ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
1. Область техники, к которой относится изобретение
Настоящее раскрытие относится к области фармацевтики; более конкретно, к многофункциональным молекулярным конструкциям, например, имеющим нацеливающие и эффекторные элементы для доставки эффектора (например, терапевтического лекарственного средства) в участки-мишени.
2. Описание предшествующего уровня техники
Непрерывное усовершенствование широкого спектра методов скрининга и отбора моноклональных антител (mAb) для антигенов-мишеней способствовало разработке большого количества терапевтических антител против многих заболеваний, которые считались неизлечимыми всего несколько лет назад. Согласно Therapeutic Antibody Database, примерно 2800 антител были исследованы или планируется исследовать в клинических испытаниях с участием людей, и примерно 80 антител были одобрены государственными органами по регулированию оборота лекарственных средств для клинического применения. Большое количество данных о терапевтическом воздействии антител обеспечило информацию, касающуюся фармакологических механизмов действия антител в качестве терапевтических средств.
Один из основных фармакологических механизмов действия антител в качестве терапевтических средств заключается в том, что антитела могут нейтрализовать или захватывать болезнетворные медиаторы, которые могут представлять собой цитокины или компоненты иммунной системы, присутствующие в кровотоке, интерстициальном пространстве или в лимфатических узлах. Нейтрализующая активность ингибирует взаимодействие болезнетворных медиаторов с их рецепторами. Следует отметить, что в качестве терапевтических средств также были разработаны белки слияния на основе растворимых рецепторов или внеклеточных частей рецепторов цитокинов и Fc-части IgG, которые действуют путем нейтрализации цитокинов или иммунных факторов подобно нейтрализующим антителам.
Некоторые терапевтические антитела, которые были одобрены для клинического применения или подвергались клиническим разработкам, опосредуют свои фармакологические эффекты путем связывания с рецепторами, блокируя таким образом взаимодействие рецепторов с их лигандами. Для этих лекарственных средств на основе антител Fc-опосредованные механизмы, такие как антителозависимая клеточная цитотоксичность (ADCC) и комплемент-опосредованный цитолиз (CMC), не являются намеченными механизмами действия антител.
Некоторые терапевтические антитела связываются с определенными поверхностными антигенами на клетках-мишенях и оказывают воздействие на клетки-мишени посредством Fc-опосредованных функций и других механизмов. Наиболее важными Fc-опосредованными механизмами являются: антителозависимая клеточная цитотоксичность (ADCC) и комплемент-опосредованный цитолиз (CMC), которые вызывают лизис клеток-мишеней, связанных с антителами. Некоторые антитела, связывающиеся с определенными антигенами клеточной поверхности, могут индуцировать апоптоз связанных клеток-мишеней.
Идея и метод получения антител с двойной специфичностью созрели более чем три десятилетия назад. В последний год усовершенствование методов создания рекомбинантных антител и стремление к разработке улучшенных медицинских препаратов стимулировали разработку биспецифических антител, демонстрирующих большое разнообразие структурных форм.
Например, двухвалентные или поливалентные антитела могут содержать два или более антиген-связывающих участка. Были сообщения о ряде способов получения поливалентных антител путем ковалентного связывания трех или четырех Fab-фрагментов с помощью присоединяющей структуры. Например, были созданы антитела, в которых представлены тандемные повторы из трех или четырех Fab.
Были раскрыты некоторые способы получения поливалентных антител путем использования синтетических сшивающих средств для объединения различных антител или связывающих фрагментов химическим путем. Один подход включает химическую сшивку трех, четырех и более отдельных Fab-фрагментов с помощью различных линкеров. Был представлен другой способ получения конструкции с несколькими Fab, собранными в одномерный ДНК-каркас. Эти различные поливалентные конструкции Ab, предназначенные для связывания с молекулами-мишенями, отличаются друг от друга по размеру, показателям периода полувыведения, гибкости конформации и способности модулировать иммунную систему. Ввиду вышеизложенного было сделано несколько сообщений о получении молекулярных конструкций с фиксированным количеством эффекторных элементов, или с двумя или более различными видами функциональных элементов (например, по меньшей мере одним нацеливающим элементом и по меньшей мере одним эффекторным элементом). Однако построение молекулярной конструкции с конкретной комбинацией нацеливающих и эффекторных элементов с применением химического синтеза либо рекомбинантной технологии часто является затруднительным. Соответственно, в уровне техники существует необходимость в обеспечении новых молекулярных платформ для построения более универсальной молекулы, подходящей для применения в широком диапазоне заболеваний.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее представлено упрощенное краткое описание настоящего раскрытия в целях обеспечения базового понимания читателем. Данное краткое описание не является широким обзором настоящего раскрытия, и в нем не определены ключевые/критически важные элементы настоящего изобретения и не очерчен объем настоящего изобретения. Его единственной целью является представление некоторых идей, раскрытых в данном документе, в упрощенной форме в качестве вступления к более подробному описанию, представленному далее.
<I> Разветвленные линкеры на основе пептидной сердцевины
В первом аспекте настоящее раскрытие направлено на линкерное звено, с которым связано по меньшей мере два функциональных элемента. Например, с линкерным звеном могут быть связаны два различных эффекторных элемента, один нацеливающий элемент и один эффекторный элемент, или один эффекторный элемент и цепь полиэтиленгликоля (PEG) для увеличения времени циркуляции в крови линкерного звена. Линкерное звено по настоящему изобретению сконструировано таким образом, чтобы оно имело по меньшей мере две различные функциональные группы, так чтобы функциональные элементы могли связываться с ним посредством реакции с соответствующими функциональными группами. Соответственно, линкерное звено по настоящему изобретению может служить в качестве платформы для получения молекулярной конструкции с двумя или более функциональными элементами.
В соответствии с различными вариантами осуществления настоящего раскрытия линкерное звено содержит центральную сердцевину и множество связывающих ветвей. Центральная сердцевина представляет собой полипептидную сердцевину, содержащую (1) множество лизиновых (K) остатков, при этом каждый остаток K отделен от следующего остатка K вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки, и количество остатков K находится в диапазоне от 2 до 15; или (2) последовательность (Хаа-K)n, где Хаа представляет собой пегилированную аминокислоту, имеющую 2-12 повторяющихся этиленгликолевых (EG) звеньев, а п представляет собой целое число от 2 до 15. Вставочная последовательность необязательно состоит из 2-20 аминокислотных остатков. В различных вариантах осуществления вставочная последовательность может иметь последовательность GS, GGS, GSG или SEQ ID NO: 1-16. В соответствии с некоторыми вариантами осуществления настоящего раскрытия центральная сердцевина содержит от 2 до 15 звеньев с последовательностью G1-5SK; центральная сердцевина предпочтительно содержит последовательность (GSK)2-15. Каждая из связывающих ветвей связана с остатками K центральной сердцевины посредством образования амидной связи между остатком K и связывающей ветвью. Связывающая ветвь, связанная с центральной сердцевиной, имеет малеимидную, N-гидроксисукцинимидильную (NHS), азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце. Кроме того, аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу; в качестве альтернативы или дополнительно, аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый (С) остаток, в котором тиольная группа аминокислотного остатка связана с соединяющей ветвью, имеющей азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на свободном конце соединяющей ветви.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия, если свободным концом связывающей ветви является азидная, алкиновая или циклооктиновая группа, то аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а свободный конец соединяющей ветви представляет собой тетразиновую или циклооктеновую группу. В соответствии с другими вариантами осуществления настоящего раскрытия, если свободный конец связывающей ветви представляет собой тетразиновую или циклооктеновую группу, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу, или аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а свободный конец соединяющей ветви представляет собой азидную, алкиновую или циклооктиновую группу.
В некоторых вариантах осуществления связывающая ветвь представляет собой цепь PEG, предпочтительно имеющую 2-20 повторяющихся EG-звеньев. В качестве альтернативы, связывающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на ее свободном конце (т.е. на конце, не связанном с остатком K центральной сердцевины). В некоторых вариантах осуществления соединяющая связывающая ветвь представляет собой цепь PEG, предпочтительно имеющую 2-12 повторяющихся EG-звеньев.
Что касается аминокислотных остатков, имеющих азидную группу, неограничивающие примеры указанных аминокислотных остатков включают L-азидогомоаланин (AHA), 4-азидо-L-фенилаланин, 4-азидо-D-фенилаланин, 3-азидо-L-аланин, 3-азидо-D-аланин, 4-азидо-L-гомоаланин, 4-азидо-D-гомоаланин, 5-азидо-L-орнитин, 5-азидо-D-орнитин, 6-азидо-L-лизин и 6-азидо-D-лизин. Что касается аминокислотных остатков, имеющих алкиновую группу, их иллюстративные примеры включают L-гомопропаргилглицин (L-HPG), D-гомопропаргилглицин (D-HPG) и бета-гомопропаргилглицин (β-HPG).
Если аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, то циклооктеновая группа на свободном конце соединяющей ветви может представлять собой транс-циклооктеновую (ТСО) группу, а циклооктиновая группа на свободном конце соединяющей ветви может представлять собой дибензоциклооктиновую (DBCO), дифторциклооктиновую (DIFO), бициклонониновую (BCN) или дибензоциклооктиновую (DICO) группу. В качестве альтернативы, тетразиновая группа на свободном конце соединяющей ветви включает без ограничения 1,2,3,4-тетразиновую, 1,2,3,5-тетразиновую и 1,2,4,5-тетразиновую группы, а также их производные, такие как 6-метилтетразиновая группа.
В соответствии с различными вариантами осуществления настоящего раскрытия линкерное звено дополнительно содержит множество первых элементов. В некоторых вариантах осуществления каждый из первых элементов связан с одной из связывающих ветвей посредством образования амидной связи между связывающей ветвью и первым элементом. В других вариантах осуществления каждый из первых элементов связан с одной из связывающих ветвей посредством реакции азид-алкинового циклоприсоединения, катализируемого медью (CuAAC), клик-реакции азид-алкинового циклоприсоединения, промотируемого напряжением (SPAAC), или реакции Дильса-Альдера с обратными электронными требованиями (iEDDA) между связывающей ветвью и первым элементом.
Необязательно, настоящее линкерное звено дополнительно содержит множество соединяющих ветвей, которые соответственно связаны со множеством связывающих ветвей посредством реакции CuAAC, реакции SPAAC или реакции iEDDA. В соответствии с вариантами осуществления настоящего раскрытия каждая из множества сопрягающих ветвей имеет малеимидную или NHS-группу на ее конце связывания элементов (т.е. на конце, который не связан со связывающей ветвью). Соответственно, каждый из первых элементов связан с одной из сопрягающих ветвей посредством тиол-малеимидной реакции между сопрягающей ветвью и первым элементом; или каждый из первых элементов соединен с одной из сопрягающих ветвей посредством образования амидной связи между сопрягающей ветвью и первым элементом. В некоторых вариантах осуществления каждая из сопрягающих ветвей представляет собой цепь PEG, предпочтительно имеющую 2-20 повторяющихся EG-звеньев. В других вариантах осуществления каждая из сопрягающих ветвей представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на конце связывания элементов.
В соответствии с различными необязательными вариантами осуществления настоящего раскрытия первый элемент представляет собой эффекторный элемент, подходящий для вызывания намеченного эффекта (например, терапевтического эффекта) у субъекта. В качестве альтернативы, первый элемент может представлять собой нацеливающий элемент для направления линкерного звена в участок, представляющий интерес. В соответствии с вариантами осуществления настоящего раскрытия первым элементом является финголимод, финголимод-фосфат, интерферон-β или одноцепочечный вариабельный фрагмент (scFv), специфичный к интегрину α4, β-амилоиду, вирусному белку, бактериальному белку.
Опять же необязательно, линкерное звено дополнительно содержит второй элемент, отличный от первых элементов. В некоторых вариантах осуществления второй элемент имеет азидную или алкиновую группу, так что он связывается с центральной сердцевиной или соединяющей ветвью посредством реакции CuAAC. В качестве альтернативы, в некоторых вариантах осуществления второй элемент, имеющий азидную или циклооктиновую группу, связан с центральной сердцевиной или соединяющей ветвью путем соединения с соответствующей циклооктиновой или азидной группой центральной сердцевины или соединяющей ветви посредством реакции SPAAC. Опять же в качестве альтернативы, в определенных вариантах осуществления второй элемент, имеющий тетразиновую или циклооктеновую группу, связан с центральной сердцевиной или соединяющей ветвью путем соединения с соответствующей циклооктеновой или тетразиновой группой центральной сердцевины или соединяющей ветви посредством реакции iEDDA. В соответствии с некоторыми вариантами осуществления линкерное звено содержит сопрягающую ветвь, которая связана со связывающей ветвью посредством реакции CuAAC, реакции SPAAC или реакции iEDDA; в этих вариантах осуществления N- или С-конец центральной сердцевины или свободный конец соединяющей ветви имеет тетразиновую или циклооктиновую группу, так что второй элемент, имеющий соответствующую циклооктеновую или тетразиновую группу, связан с центральной сердцевиной или соединяющей ветвью посредством реакции iEDDA. В соответствии с другими вариантами осуществления, линкерное звено содержит сопрягающую ветвь, которая связана со связывающей ветвью посредством реакции iEDDA; в этих условиях N- или С-конец центральной сердцевины или свободный конец соединяющей ветви имеет азидную, алкиновую или циклооктиновую группу, так что второй элемент, имеющий соответствующие химические группы, связан с центральной сердцевиной или соединяющей ветвью посредством реакции CuAAC или реакции SPAAC.
В необязательных вариантах осуществления настоящего раскрытия в тех случаях, когда первый элемент представляет собой эффекторный элемент, второй элемент может представлять собой другой эффекторный элемент, который действует аддитивно или синергично с первым элементом или независимо от него; в качестве альтернативы, второй элемент может представлять собой нацеливающий элемент или элемент для улучшения фармакокинетических свойств линкерного звена, такого как растворимость, скорость выведения, период полувыведения и биодоступность. В некоторых других необязательных вариантах осуществления в тех случаях, когда первый элемент представляет собой нацеливающий элемент, второй элемент предпочтительно представляет собой эффекторный элемент или элемент для улучшения фармакокинетических свойств линкерного звена.
В определенных вариантах осуществления линкерное звено дополнительно содержит необязательный третий элемент, отличный от первого и второго элементов. В случае, когда второй элемент непосредственно связан с центральной сердцевиной, другой конец (т.е. свободный конец, не связанный со вторым элементом) центральной сердцевины необязательно представляет собой цистеиновый остаток, который можно применять для введения необязательного третьего элемента. В частности, тиольную группу цистеинового остатка подвергают реакции с малеимидной группой цепи PEG и связываемую таким образом цепь PEG обозначают как соединяющую ветвь, которая имеет тетразиновую или циклооктеновую группу на своем свободном конце. Соответственно, третий элемент затем связывается с соединяющей ветвью посредством реакции iEDDA. В случае, когда линкерное звено содержит как второй, так и третий элементы, предпочтительно, чтобы по меньшей мере один из первого и второго элементов представлял собой эффектор, описанный выше, тогда как третий элемент может представлять собой элемент для улучшения фармакокинетических свойств линкерного звена. Одним примером элемента для улучшения фармакокинетических свойств является длинная цепь PEG, имеющая молекулярную массу приблизительно в 20000-50000 дальтон.
<II> Применение разветвленных линкеров на основе пептидной сердцевины
Линкерное звено в соответствии с первым аспектом настоящего раскрытия может оказываться полезным в клинической медицине для лечения различных заболеваний. Следовательно, второй аспект настоящего раскрытия направлен на способ лечения этих заболеваний. В соответствии с различными вариантами осуществления настоящего раскрытия способ лечения конкретного заболевания включает этап введения субъекту, нуждающемуся в этом, терапевтически эффективного количества линкерного звена в соответствии с вышеупомянутым аспектом и вариантами осуществления настоящего раскрытия. Очевидно, что указанное линкерное звено можно вводить в фармацевтическом составе, который содержит фармацевтически приемлемый наполнитель, подходящий для намеченного или желаемого пути введения, в дополнение к линкерному звену по настоящему изобретению.
Различные иллюстративные комбинации первого и второго элементов линкерного звена по настоящему изобретению для лечения некоторых конкретных заболеваний раскрыты ниже для облегчения понимания некоторых вариантов осуществления настоящего раскрытия.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия, такое линкерное звено применимо для лечения заболевания центральной нервной системы (ЦНС), например, рассеянного склероза и болезни Альцгеймера. Для лечения рассеянного склероза первым элементом может быть финголимод, финголимод-фосфат, интерферон-β или scFv, специфичный к интегрину α4. Для лечения болезни Альцгеймера этим элементом является scFv, специфичный к β-амилоиду.
В соответствии с другими вариантами осуществления настоящего раскрытия, линкерные звенья, подходящие для лечения инфекционного заболевания, в качестве первого элемента содержат scFv, специфичный к вирусному или бактериальному белку. В одном предпочтительном варианте осуществления вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; а бактериальный белок представляет собой эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
<III> Молекулярные конструкции с нацеливающими и эффекторными элементами
В третьем аспекте настоящее раскрытие направлено на молекулярную конструкцию, содержащую два линкерных звена, соединенных друг с другом непосредственно либо опосредованно, в которой сердцевина одного линкерного звена способна связываться по меньшей мере с одним нацеливающим элементом, а сердцевина другого линкерного звена способна связываться по меньшей мере с одним эффекторным элементом. Молекулярная конструкция по настоящему изобретению имеет преимущество, заключающееся в том, что два линкерных звена соединены друг с другом посредством реакции iEDDA, реакции SPAAC или реакции CuAAC. Данная архитектура обеспечивает возможность проведения легкого синтеза молекулярной конструкции со сложной структурой. В соответствии с принципами и сущностью настоящего раскрытия два линкерных звена, соответственно, несущих различные количества и (или) типы функциональных элементов, можно получить независимо и затем конъюгировать друг с другом. Таким образом, для специалиста в данной области становится практически осуществимым конструирование библиотек молекулярных конструкций, соответственно, несущих различные функциональные элементы, и последующие отбор и объединение двух молекулярных конструкций (или линкерных звеньев) из библиотек с образованием желаемых конструкций в зависимости от потребностей и (или) намеченных способов применения. Кроме того, можно контролировать количество функциональных элементов на одно линкерное звено путем регулирования количества конкретных функциональных групп в сердцевине.
В соответствии с одним вариантом осуществления настоящего раскрытия молекулярная конструкция содержит первое линкерное звено и второе линкерное звено. В частности, первое линкерное звено содержит (1) первую центральную сердцевину, (2) одну или несколько связывающих ветвей (далее - первые связывающие ветви), связанные с первой центральной сердцевиной, (3) необязательно, соединяющую ветвь (далее - первую соединяющую ветвь), связанную с первой центральной сердцевиной, и (4), необязательно, одну или несколько сопрягающих ветвей (далее - первые сопрягающие ветви), соответственно связанные с одним или несколькими первыми связывающими ветвями; второе линкерное звено содержит (1) вторую центральную сердцевину, (2) одну или несколько связывающих ветвей (далее - вторые связывающие ветви), связанные со второй центральной сердцевиной, (3) необязательно, соединяющую ветвь (далее - вторая соединяющая ветвь), связанную со второй центральной сердцевиной, и (4), необязательно, одну или несколько сопрягающих ветвей (далее - вторые сопрягающие ветви), соответственно связанные с одной или несколькими вторыми связывающими ветвями.
Первое и второе линкерные звенья соединяются друг с другом посредством реакции iEDDA, SPAAC или CuAAC, происходящей между любыми из следующих: первой и второй центральными сердцевинами, первой соединяющей ветвью и второй центральной сердцевиной, первой и второй соединяющими ветвями или первой центральной сердцевиной и второй соединяющей ветвью.
В соответствии с вариантами осуществления настоящего раскрытия как первая, так и вторая центральные сердцевины имеют множество аминогрупп. Каждая из связывающих ветвей связывается с центральной сердцевиной посредством образования амидной связи между ними, например, между N-гидроксисукцинимидильной (NHS) группой и аминогруппой. После соединения с центральной сердцевиной связывающая ветвь, таким образом, имеет NHS-группу, малеимидную, азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на ее свободном конце.
При наличии NHS-группы первый нацеливающий элемент и первый эффекторный элемент соответственно связаны с первой и второй связывающей ветвью посредством образования амидной связи между элементом (т.е. первым нацеливающим элементом или первым эффекторным элементом) и связывающей ветвью (т.е. первой связывающей ветвью или второй связывающей ветвью). В случае, когда связывающая ветвь имеет малеимидную, азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце, первый нацеливающий элемент и первый эффекторный элемент соответственно связаны с первой и второй связывающей ветвью посредством тиол-малеимидной реакции, реакции CuAAC, реакции iEDDA или реакции SPAAC между этим элементом (т.е. первым нацеливающим элементом или первым эффекторным элементом) и связывающей ветвью (т.е. первой связывающей ветвью или второй связывающей ветвью).
Необязательно, линкерное звено (т.е. первое или второе линкерное звено) настоящей молекулярной конструкции дополнительно содержит одну или несколько сопрягающих ветвей (т.е. первую или вторую сопрягающую ветвь), которые соответственно связаны с одной или несколькими связывающими ветвями посредством реакций CuAAC, iEDDA или SPAAC. В соответствии с вариантами осуществления каждая из сопрягающих ветвей имеет NHS- или малеимидную группу на свободном конце. Таким образом, каждый из элементов (т.е. первые нацеливающие элементы или первые эффекторные элементы) связан с каждой из сопрягающих ветвей либо путем образования амидной связи между этим элементом и сопрягающей ветвью, либо посредством тиол-малеимидной реакции, реакции CuAAC, реакции iEDDA или реакции SPAAC между элементом и сопрягающей ветвью.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия каждая из связывающих ветвей представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев. В качестве альтернативы, каждая из связывающих ветвей представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на ее свободном конце (т.е. конце, не связанном с центральной сердцевиной). В соответствии с некоторыми вариантами осуществления настоящего раскрытия каждая из соединяющих ветвей представляет собой цепь PEG, имеющую 2-12 повторяющихся EG-звеньев. В соответствии с некоторыми вариантами осуществления настоящего раскрытия каждая из сопрягающих ветвей представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев. В качестве альтернативы, каждая из сопрягающих ветвей представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на ее конце для связывания элементов (т.е. на конце, который не связан со связывающей ветвью).
В соответствии с различными вариантами осуществления настоящего раскрытия каждая из первой и второй центральных сердцевин может представлять собой сердцевину на основе химического соединения или полипептидную сердцевину. В некоторых примерах как первая, так и вторая центральные сердцевины представляют собой сердцевины на основе химических соединений из одинаковых или разных химических соединений. В определенных предпочтительных вариантах осуществления как первая, так и вторая центральные сердцевины представляют собой полипептидные сердцевины, имеющие одинаковые или разные последовательности. В качестве альтернативы, одна из двух сердцевин представляет собой сердцевину на основе химического соединения, а другая представляет собой полипептидную сердцевину.
[51] Неограничивающие примеры химического соединения, подходящего для применения в качестве сердцевины на основе химического соединения по настоящему изобретению, включают бензол-1,3,5-триамин, 2-(аминометил)-2-метилпропан-1,3-диамин, трис(2-аминоэтил)амин, бензол-1,2,4,5-тетраамин, 3,3',5,5'-тетраамин-1,1'-бифенил, тетракис(2-аминоэтил)метан, тетракис(этиламин)гидразин, N,N,N',N'-тетракис(аминоэтил)этилендиамин, бензол-1,2,3,4,5,6-гексаамин, 1-N,1-N,3-N,3-N,5-N,5-N-гексакис(метиламин)бензол-1,3,5-триамин, 1-N,1-N,2-N,2-N,4-N,4-N,5-N,5-N-октакис(метиламин)бензол-1,2,4,5-триамин и N,N-бис[(1-амино-3,3-диаминоэтил)пентил]метандиамин.
В случае, когда центральная сердцевина представляет собой сердцевину на основе химического соединения, соединяющая ветвь связана с одной из множества аминогрупп центральной сердцевины путем образования амидной связи между соединяющей ветвью и центральной сердцевиной. В то же время свободный конец соединяющей ветви имеет азидную, алкиновую, циклооктеновую, циклооктиновую или тетразиновую группу.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия полипептид, подходящий для применения в качестве полипептидной сердцевины по настоящему изобретению, содержит множество лизиновых (K) остатков; необязательно, 2-15 остатков K. Кроме того, каждый остаток K отделен от следующего остатка K вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки; вставочная последовательность необязательно состоит из 2-20 аминокислотных остатков. В различных вариантах осуществления вставочная последовательность может иметь последовательность GS, GGS, GSG или SEQ ID NO: 1-16. В некоторых вариантах осуществления полипептид содержит 2-15 звеньев с последовательностью G1-5SK, например, (GSK)2-15. В одном варианте осуществления полипептидная сердцевина имеет последовательность SEQ ID NO: 17, 18, 19, 21, 22, 23 или 24.
В качестве альтернативы, полипептидная сердцевина может содержать последовательность (Хаа-K)n, где Хаа представляет собой пегилированную аминокислоту, имеющую 2-12 повторяющихся этиленгликолевых (EG) звеньев, а n представляет собой целое число от 2 до 15. В одном варианте осуществления полипептидная сердцевина имеет последовательность SEQ ID NO: 25 или 26.
В случае, когда центральная сердцевина представляет собой полипептидную сердцевину, она может содержать цистеиновый остаток на своем N- или С-конце. В этих случаях соединяющая ветвь связана с цистеиновым остатком центральной сердцевины посредством реакции тиольной и малеимидной групп. Соединяющая ветвь, связанная с цистеиновым остатком, имеет азидную, алкиновую, циклооктеновую, циклооктиновую или тетразиновую группу на своем свободном конце.
Первое и второе линкерные звенья могут соединяться различными способами, которые подробно описаны ниже, в зависимости от наличия или отсутствия первой и второй соединяющих ветвей. Для линкерного звена, имеющего сердцевину на основе химического соединения, предпочтительно, чтобы оно было связано с другим линкерным звеном посредством соединяющей ветви (т.е. первой или второй соединяющей ветви), а для линкерного звена, имеющего полипептидную сердцевину, необходимость в соединяющей ветви становится необязательной.
Если первое и второе линкерные звенья, соответственно, содержат соединяющие ветви, то одна из соединяющих ветвей (допустим, например, первая соединяющая ветвь) имеет тетразиновую группу на своем свободном конце, а другая соединяющая ветвь (в данном случае вторая соединяющая ветвь) имеет циклооктеновую группу на своем свободном конце, так что два линкерных звена соединяются посредством реакции iEDDA, происходящей между двумя соединяющими ветвями (т.е. между первой и второй соединяющими ветвями). Тетразиновая группа предпочтительно представляет собой 1,2,3,4-тетразиновую, 1,2,3,5-тетразиновую и 1,2,4,5-тетразиновую группы или их производные, такие как 6-метилтетразиновая группа; а циклооктеновая группа представляет собой ТСО-группу. Это же правило также применимо в случае, когда свободные концы обеих соединяющих ветвей имеют, соответственно, азидную группу и алкиновую группу; в этом случае два линкерных звена соединяются посредством реакции CuAAC, происходящей между двумя соединяющими ветвями (т.е. первой и второй соединяющими ветвями). В качестве альтернативы, одна из соединяющих ветвей (например, первая соединяющая ветвь) имеет азидную группу, а другая соединяющая ветвь (в данном случае, вторая соединяющая ветвь) имеет циклооктиновую группу (предпочтительно DBCO, DIFO, BCN или DICO); соответственно, две соединяющие ветви могут соединяться посредством реакции SPAAC. Эти конфигурации могут иметь место между двумя линкерными звеньями в случаях, когда оба звена имеют сердцевины на основе химического соединения либо полипептидные сердцевины, а также в ситуациях, когда одно линкерное звено имеет сердцевину на основе химического соединения, а другое имеет полипептидную сердцевину.
Если только одно линкерное звено имеет соединяющую ветвь (в качестве примера, первое линкерное звено с первой соединяющей ветвью), то центральная сердцевина другого линкерного звена (например, вторая центральная сердцевина) представляет собой полипептидную сердцевину. В этом случае первый аминокислотный остаток на N- или С-конце одной второй центральной сердцевины представляет собой аминокислотный остаток, имеющий азидную или алкиновую группу. В некоторых вариантах осуществления аминокислотный остаток, имеющий азидную или алкиновую группу, будет подвергаться реакции CuAAC с соответствующей алкиновой или азидной группой первой соединяющей ветви первого линкерного звена с соединением таким образом первого и второго линкерных звеньев. В качестве альтернативы, первый аминокислотный остаток на N- или С-конце одной второй центральной сердцевины представляет собой аминокислотный остаток, имеющий азидную группу, которая может связываться с соединяющей ветвью первого линкерного звена, имеющего циклооктиновую группу (предпочтительно DBCO, DIFO, BCN или DICO) на свободном конце, посредством реакции SPAAC. Эта конфигурация может иметь место между двумя линкерными звеньями в случаях, когда оба звена имеют полипептидные сердцевины, или в ситуациях, когда одно линкерное звено имеет сердцевину на основе химического соединения, а другое имеет полипептидную сердцевину.
Также возможно, чтобы первое и второе линкерные звенья были соединены без наличия каких-либо соединяющих ветвей (т.е. первой и второй соединяющих ветвей). Иными словами, первая и вторая соединяющие ветви могут быть непосредственно связаны друг с другом. Эта конфигурация чаще всего имеет место между двумя полипептидными сердцевинами. В частности, одна из двух центральных сердцевин (допустим, например, первая центральная сердцевина) имеет аминокислотный остаток, имеющий азидную группу, на своем N- или С-конце, а другая центральная сердцевина (как, например, вторая центральная сердцевина) имеет аминокислотный остаток, имеющий алкиновую группу, на своем N- или С-конце. Таким образом, азидная группа первой центральной сердцевины реагирует с алкиновой группой второй центральной сердцевины с соединением таким образом первого и второго линкерных звеньев.
Неограничивающие примеры аминокислотных остатков, имеющих азидную группу, включают L-азидогомоаланин (AHA), 4-азидо-L-фенилаланин, 4-азидо-D-фенилаланин, 3-азидо-L-аланин, 3-азидо-D-аланин, 4-азидо-L-гомоаланин, 4-азидо-D-гомоаланин, 5-азидо-L-орнитин, 5-азидо-D-орнитин, 6-азидо-L-лизин и 6-азидо-D-лизин. Иллюстративные примеры аминокислотных остатков, имеющих алкиновую группу, включают без ограничения L-гомопропаргилглицин (L-HPG), D-гомопропаргилглицин (D-HPG) и бета-гомопропаргилглицин (β-HPG).
В соответствии с некоторыми вариантами осуществления настоящего раскрытия одно из первого и второго линкерных звеньев молекулярной конструкции дополнительно содержит дополнительную связывающую ветвь (далее - третья связывающая ветвь), связанную с первым или вторым линкерным звеном.
Подобно первой и второй связывающим ветвям, третья связывающая ветвь способна связываться с элементом либо путем образования между ними амидной связи, либо посредством тиол-малеимидной реакции, реакции CuAAC, реакции iEDDA или реакции SPAAC. В некоторых вариантах осуществления дополнительный элемент представляет собой второй нацеливающий элемент или второй эффекторный элемент, который используется для усиления нацеливающего или терапевтического эффекта данной молекулярной конструкции. В качестве альтернативы, длинная цепь PEG с молекулярной массой приблизительно 20000-50000 дальтон может использоваться как дополнительный элемент для повышения стабильности настоящей молекулярной конструкции.
В других вариантах осуществления молекулярная конструкция по настоящему изобретению дополнительно содержит третье линкерное звено. Третье линкерное звено содержит (1) третью центральную сердцевину, (2) одну или несколько связывающих ветвей (далее - третьи связывающие ветви), связанные с третьей центральной сердцевиной, (3) необязательно, соединяющую ветвь (далее - третья соединяющая ветвь), связанную с третьей центральной сердцевиной, и (4), необязательно, одну или несколько сопрягающих ветвей (далее - третьи сопрягающие ветви). В этом случае третье линкерное звено связано с первым или вторым линкерным звеном посредством реакции CuAAC, реакции iEDDA или реакции SPAAC, происходящей между любыми из следующих: первой или второй соединяющей ветвью и третьей соединяющей ветвью, первой или второй центральной сердцевиной и третьей соединяющей ветвью, первой или второй центральной соединяющей ветвью и третьей центральной сердцевиной или первой или второй центральной сердцевиной и третьей центральной сердцевиной.
Что касается третьей связывающей ветви третьего линкерного звена, она может иметь NHS-группу, малеимидную, азидную, алкиновую, циклооктеновую, циклооктиновую или тетразиновую группу на своем свободном конце. Соответственно, третья связывающая ветвь может быть непосредственно связана со вторым эффекторным элементом или с нацеливающим элементом либо путем образования между ними амидной связи, либо посредством тиол-малеимидной реакции, реакции CuAAC, реакции iEDDA или реакции SPAAC. В качестве альтернативы, третья связывающая ветвь связывается с третьей сопрягающей ветвью посредством реакции CuAAC, реакции iEDDA или реакции SPAAC; а второй эффекторный элемент или нацеливающий элемент связывается с третьей сопрягающей ветвью посредством образования между ними амидной связи или посредством тиол-малеимидной реакции.
Очевидно, что нацеливающий/эффекторный элемент (такой как лекарственное средство), имеющий NHS-группу, может быть непосредственно связан с остатком K первой, второй и (или) третьей центральной сердцевины посредством образования амидной связи между группой NHS и остатком K без наличия связывающей ветви (т.е. первой, второй или третьей связывающей ветви).
В соответствии с различными вариантами осуществления настоящего раскрытия первая, вторая и, необязательно, третья центральные сердцевины могут быть одинаковыми или разными.
<IV> Применение молекулярных конструкций с нацеливающими и эффекторными элементами
Молекулярная конструкция в соответствии с третьим аспектом настоящего раскрытия может найти применение в клинической медицине для лечения различных заболеваний. Следовательно, четвертый аспект настоящего раскрытия направлен на способ лечения этих заболеваний. В соответствии с различными вариантами осуществления настоящего раскрытия способ лечения конкретного заболевания включает этап введения субъекту, нуждающемуся в этом, молекулярной конструкции в соответствии с третьим аспектом настоящего раскрытия и его вариантами осуществления в терапевтически эффективном количестве. Очевидно, что указанную молекулярную конструкцию можно вводить в фармацевтическом составе, который содержит фармацевтически приемлемый наполнитель, подходящий для намеченного или желаемого пути введения, в дополнение к молекулярной конструкции по настоящему изобретению.
Различные иллюстративные комбинации первого и второго элементов молекулярной конструкции по настоящему изобретению для лечения некоторых конкретных заболеваний раскрыты ниже для облегчения понимания некоторых вариантов осуществления настоящего раскрытия.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия, настоящая молекулярная конструкция применима для лечения заболевания ЦНС, например, рассеянного склероза и болезни Альцгеймера. Для лечения рассеянного склероза первым элементом является scFv, специфичный к рецептору трансферрина, а вторым элементом является IFN-β, финголимод, финголимод-фосфат или scFv, специфичный к интегрину α4. Что касается лечения болезни Альцгеймера, то в качестве первого и второго элементов используются, соответственно, scFv, специфичный к рецептору трансферрина, и scFv, специфичный к β-амилоиду.
В соответствии с другими вариантами осуществления настоящего раскрытия, настоящая молекулярная конструкция применима для лечения инфекционного заболевания. В одном варианте осуществления инфекционное заболевание вызвано вирусом, а первый элемент представляет собой scFv, специфичный к вирусному белку, тогда как второй элемент представляет собой scFv, специфичный к CD16b или к CD32. Неограничивающие примеры вирусных белков включают в себя белок F вируса RSV, белок gp120 вируса HIV-1, НА белка вируса гриппа А и гликопротеин цитомегаловируса. В другом варианте осуществления инфекционное заболевание вызвано бактерией, а первый элемент представляет собой scFv, специфичный к бактериальному белку, тогда как второй элемент представляет собой scFv, специфичный к CD16b или к CD32. Примеры бактериального белка включают без ограничения эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
<V> Молекулярная конструкция для лечения заболеваний центральной нервной системы и ее применение
В пятом аспекте настоящее раскрытие направлено на молекулярную конструкцию на основе кристаллизующегося фрагмента (Fc), которая имеет по меньшей мере один нацеливающий элемент и по меньшей мере один эффекторный элемент, непосредственно или опосредованно связанные с доменом СН2-СН3 иммуноглобулина. Нацеливающие и эффекторные элементы данных молекулярных конструкций на основе Fc специально выбраны таким образом, чтобы эти молекулярные конструкции на основе Fc подходили для применения при лечении заболеваний центральной нервной системы (ЦНС) или для использования в производстве лекарственного средства для лечения заболевания ЦНС. Очевидно, что способы лечения заболеваний ЦНС с использованием таких молекулярных конструкций на основе Fc также относятся к аспекту настоящего раскрытия.
В соответствии с определенными вариантами осуществления настоящего раскрытия молекулярная конструкция на основе Fc содержит пару сегментов СН2-СН3 из IgG. Fc, пару эффекторных элементов и пару нацеливающих элементов. Пара эффекторных элементов представляет собой интерферон β1a (INF-β1a), или интерферон β1b (INF-β1b), или фрагмент антитела, специфичный к интегрину α4 или к β-амилоиду, в то время как пара нацеливающих элементов является фрагментом антитела, специфичным к рецептору трансферрина человека или к рецептору инсулина человека.
Если пара эффекторных элементов связана с N-концами пары сегментов СН2-СН3, то пара нацеливающих элементов связана с С-концами пары сегментов СН2-СН3, и наоборот. В качестве альтернативы, если как пара эффекторных элементов, так и пара нацеливающих элементов находятся в форме одноцепочечных вариабельных фрагментов (scFv), то пара нацеливающих элементов связана с N-концами пары эффекторных элементов в тандемной конфигурации или в конфигурации диатела с образованием таким образом пары биспецифических scFv, связанной с N-концами пары сегментов СН2-СН3.
В определенных вариантах осуществления пара сегментов СН2-СН3 получена из тяжелой цепи γ4 человеческого IgG или тяжелой цепи γ1 человеческого IgG.
В некоторых примерах пара эффекторных элементов или пара нацеливающих элементов имеет конфигурацию Fab (т.е. состоит из домена VH-CH1 и домена VL-Cκ); этот Fab-фрагмент связан с N-концами первой и второй тяжелых цепей, так что в молекулярной конструкции на основе Fc используется конфигурация IgG. В этих случаях пара элементов, которые находятся вне конфигурации Fab, связана с С-концами пары сегментов СН2-СН3.
В соответствии с некоторыми необязательными вариантами осуществления, эффекторный элемент представляет собой INF-β1a, INF-β1b или scFv, специфичный к интегрину α4, тогда как нацеливающий элемент представляет собой scFv, специфичный к рецептору трансферрина человека. В частности, эта молекулярная конструкция подходит для лечения рассеянного склероза.
В соответствии с другими необязательными вариантами осуществления, эффекторный элемент представляет собой scFv, специфичный к β-амилоиду, тогда как нацеливающий элемент представляет собой scFv, специфичный к рецептору трансферрина человека. В частности, эта молекулярная конструкция подходит для лечения болезни Альцгеймера.
Способы лечения заболеваний ЦНС у субъекта, нуждающегося в этом, включают стадию введения субъекту эффективного количества молекулярной конструкции согласно этому аспекту. Заболевания ЦНС, поддающиеся лечению этим методом, включают рассеянный склероз и болезнь Альцгеймера.
<VI> Молекулярные конструкции для лечения инфекционных заболеваний
В шестом аспекте настоящее раскрытие направлено на молекулярную конструкцию на основе кристаллизуемого фрагмента (Fc), которая имеет по меньшей мере один нацеливающий элемент и по меньшей мере один эффекторный элемент, прямо или косвенно связанные с доменом СН2-СН3 иммуноглобулина. Нацеливающие и эффекторные элементы настоящих молекулярных конструкций на основе Fc специально выбраны таким образом, чтобы эти молекулярные конструкции на основе Fc подходили для применения при лечении заболеваний/состояний, связанных с вирусной или бактериальной инфекцией, или для использования в производстве лекарственного средства для лечения таких заболеваний/состояний. Очевидно, что способы лечения заболеваний/состояний, связанных с вирусной или бактериальной инфекцией, с использованием таких молекулярных конструкций на основе Fc также относятся к аспекту настоящего раскрытия.
В соответствии с определенными вариантами осуществления настоящего раскрытия молекулярная конструкция на основе Fc содержит пару сегментов СН2-СН3 из IgG. Fc, пару эффекторных элементов и пару нацеливающих элементов. Пара эффекторных элементов представляет собой фрагмент антитела, специфичный к CD32 или к CD16b, тогда как пара нацеливающих элементов представляет собой фрагмент антитела, специфичный к вирусному или бактериальному белку.
Если пара эффекторных элементов связана с N-концами пары сегментов СН2-СН3, то пара нацеливающих элементов связана с С-концами пары сегментов СН2-СН3, и наоборот. В качестве альтернативы, если пара эффекторных элементов и пара нацеливающих элементов находятся в форме одноцепочечных вариабельных фрагментов (scFv), то пара нацеливающих элементов связана с N-концами пары эффекторных элементов в тандемной конфигурации или в конфигурации диатела с образованием, таким образом, пары биспецифических scFv, связанных с N-концами пары сегментов СН2-СН3.
В определенных вариантах осуществления пара сегментов СН2-СН3 получена из тяжелой цепи γ4 человеческого IgG или тяжелой цепи γ1 человеческого IgG.
В некоторых примерах пара эффекторных элементов или пара нацеливающих элементов имеет конфигурацию Fab (т.е. состоит из домена VH-CH1 и домена VL-Cκ); этот Fab-фрагмент связан с N-концами первой и второй тяжелых цепей, так что молекулярная конструкция на основе Fc принимает конфигурацию IgG. В этих случаях пара элементов, которая находится вне конфигурации Fab, связана с С-концами пары сегментов СН2-СН3.
В соответствии с некоторыми необязательными вариантами осуществления, эффекторный элемент представляет собой scFv, специфичный к CD32 или к CD16b, тогда как нацеливающий элемент представляет собой scFv, специфичный к вирусному белку. Например, вирусный белок может представлять собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса. В частности, такая молекулярная конструкция подходит для лечения вирусных инфекций.
В соответствии с другими необязательными вариантами осуществления эффекторный элемент представляет собой scFv, специфичный к CD32 или CD16b, тогда как нацеливающий элемент представляет собой scFv, специфичный к бактериальному белку. Примеры бактериального белка включают без ограничения эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli. В частности, такая молекулярная конструкция подходит для лечения бактериальных инфекций.
Способы лечения заболеваний/состояний, связанных с инфекциями (например, вирусными или бактериальными инфекциями) у субъекта, нуждающегося в этом, включают стадию введения субъекту эффективного количества молекулярной конструкции согласно этому аспекту.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Настоящее описание будет более понятным из следующего подробного описания, читаемого с учетом сопровождающих графических материалов, коротко обсуждаемых ниже.
Фигуры 1A-1N представляют собой схематические изображения, иллюстрирующие линкерные звенья в соответствии с определенными вариантами осуществления настоящего раскрытия.
Фигура 2 представляет собой схематическое изображение, иллюстрирующее линкерное звено, имеющее сердцевину на основе химического соединения.
Фигуры 3A-3D представляют собой схематические изображения, иллюстрирующие молекулярные конструкции Т-Е в соответствии с некоторыми вариантами осуществления настоящего раскрытия.
Фигура 4 представляет собой схематическое изображение, иллюстрирующее библиотеки для конструирования молекулярных конструкций в соответствии с некоторыми вариантами осуществления настоящего раскрытия.
Фигура 5А и фигура 5В представляют собой схематические изображения, иллюстрирующие молекулярные конструкции в соответствии с некоторыми вариантами осуществления настоящего раскрытия.
Фигура 6 представляет собой схематическое изображение, иллюстрирующее молекулярную конструкцию в соответствии с некоторыми вариантами осуществления настоящего раскрытия.
Фигура 7А и фигура 7В представляют собой схематические изображения, иллюстрирующие молекулярные конструкции в соответствии с различными вариантами осуществления настоящего раскрытия.
Фигуры 8А-8С представляют собой схематические изображения, иллюстрирующие молекулярные конструкции на основе Fc в соответствии с различными вариантами осуществления настоящего раскрытия.
Фигура 9 представляет собой схематическое изображение, иллюстрирующее молекулярную конструкцию на основе Fc в соответствии с различными вариантами осуществления настоящего раскрытия.
Фигуры 10А и 10В представляют собой схематические изображения, иллюстрирующие молекулярные конструкции на основе Fc в соответствии с различными вариантами осуществления настоящего раскрытия.
На фигуре 11 показан результат масс-спектрометрии методом MALDI-TOF линкерного звена на основе пептидной сердцевины, несущего одну связывающую ветвь с тетразиновой группой и три связывающих ветви PEG с малеимидными группами.
На фигуре 12 показан результат масс-спектрометрии MALDI-TOF финголимода, конъюгированного с NHS-PEG5.
На фигуре 13 показан результат анализа методом масс-спектрометрии MALDI-TOF «связки» лекарственных средств, состоящей из линкерного звена со свободной функциональной группой ТСО и набора из 5 молекул финголимода.
На фигуре 14 показан результат анализа методом масс-спектрометрии MALDI-TOF «связки» лекарственных средств, состоящей из линкерного звена со свободной функциональной группой ТСО и набора из 10 молекул финголимода.
На фигуре 15 показан результат анализа методом масс-спектрометрии MALDI-TOF «связки» лекарственных средств, состоящей из линкерного звена со свободной функциональной группой ТСО и набора из пяти молекул финголимод-фосфата.
На фигуре 16 показан результат анализа методом SDS-PAGE очищенного эктодомена человеческого CD32a.
На фигуре 17 показан результат анализа методом SDS-PAGE очищенного эктодомена человеческого TfR1.
На фигуре 18А показан результат анализа методом SDS-PAGE очищенного scFv, специфичного к белку F вируса RSV; на фигуре 18В показан результат анализа методом ELISА очищенного scFv, специфичного к белку F вируса RSV; на фигуре 18С показан результат анализа методом SDS-PAGE очищенного scFv, специфичного к эндотоксину; на фигуре 18D показан анализ методом ELISА очищенного scFv, специфичного к эндотоксину; на фигуре 18Е показан результат анализа методом SDS-PAGE очищенного scFv, специфичного к эктодомену CD32a; на фигуре 18F показан результат анализа методом ELISA очищенного scFv, специфичного к эктодомену CD32a.
На фигуре 19А показан результат анализа методом SDS-PAGE очищенного scFv, специфичного к эктодомену TfR1 крысы; на фигуре 19В показан результат анализа методом ELISА очищенного scFv, специфичного к эктодомену TfR1 крысы; на фигуре 19С показан результат анализа методом SDS-PAGE очищенного scFv, специфичного к β-амилоиду; на фигуре 19D показан анализ методом ELISА очищенного scFv, специфичного к β-амилоиду.
На фигуре 20А показаны данные титров фагов, несущих scFv, специфичные к эктодомену человеческого CD32a; на фигуре 20В показан результат анализа ELISA с одной колонией scFv на фаговых носителях, специфичных к эктодомену человеческого CD32a.
На фигуре 21А показаны данные титров фагов, несущих scFv, специфичных к эктодомену человеческого TfR1; на фигуре 21В показан результат анализа методом ELISA с одной колонией scFv на фаговых носителях, специфичных к эктодомену человеческого TfR1.
На фигуре 22А и фигуре 22В, соответственно, показаны результаты анализа методом ELISA и масс-спектрометрического анализа scFv, специфичных к CD32a и конъюгированных с ТСО.
На фигуре 23А и фигуре 23В, соответственно, показаны результаты анализа методом ELISA и масс-спектрометрического анализа scFv, специфичных к TfR1 и конъюгированных с тетразином.
На фигуре 24А показан профиль элюирования эксклюзионной колонки S75 при анализе методом FPLC для синтезированного нацеливающего линкерного звена, состоящего из линкерного звена со свободной тетразиновой функциональной группой и набора из трех scFv, специфичных к эндотоксину, в качестве нацеливающих элементов; на фигуре 24В и фигуре 24С, соответственно, показаны результаты анализа методом SDS-PAGE и результаты масс-спектрометрического анализа синтезированного нацеливающего линкерного звена с фигуры 24А.
На фигуре 25 показан результат масс-спектрометрического анализа синтезированного нацеливающего линкерного звена, состоящего из линкерного звена со свободной тетразиновой функциональной группой и набора из трех scFv, специфичных к белку F вируса RSV, в качестве нацеливающих элементов.
На фигуре 26А показан профиль элюирования катионообменной колонки при анализе методом FPLC для синтезированного нацеливающего линкерного звена, состоящего из линкерного звена со свободной функциональной группой ТСО и набора из трех scFv, специфичных к β-амилоиду, в качестве нацеливающих элементов; на фигуре 26В и фигуре 26С, соответственно, показаны результаты анализа методом SDS-PAGE и результаты анализа методом ELISА синтезированного нацеливающего линкерного звена с фигуры 26А.
На фигуре 27А показан результат масс-спектрометрического анализа молекулярной конструкции одного линкерного звена с тремя scFv, специфичными к эндотоксину, в качестве нацеливающих элементов и одним scFv, специфичным к эктодомену CD32a, в качестве эффекторного элемента; на фигуре 27В показан результат масс-спектрометрического анализа молекулярной конструкции одного линкерного звена с тремя scFv, специфичными к эндотоксину, в качестве нацеливающих элементов и одним scFv, специфичным к эктодомену CD32a, в качестве эффекторного элемента; на фигуре 27С показан результат масс-спектрометрического анализа молекулярной конструкции одного линкерного звена с одним scFv, специфичным к эктодомену TfR1, в качестве нацеливающего элемента, и тремя scFv, специфичными к β-амилоиду, в качестве эффекторных элементов.
На фигуре 28А и фигуре 28В, соответственно, показаны результаты анализа методом SDS-PAGE и масс-спектрометрического анализа молекулярной конструкции с одним scFv, специфичным к эктодомену TfR1, и одной «связки» лекарственных средств, несущей пять молекул финголимода.
На фигуре 29А показаны результаты анализа методом окрашивания В-клеток человека с экспрессией рецептора S1P1; на фигуре 29В показан результат анализа миграции финголимода в системе Трансвелл при конъюгировании с пептидной сердцевиной через связывающие ветви.
На фигуре 30А показан результат анализа методом SDS-PAGE очищенного рекомбинантного двухцепочечного (scFv α RSV)-hIgG1.Fc-(scFv α CD32) белка слияния; на фигуре 30В и фигуре 30С представлены результаты анализа методом ELISA, соответственно, иллюстрирующие активность связывания очищенного рекомбинантного белка слияния с фигуры 30А с белком F вируса RSV (фигура 30В) и эктодоменом CD32a (фигура 30С).
На фигуре 31А показан результат анализа методом SDS-PAGE очищенного рекомбинантного двухцепочечного (эндотоксин scFv α)-hIgG1.Fc-(scFv α CD32) белка слияния; на фигуре 31В и фигуре 31С представлены результаты анализов методом ELISA, соответственно, иллюстрирующие аффинность связывания очищенного рекомбинантного белка слияния с фигуры 31А с эндотоксином (фигура 31В) и эктодоменом CD32a (фигура 31С).
На фигуре 32А и фигуре 32В, соответственно, показаны результаты анализа методом SDS-PAGE и анализа методом ELISA очищенного рекомбинантного двухцепочечного (интерферон-β-1а)-hIgG.Fc-(scFv α TfR1) белка слияния.
На фигуре 33А и фигуре 33В, соответственно, показаны результаты анализа методом SDS-PAGE и анализа методом окрашивания очищенного рекомбинантного двухцепочечного (scFv α интегрин α4)-hIgG4.Fc-(scFv α TfR1) белка слияния.
На фигуре 34 показан результат анализа методом ELISA влияния очищенного рекомбинантного двухцепочечного (эндотоксин scFv α)-hIgG1.Fc-(scFv α CD32a) белка слияния на ингибирование секреции TNF-α.
В соответствии с принятой практикой, различные описанные признаки/элементы показаны с отклонением от масштаба для наилучшей иллюстрации конкретных признаков/элементов, относящихся к настоящему изобретению. Кроме того, одинаковые номера позиций и обозначения на различных графических материалах используются для указания одинаковых элементов/частей там, где это возможно.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Подразумевается, что подробное описание, представленное ниже вместе с прилагаемыми графическими материалами, является описанием примеров настоящего изобретения, и не подразумевается, что оно представляет единственные формы, в которых пример настоящего изобретения может быть сконструирован или использован. В настоящем описании изложены функции примера и последовательность этапов конструирования и эксплуатации примера. Однако, одинаковые или эквивалентные функции и последовательности могут реализовываться в различных примерах.
Для удобства здесь собраны некоторые термины, используемые в описании, примерах и прилагаемой формуле изобретения. Если в данном документе не определено иное, научные и технические термины, используемые в настоящем раскрытии, будут иметь значения, обычно понимаемые и используемые средним специалистом в данной области.
Если иное не требуется по контексту, будет понятно, что термины в единственном числе будут включать их формы множественного числа, а термины во множественном числе будут включать единственное число. В частности, используемые в данном документе и в формуле изобретения формы единственного числа включают ссылку на множественное число, если в контексте четко не указано иное. Кроме того, используемые в данном документе и в формуле изобретения термины «по меньшей мере один» и «один или более» имеют одинаковое значение и включают один, два, три или более. Более того, подразумевается, что фразы «по меньшей мере один из А, В и С», «по меньшей мере один из А, В или С» и «по меньшей мере один из А, В и (или) С», используемые во всем настоящем описании и в прилагаемой формуле изобретения, охватывают А в отдельности, В в отдельности, С в отдельности, А и В вместе, В и С вместе, А и С вместе, а также А, В и С вместе.
Несмотря на то, что числовые диапазоны и параметры, отражающие широкий объем настоящего изобретения, являются приблизительными величинами, числовые значения, изложенные в конкретных примерах, представлены как можно более точно. Любое числовое значение, однако, по своей сути заключает в себе определенные погрешности, неизбежно возникающие вследствие обычного отклонения, обнаруживаемого в соответствующих тестовых измерениях. Кроме того, используемый в данном документе термин «приблизительно», как правило, означает в пределах 10%, 5%, 1% или 0,5% от указанного значения или диапазона. В качестве альтернативы, термин «приблизительно» означает в пределах допустимой стандартной ошибки среднего при рассмотрении средним специалистом в данной области. За исключением примеров эксплуатации/рабочих примеров или если в явной форме не указано иное, все числовые диапазоны, количества, значения и процентные доли, как, например, в отношении количества материалов, продолжительности периодов времени, температур, условий эксплуатации, соотношений количеств и т.п., раскрытых в данном документе, следует понимать как модифицированные во всех случаях термином «приблизительно». Соответственно, если не указано противоположное, числовые параметры, изложенные в настоящем раскрытии и прилагаемой формуле изобретения, являются приблизительными величинами, которые при желании можно изменять. Как минимум, каждый числовой параметр следует интерпретировать по меньшей мере с учетом количества приводимых значащих цифр и путем применения стандартных методик округления. Диапазоны могут быть выражены в данном документе от одной конечной точки до другой конечной точки или между двумя конечными точками. Все диапазоны, раскрытые в данном документе, включают конечные точки, если не указано иное.
Настоящее раскрытие в целом относится к молекулярным конструкциям, при этом каждая молекулярная конструкция содержит нацеливающий элемент (Т) и эффекторный элемент (Е), и эти молекулярные конструкции иногда в данном документе называют «молекулами Т-Е», «фармацевтическими средствами Т-Е» или «лекарственными средствами Т-Е».
Используемый в данном документе термин «нацеливающий элемент» относится к части молекулярной конструкции, которая непосредственно или опосредованно связывается с мишенью, представляющей интерес (например, с рецептором на клеточной поверхности или белком в ткани), облегчая таким образом транспортировку молекулярной конструкции по настоящему изобретению к мишени, представляющей интерес. В некоторых примерах нацеливающий элемент может направлять молекулярную конструкцию в окрестность клетки-мишени. В других случаях нацеливающий элемент специфично связывается с молекулой, присутствующей на поверхности клеток-мишеней, или со второй молекулой, которая специфично связывается с молекулой, присутствующей на клеточной поверхности. В некоторых случаях нацеливающий элемент может интернализироваться вместе с молекулярной конструкцией по настоящему изобретению после его связывания с мишенью, представляющей интерес, а значит, перемещаться в цитозоль клетки-мишени. Нацеливающий элемент может представлять собой антитело или лиганд рецептора клеточной поверхности или он может представлять собой молекулу, которая связывается с таким антителом или лигандом, опосредованно нацеливая таким образом молекулярную конструкцию по настоящему изобретению на участок-мишень (например, поверхность выбранной клетки). Молекулярная конструкция по настоящему изобретению будет повышать степень или содействовать локализации эффектора (терапевтического средства) в пораженном участке по сравнению с терапевтическим средством без нацеливающей функции. Локализация относится к степени или относительной доле; она не означает абсолютную или полную локализацию эффектора в пораженном участке.
В соответствии с настоящим изобретением термин «эффекторный элемент» относится к части молекулярной конструкции, которая вызывает биологическую активность (например, индуцирует иммунные ответы, оказывает цитотоксические эффекты и т.п.) или другую функциональную активность (например, привлекает другие терапевтические молекулы, меченные гаптенами) после направления молекулярной конструкции к ее участку-мишени. «Эффект» может быть терапевтическим или диагностическим. Эффекторные элементы включают в себя те элементы, которые связываются с клетками и (или) внеклеточными иммунорегуляторными факторами. Эффекторный элемент включает в себя такие средства, как белки, нуклеиновые кислоты, липиды, углеводы, гликопептиды, компоненты-лекарственные средства (как низкомолекулярные лекарственные средства, так и биологические лекарственные средства), химические соединения, элементы и изотопы, а также их фрагменты.
Хотя термины первый, второй, третий и т.д. могут использоваться в данном документе для описания различных элементов, компонентов, областей и (или) частей, эти элементы (а также компоненты, области и (или) части) не должны ограничиваться данными терминами. Кроме того, использование таких порядковых числительных не предполагает наличие последовательности или порядка, если это четко не указано в контексте. Данные термины вместо этого используются лишь для проведения различия между одним и другим элементами. Таким образом, первый элемент, обсуждаемый ниже, можно называть вторым элементом без отступления от идей типичных вариантов осуществления.
В данной работе термины «связывать», «соединять» и «конъюгировать» используются взаимозаменяемо для обозначения любого способа присоединения двух элементов друг к другу с помощью непосредственной связи или с помощью опосредованной связи между двумя элементами.
Термин «полипептид», используемый в данном документе, относится к полимеру, имеющему по меньшей мере два аминокислотных остатка. Как правило, полипептид содержит аминокислотные остатки в диапазоне длины от 2 до приблизительно 200 остатков; предпочтительно от 2 до 50 остатков. Если в данном документе представлена аминокислотная последовательность, то также предусматриваются варианты данной последовательности, содержащие L-, D- или бета-аминокислоты. Полипептиды также включают полимеры аминокислот, в которых один или более аминокислотных остатков представляют собой искусственный химический аналог соответствующей встречающейся в природе аминокислоты, а также встречающиеся в природе полимеры аминокислот. В дополнение, данный термин применяется в отношении аминокислот, сочлененных друг с другом пептидной связью или другими «модифицированными связями» (например, в случаях, когда пептидная связь заменена α-сложноэфирной, β-сложноэфирной, тиоамидной, фосфорамидной, карбаматной, гидроксилатной и т.п.).
В определенных вариантах осуществления предусматриваются консервативные замещения аминокислот, образующих любую из последовательностей, описанных в данном документе. В различных вариантах осуществления замещены один, два, три, четыре или пять различных остатков. Термин «консервативное замещение» используется для отражения аминокислотных замещений, которые значительно не изменяют активность (например, биологическую или функциональную активность и (или) специфичность) молекулы. Как правило, консервативные аминокислотные замещения включают замещение одной аминокислоты другой аминокислотой со сходными химическими свойствами (например, зарядом или гидрофобностью). Некоторые консервативные замещения включают «замещения аналогом», при которых стандартная аминокислота заменяется нестандартной (например, редкой, синтетической и т.д.) аминокислотой, минимально отличающейся от исходного остатка. Аналоги аминокислот считаются получаемыми синтетическим методом из стандартных аминокислот без значительного изменения структуры исходной молекулы, являются изомерами или являются метаболитами-предшественниками.
В определенных вариантах осуществления также предусматриваются полипептиды, характеризующиеся по меньшей мере 80%, предпочтительно по меньшей мере 85% или 90% и более предпочтительно по меньшей мере 95% или 98% идентичностью последовательности с любой из последовательностей, описанных в данном документе.
«Процентная (%) идентичность аминокислотных последовательностей» по отношению к полипептидным последовательностям, идентифицированным в данном документе, определяется как процентная доля остатков полипептида в последовательности-кандидате, являющихся идентичными аминокислотным остаткам в конкретной полипептидной последовательности, после выравнивания последовательностей и, при необходимости, введения гэпов для достижения максимальной процентной идентичности последовательностей и без рассмотрения каких-либо консервативных замещений в качестве части идентичности последовательностей. Выравнивания в целях определения процентной идентичности последовательностей можно достичь различными способами, которые находятся в пределах компетенции специалиста в данной области, например, с помощью общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для измерения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Для целей, изложенных в данном документе, сравнение последовательностей между двумя полипептидными последовательностями проводили с помощью компьютерной программы Blastp (BLAST для сравнения последовательностей белков), предоставляемой в режиме онлайн Национальным центром биотехнологической информации (NCBI). Процентную идентичность аминокислотных последовательностей для указанной полипептидной последовательности А и указанной полипептидной последовательности В (что можно в качестве альтернативы сформулировать как определенное процентное значение идентичности аминокислотной последовательности А и указанной полипептидной последовательности В, которым характеризуется указанная полипептидная последовательность А) рассчитывают по следующей формуле:
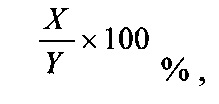
где X представляет собой количество аминокислотных остатков, подсчитанных как идентичные совпадения в программе для выравнивания последовательностей BLAST при выравнивании А и В в данной программе, и где Y представляет собой общее количество аминокислотных остатков в А или В в зависимости от того, какая из них короче.
Термин «пегилированная аминокислота», используемый в данном документе, относится к цепи полиэтиленгликоля (PEG) с одной аминогруппой и одной карбоксильной группой. Как правило, пегилированная аминокислота имеет формулу NH2-(CH2CH2O)n-COOH. В настоящем раскрытии значение n находится в диапазоне от 1 до 20; предпочтительно находится в диапазоне от 2 до 12.
Используемый в данном документе термин «концевой» по отношению к полипептиду относится к аминокислотному остатку на N- или С-конце полипептида. Применительно к полимеру термин «концевой» относится к составному звену полимера (например, полиэтиленгликоля по настоящему раскрытию), расположенному на конце основной цепи полимера. В настоящем описании и формуле изобретения термин «свободный конец» используется для обозначения концевого аминокислотного остатка или составного звена, химически не связанного с какой-либо другой молекулой.
Термин «антиген» или «Ag», используемый в данном документе, определяется как молекула, вызывающая иммунный ответ. Этот иммунный ответ может включать секреторный, гуморальный и (или) клеточный антиген-специфический ответ. В настоящем раскрытии термин «антиген» может означать любое из белка, полипептида (включая его мутантные формы или биологически активные фрагменты), полисахарида, гликопротеина, гликолипида, нуклеиновой кислоты или их комбинации.
В настоящем описании и формуле изобретения термин «антитело» используется в наиболее широком смысле и охватывает полностью собранные антитела, фрагменты антител, которые связываются с антигенами, такие как антиген-связывающий фрагмент (Fab/Fab'), F(аb')2-фрагмент (имеющий две антиген-связывающие Fab-части, связанные друг с другом дисульфидными связями), вариабельный фрагмент (Fv), одноцепочечный вариабельный фрагмент (scFv), биспецифический одноцепочечный вариабельный фрагмент (bi-scFv), нанотела, одновалентные антитела и диатела. «Фрагменты антител» включают в себя часть интактного антитела, предпочтительно антиген-связывающую область или вариабельную область интактного антитела. Как правило, «антитело» относится к белку, состоящему из одного или более полипептидов, кодируемых главным образом генами иммуноглобулинов или фрагментами генов иммуноглобулинов. Широко известные гены иммуноглобулинов включают гены константных областей каппа-, лямбда-, альфа-, гамма-, дельта-, эпсилон- и мю-цепей, а также большое число генов вариабельных областей иммуноглобулинов. Легкие цепи классифицируются как каппа либо лямбда. Тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон, что, в свою очередь, соответственно, определяет классы иммуноглобулинов IgG, IgM, IgA, IgD, и IgE. Как известно, типичная структурная единица иммуноглобулина (антитела) образует тетрамер. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, при этом каждая пара имеет одну «легкую» цепь (приблизительно 25 кДа) и одну «тяжелую» цепь (приблизительно 50-70 кДа). N-конец каждой цепи определяет вариабельную область из приблизительно 100-110 или более аминокислот, преимущественно отвечающих за распознавание антигена. Термины «вариабельная область легкой цепи (VL)» и «вариабельная область тяжелой цепи (VH)» относятся, соответственно, к этим легким и тяжелым цепям. В соответствии с вариантами осуществления настоящего раскрытия фрагмент антитела можно получить путем модификации природы антитела или путем синтеза de novo с применением методов рекомбинантных ДНК. В определенных вариантах осуществления настоящего раскрытия антитело и (или) фрагмент антитела могут быть биспецифическими и могут находиться в различных конфигурациях. Например, биспецифические антитела могут содержать два различных антиген-связывающих участка (вариабельные области). В различных вариантах осуществления биспецифические антитела можно получить с помощью гибридомной методики или методики рекомбинантных ДНК. В определенных вариантах осуществления биспецифические антитела обладают специфичностью связывания по меньшей мере с двумя различными эпитопами.
Термин «специфично связывается», используемый в данном документе, относится к способности антитела или его антиген-связывающего фрагмента к связыванию с антигеном с константой диссоциации (Kd), составляющей не более чем приблизительно 1×10-6 М, 1×10-7 М, 1×10-8 М, 1×10-9 М, 1×10-10 М, 1×10-11 М, 1×10-12 М, и (или) к связыванию с антигеном с аффинностью, по меньшей мере в два раза превышающей его аффинность к неспецифическому антигену.
Термин «лечение» используемый в данном документе, включает предупреждающее (например, профилактическое), радикальное или паллиативное лечение; и «осуществление лечения», как используется в данном документе, также включает предупреждающее (например, профилактическое), радикальное или паллиативное лечение. В частности, термин «осуществление лечения», используемый в данном документе, относится к применению молекулярной конструкции или фармацевтической композиции, содержащей ее, по отношению к субъекту, имеющему медицинское состояние или симптом, ассоциированный с медицинским состоянием, заболевание или нарушение вследствие медицинского состояния или предрасположенность к медицинскому состоянию, или ее введению ему с целью частичного или полного уменьшения интенсивности, ослабления, облегчения, задерживания начала проявления, ингибирования прогрессирования, уменьшения тяжести и (или) снижения частоты возникновения одного или более симптомов или характерных особенностей указанного конкретного заболевания, нарушения и (или) состояния. Лечение можно проводить субъекту, у которого не проявляются признаки заболевания, нарушения и (или) состояния, и (или) субъекту, у которого проявляются только ранние признаки заболевания, нарушения и (или) состояния, в целях снижения риска развития патологии, ассоциированной с заболеванием, нарушением и (или) состоянием.
Термин «эффективное количество», используемый в данном документе, относится к количеству молекулярной конструкции согласно настоящему изобретению, достаточному для достижения желаемого терапевтического ответа. Эффективное количество средства не требуется для излечения заболевания или состояния, но будет обеспечивать такое лечение заболевания или состояния, при котором начало проявления заболевания или состояния задерживается, приостанавливается или предупреждается, или симптомы заболевания или состояния ослабляются. Эффективное количество можно разделить на одну, две или более доз в форме, подходящей для введения один, два или более раз в течение всего указанного периода времени. Конкретное эффективное или достаточное количество будет изменяться в зависимости от таких факторов, как конкретное состояние, подвергаемое лечению, физическое состояние пациента (например, масса тела, возраст или пол пациента), тип субъекта, подвергаемого лечению, продолжительность лечения, характер сопутствующей терапии (если таковая имеется), а также конкретные используемые составы и структура химических соединений или их производных. Эффективное количество может быть выражено, например, в виде общей массы активного компонента (например, в граммах, миллиграммах или микрограммах) или в виде соотношения массы активного компонента и массы тела, например, в виде миллиграммов на килограмм (мг/кг).
Термины «применение» и «введение» используются в данном документе взаимозаменяемо для обозначения применения молекулярной конструкции или фармацевтической композиции по настоящему изобретению по отношению к субъекту, нуждающемуся в лечении.
Термины «субъект» и «пациент» используются в данном документе взаимозаменяемо, и подразумевается, что они означают животное, в том числе человека, которое поддается лечению с помощью молекулярной конструкции, фармацевтической композиции и (или) способа по настоящему изобретению. Подразумевается, что термин «субъект» или «пациент» относится как к мужскому, так и к женскому полу, если конкретно не указан один пол. Соответственно, термин «субъект» или «пациент» включает любое млекопитающее, для которого способ лечения по настоящему раскрытию может иметь благоприятный эффект. Примеры «субъекта» или «пациента» включают без ограничения человека, крысу, мышь, морскую свинку, обезьяну, свинью, козу, корову, лошадь, собаку, кошку, птицу и домашнюю птицу. В типичном варианте осуществления пациентом является человек. Термин «млекопитающее» относится ко всем представителям класса млекопитающих, в том числе к людям, приматам, домашним и сельскохозяйственным животным, таким как кролик, свинья, овца и крупный рогатый скот; а также к зоопарковым животным, животным, используемым в спорте, или животным-компаньонам; и грызунам, таким как мышь и крыса. Термин «млекопитающее, отличное от человека» относится ко всем представителям класса млекопитающих, за исключением человека.
Настоящее раскрытие основано по меньшей мере на конструировании фармацевтических средств Т-Е, которые можно доставлять в клетки-мишени, ткани- или органы-мишени в увеличенных долях по сравнению с кровотоком, лимфоидной системой и другими клетками, тканями или органами. При достижении этого терапевтический эффект фармацевтических средств увеличивается, а масштаб и тяжесть побочных эффектов и токсичности уменьшаются. Также возможно вводить терапевтический эффектор в форме молекулы Т-Е в более низкой дозе, чем в форме без нацеливающего элемента. Таким образом, терапевтический эффектор можно вводить в более низких дозах без утраты действенности, при этом побочные эффекты и токсичность уменьшаются.
Заболевания, в отношении которых лучшее нацеливание лекарственных средств может производить благоприятный эффект
Лекарственные средства, применяемые против многих заболеваний, можно улучшить для достижения лучшей эффективности и безопасности, если их можно нацелить на пораженные участки, т.е. если их можно локализовать в пораженных участках или разделить между ними более эффективным образом, чем в нормальных тканях или органах. Определенные лекарственные средства на основе антител, нацеленные на инфекционные микроорганизмы или их токсичные продукты, можно улучшить, если они обладают способностью задействовать иммуноциты, которые фагоцитируют и обезвреживают частицы, связанные с антителом. Далее приведены основные примеры заболеваний, лекарственные средства против которых можно улучшить, если они могут преимущественно распределяться в пораженные участки или клетки или если они могут задействовать фагоцитные иммуноциты.
I. Заболевания центральной нервной системы
Для лечения заболеваний центральной нервной системы (ЦНС) терапевтическим агентам часто необходимо преодолеть гематоэнцефалический барьер (ГЭБ), чтобы попасть в ЦНС. Некоторые терапевтические агенты не попадают в ЦНС; они регулируют определенные действия, такие как иммунная активность, в периферической нервной системе, которые затем модулируют заболевания в ЦНС. ГЭБ образован эндотелиальными клетками, выстилающими капилляры кровеносных сосудов в ЦНС. В отличие от капилляров в периферических тканях и органах, капиллярные эндотелиальные клетки в ГЭБ связаны плотными соединениями, образованными окклюдином, клоудинами и молекулами, обеспечивающими адгезию в таких соединениях.
По меньшей мере шесть антител, а именно: адуканумаб, бапинерумаб, кренезумаб, гантенерумаб, понезумаб и соланезумаб, специфичные к β-амилоиду, который вызывает болезнь Альцгеймера, были разработаны и введены в клиническую разработку. Эти антитела обычно не обладают удовлетворительной терапевтической эффективностью при лечении болезни Альцгеймера. Общим убеждением является то, что для достижения этими антителами терапевтической эффективности значительная их часть должна преодолеть ГЭБ, чтобы попасть в поврежденные участки ЦНС. Тем не менее, лишь ничтожное количество этих антител приникает сквозь ГЭБ.
Интерферон-β-1a (IFN-β-1a) и интерферон-β-1b (IFN-β-1b) использовали для лечения рассеянного склероза (PC). Фармацевтические препараты, IFN-β-1a, вырабатываемый клетками млекопитающих и IFN-β-1b, вырабатываемый бактерией Е. coli, представляют собой одноцепочечный белок из 166 аминокислотных остатков, содержащий одну дисульфидную связь. Утверждается, что эти лекарственные препараты снижают возможность рецидива PC у 18-38% получавших его пациентов. Механизмы действия IFN-β-1a и IFN-β-1b очень сложны и не полностью поняты, включая увеличение образования противовоспалительных иммунных клеток и факторов и подавление провоспалительных клеток и факторов. Лечение пациентов с PC с использованием IFN-β также снижает проникновение провоспалительных Т-клеток сквозь ГЭБ. Еще остается без ответа вопрос, опосредуют ли IFN-β-1a и IFN-β-1b частично свое фармакологическое действие попаданием на поврежденные участки ЦНС.
Когда лекарственное средство на основе антител или белков вводят в периферическую систему организма, только очень небольшое количество (около 0,1%) достигает ЦНС, поскольку лишь очень небольшая часть лекарственного средства на основе белков проходит сквозь ГЭБ. Однако также было обнаружено, что при многих заболеваниях ЦНС, включая болезнь Альцгеймера и рассеянный склероз, воспаление на пораженных участках вызывает разрушение ГЭБ, что приводит к его повышенной проницаемости. Этим мы обосновываем тот факт, что если к ГЭБ направить большее количество вводимого антитела, специфичного к β-амилоиду или IFN-β-1a и IFN-β-1b, то сквозь ГЭБ сможет пройти большая доля лекарственных средств и станет возможным достижение лучшего терапевтического эффекта.
Кроме того, были разработаны некоторые терапевтические агенты для ингибирования проникновения воспалительных иммуноцитов сквозь ГЭБ. Примечательным примером является натализумаб, специфичный к молекулам клеточной адгезии интегрина α4. Антитело функционирует, ингибируя воспалительные иммунные клетки, чтобы прикрепляться к эпителиальному слою, выстилающему ГЭБ, и проникать через него. В то время как натализумаб продемонстрировал терапевтическую эффективность, он обладает серьезным иммунодепрессивным побочным эффектом. В частности, он вызывает прогрессирующую мультифокальную лейкоэнцефалопатию, оппортунистическую инфекцию, вызванную вирусом Джона Каннингема (вирус JC). Этим мы обосновываем тот факт, что при доставке к ГЭБ большей доли антител, специфичных к интегрину α4, потребуется меньшая доза лекарственного средства, и можно будет достичь лучшего терапевтического эффекта с меньшими побочными эффектами.
Эндотелиальные клетки в капиллярах, образующих ГЭБ, экспрессируют рецепторы трансферрина и рецепторы инсулина, которые опосредуют трансцитоз молекул трансферрина и инсулина, соответственно, в паренхиму головного мозга. При использовании рецептора трансферрина в качестве транспорта сквозь барьер проникает лишь небольшая часть лекарственного средства, в то время как оставшаяся масса задерживается или разрушается. Поскольку эндотелиальные клетки, выстилающие капилляры в других частях сосудистой сети, не экспрессируют рецепторы трансферрина, эти рецепторы в эндотелиальных клетках в ГЭБ могут служить в качестве сайт-специфического антигена для доставки вводимых лекарственных средств. Как только лекарственное средство сконцентрировалось у ГЭБ, увеличенная его доля пройдет через капилляры.
Мы также обосновываем тот факт, что при установлении механизмов для направления фармацевтических препаратов к ГЭБ необходимо исследовать терапевтическое действие противовоспалительных лекарственных средств, таких как антитела к TNF-α, IL12/IL-23, IL17 и CD3, на многие виды заболеваний ЦНС.
Для лекарственного средства на основе антител, специфичного к интегрину α4, рецептор трансферрина используется в качестве рекрутера для целевого участка. Для болезни Альцгеймера эффекторный элемент может состоять из нескольких копий scFv, специфичных к β-амилоиду; для лечения рассеянного склероза эффекторный элемент может представлять собой несколько копий IFN-β-1a или IFN-β-1b или несколько копий scFv, специфичных к интегрину α4.
Варианты осуществления настоящего раскрытия указывают на тот факт, что несколько молекул Т-Е, соответственно, присутствуют в отдельных разветвленных линкерных звеньях или конфигурациях с сочлененными линкерами, каждый из которых содержит scFv, специфичный к рецептору трансферрина, в качестве нацеливающего элемента, и IFN-β-1a, или IFN-β-1b, или scFv, специфичный к интегрину α-4, в качестве эффекторного элемента. Альтернативные варианты осуществления указывают на тот факт, что молекулы Т-Е, соответственно, присутствуют в отдельных линкерных звеньях или в конфигурациях с сочлененными линкерами, каждый из которых содержит scFv, специфичный к рецептору трансферрина, в качестве нацеливающего элемента, и scFv, специфичный к β-амилоиду, в качестве эффекторного элемента.
Препарат «Финголимод» представляет собой иммунодепрессант, получаемый из природного мириоцина, первоначально выделенного из определенных грибов. Финголимод был одобрен для снижения возможности рецидива рассеянного склероза рецидивирующе-ремиттирующего типа. Финголимод фосфорилируется in vivo с образованием финголимод-фосфата, который напоминает природный сфингозин-1-фосфат (S1P), внеклеточный липидный медиатор, и может связываться с 4 из 5 рецепторов S1P. Рецепторы S1P экспрессируются на лимфоцитах и участвуют в миграции лимфоцитов. Обычно фармакологический механизм действия финголимода заключается в том, что он ингибирует выход лимфоцитов из лимфоидных тканей в кровоток и, следовательно, в ЦНС. Финголимод может проходить сквозь ГЭБ для попадания в ЦНС и многие типы клеток ЦНС экспрессируют рецепторы S1P, которые играют роль в пролиферации, морфологии и миграции клеток. Считается, что финголимод может оказывать прямое воздействие на ЦНС. Применение финголимода вызывает обычные побочные эффекты в виде головной боли и усталости, а также серьезные побочные эффекты в виде рака кожи, макулярного отека и смертельных инфекций, таких как геморрагический очаговый энцефалит.
Молекула финголимода имеет NH2-rpynny и, таким образом, обеспечивает функциональную группу для соединения бифункциональным линкером с NHS-группой. Одним из предпочтительных вариантов осуществления настоящего изобретения является обеспечение конструкции Т-Е, содержащей нацеливающий элемент для доставки к ГЭБ и «связку» с лекарственным средством финголимод в качестве эффекторного элемента. Для получения такой «связки» 5-10 молекул финголимода вводятся в линкерное звено с помощью расщепляемого линкера или нерасщепляемого линкера для конъюгирования молекул финголимода со связывающими ветвями линкерного звена. Так как финголимод после поглощения организмом пациента модифицируется в финголимод-фосфат, подобный сфингозин-1-фосфату, и становится активным, в качестве альтернативы готовят «связку» с лекарственным средством финголимод-фосфат. Линкерное звено со «связкой» финголимода или финголимод-фосфата конъюгируется с одним или двумя scFv, специфичными к рецептору трансферрина I. После введения молекулярной конструкции часть ее переносится к ГЭБ. Молекулы финголимода, высвобождаемые из расщепляющихся линкеров, проникают сквозь ГЭБ и попадают в ЦНС или же часть всей конструкции попадает в ЦНС. Расщепляемые линкеры могут быть созданы с использованием нескольких механизмов расщепления. Часто используется связь S-S, поскольку дисульфидную связь S-S можно расщепить реакцией восстановления на участке ткани-мишени. Пептидная связь между аминокислотами, чувствительная к протеазам, таким как матричные металлопротеиназы во многих тканях и катепсины в эндосомах клеткок-мишеней, также широко используется в качестве расщепляемой связи во многих конструкциях линкеров.
Некоторые примеры молекул Т-Е, соответственно, существуют в одиночных линкерных звеньях или в конфигурациях с сочлененными линкерами, каждый из которых содержит один или два scFv, специфичных к рецептору трансферрина, в качестве нацеливающего элемента и финголимод в качестве эффекторного элемента. Во всех этих молекулярных конструкциях связь между связывающими ветвями и эффекторными элементами может быть расщепляемой или нерасщепляемой. При применении платформ молекулярных конструкций по настоящему изобретению для различных целей при лечении заболеваний ЦНС нацеливающий элемент может быть собран с одной или двумя копиями scFv, специфичными к рецептору трансферрина или рецептору инсулина. Используют один или два scFv, специфичных к рецептору трансферрина или рецептору инсулина. Если scFv имеет относительно высокую аффинность (Kd<1×10-9), то используется один scFv; если scFv имеет умеренную аффинность (1×10-9<Kd<5×10-8), то используются два scFv. Предпочтительно, чтобы присутствовало не более двух копий scFv, специфичных к рецептору трансферрина или рецептору инсулина, во избежание сшивки рецепторов и эндоцитоза связанного лекарственного средства.
II. Инфекционные заболевания
Несмотря на то, что создано большое количество моноклональных антител против компонентов различных вирусов, бактерий и грибов, которые вызывают серьезные инфекции у людей и животных, было разработано лишь несколько моноклональных антител в качестве профилактических средств или терапевтических агентов для борьбы с инфекциями. Их недостатки можно связать с несколькими основными факторами. Одним из основных факторов является тот факт, что инфекционные микроорганизмы и их продукты имеют разные серотипы и переменную реактивность по отношению к конкретному антителу. Другая причина заключается в том, что микроорганизмы-мишени мутируют и избегают нацеливания конкретного антитела.
Молекулярная конструкция Т-Е по настоящему изобретению также может применяться для профилактики и лечения инфекционных заболеваний. Множество связывающих ветвей может повысить авидность и специфичность связывания с целевыми инфекционными микроорганизмами или их продуктами и вызывать иммунные реакци для облегчения обезвреживания микроорганизмов и их продуктов. Мы считаем, что повышение авидности и задействование функции иммунного клиренса могут каким-то образом преодолеть серотипические различия и решить проблемы, связанные с мутацией. Такие улучшения должны повысить эффективность испытываемых антител для профилактики и лечения инфекционных заболеваний. Многие антитела, не продемонстрировавшие ожидаемого результата в клинических испытаниях, можно сконфигурировать согласно настоящему изобретению и исследовать повторно.
Предпочтительным вариантом осуществления настоящего изобретения является использование конфигурации с сочлененными линкерами - с одним линкерным звеном для нацеливания и одним линкерным звеном для задействования эффекторной функции. Альтернативный вариант предпочтительного варианта осуществления заключается в использовании отдельных линкерных звеньев с несколькими связывающими ветвями для нацеливающих элементов и соединяющей ветвью для эффекторного элемента. Нацеливающие элементы могут относиться к одной из двух категорий: (1) scFv или sdAb, специфичные к поверхностному элементу микроорганизма или его продукта, например, к обол очечному белку gp120 вируса иммунодефицита человека 1 типа (HIV-1), белку F респираторно-синцитиального вируса (RSV), мембранному антигену бактерий Clostridium difficile или Staphylococcus aureus, эндотоксину грамотрицательных бактерий или шигаподобному токсину I или II типа бактерии Escherichia coli, или (2) внеклеточные части рецепторов клеточной поверхности вирусов, такие как домен CD4, связывающийся с белком gp120 вируса HIV-1.
Эффекторные элементы представляют собой один или два scFv или sdAb, специфичные к одному Fc-рецептору IgG, например, FcγRIIA (CD32), FcγRIIIB (CD16b) или FcγRI (CD64). Эти рецепторы экспрессируются на нейтрофилах, макрофагах и эозинофилах и являются ключевыми молекулами, опосредующими фагоцитоз связанных с антителом микроорганизмов. FcγRIIA и FcγRIIIB связываются с IgG, имея низкую аффинность (Kd в диапазоне от 10-6 до 10-7), a FcγRI связывается с IgG1 и IgG3, имея высокую аффинность (Kd порядка 10-9). Предпочтительно использование scFv или sdAb, специфичных к FcγRIIA или FcγRIIIB, поскольку они могут успешно конкурировать с IgG при связывании с рецепторами.
Антитела, специфичные к углеводным антигенам на поверхности бактерий, обычно обладают слабой аффинностью связывания и экспрессируются в IgM, а не в IgG. Молекула IgM имеет десять Fv (антиген-связывающих участков). Однако молекула IgM, имеющая молекулярную массу приблизительно 1000 кДа, не может проходить через капилляры и достигать экстраваскулярного пространства. В соответствии с конфигурацией настоящего изобретения, молекулярная конструкция, несущая шесть scFv или десять sdAb, будет иметь молекулярную массу приблизительно 150 кДа.
При использовании лекарственных средств на основе антител для обезвреживания вирусов важно, чтобы терапевтический эффект не приводил к усилению вирусной инфекции, опосредованной FcR. В этих случаях связанные вирусные частицы не фагоцитируются и не перевариваются. Некоторые вирусы, такие как вирус Денге, могут размножаться в фагоцитах. Таким образом, если вирусные частицы получают доступ к клетке и попадают в связанные клетки без разрушения, вирус может размножаться в инфицированных клетках. Поэтому набор предпочтительных вариантов осуществления данного изобретения состоит в том, чтобы молекулярная конструкция содержала два или более scFv, специфичных к рецептору Fcγ, и могла связываться с несколькими молекулами рецептора Fcγ на поверхности клеток фагоцитов таким образом, чтобы связанные вирусные частицы оказывались на пути фагоцитоза.
Среди многих антител, специфичных к вирусам, бактериям или их продуктам и подвергнутым клиническим испытаниям, только антитела, специфичные к RSV, были одобрены для клинического использования. При этом даже антитела к RSV одобрены лишь для профилактики, а не для лечения активной инфекции. Желательно разработать антитело к RSV для лечения уже инфицированных субъектов. Другие антитела все еще находятся на этапе клинических испытаний или не смогли пройти клинические испытания. На платформах молекулярной конструкции по настоящему изобретению все эти антитела могут быть использованы с повышением эффективности. Частичный список этих антител приведен ниже:
(1) Паливизумаб и фелвизумаб, специфичные к белку F вируса RSV;
(2) Сувизумаб, специфичный к белку gp120 вируса HIV-1;
(3) Либивирумаб, эксбивирумаб, тувирумаб, специфичные к мембранному антигену гепатита В (HBsAg) вируса HBV;
(4) mAb CR6261, диридавумаб и фиривумаб, специфичные к гемагглютинину А вируса гриппа А;
(5) Регавирумаб и севирумаб, специфичные к гликопротеину цитомегаловируса;
(6) Рафивирумаб, специфичный к гликопротеину вируса бешенства;
(7) Актоксумаб и безлотоксумаб, специфичные к мембранному антигену бактерии Clostridium difficile;
(8) Обильтоксаксимаб и раксибакумаб, специфичные к сибирской язве Bacillus anthracis;
(9) Панобакумаб (человеческое IgM моноклональное антитело), специфичное к серотипу IATS О11 бактерии Pseudomonas aeruginosa;
(10) Тефибазумаб и тозатоксумаб, специфичные к фактору агглютинации А бактерии Staphylococcus aureus;
(11) Эдобакомаб, специфичный к эндотоксину грамотрицательных бактерий, при лечении сепсиса;
(12) Пагибаксимаб, специфичный к липотейхоевой кислоте бактерии Staphylococcus aureus, при лечении стафилококкового сепсиса;
(13) Раксибакумаб (моноклональное человеческое антитело), специфичный к сибиреязвенному токсину;
(14) Притоксаксимаб, сетоксаксимаб и уртоксазумаб, специфичные к шигаподобному токсину I или II типа бактерии Escherichia coli.
В соответствии с несколькими вариантами осуществления настоящего раскрытия, молекулы Т-Е в конфигурациях с сочлененными линкерами для лечения инфекционных заболеваний включают scFv, специфичный к белку F респираторно-синцитиального вируса (RSV) или к белку gp120 вируса иммунодефицита человека 1 типа (HIV-1), в качестве нацеливающих/захватывающих элементов, и scFv, специфичный к FcγRIIA (CD32) или FcγRIIIB (CD16b), в качестве эффекторных/обезвреживающих элементов. В соответствии с вариантами осуществления настоящего раскрытия, некоторые молекулы Т-Е в отдельным линкерных звеньях или в конфигурации с сочлененными линкерами включают scFv, специфичный к эндотоксину грамотрицательных бактерий или липотейхоевой кислоте бактерии Staphylococcus aureus, в качестве нацеливающих/захватывающих элементов и scFv, специфичный к CD32 или к CD16b, в качестве эффекторных/очищающих элементов. Ускоренное выведение эндотоксина при инфекции грамотрицательными бактериями должно уменьшать амплитуду высвобождения цитокинов, таких как TNF-α, IL-1 и т.д., т.е. цитокиновую бурю, при опасном для жизни состоянии сепсиса. При применении платформы молекулярной конструкции по настоящему изобретению для различных целей при лечении инфекционных заболеваний эффекторный элемент может быть собран с одной или двумя копиями scFv, специфичными kCD32 или CD16b. Используются один или два scFv, специфичных к CD32 (CD32a или CD32b) или к CD16b. Если scFv обладает относительно высокой аффинностью (Kd<1×10-9), то используется один scFv; если scFv обладает умеренной аффинностью (1×10-9<Kd<5×10-8), то используются два scFv. Предпочтительно, чтобы присутствовало не более двух копий scFv, специфичных к CD32 или к CD16b, во избежание сшивки рецепторов и эндоцитоза связанного лекарственного средства.
ЧАСТЬ I. Разветвленные линкеры для лечения определенных заболеваний
I-(i) Пептидная сердцевина для использования в разветвленном линкере
Первый аспект настоящего раскрытия относится к линкерному звену, которое содержит (1) центральную сердцевину, содержащую 2-15 лизиновых (K) остатков, и (2) 2-15 связывающих ветвей, соответственно, связанных с остатками K центральной сердцевины. Центральная сердцевина по настоящему изобретению характеризуется тем, что она имеет азидную, алкиновую, тетразиновую или напряженную алкиновую группу или связана с ней на своем N- или С-конце.
При получении линкерного звена по настоящему изобретению цепь PEG, имеющая N-гидроксисукцинимидильную (NHS) группу на одном конце и функциональную группу (например, NHS, малеимидную, азидную, алкиновую, тетразиновую или напряженную алкиновую группу) на другом конце, связывается с остатком K центральной сердцевины путем образования амидной связи между NHS-группой цепи PEG и аминогруппой остатка K. В настоящем раскрытии цепь PEG, связанную с остатком K, называют связывающей ветвью, которая имеет функциональную группу на своем свободном конце.
В соответствии с вариантами осуществления настоящего раскрытия центральная сердцевина представляет собой полипептид, имеющий длину 8-120 аминокислотных остатков и содержащий 2-15 лизиновых (K) остатков, в котором каждый остаток К отделен от следующего остатка K вставочной последовательностью.
В соответствии с вариантами осуществления настоящего раскрытия вставочная последовательность содержит глициновые (G) и сериновые (S) остатки; вставочная последовательность предпочтительно состоит из 2-15 остатков, выбранных из G, S и их комбинации. Например, вставочная последовательность может представлять собой

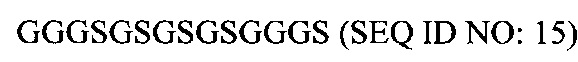 или
или

Вставочная последовательность, расположенная между двумя лизиновыми остатками, может представлять собой варианты из глициновых и сериновых остатков, имеющие в некоторой степени произвольные последовательности и (или) значения длины. Можно применять более длинные вставочные последовательности для полипептида с меньшим количеством лизиновых остатков и более короткие вставочные последовательности для полипептида с большим количеством лизиновых остатков. Во вставочные последовательности наряду с глицином и серином можно внедрять гидрофильные аминокислотные остатки, такие как аспарагиновая кислота и гистидин. В качестве альтернатив для вставочных последовательностей, образованных из глициновых и сериновых остатков, вставочные последовательности также можно взять из гибких растворимых петель широко распространенных человеческих сывороточных белков, таких как альбумин и иммуноглобулины.
В соответствии с определенными предпочтительными вариантами осуществления настоящего раскрытия центральная сердцевина содержит 2-15 звеньев с последовательностью G1-5SK. В качестве альтернативы, полипептид содержит последовательность (GSK)2-15; т.е. полипептид содержит по меньшей мере два последовательных звена с последовательностью GSK. Например, центральная сердцевина по настоящему изобретению может содержать следующую аминокислотную последовательность:
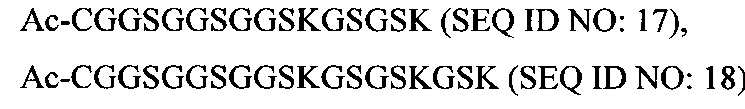 или
или
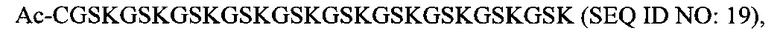
в которой Ас представляет собой ацетильную группу.
В соответствии с определенными вариантами осуществления настоящего раскрытия центральная сердцевина представляет собой полипептид, который содержит последовательность (Хаа-K)n, в которой Хаа представляет собой пегилированную аминокислоту, имеющую 2-12 повторяющихся этиленгликолевых (EG) звеньев, а n представляет собой целое число от 2 до 15.
Как описано выше, центральная сердцевина по настоящему изобретению характеризуется тем, что она имеет азидную, алкиновую, тетразиновую или напряженную алкиновую группу или связана с ней на своем N- или С-конце. В соответствии с некоторыми вариантами осуществления настоящего раскрытия центральная сердцевина по настоящему изобретению содержит на своем N- или С-конце аминокислотный остаток, имеющий азидную или алкиновую группу. Аминокислотный остаток, имеющий азидную группу, может представлять собой L-азидогомоаланин (AHA), 4-азидо-L-фенилаланин, 4-азидо-D-фенилаланин, 3-азидо-L-аланин, 3-азидо-D-аланин, 4-азидо-L-гомоаланин, 4-азидо-D-гомоаланин, 5-азидо-L-орнитин, 5-азидо-D-орнитин, 6-азидо-L-лизин или 6-азидо-D-лизин. Например, центральная сердцевина по настоящему изобретению может иметь последовательность
Ac-(GSK)2-7-(G2-4S)1-8-AAH,
Ac-AAH-(SG2-4)1-8-(GSK)2-7,
Ac-AAH-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-C,
Ac-C-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-AAH,
Ac-K-(Xaa2-12-K)2-4-Xaa2-12-AAH,
Ac-AAH-Xaa2-12-K-(Xaa2-12-K)2-4,
Ac-AAH-Xaa2-12-K-(Xaa2-12-K)1-3-Xaa2-12-C или
Ac-C-Xaa2-12-K-(Xaa2-12-K)1-3-Xaa2-12-AAH,
в которой Хаа представляет собой пегилированную аминокислоту, имеющую определенное количество повторяющихся EG-звеньев, Ас представляет собой ацетильную группу, а ААН представляет собой остаток AHA.
Типичная аминокислота, имеющая алкиновую группу, включает без ограничения L-гомопропаргилглицин (L-HPG), D-гомопропаргилглицин (D-HPG) или бета-гомопропаргилглицин (β-HPG). В этом случае центральная сердцевина по настоящему изобретению может иметь последовательность
Ac-(GSK)2-7-(G2-4S)1-8-GHP,
Ac-GHP-(SG2-4)1-8-(GSK)2-7,
Ac-GHP-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-C,
Ac-C-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-GHP,
Ac-K-(Xaa2-12-K)2-4-Xaa2-12-GHP,
Ac-GHP-Xaa2-12-K-(Xaa2-12-K)2-4,
Ac-GHP-Xaa2-12-K-(Xaa2-12-K)1-3-Xaa2-12-C или
Ac-C-Xaa2-12-K-(Xaa2-12-K)1-3-Xaa2-12-GHP,
в которой Xaa представляет собой пегилированную аминокислоту, имеющую определенное количество повторяющихся EG-звеньев, Ас представляет собой ацетильную группу, a GHP представляет собой остаток HPG.
Следует отметить, что многие аминокислоты, содержащие азидную или алкиновую группу в своих боковых цепях, и пегилированные аминокислоты доступны коммерчески в защищенных t-boc (трет-бутилоксикарбонилом) или Fmoc (9-флуоренилметоксикарбонилом) формах, легко применимых в твердофазном синтезе пептидов.
В соответствии с некоторыми рабочими примерами настоящего раскрытия центральная сердцевина может содержать последовательность
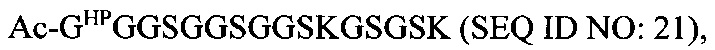
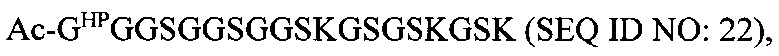
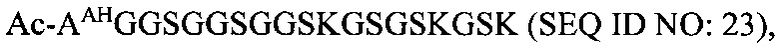
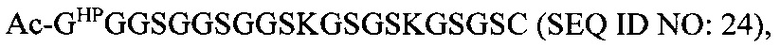
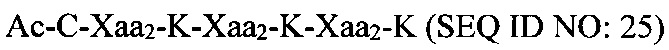 или
или
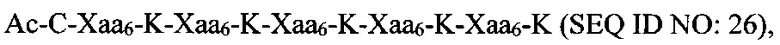
в которой Хаа представляет собой пегилированную аминокислоту, имеющую определенное количество повторяющихся EG-звеньев, Ас представляет собой ацетильную группу, AAH представляет собой остаток AHA, a GHP представляет собой остаток HPG.
В качестве альтернативы, центральная сердцевина по настоящему изобретению связана с соединяющей ветвью, которая имеет функциональную группу (например, азидную группу, алкиновую группу, тетразиновую или напряженную алкиновую группу) на своем свободном конце (т.е. на конце, не связанном с центральной сердцевиной). В этих случаях центральная сердцевина по настоящему изобретению содержит цистеиновый остаток на своем N- или С-конце. Для получения линкерного звена, связанного с соединяющей ветвью, цепь PEG, имеющая малеимидную группу на одном конце и функциональную группу на другом конце, связывается с цистеиновым остатком центральной сердцевины посредством реакции тиольной и малеимидной групп, происходящей между малеимидной группой цепи PEG и тиольной группой цистеинового остатка. В настоящем раскрытии цепь PEG, связанную с цистеиновым остатком центральной сердцевины, называют соединяющей ветвью, которая имеет функциональную группу на своем свободном конце.
Соединяющая ветвь предпочтительно имеет тетразиновую или напряженную алкиновую группу (например, циклооктеновую или циклооктиновую группу) на своем свободном конце. Эти соединяющие ветви имеют 2-12 EG-звеньев. В соответствии с вариантами осуществления настоящего раскрытия тетразиновая группа представляет собой 1,2,3,4-тетразиновую, 1,2,3,5-тетразиновую, 1,2,4,5-тетразиновую группы или их производные. Напряженная алкиновая группа может быть циклооктеновой или циклооктиновой группой. В соответствии с рабочими примерами в настоящем раскрытии, циклооктеновая группа представляет собой транс-циклооктеновую (ТСО) группу, а пример напряженной алкиновой группы включает без ограничения дибензоциклооктиновую (DBCO), дифторциклооктиновую (DIFO), бициклонониновую (BCN) и дибензоциклооктиновую (DICO) группы. В соответствии с некоторыми вариантами осуществления настоящего раскрытия тетразиновая группа представляет собой 6-метилтетразин.
Пример центральной сердцевины по настоящему изобретению, способной связываться с соединяющей ветвью, включает без ограничения
Ac-(GSK)2-7-(G2-4S)1-8-C-Xaa2-12-тетразин,
Ас-(GSK)2-7-(G2-4S)1-8-С-Хаа2-12-напряженный алкин,
Ас-K-(Хаа2-12-K)2-4-Хаа2-12-С-Хаа2-12-тетразин,
Ас-K-(Хаа2-12-K)2-4-Хаа2-12-С-Хаа2-12-напряженный алкин,
Тетразин-Xaa2-12-C(Ac)-(SG2-4)1-8-(GSK)2-7,
Напряженный алкин-Хаа2-12-С(Ас)-(SG2-4)1-8-(GSK)2-7,
Тетразин-Хаа2-12-С(Ас)-Хаа2-12-K-(Хаа2-12-K)2-4, и
Напряженный алкин-Хаа2-12-С(Ас)-Хаа2-12-K-(Хаа2-12-K)2-4.
В качестве альтернативы, центральная сердцевина имеет азидную или алкиновую группу на одном конце и соединяющую ветвь с тетразиновой или напряженной алкиновой группой на другом конце. Примерами являются следующие:
Ac-AAH-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-C-Xaa2-12-тетразин,
Ас-ААН-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-С-Хаа2-12-напряженный алкин,
Тетразин-Xaa2-12-C(Ac)-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-AAH,
Напряженный алкин-Хаа2-12-С(Ас)-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-ААН,
Ас-ААН-Хаа2-12-K-(Хаа2-12-K)1-3-Хаа2-12-С-Хаа2-12-тетразин,
Ас-ААН-Хаа2-12-K-(Хаа2-12-K)1-3-Хаа2-12-С-Хаа2-12-напряженный алкин,
Тетразин-Хаа2-12-С(Ас)-Хаа2-12-K-(Хаа2-12-K)1-3-Хаа2-12-ААН,
Напряженный алкин-Хаа2-12-С(Ас)-Хаа2-12-K-(Хаа2-12-K)1-3-Хаа2-12-ААН,
Ac-GHP-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-C-Xaa2-12-тетразин,
Ас-GHP-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-С-Хаа2-12-напряженный алкин,
Тетразин-Xaa2-12-C(Ac)-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-GHP,
Напряженный алкин-Xaa2-12-C(AC)-(SG2-4)0-7-(GSK)2-6-(G2-4S)1-8-GHP,
Ас-GHP-Хаа2-12-K-(Хаа2-12-K)1-3-Хаа2-12-С-Хаа2-12-тетразин,
Ас-GHP-Хаа2-12-K-(Хаа2-12-K)1-3-Хаа2-12-С-Хаа2-12-напряженный алкин,
Тетразин-Xaa2-12-C(Ac)-Xaa2-12-K-(Xaa2-12-K)1-3-Xaa2-12-GHP, и
Напряженный алкин-Хаа2-12-С(Ас)-Хаа2-12-K-(Хаа2-12-K)1-3-Xaa2-12-GHP.
Полипептид также можно синтезировать с помощью рекомбинантной технологии путем экспрессии сконструированных сегментов генов в клетках-хозяевах бактерий или млекопитающих. Если сердцевина имеет большое количество лизиновых остатков и существенную длину, то предпочтительно получать полипептид в виде рекомбинантных белков. При использовании твердофазного синтеза по мере увеличения длины полипептида возрастает количество ошибок, а степень чистоты и (или) выход продукта снижаются. Для получения полипептида в клетках-хозяевах бактерий или млекопитающих вставочная последовательность в диапазоне от нескольких аминокислотных остатков до 10-20 остатков может располагаться между двумя остатками K. Кроме того, поскольку AHA и HPG не являются природными аминокислотами, кодируемыми генетическими кодами, N- или С-концевой остаток в этих рекомбинантных полипептидах представляет собой цистеин. После экспрессии и очистки рекомбинантных белков концевой цистеиновый остаток затем подвергают реакции с короткими бифункциональными сшивающими средствами, которые имеют малеимидную группу на одном конце, реагирующую с SH-группой цистеинового остатка, и алкиновую, азидную, тетразиновую или напряженную алкиновую группу на другом конце.
Синтез полипептида с применением пегилированных аминокислот включает меньшее количество этапов, чем в случае применения обычных аминокислот, таких как глициновые и сериновые остатки. В дополнение, можно использовать пегилированные аминокислоты с различной длиной (т.е. количеством повторяющихся этиленгликолевых звеньев), что обеспечивает гибкость для целей растворимости и некоторое расстояние между соседними аминогруппами лизиновых остатков. Центральные сердцевины также могут быть сконструированы таким образом, чтобы они содержали, помимо пегилированных аминокислот, искусственные аминокислоты, такие как D-формы аминокислот, гомоаминокислоты, N-метилированные аминокислоты и т.д. Для конструирования центральной сердцевины предпочтительно применяют пегилированные аминокислоты с различной длиной цепи полиэтиленгликоля (PEG), поскольку PEG-компоненты, содержащиеся в молекулах аминокислот, обеспечивают конформационную гибкость и надлежащее расстояние между конъюгирующими группами, повышают растворимость в воде и, как правило, являются слабоиммуногенными. Синтез центральной сердцевины, содержащей пегилированные аминокислоты, аналогичен процедурам синтеза обычных полипептидов.
Центральная сердцевина по настоящему изобретению для целей стабильности необязательно имеет на своем N-конце ацетильную группу, блокирующую аминогруппу.
Как может быть понятно, количество связывающих ветвей, связанных с центральной сердцевиной, определяется главным образом количеством лизиновых остатков, содержащихся в центральной сердцевине. Поскольку в центральной сердцевине по настоящему изобретению содержится по меньшей мере два лизиновых остатка, линкерное звено по настоящему изобретению может содержать множество связывающих ветвей.
Обратимся теперь к фигуре 1А. Согласно иллюстрации, линкерное звено 10А содержит центральную сердцевину 11а, содержащую один остаток HPG (GHP) и четыре лизиновых (K) остатка, соответственно, разделенных вставочными последовательностями (обозначенными многоточием во всех графических материалах). Вставочные последовательности между остатком HPG и остатком K или между любыми двумя остатками K могут иметь одинаковые или разные аминокислотные последовательности. В этом примере четыре связывающие ветви 20a-20d связаны с лизиновыми остатками путем образования амидной связи между NHS-группой и аминогруппой лизинового остатка соответственно. Как может быть понятно, определенные признаки, обсуждаемые в данном документе в отношении линкерного звена 10A или любых других последующих линкерных звеньев, являются общими для других линкерных звеньев, раскрытых в данном документе, и, следовательно, некоторые или все из этих признаков также являются применимыми в следующих примерах, если это не противоречит контексту конкретного варианта осуществления. Однако, в целях краткости эти общие признаки могут не повторяться явным образом ниже.
На фигуре 1В представлено линкерное звено 10В в соответствии с другим вариантом осуществления настоящего раскрытия. Центральная сердцевина 11b содержит один цистеиновый (С) остаток и шесть лизиновых (K) остатков, соответственно, разделенных вставочными последовательностями. В этом примере линкерное звено 10А содержит шесть связывающих ветвей 20a-20f, соответственно связанных с лизиновыми остатками. В соответствии с вариантами осуществления настоящего раскрытия связывающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев.
В отличие от линкерного звена 10А на фигуре 1А, линкерное звено 1В дополнительно содержит соединяющую ветвь 60. Как обсуждается выше, цепь PEG, имеющая малеимидную группу на одном конце и функциональную группу на другом конце, применяется для образования соединяющей ветви 60. Таким образом, соединяющая ветвь 60 связана с цистеиновым остатком центральной сердцевины 11b посредством реакции тиольной и малеимидной групп. В этом примере функциональная группа на свободном конце соединяющей ветви 60 представляет собой тетразиновую группу 72. В соответствии с вариантами осуществления настоящего раскрытия соединяющая ветвь представляет собой цепь PEG, имеющую 2-12 повторяющихся EG-звеньев.
Если требуется высвобождение эффекторных элементов в участке-мишени, в связывающую ветвь можно вставить расщепляемую связь. Такая связь расщепляется путем кислотного/щелочного гидролиза, восстановления/окисления или под действием ферментов. Одним вариантом осуществления класса расщепляемых цепей PEG, которые можно применять для образования соединяющей ветви, является NHS-PEG2-20-S-S-малеимид, где S-S представляет собой дисульфидную связь, которую можно подвергнуть медленному восстановлению, а NHS-группу применяют для конъюгирования с аминогруппой центральной сердцевины, связывая таким образом цепь PEG с центральной сердцевиной. Малеимидная группа на свободном конце связывающей ветви может быть замещена азидной, алкиновой, тетразиновой или напряженной алкиновой группой. Согласно некоторым вариантам осуществления настоящего раскрытия, связывающая ветвь представляет собой цепь PEG, которая имеет 2-20 повторяющихся EG-звеньев с дисульфидной связью на ее свободном конце (т.е. на конце, не связанном с центральной сердцевиной). Обратимся теперь к фигуре 1С, на которой каждая из пяти связывающих ветвей 21a-21f, соответственно связанных с остатками К центральной сердцевины 11b, представляет собой цепь PEG с дисульфидной связью на свободном конце связывающей ветви.
В соответствии с вариантами осуществления настоящего раскрытия, связывающая ветвь, связанная с остатком К центральной сердцевины, имеет функциональную группу (т.е. малеимидную, NHS, азидную, алкиновую, тетразиновую или напряженную алкиновую группу) на своем свободном конце. Если свободный конец связывающей ветви представляет собой азидную, алкиновую или циклооктиновую группу, то аминокислотный остаток на N- или С-конце центральной сердцевины предпочтительно представляет собой цистеиновый остаток, а свободный конец соединяющей ветви представляет собой тетразиновую или циклооктеновую группу. В качестве альтернативы, если свободный конец связывающей ветви представляет собой тетразиновую или циклооктеновую группу, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу, или аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а свободный конец соединяющей ветви представляет собой азидную, алкиновую или циклооктиновую группу.
В зависимости от функциональной группы (т.е. малеимидной, NHS, азидной, алкиновой, тетразиновой или напряженной алкиновой группы), присутствующих на свободном конце связывающей ветви, с помощью соответствующей функциональной группы можно разработать функциональный элемент (например, нацеливающий элемент, эффекторный элемент или элемент для улучшения фармакокинетических свойств) таким образом, что функциональный элемент может быть связан со свободным концом связывающей ветви посредством любой из следующих химических реакций:
(1) образование между ними амидной связи: в этом случае связывающая ветвь имеет NHS-группу на свободном конце, а функциональный элемент имеет аминогруппу;
(2) тиол-малеимидная реакция: в этом случае связывающая ветвь имеет малеимидную группу на свободном конце, а функциональный элемент имеет тиольную группу;
(3) реакция азид-алкинового циклоприсоединения, катализируемого медью (I) (CuAAC или клик-реакция для краткости): один из свободных концов связывающей ветви и функционального элемента имеет азидную группу, а другой имеет алкиновую группу; реакция CuAAC проиллюстрирована на схеме 1;
(4) реакция Дильса-Альдера с обратными электронными требованиями (iEDDA): один из свободных концов связывающей ветви и функционального элемента имеет тетразиновую группу, а другой имеет циклооктеновую группу; реакция iEDDA проиллюстрирована на схеме 2; или
(5) клик-реакция азид-алкинового циклоприсоединения, промотируемого напряжением (SPAAC): один из свободных концов связывающей ветви и функционального элемента имеет азидную группу, а другой имеет циклооктиновую группу; реакция SPAAC проиллюстрирована на схеме 3.
«Схема 1. Реакция CuAAc»
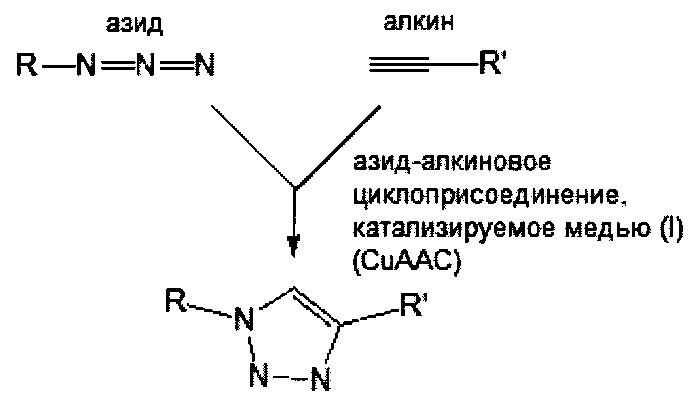
«Схема 2. Реакция iEDDA»
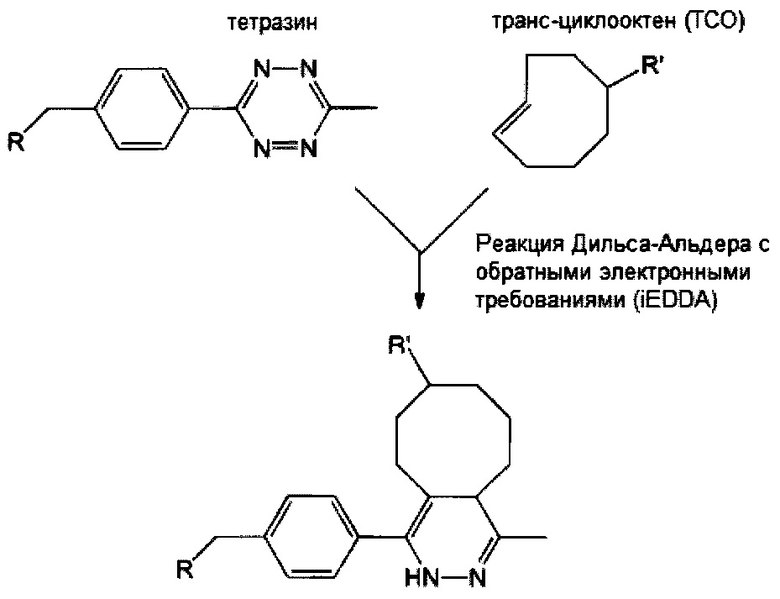
«Схема 3. Реакция SPAAC»
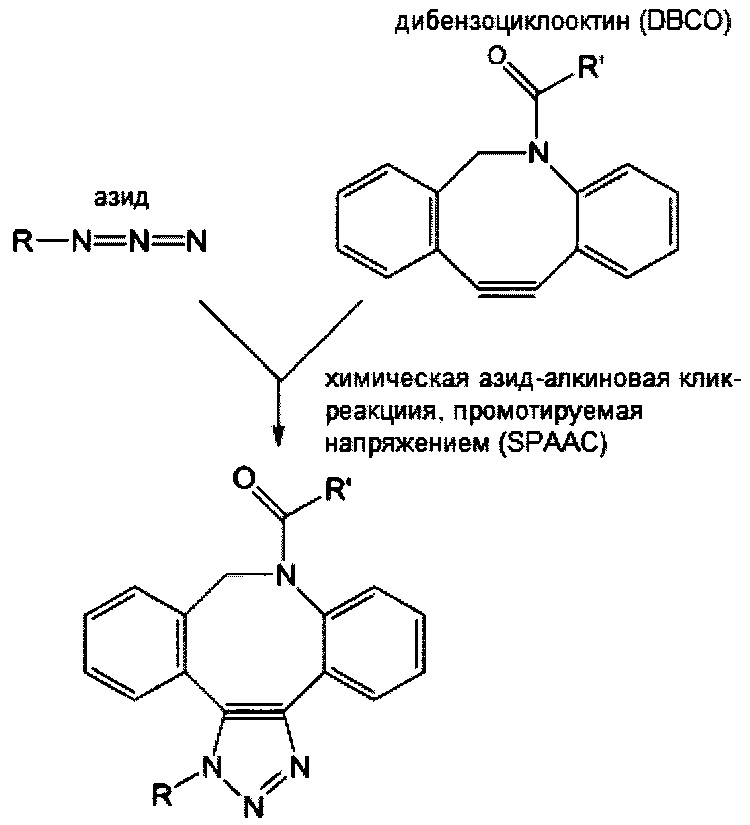
В результате реакции CuAAC получают 1,5-дизамещенный 1,2,3-триазол. Реакция между алкином и азидом является весьма селективной, и в природных биомолекулах отсутствуют алкиновые и азидные группы. Более того, реакция является быстрой и нечувствительной к рН. Было выдвинуто предположение, что вместо применения меди (I), как, например, бромида меди или йодида меди, для катализа клик-реакции лучше применять смесь меди (II) и восстановителя, такого как аскорбат натрия, для получения меди (I) в реакционной смеси in situ. В качестве альтернативы, второй элемент может быть связан с N- или С-концом центральной сердцевины по настоящему изобретению посредством реакции без использования меди, в которой в качестве катализатора для катализа азид-алкинового циклоприсоединения применяется пентаметилциклопентадиенильный комплекс хлорида рутения.
В целях иллюстрации функциональные элементы, связанные со связывающими ветвями, называют первыми элементами. Очевидно, что количество первых элементов, которые несет линкерное звено по настоящему изобретению, зависит от количества остатков K в центральной сердцевине (и, таким образом, от количества связывающих ветвей). Соответственно, средний специалист в данной области при необходимости может регулировать количество первых элементов линкерного звена, например, для достижения желаемого нацеливания или терапевтического эффекта.
Пример линкерного звена 10D, имеющего первые элементы, проиллюстрирован на фигуре 1D. За исключением признаков, обсуждаемых в данном документе ниже, фигура 1D довольно сходна с фигурой 1В. Во-первых, в центральной сердцевине 11d имеется пять остатков K и соответственно с ними связаны, соответственно, пять связывающих ветвей 20а-20е. Во-вторых, линкерное звено 10D имеет пять первых элементов 30а-30е, связанных с каждой из связывающих ветвей 20а-20е. Как обсуждается ниже, необязательная тетразиновая группа 72 обеспечивает возможность конъюгирования с дополнительным функциональным элементом, другой молекулярной конструкцией (см. часть II или часть III ниже).
На фигуре 1Е представлен альтернативный пример, в котором линкерное звено 10Е имеет структуру, аналогичную структуре линкерного звена 1D, за исключением того, что каждая из связывающих ветвей 21а-21е линкера имеет дисульфидную связь на ее концевом элементе (т.е. на конце, связанном с каждым из первых элементов 30а-30е).
В качестве альтернативы, настоящее линкерное звено дополнительно содержит множество сопрягающих ветвей, каждая из которых имеет функциональную группу (т.е. малеимидную, NHS, азидную, алкиновую, тетразиновую или напряженную алкиновую группу) на одном конце и NHS или малеимидную группу на другом конце. С использованием реакции, аналогичной реакции между первым элементом и связывающей ветвью, сопрягающая ветвь может быть связана со связывающей ветвью с соответствующей функциональной группой - либо путем образования между ними амидной связи, либо посредством тиол-малеимидной реакции, реакции CuAAC, реакции iEDDA или реакции SPAAC. Таким образом, сопрягающая ветвь, связанная со связывающей ветвью, имеет NHS или малеимидную группу на своем свободном конце (или на конце связывания элементов, т.е. на конце, не связанном со связывающей ветвью); а первый элемент, в свою очередь, связан с концом связывания элементов сопрягающей ветви путем образования между ними амидной связи или посредством тиол-малеимидной реакции.
Обратимся теперь к фигуре 1F, на которой связывающая ветвь связана с остатком K центральной сердцевины 11d, как показано на фигуре 1D. По сравнению с линкерным звеном 10D, линкерное звено 10F дополнительно содержит сопрягающую ветвь 25, связанную со связывающими ветвями 22 посредством реакции SPAAC. Первый элемент 30, в свою очередь, связан с сопрягающей ветвью 25 - либо путем образования между ними амидной связи, либо посредством тиол-малеимидной реакции. Ромб 90, изображенный на фигуре 1F, обозначает собой химическую связь, полученную в результате реакции SPAAC между связывающей ветвью 22 и сопрягающей ветвью 25.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия, сопрягающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев. В качестве альтернативы, сопрягающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на ее концевом элементе (т.е. на свободном конце, не связанном со связывающей ветвью).
В одном рабочем примере сопрягающая ветвь имеет три повторяющихся EG-звена, а также дисульфидную связь на свободном конце (иначе называемом концом связывания элементов) сопрягающей ветви. В этом случае первый элемент, связанный с концом связывания элементов сопрягающей ветви, можно эффективно высвободить из настоящего линкерного звена посредством обработки восстановителем.
В соответствии с некоторыми предпочтительными вариантами осуществления настоящего раскрытия, первыми элементами являются финголимод, финголимод-фосфат, интерферон-β или одноцепочечный вариабельный фрагмент (scFv), специфичный к интегрину α4, β-амилоиду, вирусному белку, бактериальному белку.
Неограничивающие примеры вирусных белков включают в себя белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса.
Примеры бактериального белка включают без ограничения эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигалодобный токсин I или II типа бактерии Escherichia coli.
В целях усиления намеченного или желаемого эффекта (например, терапевтического эффекта) линкерное звено по настоящему изобретению может дополнительно содержать второй элемент в дополнение к первому элементу. Например, второй элемент может представлять собой нацеливающий элемент либо эффекторный элемент. В необязательных вариантах осуществления настоящего раскрытия первый элемент представляет собой эффекторный элемент, а второй элемент может представлять собой другой эффекторный элемент, который действует аддитивно или синергично с первым элементом или независимо от него. Опять же необязательно, первый и второй элементы проявляют разные свойства; например, первый элемент представляет собой нацеливающий элемент, а второй элемент представляет собой эффекторный элемент, и наоборот. В качестве альтернативы, первый элемент представляет собой эффекторный элемент, а второй элемент представляет собой элемент, способный к улучшению фармакокинетических свойств линкерного звена, такого как растворимость, скорость выведения, период полувыведения и биодоступность. Выбор конкретного первого элемента и (или) второго элемента зависит от предполагаемого применения, при котором должно использоваться данное линкерное звено (или разветвленное линкерное звено). Примеры этих функциональных элементов обсуждаются ниже в части I-(iii) настоящего описания.
В структурном плане второй элемент связан с азидной, алкиновой, тетразиновой или напряженной алкиновой группой на N- или С-конце центральной сердцевины. В частности, второй элемент может быть необязательно конъюгирован с короткой цепью PEG (предпочтительно имеющей 2-12 повторяющихся EG-звеньев) и далее связан с N- или С-концевым аминокислотным остатком, имеющим азидную или алкиновую группу (например, остатком AHA или остатком HPG). В качестве альтернативы, второй элемент может быть необязательно конъюгирован с короткой цепью PEG и далее связан с соединяющей ветвью от центральной сердцевины.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия центральная сердцевина содержит аминокислоту, имеющую азидную группу (например, остаток AHA), на своем N- или С-конце; и, соответственно, второй элемент, имеющий алкиновую группу, связан с N- или С-концом центральной сердцевины посредством реакции CuAAC. В соответствии с другими вариантами осуществления настоящего раскрытия центральная сердцевина содержит аминокислоту, имеющую алкиновую группу (например, остаток HPG), на своем N- или С-конце; а второй элемент, имеющий азидную группу, способен, таким образом, связываться с N- или С-концом центральной сердцевины посредством реакции CuAAC.
На фигуре 1G представлен пример линкерного звена 10G по настоящему изобретению, несущего множество первых элементов и один второй элемент. В этом примере центральная сердцевина 11с содержит один остаток HPG (GHP) и пять лизиновых (K) остатков. Пять связывающих ветвей 20а-20е соответственно связаны с пятью остатками К центральной сердцевины 11с; а пять первых элементов 30а-30е соответственно связаны с указанными пятью связывающими ветвями 20а-20е посредством реакции тиольной и малеимидной групп. В дополнение к первым элементам, линкерное звено 10G дополнительно содержит один второй элемент 50, связанный с одним концом короткой цепи 62 PEG. Другой конец короткой цепи 62 PEG до конъюгирования с центральной сердцевиной 11с имеет азидную группу. Таким образом, азидная группа может реагировать с остатком HPG, имеющим алкиновую группу, посредством реакции CuAAC, так чтобы второй элемент 50 связывался с центральной сердцевиной 11с. Жирная точка 40, показанная на фигуре 1G, представляет химическую связь, образующуюся в результате реакции CuAAC, происходящей между остатком HPG и азидной группой.
В качестве альтернативы, второй элемент связан с центральной сердцевиной посредством соединяющей ветви. В соответствии с определенными вариантами осуществления настоящего раскрытия соединяющая ветвь имеет тетразиновую группу, которая может эффективно связываться со вторым элементом, имеющим ТСО-группу, посредством реакции iEDDA. В соответствии с другими вариантами осуществления настоящего раскрытия соединяющая ветвь имеет ТСО-группу, которая способна связываться со вторым элементом, имеющим тетразиновую группу, посредством реакции iEDDA. В реакции iEDDA используются напряженные циклооктены, которые характеризуются значительно сниженной энергией активации в противоположность терминальным алкинам, и поэтому устраняется необходимость в экзогенном катализаторе.
Обратимся теперь к фигуре 1Н, на которой центральная сердцевина 11d линкерного звена 10Н содержит концевой цистеиновый (С) остаток и пять лизиновых (K) остатков. Как показано на фигуре 1Н, пять связывающих ветвей 20а-20е соответственно связаны с пятью остатками K центральной сердцевины 11d, и далее пять первых элементов 30а-30е соответственно связаны с пятью связывающими ветвями 20а-20е посредством реакций тиольной и малеимидной групп. Цистеиновый остаток связан с соединяющей ветвью 60, которая до конъюгирования со вторым элементом содержит тетразиновую или ТСО-группу на своем свободном конце. В этом примере второй элемент 50, связанный с короткой цепью 62 PEG, имеющей соответствующую ТСО- или тетразиновую группу, может быть связан с соединяющей ветвью 60 посредством реакции iEDDA. Эллипс 70, показанный на фигуре 1Н, представляет химическую связь, образующуюся в результате реакции iEDDA, происходящей между соединяющей ветвью 60 и короткой цепью 62 PEG.
В соответствии с другими вариантами осуществления настоящего раскрытия соединяющая ветвь до конъюгирования со вторым элементом имеет азидную группу. В связи с этим соединяющая ветвь может быть связана со вторым элементом, имеющим циклооктиновую группу (например, DBCO-, DIFO-, BCN- или DICO-группу) на свободном конце короткой цепи PEG, посредством реакции SPAAC, и наоборот.
Обратимся теперь к фигуре 1I, на которой линкерное звено 10I имеет сходную структуру с линкерным звеном 10Н на фигуре 1Н, за исключением того, что соединяющая ветвь 60 содержит азидную или циклооктиновую группу (например, DBCO-, DIFO-, BCN- или DICO-группу) вместо тетразиновой или ТСО-группы. Соответственно, второй элемент 50, связанный с короткой цепью 62 PEG, может иметь соответствующую циклооктиновую (например, DBCO, DIFO, BCN или DICO) или азидную группу, так что он может связываться с соединяющей ветвью 60 посредством реакции SPAAC. Ромб 90, показанный на фигуре 1I, представляет химическую связь, образующуюся в результате реакции SPAAC, происходящей между соединяющей ветвью 60 и короткой цепью 62 PEG.
Схема 4 представляет собой типичную иллюстрацию способа получения линкерного звена по настоящему изобретению. На этапе 1 получают центральную сердцевину, содержащую аминокислотную последовательность (GSK)3 и L-азидогомоаланиновый (AHA) остаток на своем С-конце. На этапе 2 три связывающие ветви соответственно связываются с лизиновыми (K) остатками центральной сердцевины посредством образования амидной связи между NHS-группой и аминогруппой; связывающая ветвь, связанная с центральной сердцевиной, имеет малеимидную (Mal) группу на своем свободном конце. На этапе 3 три scFv антитела к антигену A (scFv α А) в качестве первого элемента соответственно связываются со связывающими ветвями посредством реакции тиольной и малеимидной групп. В то же время на этапе 4 один scFv антитела к антигену В (scFv α В) в качестве второго элемента связывается с короткой цепью PEG, имеющей 4 повторяющихся EG-звена и DBCO-группу на свободном конце. Наконец, на этапе 5 второй элемент связывается с остатком AHA центральной сердцевины посредством реакции SPAAC.
«Схема 4. Получение линкерного звена, связанного с двумя различными scFv посредством связывающей ветви и С-концевого аминокислотного остатка»
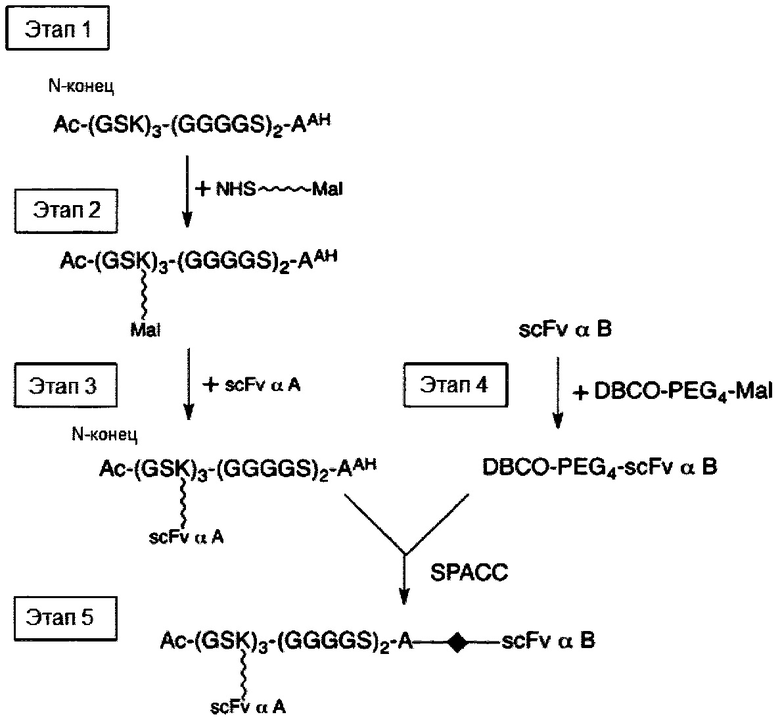
На схеме 5 проиллюстрирован другой пример способа получения линкерного звена по настоящему изобретению. На этапе 1 получают центральную сердцевину, содержащую аминокислотную последовательность (K-Хаа)3 и цистеиновый остаток на своем С-конце. На этапе 2 цепь PEG (в качестве соединяющей ветви), имеющая малеимидную (Mal) группу на одном конце и тетразиновую группу на другом конце, связывается с цистеиновым остатком посредством реакции тиольной и малеимидной групп. Далее на этапе 3 три связывающие ветви соответственно связываются с лизиновыми (K) остатками центральной сердцевины. Далее три scFv антитела к антигену A (scFv α А) в качестве первых элементов соответственно связываются со связывающими ветвями посредством реакции тиольной и малеимидной групп, как описано на этапе 4. В то же время на этапе 5 один scFv антитела к антигену В (scFv α В) в качестве второго элемента связывается с короткой цепью PEG, имеющей 3 повторяющихся EG-звена и ТСО-группу на свободном конце. Наконец, на этапе 6 второй элемент связывается с соединяющей ветвью посредством реакции iEDDA.
«Схема 5. Получение линкерного звена, связанного с двумя различными scFv посредством связывающей ветви и соединяющей ветви»
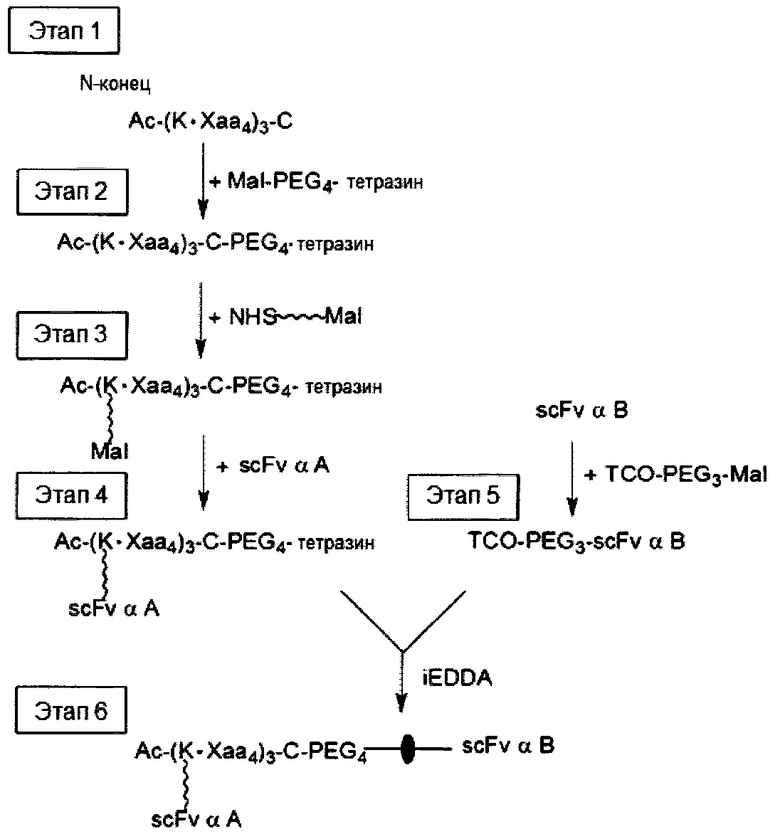
Пегилирование представляет собой процесс, при котором цепь PEG прикрепляется к молекуле (например, лекарственному средству или белку) или связывается с ней. Известно, что пегилирование придает некоторые значительные фармакологические преимущества по сравнению с немодифицированной формой, такие как улучшенная растворимость, повышенная стабильность, увеличенная продолжительность циркуляции в крови и сниженная интенсивность протеолитического распада. В соответствии с одним вариантом осуществления настоящего раскрытия второй элемент представляет собой цепь PEG, имеющую молекулярную массу приблизительно 20000-50000 дальтон.
На фигуре 1J представлен альтернативный пример линкерного звена по настоящему изобретению (линкерное звено 10J), в котором пять первых элементов 30 соответственно связаны с лизиновыми остатками посредством связывающих ветвей 20, а остаток HPG (GHP) центральной сердцевины 11е связан с цепью 80 PEG посредством реакции CuAAC. Жирная точка 40, показанная на фигуре 1J, представляет химическую связь, образующуюся в результате реакции CuAAC, происходящей между остатком HPG и цепью 80 PEG.
На фигуре 1K представлен другой пример настоящего раскрытия, в котором N-конец центральной сердцевины 11d представляет собой цистеиновый остаток, связанный с соединяющей ветвью 60. Цепь 80 PEG может эффективно связываться с соединяющей ветвью 60 посредством реакции iEDDA. Эллипс 70 линкерного звена 10K представляет химическую связь, образующуюся в результате реакции iEDDA, происходящей между соединяющей ветвью 60 и цепью 80 PEG.
На фигуре 1L представлен альтернативный пример линкерного звена по настоящему изобретению, в котором линкерное звено 10L имеет сходную структуру с линкерным звеном 10J на фигуре 1J, за исключением того, что цепь 80 PEG связана с соединяющей ветвью 60 посредством реакции SPAAC. Ромб 90, показанный на фигуре 1L, представляет химическую связь, образующуюся в результате реакции SPAAC, происходящей между соединяющей ветвью 60 и цепью 80 PEG.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия линкерное звено по настоящему изобретению в дополнение к первому и второму элементам дополнительно содержит третий элемент. В этом случае один из N- и С-конца центральной сердцевины представляет собой аминокислоту, имеющую азидную или алкиновую группу, а другой из N- и С-конца центральной сердцевины представляет собой цистеиновый остаток. Лизиновые остатки центральной сердцевины соответственно связаны со связывающими ветвями, каждая из которых имеет малеимидную группу на своем свободном конце; тогда как цистеиновый остаток центральной сердцевины связан с соединяющей ветвью, которая имеет тетразиновую или напряженную алкиновую группу на своем свободном конце. Как описано выше, первый элемент, таким образом, связан со связывающей ветвью посредством реакции тиольной и малеимидной групп, а второй элемент связан с соединяющей ветвью посредством реакции iEDDA. Кроме того, третий элемент связан с концевой аминокислотой, имеющей азидную или алкиновую группу, посредством реакции CuAAC или реакции SPAAC.
Обратимся теперь к линкерному звену 10М на фигуре 1М, в котором центральная сердцевина 11f имеет остаток HPG (GHP) на своем N-конце и цистеиновый остаток на своем С-конце. Связывающие ветви 20 и соединяющая ветвь 60 связаны соответственно с лизиновыми (K) остатками и цистеиновым (С) остатком центральной сердцевины 11f. Кроме того, пять первых элементов 30 соответственно связаны с пятью связывающими ветвями 20, второй элемент (т.е. цепь 80 PEG) связан с соединяющей ветвью 60, а третий элемент 50 связан с остатком HPG посредством короткой цепи 62 PEG. Жирная точка 40 указывает на химическую связь, образующуюся в результате реакции CuAAC, происходящей между остатком HPG и короткой цепью 62 PEG; а эллипс 70 представляет химическую связь, образующуюся в результате реакции iEDDA, происходящей между соединяющей ветвью 60 и цепью 80 PEG.
На фигуре 1N представлен другой вариант осуществления настоящего раскрытия, в котором линкерное звено 10N имеет сходную структуру с линкерным звеном 10М на фигуре 1М, за исключением того, что короткая цепь 62 PEG связана с остатком HPG посредством реакции SPAAC вместо реакции iEDDA. Ромб 90 на фигуре 1N представляет химическую связь, образующуюся в результате реакции SPAAC, происходящей между короткой цепью 62 PEG и остатком HPG.
В предпочтительных вариантах осуществления настоящего раскрытия связывающие ветви имеют малеимидную группу на свободном конце для конъюгирования с первыми элементами, имеющими сульфогидрильную группу, посредством реакции тиольной и малеимидной групп. Кроме того, на конце пептидной сердцевины существует один цистеиновый или аминокислотный остаток с азидной или алкиновой группой для прикрепления соединяющей ветви для связывания со вторым элементом.
Для специалистов в данной области понятно, что можно осуществлять изменения. На свободном конце связывающих ветвей можно применять конъюгирующую группу, отличную от малеимидной, такую как азидная, алкиновая, тетразиновая или напряженная алкиновая, для связывания с первыми элементами с помощью реакции CuAAC, реакции iEDDA или реакции SPAAC. Кроме того, не является необходимым наличие цистеинового остатка (или аминокислотного остатка с азидной или алкиновой группой) на N- или С-конце пептидной сердцевины. Более того, в состав пептидной сердцевины можно включить два или более таких остатка для прикрепления нескольких соединяющих ветвей для связывания со множеством вторых элементов.
I-(ii) Сердцевина на основе химического соединения для использования в разветвленном линкере
В дополнение к линкерному звену, описанному в части I-(i) настоящего раскрытия, в данном документе также раскрыто другое линкерное звено, в котором в качестве центральной сердцевины вместо полипептида используется химическое соединение. В частности, химическое соединение представляет собой бензол-1,3,5-триамин, 2-(аминометил)-2-метилпропан-1,3-диамин, трис(2-аминоэтил)амин, бензол-1,2,4,5-тетраамин, 3,3',5,5'-тетраамин-1,1'-бифенил, тетракис(2-аминоэтил)метан, тетракис(этиламин)гидразин, N,N,N',N'-тетракис(аминоэтил)этилендиамин, бензол-1,2,3,4,5,6-гексаамин, 1-N,1-N,3-N,3-N,5-N,5-N-гексакис(метиламин)бензол-1,3,5-триамин, 1-N,1-N,2-N,2-N,4-N,4-N,5-N,5-N-октакис(метиламин)бензол-1,2,4,5-триамин, бензол-1,2,3,4,5,6-гексаамин или N,N-бис[(1-амино-3,3-диаминоэтил)пентил]метандиамин. Каждое из этих химических соединений имеет 3 или более аминогрупп в идентичной или симметричной конфигурации. Таким образом, если одна из аминогрупп химического соединения конъюгирована с соединяющей ветвью, то все молекулы химического соединения имеют одинаковую конфигурацию.
Каждое химическое соединение, перечисленное выше, содержит множество аминогрупп, и поэтому с химическим соединением можно связать множество цепей PEG, имеющих NHS-группы, посредством образования амидной связи между аминогруппой и NHS-группой, что аналогично механизму связывания, описанному в части I-(i) настоящего раскрытия; связываемую таким образом цепь PEG обозначают как связывающую ветвь, которая имеет функциональную группу (например, NHS, малеимидную, азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу) на своем свободном конце. В то же время по меньшей мере одна из аминогрупп сердцевины на основе химического соединения связывается с другой цепью PEG, которая имеет NHS-группу на одном конце и функциональную группу (например, азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу) на другом конце; связываемую таким образом цепь PEG обозначают как соединяющую ветвь, которая имеет функциональную группу на своем свободном конце.
Соответственно, первый элемент может быть связан со связывающей ветвью посредством (1) образования между ними амидной связи, (2) тиол-малеимидной реакции, (3) реакции CuAAC, (4) реакции iEDDA или (5) реакции SPAAC. Между тем, второй элемент может быть связан с соединяющей ветвью посредством реакции CuAAC, реакции iEDDA или реакции SPAAC.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия связывающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев; предпочтительно связывающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на ее свободном конце (т.е. конце, не связанном с центральной сердцевиной). Соединяющая ветвь представляет собой цепь PEG, имеющую 2-12 повторяющихся EG-звеньев. В одном варианте осуществления как связывающая, так и соединяющая ветви имеют 12 повторяющихся EG-звеньев, при этом один конец соединяющей ветви представляет собой NHS-группу, а другой конец соединяющей ветви представляет собой алкиновую группу.
Согласно альтернативному варианту осуществления настоящего раскрытия, линкерное звено дополнительно содержит множество сопрягающих ветвей, каждая из которых связана с каждой связывающей ветвью. Множество первых элементов соответственно связано со множеством сопрягающих ветвей. В одном варианте осуществления сопрягающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев. В другом варианте осуществления сопрягающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на конце связывания элементов, не связанном со связывающей ветвью.
На схемах 6 и 7, соответственно, показано связывание между центральной сердцевиной на основе химического соединения и связывающей ветвью, а также соединяющей ветвью. В схемах 6 и 7 «NHS» представляет сложный NHS-эфир, «Mal» представляет малеимидную группу, «азид» представляет азидную группу, а «алкин» представляет алкиновую группу.
Требование наличия нескольких NH2-групп, существующих в симметричной и идентичной ориентации в химическом соединении, служащем в качестве центральной сердцевины, выдвигается по следующей причине: если одна NH2-группа применяется для присоединения бифункциональной связывающей ветви с группой N-гидроксисукцинимидильного (NHS) сложного эфира и алкиновой, азидной, тетразиновой или напряженной алкиновой группой, то продукт, а именно сердцевина с соединяющей ветвью, имеющей алкиновую, азидную, тетразиновую или напряженную алкиновую группу, является однородным и может быть очищен. Такой продукт можно затем применять для получения разветвленных линкерных звеньев, в которых все остальные NH2-группы присоединены к связывающим ветвям с малеимидными или другими соединяющими группами на других концах. Если химическое соединение имеет несколько NH2-групп в несимметричных ориентациях, то продукт с одной бифункциональной связывающей ветвью/соединяющей ветвью не является однородным.
«Схема 6. Связывание связывающей и соединяющей ветвей, имеющих, соответственно, малеимидную группу и азидную группу, с центральной сердцевиной»
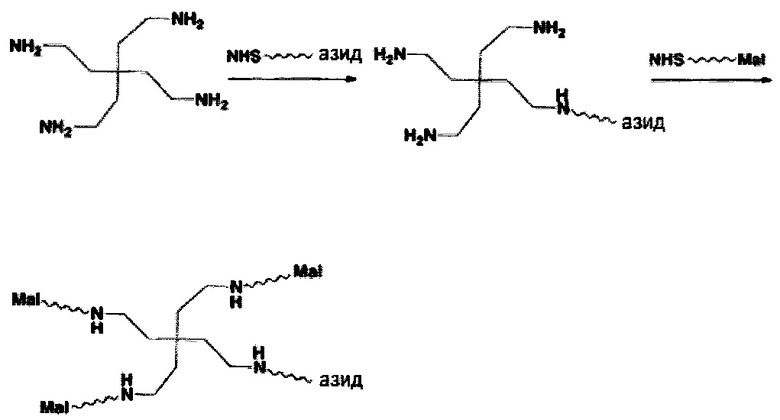
«Схема 7. Связывание связывающей и соединяющей ветвей, имеющих, соответственно, малеимидную группу и алкиновую группу, с центральной сердцевиной»
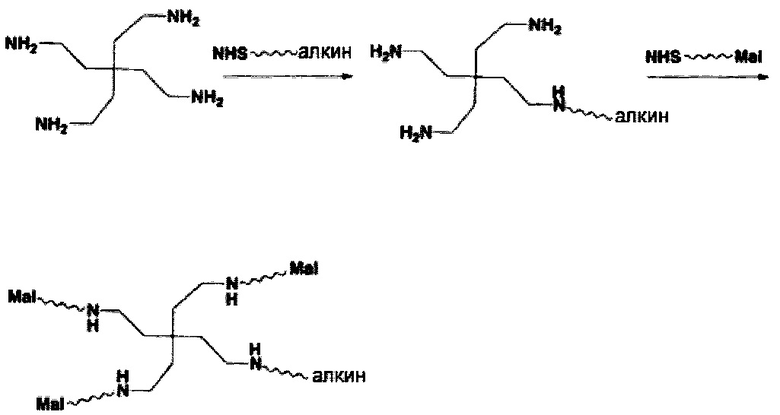
Некоторые из этих симметричных химических соединений можно дополнительно модифицировать с получением центральных сердцевин с большим количеством связывающих ветвей/соединяющих ветвей. Например, тетракис(2-аминоэтил)метан, который можно синтезировать из широко распространенных химических соединений или получить коммерческим путем, можно применять в качестве сердцевины для конструирования линкерных звеньев с четырьмя связывающими ветвями/соединяющими ветвями. Тетракис(2-аминоэтил)метан может реагировать с бис(сульфосукцинимидил)субератом с получением продукта конденсации двух молекул тетракис(2-аминоэтил)метана, который можно применять в качестве сердцевины для конструирования линкерных звеньев, имеющих шесть связывающих ветвей/соединяющих ветвей. Линкерные звенья, соответственно, имеющие 3 связывающие ветви/соединяющие ветви, 4 связывающие ветви/соединяющие ветви и 6 связывающих ветвей/соединяющих ветвей, могут удовлетворять большинство потребностей в конструировании нацеливающих/эффекторных молекул, имеющих конфигурацию с сочлененными линкерами.
Очевидно, что количество связывающих ветвей и (или) соединяющих ветвей и элементов, связанных с ними, может изменяться в зависимости от количества аминогрупп, содержащихся в центральной сердцевине. В некоторых предпочтительных вариантах осуществления количество связывающих ветвей/соединяющих ветвей и соответствующих связывающихся элементов, связанных с ними, находится в диапазоне, составляющем приблизительно 1-7.
Обратимся теперь к фигуре 2, на которой показан бензол-1,2,4,5-тетраамин, имеющий 4 аминогруппы. Три аминогруппы соответственно связаны со связывающими ветвями 20, а одна аминогруппа связана с соединяющей ветвью 60, которая имеет азидную группу на своем свободном конце. С тремя связывающими ветвями 20 далее соответственно связаны три первых элемента 30 посредством реакции тиольной и малеимидной групп, и с соединяющей ветвью 60 связан один второй элемент 50 посредством реакции CuAAC. Жирная точка 40, показанная на фигуре 2, представляет химическую связь, образующуюся в результате реакции CuAAC, происходящей между соединяющей ветвью 60 и вторым элементом 50.
I-(iii) Функциональные элементы, подходящие для использования в разветвленном линкере
В случае, если линкерное звено (или разветвленной линкер) содержит только первый элемент, но не второй и (или) третий элемент, первый элемент является эффекторным элементом, который может оказывать терапевтическое воздействие на субъект. С другой стороны, если это линкерное звено содержит другие элементы в дополнение к первому элементу, то по меньшей мере один из таких элементов представляет собой эффекторный элемент, тогда как другой может быть другим эффекторным элементом, нацеливающим элементом или элементом, способным улучшать одно или несколько фармакокинетических свойств линкерного звена (например, растворимость, скорость выведения, период полувыведения и биодоступность). Например, линкерное звено может иметь два различных типа эффекторных элементов, один эффекторный элемент и один нацеливающий элемент или один элемент, улучшающий фармакокинетические свойства, два разных типа нацеливающих элементов и один вид эффекторного элемента, два разных типа эффекторных элементов и один вид нацеливающего элемента или один вид нацеливающих элементов, один вид эффекторного элемента и один элемент, способный улучшать фармакокинетическое свойство линкерного звена.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия, нацеливающим элементом или эффекторным элементом является финголимод, финголимод-фосфат, интерферон-β или одноцепочечный вариабельный фрагмент (scFv), специфичный к интегрину α4, β-амилоиду, вирусному или бактериальному белку.
Примеры вирусных белков включают, без ограничения, белок F вируса респираторной синцитии (RSV), белок gp120 вируса иммунодефицита человека типа 1 (HIV-1), гемагглютинин А (НА) белка вируса гриппа А и гликопротеин цитомегаловируса.
Иллюстративные примеры бактериальных белков включают эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
Элементом, улучшающим фармакокинетические свойства, может являться длинная цепь PEG, имеющая молекулярную массу приблизительно 20000-50000 дальтон.
Конкретные примеры функциональных элементов, включенных в данный разветвленный линкер для лечения конкретного заболевания, обсуждаются ниже.
Для лечения заболевания ЦНС, такого как рассеянный склероз, в одном примере линкерного звена может использоваться финголимод, финголимод-фосфат, интерферон-β или scFv, специфичный к интегрину α4, в качестве первого элемента (эффекторного элемента). Для лечения болезни Альцгеймера в данном линкерном звене может использоваться scFv, специфичный к β-амилоиду, в качестве первого элемента (эффекторного элемента). Необязательно, линкерное звено для лечения рассеянного склероза, болезни Альцгеймера или других заболеваний ЦНС может дополнительно содержать второй элемент - scFv, специфичный к рецептору трансферрина, в качестве нацеливающего элемента.
Аналогично, scFv, специфичный к вирусному или бактериальному белку, может использоваться в качестве нацеливающего элемента (первого элемента) для лечения инфекции, вызванной вирусом или бактерией. В этих случаях линкерное звено может содержать необязательный второй элемент, такой как scFv, специфичный к CD32 или к CD16b, в качестве эффекторного элемента.
I-(iv) Использование разветвленного линкера
Настоящее раскрытие также относится к способу лечения различных заболеваний с использованием подходящего линкерного звена. В целом, способ включает стадию введения субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества линкерного звена в соответствии с вариантами осуществления настоящего раскрытия.
По сравнению с ранее известными терапевтическими конструкциями линкерное звено по настоящему изобретению, обсуждаемое в части I, является преимущественным по двум пунктам.
(1) Количество функциональных элементов можно регулировать в соответствии с потребностями и (или) способами применения. Линкерное звено по настоящему изобретению может содержать два элемента (т.е. первый и второй элементы) или три элемента (т.е. первый, второй и третий элементы) в соответствии с требованиями способа применения (например, заболеванием, подвергаемым лечению, путем введения линкерного звена по настоящему изобретению и авидностью и (или) аффинностью связывания антитела, переносимого линкерным звеном по настоящему изобретению). Например, в случаях прямой доставки линкерного звена по настоящему изобретению в ткань/орган (например, при обработке глаза), один элемент, выступающий в качестве эффекторного элемента, может быть достаточным, что, таким образом, будет устранять необходимость во втором элементе, выступающем в качестве нацеливающего элемента. Однако в случаях периферической доставки линкерного звена по настоящему изобретению (например, при пероральном, энтеральном, назальном, местном, чресслизистом введении, внутримышечной, внутривенной или внутрибрюшинной инъекции) может быть необходимым, чтобы линкерное звено по настоящему изобретению одновременно содержало нацеливающий элемент, специфично нацеливающий линкерное звено по настоящему изобретению на очаг поражения; и эффекторный элемент, проявляющий терапевтический эффект в очаге поражения. В целях повышения эффективности нацеливания или лечения, или повышения стабильности линкерного звена по настоящему изобретению в линкерное звено по настоящему изобретению можно дополнительно включить третий элемент (например, второй нацеливающий элемент, второй эффекторный элемент или цепь PEG).
(2) Первый элемент представлен в форме «связки». Как описано ранее, количество первых элементов может изменяться в зависимости от количества лизиновых остатков, содержащихся в центральной сердцевине. Если количество лизиновых остатков в центральной сердцевине находится в диапазоне от 2 до 15, то в каждом линкерном звене может содержаться по меньшей мере два первых элемента. Таким образом, вместо предоставления всего одной молекулы (например, цитотоксического лекарственного средства и антитела), что может выполняться в рамках традиционных терапевтических конструкции или способа, линкерное звено по настоящему изобретению способно предоставлять большее количество функциональных элементов (в качестве нацеливающих элементов либо в качестве эффекторных элементов) одновременно, что, таким образом, существенно улучшает терапевтический эффект.
В определенных способах терапевтического применения желательным является наличие одной копии нацеливающего или эффекторного элемента. Например, одна копия нацеливающего элемента может использоваться для предотвращения нежелательных эффектов из-за чрезмерно жесткой связки. Это соображение относится к случаю, если scFv обладает относительно высокой аффинностью к антигену-мишени, а также если антиген-мишень является мембранным антигеном на нормальных клетках, которые не являются больными клетками-мишенями. В качестве примера, при использовании scFv, специфичного к CD3 или CD16a, для задействования Т-клеток или NK-клеток для уничтожения клеток-мишеней, таких как клетки щитовидной железы у пациентов, пораженных болезнью Грейвса, желательна одна копия scFv, специфичная к CD3 или CD16a, чтобы можно было избежать нежелательных эффектов из-за сшивки CD3 или CD16a. Аналогично, при использовании scFv, специфичного к CD32 или к CD16b, для задействования фагоцитарных нейтрофилов и макрофагов для обезвреживания вирусных или бактериальных частиц, связанных с антителом, или их продуктов, желательна одна копия scFv. Кроме того, при использовании scFv, специфичного к рецептору трансферрина, для переноса эффекторных молекул лекарственного средства к ГЭБ при лечении заболеваний ЦНС, желательна одна копия scFv, специфичного к рецептору трансферрина. В еще одном примере желательным является наличие только одной копии длинноцепочечного PEG для усиления фармакокинетических свойств. Две или более длинных цепей PEG могут обуславливать образование сплетений и влиять на свойства связывания нацеливающих или эффекторных элементов.
ЧАСТЬ II. Молекулярные конструкции с сочлененными линкерами для лечения определенных заболеваний
Другой аспект настоящего раскрытия относится к молекулярной конструкции, содержащей по меньшей мере два линкерных звена, в которой одно линкерное звено несет один или несколько нацеливающих элементов, а другое линкерное звено несет один или несколько эффекторных элементов или элементов, улучшающих фармакокинетические свойства. В настоящем описании молекулярные конструкции как с нацеливающими, так и с эффекторными элементами (терапевтическими или фармакокинетическими) называются молекулярными конструкциями с сочлененными линкерами. В соответствии с различными вариантами осуществления настоящего раскрытия каждое линкерное звено, содержащееся в таких молекулярных конструкциях с сочлененными линкерами, может представлять собой разветвленные линкеры на основе пептидной сердцевины или сердцевины на основе химического соединения, рассмотренные выше в части I настоящего раскрытия. В соответствии с определенными вариантами осуществления настоящего раскрытия, по меньшей мере одно из линкерных звеньев молекулярной конструкции по настоящему изобретению содержит полипептидную сердцевину. Предпочтительно, по меньшей мере два линкерных звена молекулярной конструкции по настоящему изобретению содержат полипептидные сердцевины. Более предпочтительно, все линкерные звенья молекулярной конструкции по настоящему изобретению, соответственно, содержат полипептидные сердцевины.
II-(i) Структура молекулярной конструкции с сочлененными линкерами
В соответствии с некоторыми вариантами осуществления настоящего раскрытия молекулярная конструкция содержит два линкерных звена, и линкерные звенья соединены друг с другом посредством реакции CuAAC (с применением меди или пентаметилциклопентадиенильного комплекса хлорида рутения в качестве катализатора), реакции SPAAC либо реакции iEDDA. В вариантах осуществления одно линкерное звено связано со множеством первых элементов, выступающих в качестве нацеливающих элементов, а другое линкерное звено связано со множеством вторых элементов, выступающих в качестве эффекторных элементов.
В соответствии с другими вариантами осуществления настоящего раскрытия молекулярная конструкция содержит три линкерных звена, при этом первое и второе линкерные звенья соединены друг с другом посредством реакции iEDDA, и далее третье линкерное звено соединено с первым или вторым линкерными звеньями посредством реакции CuAAC. В качестве альтернативы, первое и второе линкерные звенья соединены друг с другом посредством реакции iEDDA, и третье линкерное звено соединено с первым или вторым линкерными звеньями посредством реакции SPAAC. В вариантах осуществления первое, второе и третье линкерные звенья, соответственно, несут множество первых, вторых и третьих элементов, при этом первый, второй и третий элементы являются разными. В соответствии с одним вариантом осуществления два из трех элементов (т.е. первого, второго и третьего элементов) представляют собой нацеливающие элементы, а один из трех элементов представляет собой эффекторный элемент. В соответствии с другим вариантом осуществления два из трех элементов представляют собой эффекторные элементы, а один из трех элементов представляет собой нацеливающий элемент. В соответствии с еще одним вариантом осуществления один из трех элементов представляет собой нацеливающий элемент, другой из трех элементов представляет собой эффекторный элемент, а оставшийся из трех элементов представляет собой элемент, способный к улучшению фармакокинетических свойств молекулярной конструкции, такого как растворимость, скорость выведения, период полувыведения и биодоступность.
Сначала обратимся к фигурам 3A-3D, на которых, соответственно, показано связывание между двумя линкерными звеньями. На фигуре 3А показана молекулярная конструкция, содержащая два линкерных звена (100А, 200А), соединяющихся друг с другом посредством реакции iEDDA. Первое линкерное звено 100А содержит первую центральную сердцевину 110а, связывающую ветвь 120 (в качестве первой связывающей ветви) и соединяющую ветвь 130а (в качестве первой соединяющей ветви), при этом связывающая и соединяющая ветви соответственно связаны с первой центральной сердцевиной 110а на одном из концов. Аналогично, второе линкерное звено 200А содержит вторую центральную сердцевину 210а, связывающую ветвь 220 (в качестве второй связывающей ветви) и соединяющую ветвь 230а (в качестве второй соединяющей ветви), при этом связывающая и соединяющая ветви соответственно связаны со второй центральной сердцевиной 210а на одном из концов. Одна из соединяющих ветвей 130а и 230а имеет тетразиновую группу на своем свободном конце, а другая из соединяющих ветвей 130а и 230а имеет ТСО-группу. В частности, если соединяющая ветвь 130а имеет тетразиновую группу 152 на своем свободном конце (т.е. на конце, не присоединенном к первой центральной сердцевине 110а), то соединяющая ветвь 230а будет иметь ТСО-группу 154 на своем свободном конце (т.е. на конце, не присоединенном ко второй центральной сердцевине 210а), и наоборот. Соответственно, два линкерных звена (100А, 200А) соединяются друг с другом посредством реакции iEDDA, происходящей между соответствующими свободными концами соединяющих ветвей 130а и 230а. Эллипс 156, показанный на фигуре 3А, представляет химическую связь, образующуюся в результате реакции iEDDA, происходящей между соединяющими ветвями 130а и 230а.
В показанном варианте осуществления каждая из связывающих ветвей 120 и 220 имеет малеимидную группу на своем свободном конце. Соответственно, первый нацеливающий элемент 140 и первый эффекторный элемент 240, каждый из которых имеет тиольную группу, связаны соответственно со связывающими ветвями 120 и 220 посредством реакции тиольной и малеимидной групп.
В соответствии с одним вариантом осуществления как первая, так и вторая центральные сердцевины 110а и 210а, показанные на фигуре 3А, представляют собой полипептидные сердцевины. В соответствии с другим вариантом осуществления как первая, так и вторая центральные сердцевины 110а и 210а, показанные на фигуре 3А, представляют собой сердцевины на основе химического соединения. В соответствии с еще одним вариантом осуществления одна из первой и второй центральных сердцевин 110а и 210а, показанных на фигуре 3А, представляет собой полипептидную сердцевину, а другая из первой и второй центральных сердцевин 110а и 210а, показанных на фигуре 3А, представляет собой сердцевину на основе химического соединения.
На фигуре 3В представлен альтернативный вариант осуществления настоящего раскрытия, в котором как первая, так и вторая центральные сердцевины 110b и 210b представляют собой полипептидные сердцевины и связаны соответственно с первым нацеливающим элементом 140 и первым эффекторным элементом 240 посредством связывающих ветвей 120 и 220. Уникальным признаком данного варианта осуществления является то, что одна из центральных сердцевин 110b и 210b содержит аминокислотный остаток, имеющий азидную группу (например, остаток AHA), на своем N- или С-конце, а другая из центральных сердцевин 110b и 210b содержит аминокислотный остаток, имеющий алкиновую группу (например, остаток HPG), на своем N- или С-конце, и такая конфигурация обеспечивает возможность непосредственного связывания центральных сердцевин 110а и 210а друг с другом, т.е. без присоединения с помощью каких-либо соединяющих ветвей, как показано на фигуре 3А.. В частности, если центральная сердцевина 110b содержит аминокислотный остаток, имеющий азидную группу 162, на своем N- или С-конце, то центральная сердцевина 210b будет содержать аминокислотный остаток, имеющий алкиновую группу 164, на своем N- или С-конце, и наоборот. Соответственно, линкерные звенья 100В и 200В могут непосредственно соединяться друг с другом посредством реакции CuAAC, происходящей между N- или С-концевыми аминокислотными остатками центральных сердцевин 110b и 210b. Жирная точка 166, показанная на фигуре 3В, представляет химическую связь, образующуюся между N- или С-концевыми аминокислотными остатками.
Фигура 3С представляет собой другой вариант осуществления настоящего раскрытия. Линкерные звенья 100С и 200С имеют сходные структуры с линкерными звеньями 100А и 200А, за исключением того, что соединяющие ветви 130b и 230b, соответственно, имеют азидную группу 162 и DBCO-группу 172 вместо азидной группы 152 и алкиновой группы 154, показанных в линкерных звеньях 100А и 200А на фигуре 3А. В частности, центральная сердцевина 110а связана с соединяющей ветвью 130b (в качестве первой соединяющей ветви), имеющей азидную группу 162 на своем свободном конце; а центральная сердцевина 210а связана с соединяющей ветвью 230b (в качестве второй соединяющей ветви), имеющей DBCO-группу 172 на своем свободном конце. Линкерные звенья 100С и 200С далее соединяются друг с другом посредством реакции SPAAC, происходящей между соединяющими ветвями 130b и 230b; с образованием при этом химической связи 182, показанной в виде ромба.
В одном варианте осуществления как первая, так и вторая центральные сердцевины 110а и 210а, показанные на фигуре 3С, представляют собой полипептидные сердцевины. В другом варианте осуществления как первая, так и вторая центральные сердцевины 110а и 210а, показанные на фигуре 3С, представляют собой сердцевины на основе химического соединения. В еще одном варианте осуществления одна из первой и второй центральных сердцевин 110а и 210а, показанных на фигуре 3С, представляет собой полипептидную сердцевину, а другая из первой и второй центральных сердцевин 110а и 210а, показанных на фигуре 3С, представляет собой сердцевину на основе химического соединения.
Очевидно, что два линкерных звена могут соединяться друг с другом посредством реакции CuAAC, происходящей между центральной сердцевиной и соединяющей ветвью. Обратимся теперь к фигуре 3D, на которой центральная сердцевина 110b содержит N- или С-концевой аминокислотный остаток, имеющий азидную группу 162 (например, остаток AHA), а центральная сердцевина 210а связана с соединяющей ветвью 230b, имеющей ТСО-группу 172 на своем свободном конце. Соответственно, линкерные звенья 100В и 200С могут соединяться посредством реакции SPAAC, происходящей между центральной сердцевиной 110b и соединяющей ветвью 230b; с образованием при этом химической связи 182.
В соответствии с одним вариантом осуществления линкерные звенья 100В и 200С, показанные на фигуре 3D, соответственно, содержат полипептидные сердцевины. В соответствии с другим вариантом осуществления центральная сердцевина 100В, показанная на фигуре 3D, представляет собой полипептидную сердцевину, а центральная сердцевина 200С, показанная на фигуре 3D, представляет собой сердцевину на основе химического соединения.
В качестве альтернативы, линкерное звено, которое содержит N- или С-концевой аминокислотный остаток, имеющий алкиновую группу (например, остаток HPG), и линкерное звено, содержащее соединяющую ветвь с азидной группой на своем свободном конце, могут соединяться друг с другом посредством азид-алкинового циклоприсоединения, происходящего между центральной сердцевиной и соединяющей ветвью.
Следует понимать, что по меньшей мере одно из линкерных звеньев данной молекулярной конструкции может дополнительно содержать сопрягающую ветвь, в которой один конец сопрягающей ветви связан со связывающей ветвью, а другой конец связан с функциональным элементом (нацеливающим элементом или эффекторным элементом), как показано в части I. Например, настоящая молекулярная конструкция может содержать два линкерных звена, в которых первый элемент непосредственно связан с первой связывающей ветвью, а второй элемент связан со второй связывающей ветвью через связь сопрягающей ветви. В качестве альтернативы, настоящая молекулярная конструкция может содержать два линкерных звена, где первый и второй элементы, соответственно, связаны с первой и второй связывающими ветвями через связи первой и второй сопрягающих ветвей.
Предпочтительно, если по меньшей мере одна из первой и второй связывающих ветвей связана с сопрягающей ветвью или с функциональным элементом посредством реакции CuAAC или реакции SPAAC, то первый и второй линкерные звенья соединены друг с другом посредством реакции iEDDA. В качестве альтернативы, если по меньшей мере одна из первой и второй связывающих ветвей связана с сопрягающей ветвью или с функциональным элементом посредством реакции iEDDA, то первое и второе линкерные звенья соединены друг с другом посредством реакции CuAAC или реакции SPAAC.
В соответствии с некоторыми вариантами осуществления сопрягающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев. В соответствии с другими вариантами осуществления сопрягающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на конце связывания элементов, не связанной со связывающей ветвью.
В соответствии с одним из вариантов осуществления настоящего раскрытия, первый элемент представляет собой scFv, специфичный к рецептору трансферрина, а второй элемент представляет собой интерферон-β (IFN-β), финголимод, финголимод-фосфат или scFv, специфичный к интегрину α4 или к β-амилоиду. Согласно другому варианту осуществления настоящего раскрытия, первый элемент представляет собой scFv, специфичный к вирусному или бактериальному белку, а второй элемент представляет собой scFv, специфичный к CD16b или к CD32.
По сравнению с другими терапевтическими конструкциями, молекулярная конструкция по настоящему изобретению является преимущественной по меньшей мере по трем следующим аспектам:
(1) линкерные звенья, содержащие определенные количество и (или) тип нацеливающих/эффекторных элементов, можно получать независимо, а затем переходить к их соединению друг с другом посредством реакции CuAAC, реакции iEDDA или реакции SPAAC;
(2) количество и вид нацеливающих и (или) эффекторных элементов можно изменять в соответствии с требованиями способа применения (например, с заболеванием, подвергаемым лечению, и авидностью и (или) аффинностью связывания нацеливающего и (или) эффекторного элемента). Комбинацию нацеливающих и эффекторных элементов можно регулировать в соответствии с конкретными потребностями и (или) способами применения. Каждый из нацеливающих и эффекторных элементов по настоящему изобретению можно изменять в зависимости от таких факторов, как конкретное состояние, подвергаемое лечению, физическое состояние пациента и (или) тип заболевания, подвергаемого лечению. Практикующий клиницист может объединить наиболее подходящий нацеливающий элемент и наиболее подходящий эффекторный элемент, чтобы достичь наилучшего терапевтического эффекта. В соответствии с вариантами осуществления настоящего раскрытия нацеливающий элемент может представлять собой фактор роста, пептидный гормон, цитокин или фрагмент антитела; а эффекторный элемент может представлять собой иммуномодулятор, хелатообразователь в комплексе с радиоактивным нуклидом, цитотоксическое лекарственное средство, цитокин, растворимый рецептор или антитело; и
(3) по сравнению с другими реакциями присоединения реакция CuAAC, реакция iEDDA или реакция SPAAC является более эффективной в отношении соединения любых двух линкерных звеньев.
Обратимся теперь к фигуре 4, на которой проиллюстрированы шесть библиотек, получаемых независимо. В этом варианте осуществления библиотеки 1-6, соответственно, содержат множество линкерных звеньев 300А, 300В, 300С, 400А, 400В и 400С, связанных с функциональными элементами. Все линкерные звенья 300А, 300В и 300С сходны по структуре; при этом каждое из линкерных звеньев 300А, 300В и 300С содержит одну центральную сердцевину 310, одну соединяющую ветвь 330, связанную с ней и имеющую тетразиновую группу 350 на своем свободном конце, и определенное количество связывающих ветвей 320. Например, линкерное звено 300А содержит четыре связывающие ветви 320, и, соответственно, с четырьмя связывающими ветвями 320 могут быть соответственно связаны четыре нацеливающих элемента 340а. Аналогично, два нацеливающих элемента 340b и пять нацеливающих элементов 340с могут быть соответственно связаны с линкерными звеньями 300В и 300С. Нацеливающие элементы 340а, 340b и 340с могут быть одинаковыми или разными. Что касается линкерных звеньев 400А, 400В и 400С, каждое из этих линкерных звеньев содержит одну центральную сердцевину 410, одну соединяющую ветвь 430, связанную с ней и имеющую напряженную алкиновую группу 450 на своем свободном конце, и определенное количество связывающих ветвей 420. Как показано, три эффекторных элемента 440а, пять эффекторных элементов 440b и восемь эффекторных элементов 440с могут быть соответственно связаны с линкерными звеньями 400А, 400В и 400С. Эффекторные элементы 440а, 440b и 440с могут быть одинаковыми или разными. Библиотеки 1-6 можно получать независимо. Специалист в данной области может выбрать первое линкерное звено из библиотек 1, 2 и 3 и второе линкерное звено из библиотек 4, 5 и 6, затем перейти к соединению первого и второго линкерных звеньев посредством реакции iEDDA, происходящей между тетразиновой группой 350 и напряженной алкиновой группой 450, чтобы получить молекулярную конструкцию с определенным количеством нацеливающих и эффекторных элементов.
На основании идеи библиотек можно получать молекулярные конструкции по настоящему изобретению с различными конфигурациями в зависимости от выбранных библиотек. На фигуре 5А представлен пример молекулярной конструкции по настоящему изобретению, в которой каждая из первой и второй центральных сердцевин (310, 410) связана с тремя связывающими ветвями (320, 420) и одной соединяющей ветвью (330, 430). Три первых нацеливающих элемента 340 соответственно связаны со связывающими ветвями 320; и три первых эффекторных элемента 440 соответственно связаны со связывающими ветвями 420. Два линкерных звена соединяются друг с другом посредством реакции iEDDA, происходящей между двумя соединяющими ветвями 330 и 430, с образованием при этом химической связи 356. В этой конфигурации в одной молекулярной конструкции может переноситься равное количество нескольких нацеливающих и (или) эффекторных элементов.
На фигуре 5В представлен другой пример молекулярной конструкции по настоящему изобретению, в которой первая и вторая центральные сердцевины, соответственно, содержат разные количества аминогрупп (например, лизиновых остатков), и, соответственно, молекулярная конструкция содержит неравное количество нацеливающих и эффекторных элементов. В показанном примере первая центральная сердцевина 310 связана с одной соединяющей ветвью 330 и двумя связывающими ветвями 320. Вторая центральная сердцевина 410 связана с одной соединяющей ветвью 430 и пятью связывающими ветвями 420. Соответственно, два нацеливающих элемента 340 соответственно связаны со связывающими ветвями 320; и пять эффекторных элементов 440 соответственно связаны со связывающими ветвями 420. Эллипс 356 на фигуре 5В представляет связь между двумя соединяющими ветвями 330 и 430.
В необязательных вариантах осуществления молекулярная конструкция по настоящему изобретению может дополнительно содержать относительно длинную цепь PEG, присоединенную к первой либо второй центральной сердцевине, так чтобы молекулярная конструкция по настоящему изобретению могла распадаться вдали от ретикулоэндотелиальной системы и достигала более длительного периода полувыведения после введения субъекту. В случае, когда белок модифицируют цепью PEG, чтобы улучшить его фармакокинетические свойства и (или) снизить иммуногенность, предпочтительным является PEG с длиной до 20000-50000 дальтон. Соответственно, в одном предпочтительном варианте осуществления настоящего изобретения для присоединения нацеливающих и эффекторных элементов применяются связывающие ветви относительно более короткой длины, а цепь PEG весом 20000-50000 дальтон присоединяют к любому из линкерных звеньев с целью увеличения периода полувыведения in vivo молекулярной конструкции по настоящему изобретению.
В некоторых вариантах осуществления для конструирования молекулярной конструкции по настоящему изобретению применяют несколько scFv-фрагментов в качестве нацеливающих и (или) эффекторных элементов. Фармацевтические средства с конфигурацией нацеливающий элемент/эффекторный элемент на основе молекулярных конструкций, содержащих scFv-фрагменты, должны иметь более длительные периоды полувыведения in vivo, чем отдельные фрагменты антител. Для некоторых способов клинического применения желательно значительно увеличить период полувыведения фармацевтических средств, чтобы устранить необходимость в частом введении лекарственных средств; в этих случаях цепи PEG массой 20000-50000 дальтон можно применять в качестве связывающих ветвей для связывания с scFv-фрагментами, которые служат в качестве нацеливающих или эффекторных элементов. PEG с такими значениями длины применяли для модификации большого количества терапевтических белков для увеличения их периодов полувыведения.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия линкерное звено может содержать две связывающие ветви, соответственно связанные с различными функциональными элементами. Обратимся теперь к фигуре 6, на которой молекулярная конструкция содержит два линкерных звена 100А и 200D. Первый и второй функциональные элементы 140 и 240 (один служит в качестве нацеливающего элемента, а другой служит в качестве эффекторного элемента) связаны соответственно с первой центральной сердцевиной 110а и второй центральной сердцевиной 210с посредством связывающих ветвей 120 и 220; и две центральные сердцевины 110а и 210с соединены друг с другом посредством реакции iEDDA, происходящей между соединяющими ветвями 130а и 230а, при этом эллипс 156 представляет химическую связь, образующуюся между ними. В дополнение к функциональному элементу 240, вторая центральная сердцевина 210с дополнительно связана с цепью PEG 260. В частности, вторая центральная сердцевина 210с содержит остаток AHA, который можно подвергнуть реакции и связать с цепью PEG 260, имеющей напряженную алкиновую группу, посредством реакции SPAAC, при этом ромб 182 представляет химическую связь, образующуюся в результате реакции SPAAC. В зависимости от намеченного и желаемого применения третий элемент может представлять собой второй нацеливающий элемент, второй эффекторный элемент или элемент, способный к улучшению фармацевтического свойства молекулярной конструкции. В соответствии с одним вариантом осуществления настоящего раскрытия цепь PEG 260 имеет молекулярную массу приблизительно 20000-50000 дальтон.
На основании данной идеи линкерное звено может содержать множество связывающих ветвей, которые могут быть связаны со множеством функциональных элементов. Например, линкерное звено может содержать 5-12 связывающих ветвей, которые могут быть связаны с 5-12 функциональными элементами. Это является особенно применимым в случаях, когда функциональные элементы представляют собой малые молекулы, такие как терапевтические лекарственные средства или агонисты Toll-подобных рецепторов. Линкерное звено, несущее несколько молекул терапевтических лекарственных средств, называется в данном документе «связкой» лекарственных средств.
Кроме того, полипептидные сердцевины можно использовать для получения молекулярной конструкции, содержащей три линкерных звена. Соответственно, другой аспект настоящего раскрытия направлен на молекулярную конструкцию, содержащую три линкерных звена. Среди трех линкерных звеньев два из них могут быть присоединены друг к другу посредством реакции iEDDA, а третье линкерное звено присоединяется к любому из двух линкерных звеньев посредством реакции SPAAC или реакции CuAAC. Логическое обоснование получения конструкций с несколькими линкерными звеньями (например, тремя линкерными звеньями) заключается в том, что в их состав можно включить два различных набора нацеливающих элементов или два различных набора эффекторных элементов.
Обратимся теперь к фигуре 7, на которой молекулярная конструкция содержит три линкерных звена (500, 600, 700А). Линкерные звенья 500, 600, 700А, соответственно, содержат центральную сердцевину (510, 610, 710) и связывающую ветвь (520, 620, 720) с функциональным элементом (540, 640, 740), связанным с ней. Линкерное звено 600 характеризуется тем, что оно содержит цистеиновый остаток на одном из его N- или С-конца, связанный с соединяющей ветвью 630; и аминокислотный остаток, имеющий азидную или алкиновую группу, на другом из его N- или С-конца. Одна из соединяющих ветвей 530 и 630 имеет тетразиновую группу на своем свободном конце, а другая из соединяющих ветвей 530 и 630 имеет напряженную алкиновую группу на своем свободном конце. Соответственно, линкерные звенья 500 и 600 могут соединяться друг с другом посредством реакции iEDDA, происходящей между соединяющими ветвями 530 и 630, согласно способу связывания, описанному на фигуре 3А. Что касается связывания с линкерным звеном 700, если N- или С-концевой аминокислотный остаток центральной сердцевины 610 имеет азидную группу (например, остаток AHA), то центральная сердцевина 710 содержит аминокислоту, имеющую алкиновую группу (например, остаток HPG), на своем N- или С-конце; или если N- или С-концевой аминокислотный остаток центральной сердцевины 610 имеет алкиновую группу (например, остаток HPG), то центральная сердцевина 710 содержит аминокислоту, имеющую азидную группу (например, остаток AHA), на своем N- или С-конце. Таким образом, согласно способу связывания, описанному на фигуре 3В, линкерные звенья 600 и 700А могут непосредственно соединяться друг с другом посредством реакции CuAAC, происходящей между N- или С-концевыми аминокислотными остатками центральных сердцевин 610 и 710, без наличия соединяющих ветвей. Эллипс 560 и жирная точка 670 на фигуре 7, соответственно, представляют химические связи, образующиеся в результате реакции iEDDA и реакции CuAAC.
В качестве альтернативы, два из трех линкерных звеньев могут быть присоединены друг к другу посредством реакции iEDDA, а третье линкерное звено присоединяется к любому из двух линкерных звеньев посредством реакции SPAAC. Обратимся теперь к фигуре 7В, на которой линкерные звенья 500 и 600 соединены друг с другом посредством реакции iEDDA, описанной на фигуре 7А, тогда как линкерное звено 700В связано с линкерным звеном 600 посредством реакции SPAAC, происходящей между центральной сердцевиной 610 и соединяющей ветвью 730. Ромб 672 на фигуре 7В представляет химическую связь, образующуюся в результате реакции SPAAC.
Очевидно, что количество функциональных элементов 540, 640 и 740, соответственно связанных с линкерными звеньями 500, 600, 700А или 700В в каждом случае является разным в зависимости от намеченного применения. Согласно идее библиотек, показанной на фигуре 4, линкерные звенья, соответственно, несущие различные количества и (или) типы функциональных элементов, можно получать по отдельности в виде различных библиотек, и специалист в данной области может выбрать и объединить желаемые линкерные звенья из библиотек в соответствии с различными способами применения.
В целом, соединяющую ветвь молекулярной конструкции по настоящему изобретению, описанную в вышеприведенных аспектах и (или) вариантах осуществления настоящего раскрытия, которая имеет азидную, алкиновую, тетразиновую или напряженную алкиновую группу на конце, конструируют как цепь PEG, имеющую 2-12 повторяющихся EG-звеньев. Связывающую ветвь конструируют как цепь PEG, имеющую 2-20 повторяющихся EG-звеньев; предпочтительно связывающая ветвь представляет собой цепь PEG, имеющую 2-20 повторяющихся EG-звеньев с дисульфидной связью на свободном конце, не связанном с центральной сердцевиной.
Использование полипептида в качестве центральной сердцевины обеспечивает универсальность молекулярной конструкции по настоящему изобретению, при которой несколько копий или типов нацеливающих/эффекторных элементов могут присутствовать в одной конструкции и, соответственно, достигается повышение специфичности доставки лекарственных средств и действенности в намеченных участках-мишенях. Используемая молекулярная конструкция, содержащая несколько линкерных звеньев, может принимать большое количество конфигураций. Несколькими примерами являются: первое линкерное звено, несущее три scFv в качестве нацеливающих элементов, и второе линкерное звено, несущее 5 терапевтических лекарственных средств; первое линкерное звено, несущее три scFv в качестве нацеливающих элементов, и второе линкерное звено, несущее три scFv в качестве эффекторных элементов; первое линкерное звено, несущее два scFv из первого набора нацеливающих элементов, второе линкерное звено, несущее два scFv из второго набора нацеливающих элементов, и третье линкерное звено, несущее 5 терапевтических лекарственных средств; первое линкерное звено, несущее 2 bi-scFv в качестве нацеливающих элементов, и второе линкерное звено, несущее два scFv в качестве эффекторных элементов; или первое линкерное звено, несущее три scFv в качестве нацеливающих элементов, второе линкерное звено, несущее два scFv в качестве эффекторных элементов, со связывающей ветвью с прикрепленным длинноцепочечным PEG массой 20000-50000 дальтон в целях увеличения фармакокинетических свойств.
В некоторых вариантах осуществления настоящего изобретения бифункциональный PEG, выступающий в качестве связывающей ветви, применяют для связывания антиген-связывающих фрагментов антител, которые служат в качестве нацеливающих или эффекторных элементов, с аминогруппами, расположенными в полипептидной сердцевине. Каждый PEG может иметь NHS-группу на одном конце и малеимидную группу на другом конце. NHS-группа может соединяться с аминогруппой в полипептидной сердцевине, а малеимидная группа может соединяться с сульфогидрильной группой цистеинового остатка scFv, bi-scFv или Fab-фрагмента антитела. scFv и bi-scFv конструируют таким образом, чтобы они имели полипептидный линкер с концевым цистеиновым остатком на С-конце. Fab можно получить из целого IgG путем расщепления пепсином, и свободные сульфогидрильные группы получают из межцепочечной дисульфидной связи посредством реакции мягкого восстановления.
На схемах 8-12 представлены некоторые рабочие примеры, соответственно, показывающие соединение и получение определенных линкерных звеньев.
Схема 8 представляет собой схематическое изображение, на котором показано получение молекулярной конструкции по настоящему изобретению в соответствии с одним вариантом осуществления настоящего раскрытия, в которой NHS представляет сложный NHS-эфир, Mal представляет малеимидную группу, AAH представляет L-азидогомоаланиновый (AHA) остаток, GHP представляет гомопропаргилглициновый (HPG) остаток, Ас представляет ацетильную группу, a scFv представляет одноцепочечный вариабельный фрагмент. На этапе 1, соответственно, получают первую центральную сердцевину, содержащую аминокислотную последовательность (GSK)3 и L-азидогомоаланиновый (AHA) остаток на своем С-конце; и вторую центральную сердцевину, содержащую аминокислотную последовательность (GSK)5 и гомопропаргилглициновый (HPG) остаток на своем С-конце. В целях стабилизации полипептида N-концы первой и второй центральных сердцевин, соответственно, модифицируют ацетильной группой. На этапе 2 связывающие ветви соответственно связываются с лизиновыми остатками в первой и второй центральных сердцевинах посредством образования амидной связи между ними; связывающая ветвь, связанная с центральной сердцевиной, имеет малеимидную группу на свободном конце. На этапе 3 первый нацеливающий элемент (т.е. антитело), имеющий тиольную группу (например, цистеиновый остаток), связывается со связывающей ветвью, связанной с первой центральной сердцевиной, посредством реакции тиольной и малеимидной групп; аналогично, эффекторный элемент (т.е. лекарственное средство), имеющий тиольную группу, связывается со связывающей ветвью, связанной со второй центральной сердцевиной, посредством реакции тиольной и малеимидной групп. На этапе 4 два линкерных звена соединяются посредством реакции CuAAC, происходящей между остатками AHA и HPG.
«Схема 8. Соединение линкерных звеньев посредством С-концевых аминокислотных остатков»
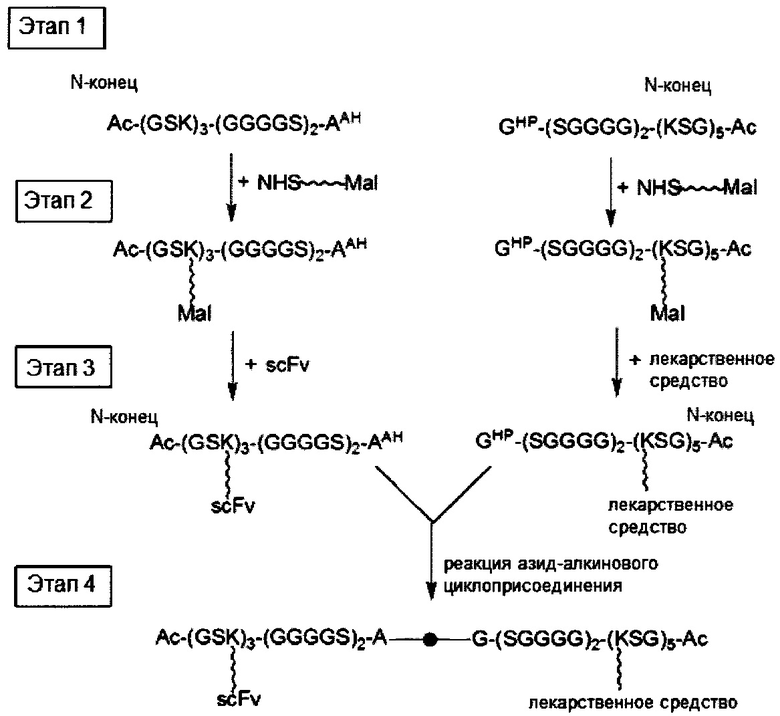
Нацеливающий/эффекторный элемент необязательно может связываться с центральной сердцевиной альтернативным способом. Схема 9 представляет собой схему, иллюстрирующую соединение эффекторного элемента с полипептидной сердцевиной, при котором сперва связывающая ветвь связывается с центральной сердцевиной, а затем эффекторный элемент (т.е. лекарственное средство) связывается со связывающей ветвью посредством реакции тиольной и малеимидной групп.
«Схема 9. Способ соединения эффекторного элемента с полипептидной сердцевиной посредством связывания со связывающими ветвями»
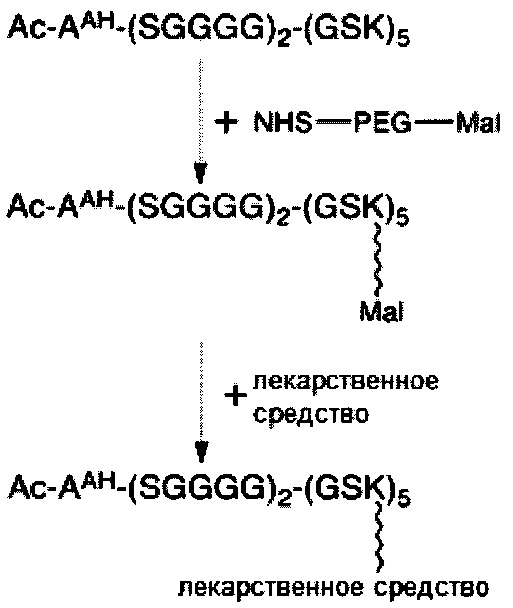
В альтернативном способе на схеме 10 эффекторный элемент (т.е. лекарственное средство) соединяется со связывающей ветвью с получением конъюгата связывающая ветвь-эффектор (т.е. PEG-лекарственное средство); далее конъюгат связывающая ветвь-эффектор связывается с центральной сердцевиной посредством образования амидной связи между лизиновыми остатками и группами сложного NHS-эфира.
«Схема 10. Альтернативный способ соединения эффекторного элемента с полипептидной сердцевиной сначала путем конъюгирования с цепью PEG, а затем связывания с аминогруппами лизиновых остатков»
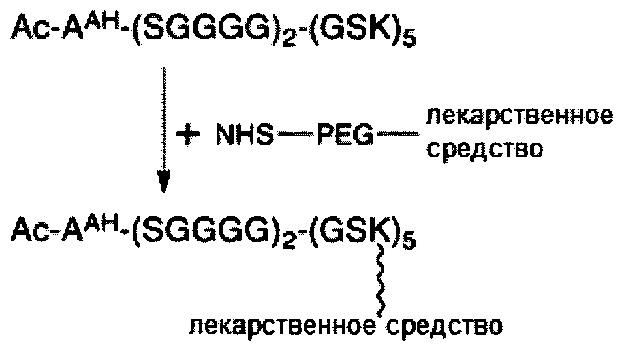
В качестве альтернативы, связывающие ветви в конфигурации с сочлененными линкерами также можно применять для связывания с биспецифическими scFv, которые выступают в качестве нацеливающих элементов или эффекторных элементов. Эти конфигурации будут увеличивать специфичность нацеливания и (или) действенность эффекторных механизмов.
На схеме 11 представлен пример получения молекулярной конструкции по настоящему изобретению, которая содержит два линкерных звена; оба линкерных звена содержат аминокислотную последовательность (K-Хаа4)3 и цистеиновый (С) остаток на своем С-конце.
«Схема 11. Получение молекулярной конструкции посредством реакции iEDDA, происходящей между соединяющими ветвями»
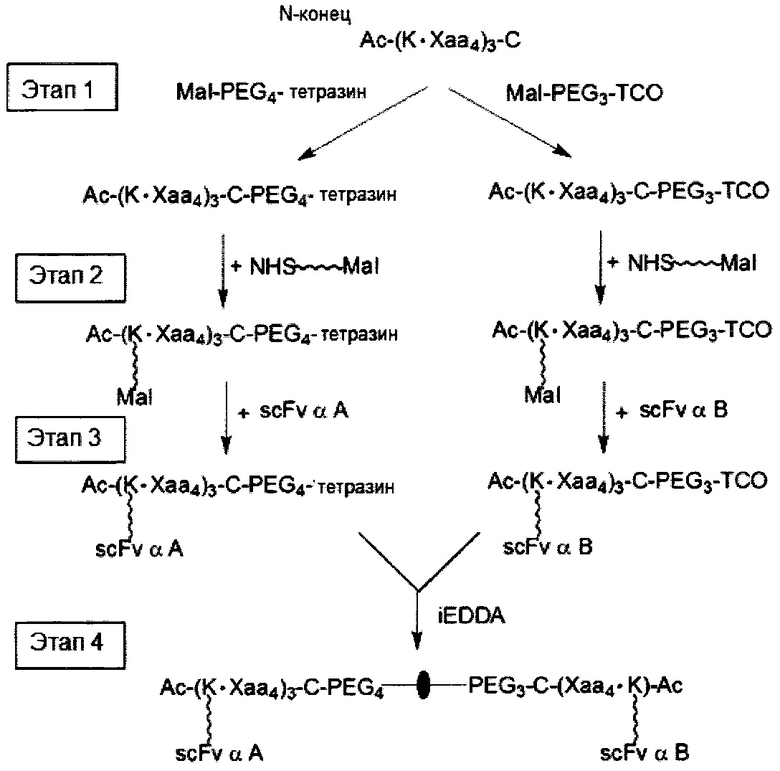
На этапе 1 две соединяющие ветви соответственно связываются с остатками С линкерных звеньев, при этом одна из соединяющих ветвей имеет малеимидную (Mal) группу на одном конце и тетразиновую группу на другом конце, а другая соединяющая ветвь имеет Mal-группу на одном конце и ТСО-группу на другом конце. На этапе 2 связывающие ветви соответственно связываются с лизиновыми (K) остатками посредством образования амидной связи между связывающей ветвью и остатком K. Далее на этапе 3 три scFv антитела к антигену A (scFv α А) и три scFv антитела к антигену В (scFv α В) соответственно связываются со связывающими ветвями линкерных звеньев посредством реакции тиольной и малеимидной групп. Наконец, на этапе 4 два линкерных звена соединяются друг с другом посредством реакции iEDDA, происходящей между тетразиновой группой и ТСО-группой.
На схеме 12 представлен пример получения молекулярной конструкции, содержащей три линкерных звена, в котором два линкерных звена, соответственно связанных с scFv α А и scFv α В, соединяются друг с другом посредством реакции iEDDA, описанной на схеме 11, а третье линкерное звено соединяется со вторым линкерным звеном посредством реакции CuAAC.
«Схема 12. Получение молекулярной конструкции, имеющей три линкерных звена с тремя функциональными элементами»
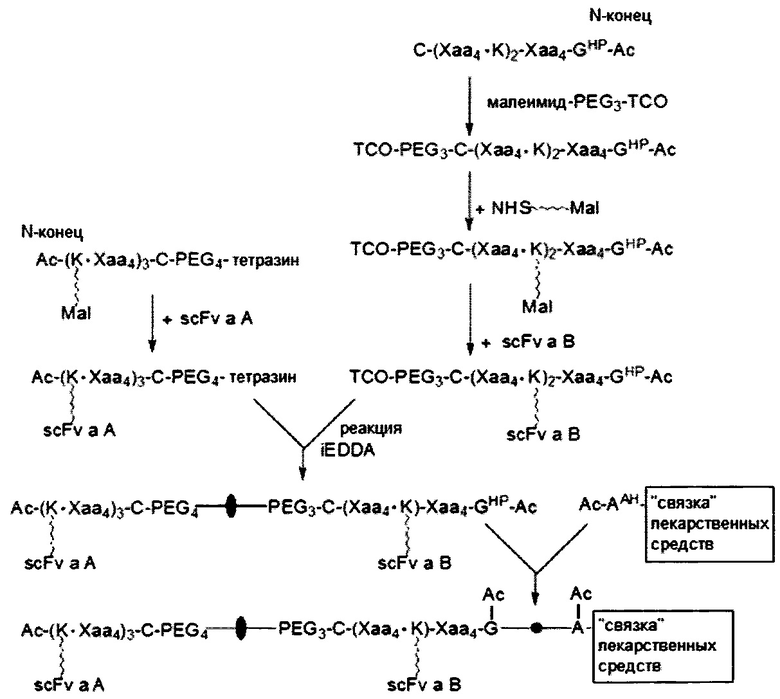
В этом примере третье линкерное звено представляет собой «связку» лекарственных средств. Эту схему реакций, однако, можно применять к третьему линкерному звену с другими элементами, такими как scFv. В примере настоящего изобретения центральное линкерное звено (т.е. второе линкерное звено) содержит остаток HPG (GHP) на своем N-конце, и, соответственно, «связка» лекарственных средств, конъюгированная с остатком AHA (ААН), может связываться со вторым линкерным звеном посредством реакции CuAAC, происходящей между остатками HPG и AHA. В качестве альтернативы, центральное линкерное звено может содержать остаток AHA на своем N- или С-конце и может связываться с третьим линкерным звеном, несущим соединяющую ветвь с DBCO или другой напряженной алкиновой группой, посредством реакции SPAAC. Образованная таким образом молекулярная конструкция на схеме 12 имеет три функциональных элемента: scFv α A, scFv α В и молекулу лекарственного средства. Молекулярные конструкции с тремя линкерными звеньями могут нести три набора scFv, из них два набора в качестве нацеливающих элементов и один набор в качестве эффекторных элементов или один набор в качестве нацеливающих элементов и два набора в качестве эффекторных элементов.
Если все нацеливающие и эффекторные элементы представляют собой scFv и применяются связывающие ветви весом 600 дальтон (12 EG-звеньев), то молекулярная конструкция в общей сложности с шестью scFv имеет молекулярную массу приблизительно 170000 дальтон. Молекулярная конструкция с семью scFv имеет молекулярную массу приблизительно 200000 дальтон, а молекулярная конструкция с восемью scFv имеет молекулярную массу приблизительно 230000 дальтон. Большинство молекулярных конструкций по настоящему изобретению имеют значения молекулярной массы, составляющие менее 200000 дальтон, и несколько молекулярных конструкций имеют значения молекулярного веса, составляющие 200000-250000 дальтон.
Если в одной молекулярной конструкции необходимо перенести четыре различных набора scFv, то предпочтительным является наличие одного линкерного звена, несущего сочлененный одноцепочечный биспецифический scFv (bi-scFv), такой как scFv1-scFv2 (например, специфичный к HER2 и HER3), и двух других линкерных звеньев, каждое из которых несет один scFv (т.е., соответственно, scFv3 и scFv4). Существует два способа конструирования биспецифического scFv1-scFv2. В «тандемной» конфигурации расположение представлено как VL1-VH1-VL2-VH2 или VH1-VL1-VH2-VL2; в конфигурации «диатела» расположение представлено как VL2-VL1-VH1-VH2 или VH2-VH1-VL1-VL2. Надлежащие линкеры с повторами GGGGS (SEQ ID NO: 6) или других последовательностей расположены между доменами иммуноглобулинов.
Как показывает практика, пептидный или PEG-линкер, который содержит малеимидную и азидную группы, может полимеризоваться при длительном хранении вследствие спонтанной реакции присоединения между малеимидной и азидной группами. Таким образом, предпочтительно, чтобы каждое линкерное звено получали заново и независимо и подвергали процессу присоединения к нацеливающим или эффекторным элементам в линкерных звеньях и незамедлительно проводили соединение линкерных звеньев посредством клик-реакции. Альтернативный предпочтительный вариант осуществления заключается в том, что как нацеливающие элементы, так и эффекторные элементы конъюгируют с линкерными звеньями с алкиновыми группами, и алкиновую группу в одном из линкерных звеньев далее превращают в азидную группу с помощью короткого гомобифункционального линкера с азидными группами на обоих концах. Линкерные звенья, одно с алкиновой группой, а другое с азидной группой, далее соединяют посредством клик-реакции.
Предпочтительные связывающие ветви для настоящего изобретения представляют собой PEG. Длина связывающих ветвей важна по нескольким соображениям. Они должны быть достаточно длинными, чтобы обеспечивать гибкость связанных scFv или других типов функциональных элементов для достижения антигенных участков-мишеней на поверхности клеток-мишеней без стерических ограничений; но недостаточно длинными, чтобы обуславливать образование внутримолекулярных и межмолекулярных сплетений связывающих ветвей и связанных с ними scFv-фрагментов или функциональных элементов или излишне увеличивать размер целой молекулярной конструкции, препятствуя проникновению в ткани. Связывающие ветви, которые являются слишком длинными, также могут быть неспособны обеспечивать сближение молекул антигенов с образованием компактных кластеров, если такие кластеры необходимы для инициирования процесса передачи сигнала для осуществления апоптоза или других клеточных эффектов. Оптимальную длину связывающих ветвей для различных типов комбинаций антигенов-мишеней и связывающихся с ними средств может определить любой специалист в соответствующей области техники без проведения излишних экспериментов. Связывающая ветвь NHS-(PEG)12-малеимид (примерно 500 дальтон) является предпочтительной в ряде молекулярных конструкций по настоящему изобретению. Полностью растянутый (PEG)12 имеет длину 40-50  .
.
Применимые связывающие ветви и соединяющие ветви не ограничены цепями PEG. Можно применять пептиды, содержащие глицин, серии и другие гидрофильные аминокислотные остатки, а также полисахариды и другие биосовместимые линейные полимеры, модифицированные таким образом, что они содержат NHS- и малеимидные группы.
Для определенных способов терапевтического применения желательно, чтобы эффекторные элементы в молекулярных конструкциях по настоящему раскрытию высвобождались из связывающих ветвей, так чтобы они могли поступать в клетки в участке-мишени, в том числе в клетки, связанные с нацеливающими элементами, или окружающие клетки, вызывая фармакологические эффекты. В этих случаях в связывающую ветвь встраивают расщепляемую связь. Были разработаны расщепляемые связи, восприимчивые к расщеплению путем гидролиза, воздействия кислот, восстановления и под действием ферментов. Например, в конструировании терапевтических конструкций применяли пептидные сегменты, восприимчивые к действию матриксных металлопротеиназ, присутствующих в воспаленных тканях. Один вариант осуществления настоящего изобретения заключается в применении PEG-линкеров NHS-PEG2-12S-S-малеимид со связью S-S рядом с малеимидной группой, где S-S представляет собой дисульфидную связь, которую можно подвергнуть медленному восстановлению.
В соответствии с некоторыми вариантами осуществления настоящего раскрытия нацеливающий элемент, описанный в вышеупомянутых вариантах осуществления, выбран из группы, состоящей из фактора роста, пептидного гормона, цитокина и фрагмента антитела; а эффекторный элемент представляет собой иммуномодулятор, хелатообразователь в комплексе с радиоактивным нуклидом, терапевтическое лекарственное средство, цитокин, растворимый рецептор или антитело.
В вариантах осуществления антитело находится в форме антиген-связывающего фрагмента (Fab), вариабельного фрагмента (Fv), одноцепочечного вариабельного фрагмента (scFv), однодоменного антитела (sdAb) или биспецифического одноцепочечного вариабельного фрагмента (bi-scFv). В соответствии с одним вариантом осуществления bi-scFv представляет собой биспецифический тандемный scFv или биспецифический scFv в форме диатела.
Для сохранения диффундирующей способности молекулярных конструкций предпочтительным является размер молекулы, составляющий менее 250000 дальтон. Таким образом, scFv-фрагменты являются предпочтительными для большинства вариантов осуществления. На уровне ДНК гены конструируют так, чтобы VL и VH были связаны в виде единого полипептида в любом порядке (VL и VH или VH и VL) пептидным линкером из 10-25 аминокислотных остатков, преобладающими остатками в котором являются глицин и серин. На С-конце встраивают короткий фрагмент с глициновым и сериновым и концевым цистеиновым остатками. Рекомбинантные scFv и bi-scFv можно получать в бактериях, таких как Е. coli и Pseudomonas putida, в дрожжах, таких как Pichia pastoris, или в клетках млекопитающих, таких как линии клеток СНО и HEK293.
В лаборатории авторов настоящего изобретения было получено большое количество антител IgG, Fab, scFv и различных фрагментов антител, белков на основе Fc и других рекомбинантных антител в линиях клеток HEK293 и СНО для проведения экспериментов в системах in vitro и в животных моделях. В лаборатории также были разработаны линии клеток для получения антител для клинических испытаний с участием людей. Для удобства можно использовать систему транзиентной экспрессии HEK293 для получения до 1 г IgG или фрагментов антител с применением нескольких колб на 1-2 литра в научно-исследовательской лаборатории. scFv-фрагменты, подлежащие применению в молекулярных конструкциях по настоящему изобретению, как правило, не имеют углеводной модификации, и углеводная модификация не требуется для активности связывания scFv с их антигенными мишенями. Более того, в scFv-фрагменте присутствует только одна дисульфидная связь и один концевой цистеин. Поэтому для получения scFv были разработаны маломасштабные бактериальные системы экспрессии в качестве производственной альтернативы. В случае с Е. coli использовали системы экспрессии для извлечения scFv из внутриклеточных телец-включений, из периплазмы и в секретируемой форме. scFv в большинстве случаев можно очищать с помощью аффинной колонки с белком L, который взаимодействует с VH большинства легких κ-цепей, или, в других случаях, с помощью ионообменных колонок.
В основе примеров настоящего изобретения лежит платформа с сочлененными линкерами, в которой в качестве нацеливающих и (или) эффекторных элементов используются главным образом scFv и Fab. Однако, также можно подвергать скринингу конкретные связывающие молекулы из больших библиотек связывающих молекул на основе sdAb или других фрагментов антител. Библиотеки связывающих молекул, в основе которых не лежат домены иммуноглобулинов, но которые напоминают антитела тем, что они характеризуются аффинностью специфичного связывания с выбранными молекулами-мишенями, включают (1) аптамеры, которые представляют собой олигонуклеотиды или короткие пептиды, выбранные для связывания с молекулами-мишенями, (2) финомеры, которые представляют собой небольшие связывающие белки, полученные из домена SH3 человеческого Fyn, (3) аффимеры, которые представляют собой связывающие белки, полученные из семейства цистатинов-ингибиторов цистеиновых протеиназ, и (4) дарпины (искусственные белки с анкириновыми повторами), которые представляют собой белки, полученные методами генной инженерии, со структурами, полученными из природных белков-анкиринов, и состоят из 3, 4 или 5 повторяющихся мотивов этих белков. Эти миметики антител имеют молекулярные массы, составляющие приблизительно 10000-20000 дальтон.
II-(ii) Функциональные элементы, подходящие для использования с молекулярной конструкцией с сочлененными линкерами
Как обсуждалось выше, данный сочлененный линкер содержит по меньшей мере два линкерных звена, в которых первое линкерное звено несет один или несколько целевых элементов, а второе линкерное звено несет один или несколько эффекторных элементов или элементов, повышающих фармакокинетические свойства, и наоборот. Конкретные примеры функциональных элементов, включенных в данную молекулярную конструкцию с сочлененными линкерами для лечения конкретного заболевания, обсуждаются ниже.
Для лечения заболеваний центральной нервной системы (ЦНС) scFv, специфичный к рецептору трансферрина, может использоваться в качестве нацеливающего элемента данных молекулярных конструкций с сочлененными линкерами в связи с эффекторными элементами, подходящими для конкретного заболевания ЦНС. Например, в молекулярных конструкциях с сочлененными линкерами для лечения рассеянного склероза в качестве эффекторного элемента могут использоваться финголимод, финголимод-фосфат, интерферон-β или scFv, специфичный к интегрину α4. В случае болезни Альцгеймера в иллюстративных молекулярных конструкциях с сочлененными линкерами в качестве эффекторного элемента может использоваться scFv, специфичный к β-амилоиду.
При заболеваниях или состояниях, связанных с инфекцией, scFv, специфичный к CD16b или к CD32, может использоваться в качестве эффекторного элемента, тогда как нацеливающий элемент зависит от причины инфекции. В частности, если инфекция вызвана вирусом, в качестве нацеливающего элемента используется scFv, специфичный к вирусному белку, а если инфекция вызвана бактерией, то в качестве нацеливающего элемента используется scFv, специфичный к бактериальному белку. Иллюстративные примеры вирусных белков включают, без ограничения, белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса. Примеры бактериального белка включают без ограничения эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
II-(iii) Использование молекулярной конструкции с сочлененными линкерами
Настоящее раскрытие также относится к способу лечения различных заболеваний с использованием подходящей молекулярной конструкции с сочлененными линкерами. Как правило, способ включает стадию введения субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества молекулярной конструкции с сочлененными линкерами в соответствии с вариантами осуществления настоящего раскрытия.
ЧАСТЬ III. Молекулярные конструкции на основе Fc для лечения заболеваний центральной нервной системы или инфекционных заболеваний
III-(i) Структура молекулярной конструкции на основе Fc
Иммуноглобулиновое антитело в широком смысле конфигурации на основе Fc может служить в качестве основы для нацеливающего или эффекторного элемента, и его соответствующий эффекторный или нацеливающий элемент может быть включен в его состав на С-конце его двух тяжелых γ-цепей в форме доменов scFv. В типичной конфигурации «на основе Fc» двухцепочечный IgG.Fc применяют в качестве основы для молекулярной платформы. Каждая из полипептидных цепей слита с одним или двумя нацеливающими и одним или двумя эффекторными элементами, так что на каждой цепи находится в общей сложности от двух до трех элементов. Молекула Т-Е с конфигурацией на основе Fc будет иметь в общей сложности от четырех до шести элементов (например, scFv или любых других фрагментов антител). Fc-часть молекулярных конструкций необязательно также заключает в себе Fc-опосредованные эффекторные функции, ADCC и (или) комплемент-опосредованную активацию. Хотя в некоторых других способах применения такие Fc-опосредованные эффекторные функции не рассматриваются.
При конструировании молекулярных конструкций на основе Fc нацеливающие элементы располагают на N- или С-конце. Если эффекторные элементы функционируют посредством связывания с компонентом клеточной поверхности, то они также должны размещаться на данном конце. Если эффекторные элементы функционируют путем связывания с растворимыми факторами и их нейтрализации, то они могут располагаться между концевым нацеливающим или эффекторным элементом и СН2-СН3.
Путем выбора элементов Т-Е данной молекулярной конструкции на основе Fc молекулярная конструкция может использоваться для лечения заболеваний центральной нервной системы (ЦНС) или инфекционных заболеваний. Настоящее раскрытие также полезно тем, что в некоторых вариантах осуществления в нем используется линкерное звено в соответствии с первым аспектом настоящего раскрытия, которое представляет собой легкое средство для управления количеством нацеливающих и эффекторных элементов настоящих молекулярных конструкций на основе Fc. В зависимости от выбранных нацеливающих и (или) эффекторных элементов настоящая молекулярная конструкция на основе Fc может принимать различные конфигурации, которые обсуждаются ниже, соответственно.
В настоящих молекулярных конструкциях на основе Fc как нацеливающий элемент, так и эффекторный элемент представляют собой антитела или их фрагменты.
Фигура 8А представляет собой схематическое изображение, иллюстрирующее молекулярную конструкцию 800А на основе Fc в соответствии с определенными вариантами осуществления настоящего раскрытия. Как проиллюстрировано, молекулярная конструкция 800А на основе Fc содержит две идентичные цепи 810 СН2-СН3, пару эффекторных элементов Е1, связанных с N-концами цепей 810 СН2-СН3, и пару нацеливающих элементов Т1, связанных с С-концами цепей 810 СН2-СН3. В этой иллюстративной конфигурации как нацеливающий элемент Т1, так и эффекторный элемент Е1 представляют собой scFv.
Молекулярная конструкция 800В на основе Fc, проиллюстрированная на фигуре 8В, довольно сходна по структуре с молекулярной конструкцией 800А на основе Fc на фигуре 8А, за исключением того, что два эффекторных элемента Е1 соответственно связаны с С-концами цепей 810 СН2-СН3, а два нацеливающих эффектора соответственно связаны с С-концами цепей 810 СН2-СН3.
В соответствии с определенными вариантами осуществления как эффекторные элементы, так и нацеливающие элементы связаны с N-концами цепей СН2-СН3. Например, если как эффекторный элемент, так и нацеливающий элемент находятся в форме одноцепочечных вариабельных фрагментов (scFv), то эффекторный элемент и нацеливающий элемент могут быть связаны в тандемной конфигурации или в конфигурации диатела с образованием, таким образом, биспецифического scFv, связанного с N-концом цепи СН2-СН3.
Молекулярная конструкция 800С на основе Fc (фигура 8С) содержит Fc-часть, и, соответственно, с N-концом каждой цепи 810 СН2-СН3 связан биспецифический scFv Т1-Е1.
В некоторых примерах пара эффекторных элементов или пара нацеливающих элементов имеет конфигурацию Fab (т.е. состоит из домена VH-CH1 и домена VL-Cκ); этот Fab-фрагмент связан с N-концами цепей СН2-СН3, так что молекулярная конструкция на основе Fc принимает конфигурацию IgG. В этих случаях пара элементов, которая не находится в конфигурации Fab, может быть связана с С-концами пары сегментов СН2-СН3.
Например, в молекулярной конструкции 900 на основе Fc на фигуре 9 каждый из двух нацеливающих элементов Т1 содержит домен 820 VH-CH1 и домен 825VL-Cκ с образованием, таким образом, конфигурации Fab 830, связанной с N-концами цепей 810 СН2-СН3, так что молекулярная конструкция 900 на основе Fc принимает конфигурацию IgG. В этом случае пара эффекторных элементов Е1 связана с С-концами пары цепей 810 СН2-СН3.
Согласно некоторым вариантам осуществления, данная молекулярная конструкция на основе Fc имеет эффекторный элемент, являющийся пептидом.
Например, в соответствии с некоторыми вариантами осуществления настоящего раскрытия, эффекторный элемент может представлять собой пептид с определенным терапевтическим эффектом, тогда как нацеливающий элемент представляет собой антитело или его фрагмент (см. фигуры 10А и 10В). Как показано на фигуре 10А, молекулярная конструкция 1000А на основе Fc содержит пару нацеливающих элементов Т1 (в виде scFv), связанных с N-концами пары сегментов 1210 СН2-СН3, и пару эффекторных элементов Е1 (в форме терапевтических пептидов), связанную с С-концами пары сегментов 1210 СН2-СН3.
Аналогично, в молекулярной конструкции 1000В на основе Fc на фигуре 10В пара нацеливающих элементов Т1 (в виде scFv) связана с С-концами пары сегментов 1210 СН2-СН3, а пара эффекторных элементов Е1 (в форме терапевтических пептидов) связана с N-концами пары сегментов 1210 СН2-СН3.
Следует понимать, что для молекулярных конструкций на основе Fc, в которых в качестве эффекторного элемента используется пептид, нацеливающий элемент может быть встроен в Fab-фрагмент, так что молекулярные конструкции принимают конфигурацию IgG.
В конфигурации, изображенной на фигурах 8А-10В, цепи СН2-СН3 унаследованы от человеческих иммуноглобулинов γ1 или γ4. Как правило, выбирают γ1, если желательны Fc-опосредованные функции, такие как антителозависимая клеточная цитотоксичность (ADCC) и комплемент-опосредованная активность (воспалительная активация или лизис клеток-мишеней). В случае если Fc-опосредованных функций следует избегать, для создания настоящих молекулярных конструкций на основе Fc выбирается γ4.
III-(ii) Функциональные элементы, подходящие для использования с молекулярной конструкцией на основе Fc
Теперь, когда основные типы структурной организации молекулярных конструкций на основе Fc были обсуждены выше, определенные комбинации конкретного (конкретных) эффекторного (эффекторных) элемента (элементов) и нацеливающего (нацеливающих) элемента (элементов) представлены ниже для целей иллюстрации.
Для лечения заболеваний центральной нервной системы (ЦНС) антитело (или его фрагмент), специфичное к рецептору трансферрина, можно использовать в качестве нацеливающего элемента в связи с эффекторными элементами, подходящими для конкретного заболевания ЦНС. Например, в молекулярных конструкциях на основе Fc для лечения рассеянного склероза в качестве эффекторного элемента может использоваться scFv, специфичный к интегрину α4. В случае болезни Альцгеймера в иллюстративных молекулярных конструкциях на основе Fc в качестве эффекторного элемента может использоваться scFv, специфичный к β-амилоиду. Вышеупомянутые молекулярные конструкции на основе Fc для лечения заболеваний ЦНС могут принимать конфигурацию, описанную в связи с любой из фигур 8А-8С и с фигурой 9.
В молекулярных конструкциях на основе Fc для лечения рассеянного склероза в качестве эффекторных элементов могут также использоваться INF-β1a или INF-β1b. В этом случае молекулярные конструкции на основе Fc могут принимать конфигурацию, описанную в связи с фигурами 10А или 10В.
При создании молекулярных конструкций на основе Fc для лечения заболеваний/состояний, связанных с инфекцией (таких как вирусные инфекции или бактериальные инфекции), в качестве нацеливающего элемента можно использовать антитело (или его фрагмент), специфичное к вирусному белку или к бактериальному белку. Что касается эффекторных элементов для лечения инфекций, можно использовать антитело (или его фрагмент), специфичное к CD32 или к CD16b. Эти молекулярные конструкции на основе Fc могут принимать конфигурацию, описанную в связи с любой из фигур 8А-8С и с фигурой 9.
Сущностью настоящего изобретения являются логическое обоснование и идея использования конкретной комбинации или составления пар нацеливающих и эффекторных элементов. Использование конфигурации слияния с Fc в молекулярных конструкциях является предпочтительным вариантом осуществления. Для специалистов в данной области возможным является связывание пар нацеливающих и эффекторных элементов по настоящему изобретению с использованием других молекулярных платформ, таких как пептиды, белки (например, альбумин), полисахариды, полиэтиленгликоль и другие типы полимеров, которые служат в качестве структурной основы для прикрепления нескольких молекулярных элементов.
III-(iii) Использование молекулярной конструкции на основе Fc
Настоящее раскрытие также относится к способу лечения заболеваний ЦНС с использованием подходящей молекулярной конструкции на основе Fc. Как правило, способ включает стадию введения субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества молекулярной конструкции на основе Fc в соответствии с вариантами осуществления настоящего раскрытия.
Настоящее раскрытие также относится к способу лечения инфекций с использованием подходящей молекулярной конструкции на основе Fc. Как правило, способ включает стадию введения субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества молекулярной конструкции на основе Fc в соответствии с вариантами осуществления настоящего раскрытия.
ЭКСПЕРИМЕНТАЛЬНЫЕ ПРИМЕРЫ
Пример 1. Синтез пептида 1 (SEQ ID NO: 18), пептида 2 (SEQ ID NO: 27) и пептида 3 (SEQ ID NO: 19) в качестве пептидных сердцевин и конъюгирование SH-группы их цистеинового остатка с малеимид-PEG3-транс-циклооктеном (ТСО) в качестве соединяющей ветви
Синтезированные пептиды 1, 2 и 3 (Chinapeptide Inc., Shanghai, China) обрабатывались сходным образом. Очищенный пептид растворяли в 100 мМ натрий-фосфатном буфере (рН 7,0), содержащем 50 мМ NaCl и 5 мМ EDTA, в конечной концентрации 2 мМ. Растворенный пептид восстанавливали с помощью 1 мМ трис(2-карбоксиэтил)фосфина (ТСЕР) при 25°С в течение 2 часов. Для конъюгирования SH-группы цистеинового остатка с малеимид-PEG3-ТСО (Conju-Probe Inc.) с получением функциональной связывающей ТСО-группы пептид и малеимид-PEG3-ТСО смешивали в молярном соотношении 1/7,5 и инкубировали при рН 7 и 25°С в течение 18 часов. Пептид, конъюгированный с ТСО, очищали с помощью обращенно-фазовой HPLC на колонке Supelco С18 (250 мм × 10 мм; 5 мкм) с применением подвижной фазы в виде ацетонитрила и 0,1% трифторуксусной кислоты, линейного градиента 0-100% ацетонитрила в течение 30 минут при скорости потока 1,0 мл/мин и температуре колонки 25°С.
Идентификацию синтезированных ТСО-пептидов (см. ниже) проводили методом масс-спектрометрии MALDI-TOF. Масс-спектрометрический анализ проводили в центре спектрометрии (Mass Core Facility) при Институте молекулярной биологии (Institute of Molecular Biology (IMB), Academia Sinica, Taipei, Taiwan). Измерения осуществляли на масс-спектрометре Bruker Autoflex III для MALDI-TOF/TOF (Bruker Daltonics, Bremen, Germany).
Синтезированный ТСО-функционализированный пептид 1, как показано ниже, имел молекулярную массу 2078,9 дальтон.
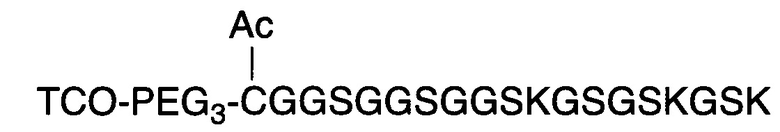
Синтезированный ТСО-функционализированный пептид 2, как показано ниже, имел молекулярную массу 2020,09 дальтон.
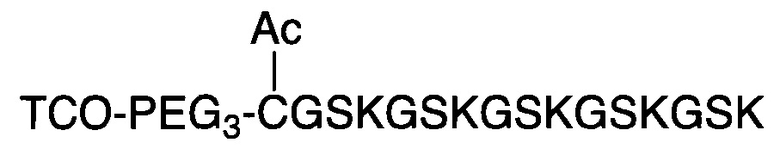
ТСО-функционализированный пептид 3, как показано ниже, имел молекулярную массу 3381,85 дальтон.
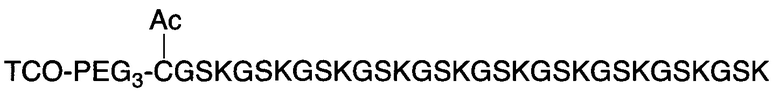
Пример 2. Синтез пептида 1 в качестве пептидной сердцевины и конъюгирование SH-группы его цистеинового остатка с малеимид-PEG4-тетразином в качестве соединяющей ветви
Синтезированный пептид 1 (Chinapeptide Inc., Shanghai, China) растворяли в 100 мМ натрий-фосфатном буфере (рН 7,0), содержащем 50 мМ NaCl и 5 мМ EDTA, в конечной концентрации 2 мМ. Растворенный пептид восстанавливали с помощью 1 мМ трис(2-карбоксиэтил)фосфина (ТСЕР) при 25°С в течение 2 часов. Для конъюгирования SH-группы цистеинового остатка с малеимид-PEG4-тетразином (Conju-probe Inc., San Diego, USA) с получением функциональной связывающей группы тетразин, пептид и малеимид-PEG4-тетразин смешивали в соотношении 1/5 и инкубировали при рН 7 и температуре 4°С в течение 18 часов. Пептид, конъюгированный с тетразином, очищали с помощью обращенно-фазовой HPLC на колонке Supelco С18 (250 мм × 10 мм; 5 мкм) с применением подвижной фазы в виде ацетонитрила и 0,1% трифторуксусной кислоты, линейного градиента 0-100% ацетонитрила в течение 30 минут при скорости потока 1,0 мл/мин, и температуры колонки 25°С.
Синтезированный тетразин-функционализированный пептид 1, как показано ниже, имел молекулярную массу 2185,2 дальтон.
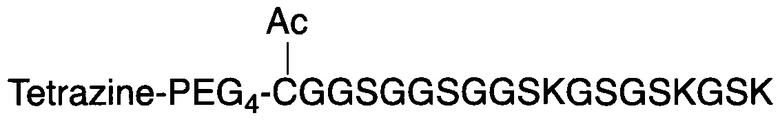
Пример 3. Синтез линкерного звена путем конъюгирования NHS-PEG12-Mal с NH2-группами ТСО-функционализированного пептида 1 в качестве связывающих ветвей
Три связывающих ветви PEG12-малеимида прикрепляли к пептидной сердцевине, ТСО-функционализированному пептиду 1. Сшивающее средство, NHS-PEG12-малеимид (сукцинимидиловый сложный эфир [(N-малеимидопропионамидо)додекаэтиленгликоля]), приобретали у Thermo Fisher Scientific Inc. (Waltham, USA). Процедуру конъюгирования осуществляли согласно инструкции изготовителя. Вкратце, пептид с лизиновыми остатками растворяли в буфере для конъюгирования, представляющем собой фосфатно-солевой буферный раствор (рН 7,5) при 100 мМ. Затем сшивающее средство NHS-PEG12-малеимид добавляли к растворенному пептиду с конечной концентрацией 1 мМ (в 10-кратном молярном избытке по отношению к 0,1 мМ раствору пептида). Реакционные смеси инкубировали в течение 18 часов при комнатной температуре. ТСО-функционализированный пептид 1, конъюгированный с малеимидом-PEG12, очищали с помощью обращенно-фазовой ВЭЖХ на колонке Supelco С18 (250 мм × 4,6 мм, 5 мкм) с применением подвижной фазы в виде ацетонитрила и 0,1% трифторуксусной кислоты с линейным градиентом 0-100% ацетонитрила в течение 30 минут при скорости потока 1,0 мл/мин и температуре колонки 25°С.
Идентификацию ТСО-функционализированного пептида 1, конъюгированного с малеимидом-PEG12, проводили методом масс-спектрометрии MALDI-TOF.
Синтезированный ТСО-функционализированный пептид 1, конъюгированный с малеимидом-PEG12, имел молекулярную массу 4332 дальтон. Как проиллюстрировано ниже, ТСО-пептид 1, конъюгированный с малеимидом-PEG12, представлял собой линкерное звено на основе пептидной сердцевины, несущее одну ТСО-группу и три связывающих ветви PEG с малеимидными группами.
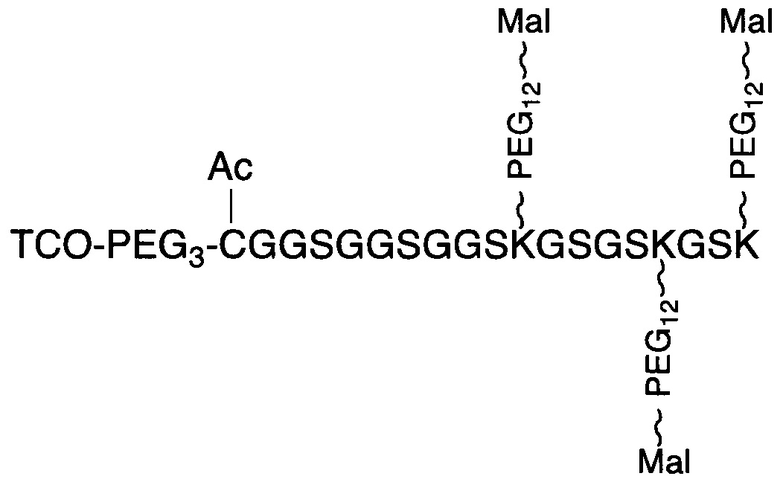
Пример 4: Синтез линкерного звена путем конъюгирования NHS-PEG12-Mal с NH2-группами тетразин-функционализированного пептида 1 в качестве связывающих ветвей
К пептидной сердцевине на основе тетразин-функционализированного пептида 1 прикрепляли три PEG12-малеимидные связывающие ветви. Сшивающее средство NHS-PEG12-малеимид (сукцинимидиловый сложный эфир [(N-малеимидопропионамидо)додекаэтиленгликоля]) приобретали у Thermo Fisher Scientific Inc. (Waltham, USA). Процедуру конъюгирования осуществляли согласно инструкции производителя. Вкратце, пептид с лизиновыми остатками растворяли в буфере для конъюгирования, представляющем собой фосфатно-солевой буферный раствор (рН 7,5), при 100 мМ. Затем к растворенному пептиду добавляли сшивающее средство MHS-PEG12-малеимид в конечной концентрации 1 мМ (в 10-кратном молярном избытке по отношению к 0,1 мМ раствору пептида). Реакционные смеси инкубировали в течение 18 часов при комнатной температуре. Тетразин-функционализированный пептид 1, конъюгированный с PEG12-малеимидом, очищали с помощью обращенно-фазовой HPLC на колонке Supelco С18 (250 мм × 4,6 мм; 5 мкм) с применением подвижной фазы в виде ацетонитрила и 0,1% трифторуксусной кислоты с линейным градиентом 0-100% ацетонитрила в течение 30 минут при скорости потока 1,0 мл/мин и температуре колонки 25°С.
Синтезированный тетразин-функционализированный пептид 1, конъюгированный с PEG12-малеимидом, проиллюстрированный ниже, представлял собой линкерное звено на основе пептидной сердцевины, несущее одну соединяющую ветвь с тетразиновой группой и две связывающие ветви PEG с малеимидными группами. Результат масс-спектрометрии MALDI-TOF на фигуре 11 показывает, что молекулярная конструкция имела молекулярную массу 4461 дальтон.
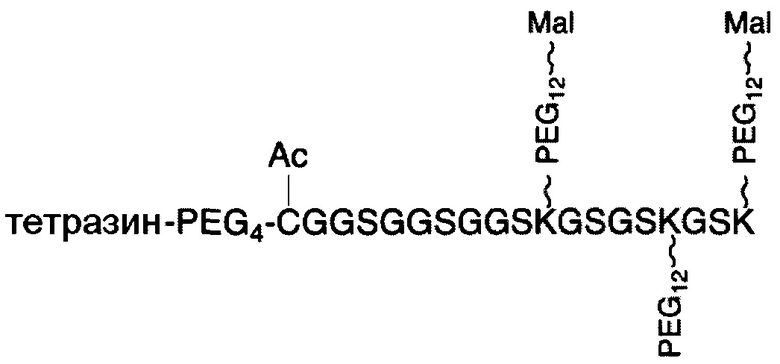
Пример 5: Конъюгирование молекул финголимода и финголимод-фосфата с помощью сшивающего средства NHS-PEG5-NHS
Финголимод был приобретен у Biotang Inc. (Lexington, USA), а финголимод-фосфат - у KM3 Scientific Corporation (New Taipei City, Taiwan). NH2-группу молекулы финголимода подвергали взаимодействию с гомо-бифункциональным сшивающим средством, NHS-PEG5-NHS, как показано на схеме 13. Финголимод растворяли в 100% ДМСО при конечной концентрации 10 мМ; NHS-PEG5-NHS растворяли в 100% ДМСО при конечной концентрации 250 мМ. Для активации NH2-группы финголимода, 6% (об./об.) к раствору финголимода добавляли основной фосфатно-солевой буферный раствор (рН 12,7) и затем инкубировали в течение 10 минут. Сшивающее средство NHS-PEG5-NHS добавляли к раствору финголимода при конечной концентрации 30 мМ (в 3-кратном молярном избытке по отношению к 10 мМ раствора финголимода). Реакционную смесь инкубировали в течение 3 часов при комнатной температуре.
«Схема 13. Конъюгирование молекул финголимода с помощью сшивающего средства NHS-PEG5-NHS»
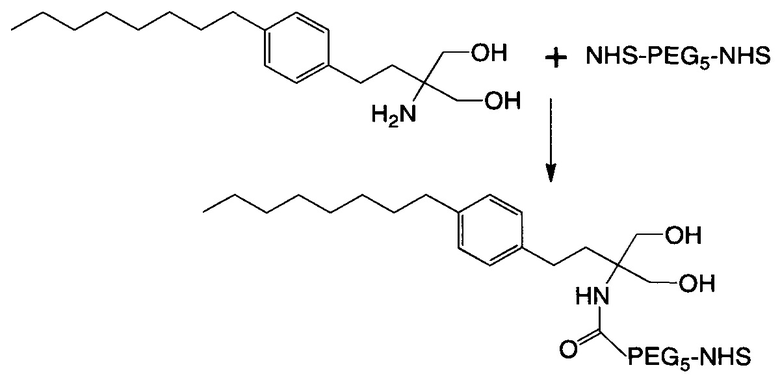
Финголимод-фосфат растворяли в 100% ДМСО при конечной концентрации 5 мМ, а сшивающее средство NHS-PEG5-NHS растворяли в 100% ДМСО при конечной концентрации 250 мМ. Сшивающее средство NHS-PEG5-NHS добавляли к раствору финголимод-фосфата с конечной концентрацией 15 мМ (в 3-кратном молярном избытке по отношению к 5 мМ раствора финголимод-фосфата). Реакционную смесь инкубировали в течение 3 часов при комнатной температуре, затем добавляли 18% (об./об.) кислотный буфер (рН=0,88) для остановки реакции. Растворитель выпаривали в вакууме.
Финголимод, конъюгированный с NHS-PEG5, и финголимод-фосфат, конъюгированный с NHS-PEG5, растворяли в 30% ацетонитриле и затем очищали с использованием обращенно-фазовой ВЭЖХ с на колонке Supelco С18 (250 мм × 4,6 мм, 5 мкм) с применением подвижной фазы в виде ацетонитрила и 0,1% трифторуксусной кислоты с линейным градиентом 30-100% ацетонитрила в течение 30 минут при скорости потока 1,0 мл/мин и температуре колонки 25°С.
На фигуре 12 показано, что синтезированный финголимод, конъюгированный с NHS-PEG5, как показано на схеме 13, имел молекулярную массу 725,41 дальтон.
Синтезированный финголимод-фосфат, конъюгированный с NHS-PEG5, как проиллюстрировано ниже, имел молекулярную массу 803,3 дальтона.
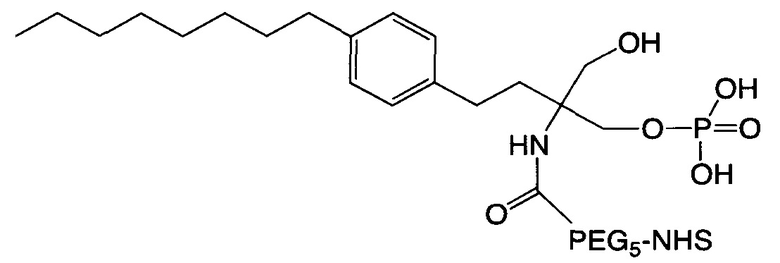
Пример 6: Конъюгирование молекул финголимода с помощью связывающей ветви азидо-NHS-S-S-PEG3
NH2-группу молекулы финголимода подвергали взаимодействию гетеро-бифункционального расщепляемого линкера NHS-S-S-PEG3-азидо (Conju-probe Inc.) с молярным соотношением 1:3. Продукт, азидо-PEG3-S-S-финголимод, очищали с помощью ВЭЖХ для удаления избыточных непрореагировавших молекул финголимода. Процедуры конъюгирования и очистки были аналогичны процедурам, описанным в предыдущем примере.
Синтезированный финголимод, конъюгированный с азидо-PEG3-S-S, как показано ниже, имел молекулярную массу 629,33 дальтона.
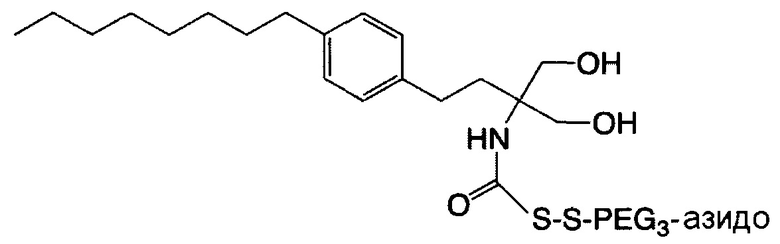
Пример 7: Конъюгирование молекул финголимода, конъюгированного с азидо-PEG3-S-S, с помощью сшивающего средства NHS-PEG4-дибензилциклооктина (DBCO)
Молекулы финголимода, конъюгированного с азидо-PEG3-S-S, растворяли в 100% ДМСО при конечной концентрации 10 мМ, а сшивающее средство NHS-PEG4-DBCO растворяли в 100% ДМСО при конечной концентрации 250 мМ. Сшивающее средство NHS-PEG4-DBCO в количестве 5 мкл добавляли к 400 мкл раствора финголимода, конъюгированного с азидо-PEG3-S-S, до конечного молярного соотношения 1/3,2 (NHS-PEG4-DBCO: финголимод, конъюгированный с азидо-PEG3-S-S) в 100 мМ фосфатно-солевого буферного раствора при рН 7,5. Реакционную смесь инкубировали в течение 3 часов при комнатной температуре.
Синтезированный финголимод, конъюгированный с NHS-PEG4-PEG3-S-S, как показано ниже, имел молекулярную массу 1278,61 дальтона. Два изотопических пика также наблюдались в спектре PC при 1279,64 и 1280,635, что соответствует [М+Н+1]+ и [М+Н+2]+.
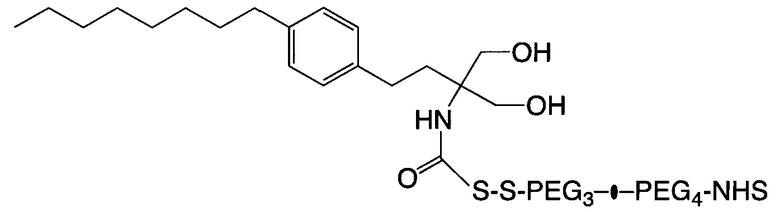
Пример 8: Конъюгирование молекул финголимода, конъюгированного с NHS-PEG5, с ТСО-функционализированным пептидом 2 и 3
ТСО-функционализированный пептид 2 растворяли в 100 мМ фосфатно-солевого буферного раствора при рН 7,5 до концентрации 20 мМ, а финголимод, конъюгированный с NHS-PEG5, растворяли в 100% ДМСО до концентрации 50 мМ.
ТСО-функционализированный пептид 2 и финголимод, конъюгированный с NHS-PEG5, смешивали при молярном соотношении 1/42 в 100% ДМСО и инкубировали в течение 3 часов при комнатной температуре. Затем к реакционному раствору вновь добавляли ТСО-функционализированный пептид 2 до конечного молярного соотношения 1/13,5 (ТСО-функционализированный пептид 2: финголимод, конъюгированный с NHS-PEG5) в 100% ДМСО. Смесь дополнительно инкубировали в течение 3 часов при комнатной температуре. На фигуре 13 показано, что «связка» лекарственных средств ТСО-функционализированного пептида 2 и финголимода имела молекулярную массу 5069 дальтон.
ТСО-функционализированный пептид 3 растворяли в 100 мМ фосфатно-солевого буферного раствора при рН 7,5 до концентрации 10 мМ, а финголимод, конъюгированный с NHS-PEG5, растворяли в 100% ДМСО до концентрации 50 мМ.
ТСО-функционализированный пептид 3 и финголимод, конъюгированный с NHS-PEG5, смешивали при молярном соотношении 1/4 при комнатной температуре в течение ночи. На фигуре 14 показано, что «связка» лекарственных средств ТСО-функционализированного пептида 3 и финголимода имела молекулярную массу 9479 дальтон, что указывает на факт конъюгирования десяти молекул финголимода в линкерное звено ТСО-функционализированного пептида 3.
Синтезированная «связка» лекарственных средств, проиллюстрированная ниже, состояла из линкерного звена со свободной функциональной ТСО-группой и набора из пяти молекул финголимода.
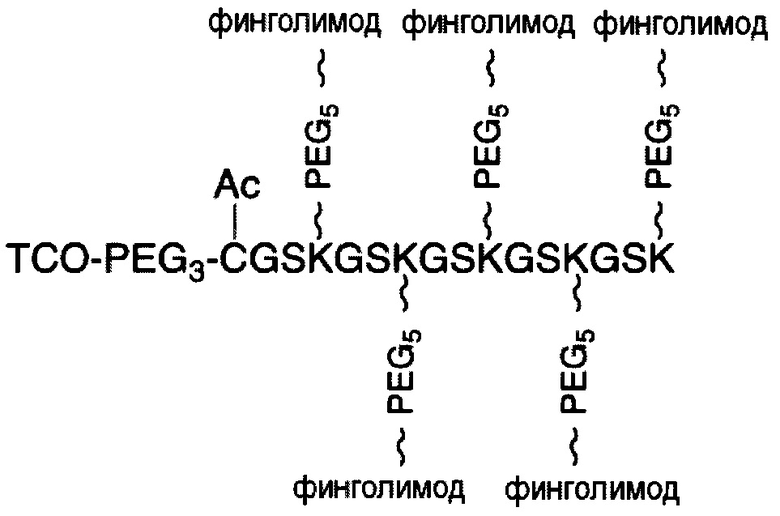
Вторая синтезированная «связка» лекарственных средств, проиллюстрированная ниже, состояла из линкерного звена со свободной функциональной ТСО-группой и набора из десяти молекул финголимода.
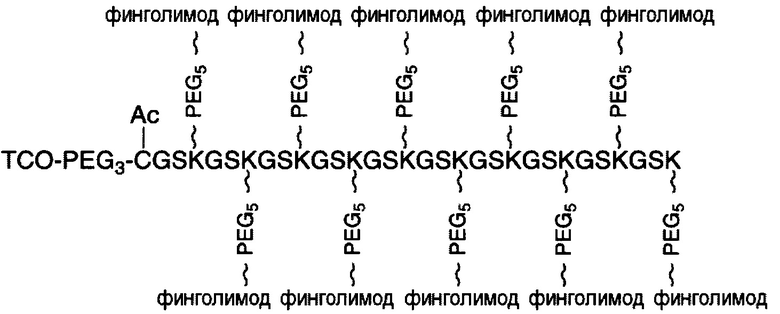
Пример 9. Конъюгирование молекул финголимод-фосфата, конъюгированного с NHS-PEG5, с ТСО-функционализированным пептидом 2.
ТСО-функционализированный пептид 2 и финголимод-фосфат, конъюгированный с NHS-PEG5, смешивали при молярном соотношении 1/42 в 100 мМ фосфатно-солевого буферного раствора при рН 7,5 при комнатной температуре в течение 3 часов. Масс-спектрометрический анализ продемонстрировал, что «связка» лекарственных средств ТСО-функционализированного пептида 2 и финголимод-фосфата имела молекулярную массу 5 379,16 дальтона (фигура 15).
Синтезированная «связка» лекарственных средств, проиллюстрированная ниже, состояла из линкерного звена со свободной функциональной ТСО-группой и набора из пяти молекул финголимод-фосфата в качестве эффекторных элементов.
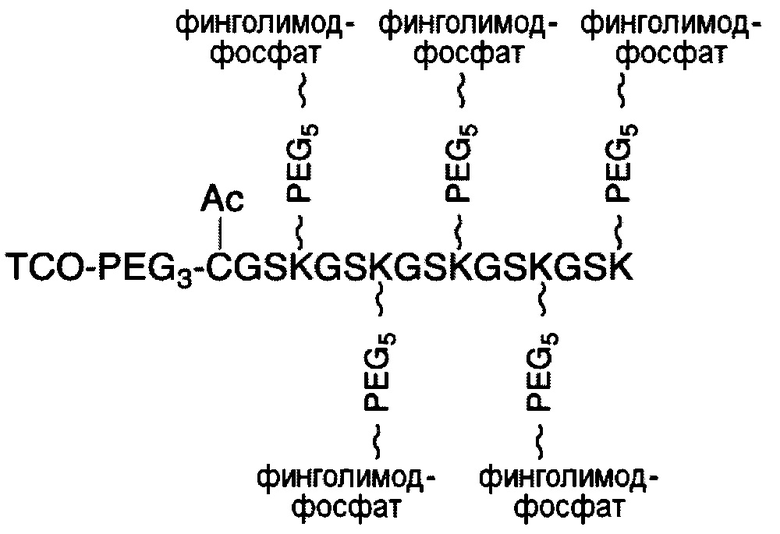
Пример 10: Конъюгирование молекул финголимода, конъюгированного с NHS-PEG4-PEG3-S-S, с ТСО-функционализированным пептидом 2
Пять молекул финголимода, конъюгированного с NHS-PEG4-PEG3-S-S, прикрепляли к ТСО-функционализированному пептиду 2. Конъюгирование молекул финголимода, конъюгированного с NHS-PEG4-PEG3-S-S, с NH2-группами лизиновых остатков ТСО-функционализированного пептида 2 было выполнено аналогично предыдущему примеру. Идентификацию проводили методом масс-спектрометрии MALDI-TOF.
Синтезированная «связка» лекарственных средств, как показано ниже, имела молекулярную массу 7815 дальтон; она состояла из линкерного звена со свободной функциональной ТСО-группой и набора из пяти молекул финголимода.
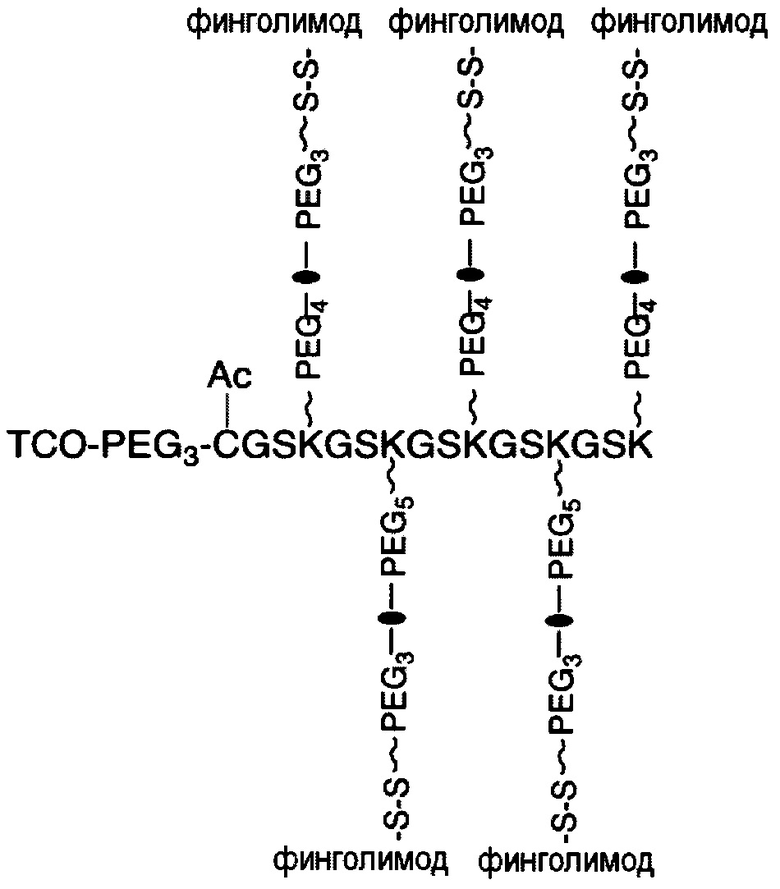
Пример 11: Получение рекомбинантного эктодомена человеческого CD32a с помощью системы сверхэкспрессии HEK293F
Последовательность кодирования генов помещали в экспрессионную кассету pG1K. Аминокислотная последовательность внеклеточной части человеческого CD32a, которая экспрессировалась в виде рекомбинантного белка с гистидиновой меткой, представлена под номером SEQ ID NO: 28. Рекомбинантный эктодомен человеческого CD32a экспрессировали в экспрессирующей системе суспензионной культуры клеток FreeStyle 293F и соответствующей среде (Invitrogen, Carlsbad, USA). Клетки FreeStyle 293F высевали при плотности 1,0×106 жизнеспособных клеток/мл в культуре объемом 600 мл и выдерживали в течение 18-24 часов перед трансфекцией, чтобы гарантировать активное деление клеток во время трансфекции. В процессе трансфекции трансфицировали 1,0×107 клеток в среде объемом 96 мл в колбе Эрленмейера объемом 2 литра с использованием линейного полиэтиленимина со средней молекулярной массой 25 кДа (Polysciences, Warrington, USA) в качестве трансфекционного реагента. Трансфицированные клетки инкубировали при температуре 37°С в течение 4 ч после трансфекции в орбитальном шейкере (125 об/мин), затем их плотность доводили свежей средой до 2,5×106 клеток/мл и инкубировали в течение 4-5 дней. Надосадочную жидкость культуры собирали и белок в среде очищали с использованием аффинной хроматографии на никеле. На фигуре 16 показан анализ SDS-PAGE очищенного белка эктодомена человеческого CD32a.
Пример 12: Получение рекомбинантного эктодомена человеческого рецептора трансферрина-1 (TfR1) с помощью системы сверхэкспрессии HEK293F
Последовательность кодирования генов помещали в экспрессионную кассету pG1K. Аминокислотная последовательность эктодомена человеческого TfR1, которая экспрессировалась в виде рекомбинантного белка с гистидиновой меткой, представлена под номером SEQ ID NO: 29. Рекомбинантный эктодомен человеческого TfR1 экспрессировали в экспрессирующей системе суспензионной культуры клеток FreeStyle 293F и соответствующей среде (Invitrogen, Carlsbad, USA). Клетки FreeStyle 293F высевали при плотности 1,0×106 жизнеспособных клеток/мл в культуре объемом 600 мл и выдерживали в течение 18-24 часов перед трансфекцией, чтобы гарантировать активное деление клеток во время трансфекции. В процессе трансфекции 1,0×107 клеток в 96 мл среды в 2-литровой смесительной колбе Эрленмейера трансфицировали с использованием линейного полиэтиленимина со средней молекулярной массой 25 кДа (Polysciences, Wamngton, USA) в качестве трансфекционного реагента. Трансфицированные клетки инкубировали при температуре 37°С в течение 4 ч после трансфекции в орбитальном шейкере (125 об/мин), затем их плотность доводили свежей средой до 2,5×106 клеток/мл и инкубировали в течение 4-5 дней. Надосадочную жидкость культуры собирали и белок в среде очищали с использованием аффинной хроматографии на никеле. На фигуре 17 показан анализ SDS-PAGE очищенного белка эктодомена человеческого TfR1.
Пример 13: Получение scFv mAb, специфичного к белку F вируса RSV, mAb, специфичного к эндотоксину, и mAb, специфичного к эктодомену CD32a, с помощью системы сверхэкспрессии Expi293F
VL и VH scFv, специфичные к белку F вируса RSV, были получены из моноклонального антитела паливизумаба; VL и VH scFv, специфичные к эндотоксину, были получены из моноклонального антитела WN1 222-5 (патент США 5858728); VL и VH scFv, специфичные к эктодомену CD32a, были получены из MDE-8 (публикация патентной заявки США US 2007/3253958). ScFv, полученный из этих антител, был сконструирован таким образом, чтобы содержать гибкий линкер GGGGSGGGGS и концевой цистеиновый остаток на С-конце. Цистеиновый остаток обеспечивает сульфогидрильную группу для конъюгирования с малеимидной группой, присутствующей на свободных концах связывающих ветвей в различных линкерных звеньях. Для получения scFv mAb, специфичного к белку F вируса RSV, mAb, специфичного к эндотоксину, и mAb, специфичного к внеклеточному компоненту CD32a, использовали последовательности ДНК VL и VH трех антител с дальнейшей оптимизацией кодонов. Были синтезированы последовательности ДНК, кодирующие VL-GSTSGSGKPGSGEGSTKG-VH-(GGGGS)2-C. Аминокислотные последовательности scFv mAb, специфичные к белку F вируса RSV, mAb, специфичные к эндотоксину, и mAb, специфичные к эктодомену CD32a, полученные для экспериментов по изобретению, приведены под номерами SEQ ID NO: 30-32, соответственно.
Для получения белков scFv с использованием системы экспрессии клеток млекопитающих для экспериментов использовалась система сверхэкспрессии, основанная на клеточной линии Expi293F™. В системе использовался набор для трансфекции ExpiFectamine™ 293 (Life Technologies, Carlsbad, USA), состоящий из линии клеток Expi293F™, реагента ExpiFectamine™ 293 на основе катионных липидов и усилителей трансфекции 1 и 2 ExpiFectamine™ 293, а также среды, которая была частью системы экспрессии (Gibco, New York, USA).
Последовательность, кодирующую scFv, помещали в кассету экспрессии pG1K. Клетки Expi293F высевали при плотности 2,0×106 жизнеспособных клеток/мл в среду для экспрессии Expi293F и выдерживали в течение 18-24 часов перед трансфекцией, чтобы убедиться в том, что клетки на момент трансфекции активно делились. В процессе трансфекции 7,5×108 клеток в 255 мл среды в 2-литровой смесительной колбе Эрленмейера трансфицировали с помощью реагента для трансфекции ExpiFectamine™ 293. Трансфицированные клетки инкубировали при 37°С в течение 16-18 часов после трансфекции в орбитальном шейкере (125 об/мин), и к клеткам в смесительную колбу добавляли усилитель трансфекции 1 и усилитель трансфекции 2 ExpiFectamine™ 293, и их инкубировали в течение 5-6 дней. Образцы надосадочной жидкости культуры собирали, и белки scFv в среде очищали с помощью аффинной хроматографии на белке L.
На фигурах 18А и 18В показаны результаты анализов SDS-PAGE и ELISA очищенного scFv mAb, специфичного к белку F вируса RSV; на фигурах 18С и 18D показаны результаты анализов SDS-PAGE и ELISA очищенного scFv mAb, специфичного к эндотоксину; на фигурах 18Е и 18F показаны результаты анализов SDS-PAGE и ELISA очищенного scFv mAb, специфичного к эктодомену CD32a. 96-луночные планшеты для анализа ELISA (Greiner Bio-one) покрывали белком F вируса RSV при концентрации 5 мкг/мл, эндотоксином при концентрации 10 мкг/мл и эктодоменом CD32a при концентрации 5 мкг/мл, соответственно. Очищенные scFv обнаруживали с помощью белка L, конъюгированного с HRP, в соотношении 1:5000.
Результаты анализа ELISA показывают, что каждый очищенный белок scFv специфически связан со своим антигеном (белок F вируса RSV, эндотоксин или эктодомен CD32a), при этом scFv адалимумаба (scFv антитела к TNF-α) использовали в качестве отрицательного контроля.
Пример 14: Получение scFv mAb, специфичного к эктодомену TfR1, и mAb, специфичного к β-амилоиду, с помощью системы сверхэкспрессии Expi293F
VL и VH scFv, специфичные к эктодомену TfR1, были получены из моноклонального антитела OX26; VL и VH scFv, специфичные к β-амилоиду, были получены из моноклонального антитела бапинеузумаба. ScFv, полученный из этих антител, был сконструирован таким образом, чтобы содержать гибкий линкер GGGGSGGGGS и концевой цистеиновый остаток на С-конце. Цистеиновый остаток обеспечивает сульфогидрильную группу для конъюгирования с малеимидной группой, присутствующей на свободных концах связывающих ветвей в различных линкерных звеньях. Для получения scFv mAb, специфичного к эктодомену TfR1, и mAb, специфичного к β-амилоиду, использовали последовательности ДНК VL и VH двух антител с дальнейшей оптимизацией кодонов. Были синтезированы последовательности ДНК, кодирующие VL-GSTSGSGKPGSGEGSTKG-VH-(GGGGS)2-C. Аминокислотные последовательности scFv mAb, специфичные к эктодомену TfR1, и mAb, специфичные к β-амилоиду, полученные для экспериментов по изобретению, приведены под номерами SEQ ID NO: 33 и 34, соответственно.
Для получения белков scFv с помощью систем экспрессии в клетках млекопитающих применяли систему сверхэкспрессии на основе линии клеток Expi293F™. В системе использовался набор для трансфекции ExpiFectamine™ 293 (Life Technologies, Карлсбад, США), состоящий из линии клеток Expi293F™, реагента ExpiFectamine™ 293 на основе катионных липидов и усилителей трансфекции 1 и 2 ExpiFectamine™ 293, а также среды (Gibco, Нью-Йорк, США).
Последовательность, кодирующую scFv, помещали в кассету экспрессии pG1K. Клетки Expi293F высевали при плотности 2,0×106 жизнеспособных клеток/мл в среду для экспрессии Expi293F и выдерживали в течение 18-24 часов перед трансфекцией, чтобы убедиться в том, что клетки на момент трансфекции активно делились. В процессе трансфекции 7,5×108 клеток в 255 мл среды в 2-литровой смесительной колбе Эрленмейера трансфицировали с помощью реагента для трансфекции ExpiFectamine™ 293. Трансфицированные клетки инкубировали при 37°С в течение 16-18 часов после трансфекции в орбитальном шейкере (125 об/мин), и к клеткам в смесительную колбу добавляли усилитель трансфекции 1 и усилитель трансфекции 2 ExpiFectamine™ 293, и их инкубировали в течение 5-6 дней. Образцы надосадочной жидкости культуры собирали, и белки scFv в среде очищали с помощью аффинной хроматографии на белке L. На фигурах 19А и 19В показаны анализы очищенных scFv для mAb, специфичных к эктодомену TfR1, по методам SDS-PAGE и ELISA, соответственно. На фигурах 19С и 19D показаны анализы очищенного scFv для mAb, специфичных к β-амилоиду, по методам SDS-PAGE и ELISA, соответственно. Планшеты для анализа ELISA покрывали эктодоменом TfR1 при концентрации 5 мкг/мл и β-амилоидом при концентрации 5 мкг/мл, соответственно. Очищенные scFv обнаруживали с помощью белка L, конъюгированного с HRP, в соотношении 1:5000.
Результаты анализа ELISA показывают, что каждый очищенный scFv-белок специфически связан со своим антигеном (эктодоменом TfR1 или β-амилоидом), при этом только белок L, конъюгированный с HRP, использовали в качестве отрицательного контроля.
Пример 15: Конструирование и селекция scFv на фаговых носителях, специфичных к эктодомену человеческого CD32a
Клоны фагов, несущие scFv, специфичные к эктодомену человеческого CD32a, были получены по контракту с лабораторией доктора An-Suei Yang (Genomics Research Center, Academia Sinica, Taipei, Taiwan). Рамочная последовательность библиотеки scFv GH2 была получена из антител G6 к VEGF Fab (код в белковом банке 2FJG) и клонирована в сайты рестрикции SfiI и NotI вектора фагмида pGANTAB5E (GE Healthcare), несущие устойчивость к ампициллину, промотор lacZ, лидерную последовательность ре1В для секреции scFv-фрагментов в надосадочную жидкость культуры, Е-метку, применимую для обнаружения. Домены VH и VL scFv-матрицы были диверсифицированы отдельно на основе процедуры олигонуклеотид-направленного мутагенеза; три CDR в каждом из вариабельных доменов были диверсифицированы одновременно. Использовали библиотеку scFv, содержащую более 109 клонов для селекции по эктодомену CD32a.
96-луночные планшеты Maxisorp (Nunc), покрытые рекомбинантньши белками CD32a (1 мкг/100 мкл PBS на лунку), использовали для пэннинга антител к CD32a. Вкратце, лунки покрывали человеческим CD32a, встряхивая покрывающий раствор в лунках в течение 2 часов при комнатной температуре. Затем лунки, покрытые CD32a, обрабатывали блокирующим буфером (5% снятого молока в PBST (фосфатно-солевой буферный раствор с 0,1% твин-20)) в течение 1 часа при комнатной температуре. Рекомбинантные фаги в блокирующем буфере, разведенные до 8×1011 КОЕ/мл, добавляли в лунки, покрытые CD32a, в течение 1 часа с осторожным встряхиванием; КОЕ обозначает колониеобразующую единицу. Затем лунки энергично промывали 10 раз PBST, а затем 6 раз PBS для удаления неспецифических связывающих фагов. Связанные фаги элюировали, используя 0,1 М HCl/глицинового буфера при рН 2,2, и элюированный раствор немедленно нейтрализовали 2 М трис-базовым буфером при рН 9,0. Для фаговой инфекции использовали штамм Е. coli ER2738 (OD600 = ~0,6) при температуре 37°С в течение 30 минут; неинфицированные бактерии Е. coli удаляли путем обработки ампициллином в течение 30 минут. После обработки ампициллином добавляли вспомогательный фаг M13KO7, несущий устойчивость к канамицину, с еще одной инкубацией в течение одного часа. Отобранные фаги, полученные с помощью вспомогательного фага в культуре Е. coli, амплифицировали, энергично встряхивая в течение ночи при температуре 37°С в присутствии канамицина. Амплифицированные фаги осаждали в PEG/NaCl и затем ресуспендировали в PBS для следующих циклов селекции/амплификации. В общей сложности на эктодомене CD32a выполняли три последовательных раунда пэннинга, повторяя эту процедуру селекции/амплификации.
Колонии ER2738, инфицированные фагами, были подсчитаны на планшетах с сериями разведении и рассчитаны титры фагов с получением выходного титра/мл (КОЕ/мл) за один раунд пэннинга. После трех раундов пэннинга было получено 1000-кратное увеличение выходного титра фагов от 1,6Е+04 КОЕ/лунка до 2,2Е+07 КОЕ/лунка. Коэффициенты отношения выходного/входного титра фагов в результате каждого раунда показаны на фигуре 20А. Для каждого раунда пэннинга коэффициенты отношения выходного/входного титра фагов показаны по оси у. Наблюдалось четкое обогащение положительных клонов в течение трех раундов пэннинга. Третий раунд пэннинга привел к 100-кратному соотношению выходного/входного титра фагов, полученного в первом раунде, поскольку связывающие клоны стали доминирующей популяцией в библиотеке.
В типичной процедуре отбора после трех раундов антиген-пэннинга на лунках с покрытием человеческого CD32a на планшетах для анализа ELISA приблизительно 80% связанных фаговых частиц были связаны с CD32a, особенно на планшетах для анализа ELISA, покрытых CD32a.
Пример 16: Анализ ELISA с одной колонией scFv на фаговых носителях, специфичных к эктодомену человеческого CD32a
Штамм бактерии Е. coli ER2738, инфицированный одноклональными фагами, каждый из которых в своем фагмиде содержал выбранный ген scFv, выращивали в середине логарифмического роста в бульоне 2YT (16 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л NaCl, pH 7,0) с ампициллином 100 мкг/мл в глубокой лунке при температуре 37°С, сопровождая встряхиванием. После того как бульон достигал OD600 1,0, добавляли ИПТГ до конечной концентрации 1 мкг/мл. Планшеты инкубировали при температуре 37°С в течение ночи при энергичном встряхивании; после этого планшеты центрифугировали при 4000 g в течение 15 минут при температуре 4°С.
Для проверки связывания растворимого scFv проводили анализ ELISA. Вкратце, 96-луночный планшет Maxisorp (Nunc) покрывали эктодоменом CD32a (0,5 мкг/100 мкл PBS на лунку) или человеческим рецептором трансферрина-1 антигена для отрицательного контроля в течение 18 часов со встряхиванием при температуре 4°С. После обработки 300 мкл блокирующего буфера в течение 1 часа 100 мкл секретируемого scFv в надосадочной жидкости смешивали с 100 мкл блокирующего буфера и затем добавляли на покрытый планшет еще на 1 час. В течение 1 часа на планшет добавляли антитело козы к Е-метке (конъюгированное с HRP, 1:4000, кат. № АВ19400, Abcam). ТМБ субстрат (50 мкл на лунку) добавляли в лунки и измеряли поглощение на 450 нм после прекращения реакции добавлением 1N HCl (50 мкл на лунку).
В общей сложности 192 фаговых клона после третьего раунда пэннинга было подвергнуто настоящему анализу. Среди них 12 scFv-клонов, которые связывались с CD32a с дифференциалом OD450, превышающим 10, дополнительно характеризовались секвенирующими генами, кодирующими эти scFv. Было идентифицировано шесть различных последовательностей ДНК. На фигуре 20В показан результат анализа ELISA scFv-клона 22D1. Аминокислотная последовательность scFV-клона 22D1, которая связывается с человеческим CD32a при OD450 0,8, приведена под номером SEQ ID NO: 35.
Пример 17: Конструирование и селекция scFv на фаговых носителях, специфичных к эктодомену человеческого TfR1
Клоны фагов, несущие scFv, специфичные к эктодомену человеческого TfR1, были получены по контракту с лабораторией доктора An-Suei Yang (Genomics Research Center, Academia Sinica, Taipei, Taiwan). Рамочная последовательность библиотеки scFv GH2 была получена из антител G6 к VEGF Fab (код в белковом банке 2FJG) и клонирована в сайты рестрикции SfiI и NotI вектора фагмида pGANTAB5E (GE Healthcare), несущие устойчивость к ампициллину, промотор lacZ, лидерную последовательность pelB для секреции scFv-фрагментов в надосадочную жидкость культуры, Е-метку, применимую для обнаружения. Домены VH и VL scFv-матрицы были диверсифицированы отдельно на основе процедуры олигонуклеотид-направленного мутагенеза; три CDR в каждом из вариабельных доменов были диверсифицированы одновременно. Использовали библиотеку scFv, содержащую более 109 клонов для селекции по эктодомену CD32a.
96-луночные планшеты Maxisorp (Nunc), покрытые рекомбинантными белками TfR1 (1 мкг/100 мкл PBS на лунку), использовали для пэннинга антител к TfR1. Вкратце, лунки покрывали человеческим TfR1, встряхивая покрывающий раствор в лунках в течение 2 часов при комнатной температуре. Затем лунки, покрытые TfR1, обрабатывали блокирующим буфером (5% снятого молока в PBST (фосфатно-солевой буферный раствор с 0,1% твин-20)) в течение 1 часа при комнатной температуре. Рекомбинантные фаги в блокирующем буфере, разведенные до 8×1011 КОЕ/мл, добавляли в лунки, покрытые TfR1, в течение 1 часа с осторожным встряхиванием; КОЕ обозначает колониеобразующую единицу. Затем лунки энергично промывали 10 раз PBST, а затем 6 раз PBS для удаления неспецифических связывающих фагов. Связанные фаги элюировали, используя 0,1 М HCl/глицинового буфера при рН 2,2, и элюированный раствор немедленно нейтрализовали 2 М трис-базовым буфером при рН 9,0. Для фаговой инфекции использовали штамм Е. coli ER2738 (OD600 = ~0,6) при температуре 37°С в течение 30 минут; неинфицированные бактерии Е. coli удаляли путем обработки ампициллином в течение 30 минут. После обработки ампициллином добавляли вспомогательный фаг M13KO7, несущий устойчивость к канамицину, с еще одной инкубацией в течение одного часа. Отобранные фаги, полученные с помощью вспомогательного фага в культуре E. coli, амплифицировали, энергично встряхивая в течение ночи при температуре 37°С в присутствии канамицина. Амплифицированные фаги осаждали в PEG/NaCl и затем ресуспендировали в PBS для следующих циклов селекции/амплификации. В общей сложности на эктодомене TfR1 выполняли три последовательных раунда пэннинга, повторяя эту процедуру селекции/амплификации.
Колонии ER2738, инфицированные фагами, были подсчитаны на планшетах с сериями разведении и рассчитаны титры фагов с получением выходного титра/мл (КОЕ/мл) за один раунд пэннинга. После трех раундов пэннинга было получено 104-кратное увеличение выходного титра фагов от 3,74Е+03 КОЕ/лунка до 1,5Е+08 КОЕ/лунка. Коэффициенты соотношения выходного/входного титра фагов в результате каждого раунда показаны на фигуре 21А. Для каждого раунда пэннинга коэффициенты отношения выходного/входного титра фагов показаны по оси у. Наблюдалось четкое обогащение положительных клонов в течение трех раундов пэннинга. Третий раунд пэннинга привел к 104-кратному соотношению выходного/входного титра фагов, полученного в первом раунде, поскольку связывающие клоны стали доминирующей популяцией в библиотеке.
В типичной процедуре отбора после трех раундов антиген-пэннинга на лунках с покрытием человеческим TfR1 на планшетах для анализа ELISA приблизительно 80% связанных фаговых частиц были связаны с TfR1, особенно на планшетах для анализа ELISA, покрытых TfR1.
Пример 18: Анализ ELISA с одной колонией scFv на фаговых носителях, специфичных к эктодомену человеческого TfR1
Штамм бактерии Е. coli ER2738, инфицированный одноклональными фагами, каждый из которых в своем фагмиде содержал выбранный ген scFv, выращивали в середине логарифмического роста в бульоне 2YT (16 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л NaCl, pH 7,0) с ампициллином 100 мкг/мл в глубокой лунке при температуре 37°С, сопровождая встряхиванием. После того как бульон достигал OD600 1,0, добавляли ИПТГ до конечной концентрации 1 мкг/мл. Планшеты инкубировали при температуре 37°С в течение ночи при энергичном встряхивании; после этого планшеты центрифугировали при 4000 g в течение 15 минут при температуре 4°С.
Для проверки связывания растворимого scFv проводили анализ ELISA. Вкратце, 96-луночный планшет Maxisorp (Nunc) покрывали эктодоменом TfR1 (0,5 мкг/100 мкл PBS на лунку) или антигена CD16b для отрицательного контроля в течение 18 часов со встряхиванием при температуре 4°С. После обработки 300 мкл блокирующего буфера в течение 1 часа 100 мкл секретируемого scFv в надосадочной жидкости смешивали с 100 мкл блокирующего буфера и затем добавляли на покрытый планшет еще на 1 час. В течение 1 часа на планшет добавляли антитело козы к Е-метке (конъюгированное с HRP, 1:4000, кат. № АВ19400, Abcam). ТМБ субстрат (50 мкл на лунку) добавляли в лунки и измеряли поглощение на 450 нм после прекращения реакции добавлением 1N HCl (50 мкл на лунку).
В общей сложности 192 фаговых клона после третьего раунда пэннинга было подвергнуто этому анализу. Среди них 12 scFv-клонов, которые связывались с TfR1 с дифференциалом OD450, превышающим 10, дополнительно характеризовались секвенирующими генами, кодирующими эти scFv. Было идентифицировано шесть различных последовательностей ДНК. На фигуре 21В показан результат анализа ELISA scFv-клона 12А1. Аминокислотная последовательность scFV-клона 12А1, которая связывается с человеческим TfR1 при OD450 1,7, приведена под номером SEQ ID NO: 36.
Пример 19. Получение TCO-scFv, специфичных к эктодомену CD32a
Последовательность ДНК, кодирующую SEQ ID NO: 32, синтезировали и экспрессировали в соответствии с вышеприведенными примерами. Для конъюгирования с Mal-PEG3-TCO (Conju-Probe, Inc.) цистеиновый остаток на С-конце очищенного scFv aHTH-CD32a mAb восстанавливали путем инкубирования с 5 мМ дитиотреитола (DTT) при комнатной температуре в течение 4 часов при осторожном встряхивании. Буфер, в котором находились восстановленные белки scFv, заменяли на натрий-фосфатный буфер (100 мМ фосфат натрия, рН 7,0, 50 мМ NaCl и 5 мМ EDTA) путем применения колонки NAP-10 с Sephadex G-25. После реакции восстановления и замены буфера проводили конъюгирование в течение ночи при комнатной температуре в реакционном молярном соотношении 10:1 ([Mal-PEG3-TCO]:[scFv]). Избыток сшивающего средства удаляли с помощью колонки для обессоливания, и анализировали scFv-продукты, конъюгированные с ТСО.
Результаты анализа методом масс-спектроскопии MALDI-TOF указывали на то, что образец scFv, конъюгированного с ТСО, специфичного к CD32a, имел молекулярную массу 27337 дальтон. Чистоту scFv, конъюгированных с ТСО, специфичных к CD32a, определяли посредством окрашивания кумасси синим 12% геля для SDS-PAGE. На фигуре 22А и фигуре 22В показаны анализы scFv, конъюгированного с ТСО, специфичного к CD32a, методом ELISA и мас-спектрометрическим методом, соответственно, в котором немодифицированные scFv антитела, специфичные к CD32a, применяли в качестве положительного контроля. В соответствии с результатами ELISA scFv, конъюгированный с ТСО, специфичный к CD32a, связывался с рекомбинантным эктодоменом человеческого CD32a.
Пример 20: Получение тетразин-scFv, специфичного к эктодомену TfR1
Последовательность ДНК под номером SEQ ID NO: 33 была синтезирована и экспрессирована, как в приведенных выше примерах. Для конъюгирования с Mal-PEG4-тетразином (Conju-probe, Inc.) цистеиновый остаток на С-конце очищенного scFv mAb, специфичного к TfR1, восстанавливали путем инкубации с 5 мМ DTT при комнатной температуре в течение 4 часов при легком встряхивании. Буфер восстановленных белков scFv заменяли на натрий-фосфатный буфер (100 мМ натрия фосфата, рН 7,0, 50 мМ NaCl и 5 мМ EDTA) с использованием колонки NAP-10 Sephadex G-25. После реакции восстановления и буферного обмена конъюгирование проводили в течение ночи при температуре 4°С при молярном соотношении реакции 10:1 ([Mal-PEG4-тетразин]:[scFv]). Избыток сшивающего средства удаляли с помощью обессоливающей колонки и анализировали продукт scFv, конъюгированный с тетразином.
Результаты анализа методом масс-спектроскопии MALDI-TOF показали, что образец scFv, конъюгированного с тетразином, специфичного к TfR1, имел молекулярную массу 27086 дальтон. Чистоту scFv, конъюгированного с тетразином, специфичного к TfR1, идентифицировали с помощью анализа в 10% геле для SDS-PAGE методом окрашивания кумасси синим. На фигуре 23А и фигуре 23В показаны, соответственно, результаты анализа ELISA и масс-спектрометрии scFv, конъюгированного с тетразином, специфичного к TfR1, в котором немодифицированный scFv, специфичный к TfR1, использовался в качестве положительного контроля. Согласно результатам анализа ELISA, scFv, конъюгированный с тетразином, специфичный к TfR1, связался с рекомбинантным эктодоменом TfR1.
Пример 21. Конъюгирование трех scFv, специфичных к эндотоксину, с тремя малеимидно-PEG12 связывающими ветвями на основе из тетразин-функционализированного пептида 1
Этот пример демонстрирует, что три scFv можно конъюгировать с тремя РЕП12-малеимидными связывающими ветвями, размещенными на основе из тетразин-функционализированного пептида 1. Перед конъюгированием с тетразин-функционализированным пептидом 1, имеющим три PEG12-малеимидные связывающие ветви, scFv, специфичный к эндотоксину, инкубировали с DTT в молярном соотношении 2:1 ([DTT]:[scFv]) при 25°С в течение 4 часов при осторожном встряхивании для сохранения его С-концевого цистеина в восстановленной форме. После этого буфер, в котором находился восстановленный scFv, специфичный к эндотоксину, заменяли на буфер для реакции присоединения между малеимидной группой и SH-группой (100 мМ натрия фосфата, рН 7,0, 50 мМ NaCl и 5 мМ EDTA) путем применения колонки NAP-10 Sephadex G-25 (GE Healthcare). После восстановления и замены буфера проводили конъюгирование с тетразин-функционализированным пептидом 1, имеющим три малеимидно-PEG12 связывающих ветви, в течение ночи при 4°С в молярном соотношении 1:4 ([линкер]: [белок]).
Тетразин-функционализированный пептид 1, конъюгированный с PEG12-малеимидом и конъюгированный с тремя scFv, специфичными к эндотоксину, отделяли от свободного scFv, свободного тетразин-функционализированного пептида 1, конъюгированного с PEG12-малеимидом, и тетразин-функционализированного пептида 1, конъюгированного с одним и двумя scFv, специфичными к эндотоксину, с помощью колонки для эксклюзионной хроматографии S75.
Фигура 24А представляет собой профиль элюирования В процессе FPLC синтезированного нацеливающего линкерного звена, состоящего из линкерного звена со свободной тетразиновой функциональной группой и набора из трех scFv, специфичных к человеческому эндотоксину, в качестве нацеливающих элементов, с объемом удержания 9,5 мл. Продукт (т.е. тетразин-функционализированный пептид 1, конъюгированный с PEG12-малеимидом, имеющий свободную тетразиновую функциональную группу и конъюгированный с набором из трех scFv, специфичных к эндотоксину) был очищен в виде элюируемой фракции и показан на дорожке 4 (обозначено стрелкой) в анализе в 10% геле для SDS-PAGE, показанном на фигуре 24В.
Пример 22. Анализ нацеливающего линкерного звена, содержащего три scFv, специфичных к эндотоксину, связанных с тремя малеимидно-PEG12 связывающими ветвями, размещенными на основе из тетразин-функционализированного пептида 1, с помощью MALDI-TOF
Состав образца нацеливающего линкерного звена с тремя scFv, специфичными к эндотоксину, связанных с тремя малеимидно-PEG12 связывающими ветвями, размещенными на основе из тетразин-функционализированного пептида 1, анализировали с помощью MALDI-TOF. Экспериментальное медианное значение молекулярного веса согласовывалось с теоретическим медианным значением молекулярного веса трех scFv специфичных к эндотоксину, конъюгированных с тетразин-функционализированным пептидом 1 с тремя малеимидно-PEG12 связывающими ветвями. В соответствии с масс-спектрометрическим профилем на фигуре 24С нацеливающее линкерное звено по настоящему изобретению имело медианный молекулярную массу, составляющую 81727 дальтон.
Ниже проиллюстрировано нацеливающее линкерное звено по настоящему изобретению, состоящее из линкерного звена со свободной тетразиновой функциональной группой и набора из трех scFv, специфичных к эндотоксину, в качестве нацеливающих элементов.
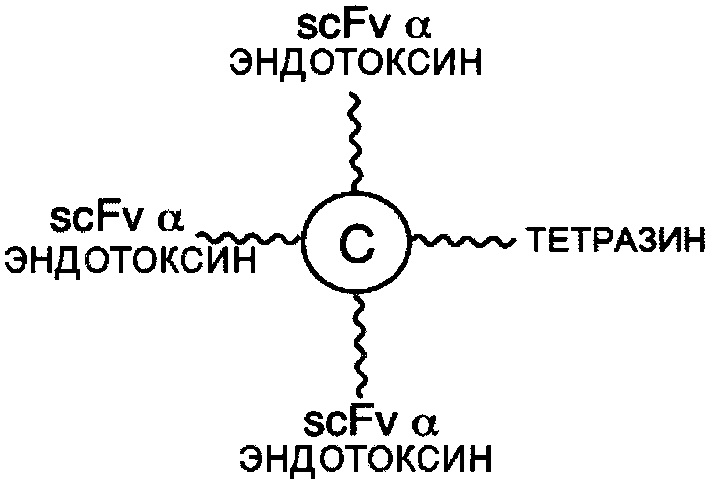
Пример 23: Получение нацеливающего линкерного звена на основе тетразин-функционализированного пептида 1 с тремя scFv, специфичными к белку F вируса RSV
Процедура конъюгирования scFv с линкерным звеном, а также очистка и анализ продукта была такой же, как в предыдущих примерах.
На фигуре 25 показан масс-спектрометрический анализ синтезированного нацеливающего линкерного звена, состоящего из линкерного звена со свободной тетразиновой функциональной группой и набора из трех scFv, специфичных к белку F вируса RSV, в качестве нацеливающих элементов (см. ниже). Как показано на фигуре 25, это нацеливающее линкерное звено имело молекулярную массу 81978 дальтон.
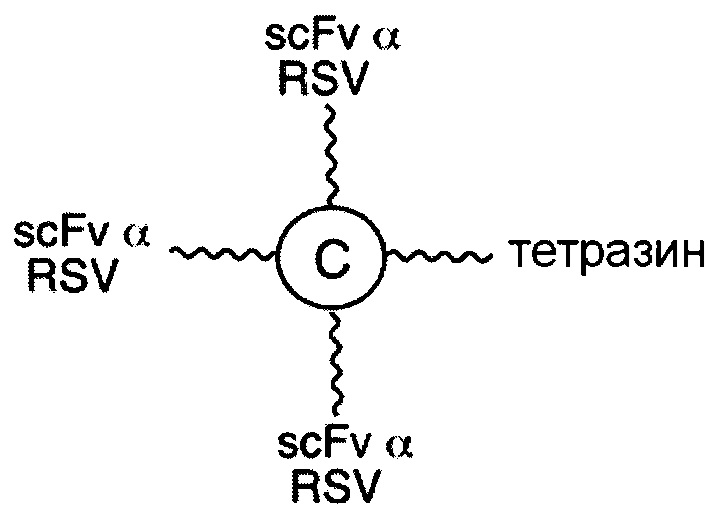
Пример 24. Коньюгирование трех scFv, специфичных к β-амилоиду, с тремя малеимидно-PEG12-связывающими ветвями на основе ТСО-функционализированного пептида 1
Этот пример был выполнен для демонстрации того, что три scFv могут быть конъюгированы с тремя малеимидно-PEG12-связывающими ветвями на основе ТСО-функционализированного пептида 1. До конъюгирования с ТСО-функционализированный пептидом 1, который имел три малеимидно-PEG12-связывающих ветви, scFv, специфичный к β-амилоиду, инкубировали с DTT при молярном соотношении 2:1 ([DTT]:[scFv]) при комнатной температуре в течение 4 часов с легким встряхиванием для сохранения С-концевого цистеина в восстановленной форме. Затем буфер восстановленного scFv, специфичного к β-амилоиду, заменяли на буфер реакции связывания малеимид-SH (100 мМ натрия фосфата, рН 7,0, 50 мМ NaCl и 5 мМ EDTA) с использованием колонки NAP-10 Sephadex G-25 (GE Healthcare). После восстановления и буферного обмена конъюгирование с ТСО-функционализированным пептидом 1, имеющим три малеимидно-PEG12-связывающих ветви, проводили в течение ночи при комнатной температуре при молярным соотношении 1:4 ([линкер]:[белок]).
Реакционную смесь из предыдущих примеров доводили до рН 5,0 и затем вносили в предварительно уравновешенную (5 мМ EDTA и 50 мМ натрия ацетата при рН 5,0) ионообменную колонку SP Sepharose FF (GE Healthcare). ТСО-функционализированный пептид 1, конъюгированный с малеимид-PEG12, конъюгированный с тремя scFv, специфичными к β-амилоиду, элюировали с линейным градиентом 0-500 мМ натрия хлорида со скоростью потока 0,5 мл/мин в течение 100 минут. ТСО-функционализированный пептид 1, конъюгированный с малеимид-PEG12, конъюгированный с тремя scFv, специфичными к β-амилоиду, отделяли от свободного scFv, свободного ТСО-функционализированного пептида 1, конъюгированного с малеимид-PEG12, и ТСО-функционализированного пептида 1, конъюгированного с малеимид-PEG12, конъюгированного с одним и двумя scFv, специфичными к β-амилоиду, с помощью ионообменной колонки SP Sepharose FF. Очищенный продукт, ТСО-функционализированный пептид 1, конъюгированный с малеимид-PEG12, конъюгированный с тремя scFv, специфичными к β-амилоиду, концентрировали и проводили замену буфера на буфер клик-реакции, 100 мМ калия фосфата при рН 7,0.
На фигуре 26А представлен профиль элюирования ионообменной колонки SP Sepharose FF при FPLC на синтезированном эффекторном линкерном звене, состоящем из линкерного звена со свободной функциональной ТСО-группой и набора из трех scFv, специфичных к β-амилоиду, в качестве эффекторных элементов. Символы #1 и #2, соответственно, указывают на пики элюирования ТСО-функционализированного пептида 1, конъюгированного с малеимидом-PEG12, конъюгированного с двумя scFv и тремя scFv, специфичными к β-амилоиду. Продукт, ТСО-функционализированный пептид 1, конъюгированный с малеимидом-PEG12, несущий свободную функциональную ТСО-группу и три scFv, специфичных к β-амилоиду, очищали и выявляли на дорожке 2 анализа SDS-PAGE в 8% геле, показанного на фигуре 26В.
Пример 25: Анализ эффекторного линкерного звена, содержащего три scFv, специфичных к β-амилоиду, связанных с тремя малеимидно-PEG12-связывающими ветвями на основе ТСО-функционализированного пептида 1 методом масс-спектрометрии MALDI-TOF
Состав образца эффекторного линкерного звена из трех scFv, специфичных к β-амилоиду, связанного с тремя малеимидно-PEG12-связывающими ветвями на основе ТСО-функционализированного пептида 1, анализировали методом масс-спектрометрии MALDI-TOF. Медиана экспериментальной молекулярной массы согласуется с медианой теоретической молекулярной массы трех scFv, специфичных к β-амилоиду, конъюгированных с ТСО-функционализированным пептидом 1, с тремя малеимидно-PEG12-связывающими ветвями. В соответствии с масс-спектрометрическим профилем на фигуре 26С, синтезированное эффекторное линкерное звено имело среднюю молекулярную массу 87160 дальтон.
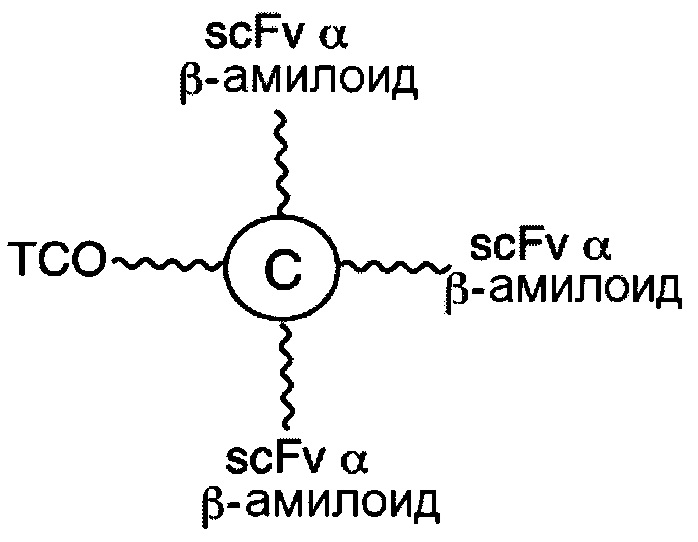
Здесь представлено синтезированное эффекторное линкерное звено, состоящее из линкерного звена со свободной функциональной ТСО-группой и набора из трех scFv, специфичных к β-амилоиду, в качестве нацеливающих элементов.
Пример 26: Получение молекулярной конструкции с тремя scFv, специфичными к белку F вируса RSV, в качестве нацеливающих элементов, и одним scFv, специфичным к эктодомену CD32a, в качестве эффекторного элемента
В этом примере нацеливающее линкерное звено из предыдущих примеров и TCO-scFv, специфичный к эктодомену CD32a, соединяли посредством реакции iEDDA между тетразиновой группой и ТСО-группой. В частности, нацеливающее линкерное звено имело три scFv, специфичных к белку F вируса RSV, и одну свободную тетразиновую группу.
Процедуру дотирования тетразиновой группы и ТСО-группы осуществляли в соответствии с инструкциями производителя (Jena Bioscience GmbH, Йена, Германия). Коротко говоря, к раствору, содержащему эффекторный элемент, добавляли 100 мкл нацеливающего линкерного звена (0,3 мг/мл) в молярном соотношении 1:1,2 ([тетразин]:[ТСО]). Реакционную смесь инкубировали в течение одного часа при комнатной температуре. Продукт подвергали масс-спектрометрическому анализу и результат указывал на молекулярную массу, составляющую 113036 дальтон (фигура 27А).
Продукт в виде молекулярной конструкции из одного линкерного звена с тремя scFv, специфичными к к белку F вируса RSV, в качестве нацеливающих элементов и одним scFv, специфичным к эктодомену CD32a, в качестве эффекторного элемента проиллюстрирован ниже.
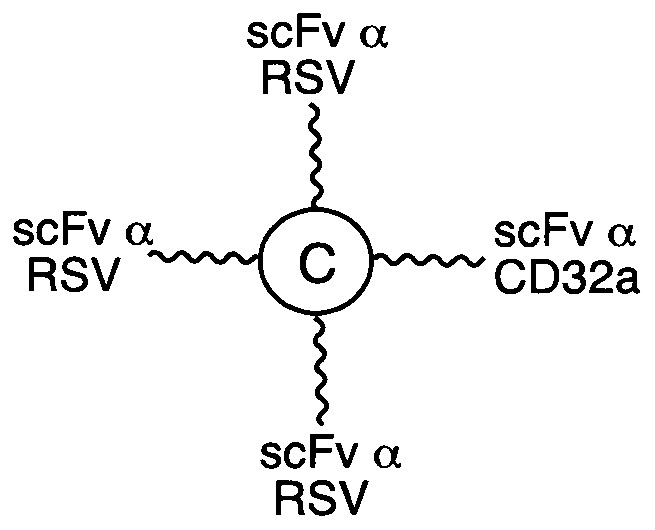
Пример 27. Получение молекулярной конструкции с тремя scFv, специфичными к эндотоксину, в качестве нацеливающих элементов и одним scFv, специфичным к эктодомену CD32a, в качестве эффекторного элемента
Нацеливающее линкерное звено, полученное в более раннем примере, и TCO-scFv, специфичный к эктодомену CD32a, соединяли посредством реакции iEDDA между тетразиновой группой и ТСО-группой.
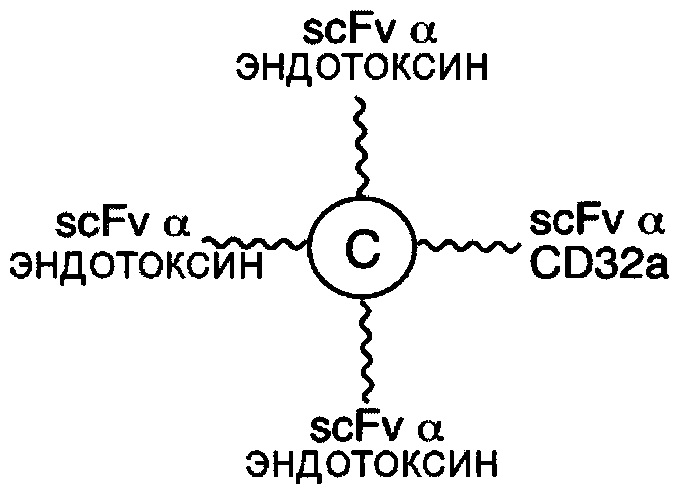
Пример 28: Получение молекулярной конструкции с одним scFv, специфичным к эктодомену TfR1, в качестве нацеливающего элемента и трех scFv, специфичных к β-амилоиду, в качестве эффекторных элементов
Нацеливающее линкерное звено, полученное в примере выше, и тетразин-функционализированный scFv, специфичный к эктодомену TfR1, связывали посредством реакции iEDDA между тетразиновой группой и ТСО-группой.
Процедуру лигирования тетразиновой группы и ТСО-группы осуществляли согласно инструкциям изготовителя (Jena Bioscience GmbH, Jena, Germany). Вкратце, 12,6 мкл нацеливающего элемента (5,65 мг/мл) добавляли к раствору, содержащему линкерное звено с эффекторными элементами при молярном соотношении 10:1 ([тетразин]:[ТСО]). Реакционную смесь инкубировали в течение 3 часов при комнатной температуре. Продукт подвергали масс-спектрометрическому анализу, результат которого показал молекулярную массу 114248 дальтон (фигура 27С).

Продукт, как проиллюстрировано здесь, представлял собой молекулярную конструкцию с одним линкерным звеном, содержащим один scFv, специфичный к эктодомену TfR1, в качестве нацеливающего элемента, и три scFv, специфичные к β-амилоиду, в качестве эффекторных элементов.
Пример 29: Получение молекулярной конструкции с одним scFv, специфичным к эктодомену TfR1, в качестве нацеливающего элемента и пяти молекул финголимода в качестве эффекторных элементов
В этом примере была построена молекулярная конструкция с одним scFv, специфичным к эктодомену TfR1, и «связкой» лекарственных средств из пяти молекул финголимода. Молекулярную конструкцию создавали посредством реакции iEDDA между тетразиновой группой и ТСО-группой. Процедуру лигирования тетразиновой группы и ТСО-группы осуществляли в соответствии с инструкциями производителя (Jena Bioscience GmbH, Йена, Германия). Вкратце, 277 мкл эффекторного линкерного звена (0,126 мкмоль) добавляли к раствору, содержащему нацеливающий элемент из одного scFv, специфичного к эктодомену TfR1, при молярном отношении 5:1 ([ТСО]:[тетразин]). Реакционную смесь инкубировали в течение 3 часов при комнатной температуре.
Продукт, как проиллюстрировано ниже, представлял собой молекулярную конструкцию с одним scFv, специфичным к эктодомену TfR1, и одной «связкой» лекарственных средств, несущей пять молекул финголимода. На фигуре 28А и фигуре 28В показаны, соответственно, результаты анализов SDS-PAGE и масс-спектрометрии по настоящему изобретению. Основная полоса, стрелка #1, указывает на молекулярную конструкцию с одним scFv, специфичным к эктодомену TfR1, и «связкой» лекарственных средств из пяти молекул финголимода, а стрелка #2 указывает на неконъюгированный scFv, специфичный к эктодомену TfR1.
Масс-спектрометрический анализ показал, что молекулярная конструкция с одним scFv, специфичным к эктодомену TfR1, и «связкой» лекарственных средств из пяти молекул финголимода имела молекулярную массу 32105 дальтон.
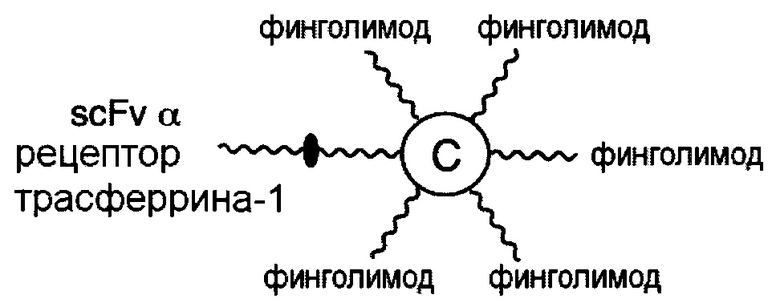
Пример 30: Анализ биологической активности финголимода при коньюгировании с пептидной сердцевиной через связывающие ветви
Модифицированные молекулы финголимода (финголимод, конъюгированный с NHC-PEG5, и «связка» лекарственных средств со свободной функциональной ТСО-группой и пятью молекулами финголимода) были синтезированы, как описано в предыдущих примерах. Для исследования биологической активности этих трех соединений выполняли анализ миграции под влиянием S1P в системе Трансвелл с первичными В-клетками, выделенными из РВМС (мононуклеары периферической крови) человека.
При получении первичных В-клеток человека В-клетки человека были выделены из РВМС (мононуклеары периферической крови) человека с помощью набора для изоляции В-клеток (Myttenyi Biotech). Затем выделенные В-клетки высевали и выдерживали в 15-см чашке в среде IMDM, дополненной 10% фетальной бычьей сывороткой (Gibco) и 20 нг/мл IL2 (Peprotech Inc.).
На фигуре 29А показан анализ выделенных S1P1-экспрессирующих человеческих В-клеток, 2×105 В-клеток инкубировали с 10 мкг/мл антитела рецептора S1P1 (AbD Serotec) в PBS, содержащем 1% BSA, на льду в течение 30 минут. Клетки промывали и инкубировали с козьими антимышиными антителами IgG, конъюгированными с FITC, разведенными 1:200 в PBS/BSA, на льду в течение 30 минут в темноте. Затем клетки анализировали методом FACS (FACSCanto II, BD Biosciences).
Для хемотаксического анализа 100 мкл исследуемых В-клеток человека (4×105 клеток) переносили в 1,5-мл пробирку Эппендорфа и добавляли финголимод, финголимод-фосфат, финголимод, конъюгированный с NHS-PEG5, и «связку» лекарственных средств со свободной функциональной ТСО-группой и пятью молекулами финголимода, соответственно, при конечной концентрации 1 и 10 мкМ при температуре 37°С в течение 4 часов. Затем в верхнюю 6,5-мм камеру системы Трансвелл добавляли 100 мкл обработанных В-клеток с 5-микронной пористой полиэфирной мембранной вставкой (Corning), а нижняя камера системы Трансвелл содержала 500 мкл среды IMDM с молекулами сфингозин-1-фосфата при конечной концентрации 10 нМ. Через 3 часа мигрировавшие клетки в нижних камерах собирали и затем окрашивали трипановым синим и подсчитывали с помощью гемоцитометра. Для каждого измерения удельная миграция рассчитывалась следующим образом: [(Количество клеток в нижней камере)/(количество клеток в нижней + верхней камере)×100] - (процентная доля миграции клеток при 0 нМ аттрактанте)]. Результат в виде процентной доли конкретных мигрировавших клеток показан на фигуре 29В.
На фигуре 29В показаны результаты анализа биологической активности финголимода, конъюгированного с NHS-PEG5, и «связки» лекарственных средств со свободной функциональной ТСО-группой и с пятью молекулами финголимода. Результат показывает, что молекула финголимода, конъюгированная со связывающей ветвью, имела биологическую активность в отношении блокировки миграции В-клеток, сходную с активностью немодифицированного финголимода.
Пример 31: Конструирование сегмента гена, кодирующего 2-цепочечный белок слияния IgG1.Fc, содержащий scFv, специфичный к белку F вируса RSV, и scFv, специфичный к эктодомену CD32a
Рекомбинантная цепь scFv1-CH2-CH3-scFv2 (человеческого γ1) была сконфигурирована путем слияния двух scFv, из которых первый, специфичный к белку F вируса RSV, сливали с N-концом домена СН2 IgG1.Fc через гибкую шарнирную область, а второй, специфичный к эктодомену CD32a, сливали с С-концом домена CH3 через гибкий линкер (GGGGS)3.
Оба scFv находились в ориентации «Vl-линкер-VH». VL и VH в каждом из двух scFv были связаны гидрофильным линкером GSTSGSGKPGSGEGSTKG. Последовательность рекомбинантной цепи в молекулярной конструкции белка слияния IgG1.Fc представлена под номером SEQ ID NO: 37.
Ниже проиллюстрирована конфигурация полученной 2-цепочечной молекулярной конструкции (scFv α RSV)-hlgG1.Fc-(scFv α CD32a).
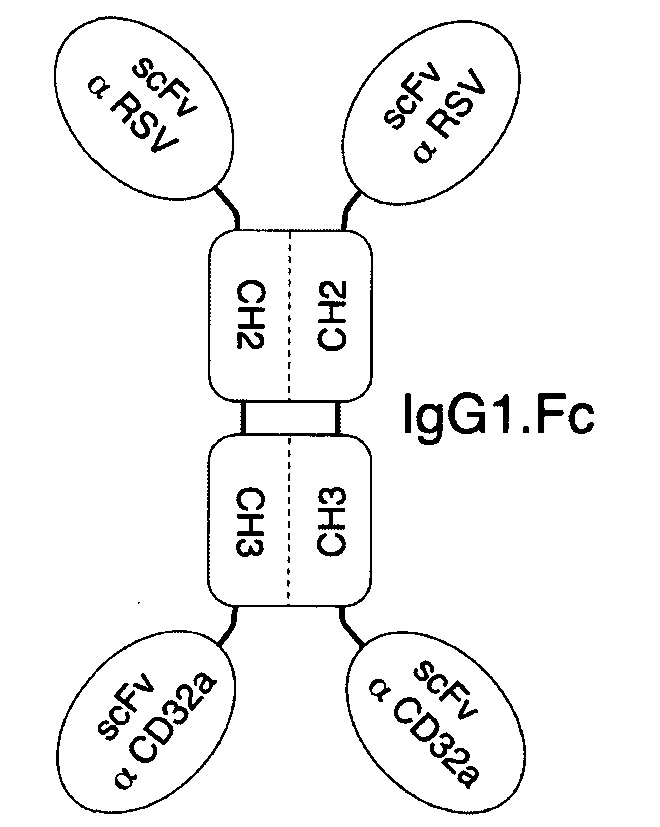
Пример 32. Экспрессия и очистка рекомбинантного 2-цепочечного белка слияния (scFv α RSV)-hIgG1.Fc-(scFv α CD32)
В этом примере последовательность, кодирующую ген, помещали в кассету экспрессии pcDNA3. Клетки Expi293F высевали при плотности 2,0×106 жизнеспособных клеток/мл в среду для экспрессии Expi293F и выдерживали в течение 18-24 часов перед трансфекцией, чтобы убедиться в том, что клетки на момент трансфекции активно делились. В процессе трансфекции 7,5×108 клеток в 255 мл среды в 2-литровой смесительной колбе Эрленмейера трансфицировали с помощью реагента для трансфекции ExpiFectamine™ 293. Трансфицированные клетки инкубировали при 37°С в течение 16-18 часов после трансфекции в орбитальном шейкере (125 об/мин), к клеткам в смесительную колбу добавляли усилитель трансфекции 1 и усилитель трансфекции 2 ExpiFectamine™ 293, и инкубировали их в течение 7 дней. Надосадочную жидкость культуры собирали, и рекомбинантный 2-цепочечный белок слияния (scFv α RSV)-hIgG1.Fc-(scFv α CD32a) в среде очищали с помощью хроматографии на белке А. После замены буфера на PBS концентрацию белка (scFv α RSV)-hIgG1.Fc-(scFv α CD32a) определяли и анализировали с помощью 12% SDS-PAGE, как показано на фигуре 30А. Молекулярная конструкция в конфигурации слияния с Fc была выявлена как основная полоса, соответствующая приблизительно 85 кДа, что согласовывалось с ожидаемым размером.
Пример 33. Анализ связывания рекомбинантного 2-цепочечного белка слияния (scFv α RSV)-hIgG1.Fc-(scFv α CD32a) методом ELISA
Для исследования связывающей способности рекомбинантного 2-цепочечного белка слияния (scFv α RSV)-hIgG1.Fc-(scFv α CD32a) как с белком F вируса RSV, так и с эктодоменом CD32a, был проведен анализ ELISA. Планшеты для анализа ELISA покрывали белком F вируса RSV (Sino Biological Inc.) при концентрации 2 мкг/мл. Рекомбинантный 2-цепочечный белок слияния (scFv α RSV)-hIgG1.Fc-(scFv α CD32a) обнаруживали козьим античеловеческим антителом IgG1.Fc, конъюгированным с HRP. Результаты анализа ELISA на фигуре 30В показывают, что рекомбинантный 2-цепочечный белок слияния (scFv α RSV)-hIgG1.Fc-(scFv а CD32a) связывался с белком F вируса RSV, в качестве контрольного scFv использовался scFv адалимумаба.
На фигуре 30С показана активность связывания рекомбинантного Fc-белка слияния с эктодоменом CD32a. Планшеты для анализа ELISA покрывали рекомбинантным эктодоменом CD32a при концентрации 5 мкг/мл. Рекомбинантный 2-цепочечный белок слияния(scFv a RSV)-hIgG1.Fc-(scFv α CD32a) обнаруживали козьим античеловеческим антителом IgG1.Fc, конъюгированным с HRP. На фигуре 30С показано, что рекомбинантный 2-цепочечный Fc-белок слияния (scFv α RSV)-hIgG1.Fc-(scFv α CD32a) обладал активностью связывания с рекомбинантным эктодоменом CD32a. Рекомбинантный 2-цепочечный белок (scFv α-эндотоксин)-IgG1.Fc использовали в качестве контрольного антитела.
Пример 34. Получение 2-цепочечного белка слияния на основе IgG1.Fc, содержащего scFv, специфичный к эндотоксину, и scFv, специфичный к эктодомену CD32a
Конфигурацию рекомбинантной цепи scFv1-CH2-CH3-scFv2 (на основе человеческого γ1) создавали путем слияния двух scFv, при этом первый из них, специфичный к эндотоксину, сливали с N-концом домена СН2 в IgG1.Fc с помощью гибкой шарнирной области, а второй из них, специфичный к эктодомену, сливали с С-концом домена СН3 с помощью гибкого линкера (GGGGS)3.
Оба scFv находились в ориентации VL-линкер-VH. VL и VH в каждом из этих двух scFv были соединены гидрофильным линкером GSTSGSGKPGSGEGSTKG.
Последовательность рекомбинантной цепи в молекулярной конструкции белка слияния на основе IgG1.Fc показана под SEQ ID NO: 38. Экспрессию сконструированного гена в клетках Expi293F и очистку экспрессируемого белка слияния осуществляли в соответствии с предыдущими примерами. Определение характеристик новой конструкции осуществляли с помощью SDS-PAGE и ELISA. Результаты SDS-PAGE, представленные на фигуре 31А, указывают на то, что рекомбинантная цепь новой конструкции имела размер, составляющий приблизительно 85 кДа, что согласуется с ожидаемым размером.
На фигуре 31В показаны результаты анализа ELISA рекомбинантного 2-цепочечного белка (scFv α эндотоксина)-(scFv α CD32a)-hIgG1.Fc, связывающегося с бактериями Е. coli LPS 0111:В4 (Sigma Aldrich). Планшеты для анализа ELISA покрывали поли-L-лизином при концентрации 50 мкг/мл. Затем покрытые поли-L-лизином планшеты дополнительно покрывали бактериями Е. coli LPS 0111:В4 в концентрации 10 мкг/мл. Рекомбинантный белок слияния детектировали козьим античеловеческим антителом IgG.Fc, конъюгированным с HRP. Результаты анализа ELISA показывают, что этот рекомбинантный Fc-белок слияния обладал связывающей активностью по отношению к E.coli LPS 0111:В4 (Sigma Aldrich); фигура 31С показывает, что рекомбинантный Fc-белок слияния обладал связывающей активностью по отношению к эктодомену CD32a.
Ниже проиллюстрирована конфигурация полученной таким образом молекулярной 2-цепочечной конструкции (scFv α эндотоксин)-(scFv α CD32)-hIgG1.Fc.
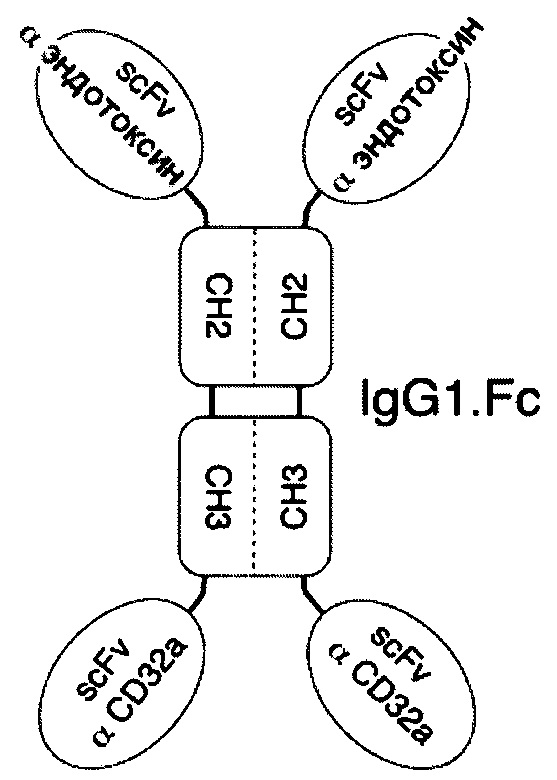
Пример 35: Конструирование сегмента гена, кодирующего 2-цепочечный белок слияния IgG4.Fc, содержащий интерферон-β-1a, и scFv, специфичный к эктодомену TfR1
2-цепочечный белок слияния IgG.Fc получали путем конфигурирования (интерферон-β-1a)-CH2-CH3-(scFv α TfR1) (человеческого γ4) в рекомбинантной цепи. С-конец интерферона-β-1а сливали с N-концом СН2 через линкер GGGGSGGGASGGS. Фрагмент scFv, специфичный к эктодомену TfR1, сливали с С-концом домена СН3 через гибкий линкер (GGGGS)3.
Фрагмент scFv (специфичный к эктодомену TfR1) находился в ориентации «VL-линкер-VH». VL и VH в scFv были связаны гидрофильным линкером GSTSGSGKPGSGEGSTKG. Последовательность рекомбинантной цепи в молекулярной конструкции белка слияния IgG4.Fc представлена под номером SEQ ID NO: 39.
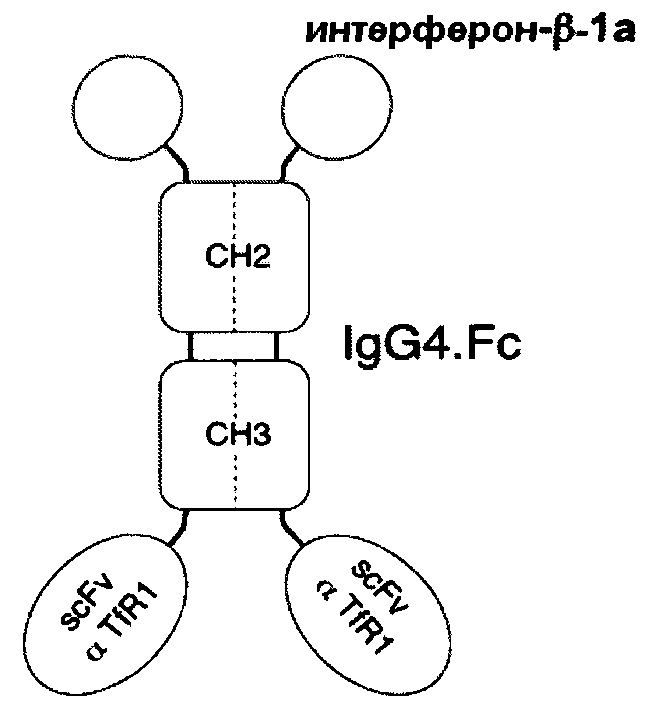
Здесь проиллюстрирована конфигурация полученной 2-цепочечной молекулярной конструкции (интерферон-β-1a)-IgG4.Fc-(scFv α TfR1).
Пример 36: Экспрессия и очистка рекомбинантного 2-цепочечного белка слияния (интерферон-β-1a)-hIgG4.Fc-(scFv α TfR1)
В этом примере последовательность, кодирующую ген, помещали в кассету экспрессии pcDNA3. Клетки Expi293F высевали при плотности 2,0×106 жизнеспособных клеток/мл в среду для экспрессии Expi293F и выдерживали в течение 18-24 часов перед трансфекцией, чтобы убедиться в том, что клетки на момент трансфекции активно делились. В процессе трансфекции 7,5×108 клеток в 255 мл среды в 2-литровой смесительной колбе Эрленмейера трансфицировали с помощью реагента для трансфекции ExpiFectamine™ 293. Трансфицированные клетки инкубировали при 37°С в течение 16-18 часов после трансфекции в орбитальном шейкере (125 об/мин) к клеткам в смесительной колбе добавляли усилитель трансфекции 1 и усилитель трансфекции 2 ExpiFectamine™ 293 и инкубировали в течение 7 дней. Надосадочную жидкость культуры собирали и рекомбинантный 2-цепочечный белок слияния (интерферон-β-1a)-hIgG4.Fc-(scFv α TfR1) в среде очищали с помощью хроматографии на белке А. После замены буфера на PBS концентрацию белка (интерферон-β-1а)-hIgG4.Fc-( scFv α TfR1) определяли и анализировали с помощью 8% SDS-PAGE, как показано на фигуре 32А. Молекулярная конструкция в конфигурации слияния с Fc была выявлена как основная полоса, соответствующая приблизительно 80 кДа, что согласовывалось с ожидаемым размером.
Пример 37. Анализ связывания рекомбинантного 2-цепочечного белка слияния (интерферон-β-1а)-hIgG1.Fc-(scFv α TfR1) методом ELISA и проточной цитометрии
Связывающую активность рекомбинантного (интерферон-β-1а)-hIgG4.Fc-(scFv α TfR1) анализировали с помощью анализа ELISA с использованием 96-луночного планшета, покрытого рекомбинантным белком (интерферон-β-1а)-hIgG4.Fc-(scFv α TfR1) в концентрации 5 мкг/мл, 100 мкл на лунку. Фрагмент scFv, специфичный к эктодомену TfR1, являлся отрицательным контролем. Рекомбинантный 2-цепочечный белок (интерферон-β-1а)-hIgG4.Fc-(scFv α TfR1) обнаруживали поликлональным кроличьим античеловеческим антителом к интерферону-β, конъюгированным с HRP (Santa Cruz Biotechnology, Dallas, USA). Затем для проявления цвета добавляли 50 мкл ТМВ-субстрата. Реакцию останавливали добавлением 50 мкл 1М HCl. Абсорбцию на 450 нм измеряли с помощью планшетного ридера. Каждый столбик представляет собой среднее значение OD450 для повторяющихся образцов.
На фигуре 32В показан результат анализа ELISA данной молекулярной конструкции. Результаты показывают, что белок слияния (интерферон-β-1а)-hIgG4.Fc-(scFv α TfR1) специфически связан с рекомбинантным эктодоменом белка TfR1.
Пример 38: Получение 2-цепочечного белка слияния IgG4.Fc, содержащего scFv, специфичный к интегрину α4, и scFv, специфичный к эктодомену TfR1
VL и VH фрагмента scFv, специфичного к интегрину α4, были получены из моноклонального антитела натализумаба. Двухцепочечный белок слияния IgG.Fc получали путем конфигурирования (scFv α интегрин α4)-CH2-CH3-(scFv α TfR1) (человеческого γ4) в рекомбинантной цепи. С-конец scFv, специфичного к интегрину α4, сливали с N-концом СН2 через линкер GGGGSGGGASGGS. Фрагмент scFv, специфичный к эктодомену TfR1, сливали с С-концом домена СН3 через гибкий линкер (GGGGS)3. Результат анализа 8% SDA-PAGE на фигуре 33А показывает, что рекомбинантная цепь новой конструкции имела размер около 85 кДа (показано стрелкой), что соответствовало ожидаемому размеру.
Оба scFv находились в ориентации «VL-линкер-VH». VL и VH в каждом из двух scFv были связаны гидрофильным линкером GSTSGSGKPGSGEGSTKG. Последовательность рекомбинантной цепи в молекулярной конструкции белка слияния IgG4.Fc представлена под номером SEQ ID NO: 40.
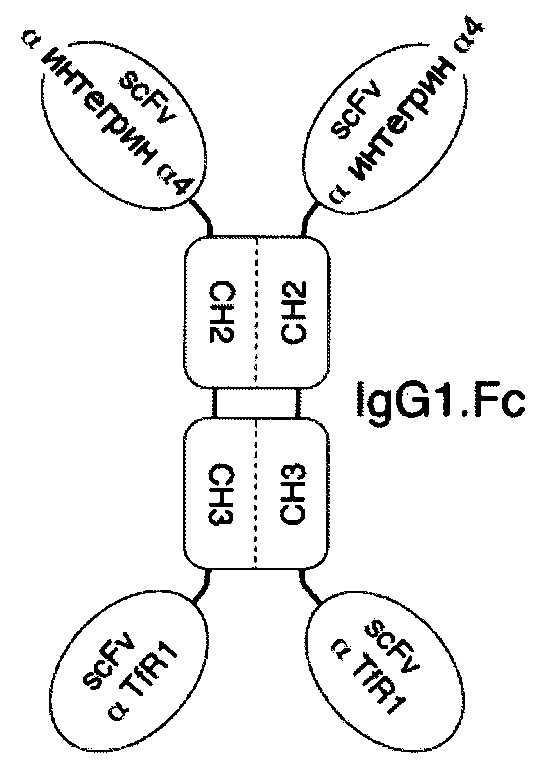
Здесь проиллюстрирована конфигурация полученной 2-цепочечной молекулярной конструкции (scFv α интегрин α4)-IgG4.Fc-(scFv α TfR1).
Для исследования связывающей способности 2-цепочечного белка рекомбинантной цепи (scFv α интегрин α4)-IgG4.Fc-(scFv α TfR1) по отношению к Т-клеткам Jurkat, экспрессирующим интегрин α4, проводили анализ клеточного связывания методом проточной цитометрии.
1×106 Т-клеток Jurkat выдерживали в среде RPMI1640, дополненной 10% FBS, с плотностью 1×105. Клетки выдерживали при температуре 37°С с 5% CO2 во влажной камере. 1×106 Т-клеток Jurkat дважды промывали связывающим буфером (фосфатно-солевой буферный раствор с 0,1% FBS, 2 мМ EDTA и 20 нг/мл NaN3). К промытым Т-клеткам Jurkat добавляли 10 мкг/мл Human BD Fc block™ (BD Biosciences, San Jose, US) для блокирования опосредованного рецептора Fc. Клетки промывали и инкубировали с 10 мкг/мл рекомбинантного белка (scFv α интегрин α4)-IgG4.Fc-(scFv α TfR1) на льду в течение 15 минут с использованием рекомбинантного 2-цепочечного белка (интерферон-β-1а)-IgG4. Fc-(scFv α TfR1) в качестве отрицательного контроля. Клетки снова промывали и инкубировали с козьим античеловеческим антителом IgG.Fc, конъюгированным с FITC (Caltag, Buckingham, UK), разбавленным в соотношении 1:200 в блокирующем буфере, на льду в течение 15 мин в темноте. Окрашенные клетки анализировали на проточном цитометре FACSCanto II (BD Biosciences).
На фигуре 33В показаны результаты анализа клеток рекомбинантного 2-цепочечного белка (scFv α integrin α4)-IgG4.Fc-(scFv α TfR1) методом окрашивания на Т-клетках Jurkat, экспрессирующих интегрин α4. Конструкция связывалась с Т-клетками Jurkat, в целом, положительно.
Пример 39: Анализ биологической активности 2-цепочечного белка IgG1.Fc, содержащего scFv, специфичный к эндотоксину, и scFv, специфичный к эктодомену CD32a, на макрофагоподобных клетках U937
Для проверки влияния рекомбинантного 2-цепочечного белка слияния (scFv α эндотоксина)-hIgG1.Fc-(scFv α CD32a) на ингибирование секреции TNF-α проводили анализ ELISA с определением количества секретируемого TNF-α в надосадочной жидкости с помощью макрофагоподобных клеток U937.
Клетки U937 выдерживали в RPMI1640, дополненном 10% фетальной бычьей сывороткой (Gibco) и 100 ед./мл пенициллин-стрептомицина (Gibco), при плотности от 3×105 до 2×106 клеток/мл. Клетки выдерживали при температуре 37°С с 5% CO2 во влажной камере. Для дифференцирования U937 на макрофагоподобные клетки, 1×106 клеток/мл U937 инкубировали с 10 нг/мл форбол-12-миристат-13-ацетатом (РМА, Sigma Aldrich). Через 48 часов неприкрепившиеся клетки удаляли, а прикрепившиеся промывали и высевали в 96-луночные планшеты.
Дифференцированные клетки U937 в количестве 5×104 клеток на лунку высевали в 96-луночные планшеты за день до анализа. Клетки стимулировали только добавлением 1 мкг/мл бактерий Е. coli LPS 0111: В4 (Sigma Aldrich) или предварительно приготовленной смеси LPS и 10 мкг/мл (scFv α эндотоксин)-hIgG1.Fc, 15 мкг/мл (scFv α эндотоксин)-hIgG1.Fc-(scFv α CD32a) или 2,5 мкг/мл анти-CD32a scFv. Стимуляция продолжалась в течение 2 часов до сбора надосадочной жидкости. Выработку TNF-α измеряли коммерчески доступным набором ELISA (Biolegend).
Уровни TNF-α в надосадочной жидкости с клетками U937 измеряли с использованием набора ELISA от R&D Systems. Лунки планшетов ELISA (Greiner Bio-One) покрывали 4 мкг/мл иммобилизованного антитела в PBS при температуре 4°С на ночь. Затем лунки блокировали добавлением 0,5% PBS в течение 1 часа и инкубировали с разбавленной надосадочной жидкостью культуры в течение 2 часов. Для обнаружения связанного TNF-α использовали 400 нг/мл меченого биотином детекторного антитела, а затем Streptavidin-HRP. Хромогенную реакцию проводили с использованием ТМВ-субстрата (Clinical Science Products) и останавливали добавлением 1N HCl. Планшеты считывали при поглощении на волне длиной 450 нм. Концентрации TNF-α определяли путем экстраполяции из 4-параметрических стандартных логистических кривых, полученных в результате разбавлений стандартного белка, поставляемого производителем.
На фигуре 34 показано, что рекомбинантный 2-цепочечный белок (scFv α эндотоксин)-hIgG.Fc-(scFv α CD32a) значительно снижал секрецию TNF-α, стимулированную бактериями Е. coli LPS 0111: В4, по сравнению с 2-цепочечным белком (scFv α эндотоксин)-hIgG1.Fc контрольных антител и scFv антител к CD32a или только со средой.
Очевидно, что вышеизложенное описание вариантов осуществления приведено только в качестве примера, и что специалисты в данной области способны выполнить их различные модификации. В вышеизложенном описании, примерах и данных представлено полное описание структуры и применения иллюстративных вариантов осуществления настоящего изобретения. Хотя различные варианты осуществления настоящего изобретения описаны выше с определенной степенью конкретности или со ссылкой на один или несколько отдельных вариантов осуществления, специалисты в данной области способны выполнить многочисленные изменения раскрытых вариантов осуществления без отступления от сущности или объема настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЯРНЫЕ КОНСТРУКЦИИ С НАЦЕЛИВАЮЩИМИ И ЭФФЕКТОРНЫМИ ЭЛЕМЕНТАМИ | 2016 |
|
RU2696378C2 |
| Линкерные звенья и содержащие их молекулярные конструкции | 2018 |
|
RU2752671C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОНСТРУКЦИИ С ПОВЫШЕННОЙ АФФИННОСТЬЮ СВЯЗЫВАНИЯ С АЛЬБУМИНОМ | 2018 |
|
RU2753880C1 |
| СОСТАВНОЙ ПОЛИПЕПТИД, ИМЕЮЩИЙ МЕТАЛЛСВЯЗЫВАЮЩИЙ МОТИВ, И СОДЕРЖАЩАЯ ИХ МОЛЕКУЛЯРНАЯ КОНСТРУКЦИЯ | 2020 |
|
RU2814475C2 |
| НУКЛЕОТИДЫ, МЕЧЕННЫЕ ЗАРЯДОМ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2751359C2 |
| СПОСОБЫ КОНЪЮГАЦИИ | 2010 |
|
RU2765240C2 |
| СПОСОБЫ КОНЪЮГАЦИИ | 2010 |
|
RU2595424C2 |
| СПОСОБ ОТБОРА И ПОЛУЧЕНИЯ СЕЛЕКТИВНЫХ И МУЛЬТИСПЕЦИФИЧЕСКИХ ТЕРАПЕВТИЧЕСКИХ МОЛЕКУЛ С ЗАДАННЫМИ СВОЙСТВАМИ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ НАЦЕЛИВАЮЩИЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2644263C2 |
| ГИПЕРГЛИКОЗИЛИРОВАННЫЕ СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2708314C2 |
| САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПОСРЕДСТВОМ ГЛИКОИНЖЕНЕРИИ | 2014 |
|
RU2711935C2 |



















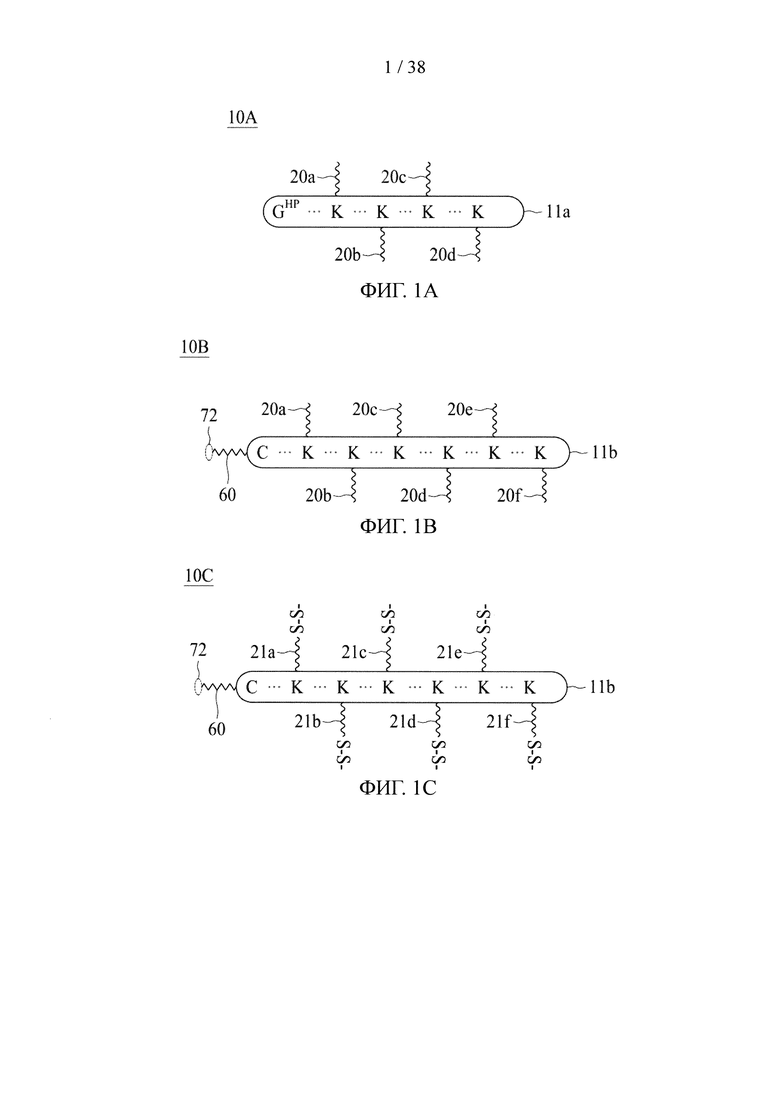
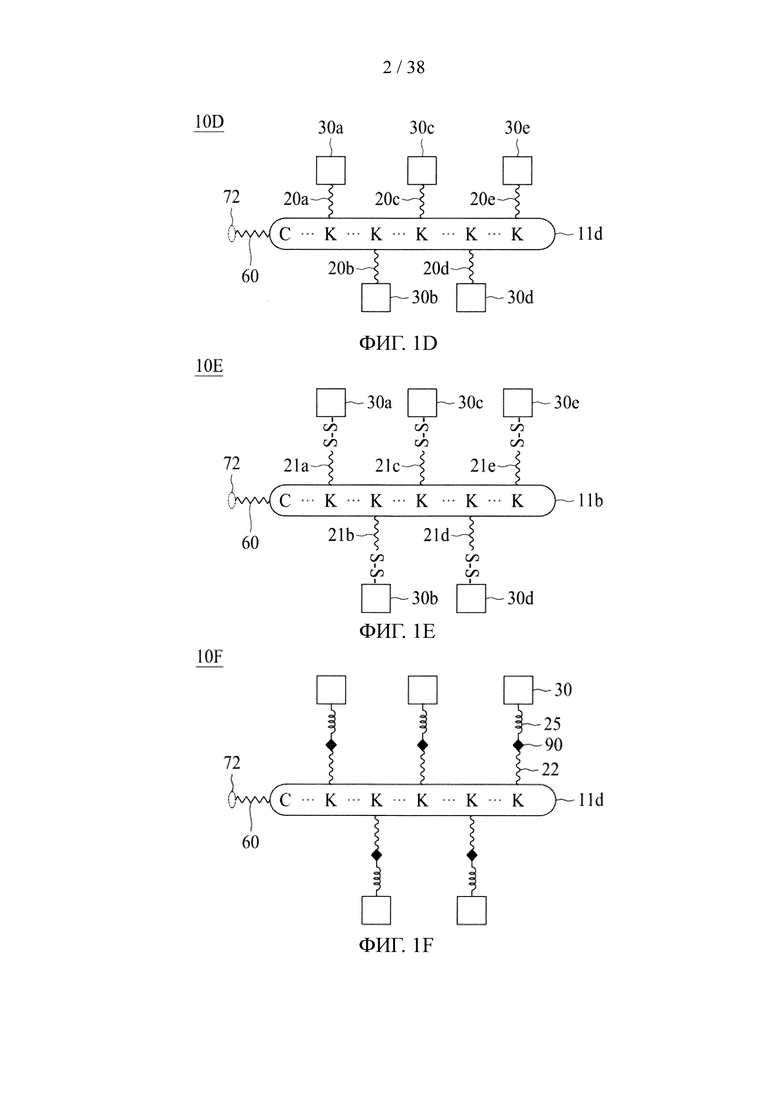
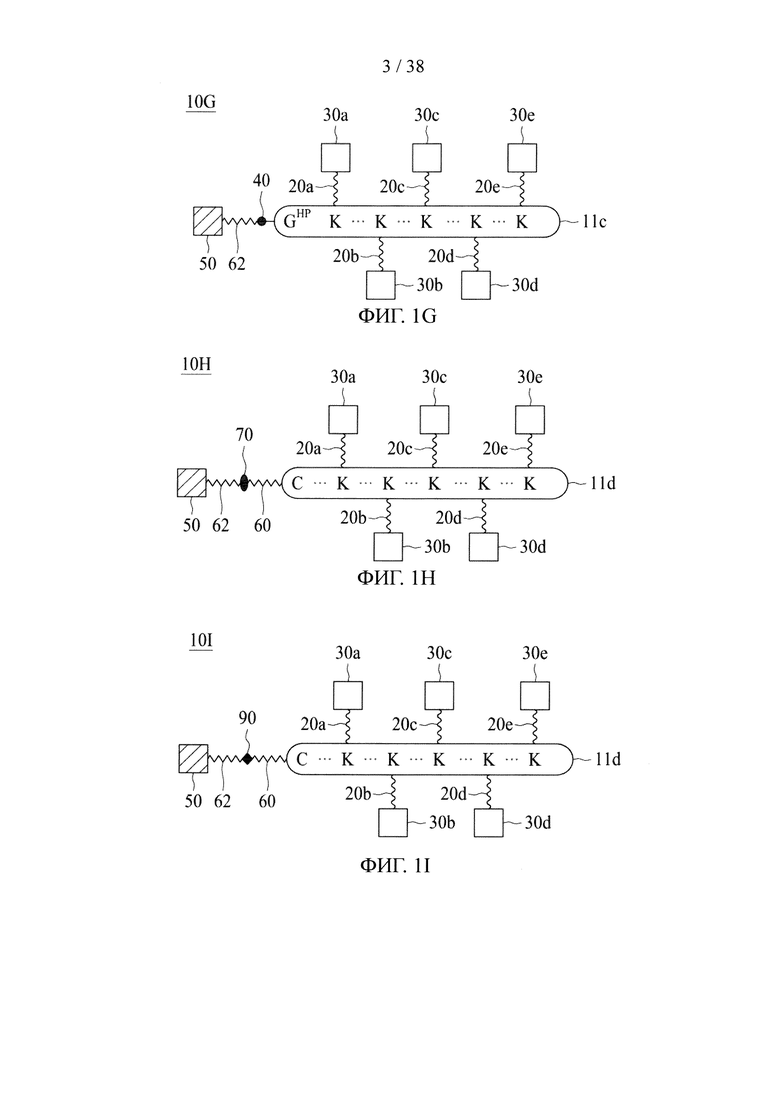
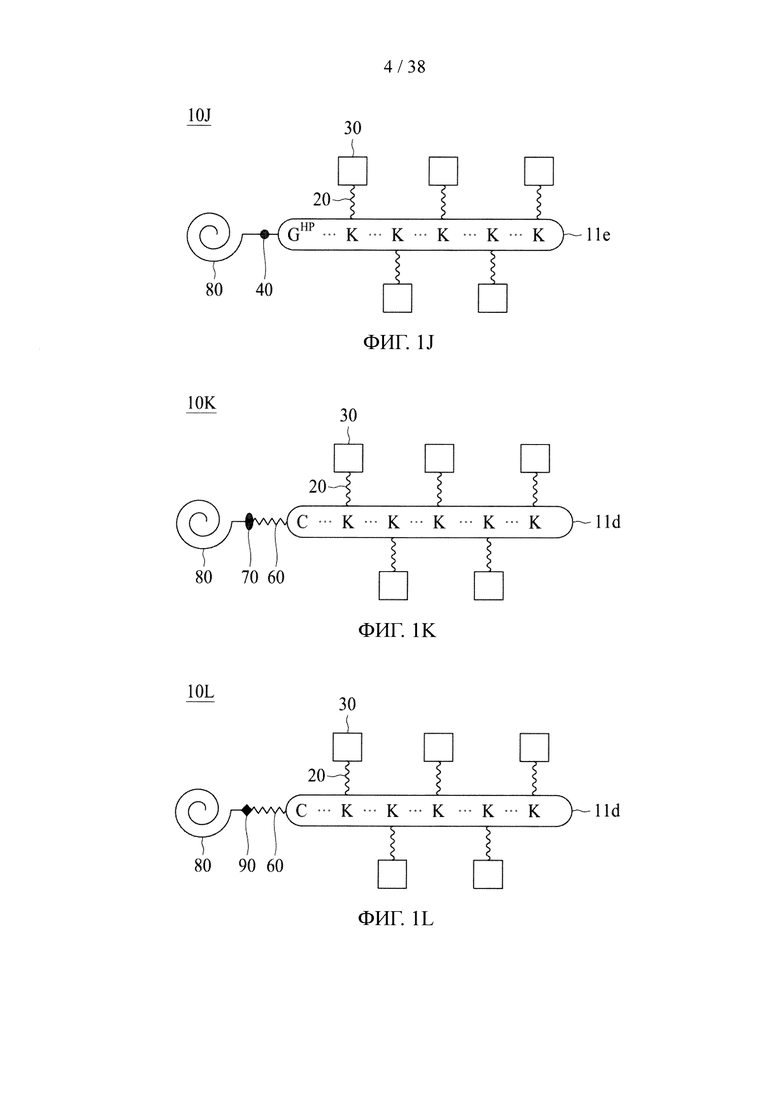
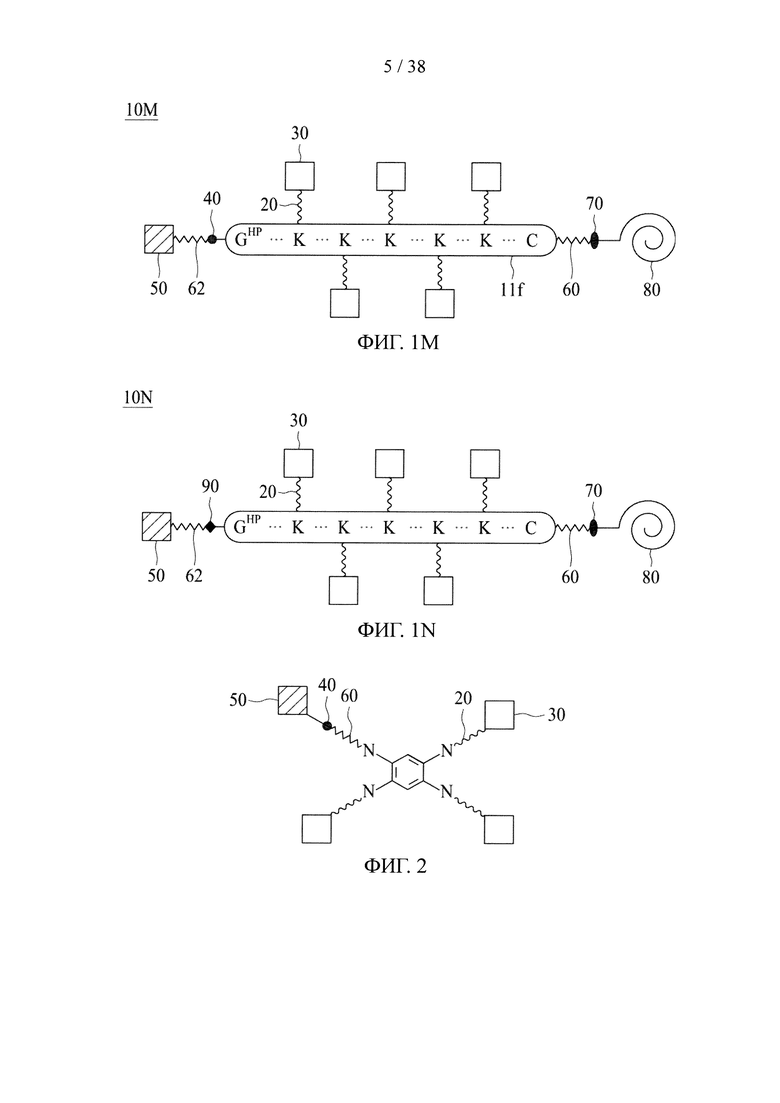
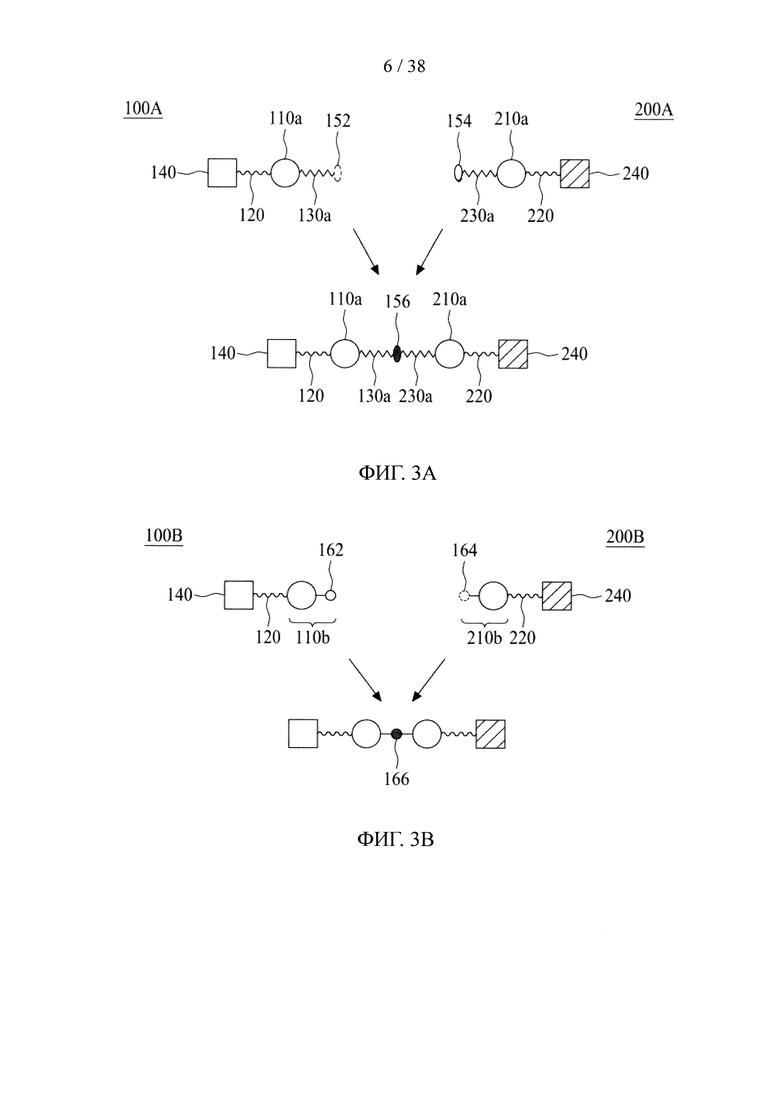
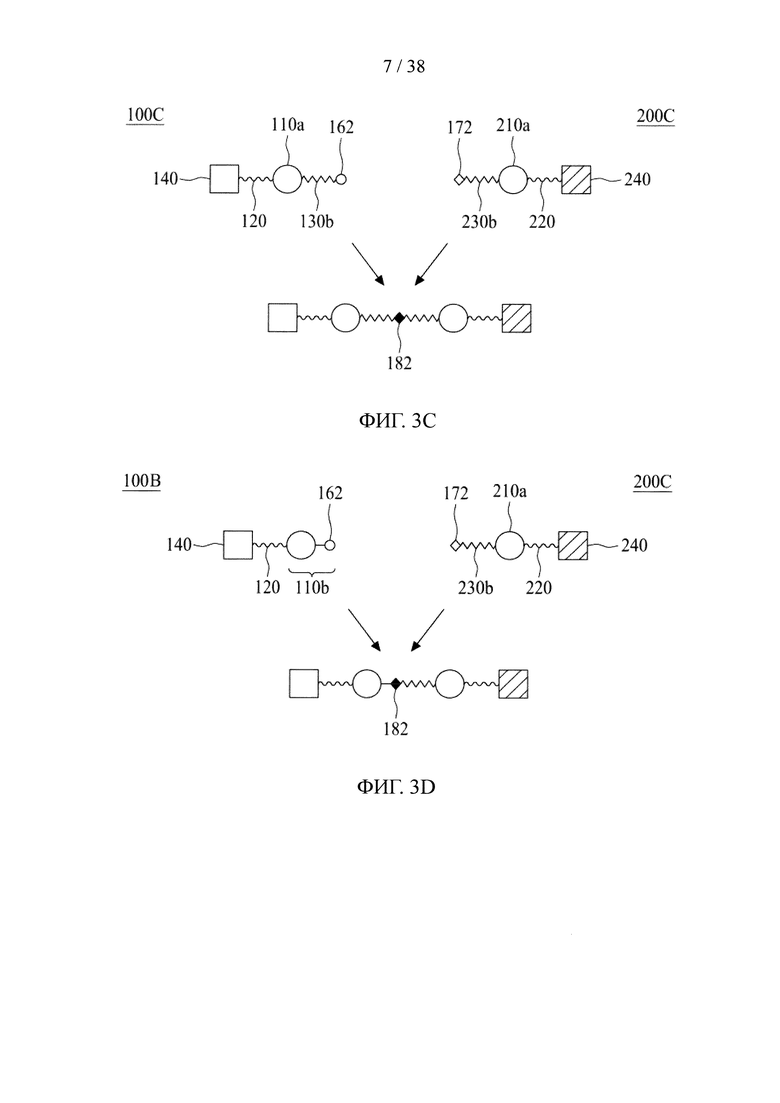
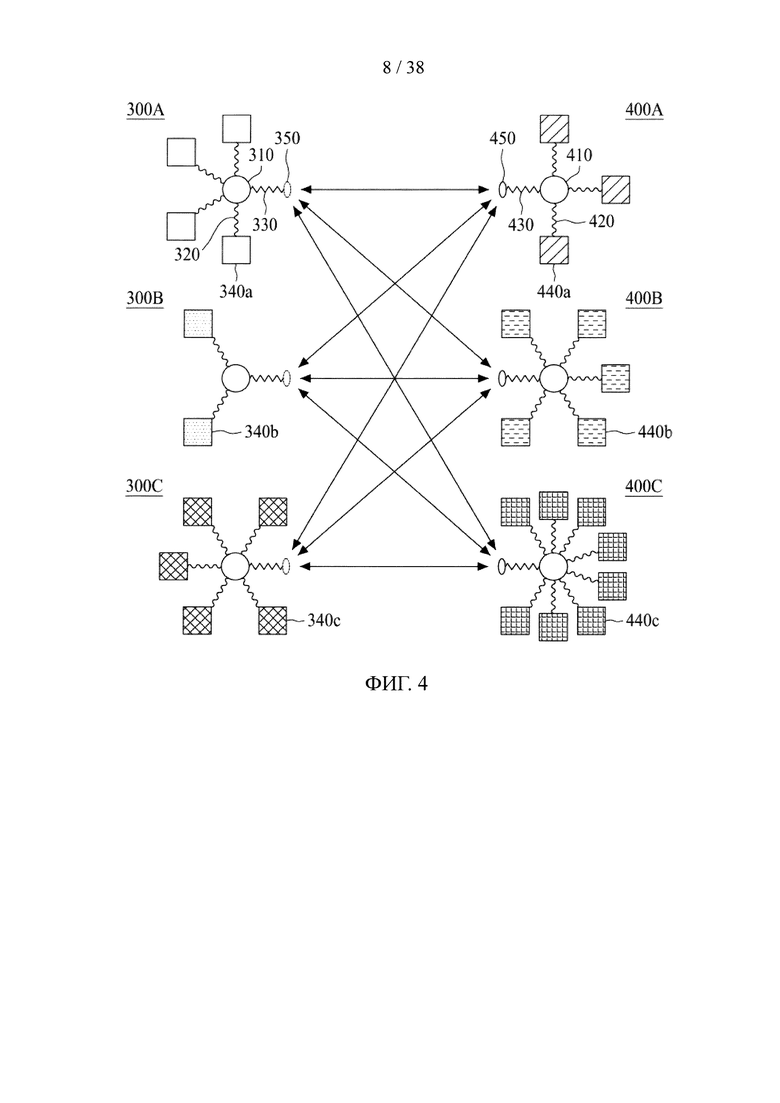
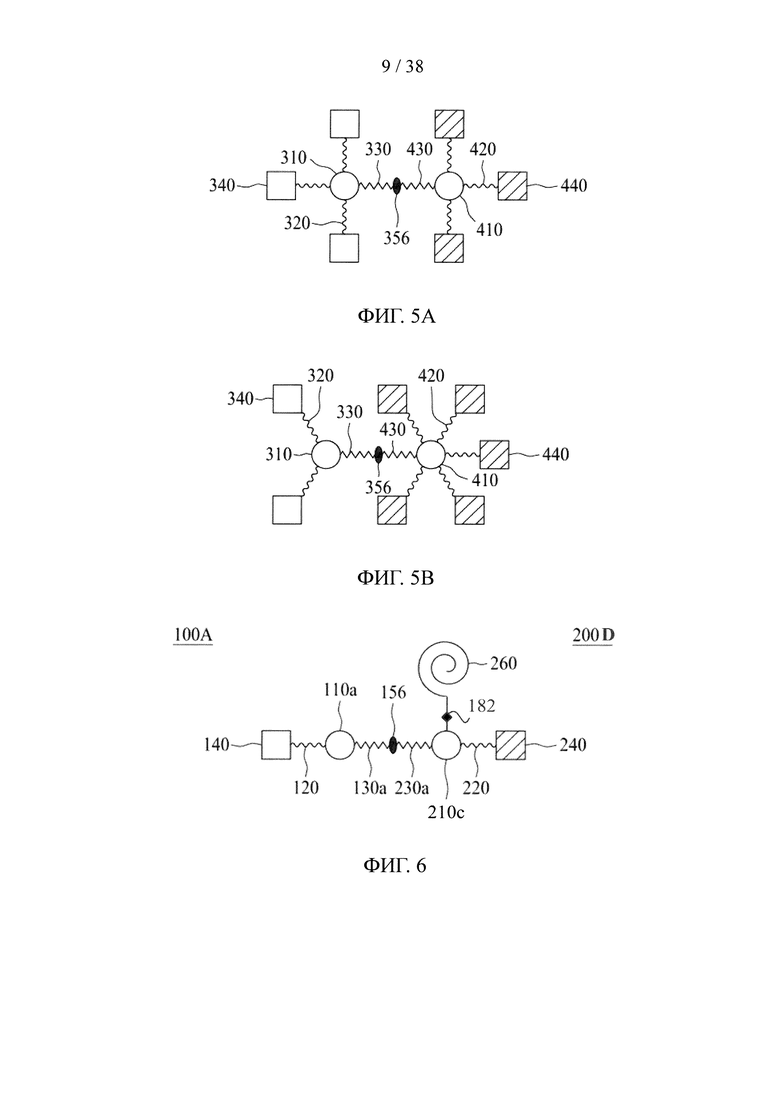
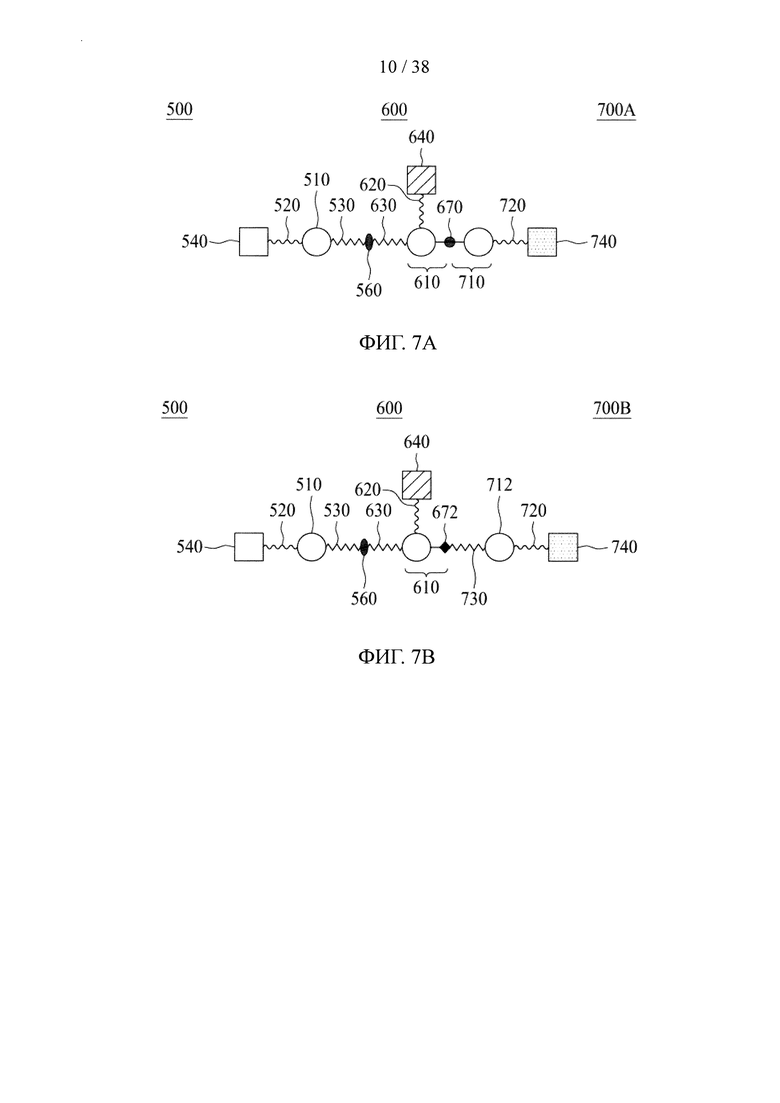
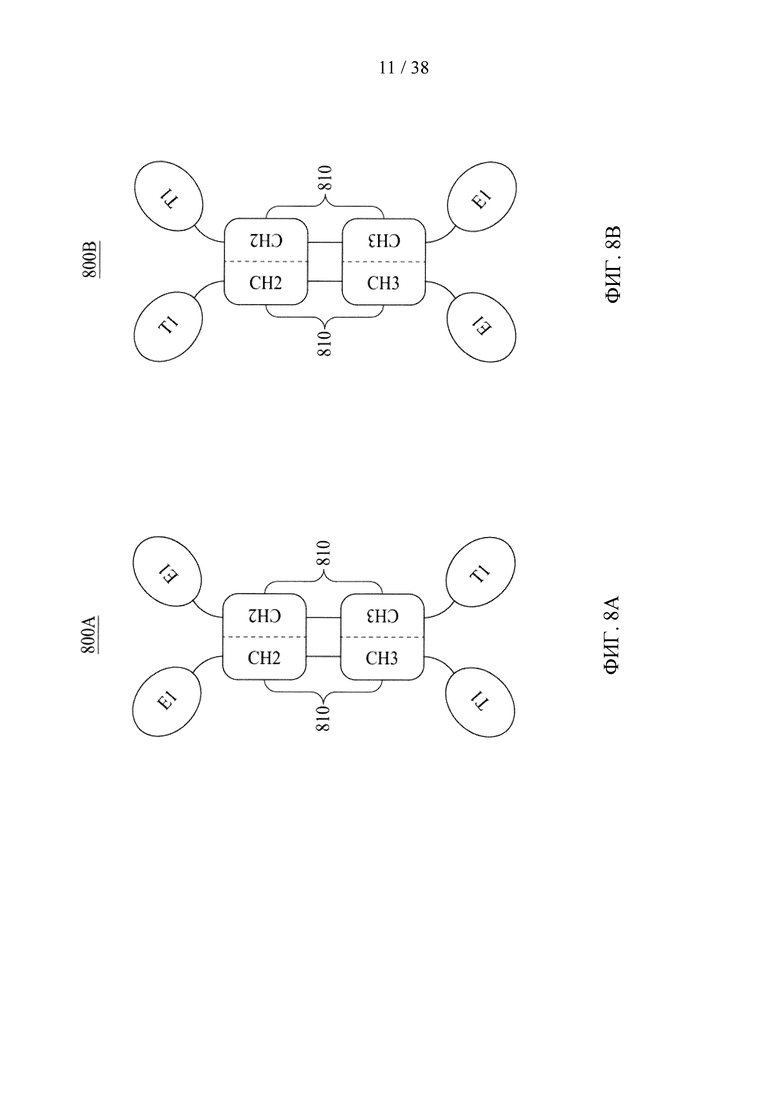
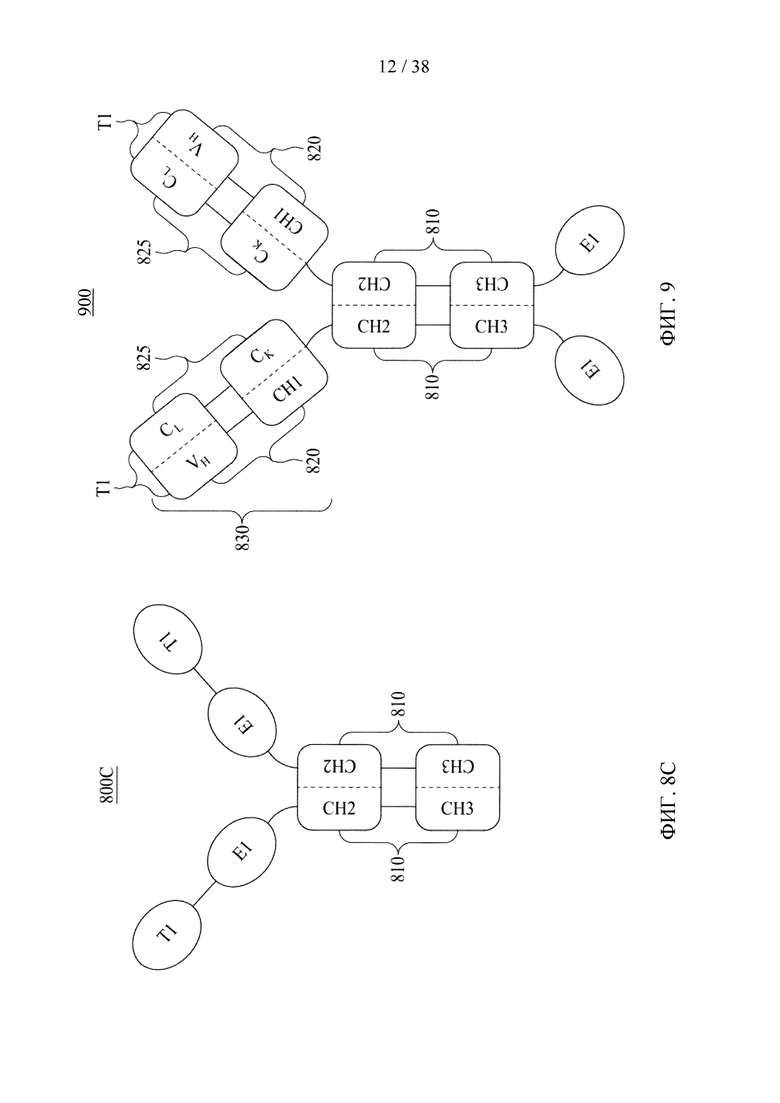
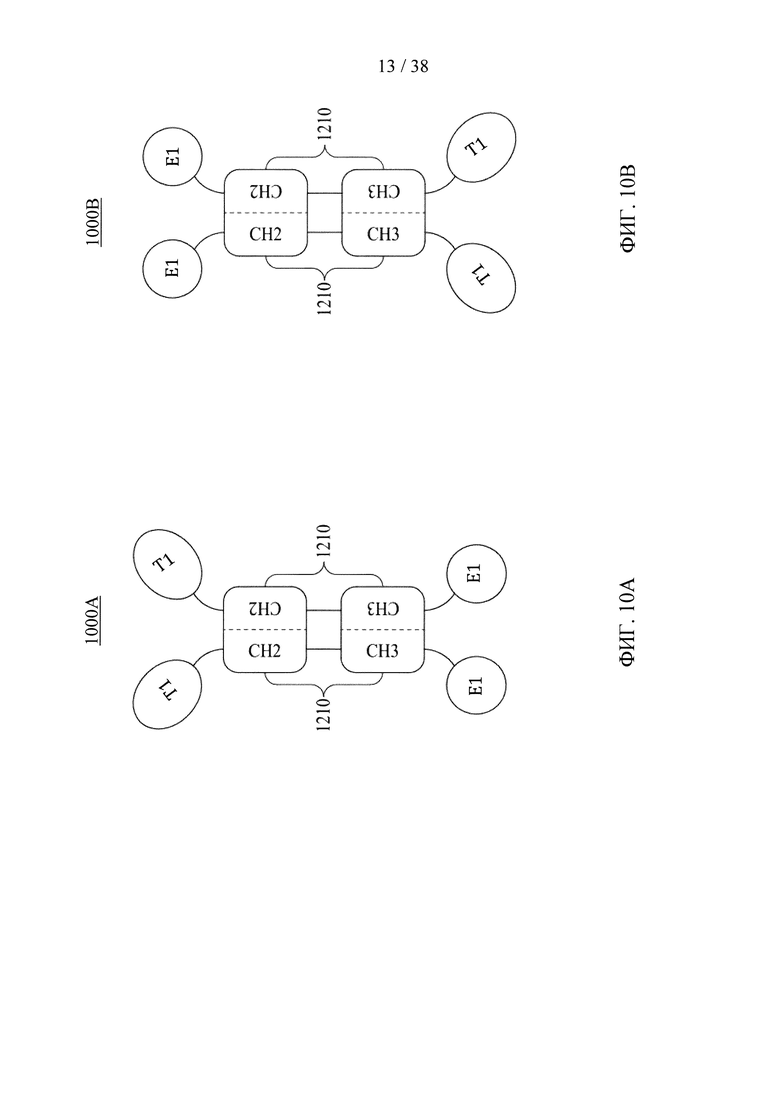
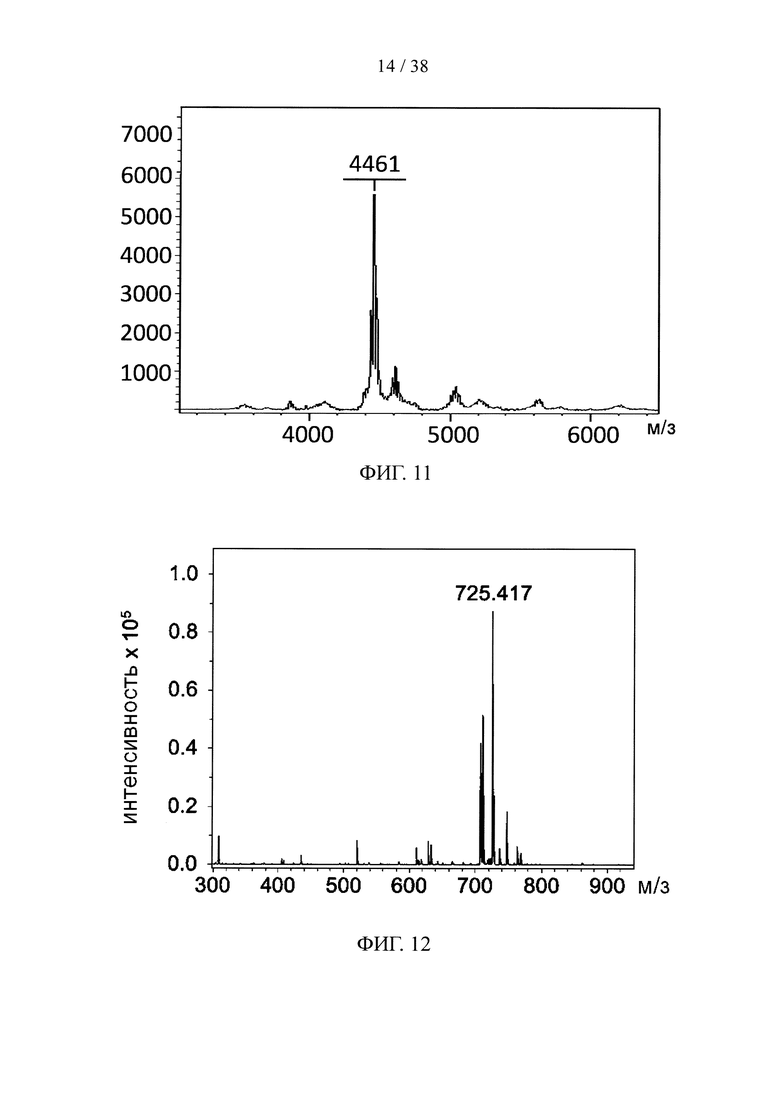
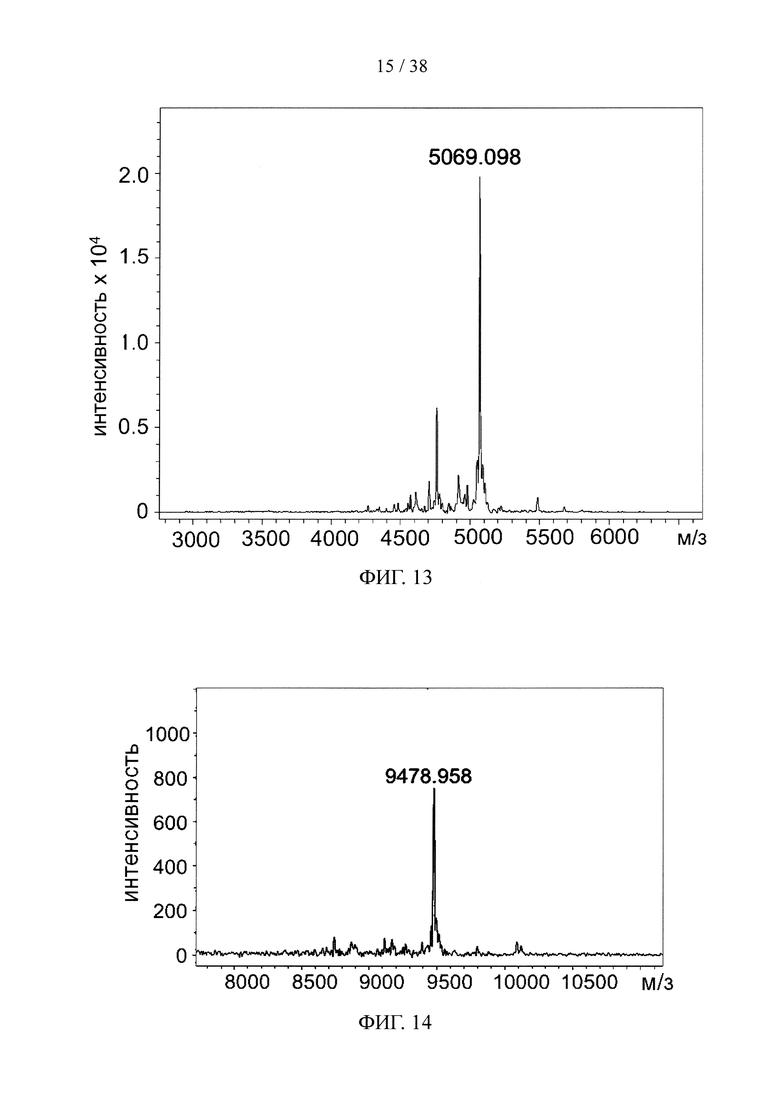
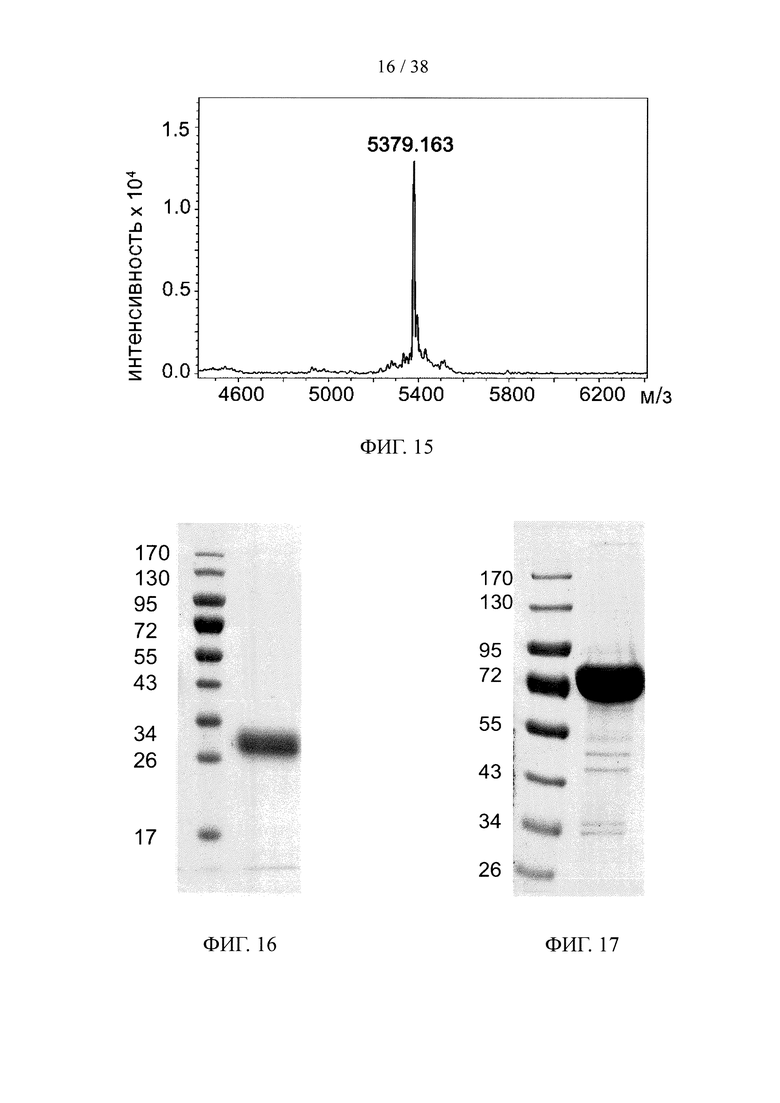
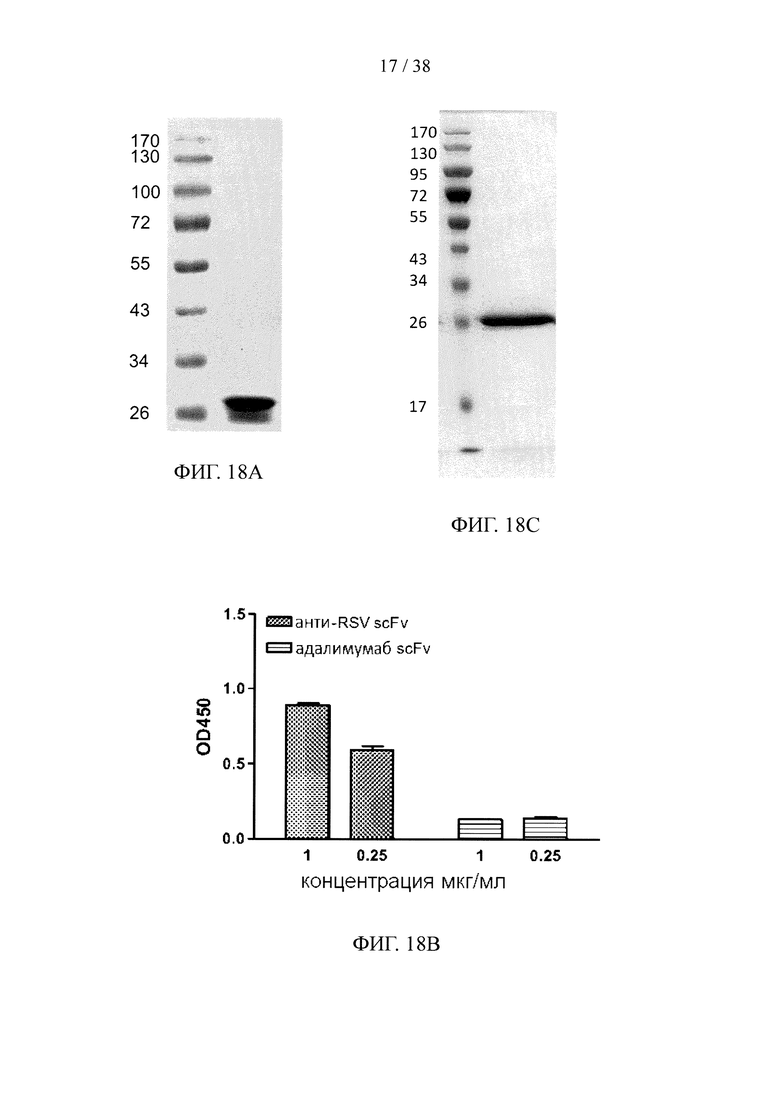
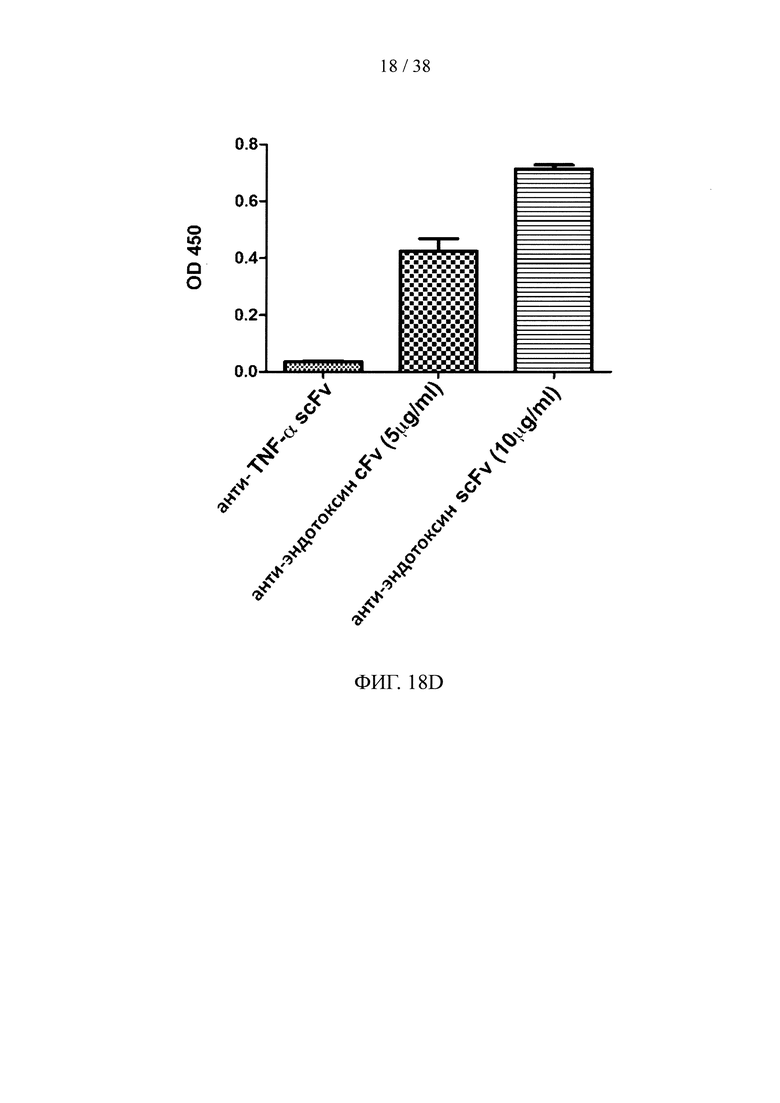
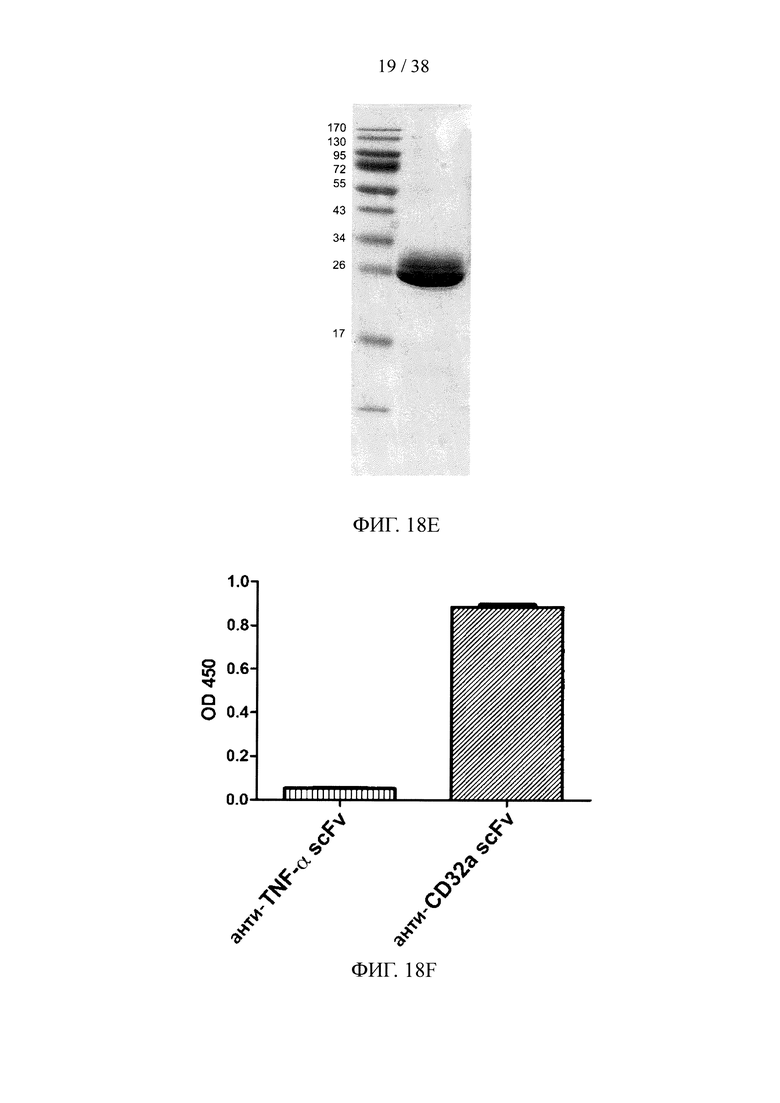
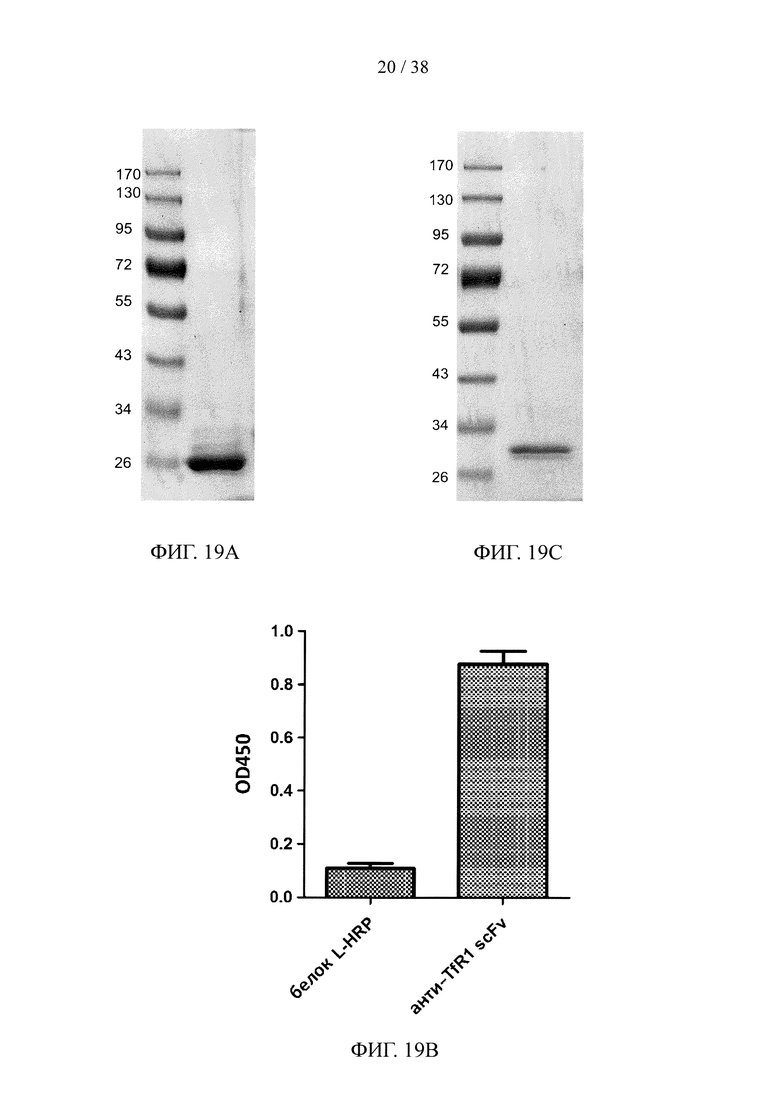
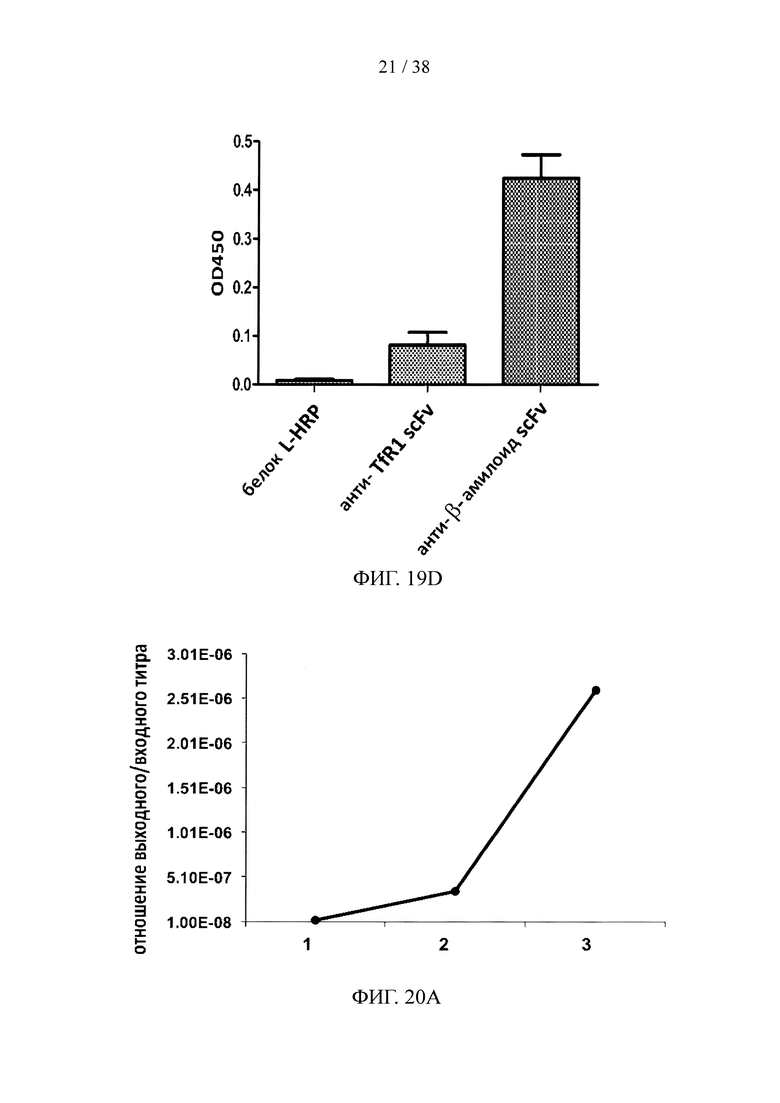
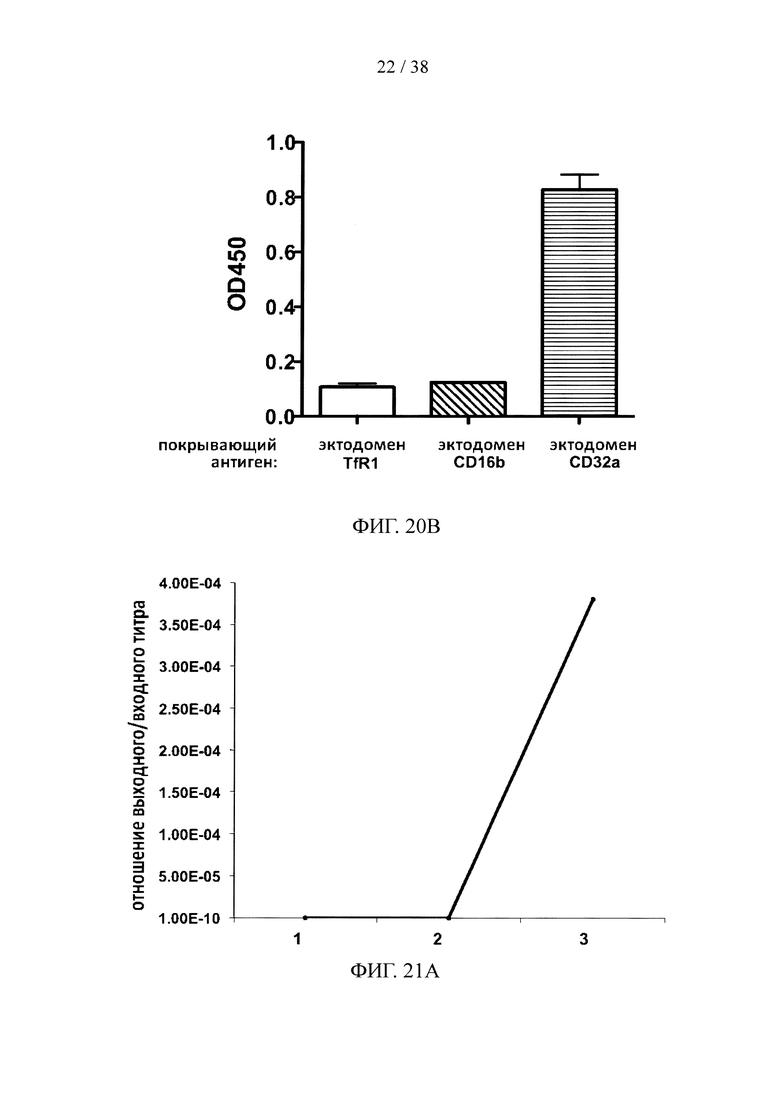
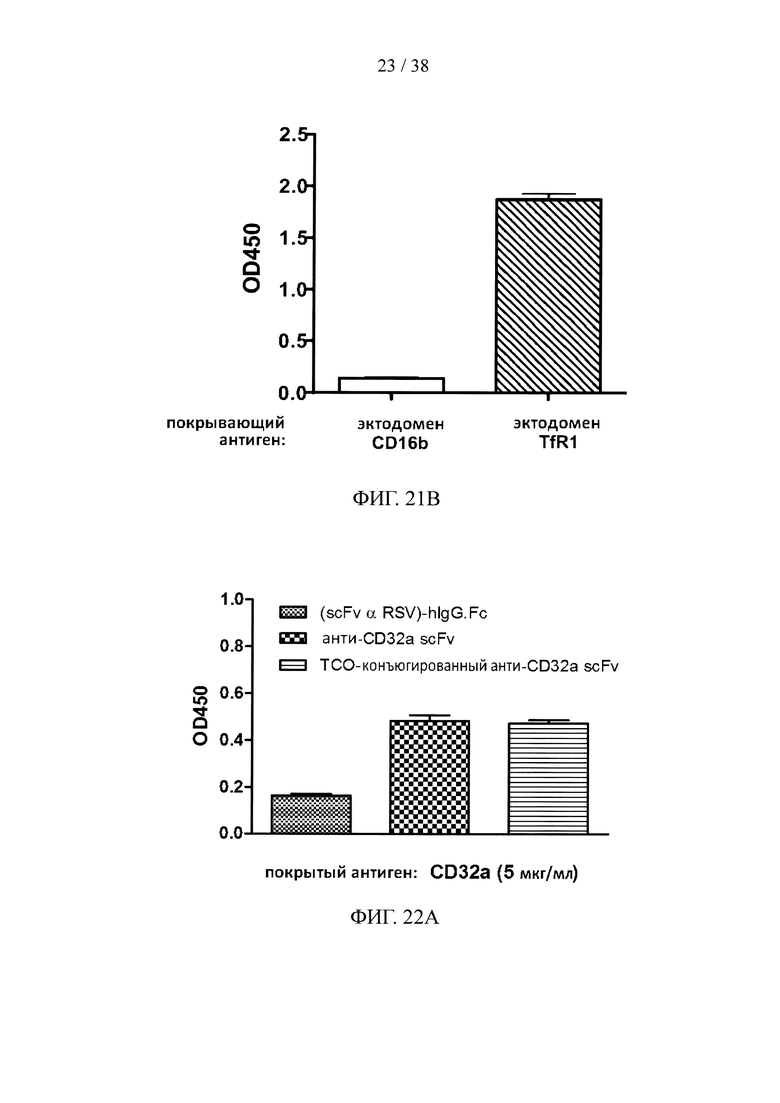
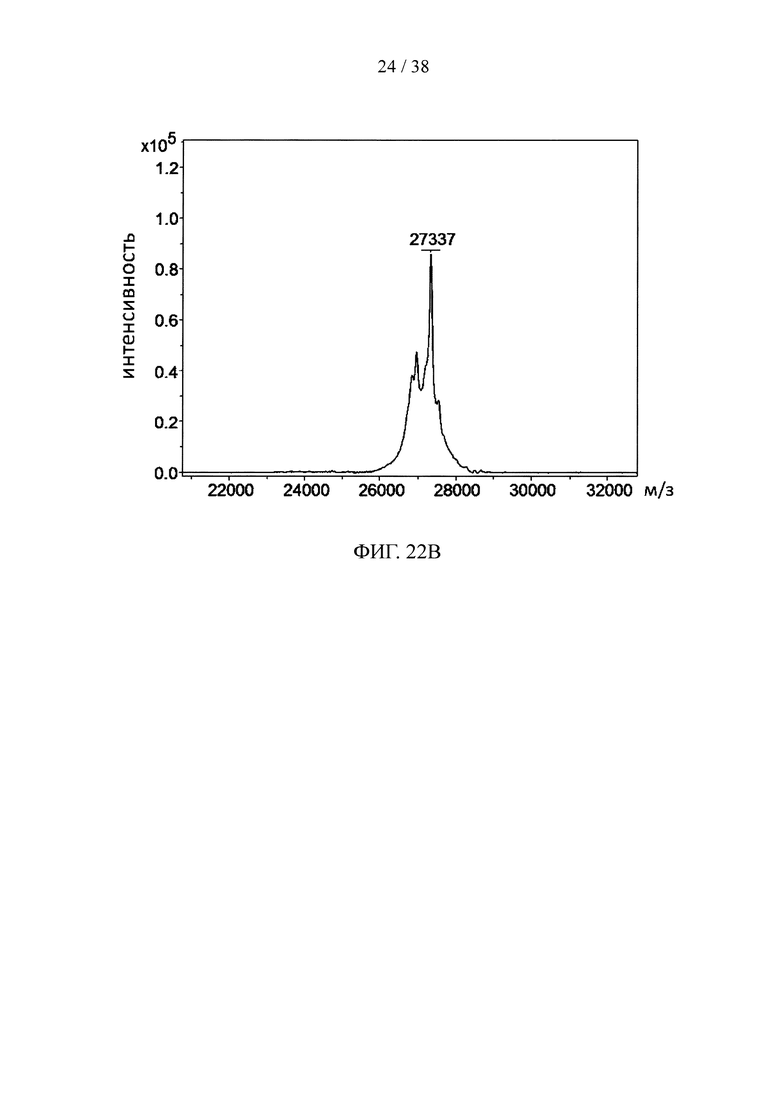
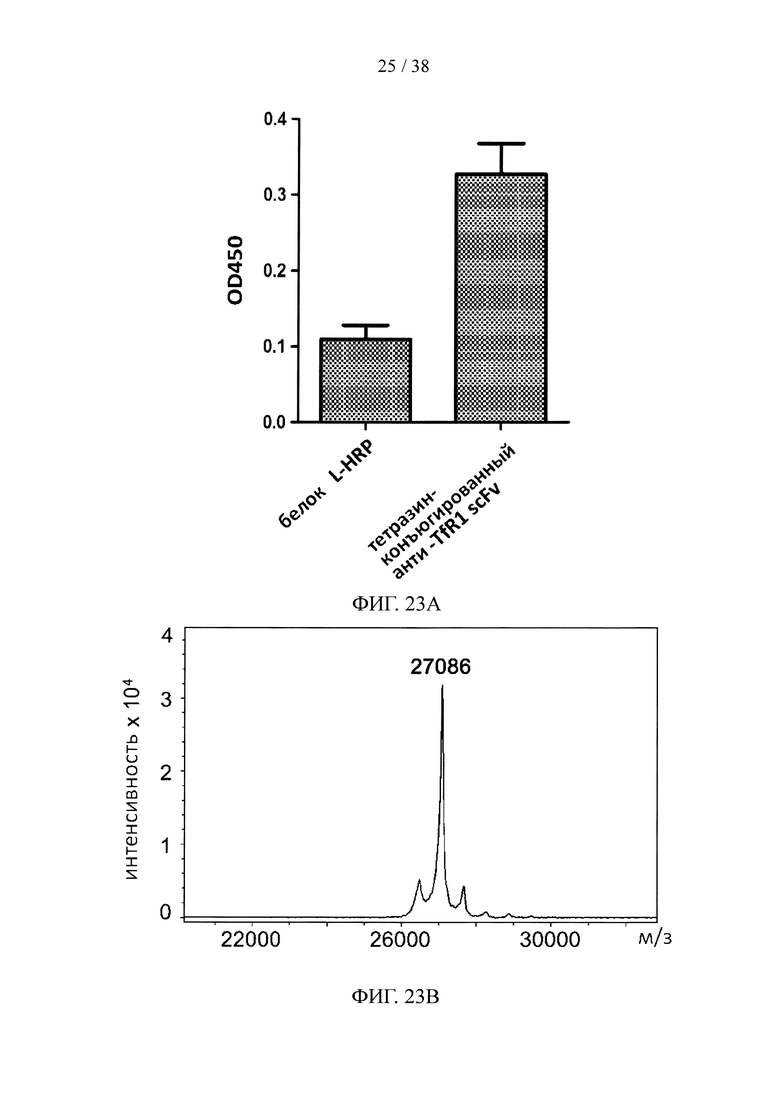
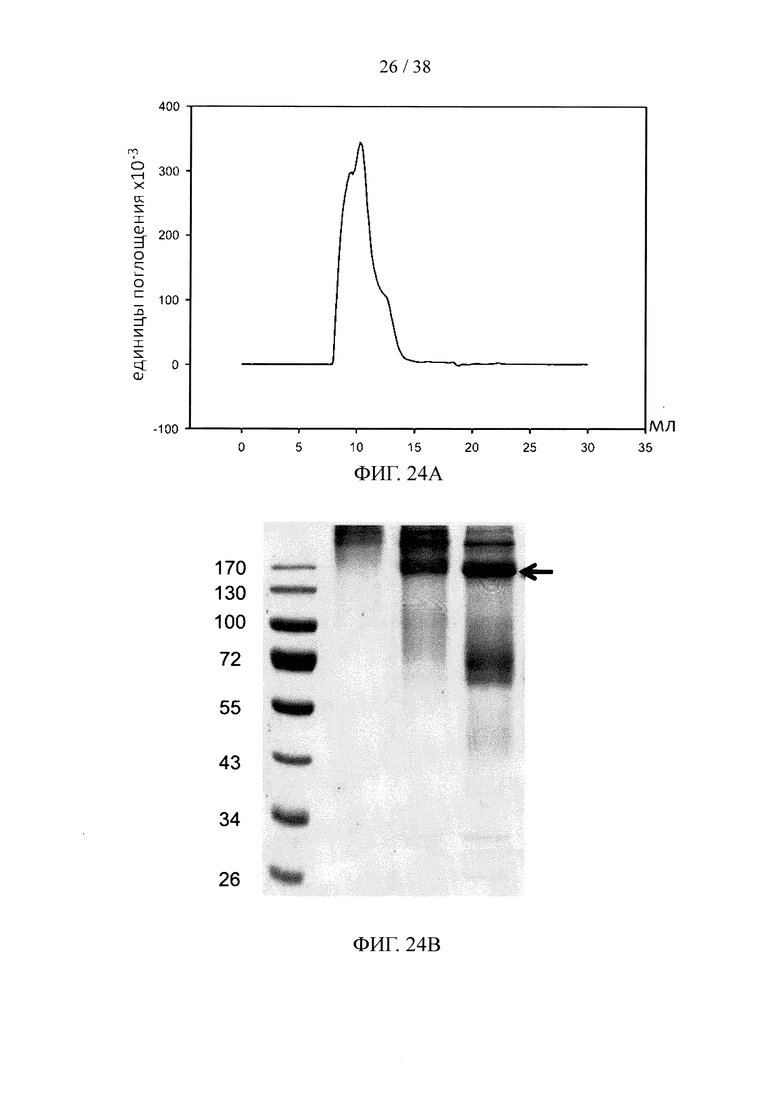
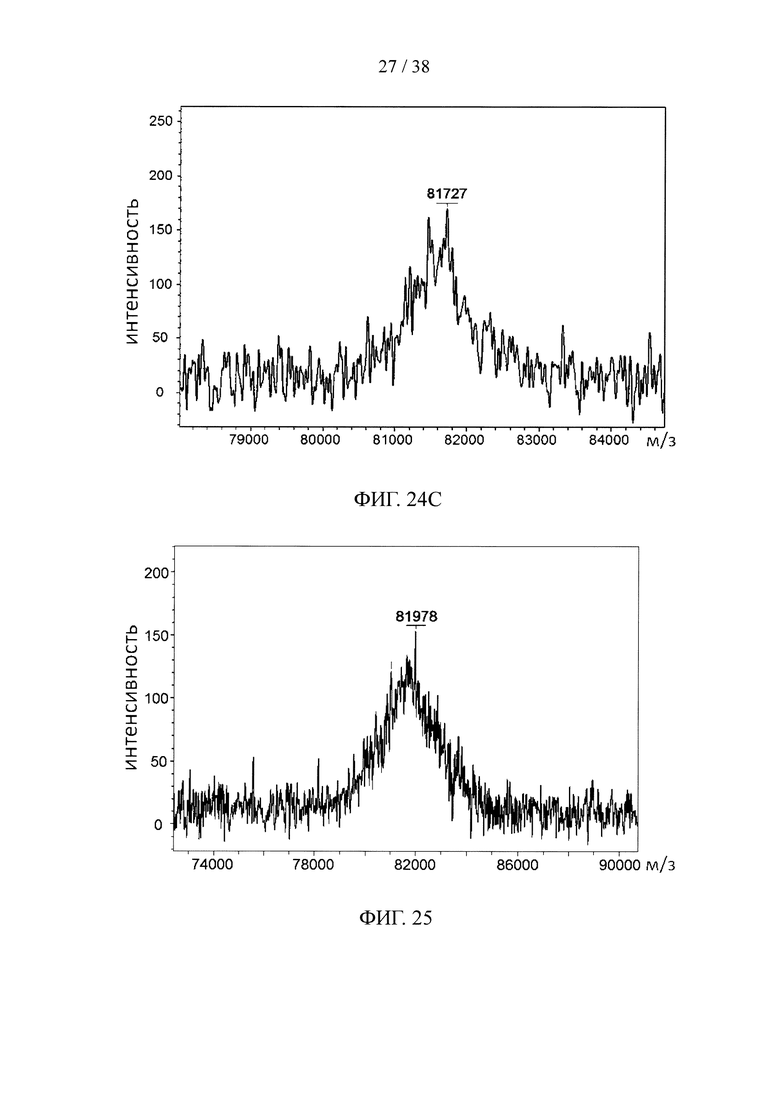
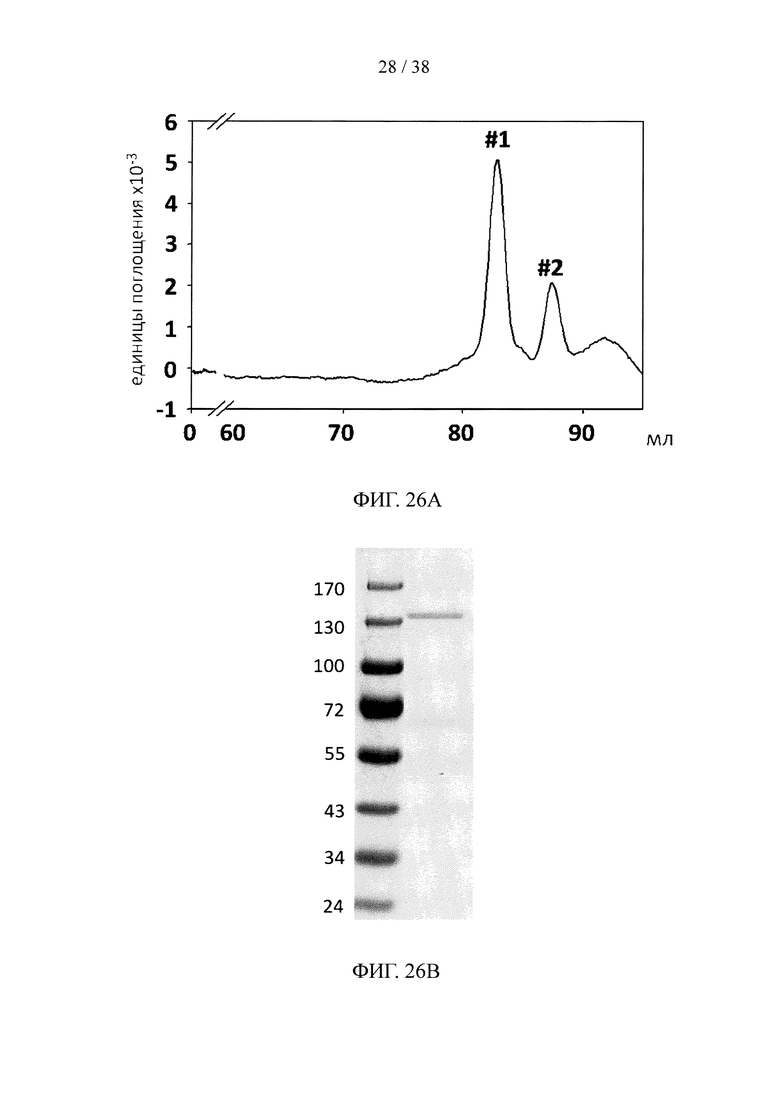
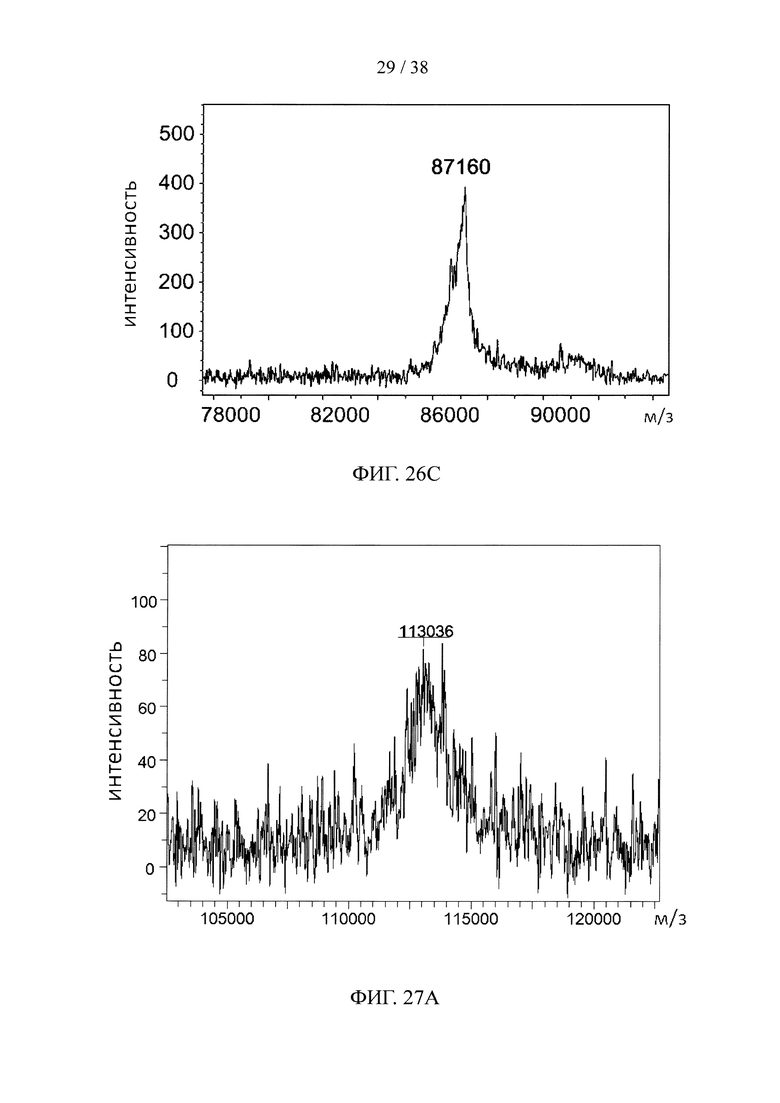

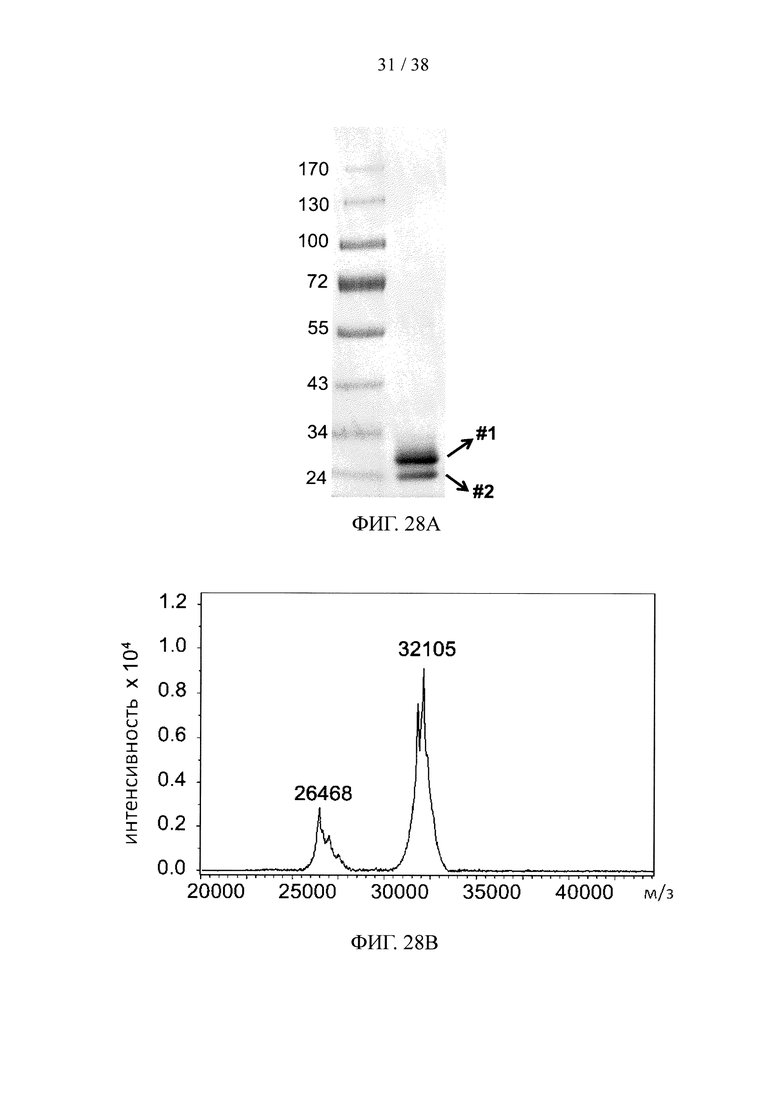
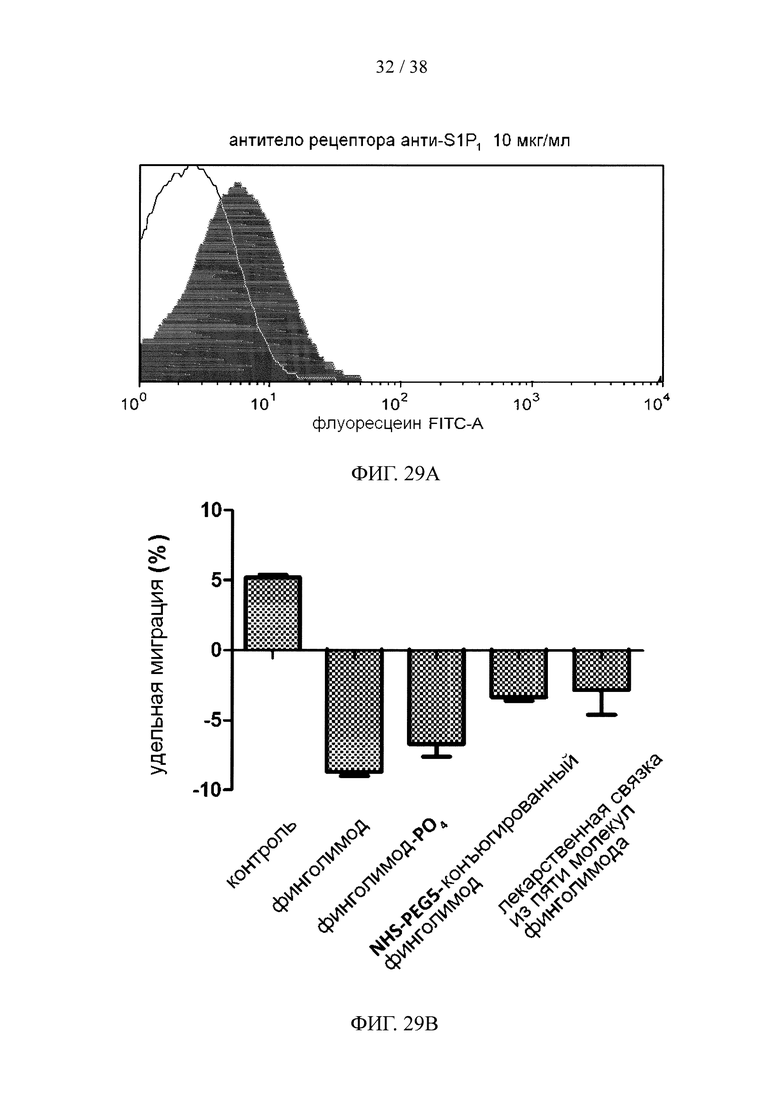
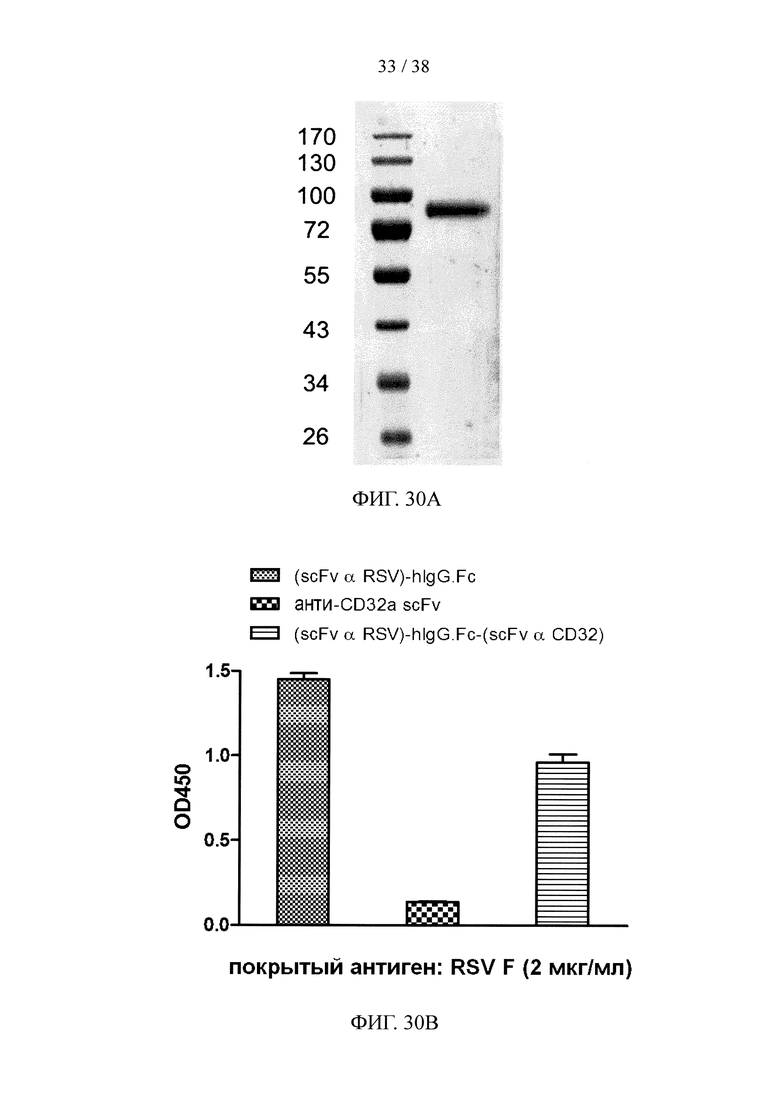
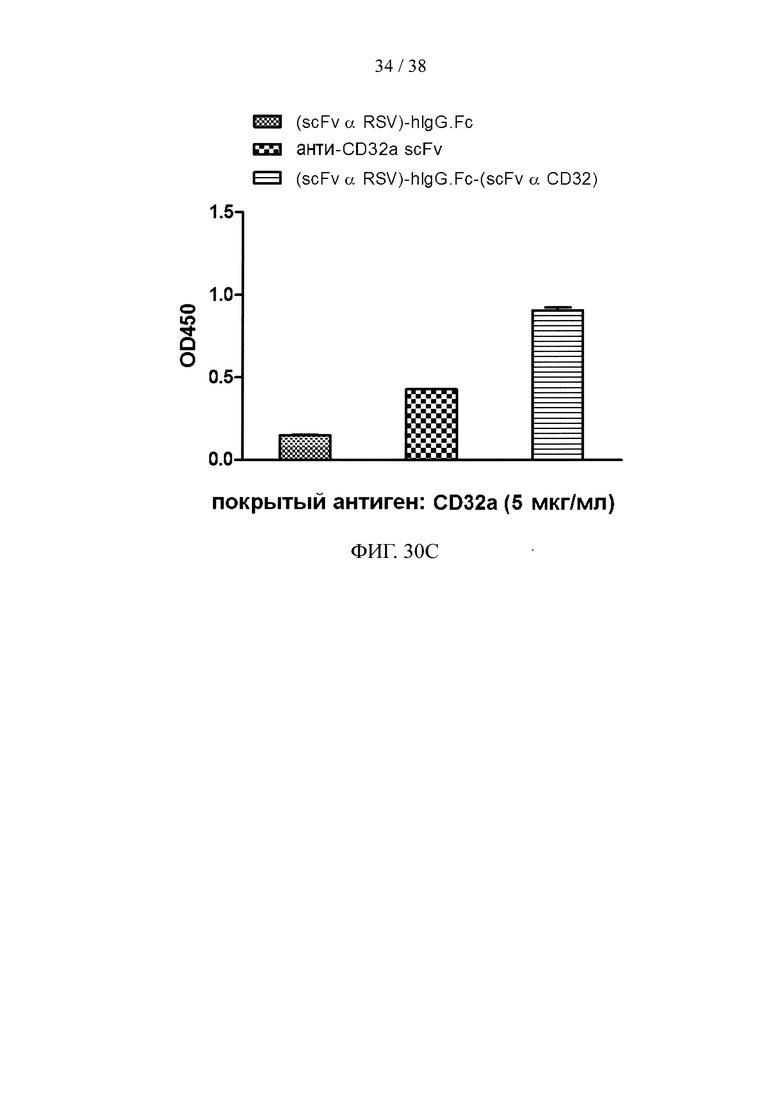
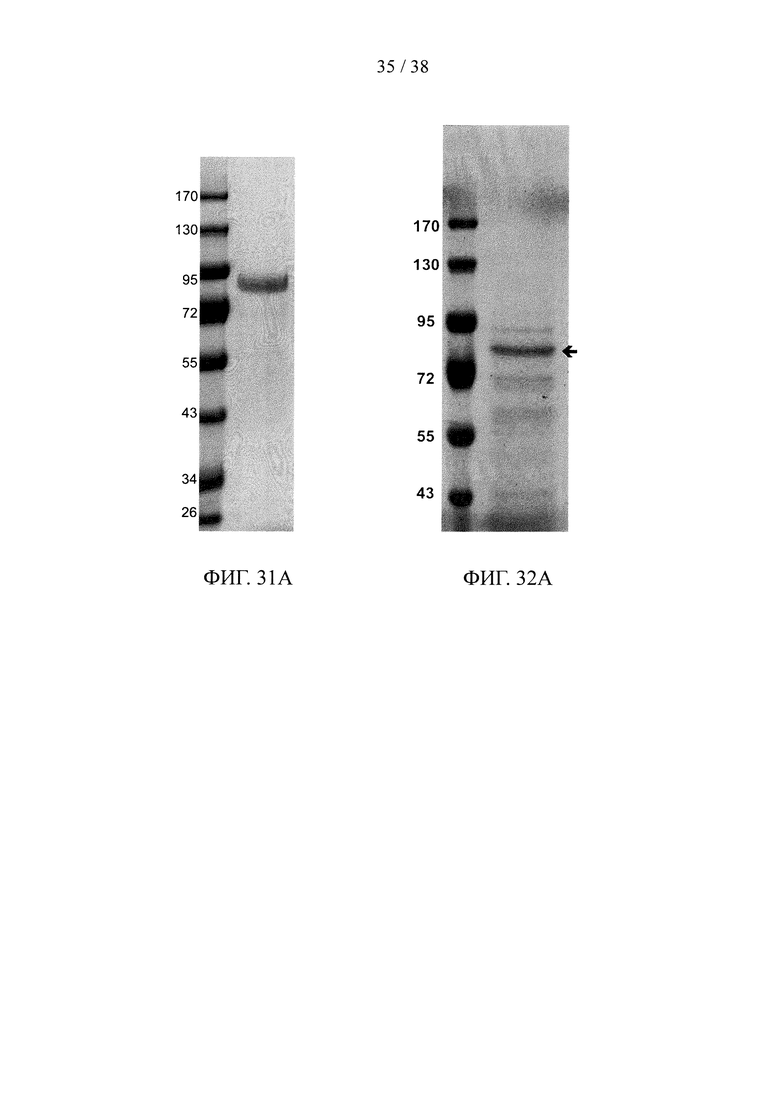
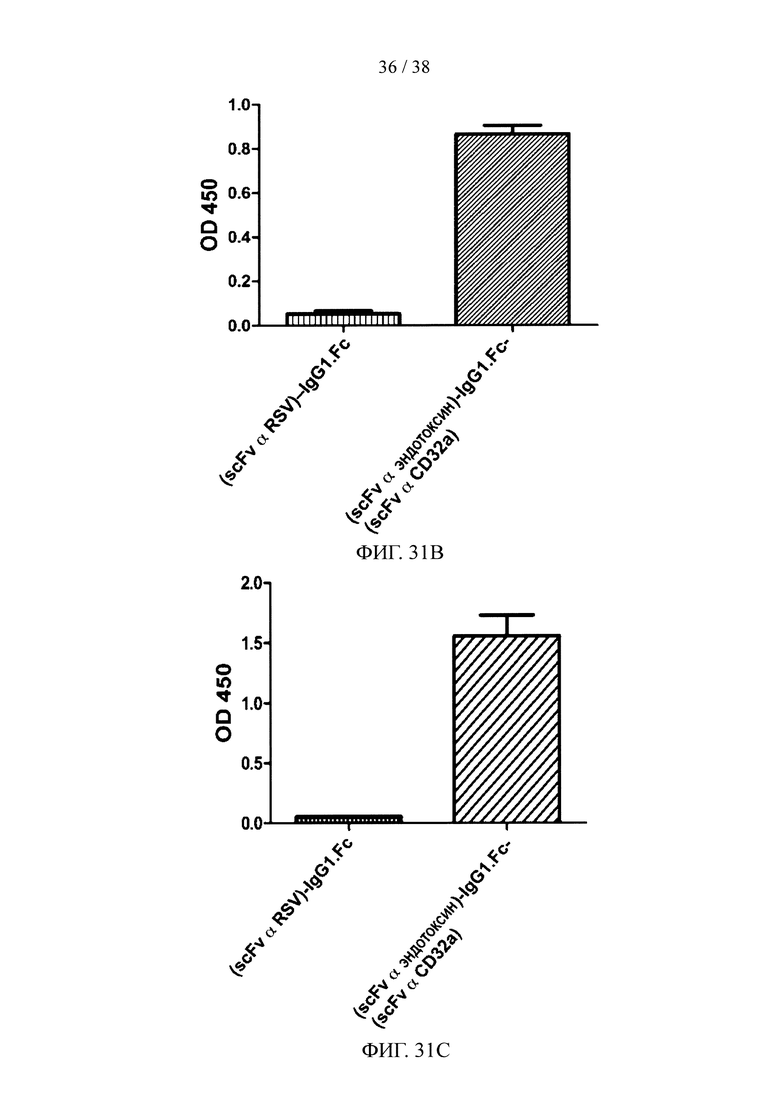
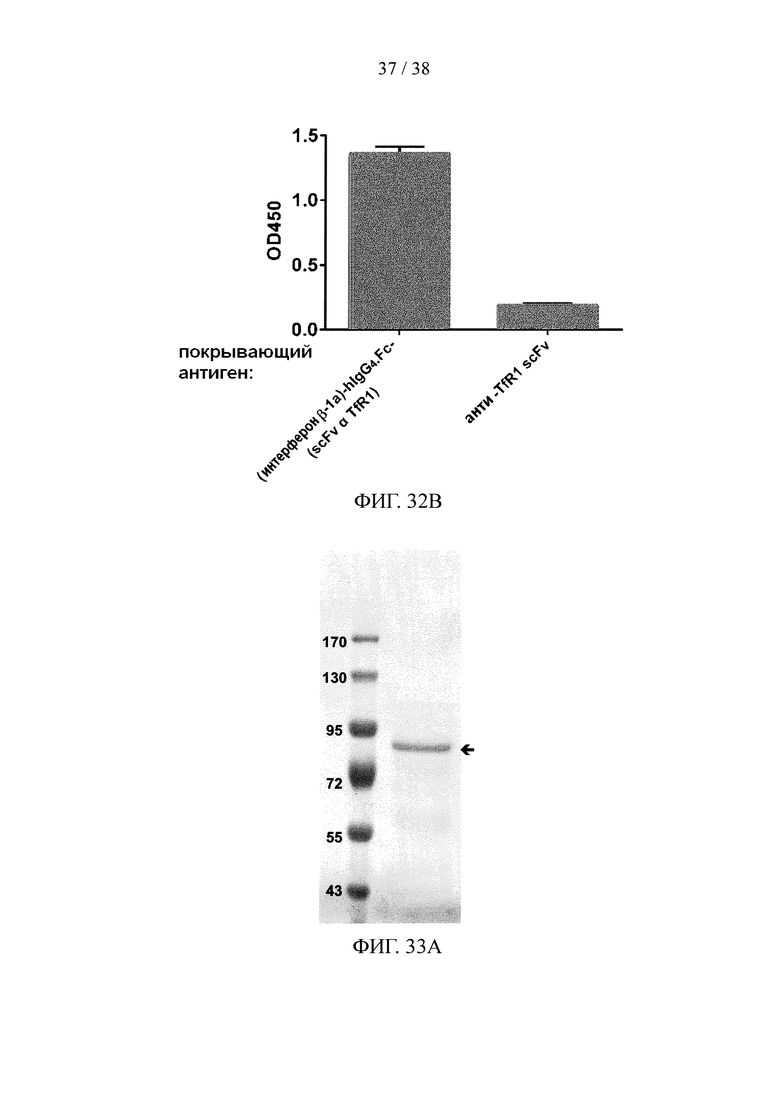

Изобретение относится к области биотехнологии. Описана группа изобретений, включающая линкерное звено, содержащее центральную сердцевину, связывающие ветви и, необязательно, соединяющую ветвь (варианты). В одном из вариантов линкерное звено содержит центральную сердцевину, которая содержит первый полипептид, содержащий лизиновые (K) остатки, где каждый и следующий за ним остаток K разделены вставочной последовательностью, содержащей глициновые и сериновые остатки, и полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K находится в диапазоне от 2 до 15; или второй полипептид, содержащий последовательность (Хаа-K)n, где каждый Хаа представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся этиленгликолевых звеньев, а n является целым числом от 2 до 15; связывающие ветви связаны с остатками K центральной сердцевины, где каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев. Изобретение расширяет арсенал линкерных звеньев. 7 н. и 9 з.п. ф-лы, 81 ил., 39 пр.
1. Линкерное звено, содержащее центральную сердцевину, множество связывающих ветвей и, необязательно, соединяющую ветвь, в котором:
центральная сердцевина содержит (1) первый полипептид, содержащий множество лизиновых (K) остатков, где каждый остаток K и следующий за ним остаток K разделены вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки, где первый полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K находится в диапазоне от 2 до 15; или (2) второй полипептид, содержащий последовательность (Хаа-K)n, где каждый Хаа представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся этиленгликолевых (EG) звеньев, а n является целым числом от 2 до 15;
множество связывающих ветвей соответственно связано с остатками K центральной сердцевины, где количество связывающих ветвей находится в диапазоне от 2 до 15, каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев, и имеет N-гидроксисукцинимидильную (NHS), азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце; и
аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу; или аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а тиольная группа цистеинового остатка связана с соединяющей ветвью, где соединяющая ветвь представляет собой цепь PEG, имеющую от 2 до 12 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем конце, при этом,
если свободный конец связывающей ветви является азидной, алкиновой или циклооктиновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является тетразиновой или циклооктеновой группой; или,
если свободный конец связывающей ветви является тетразиновой группой или циклооктеновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу или аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является азидной, алкиновой или циклооктиновой группой.
2. Линкерное звено по п. 1, в котором вставочная последовательность содержит последовательность GS, GGS, GSG или SEQ ID NO: 1-16.
3. Линкерное звено по п. 1, в котором EG-звено цепи PEG связывающей ветви имеет дисульфидную связь на ее свободном конце.
4. Линкерное звено по п. 1, в котором:
аминокислотный остаток, имеющий азидную группу, представляет собой L-азидогомоаланин (AHA), 4-азидо-L-фенилаланин, 4-азидо-D-фенилаланин, 3-азидо-L-аланин, 3-азидо-D-аланин, 4-азидо-L-гомоаланин, 4-азидо-D-гомоаланин, 5-азидо-L-орнитин, 5-азидо-D-орнитин, 6-азидо-L-лизин или 6-азидо-D-лизин;
аминокислотный остаток, имеющий алкиновую группу, представляет собой L-гомопропаргилглицин (L-HPG), D-гомопропаргилглицин (D-HPG) или бета-гомопропаргилглицин (β-HPG);
циклооктеновая группа представляет собой транс-циклооктеновую (ТСО) группу, а циклооктиновая группа представляет собой дибензоциклооктиновую (DBCO), дифторциклооктиновую (DIFO), бициклонониновую (BCN) или дибензоциклооктиновую (DICO) группу; или
тетразиновая группа представляет собой 1,2,3,4-тетразиновую, 1,2,3,5-тетразиновую или 1,2,4,5-тетразиновую группу или их производные.
5. Линкерное звено, содержащее центральную сердцевину, множество связывающих ветвей, множество первых элементов, необязательно, второй элемент и, необязательно, соединяющую ветвь, в котором:
центральная сердцевина содержит (1) первый полипептид, содержащий множество лизиновых (K) остатков, где каждый остаток K и следующий за ним остаток K разделены вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки, где первый полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K находится в диапазоне от 2 до 15; или (2) второй полипептид, содержащий последовательность (Хаа-K)n, где каждый Хаа представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся этиленгликолевых (EG) звеньев, а n является целым числом от 2 до 15;
множество связывающих ветвей соответственно связано с остатками K центральной сердцевины, где количество связывающих ветвей находится в диапазоне от 2 до 15, каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев, и имеет N-гидроксисукцинимидильную (NHS), азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце;
множество первых элементов соответственно связано со множеством связывающих ветвей путем формирования амидной связи между ними или посредством реакции азид-алкинового циклоприсоединения, катализируемого медью (CuAAC), химической азид-алкиновой клик-реакции, промотируемой напряжением (SPAAC), или реакции Дильса-Альдера с обратными электронными требованиями (iEDDA), где каждым из первых элементов является финголимод, финголимод-фосфат, интерферон-β или scFv, специфичный к интегрину α4, к β-амилоиду, к вирусному белку или к бактериальному белку, и количество первых элементов находится в диапазоне от 2 до 15;
аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу; или аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а тиольная группа цистеинового остатка связана с соединяющей ветвью, где соединяющая ветвь представляет собой цепь PEG, имеющую от 2 до 12 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на свободном конце соединяющей ветви; и,
необязательно, второй элемент связан с центральной сердцевиной посредством любой из следующих реакций: реакции CuAAC между азидной или алкиновой группой и вторым элементом; реакции SPAAC между азидной или циклооктиновой группой и вторым элементом; и реакции iEDDA между циклооктеновой группой или тетразиновой группой и вторым элементом, где вторым элементом является scFv, специфичный к рецептору трансферрина, к CD32 или к CD16b; при этом,
если свободный конец связывающей ветви является азидной, алкиновой или циклооктиновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является тетразиновой или циклооктеновой группой; или,
если свободный конец связывающей ветви является тетразиновой группой или циклооктеновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу или аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является азидной, алкиновой или циклооктиновой группой;
вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; а бактериальный белок представляет собой эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
6. Линкерное звено, содержащее центральную сердцевину, множество связывающих ветвей, множество первых элементов, соединяющую ветвь, второй элемент и, необязательно, третий элемент, в котором:
центральная сердцевина содержит (1) первый полипептид, содержащий множество лизиновых (K) остатков, где каждый остаток K и следующий за ним остаток K разделены вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки, где первый полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K находится в диапазоне от 2 до 15; или (2) второй полипептид, содержащий последовательность (Хаа-K)n, где каждый Xaa представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся этиленгликолевых (EG) звеньев, а n является целым числом от 2 до 15;
множество связывающих ветвей соответственно связано с остатками K центральной сердцевины, где количество связывающих ветвей находится в диапазоне от 2 до 15, каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев, и имеет N-гидроксисукцинимидильную (NHS) группу на своем свободном конце;
множество первых элементов соответственно связано со множеством связывающих ветвей путем формирования амидной связи между ними, где каждым из первых элементов является финголимод, финголимод-фосфат, интерферон-β или scFv, специфичный к интегрину α4, к β-амилоиду, к вирусному белку или к бактериальному белку, и количество первых элементов находится в диапазоне от 2 до 15;
аминокислотный остаток на одном из N- и С-концов центральной сердцевины имеет азидную или алкиновую группу; а аминокислотный остаток на другом из N- и С-концов центральной сердцевины представляет собой цистеиновый остаток, а тиольная группа цистеинового остатка связана с соединяющей ветвью, где соединяющая ветвь представляет собой цепь PEG, имеющую от 2 до 12 повторяющихся EG-звеньев, и имеет тетразиновую или циклооктеновую группу на своем свободном конце;
второй элемент (1) связан с азидной или алкиновой группой на N- или С-конце центральной сердцевины посредством реакции азид-алкинового циклоприсоединения, катализируемого медью (CuAAC) или (2) связан с азидной группой на N- или С-конце центральной сердцевины посредством химической азид-алкиновой клик-реакции, промотируемой напряжением (SPAAC); где вторым элементом является scFv, специфичный к рецептору трансферрина, к CD32 или к CD16b; и,
необязательно, третий элемент представляет собой длинную цепь PEG, имеющую молекулярную массу приблизительно 20000-50000 дальтон и связан с соединяющей ветвью посредством реакции Дильса-Альдера с обратными электронными требованиями (iEDDA);
вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gpl20 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; а бактериальный белок представляет собой эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
7. Линкерное звено, содержащее центральную сердцевину, множество связывающих ветвей, множество сопрягающих ветвей и, необязательно, соединяющую ветвь, в котором:
центральная сердцевина содержит (1) первый полипептид, содержащий множество лизиновых (K) остатков, где каждый остаток K и следующий за ним остаток K разделены вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки, где первый полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K находится в диапазоне от 2 до 15; или (2) второй полипептид, содержащий последовательность (Хаа-K)n, где каждый Хаа представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся этиленгликолевых (EG) звеньев, а n является целым числом от 2 до 15;
множество связывающих ветвей соответственно связано с остатками K центральной сердцевины, где количество связывающих ветвей находится в диапазоне от 2 до 15, каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце;
множество сопрягающих ветвей соответственно связано со множеством связывающих ветвей посредством реакции азид-алкинового циклоприсоединения, катализируемого медью (CuAAC), химической азид-алкиновой клик-реакции, промотируемой напряжением (SPAAC), или реакции Дильса-Альдера с обратными электронными требованиями (iEDDA), где количество сопрягающих ветвей находится в диапазоне от 2 до 15, каждая из множества сопрягающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев, и имеет малеимидную или N-гидроксисукцинимидильную (NHS) группу на своем свободном конце; и
аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу; или аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а тиольная группа цистеинового остатка связана с соединяющей ветвью, где соединяющая ветвь представляет собой цепь PEG, имеющую от 2 до 12 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на свободном конце соединяющей ветви; где,
если свободный конец связывающей ветви является азидной, алкиновой или циклооктиновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является тетразиновой или циклооктеновой группой; или,
если свободный конец связывающей ветви является тетразиновой группой или циклооктеновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу или аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является азидной, алкиновой или циклооктиновой группой.
8. Линкерное звено, содержащее центральную сердцевину, множество связывающих ветвей, множество сопрягающих ветвей, множество первых элементов, необязательно, второй элемент и, необязательно, соединяющую ветвь, в котором:
центральная сердцевина содержит (1) первый полипептид, содержащий множество лизиновых (K) остатков, где каждый остаток K и следующий за ним остаток K разделены вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки, где первый полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K находится в диапазоне от 2 до 15; или (2) второй полипептид, содержащий последовательность (Хаа-K)n, где каждый Хаа представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся этиленгликолевых (EG) звеньев, а n является целым числом от 2 до 15;
множество связывающих ветвей соответственно связано с остатками K центральной сердцевины, где количество связывающих ветвей находится в диапазоне от 2 до 15, каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце;
множество сопрягающих ветвей соответственно связано со множеством связывающих ветвей посредством реакции азид-алкинового циклоприсоединения, катализируемого медью (CuAAC), химической азид-алкиновой клик-реакции, промотируемой напряжением (SPAAC), или реакции Дильса-Альдера с обратными электронными требованиями (iEDDA), где количество сопрягающих ветвей находится в диапазоне от 2 до 15, каждая из множества сопрягающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев, и имеет малеимидную или N-гидроксисукцинимидильную (NHS) группу на своем свободном конце;
множество первых элементов соответственно связано со множеством сопрягающих ветвей посредством тиол-малеимидной реакции или путем формирования амидной связи между ними, где каждым из первых элементов является финголимод, финголимод-фосфат, интерферон-β или scFv, специфичный к интегрину α4, к β-амилоиду, к вирусному белку или к бактериальному белку, и количество первых элементов находится в диапазоне от 2 до 15;
аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу; или аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а тиольная группа цистеинового остатка связана с соединяющей ветвью, где соединяющая ветвь представляет собой цепь PEG, имеющую от 2 до 12 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на свободном конце соединяющей ветви; и,
необязательно, второй элемент связан с центральной сердцевиной посредством любой из следующих реакций: реакции CuAAC между азидной или алкиновой группой и вторым элементом; реакции SPAAC между азидной или циклооктиновой группой и вторым элементом; и реакции iEDDA между циклооктеновой группой или тетразиновой группой и вторым элементом, где вторым элементом является scFv, специфичный к рецептору трансферрина, к CD32 или к CD16b; при этом,
если свободный конец связывающей ветви является азидной, алкиновой или циклооктиновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является тетразиновой или циклооктеновой группой; или,
если свободный конец связывающей ветви является тетразиновой группой или циклооктеновой группой, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу или аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является азидной, алкиновой или циклооктиновой группой;
вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; а бактериальный белок представляет собой эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
9. Линкерное звено по любому из пп. 5, 6 и 8, в котором,
вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; и
бактериальный белок представляет собой эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
10. Линкерное звено, содержащее:
центральную сердцевину, которая содержит (1) первый полипептид, содержащий множество остатков лизина (K), при этом каждый остаток K и следующий за ним остаток K разделены вставочной последовательностью, содержащей остатки глицина (G) и серина (S), где первый полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K составляет от 2 до 15; или (2) второй полипептид, содержащий последовательность (Хаа-K)n, при этом каждый Хаа представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся звеньев этиленгликоля (EG), а n является целым числом от 2 до 15; при этом по меньшей мере один из N- и С-концевых аминокислотных остатков центральной сердцевины представляет собой аминокислоту, имеющую азидную или алкиновую группу, или является цистеиновым остатком, причем, если один из N- и С-концевых аминокислотных остатков является цистеиновым остатком, то линкерное звено дополнительно содержит, необязательно, соединяющую ветвь, которая связана с цистеиновым остатком через тиольную группу цистеинового остатка, где соединяющая ветвь представляет собой цепь PEG, имеющую от 2 до 12 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце,
множество связывающих ветвей, соответственно связанных с остатками K центральной сердцевины, где количество связывающих ветвей находится в диапазоне от 2 до 15 и каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев;
необязательно, множество сопрягающих ветвей, соответственно связанных со множеством связывающих ветвей посредством реакции CuAAC, реакции SPAAC или реакции iEDDA, где количество сопрягающих ветвей находится в диапазоне от 2 до 15 и каждая из множества сопрягающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев;
множество первых элементов, которые соответственно связаны со множеством связывающих ветвей путем образования между ними амидной связи или посредством тиол-малеимидной реакции, реакции CuAAC, реакции SPAAC или реакции iEDDA; или соответственно связаны со множеством сопрягающих ветвей путем образования между ними амидной связи или посредством тиол-малеимидной реакции; причем
количество первых элементов находится в диапазоне от 2 до 15;
каждый из первых элементов представляет собой финголимод, финголимод-фосфат, интерферон-β или одноцепочечный вариабельный фрагмент (scFv), специфичный к интегрину α4, β-амилоиду, вирусному белку, бактериальному белку; и,
если множество связывающих ветвей связано со множеством сопрягающих ветвей или со множеством первых элементов посредством реакции CuAAC или реакции SPAAC, то аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является тетразиновой или циклооктеновой группой; или,
если множество связывающих ветвей связано со множеством сопрягающих ветвей или со множеством первых элементов посредством реакции iEDDA, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу, или аминокислотный остаток на N- или С-конце центральной сердцевины является цистеиновым остатком, а свободный конец соединяющей ветви является азидной, алкиновой или циклооктиновой группой; и,
необязательно, второй элемент, представляющий собой scFv, специфичный к рецептору трансферрина, к CD32 или к CD16b и связанный с центральной сердцевиной посредством любой из следующих реакций: реакции CuAAC между азидной или алкиновой группой и вторым элементом; реакции SPAAC между азидной или циклооктиновой группой и вторым элементом; и реакции iEDDA между циклооктеновой группой или тетразиновой группой и вторым элементом;
вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; а бактериальный белок представляет собой эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
11. Линкерное звено по п. 10, в котором вставочная последовательность содержит последовательность GS, GGS, GSG или SEQ ID NO: 1-16.
12. Линкерное звено по п. 10, в котором EG-звено цепи PEG связывающей ветви или сопрягающей ветви имеет дисульфидную связь на ее свободном конце.
13. Линкерное звено по п. 10, в котором аминокислотный остаток, имеющий азидную группу, представляет собой L-азидогомоаланин (AHA), 4-азидо-L-фенилаланин, 4-азидо-D-фенилаланин, 3-азидо-L-аланин, 3-азидо-D-аланин, 4-азидо-L-гомоаланин, 4-азидо-D-гомоаланин, 5-азидо-L-орнитин, 5-азидо-D-орнитин, 6-азидо-L-лизин или 6-азидо-D-лизин;
аминокислотный остаток, имеющий алкиновую группу, представляет собой L-гомопропаргилглицин (L-HPG), D-гомопропаргилглицин (D-HPG) или бета-гомопропаргилглицин (P-HPG);
циклооктеновая группа представляет собой транс-циклооктеновую (ТСО) группу; а циклооктиновая группа представляет собой дибензоциклооктиновую (DBCO), дифторциклооктиновую (DIFO), бициклонониновую (BCN) или дибензоциклооктиновую (DICO) группу; или
тетразиновая группа представляет собой 1,2,3,4-тетразиновую, 1,2,3,5-тетразиновую или 1,2,4,5-тетразиновую группу или их производные.
14. Линкерное звено по п. 10, в котором:
первым элементом является scFv, специфичный к вирусному белку или к бактериальному белку, а вторым элементом является scFv, специфичный к CD32 или к CD16b; где
вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; и
бактериальный белок представляет собой эндотоксин грамотрицательных бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
15. Линкерное звено по п. 10, в котором первым элементом является финголимод, финголимод-фосфат, интерферон-β или scFv, специфичный к интегрину α4 или к β-амилоиду, а вторым элементом является scFv, специфичный к рецептору трансферрина.
16. Линкерное звено, содержащее:
центральную сердцевину, содержащую (1) первый полипептид, содержащий множество лизиновых (K) остатков, где каждый остаток K и следующий за ним остаток К разделены вставочной последовательностью, содержащей глициновые (G) и сериновые (S) остатки, где первый полипептид имеет длину от 8 до 120 аминокислотных остатков и количество остатков K находится в диапазоне от 2 до 15; или (2) второй полипептид, содержащий последовательность (Хаа-K)n, где каждый Хаа представляет собой пегилированную аминокислоту, имеющую от 2 до 12 повторяющихся этиленгликолевых (EG) звеньев, а n является целым числом от 2 до 15; где по меньшей мере один из N- и С-концевых аминокислотных остатков центральной сердцевины представляет собой аминокислоту, имеющую азидную или алкиновую группу, или представляет собой цистеиновый остаток, где, если один из N- и С-концевых аминокислотных остатков представляет собой цистеиновый остаток, линкерное звено дополнительно содержит, необязательно, соединяющую ветвь, связанную с цистеиновым остатком через тиольную группу цистеинового остатка, где соединяющая ветвь представляет собой цепь PEG, имеющую от 2 до 12 повторяющихся EG-звеньев, и имеет азидную, алкиновую, тетразиновую, циклооктеновую или циклооктиновую группу на своем свободном конце;
множество связывающих ветвей, соответственно связаных с остатками K центральной сердцевины, где количество связывающих ветвей находится в диапазоне от 2 до 15 и каждая из связывающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев;
необязательно, множество сопрягающих ветвей, соответственно связанных со множеством связывающих ветвей посредством реакции азид-алкинового циклоприсоединения, катализируемого медью (CuAAC), химической азид-алкиновой клик-реакции, промотируемой напряжением (SPAAC), или реакции Дильса-Альдера с обратными электронными требованиями (iEDDA), где количество сопрягающих ветвей находится в диапазоне от 2 до 15 и каждая из множества сопрягающих ветвей представляет собой цепь PEG, имеющую от 2 до 20 повторяющихся EG-звеньев;
множество первых элементов, соответственно связанных со множеством связывающих ветвей или со множеством сопрягающих ветвей посредством формирования амидной связи между ними или посредством тиол-малеимидной реакции; где
количество первых элементов находится в диапазоне от 2 до 15;
каждым из первых элементов является финголимод, финголимод-фосфат, интерферон-β или одноцепочечный вариабельный фрагмент (scFv), специфичный к интегрину α4, к β-амилоиду, к вирусному белку или к бактериальному белку; и,
если множество связывающих ветвей связано со множеством сопрягающих ветвей или со множеством первых элементов посредством реакции CuAAC или реакции SPAAC, то аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а свободный конец соединяющей ветви представляет собой тетразиновую или циклооктеновую группу; или,
если можество связывающих ветвей связано со множеством сопрягающих ветвей или со множеством первых элементов посредством реакции iEDDA, то аминокислотный остаток на N- или С-конце центральной сердцевины имеет азидную или алкиновую группу, или аминокислотный остаток на N- или С-конце центральной сердцевины представляет собой цистеиновый остаток, а свободный конец соединяющей ветви представляет собой азидную, алкиновую или циклооктиновую группу;
второй элемент, (1) связанный с азидной или алкиновой группой N- или С-концевого аминокислотного остатка центральной сердцевины посредством реакции CuAAC или (2) связанный с азидной или циклооктиновой группой N- или С-концевого аминокислотного остатка центральной сердцевины посредством реакции SPAAC; где вторым элементом является scFv, специфичный к рецептору трансферрина, к CD32 или к CD16b; и,
необязательно, третий элемент, представляющий собой длинную цепь PEG, имеющую молекулярную массу приблизительно 20000-50000 дальтон, и связанный с соединяющей ветвью посредством реакции iEDDA;
вирусный белок представляет собой белок F респираторно-синцитиального вируса (RSV), белок gp120 вируса иммунодефицита человека 1 типа (HIV-1), гемагглютинин А (НА) белка вируса гриппа А или гликопротеин цитомегаловируса; а бактериальный белок представляет собой эндотоксин грамотрицательньгх бактерий, мембранный антиген бактерии Clostridium difficile, липотейхоевую кислоту бактерии Staphylococcus aureus, сибиреязвенный токсин бактерии Bacillus anthracis или шигаподобный токсин I или II типа бактерии Escherichia coli.
| СПОСОБ ВОЗДЕЙСТВИЯ НА ОРГАНИЗМ ПУТЕМ АДРЕСНОЙ ДОСТАВКИ БИОЛОГИЧЕСКИ АКТИВНЫХ ВЕЩЕСТВ В МИТОХОНДРИИ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ И СОЕДИНЕНИЕ, ПРИМЕНЯЕМОЕ ДЛЯ ЭТОЙ ЦЕЛИ | 2005 |
|
RU2318500C2 |
| WO 2010093395 A1, 19.08.2010 | |||
| WO 2008127642 A1, 23.10.2008 | |||
| WO 2014153002 A1, 25.09.2014. | |||

