению к конформеру антитела. Указанный эксперимент описан в примере 2.1 ниже.
Во втором примере, описанном в разделе 2.2 ниже, избирательность к олигомеру по сравнению с мономером Aβ(1-42) и мономеру Aβ(1-40) исследовали с помощью дот-блот-анализа. В данном эксперименте 8F5 предпочтительно связывается с глобуломером Aβ(1-42) (по сравнению с известным антителом 4G8, картированным в области, сходной с областью для 8F5, но полученным в результате иммунизации линейным пептидом Aβ(17-24) (Abeam Ltd., Cambridge, MA)), по сравнению с мономером Aβ(1-42), а также по сравнению с мономером Aβ(1-40). 8C5 тестировали, используя протокол, идентичный протоколу для 8F5.
ПРИМЕР 2.1: ИЗБИРАТЕЛЬНОСТЬ МОНОКЛОНАЛЬНОГО АНТИТЕЛА 8F5 И 8C5 ПО ОТНОШЕНИЮ К ОЛИГОМЕРУ
a) Получение глобуломера Aβ(1-42):
9 мг Aβ(1-42) Fa. Bachem растворяли в 1,5 мл HFIP (1.1.1.3.3.3-гексафтор-2-пропаноле) и инкубировали в течение 1,5 ч при 37ºC. Раствор упаривали в SpeedVac и суспендировали в 396 мкл ДМСО (5 мМ исходный раствор Aβ). Образец обрабатывали ультразвуком в течение 20 секунд на водяной бане для озвучивания, встряхивали в течение 10 минут и хранили в течение ночи при -20ºC.
Образец разбавляли 4,5 мл PBS (20 мМ NaH2PO4; 140 мМ NaCl; pH 7,4) и добавляли 0,5 мл 2% водный раствор SDS (содержание SDS 0,2%). Смесь инкубировали в течение 7 ч при 37ºC, разбавляли 16 мл H2O и дополнительно инкубировали в течение 16 часов при 37ºC. После этого раствор глобуломера Aβ(1-42) центрифугировали в течение 20 мин при 3000 g. Надосадок концентрировали до 0,5 мл, используя концентратор Centriprep с пределом отсечения 30 кД. Концентрат диализовали против 5 мМ NaH2PO4; 35 мМ NaCl; pH 7,4 в течение ночи при 6ºC. Затем концентрат глобуломера Aβ(1-42) центрифугировали в течение 10 мин при 10000 g. Затем надосадок делили на аликвоты и хранили при -20ºC.
b) Получение мономера Aβ(1-42), предварительно обработанного HFIP:
3 мг Aβ(1-42) человека (Bachem Inc.) No. в каталоге H-1368 растворяли в 0,5 мл HFIP (суспензия 6 мг/мл) в пробирке Eppendorff объемом 1,7 мл и встряхивали (встряхиватель Eppendorff Thermo, 1400 об/мин) в течение 1,5 ч при 37ºC вплоть до получения прозрачного раствора. Образец сушили в центрифужном вакуумном концентраторе (1,5 ч) и ресуспендировали в 13,2 мкл ДМСО, встряхивали в течение 10 с с последующей обработкой на ультразвуковой бане (20 с) и встряхивали (например, используя встряхиватель Eppendorff Thermo, 1400 об./мин) в течение 10 мин.
Добавляли 6 мл 20 мМ NaH2PO4; 140 мМ NaCl; 0,1% плюроник F68; pH 7,4 и перемешивали в течение 1 часа при комнатной температуре. Образец центрифугировали в течение 20 мин при 3000 g. Надосадок отбрасывали и к осадку, растворенному в 0,6 мл 20 мМ NaH2PO4; 140 мМ NaCl; 1% плюроник F68; pH 7,4, добавляли 3,4 мл воды и перемешивали в течение 1 часа при комнатной температуре с последующим центрифугированием в течение 20 мин при 3000 g. Аликвоты надосадка 8×0,5 мл хранили при -20ºC.
c) Получение мономера Aβ(1-42) в NH 4 OH:
1 мг твердого порошка Aβ(1-42) (Bachem Inc. No. в каталоге H-1368) растворяли в 0,5 мл 0,1% NH4OH в воде (свеже приготовленный) (2 мг/мл) и сразу встряхивали в течение 30 с при комнатной температуре, получая прозрачный раствор. Образец хранили при -20ºC для дальнейшего применения.
d) Получение мономера Aβ(1-40):
1 мг Aβ(1-40) человека (Bachem Inc, No. в каталоге H-1194) суспендировали в 0,25 мл HFIP (суспензия 4 мг/мл) в пробирке Eppendorff. Пробирку встряхивали (например, на встряхивателе Eppendorff Thermo, 1400 об/мин) в течение 1,5 ч при 37ºC, получая прозрачный раствор, и затем сушили, используя центрифужный вакуумный концентратор (1,5 ч). Образец перерастворяли в 46 мкл ДМСО (раствор 21,7 мг/мл), встряхивали в течение 10 с с последующей обработкой ультразвуком в течение 20 с на ультразвуковой бане. После 10 мин встряхивания (например, на встряхивателе Eppendorff Thermo, 1400 об/мин) образец хранили при -20ºC для последующего применения.
e) Биотинилирование мышиного мАт 8F5 против Aβ:
500 мкл мышиного мАт 8F5 против Aβ (0,64 мг/мл) в PBS добавляли к 2 мкл 20 мг/мл сульфо-NHS-биотина (Pierce Inc., No. в каталоге 21420), только что растворенного в воде, и встряхивали (например, на встряхивателе Eppendorff Thermo, 1400 об/мин) в течение 30 мин, диализовали в течение 16 часов при 6ºC в диализной трубке против 500 мл 20 мМ Na Pi; 140 мМ NaCl; pH 7,4. Диализат хранили при -20ºC для дальнейшего применения. 8C5 биотинилировали соответствующим образом.
f) ELISA типа «сэндвич» образцов Aβ:
g) Перечень реагентов:
1. Планшет F96 Cert. Maxisorp NUNC-Immuno Cat. No.: 439454.
2. Связывающее антитело: мышиное мАт 6G1 против Aβ, растворенное в PBS; концентрация: 0,4 мг/мл; хранение при -20ºC.
3. Буфер для покрывания:
100 мМ гидрокарбонат натрия; pH 9,6.
4. Блокирующий реагент для ELISA; Roche Diagnostics GmbH, No. в каталоге: 1112589.
5. PBST-буфер:
20 мМ NaH2PO4; 140 мМ NaCl; 0,05% твин-20; pH 7,4.
6. Бычий альбумин, фракция V, не содержащий протеаз; Serva, No. в каталоге: 11926,03; хранение при 4ºC.
7. PBST + 0,5% БСА-буфер:
20 мМ NaH2PO4; 140 мМ NaCl; 0,05% твин-20; pH 7,4 + 0,5% БСА.
8. Стандартный раствор глобуломера Aβ(1-42):
раствор в 5 мМ NaH2PO4; 35 мМ NaCl; pH 7,4; концентрация: 10,77 мг/мл; хранение при -20ºC.
9. Стандартный исходный раствор мономера Aβ(1-42), обработанного HFIP:
раствор в 3 мМ NaH2PO4; 21 мМ NaCl; 0,15% плюроник F68; pH 7,4; концентрация: 0,45 мг/мл; хранение при -20ºC.
10. Стандартный исходный раствор мономера Aβ(1-42) в NH4OH; раствор в 0,1% NH4OH, концентрация: 2мг/мл; хранение при -20ºC.
11. Стандартный исходный раствор мономера Aβ(1-40), обработанного HFIP; раствор в ДМСО; концентрация: 21,7 мг/мл; хранение при -20ºC.
12. Биотинилированное мышиное мАт против Aβ, клон 8F5; раствор в PBS; концентрация: 0,24 мг/мл; хранение при -80ºC.
13. Конъюгат стрептавидин-POD; Fa. Roche No. в каталоге: 1089153.
14. Окрашивание:
TMB; Roche Diagnostics GmbH, No. в каталоге: 92817060; 42 мМ в ДМСО; 3% H2O2 в воде; 100 мМ ацетат натрия, pH 4,9.
15. Остановка окрашивания добавлением 2 М раствора сульфоновой кислоты.
Приготовление реагентов:
Использовали следующий протокол:
1. Связывающее антитело
Размораживали исходный раствор мАт 6G1 и разбавляли 1:400 в буфере для покрывания.
2. Блокирующий реагент:
Растворяли блокирующий реагент в 100 мл воды, чтобы получить исходный блокирующий раствор, и аликвоты по 10 ил хранили при -20ºC. Разбавляли 3 мл исходного блокирующего раствора 27 мл воды для каждого планшета, который необходимо блокировать.
3. Стандартные растворы Aβ:
a) Aβ(1-42)-глобуломер
- добавляли 1 мкл стандартного исходного раствора Aβ(1-42)-глобуломера к 1076 мкл PBST + 0,5% БСА = 10 мкг/мл;
- добавляли 50 мкл стандартного раствора 10 мкг/мл Aβ(1-42)-глобуломера к 4950 мкл PBST + 0,5% БСА = 100 нг/мл.
b) Мономер Aβ(1-42), обработанный HFIP
- добавляли 10 мкл стандартного исходного раствора мономера Aβ(1-42), предварительно обработанного HFIP, к 440 мкл PBST + 0,5% БСА = 10 мкг/мл;
- добавляли 50 мкл стандартного раствора 10 мкг/мл мономера Aβ(1-42), предварительно обработанного HFIP, к 4950 мкл PBST + 0,5% БСА = 100 нг/мл.
c) Мономер Aβ(1-42) в NH4OH
- добавляли 5 мкл стандартного исходного раствора мономера Aβ(1-42) в NH4OH к 995 мкл PBST + 0,5% БСА = 10 мкг/мл;
- добавляли 50 мкл стандартного раствора 10 мкг/мл мономера Aβ(1-42) в NH4OH к 4950 мкл PBST + 0,5% БСА = 100 нг/мл.
d) Мономер Aβ(1-40), предварительно обработанный HFIP
- добавляли 1 мкл стандартного исходного раствора мономера Aβ(1-40), предварительно обработанного HFIP, к 49 мкл PBST + 0,5% БСА = 430 мкг/мл;
- добавляли 10 мкл стандартного раствора 430 мкг/мл мономера Aβ(1-40), предварительно обработанного HFIP, к 420 мкл PBST + 0,5% БСА = 10 мкг/мл;
- добавляли 50 мкл стандартного раствора 10 мкг/мл мономера Aβ(1-40), предварительно обработанного HFIP, к 4950 мкл PBST + 0,5% БСА = 100 нг/мл.
Стандартные кривые:
1. Первое антитело: биотинилированное мАт 8F5:
Концентрированное биотинилированное анти-Aβ-мАт 8F5 разбавляли в буфере PBST + 0,5% БСА. Коэффициент разбавления 1/1200 = 0,2 мкг/мл. Антитело использовали немедленно.
2. Реагент для мечения:
Перерастворяли лиофилизат конъюгата стрептавидин-POD в 0,5 мл воды. Добавляли 500 мкл глицерина и аликвоты по 100 мкл хранили при -20ºC для последующего использования. Разбавляли концентрированный реагент для мечения в PBST-буфере. Коэффициент разбавления 1/10000. Использовали немедленно.
3. Раствор TMB для окрашивания:
Смешивали 20 мл 100 мМ раствора ацетата натрия, pH 4,9,с 200 мкл раствора TMB и 29,5 мкл 3% раствора пероксида. Использовали немедленно.
Размещение образцов в планшете (обратите внимание, что все стандарты анализировали в двух повторах):
Применяемый способ:
1. Вносили по 100 мкл раствора анти-Aβ-мАт 6G1 на лунку и инкубировали в течение ночи при 4ºC.
2. Удаляли раствор антитела и лунки три раза промывали 250 мкл PBST-буфера.
3. Добавляли по 260 мкл блокирующего раствора на лунку и инкубировали 2 часа при комнатной температуре.
4. Удаляли блокирующий раствор и лунки три раза промывали 250 мкл PBST-буфера.
5. После приготовления стандартов в планшет вносили по 100 мкл стандартов на лунку. Инкубировали 2 часа при комнатной температуре и в течение ночи при 4ºC.
6. Удаляли стандартный раствор и лунки три раза промывали 250 мкл PBST-буфера.
7. Добавляли по 200 мкл раствора первого биотинилированного антитела 8F5 на лунку и инкубировали 1,5 часа при комнатной температуре.
8. Удаляли раствор антитела и лунки три раза промывали 250 мкл PBST-буфера.
9. Добавляли по 200 мкл раствора метки на лунку и инкубировали 1 час при комнатной температуре.
10. Удаляли раствор метки и лунки три раза промывали 250 мкл PBST-буфера.
11. Добавляли по 100 мкл раствора TMB в каждую лунку и инкубировали при комнатной температуре (5-15 мин).
12. Наблюдали окрашивание и вносили по 50 мкл раствора для остановки на лунку после начала окрашивания фона.
13. Регистрация в УФ при 450 нм.
14. Рассчитывали результаты на основании стандартной кривой.
15. Оценка.
Результаты показаны на фиг. 1 для антитела 8F5 и на фиг. 8 для антитела 8C5. Значения Log EC50 значимо являются наиболее низкими для антигена глобуломера Aβ(1-42) (1,958) по сравнению с приведенными значениями для двух различным образом подготовленных мономеров Aβ(1-42) (2,745 и 3,003 соответственно) и мономера Aβ(1-40) (2,825). Приведенные данные свидетельствуют примерно о 10-кратной избирательности антитела 8F5 по отношению к глобуломеру Aβ(1-42) по сравнению с мономером Aβ(1-42).
Почти идентичные результаты получали с использованием антитела 8C5, которые показаны на фиг. 8.
ПРИМЕР 2.2: ИЗБИРАТЕЛЬНОСТЬ МОНОКЛОНАЛЬНОГО АНТИТЕЛА 8F5 И 8C5 ПО ОТНОШЕНИЮ К ОЛИГОМЕРУ
- Способ дот-блот-анализа, позволяющий отличить мономер Aβ от глобуломера Aβ: сравнение 8F5 и 8C5 с 4G8.
Получали серийные разведения глобуломера Aβ(1-42), мономера Aβ1-42 и мономера Aβ1-40 в диапазоне от 100 пмоль/мкл до 0,01 пмоль/мкл в PBS. Из каждого образца 1 мкл наносили в виде пятна на нитроцеллюлозную мембрану. Для выявления использовали мышиные моноклональные антитела 4G8 и 8F5 (0,2 мкг/мл), при этом в качестве второго антитела использовали антитело против IgG мыши, связанное со щелочной фосфатазой, а также реагент для окрашивания NBT/BCIP (Roche Diagnostics, Mannheim). Регистрируемый сигнал анализировали по его интенсивности (плотность по отражению = RD) с использованием денситометра (GS 800, Biorad, Hercules, CA, USA) при концентрации антигена 10 пмоль. При указанной концентрации для каждой Aβ-формы измеряемая плотность по отражению была в линейном диапазоне регистрации денситометра. Другое антитело 8C5 использовали в аналогичном протоколе. Результаты показаны в таблице ниже:
В частности, приведенные выше результаты свидетельствуют, что 8F5 и 8C5 имеют другие профили связывания по сравнению с коммерчески доступным анти-Aβ-(1-42)-антителом к 4G8, который картирован к Aβ(17- 24) (т.е. к линейной последовательности). Более конкретно 8F5 и 8C5 предпочтительно связываются с глобуломером по сравнению с мономером Aβ42 (см. колонку 4; 1,4 по сравнению с 1), а также предпочтительно связываются с глобуломером по сравнению с Aβ40 (колонка 5; 16,9 по сравнению с 4,2). Указанные повышенные избирательности связывания по сравнению со стандартным 4G8 должны приводить к меньшим побочным эффектам при применении 8F5 и/или 8C5, как описано выше (например, связывание с бляшками).
ПРИМЕР III
СВЯЗЫВАНИЕ 8F5 И 8C5 С ФИБРИЛЛАМИ Aβ(1-42)
Так как антитело 8F5 создавали против растворимых глобуломеров, то была выдвинута гипотеза, что 8F5 не должно связываться с материалом депонированных бляшек или фибрилл. Поэтому тестировали связывание 8F5 с суспензиями полимеризованных Aβ-фибрилл, как описано в следующем примере.
Получение фибрилл Aβ(1-42):
1 мг Aβ(1-42) (Bachem Inc., No. в каталоге: H-1368) растворяли в 500 мкл водного 0,1% раствора NH4OH (пробирка Eppendorff) и образец перемешивали в течение 1 мин при комнатной температуре с последующим 5-минутным центрифугированием при 10000 g. Надосадок собирали пипеткой в новую пробирку Eppendorff и измеряли концентрацию Aβ(1-42) с помощью анализа концентрации белка по Бредфорду (способ анализа BIO-RAD Inc.).
100 мкл такого свежеприготовленного раствора Aβ(1-42) нейтрализовали 300 мкл 20 мМ NaH2PO4; 140 мМ NaCl; pH 7,4, затем 2% HCl, чтобы довести pH до 7,4. Образец инкубировали еще в течение 20 часов при 37ºC и центрифугировали (10 мин, 10000 g). Надосадок отбрасывали, а осадок фибрилл ресуспендировали в 400 мкл 20 мМ NaH2PO4; 140 мМ NaCl; pH 7,4, перемешивая в течение 1 мин на мешалке Vortex, с последующим центрифугированием (10 мин, 10000 g . После удаления надосадка повторяли ресуспендирование и конечную суспензию фибрилл снова осаждали центрифугированием (10 мин, 10000 g). Надосадок снова отбрасывали и конечный осадок ресуспендировали в 380 мкл 20 мМ NaH2PO4; 140 мМ NaCl; pH 7,4, перемешивая в течение 1 мин на мешалке Vortex. Аликвоты образца хранили при -20ºC в морозильной камере.
80 мкл суспензии фибрилл смешивали с 320 мкл буфера, содержащего 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% твин-20, pH 7,4, и перемешивали в течение 5 мин при комнатной температуре с последующей обработкой ультразвуком (20 с). После центрифугирования (10 мин, 10000 g) осадок ресуспендировали в 190 мкл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% твин-20, pH 7,4, перемешивая на мешалке Vortex.
Связывание антител с фибриллами Aβ(1-42)
Аликвоты суспензии фибрилл объемом 10 мкл инкубировали с:
a) 10 мкл 20 мМ Na Pi; 140 мМ NaCl; pH 7,4;
b) 10 мкл 0,1 мкг/мкл мАт 6E10 Signet Inc., № в каталоге 9320 в 20 мМ;
c) NaH2PO4, 140 мМ NaCl; pH 7,4;
d) 10 мкл 0,1 мкг/мкл мАт 4G8 SignetInc., № в каталоге 9220, в 20 мМ Na Pi, 140 мМ NaCl; pH 7,4;
e) 10 мкл 0,1 мкг/мкл мАт 8F5 (8C5) в 20 мМ Na Pi; 140 мМ NaCl; pH 7,4.
Образцы инкубировали в течение 20 часов при 37ºC. Наконец образцы центрифугировали (10 мин при 10000 g). Надосадки, содержащие фракцию несвязанных антител, собирали и смешивали с 20 мкл буфера для образцов, анализируемых в SDS-ПААГ. Фракции осадка промывали 50 мкл буфера, содержащего 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4, перемешивая в течение 1 мин на мешалке Vortex с последующим центрифугированием (10 мин, 10000 g). Конечные осадки ресуспендировали в 20 мкл буфера, содержащего 20 мМ Na Pi, 140 мМ NaCl, 0,025% твин-20, pH 7,4, и растворяли в 20 мкл буфера для SDS-ПААГ.
Анализ в SDS-ПААГ
Надосадки и ресуспендированные образцы осадков нагревали в течение 5 мин при 98ºC и наносили на 4-20% гель, содержащий трис/глицин в следующих условиях:
SDS-буфер для образцов: 0,3 г SDS; 0,77 г DTT; 4 мл 1 М трис/HCl, pH 6,8; 8 мл глицерина; 1 мл 1% бромфенолового синего в этаноле; добавляли воду до 50 мл 4-20% геля с три/глицином: Invitrogen Inc., No: EC6025BOX.
Рабочий буфер: 7,5 г трис; 36 г глицина; 2,5 г SDS; добавляли воду до 2,5 л.
Электрофорез в ПААГ проводили при 20 мА. Гели красили Кумасси синим R250.
Результаты:
Окрашивание Кумасси SDS-ПААГ показало присутствие тяжелых и легких цепей антител, главным образом в надосадке суспензии фибрилл (дорожка 7, фигура 2), в остальной части суспензии фибрилл наблюдали очень небольшое количество материала антител, при этом также наблюдали частично деполимеризованный A-бета с молекулярной массой 4,5 кД. В отличие от 8F5 и 8C5 другие анти-Aβ-антитела не обнаруживались в растворимой фракции (6G10, дорожка 3, фигура 2) или обнаруживались только частично (4G8, дорожка 5, фигура 2) по сравнению со связанной с фибриллами фракцией (дорожка 6, фигура 2) .
Относительное связывание с фибриллярным типом A-бета оценивали на основе SDS-ПААГ-анализа, измеряя значения плотности по отражению для тяжелой цепи антител в связанной с фибриллами и надосадочной фракциях, и рассчитывали согласно следующей формуле:
Связанная с фибриллами фракция Ат = RDфибриллярная фракция × 100%/(RDфибриллярная фракция + RD надосадочная фракция).
Получили следующие значения:
Полученные данные свидетельствуют о значительном снижении связанных 8F5 и 8C5 по сравнению со стандартным антителом 6E10.
ПРИМЕР IV
ПРЕДПОЧТИТЕЛЬНОЕ СВЯЗЫВАНИЕ ЭНДОГЕННЫХ ГЛОБУЛОМЕРОВ Aβ(1-42) ПО СРАВНЕНИЮ С Aβ(1-40)
На основании представлений об олигомерах Aβ важно то, что антитела против олигомеров Aβ также могут проявлять предпочтительное связывание по отношению к олигомерам Aβ(1-42) in vivo, в частности по сравнению с Aβ(1-40) у пациентов с умеренным когнитивным нарушением и пациентов с AD. Идею уменьшения количества вида Aβ(1-42) по сравнению с Aβ(1-40) используют в терапевтическом способе лечения AD с применением НПВС (Weggen et al., Nature 414, 212-216 (2001)). Предполагается, что такие НПВС, которые снижают количество Aβ(1-42) по сравнению с Aβ(1-40), обладают наилучшей эффективностью при лечении болезни Альцгеймера. Отношение Aβ(1-42)/Aβ(1-40) имеет важное значение для избирательной терапии, а также для диагностических целей.
Анализ проводили на образцах СМЖ пациентов с болезнью Альцгеймера и пациентов с MCI. На основании результатов, показанных на фигуре 3 и описанных ниже, можно сделать вывод, что 8F5 имеет большое преимущество по сравнению с антителами к Aβ, подобными 6E10, так как 8F5 выявляет более высокую долю Aβ(1-42) по сравнению с менее подвергающимися агрегации Aβ(1-40). Такое преимущество позволяет более избирательно диагностировать и нейтрализовать олигомеры Aβ(1-42) типа у пациентов с MCI и AD.
A) ЭНДОГЕННЫЕ УРОВНИ АМИЛОИДА β(1-42) И АМИЛОИДА β(1-40) В СМЖ ПАЦИЕНТОВ С MCI И AD ПОСЛЕ ИММУНОПРЕЦИПИТАЦИИ ИЗБИРАТЕЛЬНЫМИ ПО ОТНОШЕНИЮ К ОЛИГОМЕРАМ МЫШИНЫМИ АНТИ-Aβ-мАт 8F5:
Иммобилизация анти-Aβ-мАт на CNBr-активированной сефарозе 4B:
a) мАт 6E10 Signet Inc., No. в каталоге 9320;
b) мАт 8F5.
0,4 г CNBr-активированной сефарозы 4B (Amersham Pharmacia Bio-tech AB, Uppsala, Sweden, Inc., No: 17-0430-01) добавляли к 10 мл водного 1 мМ раствора HCl и инкубировали в течение 30 мин при комнатной температуре. CNBr-активированную сефарозу 4B три раза промывали 10 мл 1 мМ HCl и два раза 10 мл 100 мМ NaHCO3, 500 мМ NaCl, pH 8,3. Для каждого иммобилизованного антитела 100 мкл матрицы CNBr-активированной сефарозы 4B добавляли к 950 мкл 0,5 мг/мл раствора анти-Aβ-мАт в 100 мМ NaHCO3, 500 мМ NaCl, pH 8,3. После 2-часового встряхивания при комнатной температуре образцы центрифугировали в течение 5 мин при 10000 g. Затем 500 мкл буфера, содержащего 100 мМ этаноламин, 100 мМ NaHCO3, 500 мМ NaCl, pH 8,3, добавляли к шарикам и образцы встряхивали в течение 1 часа при комнатной температуре. Образцы анти-Aβ-мАт-сефароза центрифугировали в течение 5 мин при 10000 g и 5 раз промывали 500 мкл 20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4. Перед хранением при 6ºC образцы стабилизировали добавлением азида натрия до конечной концентрации 0,02%.
Иммунопреципитация:
a) мАт 6E10-сефароза;
b) мАт 8P5-сефароза.
200 мкл образцов спинномозговой жидкости человека разбавляли 200 мкл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% твин-20, pH 7,4. Указанные образцы добавляли к 2 мкл анти-Aβ-мАт-сефарозная матрица и перемешивали в течение 2 часов при комнатной температуре. Образцы центрифугировали в течение 5 мин при 10000 g. Надосадки отбрасывали, а анти-Aβ-мАт-сефарозу дважды промывали 50 мкл PBS, перемешивали в течение 1 мин и центрифугировали (5 мин при 10000 g). Надосадки отбрасывали, а сефарозные шарики суспендировали в 50 мкл 2 мМ NaH2PO4, 14 мМ NaCl, pH 7,4, с последующим 1-минутным перемешиванием при комнатной температуре и 5-минутным центрифугированием при 10000 g. На следующей стадии анти-Aβ-мАт-сефарозные шарики обрабатывали 50 мкл 50% CH3CN, 0,2% ТФУ в воде. После 10-минутного встряхивания при комнатной температуре образцы центрифугировали в течение 5 мин при 10000 g. Надосадки собирали и переносили в пробирки Eppendorf объемом 1,5 мл. Образцы смешивали с 50 мкл воды и упаривали в концентраторе Speed Vac. Осадок перерастворяли в 4 мкл 70% HCOOH, встряхивали в течение 10 мин при комнатной температуре и нейтрализовали 76 мкл 1 М раствора трис и 720 мкл 20 мМ NaH2PO4, 140 мМ NaCl; 0,05% твин-20, pH 7,4.
Образцы для определения мономерных форм Aβ(1-40), (1-42) в СМЖ:
a) Содержание Aβ в образцах СМЖ без иммунопреципитации:
158 мкл СМЖ разбавляли 342 мкл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% твин-20, pH 7,4. Указанное разведение 1:3,16 использовали для ELISA типа «сэндвич» и учитывали при оценке.
b) Содержание Aβ в образцах СМЖ после иммунопреципитации:
Для анализа брали образцы, используемые в указанном выше способе.
Протокол ELISA типа «сэндвич», используемый для определения Aβ(1-40) в СМЖ
Список реагентов:
1. Планшет F96 Cert. Maxisorp NUNC-Immuno, No. в каталоге: 439454.
2. Связывающее антитело
Анти-Aβ-мАт, клон 6E10; Signet, No. в каталоге 9320; концентрация: 0,4 мг/мл, по Бредфорду (BioRad); хранение при -20ºC.
3. Буфер для связывания
100 мМ гидрокарбонат натрия; pH 9,6.
4. Блокирующий реагент для ELISA; Roche Diagnostics GmbH, No. в каталоге: 1112589.
5. Буфер PBST
20 мМ NaH2PO4, 140 мМ NaCl, 0,05% твин-20, pH 7,4.
6. Стандарт Aβ(1-40):
Твердый порошок Aβ(1-40); Bachem, No. в каталоге: H-1194; хранение при -20ºC.
7. Первое антитело: кроличье поликлональное Ат против Aβ(1-40); аффинно очищенное; раствор в PBS; концентрация: 0,039 мг/мл;
Signet, No. в каталоге 9130-005; хранение при -20ºC.
8. Реагент для мечения:
Конъюгат антикроличье Ат-POD; Fa. Jackson ImmunoResearch, No. в каталоге: 111-036-045.
9. Окрашивание:
TMB; Roche Diagnostics GmbH, No. в каталоге: 92817060; 42 мМ в ДМСО; 3% H2O2 в воде; 100 мМ ацетат натрия, pH 4,9.
10. Раствор для остановки: 2 М сульфоновая кислота.
Протоколы, используемые для получения реагентов:
1. Связывающее антитело:
анти-Aβ-мАт 6E10 (Signet Inc, No. в каталоге 9320) разбавляли до конечной концентрации 0,7 мкг/мл.
2. Блокирующий реагент:
Для получения исходного блокирующего раствора блокирующий реагент растворяли в 100 мл H2O и хранили при -20ºC в аликвотах по 10 мл каждая. 3 мл исходного блокирующего раствора разбавляли 27 мл H2O для блокирования одного планшета для ELISA.
3. Стандартное разведение мономерной формы Aβ(1-40):
A) Стандартный исходный раствор мономера Aβ(1-40): растворяли 0,5 мг Aβ(1-40) в 250 мкл 0,1% NH4OH, концентрация: 2 мг/мл; свежеприготовленный; немедленное использование.
B) Добавляли 5 мкл стандартного исходного раствора мономера Aβ(1-40) к 995 мкл PBST = 10 мкг/мл.
C) Добавляли 5 мкл стандартного раствора Aβ(1-40) в концентрации 10 мкг/мл к 4995 мкл PBST = 10 нг/мл.
Стандартная кривая:
Образцы:
IP: образцы иммунопреципитатов:
4. Первое антитело:
Разбавляли концентрированное поликлональное Ат против Aβ(1-40) в буфере PBST. Коэффициент разбавления 1/200 = 0,2 мкг/мл. Использовали немедленно.
5. Второе антитело:
Лиофилизованный конъюгат антикроличье антитело-POD растворяли в 0,5 мл H2O и смешивали с 500 мкл глицерина. Концентрат антитела затем хранили при -20ºC в аликвотах по 100 мкл. Концентрат разбавляли 1:10000 в буфере PBST. Раствор антител использовали немедленно.
6. Раствор TMB: 20 мл 100 мМ ацетата натрия, pH 4,9, смешивали с 200 мкл раствора TMB и 29,5 мкл 3% пероксида водорода. Полученный раствор использовали немедленно.
Размещение образцов в планшете (обратите внимание, что все стандарты и образцы анализировали в двух повторах):
Применяемый способ:
1. Вносили по 100 мкл раствора связывающего антитела на лунку и инкубировали в течение ночи при 4ºC.
2. Удаляли раствор антитела и лунки три раза промывали 250 мкл PBST-буфера.
3. Добавляли по 260 мкл блокирующего раствора на лунку и инкубировали 2 часа при комнатной температуре.
4. Удаляли блокирующий раствор и лунки три раза промывали 250 мкл PBST-буфера.
5. После приготовления стандартов и образцов в планшет вносили по 100 мкл стандартов и образцов на лунку и инкубировали 2 часа при комнатной температуре и в течение ночи при 4ºC.
6. Удаляли раствор стандарта/образца и лунки три раза промывали 250 мкл PBST-буфера.
7. Добавляли по 200 мкл раствора первого антитела на лунку и инкубировали 1,5 часа при комнатной температуре.
8. Удаляли раствор антитела и лунки три раза промывали 250 мкл PBST-буфера.
9. Добавляли по 200 мкл раствора метки на лунку и инкубировали 1 час при комнатной температуре.
10. Удаляли раствор метки и лунки три раза промывали 250 мкл PBST-буфера.
11. Добавляли по 100 мкл раствора TMB в каждую лунку и инкубировали при комнатной температуре (5-15 мин).
12. Наблюдали окрашивание и вносили по 50 мкл раствора для остановки на лунку.
13. Регистрацию проводили при 450 нм.
14. Рассчитывали результаты на основании стандартной кривой.
15. Оценка.
Если экстинкция от неизвестных образцов не была в линейном диапазоне калибровочной кривой, то ELISA повторяли с соответствующим разведением образца.
Протокол ELISA типа «сэндвич», используемый для определения мономерной формы Aβ(1-42) в СМЖ
Список реагентов:
1. Планшет F96 Cert. Maxisorp NUNC-Immuno, No. в каталоге: 439454.
2. Связывающее антитело
Анти-Aβ-мАт, клон 6E10; Signet, No. в каталоге 9320; концентрация: 0,4 мг/мл, по Бредфорду (BioRad); хранение при -20ºC.
3. Буфер для покрывания
100 мМ гидрокарбонат натрия; pH 9,6.
4. Блокирующий реагент для ELISA; Roche Diagnostics GmbH, No. в каталоге: 1112589.
5. Буфер PBST
20 мМ NaH2PO4, 140 мМ NaCl, 0,05% твин-20, pH 7,4.
6. Стандарт Aβ(1-42):
Твердый порошок Aβ(1-42); Bachem, No. в каталоге: H-1368; хранение при -20ºC.
7. Первое антитело:
кроличье поликлональное Ат против Aβ(1-42); аффинно очищенное; биотинилированное; раствор в PBS с 50% глицерином; концентрация: 0,25 мг/мл; Signet, No. в каталоге 9137-005; хранение при -20ºC.
8. Реагент для мечения:
Конъюгат антикроличье Ат-POD; Fa. Jackson ImmunoResearch, No. в каталоге: 111-036-045.
9. Окрашивание:
TMB; Roche Diagnostics GmbH, No. в каталоге: 92817060;
42 мМ в ДМСО;
3% H2O2 в воде;
100 мМ ацетат натрия, pH 4,9.
10. Раствор для остановки: 2 М сульфоновая кислота.
Способ, используемый для получения реагентов:
1. Связывающее антитело:
анти-Aβ-мАт, клон 6E10, разбавляли 1:400 в буфере для покрывания.
2. Блокирующий реагент:
Для получения исходного блокирующего раствора блокирующий реагент растворяли в 100 мл воды и аликвоты по 10 мл хранили при -20ºC. 3 мл исходного блокирующего раствора разбавляли 27 мл воды для блокирования каждого планшета.
3. Стандартное разведение мономерной формы Aβ(1-42):
Стандартный исходный раствор мономера Aβ(1-42): растворяли 0,5 мг Aβ(1-42) в 250 мкл 0,1% NH4OH, концентрация: 2 мг/мл; свежеприготовленный; использовали немедленно.
Добавляли 5 мкл стандартного исходного раствора мономера Aβ(1-42) к 995 мкл PBST = 10 мкг/мл.
Добавляли 5 мкл стандартного раствора Aβ(1-40) в концентрации 10 мкг/мл к 4995 мкл PBST = 10 нг/мл.
Стандартная кривая:
Образцы:
IP: образцы иммунопреципитатов:
1. Первое антитело:
Разбавляли концентрированное поликлональное Ат против Aβ(1-42) в буфере PBST. Коэффициент разбавления 1/1250 = 0,2 мкг/мл. Использовали немедленно.
2. Реагент для мечения:
Перерастворяли лиофилизат конъюгата антикроличье антитело-POD в 0,5 мл воды. Добавляли 500 мкл глицерина и аликвоты по 100 мкл хранили при -20ºC для последующего применения.
Концентрированный реагент для мечения разбавляли в PBST-буфере. Коэффициент разбавления 1/5000. Использовали немедленно.
3. Раствор TMB:
20 мл 100 мМ ацетата натрия, pH 4,9, смешивали с 200 мкл раствора TMB и 29,5 мкл 3% пероксида водорода. Использовали немедленно.
Размещение образцов в планшете (обратите внимание, что все стандарты и образцы анализировали в двух повторах):
Применяемый способ:
1. Вносили по 100 мкл раствора связывающего антитела на лунку и инкубировали в течение ночи при 4ºC.
2. Удаляли раствор антитела и лунки три раза промывали 250 мкл PBST-буфера.
3. Добавляли по 260 мкл блокирующего раствора на лунку и инкубировали 2 часа при комнатной температуре.
4. Удаляли блокирующий раствор и лунки три раза промывали 250 мкл PBST-буфера.
5. После приготовления стандартов и образцов в планшет вносили по 100 мкл стандартов и образцов на лунку. Инкубировали 2 часа при комнатной температуре и в течение ночи при 4ºC.
6. Удаляли раствор стандарта/образца и лунки три раза промывали 250 мкл PBST-буфера.
7. Добавляли по 200 мкл раствора первого антитела на лунку и инкубировали 1,5 часа при комнатной температуре.
8. Удаляли раствор антитела и лунки три раза промывали 250 мкл PBST-буфера.
9. Добавляли по 200 мкл раствора метки на лунку и инкубировали 1 час при комнатной температуре.
10. Удаляли раствор метки и лунки три раза промывали 250 мкл PBST-буфера.
11. Добавляли по 100 мкл раствора TMB в каждую лунку и инкубировали при комнатной температуре (5-15 мин).
12. Наблюдали окрашивание и вносили по 50 мкл раствора для остановки на лунку.
13. Регистрацию проводили при 450 нм.
14. Рассчитывали результаты на основании стандартной кривой.
15. Оценка:
Если экстинкция от неизвестных образцов не была в линейном диапазоне калибровочной кривой, то ELISA повторяли с соответствующим разведением образца.
РЕЗУЛЬТАТЫ:
Приведенные выше результаты показывают следующее:
a. Избирательное по отношению к глобуломеру антитело, подобное 8F5 (или 8C5), по сравнению с неизбирательным по отношению к глобуломеру антителом, подобным 6E10, предпочтительно связывается с Aβ42 по сравнению с Aβ40, независимо от патологического состояния. Полученный результат свидетельствует об успешном лечении болезни Альцгеймера, поскольку преимущественное элиминирование Aβ42 по сравнению с Aβ40 согласуется с концепцией лечения AD, например с применением R-флубипрофена, флуризана, который проявляет эффективность в лечении AD в клиническом испытании, результаты которого опубликованы Myriad Inc. Указанная концепция опубликована S. Weggen et al. (J. Biol. Chem. (2003) 228 (34): 31831-7). Результаты показаны на фигуре 3.
b. Избирательное по отношению к глобуломеру антитело, подобное 8F5 (или 8C5), связывается еще более эффективно с Aβ42, чем с Aβ40, у пациентов по сравнению со здоровыми людьми в контроле. Полученный результат еще более убедительно свидетельствует об успешном лечении болезни Альцгеймера, поскольку, как отмечено выше, преимущественное элиминирование Aβ42 по сравнению с Aβ40 согласуется с концепцией лечения AD (например, посредством применения нестероидных противовоспалительных лекарственных средств, подобных R-флубипрофену). (См. фигуру 3).
B) УРОВНИ ЭНДОГЕННОГО АМИЛОИДА β(1-42) и АМИЛОИДА β(1-40) В СМЖ ЧЕЛОВЕКА ПОСЛЕ ИММУНОПРЕЦИПИТАЦИИ ИЗБИРАТЕЛЬНЫМ ПО ОТНОШЕНИЮ К ГЛОБУЛОМЕРУ МЫШИНЫМ АНТИ-Aβ-мАт 8F5 ИЛИ 8C5 ПО СРАВНЕНИЮ С НЕИЗБИРАТЕЛЬНЫМ ПО ОТНОШЕНИЮ К ГЛОБУЛОМЕРУ АНТИТЕЛОМ 6E10:
b1) Иммунопреципитация (IP) шарики Dynabeads M-280, покрытые овечьими антителами против мышиного IgG
Растворы A-бета-антител
Следующие очищенные антитела получали из гибридом согласно стандартным способам очистки:
- мАт 6E10; Fa. Signet No.: 9320; 1 мг/мл в PBS-буфере;
- мАт 8F5; 1,65 мг/мл в PBS-буфере;
- мАт 8C5; 1,44 мг/мл в PBS-буфере.
Шарики Dynabeads M-280, покрытые овечьими антителами против мышиного IgG:
Овечьи антитела против мышиного IgG (Invitrogen Inc., № в каталоге: 112.02) ковалентно связаны с магнитными шариками (Dynabeads).
Активация Dynabeads моноклональными мышиными антителами:
- Исходную суспензию Dynabeads (Dynabeads M-280, покрытые овечьим антителами против мышиного IgG, Invitrogen; Prod. No. 112.02) осторожно встряхивали, чтобы не допустить образования пены.
- 1 мл асептически извлекали и переносили в реакционный сосуд объемом 1,5 мл;
- Шарики dynabeads промывали 3 раза по 5 мин, используя 1 мл буфера для промывки при иммунопреципитации (IP) (буфер для промывки при IP: PBS (20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4), 0,1% (масс./об.) БСА).
Во время процедуры промывки надосадок удаляли осторожно, при этом шарики dynabeads иммобилизовали на стенках реакционного сосуда, используя подставку магнитного сепаратора (MSS).
- Промытые шарики dynabeads инкубировали с 40 мкг A-бета-антител в 1 мл PBS, 0,1% (мас./об.) БСА.
- Активацию осуществляли посредством инкубации в течение ночи при встряхивании при 4ºC.
- Активированные шарики dynabeads промывали 4 раза по 30 мин (снова используя MSS) 1 мл буфера для промывки при IP (PBS (20 мМ NaH2PO4, 140 мМ NaCl, pH 7,4), 0,1% (мас./об.) БСА).
- Активированные шарики dynabeads ресуспендировали в 1 мл PBS, 0,1% (мас./об.) БСА, 0,02% (мас./об.) Na-азида; встряхивали и непродолжительно центрифугировали.
- Активированные антителами шарики dynabeads хранили при 4ºC вплоть до применения.
Приготовление образцов СМЖ:
400 мкл СМЖ пациента с болезнью Альцгеймера добавляли к 4 мкл полной смеси ингибиторов протеаз (Roche Inc. No. в каталоге: 1697498, 1 таблетку растворяли в 1 мл воды) и 0,8 мкл 500 мМ PMSF, растворенного в метаноле. Через 10 мин добавляли 1,6 мл 20 мМ NaH2PO4, 140 мМ NaCl, 0,05% твин-20, pH 7,4 (PBST).
Иммунопреципитация формы A-бета из СМЖ человека с AD:
- Аликвоту объемом 250 мкл приготовленного образца СМЖ добавляли к 25 мкл суспензии анти-Aβ-Dynabeads.
- Иммунопреципитацию осуществляли при перемешивании при 6ºC в течение 16 часов. Последующую промывку шариков осуществляли 3 раза по мин 1 мл PBS/0,1% (мас./об.) БСА и, наконец, один раз в течение 3 мин 1 мл 10 мМ трис/HCl-буфером, pH 7,5. Во время процедуры промывки надосадок удаляли осторожно, при этом шарики dynabeads иммобилизовали на стенках реакционного сосуда, используя подставку магнитного сепаратора (MSS).
Оставшийся надосадок тщательно удаляли после конечной стадии промывки. Пептиды A-бета и соответствующее антитело удаляли с шариков Dynabeads добавлением 25 мкл буфера для образцов без β-меркаптоэтанола (0,36 М бистрис, 0,16 М бицин, 1% SDS (мас./об.), 15% (мас./об.) сахароза, 0,004% (мас./об.) бромфеноловый синий), перенося их в пробирку Eppendorff и нагревая в течение 5 мин при 95ºC в нагревательном устройстве. После охлаждения до комнатной температуры шарики dynabeads иммобилизовали на стенках реакционного сосуда, используя подставку магнитного сепаратора (MSS), и надосадок переносили в другую пробирку Eppendorff (IP-элюат).
Анализ иммунопреципитатов A-бета в ПААГ с мочевиной с последующим Вестерн-блот-анализом:
Количественную оценку форм Aβ1-40 и Aβ1-42 осуществляли с помощью системы для электрофореза в полиакриламидном геле с 8 М мочевиной и последующим Вестерн-блот-анализом согласно способу, впервые описанному H. W. Klafki et al., Analytical Biochemistry 237, 24-29 (1996) и позднее также использованному J. Wiltfang et al., J. of Neurochemistry 81: 481-496, 2002. Были осуществлены только два небольших изменения экспериментальной процедуры:
1) Концентрацию SD в концентрирующем геле доводили до 0,25% (мас./об.), вместо 0,1% (мас./об.).
2) В случае Вестерн-блота антитело 1E8 (Senetek Drug Delivery Technologies Inc. St. Louis, MO, USA) заменяли мышиным IgG-мАт против амилоида β человека (N) (82E1) (IBL, No. в каталоге: 10323).
Аликвоты IP-элюата объемом 15 мкл из иммунопреципитированных образцов наносили на ПААГ с 8 М мочевиной. Электрофорез проводили при 100 В (15 мин) и продолжали при 60 В. Электрофорез останавливали, когда движущийся фронт синего красителя нанесенного образца был еще на расстоянии 0,5 см от конца геля.
Способ Вестерн-блоттинга:
Вестерн-блот-анализ осуществляли в камере для полусухого блоттинга (BioRad Inc., 45 мин при 75 мА) на нитроцеллюлозу 7,5 см x 9 см, 0,45 мкм (BioRad Inc.).
Буфер для блоттинга: 6 г трис; 28,1 г глицина; 500 мл метанола; доводят до 2,5 л водой.
Нитроцеллюлозный блот кипятили в течение 10 мин в PBS при 100ºC. Блот насыщали обработкой 50 мл 5% (мас./об.) БСА в PBST в течение 1 ч при комнатной температуре. После удаления жидкой фазы последующую стадию промывки осуществляли дважды: 50 мл TTBS (буфер 25 мМ трис/HCl; 150 мМ NaCl; 0,05% твин-20; pH 7,5), в течение 10 мин при комнатной температуре и затем 50 мл TBS (буфер 25 мМ трис/HCl; 150 мМ NaCl; pH 7,5) в течение 10 мин при комнатной температуре.
Для последующего проявления конечный буфер для промывки удаляли с блота и добавляли 15 мл раствора антитела I (0,2 мкг/мл 82E1 = 1:500 в растворе 3% (мас./об.) порошка обезжиренного молока (Lasana Inc.) в 15 мл TBS) в течение 20 часов при 6ºC. После удаления буфера осуществляли три стадии промывки, как описано выше. Блот инкубировали с раствором антитела II (разведение 1:10000 анти-мышиное антитело-POD в 15 мл 3% (мас./об.) порошка обезжиренного молока в 15 мл TBS) в течение 1 часа при комнатной температуре. После удаления буфера осуществляли три стадии промывки, как описано выше.
После удаления последнего буфера для промывки смешивали 2 мл субстрата с усилителем (Super Signal West Femto Maximum Sensitivity Substrate Enhancer) и 2 мл раствора пероксида. Свежеприготовленный раствор выливали на блот, который предварительно инкубировали в темноте в течение 5 мин. Хемилюминесценцию регистрировали, используя систему визуализации VersaDoc (BioRad).
Параметры визуализации:
- время экспонирования 180 с.
Регистрация картины через 30 с, 60 с, 120 с и 180 с.
Результаты получали на основании картины, соответствующей экспонированию в течение 180 с.
Приведенные выше результаты свидетельствуют, что избирательное по отношению к глобуломеру антитело, подобное 8F5 или 8C5, по сравнению с неизбирательным по отношению к глобуломеру антителом, подобным 6E10, связывается с большим количеством Aβ42, чем с Aβ40, в СМЖ человека. Полученный результат свидетельствует об успешном лечении болезни Альцгеймера, поскольку, как отмечено выше, преимущественное элиминирование Aβ42 по сравнению с Aβ40 согласуется с концепцией лечения AD (например, с применением R-флубипрофена (см. выше)).
ПРИМЕР V
8F5 УЛУЧШАЕТ РАСПОЗНАВАНИЕ НОВОГО ОБЪЕКТА У ТРАНСГЕННЫХ МЫШЕЙ APP
Чтобы проверить положительное влияние на когнитивную способность посредством нейтрализации внутреннего эпитопа глобуломера Aβ(1-42) антителом 8F5 осуществляли эксперимент по пассивной иммунизации трансгенных мышей APP, в котором мышей тестировали в отношении их способности запоминать объекты, которые они исследовали раньше. После некоторого периода задержки между первой и второй встречей с объектами трансгенные мыши APP не способны распознавать уже исследованный объект. Указанный эксперимент основан на природном любопытстве животных и значительное отсутствие интереса к уже исследованному объекту свидетельствует об узнавании объекта.
ПРИМЕР V.1: ПОВЫШЕНИЕ ИНДЕКСА РАСПОЗНАВАНИЯ МОНОКЛОНАЛЬНЫМ АНТИТЕЛОМ 8F5:
Животные:
Использовали самок мышей одной трансгенной мышиной модели болезни Альцгеймера на генетическом фоне FVB x C57Bl (APP/L, ReMYND, Leuven, Belgium) и негативное потомство одного помета в качестве контролей дикого типа от скрещивания FVB x C57Bl в возрасте 3 месяцев. Всех мышей генотипировали, используя полимеразную цепную реакцию (ПЦР), в возрасте 3 недель и давали уникальные идентификационные номера после того, как результаты ПЦР были известны и дважды проверены с использованием второй ПЦР перед началом исследования. Всех мышей рандомизировали и подбирали по возрасту, т.е. им давали случайный номер с помощью компьютера и случайным образом распределяли для обработки. Животных содержали в группах обработки 18 дней до начала исследования, чтобы позволить им ознакомиться с новым окружением в клетке. Мыши имели свободный доступ к предварительно профильтрованной и стерильной воде (УФ-лампа) и стандартному корму для мышей. Корм хранили в сухих и прохладных условиях в хорошо проветриваемом помещении для хранения. Количество воды и корма проверяли ежедневно, вносили при необходимости и пополняли дважды в неделю. Мышей содержали в обратном ритме день-ночь: 14 часов освещения/10 часов темноты, начиная в 7 часов вечера, в стандартных металлических клетках типа RVS T2 (площадь 540 см2). Клетки оборудовали твердым дном и стелили слой подстилки. Количество мышей в клетке ограничивали согласно законодательству об охране животных. За пять дней до начала тестирования поведения мышей помещали в клетки типа 2 Macrolon и переносили в лабораторию, чтобы адаптировать к лабораторным условиям при подготовке к тестированию поведения.
Обработка (пассивная иммунизация):
Осуществляли три отдельных эксперимента, в которых мыши (по меньшей мере, 9 в группе) получали внутрибрюшинные инъекции (500 мкг в 240 мкл/мышь) в 1, 8 и 15 дни. Мышей обрабатывали моноклональными антителами 6G1, 8P5 и другими неописанными антителами, которые все растворяли в фосфатно-солевом буфере, или обрабатывали 320 мкл фосфатно-солевого буфера.
Тест распознавания нового объекта:
Тест распознавания нового объекта осуществляли в день третьей обработки. В применяемом протоколе следовали способу, описанному Dewachter et al. (Journal of Neuroscience, 2002, 22(9): 3445-3453). Мышей знакомили в течение одного часа с камерой «открытое поле» из плексигласа (52 × 52 × 40 см) с черными вертикальными стенками и полупрозрачным полом, тускло освещенной лампой, помещенной снизу камеры. На следующий день животных помещали в ту же камеру и подвергали 10-минутному испытанию по обнаружению объектов. Во время указанного испытания мышей помещали по отдельности в отрытое поле в присутствии 2 идентичных объектов A (оранжевый цилиндр или зеленый куб сходного размера ± 4 см) и регистрировали продолжительность (времяAA) и частоту (частотаAA) исследования объекта A (когда морда животного была направлена к объекту на расстоянии <1 см и мыши активно обнюхивали объект) с помощью компьютеризованной системы (Ethovision, Noldus information Technology, Wageningen, Netherlands). В течение 10-минутного испытания памяти (второе испытание), осуществляемого спустя 2,5 часа, помещали новый объект (объект B, зеленый куб или оранжевый цилиндр) вместе со знакомым объектом (объект A) в открытое поле (частотаA и частотаB и времяA и времяB соответственно). Индекс распознавания (RI), определяемый в виде отношения продолжительности исследования нового объекта по сравнению с продолжительностью исследования обоих объектов [времяB/(времяA + времяB) × 100], использовали для измерения непространственной памяти. Продолжительность и частоту исследования объекта A в ходе испытания памяти (времяAA и частотаAA) использовали для измерения любопытства.
Анализ данных осуществляли, объединяя данные для трансгенных мышей APP, которые получали моноклональные антитела 6G1 или 8F5 или фосфатно-солевой буфер, и нетрансгенного потомства одного помета, которые получали фосфатно-солевой буфер, из всех трех исследований (фиг. 4). Мыши, которые не различали старый объект и новый объект, имели индекс распознавания 50. Мыши, которые узнают старый объект, предпочтительно будут исследовать новый объект, и поэтому индекс распознавания становится > 50. Мыши, которые исследуют только новый объект, имеют индекс распознавания, равный 100. Средний индекс распознавания в группе сравнивали со случайным уровнем, т.е. 50, используя t-критерий. Средний индекс распознавания всех групп также сравнивали посредством ANOVA с последующим использованием апостериорного t-критерия. Различие между группой, обработанной PBS, и группой дикого типа свидетельствует о нарушении когнитивной способности у трансгенных мышей APP в данной модели. Мыши, которым инъецировали PBS, выполняли задачу на уровне случайности (т.е. не значимое отличие от 50), тогда как все другие мыши показали распознавание объектов (фиг. 4: звездочки). В том случае, когда выполнение задачи трансгенными мышами APP, обработанными антителами, сравнивали с контрольными группами, обнаружили значимое отличие по сравнению с мышами, обработанными PBS, но не по сравнению с мышами дикого типа (фиг. 4: кружки), что свидетельствует о том, что обработка антителами восстанавливала нарушенную когнитивную способность у указанных трансгенных мышей APP.
ПРИМЕР VI
АНАЛИЗ IN SITU СПЕЦИФИЧНОГО ВЗАИМОДЕЙСТВИЯ АНТИТЕЛ 8F5 И 8C5 С ФИБРИЛЛЯРНЫМ ПЕПТИДОМ АМИЛОИДА БЕТА В ФОРМЕ АМИЛОИДНЫХ БЛЯШЕК И АМИЛОИДА В МЕНИНГЕАЛЬНЫХ СОСУДАХ У СТАРЫХ ТРАНСГЕННЫХ МЫШЕЙ APP И У ПАЦИЕНТОВ С БОЛЕЗНЬЮ АЛЬЦГЕЙМЕРА
Антитела 8F5 и 8C5 давали пониженное окрашивание отложений фибриллярного пептида Aβ, что свидетельствует о том, что их терапевтическое влияние опосредовано связыванием с растворимыми глобуломерными формами, а не с фибриллярными формами отложений пептида Aβ. Так как связывание антител с фибриллярным пептидом Aβ может приводить к быстрому растворению агрегатов и последующему увеличению концентрации растворимого Aβ, который в свою очередь, как полагают, является нейротоксичным и может приводить к микрогеморрагиям, то предпочтительной является терапия антителами, которая влияет на растворимый глобуломер, а не на мономер.
Способы:
Для указанных экспериментов использовали несколько образцов материала головного мозга: ткань коры головного мозга 2 пациентов с AD (RZ16 и RZ55) и ткань коры головного мозга 19-месячных мышей Tg2576 (APPSWE № 001349, Taconic, Hudson, NY, USA) или 12-месячных мышей APP/L (ReMYND, Leuven, Belgium).
Мыши сверхэкспрессируют APP человека с семейной мутацией болезни Альцгеймера и отложениями формы β-амилоида в паренхиме головного мозга примерно в 11-месячном возрасте и отложениями β-амилоида в более крупных сосудах головного мозга примерно в 18-месячном возрасте. Животных глубоко анестезировали и проводили транскардиальную перфузию 0,1 М фосфатно-солевым буфером (PBS), чтобы промыть от крови. Затем головной мозг извлекали из черепа и разделяли в продольном направлении. Одну полусферу головного мозга мгновенно замораживали, а другую фиксировали погружением в 4% параформальдегид. Фиксированную погружением полусферу подвергали криопротекции вымачиванием в 30% сахарозе в PBS и помещали на замораживающий микротом. Делали поперечные срезы всего переднего мозга толщиной 40 мкм, которые собирали в PBS и использовали для последующей процедуры окрашивания. Образцы неокортекса пациентов с болезнью Альцгеймера получали из Brain-Net, Munich, Germany в виде замороженной ткани, фиксировали погружением в 4% параформальдегид во время размораживания и затем обрабатывали подобно обработке тканей мышей.
Отдельные срезы красили конго красным, используя следующий протокол:
Материалы:
- набор с красителем амилоида конго красным (Sigma-Aldrich; HT-60), состоящим из спиртового раствора NaCl, раствора NaOH и раствора конго красного;
- кюветы для окрашивания;
- предметные стекла SuperfrostPlus и покровные стекла;
- этанол, ксилол, среда для заливки.
Реагенты:
- разбавление NaOH 1:100 раствором NaCl дает щелочной раствор соли;
- разбавление щелочного раствора соли 1:100 раствором конго красного дает щелочной раствор конго красного (готовят не более чем за 15 мин до применения, фильтруют);
- срезы помещают на стекла и дают им возможность высохнуть;
- инкубируют стекло в кювете для окрашивания, сначала в течение 30-40 минут в щелочном растворе соли, затем в течение 30-40 минут в щелочном растворе конго красного;
- три раза промывают свежим этанолом и заливают ксилолом.
Окрашивание сначала фотографировали, используя микроскоп Zeiss Axioplan (Zeiss, Jena, Germany), и оценивали количественно. Красная окраска свидетельствовала об отложениях амилоида и в форме бляшек, и в более крупных менингеальных сосудах. Затем оценку окрашивания антителами фокусировали на указанных структурах.
Окрашивание осуществляли посредством инкубации срезов в растворе, содержащем 0,07 - 0,7 мкг/мл соответствующего антитела, согласно следующему протоколу:
Материалы:
- раствор для промывки TBST (трис-забуференный физиологический раствор, содержащий твин-20; 10x-концентрат; DakoCytomation S3306, DAKO, Hamburg, Germany) 1:10 в очищенной воде Aqua bidest);
- 0,3% H2O2 в смеси метанол-сыворотка осла (Serotec, Dusseldorf, Germany), 5% в TBST, в качестве блокирующей сыворотки;
- моноклональные мышиные антитела против глобуломера, разбавленные в заданных концентрациях в TBST;
- второе антитело: биотинилированное антитело осла против Ig мыши (Jackson Immuno/Dianova, Hamburg, Germany; 715-065-150; разбавленное 1:500 в TBST);
- StreptABComplex (DakoCytomation K 0377, DAKO, Hamburg, Germany);
- диаминобензидин из набора для субстрата пероксидазы (=DAB; SK-4100; Vector Laboratories, Burlingame, CA, USA);
- предметные стекла SuperFrost Plus и покровные стекла;
- не содержащая ксилола среда для заливки (Medite, Burgdorf, Germany; X-tra Kitt).
Способ:
- переносили плавающие срезы в ледяную 0,3% H2O2 и инкубировали в течение 30 мин;
- промывали в течение 5 мин TBST-буфером;
- инкубировали со смесью сыворотка осла/TBST в течение 20 минут;
- инкубировали с первым антителом в течение 24 часов при комнатной температуре;
- промывали в TBST-буфере в течение 5 минут;
- инкубировали с блокирующей сывороткой в течение 20 минут;
- промывали в TBST-буфере в течение 5 минут;
- инкубировали со вторым антителом в течение 60 минут при температуре окружающей среды;
- промывали в TBST-буфере в течение 5 минут;
- инкубировали с StreptABComplex в течение 60 минут при температуре окружающей среды;
- промывали в TBST-буфере в течение 5 минут;
- инкубировали с DAB в течение 20 минут;
- срезы помещали на предметные стекла, стекла сушили на воздухе, стекла дегидратировали спиртом, и осуществляли заливку стекол.
Кроме визуального исследования срезов в микроскопе окрашивание амилоида дополнительно количественно оценивали, оптически вырезая 10 случайно выбранных бляшек из гистологических изображений с использованием системы анализа изображения ImagePro 5.0 и определяя их среднее значение по серой шкале. Значения оптической плотности рассчитывали на основании значений по серой шкале посредством вычитания средней фоновой плотности стандартного материала из плотности амилоидных бляшек (0% - не окрашивания бляшек выше окружающего фона, 100% - нет пропускания/максимальное окрашивание). Проверяли статистическую значимость различий между антителами 6E10/4G8 и 6G1, 8C5 и 8F5, соответственно, используя ANOVA.
Результаты:
Доказано, что весь описанный окрашенный антителами материал представляет собой конгофильные амилоидные отложения (фиг. 7(A)). Избирательные по отношению к глобуломеру антитела 8F5 и 8C5 окрашивали паренхимные и менингиальные конгофильные отложения Aβ-пептида значительно меньше, чем антитела 6G1 и 6E10 (фиг. 7 (B)-(C), (H)). Количественный анализ окрашивания паренхимных амилоидных бляшек выявил связывание всех антител с бляшками (статистически значимая плотность выше контроля), но связывание антител 8F5 и 8C5 было значительно меньше, чем связывание эталонного антитела 6E10 (продуцируемого к N-концевой последовательности Aβ), и такое же или ниже, чем эталонного антитела 4G8 (продуцируемого к N-концевой последовательности Aβ) (фиг. 7(D) -фиг. (G)). Антитела 8F5 и 8C5 меньше связываются с амилоидными отложениями, чем антитела, которые узнают мономер Aβ или часть последовательности Aβ. Обработка антителами, связывающимися с фибриллярным пептидом Aβ, может приводить к быстрому растворению амилоидных бляшек в ткани головного мозга и последующему увеличению концентрации растворимого Aβ, который в свою очередь, как полагают, является нейротоксичным и может приводить к микрогеморрагиям и/или к быстрому растворению амилоида в сосудах, что также может приводить к микрогеморрагиям. Поэтому терапия антителами, которые влияют на растворимый глобуломер, а не на мономер, является предпочтительной.






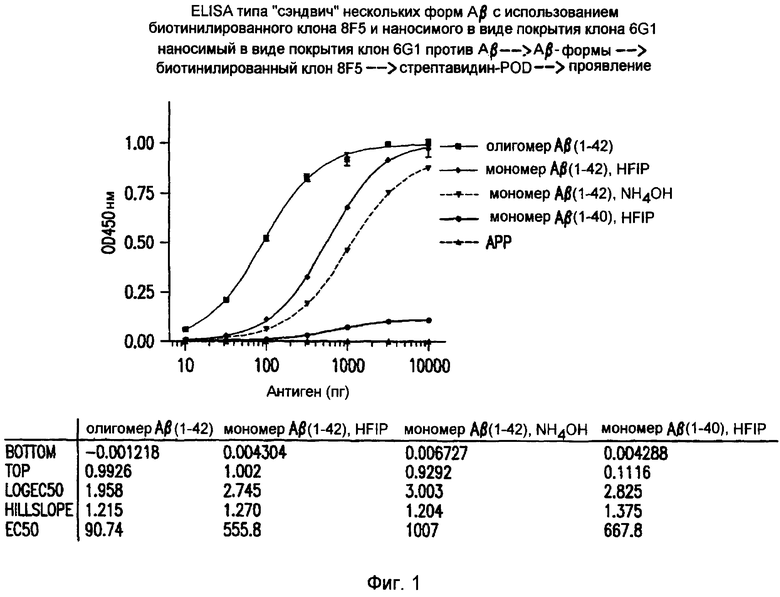

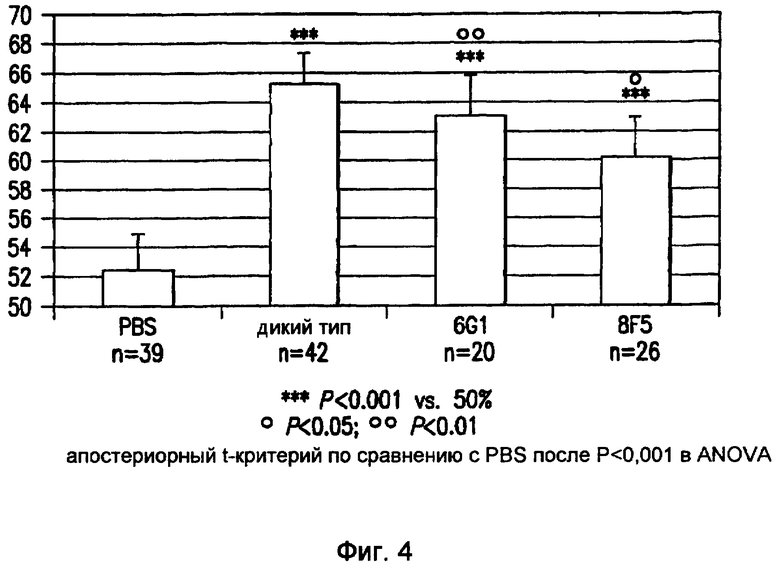




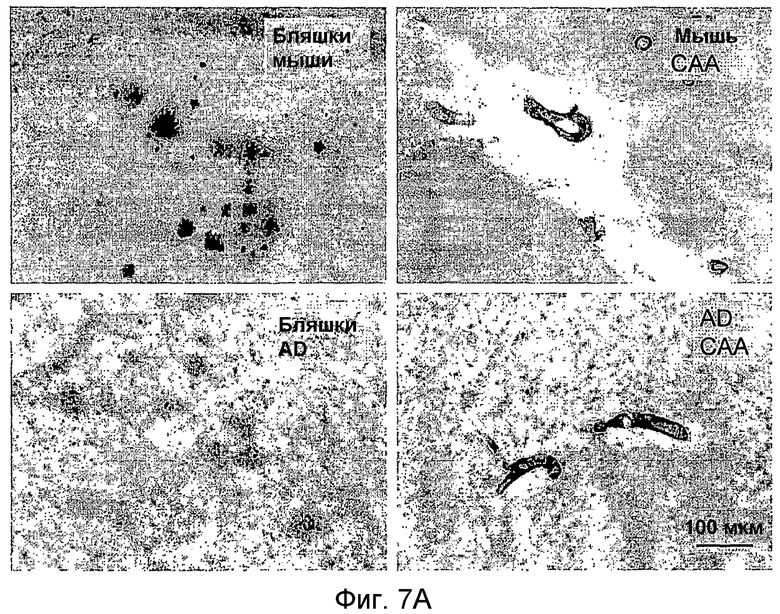
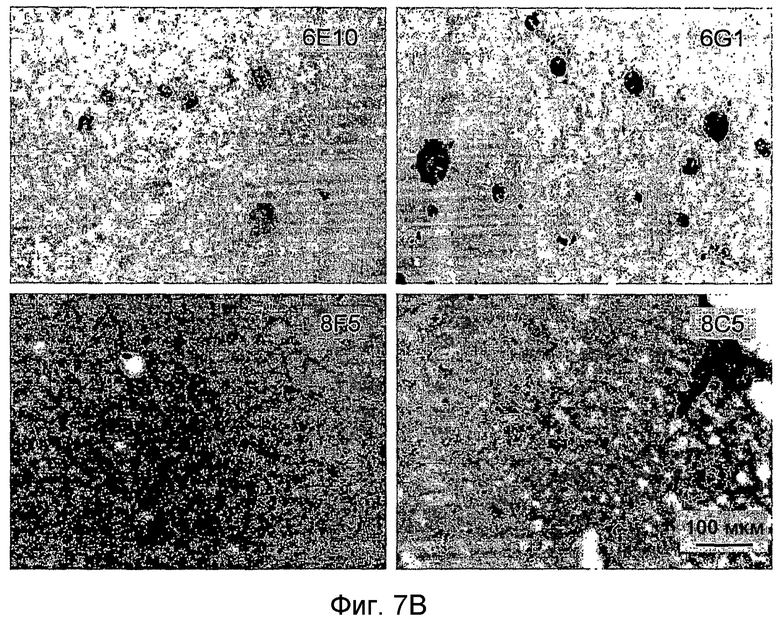
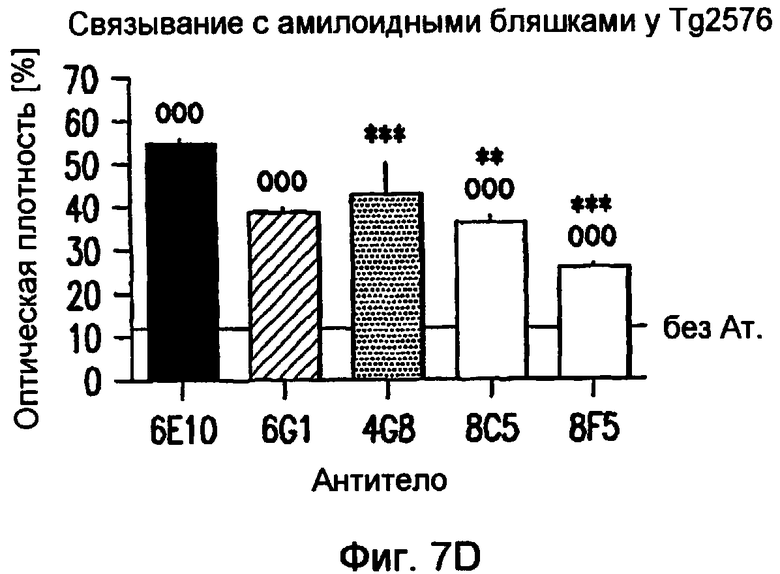
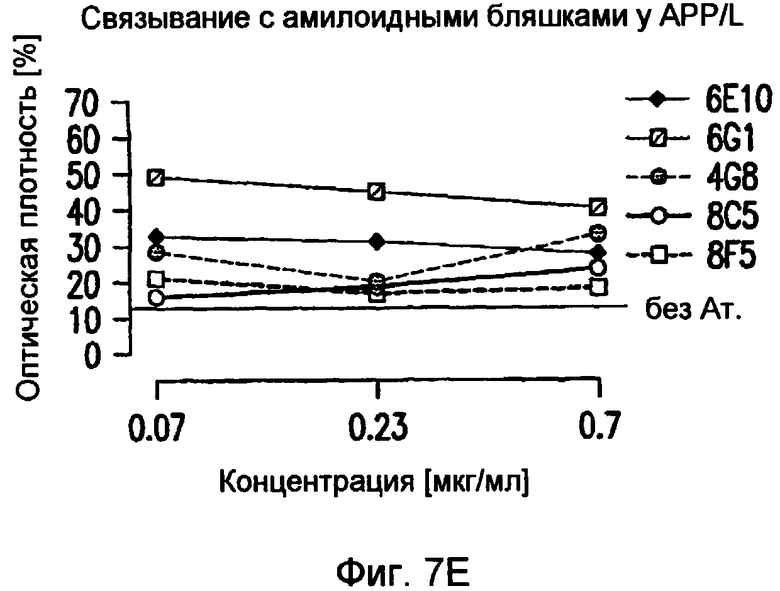
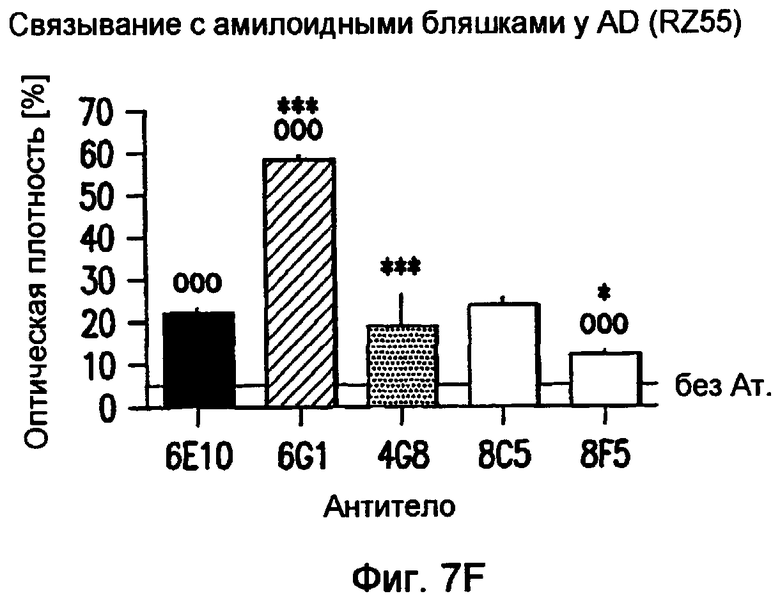
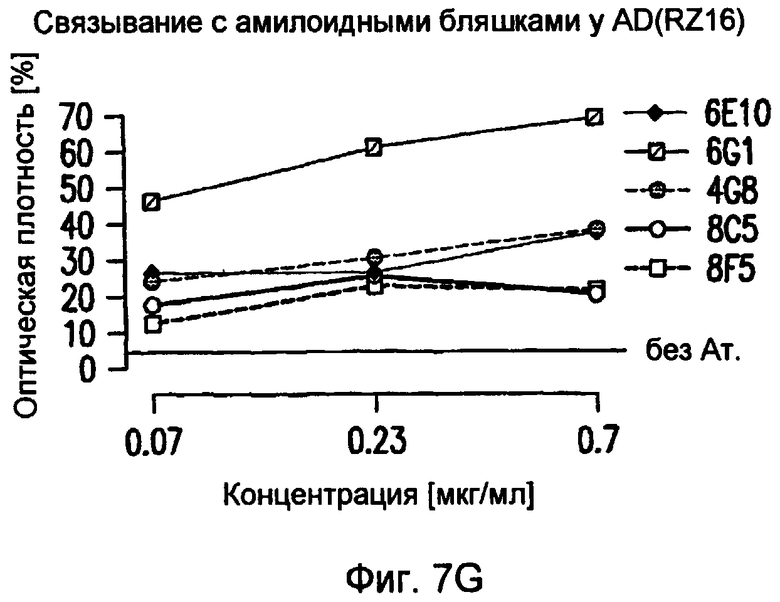
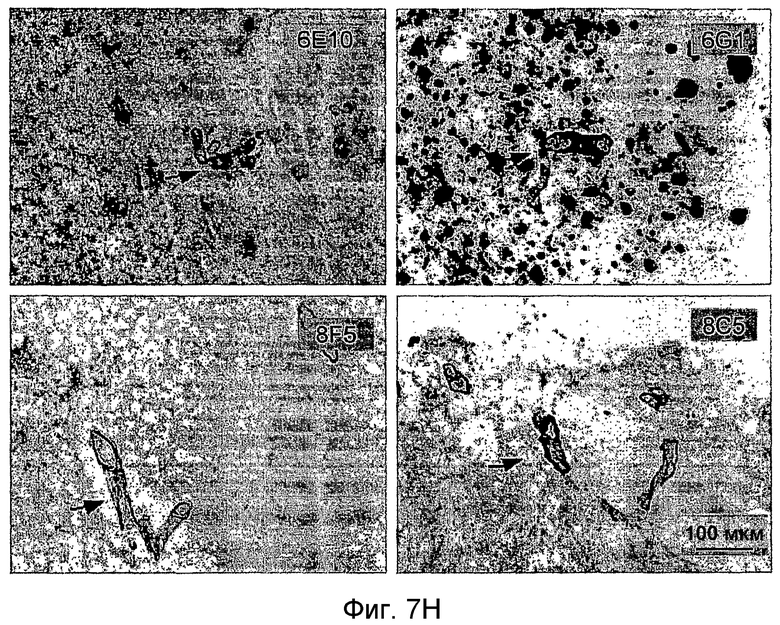
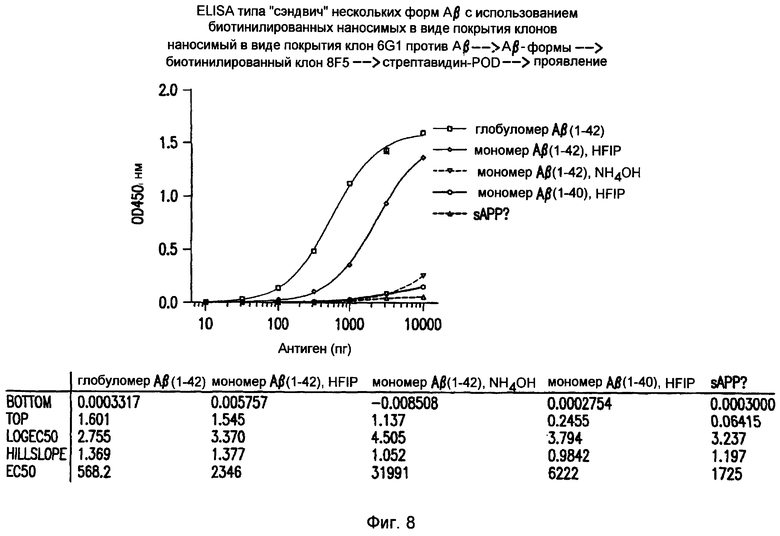
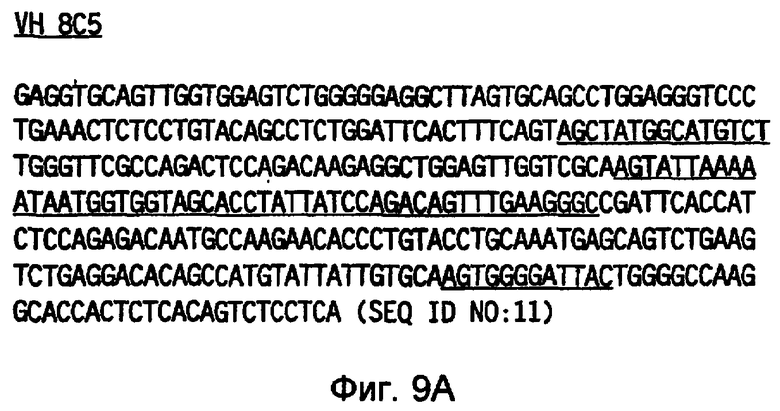
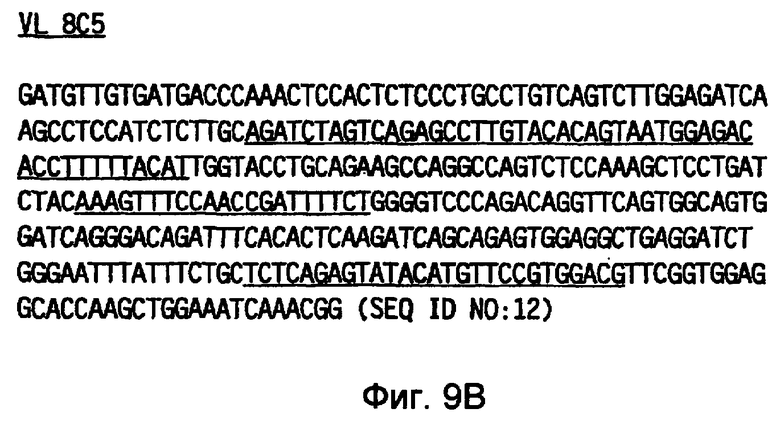


Изобретение относится к биотехнологии и иммунологии. Предложено моноклональное антитело, которое связывается с глобуломером Аβ, содержащее CDRs тяжелой и легкой цепи. Описаны композиция для диагностики болезни Альцгеймера, а также композиция и вакцина для профилактики или лечения болезни Альцгеймера на основе антитела. Раскрыто применение антитела для получения лекарственного средства по тому же назначению. Описаны варианты способа диагностики болезни Альцгеймера, набор для диагностики болезни Альцгеймера на основе антитела. Использование изобретения обеспечивает новые антитела к глобуломеру Аβ, которые выявляют более избирательно Аβ (1-42), чем известные антитела. 8 н. и 8 з.п. ф-лы, 10 ил., 1 табл.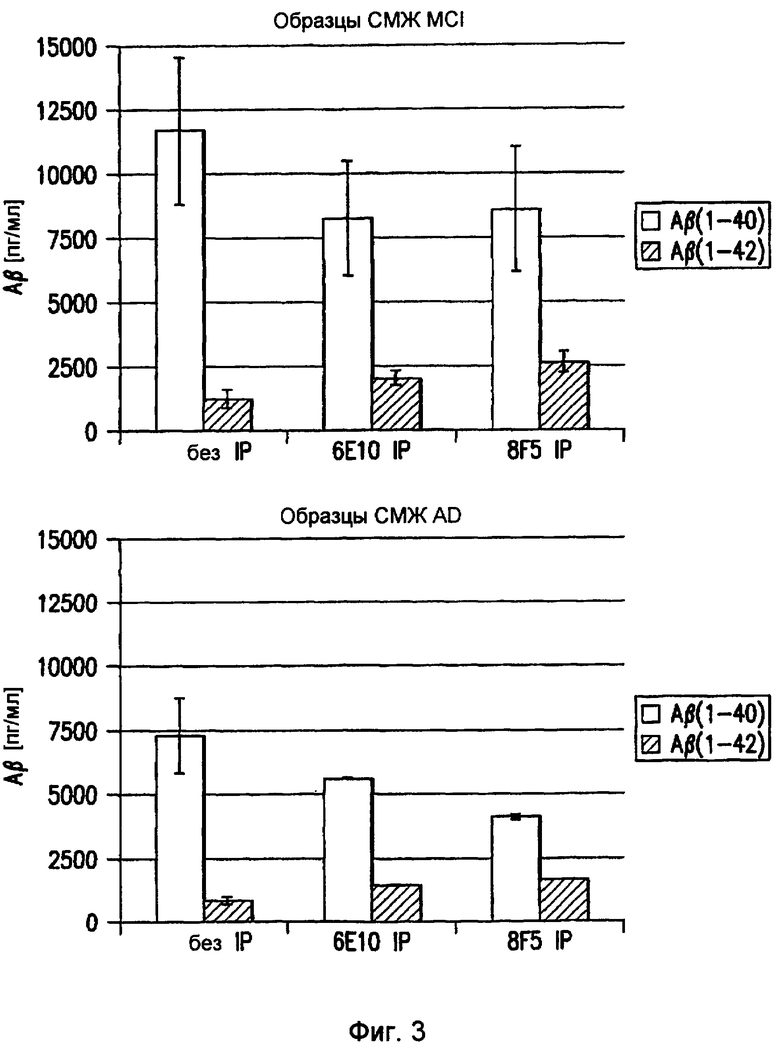
1. Моноклональное антитело, которое связывается с глобуломером Аβ, содержащее:
а) вариабельную тяжелую цепь, в которой три определяющие комплементарность области (CDR) указанной вариабельной тяжелой цепи представляют собой SEQ ID NO:5, SEQ ID NO:6 и SEQ ID NO:7; и
б) вариабельную легкую цепь, в которой три CDR указанной вариабельной легкой цепи представляют собой SEQ ID NO:8, SEQ ID NO:9 и SEQ ID NO:10.
2. Моноклональное антитело по п.1, содержащее вариабельную тяжелую и легкую цепь как определено в п.1.
3. Моноклональное антитело по п.1 или 2, содержащее:
а) вариабельную тяжелую цепь, кодируемую последовательностью SEQ ID NO:1, или вариабельную тяжелую цепь, которая по меньшей мере на 90% идентична ей; или
б) вариабельную легкую цепь, кодируемую последовательностью SEQ ID NO:2, которая по меньшей мере на 90% идентична ей; или
c) SEQ ID NO:3 или последовательность, которая по меньшей мере на 90% идентична ей; или
d) SEQ ID NO:4 или последовательность, которая по меньшей мере на 90% идентична ей.
4. Моноклональное антитело по п.3, содержащее вариабельную тяжелую и легкую цепь, как определено в п.3.
5. Моноклональное антитело по п.3, содержащее SEQ ID NO:3 или последовательность, которая по меньшей мере на 90% идентична ей, и SEQ ID NO:4 или последовательность, которая по меньшей мере на 90% идентична ей.
6. Моноклональное антитело по пп.1-5, которое связывается с большей специфичностью с глобуломером белка амилоида-бета (Аβ), чем с мономером белка амилоида-бета.
7. Моноклональное антитело по пп.1-5, где указанное антитело продуцируется гибридомой с номером РТА-7238 или РТА-7407 Американской коллекции типовых культур.
8. Моноклональное антитело по любому из пп.1-5, где указанное антитело является антителом человека или гуманизированным антителом.
9. Моноклональное антитело по любому пп.1-5, которое используется для лечения или профилактики болезни Альцгеймера.
10. Композиция для диагностики болезни Альцгеймера, содержащая эффективное количество антитела по любому из пп.1-8.
11. Композиция для профилактики или лечения болезни Альцгеймера, содержащая эффективное количество антитела по любому из пп.1-8.
12. Вакцина для профилактики или лечения болезни Альцгеймера, содержащая эффективное количество антитела по любому из пп.1-8 и фармацевтически приемлемый адъювант.
13. Применение антитела по любому из пп.1-8 для получения лекарственного средства для лечения или профилактики болезни Альцгеймера.
14. Способ диагностики болезни Альцгеймера у пациента с подозрением на наличие указанной болезни, включающий стадии:
а) приведения в контакт биологического образца, взятого у пациента, с антителом по любому из пп.1-8 в течение периода и при условиях, достаточных для образования комплексов антиген/антитело; и
b) выявления указанных комплексов антиген/антитело в указанном образце, при этом наличие указанных комплексов свидетельствует о наличии болезни Альцгеймера у указанного пациента.
15. Способ диагностики болезни Альцгеймера у пациента с подозрением на наличие указанной болезни, включающий стадии:
a) приведения в контакт биологического образца, взятого у пациента, с антигеном в течение периода и в условиях, достаточных для образования комплексов антиген/антитело;
b) добавления конъюгата к полученным комплексам антиген/антитело в течение периода и при условиях, достаточных для обеспечения связывания указанного конъюгата со связанным антителом, при этом указанный конъюгат содержит антитело по любому из пп.1-8, при этом указанный конъюгат связан с генерирующим сигнал соединением, способным генерировать регистрируемый сигнал; и
c) выявления указанных комплексов антитело/антиген в указанном образце, при этом наличие указанных комплексов свидетельствует о наличии болезни Альцгеймера у указанного пациента.
16. Набор для диагностики болезни Альцгеймера, содержащий:
a) антитело по любому из пп.1-8 и
b) конъюгат, содержащий антитело, связанное с генерирующим сигнал соединением, при этом указанное антитело указанного конъюгата отличается от указанного антитела по любому из пп.1-8 или представляет собой антиантитело против антитела по любому из пп.1-8.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| BARGHORN ет al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| - Journal of neurochemistry, 2005; 95(3), стр.834-847 | |||
| СПОСОБ ЛЕЧЕНИЯ БОЛЕЗНИ АЛЬЦГЕЙМЕРА И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ КЛИОХИНОЛА ДЛЯ ЕЕ ЛЕЧЕНИЯ | 1997 |
|
RU2193406C2 |

